
細菌輸送およびCNS侵襲を低減させるための組成物およびこの組成物を使用する方法
【課題】細菌感染、鼻輸送、鼻腔内コロニー形成およびCNS浸潤を低減または防止するように設計された組成物を提供すること。
【解決手段】本明細書では、細菌感染(例えば肺炎球菌感染)、鼻輸送、鼻腔内コロニー形成および中枢神経系侵襲を低減または防止するように設計された組成物を提供する。肺炎球菌ノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸、またはノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸のいずれかの抗原部分を含む組成物も提供する。さらに、本明細書で開示した組成物を作成および使用する方法も提供する。特に、本明細書で教示した作用因子または組成物を被験体に投与する工程を含む、被験体内で抗体を産生する方法も提供する。本明細書で教示した組成物を被験体に投与する工程を含む、被験体における鼻輸送または肺炎球菌感染を低減および防止する方法も提供する。
【解決手段】本明細書では、細菌感染(例えば肺炎球菌感染)、鼻輸送、鼻腔内コロニー形成および中枢神経系侵襲を低減または防止するように設計された組成物を提供する。肺炎球菌ノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸、またはノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸のいずれかの抗原部分を含む組成物も提供する。さらに、本明細書で開示した組成物を作成および使用する方法も提供する。特に、本明細書で教示した作用因子または組成物を被験体に投与する工程を含む、被験体内で抗体を産生する方法も提供する。本明細書で教示した組成物を被験体に投与する工程を含む、被験体における鼻輸送または肺炎球菌感染を低減および防止する方法も提供する。
【発明の詳細な説明】
【技術分野】
【0001】
(関連出願への相互参照)
本出願は、2003年11月10日に出願された米国仮出願第60/518,799号の利益を主張し、米国仮出願第60/518,799号は、本明細書においてその全体が参考として援用される。
【0002】
(謝辞)
本発明は、National Institutes of Healthからの助成金DC 04976、AI 21548、およびP30 DK 54781の下での、そしてNational Institute of Allergy and Infectious Diseasesとの契約NO1 AI 65299の下での、政府支援によって実施された。政府は本発明に一定の権利を有する。
【背景技術】
【0003】
(背景)
ストレプトコッカス・ニューモニエは、肺、中枢神経系(CNS)、中耳、および鼻道を含む複数の器官に感染しうる、かなり遍在性のヒト病原体である。これらの組織の感染は、気管支炎、肺炎、髄膜炎、および副鼻腔感染症などの様々な症状を引き起こす。S.ニューモニエは、ヒトの細菌性髄膜炎の主因であり、抗生剤処置にも関わらず、著しい死亡率および罹患率に関係している。非特許文献1。S.ニューモニエ髄膜炎は、永続的な神経性続発症を引き起こすことがある。先進国と開発途上国でのS.ニューモニエ髄膜炎の発生率は、100,000人に付きそれぞれ1〜2人および20人である。非特許文献2。米国での肺炎球菌性髄膜炎の致死率は、約18%である。非特許文献3。肺炎球菌性髄膜炎の最高の出現率は、1〜4歳の子供(すべての細菌性髄膜炎の30%)で、続いて15〜19歳(14%)および1〜11月齢の乳児(13%)で発生する。非特許文献2。高齢者も先進国と発展途上国の両方で、連鎖球菌性髄膜炎によって深刻に冒されている。非特許文献4;非特許文献5。
【0004】
世界での肺炎球菌の主な貯蔵場所は、ヒト鼻輸送(nasal carriage)である。感染の獲得は一般にキャリアからであり、感染は常に鼻輸送が先に起こる。上咽頭のコロニー形成は、肺炎球菌の下気道、副鼻腔、および中耳への蔓延の前提条件と見なされる。それゆえ輸送を防止したいずれの医学的介入も、処置を受けた人々における疾患のリスクを除去するだけではなく、集団免疫ももたらし、コミュニティにおける未処置の構成員の感染リスクすら低下させる。S.ニューモニエは重要なヒト病原体であるが、S.ニューモニエが鼻輸送または髄膜炎を引き起こす機構に関しては比較的わずかな事しか知られていない。
【0005】
ノイラミニダーゼが鼻道に独自の毒性因子であることを示唆する多少のデータが存在する。そのような観察の1つは、その親菌株よりも上咽頭から速く除去される、NanA欠損S.ニューモニエ菌株D39の研究によるものである。非特許文献6。ノイラミニダーゼは、宿主表面および体液中の広範囲に亘る糖脂質、糖タンパク質、およびオリゴ糖から末端シアル酸残基を開裂させる。肺炎球菌性髄膜炎患者の脳脊髄液(CSF)中の遊離シアル酸のレベル上昇は、予後不良に関連している。非特許文献7。ヒトにおけるS.ニューモニエ毒性に対するこの酵素の重要性は、S.ニューモニエそれぞれの新たな臨床単離物がノイラミニダーゼ活性を有する、2つの独立した研究の成果によってさらに説明される。非特許文献7;非特許文献8。その上、その毒性を上昇させることの多い肺炎球菌の単離物のマウス継代は、ノイラミニダーゼ活性の2〜5倍の上昇を引き起こすことも報告されている。非特許文献9。肺炎球菌性Cポリサッカライドは、テイコ酸としても既知であり、リポテイコ酸としても既知である肺炎球菌性F抗原のポリサッカライド部分と構造的に同一である。非特許文献10。これらの分子は、グラム陽性細菌の中でS.ニューモニエ独自の特徴である。これらの分子の主要抗原決定基は、ホスホリルコリン(PC)残基であり、PCに対するAbsは、腹腔内、静脈内、または経鼻肺炎球菌性攻撃に対して防御性である。非特許文献11;非特許文献12;非特許文献13;非特許文献14。しかしながら、これらの研究のすべてが血清によって投薬された全身感染に対する保護を評価したときに、これらのAbsが鼻腔内コロニー形成を防御する能力に関する情報は得られていない。しかしながら表面ホスホコリン残基は、呼吸細菌の表面でよく見られる。非特許文献15。
S.ニューモニエが鼻輸送と、それに続く疾患を引き起こす機構は、比較的知られていない。今日までいずれの研究も、鼻輸送を低減または防止する機構を確定していない。上咽頭のコロニー形成は、下気道、副鼻腔、全身、および脳への肺炎球菌の蔓延への前提条件と見なされており、当業界で必要なものは、粘膜免疫を初期肺炎球菌性コロニー形成の部位に供給する手段である。上咽頭での初期肺炎球菌性コロニー形成の防止は、鼻輸送を防止し、個人間でのS.ニューモニエの蔓延を低減するであろう。その上、上咽頭の粘膜表面へ免疫を提供することは、S.ニューモニエによって続いて引き起こされる疾患を防止または低減する。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Quagliarelloら,「N.Eng.J.Med.」1992年,第327巻:869−872
【非特許文献2】Anon,「CDSC European Bacterial Meningitis Surveillance Project」PHLS,London,2000年
【非特許文献3】Fedsonら,「Arch.Intern.Med.」1994年,第154巻:2531−2535
【非特許文献4】Butlerら,「Drugs Aging」1999年,第15巻(補遺1):11−19
【非特許文献5】Fedsonら,「Vaccine」1999年,第17巻補遺1:S 11−18
【非特許文献6】Tongら,「Infect.Immun.」2002年,第68巻:921−924
【非特許文献7】O’Tooleら,「J.Clin.Invest.」1971年,第50巻:979−985
【非特許文献8】Kellyら,「J.Bacteriol.」1967年,第94巻:272−273
【非特許文献9】Vishniakovaら,「Zhurnal Mikrobiologii,Epidemiologii i Immunobiologii」1992年,9−10:26−9
【非特許文献10】Fischerら,「Eur.J.Biochem」1993年,第215巻:851−857
【非特許文献11】Brilesら,「Eur.J.Immunol.」1984年,第14巻:1027−1030
【非特許文献12】Brilesら,「Nature」1981年,第294巻:88−90
【非特許文献13】Yotherら,「Infect.Immun.」1982年,第36巻:184−188
【非特許文献14】Brilesら,「J.Mol.Cell.Immunol.」1984年,第1巻:305−309
【非特許文献15】Lysenkoら,「Infect.Immun.」2000年,第68巻:1664−71
【発明の概要】
【課題を解決するための手段】
【0007】
(発明の要旨)
本明細書では、細菌感染(たとえば肺炎球菌感染)、鼻輸送、鼻腔内コロニー形成、およびCNS浸潤を低減または防止するように設計された組成物を提供する。必要に応じて、組成物は、粘膜投与のために設計されている。本明細書では、無毒化肺炎球菌ノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸(pneumococcal teichoic acid)、肺炎球菌性リポテイコ酸(pneumococcal lipoteichoic acid)、これらのいずれか1つの抗原部分およびこれらの無毒化作用因子を含有する組成物が提供される。
【0008】
肺炎球菌ノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸、またはこれらのいずれか1つの抗原部分および薬学的に受容可能なキャリアを含む組成物も提供される。必要に応じて組成物は、肺炎球菌ノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸またはこれらのいずれか1つの抗原部分のいずれかの組合せを含むことができる。無毒化肺炎球菌ノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸、またはこれらのいずれか1つの抗原部分はもちろんのこと、無毒化作用因子を含有する組成物および作用因子を使用する方法も提供される。
【0009】
被験体に作用因子を含む組成物を投与する工程を含む、被験体において肺炎球菌ノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸、あるいは肺炎球菌ノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、または肺炎球菌性リポテイコ酸のいずれか1つの抗原部分に対する抗体を産生させる方法も提供される。必要に応じて組成物は、粘膜表面への投与または全身投与に適している。
【0010】
肺炎球菌ノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸、またはこれらのいずれか1つの抗原部分に対する抗体を、薬学的に受容可能なキャリアと共に含む組成物もさらに提供される。必要に応じて組成物は、粘膜表面への投与または全身投与に適している。
【0011】
被験体の鼻粘膜に本明細書で教示した組成物を接触させる工程を含む、被験体において鼻輸送、鼻腔内コロニー形成、または細菌感染(たとえば肺炎球菌感染)を低減または防止する方法がさらに提供される。
本発明は、例えば、以下の項目を提供する:
(項目1)
無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目2)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの活性のおよそ60%を有する、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目3)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの活性のおよそ70%を有する、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目4)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの活性のおよそ80%を有する、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目5)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの活性のおよそ90%を有する、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目6)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの天然に存在するアミノ酸の少なくとも7%の欠失を含む、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目7)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの天然に存在するアミノ酸の少なくとも7%の欠失を含み、非無毒化ノイラミニダーゼの活性のおよそ60%を示す、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目8)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの天然に存在するアミノ酸の少なくとも7%の欠失を含み、非無毒化ノイラミニダーゼの活性のおよそ70%を示す、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目9)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの天然に存在するアミノ酸の少なくとも7%の欠失を含み、非無毒化ノイラミニダーゼの活性のおよそ80%を示す、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目10)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの天然に存在するアミノ酸の少なくとも7%の欠失を含み、非無毒化ノイラミニダーゼの活性のおよそ90%を示す、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目11)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの少なくとも5個のN末端アミノ酸の欠失を含む、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目12)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの少なくとも10個のN末端アミノ酸の欠失を含む、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目13)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの少なくとも15個のN末端アミノ酸の欠失を含む、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目14)
ノイラミニダーゼが、非無毒化ノイラミニダーゼのC末端アミノ酸の少なくとも10%の欠失を含む、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目15)
ノイラミニダーゼが、非無毒化ノイラミニダーゼのC末端アミノ酸の少なくとも20%の欠失を含む、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目16)
ノイラミニダーゼが、非無毒化ノイラミニダーゼのC末端アミノ酸の少なくとも30%の欠失を含む、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目17)
ノイラミニダーゼが、非無毒化ノイラミニダーゼのC末端アミノ酸の少なくとも35%の欠失を含む、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目18)
無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分および薬学的に受容可能なキャリアを含む、組成物。
(項目19)
アジュバントをさらに含む、項目18に記載の組成物。
(項目20)
被験体において肺炎球菌ノイラミニダーゼに特異的な抗体を産生する方法であって、該方法は、
該被験体に無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分を投与する工程
を包含する、方法。
(項目21)
被験体において肺炎球菌の鼻輸送を低減する方法であって、該方法は、
該被験体に無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分を投与する工程
を包含する、方法。
(項目22)
被験体において肺炎球菌感染を防止する方法であって、該方法は、
該被験体に無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分を投与する工程
を包含する、方法。
(項目23)
前記肺炎球菌感染が髄膜炎である、項目22に記載の方法。
(項目24)
前記肺炎球菌感染が中耳炎である、項目22に記載の方法。
(項目25)
前記肺炎球菌感染が肺炎である、項目22に記載の方法。
(項目26)
前記肺炎球菌感染が溶血性尿毒症である、項目22に記載の方法。
(項目27)
被験体において肺炎球菌の鼻輸送を低減または防止する方法であって、該方法は、
該鼻輸送を低減または防止する条件下で、該被験体に肺炎球菌ノイラミニダーゼまたはその抗原性フラグメントを投与する工程
を包含する、方法。
(項目28)
被験体において肺炎球菌感染を低減または防止する方法であって、該方法は、
該感染を低減または防止する条件下で、該被験体に肺炎球菌ノイラミニダーゼまたはその抗原性フラグメントを投与する工程
を包含する、方法。
(項目29)
前記肺炎球菌感染が髄膜炎である、項目28に記載の方法。
(項目30)
前記肺炎球菌感染が中耳炎である、項目28に記載の方法。
(項目31)
前記肺炎球菌感染が肺炎である、項目28に記載の方法。
(項目32)
前記肺炎球菌感染が溶血性尿毒症である、項目28に記載の方法。
(項目33)
被験体において肺炎球菌感染を低減または防止する方法であって、該方法は、
該感染を低減または防止する条件下で、該被験体に肺炎球菌ノイラミニダーゼ抗体またはそのフラグメントを投与する工程
を包含し、該投与工程は、
該被験体の粘膜表面に該抗体を接触させる工程
を包含する、方法。
(項目34)
前記肺炎球菌感染が髄膜炎である、項目33に記載の方法。
(項目35)
前記肺炎球菌感染が中耳炎である、項目33に記載の方法。
(項目36)
前記肺炎球菌感染が肺炎である、項目33に記載の方法。
(項目37)
前記肺炎球菌感染が溶血性尿毒症である、項目33に記載の方法。
(項目38)
肺炎球菌ノイラミニダーゼまたはその抗原部分および薬学的に受容可能なキャリアを含む組成物であって、該組成物は、粘膜表面への投与に適切である、組成物。
(項目39)
前記組成物が鼻スプレーである、項目38に記載の組成物。
(項目40)
前記組成物がネブライザ溶液である、項目38に記載の組成物。
(項目41)
前記組成物がエアゾール吸入薬である、項目38に記載の組成物。
(項目42)
項目38に記載の組成物を含む、容器。
(項目43)
前記容器が鼻スプレー器である、項目42に記載の容器。
(項目44)
前記容器がネブライザである、項目42に記載の容器。
(項目45)
前記容器が吸入器である、項目42に記載の容器。
(項目46)
被験体において肺炎球菌ノイラミニダーゼに特異的な抗体を産生する方法であって、該方法は、
該被験体の鼻粘膜に無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分を接触させる工程
を包含する、方法。
(項目47)
被験体において肺炎球菌の鼻輸送を低減する方法であって、該方法は、
該被験体の鼻粘膜に無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分を接触させる工程
を包含する、方法。
(項目48)
被験体において肺炎球菌感染を防止する方法であって、該方法は、
該被験体の粘膜表面に無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分を接触させる工程
を包含する、方法。
(項目49)
前記肺炎球菌感染が髄膜炎である、項目48に記載の方法。
(項目50)
前記肺炎球菌感染が中耳炎である、項目48に記載の方法。
(項目51)
前記肺炎球菌感染が肺炎である、項目48に記載の方法。
(項目52)
前記肺炎球菌感染が溶血性尿毒症である、項目48に記載の方法。
(項目53)
肺炎球菌性テイコ酸または肺炎球菌性リポテイコ酸からのホスホコリンまたはその抗原部分、および薬学的に受容可能なキャリアを含む組成物であって、該組成物は、粘膜表面への投与に適切である、組成物。
(項目54)
前記組成物が鼻スプレーである、項目53に記載の組成物。
(項目55)
前記組成物がネブライザ溶液である、項目53に記載の組成物。
(項目56)
前記組成物がエアゾール吸入薬である、項目53に記載の組成物。
(項目57)
項目53に記載の組成物を含む、容器。
(項目58)
前記容器が鼻スプレー器である、項目57に記載の容器。
(項目59)
前記容器がネブライザである、項目57に記載の容器。
(項目60)
前記容器が吸入器である、項目57に記載の容器。
(項目61)
被験体において肺炎球菌ノイラミニダーゼに特異的な抗体を産生する方法であって、該方法は、
該被験体の鼻粘膜に項目53に記載の組成物を接触させる工程
を包含する、方法。
(項目62)
被験体において肺炎球菌の鼻輸送を低減する方法であって、該方法は、
該被験体の鼻粘膜に項目53に記載の組成物を接触させる工程
を包含する、方法。
(項目63)
被験体において肺炎球菌感染を低減または防止する方法であって、該方法は、
該被験体の粘膜表面に項目53に記載の組成物を接触させる工程
を包含する、方法。
(項目64)
前記肺炎球菌感染が髄膜炎である、項目63に記載の方法。
(項目65)
前記肺炎球菌感染が中耳炎である、項目63に記載の方法。
(項目66)
前記肺炎球菌感染が肺炎である、項目63に記載の方法。
(項目67)
前記肺炎球菌感染が溶血性尿毒症である、項目63に記載の方法。
(項目68)
肺炎球菌ノイラミニダーゼまたはその抗原部分、肺炎球菌性テイコ酸または肺炎球菌性リポテイコ酸からのホスホコリンまたはその抗原部分、および薬学的に受容可能なキャリアを含む組成物であって、該組成物は、粘膜表面への投与に適切である、組成物。
(項目69)
前記組成物が鼻スプレーである、項目68の組成物。
(項目70)
前記組成物がネブライザ溶液である、項目68の組成物。
(項目71)
前記組成物がエアゾール吸入薬である、項目68の組成物。
(項目72)
項目68に記載の組成物を含む、容器。
(項目73)
前記容器が鼻スプレー器である、項目72に記載の容器。
(項目74)
前記容器がネブライザである、項目72に記載の容器。
(項目75)
前記容器が吸入器である、項目72に記載の容器。
(項目76)
被験体において肺炎球菌ノイラミニダーゼに特異性な抗体を産生する方法であって、該方法は、
該被験体の鼻粘膜に項目68に記載の組成物を接触させる工程
を包含する、方法。
(項目77)
被験体において肺炎球菌の鼻輸送を低減する方法であって、該方法は、
該被験体の鼻粘膜に項目68に記載の組成物を接触させる工程
を包含する、方法。
(項目78)
被験体において肺炎球菌感染を防止する方法であって、該方法は、
該被験体の粘膜表面に項目68に記載の組成物を接触させる工程
を包含する、方法。
(項目79)
前記肺炎球菌感染が髄膜炎である、項目78に記載の方法。
(項目80)
前記肺炎球菌感染が中耳炎である、項目78に記載の方法。
(項目81)
前記肺炎球菌感染が肺炎である、項目78に記載の方法。
(項目82)
前記肺炎球菌感染が溶血性尿毒症である、項目78に記載の方法。
(項目83)
肺炎球菌性テイコ酸または肺炎球菌性リポテイコ酸の非ホスホコリン抗原部分および薬学的に受容可能なキャリアを含む組成物であって、該組成物は、粘膜表面への投与に適切である、組成物。
(項目84)
前記組成物が鼻スプレーである、項目83に記載の組成物。
(項目85)
前記組成物がネブライザ溶液である、項目83に記載の組成物。
(項目86)
前記組成物がエアゾール吸入薬である、項目83に記載の組成物。
(項目87)
項目83に記載の組成物を含む、容器。
(項目88)
前記容器が鼻スプレー器である、項目87に記載の容器。
(項目89)
前記容器がネブライザである、項目87に記載の容器。
(項目90)
前記容器が吸入器である、項目87に記載の容器。
(項目91)
被験体において肺炎球菌ノイラミニダーゼに特異的な抗体を産生する方法であって、該方法は、
該被験体の鼻粘膜に項目83に記載の組成物を接触させる工程
を包含する、方法。
(項目92)
被験体において肺炎球菌の鼻輸送を低減する方法であって、該方法は、
該被験体の鼻粘膜に項目83に記載の組成物を接触させる工程
を包含する、方法。
(項目93)
被験体において肺炎球菌感染を防止する方法であって、該方法は、
該被験体の粘膜表面に項目83に記載の組成物を接触させる工程
を包含する、方法。
(項目94)
前記肺炎球菌感染が髄膜炎である、項目93に記載の方法。
(項目95)
前記肺炎球菌感染が中耳炎である、項目93に記載の方法。
(項目96)
前記肺炎球菌感染が肺炎である、項目93に記載の方法。
(項目97)
前記肺炎球菌感染が溶血性尿毒症である、項目93に記載の方法。
(項目98)
肺炎球菌ノイラミニダーゼまたはその抗原部分、肺炎球菌性テイコ酸または肺炎球菌性リポテイコ酸の非ホスホコリン抗原部分、および薬学的に受容可能なキャリアを含む組成物であって、該組成物は、粘膜表面への投与に適切である、組成物。
(項目99)
前記組成物が鼻スプレーである、項目98に記載の組成物。
(項目100)
前記組成物がネブライザ溶液である、項目98に記載の組成物。
(項目101)
前記組成物がエアゾール吸入薬である、項目98に記載の組成物。
(項目102)
項目98に記載の組成物を含む、容器。
(項目103)
前記容器が鼻スプレー器である、項目102に記載の容器。
(項目104)
前記容器がネブライザである、項目102に記載の容器。
(項目105)
前記容器が吸入器である、項目102に記載の容器。
(項目106)
被験体において肺炎球菌ノイラミニダーゼに特異的な抗体を産生する方法であって、該方法は、
該被験体の鼻粘膜に項目98に記載の組成物を接触させる工程
を包含する、方法。
(項目107)
被験体において肺炎球菌の鼻輸送を低減する方法であって、該方法は、
該被験体の鼻粘膜に項目98に記載の組成物を接触させる工程
を包含する、方法。
(項目108)
被験体において肺炎球菌感染を防止する方法であって、該方法は、
該被験体の粘膜表面に項目98に記載の組成物を接触させる工程
を包含する、方法。
(項目109)
前記肺炎球菌感染が髄膜炎である、項目108に記載の方法。
(項目110)
前記肺炎球菌感染が中耳炎である、項目108に記載の方法。
(項目111)
前記肺炎球菌感染が肺炎である、項目108に記載の方法。
(項目112)
前記肺炎球菌感染が溶血性尿毒症である、項目108に記載の方法。
(項目113)
ホスホコリン抗体またはそのフラグメントおよび薬学的に受容可能なキャリアを含む、組成物。
(項目114)
前記組成物が粘膜表面への投与に適切である、項目113に記載の組成物。
(項目115)
前記組成物が鼻スプレーである、項目113に記載の組成物。
(項目116)
前記組成物がネブライザ溶液である、項目113に記載の組成物。
(項目117)
前記組成物がエアゾール吸入薬である、項目113に記載の組成物。
(項目118)
項目113に記載の組成物を含む、容器。
(項目119)
前記容器が鼻スプレー器である、項目118に記載の容器。
(項目120)
前記容器がネブライザである、項目118に記載の容器。
(項目121)
前記容器が吸入器である、項目118に記載の容器。
(項目122)
被験体において肺炎球菌の鼻輸送を低減する方法であって、該方法は、
該被験体にホスホコリン抗体またはそのフラグメントを投与する工程
を包含する、方法。
(項目123)
前記投与が、前記被験体の鼻粘膜に前記抗体またはそのフラグメントを接触させる工程を包含する、項目122に記載の方法。
(項目124)
被験体において肺炎球菌感染を防止する方法であって、該方法は、
該被験体にホスホコリン抗体またはそのフラグメントを投与する工程
を包含する、方法。
(項目125)
前記投与が、前記被験体の鼻粘膜に前記抗体またはそのフラグメントを接触させる工程を包含する、項目124に記載の方法。
【0012】
さらなる利点は、以下に続く説明で一部が述べられ、一部は説明から明らかになるか、または以下で述べる態様の実施によって知られるであろう。以下で述べる利点は、添付請求項で特に指摘される要素および組合せによって実現および達成されるであろう。上述の一般的な説明および以下の詳細な説明はどちらも例示および説明のためだけであり、制限的でないことが理解されるべきである。
本明細書に含まれ、その一部を構成する添付図面は、以下で述べる複数の態様を説明する。
【図面の簡単な説明】
【0013】
【図1】xidマウスへのS.ニューモニエの非カプセル化R36A菌株または毒性EF3030菌株のどちらかの3X106CFUの経鼻送達を示す。神経組織ON/E、OB、脳組織およびリンパ系組織(NALT、CLNおよび肺)を鼻腔内攻撃の第1日〜第4日後に収集して、切り刻み、生肺炎球菌の存在を分析した。論理コロニー形成単位(CFU)の平均+1標準誤差(SE)が示されている。Y軸の0値は、検出可能なCFUの非存在を表す。1グループ当たり5匹のマウスの平均CFU+SEが示され、3つの異なる実験を表している。
【図2A】鼻腔内攻撃後のS.ニューモニエ菌株EF3030 CFUの器官分布の動力学を示す。ON/E、OB、脳、血液、NW、NALT、CLN、および肺組織を4日、11日、18日、25日、および39日に収集して、S.ニューモニエの存在を分析した。S.ニューモニエ3X106CFUの分割量は、鼻道のコロニー形成と、それに続くOBの感染を生じた。Y軸の0値は、検出可能なCFUの非存在を表す。3つの独立した実験の平均CFU+SEを示す。各時点は、マウス5匹を表す第39日を除いて、マウス10匹を表す。
【図2B】鼻腔内攻撃後のS.ニューモニエ菌株EF3030 CFUの器官分布の動力学を示す。ON/E、OB、脳、血液、NW、NALT、CLN、および肺組織を4日、11日、18日、25日、および39日に収集して、S.ニューモニエの存在を分析した。S.ニューモニエ3X106CFUの分割量は、鼻道のコロニー形成と、それに続くOBの感染を生じた。Y軸の0値は、検出可能なCFUの非存在を表す。3つの独立した実験の平均CFU+SEを示す。各時点は、マウス5匹を表す第39日を除いて、マウス10匹を表す。
【図3A】GLSによる予備インキュベーション後のS.ニューモニエ菌株EF3030の分布を示す。S.ニューモニエの分割量(3X107CFU)を、鼻への利用前に、アシアロ−GM1(a−GM1)20μgまたは混合GLS(MG)125μgを用いて30分間インキュベートした。ON/E、OB、脳およびNW、NALTおよび肺を4日後に収集して、S.ニューモニエの数を分析した。Y軸の0値は、検出可能なCFUの非存在を表す。マウス5匹の平均+1SEを示し、以下の統計解析からP値を得た。データは2つの独立した実験を表している。
【図3B】GLSによる予備インキュベーション後のS.ニューモニエ菌株EF3030の分布を示す。S.ニューモニエの分割量(3X107CFU)を、鼻への利用前に、アシアロ−GM1(a−GM1)20μgまたは混合GLS(MG)125μgを用いて30分間インキュベートした。ON/E、OB、脳およびNW、NALTおよび肺を4日後に収集して、S.ニューモニエの数を分析した。Y軸の0値は、検出可能なCFUの非存在を表す。マウス5匹の平均+1SEを示し、以下の統計解析からP値を得た。データは2つの独立した実験を表している。
【図4−1】鼻腔内攻撃後のOB中のS.ニューモニエのTIGR4菌株の検出を示す。5X105CFUの分割量を鼻腔内投与し、血液、NW、ON/E、OBおよび脳組織を攻撃1週間後にコロニー形成について分析した(パネルAおよびB)。これらの組織(DNA 10μg)もニューモリシン遺伝子の存在についてPCRによって分析した(パネルC)。加えてコントロール(D)またはS.ニューモニエ攻撃マウスのOB中のS.ニューモニエを、PspA特異性Absを用いた免疫蛍光によって描出した(パネルEおよびF)。平均+1SEを示す。データは3つの独立した実験を表している。
【図4−2】鼻腔内攻撃後のOB中のS.ニューモニエのTIGR4菌株の検出を示す。5X105CFUの分割量を鼻腔内投与し、血液、NW、ON/E、OBおよび脳組織を攻撃1週間後にコロニー形成について分析した(パネルAおよびB)。これらの組織(DNA 10μg)もニューモリシン遺伝子の存在についてPCRによって分析した(パネルC)。加えてコントロール(D)またはS.ニューモニエ攻撃マウスのOB中のS.ニューモニエを、PspA特異性Absを用いた免疫蛍光によって描出した(パネルEおよびF)。平均+1SEを示す。データは3つの独立した実験を表している。
【図5】分泌NanA、TIGR4の、および細胞壁への結合のためのLPXTG(配列番号14)モチーフを有するR6(2型)のモチーフにおける比較を示す。TIGR4遺伝子は、LPETG(配列番号13)モチーフをコード化する配列の前に停止コドンを含む。このモチーフがないと、NanAはTIGR4によって環境中へ分泌される。
【図6A】S.ニューモニエ親菌株TIGR4(黒四角)またはNanA同系変異体TIGR4/nanA−(○)によって鼻腔内感染させたCBA/Nマウスの鼻洗浄液中の生肺炎球菌の動力学を示す。各点は、各マウスの鼻洗浄液1ml当たりの細菌総数を表す。*P<0.05;**P<0.01;***P<0.005、TIGR4を接種したマウスと比較。
【図6B】S.ニューモニエ親菌株TIGR4(黒四角)またはNanA同系変異体TIGR4/nanA−(○)によって鼻腔内感染させたCBA/Nマウスでの鼻腔内コロニー形成動力学を示す。各点は、各マウスの組織1グラム当たりの細菌総数を示す。*P<0.05;**P<0.01;***P<0.005、TIGR4を接種したマウスと比較。
【図6C】S.ニューモニエ親菌株TIGR4(黒四角)またはTIGR4/nanA−(○)によって鼻腔内感染させたCBA/Nマウスの嗅球におけるCFUの動力学を示す。各点は、各マウスの組織1グラム当たりの細菌総数を示す。*P<0.05;**P<0.01;***P<0.005、TIGR4を接種したマウスと比較。
【図7A】S.ニューモニエ親菌株EF3030(黒四角)またはEF3030/nanA−(○)によって鼻腔内感染させたCBA/Nマウスの鼻腔内コロニー形成動力学を示す。各点は、各マウスの鼻洗浄液1ml当たりの細菌総数を表す。*P<0.05;**P<0.01;***P<0.005、EF3030を接種したマウスと比較。
【図7B】S.ニューモニエ親菌株EF3030(黒四角)またはEF3030/nanA−(○)によって鼻腔内感染させたCBA/Nマウスの鼻腔内コロニー形成動力学を示す。各点は、各マウスの組織1グラム当たりの細菌総数を示す。*P<0.05;**P<0.01;***P<0.005、EF3030を接種したマウスと比較。
【図7C】S.ニューモニエ親菌株EF3030(黒四角)またはNanA同系変異体EF3030/nanA−(○)によって鼻腔内感染させたCBA/Nマウスの鼻腔内コロニー形成動力学を示す。各点は、各マウスの組織1グラム当たりの細菌総数を示す。*P<0.05;**P<0.01;***P<0.005、EF3030を接種したマウスと比較。野生種および変異体データをすべての時点でプールすると、EF3030とEF3030 NanB−との比較はP=0.001であった。
【図8】S.ニューモニエ親菌株TIGR4(黒四角)またはTIGR4/nanB−(○)によって鼻腔内感染させた接種4日後のCBA/Nマウスの鼻腔内コロニー形成動力学を示す。各点は、各マウスの鼻洗浄液1mlまたは組織1グラム当たりの細菌の総数を示す。いずれの場合も、TIGR4とTIGR4/nanB−との差は統計的に有意でなかった。
【図9】S.ニューモニエ親菌株TIGR4(黒四角)、TIGR4/nanA−(○)またはTIGR4/AB−によって鼻腔内感染させた接種4日後のCBA/Nマウスの鼻腔内コロニー形成動力学を示す。各点は、各マウスの鼻洗浄液1mlまたは組織1グラム当たりの細菌の総数を示す。いずれの場合も、TIGR4/nanA−とダブル変異体TIGR4/nanAB−との差は統計的に有意でなかった。
【図10】鼻腔内攻撃後の、抗ホスホコリン特異性モノクローナル抗体によるS.ニューモニエの鼻腔内コロニー形成の阻害を示す。鼻腔内攻撃後の抗PC特異性mAbsによるS.ニューモニエの鼻腔内コロニー形成の阻害。総量1X106CFUのTIGR4菌株をIgG3サブクラスまたはIgMアイソタイプのどちらかの抗PCmAbs 5μgを用いてインキュベートした。鼻孔ごとに総量5μgを投与した。利用9および12時間後の鼻洗浄液500μl中のCFUを示す。1グループ当たりマウス5匹の平均+SDを示す。
【発明を実施するための形態】
【0014】
(詳細な説明)
本化合物,組成物,物品、器具、および/または方法を開示および説明する前に、以下で述べる態様が、具体的な合成方法または具体的な投与方法に制限されず、そのためもちろん変化することが理解されるべきである。本明細書で使用する用語が、特定の態様のみを説明する目的で使用され、制限的でないことも理解されるべきである。
【0015】
明細書および添付請求項で使用するように、単数形「a」、「an」および「the」は、文脈が明確に別途規定しない限り、複数形指示対象も含むことに注意する必要がある。それゆえたとえば、「抗原性フラグメント」への言及は、抗原性フラグメントの混合物を含み、「製薬的キャリア」または「アジュバント」への言及は、2つ以上のそのようなキャリアまたはアジュバントの混合物を含むなどである。
【0016】
全体を通じて使用するように、「被験体」は個体を意味する。それゆえ「被験体」は、たとえばネコ、イヌなどの飼い慣らされた動物、家畜(たとえば畜牛、ウマ、ブタ、ヒツジ、ヤギなど)、実験用動物(たとえばマウス、ウサギ、ラット、モルモットなど)および鳥類を含むことができる。1つの態様において、被験体は、霊長類またはヒトなどの哺乳類である。
【0017】
「任意の」または「必要に応じて」は、続いて説明される事象または状況が起こりうる、または起こらないことを、そして説明が事象または状況が起こる例および起こらない例を含むことを意味する。たとえば、「必要に応じて、組成物は組合せを含むことができる」という表現は、説明が組合せおよび組合せの非存在(すなわち組合せの個々の構成要素)を含むように、組成物が異なる分子の組合せを含む、または組合せを含まないことを意味する。
【0018】
範囲は本明細書では、「約」1つの特定の値から、および/または「約」別の特定の値までとして表現される。そのような範囲が表現されるとき、別の態様は、一方の特定の値から、および/または他方の特定の値までを含む。同様に、値が先行する「約」を用いて概数として表現されるとき、特定の値が別の態様を形成することが理解されるであろう。各範囲の終点は、他方の終点に関連しても、および他方の終点から独立しても重要であることがさらに理解されるであろう。
【0019】
本明細書では、細菌感染(たとえば肺炎球菌感染)、鼻輸送、鼻腔内コロニー形成、およびCNS浸潤を低減または防止するように設計された組成物および方法を提供する。S.ニューモニエは、Cポリサッカライド−ガングリオシド相互作用によって上皮バリアを通過して、続いて上皮細胞中へエンドサイトーシスすることによって、一部は鼻道をコロニー形成する。Cポリサッカライドは、ガングリオシド中の末端または内部GaINAcβ1−4Gal配列への結合を介して、アシアロ−GM1、アシアロ−GM2、およびフコシル−アシアロ−GM1に結合する。ヒト肺を除いて細胞原形質膜中のこれらのアシアロガングリオシドは通常は少ないが、S.ニューモニエは、2つのノイラミニダーゼNanAおよびNanBを有し(Berry et al.,(1996)J.Bacteriol.178:4854−4860)、これはそれぞれα2,3−およびα2,6結合N−アセチルノイラミン酸をガラクトースへ、そしてα2,6−結合をN−アセチル−ガラクトサミンへ開裂できる。Scanlon et al.,(1989)Enzyme 41:143−150。ガングリオシドのシアル酸残基は、ガラクトースに結合したα2,3である。S.ニューモニエのノイラミニダーゼは、すべてのモノシアロガングリオシドに存在する末端シアル酸残基、およびジ−およびトリ−シアロガングリオシドで見られるように、ガラクトース結合した複数のシアル酸残基を除去する。それゆえそれらは、最も一般的な哺乳類細胞表面ガングリオシド中のGaINAcβ1−4Ga1配列を露出できるはずである。これらの残基は、細胞表面上の推定Cポリサッカライド結合部位である。通常はさらに細胞壁に結合しているそのNanA、および分泌されると考えられるNanBを使用して、S.ニューモニエは、気道の上皮細胞上にその独自の結合部位を生成する。それゆえ肺炎球菌性Cポリサッカライドは、アシアロガングリオシド、特にアシアロ−GM1に結合し、かなり豊富なGM1をアシアロ−GM1に変換できるノイラミニダーゼは、CポリサッカライドのためにON/E上に豊富な結合部位を作成する。本機構は、鼻輸送を促進し、鼻嗅神経および鼻甲介を被覆する上皮(ON/E)、嗅球(OB)を介してS.ニューモニエのCNSへのアクセスを提供する。同様に、中耳炎およびS.ニューモニエを含む他の感染症は、中耳に分布する神経を介してCNSへのアクセスを同様に得ることができる。S.ニューモニエに加えて他の細菌は、匹敵するノイラミニダーゼを有し、それゆえ同じ機構が他の細菌でも同様に発生する。それゆえ本明細書では、各種の細菌におけるこの機構を標的とする組成物および方法を開示する。本明細書で教示される作用因子、組成物、および方法は、輸送を低減させ、CNS浸潤を防止するために本機構を妨害することに関する。
【0020】
必要に応じて組成物は、粘膜投与用に設計される。たとえば本明細書では、肺炎球菌ノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸またはこれらのいずれか1つの抗原部分および薬学的に受容可能なキャリアを含む組成物であって、粘膜表面への投与に適した組成物が提供される。必要に応じて組成物は、肺炎球菌ノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸またはこれらのいずれか1つの抗原部分のいずれかの組合せを含むことができる。
【0021】
必要に応じて組成物は、エアゾール、鼻腔用ミスト、鼻スプレー、点鼻薬、ネブライザ溶液、エアゾール吸入薬、坐剤の形、または粘膜投与(経口投与を含む)に適したいずれかの形である。必要に応じて組成物は、送達のために微小球、またはリポソームに入れることができる。「粘膜表面への投与」とは、呼吸系、胃腸系、または泌尿生殖器系を含むいずれかの粘膜表面への投与を意味する。粘膜表面の例は、これに限定されるわけではないが、鼻腔(嗅覚神経上皮を含む)、上咽頭、直腸、膣、咽頭、口、エウスタキー管、気管、気管支および他の気道、および腸粘膜を含む。
【0022】
粘膜表面への投与では、粘膜アジュバントが使用できる。アジュバントは、本発明の組成物と同時に、その直前に、またはその後に投与できる。必要に応じて、組成物はさらにアジュバントを含む。粘膜アジュバント処方物はたとえば、粘膜誘導部位を標的とする作用因子を含む。アジュバントは必要に応じて、これに限定されるわけではないが、サイトカイン、ケモカイン、成長因子、血管由来因子、アポトーシス阻害物質、およびその組合せを含む群より選択される。サイトカインがアジュバントして選択されるとき、サイトカインはこれに限定されるわけではないが、IL−1、IL−1γ、IL−1β、IL−2、IL−5、IL−6、IL−12、IL−15およびIL−18を含むインターロイキン;形質転換成長因子−β(TGF−β);顆粒球マクロファージコロニー刺激因子(GM−CSF);インターフェロン−ガンマ(IFN−γ);またはアジュバント活性を有する他のサイトカインを含む群より選択される。サイトカインの部分、あるいはアジュバント活性または他の生物活性を有するサイトカインの変異体またはミミック(またはその組合せ)も、本発明の組成物および方法で使用できる。
【0023】
ケモカインがアジュバントとして選択されるとき、ケモカインは必要に応じて、これに限定されるわけではないが、リンホタクチン、RANTES、LARC、PARC、MDC、TARC、SLCおよびFKNを含む群より選択される。アポトーシス阻害物質がアジュバントとして選択される場合、アポトーシス阻害物質は必要に応じて、これに限定されるわけではないが、カスパーゼ−8の阻害物質、およびその組合せを含む群より選択される。血管由来因子がアジュバントとして選択される場合、血管由来因子は必要に応じて、これに限定されるわけではないが、塩基性繊維芽細胞成長因子(FGF)、血管内皮成長因子(VEGF)、ヒアルロナン(HA)フラグメント、およびその組合せを含む群より選択される。実際に、プラス(+)およびマイナス(−)血管由来因子をアジュバントとして選択できる。
【0024】
本発明の実質的に非毒性の生物活性粘膜アジュバントの他の例は、ホルモン、酵素、成長因子、またはその生物活性部分を含む。そのようなホルモン、酵素、成長因子、またはその生物活性部分は、たとえばヒト、ウシ、ブタ、ヒツジ、イヌ、ネコ、ウマ、または鳥類起源であり、腫瘍壊死因子(TNF)、プロラクチン、上皮成長因子(EGF)、顆粒球コロニー刺激因子(GCSF)、インスリン様成長因子(IGF−1)、ソマトトロピン(成長ホルモン)またはインスリン、あるいはレセプタが免疫系の細胞上で発現される他のホルモンまたは成長因子でありうる。
【0025】
粘膜投与のためのアジュバントは、細菌性毒素、たとえばコレラ毒素(CT)、E.コリ易熱性毒素(LT)、クロストリジウムディフィシル毒素Aおよび百日咳毒素(PT)、あるいはその組合せ、サブユニット、トキソイド、キメラ、または変異体も含む。たとえば未変性コレラ毒素サブユニットB(CTB)の精製調製物が使用できる。フラグメント、ホモログ、誘導体、およびこれらの毒素のいずれかへの融合物も、それらがアジュバント活性を保持するという条件ならば適切である。好ましくは低下した毒性を有する変異体が使用される。適切な変異体は、たとえばWO 95/17211(Arg−7−Lys CT変異体)、WO 96/6627(Arg−192−Gly LT変異体)、およびWO 95/34323(Arg−9−LysおよびGlu−129−Gly PT変異体)に述べられている。本発明の方法および組成物で使用できる、その他のLT変異体はたとえば、Ser−63−Lys、Ala−69−Gly、Glu−110−Asp、およびGlu−112−Asp変異体を含む。他のアジュバント、たとえばRH3−リガンド;CpG−モチーフオリゴヌクレオチド;たとえばE.コリ、サルモネラ・ミネソタ、サルモネラ・チフィムリウム、またはシゲラ・フレキシネリの細菌性モノホスホリル脂質A(MPLA);サポニン(たとえばQS21)、またはポリラクチドグリコリド(PLGA)微小球も粘膜投与で使用できる。考えられる他の粘膜アジュバントは、デフェンシンおよびオリゴヌクレオチドを含有するCpGモチーフである。
【0026】
本明細書を通じて使用するように、「薬学的に受容可能なキャリア」は、生物学的にまたはそれ以外で望ましくないわけではない材料を意味し、すなわち該材料は、望ましくない生物的影響を引き起こすことなく、またはそれが含有される薬学的組成物の他の成分のいずれかと有害な方式で相互作用することなく、選択した化合物と共に個体に投与できる。
【0027】
本明細書で述べる組成物のいずれも、薬学的に受容可能なキャリア共に治療に使用できる。本明細書で述べる化合物は、薬学的に受容可能なキャリアと組み合せて、1つ以上の化合物より成る薬学的組成物へと好都合に処方できる。たとえば、代表的なキャリアおよび本明細書で述べた化合物の処方物の調製と併せて使用可能である薬学的組成物を調製する通常の方法を開示し、参照により本明細書に組み入れられている、ペンシルベニア州イーストンのE.W.Martin Mack Pub.Co.によるRemington’s Pharmaceutical Sciences、最新版を参照。最も代表的には、これらは、ヒトへの組成物投与のための標準キャリアである。1つの態様においては、ヒトおよび非ヒトであり、滅菌水、食塩水、および生理的pHの緩衝溶液などの溶液を含む。他の化合物は、当業者に使用される標準手順に従って投与されるであろう。
【0028】
本明細書で述べる薬学的組成物は、これに限定されるわけではないが、選択した分子に加えて、キャリア、増粘剤、希釈剤、緩衝剤、保存料、界面活性剤などを含むことができる。薬学的組成物は、1つ以上の活性成分、たとえば抗菌剤、抗炎症剤、麻酔剤なども含むことができる。
【0029】
肺炎球菌ノイラミニダーゼは、肺炎球菌性細菌で見られるいずれかのノイラミニダーゼ分子を意味する。表1は、複数の種からのノイラミニダーゼのアライメントを示す。ノイラミニダーゼ分子は、たとえばSP1326を含む。SP1326アミノ酸配列は、GenBankアクセッション番号AAK75424によって入手できる。Tettelin,H.,et al.,(2001)Science 293:498−506。GenBankアクセッション番号AAK75424の下でSP1326に提供されたアミノ酸および核酸配列を含むすべての情報は、この参照によりその全体が本明細書に組み入れられている。本明細書を通じて示すように、すべてのアミノ酸配列のアミノ酸残基を、肺炎球菌性菌株R6のアミノ酸配列に従って表1に示すように番号付けする。
【0030】
【表1−1】

【0031】
【表1−2】
ノイラミニダーゼのいずれの抗原性改変体も、本明細書で教示した組成物または方法で使用できる。それゆえ天然型ノイラミニダーゼは、本発明で教示した方法に従ったアミノ酸残基の置換、欠失、または変更によって修飾できる。必要に応じて、そのような修飾は、ノイラミニダーゼを無毒化するように設計されるであろう。「無毒化」とは、抗原性または免疫原性を維持しながら、酵素活性を低減または除去することを意味する。これは部位特異性突然変異誘発を使用する、ノイラミニダーゼの活性部位におけるアミノ酸の置換、欠失、または変更によって実施される。好ましくは、そのような置換、欠失、または変更は、Aspボックスの範囲内となるであろう(すなわちアミノ酸残基460〜480、530〜560、または600〜620内)。本明細書で教示されたノイラミニダーゼ構造については、その全体が参照により本明細書に組み入れられているCrennell et al.,PNAS 90:9852−9856を参照。そのような置換、欠失、または変更は、アミノ酸残基383〜387、467〜473、541〜546、または610〜616内のAspボックス内で発生しうる。Aspボックスにおける変更はたとえば、アスパラギン酸の、グルタミン酸またはトレオニンの置換を含むことができる。他の保存的または非保存的アミノ酸置換も、毒性を低下させるために、Aspボックス内のアスパラギン酸残基または他のいずれかの残基にて使用できる。ノイラミニダーゼの他の領域は必要に応じて、部位特異性突然変異誘発を標的とする。たとえば位置572におけるバリンまたはグルタミンの保存的および非保存的アミノ酸置換を含む、たとえば残基570〜580に該当する領域内での修飾が開示される。残基750〜760に該当する領域に修飾を、さらに詳細には位置754にチロシンを有するノイラミニダーゼも開示される。チロシン残基の保存的アミノ酸置換は、たとえばセリンまたはトレオニンを含む。アミノ酸残基340〜350、600〜610、または360〜370該当する領域に修飾を有するノイラミニダーゼも提供される。さらに詳細には、位置347、605、366、または367のアルギニンは、リジンまたはグルタミン、あるいは他のいずれかの保存的または非保存的アミノ酸によって置換できる。本明細書で教示した各種の修飾を組み合せて使用できる。それゆえ1つ以上の保存的または非保存的アミノ酸置換が必要に応じて、同じノイラミニダーゼ中に存在する。
【0032】
上述のように、無毒化ノイラミニダーゼは、当業界で周知であるLockら(Microb.Pathog.4:33−43,1988)のアッセイで測定されたように、非無毒化ノイラミニダーゼと比較して低下した活性を示すノイラミニダーゼである。Lockのアッセイを使用して、溶解物、血清、または血液中のNanA活性は、酵素アッセイで2’−(4−メチル−ウンベリフェリル)−α−D−N−アセチルノイラミン酸(acetlyneuraminic acid)を基質として用いて、測定される(Lock et al.1988)。基質10マイクロリットルを血清10μlと合せて、37℃にて5分間インキュベートする。0.5M炭酸ナトリウムを使用して反応を停止させる。ノイラミニダーゼ活性を1分間当たりに放出される4−メチルウンベリフェロン(MU)の量で測定する。MUは、励起波長366nMおよび放出波長445nmを有する。無毒化ノイラミニダーゼは、免疫組成物を作成するために薬学的に受容可能なキャリアと組合されるように、非無毒化ノイラミニダーゼと匹敵する抗原性または免疫原性を保持することが好ましい。比較の目的で、非無毒化ノイラミニダーゼはこれに限定されるわけではないが、表1に示すR6 NanAを含む。好ましい実施形態において、無毒化ノイラミニダーゼは、非無毒化ノイラミニダーゼの活性の少なくとも60%、70%、80%、または90%を示す。
【0033】
無毒化ノイラミニダーゼは、非無毒化ノイラミニダーゼと比較して、アミノ酸配列内に変更(すなわち置換、修飾、または欠失)を含む。好ましい実施形態において、無毒化ノイラミニダーゼは、非無毒化ノイラミニダーゼ内に見られるアミノ酸のおよそ7%、10%、15%または20%の変更を含む。好ましいアミノ酸欠失は、非無毒化ノイラミニダーゼのN末端からのアミノ酸ほぼ5、10または15個の欠失を含む。他の好ましい実施形態は、非無毒化ノイラミニダーゼのC末端のアミノ酸ほぼ60、50、40、30、20、10または5個の欠失を含む(本出願では、C末端は表1に示すように、R6 NanAのアミノ酸800にて始まる)。なお他の好ましい実施形態では無毒化ノイラミニダーゼは、非無毒化ノイラミニダーゼのC末端の17、9、8、7、4または2のアミノ酸の欠失を含む。好ましい欠失のある例を表1に示す(すなわちTIGR4 NanAアミノ酸配列)。これらの変更のいずれも、1つ以上の他の変更と組み合せることができる。そのような無毒化ノイラミニダーゼ種は、非無毒化ノイラミニダーゼの活性のおよそ60%、70%、80%または90%を示すことが好ましい。
【0034】
ノイラミニダーゼ(neuramindiase)の他の保存的および非保存的置換(substation)は、ノイラミニダーゼがその抗原性または免疫原性を維持している限り使用できる。これらの保存的置換は、天然に存在するアミノ酸が同じ特性を有する天然に存在するアミノ酸によって置換されるようになっている。そのような保存的および非保存的置換は必要に応じて、ポリペプチドの酵素機能を変化させる。たとえば保存的置換は、表2に従って行える。
【0035】
【表2】
所望の場合、修飾および変化は、本発明のポリペプチドをコード化する核酸および/または本発明のポリペプチドのアミノ酸配列において行われ、同様のまたは他の所望の特徴(たとえば抗原性または免疫原性)を有するポリペプチドがなお得られることが理解される。そのような変化は、天然単離物で発生するか、または部位特異性突然変異誘発を使用して合成により導入され、ミスマッチポリメラーゼ連鎖反応(PCR)などの手順が当業界で周知である。たとえばあるアミノ酸は、機能活性の認識できる損失なしに、ポリペプチド中の他のアミノ酸と置換される。それゆえ生物有用性または活性の認識できる損失なしに、そしておそらくはそのような有用性または活性の上昇を伴って、様々な変更が本発明のポリペプチドのアミノ酸配列(または基礎を成す核酸配列)中で行われることが検討される。
【0036】
nanA遺伝子またはnanA遺伝子のいずれかの部分の欠失は、その中で教示された方法について、参照によりその全体が本明細書に組み入れられているSung et al.,(2001)Appl Environ Microbiol 67:5190−5196によって述べられている方法を使用して実施される。試薬2,3ブタジオンは、タンパク質のArg残基と特異的に反応し、NanA分子の折畳みに対するArg残基の重要性を評価するために使用される。部位特異的突然変異誘発は、特異的アミノ酸を変更するために使用される。
【0037】
ノイラミニダーゼは、たとえば変性を含む化学処理によっても無毒化できる。化学処理は、さらに負の副作用を低減し、抗原性または免疫原性を改善するために、部位特異的突然変異誘発と組み合せることもできる。無毒化ノイラミニダーゼは、無毒化ノイラミニダーゼによる被験体の免疫化の前に、ホルマリン、グルタルアルデヒド、熱などの作用因子によって、または当業者に既知の他の作用因子によって処理できる。
【0038】
それゆえ本明細書では、無毒化肺炎球菌ノイラミニダーゼあるいはその抗原性または免疫原性部分が提供される。無毒化肺炎球菌ノイラミニダーゼおよび薬学的に受容可能なキャリアを含む組成物も提供される。必要に応じて組成物は、アジュバント(たとえば粘膜アジュバントを含む)をさらに含む。
【0039】
さらに、たとえば抗原性または免疫原性を上昇させる部分を含む、部分をノイラミニダーゼに添加できる。そのような部分はたとえば、サイトカイン、ケモカイン、成長因子、血管由来因子、アポトーシス阻害物質、ホルモン、毒素、またはアジュバントとして使用するための本明細書で述べた他の部分を含む。部分は必要に応じて、変更分子中で使用するために修飾または切断できる。それゆえ本明細書では、ノイラミニダーゼあるいはその抗原性または免疫原性フラグメントおよび抗原性または免疫原性を向上させる部分を含む、肺炎球菌ノイラミニダーゼキメラが提供される。肺炎球菌ノイラミニダーゼ誘導体および薬学的に受容可能なキャリアを含む組成物も提供される。必要に応じて組成物は、アジュバント(たとえば粘膜アジュバントを含む)をさらに含む。
【0040】
必要に応じて、本発明の修飾ノイラミニダーゼフラグメントまたはその部分は、天然型肺炎球菌ノイラミニダーゼまたはそのフラグメントと少なくとも約70%の相同性を備えたアミノ酸配列を有する。修飾ノイラミニダーゼまたはそのフラグメントをコード化する核酸がさらに提供される。いずれかの既知の改変体および誘導体、あるいは本明細書で開示された核酸およびタンパク質の発生する改変体および誘導体を定義する1つの方法は、特定の既知配列への相同性に関する改変体および誘導体の定義を通じてであることが理解される。たとえば表1に示すようにR6肺炎球菌性菌株のnanA遺伝子によってコード化されたアミノ酸配列は、肺炎球菌ノイラミニダーゼの特定の配列を説明し、タンパク質の特定のアミノ酸配列を説明する。示された配列に対して少なくとも70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99パーセントの相同性を有する、本発明で開示されたこの配列の改変体が特に開示される。当業者は、2つのタンパク質または核酸の相同性を決定する方法をただちに理解するであろう。たとえば相同性は、相同性がその最高レベルとなるように2つの配列を整列させた後に、計算できる。
【0041】
相同性を計算する別の方法は、公開されたアルゴリズムによって実施できる。比較のための配列の最適アラインメントは、Smith and Waterman Adv.Appl.Math.2:482(1981)の局所相同性アルゴリズムによって、Needleman and Wunsch,J.Mol Biol.48:443(1970)の相同性アラインメントアルゴリズムによって、Pearson and Lipman,Proc.Natl.Acad.Sci.U.S.A.85:2444(1988)の類似度方法の検索によって、これらのアルゴリズムのコンピュータによる実施によって(Wisconsin Genetics Software PackageのGAP、BESTFIT、FASTA、およびTFASTA、Genetics Computer Group, 575 Science Dr.,マジソン、ウィスコンシン州)、または検査によって実施される。核酸について同じタイプの相同性が、核酸アラインメントに関する少なくとも物質について、たとえば参照により本明細書に組み入れられている、Zuker,M.Science 244:48−52,1989,Jaeger et al.Proc.Natl.Acad.Sci.USA 86:7706−7710,1989,Jaeger et al.Methods Enzymol.183: 281−306,1989で開示されたアルゴリズムによって得られる。
【0042】
「肺炎球菌性ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸」は、肺炎球菌性細菌中に存在するホスホコリン、テイコ酸、またはリポテイコ酸を意味する。これらの化合物は、ノイラミニダーゼについて上述したように、修飾、無毒化または強化することができる。本明細書では、修飾、無毒化、および強化された化合物を含む組成物が提供される。
【0043】
「その抗原部分」とは、抗体産生を誘発する分子または化合物(たとえばノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸)のいずれかのエピトープを意味し、該抗体は該分子に向けられている。好ましくは、抗原部分は分子またはS.ニューモニエに対する免疫を誘発する。好ましくは、抗体はノイラミニダーゼの活性部位に向けられているか、または活性部位を妨害する。抗原性フラグメントの例は、これに限定されるわけではないが、保存的アミノ酸置換または修飾の存在下または非存在下でのnanA−R6アミノ酸配列の残基63〜361に該当する残基を含む。他の例は、保存的アミノ酸置換または修飾の存在下または非存在下で、ノイラミニダーゼのAsp領域(NanA−R6のアミノ酸配列の残基460〜480、530〜560、610〜620に該当する)およびNanA−R6アミノ酸配列の340〜350、360〜370、600〜610、570〜580および750〜760に該当する領域を含む。必要に応じて、ノイラミニダーゼの活性部位を妨害するように方向付けられた抗体は、配列番号15の残基347、367または605に位置するNanAのアルギニン残基に、または配列番号15の残基383〜387、467〜473、541〜546および610〜616に位置するNanAのAspボックスに結合するか、またはその結合を防止することができる。その上、抗体は、配列番号15の残基575に位置するバリン、または配列番号15の残基752に位置するチロシンに結合するか、またはその結合を防止することもできる。抗体は、配列番号15の上で示した残基のいずれかの組合せに結合するか、またはその結合を防止することもできる。
【0044】
NanAフラグメントの他の例は、アミノ酸1〜340、330〜630、620〜800、700〜1030および330〜800を含む。これらのフラグメントの2つ以上の融合物であるフラグメントもさらに提供される。融合フラグメントは、これに限定されるわけではないが、領域620〜680と融合された領域1〜340を含む。融合フラグメントは、組換えタンパク質として発現される。これらのフラグメントをコード化するフラグメントは、発現ベクター(pETベクター;Novagen,Inc.)内へクローニングされ、タンパク質が精製される。あるいはフラグメントは、ベンダーからの合成ポリペプチドとして産生される。フラグメントを使用して、動物を免疫化して抗体を産生させて、3次元構造を評価するために結晶化させる。本明細書で述べるように、保存的または非保存的アミノ酸修飾、または置換の存在下または非存在下でのフラグメントをコード化する核酸も提供される。核酸を含むベクターまたは発現系も提供される。
【0045】
本明細書では、肺炎球菌ノイラミニダーゼ、ホスホコリン、テイコ酸、リポテイコ酸、あるいはこれらの1つまたは両方の抗原部分に特異的に結合する単離抗体を含む組成物も提供される。抗体のいずれかの組合せを含む組成物も提供される。そのような抗体は、S.ニューモニエに対する受動免疫を発現させるのに有用である。抗体組成物はさらに、薬学的に受容可能なキャリアを含む。必要に応じて組成物は、粘膜表面への投与に適切であるが、上述のような全身投与を含む他の投与経路も開示されている。
【0046】
本明細書では、肺炎球菌ノイラミニダーゼ、ホスホコリン、テイコ酸、リポテイコ酸、あるいは肺炎球菌ノイラミニダーゼ、ホスホコリン、テイコ酸、またはリポテイコ酸のいずれかのエピトープに対して特異性の抗体を産生する方法も開示されている。必要に応じて抗体は、被験体の鼻粘膜に本明細書で開示された組成物の有効量を接触させることによって被験体内で(すなわち、生体内)産生される。被験体の鼻粘膜に標的分子または分子を標的とする抗体の組合せを含む組成物の有効量を接触させることによって、肺炎球菌ノイラミニダーゼ、ホスホコリン、テイコ酸、リポテイコ酸、あるいは肺炎球菌ノイラミニダーゼ、ホスホコリン、テイコ酸、またはリポテイコ酸のいずれかのエピトープのいずれかの組合せに対して特異性である抗体を産生する方法も開示される。
【0047】
必要に応じて、本明細書で述べる作用因子は、天然型であるか、無毒化または、そうでなければ修飾されたかにかかわらず、必要に応じて、たとえばベクター内の核酸として投与できる。核酸の発現は次に、被験体へのそれによって発現された所望の核酸の接触を引き起こす。それゆえたとえば、肺炎球菌ノイラミニダーゼをコード化する核酸が被験体に被験体内で発現できる形で投与する場合、次に被験体にノイラミニダーゼを接触させる。
【0048】
被験体の細胞内への外来性DNAの投与および摂取(すなわち遺伝子形質導入または形質移入)を含む本明細書で述べた方法において、開示された核酸は裸のDNAまたはRNAの形でありうるか、あるいは核酸は、核酸を細胞に送達するベクターであり、それによって抗体コード化DNAフラグメントは、当業者によって十分理解されるように、プロモータの転写調節の支配下にある。ベクターは、アデノウィルスベクター(Quantum Biotechnologies,Inc.(ラヴァル、ケベック、カナダ)などの市販の調製物でありうる。核酸またはベクターの細胞への送達は、多様な機構を介することができる。一例として、送達は、市販のリポソーム調製物、たとえばLIPOFECTIN、LIPOFECTAMINE(GIBCO−BRL,Inc.、ゲティスバーグ、メリーランド州)、SUPERFECT(Qiagen,Inc.ヒルデン、ドイツ)およびTRANSFECTAM (Promega Biotec, Inc.、マジソン、ウィスコンシン州)はもちろんのこと、当業界で標準的な手順に従って開発された他のリポソームを使用して、リポソーム経由で行われる。加えて、開示された核酸またはベクターは、遺伝子銃または他の送達方法、たとえばGenetronics,Inc.(サンディエゴ、カリフォルニア州)から入手できる技術である電気穿孔法によってはもちろんのこと、SONOPORATION装置(ImaRx Pharmaceutical Corp.、ツーソン、アリゾナ州)によっても生体内送達できる。
【0049】
一例として、ベクター送達は、ウィルス系、たとえば組換えレトロウィルスゲノムをパッケージできるレトロウィルスベクターを介して行える(たとえばPastan et al.,Proc.Natl.Acad.Sci.U.S.A.85:4486,1988;Miller et al.,Mol.Cell.Biol.6:2895,1986を参照)。次に組換えレトロウィルスを感染させるために使用して、それによってたとえば肺炎球菌ノイラミニダーゼまたは幅広い中和抗体(またはその活性フラグメント)をコード化する感染した細胞核酸に送達する。変化した核酸を哺乳類細胞に導入する正確な方法はもちろん、レトロウィルスベクターの使用に限定されない。この手順では、アデノウィルスベクター(Mitani et al.,Hum.Gene Ther.5:941−948,1994)、アデノ結合ウィルス(AAV)ベクター(Goodman et al.,Blood 84:1492−1500,1994)、レンチウィルスベクター(Naidini et al.,Science 272:263−267,1996)、シュードタイプレトロウィルスベクター(Agrawal et al.,Exper.Hematol.24:738−747,1996)を含む他の技法が広範に利用できる。物理的形質導入技法、たとえばリポソーム送達およびレセプタ仲介および他のエンドサイトーシス機構も使用できる(たとえばSchwartzenberger et al.,Blood 87:472−478,1996を参照)。このように開示された組成物および方法は、これらのまたは他の一般に使用される遺伝子導入方法のいずれかと併せて使用できる。
【0050】
一例として、抗体コード化核酸がアデノウィルスベクター内の被験体の細胞に送達される場合、ヒトへのアデノウィルス投与の投薬量は、注射1回当たり約107〜109プラーク形成単位(pfu)の範囲でありうるが、注射1回当たり1012pfuにもなりうる(Crystal,Hum.Gene Ther.8:985−1001,1997;Alvarez and Curiel,Hum.Gene Ther.8:597−613,1997)。被験体には1回の注射を投与できるか、または追加の注射が必要な場合は、それらは無期限に亘っておよび/または処置の有効性が確立されるまで、6ヶ月の間隔(または当業者によって決定されるように、他の適切な期間)で反復できる。
【0051】
核酸またはベクターの非経口投与は使用される場合、一般に注射を特徴とする。注射可能物質は、液体液剤または懸濁剤のどちらかとしての従来形で、注射前の液体への懸濁剤の溶解に適した固体形で、あるいは乳剤として調製できる。非経口投与のより最近修正された手法は、一定の投薬量が維持されるような低速放出または持続放出系の使用を包含する。たとえば参照により本明細書に組み入れられている、U.S.Patent No.3,610,795を参照。治療化合物の適切な処方および各種の投与経路のさらなる議論については、たとえばRemington: The Science and Practice of Pharmacy(19th ed.)ed.A.R.Gennaro,Mack Publishing Company,イーストン、ペンシルベニア州、1995を参照。
【0052】
被験体の鼻粘膜への本明細書で開示された組成物の有効量の接触を含む、被験体における肺炎球菌の鼻輸送を低減または防止する方法も開示される。そのような投与は、肺炎球菌感染または鼻輸送に対する能動または受動免疫あるいは防御を生成するのに有用でありうる。
【0053】
被験体の粘膜表面への本発明で開示した組成物の有効量の接触を含む、被験体における肺炎球菌感染を低減または防止する方法がさらに提供される。たとえば該方法は、肺炎球菌性髄膜炎、中耳炎、肺炎、または溶血性尿毒症を防止できる。防止または低減は、鼻輸送を低減すること、またはCNS浸潤、全身性浸潤、あるいはエウスタキー管または下気道を防止することによって行える。
【0054】
化合物の「有効量」とは本明細書で規定されるように、所望の結果を提供するための、化合物の非毒性であるが十分な量を意味する。以下で指摘されるように、正確な必要量は、被験体の種、年齢、および全身状態、処置される疾患の重症度、使用された特定の化合物、その投与方法などによって、被験体ごとに変化するであろう。それゆえ正確な「有効量」を規定することは可能ではない。しかしながら適切な有効量は、日常的な実験のみを使用して、当業者が決定できる。
【0055】
本明細書で述べる組成物の投薬量または量は、送達が行われる方法で所望の効果を生じるのに十分な多さである。投薬量は、有害な副作用、たとえば望ましくない交叉反応、アナフィラキシー反応などを引き起こすほど多くするべきではない。一般に投薬量は、年齢、状態、性別および被験体における疾患の程度によって変化し、当業者によって決定できる。投薬量は、関与する被験体の臨床状態に基づいて個々の医師が調整することができる。用量、投薬スケジュールおよび投与経路は変化しうる。抗原の鼻腔内投与の好ましい投薬量は、免疫化1回当たり約1〜1000μg、またはたとえば10〜100μgを含む、その間の任意の量を含む。
【0056】
本明細書で述べた方法による化合物または組成物の特定の用量の投与の有効性は、病歴、徴候、症状、および肺炎球菌感染した被験体または肺炎球菌性キャリアである被験体の状態を評価するのに有用であることが既知である客観的臨床試験の特定の態様を評価することによって判定できる。これらの徴候、症状、および客観的臨床試験は、そのような患者を処置する臨床医またはこの分野で実験を行う研究者に既知となるように、処置または防止される特定の疾患または状態によって変化する。たとえば適切なコントロール群および/または一般個体群または特定の個体における疾患の正常な進行の知識との比較に基づいて:1)被験体の身体的状態の改善が示される(たとえば鼻輸送が低減または除去される)、2)疾患、感染、または鼻輸送の進行が安定、減速、または後退していることが示される、あるいは3)疾患または状態を処置する他の作用因子の必要性が低下または除去される場合、特定の処置計画は有効と見なされるであろう。たとえば被験体または個体群における鼻輸送の低減または防止、CNS浸潤または他の2次肺炎球菌感染の発生の回避または低減は、有効性を示す。そのような効果は、単一の被験体(たとえば従来の粘膜表面拭取りによって検出される細菌数を減少させることによって)または個体群(たとえば疫学的研究を使用)にて決定できる。
【0057】
本明細書で述べる化合物および薬学的組成物は、局所または全身処置が望ましいかどうかによって、そして処置される部位によって多数の方法で被験体に投与することができる。それゆえたとえば、本明細書で述べる化合物または薬学的組成物は、静脈内に、皮下に、筋肉内に、リポソームまたは微小球でカプセル化して、点眼液および/または軟膏として目の表面へ、鼻スプレーとして、噴霧液として、またはエアゾールとして鼻腔または気道へ投与することができる。その上、化合物または薬学的組成物は、被験体へ経膣的に、経直腸的に、経鼻的に,経口的に、吸入により、経口的に、または挿管によって投与できる。必要に応じて組成物は、静脈内、皮下、筋肉内、または腹腔内注射によって投与される。組成物は、液体液剤または懸濁剤のどちらかとしての従来形で、液体への懸濁剤または液剤に適した固体形で、あるいは乳剤として調製できる。必要に応じて投与は、一定の投薬量が維持されるような低速放出または持続放出系による。たとえばそこで教示された方法について、参照により本明細書に組み入れられているU.S.Patent No.3,610,795を参照。
【0058】
本明細書で教示した組成物は、緩衝剤、希釈剤および他の適切な添加剤も含有できる滅菌水性または非水性液剤、懸濁剤、および乳剤を含む。非水性溶媒の例は、プロピレングリコール、ポリエチレングリコール、オリーブ油などの植物油、およびエチルオレアートなどの有機エステルである。水性キャリアは、食塩水および緩衝溶媒を含む、水、アルコール性/水性液剤、乳剤または懸濁剤である。ビヒクルは、塩化ナトリウム溶液、リンゲルデキストロース、デキストロースおよび塩化ナトリウム、乳酸リンゲル液、または不揮発性油を含む。保存料および他の添加剤、たとえば抗菌剤、抗酸化剤、キレート化剤、および不活性ガスなども存在できる。
【0059】
局所投与の処方物は、軟膏、ローション、クリーム、ゲル、滴剤、坐剤、スプレー、液剤、エアゾール、ネブライザ溶液および粉剤を含むことができる。従来の製薬キャリア、水性、粉末または油性ベース、増粘剤などは必要であるか、望ましい。
【0060】
経口投与用の組成物は、粉剤または顆粒剤、水または非水性溶媒中の懸濁剤または液剤、カプセル、サシェ、あるいは錠剤を含むことができる。増粘剤、着香剤、希釈剤、乳化剤、分散助剤または結合剤が望ましい。
【0061】
本明細書では、被験体にノイラミニダーゼインヒビタの有効量を投与する工程を含む、被験体における鼻輸送または肺炎球菌感染を低減または防止する方法が提供される。ノイラミニダーゼ阻害物質は好ましくは、被験体の内因性ノイラミニダーゼを著しく低減することなく、肺炎球菌ノイラミニダーゼ活性を阻害する。それゆえたとえば、ノイラミニダーゼがヒトに投与される場合、阻害物質は好ましくは、負の副作用がヒトに生じるように、ヒトノイラミニダーゼ活性を低減することなく、またはヒトノイラミニダーゼ活性を低減することなく、肺炎球菌ノイラミニダーゼを阻害する。既知のノイラミニダーゼインヒビタの例は、DANA、NANA、ザナミビルおよびオセルタミビルを含む。
【0062】
本明細書では、被験体に肺炎球菌ノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸に対する抗体またはそのフラグメント、あるいはこれらのいずれか1つの部分に対する抗体を含む組成物の有効量を投与する工程を含む、被験体における鼻輸送および肺炎球菌感染を低減または防止する方法である。必要に応じてこの投与は、被験体の粘膜表面に組成物を接触することを含む。組成物および抗体を含有する容器も提供される。
【0063】
「ホスホコリン抗体」という用語は本明細書で使用するように、ホスホコリンまたはその抗原性フラグメントに優先的に結合する抗体を指す。本発明の抗体は、肺炎球菌性テイコ酸または肺炎球菌性リポテイコ酸またはその抗原部分にも、あるいはノイラミニダーゼまたはそのフラグメントにも優先的に結合できる。
【0064】
「抗体」という用語は、本明細書で広範な意味で使用され、ポリクローナルおよびモノクローナル抗体の両方を含む。2重または多重抗原またはエピトープ特異性を備えたキメラ抗体、およびハイブリッド抗体、ならびにハイブリッドフラグメントを含む、F(ab’)2、Fab’、Fab、scFvなどのフラグメントも、組成物および方法で使用できる。それゆえその特異性抗原を結合する能力を維持する抗体のフラグメントが提供される。たとえばノイラミニダーゼ、ホスホコリン、テイコ酸、またはリポテイコ酸結合活性を維持する抗体のフラグメントは、用語「抗体フラグメント」の意味に含まれる。そのような抗体およびフラグメントは、当業界で既知の技法によって作成可能であり、実施例において、ならびに抗体を産生し、抗体を特異性および活性についてスクリーニングする一般的方法において述べられる方法に従って、特異性および活性についてスクリーニングできる(Harlow and Lane.Antibodies,A Laboratory Manual.Cold Spring Harbor Publications,New York,(1988)を参照)。
【0065】
本発明の組成物では、抗体フラグメントおよび抗原結合タンパク質(単鎖抗体)の結合体を使用することができる。そのような結合体は、その内容が参照により本明細書に組み入れられている、たとえばU.S.Pat.No.4,704,692に述べられている。抗体は、その所望の活性について、試験管内アッセイを使用して、または類似の方法によって試験可能であり、その後、その生体内治療および/または予防活性が、既知の臨床試験方法に従って試験される。
【0066】
「モノクローナル抗体」という用語は本明細書で使用するように、抗体の実質的に均質な個体群から得られた抗体を指し、すなわち個体群内の個々の抗体は、抗体分子の小さいサブセットに存在する考えられる天然型変異を除いて同一である。開示されたモノクローナル抗体は、モノクローナル抗体を産生するいずれかの手順を使用して作成できる。たとえば開示されたモノクローナル抗体は、Kohler and Milstein,Nature,256:495(1975)によって述べられたような、ハイブリドーマ法を使用して調製できる。ハイブリドーマ法では、マウスまたは他の適切な宿主動物は通例、免疫剤によって免疫化されて、免疫剤に特異的に結合する抗体を産生または産生できるリンパ球を発現させる。あるいはリンパ球はたとえば、本明細書で述べたHIV Env−CD4−co−レセプタ複合体を使用して、試験管内で免疫化できる。
【0067】
モノクローナル抗体は、U.S.Pat.No.4,816,567(Cabilly et al.)に述べられている方法などの、組換えDNA法によっても産生できる。開示されたモノクローナル抗体をコード化するDNAは、従来の手順を使用してただちに単離および配列決定できる(たとえばマウス抗体の重鎖および軽鎖をコード化する遺伝子に特異的に結合できるオリゴヌクレオチドプローブを使用することによって)。抗体または活性抗体フラグメントのライブラリも、たとえばBurtonらへのU.S.Patent No.5,804,440およびBarbasらへのU.S.Patent No.6,096,441に述べられているようなファージ提示技法を使用して産生およびスクリーニングできる。
【0068】
試験管内方法も、1価抗体を調製するのに適切である。そのフラグメント、特にFabフラグメントを生成するための抗体の消化は、当業界で既知の日常技法を使用して達成できる。たとえば消化は、パパインを使用して実施できる。パパイン消化の例は、1994年12月22日に公開されたWO94/29348およびU.S.Pat.No.4,342,566に述べられている。抗体のパパイン消化は通例、1個の抗原結合部位、および残りのFcフラグメントをそれぞれ備えたFabフラグメントと呼ばれる2つの同じ抗原結合フラグメントを生成する。ペプシン処置は、2つの抗原結合部位を有するフラグメントを生成し、なお抗原を架橋することができる。
【0069】
抗体フラグメントは、他の配列に結合しているかいないかにかかわらず、抗体または抗体フラグメントの活性が非修飾抗体または抗体フラグメントと比較して著しく変化または阻害されないという条件で、特定の領域または特異的アミノ酸残基の挿入、欠失、置換、あるいは他の選択された修飾を含むことができる。これらの修飾は、ジスルフィド結合可能なアミノ酸を除去および/または付加する、その生体寿命を延長する、その分泌特徴を変化させるなどの、一部の追加特性を提供できる。いずれの場合でも、抗体または抗体フラグメントは、その同種抗原への特異的結合などの生物活性特性を所有しなければならない。抗体または抗体フラグメントの機能または活性領域は、タンパク質の特異的領域の突然変異誘発と、それに続く発現ポリペプチドの発現および試験によって同定できる。そのような方法は、当業者にただちに明らかとなり、抗体または抗体フラグメントをコード化する核酸の部位特異的突然変異誘発を含むことができる(Zoller,M.J.Curr.Opin.Biotechnol.3:348−354,1992)。
【0070】
本明細書で使用するように、「抗体(antibody)」または「抗体(antibodies)」という用語は、ヒト抗体および/またはヒト化抗体を指すこともできる。多くの非ヒト抗体(たとえマウス、ラット、またはウサギに由来するもの)は当然、ヒトにおいて抗原性であり、それゆえヒトに投与したときに、望ましくない免疫反応を生じさせる可能性がある。したがって方法におけるヒトまたはヒト化抗体の使用は、ヒトに投与された抗体が望ましくない免疫反応を誘起する機会を減少させる役割を果たす。それゆえ抗体を含む組成物は必要に応じて、ヒト化または完全ヒト抗体を含む。抗体ヒト化技法は一般に抗体分子の1つ以上のポリペプチド鎖をコード化するDNA配列を操作する組換えDNA技術の使用を含む。したがって非ヒト抗体(またはそのフラグメント)のヒト化形は、キメラ抗体または抗体鎖(またはそのフラグメント、たとえばFv、Fab、Fab’、あるいはヒト(レシピエント)抗体の枠組みへ組み込まれた非ヒト(ドナー)抗体からの抗原結合部位の部分を含有する抗体の他の抗原結合部分)である。
【0071】
開示されたヒト抗体は、いずれの技法を使用しても産生できる。ヒトモノクローナル抗体産生の例は、Coleら(Monoclonal Antibodies and Cancer Therapy,Alan R.Liss,p.77,1985)によって、そしてBoernerら(J.Immunol.,147(1):86 95,1991)によって述べられているものを含む。ヒト抗体(およびそのフラグメント)も、ファージ提示ライブラリを使用して産生できる(Hoogenboom et al.,J.Mol. Biol.,227:381,1991;Marks et al.,J.Mol.Biol.,222:581,1991)。
【0072】
開示したヒト抗体は、トランスジェニック動物からも得ることができる。たとえば免疫化に反応してヒト抗体の完全レパートリーを産生できるトランスジェニックの変異体マウスが説明されている(たとえばJakobovits et al.,Proc. Natl.Acad.Sci.USA,90:2551 255(1993);Jakobovits et al.,Nature,362:255 258(1993);Bruggermann et al.,Year in Immunol.,7:33(1993)を参照)。特に、これらのキメラおよび生殖細胞系変異体マウスにおける抗体重鎖結合領域(J(H))遺伝子の同型欠失は、内因性抗体産生の完全な阻害を生じ、ヒト生殖細胞系抗体遺伝子アレイの生殖細胞系変異体マウスへの移入の成功は、抗原攻撃時にヒト抗体の産生を引き起こす。所望の活性を有する抗体は、本明細書で述べるようにEnv−CD4−co−レセプタ複合体を使用して選択される。
【0073】
ヒト化抗体を産生するためには、レシピエント(ヒト)抗体分子の1つ以上の相補性決定領域(CDR)からの残基は、所望の抗原結合特徴(たとえば標的抗原に対するあるレベルの特異性および親和性)を有することが既知であるドナー(非ヒト)抗体分子の1つ以上のCDRからの残基によって置換される。ある例において、ヒト抗体のFv枠組み(FR)残基は、対応する非ヒト残基によって置換される。ヒト化抗体は、レシピエント抗体にも、移入されたCDRまたは枠組み配列にも見出されない残基も含む。一般にヒト化抗体は、非ヒトである供給源からそれに導入された1つ以上のアミノ酸残基を有する。実際には、ヒト化抗体は通例、一部のCDR残基およびおそらく一部のFR残基がげっ歯類抗体中の類似部位からの残基によって置換されるヒト抗体である。ヒト化抗体は一般に、抗体定常領域(Fc)の少なくとも一部、通例、ヒト抗体のそれを含有する(Jones et al.,Nature,321:522 525(1986)、Reichmann et al.,Nature, 332:323 327(1988)、およびPresta,Curr.Opin.Struct.Biol.,2:593 596(1992))。
【0074】
非ヒト抗体をヒト化する方法は、当業界で周知である。たとえばヒト化抗体は、Winterおよび共同研究者(Jones et al.,Nature,321:522 525(1986),Riechmann et al.,Nature,332:323 327(1988),Verhoeyen et al.,Science,239:1534 1536(1988))の方法に従って、げっ歯類CDRまたはCDR配列をヒト抗体の対応する配列と置換することによって産生できる。ヒト化抗体の産生に使用できる方法は、U.S.Patent No.4,816,567(Cabilly et al. )、U.S. Patent No.5,565,332(Hoogenboom et al.)、U.S.Patent No.5,721,367(Kay et al.)、U.S.Patent No.5,837,243(Deo et al.)、U.S.Patent No.5,939,598(Kucherlapati et al.)、U.S.Patent No.6,130,364(Jakobovits et al.)、およびU.S.Patent No.6,180,377(Morgan et al.)でも説明されている。
【0075】
抗体の投与は、本明細書で開示するように実施できる。抗体送達のための核酸手法も存在する。抗体および抗体フラグメントも、被験体自体の細胞が核酸を吸収してコード化抗体または抗体フラグメントを産生および分泌するように、患者または被験体に、抗体または抗体フラグメントをコード化する核酸調製物(たとえばDNAまたはRNA)として投与できる。核酸の送達は、当業界で既知のいずれの手段によってもよい。
【0076】
本明細書では、本明細書で開示した作用因子および組成物を含む容器も開示する。特に容器は、鼻スプレー器、ネブライザ、吸入器、ボトル、または組成物を粘膜表面への投与形で含有する他のいずれかの手段でありうる。必要に応じて容器は、組成物の計量用量を送達できる。
【実施例】
【0077】
以下の実施例は、当業者に本明細書で説明および請求する化合物、組成物、物品、器具、および/または方法が作成および評価される方法の完全な開示および説明を提供するために与えられ、純粋な例示であるものとし、発明者がその発明と見なすものの範囲を限定するものではない。数(たとえば量、温度など)に関して正確を期するよう努力がなされたが、多少の誤差および逸脱が説明されるはずである。別途指示しない限り、部は重量部であり、温度は℃または周囲温度であり、圧力は大気圧またはその付近である。反応状態、たとえば成分濃度、所望の溶媒、溶媒混合物、温度、圧力、ならびに生成物の純度および説明した工程から得られた収量を最適化するために使用できる他の反応範囲および条件の、多数の変形および組合せがある。そのような工程状態を最適化するためには、合理的かつ日常的な実験のみが必要となるであろう。
【0078】
(実施例1)
(輸送中に嗅覚組織に浸透する鼻肺炎球菌)
(材料および方法)
(肺炎球菌性菌株)
S.ニューモニエEF3030、血清型19F、およびTIGR4菌株、血清型4の2つのカプセル化菌株、ならびに親菌株D39、血清型2に由来する無毒性の非カプセル状菌株R36Aを利用した。Avery et al.,(1944)J.Exp.Med.79:137−158。EF3030菌株は、菌血の非存在下で気道にただちにコロニー形成し(Briles et al.,(1992)Infect.Immun.60: 111−116)、静脈内接種後に菌血を持続できないため選択された。TIGR4菌株はさらに毒性であったが、中程度の鼻接種原によって、菌血なしにコロニー形成する。
【0079】
(マウス)
CBA/CAHN/xid(xid)マウス菌株をJackson Laboratory(バーハーバー、メイン州)から得た。これらマウスのBrutonのチロシンキナーゼ遺伝子における変異は、胸腺独立性II型抗原への反応をできなくするが(Amsbaugh et al.,(1972)J.Exp.Med.136:931−949;Berning et al.,(1980)J.Immunol.46:506−513)、比較的な正常なT細胞依存性免疫反応を可能にする。これらのマウスは、カプセル状ポリサッカライドに反応できず、肺炎球菌感染を高い再現性で受けやすい。xidマウスは、病原体のない状態に維持され、7〜12週齢で使用される。
【0080】
(組織収集)
血液を後腹膜神経叢からヘパリン処理毛細管内に収集した。鼻洗浄液(NW)、腎臓、脾臓、および肺の収集前にマウスを70%エタノールによって消毒した。NWの血液汚染を防止するために、気管への切開を行って、外径0.075cm、2.0cm長のTygon管(Cole−Parmer、バーノンヒルズ、イリノイ州)を上咽頭に挿入し、同時にリンゲル液を充填した注射器に取り付けた。注射器からの液体は鼻を通じて排出して、3滴を収集した。
【0081】
上咽頭関連リンパ細網組織(NALT)、ON/E、OBおよび脳の残りを上述のように得た。van Ginkel et al.,(2000)J.Immunol.165:4778−4782;Wu et al.,(1997)Scand.J.Immunol 46:506−513。脳から三叉神経節を、解剖顕微鏡を用いて慎重に切除した。ON/E、OB、三叉神経節、NALTおよび頚部リンパ節CLNそれぞれをリンゲル液0.5ml中でホモジナイズし、脳および腎臓をそれぞれリンゲル液1.0ml中でホモジナイズした。
【0082】
(組織ミンス/血液/外部排出物中の肺炎球菌の量)
組織および体液の連続3倍希釈物8個を滅菌リンゲル液中で作成し、ゲンタマイシンサルフェート4μg/mlを含有する血液寒天プレートで平板培養した。CFUは平板培養およびロウソク瓶内でのインキュベーションの24時間後に数えた。結果は、CFU/器官、NWまたは血液1ml当たりのCFUとして表した。
【0083】
(S.ニューモニエEF3030のGLSプレインキュベーション)
GLS結合部位を遮断するために、S.ニューモニエ菌株EF3030 3X107CFUを、ヒト脳からのアシアロ−GM1 20μgまたはウシ脳からの混合GLS(18% GM1、55% GD1a、15% GD1b、10% GT1b、2% 他のGLS)125μgのどちらかを用いて、氷上で30分間インキュベートした(Calbiochem−Novabiochem Corporation,Inc.,ラホーヤ、カリフォルニア州)。GLSをPBSに溶解させ、使用1日前に完璧に混合した。両親媒性GLSは、PBS中でミセルを形成し、肺炎球菌に炭水化物部分と相互作用させる。インキュベーションの後、さらに洗浄せずに鼻孔ごとに5μlをxidマウスに鼻腔内投与した。4日後に組織をCFUについて分析した。
【0084】
(PCRによるS.ニューモニエニューモリシン遺伝子の検出)
S.ニューモニエをPCRによって検出するために、組織を1% SDS中に0.1%デオキシコール酸を用いて凍結−解凍によって溶解させて、37℃で1時間インキュベートした。タンパク質は、セチルトリメチルアンモニウムブロミド/NaCl沈殿法を使用して除去した(Ausubel et al.,(1987)Current Protocols in Molecular Biology,2nd:2.4.4、セチルトリメチルアンモニウムブロミド/NaCl沈殿法の教示については、参照により本明細書に組み入れられている)。DNA 10μgをPCR増幅に使用した。ニューモリシン(ply)特異性プライマーPly1 5’−ATTTCTGTAACAGCTACCAACGA−3’(配列番号1)およびPly2 5’−GAATTCCCTGTCTTTTCAAAGTC−3’(配列番号2)をPCR混合物に添加して、400bpフラグメントを増幅した。PCR反応は、94℃での5分間の変性工程と、それに続く増幅サイクル、つまり94℃(1分)、55℃(1分)、および72℃(1分)の30サイクルを含んでいた。エチジウムブロミド染色PCRフラグメントの画像をAlpha Imager TM IS−3400(Alpha Innotech Corporation,サンリアンドロ、カリフォルニア州)で収集した。
【0085】
(PspA特異性Absを用いたOBの免疫蛍光染色)
マウスをTIGR4菌株5X105CFUで経鼻攻撃した。OBを10%緩衝ホルマリンで固定した。パラフィン画分4μm(van Ginkel(2000)J.Immunol.165:4778−4782)は、それらを室温にて加湿チャンバ内で4時間インキュベートすることによって、PspAファミリ2Abs(1:100)を染色した。スライドをPBSで洗浄して、ビオチン化ヤギF(ab’)2抗ウサギIgG(1:200)(Southern Biotechnology Associates,Inc.,バーミングハム、アラバマ州)で染色し洗浄して、ストレプトアビジン−FITC(1:100)(BD−PharMingen,サンディエゴ、カリフォルニア州)で染色した。蛍光画像は、Nikon顕微鏡によってDEI−750 CEデジタルカラービデオカメラ(Optronics,ゴレータ、カリフォルニア州)を用いて収集し、Scion Imageソフトウェア(Scion Corporation,フレデリック、メリーランド州)を用いて処理した。
【0086】
(統計)
データを平均±1標準誤差として表し、結果を統計解析により独立Mann Whitney2標本ランク検定またはStudent t検定を使用して比較して、CFUでの有意差を決定した。
【0087】
(結果)
(鼻腔内コロニー形成およびCNS浸潤での肺炎球菌性カプセルの役割)
一次感覚嗅覚ニューロンを通じた肺炎球菌の摂取を調査するために、EF3030および非カプセル化菌株R36Aが鼻道にコロニー形成して、CNSに侵入する能力を第1〜4日に測定した(図1)。第1日にON/Eでは両方の菌株で高いCFUが観察されたが、R36Aは第4日までにON/Eおよび他のすべての組織にほぼ非存在となり、長期のコロニー形成にはあるカプセルが必要であることを示した以前の結果と一致していた。Magee and Yother(2001)Infect.Immun.69:3755−3761。EF3030は、どちらの日でもOBおよび脳での明らかな存在を示し、第4日にはNWおよびNALTに多数が存在していた。これらの調査結果は、鼻腔内攻撃後のEF3030肺炎球菌のOBおよび脳への軸索輸送と一致していた。
【0088】
(鼻腔内コロニー形成およびCNS浸潤の動力学)
EF3030は、観察した39日間に亘ってすべての時点でON/E、OB、NW、およびNALTにおいて維持された(図2)。脳およびCLNでは、はるかに少ない数のCFUが見られ、存在したこれらのCFUは一般に、第18および25日に見られた。興味深いことに、肺は第1日(図1)および第18、25および39日を除いて、肺炎球菌を示さなかった(図2)。使用した経鼻用量では、菌株EF3030を用いて実施したいずれの実験の間にも、CFUがマウス血流で検出されなかったため、菌血は神経組織分布に寄与しなかった(図2)。鼻腔内投与の1、3、6、12および24時間後およびそれに続く1週間にわたって毎日、血液を菌血について監視した。血液中に細菌は検出されなかった。
【0089】
(三叉神経節のS.ニューモニエ感染)
三叉神経細胞は上咽頭を刺激し、それゆえS.ニューモニエは、鼻粘膜の感染後の三叉神経節中に予想される。これを試験するために、接種4日後に各種の組織および血液を単離し、新たな実験でのEF3030の存在について分析した。EF3030菌株は、ON/EおよびOBで、そして三叉神経節で検出された(表3)。この調査はさらに、アシアロ−GM1がS.ニューモニエによる神経標的化のレセプタとして機能することを裏付けた。他のGLSも同様に役割を果たしやすい。
【0090】
表3は、経鼻送達後の各種組織でのS.ニューモニエ菌株EF3030の分布を示す。菌株EF3030 1X107CFUの鼻腔内投与の4日後に組織を単離した。血液(50μl)、ON/E、OB、および脳組織ミンスを希釈して、次に血液寒天上で平板培養した。三叉神経節をプールして、ホモジナイズし、次にこの培地上で平板培養した。マウス5匹の平均肺炎球菌CFU±SEが示され、3つの独立した実験を表している。脳および血液では、肺炎球菌は検出されなかった。
【0091】
【表3】
(肺炎球菌性コロニー形成を阻害するガングリオシド)
EF3030菌株を鼻腔内投与前に、アシアロ−GM1または混合GLSミセルを用いてPBS中でインキュベートした。GLS混合物は、最強の阻害効果を示し、鼻腔内投与の4日後に評価したときに、NWにてCFUを10分の1に減少させた(P=0.0365)。混合GLSプレインキュベーションの結果としてのCFUの最大の減少は、ON/Eで見られた(617分の1への減少;P=0.0134)。コントロールでは平均204および166のCFUが存在する肺(P=0.0320)(図3B)およびCNS組織(P=0.0078)(図3A)の差は、まさに顕著であるが、これに対して肺炎球菌は、GLSでインキュベートしたときに検出不可能であった(検出限界=3CFU)。アシアロ−GM1プレインキュベーションは、混合GLSよりも効率的ではなかったが、それでもCNS(図3A)および肺(図3B)においてそれぞれ、コロニー形成を25分の1および63分の1に減少させた。肺は吸入した肺炎球菌によって感染し、肺に比較的豊富に存在するアシアロ−GM1へのその結合はGLSによって明らかに阻害された。このことはGLSが上皮細胞への初期結合で役割を果たすことを示した。GLS処置は、肺炎球菌の生存能力を変更させなかった。この実験の間に肺炎球菌は血液中で検出されなかった。それゆえGLSは、鼻道の神経−上皮への肺炎球菌結合ならびに肺およびCNSの感染に関して重要な標的を構成する。
【0092】
(鼻腔内攻撃後のOBへのS.ニューモニエ蓄積の検出)
OB中のEF3030の数は一般に少なすぎて、顕微鏡による細菌の描出は実現できなかった。鼻腔内投与後のOB中のS.ニューモニエを描出するために、さらに毒性の菌株TIGR4を使用した。攻撃の1、3、6、12または24時間後およびその後毎日、代表のマウスからの血液サンプルを試験した。菌血は観察されなかった。攻撃の1週間後にマウスを殺処分して、CFUについて組織を分析した(図4Aおよび4B)。わずか5X105のTIGR4 CFUの用量が、OBに〜300CFUを生じた(図3)。OB中のPspA特異性Absを染色することによって、肺炎球菌を描出した(図4D〜F)。肺炎球菌は、攻撃されたマウスのOB、すなわち糸球体層(図4F)および外部網状層(図4E)で検出された。コントロールマウスのOBには、肺炎球菌は存在しなかった(図4D)。
【0093】
TIGR4菌株は、鼻腔内投与6日後に、NW、ON/EおよびOBからのニューモリシン遺伝子のPCR増幅によっても検出された(図4C)。PCR検出可能な肺炎球菌は、この間隔で採取した血流中に、または非感染マウスによるサンプル中に存在していなかった。
【0094】
(実施例2)
(上咽頭輸送における肺炎球菌−NanAの役割およびCNSの標的化)
NanA変異体を、遺伝子的背景およびNanAの局在化の両方で異なるS.ニューモニエの3つの菌株にて産生させた。菌株EF3030(19F型)およびD39(2型)はどちらも、細胞壁に共有結合するNanAを発現するが、TIGR4菌株(4型)は、環境へ分泌される切断NanAを発現した。
【0095】
コロニー形成におけるNanBの役割も評価した。
【0096】
(細菌株および増殖条件)
本試験で使用した菌株を表4に挙げる。
【0097】
【表4】
すべての肺炎球菌株は、−80℃にて10%グリセロール中で保存して、血液寒天プレートに移して培養し、37℃にて5% CO2雰囲気中で一晩インキュベートした。肺炎球菌の培養物を、0.5%酵素抽出物をOD660 0.5まで含有するTodd−Hewitt培地中で増殖させて、−80℃にて滅菌グリセロールを用いた10%までの同じブロス添加で分割して凍結保存した。変異の安定性を確実にするために、耐抗生物質インサートを持つ変異体を適切な抗生物質中で増殖させた。
【0098】
(nanA変異体の作成)
親バックグラウンドTIGR4、EF3030およびDD39それぞれのNanA変異体菌株JW001、SAM001、およびJCP001を挿入複製突然変異誘発技法によって得た(そこで教示された技法について、参照によりその全体が本明細書に組み入れられている、Yother et al.(1992)J.Bact.174:610−618)(表4)。菌株TIGR4およびEF3030は、レシピエントとして、同質遺伝子nanA菌株D39から作成したドナー染色体DNAの形質転換に使用した(Berry et al.(2000)Infect.Immun.68:133−140)。それぞれの場合で、変異体は、親菌株への戻し交雑を3回行った。D39変異体も我々のD39親菌株への戻し交雑を3回行って、それがこれらの試験で使用した親菌株との同質遺伝子であることを確認した。D39の変異は、成熟タンパク質のアミノ酸約650個のN末端フラグメント以外のすべての欠失を可能にする挿入複製突然変異誘発によって作成した(Berry et al.(2000)Infect.Immun.68:133−140)。TIGR4/nanB同系変異体は、挿入複製突然変異誘発技法を使用して作成した(Balachandran et al.(2002)Infect.Immun.70:2536−2534;Yother et al.(1992)J.Bact.174:610−618)。nanBの461bp内部部分を、プライマー:nanBFおよびnanBR(表1)を使用して増幅し、PCRは、Taq PCR Mastermix(Invitrogen)を使用して、95℃にて1分間、45℃にて1分間、72℃にて1分間を30サイクル実施した。フラグメントは、pSF152内へクローニングした。TIGR4菌株のプラスミドDNAによる形質転換は、前と同様であった(Balachandran et al.(2002)Infect.Immun.70:2526−2534)。nanA/nanB−TIGR4ダブル変異体は、nanB/TIGR4変異体の菌株JW001から作成した染色体DNAによる形質転換によって得た。
【0099】
(マウス毒性アッセイ)
メス6〜12週齢CBA/CaHN−XID/J(CBA/N)マウスは、Jackson Laboratory(バーハーバー、マサチューセッツ州)から得た。これらマウスのBrutonのチロシンキナーゼ遺伝子における変異は、胸腺独立性II型抗原への反応をできなくするが、比較的な正常なT細胞依存性免疫反応を可能にする(Amsbaugh et al.1972 J Exp Med.136:931−949;Briles et al.1986 Curr.Top.Microbiol.Immunol.124:103−120;Potter et al.1999 Int.Immunol.11:1059−64;Wicker and Scher 1986 Curr.Top.Microbiol.Immunol.124)。これらのマウスは、カプセル状ポリサッカライドに反応できず、肺炎球菌感染を高い再現性で受けやすい(Briles et al.1986 Curr Top.Microbiol.Immunol.124:103−120;Briels et al.1981 J.Exp.Med.153:694−705)。X連鎖免疫不全(xid)マウスは、病原体のない状態に維持され、7〜12週齢で使用した。既知の濃度の生細胞を含有する凍結感染ストックを乳酸リンゲル液で希釈した。次にマウスに体積10μl中のおよそ5X105−1X106細胞を用いて、上述のように経鼻的に(I.N.)感染させた(Wu et al.1997 Microb.Pathog 23:127−137)。
【0100】
(組織収集)
すべてのマウスは、鼻洗浄および組織収集を実施する前に安楽死させた。血液を後腹膜神経叢からヘパリン処理毛細管内に収集した。鼻洗浄液(NW)、鼻組織(嗅上皮(NT)、嗅球(OB)を含む)、および脳の収集前に、マウスを70%エタノールで消毒した。これらの体液および組織を上述のように得た。NWの血液汚染を防止するために、気管への切開を行って、外径0.075cmを有する2.0cm長のTygon管(Cole−Parmer)を上咽頭に挿入し、同時にリンゲル液を充填した注射器に取り付けた。注射器からの液体は鼻を通じて排出して、3滴を収集した。ON/EおよびOBをそれぞれリンゲル液0.5ml中でホモジナイズし、脳の残りをリンゲル注射液剤1.0ml中でホモジナイズした。
【0101】
(生肺炎球菌の定量)
組織および体液の連続3倍希釈物8個を滅菌リンゲル液中で作成し、ゲンタマイシンサルフェート4μg/mlを含有する血液寒天プレートで平板培養した。コロニー形成単位(CFU)は平板培養およびロウソク瓶内でのインキュベーションの24時間後に数えた。ノイラミニダーゼ活性に使用したアッセイは、前に述べられている(アッセイ方法について、参照によりその全体が本明細書に組み入れられている、Lock et al 1988 Microb Pathog 4:33−43)。野生種肺炎球菌と変異体肺炎球菌との間の比較を行う2標本Mann−Whitneyランク検定を用いた解析によって、結果の有意性を評価した。
【0102】
(試験管内試験)
TIGR4、ならびにそのnanAおよびnanB変異体が特異性ガングリオシドに結合する能力を測定した。使用したガングリオシドは、混合ガングリオシド、アシアロ−GM1、GM1、GD1a、GD1b、GT1(Calbiochem)およびGM3ガングリオシド(Sigma)を含む。GM3ガングリオシドは、肺炎球菌性結合に関与する末端または内部GalNAcβ1−4Gal配列を欠いており、負のコントロールとして使用される。これらのガングリオシドを含有する混合、モノ−、ジ−またはトリ−シアル酸は、ELISAプレートにただちに結合する。TIGR4菌株を用いた短期インキュベーション後の初期データは、肺炎球菌がアシアロ−GM1コーティングプレートに結合するが、BSA−、GM−3、またはGM1コーティングプレートには結合しないことを示している。ガングリオシドコーティングプレートを使用して、野生種TIGR4菌株、TIGR4菌株、nanA、nanB、およびnanA/nanB変異体菌株の安定不透明および透明相改変体によるこれらのプレートへの結合能力を比較する。これらの解析は、ガングリオシドコーティングプレートへの短期インキュベーション(1時間)および長期インキュベーション(24時間)を含む。結合した肺炎球菌は、0.5%酵素抽出物を含有するTodd Hewitt培地中での短期インキュベーション(10〜15分間)によってガングリオシドプレートから除去され、放出された細菌の反復ピペッティングおよび血液寒天プレート上での平板培養が続く。あるいは41℃の、0.5%酵素抽出物を含有するTodd Hewittブロス寒天を結合した肺炎球菌の上に注入して、プレートの底からコロニーをカウントする。コントロールは、肺炎球菌のないプレートおよびガングリオシドはないが、肺炎球菌のあるプレートを含んでいる。
【0103】
ガングリオシド結合の試験に続いて、各種の細胞系は、肺炎球菌をその細胞表面に結合させて、それらを内部移行させるその能力について試験する。これらの試験は、ラット神経褐色細胞腫細胞系PC12(ATCC)およびマクロファージ細胞系P388D1に焦点を当てる。これらの2つの細胞系は、その特異的属性のために選択された。P388D1細胞系は、小膠細胞上に存在することが報告されている、高い親和性PAF−R(Valone(1988)J.Immunol.140:2389−2394)を発現する。PC12細胞系は、検出可能なPAF−Rを発現しない。Brewer et al.,(2002)J.NeuroChem 82:1502−1511。102〜105個の肺炎球菌性CFUを6ウェルまたは24ウェル組織培養プレートで増殖させたこれらの細胞系に添加して、37℃にて15分間〜6時間インキュベートし、その後、細胞を完璧に洗浄して、付着性の肺炎球菌を分析する。細胞への内部移行を判定するために、ペニシリンおよびゲンタマイシンを用いた2時間の洗浄を実施してから、血液寒天上で細胞を平板培養するか、またはそれらを41℃の0.5%酵素抽出物を含有するTodd Hewittブロス寒天で被覆する。使用した2つの細胞系は、通常はCNSにて観察されるPAF−Rの生体内発現を反映する。活性化小膠細胞は、P388D1細胞系と同様にこのレセプタを豊富に発現するが、PAF−Rレセプタは、神経細胞、たとえばPC12細胞系上に不在であるか、別個の神経小集団によって低レベルでのみ発現されるかのどちらかである。Mori et al.,(1996)J.Neurosci 16:3590−3600;Bennett et al.,(1998)Cell Dath Differ.5:867−875。肺炎球菌の両方の細胞系への付着は、PAF−Rが付着にとって必須ではなく、代わりのレセプタが存在することを示す。TIGR4不透明および透明改変体ならびにnanA−、nanB−変異体およびnanA/nanBダブル変異体を、野生種TIGR4菌株によって観察された付着と比較して、これらの細胞系への付着について試験する。肺炎球菌付着におけるPAF−R対ガングリオシドの役割をさらに分析するために、PAF−Rを欠いたCOS−7細胞系(Gerard and Gerard(1994)J.Immunol.152:793−800;Honda et al.,(1992)J.Lipid Med.5:105−107)を、1029bpのヒトPAFR読み取り枠で、pcDNA3.1/GSプラスミドを前に報告したように使用して(そこで教示した方法について、参照によりその全体が本明細書に組み入れられている、Brewer et al.,(2002)J. Neuro Chem 82: 1502−1511)形質移入し、Transfast試薬(Promega)を使用して形質移入する。プラスミド単独をコントロールとして使用し、肺炎球菌付着に影響を及ぼすパラメータをPAF−Rの存在または非存在下で解析する。本実験は、PAF−Rの付着での重要性に関する明確なデータを提供する。PAF−Rを欠いた細胞系での付着は、ガングリオシドによって仲介され、続いてガングリオシドを用いたプレインキュベーションによって遮断される。肺炎球菌が上皮細胞に付着して浸透する能力にさらに注目するために、Detroit 562ヒト咽頭上皮細胞系(ATCC)およびA549ヒト肺上皮細胞系(ATCC)を採用し、トランスウェルシステムを使用する。Millicell(登録商標)−PCF培養(Millipore,ビルリカ、マサチューセッツ州)プレートインサートを使用して、上皮細胞系をコンフルエントまで培養する。コンフルエントは、Millipore Millicell(登録商標)電気抵抗システムを使用して、経上皮耐性を測定することにより決定する。少なくとも500Ω/cm2の抵抗は、完全にコンフルエントな上皮単層が達成されたことを意味する。これらの細胞は、それらがこの上皮層に付着し、侵入して、浸透する能力を試験するために、肺炎球菌に暴露させる。結合と内部移行を区別するために、上皮細胞を洗浄して、ペニシリンおよびゲンタマイシンを含有する培地で2時間インキュベートする。最初に、TIGR4菌株、そのnanAおよびnanB変異体、ならびにダブル変異体に焦点を当てる。TIGR4菌株の安定な透明および不透明改変体は、生体内攻撃後に逆行しなかった安定な改変体が得られるまで、連続継代によって産生させた。これらのTIGR4改変体は、上皮細胞に付着、侵入、浸透する(tranverse)能力について比較する。ウェルにEMEM培地中の103〜106CFU/ウェルを装填する。0.5、1、2、4、8、および24時間後に培養物を上皮層の上下で収穫して、CFUについて解析する。細胞層を5〜6回洗浄してから、細胞に0.5%酵素抽出物および0.5%寒天を添加したTodd−Hewittブロスを重ね、41℃まで冷却し、単層と結合した肺炎球菌CFUを決定する。プレートを37℃および5% CO2にて一晩インキュベートし、その後、CFUをカウントする。細胞系をPAF−Rの発現について解析する。これらの細胞系に由来する全RNAをRT−PCRにより、2つのプライマーPAF−1(5’−CCGATACACTCTCTTCCCGA−3’(配列番号3);ヌクレオチド151〜170)およびPAF−2(5’−ACAGTTGGTGCTAAGGAGGC−3’(配列番号4);ヌクレオチド970〜951)を使用して解析し、838bpのPCR生成物を得る(そこで教示された方法について、参照によりその全体が本明細書に組み入れられている、Stengel et al.,(1997)Arterioscler.Thromb.Vasc.Biol.17:954−962)。PAFレセプタが存在する場合、PAFレセプタインヒビタ、たとえばオクチルオニウムブロミド(Biomol Research Laboratories,Inc.プリマスミーティング、ペンシルベニア州)またはPAF(Biomol)を培養物に添加して、PAF−Rの上皮付着および浸透への寄与を決定する。オクチルオニウムブロミドは、PAF−Rへ高い親和性で結合する。あるいは上述のCOS7細胞は、この目的に使用され、PAF−Rの存在または非存在下で肺炎球菌付着を比較する。
【0104】
各種の肺炎球菌株の侵襲性度は、トランスウェルシステムの頂端および基底面コンパートメントの両方での炎症性サイトカインの生成と相関している。培養物上澄みを各種の時点で上下のコンパートメント両方にて収集して、ELISA(BD PharMingen)によって解析し、炎症性サイトカインIL−1β、IL−6、IL−8、IL−10およびTNF−αの濃度を決定する。上皮単層を酢酸アルコール中で固定して、OB中での肺炎球菌の描出で以前に使用したように、PspA特異性免疫蛍光染色を使用して、肺炎球菌の細胞内存在を解析する。蛍光画像は、以前に述べたように(そこで教示された方法について、参照により本明細書に組み入れられている、Martin et al.,(1998)J.Immunol.160:3748−3758)、適切なフィルタキューブを装備したLeica/Leitz DMRB顕微鏡(Chromtechnology,バトルボロ、バージニア州)によって描出する。画像はC5810デジタルカラーカメラ(Mamamatsu Photonic System)を用いて収集し、Adobe photoshopおよびIP LAB Spectrumソフトウェアで処理する。
【0105】
(結果)
(NanAおよびNanB変異体のコロニー形成)
S.ニューモニエがCBA/Nマウスの上咽頭にコロニー形成する能力に対するNanA変異の効果を、鼻腔内(i.n.)感染させたマウスの鼻洗浄から単離された肺炎球菌細胞の数を、NanA変異体菌株で感染させたマウスの肺炎球菌細胞の数と比較することによって評価した。これらの異なる肺炎球菌菌株が含まれ、それゆえカプセル血清型および遺伝的背景が異なる菌株に対するNanA変異の効果が調査された。TIGR4/NanA−(JW001)、EF3030/NanA−(SAM001)およびD39/NanA−(JCP001)はそれぞれ、カプセル型4、19Fおよび2であった(表4)。カプセル型4臨床アイソレート、TIGR4の場合、LPETG(配列番号13)モチーフをコード化する配列の前に停止コドンがある。このモチーフがないと、NanAは、TIGR4によって環境中へ分泌されることが予想される。現在入手できる残りの4つの肺炎球菌−NanA配列、G54(19F型)、R6(2型)、Spanish 23Fおよび670(6B型)の試験(Berry et al.Gene 71:299−305;Hoskins et al.2001 J.Bacteriol 183:5709−17;Tettelin et al.2001 Science 293:498−506)は、細胞壁への共有結合のためのLPXTG(配列番号14)モチーフを有することを示した(図5)。したがってここに含まれる菌株は、NanAが分泌され、表面結合されている菌株における変異の比較を提供した。
【0106】
コロニー形成の劇的な減少は、TIGR4およびEF3030両方のNanA変異体で観察された(図6および7)。
【0107】
S.ニューモニエは別のノイラミニダーゼ、NanBを発現する。同様の相同性度がNanB間でNanAに対して共有されている。NanBは、NanAと43%の相同性を共有する(24%の同一性)。タンパク質間の共有された残基は、シアリダーゼであることを示唆した(Berry et al.1996 J.Bacteriol.178:4854−4860)。NanBは、6.5〜7であるNanAの最適pHと比較して、4.5の最適pHを有することが見出されている。その最適pHにおいてさえ、NanBは、NanAがその最適pHにあるときのシアリダーゼの約1/100の活性である。それでも、コロニー形成およびCNSの直接侵襲についてNanBの要件があるかどうか調べるために、菌株TIGR4/NanB−(JW002)、変異体を、NanBを欠いたTIGR4遺伝的背景にて、NanAおよびNanB発現の両方を欠いた菌株TIGR4/NanAB−(JW003)と同様に作成した。マウスのJW002による感染は、TIGR4菌株とほぼ同じコロニー形成のレベルを生じた(図8)。その上、NanA変異体と比較して(図9)、コロニー形成の著しい減少はダブル変異体(JW003)では起こらなかった。
【0108】
(肺炎球菌のCNSへの侵入)
S.ニューモニエの鼻組織(嗅神経を含む)への移動を追跡するために、嗅球および脳の残りをS.ニューモニエの存在について試験した。NanA変異体は、遺伝的背景とは無関係に、鼻組織および嗅球において、野生種菌株の数と比べて著しい数の減少が見られた。殺処分時に、すべてのマウスから採血したが、いずれも血液中に検出可能な肺炎球菌を示さず(<12CFU/ml血液)、肺炎球菌が鼻腔からCNS組織へ直接移動することを示している。NanB変異体は、肺炎球菌の鼻組織または嗅球内への侵入に対する影響を持たなかった(図8)。
【0109】
NanA変異体は、上咽頭およびCNSでコロニー形成および持続するその能力が明らかに弱められた。このことは、カプセル血清型およびNanAの表面への結合の両方が異なる菌株で見られた。NanAは、細菌細胞表面と宿主との間の親密性に影響を及ぼす多くの表面構造の1つに過ぎないが、肺炎球菌のCNSへの標的化と同様に、鼻輸送へのその関与は必須である。NanAの破壊は、コロニー形成およびCNSへの標的化を著しく減少させる。この結果は、TIGR4およびEF3030の両方で観察された。
【0110】
菌株EF3030(19F型)は、1ヶ月間に亘って高い効率で上咽頭にコロニー形成する。しかしながら、EF3030が持続する能力にかかわらず、NanAの変異は、鼻の肺炎球菌細胞の数を著しく減少させた。弱化は、鼻洗浄液から単離された細胞数が14日後には検出可能限界近くまで低下した、TIGR4/NanA−菌株でなおさらに劇的であった。
【0111】
自然の状況では、肺炎球菌は、他の細菌種と共存する。それゆえNanAの他の機能は、宿主タンパク質の機能を変化させることと、輸送の長期安定性に寄与することとを含む。NanAは、肺炎球菌がN.メニンギティディスまたはH.インフルエンゼを含む他の口内細菌と競合する能力も、宿主糖タンパク質を炭素源として使用できるようにすることによって向上させる。
【0112】
これらの試験の主な結果は、NanA発現がマウスの最適輸送に必要であることの証明であったが、これらのデータは、NanAを欠く肺炎球菌が嗅球でははるかに少ない数で見出されることも証明した。この時点では、活性NanAがCNS組織中でのS.ニューモニエの生存にとって重要であるかどうかを知ることは困難である。これらの組織から回収されたNanA変異体の数は親菌株よりはるかに少ないが、肺炎球菌が脳に入ったとたん、神経細胞組織でのその存在すら、NanAのさらなる毒性効果を明らかにしている。OB中のノイラミニダーゼ−変異体のレベル低下はおそらく、減少した輸送の結果であろう。この発見は、輸送がより侵襲性の疾患の前提条件であるという原則を強調し、輸送を低減できる介入、たとえばNanAを用いた免疫化がニューモニエ、髄膜炎、中耳炎および敗血症からの保護を提供するであろう。
【0113】
nanAの既知配列のうち、TIGR配列は、表面アンカーを含有しない唯一の配列であろう。本菌株では、フレームシフトがLPETG(配列番号13)モチーフの前の、分子の切断を引き起こす(Tettelin et al.2001 Science 293:498−506)。大半の菌株では、NanAのかなりの部分がソルターゼによって細胞壁に共有結合されることが予想され(Mazmanian et al.1999 Science 285:760−63)、そこで電子顕微鏡写真によって検出されている(Camara et al.Infect.Immun.62:3688−95)。これらの試験では、TIGR4はEF3030と同様に、NanA依存性輸送および嗅球での存在を示した。上澄みまたは細菌ペレットでのNanA活性の局在化の試験から、TIGR4とは異なり、EF3030のNanA活性は細胞に結合していることがわかった。それゆえNanAは、表面結合されてもされなくても、または分泌されてもされなくても、コロニー形成を促進できる。
【0114】
(実施例3)
(S.ニューモニエ病原性におけるガングリオシドの役割)
精製ノイラミニダーゼ、NanA(Calbiochem)をPBS 10μl中1、10および50μgにて、ON/E単離の15、30、および60分前に鼻腔内投与する。組織を4%パラホルムアルデヒドで固定して、パラフィン切片を作成した。GM1はビオチン化CT−Bを使用して、続いてストレプトアビジン−FITCによって染色し、染色強度を解析する。切片はローダミンに結合させたアシアロ−GM1特異性Absによっても染色して、これらの組織においてGM1染色の減少がアシアロ−GM1染色の増加と同時に起きることを確認する。同じ処置を受けるマウスの並行群をS.ニューモニエ菌株TIGR4およびEF3030によるコロニー形成について、第1日および第4日に解析して、ノイラミニダーゼ処置が鼻腔内コロニー形成レベルの上昇を引き起こしたことを確認する。マウスに菌株EF3030の高用量(1x108CFU)を鼻腔内投与して、ON/Eは、鼻腔内攻撃後の第1日および第4日に以下の間隔、すなわち、1、3、6、および12時間にて単離する。ON/Eを上で概説したように染色し、GM1およびアシアロ−GM1発現について解析する。GM1の減少およびアシアロ−GM1発現の上昇が鼻組織で観察される場合、NanAおよびNanB欠損菌株も、それらが鼻道でのGM1発現を変化させることが期待されないため、試験を行う。シアル酸残基の除去は、Thompson−Friedenreich抗原の抗原決定基に相当する、次端部ジサッカライド、β−D−ガラクトピラノシル−(1−3)N−アセチル−D−ガラクトサミンを露出させ、そのためにPNAは高い親和性を有する。それゆえON/E中のPNA−結合部位における変化は、ノイラミニダーゼ活性の別の尺度である。これらの組織から作成した凍結切片をPNA−FITCまたはPNA−HRPでただちに染色して(Medac,ハンブルク、ドイツ)、顕微鏡検査に基づいてPNA−結合部位の増加が生じたかどうかを判定する(そこで教示した方法について、参照によりその全体が本明細書に組み入れられている、Black et al.,(2000)Pediatr.Infect.Dis J.19:187−195;Klein et al.(1978)Klin.Wochenschr.56:761−765)。
【0115】
GM1部位は、菌株TIGR4またはEF3030の鼻腔内投与の前に特異的に遮断される。これは:1)CT−B対非ガングリオシドコントロールタンパク質、たとえばオボアルブミン、2)GM1に対するAbs(Calbiochem)対正常ウサギ免疫グロブリン、または3)UAB Protein AnalysisおよびPeptide Synthesis Core Facilityで合成されたGM1特異性ペプチドを使用して、3つの方法で着手される。GM1特異性ペプチドによる阻害は、CT−BおよびおそらくGM1に対するAbsが随伴性の炎症を引き起こすことが予想されるため、最良の手法である。これらの実験では、ON/EおよびOBは、肺炎球菌性CFUの投与後、第1日および第4日に解析する。GM1を遮断する代わりの手法は、GM1結合ペプチドに対して選択されるファージ提示ペンタデカペプチドライブラリの使用によって発見されたGM1結合ペプチドの使用である。このGM1結合ペプチドVWRLLAPPFSNRLLP(配列番号5)は、GM1に対する高い親和性(1010M−1)および1.0μMのIC50を有する。Matsubura et al.,(1999)FEBS Letters 456:253−256。同じ長さの、ランダムに選択された配列中の同じアミノ酸より成るペプチドをコントロールとして使用する。両方のペプチドのGM1結合能力は、生体内で使用する前にELISAによって確認する。最初に、リンゲル液またはPBS 10μl中のこれら2つのペプチド100μgを、菌株EF3030 3X106CFUの投与の10分前に鼻腔内投与する。第1日および第4日の投与後に、ON/EをCFUについて、未処置CBA/Nマウスに対するCFUの数について解析する。
【0116】
遮断実験は、PAF(Biomol Research Laboratories,Inc.プリマスミーティング、ペンシルベニア州)およびPAF−Rアンタゴニストオクチルオニウムブロミド(Biomol)を用いて実施する。本化合物は、高い親和性でPAF−Rに結合する。各ガングリオシドは個別に試験を行う。混合ガングリオシド以外に、アシアロ−GM1、GM1、GD1a、GD1b、GT1(Calbiochem)およびGM3(Sigma)を、攻撃後、第1日および第4日に評価するように鼻腔内コロニー形成を阻害するその能力について試験を行う。各種のガングリオシドは、これらのプロセスを遮断できる。GM3ガングリオシドは、公開されているCポリサッカライド結合モチーフを欠いているため、負のコントロールとして機能する。ガングリオシドプレインキュベーション後にEF3030によるコロニー形成に対して示されたデータに基づき、混合ガングリオシドは、コロニー形成の遮断ではアシアロ−GM1よりも有効である。これは、アシアロ−GM1以外の他のガングリオシドが、鼻道、肺および脳の肺炎球菌性コロニー形成のプロセスに関与することを示している。これらのガングリオシド阻害試験は、TIGR4菌株およびそのノイラミニダーゼ変異体に焦点を当てている。TIGR4菌株を用いたガングリオシドコーティングELISAプレートでの短期試験管内インキュベーションは、TIGR4菌株のアシアロ−GM1への結合を証明した。エンテロトキシンは、鼻道での肺炎球菌−ガングリオシド相互作用を異なるふうに遮断する別の手段を提供する。CTおよびLTh−1はどちらも、両方の血清型I易熱性エンテロトキシン(Pickett et al.,(1986)J.Bacteriol 165:348−352)であり、わずかに異なるが、同様のガングリオシド結合特異性を示す。Fukuta et al.,(1988)Infect Immun.56:1748−1753.CT(List Biological Laboratories,Inc.,キャンベル、カリフォルニア州)はGM1に、そしてより低い程度でGD1bに結合する。LTh−1は、GM1およびGD1bに優先的結合を示し、GM2およびアシアロ−GM1に弱く結合する。Fukuta et al.,(1988)Infect Immun.56:1748−1753。S.ニューモニエと相互作用する主なガングリオシドである場合、LTh−1の使用は GM1ガングリオシドを遮断するだけでなく、アシアロ−GM1も遮断し、このことはノイラミニダーゼ活性を必要としない、天然の頻度の低い結合を示す。血清型IIによる易熱性エンテロトキシンは、異なるガングリオシド結合特異性、特に易熱性エンテロトキシンLT−IIbを示す。この毒素は、GD1aに、より低い程度でGT1bに結合して、GM1への親和性は示さなかった。Fukuta et al.,(1988)Infect Immun.56:1748−1753。LT−IIaは、高い親和性でGD1bに、より低い親和性でGM1、GT1b、GQ1b、GD2、GD1a、およびGM2に結合する。Fukuta et al.,(1988)Infect Immun.56:1748−1753。LT−II毒素は、T.D.Connell博士のご厚意により提供された。鼻用量および最適期間を最適化してON/Eへの肺炎球菌の結合の阻害を観察するために、最初に用量反応試験(1.0または10μg)を、S.ニューモニエの鼻腔内投与に与えられる選択したエンテロトキシンに対して実施する。鼻腔内コロニー形成の阻害が第4日に観察された場合、観察は第11日まで延長され、その間にエンテロトキシンを1日おきに与える。CBA/NマウスのON/EおよびOBにおけるCFUを測定する。
【0117】
(実施例4)
(肺炎球菌性病原性におけるCポリサッカライド特異性抗体の役割)
テイコ酸としても既知の肺炎球菌性Cポリサッカライドは、リポテイコ酸としても既知の肺炎球菌F抗原と構造的に同じである。Fischer et al.(1993)Eur.J.Biochem 215:851−857。これはグラム陽性細菌の中でS.ニューモニエ独自の特徴である。これらの分子の主要抗原決定基は、ホスホリルコリン(PC)残基であり、PCに対するAbsは、i.p.または経鼻肺炎球菌攻撃に対して保護的である。Briles et al.,(1984)Eur.J.Immunol.14:1027−1030;Briles et al.,(1981)Nature 294:88−90;Yother et al.,(1982)Infect.Immun.36:184−188;Briles et al.,(1984)J.Mol.Cell.Immunol.1:305−309。それゆえ受動伝達または能動鼻腔内免疫化のどちらかによって得られたPC特異性Absの役割が調査される。保護性PC特異性Absの受動伝達では、すなわちIgG3(59.6C5)およびIgM(22.1A4)アイソタイプの両方のT15イディオタイプモノクローナルAbs(mAbs)が使用される。Briles et al.,(1981)Nature 294:88−90。T15イディオタイプは、おそらくCポリサッカライドをより効率的に結合することによって、マウスの肺炎球菌感染に対してM603またはM511イディオタイプよりも保護性であることが示されている(Briles et al.,(1984)Eur.J.Immunol.14:1027−1030)。受動Abs伝達は、鼻腔内投与された肺炎球菌を用いたT15 Abs(100μg)の直接投与を含み、鼻腔内コロニー形成を低減するためのi.v.またはi.p.投与されたAbsと比較する。コロニー形成は経時的に監視し(第4日、第11日、第18日)、これらの実験の異なる群の間で著しい相違が観察されない場合、mAbs(20μg)を1日おきに鼻腔内投与する。CBA/Nマウスは、T15イディオタイプ抗PC Absを生成しない。抗PC特異性Absの受動伝達が、鼻道中で粘膜IgAまたはPC特異性Absの他のアイソタイプを誘発することは予想されない。鼻Absを誘発するために、2つの異なる手法を取る。1つは、Cポリサッカライドに対するAb反応を誘発することが既知である、プロテアーゼ処置R36A菌株の直接鼻腔内投与である。抗PC Absの保護免疫が試験されてきたが、鼻道などの粘膜表面でのその役割に関するデータは入手できない。CBA/NマウスX染色体関連免疫不全は、T15イディオタイプの抗PC Absの産生不能を引き起こす。この不能の重要性を判定するために、CBA/Nマウスをその野生種対応物のCBA/Jマウス(Jackson Laboratories)と比較する。抗PC Abs誘発のための菌株R36Aによる免疫化は、表面タンパク質のタンパク質分解除去を含む。Krause(1970)Adv.Immunol.12:1−56。鼻腔内免疫化の代わりの手法は、上述したようなPCのタンパク質キーホール・リンペット・ヘモシアニン(KLH)へのカップリングである(そこで教示した方法について、参照により本明細書に組み入れられている、Krause(1970)Adv.Immunol 12:1−56;Chesebro and Metzger(1972)Biochemistry 11:766−771)。PC−KLHを用いた鼻腔内免疫化は、粘膜免疫反応を最適化するために、粘膜アジュバントCTを用いて実施される。マウスは、コロニー形成に対するCTの効果を防止するために、最後の免疫化の2〜3週間後に攻撃される。1週間間隔で3回の鼻腔内免疫化を実施する。当業者によって通常実施される通りにCポリサッカライドおよびPC特異性ELISAを使用して、血清Ab力価を監視する。PC特異性ELISAでは、PCは上述のようにBSAに結合される(そこで教示した方法について、参照により本明細書に組み入れられている、Chesebro and Metzger(1972)Biochemistry 11:766−771)。血清に加えて、鼻洗浄液、唾液、および気管支洗浄液中のAb力価を測定する。これらの解析は、粘膜分泌物および血清の両方でのIgA、IgM、IgG、およびIgG−サブクラス分布を含む。最適な粘膜Ab力価を誘起するプロトコルを使用して、第4日および第11日にコロニー形成が監視された後にマウスに〜5x106CFUで投与される、TIGR4菌株による粘膜攻撃試験を実施する。免疫化試験では、正常な完全免疫適格性マウス(CBA/J菌株)はもちろんのこと、CBA/Nマウスも以前の試験と同様に使用する。Wallick et al.,(1983)J.Immunol.130:2871−2875。
【0118】
(実施例5)
(S.ニューモニエ病原性におけるノイラミニダーゼ特異性抗体の役割)
鼻腔内攻撃前にマウスを鼻腔内免疫化するために、市販のS.ニューモニエ由来ノイラミニダーゼを使用する(Calbiochem)。しかしながら、提案された試験に十分な量のタンパク質を得るために、NanA遺伝子はヒスチジンタグ含有発現ベクター(Invitrogen)を使用してE.コリ内でクローニングおよび発現させる。粘膜免疫反応を最適化するために、3.4%ホルムアルデヒド処置ノイラミニダーゼの鼻腔内免疫化を対の未処置ノイラミニダーゼと、CTの存在下または非存在下で比較する。これらの免疫化は、CBA/NおよびCBA/Jマウスの両方で実施する。3回の鼻腔内免疫化を1週間おきに与え、その間に血清および唾液Abs力価をELISAによって監視する。免疫マウスをTIGR4菌株によって攻撃して、ON/E、OB、脳、血液、脾臓、および肺のコロニー形成を第4日および第11日に比較する。宿主の相互作用を遮断するために、ノイラミニダーゼおよびCポリサッカライド特異性Absの両方を同時に誘発させる。鼻腔内免疫化をノイラミニダーゼおよびT15イディオタイプmAbsの伝達による受動免疫保護と組み合せた計画を使用する。
【0119】
(実施例6)
(S.ニューモニエを用いた鼻腔内攻撃に対して保護するためのノイラミニダーゼ−PC結合体の有効性)
保護の向上を評価し、単独で使用された各抗原と比較して第11日のEF3030およびTIGR4菌株による鼻腔内コロニー形成を低下させるために、マウスを鼻腔内アジュバントとしてのCTと組み合せたノイラミニダーゼおよびPC−KLHによって免疫化する。加えて、鼻洗浄液、唾液、および血清のAb力価を上述のように解析して、免疫パラメータを鼻道内での肺炎球菌に対する保護度と関連付ける。さらに最適な免疫反応を発生させるために、ホスホコリンをノイラミニダーゼに直接結合させる。この作成物を、CBA/NおよびCBA/Jマウスの両方における鼻腔内および全身免疫化の後に、アジュバントとしてのCTを用いて、またはCTなしに送達したときに免疫原性について試験を行う。鼻洗浄液、唾液、および血漿のAb力価をELISAによって測定する。攻撃試験は、菌株EF3030 107CFUまたはTIGR4 106CFUを用いて実施する。マウスを攻撃後、第11日に殺処分して、血液、鼻洗浄液、ON/E、OB、および脳で観察されたCFUについて解析した。PCに結合させたノイラミニダーゼによる免疫化は、粘膜および全身Abレベルを上昇させることによって、これらの2つの毒性成分に対する保護を向上させる。抗原特異性IgGサブクラスの分布は、他の粘膜アジュバントを使用することによって変更される。CTは、Th2−、LT混合Th2/Th1−、およびCpGモチーフ、たとえばDNAオリゴヌクレオチド(ODN)1826 a Th1型反応をIgGサブクラス分布の関連する変化と共に引き起こす。各種のアジュバントは、ノイラミニダーゼ−Cポリサッカライド特異性免疫がS.ニューモニエによる鼻腔内コロニー形成から保護する能力をさらに向上させ、新たな肺炎球菌性ワクチン手法の考案をもたらす。
【0120】
(実施例7)
(鼻腔内攻撃後のS.ニューモニエの鼻腔内コロニー形成の、抗ホスホコリン特異性モノクローナル抗体による阻害)
合計1X106CFUのTIGR4菌株を、IgG3サブクラスまたはIgMアイソタイプのどちらかの抗ホスホコリンモノクローナル抗体5μlでインキュベートした。合計5μlを鼻孔ごとに投与した。投与9時間および12時間後の鼻洗浄液500ml中のCFUをそれぞれ示す。どちらのモノクローナル抗体でも、重大な80%超の減少が見られた。1グループ当たりマウス5匹の平均+SDを示す。データを図10に示す。
【0121】
(実施例8)
(S.ニューモニエ鼻腔内投与後の神経細胞損傷および炎症)
S.ニューモニエ菌株EF3030の鼻腔内投与後、第1日、第3日、第7日および第14日に処置マウスからON/E、OB、および脳を単離して、炎症性反応について組織学的に解析する。D39またはTIGR4菌株をそのnanA変異体菌株と、炎症性反応を発生させるその能力について比較する。殺処分時に、マウスを25℃のPBSを用いて潅流して、Zamboni固定液10ml(0.1Mリン酸緩衝液中の4%パラホルムアルデヒド、15%ピクリン酸による潅流を続ける。OBおよびON/Eを除去し、次に4℃の新しい4%パラホルムアルデヒド(PFA)中に一晩置いた。次に、切断前に凍結を防止するために、組織を4℃の30%スクロース溶液に移して48時間置いた。次に組織をOCT中で凍結させ、切片(6μm)を事前にコーティングした顕微鏡スライド上に置いた(生理的食塩水中10%BSA)。最初にヘマトキシリンおよびエオシン(H&E)染色を実施して、この期間中にOB、三叉神経節およびON/Eに浸透する(infitrate)炎症性細胞を検出する。神経細胞損傷を評価するために、神経成長因子β1(NGF−β1)を染色する。NGF−β1は、神経細胞損傷の後に生成され、アポトーシスを防止し、神経細胞の新たな成長を刺激するように機能する。三叉神経節およびOB切片をビオチン化ウサギ抗ヒトNGF−β1Abによって0.2μg/mlの濃度で染色する。Ab染色切片を4℃にて一晩インキュベートする。スライドをPBSですすぎ、次にアビジン−ビオチン複合体(ABC)のVectastain(Vector Laboratories,バーリンゲーム、カリフォルニア州)によって25℃にて30分間反応させた。組織をPBSで3回すすぎ、次に以前に報告したようにジアミノベンジデン(DAB)と5〜10分間反応させる。スライドをさらに3回すすぎ、切片をC.S.ヘマトキシリンによって30秒間対比染色する。H2Oでの洗浄後、スライドを100%アルコールおよびキシレン中で脱水する。NGF−β1の増加は、神経細胞組織の損傷度の指標を与える。神経細胞関与の別の指標は、小膠細胞の活性化である。活性化された小膠細胞は、アメーバ様球状形状を示し、これに対して静止細胞(G0/G1)は、樹枝状の分裂した外観を有する。活性化時のこのような変化によって、静止および活性化小膠細胞を区別できる。小膠細胞では、S.ニューモニエ攻撃後の活性化状態に対処するために、F4/80抗体または抗MAC−1(MI/70)が使用される。神経細胞損傷および小膠細胞活性化に加えて、OBのアポトーシスの誘発が評価される。このために、活性カスパーゼ3である、Asp−Glu−Val−Asp特異性プロテアーゼの誘発がアポトーシス経路の開始において重要であるため、それが解析される。活性カスパーゼ3(Cell Signaling Technology,Inc.,ビバリー、マサチューセッツ州)に対して特異性のAbは、アポトーシスの検出のために免疫組織化学で使用できる。カスパーゼ3活性が免疫組織化学によって神経細胞組織で検出されると、活性は、基質のタンパク質分解後の蛍光信号に基づくカスパーゼ−3アッセイキット(Molecular probes,ユージーン、オレゴン州)を用いて定量される。
【0122】
(実施例9)
(逆行性軸索輸送による、S.ニューモニエが嗅球を標的化する能力)
第一に、鼻腔内またはi.v.接種後の処置マウスの神経細胞組織、OBおよび脳への肺炎球菌の蓄積を評価する。i.v.接種後、神経細胞組織のどの肺炎球菌も血液を通じて侵入する。鼻腔内攻撃後の第1日、第4日、第11日および第18日の組織を収集する。その場合、脳およびOB1グラム当たりの細菌数は、注射後のすべての時点で同様なはずである。これに対して、鼻腔内接種後の鼻道を通じて侵入する細菌では、OBへの蓄積(組織重量当たりで表される)が先に起こり、一般に脳で観察される蓄積が待ち構えている。
【0123】
第二に、鼻腔内投与後の肺炎球菌の生体内撮像が実施される。テクネチウム−99(Tc−99m)標識TIGR4、安定不透明および透明改変体、EF3030、ならびにnanAおよび/またはnanBを欠いたTIGR4変異体を使用して、Waibel et al.(1999)Nature Biotechnol.17:897− 901によって最初に述べられた方法を使用して、アデノウィルスで以前に実施されたようにガンマカメラ撮像を使用して、マウスにおけるその存在を描出する。これは、このアイソトープの短い半減期(6時間)のために、鼻腔内投与後、最初のほぼ24時間の撮像を可能にし、鼻道で発生する早期の事象の解析を可能にする。肺炎球菌の長期撮像では、ルシフェラーゼ−またはGFP−発現肺炎球菌性EF3030(またはTIGR4)菌株を使用して、生体内での生体発光を描出する。Xenogen corporation(アラメダ、カリフォルニア州)から市販されているルシフェラーゼ発現肺炎球菌菌株EF3030を使用する。この肺炎球菌性菌株を用いた生体内撮像の成功が以前に報告されている。マウスは生体発光撮像システム(IVIS system,Xenogen,Inc.)を使用して撮像して、ルシフェラーゼ発現を検出する。同じ位置に向けたマウスで、常にルシフェリン2.5mgのi.p.注射の10分後に画像を収集する。撮像の間、マウスを37℃にてエンフルレン麻酔状態に維持する。撮像は各マウスに対して、ルシフェラーゼ−発現肺炎球菌を用いた鼻腔内攻撃後の第2日から開始して第18日まで、複数回実施する。撮像の画像取込時間は、20秒〜10分間の範囲である。データ取込ソフトウェアは、画像収集中にいずれの画素も飽和しないようにする。興味のある領域からの発光(相対光子/秒)を、Xenogen提供のソフトウェアを使用して定量する。発光の強度は、生体発光画像の擬似カラースケーリングを用いて表す。生体発光画像は通例、同時に収集されたマウスの白黒写真にオーバーレイされる。この生体内撮像は、肺炎球菌が鼻道からOBへ侵入する能力を分析することに焦点を当てている。この生体発光試験は、ルシフェラーゼ遺伝子の伝達成功後に、nanA TIGR4変異体へと拡張される。
【0124】
本出願を通じて、各種の公報が参照されている。これらの公報のその全体の開示は、本明細書で説明する化合物、組成物および方法をさらに十分に説明するために、この出願内への参照により本明細書に組み入れられている。
【0125】
各種の改良および変形を、本明細書で説明する化合物、組成物および方法に行うことができる。本明細書で説明する化合物、組成物および方法の他の態様は、本明細書で開示された化合物、組成物および方法の明細書および慣行の検討から明らかになるであろう。明細書および実施例は一例として考慮されるものとする。
【0126】
(参考文献)
【0127】
【化1】
【0128】
【化2】
【0129】
【化3】
【0130】
【化4】
【0131】
【化5】
【0132】
【化6】
【0133】
【化7】
【0134】
【化8】
【0135】
【化9】
【0136】
【化10】
【0137】
【技術分野】
【0001】
(関連出願への相互参照)
本出願は、2003年11月10日に出願された米国仮出願第60/518,799号の利益を主張し、米国仮出願第60/518,799号は、本明細書においてその全体が参考として援用される。
【0002】
(謝辞)
本発明は、National Institutes of Healthからの助成金DC 04976、AI 21548、およびP30 DK 54781の下での、そしてNational Institute of Allergy and Infectious Diseasesとの契約NO1 AI 65299の下での、政府支援によって実施された。政府は本発明に一定の権利を有する。
【背景技術】
【0003】
(背景)
ストレプトコッカス・ニューモニエは、肺、中枢神経系(CNS)、中耳、および鼻道を含む複数の器官に感染しうる、かなり遍在性のヒト病原体である。これらの組織の感染は、気管支炎、肺炎、髄膜炎、および副鼻腔感染症などの様々な症状を引き起こす。S.ニューモニエは、ヒトの細菌性髄膜炎の主因であり、抗生剤処置にも関わらず、著しい死亡率および罹患率に関係している。非特許文献1。S.ニューモニエ髄膜炎は、永続的な神経性続発症を引き起こすことがある。先進国と開発途上国でのS.ニューモニエ髄膜炎の発生率は、100,000人に付きそれぞれ1〜2人および20人である。非特許文献2。米国での肺炎球菌性髄膜炎の致死率は、約18%である。非特許文献3。肺炎球菌性髄膜炎の最高の出現率は、1〜4歳の子供(すべての細菌性髄膜炎の30%)で、続いて15〜19歳(14%)および1〜11月齢の乳児(13%)で発生する。非特許文献2。高齢者も先進国と発展途上国の両方で、連鎖球菌性髄膜炎によって深刻に冒されている。非特許文献4;非特許文献5。
【0004】
世界での肺炎球菌の主な貯蔵場所は、ヒト鼻輸送(nasal carriage)である。感染の獲得は一般にキャリアからであり、感染は常に鼻輸送が先に起こる。上咽頭のコロニー形成は、肺炎球菌の下気道、副鼻腔、および中耳への蔓延の前提条件と見なされる。それゆえ輸送を防止したいずれの医学的介入も、処置を受けた人々における疾患のリスクを除去するだけではなく、集団免疫ももたらし、コミュニティにおける未処置の構成員の感染リスクすら低下させる。S.ニューモニエは重要なヒト病原体であるが、S.ニューモニエが鼻輸送または髄膜炎を引き起こす機構に関しては比較的わずかな事しか知られていない。
【0005】
ノイラミニダーゼが鼻道に独自の毒性因子であることを示唆する多少のデータが存在する。そのような観察の1つは、その親菌株よりも上咽頭から速く除去される、NanA欠損S.ニューモニエ菌株D39の研究によるものである。非特許文献6。ノイラミニダーゼは、宿主表面および体液中の広範囲に亘る糖脂質、糖タンパク質、およびオリゴ糖から末端シアル酸残基を開裂させる。肺炎球菌性髄膜炎患者の脳脊髄液(CSF)中の遊離シアル酸のレベル上昇は、予後不良に関連している。非特許文献7。ヒトにおけるS.ニューモニエ毒性に対するこの酵素の重要性は、S.ニューモニエそれぞれの新たな臨床単離物がノイラミニダーゼ活性を有する、2つの独立した研究の成果によってさらに説明される。非特許文献7;非特許文献8。その上、その毒性を上昇させることの多い肺炎球菌の単離物のマウス継代は、ノイラミニダーゼ活性の2〜5倍の上昇を引き起こすことも報告されている。非特許文献9。肺炎球菌性Cポリサッカライドは、テイコ酸としても既知であり、リポテイコ酸としても既知である肺炎球菌性F抗原のポリサッカライド部分と構造的に同一である。非特許文献10。これらの分子は、グラム陽性細菌の中でS.ニューモニエ独自の特徴である。これらの分子の主要抗原決定基は、ホスホリルコリン(PC)残基であり、PCに対するAbsは、腹腔内、静脈内、または経鼻肺炎球菌性攻撃に対して防御性である。非特許文献11;非特許文献12;非特許文献13;非特許文献14。しかしながら、これらの研究のすべてが血清によって投薬された全身感染に対する保護を評価したときに、これらのAbsが鼻腔内コロニー形成を防御する能力に関する情報は得られていない。しかしながら表面ホスホコリン残基は、呼吸細菌の表面でよく見られる。非特許文献15。
S.ニューモニエが鼻輸送と、それに続く疾患を引き起こす機構は、比較的知られていない。今日までいずれの研究も、鼻輸送を低減または防止する機構を確定していない。上咽頭のコロニー形成は、下気道、副鼻腔、全身、および脳への肺炎球菌の蔓延への前提条件と見なされており、当業界で必要なものは、粘膜免疫を初期肺炎球菌性コロニー形成の部位に供給する手段である。上咽頭での初期肺炎球菌性コロニー形成の防止は、鼻輸送を防止し、個人間でのS.ニューモニエの蔓延を低減するであろう。その上、上咽頭の粘膜表面へ免疫を提供することは、S.ニューモニエによって続いて引き起こされる疾患を防止または低減する。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Quagliarelloら,「N.Eng.J.Med.」1992年,第327巻:869−872
【非特許文献2】Anon,「CDSC European Bacterial Meningitis Surveillance Project」PHLS,London,2000年
【非特許文献3】Fedsonら,「Arch.Intern.Med.」1994年,第154巻:2531−2535
【非特許文献4】Butlerら,「Drugs Aging」1999年,第15巻(補遺1):11−19
【非特許文献5】Fedsonら,「Vaccine」1999年,第17巻補遺1:S 11−18
【非特許文献6】Tongら,「Infect.Immun.」2002年,第68巻:921−924
【非特許文献7】O’Tooleら,「J.Clin.Invest.」1971年,第50巻:979−985
【非特許文献8】Kellyら,「J.Bacteriol.」1967年,第94巻:272−273
【非特許文献9】Vishniakovaら,「Zhurnal Mikrobiologii,Epidemiologii i Immunobiologii」1992年,9−10:26−9
【非特許文献10】Fischerら,「Eur.J.Biochem」1993年,第215巻:851−857
【非特許文献11】Brilesら,「Eur.J.Immunol.」1984年,第14巻:1027−1030
【非特許文献12】Brilesら,「Nature」1981年,第294巻:88−90
【非特許文献13】Yotherら,「Infect.Immun.」1982年,第36巻:184−188
【非特許文献14】Brilesら,「J.Mol.Cell.Immunol.」1984年,第1巻:305−309
【非特許文献15】Lysenkoら,「Infect.Immun.」2000年,第68巻:1664−71
【発明の概要】
【課題を解決するための手段】
【0007】
(発明の要旨)
本明細書では、細菌感染(たとえば肺炎球菌感染)、鼻輸送、鼻腔内コロニー形成、およびCNS浸潤を低減または防止するように設計された組成物を提供する。必要に応じて、組成物は、粘膜投与のために設計されている。本明細書では、無毒化肺炎球菌ノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸(pneumococcal teichoic acid)、肺炎球菌性リポテイコ酸(pneumococcal lipoteichoic acid)、これらのいずれか1つの抗原部分およびこれらの無毒化作用因子を含有する組成物が提供される。
【0008】
肺炎球菌ノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸、またはこれらのいずれか1つの抗原部分および薬学的に受容可能なキャリアを含む組成物も提供される。必要に応じて組成物は、肺炎球菌ノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸またはこれらのいずれか1つの抗原部分のいずれかの組合せを含むことができる。無毒化肺炎球菌ノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸、またはこれらのいずれか1つの抗原部分はもちろんのこと、無毒化作用因子を含有する組成物および作用因子を使用する方法も提供される。
【0009】
被験体に作用因子を含む組成物を投与する工程を含む、被験体において肺炎球菌ノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸、あるいは肺炎球菌ノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、または肺炎球菌性リポテイコ酸のいずれか1つの抗原部分に対する抗体を産生させる方法も提供される。必要に応じて組成物は、粘膜表面への投与または全身投与に適している。
【0010】
肺炎球菌ノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸、またはこれらのいずれか1つの抗原部分に対する抗体を、薬学的に受容可能なキャリアと共に含む組成物もさらに提供される。必要に応じて組成物は、粘膜表面への投与または全身投与に適している。
【0011】
被験体の鼻粘膜に本明細書で教示した組成物を接触させる工程を含む、被験体において鼻輸送、鼻腔内コロニー形成、または細菌感染(たとえば肺炎球菌感染)を低減または防止する方法がさらに提供される。
本発明は、例えば、以下の項目を提供する:
(項目1)
無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目2)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの活性のおよそ60%を有する、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目3)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの活性のおよそ70%を有する、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目4)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの活性のおよそ80%を有する、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目5)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの活性のおよそ90%を有する、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目6)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの天然に存在するアミノ酸の少なくとも7%の欠失を含む、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目7)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの天然に存在するアミノ酸の少なくとも7%の欠失を含み、非無毒化ノイラミニダーゼの活性のおよそ60%を示す、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目8)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの天然に存在するアミノ酸の少なくとも7%の欠失を含み、非無毒化ノイラミニダーゼの活性のおよそ70%を示す、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目9)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの天然に存在するアミノ酸の少なくとも7%の欠失を含み、非無毒化ノイラミニダーゼの活性のおよそ80%を示す、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目10)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの天然に存在するアミノ酸の少なくとも7%の欠失を含み、非無毒化ノイラミニダーゼの活性のおよそ90%を示す、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目11)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの少なくとも5個のN末端アミノ酸の欠失を含む、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目12)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの少なくとも10個のN末端アミノ酸の欠失を含む、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目13)
ノイラミニダーゼが、非無毒化ノイラミニダーゼの少なくとも15個のN末端アミノ酸の欠失を含む、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目14)
ノイラミニダーゼが、非無毒化ノイラミニダーゼのC末端アミノ酸の少なくとも10%の欠失を含む、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目15)
ノイラミニダーゼが、非無毒化ノイラミニダーゼのC末端アミノ酸の少なくとも20%の欠失を含む、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目16)
ノイラミニダーゼが、非無毒化ノイラミニダーゼのC末端アミノ酸の少なくとも30%の欠失を含む、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目17)
ノイラミニダーゼが、非無毒化ノイラミニダーゼのC末端アミノ酸の少なくとも35%の欠失を含む、無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分。
(項目18)
無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分および薬学的に受容可能なキャリアを含む、組成物。
(項目19)
アジュバントをさらに含む、項目18に記載の組成物。
(項目20)
被験体において肺炎球菌ノイラミニダーゼに特異的な抗体を産生する方法であって、該方法は、
該被験体に無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分を投与する工程
を包含する、方法。
(項目21)
被験体において肺炎球菌の鼻輸送を低減する方法であって、該方法は、
該被験体に無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分を投与する工程
を包含する、方法。
(項目22)
被験体において肺炎球菌感染を防止する方法であって、該方法は、
該被験体に無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分を投与する工程
を包含する、方法。
(項目23)
前記肺炎球菌感染が髄膜炎である、項目22に記載の方法。
(項目24)
前記肺炎球菌感染が中耳炎である、項目22に記載の方法。
(項目25)
前記肺炎球菌感染が肺炎である、項目22に記載の方法。
(項目26)
前記肺炎球菌感染が溶血性尿毒症である、項目22に記載の方法。
(項目27)
被験体において肺炎球菌の鼻輸送を低減または防止する方法であって、該方法は、
該鼻輸送を低減または防止する条件下で、該被験体に肺炎球菌ノイラミニダーゼまたはその抗原性フラグメントを投与する工程
を包含する、方法。
(項目28)
被験体において肺炎球菌感染を低減または防止する方法であって、該方法は、
該感染を低減または防止する条件下で、該被験体に肺炎球菌ノイラミニダーゼまたはその抗原性フラグメントを投与する工程
を包含する、方法。
(項目29)
前記肺炎球菌感染が髄膜炎である、項目28に記載の方法。
(項目30)
前記肺炎球菌感染が中耳炎である、項目28に記載の方法。
(項目31)
前記肺炎球菌感染が肺炎である、項目28に記載の方法。
(項目32)
前記肺炎球菌感染が溶血性尿毒症である、項目28に記載の方法。
(項目33)
被験体において肺炎球菌感染を低減または防止する方法であって、該方法は、
該感染を低減または防止する条件下で、該被験体に肺炎球菌ノイラミニダーゼ抗体またはそのフラグメントを投与する工程
を包含し、該投与工程は、
該被験体の粘膜表面に該抗体を接触させる工程
を包含する、方法。
(項目34)
前記肺炎球菌感染が髄膜炎である、項目33に記載の方法。
(項目35)
前記肺炎球菌感染が中耳炎である、項目33に記載の方法。
(項目36)
前記肺炎球菌感染が肺炎である、項目33に記載の方法。
(項目37)
前記肺炎球菌感染が溶血性尿毒症である、項目33に記載の方法。
(項目38)
肺炎球菌ノイラミニダーゼまたはその抗原部分および薬学的に受容可能なキャリアを含む組成物であって、該組成物は、粘膜表面への投与に適切である、組成物。
(項目39)
前記組成物が鼻スプレーである、項目38に記載の組成物。
(項目40)
前記組成物がネブライザ溶液である、項目38に記載の組成物。
(項目41)
前記組成物がエアゾール吸入薬である、項目38に記載の組成物。
(項目42)
項目38に記載の組成物を含む、容器。
(項目43)
前記容器が鼻スプレー器である、項目42に記載の容器。
(項目44)
前記容器がネブライザである、項目42に記載の容器。
(項目45)
前記容器が吸入器である、項目42に記載の容器。
(項目46)
被験体において肺炎球菌ノイラミニダーゼに特異的な抗体を産生する方法であって、該方法は、
該被験体の鼻粘膜に無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分を接触させる工程
を包含する、方法。
(項目47)
被験体において肺炎球菌の鼻輸送を低減する方法であって、該方法は、
該被験体の鼻粘膜に無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分を接触させる工程
を包含する、方法。
(項目48)
被験体において肺炎球菌感染を防止する方法であって、該方法は、
該被験体の粘膜表面に無毒化肺炎球菌ノイラミニダーゼまたはその抗原部分を接触させる工程
を包含する、方法。
(項目49)
前記肺炎球菌感染が髄膜炎である、項目48に記載の方法。
(項目50)
前記肺炎球菌感染が中耳炎である、項目48に記載の方法。
(項目51)
前記肺炎球菌感染が肺炎である、項目48に記載の方法。
(項目52)
前記肺炎球菌感染が溶血性尿毒症である、項目48に記載の方法。
(項目53)
肺炎球菌性テイコ酸または肺炎球菌性リポテイコ酸からのホスホコリンまたはその抗原部分、および薬学的に受容可能なキャリアを含む組成物であって、該組成物は、粘膜表面への投与に適切である、組成物。
(項目54)
前記組成物が鼻スプレーである、項目53に記載の組成物。
(項目55)
前記組成物がネブライザ溶液である、項目53に記載の組成物。
(項目56)
前記組成物がエアゾール吸入薬である、項目53に記載の組成物。
(項目57)
項目53に記載の組成物を含む、容器。
(項目58)
前記容器が鼻スプレー器である、項目57に記載の容器。
(項目59)
前記容器がネブライザである、項目57に記載の容器。
(項目60)
前記容器が吸入器である、項目57に記載の容器。
(項目61)
被験体において肺炎球菌ノイラミニダーゼに特異的な抗体を産生する方法であって、該方法は、
該被験体の鼻粘膜に項目53に記載の組成物を接触させる工程
を包含する、方法。
(項目62)
被験体において肺炎球菌の鼻輸送を低減する方法であって、該方法は、
該被験体の鼻粘膜に項目53に記載の組成物を接触させる工程
を包含する、方法。
(項目63)
被験体において肺炎球菌感染を低減または防止する方法であって、該方法は、
該被験体の粘膜表面に項目53に記載の組成物を接触させる工程
を包含する、方法。
(項目64)
前記肺炎球菌感染が髄膜炎である、項目63に記載の方法。
(項目65)
前記肺炎球菌感染が中耳炎である、項目63に記載の方法。
(項目66)
前記肺炎球菌感染が肺炎である、項目63に記載の方法。
(項目67)
前記肺炎球菌感染が溶血性尿毒症である、項目63に記載の方法。
(項目68)
肺炎球菌ノイラミニダーゼまたはその抗原部分、肺炎球菌性テイコ酸または肺炎球菌性リポテイコ酸からのホスホコリンまたはその抗原部分、および薬学的に受容可能なキャリアを含む組成物であって、該組成物は、粘膜表面への投与に適切である、組成物。
(項目69)
前記組成物が鼻スプレーである、項目68の組成物。
(項目70)
前記組成物がネブライザ溶液である、項目68の組成物。
(項目71)
前記組成物がエアゾール吸入薬である、項目68の組成物。
(項目72)
項目68に記載の組成物を含む、容器。
(項目73)
前記容器が鼻スプレー器である、項目72に記載の容器。
(項目74)
前記容器がネブライザである、項目72に記載の容器。
(項目75)
前記容器が吸入器である、項目72に記載の容器。
(項目76)
被験体において肺炎球菌ノイラミニダーゼに特異性な抗体を産生する方法であって、該方法は、
該被験体の鼻粘膜に項目68に記載の組成物を接触させる工程
を包含する、方法。
(項目77)
被験体において肺炎球菌の鼻輸送を低減する方法であって、該方法は、
該被験体の鼻粘膜に項目68に記載の組成物を接触させる工程
を包含する、方法。
(項目78)
被験体において肺炎球菌感染を防止する方法であって、該方法は、
該被験体の粘膜表面に項目68に記載の組成物を接触させる工程
を包含する、方法。
(項目79)
前記肺炎球菌感染が髄膜炎である、項目78に記載の方法。
(項目80)
前記肺炎球菌感染が中耳炎である、項目78に記載の方法。
(項目81)
前記肺炎球菌感染が肺炎である、項目78に記載の方法。
(項目82)
前記肺炎球菌感染が溶血性尿毒症である、項目78に記載の方法。
(項目83)
肺炎球菌性テイコ酸または肺炎球菌性リポテイコ酸の非ホスホコリン抗原部分および薬学的に受容可能なキャリアを含む組成物であって、該組成物は、粘膜表面への投与に適切である、組成物。
(項目84)
前記組成物が鼻スプレーである、項目83に記載の組成物。
(項目85)
前記組成物がネブライザ溶液である、項目83に記載の組成物。
(項目86)
前記組成物がエアゾール吸入薬である、項目83に記載の組成物。
(項目87)
項目83に記載の組成物を含む、容器。
(項目88)
前記容器が鼻スプレー器である、項目87に記載の容器。
(項目89)
前記容器がネブライザである、項目87に記載の容器。
(項目90)
前記容器が吸入器である、項目87に記載の容器。
(項目91)
被験体において肺炎球菌ノイラミニダーゼに特異的な抗体を産生する方法であって、該方法は、
該被験体の鼻粘膜に項目83に記載の組成物を接触させる工程
を包含する、方法。
(項目92)
被験体において肺炎球菌の鼻輸送を低減する方法であって、該方法は、
該被験体の鼻粘膜に項目83に記載の組成物を接触させる工程
を包含する、方法。
(項目93)
被験体において肺炎球菌感染を防止する方法であって、該方法は、
該被験体の粘膜表面に項目83に記載の組成物を接触させる工程
を包含する、方法。
(項目94)
前記肺炎球菌感染が髄膜炎である、項目93に記載の方法。
(項目95)
前記肺炎球菌感染が中耳炎である、項目93に記載の方法。
(項目96)
前記肺炎球菌感染が肺炎である、項目93に記載の方法。
(項目97)
前記肺炎球菌感染が溶血性尿毒症である、項目93に記載の方法。
(項目98)
肺炎球菌ノイラミニダーゼまたはその抗原部分、肺炎球菌性テイコ酸または肺炎球菌性リポテイコ酸の非ホスホコリン抗原部分、および薬学的に受容可能なキャリアを含む組成物であって、該組成物は、粘膜表面への投与に適切である、組成物。
(項目99)
前記組成物が鼻スプレーである、項目98に記載の組成物。
(項目100)
前記組成物がネブライザ溶液である、項目98に記載の組成物。
(項目101)
前記組成物がエアゾール吸入薬である、項目98に記載の組成物。
(項目102)
項目98に記載の組成物を含む、容器。
(項目103)
前記容器が鼻スプレー器である、項目102に記載の容器。
(項目104)
前記容器がネブライザである、項目102に記載の容器。
(項目105)
前記容器が吸入器である、項目102に記載の容器。
(項目106)
被験体において肺炎球菌ノイラミニダーゼに特異的な抗体を産生する方法であって、該方法は、
該被験体の鼻粘膜に項目98に記載の組成物を接触させる工程
を包含する、方法。
(項目107)
被験体において肺炎球菌の鼻輸送を低減する方法であって、該方法は、
該被験体の鼻粘膜に項目98に記載の組成物を接触させる工程
を包含する、方法。
(項目108)
被験体において肺炎球菌感染を防止する方法であって、該方法は、
該被験体の粘膜表面に項目98に記載の組成物を接触させる工程
を包含する、方法。
(項目109)
前記肺炎球菌感染が髄膜炎である、項目108に記載の方法。
(項目110)
前記肺炎球菌感染が中耳炎である、項目108に記載の方法。
(項目111)
前記肺炎球菌感染が肺炎である、項目108に記載の方法。
(項目112)
前記肺炎球菌感染が溶血性尿毒症である、項目108に記載の方法。
(項目113)
ホスホコリン抗体またはそのフラグメントおよび薬学的に受容可能なキャリアを含む、組成物。
(項目114)
前記組成物が粘膜表面への投与に適切である、項目113に記載の組成物。
(項目115)
前記組成物が鼻スプレーである、項目113に記載の組成物。
(項目116)
前記組成物がネブライザ溶液である、項目113に記載の組成物。
(項目117)
前記組成物がエアゾール吸入薬である、項目113に記載の組成物。
(項目118)
項目113に記載の組成物を含む、容器。
(項目119)
前記容器が鼻スプレー器である、項目118に記載の容器。
(項目120)
前記容器がネブライザである、項目118に記載の容器。
(項目121)
前記容器が吸入器である、項目118に記載の容器。
(項目122)
被験体において肺炎球菌の鼻輸送を低減する方法であって、該方法は、
該被験体にホスホコリン抗体またはそのフラグメントを投与する工程
を包含する、方法。
(項目123)
前記投与が、前記被験体の鼻粘膜に前記抗体またはそのフラグメントを接触させる工程を包含する、項目122に記載の方法。
(項目124)
被験体において肺炎球菌感染を防止する方法であって、該方法は、
該被験体にホスホコリン抗体またはそのフラグメントを投与する工程
を包含する、方法。
(項目125)
前記投与が、前記被験体の鼻粘膜に前記抗体またはそのフラグメントを接触させる工程を包含する、項目124に記載の方法。
【0012】
さらなる利点は、以下に続く説明で一部が述べられ、一部は説明から明らかになるか、または以下で述べる態様の実施によって知られるであろう。以下で述べる利点は、添付請求項で特に指摘される要素および組合せによって実現および達成されるであろう。上述の一般的な説明および以下の詳細な説明はどちらも例示および説明のためだけであり、制限的でないことが理解されるべきである。
本明細書に含まれ、その一部を構成する添付図面は、以下で述べる複数の態様を説明する。
【図面の簡単な説明】
【0013】
【図1】xidマウスへのS.ニューモニエの非カプセル化R36A菌株または毒性EF3030菌株のどちらかの3X106CFUの経鼻送達を示す。神経組織ON/E、OB、脳組織およびリンパ系組織(NALT、CLNおよび肺)を鼻腔内攻撃の第1日〜第4日後に収集して、切り刻み、生肺炎球菌の存在を分析した。論理コロニー形成単位(CFU)の平均+1標準誤差(SE)が示されている。Y軸の0値は、検出可能なCFUの非存在を表す。1グループ当たり5匹のマウスの平均CFU+SEが示され、3つの異なる実験を表している。
【図2A】鼻腔内攻撃後のS.ニューモニエ菌株EF3030 CFUの器官分布の動力学を示す。ON/E、OB、脳、血液、NW、NALT、CLN、および肺組織を4日、11日、18日、25日、および39日に収集して、S.ニューモニエの存在を分析した。S.ニューモニエ3X106CFUの分割量は、鼻道のコロニー形成と、それに続くOBの感染を生じた。Y軸の0値は、検出可能なCFUの非存在を表す。3つの独立した実験の平均CFU+SEを示す。各時点は、マウス5匹を表す第39日を除いて、マウス10匹を表す。
【図2B】鼻腔内攻撃後のS.ニューモニエ菌株EF3030 CFUの器官分布の動力学を示す。ON/E、OB、脳、血液、NW、NALT、CLN、および肺組織を4日、11日、18日、25日、および39日に収集して、S.ニューモニエの存在を分析した。S.ニューモニエ3X106CFUの分割量は、鼻道のコロニー形成と、それに続くOBの感染を生じた。Y軸の0値は、検出可能なCFUの非存在を表す。3つの独立した実験の平均CFU+SEを示す。各時点は、マウス5匹を表す第39日を除いて、マウス10匹を表す。
【図3A】GLSによる予備インキュベーション後のS.ニューモニエ菌株EF3030の分布を示す。S.ニューモニエの分割量(3X107CFU)を、鼻への利用前に、アシアロ−GM1(a−GM1)20μgまたは混合GLS(MG)125μgを用いて30分間インキュベートした。ON/E、OB、脳およびNW、NALTおよび肺を4日後に収集して、S.ニューモニエの数を分析した。Y軸の0値は、検出可能なCFUの非存在を表す。マウス5匹の平均+1SEを示し、以下の統計解析からP値を得た。データは2つの独立した実験を表している。
【図3B】GLSによる予備インキュベーション後のS.ニューモニエ菌株EF3030の分布を示す。S.ニューモニエの分割量(3X107CFU)を、鼻への利用前に、アシアロ−GM1(a−GM1)20μgまたは混合GLS(MG)125μgを用いて30分間インキュベートした。ON/E、OB、脳およびNW、NALTおよび肺を4日後に収集して、S.ニューモニエの数を分析した。Y軸の0値は、検出可能なCFUの非存在を表す。マウス5匹の平均+1SEを示し、以下の統計解析からP値を得た。データは2つの独立した実験を表している。
【図4−1】鼻腔内攻撃後のOB中のS.ニューモニエのTIGR4菌株の検出を示す。5X105CFUの分割量を鼻腔内投与し、血液、NW、ON/E、OBおよび脳組織を攻撃1週間後にコロニー形成について分析した(パネルAおよびB)。これらの組織(DNA 10μg)もニューモリシン遺伝子の存在についてPCRによって分析した(パネルC)。加えてコントロール(D)またはS.ニューモニエ攻撃マウスのOB中のS.ニューモニエを、PspA特異性Absを用いた免疫蛍光によって描出した(パネルEおよびF)。平均+1SEを示す。データは3つの独立した実験を表している。
【図4−2】鼻腔内攻撃後のOB中のS.ニューモニエのTIGR4菌株の検出を示す。5X105CFUの分割量を鼻腔内投与し、血液、NW、ON/E、OBおよび脳組織を攻撃1週間後にコロニー形成について分析した(パネルAおよびB)。これらの組織(DNA 10μg)もニューモリシン遺伝子の存在についてPCRによって分析した(パネルC)。加えてコントロール(D)またはS.ニューモニエ攻撃マウスのOB中のS.ニューモニエを、PspA特異性Absを用いた免疫蛍光によって描出した(パネルEおよびF)。平均+1SEを示す。データは3つの独立した実験を表している。
【図5】分泌NanA、TIGR4の、および細胞壁への結合のためのLPXTG(配列番号14)モチーフを有するR6(2型)のモチーフにおける比較を示す。TIGR4遺伝子は、LPETG(配列番号13)モチーフをコード化する配列の前に停止コドンを含む。このモチーフがないと、NanAはTIGR4によって環境中へ分泌される。
【図6A】S.ニューモニエ親菌株TIGR4(黒四角)またはNanA同系変異体TIGR4/nanA−(○)によって鼻腔内感染させたCBA/Nマウスの鼻洗浄液中の生肺炎球菌の動力学を示す。各点は、各マウスの鼻洗浄液1ml当たりの細菌総数を表す。*P<0.05;**P<0.01;***P<0.005、TIGR4を接種したマウスと比較。
【図6B】S.ニューモニエ親菌株TIGR4(黒四角)またはNanA同系変異体TIGR4/nanA−(○)によって鼻腔内感染させたCBA/Nマウスでの鼻腔内コロニー形成動力学を示す。各点は、各マウスの組織1グラム当たりの細菌総数を示す。*P<0.05;**P<0.01;***P<0.005、TIGR4を接種したマウスと比較。
【図6C】S.ニューモニエ親菌株TIGR4(黒四角)またはTIGR4/nanA−(○)によって鼻腔内感染させたCBA/Nマウスの嗅球におけるCFUの動力学を示す。各点は、各マウスの組織1グラム当たりの細菌総数を示す。*P<0.05;**P<0.01;***P<0.005、TIGR4を接種したマウスと比較。
【図7A】S.ニューモニエ親菌株EF3030(黒四角)またはEF3030/nanA−(○)によって鼻腔内感染させたCBA/Nマウスの鼻腔内コロニー形成動力学を示す。各点は、各マウスの鼻洗浄液1ml当たりの細菌総数を表す。*P<0.05;**P<0.01;***P<0.005、EF3030を接種したマウスと比較。
【図7B】S.ニューモニエ親菌株EF3030(黒四角)またはEF3030/nanA−(○)によって鼻腔内感染させたCBA/Nマウスの鼻腔内コロニー形成動力学を示す。各点は、各マウスの組織1グラム当たりの細菌総数を示す。*P<0.05;**P<0.01;***P<0.005、EF3030を接種したマウスと比較。
【図7C】S.ニューモニエ親菌株EF3030(黒四角)またはNanA同系変異体EF3030/nanA−(○)によって鼻腔内感染させたCBA/Nマウスの鼻腔内コロニー形成動力学を示す。各点は、各マウスの組織1グラム当たりの細菌総数を示す。*P<0.05;**P<0.01;***P<0.005、EF3030を接種したマウスと比較。野生種および変異体データをすべての時点でプールすると、EF3030とEF3030 NanB−との比較はP=0.001であった。
【図8】S.ニューモニエ親菌株TIGR4(黒四角)またはTIGR4/nanB−(○)によって鼻腔内感染させた接種4日後のCBA/Nマウスの鼻腔内コロニー形成動力学を示す。各点は、各マウスの鼻洗浄液1mlまたは組織1グラム当たりの細菌の総数を示す。いずれの場合も、TIGR4とTIGR4/nanB−との差は統計的に有意でなかった。
【図9】S.ニューモニエ親菌株TIGR4(黒四角)、TIGR4/nanA−(○)またはTIGR4/AB−によって鼻腔内感染させた接種4日後のCBA/Nマウスの鼻腔内コロニー形成動力学を示す。各点は、各マウスの鼻洗浄液1mlまたは組織1グラム当たりの細菌の総数を示す。いずれの場合も、TIGR4/nanA−とダブル変異体TIGR4/nanAB−との差は統計的に有意でなかった。
【図10】鼻腔内攻撃後の、抗ホスホコリン特異性モノクローナル抗体によるS.ニューモニエの鼻腔内コロニー形成の阻害を示す。鼻腔内攻撃後の抗PC特異性mAbsによるS.ニューモニエの鼻腔内コロニー形成の阻害。総量1X106CFUのTIGR4菌株をIgG3サブクラスまたはIgMアイソタイプのどちらかの抗PCmAbs 5μgを用いてインキュベートした。鼻孔ごとに総量5μgを投与した。利用9および12時間後の鼻洗浄液500μl中のCFUを示す。1グループ当たりマウス5匹の平均+SDを示す。
【発明を実施するための形態】
【0014】
(詳細な説明)
本化合物,組成物,物品、器具、および/または方法を開示および説明する前に、以下で述べる態様が、具体的な合成方法または具体的な投与方法に制限されず、そのためもちろん変化することが理解されるべきである。本明細書で使用する用語が、特定の態様のみを説明する目的で使用され、制限的でないことも理解されるべきである。
【0015】
明細書および添付請求項で使用するように、単数形「a」、「an」および「the」は、文脈が明確に別途規定しない限り、複数形指示対象も含むことに注意する必要がある。それゆえたとえば、「抗原性フラグメント」への言及は、抗原性フラグメントの混合物を含み、「製薬的キャリア」または「アジュバント」への言及は、2つ以上のそのようなキャリアまたはアジュバントの混合物を含むなどである。
【0016】
全体を通じて使用するように、「被験体」は個体を意味する。それゆえ「被験体」は、たとえばネコ、イヌなどの飼い慣らされた動物、家畜(たとえば畜牛、ウマ、ブタ、ヒツジ、ヤギなど)、実験用動物(たとえばマウス、ウサギ、ラット、モルモットなど)および鳥類を含むことができる。1つの態様において、被験体は、霊長類またはヒトなどの哺乳類である。
【0017】
「任意の」または「必要に応じて」は、続いて説明される事象または状況が起こりうる、または起こらないことを、そして説明が事象または状況が起こる例および起こらない例を含むことを意味する。たとえば、「必要に応じて、組成物は組合せを含むことができる」という表現は、説明が組合せおよび組合せの非存在(すなわち組合せの個々の構成要素)を含むように、組成物が異なる分子の組合せを含む、または組合せを含まないことを意味する。
【0018】
範囲は本明細書では、「約」1つの特定の値から、および/または「約」別の特定の値までとして表現される。そのような範囲が表現されるとき、別の態様は、一方の特定の値から、および/または他方の特定の値までを含む。同様に、値が先行する「約」を用いて概数として表現されるとき、特定の値が別の態様を形成することが理解されるであろう。各範囲の終点は、他方の終点に関連しても、および他方の終点から独立しても重要であることがさらに理解されるであろう。
【0019】
本明細書では、細菌感染(たとえば肺炎球菌感染)、鼻輸送、鼻腔内コロニー形成、およびCNS浸潤を低減または防止するように設計された組成物および方法を提供する。S.ニューモニエは、Cポリサッカライド−ガングリオシド相互作用によって上皮バリアを通過して、続いて上皮細胞中へエンドサイトーシスすることによって、一部は鼻道をコロニー形成する。Cポリサッカライドは、ガングリオシド中の末端または内部GaINAcβ1−4Gal配列への結合を介して、アシアロ−GM1、アシアロ−GM2、およびフコシル−アシアロ−GM1に結合する。ヒト肺を除いて細胞原形質膜中のこれらのアシアロガングリオシドは通常は少ないが、S.ニューモニエは、2つのノイラミニダーゼNanAおよびNanBを有し(Berry et al.,(1996)J.Bacteriol.178:4854−4860)、これはそれぞれα2,3−およびα2,6結合N−アセチルノイラミン酸をガラクトースへ、そしてα2,6−結合をN−アセチル−ガラクトサミンへ開裂できる。Scanlon et al.,(1989)Enzyme 41:143−150。ガングリオシドのシアル酸残基は、ガラクトースに結合したα2,3である。S.ニューモニエのノイラミニダーゼは、すべてのモノシアロガングリオシドに存在する末端シアル酸残基、およびジ−およびトリ−シアロガングリオシドで見られるように、ガラクトース結合した複数のシアル酸残基を除去する。それゆえそれらは、最も一般的な哺乳類細胞表面ガングリオシド中のGaINAcβ1−4Ga1配列を露出できるはずである。これらの残基は、細胞表面上の推定Cポリサッカライド結合部位である。通常はさらに細胞壁に結合しているそのNanA、および分泌されると考えられるNanBを使用して、S.ニューモニエは、気道の上皮細胞上にその独自の結合部位を生成する。それゆえ肺炎球菌性Cポリサッカライドは、アシアロガングリオシド、特にアシアロ−GM1に結合し、かなり豊富なGM1をアシアロ−GM1に変換できるノイラミニダーゼは、CポリサッカライドのためにON/E上に豊富な結合部位を作成する。本機構は、鼻輸送を促進し、鼻嗅神経および鼻甲介を被覆する上皮(ON/E)、嗅球(OB)を介してS.ニューモニエのCNSへのアクセスを提供する。同様に、中耳炎およびS.ニューモニエを含む他の感染症は、中耳に分布する神経を介してCNSへのアクセスを同様に得ることができる。S.ニューモニエに加えて他の細菌は、匹敵するノイラミニダーゼを有し、それゆえ同じ機構が他の細菌でも同様に発生する。それゆえ本明細書では、各種の細菌におけるこの機構を標的とする組成物および方法を開示する。本明細書で教示される作用因子、組成物、および方法は、輸送を低減させ、CNS浸潤を防止するために本機構を妨害することに関する。
【0020】
必要に応じて組成物は、粘膜投与用に設計される。たとえば本明細書では、肺炎球菌ノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸またはこれらのいずれか1つの抗原部分および薬学的に受容可能なキャリアを含む組成物であって、粘膜表面への投与に適した組成物が提供される。必要に応じて組成物は、肺炎球菌ノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸またはこれらのいずれか1つの抗原部分のいずれかの組合せを含むことができる。
【0021】
必要に応じて組成物は、エアゾール、鼻腔用ミスト、鼻スプレー、点鼻薬、ネブライザ溶液、エアゾール吸入薬、坐剤の形、または粘膜投与(経口投与を含む)に適したいずれかの形である。必要に応じて組成物は、送達のために微小球、またはリポソームに入れることができる。「粘膜表面への投与」とは、呼吸系、胃腸系、または泌尿生殖器系を含むいずれかの粘膜表面への投与を意味する。粘膜表面の例は、これに限定されるわけではないが、鼻腔(嗅覚神経上皮を含む)、上咽頭、直腸、膣、咽頭、口、エウスタキー管、気管、気管支および他の気道、および腸粘膜を含む。
【0022】
粘膜表面への投与では、粘膜アジュバントが使用できる。アジュバントは、本発明の組成物と同時に、その直前に、またはその後に投与できる。必要に応じて、組成物はさらにアジュバントを含む。粘膜アジュバント処方物はたとえば、粘膜誘導部位を標的とする作用因子を含む。アジュバントは必要に応じて、これに限定されるわけではないが、サイトカイン、ケモカイン、成長因子、血管由来因子、アポトーシス阻害物質、およびその組合せを含む群より選択される。サイトカインがアジュバントして選択されるとき、サイトカインはこれに限定されるわけではないが、IL−1、IL−1γ、IL−1β、IL−2、IL−5、IL−6、IL−12、IL−15およびIL−18を含むインターロイキン;形質転換成長因子−β(TGF−β);顆粒球マクロファージコロニー刺激因子(GM−CSF);インターフェロン−ガンマ(IFN−γ);またはアジュバント活性を有する他のサイトカインを含む群より選択される。サイトカインの部分、あるいはアジュバント活性または他の生物活性を有するサイトカインの変異体またはミミック(またはその組合せ)も、本発明の組成物および方法で使用できる。
【0023】
ケモカインがアジュバントとして選択されるとき、ケモカインは必要に応じて、これに限定されるわけではないが、リンホタクチン、RANTES、LARC、PARC、MDC、TARC、SLCおよびFKNを含む群より選択される。アポトーシス阻害物質がアジュバントとして選択される場合、アポトーシス阻害物質は必要に応じて、これに限定されるわけではないが、カスパーゼ−8の阻害物質、およびその組合せを含む群より選択される。血管由来因子がアジュバントとして選択される場合、血管由来因子は必要に応じて、これに限定されるわけではないが、塩基性繊維芽細胞成長因子(FGF)、血管内皮成長因子(VEGF)、ヒアルロナン(HA)フラグメント、およびその組合せを含む群より選択される。実際に、プラス(+)およびマイナス(−)血管由来因子をアジュバントとして選択できる。
【0024】
本発明の実質的に非毒性の生物活性粘膜アジュバントの他の例は、ホルモン、酵素、成長因子、またはその生物活性部分を含む。そのようなホルモン、酵素、成長因子、またはその生物活性部分は、たとえばヒト、ウシ、ブタ、ヒツジ、イヌ、ネコ、ウマ、または鳥類起源であり、腫瘍壊死因子(TNF)、プロラクチン、上皮成長因子(EGF)、顆粒球コロニー刺激因子(GCSF)、インスリン様成長因子(IGF−1)、ソマトトロピン(成長ホルモン)またはインスリン、あるいはレセプタが免疫系の細胞上で発現される他のホルモンまたは成長因子でありうる。
【0025】
粘膜投与のためのアジュバントは、細菌性毒素、たとえばコレラ毒素(CT)、E.コリ易熱性毒素(LT)、クロストリジウムディフィシル毒素Aおよび百日咳毒素(PT)、あるいはその組合せ、サブユニット、トキソイド、キメラ、または変異体も含む。たとえば未変性コレラ毒素サブユニットB(CTB)の精製調製物が使用できる。フラグメント、ホモログ、誘導体、およびこれらの毒素のいずれかへの融合物も、それらがアジュバント活性を保持するという条件ならば適切である。好ましくは低下した毒性を有する変異体が使用される。適切な変異体は、たとえばWO 95/17211(Arg−7−Lys CT変異体)、WO 96/6627(Arg−192−Gly LT変異体)、およびWO 95/34323(Arg−9−LysおよびGlu−129−Gly PT変異体)に述べられている。本発明の方法および組成物で使用できる、その他のLT変異体はたとえば、Ser−63−Lys、Ala−69−Gly、Glu−110−Asp、およびGlu−112−Asp変異体を含む。他のアジュバント、たとえばRH3−リガンド;CpG−モチーフオリゴヌクレオチド;たとえばE.コリ、サルモネラ・ミネソタ、サルモネラ・チフィムリウム、またはシゲラ・フレキシネリの細菌性モノホスホリル脂質A(MPLA);サポニン(たとえばQS21)、またはポリラクチドグリコリド(PLGA)微小球も粘膜投与で使用できる。考えられる他の粘膜アジュバントは、デフェンシンおよびオリゴヌクレオチドを含有するCpGモチーフである。
【0026】
本明細書を通じて使用するように、「薬学的に受容可能なキャリア」は、生物学的にまたはそれ以外で望ましくないわけではない材料を意味し、すなわち該材料は、望ましくない生物的影響を引き起こすことなく、またはそれが含有される薬学的組成物の他の成分のいずれかと有害な方式で相互作用することなく、選択した化合物と共に個体に投与できる。
【0027】
本明細書で述べる組成物のいずれも、薬学的に受容可能なキャリア共に治療に使用できる。本明細書で述べる化合物は、薬学的に受容可能なキャリアと組み合せて、1つ以上の化合物より成る薬学的組成物へと好都合に処方できる。たとえば、代表的なキャリアおよび本明細書で述べた化合物の処方物の調製と併せて使用可能である薬学的組成物を調製する通常の方法を開示し、参照により本明細書に組み入れられている、ペンシルベニア州イーストンのE.W.Martin Mack Pub.Co.によるRemington’s Pharmaceutical Sciences、最新版を参照。最も代表的には、これらは、ヒトへの組成物投与のための標準キャリアである。1つの態様においては、ヒトおよび非ヒトであり、滅菌水、食塩水、および生理的pHの緩衝溶液などの溶液を含む。他の化合物は、当業者に使用される標準手順に従って投与されるであろう。
【0028】
本明細書で述べる薬学的組成物は、これに限定されるわけではないが、選択した分子に加えて、キャリア、増粘剤、希釈剤、緩衝剤、保存料、界面活性剤などを含むことができる。薬学的組成物は、1つ以上の活性成分、たとえば抗菌剤、抗炎症剤、麻酔剤なども含むことができる。
【0029】
肺炎球菌ノイラミニダーゼは、肺炎球菌性細菌で見られるいずれかのノイラミニダーゼ分子を意味する。表1は、複数の種からのノイラミニダーゼのアライメントを示す。ノイラミニダーゼ分子は、たとえばSP1326を含む。SP1326アミノ酸配列は、GenBankアクセッション番号AAK75424によって入手できる。Tettelin,H.,et al.,(2001)Science 293:498−506。GenBankアクセッション番号AAK75424の下でSP1326に提供されたアミノ酸および核酸配列を含むすべての情報は、この参照によりその全体が本明細書に組み入れられている。本明細書を通じて示すように、すべてのアミノ酸配列のアミノ酸残基を、肺炎球菌性菌株R6のアミノ酸配列に従って表1に示すように番号付けする。
【0030】
【表1−1】
【0031】
【表1−2】
ノイラミニダーゼのいずれの抗原性改変体も、本明細書で教示した組成物または方法で使用できる。それゆえ天然型ノイラミニダーゼは、本発明で教示した方法に従ったアミノ酸残基の置換、欠失、または変更によって修飾できる。必要に応じて、そのような修飾は、ノイラミニダーゼを無毒化するように設計されるであろう。「無毒化」とは、抗原性または免疫原性を維持しながら、酵素活性を低減または除去することを意味する。これは部位特異性突然変異誘発を使用する、ノイラミニダーゼの活性部位におけるアミノ酸の置換、欠失、または変更によって実施される。好ましくは、そのような置換、欠失、または変更は、Aspボックスの範囲内となるであろう(すなわちアミノ酸残基460〜480、530〜560、または600〜620内)。本明細書で教示されたノイラミニダーゼ構造については、その全体が参照により本明細書に組み入れられているCrennell et al.,PNAS 90:9852−9856を参照。そのような置換、欠失、または変更は、アミノ酸残基383〜387、467〜473、541〜546、または610〜616内のAspボックス内で発生しうる。Aspボックスにおける変更はたとえば、アスパラギン酸の、グルタミン酸またはトレオニンの置換を含むことができる。他の保存的または非保存的アミノ酸置換も、毒性を低下させるために、Aspボックス内のアスパラギン酸残基または他のいずれかの残基にて使用できる。ノイラミニダーゼの他の領域は必要に応じて、部位特異性突然変異誘発を標的とする。たとえば位置572におけるバリンまたはグルタミンの保存的および非保存的アミノ酸置換を含む、たとえば残基570〜580に該当する領域内での修飾が開示される。残基750〜760に該当する領域に修飾を、さらに詳細には位置754にチロシンを有するノイラミニダーゼも開示される。チロシン残基の保存的アミノ酸置換は、たとえばセリンまたはトレオニンを含む。アミノ酸残基340〜350、600〜610、または360〜370該当する領域に修飾を有するノイラミニダーゼも提供される。さらに詳細には、位置347、605、366、または367のアルギニンは、リジンまたはグルタミン、あるいは他のいずれかの保存的または非保存的アミノ酸によって置換できる。本明細書で教示した各種の修飾を組み合せて使用できる。それゆえ1つ以上の保存的または非保存的アミノ酸置換が必要に応じて、同じノイラミニダーゼ中に存在する。
【0032】
上述のように、無毒化ノイラミニダーゼは、当業界で周知であるLockら(Microb.Pathog.4:33−43,1988)のアッセイで測定されたように、非無毒化ノイラミニダーゼと比較して低下した活性を示すノイラミニダーゼである。Lockのアッセイを使用して、溶解物、血清、または血液中のNanA活性は、酵素アッセイで2’−(4−メチル−ウンベリフェリル)−α−D−N−アセチルノイラミン酸(acetlyneuraminic acid)を基質として用いて、測定される(Lock et al.1988)。基質10マイクロリットルを血清10μlと合せて、37℃にて5分間インキュベートする。0.5M炭酸ナトリウムを使用して反応を停止させる。ノイラミニダーゼ活性を1分間当たりに放出される4−メチルウンベリフェロン(MU)の量で測定する。MUは、励起波長366nMおよび放出波長445nmを有する。無毒化ノイラミニダーゼは、免疫組成物を作成するために薬学的に受容可能なキャリアと組合されるように、非無毒化ノイラミニダーゼと匹敵する抗原性または免疫原性を保持することが好ましい。比較の目的で、非無毒化ノイラミニダーゼはこれに限定されるわけではないが、表1に示すR6 NanAを含む。好ましい実施形態において、無毒化ノイラミニダーゼは、非無毒化ノイラミニダーゼの活性の少なくとも60%、70%、80%、または90%を示す。
【0033】
無毒化ノイラミニダーゼは、非無毒化ノイラミニダーゼと比較して、アミノ酸配列内に変更(すなわち置換、修飾、または欠失)を含む。好ましい実施形態において、無毒化ノイラミニダーゼは、非無毒化ノイラミニダーゼ内に見られるアミノ酸のおよそ7%、10%、15%または20%の変更を含む。好ましいアミノ酸欠失は、非無毒化ノイラミニダーゼのN末端からのアミノ酸ほぼ5、10または15個の欠失を含む。他の好ましい実施形態は、非無毒化ノイラミニダーゼのC末端のアミノ酸ほぼ60、50、40、30、20、10または5個の欠失を含む(本出願では、C末端は表1に示すように、R6 NanAのアミノ酸800にて始まる)。なお他の好ましい実施形態では無毒化ノイラミニダーゼは、非無毒化ノイラミニダーゼのC末端の17、9、8、7、4または2のアミノ酸の欠失を含む。好ましい欠失のある例を表1に示す(すなわちTIGR4 NanAアミノ酸配列)。これらの変更のいずれも、1つ以上の他の変更と組み合せることができる。そのような無毒化ノイラミニダーゼ種は、非無毒化ノイラミニダーゼの活性のおよそ60%、70%、80%または90%を示すことが好ましい。
【0034】
ノイラミニダーゼ(neuramindiase)の他の保存的および非保存的置換(substation)は、ノイラミニダーゼがその抗原性または免疫原性を維持している限り使用できる。これらの保存的置換は、天然に存在するアミノ酸が同じ特性を有する天然に存在するアミノ酸によって置換されるようになっている。そのような保存的および非保存的置換は必要に応じて、ポリペプチドの酵素機能を変化させる。たとえば保存的置換は、表2に従って行える。
【0035】
【表2】
所望の場合、修飾および変化は、本発明のポリペプチドをコード化する核酸および/または本発明のポリペプチドのアミノ酸配列において行われ、同様のまたは他の所望の特徴(たとえば抗原性または免疫原性)を有するポリペプチドがなお得られることが理解される。そのような変化は、天然単離物で発生するか、または部位特異性突然変異誘発を使用して合成により導入され、ミスマッチポリメラーゼ連鎖反応(PCR)などの手順が当業界で周知である。たとえばあるアミノ酸は、機能活性の認識できる損失なしに、ポリペプチド中の他のアミノ酸と置換される。それゆえ生物有用性または活性の認識できる損失なしに、そしておそらくはそのような有用性または活性の上昇を伴って、様々な変更が本発明のポリペプチドのアミノ酸配列(または基礎を成す核酸配列)中で行われることが検討される。
【0036】
nanA遺伝子またはnanA遺伝子のいずれかの部分の欠失は、その中で教示された方法について、参照によりその全体が本明細書に組み入れられているSung et al.,(2001)Appl Environ Microbiol 67:5190−5196によって述べられている方法を使用して実施される。試薬2,3ブタジオンは、タンパク質のArg残基と特異的に反応し、NanA分子の折畳みに対するArg残基の重要性を評価するために使用される。部位特異的突然変異誘発は、特異的アミノ酸を変更するために使用される。
【0037】
ノイラミニダーゼは、たとえば変性を含む化学処理によっても無毒化できる。化学処理は、さらに負の副作用を低減し、抗原性または免疫原性を改善するために、部位特異的突然変異誘発と組み合せることもできる。無毒化ノイラミニダーゼは、無毒化ノイラミニダーゼによる被験体の免疫化の前に、ホルマリン、グルタルアルデヒド、熱などの作用因子によって、または当業者に既知の他の作用因子によって処理できる。
【0038】
それゆえ本明細書では、無毒化肺炎球菌ノイラミニダーゼあるいはその抗原性または免疫原性部分が提供される。無毒化肺炎球菌ノイラミニダーゼおよび薬学的に受容可能なキャリアを含む組成物も提供される。必要に応じて組成物は、アジュバント(たとえば粘膜アジュバントを含む)をさらに含む。
【0039】
さらに、たとえば抗原性または免疫原性を上昇させる部分を含む、部分をノイラミニダーゼに添加できる。そのような部分はたとえば、サイトカイン、ケモカイン、成長因子、血管由来因子、アポトーシス阻害物質、ホルモン、毒素、またはアジュバントとして使用するための本明細書で述べた他の部分を含む。部分は必要に応じて、変更分子中で使用するために修飾または切断できる。それゆえ本明細書では、ノイラミニダーゼあるいはその抗原性または免疫原性フラグメントおよび抗原性または免疫原性を向上させる部分を含む、肺炎球菌ノイラミニダーゼキメラが提供される。肺炎球菌ノイラミニダーゼ誘導体および薬学的に受容可能なキャリアを含む組成物も提供される。必要に応じて組成物は、アジュバント(たとえば粘膜アジュバントを含む)をさらに含む。
【0040】
必要に応じて、本発明の修飾ノイラミニダーゼフラグメントまたはその部分は、天然型肺炎球菌ノイラミニダーゼまたはそのフラグメントと少なくとも約70%の相同性を備えたアミノ酸配列を有する。修飾ノイラミニダーゼまたはそのフラグメントをコード化する核酸がさらに提供される。いずれかの既知の改変体および誘導体、あるいは本明細書で開示された核酸およびタンパク質の発生する改変体および誘導体を定義する1つの方法は、特定の既知配列への相同性に関する改変体および誘導体の定義を通じてであることが理解される。たとえば表1に示すようにR6肺炎球菌性菌株のnanA遺伝子によってコード化されたアミノ酸配列は、肺炎球菌ノイラミニダーゼの特定の配列を説明し、タンパク質の特定のアミノ酸配列を説明する。示された配列に対して少なくとも70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99パーセントの相同性を有する、本発明で開示されたこの配列の改変体が特に開示される。当業者は、2つのタンパク質または核酸の相同性を決定する方法をただちに理解するであろう。たとえば相同性は、相同性がその最高レベルとなるように2つの配列を整列させた後に、計算できる。
【0041】
相同性を計算する別の方法は、公開されたアルゴリズムによって実施できる。比較のための配列の最適アラインメントは、Smith and Waterman Adv.Appl.Math.2:482(1981)の局所相同性アルゴリズムによって、Needleman and Wunsch,J.Mol Biol.48:443(1970)の相同性アラインメントアルゴリズムによって、Pearson and Lipman,Proc.Natl.Acad.Sci.U.S.A.85:2444(1988)の類似度方法の検索によって、これらのアルゴリズムのコンピュータによる実施によって(Wisconsin Genetics Software PackageのGAP、BESTFIT、FASTA、およびTFASTA、Genetics Computer Group, 575 Science Dr.,マジソン、ウィスコンシン州)、または検査によって実施される。核酸について同じタイプの相同性が、核酸アラインメントに関する少なくとも物質について、たとえば参照により本明細書に組み入れられている、Zuker,M.Science 244:48−52,1989,Jaeger et al.Proc.Natl.Acad.Sci.USA 86:7706−7710,1989,Jaeger et al.Methods Enzymol.183: 281−306,1989で開示されたアルゴリズムによって得られる。
【0042】
「肺炎球菌性ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸」は、肺炎球菌性細菌中に存在するホスホコリン、テイコ酸、またはリポテイコ酸を意味する。これらの化合物は、ノイラミニダーゼについて上述したように、修飾、無毒化または強化することができる。本明細書では、修飾、無毒化、および強化された化合物を含む組成物が提供される。
【0043】
「その抗原部分」とは、抗体産生を誘発する分子または化合物(たとえばノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸)のいずれかのエピトープを意味し、該抗体は該分子に向けられている。好ましくは、抗原部分は分子またはS.ニューモニエに対する免疫を誘発する。好ましくは、抗体はノイラミニダーゼの活性部位に向けられているか、または活性部位を妨害する。抗原性フラグメントの例は、これに限定されるわけではないが、保存的アミノ酸置換または修飾の存在下または非存在下でのnanA−R6アミノ酸配列の残基63〜361に該当する残基を含む。他の例は、保存的アミノ酸置換または修飾の存在下または非存在下で、ノイラミニダーゼのAsp領域(NanA−R6のアミノ酸配列の残基460〜480、530〜560、610〜620に該当する)およびNanA−R6アミノ酸配列の340〜350、360〜370、600〜610、570〜580および750〜760に該当する領域を含む。必要に応じて、ノイラミニダーゼの活性部位を妨害するように方向付けられた抗体は、配列番号15の残基347、367または605に位置するNanAのアルギニン残基に、または配列番号15の残基383〜387、467〜473、541〜546および610〜616に位置するNanAのAspボックスに結合するか、またはその結合を防止することができる。その上、抗体は、配列番号15の残基575に位置するバリン、または配列番号15の残基752に位置するチロシンに結合するか、またはその結合を防止することもできる。抗体は、配列番号15の上で示した残基のいずれかの組合せに結合するか、またはその結合を防止することもできる。
【0044】
NanAフラグメントの他の例は、アミノ酸1〜340、330〜630、620〜800、700〜1030および330〜800を含む。これらのフラグメントの2つ以上の融合物であるフラグメントもさらに提供される。融合フラグメントは、これに限定されるわけではないが、領域620〜680と融合された領域1〜340を含む。融合フラグメントは、組換えタンパク質として発現される。これらのフラグメントをコード化するフラグメントは、発現ベクター(pETベクター;Novagen,Inc.)内へクローニングされ、タンパク質が精製される。あるいはフラグメントは、ベンダーからの合成ポリペプチドとして産生される。フラグメントを使用して、動物を免疫化して抗体を産生させて、3次元構造を評価するために結晶化させる。本明細書で述べるように、保存的または非保存的アミノ酸修飾、または置換の存在下または非存在下でのフラグメントをコード化する核酸も提供される。核酸を含むベクターまたは発現系も提供される。
【0045】
本明細書では、肺炎球菌ノイラミニダーゼ、ホスホコリン、テイコ酸、リポテイコ酸、あるいはこれらの1つまたは両方の抗原部分に特異的に結合する単離抗体を含む組成物も提供される。抗体のいずれかの組合せを含む組成物も提供される。そのような抗体は、S.ニューモニエに対する受動免疫を発現させるのに有用である。抗体組成物はさらに、薬学的に受容可能なキャリアを含む。必要に応じて組成物は、粘膜表面への投与に適切であるが、上述のような全身投与を含む他の投与経路も開示されている。
【0046】
本明細書では、肺炎球菌ノイラミニダーゼ、ホスホコリン、テイコ酸、リポテイコ酸、あるいは肺炎球菌ノイラミニダーゼ、ホスホコリン、テイコ酸、またはリポテイコ酸のいずれかのエピトープに対して特異性の抗体を産生する方法も開示されている。必要に応じて抗体は、被験体の鼻粘膜に本明細書で開示された組成物の有効量を接触させることによって被験体内で(すなわち、生体内)産生される。被験体の鼻粘膜に標的分子または分子を標的とする抗体の組合せを含む組成物の有効量を接触させることによって、肺炎球菌ノイラミニダーゼ、ホスホコリン、テイコ酸、リポテイコ酸、あるいは肺炎球菌ノイラミニダーゼ、ホスホコリン、テイコ酸、またはリポテイコ酸のいずれかのエピトープのいずれかの組合せに対して特異性である抗体を産生する方法も開示される。
【0047】
必要に応じて、本明細書で述べる作用因子は、天然型であるか、無毒化または、そうでなければ修飾されたかにかかわらず、必要に応じて、たとえばベクター内の核酸として投与できる。核酸の発現は次に、被験体へのそれによって発現された所望の核酸の接触を引き起こす。それゆえたとえば、肺炎球菌ノイラミニダーゼをコード化する核酸が被験体に被験体内で発現できる形で投与する場合、次に被験体にノイラミニダーゼを接触させる。
【0048】
被験体の細胞内への外来性DNAの投与および摂取(すなわち遺伝子形質導入または形質移入)を含む本明細書で述べた方法において、開示された核酸は裸のDNAまたはRNAの形でありうるか、あるいは核酸は、核酸を細胞に送達するベクターであり、それによって抗体コード化DNAフラグメントは、当業者によって十分理解されるように、プロモータの転写調節の支配下にある。ベクターは、アデノウィルスベクター(Quantum Biotechnologies,Inc.(ラヴァル、ケベック、カナダ)などの市販の調製物でありうる。核酸またはベクターの細胞への送達は、多様な機構を介することができる。一例として、送達は、市販のリポソーム調製物、たとえばLIPOFECTIN、LIPOFECTAMINE(GIBCO−BRL,Inc.、ゲティスバーグ、メリーランド州)、SUPERFECT(Qiagen,Inc.ヒルデン、ドイツ)およびTRANSFECTAM (Promega Biotec, Inc.、マジソン、ウィスコンシン州)はもちろんのこと、当業界で標準的な手順に従って開発された他のリポソームを使用して、リポソーム経由で行われる。加えて、開示された核酸またはベクターは、遺伝子銃または他の送達方法、たとえばGenetronics,Inc.(サンディエゴ、カリフォルニア州)から入手できる技術である電気穿孔法によってはもちろんのこと、SONOPORATION装置(ImaRx Pharmaceutical Corp.、ツーソン、アリゾナ州)によっても生体内送達できる。
【0049】
一例として、ベクター送達は、ウィルス系、たとえば組換えレトロウィルスゲノムをパッケージできるレトロウィルスベクターを介して行える(たとえばPastan et al.,Proc.Natl.Acad.Sci.U.S.A.85:4486,1988;Miller et al.,Mol.Cell.Biol.6:2895,1986を参照)。次に組換えレトロウィルスを感染させるために使用して、それによってたとえば肺炎球菌ノイラミニダーゼまたは幅広い中和抗体(またはその活性フラグメント)をコード化する感染した細胞核酸に送達する。変化した核酸を哺乳類細胞に導入する正確な方法はもちろん、レトロウィルスベクターの使用に限定されない。この手順では、アデノウィルスベクター(Mitani et al.,Hum.Gene Ther.5:941−948,1994)、アデノ結合ウィルス(AAV)ベクター(Goodman et al.,Blood 84:1492−1500,1994)、レンチウィルスベクター(Naidini et al.,Science 272:263−267,1996)、シュードタイプレトロウィルスベクター(Agrawal et al.,Exper.Hematol.24:738−747,1996)を含む他の技法が広範に利用できる。物理的形質導入技法、たとえばリポソーム送達およびレセプタ仲介および他のエンドサイトーシス機構も使用できる(たとえばSchwartzenberger et al.,Blood 87:472−478,1996を参照)。このように開示された組成物および方法は、これらのまたは他の一般に使用される遺伝子導入方法のいずれかと併せて使用できる。
【0050】
一例として、抗体コード化核酸がアデノウィルスベクター内の被験体の細胞に送達される場合、ヒトへのアデノウィルス投与の投薬量は、注射1回当たり約107〜109プラーク形成単位(pfu)の範囲でありうるが、注射1回当たり1012pfuにもなりうる(Crystal,Hum.Gene Ther.8:985−1001,1997;Alvarez and Curiel,Hum.Gene Ther.8:597−613,1997)。被験体には1回の注射を投与できるか、または追加の注射が必要な場合は、それらは無期限に亘っておよび/または処置の有効性が確立されるまで、6ヶ月の間隔(または当業者によって決定されるように、他の適切な期間)で反復できる。
【0051】
核酸またはベクターの非経口投与は使用される場合、一般に注射を特徴とする。注射可能物質は、液体液剤または懸濁剤のどちらかとしての従来形で、注射前の液体への懸濁剤の溶解に適した固体形で、あるいは乳剤として調製できる。非経口投与のより最近修正された手法は、一定の投薬量が維持されるような低速放出または持続放出系の使用を包含する。たとえば参照により本明細書に組み入れられている、U.S.Patent No.3,610,795を参照。治療化合物の適切な処方および各種の投与経路のさらなる議論については、たとえばRemington: The Science and Practice of Pharmacy(19th ed.)ed.A.R.Gennaro,Mack Publishing Company,イーストン、ペンシルベニア州、1995を参照。
【0052】
被験体の鼻粘膜への本明細書で開示された組成物の有効量の接触を含む、被験体における肺炎球菌の鼻輸送を低減または防止する方法も開示される。そのような投与は、肺炎球菌感染または鼻輸送に対する能動または受動免疫あるいは防御を生成するのに有用でありうる。
【0053】
被験体の粘膜表面への本発明で開示した組成物の有効量の接触を含む、被験体における肺炎球菌感染を低減または防止する方法がさらに提供される。たとえば該方法は、肺炎球菌性髄膜炎、中耳炎、肺炎、または溶血性尿毒症を防止できる。防止または低減は、鼻輸送を低減すること、またはCNS浸潤、全身性浸潤、あるいはエウスタキー管または下気道を防止することによって行える。
【0054】
化合物の「有効量」とは本明細書で規定されるように、所望の結果を提供するための、化合物の非毒性であるが十分な量を意味する。以下で指摘されるように、正確な必要量は、被験体の種、年齢、および全身状態、処置される疾患の重症度、使用された特定の化合物、その投与方法などによって、被験体ごとに変化するであろう。それゆえ正確な「有効量」を規定することは可能ではない。しかしながら適切な有効量は、日常的な実験のみを使用して、当業者が決定できる。
【0055】
本明細書で述べる組成物の投薬量または量は、送達が行われる方法で所望の効果を生じるのに十分な多さである。投薬量は、有害な副作用、たとえば望ましくない交叉反応、アナフィラキシー反応などを引き起こすほど多くするべきではない。一般に投薬量は、年齢、状態、性別および被験体における疾患の程度によって変化し、当業者によって決定できる。投薬量は、関与する被験体の臨床状態に基づいて個々の医師が調整することができる。用量、投薬スケジュールおよび投与経路は変化しうる。抗原の鼻腔内投与の好ましい投薬量は、免疫化1回当たり約1〜1000μg、またはたとえば10〜100μgを含む、その間の任意の量を含む。
【0056】
本明細書で述べた方法による化合物または組成物の特定の用量の投与の有効性は、病歴、徴候、症状、および肺炎球菌感染した被験体または肺炎球菌性キャリアである被験体の状態を評価するのに有用であることが既知である客観的臨床試験の特定の態様を評価することによって判定できる。これらの徴候、症状、および客観的臨床試験は、そのような患者を処置する臨床医またはこの分野で実験を行う研究者に既知となるように、処置または防止される特定の疾患または状態によって変化する。たとえば適切なコントロール群および/または一般個体群または特定の個体における疾患の正常な進行の知識との比較に基づいて:1)被験体の身体的状態の改善が示される(たとえば鼻輸送が低減または除去される)、2)疾患、感染、または鼻輸送の進行が安定、減速、または後退していることが示される、あるいは3)疾患または状態を処置する他の作用因子の必要性が低下または除去される場合、特定の処置計画は有効と見なされるであろう。たとえば被験体または個体群における鼻輸送の低減または防止、CNS浸潤または他の2次肺炎球菌感染の発生の回避または低減は、有効性を示す。そのような効果は、単一の被験体(たとえば従来の粘膜表面拭取りによって検出される細菌数を減少させることによって)または個体群(たとえば疫学的研究を使用)にて決定できる。
【0057】
本明細書で述べる化合物および薬学的組成物は、局所または全身処置が望ましいかどうかによって、そして処置される部位によって多数の方法で被験体に投与することができる。それゆえたとえば、本明細書で述べる化合物または薬学的組成物は、静脈内に、皮下に、筋肉内に、リポソームまたは微小球でカプセル化して、点眼液および/または軟膏として目の表面へ、鼻スプレーとして、噴霧液として、またはエアゾールとして鼻腔または気道へ投与することができる。その上、化合物または薬学的組成物は、被験体へ経膣的に、経直腸的に、経鼻的に,経口的に、吸入により、経口的に、または挿管によって投与できる。必要に応じて組成物は、静脈内、皮下、筋肉内、または腹腔内注射によって投与される。組成物は、液体液剤または懸濁剤のどちらかとしての従来形で、液体への懸濁剤または液剤に適した固体形で、あるいは乳剤として調製できる。必要に応じて投与は、一定の投薬量が維持されるような低速放出または持続放出系による。たとえばそこで教示された方法について、参照により本明細書に組み入れられているU.S.Patent No.3,610,795を参照。
【0058】
本明細書で教示した組成物は、緩衝剤、希釈剤および他の適切な添加剤も含有できる滅菌水性または非水性液剤、懸濁剤、および乳剤を含む。非水性溶媒の例は、プロピレングリコール、ポリエチレングリコール、オリーブ油などの植物油、およびエチルオレアートなどの有機エステルである。水性キャリアは、食塩水および緩衝溶媒を含む、水、アルコール性/水性液剤、乳剤または懸濁剤である。ビヒクルは、塩化ナトリウム溶液、リンゲルデキストロース、デキストロースおよび塩化ナトリウム、乳酸リンゲル液、または不揮発性油を含む。保存料および他の添加剤、たとえば抗菌剤、抗酸化剤、キレート化剤、および不活性ガスなども存在できる。
【0059】
局所投与の処方物は、軟膏、ローション、クリーム、ゲル、滴剤、坐剤、スプレー、液剤、エアゾール、ネブライザ溶液および粉剤を含むことができる。従来の製薬キャリア、水性、粉末または油性ベース、増粘剤などは必要であるか、望ましい。
【0060】
経口投与用の組成物は、粉剤または顆粒剤、水または非水性溶媒中の懸濁剤または液剤、カプセル、サシェ、あるいは錠剤を含むことができる。増粘剤、着香剤、希釈剤、乳化剤、分散助剤または結合剤が望ましい。
【0061】
本明細書では、被験体にノイラミニダーゼインヒビタの有効量を投与する工程を含む、被験体における鼻輸送または肺炎球菌感染を低減または防止する方法が提供される。ノイラミニダーゼ阻害物質は好ましくは、被験体の内因性ノイラミニダーゼを著しく低減することなく、肺炎球菌ノイラミニダーゼ活性を阻害する。それゆえたとえば、ノイラミニダーゼがヒトに投与される場合、阻害物質は好ましくは、負の副作用がヒトに生じるように、ヒトノイラミニダーゼ活性を低減することなく、またはヒトノイラミニダーゼ活性を低減することなく、肺炎球菌ノイラミニダーゼを阻害する。既知のノイラミニダーゼインヒビタの例は、DANA、NANA、ザナミビルおよびオセルタミビルを含む。
【0062】
本明細書では、被験体に肺炎球菌ノイラミニダーゼ、ホスホコリン、肺炎球菌性テイコ酸、肺炎球菌性リポテイコ酸に対する抗体またはそのフラグメント、あるいはこれらのいずれか1つの部分に対する抗体を含む組成物の有効量を投与する工程を含む、被験体における鼻輸送および肺炎球菌感染を低減または防止する方法である。必要に応じてこの投与は、被験体の粘膜表面に組成物を接触することを含む。組成物および抗体を含有する容器も提供される。
【0063】
「ホスホコリン抗体」という用語は本明細書で使用するように、ホスホコリンまたはその抗原性フラグメントに優先的に結合する抗体を指す。本発明の抗体は、肺炎球菌性テイコ酸または肺炎球菌性リポテイコ酸またはその抗原部分にも、あるいはノイラミニダーゼまたはそのフラグメントにも優先的に結合できる。
【0064】
「抗体」という用語は、本明細書で広範な意味で使用され、ポリクローナルおよびモノクローナル抗体の両方を含む。2重または多重抗原またはエピトープ特異性を備えたキメラ抗体、およびハイブリッド抗体、ならびにハイブリッドフラグメントを含む、F(ab’)2、Fab’、Fab、scFvなどのフラグメントも、組成物および方法で使用できる。それゆえその特異性抗原を結合する能力を維持する抗体のフラグメントが提供される。たとえばノイラミニダーゼ、ホスホコリン、テイコ酸、またはリポテイコ酸結合活性を維持する抗体のフラグメントは、用語「抗体フラグメント」の意味に含まれる。そのような抗体およびフラグメントは、当業界で既知の技法によって作成可能であり、実施例において、ならびに抗体を産生し、抗体を特異性および活性についてスクリーニングする一般的方法において述べられる方法に従って、特異性および活性についてスクリーニングできる(Harlow and Lane.Antibodies,A Laboratory Manual.Cold Spring Harbor Publications,New York,(1988)を参照)。
【0065】
本発明の組成物では、抗体フラグメントおよび抗原結合タンパク質(単鎖抗体)の結合体を使用することができる。そのような結合体は、その内容が参照により本明細書に組み入れられている、たとえばU.S.Pat.No.4,704,692に述べられている。抗体は、その所望の活性について、試験管内アッセイを使用して、または類似の方法によって試験可能であり、その後、その生体内治療および/または予防活性が、既知の臨床試験方法に従って試験される。
【0066】
「モノクローナル抗体」という用語は本明細書で使用するように、抗体の実質的に均質な個体群から得られた抗体を指し、すなわち個体群内の個々の抗体は、抗体分子の小さいサブセットに存在する考えられる天然型変異を除いて同一である。開示されたモノクローナル抗体は、モノクローナル抗体を産生するいずれかの手順を使用して作成できる。たとえば開示されたモノクローナル抗体は、Kohler and Milstein,Nature,256:495(1975)によって述べられたような、ハイブリドーマ法を使用して調製できる。ハイブリドーマ法では、マウスまたは他の適切な宿主動物は通例、免疫剤によって免疫化されて、免疫剤に特異的に結合する抗体を産生または産生できるリンパ球を発現させる。あるいはリンパ球はたとえば、本明細書で述べたHIV Env−CD4−co−レセプタ複合体を使用して、試験管内で免疫化できる。
【0067】
モノクローナル抗体は、U.S.Pat.No.4,816,567(Cabilly et al.)に述べられている方法などの、組換えDNA法によっても産生できる。開示されたモノクローナル抗体をコード化するDNAは、従来の手順を使用してただちに単離および配列決定できる(たとえばマウス抗体の重鎖および軽鎖をコード化する遺伝子に特異的に結合できるオリゴヌクレオチドプローブを使用することによって)。抗体または活性抗体フラグメントのライブラリも、たとえばBurtonらへのU.S.Patent No.5,804,440およびBarbasらへのU.S.Patent No.6,096,441に述べられているようなファージ提示技法を使用して産生およびスクリーニングできる。
【0068】
試験管内方法も、1価抗体を調製するのに適切である。そのフラグメント、特にFabフラグメントを生成するための抗体の消化は、当業界で既知の日常技法を使用して達成できる。たとえば消化は、パパインを使用して実施できる。パパイン消化の例は、1994年12月22日に公開されたWO94/29348およびU.S.Pat.No.4,342,566に述べられている。抗体のパパイン消化は通例、1個の抗原結合部位、および残りのFcフラグメントをそれぞれ備えたFabフラグメントと呼ばれる2つの同じ抗原結合フラグメントを生成する。ペプシン処置は、2つの抗原結合部位を有するフラグメントを生成し、なお抗原を架橋することができる。
【0069】
抗体フラグメントは、他の配列に結合しているかいないかにかかわらず、抗体または抗体フラグメントの活性が非修飾抗体または抗体フラグメントと比較して著しく変化または阻害されないという条件で、特定の領域または特異的アミノ酸残基の挿入、欠失、置換、あるいは他の選択された修飾を含むことができる。これらの修飾は、ジスルフィド結合可能なアミノ酸を除去および/または付加する、その生体寿命を延長する、その分泌特徴を変化させるなどの、一部の追加特性を提供できる。いずれの場合でも、抗体または抗体フラグメントは、その同種抗原への特異的結合などの生物活性特性を所有しなければならない。抗体または抗体フラグメントの機能または活性領域は、タンパク質の特異的領域の突然変異誘発と、それに続く発現ポリペプチドの発現および試験によって同定できる。そのような方法は、当業者にただちに明らかとなり、抗体または抗体フラグメントをコード化する核酸の部位特異的突然変異誘発を含むことができる(Zoller,M.J.Curr.Opin.Biotechnol.3:348−354,1992)。
【0070】
本明細書で使用するように、「抗体(antibody)」または「抗体(antibodies)」という用語は、ヒト抗体および/またはヒト化抗体を指すこともできる。多くの非ヒト抗体(たとえマウス、ラット、またはウサギに由来するもの)は当然、ヒトにおいて抗原性であり、それゆえヒトに投与したときに、望ましくない免疫反応を生じさせる可能性がある。したがって方法におけるヒトまたはヒト化抗体の使用は、ヒトに投与された抗体が望ましくない免疫反応を誘起する機会を減少させる役割を果たす。それゆえ抗体を含む組成物は必要に応じて、ヒト化または完全ヒト抗体を含む。抗体ヒト化技法は一般に抗体分子の1つ以上のポリペプチド鎖をコード化するDNA配列を操作する組換えDNA技術の使用を含む。したがって非ヒト抗体(またはそのフラグメント)のヒト化形は、キメラ抗体または抗体鎖(またはそのフラグメント、たとえばFv、Fab、Fab’、あるいはヒト(レシピエント)抗体の枠組みへ組み込まれた非ヒト(ドナー)抗体からの抗原結合部位の部分を含有する抗体の他の抗原結合部分)である。
【0071】
開示されたヒト抗体は、いずれの技法を使用しても産生できる。ヒトモノクローナル抗体産生の例は、Coleら(Monoclonal Antibodies and Cancer Therapy,Alan R.Liss,p.77,1985)によって、そしてBoernerら(J.Immunol.,147(1):86 95,1991)によって述べられているものを含む。ヒト抗体(およびそのフラグメント)も、ファージ提示ライブラリを使用して産生できる(Hoogenboom et al.,J.Mol. Biol.,227:381,1991;Marks et al.,J.Mol.Biol.,222:581,1991)。
【0072】
開示したヒト抗体は、トランスジェニック動物からも得ることができる。たとえば免疫化に反応してヒト抗体の完全レパートリーを産生できるトランスジェニックの変異体マウスが説明されている(たとえばJakobovits et al.,Proc. Natl.Acad.Sci.USA,90:2551 255(1993);Jakobovits et al.,Nature,362:255 258(1993);Bruggermann et al.,Year in Immunol.,7:33(1993)を参照)。特に、これらのキメラおよび生殖細胞系変異体マウスにおける抗体重鎖結合領域(J(H))遺伝子の同型欠失は、内因性抗体産生の完全な阻害を生じ、ヒト生殖細胞系抗体遺伝子アレイの生殖細胞系変異体マウスへの移入の成功は、抗原攻撃時にヒト抗体の産生を引き起こす。所望の活性を有する抗体は、本明細書で述べるようにEnv−CD4−co−レセプタ複合体を使用して選択される。
【0073】
ヒト化抗体を産生するためには、レシピエント(ヒト)抗体分子の1つ以上の相補性決定領域(CDR)からの残基は、所望の抗原結合特徴(たとえば標的抗原に対するあるレベルの特異性および親和性)を有することが既知であるドナー(非ヒト)抗体分子の1つ以上のCDRからの残基によって置換される。ある例において、ヒト抗体のFv枠組み(FR)残基は、対応する非ヒト残基によって置換される。ヒト化抗体は、レシピエント抗体にも、移入されたCDRまたは枠組み配列にも見出されない残基も含む。一般にヒト化抗体は、非ヒトである供給源からそれに導入された1つ以上のアミノ酸残基を有する。実際には、ヒト化抗体は通例、一部のCDR残基およびおそらく一部のFR残基がげっ歯類抗体中の類似部位からの残基によって置換されるヒト抗体である。ヒト化抗体は一般に、抗体定常領域(Fc)の少なくとも一部、通例、ヒト抗体のそれを含有する(Jones et al.,Nature,321:522 525(1986)、Reichmann et al.,Nature, 332:323 327(1988)、およびPresta,Curr.Opin.Struct.Biol.,2:593 596(1992))。
【0074】
非ヒト抗体をヒト化する方法は、当業界で周知である。たとえばヒト化抗体は、Winterおよび共同研究者(Jones et al.,Nature,321:522 525(1986),Riechmann et al.,Nature,332:323 327(1988),Verhoeyen et al.,Science,239:1534 1536(1988))の方法に従って、げっ歯類CDRまたはCDR配列をヒト抗体の対応する配列と置換することによって産生できる。ヒト化抗体の産生に使用できる方法は、U.S.Patent No.4,816,567(Cabilly et al. )、U.S. Patent No.5,565,332(Hoogenboom et al.)、U.S.Patent No.5,721,367(Kay et al.)、U.S.Patent No.5,837,243(Deo et al.)、U.S.Patent No.5,939,598(Kucherlapati et al.)、U.S.Patent No.6,130,364(Jakobovits et al.)、およびU.S.Patent No.6,180,377(Morgan et al.)でも説明されている。
【0075】
抗体の投与は、本明細書で開示するように実施できる。抗体送達のための核酸手法も存在する。抗体および抗体フラグメントも、被験体自体の細胞が核酸を吸収してコード化抗体または抗体フラグメントを産生および分泌するように、患者または被験体に、抗体または抗体フラグメントをコード化する核酸調製物(たとえばDNAまたはRNA)として投与できる。核酸の送達は、当業界で既知のいずれの手段によってもよい。
【0076】
本明細書では、本明細書で開示した作用因子および組成物を含む容器も開示する。特に容器は、鼻スプレー器、ネブライザ、吸入器、ボトル、または組成物を粘膜表面への投与形で含有する他のいずれかの手段でありうる。必要に応じて容器は、組成物の計量用量を送達できる。
【実施例】
【0077】
以下の実施例は、当業者に本明細書で説明および請求する化合物、組成物、物品、器具、および/または方法が作成および評価される方法の完全な開示および説明を提供するために与えられ、純粋な例示であるものとし、発明者がその発明と見なすものの範囲を限定するものではない。数(たとえば量、温度など)に関して正確を期するよう努力がなされたが、多少の誤差および逸脱が説明されるはずである。別途指示しない限り、部は重量部であり、温度は℃または周囲温度であり、圧力は大気圧またはその付近である。反応状態、たとえば成分濃度、所望の溶媒、溶媒混合物、温度、圧力、ならびに生成物の純度および説明した工程から得られた収量を最適化するために使用できる他の反応範囲および条件の、多数の変形および組合せがある。そのような工程状態を最適化するためには、合理的かつ日常的な実験のみが必要となるであろう。
【0078】
(実施例1)
(輸送中に嗅覚組織に浸透する鼻肺炎球菌)
(材料および方法)
(肺炎球菌性菌株)
S.ニューモニエEF3030、血清型19F、およびTIGR4菌株、血清型4の2つのカプセル化菌株、ならびに親菌株D39、血清型2に由来する無毒性の非カプセル状菌株R36Aを利用した。Avery et al.,(1944)J.Exp.Med.79:137−158。EF3030菌株は、菌血の非存在下で気道にただちにコロニー形成し(Briles et al.,(1992)Infect.Immun.60: 111−116)、静脈内接種後に菌血を持続できないため選択された。TIGR4菌株はさらに毒性であったが、中程度の鼻接種原によって、菌血なしにコロニー形成する。
【0079】
(マウス)
CBA/CAHN/xid(xid)マウス菌株をJackson Laboratory(バーハーバー、メイン州)から得た。これらマウスのBrutonのチロシンキナーゼ遺伝子における変異は、胸腺独立性II型抗原への反応をできなくするが(Amsbaugh et al.,(1972)J.Exp.Med.136:931−949;Berning et al.,(1980)J.Immunol.46:506−513)、比較的な正常なT細胞依存性免疫反応を可能にする。これらのマウスは、カプセル状ポリサッカライドに反応できず、肺炎球菌感染を高い再現性で受けやすい。xidマウスは、病原体のない状態に維持され、7〜12週齢で使用される。
【0080】
(組織収集)
血液を後腹膜神経叢からヘパリン処理毛細管内に収集した。鼻洗浄液(NW)、腎臓、脾臓、および肺の収集前にマウスを70%エタノールによって消毒した。NWの血液汚染を防止するために、気管への切開を行って、外径0.075cm、2.0cm長のTygon管(Cole−Parmer、バーノンヒルズ、イリノイ州)を上咽頭に挿入し、同時にリンゲル液を充填した注射器に取り付けた。注射器からの液体は鼻を通じて排出して、3滴を収集した。
【0081】
上咽頭関連リンパ細網組織(NALT)、ON/E、OBおよび脳の残りを上述のように得た。van Ginkel et al.,(2000)J.Immunol.165:4778−4782;Wu et al.,(1997)Scand.J.Immunol 46:506−513。脳から三叉神経節を、解剖顕微鏡を用いて慎重に切除した。ON/E、OB、三叉神経節、NALTおよび頚部リンパ節CLNそれぞれをリンゲル液0.5ml中でホモジナイズし、脳および腎臓をそれぞれリンゲル液1.0ml中でホモジナイズした。
【0082】
(組織ミンス/血液/外部排出物中の肺炎球菌の量)
組織および体液の連続3倍希釈物8個を滅菌リンゲル液中で作成し、ゲンタマイシンサルフェート4μg/mlを含有する血液寒天プレートで平板培養した。CFUは平板培養およびロウソク瓶内でのインキュベーションの24時間後に数えた。結果は、CFU/器官、NWまたは血液1ml当たりのCFUとして表した。
【0083】
(S.ニューモニエEF3030のGLSプレインキュベーション)
GLS結合部位を遮断するために、S.ニューモニエ菌株EF3030 3X107CFUを、ヒト脳からのアシアロ−GM1 20μgまたはウシ脳からの混合GLS(18% GM1、55% GD1a、15% GD1b、10% GT1b、2% 他のGLS)125μgのどちらかを用いて、氷上で30分間インキュベートした(Calbiochem−Novabiochem Corporation,Inc.,ラホーヤ、カリフォルニア州)。GLSをPBSに溶解させ、使用1日前に完璧に混合した。両親媒性GLSは、PBS中でミセルを形成し、肺炎球菌に炭水化物部分と相互作用させる。インキュベーションの後、さらに洗浄せずに鼻孔ごとに5μlをxidマウスに鼻腔内投与した。4日後に組織をCFUについて分析した。
【0084】
(PCRによるS.ニューモニエニューモリシン遺伝子の検出)
S.ニューモニエをPCRによって検出するために、組織を1% SDS中に0.1%デオキシコール酸を用いて凍結−解凍によって溶解させて、37℃で1時間インキュベートした。タンパク質は、セチルトリメチルアンモニウムブロミド/NaCl沈殿法を使用して除去した(Ausubel et al.,(1987)Current Protocols in Molecular Biology,2nd:2.4.4、セチルトリメチルアンモニウムブロミド/NaCl沈殿法の教示については、参照により本明細書に組み入れられている)。DNA 10μgをPCR増幅に使用した。ニューモリシン(ply)特異性プライマーPly1 5’−ATTTCTGTAACAGCTACCAACGA−3’(配列番号1)およびPly2 5’−GAATTCCCTGTCTTTTCAAAGTC−3’(配列番号2)をPCR混合物に添加して、400bpフラグメントを増幅した。PCR反応は、94℃での5分間の変性工程と、それに続く増幅サイクル、つまり94℃(1分)、55℃(1分)、および72℃(1分)の30サイクルを含んでいた。エチジウムブロミド染色PCRフラグメントの画像をAlpha Imager TM IS−3400(Alpha Innotech Corporation,サンリアンドロ、カリフォルニア州)で収集した。
【0085】
(PspA特異性Absを用いたOBの免疫蛍光染色)
マウスをTIGR4菌株5X105CFUで経鼻攻撃した。OBを10%緩衝ホルマリンで固定した。パラフィン画分4μm(van Ginkel(2000)J.Immunol.165:4778−4782)は、それらを室温にて加湿チャンバ内で4時間インキュベートすることによって、PspAファミリ2Abs(1:100)を染色した。スライドをPBSで洗浄して、ビオチン化ヤギF(ab’)2抗ウサギIgG(1:200)(Southern Biotechnology Associates,Inc.,バーミングハム、アラバマ州)で染色し洗浄して、ストレプトアビジン−FITC(1:100)(BD−PharMingen,サンディエゴ、カリフォルニア州)で染色した。蛍光画像は、Nikon顕微鏡によってDEI−750 CEデジタルカラービデオカメラ(Optronics,ゴレータ、カリフォルニア州)を用いて収集し、Scion Imageソフトウェア(Scion Corporation,フレデリック、メリーランド州)を用いて処理した。
【0086】
(統計)
データを平均±1標準誤差として表し、結果を統計解析により独立Mann Whitney2標本ランク検定またはStudent t検定を使用して比較して、CFUでの有意差を決定した。
【0087】
(結果)
(鼻腔内コロニー形成およびCNS浸潤での肺炎球菌性カプセルの役割)
一次感覚嗅覚ニューロンを通じた肺炎球菌の摂取を調査するために、EF3030および非カプセル化菌株R36Aが鼻道にコロニー形成して、CNSに侵入する能力を第1〜4日に測定した(図1)。第1日にON/Eでは両方の菌株で高いCFUが観察されたが、R36Aは第4日までにON/Eおよび他のすべての組織にほぼ非存在となり、長期のコロニー形成にはあるカプセルが必要であることを示した以前の結果と一致していた。Magee and Yother(2001)Infect.Immun.69:3755−3761。EF3030は、どちらの日でもOBおよび脳での明らかな存在を示し、第4日にはNWおよびNALTに多数が存在していた。これらの調査結果は、鼻腔内攻撃後のEF3030肺炎球菌のOBおよび脳への軸索輸送と一致していた。
【0088】
(鼻腔内コロニー形成およびCNS浸潤の動力学)
EF3030は、観察した39日間に亘ってすべての時点でON/E、OB、NW、およびNALTにおいて維持された(図2)。脳およびCLNでは、はるかに少ない数のCFUが見られ、存在したこれらのCFUは一般に、第18および25日に見られた。興味深いことに、肺は第1日(図1)および第18、25および39日を除いて、肺炎球菌を示さなかった(図2)。使用した経鼻用量では、菌株EF3030を用いて実施したいずれの実験の間にも、CFUがマウス血流で検出されなかったため、菌血は神経組織分布に寄与しなかった(図2)。鼻腔内投与の1、3、6、12および24時間後およびそれに続く1週間にわたって毎日、血液を菌血について監視した。血液中に細菌は検出されなかった。
【0089】
(三叉神経節のS.ニューモニエ感染)
三叉神経細胞は上咽頭を刺激し、それゆえS.ニューモニエは、鼻粘膜の感染後の三叉神経節中に予想される。これを試験するために、接種4日後に各種の組織および血液を単離し、新たな実験でのEF3030の存在について分析した。EF3030菌株は、ON/EおよびOBで、そして三叉神経節で検出された(表3)。この調査はさらに、アシアロ−GM1がS.ニューモニエによる神経標的化のレセプタとして機能することを裏付けた。他のGLSも同様に役割を果たしやすい。
【0090】
表3は、経鼻送達後の各種組織でのS.ニューモニエ菌株EF3030の分布を示す。菌株EF3030 1X107CFUの鼻腔内投与の4日後に組織を単離した。血液(50μl)、ON/E、OB、および脳組織ミンスを希釈して、次に血液寒天上で平板培養した。三叉神経節をプールして、ホモジナイズし、次にこの培地上で平板培養した。マウス5匹の平均肺炎球菌CFU±SEが示され、3つの独立した実験を表している。脳および血液では、肺炎球菌は検出されなかった。
【0091】
【表3】
(肺炎球菌性コロニー形成を阻害するガングリオシド)
EF3030菌株を鼻腔内投与前に、アシアロ−GM1または混合GLSミセルを用いてPBS中でインキュベートした。GLS混合物は、最強の阻害効果を示し、鼻腔内投与の4日後に評価したときに、NWにてCFUを10分の1に減少させた(P=0.0365)。混合GLSプレインキュベーションの結果としてのCFUの最大の減少は、ON/Eで見られた(617分の1への減少;P=0.0134)。コントロールでは平均204および166のCFUが存在する肺(P=0.0320)(図3B)およびCNS組織(P=0.0078)(図3A)の差は、まさに顕著であるが、これに対して肺炎球菌は、GLSでインキュベートしたときに検出不可能であった(検出限界=3CFU)。アシアロ−GM1プレインキュベーションは、混合GLSよりも効率的ではなかったが、それでもCNS(図3A)および肺(図3B)においてそれぞれ、コロニー形成を25分の1および63分の1に減少させた。肺は吸入した肺炎球菌によって感染し、肺に比較的豊富に存在するアシアロ−GM1へのその結合はGLSによって明らかに阻害された。このことはGLSが上皮細胞への初期結合で役割を果たすことを示した。GLS処置は、肺炎球菌の生存能力を変更させなかった。この実験の間に肺炎球菌は血液中で検出されなかった。それゆえGLSは、鼻道の神経−上皮への肺炎球菌結合ならびに肺およびCNSの感染に関して重要な標的を構成する。
【0092】
(鼻腔内攻撃後のOBへのS.ニューモニエ蓄積の検出)
OB中のEF3030の数は一般に少なすぎて、顕微鏡による細菌の描出は実現できなかった。鼻腔内投与後のOB中のS.ニューモニエを描出するために、さらに毒性の菌株TIGR4を使用した。攻撃の1、3、6、12または24時間後およびその後毎日、代表のマウスからの血液サンプルを試験した。菌血は観察されなかった。攻撃の1週間後にマウスを殺処分して、CFUについて組織を分析した(図4Aおよび4B)。わずか5X105のTIGR4 CFUの用量が、OBに〜300CFUを生じた(図3)。OB中のPspA特異性Absを染色することによって、肺炎球菌を描出した(図4D〜F)。肺炎球菌は、攻撃されたマウスのOB、すなわち糸球体層(図4F)および外部網状層(図4E)で検出された。コントロールマウスのOBには、肺炎球菌は存在しなかった(図4D)。
【0093】
TIGR4菌株は、鼻腔内投与6日後に、NW、ON/EおよびOBからのニューモリシン遺伝子のPCR増幅によっても検出された(図4C)。PCR検出可能な肺炎球菌は、この間隔で採取した血流中に、または非感染マウスによるサンプル中に存在していなかった。
【0094】
(実施例2)
(上咽頭輸送における肺炎球菌−NanAの役割およびCNSの標的化)
NanA変異体を、遺伝子的背景およびNanAの局在化の両方で異なるS.ニューモニエの3つの菌株にて産生させた。菌株EF3030(19F型)およびD39(2型)はどちらも、細胞壁に共有結合するNanAを発現するが、TIGR4菌株(4型)は、環境へ分泌される切断NanAを発現した。
【0095】
コロニー形成におけるNanBの役割も評価した。
【0096】
(細菌株および増殖条件)
本試験で使用した菌株を表4に挙げる。
【0097】
【表4】
すべての肺炎球菌株は、−80℃にて10%グリセロール中で保存して、血液寒天プレートに移して培養し、37℃にて5% CO2雰囲気中で一晩インキュベートした。肺炎球菌の培養物を、0.5%酵素抽出物をOD660 0.5まで含有するTodd−Hewitt培地中で増殖させて、−80℃にて滅菌グリセロールを用いた10%までの同じブロス添加で分割して凍結保存した。変異の安定性を確実にするために、耐抗生物質インサートを持つ変異体を適切な抗生物質中で増殖させた。
【0098】
(nanA変異体の作成)
親バックグラウンドTIGR4、EF3030およびDD39それぞれのNanA変異体菌株JW001、SAM001、およびJCP001を挿入複製突然変異誘発技法によって得た(そこで教示された技法について、参照によりその全体が本明細書に組み入れられている、Yother et al.(1992)J.Bact.174:610−618)(表4)。菌株TIGR4およびEF3030は、レシピエントとして、同質遺伝子nanA菌株D39から作成したドナー染色体DNAの形質転換に使用した(Berry et al.(2000)Infect.Immun.68:133−140)。それぞれの場合で、変異体は、親菌株への戻し交雑を3回行った。D39変異体も我々のD39親菌株への戻し交雑を3回行って、それがこれらの試験で使用した親菌株との同質遺伝子であることを確認した。D39の変異は、成熟タンパク質のアミノ酸約650個のN末端フラグメント以外のすべての欠失を可能にする挿入複製突然変異誘発によって作成した(Berry et al.(2000)Infect.Immun.68:133−140)。TIGR4/nanB同系変異体は、挿入複製突然変異誘発技法を使用して作成した(Balachandran et al.(2002)Infect.Immun.70:2536−2534;Yother et al.(1992)J.Bact.174:610−618)。nanBの461bp内部部分を、プライマー:nanBFおよびnanBR(表1)を使用して増幅し、PCRは、Taq PCR Mastermix(Invitrogen)を使用して、95℃にて1分間、45℃にて1分間、72℃にて1分間を30サイクル実施した。フラグメントは、pSF152内へクローニングした。TIGR4菌株のプラスミドDNAによる形質転換は、前と同様であった(Balachandran et al.(2002)Infect.Immun.70:2526−2534)。nanA/nanB−TIGR4ダブル変異体は、nanB/TIGR4変異体の菌株JW001から作成した染色体DNAによる形質転換によって得た。
【0099】
(マウス毒性アッセイ)
メス6〜12週齢CBA/CaHN−XID/J(CBA/N)マウスは、Jackson Laboratory(バーハーバー、マサチューセッツ州)から得た。これらマウスのBrutonのチロシンキナーゼ遺伝子における変異は、胸腺独立性II型抗原への反応をできなくするが、比較的な正常なT細胞依存性免疫反応を可能にする(Amsbaugh et al.1972 J Exp Med.136:931−949;Briles et al.1986 Curr.Top.Microbiol.Immunol.124:103−120;Potter et al.1999 Int.Immunol.11:1059−64;Wicker and Scher 1986 Curr.Top.Microbiol.Immunol.124)。これらのマウスは、カプセル状ポリサッカライドに反応できず、肺炎球菌感染を高い再現性で受けやすい(Briles et al.1986 Curr Top.Microbiol.Immunol.124:103−120;Briels et al.1981 J.Exp.Med.153:694−705)。X連鎖免疫不全(xid)マウスは、病原体のない状態に維持され、7〜12週齢で使用した。既知の濃度の生細胞を含有する凍結感染ストックを乳酸リンゲル液で希釈した。次にマウスに体積10μl中のおよそ5X105−1X106細胞を用いて、上述のように経鼻的に(I.N.)感染させた(Wu et al.1997 Microb.Pathog 23:127−137)。
【0100】
(組織収集)
すべてのマウスは、鼻洗浄および組織収集を実施する前に安楽死させた。血液を後腹膜神経叢からヘパリン処理毛細管内に収集した。鼻洗浄液(NW)、鼻組織(嗅上皮(NT)、嗅球(OB)を含む)、および脳の収集前に、マウスを70%エタノールで消毒した。これらの体液および組織を上述のように得た。NWの血液汚染を防止するために、気管への切開を行って、外径0.075cmを有する2.0cm長のTygon管(Cole−Parmer)を上咽頭に挿入し、同時にリンゲル液を充填した注射器に取り付けた。注射器からの液体は鼻を通じて排出して、3滴を収集した。ON/EおよびOBをそれぞれリンゲル液0.5ml中でホモジナイズし、脳の残りをリンゲル注射液剤1.0ml中でホモジナイズした。
【0101】
(生肺炎球菌の定量)
組織および体液の連続3倍希釈物8個を滅菌リンゲル液中で作成し、ゲンタマイシンサルフェート4μg/mlを含有する血液寒天プレートで平板培養した。コロニー形成単位(CFU)は平板培養およびロウソク瓶内でのインキュベーションの24時間後に数えた。ノイラミニダーゼ活性に使用したアッセイは、前に述べられている(アッセイ方法について、参照によりその全体が本明細書に組み入れられている、Lock et al 1988 Microb Pathog 4:33−43)。野生種肺炎球菌と変異体肺炎球菌との間の比較を行う2標本Mann−Whitneyランク検定を用いた解析によって、結果の有意性を評価した。
【0102】
(試験管内試験)
TIGR4、ならびにそのnanAおよびnanB変異体が特異性ガングリオシドに結合する能力を測定した。使用したガングリオシドは、混合ガングリオシド、アシアロ−GM1、GM1、GD1a、GD1b、GT1(Calbiochem)およびGM3ガングリオシド(Sigma)を含む。GM3ガングリオシドは、肺炎球菌性結合に関与する末端または内部GalNAcβ1−4Gal配列を欠いており、負のコントロールとして使用される。これらのガングリオシドを含有する混合、モノ−、ジ−またはトリ−シアル酸は、ELISAプレートにただちに結合する。TIGR4菌株を用いた短期インキュベーション後の初期データは、肺炎球菌がアシアロ−GM1コーティングプレートに結合するが、BSA−、GM−3、またはGM1コーティングプレートには結合しないことを示している。ガングリオシドコーティングプレートを使用して、野生種TIGR4菌株、TIGR4菌株、nanA、nanB、およびnanA/nanB変異体菌株の安定不透明および透明相改変体によるこれらのプレートへの結合能力を比較する。これらの解析は、ガングリオシドコーティングプレートへの短期インキュベーション(1時間)および長期インキュベーション(24時間)を含む。結合した肺炎球菌は、0.5%酵素抽出物を含有するTodd Hewitt培地中での短期インキュベーション(10〜15分間)によってガングリオシドプレートから除去され、放出された細菌の反復ピペッティングおよび血液寒天プレート上での平板培養が続く。あるいは41℃の、0.5%酵素抽出物を含有するTodd Hewittブロス寒天を結合した肺炎球菌の上に注入して、プレートの底からコロニーをカウントする。コントロールは、肺炎球菌のないプレートおよびガングリオシドはないが、肺炎球菌のあるプレートを含んでいる。
【0103】
ガングリオシド結合の試験に続いて、各種の細胞系は、肺炎球菌をその細胞表面に結合させて、それらを内部移行させるその能力について試験する。これらの試験は、ラット神経褐色細胞腫細胞系PC12(ATCC)およびマクロファージ細胞系P388D1に焦点を当てる。これらの2つの細胞系は、その特異的属性のために選択された。P388D1細胞系は、小膠細胞上に存在することが報告されている、高い親和性PAF−R(Valone(1988)J.Immunol.140:2389−2394)を発現する。PC12細胞系は、検出可能なPAF−Rを発現しない。Brewer et al.,(2002)J.NeuroChem 82:1502−1511。102〜105個の肺炎球菌性CFUを6ウェルまたは24ウェル組織培養プレートで増殖させたこれらの細胞系に添加して、37℃にて15分間〜6時間インキュベートし、その後、細胞を完璧に洗浄して、付着性の肺炎球菌を分析する。細胞への内部移行を判定するために、ペニシリンおよびゲンタマイシンを用いた2時間の洗浄を実施してから、血液寒天上で細胞を平板培養するか、またはそれらを41℃の0.5%酵素抽出物を含有するTodd Hewittブロス寒天で被覆する。使用した2つの細胞系は、通常はCNSにて観察されるPAF−Rの生体内発現を反映する。活性化小膠細胞は、P388D1細胞系と同様にこのレセプタを豊富に発現するが、PAF−Rレセプタは、神経細胞、たとえばPC12細胞系上に不在であるか、別個の神経小集団によって低レベルでのみ発現されるかのどちらかである。Mori et al.,(1996)J.Neurosci 16:3590−3600;Bennett et al.,(1998)Cell Dath Differ.5:867−875。肺炎球菌の両方の細胞系への付着は、PAF−Rが付着にとって必須ではなく、代わりのレセプタが存在することを示す。TIGR4不透明および透明改変体ならびにnanA−、nanB−変異体およびnanA/nanBダブル変異体を、野生種TIGR4菌株によって観察された付着と比較して、これらの細胞系への付着について試験する。肺炎球菌付着におけるPAF−R対ガングリオシドの役割をさらに分析するために、PAF−Rを欠いたCOS−7細胞系(Gerard and Gerard(1994)J.Immunol.152:793−800;Honda et al.,(1992)J.Lipid Med.5:105−107)を、1029bpのヒトPAFR読み取り枠で、pcDNA3.1/GSプラスミドを前に報告したように使用して(そこで教示した方法について、参照によりその全体が本明細書に組み入れられている、Brewer et al.,(2002)J. Neuro Chem 82: 1502−1511)形質移入し、Transfast試薬(Promega)を使用して形質移入する。プラスミド単独をコントロールとして使用し、肺炎球菌付着に影響を及ぼすパラメータをPAF−Rの存在または非存在下で解析する。本実験は、PAF−Rの付着での重要性に関する明確なデータを提供する。PAF−Rを欠いた細胞系での付着は、ガングリオシドによって仲介され、続いてガングリオシドを用いたプレインキュベーションによって遮断される。肺炎球菌が上皮細胞に付着して浸透する能力にさらに注目するために、Detroit 562ヒト咽頭上皮細胞系(ATCC)およびA549ヒト肺上皮細胞系(ATCC)を採用し、トランスウェルシステムを使用する。Millicell(登録商標)−PCF培養(Millipore,ビルリカ、マサチューセッツ州)プレートインサートを使用して、上皮細胞系をコンフルエントまで培養する。コンフルエントは、Millipore Millicell(登録商標)電気抵抗システムを使用して、経上皮耐性を測定することにより決定する。少なくとも500Ω/cm2の抵抗は、完全にコンフルエントな上皮単層が達成されたことを意味する。これらの細胞は、それらがこの上皮層に付着し、侵入して、浸透する能力を試験するために、肺炎球菌に暴露させる。結合と内部移行を区別するために、上皮細胞を洗浄して、ペニシリンおよびゲンタマイシンを含有する培地で2時間インキュベートする。最初に、TIGR4菌株、そのnanAおよびnanB変異体、ならびにダブル変異体に焦点を当てる。TIGR4菌株の安定な透明および不透明改変体は、生体内攻撃後に逆行しなかった安定な改変体が得られるまで、連続継代によって産生させた。これらのTIGR4改変体は、上皮細胞に付着、侵入、浸透する(tranverse)能力について比較する。ウェルにEMEM培地中の103〜106CFU/ウェルを装填する。0.5、1、2、4、8、および24時間後に培養物を上皮層の上下で収穫して、CFUについて解析する。細胞層を5〜6回洗浄してから、細胞に0.5%酵素抽出物および0.5%寒天を添加したTodd−Hewittブロスを重ね、41℃まで冷却し、単層と結合した肺炎球菌CFUを決定する。プレートを37℃および5% CO2にて一晩インキュベートし、その後、CFUをカウントする。細胞系をPAF−Rの発現について解析する。これらの細胞系に由来する全RNAをRT−PCRにより、2つのプライマーPAF−1(5’−CCGATACACTCTCTTCCCGA−3’(配列番号3);ヌクレオチド151〜170)およびPAF−2(5’−ACAGTTGGTGCTAAGGAGGC−3’(配列番号4);ヌクレオチド970〜951)を使用して解析し、838bpのPCR生成物を得る(そこで教示された方法について、参照によりその全体が本明細書に組み入れられている、Stengel et al.,(1997)Arterioscler.Thromb.Vasc.Biol.17:954−962)。PAFレセプタが存在する場合、PAFレセプタインヒビタ、たとえばオクチルオニウムブロミド(Biomol Research Laboratories,Inc.プリマスミーティング、ペンシルベニア州)またはPAF(Biomol)を培養物に添加して、PAF−Rの上皮付着および浸透への寄与を決定する。オクチルオニウムブロミドは、PAF−Rへ高い親和性で結合する。あるいは上述のCOS7細胞は、この目的に使用され、PAF−Rの存在または非存在下で肺炎球菌付着を比較する。
【0104】
各種の肺炎球菌株の侵襲性度は、トランスウェルシステムの頂端および基底面コンパートメントの両方での炎症性サイトカインの生成と相関している。培養物上澄みを各種の時点で上下のコンパートメント両方にて収集して、ELISA(BD PharMingen)によって解析し、炎症性サイトカインIL−1β、IL−6、IL−8、IL−10およびTNF−αの濃度を決定する。上皮単層を酢酸アルコール中で固定して、OB中での肺炎球菌の描出で以前に使用したように、PspA特異性免疫蛍光染色を使用して、肺炎球菌の細胞内存在を解析する。蛍光画像は、以前に述べたように(そこで教示された方法について、参照により本明細書に組み入れられている、Martin et al.,(1998)J.Immunol.160:3748−3758)、適切なフィルタキューブを装備したLeica/Leitz DMRB顕微鏡(Chromtechnology,バトルボロ、バージニア州)によって描出する。画像はC5810デジタルカラーカメラ(Mamamatsu Photonic System)を用いて収集し、Adobe photoshopおよびIP LAB Spectrumソフトウェアで処理する。
【0105】
(結果)
(NanAおよびNanB変異体のコロニー形成)
S.ニューモニエがCBA/Nマウスの上咽頭にコロニー形成する能力に対するNanA変異の効果を、鼻腔内(i.n.)感染させたマウスの鼻洗浄から単離された肺炎球菌細胞の数を、NanA変異体菌株で感染させたマウスの肺炎球菌細胞の数と比較することによって評価した。これらの異なる肺炎球菌菌株が含まれ、それゆえカプセル血清型および遺伝的背景が異なる菌株に対するNanA変異の効果が調査された。TIGR4/NanA−(JW001)、EF3030/NanA−(SAM001)およびD39/NanA−(JCP001)はそれぞれ、カプセル型4、19Fおよび2であった(表4)。カプセル型4臨床アイソレート、TIGR4の場合、LPETG(配列番号13)モチーフをコード化する配列の前に停止コドンがある。このモチーフがないと、NanAは、TIGR4によって環境中へ分泌されることが予想される。現在入手できる残りの4つの肺炎球菌−NanA配列、G54(19F型)、R6(2型)、Spanish 23Fおよび670(6B型)の試験(Berry et al.Gene 71:299−305;Hoskins et al.2001 J.Bacteriol 183:5709−17;Tettelin et al.2001 Science 293:498−506)は、細胞壁への共有結合のためのLPXTG(配列番号14)モチーフを有することを示した(図5)。したがってここに含まれる菌株は、NanAが分泌され、表面結合されている菌株における変異の比較を提供した。
【0106】
コロニー形成の劇的な減少は、TIGR4およびEF3030両方のNanA変異体で観察された(図6および7)。
【0107】
S.ニューモニエは別のノイラミニダーゼ、NanBを発現する。同様の相同性度がNanB間でNanAに対して共有されている。NanBは、NanAと43%の相同性を共有する(24%の同一性)。タンパク質間の共有された残基は、シアリダーゼであることを示唆した(Berry et al.1996 J.Bacteriol.178:4854−4860)。NanBは、6.5〜7であるNanAの最適pHと比較して、4.5の最適pHを有することが見出されている。その最適pHにおいてさえ、NanBは、NanAがその最適pHにあるときのシアリダーゼの約1/100の活性である。それでも、コロニー形成およびCNSの直接侵襲についてNanBの要件があるかどうか調べるために、菌株TIGR4/NanB−(JW002)、変異体を、NanBを欠いたTIGR4遺伝的背景にて、NanAおよびNanB発現の両方を欠いた菌株TIGR4/NanAB−(JW003)と同様に作成した。マウスのJW002による感染は、TIGR4菌株とほぼ同じコロニー形成のレベルを生じた(図8)。その上、NanA変異体と比較して(図9)、コロニー形成の著しい減少はダブル変異体(JW003)では起こらなかった。
【0108】
(肺炎球菌のCNSへの侵入)
S.ニューモニエの鼻組織(嗅神経を含む)への移動を追跡するために、嗅球および脳の残りをS.ニューモニエの存在について試験した。NanA変異体は、遺伝的背景とは無関係に、鼻組織および嗅球において、野生種菌株の数と比べて著しい数の減少が見られた。殺処分時に、すべてのマウスから採血したが、いずれも血液中に検出可能な肺炎球菌を示さず(<12CFU/ml血液)、肺炎球菌が鼻腔からCNS組織へ直接移動することを示している。NanB変異体は、肺炎球菌の鼻組織または嗅球内への侵入に対する影響を持たなかった(図8)。
【0109】
NanA変異体は、上咽頭およびCNSでコロニー形成および持続するその能力が明らかに弱められた。このことは、カプセル血清型およびNanAの表面への結合の両方が異なる菌株で見られた。NanAは、細菌細胞表面と宿主との間の親密性に影響を及ぼす多くの表面構造の1つに過ぎないが、肺炎球菌のCNSへの標的化と同様に、鼻輸送へのその関与は必須である。NanAの破壊は、コロニー形成およびCNSへの標的化を著しく減少させる。この結果は、TIGR4およびEF3030の両方で観察された。
【0110】
菌株EF3030(19F型)は、1ヶ月間に亘って高い効率で上咽頭にコロニー形成する。しかしながら、EF3030が持続する能力にかかわらず、NanAの変異は、鼻の肺炎球菌細胞の数を著しく減少させた。弱化は、鼻洗浄液から単離された細胞数が14日後には検出可能限界近くまで低下した、TIGR4/NanA−菌株でなおさらに劇的であった。
【0111】
自然の状況では、肺炎球菌は、他の細菌種と共存する。それゆえNanAの他の機能は、宿主タンパク質の機能を変化させることと、輸送の長期安定性に寄与することとを含む。NanAは、肺炎球菌がN.メニンギティディスまたはH.インフルエンゼを含む他の口内細菌と競合する能力も、宿主糖タンパク質を炭素源として使用できるようにすることによって向上させる。
【0112】
これらの試験の主な結果は、NanA発現がマウスの最適輸送に必要であることの証明であったが、これらのデータは、NanAを欠く肺炎球菌が嗅球でははるかに少ない数で見出されることも証明した。この時点では、活性NanAがCNS組織中でのS.ニューモニエの生存にとって重要であるかどうかを知ることは困難である。これらの組織から回収されたNanA変異体の数は親菌株よりはるかに少ないが、肺炎球菌が脳に入ったとたん、神経細胞組織でのその存在すら、NanAのさらなる毒性効果を明らかにしている。OB中のノイラミニダーゼ−変異体のレベル低下はおそらく、減少した輸送の結果であろう。この発見は、輸送がより侵襲性の疾患の前提条件であるという原則を強調し、輸送を低減できる介入、たとえばNanAを用いた免疫化がニューモニエ、髄膜炎、中耳炎および敗血症からの保護を提供するであろう。
【0113】
nanAの既知配列のうち、TIGR配列は、表面アンカーを含有しない唯一の配列であろう。本菌株では、フレームシフトがLPETG(配列番号13)モチーフの前の、分子の切断を引き起こす(Tettelin et al.2001 Science 293:498−506)。大半の菌株では、NanAのかなりの部分がソルターゼによって細胞壁に共有結合されることが予想され(Mazmanian et al.1999 Science 285:760−63)、そこで電子顕微鏡写真によって検出されている(Camara et al.Infect.Immun.62:3688−95)。これらの試験では、TIGR4はEF3030と同様に、NanA依存性輸送および嗅球での存在を示した。上澄みまたは細菌ペレットでのNanA活性の局在化の試験から、TIGR4とは異なり、EF3030のNanA活性は細胞に結合していることがわかった。それゆえNanAは、表面結合されてもされなくても、または分泌されてもされなくても、コロニー形成を促進できる。
【0114】
(実施例3)
(S.ニューモニエ病原性におけるガングリオシドの役割)
精製ノイラミニダーゼ、NanA(Calbiochem)をPBS 10μl中1、10および50μgにて、ON/E単離の15、30、および60分前に鼻腔内投与する。組織を4%パラホルムアルデヒドで固定して、パラフィン切片を作成した。GM1はビオチン化CT−Bを使用して、続いてストレプトアビジン−FITCによって染色し、染色強度を解析する。切片はローダミンに結合させたアシアロ−GM1特異性Absによっても染色して、これらの組織においてGM1染色の減少がアシアロ−GM1染色の増加と同時に起きることを確認する。同じ処置を受けるマウスの並行群をS.ニューモニエ菌株TIGR4およびEF3030によるコロニー形成について、第1日および第4日に解析して、ノイラミニダーゼ処置が鼻腔内コロニー形成レベルの上昇を引き起こしたことを確認する。マウスに菌株EF3030の高用量(1x108CFU)を鼻腔内投与して、ON/Eは、鼻腔内攻撃後の第1日および第4日に以下の間隔、すなわち、1、3、6、および12時間にて単離する。ON/Eを上で概説したように染色し、GM1およびアシアロ−GM1発現について解析する。GM1の減少およびアシアロ−GM1発現の上昇が鼻組織で観察される場合、NanAおよびNanB欠損菌株も、それらが鼻道でのGM1発現を変化させることが期待されないため、試験を行う。シアル酸残基の除去は、Thompson−Friedenreich抗原の抗原決定基に相当する、次端部ジサッカライド、β−D−ガラクトピラノシル−(1−3)N−アセチル−D−ガラクトサミンを露出させ、そのためにPNAは高い親和性を有する。それゆえON/E中のPNA−結合部位における変化は、ノイラミニダーゼ活性の別の尺度である。これらの組織から作成した凍結切片をPNA−FITCまたはPNA−HRPでただちに染色して(Medac,ハンブルク、ドイツ)、顕微鏡検査に基づいてPNA−結合部位の増加が生じたかどうかを判定する(そこで教示した方法について、参照によりその全体が本明細書に組み入れられている、Black et al.,(2000)Pediatr.Infect.Dis J.19:187−195;Klein et al.(1978)Klin.Wochenschr.56:761−765)。
【0115】
GM1部位は、菌株TIGR4またはEF3030の鼻腔内投与の前に特異的に遮断される。これは:1)CT−B対非ガングリオシドコントロールタンパク質、たとえばオボアルブミン、2)GM1に対するAbs(Calbiochem)対正常ウサギ免疫グロブリン、または3)UAB Protein AnalysisおよびPeptide Synthesis Core Facilityで合成されたGM1特異性ペプチドを使用して、3つの方法で着手される。GM1特異性ペプチドによる阻害は、CT−BおよびおそらくGM1に対するAbsが随伴性の炎症を引き起こすことが予想されるため、最良の手法である。これらの実験では、ON/EおよびOBは、肺炎球菌性CFUの投与後、第1日および第4日に解析する。GM1を遮断する代わりの手法は、GM1結合ペプチドに対して選択されるファージ提示ペンタデカペプチドライブラリの使用によって発見されたGM1結合ペプチドの使用である。このGM1結合ペプチドVWRLLAPPFSNRLLP(配列番号5)は、GM1に対する高い親和性(1010M−1)および1.0μMのIC50を有する。Matsubura et al.,(1999)FEBS Letters 456:253−256。同じ長さの、ランダムに選択された配列中の同じアミノ酸より成るペプチドをコントロールとして使用する。両方のペプチドのGM1結合能力は、生体内で使用する前にELISAによって確認する。最初に、リンゲル液またはPBS 10μl中のこれら2つのペプチド100μgを、菌株EF3030 3X106CFUの投与の10分前に鼻腔内投与する。第1日および第4日の投与後に、ON/EをCFUについて、未処置CBA/Nマウスに対するCFUの数について解析する。
【0116】
遮断実験は、PAF(Biomol Research Laboratories,Inc.プリマスミーティング、ペンシルベニア州)およびPAF−Rアンタゴニストオクチルオニウムブロミド(Biomol)を用いて実施する。本化合物は、高い親和性でPAF−Rに結合する。各ガングリオシドは個別に試験を行う。混合ガングリオシド以外に、アシアロ−GM1、GM1、GD1a、GD1b、GT1(Calbiochem)およびGM3(Sigma)を、攻撃後、第1日および第4日に評価するように鼻腔内コロニー形成を阻害するその能力について試験を行う。各種のガングリオシドは、これらのプロセスを遮断できる。GM3ガングリオシドは、公開されているCポリサッカライド結合モチーフを欠いているため、負のコントロールとして機能する。ガングリオシドプレインキュベーション後にEF3030によるコロニー形成に対して示されたデータに基づき、混合ガングリオシドは、コロニー形成の遮断ではアシアロ−GM1よりも有効である。これは、アシアロ−GM1以外の他のガングリオシドが、鼻道、肺および脳の肺炎球菌性コロニー形成のプロセスに関与することを示している。これらのガングリオシド阻害試験は、TIGR4菌株およびそのノイラミニダーゼ変異体に焦点を当てている。TIGR4菌株を用いたガングリオシドコーティングELISAプレートでの短期試験管内インキュベーションは、TIGR4菌株のアシアロ−GM1への結合を証明した。エンテロトキシンは、鼻道での肺炎球菌−ガングリオシド相互作用を異なるふうに遮断する別の手段を提供する。CTおよびLTh−1はどちらも、両方の血清型I易熱性エンテロトキシン(Pickett et al.,(1986)J.Bacteriol 165:348−352)であり、わずかに異なるが、同様のガングリオシド結合特異性を示す。Fukuta et al.,(1988)Infect Immun.56:1748−1753.CT(List Biological Laboratories,Inc.,キャンベル、カリフォルニア州)はGM1に、そしてより低い程度でGD1bに結合する。LTh−1は、GM1およびGD1bに優先的結合を示し、GM2およびアシアロ−GM1に弱く結合する。Fukuta et al.,(1988)Infect Immun.56:1748−1753。S.ニューモニエと相互作用する主なガングリオシドである場合、LTh−1の使用は GM1ガングリオシドを遮断するだけでなく、アシアロ−GM1も遮断し、このことはノイラミニダーゼ活性を必要としない、天然の頻度の低い結合を示す。血清型IIによる易熱性エンテロトキシンは、異なるガングリオシド結合特異性、特に易熱性エンテロトキシンLT−IIbを示す。この毒素は、GD1aに、より低い程度でGT1bに結合して、GM1への親和性は示さなかった。Fukuta et al.,(1988)Infect Immun.56:1748−1753。LT−IIaは、高い親和性でGD1bに、より低い親和性でGM1、GT1b、GQ1b、GD2、GD1a、およびGM2に結合する。Fukuta et al.,(1988)Infect Immun.56:1748−1753。LT−II毒素は、T.D.Connell博士のご厚意により提供された。鼻用量および最適期間を最適化してON/Eへの肺炎球菌の結合の阻害を観察するために、最初に用量反応試験(1.0または10μg)を、S.ニューモニエの鼻腔内投与に与えられる選択したエンテロトキシンに対して実施する。鼻腔内コロニー形成の阻害が第4日に観察された場合、観察は第11日まで延長され、その間にエンテロトキシンを1日おきに与える。CBA/NマウスのON/EおよびOBにおけるCFUを測定する。
【0117】
(実施例4)
(肺炎球菌性病原性におけるCポリサッカライド特異性抗体の役割)
テイコ酸としても既知の肺炎球菌性Cポリサッカライドは、リポテイコ酸としても既知の肺炎球菌F抗原と構造的に同じである。Fischer et al.(1993)Eur.J.Biochem 215:851−857。これはグラム陽性細菌の中でS.ニューモニエ独自の特徴である。これらの分子の主要抗原決定基は、ホスホリルコリン(PC)残基であり、PCに対するAbsは、i.p.または経鼻肺炎球菌攻撃に対して保護的である。Briles et al.,(1984)Eur.J.Immunol.14:1027−1030;Briles et al.,(1981)Nature 294:88−90;Yother et al.,(1982)Infect.Immun.36:184−188;Briles et al.,(1984)J.Mol.Cell.Immunol.1:305−309。それゆえ受動伝達または能動鼻腔内免疫化のどちらかによって得られたPC特異性Absの役割が調査される。保護性PC特異性Absの受動伝達では、すなわちIgG3(59.6C5)およびIgM(22.1A4)アイソタイプの両方のT15イディオタイプモノクローナルAbs(mAbs)が使用される。Briles et al.,(1981)Nature 294:88−90。T15イディオタイプは、おそらくCポリサッカライドをより効率的に結合することによって、マウスの肺炎球菌感染に対してM603またはM511イディオタイプよりも保護性であることが示されている(Briles et al.,(1984)Eur.J.Immunol.14:1027−1030)。受動Abs伝達は、鼻腔内投与された肺炎球菌を用いたT15 Abs(100μg)の直接投与を含み、鼻腔内コロニー形成を低減するためのi.v.またはi.p.投与されたAbsと比較する。コロニー形成は経時的に監視し(第4日、第11日、第18日)、これらの実験の異なる群の間で著しい相違が観察されない場合、mAbs(20μg)を1日おきに鼻腔内投与する。CBA/Nマウスは、T15イディオタイプ抗PC Absを生成しない。抗PC特異性Absの受動伝達が、鼻道中で粘膜IgAまたはPC特異性Absの他のアイソタイプを誘発することは予想されない。鼻Absを誘発するために、2つの異なる手法を取る。1つは、Cポリサッカライドに対するAb反応を誘発することが既知である、プロテアーゼ処置R36A菌株の直接鼻腔内投与である。抗PC Absの保護免疫が試験されてきたが、鼻道などの粘膜表面でのその役割に関するデータは入手できない。CBA/NマウスX染色体関連免疫不全は、T15イディオタイプの抗PC Absの産生不能を引き起こす。この不能の重要性を判定するために、CBA/Nマウスをその野生種対応物のCBA/Jマウス(Jackson Laboratories)と比較する。抗PC Abs誘発のための菌株R36Aによる免疫化は、表面タンパク質のタンパク質分解除去を含む。Krause(1970)Adv.Immunol.12:1−56。鼻腔内免疫化の代わりの手法は、上述したようなPCのタンパク質キーホール・リンペット・ヘモシアニン(KLH)へのカップリングである(そこで教示した方法について、参照により本明細書に組み入れられている、Krause(1970)Adv.Immunol 12:1−56;Chesebro and Metzger(1972)Biochemistry 11:766−771)。PC−KLHを用いた鼻腔内免疫化は、粘膜免疫反応を最適化するために、粘膜アジュバントCTを用いて実施される。マウスは、コロニー形成に対するCTの効果を防止するために、最後の免疫化の2〜3週間後に攻撃される。1週間間隔で3回の鼻腔内免疫化を実施する。当業者によって通常実施される通りにCポリサッカライドおよびPC特異性ELISAを使用して、血清Ab力価を監視する。PC特異性ELISAでは、PCは上述のようにBSAに結合される(そこで教示した方法について、参照により本明細書に組み入れられている、Chesebro and Metzger(1972)Biochemistry 11:766−771)。血清に加えて、鼻洗浄液、唾液、および気管支洗浄液中のAb力価を測定する。これらの解析は、粘膜分泌物および血清の両方でのIgA、IgM、IgG、およびIgG−サブクラス分布を含む。最適な粘膜Ab力価を誘起するプロトコルを使用して、第4日および第11日にコロニー形成が監視された後にマウスに〜5x106CFUで投与される、TIGR4菌株による粘膜攻撃試験を実施する。免疫化試験では、正常な完全免疫適格性マウス(CBA/J菌株)はもちろんのこと、CBA/Nマウスも以前の試験と同様に使用する。Wallick et al.,(1983)J.Immunol.130:2871−2875。
【0118】
(実施例5)
(S.ニューモニエ病原性におけるノイラミニダーゼ特異性抗体の役割)
鼻腔内攻撃前にマウスを鼻腔内免疫化するために、市販のS.ニューモニエ由来ノイラミニダーゼを使用する(Calbiochem)。しかしながら、提案された試験に十分な量のタンパク質を得るために、NanA遺伝子はヒスチジンタグ含有発現ベクター(Invitrogen)を使用してE.コリ内でクローニングおよび発現させる。粘膜免疫反応を最適化するために、3.4%ホルムアルデヒド処置ノイラミニダーゼの鼻腔内免疫化を対の未処置ノイラミニダーゼと、CTの存在下または非存在下で比較する。これらの免疫化は、CBA/NおよびCBA/Jマウスの両方で実施する。3回の鼻腔内免疫化を1週間おきに与え、その間に血清および唾液Abs力価をELISAによって監視する。免疫マウスをTIGR4菌株によって攻撃して、ON/E、OB、脳、血液、脾臓、および肺のコロニー形成を第4日および第11日に比較する。宿主の相互作用を遮断するために、ノイラミニダーゼおよびCポリサッカライド特異性Absの両方を同時に誘発させる。鼻腔内免疫化をノイラミニダーゼおよびT15イディオタイプmAbsの伝達による受動免疫保護と組み合せた計画を使用する。
【0119】
(実施例6)
(S.ニューモニエを用いた鼻腔内攻撃に対して保護するためのノイラミニダーゼ−PC結合体の有効性)
保護の向上を評価し、単独で使用された各抗原と比較して第11日のEF3030およびTIGR4菌株による鼻腔内コロニー形成を低下させるために、マウスを鼻腔内アジュバントとしてのCTと組み合せたノイラミニダーゼおよびPC−KLHによって免疫化する。加えて、鼻洗浄液、唾液、および血清のAb力価を上述のように解析して、免疫パラメータを鼻道内での肺炎球菌に対する保護度と関連付ける。さらに最適な免疫反応を発生させるために、ホスホコリンをノイラミニダーゼに直接結合させる。この作成物を、CBA/NおよびCBA/Jマウスの両方における鼻腔内および全身免疫化の後に、アジュバントとしてのCTを用いて、またはCTなしに送達したときに免疫原性について試験を行う。鼻洗浄液、唾液、および血漿のAb力価をELISAによって測定する。攻撃試験は、菌株EF3030 107CFUまたはTIGR4 106CFUを用いて実施する。マウスを攻撃後、第11日に殺処分して、血液、鼻洗浄液、ON/E、OB、および脳で観察されたCFUについて解析した。PCに結合させたノイラミニダーゼによる免疫化は、粘膜および全身Abレベルを上昇させることによって、これらの2つの毒性成分に対する保護を向上させる。抗原特異性IgGサブクラスの分布は、他の粘膜アジュバントを使用することによって変更される。CTは、Th2−、LT混合Th2/Th1−、およびCpGモチーフ、たとえばDNAオリゴヌクレオチド(ODN)1826 a Th1型反応をIgGサブクラス分布の関連する変化と共に引き起こす。各種のアジュバントは、ノイラミニダーゼ−Cポリサッカライド特異性免疫がS.ニューモニエによる鼻腔内コロニー形成から保護する能力をさらに向上させ、新たな肺炎球菌性ワクチン手法の考案をもたらす。
【0120】
(実施例7)
(鼻腔内攻撃後のS.ニューモニエの鼻腔内コロニー形成の、抗ホスホコリン特異性モノクローナル抗体による阻害)
合計1X106CFUのTIGR4菌株を、IgG3サブクラスまたはIgMアイソタイプのどちらかの抗ホスホコリンモノクローナル抗体5μlでインキュベートした。合計5μlを鼻孔ごとに投与した。投与9時間および12時間後の鼻洗浄液500ml中のCFUをそれぞれ示す。どちらのモノクローナル抗体でも、重大な80%超の減少が見られた。1グループ当たりマウス5匹の平均+SDを示す。データを図10に示す。
【0121】
(実施例8)
(S.ニューモニエ鼻腔内投与後の神経細胞損傷および炎症)
S.ニューモニエ菌株EF3030の鼻腔内投与後、第1日、第3日、第7日および第14日に処置マウスからON/E、OB、および脳を単離して、炎症性反応について組織学的に解析する。D39またはTIGR4菌株をそのnanA変異体菌株と、炎症性反応を発生させるその能力について比較する。殺処分時に、マウスを25℃のPBSを用いて潅流して、Zamboni固定液10ml(0.1Mリン酸緩衝液中の4%パラホルムアルデヒド、15%ピクリン酸による潅流を続ける。OBおよびON/Eを除去し、次に4℃の新しい4%パラホルムアルデヒド(PFA)中に一晩置いた。次に、切断前に凍結を防止するために、組織を4℃の30%スクロース溶液に移して48時間置いた。次に組織をOCT中で凍結させ、切片(6μm)を事前にコーティングした顕微鏡スライド上に置いた(生理的食塩水中10%BSA)。最初にヘマトキシリンおよびエオシン(H&E)染色を実施して、この期間中にOB、三叉神経節およびON/Eに浸透する(infitrate)炎症性細胞を検出する。神経細胞損傷を評価するために、神経成長因子β1(NGF−β1)を染色する。NGF−β1は、神経細胞損傷の後に生成され、アポトーシスを防止し、神経細胞の新たな成長を刺激するように機能する。三叉神経節およびOB切片をビオチン化ウサギ抗ヒトNGF−β1Abによって0.2μg/mlの濃度で染色する。Ab染色切片を4℃にて一晩インキュベートする。スライドをPBSですすぎ、次にアビジン−ビオチン複合体(ABC)のVectastain(Vector Laboratories,バーリンゲーム、カリフォルニア州)によって25℃にて30分間反応させた。組織をPBSで3回すすぎ、次に以前に報告したようにジアミノベンジデン(DAB)と5〜10分間反応させる。スライドをさらに3回すすぎ、切片をC.S.ヘマトキシリンによって30秒間対比染色する。H2Oでの洗浄後、スライドを100%アルコールおよびキシレン中で脱水する。NGF−β1の増加は、神経細胞組織の損傷度の指標を与える。神経細胞関与の別の指標は、小膠細胞の活性化である。活性化された小膠細胞は、アメーバ様球状形状を示し、これに対して静止細胞(G0/G1)は、樹枝状の分裂した外観を有する。活性化時のこのような変化によって、静止および活性化小膠細胞を区別できる。小膠細胞では、S.ニューモニエ攻撃後の活性化状態に対処するために、F4/80抗体または抗MAC−1(MI/70)が使用される。神経細胞損傷および小膠細胞活性化に加えて、OBのアポトーシスの誘発が評価される。このために、活性カスパーゼ3である、Asp−Glu−Val−Asp特異性プロテアーゼの誘発がアポトーシス経路の開始において重要であるため、それが解析される。活性カスパーゼ3(Cell Signaling Technology,Inc.,ビバリー、マサチューセッツ州)に対して特異性のAbは、アポトーシスの検出のために免疫組織化学で使用できる。カスパーゼ3活性が免疫組織化学によって神経細胞組織で検出されると、活性は、基質のタンパク質分解後の蛍光信号に基づくカスパーゼ−3アッセイキット(Molecular probes,ユージーン、オレゴン州)を用いて定量される。
【0122】
(実施例9)
(逆行性軸索輸送による、S.ニューモニエが嗅球を標的化する能力)
第一に、鼻腔内またはi.v.接種後の処置マウスの神経細胞組織、OBおよび脳への肺炎球菌の蓄積を評価する。i.v.接種後、神経細胞組織のどの肺炎球菌も血液を通じて侵入する。鼻腔内攻撃後の第1日、第4日、第11日および第18日の組織を収集する。その場合、脳およびOB1グラム当たりの細菌数は、注射後のすべての時点で同様なはずである。これに対して、鼻腔内接種後の鼻道を通じて侵入する細菌では、OBへの蓄積(組織重量当たりで表される)が先に起こり、一般に脳で観察される蓄積が待ち構えている。
【0123】
第二に、鼻腔内投与後の肺炎球菌の生体内撮像が実施される。テクネチウム−99(Tc−99m)標識TIGR4、安定不透明および透明改変体、EF3030、ならびにnanAおよび/またはnanBを欠いたTIGR4変異体を使用して、Waibel et al.(1999)Nature Biotechnol.17:897− 901によって最初に述べられた方法を使用して、アデノウィルスで以前に実施されたようにガンマカメラ撮像を使用して、マウスにおけるその存在を描出する。これは、このアイソトープの短い半減期(6時間)のために、鼻腔内投与後、最初のほぼ24時間の撮像を可能にし、鼻道で発生する早期の事象の解析を可能にする。肺炎球菌の長期撮像では、ルシフェラーゼ−またはGFP−発現肺炎球菌性EF3030(またはTIGR4)菌株を使用して、生体内での生体発光を描出する。Xenogen corporation(アラメダ、カリフォルニア州)から市販されているルシフェラーゼ発現肺炎球菌菌株EF3030を使用する。この肺炎球菌性菌株を用いた生体内撮像の成功が以前に報告されている。マウスは生体発光撮像システム(IVIS system,Xenogen,Inc.)を使用して撮像して、ルシフェラーゼ発現を検出する。同じ位置に向けたマウスで、常にルシフェリン2.5mgのi.p.注射の10分後に画像を収集する。撮像の間、マウスを37℃にてエンフルレン麻酔状態に維持する。撮像は各マウスに対して、ルシフェラーゼ−発現肺炎球菌を用いた鼻腔内攻撃後の第2日から開始して第18日まで、複数回実施する。撮像の画像取込時間は、20秒〜10分間の範囲である。データ取込ソフトウェアは、画像収集中にいずれの画素も飽和しないようにする。興味のある領域からの発光(相対光子/秒)を、Xenogen提供のソフトウェアを使用して定量する。発光の強度は、生体発光画像の擬似カラースケーリングを用いて表す。生体発光画像は通例、同時に収集されたマウスの白黒写真にオーバーレイされる。この生体内撮像は、肺炎球菌が鼻道からOBへ侵入する能力を分析することに焦点を当てている。この生体発光試験は、ルシフェラーゼ遺伝子の伝達成功後に、nanA TIGR4変異体へと拡張される。
【0124】
本出願を通じて、各種の公報が参照されている。これらの公報のその全体の開示は、本明細書で説明する化合物、組成物および方法をさらに十分に説明するために、この出願内への参照により本明細書に組み入れられている。
【0125】
各種の改良および変形を、本明細書で説明する化合物、組成物および方法に行うことができる。本明細書で説明する化合物、組成物および方法の他の態様は、本明細書で開示された化合物、組成物および方法の明細書および慣行の検討から明らかになるであろう。明細書および実施例は一例として考慮されるものとする。
【0126】
(参考文献)
【0127】
【化1】
【0128】
【化2】
【0129】
【化3】
【0130】
【化4】
【0131】
【化5】
【0132】
【化6】
【0133】
【化7】
【0134】
【化8】
【0135】
【化9】
【0136】
【化10】
【0137】
【特許請求の範囲】
【請求項1】
明細書中に記載の発明。
【請求項1】
明細書中に記載の発明。
【図1】

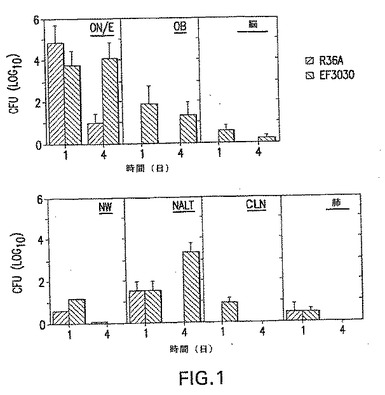
【図2A】
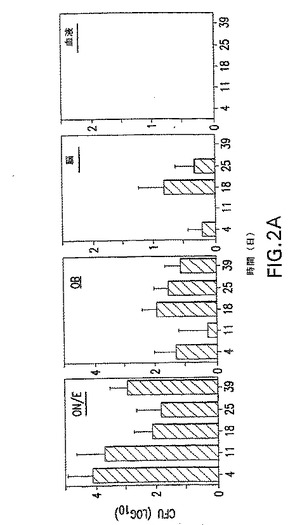
【図2B】
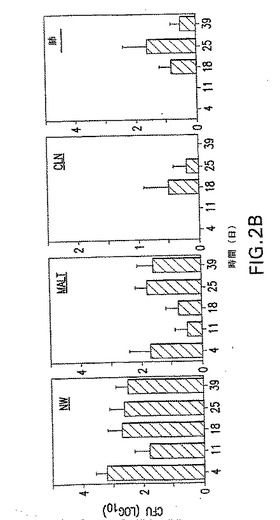
【図3A】
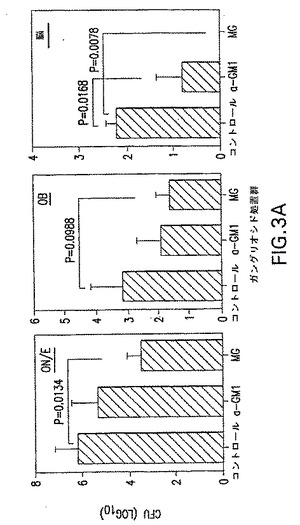
【図3B】
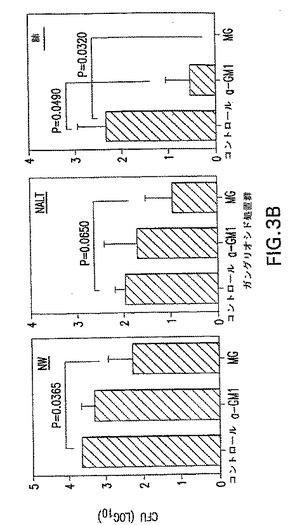
【図4−1】
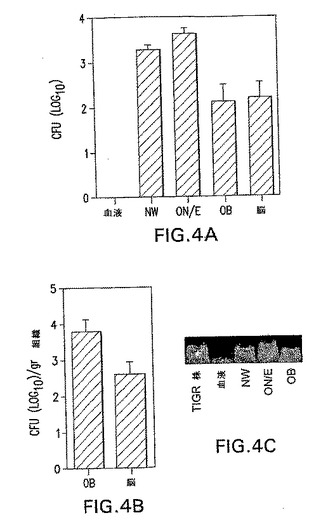
【図4−2】
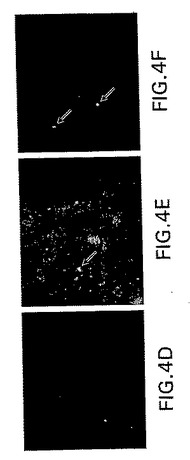
【図5】
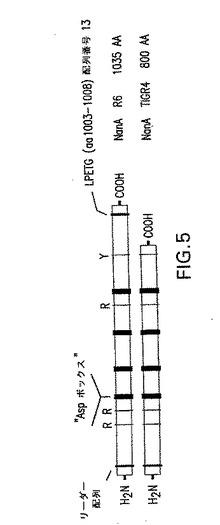
【図6A】
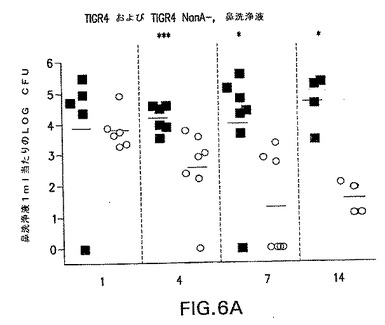
【図6B】
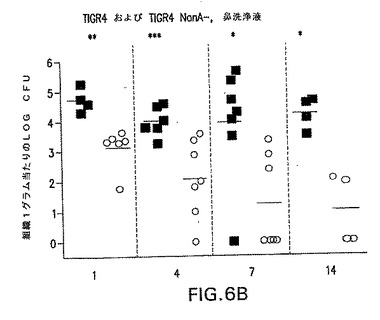
【図6C】
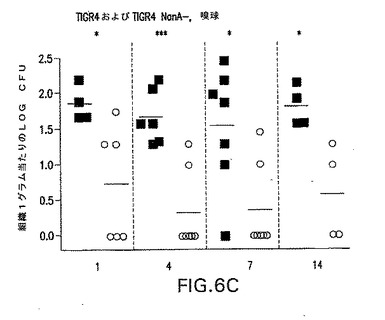
【図7A】
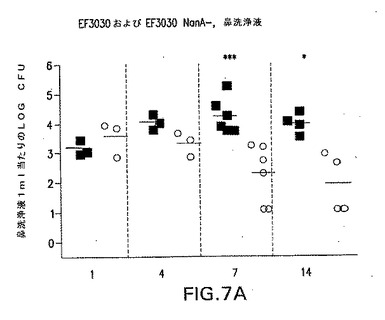
【図7B】
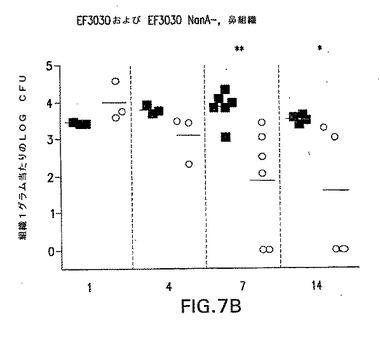
【図7C】
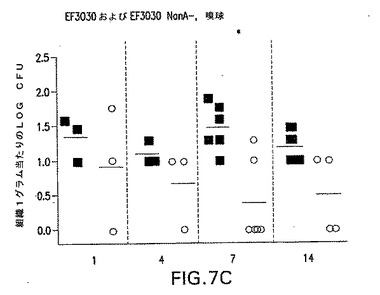
【図8】
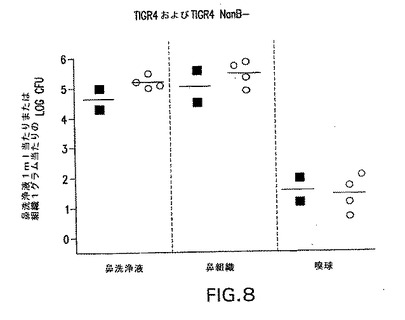
【図9】
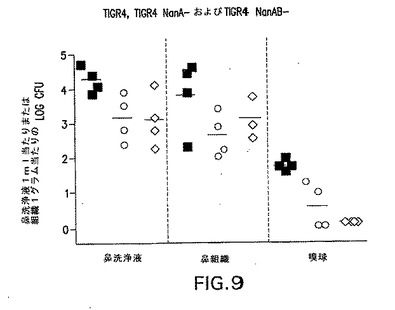
【図10】
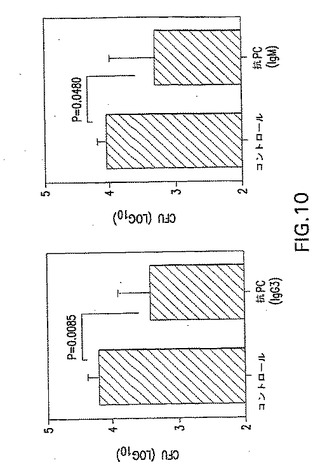
【図2A】
【図2B】
【図3A】
【図3B】
【図4−1】
【図4−2】
【図5】
【図6A】
【図6B】
【図6C】
【図7A】
【図7B】
【図7C】
【図8】
【図9】
【図10】
【公開番号】特開2011−160812(P2011−160812A)
【公開日】平成23年8月25日(2011.8.25)
【国際特許分類】
【外国語出願】
【出願番号】特願2011−113958(P2011−113958)
【出願日】平成23年5月20日(2011.5.20)
【分割の表示】特願2006−539758(P2006−539758)の分割
【原出願日】平成16年11月10日(2004.11.10)
【出願人】(504168260)ザ ユーエービー リサーチ ファウンデーション (12)
【Fターム(参考)】
【公開日】平成23年8月25日(2011.8.25)
【国際特許分類】
【出願番号】特願2011−113958(P2011−113958)
【出願日】平成23年5月20日(2011.5.20)
【分割の表示】特願2006−539758(P2006−539758)の分割
【原出願日】平成16年11月10日(2004.11.10)
【出願人】(504168260)ザ ユーエービー リサーチ ファウンデーション (12)
【Fターム(参考)】
[ Back to top ]