
組み換えタンパク質生産のための方法
本発明は、E. coliによる組み換えタンパク質の発現が誘導可能なシステムの制御下にあるようなE. coli宿主細胞培養における、上清とペリプラズムとの間での組み換えタンパク質の分画を制御するための方法を提供する。その方法は、以下の工程から構成される:a) E. coli宿主細胞培養液を提供する工程、b) E. coli宿主細胞の生育速度を変化させる工程、c) 組み換えタンパク質の発現を誘導する工程、ここで工程 (b)と (c)は、どの順序でも、あるいは同時に、行うことが可能である;そして続いて、d) 培養上清とE. coli宿主細胞ペリプラズムにおける組み換えタンパク質の収量を決定する工程、e) 工程 (d)で決定した収量と、工程 (b)で用いられた生育速度と少なくとも1つの他の場合において決定された収量とを比較する工程、f) 上清とペリプラズムの間での組み換えタンパク質の分画が、組み換えタンパク質の最初の回収に最も適しているような、工程 (e)でなされた比較の結果得られた、生育速度を選択する工程。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、一般的にはEscherichia coli宿主細胞培養液中で組み換えタンパク質を生産するための方法に関し、さらに特には、E. coli宿主細胞のペリプラズムと培養液上清との間に、前述のタンパク質の分画を制御するための方法を提供する。
【背景技術】
【0002】
E. coliは、幅広く使われ、組み換えタンパク質の生産のための便利なシステムである。このシステムの有利な点は、遺伝子操作の容易さ、遺伝子発現ベクターを含む試薬の入手可能さ、タンパク質の生産定量の容易さ、幅広く多様なタンパク質を発現するためのシステムの速度と高い適応性、などから構成される。バネイクスの、Current Opinion in Biotechnology, 1999, 10, 411-421.のレヴューを見ると、E. coliは、インシュリン、成長ホルモンそして抗体といった治療上のタンパク質のラージスケール生産に、今では幅広く使われている(Swartz, Current Opinion in Biotechnology, 2001, 12, 195−201; Humphreys and Glover, Current Opinion in Drug Discovery and Development, 2001, 4, 172-185; Verma et al., Journal of Immunological Methods, 1998, 216, 165-181; Simmons et al., Journal of Immunological Methods, 2002, 263, 133-147)。
【0003】
E. coli内での外来遺伝子の発現は、遺伝子のcDNAコピーの発現ベクターへの挿入から始まる。たくさんの形の発現ベクターが利用可能である。そうしたベクターは、大抵、DNA複製のためのプラスミドオリジン、抗生物質の選択可能なマーカー、マルチクローニングサイトによって隔てられたプロモーターと転写終結因子(発現カセット)、そして少なくとも1つのリボソーム結合サイトをコードしているDNA配列、によって構成される。外来遺伝子の転写は、本来の量で発現されたタンパク質の場合よりも高い収量になる生産物合成と、細胞増殖とが別々になることを可能にするような調節プロモーターによって、大抵制御されている。タンパク質の最高収量は、発現を誘導するための培養の間の最適なポイントを決定することによって、典型的には、一旦細胞を高密度にまで生育させることによって、達成される。
【0004】
E. coli内で発現される組み換えタンパク質は、タンパク質の性質やそのリホールディングの必要性に依存して、細胞質内か、ペリプラズムまたは培養上清に分泌されて、生産される(Makrides, Microbiological Reviews, 1996, 60, 512-538.)。例えば、タンパク質が、シグナル配列を介してペリプラズムを目標としてペリプラズムに発現することは、宿主細胞のタンパク質の数がより少ないこと、組み換えタンパク質の分解がより少ないこと、そして適当なタンパク質リホールディングを促進するような酸化的環境などを含めて、細胞質内での発現よりも、いくつかの有利な点を提供することができる。
【0005】
培養中のタンパク質収量が、E. coli内で組み換えタンパク質を生産するコストに影響を与える鍵となる因子である一方、生産されたタンパク質の下流の処理(DSP)のコストもまた、商品の最終コストを決定する上で重要である。DSPという用語は、E. coliの培養の後、最終的な精製された機能的タンパク質をもたらすまでの、すべての工程を含んでいる。生産されたタンパク質の細胞での位置に依存して、これらの工程は典型的に、遠心分離、濾過、抽出、そして濃縮・ダイアフィルトレーションなどの最初の回収工程、それに続く1つまたはより多くの精製工程を含んでいる。機能的タンパク質を高収量で得るために、これらの工程の効果を最大化することは、組み換えタンパク質のラージスケール生産の間のコストを最小化するための鍵となる。DSPの間に使われる最初の回収工程の性質は、得られる組み換えタンパク質の量と質に影響するだろうし、生産のスケール、タンパク質の性質、そして培養終了時の細胞での位置などを含むいくつかの要因に依存するだろう。例えば、ペリプラズムに生産された組み換えタンパク質に対する重要な配慮は、組み換えタンパク質が上清へと漏れる量であり、培養中に自然な量の漏れが起こったとしても、これはペリプラズムから得られた組み換えタンパク質の量と質に不利な効果があるといえる。大量の組み換えタンパク質が上清へと漏れる場合には、上清の粘度が増加し、遠心分離や濾過による細胞の最初の回収をより困難にし、そしてそれゆえに、ペリプラズムから得られるタンパク質の収量は減ってしまう。細胞自身もまた、遠心分離の間はそれほど頑丈ではなく、より溶解しやすく、結果としてタンパク質にダメージを与え、そして上清のタンパク質量が増え、再び収量が減る。さらなる問題は、抽出操作の間ペリプラズムから回収されるタンパク質の「質」もまた、正しくリホールディングされていない、または部分的に分解した、上清由来の組み換えタンパク質に汚染されることによって、低下するかもしれないということである。
【0006】
それゆえ、組み換えタンパク質の最初の回収の効率を改善すること、そして、ペリプラズムと上清の間に組み換えタンパク質の分画を制御することによって得られる、機能的タンパク質の最終的な量と質を改善することは、望ましいことである。
【発明の開示】
【発明が解決しようとする課題】
【0007】
本発明において、我々は、E. coli宿主細胞の生育速度を調節することによって、E. coli宿主細胞の培養において、上清とペリプラズムの間に組み換えタンパク質の分画を制御することができることを証明することができた。上清とペリプラズムの間でのタンパク質の分画に基づいて適切な生育速度を選択することによって、必ずしも収量だけでなく、生産されたタンパク質の、質および/または量を増加させることができる。
【課題を解決するための手段】
【0008】
本発明に従って、E. coli宿主細胞培養において、上清とペリプラズムの間に組み換えタンパク質の分画を制御するための方法を提供する。その培養において、上述の細胞による組み換えタンパク質の発現は、1つの誘導システムの下で制御される。その方法は、以下の工程から構成される:
a)E. coli宿主細胞培養液を提供する工程
b)E. coli宿主細胞の生育速度を変化させる工程
c)組み換えタンパク質の発現を誘導する工程
この点で、工程 (b)と (c)は、どの順序でも、あるいは同時に、行うことが可能である;そして続いて、
d)培養上清とE. coli宿主細胞ペリプラズムにおける組み換えタンパク質の収量を決定する工程
e)工程 (d)で決定した収量と、工程 (b)で用いられた生育速度と少なくとも1つの他の場合において決定された収量とを比較する工程
f)上清とペリプラズムの間での組み換えタンパク質の分画が、組み換えタンパク質の最初の回収に最も適しているような、工程 (e)でなされた比較の結果得られた、生育速度を選択する工程
【発明を実施するための最良の形態】
【0009】
本発明において、使用できる組み換えタンパク質は、おおよそ10以上のアミノ酸残基を持つ、ペプチド、ポリペプチド、またはタンパク質のいすれかであろう。タンパク質は、細菌を含めて、どの起源由来でも差し支えないが、より好ましくは哺乳動物であろう。1つの例では、組み換えタンパク質は抗体である。ここで使われている「抗体」という用語は、IgG, IgE, IgM, IgDそしてIgAなど、どのクラス由来の免疫グロブリン、例えばIgGクラスの1つであるIgG1、そしてまた、Fv, Fab, Fab'そしてF(ab’)2断片などの抗原が結合した免疫グロブリン断片、そして単鎖Fv断片などの誘導体、などについて言及している。これらの抗体や断片は、自然に発生し、人体に適応され、キメラであり、またはCDR移植した抗体でもよい。そして標準的な分子生物学的手法が、望みのアミノ酸残基やドメインの改変、添加、または除去に使われても差し支えない。これらの抗体分子を創るための方法は、過去の発明にてよく知られている(以下の参考文献を参照、Shrader et al., WO 92/02551; Ward et al., 1989, Nature, 341, 544; Orlandi et al., 1989, Proc. Natl. Acad. Sci. USA, 86, 3833; Riechmann et al., 1988, Nature, 322, 323; Bird et al., 1988, Science, 242, 423; Queen et al., US 5,585,089; Adair, WO 91/09967; Mountain and Admir, 1992, Biotechnol. Genet. Eng. Rev., 10, 1-142; Verna et al., 1998, Journal of Immunological Methods, 216, 165-181)。
【0010】
本発明で使用するE. coli宿主細胞は、天然に存在するE. coli菌株、または組み換えタンパク質を生産できる変異株で差し支えない。特異的な宿主E. coli株の例は、MC4100, TG1, TG2, DHB4, DH5a, DH1, BL21, XL1BlueそしてJM109が含まれる。例えば、代謝変異株やプロテアーゼ欠損株といった、改変されたE. coli株も例に含まれる。1つの好ましいE. coli宿主はE. coli W3110 (ATCC 27,325)であり、一般的に組み換えタンパク質発酵のための宿主株として使われている。
【0011】
本発明の組み換えタンパク質は、タンパク質の性質や生産のスケールに依存して、典型的に、E. coli宿主細胞のペリプラズムか、宿主細胞培養液の上清のどちらかに発現される。これらの区画へとタンパク質を目標づけるための方法は、過去の研究においてよく知られている(Makrides, Microbiological Reviews, 1996, 60, 512-538を参照)。E. coliのペリプラズムへと向かわせるための適切なシグナル配列の例は、E. coli PhoA, OmpA, OmpT, LamBそしてOmpFシグナル配列を含む。タンパク質は、天然の分泌経路によって、または、pelBリーダー、プロテインAリーダー、バクテリオシン放出タンパク質との共発現、グリシンを培地に添加することを伴ったマイトマイシン誘導バクテリオシン放出タンパク質、そして生体膜透過のためのkil遺伝子との共発現、などの使用を例とするタンパク質分泌をもたらす外膜の限定的漏えいの誘導によって、上清へと目標づけられる。
【0012】
本発明において、E. coli宿主細胞での組み換えタンパク質の発現は、誘導可能なシステムの制御下におかれる。ここで使われている「誘導可能なシステム」という用語は、E. coliにおける組み換えタンパク質の発現が誘導可能なプロモーターの制御下に置かれている、誘導可能な発現システムについて言及している。E. coliにおける使用に適切な多くの誘導可能なプロモーターは、過去の研究においてよく知られており、プロモーターに依存して、組み換えタンパク質の発現が、温度、または生育培地中の特定の物質の濃度などの、変化する要因によって誘導されることができる(Baneyx, supra; Goldstein and Doi, 1995, Biotechnol. Annu. Rev., 105-128)。誘導可能なプロモーターの例は、ラクトースまたは加水分解されていないラクトース類似物であるイソプロピル-β-D-1-チオガラクトピラノシド(IPTG)を使って誘導可能なE. coli lac, tac, そしてtrcプロモーター、そして、リン酸、トリプトファン、L-アラビノースによって、それぞれ誘導される、phoA, trpそしてaraBADプロモーターを含む。
【0013】
ここで使われている「誘導可能な発現」という用語は、例えば、誘導物質の添加、または誘導が温度依存的である場合には温度の変化によって、誘導が始まる点について言及している。組み換えタンパク質発現の誘導が、培養液への誘導物質の添加によって達成される場合は、誘導物質は、発酵システムや誘導物質に依存した適切な方法によって、例えば、単一または複数回の点添加によって、または誘導物質を与える間に逐次添加していくことによって、加えても差し支えない。誘導物質の添加と、実際のタンパク質発現の誘導との間には、遅れがあるかもしれないことも理解しなくてはならないだろう。例えば、誘導物質がラクトースの場合、元々存在している炭素源がラクトースの前に資化される間、タンパク質発現の誘導前に遅れが生じるかもしれない。
【0014】
本発明のE. coli宿主細胞培養は、E. coliの生育と組み換えタンパク質の発現に対応しているであろう、どの培地においても培養できるかもしれない。培地は、Pirt S. J. (1975) Principles of Microbe and Cell Cultivation, Blackwell Scientific Publicationsにおいて規定されている培地に、本明細書に記載されているような生育速度を制御するために適当な改変を加えた、化学的に定義づけられているどの培地でも差し支えない。適切な培地の例は、Humphreys et al., 2002, Protein Expression and Purification, 26, 309-320によって記載されている「SM6E」である。
【0015】
E. coli宿主細胞の培養は、生産に必要なスケールに依存して、振とうフラスコまたは発酵器などの適切な容器において、行うことができる。1000リットルより大きく、おおよそ10万リットルまでの容量の、様々なラージスケール発酵器が利用可能である。1000から5万リットルの好ましい発酵器が使われ、より好ましくは1000から1万リットルである。0.5と1000リットルの間の容量を持つ、より小さいスケールの発酵器もまた使われて差し支えない。
【0016】
E. coliの培養は、タンパク質と必要な収量に応じて、例えば、連続的な方法、バッチ法、またはフェドバッチ法(Thiry and Cingolani, 2002, Trends in Biotechnology, 20, 103-105)、いずれかの適したシステムにて行ってよい。バッチ法は、栄養分または誘導物質が必要な場合、その点添加とともに用いることができる。もう1つの方法として、フェドバッチ培養は使われ、そして例えば、最大の特異的生育速度でバッチ法での予備誘導において生育した培養は、発酵器内に初期から存在する栄養分や、発酵が完了するまでの誘導後の生育速度を制御するために使われた1つまたはより多くの栄養を与える状態を使い続けることができる。フェドバッチ法はまた、E. coli宿主細胞の代謝を制御するため、そして高い細胞密度に到達させるための予備誘導に用いてもよい(Lee, 1996, Tibtech, 14, 98-105)。
【0017】
本発明において、細胞は、タンパク質発現の誘導前には、どの適切な生育速度でも生育する。高い生育速度を達成するために、E. coli宿主細胞は、典型的に、栄養レベル、炭素源、温度、そして酸素などの条件が、そのような生育を許す培養において生育させる。望ましくは、本発明においてE. coli宿主細胞は、組み換えタンパク質発現の誘導の前に、高い細胞密度を得るために、高い生育速度で栄養的な制限のない条件下で生育させる。生育は、いくつかの異なった方法で測定されるが、その中で最も一般的なものは、時間ごとに繰り返して600 nmにおける光学的密度の増加を測定することによって、細胞密度の増加を測定する方法である。特異的生育速度は、時間軸に対して、OD600測定の自然対数をプロットすることによって決定することができる。生育測定は、培養の全体にわたって、例えば、オンライン測定によって、または発酵の間、規則正しい時間間隔で、培養液からサンプルを取ることによって、行われる。より好ましくは、高い細胞密度が、誘導される組み換えタンパク質の発現の前に得られるとよい。タンパク質発現での適切な密度の例は、おおよそ60から100 OD600の範囲内で誘導されるものであろう。
【0018】
本発明の方法の工程 (b)において、生育速度は、方法の工程 (a)で与えられるE. coli培養液における生育速度と比較して、変えられる。工程 (b)における生育速度の変化は、工程 (c)でのタンパク質発現の誘導の前、タンパク質発現の誘導と同時、またはタンパク質発言の誘導の後、のいずれの時にも起こるかもしれない。生育速度が工程 (c)の前に変わった1つの例では、方法の工程 (a)において、高い細胞密度に到達したその後に生育速度が変化するまでは、培養物は高い生育速度で生育され、そしてタンパク質発現がいったん誘導されると、望ましい生育速度が変化してしまうことが観測された。1つのそのような例では、生育速度が、タンパク質発現の誘導の前のおおよそ1時間から3時間の間に変化している。もう1つの例では、培養物が、方法の工程 (a)では高い生育速度で生育し、そして方法の工程 (c)ではタンパク質発現が誘導され、その後、方法の工程 (b)で生育速度が変化する。1つのそのような例では、生育速度が、工程 (c)の後のおおよそ1時間から3時間の間に変化している。もう1つの例では、細胞が、方法の工程 (a)においては高い生育速度で生育していて、そして工程 (b)と (c)が同時に起こる、すなわち、生育速度が誘導時において変化するという、おおよそ同じ時間である。
【0019】
E. coli宿主細胞の生育速度は、発酵システムに応じて、過去の研究でよく知られた多くの異なる方法によって制御されるだろう(Lee, 1996, Tibtech, 14, 98-105の例を参照)。生育速度を制御するために適した方法は、温度、摂取可能な酸素のレベル、そして栄養レベルの変化を含む。特異的な生育曲線をもたらすために必要な、適切な温度の変化、酸素レベル、または栄養分の濃度は、濃度の範囲を試したり、生育速度への影響を測定したりすることによって、当業者によって経験的に決定することができる。1つの例では、発酵開始時に存在する窒素やリン酸といった栄養素の濃度が、誘導の間、栄養分が枯渇するタイミングに応じて、生育速度を制御するために使われる。本発明の1つの特殊な例では、生育速度を制御している栄養源がリン酸である。適切なリン酸の濃度が、ここでの例において提供され、過去の研究例として知られている(Pirt, S. J. (1975) Principles of Microbe and Cell Cultivation, Blackwell Scientific Publicationsを参照)。発酵開始時に存在する適切なリン酸濃度の例は、25から40 mMの範囲内である。それゆえに、本発明での1つの例では、工程 (b)における生育速度が、発酵開始時に細胞が利用できるリン酸のレベルを調整することによって、変化する。もう1つの例では、生育速度が、例えば供給を介して、または点添加を通して培養液に加えられた炭素または窒素といった栄養素の濃度を変えることによって、調整されるだろう。
【0020】
本発明でのもう1つの例では、工程 (b)における生育速度が、E. coli宿主細胞に利用可能な炭素のレベルの調整によって変化する。細胞が利用可能な炭素源のレベルは、生育速度の必要な変化を達成するために、増加させたり減少させたりしてもよい。好ましくは、生育速度が、培養液への炭素源の供給速度を調整することによって制御される。ここで用いている「炭素源」という用語は、生育の間、E. coliのためのエネルギー源として利用可能な炭素のソースについて、そして、グルコース、ラクトース、スクロース、フルクトースなどの糖類、そして、グリセロール、コハク酸、乳酸などの他の炭素のソースについて、言及している。本発明の好ましい実施例では、炭素源はグリセロールである。好ましくは、宿主細胞はフェドバッチ発酵器で培養され、そして炭素源は、必要な生育速度を与えるような適切な供給速度で提供されたグリセロールである。適切なグリセロール供給速度は、過去の研究例にてよく知られており、そしてこの明細書に提供された例に記されたそれも含んでいる。適切なグリセロール供給速度の例は、それの0.5から11 ml/hの範囲に含まれる。
【0021】
ここで用いている「分画」という用語は、E. coli宿主細胞のペリプラズムとE. coliが生育した培養上清の間での、組み換えタンパク質の分配について言及している。組み換えタンパク質の分画を決定するために、ペリプラズムと上清の収量を、方法の工程 (d)で決定する。収量は、E. coli宿主細胞の培養の全体にわたって適切な時間間隔で一部サンプルを採取する、例えばおよそ2から4時間毎、または代わりに収穫時、例えば誘導からおよそ24から36時間後に採取することによって、決定される。タンパク質の定量の多くの方法が、過去の研究において知られており、ゲル電気泳動、HPLC、クロマトグラフィー、そしてウェスタンブロットやELISAといった免疫検出法などの方法を含んでいる。例えば、ペリプラズム抽出物と培養上清中のFab’濃度が、Humphreys et al., 2002, Protein Expression and Purification, 26, 309-320に記載されたFab’会合ELISA法によって決定できる。
【0022】
本発明の方法では、工程 (b)において2つかそれ以上の異なった生育速度で生育させたE. coli宿主細胞のペリプラズムと、上清にある組み換えタンパク質の収量は、タンパク質生産物の最初の回収に最も適したような分画をもたらす生育速度を見出すために、方法の工程 (e)において比較される。好ましくは、工程 (e)において比較された収量が、それぞれの生育速度において誘導された後、おおよそ同じ時間に、工程 (d)において決定される。これは必ずしも可能であるとは限らないだろうことは、とりわけ、より低い生育速度で生育された培養物においてよりも、最大バイオマスにより早く到達するような、より高い生育速度で生育された培養物に関して、そう理解しておくべきだろう。
【0023】
方法の工程 (b)での生育速度の変化は、生育速度の増加または減少であろう。工程 (a)で提供された培養液は、栄養分が限られていないか、栄養分が限られた条件において、適切な生育速度で生育されるだろう。好ましくは、工程 (a)で提供された培養液は、栄養分が限られていない条件下で、その条件で可能な最大の特異的な生育速度で生育している。1つの例では、工程 (a)におけるE. coli宿主細胞の生育速度は、およそ0.1と0.25/hの間であり、好ましくは0.13と0.22/hの間である。好ましくは、生育速度は、方法の工程 (b)では、減少させる。生育速度は、0/hより大きければどの適切な速度にまで減少させても差し支えなく、好ましくは、0/hと工程 (a)にて条件としていた生育速度との間である。適切な生育速度は、少なくとも2つの異なった生育速度を試し、方法の工程 (e)においてそれらを比較することによって、経験的に同定される。それぞれの生育速度は、2つまたはそれ以上に分かれた、平行して、および、または連続して生育された培養について、試してもよい。1つの例では、少なくとも2つの培養を、工程 (b)において、それぞれの培養で異なった生育速度を用いて同時に生育させる。もう1つの例では、少なくとも2つの培養を、工程 (b)において、それぞれの培養で異なった生育速度を用いて連続して生育させる。もう1つの方法として、2つまたはそれ以上の異なった生育速度が、工程 (b)での1つの生育速度を使って工程 (a)から (d)を行い、それから工程 (b)での生育速度をそれぞれの時間において少なくとももう一度変化させて工程 (b)と (d)を行うことによって、単一の培養においてテストすることができる。もし方法の工程 (e)において適切な生育速度が同定されなかった場合には、それからさらに生育速度が、方法の工程 (a)から (d)を繰り返すことによってテストされるだろうことも、理解しておかなくてはならないだろう。典型的に、異なった生育速度の範囲は、0/hと工程 (a)での培養の生育速度との間でテストされるだろう。第一の例では、テストされた異なった生育速度の範囲が、高い、そして低い生育速度の幅広い範囲にわたっていて、それは、もし必要ならば、工程 (b)においてより狭い範囲の生育速度を用いて、方法の工程 (a)から (d)を繰り返すことによって、それ以降に狭められればよい。第一の例においてテストした適切な生育速度は、例えば、栄養分の限られていない条件下での生育速度よりも、およそ2とおよそ200倍の間くらい少ないものであるかもしれない。1つの例では、生育速度は、栄養分の限られていない条件下での生育速度よりも、およそ5とおよそ200倍の間くらい少ない。もう1つの方法として、第一の例においてテストされた、異なった生育速度の範囲は、例えば、高い、または低い生育速度の狭い範囲であり、それは、もし必要ならば、工程 (b)においてより広い範囲の生育速度を用いて、方法の工程 (a)から (d)を繰り返すことによって、それ以降に広げられればよい。第一の例においてテストされた適切な生育速度は、例えば、栄養分の限られていない条件下での生育速度よりも、およそ2とおよそ50倍の間くらい、またはおよそ100とおよそ200倍の間くらい少ないものであるかもしれない。1つの例では、生育速度は、栄養分の限られていない条件下での生育速度よりも、およそ5とおよそ20倍の間くらい少ない。工程 (b)における適切な生育速度は、0.0005から0.04/hの範囲内の生育速度を含むだろう。
【0024】
一旦、2つまたはそれ以上の異なった生育速度で、組み換えタンパク質の分画が比較されると、組み換えタンパク質の最初の回収に最も適した生育速度が、方法の工程 (f)において選択される。ここで使われている「最初の回収」という用語は、無細胞の上清または、次に続く精製の工程に適したペリプラズム抽出物を生産するのに必要な、初期の回収工程について言及している。最初の回収工程の例は、遠心分離、濾過、細胞の抽出または溶解、均質化、濃縮、ダイアフィルトレーション、そして吸着流動床を含んでいる。タンパク質の最初の回収に最も適した生育速度は、最も良い収量、および/または1つまたはそれ以上の最初の回収の工程の後のタンパク質の「質」をもたらすような生育速度だろう。ここで使われている「質」という用語は、タンパク質が無傷、および/または、正確に折りたたまれているかどうか、得られたタンパク質の完全性について言及している。重要なことに、最も適切な生育速度は、E. coli宿主細胞培養液内に最大の収量の組み換えタンパク質をもたらす生育速度ではなく、その代わりに、1つまたはそれ以上の最初の回収工程の後で、より良い質のタンパク質、および/またはより高い収量をもたらすであろう、ペリプラズムと上清の間でのタンパク質の最も良い分画をもたらすような生育速度であるかもしれない。それゆえに、工程 (f)で選択した生育速度は、培養の間のタンパク質収量と、最終の収量と最終的に回収されるタンパク質の質との間で、代わる代わるにされるかもしれない。
【0025】
1つの例では、E. coli宿主細胞のペリプラズムに発現させた組み換えタンパク質は、上清に発現させたタンパク質よりも、ラージスケールにおいては、取り扱いが簡単であるかもしれない。第一に必要な、最初の回収は、遠心分離か濾過のいずれかによる、培養培地からの宿主細胞の回収である。ペリプラズムに生産されたタンパク質にとって重要な考察は、上清へと組み換えタンパク質が漏洩するレベルであり、これは最初の回収やペリプラズムのタンパク質の質に不利な効果を持ちうる。上清に存在するタンパク質の増加は、上清の粘度を増加させ、遠心分離または濾過による細胞の回収をより困難にし、そしてペリプラズムのタンパク質収量を減らす。細胞それ自身もまた、遠心分離の間はそれほど丈夫ではなく、組み換えタンパク質へのダメージをもたらすような溶解をより起こしやすく、再びペリプラズム内の組み換えタンパク質の収量を減らすような、タンパク質、DNA、そして脂質の放出のために、上清の粘度はさらに増加する。ペリプラズムから回収された組み換えタンパク質の質はまた、部分的に分解した、または正確に折りたたまれていないような、上清由来の組み換えタンパク質で汚染されることによって、低下する。これらの培養においては、それゆえ、最初の回収の後の組み換えタンパク質の収量、および/または質を上げるために、上清における組み換えタンパク質のレベルを低く保つことが重要である。この例では、後に続く最初の回収で得られた組み換えタンパク質の最終収量、および/または質が、たとえ、ペリプラズムにおける組み換えタンパク質の収量が、上清における組み換えタンパク質のレベルがより高くなるような他の生育速度のときよりも少なくなる場合でも、上清における組み換えタンパク質の量が少なくなるような生育速度を選択することによって改善されるかもしれない。それゆえ、本発明の方法においてペリプラズムに生産されたタンパク質にとって、工程 (f)で選択される生育速度は、タンパク質の55%以上がペリプラズムで発現し、タンパク質の45%以下が上清にあるような生育速度を選ぶのが好ましい。好ましくは、宿主細胞によって生産されたタンパク質の80%以上がペリプラズムに発現し、そして20%以下が上清にあるのが良い。1つの例では、宿主細胞によって生産されたタンパク質の90%以上がペリプラズムに発現し、10%以下が上清に存在している。もう1つの例では、宿主細胞によって生産されたタンパク質の95%以上がペリプラズムに発現し、5%以下が上清に存在している。
【0026】
もう1つの例では、タンパク質が上清に生産される。タンパク質は、もし抽出の工程が必要でなく、培地中に存在する宿主タンパク質はより少ないだろうならば、精製するのはより簡単であるかもしれない。タンパク質の質もまた、タンパク質の分解がより少なく、タンパク質の折りたたみが改善されているようならば、より良いかもしれない。上清に生産されたタンパク質にとって、宿主細胞によって発現され、上清へと分泌された組み換えタンパク質の量が最大化されることが重要である。宿主細胞によって生産された全ての組み換えタンパク質が、上清へと分泌されているかどうかを決定するためには、同様にペリプラズム内の組み換えタンパク質のレベルを測定することが重要である。もし、これらが依然高いのであれば、それでもさらに上清にある収量を増加させるために、組み換えタンパク質の分画を調整することは、本発明の方法を用いて可能になるかもしれない。結果として、本発明の方法において上清に生産されたタンパク質にとっては、工程 (f)で選択される生育速度は、タンパク質の55%以上が上清に発現され、タンパク質の45%以下がペリプラズムにあるような生育速度が望ましい。好ましくは、宿主細胞によって生産される組み換えタンパク質の80%以上が上清に生産され、ペリプラズムには20%以下のタンパク質が存在する。1つの例では、宿主細胞によって生産されたタンパク質の90%以上が上清に存在し、10%以下がペリプラズムに存在する。もう1つの例では、宿主細胞によって生産されたタンパク質の95%以上が上清に存在し、5%以下がペリプラズムに存在する。
【0027】
方法の工程 (f)において適切な生育速度が同定された場合、上清とペリプラズムの間での組み換えタンパク質の望ましい分画が達成されることを確認するために、同じ生育速度が、次に続くE. coli宿主細胞の発酵の間に使われうる。本発明はまた、それゆえ、当該細胞による組み換えタンパク質の発現が、誘導可能なシステムや、組み換えタンパク質の最初の回収に最も適した、上述した方法の工程 (f)において選択された生育速度でE. coli宿主細胞を生育することから成る、当該方法によってなされる上清とペリプラズムとの間の組み換えタンパク質の分画の制御下に置かれているような、E. coli宿主細胞培養における、組み換えタンパク質生産の方法を提供する。これらの後続の発酵は、上記の方法において使われたのと同じスケールか、異なったスケールであっても差し支えない。例えば、E. coli宿主細胞が、タンパク質の最初の回収に最も適した生育速度を決定するために、小さいスケールで培養され、それから、その後の発酵では、宿主細胞は、工程 (f)で選択された生育速度を用いて、より大量の組み換えタンパク質を生産するために、大きいスケールで培養されてもよい。
【0028】
本発明の方法によって生産された組み換えタンパク質は、最初の回収の直後に精製されるだろう。ここで使われる最初の回収の工程は、発酵の終了時における組み換えタンパク質の細胞内での位置に依存するだろう。例えば、ペリプラズムで発現したタンパク質は、遠心分離または濾過によって、宿主細胞内に回収されるかもしれない。タンパク質はそれから、熱処理(US 5,665,866の例を参照)または機械的な抽出といった方法を用いて、細胞内から抽出することができる。上清に発現したタンパク質は、吸着流動床、遠心分離、または濾過など細胞を除去するための方法に続いて、例えば、イオン交換、または捕捉タンパク質に対するアフィニティークロマトグラフィーといった方法を用いて回収してもよい。ペリプラズム抽出物か上清のいずれか由来のタンパク質は、その後、組み換えタンパク質の性質や、大きさ、疎水性、そして等電点といったタンパク質の物理的特性に依存して、過去の研究例に知られた方法のいずれかを用いて精製してもよい。そのような方法には、サイズ排除クロマトグラフィー、疎水性相互作用クロマトグラフィー、イオン交換、アフィニティークロマトグラフィー、そして逆相HPLCを含んでいる。従って、本発明のもう1つの様相では、当該細胞による組み換えタンパク質の発現が、誘導可能なシステムや、組み換えタンパク質の最初の回収に最も適した、上述した方法の工程 (f)において選択された生育速度でE. coli宿主細胞を生育することと、その後の組み換えタンパク質の精製から成る、当該方法によってなされる上清とペリプラズムとの間の組み換えタンパク質の分画の制御下に置かれているような、E. coli宿主細胞培養における、組み換えタンパク質生産のための方法が提供される。
【実施例1】
【0029】
誘導後の生育を制御するためにグリセロール供給の制限を用いる、Fab’ Aの分画と収量の最適化。
【0030】
材料と方法
菌株とプラスミド
この例で使われた菌株は、テトラサイクリン耐性を与え、Fab’断片であるFab’ Aの軽鎖と重鎖ポリペプチド構成成分をコードする遺伝子を持っているプラスミドを形質転換した、Escherichia coli W3110 (ATCC 27325)であった。それぞれのポリペプチドは、E. coliのOmp Aリーダーペプチドによって先行されている。単一のtacプロモーターからの発現の誘導は、ポリペプチドの一部が折りたたまれ、Fab'を形成するようなペリプラズムへと、軽鎖と重鎖ポリペプチドの分泌と合成をもたらす。
【0031】
生育培地
発酵生育培地は、3.86 g/lのリン酸二水素ナトリウム一水和物と112 g/lのグリセロールを加えた、SM6E培地(Humphreys et al., 2002, Protein Expression and Purification, 26, 309-320に記載)に基づいたものであった。
【0032】
接種材料
接種材料の培養は、10 μg/mlのテトラサイクリンを補って、同じ培地で生育させた。培養は、およそ22時間の撹拌とともに、30℃にて培養された。
【0033】
発酵
発酵器(全容量2.5リットル)は、接種材料の培養液を用いて、0.3-0.5 OD600になるまで植菌された。温度は、増殖期の間は30℃に保たれ、そして誘導の前に25℃に下げられた。溶存酸素濃度は、変化可能な撹拌と空気流によって、30%空気飽和より高く保たれた。培養pHは、15% (v/v)アンモニア水と10% (v/v)濃硫酸を用いた自動滴定によって、7.0に制御された。発泡は、10% (v/v) Struktol J673溶液(Schill and Seilacher)の添加によって制御された。
【0034】
いくらかの添加が、発酵の異なった工程においてなされた。バイオマス濃度が、おおよそ40 OD600に達した時、マグネシウム塩とリン酸二水素ナトリウム一水和物が加えられた。リン酸二水素ナトリウム一水和物のさらなる添加が、リン酸が過剰に保たれているかを確認するために、誘導期の前と誘導期の間になされた。発酵の初期に存在したグリセロールが枯渇した時(おおよそ75 OD600)には、80% (w/w)グリセロールの継続的供給が、0.5から10.9 ml/hにわたる速度で適用された。発酵中の同じポイントで、IPTGの供給が、この時点での発酵器内のIPTGの最終濃度が0.5 mMになるように、1 ml/hの速度で36時間にわたって、適用された。IPTG供給の開始が、誘導の開始として解釈された。発酵は、典型的に、70-73時間の間はより低いグリセロール供給速度で(0.5-2.5 ml/h)、そして50-60時間の間はより高いグリセロール供給速度で(5.4-10.9 ml/h)、行われた。
【0035】
バイオマス濃度と生育速度の測定
バイオマス濃度は、培養液の光学密度を600 nmで測定することによって、決定された。生育速度(μ)は、時間を通してのバイオマス濃度の変化に、このようにして関連している:
LnXt = LnX0 + μt
ここで、X0は元々のバイオマス濃度であり、Xtは時間間隔t後のバイオマス濃度である。従って、バッチ培養に関してtに対するLnXtのプロットは、μ(単位/h)と同等な傾斜の直線を与える。しかしながら、バッチ培養に対するグリセロールの直線限定的な供給の適用は、バイオマスと培養液量の増加のために、生育速度の低下をもたらす。それゆえ、誘導の時期の間の平均生育速度が、時間に対してLnOD600をプロットすることによって決定された。(最小2乗法による)適合度の線の傾斜は、供給時期の間を通しての平均生育速度を与えることによって決定された。
【0036】
ペリプラズムの抽出
細胞は、遠心分離によって培養サンプルから回収された。上清画分は、さらなる分析のために(-20℃で)保持された。細胞の塊の画分は、抽出バッファー(100 mM Tris-HCl, 10 mM EDTA; pH 7.4)中、元々の培養液の量にまで再懸濁された。続いておおよそ16時間、60℃で温めた後、抽出物は遠心分離によって浄化され、そして上清画分は、分析のために(-20℃で)保持された。
【0037】
Fab'の定量
ペリプラズム抽出物と培養上清中のFab’濃度は、Humphreys et al., 2002, Protein, Expression and Purification, 26, 309-320に記載されている、Fab’会合ELISAによって決定された。
【0038】
結果
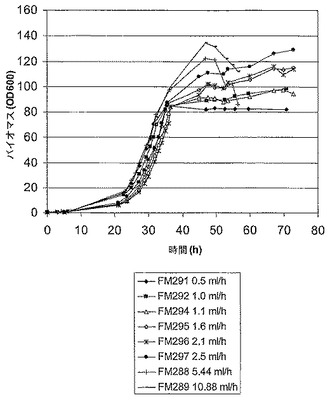
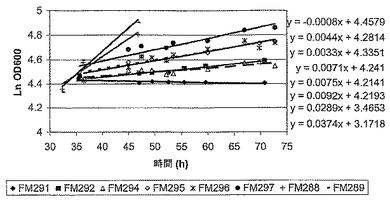
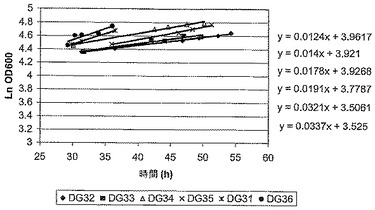
Fab’ A発酵は、生育が、誘導前は無制限で、最大の特異的速度(μmax)が生ずるような生育で、しかしそれに続く誘導後には制限されたような条件下で行われた(図1)。誘導後の生育速度は、ある範囲の流速でバッチ培養に、栄養(グリセロール)を限定した供給を適用することによって制御された。グリセロールの流速は、おおよそ0/hから上向きに生育速度の範囲を与えるように選択された。図2と表1は、グリセロールの供給速度の範囲に対して、誘導時期の間を通しての実際の平均生育速度を示している。グリセロール供給速度の上昇は、予定通り、培養の平均生育速度において増加をもたらした。
【0039】
【表1】
【0040】
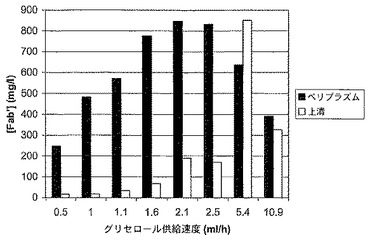
発酵は、より低いグリセロール供給速度(0.5-2.5 ml/h)では誘導後おおよそ36時間で、より高いグリセロール供給速度(5.4-10.9 ml/h)では誘導後おおよそ24時間で収穫され、そしてペリプラズム抽出物と培養上清のFab’の濃度は、Fab’会合ELISAによって決定された。図3は、Fab'の濃度と、ペリプラズムと培養培地との間での相対的分配を示している。Fab’滴定濃度に対する最適な生育速度は、おおよそ0.0075/hであることが判明した。生育速度を上げると、培養上清におけるFab’の滴定濃度が上昇するという、一般的な傾向が観測された。従って、最適な発酵能力のために選択される生育速度は、Fab’収量と、最初の回収での両立性が、最大の収量とFab'の最適な分配との間のバランスに反映するだろう。引用例では、1.6 ml/hのグリセロール供給速度(生育速度〜0.0071/h)が、発酵のスケールアップのために選択されていた。
【実施例2】
【0041】
誘導後の生育速度を制御するためのリン酸濃度の変化によるFab’ Bの収量と分画の最適化。
【0042】
材料と方法
菌株とプラスミド
この実施例で使われた菌株は、テトラサイクリンに対する耐性を与え、Fab'断片であるFab’ Bの、軽鎖と重鎖ポリペプチド構成要素をコードしている遺伝子を持っているプラスミドを形質転換したEscherichia coli W3110 (ATCC 27325)であった。単一のtacプロモーターからの発現の誘導は、ポリペプチドの一部が折りたたまれ、Fab’を形成するために会合するようなペリプラズムへの、軽鎖と重鎖ポリペプチドの分泌と、その合成をもたらす。
【0043】
生育培地
発酵生育培地は、表2に示された濃度でリン酸二水素ナトリウム一水和物を加えたSM6E培地(Humphreys et al., 2002, Protein, Expression and Purification, 26, 309-320に記載)に基づいていた。
【0044】
【表2】
【0045】
接種材料
接種材料の培養は、リン酸二水素ナトリウム一水和物の適切な濃度(表2参照)とともに、そして10 μg/mlのテトラサイクリンを補って、同じ培地で生育させた。培養は、およそ19から26時間の撹拌とともに、30℃にて培養された。
【0046】
発酵
発酵器(全容量2.5リットル)は、接種材料の培養液を用いて、0.3から0.5 OD600になるまで植菌された。温度は、増殖期の間は30℃に保たれ、そして誘導の前に27℃に下げられた。溶存酸素濃度は、変化可能な撹拌と空気流によって、30%空気飽和より高く保たれた。培養pHは、15% (v/v)アンモニア水と10% (v/v)濃硫酸を用いた自動滴定によって、7.0に制御された。発泡は、10% (v/v) Mazu溶液の添加によって制御された。
【0047】
いくらかの添加が、発酵の異なった工程においてなされた。グリセロールは、80% (w/w)溶液の45 mlを2回に分けて、最初の添加は20 OD600のバイオマス濃度で、他は40 OD600で発酵に添加された。バイオマス濃度がおおよそ40 OD600に達した時、マグネシウムとカルシウム塩もまた添加された。ラクトース(50% (w/w)溶液の60 ml)が60 OD600で添加され、誘導物質そして誘導工程の間の炭素源として供給された。誘導は、溶存酸素濃度の上昇により注目づけされたグリセロールの枯渇(およそ75 OD600と90 OD600の間)に起こった。ラクトースのさらなる添加が、0と55 g/lの間に発酵器内の濃度を保持するために必要とされて、なされた。マグネシウムのさらなる添加が、誘導後、0と2時間の間になされた。発酵は、典型的に56から61時間行われた。
【0048】
発酵の分析
ペリプラズムからの抽出、バイオマスの測定、そしてFab’の定量が、前述の通りに行われた(実施例1)。
【0049】
結果
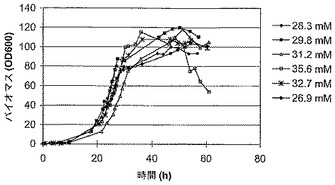
発酵培養誘導後の生育は、すべての他の開始時の栄養素を一定にした一方で、リン酸(リン酸二水素ナトリウム一水和物)の量を変化させることによって制御された。それぞれの発酵の間の生育速度は、バイオマスが増加した誘導の時期に対して決定された(図5と表3)。
【0050】
【表3】
【0051】
リン酸濃度の上昇は、誘導の期間を通した生育速度の上昇をもたらした。リン酸濃度は、誘導に関連して、様々なポイントで枯渇するように選択された(例えば、26.9 mMの場合、誘導前にはかすかに、また誘導後の様々なポイントで)。誘導が起こるポイント(バイオマス濃度)は、開始時のグリセロール濃度に関係している。望みの誘導バイオマスまでの生育を支持するために必要なリン酸の量は、Pirt, S. J. (1975) Principles of Microbe and Cell Cultivation, Blackwell Scientific Publicationsに規定されている、無機リン酸のためのバイオマス収量係数データに基づいて計算された。Fab'の収量と分配にとって最適なリン酸濃度は、ここに記載されたように経験的に決定された。
【0052】
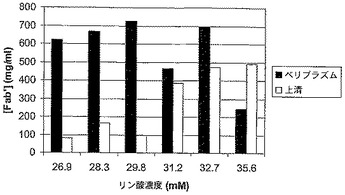
発酵は、誘導後28から31時間で収穫され、ペリプラズム抽出物と培養上清のFab’濃度はFab'会合ELISAによって決定された。図6は、収穫時でのペリプラズムと培養上清の両方のFab’濃度を示している。培養上清のFab’濃度は、リン酸濃度とともに上昇するという、一般的な傾向が観測された。ペリプラズム抽出と遠心分離または濾過を用いた、大きいスケールの最初の回収のための、リン酸の最適な値域は、29.8 mMであった。
【図面の簡単な説明】
【0053】
本発明は、以下の参考図を示す目的で記述されるだろう:
【図1】グリセロール供給速度の変動におけるE. coliの生育プロファイル
【図2】グリセロール供給速度の変動におけるE. coliの誘導後の生育速度
【図3】グリセロール供給速度の変動において生育されたE. coli培養の、ペリプラズムと上清との間でのFab’ Aの分画
【図4】無機リン酸濃度の変動におけるE. coliの生育プロファイル
【図5】無機リン酸濃度の変動におけるE. coliの誘導後の生育速度
【図6】無機リン酸濃度の変動において生育されたE. coli培養の、ペリプラズムと上清との間でのFab’ Bの分画
【技術分野】
【0001】
本発明は、一般的にはEscherichia coli宿主細胞培養液中で組み換えタンパク質を生産するための方法に関し、さらに特には、E. coli宿主細胞のペリプラズムと培養液上清との間に、前述のタンパク質の分画を制御するための方法を提供する。
【背景技術】
【0002】
E. coliは、幅広く使われ、組み換えタンパク質の生産のための便利なシステムである。このシステムの有利な点は、遺伝子操作の容易さ、遺伝子発現ベクターを含む試薬の入手可能さ、タンパク質の生産定量の容易さ、幅広く多様なタンパク質を発現するためのシステムの速度と高い適応性、などから構成される。バネイクスの、Current Opinion in Biotechnology, 1999, 10, 411-421.のレヴューを見ると、E. coliは、インシュリン、成長ホルモンそして抗体といった治療上のタンパク質のラージスケール生産に、今では幅広く使われている(Swartz, Current Opinion in Biotechnology, 2001, 12, 195−201; Humphreys and Glover, Current Opinion in Drug Discovery and Development, 2001, 4, 172-185; Verma et al., Journal of Immunological Methods, 1998, 216, 165-181; Simmons et al., Journal of Immunological Methods, 2002, 263, 133-147)。
【0003】
E. coli内での外来遺伝子の発現は、遺伝子のcDNAコピーの発現ベクターへの挿入から始まる。たくさんの形の発現ベクターが利用可能である。そうしたベクターは、大抵、DNA複製のためのプラスミドオリジン、抗生物質の選択可能なマーカー、マルチクローニングサイトによって隔てられたプロモーターと転写終結因子(発現カセット)、そして少なくとも1つのリボソーム結合サイトをコードしているDNA配列、によって構成される。外来遺伝子の転写は、本来の量で発現されたタンパク質の場合よりも高い収量になる生産物合成と、細胞増殖とが別々になることを可能にするような調節プロモーターによって、大抵制御されている。タンパク質の最高収量は、発現を誘導するための培養の間の最適なポイントを決定することによって、典型的には、一旦細胞を高密度にまで生育させることによって、達成される。
【0004】
E. coli内で発現される組み換えタンパク質は、タンパク質の性質やそのリホールディングの必要性に依存して、細胞質内か、ペリプラズムまたは培養上清に分泌されて、生産される(Makrides, Microbiological Reviews, 1996, 60, 512-538.)。例えば、タンパク質が、シグナル配列を介してペリプラズムを目標としてペリプラズムに発現することは、宿主細胞のタンパク質の数がより少ないこと、組み換えタンパク質の分解がより少ないこと、そして適当なタンパク質リホールディングを促進するような酸化的環境などを含めて、細胞質内での発現よりも、いくつかの有利な点を提供することができる。
【0005】
培養中のタンパク質収量が、E. coli内で組み換えタンパク質を生産するコストに影響を与える鍵となる因子である一方、生産されたタンパク質の下流の処理(DSP)のコストもまた、商品の最終コストを決定する上で重要である。DSPという用語は、E. coliの培養の後、最終的な精製された機能的タンパク質をもたらすまでの、すべての工程を含んでいる。生産されたタンパク質の細胞での位置に依存して、これらの工程は典型的に、遠心分離、濾過、抽出、そして濃縮・ダイアフィルトレーションなどの最初の回収工程、それに続く1つまたはより多くの精製工程を含んでいる。機能的タンパク質を高収量で得るために、これらの工程の効果を最大化することは、組み換えタンパク質のラージスケール生産の間のコストを最小化するための鍵となる。DSPの間に使われる最初の回収工程の性質は、得られる組み換えタンパク質の量と質に影響するだろうし、生産のスケール、タンパク質の性質、そして培養終了時の細胞での位置などを含むいくつかの要因に依存するだろう。例えば、ペリプラズムに生産された組み換えタンパク質に対する重要な配慮は、組み換えタンパク質が上清へと漏れる量であり、培養中に自然な量の漏れが起こったとしても、これはペリプラズムから得られた組み換えタンパク質の量と質に不利な効果があるといえる。大量の組み換えタンパク質が上清へと漏れる場合には、上清の粘度が増加し、遠心分離や濾過による細胞の最初の回収をより困難にし、そしてそれゆえに、ペリプラズムから得られるタンパク質の収量は減ってしまう。細胞自身もまた、遠心分離の間はそれほど頑丈ではなく、より溶解しやすく、結果としてタンパク質にダメージを与え、そして上清のタンパク質量が増え、再び収量が減る。さらなる問題は、抽出操作の間ペリプラズムから回収されるタンパク質の「質」もまた、正しくリホールディングされていない、または部分的に分解した、上清由来の組み換えタンパク質に汚染されることによって、低下するかもしれないということである。
【0006】
それゆえ、組み換えタンパク質の最初の回収の効率を改善すること、そして、ペリプラズムと上清の間に組み換えタンパク質の分画を制御することによって得られる、機能的タンパク質の最終的な量と質を改善することは、望ましいことである。
【発明の開示】
【発明が解決しようとする課題】
【0007】
本発明において、我々は、E. coli宿主細胞の生育速度を調節することによって、E. coli宿主細胞の培養において、上清とペリプラズムの間に組み換えタンパク質の分画を制御することができることを証明することができた。上清とペリプラズムの間でのタンパク質の分画に基づいて適切な生育速度を選択することによって、必ずしも収量だけでなく、生産されたタンパク質の、質および/または量を増加させることができる。
【課題を解決するための手段】
【0008】
本発明に従って、E. coli宿主細胞培養において、上清とペリプラズムの間に組み換えタンパク質の分画を制御するための方法を提供する。その培養において、上述の細胞による組み換えタンパク質の発現は、1つの誘導システムの下で制御される。その方法は、以下の工程から構成される:
a)E. coli宿主細胞培養液を提供する工程
b)E. coli宿主細胞の生育速度を変化させる工程
c)組み換えタンパク質の発現を誘導する工程
この点で、工程 (b)と (c)は、どの順序でも、あるいは同時に、行うことが可能である;そして続いて、
d)培養上清とE. coli宿主細胞ペリプラズムにおける組み換えタンパク質の収量を決定する工程
e)工程 (d)で決定した収量と、工程 (b)で用いられた生育速度と少なくとも1つの他の場合において決定された収量とを比較する工程
f)上清とペリプラズムの間での組み換えタンパク質の分画が、組み換えタンパク質の最初の回収に最も適しているような、工程 (e)でなされた比較の結果得られた、生育速度を選択する工程
【発明を実施するための最良の形態】
【0009】
本発明において、使用できる組み換えタンパク質は、おおよそ10以上のアミノ酸残基を持つ、ペプチド、ポリペプチド、またはタンパク質のいすれかであろう。タンパク質は、細菌を含めて、どの起源由来でも差し支えないが、より好ましくは哺乳動物であろう。1つの例では、組み換えタンパク質は抗体である。ここで使われている「抗体」という用語は、IgG, IgE, IgM, IgDそしてIgAなど、どのクラス由来の免疫グロブリン、例えばIgGクラスの1つであるIgG1、そしてまた、Fv, Fab, Fab'そしてF(ab’)2断片などの抗原が結合した免疫グロブリン断片、そして単鎖Fv断片などの誘導体、などについて言及している。これらの抗体や断片は、自然に発生し、人体に適応され、キメラであり、またはCDR移植した抗体でもよい。そして標準的な分子生物学的手法が、望みのアミノ酸残基やドメインの改変、添加、または除去に使われても差し支えない。これらの抗体分子を創るための方法は、過去の発明にてよく知られている(以下の参考文献を参照、Shrader et al., WO 92/02551; Ward et al., 1989, Nature, 341, 544; Orlandi et al., 1989, Proc. Natl. Acad. Sci. USA, 86, 3833; Riechmann et al., 1988, Nature, 322, 323; Bird et al., 1988, Science, 242, 423; Queen et al., US 5,585,089; Adair, WO 91/09967; Mountain and Admir, 1992, Biotechnol. Genet. Eng. Rev., 10, 1-142; Verna et al., 1998, Journal of Immunological Methods, 216, 165-181)。
【0010】
本発明で使用するE. coli宿主細胞は、天然に存在するE. coli菌株、または組み換えタンパク質を生産できる変異株で差し支えない。特異的な宿主E. coli株の例は、MC4100, TG1, TG2, DHB4, DH5a, DH1, BL21, XL1BlueそしてJM109が含まれる。例えば、代謝変異株やプロテアーゼ欠損株といった、改変されたE. coli株も例に含まれる。1つの好ましいE. coli宿主はE. coli W3110 (ATCC 27,325)であり、一般的に組み換えタンパク質発酵のための宿主株として使われている。
【0011】
本発明の組み換えタンパク質は、タンパク質の性質や生産のスケールに依存して、典型的に、E. coli宿主細胞のペリプラズムか、宿主細胞培養液の上清のどちらかに発現される。これらの区画へとタンパク質を目標づけるための方法は、過去の研究においてよく知られている(Makrides, Microbiological Reviews, 1996, 60, 512-538を参照)。E. coliのペリプラズムへと向かわせるための適切なシグナル配列の例は、E. coli PhoA, OmpA, OmpT, LamBそしてOmpFシグナル配列を含む。タンパク質は、天然の分泌経路によって、または、pelBリーダー、プロテインAリーダー、バクテリオシン放出タンパク質との共発現、グリシンを培地に添加することを伴ったマイトマイシン誘導バクテリオシン放出タンパク質、そして生体膜透過のためのkil遺伝子との共発現、などの使用を例とするタンパク質分泌をもたらす外膜の限定的漏えいの誘導によって、上清へと目標づけられる。
【0012】
本発明において、E. coli宿主細胞での組み換えタンパク質の発現は、誘導可能なシステムの制御下におかれる。ここで使われている「誘導可能なシステム」という用語は、E. coliにおける組み換えタンパク質の発現が誘導可能なプロモーターの制御下に置かれている、誘導可能な発現システムについて言及している。E. coliにおける使用に適切な多くの誘導可能なプロモーターは、過去の研究においてよく知られており、プロモーターに依存して、組み換えタンパク質の発現が、温度、または生育培地中の特定の物質の濃度などの、変化する要因によって誘導されることができる(Baneyx, supra; Goldstein and Doi, 1995, Biotechnol. Annu. Rev., 105-128)。誘導可能なプロモーターの例は、ラクトースまたは加水分解されていないラクトース類似物であるイソプロピル-β-D-1-チオガラクトピラノシド(IPTG)を使って誘導可能なE. coli lac, tac, そしてtrcプロモーター、そして、リン酸、トリプトファン、L-アラビノースによって、それぞれ誘導される、phoA, trpそしてaraBADプロモーターを含む。
【0013】
ここで使われている「誘導可能な発現」という用語は、例えば、誘導物質の添加、または誘導が温度依存的である場合には温度の変化によって、誘導が始まる点について言及している。組み換えタンパク質発現の誘導が、培養液への誘導物質の添加によって達成される場合は、誘導物質は、発酵システムや誘導物質に依存した適切な方法によって、例えば、単一または複数回の点添加によって、または誘導物質を与える間に逐次添加していくことによって、加えても差し支えない。誘導物質の添加と、実際のタンパク質発現の誘導との間には、遅れがあるかもしれないことも理解しなくてはならないだろう。例えば、誘導物質がラクトースの場合、元々存在している炭素源がラクトースの前に資化される間、タンパク質発現の誘導前に遅れが生じるかもしれない。
【0014】
本発明のE. coli宿主細胞培養は、E. coliの生育と組み換えタンパク質の発現に対応しているであろう、どの培地においても培養できるかもしれない。培地は、Pirt S. J. (1975) Principles of Microbe and Cell Cultivation, Blackwell Scientific Publicationsにおいて規定されている培地に、本明細書に記載されているような生育速度を制御するために適当な改変を加えた、化学的に定義づけられているどの培地でも差し支えない。適切な培地の例は、Humphreys et al., 2002, Protein Expression and Purification, 26, 309-320によって記載されている「SM6E」である。
【0015】
E. coli宿主細胞の培養は、生産に必要なスケールに依存して、振とうフラスコまたは発酵器などの適切な容器において、行うことができる。1000リットルより大きく、おおよそ10万リットルまでの容量の、様々なラージスケール発酵器が利用可能である。1000から5万リットルの好ましい発酵器が使われ、より好ましくは1000から1万リットルである。0.5と1000リットルの間の容量を持つ、より小さいスケールの発酵器もまた使われて差し支えない。
【0016】
E. coliの培養は、タンパク質と必要な収量に応じて、例えば、連続的な方法、バッチ法、またはフェドバッチ法(Thiry and Cingolani, 2002, Trends in Biotechnology, 20, 103-105)、いずれかの適したシステムにて行ってよい。バッチ法は、栄養分または誘導物質が必要な場合、その点添加とともに用いることができる。もう1つの方法として、フェドバッチ培養は使われ、そして例えば、最大の特異的生育速度でバッチ法での予備誘導において生育した培養は、発酵器内に初期から存在する栄養分や、発酵が完了するまでの誘導後の生育速度を制御するために使われた1つまたはより多くの栄養を与える状態を使い続けることができる。フェドバッチ法はまた、E. coli宿主細胞の代謝を制御するため、そして高い細胞密度に到達させるための予備誘導に用いてもよい(Lee, 1996, Tibtech, 14, 98-105)。
【0017】
本発明において、細胞は、タンパク質発現の誘導前には、どの適切な生育速度でも生育する。高い生育速度を達成するために、E. coli宿主細胞は、典型的に、栄養レベル、炭素源、温度、そして酸素などの条件が、そのような生育を許す培養において生育させる。望ましくは、本発明においてE. coli宿主細胞は、組み換えタンパク質発現の誘導の前に、高い細胞密度を得るために、高い生育速度で栄養的な制限のない条件下で生育させる。生育は、いくつかの異なった方法で測定されるが、その中で最も一般的なものは、時間ごとに繰り返して600 nmにおける光学的密度の増加を測定することによって、細胞密度の増加を測定する方法である。特異的生育速度は、時間軸に対して、OD600測定の自然対数をプロットすることによって決定することができる。生育測定は、培養の全体にわたって、例えば、オンライン測定によって、または発酵の間、規則正しい時間間隔で、培養液からサンプルを取ることによって、行われる。より好ましくは、高い細胞密度が、誘導される組み換えタンパク質の発現の前に得られるとよい。タンパク質発現での適切な密度の例は、おおよそ60から100 OD600の範囲内で誘導されるものであろう。
【0018】
本発明の方法の工程 (b)において、生育速度は、方法の工程 (a)で与えられるE. coli培養液における生育速度と比較して、変えられる。工程 (b)における生育速度の変化は、工程 (c)でのタンパク質発現の誘導の前、タンパク質発現の誘導と同時、またはタンパク質発言の誘導の後、のいずれの時にも起こるかもしれない。生育速度が工程 (c)の前に変わった1つの例では、方法の工程 (a)において、高い細胞密度に到達したその後に生育速度が変化するまでは、培養物は高い生育速度で生育され、そしてタンパク質発現がいったん誘導されると、望ましい生育速度が変化してしまうことが観測された。1つのそのような例では、生育速度が、タンパク質発現の誘導の前のおおよそ1時間から3時間の間に変化している。もう1つの例では、培養物が、方法の工程 (a)では高い生育速度で生育し、そして方法の工程 (c)ではタンパク質発現が誘導され、その後、方法の工程 (b)で生育速度が変化する。1つのそのような例では、生育速度が、工程 (c)の後のおおよそ1時間から3時間の間に変化している。もう1つの例では、細胞が、方法の工程 (a)においては高い生育速度で生育していて、そして工程 (b)と (c)が同時に起こる、すなわち、生育速度が誘導時において変化するという、おおよそ同じ時間である。
【0019】
E. coli宿主細胞の生育速度は、発酵システムに応じて、過去の研究でよく知られた多くの異なる方法によって制御されるだろう(Lee, 1996, Tibtech, 14, 98-105の例を参照)。生育速度を制御するために適した方法は、温度、摂取可能な酸素のレベル、そして栄養レベルの変化を含む。特異的な生育曲線をもたらすために必要な、適切な温度の変化、酸素レベル、または栄養分の濃度は、濃度の範囲を試したり、生育速度への影響を測定したりすることによって、当業者によって経験的に決定することができる。1つの例では、発酵開始時に存在する窒素やリン酸といった栄養素の濃度が、誘導の間、栄養分が枯渇するタイミングに応じて、生育速度を制御するために使われる。本発明の1つの特殊な例では、生育速度を制御している栄養源がリン酸である。適切なリン酸の濃度が、ここでの例において提供され、過去の研究例として知られている(Pirt, S. J. (1975) Principles of Microbe and Cell Cultivation, Blackwell Scientific Publicationsを参照)。発酵開始時に存在する適切なリン酸濃度の例は、25から40 mMの範囲内である。それゆえに、本発明での1つの例では、工程 (b)における生育速度が、発酵開始時に細胞が利用できるリン酸のレベルを調整することによって、変化する。もう1つの例では、生育速度が、例えば供給を介して、または点添加を通して培養液に加えられた炭素または窒素といった栄養素の濃度を変えることによって、調整されるだろう。
【0020】
本発明でのもう1つの例では、工程 (b)における生育速度が、E. coli宿主細胞に利用可能な炭素のレベルの調整によって変化する。細胞が利用可能な炭素源のレベルは、生育速度の必要な変化を達成するために、増加させたり減少させたりしてもよい。好ましくは、生育速度が、培養液への炭素源の供給速度を調整することによって制御される。ここで用いている「炭素源」という用語は、生育の間、E. coliのためのエネルギー源として利用可能な炭素のソースについて、そして、グルコース、ラクトース、スクロース、フルクトースなどの糖類、そして、グリセロール、コハク酸、乳酸などの他の炭素のソースについて、言及している。本発明の好ましい実施例では、炭素源はグリセロールである。好ましくは、宿主細胞はフェドバッチ発酵器で培養され、そして炭素源は、必要な生育速度を与えるような適切な供給速度で提供されたグリセロールである。適切なグリセロール供給速度は、過去の研究例にてよく知られており、そしてこの明細書に提供された例に記されたそれも含んでいる。適切なグリセロール供給速度の例は、それの0.5から11 ml/hの範囲に含まれる。
【0021】
ここで用いている「分画」という用語は、E. coli宿主細胞のペリプラズムとE. coliが生育した培養上清の間での、組み換えタンパク質の分配について言及している。組み換えタンパク質の分画を決定するために、ペリプラズムと上清の収量を、方法の工程 (d)で決定する。収量は、E. coli宿主細胞の培養の全体にわたって適切な時間間隔で一部サンプルを採取する、例えばおよそ2から4時間毎、または代わりに収穫時、例えば誘導からおよそ24から36時間後に採取することによって、決定される。タンパク質の定量の多くの方法が、過去の研究において知られており、ゲル電気泳動、HPLC、クロマトグラフィー、そしてウェスタンブロットやELISAといった免疫検出法などの方法を含んでいる。例えば、ペリプラズム抽出物と培養上清中のFab’濃度が、Humphreys et al., 2002, Protein Expression and Purification, 26, 309-320に記載されたFab’会合ELISA法によって決定できる。
【0022】
本発明の方法では、工程 (b)において2つかそれ以上の異なった生育速度で生育させたE. coli宿主細胞のペリプラズムと、上清にある組み換えタンパク質の収量は、タンパク質生産物の最初の回収に最も適したような分画をもたらす生育速度を見出すために、方法の工程 (e)において比較される。好ましくは、工程 (e)において比較された収量が、それぞれの生育速度において誘導された後、おおよそ同じ時間に、工程 (d)において決定される。これは必ずしも可能であるとは限らないだろうことは、とりわけ、より低い生育速度で生育された培養物においてよりも、最大バイオマスにより早く到達するような、より高い生育速度で生育された培養物に関して、そう理解しておくべきだろう。
【0023】
方法の工程 (b)での生育速度の変化は、生育速度の増加または減少であろう。工程 (a)で提供された培養液は、栄養分が限られていないか、栄養分が限られた条件において、適切な生育速度で生育されるだろう。好ましくは、工程 (a)で提供された培養液は、栄養分が限られていない条件下で、その条件で可能な最大の特異的な生育速度で生育している。1つの例では、工程 (a)におけるE. coli宿主細胞の生育速度は、およそ0.1と0.25/hの間であり、好ましくは0.13と0.22/hの間である。好ましくは、生育速度は、方法の工程 (b)では、減少させる。生育速度は、0/hより大きければどの適切な速度にまで減少させても差し支えなく、好ましくは、0/hと工程 (a)にて条件としていた生育速度との間である。適切な生育速度は、少なくとも2つの異なった生育速度を試し、方法の工程 (e)においてそれらを比較することによって、経験的に同定される。それぞれの生育速度は、2つまたはそれ以上に分かれた、平行して、および、または連続して生育された培養について、試してもよい。1つの例では、少なくとも2つの培養を、工程 (b)において、それぞれの培養で異なった生育速度を用いて同時に生育させる。もう1つの例では、少なくとも2つの培養を、工程 (b)において、それぞれの培養で異なった生育速度を用いて連続して生育させる。もう1つの方法として、2つまたはそれ以上の異なった生育速度が、工程 (b)での1つの生育速度を使って工程 (a)から (d)を行い、それから工程 (b)での生育速度をそれぞれの時間において少なくとももう一度変化させて工程 (b)と (d)を行うことによって、単一の培養においてテストすることができる。もし方法の工程 (e)において適切な生育速度が同定されなかった場合には、それからさらに生育速度が、方法の工程 (a)から (d)を繰り返すことによってテストされるだろうことも、理解しておかなくてはならないだろう。典型的に、異なった生育速度の範囲は、0/hと工程 (a)での培養の生育速度との間でテストされるだろう。第一の例では、テストされた異なった生育速度の範囲が、高い、そして低い生育速度の幅広い範囲にわたっていて、それは、もし必要ならば、工程 (b)においてより狭い範囲の生育速度を用いて、方法の工程 (a)から (d)を繰り返すことによって、それ以降に狭められればよい。第一の例においてテストした適切な生育速度は、例えば、栄養分の限られていない条件下での生育速度よりも、およそ2とおよそ200倍の間くらい少ないものであるかもしれない。1つの例では、生育速度は、栄養分の限られていない条件下での生育速度よりも、およそ5とおよそ200倍の間くらい少ない。もう1つの方法として、第一の例においてテストされた、異なった生育速度の範囲は、例えば、高い、または低い生育速度の狭い範囲であり、それは、もし必要ならば、工程 (b)においてより広い範囲の生育速度を用いて、方法の工程 (a)から (d)を繰り返すことによって、それ以降に広げられればよい。第一の例においてテストされた適切な生育速度は、例えば、栄養分の限られていない条件下での生育速度よりも、およそ2とおよそ50倍の間くらい、またはおよそ100とおよそ200倍の間くらい少ないものであるかもしれない。1つの例では、生育速度は、栄養分の限られていない条件下での生育速度よりも、およそ5とおよそ20倍の間くらい少ない。工程 (b)における適切な生育速度は、0.0005から0.04/hの範囲内の生育速度を含むだろう。
【0024】
一旦、2つまたはそれ以上の異なった生育速度で、組み換えタンパク質の分画が比較されると、組み換えタンパク質の最初の回収に最も適した生育速度が、方法の工程 (f)において選択される。ここで使われている「最初の回収」という用語は、無細胞の上清または、次に続く精製の工程に適したペリプラズム抽出物を生産するのに必要な、初期の回収工程について言及している。最初の回収工程の例は、遠心分離、濾過、細胞の抽出または溶解、均質化、濃縮、ダイアフィルトレーション、そして吸着流動床を含んでいる。タンパク質の最初の回収に最も適した生育速度は、最も良い収量、および/または1つまたはそれ以上の最初の回収の工程の後のタンパク質の「質」をもたらすような生育速度だろう。ここで使われている「質」という用語は、タンパク質が無傷、および/または、正確に折りたたまれているかどうか、得られたタンパク質の完全性について言及している。重要なことに、最も適切な生育速度は、E. coli宿主細胞培養液内に最大の収量の組み換えタンパク質をもたらす生育速度ではなく、その代わりに、1つまたはそれ以上の最初の回収工程の後で、より良い質のタンパク質、および/またはより高い収量をもたらすであろう、ペリプラズムと上清の間でのタンパク質の最も良い分画をもたらすような生育速度であるかもしれない。それゆえに、工程 (f)で選択した生育速度は、培養の間のタンパク質収量と、最終の収量と最終的に回収されるタンパク質の質との間で、代わる代わるにされるかもしれない。
【0025】
1つの例では、E. coli宿主細胞のペリプラズムに発現させた組み換えタンパク質は、上清に発現させたタンパク質よりも、ラージスケールにおいては、取り扱いが簡単であるかもしれない。第一に必要な、最初の回収は、遠心分離か濾過のいずれかによる、培養培地からの宿主細胞の回収である。ペリプラズムに生産されたタンパク質にとって重要な考察は、上清へと組み換えタンパク質が漏洩するレベルであり、これは最初の回収やペリプラズムのタンパク質の質に不利な効果を持ちうる。上清に存在するタンパク質の増加は、上清の粘度を増加させ、遠心分離または濾過による細胞の回収をより困難にし、そしてペリプラズムのタンパク質収量を減らす。細胞それ自身もまた、遠心分離の間はそれほど丈夫ではなく、組み換えタンパク質へのダメージをもたらすような溶解をより起こしやすく、再びペリプラズム内の組み換えタンパク質の収量を減らすような、タンパク質、DNA、そして脂質の放出のために、上清の粘度はさらに増加する。ペリプラズムから回収された組み換えタンパク質の質はまた、部分的に分解した、または正確に折りたたまれていないような、上清由来の組み換えタンパク質で汚染されることによって、低下する。これらの培養においては、それゆえ、最初の回収の後の組み換えタンパク質の収量、および/または質を上げるために、上清における組み換えタンパク質のレベルを低く保つことが重要である。この例では、後に続く最初の回収で得られた組み換えタンパク質の最終収量、および/または質が、たとえ、ペリプラズムにおける組み換えタンパク質の収量が、上清における組み換えタンパク質のレベルがより高くなるような他の生育速度のときよりも少なくなる場合でも、上清における組み換えタンパク質の量が少なくなるような生育速度を選択することによって改善されるかもしれない。それゆえ、本発明の方法においてペリプラズムに生産されたタンパク質にとって、工程 (f)で選択される生育速度は、タンパク質の55%以上がペリプラズムで発現し、タンパク質の45%以下が上清にあるような生育速度を選ぶのが好ましい。好ましくは、宿主細胞によって生産されたタンパク質の80%以上がペリプラズムに発現し、そして20%以下が上清にあるのが良い。1つの例では、宿主細胞によって生産されたタンパク質の90%以上がペリプラズムに発現し、10%以下が上清に存在している。もう1つの例では、宿主細胞によって生産されたタンパク質の95%以上がペリプラズムに発現し、5%以下が上清に存在している。
【0026】
もう1つの例では、タンパク質が上清に生産される。タンパク質は、もし抽出の工程が必要でなく、培地中に存在する宿主タンパク質はより少ないだろうならば、精製するのはより簡単であるかもしれない。タンパク質の質もまた、タンパク質の分解がより少なく、タンパク質の折りたたみが改善されているようならば、より良いかもしれない。上清に生産されたタンパク質にとって、宿主細胞によって発現され、上清へと分泌された組み換えタンパク質の量が最大化されることが重要である。宿主細胞によって生産された全ての組み換えタンパク質が、上清へと分泌されているかどうかを決定するためには、同様にペリプラズム内の組み換えタンパク質のレベルを測定することが重要である。もし、これらが依然高いのであれば、それでもさらに上清にある収量を増加させるために、組み換えタンパク質の分画を調整することは、本発明の方法を用いて可能になるかもしれない。結果として、本発明の方法において上清に生産されたタンパク質にとっては、工程 (f)で選択される生育速度は、タンパク質の55%以上が上清に発現され、タンパク質の45%以下がペリプラズムにあるような生育速度が望ましい。好ましくは、宿主細胞によって生産される組み換えタンパク質の80%以上が上清に生産され、ペリプラズムには20%以下のタンパク質が存在する。1つの例では、宿主細胞によって生産されたタンパク質の90%以上が上清に存在し、10%以下がペリプラズムに存在する。もう1つの例では、宿主細胞によって生産されたタンパク質の95%以上が上清に存在し、5%以下がペリプラズムに存在する。
【0027】
方法の工程 (f)において適切な生育速度が同定された場合、上清とペリプラズムの間での組み換えタンパク質の望ましい分画が達成されることを確認するために、同じ生育速度が、次に続くE. coli宿主細胞の発酵の間に使われうる。本発明はまた、それゆえ、当該細胞による組み換えタンパク質の発現が、誘導可能なシステムや、組み換えタンパク質の最初の回収に最も適した、上述した方法の工程 (f)において選択された生育速度でE. coli宿主細胞を生育することから成る、当該方法によってなされる上清とペリプラズムとの間の組み換えタンパク質の分画の制御下に置かれているような、E. coli宿主細胞培養における、組み換えタンパク質生産の方法を提供する。これらの後続の発酵は、上記の方法において使われたのと同じスケールか、異なったスケールであっても差し支えない。例えば、E. coli宿主細胞が、タンパク質の最初の回収に最も適した生育速度を決定するために、小さいスケールで培養され、それから、その後の発酵では、宿主細胞は、工程 (f)で選択された生育速度を用いて、より大量の組み換えタンパク質を生産するために、大きいスケールで培養されてもよい。
【0028】
本発明の方法によって生産された組み換えタンパク質は、最初の回収の直後に精製されるだろう。ここで使われる最初の回収の工程は、発酵の終了時における組み換えタンパク質の細胞内での位置に依存するだろう。例えば、ペリプラズムで発現したタンパク質は、遠心分離または濾過によって、宿主細胞内に回収されるかもしれない。タンパク質はそれから、熱処理(US 5,665,866の例を参照)または機械的な抽出といった方法を用いて、細胞内から抽出することができる。上清に発現したタンパク質は、吸着流動床、遠心分離、または濾過など細胞を除去するための方法に続いて、例えば、イオン交換、または捕捉タンパク質に対するアフィニティークロマトグラフィーといった方法を用いて回収してもよい。ペリプラズム抽出物か上清のいずれか由来のタンパク質は、その後、組み換えタンパク質の性質や、大きさ、疎水性、そして等電点といったタンパク質の物理的特性に依存して、過去の研究例に知られた方法のいずれかを用いて精製してもよい。そのような方法には、サイズ排除クロマトグラフィー、疎水性相互作用クロマトグラフィー、イオン交換、アフィニティークロマトグラフィー、そして逆相HPLCを含んでいる。従って、本発明のもう1つの様相では、当該細胞による組み換えタンパク質の発現が、誘導可能なシステムや、組み換えタンパク質の最初の回収に最も適した、上述した方法の工程 (f)において選択された生育速度でE. coli宿主細胞を生育することと、その後の組み換えタンパク質の精製から成る、当該方法によってなされる上清とペリプラズムとの間の組み換えタンパク質の分画の制御下に置かれているような、E. coli宿主細胞培養における、組み換えタンパク質生産のための方法が提供される。
【実施例1】
【0029】
誘導後の生育を制御するためにグリセロール供給の制限を用いる、Fab’ Aの分画と収量の最適化。
【0030】
材料と方法
菌株とプラスミド
この例で使われた菌株は、テトラサイクリン耐性を与え、Fab’断片であるFab’ Aの軽鎖と重鎖ポリペプチド構成成分をコードする遺伝子を持っているプラスミドを形質転換した、Escherichia coli W3110 (ATCC 27325)であった。それぞれのポリペプチドは、E. coliのOmp Aリーダーペプチドによって先行されている。単一のtacプロモーターからの発現の誘導は、ポリペプチドの一部が折りたたまれ、Fab'を形成するようなペリプラズムへと、軽鎖と重鎖ポリペプチドの分泌と合成をもたらす。
【0031】
生育培地
発酵生育培地は、3.86 g/lのリン酸二水素ナトリウム一水和物と112 g/lのグリセロールを加えた、SM6E培地(Humphreys et al., 2002, Protein Expression and Purification, 26, 309-320に記載)に基づいたものであった。
【0032】
接種材料
接種材料の培養は、10 μg/mlのテトラサイクリンを補って、同じ培地で生育させた。培養は、およそ22時間の撹拌とともに、30℃にて培養された。
【0033】
発酵
発酵器(全容量2.5リットル)は、接種材料の培養液を用いて、0.3-0.5 OD600になるまで植菌された。温度は、増殖期の間は30℃に保たれ、そして誘導の前に25℃に下げられた。溶存酸素濃度は、変化可能な撹拌と空気流によって、30%空気飽和より高く保たれた。培養pHは、15% (v/v)アンモニア水と10% (v/v)濃硫酸を用いた自動滴定によって、7.0に制御された。発泡は、10% (v/v) Struktol J673溶液(Schill and Seilacher)の添加によって制御された。
【0034】
いくらかの添加が、発酵の異なった工程においてなされた。バイオマス濃度が、おおよそ40 OD600に達した時、マグネシウム塩とリン酸二水素ナトリウム一水和物が加えられた。リン酸二水素ナトリウム一水和物のさらなる添加が、リン酸が過剰に保たれているかを確認するために、誘導期の前と誘導期の間になされた。発酵の初期に存在したグリセロールが枯渇した時(おおよそ75 OD600)には、80% (w/w)グリセロールの継続的供給が、0.5から10.9 ml/hにわたる速度で適用された。発酵中の同じポイントで、IPTGの供給が、この時点での発酵器内のIPTGの最終濃度が0.5 mMになるように、1 ml/hの速度で36時間にわたって、適用された。IPTG供給の開始が、誘導の開始として解釈された。発酵は、典型的に、70-73時間の間はより低いグリセロール供給速度で(0.5-2.5 ml/h)、そして50-60時間の間はより高いグリセロール供給速度で(5.4-10.9 ml/h)、行われた。
【0035】
バイオマス濃度と生育速度の測定
バイオマス濃度は、培養液の光学密度を600 nmで測定することによって、決定された。生育速度(μ)は、時間を通してのバイオマス濃度の変化に、このようにして関連している:
LnXt = LnX0 + μt
ここで、X0は元々のバイオマス濃度であり、Xtは時間間隔t後のバイオマス濃度である。従って、バッチ培養に関してtに対するLnXtのプロットは、μ(単位/h)と同等な傾斜の直線を与える。しかしながら、バッチ培養に対するグリセロールの直線限定的な供給の適用は、バイオマスと培養液量の増加のために、生育速度の低下をもたらす。それゆえ、誘導の時期の間の平均生育速度が、時間に対してLnOD600をプロットすることによって決定された。(最小2乗法による)適合度の線の傾斜は、供給時期の間を通しての平均生育速度を与えることによって決定された。
【0036】
ペリプラズムの抽出
細胞は、遠心分離によって培養サンプルから回収された。上清画分は、さらなる分析のために(-20℃で)保持された。細胞の塊の画分は、抽出バッファー(100 mM Tris-HCl, 10 mM EDTA; pH 7.4)中、元々の培養液の量にまで再懸濁された。続いておおよそ16時間、60℃で温めた後、抽出物は遠心分離によって浄化され、そして上清画分は、分析のために(-20℃で)保持された。
【0037】
Fab'の定量
ペリプラズム抽出物と培養上清中のFab’濃度は、Humphreys et al., 2002, Protein, Expression and Purification, 26, 309-320に記載されている、Fab’会合ELISAによって決定された。
【0038】
結果
Fab’ A発酵は、生育が、誘導前は無制限で、最大の特異的速度(μmax)が生ずるような生育で、しかしそれに続く誘導後には制限されたような条件下で行われた(図1)。誘導後の生育速度は、ある範囲の流速でバッチ培養に、栄養(グリセロール)を限定した供給を適用することによって制御された。グリセロールの流速は、おおよそ0/hから上向きに生育速度の範囲を与えるように選択された。図2と表1は、グリセロールの供給速度の範囲に対して、誘導時期の間を通しての実際の平均生育速度を示している。グリセロール供給速度の上昇は、予定通り、培養の平均生育速度において増加をもたらした。
【0039】
【表1】
【0040】
発酵は、より低いグリセロール供給速度(0.5-2.5 ml/h)では誘導後おおよそ36時間で、より高いグリセロール供給速度(5.4-10.9 ml/h)では誘導後おおよそ24時間で収穫され、そしてペリプラズム抽出物と培養上清のFab’の濃度は、Fab’会合ELISAによって決定された。図3は、Fab'の濃度と、ペリプラズムと培養培地との間での相対的分配を示している。Fab’滴定濃度に対する最適な生育速度は、おおよそ0.0075/hであることが判明した。生育速度を上げると、培養上清におけるFab’の滴定濃度が上昇するという、一般的な傾向が観測された。従って、最適な発酵能力のために選択される生育速度は、Fab’収量と、最初の回収での両立性が、最大の収量とFab'の最適な分配との間のバランスに反映するだろう。引用例では、1.6 ml/hのグリセロール供給速度(生育速度〜0.0071/h)が、発酵のスケールアップのために選択されていた。
【実施例2】
【0041】
誘導後の生育速度を制御するためのリン酸濃度の変化によるFab’ Bの収量と分画の最適化。
【0042】
材料と方法
菌株とプラスミド
この実施例で使われた菌株は、テトラサイクリンに対する耐性を与え、Fab'断片であるFab’ Bの、軽鎖と重鎖ポリペプチド構成要素をコードしている遺伝子を持っているプラスミドを形質転換したEscherichia coli W3110 (ATCC 27325)であった。単一のtacプロモーターからの発現の誘導は、ポリペプチドの一部が折りたたまれ、Fab’を形成するために会合するようなペリプラズムへの、軽鎖と重鎖ポリペプチドの分泌と、その合成をもたらす。
【0043】
生育培地
発酵生育培地は、表2に示された濃度でリン酸二水素ナトリウム一水和物を加えたSM6E培地(Humphreys et al., 2002, Protein, Expression and Purification, 26, 309-320に記載)に基づいていた。
【0044】
【表2】
【0045】
接種材料
接種材料の培養は、リン酸二水素ナトリウム一水和物の適切な濃度(表2参照)とともに、そして10 μg/mlのテトラサイクリンを補って、同じ培地で生育させた。培養は、およそ19から26時間の撹拌とともに、30℃にて培養された。
【0046】
発酵
発酵器(全容量2.5リットル)は、接種材料の培養液を用いて、0.3から0.5 OD600になるまで植菌された。温度は、増殖期の間は30℃に保たれ、そして誘導の前に27℃に下げられた。溶存酸素濃度は、変化可能な撹拌と空気流によって、30%空気飽和より高く保たれた。培養pHは、15% (v/v)アンモニア水と10% (v/v)濃硫酸を用いた自動滴定によって、7.0に制御された。発泡は、10% (v/v) Mazu溶液の添加によって制御された。
【0047】
いくらかの添加が、発酵の異なった工程においてなされた。グリセロールは、80% (w/w)溶液の45 mlを2回に分けて、最初の添加は20 OD600のバイオマス濃度で、他は40 OD600で発酵に添加された。バイオマス濃度がおおよそ40 OD600に達した時、マグネシウムとカルシウム塩もまた添加された。ラクトース(50% (w/w)溶液の60 ml)が60 OD600で添加され、誘導物質そして誘導工程の間の炭素源として供給された。誘導は、溶存酸素濃度の上昇により注目づけされたグリセロールの枯渇(およそ75 OD600と90 OD600の間)に起こった。ラクトースのさらなる添加が、0と55 g/lの間に発酵器内の濃度を保持するために必要とされて、なされた。マグネシウムのさらなる添加が、誘導後、0と2時間の間になされた。発酵は、典型的に56から61時間行われた。
【0048】
発酵の分析
ペリプラズムからの抽出、バイオマスの測定、そしてFab’の定量が、前述の通りに行われた(実施例1)。
【0049】
結果
発酵培養誘導後の生育は、すべての他の開始時の栄養素を一定にした一方で、リン酸(リン酸二水素ナトリウム一水和物)の量を変化させることによって制御された。それぞれの発酵の間の生育速度は、バイオマスが増加した誘導の時期に対して決定された(図5と表3)。
【0050】
【表3】
【0051】
リン酸濃度の上昇は、誘導の期間を通した生育速度の上昇をもたらした。リン酸濃度は、誘導に関連して、様々なポイントで枯渇するように選択された(例えば、26.9 mMの場合、誘導前にはかすかに、また誘導後の様々なポイントで)。誘導が起こるポイント(バイオマス濃度)は、開始時のグリセロール濃度に関係している。望みの誘導バイオマスまでの生育を支持するために必要なリン酸の量は、Pirt, S. J. (1975) Principles of Microbe and Cell Cultivation, Blackwell Scientific Publicationsに規定されている、無機リン酸のためのバイオマス収量係数データに基づいて計算された。Fab'の収量と分配にとって最適なリン酸濃度は、ここに記載されたように経験的に決定された。
【0052】
発酵は、誘導後28から31時間で収穫され、ペリプラズム抽出物と培養上清のFab’濃度はFab'会合ELISAによって決定された。図6は、収穫時でのペリプラズムと培養上清の両方のFab’濃度を示している。培養上清のFab’濃度は、リン酸濃度とともに上昇するという、一般的な傾向が観測された。ペリプラズム抽出と遠心分離または濾過を用いた、大きいスケールの最初の回収のための、リン酸の最適な値域は、29.8 mMであった。
【図面の簡単な説明】
【0053】
本発明は、以下の参考図を示す目的で記述されるだろう:
【図1】グリセロール供給速度の変動におけるE. coliの生育プロファイル
【図2】グリセロール供給速度の変動におけるE. coliの誘導後の生育速度
【図3】グリセロール供給速度の変動において生育されたE. coli培養の、ペリプラズムと上清との間でのFab’ Aの分画
【図4】無機リン酸濃度の変動におけるE. coliの生育プロファイル
【図5】無機リン酸濃度の変動におけるE. coliの誘導後の生育速度
【図6】無機リン酸濃度の変動において生育されたE. coli培養の、ペリプラズムと上清との間でのFab’ Bの分画
【特許請求の範囲】
【請求項1】
1つの誘導システムの下で制御されるE. coli宿主細胞による組み換えタンパク質の発現を行う培養において、上清とペリプラズムの間に組み換えタンパク質の分画を制御するための、以下の工程から構成される方法:
a)E. coli宿主細胞培養液を提供する工程
b)E. coli宿主細胞の生育速度を変化させる工程
c)組み換えタンパク質の発現を誘導する工程
この点で、工程 (b)と (c)は、どの順序でも、あるいは同時に、行うことが可能である;そして続いて、
d)培養上清とE. coli宿主細胞ペリプラズムにおける組み換えタンパク質の収量を決定する工程
e)工程 (d)で決定した収量と、工程 (b)で用いられた生育速度と少なくとも1つの他の場合において決定された収量とを比較する工程
f)上清とペリプラズムの間での組み換えタンパク質の分画が、組み換えタンパク質の最初の回収に最も適しているような、工程 (e)でなされた比較の結果得られた、生育速度を選択する工程。
【請求項2】
工程 (e)で比較された収量が、工程 (b)のそれぞれの培養において異なった生育速度を用いて、同時に生育させた少なくとも2つの培養からなる、請求項1に記載の方法。
【請求項3】
工程 (e)で比較された収量が、工程 (b)のそれぞれの培養において異なった生育速度を用いて、連続的に生育させた少なくとも2つの培養からなる、請求項1に記載の方法。
【請求項4】
工程 (e)で比較された収量が、工程 (b)の1つの生育速度を用いて工程 (a)から (d)を行う、それから工程 (b)のそれぞれの時間で少なくとも1回以上生育速度を変化させて工程 (b)と (d)を行うことによって、少なくとも2つの異なった生育速度で生育させた1つの培養からなる、請求項1に記載の方法。
【請求項5】
工程 (b)において生育速度が減少される請求項1から4のいずれかに記載の方法。
【請求項6】
工程 (b)においてE. coli宿主細胞の生育速度を変化させることが、細胞にとって利用可能な炭素のレベルを調整することから成る、請求項1から5のいずれかに記載の方法。
【請求項7】
炭素源が、グルコース、ラクトース、スクロース、フルクトース、グリセロール、コハク酸、そして乳酸から選択される、請求項6に記載の方法。
【請求項8】
炭素源がグリセロールである請求項7に記載の方法。
【請求項9】
工程 (b)においてE. coli宿主細胞の生育速度を変化させることが、細胞にとって利用可能なリン酸のレベルを調整することから成る、請求項1から8のいずれかに記載の方法。
【請求項10】
工程 (b)においてE. coli宿主細胞の生育速度を変化させることが、細胞にとって利用可能な酸素のレベルを調整することから成る、請求項1から9のいずれかに記載の方法。
【請求項11】
組み換えタンパク質がペリプラズムへと目標づけられる、請求項1から10のいずれかに記載の方法。
【請求項12】
工程 (f)で選択された生育速度が、宿主細胞によって生産された組み換えタンパク質の80%以上がペリプラズムに発現するような生育速度である、請求項11に記載の方法。
【請求項13】
組み換えタンパク質が上清へと目標づけられる、請求項1から10のいずれかに記載の方法。
【請求項14】
工程 (f)で選択された生育速度が、宿主細胞によって生産された組み換えタンパク質の80%以上が上清に生産されるような生育速度である、請求項13の方法。
【請求項15】
誘導可能なシステムが、lac由来のプロモーターから成る、請求項1から14のいずれかに記載の方法。
【請求項16】
lac由来のプロモーターが、lac、tac、またはtrcである、請求項15の方法。
【請求項17】
プロモーターが、ラクトースまたはIPTGで誘導される、請求項15または16のいずれかに記載の方法。
【請求項18】
組み換えタンパク質が、抗体またはその断片である、請求項1から17のいずれかに記載の方法。
【請求項19】
抗体がIgGである請求項18の抗体。
【請求項20】
断片が、Fab、Fab’、F(ab')2、またはscFvである、請求項18の抗体断片。
【請求項21】
当該細胞による組み換えタンパク質の発現が、誘導可能なシステムと、上清と組み換えタンパク質の最初の回収に最も適したペリプラズムとの間での組み換えタンパク質の分配が、請求項1の工程 (f)で選択された生育速度でE. coli宿主細胞を生育させることから成る上述の方法の制御下に置かれている、E. coli宿主細胞培養内で組み換えタンパク質を生産する方法。
【請求項22】
生産された組み換えタンパク質が、その後精製される、請求項21の方法。
【請求項1】
1つの誘導システムの下で制御されるE. coli宿主細胞による組み換えタンパク質の発現を行う培養において、上清とペリプラズムの間に組み換えタンパク質の分画を制御するための、以下の工程から構成される方法:
a)E. coli宿主細胞培養液を提供する工程
b)E. coli宿主細胞の生育速度を変化させる工程
c)組み換えタンパク質の発現を誘導する工程
この点で、工程 (b)と (c)は、どの順序でも、あるいは同時に、行うことが可能である;そして続いて、
d)培養上清とE. coli宿主細胞ペリプラズムにおける組み換えタンパク質の収量を決定する工程
e)工程 (d)で決定した収量と、工程 (b)で用いられた生育速度と少なくとも1つの他の場合において決定された収量とを比較する工程
f)上清とペリプラズムの間での組み換えタンパク質の分画が、組み換えタンパク質の最初の回収に最も適しているような、工程 (e)でなされた比較の結果得られた、生育速度を選択する工程。
【請求項2】
工程 (e)で比較された収量が、工程 (b)のそれぞれの培養において異なった生育速度を用いて、同時に生育させた少なくとも2つの培養からなる、請求項1に記載の方法。
【請求項3】
工程 (e)で比較された収量が、工程 (b)のそれぞれの培養において異なった生育速度を用いて、連続的に生育させた少なくとも2つの培養からなる、請求項1に記載の方法。
【請求項4】
工程 (e)で比較された収量が、工程 (b)の1つの生育速度を用いて工程 (a)から (d)を行う、それから工程 (b)のそれぞれの時間で少なくとも1回以上生育速度を変化させて工程 (b)と (d)を行うことによって、少なくとも2つの異なった生育速度で生育させた1つの培養からなる、請求項1に記載の方法。
【請求項5】
工程 (b)において生育速度が減少される請求項1から4のいずれかに記載の方法。
【請求項6】
工程 (b)においてE. coli宿主細胞の生育速度を変化させることが、細胞にとって利用可能な炭素のレベルを調整することから成る、請求項1から5のいずれかに記載の方法。
【請求項7】
炭素源が、グルコース、ラクトース、スクロース、フルクトース、グリセロール、コハク酸、そして乳酸から選択される、請求項6に記載の方法。
【請求項8】
炭素源がグリセロールである請求項7に記載の方法。
【請求項9】
工程 (b)においてE. coli宿主細胞の生育速度を変化させることが、細胞にとって利用可能なリン酸のレベルを調整することから成る、請求項1から8のいずれかに記載の方法。
【請求項10】
工程 (b)においてE. coli宿主細胞の生育速度を変化させることが、細胞にとって利用可能な酸素のレベルを調整することから成る、請求項1から9のいずれかに記載の方法。
【請求項11】
組み換えタンパク質がペリプラズムへと目標づけられる、請求項1から10のいずれかに記載の方法。
【請求項12】
工程 (f)で選択された生育速度が、宿主細胞によって生産された組み換えタンパク質の80%以上がペリプラズムに発現するような生育速度である、請求項11に記載の方法。
【請求項13】
組み換えタンパク質が上清へと目標づけられる、請求項1から10のいずれかに記載の方法。
【請求項14】
工程 (f)で選択された生育速度が、宿主細胞によって生産された組み換えタンパク質の80%以上が上清に生産されるような生育速度である、請求項13の方法。
【請求項15】
誘導可能なシステムが、lac由来のプロモーターから成る、請求項1から14のいずれかに記載の方法。
【請求項16】
lac由来のプロモーターが、lac、tac、またはtrcである、請求項15の方法。
【請求項17】
プロモーターが、ラクトースまたはIPTGで誘導される、請求項15または16のいずれかに記載の方法。
【請求項18】
組み換えタンパク質が、抗体またはその断片である、請求項1から17のいずれかに記載の方法。
【請求項19】
抗体がIgGである請求項18の抗体。
【請求項20】
断片が、Fab、Fab’、F(ab')2、またはscFvである、請求項18の抗体断片。
【請求項21】
当該細胞による組み換えタンパク質の発現が、誘導可能なシステムと、上清と組み換えタンパク質の最初の回収に最も適したペリプラズムとの間での組み換えタンパク質の分配が、請求項1の工程 (f)で選択された生育速度でE. coli宿主細胞を生育させることから成る上述の方法の制御下に置かれている、E. coli宿主細胞培養内で組み換えタンパク質を生産する方法。
【請求項22】
生産された組み換えタンパク質が、その後精製される、請求項21の方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図2】
【図3】
【図4】
【図5】
【図6】
【公表番号】特表2007−504815(P2007−504815A)
【公表日】平成19年3月8日(2007.3.8)
【国際特許分類】
【出願番号】特願2006−525876(P2006−525876)
【出願日】平成16年9月2日(2004.9.2)
【国際出願番号】PCT/GB2004/003747
【国際公開番号】WO2005/024000
【国際公開日】平成17年3月17日(2005.3.17)
【出願人】(501460693)セルテック アール アンド ディ リミテッド (29)
【Fターム(参考)】
【公表日】平成19年3月8日(2007.3.8)
【国際特許分類】
【出願日】平成16年9月2日(2004.9.2)
【国際出願番号】PCT/GB2004/003747
【国際公開番号】WO2005/024000
【国際公開日】平成17年3月17日(2005.3.17)
【出願人】(501460693)セルテック アール アンド ディ リミテッド (29)
【Fターム(参考)】
[ Back to top ]