
細胞老化に関する疾患の治療における医薬組成物を製造するための、GSE24.2誘導化剤の使用
本発明は、老化プロセスによって引き起こされた、好ましくはヒトの疾患または病状の治療における医薬または医薬組成物を製造するためのGSE24.2誘導化化合物またはGSE24.2活性化化合物の使用に関する。上記医薬組成物は、例えば上皮組織または造血細胞などの組織を再生するための治療に使用されることができ、また、バイオテクノロジーの研究またはプロセスにおいて真核細胞を使用するための、当該真核細胞の不死化に使用されることができる。
【発明の詳細な説明】
【発明の詳細な説明】
【0001】
〔技術分野〕
ヒトの健康に適用させるためのバイオテクノロジー分野、およびより具体的には、加齢および細胞老化に関する疾患を患っている人に対して治療的に適用させるための、生物学的な化合物−塩基配列、ペプチド、およびヒトの形質転換細胞。
【0002】
〔従来技術〕
TTAGGG配列(参照文献1)の多数の繰り返しにより構成されているテロメアは、DNAポリメラーゼがクロモソームの末端まで複製できないため、細胞周期のたびに短くなる。クロモソームの末端におけるテロメアの維持は、テロメラーゼ複合体によって行なわれる。この複合体のタンパク質成分であるヒトテロメラーゼ逆転写酵素(hTERT)は、触媒作用を有する逆転写酵素モチーフを含んでおり、一方、テロメアRNA成分は、内部にあるテロメアの繰り返し配列TTAGGGと相補的な鋳型を用いて、デオキシヌクレオチド三リン酸(dNTPs)の作用を導く(参照文献2〜3)。近年、ヒトテロメラーゼ酵素複合体が、2つのタンパク質成分、hTERTおよびジスケリン(dyskerin)、ならびにRNA成分であるhTRのみからどのように構成されているかについて、報告されている(参照文献7)。ジスケリンが、「H/ACA box」リボ核タンパク質に属するシュードウリジン合成酵素と推定される一方、H/ACAモチーフがhTR内に存在する。この複合体は、最終的に、hTERT、hTRおよびジスケリンのそれぞれ2つの分子により構成される。
【0003】
テロメラーゼの発現は、成人の組織における多くの体細胞内では抑制されているが、多くの癌では高いままであり、このテロメラーゼは、クロモソームの整合性を保証するため、これらの癌の細胞内において不死の表現型に寄与している(参照文献6)。テロメラーゼ(hTERT)のレベルは、最初の転写レベルで制御され(参照文献8)、またヒトでは、hTERT遺伝子の5’領域における制御領域によって制御されている。この制御領域は、最大活性化のために必要であり、またmyc結合部位(E−box)を含む(参照文献8〜9)。c−MYC転写因子は、hTERTの発現を促進させるが、mad1の発現が強化されると、hTERTを抑制する(参照文献10〜11)。c−mycの転写制御には、多数のプロモーターが含まれ、なかでもP1およびP2が最も重要である(参照文献12)。c−MYCプロモーターのヌクレアーゼ過感受性エレメントIII(NHEIII)は、c−MYC転写の85〜90%を制御し、当初はDNAseIに対して最も過感受性の部位として同定された(参照文献13)。NHEIIIは、DNA鎖上にプリン−リッチ部位を含み、この部位は、DNAにおける二重螺旋構造とnon−B形態との間のバランスを確立する能力がある(参照文献14)。プリン−リッチなDNA鎖は、異なるG−カルテット(G quartets)による2つの分子内構造を形成でき(参照文献15)、これらは転写抑制因子として機能するため、抗腫瘍性治療のターゲットとして用いられている。
【0004】
テロメアの短縮、またはその二次構造の欠失は、細胞を老化および/またはアポトーシスに向かわせる原因となる。これらの2つのプロセスは、制御不能な細胞分裂および遺伝的不安定性を防止する生物学的なコントロールポイントとして機能する(参照文献4〜5)。テロメアの長さの制御についての欠損は、早期老化症候群および癌などの様々な疾患の病状に関わる(参照文献6)。
【0005】
老化細胞は、DNA合成を停止した生細胞であり、平板化されるとともに液胞が豊富な細胞質が存在し、独特の遺伝子発現プロファイルを有する。そして、老化細胞は、酸性pHにおける、老化に付随する −ガラクトシダーゼ( −gal)活性の増加を可視化する生化学的アッセイによって、および、時間内に細胞集団が倍加できないために、同定可能である。
【0006】
ヒト細胞の老化は、テロメア侵食に関連付けられる。形質転換されていない正常な細胞は、細胞分裂プロセスごとにテロメアが短縮することによって死ぬ運命にある。一方、テロメラーゼを発現する癌細胞は、テロメアの長さを安定化させるので、不死となる。これらの知見により、テロメアが細胞の加齢を制御する生物時計として働くという仮説が提唱される。
【0007】
テロメア侵食に関連する早期老化症候群の1つは、先天性角化異常症(DC)である。これは、非常にまれな遺伝的疾患であり、骨髄機能不全および高い癌性向によって特徴付けられ、これらは先天性角化異常症の患者の死につながる最も重大な原因である(参照文献16〜18)。主にDKC遺伝子の突発性変異によって、クロモソームと結びついた形(X−DC)で引き起こされる(参照文献19〜20)。この遺伝子は、テロメラーゼ複合体の成分となる、シュードウリジン合成酵素ドメインを含む58kDのタンパク質であるジスケリンをコードする。ジスケリンは、脊椎動物において、SnoRNAs(小さい核小体のRNAs)およびhTRに存在する複数のH/ACA boxに結合する(参照文献21)。SnoRNAsは、ジスケリンを介したRNAシュードウリジル化の鋳型として機能する。H/ACA RNAは、H boxと、これに続くトリヌクレオチドACAを含む短い配列とを含む単連鎖領域によって分けられた、2つの可変長の範囲を含む(参照文献22)。SnoRNAのH/ACAにおけるDKCに結合する、3つの異なるタンパク質がある:Nap2、Gar1およびNOP10(参照文献3、22〜24)。DKC、Nap2およびNOP10は、密接に結合して、リボ核タンパク質複合体の安定性に関連する一方、Gar1による相互作用は、より緩やかでありかつ一時的である(参照文献25、26〜28)。したがって、ジスケリンは、hTR(参照文献7、24〜29)の蓄積およびリボソーマルRNAのシュードウリジル化(参照文献30)に必須であるため、テロメラーゼ活性に必要である。DCを有する患者においては、テロメラーゼ活性が減少しており、テロメア構造を維持できないため、年齢とともにクロモソームの不安定性が増加する。X−DC患者のリンパ芽球細胞および繊維芽細胞では、テロメラーゼ活性およびhTRレベルが減少し(参照文献21)、影響を受けていない細胞よりもテロメアが短い。テロメラーゼ活性の欠如は、hTERTおよびhTRの発現によって救済され得る(参照文献21、31)。hTRの変異または欠失によって、先天性角化異常症の他の形態が引き起こされ、テロメアの短縮化の加速および早死を引き起こす。この欠損は、hTRの再発現によってのみ救済され得る(参照文献31)。
【0008】
近年、ジスケリンタンパク質のシュードウリジン合成酵素ドメインをコードする、24−2として知られる遺伝的抑制因子(GSE)が報告されている。GSE24.2は、DC−Xを有する患者の細胞内においてテロメラーゼ活性を回復させるため、テロメラーゼ複合体異常によって起こるこの病気および他の病気のための新しい治療的アプローチが発展しつつある(スペイン国特許出願公開第200502511号明細書、SEQUENCE OF NUCLEOTIDES AND PEPTIDES GSE 24.2 OF DYSKERIN, WHICH CAN INDUCE TELOMERASE ACTIVITY, METHOD FOR OBTAINING SAME, THERAPEUTIC COMPOSITIONS AND APPLICATIONS THEREOF)。しかし、このGSE24.2分子の新しい活性に関する知見によって、新しい治療的ツールおよび研究ツールを発展させることができる。
【0009】
〔本発明の説明〕
<簡単な説明>
本発明の一実施形態は、老化のプロセスによって起こる、好ましくはヒトの疾患または病理学的な状態の治療に対する医薬または医薬組成物を製造するための、GSE24.2誘導化化合物またはGSE24.2活性化化合物の使用(以下本明細書において、本発明に係る使用)により構成される。
【0010】
本発明の好ましい実施形態は、本発明に係る使用からなり、GSE24.2誘導化剤が、哺乳動物の、好ましくはヒトの細胞内において老化からの回復を誘導するタンパク質またはペプチドを発現させる塩基配列(以下本明細書において、本発明に係るGSE24.2の遺伝子配列)であり、以下の群に属するGSE24.2の塩基配列の1個または数個により構成されている:
a)ヒトGSE24.2塩基配列により構成されている塩基配列(配列番号1)、
b)a)の塩基配列の変異体であって、配列番号1と少なくとも90%の同一性を示す塩基配列、
c)a)およびb)のいずれかの配列を有するフラグメント、および、
d)a)、b)およびc)のいずれかの配列を含む遺伝的構成物である、塩基配列。
【0011】
本発明の好ましい実施形態は、本発明に係る使用からなり、a)におけるGSE24.2の配列の塩基配列が、配列番号1により構成されている。
【0012】
本発明の他のより好ましい実施形態は、本発明に係る使用からなり、c)におけるGSE24.2の配列の塩基配列が、配列番号3または配列番号5により構成されており、これらはそれぞれTrubIおよびTrubIIのペプチドドメインをコードするものである。
【0013】
また、本発明の他の好ましい実施形態は、本発明に係る使用からなり、GSE24.2誘導化剤が、哺乳動物の、好ましくはヒトの細胞内において老化を逆転させる活性を表すタンパク質またはペプチド(以下本明細書において、本発明に係るGSE24.2タンパク質)であり、以下の群に属するアミノ酸配列の1個または数個により構成されている:
a)ヒトGSE24.2のアミノ酸配列により構成されているアミノ酸配列(配列番号2)、
b)a)の配列に基づくアミノ酸配列の変異体であって、配列番号2のポリペプチドと少なくとも90%の同一性を示すアミノ酸配列、
c)a)およびb)のいずれかの配列を有するフラグメント、および、
d)a)、b)およびc)に属するいずれかの配列を含むアミノ酸配列。
【0014】
本発明の他のより好ましい実施形態は、本発明に係る使用からなり、GSE24.2誘導化剤はタンパク質であり、このタンパク質におけるa)のアミノ酸配列が配列番号2により構成されている。
【0015】
本発明の他のより好ましい実施形態は、本発明に係る使用からなり、GSE24.2誘導化剤はタンパク質であり、このタンパク質におけるc)のアミノ酸配列が、配列番号4または配列番号6により構成されている。
【0016】
本発明の他の実施形態は、老化異常に関連する疾患または病状を治療するための医薬組成物または医薬(以下本明細書において、本発明に係る医薬組成物)からなり、必要に応じて1つ以上のアジュバントおよび/または医薬的に認容され得る賦形剤とともに、治療的に有効な量にてGSE24.2化合物またはGSE24.2活性化化合物を含んでいる。また、この医薬組成物または医薬は、老化のプロセスを減少または逆転させることが可能であり、かつ、本発明に係る使用により獲得される。
【0017】
本発明の他の実施形態は、老化の異常なプロセスによって引き起こされる、好ましくはヒトの疾患または病状を治療するための処置における、本発明に係る医薬的化合物の使用からなる。
【0018】
本発明の他の好ましい実施形態は、真核細胞、好ましくはヒト細胞からなり、その老化のプロセスが逆転または減少されており、かつ本発明に用いられるGSE24.2分子を含むものである。
【0019】
<本発明の詳細な説明>
本発明は、ジスケリンのフラグメントとして同定された(スペイン国特許出願公開第200502511号明細書を参照)GSE24−2タンパク質(配列番号1)の発現が、hTERTおよびhTRのRNAレベルを制御し、テロメラーゼ活性の回復を誘導できるという、本発明者らの観察結果に基づいている。この結果は、種々の細胞種;ジスケリンの変異体が発現されているDC患者(スペイン国特許出願公開第200502511号明細書)の細胞、および野生型のDKC対立遺伝子を有し、かつ低いテロメラーゼ活性を有する者(VA13)の細胞の両方の細胞において観察されている(実施例4)。また、本発明における意味において、初めてGSE24.2ペプチドを直接に(その遺伝子発現を利用せずに)使用し、その結果これがテロメラーゼ活性を再活性化できることを明らかにした(実施例1)。
【0020】
同時に、GSE24−2タンパク質が、c−myc発現を調節することによってhTERTタンパク質の転写を活性化することを示した。これは、myc/madの融合分子が、hTERTの転写活性化を完全に消滅させるからである。欠失および変異解析によって、ヒトc−myc遺伝子のプロモーターにある、GSE24−2タンパク質の活性化因子としての標的領域は、P1領域を超えて位置するNHEIIIドメインであることが示された。さらに、異常なDNA構造によく結合するCCR5およびPDGFA(参照文献61)などの他の遺伝子プロモーターにも見られるポリプリン配列(Pu27)は、c−myc転写を刺激するためにインタクトのままでなければならない(実施例3)。そのため、NHEIII部位におけるGカルテットの二次構造の異常は、GSE24−2タンパク質におけるこの配列を介した活性を変化させる。NHE部位と相互作用して活性化するタンパク質がほとんど知られていないことは、GSE24.2タンパク質が、NHEドメインを有するプロモーターを持つ遺伝子を制御するために選択されて発現され得ることを意味しており、これは、強調されるべき1つの点である。これらのタンパク質の1つはNH23−H2またはNDPキナーゼBであり、これは、特異的なDNA配列に結合し、c−mycプロモーターのNHE部位に対する親和性を有するタンパク質である。それはまた、ミエロペルオキシダーゼ、CD11b、CCR5などの他の遺伝子の発現を活性化し、PDGFAなどの他の遺伝子の発現を抑制する(参照文献61〜62)。このタンパク質はまた、ヒトにおいてc−mycプロモーターのNHEおよびテロメアに見られるようなグアニン−リッチ配列にインビトロにて結合し得る(参照文献61〜62)。そのため、GSE24−2がNHEを介して転写を活性化し得るための1つの可能性あるメカニズムは、GSE24−2がNM23H2などのタンパク質と結合することによりDNAと相互作用しやすくなり、その結果、転写を増加させるという手段によるものである。
【0021】
X−DCおよびVA13細胞において得られた結果は、GSE24−2ペプチドが、2つの異なるメカニズムの組み合わせによってテロメラーゼ活性を増加させることを示す:1)hTERT転写を活性化し、そして、2)プロモーターの活性には影響を与えることなく、RNAの安定化によってhTRレベルを増加させる。さらに、GSE24.2は、ジスケリンが欠損している場合であっても、H/ACA boxにおけるhNaf1またはNop10などの他のタンパク質の会合を促進できる(参照文献28)。
【0022】
最後に、そして驚くことに、GS24.2タンパク質によるこのテロメラーゼ活性の増加は、繊維芽細胞の繁殖能を回復させるなどの方法によって、X−DC患者におけるこれらの細胞の早期の老化を回復させ、この活性を140日以上維持することが、本発明において観察されている(実施例4、図5を参照)。これは、hTERTおよびhTRの両方のレベルの増加によって媒介され得る。この意味において、GSE24.2のタンパク質または遺伝子は、細胞老化に関連する疾患および病状のプロセスのための治療的アプローチを構成し得る、または、真核細胞を不死化させるため、もしくはその増殖能を増加させるためのツールを構成し得る。一方、GSE24.2の発現が腫瘍形成能を誘導せず、かつ細胞増殖の異常性を増加させないであろうことが、本発明の結果によって示されている(実施例3)。
【0023】
上述した全ての点に照らせば、GSE24.2は、正常なジスケリンを発現するVA13細胞などの細胞、または常染色体優性の先天性角化異常症(DC)もしくは再生不良性貧血などの変異体を発現する細胞を含む、細胞老化を異常化または加速するプロセスに関連する疾患または病状のプロセスを治療するための、医薬または実験用試薬を調製するために使用され得る。
【0024】
したがって、本発明の一実施形態は、老化異常のプロセスによって起こる、好ましくはヒトの疾患または病状の治療に対する医薬または医薬組成物を製造するための、GSE24.2誘導化化合物またはGSE24.2活性化化合物の使用(以下、本発明に係る使用)からなる。本発明において、用語「医薬組成物」は、本発明において記載され、または使用されているGSE24.2の薬剤により構成されている組成物であって、研究分野において(実験用試薬など)、または産業的なバイオテクノロジー関連の活動において使用されるものもまた含む。
【0025】
本発明において使用される場合、GSE24.2誘導化化合物またはGSE24.2活性化化合物の用語は、ジスケリンにおけるGSE24.2フラグメントの塩基配列、または当該塩基配列によってコードされるタンパク質もしくはペプチドの配列であって、哺乳動物、好ましくはヒトの細胞内における老化プロセスを減少または逆転させることが可能なものをさす。この定義はまた、GSE24.2タンパク質をコードする塩基配列を発現させる化合物または分子をも含む。活性化化合物は、ペプチド、タンパク質または塩基配列、抗体および多糖により構成されていてもよい。
【0026】
本発明において使用される場合、用語「正常な、または加速された老化プロセスによって起こる疾患または病状」は、細胞および組織が増殖能の減少(加齢による自然なものを含む)、または加速を示す疾患をさす(本発明の従来技術に記載されている老化の概念を参照)。このように、本発明において使用されるこの用語はまた、ヒト細胞、好ましくは上皮の、または高レベルの増殖を伴うヒト細胞であり、説明するためであって本発明の範囲を限定するものではないが、以下の群に属する細胞の、自然な加齢によって異常化した生理的な状態をもさす:皮膚細胞、好ましくは表皮の皮膚細胞、腸上皮組織、角膜、肝臓、肺、毛球等、または造血系細胞、好ましくはリンパ球、マクロファージおよび赤血球;精巣および卵巣における胚芽細胞等であって、それぞれ、体細胞か、幹細胞か、または胚細胞。老化のプロセスおよび/またはテロメラーゼ活性の制御による、ヒトの疾患または病状のプロセスの例は、特に限定されないが、Blasco MAの改訂版に見られる(Blasco MA, Telomere length, stem cells and aging. Nature Chemical Biology 3 (10): 640-649, 2007; Crabbe L, Jauch A, Naeger CM, Holtgreve-Grez H., and Karlseder J. Telomere dysfunction as a cause of genomic instability in Werner syndrome. PNAS 104 (7): 2205-2210, 2007)。
【0027】
細胞老化のプロセスの逆転または減少は、分裂して器官の恒常性を維持する能力を有する細胞において治療的に使用され得る。これによって、加齢か、毒素による組織の損傷か、外科的処置に続く組織の損傷か、そして先天的または遺伝的な原因、例えば肺の形成不全など、の再生不良性の組織であるか、の原因に無関係に、組織の再生能力を増加させる。さらに、この細胞老化のプロセスにおける逆転または減少は、移植治療または細胞治療において、幹細胞、体細胞または胚細胞の集団の生存能力を増加させるために使用され得る。これらの治療において、幹細胞はテロメラーゼを発現するが、それらはテロメアの長さを長期間維持できないため、GSE24.2ペプチドを用いた細胞の形質転換または処理によって、これらの細胞の生存機能を高め、そしてそれゆえこれらの細胞処理の効率性を高めることができる。このように、例えば、この医薬組成物は、高齢者の表皮を再生するために使用され得る。または、この医薬組成物は、皮膚の創傷もしくは火傷の皮膚再生を促進するため、または化学療法による治療をされた患者に移植された自家造血細胞を刺激するために使用され得る。
【0028】
したがって、本発明の好ましい実施形態は、本発明に係る使用を含み、GSE24.2誘導化剤は、哺乳動物の、好ましくはヒトの細胞内において老化からの回復を誘導するタンパク質またはペプチドを発現させる塩基配列(以下本明細書において、本発明に係るGSE24.2の遺伝子配列)であって、以下の群に属するGSE24.2の塩基配列の1個または数個により構成されているものである:
a)ヒトGSE24.2塩基配列により構成されている塩基配列(配列番号1)、
b)a)の塩基配列の変異体であって、配列番号1と少なくとも90%の同一性を示す塩基配列、
c)a)およびb)のいずれかの配列を有するフラグメント、および、
d)a)、b)およびc)のいずれかの配列を含む塩基配列である、遺伝的構成物。
【0029】
この記載に使用される意味において、用語「変異体」は、配列番号1の配列に基づいて、単離され得る塩基配列、または、例えば1つ以上の塩基の挿入を含む、塩基の保存的置換または非保存的な置換の導入、分子のいずれかの末端に対する1つ以上の塩基の付加、もしくはいずれかの末端か配列内部かからの1つ以上の塩基の欠失、によって構築され得る塩基配列のいずれかを含み、かつ、GSE24.2(配列番号2)またはそのフラグメント(配列番号4および配列番号6)の配列の活性を模倣することができるペプチドまたはタンパク質をコードし得ることを意図する。
【0030】
酵素ジスケリンは、様々な生物に存在するシュードウリジン合成酵素ファミリーに属する(図3Bを参照、Mitchel et al, 1999)。当業者は、本発明に含まれる情報、−およびスペイン国特許出願公開第200502511号明細書−に基づき、自然界に存在する種々の生物から、本明細書に記載した塩基配列のアナロガスを同定または構築することができる。
【0031】
本発明において使用される場合、用語「塩基配列」は、DNA、cDNAまたはmRNAの配列をさす。
【0032】
本発明のより好ましい実施形態は、本発明に係る使用からなり、a)におけるGSE24.2の配列の塩基配列が、配列番号1により構成されている。
【0033】
本発明の他のより好ましい実施形態は、本発明に係る使用からなり、c)におけるGSE24.2の配列の塩基配列が、配列番号3または配列番号5により構成されており、これらはそれぞれTrubIおよびTrubIIのペプチドドメインをコードするものである。
【0034】
GSE24.2の塩基配列は、GSE24.2の遺伝的構成物に相当するd)として同定される。この本発明に係るGSE24.2の遺伝的構成物はまた、もし必要であれば、発現したペプチドの細胞質からの単離、検出または分泌をよりよく行なうために、単離、検出または分泌の目的で使用され得るペプチドをコードする塩基配列を含んでいてもよい。したがって、本発明の他の好ましい実施形態は、本発明に係る使用からなり、その塩基配列は、GSE24.2の遺伝的構成物であって、GSE24.2の塩基配列に加えて、発現したペプチドを細胞質から単離、検出または分泌させるペプチドまたはペプチド配列を含む。このペプチドまたはペプチド配列は、例えば、本発明の範囲を限定するものではないが、ポリヒスチジン(6×His)配列、モノクローナル抗体により認識可能なペプチド配列(例えば、その同定のための抗体、または免疫親和性クロマトグラフィーを用いて、その結果の融合タンパク質を精製するために利用する他の抗体:c−myc、HA、E−tagなどのタグペプチド)(抗体の使用:実験用マニュアル、Ed. Harlow and David Lane (1999). Cold Spring Harbor Laboratory Press. New York. Chapter: Tagging proteins. Pp. 347-377)などである。
【0035】
上述したGSE24.2の塩基配列およびGSE24.2の遺伝的構成物は、従来技術における公知の方法により、専門家によって取得できる(Sambrook et al. “Molecular cloning, a Laboratory Manual 2nd ed., Cold Spring Harbor Laboratory Press, N.Y., 1989 vol 1-3)。この塩基配列は、細胞内にて適した条件下でその発現を制御することができる遺伝子発現ベクター内に組み込まれてもよい。
【0036】
したがって、本発明の他の好ましい目的は、本発明に係る使用からなり、GSE24.2誘導化剤またはGSE24.2活性化剤は、GSE24.2の発現ベクターであって、これは、本発明において記載されるGSE24.2の塩基配列またはGSE24.2の遺伝的構成物を含み、そして、哺乳動物、好ましくはヒトの細胞内において老化のプロセスを減少または逆転させることが可能なタンパク質またはペプチドを発現させるものである。より好ましい実施形態は、発明pLNCX24.2の発現ベクターによって表される(スペイン国特許出願公開第200502511号明細書をも参照)。
【0037】
一般的に、発現ベクターは、本発明に記載のGSE24.2の塩基配列または24.2の遺伝的構成物に加えて、その転写を導くプロモーター(例えば、pT7、plac、ptrc、ptac、pBAD、retなど)であって機能的に結びついているもの、ならびにこの転写、および適切な場所における所定の産物の翻訳を調節および制御する他の必要な、または適切な配列、例えば、転写開始シグナルおよびストップシグナル(tlt2など)、ポリアデニル化シグナル、複製点、リボソーム結合配列(RBS)、転写制御因子をコードする配列、(エンハンサー)、転写サイレンサー、リプレッサーなど、を含む。好適な発現ベクターの例は、発現プラスミド、ウイルスベクター(DNAまたはRNA)、コスミド、人工クロモソームなどの中からそれぞれの特定の場合における条件および必要に応じて選択されることができ、また、所定の1つまたは複数の遺伝子を用いて形質移入された細胞または形質転換された細胞を選択するために使用され得るマーカーをも含み得る。ベクターの選択は、宿主細胞、および要求される使用形態に依存され得る。したがって、本発明の好適な実施形態によれば、ベクターは、プラスミドまたはウイルスベクターである。微生物および真核細胞の形質転換に、異なった複数の周知の方法(化学形質転換、エレクトロポレーション、マイクロインジェクションなど)が使用され得る(様々なマニュアルに記載されている(Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989). Molecular cloning: a laboratory manual, 2nd ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.))ように同様に、ベクターは、当業者によって知られている慣用的な方法を用いて取得され得る。
【0038】
さらに、本発明の他の好ましい実施形態は、本発明に係る使用からなり、GSE24.2誘導化剤は、哺乳動物、好ましくはヒトの細胞内において老化を逆転させる活性を表すタンパク質またはペプチド(以下本明細書において、本発明に係るGSE24.2タンパク質)であって、以下の群に属するアミノ酸配列の1個または数個により構成されている:
a)ヒトGSE24.2のアミノ酸配列により構成されているアミノ酸配列(配列番号2)、
b)a)の配列に基づくアミノ酸配列の変異体であって、配列番号2のポリペプチドと少なくとも90%の同一性を示すアミノ酸配列、
c)a)およびb)のいずれかの配列を有するフラグメント、および、
d)a)、b)およびc)に属するいずれかの配列を含むアミノ酸配列。
【0039】
本発明の原理から理解されるように、配列番号2の配列に示されるポリペプチドの変異体は、当該配列から、1つ以上のアミノ酸の置換、挿入または欠失によって取得されうるいずれかのペプチドである。置換による変異体の場合には、置換は好ましくは保存的置換であり、すなわち、アミノ酸は、その側鎖の性質が似た特徴を有する他のアミノ酸に置換される。したがって、保存的置換は、表1に示すアミノ酸の群の中での置換を含む。
【0040】
【表1】

【0041】
さらに、本発明の変異体における1つ以上のアミノ酸は、特別な天然または合成アミノ酸によって置換されてもよい。このアミノ酸は、例えばベータ−アミノ酸、2−アミノアジピン酸、アルファ−アスパラギン、2−アミノブタン酸、2−アミノカプロン酸、アルファ−グルタミン、アルファ−メチルアラニン、2−アミノピメリン酸、ガンマ−アミノ−ベータ−ヒドロキシベンゼンペンタン酸、2−アミノスベリン酸、2−カルボキシアゼチジン、ベータ−アラニン、ベータ−アスパラギン酸、3,6ジアミノヘキサン酸、酪酸、4−アミノ 4−アミノ−3−ヒドロキシ酪酸、ガンマ−アミノ−ベータ−ヒドロキシシクロヘキサンペンタン酸、N5−アミノカルボニルオルニチン、3−スルホアラニン、2,4 ジアミノ酪酸、ジアミノピメリン酸、2,3ジアミノプロパン酸、2,7ジアミノスベリン酸、S−エチルチオシステイン、ガンマ−グルタミン酸、ガンマ−カルボキシグルタミン酸、ピログルタミン酸、ホモアルギニン、ホモシステイン、ホモヒスチジン、ホモセリン、2−ヒドロキシイソ吉草酸(2-hydroxyisovaleric acid)、2−ヒドロキシペンタン酸、5−ヒドロキシリジン、4−ヒドロキシプロリン、2−カルボキシオクタヒドロインドール、3−カルボキシイソキノリン、イソバリン、2−ヒドロキシプロパン酸、メルカプト酢酸、メルカプト酪酸、4−メチル−3−ヒドロキシプロリン、メルカプトプロパン酸、ノルロイシン、ノルチロシン、ノルバリン、オルニチン、ペニシラミン、2−フェニルグリシン、2−カルボキシピペリジン、サルコシン、1−アミノ−1−カルボキシシクロペンタン、スタチン(statin)、3−チエニルアラニン(3-thyenylalanine)、イプシロン−N−トリメチルリジン、3−チアゾールアラニン、アルファ−アミノ−2,4−ジオキソピリミジンプロピオン酸などである。
【0042】
さらに、本発明は、本発明に係るペプチドの変異体として、1つ以上のアミノ酸の側鎖を変化させた変異体の使用について検討する。本発明における側鎖の改変の例は、アルキル化、アミジン化、アシル化、カルボモイル化(carbomoylation)、トリニトロベンジル化、ピリドキシル化などのアミノ基の改変、複素環縮合の形成からなるアルギニン残基のグアニジン基の改変;アミド化によるカルボキシル基の改変、メトキシル化によるチロシンの改変、アルキル化またはN−カルボキシエチル化によるヒスチジンのイミダゾール環の改変、4位のヒドロキシル化によるプロリンの改変などを含む。あるいは、本発明は、本発明に係るペプチドの変異体として、グリコシル化、すなわち、セリンおよび/もしくはスレオニン(O−グリコシル化)の側鎖、またはアスパラギンおよび/またはグルタミン(N−グリコシル化)の側鎖のどちらかへのグリカン基の付加による変異体について検討する。本発明におけるポリペプチドに組み込まれ得るグリカンが含む糖ユニットの数は、変えることができる(単糖、二糖、三糖、四糖など)。グリカンを形成する単糖は、D−アロース、D−アルトロース、D−グルコース、D−マンノース、D−グロース、D−イドース、D−ガラクトース、D−タロース、D−ガラクトサミン、D−グルコサミン、D−N−アセチルグルコサミン、D−N−アセチルガラクトサミン、D−フコース、またはD−アラビノースを含む。
【0043】
あるいは、本発明は、本発明に記載されたポリペプチドの変異体として、逆反転(retro-inverse)アイソマーを生じさせるために、ペプチド鎖を構成するアミノ酸の少なくとも1つとしてD立体異性体が含まれる変異体の使用を検討する。
【0044】
他の実施形態において、本発明は、本発明に記載されたポリペプチドにおける擬態ペプチド(peptidomimetics)、すなわち1つ以上のペプチド結合が共有結合に代わるものにより置き換わっている変異体の使用を検討する。この擬態ペプチドは、プロテアーゼに対してより耐性であるため、より高い安定性を示すことによって特徴付けられる。ペプチド骨格の改変は、ペプチド結合の構成要素(−NH−、−CH−、−CO−)に対する基の置換または挿入を含み、この基とは、例えば、−NH−の代わりに−O−、−S−、−CH2など、−CHRの代わりに−N−、−C−アルキル p −BH−など、および、−CO−の代わりに−CS−、−CH2−、−SOn−、−P=O(OH)−または−B(OH)−などである。さらに、N末端をブロックする基を用いれば、本発明に係るペプチドの安定性を高めることができる。この基とは、例えば、t−ブチルオキシカルボニル基、アセチル基、スクシニル基、メトキシスクシニル基、スベリル基(suberyl)、アジピル基(adipyl)、ダンシル基(dansyl)、ベンジルオキシカルボニル基、フルオレニルメトキシカルボニル基、メトキシアジピル基、メトキシアジピル基、メトキシスベリル2,3−ジニトロフェニル基などである。この代わりに、または同時に、アミド化によって本発明に係るペプチドのC末端を改変することが可能である。
【0045】
変異体と、配列番号2の配列によって示されるポリペプチドとの間の同一性は、コンピュータアルゴリズムおよび当業者に周知の方法を用いて決定される。2つのアミノ酸配列間の相同性は、BLASTPアルゴリズム(BLASTManual, Altschul, S., et al, NCBI NLM NIH Bethesda, Md. 20894, Altschul, S., et al., J. MoI. Biol. 21 5: 403-410 (1990))を用いて決定されることが好ましい。本発明において使用されるポリペプチドは、配列番号1〜19の配列によって示されるポリペプチドに対し、少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも91%、少なくとも92%、少なくとも93%、少なくとも94%、少なくとも95%、少なくとも96%、少なくとも97%、少なくとも98%、または少なくとも99%の配列の同一性を示すことが好ましい。
【0046】
酵素ジスケリンは、種々の生物に存在するシュードウリジン合成酵素ファミリーに属する(図3B、Mitchel et al, 1999を参照)。当業者は、本明細書に記載の情報および自然界に存在する種々の生物から、本明細書に記載のアミノ酸配列と類似したアミノ酸配列を同定または構築することができる。
【0047】
本発明の他のより好ましい実施形態は、本発明に係る使用からなり、GSE24.2誘導化剤がタンパク質であって、そのa)のアミノ酸配列が配列番号2により構成されているものである。
【0048】
本発明の他のより好ましい実施形態は、本発明に係る使用からなり、GSE24.2誘導化剤がタンパク質であって、そのc)のアミノ酸配列が配列番号4または配列番号6により構成されているものである。
【0049】
一方、本発明の他のさらなる実施形態は細胞(以下本明細書において、本発明に係るGSE24.2細胞)からなり、この細胞は、真核細胞−好ましくはヒト細胞、より好ましくは単離された細胞−または原核細胞であり、遺伝的に改変されており、かつ本発明に係るGSE24.2の塩基配列、構成物および発現ベクターを含んでおり、本発明において使用されるGSE24.2のペプチドまたはタンパク質が安定的に発現され得る。これらの細胞は、当業者に公知の遺伝子工学技術を用いて、上述した塩基配列により形質転換、感染または形質移入され得る(Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989). Molecular cloning:実験用マニュアル、2nd ed. Cold Spring Harbor Laboratoryおよび本発明に含まれる)。これらの細胞は、例えば老化プロセスを逆転させ、または減少させることができるペプチドを生産するために有用であり得る。また、これらの細胞は、例えば上述した塩基配列の組み換え体を増幅するために、医薬組成物のための基材として貢献し得る。あるいは、これらの細胞は、例えば遺伝子治療において細胞自体として有用であり得る。好ましい実施形態は、上述したGSE24.2の塩基配列により形質転換された、種々の細胞系統由来のヒト細胞の使用であってもよく、この細胞は、ヒト組織の再生細胞として使用できる。
【0050】
他の実施形態において、本発明は、本発明に係る遺伝的構成物または本発明において規定されたベクターを含む細胞または宿主生物に関する。本発明には、当業者に公知の宿主生物のいかなる類型をも使用でき、例えば細菌系統(大腸菌(Escherichia coli)、枯草菌(Bacillus subtilis)など)、酵母系統(酵母(Saccharomyces cerevisiae)、ピキア・パストリス(Pichia pastoris)、クリベロマイセス・ラクティス(Kluyveromyces lactis)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)など)、遺伝子導入植物(双子葉植物または単子葉植物)、昆虫細胞、例えば、バキュロ・ウイルス、哺乳動物細胞(COS細胞、CHO細胞、C127細胞、HeLa細胞など)、および非ヒトトランスジェニック(例えば、マウス、ウシ、ヤギ、ウサギ、ブタなど)などを使用できる。
【0051】
宿主細胞のゲノム中に新しい遺伝的物質が組み込まれるか組み込まれないかは、遺伝子発現系によって決まる。このように、GSE24.2の塩基配列、遺伝的構成物または発現ベクターはいずれも、異常な老化プロセスに関連する疾患に影響を受けるヒトに対する遺伝子治療の治療法および予防法において、異常化している(増加している)か否かにかかわらず老化プロセスに影響を受ける宿主細胞、好ましくはヒト細胞を保護するための医薬として使用され得る。同様に、本発明に係るGSE24.2細胞は、ヒトの組織または細胞の再生または移植のための医薬として使用され得る。これらの生物製剤ツールおよび遺伝子治療法は、本発明に記載の情報を過度の努力なしに発展させられる技術を有する当業者に充分に知られている。さらに、このタンパク質またはペプチドおよび細胞それら自身は、バイオドラッグとなり得る。
【0052】
本発明の他の実施形態は、老化の異常に関連する疾患または病状を治療するための医薬組成物または医薬(以下本明細書において、本発明に係る医薬組成物)からなる。この医薬組成物は、必要に応じて1つ以上のアジュバントおよび/または薬剤的に認容され得る溶媒とともに、GSE24.2誘導化化合物またはGSE24.2活性化化合物を治療的に有効な量において含む。また、この医薬組成物は、老化プロセスを減少させまたは逆転させることができ、かつ本発明に係る使用を用いて取得される。
【0053】
医薬に使用するために、本発明に係る化合物を有する化合物および組合せは、医薬品の観点から認容され得る賦形剤とともに処方され得る。本発明における使用のための好ましい賦形剤は、糖、デンプン、セルロース、粘剤およびタンパク質を含む。
【0054】
この記載中に使用される意味において、用語「治療的に有効な量」とは、老化からの回復が可能な薬剤または化合物の量をさし、所望の効果を生み出すために算出されるとともに、一般に、その化合物の固有の特性に基づいて、年齢、患者の状態、異常性または障害の重症度、および投与の経路および頻度を含む他の原因から決定されてもよい。
【0055】
好ましい実施形態において、本発明に係る医薬的組成物は、固形(例えば、錠剤、カプセル剤、トローチ剤、顆粒剤、座薬など)または液体(剤剤、懸濁剤、乳剤など)にて投与する医薬的な形態として処方されてもよい。他の好ましい実施形態では、本発明に係る医薬組成物は、限定されないが、経口、静脈内、筋肉内、動脈内、髄内、くも膜下腔内、脳室内、経皮、皮下、腹腔内、鼻腔内、腸内、局所的、舌下または直腸を含むいかなる経路を解して投与されてもよい。有効成分の種々の投与形態、使用される賦形剤およびその製造方法についての総説を、Galenic Pharmacy Treatise (Tratado de Farmacia Galenica), C. Fauli i Trillo, Luzan 5, S.A. de Ediciones, 1993.にて見出すことができる。
【0056】
本発明の他の実施形態は、好ましくはヒトにおいて、異常な老化のプロセスによって引き起こされる疾患または病状を治療するための方法における、本発明に係る医薬組成物の使用からなる。
【0057】
他の好ましい実施形態が、生物医学研究に使用する真核細胞、好ましくはヒト細胞または動物細胞を不死化するための方法における本発明に係る医薬組成物の使用からなることは、非常に重要である。真核細胞の不死化は、組み換えタンパク質生産工程に関して、好ましくは生物医学的適用における組み換えタンパク質生産工程に関して、その有効寿命を増加させるため、バイオテクノロジーのR+Dの分野において大いに必要な操作である。また、治療化合物として可能性のある化合物の効果の解析に用いられる正常細胞の有効寿命を増加させ、また、そのより長い寿命のおかげで二次的な効果を長期にわたって解析するのに好都合であるため、バイオテクノロジーのR+Dの分野において大いに必要な操作である。
【0058】
本発明の他の好ましい実施形態は、老化プロセスが逆転され、または減少しており、かつ本発明に用いられるGSE24.2の構成要素を含む、真核細胞、好ましくはヒト細胞からなる。
【0059】
〔図面の説明〕
<図1−シスプラチンへの応答におけるテロメラーゼ活性に対するGSE24−2ペプチドの発現の効果>
(C)シスプラチンおよびGSE24−2ペプチドを用いた、293T細胞抽出物のインビトロでの処理に続く、テロメラーゼ活性。シスプラチン、および精製したペプチドGSE24−2の量を増加させたもの(3−5列目)、熱により失活させたGSE24−2ペプチド(7列目)またはコントロールペプチド(8列目)を用いて、293T細胞抽出物をインビトロにて処理した後、後者をTPAPアッセイに用いた。テロメラーゼアッセイを行なうために、テロメアの配列を有する特異的な鋳型を用いた。この実験を3回繰り返して同様の結果を得た。(D)GSE24−2を発現する細胞抽出物を用いたゲルシフト試験。プラスミドpcDNA3−9E1024−2またはpcDNA3−9E10を発現する細胞の核抽出物をゲルシフト試験に供した。特異的なテロメアの配列を含むTEL1プローブを使用した。特異的な抗体9E10(3列目)、およびTEL1とハイブリダイズして競合分子として機能するオリゴヌクレオチドCXext(4−5列目)を用いることにより、特異的な結合が確立された。この実験を3回繰り返して同様の結果を得た。
【0060】
<図2−GSE24−2はhTERTプロモーターの活性を増加させる>
(A)pLNCX、DXC、GSE24−2、Cbf5、モチーフIまたはモチーフII(細胞百万個あたり10μgのDNA)およびレポータープラスミドhTERT−luc(細胞百万個あたり1μg)を用いて、293T細胞を同時形質移入した。形質移入して24時間後のテロメラーゼ活性を測定した。*は統計値がp<0.05であることを示す。(B)レポーターベクターhTERT−luc(細胞百万個あたり1μg)を用いて、293T細胞(黒)および293T24−2細胞(白)を同時形質移入した。24時間後、これらの細胞をシスプラチン(3 g/ml)により8時間処理した後、ルシフェラーゼ活性を測定した。(C)提示された構築物(細胞百万個あたり10μgのDNA)およびレポーターベクターhTR−luc(細胞百万個あたり1μg)を用いて、293T細胞を同時形質移入した。形質移入して24時間後のルシフェラーゼ活性を測定した。(D)レポーターベクターhTERT−luc、および様々な量のMad/mycの発現ベクターを用いて、これらの細胞系統を同時形質移入した。形質移入して24時間後のルシフェラーゼ活性を測定した。(E)レポーターベクターHIV−lucと、発現ベクターmad/mycの量を増加させたものとを用いて、これらの細胞系統を同時形質移入した。形質移入して24時間後に、50ng/mlのTNF を用いてこれらの細胞を6時間刺激した後、ルシフェラーゼ活性を測定した。形質移入効率を制御するために、CMV−Renillaベクターを用いた(0.1μg/ml)。それぞれのグラフは、4通りにて行なった2つの実験の平均値および標準偏差を表す。
【0061】
<図3−GSE24.2はc−MYC遺伝子のプロモーターを介してその活性を誘導する>
(A)実験に使用したそれぞれの構築物を示す、c−MYCプロモーターの概略図。(B)レポーターベクターc−MYC−luc(細胞百万個あたり1μg)のそれぞれの構築物を用いて、293T pLNCXおよび293T GSE24−2の細胞を形質移入した。形質移入して24時間後にルシフェラーゼ活性を測定した。形質移入効率のコントロールとして、CMV−Renillaベクターを用いた(細胞百万個あたり0.1μg/ml)。それぞれのグラフは、4通りにて行なった2つの実験の平均値および標準偏差を表す。(C)NHEIIIにおいて作製した変異体の概略図。(D)プラスミドpx3.2由来の変異体のレポーターベクター(細胞百万個あたり1μg)を用いて、提示された細胞系統を形質移入した。形質移入して24時間後にルシフェラーゼ活性を測定した。形質移入効率のコントロールとして、CMV−Renillaベクターを用いた(細胞百万個あたり0.1μg/ml)。それぞれのグラフは、4通りにて行なった2つの実験の平均値および標準偏差を表す。(E)発現ベクターpLNCX、DKCまたはGSE24−2を用いて、MEF細胞を形質移入した。形質移入して24時間後、全RNAを単離し、RT−PCRによって、マウスのc−myc遺伝子および −アクチン遺伝子の発現を測定した。この実験を3回繰り返して同様の結果を得た。(F)発現ベクターpLNCX、DKCまたはGSE24−2を用いて、MEF細胞を形質移入した。形質移入して24時間後、全タンパク質の抽出物を単離し、特異的な抗体を用いたウェスタンブロットによって、マウスのc−mycタンパク質および −チューブリンタンパク質の発現を決定した。この実験を3回繰り返して同様の結果を得た。
【0062】
<図4−GSE24−2は、X染色体に関連する先天性角化異常症の患者の細胞、およびVA13細胞におけるテロメラーゼ活性を再活性化させる>
(A)キャリアの母親(DC−C)、および影響を受けた子供(DC−1、DC−2およびDC−3)から取得した、X染色体に関連する先天性角化異常症を患う患者のリンパ芽球において、テロメラーゼ活性を測定した。これらの細胞に対し、空のベクター(−)またはGSE24−2(+)を細胞百万個あたり3μg用いて一過的に形質移入した。その24時間後にテロメラーゼ活性を測定した。(B)細胞百万個あたり3μgの発現ベクターpLNCX、GSE24−2またはDKCを用いて、一過的にDC2細胞を形質移入した。その24時間後にテロメラーゼ活性を測定した。(C)細胞百万個あたり3μgの空のベクターまたはGSE24.2により形質移入された患者(DC−3)の1人の細胞における、hTERTおよびhTRの発現レベル。RNAレベルは、RT−PCRにより検出した。GAPDH発現をコントロールとして用いた。(D)X−DC GMO1787の患者の繊維芽細胞を、ベクターpLNCXまたはGSE24−2由来のウイルスによって感染させ、安定な細胞系統をテロメラーゼ活性測定のために用いた(タンパク質の全量を三角形にて示す)。(E)プラスミドpLNCXおよびpGSE24−2によって形質移入したGMO1787細胞におけるhTERTおよびhTRの発現レベルを、RT−PCRにより測定した。GAPDH発現をコントロールとして用いた。(F)パネルDに記載したように形質移入されたGMO1787細胞の増殖速度を、一定時間における個体数の重複の累積を用いて評価した。全ての実験について3回繰り返し、同様の結果を得た。(G)VA13細胞におけるテロメラーゼ活性。この細胞に対し、細胞百万個あたり16μgのコントロールベクターpLNCXまたはGSE24.2を用いて一過的に形質移入した。その24時間後に、タンパク質抽出液の連続した希釈液を用いて、テロメラーゼ活性を測定した(タンパク質の全量を三角形にて表す)。(H)細胞百万個あたり16μgのプラスミドpLNCX、DKCまたはGSE24−2により形質移入されたVA13細胞における、hTERTおよびhTRの発現レベル。RT−PCRによりRNAレベルを測定した。GAPDHレベルをコントロールとして用いた。
【0063】
<図5−ペプチドGSE24.2は細胞老化から回復させる>
図4Dに記載されたように形質移入されたGMO1787細胞の増殖速度を、一定時間における個体数の重複の累積を用いて評価した。全ての実験について3回繰り返し、同様の結果を得た。
【0064】
〔本発明の実施例〕
(実施例1−GSE24.2ペプチドはインビトロにてテロメラーゼ活性を救済させる)
テロメア配列TTAGGGは、シスプラチンの標的である(参照文献45)。シスプラチンがこの領域に結合することによって、テロメラーゼの阻害とテロメアの短縮化とがもたらされる(参照文献43)。したがって、ジスケリンはテロメラーゼ複合体の構成要素の1つであるため(参照文献7)、本発明者らはTRAP(参照文献37)アッセイを用いてテロメラーゼ活性におけるシスプラチンおよびGSE24−2の発現の効果についての研究を続け、GSE24−2を発現させるベクターにより形質移入された細胞が高いテロメラーゼ活性を示すことを見出した(スペイン国特許出願公開第200502511号明細書)。
【0065】
本説明では、はじめに、293T細胞からの細胞抽出物、シスプラチン、およびシスプラチンに処理されたテロメアプローブを用いたインビトロのテロメラーゼ活性阻害試験における、前述したGSE24.2ペプチドの使用について記載する。この試験では、GSE24−2ペプチドを用量依存的な様式にて加えた結果、抽出物におけるテロメラーゼ活性をシスプラチンによる阻害から保護した(図1C、3−5列)。一方、熱により失活されたGSE24−2ペプチドまたは無関係のペプチドは、シスプラチンにより誘導されたテロメラーゼ阻害を救済することができなかった(図1C、7−8列)。これらによって、GSE24−2ペプチドが、インビトロでテロメアにおいてシスプラチンにより誘導される架橋を阻害できることが示された。さらに、小さい3’の吊り下がった配列によってテロメアの二重鎖配列を再構築する、ループを形成するオリゴヌクレオチド(TEL1)(参照文献41)を用いて、GSE24−2ペプチドにおけるテロメア配列に対する結合の可能性について、ゲルシフト試験によって解析した。特異的な複合体GSE24−2/TEL1(図1D)が検出された。一本鎖の延長部分の3’にハイブリダイズするオリゴヌクレオチドCxextは、複合体GSE24−2/TEL1の形成を完全に阻害した。併せて、これらの結果は、GSE24−2がテロメアの配列に結合し得ること、およびGSE24−2のテロメアプローブに対する保護効果によってシスプラチンによるテロメラーゼ活性の阻害が部分的に欠如することを示す。
【0066】
(実施例2−hTERTのプロモーターはGSE24−2タンパク質によって活性化されるが、hTRのプロモーターは活性化されない)
従来、GSE24.2におけるシスプラチンに対して保護できる能力によって、hTERTの高いRNAレベルが維持され得ることが観察された(スペイン国特許出願公開第200502511号明細書)。この仮説により、hTERTを過剰発現する293T細胞は、コントロール細胞よりもシスプラチンに対してより耐性であった。
【0067】
シスプラチンがhTERTのmRNAの減少を誘導し、かつGSE24−2の発現がこの効果を補償しているのではないかと考え、本発明者らは、GSE24−2がhTERTの転写を変化させ得るかどうかを詳細に調べた(参照文献53)。本発明者らは、hTERTプロモーターの3402bpを含むレポーターベクターを用いて、GSE24−2細胞におけるこのプロモーター活性を調査した(参照文献33)。GSE24−2の発現は、一過的な形質移入試験においてhTERTプロモーターの転写を刺激できたが、DKCはできなかった(図2A)。また、シスプラチンは、コントロール細胞よりもGSE24−2細胞において、このプロモーター活性を刺激する(図2B)。これらの結果は、GSE24−2の発現が、シスプラチン存在下であっても転写を刺激でき、その結果hTERTのmRNAレベルが増加したことを示す。本発明者らはまた、プラスミドGSE24−2により形質移入されたMEF細胞(一次マウスの繊維芽細胞)におけるhTERTプロモーターの刺激について検出できた(データは示さない)。DKCまたはGSE24−2を発現する細胞を用いたところ、hTRプロモーター(参照文献9)に対するGSE24−2の発現の効果は観察されなかった(図2C)。しかし、以前に報告されたように、阻害剤JNK SP60125(参照文献9)の処理は、このプロモーター活性を活性化することができた(データは示さない)(参照文献11)。これらの結果は、GSE24−2活性がhTERT発現に対して特異的であることを示す。
【0068】
ジスケリンは、当初は、酵母(S. cerevisiae)のCbf5遺伝子と、タンパク質レベルで85%の同一性を示す、ヒトの相同遺伝子cbf5(参照文献55)として同定された(参照文献56)。そのため、本発明者らは、酵母遺伝子S. cerevisiae cbf5から、GSE24−2に相当する配列(配列番号2)をベクターpLNCXにクローニングした。この構築物の発現もまた、hTERTプロモーターの発現を増加させる(図2A)が、hTRプロモーターの発現は増加させない(図2C)。これは、このDKC領域の機能が高度に保存されていることを示す。
【0069】
(実施例3−c−MYCプロモーターのNHEIIIドメインはGSE24−2の標的配列である)
hTERTプロモーターは、myc/maxヘテロダイマーに結合可能な2つのE−boxを含む(参照文献32、57)。madをトランス活性化するc−mycのDNA結合ドメインを含むハイブリッド分子は、E boxに結合可能であり、そのためc−myc依存的な転写を阻害する。mad/mycの発現は、DKC細胞だけでなくコントロール細胞においても、投与量に依存する様式にて、hTERTプロモーター活性を阻害することができる(図3D)。また、myc/madの発現は、GSE24−2細胞におけるGSE24.2タンパク質によって介される転写活性化を消失させることができる。これにより、GSE24−2によって誘導されるhTERTの転写の活性化が、c−myc依存的であることが示された(図3D)。myc/mad構築物をNFκBプロモーターのためのレポーターベクター(HIVLuc)とともに形質移入しても、TNFαによって誘導される転写に影響を及ぼさないことから、この阻害は特異的である(図4E)。
【0070】
3.2Kbのc−myc遺伝子(px3.2myc)を含み、かつルシフェラーゼ遺伝子に結合されたプロモーターP0、P1およびP3を包含するレポーターベクター(図3A)を用いて、コントロール細胞およびGSE24−2細胞を形質移入した。GSE24−2細胞におけるプロモーター活性が、コントロール細胞と比べて3倍に増加したことが観察された(図3B)。レポーターベクターpx3.2mycとともにベクターpLNCXまたはGSE24−2を発現するプラスミドにより293T細胞を形質移入した場合にも、同様の結果が得られた(データは示さない)。GSE24−2が介する活性化に必要な領域を調べるために、c−mycプロモーターの3つの欠失変異体を用いた(図3A)。NHEIIIが含まれる、末端のプロモーターP1およびP0が包含される構築物のみが、GSE24−2細胞において活性化されることができたが、コントロール細胞においては活性化されなかった(図3B)。これにより、最も近いプロモーターP2は、GSEを介した活性化に関わっていないことが示された。
【0071】
G−カルテットの二次構造の維持に関わっているプリンリッチ領域(Pu27)におけるグアニン残基を置換することによって、NHEIIIの種々の変異体を取得した。4つ組のG群の2番目におけるG12A変異体、2番目の3つ組のGからG17Aの変異体、そして最後にプリンリッチ領域の末端における2つの連続したグアニン(G26A/G27A)の変異体を取得した(図3C)。この結果から、オリジナルの配列のみがGSE24−2によって活性化されることが示された(図3D)。概して、これらの結果により、GSE24−2が、Pu27領域の二次構造の変形を誘導してc−myc転写させる活性のある配置にさせることが可能であることが示唆される。
【0072】
本発明者らはまた、c−mycプロモーター活性にて観察される変化が、その発現に反映されるのかを調べた。この効果のために、空のベクター、DKCの発現ベクターまたはGSE24−2発現ベクターを用いてMEF細胞を一過的に形質移入し、次にmRNAおよびc−mycタンパク質のレベルを測定した(図3E y 3F)。予想どおり、GSE24−2の発現のみが、c−mycのmRNAとタンパク質との両方について、その発現の活性化を誘導できた。c−MYC発現の増加が細胞の形質転換を誘導し得ることは、GSE24−2発現による不要な効果の1つとなり得る。この可能性を評価するために、極小マウス(atimic mice)に、空のベクター、ヒトc−myc遺伝子、またはGSE24−2を発現しているPam212細胞を接種した。c−mycを発現している細胞を接種された動物のみが、最初の10日以内に腫瘍を発症し、GSE24−2を発現している細胞は接種後3ヶ月間でさえも腫瘍を発症させなかった。
【0073】
(実施例4−GSE24−2を用いて刺激されたVA13細胞およびX−DC患者におけるテロメラーゼ活性は、老化プロセスからそれらを回復させる)
本発明者らは、X−DCを患っている患者の細胞におけるGSE24−2発現の効果を調べた。キャリアの母親(DC、正常なdkc対立遺伝子)および臨床的に影響を受けた彼女の3人の子供(DC1、DC2およびDC3、変異体dkc対立遺伝子)から取得された市販のリンパ芽球を用いた。これらの細胞は不死化されておらず、培養中に正常に老化し始める。さらに以前観察されたように、すなわちGSE24−2の発現はこれらの細胞系統におけるテロメラーゼ活性を増加させ(図4A)、前述したように(参照文献21)、全長のジスケリン発現によるテロメラーゼ活性の増加は観察されなかった(図4B)。GSE24−2により形質移入された細胞において、hTERTおよびhTRのRNAレベルが増加した(図5C)ことは、GSE24−2の形質移入によるテロメラーゼ活性の回復が、両方の分子の増加に起因することを示唆する。
【0074】
一方、X−DC患者における本来の真皮の繊維芽細胞は、連続培養において3〜4週間以内に完全に老化するが、正常な細胞は、2ヶ月増殖した後に老化期に到達する(参照文献21)。hTERTを発現するX−DC細胞は、より長く分裂し、数ヶ月間連続して生育する能力を獲得する(参照文献21)。X−DCにおけるGMO1787細胞内でのGSE24−2の発現は、前述したように、X−DCにおける繊維芽細胞内のhTRおよびhTERTのRNAレベルの発現を増加させることによって、テロメラーゼ活性の増加を誘導することができた(図4D)。そして、驚いたことに、これらの細胞は、連続して160日以上成育することができた(図5)。ウェルナー症候群の患者における細胞(老化した真皮の繊維芽細胞)もまた、連続して150日以上生育することができた(図6)。
【0075】
さらに興味深いことに、hTERTおよびhTRの欠如を示し(参照文献59)、かつALTメカニズムを用いてテロメアを維持するVA13のテロメラーゼ欠失細胞系統(野生型のジスケリン)において、GSE24−2の発現が、テロメラーゼ活性(図4G)、およびhTERTおよびhTRのRNAレベル(図4H)を回復させることができた。
【0076】
〔材料および方法〕
(構築物および細胞系統)
GSE24−2、DKC、DKC5’、モチーフI、モチーフIIおよびGSE24−2と相同なCBF5遺伝子の領域の配列を、pLNCXベクター(BD Biosciences, Madrid)にクローニングした。hTERTプロモーターを有する構築物は、Dr. T.Kim(参照文献32)から取得し、hTRプロモーターを有する構築物は、Dr. N.Keith(参照文献9)から取得し、c−MYCプロモーター(px3.2)を有する構築物は、Dra. A. Arandaから取得し、そしてハイブリッド構築物Myc/mad、pECLおよびpECLc−mycは、Dr. J. Leonから取得した。プラスミドBASE−TERTはDra. K. Collinsから取得した(参照文献31)。プラスミドPGATEV(参照文献33)は、Dr. G. Montoyaから取得した。プラスミドPGATEV−24−2は、pGATEVプラスミドのNdeI/XhoI部位の24−2断片をサブクローニングすることにより取得した。プラスミドpCDNA3−9E10−24−2は、エピトープ9E10−mycを含むpCDNA3ベクターのEcoR1/XbaI部位の間の24−4断片をサブクローニングすることによって取得した。293T細胞(American Type Culture Collection)Phoenixを、10%FBSを添加したDMEM培地にて培養した。細胞系統VA13は、Dr. M. Serranoから取得した。ファミリー(DC、DC1、DC2およびDC3)のX−DCリンパ芽球については、臨床的に報告されており(参照文献34〜35)、そのジスケリン遺伝子は1塩基置換を示す:T66A。X−DCの皮膚の繊維芽細胞GMO1787は、10%FCSを添加したDMEMにて培養し、そしてこの細胞には、ロイシン37を欠失したジスケリンタンパク質がコードされていた(参照文献31)。マウスの胚の繊維芽細胞(MEF)は、10%FBSと非必須アミノ酸(Gibco, Carlsbad, USA)とを添加したDMEMにて培養した。安定な形質移入のために、293T細胞を、細胞百万個あたり10μgのGSE24−2またはpLNCXと1μgのプラスミドp−BabePUR(Clontech)とを用いて同時形質移入し、形質転換24時間後にピューロマイシン耐性のものを選択した。Pam212細胞は、ベクターpECL、pECLc−mycまたはGSE24−2を用いて形質移入し、G418耐性のものを選択した。X−DC繊維芽細胞の感染のために、リン酸カルシウム法によりpLNCX由来のベクターを用いてPhoenix細胞を形質移入した。形質移入48時間後、ウイルスを回収し、X−DC繊維芽細胞を感染させるために48時間使用した。安定な細胞系統は、G−418(Gibco)より選択した。
【0077】
(GSEsのライブラリーおよびシスプラチンによる選択)
基本的に、以前にRoninson et al(参照文献36)によって記載されたように、GSEsのライブラリーを構築した。
【0078】
(試薬)
シスプラチン、テロメラーゼIの阻害剤およびSP60125は、Calbiochem(San Diego, USA)から購入した。TNFαは、Upstate(Charlottesville, USA)から購入した。
【0079】
(ペプチドGSE24−2の生産および精製)
大腸菌(E.coli)のDH5α細胞を、ベクターpGATEV GSE24−2を用いて形質転換し、その溶菌液を前述されたように調製した(参照文献33)。融合タンパク質をグルタチオン−セファロース法によって精製し、その純度をゲル電気泳動によって解析した。製造者の指示にしたがってプロテアーゼTEVにより消化して、GSE24−2を取得した。通常、SDS−PAGE電気泳動にて取得されるものによれば、消化されたタンパク質の90%が取得される。このタンパク質は、Hi−Trap Ni−NTAカラムに2回通され、ポリヒスチジンテール、未消化のタンパク質、TEVプロテアーゼおよび不純物を廃棄した。
【0080】
(定方向突然変異誘発)
Quickchange X-L site-directed mutagenesis kit(Stratagene, Santa Clara, USA)を用いて、製造者の指示にしたがってc−MYCプロモーター(px3.2)のNHEIII領域におけるグアニン残基の変異導入を行なった。
【0081】
(テロメアリピート増幅プロトコール(TRAP))
TRAPeze(参照文献37) telomerase detection kit(Intergen, Purchase, USA)を用いて製造者の指示にしたがってテロメラーゼ活性を測定した。TRAPアッセイは、存在する全タンパク質の濃度によりそれぞれのタンパク質抽出物を標識して行なった。所望の実験において、テロメアのプローブをTRAPアッセイに使用した(参照文献38)。TRAP反応において直接にシスプラチンとともにインキュベートすることによってインビトロでのプラチナ化(platination)を行なった。
【0082】
(イムノブロットおよび抗体)
前述されたように細胞を溶解させ(参照文献39)、その後20μgのタンパク質を10%にてSDS−PAGEにより解析した。使用した抗体:抗pJNK(V7391、Promega, Madison, USA)、抗JNK1(C−17、Santa Cruz Biotechnologies, Santa Cruz, USA)、抗p−P38(Cell Signaling, Charlottesville, USA)、9−E10(A14−Santa Cruz Biotechnologies)および抗Flag(Invitrogen, Carlsbad, USA)。
【0083】
(cDNAの調製およびRT−PCR)
Trizol(Life Technologies, Carlsbad, USA)を用いて製造者の指示にしたがって細胞の全RNAを抽出した。逆転写酵素M−Mlv(Promega)を用いて、各反応において2μgの全RNAをcDNAに転写させた。
【0084】
(形質移入および遺伝子発現解析)
293T細胞は、前述されたようにリン酸カルシウム法を用いて一過的に形質移入した(参照文献39)。DNAの全量は、常に細胞百万個あたり10μgを維持した。X−DC細胞は、細胞百万個あたり3μgのpLNCX−GSE24−2を用いたエレクトロポレーション法により一過的に形質移入した。VA13のMEFs細胞は、100万個の細胞あたり16μgのDNAを用いてリポフェクタミン プラス(lipofectamine plus)(Invitrogen)により製造者の指示にしたがって形質移入した。タンパク質抽出液を調製し、市販のキット(Promega)を用いてルシフェラーゼを測定した。各試験は3通りにて行ない、実験は3回繰り返した。形質移入効率は、プラスミドp−CMV−Renillaを同時形質移入することによって修正した。
【0085】
(ゲル易動度シフトアッセイ)
プラスミドpCDNA3−E1024−2または空のベクターによって293T細胞を形質移入し、G−418により選択することによって安定な細胞系統を作製した。これらの細胞は、エピトープE10−myc GSE24−2の融合タンパク質を発現する。核タンパク質(参照文献40)の抽出液における結合反応を、前述されたように行なった(参照文献41)。テロメア配列を含むオリゴヌクレオチドTEL1の5’末端、及びTEL1のハンギング領域(hanging zone)にハイブリダイズするCXext(ネガティブコントロールとして)(参照文献41)の5’末端を、T4ポリヌクレオチドキナーゼ(New England Biolabs, Beverly, MA)および 32P ATP(Amersham Bioscences, Orsay, France)によって標識した。エピトープ9E10に対する抗体を用いて、GSE24−2結合と競合させた。DNAに結合した複合体は、トリス・ホウ酸塩・EDTAバッファーを用いた0.5%アクリルアミドの非変性ゲルにおいて分離させた。
【0086】
(生存度アッセイおよび累積重複数(PLD))
前述されたように、クリスタル・バイオレット染色法を用いて、細胞増殖および生存度を調べた(参照文献39)。感染後にDKC繊維芽細胞を混合し、G−418耐性細胞を選択した。細胞を選択する最初のステップにおいて、培養中のPLDsを定量化して増殖を測定した。
【0087】
〔参照文献〕
1.de Lange T. Protection of mammalian telomeres. Oncogene. 2002; 21: 532-540.
2.Blackburn EH. Switching and signaling at the telomere. Cell. 2001; 106: 661-673.
3.Collins K, Mitchell JR. Telomerase in the human organism. Oncogene. 2002; 21: 564-579.
4.Colgin LM, Reddel RR. Telomere maintenance mechanisms and cellular immortalization. Curr Opin Genet Dev. 1999; 9: 97-103.
5.Harley CB. Telomerase therapeutics for degenerative diseases. Curr Mol Med. 2005; 5: 205-211.
6.Blasco MA. Telomeres and human disease: ageing, cancer and beyond. Nat Rev Genet. 2005; 6: 611-622.
7.Cohen SB, Graham ME, Lovrec GO, et al, Protein composition of catalytically active human telomerase from immortal cells. Science, 2007; 315: 1850-1853
8.Zhao J Q, Hoare S F, McFarlane R, et al. Cloning and characterization of human and mouse telomerase RNA gene promoter sequences. Oncogene. 1998; 16: 1345-1350.
9.Takakura M, Kyo S, Kanaya T, et al. Cloning of human telomerase catalytic subunit (hTERT) gene promoter and identification of proximal core promoter sequences essential for transcriptional activation in immortalized and cancer cells. Cancer Res. 1999; 59: 551-5577.
10.Gomez D, Aouali N, Renaud A, et al. Resistance to senescence induction and telomere shortening by a G-quadruplex ligand inhibitor of telomerase. Cancer Res. 2003; 63: 6149-6153.
11.Li H, Xu D, Li J, Berndt MC, Liu JP. Transforming growth factor beta suppresses human telomerase reverse transcriptase (hTERT) by Smad3 interactions with c-Myc and the hTERT gene. J Biol Chem. 2006; 281: 25588-255600.
12.Levens D, Duncan R C, Tomonaga T, et al. DNA conformation, topology, and the regulation of c-myc expression. Curr Top Microbiol Immunol. 1997; 224: 33-46.
13.Siebenlist U, Hennighausen L, Battey J, Leder P. Chromatin structure and protein binding in the putative regulatory region of the c-myc gene in Burkitt lymphoma. Cell. 1984; 37: 381-391.
14.Mirkin S M, Frank-Kamenetskii MD. H-DNA and related structures. Annu Rev Biophys Biomol Struct. 1994; 23: 541-576
15.Siddiqui-Jain A, Grand CL, Bearss DJ, Hurley LH. Direct evidence for a G-quadruplex in a promoter region and its targeting with a small molecule to repress c-MYC transcription. Proc Natl Acad Sci U S A. 2002; 99: 11593-11598.
16.Bessler M, Wilson DB, Mason PJ. Dyskeratosis congenita and telomerase. Curr Opin Pediatr. 2004; 16: 23-28.
17.Marrone A, Mason PJ. Dyskeratosis congenita. Cell Mol Life Sci. 2003; 60: 507-517.
18.Mason PJ, Wilson DB, Bessler M. Dyskeratosis congenita -- a disease of dysfunctional telomere maintenance. In Curr Mol Med. 2005; 5: 159-170.
19.Heiss NS, Knight SW, VulliamyTJ, et al. X-linked dyskeratosis congenita is caused by mutations in a highly conserved gene with putative nucleolar functions. Nat Genet. 1998; 19: 32-38.
20.Knight SW, Heiss NS, Vulliamy TJ, et al. X-linked dyskeratosis congenita is predominantly caused by missense mutations in the DKC1 gene. Am J Hum Genet. 1999; 65: 50-58.
21.Mitchell J, Wood E, Collins K. A telomerase component is defective in the human disease dyskeratosis congenita. Nature. 1999; 402: 551-555.
22.Normand C, Capeyrou R, Quevillon-Cheruel S, Mougin A, Henry Y, Caizergues-Ferrer M. Analysis of the binding of the N-terminal conserved domain of yeast Cbf5p to a box H/ACA snoRNA. RNA. 2006; 12: 1868-82.
23.Henras A, Henry Y, Bousquet-Antonelli C, Noaillac-Depeyre J, Gelugne JP, Caizergues-Ferrer M. Nhp2p and Nop10p are essential for the function of H/ACA snoRNPs.; EMBO J. 1998; 17: 7078-7090.
24.Pogacic V, Dragon F, Filipowicz W.Human H/ACA small nucleolar RNPs and telomerase share evolutionarily conserved proteins NHP2 and NOP10.. Mol Cell Biol. 2000; 20: 9028-40.
25.Wang C, Meier UT. Architecture and assembly of mammalian H/ACA- small nucleolar and telomerase ribonucleoproteins. EMBO J. 2004; 23: 1857-1867
26.Henras AK, Capeyrou R, Henry Y, et al Cbf5p, the putative pseudouridine synthase of H/ACA-type snoRNPs, can form a complex with Gar1p and Nop10p in absence of Nhp2p and box H/ACA snoRNAs. RNA. 2004; 10: 1704-12.
27.Rashid R, Liang B, Baker D L, et al. Crystal structure of a Cbf5-Nop10-Gar1 complex and implications in RNA-guided pseudouridylation and dyskeratosis congenita.. Mol Cell.2006; 21: 249-260.
28.Darzacq X, Kittur N, Roy S, et al Stepwise RNP assembly at the site of H/ACA RNA transcription in human cells. J Cell Biol. 2006; 173: 207-18.
29.Tollervey D, Kiss T. Function and synthesis of small nucleolar RNAs. Curr Opin Cell Biol. 1997; 9: 337-342.
30.Fu D, Collins K. Distinct biogenesis pathways for human telomerase RNA and H/ACA small nucleolar RNAs. Mol Cell. 2003; 11: 1361-1372.
31.Wong JM, Collins K. Telomerase RNA level limits telomere maintenance in X-linked dyskeratosis congenita. Genes Dev. 2006; 2: 2848-2858.
32.Oh S, Song YH, Kim UJ, Yim J, Kim TK. In vivo and in vitro analyses of Myc for differential promoter activities of the human telomerase (hTERT) gene in normal and tumor cells. Biochem Biophys Res Commun. 1999; 263: 361-365
33.Kalinin A, Thoma NH, Iakovenko A, Heinemann I, Rostkova E, Constantinescu AT, Alexandrov K. Expression of mammalian geranylgeranyltransferase type-II in Escherichia coli and its application for in vitro prenylation of Rab proteins. Protein Expr Purif. 2001; 22: 84-91.
34.Sirinavin C, Trowbridge AA. Dyskeratosis congenita: clinical features and genetic aspects. Report of a family and review of the literature. J Med Genet. 1975; 12: 339-354
35.Trowbridge AA, Sirinavin C, Linman JW. Dyskeratosis congenita: hematologic evaluation of a sibship and review of the literature. Am J Hematol. 1977; 3: 143-152.
36.Roninson IB, Gudkov AV, Holzmayer TA, et al. Genetic suppressor elements: new tools for molecular oncology. Cancer Res. 1995; 55: 4023-4028.
37.Wright WE, Shay JW, Piatyszek MA. Modifications of a telomeric repeat amplification protocol (TRAP) result in increased reliability, linearity and sensitivity. Nucleic Acids Res. 1995; 23: 3794-3795.
38.Gomez D, Mergny JL, Riou JF. Detection of telomerase inhibitors based on g-quadruplex ligands by a modified telomeric repeat amplification protocol assay Cancer Res. 2002; 62: 3365-8.
39.Sanchez-Perez I, Perona R. Lack of c-Jun activity increases survival to cisplatin. FEBS Lett. 1999; 453: 151-158.
40.Perona R, Montaner S, Saniger L, Sanchez-Perez I, Bravo R, Lacal JC.1997 Activation of the nuclear factor-kappaB by Rho, CDC42, and Rac-1 proteins. Genes Dev. 1997; 11: 463-75.
41.Gomez D, O'Donohue MF, Wenner T, Douarre C, Macadre J, Koebel P, Giraud-Panis MJ, Kaplan H, Kolkes A, Shin-ya K, Riou JF.The G-quadruplex ligand telomestatin inhibits POT1 binding to telomeric sequences in vitro and induces GFP-POT1 dissociation from telomeres in human cells. Cancer Res. 2006; 66: 6908-12.
42.Redon S, Bombard S, Elizondo-Riojas MA, Chottard JC. Platination of the (T2G4) 4 telomeric sequence: a structural and cross-linking study. Biochemistry. 2001; 40: 8463-8470.
43.Ishibashi T, Lippard SJ. Telomere loss in cells treated with cisplatin. Proc Natl Acad Sci U S A. 1998; 95: 4219-4223.
44.Parkinson GN, Lee M P, Neidle S. Crystal structure of parallel quadruplexes from human telomeric DNA. Nature. 2002; 417: 876-880.
45.Bednarek A, Shilkaitis A, Green A, et al. Suppression of cell proliferation and telomerase activity in 4-(hydroxyphenyl)retinamide-treated mammary tumors. Carcinogenesis. 1999; 20: 879-883.
46.Kim JH, Kim JH, Lee GE, Kim SW, Chung IK. Identification of a quinoxaline derivative that is a potent telomerase inhibitor leading to cellular senescence of human cancer cells. Biochem J. 2003; 373: 523-529.
47.Hurley LH. DNA and its associated processes as targets for cancer therapy. Nat Rev Cancer. 2002; 2: 188-200.
48.Sun D, Thompson B, Cathers BE, et al. Inhibition of human telomerase by a G-quadruplex-interactive compound. J Med Chem. 1997; 40: 2113-2116.
49.Cuesta J, Read MA, Neidle S. The design of G-quadruplex ligands as telomerase inhibitors. Mini Rev Med Chem. 2003; 3: 11-21.
50.Chau NP, Deschatrette J, Wolfrom C Reversal of hepatoma cells resistance to anticancer drugs is correlated to cell proliferation kinetics, telomere length and telomerase activity. Anticancer Res. 2005; 25: 3279-85
51.Lin CP, Liu JD, Chow JM, Liu CR, Liu HE. Small-molecule c-Myc inhibitor, 10058-F4, inhibits proliferation, downregulates human telomerase reverse transcriptase and enhances chemosensitivity in human hepatocellular carcinoma cells. Anticancer Drugs. 2007; 18: 161-70.
52.Sun PM, Wei LH, Luo MY, Liu G, Wang JL, Mustea A, Konsgen D, Lichtenegger W, Sehouli J. The telomerase activity and expression of hTERT gene can serve as indicators in the anti-cancer treatment of human ovarian cancer. Eur J Obstet Gynecol Reprod Biol. 2007; 130: 249-57.
53.Poole JC, Andrews LG, Tollefsbol TO. Activity, function, and gene regulation of the catalytic subunit of telomerase (hTERT). Gene. 2001; 269: 1-12.
54.Meier UT, Blobel G. NAP57, a mammalian nucleolar protein with a putative homolog in yeast and bacteria.. J Cell Biol. 1994; 127: 1505-14.
55.Meier UT. The many facets of H/ACA ribonucleoproteins. Chromosoma. 2005; 114: 1-14.
56.Zucchini C, Strippoli P, Biolchi A, et al. The human TruB family of pseudouridine synthase genes, including the Dyskeratosis Congenita 1 gene and the novel member TRUB1. Int J Mol Med. 2003; 11: 697-704.
57.Wu KJ, Grandori C, Amacker M, et al. Direct activation of TERT transcription by c-MYC. Nat Genet. 1999; 21: 220-224.
58.Mochizuki Y, He J, Kulkarni S, Bessler M, Mason PJ. Mouse dyskerin mutations affect accumulation of telomerase RNA and small nucleolar RNA, telomerase activity, and ribosomal RNA processing. Proc Natl Acad Sci U S A. 2004; 101: 10756-10761.
59.Bryan TM, Marusic L, Bacchetti S, Namba M, Reddel RR The telomere lengthening mechanism in telomerase-negative immortal human cells does not involve the telomerase RNA subunit. Hum Mol Genet. 1997; 6: 921-6.
60.Kunifuji Y, Gotoh S, Abe T, Miura M, Karasaki Y. Down-regulation of telomerase activity by anticancer drugs in human ovarian cancer cells. Anticancer Drugs. 2002; 13: 595-598.
61.Postel EH, Berberich SJ, Rooney JW, Kaetzel DM. Human NM23/nucleoside diphosphate kinase regulates gene expression through DNA binding to nuclease-hypersensitive transcriptional elements. J Bioenerg Biomembr. 2000; 32: 277-284.
62.Nosaka K, Kawahara M, Masuda M, Satomi Y, Nishino H. Association of nucleoside diphosphate kinase nm23-H2 with human telomeres. Biochem Biophys Res Commun. 1998; 243: 342-348.
【図面の簡単な説明】
【0088】
【図1a】シスプラチンへの応答におけるテロメラーゼ活性に対するGSE24−2ペプチドの発現の効果。
【図1b】シスプラチンへの応答におけるテロメラーゼ活性に対するGSE24−2ペプチドの発現の効果。
【図1c】(C)シスプラチンおよびGSE24−2ペプチドを用いた、293T細胞抽出物のインビトロでの処理に続く、テロメラーゼ活性。シスプラチン、および精製したペプチドGSE24−2の量を増加させたもの(3−5列目)、熱により失活させたGSE24−2ペプチド(7列目)またはコントロールペプチド(8列目)を用いて、293T細胞抽出物をインビトロにて処理した後、後者をTPAPアッセイに用いた。テロメラーゼアッセイを行なうために、テロメアの配列を有する特異的な鋳型を用いた。この実験を3回繰り返して同様の結果を得た。
【図1d】(D)GSE24−2を発現する細胞抽出物を用いたゲルシフト試験。プラスミドpcDNA3−9E1024−2またはpcDNA3−9E10を発現する細胞の核抽出物をゲルシフト試験に供した。特異的なテロメアの配列を含むTEL1プローブを使用した。特異的な抗体9E10(3列目)、およびTEL1とハイブリダイズして競合分子として機能するオリゴヌクレオチドCXext(4−5列目)を用いることにより、特異的な結合が確立された。この実験を3回繰り返して同様の結果を得た。
【図2a】GSE24−2はhTERTプロモーターの活性を増加させる。(A)pLNCX、DXC、GSE24−2、Cbf5、モチーフIまたはモチーフII(細胞百万個あたり10μgのDNA)およびレポータープラスミドhTERT−luc(細胞百万個あたり1μg)を用いて、293T細胞を同時形質移入した。形質移入して24時間後のテロメラーゼ活性を測定した。*は統計値がp<0.05であることを示す。
【図2b】(B)レポーターベクターhTERT−luc(細胞百万個あたり1μg)を用いて、293T細胞(黒)および293T24−2細胞(白)を同時形質移入した。24時間後、これらの細胞をシスプラチン(3 g/ml)により8時間処理した後、ルシフェラーゼ活性を測定した。
【図2c】(C)提示された構築物(細胞百万個あたり10μgのDNA)およびレポーターベクターhTR−luc(細胞百万個あたり1μg)を用いて、293T細胞を同時形質移入した。形質移入して24時間後のルシフェラーゼ活性を測定した。
【図2d】(D)レポーターベクターhTERT−luc、および様々な量のMad/mycの発現ベクターを用いて、これらの細胞系統を同時形質移入した。形質移入して24時間後のルシフェラーゼ活性を測定した。
【図2e】(E)レポーターベクターHIV−lucと、発現ベクターmad/mycの量を増加させたものとを用いて、これらの細胞系統を同時形質移入した。形質移入して24時間後に、50ng/mlのTNF を用いてこれらの細胞を6時間刺激した後、ルシフェラーゼ活性を測定した。形質移入効率を制御するために、CMV−Renillaベクターを用いた(0.1μg/ml)。それぞれのグラフは、4通りにて行なった2つの実験の平均値および標準偏差を表す。
【図3a】GSE24.2はc−MYC遺伝子のプロモーターを介してその活性を誘導する。(A)実験に使用したそれぞれの構築物を示す、c−MYCプロモーターの概略図。
【図3b】(B)レポーターベクターc−MYC−luc(細胞百万個あたり1μg)のそれぞれの構築物を用いて、293T pLNCXおよび293T GSE24−2の細胞を形質移入した。形質移入して24時間後にルシフェラーゼ活性を測定した。形質移入効率のコントロールとして、CMV−Renillaベクターを用いた(細胞百万個あたり0.1μg/ml)。それぞれのグラフは、4通りにて行なった2つの実験の平均値および標準偏差を表す。
【図3c】(C)NHEIIIにおいて作製した変異体の概略図。
【図3d】(D)プラスミドpx3.2由来の変異体のレポーターベクター(細胞百万個あたり1μg)を用いて、提示された細胞系統を形質移入した。形質移入して24時間後にルシフェラーゼ活性を測定した。形質移入効率のコントロールとして、CMV−Renillaベクターを用いた(細胞百万個あたり0.1μg/ml)。それぞれのグラフは、4通りにて行なった2つの実験の平均値および標準偏差を表す。
【図3e】(E)発現ベクターpLNCX、DKCまたはGSE24−2を用いて、MEF細胞を形質移入した。形質移入して24時間後、全RNAを単離し、RT−PCRによって、マウスのc−myc遺伝子および −アクチン遺伝子の発現を測定した。この実験を3回繰り返して同様の結果を得た。
【図3f】(F)発現ベクターpLNCX、DKCまたはGSE24−2を用いて、MEF細胞を形質移入した。形質移入して24時間後、全タンパク質の抽出物を単離し、特異的な抗体を用いたウェスタンブロットによって、マウスのc−mycタンパク質および −チューブリンタンパク質の発現を決定した。この実験を3回繰り返して同様の結果を得た。
【図4a】GSE24−2は、X染色体に関連する先天性角化異常症の患者の細胞、およびVA13細胞におけるテロメラーゼ活性を再活性化させる。(A)キャリアの母親(DC−C)、および影響を受けた子供(DC−1、DC−2およびDC−3)から取得した、X染色体に関連する先天性角化異常症を患う患者のリンパ芽球において、テロメラーゼ活性を測定した。これらの細胞に対し、空のベクター(−)またはGSE24−2(+)を細胞百万個あたり3μg用いて一過的に形質移入した。その24時間後にテロメラーゼ活性を測定した。
【図4b】(B)細胞百万個あたり3μgの発現ベクターpLNCX、GSE24−2またはDKCを用いて、一過的にDC2細胞を形質移入した。その24時間後にテロメラーゼ活性を測定した。
【図4c】(C)細胞百万個あたり3μgの空のベクターまたはGSE24.2により形質移入された患者(DC−3)の1人の細胞における、hTERTおよびhTRの発現レベル。RNAレベルは、RT−PCRにより検出した。GAPDH発現をコントロールとして用いた。
【図4d】(D)X−DC GMO1787の患者の繊維芽細胞を、ベクターpLNCXまたはGSE24−2由来のウイルスによって感染させ、安定な細胞系統をテロメラーゼ活性測定のために用いた(タンパク質の全量を三角形にて示す)。
【図4e】(E)プラスミドpLNCXおよびpGSE24−2によって形質移入したGMO1787細胞におけるhTERTおよびhTRの発現レベルを、RT−PCRにより測定した。GAPDH発現をコントロールとして用いた。
【図4f】(F)パネルDに記載したように形質移入されたGMO1787細胞の増殖速度を、一定時間における個体数の重複の累積を用いて評価した。全ての実験について3回繰り返し、同様の結果を得た。
【図4g】(G)VA13細胞におけるテロメラーゼ活性。この細胞に対し、細胞百万個あたり16μgのコントロールベクターpLNCXまたはGSE24.2を用いて一過的に形質移入した。その24時間後に、タンパク質抽出液の連続した希釈液を用いて、テロメラーゼ活性を測定した(タンパク質の全量を三角形にて表す)。
【図4h】(H)細胞百万個あたり16μgのプラスミドpLNCX、DKCまたはGSE24−2により形質移入されたVA13細胞における、hTERTおよびhTRの発現レベル。RT−PCRによりRNAレベルを測定した。GAPDHレベルをコントロールとして用いた。
【図5】ペプチドGSE24.2は細胞老化から回復させる。図4Dに記載されたように形質移入されたGMO1787細胞の増殖速度を、一定時間における個体数の重複の累積を用いて評価した。全ての実験について3回繰り返し、同様の結果を得た。
【図6】ウェルナー症候群の患者における細胞(老化した真皮の繊維芽細胞)もまた、連続して150日以上生育することができたことを示す図である。
【発明の詳細な説明】
【0001】
〔技術分野〕
ヒトの健康に適用させるためのバイオテクノロジー分野、およびより具体的には、加齢および細胞老化に関する疾患を患っている人に対して治療的に適用させるための、生物学的な化合物−塩基配列、ペプチド、およびヒトの形質転換細胞。
【0002】
〔従来技術〕
TTAGGG配列(参照文献1)の多数の繰り返しにより構成されているテロメアは、DNAポリメラーゼがクロモソームの末端まで複製できないため、細胞周期のたびに短くなる。クロモソームの末端におけるテロメアの維持は、テロメラーゼ複合体によって行なわれる。この複合体のタンパク質成分であるヒトテロメラーゼ逆転写酵素(hTERT)は、触媒作用を有する逆転写酵素モチーフを含んでおり、一方、テロメアRNA成分は、内部にあるテロメアの繰り返し配列TTAGGGと相補的な鋳型を用いて、デオキシヌクレオチド三リン酸(dNTPs)の作用を導く(参照文献2〜3)。近年、ヒトテロメラーゼ酵素複合体が、2つのタンパク質成分、hTERTおよびジスケリン(dyskerin)、ならびにRNA成分であるhTRのみからどのように構成されているかについて、報告されている(参照文献7)。ジスケリンが、「H/ACA box」リボ核タンパク質に属するシュードウリジン合成酵素と推定される一方、H/ACAモチーフがhTR内に存在する。この複合体は、最終的に、hTERT、hTRおよびジスケリンのそれぞれ2つの分子により構成される。
【0003】
テロメラーゼの発現は、成人の組織における多くの体細胞内では抑制されているが、多くの癌では高いままであり、このテロメラーゼは、クロモソームの整合性を保証するため、これらの癌の細胞内において不死の表現型に寄与している(参照文献6)。テロメラーゼ(hTERT)のレベルは、最初の転写レベルで制御され(参照文献8)、またヒトでは、hTERT遺伝子の5’領域における制御領域によって制御されている。この制御領域は、最大活性化のために必要であり、またmyc結合部位(E−box)を含む(参照文献8〜9)。c−MYC転写因子は、hTERTの発現を促進させるが、mad1の発現が強化されると、hTERTを抑制する(参照文献10〜11)。c−mycの転写制御には、多数のプロモーターが含まれ、なかでもP1およびP2が最も重要である(参照文献12)。c−MYCプロモーターのヌクレアーゼ過感受性エレメントIII(NHEIII)は、c−MYC転写の85〜90%を制御し、当初はDNAseIに対して最も過感受性の部位として同定された(参照文献13)。NHEIIIは、DNA鎖上にプリン−リッチ部位を含み、この部位は、DNAにおける二重螺旋構造とnon−B形態との間のバランスを確立する能力がある(参照文献14)。プリン−リッチなDNA鎖は、異なるG−カルテット(G quartets)による2つの分子内構造を形成でき(参照文献15)、これらは転写抑制因子として機能するため、抗腫瘍性治療のターゲットとして用いられている。
【0004】
テロメアの短縮、またはその二次構造の欠失は、細胞を老化および/またはアポトーシスに向かわせる原因となる。これらの2つのプロセスは、制御不能な細胞分裂および遺伝的不安定性を防止する生物学的なコントロールポイントとして機能する(参照文献4〜5)。テロメアの長さの制御についての欠損は、早期老化症候群および癌などの様々な疾患の病状に関わる(参照文献6)。
【0005】
老化細胞は、DNA合成を停止した生細胞であり、平板化されるとともに液胞が豊富な細胞質が存在し、独特の遺伝子発現プロファイルを有する。そして、老化細胞は、酸性pHにおける、老化に付随する −ガラクトシダーゼ( −gal)活性の増加を可視化する生化学的アッセイによって、および、時間内に細胞集団が倍加できないために、同定可能である。
【0006】
ヒト細胞の老化は、テロメア侵食に関連付けられる。形質転換されていない正常な細胞は、細胞分裂プロセスごとにテロメアが短縮することによって死ぬ運命にある。一方、テロメラーゼを発現する癌細胞は、テロメアの長さを安定化させるので、不死となる。これらの知見により、テロメアが細胞の加齢を制御する生物時計として働くという仮説が提唱される。
【0007】
テロメア侵食に関連する早期老化症候群の1つは、先天性角化異常症(DC)である。これは、非常にまれな遺伝的疾患であり、骨髄機能不全および高い癌性向によって特徴付けられ、これらは先天性角化異常症の患者の死につながる最も重大な原因である(参照文献16〜18)。主にDKC遺伝子の突発性変異によって、クロモソームと結びついた形(X−DC)で引き起こされる(参照文献19〜20)。この遺伝子は、テロメラーゼ複合体の成分となる、シュードウリジン合成酵素ドメインを含む58kDのタンパク質であるジスケリンをコードする。ジスケリンは、脊椎動物において、SnoRNAs(小さい核小体のRNAs)およびhTRに存在する複数のH/ACA boxに結合する(参照文献21)。SnoRNAsは、ジスケリンを介したRNAシュードウリジル化の鋳型として機能する。H/ACA RNAは、H boxと、これに続くトリヌクレオチドACAを含む短い配列とを含む単連鎖領域によって分けられた、2つの可変長の範囲を含む(参照文献22)。SnoRNAのH/ACAにおけるDKCに結合する、3つの異なるタンパク質がある:Nap2、Gar1およびNOP10(参照文献3、22〜24)。DKC、Nap2およびNOP10は、密接に結合して、リボ核タンパク質複合体の安定性に関連する一方、Gar1による相互作用は、より緩やかでありかつ一時的である(参照文献25、26〜28)。したがって、ジスケリンは、hTR(参照文献7、24〜29)の蓄積およびリボソーマルRNAのシュードウリジル化(参照文献30)に必須であるため、テロメラーゼ活性に必要である。DCを有する患者においては、テロメラーゼ活性が減少しており、テロメア構造を維持できないため、年齢とともにクロモソームの不安定性が増加する。X−DC患者のリンパ芽球細胞および繊維芽細胞では、テロメラーゼ活性およびhTRレベルが減少し(参照文献21)、影響を受けていない細胞よりもテロメアが短い。テロメラーゼ活性の欠如は、hTERTおよびhTRの発現によって救済され得る(参照文献21、31)。hTRの変異または欠失によって、先天性角化異常症の他の形態が引き起こされ、テロメアの短縮化の加速および早死を引き起こす。この欠損は、hTRの再発現によってのみ救済され得る(参照文献31)。
【0008】
近年、ジスケリンタンパク質のシュードウリジン合成酵素ドメインをコードする、24−2として知られる遺伝的抑制因子(GSE)が報告されている。GSE24.2は、DC−Xを有する患者の細胞内においてテロメラーゼ活性を回復させるため、テロメラーゼ複合体異常によって起こるこの病気および他の病気のための新しい治療的アプローチが発展しつつある(スペイン国特許出願公開第200502511号明細書、SEQUENCE OF NUCLEOTIDES AND PEPTIDES GSE 24.2 OF DYSKERIN, WHICH CAN INDUCE TELOMERASE ACTIVITY, METHOD FOR OBTAINING SAME, THERAPEUTIC COMPOSITIONS AND APPLICATIONS THEREOF)。しかし、このGSE24.2分子の新しい活性に関する知見によって、新しい治療的ツールおよび研究ツールを発展させることができる。
【0009】
〔本発明の説明〕
<簡単な説明>
本発明の一実施形態は、老化のプロセスによって起こる、好ましくはヒトの疾患または病理学的な状態の治療に対する医薬または医薬組成物を製造するための、GSE24.2誘導化化合物またはGSE24.2活性化化合物の使用(以下本明細書において、本発明に係る使用)により構成される。
【0010】
本発明の好ましい実施形態は、本発明に係る使用からなり、GSE24.2誘導化剤が、哺乳動物の、好ましくはヒトの細胞内において老化からの回復を誘導するタンパク質またはペプチドを発現させる塩基配列(以下本明細書において、本発明に係るGSE24.2の遺伝子配列)であり、以下の群に属するGSE24.2の塩基配列の1個または数個により構成されている:
a)ヒトGSE24.2塩基配列により構成されている塩基配列(配列番号1)、
b)a)の塩基配列の変異体であって、配列番号1と少なくとも90%の同一性を示す塩基配列、
c)a)およびb)のいずれかの配列を有するフラグメント、および、
d)a)、b)およびc)のいずれかの配列を含む遺伝的構成物である、塩基配列。
【0011】
本発明の好ましい実施形態は、本発明に係る使用からなり、a)におけるGSE24.2の配列の塩基配列が、配列番号1により構成されている。
【0012】
本発明の他のより好ましい実施形態は、本発明に係る使用からなり、c)におけるGSE24.2の配列の塩基配列が、配列番号3または配列番号5により構成されており、これらはそれぞれTrubIおよびTrubIIのペプチドドメインをコードするものである。
【0013】
また、本発明の他の好ましい実施形態は、本発明に係る使用からなり、GSE24.2誘導化剤が、哺乳動物の、好ましくはヒトの細胞内において老化を逆転させる活性を表すタンパク質またはペプチド(以下本明細書において、本発明に係るGSE24.2タンパク質)であり、以下の群に属するアミノ酸配列の1個または数個により構成されている:
a)ヒトGSE24.2のアミノ酸配列により構成されているアミノ酸配列(配列番号2)、
b)a)の配列に基づくアミノ酸配列の変異体であって、配列番号2のポリペプチドと少なくとも90%の同一性を示すアミノ酸配列、
c)a)およびb)のいずれかの配列を有するフラグメント、および、
d)a)、b)およびc)に属するいずれかの配列を含むアミノ酸配列。
【0014】
本発明の他のより好ましい実施形態は、本発明に係る使用からなり、GSE24.2誘導化剤はタンパク質であり、このタンパク質におけるa)のアミノ酸配列が配列番号2により構成されている。
【0015】
本発明の他のより好ましい実施形態は、本発明に係る使用からなり、GSE24.2誘導化剤はタンパク質であり、このタンパク質におけるc)のアミノ酸配列が、配列番号4または配列番号6により構成されている。
【0016】
本発明の他の実施形態は、老化異常に関連する疾患または病状を治療するための医薬組成物または医薬(以下本明細書において、本発明に係る医薬組成物)からなり、必要に応じて1つ以上のアジュバントおよび/または医薬的に認容され得る賦形剤とともに、治療的に有効な量にてGSE24.2化合物またはGSE24.2活性化化合物を含んでいる。また、この医薬組成物または医薬は、老化のプロセスを減少または逆転させることが可能であり、かつ、本発明に係る使用により獲得される。
【0017】
本発明の他の実施形態は、老化の異常なプロセスによって引き起こされる、好ましくはヒトの疾患または病状を治療するための処置における、本発明に係る医薬的化合物の使用からなる。
【0018】
本発明の他の好ましい実施形態は、真核細胞、好ましくはヒト細胞からなり、その老化のプロセスが逆転または減少されており、かつ本発明に用いられるGSE24.2分子を含むものである。
【0019】
<本発明の詳細な説明>
本発明は、ジスケリンのフラグメントとして同定された(スペイン国特許出願公開第200502511号明細書を参照)GSE24−2タンパク質(配列番号1)の発現が、hTERTおよびhTRのRNAレベルを制御し、テロメラーゼ活性の回復を誘導できるという、本発明者らの観察結果に基づいている。この結果は、種々の細胞種;ジスケリンの変異体が発現されているDC患者(スペイン国特許出願公開第200502511号明細書)の細胞、および野生型のDKC対立遺伝子を有し、かつ低いテロメラーゼ活性を有する者(VA13)の細胞の両方の細胞において観察されている(実施例4)。また、本発明における意味において、初めてGSE24.2ペプチドを直接に(その遺伝子発現を利用せずに)使用し、その結果これがテロメラーゼ活性を再活性化できることを明らかにした(実施例1)。
【0020】
同時に、GSE24−2タンパク質が、c−myc発現を調節することによってhTERTタンパク質の転写を活性化することを示した。これは、myc/madの融合分子が、hTERTの転写活性化を完全に消滅させるからである。欠失および変異解析によって、ヒトc−myc遺伝子のプロモーターにある、GSE24−2タンパク質の活性化因子としての標的領域は、P1領域を超えて位置するNHEIIIドメインであることが示された。さらに、異常なDNA構造によく結合するCCR5およびPDGFA(参照文献61)などの他の遺伝子プロモーターにも見られるポリプリン配列(Pu27)は、c−myc転写を刺激するためにインタクトのままでなければならない(実施例3)。そのため、NHEIII部位におけるGカルテットの二次構造の異常は、GSE24−2タンパク質におけるこの配列を介した活性を変化させる。NHE部位と相互作用して活性化するタンパク質がほとんど知られていないことは、GSE24.2タンパク質が、NHEドメインを有するプロモーターを持つ遺伝子を制御するために選択されて発現され得ることを意味しており、これは、強調されるべき1つの点である。これらのタンパク質の1つはNH23−H2またはNDPキナーゼBであり、これは、特異的なDNA配列に結合し、c−mycプロモーターのNHE部位に対する親和性を有するタンパク質である。それはまた、ミエロペルオキシダーゼ、CD11b、CCR5などの他の遺伝子の発現を活性化し、PDGFAなどの他の遺伝子の発現を抑制する(参照文献61〜62)。このタンパク質はまた、ヒトにおいてc−mycプロモーターのNHEおよびテロメアに見られるようなグアニン−リッチ配列にインビトロにて結合し得る(参照文献61〜62)。そのため、GSE24−2がNHEを介して転写を活性化し得るための1つの可能性あるメカニズムは、GSE24−2がNM23H2などのタンパク質と結合することによりDNAと相互作用しやすくなり、その結果、転写を増加させるという手段によるものである。
【0021】
X−DCおよびVA13細胞において得られた結果は、GSE24−2ペプチドが、2つの異なるメカニズムの組み合わせによってテロメラーゼ活性を増加させることを示す:1)hTERT転写を活性化し、そして、2)プロモーターの活性には影響を与えることなく、RNAの安定化によってhTRレベルを増加させる。さらに、GSE24.2は、ジスケリンが欠損している場合であっても、H/ACA boxにおけるhNaf1またはNop10などの他のタンパク質の会合を促進できる(参照文献28)。
【0022】
最後に、そして驚くことに、GS24.2タンパク質によるこのテロメラーゼ活性の増加は、繊維芽細胞の繁殖能を回復させるなどの方法によって、X−DC患者におけるこれらの細胞の早期の老化を回復させ、この活性を140日以上維持することが、本発明において観察されている(実施例4、図5を参照)。これは、hTERTおよびhTRの両方のレベルの増加によって媒介され得る。この意味において、GSE24.2のタンパク質または遺伝子は、細胞老化に関連する疾患および病状のプロセスのための治療的アプローチを構成し得る、または、真核細胞を不死化させるため、もしくはその増殖能を増加させるためのツールを構成し得る。一方、GSE24.2の発現が腫瘍形成能を誘導せず、かつ細胞増殖の異常性を増加させないであろうことが、本発明の結果によって示されている(実施例3)。
【0023】
上述した全ての点に照らせば、GSE24.2は、正常なジスケリンを発現するVA13細胞などの細胞、または常染色体優性の先天性角化異常症(DC)もしくは再生不良性貧血などの変異体を発現する細胞を含む、細胞老化を異常化または加速するプロセスに関連する疾患または病状のプロセスを治療するための、医薬または実験用試薬を調製するために使用され得る。
【0024】
したがって、本発明の一実施形態は、老化異常のプロセスによって起こる、好ましくはヒトの疾患または病状の治療に対する医薬または医薬組成物を製造するための、GSE24.2誘導化化合物またはGSE24.2活性化化合物の使用(以下、本発明に係る使用)からなる。本発明において、用語「医薬組成物」は、本発明において記載され、または使用されているGSE24.2の薬剤により構成されている組成物であって、研究分野において(実験用試薬など)、または産業的なバイオテクノロジー関連の活動において使用されるものもまた含む。
【0025】
本発明において使用される場合、GSE24.2誘導化化合物またはGSE24.2活性化化合物の用語は、ジスケリンにおけるGSE24.2フラグメントの塩基配列、または当該塩基配列によってコードされるタンパク質もしくはペプチドの配列であって、哺乳動物、好ましくはヒトの細胞内における老化プロセスを減少または逆転させることが可能なものをさす。この定義はまた、GSE24.2タンパク質をコードする塩基配列を発現させる化合物または分子をも含む。活性化化合物は、ペプチド、タンパク質または塩基配列、抗体および多糖により構成されていてもよい。
【0026】
本発明において使用される場合、用語「正常な、または加速された老化プロセスによって起こる疾患または病状」は、細胞および組織が増殖能の減少(加齢による自然なものを含む)、または加速を示す疾患をさす(本発明の従来技術に記載されている老化の概念を参照)。このように、本発明において使用されるこの用語はまた、ヒト細胞、好ましくは上皮の、または高レベルの増殖を伴うヒト細胞であり、説明するためであって本発明の範囲を限定するものではないが、以下の群に属する細胞の、自然な加齢によって異常化した生理的な状態をもさす:皮膚細胞、好ましくは表皮の皮膚細胞、腸上皮組織、角膜、肝臓、肺、毛球等、または造血系細胞、好ましくはリンパ球、マクロファージおよび赤血球;精巣および卵巣における胚芽細胞等であって、それぞれ、体細胞か、幹細胞か、または胚細胞。老化のプロセスおよび/またはテロメラーゼ活性の制御による、ヒトの疾患または病状のプロセスの例は、特に限定されないが、Blasco MAの改訂版に見られる(Blasco MA, Telomere length, stem cells and aging. Nature Chemical Biology 3 (10): 640-649, 2007; Crabbe L, Jauch A, Naeger CM, Holtgreve-Grez H., and Karlseder J. Telomere dysfunction as a cause of genomic instability in Werner syndrome. PNAS 104 (7): 2205-2210, 2007)。
【0027】
細胞老化のプロセスの逆転または減少は、分裂して器官の恒常性を維持する能力を有する細胞において治療的に使用され得る。これによって、加齢か、毒素による組織の損傷か、外科的処置に続く組織の損傷か、そして先天的または遺伝的な原因、例えば肺の形成不全など、の再生不良性の組織であるか、の原因に無関係に、組織の再生能力を増加させる。さらに、この細胞老化のプロセスにおける逆転または減少は、移植治療または細胞治療において、幹細胞、体細胞または胚細胞の集団の生存能力を増加させるために使用され得る。これらの治療において、幹細胞はテロメラーゼを発現するが、それらはテロメアの長さを長期間維持できないため、GSE24.2ペプチドを用いた細胞の形質転換または処理によって、これらの細胞の生存機能を高め、そしてそれゆえこれらの細胞処理の効率性を高めることができる。このように、例えば、この医薬組成物は、高齢者の表皮を再生するために使用され得る。または、この医薬組成物は、皮膚の創傷もしくは火傷の皮膚再生を促進するため、または化学療法による治療をされた患者に移植された自家造血細胞を刺激するために使用され得る。
【0028】
したがって、本発明の好ましい実施形態は、本発明に係る使用を含み、GSE24.2誘導化剤は、哺乳動物の、好ましくはヒトの細胞内において老化からの回復を誘導するタンパク質またはペプチドを発現させる塩基配列(以下本明細書において、本発明に係るGSE24.2の遺伝子配列)であって、以下の群に属するGSE24.2の塩基配列の1個または数個により構成されているものである:
a)ヒトGSE24.2塩基配列により構成されている塩基配列(配列番号1)、
b)a)の塩基配列の変異体であって、配列番号1と少なくとも90%の同一性を示す塩基配列、
c)a)およびb)のいずれかの配列を有するフラグメント、および、
d)a)、b)およびc)のいずれかの配列を含む塩基配列である、遺伝的構成物。
【0029】
この記載に使用される意味において、用語「変異体」は、配列番号1の配列に基づいて、単離され得る塩基配列、または、例えば1つ以上の塩基の挿入を含む、塩基の保存的置換または非保存的な置換の導入、分子のいずれかの末端に対する1つ以上の塩基の付加、もしくはいずれかの末端か配列内部かからの1つ以上の塩基の欠失、によって構築され得る塩基配列のいずれかを含み、かつ、GSE24.2(配列番号2)またはそのフラグメント(配列番号4および配列番号6)の配列の活性を模倣することができるペプチドまたはタンパク質をコードし得ることを意図する。
【0030】
酵素ジスケリンは、様々な生物に存在するシュードウリジン合成酵素ファミリーに属する(図3Bを参照、Mitchel et al, 1999)。当業者は、本発明に含まれる情報、−およびスペイン国特許出願公開第200502511号明細書−に基づき、自然界に存在する種々の生物から、本明細書に記載した塩基配列のアナロガスを同定または構築することができる。
【0031】
本発明において使用される場合、用語「塩基配列」は、DNA、cDNAまたはmRNAの配列をさす。
【0032】
本発明のより好ましい実施形態は、本発明に係る使用からなり、a)におけるGSE24.2の配列の塩基配列が、配列番号1により構成されている。
【0033】
本発明の他のより好ましい実施形態は、本発明に係る使用からなり、c)におけるGSE24.2の配列の塩基配列が、配列番号3または配列番号5により構成されており、これらはそれぞれTrubIおよびTrubIIのペプチドドメインをコードするものである。
【0034】
GSE24.2の塩基配列は、GSE24.2の遺伝的構成物に相当するd)として同定される。この本発明に係るGSE24.2の遺伝的構成物はまた、もし必要であれば、発現したペプチドの細胞質からの単離、検出または分泌をよりよく行なうために、単離、検出または分泌の目的で使用され得るペプチドをコードする塩基配列を含んでいてもよい。したがって、本発明の他の好ましい実施形態は、本発明に係る使用からなり、その塩基配列は、GSE24.2の遺伝的構成物であって、GSE24.2の塩基配列に加えて、発現したペプチドを細胞質から単離、検出または分泌させるペプチドまたはペプチド配列を含む。このペプチドまたはペプチド配列は、例えば、本発明の範囲を限定するものではないが、ポリヒスチジン(6×His)配列、モノクローナル抗体により認識可能なペプチド配列(例えば、その同定のための抗体、または免疫親和性クロマトグラフィーを用いて、その結果の融合タンパク質を精製するために利用する他の抗体:c−myc、HA、E−tagなどのタグペプチド)(抗体の使用:実験用マニュアル、Ed. Harlow and David Lane (1999). Cold Spring Harbor Laboratory Press. New York. Chapter: Tagging proteins. Pp. 347-377)などである。
【0035】
上述したGSE24.2の塩基配列およびGSE24.2の遺伝的構成物は、従来技術における公知の方法により、専門家によって取得できる(Sambrook et al. “Molecular cloning, a Laboratory Manual 2nd ed., Cold Spring Harbor Laboratory Press, N.Y., 1989 vol 1-3)。この塩基配列は、細胞内にて適した条件下でその発現を制御することができる遺伝子発現ベクター内に組み込まれてもよい。
【0036】
したがって、本発明の他の好ましい目的は、本発明に係る使用からなり、GSE24.2誘導化剤またはGSE24.2活性化剤は、GSE24.2の発現ベクターであって、これは、本発明において記載されるGSE24.2の塩基配列またはGSE24.2の遺伝的構成物を含み、そして、哺乳動物、好ましくはヒトの細胞内において老化のプロセスを減少または逆転させることが可能なタンパク質またはペプチドを発現させるものである。より好ましい実施形態は、発明pLNCX24.2の発現ベクターによって表される(スペイン国特許出願公開第200502511号明細書をも参照)。
【0037】
一般的に、発現ベクターは、本発明に記載のGSE24.2の塩基配列または24.2の遺伝的構成物に加えて、その転写を導くプロモーター(例えば、pT7、plac、ptrc、ptac、pBAD、retなど)であって機能的に結びついているもの、ならびにこの転写、および適切な場所における所定の産物の翻訳を調節および制御する他の必要な、または適切な配列、例えば、転写開始シグナルおよびストップシグナル(tlt2など)、ポリアデニル化シグナル、複製点、リボソーム結合配列(RBS)、転写制御因子をコードする配列、(エンハンサー)、転写サイレンサー、リプレッサーなど、を含む。好適な発現ベクターの例は、発現プラスミド、ウイルスベクター(DNAまたはRNA)、コスミド、人工クロモソームなどの中からそれぞれの特定の場合における条件および必要に応じて選択されることができ、また、所定の1つまたは複数の遺伝子を用いて形質移入された細胞または形質転換された細胞を選択するために使用され得るマーカーをも含み得る。ベクターの選択は、宿主細胞、および要求される使用形態に依存され得る。したがって、本発明の好適な実施形態によれば、ベクターは、プラスミドまたはウイルスベクターである。微生物および真核細胞の形質転換に、異なった複数の周知の方法(化学形質転換、エレクトロポレーション、マイクロインジェクションなど)が使用され得る(様々なマニュアルに記載されている(Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989). Molecular cloning: a laboratory manual, 2nd ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.))ように同様に、ベクターは、当業者によって知られている慣用的な方法を用いて取得され得る。
【0038】
さらに、本発明の他の好ましい実施形態は、本発明に係る使用からなり、GSE24.2誘導化剤は、哺乳動物、好ましくはヒトの細胞内において老化を逆転させる活性を表すタンパク質またはペプチド(以下本明細書において、本発明に係るGSE24.2タンパク質)であって、以下の群に属するアミノ酸配列の1個または数個により構成されている:
a)ヒトGSE24.2のアミノ酸配列により構成されているアミノ酸配列(配列番号2)、
b)a)の配列に基づくアミノ酸配列の変異体であって、配列番号2のポリペプチドと少なくとも90%の同一性を示すアミノ酸配列、
c)a)およびb)のいずれかの配列を有するフラグメント、および、
d)a)、b)およびc)に属するいずれかの配列を含むアミノ酸配列。
【0039】
本発明の原理から理解されるように、配列番号2の配列に示されるポリペプチドの変異体は、当該配列から、1つ以上のアミノ酸の置換、挿入または欠失によって取得されうるいずれかのペプチドである。置換による変異体の場合には、置換は好ましくは保存的置換であり、すなわち、アミノ酸は、その側鎖の性質が似た特徴を有する他のアミノ酸に置換される。したがって、保存的置換は、表1に示すアミノ酸の群の中での置換を含む。
【0040】
【表1】
【0041】
さらに、本発明の変異体における1つ以上のアミノ酸は、特別な天然または合成アミノ酸によって置換されてもよい。このアミノ酸は、例えばベータ−アミノ酸、2−アミノアジピン酸、アルファ−アスパラギン、2−アミノブタン酸、2−アミノカプロン酸、アルファ−グルタミン、アルファ−メチルアラニン、2−アミノピメリン酸、ガンマ−アミノ−ベータ−ヒドロキシベンゼンペンタン酸、2−アミノスベリン酸、2−カルボキシアゼチジン、ベータ−アラニン、ベータ−アスパラギン酸、3,6ジアミノヘキサン酸、酪酸、4−アミノ 4−アミノ−3−ヒドロキシ酪酸、ガンマ−アミノ−ベータ−ヒドロキシシクロヘキサンペンタン酸、N5−アミノカルボニルオルニチン、3−スルホアラニン、2,4 ジアミノ酪酸、ジアミノピメリン酸、2,3ジアミノプロパン酸、2,7ジアミノスベリン酸、S−エチルチオシステイン、ガンマ−グルタミン酸、ガンマ−カルボキシグルタミン酸、ピログルタミン酸、ホモアルギニン、ホモシステイン、ホモヒスチジン、ホモセリン、2−ヒドロキシイソ吉草酸(2-hydroxyisovaleric acid)、2−ヒドロキシペンタン酸、5−ヒドロキシリジン、4−ヒドロキシプロリン、2−カルボキシオクタヒドロインドール、3−カルボキシイソキノリン、イソバリン、2−ヒドロキシプロパン酸、メルカプト酢酸、メルカプト酪酸、4−メチル−3−ヒドロキシプロリン、メルカプトプロパン酸、ノルロイシン、ノルチロシン、ノルバリン、オルニチン、ペニシラミン、2−フェニルグリシン、2−カルボキシピペリジン、サルコシン、1−アミノ−1−カルボキシシクロペンタン、スタチン(statin)、3−チエニルアラニン(3-thyenylalanine)、イプシロン−N−トリメチルリジン、3−チアゾールアラニン、アルファ−アミノ−2,4−ジオキソピリミジンプロピオン酸などである。
【0042】
さらに、本発明は、本発明に係るペプチドの変異体として、1つ以上のアミノ酸の側鎖を変化させた変異体の使用について検討する。本発明における側鎖の改変の例は、アルキル化、アミジン化、アシル化、カルボモイル化(carbomoylation)、トリニトロベンジル化、ピリドキシル化などのアミノ基の改変、複素環縮合の形成からなるアルギニン残基のグアニジン基の改変;アミド化によるカルボキシル基の改変、メトキシル化によるチロシンの改変、アルキル化またはN−カルボキシエチル化によるヒスチジンのイミダゾール環の改変、4位のヒドロキシル化によるプロリンの改変などを含む。あるいは、本発明は、本発明に係るペプチドの変異体として、グリコシル化、すなわち、セリンおよび/もしくはスレオニン(O−グリコシル化)の側鎖、またはアスパラギンおよび/またはグルタミン(N−グリコシル化)の側鎖のどちらかへのグリカン基の付加による変異体について検討する。本発明におけるポリペプチドに組み込まれ得るグリカンが含む糖ユニットの数は、変えることができる(単糖、二糖、三糖、四糖など)。グリカンを形成する単糖は、D−アロース、D−アルトロース、D−グルコース、D−マンノース、D−グロース、D−イドース、D−ガラクトース、D−タロース、D−ガラクトサミン、D−グルコサミン、D−N−アセチルグルコサミン、D−N−アセチルガラクトサミン、D−フコース、またはD−アラビノースを含む。
【0043】
あるいは、本発明は、本発明に記載されたポリペプチドの変異体として、逆反転(retro-inverse)アイソマーを生じさせるために、ペプチド鎖を構成するアミノ酸の少なくとも1つとしてD立体異性体が含まれる変異体の使用を検討する。
【0044】
他の実施形態において、本発明は、本発明に記載されたポリペプチドにおける擬態ペプチド(peptidomimetics)、すなわち1つ以上のペプチド結合が共有結合に代わるものにより置き換わっている変異体の使用を検討する。この擬態ペプチドは、プロテアーゼに対してより耐性であるため、より高い安定性を示すことによって特徴付けられる。ペプチド骨格の改変は、ペプチド結合の構成要素(−NH−、−CH−、−CO−)に対する基の置換または挿入を含み、この基とは、例えば、−NH−の代わりに−O−、−S−、−CH2など、−CHRの代わりに−N−、−C−アルキル p −BH−など、および、−CO−の代わりに−CS−、−CH2−、−SOn−、−P=O(OH)−または−B(OH)−などである。さらに、N末端をブロックする基を用いれば、本発明に係るペプチドの安定性を高めることができる。この基とは、例えば、t−ブチルオキシカルボニル基、アセチル基、スクシニル基、メトキシスクシニル基、スベリル基(suberyl)、アジピル基(adipyl)、ダンシル基(dansyl)、ベンジルオキシカルボニル基、フルオレニルメトキシカルボニル基、メトキシアジピル基、メトキシアジピル基、メトキシスベリル2,3−ジニトロフェニル基などである。この代わりに、または同時に、アミド化によって本発明に係るペプチドのC末端を改変することが可能である。
【0045】
変異体と、配列番号2の配列によって示されるポリペプチドとの間の同一性は、コンピュータアルゴリズムおよび当業者に周知の方法を用いて決定される。2つのアミノ酸配列間の相同性は、BLASTPアルゴリズム(BLASTManual, Altschul, S., et al, NCBI NLM NIH Bethesda, Md. 20894, Altschul, S., et al., J. MoI. Biol. 21 5: 403-410 (1990))を用いて決定されることが好ましい。本発明において使用されるポリペプチドは、配列番号1〜19の配列によって示されるポリペプチドに対し、少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも91%、少なくとも92%、少なくとも93%、少なくとも94%、少なくとも95%、少なくとも96%、少なくとも97%、少なくとも98%、または少なくとも99%の配列の同一性を示すことが好ましい。
【0046】
酵素ジスケリンは、種々の生物に存在するシュードウリジン合成酵素ファミリーに属する(図3B、Mitchel et al, 1999を参照)。当業者は、本明細書に記載の情報および自然界に存在する種々の生物から、本明細書に記載のアミノ酸配列と類似したアミノ酸配列を同定または構築することができる。
【0047】
本発明の他のより好ましい実施形態は、本発明に係る使用からなり、GSE24.2誘導化剤がタンパク質であって、そのa)のアミノ酸配列が配列番号2により構成されているものである。
【0048】
本発明の他のより好ましい実施形態は、本発明に係る使用からなり、GSE24.2誘導化剤がタンパク質であって、そのc)のアミノ酸配列が配列番号4または配列番号6により構成されているものである。
【0049】
一方、本発明の他のさらなる実施形態は細胞(以下本明細書において、本発明に係るGSE24.2細胞)からなり、この細胞は、真核細胞−好ましくはヒト細胞、より好ましくは単離された細胞−または原核細胞であり、遺伝的に改変されており、かつ本発明に係るGSE24.2の塩基配列、構成物および発現ベクターを含んでおり、本発明において使用されるGSE24.2のペプチドまたはタンパク質が安定的に発現され得る。これらの細胞は、当業者に公知の遺伝子工学技術を用いて、上述した塩基配列により形質転換、感染または形質移入され得る(Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989). Molecular cloning:実験用マニュアル、2nd ed. Cold Spring Harbor Laboratoryおよび本発明に含まれる)。これらの細胞は、例えば老化プロセスを逆転させ、または減少させることができるペプチドを生産するために有用であり得る。また、これらの細胞は、例えば上述した塩基配列の組み換え体を増幅するために、医薬組成物のための基材として貢献し得る。あるいは、これらの細胞は、例えば遺伝子治療において細胞自体として有用であり得る。好ましい実施形態は、上述したGSE24.2の塩基配列により形質転換された、種々の細胞系統由来のヒト細胞の使用であってもよく、この細胞は、ヒト組織の再生細胞として使用できる。
【0050】
他の実施形態において、本発明は、本発明に係る遺伝的構成物または本発明において規定されたベクターを含む細胞または宿主生物に関する。本発明には、当業者に公知の宿主生物のいかなる類型をも使用でき、例えば細菌系統(大腸菌(Escherichia coli)、枯草菌(Bacillus subtilis)など)、酵母系統(酵母(Saccharomyces cerevisiae)、ピキア・パストリス(Pichia pastoris)、クリベロマイセス・ラクティス(Kluyveromyces lactis)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)など)、遺伝子導入植物(双子葉植物または単子葉植物)、昆虫細胞、例えば、バキュロ・ウイルス、哺乳動物細胞(COS細胞、CHO細胞、C127細胞、HeLa細胞など)、および非ヒトトランスジェニック(例えば、マウス、ウシ、ヤギ、ウサギ、ブタなど)などを使用できる。
【0051】
宿主細胞のゲノム中に新しい遺伝的物質が組み込まれるか組み込まれないかは、遺伝子発現系によって決まる。このように、GSE24.2の塩基配列、遺伝的構成物または発現ベクターはいずれも、異常な老化プロセスに関連する疾患に影響を受けるヒトに対する遺伝子治療の治療法および予防法において、異常化している(増加している)か否かにかかわらず老化プロセスに影響を受ける宿主細胞、好ましくはヒト細胞を保護するための医薬として使用され得る。同様に、本発明に係るGSE24.2細胞は、ヒトの組織または細胞の再生または移植のための医薬として使用され得る。これらの生物製剤ツールおよび遺伝子治療法は、本発明に記載の情報を過度の努力なしに発展させられる技術を有する当業者に充分に知られている。さらに、このタンパク質またはペプチドおよび細胞それら自身は、バイオドラッグとなり得る。
【0052】
本発明の他の実施形態は、老化の異常に関連する疾患または病状を治療するための医薬組成物または医薬(以下本明細書において、本発明に係る医薬組成物)からなる。この医薬組成物は、必要に応じて1つ以上のアジュバントおよび/または薬剤的に認容され得る溶媒とともに、GSE24.2誘導化化合物またはGSE24.2活性化化合物を治療的に有効な量において含む。また、この医薬組成物は、老化プロセスを減少させまたは逆転させることができ、かつ本発明に係る使用を用いて取得される。
【0053】
医薬に使用するために、本発明に係る化合物を有する化合物および組合せは、医薬品の観点から認容され得る賦形剤とともに処方され得る。本発明における使用のための好ましい賦形剤は、糖、デンプン、セルロース、粘剤およびタンパク質を含む。
【0054】
この記載中に使用される意味において、用語「治療的に有効な量」とは、老化からの回復が可能な薬剤または化合物の量をさし、所望の効果を生み出すために算出されるとともに、一般に、その化合物の固有の特性に基づいて、年齢、患者の状態、異常性または障害の重症度、および投与の経路および頻度を含む他の原因から決定されてもよい。
【0055】
好ましい実施形態において、本発明に係る医薬的組成物は、固形(例えば、錠剤、カプセル剤、トローチ剤、顆粒剤、座薬など)または液体(剤剤、懸濁剤、乳剤など)にて投与する医薬的な形態として処方されてもよい。他の好ましい実施形態では、本発明に係る医薬組成物は、限定されないが、経口、静脈内、筋肉内、動脈内、髄内、くも膜下腔内、脳室内、経皮、皮下、腹腔内、鼻腔内、腸内、局所的、舌下または直腸を含むいかなる経路を解して投与されてもよい。有効成分の種々の投与形態、使用される賦形剤およびその製造方法についての総説を、Galenic Pharmacy Treatise (Tratado de Farmacia Galenica), C. Fauli i Trillo, Luzan 5, S.A. de Ediciones, 1993.にて見出すことができる。
【0056】
本発明の他の実施形態は、好ましくはヒトにおいて、異常な老化のプロセスによって引き起こされる疾患または病状を治療するための方法における、本発明に係る医薬組成物の使用からなる。
【0057】
他の好ましい実施形態が、生物医学研究に使用する真核細胞、好ましくはヒト細胞または動物細胞を不死化するための方法における本発明に係る医薬組成物の使用からなることは、非常に重要である。真核細胞の不死化は、組み換えタンパク質生産工程に関して、好ましくは生物医学的適用における組み換えタンパク質生産工程に関して、その有効寿命を増加させるため、バイオテクノロジーのR+Dの分野において大いに必要な操作である。また、治療化合物として可能性のある化合物の効果の解析に用いられる正常細胞の有効寿命を増加させ、また、そのより長い寿命のおかげで二次的な効果を長期にわたって解析するのに好都合であるため、バイオテクノロジーのR+Dの分野において大いに必要な操作である。
【0058】
本発明の他の好ましい実施形態は、老化プロセスが逆転され、または減少しており、かつ本発明に用いられるGSE24.2の構成要素を含む、真核細胞、好ましくはヒト細胞からなる。
【0059】
〔図面の説明〕
<図1−シスプラチンへの応答におけるテロメラーゼ活性に対するGSE24−2ペプチドの発現の効果>
(C)シスプラチンおよびGSE24−2ペプチドを用いた、293T細胞抽出物のインビトロでの処理に続く、テロメラーゼ活性。シスプラチン、および精製したペプチドGSE24−2の量を増加させたもの(3−5列目)、熱により失活させたGSE24−2ペプチド(7列目)またはコントロールペプチド(8列目)を用いて、293T細胞抽出物をインビトロにて処理した後、後者をTPAPアッセイに用いた。テロメラーゼアッセイを行なうために、テロメアの配列を有する特異的な鋳型を用いた。この実験を3回繰り返して同様の結果を得た。(D)GSE24−2を発現する細胞抽出物を用いたゲルシフト試験。プラスミドpcDNA3−9E1024−2またはpcDNA3−9E10を発現する細胞の核抽出物をゲルシフト試験に供した。特異的なテロメアの配列を含むTEL1プローブを使用した。特異的な抗体9E10(3列目)、およびTEL1とハイブリダイズして競合分子として機能するオリゴヌクレオチドCXext(4−5列目)を用いることにより、特異的な結合が確立された。この実験を3回繰り返して同様の結果を得た。
【0060】
<図2−GSE24−2はhTERTプロモーターの活性を増加させる>
(A)pLNCX、DXC、GSE24−2、Cbf5、モチーフIまたはモチーフII(細胞百万個あたり10μgのDNA)およびレポータープラスミドhTERT−luc(細胞百万個あたり1μg)を用いて、293T細胞を同時形質移入した。形質移入して24時間後のテロメラーゼ活性を測定した。*は統計値がp<0.05であることを示す。(B)レポーターベクターhTERT−luc(細胞百万個あたり1μg)を用いて、293T細胞(黒)および293T24−2細胞(白)を同時形質移入した。24時間後、これらの細胞をシスプラチン(3 g/ml)により8時間処理した後、ルシフェラーゼ活性を測定した。(C)提示された構築物(細胞百万個あたり10μgのDNA)およびレポーターベクターhTR−luc(細胞百万個あたり1μg)を用いて、293T細胞を同時形質移入した。形質移入して24時間後のルシフェラーゼ活性を測定した。(D)レポーターベクターhTERT−luc、および様々な量のMad/mycの発現ベクターを用いて、これらの細胞系統を同時形質移入した。形質移入して24時間後のルシフェラーゼ活性を測定した。(E)レポーターベクターHIV−lucと、発現ベクターmad/mycの量を増加させたものとを用いて、これらの細胞系統を同時形質移入した。形質移入して24時間後に、50ng/mlのTNF を用いてこれらの細胞を6時間刺激した後、ルシフェラーゼ活性を測定した。形質移入効率を制御するために、CMV−Renillaベクターを用いた(0.1μg/ml)。それぞれのグラフは、4通りにて行なった2つの実験の平均値および標準偏差を表す。
【0061】
<図3−GSE24.2はc−MYC遺伝子のプロモーターを介してその活性を誘導する>
(A)実験に使用したそれぞれの構築物を示す、c−MYCプロモーターの概略図。(B)レポーターベクターc−MYC−luc(細胞百万個あたり1μg)のそれぞれの構築物を用いて、293T pLNCXおよび293T GSE24−2の細胞を形質移入した。形質移入して24時間後にルシフェラーゼ活性を測定した。形質移入効率のコントロールとして、CMV−Renillaベクターを用いた(細胞百万個あたり0.1μg/ml)。それぞれのグラフは、4通りにて行なった2つの実験の平均値および標準偏差を表す。(C)NHEIIIにおいて作製した変異体の概略図。(D)プラスミドpx3.2由来の変異体のレポーターベクター(細胞百万個あたり1μg)を用いて、提示された細胞系統を形質移入した。形質移入して24時間後にルシフェラーゼ活性を測定した。形質移入効率のコントロールとして、CMV−Renillaベクターを用いた(細胞百万個あたり0.1μg/ml)。それぞれのグラフは、4通りにて行なった2つの実験の平均値および標準偏差を表す。(E)発現ベクターpLNCX、DKCまたはGSE24−2を用いて、MEF細胞を形質移入した。形質移入して24時間後、全RNAを単離し、RT−PCRによって、マウスのc−myc遺伝子および −アクチン遺伝子の発現を測定した。この実験を3回繰り返して同様の結果を得た。(F)発現ベクターpLNCX、DKCまたはGSE24−2を用いて、MEF細胞を形質移入した。形質移入して24時間後、全タンパク質の抽出物を単離し、特異的な抗体を用いたウェスタンブロットによって、マウスのc−mycタンパク質および −チューブリンタンパク質の発現を決定した。この実験を3回繰り返して同様の結果を得た。
【0062】
<図4−GSE24−2は、X染色体に関連する先天性角化異常症の患者の細胞、およびVA13細胞におけるテロメラーゼ活性を再活性化させる>
(A)キャリアの母親(DC−C)、および影響を受けた子供(DC−1、DC−2およびDC−3)から取得した、X染色体に関連する先天性角化異常症を患う患者のリンパ芽球において、テロメラーゼ活性を測定した。これらの細胞に対し、空のベクター(−)またはGSE24−2(+)を細胞百万個あたり3μg用いて一過的に形質移入した。その24時間後にテロメラーゼ活性を測定した。(B)細胞百万個あたり3μgの発現ベクターpLNCX、GSE24−2またはDKCを用いて、一過的にDC2細胞を形質移入した。その24時間後にテロメラーゼ活性を測定した。(C)細胞百万個あたり3μgの空のベクターまたはGSE24.2により形質移入された患者(DC−3)の1人の細胞における、hTERTおよびhTRの発現レベル。RNAレベルは、RT−PCRにより検出した。GAPDH発現をコントロールとして用いた。(D)X−DC GMO1787の患者の繊維芽細胞を、ベクターpLNCXまたはGSE24−2由来のウイルスによって感染させ、安定な細胞系統をテロメラーゼ活性測定のために用いた(タンパク質の全量を三角形にて示す)。(E)プラスミドpLNCXおよびpGSE24−2によって形質移入したGMO1787細胞におけるhTERTおよびhTRの発現レベルを、RT−PCRにより測定した。GAPDH発現をコントロールとして用いた。(F)パネルDに記載したように形質移入されたGMO1787細胞の増殖速度を、一定時間における個体数の重複の累積を用いて評価した。全ての実験について3回繰り返し、同様の結果を得た。(G)VA13細胞におけるテロメラーゼ活性。この細胞に対し、細胞百万個あたり16μgのコントロールベクターpLNCXまたはGSE24.2を用いて一過的に形質移入した。その24時間後に、タンパク質抽出液の連続した希釈液を用いて、テロメラーゼ活性を測定した(タンパク質の全量を三角形にて表す)。(H)細胞百万個あたり16μgのプラスミドpLNCX、DKCまたはGSE24−2により形質移入されたVA13細胞における、hTERTおよびhTRの発現レベル。RT−PCRによりRNAレベルを測定した。GAPDHレベルをコントロールとして用いた。
【0063】
<図5−ペプチドGSE24.2は細胞老化から回復させる>
図4Dに記載されたように形質移入されたGMO1787細胞の増殖速度を、一定時間における個体数の重複の累積を用いて評価した。全ての実験について3回繰り返し、同様の結果を得た。
【0064】
〔本発明の実施例〕
(実施例1−GSE24.2ペプチドはインビトロにてテロメラーゼ活性を救済させる)
テロメア配列TTAGGGは、シスプラチンの標的である(参照文献45)。シスプラチンがこの領域に結合することによって、テロメラーゼの阻害とテロメアの短縮化とがもたらされる(参照文献43)。したがって、ジスケリンはテロメラーゼ複合体の構成要素の1つであるため(参照文献7)、本発明者らはTRAP(参照文献37)アッセイを用いてテロメラーゼ活性におけるシスプラチンおよびGSE24−2の発現の効果についての研究を続け、GSE24−2を発現させるベクターにより形質移入された細胞が高いテロメラーゼ活性を示すことを見出した(スペイン国特許出願公開第200502511号明細書)。
【0065】
本説明では、はじめに、293T細胞からの細胞抽出物、シスプラチン、およびシスプラチンに処理されたテロメアプローブを用いたインビトロのテロメラーゼ活性阻害試験における、前述したGSE24.2ペプチドの使用について記載する。この試験では、GSE24−2ペプチドを用量依存的な様式にて加えた結果、抽出物におけるテロメラーゼ活性をシスプラチンによる阻害から保護した(図1C、3−5列)。一方、熱により失活されたGSE24−2ペプチドまたは無関係のペプチドは、シスプラチンにより誘導されたテロメラーゼ阻害を救済することができなかった(図1C、7−8列)。これらによって、GSE24−2ペプチドが、インビトロでテロメアにおいてシスプラチンにより誘導される架橋を阻害できることが示された。さらに、小さい3’の吊り下がった配列によってテロメアの二重鎖配列を再構築する、ループを形成するオリゴヌクレオチド(TEL1)(参照文献41)を用いて、GSE24−2ペプチドにおけるテロメア配列に対する結合の可能性について、ゲルシフト試験によって解析した。特異的な複合体GSE24−2/TEL1(図1D)が検出された。一本鎖の延長部分の3’にハイブリダイズするオリゴヌクレオチドCxextは、複合体GSE24−2/TEL1の形成を完全に阻害した。併せて、これらの結果は、GSE24−2がテロメアの配列に結合し得ること、およびGSE24−2のテロメアプローブに対する保護効果によってシスプラチンによるテロメラーゼ活性の阻害が部分的に欠如することを示す。
【0066】
(実施例2−hTERTのプロモーターはGSE24−2タンパク質によって活性化されるが、hTRのプロモーターは活性化されない)
従来、GSE24.2におけるシスプラチンに対して保護できる能力によって、hTERTの高いRNAレベルが維持され得ることが観察された(スペイン国特許出願公開第200502511号明細書)。この仮説により、hTERTを過剰発現する293T細胞は、コントロール細胞よりもシスプラチンに対してより耐性であった。
【0067】
シスプラチンがhTERTのmRNAの減少を誘導し、かつGSE24−2の発現がこの効果を補償しているのではないかと考え、本発明者らは、GSE24−2がhTERTの転写を変化させ得るかどうかを詳細に調べた(参照文献53)。本発明者らは、hTERTプロモーターの3402bpを含むレポーターベクターを用いて、GSE24−2細胞におけるこのプロモーター活性を調査した(参照文献33)。GSE24−2の発現は、一過的な形質移入試験においてhTERTプロモーターの転写を刺激できたが、DKCはできなかった(図2A)。また、シスプラチンは、コントロール細胞よりもGSE24−2細胞において、このプロモーター活性を刺激する(図2B)。これらの結果は、GSE24−2の発現が、シスプラチン存在下であっても転写を刺激でき、その結果hTERTのmRNAレベルが増加したことを示す。本発明者らはまた、プラスミドGSE24−2により形質移入されたMEF細胞(一次マウスの繊維芽細胞)におけるhTERTプロモーターの刺激について検出できた(データは示さない)。DKCまたはGSE24−2を発現する細胞を用いたところ、hTRプロモーター(参照文献9)に対するGSE24−2の発現の効果は観察されなかった(図2C)。しかし、以前に報告されたように、阻害剤JNK SP60125(参照文献9)の処理は、このプロモーター活性を活性化することができた(データは示さない)(参照文献11)。これらの結果は、GSE24−2活性がhTERT発現に対して特異的であることを示す。
【0068】
ジスケリンは、当初は、酵母(S. cerevisiae)のCbf5遺伝子と、タンパク質レベルで85%の同一性を示す、ヒトの相同遺伝子cbf5(参照文献55)として同定された(参照文献56)。そのため、本発明者らは、酵母遺伝子S. cerevisiae cbf5から、GSE24−2に相当する配列(配列番号2)をベクターpLNCXにクローニングした。この構築物の発現もまた、hTERTプロモーターの発現を増加させる(図2A)が、hTRプロモーターの発現は増加させない(図2C)。これは、このDKC領域の機能が高度に保存されていることを示す。
【0069】
(実施例3−c−MYCプロモーターのNHEIIIドメインはGSE24−2の標的配列である)
hTERTプロモーターは、myc/maxヘテロダイマーに結合可能な2つのE−boxを含む(参照文献32、57)。madをトランス活性化するc−mycのDNA結合ドメインを含むハイブリッド分子は、E boxに結合可能であり、そのためc−myc依存的な転写を阻害する。mad/mycの発現は、DKC細胞だけでなくコントロール細胞においても、投与量に依存する様式にて、hTERTプロモーター活性を阻害することができる(図3D)。また、myc/madの発現は、GSE24−2細胞におけるGSE24.2タンパク質によって介される転写活性化を消失させることができる。これにより、GSE24−2によって誘導されるhTERTの転写の活性化が、c−myc依存的であることが示された(図3D)。myc/mad構築物をNFκBプロモーターのためのレポーターベクター(HIVLuc)とともに形質移入しても、TNFαによって誘導される転写に影響を及ぼさないことから、この阻害は特異的である(図4E)。
【0070】
3.2Kbのc−myc遺伝子(px3.2myc)を含み、かつルシフェラーゼ遺伝子に結合されたプロモーターP0、P1およびP3を包含するレポーターベクター(図3A)を用いて、コントロール細胞およびGSE24−2細胞を形質移入した。GSE24−2細胞におけるプロモーター活性が、コントロール細胞と比べて3倍に増加したことが観察された(図3B)。レポーターベクターpx3.2mycとともにベクターpLNCXまたはGSE24−2を発現するプラスミドにより293T細胞を形質移入した場合にも、同様の結果が得られた(データは示さない)。GSE24−2が介する活性化に必要な領域を調べるために、c−mycプロモーターの3つの欠失変異体を用いた(図3A)。NHEIIIが含まれる、末端のプロモーターP1およびP0が包含される構築物のみが、GSE24−2細胞において活性化されることができたが、コントロール細胞においては活性化されなかった(図3B)。これにより、最も近いプロモーターP2は、GSEを介した活性化に関わっていないことが示された。
【0071】
G−カルテットの二次構造の維持に関わっているプリンリッチ領域(Pu27)におけるグアニン残基を置換することによって、NHEIIIの種々の変異体を取得した。4つ組のG群の2番目におけるG12A変異体、2番目の3つ組のGからG17Aの変異体、そして最後にプリンリッチ領域の末端における2つの連続したグアニン(G26A/G27A)の変異体を取得した(図3C)。この結果から、オリジナルの配列のみがGSE24−2によって活性化されることが示された(図3D)。概して、これらの結果により、GSE24−2が、Pu27領域の二次構造の変形を誘導してc−myc転写させる活性のある配置にさせることが可能であることが示唆される。
【0072】
本発明者らはまた、c−mycプロモーター活性にて観察される変化が、その発現に反映されるのかを調べた。この効果のために、空のベクター、DKCの発現ベクターまたはGSE24−2発現ベクターを用いてMEF細胞を一過的に形質移入し、次にmRNAおよびc−mycタンパク質のレベルを測定した(図3E y 3F)。予想どおり、GSE24−2の発現のみが、c−mycのmRNAとタンパク質との両方について、その発現の活性化を誘導できた。c−MYC発現の増加が細胞の形質転換を誘導し得ることは、GSE24−2発現による不要な効果の1つとなり得る。この可能性を評価するために、極小マウス(atimic mice)に、空のベクター、ヒトc−myc遺伝子、またはGSE24−2を発現しているPam212細胞を接種した。c−mycを発現している細胞を接種された動物のみが、最初の10日以内に腫瘍を発症し、GSE24−2を発現している細胞は接種後3ヶ月間でさえも腫瘍を発症させなかった。
【0073】
(実施例4−GSE24−2を用いて刺激されたVA13細胞およびX−DC患者におけるテロメラーゼ活性は、老化プロセスからそれらを回復させる)
本発明者らは、X−DCを患っている患者の細胞におけるGSE24−2発現の効果を調べた。キャリアの母親(DC、正常なdkc対立遺伝子)および臨床的に影響を受けた彼女の3人の子供(DC1、DC2およびDC3、変異体dkc対立遺伝子)から取得された市販のリンパ芽球を用いた。これらの細胞は不死化されておらず、培養中に正常に老化し始める。さらに以前観察されたように、すなわちGSE24−2の発現はこれらの細胞系統におけるテロメラーゼ活性を増加させ(図4A)、前述したように(参照文献21)、全長のジスケリン発現によるテロメラーゼ活性の増加は観察されなかった(図4B)。GSE24−2により形質移入された細胞において、hTERTおよびhTRのRNAレベルが増加した(図5C)ことは、GSE24−2の形質移入によるテロメラーゼ活性の回復が、両方の分子の増加に起因することを示唆する。
【0074】
一方、X−DC患者における本来の真皮の繊維芽細胞は、連続培養において3〜4週間以内に完全に老化するが、正常な細胞は、2ヶ月増殖した後に老化期に到達する(参照文献21)。hTERTを発現するX−DC細胞は、より長く分裂し、数ヶ月間連続して生育する能力を獲得する(参照文献21)。X−DCにおけるGMO1787細胞内でのGSE24−2の発現は、前述したように、X−DCにおける繊維芽細胞内のhTRおよびhTERTのRNAレベルの発現を増加させることによって、テロメラーゼ活性の増加を誘導することができた(図4D)。そして、驚いたことに、これらの細胞は、連続して160日以上成育することができた(図5)。ウェルナー症候群の患者における細胞(老化した真皮の繊維芽細胞)もまた、連続して150日以上生育することができた(図6)。
【0075】
さらに興味深いことに、hTERTおよびhTRの欠如を示し(参照文献59)、かつALTメカニズムを用いてテロメアを維持するVA13のテロメラーゼ欠失細胞系統(野生型のジスケリン)において、GSE24−2の発現が、テロメラーゼ活性(図4G)、およびhTERTおよびhTRのRNAレベル(図4H)を回復させることができた。
【0076】
〔材料および方法〕
(構築物および細胞系統)
GSE24−2、DKC、DKC5’、モチーフI、モチーフIIおよびGSE24−2と相同なCBF5遺伝子の領域の配列を、pLNCXベクター(BD Biosciences, Madrid)にクローニングした。hTERTプロモーターを有する構築物は、Dr. T.Kim(参照文献32)から取得し、hTRプロモーターを有する構築物は、Dr. N.Keith(参照文献9)から取得し、c−MYCプロモーター(px3.2)を有する構築物は、Dra. A. Arandaから取得し、そしてハイブリッド構築物Myc/mad、pECLおよびpECLc−mycは、Dr. J. Leonから取得した。プラスミドBASE−TERTはDra. K. Collinsから取得した(参照文献31)。プラスミドPGATEV(参照文献33)は、Dr. G. Montoyaから取得した。プラスミドPGATEV−24−2は、pGATEVプラスミドのNdeI/XhoI部位の24−2断片をサブクローニングすることにより取得した。プラスミドpCDNA3−9E10−24−2は、エピトープ9E10−mycを含むpCDNA3ベクターのEcoR1/XbaI部位の間の24−4断片をサブクローニングすることによって取得した。293T細胞(American Type Culture Collection)Phoenixを、10%FBSを添加したDMEM培地にて培養した。細胞系統VA13は、Dr. M. Serranoから取得した。ファミリー(DC、DC1、DC2およびDC3)のX−DCリンパ芽球については、臨床的に報告されており(参照文献34〜35)、そのジスケリン遺伝子は1塩基置換を示す:T66A。X−DCの皮膚の繊維芽細胞GMO1787は、10%FCSを添加したDMEMにて培養し、そしてこの細胞には、ロイシン37を欠失したジスケリンタンパク質がコードされていた(参照文献31)。マウスの胚の繊維芽細胞(MEF)は、10%FBSと非必須アミノ酸(Gibco, Carlsbad, USA)とを添加したDMEMにて培養した。安定な形質移入のために、293T細胞を、細胞百万個あたり10μgのGSE24−2またはpLNCXと1μgのプラスミドp−BabePUR(Clontech)とを用いて同時形質移入し、形質転換24時間後にピューロマイシン耐性のものを選択した。Pam212細胞は、ベクターpECL、pECLc−mycまたはGSE24−2を用いて形質移入し、G418耐性のものを選択した。X−DC繊維芽細胞の感染のために、リン酸カルシウム法によりpLNCX由来のベクターを用いてPhoenix細胞を形質移入した。形質移入48時間後、ウイルスを回収し、X−DC繊維芽細胞を感染させるために48時間使用した。安定な細胞系統は、G−418(Gibco)より選択した。
【0077】
(GSEsのライブラリーおよびシスプラチンによる選択)
基本的に、以前にRoninson et al(参照文献36)によって記載されたように、GSEsのライブラリーを構築した。
【0078】
(試薬)
シスプラチン、テロメラーゼIの阻害剤およびSP60125は、Calbiochem(San Diego, USA)から購入した。TNFαは、Upstate(Charlottesville, USA)から購入した。
【0079】
(ペプチドGSE24−2の生産および精製)
大腸菌(E.coli)のDH5α細胞を、ベクターpGATEV GSE24−2を用いて形質転換し、その溶菌液を前述されたように調製した(参照文献33)。融合タンパク質をグルタチオン−セファロース法によって精製し、その純度をゲル電気泳動によって解析した。製造者の指示にしたがってプロテアーゼTEVにより消化して、GSE24−2を取得した。通常、SDS−PAGE電気泳動にて取得されるものによれば、消化されたタンパク質の90%が取得される。このタンパク質は、Hi−Trap Ni−NTAカラムに2回通され、ポリヒスチジンテール、未消化のタンパク質、TEVプロテアーゼおよび不純物を廃棄した。
【0080】
(定方向突然変異誘発)
Quickchange X-L site-directed mutagenesis kit(Stratagene, Santa Clara, USA)を用いて、製造者の指示にしたがってc−MYCプロモーター(px3.2)のNHEIII領域におけるグアニン残基の変異導入を行なった。
【0081】
(テロメアリピート増幅プロトコール(TRAP))
TRAPeze(参照文献37) telomerase detection kit(Intergen, Purchase, USA)を用いて製造者の指示にしたがってテロメラーゼ活性を測定した。TRAPアッセイは、存在する全タンパク質の濃度によりそれぞれのタンパク質抽出物を標識して行なった。所望の実験において、テロメアのプローブをTRAPアッセイに使用した(参照文献38)。TRAP反応において直接にシスプラチンとともにインキュベートすることによってインビトロでのプラチナ化(platination)を行なった。
【0082】
(イムノブロットおよび抗体)
前述されたように細胞を溶解させ(参照文献39)、その後20μgのタンパク質を10%にてSDS−PAGEにより解析した。使用した抗体:抗pJNK(V7391、Promega, Madison, USA)、抗JNK1(C−17、Santa Cruz Biotechnologies, Santa Cruz, USA)、抗p−P38(Cell Signaling, Charlottesville, USA)、9−E10(A14−Santa Cruz Biotechnologies)および抗Flag(Invitrogen, Carlsbad, USA)。
【0083】
(cDNAの調製およびRT−PCR)
Trizol(Life Technologies, Carlsbad, USA)を用いて製造者の指示にしたがって細胞の全RNAを抽出した。逆転写酵素M−Mlv(Promega)を用いて、各反応において2μgの全RNAをcDNAに転写させた。
【0084】
(形質移入および遺伝子発現解析)
293T細胞は、前述されたようにリン酸カルシウム法を用いて一過的に形質移入した(参照文献39)。DNAの全量は、常に細胞百万個あたり10μgを維持した。X−DC細胞は、細胞百万個あたり3μgのpLNCX−GSE24−2を用いたエレクトロポレーション法により一過的に形質移入した。VA13のMEFs細胞は、100万個の細胞あたり16μgのDNAを用いてリポフェクタミン プラス(lipofectamine plus)(Invitrogen)により製造者の指示にしたがって形質移入した。タンパク質抽出液を調製し、市販のキット(Promega)を用いてルシフェラーゼを測定した。各試験は3通りにて行ない、実験は3回繰り返した。形質移入効率は、プラスミドp−CMV−Renillaを同時形質移入することによって修正した。
【0085】
(ゲル易動度シフトアッセイ)
プラスミドpCDNA3−E1024−2または空のベクターによって293T細胞を形質移入し、G−418により選択することによって安定な細胞系統を作製した。これらの細胞は、エピトープE10−myc GSE24−2の融合タンパク質を発現する。核タンパク質(参照文献40)の抽出液における結合反応を、前述されたように行なった(参照文献41)。テロメア配列を含むオリゴヌクレオチドTEL1の5’末端、及びTEL1のハンギング領域(hanging zone)にハイブリダイズするCXext(ネガティブコントロールとして)(参照文献41)の5’末端を、T4ポリヌクレオチドキナーゼ(New England Biolabs, Beverly, MA)および 32P ATP(Amersham Bioscences, Orsay, France)によって標識した。エピトープ9E10に対する抗体を用いて、GSE24−2結合と競合させた。DNAに結合した複合体は、トリス・ホウ酸塩・EDTAバッファーを用いた0.5%アクリルアミドの非変性ゲルにおいて分離させた。
【0086】
(生存度アッセイおよび累積重複数(PLD))
前述されたように、クリスタル・バイオレット染色法を用いて、細胞増殖および生存度を調べた(参照文献39)。感染後にDKC繊維芽細胞を混合し、G−418耐性細胞を選択した。細胞を選択する最初のステップにおいて、培養中のPLDsを定量化して増殖を測定した。
【0087】
〔参照文献〕
1.de Lange T. Protection of mammalian telomeres. Oncogene. 2002; 21: 532-540.
2.Blackburn EH. Switching and signaling at the telomere. Cell. 2001; 106: 661-673.
3.Collins K, Mitchell JR. Telomerase in the human organism. Oncogene. 2002; 21: 564-579.
4.Colgin LM, Reddel RR. Telomere maintenance mechanisms and cellular immortalization. Curr Opin Genet Dev. 1999; 9: 97-103.
5.Harley CB. Telomerase therapeutics for degenerative diseases. Curr Mol Med. 2005; 5: 205-211.
6.Blasco MA. Telomeres and human disease: ageing, cancer and beyond. Nat Rev Genet. 2005; 6: 611-622.
7.Cohen SB, Graham ME, Lovrec GO, et al, Protein composition of catalytically active human telomerase from immortal cells. Science, 2007; 315: 1850-1853
8.Zhao J Q, Hoare S F, McFarlane R, et al. Cloning and characterization of human and mouse telomerase RNA gene promoter sequences. Oncogene. 1998; 16: 1345-1350.
9.Takakura M, Kyo S, Kanaya T, et al. Cloning of human telomerase catalytic subunit (hTERT) gene promoter and identification of proximal core promoter sequences essential for transcriptional activation in immortalized and cancer cells. Cancer Res. 1999; 59: 551-5577.
10.Gomez D, Aouali N, Renaud A, et al. Resistance to senescence induction and telomere shortening by a G-quadruplex ligand inhibitor of telomerase. Cancer Res. 2003; 63: 6149-6153.
11.Li H, Xu D, Li J, Berndt MC, Liu JP. Transforming growth factor beta suppresses human telomerase reverse transcriptase (hTERT) by Smad3 interactions with c-Myc and the hTERT gene. J Biol Chem. 2006; 281: 25588-255600.
12.Levens D, Duncan R C, Tomonaga T, et al. DNA conformation, topology, and the regulation of c-myc expression. Curr Top Microbiol Immunol. 1997; 224: 33-46.
13.Siebenlist U, Hennighausen L, Battey J, Leder P. Chromatin structure and protein binding in the putative regulatory region of the c-myc gene in Burkitt lymphoma. Cell. 1984; 37: 381-391.
14.Mirkin S M, Frank-Kamenetskii MD. H-DNA and related structures. Annu Rev Biophys Biomol Struct. 1994; 23: 541-576
15.Siddiqui-Jain A, Grand CL, Bearss DJ, Hurley LH. Direct evidence for a G-quadruplex in a promoter region and its targeting with a small molecule to repress c-MYC transcription. Proc Natl Acad Sci U S A. 2002; 99: 11593-11598.
16.Bessler M, Wilson DB, Mason PJ. Dyskeratosis congenita and telomerase. Curr Opin Pediatr. 2004; 16: 23-28.
17.Marrone A, Mason PJ. Dyskeratosis congenita. Cell Mol Life Sci. 2003; 60: 507-517.
18.Mason PJ, Wilson DB, Bessler M. Dyskeratosis congenita -- a disease of dysfunctional telomere maintenance. In Curr Mol Med. 2005; 5: 159-170.
19.Heiss NS, Knight SW, VulliamyTJ, et al. X-linked dyskeratosis congenita is caused by mutations in a highly conserved gene with putative nucleolar functions. Nat Genet. 1998; 19: 32-38.
20.Knight SW, Heiss NS, Vulliamy TJ, et al. X-linked dyskeratosis congenita is predominantly caused by missense mutations in the DKC1 gene. Am J Hum Genet. 1999; 65: 50-58.
21.Mitchell J, Wood E, Collins K. A telomerase component is defective in the human disease dyskeratosis congenita. Nature. 1999; 402: 551-555.
22.Normand C, Capeyrou R, Quevillon-Cheruel S, Mougin A, Henry Y, Caizergues-Ferrer M. Analysis of the binding of the N-terminal conserved domain of yeast Cbf5p to a box H/ACA snoRNA. RNA. 2006; 12: 1868-82.
23.Henras A, Henry Y, Bousquet-Antonelli C, Noaillac-Depeyre J, Gelugne JP, Caizergues-Ferrer M. Nhp2p and Nop10p are essential for the function of H/ACA snoRNPs.; EMBO J. 1998; 17: 7078-7090.
24.Pogacic V, Dragon F, Filipowicz W.Human H/ACA small nucleolar RNPs and telomerase share evolutionarily conserved proteins NHP2 and NOP10.. Mol Cell Biol. 2000; 20: 9028-40.
25.Wang C, Meier UT. Architecture and assembly of mammalian H/ACA- small nucleolar and telomerase ribonucleoproteins. EMBO J. 2004; 23: 1857-1867
26.Henras AK, Capeyrou R, Henry Y, et al Cbf5p, the putative pseudouridine synthase of H/ACA-type snoRNPs, can form a complex with Gar1p and Nop10p in absence of Nhp2p and box H/ACA snoRNAs. RNA. 2004; 10: 1704-12.
27.Rashid R, Liang B, Baker D L, et al. Crystal structure of a Cbf5-Nop10-Gar1 complex and implications in RNA-guided pseudouridylation and dyskeratosis congenita.. Mol Cell.2006; 21: 249-260.
28.Darzacq X, Kittur N, Roy S, et al Stepwise RNP assembly at the site of H/ACA RNA transcription in human cells. J Cell Biol. 2006; 173: 207-18.
29.Tollervey D, Kiss T. Function and synthesis of small nucleolar RNAs. Curr Opin Cell Biol. 1997; 9: 337-342.
30.Fu D, Collins K. Distinct biogenesis pathways for human telomerase RNA and H/ACA small nucleolar RNAs. Mol Cell. 2003; 11: 1361-1372.
31.Wong JM, Collins K. Telomerase RNA level limits telomere maintenance in X-linked dyskeratosis congenita. Genes Dev. 2006; 2: 2848-2858.
32.Oh S, Song YH, Kim UJ, Yim J, Kim TK. In vivo and in vitro analyses of Myc for differential promoter activities of the human telomerase (hTERT) gene in normal and tumor cells. Biochem Biophys Res Commun. 1999; 263: 361-365
33.Kalinin A, Thoma NH, Iakovenko A, Heinemann I, Rostkova E, Constantinescu AT, Alexandrov K. Expression of mammalian geranylgeranyltransferase type-II in Escherichia coli and its application for in vitro prenylation of Rab proteins. Protein Expr Purif. 2001; 22: 84-91.
34.Sirinavin C, Trowbridge AA. Dyskeratosis congenita: clinical features and genetic aspects. Report of a family and review of the literature. J Med Genet. 1975; 12: 339-354
35.Trowbridge AA, Sirinavin C, Linman JW. Dyskeratosis congenita: hematologic evaluation of a sibship and review of the literature. Am J Hematol. 1977; 3: 143-152.
36.Roninson IB, Gudkov AV, Holzmayer TA, et al. Genetic suppressor elements: new tools for molecular oncology. Cancer Res. 1995; 55: 4023-4028.
37.Wright WE, Shay JW, Piatyszek MA. Modifications of a telomeric repeat amplification protocol (TRAP) result in increased reliability, linearity and sensitivity. Nucleic Acids Res. 1995; 23: 3794-3795.
38.Gomez D, Mergny JL, Riou JF. Detection of telomerase inhibitors based on g-quadruplex ligands by a modified telomeric repeat amplification protocol assay Cancer Res. 2002; 62: 3365-8.
39.Sanchez-Perez I, Perona R. Lack of c-Jun activity increases survival to cisplatin. FEBS Lett. 1999; 453: 151-158.
40.Perona R, Montaner S, Saniger L, Sanchez-Perez I, Bravo R, Lacal JC.1997 Activation of the nuclear factor-kappaB by Rho, CDC42, and Rac-1 proteins. Genes Dev. 1997; 11: 463-75.
41.Gomez D, O'Donohue MF, Wenner T, Douarre C, Macadre J, Koebel P, Giraud-Panis MJ, Kaplan H, Kolkes A, Shin-ya K, Riou JF.The G-quadruplex ligand telomestatin inhibits POT1 binding to telomeric sequences in vitro and induces GFP-POT1 dissociation from telomeres in human cells. Cancer Res. 2006; 66: 6908-12.
42.Redon S, Bombard S, Elizondo-Riojas MA, Chottard JC. Platination of the (T2G4) 4 telomeric sequence: a structural and cross-linking study. Biochemistry. 2001; 40: 8463-8470.
43.Ishibashi T, Lippard SJ. Telomere loss in cells treated with cisplatin. Proc Natl Acad Sci U S A. 1998; 95: 4219-4223.
44.Parkinson GN, Lee M P, Neidle S. Crystal structure of parallel quadruplexes from human telomeric DNA. Nature. 2002; 417: 876-880.
45.Bednarek A, Shilkaitis A, Green A, et al. Suppression of cell proliferation and telomerase activity in 4-(hydroxyphenyl)retinamide-treated mammary tumors. Carcinogenesis. 1999; 20: 879-883.
46.Kim JH, Kim JH, Lee GE, Kim SW, Chung IK. Identification of a quinoxaline derivative that is a potent telomerase inhibitor leading to cellular senescence of human cancer cells. Biochem J. 2003; 373: 523-529.
47.Hurley LH. DNA and its associated processes as targets for cancer therapy. Nat Rev Cancer. 2002; 2: 188-200.
48.Sun D, Thompson B, Cathers BE, et al. Inhibition of human telomerase by a G-quadruplex-interactive compound. J Med Chem. 1997; 40: 2113-2116.
49.Cuesta J, Read MA, Neidle S. The design of G-quadruplex ligands as telomerase inhibitors. Mini Rev Med Chem. 2003; 3: 11-21.
50.Chau NP, Deschatrette J, Wolfrom C Reversal of hepatoma cells resistance to anticancer drugs is correlated to cell proliferation kinetics, telomere length and telomerase activity. Anticancer Res. 2005; 25: 3279-85
51.Lin CP, Liu JD, Chow JM, Liu CR, Liu HE. Small-molecule c-Myc inhibitor, 10058-F4, inhibits proliferation, downregulates human telomerase reverse transcriptase and enhances chemosensitivity in human hepatocellular carcinoma cells. Anticancer Drugs. 2007; 18: 161-70.
52.Sun PM, Wei LH, Luo MY, Liu G, Wang JL, Mustea A, Konsgen D, Lichtenegger W, Sehouli J. The telomerase activity and expression of hTERT gene can serve as indicators in the anti-cancer treatment of human ovarian cancer. Eur J Obstet Gynecol Reprod Biol. 2007; 130: 249-57.
53.Poole JC, Andrews LG, Tollefsbol TO. Activity, function, and gene regulation of the catalytic subunit of telomerase (hTERT). Gene. 2001; 269: 1-12.
54.Meier UT, Blobel G. NAP57, a mammalian nucleolar protein with a putative homolog in yeast and bacteria.. J Cell Biol. 1994; 127: 1505-14.
55.Meier UT. The many facets of H/ACA ribonucleoproteins. Chromosoma. 2005; 114: 1-14.
56.Zucchini C, Strippoli P, Biolchi A, et al. The human TruB family of pseudouridine synthase genes, including the Dyskeratosis Congenita 1 gene and the novel member TRUB1. Int J Mol Med. 2003; 11: 697-704.
57.Wu KJ, Grandori C, Amacker M, et al. Direct activation of TERT transcription by c-MYC. Nat Genet. 1999; 21: 220-224.
58.Mochizuki Y, He J, Kulkarni S, Bessler M, Mason PJ. Mouse dyskerin mutations affect accumulation of telomerase RNA and small nucleolar RNA, telomerase activity, and ribosomal RNA processing. Proc Natl Acad Sci U S A. 2004; 101: 10756-10761.
59.Bryan TM, Marusic L, Bacchetti S, Namba M, Reddel RR The telomere lengthening mechanism in telomerase-negative immortal human cells does not involve the telomerase RNA subunit. Hum Mol Genet. 1997; 6: 921-6.
60.Kunifuji Y, Gotoh S, Abe T, Miura M, Karasaki Y. Down-regulation of telomerase activity by anticancer drugs in human ovarian cancer cells. Anticancer Drugs. 2002; 13: 595-598.
61.Postel EH, Berberich SJ, Rooney JW, Kaetzel DM. Human NM23/nucleoside diphosphate kinase regulates gene expression through DNA binding to nuclease-hypersensitive transcriptional elements. J Bioenerg Biomembr. 2000; 32: 277-284.
62.Nosaka K, Kawahara M, Masuda M, Satomi Y, Nishino H. Association of nucleoside diphosphate kinase nm23-H2 with human telomeres. Biochem Biophys Res Commun. 1998; 243: 342-348.
【図面の簡単な説明】
【0088】
【図1a】シスプラチンへの応答におけるテロメラーゼ活性に対するGSE24−2ペプチドの発現の効果。
【図1b】シスプラチンへの応答におけるテロメラーゼ活性に対するGSE24−2ペプチドの発現の効果。
【図1c】(C)シスプラチンおよびGSE24−2ペプチドを用いた、293T細胞抽出物のインビトロでの処理に続く、テロメラーゼ活性。シスプラチン、および精製したペプチドGSE24−2の量を増加させたもの(3−5列目)、熱により失活させたGSE24−2ペプチド(7列目)またはコントロールペプチド(8列目)を用いて、293T細胞抽出物をインビトロにて処理した後、後者をTPAPアッセイに用いた。テロメラーゼアッセイを行なうために、テロメアの配列を有する特異的な鋳型を用いた。この実験を3回繰り返して同様の結果を得た。
【図1d】(D)GSE24−2を発現する細胞抽出物を用いたゲルシフト試験。プラスミドpcDNA3−9E1024−2またはpcDNA3−9E10を発現する細胞の核抽出物をゲルシフト試験に供した。特異的なテロメアの配列を含むTEL1プローブを使用した。特異的な抗体9E10(3列目)、およびTEL1とハイブリダイズして競合分子として機能するオリゴヌクレオチドCXext(4−5列目)を用いることにより、特異的な結合が確立された。この実験を3回繰り返して同様の結果を得た。
【図2a】GSE24−2はhTERTプロモーターの活性を増加させる。(A)pLNCX、DXC、GSE24−2、Cbf5、モチーフIまたはモチーフII(細胞百万個あたり10μgのDNA)およびレポータープラスミドhTERT−luc(細胞百万個あたり1μg)を用いて、293T細胞を同時形質移入した。形質移入して24時間後のテロメラーゼ活性を測定した。*は統計値がp<0.05であることを示す。
【図2b】(B)レポーターベクターhTERT−luc(細胞百万個あたり1μg)を用いて、293T細胞(黒)および293T24−2細胞(白)を同時形質移入した。24時間後、これらの細胞をシスプラチン(3 g/ml)により8時間処理した後、ルシフェラーゼ活性を測定した。
【図2c】(C)提示された構築物(細胞百万個あたり10μgのDNA)およびレポーターベクターhTR−luc(細胞百万個あたり1μg)を用いて、293T細胞を同時形質移入した。形質移入して24時間後のルシフェラーゼ活性を測定した。
【図2d】(D)レポーターベクターhTERT−luc、および様々な量のMad/mycの発現ベクターを用いて、これらの細胞系統を同時形質移入した。形質移入して24時間後のルシフェラーゼ活性を測定した。
【図2e】(E)レポーターベクターHIV−lucと、発現ベクターmad/mycの量を増加させたものとを用いて、これらの細胞系統を同時形質移入した。形質移入して24時間後に、50ng/mlのTNF を用いてこれらの細胞を6時間刺激した後、ルシフェラーゼ活性を測定した。形質移入効率を制御するために、CMV−Renillaベクターを用いた(0.1μg/ml)。それぞれのグラフは、4通りにて行なった2つの実験の平均値および標準偏差を表す。
【図3a】GSE24.2はc−MYC遺伝子のプロモーターを介してその活性を誘導する。(A)実験に使用したそれぞれの構築物を示す、c−MYCプロモーターの概略図。
【図3b】(B)レポーターベクターc−MYC−luc(細胞百万個あたり1μg)のそれぞれの構築物を用いて、293T pLNCXおよび293T GSE24−2の細胞を形質移入した。形質移入して24時間後にルシフェラーゼ活性を測定した。形質移入効率のコントロールとして、CMV−Renillaベクターを用いた(細胞百万個あたり0.1μg/ml)。それぞれのグラフは、4通りにて行なった2つの実験の平均値および標準偏差を表す。
【図3c】(C)NHEIIIにおいて作製した変異体の概略図。
【図3d】(D)プラスミドpx3.2由来の変異体のレポーターベクター(細胞百万個あたり1μg)を用いて、提示された細胞系統を形質移入した。形質移入して24時間後にルシフェラーゼ活性を測定した。形質移入効率のコントロールとして、CMV−Renillaベクターを用いた(細胞百万個あたり0.1μg/ml)。それぞれのグラフは、4通りにて行なった2つの実験の平均値および標準偏差を表す。
【図3e】(E)発現ベクターpLNCX、DKCまたはGSE24−2を用いて、MEF細胞を形質移入した。形質移入して24時間後、全RNAを単離し、RT−PCRによって、マウスのc−myc遺伝子および −アクチン遺伝子の発現を測定した。この実験を3回繰り返して同様の結果を得た。
【図3f】(F)発現ベクターpLNCX、DKCまたはGSE24−2を用いて、MEF細胞を形質移入した。形質移入して24時間後、全タンパク質の抽出物を単離し、特異的な抗体を用いたウェスタンブロットによって、マウスのc−mycタンパク質および −チューブリンタンパク質の発現を決定した。この実験を3回繰り返して同様の結果を得た。
【図4a】GSE24−2は、X染色体に関連する先天性角化異常症の患者の細胞、およびVA13細胞におけるテロメラーゼ活性を再活性化させる。(A)キャリアの母親(DC−C)、および影響を受けた子供(DC−1、DC−2およびDC−3)から取得した、X染色体に関連する先天性角化異常症を患う患者のリンパ芽球において、テロメラーゼ活性を測定した。これらの細胞に対し、空のベクター(−)またはGSE24−2(+)を細胞百万個あたり3μg用いて一過的に形質移入した。その24時間後にテロメラーゼ活性を測定した。
【図4b】(B)細胞百万個あたり3μgの発現ベクターpLNCX、GSE24−2またはDKCを用いて、一過的にDC2細胞を形質移入した。その24時間後にテロメラーゼ活性を測定した。
【図4c】(C)細胞百万個あたり3μgの空のベクターまたはGSE24.2により形質移入された患者(DC−3)の1人の細胞における、hTERTおよびhTRの発現レベル。RNAレベルは、RT−PCRにより検出した。GAPDH発現をコントロールとして用いた。
【図4d】(D)X−DC GMO1787の患者の繊維芽細胞を、ベクターpLNCXまたはGSE24−2由来のウイルスによって感染させ、安定な細胞系統をテロメラーゼ活性測定のために用いた(タンパク質の全量を三角形にて示す)。
【図4e】(E)プラスミドpLNCXおよびpGSE24−2によって形質移入したGMO1787細胞におけるhTERTおよびhTRの発現レベルを、RT−PCRにより測定した。GAPDH発現をコントロールとして用いた。
【図4f】(F)パネルDに記載したように形質移入されたGMO1787細胞の増殖速度を、一定時間における個体数の重複の累積を用いて評価した。全ての実験について3回繰り返し、同様の結果を得た。
【図4g】(G)VA13細胞におけるテロメラーゼ活性。この細胞に対し、細胞百万個あたり16μgのコントロールベクターpLNCXまたはGSE24.2を用いて一過的に形質移入した。その24時間後に、タンパク質抽出液の連続した希釈液を用いて、テロメラーゼ活性を測定した(タンパク質の全量を三角形にて表す)。
【図4h】(H)細胞百万個あたり16μgのプラスミドpLNCX、DKCまたはGSE24−2により形質移入されたVA13細胞における、hTERTおよびhTRの発現レベル。RT−PCRによりRNAレベルを測定した。GAPDHレベルをコントロールとして用いた。
【図5】ペプチドGSE24.2は細胞老化から回復させる。図4Dに記載されたように形質移入されたGMO1787細胞の増殖速度を、一定時間における個体数の重複の累積を用いて評価した。全ての実験について3回繰り返し、同様の結果を得た。
【図6】ウェルナー症候群の患者における細胞(老化した真皮の繊維芽細胞)もまた、連続して150日以上生育することができたことを示す図である。
【特許請求の範囲】
【請求項1】
老化異常のプロセスによって起こる、好ましくはヒトの疾患または病状の治療に対する医薬または医薬組成物を製造するための、GSE24.2誘導化化合物またはGSE24.2活性化化合物の使用。
【請求項2】
上記疾患が、自然老化の結果としての自然な老化のプロセスに起因するものであることを特徴とする請求項1に記載の化合物の使用。
【請求項3】
上記疾患が、加速された老化における病理学的なプロセスに起因するものであることを特徴とする請求項1に記載の化合物の使用。
【請求項4】
上記自然な老化のプロセスは、上皮組織または高い増殖レベルを有する組織に影響を与えるものであることを特徴とする請求項2に記載の化合物の使用。
【請求項5】
上記上皮組織は、皮膚、好ましくは、表皮、内臓、角膜、肝臓、肺および毛球の群に属するものであることを特徴とする請求項4に記載の化合物の使用。
【請求項6】
上記高い増殖レベルを有する組織は、造血系細胞、好ましくはリンパ球、マクロファージおよび赤血球、ならびに胚芽細胞の群に属するものであることを特徴とする請求項4に記載の化合物の使用。
【請求項7】
上記細胞は、体細胞、体幹細胞または胚細胞であることを特徴とする請求項4から6のいずれか1項に記載の化合物の使用。
【請求項8】
GSE24.2誘導化剤は、哺乳動物の、好ましくはヒトの細胞内において老化からの回復を誘導するタンパク質またはペプチドを発現させる塩基配列であって、以下の群に属するGSE24.2の塩基配列の1個または数個により構成されているものであることを特徴とする、請求項1に記載の化合物の使用:
a)ヒトGSE24.2塩基配列により構成されている塩基配列(配列番号1)、
b)a)の塩基配列の変異体であって、配列番号1と少なくとも90%の同一性を示す塩基配列、
c)a)およびb)のいずれかの配列を有するフラグメント、および、
d)a)、b)およびc)のいずれかの配列を含む塩基配列である、遺伝的構成物。
【請求項9】
上記塩基配列がDNA、cDNAまたはmRNAの配列であることを特徴とする請求項8に記載の化合物の使用。
【請求項10】
a)におけるGSE24.2の配列の塩基配列が、配列番号1により構成されていることを特徴とする請求項8に記載の化合物の使用。
【請求項11】
c)におけるGSE24.2の配列の塩基配列が、配列番号3または配列番号5により構成されており、これらはそれぞれTrubIおよびTrubIIのペプチドドメインをコードするものであることを特徴とする請求項8に記載の化合物の使用。
【請求項12】
上記塩基配列は、GSE24.2の配列を含むGSE24.2の遺伝的構成物であることを特徴とする請求項8に記載の化合物の使用。
【請求項13】
上記塩基配列は、GSE24.2の塩基配列またはGSE24.2の遺伝的構成物を含んでおり、かつ老化のプロセスを減少または逆転させることが可能なタンパク質またはペプチドを発現させる、GSE24.2の発現ベクターであることを特徴とする請求項8に記載の化合物の使用。
【請求項14】
上記発現ベクターは、プラスミドpLNCX24.2であることを特徴とする請求項13に記載の化合物の使用。
【請求項15】
GSE24.2誘導化剤は、哺乳動物、好ましくはヒトの細胞内において老化を逆転させる活性を表すタンパク質またはペプチドであって、以下の群に属するアミノ酸配列の1個または数個により構成されている、請求項1に記載の化合物の使用:
a)ヒトGSE24.2のアミノ酸配列により構成されているアミノ酸配列(配列番号2)、
b)a)の配列に基づくアミノ酸配列の変異体であって、配列番号2のポリペプチドと少なくとも90%の同一性を示すアミノ酸配列、
c)a)およびb)のいずれかの配列を有するフラグメント、および、
d)a)、b)およびc)に属するいずれかの配列を含むアミノ酸配列。
【請求項16】
GSE24.2誘導化剤はタンパク質であり、
上記タンパク質におけるa)のアミノ酸配列が配列番号2により構成されていることを特徴とする請求項15に記載の化合物の使用。
【請求項17】
GSE24.2誘導化剤はタンパク質であり、
上記タンパク質におけるc)のアミノ酸配列が配列番号4または配列番号6により構成されていることを特徴とする請求項15に記載の化合物の使用。
【請求項18】
GSE24.2誘導化剤は、ヒト組織の再生細胞として用いられ得る、GSE24.2の上記塩基配列を用いて形質転換された、種々の細胞系統における真核細胞、好ましくはヒト細胞であることを特徴とする請求項1に記載の化合物の使用。
【請求項19】
老化異常に関連する疾患または病状を治療するための医薬組成物または医薬であって、
必要に応じて1つ以上のアジュバントおよび/または医薬的に認容され得る賦形剤とともに、治療的に有効な量にてGSE24.2化合物またはGSE24.2活性化化合物を含んでおり、
老化のプロセスを減少または逆転させることが可能であり、かつ、本発明に係る使用により獲得され、ならびに請求項1から18のいずれかに記載の化合物の使用により獲得される、医薬組成物または医薬。
【請求項20】
ヒトに影響を及ぼす異常な老化のプロセスによって引き起こされる疾患または病状の治療の処置における、請求項19に記載の医薬組成物の、使用。
【請求項21】
生物医学研究に用いられる真核細胞、好ましくはヒトまたは動物の細胞を不死化させる処置における、請求項19に記載の医薬組成物の、使用。
【請求項22】
老化のプロセスが逆転または減少されているとともに、GSE24.2分子を含むことを特徴とする真核細胞、好ましくはヒト細胞。
【請求項1】
老化異常のプロセスによって起こる、好ましくはヒトの疾患または病状の治療に対する医薬または医薬組成物を製造するための、GSE24.2誘導化化合物またはGSE24.2活性化化合物の使用。
【請求項2】
上記疾患が、自然老化の結果としての自然な老化のプロセスに起因するものであることを特徴とする請求項1に記載の化合物の使用。
【請求項3】
上記疾患が、加速された老化における病理学的なプロセスに起因するものであることを特徴とする請求項1に記載の化合物の使用。
【請求項4】
上記自然な老化のプロセスは、上皮組織または高い増殖レベルを有する組織に影響を与えるものであることを特徴とする請求項2に記載の化合物の使用。
【請求項5】
上記上皮組織は、皮膚、好ましくは、表皮、内臓、角膜、肝臓、肺および毛球の群に属するものであることを特徴とする請求項4に記載の化合物の使用。
【請求項6】
上記高い増殖レベルを有する組織は、造血系細胞、好ましくはリンパ球、マクロファージおよび赤血球、ならびに胚芽細胞の群に属するものであることを特徴とする請求項4に記載の化合物の使用。
【請求項7】
上記細胞は、体細胞、体幹細胞または胚細胞であることを特徴とする請求項4から6のいずれか1項に記載の化合物の使用。
【請求項8】
GSE24.2誘導化剤は、哺乳動物の、好ましくはヒトの細胞内において老化からの回復を誘導するタンパク質またはペプチドを発現させる塩基配列であって、以下の群に属するGSE24.2の塩基配列の1個または数個により構成されているものであることを特徴とする、請求項1に記載の化合物の使用:
a)ヒトGSE24.2塩基配列により構成されている塩基配列(配列番号1)、
b)a)の塩基配列の変異体であって、配列番号1と少なくとも90%の同一性を示す塩基配列、
c)a)およびb)のいずれかの配列を有するフラグメント、および、
d)a)、b)およびc)のいずれかの配列を含む塩基配列である、遺伝的構成物。
【請求項9】
上記塩基配列がDNA、cDNAまたはmRNAの配列であることを特徴とする請求項8に記載の化合物の使用。
【請求項10】
a)におけるGSE24.2の配列の塩基配列が、配列番号1により構成されていることを特徴とする請求項8に記載の化合物の使用。
【請求項11】
c)におけるGSE24.2の配列の塩基配列が、配列番号3または配列番号5により構成されており、これらはそれぞれTrubIおよびTrubIIのペプチドドメインをコードするものであることを特徴とする請求項8に記載の化合物の使用。
【請求項12】
上記塩基配列は、GSE24.2の配列を含むGSE24.2の遺伝的構成物であることを特徴とする請求項8に記載の化合物の使用。
【請求項13】
上記塩基配列は、GSE24.2の塩基配列またはGSE24.2の遺伝的構成物を含んでおり、かつ老化のプロセスを減少または逆転させることが可能なタンパク質またはペプチドを発現させる、GSE24.2の発現ベクターであることを特徴とする請求項8に記載の化合物の使用。
【請求項14】
上記発現ベクターは、プラスミドpLNCX24.2であることを特徴とする請求項13に記載の化合物の使用。
【請求項15】
GSE24.2誘導化剤は、哺乳動物、好ましくはヒトの細胞内において老化を逆転させる活性を表すタンパク質またはペプチドであって、以下の群に属するアミノ酸配列の1個または数個により構成されている、請求項1に記載の化合物の使用:
a)ヒトGSE24.2のアミノ酸配列により構成されているアミノ酸配列(配列番号2)、
b)a)の配列に基づくアミノ酸配列の変異体であって、配列番号2のポリペプチドと少なくとも90%の同一性を示すアミノ酸配列、
c)a)およびb)のいずれかの配列を有するフラグメント、および、
d)a)、b)およびc)に属するいずれかの配列を含むアミノ酸配列。
【請求項16】
GSE24.2誘導化剤はタンパク質であり、
上記タンパク質におけるa)のアミノ酸配列が配列番号2により構成されていることを特徴とする請求項15に記載の化合物の使用。
【請求項17】
GSE24.2誘導化剤はタンパク質であり、
上記タンパク質におけるc)のアミノ酸配列が配列番号4または配列番号6により構成されていることを特徴とする請求項15に記載の化合物の使用。
【請求項18】
GSE24.2誘導化剤は、ヒト組織の再生細胞として用いられ得る、GSE24.2の上記塩基配列を用いて形質転換された、種々の細胞系統における真核細胞、好ましくはヒト細胞であることを特徴とする請求項1に記載の化合物の使用。
【請求項19】
老化異常に関連する疾患または病状を治療するための医薬組成物または医薬であって、
必要に応じて1つ以上のアジュバントおよび/または医薬的に認容され得る賦形剤とともに、治療的に有効な量にてGSE24.2化合物またはGSE24.2活性化化合物を含んでおり、
老化のプロセスを減少または逆転させることが可能であり、かつ、本発明に係る使用により獲得され、ならびに請求項1から18のいずれかに記載の化合物の使用により獲得される、医薬組成物または医薬。
【請求項20】
ヒトに影響を及ぼす異常な老化のプロセスによって引き起こされる疾患または病状の治療の処置における、請求項19に記載の医薬組成物の、使用。
【請求項21】
生物医学研究に用いられる真核細胞、好ましくはヒトまたは動物の細胞を不死化させる処置における、請求項19に記載の医薬組成物の、使用。
【請求項22】
老化のプロセスが逆転または減少されているとともに、GSE24.2分子を含むことを特徴とする真核細胞、好ましくはヒト細胞。
【図1a】

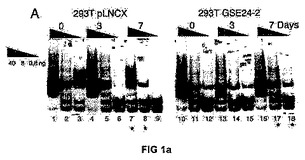
【図1b】
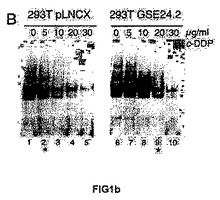
【図1c】
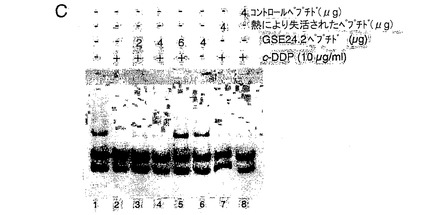
【図1d】
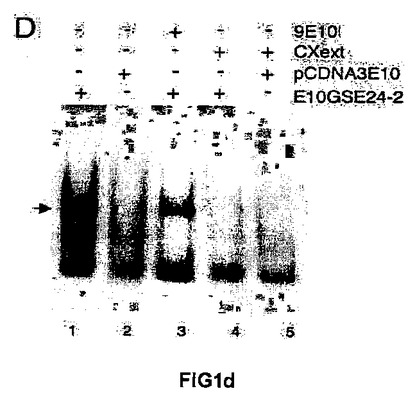
【図2a】
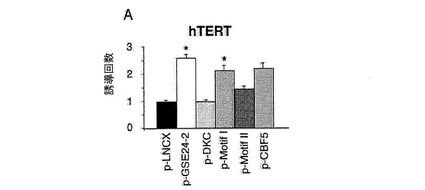
【図2b】
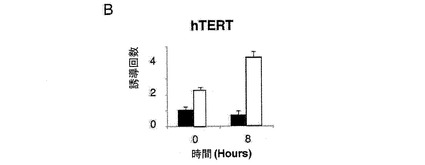
【図2c】
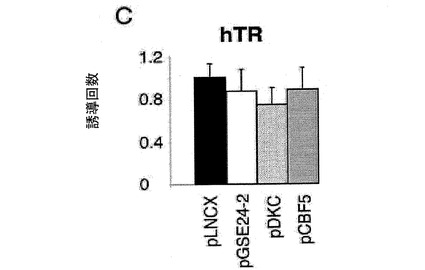
【図2d】
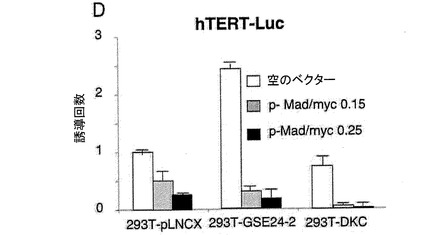
【図2e】
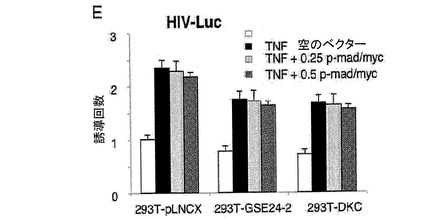
【図3a】
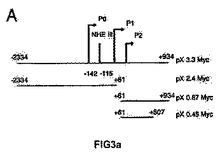
【図3b】
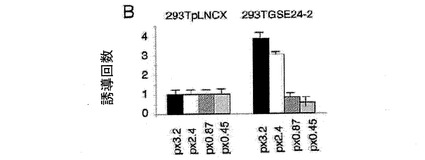
【図3c】
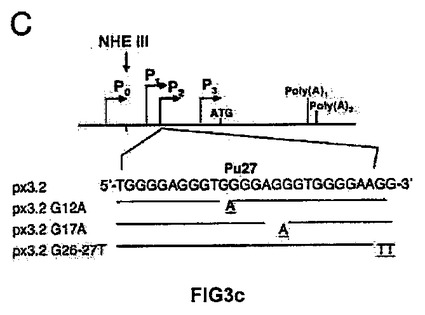
【図3d】
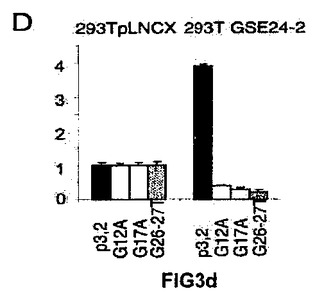
【図3e】
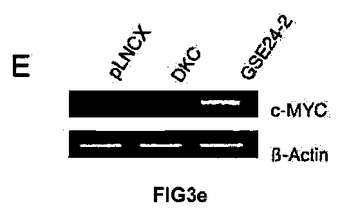
【図3f】
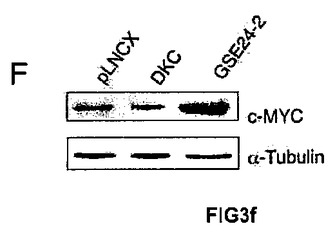
【図4a】
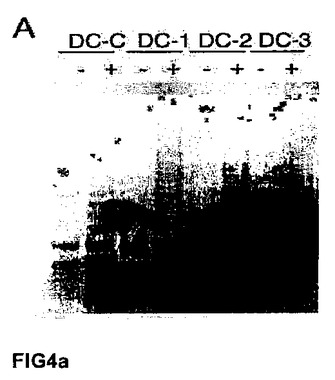
【図4b】

【図4c】
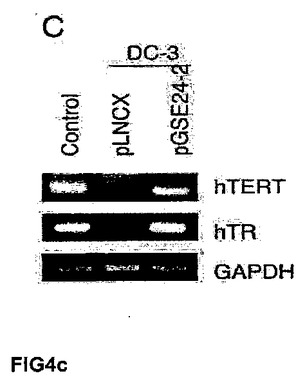
【図4d】
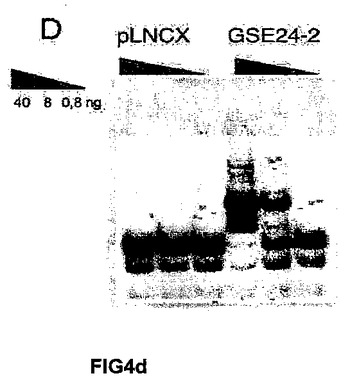
【図4e】

【図4f】
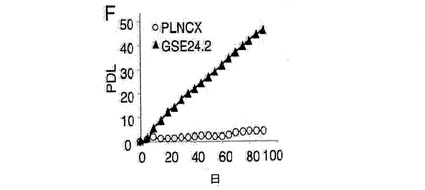
【図4g】
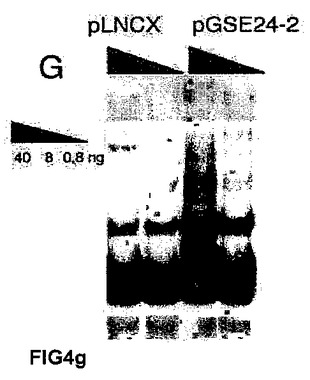
【図4h】

【図5】
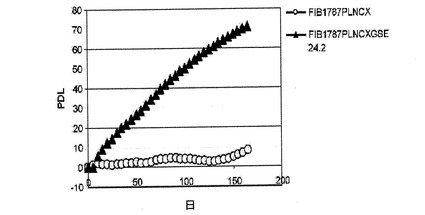
【図6】
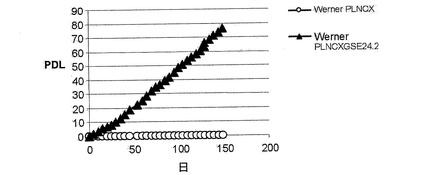
【図1b】
【図1c】
【図1d】
【図2a】
【図2b】
【図2c】
【図2d】
【図2e】
【図3a】
【図3b】
【図3c】
【図3d】
【図3e】
【図3f】
【図4a】
【図4b】
【図4c】
【図4d】
【図4e】
【図4f】
【図4g】
【図4h】
【図5】
【図6】
【公表番号】特表2011−504483(P2011−504483A)
【公表日】平成23年2月10日(2011.2.10)
【国際特許分類】
【出願番号】特願2010−534510(P2010−534510)
【出願日】平成20年11月21日(2008.11.21)
【国際出願番号】PCT/ES2008/070212
【国際公開番号】WO2009/065994
【国際公開日】平成21年5月28日(2009.5.28)
【出願人】(593005895)コンセホ・スペリオール・デ・インベスティガシオネス・シエンティフィカス (67)
【氏名又は名称原語表記】CONSEJO SUPERIOR DE INVESTIGACIONES CIENTIFICAS
【出願人】(506255603)ウニベルシダッド アウトノマ デ マドリッド (4)
【Fターム(参考)】
【公表日】平成23年2月10日(2011.2.10)
【国際特許分類】
【出願日】平成20年11月21日(2008.11.21)
【国際出願番号】PCT/ES2008/070212
【国際公開番号】WO2009/065994
【国際公開日】平成21年5月28日(2009.5.28)
【出願人】(593005895)コンセホ・スペリオール・デ・インベスティガシオネス・シエンティフィカス (67)
【氏名又は名称原語表記】CONSEJO SUPERIOR DE INVESTIGACIONES CIENTIFICAS
【出願人】(506255603)ウニベルシダッド アウトノマ デ マドリッド (4)
【Fターム(参考)】
[ Back to top ]