
細菌のATP合成酵素の結合ドメイン
【課題】 単離された変異体atpEタンパク質、および前記変異体atpEタンパク質から出発してATPアーゼ結合ドメインの同定を提供する。
【解決手段】 特定のアミノ酸配列において少なくとも1つの点突然変異を含むポリペプチドの単離された変異体atpEタンパク質を使用することにより、それに関連する核酸、ベクター、宿主細胞、製薬学的組成物および製品等も提供される。
【解決手段】 特定のアミノ酸配列において少なくとも1つの点突然変異を含むポリペプチドの単離された変異体atpEタンパク質を使用することにより、それに関連する核酸、ベクター、宿主細胞、製薬学的組成物および製品等も提供される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、単離された変異体atpEタンパク質、および前記変異体atpEタンパク質から出発してATPアーゼ結合ドメインの同定を提供する。本発明はまた、関連する核酸、ベクター、宿主細胞、製薬学的組成物および製品も提供する。本発明はさらに、試験化合物がatpEタンパク質と、すなわち本発明のATPアーゼ結合ドメインと相互作用するかどうかの決定方法、ならびに、とりわけ抗菌薬としての、より具体的には抗ミコバクテリウム薬としての、なおより具体的には被験体における結核を処置するための前記試験化合物を含んでなる製薬学的組成物を提供する。
【背景技術】
【0002】
AIDS後、結核(TB)は世界の成人の死亡の第一位の原因(年あたり2〜3百万の死亡)であり、そして世界的な貧困および被害の軽減に対する決定的な障害である(非特許文献1)。該疾患の復活に寄与する因子は、多くの国における抗TBプログラムの実行における困難、免疫抑制個体の数の劇的な増加(主としてHIV感染による)、およびTBが固有である地域を通過しかつそこからの人々の動きを包含する。TBおよびHIVの異常発生は、共感染した人々(現在1,100万人の成人)において相互に刺激し、罹患率および死亡率双方を増大させる(非特許文献2、3)。加えて、TBはHIVに感染した人々における死亡の第一位の原因である(非特許文献4)。
【0003】
第一列の抗TB薬物レジメンは90%以上の有効率を達成し得るとは言え、それらの複雑さは、十分な医学的支援およびTB処置プログラムが利用可能でない場合に乏しいコンプライアンス、そして順に耐性の発生につながり得る(非特許文献5)。TBの多剤耐性(MDR)株は処置を大きく複雑にする(非特許文献6)。結核治療薬開発のための世界同盟(Global Alliance for TB Drug Development)は、いかなる新たな処置も、既存の治療法を上回る以下の3つの利点、すなわち、TBの効果的処置を短縮若しくは単純化すること;MDR−TBに対する有効性を増大させること;および潜伏型のTB感染の処置を改良することの最低1つを提供すべきであると勧告した。こうした新薬は患者コンプライアンスを大きく改善して、それにより世界保健機関(WHO)の直接監視下短期化学療法(Directly Observed Treatment Short−course)(DOTS)戦略のようなTB処置プログラムの費用を低減させるとみられる(非特許文献7)。
【0004】
現在前臨床および臨床開発中のより新たな抗TBの候補は、既存の薬物群(モキシフロキサシンのような)、若しくはMJH−98−1−81(イソニアジドから)、オキサゾリジノンおよびリファペンチン(リファンピンの近接アナログ)のような第一列の薬物のアナログのいずれかからである傾向がある(非特許文献8)。これらの新薬は強力でありうるとは言え、アナログ化合物は耐性に対する一時的な解決策のみを提供する(非特許文献9)。それらは既存の薬物群と同一の作用機序に頼るからである。
【0005】
抗生物質は、一般に、通常、細菌の代謝を特定の機序により阻害することにより細菌の複製を阻害する。例えば、イソニアジドはミコール酸(細胞壁の必要な成分)を合成する酵素機構を妨害する一方、リファンピシンはDNAからRNAを転写するための細菌の機構を妨害する。従って、既知の剤に比較して、細胞の増殖および複製の異なったミコバクテリウム特異的局面を標的とする抗TB化合物の新規同定方法を探索することが興味深い。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Global Alliance for TB Drug Development、Developing a faster TB cure(2004;http://www.tballiance.org)
【非特許文献2】E.L.Corbettら、Arch.Invern.Med.163、1009(2003)
【非特許文献3】UNAIDS、AIDS epidemic update 2003(2003;www.unaids.org/Unaids/EN/Resources)。
【非特許文献4】World Health Organization、Tuberculosis(2004;http://www.who.int/health_topics/tuberculosis/en/)。
【非特許文献5】R.J.O’Brien、P.P.Nunn、Am.J.Respir.Crit.Care Med.163、1055(2001)。
【非特許文献6】World Health Organization、Tuberculosis Fact Sheet No 104(2004;http://www.who.int/mediacentre/factsheets/fs104/en/)。
【非特許文献7】A.J.Claxton、J.Cramer、C.Pierce、Clin.Ther.23、1296(2001)。
【非特許文献8】N.Lounisら、Antimicrob.Agents Chemother.45、3482(2001)。
【非特許文献9】A.S.Ginsburg、J.H.Grosset、W.R.Bishai、Lancet Infect.Dis.3、432(2003)。
【発明の概要】
【0007】
本発明は、とりわけ(配列番号1)、(配列番号2)、(配列番号3)、(配列番号4)および(配列番号5)から選択されるアミノ酸配列によりコードされる単離された変異体atpEタンパク質、とりわけ(配列番号6)、(配列番号7)、(配列番号8)、(配列番号9)および(配列番号10)よりなる群から選択される前記変異体atpEタンパク質をコードする単離された核酸、ならびに本核酸を含んでなるベクターを提供する。特定の一態様において、変異体atpEタンパク質は配列番号2によりコードされ、また、前記タンパク質をコードする単離された核酸配列は配列番号7よりなる。
【0008】
本発明はさらに、本発現ベクターをその中に有する宿主細胞を含んでなる宿主−ベクター系を提供する。
【0009】
本発明はさらに、変異体atpEタンパク質が該細胞中で抗菌耐性を誘導する、前記タンパク質を含んでなる単離された細胞を提供する。
【0010】
本発明はさらに、抗菌化合物の同定方法を提供し、前記方法は、
a)atpEタンパク質を発現する細胞を、生理学的条件下で試験化合物と接触させる段階、および
b)該試験化合物がatpEタンパク質と相互作用するかどうかを決定する段階
を含んでなる。
【0011】
本発明はさらに、atpEタンパク質と相互作用する試験化合物の潜在能力の評価方法を提供し、前記方法は;
(a)分子モデル化技術を使用して、atpEタンパク質の三次元構造を生成させること;
(b)コンピュータによる手段を使用して、試験化合物と、atpEタンパク質の三次元構造の間のフィッティング操作を実施すること;および
(c)前記フィッティング操作の結果を解析して、試験化合物のatpEタンパク質の三次元構造との会合を定量化すること
を含んでなる。
【0012】
1個のCサブユニットの少なくともアミノ酸Ala24、Gly27、Phe53、Val57、Gly58、Glu61、Tyr64およびPhe65;1個のAサブユニットのアミノ酸Ser182、Leu183、Ser184、Leu185およびArg186を含んでなり、かつ、前記アミノ酸は表3、4若しくは5のいずれかの原子座標を有する、ATPアーゼのF0部分の結合部位を提供することもまた、本発明の一目的である。
【0013】
さらなる一目的において、本発明は、ATPアーゼのF0部分と相互作用する化合物の同定方法、および、抗菌化合物としてのそれらの潜在能力、とりわけ抗ミコバクテリウム化合物の同定方法における、前述の結合ドメインの使用を提供する。
【0014】
従って、ATPアーゼのF0部分と、とりわけ耐性を賦与する突然変異部位でatpEタンパク質と、若しくは本発明の結合部位と相互作用する化合物を被験体に投与することを含んでなる、微生物に基づく感染症を伴う被験体の処置方法を提供することが、本発明の一目的である。本発明はさらに、前述のスクリーニング方法のいずれかを使用してatpEタンパク質と相互作用する剤を被験体に投与することを含んでなる、結核に苦しめられる被験体の処置方法を提供する。ATPアーゼのF0部分、とりわけatpEタンパク質と相互作用する化合物の使用を含んでなる処置方法において、ATPアーゼのF0部分、およびとりわけatpEタンパク質と相互作用することが以前に知られている化合物は除外されるべきである。より具体的には、開示された処置方法のいずれかにおける(11)に記述されるDARQ J化合物の使用が除外されるべきである。
【0015】
本発明はさらに、細胞中でatpEタンパク質と相互作用する剤および製薬学的に許容できる担体を含んでなる製薬学的組成物を提供する。最後に、本発明は、(a)製薬学的剤が細胞中でatpEタンパク質と相互作用し、かつ(b)包装が被験体における細菌感染症を処置するための該剤の使用を示すラベルを含んでなる、包装および製薬学的剤を含んでなる製品を提供する。特定の一態様において、本発明は抗菌医薬品の製造におけるDARQ Jの使用を提供する。
【0016】
本発明のこれおよびさらなる局面は下でより詳細に論考されるであろう。
【0017】
[図面の簡単な説明]
表1 多様なミコバクテリウム種の増殖を90%阻害した、リードDARQ化合物(J)の最小阻害濃度(MIC)。試験した株の数は別の方法で示されない限りn=1であった。
【0018】
表2 DARQ J化合物の結合部位の周囲のアミノ酸。
【0019】
表3 野性型およびDARQ J変異体ヒト結核菌(M.tuberculosis)株双方由来のDARQ J化合物の結合部位の周囲のアミノ酸の原子座標。
【0020】
表4 野性型ヒト結核菌(M.tuberculosis)のDARQ J化合物の結合部位の原子座標。
【0021】
表5 DARQ J変異体ヒト結核菌(M.tuberculosis)株のDARQ J化合物の結合部位の原子座標。
【0022】
表6 ヒト結核菌(M.tuberculosis)の変異体atpEタンパク質(配列番号2)の原子座標。
【0023】
表7 ヒト結核菌(M.tuberculosis)の野性型atpEタンパク質(配列番号1)の原子座標。
【0024】
図1 下でJ若しくはDARQ Jともまた称されるR207910の絶対配置。
【0025】
図2 ヒト結核菌(M.tuberculosis)およびスメグマ菌(M.smegmatis)変異体のatpEタンパク質配列アライメント。Mtb_S:薬物感受性株ヒト結核菌(M.tuberculosis)H37RvのatpE(1−81)。受託番号:Swiss−Prot Q10598(配列番号1)。Mtb_R:薬物耐性株ヒト結核菌(M.tuberculosis)BK12のatpE(1−81)(配列番号2)。Msm_S:薬物感受性株スメグマ菌(M.smegmatis)のatpE(1−86)。ゲノム研究所(Institute for Genome Research)により得られた配列(配列番号3)。Msm_R09(配列番号4)およびR10(配列番号5):薬物耐性株スメグマ菌(M.smegmatis)のatpE(1−86)。社内で得られた配列。Human:ヒト(Homo sapiens)、ATP5G3(66−142)。受託番号:Ensembl ENSP00000284727。上の番号付け:ヒト結核菌(M.tuberculosis)およびスメグマ菌(M.smegmatis)のatpE。下の番号付け:ヒト(H.sapiens)ATP5G3(66−142)陰影はBLOSUM62マトリックスを使用するアミノ酸類似性を示す(黒=高、灰色=中)。矢印は耐性株で観察される点突然変異の位置を示す。
【0026】
図3 DARQ J、イソニアジドおよびDCCDの存在下でのヒト結核菌(M.tuberculosis)の全細胞ATP測定。野性型ヒト結核菌(M.tuberculosis)およびDARQ J変異体ヒト結核菌(M.tuberculosis)双方で526nmで測定したオキシルシフェリンの相対発光単位。
【0027】
図4 一緒になってDARQ J化合物の結合部位を形成する3個のCサブユニット(A鎖、K鎖およびL鎖)ならびにAサブユニット(M鎖)のリボン表示。
【0028】
[発明の詳細な記述]
定義
別の方法で本明細書に明らかに提供されるところを除き、本出願で使用されるところの以下の用語のそれぞれは、下に示される意味するところを有する。
【0029】
「atpEタンパク質」は、ヒト結核菌(M.tuberculosis)についてSwissProtエントリQ10598により表されるところのATPアーゼ複合体のF0サブユニットのC鎖、または前記ヒト結核菌(M.tuberculosis)配列に対する最低70、80、90、95、97若しくは99%の配列同一性を有するタンパク質を意味している。
【0030】
ATPアーゼ、ATP合成酵素若しくはF0F1ATPアーゼともまた称される「F1F0ATPアーゼ」は、ATPの合成若しくは加水分解を触媒する大型の多サブユニット複合体を意味している。F0F1ATPアーゼは2ドメイン、すなわち、膜に対し外部でありかつ触媒部位を含有するF1部位、および二層におよびかつプロトン孔を含有するF
0部分より構成される。ATPアーゼは、細菌の形質膜、葉緑体のチラコイド膜、およびミトコンドリアの内膜で見出され、そこでそれらはプロトンの電子化学的勾配のエネルギーを使用してATP合成を駆動する。
【0031】
「投与すること」は、当業者に既知の多様な方法および送達系のいずれかを使用して遂げられ若しくは実施されるある様式で送達することを意味している。投与することは、例えば、局所で、静脈内に、心膜に、経口で、植込物を介して、経粘膜で、経皮で、筋肉内に、皮下に、腹腔内に、クモ膜下腔内に、リンパ内に(intralymphatically)、病変内に、若しくは硬膜外に実施し得る。投与することはまた、例えば1回、複数回、および/または1つ若しくはそれ以上の延長された期間にわたっても実施し得る。
【0032】
「宿主細胞」は、限定されるものでないが、細菌細胞、酵母細胞、真菌細胞、昆虫細胞および哺乳動物細胞を挙げることができる。細菌細胞は、リン酸カルシウム沈殿、電気穿孔法および微小注入法のような当該技術分野で公知の方法によりトランスフェクトし得る。
【0033】
atpEタンパク質に関して「単離された」は、atpEがその天然の機能を保持しかつその天然の環境中の他のタンパク質の数種若しくは全部を含まない、atpEタンパク質を含有する膜断片調製物若しくは他の適する調製物を意味している。それは、F0F1ATPアーゼのF0部分、とりわけ本発明の変異体atpEタンパク質を含んでなるF0部分を含んでなる膜調製物を包含することを意味している。
【0034】
「細菌細胞」はいかなる細菌細胞も意味している。細菌細胞は、正常、異常および形質転換され、かつ、ミコバクテリウム、とりわけヒト結核菌(Mycobacterium
tuberculosis)およびスメグマ菌(Mycobacterium smegmatis)、コリネバクテリウム、ノカルジア類、例えば連鎖球菌、ブドウ球菌および腸球菌のようなグラム陽性細菌、若しくは例えば大腸菌(Escherichia coli)、インフルエンザ菌(Heamophilus influenzae)およびヘリコバクター ピロリ(Helicobacter pylori)のようなグラム陰性細菌により例示される細胞を制限なしに包含する。
【0035】
「核酸」および「ポリヌクレオチド」という用語は本明細書で互換性に使用され、そしてそれぞれデオキシリボヌクレオチドおよび/若しくはリボヌクレオチドのポリマーを指す。デオキシリボヌクレオチドおよびリボヌクレオチドは天然に存在し得るか、若しくはその合成のアナログであり得る。
【0036】
「生理学的条件」という用語は、所定の細胞に関して、該細胞の生化学的環境を通常構成しうる条件を意味している。細胞の生化学的環境は、該細胞が通常曝露される数種若しくは全部のプロテアーゼを制限なしに包含する。こうした条件は、限定されるものでないがin vivo状態を挙げることができる。
【0037】
「ポリペプチド」、「ペプチド」および「タンパク質」という用語は本明細書で互換性に使用され、そしてそれぞれアミノ酸残基のポリマーを意味している。アミノ酸残基は天然に存在し得るか、若しくはそれらの化学的アナログであり得る。ポリペプチド、ペプチドおよびタンパク質はまた、グリコシル化、脂質結合、硫酸化、水酸化およびADP−リボシル化のような修飾も包含し得る。
【0038】
「被験体」は、ウシ、ウマ、ヒツジ、ブタ、イヌ、ネコ、マウス若しくはラットのようなげっ歯類、シチメンチョウ、ニワトリおよび霊長類を制限なしに包含する哺乳動物若し
くは鳥類のようないかなる動物も意味している。好ましい態様において、被験体はヒトである。
【0039】
「処置すること」は、被験体における疾患を除外すること、その経過を反転すること、その進行を遅らせること、その症状を低下させること、若しくは別の方法で改善することを制限なしに包含する。
【0040】
「ベクター」は当該技術分野で既知のいかなる核酸ベクターも意味している。こうしたベクターは、限定されるものでないが、プラスミドベクター、コスミドベクターおよびバクテリオファージベクターを挙げることができる。
【0041】
「候補物質」および「試験化合物」という用語は互換性に使用され、そして、生物学的応答修飾因子として別の部分すなわちatpEタンパク質と相互作用すると考えられている物質を指す。例えば、代表的な候補物質は、atpEタンパク質と相互作用すると考えられ、また、ATPアーゼ活性を改変すると考えられている。本発明の方法を使用して検討され得る例示的候補物質は、限定されるものでないが、ペプチド、酵素、酵素基質、補助因子、糖、オリゴヌクレオチド、化合物小分子およびモノクローナル抗体を挙げることができる。
【0042】
「調節する」は、野性型若しくは変異体atpEタンパク質のいずれかの若しくは全部の化学的および生物学的活性若しくは特性の増大、減少若しくは他の変化を意味している。
【0043】
「相互作用する」は、分子間の「結合」相互作用を包含する分子間の検出可能な相互作用を意味している。相互作用は、例えば、性質がタンパク質−タンパク質若しくはタンパク質−核酸であり得る。こうした相互作用は技術既知の手順、例えば酵母2ハイブリッドアッセイ、免疫沈降、SPAアッセイ若しくはフィルター結合アッセイを使用して検出し得る。
【0044】
本明細書で使用されるところの「原子座標」若しくは「構造座標」という用語は、各原子についてX、Y、ZおよびBを包含するタンパク質データバンク(PDB)フォーマットで原子の位置を記述する数学的座標を指す。当業者は、X線結晶学により決定される一組の構造座標が標準誤差を伴わなくはないことを理解している。本発明の目的上、表3、4、5、6若しくは7の対応する原子座標の水素以外の原子位置に重ね合わせられる場合に1.5Å未満の水素以外の原子の二乗平均偏差を有するいかなる供給源からのAPT合成酵素の構造座標のいずれの組も、実質的に同一若しくは相同と考えられる。より好ましい一態様において、表3、4、5、6若しくは7の対応する原子座標の水素以外の原子位置に重ね合わせられる場合に0.75Å未満の水素以外の原子の二乗平均偏差を有するいかなる供給源からのAPT合成酵素の構造座標のいずれの組も、実質的に同一と考えられる。
【0045】
本発明の態様
変異体atpE(または「変異atpE」という)タンパク質
本発明は、単離された変異体atpEタンパク質、とりわけ細菌のatpEタンパク質、より具体的にはミコバクテリウムのatpEタンパク質、なおより具体的にはヒト結核菌(M.tuberculosis)若しくはスメグマ菌(M.smegmatis)のatpEタンパク質を提供する。突然変異は単一の点突然変異、挿入若しくは欠失から選択される。本発明の一態様において、突然変異は、図2の配列アライメントに示されるところのアミノ酸20ないし40、とりわけ30ないし40のいずれか1つ、好ましいアミノ酸34の、若しくはアミノ酸60ないし75の、とりわけ62ないし73のいずれか1
つ、好ましくはアミノ酸69に位置する最低1個の点突然変異よりなる。さらなる一態様において、単離された変異体atpEタンパク質は、図2に示されるところのMtb_R(配列番号2)、Msm_R09(配列番号4)およびMsm_R10(配列番号5)、または前述のアミノ酸配列のいずれかに対する最低70、80、90、95、97若しくは98%の配列同一性を有するアミノ酸配列のから選択される。
【0046】
本発明はさらに、前記変異体atpEタンパク質をコードする単離された核酸を提供する。一態様において、前記核酸配列は、例えばJ.Biol.Chem.、1994、Vol.269(10)、p.7285−7289に記述されるようなF0部分をコードする全部の遺伝子よりなり、ここで前記遺伝子は単一のプロモーターから転写され、そして本発明の変異体atpEタンパク質をコードする核酸配列を含んでなる。核酸はDNA若しくはRNA、および好ましくはDNAであり得、そして、さらなる一態様において、Mtb_R(配列番号7)Msm_R09(配列番号9)、Msm_R10(配列番号10)をコードする核酸配列、または前述の核酸配列のいずれかに対する最低70、80、90、95、97若しくは98%の配列同一性を有する核酸配列から選択される。
【0047】
核酸およびポリペプチド配列の同一性パーセントは、参照配列をクエリ配列と比較する商業的に入手可能なアルゴリズムを使用して計算し得る。以下のプログラム(国立バイオテクノロジー情報センター(National Center for Biotechnology Information)により提供される)、すなわち、BLAST、gapped BLAST、BLASTNおよびPSI−BLASTを使用して相同性/同一性を決定することができ、これらはデフォルトのパラメータとともに使用しうる。
【0048】
アルゴリズムGAP(Genetics Computer Group、ウィスコンシン州マディソン)は、NeedlemanとWunschのアルゴリズムを使用して、一致の数を最大にしかつギャップの数を最少にする2種の完全な配列を整列させる。一般に、ギャップ創製ペナルティ(gap creation penalty)=12およびギャップ伸長ペナルティ(gap extension penalty)=4を伴うデフォルトのパラメータを使用する。
【0049】
核酸配列若しくはその一部分とクエリ配列の間の最良の全体的一致の別の決定方法は、Brutlagら(Comp.App.Biosci.、6:237−245(1990))のアルゴリズムに基づくFASTDBコンピュータプログラムの使用である。該プログラムは包括的配列アライメントを提供する。前記包括的配列アライメントの結果は同一性パーセントにある。同一性パーセントを計算するためにDNA配列のFASTDB検索での使用に適するパラメータは:マトリックス(Matrix)=Unitary、k−タプル(k−tuple)=4、ミスマッチペナルティ(Mismatch penalty)=1、結合ペナルティ(Joining Penalty)=30、無作為化グループ長(Randomization Group Length)=0、カットオフスコア(Cutoff Score)=1、ギャップペナルティ(Gap Penalty)=5、ギャップサイズペナルティ(Gap Size Penalty)=0.05およびウィンドウサイズ(Window Size)=500若しくはヌクレオチド塩基中のクエリ配列長さのいずれか短い方、である。アミノ酸アライメントの同一性および類似性パーセントを計算するための適するパラメータは:マトリックス(Matrix)=PAM 150、k−タプル(k−tuple)=2、ミスマッチペナルティ(Mismatch penalty)=1、結合ペナルティ(Joining Penalty)=20、無作為化グループ長(Randomization Group Length)=0、カットオフスコア(Cutoff Score)=1、ギャップペナルティ(Gap Penalty)=5、ギャップサイズペナルティ(Gap Size Penalty)=0.05およびウィンドウサイズ(Window Size)=500若しくは
ヌクレオチド塩基中のクエリ配列長さのいずれか短い方、である。
【0050】
本発明はさらに、本核酸を含んでなるベクターを提供する。一態様において、ベクターはプラスミドベクターである。
【0051】
本発明はさらに、本プラスミドベクターをその中に有する宿主細胞を含んでなる宿主−ベクター系を提供する。該細胞は原核生物若しくは真核生物であり得、一態様において、宿主細胞は細菌細胞、とりわけ例えばヒト結核菌(M.tuberculosis)若しくはスメグマ菌(M.smegmatis)のようなミコバクテリウム細胞である。
【0052】
本発明はさらに、変異体atpEタンパク質(このタンパク質は細胞中で抗菌耐性を誘導する)を含んでなる単離された細胞を提供する。一態様において、単離された細胞は、変異体のミコバクテリウムatpEタンパク質で形質転換された、とりわけ、突然変異が図2の配列アライメントに示されるところのアミノ酸20ないし40、より具体的には30ないし40のいずれか1つ、好ましいアミノ酸34に、若しくはアミノ酸60ないし75、とりわけ62ないし73のいずれか1つ、好ましくはアミノ酸69に位置する最低1個の点突然変異よりなる変異体のミコバクテリウムatpEタンパク質で形質転換されたスメグマ菌(M.smegmatis)細胞よりなる。
【0053】
スクリーニング方法
本発明はさらに、抗菌化合物の同定方法を提供し、前記方法は
(a)atpEタンパク質を発現する細胞を、生理学的条件下で試験化合物と接触させる段階;
(b)該試験化合物がatpEタンパク質と相互作用するかどうかを決定する段階
を含んでなる。
【0054】
一態様において、前述の方法で使用されるatpEタンパク質は、細菌のatpEタンパク質、とりわけミコバクテリウムのタンパク質よりなり、かつ、野性型atpEタンパク質ならびに上述されたところの変異体atpEタンパク質の双方を包含することを意味している。本発明のさらなる一態様において、前述の方法で使用されるミコバクテリウムのatpEタンパク質は、本発明の変異体のミコバクテリウムatpEタンパク質よりなる。前述のアッセイの特定の一態様において、本発明の変異体atpEタンパク質で形質転換した細胞を使用し、そして、試験化合物の前記atpEタンパク質との相互作用を、前記変異体atpEタンパク質を含んでなるF1F0−ATPアーゼの酵素活性の可能な阻害を測定することにより評価する。F1F0−ATPアーゼ活性の阻害は、例えば、ADPの産生をピルビン酸キナーゼおよび乳酸脱水素酵素の反応を介するNADHの酸化に結びつけることによる酵素活性の検出を用いる、F1F0−ATPアーゼおよび基質としてのATPを含んでなる系への物質の添加によるような技術既知の手順を使用して測定する。
【0055】
アッセイの一態様において、atpEタンパク質を結合アッセイで使用しうる。結合アッセイは競合的でも若しくは非競合的でもよい。こうしたアッセイは、どの化合物(あれば)がポリペプチドに結合することが可能であるかを決定するための多数の化合物の迅速スクリーニングを提供し得る。
【0056】
この情況内で、本発明は、試験化合物が本発明の単離されたatpEタンパク質に結合するかどうか、そして従って潜在的抗菌化合物であるかどうかの同定方法を提供し、前記方法は;
a)atpEタンパク質を発現する細胞(こうした細胞は前記atpEタンパク質を通常は発現しない)を、atpEタンパク質を結合することが既知の化合物の存在および非存
在下に試験化合物と接触させること、
b)atpEタンパク質に結合することが既知の化合物を参照として使用してatpEタンパク質への試験化合物の結合を測定すること
を含んでなる。
【0057】
試験化合物、若しくは下で参照化合物ともまた称されるatpEタンパク質に結合することが既知の化合物の結合を、タンパク質−リガンド相互作用の技術既知の試験方法を使用して評価する。例えば、こうした結合は標識した物質若しくは参照化合物を使用することにより測定し得る。試験化合物若しくは参照化合物、とりわけ化合物J(図1)を、当該技術分野で既知のいずれかの便宜的様式、例えば放射活性で、蛍光で若しくは酵素的に標識し得る。前述の方法の特定の一態様において、参照化合物としてもまた知られるatpEタンパク質に結合することが既知の化合物を、検出可能に標識し、そして。前記標識を使用して、試験化合物のatpEタンパク質への結合を測定する。前記参照化合物は、放射標識、蛍光標識若しくは酵素標識、より好ましくは放射標識を使用して標識する。
【0058】
本発明の代替の一態様において、前述の結合アッセイは、細胞組成物、すなわち上で定義されたところのatpEタンパク質を含んでなる細胞抽出物、細胞画分若しくは細胞小器官(organel)上で実施される。より具体的には、前述の結合アッセイは、細胞組成物、すなわち上で定義されたところのatpEタンパク質を含んでなる膜調節物上で実施され、前記細胞組成物すなわち膜調製物は、変異体のミコバクテリウムatpEタンパク質で形質転換された、とりわけ、突然変異が図2の配列アライメントに示されるところのアミノ酸20ないし40、とりわけ30ないし40のいずれか1つ、好ましいアミノ酸34に、またはアミノ酸60ないし75、とりわけ62ないし73のいずれか1つ、好ましくはアミノ酸69に位置する最低1個の点突然変異よりなる変異体のミコバクテリウムatpEタンパク質で形質転換されたスメグマ菌(M.smegmatis)細胞から得られる。Mtb_S(配列番号1)若しくはMtb_R(配列番号2)の番号付けを参照として採用すれば、前述の領域は、アミノ酸14ないし34、とりわけ24ないし34、好ましくはアミノ酸28中に、若しくはアミノ酸54ないし69、とりわけ56ないし67、好ましくはアミノ酸63に対応する。
【0059】
一態様において、結合アッセイは膜調製物を使用して実施される。これらの膜調製物は、放射標識atpEリガンド(3H標識DARQを包含する)の結合およびこうした放射リガンドの結合部位についての競合体による置換を検出するために、慣習的フィルター結合アッセイ(例えばBrandelフィルターアッセイ装置を使用して)若しくはハイスループットシンチレーション近接型結合アッセイ(SPAおよびCytostar−Tフラッシュプレート技術;Amersham Pharmacia Biotech)で使用し得る。放射活性は、96、384、1536マイクロタイタープレート形式から迅速測定を行うことが可能なPackard Topcount若しくは類似の装置で測定し得る。SPA/Cytostar−T技術はハイスループットスクリーニングにとりわけ従いやすく、そして、従って、この技術は標準的リガンドを置換することが可能な化合物についてのスクリーニングとして使用するのに適する。
【0060】
天然の状況に近づける環境でのatpEタンパク質へのリガンドの結合を研究するための別のアプローチは、Biacore装置(Biacore)により活用される表面プラスモン共鳴効果を利用する。膜調製物若しくは全細胞中のatpEタンパク質をBiacoreのバイオセンサーチップに結合し得、そして、リガンドの結合を化合物の存在および非存在下で検査して、結合部位の競合体を同定し得る。
【0061】
分子モデル化
本発明はさらに、atpEタンパク質と相互作用する試験化合物の潜在能力の評価方法
を提供し、前記方法は;
(a)分子モデル化技術を使用してatpEタンパク質の三次元構造を明確に表すこと;(b)コンピュータによる手段を使用して、試験化合物とatpEタンパク質の三次元構造との間のフィッティング操作を実施すること;および
(c)前記フィッティング操作の結果を解析して、atpEタンパク質の三次元構造との試験化合物の会合を定量化すること
を含んでなる。
【0062】
受容体および酵素のコンホメーションのモデルを創製かつ利用するのに適切なハードウェアおよびソフトウェア双方を包含する分子モデル化技術は当該技術分野で既知である。
【0063】
多数のコンピュータプログラムが、本明細書に記述される方法で潜在的なatpEと相互作用する化合物をコンピュータモデル化し、モデル構築しかつコンピュータにより同定、選択および評価する方法に利用可能でありかつ適する。これらは、例えば、GRID(英国オックスフォード大学から入手可能)、MCSS(Accelrys,Inc.、カリフォルニア州サンディエゴから入手可能)、AUTODOCK(Oxford Molecular Groupから入手可能)、FLEX X(Tripos、ミズーリ州セントルイスから入手可能)、DOCK(カリフォルニア大学、カリフォルニア州サンフランシスコから入手可能)、CAVEAT(カリフォルニア大学、バークレーから入手可能)、HOOK(Accelrys,Inc.、カリフォルニア州サンディエゴから入手可能)、ならびにMACCS−3D(MDL Information Systems、カリフォルニア州サンリアンドロから入手可能)、UNITY(Tripos、ミズーリ州セントルイスから入手可能)およびCATALYST(Accelrys,Inc.、カリフォルニア州サンディエゴから入手可能)のような3Dデータベースシステムを包含する。潜在的候補物質はまた、LUDI(Biosym Technologies、カリフォルニア州サンディエゴから入手可能)、LEGEND(Accelrys,Inc.、カリフォルニア州サンディエゴから入手可能)およびLEAPFROG(Tripos、ミズーリ州セントルイスから入手可能)のようなソフトウェアパッケージを使用してコンピュータで「新規に(de novo)」設計されうる。化合物の変形エネルギーおよび静電反発力は、GAUSSIAN 92、AMBER、QUANTA/CHARMMおよびINSIGHT II/DISCOVERのようなプログラムを使用して解析しうる。これらのコンピュータ評価およびモデル化技術は、例えばSilicon Graphics、Sun Microsystemsおよび他者から入手可能なワークステーションを包含するいずれの適するハードウェアでも実施しうる。これらのモデル化技術、方法、ハードウェアおよびソフトウェアパッケージは代表的なものであり、かつ、包括的列挙であることを意図していない。当該技術分野で既知の他のモデル化技術もまた本発明で使用しうる。例えば、N.C.Cohen、Molecular Modeling in Drug Design、Academic Press(1996)を参照されたい。
【0064】
本発明の一態様において、atpEタンパク質の三次元構造は、大腸菌(E.coli)(タンパク質データベース1Q01)のIle28、Glu61およびIle63の原子座標+/−10Åを超えない、好ましくは5Åを超えない前記アミノ酸のバックボーン原子の二乗平均偏差を使用して生成される。
【0065】
下の実施例で提供されるとおり、atpEタンパク質の三次元構造を提供することが本発明の一目的であった。表6および7は、配列番号2および配列番号1を伴う変異体および野性型のatpEタンパク質の原子座標を提供する。従って、一態様において、atpEタンパク質の三次元構造は表6若しくは7の原子座標を使用して生成される。特定の一態様において、atpEタンパク質の三次元構造は表7の原子座標を使用して生成される
。DARQ J化合物は、その脱プロトン化された形態のAサブユニットのArg186のCサブユニットのGlu61との相互作用を阻害する。従って、atpEタンパク質と相互作用する試験化合物の潜在能力の評価方法における表6若しくは7の原子座標の使用を提供することが、本発明の一目的である。
【0066】
結合部位
別の態様において、本発明は、ATPアーゼのF0部分の結合部位の特徴を提供する。DARQ J化合物を結合することが可能であると同定されたこの結合部位は、ヒト結核菌(M.tuberculosis)およびスメグマ菌(M.smegmatis)のatpEタンパク質中の耐性を賦与する突然変異部位として上で同定された領域と一致することが見出された(17)。これゆえに、本発明は、それがatpEタンパク質の耐性を賦与する突然変異部位を含んでなることを特徴とするATPアーゼのF0部分の結合部位を提供する。本明細書で使用されるところの耐性を賦与する突然変異部位は、Mtb_S(配列番号1)若しくはMtb_R(配列番号2)の番号付けを参照として採用して、atpEタンパク質のアミノ酸14ないし34、とりわけ24ないし34、およびアミノ酸53ないし69、とりわけ56ないし67を指す。
【0067】
さらなる一態様において、該結合部位は、少なくとも、1個のCサブユニットのアミノ酸Ala24、Gly27、Phe53、Val57、Gly58、Glu61、Tyr64およびPhe65、ならびに1個のAサブユニットのアミノ酸Ser182、Leu183、Leu185およびArg186(Aサブユニットについて表3、4および5のコードSer206−Leu207−Leu209およびArg210を有する)を含んでなり、前記アミノ酸は、表3、4若しくは5のいずれかの原子座標、または、表3、4若しくは5の対応する原子座標の水素以外の原子位置に重ね合わせられる場合に約1.5Å未満の、好ましくは0.75Åを超えない水素以外の原子の二乗平均偏差を含んでなる相同な構造座標を有する。特定の一態様において、結合部位は、第一のCサブユニットのアミノ酸Ala21、Gly25;第二のCサブユニットのアミノ酸Ala24、Gly27、Phe53、Phe54、Val57、Gly58、Glu61、Tyr64、Phe65;第三のCサブユニットのアミノ酸Met17、Gly19、Gly20、Ala21、Ile22、Gly23、Ala24、Gly25、Ile26、Gly27、Asp28、Gly29、Ala31、Phe53、Thr56、Val57、Gly58、Leu59、Val60、Glu61、Ala62、Ala63/Pro63、Tyr64、Phe65、ならびにAサブユニットのアミノ酸Leu183、Leu185およびArg186を含んでなり;前記アミノ酸は、表3、4若しくは5のいずれかの原子座標、または、表3、4若しくは5の対応する原子座標の水素以外の原子位置に重ね合わせられる場合に約1.5Å未満の、好ましくは0.75Åを超えない水素以外の原子の二乗平均偏差を含んでなる相同な構造座標を有する。なおより具体的な一態様において、結合部位は、第一のCサブユニットのアミノ酸Ala21、Gly25;第二のCサブユニットのアミノ酸Ala24、Gly27、Phe53、Phe54、Val57、Gly58、Glu61、Tyr64、Phe65;第三のCサブユニットのアミノ酸Met17、Gly19、Gly20、Ala21、Ile22、Gly23、Ala24、Gly25、Ile26、Gly27、Asp28、Gly29、Ala31、Phe53、Thr56、Val57、Gly58、Leu59、Val60、Glu61、Ala62、Ala63/Pro63、Tyr64、Phe65、ならびにAサブユニットのアミノ酸Leu183、Leu185およびArg186よりなり;前記アミノ酸は表3、4若しくは5のいずれかの原子座標を有する。最も具体的な一態様において、結合部位は、第一のCサブユニットのアミノ酸Ala21、Gly25;第二のCサブユニットのアミノ酸Ala24、Gly27、Phe53、Phe54、Val57、Gly58、Glu61、Tyr64、Phe65;第三のCサブユニットのアミノ酸Met17、Gly19、Gly20、Ala21、Ile22、Gly23、Ala24、Gly25、I
le26、Gly27、Asp28、Gly29、Ala31、Phe53、Thr56、Val57、Gly58、Leu59、Val60、Glu61、Ala62、Ala63/Pro63、Tyr64、Phe65、ならびにAサブユニットのアミノ酸Leu183、Leu185およびArg186よりなり;前記アミノ酸は表3の原子座標を有する。
【0068】
従って、コンピュータによるスクリーニングプログラムにおいて、上で概説されたところの原子座標を使用してatpEタンパク質と相互作用する試験化合物の潜在能力を評価することが、本発明の一目的である。一態様において、本発明は、atpEタンパク質と相互作用する試験化合物の潜在能力の評価方法を提供し、前記方法は;−ATPアーゼのF0部分の結合部位の三次元構造を生成させるための分子モデル化技術;−コンピュータによる手段を使用して、試験化合物と結合部位の三次元構造の間のフィッティング操作を実施すること;および−前記フィッティング操作の結果を解析して結合部位の三次元構造との試験化合物の会合を定量化することを含んでなる。本発明のさらなる一態様において、結合部位の三次元構造は、表3、4若しくは5の原子座標、または、表3、4若しくは5の対応する原子座標の水素以外の原子位置に重ね合わせられる場合に約1.5Å未満の、好ましくは0.75Åを超えない水素以外の原子の二乗平均偏差を含んでなる相同な構造座標を使用して生成される。特定の一態様において、三次元構造は、表3、4若しくは5のいずれかのA鎖のアミノ酸Ala21、Gly25;表3、4若しくは5のいずれかのK鎖のアミノ酸Ala24、Gly27、Phe53、Phe54、Val57、Gly58、Glu61、Tyr64、Phe65;表3、4若しくは5のいずれかのL鎖のアミノ酸Met17、Gly19、Gly20、Ala21、Ile22、Gly23、Ala24、Gly25、Ile26、Gly27、Asp28、Gly29、Ala31、Phe53、Thr56、Val57、Gly58、Leu59、Val60、Glu61、Ala62、Ala63/Pro63、Tyr64、Phe65;ならびに表3、4若しくは5のいずれかのM鎖のアミノ酸Ser206、Leu207、Leu207およびArg210の原子座標を使用して生成される。
【0069】
このスクリーニングにおいて、こうした化合物の結合部位へのフィッティングの質は、形状の相補性若しくは推定される相互作用エネルギーのいずれかにより判断しうる(Meng,E.C.ら、J.Coma.Chem 13:505−524(1992))。
【0070】
結合部位の使用
本発明のatpEに結合するか、その機能的活性を促進するか若しくは阻害する化合物の設計は、一般に、2種の因子の考慮を必要とする。第一に、該化合物はatpEと物理的かつ構造的に会合することが可能でなければならない。atpEの化合物との会合において重要な非共有分子相互作用は、水素結合形成、ファンデルワールスおよび疎水性相互作用を包含する。第二に、化合物は、それがatpEと会合することを可能にするコンホメーションをとることが可能でなければならない。化合物のある部分がatpEとの会合に直接参画しなくてもよいとは言え、それらの部分はなお、該分子の全体的コンホメーションに影響しうる。これは、順に、結合親和性、治療的有効性、薬物様の質および効力に対する大きな影響を有しうる。こうしたコンホメーションの要件は、atpEの活性部位若しくは他の領域の全部若しくは一部分に関しての化学的実体若しくは化合物の全体的な三次元構造および幾何学的配置、またはatpEと直接相互作用する数種の化学的実体を含んでなる化合物の官能基間の間隔を包含する。
【0071】
atpEに対するリガンド若しくは他の化合物の潜在的な予測される阻害性のアゴニスト、アンタゴニスト若しくは結合の効果は、コンピュータモデル化技術の使用により、その実際の合成および試験の前に解析しうる。所定の化合物の理論的構造がそれとatpEの間の不十分な相互作用および会合を示唆する場合は、該化合物の合成および試験を未然
に防ぎうる。しかしながら、コンピュータモデル化が強い相互作用を示す場合には、該分子を合成しかつatpEと相互作用するその能力について試験しうる。この様式で、無効の化合物の合成を回避しうる。いくつかの場合には、モデル化で予測される不活性化合物を合成しかつその後試験してatpEの特定の領域と相互作用する化合物のSAR(構造活性相関)を作成する。当業者は、いくつかの方法の1つを使用して、化学的実体のフラグメント、化合物若しくは剤をatpEおよびより具体的にはatpEの個々の結合ポケットすなわち活性部位と会合するそれらの能力についてスクリーニングしうる。この方法は、例えば、atpE若しくはリガンドと複合体形成したatpEの原子座標に基づくコンピュータスクリーニングでの活性部位の目視検査により開始しうる。選択された化学的実体、化合物若しくは剤をその後、多様な幾何学的配置に配置しうるか、若しくはatpEの個々の結合ポケット内にドッキングしうる。ドッキングは、QuantaおよびSybylのようなソフトウェア、次いでCHARMMおよびAMBERのような標準的分子の力学上の力場(mechanics forcefield)を用いるエネルギー最小化および分子動力学を使用して達成しうる。
【0072】
特化されたコンピュータプログラムもまた化学的実体の選択の方法で補助しうる。これらは、限定されるものでないが:GRID(Goodford,P.J.、“A Computational Procedure for Determining Energetically Favorable Binding Sites on Biologically Important Macromolecules,”J.Med.Chem.28:849−857(1985)、オックスフォード大学、英国オックスフォードから入手可能);MCSS(Miranker,A.とM.Karplus、“Functionality Maps of Bioding Sites:A Multiple Copy Simultaneous Search Method.”Proteins:Structure,Function and Genetics 11:29−34(1991)、Molecular Simulations、マサチューセッツ州バーリントンから入手可能);AUTODOCK(Goodsell,D.S.とA.J.Olsen、“Automated Docking of Substrates to Proteins by Simulated Aneealing”Proteins:Structure,Function and Genetics 8:195−202(1990)、Scripps Research Institute、カリフォルニア州ラホヤから入手可能);およびDOCK(Kuntz,I.D.ら、“A Geometric Approach to Macromolecule−Ligand Interactions,”J.−Mol.Biol.161:269−288(1982)、カリフォルニア大学、カリフォルニア州サンフランシスコから入手可能)を挙げることができる。
【0073】
多様な官能基の特徴をもつプローブと巨大分子表面の間のありそうな相互作用部位を決定するプログラムであるGRIDのようなソフトウェアの使用は、表面部位を分析して類似の阻害するタンパク質若しくは化合物の構造を決定するのに使用される。プローブとしての分子上の適する阻害する基(例えばプロトン化された一級アミン)を用いるGRID計算を使用して、適するエネルギー等高線(contour)の水準の到達可能な位置の周囲の潜在的ホットスポットを同定する。DOCKプログラムを使用して、活性部位若しくはリガンド結合部位を解析しかつ相補的立体構造特性をもつリガンドを示唆しうる。
【0074】
適する化学的実体、化合物若しくは剤が一旦選択されれば、それらを単一のリガンド若しくは化合物若しくは阻害剤若しくは活性化物質に集成し得る。集成は、三次元像での相互に対するフラグメントの関係の目視検査により進行しうる。これに次いで、Quanta若しくはSybylのようなソフトウェアを使用して人的にモデルを構築しうる。
【0075】
個々の化学的実体、化合物若しくは剤の結合において補助するための有用なプログラムは、限定されるものでないが:CAVEAT(Bartlett,P.A.ら、“CAVEAT:A Program to Facilitate the Structure−Derived Design of Biologically Active Molecules.” Molecular Recognition in Chemical and Biological Problems、Special Pub.、Royal Chem.Soc.中、78、pp.82−196(1989));MACCS−3D(MDL Information Systems、カリフォルニア州サンリアンドロ、およびMartin,Y.C.、“3D Database Searching in Drug Design”、J.Med.Chem.35:2145−2154(1992)のような3Dデータベースシステム;ならびにHOOK(Molecular Simulations、マサチューセッツ州バーリントンから入手可能)を挙げることができる。
【0076】
ファルマコフォア仮説を検定しかつスクリーニングのための化合物を選択するために三次元データベースを検索するためのいくつかの方法論が利用可能である。これらはCAVEATプログラム(Baconら、J.Mol.Biol.225:849−858(1992))を包含する。例えば、CAVEATは、活性部位に既に配置されているいずれかの数の化学的断片を結合するための「スペーサー」として作用し得る環状化合物のデータベースを使用する。これは、当業者が、密接な結合に必要であることが既に知られているか若しくは疑われる断片を結合するための何百もの可能な方法を迅速に生成することを可能にする。上述されたとおり一度に1種の化学的実体を段階的様式でatpEの阻害剤活性化物質、アゴニスト若しくはアンタゴニストを構築するように進行する代わりに、こうした化合物は、空の結合部位を使用すること、または既知分子のいくつかの部分(1個若しくは複数)を場合によっては包含することのいずれかを使用して、全体としてすなわち「新規に」設計しうる。これらの方法は:LUDI(Bohm,H.−J.、“The
Computer Program LUDI:A New Method for the De Novo Design of Enzyme Inhinitors”,J.ComR.Aid.Molec.Design、6、pp.61−78(1992)、Biosym Technologies、カリフォルニア州サンディエゴから入手可能);LEGEND(Nishibata,Y.とA.Itai、Tetrahedron 47:8985(1991)、Molecular Simulations、マサチューセッツ州バーリントンから入手可能);およびLeapFrog(Tripos
Associates、ミズーリ州セントルイスから入手可能)を包含する。例えば、LUDIプログラムは、水素結合形成および疎水性フラグメントの双方が位置するべき相互作用部位の一覧を決定し得る。LUDIはその後、リンカーのライブラリーを使用して、4個までの異なる相互作用部位を断片に結合する。その後、――CH2−および――COO――のようなより小さい「結合」基を使用してこれらの断片を結合する。例えば、酵素DHFRについて、公知の阻害剤メトトレキセート中の重要な官能基の配置がLUDIにより再現された。RotsteinとMurcko、J.Med.Chem.36:1700−1710(1992)もまた参照されたい。
【0077】
他の分子モデル化技術もまた本発明で使用しうる。例えば、Cohen,N.C.ら、“Molecular Modeling Software and Methods
for Medicinal Chemistry、J.Med.Chem.33:883−894(1990)を参照されたい。Navia,M.A.とM.A.Murcko、“The Use of Structural Information in Drug Design,”Current Opinions in Structural Biology、2、pp.202−210(1992)もまた参照されたい。
【0078】
化合物が上の方法により一旦設計若しくは選択されれば、その化合物がatpEを結合若しくはそれと会合する親和性を、コンピュータによる評価および/若しくは該化合物を合成した後に生物学的活性を試験することにより試験かつ最適化しうる。阻害剤若しくは化合物は、全体的な結合エネルギーが類似である1種以上のコンホメーションでatpEと相互作用しうる。それらの場合に、結合の変形エネルギーは、遊離化合物のエネルギーと、該化合物がatpEに結合する場合に観察されるコンホメーションの平均エネルギーの間の差違であると解釈される。
【0079】
atpEと結合若しくは会合するとして設計若しくは選択される化合物は、その結合された状態でそれが好ましくはatpEとの反発性静電相互作用を欠くであろうようにさらにコンピュータにより最適化しうる。こうした非相補的(例えば静電的)相互作用は、反発性の電荷−電荷、双極子−双極子および電荷−双極子相互作用を包含する。とりわけ、阻害剤が結合されている場合の阻害剤とatpEの間の全部の静電的相互作用の総和は、好ましくは、結合のエンタルピーへの中立の若しくは好ましい貢献をなす。弱い結合化合物もまた、SARを決定するようにこれらの方法により設計することができる。例えば、米国特許出願第60/275,629号;同第60/331,235号;同第60/379,617号;および同第10/097,249号明細書を参照されたい。
【0080】
特定のコンピュータソフトウェアが、化合物の変形エネルギーおよび静電的相互作用を評価するのに当該技術分野で使用可能である。こうした用途のため設計されたプログラムの例は:Gaussian 92、revision C(M.J.Frisch、Gaussian,Inc.、ペンシルバニア州ピッツバーグ、COPYRGT 1992);AMBER,version 4.0(P.A.Kollman、カリフォルニア大学サンフランシスコ校、GOPYRGT 1994);QUANTA/CHARMM(Molecular Simulations,Inc.、マサチューセッツ州バーリントン、COPYRGT 1994);およびInsight II/Discover(Biosysm Technologies Inc.、カリフォルニア州サンディエゴ、COPYRGT 1994)を包含する。他のハードウェアシステムおよびソフトウェアパッケージは当業者に既知であろう。
【0081】
atpEと会合する化合物が上述されたとおり一旦最適に選択若しくは設計されれば、その後、その結合特性を改良若しくは改変するために、その原子若しくは側基のいくつかで置換を行いうる。一般に、初期置換は保存的であり、すなわち、置換基は元の基とほぼ同一の大きさ、形状、疎水性および電荷を有することができる。もちろん、コンホメーションを変えることが当該技術分野で既知の化合物は回避されうることが理解されるべきである。こうした置換された化合物をその後、詳細に上述された同一のコンピュータ法によりatpEへのフィッティングの効率について解析しうる。
【0082】
本発明はさらに、本明細書に記述される配列および/若しくは構造座標を含有するシステム、とりわけコンピュータに基づくシステムを提供する。こうした系は、atpE若しくはATPアーゼのF0部分の結合部位の構造決定および理論的ドラッグデザインを行うよう設計される。コンピュータに基づくシステムは、詳細に上述されたコンピュータ法のいずれかで本発明の配列および/若しくは構造座標を解析するのに使用されるハードウェア手段、ソフトウェア手段およびデータ記憶手段を指す。本発明のコンピュータに基づくシステムの最小ハードウェア手段は、中央演算処理装置(CPU)、入力手段、出力手段およびデータ記憶手段を含んでなる。当業者は、現在利用可能なコンピュータに基づくシステムのどれが本発明での使用に適するかを容易に認識し得る。
【0083】
従って、本明細書に記述される構造座標を含有するコンピュータで読み取り可能なデータ記憶媒体を提供することが、本発明の一目的である。本明細書で使用されるところの「
コンピュータで読み取り可能なデータ記憶媒体」は、コンピュータにより直接読み取り若しくはアクセスし得るいかなる媒体も指す。こうした培体は、限定されるものでないが:フロッピーディスク、ハードディスク記憶媒体および磁気テープのような磁気記憶媒体;光ディスク若しくはCD−ROMのような光学式記憶媒体;RAMおよびROMのような電気的記憶媒体;ならびに磁気/光学式記憶媒体のようなこれらの範疇のハイブリッドを挙げることができる。
【0084】
処置方法
上で既に挙げられたとおり、微生物に基づく感染症を伴う被験体の処置方法において、前述のスクリーニング方法のいずれかを使用して同定される化合物の使用を提供することもまた、本発明の一目的である。一般に、細菌性病原体はグラム陽性若しくはグラム陰性いずれかの病原体に分類しうる。グラム陽性およびグラム陰性双方の病原体に対する活性をもつ抗菌化合物は、一般に広範囲の活性を有するとみなされる。本発明の化合物は、グラム陽性および/若しくはグラム陰性の細菌性病原体に対し活性とみなされる。とりわけ、本化合物は、最低1種のグラム陽性細菌、好ましくは数種のグラム陽性細菌、より好ましくは1種若しくはそれ以上のグラム陽性細菌および/または1種若しくはそれ以上のグラム陰性細菌に対し活性である。
【0085】
グラム陽性およびグラム陰性の好気性および嫌気性細菌の例は、ブドウ球菌、例えば黄色ブドウ球菌(S.aureus);腸球菌、例えばE.フェーカリス(E.faecalis);連鎖球菌、例えば肺炎連鎖球菌(S.penaumoniae)、S.ミュータンス(S.mutans)、化膿性連鎖球菌(S.pyogens);バチルス属の細菌、例えば枯草菌(Bacillus subtilis)、リステリア属(Listeria)、例えばリステリア菌(Listeria monocytogenes);ヘモフィルス属(Haemophilus)、例えばインフルエンザ菌(H.influenza);モラクセラ属(Moraxella)、例えばM.カタラリス(M.chatarrhalis);シュードモナス属(Pseudomonas)、例えば緑膿菌(Pseudomonas aeruginosa);およびエシェリキア属(Escherichia)、例えば大腸菌(E.coli)を包含する。グラム陽性病原体、例えば、ブドウ球菌、腸球菌および連鎖球菌は、処置することが困難かつ一旦確立すれば例えば病院環境から根絶することが困難の双方である耐性株の発生によりとりわけ重要である。こうした株の例は、メチシリン耐性黄色ブドウ球菌(Staphylococcus aureus)(MRSA)、メチシリン耐性凝固酵素陰性ブドウ球菌(MRCNS)、ペニシリン耐性肺炎連鎖球菌(Streptococcus pneumoniae)および多剤耐性エンテロコッカス ファシウム(Enterococcu faecium)である。
【0086】
本発明の化合物は耐性細菌株に対する活性もまた示す。
【0087】
本発明の化合物は、生存率がF1F0ATP合成酵素の適正な機能に依存する細菌に対しとりわけ活性である。いずれかの論理により束縛されずに、本化合物の活性がF1F0ATP合成酵素の阻害、とりわけF1F0ATP合成酵素のF0複合体の阻害、より具体的にはF1F0ATP合成酵素のF0複合体のAサブユニットのArg186からCサブユニットのGlu61へのプロトンの移動の阻害に存し、細菌の細胞ATP濃度の枯渇により該細菌の死滅に至ることが教示される。前述のスクリーニング方法のいずれかを使用して同定される化合物は、グラム陽性細菌、より具体的にはミコバクテリウム、および最も具体的にはM.アフリカヌム(M.africanum)、トリ結核菌(M.avium)、ウシ結核菌(M.bovis)、ウシ結核菌−BCG(M.bovis−BCG)、カメ結核菌(M.chelonae)、M.フォルツイタム(M.fortuitum)、M.ゴルドネ(M.gordonae)、M.イントラセルラレ(M.intrac
ellulare)、M.カンサシイ(M.kansasii)、M.ミクロティ(M.microti)、M.スクロフラセウム(M.scrofulaceum)、パラ結核菌(M.paratuberculosis)、らい菌(M.leprea)、ヒト結核菌(M.tuberculosis)、M.ウルセランス(M.ulcerans)およびM.ラネ(M.ranae)により引き起こされる感染症に対しとりわけ活性である。
【0088】
上記若しくは下記で使用される場合はいつも、該化合物が細菌感染症を処置し得ることは、該化合物が1種若しくはそれ以上の細菌株への感染を処置し得ることを意味している。抗菌化合物としてのDARQ Jの使用に関してしかしながら使用される場合、抗菌薬は、1種若しくはそれ以上の細菌株への感染を処置し得る化合物であることを意味しているが、但し前記細菌株はミコバクテリウム以外である。
【0089】
本化合物により処置されうる細菌感染症は、例えば、中枢神経系の感染症、外耳感染症、急性中耳炎のような中耳の感染症、硬膜静脈洞の感染症、眼の感染症、歯、歯肉および粘膜の感染症のような口腔の感染症、上気道感染症、下気道感染症、泌尿生殖器感染症、胃腸感染症、産婦人科感染症、敗血症、骨および関節の感染症、皮膚および皮膚構造の感染症、細菌性心内膜炎、火傷、手術の抗菌的予防、ならびに癌化学療法を受領している患者若しくは臓器移植患者のような免疫抑制患者における抗菌的予防を包含する。
【0090】
本発明はさらに、atpEタンパク質と相互作用する剤を被験体に投与することを含んでなる、結核に苦しめられる被験体の処置方法を提供する。
【0091】
製薬学的組成物
本発明はさらに、細胞中でatpEタンパク質と相互作用する剤および製薬学的に許容できる担体を含んでなる製薬学的組成物を提供する。こうした剤は、製薬学的に許容できる担体若しくは希釈剤と一緒になって剤を含んでなる組成物に処方しうる。剤は、エステル若しくは塩のような、酸付加塩若しくは塩基金属塩、またはN若しくはSオキシドのような生理学的に機能的な誘導体の形態にあることができる。組成物はいかなる適する投与経路および手段のためにも処方しうる。製薬学的に許容できる担体若しくは希釈剤は、経口、直腸、鼻、吸入可能、局所(頬側および舌下を包含する)、膣若しくは非経口(皮下、筋肉内、静脈内、皮内、クモ膜下腔内および硬膜外を包含する)投与に適する製剤で使用されるものを包含する。担体若しくは希釈剤の選択はもちろん、剤およびその治療目的に依存しうる提案される投与経路に依存することができる。製剤は、便宜的には単位投薬形態物で提示されることができ、また、製薬学の技術分野で公知の方法のいずれによっても製造しうる。こうした方法は、有効成分を1種若しくはそれ以上の付属成分を構成する担体との連合にもたらす段階を包含する。一般に、製剤は、有効成分を液体担体若しくは微粉化した固体担体または双方との連合に均一かつ緊密にもたらすこと、およびその後必要な場合は生成物を造形することにより製造する。
【0092】
固体の組成物について、慣習的な非毒性の固体担体は、例えば、製薬学的等級のマンニトール、乳糖、セルロース、セルロース誘導体、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、滑石、ブドウ糖、ショ糖、炭酸マグネシウムなどを包含するものを使用しうる。上記で定義されるところの有効成分を、例えばポリアルキレングリコール、アセチル化トリグリセリドなどを担体として使用して坐剤として処方しうる。液体の製薬学的に投与可能な組成物は、例えば、例えば水、生理的食塩水、水性D−ブドウ糖、グリセロール、エタノールなどのような担体中に、上で定義されたところの有効成分および任意の製薬学的補助物質を溶解、分散などしてそれにより溶液若しくは懸濁液を形成することにより製造し得る。所望の場合は、投与されるべき製薬学的組成物は、少量の湿潤剤若しくは乳化剤、pH緩衝剤など、例えば酢酸ナトリウム、ソルビタンモノラウレート、酢酸トリエタノールアミンナトリウム、ソルビタンモノラウレート、トリエタノールアミ
ンオレエートなどのような非毒性の補助物質もまた含有しうる。こうした投薬形態物の実際の製造方法は当業者に既知であるか若しくは明らかであろう;例えば、Gennaroら、Remington’s Pharmaceutical Sciences、Mack Publishing Company、ペンシルバニア州イーストン、第18版、1990を参照されたい。
【0093】
組成物若しくは製剤は、いずれにしても、処置されている被験体の症状を緩和するのに有効な量の、ある量の有効成分(1種若しくは複数)を含有することができる。
【0094】
本化合物の正確な投薬量および投与の頻度は、当業者に公知であるとおり、使用される特定の化合物、処置されている特定の状態、処置されている状態の重症度、特定の患者の齢、重量、性、食餌、投与時間および全身の健康状態、投与様式、ならびに該個体が服用しうる他の医薬品に依存する。さらに、有効1日量は、処置される被験体の応答に依存して、かつ/若しくは本発明の化合物を処方する内科医の評価に依存して減少若しくは増大させうる。
【0095】
非毒性の担体から構成されるバランス(balance)を伴う0.25ないし95%の範囲の有効成分を含有する投薬形態物若しくは組成物を製造しうる。投与様式に依存して、製薬学的組成物は、好ましくは、0.05から99重量%まで、より好ましくは0.1から70重量%までの有効成分、および1から99.95重量%まで、より好ましくは30から99.9重量%までの製薬学的に許容できる担体を含むことができ、全部のパーセンテージは全組成物に基づく。
【0096】
経口投与のため、製薬学的に許容できる非毒性の組成物は、例えば製薬学的等級のマンニトール、乳糖、セルロース、セルロース誘導体、クロスカルメロースナトリウム、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、滑石、ブドウ糖、ショ糖、マグネシウム、炭酸塩などのような通常使用される賦形剤のいずれかの組み込みにより形成される。こうした組成物は溶液、懸濁剤、錠剤、丸剤、カプセル剤、散剤、除放製剤などの形態を取る。こうした組成物は1%〜95%の有効成分、より好ましくは2〜50%、最も好ましくは5〜8%を含有しうる。
【0097】
非経口投与は、一般に皮下、筋肉内若しくは静脈内いずれかでの注入を特徴とする。注入可能物は、慣習的形態で、液体の溶液若しくは懸濁液、注入前の液体中の溶液若しくは懸濁液に適する固体の形態、または乳剤のいずれかとして製造し得る。適する賦形剤は、例えば水、生理的食塩水、D−ブドウ糖、グリセロール、エタノールなどである。加えて、所望の場合は、投与されるべき製薬学的組成物は、少量の例えば酢酸ナトリウム、ソルビタンモノラウレート、トリエタノールアミンオレエート、酢酸トリエタノールアミンナトリウムなどのような、湿潤剤若しくは乳化剤、pH緩衝剤などのような非毒性の補助物質もまた含有しうる。
【0098】
こうした非経口組成物中に含有される有効成分のパーセンテージは、その特定の性質、ならびに化合物の活性および被験体の必要性に高度に依存する。しかしながら、溶液中0.1%ないし10%という有効成分のパーセンテージが使用可能であり、そして、該組成物が上のパーセンテージに後に希釈されることができる固体である場合はより高いことができる。好ましくは、組成物は溶液中に0.2〜2%の有効成分を含むことができる。
【0099】
最後に、本発明は、(a)製薬学的剤が細胞中でatpEタンパク質と相互作用し、かつ(b)包装が被験体における細菌感染症を処置するための該剤の使用を示すラベルを含んでなる、包装および製薬学的剤を含んでなる、とりわけ、抗ミコバクテリウム医薬品としての製品を提供する。
【0100】
本記述を通じ、「標準的方法」、「標準的プロトコル」および「標準的手順」という用語は、分子生物学技術の文脈で使用される場合に、Current Protocols
in Molecular Bioloty、編者F.Ausubelら、John Wiley and Sons,Inc.1994、若しくはSambrook,J.、Fritsch,E.F.およびManiatis,T.、Molecular Cloning:A laboratory manual、第2版、Cold Spring
Harbor Laboratory Press、ニューヨーク州コールドスプリングハーバー、1989のような通常の実験室手引書に見出されるプロトコルおよび手順として理解されるべきである。
【0101】
本発明は、後に続く実験の詳細を参照してより良好に理解されるであろうが、しかし、当業者は、これらは下に続く請求の範囲により完全に記述されるところの本発明を具体的に説明するのみであることを容易に認識するであろう。加えて、本出願を通じ多様な刊行物が引用される。これらの刊行物の開示は、本発明が関する従来技術をより完全に記述するために、本出願に引用することによりここに組み込まれる。
【図面の簡単な説明】
【0102】
【図1】下でJ若しくはDARQ Jともまた称されるR207910の絶対配置。
【図2】ヒト結核菌(M.tuberculosis)およびスメグマ菌(M.smegmatis)変異体のatpEタンパク質配列アライメント。Mtb_S:薬物感受性株ヒト結核菌(M.tuberculosis)H37RvのatpE(1−81)。受託番号:Swiss−Prot Q10598(配列番号1)。Mtb_R:薬物耐性株ヒト結核菌(M.tuberculosis)BK12のatpE(1−81)(配列番号2)。Msm_S:薬物感受性株スメグマ菌(M.smegmatis)のatpE(1−86)。ゲノム研究所(Institute for Genome Research)により得られた配列(配列番号3)。Msm_R09(配列番号4)およびR10(配列番号5):薬物耐性株スメグマ菌(M.smegmatis)のatpE(1−86)。社内で得られた配列。Human:ヒト(Homo sapiens)、ATP5G3(66−142)。受託番号:Ensembl ENSP00000284727。上の番号付け:ヒト結核菌(M.tuberculosis)およびスメグマ菌(M.smegmatis)のatpE。下の番号付け:ヒト(H.sapiens)ATP5G3(66−142)陰影はBLOSUM62マトリックスを使用するアミノ酸類似性を示す(黒=高、灰色=中)。矢印は耐性株で観察される点突然変異の位置を示す。
【図3】DARQ J、イソニアジドおよびDCCDの存在下でのヒト結核菌(M.tuberculosis)の全細胞ATP測定。野性型ヒト結核菌(M.tuberculosis)およびDARQ J変異体ヒト結核菌(M.tuberculosis)双方で526nmで測定したオキシルシフェリンの相対発光単位。
【図4】一緒になってDARQ J化合物の結合部位を形成する3個のCサブユニット(A鎖、K鎖およびL鎖)ならびにAサブユニット(M鎖)のリボン表示。
【0103】
実験
スメグマ菌(Mycobacterium smegmatis)を代理物として使用して、われわれは、数種のミコバクテリウムに対する強力なin vitro活性をもつ一連のDARQを発見した(11)。今日まで、DARQ系列の20種の分子がヒト結核菌(Mycobacterium tuberculosis)H37Rvに対し0.5μg/mlより下の最小阻害濃度(MIC)を有し、また、これらの3種について、抗ミコバクテリウム活性がin vivoマウスモデルで確認された。
【0104】
構造的におよび機構的に、DARQは、フルオロキノロン(メトキシキノロンを包含す
る)、ならびにメフロキンならびにそのアナログ4−メチルキノリンおよび4−キノリルヒドラゾンを包含する他のキノロン分類の双方と非常に異なる(12−16)。DARQと他のキノロン若しくはキノリン分類の間の主要な構造上の差違の1つは、DARQ分類により担持される官能性化された側(3’)鎖の特異性である。加えて、既存の化学分類とのミコバクテリウムの交差耐性の欠如が異なる作用機序を指摘する。
【0105】
下でJ若しくはDARQ Jと称されるDARQのリード化合物(図1)が、in vitroで独特の範囲の強力かつ選択的な抗ミコバクテリウム活性を有することが見出された(表1)。実験室株H37Rvおよび6種の完全に感受性の単離物について得られたMICの中央値は、リファンピンについて1.00μg/mlに対し0.060μg/mlであった。Jは、第一列のTB薬リファンピン、ストレプトマイシン、エタンブトールおよびピラジナミド;ならびに第二列のTB薬モキシフロキサシンに対し耐性のヒト結核菌(M.tuberculosis)臨床単離物に対する類似のin vitro有効性を示した。イソニアジドに対し耐性の8種の臨床単離物について、MICの中央値は0.010μg/mlであった。現在使用されている抗TB薬との交差耐性の欠如は、JがMDR−TB株に対する活性を保持しうることを示唆した。事実、BACTECTM培養系を使用して、細菌の増殖の明瞭な濃度依存性の阻害が、MDR−TB株を固定濃度のJに曝露した場合に見られた。MDR−TBの30種の単離物のうち、13種(43%)が0.100μg/mlのJに対し感受性であることが見出され、また、17種(57%)が0.010μg/mlのJに対し感受性であった。類似の高い程度の感受性(0.010μg/mlより下のMIC)は、BACTECTM系を使用して試験した場合に10種の付加的な完全に薬物感受性の株の1種のみについて見られた一方、全部の株が0.100μg/mlのJに対し感受性であった。
【0106】
強力な活性は、ウシ結核菌(Mycobacterium bovis)およびミコバクテリウム カンサシイ(Mycobacterium kansasii)を包含する他のミコバクテリウム種、ならびに、トリ結核菌(Mycobacterium avium)群(MAC)、カメ結核菌(Mycobacterium abcessus)、ミコバクテリウム フォルツイタム(Mycobacterium fortuitum)およびミコバクテリウム マリナム(Mycobacterium marinum)のような、多くの他の抗TB薬に対し天然に耐性かつ日和見感染症に関与する種に対してもまた示された(表1)。
【0107】
驚くべきことに、Jの活性はミコバクテリウムに特異的であるようであった。Jは、コリネバクテリウム属(Corynebacterium)(MIC 4.00μg/ml)およびノカルジア属(Nocardia)(MIC>4.00μg/ml)のようなミコバクテリウムに近い種に対しほとんど活性でなく、また、グラム陽性の肺炎連鎖球菌(Streptococcus pneumoniae)、黄色ブドウ球菌(Staphylococcus aureus)(メシチリン耐性株(MIC>32μg/ml)を包含する)およびエンテロコッカス フェーカリス(Enterococcus faecalis)、若しくはグラム陰性の大腸菌(Escherichia coli)、インフルエンザ菌(Haemophilus influenzae)およびヘリコバクター
ピロリ(Helicobacter pylori)を包含する他の生物体に対し活性でなかった。対数期増殖にあるヒト結核菌(M.tuberculosis)の100×MICのJの濃度への曝露は、12日後に細菌数の103対数の減少をもたらし、Jがin vitroで殺菌活性を有することを示した。静止期の結核桿菌に対するJの影響は未だ研究されていない。
【0108】
変異体の単離、交差耐性および想定される薬物標的
ミコバクテリウム耐性を検討することにより、われわれは分子薬物標的を同定しかつ作
用機序を提案することを目的とした。ヒト結核菌(M.tuberculosis)およびスメグマ菌(M.smegmatis)の耐性変異体を、
−ミコバクテリウム中の耐性変異体の比率を定量化する(対照としてリファンピンを用いて)
−耐性変異体の耐性パターン(キノロンに対する交差/非交差耐性を包含する)を評価する
−作用機序を検討する
ため、阻害濃度のJでのin vitro選択により導き出した。
【0109】
選択実験から、Jに対する低下された感受性を伴う変異体の比率は、ヒト結核菌(M.tuberculosis)およびスメグマ菌(M.smegmatis)についてそれぞれMIC×4で5×10−7および2×10−8、ならびにMIC×8で5×10−8および1×10−8であった(サポーティングオンラインテキスト)。ヒト結核菌(M.tuberculosis)の場合、これらの比率はリファンピンに対し耐性の変異体のもの(10−7ないし10−8)に匹敵し、そして、Jに対する天然に存在する耐性がまれであることを示す。加えて、Jに対し耐性のヒト結核菌(M.tuberculosis)株の感受性は、抗TB薬イソニアジド、リファンピン、ストレプトマイシン、アミカシン、エタンブトールおよびモキシフロキサシンに対し不変のままであった。Jに対する低下された感受性を伴うヒト結核菌(M.tuberculosis)およびスメグマ菌(M.smegmatis)変異体のさらなる分析は、キノロン耐性が典型的に発生する配列すなわちDNAジャイレース領域のgyrAおよびgyrBに突然変異が存在しなかったことを示した。これは、Jの分子標的がフルオロキノロンのものと異なることを確認する。
【0110】
Jの分子標的を決定しかつ作用機序を推察することへの1アプローチは、ヒト結核菌(M.tuberculosis)およびスメグマ菌(M.smegmatis)の感受性および耐性株で耐性を賦与する突然変異を同定かつ比較することである。耐性のヒト結核菌(M.tuberculosis)株BK12ならびに2種の耐性のスメグマ菌(M.smegmatis)株R09およびR10、ならびに親スメグマ菌(M.smegmatis)のゲノムを完了近くまで配列決定した。われわれは、ヒト結核菌(M.tuberculosis)およびスメグマ菌(M.smegmatis)の感受性および耐性株のゲノム配列の比較分析により、耐性を賦与する突然変異を同定した(図2)。われわれは、対応する親野性型に対し全3種の独立の変異体中で影響を及ぼされた唯一の遺伝子がatpE(ATP合成酵素のF0サブユニットの一部)をコードすることを示した。これは、atpEが変異体株でのJに対する耐性の原因であることを示唆し、Jが新たなヒト結核菌(M.tuberculosis)の標的すなわちATP合成酵素のプロトンポンプを阻害することを示す。
【0111】
変異体atpE遺伝子がJに対する耐性の原因であること、および直接推測によりatpE遺伝子産物がミコバクテリウムでのJの標的であることを示すため、補完的研究を実施した。ATP合成酵素のオペロンの全遺伝子が協調された方法で発現されなければならない、すなわち、F0部分をコードする全遺伝子が同一の場所から発現されなければならないことが既知であるという事実を考え、われわれは、該オペロンのF0部分を耐性のスメグマ菌(M.smegmatis)株(D32V)から増幅し、そしてPCR過程により付加的な突然変異を獲得しなかったクローンを選択した。野性型スメグマ菌(M.smegmatis)を、かように選択した変異体F0フラグメントを含有するプラスミドで形質転換した。これは、細胞を、耐性株のスメグマ菌(M.smegmatis)R09(D32V)のものと実際上同一のMICを伴い、Jに対し耐性にした。加えて、該プラスミドをこれらの形質転換体から再単離しかつatpE遺伝子を配列決定した場合に、それが変異体アレル(D32V)を残存していたことが示された。
【0112】
ヒト結核菌(M.tuberculosis)でのATP産生に対するDARQ Jの実際の影響を、RocheのATP生物発光ルシフェラーゼアッセイキットHS IIを使用してミコバクテリウム中に存在する全細胞ATPに対するJの影響を測定することにより、さらに示した。このアッセイは、526nmで測定し得るD−ルシフェリンのオキシルシフェリンへのATPに駆動される変換に基づく。
【0113】
【化1】

【0114】
簡潔には、全ATPに対するDARQ Jの影響を野性型ヒト結核菌(M.tuberculosis)および変異体株双方で試験した。ATP合成酵素の公知の阻害剤であるDCCDを陽性対照として使用し、また、ある種の細胞壁成分の生合成の阻害剤であるがしかしATP産生に対し影響を有しないイソニアジドを陰性対照として使用した。
【0115】
図3に見ることができるとおり、DARQ Jでの野性型ヒト結核菌(M.tuberculosis)の処理は、これらの細菌中でのATP産生の用量依存性の減少に至る。対照的に、イソニアジドはATP産生に対する影響を有しない。既に上述されたとおり、これらの細菌を高濃度のDARQ Jに曝露することは、ヒト結核菌(M.tuberculosis)のジアリールキノリン耐性変異体を生じさせた。これらの耐性ヒト結核菌(M.tuberculosis)をDARQ Jで処理した場合、これらの細菌は、この化合物の最小阻害濃度(MIC)の100倍ででさえATP産生のいかなる減少も示さなかった。対照的に、DCCDはこれらの桿菌中でのATP産生を阻害することが可能であり、DARQ JおよびDCCDがATP合成において異なる結合ポケットを有することを示唆した。
【0116】
DARQ J結合領域のコンピュータモデル化および同定
DCCDおよびDARQ Jの異なる作用様式をさらに検討するため、野性型およびDARQ J変異体双方のヒト結核菌(M.tuberculosis)のATP合成酵素のコンピュータ生成される3Dモデルを生成した。表4および5に提供される原子座標は、公表されたアミノ酸配列P63691およびAJ865377の3D構造のモデルを作成することにより計算した。実際のDARQ J結合部位は、AおよびCサブユニットの接触領域、より具体的には、表3、4若しくは5で称されるところのAサブユニットのアミノ酸「Arg210」およびCサブユニットの「Glu61」周辺に位置することが見出された。これは、上述されたヒト結核菌(M.tuberculosis)およびスメグマ菌(M.smegmatis)の感受性および耐性株での耐性を賦与する突然変異のスクリーニングについて見られた結果とよく一致する。
【0117】
該モデルは、ATPアーゼ構造のAおよびCヘリックスの相対配置の最適化、ならびに最小のコンピュータ化した内部株(internal strain)に対するアミノ酸バックボーンおよび側鎖の幾何学的配置に基づく。該配置は、異なる生物体[E−Coli PDBエントリコード1C17−V.K.RastogiとM.E.Girvin、Nature、402、263−268(1999)]の以前に公表された全般的らせん配置から出発する多数の分子動力学シミュレーション周期および分子力学の緩和によって得た。分子動力学および配置緩和は双方ともMMFF94に基づく力場のパラメータ化[Halgren,T.A.(1996)、J.Comput.Chem.、17、490
−519]を用いて実施したが、しかし、いずれかの従来技術の分子動力学ソフトウェア[Berendsen,H.J.C.、van der Spoel,D.とvan Drunen,R.、Comp.Phys.Comm.91(1995)、43−56;Lindahl,E.、Hess,B.およびvan der Spoel,D.、J.Mol.Mod.7(2001)306−317.]、次いで適する配置最適化[J.W.PonderとF.M.Richards、J.Comput.Chem.、8、1016−1024(1987)]を使用し得た。
【0118】
表3、4および5のコンピュータ化した座標は、提案された阻害様式、および生物学的アッセイにおける点突然変異を誘導する耐性の発生に基づき、これらの酵素中のMTB ATPアーゼ活性の阻害に関連すると考えられる領域の予測された構造の一部(30オングストロームの半径量を伴う)を含んでなる。
【0119】
考察
DARQ Jは、参照化合物のMICに等しいか若しくはそれより小さいMICをもつ、抗TB薬の新たな化学的分類の1メンバーである。そのスペクトルは、ヒトで重要な異型の種;MAC、M.カンサシイ(M.kansasii)ならびに迅速増殖体M.フォルツイタム(M.fortuitum)およびカメ結核菌(M.abscessus)を包含するミコバクテリウムに対するその特異性において独特である。この抗ミコバクテリウム特異的スペクトルは、MACに対する活性を有しないイソニアジドのものと異なる。Jの臨床使用はTBおよびミコバクテリウム感染症の処置に高度に標的を定めることができる。ミコバクテリウム以外を阻害するJの不能は、より広範なスペクトルをもつ抗生物質と比較した場合に、他の細菌種で発生するより小さい選択圧および耐性のより低いリスクになるはずである(9)。
【0120】
Jの標的および作用機序は他の抗TB薬のものと異なる。多様な細菌のATP合成酵素および真核生物のATP合成酵素、ならびにとりわけATPアーゼ複合体のF0サブユニットのC鎖の配列の比較は、野性型および変異体のヒト結核菌(M.tuberculosis)のATP合成酵素の3Dモデル化と一緒になって、抗菌スペクトルの特異性、およびより少ない程度まで安全性プロファイルに理論的根拠を提供する。
【0121】
構築されたミコバクテリウム属(Mycobacterium)のATPアーゼモデルで実施される動力学的研究は、これらの構造中で空洞(表3の原子座標による結合部位)がAおよびCサブユニットの接触領域(Aサブユニットのアミノ酸「Arg210」およびCサブユニットの「Glu61」周辺)に存在することを示す。表4および5は、この部位を取り囲む原子の2種の研究されたバリアントの座標、およびそれらの平均位置を提供する。DARQ J阻害剤は、これら2アミノ酸が相互作用することを禁止することにより、これら2アミノ酸を伴う通常のプロトン移動段階を妨害することが可能である。DARQ Jの立体特異性は予測された結合部位の非対称性から理解され得;活性のキラル鏡像異性体がこの空洞を最適に提供し、該化合物の他の形態およびATPアーゼのバリアントがより少なく良好に一致する。
【0122】
われわれがここで導き出した結合部位は、ATPアーゼ系のDCCDと完全に異なる部分にある(DARQは酵素の膜部分にあり、DCCD結合は細胞の内側からおよそ90オングストローム離れて発生する;C.Gibbons、M.G.Montgomery、A.G.W.Leslie、J.E.Walker、Nat.Struct.Biol.、7、1055(2000)に公表されたウシATPアーゼ結晶「1E79」のPDB構造に基づく)。従って、MTB ATPアーゼのDCCD型の阻害と潜在的に関与し得る原子は、結合部位(酵素の膜部分にのみちょうどわたる)の座標の表に列挙される領域中になく、そして、相互に、DARQ Jの阻害様式に関与する酵素のこの部分は、公表さ
れた「1E79」構造(細胞内部分のみ示す)に存在しない。結合部位のこの差違は、in vitro ATP産生アッセイで観察されたヒト結核菌(M.tuberculosis)の異なる応答を説明しうる。
【0123】
上にもかかわらず、ミトコンドリアのATPアーゼでDCCDを用いるより古い研究は、例えばSebald W、Machleidt W、Wachter E.、Proc
Natl Acad Sci U.S.A.1980年2月;77(2):785−789により、該酵素のF0領域の親油性環境の1酸性アミノ酸の周囲に位置する別の結合部位を示唆している。ミトコンドリアのATPアーゼのこの結合位置は、ミコバクテリウム属(Mycobacterium)種についてここで記述されるものに類似とみなし得る。
【0124】
同時に、新たな機構を標的とすることは、利用可能な処置に対する耐性突然変異を伴う現在循環しているTB株がJに対し交差耐性でないことを確実にする。Jが、MDR−TB単離物に対し、なお、広範な4種の薬物耐性をもつものに対して、ヒト結核菌(M.tuberculosis)の通常の野性型汎感受性株に対すると少なくとも同じくらい高い抗菌効果を有することが、われわれのin vitro研究から明らかである。この観察結果は、既存の抗TB薬物との交差耐性が存在しないことをそれが明瞭に示すために、重要である。ATPアーゼの膜部分の結合ポケットのさらなる同定を考えれば、本研究の結果は、新たな抗菌化合物、とりわけこれらの生物体中でのATP合成を標的とする抗ミコバクテリウム化合物のさらなる開発を可能にするであろう。
【0125】
【表1】
【0126】
【表2】
【0127】
【表3】
【0128】
【表4】
【0129】
【表5】
【0130】
【表6】
【0131】
【表7】
【0132】
【表8】
【0133】
【表9】
【0134】
【表10】
【0135】
【表11】
【0136】
【表12】
【0137】
【表13】
【0138】
【表14】
【0139】
【表15】
【0140】
【表16】
【0141】
【表17】
【0142】
【表18】
【0143】
【表19】
【0144】
【表20】
【0145】
【表21】
【0146】
【表22】
【0147】
【表23】
【0148】
【表24】
【0149】
【表25】
【0150】
【表26】
【0151】
【表27】
【0152】
【表28】
【0153】
【表29】
【0154】
【表30】
【0155】
【表31】
【0156】
【表32】
【0157】
【表33】
【0158】
【表34】
【0159】
【表35】
【0160】
【表36】
【0161】
【表37】
【0162】
【表38】
【0163】
【表39】
【0164】
【表40】
【0165】
【表41】
【0166】
【表42】
【0167】
【表43】
【0168】
【表44】
【0169】
【表45】
【0170】
【表46】
【0171】
【表47】
【技術分野】
【0001】
本発明は、単離された変異体atpEタンパク質、および前記変異体atpEタンパク質から出発してATPアーゼ結合ドメインの同定を提供する。本発明はまた、関連する核酸、ベクター、宿主細胞、製薬学的組成物および製品も提供する。本発明はさらに、試験化合物がatpEタンパク質と、すなわち本発明のATPアーゼ結合ドメインと相互作用するかどうかの決定方法、ならびに、とりわけ抗菌薬としての、より具体的には抗ミコバクテリウム薬としての、なおより具体的には被験体における結核を処置するための前記試験化合物を含んでなる製薬学的組成物を提供する。
【背景技術】
【0002】
AIDS後、結核(TB)は世界の成人の死亡の第一位の原因(年あたり2〜3百万の死亡)であり、そして世界的な貧困および被害の軽減に対する決定的な障害である(非特許文献1)。該疾患の復活に寄与する因子は、多くの国における抗TBプログラムの実行における困難、免疫抑制個体の数の劇的な増加(主としてHIV感染による)、およびTBが固有である地域を通過しかつそこからの人々の動きを包含する。TBおよびHIVの異常発生は、共感染した人々(現在1,100万人の成人)において相互に刺激し、罹患率および死亡率双方を増大させる(非特許文献2、3)。加えて、TBはHIVに感染した人々における死亡の第一位の原因である(非特許文献4)。
【0003】
第一列の抗TB薬物レジメンは90%以上の有効率を達成し得るとは言え、それらの複雑さは、十分な医学的支援およびTB処置プログラムが利用可能でない場合に乏しいコンプライアンス、そして順に耐性の発生につながり得る(非特許文献5)。TBの多剤耐性(MDR)株は処置を大きく複雑にする(非特許文献6)。結核治療薬開発のための世界同盟(Global Alliance for TB Drug Development)は、いかなる新たな処置も、既存の治療法を上回る以下の3つの利点、すなわち、TBの効果的処置を短縮若しくは単純化すること;MDR−TBに対する有効性を増大させること;および潜伏型のTB感染の処置を改良することの最低1つを提供すべきであると勧告した。こうした新薬は患者コンプライアンスを大きく改善して、それにより世界保健機関(WHO)の直接監視下短期化学療法(Directly Observed Treatment Short−course)(DOTS)戦略のようなTB処置プログラムの費用を低減させるとみられる(非特許文献7)。
【0004】
現在前臨床および臨床開発中のより新たな抗TBの候補は、既存の薬物群(モキシフロキサシンのような)、若しくはMJH−98−1−81(イソニアジドから)、オキサゾリジノンおよびリファペンチン(リファンピンの近接アナログ)のような第一列の薬物のアナログのいずれかからである傾向がある(非特許文献8)。これらの新薬は強力でありうるとは言え、アナログ化合物は耐性に対する一時的な解決策のみを提供する(非特許文献9)。それらは既存の薬物群と同一の作用機序に頼るからである。
【0005】
抗生物質は、一般に、通常、細菌の代謝を特定の機序により阻害することにより細菌の複製を阻害する。例えば、イソニアジドはミコール酸(細胞壁の必要な成分)を合成する酵素機構を妨害する一方、リファンピシンはDNAからRNAを転写するための細菌の機構を妨害する。従って、既知の剤に比較して、細胞の増殖および複製の異なったミコバクテリウム特異的局面を標的とする抗TB化合物の新規同定方法を探索することが興味深い。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Global Alliance for TB Drug Development、Developing a faster TB cure(2004;http://www.tballiance.org)
【非特許文献2】E.L.Corbettら、Arch.Invern.Med.163、1009(2003)
【非特許文献3】UNAIDS、AIDS epidemic update 2003(2003;www.unaids.org/Unaids/EN/Resources)。
【非特許文献4】World Health Organization、Tuberculosis(2004;http://www.who.int/health_topics/tuberculosis/en/)。
【非特許文献5】R.J.O’Brien、P.P.Nunn、Am.J.Respir.Crit.Care Med.163、1055(2001)。
【非特許文献6】World Health Organization、Tuberculosis Fact Sheet No 104(2004;http://www.who.int/mediacentre/factsheets/fs104/en/)。
【非特許文献7】A.J.Claxton、J.Cramer、C.Pierce、Clin.Ther.23、1296(2001)。
【非特許文献8】N.Lounisら、Antimicrob.Agents Chemother.45、3482(2001)。
【非特許文献9】A.S.Ginsburg、J.H.Grosset、W.R.Bishai、Lancet Infect.Dis.3、432(2003)。
【発明の概要】
【0007】
本発明は、とりわけ(配列番号1)、(配列番号2)、(配列番号3)、(配列番号4)および(配列番号5)から選択されるアミノ酸配列によりコードされる単離された変異体atpEタンパク質、とりわけ(配列番号6)、(配列番号7)、(配列番号8)、(配列番号9)および(配列番号10)よりなる群から選択される前記変異体atpEタンパク質をコードする単離された核酸、ならびに本核酸を含んでなるベクターを提供する。特定の一態様において、変異体atpEタンパク質は配列番号2によりコードされ、また、前記タンパク質をコードする単離された核酸配列は配列番号7よりなる。
【0008】
本発明はさらに、本発現ベクターをその中に有する宿主細胞を含んでなる宿主−ベクター系を提供する。
【0009】
本発明はさらに、変異体atpEタンパク質が該細胞中で抗菌耐性を誘導する、前記タンパク質を含んでなる単離された細胞を提供する。
【0010】
本発明はさらに、抗菌化合物の同定方法を提供し、前記方法は、
a)atpEタンパク質を発現する細胞を、生理学的条件下で試験化合物と接触させる段階、および
b)該試験化合物がatpEタンパク質と相互作用するかどうかを決定する段階
を含んでなる。
【0011】
本発明はさらに、atpEタンパク質と相互作用する試験化合物の潜在能力の評価方法を提供し、前記方法は;
(a)分子モデル化技術を使用して、atpEタンパク質の三次元構造を生成させること;
(b)コンピュータによる手段を使用して、試験化合物と、atpEタンパク質の三次元構造の間のフィッティング操作を実施すること;および
(c)前記フィッティング操作の結果を解析して、試験化合物のatpEタンパク質の三次元構造との会合を定量化すること
を含んでなる。
【0012】
1個のCサブユニットの少なくともアミノ酸Ala24、Gly27、Phe53、Val57、Gly58、Glu61、Tyr64およびPhe65;1個のAサブユニットのアミノ酸Ser182、Leu183、Ser184、Leu185およびArg186を含んでなり、かつ、前記アミノ酸は表3、4若しくは5のいずれかの原子座標を有する、ATPアーゼのF0部分の結合部位を提供することもまた、本発明の一目的である。
【0013】
さらなる一目的において、本発明は、ATPアーゼのF0部分と相互作用する化合物の同定方法、および、抗菌化合物としてのそれらの潜在能力、とりわけ抗ミコバクテリウム化合物の同定方法における、前述の結合ドメインの使用を提供する。
【0014】
従って、ATPアーゼのF0部分と、とりわけ耐性を賦与する突然変異部位でatpEタンパク質と、若しくは本発明の結合部位と相互作用する化合物を被験体に投与することを含んでなる、微生物に基づく感染症を伴う被験体の処置方法を提供することが、本発明の一目的である。本発明はさらに、前述のスクリーニング方法のいずれかを使用してatpEタンパク質と相互作用する剤を被験体に投与することを含んでなる、結核に苦しめられる被験体の処置方法を提供する。ATPアーゼのF0部分、とりわけatpEタンパク質と相互作用する化合物の使用を含んでなる処置方法において、ATPアーゼのF0部分、およびとりわけatpEタンパク質と相互作用することが以前に知られている化合物は除外されるべきである。より具体的には、開示された処置方法のいずれかにおける(11)に記述されるDARQ J化合物の使用が除外されるべきである。
【0015】
本発明はさらに、細胞中でatpEタンパク質と相互作用する剤および製薬学的に許容できる担体を含んでなる製薬学的組成物を提供する。最後に、本発明は、(a)製薬学的剤が細胞中でatpEタンパク質と相互作用し、かつ(b)包装が被験体における細菌感染症を処置するための該剤の使用を示すラベルを含んでなる、包装および製薬学的剤を含んでなる製品を提供する。特定の一態様において、本発明は抗菌医薬品の製造におけるDARQ Jの使用を提供する。
【0016】
本発明のこれおよびさらなる局面は下でより詳細に論考されるであろう。
【0017】
[図面の簡単な説明]
表1 多様なミコバクテリウム種の増殖を90%阻害した、リードDARQ化合物(J)の最小阻害濃度(MIC)。試験した株の数は別の方法で示されない限りn=1であった。
【0018】
表2 DARQ J化合物の結合部位の周囲のアミノ酸。
【0019】
表3 野性型およびDARQ J変異体ヒト結核菌(M.tuberculosis)株双方由来のDARQ J化合物の結合部位の周囲のアミノ酸の原子座標。
【0020】
表4 野性型ヒト結核菌(M.tuberculosis)のDARQ J化合物の結合部位の原子座標。
【0021】
表5 DARQ J変異体ヒト結核菌(M.tuberculosis)株のDARQ J化合物の結合部位の原子座標。
【0022】
表6 ヒト結核菌(M.tuberculosis)の変異体atpEタンパク質(配列番号2)の原子座標。
【0023】
表7 ヒト結核菌(M.tuberculosis)の野性型atpEタンパク質(配列番号1)の原子座標。
【0024】
図1 下でJ若しくはDARQ Jともまた称されるR207910の絶対配置。
【0025】
図2 ヒト結核菌(M.tuberculosis)およびスメグマ菌(M.smegmatis)変異体のatpEタンパク質配列アライメント。Mtb_S:薬物感受性株ヒト結核菌(M.tuberculosis)H37RvのatpE(1−81)。受託番号:Swiss−Prot Q10598(配列番号1)。Mtb_R:薬物耐性株ヒト結核菌(M.tuberculosis)BK12のatpE(1−81)(配列番号2)。Msm_S:薬物感受性株スメグマ菌(M.smegmatis)のatpE(1−86)。ゲノム研究所(Institute for Genome Research)により得られた配列(配列番号3)。Msm_R09(配列番号4)およびR10(配列番号5):薬物耐性株スメグマ菌(M.smegmatis)のatpE(1−86)。社内で得られた配列。Human:ヒト(Homo sapiens)、ATP5G3(66−142)。受託番号:Ensembl ENSP00000284727。上の番号付け:ヒト結核菌(M.tuberculosis)およびスメグマ菌(M.smegmatis)のatpE。下の番号付け:ヒト(H.sapiens)ATP5G3(66−142)陰影はBLOSUM62マトリックスを使用するアミノ酸類似性を示す(黒=高、灰色=中)。矢印は耐性株で観察される点突然変異の位置を示す。
【0026】
図3 DARQ J、イソニアジドおよびDCCDの存在下でのヒト結核菌(M.tuberculosis)の全細胞ATP測定。野性型ヒト結核菌(M.tuberculosis)およびDARQ J変異体ヒト結核菌(M.tuberculosis)双方で526nmで測定したオキシルシフェリンの相対発光単位。
【0027】
図4 一緒になってDARQ J化合物の結合部位を形成する3個のCサブユニット(A鎖、K鎖およびL鎖)ならびにAサブユニット(M鎖)のリボン表示。
【0028】
[発明の詳細な記述]
定義
別の方法で本明細書に明らかに提供されるところを除き、本出願で使用されるところの以下の用語のそれぞれは、下に示される意味するところを有する。
【0029】
「atpEタンパク質」は、ヒト結核菌(M.tuberculosis)についてSwissProtエントリQ10598により表されるところのATPアーゼ複合体のF0サブユニットのC鎖、または前記ヒト結核菌(M.tuberculosis)配列に対する最低70、80、90、95、97若しくは99%の配列同一性を有するタンパク質を意味している。
【0030】
ATPアーゼ、ATP合成酵素若しくはF0F1ATPアーゼともまた称される「F1F0ATPアーゼ」は、ATPの合成若しくは加水分解を触媒する大型の多サブユニット複合体を意味している。F0F1ATPアーゼは2ドメイン、すなわち、膜に対し外部でありかつ触媒部位を含有するF1部位、および二層におよびかつプロトン孔を含有するF
0部分より構成される。ATPアーゼは、細菌の形質膜、葉緑体のチラコイド膜、およびミトコンドリアの内膜で見出され、そこでそれらはプロトンの電子化学的勾配のエネルギーを使用してATP合成を駆動する。
【0031】
「投与すること」は、当業者に既知の多様な方法および送達系のいずれかを使用して遂げられ若しくは実施されるある様式で送達することを意味している。投与することは、例えば、局所で、静脈内に、心膜に、経口で、植込物を介して、経粘膜で、経皮で、筋肉内に、皮下に、腹腔内に、クモ膜下腔内に、リンパ内に(intralymphatically)、病変内に、若しくは硬膜外に実施し得る。投与することはまた、例えば1回、複数回、および/または1つ若しくはそれ以上の延長された期間にわたっても実施し得る。
【0032】
「宿主細胞」は、限定されるものでないが、細菌細胞、酵母細胞、真菌細胞、昆虫細胞および哺乳動物細胞を挙げることができる。細菌細胞は、リン酸カルシウム沈殿、電気穿孔法および微小注入法のような当該技術分野で公知の方法によりトランスフェクトし得る。
【0033】
atpEタンパク質に関して「単離された」は、atpEがその天然の機能を保持しかつその天然の環境中の他のタンパク質の数種若しくは全部を含まない、atpEタンパク質を含有する膜断片調製物若しくは他の適する調製物を意味している。それは、F0F1ATPアーゼのF0部分、とりわけ本発明の変異体atpEタンパク質を含んでなるF0部分を含んでなる膜調製物を包含することを意味している。
【0034】
「細菌細胞」はいかなる細菌細胞も意味している。細菌細胞は、正常、異常および形質転換され、かつ、ミコバクテリウム、とりわけヒト結核菌(Mycobacterium
tuberculosis)およびスメグマ菌(Mycobacterium smegmatis)、コリネバクテリウム、ノカルジア類、例えば連鎖球菌、ブドウ球菌および腸球菌のようなグラム陽性細菌、若しくは例えば大腸菌(Escherichia coli)、インフルエンザ菌(Heamophilus influenzae)およびヘリコバクター ピロリ(Helicobacter pylori)のようなグラム陰性細菌により例示される細胞を制限なしに包含する。
【0035】
「核酸」および「ポリヌクレオチド」という用語は本明細書で互換性に使用され、そしてそれぞれデオキシリボヌクレオチドおよび/若しくはリボヌクレオチドのポリマーを指す。デオキシリボヌクレオチドおよびリボヌクレオチドは天然に存在し得るか、若しくはその合成のアナログであり得る。
【0036】
「生理学的条件」という用語は、所定の細胞に関して、該細胞の生化学的環境を通常構成しうる条件を意味している。細胞の生化学的環境は、該細胞が通常曝露される数種若しくは全部のプロテアーゼを制限なしに包含する。こうした条件は、限定されるものでないがin vivo状態を挙げることができる。
【0037】
「ポリペプチド」、「ペプチド」および「タンパク質」という用語は本明細書で互換性に使用され、そしてそれぞれアミノ酸残基のポリマーを意味している。アミノ酸残基は天然に存在し得るか、若しくはそれらの化学的アナログであり得る。ポリペプチド、ペプチドおよびタンパク質はまた、グリコシル化、脂質結合、硫酸化、水酸化およびADP−リボシル化のような修飾も包含し得る。
【0038】
「被験体」は、ウシ、ウマ、ヒツジ、ブタ、イヌ、ネコ、マウス若しくはラットのようなげっ歯類、シチメンチョウ、ニワトリおよび霊長類を制限なしに包含する哺乳動物若し
くは鳥類のようないかなる動物も意味している。好ましい態様において、被験体はヒトである。
【0039】
「処置すること」は、被験体における疾患を除外すること、その経過を反転すること、その進行を遅らせること、その症状を低下させること、若しくは別の方法で改善することを制限なしに包含する。
【0040】
「ベクター」は当該技術分野で既知のいかなる核酸ベクターも意味している。こうしたベクターは、限定されるものでないが、プラスミドベクター、コスミドベクターおよびバクテリオファージベクターを挙げることができる。
【0041】
「候補物質」および「試験化合物」という用語は互換性に使用され、そして、生物学的応答修飾因子として別の部分すなわちatpEタンパク質と相互作用すると考えられている物質を指す。例えば、代表的な候補物質は、atpEタンパク質と相互作用すると考えられ、また、ATPアーゼ活性を改変すると考えられている。本発明の方法を使用して検討され得る例示的候補物質は、限定されるものでないが、ペプチド、酵素、酵素基質、補助因子、糖、オリゴヌクレオチド、化合物小分子およびモノクローナル抗体を挙げることができる。
【0042】
「調節する」は、野性型若しくは変異体atpEタンパク質のいずれかの若しくは全部の化学的および生物学的活性若しくは特性の増大、減少若しくは他の変化を意味している。
【0043】
「相互作用する」は、分子間の「結合」相互作用を包含する分子間の検出可能な相互作用を意味している。相互作用は、例えば、性質がタンパク質−タンパク質若しくはタンパク質−核酸であり得る。こうした相互作用は技術既知の手順、例えば酵母2ハイブリッドアッセイ、免疫沈降、SPAアッセイ若しくはフィルター結合アッセイを使用して検出し得る。
【0044】
本明細書で使用されるところの「原子座標」若しくは「構造座標」という用語は、各原子についてX、Y、ZおよびBを包含するタンパク質データバンク(PDB)フォーマットで原子の位置を記述する数学的座標を指す。当業者は、X線結晶学により決定される一組の構造座標が標準誤差を伴わなくはないことを理解している。本発明の目的上、表3、4、5、6若しくは7の対応する原子座標の水素以外の原子位置に重ね合わせられる場合に1.5Å未満の水素以外の原子の二乗平均偏差を有するいかなる供給源からのAPT合成酵素の構造座標のいずれの組も、実質的に同一若しくは相同と考えられる。より好ましい一態様において、表3、4、5、6若しくは7の対応する原子座標の水素以外の原子位置に重ね合わせられる場合に0.75Å未満の水素以外の原子の二乗平均偏差を有するいかなる供給源からのAPT合成酵素の構造座標のいずれの組も、実質的に同一と考えられる。
【0045】
本発明の態様
変異体atpE(または「変異atpE」という)タンパク質
本発明は、単離された変異体atpEタンパク質、とりわけ細菌のatpEタンパク質、より具体的にはミコバクテリウムのatpEタンパク質、なおより具体的にはヒト結核菌(M.tuberculosis)若しくはスメグマ菌(M.smegmatis)のatpEタンパク質を提供する。突然変異は単一の点突然変異、挿入若しくは欠失から選択される。本発明の一態様において、突然変異は、図2の配列アライメントに示されるところのアミノ酸20ないし40、とりわけ30ないし40のいずれか1つ、好ましいアミノ酸34の、若しくはアミノ酸60ないし75の、とりわけ62ないし73のいずれか1
つ、好ましくはアミノ酸69に位置する最低1個の点突然変異よりなる。さらなる一態様において、単離された変異体atpEタンパク質は、図2に示されるところのMtb_R(配列番号2)、Msm_R09(配列番号4)およびMsm_R10(配列番号5)、または前述のアミノ酸配列のいずれかに対する最低70、80、90、95、97若しくは98%の配列同一性を有するアミノ酸配列のから選択される。
【0046】
本発明はさらに、前記変異体atpEタンパク質をコードする単離された核酸を提供する。一態様において、前記核酸配列は、例えばJ.Biol.Chem.、1994、Vol.269(10)、p.7285−7289に記述されるようなF0部分をコードする全部の遺伝子よりなり、ここで前記遺伝子は単一のプロモーターから転写され、そして本発明の変異体atpEタンパク質をコードする核酸配列を含んでなる。核酸はDNA若しくはRNA、および好ましくはDNAであり得、そして、さらなる一態様において、Mtb_R(配列番号7)Msm_R09(配列番号9)、Msm_R10(配列番号10)をコードする核酸配列、または前述の核酸配列のいずれかに対する最低70、80、90、95、97若しくは98%の配列同一性を有する核酸配列から選択される。
【0047】
核酸およびポリペプチド配列の同一性パーセントは、参照配列をクエリ配列と比較する商業的に入手可能なアルゴリズムを使用して計算し得る。以下のプログラム(国立バイオテクノロジー情報センター(National Center for Biotechnology Information)により提供される)、すなわち、BLAST、gapped BLAST、BLASTNおよびPSI−BLASTを使用して相同性/同一性を決定することができ、これらはデフォルトのパラメータとともに使用しうる。
【0048】
アルゴリズムGAP(Genetics Computer Group、ウィスコンシン州マディソン)は、NeedlemanとWunschのアルゴリズムを使用して、一致の数を最大にしかつギャップの数を最少にする2種の完全な配列を整列させる。一般に、ギャップ創製ペナルティ(gap creation penalty)=12およびギャップ伸長ペナルティ(gap extension penalty)=4を伴うデフォルトのパラメータを使用する。
【0049】
核酸配列若しくはその一部分とクエリ配列の間の最良の全体的一致の別の決定方法は、Brutlagら(Comp.App.Biosci.、6:237−245(1990))のアルゴリズムに基づくFASTDBコンピュータプログラムの使用である。該プログラムは包括的配列アライメントを提供する。前記包括的配列アライメントの結果は同一性パーセントにある。同一性パーセントを計算するためにDNA配列のFASTDB検索での使用に適するパラメータは:マトリックス(Matrix)=Unitary、k−タプル(k−tuple)=4、ミスマッチペナルティ(Mismatch penalty)=1、結合ペナルティ(Joining Penalty)=30、無作為化グループ長(Randomization Group Length)=0、カットオフスコア(Cutoff Score)=1、ギャップペナルティ(Gap Penalty)=5、ギャップサイズペナルティ(Gap Size Penalty)=0.05およびウィンドウサイズ(Window Size)=500若しくはヌクレオチド塩基中のクエリ配列長さのいずれか短い方、である。アミノ酸アライメントの同一性および類似性パーセントを計算するための適するパラメータは:マトリックス(Matrix)=PAM 150、k−タプル(k−tuple)=2、ミスマッチペナルティ(Mismatch penalty)=1、結合ペナルティ(Joining Penalty)=20、無作為化グループ長(Randomization Group Length)=0、カットオフスコア(Cutoff Score)=1、ギャップペナルティ(Gap Penalty)=5、ギャップサイズペナルティ(Gap Size Penalty)=0.05およびウィンドウサイズ(Window Size)=500若しくは
ヌクレオチド塩基中のクエリ配列長さのいずれか短い方、である。
【0050】
本発明はさらに、本核酸を含んでなるベクターを提供する。一態様において、ベクターはプラスミドベクターである。
【0051】
本発明はさらに、本プラスミドベクターをその中に有する宿主細胞を含んでなる宿主−ベクター系を提供する。該細胞は原核生物若しくは真核生物であり得、一態様において、宿主細胞は細菌細胞、とりわけ例えばヒト結核菌(M.tuberculosis)若しくはスメグマ菌(M.smegmatis)のようなミコバクテリウム細胞である。
【0052】
本発明はさらに、変異体atpEタンパク質(このタンパク質は細胞中で抗菌耐性を誘導する)を含んでなる単離された細胞を提供する。一態様において、単離された細胞は、変異体のミコバクテリウムatpEタンパク質で形質転換された、とりわけ、突然変異が図2の配列アライメントに示されるところのアミノ酸20ないし40、より具体的には30ないし40のいずれか1つ、好ましいアミノ酸34に、若しくはアミノ酸60ないし75、とりわけ62ないし73のいずれか1つ、好ましくはアミノ酸69に位置する最低1個の点突然変異よりなる変異体のミコバクテリウムatpEタンパク質で形質転換されたスメグマ菌(M.smegmatis)細胞よりなる。
【0053】
スクリーニング方法
本発明はさらに、抗菌化合物の同定方法を提供し、前記方法は
(a)atpEタンパク質を発現する細胞を、生理学的条件下で試験化合物と接触させる段階;
(b)該試験化合物がatpEタンパク質と相互作用するかどうかを決定する段階
を含んでなる。
【0054】
一態様において、前述の方法で使用されるatpEタンパク質は、細菌のatpEタンパク質、とりわけミコバクテリウムのタンパク質よりなり、かつ、野性型atpEタンパク質ならびに上述されたところの変異体atpEタンパク質の双方を包含することを意味している。本発明のさらなる一態様において、前述の方法で使用されるミコバクテリウムのatpEタンパク質は、本発明の変異体のミコバクテリウムatpEタンパク質よりなる。前述のアッセイの特定の一態様において、本発明の変異体atpEタンパク質で形質転換した細胞を使用し、そして、試験化合物の前記atpEタンパク質との相互作用を、前記変異体atpEタンパク質を含んでなるF1F0−ATPアーゼの酵素活性の可能な阻害を測定することにより評価する。F1F0−ATPアーゼ活性の阻害は、例えば、ADPの産生をピルビン酸キナーゼおよび乳酸脱水素酵素の反応を介するNADHの酸化に結びつけることによる酵素活性の検出を用いる、F1F0−ATPアーゼおよび基質としてのATPを含んでなる系への物質の添加によるような技術既知の手順を使用して測定する。
【0055】
アッセイの一態様において、atpEタンパク質を結合アッセイで使用しうる。結合アッセイは競合的でも若しくは非競合的でもよい。こうしたアッセイは、どの化合物(あれば)がポリペプチドに結合することが可能であるかを決定するための多数の化合物の迅速スクリーニングを提供し得る。
【0056】
この情況内で、本発明は、試験化合物が本発明の単離されたatpEタンパク質に結合するかどうか、そして従って潜在的抗菌化合物であるかどうかの同定方法を提供し、前記方法は;
a)atpEタンパク質を発現する細胞(こうした細胞は前記atpEタンパク質を通常は発現しない)を、atpEタンパク質を結合することが既知の化合物の存在および非存
在下に試験化合物と接触させること、
b)atpEタンパク質に結合することが既知の化合物を参照として使用してatpEタンパク質への試験化合物の結合を測定すること
を含んでなる。
【0057】
試験化合物、若しくは下で参照化合物ともまた称されるatpEタンパク質に結合することが既知の化合物の結合を、タンパク質−リガンド相互作用の技術既知の試験方法を使用して評価する。例えば、こうした結合は標識した物質若しくは参照化合物を使用することにより測定し得る。試験化合物若しくは参照化合物、とりわけ化合物J(図1)を、当該技術分野で既知のいずれかの便宜的様式、例えば放射活性で、蛍光で若しくは酵素的に標識し得る。前述の方法の特定の一態様において、参照化合物としてもまた知られるatpEタンパク質に結合することが既知の化合物を、検出可能に標識し、そして。前記標識を使用して、試験化合物のatpEタンパク質への結合を測定する。前記参照化合物は、放射標識、蛍光標識若しくは酵素標識、より好ましくは放射標識を使用して標識する。
【0058】
本発明の代替の一態様において、前述の結合アッセイは、細胞組成物、すなわち上で定義されたところのatpEタンパク質を含んでなる細胞抽出物、細胞画分若しくは細胞小器官(organel)上で実施される。より具体的には、前述の結合アッセイは、細胞組成物、すなわち上で定義されたところのatpEタンパク質を含んでなる膜調節物上で実施され、前記細胞組成物すなわち膜調製物は、変異体のミコバクテリウムatpEタンパク質で形質転換された、とりわけ、突然変異が図2の配列アライメントに示されるところのアミノ酸20ないし40、とりわけ30ないし40のいずれか1つ、好ましいアミノ酸34に、またはアミノ酸60ないし75、とりわけ62ないし73のいずれか1つ、好ましくはアミノ酸69に位置する最低1個の点突然変異よりなる変異体のミコバクテリウムatpEタンパク質で形質転換されたスメグマ菌(M.smegmatis)細胞から得られる。Mtb_S(配列番号1)若しくはMtb_R(配列番号2)の番号付けを参照として採用すれば、前述の領域は、アミノ酸14ないし34、とりわけ24ないし34、好ましくはアミノ酸28中に、若しくはアミノ酸54ないし69、とりわけ56ないし67、好ましくはアミノ酸63に対応する。
【0059】
一態様において、結合アッセイは膜調製物を使用して実施される。これらの膜調製物は、放射標識atpEリガンド(3H標識DARQを包含する)の結合およびこうした放射リガンドの結合部位についての競合体による置換を検出するために、慣習的フィルター結合アッセイ(例えばBrandelフィルターアッセイ装置を使用して)若しくはハイスループットシンチレーション近接型結合アッセイ(SPAおよびCytostar−Tフラッシュプレート技術;Amersham Pharmacia Biotech)で使用し得る。放射活性は、96、384、1536マイクロタイタープレート形式から迅速測定を行うことが可能なPackard Topcount若しくは類似の装置で測定し得る。SPA/Cytostar−T技術はハイスループットスクリーニングにとりわけ従いやすく、そして、従って、この技術は標準的リガンドを置換することが可能な化合物についてのスクリーニングとして使用するのに適する。
【0060】
天然の状況に近づける環境でのatpEタンパク質へのリガンドの結合を研究するための別のアプローチは、Biacore装置(Biacore)により活用される表面プラスモン共鳴効果を利用する。膜調製物若しくは全細胞中のatpEタンパク質をBiacoreのバイオセンサーチップに結合し得、そして、リガンドの結合を化合物の存在および非存在下で検査して、結合部位の競合体を同定し得る。
【0061】
分子モデル化
本発明はさらに、atpEタンパク質と相互作用する試験化合物の潜在能力の評価方法
を提供し、前記方法は;
(a)分子モデル化技術を使用してatpEタンパク質の三次元構造を明確に表すこと;(b)コンピュータによる手段を使用して、試験化合物とatpEタンパク質の三次元構造との間のフィッティング操作を実施すること;および
(c)前記フィッティング操作の結果を解析して、atpEタンパク質の三次元構造との試験化合物の会合を定量化すること
を含んでなる。
【0062】
受容体および酵素のコンホメーションのモデルを創製かつ利用するのに適切なハードウェアおよびソフトウェア双方を包含する分子モデル化技術は当該技術分野で既知である。
【0063】
多数のコンピュータプログラムが、本明細書に記述される方法で潜在的なatpEと相互作用する化合物をコンピュータモデル化し、モデル構築しかつコンピュータにより同定、選択および評価する方法に利用可能でありかつ適する。これらは、例えば、GRID(英国オックスフォード大学から入手可能)、MCSS(Accelrys,Inc.、カリフォルニア州サンディエゴから入手可能)、AUTODOCK(Oxford Molecular Groupから入手可能)、FLEX X(Tripos、ミズーリ州セントルイスから入手可能)、DOCK(カリフォルニア大学、カリフォルニア州サンフランシスコから入手可能)、CAVEAT(カリフォルニア大学、バークレーから入手可能)、HOOK(Accelrys,Inc.、カリフォルニア州サンディエゴから入手可能)、ならびにMACCS−3D(MDL Information Systems、カリフォルニア州サンリアンドロから入手可能)、UNITY(Tripos、ミズーリ州セントルイスから入手可能)およびCATALYST(Accelrys,Inc.、カリフォルニア州サンディエゴから入手可能)のような3Dデータベースシステムを包含する。潜在的候補物質はまた、LUDI(Biosym Technologies、カリフォルニア州サンディエゴから入手可能)、LEGEND(Accelrys,Inc.、カリフォルニア州サンディエゴから入手可能)およびLEAPFROG(Tripos、ミズーリ州セントルイスから入手可能)のようなソフトウェアパッケージを使用してコンピュータで「新規に(de novo)」設計されうる。化合物の変形エネルギーおよび静電反発力は、GAUSSIAN 92、AMBER、QUANTA/CHARMMおよびINSIGHT II/DISCOVERのようなプログラムを使用して解析しうる。これらのコンピュータ評価およびモデル化技術は、例えばSilicon Graphics、Sun Microsystemsおよび他者から入手可能なワークステーションを包含するいずれの適するハードウェアでも実施しうる。これらのモデル化技術、方法、ハードウェアおよびソフトウェアパッケージは代表的なものであり、かつ、包括的列挙であることを意図していない。当該技術分野で既知の他のモデル化技術もまた本発明で使用しうる。例えば、N.C.Cohen、Molecular Modeling in Drug Design、Academic Press(1996)を参照されたい。
【0064】
本発明の一態様において、atpEタンパク質の三次元構造は、大腸菌(E.coli)(タンパク質データベース1Q01)のIle28、Glu61およびIle63の原子座標+/−10Åを超えない、好ましくは5Åを超えない前記アミノ酸のバックボーン原子の二乗平均偏差を使用して生成される。
【0065】
下の実施例で提供されるとおり、atpEタンパク質の三次元構造を提供することが本発明の一目的であった。表6および7は、配列番号2および配列番号1を伴う変異体および野性型のatpEタンパク質の原子座標を提供する。従って、一態様において、atpEタンパク質の三次元構造は表6若しくは7の原子座標を使用して生成される。特定の一態様において、atpEタンパク質の三次元構造は表7の原子座標を使用して生成される
。DARQ J化合物は、その脱プロトン化された形態のAサブユニットのArg186のCサブユニットのGlu61との相互作用を阻害する。従って、atpEタンパク質と相互作用する試験化合物の潜在能力の評価方法における表6若しくは7の原子座標の使用を提供することが、本発明の一目的である。
【0066】
結合部位
別の態様において、本発明は、ATPアーゼのF0部分の結合部位の特徴を提供する。DARQ J化合物を結合することが可能であると同定されたこの結合部位は、ヒト結核菌(M.tuberculosis)およびスメグマ菌(M.smegmatis)のatpEタンパク質中の耐性を賦与する突然変異部位として上で同定された領域と一致することが見出された(17)。これゆえに、本発明は、それがatpEタンパク質の耐性を賦与する突然変異部位を含んでなることを特徴とするATPアーゼのF0部分の結合部位を提供する。本明細書で使用されるところの耐性を賦与する突然変異部位は、Mtb_S(配列番号1)若しくはMtb_R(配列番号2)の番号付けを参照として採用して、atpEタンパク質のアミノ酸14ないし34、とりわけ24ないし34、およびアミノ酸53ないし69、とりわけ56ないし67を指す。
【0067】
さらなる一態様において、該結合部位は、少なくとも、1個のCサブユニットのアミノ酸Ala24、Gly27、Phe53、Val57、Gly58、Glu61、Tyr64およびPhe65、ならびに1個のAサブユニットのアミノ酸Ser182、Leu183、Leu185およびArg186(Aサブユニットについて表3、4および5のコードSer206−Leu207−Leu209およびArg210を有する)を含んでなり、前記アミノ酸は、表3、4若しくは5のいずれかの原子座標、または、表3、4若しくは5の対応する原子座標の水素以外の原子位置に重ね合わせられる場合に約1.5Å未満の、好ましくは0.75Åを超えない水素以外の原子の二乗平均偏差を含んでなる相同な構造座標を有する。特定の一態様において、結合部位は、第一のCサブユニットのアミノ酸Ala21、Gly25;第二のCサブユニットのアミノ酸Ala24、Gly27、Phe53、Phe54、Val57、Gly58、Glu61、Tyr64、Phe65;第三のCサブユニットのアミノ酸Met17、Gly19、Gly20、Ala21、Ile22、Gly23、Ala24、Gly25、Ile26、Gly27、Asp28、Gly29、Ala31、Phe53、Thr56、Val57、Gly58、Leu59、Val60、Glu61、Ala62、Ala63/Pro63、Tyr64、Phe65、ならびにAサブユニットのアミノ酸Leu183、Leu185およびArg186を含んでなり;前記アミノ酸は、表3、4若しくは5のいずれかの原子座標、または、表3、4若しくは5の対応する原子座標の水素以外の原子位置に重ね合わせられる場合に約1.5Å未満の、好ましくは0.75Åを超えない水素以外の原子の二乗平均偏差を含んでなる相同な構造座標を有する。なおより具体的な一態様において、結合部位は、第一のCサブユニットのアミノ酸Ala21、Gly25;第二のCサブユニットのアミノ酸Ala24、Gly27、Phe53、Phe54、Val57、Gly58、Glu61、Tyr64、Phe65;第三のCサブユニットのアミノ酸Met17、Gly19、Gly20、Ala21、Ile22、Gly23、Ala24、Gly25、Ile26、Gly27、Asp28、Gly29、Ala31、Phe53、Thr56、Val57、Gly58、Leu59、Val60、Glu61、Ala62、Ala63/Pro63、Tyr64、Phe65、ならびにAサブユニットのアミノ酸Leu183、Leu185およびArg186よりなり;前記アミノ酸は表3、4若しくは5のいずれかの原子座標を有する。最も具体的な一態様において、結合部位は、第一のCサブユニットのアミノ酸Ala21、Gly25;第二のCサブユニットのアミノ酸Ala24、Gly27、Phe53、Phe54、Val57、Gly58、Glu61、Tyr64、Phe65;第三のCサブユニットのアミノ酸Met17、Gly19、Gly20、Ala21、Ile22、Gly23、Ala24、Gly25、I
le26、Gly27、Asp28、Gly29、Ala31、Phe53、Thr56、Val57、Gly58、Leu59、Val60、Glu61、Ala62、Ala63/Pro63、Tyr64、Phe65、ならびにAサブユニットのアミノ酸Leu183、Leu185およびArg186よりなり;前記アミノ酸は表3の原子座標を有する。
【0068】
従って、コンピュータによるスクリーニングプログラムにおいて、上で概説されたところの原子座標を使用してatpEタンパク質と相互作用する試験化合物の潜在能力を評価することが、本発明の一目的である。一態様において、本発明は、atpEタンパク質と相互作用する試験化合物の潜在能力の評価方法を提供し、前記方法は;−ATPアーゼのF0部分の結合部位の三次元構造を生成させるための分子モデル化技術;−コンピュータによる手段を使用して、試験化合物と結合部位の三次元構造の間のフィッティング操作を実施すること;および−前記フィッティング操作の結果を解析して結合部位の三次元構造との試験化合物の会合を定量化することを含んでなる。本発明のさらなる一態様において、結合部位の三次元構造は、表3、4若しくは5の原子座標、または、表3、4若しくは5の対応する原子座標の水素以外の原子位置に重ね合わせられる場合に約1.5Å未満の、好ましくは0.75Åを超えない水素以外の原子の二乗平均偏差を含んでなる相同な構造座標を使用して生成される。特定の一態様において、三次元構造は、表3、4若しくは5のいずれかのA鎖のアミノ酸Ala21、Gly25;表3、4若しくは5のいずれかのK鎖のアミノ酸Ala24、Gly27、Phe53、Phe54、Val57、Gly58、Glu61、Tyr64、Phe65;表3、4若しくは5のいずれかのL鎖のアミノ酸Met17、Gly19、Gly20、Ala21、Ile22、Gly23、Ala24、Gly25、Ile26、Gly27、Asp28、Gly29、Ala31、Phe53、Thr56、Val57、Gly58、Leu59、Val60、Glu61、Ala62、Ala63/Pro63、Tyr64、Phe65;ならびに表3、4若しくは5のいずれかのM鎖のアミノ酸Ser206、Leu207、Leu207およびArg210の原子座標を使用して生成される。
【0069】
このスクリーニングにおいて、こうした化合物の結合部位へのフィッティングの質は、形状の相補性若しくは推定される相互作用エネルギーのいずれかにより判断しうる(Meng,E.C.ら、J.Coma.Chem 13:505−524(1992))。
【0070】
結合部位の使用
本発明のatpEに結合するか、その機能的活性を促進するか若しくは阻害する化合物の設計は、一般に、2種の因子の考慮を必要とする。第一に、該化合物はatpEと物理的かつ構造的に会合することが可能でなければならない。atpEの化合物との会合において重要な非共有分子相互作用は、水素結合形成、ファンデルワールスおよび疎水性相互作用を包含する。第二に、化合物は、それがatpEと会合することを可能にするコンホメーションをとることが可能でなければならない。化合物のある部分がatpEとの会合に直接参画しなくてもよいとは言え、それらの部分はなお、該分子の全体的コンホメーションに影響しうる。これは、順に、結合親和性、治療的有効性、薬物様の質および効力に対する大きな影響を有しうる。こうしたコンホメーションの要件は、atpEの活性部位若しくは他の領域の全部若しくは一部分に関しての化学的実体若しくは化合物の全体的な三次元構造および幾何学的配置、またはatpEと直接相互作用する数種の化学的実体を含んでなる化合物の官能基間の間隔を包含する。
【0071】
atpEに対するリガンド若しくは他の化合物の潜在的な予測される阻害性のアゴニスト、アンタゴニスト若しくは結合の効果は、コンピュータモデル化技術の使用により、その実際の合成および試験の前に解析しうる。所定の化合物の理論的構造がそれとatpEの間の不十分な相互作用および会合を示唆する場合は、該化合物の合成および試験を未然
に防ぎうる。しかしながら、コンピュータモデル化が強い相互作用を示す場合には、該分子を合成しかつatpEと相互作用するその能力について試験しうる。この様式で、無効の化合物の合成を回避しうる。いくつかの場合には、モデル化で予測される不活性化合物を合成しかつその後試験してatpEの特定の領域と相互作用する化合物のSAR(構造活性相関)を作成する。当業者は、いくつかの方法の1つを使用して、化学的実体のフラグメント、化合物若しくは剤をatpEおよびより具体的にはatpEの個々の結合ポケットすなわち活性部位と会合するそれらの能力についてスクリーニングしうる。この方法は、例えば、atpE若しくはリガンドと複合体形成したatpEの原子座標に基づくコンピュータスクリーニングでの活性部位の目視検査により開始しうる。選択された化学的実体、化合物若しくは剤をその後、多様な幾何学的配置に配置しうるか、若しくはatpEの個々の結合ポケット内にドッキングしうる。ドッキングは、QuantaおよびSybylのようなソフトウェア、次いでCHARMMおよびAMBERのような標準的分子の力学上の力場(mechanics forcefield)を用いるエネルギー最小化および分子動力学を使用して達成しうる。
【0072】
特化されたコンピュータプログラムもまた化学的実体の選択の方法で補助しうる。これらは、限定されるものでないが:GRID(Goodford,P.J.、“A Computational Procedure for Determining Energetically Favorable Binding Sites on Biologically Important Macromolecules,”J.Med.Chem.28:849−857(1985)、オックスフォード大学、英国オックスフォードから入手可能);MCSS(Miranker,A.とM.Karplus、“Functionality Maps of Bioding Sites:A Multiple Copy Simultaneous Search Method.”Proteins:Structure,Function and Genetics 11:29−34(1991)、Molecular Simulations、マサチューセッツ州バーリントンから入手可能);AUTODOCK(Goodsell,D.S.とA.J.Olsen、“Automated Docking of Substrates to Proteins by Simulated Aneealing”Proteins:Structure,Function and Genetics 8:195−202(1990)、Scripps Research Institute、カリフォルニア州ラホヤから入手可能);およびDOCK(Kuntz,I.D.ら、“A Geometric Approach to Macromolecule−Ligand Interactions,”J.−Mol.Biol.161:269−288(1982)、カリフォルニア大学、カリフォルニア州サンフランシスコから入手可能)を挙げることができる。
【0073】
多様な官能基の特徴をもつプローブと巨大分子表面の間のありそうな相互作用部位を決定するプログラムであるGRIDのようなソフトウェアの使用は、表面部位を分析して類似の阻害するタンパク質若しくは化合物の構造を決定するのに使用される。プローブとしての分子上の適する阻害する基(例えばプロトン化された一級アミン)を用いるGRID計算を使用して、適するエネルギー等高線(contour)の水準の到達可能な位置の周囲の潜在的ホットスポットを同定する。DOCKプログラムを使用して、活性部位若しくはリガンド結合部位を解析しかつ相補的立体構造特性をもつリガンドを示唆しうる。
【0074】
適する化学的実体、化合物若しくは剤が一旦選択されれば、それらを単一のリガンド若しくは化合物若しくは阻害剤若しくは活性化物質に集成し得る。集成は、三次元像での相互に対するフラグメントの関係の目視検査により進行しうる。これに次いで、Quanta若しくはSybylのようなソフトウェアを使用して人的にモデルを構築しうる。
【0075】
個々の化学的実体、化合物若しくは剤の結合において補助するための有用なプログラムは、限定されるものでないが:CAVEAT(Bartlett,P.A.ら、“CAVEAT:A Program to Facilitate the Structure−Derived Design of Biologically Active Molecules.” Molecular Recognition in Chemical and Biological Problems、Special Pub.、Royal Chem.Soc.中、78、pp.82−196(1989));MACCS−3D(MDL Information Systems、カリフォルニア州サンリアンドロ、およびMartin,Y.C.、“3D Database Searching in Drug Design”、J.Med.Chem.35:2145−2154(1992)のような3Dデータベースシステム;ならびにHOOK(Molecular Simulations、マサチューセッツ州バーリントンから入手可能)を挙げることができる。
【0076】
ファルマコフォア仮説を検定しかつスクリーニングのための化合物を選択するために三次元データベースを検索するためのいくつかの方法論が利用可能である。これらはCAVEATプログラム(Baconら、J.Mol.Biol.225:849−858(1992))を包含する。例えば、CAVEATは、活性部位に既に配置されているいずれかの数の化学的断片を結合するための「スペーサー」として作用し得る環状化合物のデータベースを使用する。これは、当業者が、密接な結合に必要であることが既に知られているか若しくは疑われる断片を結合するための何百もの可能な方法を迅速に生成することを可能にする。上述されたとおり一度に1種の化学的実体を段階的様式でatpEの阻害剤活性化物質、アゴニスト若しくはアンタゴニストを構築するように進行する代わりに、こうした化合物は、空の結合部位を使用すること、または既知分子のいくつかの部分(1個若しくは複数)を場合によっては包含することのいずれかを使用して、全体としてすなわち「新規に」設計しうる。これらの方法は:LUDI(Bohm,H.−J.、“The
Computer Program LUDI:A New Method for the De Novo Design of Enzyme Inhinitors”,J.ComR.Aid.Molec.Design、6、pp.61−78(1992)、Biosym Technologies、カリフォルニア州サンディエゴから入手可能);LEGEND(Nishibata,Y.とA.Itai、Tetrahedron 47:8985(1991)、Molecular Simulations、マサチューセッツ州バーリントンから入手可能);およびLeapFrog(Tripos
Associates、ミズーリ州セントルイスから入手可能)を包含する。例えば、LUDIプログラムは、水素結合形成および疎水性フラグメントの双方が位置するべき相互作用部位の一覧を決定し得る。LUDIはその後、リンカーのライブラリーを使用して、4個までの異なる相互作用部位を断片に結合する。その後、――CH2−および――COO――のようなより小さい「結合」基を使用してこれらの断片を結合する。例えば、酵素DHFRについて、公知の阻害剤メトトレキセート中の重要な官能基の配置がLUDIにより再現された。RotsteinとMurcko、J.Med.Chem.36:1700−1710(1992)もまた参照されたい。
【0077】
他の分子モデル化技術もまた本発明で使用しうる。例えば、Cohen,N.C.ら、“Molecular Modeling Software and Methods
for Medicinal Chemistry、J.Med.Chem.33:883−894(1990)を参照されたい。Navia,M.A.とM.A.Murcko、“The Use of Structural Information in Drug Design,”Current Opinions in Structural Biology、2、pp.202−210(1992)もまた参照されたい。
【0078】
化合物が上の方法により一旦設計若しくは選択されれば、その化合物がatpEを結合若しくはそれと会合する親和性を、コンピュータによる評価および/若しくは該化合物を合成した後に生物学的活性を試験することにより試験かつ最適化しうる。阻害剤若しくは化合物は、全体的な結合エネルギーが類似である1種以上のコンホメーションでatpEと相互作用しうる。それらの場合に、結合の変形エネルギーは、遊離化合物のエネルギーと、該化合物がatpEに結合する場合に観察されるコンホメーションの平均エネルギーの間の差違であると解釈される。
【0079】
atpEと結合若しくは会合するとして設計若しくは選択される化合物は、その結合された状態でそれが好ましくはatpEとの反発性静電相互作用を欠くであろうようにさらにコンピュータにより最適化しうる。こうした非相補的(例えば静電的)相互作用は、反発性の電荷−電荷、双極子−双極子および電荷−双極子相互作用を包含する。とりわけ、阻害剤が結合されている場合の阻害剤とatpEの間の全部の静電的相互作用の総和は、好ましくは、結合のエンタルピーへの中立の若しくは好ましい貢献をなす。弱い結合化合物もまた、SARを決定するようにこれらの方法により設計することができる。例えば、米国特許出願第60/275,629号;同第60/331,235号;同第60/379,617号;および同第10/097,249号明細書を参照されたい。
【0080】
特定のコンピュータソフトウェアが、化合物の変形エネルギーおよび静電的相互作用を評価するのに当該技術分野で使用可能である。こうした用途のため設計されたプログラムの例は:Gaussian 92、revision C(M.J.Frisch、Gaussian,Inc.、ペンシルバニア州ピッツバーグ、COPYRGT 1992);AMBER,version 4.0(P.A.Kollman、カリフォルニア大学サンフランシスコ校、GOPYRGT 1994);QUANTA/CHARMM(Molecular Simulations,Inc.、マサチューセッツ州バーリントン、COPYRGT 1994);およびInsight II/Discover(Biosysm Technologies Inc.、カリフォルニア州サンディエゴ、COPYRGT 1994)を包含する。他のハードウェアシステムおよびソフトウェアパッケージは当業者に既知であろう。
【0081】
atpEと会合する化合物が上述されたとおり一旦最適に選択若しくは設計されれば、その後、その結合特性を改良若しくは改変するために、その原子若しくは側基のいくつかで置換を行いうる。一般に、初期置換は保存的であり、すなわち、置換基は元の基とほぼ同一の大きさ、形状、疎水性および電荷を有することができる。もちろん、コンホメーションを変えることが当該技術分野で既知の化合物は回避されうることが理解されるべきである。こうした置換された化合物をその後、詳細に上述された同一のコンピュータ法によりatpEへのフィッティングの効率について解析しうる。
【0082】
本発明はさらに、本明細書に記述される配列および/若しくは構造座標を含有するシステム、とりわけコンピュータに基づくシステムを提供する。こうした系は、atpE若しくはATPアーゼのF0部分の結合部位の構造決定および理論的ドラッグデザインを行うよう設計される。コンピュータに基づくシステムは、詳細に上述されたコンピュータ法のいずれかで本発明の配列および/若しくは構造座標を解析するのに使用されるハードウェア手段、ソフトウェア手段およびデータ記憶手段を指す。本発明のコンピュータに基づくシステムの最小ハードウェア手段は、中央演算処理装置(CPU)、入力手段、出力手段およびデータ記憶手段を含んでなる。当業者は、現在利用可能なコンピュータに基づくシステムのどれが本発明での使用に適するかを容易に認識し得る。
【0083】
従って、本明細書に記述される構造座標を含有するコンピュータで読み取り可能なデータ記憶媒体を提供することが、本発明の一目的である。本明細書で使用されるところの「
コンピュータで読み取り可能なデータ記憶媒体」は、コンピュータにより直接読み取り若しくはアクセスし得るいかなる媒体も指す。こうした培体は、限定されるものでないが:フロッピーディスク、ハードディスク記憶媒体および磁気テープのような磁気記憶媒体;光ディスク若しくはCD−ROMのような光学式記憶媒体;RAMおよびROMのような電気的記憶媒体;ならびに磁気/光学式記憶媒体のようなこれらの範疇のハイブリッドを挙げることができる。
【0084】
処置方法
上で既に挙げられたとおり、微生物に基づく感染症を伴う被験体の処置方法において、前述のスクリーニング方法のいずれかを使用して同定される化合物の使用を提供することもまた、本発明の一目的である。一般に、細菌性病原体はグラム陽性若しくはグラム陰性いずれかの病原体に分類しうる。グラム陽性およびグラム陰性双方の病原体に対する活性をもつ抗菌化合物は、一般に広範囲の活性を有するとみなされる。本発明の化合物は、グラム陽性および/若しくはグラム陰性の細菌性病原体に対し活性とみなされる。とりわけ、本化合物は、最低1種のグラム陽性細菌、好ましくは数種のグラム陽性細菌、より好ましくは1種若しくはそれ以上のグラム陽性細菌および/または1種若しくはそれ以上のグラム陰性細菌に対し活性である。
【0085】
グラム陽性およびグラム陰性の好気性および嫌気性細菌の例は、ブドウ球菌、例えば黄色ブドウ球菌(S.aureus);腸球菌、例えばE.フェーカリス(E.faecalis);連鎖球菌、例えば肺炎連鎖球菌(S.penaumoniae)、S.ミュータンス(S.mutans)、化膿性連鎖球菌(S.pyogens);バチルス属の細菌、例えば枯草菌(Bacillus subtilis)、リステリア属(Listeria)、例えばリステリア菌(Listeria monocytogenes);ヘモフィルス属(Haemophilus)、例えばインフルエンザ菌(H.influenza);モラクセラ属(Moraxella)、例えばM.カタラリス(M.chatarrhalis);シュードモナス属(Pseudomonas)、例えば緑膿菌(Pseudomonas aeruginosa);およびエシェリキア属(Escherichia)、例えば大腸菌(E.coli)を包含する。グラム陽性病原体、例えば、ブドウ球菌、腸球菌および連鎖球菌は、処置することが困難かつ一旦確立すれば例えば病院環境から根絶することが困難の双方である耐性株の発生によりとりわけ重要である。こうした株の例は、メチシリン耐性黄色ブドウ球菌(Staphylococcus aureus)(MRSA)、メチシリン耐性凝固酵素陰性ブドウ球菌(MRCNS)、ペニシリン耐性肺炎連鎖球菌(Streptococcus pneumoniae)および多剤耐性エンテロコッカス ファシウム(Enterococcu faecium)である。
【0086】
本発明の化合物は耐性細菌株に対する活性もまた示す。
【0087】
本発明の化合物は、生存率がF1F0ATP合成酵素の適正な機能に依存する細菌に対しとりわけ活性である。いずれかの論理により束縛されずに、本化合物の活性がF1F0ATP合成酵素の阻害、とりわけF1F0ATP合成酵素のF0複合体の阻害、より具体的にはF1F0ATP合成酵素のF0複合体のAサブユニットのArg186からCサブユニットのGlu61へのプロトンの移動の阻害に存し、細菌の細胞ATP濃度の枯渇により該細菌の死滅に至ることが教示される。前述のスクリーニング方法のいずれかを使用して同定される化合物は、グラム陽性細菌、より具体的にはミコバクテリウム、および最も具体的にはM.アフリカヌム(M.africanum)、トリ結核菌(M.avium)、ウシ結核菌(M.bovis)、ウシ結核菌−BCG(M.bovis−BCG)、カメ結核菌(M.chelonae)、M.フォルツイタム(M.fortuitum)、M.ゴルドネ(M.gordonae)、M.イントラセルラレ(M.intrac
ellulare)、M.カンサシイ(M.kansasii)、M.ミクロティ(M.microti)、M.スクロフラセウム(M.scrofulaceum)、パラ結核菌(M.paratuberculosis)、らい菌(M.leprea)、ヒト結核菌(M.tuberculosis)、M.ウルセランス(M.ulcerans)およびM.ラネ(M.ranae)により引き起こされる感染症に対しとりわけ活性である。
【0088】
上記若しくは下記で使用される場合はいつも、該化合物が細菌感染症を処置し得ることは、該化合物が1種若しくはそれ以上の細菌株への感染を処置し得ることを意味している。抗菌化合物としてのDARQ Jの使用に関してしかしながら使用される場合、抗菌薬は、1種若しくはそれ以上の細菌株への感染を処置し得る化合物であることを意味しているが、但し前記細菌株はミコバクテリウム以外である。
【0089】
本化合物により処置されうる細菌感染症は、例えば、中枢神経系の感染症、外耳感染症、急性中耳炎のような中耳の感染症、硬膜静脈洞の感染症、眼の感染症、歯、歯肉および粘膜の感染症のような口腔の感染症、上気道感染症、下気道感染症、泌尿生殖器感染症、胃腸感染症、産婦人科感染症、敗血症、骨および関節の感染症、皮膚および皮膚構造の感染症、細菌性心内膜炎、火傷、手術の抗菌的予防、ならびに癌化学療法を受領している患者若しくは臓器移植患者のような免疫抑制患者における抗菌的予防を包含する。
【0090】
本発明はさらに、atpEタンパク質と相互作用する剤を被験体に投与することを含んでなる、結核に苦しめられる被験体の処置方法を提供する。
【0091】
製薬学的組成物
本発明はさらに、細胞中でatpEタンパク質と相互作用する剤および製薬学的に許容できる担体を含んでなる製薬学的組成物を提供する。こうした剤は、製薬学的に許容できる担体若しくは希釈剤と一緒になって剤を含んでなる組成物に処方しうる。剤は、エステル若しくは塩のような、酸付加塩若しくは塩基金属塩、またはN若しくはSオキシドのような生理学的に機能的な誘導体の形態にあることができる。組成物はいかなる適する投与経路および手段のためにも処方しうる。製薬学的に許容できる担体若しくは希釈剤は、経口、直腸、鼻、吸入可能、局所(頬側および舌下を包含する)、膣若しくは非経口(皮下、筋肉内、静脈内、皮内、クモ膜下腔内および硬膜外を包含する)投与に適する製剤で使用されるものを包含する。担体若しくは希釈剤の選択はもちろん、剤およびその治療目的に依存しうる提案される投与経路に依存することができる。製剤は、便宜的には単位投薬形態物で提示されることができ、また、製薬学の技術分野で公知の方法のいずれによっても製造しうる。こうした方法は、有効成分を1種若しくはそれ以上の付属成分を構成する担体との連合にもたらす段階を包含する。一般に、製剤は、有効成分を液体担体若しくは微粉化した固体担体または双方との連合に均一かつ緊密にもたらすこと、およびその後必要な場合は生成物を造形することにより製造する。
【0092】
固体の組成物について、慣習的な非毒性の固体担体は、例えば、製薬学的等級のマンニトール、乳糖、セルロース、セルロース誘導体、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、滑石、ブドウ糖、ショ糖、炭酸マグネシウムなどを包含するものを使用しうる。上記で定義されるところの有効成分を、例えばポリアルキレングリコール、アセチル化トリグリセリドなどを担体として使用して坐剤として処方しうる。液体の製薬学的に投与可能な組成物は、例えば、例えば水、生理的食塩水、水性D−ブドウ糖、グリセロール、エタノールなどのような担体中に、上で定義されたところの有効成分および任意の製薬学的補助物質を溶解、分散などしてそれにより溶液若しくは懸濁液を形成することにより製造し得る。所望の場合は、投与されるべき製薬学的組成物は、少量の湿潤剤若しくは乳化剤、pH緩衝剤など、例えば酢酸ナトリウム、ソルビタンモノラウレート、酢酸トリエタノールアミンナトリウム、ソルビタンモノラウレート、トリエタノールアミ
ンオレエートなどのような非毒性の補助物質もまた含有しうる。こうした投薬形態物の実際の製造方法は当業者に既知であるか若しくは明らかであろう;例えば、Gennaroら、Remington’s Pharmaceutical Sciences、Mack Publishing Company、ペンシルバニア州イーストン、第18版、1990を参照されたい。
【0093】
組成物若しくは製剤は、いずれにしても、処置されている被験体の症状を緩和するのに有効な量の、ある量の有効成分(1種若しくは複数)を含有することができる。
【0094】
本化合物の正確な投薬量および投与の頻度は、当業者に公知であるとおり、使用される特定の化合物、処置されている特定の状態、処置されている状態の重症度、特定の患者の齢、重量、性、食餌、投与時間および全身の健康状態、投与様式、ならびに該個体が服用しうる他の医薬品に依存する。さらに、有効1日量は、処置される被験体の応答に依存して、かつ/若しくは本発明の化合物を処方する内科医の評価に依存して減少若しくは増大させうる。
【0095】
非毒性の担体から構成されるバランス(balance)を伴う0.25ないし95%の範囲の有効成分を含有する投薬形態物若しくは組成物を製造しうる。投与様式に依存して、製薬学的組成物は、好ましくは、0.05から99重量%まで、より好ましくは0.1から70重量%までの有効成分、および1から99.95重量%まで、より好ましくは30から99.9重量%までの製薬学的に許容できる担体を含むことができ、全部のパーセンテージは全組成物に基づく。
【0096】
経口投与のため、製薬学的に許容できる非毒性の組成物は、例えば製薬学的等級のマンニトール、乳糖、セルロース、セルロース誘導体、クロスカルメロースナトリウム、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、滑石、ブドウ糖、ショ糖、マグネシウム、炭酸塩などのような通常使用される賦形剤のいずれかの組み込みにより形成される。こうした組成物は溶液、懸濁剤、錠剤、丸剤、カプセル剤、散剤、除放製剤などの形態を取る。こうした組成物は1%〜95%の有効成分、より好ましくは2〜50%、最も好ましくは5〜8%を含有しうる。
【0097】
非経口投与は、一般に皮下、筋肉内若しくは静脈内いずれかでの注入を特徴とする。注入可能物は、慣習的形態で、液体の溶液若しくは懸濁液、注入前の液体中の溶液若しくは懸濁液に適する固体の形態、または乳剤のいずれかとして製造し得る。適する賦形剤は、例えば水、生理的食塩水、D−ブドウ糖、グリセロール、エタノールなどである。加えて、所望の場合は、投与されるべき製薬学的組成物は、少量の例えば酢酸ナトリウム、ソルビタンモノラウレート、トリエタノールアミンオレエート、酢酸トリエタノールアミンナトリウムなどのような、湿潤剤若しくは乳化剤、pH緩衝剤などのような非毒性の補助物質もまた含有しうる。
【0098】
こうした非経口組成物中に含有される有効成分のパーセンテージは、その特定の性質、ならびに化合物の活性および被験体の必要性に高度に依存する。しかしながら、溶液中0.1%ないし10%という有効成分のパーセンテージが使用可能であり、そして、該組成物が上のパーセンテージに後に希釈されることができる固体である場合はより高いことができる。好ましくは、組成物は溶液中に0.2〜2%の有効成分を含むことができる。
【0099】
最後に、本発明は、(a)製薬学的剤が細胞中でatpEタンパク質と相互作用し、かつ(b)包装が被験体における細菌感染症を処置するための該剤の使用を示すラベルを含んでなる、包装および製薬学的剤を含んでなる、とりわけ、抗ミコバクテリウム医薬品としての製品を提供する。
【0100】
本記述を通じ、「標準的方法」、「標準的プロトコル」および「標準的手順」という用語は、分子生物学技術の文脈で使用される場合に、Current Protocols
in Molecular Bioloty、編者F.Ausubelら、John Wiley and Sons,Inc.1994、若しくはSambrook,J.、Fritsch,E.F.およびManiatis,T.、Molecular Cloning:A laboratory manual、第2版、Cold Spring
Harbor Laboratory Press、ニューヨーク州コールドスプリングハーバー、1989のような通常の実験室手引書に見出されるプロトコルおよび手順として理解されるべきである。
【0101】
本発明は、後に続く実験の詳細を参照してより良好に理解されるであろうが、しかし、当業者は、これらは下に続く請求の範囲により完全に記述されるところの本発明を具体的に説明するのみであることを容易に認識するであろう。加えて、本出願を通じ多様な刊行物が引用される。これらの刊行物の開示は、本発明が関する従来技術をより完全に記述するために、本出願に引用することによりここに組み込まれる。
【図面の簡単な説明】
【0102】
【図1】下でJ若しくはDARQ Jともまた称されるR207910の絶対配置。
【図2】ヒト結核菌(M.tuberculosis)およびスメグマ菌(M.smegmatis)変異体のatpEタンパク質配列アライメント。Mtb_S:薬物感受性株ヒト結核菌(M.tuberculosis)H37RvのatpE(1−81)。受託番号:Swiss−Prot Q10598(配列番号1)。Mtb_R:薬物耐性株ヒト結核菌(M.tuberculosis)BK12のatpE(1−81)(配列番号2)。Msm_S:薬物感受性株スメグマ菌(M.smegmatis)のatpE(1−86)。ゲノム研究所(Institute for Genome Research)により得られた配列(配列番号3)。Msm_R09(配列番号4)およびR10(配列番号5):薬物耐性株スメグマ菌(M.smegmatis)のatpE(1−86)。社内で得られた配列。Human:ヒト(Homo sapiens)、ATP5G3(66−142)。受託番号:Ensembl ENSP00000284727。上の番号付け:ヒト結核菌(M.tuberculosis)およびスメグマ菌(M.smegmatis)のatpE。下の番号付け:ヒト(H.sapiens)ATP5G3(66−142)陰影はBLOSUM62マトリックスを使用するアミノ酸類似性を示す(黒=高、灰色=中)。矢印は耐性株で観察される点突然変異の位置を示す。
【図3】DARQ J、イソニアジドおよびDCCDの存在下でのヒト結核菌(M.tuberculosis)の全細胞ATP測定。野性型ヒト結核菌(M.tuberculosis)およびDARQ J変異体ヒト結核菌(M.tuberculosis)双方で526nmで測定したオキシルシフェリンの相対発光単位。
【図4】一緒になってDARQ J化合物の結合部位を形成する3個のCサブユニット(A鎖、K鎖およびL鎖)ならびにAサブユニット(M鎖)のリボン表示。
【0103】
実験
スメグマ菌(Mycobacterium smegmatis)を代理物として使用して、われわれは、数種のミコバクテリウムに対する強力なin vitro活性をもつ一連のDARQを発見した(11)。今日まで、DARQ系列の20種の分子がヒト結核菌(Mycobacterium tuberculosis)H37Rvに対し0.5μg/mlより下の最小阻害濃度(MIC)を有し、また、これらの3種について、抗ミコバクテリウム活性がin vivoマウスモデルで確認された。
【0104】
構造的におよび機構的に、DARQは、フルオロキノロン(メトキシキノロンを包含す
る)、ならびにメフロキンならびにそのアナログ4−メチルキノリンおよび4−キノリルヒドラゾンを包含する他のキノロン分類の双方と非常に異なる(12−16)。DARQと他のキノロン若しくはキノリン分類の間の主要な構造上の差違の1つは、DARQ分類により担持される官能性化された側(3’)鎖の特異性である。加えて、既存の化学分類とのミコバクテリウムの交差耐性の欠如が異なる作用機序を指摘する。
【0105】
下でJ若しくはDARQ Jと称されるDARQのリード化合物(図1)が、in vitroで独特の範囲の強力かつ選択的な抗ミコバクテリウム活性を有することが見出された(表1)。実験室株H37Rvおよび6種の完全に感受性の単離物について得られたMICの中央値は、リファンピンについて1.00μg/mlに対し0.060μg/mlであった。Jは、第一列のTB薬リファンピン、ストレプトマイシン、エタンブトールおよびピラジナミド;ならびに第二列のTB薬モキシフロキサシンに対し耐性のヒト結核菌(M.tuberculosis)臨床単離物に対する類似のin vitro有効性を示した。イソニアジドに対し耐性の8種の臨床単離物について、MICの中央値は0.010μg/mlであった。現在使用されている抗TB薬との交差耐性の欠如は、JがMDR−TB株に対する活性を保持しうることを示唆した。事実、BACTECTM培養系を使用して、細菌の増殖の明瞭な濃度依存性の阻害が、MDR−TB株を固定濃度のJに曝露した場合に見られた。MDR−TBの30種の単離物のうち、13種(43%)が0.100μg/mlのJに対し感受性であることが見出され、また、17種(57%)が0.010μg/mlのJに対し感受性であった。類似の高い程度の感受性(0.010μg/mlより下のMIC)は、BACTECTM系を使用して試験した場合に10種の付加的な完全に薬物感受性の株の1種のみについて見られた一方、全部の株が0.100μg/mlのJに対し感受性であった。
【0106】
強力な活性は、ウシ結核菌(Mycobacterium bovis)およびミコバクテリウム カンサシイ(Mycobacterium kansasii)を包含する他のミコバクテリウム種、ならびに、トリ結核菌(Mycobacterium avium)群(MAC)、カメ結核菌(Mycobacterium abcessus)、ミコバクテリウム フォルツイタム(Mycobacterium fortuitum)およびミコバクテリウム マリナム(Mycobacterium marinum)のような、多くの他の抗TB薬に対し天然に耐性かつ日和見感染症に関与する種に対してもまた示された(表1)。
【0107】
驚くべきことに、Jの活性はミコバクテリウムに特異的であるようであった。Jは、コリネバクテリウム属(Corynebacterium)(MIC 4.00μg/ml)およびノカルジア属(Nocardia)(MIC>4.00μg/ml)のようなミコバクテリウムに近い種に対しほとんど活性でなく、また、グラム陽性の肺炎連鎖球菌(Streptococcus pneumoniae)、黄色ブドウ球菌(Staphylococcus aureus)(メシチリン耐性株(MIC>32μg/ml)を包含する)およびエンテロコッカス フェーカリス(Enterococcus faecalis)、若しくはグラム陰性の大腸菌(Escherichia coli)、インフルエンザ菌(Haemophilus influenzae)およびヘリコバクター
ピロリ(Helicobacter pylori)を包含する他の生物体に対し活性でなかった。対数期増殖にあるヒト結核菌(M.tuberculosis)の100×MICのJの濃度への曝露は、12日後に細菌数の103対数の減少をもたらし、Jがin vitroで殺菌活性を有することを示した。静止期の結核桿菌に対するJの影響は未だ研究されていない。
【0108】
変異体の単離、交差耐性および想定される薬物標的
ミコバクテリウム耐性を検討することにより、われわれは分子薬物標的を同定しかつ作
用機序を提案することを目的とした。ヒト結核菌(M.tuberculosis)およびスメグマ菌(M.smegmatis)の耐性変異体を、
−ミコバクテリウム中の耐性変異体の比率を定量化する(対照としてリファンピンを用いて)
−耐性変異体の耐性パターン(キノロンに対する交差/非交差耐性を包含する)を評価する
−作用機序を検討する
ため、阻害濃度のJでのin vitro選択により導き出した。
【0109】
選択実験から、Jに対する低下された感受性を伴う変異体の比率は、ヒト結核菌(M.tuberculosis)およびスメグマ菌(M.smegmatis)についてそれぞれMIC×4で5×10−7および2×10−8、ならびにMIC×8で5×10−8および1×10−8であった(サポーティングオンラインテキスト)。ヒト結核菌(M.tuberculosis)の場合、これらの比率はリファンピンに対し耐性の変異体のもの(10−7ないし10−8)に匹敵し、そして、Jに対する天然に存在する耐性がまれであることを示す。加えて、Jに対し耐性のヒト結核菌(M.tuberculosis)株の感受性は、抗TB薬イソニアジド、リファンピン、ストレプトマイシン、アミカシン、エタンブトールおよびモキシフロキサシンに対し不変のままであった。Jに対する低下された感受性を伴うヒト結核菌(M.tuberculosis)およびスメグマ菌(M.smegmatis)変異体のさらなる分析は、キノロン耐性が典型的に発生する配列すなわちDNAジャイレース領域のgyrAおよびgyrBに突然変異が存在しなかったことを示した。これは、Jの分子標的がフルオロキノロンのものと異なることを確認する。
【0110】
Jの分子標的を決定しかつ作用機序を推察することへの1アプローチは、ヒト結核菌(M.tuberculosis)およびスメグマ菌(M.smegmatis)の感受性および耐性株で耐性を賦与する突然変異を同定かつ比較することである。耐性のヒト結核菌(M.tuberculosis)株BK12ならびに2種の耐性のスメグマ菌(M.smegmatis)株R09およびR10、ならびに親スメグマ菌(M.smegmatis)のゲノムを完了近くまで配列決定した。われわれは、ヒト結核菌(M.tuberculosis)およびスメグマ菌(M.smegmatis)の感受性および耐性株のゲノム配列の比較分析により、耐性を賦与する突然変異を同定した(図2)。われわれは、対応する親野性型に対し全3種の独立の変異体中で影響を及ぼされた唯一の遺伝子がatpE(ATP合成酵素のF0サブユニットの一部)をコードすることを示した。これは、atpEが変異体株でのJに対する耐性の原因であることを示唆し、Jが新たなヒト結核菌(M.tuberculosis)の標的すなわちATP合成酵素のプロトンポンプを阻害することを示す。
【0111】
変異体atpE遺伝子がJに対する耐性の原因であること、および直接推測によりatpE遺伝子産物がミコバクテリウムでのJの標的であることを示すため、補完的研究を実施した。ATP合成酵素のオペロンの全遺伝子が協調された方法で発現されなければならない、すなわち、F0部分をコードする全遺伝子が同一の場所から発現されなければならないことが既知であるという事実を考え、われわれは、該オペロンのF0部分を耐性のスメグマ菌(M.smegmatis)株(D32V)から増幅し、そしてPCR過程により付加的な突然変異を獲得しなかったクローンを選択した。野性型スメグマ菌(M.smegmatis)を、かように選択した変異体F0フラグメントを含有するプラスミドで形質転換した。これは、細胞を、耐性株のスメグマ菌(M.smegmatis)R09(D32V)のものと実際上同一のMICを伴い、Jに対し耐性にした。加えて、該プラスミドをこれらの形質転換体から再単離しかつatpE遺伝子を配列決定した場合に、それが変異体アレル(D32V)を残存していたことが示された。
【0112】
ヒト結核菌(M.tuberculosis)でのATP産生に対するDARQ Jの実際の影響を、RocheのATP生物発光ルシフェラーゼアッセイキットHS IIを使用してミコバクテリウム中に存在する全細胞ATPに対するJの影響を測定することにより、さらに示した。このアッセイは、526nmで測定し得るD−ルシフェリンのオキシルシフェリンへのATPに駆動される変換に基づく。
【0113】
【化1】
【0114】
簡潔には、全ATPに対するDARQ Jの影響を野性型ヒト結核菌(M.tuberculosis)および変異体株双方で試験した。ATP合成酵素の公知の阻害剤であるDCCDを陽性対照として使用し、また、ある種の細胞壁成分の生合成の阻害剤であるがしかしATP産生に対し影響を有しないイソニアジドを陰性対照として使用した。
【0115】
図3に見ることができるとおり、DARQ Jでの野性型ヒト結核菌(M.tuberculosis)の処理は、これらの細菌中でのATP産生の用量依存性の減少に至る。対照的に、イソニアジドはATP産生に対する影響を有しない。既に上述されたとおり、これらの細菌を高濃度のDARQ Jに曝露することは、ヒト結核菌(M.tuberculosis)のジアリールキノリン耐性変異体を生じさせた。これらの耐性ヒト結核菌(M.tuberculosis)をDARQ Jで処理した場合、これらの細菌は、この化合物の最小阻害濃度(MIC)の100倍ででさえATP産生のいかなる減少も示さなかった。対照的に、DCCDはこれらの桿菌中でのATP産生を阻害することが可能であり、DARQ JおよびDCCDがATP合成において異なる結合ポケットを有することを示唆した。
【0116】
DARQ J結合領域のコンピュータモデル化および同定
DCCDおよびDARQ Jの異なる作用様式をさらに検討するため、野性型およびDARQ J変異体双方のヒト結核菌(M.tuberculosis)のATP合成酵素のコンピュータ生成される3Dモデルを生成した。表4および5に提供される原子座標は、公表されたアミノ酸配列P63691およびAJ865377の3D構造のモデルを作成することにより計算した。実際のDARQ J結合部位は、AおよびCサブユニットの接触領域、より具体的には、表3、4若しくは5で称されるところのAサブユニットのアミノ酸「Arg210」およびCサブユニットの「Glu61」周辺に位置することが見出された。これは、上述されたヒト結核菌(M.tuberculosis)およびスメグマ菌(M.smegmatis)の感受性および耐性株での耐性を賦与する突然変異のスクリーニングについて見られた結果とよく一致する。
【0117】
該モデルは、ATPアーゼ構造のAおよびCヘリックスの相対配置の最適化、ならびに最小のコンピュータ化した内部株(internal strain)に対するアミノ酸バックボーンおよび側鎖の幾何学的配置に基づく。該配置は、異なる生物体[E−Coli PDBエントリコード1C17−V.K.RastogiとM.E.Girvin、Nature、402、263−268(1999)]の以前に公表された全般的らせん配置から出発する多数の分子動力学シミュレーション周期および分子力学の緩和によって得た。分子動力学および配置緩和は双方ともMMFF94に基づく力場のパラメータ化[Halgren,T.A.(1996)、J.Comput.Chem.、17、490
−519]を用いて実施したが、しかし、いずれかの従来技術の分子動力学ソフトウェア[Berendsen,H.J.C.、van der Spoel,D.とvan Drunen,R.、Comp.Phys.Comm.91(1995)、43−56;Lindahl,E.、Hess,B.およびvan der Spoel,D.、J.Mol.Mod.7(2001)306−317.]、次いで適する配置最適化[J.W.PonderとF.M.Richards、J.Comput.Chem.、8、1016−1024(1987)]を使用し得た。
【0118】
表3、4および5のコンピュータ化した座標は、提案された阻害様式、および生物学的アッセイにおける点突然変異を誘導する耐性の発生に基づき、これらの酵素中のMTB ATPアーゼ活性の阻害に関連すると考えられる領域の予測された構造の一部(30オングストロームの半径量を伴う)を含んでなる。
【0119】
考察
DARQ Jは、参照化合物のMICに等しいか若しくはそれより小さいMICをもつ、抗TB薬の新たな化学的分類の1メンバーである。そのスペクトルは、ヒトで重要な異型の種;MAC、M.カンサシイ(M.kansasii)ならびに迅速増殖体M.フォルツイタム(M.fortuitum)およびカメ結核菌(M.abscessus)を包含するミコバクテリウムに対するその特異性において独特である。この抗ミコバクテリウム特異的スペクトルは、MACに対する活性を有しないイソニアジドのものと異なる。Jの臨床使用はTBおよびミコバクテリウム感染症の処置に高度に標的を定めることができる。ミコバクテリウム以外を阻害するJの不能は、より広範なスペクトルをもつ抗生物質と比較した場合に、他の細菌種で発生するより小さい選択圧および耐性のより低いリスクになるはずである(9)。
【0120】
Jの標的および作用機序は他の抗TB薬のものと異なる。多様な細菌のATP合成酵素および真核生物のATP合成酵素、ならびにとりわけATPアーゼ複合体のF0サブユニットのC鎖の配列の比較は、野性型および変異体のヒト結核菌(M.tuberculosis)のATP合成酵素の3Dモデル化と一緒になって、抗菌スペクトルの特異性、およびより少ない程度まで安全性プロファイルに理論的根拠を提供する。
【0121】
構築されたミコバクテリウム属(Mycobacterium)のATPアーゼモデルで実施される動力学的研究は、これらの構造中で空洞(表3の原子座標による結合部位)がAおよびCサブユニットの接触領域(Aサブユニットのアミノ酸「Arg210」およびCサブユニットの「Glu61」周辺)に存在することを示す。表4および5は、この部位を取り囲む原子の2種の研究されたバリアントの座標、およびそれらの平均位置を提供する。DARQ J阻害剤は、これら2アミノ酸が相互作用することを禁止することにより、これら2アミノ酸を伴う通常のプロトン移動段階を妨害することが可能である。DARQ Jの立体特異性は予測された結合部位の非対称性から理解され得;活性のキラル鏡像異性体がこの空洞を最適に提供し、該化合物の他の形態およびATPアーゼのバリアントがより少なく良好に一致する。
【0122】
われわれがここで導き出した結合部位は、ATPアーゼ系のDCCDと完全に異なる部分にある(DARQは酵素の膜部分にあり、DCCD結合は細胞の内側からおよそ90オングストローム離れて発生する;C.Gibbons、M.G.Montgomery、A.G.W.Leslie、J.E.Walker、Nat.Struct.Biol.、7、1055(2000)に公表されたウシATPアーゼ結晶「1E79」のPDB構造に基づく)。従って、MTB ATPアーゼのDCCD型の阻害と潜在的に関与し得る原子は、結合部位(酵素の膜部分にのみちょうどわたる)の座標の表に列挙される領域中になく、そして、相互に、DARQ Jの阻害様式に関与する酵素のこの部分は、公表さ
れた「1E79」構造(細胞内部分のみ示す)に存在しない。結合部位のこの差違は、in vitro ATP産生アッセイで観察されたヒト結核菌(M.tuberculosis)の異なる応答を説明しうる。
【0123】
上にもかかわらず、ミトコンドリアのATPアーゼでDCCDを用いるより古い研究は、例えばSebald W、Machleidt W、Wachter E.、Proc
Natl Acad Sci U.S.A.1980年2月;77(2):785−789により、該酵素のF0領域の親油性環境の1酸性アミノ酸の周囲に位置する別の結合部位を示唆している。ミトコンドリアのATPアーゼのこの結合位置は、ミコバクテリウム属(Mycobacterium)種についてここで記述されるものに類似とみなし得る。
【0124】
同時に、新たな機構を標的とすることは、利用可能な処置に対する耐性突然変異を伴う現在循環しているTB株がJに対し交差耐性でないことを確実にする。Jが、MDR−TB単離物に対し、なお、広範な4種の薬物耐性をもつものに対して、ヒト結核菌(M.tuberculosis)の通常の野性型汎感受性株に対すると少なくとも同じくらい高い抗菌効果を有することが、われわれのin vitro研究から明らかである。この観察結果は、既存の抗TB薬物との交差耐性が存在しないことをそれが明瞭に示すために、重要である。ATPアーゼの膜部分の結合ポケットのさらなる同定を考えれば、本研究の結果は、新たな抗菌化合物、とりわけこれらの生物体中でのATP合成を標的とする抗ミコバクテリウム化合物のさらなる開発を可能にするであろう。
【0125】
【表1】
【0126】
【表2】
【0127】
【表3】
【0128】
【表4】
【0129】
【表5】
【0130】
【表6】
【0131】
【表7】
【0132】
【表8】
【0133】
【表9】
【0134】
【表10】
【0135】
【表11】
【0136】
【表12】
【0137】
【表13】
【0138】
【表14】
【0139】
【表15】
【0140】
【表16】
【0141】
【表17】
【0142】
【表18】
【0143】
【表19】
【0144】
【表20】
【0145】
【表21】
【0146】
【表22】
【0147】
【表23】
【0148】
【表24】
【0149】
【表25】
【0150】
【表26】
【0151】
【表27】
【0152】
【表28】
【0153】
【表29】
【0154】
【表30】
【0155】
【表31】
【0156】
【表32】
【0157】
【表33】
【0158】
【表34】
【0159】
【表35】
【0160】
【表36】
【0161】
【表37】
【0162】
【表38】
【0163】
【表39】
【0164】
【表40】
【0165】
【表41】
【0166】
【表42】
【0167】
【表43】
【0168】
【表44】
【0169】
【表45】
【0170】
【表46】
【0171】
【表47】
【特許請求の範囲】
【請求項1】
アミノ酸14ないし34またはアミノ酸54ないし69のいずれか一の位置に少なくとも1つの点突然変異を含む配列番号1およびアミノ酸20ないし40またはアミノ酸60ないし75のいずれか一の位置に少なくとも点突然変異を含む配列番号3から選ばれるポリペプチド配列を含んでなる単離された変異atpEタンパク質。
【請求項2】
アミノ酸28またはアミノ酸63に位置する少なくとも点突然変異を含む配列番号1からなる群より選ばれるポリペプチド配列を含んでなる請求項1に記載の単離された変異atpEタンパク質。
【請求項3】
bu配列番号2に示されるMtb_R、配列番号4に示されるMsm_R09および配列番号5に示されるMsm_R10から選ばれる請求項1または2に記載の単離された変異atpEタンパク質。
【請求項4】
ATPアーゼのF0部分と相互作用する化合物、および抗菌化合物としてのそれらの潜在能力の同定方法における、請求項1ないし3のいずれか1つに記載のタンパク質の使用。
【請求項5】
atpEタンパク質と相互作用する試験化合物の潜在能力の評価方法であって、
(a)請求項1ないし3のいずれかの1つに記載のatpEタンパク質の結合部位の三次元構造を生成させるための分子モデル化技術、
(b)計算手段を使用して試験化合物と該結合部位の三次元構造の間のフィッティング操作を実施すること、および
(c)前記フィッティング操作の結果を解析して、試験化合物の該結合部位の三次元構造との関連性を定量化すること
を含んでなる、上記方法。
【請求項6】
三次元構造が、表3、4若しくは5のいずれかの原子座標、または1.5Å未満の水素以外の原子の二乗平均偏差を含んでなる相同な構造座標を使用して、1個のCサブユニットの少なくともアミノ酸Ala24、Gly27、Phe53、Val57、Gly58、Glu61、Tyr64およびPhe65、ならびに1個のAサブユニットのアミノ酸Ser182、Leu183、Leu185およびArg186を伴い生成される、請求項5に記載の方法。
【請求項7】
三次元構造が、表3、4若しくは5のいずれかのA鎖のアミノ酸Ala21、Gly25;表3、4若しくは5のいずれかのK鎖のアミノ酸Ala24、Gly27、Phe53、Phe54、Val57、Gly58、Glu61、Tyr64、Phe65:表3、4若しくは5のいずれかのL鎖のアミノ酸Met17、Gly19、Gly20、Ala21、Ile22、Gly23、Ala24、Gly25、Ile26、Gly27、Asp28、Gly29、Ala31、Phe53、Thr56、Val57、Gly58、Leu59、Val60、Glu61、Ala62、Ala63/Pro63、Tyr64、Phe65;ならびに表3、4若しくは5のいずれかのM鎖のアミノ酸Ser182、Leu183、Leu185およびArg186の原子座標を使用して生成される、請求項5に記載の方法。
【請求項8】
請求項1ないし3のいずれか1つに記載のatpEタンパク質と耐性を賦与する突然変異部位において相互作用する化合物を活性成分とする微生物に基づく感染症を伴う被験体の処置用の製薬学的製剤。
【請求項9】
耐性を賦与する突然変異部位が、配列番号1に示されるMtb_S若しくは配列番号2に示されるMtb_Rの番号付けを参照として採用して、atpEタンパク質のアミノ酸14ないし34およびアミノ酸53ないし69を指す、請求項8に記載の製薬学的製剤。
【請求項10】
表6若しくは7のいずれかのatpEタンパク質のアミノ酸14−34若しくは54−69の構造座標、または1.5Å未満の水素以外の原子の二乗平均偏差を含んでなる相同な構造座標によって規定されるATPaseのF0部分におえる結合部位を含んでなる単離、精製されたポリペプチド。
【請求項11】
結合部位が、Cサブユニットの少なくともアミノ酸Ala24、Gly27、Phe53、Val57、Gly58、Glu61、Tyr64およびPhe65、ならびに1個のAサブユニットのアミノ酸Ser182、Leu183、Leu185、Arg186を含んでなる、請求項10記載の単離、精製されたポリペプチド。
【請求項12】
請求項10または11に記載のポリペプチドの結合部位と相互作用する化合物を活性成分として含んでなる微生物に基づく感染症を伴う被験体の処置用の製薬学的製剤。
【請求項13】
請求項5ないし7のいずれかの1に記載の方法を使用して同定された化合物を活性成分として含んでなる微生物に基づく感染症を伴う被験体の処置用の製薬学的製剤。
【請求項14】
感染症がグラム陽性細菌により引き起こされる、請求項13に記載の製薬学的製剤。
【請求項15】
感染症がミコバクテリウムにより引き起こされる、請求項13に記載の製薬学的製剤。
【請求項16】
感染症がM.アフリカヌム(M.africanum)、トリ結核菌(M.avium)、ウシ結核菌(M.bovis)、ウシ結核菌−BCG(M.bovis−BCG)、カメ結核菌(M.chelonae)、M.フォルツイタム(M.fortuitum)、M.ゴルドネ(M.gordonae)、M.イントラセルラレ(M.intracellulare)、M.カンサシイ(M.kansasii)、M.ミクロティ(M.microti)、M.スクロフラセウム(M.scrofulaceum)、パラ結核菌(M.paratuberculosis)、らい菌(M.leprea)、ヒト結核菌(M.tuberculosis)、M.ウルセランス(M.ulcerans)およびM.ラネ(M.ranae)からなる群より選択される微生物により引き起こされる、請求項13に記載の製薬学的製剤。
【請求項17】
請求項13に記載の製薬学的製剤であって、中枢神経系の感染症、外耳感染症、急性中耳炎のような中耳の感染症、硬膜静脈洞の感染症、眼の感染症、歯、歯肉および粘膜の感染症のような口腔の感染症、上気道感染症、下気道感染症、泌尿生殖器感染症、胃腸感染症、産婦人科感染症、敗血症、骨および関節の感染症、皮膚および皮膚構造の感染症、細菌性心内膜炎、火傷、手術の抗菌的予防、ならびに癌化学療法を受領している患者若しくは臓器移植患者のような免疫抑制患者における抗菌的予防を処置するための、製薬学的製剤。
【請求項18】
請求項1ないし3のいずれかの1つに記載の単離された変異atpEタンパク質をコードする単離された核酸分子。
【請求項19】
請求項18に記載の核酸分子を含んでなるベクター。
【請求項20】
請求項19に記載のベクターを担持する宿主細胞。
【請求項21】
抗菌化合物の同定方法であって、
a)atpEタンパク質を発現する細胞を、生理学的条件下で試験化合物と接触させる工程、および
b)該試験化合物がatpEタンパク質と相互作用するかどうかを決定する工程を含んでなり、かつ、atpEタンパク質が請求項1ないし3のいずれかの1つに記載の変異atpEタンパク質である、ことを特徴とする上記方法。
【請求項22】
単離されたatpEタンパク質に試験化合物が結合するかどうかの同定方法であって、a)atpEタンパク質を発現する細胞(こうした細胞は前記atpEタンパク質を通常は発現しない)を、atpEタンパク質を結合することが既知の化合物の存在および非存在下で試験化合物と接触させる工程、
b)atpEタンパク質に結合することが既知の化合物を参照として使用して、atpEタンパク質への該試験化合物の結合を測定する工程を含んでなり、かつ、atpEタンパク質が請求項10または11に記載のポリペプチドを含む変異atpEタンパク質である、ことを特徴とする上記方法。
【請求項23】
atpEタンパク質に結合することが既知の化合物が、検出可能に標識されており、図1に記載された化合物からなる、請求項22に記載の方法。
【請求項24】
工程a)が、atpEタンパク質を含む細胞組成物を、atpEタンパク質を結合することが既知の化合物の存在および非存在下で試験化合物と接触させることよりなる、請求項22に記載の方法。
【請求項25】
細胞組成物が、請求項20に記載の細胞から得られる膜調製物よりなる、請求項24に記載の方法。
【請求項26】
請求項1ないし3のいずれか1つに記載のatpEタンパク質と相互作用する試験化合物の潜在能力の評価方法であって、
a)分子モデル化技術を使用して請求項1ないし3のいずれか1つに記載のatpEタンパク質の三次元構造を明確に表すこと、
b)計算手段を使用して、試験化合物と請求項1ないし3のいずれか1つに記載のatpEタンパク質の三次元構造の間のフィッティング操作を実施すること、および
c)前記フィッティング操作の結果を解析して、請求項1ないし3のいずれか1つに記載のatpEタンパク質の三次元構造との試験化合物の会合を定量化すること
を含んでなる、上記方法。
【請求項27】
atpEタンパク質の三次元構造が、大腸菌(E.coli)(タンパク質データベース1Q01)のIle28、Glu61およびIle63の原子座標+/−10Åを超えない前記アミノ酸のバックボーン原子の二乗平均偏差を使用して生成される、請求項26に記載の方法。
【請求項28】
atpEタンパク質の三次元構造が、表6若しくは7の原子座標+/−1.5Åを超えない前記アミノ酸のバックボーン原子の二乗平均偏差を使用して生成される、請求項26に記載の方法。
【請求項1】
アミノ酸14ないし34またはアミノ酸54ないし69のいずれか一の位置に少なくとも1つの点突然変異を含む配列番号1およびアミノ酸20ないし40またはアミノ酸60ないし75のいずれか一の位置に少なくとも点突然変異を含む配列番号3から選ばれるポリペプチド配列を含んでなる単離された変異atpEタンパク質。
【請求項2】
アミノ酸28またはアミノ酸63に位置する少なくとも点突然変異を含む配列番号1からなる群より選ばれるポリペプチド配列を含んでなる請求項1に記載の単離された変異atpEタンパク質。
【請求項3】
bu配列番号2に示されるMtb_R、配列番号4に示されるMsm_R09および配列番号5に示されるMsm_R10から選ばれる請求項1または2に記載の単離された変異atpEタンパク質。
【請求項4】
ATPアーゼのF0部分と相互作用する化合物、および抗菌化合物としてのそれらの潜在能力の同定方法における、請求項1ないし3のいずれか1つに記載のタンパク質の使用。
【請求項5】
atpEタンパク質と相互作用する試験化合物の潜在能力の評価方法であって、
(a)請求項1ないし3のいずれかの1つに記載のatpEタンパク質の結合部位の三次元構造を生成させるための分子モデル化技術、
(b)計算手段を使用して試験化合物と該結合部位の三次元構造の間のフィッティング操作を実施すること、および
(c)前記フィッティング操作の結果を解析して、試験化合物の該結合部位の三次元構造との関連性を定量化すること
を含んでなる、上記方法。
【請求項6】
三次元構造が、表3、4若しくは5のいずれかの原子座標、または1.5Å未満の水素以外の原子の二乗平均偏差を含んでなる相同な構造座標を使用して、1個のCサブユニットの少なくともアミノ酸Ala24、Gly27、Phe53、Val57、Gly58、Glu61、Tyr64およびPhe65、ならびに1個のAサブユニットのアミノ酸Ser182、Leu183、Leu185およびArg186を伴い生成される、請求項5に記載の方法。
【請求項7】
三次元構造が、表3、4若しくは5のいずれかのA鎖のアミノ酸Ala21、Gly25;表3、4若しくは5のいずれかのK鎖のアミノ酸Ala24、Gly27、Phe53、Phe54、Val57、Gly58、Glu61、Tyr64、Phe65:表3、4若しくは5のいずれかのL鎖のアミノ酸Met17、Gly19、Gly20、Ala21、Ile22、Gly23、Ala24、Gly25、Ile26、Gly27、Asp28、Gly29、Ala31、Phe53、Thr56、Val57、Gly58、Leu59、Val60、Glu61、Ala62、Ala63/Pro63、Tyr64、Phe65;ならびに表3、4若しくは5のいずれかのM鎖のアミノ酸Ser182、Leu183、Leu185およびArg186の原子座標を使用して生成される、請求項5に記載の方法。
【請求項8】
請求項1ないし3のいずれか1つに記載のatpEタンパク質と耐性を賦与する突然変異部位において相互作用する化合物を活性成分とする微生物に基づく感染症を伴う被験体の処置用の製薬学的製剤。
【請求項9】
耐性を賦与する突然変異部位が、配列番号1に示されるMtb_S若しくは配列番号2に示されるMtb_Rの番号付けを参照として採用して、atpEタンパク質のアミノ酸14ないし34およびアミノ酸53ないし69を指す、請求項8に記載の製薬学的製剤。
【請求項10】
表6若しくは7のいずれかのatpEタンパク質のアミノ酸14−34若しくは54−69の構造座標、または1.5Å未満の水素以外の原子の二乗平均偏差を含んでなる相同な構造座標によって規定されるATPaseのF0部分におえる結合部位を含んでなる単離、精製されたポリペプチド。
【請求項11】
結合部位が、Cサブユニットの少なくともアミノ酸Ala24、Gly27、Phe53、Val57、Gly58、Glu61、Tyr64およびPhe65、ならびに1個のAサブユニットのアミノ酸Ser182、Leu183、Leu185、Arg186を含んでなる、請求項10記載の単離、精製されたポリペプチド。
【請求項12】
請求項10または11に記載のポリペプチドの結合部位と相互作用する化合物を活性成分として含んでなる微生物に基づく感染症を伴う被験体の処置用の製薬学的製剤。
【請求項13】
請求項5ないし7のいずれかの1に記載の方法を使用して同定された化合物を活性成分として含んでなる微生物に基づく感染症を伴う被験体の処置用の製薬学的製剤。
【請求項14】
感染症がグラム陽性細菌により引き起こされる、請求項13に記載の製薬学的製剤。
【請求項15】
感染症がミコバクテリウムにより引き起こされる、請求項13に記載の製薬学的製剤。
【請求項16】
感染症がM.アフリカヌム(M.africanum)、トリ結核菌(M.avium)、ウシ結核菌(M.bovis)、ウシ結核菌−BCG(M.bovis−BCG)、カメ結核菌(M.chelonae)、M.フォルツイタム(M.fortuitum)、M.ゴルドネ(M.gordonae)、M.イントラセルラレ(M.intracellulare)、M.カンサシイ(M.kansasii)、M.ミクロティ(M.microti)、M.スクロフラセウム(M.scrofulaceum)、パラ結核菌(M.paratuberculosis)、らい菌(M.leprea)、ヒト結核菌(M.tuberculosis)、M.ウルセランス(M.ulcerans)およびM.ラネ(M.ranae)からなる群より選択される微生物により引き起こされる、請求項13に記載の製薬学的製剤。
【請求項17】
請求項13に記載の製薬学的製剤であって、中枢神経系の感染症、外耳感染症、急性中耳炎のような中耳の感染症、硬膜静脈洞の感染症、眼の感染症、歯、歯肉および粘膜の感染症のような口腔の感染症、上気道感染症、下気道感染症、泌尿生殖器感染症、胃腸感染症、産婦人科感染症、敗血症、骨および関節の感染症、皮膚および皮膚構造の感染症、細菌性心内膜炎、火傷、手術の抗菌的予防、ならびに癌化学療法を受領している患者若しくは臓器移植患者のような免疫抑制患者における抗菌的予防を処置するための、製薬学的製剤。
【請求項18】
請求項1ないし3のいずれかの1つに記載の単離された変異atpEタンパク質をコードする単離された核酸分子。
【請求項19】
請求項18に記載の核酸分子を含んでなるベクター。
【請求項20】
請求項19に記載のベクターを担持する宿主細胞。
【請求項21】
抗菌化合物の同定方法であって、
a)atpEタンパク質を発現する細胞を、生理学的条件下で試験化合物と接触させる工程、および
b)該試験化合物がatpEタンパク質と相互作用するかどうかを決定する工程を含んでなり、かつ、atpEタンパク質が請求項1ないし3のいずれかの1つに記載の変異atpEタンパク質である、ことを特徴とする上記方法。
【請求項22】
単離されたatpEタンパク質に試験化合物が結合するかどうかの同定方法であって、a)atpEタンパク質を発現する細胞(こうした細胞は前記atpEタンパク質を通常は発現しない)を、atpEタンパク質を結合することが既知の化合物の存在および非存在下で試験化合物と接触させる工程、
b)atpEタンパク質に結合することが既知の化合物を参照として使用して、atpEタンパク質への該試験化合物の結合を測定する工程を含んでなり、かつ、atpEタンパク質が請求項10または11に記載のポリペプチドを含む変異atpEタンパク質である、ことを特徴とする上記方法。
【請求項23】
atpEタンパク質に結合することが既知の化合物が、検出可能に標識されており、図1に記載された化合物からなる、請求項22に記載の方法。
【請求項24】
工程a)が、atpEタンパク質を含む細胞組成物を、atpEタンパク質を結合することが既知の化合物の存在および非存在下で試験化合物と接触させることよりなる、請求項22に記載の方法。
【請求項25】
細胞組成物が、請求項20に記載の細胞から得られる膜調製物よりなる、請求項24に記載の方法。
【請求項26】
請求項1ないし3のいずれか1つに記載のatpEタンパク質と相互作用する試験化合物の潜在能力の評価方法であって、
a)分子モデル化技術を使用して請求項1ないし3のいずれか1つに記載のatpEタンパク質の三次元構造を明確に表すこと、
b)計算手段を使用して、試験化合物と請求項1ないし3のいずれか1つに記載のatpEタンパク質の三次元構造の間のフィッティング操作を実施すること、および
c)前記フィッティング操作の結果を解析して、請求項1ないし3のいずれか1つに記載のatpEタンパク質の三次元構造との試験化合物の会合を定量化すること
を含んでなる、上記方法。
【請求項27】
atpEタンパク質の三次元構造が、大腸菌(E.coli)(タンパク質データベース1Q01)のIle28、Glu61およびIle63の原子座標+/−10Åを超えない前記アミノ酸のバックボーン原子の二乗平均偏差を使用して生成される、請求項26に記載の方法。
【請求項28】
atpEタンパク質の三次元構造が、表6若しくは7の原子座標+/−1.5Åを超えない前記アミノ酸のバックボーン原子の二乗平均偏差を使用して生成される、請求項26に記載の方法。
【図1】

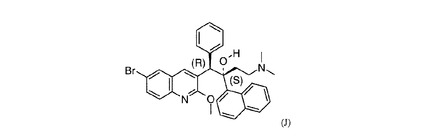
【図2】
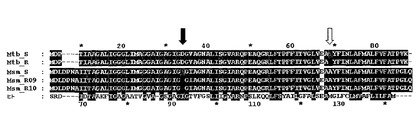
【図3】
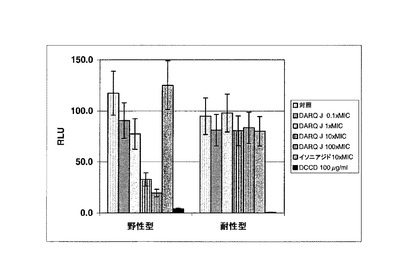
【図4】
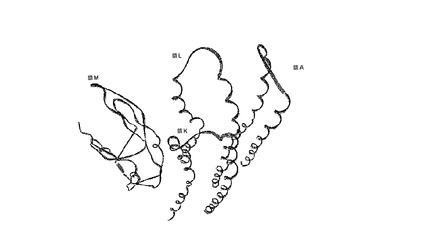
【図2】
【図3】
【図4】
【公開番号】特開2012−210210(P2012−210210A)
【公開日】平成24年11月1日(2012.11.1)
【国際特許分類】
【出願番号】特願2012−109892(P2012−109892)
【出願日】平成24年5月11日(2012.5.11)
【分割の表示】特願2007−532910(P2007−532910)の分割
【原出願日】平成17年9月28日(2005.9.28)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.フロッピー
【出願人】(390033008)ジヤンセン・フアーマシユーチカ・ナームローゼ・フエンノートシヤツプ (616)
【氏名又は名称原語表記】JANSSEN PHARMACEUTICA NAAMLOZE VENNOOTSCHAP
【Fターム(参考)】
【公開日】平成24年11月1日(2012.11.1)
【国際特許分類】
【出願日】平成24年5月11日(2012.5.11)
【分割の表示】特願2007−532910(P2007−532910)の分割
【原出願日】平成17年9月28日(2005.9.28)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.フロッピー
【出願人】(390033008)ジヤンセン・フアーマシユーチカ・ナームローゼ・フエンノートシヤツプ (616)
【氏名又は名称原語表記】JANSSEN PHARMACEUTICA NAAMLOZE VENNOOTSCHAP
【Fターム(参考)】
[ Back to top ]