
組み換え三重足場ベースポリペプチド
【課題】高い親和性、低い細胞分裂促進作用、および高いインビボ安定性を備えるタンパク質複合体ベースの結合試薬の提供。
【解決手段】足場ドメインおよび異種ドメインをそれぞれ有する融合ポリペプチド鎖を形成する三重らせんコイルを含むタンパク質複合体を開示する。また、関連する単離融合ポリペプチド、核酸、ベクター、宿主細胞、および製造方法も開示する。
【解決手段】足場ドメインおよび異種ドメインをそれぞれ有する融合ポリペプチド鎖を形成する三重らせんコイルを含むタンパク質複合体を開示する。また、関連する単離融合ポリペプチド、核酸、ベクター、宿主細胞、および製造方法も開示する。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願
本出願は、2005年12月15日に出願された米国特許出願番号60/750,746の優先権を主張するものであり、その内容はここに参照することにより組み込まれる。
【0002】
本発明は、高い親和性、低い細胞分裂促進作用、および高いインビボ安定性を備えるタンパク質複合体ベースの結合試薬に関する。該試薬は多価(multi-valent)あるいは多重特異性(multi-specific)にもなり得る。
【背景技術】
【0003】
タンパク質ベースの結合試薬は治療または診断の適用において様々な用途がある。抗体がこのような試薬の優れたパラダイムであることは証明されている。実際、多くのモノクローナル抗体(mAb)が、癌、感染病、および炎症性疾患の治療のために首尾よく使用されている(Adamsら、Nat. Biotechnol.2005 Sep;23(9):1147-57)。
【0004】
マウスmAbの有効性および安全性はいずれもキメラ化およびヒト化などの組み換えDNA技術によって高められた。しかし、CDR(相補決定領域)グラフティングを用いたマウスmAbのヒト化は、マウス対応物よりも結合親和性が低くなることが多い。抗体親和性は、治療薬としての抗体の成功の鍵となる要因である。高親和性を有する抗体は、標的レセプターに対し天然リガンドと有効に競合できるものとなり、投与量、毒性、およびコストを低減させる。親和性はまた、抗体の薬物動態、例えばその標的組織および宿主の血液循環内における分布と排出にも影響する。インビボで有効なモノクローナル抗体に要求されるのは、無関係のタンパク質に非特異的に結合することのない、標的抗原に対する強い親和性である。抗原結合部位の多量体化は、抗体親和性(機能的親和性(functional affinity))として定義される、抗原への抗体の全体的な結合の強さを向上させる効果的な手段と見られている(Millerら、J Immunol 170:4854-4861、2003; Rheinneckerら、J Immunol 157:2989-2997、1996; Shopes、J Immunol 148:2918-2922、1992; Shufordら、Science 252:724-727、1991; Wolffら、J Immunol 148:2469-2474、1992))。それらはインビボにおける抗腫瘍活性を向上させる(Liu ら、Int Immunopharmacol 6:791-799、2006; Wolffら、Cancer Res 53:2560-2565、1993)。免疫グロブリンG(IgG)の二価の性質により、従来および人工のIgGは、2つを超える異なった抗原に同時に結合させるために用いることはできない。よって、多価または多重特異性のタンパク質ベース結合試薬が必要とされる。
【0005】
場合によって、Fc領域を操作することを通し、例えば抗体依存性細胞介在性障害(ADCC)および補体依存性細胞障害(CDC)のようなエフェクター作用を回避することは、細胞分裂促進性の副作用を低減するために必要である。例として、マウスの抗ヒトCD3 mAb(オルソクローンOKT3、ムロモナブ−CD3)は、ヒトT細胞上のT細胞レセプター(TCR)/CD3複合体を標的にする強力な免疫抑制剤である。過去20年の間、それは同種移植の拒否反応を防ぐまたは治療するために用いられてきた(Cosimiら、N Engl J Med 305:308-314、1981; Group、N Engl J Med 313:337-342、1985; Kungら、Science 206:347-349、1979)。しかし、この治療を用いることの主な欠点の1つは、インフルエンザに似た症状、呼吸困難、神経学的症状および急性尿細管壊死を含む一連の有害な細胞分裂促進作用をもたらすTNF-α、IL-2およびIFN-γのようなサイトカインの全身的な放出である(Abramowiczら、Transplantation 47:606-608、1989; Chatenoudら、N Engl J Med 320:1420-1421、1989; Goldmanら、Transplantation 50:158-159、1990; Toussaintら、Transplantation 48:524-526、1989)。OKT3およびその他の抗CD3 mAbの細胞分裂促進活性は、FcR陽性細胞(例えば単球)への結合を介しての広範なTCR/CD3架橋によるため、最近は、FcRへの結合を修正することによる非細胞分裂促進型の抗CD3抗体の開発に努力が注がれている。したがって、高い親和性、低い細胞分裂促進作用、および高いインビボ安定性を備えるタンパク質ベースの結合試薬が必要とされる。
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明の目的は、高い親和性、低い細胞分裂促進作用、および高いインビボ安定性を備えるタンパク質複合体ベースの結合試薬を提供することにある。該試薬は多価あるいは多重特異性とすることもできる。
【課題を解決するための手段】
【0007】
したがって、本発明の1態様は、第1の足場ドメイン(scaffold domain)および第1の足場ドメインの1つの末端にインフレームで(in-frame)融合する第1の異種ドメイン(heterologous domain)を含む第1の融合ポリペプチド鎖と、第2の足場ドメインを含む第2の融合ポリペプチド鎖と、第3の足場ドメインを含む第3の融合ポリペプチド鎖とを含んでいる単離組み換えタンパク質複合体に特徴を有する。第1、第2および第3の足場ドメインは、三重らせんコイル(triple helix coil)を形成するように並ぶ。第1の足場ドメインおよび第1の異種ドメインはインフレームで融合し、かつ同一のペプチド鎖に存する。
【0008】
異種ドメインには酵素ドメインまたは蛍光タンパク質の配列が含まれ得る。蛍光タンパク質の例としては、GFPおよびdsRed、さらにそれらの変異体などがある。酵素ドメインの例には、グルタチオンS-トランスフェラーゼ、ルシフェラーゼ、β-ガラクトシダーゼ、およびβ-ラクタマーゼのドメインが含まれる。
【0009】
異種ドメインには、結合パートナーと結合する結合ドメイン(例えばリガンド結合ドメイン、リガンド、レセプター、またはプロテオグリカン)が含まれ得る。「結合パートナー」とは、目的とする所望の化合物(例えばタンパク質)の部分に対して特異的、共有結合的または非共有結合的な親和性を備える任意の分子を意味する。結合パートナーの例としては、抗原/抗体のペア、タンパク質/阻害剤のペア、レセプター/リガンドのペア(例えば細胞表面または核レセプター/リガンドのペア)、酵素/基質のペア(例えばキナーゼ/基質のペア)、レクチン/炭水化物のペア、オリゴマーまたはへテロオリゴマータンパク質のペア、DNA結合タンパク質/DNA結合部位のペア、およびRNA/タンパク質のペアがある。また、結合ドメインの例には、例えばヒスチジンタグ、mycタグ、またはヘマグルチン(hemagglutin)タグのような親和性タグの配列も含まれる。
【0010】
上述した第1の異種ドメインには、免疫グロブリンの1つまたはそれ以上の相補決定領域(CDR)が含まれてもよい。よって、異種ドメインには、例えばVHドメインおよびFabのような抗体の抗原結合部位が含まれ得る。1つの実施形態において、第1の異種ドメインには、抗原結合性フラグメントまたは一本鎖抗体、例えば分化抗原3(CD3)または上皮増殖因子レセプター(EGFR)に特異なものの配列が含まれる。第1のポリペプチド鎖はさらに、第1の足場ドメインのもう1つの末端にインフレームで融合する第2の異種ドメインを含んでいてもよい。第1の異種ドメインのように、第2の異種ドメインにも、結合パートナーと結合するドメインが含まれ得る。第1および第2の異種ドメインは互いに同一であってもまたは異なっていてもよい。これらは、同一の結合パートナーまたは2つの異なる結合パートナーに結合することができる。例えば、第1の異種ドメインおよび第2の異種ドメインには、CD3およびEGFRにそれぞれ特異的に結合する第1の一本鎖抗体および第2の一本鎖抗体の配列が各々含まれていてもよい。
【0011】
上述したタンパク質複合体において、第2の融合ポリペプチド鎖は、第2の足場ドメインの1つの末端にインフレームで融合する第3の異種ドメイン、第2の足場ドメインのもう1つの末端にインフレームで融合する第4の異種ドメイン、または2つの末端にインフレームで融合する両方のドメインを含んでいてもよい。同様に、第3の融合ポリペプチド鎖は、第3の足場ドメインの1つの末端にインフレームで融合する第5の異種ドメイン、または、第3の足場ドメインのもう1つの末端にインフレームで融合する第6の異種ドメイン、または、両方を含んでいてもよい。上述した第1および第2の異種ドメインのように、これら4つの異種ドメインの各々には、結合パートナーと結合する結合ドメインも含まれ得る。6つ全ての異種ドメインは互いに同一であってもよくまたは異なってもよい。よってこれらは、1、2、3、4、5または6つの結合パートナーと結合できる。言い換えると、当該タンパク質複合体は、一、二、三、四、五、または六価であり得る。
【0012】
第1、第2、および第3の足場ドメインが三重らせんコイルを形成するよう、3つの足場ドメインのそれぞれは1つまたはそれ以上の三重らせんリピートを含んでおり、各リピートは次の式:(X1−X2−X3)nの配列を含み、式中、X1はGly残基であり、X2およびX3は任意のアミノ酸残基、好ましくはイミノ酸のプロリンまたはヒドロキシプロリンであり、かつ、nは5またはこれより大きい数である。例えば、第1、第2、および第3の足場ドメインは、(GPP)10、または、1つもしくはそれ以上のヒト補体C1q、コレクチン、もしくはコラーゲンポリペプチド鎖の三重らせんリピートを含むことができる。
【0013】
1つの実施形態において、上述した第1、第2および第3の融合ポリペプチドは実質的に同じであり、互いに少なくとも75%(例えば75%から100%の間の任意の数を含む)の配列同一性を有している。3つの同じ融合ポリペプチドから形成される複合体はホモ三量体である。これら3つの融合ポリペプチドは機能的に等価であり得る。「機能的に等価」とは、例えば、1もしくはそれ以上の点変異、挿入、欠失、切断を有するタンパク質、融合タンパク質、またはそれらの組み合わせなど共通のポリペプチドのポリペプチド誘導体が、三重らせんコイルを形成する能力と、例えばリガンドに結合するといった異種ドメインの活性を実質的に保持していることをいう。
【0014】
異種ポリペプチド、核酸、または遺伝子は異種から由来するもの、または、同種からのものである場合はその原形に実質的に修飾が加えられたものである。2つの融合ドメインまたは配列は、それらが天然のタンパク質または核酸中で互いに隣り合っていない場合は、互いに異種である。例えば、天然のヒトミニコラーゲン(XXI型)、コレクチンファミリータンパク質、または補体1q(C1q)の一部でないポリペプチド配列は、ヒトタンパク質におけるドメインに対して異種である。
【0015】
本発明はまた、(i)三重らせんコイルを形成するのに用いられる足場ドメイン、および(ii)足場ドメインの1つの末端にインフレームで融合する第1の異種ドメイン、または足場ドメインのもう1つの末端にインフレームで融合する第2の異種ドメインを含む単離組み換え融合ポリペプチド(例えば上述した3つの各融合ポリペプチド)にも特徴を有する。足場ドメインは、1つまたはそれ以上の上述した三重らせんリピート、例えばヒトC1qまたはコラーゲンポリペプチド鎖のそれを含み得る。異種ドメインは、上述した結合ドメインの1つを含んでいてもよく、かつ、ファージディスプレイスクリーニングのような当該分野で知られるさまざまな方法によって得ることができる。
【0016】
単離ポリペプチドまたはタンパク質複合体とは、天然の関連分子を実質的に含まないポリペプチドまたはタンパク質複合体のことをいい、つまり、それは乾燥重量で少なくとも純度75%(すなわち75%および100%の間の任意の数を含む)である。純度は任意の適した標準的方法、例えばカラムクロマトグラフィ、ポリアクリルアミドゲル電気泳動、またはHPLC分析によって測定することができる。本発明の単離ポリペプチドまたはタンパク質複合体は、天然資源から精製でき、また組み換えDNA技術で作製することができる。
【0017】
本発明は、先ほど述べた融合ポリペプチドをコードする配列またはその配列の補体を含む単離核酸にも特徴を有する。核酸とはDNA分子(例えばcDNAもしくはゲノムDNA)、RNA分子(例えばmRNA)、またはDNAもしくはRNAアナログのことをいう。DNAまたはRNAアナログはヌクレオチドアナログから合成されたものであってよい。核酸分子は、一本鎖または二本鎖であってもよいが、好ましいのは二本鎖DNAである。「単離核酸」とは、その構造が、どの天然に存在する核酸の構造とも、またはどの天然に存在するゲノム核酸のフラグメントの構造とも同一でない核酸である。よってこの用語は、例えば(a)天然に存在するゲノムDNA分子の一部の配列を有するが、生物に天然に存在するゲノムにおいてその分子の一部の側方に位置する2つのコーディング配列の隣にはないDNA、(b)得られる分子がいかなる天然に存在するベクターまたはゲノムDNAとも同一にならないような方式で、ベクターまたは原核生物もしくは真核生物のゲノムDNAに組み込まれる核酸、(c)例えばcDNA、ゲノムフラグメント、ポリメラーゼ連鎖反応(PCR)により作製されたフラグメントまたは制限フラグメントのような分離された分子、および(d)ハイブリッド遺伝子、つまり融合タンパク質をコードする遺伝子の一部である組み換えヌクレオチド配列、を包含する。上述した核酸は本発明のポリペプチドを発現するのに用いることができる。この目的のために、核酸を適した調節配列へ動作可能に連結させて発現ベクターを生成することができる。
【0018】
ベクターとは、それに連結された別の核酸を運ぶ能力のある核酸分子のことをいう。ベクターは自己複製または宿主DNAと統一化することが可能である。ベクターの例としては、プラスミド、コスミド、またはウィルスベクターなどがある。本発明のベクターは、宿主細胞における核酸の発現に適したかたちで核酸を含む。ベクターが、発現させようとする核酸配列に操作により連結した1つまたはそれ以上の調節配列を含んでいると好ましい。「調節配列」には、プロモーター、エンハンサー、またはその他の発現制御要素(例えばポリアデニル化信号)が含まれる。調節配列には、組織特異的な調節および/または誘導(inducible)配列だけでなく、ヌクレオチド配列を直接構成的発現させるものも含まれる。発現ベクターの設計は、形質転換させるべき宿主細胞の選択、所望のタンパク質の発現レベルなどといった要素に応じて行うことができる。発現ベクターを宿主細胞に導入することにより本発明のポリペプチドが生産される。上述した核酸を含む宿主細胞もまた本発明の範囲内に含まれる。例として、E.Coli細胞、昆虫細胞(例えばショウジョウバエS2またはバキュロウイルス細胞を使用)、酵母細胞、または哺乳動物の細胞(例えばマウス骨髄腫NS0細胞)などがある。例えば、Goeddel、(1990)Gene Expression Technology:Method in Enzymology 185、Academic Press、San Diego、CAを参照されたい。
【0019】
本発明の融合ポリペプチドを作製するには、本発明の核酸によってコードされるポリペプチドの発現を可能とする条件下、培地中で宿主細胞を培養し、培養された細胞または細胞の培地からポリペプチドを精製することができる。あるいは、本発明の核酸は、例えばT7プロモーター調節配列およびT7ポリメラーゼを用いてインビトロの複写および翻訳がされてもよい。
【0020】
本発明のタンパク質複合体を作製するには、上述した第1、第2および第3の融合ポリペプチドをそれぞれコードする第1、第2および第3の核酸を含む宿主細胞を、これら3つの核酸によりコードされるポリペプチドの発現、および、発現されたポリペプチド間における三重らせんコイルの形成を可能とさせる条件下、培地中で培養し、培養された細胞または細胞の培地からタンパク質複合体を精製することができる。宿主細胞が、プロリン残基をヒドロキシル化する酵素活性を含む真核細胞であると好ましい。
【0021】
本発明の1つまたはそれ以上の実施形態の詳細を、添付の図面および以下の記載において説明する。本発明のその他の特徴、目的および特長は、当該記載および図面から、そして特許請求の範囲から明らかとなるであろう。
【図面の簡単な説明】
【0022】
【図1】ヒトXXI型コラーゲン由来のミニコラーゲン三重らせんコイル足場を有するタンパク質複合体を示す図である。三重らせんコイルの6つの末端は、一本鎖抗体のVLおよびVHドメインを有する6つのFvフラグメント(scFv)にそれぞれインフレームで融合している。
【図2】それぞれ、(A)その3つのN末端が3つの一本鎖抗体OKT3(抗CD3)、528(抗EGFR)、およびerb_scFv(抗EGFR)にそれぞれインフレームで融合する三重らせんコイル足場を有するタンパク質複合体を示す図、(B)該タンパク質複合体のウェスタンブロットの結果を示す写真である。安定にトランスフェクトされたショウジョウバエS2細胞由来の培地を、非還元条件下SDS−PAGEで電気泳動してから、XXI型コラーゲン、3E2のC末端に対するモノクローナル抗体で免疫ブロットした。Tは鎖間でジスルフィド結合した三量体であり、Mtは鎖間でジスルフィド結合した三量体を含む単量体である。
【図3】三重らせんコイル足場を有するタンパク質複合体を示す図である。三重らせんコイルの3つのN末端は3つのOKT3一本鎖抗体にインフレームで融合し、三重らせんコイルの3つのC末端は3つの528一本鎖抗体にインフレームで融合している。
【図4】それぞれ異なる形式の抗体を説明するものである。(A)コラーゲン足場抗体:アミノ末端scFv、ヒトIgGのヒンジ領域、コラーゲン様ドメイン(GPP)10、およびXXI型コラーゲンのカルボキシル末端NC1ドメインを含むscFv−Col(左パネル)、サーファクタントタンパク質D(SPD)コレクチンのアミノ末端部およびカルボキシル末端のscFvを含むNSPD−scFv(右パネル)。(B)左から右に:免疫グロブリンG(IgG)、キメラ(scFv−Fc)、および一本鎖抗体(scFv)と、これらのおおよその分子量。グレーのエリアはVHおよびVLフラグメントを表す。点線:鎖間ジスルフィド結合。
【発明を実施するための形態】
【0023】
本発明は、ヒト三重らせんコイルの足場ドメインにインフレームで融合する異種タンパク質結合ドメインが結合活性を保持する、および得られる融合ポリペプチドが三重らせんコイルを形成する、という予期せぬ発見に、少なくとも一部基づいている。図1に示されているのは、本発明のタンパク質複合体の1例である。この図に示されているように、三重らせんコイルタンパク質複合体の6つの末端は、6つの異種タンパク質結合ドメイン、すなわち一本鎖抗体(scFv)のFvフラグメントにインフレームで融合する。
【0024】
本発明のタンパク質複合体は従来の抗体よりも優れている。また、6つのドメインのうち2つまたはそれ以上が互いに同一である場合、該タンパク質複合体は1つの結合パートナー(例えば抗原)に特異な2〜6つの結合ドメインを備えることができ、これに比べて従来の抗体はそのようなドメインを2つしか備えない。言い換えると、抗原に対し二価でしかない従来の抗体とは異なり、当該タンパク質複合体は二、三、四、五または六価となり得る。その結果として、従来の抗体よりも高い各種親和性を持つようにすることができる。その高い親和性のために、従来の抗体よりも、必要とされるタンパク質複合体の量は少なく、かつインキュベーション期間は短くなり、望ましいゴールが達成される。例えば治療上の効果としては、これにより治療コストが低くなり、かつ副作用(例えば望ましくない免疫反応)が最小限に抑えられる。一方、6つのドメインのうち2つまたはそれ以上が互いに異なる場合、本発明のタンパク質複合体は、2〜6つの異なる結合パートナーに特異である2〜6つの結合ドメインを備えることができる。特異性の異なる多数の結合パートナー部位を1ユニットに統合することによって、多数の結合パートナーをまとめる能力が備わるため、治療、組織再生、および活性タンパク質機械(active protein machinery)(例えばマルチサブユニット酵素)の組立において、ナノメーターレベルでの望ましい使用が可能となる。
【0025】
ヒトのインビボでの使用において、本発明のタンパク質複合体はヒト由来であることが好ましい。例えば、それには、ヒトC1q、コレクチンファミリータンパク質、またはコラーゲンポリペプチド鎖のようなヒト由来のらせんコイル足場にインフレームで融合するヒト化一本鎖抗体配列などが含まれる。ヒトC1qおよびコラーゲンはいずれも血中で極めて安定であるので、当該タンパク質複合体は従来の抗体に比べてより安定である。
【0026】
コラーゲンは哺乳動物に存在する最も豊富なタンパク質である。それは、リピートトリプレット配列Gly−X1−X2を含むと共に、かかるトリプレットの存在が3本のコラーゲンのポリペプチド鎖(α鎖)を三重らせん構造に折り畳ませ得る細胞外基質タンパク質である。トリプレット配列Gly−X1−X2におけるX2の位置のアミノ酸はプロリンであることが多く、それはコラーゲンの三重らせん構造を安定にするためコラーゲンポリペプチド鎖の翻訳後修飾によってしばしば4−ヒドロキシル化される。プロリンのヒドロキシル化なしには、コラーゲンの主要な三重らせん構造が生理的温度以下で熱的に不安定になる(BergおよびProckop、Biochem Biophys Res Commun 52:115-120、1973;Rosenbloomら、Arch Biochem Biophys 158:478-484、1973)。ヒトの血清中には多くのコラーゲン様ドメインを有するコラーゲン様タンパク質が存在しており、感染性生物からの防御において先天性免疫システムの役目を果たす。これらには、補体タンパク質C1q、コレクチンファミリータンパク質−マンノース結合レクチン(MBL)、サーファクタントタンパク質AおよびD(SP−AおよびSP−D)が含まれる。これらコラーゲン様タンパク質間で共通な構造的特徴は、これらがいずれも、そのコラーゲン様ドメインの三重らせんが集合した後、三量体分子が堆積または鎖間ジスルフィド架橋することにより、多量体タンパク質ユニットの形をとることにある。したがって、これら「防御コラーゲン」分子の結合ドメインの機能的親和性(functional affinity)は、多量体化を通して大いに向上する。
【0027】
本発明のタンパク質複合体またはポリペプチドは組み換え技術により得ることができる。該複合体のポリペプチドをコードする適当な宿主細胞の核酸に導入してから、該核酸にコードされたポリペプチドの発現と該ポリペプチド間における三重らせんコイルの形成とを可能とする条件下でポリペプチドを発現させることができる。三重らせんコイル足場の形成を促進するために、コラーゲン生合成の鍵酵素である宿主細胞のプロリル4−ヒドロキシラーゼ(P4HA)で同時発現させることが可能である。
【0028】
異種タンパク質ドメインには抗体またはそのフラグメント(例えばその抗原結合フラグメント)が含まれ得る。ここで用いられる「抗体」という用語は、免疫グロブリン分子またはその免疫学的に活性である部位、すなわち抗原結合部位のことをいう。それは、少なくとも1つ、そして好ましくは2つの重(H)鎖可変領域(VH)と、少なくとも1つ、そして好ましくは2つの軽(L)鎖可変領域(VL)とを含むタンパク質のことをいう。VHおよびVL領域はさらに、「フレームワーク領域」(FR)と呼ばれるより保存性の高い領域の間に位置する、「相補決定領域(「CDR」)」と呼ばれる超可変の領域に細分されていてもよい。フレームワーク領域およびCDRの範囲ははっきりと定義がされている(参照することにより本明細書に組み込まれるKabatら(1991)Sequences of Proteins of Immunological Interes、Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242、およびChothiaら(1987)J. Mol. Biol. 196:901-917を参照)。各VHおよびVLはアミノ末端からカルボキシ末端まで次の順序:FR1、CDR1、FR2、CDR2、FR3、CDR3、FR4、で並ぶ3つのCDRおよび4つのFRよりなっている。抗体はさらに、重および軽鎖定常領域を含み、これにより重および軽免疫グロブリン鎖がそれぞれ形成されるようになっていてもよい。重鎖定常領域は3つのドメイン、CH1、CH2およびCH3からなる。軽鎖定常領域は1つのドメイン、CLからなる。重および軽鎖の可変領域は、抗原と作用する結合ドメインを含む。抗体の定常領域は通常、免疫システムの各種細胞(例えばエフェクター細胞)および古典的補体系の第1成分(C1q)を含む宿主組織または因子への抗体の結合を仲介する。
【0029】
本明細書で「免疫グロブリン」という用語は、実質的に免疫グロブリン遺伝子によってコードされる1つまたはそれ以上のポリペプチドからなるタンパク質のことをいう。存在が認められているヒト免疫グロブリン遺伝子には、カッパ、ラムダ、アルファ(IgA1およびIgA2)、ガンマ(IgG1、IgG2,IgG3、IgG4)、デルタ、エプシロンならびにミュー定常領域遺伝子、そしてさらに無数の免疫グロブリン可変領域遺伝子がある。完全長の免疫グロブリン「軽鎖」(約25KDaまたは214のアミノ酸)は、NH2-末端(約110のアミノ酸)の可変領域遺伝子と、COOH-末端のカッパまたはラムダ定常領域遺伝子とによってコードされる。同じように、完全長の免疫グロブリン「重鎖」(約50KDaまたは446のアミノ酸)は、可変領域遺伝子(約116のアミノ酸)と、他の上記定常領域遺伝子のうちの1つ、例えばガンマ(約330のアミノ酸をコードする)とによってコードされる。
【0030】
抗体の「抗原結合フラグメント」(または「抗体部分(antibody portion)」、または「フラグメント」)は、本明細書においては、例えばEGFRまたはCD3ポリペプチドあるいはそのフラグメントである抗原に特異的に結合する能力を保持する1つまたはそれ以上の完全長の抗体のフラグメントのことをいう。抗体の抗原結合フラグメントの例としては、限定はされないが、(i)Fabフラグメント、VL、VH、CLおよびCH1ドメインからなる一価フラグメント、(ii)F(ab’)2フラグメント、ヒンジ領域にてジスルフィドブリッジにより連結される2つのFabフラグメントを含む二価フラグメント、(iii)VHおよびCHlドメインからなるFdフラグメント、(iv)抗体の1本のアームのVLおよびVHドメインからなるFvフラグメント、(v)VHドメインからなるdAb領域(Wardら、(1987) Nature 341:544-546)、(vi)単離相補決定領域(CDR)、ならびに(vii)VLまたはVHドメインが含まれる。さらに、Fvフラグメントの2つのドメイン、VLおよびVHは、別々の遺伝子によりコードされるが、これらは組み換え法を用い、合成リンカーにより結合させることができ、この合成リンカーはこれらを、VLおよびVH領域が組になって一価分子を形成する一本のタンパク質鎖とさせることができる(一本鎖Fv(scFv)として知られる。例えばBirdら (1988) Science 242:423-426、およびHustonら (1988)PROC. Natl. Acad. Sci USA 85:5879-5883参照)。このような一本鎖抗体もまた、抗体の「抗原結合フラグメント」という用語に包含される。これらの抗体フラグメントは当業者に知られる従来の手法により得ることができ、かつ、当該フラグメントは実用に供することのできるよう、インタクト抗体と同じ方式で選別される。
【0031】
適した抗体にはモノクローナル抗体がある。別の実施形態において、抗体は、組み換え技術、例えばファージディスプレイまたはコンビナトリアル手法により作製したものであってもよい。抗体を作製するためのファージディスプレイおよびコンビナトリアル手法は当該技術分野において周知である(例えば、Ladnerら、米国特許第5,223,409号明細書;Kangら、国際公開第92/18619号パンフレット;Dowerら、国際公開第91/17271号パンフレット;Winterら、国際公開第O92/20791号パンフレット;Marklandら、国際公開第92/15679号パンフレット;Breitlingら、国際公開第93/01288号パンフレット;McCaffertyら、国際公開第92/01047号パンフレット;Garrardら、国際公開第92/09690号パンフレット;Ladnerら、国際公開第90/02809号パンフレット;Fuchsら、(1991)Bio/Technology 9:1370-1372;Hayら、(1992) Hum Antibod Hybridomas 3:81-85;Huseら、(1989)Science 246:1275-1281;Griffthsら、(1993)EMBO J 12:725-734;Hawkinsら、(1992)J Mol Biol 226:889-896;Clacksonら、(1991)Nature 352:624-628;Gramら、(1992) PNAS 89:3576-3580;Garradら、(1991)Bio/Technology 9:1373-1377;Hoogenboomら、(1991)Nuc Acid Res 19:4133-4137、およびBarbasら、(1991)PNAS 88:7978-7982を参照、これらすべての内容は参照することにより本明細書に組み込まれる)。
【0032】
1つの実施形態において、抗体は、完全ヒト抗体(例えば、ヒト免疫グロブリン配列からの抗体を産出するよう遺伝子操作を受けたマウスで作られた抗体)、または非ヒト抗体、例えば齧歯動物(マウスまたはラット)、ヤギ、霊長類の動物(例えばサル)、ラクダの抗体である。非ヒト抗体は齧歯動物(マウスまたはラット抗体)とするのが好ましい。齧歯動物抗体を作製する方法は当該技術分野において周知である。
【0033】
ヒトモノクローナル抗体は、マウスの系ではなく、ヒト免疫グロブリン遺伝子を持つトランスジェニックマウスを用いて生成することができる。目的の抗原で免疫されたこれらトランスジェニックマウスからの脾細胞は、ヒトタンパク質のエピトープに対し特異的親和性があるヒトmAbを分泌するハイブリドーマを作出するのに用いられる(例えば、Woodら、国際公開第91/00906号パンフレット;Kucherlapatiら、国際公開第91/10741号パンフレット;Lonbergら、国際公開第92/03918号パンフレット;Kayら、国際公開第92/03917号パンフレット;Lonberg,Nら、1994 Nature 368:856-859、Green,L.Lら、1994 Nature Genet. 7:13-21;Morrisonら、1994 Proc. Natl.Acad. Sci. USA 81:6851-6855;Bruggemanら、1993 Year Immunol 7:33-40;Tuaillonら、1993 PNAS 90:3720-3724;Bruggemanら、1991 Eur J Immuno 21:1323-1326を参照。)。
【0034】
抗体は、可変領域または例えばCDRなどその一部が、非ヒト生物、例えばラットまたはマウスにおいて産出されたものであってよい。キメラ、CDR−グラフト、およびヒト化抗体が使用可能である。例えばラットまたはマウスのような非ヒト生物で産出されてから、ヒトにおける抗原性を低減するために例えば可変フレームワークまたは定常領域において修飾がなされた抗体は、本発明の範囲に含まれる。
【0035】
キメラ抗体は、当該技術分野において周知である組み換えDNA技術によって作製することができる。例として、マウス(またはその他の種の)モノクローナル抗体分子のFc定常領域をコードする遺伝子を制限酵素により消化して、マウスFcをコードする領域を除去してから、ヒトFc定常領域をコードする遺伝子の相当部位で置換する(Robinsonら、国際特許出願PCT/US86/02269号;Akiraら、欧州特許出願第184,187号明細書;Taniguchi,M、欧州特許出願第171,496号明細書;Morrisonら、欧州特許出願第173,494号明細書;Neubergerら、国際公開第86/01533号パンフレット;Cabillyら、米国特許第4,816,567号明細書;Cabillyら 欧州特許出願第125,023号明細書;Betterら、(1998 Science 240:1041-1043);Liuら、(1987) PNAS 84:3439-3443;Liuら、1987,J.Immunol 139:3521-3526;Sunら、(1987) PNAS 84:214-218;Nishimuraら、1987,Canc.Res. 47:999-1005;Woodら、(1985) Nature 314:446-449;およびShawら、1998, J. Natl Cancer Inst. 89:1553-1559を参照)。
【0036】
ヒト化またはCDRグラフト抗体は、少なくとも1つまたは2つ、ただし通常は3つすべてのレシピエントの(重およびまたは軽免疫グロブリン鎖の)CDRが、ドナーCDRで置き換えられる。抗体が非ヒトCDRの少なくとも1部で置き換えられても、あるいは、CDRのいくつかだけが非ヒトCDRで置き換えられてもよい。ヒト化抗体またはそのフラグメントの結合に要されるCDRの数だけ置き換えればよいのである。ドナーが齧歯動物の抗体、例えばラットまたはマウス抗体であり、かつレシピエントがヒトフレームワークまたはヒトコンセンサスフレームワークであると好ましい。一般に、CDRを提供する免疫グロブリンは「ドナー」と呼ばれ、フレームワークを提供する免疫グロブリンは「アクセプター」と呼ばれる。1つの実施形態において、ドナー免疫グロブリンはヒト以外(例えば齧歯動物)である。アクセプターのフレームワークは、天然(例えばヒト)フレームワークまたはコンセンサスフレームワーク、または、約85%またはこれ以上、好ましくは90%、95%、99%またはこれ以上がそれと同一な配列である。本明細書で「コンセンサス配列」とは、関連配列のファミリー中にて最も頻繁に出現するアミノ酸(またはヌクレオチド)から形成される配列のことをいう(例えば、Winnaker、From Genes to Clones (Verlagsgesellschaft, Weinheim, Germany 1987) 参照)。タンパク質のファミリーにおいて、コンセンサス配列中の各位置は、該ファミリーにおけるその位置に最も頻繁に出現するアミノ酸によって占められる。2つのアミノ酸が同等に頻繁に出現する場合は、どちらかがコンセンサス配列に含まれ得る。「コンセンサスフレームワーク」とは、コンセンサス免疫グロブリン配列中のフレームワーク領域のことをいう。
【0037】
抗体は当該技術分野において周知の方法でヒト化することができる。ヒト化抗体は、抗原の結合に直接かかわりのないFv可変領域の配列を、ヒトFv可変領域からの相当する配列で置き換えることによって生成することができる。ヒト化抗体を生成する一般的な方法は、Morrison,S.L、1985,Science 229:1202-1207、Oiら、1986,BioTechniques 4:214、ならびにQueenら、米国特許第5,585,089号明細書、米国特許第5,693,761号明細書および米国特許第5,693,762号明細書により提示されており、これらのすべての内容は参照することにより本明細書に組み込まれる。それらの方法は、少なくとも1つの重または軽鎖からの免疫グロブリンFv可変領域のすべてまたは一部をコードする核酸配列を単離する工程、操作する工程、および発現させる工程を含む。かかる核酸のソースは当業者にはよく知られており、例えば、目的のポリペプチドまたはそのフラグメントに対する抗体を産出するハイブリドーマから得ることができる。よって、ヒト化抗体またはそのフラグメントをコードする組み換えDNAを、適当な発現ベクターにクローニングすることができる。
【0038】
特定のアミノ酸が置換、欠失または追加されたヒト化抗体は足場に融合することもできる。好ましいヒト化抗体は、例えば抗原への結合を改善する目的で、フレームワーク領域にアミノ酸置換基を有する。例えば、ヒト化抗体は、ドナーのフレームワークの残基またはレシピエントのフレームワーク残基以外の別なアミノ酸と同一であるフレームワークの残基を備える。このような抗体を生成するため、選択された、少数のヒト化免疫グロブリン鎖のアクセプターフレームワーク残基を、対応するドナーアミノ酸で置き換えることができる。好ましい置換基の配置としては、アミノ酸残基をCDRに隣接させる、またはCDRと作用することのできる配置などがある。ドナーからアミノ酸を選ぶ基準は、US5,585,089号に記載されており、その内容は本明細書では参照することにより組み込まれる。ヒト化抗体のその他の技術は、Padlanら、欧州特許出願公開第519596号明細書に記載されている。
【0039】
本発明は、本発明のタンパク質複合体を形成する融合ポリペプチドをコードする核酸をも含む。核酸は、ファージディスプレイライブラリーから選別された、または上記の適した抗体または抗体の誘導体を発現する細胞株から(RT−PCRによって)単離されたものとすることができる。核酸は発現ベクターに機能的にライゲートされてもよい。核酸またはベクターで形質転換された細胞を、本発明の融合ポリペプチドまたはタンパク質複合体を作製するのに用いることができる。抗体を作製するのに有用な細胞としては、昆虫細胞および哺乳動物の細胞(例えばCHOまたはリンパ細胞)などがある。
【0040】
本発明のタンパク質複合体は、細胞毒素のような治療成分(therapeutic moiety)、治療薬または放射性イオンにコンジュゲートすることができる。細胞毒素または細胞毒性薬には、細胞に有害なあらゆる薬剤が含まれる。例としては、タクソール、サイトカラシンB、グラミシジンD、エチジウムブロマイド、エメチン、マイトマイシン、エトポシド、テノポシド(tenoposide)、ビンクリスチン、ビンブラスチン、コルヒチン、ドキソルビシン、ダウノルビシン、ジヒドロキシアントラシンジオン、ミトキサントロン、ミトラマイシン、アクチノマイシンD、1−デヒドロテストステロン、グルココルチコイド、プロカイン、テトラカイン、リドカイン、プロプラノロール、ピューロマイシン、メイタンシノイド(maytansinoids)、例えばメイタンシノール(maytansinol)(米国特許第5,208,020号明細書参照)、CC−1065(米国特許第5,475,092号明細書、米国特許第5,585,499号明細書、米国特許第5,846,545号明細書参照)およびそのアナログまたはホモログがある。治療薬には、限定はされないが、代謝拮抗物質(例えばメトトレキサート、6−メルカプトプリン、6−チオグアニン、シタラビン、5−フルオロウラシルデカルバジン)、アルキル化剤(例えばメクロレタミン、チオエパクロラムブシル(thioepa chlorambucil)、CC−1065、メルファラン、カルムスチン(BSNU)およびロムスチン(CCNU)、シクロトスファミド(cyclothosphamide)、ブスルファン、ジブロモマンニトール、ストレプトゾトシン、マイトマイシンC、およびシスジクロロジアミン白金(II)(DDP)(シスプラチン)、アントラサイクリン(例えばダウノルビシン(旧ダウノマイシン)およびドクソルビシン)、抗生物質(例えばダクチノマイシン、(旧アクチノマイシン)、ブレオマイシン、ミトラマイシンおよびアントラマイシン(AMC))、ならびに抗分裂剤(例えばビンクリスチン、ビンブラスチン、タクソールおよびメイタンシノイド)が含まれる。放射性イオンには、限定はされないが、ヨード、イットリウムおよびプラセオジムが含まれる。
【0041】
コンジュゲートは所定の生物学的反応を修飾するのに用いることができ、薬物成分(drug moiety)は古典的な化学治療薬に限定されると解されるべきではない。例えば、薬物成分は、所望の生物学的活性を備えたタンパク質またはポリペプチドであり得る。かかるタンパク質には、例として、アブリン、リシンA、緑膿菌外毒素もしくはジフテリア毒素のような毒素、腫瘍壊死因子、α−インターフェロン、β−インターフェロン、神経成長因子、血小板由来成長因子、組織プラスミノーゲンアクチベータのようなタンパク質、または、例えばリンフォカイン、インターロイキン−1(「IL−1」)、インターロイキン−2(「IL−2」)、インターロイキン−6(「IL−6」)、顆粒球マクロファージコロニー刺激因子(「GM−CSF」)、 顆粒球コロニー刺激因子(「G-CSF」)もしくはその他の成長因子のような生物学的反応修飾物質が含まれる。
【0042】
異種結合ドメインの特異性をベースとしている上述のタンパク質複合体およびコンジュゲートは、ガン、炎症疾患、代謝疾患、繊維化疾患、および心臓血管疾患を含むさまざまな疾患を治療するために用いることができる。よって本発明は、例えば、それを必要とする対象に本発明のタンパク質複合体を有効量投与することによって、このような疾患を治療する方法に特徴を有する。治療を受けるべき対象は、疾患に特徴付けられる状態を持つまたは獲得する危険があるものとして確認することができる。この方法は、単独で、またはその他の薬物もしくは治療法と併用して行うことが可能である。
【0043】
その多重特異性という特徴により、本発明のタンパク質複合体は、本来は互いにつながっていない分子または細胞を架橋するのに用いられ得る。この特徴は細胞ベースの治療に対しとりわけ有用である。一例において、タンパク質複合体における1つの異種ドメインは、細胞障害性細胞上のエフェクター抗原に特異的に結合することによって細胞傷害性細胞(例えば細胞傷害性T細胞)を活性化させることができ、同時に他の異種ドメインは、破壊するべき病原細胞または悪性細胞上の標的抗原に特異的に結合する。このようにして、タンパク質複合体は病原または悪性細胞によって引き起こされる疾病を治療することができるのである。
【0044】
細胞傷害性T細胞の活性化は、本発明のタンパク質複合体による、細胞障害性T細胞表面のエフェクター抗原としてのCD3抗原の結合を通して起こり得る。その他のリンパ球様細胞結合型(lymphoid cell-associated)エフェクター抗原には、ヒトCD16抗原、NKG2D抗原、NKp46抗原、CD2抗原、CD28抗原、CD25抗原、CD64抗原、およびCD89抗原などがある。これらエフェクター抗原に結合することによって、単球、好中性顆粒球および樹枝状細胞のようなエフェクター細胞の活性化がもたらされる。よって、これら活性化された細胞は、標的細胞に対し細胞傷害またはアポトーシス効果を発揮する。
【0045】
標的抗原は、病気の状態に付随する標的細胞上にユニークに発現(uniquely express)する抗原であるが、健康な状態では発現しない、低レベルで発現する、あるいはアクセスできない(non-accessible)ものである。悪性細胞に関連するかかる標的抗原の例には、EpCAM、CCR5、CD19、HER−2neu、HER−3、HER−4、EGFR、PSMA、CEA、MUC−1(ムチン)、MUC2、MUC3、MUC4、MUC5.sub.AC、MUC5.sub.B、MUC7、ベータ.hCG、ルイス−Y、CD20、CD33、CD30、ガングリオシドGD3、9−O−アセチル−GD3、GM2、Globo H、フコシル GM1、ポリSA、GD2、カルボアンヒドラーゼ(Carboanhydrase)IX (MN/CA IX)、D44v6、ソニックヘッジホッグ(Shh)、Wue−1、形質細胞抗原、(膜結合性)IgE、メラノーマコンドロイチンサルフェートプロテオグリカン(MCSP)、CCR8、TNF−アルファ前駆体、STEAP、メソテリン(mesothelin)、A33抗原、前立腺幹細胞抗原(PSCA)、Ly−6;デスモグレイン4、E−カドヘリンネオエピトープ、胎児アセチルコリンレセプター、CD25、CA19−9マーカー、CA−125マーカーおよびミュラー管抑制物質(MIS)レセプタータイプII、sTn(シリル化Tn抗原;TAG−72)、FAP(線維芽細胞活性化抗原)、エンドシアリン(endosialin)、EGFRvIII、LG、SASならびにCD63などがある。
【0046】
「治療する(treating)」という用語は、病気、病気の症状、病気による二次的な病状、または病気にかかり易い素因を治す(cure)、緩和する(alleviate)、軽減する(relieve)、修復する(remedy)、予防する(prevent)、または改善する(ameliorate)ことを目的とし、対象に組成物を投与することとして定義される。「有効量」とは、治療を受ける対象において例えば上述したような医学的に望ましい結果を生じさせることのできる組成物の量のことである。
【0047】
1つのインビボアプローチでは、治療組成物(例えば本発明のタンパク質複合体を含む組成物)を対象に投与する。通常、該複合体は医薬的に許容される担体(例えば生理食塩水)に懸濁されており、経口でもしくは静脈内注射により投与される、または皮下、筋肉、鞘内、腹腔内、直腸内、膣内、鼻腔内、胃内、気管内もしくは肺内に注射もしくは注入される。
【0048】
必要な投与量は、投与経路の選択、剤形の性質、対象の疾病の性質、対象の大きさ、体重、表面積、年齢および性別、投与されている他の薬物、ならびに主治医の判断によって決まる。適切な投与量は0.01〜100.0mg/kgの範囲である。必要とされる投与量の増減は、利用可能な組成物の種類、および各種投与経路の異なる効率を考慮して予測される。例えば、経口投与は静脈注射による投与よりも高い投与量を要するであろうことが予想できる。これら投与量の増減の程度は、当該技術分野においてよく理解されているような、標準的な経験的ルーチンによって調整し至適化することができる。適した運搬媒体(例えば高分子微粒子または埋め込み型装置)に組成物をカプセル封入すれば、特に経口デリバリーにおいて、運搬の効率を向上させることができる。
【0049】
医薬的に許容される担体および有効量の本発明のタンパク質複合体を含む医薬組成物も、本発明の範囲内に含まれる。医薬組成物は上記した疾患を処置するのに用いることができる。医薬的に許容される担体には、溶剤、分散媒、コーティング、抗菌および抗真菌剤、ならびに等張および吸収遅延剤が含まれる。医薬組成物は、従来の方法を用い、それぞれ異なる投与経路に合った剤形に形成することができる。
【0050】
本発明の組成物の有効性はインビトロとインビボの両方において評価できる。インビボ試験では、組成物を動物(例えばマウスモデル)に注射してから、その治療上の効果を調べることができる。その結果に基づいて、適切な投与量の範囲と投与経路が決められる。
【0051】
以下の具体例は、説明にすぎず、いかなる方式においてもこの開示以外のものを限定するものではないと解されるべきである。詳述はしなくとも、当業者は、本明細書の記載に基づいて、本発明を最大限利用できると考えられる。本明細書に引用されるすべての刊行物は、参照することによりその全体が本明細書に組み込まれる。
【実施例】
【0052】
実施例1
M13ファージディスプレイライブラリーをスクリーニングして、EGFRに特異的に結合するヒト一本鎖可変フラグメント(scFv)を同定した。多数のクローンを同定した。ウェスタンブロッティングおよびELISAによる確認ののち、さらなる実験のために1つのクローン、erb_scFvを選んだ。標準方法により、erb_scFvをコードするcDNAを得てから、発現ベクターにライゲートした。以下に示すのは、erb_scFvのポリペプチド配列(配列番号1)とそれをコードするヌクレオチド配列(配列番号2)である。
【0053】
配列番号1
MetAlaGluValGlnLeuLeuGluSerGlyGlyGlyLeuValGlnProGlyGlySerLeuArgLeuSerCysAlaAlaSerGlyPheThrPheSerSerTyrAlaMetSerTrpValArgGlnAlaProGlyLysGlyLeuGluTrpValSerAspIleGlyAlaSerGlySerAlaThrSerTyrAlaAspSerValLysGlyArgPheThrIleSerArgAspAsnSerLysAsnThrLeuTyrLeuGlnMetAsnSerLeuArgAlaGluAspThrAlaValTyrTyrCysAlaLysSerThrThrThrPheAspTyrTrpGlyGlnGlyThrLeuValThrValSerSerGlyGlyGlyGlySerGlyGlyGlyGlySerGlyGlyGlyGlySerThrAspIleGlnMetThrGlnSerProSerSerLeuSerAlaSerValGlyAspArgValThrIleThrCysArgAlaSerGlnSerIleSerSerTyrLeuAsnTrpTyrGlnGlnLysProGlyLysAlaProLysLeuLeuIleTyrAspAlaSerAlaLeuGlnSerGlyValProSerArgPheSerGlySerGlySerGlyThrAspPheThrLeuThrIleSerSerLeuGlnProGluAspPheAlaThrTyrTyrCysGlnGlnTyrAlaAspTyrProThrThrPheGlyGlnGlyThrLysValGluIleLysArg
【0054】
配列番号2
ATGGCCGAGGTGCAGCTGTTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTTAGCAGCTATGCCATGAGCTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTCTCAGATATTGGTGCTTCTGGTTCTGCTACATCTTACGCAGACTCCGTGAAGGGCCGGTTCACCATCTCCAGAGACAATTCCAAGAACACGCTGTATCTGCAAATGAACAGCCTGAGAGCCGAGGACACGGCCGTATATTACTGTGCGAAATCTACTACTACTTTTGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCGAGCGGTGGAGGCGGTTCAGGCGGAGGTGGCAGCGGCGGTGGCGGGTCGACGGACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAGCAGCTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGATGCATCCGCTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAACAGTATGCTGATTATCCTACTACGTTCGGCCAAGGGACCAAGGTGGAAATCAAACGG
【0055】
次いで、発現ベクターを昆虫細胞株ショウジョウバエS2中で発現させた。抗EGFR erb_scFvを精製し、ウェスタンブロット分析およびELISAにかけて、そのEGFRに対する特異性を確認した。
【0056】
RT−PCRを行って、抗CD3モノクローナル抗体OKT3の重鎖可変領域 (VH)および軽鎖可変領域(VL)をコードするcDNAをハイブリドーマ細胞株から得た。続いて、2つのcDNAをライゲートして、OKT3のVH−VLの融合タンパク質をコードする融合配列を作製した。以下に示すのは、この融合タンパク質のポリペプチド配列(配列番号3)とそれをコードするcDNAの配列(配列番号4)である。
【0057】
配列番号3
ValGlnLeuGlnGlnSerGlyAlaGluLeuAlaArgProGlyAlaSerValLysMetSerCysLysAlaSerGlyTyrThrPheThrArgTyrThrMetHisTrpValLysGlnArgProGlyGlnGlyLeuGluTrpIleGlyTyrIleAsnProSerArgGlyTyrThrAsnTyrAsnGlnLysPheLysAspLysAlaThrLeuThrThrAspLysSerSerSerThrAlaTyrMetGlnLeuSerSerLeuThrSerGluAspSerAlaValTyrTyrCysAlaArgTyrTyrAspAspHisTyrCysLeuAspTyrTrpGlyGlnGlyThrThrValThrValSerSerGlyGlyGlyGlySerGlyGlyGlyGlySerGlyGlyGlyGlySerAspIleValLeuThrGlnSerProAlaIleMetSerAlaSerProGlyGluLysValThrMetThrCysSerAlaSerSerSerValSerTyrMetAsnTrpTyrGlnGlnLysSerGlyThrSerProLysArgTrpIleTyrAspThrSerLysLeuAlaSerGlyValProAlaHisPheArgGlySerGlySerGlyThrSerTyrSerLeuThrIleSerGlyMetGluAlaGluAspAlaAlaThrTyrTyrCysGlnGlnTrpSerSerAsnProPheThrPheGlySerGlyThrLysLeuGluLeuLysArg
【0058】
配列番号4
GTCCAGCTGCAGCAGTCAGGGGCTGAACTGGCAAGACCTGGGGCCTCAGTGAAGATGTCCTGCAAGGCTTCTGGCTACACCTTTACTAGGTACACGATGCACTGGGTAAAACAGAGGCCTGGACAGGGTCTGGAATGGATTGGATACATTAATCCTAGCCGTGGTTATACTAATTACAATCAGAAGTTCAAGGACAAGGCCACATTGACTACAGACAAATCCTCCAGCACAGCCTACATGCAACTGAGCAGCCTGACATCTGAGGACTCTGCAGTCTATTACTGTGCAAGATATTATGATGATCATTACTGCCTTGACTACTGGGGCCAAGGGACCACGGTCACCGTCTCCTCAGGTGGAGGCGGTTCAGGCGGAGGTGGCTCTGGCGGTGGCGGATCGGACATTGTGCTAACCCAGTCTCCAGCAATCATGTCTGCATCTCCAGGGGAGAAGGTCACCATGACCTGCAGTGCCAGCTCAAGTGTAAGTTACATGAACTGGTACCAGCAGAAGTCAGGCACCTCCCCCAAAAGATGGATTTATGACACATCCAAACTGGCTTCTGGAGTCCCTGCTCACTTCAGGGGCAGTGGGTCTGGGACCTCTTACTCTCTCACAATCAGCGGCATGGAGGCTGAAGATGCTGCCACTTATTACTGCCAGCAGTGGAGTAGTAACCCATTCACGTTCGGCTCGGGGACCAAGCTGGAGCTGAAACGA
【0059】
同じ手順を行って、抗EGFRモノクローナル抗体528のVHおよびVLをコードするcDNA、ならびに抗EGFR528のVH−VLの融合タンパク質をコードする融合配列を得た。528モノクローナル抗体は、細胞膜、例えばヒト類表皮癌A431細胞などの細胞膜上のEGFRに結合する。この528一本鎖抗体のポリペプチド配列(配列番号5)およびそれをコードするcDNAの配列(配列番号6)を以下に示す。
【0060】
配列番号5
ValLysLeuGlnGluSerGlySerGluMetAlaArgProGlyAlaSerValLysLeuProCysLysAlaSerGlyAspThrPheThrSerTyrTrpMetHisTrpValLysGlnArgHisGlyHisGlyProGluTrpIleGlyAsnIleTyrProGlySerGlyGlyThrAsnTyrAlaGluLysPheLysAsnLysValThrLeuThrValAspArgSerSerArgThrValTyrMetHisLeuSerArgLeuThrSerGluAspPheAlaValTyrTyrCysThrArgSerGlyGlyProTyrPhePheAspTyrTrpGlyGlnGlyThrThrValThrValSerSerGlyGlyGlyGlySerGlyGlyGlyGlySerGlyGlyGlyGlySerMetThrGlnThrProLeuSerLeuProValSerLeuGlyAspGlnAlaSerIleSerCysArgSerSerGlnAsnIleValHisAsnAsnGlyIleThrTyrLeuGluTrpTyrLeuGlnArgProGlyGlnSerProLysLeuLeuIleTyrLysValSerAspArgPheSerGlyValProAspArgPheSerGlySerGlySerGlyThrAspPheThrLeuLysIleSerArgValGluAlaGluAspLeuGlyIleTyrTyrCysPheGlnGlySerHisHisProProThrPheGlyGlyGlyThrLysLeuGlu
【0061】
配列番号6
GTCAAGCTGCAGGAGTCAGGGTCTGAGATGGCGAGGCCTGGAGCTTCAGTGAAGCTGCCCTGCAAGGCTTCTGGCGACACATTCACCAGTTACTGGATGCACTGGGTGAAGCAGAGGCATGGACATGGCCCTGAGTGGATCGGAAATATTTATCCAGGTAGTGGTGGTACTAACTACGCTGAGAAGTTCAAGAACAAGGTCACTCTGACTGTAGACAGGTCCTCCCGCACAGTCTACATGCACCTCAGCAGGCTGACATCTGAGGACTTTGCGGTCTATTATTGTACAAGATCGGGGGGTCCCTACTTCTTTGACTACTGGGGCCAAGGGACCACGGTCACCGTCTCCTCAGGTGGAGGCGGTTCAGGCGGAGGTGGCTCTGGCGGTGGCGGATCGATGACCCAAACTCCACTCTCCCTGCCTGTCAGTCTTGGAGATCAAGCCTCCATCTCTTGCAGATCTAGTCAGAACATTGTACATAATAATGGAATCACCTATTTAGAATGGTACCTGCAAAGGCCAGGCCAGTCTCCAAAGCTCCTGATCTACAAAGTTTCCGACCGATTTTCTGGGGTCCCAGACAGGTTCAGTGGCAGTGGATCAGGGACAGATTTCACACTCAAGATCAGCAGAGTAGAGGCTGAGGATCTGGGAATTTATTACTGCTTTCAAGGTTCACATCATCCTCCCACGTTCGGCGGGGGGACCAAGCTGGAA
【0062】
上述の抗EGFR scFv03、OKT3VH−VL、および抗EGFR 528VH−VLをコードするcDNAを、5’末端の短いヒンジ配列、および3’末端のヒスチジンタグ配列を含むXXI型ヒトミニコラーゲンのcDNAにそれぞれインフレームで融合した。以下に示すのは、XXI型ヒトミニコラーゲンのポリペプチドおよびcDNAの配列(それぞれ配列番号7および8)である。
【0063】
配列番号7
GlyGlyArgGluProLysSerCysAspLysThrHisThrCysProProCysProArgSerIleProGlyProProGlyProIleGlyProGluGlyProArgGlyLeuProGlyLeuProGlyArgAspGlyValProGlyLeuValGlyValProGlyArgProGlyValArgGlyLeuLysGlyLeuProGlyArgAsnGlyGluLysGlySerGlnGlyPheGlyTyrProGlyGluGlnGlyProProGlyProProGlyProGluGlyProProGlyIleSerLysGluGlyProProGlyAspProGlyLeuProGlyLysAspGlyAspHisGlyLysProGlyIleGlnGlyGlnProGlyProProGlyIleCysAspProSerLeuCysPheSerValIleAlaArgArgAspProPheArgLysGlyProAsnTyrSerLeuAspAspSerSerHisHisHisHisHisHisSerSerGly
(注:Proはプロリンまたはヒドロキシプロリンである)
【0064】
配列番号8
GGCGGCCGCGAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAAGATCTATTCCTGGGCCACCTGGTCCGATAGGCCCAGAGGGTCCCAGAGGATTACCTGGTTTGCCAGGAAGAGATGGTGTTCCTGGATTAGTGGGTGTCCCTGGACGTCCAGGTGTCAGAGGATTAAAAGGCCTACCAGGAAGAAATGGGGAAAAAGGGAGCCAAGGGTTTGGGTATCCTGGAGAACAAGGTCCTCCTGGTCCCCCAGGTCCAGAGGGCCCTCCTGGAATAAGCAAAGAAGGTCCTCCAGGAGACCCAGGTCTCCCTGGCAAAGATGGAGACCATGGAAAACCTGGAATCCAAGGGCAACCAGGCCCCCCAGGCATCTGCGACCCATCACTATGTTTTAGTGTAATTGCCAGAAGAGATCCGTTCAGAAAAGGACCAAACTATAGTCTAGACGACAGCAGCCATCATCACCATCACCATAGCAGCGGC
【0065】
得られた3つの発現ベクターを、ショウジョウバエS2細胞に同時トランスフェクト(co-transfect)した。その細胞をブラストサイジン存在下で培養し、ブラストサイジンに耐性を持つ細胞を選別した。細胞培養上清を回収してから、EGFRおよびCD3に対する抗体活性に対して、ウェスタンブロットとELISAでスクリーニングした。一部のクローンの細胞が三重らせん状の複合体(triple helix complex)を安定に発現していることが発見された。XXI型ヒトミニコラーゲンに似たこれら三重らせん状の複合体は、熱およびペプシンに耐性を示した。より重要なのは、これらがEGFRとCD3の両方に特異的に結合したということである。
【0066】
実施例2
この実施例では、3つの融合ポリペプチド、OKT3_scFv−Col、erb_scFv−Col、およびerb_NSPD−scFvを作製した。
【0067】
ファージライブラリーの選択
上皮細成長因子レセプター細胞外ドメイン(EGFR−ECD)結合可変フラグメント(scFv)を含むerbファージミドを、ヒトシングルフォールドscFvファージディスプレイライブラリー(Tomlinson I + J;I. M. Tomlinson and G. Winterより親切に提供された、MRCラボラトリー オブ モレキュラーバイオロジー、ケンブリッジ、英国)をスクリーニングすることにより単離した。精製したEGFレセプターの組み換え細胞外ドメイン(EGFR-ECD;リサーチ ダイアグノスティック社(Research Diagnostics, Inc.))10μgを塗布したイムノチューブ(Maxisorp;ヌン(Nunc)、 ロスキレ、デンマーク)を用いて選別を行った。メーカーのプロトコールにしたがって、ブロッキング、パンニング(panning)、洗浄、溶出および溶出ファージの再増幅を行った。
【0068】
組み換えプラスミドの構築
erbのscFvをコードするcDNAをPCRによりerbファージミドから増幅した。OKT3ハイブリドーマ(ATCC、CRL-8001)の逆転写産物により、ムリンIgG2a抗CD3 mAb OKT3(オルト ファーマシューティカル コーポレーション)をコードする配列を得た。公開されたヌクレオチド配列を基に、RT−PCRによってOKT3 mAbのVLおよびVHのcDNAを得た。グリシンリンカー(GGGS)3でVLおよびVH鎖をつなぐことにより、erbおよびOKT3のscFv PCRの融合を発生させた。
【0069】
scFv-Colを生成するよう、scFv-Colのコーディング領域には、N末端のscFvヌクレオチド配列と、ヒトIgGのヒンジ領域、コラーゲン様ドメイン(下線部)およびXXI型コラーゲンのNC1ドメインを含む、EPKSCDKTHTCPPCPRSIP(GPP)10GICDPSLCFSVIA-RRDPFRKGPNYのペプチド配列をコードするC末端合成コラーゲン足場遺伝子とが含まれる。以下に示すのは、合成コラーゲン足場ポリペプチドおよびcDNAの配列(それぞれ配列番号9および10)である。
【0070】
配列番号9
SEQ ID NO: 9:
GluProLysSerCysAspLysThrHisThrCysProProCysProArgSerIleProGlyProProGlyProProGlyProProGlyProProGlyProProGlyProProGlyProProGlyProProGlyProProGlyProProGlyIleCysAspProSerLeuCysPheSerValIleAlaArgArgAspProPheArgLysGlyProAsnTyr
【0071】
配列番号10
GAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAAGATCTATTCCTGGGCCACCTGGTCCCCCAGGTCCTCCAGGACCCCCAGGGCCCCCAGGCCCCCCCGGGCCGCCTGGACCCCCAGGGCCACCAGGCCCCCCAGGCATCTGCGACCCATCACTATGTTTTAGTGTAATTGCCAGAAGAGATCCGTTCAGAAAAGGACCAAACTAT
【0072】
この合成配列(配列番号10)をオーバーラップPCRにより作製し、NotIおよびXhoI部位の隣に位置する(flanking)PCR産物を、同部位の発現ベクターpSecTag2/Hygro(インビトロジェン)にクローニングした。続いて、erbおよびOKT3のscFvを、AscIおよびNotI部位で上述したC末端コラーゲン足場含有構成体にインフレームでクローニングして、erb_scFv−ColおよびOKT3_scFv−Colの発現構成体をそれぞれ作った。
【0073】
次に、erb_NSPD−scFvを作製した。NSPD−scFvのコーディング領域には、ヒトサーファクタントタンパク質D(SPD)のN末端の254のアミノ酸、コレクチンファミリーのメンバー、およびC末端のscFvが含まれていた。以下に示すのは、ヒトサーファクタントタンパク質DのポリペプチドのN末端254アミノ酸およびそのcDNAの配列である(それぞれ配列番号11および12)。
【0074】
配列番号11
MetLeuLeuPheLeuLeuSerAlaLeuValLeuLeuThrGlnProLeuGlyTyrLeuGluAlaGluMetLysThrTyrSerHisArgThrMetProSerAlaCysThrLeuValMetCysSerSerValGluSerGlyLeuProGlyArgAspGlyArgAspGlyArgGluGlyProArgGlyGluLysGlyAspProGlyLeuProGlyAlaAlaGlyGlnAlaGlyMetProGlyGlnAlaGlyProValGlyProLysGlyAspAsnGlySerValGlyGluProGlyProLysGlyAspThrGlyProSerGlyProProGlyProProGlyValProGlyProAlaGlyArgGluGlyProLeuGlyLysGlnGlyAsnIleGlyProGlnGlyLysProGlyProLysGlyGluAlaGlyProLysGlyGluValGlyAlaProGlyMetGlnGlySerAlaGlyAlaArgGlyLeuAlaGlyProLysGlyGluArgGlyValProGlyGluArgGlyValProGlyAsnThrGlyAlaAlaGlySerAlaGlyAlaMetGlyProGlnGlySerProGlyAlaArgGlyProProGlyLeuLysGlyAspLysGlyIleProGlyAspLysGlyAlaLysGlyGluSerGlyLeuProAspValAlaSerLeuArgGlnGlnValGluAlaLeuGlnGlyGlnValGlnHisLeuGlnAlaAlaPheSerGlnTyrLysLysValGluLeuPhe
【0075】
配列番号12
ATGCTGCTCTTCCTCCTCTCTGCACTGGTCCTGCTCACACAGCCCCTGGGCTACCTGGAAGCAGAAATGAAGACCTACTCCCACAGAACAATGCCCAGTGCTTGCACCCTGGTCATGTGTAGCTCAGTGGAGAGTGGCCTGCCTGGTCGCGATGGACGGGATGGGAGAGAGGGCCCTCGGGGCGAGAAGGGGGACCCAGGTTTGCCAGGAGCTGCAGGGCAAGCAGGGATGCCTGGACAAGCTGGCCCAGTTGGGCCCAAAGGGGACAATGGCTCTGTTGGAGAACCTGGACCAAAGGGAGACACTGGGCCAAGTGGACCTCCAGGACCTCCCGGTGTGCCTGGTCCAGCTGGAAGAGAAGGTCCCCTGGGGAAGCAGGGGAACATAGGACCTCAGGGCAAGCCAGGCCCAAAAGGAGAAGCTGGGCCCAAAGGAGAAGTAGGTGCCCCAGGCATGCAGGGCTCGGCAGGGGCAAGAGGCCTCGCAGGCCCTAAGGGAGAGCGAGGTGTCCCTGGTGAGCGTGGAGTCCCTGGAAACACAGGGGCAGCAGGGTCTGCTGGAGCCATGGGTCCCCAGGGAAGTCCAGGTGCCAGGGGACCCCCGGGATTGAAGGGGGACAAAGGCATTCCTGGAGACAAAGGAGCAAAGGGAGAAAGTGGGCTTCCAGATGTTGCTTCTCTGAGGCAGCAGGTTGAGGCCTTACAGGGACAAGTACAGCACCTCCAGGCTGCTTTCTCTCAGTATAAGAAAGTTGAGCTCTTC
【0076】
N末端SPD cDNAを、発現ベクター、pSecTag2/Hygro(インビトロジェン)にNheIおよびAscI部位でクローニングした。続いて、erbのscFvを、AscIおよびXhoI部位で上述したN末端SPD含有構成体にインフレームでクローニングして、erb_NSPD−scFvの発現構成体を作った。
【0077】
erb_scFv−Col、erb_NSPD−scFvおよびOKT3_scFv−Colの各読み取り枠(open reading frame)には、N末端リーダー配列と、分泌、検出および精製に利用されるC末端mycエピトープ/ポリヒスチジンタグとをコードする配列が含まれていた。下表にまとめたのは、上述の発現構成体によりコードされる各種組み換えタンパク質/抗体である。
【0078】
【表1】

【0079】
抗体の発現および精製
組み換えタンパク質複合体/抗体を作製するため、Effectene(Qiagen)を用い、メーカーの取扱説明書にしたがって上述の構成体をマウス骨髄腫NS0細胞に導入した。ハイグロマイシン(400μg/ml)で4週間選別したのち、各安定クローンを、振盪フラスコ中の2%のウシ胎仔血清を含む既知組成培地HyQCDM4NS0(ハイクローン(Hyclone))にて、初播種密度2×105細胞/mlとして培養した。該培養は37℃で5日間、150rpmに保って行った。それら細胞が、上述の抗体ドメインと、コラーゲン足場ドメインつまりコラーゲン足場抗体(CSA)と、を含むタンパク質をコードする発現構成体を保持できるように、培地にアスコルビン酸ナトリウム(80μg/ml)を毎日加えた。
【0080】
erb_scFv、erb_scFv−Fc、erb_scFv−Col、またはOKT3_scFv−Colタンパク質あるいはタンパク質複合体を精製するべく、ろ過した各培地約2Lを、流速60ml/時間で、pH8.0の50mMトリスHClバッファーで平衡化したTゲルカラム(1.5×8cm、Pierce)にかけた。同バッファーで洗浄したのち、その組み換えタンパク質またはタンパク質複合体をpH4.0の50mM酢酸ナトリウムバッファーで溶出した。それらのUV吸収度を280nmで測定し、そのピーク画分を、流速60ml/時間で、pH8.0の0.5M NaCl含有50mMトリスHClバッファーにより平衡化したZnSO4チャージキレーティングセファロースハイトラップカラム(ZnSO4-charged chelating Sepharose HighTrap column)(ベッドボリューム1−ml、GE ヘルスケア)にかけた。そのカラムを先ず20mMのイミダゾールで洗浄してから、結合タンパク質またはタンパク質複合体を同バッファー中にて0.25Mのイミダゾールで溶出した。最終的な調製液をpH7.0の50mM Hepesバッファーに透析した。
【0081】
次に、MOPSを用いる10%NuPAGEビス−トリスポリアクリルアミドゲルか、酢酸ナトリウムをランニングバッファー(インビトロジェン)に用いる7%SDS/トリスアセテートポリアクリルアミドゲルのいずれかにより、SDS−PAGEを実行した。続いてクマシーブリリアントブルーR−250でタンパク質を染色した。ChemiImager5500(アルファ イノテック(Alpha Innotech)、サンリアンドロ、カリフォルニア)とソフトウェアAlpha EaseFC(v.4.0; アルファ イノテック)を用い濃度測定を行うことによってタンパク質バンドの濃度を定量化した。
【0082】
三重らせんの性質を調べるため、精製されたerb_scFv−Col(1mg/ml)を10mM DTTの非存在または存在下、37℃で1時間インキュベートした。DDT処理サンプルのアリコートをさらに常温で50mM N−エチル−マレイミド(NEM)と30分間反応させて、遊離スルフヒドリル基および三量体の再編成を完全にブロックした。各サンプルからの等量のタンパク質を、酢酸ナトリウムをランニングバッファーとする7% SDS/トリスアセテートポリアクリルアミドゲル上で電気泳動した。そのゲルをクマシーブルーで染色した。精製されたCSAは、ホモ三量体、または、マイルドな還元条件下で2つの三量体に解離可能な鎖間ジスルフィド結合六量体であることが発見された。
【0083】
erb_scFv−Colの三量体構造の熱安定性を調べた。2M尿素含有50mMトリス−HCl(pH8.0)中の精製されたerb_scFv−Colを10mMトリス(2−カルボキシエチル)ホスフィン(TCEP)の非存在または存在下常温で処理した。次いで、還元されたサンプルを常温下50mM NEMでアルキル化した。SDSローディングバッファーを混合する前に、等量のタンパク質を含む各サンプルを35、45、55、65、75および85℃で10分間加熱した。そのサンプルを、非還元条件下、MOPSバッファーを用い10%SDS/ビス−トリスポリアクリルアミドゲル上で電気泳動した。そのゲルをクマシーブルーで染色した。その結果、erb_scFv−Col三量体が高い熱安定性を有することが示された。実際には、65℃で10分間処理したのち、50%を超える三量体が残っていた。また、erb_scFv−Colのコラーゲン様ドメインの三量体構造は、プロリルのヒドロキシル化がなされていることもわかった。
【0084】
結合試験
各種の型のerb抗体のEGFR−ECDに対する結合動態(binding kinetics)を、ランニングバッファーHBS-EP(pH7.4の10mM HEPES、150mM NaCl、3mM EDTA、0.005%界面活性剤P20)中で、BIAcore Xバイオセンサー(ビアコア社(BIACORE, Inc.)、ウプサラ、スウェーデン)を用いて測定した。簡単に言うと、EGFR−ECDをアミンカップリングにより、レスポンス単位(RU)が1700程度となるまで、Clセンサーチップ上に固定化し、各種濃度で精製した抗体を流速10μl/分で注入した。pH 3.5の10mMグリシン−HClを5μl注入することによってその表面を再生した。各濃度におけるセンサーグラムを得ると共に、プログラム、BIA Evaluation3.2を用いて評価した。結合データを1:1ラングミュア結合モデルでフィッティングし、解離速度(kdiss)/会合速度(kass)の比として定義される親和定数KDを計算した。その結果を以下の表2に示す。
【0085】
【表2】
【0086】
表2に示されるように、EGFR−ECDに対するerb_scFv_Colの結合親和性は、二価(erb_scFv−Fc)および一価 (erb_scFv) mAbカウンターパートよりもそれぞれ約20倍および1000倍強かった。
【0087】
安定性および薬物動態アッセイ
血清安定性アッセイでは、各種形態のerb抗体、erb_scFv_Col、erb_scFv−Fcまたはerb_scFvの安定性を37℃にてヒト血清でインキュベートすることにより測定した。各インキュベーション期間経過後に残った活性抗EGFRの量を定量的ELISAにより測定した。ELISAは、(捕捉試薬としての)組み換えEGFR−ECDと抗c-myc mAb(9E10、シグマケミカル社)、続いて、HRPコンジュゲートアフィニティ精製ポリクローナルヤギ抗マウスIgGと化学発光基質(ピアース バイオテクノロジー社(Pierce Biotechnology, Inc.))を用いて行った。薬物動態アッセイでは、3匹のBALB/cヌードマウスを用いてerb_scFv−Colのクリアランスを分析した。簡単に言うと、事前の採血(pre-bleed)を行ってから、各マウスに25μg(2mg/体重キログラム )のerb_scFv−Colを皮下注射(s.c.)した。続く70時間の間に、定期的に血液サンプルを回収し、ELISAによってそれらのerb_scFv−Colの含有量を評価した。タンパク質はきわめて安定しているということがわかった。
【0088】
T細胞増殖アッセイおよび混合リンパ球反応(MLR)
5−ブロモ-2'−デオキシウリジン(BrdU)細胞増殖アッセイを行った。簡単に言うと、ヒト末梢血単核細胞(PBMC)を37℃で66時間、10倍段階希釈したOKT3(イーバイオサイエンス社(eBioscience, Inc.))またはOLT3_scFv−Colの存在下、ブラック96ウェル平底組織培養プレート中に2×105細胞/ウェルとなるように加えた10%FBS含有100μl RPMI−1640培地にプレートした。次いでその細胞を10μMのBrdUにより5時間パルス処理した。培地を除去したのち、FixDenatによりワンステップで細胞の固定とDNAの変性を行った。その後、その細胞をペルオキシダーゼ標識抗BrdU抗体(抗BrdU POD、Fagフラグメント)で1.5時間室温にてインキュベートした。マイクロプレートルミノメーター(Hidex, CHAMELEON detection platform、フィンランド)を用いて化学発光検出および定量化を行った。
【0089】
一方向性混合リンパ球反応におけるT細胞増殖および免疫抑制を以下のようにして評価した。ヒトPBMCを2人の健康なドナーから採取した(刺激および応答)。刺激または応答細胞を、37℃、5%のCO2を含んだ加湿空気中、完全培地(10%ヒトAB血清、2mMグルタミン、50nM 2−メルカプトエタノール、ならびにそれぞれ100単位/mlのペニシリンおよびストレプトマイシンを加えたRPMI 1640)にて25μg/mlのマイトマイシンC(シグマ−アルドリッチ)で30分間処理してから、RPMI 1640培地中で3回洗浄した。応答細胞を単独で、またはマイトマイシンC処理した刺激細胞もしくはマイトマイシンC応答細胞と1:1で混合し、2×105細胞/ウェルとして200μlの完全培地で培養した。応答細胞をプレートしたのち、精製したOKT3_scFv−ColまたはOKT3を異なる濃度で培養物に速やかに加えた。5日後、培養細胞を10μMのBrdUでパルス処理し、24時間後に回収した。次いで、細胞増殖アッセイを上述の手法で行った。
【0090】
OKT3_scFv−Colは、T細胞増殖の免疫抑制においてより効果が高く、かつ、T細胞増殖の刺激においては無視できる程度の細胞分裂促進活性を示すことが見出された。
【0091】
サイトカイン測定
ヒトPBMCを、37℃、10倍段階希釈したOKT3またはOKT3_scFv−Colの存在下、2×105細胞/ウェルとなるように、10%FBS含有0.1ml RPMI−1640培地にプレートした。その上清を各時点で回収し、ヒトサイトカインイムノアッセイキット(イーバイオサイエンス社)を用いて多数のサイトカインを測定した。その結果、OKT3_scFv_Colの投与は、マウスOKT3mAbに比較して、無視できる程度のサイトカイン放出しか生じさせないことが示された。
【0092】
抗体変位(Antibody Displacement)アッセイ
下記する全ての手順は4℃下で行った。ヒトT細胞を1×106細胞/mlの密度でFCMバッファー(2%FBSおよび0.1%アジ化ナトリウム含有リン酸緩衝食塩水)に懸濁させた。その細胞をマウス総IgG(2μg/ml、ジャクソン イムノリサーチ ラボラトリーズ)で30分処理してから、段階希釈したOKT3_scFv−ColまたはOKT3抗体で1時間インキュベートした。固定した、飽和量(フローサイトメトリーにより判断)のFITCコンジュゲートOKT3(0.25μg/ml、イーバイオサイエンス社より購入) を直接加えた。1時間のインキュベーション後、その細胞をFCMバッファーで洗浄し、FACScan(ベクトンデッキンソン、サンジョゼ、カリフォルニア)を用いフローサイトメトリー法により免疫発光解析を行った。そのデータを、遮断抗体の非存在下OKT3−FITCでT細胞を染色することにより得た平均蛍光強度として定義される最大蛍光強度の抑制率(percent inhibition)として表した。
【0093】
その結果、OKT3_scFv_Colが、天然マウスOKT3 mAbよりも、強力にヒトCD3+T細胞に結合することがわかった。
【0094】
ヒトIgGを標準として、Bradfordアッセイ(ピアース バイオテクノロジー社製、クーマシープラス試薬(Coomassie plus reagent))によりタンパク質濃度を測定した。アミノ酸分析については、精製したerb_scFv−Colを50mM酢酸に透析し、110℃で24時間、6NのHCl中で加水分解し、Waters PicoTag(登録商標)システムでアミノ酸分析を行った。
【0095】
これら結果によって、コラーゲン足場抗体が、抗腫瘍および免疫調節いずれの用途においても治療抗体設計に理想的な構造であるということが示される。
【0096】
本明細書に開示されたすべての特徴は、どのような組合せにも組み合わせることができる。本明細書に開示された各特徴は、同一、均等、または類似の目的を果たす他の特徴で置き換えられてもよい。したがって、特に明示する場合を除いては、開示された各特徴は、単に、包括的な一連の均等または同様の特徴の例に過ぎない。
【0097】
上述の記載から、当業者は、本発明の主要な特徴を容易に確認することができると共に、その精神と範囲を逸脱することなく、各種使用および条件に適合するように本発明をさまざまに変更および修飾することができる。よって、その他の実施形態もまた、以下の特許請求の範囲の範囲内に含まれる。
【技術分野】
【0001】
関連出願
本出願は、2005年12月15日に出願された米国特許出願番号60/750,746の優先権を主張するものであり、その内容はここに参照することにより組み込まれる。
【0002】
本発明は、高い親和性、低い細胞分裂促進作用、および高いインビボ安定性を備えるタンパク質複合体ベースの結合試薬に関する。該試薬は多価(multi-valent)あるいは多重特異性(multi-specific)にもなり得る。
【背景技術】
【0003】
タンパク質ベースの結合試薬は治療または診断の適用において様々な用途がある。抗体がこのような試薬の優れたパラダイムであることは証明されている。実際、多くのモノクローナル抗体(mAb)が、癌、感染病、および炎症性疾患の治療のために首尾よく使用されている(Adamsら、Nat. Biotechnol.2005 Sep;23(9):1147-57)。
【0004】
マウスmAbの有効性および安全性はいずれもキメラ化およびヒト化などの組み換えDNA技術によって高められた。しかし、CDR(相補決定領域)グラフティングを用いたマウスmAbのヒト化は、マウス対応物よりも結合親和性が低くなることが多い。抗体親和性は、治療薬としての抗体の成功の鍵となる要因である。高親和性を有する抗体は、標的レセプターに対し天然リガンドと有効に競合できるものとなり、投与量、毒性、およびコストを低減させる。親和性はまた、抗体の薬物動態、例えばその標的組織および宿主の血液循環内における分布と排出にも影響する。インビボで有効なモノクローナル抗体に要求されるのは、無関係のタンパク質に非特異的に結合することのない、標的抗原に対する強い親和性である。抗原結合部位の多量体化は、抗体親和性(機能的親和性(functional affinity))として定義される、抗原への抗体の全体的な結合の強さを向上させる効果的な手段と見られている(Millerら、J Immunol 170:4854-4861、2003; Rheinneckerら、J Immunol 157:2989-2997、1996; Shopes、J Immunol 148:2918-2922、1992; Shufordら、Science 252:724-727、1991; Wolffら、J Immunol 148:2469-2474、1992))。それらはインビボにおける抗腫瘍活性を向上させる(Liu ら、Int Immunopharmacol 6:791-799、2006; Wolffら、Cancer Res 53:2560-2565、1993)。免疫グロブリンG(IgG)の二価の性質により、従来および人工のIgGは、2つを超える異なった抗原に同時に結合させるために用いることはできない。よって、多価または多重特異性のタンパク質ベース結合試薬が必要とされる。
【0005】
場合によって、Fc領域を操作することを通し、例えば抗体依存性細胞介在性障害(ADCC)および補体依存性細胞障害(CDC)のようなエフェクター作用を回避することは、細胞分裂促進性の副作用を低減するために必要である。例として、マウスの抗ヒトCD3 mAb(オルソクローンOKT3、ムロモナブ−CD3)は、ヒトT細胞上のT細胞レセプター(TCR)/CD3複合体を標的にする強力な免疫抑制剤である。過去20年の間、それは同種移植の拒否反応を防ぐまたは治療するために用いられてきた(Cosimiら、N Engl J Med 305:308-314、1981; Group、N Engl J Med 313:337-342、1985; Kungら、Science 206:347-349、1979)。しかし、この治療を用いることの主な欠点の1つは、インフルエンザに似た症状、呼吸困難、神経学的症状および急性尿細管壊死を含む一連の有害な細胞分裂促進作用をもたらすTNF-α、IL-2およびIFN-γのようなサイトカインの全身的な放出である(Abramowiczら、Transplantation 47:606-608、1989; Chatenoudら、N Engl J Med 320:1420-1421、1989; Goldmanら、Transplantation 50:158-159、1990; Toussaintら、Transplantation 48:524-526、1989)。OKT3およびその他の抗CD3 mAbの細胞分裂促進活性は、FcR陽性細胞(例えば単球)への結合を介しての広範なTCR/CD3架橋によるため、最近は、FcRへの結合を修正することによる非細胞分裂促進型の抗CD3抗体の開発に努力が注がれている。したがって、高い親和性、低い細胞分裂促進作用、および高いインビボ安定性を備えるタンパク質ベースの結合試薬が必要とされる。
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明の目的は、高い親和性、低い細胞分裂促進作用、および高いインビボ安定性を備えるタンパク質複合体ベースの結合試薬を提供することにある。該試薬は多価あるいは多重特異性とすることもできる。
【課題を解決するための手段】
【0007】
したがって、本発明の1態様は、第1の足場ドメイン(scaffold domain)および第1の足場ドメインの1つの末端にインフレームで(in-frame)融合する第1の異種ドメイン(heterologous domain)を含む第1の融合ポリペプチド鎖と、第2の足場ドメインを含む第2の融合ポリペプチド鎖と、第3の足場ドメインを含む第3の融合ポリペプチド鎖とを含んでいる単離組み換えタンパク質複合体に特徴を有する。第1、第2および第3の足場ドメインは、三重らせんコイル(triple helix coil)を形成するように並ぶ。第1の足場ドメインおよび第1の異種ドメインはインフレームで融合し、かつ同一のペプチド鎖に存する。
【0008】
異種ドメインには酵素ドメインまたは蛍光タンパク質の配列が含まれ得る。蛍光タンパク質の例としては、GFPおよびdsRed、さらにそれらの変異体などがある。酵素ドメインの例には、グルタチオンS-トランスフェラーゼ、ルシフェラーゼ、β-ガラクトシダーゼ、およびβ-ラクタマーゼのドメインが含まれる。
【0009】
異種ドメインには、結合パートナーと結合する結合ドメイン(例えばリガンド結合ドメイン、リガンド、レセプター、またはプロテオグリカン)が含まれ得る。「結合パートナー」とは、目的とする所望の化合物(例えばタンパク質)の部分に対して特異的、共有結合的または非共有結合的な親和性を備える任意の分子を意味する。結合パートナーの例としては、抗原/抗体のペア、タンパク質/阻害剤のペア、レセプター/リガンドのペア(例えば細胞表面または核レセプター/リガンドのペア)、酵素/基質のペア(例えばキナーゼ/基質のペア)、レクチン/炭水化物のペア、オリゴマーまたはへテロオリゴマータンパク質のペア、DNA結合タンパク質/DNA結合部位のペア、およびRNA/タンパク質のペアがある。また、結合ドメインの例には、例えばヒスチジンタグ、mycタグ、またはヘマグルチン(hemagglutin)タグのような親和性タグの配列も含まれる。
【0010】
上述した第1の異種ドメインには、免疫グロブリンの1つまたはそれ以上の相補決定領域(CDR)が含まれてもよい。よって、異種ドメインには、例えばVHドメインおよびFabのような抗体の抗原結合部位が含まれ得る。1つの実施形態において、第1の異種ドメインには、抗原結合性フラグメントまたは一本鎖抗体、例えば分化抗原3(CD3)または上皮増殖因子レセプター(EGFR)に特異なものの配列が含まれる。第1のポリペプチド鎖はさらに、第1の足場ドメインのもう1つの末端にインフレームで融合する第2の異種ドメインを含んでいてもよい。第1の異種ドメインのように、第2の異種ドメインにも、結合パートナーと結合するドメインが含まれ得る。第1および第2の異種ドメインは互いに同一であってもまたは異なっていてもよい。これらは、同一の結合パートナーまたは2つの異なる結合パートナーに結合することができる。例えば、第1の異種ドメインおよび第2の異種ドメインには、CD3およびEGFRにそれぞれ特異的に結合する第1の一本鎖抗体および第2の一本鎖抗体の配列が各々含まれていてもよい。
【0011】
上述したタンパク質複合体において、第2の融合ポリペプチド鎖は、第2の足場ドメインの1つの末端にインフレームで融合する第3の異種ドメイン、第2の足場ドメインのもう1つの末端にインフレームで融合する第4の異種ドメイン、または2つの末端にインフレームで融合する両方のドメインを含んでいてもよい。同様に、第3の融合ポリペプチド鎖は、第3の足場ドメインの1つの末端にインフレームで融合する第5の異種ドメイン、または、第3の足場ドメインのもう1つの末端にインフレームで融合する第6の異種ドメイン、または、両方を含んでいてもよい。上述した第1および第2の異種ドメインのように、これら4つの異種ドメインの各々には、結合パートナーと結合する結合ドメインも含まれ得る。6つ全ての異種ドメインは互いに同一であってもよくまたは異なってもよい。よってこれらは、1、2、3、4、5または6つの結合パートナーと結合できる。言い換えると、当該タンパク質複合体は、一、二、三、四、五、または六価であり得る。
【0012】
第1、第2、および第3の足場ドメインが三重らせんコイルを形成するよう、3つの足場ドメインのそれぞれは1つまたはそれ以上の三重らせんリピートを含んでおり、各リピートは次の式:(X1−X2−X3)nの配列を含み、式中、X1はGly残基であり、X2およびX3は任意のアミノ酸残基、好ましくはイミノ酸のプロリンまたはヒドロキシプロリンであり、かつ、nは5またはこれより大きい数である。例えば、第1、第2、および第3の足場ドメインは、(GPP)10、または、1つもしくはそれ以上のヒト補体C1q、コレクチン、もしくはコラーゲンポリペプチド鎖の三重らせんリピートを含むことができる。
【0013】
1つの実施形態において、上述した第1、第2および第3の融合ポリペプチドは実質的に同じであり、互いに少なくとも75%(例えば75%から100%の間の任意の数を含む)の配列同一性を有している。3つの同じ融合ポリペプチドから形成される複合体はホモ三量体である。これら3つの融合ポリペプチドは機能的に等価であり得る。「機能的に等価」とは、例えば、1もしくはそれ以上の点変異、挿入、欠失、切断を有するタンパク質、融合タンパク質、またはそれらの組み合わせなど共通のポリペプチドのポリペプチド誘導体が、三重らせんコイルを形成する能力と、例えばリガンドに結合するといった異種ドメインの活性を実質的に保持していることをいう。
【0014】
異種ポリペプチド、核酸、または遺伝子は異種から由来するもの、または、同種からのものである場合はその原形に実質的に修飾が加えられたものである。2つの融合ドメインまたは配列は、それらが天然のタンパク質または核酸中で互いに隣り合っていない場合は、互いに異種である。例えば、天然のヒトミニコラーゲン(XXI型)、コレクチンファミリータンパク質、または補体1q(C1q)の一部でないポリペプチド配列は、ヒトタンパク質におけるドメインに対して異種である。
【0015】
本発明はまた、(i)三重らせんコイルを形成するのに用いられる足場ドメイン、および(ii)足場ドメインの1つの末端にインフレームで融合する第1の異種ドメイン、または足場ドメインのもう1つの末端にインフレームで融合する第2の異種ドメインを含む単離組み換え融合ポリペプチド(例えば上述した3つの各融合ポリペプチド)にも特徴を有する。足場ドメインは、1つまたはそれ以上の上述した三重らせんリピート、例えばヒトC1qまたはコラーゲンポリペプチド鎖のそれを含み得る。異種ドメインは、上述した結合ドメインの1つを含んでいてもよく、かつ、ファージディスプレイスクリーニングのような当該分野で知られるさまざまな方法によって得ることができる。
【0016】
単離ポリペプチドまたはタンパク質複合体とは、天然の関連分子を実質的に含まないポリペプチドまたはタンパク質複合体のことをいい、つまり、それは乾燥重量で少なくとも純度75%(すなわち75%および100%の間の任意の数を含む)である。純度は任意の適した標準的方法、例えばカラムクロマトグラフィ、ポリアクリルアミドゲル電気泳動、またはHPLC分析によって測定することができる。本発明の単離ポリペプチドまたはタンパク質複合体は、天然資源から精製でき、また組み換えDNA技術で作製することができる。
【0017】
本発明は、先ほど述べた融合ポリペプチドをコードする配列またはその配列の補体を含む単離核酸にも特徴を有する。核酸とはDNA分子(例えばcDNAもしくはゲノムDNA)、RNA分子(例えばmRNA)、またはDNAもしくはRNAアナログのことをいう。DNAまたはRNAアナログはヌクレオチドアナログから合成されたものであってよい。核酸分子は、一本鎖または二本鎖であってもよいが、好ましいのは二本鎖DNAである。「単離核酸」とは、その構造が、どの天然に存在する核酸の構造とも、またはどの天然に存在するゲノム核酸のフラグメントの構造とも同一でない核酸である。よってこの用語は、例えば(a)天然に存在するゲノムDNA分子の一部の配列を有するが、生物に天然に存在するゲノムにおいてその分子の一部の側方に位置する2つのコーディング配列の隣にはないDNA、(b)得られる分子がいかなる天然に存在するベクターまたはゲノムDNAとも同一にならないような方式で、ベクターまたは原核生物もしくは真核生物のゲノムDNAに組み込まれる核酸、(c)例えばcDNA、ゲノムフラグメント、ポリメラーゼ連鎖反応(PCR)により作製されたフラグメントまたは制限フラグメントのような分離された分子、および(d)ハイブリッド遺伝子、つまり融合タンパク質をコードする遺伝子の一部である組み換えヌクレオチド配列、を包含する。上述した核酸は本発明のポリペプチドを発現するのに用いることができる。この目的のために、核酸を適した調節配列へ動作可能に連結させて発現ベクターを生成することができる。
【0018】
ベクターとは、それに連結された別の核酸を運ぶ能力のある核酸分子のことをいう。ベクターは自己複製または宿主DNAと統一化することが可能である。ベクターの例としては、プラスミド、コスミド、またはウィルスベクターなどがある。本発明のベクターは、宿主細胞における核酸の発現に適したかたちで核酸を含む。ベクターが、発現させようとする核酸配列に操作により連結した1つまたはそれ以上の調節配列を含んでいると好ましい。「調節配列」には、プロモーター、エンハンサー、またはその他の発現制御要素(例えばポリアデニル化信号)が含まれる。調節配列には、組織特異的な調節および/または誘導(inducible)配列だけでなく、ヌクレオチド配列を直接構成的発現させるものも含まれる。発現ベクターの設計は、形質転換させるべき宿主細胞の選択、所望のタンパク質の発現レベルなどといった要素に応じて行うことができる。発現ベクターを宿主細胞に導入することにより本発明のポリペプチドが生産される。上述した核酸を含む宿主細胞もまた本発明の範囲内に含まれる。例として、E.Coli細胞、昆虫細胞(例えばショウジョウバエS2またはバキュロウイルス細胞を使用)、酵母細胞、または哺乳動物の細胞(例えばマウス骨髄腫NS0細胞)などがある。例えば、Goeddel、(1990)Gene Expression Technology:Method in Enzymology 185、Academic Press、San Diego、CAを参照されたい。
【0019】
本発明の融合ポリペプチドを作製するには、本発明の核酸によってコードされるポリペプチドの発現を可能とする条件下、培地中で宿主細胞を培養し、培養された細胞または細胞の培地からポリペプチドを精製することができる。あるいは、本発明の核酸は、例えばT7プロモーター調節配列およびT7ポリメラーゼを用いてインビトロの複写および翻訳がされてもよい。
【0020】
本発明のタンパク質複合体を作製するには、上述した第1、第2および第3の融合ポリペプチドをそれぞれコードする第1、第2および第3の核酸を含む宿主細胞を、これら3つの核酸によりコードされるポリペプチドの発現、および、発現されたポリペプチド間における三重らせんコイルの形成を可能とさせる条件下、培地中で培養し、培養された細胞または細胞の培地からタンパク質複合体を精製することができる。宿主細胞が、プロリン残基をヒドロキシル化する酵素活性を含む真核細胞であると好ましい。
【0021】
本発明の1つまたはそれ以上の実施形態の詳細を、添付の図面および以下の記載において説明する。本発明のその他の特徴、目的および特長は、当該記載および図面から、そして特許請求の範囲から明らかとなるであろう。
【図面の簡単な説明】
【0022】
【図1】ヒトXXI型コラーゲン由来のミニコラーゲン三重らせんコイル足場を有するタンパク質複合体を示す図である。三重らせんコイルの6つの末端は、一本鎖抗体のVLおよびVHドメインを有する6つのFvフラグメント(scFv)にそれぞれインフレームで融合している。
【図2】それぞれ、(A)その3つのN末端が3つの一本鎖抗体OKT3(抗CD3)、528(抗EGFR)、およびerb_scFv(抗EGFR)にそれぞれインフレームで融合する三重らせんコイル足場を有するタンパク質複合体を示す図、(B)該タンパク質複合体のウェスタンブロットの結果を示す写真である。安定にトランスフェクトされたショウジョウバエS2細胞由来の培地を、非還元条件下SDS−PAGEで電気泳動してから、XXI型コラーゲン、3E2のC末端に対するモノクローナル抗体で免疫ブロットした。Tは鎖間でジスルフィド結合した三量体であり、Mtは鎖間でジスルフィド結合した三量体を含む単量体である。
【図3】三重らせんコイル足場を有するタンパク質複合体を示す図である。三重らせんコイルの3つのN末端は3つのOKT3一本鎖抗体にインフレームで融合し、三重らせんコイルの3つのC末端は3つの528一本鎖抗体にインフレームで融合している。
【図4】それぞれ異なる形式の抗体を説明するものである。(A)コラーゲン足場抗体:アミノ末端scFv、ヒトIgGのヒンジ領域、コラーゲン様ドメイン(GPP)10、およびXXI型コラーゲンのカルボキシル末端NC1ドメインを含むscFv−Col(左パネル)、サーファクタントタンパク質D(SPD)コレクチンのアミノ末端部およびカルボキシル末端のscFvを含むNSPD−scFv(右パネル)。(B)左から右に:免疫グロブリンG(IgG)、キメラ(scFv−Fc)、および一本鎖抗体(scFv)と、これらのおおよその分子量。グレーのエリアはVHおよびVLフラグメントを表す。点線:鎖間ジスルフィド結合。
【発明を実施するための形態】
【0023】
本発明は、ヒト三重らせんコイルの足場ドメインにインフレームで融合する異種タンパク質結合ドメインが結合活性を保持する、および得られる融合ポリペプチドが三重らせんコイルを形成する、という予期せぬ発見に、少なくとも一部基づいている。図1に示されているのは、本発明のタンパク質複合体の1例である。この図に示されているように、三重らせんコイルタンパク質複合体の6つの末端は、6つの異種タンパク質結合ドメイン、すなわち一本鎖抗体(scFv)のFvフラグメントにインフレームで融合する。
【0024】
本発明のタンパク質複合体は従来の抗体よりも優れている。また、6つのドメインのうち2つまたはそれ以上が互いに同一である場合、該タンパク質複合体は1つの結合パートナー(例えば抗原)に特異な2〜6つの結合ドメインを備えることができ、これに比べて従来の抗体はそのようなドメインを2つしか備えない。言い換えると、抗原に対し二価でしかない従来の抗体とは異なり、当該タンパク質複合体は二、三、四、五または六価となり得る。その結果として、従来の抗体よりも高い各種親和性を持つようにすることができる。その高い親和性のために、従来の抗体よりも、必要とされるタンパク質複合体の量は少なく、かつインキュベーション期間は短くなり、望ましいゴールが達成される。例えば治療上の効果としては、これにより治療コストが低くなり、かつ副作用(例えば望ましくない免疫反応)が最小限に抑えられる。一方、6つのドメインのうち2つまたはそれ以上が互いに異なる場合、本発明のタンパク質複合体は、2〜6つの異なる結合パートナーに特異である2〜6つの結合ドメインを備えることができる。特異性の異なる多数の結合パートナー部位を1ユニットに統合することによって、多数の結合パートナーをまとめる能力が備わるため、治療、組織再生、および活性タンパク質機械(active protein machinery)(例えばマルチサブユニット酵素)の組立において、ナノメーターレベルでの望ましい使用が可能となる。
【0025】
ヒトのインビボでの使用において、本発明のタンパク質複合体はヒト由来であることが好ましい。例えば、それには、ヒトC1q、コレクチンファミリータンパク質、またはコラーゲンポリペプチド鎖のようなヒト由来のらせんコイル足場にインフレームで融合するヒト化一本鎖抗体配列などが含まれる。ヒトC1qおよびコラーゲンはいずれも血中で極めて安定であるので、当該タンパク質複合体は従来の抗体に比べてより安定である。
【0026】
コラーゲンは哺乳動物に存在する最も豊富なタンパク質である。それは、リピートトリプレット配列Gly−X1−X2を含むと共に、かかるトリプレットの存在が3本のコラーゲンのポリペプチド鎖(α鎖)を三重らせん構造に折り畳ませ得る細胞外基質タンパク質である。トリプレット配列Gly−X1−X2におけるX2の位置のアミノ酸はプロリンであることが多く、それはコラーゲンの三重らせん構造を安定にするためコラーゲンポリペプチド鎖の翻訳後修飾によってしばしば4−ヒドロキシル化される。プロリンのヒドロキシル化なしには、コラーゲンの主要な三重らせん構造が生理的温度以下で熱的に不安定になる(BergおよびProckop、Biochem Biophys Res Commun 52:115-120、1973;Rosenbloomら、Arch Biochem Biophys 158:478-484、1973)。ヒトの血清中には多くのコラーゲン様ドメインを有するコラーゲン様タンパク質が存在しており、感染性生物からの防御において先天性免疫システムの役目を果たす。これらには、補体タンパク質C1q、コレクチンファミリータンパク質−マンノース結合レクチン(MBL)、サーファクタントタンパク質AおよびD(SP−AおよびSP−D)が含まれる。これらコラーゲン様タンパク質間で共通な構造的特徴は、これらがいずれも、そのコラーゲン様ドメインの三重らせんが集合した後、三量体分子が堆積または鎖間ジスルフィド架橋することにより、多量体タンパク質ユニットの形をとることにある。したがって、これら「防御コラーゲン」分子の結合ドメインの機能的親和性(functional affinity)は、多量体化を通して大いに向上する。
【0027】
本発明のタンパク質複合体またはポリペプチドは組み換え技術により得ることができる。該複合体のポリペプチドをコードする適当な宿主細胞の核酸に導入してから、該核酸にコードされたポリペプチドの発現と該ポリペプチド間における三重らせんコイルの形成とを可能とする条件下でポリペプチドを発現させることができる。三重らせんコイル足場の形成を促進するために、コラーゲン生合成の鍵酵素である宿主細胞のプロリル4−ヒドロキシラーゼ(P4HA)で同時発現させることが可能である。
【0028】
異種タンパク質ドメインには抗体またはそのフラグメント(例えばその抗原結合フラグメント)が含まれ得る。ここで用いられる「抗体」という用語は、免疫グロブリン分子またはその免疫学的に活性である部位、すなわち抗原結合部位のことをいう。それは、少なくとも1つ、そして好ましくは2つの重(H)鎖可変領域(VH)と、少なくとも1つ、そして好ましくは2つの軽(L)鎖可変領域(VL)とを含むタンパク質のことをいう。VHおよびVL領域はさらに、「フレームワーク領域」(FR)と呼ばれるより保存性の高い領域の間に位置する、「相補決定領域(「CDR」)」と呼ばれる超可変の領域に細分されていてもよい。フレームワーク領域およびCDRの範囲ははっきりと定義がされている(参照することにより本明細書に組み込まれるKabatら(1991)Sequences of Proteins of Immunological Interes、Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242、およびChothiaら(1987)J. Mol. Biol. 196:901-917を参照)。各VHおよびVLはアミノ末端からカルボキシ末端まで次の順序:FR1、CDR1、FR2、CDR2、FR3、CDR3、FR4、で並ぶ3つのCDRおよび4つのFRよりなっている。抗体はさらに、重および軽鎖定常領域を含み、これにより重および軽免疫グロブリン鎖がそれぞれ形成されるようになっていてもよい。重鎖定常領域は3つのドメイン、CH1、CH2およびCH3からなる。軽鎖定常領域は1つのドメイン、CLからなる。重および軽鎖の可変領域は、抗原と作用する結合ドメインを含む。抗体の定常領域は通常、免疫システムの各種細胞(例えばエフェクター細胞)および古典的補体系の第1成分(C1q)を含む宿主組織または因子への抗体の結合を仲介する。
【0029】
本明細書で「免疫グロブリン」という用語は、実質的に免疫グロブリン遺伝子によってコードされる1つまたはそれ以上のポリペプチドからなるタンパク質のことをいう。存在が認められているヒト免疫グロブリン遺伝子には、カッパ、ラムダ、アルファ(IgA1およびIgA2)、ガンマ(IgG1、IgG2,IgG3、IgG4)、デルタ、エプシロンならびにミュー定常領域遺伝子、そしてさらに無数の免疫グロブリン可変領域遺伝子がある。完全長の免疫グロブリン「軽鎖」(約25KDaまたは214のアミノ酸)は、NH2-末端(約110のアミノ酸)の可変領域遺伝子と、COOH-末端のカッパまたはラムダ定常領域遺伝子とによってコードされる。同じように、完全長の免疫グロブリン「重鎖」(約50KDaまたは446のアミノ酸)は、可変領域遺伝子(約116のアミノ酸)と、他の上記定常領域遺伝子のうちの1つ、例えばガンマ(約330のアミノ酸をコードする)とによってコードされる。
【0030】
抗体の「抗原結合フラグメント」(または「抗体部分(antibody portion)」、または「フラグメント」)は、本明細書においては、例えばEGFRまたはCD3ポリペプチドあるいはそのフラグメントである抗原に特異的に結合する能力を保持する1つまたはそれ以上の完全長の抗体のフラグメントのことをいう。抗体の抗原結合フラグメントの例としては、限定はされないが、(i)Fabフラグメント、VL、VH、CLおよびCH1ドメインからなる一価フラグメント、(ii)F(ab’)2フラグメント、ヒンジ領域にてジスルフィドブリッジにより連結される2つのFabフラグメントを含む二価フラグメント、(iii)VHおよびCHlドメインからなるFdフラグメント、(iv)抗体の1本のアームのVLおよびVHドメインからなるFvフラグメント、(v)VHドメインからなるdAb領域(Wardら、(1987) Nature 341:544-546)、(vi)単離相補決定領域(CDR)、ならびに(vii)VLまたはVHドメインが含まれる。さらに、Fvフラグメントの2つのドメイン、VLおよびVHは、別々の遺伝子によりコードされるが、これらは組み換え法を用い、合成リンカーにより結合させることができ、この合成リンカーはこれらを、VLおよびVH領域が組になって一価分子を形成する一本のタンパク質鎖とさせることができる(一本鎖Fv(scFv)として知られる。例えばBirdら (1988) Science 242:423-426、およびHustonら (1988)PROC. Natl. Acad. Sci USA 85:5879-5883参照)。このような一本鎖抗体もまた、抗体の「抗原結合フラグメント」という用語に包含される。これらの抗体フラグメントは当業者に知られる従来の手法により得ることができ、かつ、当該フラグメントは実用に供することのできるよう、インタクト抗体と同じ方式で選別される。
【0031】
適した抗体にはモノクローナル抗体がある。別の実施形態において、抗体は、組み換え技術、例えばファージディスプレイまたはコンビナトリアル手法により作製したものであってもよい。抗体を作製するためのファージディスプレイおよびコンビナトリアル手法は当該技術分野において周知である(例えば、Ladnerら、米国特許第5,223,409号明細書;Kangら、国際公開第92/18619号パンフレット;Dowerら、国際公開第91/17271号パンフレット;Winterら、国際公開第O92/20791号パンフレット;Marklandら、国際公開第92/15679号パンフレット;Breitlingら、国際公開第93/01288号パンフレット;McCaffertyら、国際公開第92/01047号パンフレット;Garrardら、国際公開第92/09690号パンフレット;Ladnerら、国際公開第90/02809号パンフレット;Fuchsら、(1991)Bio/Technology 9:1370-1372;Hayら、(1992) Hum Antibod Hybridomas 3:81-85;Huseら、(1989)Science 246:1275-1281;Griffthsら、(1993)EMBO J 12:725-734;Hawkinsら、(1992)J Mol Biol 226:889-896;Clacksonら、(1991)Nature 352:624-628;Gramら、(1992) PNAS 89:3576-3580;Garradら、(1991)Bio/Technology 9:1373-1377;Hoogenboomら、(1991)Nuc Acid Res 19:4133-4137、およびBarbasら、(1991)PNAS 88:7978-7982を参照、これらすべての内容は参照することにより本明細書に組み込まれる)。
【0032】
1つの実施形態において、抗体は、完全ヒト抗体(例えば、ヒト免疫グロブリン配列からの抗体を産出するよう遺伝子操作を受けたマウスで作られた抗体)、または非ヒト抗体、例えば齧歯動物(マウスまたはラット)、ヤギ、霊長類の動物(例えばサル)、ラクダの抗体である。非ヒト抗体は齧歯動物(マウスまたはラット抗体)とするのが好ましい。齧歯動物抗体を作製する方法は当該技術分野において周知である。
【0033】
ヒトモノクローナル抗体は、マウスの系ではなく、ヒト免疫グロブリン遺伝子を持つトランスジェニックマウスを用いて生成することができる。目的の抗原で免疫されたこれらトランスジェニックマウスからの脾細胞は、ヒトタンパク質のエピトープに対し特異的親和性があるヒトmAbを分泌するハイブリドーマを作出するのに用いられる(例えば、Woodら、国際公開第91/00906号パンフレット;Kucherlapatiら、国際公開第91/10741号パンフレット;Lonbergら、国際公開第92/03918号パンフレット;Kayら、国際公開第92/03917号パンフレット;Lonberg,Nら、1994 Nature 368:856-859、Green,L.Lら、1994 Nature Genet. 7:13-21;Morrisonら、1994 Proc. Natl.Acad. Sci. USA 81:6851-6855;Bruggemanら、1993 Year Immunol 7:33-40;Tuaillonら、1993 PNAS 90:3720-3724;Bruggemanら、1991 Eur J Immuno 21:1323-1326を参照。)。
【0034】
抗体は、可変領域または例えばCDRなどその一部が、非ヒト生物、例えばラットまたはマウスにおいて産出されたものであってよい。キメラ、CDR−グラフト、およびヒト化抗体が使用可能である。例えばラットまたはマウスのような非ヒト生物で産出されてから、ヒトにおける抗原性を低減するために例えば可変フレームワークまたは定常領域において修飾がなされた抗体は、本発明の範囲に含まれる。
【0035】
キメラ抗体は、当該技術分野において周知である組み換えDNA技術によって作製することができる。例として、マウス(またはその他の種の)モノクローナル抗体分子のFc定常領域をコードする遺伝子を制限酵素により消化して、マウスFcをコードする領域を除去してから、ヒトFc定常領域をコードする遺伝子の相当部位で置換する(Robinsonら、国際特許出願PCT/US86/02269号;Akiraら、欧州特許出願第184,187号明細書;Taniguchi,M、欧州特許出願第171,496号明細書;Morrisonら、欧州特許出願第173,494号明細書;Neubergerら、国際公開第86/01533号パンフレット;Cabillyら、米国特許第4,816,567号明細書;Cabillyら 欧州特許出願第125,023号明細書;Betterら、(1998 Science 240:1041-1043);Liuら、(1987) PNAS 84:3439-3443;Liuら、1987,J.Immunol 139:3521-3526;Sunら、(1987) PNAS 84:214-218;Nishimuraら、1987,Canc.Res. 47:999-1005;Woodら、(1985) Nature 314:446-449;およびShawら、1998, J. Natl Cancer Inst. 89:1553-1559を参照)。
【0036】
ヒト化またはCDRグラフト抗体は、少なくとも1つまたは2つ、ただし通常は3つすべてのレシピエントの(重およびまたは軽免疫グロブリン鎖の)CDRが、ドナーCDRで置き換えられる。抗体が非ヒトCDRの少なくとも1部で置き換えられても、あるいは、CDRのいくつかだけが非ヒトCDRで置き換えられてもよい。ヒト化抗体またはそのフラグメントの結合に要されるCDRの数だけ置き換えればよいのである。ドナーが齧歯動物の抗体、例えばラットまたはマウス抗体であり、かつレシピエントがヒトフレームワークまたはヒトコンセンサスフレームワークであると好ましい。一般に、CDRを提供する免疫グロブリンは「ドナー」と呼ばれ、フレームワークを提供する免疫グロブリンは「アクセプター」と呼ばれる。1つの実施形態において、ドナー免疫グロブリンはヒト以外(例えば齧歯動物)である。アクセプターのフレームワークは、天然(例えばヒト)フレームワークまたはコンセンサスフレームワーク、または、約85%またはこれ以上、好ましくは90%、95%、99%またはこれ以上がそれと同一な配列である。本明細書で「コンセンサス配列」とは、関連配列のファミリー中にて最も頻繁に出現するアミノ酸(またはヌクレオチド)から形成される配列のことをいう(例えば、Winnaker、From Genes to Clones (Verlagsgesellschaft, Weinheim, Germany 1987) 参照)。タンパク質のファミリーにおいて、コンセンサス配列中の各位置は、該ファミリーにおけるその位置に最も頻繁に出現するアミノ酸によって占められる。2つのアミノ酸が同等に頻繁に出現する場合は、どちらかがコンセンサス配列に含まれ得る。「コンセンサスフレームワーク」とは、コンセンサス免疫グロブリン配列中のフレームワーク領域のことをいう。
【0037】
抗体は当該技術分野において周知の方法でヒト化することができる。ヒト化抗体は、抗原の結合に直接かかわりのないFv可変領域の配列を、ヒトFv可変領域からの相当する配列で置き換えることによって生成することができる。ヒト化抗体を生成する一般的な方法は、Morrison,S.L、1985,Science 229:1202-1207、Oiら、1986,BioTechniques 4:214、ならびにQueenら、米国特許第5,585,089号明細書、米国特許第5,693,761号明細書および米国特許第5,693,762号明細書により提示されており、これらのすべての内容は参照することにより本明細書に組み込まれる。それらの方法は、少なくとも1つの重または軽鎖からの免疫グロブリンFv可変領域のすべてまたは一部をコードする核酸配列を単離する工程、操作する工程、および発現させる工程を含む。かかる核酸のソースは当業者にはよく知られており、例えば、目的のポリペプチドまたはそのフラグメントに対する抗体を産出するハイブリドーマから得ることができる。よって、ヒト化抗体またはそのフラグメントをコードする組み換えDNAを、適当な発現ベクターにクローニングすることができる。
【0038】
特定のアミノ酸が置換、欠失または追加されたヒト化抗体は足場に融合することもできる。好ましいヒト化抗体は、例えば抗原への結合を改善する目的で、フレームワーク領域にアミノ酸置換基を有する。例えば、ヒト化抗体は、ドナーのフレームワークの残基またはレシピエントのフレームワーク残基以外の別なアミノ酸と同一であるフレームワークの残基を備える。このような抗体を生成するため、選択された、少数のヒト化免疫グロブリン鎖のアクセプターフレームワーク残基を、対応するドナーアミノ酸で置き換えることができる。好ましい置換基の配置としては、アミノ酸残基をCDRに隣接させる、またはCDRと作用することのできる配置などがある。ドナーからアミノ酸を選ぶ基準は、US5,585,089号に記載されており、その内容は本明細書では参照することにより組み込まれる。ヒト化抗体のその他の技術は、Padlanら、欧州特許出願公開第519596号明細書に記載されている。
【0039】
本発明は、本発明のタンパク質複合体を形成する融合ポリペプチドをコードする核酸をも含む。核酸は、ファージディスプレイライブラリーから選別された、または上記の適した抗体または抗体の誘導体を発現する細胞株から(RT−PCRによって)単離されたものとすることができる。核酸は発現ベクターに機能的にライゲートされてもよい。核酸またはベクターで形質転換された細胞を、本発明の融合ポリペプチドまたはタンパク質複合体を作製するのに用いることができる。抗体を作製するのに有用な細胞としては、昆虫細胞および哺乳動物の細胞(例えばCHOまたはリンパ細胞)などがある。
【0040】
本発明のタンパク質複合体は、細胞毒素のような治療成分(therapeutic moiety)、治療薬または放射性イオンにコンジュゲートすることができる。細胞毒素または細胞毒性薬には、細胞に有害なあらゆる薬剤が含まれる。例としては、タクソール、サイトカラシンB、グラミシジンD、エチジウムブロマイド、エメチン、マイトマイシン、エトポシド、テノポシド(tenoposide)、ビンクリスチン、ビンブラスチン、コルヒチン、ドキソルビシン、ダウノルビシン、ジヒドロキシアントラシンジオン、ミトキサントロン、ミトラマイシン、アクチノマイシンD、1−デヒドロテストステロン、グルココルチコイド、プロカイン、テトラカイン、リドカイン、プロプラノロール、ピューロマイシン、メイタンシノイド(maytansinoids)、例えばメイタンシノール(maytansinol)(米国特許第5,208,020号明細書参照)、CC−1065(米国特許第5,475,092号明細書、米国特許第5,585,499号明細書、米国特許第5,846,545号明細書参照)およびそのアナログまたはホモログがある。治療薬には、限定はされないが、代謝拮抗物質(例えばメトトレキサート、6−メルカプトプリン、6−チオグアニン、シタラビン、5−フルオロウラシルデカルバジン)、アルキル化剤(例えばメクロレタミン、チオエパクロラムブシル(thioepa chlorambucil)、CC−1065、メルファラン、カルムスチン(BSNU)およびロムスチン(CCNU)、シクロトスファミド(cyclothosphamide)、ブスルファン、ジブロモマンニトール、ストレプトゾトシン、マイトマイシンC、およびシスジクロロジアミン白金(II)(DDP)(シスプラチン)、アントラサイクリン(例えばダウノルビシン(旧ダウノマイシン)およびドクソルビシン)、抗生物質(例えばダクチノマイシン、(旧アクチノマイシン)、ブレオマイシン、ミトラマイシンおよびアントラマイシン(AMC))、ならびに抗分裂剤(例えばビンクリスチン、ビンブラスチン、タクソールおよびメイタンシノイド)が含まれる。放射性イオンには、限定はされないが、ヨード、イットリウムおよびプラセオジムが含まれる。
【0041】
コンジュゲートは所定の生物学的反応を修飾するのに用いることができ、薬物成分(drug moiety)は古典的な化学治療薬に限定されると解されるべきではない。例えば、薬物成分は、所望の生物学的活性を備えたタンパク質またはポリペプチドであり得る。かかるタンパク質には、例として、アブリン、リシンA、緑膿菌外毒素もしくはジフテリア毒素のような毒素、腫瘍壊死因子、α−インターフェロン、β−インターフェロン、神経成長因子、血小板由来成長因子、組織プラスミノーゲンアクチベータのようなタンパク質、または、例えばリンフォカイン、インターロイキン−1(「IL−1」)、インターロイキン−2(「IL−2」)、インターロイキン−6(「IL−6」)、顆粒球マクロファージコロニー刺激因子(「GM−CSF」)、 顆粒球コロニー刺激因子(「G-CSF」)もしくはその他の成長因子のような生物学的反応修飾物質が含まれる。
【0042】
異種結合ドメインの特異性をベースとしている上述のタンパク質複合体およびコンジュゲートは、ガン、炎症疾患、代謝疾患、繊維化疾患、および心臓血管疾患を含むさまざまな疾患を治療するために用いることができる。よって本発明は、例えば、それを必要とする対象に本発明のタンパク質複合体を有効量投与することによって、このような疾患を治療する方法に特徴を有する。治療を受けるべき対象は、疾患に特徴付けられる状態を持つまたは獲得する危険があるものとして確認することができる。この方法は、単独で、またはその他の薬物もしくは治療法と併用して行うことが可能である。
【0043】
その多重特異性という特徴により、本発明のタンパク質複合体は、本来は互いにつながっていない分子または細胞を架橋するのに用いられ得る。この特徴は細胞ベースの治療に対しとりわけ有用である。一例において、タンパク質複合体における1つの異種ドメインは、細胞障害性細胞上のエフェクター抗原に特異的に結合することによって細胞傷害性細胞(例えば細胞傷害性T細胞)を活性化させることができ、同時に他の異種ドメインは、破壊するべき病原細胞または悪性細胞上の標的抗原に特異的に結合する。このようにして、タンパク質複合体は病原または悪性細胞によって引き起こされる疾病を治療することができるのである。
【0044】
細胞傷害性T細胞の活性化は、本発明のタンパク質複合体による、細胞障害性T細胞表面のエフェクター抗原としてのCD3抗原の結合を通して起こり得る。その他のリンパ球様細胞結合型(lymphoid cell-associated)エフェクター抗原には、ヒトCD16抗原、NKG2D抗原、NKp46抗原、CD2抗原、CD28抗原、CD25抗原、CD64抗原、およびCD89抗原などがある。これらエフェクター抗原に結合することによって、単球、好中性顆粒球および樹枝状細胞のようなエフェクター細胞の活性化がもたらされる。よって、これら活性化された細胞は、標的細胞に対し細胞傷害またはアポトーシス効果を発揮する。
【0045】
標的抗原は、病気の状態に付随する標的細胞上にユニークに発現(uniquely express)する抗原であるが、健康な状態では発現しない、低レベルで発現する、あるいはアクセスできない(non-accessible)ものである。悪性細胞に関連するかかる標的抗原の例には、EpCAM、CCR5、CD19、HER−2neu、HER−3、HER−4、EGFR、PSMA、CEA、MUC−1(ムチン)、MUC2、MUC3、MUC4、MUC5.sub.AC、MUC5.sub.B、MUC7、ベータ.hCG、ルイス−Y、CD20、CD33、CD30、ガングリオシドGD3、9−O−アセチル−GD3、GM2、Globo H、フコシル GM1、ポリSA、GD2、カルボアンヒドラーゼ(Carboanhydrase)IX (MN/CA IX)、D44v6、ソニックヘッジホッグ(Shh)、Wue−1、形質細胞抗原、(膜結合性)IgE、メラノーマコンドロイチンサルフェートプロテオグリカン(MCSP)、CCR8、TNF−アルファ前駆体、STEAP、メソテリン(mesothelin)、A33抗原、前立腺幹細胞抗原(PSCA)、Ly−6;デスモグレイン4、E−カドヘリンネオエピトープ、胎児アセチルコリンレセプター、CD25、CA19−9マーカー、CA−125マーカーおよびミュラー管抑制物質(MIS)レセプタータイプII、sTn(シリル化Tn抗原;TAG−72)、FAP(線維芽細胞活性化抗原)、エンドシアリン(endosialin)、EGFRvIII、LG、SASならびにCD63などがある。
【0046】
「治療する(treating)」という用語は、病気、病気の症状、病気による二次的な病状、または病気にかかり易い素因を治す(cure)、緩和する(alleviate)、軽減する(relieve)、修復する(remedy)、予防する(prevent)、または改善する(ameliorate)ことを目的とし、対象に組成物を投与することとして定義される。「有効量」とは、治療を受ける対象において例えば上述したような医学的に望ましい結果を生じさせることのできる組成物の量のことである。
【0047】
1つのインビボアプローチでは、治療組成物(例えば本発明のタンパク質複合体を含む組成物)を対象に投与する。通常、該複合体は医薬的に許容される担体(例えば生理食塩水)に懸濁されており、経口でもしくは静脈内注射により投与される、または皮下、筋肉、鞘内、腹腔内、直腸内、膣内、鼻腔内、胃内、気管内もしくは肺内に注射もしくは注入される。
【0048】
必要な投与量は、投与経路の選択、剤形の性質、対象の疾病の性質、対象の大きさ、体重、表面積、年齢および性別、投与されている他の薬物、ならびに主治医の判断によって決まる。適切な投与量は0.01〜100.0mg/kgの範囲である。必要とされる投与量の増減は、利用可能な組成物の種類、および各種投与経路の異なる効率を考慮して予測される。例えば、経口投与は静脈注射による投与よりも高い投与量を要するであろうことが予想できる。これら投与量の増減の程度は、当該技術分野においてよく理解されているような、標準的な経験的ルーチンによって調整し至適化することができる。適した運搬媒体(例えば高分子微粒子または埋め込み型装置)に組成物をカプセル封入すれば、特に経口デリバリーにおいて、運搬の効率を向上させることができる。
【0049】
医薬的に許容される担体および有効量の本発明のタンパク質複合体を含む医薬組成物も、本発明の範囲内に含まれる。医薬組成物は上記した疾患を処置するのに用いることができる。医薬的に許容される担体には、溶剤、分散媒、コーティング、抗菌および抗真菌剤、ならびに等張および吸収遅延剤が含まれる。医薬組成物は、従来の方法を用い、それぞれ異なる投与経路に合った剤形に形成することができる。
【0050】
本発明の組成物の有効性はインビトロとインビボの両方において評価できる。インビボ試験では、組成物を動物(例えばマウスモデル)に注射してから、その治療上の効果を調べることができる。その結果に基づいて、適切な投与量の範囲と投与経路が決められる。
【0051】
以下の具体例は、説明にすぎず、いかなる方式においてもこの開示以外のものを限定するものではないと解されるべきである。詳述はしなくとも、当業者は、本明細書の記載に基づいて、本発明を最大限利用できると考えられる。本明細書に引用されるすべての刊行物は、参照することによりその全体が本明細書に組み込まれる。
【実施例】
【0052】
実施例1
M13ファージディスプレイライブラリーをスクリーニングして、EGFRに特異的に結合するヒト一本鎖可変フラグメント(scFv)を同定した。多数のクローンを同定した。ウェスタンブロッティングおよびELISAによる確認ののち、さらなる実験のために1つのクローン、erb_scFvを選んだ。標準方法により、erb_scFvをコードするcDNAを得てから、発現ベクターにライゲートした。以下に示すのは、erb_scFvのポリペプチド配列(配列番号1)とそれをコードするヌクレオチド配列(配列番号2)である。
【0053】
配列番号1
MetAlaGluValGlnLeuLeuGluSerGlyGlyGlyLeuValGlnProGlyGlySerLeuArgLeuSerCysAlaAlaSerGlyPheThrPheSerSerTyrAlaMetSerTrpValArgGlnAlaProGlyLysGlyLeuGluTrpValSerAspIleGlyAlaSerGlySerAlaThrSerTyrAlaAspSerValLysGlyArgPheThrIleSerArgAspAsnSerLysAsnThrLeuTyrLeuGlnMetAsnSerLeuArgAlaGluAspThrAlaValTyrTyrCysAlaLysSerThrThrThrPheAspTyrTrpGlyGlnGlyThrLeuValThrValSerSerGlyGlyGlyGlySerGlyGlyGlyGlySerGlyGlyGlyGlySerThrAspIleGlnMetThrGlnSerProSerSerLeuSerAlaSerValGlyAspArgValThrIleThrCysArgAlaSerGlnSerIleSerSerTyrLeuAsnTrpTyrGlnGlnLysProGlyLysAlaProLysLeuLeuIleTyrAspAlaSerAlaLeuGlnSerGlyValProSerArgPheSerGlySerGlySerGlyThrAspPheThrLeuThrIleSerSerLeuGlnProGluAspPheAlaThrTyrTyrCysGlnGlnTyrAlaAspTyrProThrThrPheGlyGlnGlyThrLysValGluIleLysArg
【0054】
配列番号2
ATGGCCGAGGTGCAGCTGTTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTTAGCAGCTATGCCATGAGCTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTCTCAGATATTGGTGCTTCTGGTTCTGCTACATCTTACGCAGACTCCGTGAAGGGCCGGTTCACCATCTCCAGAGACAATTCCAAGAACACGCTGTATCTGCAAATGAACAGCCTGAGAGCCGAGGACACGGCCGTATATTACTGTGCGAAATCTACTACTACTTTTGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCGAGCGGTGGAGGCGGTTCAGGCGGAGGTGGCAGCGGCGGTGGCGGGTCGACGGACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAGCAGCTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGATGCATCCGCTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAACAGTATGCTGATTATCCTACTACGTTCGGCCAAGGGACCAAGGTGGAAATCAAACGG
【0055】
次いで、発現ベクターを昆虫細胞株ショウジョウバエS2中で発現させた。抗EGFR erb_scFvを精製し、ウェスタンブロット分析およびELISAにかけて、そのEGFRに対する特異性を確認した。
【0056】
RT−PCRを行って、抗CD3モノクローナル抗体OKT3の重鎖可変領域 (VH)および軽鎖可変領域(VL)をコードするcDNAをハイブリドーマ細胞株から得た。続いて、2つのcDNAをライゲートして、OKT3のVH−VLの融合タンパク質をコードする融合配列を作製した。以下に示すのは、この融合タンパク質のポリペプチド配列(配列番号3)とそれをコードするcDNAの配列(配列番号4)である。
【0057】
配列番号3
ValGlnLeuGlnGlnSerGlyAlaGluLeuAlaArgProGlyAlaSerValLysMetSerCysLysAlaSerGlyTyrThrPheThrArgTyrThrMetHisTrpValLysGlnArgProGlyGlnGlyLeuGluTrpIleGlyTyrIleAsnProSerArgGlyTyrThrAsnTyrAsnGlnLysPheLysAspLysAlaThrLeuThrThrAspLysSerSerSerThrAlaTyrMetGlnLeuSerSerLeuThrSerGluAspSerAlaValTyrTyrCysAlaArgTyrTyrAspAspHisTyrCysLeuAspTyrTrpGlyGlnGlyThrThrValThrValSerSerGlyGlyGlyGlySerGlyGlyGlyGlySerGlyGlyGlyGlySerAspIleValLeuThrGlnSerProAlaIleMetSerAlaSerProGlyGluLysValThrMetThrCysSerAlaSerSerSerValSerTyrMetAsnTrpTyrGlnGlnLysSerGlyThrSerProLysArgTrpIleTyrAspThrSerLysLeuAlaSerGlyValProAlaHisPheArgGlySerGlySerGlyThrSerTyrSerLeuThrIleSerGlyMetGluAlaGluAspAlaAlaThrTyrTyrCysGlnGlnTrpSerSerAsnProPheThrPheGlySerGlyThrLysLeuGluLeuLysArg
【0058】
配列番号4
GTCCAGCTGCAGCAGTCAGGGGCTGAACTGGCAAGACCTGGGGCCTCAGTGAAGATGTCCTGCAAGGCTTCTGGCTACACCTTTACTAGGTACACGATGCACTGGGTAAAACAGAGGCCTGGACAGGGTCTGGAATGGATTGGATACATTAATCCTAGCCGTGGTTATACTAATTACAATCAGAAGTTCAAGGACAAGGCCACATTGACTACAGACAAATCCTCCAGCACAGCCTACATGCAACTGAGCAGCCTGACATCTGAGGACTCTGCAGTCTATTACTGTGCAAGATATTATGATGATCATTACTGCCTTGACTACTGGGGCCAAGGGACCACGGTCACCGTCTCCTCAGGTGGAGGCGGTTCAGGCGGAGGTGGCTCTGGCGGTGGCGGATCGGACATTGTGCTAACCCAGTCTCCAGCAATCATGTCTGCATCTCCAGGGGAGAAGGTCACCATGACCTGCAGTGCCAGCTCAAGTGTAAGTTACATGAACTGGTACCAGCAGAAGTCAGGCACCTCCCCCAAAAGATGGATTTATGACACATCCAAACTGGCTTCTGGAGTCCCTGCTCACTTCAGGGGCAGTGGGTCTGGGACCTCTTACTCTCTCACAATCAGCGGCATGGAGGCTGAAGATGCTGCCACTTATTACTGCCAGCAGTGGAGTAGTAACCCATTCACGTTCGGCTCGGGGACCAAGCTGGAGCTGAAACGA
【0059】
同じ手順を行って、抗EGFRモノクローナル抗体528のVHおよびVLをコードするcDNA、ならびに抗EGFR528のVH−VLの融合タンパク質をコードする融合配列を得た。528モノクローナル抗体は、細胞膜、例えばヒト類表皮癌A431細胞などの細胞膜上のEGFRに結合する。この528一本鎖抗体のポリペプチド配列(配列番号5)およびそれをコードするcDNAの配列(配列番号6)を以下に示す。
【0060】
配列番号5
ValLysLeuGlnGluSerGlySerGluMetAlaArgProGlyAlaSerValLysLeuProCysLysAlaSerGlyAspThrPheThrSerTyrTrpMetHisTrpValLysGlnArgHisGlyHisGlyProGluTrpIleGlyAsnIleTyrProGlySerGlyGlyThrAsnTyrAlaGluLysPheLysAsnLysValThrLeuThrValAspArgSerSerArgThrValTyrMetHisLeuSerArgLeuThrSerGluAspPheAlaValTyrTyrCysThrArgSerGlyGlyProTyrPhePheAspTyrTrpGlyGlnGlyThrThrValThrValSerSerGlyGlyGlyGlySerGlyGlyGlyGlySerGlyGlyGlyGlySerMetThrGlnThrProLeuSerLeuProValSerLeuGlyAspGlnAlaSerIleSerCysArgSerSerGlnAsnIleValHisAsnAsnGlyIleThrTyrLeuGluTrpTyrLeuGlnArgProGlyGlnSerProLysLeuLeuIleTyrLysValSerAspArgPheSerGlyValProAspArgPheSerGlySerGlySerGlyThrAspPheThrLeuLysIleSerArgValGluAlaGluAspLeuGlyIleTyrTyrCysPheGlnGlySerHisHisProProThrPheGlyGlyGlyThrLysLeuGlu
【0061】
配列番号6
GTCAAGCTGCAGGAGTCAGGGTCTGAGATGGCGAGGCCTGGAGCTTCAGTGAAGCTGCCCTGCAAGGCTTCTGGCGACACATTCACCAGTTACTGGATGCACTGGGTGAAGCAGAGGCATGGACATGGCCCTGAGTGGATCGGAAATATTTATCCAGGTAGTGGTGGTACTAACTACGCTGAGAAGTTCAAGAACAAGGTCACTCTGACTGTAGACAGGTCCTCCCGCACAGTCTACATGCACCTCAGCAGGCTGACATCTGAGGACTTTGCGGTCTATTATTGTACAAGATCGGGGGGTCCCTACTTCTTTGACTACTGGGGCCAAGGGACCACGGTCACCGTCTCCTCAGGTGGAGGCGGTTCAGGCGGAGGTGGCTCTGGCGGTGGCGGATCGATGACCCAAACTCCACTCTCCCTGCCTGTCAGTCTTGGAGATCAAGCCTCCATCTCTTGCAGATCTAGTCAGAACATTGTACATAATAATGGAATCACCTATTTAGAATGGTACCTGCAAAGGCCAGGCCAGTCTCCAAAGCTCCTGATCTACAAAGTTTCCGACCGATTTTCTGGGGTCCCAGACAGGTTCAGTGGCAGTGGATCAGGGACAGATTTCACACTCAAGATCAGCAGAGTAGAGGCTGAGGATCTGGGAATTTATTACTGCTTTCAAGGTTCACATCATCCTCCCACGTTCGGCGGGGGGACCAAGCTGGAA
【0062】
上述の抗EGFR scFv03、OKT3VH−VL、および抗EGFR 528VH−VLをコードするcDNAを、5’末端の短いヒンジ配列、および3’末端のヒスチジンタグ配列を含むXXI型ヒトミニコラーゲンのcDNAにそれぞれインフレームで融合した。以下に示すのは、XXI型ヒトミニコラーゲンのポリペプチドおよびcDNAの配列(それぞれ配列番号7および8)である。
【0063】
配列番号7
GlyGlyArgGluProLysSerCysAspLysThrHisThrCysProProCysProArgSerIleProGlyProProGlyProIleGlyProGluGlyProArgGlyLeuProGlyLeuProGlyArgAspGlyValProGlyLeuValGlyValProGlyArgProGlyValArgGlyLeuLysGlyLeuProGlyArgAsnGlyGluLysGlySerGlnGlyPheGlyTyrProGlyGluGlnGlyProProGlyProProGlyProGluGlyProProGlyIleSerLysGluGlyProProGlyAspProGlyLeuProGlyLysAspGlyAspHisGlyLysProGlyIleGlnGlyGlnProGlyProProGlyIleCysAspProSerLeuCysPheSerValIleAlaArgArgAspProPheArgLysGlyProAsnTyrSerLeuAspAspSerSerHisHisHisHisHisHisSerSerGly
(注:Proはプロリンまたはヒドロキシプロリンである)
【0064】
配列番号8
GGCGGCCGCGAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAAGATCTATTCCTGGGCCACCTGGTCCGATAGGCCCAGAGGGTCCCAGAGGATTACCTGGTTTGCCAGGAAGAGATGGTGTTCCTGGATTAGTGGGTGTCCCTGGACGTCCAGGTGTCAGAGGATTAAAAGGCCTACCAGGAAGAAATGGGGAAAAAGGGAGCCAAGGGTTTGGGTATCCTGGAGAACAAGGTCCTCCTGGTCCCCCAGGTCCAGAGGGCCCTCCTGGAATAAGCAAAGAAGGTCCTCCAGGAGACCCAGGTCTCCCTGGCAAAGATGGAGACCATGGAAAACCTGGAATCCAAGGGCAACCAGGCCCCCCAGGCATCTGCGACCCATCACTATGTTTTAGTGTAATTGCCAGAAGAGATCCGTTCAGAAAAGGACCAAACTATAGTCTAGACGACAGCAGCCATCATCACCATCACCATAGCAGCGGC
【0065】
得られた3つの発現ベクターを、ショウジョウバエS2細胞に同時トランスフェクト(co-transfect)した。その細胞をブラストサイジン存在下で培養し、ブラストサイジンに耐性を持つ細胞を選別した。細胞培養上清を回収してから、EGFRおよびCD3に対する抗体活性に対して、ウェスタンブロットとELISAでスクリーニングした。一部のクローンの細胞が三重らせん状の複合体(triple helix complex)を安定に発現していることが発見された。XXI型ヒトミニコラーゲンに似たこれら三重らせん状の複合体は、熱およびペプシンに耐性を示した。より重要なのは、これらがEGFRとCD3の両方に特異的に結合したということである。
【0066】
実施例2
この実施例では、3つの融合ポリペプチド、OKT3_scFv−Col、erb_scFv−Col、およびerb_NSPD−scFvを作製した。
【0067】
ファージライブラリーの選択
上皮細成長因子レセプター細胞外ドメイン(EGFR−ECD)結合可変フラグメント(scFv)を含むerbファージミドを、ヒトシングルフォールドscFvファージディスプレイライブラリー(Tomlinson I + J;I. M. Tomlinson and G. Winterより親切に提供された、MRCラボラトリー オブ モレキュラーバイオロジー、ケンブリッジ、英国)をスクリーニングすることにより単離した。精製したEGFレセプターの組み換え細胞外ドメイン(EGFR-ECD;リサーチ ダイアグノスティック社(Research Diagnostics, Inc.))10μgを塗布したイムノチューブ(Maxisorp;ヌン(Nunc)、 ロスキレ、デンマーク)を用いて選別を行った。メーカーのプロトコールにしたがって、ブロッキング、パンニング(panning)、洗浄、溶出および溶出ファージの再増幅を行った。
【0068】
組み換えプラスミドの構築
erbのscFvをコードするcDNAをPCRによりerbファージミドから増幅した。OKT3ハイブリドーマ(ATCC、CRL-8001)の逆転写産物により、ムリンIgG2a抗CD3 mAb OKT3(オルト ファーマシューティカル コーポレーション)をコードする配列を得た。公開されたヌクレオチド配列を基に、RT−PCRによってOKT3 mAbのVLおよびVHのcDNAを得た。グリシンリンカー(GGGS)3でVLおよびVH鎖をつなぐことにより、erbおよびOKT3のscFv PCRの融合を発生させた。
【0069】
scFv-Colを生成するよう、scFv-Colのコーディング領域には、N末端のscFvヌクレオチド配列と、ヒトIgGのヒンジ領域、コラーゲン様ドメイン(下線部)およびXXI型コラーゲンのNC1ドメインを含む、EPKSCDKTHTCPPCPRSIP(GPP)10GICDPSLCFSVIA-RRDPFRKGPNYのペプチド配列をコードするC末端合成コラーゲン足場遺伝子とが含まれる。以下に示すのは、合成コラーゲン足場ポリペプチドおよびcDNAの配列(それぞれ配列番号9および10)である。
【0070】
配列番号9
SEQ ID NO: 9:
GluProLysSerCysAspLysThrHisThrCysProProCysProArgSerIleProGlyProProGlyProProGlyProProGlyProProGlyProProGlyProProGlyProProGlyProProGlyProProGlyProProGlyIleCysAspProSerLeuCysPheSerValIleAlaArgArgAspProPheArgLysGlyProAsnTyr
【0071】
配列番号10
GAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAAGATCTATTCCTGGGCCACCTGGTCCCCCAGGTCCTCCAGGACCCCCAGGGCCCCCAGGCCCCCCCGGGCCGCCTGGACCCCCAGGGCCACCAGGCCCCCCAGGCATCTGCGACCCATCACTATGTTTTAGTGTAATTGCCAGAAGAGATCCGTTCAGAAAAGGACCAAACTAT
【0072】
この合成配列(配列番号10)をオーバーラップPCRにより作製し、NotIおよびXhoI部位の隣に位置する(flanking)PCR産物を、同部位の発現ベクターpSecTag2/Hygro(インビトロジェン)にクローニングした。続いて、erbおよびOKT3のscFvを、AscIおよびNotI部位で上述したC末端コラーゲン足場含有構成体にインフレームでクローニングして、erb_scFv−ColおよびOKT3_scFv−Colの発現構成体をそれぞれ作った。
【0073】
次に、erb_NSPD−scFvを作製した。NSPD−scFvのコーディング領域には、ヒトサーファクタントタンパク質D(SPD)のN末端の254のアミノ酸、コレクチンファミリーのメンバー、およびC末端のscFvが含まれていた。以下に示すのは、ヒトサーファクタントタンパク質DのポリペプチドのN末端254アミノ酸およびそのcDNAの配列である(それぞれ配列番号11および12)。
【0074】
配列番号11
MetLeuLeuPheLeuLeuSerAlaLeuValLeuLeuThrGlnProLeuGlyTyrLeuGluAlaGluMetLysThrTyrSerHisArgThrMetProSerAlaCysThrLeuValMetCysSerSerValGluSerGlyLeuProGlyArgAspGlyArgAspGlyArgGluGlyProArgGlyGluLysGlyAspProGlyLeuProGlyAlaAlaGlyGlnAlaGlyMetProGlyGlnAlaGlyProValGlyProLysGlyAspAsnGlySerValGlyGluProGlyProLysGlyAspThrGlyProSerGlyProProGlyProProGlyValProGlyProAlaGlyArgGluGlyProLeuGlyLysGlnGlyAsnIleGlyProGlnGlyLysProGlyProLysGlyGluAlaGlyProLysGlyGluValGlyAlaProGlyMetGlnGlySerAlaGlyAlaArgGlyLeuAlaGlyProLysGlyGluArgGlyValProGlyGluArgGlyValProGlyAsnThrGlyAlaAlaGlySerAlaGlyAlaMetGlyProGlnGlySerProGlyAlaArgGlyProProGlyLeuLysGlyAspLysGlyIleProGlyAspLysGlyAlaLysGlyGluSerGlyLeuProAspValAlaSerLeuArgGlnGlnValGluAlaLeuGlnGlyGlnValGlnHisLeuGlnAlaAlaPheSerGlnTyrLysLysValGluLeuPhe
【0075】
配列番号12
ATGCTGCTCTTCCTCCTCTCTGCACTGGTCCTGCTCACACAGCCCCTGGGCTACCTGGAAGCAGAAATGAAGACCTACTCCCACAGAACAATGCCCAGTGCTTGCACCCTGGTCATGTGTAGCTCAGTGGAGAGTGGCCTGCCTGGTCGCGATGGACGGGATGGGAGAGAGGGCCCTCGGGGCGAGAAGGGGGACCCAGGTTTGCCAGGAGCTGCAGGGCAAGCAGGGATGCCTGGACAAGCTGGCCCAGTTGGGCCCAAAGGGGACAATGGCTCTGTTGGAGAACCTGGACCAAAGGGAGACACTGGGCCAAGTGGACCTCCAGGACCTCCCGGTGTGCCTGGTCCAGCTGGAAGAGAAGGTCCCCTGGGGAAGCAGGGGAACATAGGACCTCAGGGCAAGCCAGGCCCAAAAGGAGAAGCTGGGCCCAAAGGAGAAGTAGGTGCCCCAGGCATGCAGGGCTCGGCAGGGGCAAGAGGCCTCGCAGGCCCTAAGGGAGAGCGAGGTGTCCCTGGTGAGCGTGGAGTCCCTGGAAACACAGGGGCAGCAGGGTCTGCTGGAGCCATGGGTCCCCAGGGAAGTCCAGGTGCCAGGGGACCCCCGGGATTGAAGGGGGACAAAGGCATTCCTGGAGACAAAGGAGCAAAGGGAGAAAGTGGGCTTCCAGATGTTGCTTCTCTGAGGCAGCAGGTTGAGGCCTTACAGGGACAAGTACAGCACCTCCAGGCTGCTTTCTCTCAGTATAAGAAAGTTGAGCTCTTC
【0076】
N末端SPD cDNAを、発現ベクター、pSecTag2/Hygro(インビトロジェン)にNheIおよびAscI部位でクローニングした。続いて、erbのscFvを、AscIおよびXhoI部位で上述したN末端SPD含有構成体にインフレームでクローニングして、erb_NSPD−scFvの発現構成体を作った。
【0077】
erb_scFv−Col、erb_NSPD−scFvおよびOKT3_scFv−Colの各読み取り枠(open reading frame)には、N末端リーダー配列と、分泌、検出および精製に利用されるC末端mycエピトープ/ポリヒスチジンタグとをコードする配列が含まれていた。下表にまとめたのは、上述の発現構成体によりコードされる各種組み換えタンパク質/抗体である。
【0078】
【表1】
【0079】
抗体の発現および精製
組み換えタンパク質複合体/抗体を作製するため、Effectene(Qiagen)を用い、メーカーの取扱説明書にしたがって上述の構成体をマウス骨髄腫NS0細胞に導入した。ハイグロマイシン(400μg/ml)で4週間選別したのち、各安定クローンを、振盪フラスコ中の2%のウシ胎仔血清を含む既知組成培地HyQCDM4NS0(ハイクローン(Hyclone))にて、初播種密度2×105細胞/mlとして培養した。該培養は37℃で5日間、150rpmに保って行った。それら細胞が、上述の抗体ドメインと、コラーゲン足場ドメインつまりコラーゲン足場抗体(CSA)と、を含むタンパク質をコードする発現構成体を保持できるように、培地にアスコルビン酸ナトリウム(80μg/ml)を毎日加えた。
【0080】
erb_scFv、erb_scFv−Fc、erb_scFv−Col、またはOKT3_scFv−Colタンパク質あるいはタンパク質複合体を精製するべく、ろ過した各培地約2Lを、流速60ml/時間で、pH8.0の50mMトリスHClバッファーで平衡化したTゲルカラム(1.5×8cm、Pierce)にかけた。同バッファーで洗浄したのち、その組み換えタンパク質またはタンパク質複合体をpH4.0の50mM酢酸ナトリウムバッファーで溶出した。それらのUV吸収度を280nmで測定し、そのピーク画分を、流速60ml/時間で、pH8.0の0.5M NaCl含有50mMトリスHClバッファーにより平衡化したZnSO4チャージキレーティングセファロースハイトラップカラム(ZnSO4-charged chelating Sepharose HighTrap column)(ベッドボリューム1−ml、GE ヘルスケア)にかけた。そのカラムを先ず20mMのイミダゾールで洗浄してから、結合タンパク質またはタンパク質複合体を同バッファー中にて0.25Mのイミダゾールで溶出した。最終的な調製液をpH7.0の50mM Hepesバッファーに透析した。
【0081】
次に、MOPSを用いる10%NuPAGEビス−トリスポリアクリルアミドゲルか、酢酸ナトリウムをランニングバッファー(インビトロジェン)に用いる7%SDS/トリスアセテートポリアクリルアミドゲルのいずれかにより、SDS−PAGEを実行した。続いてクマシーブリリアントブルーR−250でタンパク質を染色した。ChemiImager5500(アルファ イノテック(Alpha Innotech)、サンリアンドロ、カリフォルニア)とソフトウェアAlpha EaseFC(v.4.0; アルファ イノテック)を用い濃度測定を行うことによってタンパク質バンドの濃度を定量化した。
【0082】
三重らせんの性質を調べるため、精製されたerb_scFv−Col(1mg/ml)を10mM DTTの非存在または存在下、37℃で1時間インキュベートした。DDT処理サンプルのアリコートをさらに常温で50mM N−エチル−マレイミド(NEM)と30分間反応させて、遊離スルフヒドリル基および三量体の再編成を完全にブロックした。各サンプルからの等量のタンパク質を、酢酸ナトリウムをランニングバッファーとする7% SDS/トリスアセテートポリアクリルアミドゲル上で電気泳動した。そのゲルをクマシーブルーで染色した。精製されたCSAは、ホモ三量体、または、マイルドな還元条件下で2つの三量体に解離可能な鎖間ジスルフィド結合六量体であることが発見された。
【0083】
erb_scFv−Colの三量体構造の熱安定性を調べた。2M尿素含有50mMトリス−HCl(pH8.0)中の精製されたerb_scFv−Colを10mMトリス(2−カルボキシエチル)ホスフィン(TCEP)の非存在または存在下常温で処理した。次いで、還元されたサンプルを常温下50mM NEMでアルキル化した。SDSローディングバッファーを混合する前に、等量のタンパク質を含む各サンプルを35、45、55、65、75および85℃で10分間加熱した。そのサンプルを、非還元条件下、MOPSバッファーを用い10%SDS/ビス−トリスポリアクリルアミドゲル上で電気泳動した。そのゲルをクマシーブルーで染色した。その結果、erb_scFv−Col三量体が高い熱安定性を有することが示された。実際には、65℃で10分間処理したのち、50%を超える三量体が残っていた。また、erb_scFv−Colのコラーゲン様ドメインの三量体構造は、プロリルのヒドロキシル化がなされていることもわかった。
【0084】
結合試験
各種の型のerb抗体のEGFR−ECDに対する結合動態(binding kinetics)を、ランニングバッファーHBS-EP(pH7.4の10mM HEPES、150mM NaCl、3mM EDTA、0.005%界面活性剤P20)中で、BIAcore Xバイオセンサー(ビアコア社(BIACORE, Inc.)、ウプサラ、スウェーデン)を用いて測定した。簡単に言うと、EGFR−ECDをアミンカップリングにより、レスポンス単位(RU)が1700程度となるまで、Clセンサーチップ上に固定化し、各種濃度で精製した抗体を流速10μl/分で注入した。pH 3.5の10mMグリシン−HClを5μl注入することによってその表面を再生した。各濃度におけるセンサーグラムを得ると共に、プログラム、BIA Evaluation3.2を用いて評価した。結合データを1:1ラングミュア結合モデルでフィッティングし、解離速度(kdiss)/会合速度(kass)の比として定義される親和定数KDを計算した。その結果を以下の表2に示す。
【0085】
【表2】
【0086】
表2に示されるように、EGFR−ECDに対するerb_scFv_Colの結合親和性は、二価(erb_scFv−Fc)および一価 (erb_scFv) mAbカウンターパートよりもそれぞれ約20倍および1000倍強かった。
【0087】
安定性および薬物動態アッセイ
血清安定性アッセイでは、各種形態のerb抗体、erb_scFv_Col、erb_scFv−Fcまたはerb_scFvの安定性を37℃にてヒト血清でインキュベートすることにより測定した。各インキュベーション期間経過後に残った活性抗EGFRの量を定量的ELISAにより測定した。ELISAは、(捕捉試薬としての)組み換えEGFR−ECDと抗c-myc mAb(9E10、シグマケミカル社)、続いて、HRPコンジュゲートアフィニティ精製ポリクローナルヤギ抗マウスIgGと化学発光基質(ピアース バイオテクノロジー社(Pierce Biotechnology, Inc.))を用いて行った。薬物動態アッセイでは、3匹のBALB/cヌードマウスを用いてerb_scFv−Colのクリアランスを分析した。簡単に言うと、事前の採血(pre-bleed)を行ってから、各マウスに25μg(2mg/体重キログラム )のerb_scFv−Colを皮下注射(s.c.)した。続く70時間の間に、定期的に血液サンプルを回収し、ELISAによってそれらのerb_scFv−Colの含有量を評価した。タンパク質はきわめて安定しているということがわかった。
【0088】
T細胞増殖アッセイおよび混合リンパ球反応(MLR)
5−ブロモ-2'−デオキシウリジン(BrdU)細胞増殖アッセイを行った。簡単に言うと、ヒト末梢血単核細胞(PBMC)を37℃で66時間、10倍段階希釈したOKT3(イーバイオサイエンス社(eBioscience, Inc.))またはOLT3_scFv−Colの存在下、ブラック96ウェル平底組織培養プレート中に2×105細胞/ウェルとなるように加えた10%FBS含有100μl RPMI−1640培地にプレートした。次いでその細胞を10μMのBrdUにより5時間パルス処理した。培地を除去したのち、FixDenatによりワンステップで細胞の固定とDNAの変性を行った。その後、その細胞をペルオキシダーゼ標識抗BrdU抗体(抗BrdU POD、Fagフラグメント)で1.5時間室温にてインキュベートした。マイクロプレートルミノメーター(Hidex, CHAMELEON detection platform、フィンランド)を用いて化学発光検出および定量化を行った。
【0089】
一方向性混合リンパ球反応におけるT細胞増殖および免疫抑制を以下のようにして評価した。ヒトPBMCを2人の健康なドナーから採取した(刺激および応答)。刺激または応答細胞を、37℃、5%のCO2を含んだ加湿空気中、完全培地(10%ヒトAB血清、2mMグルタミン、50nM 2−メルカプトエタノール、ならびにそれぞれ100単位/mlのペニシリンおよびストレプトマイシンを加えたRPMI 1640)にて25μg/mlのマイトマイシンC(シグマ−アルドリッチ)で30分間処理してから、RPMI 1640培地中で3回洗浄した。応答細胞を単独で、またはマイトマイシンC処理した刺激細胞もしくはマイトマイシンC応答細胞と1:1で混合し、2×105細胞/ウェルとして200μlの完全培地で培養した。応答細胞をプレートしたのち、精製したOKT3_scFv−ColまたはOKT3を異なる濃度で培養物に速やかに加えた。5日後、培養細胞を10μMのBrdUでパルス処理し、24時間後に回収した。次いで、細胞増殖アッセイを上述の手法で行った。
【0090】
OKT3_scFv−Colは、T細胞増殖の免疫抑制においてより効果が高く、かつ、T細胞増殖の刺激においては無視できる程度の細胞分裂促進活性を示すことが見出された。
【0091】
サイトカイン測定
ヒトPBMCを、37℃、10倍段階希釈したOKT3またはOKT3_scFv−Colの存在下、2×105細胞/ウェルとなるように、10%FBS含有0.1ml RPMI−1640培地にプレートした。その上清を各時点で回収し、ヒトサイトカインイムノアッセイキット(イーバイオサイエンス社)を用いて多数のサイトカインを測定した。その結果、OKT3_scFv_Colの投与は、マウスOKT3mAbに比較して、無視できる程度のサイトカイン放出しか生じさせないことが示された。
【0092】
抗体変位(Antibody Displacement)アッセイ
下記する全ての手順は4℃下で行った。ヒトT細胞を1×106細胞/mlの密度でFCMバッファー(2%FBSおよび0.1%アジ化ナトリウム含有リン酸緩衝食塩水)に懸濁させた。その細胞をマウス総IgG(2μg/ml、ジャクソン イムノリサーチ ラボラトリーズ)で30分処理してから、段階希釈したOKT3_scFv−ColまたはOKT3抗体で1時間インキュベートした。固定した、飽和量(フローサイトメトリーにより判断)のFITCコンジュゲートOKT3(0.25μg/ml、イーバイオサイエンス社より購入) を直接加えた。1時間のインキュベーション後、その細胞をFCMバッファーで洗浄し、FACScan(ベクトンデッキンソン、サンジョゼ、カリフォルニア)を用いフローサイトメトリー法により免疫発光解析を行った。そのデータを、遮断抗体の非存在下OKT3−FITCでT細胞を染色することにより得た平均蛍光強度として定義される最大蛍光強度の抑制率(percent inhibition)として表した。
【0093】
その結果、OKT3_scFv_Colが、天然マウスOKT3 mAbよりも、強力にヒトCD3+T細胞に結合することがわかった。
【0094】
ヒトIgGを標準として、Bradfordアッセイ(ピアース バイオテクノロジー社製、クーマシープラス試薬(Coomassie plus reagent))によりタンパク質濃度を測定した。アミノ酸分析については、精製したerb_scFv−Colを50mM酢酸に透析し、110℃で24時間、6NのHCl中で加水分解し、Waters PicoTag(登録商標)システムでアミノ酸分析を行った。
【0095】
これら結果によって、コラーゲン足場抗体が、抗腫瘍および免疫調節いずれの用途においても治療抗体設計に理想的な構造であるということが示される。
【0096】
本明細書に開示されたすべての特徴は、どのような組合せにも組み合わせることができる。本明細書に開示された各特徴は、同一、均等、または類似の目的を果たす他の特徴で置き換えられてもよい。したがって、特に明示する場合を除いては、開示された各特徴は、単に、包括的な一連の均等または同様の特徴の例に過ぎない。
【0097】
上述の記載から、当業者は、本発明の主要な特徴を容易に確認することができると共に、その精神と範囲を逸脱することなく、各種使用および条件に適合するように本発明をさまざまに変更および修飾することができる。よって、その他の実施形態もまた、以下の特許請求の範囲の範囲内に含まれる。
【特許請求の範囲】
【請求項1】
第1の足場ドメインおよび該第1の足場ドメインの1つの末端に融合する第1の異種ドメインを含む第1の融合ポリペプチド鎖と、
第2の足場ドメインを含む第2の融合ポリペプチド鎖と、
第3の足場ドメインを含む第3の融合ポリペプチド鎖と、を含み、
該第1、第2および第3の足場ドメインが、三重らせんコイルを形成するよう並んでいる組み換えタンパク質複合体。
【請求項2】
前記異種ドメインが結合ドメインを含む請求項1記載のタンパク質複合体。
【請求項3】
前記結合ドメインが、リガンド結合ドメイン、リガンド、レセプター、親和性タグ、またはプロテオグリカンを含む請求項2記載のタンパク質複合体。
【請求項4】
前記結合ドメインが、免疫グロブリンの1つまたはそれ以上の相補決定領域を含む請求項2記載のタンパク質複合体。
【請求項5】
前記結合ドメインが、抗原結合フラグメントの配列を含む請求項4記載のタンパク質複合体。
【請求項6】
前記抗原結合フラグメントがCD3またはEGFRに特異的に結合する請求項5記載のタンパク質複合体。
【請求項7】
前記抗原結合フラグメントが、一本鎖抗体の配列を含む請求項5記載のタンパク質複合体。
【請求項8】
前記第1の融合ポリペプチド鎖が、前記第1の足場ドメインのもう1つの末端に融合する第2の異種ドメインをさらに含む請求項1記載のタンパク質複合体。
【請求項9】
前記第1の異種ドメインが、CD3に特異的に結合する第1の一本鎖抗体の配列を含む請求項8記載のタンパク質複合体。
【請求項10】
前記第2の異種ドメインが、EGFRに特異的に結合する第2の一本鎖抗体の配列を含む請求項8記載のタンパク質複合体。
【請求項11】
前記第2の融合ポリペプチド鎖が、前記第2の足場ドメインの1つの末端に融合する第3の異種ドメインを含む請求項8記載のタンパク質複合体。
【請求項12】
前記第2の融合ポリペプチド鎖が、前記第2の足場ドメインのもう1つの末端に融合する第4の異種ドメインをさらに含む請求項11記載のタンパク質複合体。
【請求項13】
前記第3の融合ポリペプチド鎖が、前記第3の足場ドメインの1つの末端に融合する第5の異種ドメインを含む請求項12記載のタンパク質複合体。
【請求項14】
前記第3の融合ポリペプチド鎖が、前記第3の足場ドメインのもう1つの末端に融合する第6の異種ドメインを含む請求項13記載のタンパク質複合体。
【請求項15】
前記第1、第2または第3の足場ドメインが、1またはそれ以上の三重らせんリピートを含み、各リピートは次の式:(X1−X2−X3)nの配列を含んでおり、式中、X1はGly残基、X2またはX3は任意のアミノ酸残基であり、かつnは5またはこれより大きい数である請求項1記載のタンパク質複合体。
【請求項16】
前記第1、第2または第3の足場ドメインが、1つまたはそれ以上のC1q、コレクチン、またはコラーゲンポリペプチド鎖の三重らせんリピートを含む請求項15記載のタンパク質複合体。
【請求項17】
X3がプロリンまたはヒドロキシプロリン残基である請求項15記載のタンパク質複合体。
【請求項18】
各リピートが(GPP)10の配列を含んでいる請求項15記載のタンパク質複合体。
【請求項19】
前記異種ドメインが酵素ドメインまたは蛍光タンパク質の配列を含む請求項1記載のタンパク質複合体。
【請求項20】
前記第1、第2および第3の融合ポリペプチドが実質的に同一である請求項1記載のタンパク質複合体。
【請求項21】
三重らせんコイルを形成するための足場ドメインと、該足場ドメインの1つの末端に融合する第1の異種ドメインとを含む組み換え融合ポリペプチド。
【請求項22】
前記足場ドメインが、1またはそれ以上の三重らせんリピートを含み、各リピートが次の式:(X1−X2−X3)nの配列を含んでおり、式中、X1はGly残基、X2またはX3は任意のアミノ酸残基であり、かつnは5またはこれより大きい数である請求項21記載の融合ポリペプチド。
【請求項23】
X3がプロリンまたはヒドロキシプロリン残基である請求項22記載の融合ポリペプチド。
【請求項24】
前記足場ドメインのもう1つの末端に融合する第2の異種ドメインをさらに含む請求項21記載の融合ポリペプチド。
【請求項25】
前記足場ドメインが、C1q、コレクチン、またはコラーゲンポリペプチド鎖の、1つまたはそれ以上の三重らせんリピートを含む請求項21記載の融合ポリペプチド。
【請求項26】
前記異種ドメインが、リガンド結合ドメイン、リガンド、レセプターまたは多糖類を含む請求項21記載の融合ポリペプチド。
【請求項27】
前記異種ドメインがファージディスプレイスクリーニングによって得られる請求項26記載の融合ポリペプチド。
【請求項28】
請求項21記載の融合ポリペプチドをコードする配列またはその補体を含む単離核酸。
【請求項29】
請求項28記載の核酸を含む宿主細胞。
【請求項30】
前記細胞が哺乳類または昆虫の細胞である請求項29記載の宿主細胞。
【請求項31】
前記哺乳類の細胞がマウス骨髄腫NS0細胞である請求項30記載の宿主細胞。
【請求項32】
請求項28記載の核酸を含む発現ベクター。
【請求項33】
請求項29記載の宿主細胞を、前記核酸によりコードされるポリペプチドの発現を可能とする条件下、培地中で培養する工程と、前記培養された細胞または前記細胞の前記培地から前記ポリペプチドを精製する工程とを含む融合ポリペプチドの製造方法。
【請求項34】
請求項1記載のタンパク質複合体の製造方法であって、
第1の足場ドメインおよび該第1の足場ドメインの1つの末端に融合する第1の異種ドメインを含む第1の融合ポリペプチド鎖をコードする第1の核酸と、第2の足場ドメインを含む第2の融合ポリペプチド鎖をコードする第2の核酸と、第3の足場ドメインを含む第3の融合ポリペプチド鎖をコードする第3の核酸とを含有する宿主細胞を、該3つの核酸にコードされるポリペプチドの発現およびそれらの間における三重らせんコイルの形成を可能とする条件下、培地で培養する工程、ならびに
培養された細胞または細胞の培地からタンパク質複合体を精製する工程
を含む方法。
【請求項35】
前記宿主細胞が、プロリン残基をヒドロキシル化する酵素活性を含む真核細胞である請求項34記載の方法。
【請求項1】
第1の足場ドメインおよび該第1の足場ドメインの1つの末端に融合する第1の異種ドメインを含む第1の融合ポリペプチド鎖と、
第2の足場ドメインを含む第2の融合ポリペプチド鎖と、
第3の足場ドメインを含む第3の融合ポリペプチド鎖と、を含み、
該第1、第2および第3の足場ドメインが、三重らせんコイルを形成するよう並んでいる組み換えタンパク質複合体。
【請求項2】
前記異種ドメインが結合ドメインを含む請求項1記載のタンパク質複合体。
【請求項3】
前記結合ドメインが、リガンド結合ドメイン、リガンド、レセプター、親和性タグ、またはプロテオグリカンを含む請求項2記載のタンパク質複合体。
【請求項4】
前記結合ドメインが、免疫グロブリンの1つまたはそれ以上の相補決定領域を含む請求項2記載のタンパク質複合体。
【請求項5】
前記結合ドメインが、抗原結合フラグメントの配列を含む請求項4記載のタンパク質複合体。
【請求項6】
前記抗原結合フラグメントがCD3またはEGFRに特異的に結合する請求項5記載のタンパク質複合体。
【請求項7】
前記抗原結合フラグメントが、一本鎖抗体の配列を含む請求項5記載のタンパク質複合体。
【請求項8】
前記第1の融合ポリペプチド鎖が、前記第1の足場ドメインのもう1つの末端に融合する第2の異種ドメインをさらに含む請求項1記載のタンパク質複合体。
【請求項9】
前記第1の異種ドメインが、CD3に特異的に結合する第1の一本鎖抗体の配列を含む請求項8記載のタンパク質複合体。
【請求項10】
前記第2の異種ドメインが、EGFRに特異的に結合する第2の一本鎖抗体の配列を含む請求項8記載のタンパク質複合体。
【請求項11】
前記第2の融合ポリペプチド鎖が、前記第2の足場ドメインの1つの末端に融合する第3の異種ドメインを含む請求項8記載のタンパク質複合体。
【請求項12】
前記第2の融合ポリペプチド鎖が、前記第2の足場ドメインのもう1つの末端に融合する第4の異種ドメインをさらに含む請求項11記載のタンパク質複合体。
【請求項13】
前記第3の融合ポリペプチド鎖が、前記第3の足場ドメインの1つの末端に融合する第5の異種ドメインを含む請求項12記載のタンパク質複合体。
【請求項14】
前記第3の融合ポリペプチド鎖が、前記第3の足場ドメインのもう1つの末端に融合する第6の異種ドメインを含む請求項13記載のタンパク質複合体。
【請求項15】
前記第1、第2または第3の足場ドメインが、1またはそれ以上の三重らせんリピートを含み、各リピートは次の式:(X1−X2−X3)nの配列を含んでおり、式中、X1はGly残基、X2またはX3は任意のアミノ酸残基であり、かつnは5またはこれより大きい数である請求項1記載のタンパク質複合体。
【請求項16】
前記第1、第2または第3の足場ドメインが、1つまたはそれ以上のC1q、コレクチン、またはコラーゲンポリペプチド鎖の三重らせんリピートを含む請求項15記載のタンパク質複合体。
【請求項17】
X3がプロリンまたはヒドロキシプロリン残基である請求項15記載のタンパク質複合体。
【請求項18】
各リピートが(GPP)10の配列を含んでいる請求項15記載のタンパク質複合体。
【請求項19】
前記異種ドメインが酵素ドメインまたは蛍光タンパク質の配列を含む請求項1記載のタンパク質複合体。
【請求項20】
前記第1、第2および第3の融合ポリペプチドが実質的に同一である請求項1記載のタンパク質複合体。
【請求項21】
三重らせんコイルを形成するための足場ドメインと、該足場ドメインの1つの末端に融合する第1の異種ドメインとを含む組み換え融合ポリペプチド。
【請求項22】
前記足場ドメインが、1またはそれ以上の三重らせんリピートを含み、各リピートが次の式:(X1−X2−X3)nの配列を含んでおり、式中、X1はGly残基、X2またはX3は任意のアミノ酸残基であり、かつnは5またはこれより大きい数である請求項21記載の融合ポリペプチド。
【請求項23】
X3がプロリンまたはヒドロキシプロリン残基である請求項22記載の融合ポリペプチド。
【請求項24】
前記足場ドメインのもう1つの末端に融合する第2の異種ドメインをさらに含む請求項21記載の融合ポリペプチド。
【請求項25】
前記足場ドメインが、C1q、コレクチン、またはコラーゲンポリペプチド鎖の、1つまたはそれ以上の三重らせんリピートを含む請求項21記載の融合ポリペプチド。
【請求項26】
前記異種ドメインが、リガンド結合ドメイン、リガンド、レセプターまたは多糖類を含む請求項21記載の融合ポリペプチド。
【請求項27】
前記異種ドメインがファージディスプレイスクリーニングによって得られる請求項26記載の融合ポリペプチド。
【請求項28】
請求項21記載の融合ポリペプチドをコードする配列またはその補体を含む単離核酸。
【請求項29】
請求項28記載の核酸を含む宿主細胞。
【請求項30】
前記細胞が哺乳類または昆虫の細胞である請求項29記載の宿主細胞。
【請求項31】
前記哺乳類の細胞がマウス骨髄腫NS0細胞である請求項30記載の宿主細胞。
【請求項32】
請求項28記載の核酸を含む発現ベクター。
【請求項33】
請求項29記載の宿主細胞を、前記核酸によりコードされるポリペプチドの発現を可能とする条件下、培地中で培養する工程と、前記培養された細胞または前記細胞の前記培地から前記ポリペプチドを精製する工程とを含む融合ポリペプチドの製造方法。
【請求項34】
請求項1記載のタンパク質複合体の製造方法であって、
第1の足場ドメインおよび該第1の足場ドメインの1つの末端に融合する第1の異種ドメインを含む第1の融合ポリペプチド鎖をコードする第1の核酸と、第2の足場ドメインを含む第2の融合ポリペプチド鎖をコードする第2の核酸と、第3の足場ドメインを含む第3の融合ポリペプチド鎖をコードする第3の核酸とを含有する宿主細胞を、該3つの核酸にコードされるポリペプチドの発現およびそれらの間における三重らせんコイルの形成を可能とする条件下、培地で培養する工程、ならびに
培養された細胞または細胞の培地からタンパク質複合体を精製する工程
を含む方法。
【請求項35】
前記宿主細胞が、プロリン残基をヒドロキシル化する酵素活性を含む真核細胞である請求項34記載の方法。
【図1】

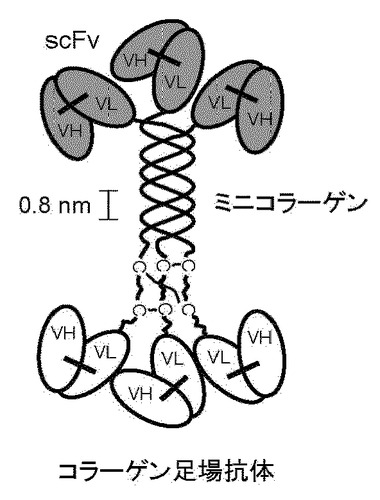
【図2】
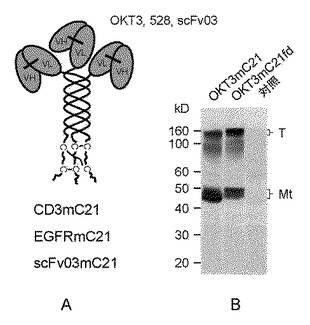
【図3】
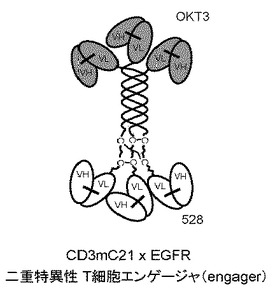
【図4】
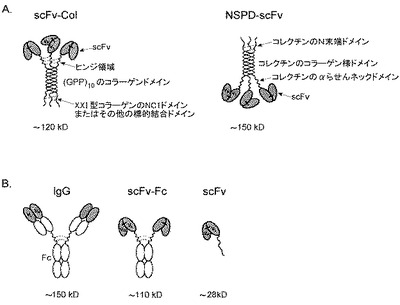
【図2】
【図3】
【図4】
【公開番号】特開2011−217760(P2011−217760A)
【公開日】平成23年11月4日(2011.11.4)
【国際特許分類】
【外国語出願】
【出願番号】特願2011−165511(P2011−165511)
【出願日】平成23年7月28日(2011.7.28)
【分割の表示】特願2006−338520(P2006−338520)の分割
【原出願日】平成18年12月15日(2006.12.15)
【出願人】(390023582)財団法人工業技術研究院 (524)
【氏名又は名称原語表記】INDUSTRIAL TECHNOLOGY RESEARCH INSTITUTE
【住所又は居所原語表記】195 Chung Hsing Rd.,Sec.4,Chutung,Hsin−Chu,Taiwan R.O.C
【Fターム(参考)】
【公開日】平成23年11月4日(2011.11.4)
【国際特許分類】
【出願番号】特願2011−165511(P2011−165511)
【出願日】平成23年7月28日(2011.7.28)
【分割の表示】特願2006−338520(P2006−338520)の分割
【原出願日】平成18年12月15日(2006.12.15)
【出願人】(390023582)財団法人工業技術研究院 (524)
【氏名又は名称原語表記】INDUSTRIAL TECHNOLOGY RESEARCH INSTITUTE
【住所又は居所原語表記】195 Chung Hsing Rd.,Sec.4,Chutung,Hsin−Chu,Taiwan R.O.C
【Fターム(参考)】
[ Back to top ]