
細胞機能を調節する能力が増強された治療産物
本発明は、ヒトIgGのIIB型受容体(FcγRIIB/CD32)を介した阻害機能誘発などの1つまたはいくつかの特定のFc受容体の活性を調節し得る、ヒトIgGのFc領域を含む、ヒトの、またはキメラの、またはヒト化された、抗体または分子を作製および選択する方法に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、IgGのヒトII型受容体(FcγRII/CD32)を介して阻害機能を誘発するような、1つまたはいくつかの特定のFc受容体の活性を調節し得る、ヒトIgGのFc領域を含むヒトもしくはキメラもしくはヒト化抗体または分子を作製および選択する方法に関する。
【背景技術】
【0002】
導入
ヒトにおける抗体に基づく治療法は、部分的ヒト抗体(キメラ抗体)、つい最近ではヒト化抗体を作製できる可能性と共に出現した。ヒト化組み換え抗体またはマウス/ヒトキメラ組み換え抗体は、マウス抗体で処置した患者におけるヒト抗マウス抗体(「HAMA」)の出現を防ぎ、HAMAと注入された抗体との免疫複合体の形成による副作用を回避するために開発された。さらに、そのような操作された抗体は、ヒトFcγRに対する結合が向上するために、マウス対応物よりも強力なエフェクター機能を示す。現在、数多くの抗体が例えば癌治療の臨床試験において試験されている。しかし、今日現在、ほんのわずかな抗体しか低用量で有効性を示しておらず、いくつかの治験は時期尚早に打ち切られている。高用量の抗体投与は、主に2つの要因から制限される。第一に、それにより同様に多くの副作用が増強される。第二に、大量の抗体を作製することはやはり難題であり、拡大に伴うコストは必ずしも経済的に存立できるとは限らない。
【0003】
様々な疾患に苦しむ多くの人々に抗体療法を適用する前に、これらの問題に取り組む必要がある。
【0004】
IgGのFc領域は抗体の機能に必須であることが示されている。IgGのFc領域を介したC1qおよび膜FcγRの補充により、細胞傷害性、サイトカインの放出、またはエンドサイトーシス等の種々のエフェクター機能が誘発され得る。例えば、IgG1、IgG2、IgG3、およびIgG4を含むヒトIgGサブクラスのうち最もよく説明されているヒトIgG1は、最も高い抗体依存性細胞傷害(ADCC)を誘発することが示されている。それは、ヒトIgG1が、NK細胞および単球/マクロファージ上に発現されるFcγRと効率的に結合する能力に起因する。さらに、FcγRは、阻害機能の誘発により、免疫調節において重要な役割を担う。
【0005】
FcγRの3つの異なるクラスが、ヒトおよびマウスにおいて明らかにされている。ヒトFcγRには、高親和性FcγRI(CD64)、ならびに低親和性FcγRであるFcγRII(CD32)およびFcγRIII(CD16)が含まれる。3つの遺伝子がFcγRIアイソフォーム(A、B、C)をコードし、3つの遺伝子がFcγRIIアイソフォーム(A1、B1、B2)をコードし、2つの遺伝子がFcγRIIIアイソフォーム(A、B)をコードしている。Fc領域とFcγRとの相互作用の調節を達成できるようにFcドメイン内のアミノ酸配列を改変することにより、IgG抗体の有効性を改善する試みが行われている(例えば、WO99/54572を参照のこと)。
【0006】
生物物理学的研究および分子的研究から、CH1ドメインとCH2ドメインの間のヒンジ領域に位置する、およびIgG1のCH2ドメインのN末端に直接隣接するいくつかのアミノ酸残基、ならびにAsn297位においてCH2ドメインに結合している糖鎖が、FcγR結合において重要な役割を果たすことが示されている。一方では、IgG1残基の共通のセットがすべてのFcγR(I、II、III)に対する結合に関与しているが、FcγRIIおよびFcγRIIIとヒトFcγ1との相互作用を詳細に研究した際に、この共通セットの外側の残基が同定された(Shields et al., J. Biol. Chem., 276, 6591-6604, 2001)。他方では、FcγRとの直接的接触ではないが、Fc上の保存された残基Asn297に結合している糖鎖が、Fc上の受容体結合エピトープの高次構造を安定化している可能性が高い(Radaev et al., J. Biol. Chem., 276, 16469-16477, 2001)。脱グリコシル化が、Fcが開いた高次構造から閉じた高次構造に移行するような、2つのCH2ドメインの相対配向の高次構造変化を引き起こし、FcγRへの結合を妨げることが仮定されている(Redaev & Sun, J. Biol. Chem., 276, 16478-16483, 2001)。継続的に切断されたオリゴ糖を有するIgG1グリコフォームを解析することにより、糖残基の除去によって、結果的に閉じた高次構造を生じるCH2ドメインの相互の接近が可能になることが確認された(Krapp et al., J. Mol. Biol., 325, 979-989, 2003)。
【0007】
操作したIgGグリコフォームが、FcγRIIIの補充を介して最適なADCCを誘発することが示された。最初に、Umanaらは、二分岐複合オリゴ糖(二分岐N-アセチルグルコサミン、GlcNAC)の量を多く含むように操作したIgG1抗体が、親の対応物と比較して強力なADCCを誘発し得ることを提唱した(Umana et al., Nature Biotechnol., 17, 176-180, 1999)が、この主張は意義を唱えられた(Shinkawa et al., J.Biol. Chem, 2002)。
【0008】
そのような矛盾する結果から、所与のFcRの活性化または阻害に関与する実際に最適なオリゴ糖構造を同定することは困難であることが示される。本発明者らは、独特の構造に焦点を当てるよりもむしろ結合の改善に関与するパターンを同定する方がより適切であることを見出した。実際に本発明者らは、類似した構造の範囲内で結合が起こることを認めた。
【0009】
この点において、本発明者らは、Fcドメインに結合した特定のグリコシル化パターンがFc領域-FcRIII受容体相互作用の増強に関与することを実証した。この関連で、本発明者らはYB2/0等の特定の細胞株から改善されたADCC特性を有する抗体の組成物を調製することが可能であることを示し、特定のグリコシル化パターンの役割を実証するWO 01/77181を出願した。そのような抗体は、短いオリゴ糖鎖、弱いシアリル化、ゼロまたは弱いレベルの二分岐GlcNac、および20%〜50%という低レベルのフコースを示す。
【0010】
WO 00/61739においても、フコースの存在または非存在が抗体の活性を調節すると仮定された。ヒトIgG1 N結合型オリゴ糖におけるフコースの欠如は、FcγRIII結合およびADCCを改善することが示された。野生型CHO細胞で産生される同じIgG1と比較して低いフコース含量を示すかまたはフコースが欠如した、YB2/0細胞(Shinkawa et al., J. Biol. Chem., 2002)またはCHO-Lec13細胞(Shields et al., J. Biol. Chem., 277, 26733-26740, 2002)で産生される組み換えヒトIgG1は、細胞傷害性を誘発する能力の亢進を示した。それに反して、ガラクトースとADCCとの相関関係は認められず、二分岐GlcNACの含量はわずかにADCCに影響を及ぼしたに過ぎなかった(Shinkawa et al., J. Biol. Chem., 2002)。
【0011】
本発明者らが出願WO 01/77181に以前に記載したことが上記の結果によって確認されるが、本発明者らは現在、抗体活性の変化は単にフコースレベルで要約し得ないことを示す証拠を有する。GlcNac、マンノース、シアル酸、ガラクトース、およびそれらのそれぞれの位置の関係は、異なる細胞株から産生される抗体において非常に変化しやすい。この知見から、異なるグリコシル化パターンはFc領域-Fc受容体の相互関係に関与するばかりでなく、抗体の機能にも関与することが裏付けられる。
【0012】
したがって、本発明の全体的な目的は、異なるFc受容体に関して所与の抗体の機能を微調整する方法を提供することにある。異なる規定のパターンにより、異なるFcRの調節が可能になる。
【0013】
IgGのFc領域の受容体の中でも、IIB型Fcγ受容体が特に関心の対象である。FcRIIは、免疫複合体中に存在するかまたは細胞表面抗原に結合したIgGと親和性を増して結合する一本鎖、低親和性受容体である。遺伝子解析およびタンパク質解析から、FcγRIIBは、ヒトでは、受容体の細胞質内ドメインをコードする3つのエキソンの選択的スプライシングによって生じる2つのアイソフォーム(FcγRIIB1およびFcγRIIB2)で存在する。FcγRIIBは、B細胞、単球、樹状細胞、肥満細胞、および好塩基球上に発現される。他のすべての既知FcγRとは対照的に、FcγRIIBアイソフォームは、活性化受容体を介して誘導される細胞活性化を阻害する(Amigorena et al., Science, 256, 1808-1812, 1992)。分子的解析により、FcγRIIBの細胞内ドメイン内の「免疫受容体チロシンに基づく阻害性モチーフ」(ITIM)と称されるYxxLモチーフが、受容体の阻害機能に直接関与していることが示された。チロシンがリン酸化されると、ITIMは、ホスファターゼ、すなわちSHP-1、SHP-2、およびホスファチジルイノシトールポリリン酸5-ホスファターゼ、SHIPのSH2ドメインと結合する。SHIPは、PIP3を脱リン酸化してPI(3,4)P2にし、ホスホリパーゼC-γ(PLCγ)をリン酸化するキナーゼbtkの補充を妨げ得る。次いで、イノシトール(1,4,5)三リン酸(IP3)の生成が阻止される。これにより、活性化受容体の補充によって誘発される初期事象である、細胞外カルシウムの流入が阻害される。サイトカインの産生または細胞の増殖等の後期事象もまた阻害される(O. Malbec et al., 「Immunoreceptor Tyrosine-based Inhibition Motifs」、M. Daeron & E, Vivier eds, 1999, Springer-Verlag. Berlin, pp. 13-27)。
【0014】
FcγRIIに依存した負の制御の生物学的重要性は、多くの著者によって検討されている。抗体応答の負の制御においては他の機構も同時に起こる可能性が高いものの、抗体応答の負の制御はFcγRIIBの補充を介して達成され得ることが示されている。したがって、抗体に基づく自己免疫疾患または病原性抗体の出現をもたらす望ましくない同種免疫が、FcγRIIBに結合し得る抗体または他の分子によるFcγRIIBの最適な補充を介して回避され得ることが示唆され得る。
【0015】
さらに、アレルゲン複合化IgGによる肥満細胞および好塩基球上に存在するFcγRIIBの補充、およびIgEが同じアレルゲンに複合化した時点での、次に起こるFcγRIIBとFcεRIとの共凝集は、これらの細胞によるアナフィラキシー反応の遮断に関与する。したがって、FcγRIIBと結合し、それ故にFcγRIIB阻害機能を誘発する最適な能力を有するIgG抗体は、アレルギーの治療における有効な治療手段を表し得る。
【0016】
最終的には、FcγRIIBの補充によって誘発される阻害機能は、癌患者において利用され得る。一方では、免疫不全FcγRIIB-/-マウスに腫瘍を移植し、それぞれ抗CD-20もしくは抗HER2/Neuのキメラ抗体またはヒト化抗体等の組み換えモノクローナル抗体で処置したモデルにおいて、FcγRIIBノックアウトマウスはより優れた抗腫瘍反応を示すことが示された。したがって、強力なFcγRII阻害機能を誘発し得ないモノクローナルIgG抗体は、腫瘍治療における最適な手段となる可能性がある。他方では、FcγRIIbは受容体チロシンキナーゼ(RTK)に依存して造血細胞増殖を負に制御する。したがって、RTKとFcγRIIBを効率的に共凝集させ得る最適なFc領域を有する抗RTK IgG抗体等の二機能性分子は、FcγRIIB+腫瘍細胞の増殖を阻止する。
【0017】
FcγRII/ヒトIgG相互作用に及ぼすグリコシル化の影響はまた、可溶性FcγR外部ドメインを用いても実証されている。マウスFcγRIIおよびヒトFcγRIIBのどちらに関しても、それぞれ超遠心および等温滴定熱量計(ITC)を用いた沈降平衡法から1:1化学量論が推定された(Mimura et al., J. Biol. Chem., 276, 45539-45547, 2001;Kato et al., J. Mol. Biol., 295, 213-224, 2001)。Asn297と主要なGlcNAC残基との相互作用が、ヒトFcγRIIによるヒトキメラIgG3の認識に重要であることが示された(Lund et al., FASEB J., 116, 115-119, 1995)。α(1-6)マンノースアーム上のガラクトースの接触残基の置換はFcγRII認識に影響を及ぼさないのに対し、コアGlcNAC残基との接点であるAsp265の置換はFcγRII認識の喪失をもたらす(Jefferis et al., Immunol. Letters, 44, 111-117, 1995)。さらに別の研究から、GlcNAC残基はヒトIgG1の受容体結合にわずかに寄与するのみである一方、α(1-3)およびα(1-6)アームマンノース残基を除去することによって親和性が有意に減少することが示された(Mimura et al., J. Biol. Chem., 276, 45539-45547, 2001)。同じ研究では、オリゴ糖の切断によって2つのCH2ドメインの閉じた配置がもたらされ、それに伴ってFcγIIBへの結合が減少すると結論づけた。FcγRIIIとは対照的に、CHO-Lec13細胞で産生された、故にフコースを欠く抗体は、可溶性固定化Arg131-FcγRIIA多型および可溶性FcγRIIB型に対する結合においてわずかな改善を示したが、可溶性His131-FcγRIIA多型に対する結合では改善は認められなかった(それらはいずれも細胞外ドメインおよび膜貫通ドメインに相当する)。前者の受容体は131位にアルギニンを有するため、フコースがこの位置のFcγII残基と直接結合するか、またはその存在によってFcγRII結合に対するわずかな弊害が誘導されるようにIgG1高次構造を変化させると仮定された(Shields et al., J. Biol. Chem., 277, 26733-26740, 2002)。同じ研究から、ガラクトース含量はFcγRIIに対する結合に影響を及ぼさないことが示唆された。
【0018】
本明細書において、本発明者らは、標的および疾患に応じて微調整し得る薬剤を作製するために、異なる結合プロファイルを示す種々のクラスの抗体を調製することを提案する。そのような抗体の調製法により、有効性が増強された、ならびに/または免疫調節機能およびFcγRに対する特異性が増強された抗体を産生するのに適した細胞株を同定することが初めて可能になる。本発明はまた、特定の結合プロファイルに関与する抗体のグリカン構造、およびそのような特定の抗体の作製法を提供する。
【発明の開示】
【0019】
本発明は、異なるFcR、特にFcγRIIB、FcγRIIA、およびFcγRIIIAを使用する結合アッセイ法-免疫蛍光法-による、規定されたグリコシル化パターンを有するヒトIgGのFc領域を含むヒト組み換え抗体またはキメラ抗体またはヒト化抗体もしくは分子を作製および選択する方法を提供する。例えば、本方法は、カルシウム動員アッセイ法およびサイトカイン分泌アッセイ法等の機能アッセイ法による、IgGのFc領域に対するヒトIIB型受容体(FcγRIIB/CD32B)を介して阻害機能を特異的に誘発し得る規定のグリコシル化パターンを有するヒト組み換え抗体または抗体由来分子の選択に適用し得る。
【0020】
本発明は、特殊化したかつ微調整された抗体をもたらす規定の培地中で培養した種々の細胞株による、異なるFcグリコシル化パターンを有する組み換えヒト抗体、ヒト化抗体、またはキメラ抗体の産生に基づく。
【0021】
より具体的には、本発明は、B細胞、単球もしくは他の抗原提示細胞(APC)、肥満細胞、好塩基球、または形質転換細胞および腫瘍細胞等の任意の他のFcγRIIB+細胞等のFcγRIIB+細胞(すなわち、FcγRIIBを発現している細胞)上のFcγRIIBによってもたらされる負の制御のオンまたはオフを可能にする、異なるFcグリコシル化パターンによって決定される特定の構造特性を有する抗体の作製および選択に関する。
【0022】
本発明は、抗体応答を調節するための有用な治療手段となると考えられる最適な抗体に関する。これにより、抗体に基づく自己免疫疾患または特に注入された組み換え抗体に対する病原性抗体の出現をもたらす望ましくない同種免疫において、FcγRIIBの補充を介して免疫調節阻害機能を誘発し得る、FcγRIIBに対する結合に関して最適化された任意の特異性の抗体または他の分子の作製および選択が可能になる。
【0023】
本発明は、抗体に基づいた治療において抗体の有効性を調節する有用な治療手段となると考えられる任意の特異性の最適な抗体に関する。それにより、弱いFcγRIIB結合を有するかまたはFcγRIIB結合のない抗体を作製および選択し、治療抗腫瘍抗体による癌治療等の抗体に基づく治療処置過程において、FcγRIIBを介した免疫調節阻害機能の誘発を妨げることが可能になる。
【0024】
第1の局面において、本発明は、ヒトIgGのFc領域を含みかつ異なる結合プロファイルを有する、ヒトの、ヒト化された、もしくはキメラの抗体またはポリペプチドを調製する方法であって、以下からなる段階を含む方法を対象とする:
(a) ハイブリドーマ、ヘテロハイブリドーマ、EBV形質転換ヒトB細胞株を含む動物細胞株に由来もしくは真核微生物に由来する細胞により天然に産生される、または、該抗体もしくはポリペプチドのコード配列を含むベクターによるそのトランスフェクション後に産生される、ヒトIgGのFc領域を含む、ヒトの、ヒト化された、もしくはキメラの抗体候補またはポリペプチド候補を提供する段階;
(b) FcγRIIIA、FcγRIIA、およびFcγRIIBを含むFcγ受容体において該抗体またはポリペプチドの結合を試験する段階;
(c)
i) FcγRIIIA、FcγRIIA、およびFcγRIIBのいずれにも結合する、または
ii) FcγRIIAおよびFcγRIIBには結合するが、FcγRIIIAには結合しないかまたはごく弱く結合する、または
iii) FcγRIIIA、FcγRIIA、およびFcγRIIBのいずれにも結合しないかまたはごく弱く結合する
抗体またはポリペプチドを選択する段階。
【0025】
FcγRIIIA、FcγRIIA、およびFcγRIIBに対するこれらの抗体の結合は、それらの定常領域(すなわち、CH1、CH2、CH3)を介する、より具体的にはそれらのFc領域を介する。これらの抗体は、それらの可変領域を介して、すなわちそれらの抗原結合領域を介してまたはそれらの相補性決定領域(CDR)を介して任意の種類の特異性を示し得る。
【0026】
本明細書において規定する手段によって調製される抗体は、それぞれ特定の結合プロファイルを有し、結果的に異なる特性を有する3つクラスのうちの1つに属し、各クラスを特定の明確な治療目的のために使用することが可能になる。
【0027】
第1クラスに属する抗体またはポリペプチドは、上記の方法の段階c) i)を用いて調製し得る。それらはFcγRIIIAに結合し得るため、この受容体を介してADCCを誘導し得る。さらに、それらはFcγRIIAに結合し得るため、FcγRIIA陽性単球およびマクロファージを介してADCCおよび食作用を誘導し得る。さらに、このクラスの抗体またはポリペプチドはまた、それらが結合し得るFcγRIIBを介して阻害機能を誘発し得る。したがって、これらの抗体はFcγRIIBを介して免疫調節機能を発揮し得る。これは、そのような抗体が免疫系の細胞または腫瘍細胞を負に免疫調節する能力に関する。そのような能力は、多くの疾患においておよび移植、同種免疫、または抗体に基づく処置等の種々の臨床状況において主に重要である。
【0028】
したがって、このクラスに属する抗体またはポリペプチドは、細胞傷害性が高い抗体(またはポリペプチド)であり、かつ強力な免疫調節抗体(またはポリペプチド)である。
【0029】
第2クラスの抗体またはポリペプチドは、上記の方法の段階c) ii)を用いて調製し得る。それらは、FcγRIII、特にFcγRIIIAに対する結合が弱いために第1クラスに属する抗体と比較して細胞傷害性が乏しいことから、関心対象となる。より詳細には、それらはポリクローナル多価IgG(静注用IgG、IVIg)の存在下において細胞傷害性を示さない。それらは、FcγRIIAに結合する能力により、FcγRIIA陽性単球およびマクロファージを介してADCCおよび食作用を誘導し得る。さらに、それらはFcγRIIBと結合する能力のために免疫調節抗体である。
【0030】
最後に、第3クラスの抗体は上記の方法の段階c) iii)を用いて調製し得る。このクラスの抗体は細胞傷害性を示さず、また免疫調節活性を全く提供しない。より詳細には、それらはポリクローナル多価IgG(静注用IgG、IVIg)の存在下において細胞傷害性を示さない。それらは、FcγRIIIA、FcγRIIA、およびFcγRIIBに結合できないかまたは弱く結合するために、FcγR補充を介して達成されるエフェクター機能の誘発を起こさずインビボで使用し得ることから、関心対象となる。
【0031】
本発明の方法により、どのFcγR(IIIA、IIA、IIB)を標的するかに応じて、特定のまたは所望の治療プロファイルを有する抗体を調製することが可能となる。
【0032】
有利には、本発明による方法は、競合物として単量体抗体を用いる競合アッセイ法に基づく。
【0033】
競合物として単量体抗体を用い得る競合アッセイ法は、免疫蛍光競合アッセイ法である。
【0034】
別の態様において、競合物として単量体抗体を用い得る競合アッセイ法は、直接的または間接的免疫蛍光競合アッセイ法である。
【0035】
本アッセイ法において、ID50とは、蛍光色素と結合させた抗FcγR抗体を参照して、FcγR陽性指標細胞の染色の50%減少をもたらす被験抗体の用量(マイクロMで表す)である。
【0036】
本アッセイ法において、ID30とは、蛍光色素と結合させた抗FcγR抗体を参照して、FcγR陽性指標細胞の染色の30%減少をもたらす被験抗体の用量(マイクロMで表す)である。
【0037】
本アッセイ法において、ID10とは、蛍光色素と結合させた抗FcγR抗体を参照して、FcγR陽性指標細胞の染色の10%減少をもたらす被験抗体の用量(マイクロMで表す)である。
【0038】
「結合する」という表現は、マウスモノクローナル抗体蛍光色素標識3G8を用いた競合アッセイにおいて0.2マイクロMより低いID50、または0.1マイクロMより低い、有利には0.05マイクロMより低い、さらにより有意には0.02マイクロM未満のID50を有する、FcγRIIIAに結合する単量体型の抗体を意味する。3G8とはFcγRIIIに対する抗体であり、当技術分野において周知である(3G8-PE、Immunotech、フランス、カタログ番号A07766)。
【0039】
「結合する」という表現は、マウスモノクローナル抗体蛍光色素標識AT10を用いた競合アッセイにおいて6マイクロMより低いID50、または2マイクロMより低い、有利には1マイクロMより低い、さらにより有意には0.85マイクロM未満のID50を有する、FcγRIIAに結合する単量体型の抗体を意味する。AT10とはFcγRIIAおよびFcγRIIBに対する抗体であり、当技術分野において周知である(AT10-FITC、Serotec、カタログ番号MCA 1075F)。
【0040】
「結合する」という表現は、マウスモノクローナル抗体蛍光色素標識AT10を用いた競合アッセイにおいて6マイクロMより低い、またはより有利には2マイクロMより低い、さらにより有意には1マイクロMより低いID50を有する、FcγRIIBに結合する単量体型の抗体を意味する。AT10とはFcγRIIAおよびFcγRIIBに対する抗体であり、当技術分野において周知である(AT10-FITC、Serotec、英国、カタログ番号MCA 1075F)。
【0041】
「結合しないかまたはごく弱く結合する」という表現は、マウスモノクローナル抗体蛍光色素標識3G8を用いた競合アッセイにおいて0.2マイクロM以上の、有利には0.5マイクロMを超える、より有利には1マイクロMを超えるもしくは2マイクロMを超える、最も有利には10マイクロMを超えるID50を有する、FcγRIIIAに結合する単量体型の抗体を意味する。
【0042】
「結合しないかまたはごく弱く結合する」という表現は、マウスモノクローナル抗体蛍光色素標識AT10を用いた競合アッセイにおいて6マイクロM以上の、有利には9マイクロMを超える、より有利には15マイクロMを超えるもしくは20マイクロMを超える、最も有利には50マイクロMを超えるID50を有する、FcγRIIAに結合する単量体型の抗体を意味する。
【0043】
「結合しないかまたはごく弱く結合する」という表現は、マウスモノクローナル抗体蛍光色素標識AT10を用いた競合アッセイにおいて6マイクロM以上の、有利には9マイクロMを超える、より有利には15マイクロを超えるもしくは20マイクロMを超える、最も有利には50マイクロMを超えるID50を有する、FcγRIIBに結合する単量体型の抗体を意味する。
【0044】
本明細書で用いるFcγRIIIA、FcγRIIA、およびFcγRIIBに対する結合とは、抗体調製物中に低い割合の凝集物(<2%)しか存在せず、抗体が熱もしくは化学的凝集等の任意の手段により、または関連抗原もしくはF(ab')2抗ヒトIgGとの免疫複合体の形成により故意に複合化されていない場合の抗体の結合を指す。
【0045】
別の態様において、本方法は複合化抗体を用いた結合アッセイ法に基づく。
【0046】
結合アッセイ法は、複合化抗体を用いた免疫蛍光結合アッセイ法であってよい。
【0047】
有利には、結合アッセイ法は、複合化抗体を用いる直接的または間接的免疫蛍光結合アッセイ法である。
【0048】
本明細書で用いるFcγRIIIA、FcγRIIA、およびFcγRIIBに対する結合とは、F(ab')2抗ヒトIgGと複合化した抗体の結合を指す。
【0049】
本発明によると、複合化抗体を用いる免疫蛍光結合アッセイ法を用いる方法における「結合する」という表現は、FcγRIIIA陽性指標細胞に対する複合化抗体(それぞれ500、250、および100 ng/mlで使用)の結合を測定する免疫蛍光アッセイ法において、少なくとも30%、15%、5%の割合の陽性指標細胞をもたらす抗体を意味する。
【0050】
本発明によると、複合化抗体を用いる免疫蛍光結合アッセイ法を用いる方法における「結合する」という表現は、FcγRIIA陽性指標細胞に対する抗体(それぞれ10、5、および1マイクロg/mlで使用)の結合を測定する免疫蛍光アッセイ法において、少なくとも70%、60%、30%の割合の陽性指標細胞をもたらす抗体を意味する。
【0051】
本発明によると、複合化抗体を用いる免疫蛍光結合アッセイ法を用いる方法における「結合する」という表現は、FcγRIIB陽性指標細胞に対する抗体(それぞれ50および25マイクロg/mlで使用)の結合を測定する免疫蛍光アッセイ法において、少なくとも50%、25%の割合の陽性指標細胞をもたらす抗体を意味する。
【0052】
複合化抗体を用いる免疫蛍光結合アッセイ法を用いる方法における「結合しないかまたはごく弱く結合する」という表現は、FcγRIIIA陽性指標細胞に対する複合化抗体(それぞれ500、250、および100 ng/mlで使用)の結合を測定する免疫蛍光アッセイ法において、30%、15%、5%よりも低い割合の陽性指標細胞をもたらす抗体を意味する。
【0053】
複合化抗体を用いる免疫蛍光結合アッセイ法を用いる方法における「結合しないかまたはごく弱く結合する」という表現は、FcγRIIA陽性指標細胞に対する抗体(それぞれ10、5、および1マイクロg/mlで使用)の結合を測定する免疫蛍光アッセイ法において、70%、60%、30%よりも低い割合の陽性指標細胞をもたらす抗体を意味する。
【0054】
複合化抗体を用いる免疫蛍光結合アッセイ法を用いる方法における「結合しないかまたはごく弱く結合する」という表現は、FcγRIIB陽性指標細胞に対する抗体(それぞれ50および25マイクロg/mlで使用)の結合を測定する免疫蛍光アッセイ法において、少なくとも30%、10%よりも低い割合の陽性指標細胞をもたらす抗体を意味する。
【0055】
有利には、段階c) i)で選択される抗体またはポリペプチドは、リンパ球系(lymphoid)細胞株またはリンパ球系細胞由来(lymphoid-derived)細胞株またはハイブリドーマまたは腎臓上皮細胞株由来の細胞により産生され、段階c) ii)で選択される抗体またはポリペプチドは非リンパ球系(non-lymphoid)細胞株由来の細胞により産生され、段階c) iii)で選択される抗体またはポリペプチドは、EBV形質転換細胞株由来の細胞にまたはヒトドナー由来のBリンパ球に融合させたヘテロハイブリドーマ由来の細胞により産生される。
【0056】
「リンパ球系細胞株」という用語は、任意の手段により長期インビトロ培養で培養したヒトリンパ球に由来する細胞を指す。具体的には、そのような細胞株はEBV形質転換により取得することができる。「リンパ球系細胞由来細胞株」という用語は、リンパ球系細胞株由来の細胞とパートナー細胞との融合のような任意の手段によるリンパ球系細胞株に由来する細胞を指す。そのようなパートナー細胞は、ヒトドナーに由来するBリンパ球であってよい。「ハイブリドーマ」という用語は、B細胞と骨髄腫細胞との融合に由来する細胞を指す。「ヘテロハイブリドーマ」という用語は、マウス骨髄腫細胞由来の細胞と、結節型リンパ腫患者によるヒト悪性リンパ球との融合によって得られる細胞を指す。
【0057】
本明細書に述べるように、同定された抗体またはポリペプチドは、上記の関連特性を示す抗体または抗体由来分子を産生し得る同定された細胞株を包含する。
【0058】
本発明の特定の局面において、リンパ球系細胞由来細胞株はラット骨髄腫細胞株もしくはハイブリドーマYB2/0細胞株(ATCC番号CRL-1662)もしくはそれらに由来する細胞株であり、かつ/または、腎臓上皮細胞株はVERO(ATCC番号CCL-81)またはそれに由来する細胞であり、かつ/または、非リンパ球系細胞株はCHO(ATCC番号CCL-61)またはそれに由来する細胞であり、かつ/または、ヘテロハイブリドーマはK6H6B5(ATCC番号CRL-1823)またはそれに由来する細胞株である。
【0059】
CHO細胞株はCHO dhfr-細胞株(コロンビア大学、米国、ニューヨーク州)であってよい。
【0060】
本発明の好ましい態様において、結合アッセイは以下を用いて行われ得る:
i) 細胞表面上に異なるFc受容体を発現する細胞株由来の指標細胞、
ii) FcγR外部ドメインを含む組み換えFc受容体、Fc受容体由来ペプチド。
【0061】
指標細胞は、例えば(FcγRIIIAを発現している)Jurkat-CD16、(FcγRIIAを発現している)CD32陽性K562(ATCC番号CCL-243)、または(FcγRIIBを発現している)CD32陽性IIA1.6であってよい。
【0062】
「FcγR外部ドメインを含むFc受容体」という用語は、2つの外部ドメイン(D1およびD2)からなるFcγ受容体IIA、IIB、およびIIIA型の細胞外領域を指す。
【0063】
好ましい態様においては、FcγRIIIA、FcγRIIA、およびFcγRIIBのいずれにも結合する能力に関して選択された抗体またはポリペプチドのサブセット、ならびにFcγRIIAおよびFcγRIIBの両方に結合する能力に関して選択された抗体またはポリペプチドのサブセットは、i) FcγRIIIAを誘発してADCCの改善、インターロイキン-2(IL-2)等のサイトカインおよび腫瘍壊死因子α(TNFα)等の炎症促進性分子の産生の増加をもたらす能力;およびii) FcγRIIBを誘発してB細胞および単球等のFcγRIIBを発現する細胞によるカルシウム動員の阻害およびIL-2等のサイトカイン産生の阻害をもたらす能力に関する機能アッセイにより、さらに試験および選択される。
【0064】
有利には、機能アッセイ法は、カルシウム動員阻害アッセイ法および/またはサイトカイン分泌阻害アッセイ法からなる。
【0065】
本発明に準じて第1および第2クラスに属する抗体の活性には、表面FcγRIIBに結合したそのような抗体とBCR、TCR、FcεRI等の活性化表面受容体との共凝集によるカルシウム動員の阻害が含まれる。本明細書で使用するカルシウム動員とは、BCR等の活性化受容体の補充後の細胞内への細胞外カルシウムの流入を指す。
【0066】
本発明の抗体の活性には、表面FcγRIIBに結合したそのような抗体とBCR等の活性化表面分子との共凝集によるサイトカイン産生の阻害が含まれる。
【0067】
本明細書において使用するサイトカイン産生とは、BCR等の活性化受容体の補充後の細胞によるサイトカインの放出を指す。
【0068】
本発明の分子によるサイトカイン産生の阻害には、インターロイキン-2(IL-2)産生の阻害が含まれる。
【0069】
別の態様において、機能アッセイ法には特異的FcγRIIIA ADCCアッセイ法がさらに含まれ得る。
【0070】
基本的に、この特異的FcγRIIIA ADCCアッセイ法は、各抗体またはポリペプチドを、標的細胞、FcγRIIIAを発現しているエフェクター細胞、および多価IgGを含む異なる反応混合物に添加する段階、および標的細胞の溶解の割合を決定する段階を含む。
【0071】
本発明の別の目的は、FcγRIIIA、FcγRIIA、およびFcγRIIBに結合し得る抗体を産生させるための、リンパ球系細胞株またはリンパ球系細胞由来細胞株またはハイブリドーマまたは腎臓上皮細胞株に由来する細胞の使用である。
【0072】
「ハイブリドーマ」という用語は、上記のハイブリドーマと同じ意味を有する。有利には、ハイブリドーマはリンパ球に融合させたラット骨髄腫細胞に由来し得る。
【0073】
YB2/0ハイブリドーマ細胞により産生される抗体の、FcγRIIIA、FcγRIIA、およびFcγRIIBに対する結合に対するID10、ID30、およびID50の値を図9に要約するが、これにより3つの型の受容体に関するこれらの抗体の強力な結合が示される。
【0074】
本発明の特定の態様において、リンパ球系細胞株またはリンパ球系細胞由来細胞株またはハイブリドーマまたは腎臓上皮細胞株に由来する細胞によって産生される抗体は免疫調節抗体である。
【0075】
ハイブリドーマ細胞、より詳細にはB細胞とラット骨髄腫細胞との融合によって生じるハイブリドーマ細胞により産生される抗体はFcγRIIBに結合し、ひいてはFcγRIIBを介して細胞機能の阻害を誘発し得る。
【0076】
本発明によれば、ハイブリドーマ細胞、好ましくはB細胞とラット骨髄腫細胞との融合によって生じるハイブリドーマ細胞により産生される抗体は、上記の第1クラスの抗体に属する。結果として、それらはまた上記の方法の段階c) i)によって選択され得る。
【0077】
リンパ球系細胞株またはリンパ球系細胞由来細胞株またはハイブリドーマまたは腎臓上皮細胞株に由来する細胞によって産生されるこれらの抗体は、免疫調節かつ細胞傷害性の抗体である。
【0078】
ハイブリドーマ細胞、好ましくはB細胞とラット骨髄腫細胞との融合によって生じるハイブリドーマ細胞により産生されるこれらの免疫調節抗体は、FcγRIIIAおよびFcγRIIAに結合し得る。これらの受容体は活性化受容体であり、細胞傷害活性を誘発する。
【0079】
好ましい態様において、ハイブリドーマ細胞株はYB2/0である。
【0080】
別の好ましい態様において、腎臓上皮細胞株はVEROである。
【0081】
本発明の別の目的は、FcγRIIIAには結合しないかまたはごく弱く結合するが、FcγRIIAおよびFcγRIIBの両方には結合する抗体を産生させるための、非リンパ球系細胞株由来の細胞の使用である。
【0082】
本出願人らは本出願において、非リンパ球系細胞株によって産生される抗体は、FcγRIIIAには結合しないとは言えないにしても弱く結合するが、FcγRIIAおよびFcγRIIBには結合することを示す。図9に細胞株CHOによって産生される抗体のID10、ID30、およびID50の値を要約するが、これによりFcγRIIIAに対するこれらの抗体の弱い結合ならびにFcγRIIAおよびFcγRIIBに対するこれらの抗体の結合が示される。
【0083】
非リンパ球系細胞株由来の細胞により産生されるこれらの抗体は、免疫調節抗体である。
【0084】
この免疫調節活性は、FcγRIIBに対するこれらの抗体の結合に起因する。
【0085】
さらに、非リンパ球系細胞株により産生されるこれらの抗体は、FcγRIIAを発現している単球およびマクロファージによるADCCおよび食作用を誘導する。
【0086】
上記の第2クラスの抗体に属するこれらの抗体はまた、上記の方法の段階c) ii)によって調製または選択され得る。
【0087】
好ましい態様において、非リンパ球系細胞株はCHO(ATCC番号CCL-61)である。
【0088】
別の態様において、CHO細胞株はCHO dhfr-(コロンビア大学、米国、ニューヨーク州)であってよい。
【0089】
本発明のさらに別の目的は、FcγRIIIA、FcγRIIA、およびFcγRIIBのいずれにも結合しないかまたは弱く結合する抗体を産生させるための、EBV形質転換細胞株由来の細胞とまたはB細胞と融合させたヘテロハイブリドーマ由来の細胞の使用である。
【0090】
本出願人らは本出願において、B細胞と融合させたヘテロハイブリドーマによって産生される抗体が、FcγRIIIA、FcγRIIA、およびFcγRIIBに弱く結合することを示す。図9にB細胞と融合させたヘテロハイブリドーマK6H6B5由来の細胞によって産生される抗体のID10、ID30、およびID50の値を要約するが、これによりFcγRIIIA、FcγRIIA、およびFcγRIIBに対するこれらの抗体の弱い結合が示される。
【0091】
有利には、EBV形質転換細胞株由来の細胞とまたはB細胞と融合させたヘテロハイブリドーマ由来の細胞によって産生されるこれらの抗体は、IgG4の使用に代わる治療法として用いられる。
【0092】
IgG4は、FcγRIIIA、FcγRIIA、およびFcγRIIBを介したエフェクター機能の補充を回避するために用いられる。本発明に従って、B細胞またはEBV形質転換B細胞に融合させたヘテロハイブリドーマから産生される抗体は、IgG1アイソタイプに属する。これらは、特定の疾患の治療過程において、FcγRIIIA、FcγRIIA、およびFcγRIIBの補充を介して達成されるエフェクター機能を誘発することなく、FcγRおよびFcεRI結合によって誘発される重篤な副作用の発症を妨げてインビボで使用し得る。
【0093】
好ましい態様において、これらの細胞はヒトB細胞に融合させたヘテロハイブリドーマK6H6B5(ATCC番号CRL-1823)に由来する細胞である。
【0094】
本発明のさらに別の目的は、10%〜55%のフコース、および60%〜98%のガラクトースを含む、ヒトIgGのFc領域を含む抗体またはポリペプチドである。
【0095】
10%〜55%のフコース、および60%〜98%のガラクトースを含むこの抗体またはポリペプチドは、FcγRIIIA、FcγRIIA、およびFcγRIIBのいずれにも結合する。
【0096】
これらの2つの特性(Fcグリコシル化プロファイルおよびFcγR結合プロファイル)は共に関連している。本出願人らは、これら2つの特性がまた、このクラスの抗体がハイブリドーマ細胞株によって産生され得るという事実にも関連していることを示す。
【0097】
この抗体またはポリペプチドは、リンパ球系細胞株またはリンパ球系細胞由来細胞株またはハイブリドーマまたは腎臓上皮細胞株に由来する細胞によって産生され得る。
【0098】
有利には、このハイブリドーマ細胞株は、AOラットの脾臓細胞と融合させた骨髄腫細胞株、LOU/C Y3/Ag 1.2.3に由来するハイブリドーマ細胞株、YB2/3HLに由来するYB2/0細胞株(ATCC番号CRL-1662)である。
【0099】
別の態様において、この抗体またはポリペプチドは腎臓上皮細胞株VERO(ATCC番号CCL-81)によって産生される。
【0100】
さらに、10%〜55%のフコース、および60%〜98%のガラクトースを含むこの抗体またはポリペプチドは、上記の調製過程の段階c) i)により取得することができる。
【0101】
これらの抗体またはポリペプチドは上記の第1クラスに属する。
【0102】
本発明はまた、この第1クラスの抗体またはポリペプチドを少なくとも80%、好ましくは少なくとも95%含む組成物を対象とする。
【0103】
好ましい態様において、組成物は上記の抗体またはポリペプチドを少なくとも80%ないし90%、好ましくは少なくとも95%ないし99%含み得る。
【0104】
白血病、リンパ腫、骨髄腫、セザリー症候群、もしくは固形腫瘍等の癌、自己免疫疾患、アレルギー、移植後の同種免疫、母子間同種免疫、移植片対宿主(GVH)反応、または感染症の治療用の医薬品を製造するためにこの組成物を使用することも本発明のさらなる対象である。
【0105】
第1クラスの抗体は、標的細胞、例えば癌細胞を死滅させるために、(FcγRIIIAおよびFcγRIIAに結合する能力に起因して)細胞傷害性細胞を活性化し得る。
【0106】
そのような抗体は、FcγRIIBにより受容体チロシンキナーゼ(RTK)に依存して、造血細胞増殖の制御を誘発するための有用な治療手段になると考えられる。RTKとFcγRIIBを効率的に共凝集させ得る最適なFc領域を有する抗RTK IgG抗体のような二機能性分子は、FcγRIIB+腫瘍細胞の増殖を阻止し得る。
【0107】
本発明の特定の局面において、この組成物を用いて治療する母子間同種免疫は新生児溶血性疾患(HDNB)である。
【0108】
このクラスの抗体は赤血球を死滅させ得り、それと同時に赤血球に対する抗体を産生するB細胞を免疫調節し得る。
【0109】
本発明の別の特定の局面において、自己免疫疾患は、全身性エリテマトーデス(SLE)、特発性血小板減少性紫斑病(ITP)、川崎症候群等の自己抗体を産生するB細胞関連自己免疫疾患である。
【0110】
別の好ましい局面において、治療するアレルギーは、喘息、アレルギー性鼻炎、アレルギー性副鼻腔炎、アナフィラキシー症候群、じんま疹、血管性水腫、アトピー性皮膚炎、アレルギー性接触皮膚炎、および紅斑である。
【0111】
本発明のさらに別の目的は、70%〜100%、例えば96%〜99%のフコース、および60%〜98%、例えば74%〜89%のガラクトースを含むことを特徴とする、ヒトIgGのFc領域を含む抗体またはポリペプチドである。
【0112】
70%〜100%のフコースおよび60%〜98%のガラクトースを含むこの抗体またはポリペプチドは、FcγRIIAおよびFcγRIIBの両方に結合するが、FcγRIIIAには結合しないかまたはごく弱く結合する。
【0113】
本出願人らは、結合プロファイルがFcグリコシル化プロファイルと関連していることを示す。さらに本出願人らは、この抗体またはポリペプチドが非リンパ球系細胞株由来の細胞によって産生されることを示す;よって、結合プロファイル、グリコシル化プロファイル、および抗体産生細胞株は関連している。
【0114】
より詳細には、この非リンパ球系細胞株はCHO細胞株(ATCC番号CCL-61)である。
【0115】
特に関心対象である局面において、この抗体またはポリペプチドは上記の調製過程の段階c) ii)によって取得することができる。
【0116】
これらの抗体またはポリペプチドは、上記の第2クラスの抗体に属する。
【0117】
本発明はまた、上記の第2クラスの抗体を少なくとも80%、好ましくは少なくとも95%含む組成物を包含する。
【0118】
好ましい態様において、組成物は上記の第2クラスに属する抗体またはポリペプチドを少なくとも80%〜90%、好ましくは少なくとも95%ないし99%含み得る。
【0119】
本発明はまた、自己免疫疾患、母子間同種免疫、および炎症性疾患の治療用の医薬品を製造するためのこの組成物の使用に関する。
【0120】
これらの最適化された抗体は、アレルゲン複合化IgGにより、肥満細胞および好塩基球上に存在するFcγRIIBの補充を調節するための有用な治療手段となると考えられる。
【0121】
例として、FcγRIIBと結合し、IgEが同じアレルゲンに複合化した時点での、次に起こるFcγRIIBとFcεRIとの共凝集を介してFcγRIIBの阻害機能を誘発する最適な能力を有するそのような抗体が挙げられる。FcγRIIBと結合し、それ故にFcγRIIB阻害機能を誘発する最適な能力を有する抗アレルゲンIgG抗体は、アレルギーの治療における有効な治療手段を表し得る。例として、本発明により、FcγRIIBと結合し、次に起こるFcγRIIBとFcεRIとの共凝集を介してFcγRIIB阻害機能を誘発する最適な能力を有する抗FcεRI IgG抗体を作製および選択することが可能になる。FcγRIIBと結合し、それ故にFcγRIIB阻害機能を誘発する最適な能力を有する抗FcεRI IgG抗体は、アレルギーの治療における有効な治療手段を表し得る。
【0122】
本発明は、APC(抗原提示細胞)によって発現されるFcγRIIBの効果を調節するための有用な治療手段となると考えられる最適な抗体に関する。そのような効果の例には、抗原提示およびT細胞動員の制御ならびに獲得免疫の制御が含まれる。
【0123】
さらに別の局面において、本発明は、80%〜100%のフコース、および60%〜98%のガラクトースを含み、ガラクトースが30%〜80%のシアリル化型を含む、ヒトIgGのFc領域を含む抗体またはポリペプチドを提供する。
【0124】
本発明によれば、この抗体またはポリペプチドは、FcγRIIIA、FcγRIIA、およびFcγRIIBに結合しないかまたはごく弱く結合する。
【0125】
本出願人らは、結合プロファイルがFcグリコシル化プロファイルと関連していることを示す。本出願人らはまた、このクラスの抗体(上記の第3クラス)が、B細胞またはEBV形質転換B細胞に融合させたヘテロハイブリドーマ由来の細胞によって産生されることを示す。
【0126】
特に関心対象である局面において、この抗体またはポリペプチドはB細胞に融合させたヘテロハイブリドーマ由来の細胞によって産生される。
【0127】
特定の態様において、このヘテロハイブリドーマはEBV形質転換細胞またはヒトドナー由来のBリンパ球に融合させたK6H6B5(ATCC番号CRL-1823)である。
【0128】
この抗体またはポリペプチドは、上記の調製過程の段階c) iii)によって取得するとができる。
【0129】
この抗体またはポリペプチドは上記の第3クラスに属する。
【0130】
本発明はまた、第3クラスに属するこの抗体またはポリペプチドを少なくとも80%、好ましくは少なくとも95%含む組成物を対象とする。
【0131】
好ましい態様において、組成物は、上記の抗体またはポリペプチドを少なくとも80%〜90%、好ましくは少なくとも95%ないし99%含み得る。
【0132】
特に関心対象である局面において、この組成物は、炎症性疾患、クローン病、または関節リウマチの治療用の医薬品を製造するために用いられる。
【0133】
この第3クラスの抗体またはポリペプチドは、FcγRIIBを介して媒介される阻害機能を誘発すること、およびADCCまたは別の細胞傷害活性を誘発することができない。これらの抗体またはポリペプチドは、本発明において、免疫エフェクター機能の補充を妨げるためにい用いられる。
【0134】
本発明による特に関心対象の抗体またはポリペプチドは、IgG1サブクラスに属する。
【0135】
別の態様において、特に関心対象の抗体またはポリペプチドはIgG3サブクラスに属する。
【0136】
本発明はさらに、抗体が標的細胞の細胞表面上に発現されるまたは細胞表面に結合した抗原を認識する医薬品を製造するための上記組成物の使用に関する。そのような抗原には、これらに限定されないが、同種抗原、移植抗原、クラスIおよびクラスII主要組織適合抗原(HLA)等の自己抗原、FcγRI、FcγRIIA、FcγRIIIA分子、FcαR、FcεRI、B細胞受容体、T細胞受容体、CD20、Her2/NEU、CEA、GD2等の腫瘍抗原、ホスホリパーゼA2等のアレルゲン、およびIgE等が含まれる。
【0137】
より詳細には、本発明の抗体は、新生児溶血性疾患(HDNB)をもたらすRh陰性患者のRh同種免疫を治療または予防する医薬品を製造するためにRhD抗原を認識する。
【0138】
より詳細には、本発明の抗体は、特発性血小板減少性紫斑病(ITP)を治療または予防する医薬品を製造するためにRhD抗原を認識する。
【0139】
より詳細には、本発明の抗体は、癌、自己免疫疾患、または移植片拒絶を治療または予防する医薬品を製造するためにHLAクラスII分子を認識する。
【0140】
さらなる詳細を以下の実施例1に提供する。これらの結合および機能試験の結果を図に示す。
【0141】
実施例1:FcγRIIB媒介性阻害を誘発する改善された特性を有する抗RhD抗体の選択
材料および方法
免疫学的に活性のある分子を選択する本発明の方法を、RhD抗原に対するヒトIgG抗体を用いて以下に説明する。例として、RhDに対するLFB1、LFB2、およびLFB3ヒトモノクローナル抗体を使用している。
【0142】
F(ab)'2抗ヒトIgと複合化したまたはしていない精製ヒト抗体が指標細胞(IIA1.6-huFcγRIIB1と命名された、機能的なマウスBCRおよびヒト組み換えFcγRIIB1を発現するマウスIIA1.6リンパ腫B細胞)上に発現されるFcγRIIB1に結合する能力を、それぞれ蛍光抗体アッセイ法(複合化)または免疫蛍光競合アッセイ法(単量体)により測定する。
【0143】
精製ヒト抗体がカルシウム動員の阻害を誘導する能力は、カルシウム測定アッセイ法により評価し得る。カルシウム動員の阻害は、指標細胞(機能的なマウスBCRおよびヒト組み換えFcγRIIB1を発現するマウスIIA1.6リンパ腫B細胞)上に発現されるFcγRIIBに結合した精製ヒト抗体とBCRを共凝集させることにより評価する。これを、マウスIgGと交差反応するF(ab)'2ウサギ抗ヤギIgG(RAG)を用いて誘導されるBCR媒介性カルシウム動員と比較する。
【0144】
精製ヒト抗体がインターロイキン-2(IL-2)産生の阻害を誘導する能力は、IL-2を検出し定量化する酵素結合免疫吸着測定法(ELISA)により測定し得る。IL-2産生の阻害は、指標細胞(機能的なマウスBCRおよびヒト組み換えFcγRIIB1を発現するマウスIIA1.6リンパ腫B細胞)上に発現されるFcγRIIBに結合した精製ヒトまたはキメラ抗体とBCRを共凝集させることにより評価する。これを、F(ab)'2ヤギ抗マウスIgG(H+L)を用いて誘導されるBCR媒介性IL-2産生と比較する。
【0145】
細胞株および抗体
マウスIIA1.6 B細胞リンパ腫は、FcγRII遺伝子の5'末端の欠失を有する(Lewis et al., 1986;Bonnerot et al., 1991)A20 B細胞のFcγR欠損変種(Jones et al., 1986)であり、マウスFcγRIIIのαおよびγ鎖をコードする遺伝子を転写しない(Bonnerot et al., 1991)。M. Hogarth博士(メルボルン大学、オーストラリア、ビクトリア州、パークビル)の供与によるFcγRIIB1をコードするcDNA(Hogarth et al., 1987)をIIA.1.6細胞にトランスフェクションした。IIA1.6およびIIA1.6-huFcγRIIB1細胞を、Click培地[10%熱非働化FCS(Hyclone Laboratories Inc.、米国、ユタ州、ローガン)、100 U/mlペニシリン、100μg/mlストレプトマイシン、2 mM L-グルタミン、5 mMピルビン酸ナトリウム、0.5μM 2-β-メルカプトエタノールを添加したRPMI 1640(GIBCO、スコットランド、ペーズリー)]中で培養した。
【0146】
ヒト組み換え抗Rh D(LFB1)モノクローナル抗体を産生させるには、YB2/0細胞株(ATCC番号CRL-1662)を使用した。YB2/0トランスフェクション細胞は、5% FCSを添加した特別なEM培地中で培養した。
【0147】
LFB2抗Rh Dヒト組み換え抗体を産生させるため、CHO dhfr-(コロンビア大学、米国、ニューヨーク州)細胞株を5% FCSを添加した特別なEM培地中で培養した。
【0148】
LFB1/LFB2抗体の重鎖の定常領域の配列(Kabatにより規定されるCH1の5'末端から開始)を以下に示す(配列番号:5)。

【0149】
抗Rh D LFB3抗体を産生するB細胞株は、ヒトRh陽性赤血球で免疫したドナーによるヒトBリンパ球をヘテロミエローマ細胞株K6H6/B5(ATCC番号CRL-1823)と融合することによって取得し、次いで限界希釈によりクローニングを行った。LFB3細胞は、5% FCSを添加したRPMI 1640培地中で培養した。
【0150】
LFB3抗体の重鎖の定常領域の配列(Kabatにより規定されるCH1の5'末端から開始)を以下に示す(配列番号:6)。
【0151】
産生細胞株はすべて無タンパク質培地に適応化させてそれにより培養し、産生はローラーボトルまたは発酵槽で行った。産生後、プロテイン-Aアフィニティークロマトグラフィーによりモノクローナル抗体を精製した。次いで、精製抗体を-20℃で保存した。
【0152】
リツキサン(Rituxan)(リツキシマブ(Rituximab))(Genentech、米国、カリフォルニア州、サウスサンフランシスコ)は、再発性または難治性低悪性度またはろ胞性、CD20+、B細胞非ホジキンリンパ腫(NHL)の治療において用いられるキメラマウス/ヒト抗CD20モノクローナル抗体である。市販のリツキサンバイアルは、無菌の防腐剤の入っていない10 mg/ml濃度溶液で2℃〜8℃にて保存する。
【0153】
AT10-FITCは、FcγRIIAおよびFcγRIIBの結合部位に対するフルオレセインイソチオシアネート標識マウスモノクローナル抗体である(AT10-FITC、Serotec、英国、カタログ番号MCA 1075F)。
【0154】
免疫蛍光結合アッセイ法
まず、組み換えヒト抗体を、アフィニティー精製したヤギF(ab)'2抗ヒトIgG(Jackson ImmunoResearch Laboratories Inc.、米国、ペンシルベニア州、ウェストグローブ)を用いて30分間、重合型に複合化した。次いで、指標細胞5×105個を、0.5%ウシ血清アルブミンを含む氷冷リン酸緩衝食塩水(PBS-BSA)中で複合化抗体と共に30分間インキュベートした。その後、細胞をPBS-BSAで洗浄し、FITC標識マウスF(ab)'2抗ヒトIgG(H+L)(Jackson ImmunoResearch)と共に氷上で30分間インキュベートすることにより、抗体結合を検出した。さらに洗浄した後、Cell Quest Proソフトウェアを用いてFACScalibur 4CA(Becton Dickinson、米国、カリフォルニア州、マウンテンビュー)によりフローサイトメトリー解析を行った。
【0155】
免疫蛍光競合アッセイ法
IIA1.6-huFcγRIIB細胞をまず種々の濃度の単量体LFB1または単量体LFB2または単量体LFB3または単量体リツキシマブと共に、次いでAT10-FITCと共にインキュベートした。続いて、競合物としての単量体抗体の存在下または非存在下におけるヒトFcγRIIBに対するFITC-AT10モノクローナル抗体の結合を、直接免疫蛍光法により評価した。フローサイトメトリー解析は、Cell Quest Proソフトウェアを用いてFACScalibur 4CA(Becton Dickinson、米国、カリフォルニア州、マウンテンビュー)により行った。
【0156】
Facscalibur 4CAの技術仕様
FITC蛍光色素の獲得には、15 mW、488 nm空冷アルゴンイオンレーザーを用いる。推定検出限界は、粒子当たり200 FITC相当分子である。FL1(FITC)の対数増幅器は、4 log10(log decade)範囲を提供する。ソーティング速度は300細胞/秒であり、ソーティング精度は95%を超える。
【0157】
カルシウム動員の阻害
IIA1.6-huFcγRIIB1細胞をLFB1抗体と共にまたはなしで氷上で30分間インキュベートした。0.2% Pluronic F-127(Sigma Cnemicals Co.,米国、ミズーリ州、セントルイス)を含むRPMI 1640中、106個細胞を5 mM Fluo-3 AM(Molecular Probes、米国、オレゴン州、ユージーン)と共に室温で30分間インキュベートした。次いで、負荷された細胞をRPMI 1640で3回洗浄し、106個/mlになるよう調整した。1 mM EGTAを含むRPMI 1640中、ウサギF(ab)'2抗ヤギIgG(H+L)(RAG)(Jackson ImmunoResearch)単独またはウサギF(ab)'2抗ヤギIgG(H+L)(RAG)およびヤギF(ab)'2抗ヒトIgG(H+L)(GAH)(Jackson ImmunoResearch)で細胞を刺激した。刺激してから150秒後に、6 mM CaCl2を添加した。Cell Quest Proソフトウェアを用いてFACScalibur 4CAにより実施するフローサイトメトリーによって、細胞内カルシウム動員を検出した。次いで、FCS assistant 1.2.9βソフトウェア(Becton Dickinson)により、細胞内カルシウム濃度平均値を算出した。
【0158】
IL-2産生の阻害
IIA1.6-huFcγRIIB1細胞5×105個を、F(ab)'2ウサギ抗ヤギ(Jackson ImmunoResearch)単独またはF(ab)'2ウサギ抗ヤギおよびLFB1、LFB2、またはLFB3で22時間刺激した。1μg/mlラット抗マウスIL-2(Pharmingen)を捕獲抗体として、および1μg/mlビオチン化ラット抗マウスIL-2(Pharmingen)を検出抗体として使用するELISAアッセイにより、マウスIL-2の存在を測定した。
【図面の簡単な説明】
【0159】
【図1】純度および分子の完全性および重合型が存在しないことを管理するための、モノクローナル抗体[RhDに対する(抗RhD)LFB1抗体]のHPLCプロファイルを表す:LFB1モノクローナル抗体は99%の単量体から構成される。
【図2】キャピラリー電気泳動によって決定される、RhDに対する種々の抗体(LFB1、LFB2、LFB3)のグリコシル化パターンを表す。結果は同定された種々の構造の割合として表す。抗体は異なる細胞株によって産生された。LFB1はYB2/0細胞により産生され、LFB2はCHO細胞によって産生され、LFB3はヒトB細胞に融合させたヘテロハイブリドーマK6H6B5によって産生される。
【図3】Jurkat-FcγRIIIA細胞上に発現されるヒトFcγRIIIAに対する、F(ab)'2ヤギ抗ヒトIgG(H+L)複合化LFB1、LFB2、およびLFB3抗体の結合を表す。ヒトFcγRIIIAに対するLFB1、LFB2、およびLFB3ヒト抗RhD抗体の結合は、間接的免疫蛍光法により評価する。まずヒト抗体(100 ng/ml;250 ng/ml;500 ng/ml)をF(ab)'2ヤギ抗ヒトIgG(H+L)(それぞれ150 ng/ml;375 ng/ml;750 ng/mlの用量)により複合化し、次いでJurkat-FcγRIIIA細胞と共にインキュベートする。続いて、FITC標識F(ab)'2マウス抗ヒトIgG(H+L)を用いて、LFB1、LFB2、およびLFB3抗体の結合を明らかにする(黒い曲線)。点線の曲線は、FITC-F(ab)'2マウス抗ヒトIgG(H+L)とのみインキュベートした細胞のバックグラウンド蛍光を示す。結果およびコメント:Jurkat-FcγRIIIA細胞に対するLFB1の結合は、100 ng/ml〜500 ng/mlにおいて常に検出される。最も高い濃度では、ほぼ50%のJurkat-FcγRIIIA細胞が染色される。それに反して、Jurkat-FcγRIIIA細胞によって発現されるヒトFcγRIIIAに対するLFB2抗体の結合は100 ng/mlにおいて認められず、また250 ng/mlおよび500 ng/mlにおいては3%のJurkat-FcγRIIIA細胞が弱く標識されるにすぎない。最後に、Jurkat-FcγRIIIA細胞によって発現されるヒトFcγRIIIAに対するLFB3抗体の結合は、試験したいずれの濃度においても認められない。したがって、YB2/0細胞によって産生される規定のグリコシル化パターンを有するLFB1は、FcγRIIIAに結合するモノクローナル抗体の例である。さらに、異なるが規定のグリコシル化パターンを有する、CHO細胞によって産生されるLFB2およびB細胞に融合させたヘテロハイブリドーマK6H6B5由来の細胞によって産生されるLFB3は、FcγRIIIAに結合しないかまたはごく弱く結合するモノクローナル抗体の例である。
【図4】単量体LFB1、LFB2、LFB3、またはリツキシマブ抗体による、Jurkat-FcγRIIIA細胞上に発現されるヒトFcγRIIIAに対する3G8-PE結合の阻害を表す。まずJurkat-FcγRIIIA細胞を種々の濃度の単量体LFB1(1マイクロg/ml〜1000マイクロg/ml)または単量体LFB2(1マイクロg/ml〜100マイクロg/ml)または単量体LFB3(1マイクロg/ml〜1000マイクロg/ml)または単量体リツキシマブ(1マイクロg/ml〜1000マイクロg/ml)と共に、次いで0.01マイクロg/ml 3G8-PEと共にインキュベートする。続いて、競合物としての単量体抗体の存在下または非存在下におけるヒトFcγRIIIAに対するPE標識マウス3G8モノクローナル抗体の結合を、直接免疫蛍光法により評価する。結果およびコメント:単量体LFB1は、Jurkat-FcγRIIIA細胞上に発現されるヒトFcγRIIIAに対する3G8-PE結合の用量依存的阻害を誘導する。10マイクロg/mlのLFB1は3G8-PEによって標識される細胞の96%という割合の減少を誘導するのに対し、10マイクロg/mlのリツキシマブでは2%の減少しか達成されない。用いたうち最も高い用量(1000マイクロg/ml)では、LFB1は100%阻害を誘導する(1000マイクロg/mlのリツキシマブでは72%)。単量体LFB2は、LFB1によって得られる阻害よりも低い、FcγRIIIAに対する3G8-PE結合の阻害を誘導する。10マイクロg/mlのLFB2は、3G8-PEによって標識される細胞の5%未満という割合の減少を誘導する。しかし、高濃度(100マイクロg/ml)では、(同じ用量のリツキシマブでのたった15%と比較して)染色されたJurkat-FcγRIIIA細胞の約95%という割合の減少が認められる。それに反して、単量体LFB3によって誘導される阻害は、用いたどの用量においても非常に弱い。1000マイクロg/mlのLFB3を用いた場合、阻害は20%よりも低い。 したがって、このアッセイにより、単量体型の抗体は、それらを産生する細胞株およびそれらのグリコシル化プロファイルに依存して、FcγRIIIAに対して異なる結合能を示すことが示される。
【図5】K562細胞上に発現されるヒトFcγRIIAに対する、F(ab)'2ヤギ抗ヒトIgG(H+L)複合化LFB1、LFB2、およびLFB3抗体の結合を表す。ヒトFcγRIIAに対するLFB1、LFB2、およびLFB3ヒト抗RhD抗体の結合は、間接的免疫蛍光法により評価する。まずヒト抗体(1μg/ml;5μg/ml;10μg/ml)をF(ab)'2ヤギ抗ヒトIgG(H+L)(それぞれ1.5μg/ml;7.5μg/ml;15μg/mlの用量)により複合化し、次いでK562細胞と共にインキュベートする。続いて、FITC標識F(ab)'2マウス抗ヒトIgG(H+L)を用いて、LFB1、LFB2、およびLFB3抗体の結合を明らかにする(黒い曲線)。点線の曲線は、FITC-F(ab)'2マウス抗ヒトIgG(H+L)とのみインキュベートした細胞のバックグラウンド蛍光を示す。結果およびコメント:LFB1抗体は、1μg/mlというような低い用量においてさえもK562細胞に結合する(この低濃度において、50%を超えるK562細胞が染色される)。最も高い用量のLFB1抗体(10μg/ml)では、ほぼ100%のK562細胞が標識される。LFB2は、K562細胞上に発現されるヒトFcγRIIAに対してLFB1と同様の結合パターンを有する。LFB2は使用したいずれの用量においても(1μg/ml〜10μg/ml)ヒトFcγRIIAに結合し、10μg/mlではほぼ100%のK562細胞が染色される。それに反して、K562細胞は、1μg/ml、5μg/ml、または10μg/mlのLFB3により弱く標識される。したがって、異なるが規定のグリコシル化パターンを有する、YB2/0細胞によって産生されるLFB1およびCHO細胞によって産生されるLFB2は、FcγRIIAに結合するモノクローナル抗体の例である。さらに、規定のグリコシル化パターンを有する、B細胞に融合させたヘテロハイブリドーマK6H6B5由来の細胞によって産生されるLFB3は、FcγRIIAに結合しないかまたはごく弱く結合するモノクローナル抗体の例である。
【図6】単量体LFB1、LFB2、LFB3、またはリツキシマブ抗体による、K562細胞上に発現されるヒトFcγRIIAに対する(FcγRIIAおよびFcγRIIBに対する)AT10-FITC結合の阻害を表す。まずK562細胞を種々の濃度の単量体LFB1(50μg/ml〜1000μg/ml)または単量体LFB2(50μg/ml〜250μg/ml)または単量体LFB3(50μg/ml〜1000μg/ml)または単量体リツキシマブ(50μg/ml〜1000μg/ml)と共に、次いで0.25μg/ml AT10-FITCと共にインキュベートする。競合物としての単量体抗体の存在下または非存在下におけるヒトFcγRIIAに対するFITC標識マウスAT10モノクローナル抗体の結合を、直接免疫蛍光法により評価する。結果およびコメント:単量体LFB1は、K562細胞上に発現されるヒトFcγRIIAに対するAT10-FITC結合の用量依存的阻害を誘導する。100μg/mlのLFB1はAT10-FITCによって標識される細胞のほぼ50%という割合の減少を誘導するのに対し、100μg/mlのリツキシマブでは19%の減少しか達成されない。用いたうち最も高い用量(1000μg/ml)では、(1000μg/mlのリツキシマブでの43%と比較して)90%に達する。単量体LFB2もまた、FcγRIIAに対するAT10-FITC結合の用量依存的阻害を誘導する。用いうち最も高濃度(250μg/ml)のLFB2では、(同じ用量のリツキシマブでの24%と比較して)染色されたK562細胞の約40%という割合の減少が認められる。それに反して、単量体LFB3によって誘導される阻害は弱い。1000μg/mlのLFB3を用いた場合、阻害は30%よりも低い。したがって、このアッセイにより、単量体型の抗体は、それらを産生する細胞株およびそれらのグリコシル化プロファイルに依存して、FcγRIIAに対して異なる結合能を示すことが示される。
【図7】IIA1.6-huFcγRIIB細胞上に発現されるヒトFcγRIIBに対する、F(ab)'2ヤギ抗ヒトIgG(H+L)複合化LFB1、LFB2、およびLFB3抗体の結合を表す。ヒトFcγRIIBに対するLFB1、LFB2、およびLFB3ヒト抗RhD抗体の結合は、間接的免疫蛍光法により評価する。まずヒト抗体(25μg/ml;50μg/ml)をF(ab)'2ヤギ抗ヒトIgG(H+L)(それぞれ37.5μg/ml;75μg/mlの用量)により複合化し、次いでIIA1.6-huFcγRIIB細胞と共にインキュベートする。続いて、FITC標識F(ab)'2マウス抗ヒトIgG(H+L)を用いて、LFB1、LFB2、およびLFB3抗体の結合を明らかにする(黒い曲線)。点線の曲線は、FITC-F(ab)'2マウス抗ヒトIgG(H+L)とのみインキュベートした細胞のバックグラウンド蛍光を示す。結果およびコメント:LFB1抗体は、25μg/mlおよび50μg/mlおいてIIA1.6-huFcγRIIB細胞に結合する(50μg/mlおいて、50%を超えるIIA1.6-huFcγRIIB細胞が染色される)。LFB2は、ヒトFcγRIIBに対してLFB1と同様の結合パターンを有する。LFB2は使用したいずれの用量においてもヒトFcγRIIBに結合し、50μg/mlでは50%を超えるIIA1.6-huFcγRIIB細胞が標識される。それに反して、ヒトFcγRIIBに対するLFB3の結合は、試験したいずれの濃度においても認められない。したがって、異なるが規定のグリコシル化パターンを有する、YB2/0細胞によって産生されるLFB1およびCHO細胞によって産生されるLFB2は、FcγRIIBに結合するモノクローナル抗体の例である。さらに、規定のグリコシル化パターンを有する、B細胞に融合させたヘテロハイブリドーマK6H6B5由来の細胞によって産生されるLFB3は、FcγRIIBに結合しないかまたはごく弱く結合するモノクローナル抗体の例である。
【図8】単量体LFB1、LFB2、LFB3、およびリツキシマブ抗体による、IIA1.6-huFcγRIIB細胞上に発現されるヒトFcγRIIBに対するAT10-FITC結合の阻害を表す。まずIIA1.6-huFcγRIIB細胞を種々の濃度の単量体LFB1(50μg/ml〜1000μg/ml)または単量体LFB2(50μg/ml〜250μg/ml)または単量体LFB3(50μg/ml〜1000μg/ml)または単量体リツキシマブ(50μg/ml〜1000μg/ml)と共に、次いで0.25μg/ml AT10-FITCと共にインキュベートする。続いて、競合物としての単量体抗体の存在下または非存在下におけるヒトFcγRIIBに対するFITC標識マウスAT10モノクローナル抗体の結合を、直接免疫蛍光法により評価する。結果およびコメント:単量体LFB1は、IIA1.6-huFcγRIIB細胞上に発現されるヒトFcγRIIBに対するAT10-FITC結合の用量依存的阻害を誘導する。100μg/mlのLFB1はAT10-FITCによって標識される細胞のほぼ40%という割合の減少を誘導するのに対し、100μg/mlのリツキシマブでは11%の減少しか達成されない。用いたうち最も高い用量(1000μg/ml)では、(1000μg/mlのリツキシマブでの23%と比較して)80%に達する。単量体LFB2は、LFB1によって得られる阻害よりも低い、FcγRIIBに対するAT10-FITC結合の阻害を誘導し、これは単量体リツキシマブによって誘発される阻害と類似している(100μg/mlのこれら抗体を用いた場合、AT10-FITCによって標識される細胞の約10%の減少が認められる)。1000μg/mlのLFB3を用いた場合、AT10-FITCによって標識される細胞の減少の割合は20%未満であり、単量体LFB3によって誘導される阻害は弱い。 したがって、このアッセイにより、単量体型の抗体は、それらを産生する細胞株およびそれらのグリコシル化プロファイルに依存して、FcγRIIBに対して異なる結合能を示すことが示される。
【図9】グリコシル化プロファイル、および競合アッセイ法によって評価したLFB1、LFB2、LFB3、およびリツキシマブのID10、ID30、OD50を要約する。FCγRIIIAを試験した場合に0.02マイクロMというID50、FCγRIIAを試験した場合に0.85マイクロMというID50、およびFCγRIIBを試験した場合に1マイクロMというID50を示すLFB1は、FcγRIIIA、FcγRIIA、およびFcγRIIBに「結合する」モノクローナル抗体の例である。 FCγRIIIAを試験した場合にID50>6マイクロM、FCγRIIAを試験した場合にID50>6マイクロM、およびFCγRIIBを試験した場合にID50>6マイクロMを示すLFB3は、FcγRIIIA、FcγRIIA、およびFcγRIIBに「結合しないかまたはごく弱く結合する」モノクローナル抗体の例である。
【図10】LFB1ヒト抗RhD抗体によるカルシウム動員の阻害を表す。IIA1.6-huFcγRIIB細胞をマウスIgと交差反応するF(ab)'2ウサギ抗ヤギIgG(RAG)(10μg/ml)により、またはF(ab)'2ヤギ抗ヒトIgG(H+L)(GAH)(10μg/ml)およびF(ab)'2ウサギ抗ヤギIgG(RAG)(10μg/ml)の存在下においてLFB1抗体により刺激する。この後者のアッセイにより、FcγRIIB受容体に結合したヒトモノクローナルIgG抗体を介して、IIA1.6-huFcγRIIB細胞によって発現されるマウス表面IgGとヒトFcγRIIBが効率的に架橋され得る。F(ab)'2 RAG(10μg/ml)によって誘導されるBCR媒介性Ca2+流入を、BCR/FcγRIIB架橋後に観察されるCa2+流入と比較する。Fluo-3 AM負荷細胞における細胞内遊離Ca2+レベルを、フローサイトメトリーによってモニターする。1 mM EGTAの存在下において細胞内Ca2+貯蔵からのCa2+放出を測定し(曲線の左部分)、6 mM CaCl2を添加した後に細胞内に流入するCa2+を測定する(曲線の右部分)。結果およびコメント:F(ab)'2ウサギ抗ヤギIgG(RAG)抗体を添加することにより、細胞内貯蔵からのカルシウム放出(曲線の左側のピーク、点線)およびIIA1.6-huFcγRIIB細胞へのカルシウム流入(曲線の右側のピーク、点線)が誘発される。F(ab)'2ヤギ抗ヒトIgG(H+L)(GAH)(10μg/ml)およびF(ab)'2ウサギ抗ヤギIgG(RAG)(10μg/ml)の存在下において、IIA1.6-huFcγRIIB細胞をLFB1抗体(100μg/ml)と共にインキュベートすることにより、B細胞上のFcγRIIBの結合の特徴であるカルシウム流入の強い阻害が得られる(曲線の右側のピーク、黒線)。FcγRIIB阻害機能が誘発される場合に予想される通り、細胞内貯蔵からのカルシウム放出の阻害は認められない(曲線の左側のピーク、黒線)。したがって、この実験により、LFB1モノクローナル抗体は、B細胞活性化および抗体産生細胞への分化の遮断を起こす要因である、リンパ腫B細胞上に発現されるFcγRIIBによって媒介される阻害機能を誘発し得ることが示される。
【図11】LFB1およびLFB2ヒト抗RhD抗体によるIL-2産生の阻害を表す。IIA1.6-huFcγRIIB細胞5×105個を、50μg/ml F(ab)'2ウサギ抗ヤギIgG(RAG)単独で、または50μg/ml F(ab)'2ウサギ抗ヤギIgGおよび横軸に表示した種々の濃度の(10〜100μg/ml)のLFB1またはLFB2抗体で22時間刺激する。RAG F(ab)'2断片はマウスIgGおよびヒトIgGの両方と交差反応し、IIA1.6-huFcγRIIB陽性細胞によって発現されるマウス表面IgGとFcγRIIBに結合したヒトモノクローナルIgG抗体とを架橋させ得る。培養上清中のIIA1.6-huFcγRIIB細胞によるマウスIL-2の分泌を、1μg/mlラット抗マウスIL-2を捕獲抗体としておよび1μg/mlビオチン化ラット抗マウスIL-2を検出抗体として使用するELISAアッセイにより測定する。結果およびコメント:表面IgGを発現するIIA1.6-huFcγRIIB陽性細胞をマウスIgGと交差反応するF(ab)'2ウサギ抗ヤギIgG(RAG)により刺激すると、IL-2の分泌が誘発される(IIA1.6-huFcγRIIB細胞5×105個を22時間培養した後、約380 pg/ml)。マウスIgGおよびヒトIgGの両方と交差反応するRAG F(ab)'2断片の存在下では、種々の用量のLFB1抗体(10μg/ml〜100μg/ml)の存在により、IL-2分泌の用量依存的阻害が誘導される。10μg/mlという低いLFB1抗体により、IL-2産生の約40%の阻害が誘導される。用いたうち最も高い用量のLFB1(100μg/ml)では、IL-2産生の阻害は60%に達する。さらに、LFB2抗体もまた、RAG F(ab)'2の存在下で培養したIIA1.6-huFcγRIIB1細胞によるIL-2産生を阻害する。この抗体10μ/mlの存在によりIL-2の30%阻害が誘導され、100μg/mlのLFB2によりIL-2産生の約50%阻害が誘導されることから、LFB2はIL-2産生の用量依存的阻害を誘導する。 したがってこの実験により、IIA1.6-huFcγRIIB細胞によって発現されるFcγRIIBに結合する、YB2/0細胞によって産生されるLFB1およびCHO細胞によって産生されるLFB2は、B細胞上に発現されるFcγRIIBによって媒介される阻害機能を誘発し得るモノクローナル抗体の例であることが示される。特に、これらの抗体によって、これらの細胞によるサイトカイン産生の遮断がもたらされる。
【図12】LFB3ヒト抗RhD抗体によってではなくLFB1ヒト抗RhD抗体によって起こる、IL-2産生の阻害を表す。IIA1.6-huFcγRIIB細胞5×105個を、50μg/ml F(ab)'2ウサギ抗ヤギIgG(RAG)単独で、または50μg/ml F(ab)'2ウサギ抗ヤギIgGおよび横軸に表示した種々の濃度の(10〜200μg/ml)のLFB1またはLFB3抗体で22時間刺激する。RAG F(ab)'2断片はマウスIgGおよびヒトIgGの両方と交差反応し、IIA1.6-huFcγRIIB陽性細胞によって発現されるマウス表面IgGとFcγRIIBに結合したヒトモノクローナルIgG抗体とを架橋させ得る。培養上清中のIIA1.6-huFcγRIIB細胞によるマウスIL-2の分泌を、1μg/mlラット抗マウスIL-2を捕獲抗体としておよび1μg/mlビオチン化ラット抗マウスIL-2を検出抗体として使用するELISAアッセイにより測定する。結果およびコメント:LFB1抗体(10μg/ml〜200μg/ml)はIL-2分泌の用量依存的阻害を誘導する。10μg/mlという低いLFB1抗体によりIL-2産生の約20%の阻害が誘導され、100μg/mlにより50%に達する阻害が誘導される。用いたうち最も高い用量のLFB1(200μg/ml)では、IL-2産生の阻害は70%に達する。これに反してLFB3抗体は、200μg/mlというような高い用量で使用した場合にでさえ、RAG F(ab)'2断片の存在下で培養したIIA1.6-huFcγRIIB細胞によるIL-2産生を阻害しない(<3%減少)。したがってこの実験により、IIA1.6-huFcγRIIB1細胞によって発現されるFcγRIIBに結合する、YB2/0細胞によって産生されるLFB1モノクローナル抗体は、B細胞上に発現されるFcγRIIBによって媒介される阻害機能を誘発し得るモノクローナル抗体の例であることが示される。特に、これによりB細胞によるIL-2産生の遮断がもたらされる。さらにこの実験により、IIA1.6-huFcγRIIB1細胞によって発現されるFcγRIIBに結合しない、B細胞に融合させたヘテロハイブリドーマK6H6B5由来の細胞によって産生されるLFB3モノクローナル抗体は、B細胞上に発現されるFcγRIIBによって媒介される阻害機能を誘発し得ないモノクローナル抗体の例であることが示される。
【技術分野】
【0001】
本発明は、IgGのヒトII型受容体(FcγRII/CD32)を介して阻害機能を誘発するような、1つまたはいくつかの特定のFc受容体の活性を調節し得る、ヒトIgGのFc領域を含むヒトもしくはキメラもしくはヒト化抗体または分子を作製および選択する方法に関する。
【背景技術】
【0002】
導入
ヒトにおける抗体に基づく治療法は、部分的ヒト抗体(キメラ抗体)、つい最近ではヒト化抗体を作製できる可能性と共に出現した。ヒト化組み換え抗体またはマウス/ヒトキメラ組み換え抗体は、マウス抗体で処置した患者におけるヒト抗マウス抗体(「HAMA」)の出現を防ぎ、HAMAと注入された抗体との免疫複合体の形成による副作用を回避するために開発された。さらに、そのような操作された抗体は、ヒトFcγRに対する結合が向上するために、マウス対応物よりも強力なエフェクター機能を示す。現在、数多くの抗体が例えば癌治療の臨床試験において試験されている。しかし、今日現在、ほんのわずかな抗体しか低用量で有効性を示しておらず、いくつかの治験は時期尚早に打ち切られている。高用量の抗体投与は、主に2つの要因から制限される。第一に、それにより同様に多くの副作用が増強される。第二に、大量の抗体を作製することはやはり難題であり、拡大に伴うコストは必ずしも経済的に存立できるとは限らない。
【0003】
様々な疾患に苦しむ多くの人々に抗体療法を適用する前に、これらの問題に取り組む必要がある。
【0004】
IgGのFc領域は抗体の機能に必須であることが示されている。IgGのFc領域を介したC1qおよび膜FcγRの補充により、細胞傷害性、サイトカインの放出、またはエンドサイトーシス等の種々のエフェクター機能が誘発され得る。例えば、IgG1、IgG2、IgG3、およびIgG4を含むヒトIgGサブクラスのうち最もよく説明されているヒトIgG1は、最も高い抗体依存性細胞傷害(ADCC)を誘発することが示されている。それは、ヒトIgG1が、NK細胞および単球/マクロファージ上に発現されるFcγRと効率的に結合する能力に起因する。さらに、FcγRは、阻害機能の誘発により、免疫調節において重要な役割を担う。
【0005】
FcγRの3つの異なるクラスが、ヒトおよびマウスにおいて明らかにされている。ヒトFcγRには、高親和性FcγRI(CD64)、ならびに低親和性FcγRであるFcγRII(CD32)およびFcγRIII(CD16)が含まれる。3つの遺伝子がFcγRIアイソフォーム(A、B、C)をコードし、3つの遺伝子がFcγRIIアイソフォーム(A1、B1、B2)をコードし、2つの遺伝子がFcγRIIIアイソフォーム(A、B)をコードしている。Fc領域とFcγRとの相互作用の調節を達成できるようにFcドメイン内のアミノ酸配列を改変することにより、IgG抗体の有効性を改善する試みが行われている(例えば、WO99/54572を参照のこと)。
【0006】
生物物理学的研究および分子的研究から、CH1ドメインとCH2ドメインの間のヒンジ領域に位置する、およびIgG1のCH2ドメインのN末端に直接隣接するいくつかのアミノ酸残基、ならびにAsn297位においてCH2ドメインに結合している糖鎖が、FcγR結合において重要な役割を果たすことが示されている。一方では、IgG1残基の共通のセットがすべてのFcγR(I、II、III)に対する結合に関与しているが、FcγRIIおよびFcγRIIIとヒトFcγ1との相互作用を詳細に研究した際に、この共通セットの外側の残基が同定された(Shields et al., J. Biol. Chem., 276, 6591-6604, 2001)。他方では、FcγRとの直接的接触ではないが、Fc上の保存された残基Asn297に結合している糖鎖が、Fc上の受容体結合エピトープの高次構造を安定化している可能性が高い(Radaev et al., J. Biol. Chem., 276, 16469-16477, 2001)。脱グリコシル化が、Fcが開いた高次構造から閉じた高次構造に移行するような、2つのCH2ドメインの相対配向の高次構造変化を引き起こし、FcγRへの結合を妨げることが仮定されている(Redaev & Sun, J. Biol. Chem., 276, 16478-16483, 2001)。継続的に切断されたオリゴ糖を有するIgG1グリコフォームを解析することにより、糖残基の除去によって、結果的に閉じた高次構造を生じるCH2ドメインの相互の接近が可能になることが確認された(Krapp et al., J. Mol. Biol., 325, 979-989, 2003)。
【0007】
操作したIgGグリコフォームが、FcγRIIIの補充を介して最適なADCCを誘発することが示された。最初に、Umanaらは、二分岐複合オリゴ糖(二分岐N-アセチルグルコサミン、GlcNAC)の量を多く含むように操作したIgG1抗体が、親の対応物と比較して強力なADCCを誘発し得ることを提唱した(Umana et al., Nature Biotechnol., 17, 176-180, 1999)が、この主張は意義を唱えられた(Shinkawa et al., J.Biol. Chem, 2002)。
【0008】
そのような矛盾する結果から、所与のFcRの活性化または阻害に関与する実際に最適なオリゴ糖構造を同定することは困難であることが示される。本発明者らは、独特の構造に焦点を当てるよりもむしろ結合の改善に関与するパターンを同定する方がより適切であることを見出した。実際に本発明者らは、類似した構造の範囲内で結合が起こることを認めた。
【0009】
この点において、本発明者らは、Fcドメインに結合した特定のグリコシル化パターンがFc領域-FcRIII受容体相互作用の増強に関与することを実証した。この関連で、本発明者らはYB2/0等の特定の細胞株から改善されたADCC特性を有する抗体の組成物を調製することが可能であることを示し、特定のグリコシル化パターンの役割を実証するWO 01/77181を出願した。そのような抗体は、短いオリゴ糖鎖、弱いシアリル化、ゼロまたは弱いレベルの二分岐GlcNac、および20%〜50%という低レベルのフコースを示す。
【0010】
WO 00/61739においても、フコースの存在または非存在が抗体の活性を調節すると仮定された。ヒトIgG1 N結合型オリゴ糖におけるフコースの欠如は、FcγRIII結合およびADCCを改善することが示された。野生型CHO細胞で産生される同じIgG1と比較して低いフコース含量を示すかまたはフコースが欠如した、YB2/0細胞(Shinkawa et al., J. Biol. Chem., 2002)またはCHO-Lec13細胞(Shields et al., J. Biol. Chem., 277, 26733-26740, 2002)で産生される組み換えヒトIgG1は、細胞傷害性を誘発する能力の亢進を示した。それに反して、ガラクトースとADCCとの相関関係は認められず、二分岐GlcNACの含量はわずかにADCCに影響を及ぼしたに過ぎなかった(Shinkawa et al., J. Biol. Chem., 2002)。
【0011】
本発明者らが出願WO 01/77181に以前に記載したことが上記の結果によって確認されるが、本発明者らは現在、抗体活性の変化は単にフコースレベルで要約し得ないことを示す証拠を有する。GlcNac、マンノース、シアル酸、ガラクトース、およびそれらのそれぞれの位置の関係は、異なる細胞株から産生される抗体において非常に変化しやすい。この知見から、異なるグリコシル化パターンはFc領域-Fc受容体の相互関係に関与するばかりでなく、抗体の機能にも関与することが裏付けられる。
【0012】
したがって、本発明の全体的な目的は、異なるFc受容体に関して所与の抗体の機能を微調整する方法を提供することにある。異なる規定のパターンにより、異なるFcRの調節が可能になる。
【0013】
IgGのFc領域の受容体の中でも、IIB型Fcγ受容体が特に関心の対象である。FcRIIは、免疫複合体中に存在するかまたは細胞表面抗原に結合したIgGと親和性を増して結合する一本鎖、低親和性受容体である。遺伝子解析およびタンパク質解析から、FcγRIIBは、ヒトでは、受容体の細胞質内ドメインをコードする3つのエキソンの選択的スプライシングによって生じる2つのアイソフォーム(FcγRIIB1およびFcγRIIB2)で存在する。FcγRIIBは、B細胞、単球、樹状細胞、肥満細胞、および好塩基球上に発現される。他のすべての既知FcγRとは対照的に、FcγRIIBアイソフォームは、活性化受容体を介して誘導される細胞活性化を阻害する(Amigorena et al., Science, 256, 1808-1812, 1992)。分子的解析により、FcγRIIBの細胞内ドメイン内の「免疫受容体チロシンに基づく阻害性モチーフ」(ITIM)と称されるYxxLモチーフが、受容体の阻害機能に直接関与していることが示された。チロシンがリン酸化されると、ITIMは、ホスファターゼ、すなわちSHP-1、SHP-2、およびホスファチジルイノシトールポリリン酸5-ホスファターゼ、SHIPのSH2ドメインと結合する。SHIPは、PIP3を脱リン酸化してPI(3,4)P2にし、ホスホリパーゼC-γ(PLCγ)をリン酸化するキナーゼbtkの補充を妨げ得る。次いで、イノシトール(1,4,5)三リン酸(IP3)の生成が阻止される。これにより、活性化受容体の補充によって誘発される初期事象である、細胞外カルシウムの流入が阻害される。サイトカインの産生または細胞の増殖等の後期事象もまた阻害される(O. Malbec et al., 「Immunoreceptor Tyrosine-based Inhibition Motifs」、M. Daeron & E, Vivier eds, 1999, Springer-Verlag. Berlin, pp. 13-27)。
【0014】
FcγRIIに依存した負の制御の生物学的重要性は、多くの著者によって検討されている。抗体応答の負の制御においては他の機構も同時に起こる可能性が高いものの、抗体応答の負の制御はFcγRIIBの補充を介して達成され得ることが示されている。したがって、抗体に基づく自己免疫疾患または病原性抗体の出現をもたらす望ましくない同種免疫が、FcγRIIBに結合し得る抗体または他の分子によるFcγRIIBの最適な補充を介して回避され得ることが示唆され得る。
【0015】
さらに、アレルゲン複合化IgGによる肥満細胞および好塩基球上に存在するFcγRIIBの補充、およびIgEが同じアレルゲンに複合化した時点での、次に起こるFcγRIIBとFcεRIとの共凝集は、これらの細胞によるアナフィラキシー反応の遮断に関与する。したがって、FcγRIIBと結合し、それ故にFcγRIIB阻害機能を誘発する最適な能力を有するIgG抗体は、アレルギーの治療における有効な治療手段を表し得る。
【0016】
最終的には、FcγRIIBの補充によって誘発される阻害機能は、癌患者において利用され得る。一方では、免疫不全FcγRIIB-/-マウスに腫瘍を移植し、それぞれ抗CD-20もしくは抗HER2/Neuのキメラ抗体またはヒト化抗体等の組み換えモノクローナル抗体で処置したモデルにおいて、FcγRIIBノックアウトマウスはより優れた抗腫瘍反応を示すことが示された。したがって、強力なFcγRII阻害機能を誘発し得ないモノクローナルIgG抗体は、腫瘍治療における最適な手段となる可能性がある。他方では、FcγRIIbは受容体チロシンキナーゼ(RTK)に依存して造血細胞増殖を負に制御する。したがって、RTKとFcγRIIBを効率的に共凝集させ得る最適なFc領域を有する抗RTK IgG抗体等の二機能性分子は、FcγRIIB+腫瘍細胞の増殖を阻止する。
【0017】
FcγRII/ヒトIgG相互作用に及ぼすグリコシル化の影響はまた、可溶性FcγR外部ドメインを用いても実証されている。マウスFcγRIIおよびヒトFcγRIIBのどちらに関しても、それぞれ超遠心および等温滴定熱量計(ITC)を用いた沈降平衡法から1:1化学量論が推定された(Mimura et al., J. Biol. Chem., 276, 45539-45547, 2001;Kato et al., J. Mol. Biol., 295, 213-224, 2001)。Asn297と主要なGlcNAC残基との相互作用が、ヒトFcγRIIによるヒトキメラIgG3の認識に重要であることが示された(Lund et al., FASEB J., 116, 115-119, 1995)。α(1-6)マンノースアーム上のガラクトースの接触残基の置換はFcγRII認識に影響を及ぼさないのに対し、コアGlcNAC残基との接点であるAsp265の置換はFcγRII認識の喪失をもたらす(Jefferis et al., Immunol. Letters, 44, 111-117, 1995)。さらに別の研究から、GlcNAC残基はヒトIgG1の受容体結合にわずかに寄与するのみである一方、α(1-3)およびα(1-6)アームマンノース残基を除去することによって親和性が有意に減少することが示された(Mimura et al., J. Biol. Chem., 276, 45539-45547, 2001)。同じ研究では、オリゴ糖の切断によって2つのCH2ドメインの閉じた配置がもたらされ、それに伴ってFcγIIBへの結合が減少すると結論づけた。FcγRIIIとは対照的に、CHO-Lec13細胞で産生された、故にフコースを欠く抗体は、可溶性固定化Arg131-FcγRIIA多型および可溶性FcγRIIB型に対する結合においてわずかな改善を示したが、可溶性His131-FcγRIIA多型に対する結合では改善は認められなかった(それらはいずれも細胞外ドメインおよび膜貫通ドメインに相当する)。前者の受容体は131位にアルギニンを有するため、フコースがこの位置のFcγII残基と直接結合するか、またはその存在によってFcγRII結合に対するわずかな弊害が誘導されるようにIgG1高次構造を変化させると仮定された(Shields et al., J. Biol. Chem., 277, 26733-26740, 2002)。同じ研究から、ガラクトース含量はFcγRIIに対する結合に影響を及ぼさないことが示唆された。
【0018】
本明細書において、本発明者らは、標的および疾患に応じて微調整し得る薬剤を作製するために、異なる結合プロファイルを示す種々のクラスの抗体を調製することを提案する。そのような抗体の調製法により、有効性が増強された、ならびに/または免疫調節機能およびFcγRに対する特異性が増強された抗体を産生するのに適した細胞株を同定することが初めて可能になる。本発明はまた、特定の結合プロファイルに関与する抗体のグリカン構造、およびそのような特定の抗体の作製法を提供する。
【発明の開示】
【0019】
本発明は、異なるFcR、特にFcγRIIB、FcγRIIA、およびFcγRIIIAを使用する結合アッセイ法-免疫蛍光法-による、規定されたグリコシル化パターンを有するヒトIgGのFc領域を含むヒト組み換え抗体またはキメラ抗体またはヒト化抗体もしくは分子を作製および選択する方法を提供する。例えば、本方法は、カルシウム動員アッセイ法およびサイトカイン分泌アッセイ法等の機能アッセイ法による、IgGのFc領域に対するヒトIIB型受容体(FcγRIIB/CD32B)を介して阻害機能を特異的に誘発し得る規定のグリコシル化パターンを有するヒト組み換え抗体または抗体由来分子の選択に適用し得る。
【0020】
本発明は、特殊化したかつ微調整された抗体をもたらす規定の培地中で培養した種々の細胞株による、異なるFcグリコシル化パターンを有する組み換えヒト抗体、ヒト化抗体、またはキメラ抗体の産生に基づく。
【0021】
より具体的には、本発明は、B細胞、単球もしくは他の抗原提示細胞(APC)、肥満細胞、好塩基球、または形質転換細胞および腫瘍細胞等の任意の他のFcγRIIB+細胞等のFcγRIIB+細胞(すなわち、FcγRIIBを発現している細胞)上のFcγRIIBによってもたらされる負の制御のオンまたはオフを可能にする、異なるFcグリコシル化パターンによって決定される特定の構造特性を有する抗体の作製および選択に関する。
【0022】
本発明は、抗体応答を調節するための有用な治療手段となると考えられる最適な抗体に関する。これにより、抗体に基づく自己免疫疾患または特に注入された組み換え抗体に対する病原性抗体の出現をもたらす望ましくない同種免疫において、FcγRIIBの補充を介して免疫調節阻害機能を誘発し得る、FcγRIIBに対する結合に関して最適化された任意の特異性の抗体または他の分子の作製および選択が可能になる。
【0023】
本発明は、抗体に基づいた治療において抗体の有効性を調節する有用な治療手段となると考えられる任意の特異性の最適な抗体に関する。それにより、弱いFcγRIIB結合を有するかまたはFcγRIIB結合のない抗体を作製および選択し、治療抗腫瘍抗体による癌治療等の抗体に基づく治療処置過程において、FcγRIIBを介した免疫調節阻害機能の誘発を妨げることが可能になる。
【0024】
第1の局面において、本発明は、ヒトIgGのFc領域を含みかつ異なる結合プロファイルを有する、ヒトの、ヒト化された、もしくはキメラの抗体またはポリペプチドを調製する方法であって、以下からなる段階を含む方法を対象とする:
(a) ハイブリドーマ、ヘテロハイブリドーマ、EBV形質転換ヒトB細胞株を含む動物細胞株に由来もしくは真核微生物に由来する細胞により天然に産生される、または、該抗体もしくはポリペプチドのコード配列を含むベクターによるそのトランスフェクション後に産生される、ヒトIgGのFc領域を含む、ヒトの、ヒト化された、もしくはキメラの抗体候補またはポリペプチド候補を提供する段階;
(b) FcγRIIIA、FcγRIIA、およびFcγRIIBを含むFcγ受容体において該抗体またはポリペプチドの結合を試験する段階;
(c)
i) FcγRIIIA、FcγRIIA、およびFcγRIIBのいずれにも結合する、または
ii) FcγRIIAおよびFcγRIIBには結合するが、FcγRIIIAには結合しないかまたはごく弱く結合する、または
iii) FcγRIIIA、FcγRIIA、およびFcγRIIBのいずれにも結合しないかまたはごく弱く結合する
抗体またはポリペプチドを選択する段階。
【0025】
FcγRIIIA、FcγRIIA、およびFcγRIIBに対するこれらの抗体の結合は、それらの定常領域(すなわち、CH1、CH2、CH3)を介する、より具体的にはそれらのFc領域を介する。これらの抗体は、それらの可変領域を介して、すなわちそれらの抗原結合領域を介してまたはそれらの相補性決定領域(CDR)を介して任意の種類の特異性を示し得る。
【0026】
本明細書において規定する手段によって調製される抗体は、それぞれ特定の結合プロファイルを有し、結果的に異なる特性を有する3つクラスのうちの1つに属し、各クラスを特定の明確な治療目的のために使用することが可能になる。
【0027】
第1クラスに属する抗体またはポリペプチドは、上記の方法の段階c) i)を用いて調製し得る。それらはFcγRIIIAに結合し得るため、この受容体を介してADCCを誘導し得る。さらに、それらはFcγRIIAに結合し得るため、FcγRIIA陽性単球およびマクロファージを介してADCCおよび食作用を誘導し得る。さらに、このクラスの抗体またはポリペプチドはまた、それらが結合し得るFcγRIIBを介して阻害機能を誘発し得る。したがって、これらの抗体はFcγRIIBを介して免疫調節機能を発揮し得る。これは、そのような抗体が免疫系の細胞または腫瘍細胞を負に免疫調節する能力に関する。そのような能力は、多くの疾患においておよび移植、同種免疫、または抗体に基づく処置等の種々の臨床状況において主に重要である。
【0028】
したがって、このクラスに属する抗体またはポリペプチドは、細胞傷害性が高い抗体(またはポリペプチド)であり、かつ強力な免疫調節抗体(またはポリペプチド)である。
【0029】
第2クラスの抗体またはポリペプチドは、上記の方法の段階c) ii)を用いて調製し得る。それらは、FcγRIII、特にFcγRIIIAに対する結合が弱いために第1クラスに属する抗体と比較して細胞傷害性が乏しいことから、関心対象となる。より詳細には、それらはポリクローナル多価IgG(静注用IgG、IVIg)の存在下において細胞傷害性を示さない。それらは、FcγRIIAに結合する能力により、FcγRIIA陽性単球およびマクロファージを介してADCCおよび食作用を誘導し得る。さらに、それらはFcγRIIBと結合する能力のために免疫調節抗体である。
【0030】
最後に、第3クラスの抗体は上記の方法の段階c) iii)を用いて調製し得る。このクラスの抗体は細胞傷害性を示さず、また免疫調節活性を全く提供しない。より詳細には、それらはポリクローナル多価IgG(静注用IgG、IVIg)の存在下において細胞傷害性を示さない。それらは、FcγRIIIA、FcγRIIA、およびFcγRIIBに結合できないかまたは弱く結合するために、FcγR補充を介して達成されるエフェクター機能の誘発を起こさずインビボで使用し得ることから、関心対象となる。
【0031】
本発明の方法により、どのFcγR(IIIA、IIA、IIB)を標的するかに応じて、特定のまたは所望の治療プロファイルを有する抗体を調製することが可能となる。
【0032】
有利には、本発明による方法は、競合物として単量体抗体を用いる競合アッセイ法に基づく。
【0033】
競合物として単量体抗体を用い得る競合アッセイ法は、免疫蛍光競合アッセイ法である。
【0034】
別の態様において、競合物として単量体抗体を用い得る競合アッセイ法は、直接的または間接的免疫蛍光競合アッセイ法である。
【0035】
本アッセイ法において、ID50とは、蛍光色素と結合させた抗FcγR抗体を参照して、FcγR陽性指標細胞の染色の50%減少をもたらす被験抗体の用量(マイクロMで表す)である。
【0036】
本アッセイ法において、ID30とは、蛍光色素と結合させた抗FcγR抗体を参照して、FcγR陽性指標細胞の染色の30%減少をもたらす被験抗体の用量(マイクロMで表す)である。
【0037】
本アッセイ法において、ID10とは、蛍光色素と結合させた抗FcγR抗体を参照して、FcγR陽性指標細胞の染色の10%減少をもたらす被験抗体の用量(マイクロMで表す)である。
【0038】
「結合する」という表現は、マウスモノクローナル抗体蛍光色素標識3G8を用いた競合アッセイにおいて0.2マイクロMより低いID50、または0.1マイクロMより低い、有利には0.05マイクロMより低い、さらにより有意には0.02マイクロM未満のID50を有する、FcγRIIIAに結合する単量体型の抗体を意味する。3G8とはFcγRIIIに対する抗体であり、当技術分野において周知である(3G8-PE、Immunotech、フランス、カタログ番号A07766)。
【0039】
「結合する」という表現は、マウスモノクローナル抗体蛍光色素標識AT10を用いた競合アッセイにおいて6マイクロMより低いID50、または2マイクロMより低い、有利には1マイクロMより低い、さらにより有意には0.85マイクロM未満のID50を有する、FcγRIIAに結合する単量体型の抗体を意味する。AT10とはFcγRIIAおよびFcγRIIBに対する抗体であり、当技術分野において周知である(AT10-FITC、Serotec、カタログ番号MCA 1075F)。
【0040】
「結合する」という表現は、マウスモノクローナル抗体蛍光色素標識AT10を用いた競合アッセイにおいて6マイクロMより低い、またはより有利には2マイクロMより低い、さらにより有意には1マイクロMより低いID50を有する、FcγRIIBに結合する単量体型の抗体を意味する。AT10とはFcγRIIAおよびFcγRIIBに対する抗体であり、当技術分野において周知である(AT10-FITC、Serotec、英国、カタログ番号MCA 1075F)。
【0041】
「結合しないかまたはごく弱く結合する」という表現は、マウスモノクローナル抗体蛍光色素標識3G8を用いた競合アッセイにおいて0.2マイクロM以上の、有利には0.5マイクロMを超える、より有利には1マイクロMを超えるもしくは2マイクロMを超える、最も有利には10マイクロMを超えるID50を有する、FcγRIIIAに結合する単量体型の抗体を意味する。
【0042】
「結合しないかまたはごく弱く結合する」という表現は、マウスモノクローナル抗体蛍光色素標識AT10を用いた競合アッセイにおいて6マイクロM以上の、有利には9マイクロMを超える、より有利には15マイクロMを超えるもしくは20マイクロMを超える、最も有利には50マイクロMを超えるID50を有する、FcγRIIAに結合する単量体型の抗体を意味する。
【0043】
「結合しないかまたはごく弱く結合する」という表現は、マウスモノクローナル抗体蛍光色素標識AT10を用いた競合アッセイにおいて6マイクロM以上の、有利には9マイクロMを超える、より有利には15マイクロを超えるもしくは20マイクロMを超える、最も有利には50マイクロMを超えるID50を有する、FcγRIIBに結合する単量体型の抗体を意味する。
【0044】
本明細書で用いるFcγRIIIA、FcγRIIA、およびFcγRIIBに対する結合とは、抗体調製物中に低い割合の凝集物(<2%)しか存在せず、抗体が熱もしくは化学的凝集等の任意の手段により、または関連抗原もしくはF(ab')2抗ヒトIgGとの免疫複合体の形成により故意に複合化されていない場合の抗体の結合を指す。
【0045】
別の態様において、本方法は複合化抗体を用いた結合アッセイ法に基づく。
【0046】
結合アッセイ法は、複合化抗体を用いた免疫蛍光結合アッセイ法であってよい。
【0047】
有利には、結合アッセイ法は、複合化抗体を用いる直接的または間接的免疫蛍光結合アッセイ法である。
【0048】
本明細書で用いるFcγRIIIA、FcγRIIA、およびFcγRIIBに対する結合とは、F(ab')2抗ヒトIgGと複合化した抗体の結合を指す。
【0049】
本発明によると、複合化抗体を用いる免疫蛍光結合アッセイ法を用いる方法における「結合する」という表現は、FcγRIIIA陽性指標細胞に対する複合化抗体(それぞれ500、250、および100 ng/mlで使用)の結合を測定する免疫蛍光アッセイ法において、少なくとも30%、15%、5%の割合の陽性指標細胞をもたらす抗体を意味する。
【0050】
本発明によると、複合化抗体を用いる免疫蛍光結合アッセイ法を用いる方法における「結合する」という表現は、FcγRIIA陽性指標細胞に対する抗体(それぞれ10、5、および1マイクロg/mlで使用)の結合を測定する免疫蛍光アッセイ法において、少なくとも70%、60%、30%の割合の陽性指標細胞をもたらす抗体を意味する。
【0051】
本発明によると、複合化抗体を用いる免疫蛍光結合アッセイ法を用いる方法における「結合する」という表現は、FcγRIIB陽性指標細胞に対する抗体(それぞれ50および25マイクロg/mlで使用)の結合を測定する免疫蛍光アッセイ法において、少なくとも50%、25%の割合の陽性指標細胞をもたらす抗体を意味する。
【0052】
複合化抗体を用いる免疫蛍光結合アッセイ法を用いる方法における「結合しないかまたはごく弱く結合する」という表現は、FcγRIIIA陽性指標細胞に対する複合化抗体(それぞれ500、250、および100 ng/mlで使用)の結合を測定する免疫蛍光アッセイ法において、30%、15%、5%よりも低い割合の陽性指標細胞をもたらす抗体を意味する。
【0053】
複合化抗体を用いる免疫蛍光結合アッセイ法を用いる方法における「結合しないかまたはごく弱く結合する」という表現は、FcγRIIA陽性指標細胞に対する抗体(それぞれ10、5、および1マイクロg/mlで使用)の結合を測定する免疫蛍光アッセイ法において、70%、60%、30%よりも低い割合の陽性指標細胞をもたらす抗体を意味する。
【0054】
複合化抗体を用いる免疫蛍光結合アッセイ法を用いる方法における「結合しないかまたはごく弱く結合する」という表現は、FcγRIIB陽性指標細胞に対する抗体(それぞれ50および25マイクロg/mlで使用)の結合を測定する免疫蛍光アッセイ法において、少なくとも30%、10%よりも低い割合の陽性指標細胞をもたらす抗体を意味する。
【0055】
有利には、段階c) i)で選択される抗体またはポリペプチドは、リンパ球系(lymphoid)細胞株またはリンパ球系細胞由来(lymphoid-derived)細胞株またはハイブリドーマまたは腎臓上皮細胞株由来の細胞により産生され、段階c) ii)で選択される抗体またはポリペプチドは非リンパ球系(non-lymphoid)細胞株由来の細胞により産生され、段階c) iii)で選択される抗体またはポリペプチドは、EBV形質転換細胞株由来の細胞にまたはヒトドナー由来のBリンパ球に融合させたヘテロハイブリドーマ由来の細胞により産生される。
【0056】
「リンパ球系細胞株」という用語は、任意の手段により長期インビトロ培養で培養したヒトリンパ球に由来する細胞を指す。具体的には、そのような細胞株はEBV形質転換により取得することができる。「リンパ球系細胞由来細胞株」という用語は、リンパ球系細胞株由来の細胞とパートナー細胞との融合のような任意の手段によるリンパ球系細胞株に由来する細胞を指す。そのようなパートナー細胞は、ヒトドナーに由来するBリンパ球であってよい。「ハイブリドーマ」という用語は、B細胞と骨髄腫細胞との融合に由来する細胞を指す。「ヘテロハイブリドーマ」という用語は、マウス骨髄腫細胞由来の細胞と、結節型リンパ腫患者によるヒト悪性リンパ球との融合によって得られる細胞を指す。
【0057】
本明細書に述べるように、同定された抗体またはポリペプチドは、上記の関連特性を示す抗体または抗体由来分子を産生し得る同定された細胞株を包含する。
【0058】
本発明の特定の局面において、リンパ球系細胞由来細胞株はラット骨髄腫細胞株もしくはハイブリドーマYB2/0細胞株(ATCC番号CRL-1662)もしくはそれらに由来する細胞株であり、かつ/または、腎臓上皮細胞株はVERO(ATCC番号CCL-81)またはそれに由来する細胞であり、かつ/または、非リンパ球系細胞株はCHO(ATCC番号CCL-61)またはそれに由来する細胞であり、かつ/または、ヘテロハイブリドーマはK6H6B5(ATCC番号CRL-1823)またはそれに由来する細胞株である。
【0059】
CHO細胞株はCHO dhfr-細胞株(コロンビア大学、米国、ニューヨーク州)であってよい。
【0060】
本発明の好ましい態様において、結合アッセイは以下を用いて行われ得る:
i) 細胞表面上に異なるFc受容体を発現する細胞株由来の指標細胞、
ii) FcγR外部ドメインを含む組み換えFc受容体、Fc受容体由来ペプチド。
【0061】
指標細胞は、例えば(FcγRIIIAを発現している)Jurkat-CD16、(FcγRIIAを発現している)CD32陽性K562(ATCC番号CCL-243)、または(FcγRIIBを発現している)CD32陽性IIA1.6であってよい。
【0062】
「FcγR外部ドメインを含むFc受容体」という用語は、2つの外部ドメイン(D1およびD2)からなるFcγ受容体IIA、IIB、およびIIIA型の細胞外領域を指す。
【0063】
好ましい態様においては、FcγRIIIA、FcγRIIA、およびFcγRIIBのいずれにも結合する能力に関して選択された抗体またはポリペプチドのサブセット、ならびにFcγRIIAおよびFcγRIIBの両方に結合する能力に関して選択された抗体またはポリペプチドのサブセットは、i) FcγRIIIAを誘発してADCCの改善、インターロイキン-2(IL-2)等のサイトカインおよび腫瘍壊死因子α(TNFα)等の炎症促進性分子の産生の増加をもたらす能力;およびii) FcγRIIBを誘発してB細胞および単球等のFcγRIIBを発現する細胞によるカルシウム動員の阻害およびIL-2等のサイトカイン産生の阻害をもたらす能力に関する機能アッセイにより、さらに試験および選択される。
【0064】
有利には、機能アッセイ法は、カルシウム動員阻害アッセイ法および/またはサイトカイン分泌阻害アッセイ法からなる。
【0065】
本発明に準じて第1および第2クラスに属する抗体の活性には、表面FcγRIIBに結合したそのような抗体とBCR、TCR、FcεRI等の活性化表面受容体との共凝集によるカルシウム動員の阻害が含まれる。本明細書で使用するカルシウム動員とは、BCR等の活性化受容体の補充後の細胞内への細胞外カルシウムの流入を指す。
【0066】
本発明の抗体の活性には、表面FcγRIIBに結合したそのような抗体とBCR等の活性化表面分子との共凝集によるサイトカイン産生の阻害が含まれる。
【0067】
本明細書において使用するサイトカイン産生とは、BCR等の活性化受容体の補充後の細胞によるサイトカインの放出を指す。
【0068】
本発明の分子によるサイトカイン産生の阻害には、インターロイキン-2(IL-2)産生の阻害が含まれる。
【0069】
別の態様において、機能アッセイ法には特異的FcγRIIIA ADCCアッセイ法がさらに含まれ得る。
【0070】
基本的に、この特異的FcγRIIIA ADCCアッセイ法は、各抗体またはポリペプチドを、標的細胞、FcγRIIIAを発現しているエフェクター細胞、および多価IgGを含む異なる反応混合物に添加する段階、および標的細胞の溶解の割合を決定する段階を含む。
【0071】
本発明の別の目的は、FcγRIIIA、FcγRIIA、およびFcγRIIBに結合し得る抗体を産生させるための、リンパ球系細胞株またはリンパ球系細胞由来細胞株またはハイブリドーマまたは腎臓上皮細胞株に由来する細胞の使用である。
【0072】
「ハイブリドーマ」という用語は、上記のハイブリドーマと同じ意味を有する。有利には、ハイブリドーマはリンパ球に融合させたラット骨髄腫細胞に由来し得る。
【0073】
YB2/0ハイブリドーマ細胞により産生される抗体の、FcγRIIIA、FcγRIIA、およびFcγRIIBに対する結合に対するID10、ID30、およびID50の値を図9に要約するが、これにより3つの型の受容体に関するこれらの抗体の強力な結合が示される。
【0074】
本発明の特定の態様において、リンパ球系細胞株またはリンパ球系細胞由来細胞株またはハイブリドーマまたは腎臓上皮細胞株に由来する細胞によって産生される抗体は免疫調節抗体である。
【0075】
ハイブリドーマ細胞、より詳細にはB細胞とラット骨髄腫細胞との融合によって生じるハイブリドーマ細胞により産生される抗体はFcγRIIBに結合し、ひいてはFcγRIIBを介して細胞機能の阻害を誘発し得る。
【0076】
本発明によれば、ハイブリドーマ細胞、好ましくはB細胞とラット骨髄腫細胞との融合によって生じるハイブリドーマ細胞により産生される抗体は、上記の第1クラスの抗体に属する。結果として、それらはまた上記の方法の段階c) i)によって選択され得る。
【0077】
リンパ球系細胞株またはリンパ球系細胞由来細胞株またはハイブリドーマまたは腎臓上皮細胞株に由来する細胞によって産生されるこれらの抗体は、免疫調節かつ細胞傷害性の抗体である。
【0078】
ハイブリドーマ細胞、好ましくはB細胞とラット骨髄腫細胞との融合によって生じるハイブリドーマ細胞により産生されるこれらの免疫調節抗体は、FcγRIIIAおよびFcγRIIAに結合し得る。これらの受容体は活性化受容体であり、細胞傷害活性を誘発する。
【0079】
好ましい態様において、ハイブリドーマ細胞株はYB2/0である。
【0080】
別の好ましい態様において、腎臓上皮細胞株はVEROである。
【0081】
本発明の別の目的は、FcγRIIIAには結合しないかまたはごく弱く結合するが、FcγRIIAおよびFcγRIIBの両方には結合する抗体を産生させるための、非リンパ球系細胞株由来の細胞の使用である。
【0082】
本出願人らは本出願において、非リンパ球系細胞株によって産生される抗体は、FcγRIIIAには結合しないとは言えないにしても弱く結合するが、FcγRIIAおよびFcγRIIBには結合することを示す。図9に細胞株CHOによって産生される抗体のID10、ID30、およびID50の値を要約するが、これによりFcγRIIIAに対するこれらの抗体の弱い結合ならびにFcγRIIAおよびFcγRIIBに対するこれらの抗体の結合が示される。
【0083】
非リンパ球系細胞株由来の細胞により産生されるこれらの抗体は、免疫調節抗体である。
【0084】
この免疫調節活性は、FcγRIIBに対するこれらの抗体の結合に起因する。
【0085】
さらに、非リンパ球系細胞株により産生されるこれらの抗体は、FcγRIIAを発現している単球およびマクロファージによるADCCおよび食作用を誘導する。
【0086】
上記の第2クラスの抗体に属するこれらの抗体はまた、上記の方法の段階c) ii)によって調製または選択され得る。
【0087】
好ましい態様において、非リンパ球系細胞株はCHO(ATCC番号CCL-61)である。
【0088】
別の態様において、CHO細胞株はCHO dhfr-(コロンビア大学、米国、ニューヨーク州)であってよい。
【0089】
本発明のさらに別の目的は、FcγRIIIA、FcγRIIA、およびFcγRIIBのいずれにも結合しないかまたは弱く結合する抗体を産生させるための、EBV形質転換細胞株由来の細胞とまたはB細胞と融合させたヘテロハイブリドーマ由来の細胞の使用である。
【0090】
本出願人らは本出願において、B細胞と融合させたヘテロハイブリドーマによって産生される抗体が、FcγRIIIA、FcγRIIA、およびFcγRIIBに弱く結合することを示す。図9にB細胞と融合させたヘテロハイブリドーマK6H6B5由来の細胞によって産生される抗体のID10、ID30、およびID50の値を要約するが、これによりFcγRIIIA、FcγRIIA、およびFcγRIIBに対するこれらの抗体の弱い結合が示される。
【0091】
有利には、EBV形質転換細胞株由来の細胞とまたはB細胞と融合させたヘテロハイブリドーマ由来の細胞によって産生されるこれらの抗体は、IgG4の使用に代わる治療法として用いられる。
【0092】
IgG4は、FcγRIIIA、FcγRIIA、およびFcγRIIBを介したエフェクター機能の補充を回避するために用いられる。本発明に従って、B細胞またはEBV形質転換B細胞に融合させたヘテロハイブリドーマから産生される抗体は、IgG1アイソタイプに属する。これらは、特定の疾患の治療過程において、FcγRIIIA、FcγRIIA、およびFcγRIIBの補充を介して達成されるエフェクター機能を誘発することなく、FcγRおよびFcεRI結合によって誘発される重篤な副作用の発症を妨げてインビボで使用し得る。
【0093】
好ましい態様において、これらの細胞はヒトB細胞に融合させたヘテロハイブリドーマK6H6B5(ATCC番号CRL-1823)に由来する細胞である。
【0094】
本発明のさらに別の目的は、10%〜55%のフコース、および60%〜98%のガラクトースを含む、ヒトIgGのFc領域を含む抗体またはポリペプチドである。
【0095】
10%〜55%のフコース、および60%〜98%のガラクトースを含むこの抗体またはポリペプチドは、FcγRIIIA、FcγRIIA、およびFcγRIIBのいずれにも結合する。
【0096】
これらの2つの特性(Fcグリコシル化プロファイルおよびFcγR結合プロファイル)は共に関連している。本出願人らは、これら2つの特性がまた、このクラスの抗体がハイブリドーマ細胞株によって産生され得るという事実にも関連していることを示す。
【0097】
この抗体またはポリペプチドは、リンパ球系細胞株またはリンパ球系細胞由来細胞株またはハイブリドーマまたは腎臓上皮細胞株に由来する細胞によって産生され得る。
【0098】
有利には、このハイブリドーマ細胞株は、AOラットの脾臓細胞と融合させた骨髄腫細胞株、LOU/C Y3/Ag 1.2.3に由来するハイブリドーマ細胞株、YB2/3HLに由来するYB2/0細胞株(ATCC番号CRL-1662)である。
【0099】
別の態様において、この抗体またはポリペプチドは腎臓上皮細胞株VERO(ATCC番号CCL-81)によって産生される。
【0100】
さらに、10%〜55%のフコース、および60%〜98%のガラクトースを含むこの抗体またはポリペプチドは、上記の調製過程の段階c) i)により取得することができる。
【0101】
これらの抗体またはポリペプチドは上記の第1クラスに属する。
【0102】
本発明はまた、この第1クラスの抗体またはポリペプチドを少なくとも80%、好ましくは少なくとも95%含む組成物を対象とする。
【0103】
好ましい態様において、組成物は上記の抗体またはポリペプチドを少なくとも80%ないし90%、好ましくは少なくとも95%ないし99%含み得る。
【0104】
白血病、リンパ腫、骨髄腫、セザリー症候群、もしくは固形腫瘍等の癌、自己免疫疾患、アレルギー、移植後の同種免疫、母子間同種免疫、移植片対宿主(GVH)反応、または感染症の治療用の医薬品を製造するためにこの組成物を使用することも本発明のさらなる対象である。
【0105】
第1クラスの抗体は、標的細胞、例えば癌細胞を死滅させるために、(FcγRIIIAおよびFcγRIIAに結合する能力に起因して)細胞傷害性細胞を活性化し得る。
【0106】
そのような抗体は、FcγRIIBにより受容体チロシンキナーゼ(RTK)に依存して、造血細胞増殖の制御を誘発するための有用な治療手段になると考えられる。RTKとFcγRIIBを効率的に共凝集させ得る最適なFc領域を有する抗RTK IgG抗体のような二機能性分子は、FcγRIIB+腫瘍細胞の増殖を阻止し得る。
【0107】
本発明の特定の局面において、この組成物を用いて治療する母子間同種免疫は新生児溶血性疾患(HDNB)である。
【0108】
このクラスの抗体は赤血球を死滅させ得り、それと同時に赤血球に対する抗体を産生するB細胞を免疫調節し得る。
【0109】
本発明の別の特定の局面において、自己免疫疾患は、全身性エリテマトーデス(SLE)、特発性血小板減少性紫斑病(ITP)、川崎症候群等の自己抗体を産生するB細胞関連自己免疫疾患である。
【0110】
別の好ましい局面において、治療するアレルギーは、喘息、アレルギー性鼻炎、アレルギー性副鼻腔炎、アナフィラキシー症候群、じんま疹、血管性水腫、アトピー性皮膚炎、アレルギー性接触皮膚炎、および紅斑である。
【0111】
本発明のさらに別の目的は、70%〜100%、例えば96%〜99%のフコース、および60%〜98%、例えば74%〜89%のガラクトースを含むことを特徴とする、ヒトIgGのFc領域を含む抗体またはポリペプチドである。
【0112】
70%〜100%のフコースおよび60%〜98%のガラクトースを含むこの抗体またはポリペプチドは、FcγRIIAおよびFcγRIIBの両方に結合するが、FcγRIIIAには結合しないかまたはごく弱く結合する。
【0113】
本出願人らは、結合プロファイルがFcグリコシル化プロファイルと関連していることを示す。さらに本出願人らは、この抗体またはポリペプチドが非リンパ球系細胞株由来の細胞によって産生されることを示す;よって、結合プロファイル、グリコシル化プロファイル、および抗体産生細胞株は関連している。
【0114】
より詳細には、この非リンパ球系細胞株はCHO細胞株(ATCC番号CCL-61)である。
【0115】
特に関心対象である局面において、この抗体またはポリペプチドは上記の調製過程の段階c) ii)によって取得することができる。
【0116】
これらの抗体またはポリペプチドは、上記の第2クラスの抗体に属する。
【0117】
本発明はまた、上記の第2クラスの抗体を少なくとも80%、好ましくは少なくとも95%含む組成物を包含する。
【0118】
好ましい態様において、組成物は上記の第2クラスに属する抗体またはポリペプチドを少なくとも80%〜90%、好ましくは少なくとも95%ないし99%含み得る。
【0119】
本発明はまた、自己免疫疾患、母子間同種免疫、および炎症性疾患の治療用の医薬品を製造するためのこの組成物の使用に関する。
【0120】
これらの最適化された抗体は、アレルゲン複合化IgGにより、肥満細胞および好塩基球上に存在するFcγRIIBの補充を調節するための有用な治療手段となると考えられる。
【0121】
例として、FcγRIIBと結合し、IgEが同じアレルゲンに複合化した時点での、次に起こるFcγRIIBとFcεRIとの共凝集を介してFcγRIIBの阻害機能を誘発する最適な能力を有するそのような抗体が挙げられる。FcγRIIBと結合し、それ故にFcγRIIB阻害機能を誘発する最適な能力を有する抗アレルゲンIgG抗体は、アレルギーの治療における有効な治療手段を表し得る。例として、本発明により、FcγRIIBと結合し、次に起こるFcγRIIBとFcεRIとの共凝集を介してFcγRIIB阻害機能を誘発する最適な能力を有する抗FcεRI IgG抗体を作製および選択することが可能になる。FcγRIIBと結合し、それ故にFcγRIIB阻害機能を誘発する最適な能力を有する抗FcεRI IgG抗体は、アレルギーの治療における有効な治療手段を表し得る。
【0122】
本発明は、APC(抗原提示細胞)によって発現されるFcγRIIBの効果を調節するための有用な治療手段となると考えられる最適な抗体に関する。そのような効果の例には、抗原提示およびT細胞動員の制御ならびに獲得免疫の制御が含まれる。
【0123】
さらに別の局面において、本発明は、80%〜100%のフコース、および60%〜98%のガラクトースを含み、ガラクトースが30%〜80%のシアリル化型を含む、ヒトIgGのFc領域を含む抗体またはポリペプチドを提供する。
【0124】
本発明によれば、この抗体またはポリペプチドは、FcγRIIIA、FcγRIIA、およびFcγRIIBに結合しないかまたはごく弱く結合する。
【0125】
本出願人らは、結合プロファイルがFcグリコシル化プロファイルと関連していることを示す。本出願人らはまた、このクラスの抗体(上記の第3クラス)が、B細胞またはEBV形質転換B細胞に融合させたヘテロハイブリドーマ由来の細胞によって産生されることを示す。
【0126】
特に関心対象である局面において、この抗体またはポリペプチドはB細胞に融合させたヘテロハイブリドーマ由来の細胞によって産生される。
【0127】
特定の態様において、このヘテロハイブリドーマはEBV形質転換細胞またはヒトドナー由来のBリンパ球に融合させたK6H6B5(ATCC番号CRL-1823)である。
【0128】
この抗体またはポリペプチドは、上記の調製過程の段階c) iii)によって取得するとができる。
【0129】
この抗体またはポリペプチドは上記の第3クラスに属する。
【0130】
本発明はまた、第3クラスに属するこの抗体またはポリペプチドを少なくとも80%、好ましくは少なくとも95%含む組成物を対象とする。
【0131】
好ましい態様において、組成物は、上記の抗体またはポリペプチドを少なくとも80%〜90%、好ましくは少なくとも95%ないし99%含み得る。
【0132】
特に関心対象である局面において、この組成物は、炎症性疾患、クローン病、または関節リウマチの治療用の医薬品を製造するために用いられる。
【0133】
この第3クラスの抗体またはポリペプチドは、FcγRIIBを介して媒介される阻害機能を誘発すること、およびADCCまたは別の細胞傷害活性を誘発することができない。これらの抗体またはポリペプチドは、本発明において、免疫エフェクター機能の補充を妨げるためにい用いられる。
【0134】
本発明による特に関心対象の抗体またはポリペプチドは、IgG1サブクラスに属する。
【0135】
別の態様において、特に関心対象の抗体またはポリペプチドはIgG3サブクラスに属する。
【0136】
本発明はさらに、抗体が標的細胞の細胞表面上に発現されるまたは細胞表面に結合した抗原を認識する医薬品を製造するための上記組成物の使用に関する。そのような抗原には、これらに限定されないが、同種抗原、移植抗原、クラスIおよびクラスII主要組織適合抗原(HLA)等の自己抗原、FcγRI、FcγRIIA、FcγRIIIA分子、FcαR、FcεRI、B細胞受容体、T細胞受容体、CD20、Her2/NEU、CEA、GD2等の腫瘍抗原、ホスホリパーゼA2等のアレルゲン、およびIgE等が含まれる。
【0137】
より詳細には、本発明の抗体は、新生児溶血性疾患(HDNB)をもたらすRh陰性患者のRh同種免疫を治療または予防する医薬品を製造するためにRhD抗原を認識する。
【0138】
より詳細には、本発明の抗体は、特発性血小板減少性紫斑病(ITP)を治療または予防する医薬品を製造するためにRhD抗原を認識する。
【0139】
より詳細には、本発明の抗体は、癌、自己免疫疾患、または移植片拒絶を治療または予防する医薬品を製造するためにHLAクラスII分子を認識する。
【0140】
さらなる詳細を以下の実施例1に提供する。これらの結合および機能試験の結果を図に示す。
【0141】
実施例1:FcγRIIB媒介性阻害を誘発する改善された特性を有する抗RhD抗体の選択
材料および方法
免疫学的に活性のある分子を選択する本発明の方法を、RhD抗原に対するヒトIgG抗体を用いて以下に説明する。例として、RhDに対するLFB1、LFB2、およびLFB3ヒトモノクローナル抗体を使用している。
【0142】
F(ab)'2抗ヒトIgと複合化したまたはしていない精製ヒト抗体が指標細胞(IIA1.6-huFcγRIIB1と命名された、機能的なマウスBCRおよびヒト組み換えFcγRIIB1を発現するマウスIIA1.6リンパ腫B細胞)上に発現されるFcγRIIB1に結合する能力を、それぞれ蛍光抗体アッセイ法(複合化)または免疫蛍光競合アッセイ法(単量体)により測定する。
【0143】
精製ヒト抗体がカルシウム動員の阻害を誘導する能力は、カルシウム測定アッセイ法により評価し得る。カルシウム動員の阻害は、指標細胞(機能的なマウスBCRおよびヒト組み換えFcγRIIB1を発現するマウスIIA1.6リンパ腫B細胞)上に発現されるFcγRIIBに結合した精製ヒト抗体とBCRを共凝集させることにより評価する。これを、マウスIgGと交差反応するF(ab)'2ウサギ抗ヤギIgG(RAG)を用いて誘導されるBCR媒介性カルシウム動員と比較する。
【0144】
精製ヒト抗体がインターロイキン-2(IL-2)産生の阻害を誘導する能力は、IL-2を検出し定量化する酵素結合免疫吸着測定法(ELISA)により測定し得る。IL-2産生の阻害は、指標細胞(機能的なマウスBCRおよびヒト組み換えFcγRIIB1を発現するマウスIIA1.6リンパ腫B細胞)上に発現されるFcγRIIBに結合した精製ヒトまたはキメラ抗体とBCRを共凝集させることにより評価する。これを、F(ab)'2ヤギ抗マウスIgG(H+L)を用いて誘導されるBCR媒介性IL-2産生と比較する。
【0145】
細胞株および抗体
マウスIIA1.6 B細胞リンパ腫は、FcγRII遺伝子の5'末端の欠失を有する(Lewis et al., 1986;Bonnerot et al., 1991)A20 B細胞のFcγR欠損変種(Jones et al., 1986)であり、マウスFcγRIIIのαおよびγ鎖をコードする遺伝子を転写しない(Bonnerot et al., 1991)。M. Hogarth博士(メルボルン大学、オーストラリア、ビクトリア州、パークビル)の供与によるFcγRIIB1をコードするcDNA(Hogarth et al., 1987)をIIA.1.6細胞にトランスフェクションした。IIA1.6およびIIA1.6-huFcγRIIB1細胞を、Click培地[10%熱非働化FCS(Hyclone Laboratories Inc.、米国、ユタ州、ローガン)、100 U/mlペニシリン、100μg/mlストレプトマイシン、2 mM L-グルタミン、5 mMピルビン酸ナトリウム、0.5μM 2-β-メルカプトエタノールを添加したRPMI 1640(GIBCO、スコットランド、ペーズリー)]中で培養した。
【0146】
ヒト組み換え抗Rh D(LFB1)モノクローナル抗体を産生させるには、YB2/0細胞株(ATCC番号CRL-1662)を使用した。YB2/0トランスフェクション細胞は、5% FCSを添加した特別なEM培地中で培養した。
【0147】
LFB2抗Rh Dヒト組み換え抗体を産生させるため、CHO dhfr-(コロンビア大学、米国、ニューヨーク州)細胞株を5% FCSを添加した特別なEM培地中で培養した。
【0148】
LFB1/LFB2抗体の重鎖の定常領域の配列(Kabatにより規定されるCH1の5'末端から開始)を以下に示す(配列番号:5)。
【0149】
抗Rh D LFB3抗体を産生するB細胞株は、ヒトRh陽性赤血球で免疫したドナーによるヒトBリンパ球をヘテロミエローマ細胞株K6H6/B5(ATCC番号CRL-1823)と融合することによって取得し、次いで限界希釈によりクローニングを行った。LFB3細胞は、5% FCSを添加したRPMI 1640培地中で培養した。
【0150】
LFB3抗体の重鎖の定常領域の配列(Kabatにより規定されるCH1の5'末端から開始)を以下に示す(配列番号:6)。
【0151】
産生細胞株はすべて無タンパク質培地に適応化させてそれにより培養し、産生はローラーボトルまたは発酵槽で行った。産生後、プロテイン-Aアフィニティークロマトグラフィーによりモノクローナル抗体を精製した。次いで、精製抗体を-20℃で保存した。
【0152】
リツキサン(Rituxan)(リツキシマブ(Rituximab))(Genentech、米国、カリフォルニア州、サウスサンフランシスコ)は、再発性または難治性低悪性度またはろ胞性、CD20+、B細胞非ホジキンリンパ腫(NHL)の治療において用いられるキメラマウス/ヒト抗CD20モノクローナル抗体である。市販のリツキサンバイアルは、無菌の防腐剤の入っていない10 mg/ml濃度溶液で2℃〜8℃にて保存する。
【0153】
AT10-FITCは、FcγRIIAおよびFcγRIIBの結合部位に対するフルオレセインイソチオシアネート標識マウスモノクローナル抗体である(AT10-FITC、Serotec、英国、カタログ番号MCA 1075F)。
【0154】
免疫蛍光結合アッセイ法
まず、組み換えヒト抗体を、アフィニティー精製したヤギF(ab)'2抗ヒトIgG(Jackson ImmunoResearch Laboratories Inc.、米国、ペンシルベニア州、ウェストグローブ)を用いて30分間、重合型に複合化した。次いで、指標細胞5×105個を、0.5%ウシ血清アルブミンを含む氷冷リン酸緩衝食塩水(PBS-BSA)中で複合化抗体と共に30分間インキュベートした。その後、細胞をPBS-BSAで洗浄し、FITC標識マウスF(ab)'2抗ヒトIgG(H+L)(Jackson ImmunoResearch)と共に氷上で30分間インキュベートすることにより、抗体結合を検出した。さらに洗浄した後、Cell Quest Proソフトウェアを用いてFACScalibur 4CA(Becton Dickinson、米国、カリフォルニア州、マウンテンビュー)によりフローサイトメトリー解析を行った。
【0155】
免疫蛍光競合アッセイ法
IIA1.6-huFcγRIIB細胞をまず種々の濃度の単量体LFB1または単量体LFB2または単量体LFB3または単量体リツキシマブと共に、次いでAT10-FITCと共にインキュベートした。続いて、競合物としての単量体抗体の存在下または非存在下におけるヒトFcγRIIBに対するFITC-AT10モノクローナル抗体の結合を、直接免疫蛍光法により評価した。フローサイトメトリー解析は、Cell Quest Proソフトウェアを用いてFACScalibur 4CA(Becton Dickinson、米国、カリフォルニア州、マウンテンビュー)により行った。
【0156】
Facscalibur 4CAの技術仕様
FITC蛍光色素の獲得には、15 mW、488 nm空冷アルゴンイオンレーザーを用いる。推定検出限界は、粒子当たり200 FITC相当分子である。FL1(FITC)の対数増幅器は、4 log10(log decade)範囲を提供する。ソーティング速度は300細胞/秒であり、ソーティング精度は95%を超える。
【0157】
カルシウム動員の阻害
IIA1.6-huFcγRIIB1細胞をLFB1抗体と共にまたはなしで氷上で30分間インキュベートした。0.2% Pluronic F-127(Sigma Cnemicals Co.,米国、ミズーリ州、セントルイス)を含むRPMI 1640中、106個細胞を5 mM Fluo-3 AM(Molecular Probes、米国、オレゴン州、ユージーン)と共に室温で30分間インキュベートした。次いで、負荷された細胞をRPMI 1640で3回洗浄し、106個/mlになるよう調整した。1 mM EGTAを含むRPMI 1640中、ウサギF(ab)'2抗ヤギIgG(H+L)(RAG)(Jackson ImmunoResearch)単独またはウサギF(ab)'2抗ヤギIgG(H+L)(RAG)およびヤギF(ab)'2抗ヒトIgG(H+L)(GAH)(Jackson ImmunoResearch)で細胞を刺激した。刺激してから150秒後に、6 mM CaCl2を添加した。Cell Quest Proソフトウェアを用いてFACScalibur 4CAにより実施するフローサイトメトリーによって、細胞内カルシウム動員を検出した。次いで、FCS assistant 1.2.9βソフトウェア(Becton Dickinson)により、細胞内カルシウム濃度平均値を算出した。
【0158】
IL-2産生の阻害
IIA1.6-huFcγRIIB1細胞5×105個を、F(ab)'2ウサギ抗ヤギ(Jackson ImmunoResearch)単独またはF(ab)'2ウサギ抗ヤギおよびLFB1、LFB2、またはLFB3で22時間刺激した。1μg/mlラット抗マウスIL-2(Pharmingen)を捕獲抗体として、および1μg/mlビオチン化ラット抗マウスIL-2(Pharmingen)を検出抗体として使用するELISAアッセイにより、マウスIL-2の存在を測定した。
【図面の簡単な説明】
【0159】
【図1】純度および分子の完全性および重合型が存在しないことを管理するための、モノクローナル抗体[RhDに対する(抗RhD)LFB1抗体]のHPLCプロファイルを表す:LFB1モノクローナル抗体は99%の単量体から構成される。
【図2】キャピラリー電気泳動によって決定される、RhDに対する種々の抗体(LFB1、LFB2、LFB3)のグリコシル化パターンを表す。結果は同定された種々の構造の割合として表す。抗体は異なる細胞株によって産生された。LFB1はYB2/0細胞により産生され、LFB2はCHO細胞によって産生され、LFB3はヒトB細胞に融合させたヘテロハイブリドーマK6H6B5によって産生される。
【図3】Jurkat-FcγRIIIA細胞上に発現されるヒトFcγRIIIAに対する、F(ab)'2ヤギ抗ヒトIgG(H+L)複合化LFB1、LFB2、およびLFB3抗体の結合を表す。ヒトFcγRIIIAに対するLFB1、LFB2、およびLFB3ヒト抗RhD抗体の結合は、間接的免疫蛍光法により評価する。まずヒト抗体(100 ng/ml;250 ng/ml;500 ng/ml)をF(ab)'2ヤギ抗ヒトIgG(H+L)(それぞれ150 ng/ml;375 ng/ml;750 ng/mlの用量)により複合化し、次いでJurkat-FcγRIIIA細胞と共にインキュベートする。続いて、FITC標識F(ab)'2マウス抗ヒトIgG(H+L)を用いて、LFB1、LFB2、およびLFB3抗体の結合を明らかにする(黒い曲線)。点線の曲線は、FITC-F(ab)'2マウス抗ヒトIgG(H+L)とのみインキュベートした細胞のバックグラウンド蛍光を示す。結果およびコメント:Jurkat-FcγRIIIA細胞に対するLFB1の結合は、100 ng/ml〜500 ng/mlにおいて常に検出される。最も高い濃度では、ほぼ50%のJurkat-FcγRIIIA細胞が染色される。それに反して、Jurkat-FcγRIIIA細胞によって発現されるヒトFcγRIIIAに対するLFB2抗体の結合は100 ng/mlにおいて認められず、また250 ng/mlおよび500 ng/mlにおいては3%のJurkat-FcγRIIIA細胞が弱く標識されるにすぎない。最後に、Jurkat-FcγRIIIA細胞によって発現されるヒトFcγRIIIAに対するLFB3抗体の結合は、試験したいずれの濃度においても認められない。したがって、YB2/0細胞によって産生される規定のグリコシル化パターンを有するLFB1は、FcγRIIIAに結合するモノクローナル抗体の例である。さらに、異なるが規定のグリコシル化パターンを有する、CHO細胞によって産生されるLFB2およびB細胞に融合させたヘテロハイブリドーマK6H6B5由来の細胞によって産生されるLFB3は、FcγRIIIAに結合しないかまたはごく弱く結合するモノクローナル抗体の例である。
【図4】単量体LFB1、LFB2、LFB3、またはリツキシマブ抗体による、Jurkat-FcγRIIIA細胞上に発現されるヒトFcγRIIIAに対する3G8-PE結合の阻害を表す。まずJurkat-FcγRIIIA細胞を種々の濃度の単量体LFB1(1マイクロg/ml〜1000マイクロg/ml)または単量体LFB2(1マイクロg/ml〜100マイクロg/ml)または単量体LFB3(1マイクロg/ml〜1000マイクロg/ml)または単量体リツキシマブ(1マイクロg/ml〜1000マイクロg/ml)と共に、次いで0.01マイクロg/ml 3G8-PEと共にインキュベートする。続いて、競合物としての単量体抗体の存在下または非存在下におけるヒトFcγRIIIAに対するPE標識マウス3G8モノクローナル抗体の結合を、直接免疫蛍光法により評価する。結果およびコメント:単量体LFB1は、Jurkat-FcγRIIIA細胞上に発現されるヒトFcγRIIIAに対する3G8-PE結合の用量依存的阻害を誘導する。10マイクロg/mlのLFB1は3G8-PEによって標識される細胞の96%という割合の減少を誘導するのに対し、10マイクロg/mlのリツキシマブでは2%の減少しか達成されない。用いたうち最も高い用量(1000マイクロg/ml)では、LFB1は100%阻害を誘導する(1000マイクロg/mlのリツキシマブでは72%)。単量体LFB2は、LFB1によって得られる阻害よりも低い、FcγRIIIAに対する3G8-PE結合の阻害を誘導する。10マイクロg/mlのLFB2は、3G8-PEによって標識される細胞の5%未満という割合の減少を誘導する。しかし、高濃度(100マイクロg/ml)では、(同じ用量のリツキシマブでのたった15%と比較して)染色されたJurkat-FcγRIIIA細胞の約95%という割合の減少が認められる。それに反して、単量体LFB3によって誘導される阻害は、用いたどの用量においても非常に弱い。1000マイクロg/mlのLFB3を用いた場合、阻害は20%よりも低い。 したがって、このアッセイにより、単量体型の抗体は、それらを産生する細胞株およびそれらのグリコシル化プロファイルに依存して、FcγRIIIAに対して異なる結合能を示すことが示される。
【図5】K562細胞上に発現されるヒトFcγRIIAに対する、F(ab)'2ヤギ抗ヒトIgG(H+L)複合化LFB1、LFB2、およびLFB3抗体の結合を表す。ヒトFcγRIIAに対するLFB1、LFB2、およびLFB3ヒト抗RhD抗体の結合は、間接的免疫蛍光法により評価する。まずヒト抗体(1μg/ml;5μg/ml;10μg/ml)をF(ab)'2ヤギ抗ヒトIgG(H+L)(それぞれ1.5μg/ml;7.5μg/ml;15μg/mlの用量)により複合化し、次いでK562細胞と共にインキュベートする。続いて、FITC標識F(ab)'2マウス抗ヒトIgG(H+L)を用いて、LFB1、LFB2、およびLFB3抗体の結合を明らかにする(黒い曲線)。点線の曲線は、FITC-F(ab)'2マウス抗ヒトIgG(H+L)とのみインキュベートした細胞のバックグラウンド蛍光を示す。結果およびコメント:LFB1抗体は、1μg/mlというような低い用量においてさえもK562細胞に結合する(この低濃度において、50%を超えるK562細胞が染色される)。最も高い用量のLFB1抗体(10μg/ml)では、ほぼ100%のK562細胞が標識される。LFB2は、K562細胞上に発現されるヒトFcγRIIAに対してLFB1と同様の結合パターンを有する。LFB2は使用したいずれの用量においても(1μg/ml〜10μg/ml)ヒトFcγRIIAに結合し、10μg/mlではほぼ100%のK562細胞が染色される。それに反して、K562細胞は、1μg/ml、5μg/ml、または10μg/mlのLFB3により弱く標識される。したがって、異なるが規定のグリコシル化パターンを有する、YB2/0細胞によって産生されるLFB1およびCHO細胞によって産生されるLFB2は、FcγRIIAに結合するモノクローナル抗体の例である。さらに、規定のグリコシル化パターンを有する、B細胞に融合させたヘテロハイブリドーマK6H6B5由来の細胞によって産生されるLFB3は、FcγRIIAに結合しないかまたはごく弱く結合するモノクローナル抗体の例である。
【図6】単量体LFB1、LFB2、LFB3、またはリツキシマブ抗体による、K562細胞上に発現されるヒトFcγRIIAに対する(FcγRIIAおよびFcγRIIBに対する)AT10-FITC結合の阻害を表す。まずK562細胞を種々の濃度の単量体LFB1(50μg/ml〜1000μg/ml)または単量体LFB2(50μg/ml〜250μg/ml)または単量体LFB3(50μg/ml〜1000μg/ml)または単量体リツキシマブ(50μg/ml〜1000μg/ml)と共に、次いで0.25μg/ml AT10-FITCと共にインキュベートする。競合物としての単量体抗体の存在下または非存在下におけるヒトFcγRIIAに対するFITC標識マウスAT10モノクローナル抗体の結合を、直接免疫蛍光法により評価する。結果およびコメント:単量体LFB1は、K562細胞上に発現されるヒトFcγRIIAに対するAT10-FITC結合の用量依存的阻害を誘導する。100μg/mlのLFB1はAT10-FITCによって標識される細胞のほぼ50%という割合の減少を誘導するのに対し、100μg/mlのリツキシマブでは19%の減少しか達成されない。用いたうち最も高い用量(1000μg/ml)では、(1000μg/mlのリツキシマブでの43%と比較して)90%に達する。単量体LFB2もまた、FcγRIIAに対するAT10-FITC結合の用量依存的阻害を誘導する。用いうち最も高濃度(250μg/ml)のLFB2では、(同じ用量のリツキシマブでの24%と比較して)染色されたK562細胞の約40%という割合の減少が認められる。それに反して、単量体LFB3によって誘導される阻害は弱い。1000μg/mlのLFB3を用いた場合、阻害は30%よりも低い。したがって、このアッセイにより、単量体型の抗体は、それらを産生する細胞株およびそれらのグリコシル化プロファイルに依存して、FcγRIIAに対して異なる結合能を示すことが示される。
【図7】IIA1.6-huFcγRIIB細胞上に発現されるヒトFcγRIIBに対する、F(ab)'2ヤギ抗ヒトIgG(H+L)複合化LFB1、LFB2、およびLFB3抗体の結合を表す。ヒトFcγRIIBに対するLFB1、LFB2、およびLFB3ヒト抗RhD抗体の結合は、間接的免疫蛍光法により評価する。まずヒト抗体(25μg/ml;50μg/ml)をF(ab)'2ヤギ抗ヒトIgG(H+L)(それぞれ37.5μg/ml;75μg/mlの用量)により複合化し、次いでIIA1.6-huFcγRIIB細胞と共にインキュベートする。続いて、FITC標識F(ab)'2マウス抗ヒトIgG(H+L)を用いて、LFB1、LFB2、およびLFB3抗体の結合を明らかにする(黒い曲線)。点線の曲線は、FITC-F(ab)'2マウス抗ヒトIgG(H+L)とのみインキュベートした細胞のバックグラウンド蛍光を示す。結果およびコメント:LFB1抗体は、25μg/mlおよび50μg/mlおいてIIA1.6-huFcγRIIB細胞に結合する(50μg/mlおいて、50%を超えるIIA1.6-huFcγRIIB細胞が染色される)。LFB2は、ヒトFcγRIIBに対してLFB1と同様の結合パターンを有する。LFB2は使用したいずれの用量においてもヒトFcγRIIBに結合し、50μg/mlでは50%を超えるIIA1.6-huFcγRIIB細胞が標識される。それに反して、ヒトFcγRIIBに対するLFB3の結合は、試験したいずれの濃度においても認められない。したがって、異なるが規定のグリコシル化パターンを有する、YB2/0細胞によって産生されるLFB1およびCHO細胞によって産生されるLFB2は、FcγRIIBに結合するモノクローナル抗体の例である。さらに、規定のグリコシル化パターンを有する、B細胞に融合させたヘテロハイブリドーマK6H6B5由来の細胞によって産生されるLFB3は、FcγRIIBに結合しないかまたはごく弱く結合するモノクローナル抗体の例である。
【図8】単量体LFB1、LFB2、LFB3、およびリツキシマブ抗体による、IIA1.6-huFcγRIIB細胞上に発現されるヒトFcγRIIBに対するAT10-FITC結合の阻害を表す。まずIIA1.6-huFcγRIIB細胞を種々の濃度の単量体LFB1(50μg/ml〜1000μg/ml)または単量体LFB2(50μg/ml〜250μg/ml)または単量体LFB3(50μg/ml〜1000μg/ml)または単量体リツキシマブ(50μg/ml〜1000μg/ml)と共に、次いで0.25μg/ml AT10-FITCと共にインキュベートする。続いて、競合物としての単量体抗体の存在下または非存在下におけるヒトFcγRIIBに対するFITC標識マウスAT10モノクローナル抗体の結合を、直接免疫蛍光法により評価する。結果およびコメント:単量体LFB1は、IIA1.6-huFcγRIIB細胞上に発現されるヒトFcγRIIBに対するAT10-FITC結合の用量依存的阻害を誘導する。100μg/mlのLFB1はAT10-FITCによって標識される細胞のほぼ40%という割合の減少を誘導するのに対し、100μg/mlのリツキシマブでは11%の減少しか達成されない。用いたうち最も高い用量(1000μg/ml)では、(1000μg/mlのリツキシマブでの23%と比較して)80%に達する。単量体LFB2は、LFB1によって得られる阻害よりも低い、FcγRIIBに対するAT10-FITC結合の阻害を誘導し、これは単量体リツキシマブによって誘発される阻害と類似している(100μg/mlのこれら抗体を用いた場合、AT10-FITCによって標識される細胞の約10%の減少が認められる)。1000μg/mlのLFB3を用いた場合、AT10-FITCによって標識される細胞の減少の割合は20%未満であり、単量体LFB3によって誘導される阻害は弱い。 したがって、このアッセイにより、単量体型の抗体は、それらを産生する細胞株およびそれらのグリコシル化プロファイルに依存して、FcγRIIBに対して異なる結合能を示すことが示される。
【図9】グリコシル化プロファイル、および競合アッセイ法によって評価したLFB1、LFB2、LFB3、およびリツキシマブのID10、ID30、OD50を要約する。FCγRIIIAを試験した場合に0.02マイクロMというID50、FCγRIIAを試験した場合に0.85マイクロMというID50、およびFCγRIIBを試験した場合に1マイクロMというID50を示すLFB1は、FcγRIIIA、FcγRIIA、およびFcγRIIBに「結合する」モノクローナル抗体の例である。 FCγRIIIAを試験した場合にID50>6マイクロM、FCγRIIAを試験した場合にID50>6マイクロM、およびFCγRIIBを試験した場合にID50>6マイクロMを示すLFB3は、FcγRIIIA、FcγRIIA、およびFcγRIIBに「結合しないかまたはごく弱く結合する」モノクローナル抗体の例である。
【図10】LFB1ヒト抗RhD抗体によるカルシウム動員の阻害を表す。IIA1.6-huFcγRIIB細胞をマウスIgと交差反応するF(ab)'2ウサギ抗ヤギIgG(RAG)(10μg/ml)により、またはF(ab)'2ヤギ抗ヒトIgG(H+L)(GAH)(10μg/ml)およびF(ab)'2ウサギ抗ヤギIgG(RAG)(10μg/ml)の存在下においてLFB1抗体により刺激する。この後者のアッセイにより、FcγRIIB受容体に結合したヒトモノクローナルIgG抗体を介して、IIA1.6-huFcγRIIB細胞によって発現されるマウス表面IgGとヒトFcγRIIBが効率的に架橋され得る。F(ab)'2 RAG(10μg/ml)によって誘導されるBCR媒介性Ca2+流入を、BCR/FcγRIIB架橋後に観察されるCa2+流入と比較する。Fluo-3 AM負荷細胞における細胞内遊離Ca2+レベルを、フローサイトメトリーによってモニターする。1 mM EGTAの存在下において細胞内Ca2+貯蔵からのCa2+放出を測定し(曲線の左部分)、6 mM CaCl2を添加した後に細胞内に流入するCa2+を測定する(曲線の右部分)。結果およびコメント:F(ab)'2ウサギ抗ヤギIgG(RAG)抗体を添加することにより、細胞内貯蔵からのカルシウム放出(曲線の左側のピーク、点線)およびIIA1.6-huFcγRIIB細胞へのカルシウム流入(曲線の右側のピーク、点線)が誘発される。F(ab)'2ヤギ抗ヒトIgG(H+L)(GAH)(10μg/ml)およびF(ab)'2ウサギ抗ヤギIgG(RAG)(10μg/ml)の存在下において、IIA1.6-huFcγRIIB細胞をLFB1抗体(100μg/ml)と共にインキュベートすることにより、B細胞上のFcγRIIBの結合の特徴であるカルシウム流入の強い阻害が得られる(曲線の右側のピーク、黒線)。FcγRIIB阻害機能が誘発される場合に予想される通り、細胞内貯蔵からのカルシウム放出の阻害は認められない(曲線の左側のピーク、黒線)。したがって、この実験により、LFB1モノクローナル抗体は、B細胞活性化および抗体産生細胞への分化の遮断を起こす要因である、リンパ腫B細胞上に発現されるFcγRIIBによって媒介される阻害機能を誘発し得ることが示される。
【図11】LFB1およびLFB2ヒト抗RhD抗体によるIL-2産生の阻害を表す。IIA1.6-huFcγRIIB細胞5×105個を、50μg/ml F(ab)'2ウサギ抗ヤギIgG(RAG)単独で、または50μg/ml F(ab)'2ウサギ抗ヤギIgGおよび横軸に表示した種々の濃度の(10〜100μg/ml)のLFB1またはLFB2抗体で22時間刺激する。RAG F(ab)'2断片はマウスIgGおよびヒトIgGの両方と交差反応し、IIA1.6-huFcγRIIB陽性細胞によって発現されるマウス表面IgGとFcγRIIBに結合したヒトモノクローナルIgG抗体とを架橋させ得る。培養上清中のIIA1.6-huFcγRIIB細胞によるマウスIL-2の分泌を、1μg/mlラット抗マウスIL-2を捕獲抗体としておよび1μg/mlビオチン化ラット抗マウスIL-2を検出抗体として使用するELISAアッセイにより測定する。結果およびコメント:表面IgGを発現するIIA1.6-huFcγRIIB陽性細胞をマウスIgGと交差反応するF(ab)'2ウサギ抗ヤギIgG(RAG)により刺激すると、IL-2の分泌が誘発される(IIA1.6-huFcγRIIB細胞5×105個を22時間培養した後、約380 pg/ml)。マウスIgGおよびヒトIgGの両方と交差反応するRAG F(ab)'2断片の存在下では、種々の用量のLFB1抗体(10μg/ml〜100μg/ml)の存在により、IL-2分泌の用量依存的阻害が誘導される。10μg/mlという低いLFB1抗体により、IL-2産生の約40%の阻害が誘導される。用いたうち最も高い用量のLFB1(100μg/ml)では、IL-2産生の阻害は60%に達する。さらに、LFB2抗体もまた、RAG F(ab)'2の存在下で培養したIIA1.6-huFcγRIIB1細胞によるIL-2産生を阻害する。この抗体10μ/mlの存在によりIL-2の30%阻害が誘導され、100μg/mlのLFB2によりIL-2産生の約50%阻害が誘導されることから、LFB2はIL-2産生の用量依存的阻害を誘導する。 したがってこの実験により、IIA1.6-huFcγRIIB細胞によって発現されるFcγRIIBに結合する、YB2/0細胞によって産生されるLFB1およびCHO細胞によって産生されるLFB2は、B細胞上に発現されるFcγRIIBによって媒介される阻害機能を誘発し得るモノクローナル抗体の例であることが示される。特に、これらの抗体によって、これらの細胞によるサイトカイン産生の遮断がもたらされる。
【図12】LFB3ヒト抗RhD抗体によってではなくLFB1ヒト抗RhD抗体によって起こる、IL-2産生の阻害を表す。IIA1.6-huFcγRIIB細胞5×105個を、50μg/ml F(ab)'2ウサギ抗ヤギIgG(RAG)単独で、または50μg/ml F(ab)'2ウサギ抗ヤギIgGおよび横軸に表示した種々の濃度の(10〜200μg/ml)のLFB1またはLFB3抗体で22時間刺激する。RAG F(ab)'2断片はマウスIgGおよびヒトIgGの両方と交差反応し、IIA1.6-huFcγRIIB陽性細胞によって発現されるマウス表面IgGとFcγRIIBに結合したヒトモノクローナルIgG抗体とを架橋させ得る。培養上清中のIIA1.6-huFcγRIIB細胞によるマウスIL-2の分泌を、1μg/mlラット抗マウスIL-2を捕獲抗体としておよび1μg/mlビオチン化ラット抗マウスIL-2を検出抗体として使用するELISAアッセイにより測定する。結果およびコメント:LFB1抗体(10μg/ml〜200μg/ml)はIL-2分泌の用量依存的阻害を誘導する。10μg/mlという低いLFB1抗体によりIL-2産生の約20%の阻害が誘導され、100μg/mlにより50%に達する阻害が誘導される。用いたうち最も高い用量のLFB1(200μg/ml)では、IL-2産生の阻害は70%に達する。これに反してLFB3抗体は、200μg/mlというような高い用量で使用した場合にでさえ、RAG F(ab)'2断片の存在下で培養したIIA1.6-huFcγRIIB細胞によるIL-2産生を阻害しない(<3%減少)。したがってこの実験により、IIA1.6-huFcγRIIB1細胞によって発現されるFcγRIIBに結合する、YB2/0細胞によって産生されるLFB1モノクローナル抗体は、B細胞上に発現されるFcγRIIBによって媒介される阻害機能を誘発し得るモノクローナル抗体の例であることが示される。特に、これによりB細胞によるIL-2産生の遮断がもたらされる。さらにこの実験により、IIA1.6-huFcγRIIB1細胞によって発現されるFcγRIIBに結合しない、B細胞に融合させたヘテロハイブリドーマK6H6B5由来の細胞によって産生されるLFB3モノクローナル抗体は、B細胞上に発現されるFcγRIIBによって媒介される阻害機能を誘発し得ないモノクローナル抗体の例であることが示される。
【特許請求の範囲】
【請求項1】
ヒトIgGのFc領域を含みかつ異なる結合プロファイルを有する、ヒトの、ヒト化された、またはキメラの抗体またはポリペプチドを調製する方法であって、以下からなる段階を含む方法:
(a) ハイブリドーマ、ヘテロハイブリドーマ、EBV形質転換ヒトB細胞株を含む動物細胞株に由来もしくは真核微生物に由来する細胞により天然に産生される、または、該抗体もしくはポリペプチドのコード配列を含むベクターによるそのトランスフェクション後に産生される、ヒトIgGのFc領域を含む、ヒトの、ヒト化された、またはキメラの抗体候補またはポリペプチド候補を提供する段階;
(b) FcγRIIIA、FcγRIIA、およびFcγRIIBを含むFcγ受容体において該抗体またはポリペプチドの結合を試験する段階;
(c)
i) FcγRIIIA、FcγRIIA、およびFcγRIIBのいずれにも結合する、または
ii) FcγRIIAおよびFcγRIIBの両方に結合するが、FcγRIIIAには結合しないかまたはごく弱く結合する、または
iii) FcγRIIIA、FcγRIIA、およびFcγRIIBのいずれにも結合しないかまたはごく弱く結合する
抗体またはポリペプチドを選択する段階。
【請求項2】
段階c) i)で選択される抗体またはポリペプチドが、リンパ球系(lymphoid)細胞株またはリンパ球系細胞由来(lymphoid-derived)細胞株またはハイブリドーマまたは腎臓上皮細胞株由来の細胞により産生され、
段階c) ii)で選択される抗体またはポリペプチドが非リンパ球系(non-lymphoid)細胞株由来の細胞により産生され、かつ、
段階c) iii)で選択される抗体またはポリペプチドが、EBV形質転換細胞株由来の細胞にまたはヒトドナー由来のBリンパ球に融合させたヘテロハイブリドーマ由来の細胞により産生される、請求項1記載の方法。
【請求項3】
リンパ球系細胞由来細胞株が、ラット骨髄腫細胞株もしくはハイブリドーマYB2/0細胞株(ATCC番号CRL-1662)もしくはそれらに由来する細胞株であり、かつ/または、腎臓上皮細胞株がVERO(ATCC番号CCL-81)もしくはそれに由来する細胞であり、かつ/または、非リンパ球系細胞株がCHO(ATCC番号CCL-61)もしくはそれに由来する細胞であり、かつ/または、ヘテロハイブリドーマがK6H6B5(ATCC番号CRL-1823)もしくはそれに由来する細胞株である、請求項2記載の方法。
【請求項4】
結合アッセイが以下を用いて行われ得る、前期請求項のいずれか一項記載の方法:
i) 細胞表面上に異なるFc受容体を発現する細胞株由来の指標細胞;
ii) FcγR外部ドメイン(ectodomain)であるFc受容体由来ペプチドを含む、組み換えFc受容体。
【請求項5】
FcγRIIIA、FcγRIIA、およびFcγRIIBのいずれにも結合する能力に関して選択された抗体またはポリペプチドのサブセット、ならびに、FcγRIIAおよびFcγRIIBの両方に結合する能力に関して選択された抗体またはポリペプチドのサブセットが、i) FcγRIIIAを誘発してADCCの改善、インターロイキン-2(IL-2)等のサイトカインおよび腫瘍壊死因子α(TNFα)等の炎症促進性分子の産生の増加をもたらす能力;ならびにii) FcγRIIBを誘発して、B細胞および単球等のFcγRIIBを発現する細胞による、カルシウム動員の阻害およびIL-2等のサイトカイン産生の阻害をもたらす能力に関する機能アッセイにより、さらに試験および選択される、前期請求項のいずれか一項記載の方法。
【請求項6】
機能アッセイ法が、カルシウム動員阻害アッセイ法および/またはサイトカイン分泌阻害アッセイ法からなる、前期請求項のいずれか一項記載の方法。
【請求項7】
機能アッセイ法が特異的FcγRIIIA ADCCアッセイ法をさらに含み得る、請求項5または6記載の方法。
【請求項8】
FcγRIIIA、FcγRIIA、およびFcγRIIBに結合し得る抗体を産生させるための、リンパ球系細胞株またはリンパ球系細胞由来細胞株またはハイブリドーマまたは腎臓上皮細胞株に由来する細胞の使用。
【請求項9】
抗体が免疫調節抗体である、請求項8記載の使用。
【請求項10】
抗体が免疫調節かつ細胞傷害性の抗体である、請求項8または9記載の方法。
【請求項11】
ハイブリドーマ細胞株がYB2/0である、請求項8〜10のいずれか一項記載の方法。
【請求項12】
腎臓上皮細胞株がVERO(ATCC番号CCL-81)である、請求項8〜10のいずれか一項記載の方法。
【請求項13】
FcγRIIIAには結合しないかまたはごく弱く結合するが、FcγRIIAおよびFcγRIIBの両方には結合する抗体を産生させるための、非リンパ球系細胞株由来の細胞の使用。
【請求項14】
抗体が免疫調節抗体である、請求項13記載の使用。
【請求項15】
抗体が、FcγRIIAを発現している単球およびマクロファージによるADCCおよび食作用を誘導する、請求項13または14記載の方法。
【請求項16】
非リンパ球系細胞株由来の細胞がCHO(ATCC番号CCL-61)である、請求項13〜15のいずれか一項記載の方法。
【請求項17】
FcγRIIIA、FcγRIIA、およびFcγRIIBのいずれにも結合しないかまたは弱く結合する抗体を産生させるための、EBV形質転換細胞株由来の細胞とまたはB細胞と融合させたヘテロハイブリドーマ由来の細胞の使用。
【請求項18】
抗体が、IgG4の使用に代わる治療法として用いられる、請求項17記載の使用。
【請求項19】
細胞が、ヒトB細胞に融合させたヘテロハイブリドーマK6H6B5(ATCC番号CRL-1823)に由来する、請求項17または18記載の使用。
【請求項20】
10%〜55%のフコース、および60%〜98%のガラクトースを含むことを特徴とする、ヒトIgGのFc領域を含む抗体またはポリペプチド。
【請求項21】
リンパ球系細胞株またはリンパ球系細胞由来細胞株またはハイブリドーマまたは腎臓上皮細胞株に由来する細胞によって産生されることを特徴とする、請求項20記載の抗体またはポリペプチド。
【請求項22】
ハイブリドーマ細胞株がYB2/0細胞株(ATCC番号CRL-1622)である、請求項21記載の抗体またはポリペプチド。
【請求項23】
腎臓上皮細胞株がVERO(ATCC番号CCL-81)である、請求項21記載の抗体またはポリペプチド。
【請求項24】
FcγRIIIA、FcγRIIA、およびFcγRIIBのいずれにも結合することを特徴とする、請求項20〜23のいずれか一項記載の抗体またはポリペプチド。
【請求項25】
請求項1〜7のいずれか一項記載の方法における段階c) i)によって取得できることを特徴とする、請求項20〜24のいずれか一項記載の抗体またはポリペプチド。
【請求項26】
70%〜100%のフコースおよび60%〜98%のガラクトースを含むことを特徴とする、ヒトIgGのFc領域を含む抗体またはポリペプチド。
【請求項27】
非リンパ球系細胞株由来の細胞によって産生されることを特徴とする、請求項26記載の抗体またはポリペプチド。
【請求項28】
非リンパ球系細胞株がCHO細胞株(ATCC番号CCL-61)であることを特徴とする、請求項27記載の抗体またはポリペプチド。
【請求項29】
FcγRIIAおよびFcγRIIBには結合するが、FcγRIIIAには結合しないかまたはごく弱く結合することを特徴とする、請求項26〜28のいずれか一項記載の抗体またはポリペプチド。
【請求項30】
請求項1〜7のいずれか一項記載の方法における段階c) ii)によって取得できることを特徴とする、請求項26〜29のいずれか一項記載の抗体またはポリペプチド。
【請求項31】
80%〜100%のフコース、および60%〜98%のガラクトースを含むこと、ならびに、ガラクトースが30%〜80%のシアリル化型を含むことを特徴とする、ヒトIgGのFc領域を含む抗体またはポリペプチド。
【請求項32】
EBV形質転換B細胞にまたはヒトドナー由来のBリンパ球に融合させたヘテロハイブリドーマ由来の細胞によって産生されることを特徴とする、請求項31記載の抗体またはポリペプチド。
【請求項33】
ヘテロハイブリドーマが、EBV形質転換細胞株由来の細胞に融合させたK6H6B5(ATCC番号CRL-1823)であることを特徴とする、請求項32記載の抗体またはポリペプチド。
【請求項34】
FcγRIIIA、FcγRIIA、およびFcγRIIBのいずれにも結合しないかまたはごく弱く結合する、請求項31〜33のいずれか一項記載の抗体またはポリペプチド。
【請求項35】
請求項1〜7のいずれか一項記載の方法における段階c) iii)によって取得できることを特徴とする、請求項31〜34のいずれか一項記載の抗体またはポリペプチド。
【請求項36】
IgG1サブクラスに属することを特徴とする、請求項1〜35のいずれか一項記載の抗体またはポリペプチド。
【請求項37】
IgG3サブクラスに属することを特徴とする、請求項1〜35のいずれか一項記載の抗体またはポリペプチド。
【請求項38】
請求項20〜25のいずれか一項記載の抗体またはポリペプチドを少なくとも80%、好ましくは少なくとも95%含む、組成物。
【請求項39】
癌、自己免疫疾患、アレルギー、移植後の同種免疫、母子間(materno-fetal)同種免疫、移植片対宿主(GVH)反応、または感染症の治療用の医薬品を製造するための、請求項38記載の組成物の使用。
【請求項40】
癌が白血病、リンパ腫、骨髄腫、セザリー症候群、または固形腫瘍である、請求項39記載の使用。
【請求項41】
母子間同種免疫が新生児溶血性疾患(HDNB)である、請求項39記載の使用。
【請求項42】
自己免疫疾患が、全身性エリテマトーデス(SLE)、特発性血小板減少性紫斑病(ITP)、川崎症候群等の自己抗体を産生するB細胞関連自己免疫疾患である、請求項39記載の使用。
【請求項43】
アレルギーが、喘息、アレルギー性鼻炎、アレルギー性副鼻腔炎、アナフィラキシー症候群、じんま疹、血管性水腫、アトピー性皮膚炎、アレルギー性接触皮膚炎、および紅斑である、請求項39記載の使用。
【請求項44】
請求項26〜30のいずれか一項記載の抗体またはポリペプチドを少なくとも80%、好ましくは少なくとも95%含む、組成物。
【請求項45】
自己免疫疾患、母子間同種免疫、および炎症性疾患の治療用の医薬品を製造するための、請求項44記載の組成物の使用。
【請求項46】
請求項31〜35のいずれか一項記載の抗体またはポリペプチドを少なくとも80%、好ましくは少なくとも95%含む、組成物。
【請求項47】
炎症性疾患、クローン病、または関節リウマチの治療用の医薬品を製造するための、請求項46記載の組成物の使用。
【請求項1】
ヒトIgGのFc領域を含みかつ異なる結合プロファイルを有する、ヒトの、ヒト化された、またはキメラの抗体またはポリペプチドを調製する方法であって、以下からなる段階を含む方法:
(a) ハイブリドーマ、ヘテロハイブリドーマ、EBV形質転換ヒトB細胞株を含む動物細胞株に由来もしくは真核微生物に由来する細胞により天然に産生される、または、該抗体もしくはポリペプチドのコード配列を含むベクターによるそのトランスフェクション後に産生される、ヒトIgGのFc領域を含む、ヒトの、ヒト化された、またはキメラの抗体候補またはポリペプチド候補を提供する段階;
(b) FcγRIIIA、FcγRIIA、およびFcγRIIBを含むFcγ受容体において該抗体またはポリペプチドの結合を試験する段階;
(c)
i) FcγRIIIA、FcγRIIA、およびFcγRIIBのいずれにも結合する、または
ii) FcγRIIAおよびFcγRIIBの両方に結合するが、FcγRIIIAには結合しないかまたはごく弱く結合する、または
iii) FcγRIIIA、FcγRIIA、およびFcγRIIBのいずれにも結合しないかまたはごく弱く結合する
抗体またはポリペプチドを選択する段階。
【請求項2】
段階c) i)で選択される抗体またはポリペプチドが、リンパ球系(lymphoid)細胞株またはリンパ球系細胞由来(lymphoid-derived)細胞株またはハイブリドーマまたは腎臓上皮細胞株由来の細胞により産生され、
段階c) ii)で選択される抗体またはポリペプチドが非リンパ球系(non-lymphoid)細胞株由来の細胞により産生され、かつ、
段階c) iii)で選択される抗体またはポリペプチドが、EBV形質転換細胞株由来の細胞にまたはヒトドナー由来のBリンパ球に融合させたヘテロハイブリドーマ由来の細胞により産生される、請求項1記載の方法。
【請求項3】
リンパ球系細胞由来細胞株が、ラット骨髄腫細胞株もしくはハイブリドーマYB2/0細胞株(ATCC番号CRL-1662)もしくはそれらに由来する細胞株であり、かつ/または、腎臓上皮細胞株がVERO(ATCC番号CCL-81)もしくはそれに由来する細胞であり、かつ/または、非リンパ球系細胞株がCHO(ATCC番号CCL-61)もしくはそれに由来する細胞であり、かつ/または、ヘテロハイブリドーマがK6H6B5(ATCC番号CRL-1823)もしくはそれに由来する細胞株である、請求項2記載の方法。
【請求項4】
結合アッセイが以下を用いて行われ得る、前期請求項のいずれか一項記載の方法:
i) 細胞表面上に異なるFc受容体を発現する細胞株由来の指標細胞;
ii) FcγR外部ドメイン(ectodomain)であるFc受容体由来ペプチドを含む、組み換えFc受容体。
【請求項5】
FcγRIIIA、FcγRIIA、およびFcγRIIBのいずれにも結合する能力に関して選択された抗体またはポリペプチドのサブセット、ならびに、FcγRIIAおよびFcγRIIBの両方に結合する能力に関して選択された抗体またはポリペプチドのサブセットが、i) FcγRIIIAを誘発してADCCの改善、インターロイキン-2(IL-2)等のサイトカインおよび腫瘍壊死因子α(TNFα)等の炎症促進性分子の産生の増加をもたらす能力;ならびにii) FcγRIIBを誘発して、B細胞および単球等のFcγRIIBを発現する細胞による、カルシウム動員の阻害およびIL-2等のサイトカイン産生の阻害をもたらす能力に関する機能アッセイにより、さらに試験および選択される、前期請求項のいずれか一項記載の方法。
【請求項6】
機能アッセイ法が、カルシウム動員阻害アッセイ法および/またはサイトカイン分泌阻害アッセイ法からなる、前期請求項のいずれか一項記載の方法。
【請求項7】
機能アッセイ法が特異的FcγRIIIA ADCCアッセイ法をさらに含み得る、請求項5または6記載の方法。
【請求項8】
FcγRIIIA、FcγRIIA、およびFcγRIIBに結合し得る抗体を産生させるための、リンパ球系細胞株またはリンパ球系細胞由来細胞株またはハイブリドーマまたは腎臓上皮細胞株に由来する細胞の使用。
【請求項9】
抗体が免疫調節抗体である、請求項8記載の使用。
【請求項10】
抗体が免疫調節かつ細胞傷害性の抗体である、請求項8または9記載の方法。
【請求項11】
ハイブリドーマ細胞株がYB2/0である、請求項8〜10のいずれか一項記載の方法。
【請求項12】
腎臓上皮細胞株がVERO(ATCC番号CCL-81)である、請求項8〜10のいずれか一項記載の方法。
【請求項13】
FcγRIIIAには結合しないかまたはごく弱く結合するが、FcγRIIAおよびFcγRIIBの両方には結合する抗体を産生させるための、非リンパ球系細胞株由来の細胞の使用。
【請求項14】
抗体が免疫調節抗体である、請求項13記載の使用。
【請求項15】
抗体が、FcγRIIAを発現している単球およびマクロファージによるADCCおよび食作用を誘導する、請求項13または14記載の方法。
【請求項16】
非リンパ球系細胞株由来の細胞がCHO(ATCC番号CCL-61)である、請求項13〜15のいずれか一項記載の方法。
【請求項17】
FcγRIIIA、FcγRIIA、およびFcγRIIBのいずれにも結合しないかまたは弱く結合する抗体を産生させるための、EBV形質転換細胞株由来の細胞とまたはB細胞と融合させたヘテロハイブリドーマ由来の細胞の使用。
【請求項18】
抗体が、IgG4の使用に代わる治療法として用いられる、請求項17記載の使用。
【請求項19】
細胞が、ヒトB細胞に融合させたヘテロハイブリドーマK6H6B5(ATCC番号CRL-1823)に由来する、請求項17または18記載の使用。
【請求項20】
10%〜55%のフコース、および60%〜98%のガラクトースを含むことを特徴とする、ヒトIgGのFc領域を含む抗体またはポリペプチド。
【請求項21】
リンパ球系細胞株またはリンパ球系細胞由来細胞株またはハイブリドーマまたは腎臓上皮細胞株に由来する細胞によって産生されることを特徴とする、請求項20記載の抗体またはポリペプチド。
【請求項22】
ハイブリドーマ細胞株がYB2/0細胞株(ATCC番号CRL-1622)である、請求項21記載の抗体またはポリペプチド。
【請求項23】
腎臓上皮細胞株がVERO(ATCC番号CCL-81)である、請求項21記載の抗体またはポリペプチド。
【請求項24】
FcγRIIIA、FcγRIIA、およびFcγRIIBのいずれにも結合することを特徴とする、請求項20〜23のいずれか一項記載の抗体またはポリペプチド。
【請求項25】
請求項1〜7のいずれか一項記載の方法における段階c) i)によって取得できることを特徴とする、請求項20〜24のいずれか一項記載の抗体またはポリペプチド。
【請求項26】
70%〜100%のフコースおよび60%〜98%のガラクトースを含むことを特徴とする、ヒトIgGのFc領域を含む抗体またはポリペプチド。
【請求項27】
非リンパ球系細胞株由来の細胞によって産生されることを特徴とする、請求項26記載の抗体またはポリペプチド。
【請求項28】
非リンパ球系細胞株がCHO細胞株(ATCC番号CCL-61)であることを特徴とする、請求項27記載の抗体またはポリペプチド。
【請求項29】
FcγRIIAおよびFcγRIIBには結合するが、FcγRIIIAには結合しないかまたはごく弱く結合することを特徴とする、請求項26〜28のいずれか一項記載の抗体またはポリペプチド。
【請求項30】
請求項1〜7のいずれか一項記載の方法における段階c) ii)によって取得できることを特徴とする、請求項26〜29のいずれか一項記載の抗体またはポリペプチド。
【請求項31】
80%〜100%のフコース、および60%〜98%のガラクトースを含むこと、ならびに、ガラクトースが30%〜80%のシアリル化型を含むことを特徴とする、ヒトIgGのFc領域を含む抗体またはポリペプチド。
【請求項32】
EBV形質転換B細胞にまたはヒトドナー由来のBリンパ球に融合させたヘテロハイブリドーマ由来の細胞によって産生されることを特徴とする、請求項31記載の抗体またはポリペプチド。
【請求項33】
ヘテロハイブリドーマが、EBV形質転換細胞株由来の細胞に融合させたK6H6B5(ATCC番号CRL-1823)であることを特徴とする、請求項32記載の抗体またはポリペプチド。
【請求項34】
FcγRIIIA、FcγRIIA、およびFcγRIIBのいずれにも結合しないかまたはごく弱く結合する、請求項31〜33のいずれか一項記載の抗体またはポリペプチド。
【請求項35】
請求項1〜7のいずれか一項記載の方法における段階c) iii)によって取得できることを特徴とする、請求項31〜34のいずれか一項記載の抗体またはポリペプチド。
【請求項36】
IgG1サブクラスに属することを特徴とする、請求項1〜35のいずれか一項記載の抗体またはポリペプチド。
【請求項37】
IgG3サブクラスに属することを特徴とする、請求項1〜35のいずれか一項記載の抗体またはポリペプチド。
【請求項38】
請求項20〜25のいずれか一項記載の抗体またはポリペプチドを少なくとも80%、好ましくは少なくとも95%含む、組成物。
【請求項39】
癌、自己免疫疾患、アレルギー、移植後の同種免疫、母子間(materno-fetal)同種免疫、移植片対宿主(GVH)反応、または感染症の治療用の医薬品を製造するための、請求項38記載の組成物の使用。
【請求項40】
癌が白血病、リンパ腫、骨髄腫、セザリー症候群、または固形腫瘍である、請求項39記載の使用。
【請求項41】
母子間同種免疫が新生児溶血性疾患(HDNB)である、請求項39記載の使用。
【請求項42】
自己免疫疾患が、全身性エリテマトーデス(SLE)、特発性血小板減少性紫斑病(ITP)、川崎症候群等の自己抗体を産生するB細胞関連自己免疫疾患である、請求項39記載の使用。
【請求項43】
アレルギーが、喘息、アレルギー性鼻炎、アレルギー性副鼻腔炎、アナフィラキシー症候群、じんま疹、血管性水腫、アトピー性皮膚炎、アレルギー性接触皮膚炎、および紅斑である、請求項39記載の使用。
【請求項44】
請求項26〜30のいずれか一項記載の抗体またはポリペプチドを少なくとも80%、好ましくは少なくとも95%含む、組成物。
【請求項45】
自己免疫疾患、母子間同種免疫、および炎症性疾患の治療用の医薬品を製造するための、請求項44記載の組成物の使用。
【請求項46】
請求項31〜35のいずれか一項記載の抗体またはポリペプチドを少なくとも80%、好ましくは少なくとも95%含む、組成物。
【請求項47】
炎症性疾患、クローン病、または関節リウマチの治療用の医薬品を製造するための、請求項46記載の組成物の使用。
【図1】

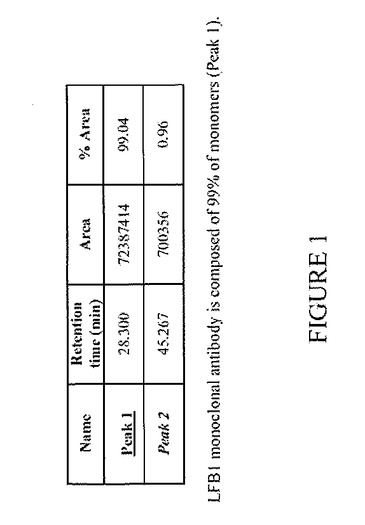
【図2】

【図3】
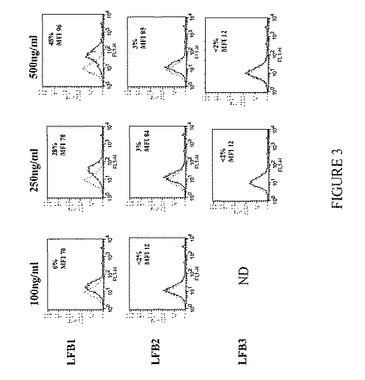
【図4】
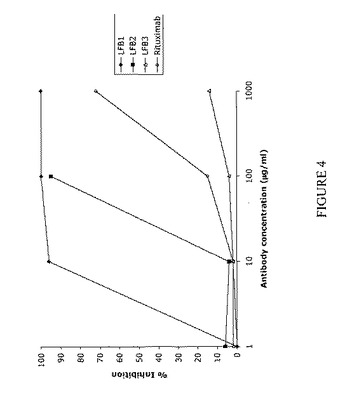
【図5】
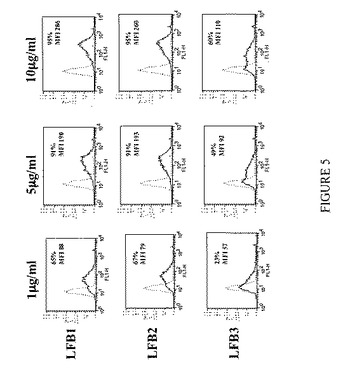
【図6】
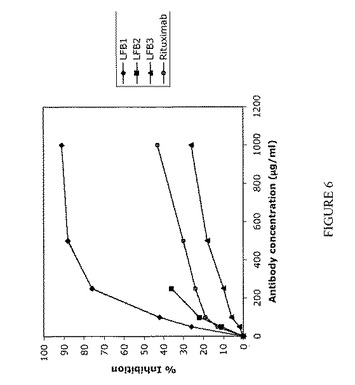
【図7】
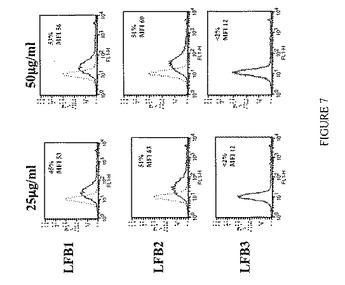
【図8】
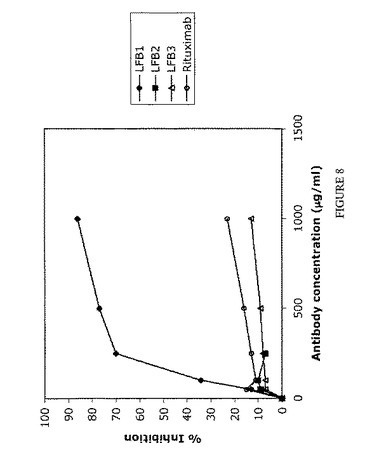
【図9】
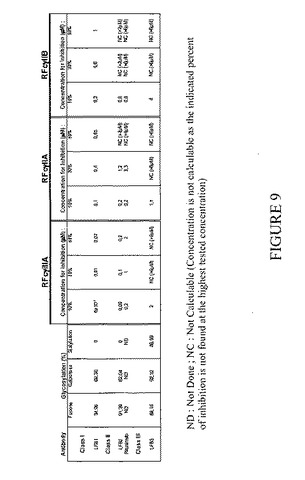
【図10】
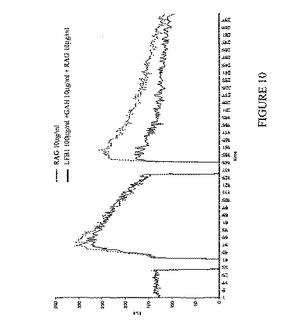
【図11】
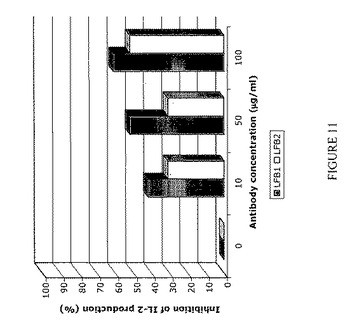
【図12】
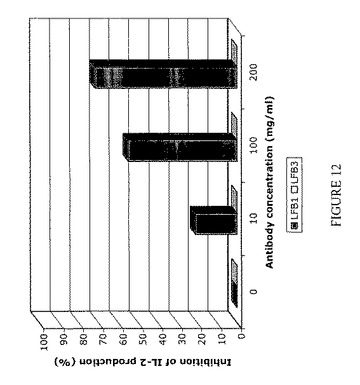
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【公表番号】特表2007−527696(P2007−527696A)
【公表日】平成19年10月4日(2007.10.4)
【国際特許分類】
【出願番号】特願2006−506574(P2006−506574)
【出願日】平成16年4月5日(2004.4.5)
【国際出願番号】PCT/IB2004/001390
【国際公開番号】WO2004/087757
【国際公開日】平成16年10月14日(2004.10.14)
【出願人】(505093378)ラボラトワール フランセ デュ フラクションヌメント エ デ バイオテクノロジーズ (6)
【出願人】(504006489)インスティチュート ナショナル デ ラ サンテ エ デ ラ ルシェルシュ メディカル (インセルム) (5)
【Fターム(参考)】
【公表日】平成19年10月4日(2007.10.4)
【国際特許分類】
【出願日】平成16年4月5日(2004.4.5)
【国際出願番号】PCT/IB2004/001390
【国際公開番号】WO2004/087757
【国際公開日】平成16年10月14日(2004.10.14)
【出願人】(505093378)ラボラトワール フランセ デュ フラクションヌメント エ デ バイオテクノロジーズ (6)
【出願人】(504006489)インスティチュート ナショナル デ ラ サンテ エ デ ラ ルシェルシュ メディカル (インセルム) (5)
【Fターム(参考)】
[ Back to top ]