
組み換え細胞毒性RNAseを含む融合タンパク質
【課題】非哺乳動物の細胞毒性RNAseおよび免疫グロブリンおよび免疫グロブリンフラグメントの融合タンパク質を含む免疫毒素を提供する。
【解決手段】グリコシル化された免疫毒素であって、(a)第一の免疫グロブリン可変ドメインに融合された非哺乳動物リボヌクレアーゼを含む融合ポリペプチドと、(b)第二の免疫グロブリン可変ドメインを含む第二のポリペプチドとを含み、前記第一および第二の免疫グロブリン可変ドメインが、一緒になって抗原結合部位を形成する抗体の免疫グロブリン重鎖および免疫グロブリン軽鎖のCDRを含み、前記非哺乳動物リボヌクレアーゼが、前記第一の免疫グロブリン可変ドメインのN末端に融合されており、前記非哺乳動物リボヌクレアーゼが、N末端ピログルタミン酸残基を保有している、免疫毒素。
【解決手段】グリコシル化された免疫毒素であって、(a)第一の免疫グロブリン可変ドメインに融合された非哺乳動物リボヌクレアーゼを含む融合ポリペプチドと、(b)第二の免疫グロブリン可変ドメインを含む第二のポリペプチドとを含み、前記第一および第二の免疫グロブリン可変ドメインが、一緒になって抗原結合部位を形成する抗体の免疫グロブリン重鎖および免疫グロブリン軽鎖のCDRを含み、前記非哺乳動物リボヌクレアーゼが、前記第一の免疫グロブリン可変ドメインのN末端に融合されており、前記非哺乳動物リボヌクレアーゼが、N末端ピログルタミン酸残基を保有している、免疫毒素。
【発明の詳細な説明】
【技術分野】
【0001】
本願は、その内容が全体として引用することにより本明細書に組み込まれる、2004年2
月13日出願の米国仮特許出願第60/544,227号に対する優先権を主張する。
【0002】
本発明は、非哺乳動物の細胞毒性RNAseおよび免疫グロブリンおよび免疫グロブリ
ンフラグメントの融合タンパク質を含む免疫毒素を提供する。
【背景技術】
【0003】
ランピルナーゼ(ranpirnase)は、疾患の処置のための細胞毒性因子としての見込みを示
す非哺乳動物リボヌクレアーゼ(RNAse)のあるクラスのメンバーである。これらの
RNAseの細胞毒性は、細胞表面レセプターに対する当初の結合および細胞のサイトゾ
ルへの内部移行(リボソームRNAの付随する分解および細胞タンパク質合成の阻害を伴
う)に関与する機構に起因している。Wu et al.,J.Biol.Chem.,268:10686-10693(1993)を
参照のこと。当該RNAse活性は、哺乳動物のRNAseインヒビターに対して耐性で
あり、これによって哺乳動物の酵素に比較して観察される細胞毒性の増強が説明され得る
。同上。ランピルナーゼは、このファミリーのRNAseの原型的なメンバーであり、ヒ
ョウガエルの卵母細胞および初期の胚から精製され得る。ランピルナーゼは、12,00
0の分子量を有し、そしてウサギの網状赤血球溶解物において(IC50 10-11M)、
そしてアフリカツメガエル(Xenopus)の卵母細胞に微量注入された場合(IC50 10-10
M)、タンパク質合成の強力な阻害を生じる。RNase Aスーパーファミリーの他の
メンバーとは異なり、これらの細胞毒性リボヌクレアーゼは、卵母細胞rRNAを分解し
ない。Lin et al.,Biochem Biophys Res Commun.204:156-62(1994)。これらの分子は、酵
素のリン酸結合ポケットの一部を形成し、RNAseおよび抗腫瘍活性に必須である必須
のN末端ピログルタミル残基を含む。
【0004】
動物の毒性研究によって、ランピルナーゼが予測可能な用量依存性でかつ可逆性の毒性
を、ラット(用量範囲0.01〜0.02mg/kg)およびイヌ(0.005〜0.1
5mg/kg)の両方で示すということが示される。侵襲性のM109 Madison肺癌腫を接種
されて、かつ毎日および毎週の両方のスケジュールでランピルナーゼの腹腔内投与の処置
をされたマウスは、有意に延長された生存を示した。ほとんどの著しい結果は、ランピル
ナーゼの毎週のスケジュールで処置されたマウスの群でみられ、ここでは、18匹の動物
のうち6匹が、長期間生存して、明らかに癌が治癒した。Mikulski et al.,J Natl Cance
r Inst.82:151-3(1990)。
【0005】
天然のランピルナーゼは、臨床試験において、種々の固体腫瘍に対して抗腫瘍活性を有
することが示されている。これに関しては、ランピルナーゼは、例えば、膵臓癌を有する
患者を処置する場合、単独で、そしてタモキシフェンのような他の抗腫瘍因子と組み合わ
せての両方で用いられている。抗腫瘍因子として用いられる場合、これらの細胞毒性RN
Aseは、特定の細胞タイプに対する標的を可能にするためのマーカーに複合体化され得
る。
【0006】
第I相試験では、種々の再発性および耐性の腫瘍に罹患している患者を、ランピルナー
ゼ(用量範囲60〜960μg/m2)を用いて毎週静脈内処置した。観察される副作用
としては、一般に顔面紅潮、筋肉痛、一過性の眩暈および食欲減退が含まれた。タンパク
尿の増大、末梢浮腫、高窒素血症、クレアチニンクリアランスの低下、および疲労によっ
て呈示される、用量を制限する腎臓毒性を含む、観察される毒性は、用量依存性でかつ可
逆性であり、これは、動物での毒物学的研究と一致している。真の免疫学的感作の臨床的
な顕在化は、ランピルナーゼの反復された毎週の静脈内用量後でさえ証明されていない。
最大耐容量は、主に腎毒性に起因して、960μg/m2であることが見出された。非小
細胞肺癌、食道癌および結腸直腸癌にはなんらかの他覚的な応答があった。Mikulski et
al.,Int J Oncol 3:57-64,(1993);Mikulski et al.,J Clin Oncol.20:274-81(2002)を参
照のこと。それにもかかわらず、ランピルナーゼは、試験された動物およびほとんどのヒ
ト患者では十分耐容されて、一貫してかつ可逆性の臨床毒性パターンを示し、そしてほと
んどの従来の化学療法剤に伴うほとんどの毒性、例えば、骨髄抑制および脱毛症は誘発し
なかった。
【0007】
WO97/31116は、(a)N末端メチオニンに続いてグルタミン酸以外のアミノ酸、(b)
システインを26位、40位、58位、84位、95位、および110位に、リジンを4
1位に、そしてヒスチジンを119位に(ウシRNAseAとの最大一致について整列さ
せた場合)を有する組み換えリボヌクレアーゼ、ならびに天然のランピルナーゼ由来のア
ミノ酸配列を開示している。しかし、RNAse改変体(variant)を含む組み換え細胞毒
性RNAseは、多段階のプロセスによって細菌で調製されており、このプロセスには、
細菌によって挿入されたN末端ホルミルメチオニン残基を除去するため、そして必須のN
末端ピログルタミル残基を生成するためのさらなる工程を要する。それにもかかわらず、
細菌中での産生は、グリコシル化された細胞毒性RNAse含有融合タンパク質の調製を
妨げる。従って、真核生物細胞において組み換え細胞毒性RNAse融合タンパク質を産
生することが有利である。真核生物細胞では、細胞毒性RNAseは、両生類の供給源か
ら精製されたランピルナーゼの細胞毒性特性を保持するが、ヒトにおける所望されない免
疫応答は少ないか、またはない。しかし、真核生物細胞における細胞毒性RNAseの発
現は、RNAseの細胞毒性活性に起因して細胞死を生じるということも予想される。
【0008】
従って、グリコシル化RNAse融合タンパク質を含む、細胞性RNAse含有融合タ
ンパク質を調製する改良方法が極めて望ましいということが明らかである。グリコシル化
細胞毒性RNAse融合タンパク質自体が極めて所望される分子であるということも明ら
かである。
【発明の開示】
【0009】
従って、本発明の目的は、(a)第一の免疫グロブリン可変ドメインに融合された非哺
乳動物の細胞毒性リボヌクレアーゼを含む融合ポリペプチドと、(b)第二の免疫グロブ
リン可変ドメインを含む第二のポリペプチドとを含み、この免疫グロブリン可変ドメイン
の1つが軽鎖可変ドメインであり、かつもう一方の免疫グロブリン可変ドメインが重鎖可
変ドメインであり、この第一および第二の免疫グロブリン可変ドメインが一緒になって抗
原結合部位を形成し、そしてこの免疫毒素がグリコシル化されている、免疫毒素を提供す
ることである。
【0010】
本発明の別の目的は、哺乳動物宿主細胞を培養することによってこれらおよび他の免疫
毒素を作製する方法を提供することであって、この宿主細胞は、(a)第一の免疫グロブ
リン可変ドメインに融合された非哺乳動物細胞毒性リボヌクレアーゼを含む融合ポリペプ
チドをコードする核酸配列と、(b)第二の免疫グロブリン可変ドメインを含む第二のポ
リペプチドをコードする核酸配列とで形質転換され、この第一および第二の免疫グロブリ
ンの可変ドメインは一緒になって抗原結合部位を形成する。
【0011】
本発明のさらに別の目的は、細胞毒性RNAse部分に融合された内部移行抗体(inter
nalizing antibody)または抗体フラグメントを含む免疫毒素であって、このRNAse部
分がN末端ピログルタミン酸残基を保有し、かつこのRNAse部分がそのC末端におい
て、この抗体または抗体フラグメントの軽鎖を含むポリペプチドのN末端、あるいはこの
抗体またはフラグメントの重鎖を含むポリペプチドのN末端、あるいはこの抗体またはフ
ラグメントの軽鎖および重鎖の両方のN末端に融合されている免疫毒素を提供することで
ある。
【0012】
例えば、一実施形態では、非哺乳動物細胞毒性リボヌクレアーゼは、第一の免疫グロブ
リン可変ドメイン、例えば、軽鎖可変ドメインのN末端に融合されてもよい。この非哺乳
動物リボヌクレアーゼは、N末端ピログルタミン酸残基を有してもよい。
【0013】
上記融合ポリペプチドおよび第二の免疫グロブリン可変ドメインは、別の分子であって
もよく、そして宿主細胞中で別の分子として産生されてもよい。
【0014】
上記融合ポリペプチドはさらに、CLドメインを含んでもよく、そして上記第二のポリ
ペプチドはさらにCH1ドメインを含んでもよい。
【0015】
上記第二のポリペプチドはさらに、CH2ドメインおよびCH3ドメインを含んでもよ
い。
【0016】
別の実施形態では、非哺乳動物リボヌクレアーゼは、重鎖可変ドメインのN末端に融合
されてもよい。この非哺乳動物リボヌクレアーゼは、N末端ピログルタミン酸残基を有し
てもよい。この融合ポリペプチドおよび第二の免疫グロブリン可変ドメインは、別の分子
であってもよく、そして宿主細胞中で別の分子として産生されてもよい。
【0017】
この融合ポリペプチドはさらに、CH1ドメインを含んでもよく、そして第二のポリペ
プチドはさらにCLドメインを含んでもよい。
【0018】
この融合ポリペプチドはさらに、CH2ドメインおよびCH3ドメインを含んでもよい
。
【0019】
さらに別の実施形態では、非哺乳動物リボヌクレアーゼは、第一および第二の免疫グロ
ブリン可変ドメインの両方のN末端に融合されてもよい。この非哺乳動物リボヌクレアー
ゼは、N末端ピログルタミン酸残基を有してもよい。この第一の免疫グロブリン可変ドメ
インは、重鎖可変ドメインであってもよく、そして第二の免疫グロブリン可変ドメインは
、軽鎖可変ドメインであってもよい。
【0020】
この第一および第二の融合ポリペプチドは、別の分子であってもよく、そして宿主細胞
中で別の分子として産生されてもよい。
【0021】
この第一の融合ポリペプチドはさらに、CH1ドメインを含んでもよく、そして第二の
融合ポリペプチドはさらにCLドメインを含んでもよい。
【0022】
この第一の融合ポリペプチドはさらに、CH2ドメインおよびCH3ドメインを含んで
もよい。
【0023】
免疫毒素は、例えば、CH2ドメイン上で、および/またはRNAse上でグリコシル
化されてもよい。
【0024】
上記の非非哺乳動物リボヌクレアーゼは、例えば、ヒョウガエルから精製されたランピ
ルナーゼの配列および構造を有する酵素であってもよい。
【0025】
上記抗原結合部位は、細胞表面分子に対して特異的に結合してもよく、そして上記免疫
毒素は、この細胞表面分子を保有する細胞結合した際に内部移行されてもよい。
【0026】
上記免疫毒素は、ガン細胞、感染/炎症の部位、自己免疫疾患に対するか、または微生
物および寄生生物に対するものであり得る。
【0027】
上記免疫毒素は、B細胞抗原、T細胞抗原、形質細胞抗原、HLA−DR系統抗原、C
EA,NCA、MUC1、MUC2、MUC3、MUC4およびMUC16抗原、EGP
−1抗原、GEP−2抗原、胎盤アルカリホスファターゼ抗原、IL−6、VEGF、P
1GF、ILGF、EGFR、Her2/neu、テネイシン、CD33、CD74、P
SMA、PSA、PAP、Leyを含む腫瘍関連抗原、自己免疫疾患、感染/炎症および
感染性疾患に関連する抗原からなる群より選択される抗原に対するものであってもよい。
【0028】
上記抗原は、B細胞もしくはT細胞リンパ腫に関連する標的抗原、または自己免疫疾患
に関連するB細胞もしくはT細胞であってもよい。
【0029】
上記抗原は、悪性疾患によって発現されるCD19、CD22、CD40、CD74、
CEA、NCA、MUC1、MUC2、MUC3、MUC4、HLA−DR、EGP−1
、EGP−2、EGFR、Her2/neu、IL−15およびHLA−DRからなる群
より選択される抗原であってもよい。
【0030】
上記免疫毒素の免疫グロブリン可変ドメインは、悪性疾患を標的する場合、例えば、R
S11、17−1A、RS7、LL1、LL2、MN−3、MN−14、MN−15もし
くはPAM4,またはそのヒト化バージョンに由来し得る。
【0031】
免疫毒素によって結合される抗原は、特定の悪性疾患に関しては、例えば、EGP−2
,EGP−1、CD22、CD74、CEA、炭酸脱水素酵素IX、またはMUC1であ
ってもよい。
【0032】
標的化抗原は、細菌、ウイルス、真菌、マイコプラズマ、寄生生物、または他の微生物
によって発現されてもよい。
【0033】
標的化合物抗原は、活性化顆粒球のような、感染の部位での宿主細胞蓄積によって発現
されてもよい(例えば、CD15、CD33、CD66a、CD66b、およびCD66
c(NCA)など)。適切なこのような顆粒球抗体は、MN−3であり、そのFab’は
LeukoScan(商標)で用いられる。
【0034】
上記の方法および/または組成物のいずれかにおいて、可変ドメインはヒト化されてよ
いしまたはヒトドメインであってもよい。
【0035】
本発明の別の実施形態では、薬学的に受容可能なキャリアとともに上記されたような免
疫毒素を含む薬学的組成物が提供される。
【0036】
本発明のさらに別の実施形態では、被験体に対して上記のような組成物を、疾患または
症候群の症状を処置または改善するために有効な量で投与することによって、この被験体
におけるこの疾患または症候群を処置する方法が提供される。この疾患または症候群は、
この被験体における細胞、例えばT細胞、骨髄細胞または形質細胞の望ましくないまたは
不適切な増殖または活性化によって特徴付けられ得る。あるいは、この疾患は、感染性微
生物または寄生生物の存在によって特徴付けられ得る。
【0037】
この疾患は、例えば、癌腫、肉腫、神経膠腫、白血病、リンパ腫、黒色腫、神経芽細胞
腫、および骨髄腫からなるガンの群より選択され得る。
【0038】
この疾患または症候群は、固形腫瘍、例えば、神経芽細胞腫、悪性黒色腫、または癌腫
、例えば、乳房、藍藻、前立腺、肺、腎臓、胃、結腸直腸、肝臓、または膵臓の癌腫の存
在によって特徴付けられ得る。
【0039】
これらの方法では、薬学的組成物は、被験体に対して、1日あたり2回以上、例えば、
0.1〜約1000mgの量で投与されてもよいが、それより多く用いられても少なく用
いられてもよい。
【0040】
望ましくないまたは不適切な増殖または活性化を受けている細胞は、例えば、関節リウ
マチ、シェーグレン症候群、急性特発性血小板減少性紫斑病、慢性特発性血小板減少性紫
斑病、皮膚筋炎、シデナム舞踏病、重症筋無力症、全身性エリテマトーデス、ループス腎
炎、リウマチ熱、多腺性症候群、水疱性類天疱瘡、糖尿病、ヘノッホ・シェーンライン紫
斑病、連鎖球菌感染後腎炎、結節性紅斑、高安動脈炎、アジソン病、多発性硬化症、サル
コイドーシス、潰瘍性大腸炎、多形性紅斑、IgA腎症、結節性多発性動脈炎、強直性脊
椎炎、グッドパスチャー症候群、血栓血管炎(thromboangitis ubiterans)、原発性胆汁性
肝硬変、橋本甲状腺炎、甲状腺中毒症、強皮症、慢性活動性肝炎、多発性筋炎/皮膚筋炎
、多発性軟骨炎、尋常性天疱瘡、ヴェーゲナー肉芽腫症、膜性腎症、筋萎縮性側索硬化症
、脊髄癆、巨細胞性動脈炎/多発性筋痛、悪性貧血、急速進行性糸球体腎炎および線維化
性肺胞炎のような1つ以上の自己免疫疾患の発症および進行に関与し得る。
【0041】
本発明の組成物が、感染の治療的処置のために用いられる場合、免疫毒素の免疫グロブ
リン構成要素は、疾患原因の微生物、例えば、病原性細菌、ウイルス、真菌および多様な
寄生生物に特異的に結合し、そして抗体は、これらの微生物、それらの産物もしくは表面
抗原、またはそれらの病変に関連する抗原を標的し得る。微生物の例としては、限定はし
ないが、
ストレプトコッカス・アガラクティエ、在郷軍人病菌、化膿連鎖球菌、大腸菌、淋菌、
髄膜炎菌、肺炎球菌、Hemophilis influenzae B、梅毒トレポネーマ、ライム病スピロヘ
ータ、緑膿菌、ハンセン菌、ウシ流産菌、ヒト結核菌、破傷風毒素、抗ウイルスモノクロ
ーナル抗体、HIV−1、HIV−2、HIV−3、A型肝炎、B型肝炎、C型肝炎、D
型肝炎、狂犬病ウイルス、インフルエンザウイルス、サイトメガロウイルス、単純疱疹I
およびII、ヒト血清パルボ様ウイルス、呼吸器合胞体ウイルス、水痘帯状疱疹ウイルス
、B型肝炎ウイルス、麻疹ウイルス、アデノウイルス、ヒトT細胞白血病ウイルス、エプ
スタイン・バーウイルス、マウス白血病ウイルス、流行性耳下腺炎ウイルス、水疱性口内
炎ウイルス、シンドビスウイルス、リンパ球性脈絡髄膜炎ウイルス、疣ウイルス、ブルー
・タングウイルス、センダイウイルス、猫白血病ウイルス、レオウイルス、ポリオウイル
ス、シミアン・ウイルス40、マウス乳癌ウイルス、デング熱ウイルス、ポリオーマウイ
ルス、パピローマウイルス、SARSウイルス、種々のインフルエンザウイルス、風疹ウ
イルス、原生動物、熱帯熱マラリア原虫(Plasmodium falciparum)、三日熱マラリア原虫(
Plasmodium vivax)、トキソプラズマ原虫、Trypanosoma rangeli、クルーズ・トリパノソ
ーマ、Trypanosoma rhodesiensei、トリパノソーマ・ブルーセイ、マンソン住血吸虫,Sc
histosoma japanicum、Babesia bovis、Elmeria tenella、回旋糸状虫、熱帯リーシュマ
ニア、旋毛虫、Theileria parva、Taenia hydatigena、Taenia ovis、無鉤条虫、単包条
虫、Mesocestoides corti、マイコプラズマ、Mycoplasma arthritidis、M.hyorhinis、M.
orale、M.arginini、Acholeplasma laidlawii、M.salivarium、およびM肺炎が挙げられ
る。
【0042】
本発明の他の目的、特徴および利点は、以下の詳細な説明から明らかになる。しかし、
この詳細な説明および特定の実施例は、本発明の好ましい実施形態を示すが、例示のため
だけに示していることが理解されるべきである。なぜなら、本発明の趣旨および範囲内の
種々の変化および改変が、この詳細な説明から当業者には明白になるからである。
【図面の簡単な説明】
【0043】
【図1】ベクターpdHL2−rap−hLL1−γ4Pの構成を示す。構築物である、リーダーペプチド−RNAse、−リンカー−hLL1のVk領域を含むXbaI−BamHIフラグメントを、pBSベクターに挿入して、pdHL2ベクターに連結した。注記:本発明においては、rpRNAseおよびrapという用語は、交換可能である。
【図2】宿主細胞から分泌された融合タンパク質のHPLCプロフィールを示す。単一のピークが7.7分で観察された。
【図3】分泌された融合タンパク質のSDS−PAGEを示す。2つの接近して移動する軽鎖バンドをみることができるが、これはhLL1単独の軽鎖よりも長い(25kD)。
【図4a】天然の条件下で分泌された融合タンパク質の質量スペクトルを示す:177150の質量を有するrap−hLL1−γ4P融合タンパク質に相当する、1つの主要なピークが観察された。
【図4b】変性条件下の質量スペクトルを示す。重鎖、およびrap−hLL1−γ4P融合タンパク質の糖分子の有無の2つの軽鎖に相当する3つの主要なピークが見出され得る。
【図5】融合タンパク質のウエスタンブロットを示す。両方の分子ともrapを含むことを示している、糖有の軽鎖および糖無の軽鎖が示され得る。
【図6】N−グリコシダーゼ処理による軽鎖糖分子の除去を示すSDS−PAGEゲルを示す:レーン1は、未処理のサンプルに相当し、ここでは2つの軽鎖バンドを見ることができる。レーン2〜5は、Nグリコシダーゼ酵素で処理されたサンプルである。2つの軽鎖に相当するバンドは、糖の除去の際に1つのバンドに集束した。
【図7】インビトロ転写翻訳アッセイによるRNAse活性を示す。
【図8】競合結合アッセイを示しており、これは、hLL1およびrap−hLL1融合タンパク質の両方が、hLL1の抗イディオタイプ抗体であるWPについて同じ親和性を有することを示している。
【図9】Daudi細胞における融合タンパク質のインビトロ細胞毒性を示す:A)MTSアッセイによって測定した細胞毒性;B)BRdUアッセイ法によって測定した細胞毒性。
【図10】MTSアッセイによるMC/CAR細胞における融合タンパク質のインビトロ細胞毒性を示す。
【図11】ナイーブなSCIDマウスにおける2L−Rap−hLL1−γ4Pの血液クリアランスを示す。ナイーブなSCIDマウスに、88Y−DTPA−hLL1(O)および111In−DTPA−2L−Rap−hLL1−γ4P(□)を静脈内に同時注射した。投与後選択された時間で、心臓穿刺によってマウスを放血させて、血液サンプルを放射能についてカウントした。データは、血液中の注射用量の平均±S.D.である(n=3)。
【図12】2L−Rap−hLL1−γ4Pまたは成分のタンパク質を用いた侵襲最小ダウディリンパ腫(aggressive minimal Daudi lymphoma)の処置を示す。SCIDマウス(1群あたり8〜10匹のマウス)に、1.5×107個のDaudi細胞を静脈内に接種した。1日後、マウスを、2L−Rap−hLL1−γ4Pの1μg(×)、5μg(黒四角)、15μg(黒三角)、30μg(下向き黒三角)、40μg(◆)、または50μg(●)の単回ボーラス注射で処置した。コントロール群には、50μgの免疫毒素(*)に等価な構成要素のタンパク質またはPBS(□)のみを注射した。
【図13】インビトロの転写/翻訳アッセイによって測定されたRNase活性を示す。rRap(黒四角)、2L−Rap−hLL1−γ4P(黒上向き三角)およびhLL1−γ4P(◆)の濃度を、相対発光単位(relative luminescence units)(RLU)に対してプロットした。
【発明を実施するための形態】
【0044】
本発明者らは、驚くべき事に、細胞毒性RNAse分子を含む融合分子が哺乳動物細胞
において調製され得ることを見出した。この結果は、驚くべきである。なぜなら、酵素的
に活性な細胞毒性RNAse分子を含む分子は、細胞増殖を妨害して、これによって融合
タンパク質の組み換え産生を妨げる程度まで細胞毒性であることが予想されるからである
。
【0045】
この結果によって、原核生物宿主細胞を用いて組み換え方法によって調製することが以
前には困難であるかまたは不能であった哺乳動物細胞において、細胞毒性RNAse融合
タンパク質を調製する可能性がもたらされる。例えば、哺乳動物宿主細胞における産生に
よって、原核生物宿主細胞を用いる場合、必須のN末端ピログルタミル残基を生成するこ
とに関連する困難性が回避される。さらに、哺乳動物宿主細胞の使用によって、マルチサ
ブユニットタンパク質、例えば、インタクトなFc領域を含むIgG構造の容易な調製が
可能になり、そしてまたグリコシル化融合タンパク質、例えば、Fc領域上でグリコシル
化されるインタクトな抗体の調製が可能になる。細胞毒性RNAseと免疫グロブリン、
例えば抗体または抗体フラグメントとの間の融合タンパク質は、本明細書において以降で
は免疫毒素と呼ばれる。この方法はまた、任意の適切な抗体フラグメント、例えば、F(
ab’)2、F(ab)2、Fab’、Fab、FvおよびscFvを含む免疫毒素を調製
するために用いられ得る。
【0046】
本発明はまた、RNAseが、Ig可変ドメイン(例えばVHまたはVLドメイン)の
1つのN末端に融合される、細胞毒性RNAse含有免疫グロブリン融合タンパク質を提
供する。これは驚くべきことに、細胞毒性RNAse活性を保持するだけでなく、抗原結
合部位でIg結合特異性を保持しており、N末端融合の調製によって立体的にブロックさ
れることが期待されている融合タンパク質を提供する。
【0047】
さらに、本発明者らは、驚くべきことに、本発明の細胞毒性RNAse融合タンパク質
が細胞および微生物/寄生生物の殺傷に有効であることを見出している。詳細には、迅速
な内部移行抗体成分を含む免疫毒素は細胞特異的な様式で極めて細胞毒性であることが見
出されている。例えば、ヒト化LL1抗体のN末端に対する細胞毒性RNAseの融合に
よって、CD74(LL−1の標的)の不変鎖を発現する細胞に対して特異的に結合して
、結合の際その細胞集団に対して細胞毒性である免疫毒素が産生された。この結果は、驚
くべきことである。なぜなら、融合タンパク質は、融合タンパク質の細胞毒性を大きく低
下させるか、またはさらに排除する可能性が高いことが予期される、内部移行の際に細胞
のリソソームに送達されて、そこで分解されることが予想されるからである。
【0048】
本発明はまた、疾患または症候群に罹患している被験体を、上記で記載されるタイプの
免疫毒素の有効量をこの被験体に投与することによって処置する方法を提供する。
【0049】
(定義)
他に特定しない限り、用いられる全ての技術および科学的用語は、当業者によって通常
理解されるのと同じ意味を有する。さらに、本明細書に引用される全ての特許および他の
引用文献の内容は、その全体が引用することにより組み込まれる。本発明の目的のために
は、以下の用語は以下のとおり規定される:
【0050】
アミノ酸は、名称によるか、またはその一般に公知の3文字記号か、もしくは1文字の
IUPAC記号のいずれかによって言及される。ヌクレオチドは、その通常受容される一
文字コードによって呼ばれる。
【0051】
特定の核酸配列の「保存的に改変されたバリエーション(conservatively modified var
iations)」とは、同一であるかまたは本質的に同一であるアミノ酸配列をコードする核酸
をいうか、またはこの核酸が、アミノ酸配列をコードしない場合は、本質的に同一の配列
をいう。遺伝子コードの縮重の理由で、多数の機能的に同一の核酸が、任意の所定のポリ
ペプチドをコードする。例えば、コドンGCA、GCC、GCGおよびGCUは全てがア
ラニンというアミノ酸をコードする。従って、アラニンがコドンによって特定されるあら
ゆる位置では、このコドンは、コードされたポリペプチドを変化させることなく、記載さ
れる任意の対応するコドンに変化され得る。メチオニンをコードするAUG以外の核酸中
の各々のコドンが、機能的に同一の分子を生じるように改変され得る。本明細書に記載さ
れる核酸配列はまた、これらの変更を包含する。
【0052】
アミノ酸配列の「保存的に改変されたバリエーション」とは、コードされた配列中の単
独のアミノ酸または小さい割合のアミノ酸を変更させる個々の置換であって、この変更に
よって、化学的に類似のアミノ酸でのアミノ酸の置換が生じる置換を包含する。保存的置
換は当業者に周知である。以下の6つの群の各々は、お互いについて保存的置換であるア
ミノ酸を含む:
1.アラニン、セリン、トレオニン
2.アスパラギン酸、グルタミン酸
3.アスパラギン、グルタミン
4.アルギニン、リジン
5.イソロイシン、ロイシン、メチオニン、バリン、そして
6.フェニルアラニン、チロシン、トリプトファン。
【0053】
アミノ酸配列の「保存的に改変されたバリエーション」はまた、コードされた配列中の
単独のアミノ酸または小さい割合のアミノ酸の欠失または付加であって、この付加および
欠失が、化学的に類似のアミノ酸でのアミノ酸の置換を生じる欠失または付加を包含する
。本明細書に記載されるアミノ酸配列はまた、これらのバリエーションを包含する。
【0054】
「単離された(isolated)」または「生物学的に純粋な(biologically pure)」という用
語は、その天然に存在する環境において見出されるような通常伴う成分を実質的にまたは
本質的に含まない物質をいう。この単離された物質は状況に応じて、その天然の環境にお
いてこの物質とともに見出されない物質を含む。
【0055】
「核酸(nucleic acid)」という用語は、デオキシリボヌクレアーゼまたはリボヌクレオ
チドポリマーであって、一本鎖型または二本鎖型のいずれかをいい、他に限定しない限り
、天然に存在するヌクレオチドに対して同様の様式で核酸にハイブリダイズする天然のヌ
クレオチドの公知のアナログを包含する。他に示さない限り、特定の核酸配列とは、その
相補的な配列を含む。
【0056】
「発現ベクター(expression vector)」とは、組み換え発現カセットを含み、この組み
換え発現カセットは、細胞によって転写および翻訳され得る、本発明によるポリペプチド
をコードする核酸を包含する。組み換え発現カセットとは、組み換え的にまたは合成的に
生成された核酸構築物であり、標的細胞において特定の核酸の転写を可能にする一連の特
定の核酸エレメントを伴う。発現ベクターは、プラスミド、ウイルスまたは核酸フラグメ
ントの一部であってもよい。代表的には、この発現ベクターのこの組み換え発現カセット
部分は、転写されるべき核酸およびそれに作動可能に連結されるプロモーターを包含する
。
【0057】
「組み換え体(recombinant)」という用語は、タンパク質に関して用いられる場合、あ
る細胞がその細胞には外因性である起源の核酸によってコードされるペプチドまたはタン
パク質を発現することを示す。組み換え細胞は、その細胞の天然の(非組み換え体)型内
では見出されない遺伝子を発現し得る。組み換え細胞はまた、細胞の天然の形態で見出さ
れる遺伝子を発現し得、この遺伝子は、人工的な方法によって、例えば、異種プロモータ
ーの制御下で細胞に再導入される。
【0058】
「実質的な同一性(substantial identity)」または「実質的な類似性(substantial sim
ilarity)」という用語は、ポリペプチドの文脈では、あるポリペプチドが参照配列と少な
くとも80%、さらに好ましくは90%、そして最も好ましくは少なくとも95%の同一
性を有する配列を含むことを示す。実質的に同一である2つのポリペプチドとは、このポ
リペプチドの1つが第二のペプチドに対して惹起された抗体と免疫学的に反応性であるこ
とを意味する。2つの核酸が実質的に同一であるとは、この2つの分子がストリンジェン
トな条件下でお互いにハイブリダイズすることである。一般に、ストリンジェントな条件
とは、所定のイオン強度およびpHにおいて特定の配列についての融点(Tm)よりも約
5℃〜20℃低いように選択される。このTmは、標的配列の50%が完全にマッチした
プローブにハイブリダイズする温度(所定のイオン強度およびpH下で)である。しかし
、ストリンジェントな条件下でお互いに対してハイブリダイズしない核酸は、それらがコ
ードするポリペプチドが実質的に同一であるならば、やはり実質的に同一である。
【0059】
「抗体(antibody)」とは、当分野で周知の種々の免疫グロブリン種(IgG1、IgG4
、IgMなど)の抗体全体および抗体フラグメント、例えば、F(ab’)2、F(ab
)2、Fab’Fab、Fvなどを含み、このフラグメントとしては、標的抗原に結合す
る能力を保持するハイブリッドフラグメントが挙げられる。また、免疫グロブリンの超可
変性の抗原結合領域を保持する任意の小フラグメントも有用である。抗体全体から、抗原
に特異的に結合する能力を保持している抗体フラグメントを作成する方法は、当分野で周
知であり、そして例えば、Harlow and Lane「Antibodies:A Laboratory Manual」CSHL Pr
ess(1988)に記載されている。組み換え抗体または抗体フラグメントを作成する方法は、
当分野で周知であり、例えば、McCafferty et al.,「Antibody Engineering:A Practical
Approach」IRL Press(1996)に記載されている。Fabフラグメントは、代表的には鎖間
ジスルフィド結合を含む(VL−CL)−(VH−CH1)二量体を含む。Fab’フラ
グメントは、類似であるが、また重鎖ヒンジドメインの一部を含む。F(ab’)2フラ
グメントとは、従来、2つのFab’フラグメントのジスルフィド結合二量体を指すこと
が理解される。本発明の抗体は、任意の種の抗体であってもよいが、この抗体はヒト抗体
またはヒト化抗体であることが有利である。抗体はまた、任意の周知の形態または組み換
え抗体、例えば、単鎖Fv(VH鎖に対する任意のリンカーによって結合されたVL鎖を
含む)であってもよい。このリンカーが短いかまたは存在しない場合、この単鎖Fvは二
量体または多量体に二量化し、この二量体とは二重特異性抗体として公知である。特定の
環境下では、本発明の抗体はまた、単結合ドメイン抗体、例えば、ラクダ科抗体(Dumoul
in et al.,「Single-domain antibody fragments with high conformational stability.
」Protein Sci. 11,500-515(2002);Hamers-Casterman et al.,「Naturally occurring an
tibodies devoid of light chains」Nature 363,446-448(1993);ならびにMuyldermans.「
Single domain camel antibodies:current status」J Biotechnol.74,277-302(2001)を参
照のこと)、または「ドメイン抗体(domain antibody)」を含み得る。WO92/01787および
そこに引用される参考文献を参照のこと。ヒト抗体とは、ヒト被験体において産生される
抗体由来であるか、またはヒトにおいて産生される抗体の特徴である配列を有するかのい
ずれかの配列を有する、抗体または抗体フラグメントである。このような抗体は、例えば
、米国特許第6,300,064号に記載される。ヒト抗体はまた、ヒト抗体ライブラリーから単
離され得る。例えば、米国特許第6,300,064号および同第6,172,197号、ならびにそこに記
載される参考文献を参照のこと。ヒト化抗体は、非ヒト抗体からヒトフレームワークへC
RD領域を挿入することによって、続いて必要に応じて抗原結合を回復または最適化させ
るための特定のフレームワーク残基の変異によって調製される抗体である。ヒト化抗体は
当分野で周知である、そして例えば、Jones et al.,Nature 321:522(1988);Riechmann et
al.,Nature 332:323(1988);およびWinter&Milstein,Nature,349:293(1991)に記載される
。
【0060】
「標的化部分(targeting moiety)」とは、所定の細胞タイプの上のマーカーまたはレセ
プターに特異的である抗体、ペプチド、サイトカイン、オリゴヌクレオチド、または増殖
因子である。標的化部分は、所定の細胞タイプに関連するマーカーと優先的に関連するこ
とによって、その細胞タイプに対して、結合された分子を特異的に送達するために用いら
れ得る。
【0061】
「融合タンパク質(fusion protein)」とは、2つ以上のポリペプチド、さらに詳細には
、細胞毒性RNAseおよび標的化部分を結合することによって形成されるキメラ分子で
ある。細胞毒性RNAseおよび標的化部分は、標的化部分のアミノ末端とRNAseの
カルボキシル末端との間で形成されたペプチド結合を通じて結合され、そして融合タンパ
ク質をコードする核酸配列によって組み換え的に発現される。単鎖融合タンパク質は、単
一の連続するポリペプチド骨格を有する融合タンパク質である。
【0062】
「化学的複合体(chemical conjugate)」とは、細胞毒性RNAseおよび標的化部分の
化学的カップリングによって形成される複合体である。
【0063】
「薬学的に受容可能なキャリア(pharmaceutically acceptable carrier)」とは、免疫
毒素を投与するためのビヒクルとして用いられ得る物質であって、この物質は、不活性で
あるかそうでなければ医学的に受容可能であり、そして融合タンパク質またはアームのリ
ガンドと適合性であるという理由でビヒクルとして用いられ得る物質である。
【0064】
(細胞毒性RNAseをコードする核酸の調製)
当業者は、本発明における使用に適切な細胞毒性RNAse部分が、天然のランピルナ
ーゼ構造を有するポリペプチドおよび全ての酵素的に活性なその改変体を含むことを認識
する。これらの分子は有利には、RNAse活性に必須であると考えられるN末端ピログ
ルタミン酸残基を有し、哺乳動物RNAseインヒビターによって実質的に阻害されない
。天然の細胞毒性RNAseをコードする核酸は、適切な配列のクローニングおよび制限
によって、またはポリメラーゼ連鎖反応(PCR)によるDNA増幅を用いることによっ
て調製され得る。ヒョウガエルのランピルナーゼのアミノ酸配列は、Ardelt et al.,J.Bi
ol.Chem.,256:245(1991)から得ることが可能であり、そして天然のランピルナーゼをコー
ドするcDNA配列、またはその保存的に改変された改変体は、Leung et al.,Mol.Immun
ol.,32:1413(1995)のhLL2ヒト化に用いられるen bloc V-geneアセンブリ法と同様の
方法によって遺伝子合成され得る。細胞毒性RNAse改変体を作成する方法は、当分野
で公知であり、そして慣用的な技術の範囲内である。
【0065】
あるいは、細胞毒性RNAseまたはその改変体をコードする核酸は、インビトロで合
成され得る。化学的合成によって一本鎖オリゴヌクレオチドが生成される。相補的な配列
とのハイブリダイゼーションによって、または短いプライマーおよび一本鎖をテンプレー
トとして用いるDNAポリメラーゼでの重合によって、これは二本鎖DNAに変換され得
る。化学的合成は、約100塩基の配列に最も適合するが、より短い配列を連結すること
によってそれより長い配列を得てもよい。下の実施例2は、細胞毒性RNAse遺伝子を
得るための1つの例示的な方法を提供する。
【0066】
(免疫グロブリン遺伝子の調製)
抗体をコードする遺伝子を調製する方法は、当分野で周知である。例えば、変性プライ
マーセットを用いるPCRを用いて、抗体産生細胞、例えばハイブリドーマから得られる
可変鎖配列cDNAを増幅してもよい(または周知の方法を用いる直接RT−PCRが、
細胞から得られるmRNAに対して直接用いられ得る)。例えば、Marks et al.,J Mol B
iol.(1991)222:581-97、およびそこに引用される参考文献を参照のこと。非ヒト抗体は、
当分野で周知である方法によってヒト化されてもよい。Leung、前出およびそこに引用さ
れる方法を参照のこと。あるいは、ヒト(または他の種)の抗体可変領域ドメインをコー
ドする遺伝子は、MorphoSys(Martinsried,Germany)またはCambridge Antibody Technolog
y(Cambridge,UK)から市販されているタイプのファージ・ディスプレイ・ライブラリーか
ら得てもよい。下の実施例1および実施例3はまた、抗体可変ドメインをコードする核酸
配列を得るための例示的な方法を示す。
【0067】
本質的に、所望の特異性を有する任意の抗体配列が、本発明による免疫毒素を調製する
ために用いられ得る。特に好ましいのは、治療上有用であることが示されている特異性を
有する抗体である。適切な例としては、限定はしないが、腫瘍に結合する抗体、例えば、
B細胞抗原、T細胞抗原、形質細胞抗原、HLA−DR系統抗原、MUC1、MUC2、
MUC3、およびMUC4およびMUC16抗原、EGP−1抗原、EGP−2抗原、胎
盤アルカリホスファターゼ抗原、炭酸脱水素酵素IX、IL−6、VEGF、P1GF、
ILGF、Her2/neu、テネイシン、CD33、CD40、CD74、CD80、
PSMA、PSAおよびPAP;ならびにHLA−DR、CD27、CD40、B細胞お
よびT細胞抗原(例えば、CD15、CD19、CD20、CD21、CD25およびC
D22抗原)を含む自己免疫疾患に関連する抗原が挙げられる。この抗原は、CD19、
CD21、CD22、CD40、MUC1、HLA−DRおよびIL−15のようなB細
胞またはT細胞リンパ腫または白血病に関連する抗原であってもよい。この抗原がCD7
4またはCD74−HLA−DR複合体である場合、適切な抗体はLL1またはそのヒト
化バージョンである。抗原がCD22である場合、適切な抗体はLL2もしくはRBF4
であるか、またはそのヒト化バージョンである。抗原がMUC1である場合、適切な抗体
はPAM4、またはそのヒト化バージョンである。抗原がCD16である場合、適切な抗
体は、CA125またはMu−9のいずれかである。抗原がEGP−1である場合、適切
な抗体はRS7またはそのヒト化バージョンである。抗原がEGP−2である場合、適切
な抗体としてはRS11もしくは17−1Aまたはそのヒト化バージョンが挙げられる。
この抗体は急速に内部移行する抗体であることが有利である。
【0068】
可変ドメイン遺伝子の挿入に適切な抗体定常ドメインを含むか、または含むように操作
され得るベクターは、当分野で公知であり、そしてこのようなベクターはまた、本発明の
免疫毒素の発現のために適切である。このようなベクターは、哺乳動物宿主細胞において
、コードされた遺伝子の転写を開始するのに必須である必須の制御配列を含む。このよう
なコントロール配列は、当分野で周知である。例えば、その全体において引用することに
より本明細書に組み込まれる、米国特許第5,650,150号および同第6,472,511号のpdHL
2ベクターの説明を参照のこと。あるいは、定常ドメイン遺伝子(CH1、CH2、CH
3およびCL)は、抗体産生細胞から得られてもよいし、当分野で周知の方法を用いて適
切な発現ベクターにクローニングされてもよい。
【0069】
(免疫毒素遺伝子の調製)
本発明による免疫毒素コード遺伝子の調製は、例示の目的でのみ提供されて、限定され
るものではない、下の実施例1および4に詳細に例示される。要するに、PCRを用いて
例えば、抗体軽鎖可変ドメイン(または必要に応じて重鎖可変ドメイン)をコードする遺
伝子を得てもよく、当分野で公知のPCR法を用いて得てもよい。適切な制限部位が、P
CRプライマーに導入されてもよく、それによってこの遺伝子とさらなる配列とのその後
の結合が容易になる。同様に、細胞毒性RNAseコード遺伝子は、都合のよい制限部位
を提供するプライマーをここでも用いて、適切なテンプレートからPCRによって得られ
てもよい。次いで、細胞毒性RNAse遺伝子および可変ドメイン遺伝子は、適切な制限
エンドヌクレアーゼで消化されて、可変鎖RNAse融合タンパク質をコードする遺伝子
を提供するために直接一緒に結合されてもよい。細胞毒性RNAse遺伝子は、有利には
、可変ドメイン遺伝子のN末端に結合され、さらに有利には軽鎖可変鎖遺伝子のN末端に
結合されるが、RNAse遺伝子は、あるいは、重鎖可変ドメインのN末端に、または重
鎖および軽鎖可変ドメインの両方のN末端に結合されてもよい。
【0070】
あるいは、適切なリンカーは、合成オリゴヌクレオチド(例えば、細胞毒性RNAse
および免疫グロブリン遺伝子に含まれる制限部位と適合しており、そして細胞毒性RNA
seと可変ドメイン遺伝子とリンカー配列との直接の結合をもたらす制限部位を含むよう
に調製され得る)を用いて、可変ドメイン遺伝子と細胞毒性RNAse遺伝子との間に導
入され得る。当業者は、種々のリンカーが本発明における使用に適切であることを認識す
る。しかし、リンカーは有利には、10〜30アミノ酸長であり、そして二次構造も三次
構造もほとんどまたは全く有さない、親水性アミノ酸を含むリンカーである。このような
リンカーは当分野で周知であり、そして例えば、単鎖Fv分子を構築するのに用いられる
。例えば、米国特許第4,946,778号を参照のこと。適切なリンカーとしては、配列GGG
GSなどの多量体が挙げられる。免疫毒素遺伝子を調製する他の方法は、その全体が引用
することにより本明細書に組み込まれる、米国特許出願第20030099629号に記載される。
【0071】
インタクトな抗体構造を含む免疫毒素(すなわち、軽鎖VLおよびCLドメイン、なら
びに重鎖VHおよびCH1−3ドメインを含む免疫毒素)が調製されるべき場合、上記の
可変ドメイン鎖は、これらのさらなる定常ドメインをコードする配列に結合されなければ
ならない。有利には、適切な定常ドメインを既に含むベクター、例えば、上記のpdHL
2を用いる。適切な定常ドメインを調製して、それらを可変ドメインに結合させる別の方
法が当分野で周知である。Fabフラグメントについては、CLおよびCH1ドメインの
みが必要である。一旦、適切な形態に結合されれば、次にこれらの配列は、宿主細胞のト
ランスフェクションの前に適切な発現ベクターに挿入され得る。哺乳動物発現に適切であ
る発現ベクターは、当分野で周知であり、そしてまた例えば、Invitrogen,Carlsbad,CAか
らも市販されている。
【0072】
(哺乳動物細胞における免疫毒素の発現)
一旦適切な発現ベクターが調製されれば、適切な宿主細胞にトランスフェクトされる。
組み換え抗体の発現に適切な宿主細胞は、当分野で周知であり、そしてこれには例えば、
CHO細胞、および有利には、骨髄細胞株、例えば、NSO細胞が挙げられる。CHOお
よびNSO細胞の両方がATCC(Manassas,VA)から入手可能である。ヒト抗体配列を含
む免疫毒素の産生のために特に適切な別の宿主細胞は、Karpas et al.,Proc.Natl.Acad.S
ci USA 98:1799(2001)に記載されるKarpas 707H系統である。トランスフェクションは、
周知の方法、例えば、エレクトロポレーション、リポフェクション、およびDEAEデキ
ストラン媒介性トランスフェクションによって達成され得る。有利には、発現ベクターは
、選択マーカー、例えば、DHFRを含み、そして宿主細胞はDHFR-である。この細
胞は、選択因子(例えば、選択マーカーがDHFRである場合はメトトレキセート)の存
在下で培養される。陽性のクローンは、例えば、免疫毒素の細胞毒性RNAse部分に特
異的に結合する抗体を用いてELISA法によって確認され得る。陽性クローンは、後の
増殖のために凍結されてもよいし、または直接増殖されてもよい。陽性クローンの培養物
を、適切な培地、例えば完全HSFM培地中で増殖させる場合、免疫毒素を含む培養上清
を収集する。
【0073】
(免疫毒素の精製)
免疫毒素は、当分野で周知である方法を用いて容易に精製され得る。例えば、アフィニ
ティー精製が用いられてもよい。完全な抗体構造を含む免疫毒素については、プロテイン
Aアフィニティー精製を用いて、純粋な免疫毒素を単独の精製工程で得てもよい。さらな
る精製工程は、必要に応じて、イオン交換クロマトグラフィー、サイズ排除クロマトグラ
フィーなどのような従来の方法を用いて行なわれ得る。免疫毒素は、従来の方法、例えば
、RP−HPLC、SDS−PAGE、質量分析、および例えば抗細胞毒性RNAse抗
体を用いるウエスタンブロットによって特徴付けられ得る。ヒョウガエル由来のランピル
ナーゼは、潜在的なグリコシル化部位を含む(実施例1を参照のこと)。この部位のグリ
コシル化が所望されない事象では、免疫毒素は、オンコナーゼ(onconase)上に存在する炭
水化物部分を除去するためにグリコシダーゼ酵素で処理されてもよい。あるいは、RNA
seをコードする遺伝子配列は、保存的アミノ酸置換を介してAsn残基を置換すること
によってグリコシル化部位を欠失するために、部位特異的変異誘発法によって変異されて
もよい。例えば、69位置のAsnは、グルタミンで(変異体N69Q)、または本明細
書に記載される任意の構築物中の他の適切なアミノ酸で置換されてもよい。このグリコシ
ル化変異体の適切な例としては、2L−Rap(N69Q)−hLL1−r4P、2L−
Rap(N69Q)−hRS7および2L−Rap(N69Q)−hLL2が挙げられる
。
【0074】
(細胞毒性RNAse免疫毒性を用いる治療方法)
本明細書に記載の免疫毒素は、広範な種々の細胞毒性の治療様式のための薬学的組成物
に処方され得る。これらの組成物は、任意の欲されない細胞タイプ、例えば、不適切な増
殖または活性化を受けている細胞を標的するために用いられてもよいし、または病原性微
生物を標的することによって感染性疾患を処置するために用いられてもよい。詳細には、
この組成物は、種々のガンおよび自己免疫疾患を処置するために有用である。処置され得
るガンとしては、例えば、リンパ腫、白血病、黒色腫、神経芽細胞腫、および骨髄腫、肉
腫、神経膠腫、または癌腫、例えば、乳癌、結腸直腸癌、卵巣癌、肝臓癌、胃癌、結腸直
腸癌、前立腺癌、肺癌、腎臓癌または膵臓癌が挙げられる。処置され得る自己免疫疾患と
しては、例えば、関節リウマチ、全身性エリテマトーデス、シェーグレン症候群、急性特
発性血小板減少性紫斑病、慢性特発性血小板減少性紫斑病、皮膚筋炎、シデナム舞踏病、
重症筋無力症、ループス腎炎、リウマチ熱、多腺性症候群、水疱性類天疱瘡、糖尿病、ヘ
ノッホ・シェーンライン紫斑病、連鎖球菌感染後腎炎、結節性紅斑、高安動脈炎、アジソ
ン病、多発性硬化症、サルコイドーシス、潰瘍性大腸炎、多形性紅斑、IgA腎症、結節
性多発性動脈炎、強直性脊椎炎、グッドパスチャー症候群、血栓血管炎(thromboangitis
ubiterans)、原発性胆汁性肝硬変、橋本甲状腺炎、甲状腺中毒症、強皮症、慢性活動性肝
炎、多発性筋炎/皮膚筋炎、多発性軟骨炎、尋常性天疱瘡、ヴェーゲナー肉芽腫症、膜性
腎症、筋萎縮性側索硬化症、脊髄癆、巨細胞性動脈炎/多発性筋痛、悪性貧血、急速進行
性糸球体腎炎および線維化性肺胞炎が挙げられる。
【0075】
本発明の組成物はまた、感染の治療的処置のためにも有用であり、ここでは免疫毒素の
免疫グロブリン成分は、疾患を生じる微生物に特異的に結合する。本発明の状況では、疾
患を生じる微生物としては、病原性細菌、ウイルス、真菌および多様な寄生生物が挙げら
れ、そしてこの抗体は、これらの微生物、それらの産物またはそれらの病変に関連する抗
原を標的し得る。微生物の例としては、限定はしないが以下が挙げられる:
ストレプトコッカス・アガラクティエ、在郷軍人病菌、化膿連鎖球菌、大腸菌、淋菌、
髄膜炎菌、肺炎球菌、Hemophilis influenzae B、梅毒トレポネーマ、ライム病スピロヘ
ータ、緑膿菌、ハンセン菌、ウシ流産菌、ヒト結核菌、破傷風毒素、HIV−1、HIV
−2、HIV−3、A型肝炎、B型肝炎、C型肝炎、D型肝炎、狂犬病ウイルス、インフ
ルエンザウイルス、サイトメガロウイルス、単純疱疹IおよびII、ヒト血清パルボ様ウ
イルス、パピローマウイルス、ポリオーマウイルス、呼吸器合胞体ウイルス、水痘帯状疱
疹ウイルス、B型肝炎ウイルス、パピローマウイルス、麻疹ウイルス、アデノウイルス、
ヒトT細胞白血病ウイルス、エプスタイン・バーウイルス、マウス白血病ウイルス、流行
性耳下腺炎ウイルス、水疱性口内炎ウイルス、シンドビスウイルス、リンパ球性脈絡髄膜
炎ウイルス、疣ウイルス、ブルー・タングウイルス、センダイウイルス、猫白血病ウイル
ス、レオウイルス、ポリオウイルス、シミアン・ウイルス40、マウス乳癌ウイルス、デ
ング熱ウイルス、風疹ウイルス、原生動物、熱帯熱マラリア原虫、三日熱マラリア原虫、
トキソプラズマ原虫、Trypanosoma rangeli、クルーズ・トリパノソーマ、Trypanosoma r
hodesiensei、トリパノソーマ・ブルーセイ、マンソン住血吸虫,Schistosoma japanicum
、Babesia bovis、Elmeria tenella、回旋糸状虫、熱帯リーシュマニア、旋毛虫、Theile
ria parva、Taenia hydatigena、Taenia ovis、無鉤条虫、単包条虫、Mesocestoides cor
ti、抗マイコプラズマモノクローナル抗体、Mycoplasma arthritidis、M.hyorhinis、M.o
rale、M.arginini、Acholeplasma laidlawii、M.salivarium、およびM肺炎。これらの病
原性微生物に結合するモノクローナル抗体は当分野で周知である。
【0076】
当業者は、これらの実施例が例示的であって、本発明を限定するものではないことを理
解する。
【0077】
これらの方法では、薬学的組成物は、被験体に1日あたり2回以上、例えば、0.1〜
約1000mgの量で投与されてもよいが、それより多くてもまたは少なくても適切とみ
なして用いられ得る。
【0078】
この組成物のタンパク質性の性質に起因して、それらの組成物は特に、非経口投与、例
えば静脈内投与または皮下投与(または皮内投与)に適切であるが、当業者は、他の送達
方法が用いられ得ることを理解する。この方法は、現在開発中であるか、または将来開発
されるタンパク質送達のための方法を包含し、そしてまた例えば、腹腔、頭蓋、胸膜など
のような局所投与を包含し得る。非経口投与のためには、組成物は、薬学的に受容可能な
キャリア、好ましくは、緩衝化生理食塩水のような水性キャリアに溶解されるかまたは懸
濁された免疫毒素の溶液を含んでもよい。これらの溶液は、無菌であって、補助物質、例
えば、pH調節剤および緩衝化剤、ならびに毒性および等張性の調節剤を含んでもよい。
【0079】
免疫毒素の用量は、1日あたり1患者あたり約0.1〜10mgであるが、特にこの薬
物が局所的に投与され、血流中ではない場合には、1日あたり1患者あたり最大100m
gの用量が用いられてもよい。この用量は、より少量のRNAse成分を含む、抗体融合
タンパク質に基づく。
【0080】
治療適用では、この組成物は、疾患に罹患している被験体に対して、目的の細胞を殺傷
するのに十分な量として規定される、治療上細胞毒性の量で投与される。これを達成する
のに十分な量は、「薬学的に有効な量(therapeutically effective amount)」として規定
される。任意の所定の環境における正確な量は、疾患の重篤度および患者の健康の一般的
状態を含む種々の要因に依存する。この組成物の単回投与または複数回投与は、必要な用
量に依存して投与され得る。適切な投薬レジメンの決定は、当分野で周知の慣用的方法を
用いて行なわれる。
【0081】
免疫毒素はまた、インビトロにおいて細胞の集団を処置するために用いられ得る。例え
ば、それらは、骨髄アブレーションを受けている患者への移植の前に骨髄中の欲されない
細胞タイプを選択的に殺傷するために用いられ得る。
【0082】
このように一般的に記載された本発明は、例示のために記載されており、本発明の限定
は意図していない以下の実施例を参照してさらに容易に理解される。
【実施例1】
【0083】
(2L−rap−hLL1−γ4Pの発現および特徴付け)
実施例1では、rapはランピルナーゼである。
【0084】
(pdHL−IgG4P改変体の構築)
IgG4遺伝子を含むB13−24細胞は、ATCC(ATCC番号 CRL−113
97)から購入して、ゲノムDNAを単離した。要するに、細胞をPBSで洗浄して、消
化緩衝液(100mM NaCl,10mMのTris−HCl pH8.0、25mM
のEDTA pH8.0、0.5%のSDS、0.1mg/mlのプロテイナーゼK)に
再懸濁して、50℃で18時間インキュベートさせた。等容積のフェノール/クロロホル
ム/イソアミルアルコールを用いてこのサンプルを抽出して、7.5MのNH4Ac/1
00% EtOHで沈殿させた。ゲノムDNAは、遠心分離によって回収して、TE緩衝
液に溶解した。テンプレートとしてゲノムDNAを用い、IgG4遺伝子を、以下のプラ
イマーを用いてPCRによって増幅させた。
プライマー−SacII:5'CCGCGGTCAC ATGGCACCAC CTCTCTTGCA GCTTCCACCA AGGGCCC 3'
(47マー);
プライマー−EagI:5'CCGGCCGTCG CACTCAT TTA CCCAGAGACA GGG 3'(33マー)
【0085】
増幅されたPCR産物を、TOPO−TA配列決定ベクター(Invitrogen)中にクローニ
ングして、DNA配列決定によって確認した。pdHL−hLL2中のIgG1の重鎖定
常領域を含むSacII−EagIフラグメントを、TOPO−TA−IgG4プラスミ
ドのSacII−EagIで置換して、pdHL2−hLL2−IgG4(pdHL2−
hLL2−γ4)ベクターを生成した。
【0086】
(IgG4−プロリン変異)
Ser228Proの変異をIgG4のヒンジ領域に導入して、半分子の形成を回避し
た。変異したヒンジ領域の56bpフラグメント(PstI−StuI)を合成し(トッ
プ:5'GAG TCC AAA TAT GGT CCC CCA TGC CCA CCG TGC CCA GGT AAG CCA ACC CAG G3';ホ
゛トム:5'C CTG GGT TGG CTT ACC TGG GCA CGG TGG GCA TGG GGG ACC ATA TTT GGA CTC T
GC A 3')、アニーリングして、IgG4のPstI−StuIフラグメントで置換した。
この構築で、最終ベクターpdHL2−hLL2−γ4Pを得た。
【0087】
(pdHL2−hLL1−γ4Pの構築)
pdHL2−hLL2−γ4PのXbaI−HindIIIフラグメントを、Vkおよ
びVH領域を含むpdHL2−hLL1のXba−HindIIIフラグメントで置換し
て、hLL1−γ4P構築物を生成した。
【0088】
(pdHL2−2L−rap−hLL1−γ4Pの構築)
可塑性リンカー(GGGGS)3を用いて、hLL1のVkのN末端に対してRapの
C末端を結合させた。各々の軽鎖のN末端に1つのrap分子を結合させた。この分子に
ついてのDNAの構築は、以下のプライマーを用いてPCRによって行なった:
P1;リーダー5’(XbaI):5'CTC TAG ACA CAG GAC CTC ACC ATG GGA TGG 3'(
30マー)
P2;リーダー3’:5'TGA AAC GTT AGC CAA TCC TGG GAG TGG ACA CCT GTG GA 3'(
38マー)
P3;Onc 5’:5'TCC ACA GGT GTC CAC TCC CAG GAT TGG CTA ACG TTT CA 3'(
38マー)
P4;Onc 3’(EcoRV):5'AGT CAG CTG GAT ATC GGA GCC ACC GCC TCC AG
A TC 3'(35マー)
P5;LL1−Vk 5’(EcoRV):5'GAT CTG GAG GCG GTG GCT CCG ATA TCC
AGC TGA CT 3'(35マー)
P6;LL1−Vk 3’(BamHI):5'GGG ATC CAA CTG AGG AAG CAA AGT TTA
A 3'(28マー)
【0089】
図1に図示されるように、pdHL2−hLL1−γ4PのXba−BamHIフラグ
メントを、pBS−2L−rap−hLL1のXba−BamHI(Xba−Leade
r−rap−Linker−Vk−BamHI)フラグメントで置換して、最終のベクタ
ーpdHL2−2L−rap−hLL1−γ4Pを完成させた。
【0090】
(トランスフェクション)
ベクターDNA(30μg)をSalI酵素で直線化して、エレクトロポレーション(
450V)によってNS0(4×106細胞/mL)またはSp2/0−Ag14(5×
106個の細胞/mL)の骨髄腫細胞にトランスフェクトした。低IgG FBS(10
%)、ペニシリン(100単位/mL)、ストレプトマイシン(100μg/mL)、L
−グルタミン(2mM)、ピルビン酸ナトリウム(1mM)、非必須アミノ酸(100μ
M)およびメトトレキセート(0.1μM)を補充した完全なハイブリドーマ−SFM培
地中で細胞を増殖させた。陽性のクローンをELISAによってスクリーニングした。要
するに、プレートを、PBS培地中に含まれる5μg/mLの50μlの抗rap抗体を
用いてコーティングして、4℃で一晩インキュベートさせた。PBSでプレートを洗浄し
、2%BSAでブロックした後に細胞培養上清を添加した。HRP複合体化ヤギ抗ヒトI
gG4抗体を検出のために用いて、OPDを発色のための基質として用いた。プレートを
490nmで読み取った。陽性のクローンを増殖させて、さらなる使用のために凍結させ
た。クローンC6を、最適のプロデューサーとして特定して、さらなる開発のために用い
た。
【0091】
(発現および精製)
各々に500mlの培地を有する2つのローラーボトル中で細胞を最終培養物まで増殖
させて(10〜20%の生存度)、その細胞を遠心分離によって取り出した。培養上清を
充填して、プロテインAカラムに加え、20mM Tris−HCl/100mM Na
Cl緩衝液(pH8.5)で平衡化した。ローディング後、このカラムを100mMのク
エン酸ナトリウム緩衝液(pH7.0)で洗浄して、100mMのクエン酸ナトリウム緩
衝液(pH3.5)で溶出させ、融合タンパク質を得た。生成物を含有するピークを、3
M Tris−HCl、pH8.0を用いてpH7.0に調節して、10mMのPBS緩
衝液に対して透析した。濃縮後、この生成物を0.22μmのフィルターを通して濾過し
、2〜8℃で保管した。1Lの培養物から、精製後に16mgを回収した。
【0092】
(2L−rap−hLL1−γ4Pの特徴付け)
HPLC:タンパク質の純度および濃度は、HPLCでチエックした。先鋭な単独のピ
ークが、図2に示されるように7.7分で観察され、この保持時間によって、この分子が
IgGより大きいことが示された。
【0093】
SDS−PAGE:4〜20% Tris−Glycineゲル(PAGEr(商標)
Gold Precast Gels,Cambrex)を用いて、SDS−PAGEを還元条件下で行なった。図
3に示されるように、予想されるサイズ約50kDの重鎖に関連するバンド、ならびにh
LL1の軽鎖(約25kD)よりも両方とも大きい、約37kDおよび39kDの分子量
の2つのバンドが観察された。2つの軽鎖の存在は、融合タンパク質上のrapのグリコ
シル化に起因することが示された(以下参照)。
【0094】
質量分析法:The Scripps Research Institute,CAにおいて、MALDI−TOF法に
よって質量分析を行った。2つのサンプル、その1つは天然の状態(1.6mg/mLを
10mMのPBSに含有)、そしてもう一方は、還元状態(1.6mg/mLを1mMの
HEPES/10mMのDTT、pH7.5緩衝液)を、分析のために送った。天然のサ
ンプル(図4a)は、質量177150の1つの主要なピークを示し、これは、IgGに
加えて2つのrapのMWとよく一致している。この還元サンプルは、50560(重鎖
に相当する)、38526および36700(rapを含有する2つの軽鎖に相当する)
において3つの主なピークを示した(図4b)。
【0095】
ウエスタンブロッティング:精製されたタンパク質におけるrapの存在を確認するた
めに、ウエスタンブロッティングを行った。還元条件下のSDS−PAGEゲル由来のサ
ンプルを、PVDF-Plus(Osmonics,Inc.,)メンブレン上に電気的に転写した。5% BSA
でのブロッキング後、マウスの抗rap抗体を、1:10,000希釈または100ng
/mlで添加して、1時間インキュベートさせた。洗浄後、HRP複合体化ヤギ抗マウス
Fc抗体を添加して1時間インキュベートさせた。6回の洗浄後、LumiGloTM(Kirkegaard
&Perry Laboratories)基質を添加して、コダックのフィルムを現像した。図5に示される
ように、融合された軽鎖に相当する両方のバンドをフィルム上で検出し、これによって両
方の軽鎖上のrapの存在を確認した。
【0096】
N−グリコシダーゼでの処理:rapは、潜在的なNグリコシル化部位、Asn−X−
Thr/Ser、Asn69−Val70−Thr71を有するので、分子量で2kDの
相違があるという2つの軽鎖の観察は、rapの一様でないグリコシル化の結果であり得
る。この可能性を検討するために、rap−hLL1抗体を、供給業者の推奨に従って変
性条件下でNグリコシダーゼ(New England Biolabs)とともにインキュベートさせた。図
6に示されるとおり、N−グリコシダーゼ処理後、2つの軽鎖に相当する2つのバンドが
1つ(より早く移動するバンド)に集束し、これによってカルボヒドラーゼの一様でない
分解が、SDS−PAGEでの2つのバンドという観察の原因であったことが確認された
。除去されたグリコシル化部位を有するRapの改変体であるRap(N69Q)が組み
換え構築物中でRapについて置換される場合、唯一のrap融合軽鎖の観察によってさ
らなる支持が得られた(データ示さず)。
【0097】
rapの活性:RNAse活性は、供給業者の推奨に従ってBright-GloTM Luciferase
Reporter Assayシステム(Promega)を用いてTNT(商標)Quick Coupled Transcription
/Translation System(Promega)によって試験した。このアッセイの原理は、ルシフェラー
ゼレポーター系を用いるRNase活性の結果としてのタンパク質合成の阻害(mRNA
分解)の測定であって。サンプルは、種々の希釈中で遊離のrap(0.001〜2.5
nM)、hLL1−rap(0.01−20nM)またはPKI−LL2−Oncおよび
PKII−LL2−Onc(0.01〜20nM)として表されるhLL2−rapの化
学的複合体を調製した。各々のサンプル(5μL)を、20μlのTNTマスター混合物
と混合して、96ウェルプレート中で30℃で2時間インキュベートして、それから1μ
lを、50μlのBright-GloTM基質での分析のために取り出した。その結果を、Excelま
たはPrism Padソフトウェアを用いて、図7に示した。EC50値は、rap−hLL1お
よびhLL2−Oncの化学的複合体については約300pM、そして遊離のrapにつ
いては30pMであった。
【0098】
WPについての競合結合:WPはhLL1の抗イディオタイプ抗体である。WPに対す
るhLL1抗体と比較したrap−hLL1抗体の親和性は、競合結合アッセイによって
評価した。要するに、96ウェルプレートを50μlのWPを5μg/mLで用いてコー
ティングして、4℃で一晩インキュベートさせた。タンパク質サンプルの3つのタイプで
あるhLL1、rap−hLL、またはhA20は、種々の2倍希釈(最終濃度は0.4
9〜1000nMに及ぶ)中で調製して、等容積の2×HRP複合体化mLL1抗体(最
終希釈は1/20,000)と混合した。上記のようなHRP複合体化mLL1と混合し
た50μLのタンパク質サンプルを、各々のウェルに添加して、1時間インキュベートさ
せた。洗浄後、H2O2を含有するOPD基質を添加してプレートを490nmで読んだ。
図8に示されるように、吸光度に対するタンパク質濃度を、ExcelまたはPrism Pad graph
ソフトウェアを用いてプロットした。hA20(ヒト化抗−CD20抗体)を、陰性のコ
ントロールとして用いた。図8から、rap−hLL1は、hLL1と同様の結合親和性
を有し、そして陰性コントロールhA20は、親和性を全く有さないことが明らかである
。抗原の供給源としてRaji細胞を用いて同様の結果を得た。
【0099】
インビトロ細胞毒性:インビトロ細胞毒性は、B細胞リンパ腫細胞株(Daudi)および複
数の黒色腫細胞株(MC/CAR)で測定した。細胞(0.1ml中に10,000個)
を、96ウェルプレートの各々のウェルに入れた。24時間後、遊離のhLL1、遊離の
rapまたはrap−hLL1(10μl)を適切なウェルに添加して、その細胞をイン
キュベーター中で37℃で3日間インキュベートした。細胞増殖は、MTSテトラゾリウ
ム色素還元アッセイまたはBrDU比色定量アッセイを用いて決定した。結果は、Prism
Padソフトウェアを用いてグラフとして得られたEC50で表す。図(図9〜10)から、
rap−hLL1が、B−細胞リンパ腫細胞株(Daudi)および多発性骨髄腫細胞株(MC
/CAR)の両方に対して感受性であることが証明される。rap−hLL1は、EC50
値によって反映されるように、MC/CAR細胞に比較してDaudi細胞に対して有意によ
り強力(細胞毒性)であった(図9および図10)。MC/CAR細胞については、EC
50値は、試験した濃度では達成されなかった。最高濃度(56nM)では、細胞の生存度
は57%であった。将来の実験は、Daudi細胞上のrap−hLL1について正確なEC5
0値を測定することが目的である。hLL1または遊離のrapは、それ自体では、いず
れの細胞株でも細胞毒性を実証しなかった。
【0100】
薬物動態および体内分布:hLL1または2L−Rap−hLL1−γ4Pを、Sharke
y et al.,(Int J Cancer.1990;46:79-85)に記載のように、2−(4−イソチオシアナー
トベンジル)DTPA(Macrocyclics,Dallas,TX)を用いてジエチレントリアミノペンタ酢
酸(DTPA)と複合体化させて、DTPA−hLL1またはDTPA−2L−Rap−
hLL1−γ4Pを得た。これはそれぞれ、薬物動態学的研究および生体分布の研究のた
めに、88Y塩化物(Los Alamos National Laboratory(Los Alamos,NM)または111In塩化
物(Perkin Elmer Life Sciences,Boston,MA)で標識された。ナイーブな雌性SCIDマウ
ス(8週齢、18〜22g)に対して各々の動物に、hLL1および2L−Rap−hL
L1−γ4Pの各々10μgの総用量を投与するように、未標識の、hLL1または2L
−Rap−hLL1−γ4PのDTPA複合体を補充した0.001mCi88Y−DTP
A−hLL1および0.02mCiの111In−DTPA−2L−Rap−hLL1−γ
4Pの混合物を静脈内注射した。投与後選択された時点(1、2、4、16、48、72
、168時間)後に、5匹のマウスの群を麻酔して、血液サンプルを心臓穿刺によって採
取した。主な組織を取り出して、秤量して、容器中に入れた。血液サンプルおよび組織を
、目盛り付きのガンマカウンターであるMinaxiλ Auto-Gamma(商標)5000シリーズ
のガンマカウンター(Packard Instrument Company;Downers Grove,IL)で、111In(チャ
ネル120〜480)および88Y(チャネル600〜2000)についてカウントした。
111Inカウンティングウインドウへの88Yエネルギーの後方散乱について補正するため
に交差曲線を作成した。
【0101】
インビトロの毒性:ナイーブなSCIDまたはBALB/cマウスに、マウス1匹あた
り25〜400μgに及ぶ種々の用量の2L−Rap−hLL1−γ4Pを静脈内注射し
て、毒性および体重の損失の可視的な徴候について毎日モニターした。最大耐容用量(M
TD)は、死亡が生じず、かつ体重減少が処置前の動物の重量(約20g)の20%以下
である最高用量として規定した。毒性効果を経験した動物は屠殺して、回収して、組織病
理学的分析に供した。ナイーブなSCIDマウスでは、2L−Rap−hLL1−γ4P
の100、150、200、250、300または400μgの単回静脈内用量によって
、動物の重篤な体重減少および死亡が生じたが、全てのマウスは、25または50μgの
用量で生残した(表2)。BALB/cマウスでは、全てのマウスが2L−Rap−hL
L1−γ4Pの30または50μgの単回静脈内用量で生残したが、100または200
μgでは生残しなかった。別の実験では、2L−Rap−hLL1−γ4Pの75μgの
用量は、SCIDマウスに対して毒性であることが見いだされた(データ示さず)。従っ
て、単回ボーラス注射として与えられた2L−Rap−hLL1−γ4PのMTDは、S
CIDマウスでは50〜75μgであって、BALB/cマウスでは50〜100μgで
ある。死亡マウスおよび屠殺されたマウスの全体の病理学的な検査では、重篤な肝臓およ
び脾臓の毒性が示された。肝臓は、色が青ざめており、脾臓は縮んで、通常のサイズより
小さかった。組織病理学的検査によって、肝臓および脾臓の壊死が明らかになった。代表
的なマウスの血清サンプルでは、アラニンアミノトランスフェラーゼ(ALT)、アアス
パルテートアミノトランスフェラーゼ(AST)および総ビリルビンのレベル上昇があり
、このことはこれらの高用量での有意な肝臓毒性を示唆していた。
【0102】
データ分析:インビトロの細胞毒性研究のために、三連の測定値の平均から用量応答曲
線を作成して、GraphPad Prismソフトウェア(Advanced Graphics Software,Encinitas,CA
)を用いて50%阻害濃度(IC50)値を得た。薬物動態学的データは、ノンコンパート
メント分析プログラムWinNonlin,バージョン4.1(Pharsight,Mountain View,CA)の標
準的アルゴリズムを用いて分析した。このプログラムは、線形補間による線形台形公式を
用いて曲線下免疫(AUC)を算出する。排出速度定数(kβ)は、一次速度式を仮定し
て終末半減期(t1/2β)から算出した。生残研究は、GraphPad Prismソフトウェアを用
いるKaplan-Meierプロット(ログランク分析)を用いて分析した。相違はP<0.05で
有意とみなした。
【0103】
薬物動態および生体分布のデータ:放射性標識したhLL1および2L−Rap−hL
L1−γ4Pの薬物動態および生体分布を、ナイーブなSCIDマウスで測定した。hL
L1および2L−Rap−hLL1−γ4Pを、DTPAと結合体化して、それぞれ標識
した88Yおよび111Inで追跡した。図11に示されるように、111IIn−標識した2L
−Rap−hLL1−γ4Pは、初期の急速な再分布相(α)および後期の緩徐な排出相
(β)によって特徴付けられる、88Y−標識されたhLL1と同様の血液からの二相性の
クリアランスを示す。わずかに短いα半減期が、hLL1(4時間)に比較して、2L−
Rap−hLL1−γ4P(5.1時間)について観察された。5時間を超えるデータポ
イントを用いて、t1/2β、kβ、AUC、平均滞留時間(mean residence time)(MRT
)、見かけの分布容積(Vd)、およびクリアランスの速度(Cl)を算出して、これら
のパラメーターの値を表1に示す。111In−標識された2L−Rap−hLL1−γ4
Pの組織取り込みは、88Y−標識hLL1の取り込みと同様であった(データ示さず)。
【表1】

【0104】
インビトロの毒性:ナイーブなSCIDマウスでは、100、150、200、250
、300または400μgの単回静脈内用量の2L−Rap−hLL1−γ4Pは、動物
の重篤な体重減少および死亡を生じたが、全てのマウスは、25または50μgの用量で
生残した(表2)。BALB/cマウスでは、全てのマウスが2L−Rap−hLL1−
γ4Pの30または50μgの単回静脈内用量では生残したが、100または200μg
では生残しなかった(表2)。別の実験では、2L−Rap−hLL1−γ4Pの75μ
gの用量は、SCIDマウスに対して毒性であることが見いだされた(データ示さず)。
従って、単回ボーラス注射として与えられた2L−Rap−hLL1−γ4PのMTDは
、SCIDマウスでは50〜75μgであって、BALB/cマウスでは50〜100μ
gである。死亡マウスおよび屠殺されたマウスの全体的な病理学的な検査では、重篤な肝
臓および脾臓の毒性が示された。肝臓は、色が青ざめており、脾臓は縮んで、通常のサイ
ズより小さかった。組織病理学的検査によって、肝臓および脾臓の壊死が明らかになった
。代表的なマウスの血清サンプルでは、アラニンアミノトランスフェラーゼ(ALT)、
アアスパルテートアミノトランスフェラーゼ(AST)および総ビリルビンのレベル上昇
があり、このことはこれらの高用量での有意な肝臓毒性を示唆していた。
【表2】
【0105】
(腫瘍保有マウスでの治療有効性)
腫瘍保有マウスでの治療有効性:1群あたり8〜9匹の雌性SCIDマウス(8週齢、
18〜22g)に、1.5×107個のDaudi細胞を静脈内注射して、1日後に処置した。
後肢の麻痺についてマウスを毎日検査して、毎週秤量した。動物が後肢の麻痺を発症する
かまたはその処置前の体重の20%が低下した場合、それらの動物を安楽死させた。各々
のセットの治療実験は180日後に終えた。
【0106】
図12に示されるように、未処置のマウス(PBSのみ)は、30日内に全て死亡し、
平均生残時間(median survival time)(MST)は28日であった。50μgの2L−R
ap−hLL1−γ4P中の成分タンパク質の組成に相当する、hLL1−γ4P(43
.2μg)およびRap(6.6μg)の混合物を投与されたコントロール群のMSTは
、70日であった(P<0.0001対PBS群)。対照的に、5μgまたは15μgの
いずれかの2L−Rap−hLL1−γ4Pの単回注射を受けた全てのマウスでは、10
0日より長く生きており(MST>180日;P=0.0005対成分処置群)、そして
この研究の終わり近くで各々の群から欠けたのは1匹のマウスだけであった。180日後
に研究が終わったとき、5、15、30、40または50μgの2L−Rap−hLL1
−γ4Pの単回注射を投与されたマウスの90%が治癒した。1μgの単回注射を受けた
マウスのMSTは、230%の増大に相当する、未処置の群の28日に比較して92日で
あった(P<0.0001)ことに注目すべきである。
【実施例2】
【0107】
(細胞毒性RNAseをコードするPCR増幅したDNAの合成)
組み換え細胞毒性RNAseのN末端配列(46アミノ酸)をコードするセンス鎖配列
[5'-TGG CTA ACG TTT CAG AAG AAA CAT ATC ACG AAT ACA CGA GAT GTA GAC TGG GAC AAT
ATA ATG TCT ACG AAT CTG TTT CAC TGT AAG GAT AAG AAT ACC TTT ATA TAC AGT CGC CCA
GAG CCT GTA AAG GCT ATC TGT A-3']を有する139マーのDNAヌクレオチド、ON
CO−Nを、自動DNAシンセサイザー(Applied Biosystem 392 DNA/RNA Synthesizer)
によって合成して、隣接するプライマーONNBACK[5'-AAG CTT CAT ATG CAG GAT T
GG CTA ACG TTT CAG AAG AAA-3'、およびONNFOR[5'-CTT ACT CGC GAT AAT GCC TT
T ACA GAT AGC CTT TAC AGG CTC TG-3']を用いるPCR増幅のためのテンプレートとし
て用いた。得られた二本鎖PCR産物は、細胞毒性RNAseのN末端の半分の54アミ
ノ酸残基をコードするcDNA配列を含む。ONNBACKは、ステージングベクターに
、または細菌発現ベクターへのインフレーム結合のために(NdeI部位)サブクローニ
ングを容易にするために、制限部位HindIII(AAAGCTT)およびNdeI(
CATATG)を含む。NruI部位(TCGCGA)を、ONNFORプライマーに組
み込んで、細胞毒性RNAseのC末端半分をコードするcDNAとのインフレーム結合
を容易にする。
【0108】
同様に、細胞毒性DNAseのC末端配列(46アミノ酸)をコードするセンス鎖配列
[TGC TGA CTA CTT CCG AGT TCT ATC TGT CCG ATT GCA ATG TGA CTT CAC GGC CCT GCA AA
T ATA AGC TGA AGA AAA GCA CTA ACA AAT TTT GCG TAA CTT GCG AGA ACC AGG CTC CTG TA
C ATT TCG TTG GAG TCG GG-3']を有する137マーのDNAヌクレオチド、ONCO−
Cを合成して、プライマーONCBACK[5'-ATT ATC GCG AGT AAG AAC GTG CTG ACT A
CT TCC GAG TTC TAT-、およびONCFOR[5'-TTA GGA TCC TTA GCA GCT CCC GAC TCC AAC GAA
ATG TAC-3']によってPCR増幅させる。最終の二本鎖PCR産物は、細胞毒性RNA
seのC末端の半分の残りの51アミノ酸をコードするcDNA配列を含んだ。NruI
部位によって、ONCBACKに組み込まれたPCR増幅されたDNAのN末端の半分と
のインフレームの結合が可能になった。ステージングベクターへ、または細菌発現ベクタ
ーへのサブクローニングのために、終止コドン(太字で示される)およびBamHI制限
部位(下線)をONCFOR配列に入れた。
【0109】
適切な制限酵素での処理後に細胞毒性RNAseのN末端およびC末端の半分をコード
する、PCR増幅されたDNAをNruI部位に結合して、ステージングベクター、例え
ばStratageneのpBluescript中にサブクローニングした。この結合された配列は、N末端
のMetを有する105アミノ酸のポリペプチドをコードしなければならない。
【実施例3】
【0110】
(LL2およびMN−14のV−領域配列のクローニング、ならびにLL2およびMN−
14のヒト化)
hLL2およびhMN−14のV領域配列は公開されている。Leung et al.,Mol.Immun
ol.,32:1413(1995);米国特許第5,874,540号。LL2およびMN−14のVKおよびVH
の配列は、公開された方法およびプライマーを用いてPCR増幅させた。
【0111】
PCR増幅されたDNAの配列分析によって、それらが抗体のVKおよびVHドメイン
の代表的なタンパク質をコードすることが示された。PCR−増幅されたLL2およびM
N−14の配列に基づいて構築されたキメラ抗体は、その親抗体に匹敵する免疫反応性を
示しており、これによって得られた配列の信頼性が確認された。
【0112】
LL2抗体の配列分析によって、フレームワーク−1領域におけるVK付加されたN結
合グリコシル化部位の存在が明らかになった。突然変異研究によって、VK付加部位での
グリコシル化は、抗体の免疫反応性を維持するには必要ないことが示された。FR−1グ
リコシル化部位の包含なしに、REIフレームワーク配列を、軽鎖CDRを接合するため
、そしてEU/NEWMをLL2の重鎖CDRを接合するための足場として用いた。ヒト
化LL2(hLL2)の免疫反応性は、マウスおよびキメラのLL2の免疫反応性に匹敵
することが示された。LL2の内部移行の速度は、抗体のキメラ化またはヒト化によって
は影響されなかった。
【実施例4】
【0113】
(ヒト化LL2および細胞毒性RNAseの融合タンパク質をコードする遺伝子の構築)
hLL2のVHおよびVKの配列をテンプレートとして用いて、標準的なPCR手順に
よってhLL2−scFv遺伝子をアセンブルした。遺伝子の構成は、Met(−1)−
VL−(GGGS)4−VH−(His)6であった。VHドメインに対して16アミノ酸
のリンカー(GGGS)6を介して結合されたVL遺伝子のN末端に、Met(ATG)
開始コドンを−1位置で組み込んだ。6つのヒスチジル残基からなるテールを、VH鎖の
カルボキシル末端に入れて、金属キレートクロマトグラフィーを介した融合タンパク質の
精製を容易にする。
【0114】
ランピルナーゼ−hLL2scFvの免疫毒素融合タンパク質遺伝子を、制限消化およ
び結合方法によって同様の方式で構築した。cDNA配列は発現された場合、以下の構築
物:
ランピルナーゼ−[リンカー]−VL−(GGGS)4−VH−(His)6の融合タン
パク質をコードした。
【0115】
細胞毒性RNAseのC末端とVLドメインのN末端との間に挿入され得る種々のリン
カーが存在する。好ましいリンカーは、シュードモナスの外毒素(PE)のC末端位置2
73〜281由来のアミノ酸配列TRHRQPRGWである。この配列は、配列のG残基
とW残基との間に存在する切断でのサブチリシンによる、活性フラグメントへのPEの細
胞内切断の認識部位であることが示されている。Chiron et al.,J.Biol Chem.,269:18167
(1994)。この配列の組み込みによって、融合免疫毒素の内部移行後の活性細胞毒性RNA
seの放出が容易になる。あるいは、細胞毒性RNAseとscFvとの間の可塑性の結
合を可能にするために、EDN−scFvの構築に用いられるStaphylococcalのプロテイ
ンAのフラグメントBのアミノ酸残基48〜60からなる13アミノ酸残基のスペーサー
を代わりに用いてもよい。Tai et al.,Biochemistry,29:8024(1990)、およびRybak et al
.,Tumor Targeting,1:141(1995)。
【実施例5】
【0116】
(ヒト化MN−14およびランピルナーゼの融合タンパク質をコードする遺伝子の構築)
MN−14scFvは、ヒト化MN−14トランスフェクトーマ(transfectoma)由来の
cDNAのPCR増幅によって生成した。MN−14 scFvのために用いられるリン
カーは、15アミノ酸リンカー(GGSGS)3であり、そして方向はVL−リンカー−V
Hであった。DNA配列の確認後、単鎖構築物を、実施例1に記載のような真核生物発現
ベクター中にサブクローニングして、発現のために適切な哺乳動物宿主細胞中にトランス
フェクトする。
【0117】
別の単鎖構築物も作成した。これは、重鎖および軽鎖の反対の5’−3’方向で作成し
、pCANTABE5E(Pharmacia Biotech,Piscataway,N.J.)中でアセンブルして、フ
ァージ中で発現させた。このscFvを発現する組み換えファージの特異的結合はELI
SAによって実証された。
【0118】
VL−リンカー−V配列を、以下に図示されるように、ランピルナーゼ−MN−14融
合タンパク質の構築のために用いた。ランピルナーゼをコードするDNAフラグメントを
実施例1に記載のように得た。23アミノ酸リンカーをランピルナーゼ配列とscFvと
の間で用いた。Kurucz et al.(1995)。あるいは、上記のMN−14 scFvの構築に
用いられる(GGSGS)3リンカーを用いた。融合タンパク質の好ましい構成は以下:
ランピルナーゼ−リンカー−−VL−−(GGSGS)3−−VH
であった。
【0119】
本明細書に引用される全ての特許および他の引用文献の内容は、その全体が引用するこ
とにより本明細書に組み込まれる。
【技術分野】
【0001】
本願は、その内容が全体として引用することにより本明細書に組み込まれる、2004年2
月13日出願の米国仮特許出願第60/544,227号に対する優先権を主張する。
【0002】
本発明は、非哺乳動物の細胞毒性RNAseおよび免疫グロブリンおよび免疫グロブリ
ンフラグメントの融合タンパク質を含む免疫毒素を提供する。
【背景技術】
【0003】
ランピルナーゼ(ranpirnase)は、疾患の処置のための細胞毒性因子としての見込みを示
す非哺乳動物リボヌクレアーゼ(RNAse)のあるクラスのメンバーである。これらの
RNAseの細胞毒性は、細胞表面レセプターに対する当初の結合および細胞のサイトゾ
ルへの内部移行(リボソームRNAの付随する分解および細胞タンパク質合成の阻害を伴
う)に関与する機構に起因している。Wu et al.,J.Biol.Chem.,268:10686-10693(1993)を
参照のこと。当該RNAse活性は、哺乳動物のRNAseインヒビターに対して耐性で
あり、これによって哺乳動物の酵素に比較して観察される細胞毒性の増強が説明され得る
。同上。ランピルナーゼは、このファミリーのRNAseの原型的なメンバーであり、ヒ
ョウガエルの卵母細胞および初期の胚から精製され得る。ランピルナーゼは、12,00
0の分子量を有し、そしてウサギの網状赤血球溶解物において(IC50 10-11M)、
そしてアフリカツメガエル(Xenopus)の卵母細胞に微量注入された場合(IC50 10-10
M)、タンパク質合成の強力な阻害を生じる。RNase Aスーパーファミリーの他の
メンバーとは異なり、これらの細胞毒性リボヌクレアーゼは、卵母細胞rRNAを分解し
ない。Lin et al.,Biochem Biophys Res Commun.204:156-62(1994)。これらの分子は、酵
素のリン酸結合ポケットの一部を形成し、RNAseおよび抗腫瘍活性に必須である必須
のN末端ピログルタミル残基を含む。
【0004】
動物の毒性研究によって、ランピルナーゼが予測可能な用量依存性でかつ可逆性の毒性
を、ラット(用量範囲0.01〜0.02mg/kg)およびイヌ(0.005〜0.1
5mg/kg)の両方で示すということが示される。侵襲性のM109 Madison肺癌腫を接種
されて、かつ毎日および毎週の両方のスケジュールでランピルナーゼの腹腔内投与の処置
をされたマウスは、有意に延長された生存を示した。ほとんどの著しい結果は、ランピル
ナーゼの毎週のスケジュールで処置されたマウスの群でみられ、ここでは、18匹の動物
のうち6匹が、長期間生存して、明らかに癌が治癒した。Mikulski et al.,J Natl Cance
r Inst.82:151-3(1990)。
【0005】
天然のランピルナーゼは、臨床試験において、種々の固体腫瘍に対して抗腫瘍活性を有
することが示されている。これに関しては、ランピルナーゼは、例えば、膵臓癌を有する
患者を処置する場合、単独で、そしてタモキシフェンのような他の抗腫瘍因子と組み合わ
せての両方で用いられている。抗腫瘍因子として用いられる場合、これらの細胞毒性RN
Aseは、特定の細胞タイプに対する標的を可能にするためのマーカーに複合体化され得
る。
【0006】
第I相試験では、種々の再発性および耐性の腫瘍に罹患している患者を、ランピルナー
ゼ(用量範囲60〜960μg/m2)を用いて毎週静脈内処置した。観察される副作用
としては、一般に顔面紅潮、筋肉痛、一過性の眩暈および食欲減退が含まれた。タンパク
尿の増大、末梢浮腫、高窒素血症、クレアチニンクリアランスの低下、および疲労によっ
て呈示される、用量を制限する腎臓毒性を含む、観察される毒性は、用量依存性でかつ可
逆性であり、これは、動物での毒物学的研究と一致している。真の免疫学的感作の臨床的
な顕在化は、ランピルナーゼの反復された毎週の静脈内用量後でさえ証明されていない。
最大耐容量は、主に腎毒性に起因して、960μg/m2であることが見出された。非小
細胞肺癌、食道癌および結腸直腸癌にはなんらかの他覚的な応答があった。Mikulski et
al.,Int J Oncol 3:57-64,(1993);Mikulski et al.,J Clin Oncol.20:274-81(2002)を参
照のこと。それにもかかわらず、ランピルナーゼは、試験された動物およびほとんどのヒ
ト患者では十分耐容されて、一貫してかつ可逆性の臨床毒性パターンを示し、そしてほと
んどの従来の化学療法剤に伴うほとんどの毒性、例えば、骨髄抑制および脱毛症は誘発し
なかった。
【0007】
WO97/31116は、(a)N末端メチオニンに続いてグルタミン酸以外のアミノ酸、(b)
システインを26位、40位、58位、84位、95位、および110位に、リジンを4
1位に、そしてヒスチジンを119位に(ウシRNAseAとの最大一致について整列さ
せた場合)を有する組み換えリボヌクレアーゼ、ならびに天然のランピルナーゼ由来のア
ミノ酸配列を開示している。しかし、RNAse改変体(variant)を含む組み換え細胞毒
性RNAseは、多段階のプロセスによって細菌で調製されており、このプロセスには、
細菌によって挿入されたN末端ホルミルメチオニン残基を除去するため、そして必須のN
末端ピログルタミル残基を生成するためのさらなる工程を要する。それにもかかわらず、
細菌中での産生は、グリコシル化された細胞毒性RNAse含有融合タンパク質の調製を
妨げる。従って、真核生物細胞において組み換え細胞毒性RNAse融合タンパク質を産
生することが有利である。真核生物細胞では、細胞毒性RNAseは、両生類の供給源か
ら精製されたランピルナーゼの細胞毒性特性を保持するが、ヒトにおける所望されない免
疫応答は少ないか、またはない。しかし、真核生物細胞における細胞毒性RNAseの発
現は、RNAseの細胞毒性活性に起因して細胞死を生じるということも予想される。
【0008】
従って、グリコシル化RNAse融合タンパク質を含む、細胞性RNAse含有融合タ
ンパク質を調製する改良方法が極めて望ましいということが明らかである。グリコシル化
細胞毒性RNAse融合タンパク質自体が極めて所望される分子であるということも明ら
かである。
【発明の開示】
【0009】
従って、本発明の目的は、(a)第一の免疫グロブリン可変ドメインに融合された非哺
乳動物の細胞毒性リボヌクレアーゼを含む融合ポリペプチドと、(b)第二の免疫グロブ
リン可変ドメインを含む第二のポリペプチドとを含み、この免疫グロブリン可変ドメイン
の1つが軽鎖可変ドメインであり、かつもう一方の免疫グロブリン可変ドメインが重鎖可
変ドメインであり、この第一および第二の免疫グロブリン可変ドメインが一緒になって抗
原結合部位を形成し、そしてこの免疫毒素がグリコシル化されている、免疫毒素を提供す
ることである。
【0010】
本発明の別の目的は、哺乳動物宿主細胞を培養することによってこれらおよび他の免疫
毒素を作製する方法を提供することであって、この宿主細胞は、(a)第一の免疫グロブ
リン可変ドメインに融合された非哺乳動物細胞毒性リボヌクレアーゼを含む融合ポリペプ
チドをコードする核酸配列と、(b)第二の免疫グロブリン可変ドメインを含む第二のポ
リペプチドをコードする核酸配列とで形質転換され、この第一および第二の免疫グロブリ
ンの可変ドメインは一緒になって抗原結合部位を形成する。
【0011】
本発明のさらに別の目的は、細胞毒性RNAse部分に融合された内部移行抗体(inter
nalizing antibody)または抗体フラグメントを含む免疫毒素であって、このRNAse部
分がN末端ピログルタミン酸残基を保有し、かつこのRNAse部分がそのC末端におい
て、この抗体または抗体フラグメントの軽鎖を含むポリペプチドのN末端、あるいはこの
抗体またはフラグメントの重鎖を含むポリペプチドのN末端、あるいはこの抗体またはフ
ラグメントの軽鎖および重鎖の両方のN末端に融合されている免疫毒素を提供することで
ある。
【0012】
例えば、一実施形態では、非哺乳動物細胞毒性リボヌクレアーゼは、第一の免疫グロブ
リン可変ドメイン、例えば、軽鎖可変ドメインのN末端に融合されてもよい。この非哺乳
動物リボヌクレアーゼは、N末端ピログルタミン酸残基を有してもよい。
【0013】
上記融合ポリペプチドおよび第二の免疫グロブリン可変ドメインは、別の分子であって
もよく、そして宿主細胞中で別の分子として産生されてもよい。
【0014】
上記融合ポリペプチドはさらに、CLドメインを含んでもよく、そして上記第二のポリ
ペプチドはさらにCH1ドメインを含んでもよい。
【0015】
上記第二のポリペプチドはさらに、CH2ドメインおよびCH3ドメインを含んでもよ
い。
【0016】
別の実施形態では、非哺乳動物リボヌクレアーゼは、重鎖可変ドメインのN末端に融合
されてもよい。この非哺乳動物リボヌクレアーゼは、N末端ピログルタミン酸残基を有し
てもよい。この融合ポリペプチドおよび第二の免疫グロブリン可変ドメインは、別の分子
であってもよく、そして宿主細胞中で別の分子として産生されてもよい。
【0017】
この融合ポリペプチドはさらに、CH1ドメインを含んでもよく、そして第二のポリペ
プチドはさらにCLドメインを含んでもよい。
【0018】
この融合ポリペプチドはさらに、CH2ドメインおよびCH3ドメインを含んでもよい
。
【0019】
さらに別の実施形態では、非哺乳動物リボヌクレアーゼは、第一および第二の免疫グロ
ブリン可変ドメインの両方のN末端に融合されてもよい。この非哺乳動物リボヌクレアー
ゼは、N末端ピログルタミン酸残基を有してもよい。この第一の免疫グロブリン可変ドメ
インは、重鎖可変ドメインであってもよく、そして第二の免疫グロブリン可変ドメインは
、軽鎖可変ドメインであってもよい。
【0020】
この第一および第二の融合ポリペプチドは、別の分子であってもよく、そして宿主細胞
中で別の分子として産生されてもよい。
【0021】
この第一の融合ポリペプチドはさらに、CH1ドメインを含んでもよく、そして第二の
融合ポリペプチドはさらにCLドメインを含んでもよい。
【0022】
この第一の融合ポリペプチドはさらに、CH2ドメインおよびCH3ドメインを含んで
もよい。
【0023】
免疫毒素は、例えば、CH2ドメイン上で、および/またはRNAse上でグリコシル
化されてもよい。
【0024】
上記の非非哺乳動物リボヌクレアーゼは、例えば、ヒョウガエルから精製されたランピ
ルナーゼの配列および構造を有する酵素であってもよい。
【0025】
上記抗原結合部位は、細胞表面分子に対して特異的に結合してもよく、そして上記免疫
毒素は、この細胞表面分子を保有する細胞結合した際に内部移行されてもよい。
【0026】
上記免疫毒素は、ガン細胞、感染/炎症の部位、自己免疫疾患に対するか、または微生
物および寄生生物に対するものであり得る。
【0027】
上記免疫毒素は、B細胞抗原、T細胞抗原、形質細胞抗原、HLA−DR系統抗原、C
EA,NCA、MUC1、MUC2、MUC3、MUC4およびMUC16抗原、EGP
−1抗原、GEP−2抗原、胎盤アルカリホスファターゼ抗原、IL−6、VEGF、P
1GF、ILGF、EGFR、Her2/neu、テネイシン、CD33、CD74、P
SMA、PSA、PAP、Leyを含む腫瘍関連抗原、自己免疫疾患、感染/炎症および
感染性疾患に関連する抗原からなる群より選択される抗原に対するものであってもよい。
【0028】
上記抗原は、B細胞もしくはT細胞リンパ腫に関連する標的抗原、または自己免疫疾患
に関連するB細胞もしくはT細胞であってもよい。
【0029】
上記抗原は、悪性疾患によって発現されるCD19、CD22、CD40、CD74、
CEA、NCA、MUC1、MUC2、MUC3、MUC4、HLA−DR、EGP−1
、EGP−2、EGFR、Her2/neu、IL−15およびHLA−DRからなる群
より選択される抗原であってもよい。
【0030】
上記免疫毒素の免疫グロブリン可変ドメインは、悪性疾患を標的する場合、例えば、R
S11、17−1A、RS7、LL1、LL2、MN−3、MN−14、MN−15もし
くはPAM4,またはそのヒト化バージョンに由来し得る。
【0031】
免疫毒素によって結合される抗原は、特定の悪性疾患に関しては、例えば、EGP−2
,EGP−1、CD22、CD74、CEA、炭酸脱水素酵素IX、またはMUC1であ
ってもよい。
【0032】
標的化抗原は、細菌、ウイルス、真菌、マイコプラズマ、寄生生物、または他の微生物
によって発現されてもよい。
【0033】
標的化合物抗原は、活性化顆粒球のような、感染の部位での宿主細胞蓄積によって発現
されてもよい(例えば、CD15、CD33、CD66a、CD66b、およびCD66
c(NCA)など)。適切なこのような顆粒球抗体は、MN−3であり、そのFab’は
LeukoScan(商標)で用いられる。
【0034】
上記の方法および/または組成物のいずれかにおいて、可変ドメインはヒト化されてよ
いしまたはヒトドメインであってもよい。
【0035】
本発明の別の実施形態では、薬学的に受容可能なキャリアとともに上記されたような免
疫毒素を含む薬学的組成物が提供される。
【0036】
本発明のさらに別の実施形態では、被験体に対して上記のような組成物を、疾患または
症候群の症状を処置または改善するために有効な量で投与することによって、この被験体
におけるこの疾患または症候群を処置する方法が提供される。この疾患または症候群は、
この被験体における細胞、例えばT細胞、骨髄細胞または形質細胞の望ましくないまたは
不適切な増殖または活性化によって特徴付けられ得る。あるいは、この疾患は、感染性微
生物または寄生生物の存在によって特徴付けられ得る。
【0037】
この疾患は、例えば、癌腫、肉腫、神経膠腫、白血病、リンパ腫、黒色腫、神経芽細胞
腫、および骨髄腫からなるガンの群より選択され得る。
【0038】
この疾患または症候群は、固形腫瘍、例えば、神経芽細胞腫、悪性黒色腫、または癌腫
、例えば、乳房、藍藻、前立腺、肺、腎臓、胃、結腸直腸、肝臓、または膵臓の癌腫の存
在によって特徴付けられ得る。
【0039】
これらの方法では、薬学的組成物は、被験体に対して、1日あたり2回以上、例えば、
0.1〜約1000mgの量で投与されてもよいが、それより多く用いられても少なく用
いられてもよい。
【0040】
望ましくないまたは不適切な増殖または活性化を受けている細胞は、例えば、関節リウ
マチ、シェーグレン症候群、急性特発性血小板減少性紫斑病、慢性特発性血小板減少性紫
斑病、皮膚筋炎、シデナム舞踏病、重症筋無力症、全身性エリテマトーデス、ループス腎
炎、リウマチ熱、多腺性症候群、水疱性類天疱瘡、糖尿病、ヘノッホ・シェーンライン紫
斑病、連鎖球菌感染後腎炎、結節性紅斑、高安動脈炎、アジソン病、多発性硬化症、サル
コイドーシス、潰瘍性大腸炎、多形性紅斑、IgA腎症、結節性多発性動脈炎、強直性脊
椎炎、グッドパスチャー症候群、血栓血管炎(thromboangitis ubiterans)、原発性胆汁性
肝硬変、橋本甲状腺炎、甲状腺中毒症、強皮症、慢性活動性肝炎、多発性筋炎/皮膚筋炎
、多発性軟骨炎、尋常性天疱瘡、ヴェーゲナー肉芽腫症、膜性腎症、筋萎縮性側索硬化症
、脊髄癆、巨細胞性動脈炎/多発性筋痛、悪性貧血、急速進行性糸球体腎炎および線維化
性肺胞炎のような1つ以上の自己免疫疾患の発症および進行に関与し得る。
【0041】
本発明の組成物が、感染の治療的処置のために用いられる場合、免疫毒素の免疫グロブ
リン構成要素は、疾患原因の微生物、例えば、病原性細菌、ウイルス、真菌および多様な
寄生生物に特異的に結合し、そして抗体は、これらの微生物、それらの産物もしくは表面
抗原、またはそれらの病変に関連する抗原を標的し得る。微生物の例としては、限定はし
ないが、
ストレプトコッカス・アガラクティエ、在郷軍人病菌、化膿連鎖球菌、大腸菌、淋菌、
髄膜炎菌、肺炎球菌、Hemophilis influenzae B、梅毒トレポネーマ、ライム病スピロヘ
ータ、緑膿菌、ハンセン菌、ウシ流産菌、ヒト結核菌、破傷風毒素、抗ウイルスモノクロ
ーナル抗体、HIV−1、HIV−2、HIV−3、A型肝炎、B型肝炎、C型肝炎、D
型肝炎、狂犬病ウイルス、インフルエンザウイルス、サイトメガロウイルス、単純疱疹I
およびII、ヒト血清パルボ様ウイルス、呼吸器合胞体ウイルス、水痘帯状疱疹ウイルス
、B型肝炎ウイルス、麻疹ウイルス、アデノウイルス、ヒトT細胞白血病ウイルス、エプ
スタイン・バーウイルス、マウス白血病ウイルス、流行性耳下腺炎ウイルス、水疱性口内
炎ウイルス、シンドビスウイルス、リンパ球性脈絡髄膜炎ウイルス、疣ウイルス、ブルー
・タングウイルス、センダイウイルス、猫白血病ウイルス、レオウイルス、ポリオウイル
ス、シミアン・ウイルス40、マウス乳癌ウイルス、デング熱ウイルス、ポリオーマウイ
ルス、パピローマウイルス、SARSウイルス、種々のインフルエンザウイルス、風疹ウ
イルス、原生動物、熱帯熱マラリア原虫(Plasmodium falciparum)、三日熱マラリア原虫(
Plasmodium vivax)、トキソプラズマ原虫、Trypanosoma rangeli、クルーズ・トリパノソ
ーマ、Trypanosoma rhodesiensei、トリパノソーマ・ブルーセイ、マンソン住血吸虫,Sc
histosoma japanicum、Babesia bovis、Elmeria tenella、回旋糸状虫、熱帯リーシュマ
ニア、旋毛虫、Theileria parva、Taenia hydatigena、Taenia ovis、無鉤条虫、単包条
虫、Mesocestoides corti、マイコプラズマ、Mycoplasma arthritidis、M.hyorhinis、M.
orale、M.arginini、Acholeplasma laidlawii、M.salivarium、およびM肺炎が挙げられ
る。
【0042】
本発明の他の目的、特徴および利点は、以下の詳細な説明から明らかになる。しかし、
この詳細な説明および特定の実施例は、本発明の好ましい実施形態を示すが、例示のため
だけに示していることが理解されるべきである。なぜなら、本発明の趣旨および範囲内の
種々の変化および改変が、この詳細な説明から当業者には明白になるからである。
【図面の簡単な説明】
【0043】
【図1】ベクターpdHL2−rap−hLL1−γ4Pの構成を示す。構築物である、リーダーペプチド−RNAse、−リンカー−hLL1のVk領域を含むXbaI−BamHIフラグメントを、pBSベクターに挿入して、pdHL2ベクターに連結した。注記:本発明においては、rpRNAseおよびrapという用語は、交換可能である。
【図2】宿主細胞から分泌された融合タンパク質のHPLCプロフィールを示す。単一のピークが7.7分で観察された。
【図3】分泌された融合タンパク質のSDS−PAGEを示す。2つの接近して移動する軽鎖バンドをみることができるが、これはhLL1単独の軽鎖よりも長い(25kD)。
【図4a】天然の条件下で分泌された融合タンパク質の質量スペクトルを示す:177150の質量を有するrap−hLL1−γ4P融合タンパク質に相当する、1つの主要なピークが観察された。
【図4b】変性条件下の質量スペクトルを示す。重鎖、およびrap−hLL1−γ4P融合タンパク質の糖分子の有無の2つの軽鎖に相当する3つの主要なピークが見出され得る。
【図5】融合タンパク質のウエスタンブロットを示す。両方の分子ともrapを含むことを示している、糖有の軽鎖および糖無の軽鎖が示され得る。
【図6】N−グリコシダーゼ処理による軽鎖糖分子の除去を示すSDS−PAGEゲルを示す:レーン1は、未処理のサンプルに相当し、ここでは2つの軽鎖バンドを見ることができる。レーン2〜5は、Nグリコシダーゼ酵素で処理されたサンプルである。2つの軽鎖に相当するバンドは、糖の除去の際に1つのバンドに集束した。
【図7】インビトロ転写翻訳アッセイによるRNAse活性を示す。
【図8】競合結合アッセイを示しており、これは、hLL1およびrap−hLL1融合タンパク質の両方が、hLL1の抗イディオタイプ抗体であるWPについて同じ親和性を有することを示している。
【図9】Daudi細胞における融合タンパク質のインビトロ細胞毒性を示す:A)MTSアッセイによって測定した細胞毒性;B)BRdUアッセイ法によって測定した細胞毒性。
【図10】MTSアッセイによるMC/CAR細胞における融合タンパク質のインビトロ細胞毒性を示す。
【図11】ナイーブなSCIDマウスにおける2L−Rap−hLL1−γ4Pの血液クリアランスを示す。ナイーブなSCIDマウスに、88Y−DTPA−hLL1(O)および111In−DTPA−2L−Rap−hLL1−γ4P(□)を静脈内に同時注射した。投与後選択された時間で、心臓穿刺によってマウスを放血させて、血液サンプルを放射能についてカウントした。データは、血液中の注射用量の平均±S.D.である(n=3)。
【図12】2L−Rap−hLL1−γ4Pまたは成分のタンパク質を用いた侵襲最小ダウディリンパ腫(aggressive minimal Daudi lymphoma)の処置を示す。SCIDマウス(1群あたり8〜10匹のマウス)に、1.5×107個のDaudi細胞を静脈内に接種した。1日後、マウスを、2L−Rap−hLL1−γ4Pの1μg(×)、5μg(黒四角)、15μg(黒三角)、30μg(下向き黒三角)、40μg(◆)、または50μg(●)の単回ボーラス注射で処置した。コントロール群には、50μgの免疫毒素(*)に等価な構成要素のタンパク質またはPBS(□)のみを注射した。
【図13】インビトロの転写/翻訳アッセイによって測定されたRNase活性を示す。rRap(黒四角)、2L−Rap−hLL1−γ4P(黒上向き三角)およびhLL1−γ4P(◆)の濃度を、相対発光単位(relative luminescence units)(RLU)に対してプロットした。
【発明を実施するための形態】
【0044】
本発明者らは、驚くべき事に、細胞毒性RNAse分子を含む融合分子が哺乳動物細胞
において調製され得ることを見出した。この結果は、驚くべきである。なぜなら、酵素的
に活性な細胞毒性RNAse分子を含む分子は、細胞増殖を妨害して、これによって融合
タンパク質の組み換え産生を妨げる程度まで細胞毒性であることが予想されるからである
。
【0045】
この結果によって、原核生物宿主細胞を用いて組み換え方法によって調製することが以
前には困難であるかまたは不能であった哺乳動物細胞において、細胞毒性RNAse融合
タンパク質を調製する可能性がもたらされる。例えば、哺乳動物宿主細胞における産生に
よって、原核生物宿主細胞を用いる場合、必須のN末端ピログルタミル残基を生成するこ
とに関連する困難性が回避される。さらに、哺乳動物宿主細胞の使用によって、マルチサ
ブユニットタンパク質、例えば、インタクトなFc領域を含むIgG構造の容易な調製が
可能になり、そしてまたグリコシル化融合タンパク質、例えば、Fc領域上でグリコシル
化されるインタクトな抗体の調製が可能になる。細胞毒性RNAseと免疫グロブリン、
例えば抗体または抗体フラグメントとの間の融合タンパク質は、本明細書において以降で
は免疫毒素と呼ばれる。この方法はまた、任意の適切な抗体フラグメント、例えば、F(
ab’)2、F(ab)2、Fab’、Fab、FvおよびscFvを含む免疫毒素を調製
するために用いられ得る。
【0046】
本発明はまた、RNAseが、Ig可変ドメイン(例えばVHまたはVLドメイン)の
1つのN末端に融合される、細胞毒性RNAse含有免疫グロブリン融合タンパク質を提
供する。これは驚くべきことに、細胞毒性RNAse活性を保持するだけでなく、抗原結
合部位でIg結合特異性を保持しており、N末端融合の調製によって立体的にブロックさ
れることが期待されている融合タンパク質を提供する。
【0047】
さらに、本発明者らは、驚くべきことに、本発明の細胞毒性RNAse融合タンパク質
が細胞および微生物/寄生生物の殺傷に有効であることを見出している。詳細には、迅速
な内部移行抗体成分を含む免疫毒素は細胞特異的な様式で極めて細胞毒性であることが見
出されている。例えば、ヒト化LL1抗体のN末端に対する細胞毒性RNAseの融合に
よって、CD74(LL−1の標的)の不変鎖を発現する細胞に対して特異的に結合して
、結合の際その細胞集団に対して細胞毒性である免疫毒素が産生された。この結果は、驚
くべきことである。なぜなら、融合タンパク質は、融合タンパク質の細胞毒性を大きく低
下させるか、またはさらに排除する可能性が高いことが予期される、内部移行の際に細胞
のリソソームに送達されて、そこで分解されることが予想されるからである。
【0048】
本発明はまた、疾患または症候群に罹患している被験体を、上記で記載されるタイプの
免疫毒素の有効量をこの被験体に投与することによって処置する方法を提供する。
【0049】
(定義)
他に特定しない限り、用いられる全ての技術および科学的用語は、当業者によって通常
理解されるのと同じ意味を有する。さらに、本明細書に引用される全ての特許および他の
引用文献の内容は、その全体が引用することにより組み込まれる。本発明の目的のために
は、以下の用語は以下のとおり規定される:
【0050】
アミノ酸は、名称によるか、またはその一般に公知の3文字記号か、もしくは1文字の
IUPAC記号のいずれかによって言及される。ヌクレオチドは、その通常受容される一
文字コードによって呼ばれる。
【0051】
特定の核酸配列の「保存的に改変されたバリエーション(conservatively modified var
iations)」とは、同一であるかまたは本質的に同一であるアミノ酸配列をコードする核酸
をいうか、またはこの核酸が、アミノ酸配列をコードしない場合は、本質的に同一の配列
をいう。遺伝子コードの縮重の理由で、多数の機能的に同一の核酸が、任意の所定のポリ
ペプチドをコードする。例えば、コドンGCA、GCC、GCGおよびGCUは全てがア
ラニンというアミノ酸をコードする。従って、アラニンがコドンによって特定されるあら
ゆる位置では、このコドンは、コードされたポリペプチドを変化させることなく、記載さ
れる任意の対応するコドンに変化され得る。メチオニンをコードするAUG以外の核酸中
の各々のコドンが、機能的に同一の分子を生じるように改変され得る。本明細書に記載さ
れる核酸配列はまた、これらの変更を包含する。
【0052】
アミノ酸配列の「保存的に改変されたバリエーション」とは、コードされた配列中の単
独のアミノ酸または小さい割合のアミノ酸を変更させる個々の置換であって、この変更に
よって、化学的に類似のアミノ酸でのアミノ酸の置換が生じる置換を包含する。保存的置
換は当業者に周知である。以下の6つの群の各々は、お互いについて保存的置換であるア
ミノ酸を含む:
1.アラニン、セリン、トレオニン
2.アスパラギン酸、グルタミン酸
3.アスパラギン、グルタミン
4.アルギニン、リジン
5.イソロイシン、ロイシン、メチオニン、バリン、そして
6.フェニルアラニン、チロシン、トリプトファン。
【0053】
アミノ酸配列の「保存的に改変されたバリエーション」はまた、コードされた配列中の
単独のアミノ酸または小さい割合のアミノ酸の欠失または付加であって、この付加および
欠失が、化学的に類似のアミノ酸でのアミノ酸の置換を生じる欠失または付加を包含する
。本明細書に記載されるアミノ酸配列はまた、これらのバリエーションを包含する。
【0054】
「単離された(isolated)」または「生物学的に純粋な(biologically pure)」という用
語は、その天然に存在する環境において見出されるような通常伴う成分を実質的にまたは
本質的に含まない物質をいう。この単離された物質は状況に応じて、その天然の環境にお
いてこの物質とともに見出されない物質を含む。
【0055】
「核酸(nucleic acid)」という用語は、デオキシリボヌクレアーゼまたはリボヌクレオ
チドポリマーであって、一本鎖型または二本鎖型のいずれかをいい、他に限定しない限り
、天然に存在するヌクレオチドに対して同様の様式で核酸にハイブリダイズする天然のヌ
クレオチドの公知のアナログを包含する。他に示さない限り、特定の核酸配列とは、その
相補的な配列を含む。
【0056】
「発現ベクター(expression vector)」とは、組み換え発現カセットを含み、この組み
換え発現カセットは、細胞によって転写および翻訳され得る、本発明によるポリペプチド
をコードする核酸を包含する。組み換え発現カセットとは、組み換え的にまたは合成的に
生成された核酸構築物であり、標的細胞において特定の核酸の転写を可能にする一連の特
定の核酸エレメントを伴う。発現ベクターは、プラスミド、ウイルスまたは核酸フラグメ
ントの一部であってもよい。代表的には、この発現ベクターのこの組み換え発現カセット
部分は、転写されるべき核酸およびそれに作動可能に連結されるプロモーターを包含する
。
【0057】
「組み換え体(recombinant)」という用語は、タンパク質に関して用いられる場合、あ
る細胞がその細胞には外因性である起源の核酸によってコードされるペプチドまたはタン
パク質を発現することを示す。組み換え細胞は、その細胞の天然の(非組み換え体)型内
では見出されない遺伝子を発現し得る。組み換え細胞はまた、細胞の天然の形態で見出さ
れる遺伝子を発現し得、この遺伝子は、人工的な方法によって、例えば、異種プロモータ
ーの制御下で細胞に再導入される。
【0058】
「実質的な同一性(substantial identity)」または「実質的な類似性(substantial sim
ilarity)」という用語は、ポリペプチドの文脈では、あるポリペプチドが参照配列と少な
くとも80%、さらに好ましくは90%、そして最も好ましくは少なくとも95%の同一
性を有する配列を含むことを示す。実質的に同一である2つのポリペプチドとは、このポ
リペプチドの1つが第二のペプチドに対して惹起された抗体と免疫学的に反応性であるこ
とを意味する。2つの核酸が実質的に同一であるとは、この2つの分子がストリンジェン
トな条件下でお互いにハイブリダイズすることである。一般に、ストリンジェントな条件
とは、所定のイオン強度およびpHにおいて特定の配列についての融点(Tm)よりも約
5℃〜20℃低いように選択される。このTmは、標的配列の50%が完全にマッチした
プローブにハイブリダイズする温度(所定のイオン強度およびpH下で)である。しかし
、ストリンジェントな条件下でお互いに対してハイブリダイズしない核酸は、それらがコ
ードするポリペプチドが実質的に同一であるならば、やはり実質的に同一である。
【0059】
「抗体(antibody)」とは、当分野で周知の種々の免疫グロブリン種(IgG1、IgG4
、IgMなど)の抗体全体および抗体フラグメント、例えば、F(ab’)2、F(ab
)2、Fab’Fab、Fvなどを含み、このフラグメントとしては、標的抗原に結合す
る能力を保持するハイブリッドフラグメントが挙げられる。また、免疫グロブリンの超可
変性の抗原結合領域を保持する任意の小フラグメントも有用である。抗体全体から、抗原
に特異的に結合する能力を保持している抗体フラグメントを作成する方法は、当分野で周
知であり、そして例えば、Harlow and Lane「Antibodies:A Laboratory Manual」CSHL Pr
ess(1988)に記載されている。組み換え抗体または抗体フラグメントを作成する方法は、
当分野で周知であり、例えば、McCafferty et al.,「Antibody Engineering:A Practical
Approach」IRL Press(1996)に記載されている。Fabフラグメントは、代表的には鎖間
ジスルフィド結合を含む(VL−CL)−(VH−CH1)二量体を含む。Fab’フラ
グメントは、類似であるが、また重鎖ヒンジドメインの一部を含む。F(ab’)2フラ
グメントとは、従来、2つのFab’フラグメントのジスルフィド結合二量体を指すこと
が理解される。本発明の抗体は、任意の種の抗体であってもよいが、この抗体はヒト抗体
またはヒト化抗体であることが有利である。抗体はまた、任意の周知の形態または組み換
え抗体、例えば、単鎖Fv(VH鎖に対する任意のリンカーによって結合されたVL鎖を
含む)であってもよい。このリンカーが短いかまたは存在しない場合、この単鎖Fvは二
量体または多量体に二量化し、この二量体とは二重特異性抗体として公知である。特定の
環境下では、本発明の抗体はまた、単結合ドメイン抗体、例えば、ラクダ科抗体(Dumoul
in et al.,「Single-domain antibody fragments with high conformational stability.
」Protein Sci. 11,500-515(2002);Hamers-Casterman et al.,「Naturally occurring an
tibodies devoid of light chains」Nature 363,446-448(1993);ならびにMuyldermans.「
Single domain camel antibodies:current status」J Biotechnol.74,277-302(2001)を参
照のこと)、または「ドメイン抗体(domain antibody)」を含み得る。WO92/01787および
そこに引用される参考文献を参照のこと。ヒト抗体とは、ヒト被験体において産生される
抗体由来であるか、またはヒトにおいて産生される抗体の特徴である配列を有するかのい
ずれかの配列を有する、抗体または抗体フラグメントである。このような抗体は、例えば
、米国特許第6,300,064号に記載される。ヒト抗体はまた、ヒト抗体ライブラリーから単
離され得る。例えば、米国特許第6,300,064号および同第6,172,197号、ならびにそこに記
載される参考文献を参照のこと。ヒト化抗体は、非ヒト抗体からヒトフレームワークへC
RD領域を挿入することによって、続いて必要に応じて抗原結合を回復または最適化させ
るための特定のフレームワーク残基の変異によって調製される抗体である。ヒト化抗体は
当分野で周知である、そして例えば、Jones et al.,Nature 321:522(1988);Riechmann et
al.,Nature 332:323(1988);およびWinter&Milstein,Nature,349:293(1991)に記載される
。
【0060】
「標的化部分(targeting moiety)」とは、所定の細胞タイプの上のマーカーまたはレセ
プターに特異的である抗体、ペプチド、サイトカイン、オリゴヌクレオチド、または増殖
因子である。標的化部分は、所定の細胞タイプに関連するマーカーと優先的に関連するこ
とによって、その細胞タイプに対して、結合された分子を特異的に送達するために用いら
れ得る。
【0061】
「融合タンパク質(fusion protein)」とは、2つ以上のポリペプチド、さらに詳細には
、細胞毒性RNAseおよび標的化部分を結合することによって形成されるキメラ分子で
ある。細胞毒性RNAseおよび標的化部分は、標的化部分のアミノ末端とRNAseの
カルボキシル末端との間で形成されたペプチド結合を通じて結合され、そして融合タンパ
ク質をコードする核酸配列によって組み換え的に発現される。単鎖融合タンパク質は、単
一の連続するポリペプチド骨格を有する融合タンパク質である。
【0062】
「化学的複合体(chemical conjugate)」とは、細胞毒性RNAseおよび標的化部分の
化学的カップリングによって形成される複合体である。
【0063】
「薬学的に受容可能なキャリア(pharmaceutically acceptable carrier)」とは、免疫
毒素を投与するためのビヒクルとして用いられ得る物質であって、この物質は、不活性で
あるかそうでなければ医学的に受容可能であり、そして融合タンパク質またはアームのリ
ガンドと適合性であるという理由でビヒクルとして用いられ得る物質である。
【0064】
(細胞毒性RNAseをコードする核酸の調製)
当業者は、本発明における使用に適切な細胞毒性RNAse部分が、天然のランピルナ
ーゼ構造を有するポリペプチドおよび全ての酵素的に活性なその改変体を含むことを認識
する。これらの分子は有利には、RNAse活性に必須であると考えられるN末端ピログ
ルタミン酸残基を有し、哺乳動物RNAseインヒビターによって実質的に阻害されない
。天然の細胞毒性RNAseをコードする核酸は、適切な配列のクローニングおよび制限
によって、またはポリメラーゼ連鎖反応(PCR)によるDNA増幅を用いることによっ
て調製され得る。ヒョウガエルのランピルナーゼのアミノ酸配列は、Ardelt et al.,J.Bi
ol.Chem.,256:245(1991)から得ることが可能であり、そして天然のランピルナーゼをコー
ドするcDNA配列、またはその保存的に改変された改変体は、Leung et al.,Mol.Immun
ol.,32:1413(1995)のhLL2ヒト化に用いられるen bloc V-geneアセンブリ法と同様の
方法によって遺伝子合成され得る。細胞毒性RNAse改変体を作成する方法は、当分野
で公知であり、そして慣用的な技術の範囲内である。
【0065】
あるいは、細胞毒性RNAseまたはその改変体をコードする核酸は、インビトロで合
成され得る。化学的合成によって一本鎖オリゴヌクレオチドが生成される。相補的な配列
とのハイブリダイゼーションによって、または短いプライマーおよび一本鎖をテンプレー
トとして用いるDNAポリメラーゼでの重合によって、これは二本鎖DNAに変換され得
る。化学的合成は、約100塩基の配列に最も適合するが、より短い配列を連結すること
によってそれより長い配列を得てもよい。下の実施例2は、細胞毒性RNAse遺伝子を
得るための1つの例示的な方法を提供する。
【0066】
(免疫グロブリン遺伝子の調製)
抗体をコードする遺伝子を調製する方法は、当分野で周知である。例えば、変性プライ
マーセットを用いるPCRを用いて、抗体産生細胞、例えばハイブリドーマから得られる
可変鎖配列cDNAを増幅してもよい(または周知の方法を用いる直接RT−PCRが、
細胞から得られるmRNAに対して直接用いられ得る)。例えば、Marks et al.,J Mol B
iol.(1991)222:581-97、およびそこに引用される参考文献を参照のこと。非ヒト抗体は、
当分野で周知である方法によってヒト化されてもよい。Leung、前出およびそこに引用さ
れる方法を参照のこと。あるいは、ヒト(または他の種)の抗体可変領域ドメインをコー
ドする遺伝子は、MorphoSys(Martinsried,Germany)またはCambridge Antibody Technolog
y(Cambridge,UK)から市販されているタイプのファージ・ディスプレイ・ライブラリーか
ら得てもよい。下の実施例1および実施例3はまた、抗体可変ドメインをコードする核酸
配列を得るための例示的な方法を示す。
【0067】
本質的に、所望の特異性を有する任意の抗体配列が、本発明による免疫毒素を調製する
ために用いられ得る。特に好ましいのは、治療上有用であることが示されている特異性を
有する抗体である。適切な例としては、限定はしないが、腫瘍に結合する抗体、例えば、
B細胞抗原、T細胞抗原、形質細胞抗原、HLA−DR系統抗原、MUC1、MUC2、
MUC3、およびMUC4およびMUC16抗原、EGP−1抗原、EGP−2抗原、胎
盤アルカリホスファターゼ抗原、炭酸脱水素酵素IX、IL−6、VEGF、P1GF、
ILGF、Her2/neu、テネイシン、CD33、CD40、CD74、CD80、
PSMA、PSAおよびPAP;ならびにHLA−DR、CD27、CD40、B細胞お
よびT細胞抗原(例えば、CD15、CD19、CD20、CD21、CD25およびC
D22抗原)を含む自己免疫疾患に関連する抗原が挙げられる。この抗原は、CD19、
CD21、CD22、CD40、MUC1、HLA−DRおよびIL−15のようなB細
胞またはT細胞リンパ腫または白血病に関連する抗原であってもよい。この抗原がCD7
4またはCD74−HLA−DR複合体である場合、適切な抗体はLL1またはそのヒト
化バージョンである。抗原がCD22である場合、適切な抗体はLL2もしくはRBF4
であるか、またはそのヒト化バージョンである。抗原がMUC1である場合、適切な抗体
はPAM4、またはそのヒト化バージョンである。抗原がCD16である場合、適切な抗
体は、CA125またはMu−9のいずれかである。抗原がEGP−1である場合、適切
な抗体はRS7またはそのヒト化バージョンである。抗原がEGP−2である場合、適切
な抗体としてはRS11もしくは17−1Aまたはそのヒト化バージョンが挙げられる。
この抗体は急速に内部移行する抗体であることが有利である。
【0068】
可変ドメイン遺伝子の挿入に適切な抗体定常ドメインを含むか、または含むように操作
され得るベクターは、当分野で公知であり、そしてこのようなベクターはまた、本発明の
免疫毒素の発現のために適切である。このようなベクターは、哺乳動物宿主細胞において
、コードされた遺伝子の転写を開始するのに必須である必須の制御配列を含む。このよう
なコントロール配列は、当分野で周知である。例えば、その全体において引用することに
より本明細書に組み込まれる、米国特許第5,650,150号および同第6,472,511号のpdHL
2ベクターの説明を参照のこと。あるいは、定常ドメイン遺伝子(CH1、CH2、CH
3およびCL)は、抗体産生細胞から得られてもよいし、当分野で周知の方法を用いて適
切な発現ベクターにクローニングされてもよい。
【0069】
(免疫毒素遺伝子の調製)
本発明による免疫毒素コード遺伝子の調製は、例示の目的でのみ提供されて、限定され
るものではない、下の実施例1および4に詳細に例示される。要するに、PCRを用いて
例えば、抗体軽鎖可変ドメイン(または必要に応じて重鎖可変ドメイン)をコードする遺
伝子を得てもよく、当分野で公知のPCR法を用いて得てもよい。適切な制限部位が、P
CRプライマーに導入されてもよく、それによってこの遺伝子とさらなる配列とのその後
の結合が容易になる。同様に、細胞毒性RNAseコード遺伝子は、都合のよい制限部位
を提供するプライマーをここでも用いて、適切なテンプレートからPCRによって得られ
てもよい。次いで、細胞毒性RNAse遺伝子および可変ドメイン遺伝子は、適切な制限
エンドヌクレアーゼで消化されて、可変鎖RNAse融合タンパク質をコードする遺伝子
を提供するために直接一緒に結合されてもよい。細胞毒性RNAse遺伝子は、有利には
、可変ドメイン遺伝子のN末端に結合され、さらに有利には軽鎖可変鎖遺伝子のN末端に
結合されるが、RNAse遺伝子は、あるいは、重鎖可変ドメインのN末端に、または重
鎖および軽鎖可変ドメインの両方のN末端に結合されてもよい。
【0070】
あるいは、適切なリンカーは、合成オリゴヌクレオチド(例えば、細胞毒性RNAse
および免疫グロブリン遺伝子に含まれる制限部位と適合しており、そして細胞毒性RNA
seと可変ドメイン遺伝子とリンカー配列との直接の結合をもたらす制限部位を含むよう
に調製され得る)を用いて、可変ドメイン遺伝子と細胞毒性RNAse遺伝子との間に導
入され得る。当業者は、種々のリンカーが本発明における使用に適切であることを認識す
る。しかし、リンカーは有利には、10〜30アミノ酸長であり、そして二次構造も三次
構造もほとんどまたは全く有さない、親水性アミノ酸を含むリンカーである。このような
リンカーは当分野で周知であり、そして例えば、単鎖Fv分子を構築するのに用いられる
。例えば、米国特許第4,946,778号を参照のこと。適切なリンカーとしては、配列GGG
GSなどの多量体が挙げられる。免疫毒素遺伝子を調製する他の方法は、その全体が引用
することにより本明細書に組み込まれる、米国特許出願第20030099629号に記載される。
【0071】
インタクトな抗体構造を含む免疫毒素(すなわち、軽鎖VLおよびCLドメイン、なら
びに重鎖VHおよびCH1−3ドメインを含む免疫毒素)が調製されるべき場合、上記の
可変ドメイン鎖は、これらのさらなる定常ドメインをコードする配列に結合されなければ
ならない。有利には、適切な定常ドメインを既に含むベクター、例えば、上記のpdHL
2を用いる。適切な定常ドメインを調製して、それらを可変ドメインに結合させる別の方
法が当分野で周知である。Fabフラグメントについては、CLおよびCH1ドメインの
みが必要である。一旦、適切な形態に結合されれば、次にこれらの配列は、宿主細胞のト
ランスフェクションの前に適切な発現ベクターに挿入され得る。哺乳動物発現に適切であ
る発現ベクターは、当分野で周知であり、そしてまた例えば、Invitrogen,Carlsbad,CAか
らも市販されている。
【0072】
(哺乳動物細胞における免疫毒素の発現)
一旦適切な発現ベクターが調製されれば、適切な宿主細胞にトランスフェクトされる。
組み換え抗体の発現に適切な宿主細胞は、当分野で周知であり、そしてこれには例えば、
CHO細胞、および有利には、骨髄細胞株、例えば、NSO細胞が挙げられる。CHOお
よびNSO細胞の両方がATCC(Manassas,VA)から入手可能である。ヒト抗体配列を含
む免疫毒素の産生のために特に適切な別の宿主細胞は、Karpas et al.,Proc.Natl.Acad.S
ci USA 98:1799(2001)に記載されるKarpas 707H系統である。トランスフェクションは、
周知の方法、例えば、エレクトロポレーション、リポフェクション、およびDEAEデキ
ストラン媒介性トランスフェクションによって達成され得る。有利には、発現ベクターは
、選択マーカー、例えば、DHFRを含み、そして宿主細胞はDHFR-である。この細
胞は、選択因子(例えば、選択マーカーがDHFRである場合はメトトレキセート)の存
在下で培養される。陽性のクローンは、例えば、免疫毒素の細胞毒性RNAse部分に特
異的に結合する抗体を用いてELISA法によって確認され得る。陽性クローンは、後の
増殖のために凍結されてもよいし、または直接増殖されてもよい。陽性クローンの培養物
を、適切な培地、例えば完全HSFM培地中で増殖させる場合、免疫毒素を含む培養上清
を収集する。
【0073】
(免疫毒素の精製)
免疫毒素は、当分野で周知である方法を用いて容易に精製され得る。例えば、アフィニ
ティー精製が用いられてもよい。完全な抗体構造を含む免疫毒素については、プロテイン
Aアフィニティー精製を用いて、純粋な免疫毒素を単独の精製工程で得てもよい。さらな
る精製工程は、必要に応じて、イオン交換クロマトグラフィー、サイズ排除クロマトグラ
フィーなどのような従来の方法を用いて行なわれ得る。免疫毒素は、従来の方法、例えば
、RP−HPLC、SDS−PAGE、質量分析、および例えば抗細胞毒性RNAse抗
体を用いるウエスタンブロットによって特徴付けられ得る。ヒョウガエル由来のランピル
ナーゼは、潜在的なグリコシル化部位を含む(実施例1を参照のこと)。この部位のグリ
コシル化が所望されない事象では、免疫毒素は、オンコナーゼ(onconase)上に存在する炭
水化物部分を除去するためにグリコシダーゼ酵素で処理されてもよい。あるいは、RNA
seをコードする遺伝子配列は、保存的アミノ酸置換を介してAsn残基を置換すること
によってグリコシル化部位を欠失するために、部位特異的変異誘発法によって変異されて
もよい。例えば、69位置のAsnは、グルタミンで(変異体N69Q)、または本明細
書に記載される任意の構築物中の他の適切なアミノ酸で置換されてもよい。このグリコシ
ル化変異体の適切な例としては、2L−Rap(N69Q)−hLL1−r4P、2L−
Rap(N69Q)−hRS7および2L−Rap(N69Q)−hLL2が挙げられる
。
【0074】
(細胞毒性RNAse免疫毒性を用いる治療方法)
本明細書に記載の免疫毒素は、広範な種々の細胞毒性の治療様式のための薬学的組成物
に処方され得る。これらの組成物は、任意の欲されない細胞タイプ、例えば、不適切な増
殖または活性化を受けている細胞を標的するために用いられてもよいし、または病原性微
生物を標的することによって感染性疾患を処置するために用いられてもよい。詳細には、
この組成物は、種々のガンおよび自己免疫疾患を処置するために有用である。処置され得
るガンとしては、例えば、リンパ腫、白血病、黒色腫、神経芽細胞腫、および骨髄腫、肉
腫、神経膠腫、または癌腫、例えば、乳癌、結腸直腸癌、卵巣癌、肝臓癌、胃癌、結腸直
腸癌、前立腺癌、肺癌、腎臓癌または膵臓癌が挙げられる。処置され得る自己免疫疾患と
しては、例えば、関節リウマチ、全身性エリテマトーデス、シェーグレン症候群、急性特
発性血小板減少性紫斑病、慢性特発性血小板減少性紫斑病、皮膚筋炎、シデナム舞踏病、
重症筋無力症、ループス腎炎、リウマチ熱、多腺性症候群、水疱性類天疱瘡、糖尿病、ヘ
ノッホ・シェーンライン紫斑病、連鎖球菌感染後腎炎、結節性紅斑、高安動脈炎、アジソ
ン病、多発性硬化症、サルコイドーシス、潰瘍性大腸炎、多形性紅斑、IgA腎症、結節
性多発性動脈炎、強直性脊椎炎、グッドパスチャー症候群、血栓血管炎(thromboangitis
ubiterans)、原発性胆汁性肝硬変、橋本甲状腺炎、甲状腺中毒症、強皮症、慢性活動性肝
炎、多発性筋炎/皮膚筋炎、多発性軟骨炎、尋常性天疱瘡、ヴェーゲナー肉芽腫症、膜性
腎症、筋萎縮性側索硬化症、脊髄癆、巨細胞性動脈炎/多発性筋痛、悪性貧血、急速進行
性糸球体腎炎および線維化性肺胞炎が挙げられる。
【0075】
本発明の組成物はまた、感染の治療的処置のためにも有用であり、ここでは免疫毒素の
免疫グロブリン成分は、疾患を生じる微生物に特異的に結合する。本発明の状況では、疾
患を生じる微生物としては、病原性細菌、ウイルス、真菌および多様な寄生生物が挙げら
れ、そしてこの抗体は、これらの微生物、それらの産物またはそれらの病変に関連する抗
原を標的し得る。微生物の例としては、限定はしないが以下が挙げられる:
ストレプトコッカス・アガラクティエ、在郷軍人病菌、化膿連鎖球菌、大腸菌、淋菌、
髄膜炎菌、肺炎球菌、Hemophilis influenzae B、梅毒トレポネーマ、ライム病スピロヘ
ータ、緑膿菌、ハンセン菌、ウシ流産菌、ヒト結核菌、破傷風毒素、HIV−1、HIV
−2、HIV−3、A型肝炎、B型肝炎、C型肝炎、D型肝炎、狂犬病ウイルス、インフ
ルエンザウイルス、サイトメガロウイルス、単純疱疹IおよびII、ヒト血清パルボ様ウ
イルス、パピローマウイルス、ポリオーマウイルス、呼吸器合胞体ウイルス、水痘帯状疱
疹ウイルス、B型肝炎ウイルス、パピローマウイルス、麻疹ウイルス、アデノウイルス、
ヒトT細胞白血病ウイルス、エプスタイン・バーウイルス、マウス白血病ウイルス、流行
性耳下腺炎ウイルス、水疱性口内炎ウイルス、シンドビスウイルス、リンパ球性脈絡髄膜
炎ウイルス、疣ウイルス、ブルー・タングウイルス、センダイウイルス、猫白血病ウイル
ス、レオウイルス、ポリオウイルス、シミアン・ウイルス40、マウス乳癌ウイルス、デ
ング熱ウイルス、風疹ウイルス、原生動物、熱帯熱マラリア原虫、三日熱マラリア原虫、
トキソプラズマ原虫、Trypanosoma rangeli、クルーズ・トリパノソーマ、Trypanosoma r
hodesiensei、トリパノソーマ・ブルーセイ、マンソン住血吸虫,Schistosoma japanicum
、Babesia bovis、Elmeria tenella、回旋糸状虫、熱帯リーシュマニア、旋毛虫、Theile
ria parva、Taenia hydatigena、Taenia ovis、無鉤条虫、単包条虫、Mesocestoides cor
ti、抗マイコプラズマモノクローナル抗体、Mycoplasma arthritidis、M.hyorhinis、M.o
rale、M.arginini、Acholeplasma laidlawii、M.salivarium、およびM肺炎。これらの病
原性微生物に結合するモノクローナル抗体は当分野で周知である。
【0076】
当業者は、これらの実施例が例示的であって、本発明を限定するものではないことを理
解する。
【0077】
これらの方法では、薬学的組成物は、被験体に1日あたり2回以上、例えば、0.1〜
約1000mgの量で投与されてもよいが、それより多くてもまたは少なくても適切とみ
なして用いられ得る。
【0078】
この組成物のタンパク質性の性質に起因して、それらの組成物は特に、非経口投与、例
えば静脈内投与または皮下投与(または皮内投与)に適切であるが、当業者は、他の送達
方法が用いられ得ることを理解する。この方法は、現在開発中であるか、または将来開発
されるタンパク質送達のための方法を包含し、そしてまた例えば、腹腔、頭蓋、胸膜など
のような局所投与を包含し得る。非経口投与のためには、組成物は、薬学的に受容可能な
キャリア、好ましくは、緩衝化生理食塩水のような水性キャリアに溶解されるかまたは懸
濁された免疫毒素の溶液を含んでもよい。これらの溶液は、無菌であって、補助物質、例
えば、pH調節剤および緩衝化剤、ならびに毒性および等張性の調節剤を含んでもよい。
【0079】
免疫毒素の用量は、1日あたり1患者あたり約0.1〜10mgであるが、特にこの薬
物が局所的に投与され、血流中ではない場合には、1日あたり1患者あたり最大100m
gの用量が用いられてもよい。この用量は、より少量のRNAse成分を含む、抗体融合
タンパク質に基づく。
【0080】
治療適用では、この組成物は、疾患に罹患している被験体に対して、目的の細胞を殺傷
するのに十分な量として規定される、治療上細胞毒性の量で投与される。これを達成する
のに十分な量は、「薬学的に有効な量(therapeutically effective amount)」として規定
される。任意の所定の環境における正確な量は、疾患の重篤度および患者の健康の一般的
状態を含む種々の要因に依存する。この組成物の単回投与または複数回投与は、必要な用
量に依存して投与され得る。適切な投薬レジメンの決定は、当分野で周知の慣用的方法を
用いて行なわれる。
【0081】
免疫毒素はまた、インビトロにおいて細胞の集団を処置するために用いられ得る。例え
ば、それらは、骨髄アブレーションを受けている患者への移植の前に骨髄中の欲されない
細胞タイプを選択的に殺傷するために用いられ得る。
【0082】
このように一般的に記載された本発明は、例示のために記載されており、本発明の限定
は意図していない以下の実施例を参照してさらに容易に理解される。
【実施例1】
【0083】
(2L−rap−hLL1−γ4Pの発現および特徴付け)
実施例1では、rapはランピルナーゼである。
【0084】
(pdHL−IgG4P改変体の構築)
IgG4遺伝子を含むB13−24細胞は、ATCC(ATCC番号 CRL−113
97)から購入して、ゲノムDNAを単離した。要するに、細胞をPBSで洗浄して、消
化緩衝液(100mM NaCl,10mMのTris−HCl pH8.0、25mM
のEDTA pH8.0、0.5%のSDS、0.1mg/mlのプロテイナーゼK)に
再懸濁して、50℃で18時間インキュベートさせた。等容積のフェノール/クロロホル
ム/イソアミルアルコールを用いてこのサンプルを抽出して、7.5MのNH4Ac/1
00% EtOHで沈殿させた。ゲノムDNAは、遠心分離によって回収して、TE緩衝
液に溶解した。テンプレートとしてゲノムDNAを用い、IgG4遺伝子を、以下のプラ
イマーを用いてPCRによって増幅させた。
プライマー−SacII:5'CCGCGGTCAC ATGGCACCAC CTCTCTTGCA GCTTCCACCA AGGGCCC 3'
(47マー);
プライマー−EagI:5'CCGGCCGTCG CACTCAT TTA CCCAGAGACA GGG 3'(33マー)
【0085】
増幅されたPCR産物を、TOPO−TA配列決定ベクター(Invitrogen)中にクローニ
ングして、DNA配列決定によって確認した。pdHL−hLL2中のIgG1の重鎖定
常領域を含むSacII−EagIフラグメントを、TOPO−TA−IgG4プラスミ
ドのSacII−EagIで置換して、pdHL2−hLL2−IgG4(pdHL2−
hLL2−γ4)ベクターを生成した。
【0086】
(IgG4−プロリン変異)
Ser228Proの変異をIgG4のヒンジ領域に導入して、半分子の形成を回避し
た。変異したヒンジ領域の56bpフラグメント(PstI−StuI)を合成し(トッ
プ:5'GAG TCC AAA TAT GGT CCC CCA TGC CCA CCG TGC CCA GGT AAG CCA ACC CAG G3';ホ
゛トム:5'C CTG GGT TGG CTT ACC TGG GCA CGG TGG GCA TGG GGG ACC ATA TTT GGA CTC T
GC A 3')、アニーリングして、IgG4のPstI−StuIフラグメントで置換した。
この構築で、最終ベクターpdHL2−hLL2−γ4Pを得た。
【0087】
(pdHL2−hLL1−γ4Pの構築)
pdHL2−hLL2−γ4PのXbaI−HindIIIフラグメントを、Vkおよ
びVH領域を含むpdHL2−hLL1のXba−HindIIIフラグメントで置換し
て、hLL1−γ4P構築物を生成した。
【0088】
(pdHL2−2L−rap−hLL1−γ4Pの構築)
可塑性リンカー(GGGGS)3を用いて、hLL1のVkのN末端に対してRapの
C末端を結合させた。各々の軽鎖のN末端に1つのrap分子を結合させた。この分子に
ついてのDNAの構築は、以下のプライマーを用いてPCRによって行なった:
P1;リーダー5’(XbaI):5'CTC TAG ACA CAG GAC CTC ACC ATG GGA TGG 3'(
30マー)
P2;リーダー3’:5'TGA AAC GTT AGC CAA TCC TGG GAG TGG ACA CCT GTG GA 3'(
38マー)
P3;Onc 5’:5'TCC ACA GGT GTC CAC TCC CAG GAT TGG CTA ACG TTT CA 3'(
38マー)
P4;Onc 3’(EcoRV):5'AGT CAG CTG GAT ATC GGA GCC ACC GCC TCC AG
A TC 3'(35マー)
P5;LL1−Vk 5’(EcoRV):5'GAT CTG GAG GCG GTG GCT CCG ATA TCC
AGC TGA CT 3'(35マー)
P6;LL1−Vk 3’(BamHI):5'GGG ATC CAA CTG AGG AAG CAA AGT TTA
A 3'(28マー)
【0089】
図1に図示されるように、pdHL2−hLL1−γ4PのXba−BamHIフラグ
メントを、pBS−2L−rap−hLL1のXba−BamHI(Xba−Leade
r−rap−Linker−Vk−BamHI)フラグメントで置換して、最終のベクタ
ーpdHL2−2L−rap−hLL1−γ4Pを完成させた。
【0090】
(トランスフェクション)
ベクターDNA(30μg)をSalI酵素で直線化して、エレクトロポレーション(
450V)によってNS0(4×106細胞/mL)またはSp2/0−Ag14(5×
106個の細胞/mL)の骨髄腫細胞にトランスフェクトした。低IgG FBS(10
%)、ペニシリン(100単位/mL)、ストレプトマイシン(100μg/mL)、L
−グルタミン(2mM)、ピルビン酸ナトリウム(1mM)、非必須アミノ酸(100μ
M)およびメトトレキセート(0.1μM)を補充した完全なハイブリドーマ−SFM培
地中で細胞を増殖させた。陽性のクローンをELISAによってスクリーニングした。要
するに、プレートを、PBS培地中に含まれる5μg/mLの50μlの抗rap抗体を
用いてコーティングして、4℃で一晩インキュベートさせた。PBSでプレートを洗浄し
、2%BSAでブロックした後に細胞培養上清を添加した。HRP複合体化ヤギ抗ヒトI
gG4抗体を検出のために用いて、OPDを発色のための基質として用いた。プレートを
490nmで読み取った。陽性のクローンを増殖させて、さらなる使用のために凍結させ
た。クローンC6を、最適のプロデューサーとして特定して、さらなる開発のために用い
た。
【0091】
(発現および精製)
各々に500mlの培地を有する2つのローラーボトル中で細胞を最終培養物まで増殖
させて(10〜20%の生存度)、その細胞を遠心分離によって取り出した。培養上清を
充填して、プロテインAカラムに加え、20mM Tris−HCl/100mM Na
Cl緩衝液(pH8.5)で平衡化した。ローディング後、このカラムを100mMのク
エン酸ナトリウム緩衝液(pH7.0)で洗浄して、100mMのクエン酸ナトリウム緩
衝液(pH3.5)で溶出させ、融合タンパク質を得た。生成物を含有するピークを、3
M Tris−HCl、pH8.0を用いてpH7.0に調節して、10mMのPBS緩
衝液に対して透析した。濃縮後、この生成物を0.22μmのフィルターを通して濾過し
、2〜8℃で保管した。1Lの培養物から、精製後に16mgを回収した。
【0092】
(2L−rap−hLL1−γ4Pの特徴付け)
HPLC:タンパク質の純度および濃度は、HPLCでチエックした。先鋭な単独のピ
ークが、図2に示されるように7.7分で観察され、この保持時間によって、この分子が
IgGより大きいことが示された。
【0093】
SDS−PAGE:4〜20% Tris−Glycineゲル(PAGEr(商標)
Gold Precast Gels,Cambrex)を用いて、SDS−PAGEを還元条件下で行なった。図
3に示されるように、予想されるサイズ約50kDの重鎖に関連するバンド、ならびにh
LL1の軽鎖(約25kD)よりも両方とも大きい、約37kDおよび39kDの分子量
の2つのバンドが観察された。2つの軽鎖の存在は、融合タンパク質上のrapのグリコ
シル化に起因することが示された(以下参照)。
【0094】
質量分析法:The Scripps Research Institute,CAにおいて、MALDI−TOF法に
よって質量分析を行った。2つのサンプル、その1つは天然の状態(1.6mg/mLを
10mMのPBSに含有)、そしてもう一方は、還元状態(1.6mg/mLを1mMの
HEPES/10mMのDTT、pH7.5緩衝液)を、分析のために送った。天然のサ
ンプル(図4a)は、質量177150の1つの主要なピークを示し、これは、IgGに
加えて2つのrapのMWとよく一致している。この還元サンプルは、50560(重鎖
に相当する)、38526および36700(rapを含有する2つの軽鎖に相当する)
において3つの主なピークを示した(図4b)。
【0095】
ウエスタンブロッティング:精製されたタンパク質におけるrapの存在を確認するた
めに、ウエスタンブロッティングを行った。還元条件下のSDS−PAGEゲル由来のサ
ンプルを、PVDF-Plus(Osmonics,Inc.,)メンブレン上に電気的に転写した。5% BSA
でのブロッキング後、マウスの抗rap抗体を、1:10,000希釈または100ng
/mlで添加して、1時間インキュベートさせた。洗浄後、HRP複合体化ヤギ抗マウス
Fc抗体を添加して1時間インキュベートさせた。6回の洗浄後、LumiGloTM(Kirkegaard
&Perry Laboratories)基質を添加して、コダックのフィルムを現像した。図5に示される
ように、融合された軽鎖に相当する両方のバンドをフィルム上で検出し、これによって両
方の軽鎖上のrapの存在を確認した。
【0096】
N−グリコシダーゼでの処理:rapは、潜在的なNグリコシル化部位、Asn−X−
Thr/Ser、Asn69−Val70−Thr71を有するので、分子量で2kDの
相違があるという2つの軽鎖の観察は、rapの一様でないグリコシル化の結果であり得
る。この可能性を検討するために、rap−hLL1抗体を、供給業者の推奨に従って変
性条件下でNグリコシダーゼ(New England Biolabs)とともにインキュベートさせた。図
6に示されるとおり、N−グリコシダーゼ処理後、2つの軽鎖に相当する2つのバンドが
1つ(より早く移動するバンド)に集束し、これによってカルボヒドラーゼの一様でない
分解が、SDS−PAGEでの2つのバンドという観察の原因であったことが確認された
。除去されたグリコシル化部位を有するRapの改変体であるRap(N69Q)が組み
換え構築物中でRapについて置換される場合、唯一のrap融合軽鎖の観察によってさ
らなる支持が得られた(データ示さず)。
【0097】
rapの活性:RNAse活性は、供給業者の推奨に従ってBright-GloTM Luciferase
Reporter Assayシステム(Promega)を用いてTNT(商標)Quick Coupled Transcription
/Translation System(Promega)によって試験した。このアッセイの原理は、ルシフェラー
ゼレポーター系を用いるRNase活性の結果としてのタンパク質合成の阻害(mRNA
分解)の測定であって。サンプルは、種々の希釈中で遊離のrap(0.001〜2.5
nM)、hLL1−rap(0.01−20nM)またはPKI−LL2−Oncおよび
PKII−LL2−Onc(0.01〜20nM)として表されるhLL2−rapの化
学的複合体を調製した。各々のサンプル(5μL)を、20μlのTNTマスター混合物
と混合して、96ウェルプレート中で30℃で2時間インキュベートして、それから1μ
lを、50μlのBright-GloTM基質での分析のために取り出した。その結果を、Excelま
たはPrism Padソフトウェアを用いて、図7に示した。EC50値は、rap−hLL1お
よびhLL2−Oncの化学的複合体については約300pM、そして遊離のrapにつ
いては30pMであった。
【0098】
WPについての競合結合:WPはhLL1の抗イディオタイプ抗体である。WPに対す
るhLL1抗体と比較したrap−hLL1抗体の親和性は、競合結合アッセイによって
評価した。要するに、96ウェルプレートを50μlのWPを5μg/mLで用いてコー
ティングして、4℃で一晩インキュベートさせた。タンパク質サンプルの3つのタイプで
あるhLL1、rap−hLL、またはhA20は、種々の2倍希釈(最終濃度は0.4
9〜1000nMに及ぶ)中で調製して、等容積の2×HRP複合体化mLL1抗体(最
終希釈は1/20,000)と混合した。上記のようなHRP複合体化mLL1と混合し
た50μLのタンパク質サンプルを、各々のウェルに添加して、1時間インキュベートさ
せた。洗浄後、H2O2を含有するOPD基質を添加してプレートを490nmで読んだ。
図8に示されるように、吸光度に対するタンパク質濃度を、ExcelまたはPrism Pad graph
ソフトウェアを用いてプロットした。hA20(ヒト化抗−CD20抗体)を、陰性のコ
ントロールとして用いた。図8から、rap−hLL1は、hLL1と同様の結合親和性
を有し、そして陰性コントロールhA20は、親和性を全く有さないことが明らかである
。抗原の供給源としてRaji細胞を用いて同様の結果を得た。
【0099】
インビトロ細胞毒性:インビトロ細胞毒性は、B細胞リンパ腫細胞株(Daudi)および複
数の黒色腫細胞株(MC/CAR)で測定した。細胞(0.1ml中に10,000個)
を、96ウェルプレートの各々のウェルに入れた。24時間後、遊離のhLL1、遊離の
rapまたはrap−hLL1(10μl)を適切なウェルに添加して、その細胞をイン
キュベーター中で37℃で3日間インキュベートした。細胞増殖は、MTSテトラゾリウ
ム色素還元アッセイまたはBrDU比色定量アッセイを用いて決定した。結果は、Prism
Padソフトウェアを用いてグラフとして得られたEC50で表す。図(図9〜10)から、
rap−hLL1が、B−細胞リンパ腫細胞株(Daudi)および多発性骨髄腫細胞株(MC
/CAR)の両方に対して感受性であることが証明される。rap−hLL1は、EC50
値によって反映されるように、MC/CAR細胞に比較してDaudi細胞に対して有意によ
り強力(細胞毒性)であった(図9および図10)。MC/CAR細胞については、EC
50値は、試験した濃度では達成されなかった。最高濃度(56nM)では、細胞の生存度
は57%であった。将来の実験は、Daudi細胞上のrap−hLL1について正確なEC5
0値を測定することが目的である。hLL1または遊離のrapは、それ自体では、いず
れの細胞株でも細胞毒性を実証しなかった。
【0100】
薬物動態および体内分布:hLL1または2L−Rap−hLL1−γ4Pを、Sharke
y et al.,(Int J Cancer.1990;46:79-85)に記載のように、2−(4−イソチオシアナー
トベンジル)DTPA(Macrocyclics,Dallas,TX)を用いてジエチレントリアミノペンタ酢
酸(DTPA)と複合体化させて、DTPA−hLL1またはDTPA−2L−Rap−
hLL1−γ4Pを得た。これはそれぞれ、薬物動態学的研究および生体分布の研究のた
めに、88Y塩化物(Los Alamos National Laboratory(Los Alamos,NM)または111In塩化
物(Perkin Elmer Life Sciences,Boston,MA)で標識された。ナイーブな雌性SCIDマウ
ス(8週齢、18〜22g)に対して各々の動物に、hLL1および2L−Rap−hL
L1−γ4Pの各々10μgの総用量を投与するように、未標識の、hLL1または2L
−Rap−hLL1−γ4PのDTPA複合体を補充した0.001mCi88Y−DTP
A−hLL1および0.02mCiの111In−DTPA−2L−Rap−hLL1−γ
4Pの混合物を静脈内注射した。投与後選択された時点(1、2、4、16、48、72
、168時間)後に、5匹のマウスの群を麻酔して、血液サンプルを心臓穿刺によって採
取した。主な組織を取り出して、秤量して、容器中に入れた。血液サンプルおよび組織を
、目盛り付きのガンマカウンターであるMinaxiλ Auto-Gamma(商標)5000シリーズ
のガンマカウンター(Packard Instrument Company;Downers Grove,IL)で、111In(チャ
ネル120〜480)および88Y(チャネル600〜2000)についてカウントした。
111Inカウンティングウインドウへの88Yエネルギーの後方散乱について補正するため
に交差曲線を作成した。
【0101】
インビトロの毒性:ナイーブなSCIDまたはBALB/cマウスに、マウス1匹あた
り25〜400μgに及ぶ種々の用量の2L−Rap−hLL1−γ4Pを静脈内注射し
て、毒性および体重の損失の可視的な徴候について毎日モニターした。最大耐容用量(M
TD)は、死亡が生じず、かつ体重減少が処置前の動物の重量(約20g)の20%以下
である最高用量として規定した。毒性効果を経験した動物は屠殺して、回収して、組織病
理学的分析に供した。ナイーブなSCIDマウスでは、2L−Rap−hLL1−γ4P
の100、150、200、250、300または400μgの単回静脈内用量によって
、動物の重篤な体重減少および死亡が生じたが、全てのマウスは、25または50μgの
用量で生残した(表2)。BALB/cマウスでは、全てのマウスが2L−Rap−hL
L1−γ4Pの30または50μgの単回静脈内用量で生残したが、100または200
μgでは生残しなかった。別の実験では、2L−Rap−hLL1−γ4Pの75μgの
用量は、SCIDマウスに対して毒性であることが見いだされた(データ示さず)。従っ
て、単回ボーラス注射として与えられた2L−Rap−hLL1−γ4PのMTDは、S
CIDマウスでは50〜75μgであって、BALB/cマウスでは50〜100μgで
ある。死亡マウスおよび屠殺されたマウスの全体の病理学的な検査では、重篤な肝臓およ
び脾臓の毒性が示された。肝臓は、色が青ざめており、脾臓は縮んで、通常のサイズより
小さかった。組織病理学的検査によって、肝臓および脾臓の壊死が明らかになった。代表
的なマウスの血清サンプルでは、アラニンアミノトランスフェラーゼ(ALT)、アアス
パルテートアミノトランスフェラーゼ(AST)および総ビリルビンのレベル上昇があり
、このことはこれらの高用量での有意な肝臓毒性を示唆していた。
【0102】
データ分析:インビトロの細胞毒性研究のために、三連の測定値の平均から用量応答曲
線を作成して、GraphPad Prismソフトウェア(Advanced Graphics Software,Encinitas,CA
)を用いて50%阻害濃度(IC50)値を得た。薬物動態学的データは、ノンコンパート
メント分析プログラムWinNonlin,バージョン4.1(Pharsight,Mountain View,CA)の標
準的アルゴリズムを用いて分析した。このプログラムは、線形補間による線形台形公式を
用いて曲線下免疫(AUC)を算出する。排出速度定数(kβ)は、一次速度式を仮定し
て終末半減期(t1/2β)から算出した。生残研究は、GraphPad Prismソフトウェアを用
いるKaplan-Meierプロット(ログランク分析)を用いて分析した。相違はP<0.05で
有意とみなした。
【0103】
薬物動態および生体分布のデータ:放射性標識したhLL1および2L−Rap−hL
L1−γ4Pの薬物動態および生体分布を、ナイーブなSCIDマウスで測定した。hL
L1および2L−Rap−hLL1−γ4Pを、DTPAと結合体化して、それぞれ標識
した88Yおよび111Inで追跡した。図11に示されるように、111IIn−標識した2L
−Rap−hLL1−γ4Pは、初期の急速な再分布相(α)および後期の緩徐な排出相
(β)によって特徴付けられる、88Y−標識されたhLL1と同様の血液からの二相性の
クリアランスを示す。わずかに短いα半減期が、hLL1(4時間)に比較して、2L−
Rap−hLL1−γ4P(5.1時間)について観察された。5時間を超えるデータポ
イントを用いて、t1/2β、kβ、AUC、平均滞留時間(mean residence time)(MRT
)、見かけの分布容積(Vd)、およびクリアランスの速度(Cl)を算出して、これら
のパラメーターの値を表1に示す。111In−標識された2L−Rap−hLL1−γ4
Pの組織取り込みは、88Y−標識hLL1の取り込みと同様であった(データ示さず)。
【表1】
【0104】
インビトロの毒性:ナイーブなSCIDマウスでは、100、150、200、250
、300または400μgの単回静脈内用量の2L−Rap−hLL1−γ4Pは、動物
の重篤な体重減少および死亡を生じたが、全てのマウスは、25または50μgの用量で
生残した(表2)。BALB/cマウスでは、全てのマウスが2L−Rap−hLL1−
γ4Pの30または50μgの単回静脈内用量では生残したが、100または200μg
では生残しなかった(表2)。別の実験では、2L−Rap−hLL1−γ4Pの75μ
gの用量は、SCIDマウスに対して毒性であることが見いだされた(データ示さず)。
従って、単回ボーラス注射として与えられた2L−Rap−hLL1−γ4PのMTDは
、SCIDマウスでは50〜75μgであって、BALB/cマウスでは50〜100μ
gである。死亡マウスおよび屠殺されたマウスの全体的な病理学的な検査では、重篤な肝
臓および脾臓の毒性が示された。肝臓は、色が青ざめており、脾臓は縮んで、通常のサイ
ズより小さかった。組織病理学的検査によって、肝臓および脾臓の壊死が明らかになった
。代表的なマウスの血清サンプルでは、アラニンアミノトランスフェラーゼ(ALT)、
アアスパルテートアミノトランスフェラーゼ(AST)および総ビリルビンのレベル上昇
があり、このことはこれらの高用量での有意な肝臓毒性を示唆していた。
【表2】
【0105】
(腫瘍保有マウスでの治療有効性)
腫瘍保有マウスでの治療有効性:1群あたり8〜9匹の雌性SCIDマウス(8週齢、
18〜22g)に、1.5×107個のDaudi細胞を静脈内注射して、1日後に処置した。
後肢の麻痺についてマウスを毎日検査して、毎週秤量した。動物が後肢の麻痺を発症する
かまたはその処置前の体重の20%が低下した場合、それらの動物を安楽死させた。各々
のセットの治療実験は180日後に終えた。
【0106】
図12に示されるように、未処置のマウス(PBSのみ)は、30日内に全て死亡し、
平均生残時間(median survival time)(MST)は28日であった。50μgの2L−R
ap−hLL1−γ4P中の成分タンパク質の組成に相当する、hLL1−γ4P(43
.2μg)およびRap(6.6μg)の混合物を投与されたコントロール群のMSTは
、70日であった(P<0.0001対PBS群)。対照的に、5μgまたは15μgの
いずれかの2L−Rap−hLL1−γ4Pの単回注射を受けた全てのマウスでは、10
0日より長く生きており(MST>180日;P=0.0005対成分処置群)、そして
この研究の終わり近くで各々の群から欠けたのは1匹のマウスだけであった。180日後
に研究が終わったとき、5、15、30、40または50μgの2L−Rap−hLL1
−γ4Pの単回注射を投与されたマウスの90%が治癒した。1μgの単回注射を受けた
マウスのMSTは、230%の増大に相当する、未処置の群の28日に比較して92日で
あった(P<0.0001)ことに注目すべきである。
【実施例2】
【0107】
(細胞毒性RNAseをコードするPCR増幅したDNAの合成)
組み換え細胞毒性RNAseのN末端配列(46アミノ酸)をコードするセンス鎖配列
[5'-TGG CTA ACG TTT CAG AAG AAA CAT ATC ACG AAT ACA CGA GAT GTA GAC TGG GAC AAT
ATA ATG TCT ACG AAT CTG TTT CAC TGT AAG GAT AAG AAT ACC TTT ATA TAC AGT CGC CCA
GAG CCT GTA AAG GCT ATC TGT A-3']を有する139マーのDNAヌクレオチド、ON
CO−Nを、自動DNAシンセサイザー(Applied Biosystem 392 DNA/RNA Synthesizer)
によって合成して、隣接するプライマーONNBACK[5'-AAG CTT CAT ATG CAG GAT T
GG CTA ACG TTT CAG AAG AAA-3'、およびONNFOR[5'-CTT ACT CGC GAT AAT GCC TT
T ACA GAT AGC CTT TAC AGG CTC TG-3']を用いるPCR増幅のためのテンプレートとし
て用いた。得られた二本鎖PCR産物は、細胞毒性RNAseのN末端の半分の54アミ
ノ酸残基をコードするcDNA配列を含む。ONNBACKは、ステージングベクターに
、または細菌発現ベクターへのインフレーム結合のために(NdeI部位)サブクローニ
ングを容易にするために、制限部位HindIII(AAAGCTT)およびNdeI(
CATATG)を含む。NruI部位(TCGCGA)を、ONNFORプライマーに組
み込んで、細胞毒性RNAseのC末端半分をコードするcDNAとのインフレーム結合
を容易にする。
【0108】
同様に、細胞毒性DNAseのC末端配列(46アミノ酸)をコードするセンス鎖配列
[TGC TGA CTA CTT CCG AGT TCT ATC TGT CCG ATT GCA ATG TGA CTT CAC GGC CCT GCA AA
T ATA AGC TGA AGA AAA GCA CTA ACA AAT TTT GCG TAA CTT GCG AGA ACC AGG CTC CTG TA
C ATT TCG TTG GAG TCG GG-3']を有する137マーのDNAヌクレオチド、ONCO−
Cを合成して、プライマーONCBACK[5'-ATT ATC GCG AGT AAG AAC GTG CTG ACT A
CT TCC GAG TTC TAT-、およびONCFOR[5'-TTA GGA TCC TTA GCA GCT CCC GAC TCC AAC GAA
ATG TAC-3']によってPCR増幅させる。最終の二本鎖PCR産物は、細胞毒性RNA
seのC末端の半分の残りの51アミノ酸をコードするcDNA配列を含んだ。NruI
部位によって、ONCBACKに組み込まれたPCR増幅されたDNAのN末端の半分と
のインフレームの結合が可能になった。ステージングベクターへ、または細菌発現ベクタ
ーへのサブクローニングのために、終止コドン(太字で示される)およびBamHI制限
部位(下線)をONCFOR配列に入れた。
【0109】
適切な制限酵素での処理後に細胞毒性RNAseのN末端およびC末端の半分をコード
する、PCR増幅されたDNAをNruI部位に結合して、ステージングベクター、例え
ばStratageneのpBluescript中にサブクローニングした。この結合された配列は、N末端
のMetを有する105アミノ酸のポリペプチドをコードしなければならない。
【実施例3】
【0110】
(LL2およびMN−14のV−領域配列のクローニング、ならびにLL2およびMN−
14のヒト化)
hLL2およびhMN−14のV領域配列は公開されている。Leung et al.,Mol.Immun
ol.,32:1413(1995);米国特許第5,874,540号。LL2およびMN−14のVKおよびVH
の配列は、公開された方法およびプライマーを用いてPCR増幅させた。
【0111】
PCR増幅されたDNAの配列分析によって、それらが抗体のVKおよびVHドメイン
の代表的なタンパク質をコードすることが示された。PCR−増幅されたLL2およびM
N−14の配列に基づいて構築されたキメラ抗体は、その親抗体に匹敵する免疫反応性を
示しており、これによって得られた配列の信頼性が確認された。
【0112】
LL2抗体の配列分析によって、フレームワーク−1領域におけるVK付加されたN結
合グリコシル化部位の存在が明らかになった。突然変異研究によって、VK付加部位での
グリコシル化は、抗体の免疫反応性を維持するには必要ないことが示された。FR−1グ
リコシル化部位の包含なしに、REIフレームワーク配列を、軽鎖CDRを接合するため
、そしてEU/NEWMをLL2の重鎖CDRを接合するための足場として用いた。ヒト
化LL2(hLL2)の免疫反応性は、マウスおよびキメラのLL2の免疫反応性に匹敵
することが示された。LL2の内部移行の速度は、抗体のキメラ化またはヒト化によって
は影響されなかった。
【実施例4】
【0113】
(ヒト化LL2および細胞毒性RNAseの融合タンパク質をコードする遺伝子の構築)
hLL2のVHおよびVKの配列をテンプレートとして用いて、標準的なPCR手順に
よってhLL2−scFv遺伝子をアセンブルした。遺伝子の構成は、Met(−1)−
VL−(GGGS)4−VH−(His)6であった。VHドメインに対して16アミノ酸
のリンカー(GGGS)6を介して結合されたVL遺伝子のN末端に、Met(ATG)
開始コドンを−1位置で組み込んだ。6つのヒスチジル残基からなるテールを、VH鎖の
カルボキシル末端に入れて、金属キレートクロマトグラフィーを介した融合タンパク質の
精製を容易にする。
【0114】
ランピルナーゼ−hLL2scFvの免疫毒素融合タンパク質遺伝子を、制限消化およ
び結合方法によって同様の方式で構築した。cDNA配列は発現された場合、以下の構築
物:
ランピルナーゼ−[リンカー]−VL−(GGGS)4−VH−(His)6の融合タン
パク質をコードした。
【0115】
細胞毒性RNAseのC末端とVLドメインのN末端との間に挿入され得る種々のリン
カーが存在する。好ましいリンカーは、シュードモナスの外毒素(PE)のC末端位置2
73〜281由来のアミノ酸配列TRHRQPRGWである。この配列は、配列のG残基
とW残基との間に存在する切断でのサブチリシンによる、活性フラグメントへのPEの細
胞内切断の認識部位であることが示されている。Chiron et al.,J.Biol Chem.,269:18167
(1994)。この配列の組み込みによって、融合免疫毒素の内部移行後の活性細胞毒性RNA
seの放出が容易になる。あるいは、細胞毒性RNAseとscFvとの間の可塑性の結
合を可能にするために、EDN−scFvの構築に用いられるStaphylococcalのプロテイ
ンAのフラグメントBのアミノ酸残基48〜60からなる13アミノ酸残基のスペーサー
を代わりに用いてもよい。Tai et al.,Biochemistry,29:8024(1990)、およびRybak et al
.,Tumor Targeting,1:141(1995)。
【実施例5】
【0116】
(ヒト化MN−14およびランピルナーゼの融合タンパク質をコードする遺伝子の構築)
MN−14scFvは、ヒト化MN−14トランスフェクトーマ(transfectoma)由来の
cDNAのPCR増幅によって生成した。MN−14 scFvのために用いられるリン
カーは、15アミノ酸リンカー(GGSGS)3であり、そして方向はVL−リンカー−V
Hであった。DNA配列の確認後、単鎖構築物を、実施例1に記載のような真核生物発現
ベクター中にサブクローニングして、発現のために適切な哺乳動物宿主細胞中にトランス
フェクトする。
【0117】
別の単鎖構築物も作成した。これは、重鎖および軽鎖の反対の5’−3’方向で作成し
、pCANTABE5E(Pharmacia Biotech,Piscataway,N.J.)中でアセンブルして、フ
ァージ中で発現させた。このscFvを発現する組み換えファージの特異的結合はELI
SAによって実証された。
【0118】
VL−リンカー−V配列を、以下に図示されるように、ランピルナーゼ−MN−14融
合タンパク質の構築のために用いた。ランピルナーゼをコードするDNAフラグメントを
実施例1に記載のように得た。23アミノ酸リンカーをランピルナーゼ配列とscFvと
の間で用いた。Kurucz et al.(1995)。あるいは、上記のMN−14 scFvの構築に
用いられる(GGSGS)3リンカーを用いた。融合タンパク質の好ましい構成は以下:
ランピルナーゼ−リンカー−−VL−−(GGSGS)3−−VH
であった。
【0119】
本明細書に引用される全ての特許および他の引用文献の内容は、その全体が引用するこ
とにより本明細書に組み込まれる。
【特許請求の範囲】
【請求項1】
哺乳動物宿主細胞を培養するステップを含む、免疫毒素を調製する方法であって、
前記宿主細胞が、(a)第一の免疫グロブリン可変ドメインに融合された非哺乳動物リ
ボヌクレアーゼを含む融合ポリペプチドをコードする第一の核酸配列と、(b)第二の免
疫グロブリン可変ドメインを含む第二のポリペプチドをコードする第二の核酸配列とで形
質転換されており、前記第一および第二の免疫グロブリンの可変ドメインが一緒になって
抗原結合部位を形成する、方法。
【請求項2】
前記非哺乳動物リボヌクレアーゼが、前記第一の免疫グロブリン可変ドメインのN末端
に融合されている、請求項1に記載の方法。
【請求項3】
前記非哺乳動物リボヌクレアーゼが、N末端ピログルタミン酸残基を保有する、請求項
2に記載の方法。
【請求項4】
前記第一の免疫グロブリン可変ドメインが軽鎖可変ドメインである、請求項3に記載の
方法。
【請求項5】
前記融合ポリペプチドと前記第二の免疫グロブリン可変ドメインとが、前記宿主細胞中
で別の分子として産生される、請求項1〜4のいずれかに記載の方法。
【請求項6】
前記融合ポリペプチドがCLドメインをさらに含み、前記第二のポリペプチドがCH1
ドメインをさらに含む、請求項4に記載の方法。
【請求項7】
前記融合ポリペプチドがCH1ドメインをさらに含み、前記第二のポリペプチドがCL
ドメインをさらに含む、請求項2に記載の方法。
【請求項8】
前記第二のポリペプチドが、CH2ドメインおよびCH3ドメインをさらに含む、請求
項6に記載の方法。
【請求項9】
前記融合ポリペプチドが、CH2ドメインおよびCH3ドメインをさらに含む、請求項
6に記載の方法。
【請求項10】
前記免疫毒素がグリコシル化されている、請求項1に記載の方法。
【請求項11】
前記免疫毒素が、CH2ドメイン上でグリコシル化されている、請求項8または9に記
載の方法。
【請求項12】
前記非哺乳動物のリボヌクレアーゼが、ランピルナーゼまたはその改変体である、請求
項1〜11のいずれかに記載の方法。
【請求項13】
前記細胞毒性RNAseが、ヒョウガエルから精製されたランピルナーゼの配列および
構造を有する、請求項12に記載の方法。
【請求項14】
免疫毒素であって、(a)第一の免疫グロブリン可変ドメインに融合された非哺乳動物
リボヌクレアーゼを含む融合ポリペプチドと、(b)第二の免疫グロブリン可変ドメイン
を含む第二のポリペプチドとを含み、前記免疫グロブリン可変ドメインの1つが軽鎖可変
ドメインであり、もう一方の免疫グロブリン可変ドメインが重鎖可変ドメインであり、前
記第一および第二の免疫グロブリン可変ドメインが一緒になって抗原結合部位を形成し、
前記免疫毒素がグリコシル化されている、免疫毒素。
【請求項15】
前記非哺乳動物リボヌクレアーゼが、前記第一の免疫グロブリン可変ドメインのN末端
に融合されている、請求項14に記載の免疫毒素。
【請求項16】
前記非哺乳動物リボヌクレアーゼが、N末端ピログルタミン酸残基を保有する、請求項
15に記載の免疫毒素。
【請求項17】
前記第一の免疫グロブリン可変ドメインが軽鎖可変ドメインである、請求項16に記載
の免疫毒素。
【請求項18】
前記融合ポリペプチドと前記第二の免疫グロブリン可変ドメインとが、別の分子である
、請求項14〜17のいずれかに記載の免疫毒素。
【請求項19】
前記融合ポリペプチドがCLドメインをさらに含み、前記第二のポリペプチドがCH1
ドメインをさらに含む、請求項17に記載の免疫毒素。
【請求項20】
前記融合ポリペプチドがCH1ドメインをさらに含み、前記第二のポリペプチドがCL
ドメインをさらに含む、請求項17に記載の免疫毒素。
【請求項21】
前記第二のポリペプチドが、CH2ドメインおよびCH3ドメインをさらに含む、請求
項19に記載の免疫毒素。
【請求項22】
前記融合ポリペプチドが、CH2ドメインおよびCH3ドメインをさらに含む、請求項
20に記載の免疫毒素。
【請求項23】
前記免疫毒素が、VLドメイン上でグリコシル化されている、請求項14〜22のいず
れかに記載の免疫毒素。
【請求項24】
前記免疫毒素が、CH2ドメイン上でグリコシル化されている、請求項19〜23に記
載の免疫毒素。
【請求項25】
前記非哺乳動物のリボヌクレアーゼが、ランピルナーゼまたはその改変体である、請求
項14〜25のいずれかに記載の免疫毒素。
【請求項26】
前記細胞毒性RNAseが、ヒョウガエルから精製されたランピルナーゼの配列および
構造を有する、請求項25に記載の免疫毒素。
【請求項27】
前記抗原結合部位が細胞表面分子に対して特異的に結合し、前記免疫毒素が前記細胞表
面分子を保有する細胞に結合した際に内部移行されるという特性を有する、請求項14〜
26のいずれかに記載の免疫毒素。
【請求項28】
細胞毒性RNAse部分に融合された内部移行抗体または抗体フラグメントを含む免疫
毒素であって、前記細胞毒性RNAse部分がN末端ピログルタミン酸残基を保有し、前
記細胞毒性RNAse部分がそのC末端において前記抗体または抗体フラグメントの軽鎖
を含むポリペプチドのN末端に融合されており、前記抗体または抗体フラグメントが別の
軽鎖および重鎖を含む、免疫毒素。
【請求項29】
腫瘍関連抗原、B細胞抗原、T細胞抗原、形質細胞抗原、HLA−DR系統抗原、CE
A,NCA、MUC1、MUC2、MUC3、MUC4およびMUC16抗原、EGP−
1抗原、GEP−2抗原、胎盤アルカリホスファターゼ抗原、IL−6、EGFR、VE
GF、ILGF、P1GF、テネイシン、CD33、PSMA、PSA、PAP、Her
2/neu、炭酸脱水素酵素IX、および自己免疫疾患に関連する抗原からなる群より選
択される抗原に対する、請求項14〜28のいずれかに記載の免疫毒素。
【請求項30】
前記抗原が、B細胞またはT細胞リンパ腫または白血病に関連する標的抗原である、請
求項29に記載の免疫毒素。
【請求項31】
前記抗原が、CD15、CD19、CD21、CD22、CD25、CD40、CD8
0、MUC1、HLA−DR、IL−6、P1GF、VEGF、CD66a、CD66b
、CD66c、およびCD66dからなる群より選択される抗原である、請求項30に記
載の免疫毒素。
【請求項32】
前記抗原がHLA−DRである、請求項31に記載の免疫毒素。
【請求項33】
前記免疫グロブリン可変ドメインが、LL1に由来する、請求項32に記載の免疫毒素
。
【請求項34】
前記抗原がCD22である、請求項31に記載の免疫毒素。
【請求項35】
前記免疫グロブリン可変ドメインが、LL2に由来する、請求項34に記載の免疫毒素
。
【請求項36】
前記抗原がMUC1である、請求項31に記載の免疫毒素。
【請求項37】
前記免疫グロブリン可変ドメインがPAM4由来である、請求項36に記載の免疫毒素
。
【請求項38】
前記抗原がEGP−1である、請求項31に記載の免疫毒素。
【請求項39】
前記免疫グロブリン可変ドメインが、RS7に由来する、請求項38に記載の免疫毒素
。
【請求項40】
前記抗原がEGP−2である、請求項31に記載の免疫毒素。
【請求項41】
前記免疫グロブリン可変ドメインが、RS11または17−1Aに由来する、請求項4
0に記載の免疫毒素。
【請求項42】
前記抗原がCEAである、請求項31に記載の免疫毒素。
【請求項43】
前記免疫グロブリン可変ドメインがMN−14由来である、請求項42に記載の免疫毒
素。
【請求項44】
前記抗原がNCAである、請求項31に記載の免疫毒素。
【請求項45】
前記免疫グロブリン可変ドメインが、MN−3またはMN−15に由来する、請求項4
4に記載の免疫毒素。
【請求項46】
前記可変ドメインが、ヒト化ドメインまたはヒトドメインである、請求項1〜13のい
ずれかに記載の方法。
【請求項47】
前記可変ドメインが、ヒト化ドメインまたはヒトドメインである、請求項14〜46の
いずれかに記載の免疫毒素。
【請求項48】
請求項1〜47のいずれかに記載の免疫毒素、および薬学的に受容可能なキャリアを含
む薬学的組成物。
【請求項49】
被験体における疾患または症候群を処置する方法であって、前記被験体に対して、請求
項48に記載の組成物を、前記疾患または症候群の症状を処置または改善するのに有効な
量で投与するステップを含む、方法。
【請求項50】
前記疾患またな症候群が、前記被験体における細胞の望ましくないまたは不適切な増殖
または活性化によって特徴付けられる、請求項49に記載の方法。
【請求項51】
前記疾患または症候群が、前記被験体におけるB細胞、T細胞、骨髄細胞または形質細
胞の望ましくないまたは不適切な増殖または活性化によって特徴付けられる、請求項50
に記載の方法。
【請求項52】
前記疾患が、癌腫、肉腫、神経膠腫、リンパ腫、白血病、黒色腫、神経芽細胞腫および
骨髄腫からなるガンの群より選択される、請求項50に記載の方法。
【請求項53】
前記疾患または症候群が、固形腫瘍の存在によって特徴付けられる、請求項50に記載
の方法。
【請求項54】
前記固形腫瘍が、神経芽細胞腫、神経膠腫、悪性黒色腫、乳房、結腸直腸、胃、卵巣、
前立腺、膀胱、肝臓、肺、腎臓、および膵臓の腫瘍からなる群より選択される腫瘍である
、請求項53に記載の方法。
【請求項55】
前記薬学的組成物が、前記被験体に対して2回以上投与される、請求項48〜54のい
ずれかに記載の方法。
【請求項56】
1日あたり0.1〜約1000mgの前記薬学的組成物が、前記被験体に投与される、
請求項48〜55のいずれかに記載の方法。
【請求項57】
前記組成物が非経口的に投与される、請求項56に記載の方法。
【請求項58】
前記組成物が、静脈内に、動脈内に、皮内にまたは皮下に投与される、請求項56に記
載の方法。
【請求項59】
前記細胞が、1つ以上の自己免疫疾患の発生および進行に関与する、請求項50に記載
の方法。
【請求項60】
前記自己免疫疾患が、関節リウマチ、全身性エリテマトーデス、シェーグレン症候群、
急性特発性血小板減少性紫斑病、慢性特発性血小板減少性紫斑病、皮膚筋炎、シデナム舞
踏病、重症筋無力症、全身性エリテマトーデス、ループス腎炎、リウマチ熱、多腺性症候
群、水疱性類天疱瘡、糖尿病、ヘノッホ・シェーンライン紫斑病、連鎖球菌感染後腎炎、
結節性紅斑、高安動脈炎、アジソン病、多発性硬化症、サルコイドーシス、潰瘍性大腸炎
、多形性紅斑、IgA腎症、結節性多発性動脈炎、強直性脊椎炎、グッドパスチャー症候
群、血栓血管炎(thromboangitis ubiterans)、原発性胆汁性肝硬変、橋本甲状腺炎、甲状
腺中毒症、強皮症、慢性活動性肝炎、多発性筋炎/皮膚筋炎、多発性軟骨炎、尋常性天疱
瘡、ヴェーゲナー肉芽腫症、膜性腎症、筋萎縮性側索硬化症、脊髄癆、巨細胞性動脈炎/
多発性筋痛、悪性貧血、急速進行性糸球体腎炎および線維化性肺胞炎からなる群より選択
される、請求項59に記載の方法。
【請求項61】
感染性の生物によって引き起こされる疾患を処置する方法であって、前記疾患に罹患し
ている被験体に対して、請求項14に記載の免疫毒素の有効量を投与するステップを含み
、前記免疫毒素の免疫グロブリン成分が、疾患を引き起こす微生物または寄生生物に特異
的に結合する、方法。
【請求項62】
前記疾患を引き起こす微生物が、
ストレプトコッカス・アガラクティエ、在郷軍人病菌、化膿連鎖球菌、大腸菌、淋菌、
髄膜炎菌、肺炎球菌、Hemophilis influenzae B、梅毒トレポネーマ、ライム病スピロヘ
ータ、緑膿菌、ハンセン菌、ウシ流産菌、ヒト結核菌、破傷風毒素、抗ウイルスモノクロ
ーナル抗体、HIV−1、HIV−2、HIV−3、A型肝炎、B型肝炎、C型肝炎、D
型肝炎、パピローマウイルス、狂犬病ウイルス、インフルエンザウイルス、サイトメガロ
ウイルス、単純疱疹IおよびII、ヒト血清パルボ様ウイルス、ポリオーマウイルス、パ
ピローマウイルス、呼吸器合胞体ウイルス、水痘帯状疱疹ウイルス、B型肝炎ウイルス、
麻疹ウイルス、アデノウイルス、ヒトT細胞白血病ウイルス、エプスタイン・バーウイル
ス、マウス白血病ウイルス、流行性耳下腺炎ウイルス、水疱性口内炎ウイルス、シンドビ
スウイルス、リンパ球性脈絡髄膜炎ウイルス、疣ウイルス、ブルー・タングウイルス、セ
ンダイウイルス、猫白血病ウイルス、レオウイルス、ポリオウイルス、シミアン・ウイル
ス40、マウス乳癌ウイルス、デング熱ウイルス、風疹ウイルス、原生動物、熱帯熱マラ
リア原虫、三日熱マラリア原虫、トキソプラズマ原虫、Trypanosoma rangeli、クルーズ
・トリパノソーマ、Trypanosoma rhodesiensei、トリパノソーマ・ブルーセイ、マンソン
住血吸虫、Schistosoma japanicum、Babesia bovis、Elmeria tenella、回旋糸状虫、熱
帯リーシュマニア、旋毛虫、Theileria parva、Taenia hydatigena、Taenia ovis、無鉤
条虫、単包条虫、Mesocestoides corti、抗マイコプラズマMAbs、Mycoplasma arthri
tidis、M.hyorhinis、M.orale、M.arginini、Acholeplasma laidlawii、M.salivarium、
およびM肺炎からなる群より選択される、請求項61に記載の方法。
【請求項63】
前記第一および第二の核酸配列が、pdHL2ベクター内に含まれる、請求項1〜13
のいずれかに記載の方法。
【請求項64】
前記第一および第二の免疫グロブリンドメインが、半分子の形成を阻害する変異を含む
IgG4ドメインである、請求項8または9に記載の方法。
【請求項65】
前記第一の免疫グロブリン可変ドメインが重鎖可変ドメインである、請求項3に記載の
方法。
【請求項66】
前記第一の免疫グロブリン可変ドメインが軽鎖可変ドメインであり、前記第二の免疫グ
ロブリン可変ドメインが重鎖可変ドメインであり、前記免疫グロブリン重鎖可変ドメイン
が、前記第二の免疫グロブリン重鎖可変ドメインのN末端に融合された非哺乳動物リボヌ
クレアーゼを含む、請求項3に記載の方法。
【請求項1】
哺乳動物宿主細胞を培養するステップを含む、免疫毒素を調製する方法であって、
前記宿主細胞が、(a)第一の免疫グロブリン可変ドメインに融合された非哺乳動物リ
ボヌクレアーゼを含む融合ポリペプチドをコードする第一の核酸配列と、(b)第二の免
疫グロブリン可変ドメインを含む第二のポリペプチドをコードする第二の核酸配列とで形
質転換されており、前記第一および第二の免疫グロブリンの可変ドメインが一緒になって
抗原結合部位を形成する、方法。
【請求項2】
前記非哺乳動物リボヌクレアーゼが、前記第一の免疫グロブリン可変ドメインのN末端
に融合されている、請求項1に記載の方法。
【請求項3】
前記非哺乳動物リボヌクレアーゼが、N末端ピログルタミン酸残基を保有する、請求項
2に記載の方法。
【請求項4】
前記第一の免疫グロブリン可変ドメインが軽鎖可変ドメインである、請求項3に記載の
方法。
【請求項5】
前記融合ポリペプチドと前記第二の免疫グロブリン可変ドメインとが、前記宿主細胞中
で別の分子として産生される、請求項1〜4のいずれかに記載の方法。
【請求項6】
前記融合ポリペプチドがCLドメインをさらに含み、前記第二のポリペプチドがCH1
ドメインをさらに含む、請求項4に記載の方法。
【請求項7】
前記融合ポリペプチドがCH1ドメインをさらに含み、前記第二のポリペプチドがCL
ドメインをさらに含む、請求項2に記載の方法。
【請求項8】
前記第二のポリペプチドが、CH2ドメインおよびCH3ドメインをさらに含む、請求
項6に記載の方法。
【請求項9】
前記融合ポリペプチドが、CH2ドメインおよびCH3ドメインをさらに含む、請求項
6に記載の方法。
【請求項10】
前記免疫毒素がグリコシル化されている、請求項1に記載の方法。
【請求項11】
前記免疫毒素が、CH2ドメイン上でグリコシル化されている、請求項8または9に記
載の方法。
【請求項12】
前記非哺乳動物のリボヌクレアーゼが、ランピルナーゼまたはその改変体である、請求
項1〜11のいずれかに記載の方法。
【請求項13】
前記細胞毒性RNAseが、ヒョウガエルから精製されたランピルナーゼの配列および
構造を有する、請求項12に記載の方法。
【請求項14】
免疫毒素であって、(a)第一の免疫グロブリン可変ドメインに融合された非哺乳動物
リボヌクレアーゼを含む融合ポリペプチドと、(b)第二の免疫グロブリン可変ドメイン
を含む第二のポリペプチドとを含み、前記免疫グロブリン可変ドメインの1つが軽鎖可変
ドメインであり、もう一方の免疫グロブリン可変ドメインが重鎖可変ドメインであり、前
記第一および第二の免疫グロブリン可変ドメインが一緒になって抗原結合部位を形成し、
前記免疫毒素がグリコシル化されている、免疫毒素。
【請求項15】
前記非哺乳動物リボヌクレアーゼが、前記第一の免疫グロブリン可変ドメインのN末端
に融合されている、請求項14に記載の免疫毒素。
【請求項16】
前記非哺乳動物リボヌクレアーゼが、N末端ピログルタミン酸残基を保有する、請求項
15に記載の免疫毒素。
【請求項17】
前記第一の免疫グロブリン可変ドメインが軽鎖可変ドメインである、請求項16に記載
の免疫毒素。
【請求項18】
前記融合ポリペプチドと前記第二の免疫グロブリン可変ドメインとが、別の分子である
、請求項14〜17のいずれかに記載の免疫毒素。
【請求項19】
前記融合ポリペプチドがCLドメインをさらに含み、前記第二のポリペプチドがCH1
ドメインをさらに含む、請求項17に記載の免疫毒素。
【請求項20】
前記融合ポリペプチドがCH1ドメインをさらに含み、前記第二のポリペプチドがCL
ドメインをさらに含む、請求項17に記載の免疫毒素。
【請求項21】
前記第二のポリペプチドが、CH2ドメインおよびCH3ドメインをさらに含む、請求
項19に記載の免疫毒素。
【請求項22】
前記融合ポリペプチドが、CH2ドメインおよびCH3ドメインをさらに含む、請求項
20に記載の免疫毒素。
【請求項23】
前記免疫毒素が、VLドメイン上でグリコシル化されている、請求項14〜22のいず
れかに記載の免疫毒素。
【請求項24】
前記免疫毒素が、CH2ドメイン上でグリコシル化されている、請求項19〜23に記
載の免疫毒素。
【請求項25】
前記非哺乳動物のリボヌクレアーゼが、ランピルナーゼまたはその改変体である、請求
項14〜25のいずれかに記載の免疫毒素。
【請求項26】
前記細胞毒性RNAseが、ヒョウガエルから精製されたランピルナーゼの配列および
構造を有する、請求項25に記載の免疫毒素。
【請求項27】
前記抗原結合部位が細胞表面分子に対して特異的に結合し、前記免疫毒素が前記細胞表
面分子を保有する細胞に結合した際に内部移行されるという特性を有する、請求項14〜
26のいずれかに記載の免疫毒素。
【請求項28】
細胞毒性RNAse部分に融合された内部移行抗体または抗体フラグメントを含む免疫
毒素であって、前記細胞毒性RNAse部分がN末端ピログルタミン酸残基を保有し、前
記細胞毒性RNAse部分がそのC末端において前記抗体または抗体フラグメントの軽鎖
を含むポリペプチドのN末端に融合されており、前記抗体または抗体フラグメントが別の
軽鎖および重鎖を含む、免疫毒素。
【請求項29】
腫瘍関連抗原、B細胞抗原、T細胞抗原、形質細胞抗原、HLA−DR系統抗原、CE
A,NCA、MUC1、MUC2、MUC3、MUC4およびMUC16抗原、EGP−
1抗原、GEP−2抗原、胎盤アルカリホスファターゼ抗原、IL−6、EGFR、VE
GF、ILGF、P1GF、テネイシン、CD33、PSMA、PSA、PAP、Her
2/neu、炭酸脱水素酵素IX、および自己免疫疾患に関連する抗原からなる群より選
択される抗原に対する、請求項14〜28のいずれかに記載の免疫毒素。
【請求項30】
前記抗原が、B細胞またはT細胞リンパ腫または白血病に関連する標的抗原である、請
求項29に記載の免疫毒素。
【請求項31】
前記抗原が、CD15、CD19、CD21、CD22、CD25、CD40、CD8
0、MUC1、HLA−DR、IL−6、P1GF、VEGF、CD66a、CD66b
、CD66c、およびCD66dからなる群より選択される抗原である、請求項30に記
載の免疫毒素。
【請求項32】
前記抗原がHLA−DRである、請求項31に記載の免疫毒素。
【請求項33】
前記免疫グロブリン可変ドメインが、LL1に由来する、請求項32に記載の免疫毒素
。
【請求項34】
前記抗原がCD22である、請求項31に記載の免疫毒素。
【請求項35】
前記免疫グロブリン可変ドメインが、LL2に由来する、請求項34に記載の免疫毒素
。
【請求項36】
前記抗原がMUC1である、請求項31に記載の免疫毒素。
【請求項37】
前記免疫グロブリン可変ドメインがPAM4由来である、請求項36に記載の免疫毒素
。
【請求項38】
前記抗原がEGP−1である、請求項31に記載の免疫毒素。
【請求項39】
前記免疫グロブリン可変ドメインが、RS7に由来する、請求項38に記載の免疫毒素
。
【請求項40】
前記抗原がEGP−2である、請求項31に記載の免疫毒素。
【請求項41】
前記免疫グロブリン可変ドメインが、RS11または17−1Aに由来する、請求項4
0に記載の免疫毒素。
【請求項42】
前記抗原がCEAである、請求項31に記載の免疫毒素。
【請求項43】
前記免疫グロブリン可変ドメインがMN−14由来である、請求項42に記載の免疫毒
素。
【請求項44】
前記抗原がNCAである、請求項31に記載の免疫毒素。
【請求項45】
前記免疫グロブリン可変ドメインが、MN−3またはMN−15に由来する、請求項4
4に記載の免疫毒素。
【請求項46】
前記可変ドメインが、ヒト化ドメインまたはヒトドメインである、請求項1〜13のい
ずれかに記載の方法。
【請求項47】
前記可変ドメインが、ヒト化ドメインまたはヒトドメインである、請求項14〜46の
いずれかに記載の免疫毒素。
【請求項48】
請求項1〜47のいずれかに記載の免疫毒素、および薬学的に受容可能なキャリアを含
む薬学的組成物。
【請求項49】
被験体における疾患または症候群を処置する方法であって、前記被験体に対して、請求
項48に記載の組成物を、前記疾患または症候群の症状を処置または改善するのに有効な
量で投与するステップを含む、方法。
【請求項50】
前記疾患またな症候群が、前記被験体における細胞の望ましくないまたは不適切な増殖
または活性化によって特徴付けられる、請求項49に記載の方法。
【請求項51】
前記疾患または症候群が、前記被験体におけるB細胞、T細胞、骨髄細胞または形質細
胞の望ましくないまたは不適切な増殖または活性化によって特徴付けられる、請求項50
に記載の方法。
【請求項52】
前記疾患が、癌腫、肉腫、神経膠腫、リンパ腫、白血病、黒色腫、神経芽細胞腫および
骨髄腫からなるガンの群より選択される、請求項50に記載の方法。
【請求項53】
前記疾患または症候群が、固形腫瘍の存在によって特徴付けられる、請求項50に記載
の方法。
【請求項54】
前記固形腫瘍が、神経芽細胞腫、神経膠腫、悪性黒色腫、乳房、結腸直腸、胃、卵巣、
前立腺、膀胱、肝臓、肺、腎臓、および膵臓の腫瘍からなる群より選択される腫瘍である
、請求項53に記載の方法。
【請求項55】
前記薬学的組成物が、前記被験体に対して2回以上投与される、請求項48〜54のい
ずれかに記載の方法。
【請求項56】
1日あたり0.1〜約1000mgの前記薬学的組成物が、前記被験体に投与される、
請求項48〜55のいずれかに記載の方法。
【請求項57】
前記組成物が非経口的に投与される、請求項56に記載の方法。
【請求項58】
前記組成物が、静脈内に、動脈内に、皮内にまたは皮下に投与される、請求項56に記
載の方法。
【請求項59】
前記細胞が、1つ以上の自己免疫疾患の発生および進行に関与する、請求項50に記載
の方法。
【請求項60】
前記自己免疫疾患が、関節リウマチ、全身性エリテマトーデス、シェーグレン症候群、
急性特発性血小板減少性紫斑病、慢性特発性血小板減少性紫斑病、皮膚筋炎、シデナム舞
踏病、重症筋無力症、全身性エリテマトーデス、ループス腎炎、リウマチ熱、多腺性症候
群、水疱性類天疱瘡、糖尿病、ヘノッホ・シェーンライン紫斑病、連鎖球菌感染後腎炎、
結節性紅斑、高安動脈炎、アジソン病、多発性硬化症、サルコイドーシス、潰瘍性大腸炎
、多形性紅斑、IgA腎症、結節性多発性動脈炎、強直性脊椎炎、グッドパスチャー症候
群、血栓血管炎(thromboangitis ubiterans)、原発性胆汁性肝硬変、橋本甲状腺炎、甲状
腺中毒症、強皮症、慢性活動性肝炎、多発性筋炎/皮膚筋炎、多発性軟骨炎、尋常性天疱
瘡、ヴェーゲナー肉芽腫症、膜性腎症、筋萎縮性側索硬化症、脊髄癆、巨細胞性動脈炎/
多発性筋痛、悪性貧血、急速進行性糸球体腎炎および線維化性肺胞炎からなる群より選択
される、請求項59に記載の方法。
【請求項61】
感染性の生物によって引き起こされる疾患を処置する方法であって、前記疾患に罹患し
ている被験体に対して、請求項14に記載の免疫毒素の有効量を投与するステップを含み
、前記免疫毒素の免疫グロブリン成分が、疾患を引き起こす微生物または寄生生物に特異
的に結合する、方法。
【請求項62】
前記疾患を引き起こす微生物が、
ストレプトコッカス・アガラクティエ、在郷軍人病菌、化膿連鎖球菌、大腸菌、淋菌、
髄膜炎菌、肺炎球菌、Hemophilis influenzae B、梅毒トレポネーマ、ライム病スピロヘ
ータ、緑膿菌、ハンセン菌、ウシ流産菌、ヒト結核菌、破傷風毒素、抗ウイルスモノクロ
ーナル抗体、HIV−1、HIV−2、HIV−3、A型肝炎、B型肝炎、C型肝炎、D
型肝炎、パピローマウイルス、狂犬病ウイルス、インフルエンザウイルス、サイトメガロ
ウイルス、単純疱疹IおよびII、ヒト血清パルボ様ウイルス、ポリオーマウイルス、パ
ピローマウイルス、呼吸器合胞体ウイルス、水痘帯状疱疹ウイルス、B型肝炎ウイルス、
麻疹ウイルス、アデノウイルス、ヒトT細胞白血病ウイルス、エプスタイン・バーウイル
ス、マウス白血病ウイルス、流行性耳下腺炎ウイルス、水疱性口内炎ウイルス、シンドビ
スウイルス、リンパ球性脈絡髄膜炎ウイルス、疣ウイルス、ブルー・タングウイルス、セ
ンダイウイルス、猫白血病ウイルス、レオウイルス、ポリオウイルス、シミアン・ウイル
ス40、マウス乳癌ウイルス、デング熱ウイルス、風疹ウイルス、原生動物、熱帯熱マラ
リア原虫、三日熱マラリア原虫、トキソプラズマ原虫、Trypanosoma rangeli、クルーズ
・トリパノソーマ、Trypanosoma rhodesiensei、トリパノソーマ・ブルーセイ、マンソン
住血吸虫、Schistosoma japanicum、Babesia bovis、Elmeria tenella、回旋糸状虫、熱
帯リーシュマニア、旋毛虫、Theileria parva、Taenia hydatigena、Taenia ovis、無鉤
条虫、単包条虫、Mesocestoides corti、抗マイコプラズマMAbs、Mycoplasma arthri
tidis、M.hyorhinis、M.orale、M.arginini、Acholeplasma laidlawii、M.salivarium、
およびM肺炎からなる群より選択される、請求項61に記載の方法。
【請求項63】
前記第一および第二の核酸配列が、pdHL2ベクター内に含まれる、請求項1〜13
のいずれかに記載の方法。
【請求項64】
前記第一および第二の免疫グロブリンドメインが、半分子の形成を阻害する変異を含む
IgG4ドメインである、請求項8または9に記載の方法。
【請求項65】
前記第一の免疫グロブリン可変ドメインが重鎖可変ドメインである、請求項3に記載の
方法。
【請求項66】
前記第一の免疫グロブリン可変ドメインが軽鎖可変ドメインであり、前記第二の免疫グ
ロブリン可変ドメインが重鎖可変ドメインであり、前記免疫グロブリン重鎖可変ドメイン
が、前記第二の免疫グロブリン重鎖可変ドメインのN末端に融合された非哺乳動物リボヌ
クレアーゼを含む、請求項3に記載の方法。
【図1】

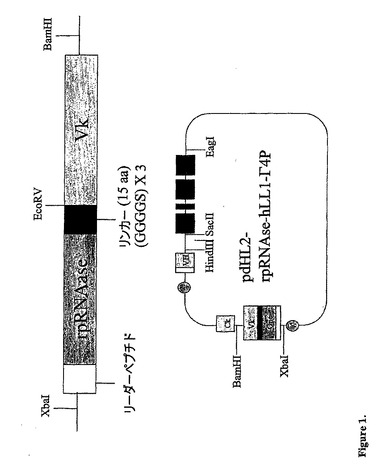
【図2】
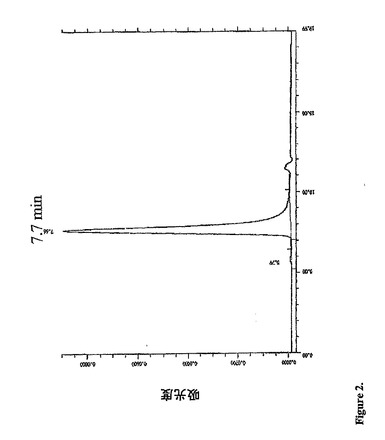
【図3】
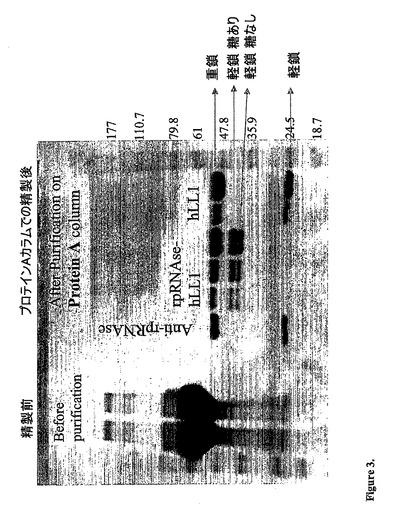
【図4a】
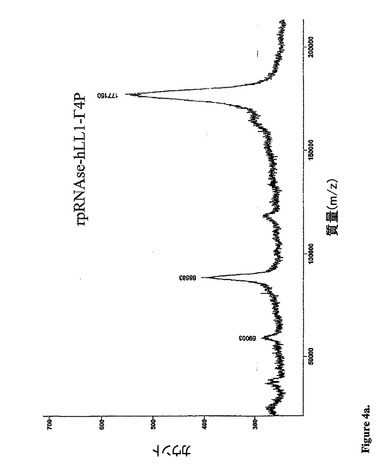
【図4b】
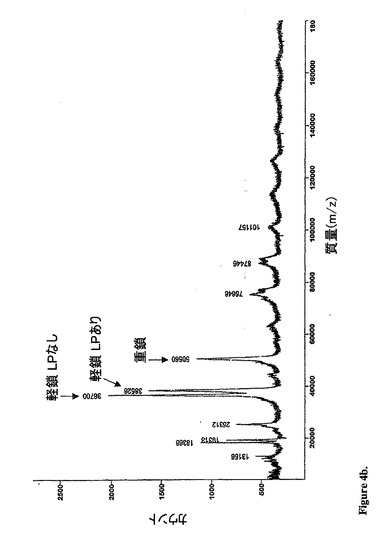
【図5】
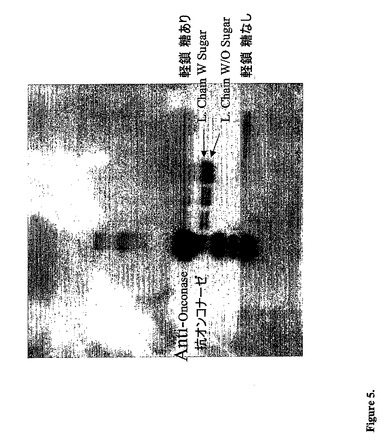
【図6】
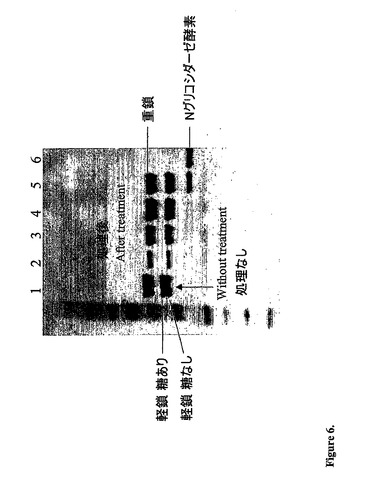
【図7】
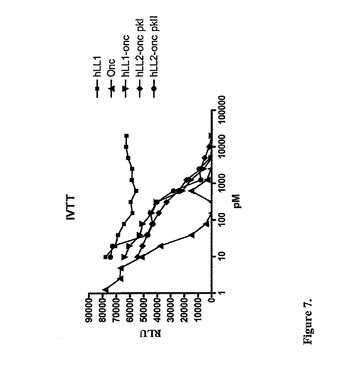
【図8】
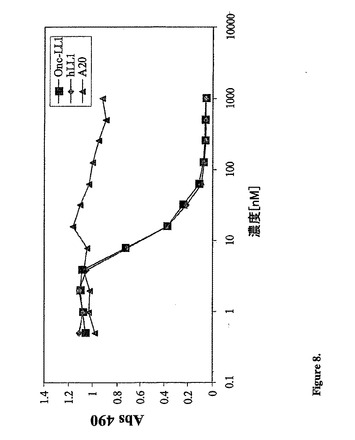
【図9】
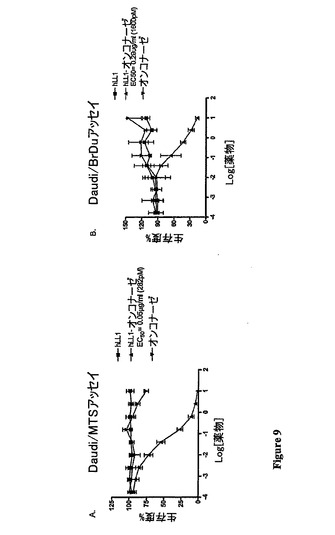
【図10】
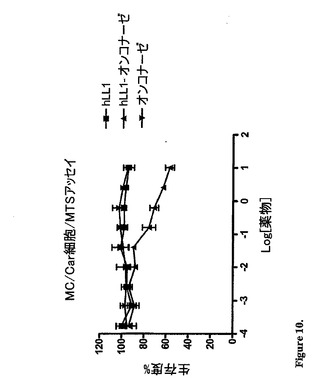
【図11】
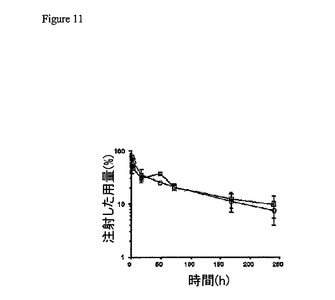
【図12】
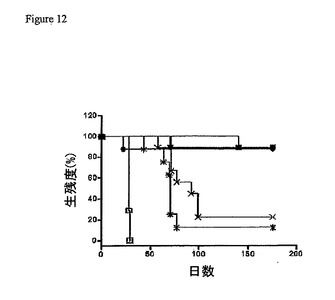
【図13】

【図2】
【図3】
【図4a】
【図4b】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【公開番号】特開2012−197286(P2012−197286A)
【公開日】平成24年10月18日(2012.10.18)
【国際特許分類】
【出願番号】特願2012−116401(P2012−116401)
【出願日】平成24年5月22日(2012.5.22)
【分割の表示】特願2006−553352(P2006−553352)の分割
【原出願日】平成17年2月14日(2005.2.14)
【出願人】(504149971)イミューノメディクス、インコーポレイテッド (48)
【氏名又は名称原語表記】IMMUNOMEDICS, INC.
【Fターム(参考)】
【公開日】平成24年10月18日(2012.10.18)
【国際特許分類】
【出願日】平成24年5月22日(2012.5.22)
【分割の表示】特願2006−553352(P2006−553352)の分割
【原出願日】平成17年2月14日(2005.2.14)
【出願人】(504149971)イミューノメディクス、インコーポレイテッド (48)
【氏名又は名称原語表記】IMMUNOMEDICS, INC.
【Fターム(参考)】
[ Back to top ]