
組み換え酵母株を用いたグルコース、ガラクトース、およびアラビノースからのエタノールの発酵生産
本発明は、
a)糖組成物をサッカロミセス(Saccharomyces)、クリヴェロミセス(Kluyveromyces)、カンジダ(Candida)、ピチア(Pichia)、分裂酵母(SchizoSaccharomyces)、ハンゼヌラ(Hansenula)、クロエケラ(Kloeckera)、シュワニオミセス(Schwanniomyces)またはヤロウィア(Yarrowia)属に属する酵母の存在下で発酵させるステップと、b)発酵産物を回収するステップを含んでなり、酵母が遺伝子araA、araB、およびaraDを含んでなり、糖組成物がグルコース、ガラクトース、およびアラビノースを含んでなる、糖組成物から1つ以上の発酵産物を生成する方法に関する。

a)糖組成物をサッカロミセス(Saccharomyces)、クリヴェロミセス(Kluyveromyces)、カンジダ(Candida)、ピチア(Pichia)、分裂酵母(SchizoSaccharomyces)、ハンゼヌラ(Hansenula)、クロエケラ(Kloeckera)、シュワニオミセス(Schwanniomyces)またはヤロウィア(Yarrowia)属に属する酵母の存在下で発酵させるステップと、b)発酵産物を回収するステップを含んでなり、酵母が遺伝子araA、araB、およびaraDを含んでなり、糖組成物がグルコース、ガラクトース、およびアラビノースを含んでなる、糖組成物から1つ以上の発酵産物を生成する方法に関する。
【発明の詳細な説明】
【発明の詳細な説明】
【0001】
[技術分野]
本発明は、混合糖の発酵、特にグルコース、ガラクトース、およびアラビノースを含んでなる糖組成物の発酵に関する。糖組成は、リグノセルロース系材料が起源であってもよい。
【0002】
[背景技術]
化石燃料代替物として生産されるエタノールのほとんどは、目下、コーンスターチおよびサトウキビベースのスクロースの発酵に由来する。再生可能燃料を生産するという意欲的目標に到達するために、非食物バイオマスをエタノールなどの発酵産物に転換する新たな技術が開発されている。サッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)は、エタノール産業において一般に好まれる生物であるが、それはバイオマス原材料のヘミセルロース構成要素に含有される五炭素糖を利用し得ない。ヘミセルロースは、バイオマスの20〜30%を構成し得て、キシロースおよびアラビノースが最も豊富なC5糖である。キシロースイソメラーゼ(XI)の異種性発現は、酵母細胞がキシロースを代謝して発酵できるようにするための一つの選択肢である。同様にS.セレヴィシエ(S.cerevisiae)株中における細菌遺伝子araA、araB、およびaraDの発現は、アラビノースの利用と効率的なアルコール発酵をもたらす。ガラクトースはC6糖であり、それは頻繁に経済的理由のために無視できない量(全糖の約4%)でリグノセルロース中に存在することが多い糖でもある。
【0003】
J. van den Brink et al,Microbiology(2009)155,1340−1350は、グルコースがサッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)の好む炭素源であり、嫌気的条件下において、グルコース限定的発酵条件からガラクトース過剰条件への切り替えに際して、ガラクトースが消費されなかったことを開示する。
【0004】
今までのところ、グルコースおよび1つ以上のC5糖を用いるのと同じ方法で、ガラクトースを発酵産物に転換する方法は開示されていない。したがって本発明の目的は、グルコースおよび1つ以上のC5糖を用いるのと同じ方法で、ガラクトースを発酵産物に転換する方法を提供することである。
【0005】
[発明の概要]
本発明は、
a)サッカロミセス(Saccharomyces)、クリヴェロミセス(Kluyveromyces)、カンジダ(Candida)、ピチア(Pichia)、分裂酵母(Schizosaccharomyces)、ハンゼヌラ(Hansenula)、クロエケラ(Kloeckera)、シュワニオミセス(Schwanniomyces)またはヤロウィア(Yarrowia)属に属する酵母の存在下で糖組成物を発酵させるステップと、
b)発酵産物を回収するステップ
を含んでなる、糖組成物から1つ以上の発酵産物を生産する方法を提供し、酵母は遺伝子araA、araB、およびaraDを含んでなり、糖組成物はグルコース、ガラクトース、およびアラビノースを含んでなる。
【0006】
有利には糖グルコース、ガラクトース、およびアラビノースは、発酵産物に変換される。
【0007】
好ましくは混合糖細胞は、サッカロミセス(Saccharomyces)より好ましくはサッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)属に属する。
【0008】
本発明は、グルコース発酵株上における、これらの遺伝子の発現を通じて、アラビノース存在下でガラクトースを嫌気的に発酵する能力をもたらす、遺伝子araA、araB、およびaraDの使用にさらに関する。
【図面の簡単な説明】
【0009】
【図1】プラスミドpPWT006の物理的地図を示す。
【図2】プラスミドpPWT018の物理的地図を示す。
【図3】サザンブロット法オートラジオグラムを示す。野生型株CEN.PK113−7D(レーン1)およびBIE104A2(レーン2)の染色体DNAをEcoRIおよびHindIIIの双方で消化する。ブロットを特異的SIT2プローブとハイブリダイズする。
【図4】野生型SIT2遺伝子座の物理的地図(パネルa)、およびプラスミドpPWT018組み込みによるara遺伝子導入と、それに続く、ベクター損失と選択可能なマーカー配列をもたらす分子内遺伝子組み換え後の物理的地図(パネルb)を示す。プローブのハイブリダイゼーションが表示される。
【図5】配列が配列番号4に記載される、プラスミドpPWT080の物理的地図を示す。
【図6−1】野生型GRE3遺伝子座の物理的地図(パネルa)、およびGRE3遺伝子座への1コピーのPWT080の組み込みの物理的地図を示す(パネルbはプライマー結合部位を示し、パネルcはRKI1プローブ結合部位を示す)。
【図6−2】野生型GRE3遺伝子座の物理的地図(パネルa)、およびGRE3遺伝子座への1コピーのPWT080の組み込みの物理的地図を示す(パネルbはプライマー結合部位を示し、パネルcはRKI1プローブ結合部位を示す)。
【図7】GRE3遺伝子のコード領域がPPP遺伝子TAL1、TKL1、RKI1、およびRPE1の組み込みによって置換されたGRE3遺伝子座の物理的地図を示す。パネルaは配列番号5および6のプライマー結合部位を示し、パネルbはRKI1プローブ結合部位を示す。
【図8】異なる培地上でのBIE104P1A2の好気的条件下の成長曲線を示す。BIE104A2P1株は、YNB2%ガラクトース上で前培養された。成長曲線は2%ガラクトースおよび1%アラビノースで開始され、グラフで番号(1)で示される事象である2%アラビノースを唯一の炭素源とするYNBへの移動がそれに続いた。OD600が1を超えた後、培養物を0.2の開始OD600で新鮮な培地に移した。純粋アラビノース培地上での3回の移動後、得られた株をBIE104P1A2cと命名した。
【図9】嫌気的条件下における、2%アラビノースを唯一の炭素源とするYNB上でのBIE104P1A2cの成長曲線を示す。OD600が1を超えた後、培養物を0.2の開始OD600で新鮮な培地に移した。数回の移動後、得られた株をBIE104P1A2d(=BIE201)と命名した。
【図10】合成トウモロコシ繊維モデル培地上でのBIE104の糖転換および生成物形成を示す。CO2生成を絶えず測定した。成長は培養物の光学濃度を追跡してモニターした。前培養は2%グルコース上で成長させた。
【図11】合成トウモロコシ繊維モデル培地上でのBIE104P1A2cの糖転換および生成物形成を示す。CO2生成を絶えず測定した。成長は培養物の光学濃度を追跡してモニターした。前培養は2%グルコース上で成長させた。
【図12】合成トウモロコシ繊維モデル培地上でのBIE201の糖転換および生成物形成を示す。CO2生成を絶えず測定した。成長は培養物の光学濃度を追跡してモニターした。前培養は2%グルコース上で成長させた。
【図13】合成トウモロコシ繊維モデル培地上でのBIE104A2の糖転換および生成物形成を示す。CO2生成を絶えず測定した。成長は培養物の光学濃度を追跡してモニターした。前培養は2%グルコース上で成長させた。
【図14】合成トウモロコシ繊維モデル培地上でのBIE105A2の糖転換および生成物形成を示す。CO2生成を絶えず測定した。成長は培養物の光学濃度を追跡してモニターした。前培養は2%グルコース上で成長させた。
【図15】プラスミドpPWT007の物理的地図を示す。
【図16】プラスミドpPWT042の物理的地図を示す。
【図17】野生型SIT4遺伝子座の物理的地図を示す(パネルa)、およびSIT4遺伝子座への1コピーのPWT080組み込み(パネルbはプライマー結合部位を示す)。
【図18−1】異なる培地上でのBIE104A2P1Y9株の成長曲線グラフである。パネルa:BIE104A2P1Y9株をグルコース上で成長させて、グラフで番号(1)で示される事象である1%アラビノース+1%キシロースへの移動、(2)2%キシロース+0.2%アラビノースへの移動がそれに続いた。パネルb:BIE104A2P1Y9株をガラクトース上で成長させて、(1)1%アラビノース+1%キシロースへの移動、および(2)2%キシロース+0.2%アラビノースへの移動がそれに続いた。
【図18−2】異なる培地上でのBIE104A2P1Y9株の成長曲線グラフである。パネルa:BIE104A2P1Y9株をグルコース上で成長させて、グラフで番号(1)で示される事象である1%アラビノース+1%キシロースへの移動、(2)2%キシロース+0.2%アラビノースへの移動がそれに続いた。パネルb:BIE104A2P1Y9株をガラクトース上で成長させて、(1)1%アラビノース+1%キシロースへの移動、および(2)2%キシロース+0.2%アラビノースへの移動がそれに続いた。
【図19】2%キシロースを添加したVerduyn培地上でのBIE104A2P1Y9株の成長を示す。2つの独立したコロニーを試験した。OD600が2に達した後、株を新鮮な培地に移して、即座にキシロース上での生育を再開した。
【図20】プラスミドpGBS416ARAABDの物理的地図を示す。
【0010】
[配列表の簡単な説明]
配列番号1は、バクテロイデス・ユニフォルミス(Bacteroides uniformis)ATCC8492からの野生型キシロースイソメラーゼ配列を示す。Genbank登録番号AAYH02000036。
配列番号2は、配列番号1に由来するコドン最適化された配列を示す。
配列番号3は、バクテロイデス・ユニフォルミス(Bacteroides uniformis)ATCC8492からのキシロースイソメラーゼのアミノ酸配列を示す。
配列番号4は、プラスミドpPWT080の配列を示す。
配列番号5は、順方向プライマーの配列を示す。
配列番号6は、逆方向プライマーの配列を示す。
配列番号7は、診断PCRのための順方向多官能性プライマーの配列を示す。
配列番号8は、診断PCRのための逆方向多官能性プライマーの配列を示す。
配列番号9は、順方向プライマーRKI1プローブの配列を示す。
配列番号10は、逆方向プライマーRKI1プローブの配列を示す。
配列番号11は、順方向プライマーkanMX−カセットの配列を示す。
配列番号12は、逆方向プライマーkanMX−カセットの配列を示す。
配列番号13は、順方向プライマーの配列を示す。
配列番号14は、逆方向プライマーの配列を示す。
配列番号15は、診断PCRのための順方向多官能性プライマーの配列を示す。
配列番号16は、診断PCRのための逆方向多官能性プライマーの配列を示す。
配列番号17は、プラスミドpPWT018の配列を示す。
配列番号18は、順方向プライマー組み込みpPWT018の配列を示す。
配列番号19は、逆方向プライマー組み込みpPWT018の配列を示す。
配列番号20は、順方向プライマーSIT2プローブの配列を示す。
配列番号21は、逆方向プライマーSIT2プローブの配列を示す。
配列番号22は、araABD発現カセットを増幅する順方向プライマーの配列を示す。
配列番号23は、araABD発現カセットを増幅する逆方向プライマーの配列を示す。
【0011】
[発明の詳細な説明]
本明細書および添付の特許請求の範囲全体を通じて、語句「含んでなる(comprise)」および「含む(include)」、および「含んでなる(comprises)」、「含んでなる(comprising)」、「含む(includes)」、および「含む(including)」などのバリエーションは、包括的に解釈されるものとする。すなわちこれらの語句は、文脈が許せば、具体的に列挙されないその他の要素または整数の可能な包含を伝えることが意図される。
【0012】
冠詞「a」および「an」は、本明細書では、1つまたは1つを超える(すなわち1つのまたは少なくとも1つの)冠詞の文法的目的語に言及するために使用される。一例として、「要素(an element)」は、1つの要素または1つを超える要素を意味してもよい。
【0013】
本明細書に記載される本発明の様々な実施態様は、交差して組み合わせされてもよい。
【0014】
[糖組成物]
本発明に従った糖組成物は、グルコース、アラビノース、およびガラクトースを含んでなる。本発明の方法では、有利には、糖グルコース、ガラクトース、およびアラビノースは、発酵産物に変換される。
【0015】
本発明では、これらの基準を満たすあらゆる糖組成物を使用してもよい。好ましい実施態様では、糖組成物は1つ以上のリグノセルロース系材料の加水分解産物である。本明細書でリグノセルロースは、ヘミセルロースおよびバイオマスのヘミセルロース部分を含む。またリグノセルロースは、バイオマスのリグノセルロース性画分も含む。適切なリグノセルロース系材料は、以下の一覧にあってもよい:果樹園の剪定物、シャパラル、製材廃棄物、都市木材廃棄物、一般廃棄物、伐採廃棄物、森林間伐材、短期輪作木質作物、産業廃棄物、麦わら、オート麦わら、稲わら、大麦わら、ライ麦わら、亜麻のわら、大豆殻、もみ殻、稲わら、コーングルテン飼料、オート麦外皮、サトウキビ、トウモロコシ茎葉、トウモロコシ茎、トウモロコシ穂軸、トウモロコシ苞葉、スイッチグラス、ススキ、サトウモロコシ、菜種茎、大豆茎、プレリーグラス、ガマグラス、エノコログサ;甜菜パルプ、柑橘類パルプ、種子殻、セルロース性家畜排泄物、刈り取った芝草、綿、海藻、樹木、針葉樹、広葉樹、ポプラ、松、灌木、草、小麦、麦わら、サトウキビバガス、トウモロコシ、トウモロコシ苞葉、トウモロコシ穂軸、トウモロコシ穀粒、穀物の湿式または乾式製粉からの製品または副産物、都市固形廃棄物、古紙、庭ごみ、草質材料、農業残渣、林業残渣、都市固体廃棄物、古紙、パルプ、製紙工場残渣、枝、茂み、籐類、トウモロコシ、トウモロコシ苞葉、エネルギー作物、森林、果物、花、穀物、草、草本作物、葉、樹皮、針葉、原木、根、苗木、灌木、スイッチグラス、木、野菜、果皮、つる植物、甜菜パルプ、ミドリング粉、オート麦外皮、硬材または軟材、農産加工から発生する有機廃棄物、林業木材廃棄物、またはそれらの任意の2つ以上の組み合わせ。
【0016】
リグノセルロースに由来するいくつかの適切な糖組成物、およびそれらの加水分解産物の糖組成の概要を表1に示す。列挙したリグノセルロースには、トウモロコシ穂軸、トウモロコシ繊維、もみ殻、メロンの皮、甜菜パルプ、麦わら、サトウキビバガス、木材、草、およびオリブ絞りかすが含まれる。
【0017】
【表1】
【0018】
表1から、これらのリグノセルロース中では、かなりの量の糖(平均して3.8%)がガラクトースであることが明らかである。したがってガラクトースの発酵産物への転換は、経済上重大である。
【0019】
[混合糖細胞]
混合糖細胞は、以下で定義されるように、遺伝子araA、araB、およびaraDを含んでなる。これはグルコース、アラビノース、およびガラクトースを発酵できる。本発明の一実施態様では、混合糖細胞は、好ましくはC5および/またはC6糖である1つ以上の追加的糖を発酵できる。本発明の実施態様では、混合糖細胞は、以下の1つ以上を含んでなる:混合糖細胞がキシロースを発酵できるようにするxylA遺伝子および/またはXKS1遺伝子;アルドース還元酵素(GRE3)遺伝子の欠失;細胞中のペントースリン酸経路を通じた流量を増大させるPPP遺伝子TAL1、TKL1、RPE1およびRKI1の過剰発現。
【0020】
本発明の一実施態様では、混合糖細胞は好ましくはC5および/またはC6糖である1つ以上の追加的糖を発酵できる。本発明の一実施態様では、混合糖細胞は以下の1つ以上を含んでなる:混合糖細胞にキシロースを発酵させるxylA遺伝子、XYL1遺伝子、およびXYL2遺伝子および/またはXKS1遺伝子;アルドース還元酵素(GRE3)遺伝子の欠失;細胞中のペントースリン酸経路を通じた流量を増大させるPPP遺伝子TAL1、TKL1、RPE1、およびRKI1の過剰発現。
【0021】
一実施態様では、混合糖細胞は工業用細胞であり、より好ましくは工業用酵母である。工業用細胞および工業用酵母細胞は、次のように定義されてもよい。工業工程における(酵母)細胞の生存環境が、実験室の環境とは顕著に異なる。工業用酵母細胞は、工程中に変動し得る複数の環境条件下で良好に機能できなくてはならない。このような変動としては、一緒になってサッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)細胞の成長とエタノール産生に潜在的影響を及ぼす栄養源、pH、エタノール濃度、温度、酸素濃度の変化などが挙げられる。有害工業条件の下で、環境耐性株は、頑強な成長および生産が可能であるべきである。工業酵母株は、一般に製パン工業、醸造業、ぶどう酒醸造、およびエタノール産業などのそれらが使用される用途において起こり得るこれらの環境条件の変化に対してより頑強である。一実施態様では、工業混合糖細胞は工業宿主細胞に基づいて構築され、構築は下述するように実施される。工業酵母(S.セレヴィシエ(S.cerevisiae))の例は、Ethanol Red(登録商標)(Fermentis)、Fermiol(登録商標)(DSM)、およびThermosacc(登録商標)(Lallemand)である。
【0022】
一実施態様では、混合糖細胞は阻害物質耐性である。阻害物質耐性は、阻害性化合物に対する抵抗性である。リグノセルロース中の阻害性化合物の存在およびレベルは、原材料、前処理方法、加水分解処理の変動にともなって、変化に富んでもよい。阻害物質カテゴリーの例は、カルボン酸、フランおよび/またはフェノール化合物である。カルボン酸の例は、乳酸、酢酸またはギ酸である。フランの例は、フルフラールおよびヒドロキシメチルフルフラールである。フェノール化合物の例は、バニリン、シリング酸、フェルラ酸、およびクマル酸である。阻害物質の典型的量は次のとおりである。カルボン酸では、原材料、前処理、および加水分解条件次第で、1リットル当たり数グラム、1リットル当たり20グラム以上まで。フランでは、原材料、前処理、および加水分解条件次第で、1リットル当たり数百ミリグラム、1リットル当たり数グラムまで。フェノール類では、原材料、前処理、および加水分解条件次第で、1リットル当たり数十ミリグラム、1リットル当たり1グラムまで。
【0023】
本発明に従った混合糖株は、阻害物質耐性であり、すなわちそれらは、混合糖株が幅広い用途に適用することができ、すなわちそれが異なる原材料、異なる前処理方法、および異なる加水分解条件に対して高い適用性を有するように、一般的な前処理および加水分解条件レベルで、それらが典型的に有する一般的な阻害物質に耐え得る。
【0024】
一実施態様では、工業混合糖細胞は、阻害物質耐性宿主細胞に基づいて構築され、構築は下述するように実施される。阻害物質耐性宿主細胞は、阻害物質耐性S.セレヴィシエ(S.cerevisiae)ATCC26602株が選択された、Kadar et al,Appl.Biochem.Biotechnol.(2007),Vol.136−140,847−858で示されるような阻害物質含有材料上での成長について、株をスクリーニングすることで選択されてもよい。
【0025】
一実施態様では、混合糖細胞はマーカーフリーである。本明細書での用法では、「マーカー」という用語は、マーカーを含有する宿主細胞を選択またはスクリーニングできるようにする形質または表現型をコードする遺伝子を指す。マーカーフリーとは、混合糖細胞内にマーカーが本質的に不在であることを意味する。マーカーフリーであることは、混合糖細胞の構築において抗生物質マーカーが使用され、その後除去される場合に特に有利である。マーカーの除去は、例えば分子内組み換えなどのあらゆる適切な先行技術を使用して実施されてもよい。マーカーを除去する適切な方法は、実施例で示される。
【0026】
混合糖細胞は、植物バイオマス、セルロース、ヘミセルロース、ペクチン、ラムノース、ガラクトース、フルクトース、マルトース、マルトデキストリン、リボース、リブロース、またはデンプン、デンプン誘導体、スクロース、乳糖、およびグリセロールを例えば発酵性糖に転換できてもよい。したがって混合糖細胞は、セルロースのグルコースモノマーへの、そしてヘミセルロースのキシロースおよびアラビノースモノマーへの転換に必要なセルラーゼ(エンドセルラーゼまたはエキソセルラーゼ)、ヘミセルラーゼ(エンドまたはエキソキシラナーゼまたはアラビナーゼ);ペクチンをグルクロン酸およびガラクツロン酸に転換できるペクチナーゼ;またはデンプンをグルコースモノマーに転換するアミラーゼなどの1つ以上の酵素を発現してもよい。
【0027】
混合糖細胞は、ピルビン酸から、エタノール、ブタノール、乳酸、3−ヒドロキシ−プロピオン酸、アクリル酸、酢酸、コハク酸、クエン酸、フマル酸、リンゴ酸、イタコン酸、アミノ酸、1,3−プロパン−ジオール、エチレン、グリセロール、β−ラクタム抗生物質またはセファロスポリンなどの所望の発酵産物への転換に必要とされる酵素活性をさらに含んでなってもよい。
【0028】
一実施態様では、混合糖細胞は、天然でアルコール発酵、好ましくは嫌気的アルコール発酵できる細胞である。混合糖細胞は、好ましくは高いエタノール耐性、高い低pH耐性(すなわち約5、約4、約3、または約2.5よりも低いpHで成長できる)、および有機物に対する高い耐性、および/または高い高温耐性を有する。
【0029】
上記のいずれかの混合糖細胞の特徴または活性は、細胞内に天然に存在してもよく、または遺伝子改変によって導入または修飾されてもよい。
【0030】
[混合糖株の構築]
遺伝子は、
a)強力なプロモーター制御下のPPP遺伝子、TAL1、TKL1、RPE1、およびRKI1からなるクラスター;
b)どちらも構成的プロモーター制御下のxylA遺伝子およびXKS1遺伝子からなるクラスター;
c)遺伝子araA、araB、およびaraDからなるクラスターおよび/またはxylA遺伝子および/またはXKS1遺伝子のクラスター;および
d)アルドース還元酵素遺伝子の欠失
の宿主細胞への導入と、混合糖細胞を生じる適応進化によって、混合糖細胞に導入されてもよい。上記細胞は、組み換え発現技術を使用して構築されてもよい。
【0031】
[組み換え発現]
本発明の細胞は、組み換え細胞である。すなわち本発明の細胞は、問題となっている細胞中では天然では生じないヌクレオチド配列を含んでなり、またはそれによって形質転換され、またはそれによって遺伝子改変されている。
【0032】
細胞中の酵素組み換え発現技術、ならびに本発明の細胞の追加的遺伝子改変は、当業者に良く知られている。典型的に、このような技術は、妥当な配列を含んでなる核酸コンストラクトによる細胞の形質転換を伴う。このような方法は、例えばSambrook and Russel(2001)”Molecular Cloning:A Laboratory Manual”(3rd edition),Cold Spring Harbor Laboratory,Cold Spring Harbor Laboratory Press、またはF.Ausubel etal.,eds.,”Current protocols in molecular biology”,Green Publishing and Wiley Interscience,New York(1987)などの標準的手引き書から知られている。真菌宿主細胞の形質転換および遺伝子改変方法は、例えば欧州特許出願公開第A−0635574号明細書、国際公開第98/46772号パンフレット、国際公開第99/60102号パンフレット、国際公開第00/37671号パンフレット、国際公開第90/14423号パンフレット、欧州特許出願公開第A−0481008号明細書、欧州特許出願公開第A−0635574号明細書、および米国特許第6,265,186号明細書から知られている。
【0033】
典型的に、核酸コンストラクトは、プラスミド、例えば低コピー数プラスミドまたは高コピー数プラスミドであってもよい。本発明に従った細胞は、例えばヌクレオチドコンストラクトの複数コピーによって、または酵素配列の複数コピーを有するコンストラクトの使用によって、酵素をコードするヌクレオチド配列の単数または複数コピーを含んでなってもよい。
【0034】
核酸コンストラクトはエピソームに保持されてもよく、したがって常染色体性複製配列などの自律複製配列を含んでなってもよい。適切なエピソームの核酸コンストラクトは、例えば酵母2μまたはpKD1プラスミド(Gleer et al.,1991,Biotechnology 9:968−975)に、またはAMAプラスミド(Fierro et al.,1995,Curr Genet.29:482−489)に基づいてもよい。代案としては、各核酸コンストラクトが1つ以上のコピーで、細胞ゲノムに組み込まれてもよい。細胞ゲノムへの組み込みは非相同的組換えによってランダムに生じてもよいが、好ましくは核酸コンストラクトは、当該技術分野で良く知られている相同的組換えによって細胞のゲノムに組み込まれてもよい(例えば国際公開第90/14423号パンフレット、欧州特許出願公開A−0481008号明細書、欧州特許出願公開A−0635574号明細書、および米国特許第6,265,186号明細書参照)。
【0035】
ほとんどのエピソームのプラスミドまたは2μプラスミドは比較的不安定であり、各世代毎におよそ10ー2個以上の細胞内で失われる。選択的成長条件下でさえも、細胞の60%〜95%のみがエピソームのプラスミドを保持する。ほとんどのエピソームのプラスミドのコピー数は、cir+宿主の細胞当たり10〜40個の範囲である。しかしプラスミドは細胞間で均等に分布しておらず、集団中の細胞当たりのコピー数には大きな相違がある。組み込みプラスミドで形質転換された株は、選択的圧力の不在下でさえも極めて安定している。しかしプラスミドの損失は、縦列反復DNA間の相同的組換えによっておよそ10ー3〜10ー4の頻度で起こり得て、ベクター配列のループアウトにつながる。したがって、好ましくは安定した組み込みの場合のベクターデザインは、(分子内相同的組換えによっても起きる)選択マーカー遺伝子の損失に際して、組み込みコンストラクトのループアウトはもはや可能でない。好ましくは遺伝子はこのようにして安定して組み込まれる。安定した組み込みは、本明細書では、組み込みコンストラクトのループアウトがもはや可能でない、ゲノムへの組み込みと定義される。好ましくは選択マーカーは不在である。典型的に酵素をコードする配列は、酵素配列の転写および/または翻訳を提供でき、またはそれを助けることができる、1つ以上の核酸配列と作動的に連結する。
【0036】
「作動的に連結する」という用語は、記載される構成要素がそれらの意図される様式で機能できる関係にある並置を指す。例えばプロモーターまたはエンハンサーは、コード配列と作動的に連結し、上記プロモーターまたはエンハンサーはコード配列の転写に影響を及ぼす。
【0037】
本明細書での用法では、「プロモーター」という用語は、1つ以上の遺伝子の転写を制御するよう機能する核酸断片を指し、それは遺伝子の転写開始部位の転写方向に対して上流に位置し、DNA依存RNAポリメラーゼ結合部位、転写開始部位、およびあらゆるその他の当業者に知られているDNA配列の存在によって、構造的に同定される。「構成的」プロモーターは、ほとんどの環境および発生的条件の下で活性のプロモーターである。「誘導性」プロモーターは、環境または発生的調節下で活性のプロモーターである。
【0038】
本発明に従った酵素をコードするヌクレオチド配列の発現を達成するのに使用できるプロモーターは、発現される酵素をコードするヌクレオチド配列にもともと備わっていなくてもよく、すなわちそれが作動的に連結するヌクレオチド配列(コード配列)に異種性のプロモーターであってもよい。しかしプロモーターは、宿主細胞に相同的、すなわち内在性であってもよい。
【0039】
プロモーターは広く入手でき、当業者に知られている。このようなプロモーターの適切な例としては、例えば酵母または糸状菌からのホスホフルクトキナーゼ(PFK)、トリオースリン酸イソメラーゼ(TPI)、グリセルアルデヒド−3−リン酸脱水素酵素(GPD、TDH3またはGAPDH)、ピルビン酸キナーゼ(PYK)、ホスホグリセリン酸キナーゼ(PGK)プロモーターなどの解糖作用遺伝子からのプロモーターが挙げられ、このような酵母からのプロモーターについてより詳しくは、(国際公開第93/03159号パンフレット)に見つかる。その他の別の有用なプロモーターは、リボソームタンパク質コーディング遺伝子プロモーター、ラクターゼ遺伝子プロモーター(LAC4)、アルコールデヒドロゲナーゼプロモーター(ADHI、ADH4など)、およびエノラーゼプロモーター(ENO)である。構成および誘導性の双方のその他のプロモーター、およびエンハンサーまたは上流活性化配列については、当業者に知られている。本発明の宿主細胞で使用されるプロモーターは、所望ならば修飾して、それらの制御特性に影響を及ぼしてもよい。本明細書の文脈で適切なプロモーターとしては、構成および誘導性双方の天然プロモーターならびに当業者に良く知られている遺伝子操作されたプロモーターが挙げられる。真核生物の宿主細胞中の適切なプロモーターは、GAL7、GAL10、またはGAL1、CYC1、HIS3、ADH1、PGL、PH05、GAPDH、ADC1、TRPI、URA3、LEU2、EN01、TPI1、およびAOX1であってもよい。その他の適切なプロモーターとしては、PDC1、GPD1、PGK1、TEF1、およびTDH3が挙げられる。
【0040】
本発明の細胞中では、酵素をコードするヌクレオチド酸配列の3’末端は、好ましくは転写ターミネーター配列と作動的に連結する。好ましくはターミネーター配列は、例えば選択された酵母種などの選択された宿主細胞中で作動可能である。いずれの場合もターミネーターの選択は重要でなく、それは例えばあらゆる酵母遺伝子からのものであってもよいが、非酵母、真核生物遺伝子からのものであれば、ターミネーターが時に機能することもある。通常酵素をコードするヌクレオチド配列は、ターミネーターを含んでなる。好ましくは、このようなターミネーターは、本発明の宿主細胞中のナンセンス仲介mRNA崩壊を妨げる変異と組み合わせられる(例えばShirley et al.,2002,Genetics 161:1465−1482を参照されたい)。
【0041】
転写終結配列は、好ましくはさらにポリアデニル化シグナルを含んでなる。
【0042】
任意選択的に、選択可能なマーカーは、本発明で使用するのに適した核酸コンストラクト中に存在してもよい。本明細書での用法では、「マーカー」という用語は、マーカーを含有する宿主細胞を選択またはスクリーニングできるようにする、形質または表現型をコードする遺伝子を指す。マーカー遺伝子は、それによって適切な抗生物質を使用して、形質転換されていない細胞から形質転換された細胞を選択し得る、抗生物質耐性遺伝子であってもよい。適切な抗生物質耐性マーカーの例としては、例えばジヒドロ葉酸還元酵素、ハイグロマイシン−B−リン酸転移酵素、3’−O−リン酸転移酵素Il(カナマイシン、ネオマイシン、およびG418耐性)が挙げられる。抗生物質耐性マーカーは、倍数体宿主細胞の形質転換のために最も都合よいかもしれず、栄養要求性マーカー(URA3、TRPI、LEU2)またはS.ポンベ(S.pombe)TPI遺伝子などの非抗生物質耐性マーカーもまた使用してもよい(Russell P R,1985,Gene 40:125−130によって記載される)。好ましい実施態様では、核酸コンストラクトによって形質転換される宿主細胞は、マーカー遺伝子フリーである。組み換えマーカー遺伝子フリー微生物宿主細胞を構築する方法は、欧州特許出願公開A−0635574号明細書で開示され、A.ニデュランス(A.nidulans)amdS(アセトアミダーゼ)遺伝子または酵母URA3、およびLYS2遺伝子などの二方向性マーカーの使用に基づく。代案としては、緑色蛍光タンパク質、lacL、ルシフェラーゼ、クロラムフェニコールアセチルトランスフェラーゼ、βグルクロニダーゼなどのスクリーニング可能マーカーを本発明の核酸コンストラクトに組み込んで、形質転換された細胞をスクリーニングできるようにしてもよい。
【0043】
本発明で使用するのに適した中核酸コンストラクトに存在してもよい任意のさらなる要素としては、1つ以上のリーダー配列、エンハンサー、組み込み因子、および/またはレポーター遺伝子、イントロン配列、セントロメア、テロマーおよび/またはマトリックス付着(MAR)配列が挙げられるが、これに限定されるものではない。本発明の核酸コンストラクトは、ARS配列などの自律複製のための配列をさらに含んでなってもよい。
【0044】
したがって遺伝子組み換え過程は、既知の遺伝子組み換え技術を使用して実施されてもよい。本発明の細胞中での酵素発現および過剰発現のための様々な手段が、当業者に知られている。特に、例えば遺伝子の追加的コピーを宿主細胞のゲノム中に組み込むことによって、エピソームのマルチコピー発現ベクターからの遺伝子を発現することによって、または遺伝子の複数コピーを含んでなるエピソームの発現ベクターを導入することによって、宿主細胞中で酵素をコードする遺伝子のコピー数を増大させ酵素を過剰発現させてもよい。
【0045】
代案としては、本発明の宿主細胞中の酵素の過剰発現は、過剰発現させる酵素をコードする配列に天然でないプロモーター、すなわちそれが作動的に連結するコード配列に異種性のプロモーターを使用することによって達成してもよい。プロモーターは、好ましくはそれが作動的に連結するコード配列に異種性であるが、プロモーターが相同的である、すなわち宿主細胞に内在性であることもまた好ましい。好ましくは異種性プロモーターは、コード配列に天然のプロモーターよりも、コード配列を含んでなる転写物のより高い定常状態レベルを生じることができる(または単位時間当たりより多くの転写物分子、すなわちmRNA分子を生成できる)。この文脈で適切なプロモーターとしては、構成および誘導性双方の天然プロモーターならびに改変プロモーターが挙げられる。
【0046】
一実施態様では、混合糖細胞はマーカーフリーであり、それは栄養要求性または優勢マーカー、特に抗生物質耐性マーカーが、ゲノム中または染色体外に存在しないことを意味する。
【0047】
上述の酵素の過剰発現で使用されるコード配列は、好ましくは本発明の宿主細胞と相同的であってもよい。しかし本発明の宿主細胞と異種性のコード配列を使用してもよい。
【0048】
酵素の過剰発現は、遺伝子改変細胞中での酵素生成について言及する場合、同一条件下の未改変宿主細胞と比較して、特定の酵素活性がより高レベルで酵素が生成されることを意味する。通常これは、酵素的に活性なタンパク質(またはマルチサブユニット酵素の場合は複数のタンパク質)が、同一条件下の未改変宿主細胞と比較してより多量に、またはかなりより高い定常状態レベルで生成されることを意味する。同様にこれは通常、酵素的に活性なタンパク質をコードするmRNAが、同一条件下の未改変宿主細胞と比較してより多量に、またはかなりより高い定常状態レベルで生成されることを意味する。好ましくは本発明の宿主細胞中では、過剰発現される酵素は、過剰発現を引き起こす遺伝子改変以外は遺伝的に同一の株と比較して、少なくとも約1.1、約1.2、約1.5、約2、約5、約10または約20倍過剰発現される。これらの過剰発現レベルは、酵素活性の定常状態レベルに、酵素タンパク質の定常状態レベルに、ならびに酵素をコードする転写物の定常状態レベルに当てはめられてもよいものと理解されるものとする。
【0049】
[適応]
適応は、それによって集団がその生息環境または複数の生息環境により良く適する(順応する)進化の過程である。この過程は、数世代から多世代にわたって起き、基本的な生物学的現象の1つである。
【0050】
適応という用語は、生物の生存に特に重要な特徴もまた指してもよい。このような適応は、より良く適した形態がより成功裏に繁殖することによって、自然淘汰によって可変集団中で生じる。
【0051】
環境条件の変化は、自然淘汰の結果を変化させ、新しい条件下における生物の適応度を改善する引き続く適応の選択的利点に影響を及ぼす。極端な環境変化の場合、有益な適応の出現と固定化が生存のために必須であり得る。例えば養分利用性、温度、酸素利用性などの多数の異なる要因が、適応進化を駆動し得る。
【0052】
[適合]
適応(生物が特定の生息環境内で生きて繁殖できる程度)と適合の間には、明らかな関係がある。適合は、自然淘汰率の推定および予測である。自然淘汰の適用によって、代替の表現型の相対頻度は、それらが遺伝性であるならば経時的に変動する。
【0053】
[遺伝的変化]
自然淘汰が集団の遺伝的変動に作用する場合、遺伝的変化が根底にある機序である。これは集団が、その状況に遺伝的に適応することを意味する。遺伝的変化は、生息環境の変化に適するように、可視的構造をもたらしてもよく、または生物の生理学的活性を調節してもよい。
【0054】
[適応進化]
混合糖細胞は、それらの調製中に適応進化を被る。本発明の細胞は、好ましくは唯一の炭素源としての所望の糖の上での、より好ましくは嫌気的条件下における成長について、自然発生または誘発的(例えば放射線または化学薬品による)いずれかの変異体を選択することで、糖利用のために適応させてもよい。変異体の選択は、例えばKuyperら(2004,FEMS Yeast Res.4:655−664)によって記述される継代培養をはじめとする技術によって、またはケモスタット培養中の選択的圧力下における培養によって実施されてもよい。例えば本発明の好ましい宿主細胞中で、変異体の選択によって得られる改変をはじめとする上述の遺伝子改変の少なくとも1つは、炭素源として、好ましくは唯一の炭素源としてキシロース上で、好ましくは嫌気的条件下で、生育する能力を宿主細胞にもたらす。好ましくは細胞は本質的にキシリトールを生成せず、例えば生成するキシリトールが検出限界未満であり、または例えばモル基準で消費される炭素の約5、約2、約1、約0.5、または約0.3%未満である。
【0055】
適応進化についてはまた、例えばH.W. et al,Applied and Environmental Microbiology Aug.2007,p.4881−4891でも記載される。
【0056】
適応進化の一実施態様では、異なる培地、例えば組成の異なる3種類の培地中(グルコース、キシロース、およびアラビノース;キシロースおよびアラビノース)における、連続成長を用いた反復バッチ培養からなる投与計画が適用されるWisselink et al.(2009)Applied and Environmental Microbiology,Feb.2009,p.907−914参照。
【0057】
[酵母形質転換および遺伝的安定性]
組み換えDNAを用いた酵母細胞の遺伝子操作、すなわち形質転換は、1978年に初めて実行可能になった[Beggs,1978;Hinnen et al.,1978]。酵母中の組み換えDNA技術は、それ以来地位を確立した。多数の異なるベクターコンストラクトが利用可能である。一般にシャトルベクターと称されるこれらのプラスミドベクターは、酵母細胞への形質転換に先だって、それらが大腸菌(E.coli)中で増殖できるようにする、複製起点と選択可能なマーカー(βラクタマーゼ遺伝子、ampRであることが多い)からなる、大腸菌(E.coli)ベクターに由来する遺伝物質を含有する。さらにシャトルベクターは酵母中における選択のための選択可能なマーカーを含有する。マーカーは、対応するゲノムの欠失(または突然変異)を保有する細胞が、栄養要求性または独立栄養性について補足されるような、特定のアミノ酸またはヌクレオチド合成のための酵素をコードする遺伝子であり得る。代案としては、これらのベクターは、g418(Geneticin)、ハイグロマイシンBまたはフレオマイシンのような特定の抗生物質に対する耐性を組み換え酵母細胞(すなわちDNAを取り込んでマーカー遺伝子を発現する細胞)に提供する、異種性顕性耐性マーカーを含有する。さらにこれらのベクターは、外来性DNAをこれらの部位にクローンできるようにする(組み合わされた)制限部位(多重クローニング部位またはMCS)配列を含有してもよいが、代案の方法もまた存在する。
【0058】
伝統的に、追加的遺伝的要素の不在または存在によって、4種のシャトルベクターを区別し得る。
●組み込みプラスミド(YIp)。限定酵素によって開裂され、酵母細胞の形質転換のために直線化DNAが使用される場合は、これはマーカーまたは別の遺伝子の遺伝子座において、相同的組換えにより宿主ゲノムに組み込まれる。これは一般に、ゲノム中のこの特定部位に挿入された外来性DNAの1コピーの存在をもたらす。
●エピソームプラスミド(YEp)。これは酵母細胞中の自律複製に必要な2μプラスミドDNA配列の部分を保有する。複数コピーの形質転換プラスミドが酵母細胞中で増殖され、エピソームとして維持される。
●自己複製プラスミド(YRp)。これは形質転換プラスミドを数百倍に増殖できるようにする、酵母複製起点(ARS、自律的複製配列)を保有する。
●CENプラスミド(YCp)。これはARS配列に加えて、常態では安定した有糸分裂分離を保証し、通常は自己複製プラスミドのコピー数を1つだけに減少させるセントロメア配列(核染色体の1つに由来する)を保有する。
【0059】
これらのプラスミドは、形質転換によって酵母細胞に導入される。酵母細胞の形質転換は、酢酸リチウムによる細胞透過性上昇(Ito et al,1983)、および電気穿孔方法などのいくつかの異なる技術によって達成されてもよい。
【0060】
組み換え微生物の商業的用途では、プラスミドの不安定性が最重要課題である。不安定性は、形質転換された細胞が、プラスミドの変化または損失のためにそれらの改変された特性を失う傾向である。この問題は、Zhangら(Plasmid stability in recombinant Saccharomyces cerevisiae.Biotechnology Advances,Vol.14,No.4,pp.401−435,1996)で、詳細に考察されている。組み込みプラスミドで形質転換された株は、選択圧力の不在下であってさえも極めて安定している(Sherman,F.http://dbb.urmc.rochester.edu/labs/sherman_f/yeast/9.htmlおよびその中の参考文献)。
【0061】
異種性DNAは、通常、染色体外プラスミドの形態(YEp、YCp、およびYRp)で生物に導入される。残念ながら、特に淘汰圧が連続的にかからない場合は、細菌および酵母のどちらでも新しい特徴が保持されないこともあり得ることが分かった。これは組み換え細胞が長期にわたって成長した際のハイブリッドプラスミドの分離不安定性のためである。これは、集団不均一性、クローン変動性、最終的にはその中で大部分の細胞が形質転換により導入された特性を失う細胞集団をもたらす。栄養要求性マーカーを持つベクターが使用される場合、ベクターは最少培地中でのみ維持されるので、富栄養培地中の培養はベクターの迅速な損失をもたらすことが多い。代案である顕性抗生物質耐性マーカーの使用は、製造工程に適合しないことが多い。抗生物質の使用はまた、登録の観点(微量の抗生物質が最終製品中に入る可能性)から、または経済的理由(工業規模での抗生物質の使用コスト)のために、所望されないこともあり得る。
【0062】
ベクターの損失は、大量生産状況において問題をもたらす。組み込みプラスミド(YIp)の使用などのDNAを導入する代案の方法が、酵母に対して存在する。DNAは遺伝子組み換えによって宿主ゲノムに組み込まれ、高い安定性がもたらされる(Caunt,P.Stability of recombinant plasmids in yeast.Journal of Biotechnology 9(1988)173−192)。我々は、宿主トランスポゾンを使用した組み込み法が、良い代案であることを発見した。
【0063】
[トランスポゾン]
本発明の実施態様では、細胞は、所望の遺伝子の1つを超えるコピーを含んでなってもよい。例えば2つ以上のキシロースイソメラーゼ遺伝子またはキシロース還元酵素遺伝子およびキシリトールデヒドロゲナーゼが、混合糖細胞ゲノムに組み込まれてもよい。これは遺伝子の導入をもたらすことが当該技術分野で知られているあらゆる方法によって、実施されてもよい。好ましい実施態様では、これは宿主細胞の反復配列(トランスポゾン)と相同的な部分がある、ベクターを使用して達成されてもよい。宿主細胞が酵母細胞である場合、適切な反復配列はδ配列として知られているTy因子の長い末端反復(LTR)である。
【0064】
Ty因子は、Ty1およびTy2と称される、2つのかなり類似した亜科に分類される。これらの因子は、長さが約6キロベース(kb)であり、約335塩基対の配列である長い末端反復(LTR)と境を接する(Boeke JD et al,The Saccharomyces cerevisiae Genome Contains Functional and Nonfunctional Copies of Transposon Ty1.Molecular and Cellular Biology,Apr.1988,p.1432−1442 Vol.8,No.4)。完全に配列決定されたS.セレヴィシエ(S.cerevisiae)S288c株中で最も豊富なトランスポゾンは、Ty1(31コピー)およびTy2(13コピー)である(Gabriel A,Dapprich J,Kunkel M,Gresham D,Pratt SC,et al.(2006)Global mapping of transposon location.PLoS Genet 2(12):e212.doi:10.1371/journal.pgen.0020212)。これらのトランスポゾンは、そのそれぞれがいくつかのタンパク質をコードする、2つの重複する読み取り枠(ORF)から構成される。コード領域は、前述のほぼ同一のLTRで挟まれる。S.セレヴィシエ(S.cerevisiae)中の別のものであるが、より少なくより特徴的であるTy因子は、Ty3、Ty4、およびTy5からなる。全長Ty因子の各ファミリーで、一桁多い単独LTR因子がゲノム全体に分散する。これらは、内部タンパク質コード領域のループアウトと共に、全長因子のLTR−LTR組み換えによって生じると考えられる。
【0065】
Tyレトロトランスポゾンのレトロ転位機序は、ゲノム全体に複数コピーを組み込むのに利用されている(Boeke et al.,1988;Jacobs et al.,1988)。δ配列として知られているTy因子の長い末端反復(LTR)はまた、Ty結合または単独部位のどちらかである約150〜200コピーで存在するので、相同的組換えによる組み込みの良好な標的でもある。(Boeke,1989;Kingsman and Kingsman,1988)(Parekh R.N.(1996).An Integrating Vector for Tunable, High Copy,Stable Integration into the Dispersed Ty DELTA Sites of Saccharomyces cerevisiae.Biotechnol.Prog.1996,12,16−21)。
【0066】
[宿主細胞]
宿主細胞は、有用な生成物の生産に適したあらゆる宿主細胞であってもよい。本発明の細胞は、細菌などの原核細胞、または真核細胞などのあらゆる適切な細胞であってもよい。典型的に細胞は、例えば酵母または糸状菌などの真核細胞である。
【0067】
酵母は、本明細書で真核微生物と定義され、大部分は単細胞形態で成長する真菌植物亜門(Alexopoulos,C.J.,1962,In:Introductory Mycology,John Wiley & Sons,Inc.,New York)の全ての種を含む。
【0068】
酵母は単細胞性葉状体の出芽によって成長してもよく、または生物体の分裂によって成長してもよい。本発明の細胞として好ましい酵母は、サッカロミセス(Saccharomyces)、クリヴェロミセス(Kluyveromyces)、カンジダ(Candida)、ピチア(Pichia)、分裂酵母(Schizosaccharomyces)、ハンゼヌラ(Hansenula)、クロエケラ(Kloeckera)、シュワニオミセス(Schwanniomyces)またはヤロウィア(Yarrowia)属に属してもよい。好ましくは酵母は、嫌気的または酸素限定発酵ができるものであり、より好ましくは嫌気的アルコール発酵ができるものである。
【0069】
糸状菌は、本明細書で、真菌植物亜門の全ての糸状体を含む真核微生物と定義される。これらの真菌は、キチン、セルロース、および複合多糖からなる栄養菌糸体によって特徴付けられる。本発明の細胞として使用するのに適した糸状菌は、形態的、生理的、および遺伝的に酵母と異なる。ほとんどの真菌は増殖のために無菌条件を必要とせず、バクテリオファージ感染に感受性がないので、有利には糸状菌細胞を使用してもよい。糸状菌による栄養成長は菌糸伸長と炭素異化作用により、ほとんどの糸状菌で偏性好気性である。本発明の宿主細胞として好ましい糸状菌は、アスペルギルス(Aspergillus)、トリコデルマ(Trichoderma)、フミコラ(Humicola)、アクレモニウム(Acremonium)、フザリウム(Fusarium)またはペニシリウム(Penicillium)属に属してもよい。より好ましくは、糸状菌細胞は、アスペルギルス・ニガー(Aspergillus niger)、コウジカビ(Aspergillus oryzae)、ペニシリウム・クリソゲヌム(Penicillium chrysogenum)、またはリゾプス・オリゼー(Rhizopus oryzae)細胞であってもよい。
【0070】
一実施態様では宿主細胞は、酵母であってもよい。
【0071】
好ましくは宿主は、工業宿主、より好ましくは工業酵母である。工業宿主および工業酵母細胞は、以下のように定義されてもよい。工業工程中の酵母細胞の生息環境は、実験室のそれと顕著に異なる。工業用酵母細胞は、工程中に変動し得る複数の環境条件下で良好に機能できなくてはならない。このような変動としては、一緒になってサッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)細胞の成長とエタノール産生に潜在的影響を及ぼす栄養源、pH、エタノール濃度、温度、酸素濃度の変化などが挙げられる。有害工業条件の下で、環境耐性株は、頑強な成長および生産が可能であるべきである。工業酵母株は、一般に、製パン工業、醸造業、ぶどう酒醸造、およびエタノール産業などのそれらが使用される用途において起こり得るこれらの環境条件の変化に対してより頑強である。工業酵母(S.セレヴィシエ(S.cerevisiae))の例は、Ethanol Red(登録商標)(Fermentis)、Fermiol(登録商標)(DSM)、およびThermosacc(登録商標)(Lallemand)である。
【0072】
一実施態様では宿主は、阻害物質耐性である。阻害物質耐性宿主細胞は、阻害物質耐性S.セレヴィシエ(S.cerevisiae)ATCC26602株が選択された、Kadar et al,Appl.Biochem.Biotechnol.(2007),Vol.136−140,847−858で示されるような阻害物質含有材料上での成長について、株をスクリーニングすることで選択されてもよい。
【0073】
好ましくは、宿主細胞は工業用および阻害物質耐性である。
【0074】
[AraA、AraB、およびAraD遺伝子]
本発明の細胞は、アラビノースを使用できる。したがって本発明の細胞は、L−アラビノースをL−リブロースおよび/またはキシルロース−5−リン酸、および/または例えば本明細書で言及されるものなどの所望の発酵産物に転換できる。
【0075】
L−アラビノースからエタノールを生成できる生物、例えばS.セレヴィシエ(S.cerevisiae)株は、好適な起源からのaraA(L−アラビノースイソメラーゼ)、araB(L−リブロキナーゼ)、およびaraD(L−リブロース−5−P4−エピメラーゼ)遺伝子を導入して細胞を改変することにより、生成されてもよい。このような遺伝子は、本発明の細胞がアラビノースを使用できるように導入されてもよい。このようなアプローチについては、国際公開第2003/095627号パンフレットに記載される。ラクトバチルス・プランタルム(Lactobacillus plantarum)からのaraA、araB、およびaraD遺伝子を使用してもよく、国際公開第2008/041840号パンフレットで開示される。枯草菌(Bacillus subtilis)からのaraA遺伝子、および大腸菌(Escherichia coli)からのaraBおよびaraD遺伝子を使用してもよく、欧州特許第1499708号明細書で開示される。別の実施態様では、araA、araB、およびaraD遺伝子は、クラビバクター(Clavibacter)、アルスロバクター(Arthrobacter)および/またはグラメラ(Gramella)属の少なくとも1つ、特に国際公開第2009011591号パンフレットで開示されるようなクラビバクター・ミシガンエンシス(Clavibacter michiganensis)、アルスロバクター・アウレセンス(Arthrobacter aurescens)、および/またはグラメラ・フォルセティ(Gramella forsetii)の1つに由来してもよい。
【0076】
[PPP遺伝子]
本発明の細胞は、ペントースリン酸経路の流量を増大させる1つ以上の遺伝子改変を含んでなってもよい。特に遺伝子改変は、非酸化的部分ペントースリン酸経路を通じて、流量の増大をもたらしてもよい。ペントースリン酸経路の非酸化的部分の流量の増大を引き起こす遺伝子改変は、本明細書では、流量の増大を引き起こす遺伝子改変以外は、遺伝的に同一の株中の流量と比較して、流量を少なくとも約1.1、約1.2、約1.5、約2、約5、約10または約20倍増大させる改変を意味するものと理解される。ペントースリン酸経路の非酸化的部分の流量は、いくらかでもキシリトールが生成される場合は、唯一の炭素源としてのキシロース上で改変宿主を成長させてキシロース特異的消費速度を判定し、キシロース特異的消費速度からキシリトール特異的生成速度を差し引くことによって測定されてもよい。しかしペントースリン酸経路の非酸化的部分の流量は、唯一の炭素源としてのキシロース上での成長速度、好ましくは唯一の炭素源としてのキシロース上での嫌気的成長速度と比例する。唯一の炭素源としてのキシロース上での成長速度(μmax)とペントースリン酸経路の非酸化的部分の流量の間には直線関係がある。(嫌気性、増殖培地、pH、株の遺伝的背景などの所定の条件の組の下では)糖の上でのバイオマス収率は一定であるので、キシロースの比消費速度(Qs)は、糖の上でのバイオマス収率(Yxs)で除した成長速度(μ)に等しい(すなわちQs=μ/Yxs)。したがってペントースリン酸経路の非酸化的部分の流量の増大は、輸送(取り込)みが制限されない限り、これらの条件下の最大成長速度の増大から推定されてもよい。
【0077】
ペントースリン酸経路流量を増大させる1つ以上の遺伝子改変は、様々なやり方で宿主細胞に導入されてもよい。これらとしては、例えばキシルロースキナーゼの、および/または非酸化的部分ペントースリン酸経路酵素の1つ以上の、より高い定常状態活性レベルを達成すること、および/または非特異的アルドース還元酵素の低下した定常状態活性レベルを達成することが挙げられる。定常状態活性レベルにおけるこれらの変化は、(自然発生または化学薬品または放射線により誘発される)変異体の選択によって、および/または組み換えDNA技術によって、例えばそれぞれ遺伝子をコードする酵素またはこれらの遺伝子を制御する因子の過剰発現または不活性化によって、達成されてもよい。
【0078】
好ましい宿主細胞では、遺伝子改変は、(非酸化的部分)ペントースリン酸経路の少なくとも1つの酵素の過剰発現を含んでなる。好ましくは酵素は、リブロース−5−リン酸イソメラーゼ、リブロース−5−リン酸エピメラーゼ、トランスケトラーゼ、およびトランスアルドラーゼをコードする酵素からなる群から選択される。(非酸化的部分)ペントースリン酸経路の酵素の様々な組み合わせが過剰発現されてもよい。例えば過剰発現される酵素は、少なくとも酵素リブロース−5−リン酸イソメラーゼおよびリブロース−5−リン酸エピメラーゼ;または少なくとも酵素リブロース−5−リン酸イソメラーゼおよびトランスケトラーゼ;または少なくとも酵素リブロース−5−リン酸イソメラーゼおよびトランスアルドラーゼ;または少なくとも酵素リブロース−5−リン酸エピメラーゼおよびトランスケトラーゼ;または少なくとも酵素リブロース−5ホスフェートエピメラーゼおよびトランスアルドラーゼ;または少なくとも酵素トランスケトラーゼおよびトランスアルドラーゼ;または少なくとも酵素リブロース−5−リン酸エピメラーゼ、トランスケトラーゼおよびトランスアルドラーゼ;または少なくとも酵素リブロース−5−リン酸イソメラーゼ、トランスケトラーゼおよびトランスアルドラーゼ;または少なくとも酵素リブロース−5−リン酸イソメラーゼ、リブロース−5−リン酸エピメラーゼ、およびトランスアルドラーゼ;または少なくとも酵素リブロース−5−リン酸イソメラーゼ、リブロース−5−リン酸エピメラーゼ、およびトランスケトラーゼであってもよい。本発明の一実施態様では、リブロース−5−リン酸イソメラーゼ、リブロース−5−リン酸エピメラーゼ、トランスケトラーゼ、およびトランスアルドラーゼ酵素のそれぞれが、宿主細胞中で過剰発現される。より好ましいのは、その中の遺伝子改変が、少なくともトランスケトラーゼおよびトランスアルドラーゼ酵素双方の過剰発現を含んでなる宿主細胞であり、それはこのような宿主細胞が既にキシロース上で嫌気的成長できるためである。実際には、条件によっては、トランスケトラーゼおよびトランスアルドラーゼのみを過剰に発現する宿主細胞は、4つの酵素全て、すなわちリブロース−5−リン酸イソメラーゼ、リブロース−5−リン酸エピメラーゼ、トランスケトラーゼ、およびトランスアルドラーゼを過剰に発現する宿主細胞と同一の嫌気的成長速度をキシロース上で既に有する。さらに酵素リブロース−5−リン酸イソメラーゼおよびリブロース−5−リン酸エピメラーゼの双方を過剰に発現する宿主細胞が、イソメラーゼのみまたはエピメラーゼのみを過剰に発現する宿主細胞よりも好ましいが、それはこれらの酵素の1つのみの過剰発現が代謝不均衡を生じ得るためである。
【0079】
本明細書で酵素「リブロース−5−リン酸エピメラーゼ」(EC5.1.3.1)は、D−キシルロース−5−リン酸のD−リブロース−5−リン酸へのエピマー化を触媒する酵素と定義され、逆の場合も同じである。酵素は、ホスホリブロースエピメラーゼ;エリスロース−4−ホスフェートイソメラーゼ;ホスホケトペントース3−エピメラーゼ;キシルロースホスフェート3−エピメラーゼ;ホスホケトペントースエピメラーゼ;リブロース−5−リン酸3−エピメラーゼ;D−リブロースホスフェート−3−エピメラーゼ;D−リブロース−5−リン酸エピメラーゼ;D−リブロース−5−P3−エピメラーゼ;D−キシルロース−5−リン酸3−エピメラーゼ;ペントースー5−ホスフェート3−エピメラーゼ;またはD−リブロース−5−リン酸3−エピメラーゼとしてもまた知られている。リブロース−5−リン酸エピメラーゼは、そのアミノ酸配列によってさらに定義されてもよい。同様にリブロース−5−リン酸エピメラーゼは、酵素をコードするヌクレオチド配列によって、ならびにリブロース−5−リン酸エピメラーゼをコードする参照ヌクレオチド配列とハイブリッドするヌクレオチド配列によって、定義されてもよい。リブロース−5−リン酸エピメラーゼをコードするヌクレオチド配列は、本明細書でRPE1と命名される。
【0080】
本明細書で酵素「リブロース−5−リン酸イソメラーゼ」(EC5.3.1.6)は、D−リボース5−ホスフェートのD−リブロース5−ホスフェートへの直接異性化を触媒する酵素と定義され、逆の場合も同じである。酵素は、ホスホペントースイソメラーゼ;ホスホリボイソメラーゼ;リボースホスフェートイソメラーゼ;5−ホスホリボースイソメラーゼ;D−リボース5−ホスフェートイソメラーゼ;D−リボース−5−ホスフェートケトール−イソメラーゼ;またはD−リボース−5−ホスフェートアルドース−ケトース−イソメラーゼとしてもまた知られている。リブロース−5−リン酸イソメラーゼは、そのアミノ酸配列によってさらに定義されてもよい。同様にリブロース−5−リン酸イソメラーゼは、酵素をコードするヌクレオチド配列によって、ならびリブロース−5−リン酸イソメラーゼをコードする参照ヌクレオチド配列とハイブリッドするヌクレオチド配列によって定義されてもよい。リブロース−5−リン酸イソメラーゼをコードするヌクレオチド配列は、本明細書でRKI1と命名される。
【0081】
本明細書において酵素「トランスケトラーゼ」(EC2.2.1.1)は、D−リボース5−ホスフェート+D−キシルロース−5−リン酸←→セドヘプツロース7−ホスフェート+D−グリセルアルデヒド3−ホスフェートの反応を触媒する酵素と定義され、逆の場合も同じである。この酵素は、グリコールアルデヒドトランスフェラーゼまたはセドヘプツロース−7−ホスフェート:D−グリセルアルデヒド−3−リン酸グリコールアルデヒドトランスフェラーゼとしてもまた知られている。トランスケトラーゼは、そのアミノ酸によってさらに定義されてもよい。同様にトランスケトラーゼは、酵素をコードするヌクレオチド配列によって、ならびにトランスケトラーゼをコードする参照ヌクレオチド配列とハイブリッドするヌクレオチド配列によって定義されてもよい。トランスケトラーゼをコードするヌクレオチド配列は、本明細書でTKL1と命名される。
【0082】
本明細書で酵素「トランスアルドラーゼ」(EC2.2.1.2)は、セドヘプツロース7−ホスフェート+D−グリセルアルデヒド3−ホスフェート←→D−エリスロース4−ホスフェート+D−果糖6−ホスフェートの反応を触媒する酵素と定義され、逆の場合も同じである。酵素は、ジヒドロキシアセトントランスフェラーゼ;ジヒドロキシアセトンシンターゼ;ホルムアルデヒドトランスケトラーゼ;またはセドヘプツロース−7−ホスフェート:D−グリセルアルデヒド−3−ホスフェートグリセロントランスフェラーゼとしてもまた知られている。トランスアルドラーゼは、そのアミノ酸配列によってさらに定義されてもよい。同様にトランスアルドラーゼは、酵素をコードするヌクレオチド配列によって、ならびにトランスアルドラーゼをコードする参照ヌクレオチド配列とハイブリッドするヌクレオチド配列によって、定義されてもよい。トランスケトラーゼをコードするヌクレオチド配列は、本明細書でTAL1と命名される。
【0083】
[キシロースイソメラーゼまたはキシロース還元酵素およびキシリトールデヒドロゲナーゼ遺伝子]
本発明に従って、1つ以上のキシロースイソメラーゼ遺伝子および/または1つ以上のキシロース還元酵素およびキシリトールデヒドロゲナーゼのコピーの1つ、2つまたはそれ以上のコピーが、宿主細胞のゲノムに導入される。これらの2つ以上の遺伝要素の存在は、細胞に異性化または還元によってキシロースを転換する能力を与える。
【0084】
一実施態様では1つ以上のキシロースイソメラーゼ遺伝子の1つ、2つまたはそれ以上のコピーが宿主細胞のゲノムに導入される。
【0085】
本明細書で「キシロースイソメラーゼ」(EC5.3.1.5)は、D−キシロースのD−キシルロースへの直接異性化を触媒する酵素と定義され、逆もまた同様である。酵素は、D−キシロースケトイソメラーゼとしてもまた知られている。本明細書でキシロースイソメラーゼはまた、D−グルコースとD−果糖の間の転換を触媒できてもよい(したがって適宜にグルコースイソメラーゼと称されてもよい)。本明細書でキシロースイソメラーゼは、補助因子としてマグネシウム、マンガンまたはコバルトなどの二価のカチオンを必要としてもよい。
【0086】
したがってこのような混合糖細胞は、キシロースをキシルロースに異性化できる。キシロースをキシルロースに異性化する能力は、定義されたキシロースイソメラーゼをコードするヌクレオチド配列を含んでなる核酸コンストラクトで宿主細胞を形質転換することによって、宿主細胞に与えられる。混合糖細胞は、キシロースのキシルロースへの直接異性化によって、キシロースをキシルロースに異性化する。
【0087】
本明細書で、キシロースイソメラーゼ活性単位(U)は、Kuyper et al.(2003,FEMS Yeast Res.4:69−78)によって記述される条件下で、1分当たり1nmolのキシルロースを生成する酵素量と定義されてもよい。キシロース異性化遺伝子は、例えば国際公開第2006/009434号パンフレットで開示されるようなピロミセス(Piromyces)種などの様々な起源を有してもよい。その他の適切な起源は、バクテロイデス(Bacteroides)、特にPCT欧州特許第2009/52623号明細書に記載されるようなバクテロイデス・ユニフォルミス(Bacteroides uniformis)、バチルス(Bacillus)、特にPCT欧州特許第2009/052625号明細書に記載されるようなバチルス・ステアロサーモフィラス(Bacillus stearothermophilus)である。
【0088】
別の実施態様では、1つ以上のキシロース還元酵素およびキシリトールデヒドロゲナーゼ遺伝子の2つ以上のコピーが宿主細胞ゲノムに導入される。この実施態様では、キシロースの転換は、それぞれキシロース還元酵素およびキシリトールデヒドロゲナーゼによって触媒される、キシロースからキシリトール中間体を経由するキシルロースへの転換の二段階で実施される。一実施態様では、国際公開第2004085627号パンフレットで開示されるように、キシロース還元酵素(XR)、キシリトールデヒドロゲナーゼ(XDH)、およびキシルロキナーゼ(XK)が過剰発現されてもよく、任意にNADPH生成酵素をコードする遺伝子の1つ以上が上方制御されて、NADH消費酵素をコードする遺伝子の1つ以上が上方制御される。
【0089】
[XKS1遺伝子]
本発明の細胞は、特異的キシルロースキナーゼ活性を増大させる、1つ以上の遺伝子改変を含んでなってもよい。好ましくは1つまたは複数の遺伝子改変は、例えばキシルロースキナーゼをコードするヌクレオチド配列の過剰発現によってキシルロースキナーゼの過剰発現を引き起こす。キシルロースキナーゼをコードする遺伝子は宿主細胞に内在性であってもよく、または宿主細胞に異種性のキシルロースキナーゼであってもよい。本発明の宿主細胞中におけるキシルロースキナーゼ過剰発現のために使用されるヌクレオチド配列は、キシルロースキナーゼ活性があるポリペプチドをコードするヌクレオチド配列である。
【0090】
本明細書で酵素「キシルロースキナーゼ」(EC2.7.1.17)は、ATP+D−キシルロース=ADP+D−キシルロース−5−リン酸の反応を触媒する酵素と定義される。酵素はホスホリル化キシルロキナーゼ、D−キシルロキナーゼまたはATP:D−キシルロース5−リン酸転移酵素としてもまた知られている。本発明のキシルロースキナーゼは、そのアミノ酸配列によってさらに定義されてもよい。同様にキシルロースキナーゼは、酵素をコードするヌクレオチド配列によって、ならびにキシルロースキナーゼをコードする参照ヌクレオチド配列とハイブリッドするヌクレオチド配列によって定義されてもよい。
【0091】
本発明の細胞中では、特異的キシルロースキナーゼ活性を増大させる1つまたは複数の遺伝子改変は、上述のようなペントースリン酸経路の流量を増大させる修飾のいずれかと組み合わされてもよい。しかしこれは必須でない。
【0092】
したがって本発明の宿主細胞は、特異的キシルロースキナーゼ活性を増大させる遺伝子改変修飾のみを含んでなってもよい。本発明宿主細胞中でのキシルロースキナーゼの過剰発現を達成し分析するための当該技術分野で利用できる様々な手段は、ペントースリン酸経路酵素について上述したのと同じである。好ましくは本発明の宿主細胞中では、過剰発現されるキシルロースキナーゼは、過剰発現を引き起こす遺伝子改変以外は遺伝的に同一の株と比較して、少なくとも約1.1、約1.2、約1.5、約2、約5、約10または約20倍過剰発現される。これらの過剰発現レベルは、酵素活性の定常状態レベルに、酵素タンパク質の定常状態レベルに、ならびに酵素をコードする転写物の定常状態レベルに当てはまってもよいと理解されるものとする。
【0093】
[アルドース還元酵素(GRE3)遺伝子欠失]
XIがキシロースを転換する遺伝子として使用される実施態様では、アルドース還元酵素活性を低下させることが都合良いこともあり得る。したがって本発明の細胞は、宿主細胞の非特異的アルドース還元酵素活性を低下させる、1つ以上の遺伝子改変を含んでなってもよい。好ましくは、非特異的アルドース還元酵素をコードする遺伝子の発現を低下させまたは不活性化する1つ以上の遺伝子改変によって、宿主細胞中で非特異的アルドース還元酵素活性が低下する。好ましくは遺伝子改変は、宿主細胞中で非特異的アルドース還元酵素をコードする遺伝子の各内在性コピーの発現を低下させまたは不活性化する(本明細書中でGRE3欠失と称する)。宿主細胞は二倍性、倍数性または異数性の結果として、非特異的アルドース還元酵素をコードする遺伝子の複数コピーを含んでなってもよく、および/または宿主細胞はアミノ酸配列が異なり、それぞれ異なる遺伝子によってコードされるアルドース還元酵素活性があるいくつかの異なる酵素(アイソザイム)を含有してもよい。またこのような場合、好ましくは、非特異的アルドース還元酵素をコードする各遺伝子の発現が低下し、または不活性化される。好ましくは、遺伝子は、遺伝子の少なくとも一部の欠失によって、または遺伝子中断によって不活性化され、この文脈では、遺伝子という用語は、その(部分的)欠失または不活性化が宿主細胞中における非特異的アルドース還元酵素活性の発現低下をもたらすコード配列の上流または下流にある、いかなる非コード配列もまた含む。
【0094】
本発明の宿主細胞中でその活性が低下されるアルドース還元酵素をコードするヌクレオチド配列は、アルドース還元酵素活性があるポリペプチドをコードするヌクレオチド配列である。
【0095】
したがって宿主細胞中で非特異的アルドース還元酵素活性を低下させる遺伝子改変のみを含んでなる本発明の宿主細胞は、明確に本発明に含まれる。
【0096】
酵素「アルドース還元酵素」(EC1.1.1.21)は、本明細書でキシロースまたはキシルロースをキシリトールに還元できるあらゆる酵素と定義される。本発明の文脈でアルドース還元酵素は、本発明の宿主細胞に天然(内在性)であり、キシロースまたはキシルロースをキシリトールに還元できるあらゆる非特異的アルドース還元酵素であってもよい。非特異的アルドース還元酵素は、次の反応を触媒する。
アルドース+NAD(P)H+H+←→アルジトール+NAD(P)+
【0097】
酵素は広い特異性を有し、アルドース還元酵素;ポリオールデヒドロゲナーゼ(NADP+);アルジトール:NADPオキシド還元酵素;アルジトール:NADP+1−オキシド還元酵素;NADPH−アルドペントース還元酵素;またはNADPH−アルドース還元酵素としてもまた知られている。
【0098】
S.セレヴィシエ(S.cerevisiae)に内在性であるこのような非特異的アルドース還元酵素の特定の例は、GRE3遺伝子によってコードされる(Traff et al.,2001,Appl.Environ.Microbiol.67:5668−74)。したがって本発明のアルドース還元酵素は、そのアミノ酸配列によってさらに定義されてもよい。同様にアルドース還元酵素は、酵素をコードするヌクレオチド配列によって、ならびに参照ヌクレオチド配列とハイブリッドする、アルドース還元酵素をコードするヌクレオチド配列によって定義されてもよい。
【0099】
[配列同一性]
本明細書で配列同一性(または配列類似性)とは、2つ以上のアミノ酸(ポリペプチドまたはタンパク質)配列または2つ以上の核酸(ポリヌクレオチド)配列を比較して判定される、配列間の関係と定義される。通常、配列アイデンティティまたは類似性が比較され、典型的に全長にわたって配列が比較される。しかし配列は、より短い比較ウィンドウにわたって比較されてもよい。当該技術分野で「アイデンティティ」はまた、場合によってはそのような配列のストリング間のマッチによって判定される。アミノ酸または核酸配列間の配列関連性の程度を意味する。
【0100】
アイデンティティを判定する好ましい方法は、試験される配列間に最大のマッチを与えるようにデザインされる。アイデンティティおよび類似性を判定する方法は、公的に入手可能なコンピュータプログラムにおいて体系化される。2つの配列間のアイデンティティおよび類似性を判定する好ましいコンピュータプログラム法としては、例えばBestFit、BLASTP、BLASTN、およびFASTA(Altschul,S.F.et al.,J.Mol.Biol.215:403−410(1990)、NCBIおよびその他の情報源から公的に入手可能(BLAST Manual,Altschul,S.,et al.,NCBI NLM NIH Bethesda,MD 20894)が挙げられる。BLASTPを使用したアミノ酸配列比較の好ましいパラメーターは、gap open 11.0、gap extend 1、Blosum62 matrixである。BLASTPを使用した核酸配列比較の好ましいパラメーターは、gap open 11.0、gap extend 1、DNA full matrix(DNA identity matrix)である。
【0101】
任意選択的に、アミノ酸の類似度を判定する上で、当業者はまた、当業者には明らかであるように、いわゆる「保存的」アミノ酸置換を考慮してもよい。
【0102】
保存的アミノ酸置換とは、同様の側鎖を有する残基の互換性を指す。例えば脂肪族側鎖を有する一群のアミノ酸はグリシン、アラニン、バリン、ロイシン、およびイソロイシンであり;脂肪族ヒドロキシル側鎖を有する一群のアミノ酸はセリンおよびスレオニンであり;アミド含有側鎖を有する一群のアミノ酸はアスパラギンおよびグルタミンであり;芳香族側鎖を有する一群のアミノ酸はフェニルアラニン、チロシン、およびトリプトファンであり;塩基性側鎖を有する一群のアミノ酸はリジン、アルギニン、およびヒスチジンであり;イオウ含有側鎖を有する一群のアミノ酸はシステインおよびメチオニンである。
【0103】
好ましい保存的アミノ酸置換基は、バリン−ロイシン−イソロイシン、フェニルアラニン−チロシン、リジン−アルギニン、アラニン−バリン、およびアスパラギン−グルタミンである。本明細書で開示される置換変異型アミノ酸配列とは、その中で開示される配列の少なくとも1つの残基が除去され、その場所に異なる残基が挿入されるものである。好ましくは、アミノ酸変化は保存的である。天然アミノ酸のそれぞれについて、好ましい保存的置換を下に示す。Alaからser;Argからlys;Asnからglnまたはhis;Aspからglu;Cysからserまたはala;Glnからasn;Gluからasp;Glyからpro;Hisからasnまたはgln;Heからleuまたはval;Leuからileまたはval;Lysからarg;glnまたはglu;Metからleuまたはile;Pheからmet、leuまたはtyr;Serからthr;Thrからser;Trpからtyr;Tyrからtrpまたはphe;およびValからileまたはleu。
【0104】
ストリンジェントなハイブリダイゼーション条件は、本明細書で、少なくとも約25個、好ましくは約50個のヌクレオチド、75個または100個および最も好ましくは約200個以上のヌクレオチドの核酸配列を約1Mの塩、好ましくは6×SSC(塩化ナトリウム、クエン酸ナトリウム)を含んでなる溶液中、または比較できるイオン強度を有するあらゆるその他の溶液中において、約65℃の温度でハイブリッド形成させ、約0.1M以下の塩、好ましくは0.2×SSCを含んでなる溶液、または比較できるイオン強度を有するあらゆるその他の溶液中において、65℃で洗浄する条件と定義される。好ましくはハイブリダイゼーションは、一晩、すなわち少なくとも10時間にわたり実施され、好ましくは洗浄は、洗浄溶液を少なくとも2回取り替えて少なくとも1時間にわたり実施される。これらの条件は、通常、約90%以上の配列同一性を有する配列が特異的にハイブリダイゼーションできるようにする。
【0105】
本明細書で中程度の条件は、少なくとも50個のヌクレオチド、好ましくは約200個以上のヌクレオチドの核酸配列を約1Mの塩、好ましくは6×SSCを含んでなる溶液中、または比較できるイオン強度を有するあらゆるその他の溶液中において、約45℃の温度でハイブリッド形成させ、約1M塩、好ましくは6×SSCを含んでなる溶液、または比較できるイオン強度を有するあらゆるその他の溶液中において、室温で洗浄する条件と定義される。好ましくはハイブリダイゼーションは、一晩、すなわち少なくとも10時間にわたり実施され、好ましくは洗浄は、洗浄溶液を少なくとも2回取り替えて少なくとも1時間にわたり実施される。これらの条件は、通常、50%以下の配列同一性を有する配列が特異的にハイブリダイゼーションできるようにする。当業者は、アイデンティティが50%〜90%で変動する配列を特異的に同定するために、これらのハイブリダイゼーション条件を変更できる。
【0106】
導入された酵素が本発明の細胞中において活性形態で発現する可能性を増大させるために、対応するコードディングヌクレオチド配列を適応させて、そのコドン使用頻度を選択された酵母細胞にとって最適化させてもよい。いくつかのコドン最適化法が、当該技術分野で知られている。酵母のヌクレオチド配列のコドン使用頻度を最適化する好ましい方法は、国際公開第2006/077258号パンフレットおよび/または国際公開第2008/000632号パンフレットで開示されるようなコドンペア最適化技術である。国際公開第2008/000632号パンフレットは、コドンペア最適化に対処する。コドンペア最適化は、ポリペプチドコードするヌクレオチド配列をそれらのコドン、特に使用されるコドンペアの使用頻度について改変し、ポリペプチドをコードするヌクレオチド配列の改善された発現および/またはコードされたポリペプチドの改善された生成を得る方法である。コドンペアは、コード配中の2個の引き続く三塩基(コドン)の組と定義される。
【0107】
本明細書では、遺伝子発現および翻訳効率の簡単な尺度として、Xuhua Xia,Evolutionary Bioinformatics 2007,:3 53−58で記載されるコドン適応指数(CAI)を使用する。指数は、高度に発現される種からの遺伝子の参照集合を使用して各コドンの優劣を評価し、遺伝子のスコアをその遺伝子中の全コドンの使用頻度から計算する。指数は、コドン使用頻度パターンを形成する上で選択が効果的である程度を評価する。その点において、それらの宿主に対するウィルス性遺伝子の適応を評価するために、異なる生物中のコドン使用頻度の比較を行うために、遺伝子発現レベルを予測することが有用である。指数はまた、異種遺伝子発現の成功可能性のおよその指標も与えてもよい。本発明に従ったコドンペア最適化された遺伝子中では、CAIは0.6以上、0.7以上、0.8以上、0.85以上、0.87以上、0.90以上、0.95以上、または約1.0である。
【0108】
したがって本発明の細胞は、上で定義されるaraA、araB、およびaraD遺伝子をコードするヌクレオチド配列を含んでなる核酸コンストラクトを含んでなる、すなわちそれで形質転換された細胞である。araAコード配列を含んでなる核酸コンストラクトは、好ましくは宿主細胞中でaraA遺伝子を発現できる。
【0109】
好ましくは遺伝子は、細胞質ゾル中で発現される。細胞質性発現は、ミトコンドリアまたはペルオキシソーム標的シグナルの欠失または修飾によって達成されてもよい。
【0110】
[バイオ製品生産]
長年にわたって、糖科作物からバイオエタノールを生産するために、様々な生物の導入が提案されてきた。しかし実際には、全ての主要なバイオエタノール製造工程は、エタノール生産菌としてサッカロミセス(Saccharomyces)属の酵母を使用し続けている。これは工業的方法のための、サッカロミセス(Saccharomyces)種の数多くの魅力的な特徴、すなわち高い酸、エタノール、および浸透圧耐性、嫌気的成長能力、そしてもちろんその高いアルコール発酵能力のためである。宿主細胞として好ましい酵母種としては、S.セレヴィシエ(S.cerevisiae)、S.ブルデリ(S.bulderi)、S.バルネッチ(S.barnetti)、S.エクシグウス(S.exiguus)、S.ウバラム(S.uvarum)、S.ディアスタティカス(S.diastaticus)、K.ラクチス(K.lactis)、K.マルキシアヌス(K.marxianus)、またはK.フラギリス(K.fragilis)が挙げられる。
【0111】
本発明の細胞は、植物バイオマス、セルロース、ヘミセルロース、ペクチン、ラムノース、ガラクトース、果糖、マルトース、マルトデキストリン、リボース、リブロース、またはデンプン、デンプン誘導体、スクロース、乳糖およびグリセロールを例えば発酵性糖に転換できてもよい。したがって本発明の細胞は、セルロースのグルコースモノマーへの、そしてヘミセルロースのキシロースおよびアラビノースモノマーへの転換に必要なセルラーゼ(エンドセルラーゼまたはエキソセルラーゼ)、ヘミセルラーゼ(エンドまたはエキソキシラナーゼまたはアラビナーゼ);ペクチンをグルクロン酸およびガラクツロン酸に転換できるペクチナーゼ;またはデンプンをグルコースモノマーに転換するアミラーゼなどの1つ以上の酵素を発現してもよい。
【0112】
本細胞は、さらに好ましくは、ピルビン酸から、エタノール、ブタノール、乳酸、3−ヒドロキシ−プロピオン酸、アクリル酸、酢酸、コハク酸、クエン酸、フマル酸、リンゴ酸、イタコン酸、アミノ酸、1,3−プロパン−ジオール、エチレン、グリセロール、β−ラクタム抗生物質またはセファロスポリンなどの所望の発酵産物への転換に必要とされる酵素活性をさらに含んでなる。
【0113】
好ましい本発明の細胞は、天然でアルコール発酵、好ましくは嫌気性アルコール発酵できる細胞である。本発明の細胞は好ましくは、高いエタノール耐性、高い低pH耐性(すなわち約5、約4、約3、または約2.5よりも低いpHで成長できる)、および乳酸、酢酸またはギ酸のような有機酸、および/またはフルフラールおよびヒドロキシメチルフルフラールなどの糖分解生成物に対する高い耐性、および/または高い高温耐性を有する。
【0114】
上記のいずれかの本発明の細胞の特徴または活性は、細胞内に天然に存在してもよく、または遺伝子改変によって導入または改変されてもよい。
【0115】
本発明の細胞は、エタノール生産に適した細胞であってもよい。しかし本発明の細胞は、エタノール以外の発酵産物生産に適していてもよい。このような非エタノール発酵産物としては、原則的には、酵母または糸状菌などの真核微生物が生成可能なあらゆるバルクまたはファインケミカルが挙げられる。
【0116】
このような発酵産物は、例えばブタノール、乳酸、3−ヒドロキシ−プロピオン酸、アクリル酸、酢酸、コハク酸、クエン酸、リンゴ酸、フマル酸、イタコン酸、アミノ酸、1,3−プロパン−ジオール、エチレン、グリセロール、β−ラクタム抗生物質またはセファロスポリンであってもよい。非エタノール発酵産物のための好ましい本発明の細胞は、アルコールデヒドロゲナーゼ活性の低下をもたらす遺伝子改変を含有する宿主細胞である。
【0117】
さらなる態様では、本発明は、その中で本発明細胞が、キシロースなどのキシロース源を含んでなる炭素源発酵のために使用される発酵工程に関する。キシロース源に加えて発酵培地中の炭素源は、グルコース源もまた含んでなってもよい。キシロースまたはグルコース源は、キシロースまたはグルコースそれ自体であってもよく、または例えばリグノセルロース、キシラン、セルロース、デンプンなどのキシロースまたはグルコース単位を含んでなる、あらゆる炭水化物オリゴマーまたはポリマーであってもよい。このような炭水化物からのキシロースまたはグルコース単位の放出のために、適切なカルボヒドラーゼ(キシラナーゼ、グルカナーゼ、アミラーゼなど)が発酵培地に添加されてもよく、または細胞によって生成されてもよい。後者の場合、細胞は遺伝子操作されて、このようなカルボヒドラーゼを生成し分泌してもよい。オリゴまたはポリマーグルコース源を使用することの追加的利点は、例えば律速量のカルボヒドラーゼを使用することにより、それが発酵中に(より)低い濃度の遊離グルコースを維持できるようにすることである。これは、次に、キシロースなどの非グルコース糖の代謝および輸送に必要とされるシステムの抑制を防止する。
【0118】
好ましい方法では、細胞はキシロースおよびグルコースの双方を好ましくは同時に発酵し、その場合、好ましくはジオーキシー成長を妨げるグルコース抑制に非感受性の細胞が使用される。炭素源としてのキシロース(およびグルコース)源に加えて、発酵培地は、細胞成長に必要とされる適切な成分をさらに含んでなる。酵母などの微生物の成長のための発酵培地組成物は、当該技術分野で良く知られている。発酵過程は、例えばエタノール、ブタノール、乳酸、3−ヒドロキシ−プロピオン酸、アクリル酸、酢酸、コハク酸、クエン酸、リンゴ酸、フマル酸、イタコン酸、アミノ酸、1,3−プロパン−ジオール、エチレン、グリセロール、ペニシリンGまたはペニシリンVとその発酵誘導体などのβ−ラクタム抗生物質、およびセファロスポリンなどの発酵産物生産のための過程である。
【0119】
[リグノセルロース]
潜在的な再生可能原材料とみなされてもよいリグノセルロースは、一般に多糖類セルロース(グルカン)およびヘミセルロース(キシラン、ヘテロキシラン、およびキシログルカン)を含んでなる。これに加えて例えば木材由来原材料の中に、ある種のヘミセルロースがグルコマンナンとして存在してもよい。例えばこれらの多糖類のグルコース、セロビオース、キシロース、アラビノース、ガラクトース、果糖、マンノース、ラムノース、リボース、ガラクツロン酸、グルクロン酸、およびその他のヘキソースおよび五炭糖などのモノマーおよび多量体の双方をはじめとする可溶性糖への酵素加水分解は、協調して作用する異なる酵素作用の下で起きる。
【0120】
さらにペクチン、およびアラビナンなどのその他のペクチン質が、典型的な非木本組織からの細胞壁の乾燥質量のかなりの部分を構成してもよい(乾燥質量の約4分の1から2分の1がペクチンであってもよい)。
【0121】
[前処理]
(ヘミセルロース系をはじめとする)リグノセルロース系材料から、本発明に従って発酵させてもよい糖を放出させるために前処理が望ましいこともあり得る。このステップは、例えば以下のような従来の方法で実施してもよい。
【0122】
[酵素加水分解]
酵素加水分解は、従来の方法で実施してもよい。
【0123】
[発酵]
発酵工程は、好気的または嫌気的発酵工程であってもよい。嫌気的発酵工程は、本明細書で、酸素不在下で進行する発酵工程、またはその中で実質的に酸素が消費されず、好ましくは約5、約2.5または約1mmol/L/h未満、より好ましくは0mmol/L/h未満が消費され(すなわち酸素消費が検出可能でない)、その中で有機分子が電子供与体および電子受容体の双方の役割を果たす発酵工程と定義される。酸素の不在下で、解糖およびバイオマス形成中に生じるNADHは、酸化的リン酸化によって酸化され得ない。この問題を解決するために、多数の微生物は、電子および水素受容体としてピルビン酸またはその誘導体の1つを使用し、それによってNAD+を再生する。
【0124】
したがって好ましい嫌気的発酵工程では、ピルビン酸が電子(および水素受容体)として使用され、還元されて、エタノール、ブタノール、乳酸、3−ヒドロキシ−プロピオン酸、アクリル酸、酢酸、コハク酸、クエン酸、リンゴ酸、フマル酸、アミノ酸、1,3−プロパン−ジオール、エチレン、グリセロール、β−ラクタム抗生物質、およびセファロスポリンなどの発酵産物になる。
【0125】
発酵工程は、好ましくは細胞にとって最適の温度で実施される。したがってほとんどの酵母または真菌宿主細胞では、発酵工程は、約42℃未満、好ましくは約38℃未満の温度で実施される。酵母または糸状菌宿主細胞では、発酵工程は好ましくは約35、約33、約30または約28℃よりも低い温度、および約20、約22、または約25℃よりも高い温度で実施される。
【0126】
工程におけるキシロースおよび/またはグルコース上のエタノール収率は、好ましくは少なくとも約50、約60、約70、約80、約90、約95または約98%である。エタノール収率は、本明細書で理論的最大収率の百分率と定義される。
【0127】
本発明はまた、発酵産物を製造する方法にも関する。
【0128】
発酵工程は、バッチ、流加または連続様式で実施してもよい。別個の加水分解および発酵(SHF)工程、または同時の糖化および発酵(SSF)工程もまた適用してもよい。最適生産性のために、これらの発酵工程様式の併用もまた可能であってもよい。
【0129】
本発明に従った発酵工程は、好気的および嫌気的条件下で進行してもよい。好ましくは工程は、微好気的または酸素限定条件下で実施される。
【0130】
嫌気的発酵工程は、本明細書では、酸素不在下で進行し、またはその中で実質的に酸素が消費されず、好ましくは約5、約2.5または約1mmol/L/h未満が消費され、有機分子が電子供与体および電子受容体の双方の役割を果たす発酵工程、と定義される。
【0131】
酸素限定発酵工程は、気体から液体への酸素移動によって、酸素消費が制限される工程である。酸素制限の程度は、流入ガス流量および組成、ならびに使用される発酵装置の実際の混合/質量移動特性によって定まる。好ましくは、酸素限定条件下の工程では、酸素消費速度は、少なくとも約5.5、より好ましくは少なくとも約6、少なくとも7mmol/L/hなどである。本発明の方法は、発酵産物の回収を含んでなる。
【0132】
好ましい方法では、細胞はキシロースおよびグルコースの双方を好ましくは同時に発酵し、その場合、好ましくはジオーキシー成長を妨げるグルコース抑制に非感受性の細胞が使用される。炭素源としてのキシロース(およびグルコース)源に加えて、発酵培地は、細胞成長に必要とされる適切な成分をさらに含んでなる。酵母などの微生物の成長のための発酵培地組成物は、当該技術分野で周知である。
【0133】
発酵工程は、バッチ、流加または連続様式で実施してもよい。別個の加水分解および発酵(SHF)工程、または同時の糖化および発酵(SSF)工程もまた応用してもよい。最適生産性のために、これらの発酵工程様式の併用もまた可能であり得る。これらの工程については、以下でより詳細に説明する。
【0134】
[SSF様式]
同時糖化および発酵(SSF)様式では、液化/加水分解または前糖化工程のための反応時間は、所望の収率、すなわちセルロースからグルコースへの収率を実現する時間に左右される。このような収率は、好ましくは可能な限り高く、好ましくは60%以上、65%以上、70%以上、75%以上80%以上、85%以上、90%以上、95%以上、96%以上、97%以上、98%以上、99%以上、99.5%以上または99.9%以上でさえある。
【0135】
本発明に従って、SHF様式における非常に高い糖濃度力、およびSSF様式における非常に高い生成物(例えばエタノール)濃度が実現する。SHF操作では、グルコース濃度は、25g/L以上、30g/L以上、35g/L以上、40g/L以上、45g/L以上、50g/L以上、55g/L以上、60g/L以上、65g/L以上、70g/L以上、75g/L以上、80g/L以上、85g/L以上、90g/L以上、95g/L以上、100g/L以上、110g/L以上、120g/L以上であり、または例えば25g/L〜250g/L、30gl/L〜200g/L、40g/L〜200g/L、50g/L〜200g/L、60g/L〜200g/L、70g/L〜200g/L、80g/L〜200g/L、90g/L、80g/L〜200g/Lであってもよい。
【0136】
[SSF様式における生成物濃度]
SSF操作では、生成物濃度(g/L)は生成されるグルコース量に左右されるが、糖はSSF中で生成物に転換され、生成物濃度は、理論的な最小収率の増大により根底にあるグルコース濃度に関連し得る(Yps maxは、グルコース1グラム当たりの生成物グラムである)ので、これは目に見えない。
【0137】
発酵産物の理論的最大収率(Yps maxは、グルコース1グラム当たりの生成物グラムである)は、教科書通りの生化学から導かれ得る。エタノールでは、1モルのグルコース(180g)は、酵母中の通常解糖発酵経路に従って、2モルのエタノール(=2×46=92グラムのエタノール)を生じる。したがってグルコース上のエタノールの理論的最大収率は、92/180=0.511gのエタノール/グルコース1グラムである。
【0138】
ブタノール(MW74g/モル)またはイソブタノールでは、理論的最大収率はグルコース1グラム当たり1モルのブタノールである。したがって(イソ−)ブタノールのYps max=74/180=0.411g(イソ−)ブタノール/グルコース1グラムである。
【0139】
乳酸では、ホモ乳酸発酵の発酵収率は、1モルのグルコース当たり2モルの乳酸(MW=90g/モル)である。この化学量論によれば、Yps max=乳酸1g/グルコース1gである。
【0140】
その他の発酵産物についても、同様の計算を実施してもよい。
【0141】
[SSF様式]
SSF操作では、生成物濃度は、25g*Yps g/L/L以上、30*Yps g/L以上、35g*Yps/L以上、40*Yps g/L以上、45*Yps g/L以上、50*Yps g/L以上、55*Yps g/L以上、60*Yps g/L以上、65*Yps g/L以上、70*Yps g/L以上、75*Yps g/L以上、80*Yps g/L以上、85*Yps g/L以上、90*Yps g/L以上、95*Yps g/L以上、100*Yps g/L以上、110*Yps g/L以上、120g/L*Yps以上であり、または例えば25*Yps g/L〜250*Yps g/L、30*Yps gl/L〜200*Yps g/L、40*Yps g/L〜200*Yps g/L、50*Yps g/L〜200*Yps g/L、60*Yps g/L〜200*Yps g/L、70*Yps g/L〜200*Yps g/L、80*Yps g/L〜200*Yps g/L、90*Yps g/L、80*Yps g/L〜200*Yps g/Lであってもよい。
【0142】
したがって本発明は発酵産物を調製する方法を提供し、方法は、
a.本明細書に記載される方法を使用してリグノセルロースを分解するステップと;
b.得られた物質を発酵し、
それによって発酵産物を調製するステップ
を含んでなる。
【0143】
[発酵産物]
本発明の発酵産物は、いかなる有用な生成物であってもよい。一実施態様では、これは、エタノール;n−ブタノール;イソブタノール;乳酸;3−ヒドロキシ−プロピオン酸;アクリル酸;酢酸;コハク酸;フマル酸;リンゴ酸;イタコン酸;マレイン酸;クエン酸;アジピン酸;リジン、メチオニン、トリプトファン、スレオニン、およびアスパラギン酸などのアミノ酸;1,3−プロパン−ジオール;エチレン;グリセロール;β−ラクタム抗生物質およびセファロスポリン;ビタミン;医薬品;動物飼料補給剤;特殊化学薬品;化学原料;プラスチック;溶剤;生物燃料およびバイオガスまたは有機ポリマーをはじめとする燃料;およびプロテアーゼ、セルラーゼ、アミラーゼ、グルカナーゼ、ラクターゼ、リパーゼ、リアーゼ、オキシド還元酵素、トランスフェラーゼまたはキシラナーゼなどの工業用酵素からなる群から選択される生成物である。
【0144】
[発酵産物の回収]
発酵生成物の回収のためには、既存の技術が使用される。異なる発酵産物では、異なる回収工程が適切である。水性混合物からエタノールを回収する既存の方法は、一般に分留および吸着技術を使用する。例えばビール蒸留缶を使用して、エタノールを水性混合物中に含有する発酵産物を処理して濃縮エタノール含有混合物を生成し、次に分留(例えば分別蒸留またはその他の類似技術)を実施し得る。次に最高濃度のエタノールを含有する留分を吸収材に通過させ、エタノールから残りの水の全部ではないが大部分を除去し得る。
【0145】
以下の実施例は、本発明を例証する。
【0146】
[実施例]
特に断りのない限り、使用された方法は標準生化学的技術である。適切な一般方法の教科書は例えば、Sambrook et al.,Molecular Cloning,a Laboratory Manual(1989)and Ausubel et al.,Current Protocols in Molecular Biology(1995),John Wiley & Sons,Inc.を含む。
【0147】
[S.セレヴィシエ(S.cerevisiae)の形質転換]
S.セレヴィシエ(S.cerevisiae)の形質転換は、Gietz and Woods(2002;Transformation of the yeast by the LiAc/SS carrier DNA/PEG method.Methods in Enzymology 350:87−96)に記載されるようにして実施した。
【0148】
[コロニーPCR]
単一コロニー分離株をプラスチック爪楊枝で拾って50μlミリQ水に再懸濁した。サンプルを99℃で10分間インキュベートした。5μlのインキュベートしたサンプルをPhusion(登録商標)DNAポリメラーゼ(Finnzymes)を使用したPCR反応のためのテンプレートとして、供給元によって提供される使用説明書に従って使用した。
【0149】
【表2】
【0150】
[培地組成]
成長実験:サッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)株を以下の組成を有する培地上で成長させた。0.67%(w/v)酵母窒素ベースまたは合成培地(Verduyn et al.,Yeast 8:501−517,1992)およびグルコース、アラビノース、ガラクトースまたはキシロースのいずれか、またはこれらの基質の組み合わせ(下記参照)。寒天プレートでは、培地に2%(w/v)細菌用寒天を添加した。
【0151】
[エタノール生成]
培養は、30℃でBAM(Biological Activity Monitor;オランダ国(The Netherlands)のHalotec)内において、100mlの合成モデル培地(5%グルコース、5%キシロース、3.5%アラビノースおよび1〜1.5%ガラクトースを添加したVerduyn培地(Verduyn et al.,Yeast 8:501−517,1992))中で実施した。滅菌に先だって、培地のpHを2MのNaOH/H2SO4で4.2に調節した。嫌気的培養のための合成培地に、エタノールに溶解させた0.01g/Lのエルゴステロールおよび0.42g/LのTween80を添加した(Andreasen and Stier.J.Cell Physiol.41:23−36,1953;およびAndreasen and Stier.J.Cell Physiol.43:271−281,1954)。培養物を磁気撹拌機によって撹拌した。培養物に通気しなかったために、嫌気的条件は発酵中に迅速に発生した。CO2の生成を絶えずモニターした。糖転換および生成物形成をNMRによって分析した。培養物の光学濃度をLKB Ultrospec K分光光度計上で600nmで追跡することにより、成長をモニターした。
【0152】
100ml振盪フラスコ内の2%グルコースを添加した25mlのVerduyn培地(Verduyn et al.,Yeast 8:501−517,1992)に、冷凍保存培養または寒天プレートからの単一コロニーを接種して、前培養を調製した。軌道振盪機(200rpm)内における30℃でおよそ24時間のインキュベーション後にこの培養物を収集し、およそ2のOD600でBAM接種のために使用した。
【0153】
[実施例1]
「S.セレヴィシエ(S.cerevisiae)ゲノムへの遺伝子araA、araB、およびaraDの導入]
[1.1 アラビノース経路の遺伝子を含有する発現ベクターの構築]
図2に記載されるプラスミドpPWT018を次のようにして構築した。ベクターpPWT006(図1、SIT2遺伝子座からなる(Gottlin−Ninfa and Kaback(1986)Molecular and Cell Biology vol.6,no.6,2185−2197))と、抗生物質G418上およびアセトアミド上で成長する能力(上記参照)により形質転換体を選択できるようにするマーカーとを制限酵素BsiWIおよびMluIで消化した。G418耐性をもたらすkanMX−マーカーは、p427TEF(Dualsystems Biotech)から単離され、amdSマーカーを含有する断片については文献で記載されている(Swinkels,B.W.,Noordermeer,A.C.M.and Renniers,A.C.H.M(1995)The use of the amdS cDNA of Aspergillus nidulans as a dominant,bidirectional selectable marker for yeast transformation.Yeast Volume 11,Issue 1995A,page S579;および米国特許第6051431号明細書)。国際公開第2008/041840号パンフレットで開示されるような、ラクトバチルス・プランタルム(Lactobacillus plantarum)からのアラビノースイソメラーゼ(araA)、L−リブロキナーゼ(araB)、およびL−リブロース−5−リン酸−4−エピメラーゼ(araD)をコードする遺伝子は、のBaseClear(オランダ国ライデン(Leiden,The Netherlands))によって合成された。S.セレヴィシエ(S.cerevisiae)からの強力なプロモーター、すなわちaraA遺伝子の発現を制御するTDH3プロモーター、araB遺伝子を制御するENO1プロモーター、およびaraD遺伝子を制御するPGI1プロモーターの制御下にある(またはそれらと作動可能に結合している)上述の3つのアラビノース遺伝子を含む、1個の大きな断片が合成された。この断片は、ユニークな制限酵素Acc65lおよびMluIで取り囲まれた。この断片のpPWT006へのクローニング、BsiWIおよびMluIによる消化は、プラスミドpPWT018をもたらした(図2)。プラスミドpPWT018の配列を配列番号17で記載する。
【0154】
[1.2 酵母形質転換]
供給元によって提供される使用説明書に従って、あらかじめSfiI(New England Biolabs)で直線化したプラスミドpPWT018でCEN.PK113−7D[MATa URA3 HIS3 LEU2 TRP1 MAL2−8 SUC2)を形質転換した。合成SfiI部位をSIT2遺伝子の5’−flank中にデザインした(図2参照)。形質転換混合物を1ml当たり100μgのG418(シグマアルドリッチ(Sigma Aldrich))を含有するYPD−寒天(1リットル当たり10グラムの酵母抽出物、1リットル当たり20グラムのペプトン、1リットル当たり20グラムのデキストロース、20グラムの寒天)上に播種した。2〜4日後にはコロニーがプレート上に出現したが、陰性対照(すなわち形質転換実験中でDNA添加なし)はブランクのYPD/G418プレートをもたらした。プラスミドpPWT018の組み込みは、SIT2遺伝子座を対象とする。質転換体は、PCRおよびサザンブロット技術を使用して特性決定した。
【0155】
配列番号18および15と、15および14で示されるプライマーを用いて、プラスミドpPWT018の1コピーの正しい組み込みの指標となるPCR反応を実施した(図4参照)。配列番号18および15のプライマー対によって、SIT2遺伝子座での正しい組み込みをチェックした。プラスミドpPWT018が複数コピー組み込まれれば(頭尾組み込み)、配列番号15および14のプライマー対はPCR生成物を与える。後者のPCR産物が不在であれば、これはpPWT018の1コピー組み込みの徴候である。プラスミドpPWT018の1コピーがSIT2遺伝子座中に組み込まれた株をBIE104R2と命名した。
【0156】
[1.3 マーカーレスキュー]
同一選択マーカーを使用して、酵母株をその他のコンストラクトで形質転換できるようにするためには、選択可能なマーカーを除去することが必要である。プラスミドpPWT018のデザインによって、染色体へのpPWT018組み込みに際し、相同的な配列が互いにごく接近するようになる。このデザインは、これらの相同領域の突発性分子内組み換えによって、選択可能なマーカーが喪失できるようにする。
【0157】
栄養成長に際して、低頻度ではあるが分子内組み換えが起きる。この遺伝子組み換え頻度は、相同性の長さおよびゲノム中の遺伝子座(結果未公開)に左右される。培養物副画分の新鮮培地への連続的移動に際して、分子内組み換えは経時的に蓄積する。
【0158】
この目的を達成するために、単一コロニー分離株から開始して、YPD培地(1リットル当たり10グラムの酵母抽出物、1リットル当たり20グラムのペプトン、1リットル当たり20グラムのデキストロース)中でBIE104R2株を培養した。25μlの一晩培養物を使用して、新鮮なYPD培地に接種した。少なくとも5回のこのような連続的移動後、培養物の光学濃度を測定し、1mlあたりおよそ5000個の濃度に細胞を希釈した。100μlの細胞懸濁液を30mMのKPi(pH6.8)、0.1%(NH4)2SO4、40mMのフルオロ−アセトアミド(アマシャム(Amersham))、および1.8%の寒天(ディフコ(Difco))を含有する酵母炭素ベース培地(ディフコ)に播種した。BIE104R2株の細胞と同一である、すなわち細胞内組み換えのない細胞は、依然としてamdS遺伝子を含有した。これらの細胞にとって、フルオロ−アセトアミドは有毒である。これらの細胞はフルオロアセトアミド含有培地上で成長できず、コロニーを形成しない。しかし分子内組み換えが起きた場合、選択可能なマーカーを失ったBIE104R2変異株はフルオロ−アセトアミドを成長阻害化合物に転換できないので、フルオロアセトアミド培地上で成長できる。これらの細胞はこの寒天培地上でコロニーを形成する。
【0159】
このようにして得られたフルオロ−アセトアミド抵抗性コロニーに、配列番号18および15と、14および19のプライマーを使用したPCR分析を実施した。配列番号18および5のプライマーは、選択可能なマーカーの組み換えが意図したように起きていればバンドを与える。その結果、強力酵母プロモーター制御下の遺伝子araA、araB、およびaraDのあるカセットが、宿主株のゲノムSIT2遺伝子座に組み込まれた。この場合、プライマー14は組み換えのために失われたはずの領域内でプライムするので、配列番号14および19のプライマーを使用したPCR反応は、PCR産物をもたらさないはずである。後者のプライマーによってバンドが得られた場合、これはゲノム中の完全なプラスミドpPWT018の存在の徴候であり、したがって組み換えは起きていない。
【0160】
配列番号18および15のプライマーがPCR産物をもたらさなければ、組み換えは起きているが、完全なプラスミドpPWT018は組み換えによってゲノムの外にある。選択可能なマーカーだけでなく、アラビノース遺伝子もまた失われた。実際、野生型酵母が回復されていた。
【0161】
pPWT018の1コピーの組み込みに従ってPCR結果を示す分離株に、サザンブロット分析を実施した。CEN.PK113−7D株の染色体DNAと正確な組み換体えをEcoRIおよびHindIIIで消化した(二重消化)。テンプレートとしてCEN.PK113−7Dの染色体DNAを使用して、配列番号20および21プライマーによりSIT2プローブを調製した。ハイブリダイゼーション実験の結果を図3に示す。予期されるハイブリダイゼーションパターンは、図4(パネルaおよびb)に示すような物理的地図から推定されてもよい。
【0162】
野生型株では、2.35kbのバンドが観察され、それは野性型遺伝子(図4、パネルa)の予期されるサイズに一致した。プラスミドpPWT018の組み換えによる組み込みおよび部分的損失に際して、1.06kbのバンドが予期された(図4、パネルb)。実際、このバンドは図3(レーン2)に示すように観察された。
【0163】
サザンブロット法でバンドの正確なパターン(図3から推定し得るような)を示した株の1つは、BIE104A2と命名された株である。
【0164】
[1.4 非酸化的ペントースリン酸経路の4つの構成的に発現される遺伝子の導入]
遺伝子araA、araB、およびaraDを構成的に発現するサッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)BIE104A2をプラスミドpPWT080で形質転換した(図5)。プラスミドpPWT080の配列は、配列番号4に記載される。1コピーの形質転換体を選択した後の形質転換および選択手順は、1.1、1.2、および1.3節で上述したのと同じであった。手短に言えば、BIE104A2をSfiI消化pPWT080で形質転換した。形質転換混合物を1mlあたり100μgのG418(シグマアルドリッチ)を含有するYPD寒天(1リットル当たり10グラムの酵母抽出物、1リットル当たり20グラムのペプトン、1リットル当たり20グラムのデキストロース、20グラムの寒天)上に播種した。
【0165】
2〜4日後にはコロニーがプレート上に出現したが、陰性対照(すなわち形質転換実験中でDNA添加なし)はブランクのYPD/G418プレートをもたらした。
【0166】
プラスミドpPWT080の組み込みは、GRE3遺伝子座を対象とする。形質転換体は、PCRおよびサザンブロット技術を使用して特性決定した。
【0167】
予期されるハイブリダイゼーションパターンに従って、プラスミドpPWT080の1コピーの正しい組み込みを示す形質転換体をBIE104A2F1と命名した。
【0168】
キシロースイソメラーゼおよびキシルロキナーゼをコードする遺伝子を導入できるように(実施例5)、プラスミドpPWT080の組み込みによって導入された選択マーカーを除去することが必要である。この目的を達成するために、コロニー分離株から開始して、BIE104A2F1株をYPD培地中で培養した。25μlの一晩培養物を使用して新鮮なYPD培地に接種した。5回の連続的移動後、培養物の光学濃度を測定し、1ml当たりおよそ5000個の濃度に細胞を希釈した。100μlの細胞懸濁液を30mMのKPi(pH6.8)、0.1%のH2SO4、40mMのフルオロ−アセトアミド(アマシャム)、および1.8%の寒天(ディフコ)を含有する酵母炭素ベース培地(ディフコ)に播種した。フルオロアセトアミド抵抗性コロニーにPCR分析を実施して、正確なPCRプロフィールの場合は、サザンブロット分析を実施した(実施例1の1.3節)。サザンブロット法で正確なバンドパターンを示す株の1つは、BIE104A2P1と命名された株である。
【0169】
[実施例2]
[適応進化]
[2.1 適応進化(好気的)]
BIE104A2P1株の単一コロニー分離株を使用して、2%ガラクトースを添加したYNB培地(ディフコ)に接種した。前培養を30℃および280rpmでおよそ24時間インキュベートした。細胞を収集して、0.2の開始OD600で1%ガラクトースおよび1%アラビノースを含有するYNB培地に接種した(図8)。細胞を30℃および280rpmで成長させた。600nmでの光学濃度を定期的にモニターした。
【0170】
光学濃度の値が5に達した際、培養物のアリコートを同一培地を含有する新鮮なYNB培地に移した。添加された細胞量は、培養物の開始OD600が0.2になるような量であった。OD600が再度5に達したら、培養物のアリコートを唯一の炭素源として2%アラビノースを含有するYNB培地に移した(図8の(1)で示される事象)。
【0171】
唯一の炭素源として2%アラビノース添加YNBに移すと、およそ2週間後に成長が観察できた。600nmでの光学濃度の値が少なくとも1に達した際、細胞を0.2の開始OD600で、2%アラビノースを添加した新鮮なYNB培地を入れた振盪フラスコに移した(図8)。
【0172】
図8に示すように連続的移動を3回繰り返した。得られたアラビノースで迅速に成長できる株をBIE104A2P1cと命名した。
【0173】
[2.2適応進化(嫌気的)]
好気的条件下におけるアラビノース上での成長適応後、BIE104A2P1c株からの単一コロニーを2%グルコースを添加したYNB培地上に接種した。前培養を30℃および280rpmでおよそ24時間インキュベートした。
【0174】
細胞を収集して2%アラビノースを含有するYNB培地中に、0.2の光学濃度OD600で接種した。フラスコをウォーターロックで閉鎖して、培地およびヘッドスペースから酸素が枯渇した後の嫌気的成長条件を確実にした。OD600が最低3に達したら、培養物のアリコートを2%アラビノースを含有する新鮮なYNB培地にそれぞれ0.2の光学濃度OD600で移した(図9)。
【0175】
数回の移動後、得られた株をBIE104A2P1d(=BIE201)と命名した。
【0176】
[実施例3]
[発酵能力測定]
BIE104、BIE104A2P1c、およびBIE201株の単一コロニー分離株を使用して、2%グルコースを添加したYNB培地(ディフコ)に接種した。前培養を30℃および280rpmでおよそ24時間インキュベートした。細胞を収集し、BAM内で合成モデル培地(Verduyn et al.,Yeast 8:501−517、1992;5%グルコース、5%キシロース、3.5%アラビノース、1%ガラクトース)におよそ2の開始OD600で接種した。CO2生成を絶えずモニターした。糖転換および生成物形成をNMRによって分析した。データは指定の糖の残留量(1リットルあたりのグルコース、アラビノース、ガラクトースおよびキシロースのグラム数)および(副)生成物形成(エタノール、グリセロール)を表す。600nmで培養物の光学濃度を追跡して成長をモニターした(図10、11、12)。実験はおよそ140時間実施した。
【0177】
実験は参照株BIE104がグルコースを迅速に転換したが、アラビノースまたはガラクトースのどちらも140時間以内に転換できなかったことを明らかに示した(図10)。しかしBIE104A2P1cおよびBIE201株は、アラビノースおよびガラクトースを転換できた(それぞれ図11および12)。ガラクトースおよびアラビノース利用は、20時間以内にグルコース枯渇の直後に始まった。どちらの糖も同時に転換された。しかし嫌気的条件下におけるアラビノース成長のために改善されたBIE201株は、双方の糖をより迅速に消費した(図12)。全ての発酵で、副産物としてはグリセロールのみが生じた。BIE201の発酵のデータは、本明細書で表2に示す。
【0178】
【表3】
【0179】
以下からの最大エタノール濃度(g/l)
グルコース 21.8
アラビノース 16.1
ガラクトース 6.6
合計 44.5
実験的エタノール収率0.45gエタノール/g糖
【0180】
この計算から、糖グルコース、ガラクトース、およびアラビノースが、それぞれエタノールに変換されることが明らかである。
【0181】
[実施例4]
[糖転換に対するPPP遺伝子の影響]
PPP遺伝子が糖転換におよぼす影響を試験するために、BIE104A2およびBIE105A2株からの単一コロニーを2%グルコースを添加したYNB培地(ディフコ)に接種した。どちらの株もアラビノース遺伝子を含有し、アラビノース上での成長のために進化させた(実施例2、2.1節に記載されるように)。BIE105A2株は工業株のバックグラウンドを有する。しかしこれは前述したのと同じ方法およびコンストラクトで形質転換された(実施例1、1.2節)。
【0182】
前培養を収集し、BAM内で合成トウモロコシ繊維モデル培地(Verduyn et al.,Yeast 8:501−517、1992;5%グルコース、5%キシロース、3.5%アラビノース、1.5%ガラクトース)におよそ2の開始OD600で接種した。CO2生成を絶えずモニターした。糖転換および生成物形成をNMRによって分析した。データは指定の糖の残留量(1リットルあたりのグルコース、アラビノース、ガラクトースおよびキシロースのグラム数)および(副)生成物形成(エタノール、グリセロール)を表す。600nmで培養物の光学濃度を追跡して成長をモニターした。実験はおよそ160時間実施した。
【0183】
実験はどちらの株もグルコース枯渇直後に、PPP遺伝子を過剰発現することなく、アラビノースおよびガラクトースを転換できることを示す(図13および14)。
【0184】
[実施例5]
[キシロースイソメラーゼおよびキシルロキナーゼをコードする構成的に発現される遺伝子の導入]
[5.1 酵母形質転換]
BIE104A2P1株(MATa URA3 HIS3 LEU2 TRP1 MAL2−8 SUC2 SIT2::[TDH3−araA、ENO1−araB、PGI1−araD] ΔGRE3::[TPI1p−TAL1、ADH1p−TKL1、PGI1p−RPE1、ENO1 P−RKI1])をプラスミドpPWT042で形質転換した(図16)。プラスミドpPWT042は、ベクターpPWT007に由来する(図15)。それはPCT欧州特許第2009/52623号明細書で開示されるように、S.セレヴィシエ(S.cerevisiae)からのコドンペア最適化されたキシルロキナーゼ、およびバクテロイデス・ユニフォルミス(Bacteroides uniformis)からのコドンペア最適化されたキシロースイソメラーゼ(配列番号Q2)を含有する。BIE104A2P1の形質転換に先だって、供給元が提供する使用説明書に従って、制限酵素SfiIを使用してpPWT042を直線化した。形質転換混合物を1mlあたり100μgのG418(シグマアルドリッチ)を含有するYPD寒天(1リットル当たり10グラムの酵母抽出物、1リットル当たり20グラムのペプトン、1リットル当たり20グラムのデキストロース、20グラムの寒天)上に播種した。
【0185】
2〜4日後にはコロニーがプレート上に出現したが、陰性対照(すなわち形質転換実験中でDNA添加なし)はブランクのYPD/G418プレートをもたらした。
【0186】
プラスミドをSfiIでpPWT042消化すると、その組み込みはゲノム中のSIT4遺伝子座を対象とする(Gottlin−Ninfa and Kaback(1986)Molecular and Cellular Biology Vol.6,No.6,2185−2197)(図17)。形質転換体は、実施例1(1.2節)に記載されるようにして、PCRおよびサザンブロティング技術を使用して特性決定した。
【0187】
1コピーのpPWT042プラスミドがゲノムに組み込まれた株をBIE104A2P1Y9と命名した。
【0188】
[5.2 成長実験]
BIE104A2P1Y9株の単一コロニー分離株を使用して、2%グルコースまたは2%ガラクトースを添加したYNB培地(ディフコ)に接種した。接種されたフラスコは、600nmでの光学濃度が少なくとも2.0の値に達するまで、30℃および280rpmでインキュベートした。
【0189】
1%アラビノースおよび1%キシロースを添加したYNB培地に、0.2の開始OD600で一晩培養物を接種した。細胞を30℃および280rpmで成長させた。600nmの光学濃度を定期的にモニターした。光学濃度が2.0を超える値に達した際、培養物のアリコートを2%キシロースおよび0.2%アラビノースを含有する新鮮なYNB培地に移した。添加された細胞量は、培養物の開始OD600が0.2になるような量であった。
【0190】
光学濃度を定期的にモニターした。結果を図18のパネルa(ガラクトース上の前培養)およびパネルb(グルコース上の前培養)に示す。
【0191】
結果は株が、グルコース、ガラクトース、アラビノース、およびキシロースを利用できることを明らかに示す。
【0192】
[5.3 マーカーレスキュー]
プラスミドpPWT042の組み込みによって導入された選択マーカー除去するために、コロニー分離株から開始して、BIE104A2P1Y9株をYPD培地中で培養した。25μlの一晩培養物を使用して新鮮なYPD培地に接種した。連続的移動後、培養物の光学濃度を測定し、1ml当たりおよそ5000個の濃度に細胞を希釈した。100μlの細胞懸濁液を30mMのKPi(pH6.8)、0.1%の(NH4)2SO4、40mMのフルオロ−アセトアミド(アマシャム)、および1.8%の寒天(ディフコ)を含有する酵母炭素ベース培地(ディフコ)に播種した。フルオロアセトアミド抵抗性コロニーにPCR分析を実施して、正確なPCRプロフィールの場合は、サザンブロット分析を実施した(実施例1、1.3節)。サザンブロット法で正確なバンドパターンを示す株の1つは、BIE104A2P1X9と命名された株である。
【0193】
[5.4 成長実験]
BIE104A2P1X9(BIE104A2P1X9a1およびBIE104A2P1X9a2)株の単一コロニー分離株を使用して、2%グルコースを添加したVerduyn培地(ディフコ)に接種した。接種したフラスコを30℃および280rpmでおよそ24時間インキュベートした。
【0194】
2%キシロースを添加したVerduyn培地に一晩培養物を0.2の開始OD600で接種した。細胞を30℃および280rpmで成長させた。600nmでの光学濃度を定期的にモニターした。結果を図19に示す。結果は、BIE104A2P1X9株の独立したコロニーの双方が、マーカーレスキュー後に依然としてキシロースを利用できることを明らかに示す。実施例3で既に示されたように、株はグルコース、アラビノース、およびガラクトースを利用できる(図11および図12)。
【0195】
[実施例6]
[アラビノースおよびガラクトース上のコハク酸生成のためのS.セレヴィシエ(S.cerevisiae)形質転換]
[6.1. 発現コンストラクト]
アクチノバチルス・サクシノジェネス(Actinobacillus succinogenes)からのホスホエノールピルビン酸カルボキシキナーゼPCKa(E.C.4.1.1.49)と、トリパノソーマ・ブルセイ(Trypanosoma brucei)からのグリコソームフマル酸還元酵素FRDg(E.C.1.3.1.6)とを含んでなる発現コンストラクトpGBS414PPK−3、およびリゾプス・オリゼー(Rhizopus oryzae)からのフマラーゼ(E.C.4.2.1.2.)と、ペルオキシソームリンゴ酸デヒドロゲナーゼMDH3(E.C.1.1.1.37)を含んでなる発現コンストラクトpGBS415FUM3を本明細書に図および配列表を含めて参照として組み込んだ国際公開第2009/065778号パンフレットの19〜20および22〜30頁で既述されたようにして作成する。
【0196】
プラスミドpPWT018からのaraABD発現カセットを含んでなるPCR産物をプラスミドpRS416にクローニングすることで、ラクトバチルス・プランタルム(Lactobacillus plantarum)に由来する遺伝子araA、araB、およびaraDを含んでなる発現コンストラクトpGBS416ARAABDを構築する。Phusion(登録商標)DNAポリメラーゼ(Finnzymes)と、本明細書で配列番号22および配列番号23で定義されるPCRプライマーとを使用してPCR断片を作成する。PCR産物を制限酵素SalIおよびNotIで切断し、プラスミドpRS416も同様にする。ライゲーションおよび大腸菌(E.coli)TOP10への形質転換後、制限酵素分析に基づいて正しい組み換え体を選択する。プラスミドpGBS416ARAABDの物理的地図を図20に記載する。
【0197】
[6.2. S.セレヴィシエ(S.cerevisiae)株]
プラスミドpGBS414PPK−3、pGBS415−FUM−3をS.セレヴィシエ(S.cerevisiae)株CEN.PK113−6B[MATA ura3−52 Ieu2−112 trp1−289)に形質転換する。さらにプラスミドpGBS416ARAABDをこの酵母に形質転換して、原栄養酵母株を作り出す。発現ベクターを電気穿孔によって酵母に形質転換する。形質転換混合物を酵母窒素ベース(YNB)w/o AA(ディフコ)+2%グルコース上に播種する。
【0198】
株に、唯一の炭素源としてのアラビノース上での成長のための適応進化(2節参照)を実施する。
【0199】
[6.3. 成長実験およびコハク酸生成]
形質転換体を2%ガラクトース(w/v)を含んでなるVerduyn培地(Verduyn et al.,1992,Yeast.Jul;8(7):501−17)からなる20mlの前培養液に接種して、好気的条件下で振盪恒温器内の100ml振盪フラスコ内で30℃および250rpmで成長させる。およそ24時間後、細胞を2%グルコース、2%ガラクトースまたは2%アラビノース、またはそれらの混合物のいずれかを含有する新鮮なVerduyn培地に4連で移す。2本のフラスコは好気的条件下でインキュベートし、2本のフラスコは、例えばウォーターロックを使用してフラスコを閉鎖して、または嫌気性軌道振盪機内でのインキュベーションによって、嫌気的条件下でインキュベートする。周期的に培養サンプルを採取する。サンプルを4750rpmで5分間遠心分離する。1mlの上清を使用し、6.4.節に記載されるようにしてHPLCによりコハク酸レベルを測定する。
【0200】
[6.4. HPLC分析]
有機酸および糖の定量のためにHPLCを実施する。Phenomenex Rezex−RHM−単糖類カラムの分離の原理は、逆相機序を使用した、サイズ排除、イオン排除、およびイオン交換に基づく。検出を示差屈折率および紫外線検出器によって実施する。
【0201】
[文献]
文献番号 出典
(1)Bioresource Technology 1994 Vol.47 283−284頁
(2)Micard,Enzyme Microbiol Technology 1996 Vol 19 163−170頁
(3)DOE Radke,Idaho wheat straw composition
(4)Grohman and Botast Process Biochemistry 1997 Vol.32 No 5 405−415
(5)Saska B & B 1995 517−523
(6)PCT/欧州特許第2009/52623号明細書
(7)Zheng Appl.Biochem.Microbiol.2007,Vol.136−140 423−436頁
(8)Bradshaw Appl Biochem.Microbiol.2007 Vol 136−140 395−406頁
(9)Cara Appl Biochem.Microbiol.2007 Vol 136−140 379−394頁
【発明の詳細な説明】
【0001】
[技術分野]
本発明は、混合糖の発酵、特にグルコース、ガラクトース、およびアラビノースを含んでなる糖組成物の発酵に関する。糖組成は、リグノセルロース系材料が起源であってもよい。
【0002】
[背景技術]
化石燃料代替物として生産されるエタノールのほとんどは、目下、コーンスターチおよびサトウキビベースのスクロースの発酵に由来する。再生可能燃料を生産するという意欲的目標に到達するために、非食物バイオマスをエタノールなどの発酵産物に転換する新たな技術が開発されている。サッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)は、エタノール産業において一般に好まれる生物であるが、それはバイオマス原材料のヘミセルロース構成要素に含有される五炭素糖を利用し得ない。ヘミセルロースは、バイオマスの20〜30%を構成し得て、キシロースおよびアラビノースが最も豊富なC5糖である。キシロースイソメラーゼ(XI)の異種性発現は、酵母細胞がキシロースを代謝して発酵できるようにするための一つの選択肢である。同様にS.セレヴィシエ(S.cerevisiae)株中における細菌遺伝子araA、araB、およびaraDの発現は、アラビノースの利用と効率的なアルコール発酵をもたらす。ガラクトースはC6糖であり、それは頻繁に経済的理由のために無視できない量(全糖の約4%)でリグノセルロース中に存在することが多い糖でもある。
【0003】
J. van den Brink et al,Microbiology(2009)155,1340−1350は、グルコースがサッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)の好む炭素源であり、嫌気的条件下において、グルコース限定的発酵条件からガラクトース過剰条件への切り替えに際して、ガラクトースが消費されなかったことを開示する。
【0004】
今までのところ、グルコースおよび1つ以上のC5糖を用いるのと同じ方法で、ガラクトースを発酵産物に転換する方法は開示されていない。したがって本発明の目的は、グルコースおよび1つ以上のC5糖を用いるのと同じ方法で、ガラクトースを発酵産物に転換する方法を提供することである。
【0005】
[発明の概要]
本発明は、
a)サッカロミセス(Saccharomyces)、クリヴェロミセス(Kluyveromyces)、カンジダ(Candida)、ピチア(Pichia)、分裂酵母(Schizosaccharomyces)、ハンゼヌラ(Hansenula)、クロエケラ(Kloeckera)、シュワニオミセス(Schwanniomyces)またはヤロウィア(Yarrowia)属に属する酵母の存在下で糖組成物を発酵させるステップと、
b)発酵産物を回収するステップ
を含んでなる、糖組成物から1つ以上の発酵産物を生産する方法を提供し、酵母は遺伝子araA、araB、およびaraDを含んでなり、糖組成物はグルコース、ガラクトース、およびアラビノースを含んでなる。
【0006】
有利には糖グルコース、ガラクトース、およびアラビノースは、発酵産物に変換される。
【0007】
好ましくは混合糖細胞は、サッカロミセス(Saccharomyces)より好ましくはサッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)属に属する。
【0008】
本発明は、グルコース発酵株上における、これらの遺伝子の発現を通じて、アラビノース存在下でガラクトースを嫌気的に発酵する能力をもたらす、遺伝子araA、araB、およびaraDの使用にさらに関する。
【図面の簡単な説明】
【0009】
【図1】プラスミドpPWT006の物理的地図を示す。
【図2】プラスミドpPWT018の物理的地図を示す。
【図3】サザンブロット法オートラジオグラムを示す。野生型株CEN.PK113−7D(レーン1)およびBIE104A2(レーン2)の染色体DNAをEcoRIおよびHindIIIの双方で消化する。ブロットを特異的SIT2プローブとハイブリダイズする。
【図4】野生型SIT2遺伝子座の物理的地図(パネルa)、およびプラスミドpPWT018組み込みによるara遺伝子導入と、それに続く、ベクター損失と選択可能なマーカー配列をもたらす分子内遺伝子組み換え後の物理的地図(パネルb)を示す。プローブのハイブリダイゼーションが表示される。
【図5】配列が配列番号4に記載される、プラスミドpPWT080の物理的地図を示す。
【図6−1】野生型GRE3遺伝子座の物理的地図(パネルa)、およびGRE3遺伝子座への1コピーのPWT080の組み込みの物理的地図を示す(パネルbはプライマー結合部位を示し、パネルcはRKI1プローブ結合部位を示す)。
【図6−2】野生型GRE3遺伝子座の物理的地図(パネルa)、およびGRE3遺伝子座への1コピーのPWT080の組み込みの物理的地図を示す(パネルbはプライマー結合部位を示し、パネルcはRKI1プローブ結合部位を示す)。
【図7】GRE3遺伝子のコード領域がPPP遺伝子TAL1、TKL1、RKI1、およびRPE1の組み込みによって置換されたGRE3遺伝子座の物理的地図を示す。パネルaは配列番号5および6のプライマー結合部位を示し、パネルbはRKI1プローブ結合部位を示す。
【図8】異なる培地上でのBIE104P1A2の好気的条件下の成長曲線を示す。BIE104A2P1株は、YNB2%ガラクトース上で前培養された。成長曲線は2%ガラクトースおよび1%アラビノースで開始され、グラフで番号(1)で示される事象である2%アラビノースを唯一の炭素源とするYNBへの移動がそれに続いた。OD600が1を超えた後、培養物を0.2の開始OD600で新鮮な培地に移した。純粋アラビノース培地上での3回の移動後、得られた株をBIE104P1A2cと命名した。
【図9】嫌気的条件下における、2%アラビノースを唯一の炭素源とするYNB上でのBIE104P1A2cの成長曲線を示す。OD600が1を超えた後、培養物を0.2の開始OD600で新鮮な培地に移した。数回の移動後、得られた株をBIE104P1A2d(=BIE201)と命名した。
【図10】合成トウモロコシ繊維モデル培地上でのBIE104の糖転換および生成物形成を示す。CO2生成を絶えず測定した。成長は培養物の光学濃度を追跡してモニターした。前培養は2%グルコース上で成長させた。
【図11】合成トウモロコシ繊維モデル培地上でのBIE104P1A2cの糖転換および生成物形成を示す。CO2生成を絶えず測定した。成長は培養物の光学濃度を追跡してモニターした。前培養は2%グルコース上で成長させた。
【図12】合成トウモロコシ繊維モデル培地上でのBIE201の糖転換および生成物形成を示す。CO2生成を絶えず測定した。成長は培養物の光学濃度を追跡してモニターした。前培養は2%グルコース上で成長させた。
【図13】合成トウモロコシ繊維モデル培地上でのBIE104A2の糖転換および生成物形成を示す。CO2生成を絶えず測定した。成長は培養物の光学濃度を追跡してモニターした。前培養は2%グルコース上で成長させた。
【図14】合成トウモロコシ繊維モデル培地上でのBIE105A2の糖転換および生成物形成を示す。CO2生成を絶えず測定した。成長は培養物の光学濃度を追跡してモニターした。前培養は2%グルコース上で成長させた。
【図15】プラスミドpPWT007の物理的地図を示す。
【図16】プラスミドpPWT042の物理的地図を示す。
【図17】野生型SIT4遺伝子座の物理的地図を示す(パネルa)、およびSIT4遺伝子座への1コピーのPWT080組み込み(パネルbはプライマー結合部位を示す)。
【図18−1】異なる培地上でのBIE104A2P1Y9株の成長曲線グラフである。パネルa:BIE104A2P1Y9株をグルコース上で成長させて、グラフで番号(1)で示される事象である1%アラビノース+1%キシロースへの移動、(2)2%キシロース+0.2%アラビノースへの移動がそれに続いた。パネルb:BIE104A2P1Y9株をガラクトース上で成長させて、(1)1%アラビノース+1%キシロースへの移動、および(2)2%キシロース+0.2%アラビノースへの移動がそれに続いた。
【図18−2】異なる培地上でのBIE104A2P1Y9株の成長曲線グラフである。パネルa:BIE104A2P1Y9株をグルコース上で成長させて、グラフで番号(1)で示される事象である1%アラビノース+1%キシロースへの移動、(2)2%キシロース+0.2%アラビノースへの移動がそれに続いた。パネルb:BIE104A2P1Y9株をガラクトース上で成長させて、(1)1%アラビノース+1%キシロースへの移動、および(2)2%キシロース+0.2%アラビノースへの移動がそれに続いた。
【図19】2%キシロースを添加したVerduyn培地上でのBIE104A2P1Y9株の成長を示す。2つの独立したコロニーを試験した。OD600が2に達した後、株を新鮮な培地に移して、即座にキシロース上での生育を再開した。
【図20】プラスミドpGBS416ARAABDの物理的地図を示す。
【0010】
[配列表の簡単な説明]
配列番号1は、バクテロイデス・ユニフォルミス(Bacteroides uniformis)ATCC8492からの野生型キシロースイソメラーゼ配列を示す。Genbank登録番号AAYH02000036。
配列番号2は、配列番号1に由来するコドン最適化された配列を示す。
配列番号3は、バクテロイデス・ユニフォルミス(Bacteroides uniformis)ATCC8492からのキシロースイソメラーゼのアミノ酸配列を示す。
配列番号4は、プラスミドpPWT080の配列を示す。
配列番号5は、順方向プライマーの配列を示す。
配列番号6は、逆方向プライマーの配列を示す。
配列番号7は、診断PCRのための順方向多官能性プライマーの配列を示す。
配列番号8は、診断PCRのための逆方向多官能性プライマーの配列を示す。
配列番号9は、順方向プライマーRKI1プローブの配列を示す。
配列番号10は、逆方向プライマーRKI1プローブの配列を示す。
配列番号11は、順方向プライマーkanMX−カセットの配列を示す。
配列番号12は、逆方向プライマーkanMX−カセットの配列を示す。
配列番号13は、順方向プライマーの配列を示す。
配列番号14は、逆方向プライマーの配列を示す。
配列番号15は、診断PCRのための順方向多官能性プライマーの配列を示す。
配列番号16は、診断PCRのための逆方向多官能性プライマーの配列を示す。
配列番号17は、プラスミドpPWT018の配列を示す。
配列番号18は、順方向プライマー組み込みpPWT018の配列を示す。
配列番号19は、逆方向プライマー組み込みpPWT018の配列を示す。
配列番号20は、順方向プライマーSIT2プローブの配列を示す。
配列番号21は、逆方向プライマーSIT2プローブの配列を示す。
配列番号22は、araABD発現カセットを増幅する順方向プライマーの配列を示す。
配列番号23は、araABD発現カセットを増幅する逆方向プライマーの配列を示す。
【0011】
[発明の詳細な説明]
本明細書および添付の特許請求の範囲全体を通じて、語句「含んでなる(comprise)」および「含む(include)」、および「含んでなる(comprises)」、「含んでなる(comprising)」、「含む(includes)」、および「含む(including)」などのバリエーションは、包括的に解釈されるものとする。すなわちこれらの語句は、文脈が許せば、具体的に列挙されないその他の要素または整数の可能な包含を伝えることが意図される。
【0012】
冠詞「a」および「an」は、本明細書では、1つまたは1つを超える(すなわち1つのまたは少なくとも1つの)冠詞の文法的目的語に言及するために使用される。一例として、「要素(an element)」は、1つの要素または1つを超える要素を意味してもよい。
【0013】
本明細書に記載される本発明の様々な実施態様は、交差して組み合わせされてもよい。
【0014】
[糖組成物]
本発明に従った糖組成物は、グルコース、アラビノース、およびガラクトースを含んでなる。本発明の方法では、有利には、糖グルコース、ガラクトース、およびアラビノースは、発酵産物に変換される。
【0015】
本発明では、これらの基準を満たすあらゆる糖組成物を使用してもよい。好ましい実施態様では、糖組成物は1つ以上のリグノセルロース系材料の加水分解産物である。本明細書でリグノセルロースは、ヘミセルロースおよびバイオマスのヘミセルロース部分を含む。またリグノセルロースは、バイオマスのリグノセルロース性画分も含む。適切なリグノセルロース系材料は、以下の一覧にあってもよい:果樹園の剪定物、シャパラル、製材廃棄物、都市木材廃棄物、一般廃棄物、伐採廃棄物、森林間伐材、短期輪作木質作物、産業廃棄物、麦わら、オート麦わら、稲わら、大麦わら、ライ麦わら、亜麻のわら、大豆殻、もみ殻、稲わら、コーングルテン飼料、オート麦外皮、サトウキビ、トウモロコシ茎葉、トウモロコシ茎、トウモロコシ穂軸、トウモロコシ苞葉、スイッチグラス、ススキ、サトウモロコシ、菜種茎、大豆茎、プレリーグラス、ガマグラス、エノコログサ;甜菜パルプ、柑橘類パルプ、種子殻、セルロース性家畜排泄物、刈り取った芝草、綿、海藻、樹木、針葉樹、広葉樹、ポプラ、松、灌木、草、小麦、麦わら、サトウキビバガス、トウモロコシ、トウモロコシ苞葉、トウモロコシ穂軸、トウモロコシ穀粒、穀物の湿式または乾式製粉からの製品または副産物、都市固形廃棄物、古紙、庭ごみ、草質材料、農業残渣、林業残渣、都市固体廃棄物、古紙、パルプ、製紙工場残渣、枝、茂み、籐類、トウモロコシ、トウモロコシ苞葉、エネルギー作物、森林、果物、花、穀物、草、草本作物、葉、樹皮、針葉、原木、根、苗木、灌木、スイッチグラス、木、野菜、果皮、つる植物、甜菜パルプ、ミドリング粉、オート麦外皮、硬材または軟材、農産加工から発生する有機廃棄物、林業木材廃棄物、またはそれらの任意の2つ以上の組み合わせ。
【0016】
リグノセルロースに由来するいくつかの適切な糖組成物、およびそれらの加水分解産物の糖組成の概要を表1に示す。列挙したリグノセルロースには、トウモロコシ穂軸、トウモロコシ繊維、もみ殻、メロンの皮、甜菜パルプ、麦わら、サトウキビバガス、木材、草、およびオリブ絞りかすが含まれる。
【0017】
【表1】
【0018】
表1から、これらのリグノセルロース中では、かなりの量の糖(平均して3.8%)がガラクトースであることが明らかである。したがってガラクトースの発酵産物への転換は、経済上重大である。
【0019】
[混合糖細胞]
混合糖細胞は、以下で定義されるように、遺伝子araA、araB、およびaraDを含んでなる。これはグルコース、アラビノース、およびガラクトースを発酵できる。本発明の一実施態様では、混合糖細胞は、好ましくはC5および/またはC6糖である1つ以上の追加的糖を発酵できる。本発明の実施態様では、混合糖細胞は、以下の1つ以上を含んでなる:混合糖細胞がキシロースを発酵できるようにするxylA遺伝子および/またはXKS1遺伝子;アルドース還元酵素(GRE3)遺伝子の欠失;細胞中のペントースリン酸経路を通じた流量を増大させるPPP遺伝子TAL1、TKL1、RPE1およびRKI1の過剰発現。
【0020】
本発明の一実施態様では、混合糖細胞は好ましくはC5および/またはC6糖である1つ以上の追加的糖を発酵できる。本発明の一実施態様では、混合糖細胞は以下の1つ以上を含んでなる:混合糖細胞にキシロースを発酵させるxylA遺伝子、XYL1遺伝子、およびXYL2遺伝子および/またはXKS1遺伝子;アルドース還元酵素(GRE3)遺伝子の欠失;細胞中のペントースリン酸経路を通じた流量を増大させるPPP遺伝子TAL1、TKL1、RPE1、およびRKI1の過剰発現。
【0021】
一実施態様では、混合糖細胞は工業用細胞であり、より好ましくは工業用酵母である。工業用細胞および工業用酵母細胞は、次のように定義されてもよい。工業工程における(酵母)細胞の生存環境が、実験室の環境とは顕著に異なる。工業用酵母細胞は、工程中に変動し得る複数の環境条件下で良好に機能できなくてはならない。このような変動としては、一緒になってサッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)細胞の成長とエタノール産生に潜在的影響を及ぼす栄養源、pH、エタノール濃度、温度、酸素濃度の変化などが挙げられる。有害工業条件の下で、環境耐性株は、頑強な成長および生産が可能であるべきである。工業酵母株は、一般に製パン工業、醸造業、ぶどう酒醸造、およびエタノール産業などのそれらが使用される用途において起こり得るこれらの環境条件の変化に対してより頑強である。一実施態様では、工業混合糖細胞は工業宿主細胞に基づいて構築され、構築は下述するように実施される。工業酵母(S.セレヴィシエ(S.cerevisiae))の例は、Ethanol Red(登録商標)(Fermentis)、Fermiol(登録商標)(DSM)、およびThermosacc(登録商標)(Lallemand)である。
【0022】
一実施態様では、混合糖細胞は阻害物質耐性である。阻害物質耐性は、阻害性化合物に対する抵抗性である。リグノセルロース中の阻害性化合物の存在およびレベルは、原材料、前処理方法、加水分解処理の変動にともなって、変化に富んでもよい。阻害物質カテゴリーの例は、カルボン酸、フランおよび/またはフェノール化合物である。カルボン酸の例は、乳酸、酢酸またはギ酸である。フランの例は、フルフラールおよびヒドロキシメチルフルフラールである。フェノール化合物の例は、バニリン、シリング酸、フェルラ酸、およびクマル酸である。阻害物質の典型的量は次のとおりである。カルボン酸では、原材料、前処理、および加水分解条件次第で、1リットル当たり数グラム、1リットル当たり20グラム以上まで。フランでは、原材料、前処理、および加水分解条件次第で、1リットル当たり数百ミリグラム、1リットル当たり数グラムまで。フェノール類では、原材料、前処理、および加水分解条件次第で、1リットル当たり数十ミリグラム、1リットル当たり1グラムまで。
【0023】
本発明に従った混合糖株は、阻害物質耐性であり、すなわちそれらは、混合糖株が幅広い用途に適用することができ、すなわちそれが異なる原材料、異なる前処理方法、および異なる加水分解条件に対して高い適用性を有するように、一般的な前処理および加水分解条件レベルで、それらが典型的に有する一般的な阻害物質に耐え得る。
【0024】
一実施態様では、工業混合糖細胞は、阻害物質耐性宿主細胞に基づいて構築され、構築は下述するように実施される。阻害物質耐性宿主細胞は、阻害物質耐性S.セレヴィシエ(S.cerevisiae)ATCC26602株が選択された、Kadar et al,Appl.Biochem.Biotechnol.(2007),Vol.136−140,847−858で示されるような阻害物質含有材料上での成長について、株をスクリーニングすることで選択されてもよい。
【0025】
一実施態様では、混合糖細胞はマーカーフリーである。本明細書での用法では、「マーカー」という用語は、マーカーを含有する宿主細胞を選択またはスクリーニングできるようにする形質または表現型をコードする遺伝子を指す。マーカーフリーとは、混合糖細胞内にマーカーが本質的に不在であることを意味する。マーカーフリーであることは、混合糖細胞の構築において抗生物質マーカーが使用され、その後除去される場合に特に有利である。マーカーの除去は、例えば分子内組み換えなどのあらゆる適切な先行技術を使用して実施されてもよい。マーカーを除去する適切な方法は、実施例で示される。
【0026】
混合糖細胞は、植物バイオマス、セルロース、ヘミセルロース、ペクチン、ラムノース、ガラクトース、フルクトース、マルトース、マルトデキストリン、リボース、リブロース、またはデンプン、デンプン誘導体、スクロース、乳糖、およびグリセロールを例えば発酵性糖に転換できてもよい。したがって混合糖細胞は、セルロースのグルコースモノマーへの、そしてヘミセルロースのキシロースおよびアラビノースモノマーへの転換に必要なセルラーゼ(エンドセルラーゼまたはエキソセルラーゼ)、ヘミセルラーゼ(エンドまたはエキソキシラナーゼまたはアラビナーゼ);ペクチンをグルクロン酸およびガラクツロン酸に転換できるペクチナーゼ;またはデンプンをグルコースモノマーに転換するアミラーゼなどの1つ以上の酵素を発現してもよい。
【0027】
混合糖細胞は、ピルビン酸から、エタノール、ブタノール、乳酸、3−ヒドロキシ−プロピオン酸、アクリル酸、酢酸、コハク酸、クエン酸、フマル酸、リンゴ酸、イタコン酸、アミノ酸、1,3−プロパン−ジオール、エチレン、グリセロール、β−ラクタム抗生物質またはセファロスポリンなどの所望の発酵産物への転換に必要とされる酵素活性をさらに含んでなってもよい。
【0028】
一実施態様では、混合糖細胞は、天然でアルコール発酵、好ましくは嫌気的アルコール発酵できる細胞である。混合糖細胞は、好ましくは高いエタノール耐性、高い低pH耐性(すなわち約5、約4、約3、または約2.5よりも低いpHで成長できる)、および有機物に対する高い耐性、および/または高い高温耐性を有する。
【0029】
上記のいずれかの混合糖細胞の特徴または活性は、細胞内に天然に存在してもよく、または遺伝子改変によって導入または修飾されてもよい。
【0030】
[混合糖株の構築]
遺伝子は、
a)強力なプロモーター制御下のPPP遺伝子、TAL1、TKL1、RPE1、およびRKI1からなるクラスター;
b)どちらも構成的プロモーター制御下のxylA遺伝子およびXKS1遺伝子からなるクラスター;
c)遺伝子araA、araB、およびaraDからなるクラスターおよび/またはxylA遺伝子および/またはXKS1遺伝子のクラスター;および
d)アルドース還元酵素遺伝子の欠失
の宿主細胞への導入と、混合糖細胞を生じる適応進化によって、混合糖細胞に導入されてもよい。上記細胞は、組み換え発現技術を使用して構築されてもよい。
【0031】
[組み換え発現]
本発明の細胞は、組み換え細胞である。すなわち本発明の細胞は、問題となっている細胞中では天然では生じないヌクレオチド配列を含んでなり、またはそれによって形質転換され、またはそれによって遺伝子改変されている。
【0032】
細胞中の酵素組み換え発現技術、ならびに本発明の細胞の追加的遺伝子改変は、当業者に良く知られている。典型的に、このような技術は、妥当な配列を含んでなる核酸コンストラクトによる細胞の形質転換を伴う。このような方法は、例えばSambrook and Russel(2001)”Molecular Cloning:A Laboratory Manual”(3rd edition),Cold Spring Harbor Laboratory,Cold Spring Harbor Laboratory Press、またはF.Ausubel etal.,eds.,”Current protocols in molecular biology”,Green Publishing and Wiley Interscience,New York(1987)などの標準的手引き書から知られている。真菌宿主細胞の形質転換および遺伝子改変方法は、例えば欧州特許出願公開第A−0635574号明細書、国際公開第98/46772号パンフレット、国際公開第99/60102号パンフレット、国際公開第00/37671号パンフレット、国際公開第90/14423号パンフレット、欧州特許出願公開第A−0481008号明細書、欧州特許出願公開第A−0635574号明細書、および米国特許第6,265,186号明細書から知られている。
【0033】
典型的に、核酸コンストラクトは、プラスミド、例えば低コピー数プラスミドまたは高コピー数プラスミドであってもよい。本発明に従った細胞は、例えばヌクレオチドコンストラクトの複数コピーによって、または酵素配列の複数コピーを有するコンストラクトの使用によって、酵素をコードするヌクレオチド配列の単数または複数コピーを含んでなってもよい。
【0034】
核酸コンストラクトはエピソームに保持されてもよく、したがって常染色体性複製配列などの自律複製配列を含んでなってもよい。適切なエピソームの核酸コンストラクトは、例えば酵母2μまたはpKD1プラスミド(Gleer et al.,1991,Biotechnology 9:968−975)に、またはAMAプラスミド(Fierro et al.,1995,Curr Genet.29:482−489)に基づいてもよい。代案としては、各核酸コンストラクトが1つ以上のコピーで、細胞ゲノムに組み込まれてもよい。細胞ゲノムへの組み込みは非相同的組換えによってランダムに生じてもよいが、好ましくは核酸コンストラクトは、当該技術分野で良く知られている相同的組換えによって細胞のゲノムに組み込まれてもよい(例えば国際公開第90/14423号パンフレット、欧州特許出願公開A−0481008号明細書、欧州特許出願公開A−0635574号明細書、および米国特許第6,265,186号明細書参照)。
【0035】
ほとんどのエピソームのプラスミドまたは2μプラスミドは比較的不安定であり、各世代毎におよそ10ー2個以上の細胞内で失われる。選択的成長条件下でさえも、細胞の60%〜95%のみがエピソームのプラスミドを保持する。ほとんどのエピソームのプラスミドのコピー数は、cir+宿主の細胞当たり10〜40個の範囲である。しかしプラスミドは細胞間で均等に分布しておらず、集団中の細胞当たりのコピー数には大きな相違がある。組み込みプラスミドで形質転換された株は、選択的圧力の不在下でさえも極めて安定している。しかしプラスミドの損失は、縦列反復DNA間の相同的組換えによっておよそ10ー3〜10ー4の頻度で起こり得て、ベクター配列のループアウトにつながる。したがって、好ましくは安定した組み込みの場合のベクターデザインは、(分子内相同的組換えによっても起きる)選択マーカー遺伝子の損失に際して、組み込みコンストラクトのループアウトはもはや可能でない。好ましくは遺伝子はこのようにして安定して組み込まれる。安定した組み込みは、本明細書では、組み込みコンストラクトのループアウトがもはや可能でない、ゲノムへの組み込みと定義される。好ましくは選択マーカーは不在である。典型的に酵素をコードする配列は、酵素配列の転写および/または翻訳を提供でき、またはそれを助けることができる、1つ以上の核酸配列と作動的に連結する。
【0036】
「作動的に連結する」という用語は、記載される構成要素がそれらの意図される様式で機能できる関係にある並置を指す。例えばプロモーターまたはエンハンサーは、コード配列と作動的に連結し、上記プロモーターまたはエンハンサーはコード配列の転写に影響を及ぼす。
【0037】
本明細書での用法では、「プロモーター」という用語は、1つ以上の遺伝子の転写を制御するよう機能する核酸断片を指し、それは遺伝子の転写開始部位の転写方向に対して上流に位置し、DNA依存RNAポリメラーゼ結合部位、転写開始部位、およびあらゆるその他の当業者に知られているDNA配列の存在によって、構造的に同定される。「構成的」プロモーターは、ほとんどの環境および発生的条件の下で活性のプロモーターである。「誘導性」プロモーターは、環境または発生的調節下で活性のプロモーターである。
【0038】
本発明に従った酵素をコードするヌクレオチド配列の発現を達成するのに使用できるプロモーターは、発現される酵素をコードするヌクレオチド配列にもともと備わっていなくてもよく、すなわちそれが作動的に連結するヌクレオチド配列(コード配列)に異種性のプロモーターであってもよい。しかしプロモーターは、宿主細胞に相同的、すなわち内在性であってもよい。
【0039】
プロモーターは広く入手でき、当業者に知られている。このようなプロモーターの適切な例としては、例えば酵母または糸状菌からのホスホフルクトキナーゼ(PFK)、トリオースリン酸イソメラーゼ(TPI)、グリセルアルデヒド−3−リン酸脱水素酵素(GPD、TDH3またはGAPDH)、ピルビン酸キナーゼ(PYK)、ホスホグリセリン酸キナーゼ(PGK)プロモーターなどの解糖作用遺伝子からのプロモーターが挙げられ、このような酵母からのプロモーターについてより詳しくは、(国際公開第93/03159号パンフレット)に見つかる。その他の別の有用なプロモーターは、リボソームタンパク質コーディング遺伝子プロモーター、ラクターゼ遺伝子プロモーター(LAC4)、アルコールデヒドロゲナーゼプロモーター(ADHI、ADH4など)、およびエノラーゼプロモーター(ENO)である。構成および誘導性の双方のその他のプロモーター、およびエンハンサーまたは上流活性化配列については、当業者に知られている。本発明の宿主細胞で使用されるプロモーターは、所望ならば修飾して、それらの制御特性に影響を及ぼしてもよい。本明細書の文脈で適切なプロモーターとしては、構成および誘導性双方の天然プロモーターならびに当業者に良く知られている遺伝子操作されたプロモーターが挙げられる。真核生物の宿主細胞中の適切なプロモーターは、GAL7、GAL10、またはGAL1、CYC1、HIS3、ADH1、PGL、PH05、GAPDH、ADC1、TRPI、URA3、LEU2、EN01、TPI1、およびAOX1であってもよい。その他の適切なプロモーターとしては、PDC1、GPD1、PGK1、TEF1、およびTDH3が挙げられる。
【0040】
本発明の細胞中では、酵素をコードするヌクレオチド酸配列の3’末端は、好ましくは転写ターミネーター配列と作動的に連結する。好ましくはターミネーター配列は、例えば選択された酵母種などの選択された宿主細胞中で作動可能である。いずれの場合もターミネーターの選択は重要でなく、それは例えばあらゆる酵母遺伝子からのものであってもよいが、非酵母、真核生物遺伝子からのものであれば、ターミネーターが時に機能することもある。通常酵素をコードするヌクレオチド配列は、ターミネーターを含んでなる。好ましくは、このようなターミネーターは、本発明の宿主細胞中のナンセンス仲介mRNA崩壊を妨げる変異と組み合わせられる(例えばShirley et al.,2002,Genetics 161:1465−1482を参照されたい)。
【0041】
転写終結配列は、好ましくはさらにポリアデニル化シグナルを含んでなる。
【0042】
任意選択的に、選択可能なマーカーは、本発明で使用するのに適した核酸コンストラクト中に存在してもよい。本明細書での用法では、「マーカー」という用語は、マーカーを含有する宿主細胞を選択またはスクリーニングできるようにする、形質または表現型をコードする遺伝子を指す。マーカー遺伝子は、それによって適切な抗生物質を使用して、形質転換されていない細胞から形質転換された細胞を選択し得る、抗生物質耐性遺伝子であってもよい。適切な抗生物質耐性マーカーの例としては、例えばジヒドロ葉酸還元酵素、ハイグロマイシン−B−リン酸転移酵素、3’−O−リン酸転移酵素Il(カナマイシン、ネオマイシン、およびG418耐性)が挙げられる。抗生物質耐性マーカーは、倍数体宿主細胞の形質転換のために最も都合よいかもしれず、栄養要求性マーカー(URA3、TRPI、LEU2)またはS.ポンベ(S.pombe)TPI遺伝子などの非抗生物質耐性マーカーもまた使用してもよい(Russell P R,1985,Gene 40:125−130によって記載される)。好ましい実施態様では、核酸コンストラクトによって形質転換される宿主細胞は、マーカー遺伝子フリーである。組み換えマーカー遺伝子フリー微生物宿主細胞を構築する方法は、欧州特許出願公開A−0635574号明細書で開示され、A.ニデュランス(A.nidulans)amdS(アセトアミダーゼ)遺伝子または酵母URA3、およびLYS2遺伝子などの二方向性マーカーの使用に基づく。代案としては、緑色蛍光タンパク質、lacL、ルシフェラーゼ、クロラムフェニコールアセチルトランスフェラーゼ、βグルクロニダーゼなどのスクリーニング可能マーカーを本発明の核酸コンストラクトに組み込んで、形質転換された細胞をスクリーニングできるようにしてもよい。
【0043】
本発明で使用するのに適した中核酸コンストラクトに存在してもよい任意のさらなる要素としては、1つ以上のリーダー配列、エンハンサー、組み込み因子、および/またはレポーター遺伝子、イントロン配列、セントロメア、テロマーおよび/またはマトリックス付着(MAR)配列が挙げられるが、これに限定されるものではない。本発明の核酸コンストラクトは、ARS配列などの自律複製のための配列をさらに含んでなってもよい。
【0044】
したがって遺伝子組み換え過程は、既知の遺伝子組み換え技術を使用して実施されてもよい。本発明の細胞中での酵素発現および過剰発現のための様々な手段が、当業者に知られている。特に、例えば遺伝子の追加的コピーを宿主細胞のゲノム中に組み込むことによって、エピソームのマルチコピー発現ベクターからの遺伝子を発現することによって、または遺伝子の複数コピーを含んでなるエピソームの発現ベクターを導入することによって、宿主細胞中で酵素をコードする遺伝子のコピー数を増大させ酵素を過剰発現させてもよい。
【0045】
代案としては、本発明の宿主細胞中の酵素の過剰発現は、過剰発現させる酵素をコードする配列に天然でないプロモーター、すなわちそれが作動的に連結するコード配列に異種性のプロモーターを使用することによって達成してもよい。プロモーターは、好ましくはそれが作動的に連結するコード配列に異種性であるが、プロモーターが相同的である、すなわち宿主細胞に内在性であることもまた好ましい。好ましくは異種性プロモーターは、コード配列に天然のプロモーターよりも、コード配列を含んでなる転写物のより高い定常状態レベルを生じることができる(または単位時間当たりより多くの転写物分子、すなわちmRNA分子を生成できる)。この文脈で適切なプロモーターとしては、構成および誘導性双方の天然プロモーターならびに改変プロモーターが挙げられる。
【0046】
一実施態様では、混合糖細胞はマーカーフリーであり、それは栄養要求性または優勢マーカー、特に抗生物質耐性マーカーが、ゲノム中または染色体外に存在しないことを意味する。
【0047】
上述の酵素の過剰発現で使用されるコード配列は、好ましくは本発明の宿主細胞と相同的であってもよい。しかし本発明の宿主細胞と異種性のコード配列を使用してもよい。
【0048】
酵素の過剰発現は、遺伝子改変細胞中での酵素生成について言及する場合、同一条件下の未改変宿主細胞と比較して、特定の酵素活性がより高レベルで酵素が生成されることを意味する。通常これは、酵素的に活性なタンパク質(またはマルチサブユニット酵素の場合は複数のタンパク質)が、同一条件下の未改変宿主細胞と比較してより多量に、またはかなりより高い定常状態レベルで生成されることを意味する。同様にこれは通常、酵素的に活性なタンパク質をコードするmRNAが、同一条件下の未改変宿主細胞と比較してより多量に、またはかなりより高い定常状態レベルで生成されることを意味する。好ましくは本発明の宿主細胞中では、過剰発現される酵素は、過剰発現を引き起こす遺伝子改変以外は遺伝的に同一の株と比較して、少なくとも約1.1、約1.2、約1.5、約2、約5、約10または約20倍過剰発現される。これらの過剰発現レベルは、酵素活性の定常状態レベルに、酵素タンパク質の定常状態レベルに、ならびに酵素をコードする転写物の定常状態レベルに当てはめられてもよいものと理解されるものとする。
【0049】
[適応]
適応は、それによって集団がその生息環境または複数の生息環境により良く適する(順応する)進化の過程である。この過程は、数世代から多世代にわたって起き、基本的な生物学的現象の1つである。
【0050】
適応という用語は、生物の生存に特に重要な特徴もまた指してもよい。このような適応は、より良く適した形態がより成功裏に繁殖することによって、自然淘汰によって可変集団中で生じる。
【0051】
環境条件の変化は、自然淘汰の結果を変化させ、新しい条件下における生物の適応度を改善する引き続く適応の選択的利点に影響を及ぼす。極端な環境変化の場合、有益な適応の出現と固定化が生存のために必須であり得る。例えば養分利用性、温度、酸素利用性などの多数の異なる要因が、適応進化を駆動し得る。
【0052】
[適合]
適応(生物が特定の生息環境内で生きて繁殖できる程度)と適合の間には、明らかな関係がある。適合は、自然淘汰率の推定および予測である。自然淘汰の適用によって、代替の表現型の相対頻度は、それらが遺伝性であるならば経時的に変動する。
【0053】
[遺伝的変化]
自然淘汰が集団の遺伝的変動に作用する場合、遺伝的変化が根底にある機序である。これは集団が、その状況に遺伝的に適応することを意味する。遺伝的変化は、生息環境の変化に適するように、可視的構造をもたらしてもよく、または生物の生理学的活性を調節してもよい。
【0054】
[適応進化]
混合糖細胞は、それらの調製中に適応進化を被る。本発明の細胞は、好ましくは唯一の炭素源としての所望の糖の上での、より好ましくは嫌気的条件下における成長について、自然発生または誘発的(例えば放射線または化学薬品による)いずれかの変異体を選択することで、糖利用のために適応させてもよい。変異体の選択は、例えばKuyperら(2004,FEMS Yeast Res.4:655−664)によって記述される継代培養をはじめとする技術によって、またはケモスタット培養中の選択的圧力下における培養によって実施されてもよい。例えば本発明の好ましい宿主細胞中で、変異体の選択によって得られる改変をはじめとする上述の遺伝子改変の少なくとも1つは、炭素源として、好ましくは唯一の炭素源としてキシロース上で、好ましくは嫌気的条件下で、生育する能力を宿主細胞にもたらす。好ましくは細胞は本質的にキシリトールを生成せず、例えば生成するキシリトールが検出限界未満であり、または例えばモル基準で消費される炭素の約5、約2、約1、約0.5、または約0.3%未満である。
【0055】
適応進化についてはまた、例えばH.W. et al,Applied and Environmental Microbiology Aug.2007,p.4881−4891でも記載される。
【0056】
適応進化の一実施態様では、異なる培地、例えば組成の異なる3種類の培地中(グルコース、キシロース、およびアラビノース;キシロースおよびアラビノース)における、連続成長を用いた反復バッチ培養からなる投与計画が適用されるWisselink et al.(2009)Applied and Environmental Microbiology,Feb.2009,p.907−914参照。
【0057】
[酵母形質転換および遺伝的安定性]
組み換えDNAを用いた酵母細胞の遺伝子操作、すなわち形質転換は、1978年に初めて実行可能になった[Beggs,1978;Hinnen et al.,1978]。酵母中の組み換えDNA技術は、それ以来地位を確立した。多数の異なるベクターコンストラクトが利用可能である。一般にシャトルベクターと称されるこれらのプラスミドベクターは、酵母細胞への形質転換に先だって、それらが大腸菌(E.coli)中で増殖できるようにする、複製起点と選択可能なマーカー(βラクタマーゼ遺伝子、ampRであることが多い)からなる、大腸菌(E.coli)ベクターに由来する遺伝物質を含有する。さらにシャトルベクターは酵母中における選択のための選択可能なマーカーを含有する。マーカーは、対応するゲノムの欠失(または突然変異)を保有する細胞が、栄養要求性または独立栄養性について補足されるような、特定のアミノ酸またはヌクレオチド合成のための酵素をコードする遺伝子であり得る。代案としては、これらのベクターは、g418(Geneticin)、ハイグロマイシンBまたはフレオマイシンのような特定の抗生物質に対する耐性を組み換え酵母細胞(すなわちDNAを取り込んでマーカー遺伝子を発現する細胞)に提供する、異種性顕性耐性マーカーを含有する。さらにこれらのベクターは、外来性DNAをこれらの部位にクローンできるようにする(組み合わされた)制限部位(多重クローニング部位またはMCS)配列を含有してもよいが、代案の方法もまた存在する。
【0058】
伝統的に、追加的遺伝的要素の不在または存在によって、4種のシャトルベクターを区別し得る。
●組み込みプラスミド(YIp)。限定酵素によって開裂され、酵母細胞の形質転換のために直線化DNAが使用される場合は、これはマーカーまたは別の遺伝子の遺伝子座において、相同的組換えにより宿主ゲノムに組み込まれる。これは一般に、ゲノム中のこの特定部位に挿入された外来性DNAの1コピーの存在をもたらす。
●エピソームプラスミド(YEp)。これは酵母細胞中の自律複製に必要な2μプラスミドDNA配列の部分を保有する。複数コピーの形質転換プラスミドが酵母細胞中で増殖され、エピソームとして維持される。
●自己複製プラスミド(YRp)。これは形質転換プラスミドを数百倍に増殖できるようにする、酵母複製起点(ARS、自律的複製配列)を保有する。
●CENプラスミド(YCp)。これはARS配列に加えて、常態では安定した有糸分裂分離を保証し、通常は自己複製プラスミドのコピー数を1つだけに減少させるセントロメア配列(核染色体の1つに由来する)を保有する。
【0059】
これらのプラスミドは、形質転換によって酵母細胞に導入される。酵母細胞の形質転換は、酢酸リチウムによる細胞透過性上昇(Ito et al,1983)、および電気穿孔方法などのいくつかの異なる技術によって達成されてもよい。
【0060】
組み換え微生物の商業的用途では、プラスミドの不安定性が最重要課題である。不安定性は、形質転換された細胞が、プラスミドの変化または損失のためにそれらの改変された特性を失う傾向である。この問題は、Zhangら(Plasmid stability in recombinant Saccharomyces cerevisiae.Biotechnology Advances,Vol.14,No.4,pp.401−435,1996)で、詳細に考察されている。組み込みプラスミドで形質転換された株は、選択圧力の不在下であってさえも極めて安定している(Sherman,F.http://dbb.urmc.rochester.edu/labs/sherman_f/yeast/9.htmlおよびその中の参考文献)。
【0061】
異種性DNAは、通常、染色体外プラスミドの形態(YEp、YCp、およびYRp)で生物に導入される。残念ながら、特に淘汰圧が連続的にかからない場合は、細菌および酵母のどちらでも新しい特徴が保持されないこともあり得ることが分かった。これは組み換え細胞が長期にわたって成長した際のハイブリッドプラスミドの分離不安定性のためである。これは、集団不均一性、クローン変動性、最終的にはその中で大部分の細胞が形質転換により導入された特性を失う細胞集団をもたらす。栄養要求性マーカーを持つベクターが使用される場合、ベクターは最少培地中でのみ維持されるので、富栄養培地中の培養はベクターの迅速な損失をもたらすことが多い。代案である顕性抗生物質耐性マーカーの使用は、製造工程に適合しないことが多い。抗生物質の使用はまた、登録の観点(微量の抗生物質が最終製品中に入る可能性)から、または経済的理由(工業規模での抗生物質の使用コスト)のために、所望されないこともあり得る。
【0062】
ベクターの損失は、大量生産状況において問題をもたらす。組み込みプラスミド(YIp)の使用などのDNAを導入する代案の方法が、酵母に対して存在する。DNAは遺伝子組み換えによって宿主ゲノムに組み込まれ、高い安定性がもたらされる(Caunt,P.Stability of recombinant plasmids in yeast.Journal of Biotechnology 9(1988)173−192)。我々は、宿主トランスポゾンを使用した組み込み法が、良い代案であることを発見した。
【0063】
[トランスポゾン]
本発明の実施態様では、細胞は、所望の遺伝子の1つを超えるコピーを含んでなってもよい。例えば2つ以上のキシロースイソメラーゼ遺伝子またはキシロース還元酵素遺伝子およびキシリトールデヒドロゲナーゼが、混合糖細胞ゲノムに組み込まれてもよい。これは遺伝子の導入をもたらすことが当該技術分野で知られているあらゆる方法によって、実施されてもよい。好ましい実施態様では、これは宿主細胞の反復配列(トランスポゾン)と相同的な部分がある、ベクターを使用して達成されてもよい。宿主細胞が酵母細胞である場合、適切な反復配列はδ配列として知られているTy因子の長い末端反復(LTR)である。
【0064】
Ty因子は、Ty1およびTy2と称される、2つのかなり類似した亜科に分類される。これらの因子は、長さが約6キロベース(kb)であり、約335塩基対の配列である長い末端反復(LTR)と境を接する(Boeke JD et al,The Saccharomyces cerevisiae Genome Contains Functional and Nonfunctional Copies of Transposon Ty1.Molecular and Cellular Biology,Apr.1988,p.1432−1442 Vol.8,No.4)。完全に配列決定されたS.セレヴィシエ(S.cerevisiae)S288c株中で最も豊富なトランスポゾンは、Ty1(31コピー)およびTy2(13コピー)である(Gabriel A,Dapprich J,Kunkel M,Gresham D,Pratt SC,et al.(2006)Global mapping of transposon location.PLoS Genet 2(12):e212.doi:10.1371/journal.pgen.0020212)。これらのトランスポゾンは、そのそれぞれがいくつかのタンパク質をコードする、2つの重複する読み取り枠(ORF)から構成される。コード領域は、前述のほぼ同一のLTRで挟まれる。S.セレヴィシエ(S.cerevisiae)中の別のものであるが、より少なくより特徴的であるTy因子は、Ty3、Ty4、およびTy5からなる。全長Ty因子の各ファミリーで、一桁多い単独LTR因子がゲノム全体に分散する。これらは、内部タンパク質コード領域のループアウトと共に、全長因子のLTR−LTR組み換えによって生じると考えられる。
【0065】
Tyレトロトランスポゾンのレトロ転位機序は、ゲノム全体に複数コピーを組み込むのに利用されている(Boeke et al.,1988;Jacobs et al.,1988)。δ配列として知られているTy因子の長い末端反復(LTR)はまた、Ty結合または単独部位のどちらかである約150〜200コピーで存在するので、相同的組換えによる組み込みの良好な標的でもある。(Boeke,1989;Kingsman and Kingsman,1988)(Parekh R.N.(1996).An Integrating Vector for Tunable, High Copy,Stable Integration into the Dispersed Ty DELTA Sites of Saccharomyces cerevisiae.Biotechnol.Prog.1996,12,16−21)。
【0066】
[宿主細胞]
宿主細胞は、有用な生成物の生産に適したあらゆる宿主細胞であってもよい。本発明の細胞は、細菌などの原核細胞、または真核細胞などのあらゆる適切な細胞であってもよい。典型的に細胞は、例えば酵母または糸状菌などの真核細胞である。
【0067】
酵母は、本明細書で真核微生物と定義され、大部分は単細胞形態で成長する真菌植物亜門(Alexopoulos,C.J.,1962,In:Introductory Mycology,John Wiley & Sons,Inc.,New York)の全ての種を含む。
【0068】
酵母は単細胞性葉状体の出芽によって成長してもよく、または生物体の分裂によって成長してもよい。本発明の細胞として好ましい酵母は、サッカロミセス(Saccharomyces)、クリヴェロミセス(Kluyveromyces)、カンジダ(Candida)、ピチア(Pichia)、分裂酵母(Schizosaccharomyces)、ハンゼヌラ(Hansenula)、クロエケラ(Kloeckera)、シュワニオミセス(Schwanniomyces)またはヤロウィア(Yarrowia)属に属してもよい。好ましくは酵母は、嫌気的または酸素限定発酵ができるものであり、より好ましくは嫌気的アルコール発酵ができるものである。
【0069】
糸状菌は、本明細書で、真菌植物亜門の全ての糸状体を含む真核微生物と定義される。これらの真菌は、キチン、セルロース、および複合多糖からなる栄養菌糸体によって特徴付けられる。本発明の細胞として使用するのに適した糸状菌は、形態的、生理的、および遺伝的に酵母と異なる。ほとんどの真菌は増殖のために無菌条件を必要とせず、バクテリオファージ感染に感受性がないので、有利には糸状菌細胞を使用してもよい。糸状菌による栄養成長は菌糸伸長と炭素異化作用により、ほとんどの糸状菌で偏性好気性である。本発明の宿主細胞として好ましい糸状菌は、アスペルギルス(Aspergillus)、トリコデルマ(Trichoderma)、フミコラ(Humicola)、アクレモニウム(Acremonium)、フザリウム(Fusarium)またはペニシリウム(Penicillium)属に属してもよい。より好ましくは、糸状菌細胞は、アスペルギルス・ニガー(Aspergillus niger)、コウジカビ(Aspergillus oryzae)、ペニシリウム・クリソゲヌム(Penicillium chrysogenum)、またはリゾプス・オリゼー(Rhizopus oryzae)細胞であってもよい。
【0070】
一実施態様では宿主細胞は、酵母であってもよい。
【0071】
好ましくは宿主は、工業宿主、より好ましくは工業酵母である。工業宿主および工業酵母細胞は、以下のように定義されてもよい。工業工程中の酵母細胞の生息環境は、実験室のそれと顕著に異なる。工業用酵母細胞は、工程中に変動し得る複数の環境条件下で良好に機能できなくてはならない。このような変動としては、一緒になってサッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)細胞の成長とエタノール産生に潜在的影響を及ぼす栄養源、pH、エタノール濃度、温度、酸素濃度の変化などが挙げられる。有害工業条件の下で、環境耐性株は、頑強な成長および生産が可能であるべきである。工業酵母株は、一般に、製パン工業、醸造業、ぶどう酒醸造、およびエタノール産業などのそれらが使用される用途において起こり得るこれらの環境条件の変化に対してより頑強である。工業酵母(S.セレヴィシエ(S.cerevisiae))の例は、Ethanol Red(登録商標)(Fermentis)、Fermiol(登録商標)(DSM)、およびThermosacc(登録商標)(Lallemand)である。
【0072】
一実施態様では宿主は、阻害物質耐性である。阻害物質耐性宿主細胞は、阻害物質耐性S.セレヴィシエ(S.cerevisiae)ATCC26602株が選択された、Kadar et al,Appl.Biochem.Biotechnol.(2007),Vol.136−140,847−858で示されるような阻害物質含有材料上での成長について、株をスクリーニングすることで選択されてもよい。
【0073】
好ましくは、宿主細胞は工業用および阻害物質耐性である。
【0074】
[AraA、AraB、およびAraD遺伝子]
本発明の細胞は、アラビノースを使用できる。したがって本発明の細胞は、L−アラビノースをL−リブロースおよび/またはキシルロース−5−リン酸、および/または例えば本明細書で言及されるものなどの所望の発酵産物に転換できる。
【0075】
L−アラビノースからエタノールを生成できる生物、例えばS.セレヴィシエ(S.cerevisiae)株は、好適な起源からのaraA(L−アラビノースイソメラーゼ)、araB(L−リブロキナーゼ)、およびaraD(L−リブロース−5−P4−エピメラーゼ)遺伝子を導入して細胞を改変することにより、生成されてもよい。このような遺伝子は、本発明の細胞がアラビノースを使用できるように導入されてもよい。このようなアプローチについては、国際公開第2003/095627号パンフレットに記載される。ラクトバチルス・プランタルム(Lactobacillus plantarum)からのaraA、araB、およびaraD遺伝子を使用してもよく、国際公開第2008/041840号パンフレットで開示される。枯草菌(Bacillus subtilis)からのaraA遺伝子、および大腸菌(Escherichia coli)からのaraBおよびaraD遺伝子を使用してもよく、欧州特許第1499708号明細書で開示される。別の実施態様では、araA、araB、およびaraD遺伝子は、クラビバクター(Clavibacter)、アルスロバクター(Arthrobacter)および/またはグラメラ(Gramella)属の少なくとも1つ、特に国際公開第2009011591号パンフレットで開示されるようなクラビバクター・ミシガンエンシス(Clavibacter michiganensis)、アルスロバクター・アウレセンス(Arthrobacter aurescens)、および/またはグラメラ・フォルセティ(Gramella forsetii)の1つに由来してもよい。
【0076】
[PPP遺伝子]
本発明の細胞は、ペントースリン酸経路の流量を増大させる1つ以上の遺伝子改変を含んでなってもよい。特に遺伝子改変は、非酸化的部分ペントースリン酸経路を通じて、流量の増大をもたらしてもよい。ペントースリン酸経路の非酸化的部分の流量の増大を引き起こす遺伝子改変は、本明細書では、流量の増大を引き起こす遺伝子改変以外は、遺伝的に同一の株中の流量と比較して、流量を少なくとも約1.1、約1.2、約1.5、約2、約5、約10または約20倍増大させる改変を意味するものと理解される。ペントースリン酸経路の非酸化的部分の流量は、いくらかでもキシリトールが生成される場合は、唯一の炭素源としてのキシロース上で改変宿主を成長させてキシロース特異的消費速度を判定し、キシロース特異的消費速度からキシリトール特異的生成速度を差し引くことによって測定されてもよい。しかしペントースリン酸経路の非酸化的部分の流量は、唯一の炭素源としてのキシロース上での成長速度、好ましくは唯一の炭素源としてのキシロース上での嫌気的成長速度と比例する。唯一の炭素源としてのキシロース上での成長速度(μmax)とペントースリン酸経路の非酸化的部分の流量の間には直線関係がある。(嫌気性、増殖培地、pH、株の遺伝的背景などの所定の条件の組の下では)糖の上でのバイオマス収率は一定であるので、キシロースの比消費速度(Qs)は、糖の上でのバイオマス収率(Yxs)で除した成長速度(μ)に等しい(すなわちQs=μ/Yxs)。したがってペントースリン酸経路の非酸化的部分の流量の増大は、輸送(取り込)みが制限されない限り、これらの条件下の最大成長速度の増大から推定されてもよい。
【0077】
ペントースリン酸経路流量を増大させる1つ以上の遺伝子改変は、様々なやり方で宿主細胞に導入されてもよい。これらとしては、例えばキシルロースキナーゼの、および/または非酸化的部分ペントースリン酸経路酵素の1つ以上の、より高い定常状態活性レベルを達成すること、および/または非特異的アルドース還元酵素の低下した定常状態活性レベルを達成することが挙げられる。定常状態活性レベルにおけるこれらの変化は、(自然発生または化学薬品または放射線により誘発される)変異体の選択によって、および/または組み換えDNA技術によって、例えばそれぞれ遺伝子をコードする酵素またはこれらの遺伝子を制御する因子の過剰発現または不活性化によって、達成されてもよい。
【0078】
好ましい宿主細胞では、遺伝子改変は、(非酸化的部分)ペントースリン酸経路の少なくとも1つの酵素の過剰発現を含んでなる。好ましくは酵素は、リブロース−5−リン酸イソメラーゼ、リブロース−5−リン酸エピメラーゼ、トランスケトラーゼ、およびトランスアルドラーゼをコードする酵素からなる群から選択される。(非酸化的部分)ペントースリン酸経路の酵素の様々な組み合わせが過剰発現されてもよい。例えば過剰発現される酵素は、少なくとも酵素リブロース−5−リン酸イソメラーゼおよびリブロース−5−リン酸エピメラーゼ;または少なくとも酵素リブロース−5−リン酸イソメラーゼおよびトランスケトラーゼ;または少なくとも酵素リブロース−5−リン酸イソメラーゼおよびトランスアルドラーゼ;または少なくとも酵素リブロース−5−リン酸エピメラーゼおよびトランスケトラーゼ;または少なくとも酵素リブロース−5ホスフェートエピメラーゼおよびトランスアルドラーゼ;または少なくとも酵素トランスケトラーゼおよびトランスアルドラーゼ;または少なくとも酵素リブロース−5−リン酸エピメラーゼ、トランスケトラーゼおよびトランスアルドラーゼ;または少なくとも酵素リブロース−5−リン酸イソメラーゼ、トランスケトラーゼおよびトランスアルドラーゼ;または少なくとも酵素リブロース−5−リン酸イソメラーゼ、リブロース−5−リン酸エピメラーゼ、およびトランスアルドラーゼ;または少なくとも酵素リブロース−5−リン酸イソメラーゼ、リブロース−5−リン酸エピメラーゼ、およびトランスケトラーゼであってもよい。本発明の一実施態様では、リブロース−5−リン酸イソメラーゼ、リブロース−5−リン酸エピメラーゼ、トランスケトラーゼ、およびトランスアルドラーゼ酵素のそれぞれが、宿主細胞中で過剰発現される。より好ましいのは、その中の遺伝子改変が、少なくともトランスケトラーゼおよびトランスアルドラーゼ酵素双方の過剰発現を含んでなる宿主細胞であり、それはこのような宿主細胞が既にキシロース上で嫌気的成長できるためである。実際には、条件によっては、トランスケトラーゼおよびトランスアルドラーゼのみを過剰に発現する宿主細胞は、4つの酵素全て、すなわちリブロース−5−リン酸イソメラーゼ、リブロース−5−リン酸エピメラーゼ、トランスケトラーゼ、およびトランスアルドラーゼを過剰に発現する宿主細胞と同一の嫌気的成長速度をキシロース上で既に有する。さらに酵素リブロース−5−リン酸イソメラーゼおよびリブロース−5−リン酸エピメラーゼの双方を過剰に発現する宿主細胞が、イソメラーゼのみまたはエピメラーゼのみを過剰に発現する宿主細胞よりも好ましいが、それはこれらの酵素の1つのみの過剰発現が代謝不均衡を生じ得るためである。
【0079】
本明細書で酵素「リブロース−5−リン酸エピメラーゼ」(EC5.1.3.1)は、D−キシルロース−5−リン酸のD−リブロース−5−リン酸へのエピマー化を触媒する酵素と定義され、逆の場合も同じである。酵素は、ホスホリブロースエピメラーゼ;エリスロース−4−ホスフェートイソメラーゼ;ホスホケトペントース3−エピメラーゼ;キシルロースホスフェート3−エピメラーゼ;ホスホケトペントースエピメラーゼ;リブロース−5−リン酸3−エピメラーゼ;D−リブロースホスフェート−3−エピメラーゼ;D−リブロース−5−リン酸エピメラーゼ;D−リブロース−5−P3−エピメラーゼ;D−キシルロース−5−リン酸3−エピメラーゼ;ペントースー5−ホスフェート3−エピメラーゼ;またはD−リブロース−5−リン酸3−エピメラーゼとしてもまた知られている。リブロース−5−リン酸エピメラーゼは、そのアミノ酸配列によってさらに定義されてもよい。同様にリブロース−5−リン酸エピメラーゼは、酵素をコードするヌクレオチド配列によって、ならびにリブロース−5−リン酸エピメラーゼをコードする参照ヌクレオチド配列とハイブリッドするヌクレオチド配列によって、定義されてもよい。リブロース−5−リン酸エピメラーゼをコードするヌクレオチド配列は、本明細書でRPE1と命名される。
【0080】
本明細書で酵素「リブロース−5−リン酸イソメラーゼ」(EC5.3.1.6)は、D−リボース5−ホスフェートのD−リブロース5−ホスフェートへの直接異性化を触媒する酵素と定義され、逆の場合も同じである。酵素は、ホスホペントースイソメラーゼ;ホスホリボイソメラーゼ;リボースホスフェートイソメラーゼ;5−ホスホリボースイソメラーゼ;D−リボース5−ホスフェートイソメラーゼ;D−リボース−5−ホスフェートケトール−イソメラーゼ;またはD−リボース−5−ホスフェートアルドース−ケトース−イソメラーゼとしてもまた知られている。リブロース−5−リン酸イソメラーゼは、そのアミノ酸配列によってさらに定義されてもよい。同様にリブロース−5−リン酸イソメラーゼは、酵素をコードするヌクレオチド配列によって、ならびリブロース−5−リン酸イソメラーゼをコードする参照ヌクレオチド配列とハイブリッドするヌクレオチド配列によって定義されてもよい。リブロース−5−リン酸イソメラーゼをコードするヌクレオチド配列は、本明細書でRKI1と命名される。
【0081】
本明細書において酵素「トランスケトラーゼ」(EC2.2.1.1)は、D−リボース5−ホスフェート+D−キシルロース−5−リン酸←→セドヘプツロース7−ホスフェート+D−グリセルアルデヒド3−ホスフェートの反応を触媒する酵素と定義され、逆の場合も同じである。この酵素は、グリコールアルデヒドトランスフェラーゼまたはセドヘプツロース−7−ホスフェート:D−グリセルアルデヒド−3−リン酸グリコールアルデヒドトランスフェラーゼとしてもまた知られている。トランスケトラーゼは、そのアミノ酸によってさらに定義されてもよい。同様にトランスケトラーゼは、酵素をコードするヌクレオチド配列によって、ならびにトランスケトラーゼをコードする参照ヌクレオチド配列とハイブリッドするヌクレオチド配列によって定義されてもよい。トランスケトラーゼをコードするヌクレオチド配列は、本明細書でTKL1と命名される。
【0082】
本明細書で酵素「トランスアルドラーゼ」(EC2.2.1.2)は、セドヘプツロース7−ホスフェート+D−グリセルアルデヒド3−ホスフェート←→D−エリスロース4−ホスフェート+D−果糖6−ホスフェートの反応を触媒する酵素と定義され、逆の場合も同じである。酵素は、ジヒドロキシアセトントランスフェラーゼ;ジヒドロキシアセトンシンターゼ;ホルムアルデヒドトランスケトラーゼ;またはセドヘプツロース−7−ホスフェート:D−グリセルアルデヒド−3−ホスフェートグリセロントランスフェラーゼとしてもまた知られている。トランスアルドラーゼは、そのアミノ酸配列によってさらに定義されてもよい。同様にトランスアルドラーゼは、酵素をコードするヌクレオチド配列によって、ならびにトランスアルドラーゼをコードする参照ヌクレオチド配列とハイブリッドするヌクレオチド配列によって、定義されてもよい。トランスケトラーゼをコードするヌクレオチド配列は、本明細書でTAL1と命名される。
【0083】
[キシロースイソメラーゼまたはキシロース還元酵素およびキシリトールデヒドロゲナーゼ遺伝子]
本発明に従って、1つ以上のキシロースイソメラーゼ遺伝子および/または1つ以上のキシロース還元酵素およびキシリトールデヒドロゲナーゼのコピーの1つ、2つまたはそれ以上のコピーが、宿主細胞のゲノムに導入される。これらの2つ以上の遺伝要素の存在は、細胞に異性化または還元によってキシロースを転換する能力を与える。
【0084】
一実施態様では1つ以上のキシロースイソメラーゼ遺伝子の1つ、2つまたはそれ以上のコピーが宿主細胞のゲノムに導入される。
【0085】
本明細書で「キシロースイソメラーゼ」(EC5.3.1.5)は、D−キシロースのD−キシルロースへの直接異性化を触媒する酵素と定義され、逆もまた同様である。酵素は、D−キシロースケトイソメラーゼとしてもまた知られている。本明細書でキシロースイソメラーゼはまた、D−グルコースとD−果糖の間の転換を触媒できてもよい(したがって適宜にグルコースイソメラーゼと称されてもよい)。本明細書でキシロースイソメラーゼは、補助因子としてマグネシウム、マンガンまたはコバルトなどの二価のカチオンを必要としてもよい。
【0086】
したがってこのような混合糖細胞は、キシロースをキシルロースに異性化できる。キシロースをキシルロースに異性化する能力は、定義されたキシロースイソメラーゼをコードするヌクレオチド配列を含んでなる核酸コンストラクトで宿主細胞を形質転換することによって、宿主細胞に与えられる。混合糖細胞は、キシロースのキシルロースへの直接異性化によって、キシロースをキシルロースに異性化する。
【0087】
本明細書で、キシロースイソメラーゼ活性単位(U)は、Kuyper et al.(2003,FEMS Yeast Res.4:69−78)によって記述される条件下で、1分当たり1nmolのキシルロースを生成する酵素量と定義されてもよい。キシロース異性化遺伝子は、例えば国際公開第2006/009434号パンフレットで開示されるようなピロミセス(Piromyces)種などの様々な起源を有してもよい。その他の適切な起源は、バクテロイデス(Bacteroides)、特にPCT欧州特許第2009/52623号明細書に記載されるようなバクテロイデス・ユニフォルミス(Bacteroides uniformis)、バチルス(Bacillus)、特にPCT欧州特許第2009/052625号明細書に記載されるようなバチルス・ステアロサーモフィラス(Bacillus stearothermophilus)である。
【0088】
別の実施態様では、1つ以上のキシロース還元酵素およびキシリトールデヒドロゲナーゼ遺伝子の2つ以上のコピーが宿主細胞ゲノムに導入される。この実施態様では、キシロースの転換は、それぞれキシロース還元酵素およびキシリトールデヒドロゲナーゼによって触媒される、キシロースからキシリトール中間体を経由するキシルロースへの転換の二段階で実施される。一実施態様では、国際公開第2004085627号パンフレットで開示されるように、キシロース還元酵素(XR)、キシリトールデヒドロゲナーゼ(XDH)、およびキシルロキナーゼ(XK)が過剰発現されてもよく、任意にNADPH生成酵素をコードする遺伝子の1つ以上が上方制御されて、NADH消費酵素をコードする遺伝子の1つ以上が上方制御される。
【0089】
[XKS1遺伝子]
本発明の細胞は、特異的キシルロースキナーゼ活性を増大させる、1つ以上の遺伝子改変を含んでなってもよい。好ましくは1つまたは複数の遺伝子改変は、例えばキシルロースキナーゼをコードするヌクレオチド配列の過剰発現によってキシルロースキナーゼの過剰発現を引き起こす。キシルロースキナーゼをコードする遺伝子は宿主細胞に内在性であってもよく、または宿主細胞に異種性のキシルロースキナーゼであってもよい。本発明の宿主細胞中におけるキシルロースキナーゼ過剰発現のために使用されるヌクレオチド配列は、キシルロースキナーゼ活性があるポリペプチドをコードするヌクレオチド配列である。
【0090】
本明細書で酵素「キシルロースキナーゼ」(EC2.7.1.17)は、ATP+D−キシルロース=ADP+D−キシルロース−5−リン酸の反応を触媒する酵素と定義される。酵素はホスホリル化キシルロキナーゼ、D−キシルロキナーゼまたはATP:D−キシルロース5−リン酸転移酵素としてもまた知られている。本発明のキシルロースキナーゼは、そのアミノ酸配列によってさらに定義されてもよい。同様にキシルロースキナーゼは、酵素をコードするヌクレオチド配列によって、ならびにキシルロースキナーゼをコードする参照ヌクレオチド配列とハイブリッドするヌクレオチド配列によって定義されてもよい。
【0091】
本発明の細胞中では、特異的キシルロースキナーゼ活性を増大させる1つまたは複数の遺伝子改変は、上述のようなペントースリン酸経路の流量を増大させる修飾のいずれかと組み合わされてもよい。しかしこれは必須でない。
【0092】
したがって本発明の宿主細胞は、特異的キシルロースキナーゼ活性を増大させる遺伝子改変修飾のみを含んでなってもよい。本発明宿主細胞中でのキシルロースキナーゼの過剰発現を達成し分析するための当該技術分野で利用できる様々な手段は、ペントースリン酸経路酵素について上述したのと同じである。好ましくは本発明の宿主細胞中では、過剰発現されるキシルロースキナーゼは、過剰発現を引き起こす遺伝子改変以外は遺伝的に同一の株と比較して、少なくとも約1.1、約1.2、約1.5、約2、約5、約10または約20倍過剰発現される。これらの過剰発現レベルは、酵素活性の定常状態レベルに、酵素タンパク質の定常状態レベルに、ならびに酵素をコードする転写物の定常状態レベルに当てはまってもよいと理解されるものとする。
【0093】
[アルドース還元酵素(GRE3)遺伝子欠失]
XIがキシロースを転換する遺伝子として使用される実施態様では、アルドース還元酵素活性を低下させることが都合良いこともあり得る。したがって本発明の細胞は、宿主細胞の非特異的アルドース還元酵素活性を低下させる、1つ以上の遺伝子改変を含んでなってもよい。好ましくは、非特異的アルドース還元酵素をコードする遺伝子の発現を低下させまたは不活性化する1つ以上の遺伝子改変によって、宿主細胞中で非特異的アルドース還元酵素活性が低下する。好ましくは遺伝子改変は、宿主細胞中で非特異的アルドース還元酵素をコードする遺伝子の各内在性コピーの発現を低下させまたは不活性化する(本明細書中でGRE3欠失と称する)。宿主細胞は二倍性、倍数性または異数性の結果として、非特異的アルドース還元酵素をコードする遺伝子の複数コピーを含んでなってもよく、および/または宿主細胞はアミノ酸配列が異なり、それぞれ異なる遺伝子によってコードされるアルドース還元酵素活性があるいくつかの異なる酵素(アイソザイム)を含有してもよい。またこのような場合、好ましくは、非特異的アルドース還元酵素をコードする各遺伝子の発現が低下し、または不活性化される。好ましくは、遺伝子は、遺伝子の少なくとも一部の欠失によって、または遺伝子中断によって不活性化され、この文脈では、遺伝子という用語は、その(部分的)欠失または不活性化が宿主細胞中における非特異的アルドース還元酵素活性の発現低下をもたらすコード配列の上流または下流にある、いかなる非コード配列もまた含む。
【0094】
本発明の宿主細胞中でその活性が低下されるアルドース還元酵素をコードするヌクレオチド配列は、アルドース還元酵素活性があるポリペプチドをコードするヌクレオチド配列である。
【0095】
したがって宿主細胞中で非特異的アルドース還元酵素活性を低下させる遺伝子改変のみを含んでなる本発明の宿主細胞は、明確に本発明に含まれる。
【0096】
酵素「アルドース還元酵素」(EC1.1.1.21)は、本明細書でキシロースまたはキシルロースをキシリトールに還元できるあらゆる酵素と定義される。本発明の文脈でアルドース還元酵素は、本発明の宿主細胞に天然(内在性)であり、キシロースまたはキシルロースをキシリトールに還元できるあらゆる非特異的アルドース還元酵素であってもよい。非特異的アルドース還元酵素は、次の反応を触媒する。
アルドース+NAD(P)H+H+←→アルジトール+NAD(P)+
【0097】
酵素は広い特異性を有し、アルドース還元酵素;ポリオールデヒドロゲナーゼ(NADP+);アルジトール:NADPオキシド還元酵素;アルジトール:NADP+1−オキシド還元酵素;NADPH−アルドペントース還元酵素;またはNADPH−アルドース還元酵素としてもまた知られている。
【0098】
S.セレヴィシエ(S.cerevisiae)に内在性であるこのような非特異的アルドース還元酵素の特定の例は、GRE3遺伝子によってコードされる(Traff et al.,2001,Appl.Environ.Microbiol.67:5668−74)。したがって本発明のアルドース還元酵素は、そのアミノ酸配列によってさらに定義されてもよい。同様にアルドース還元酵素は、酵素をコードするヌクレオチド配列によって、ならびに参照ヌクレオチド配列とハイブリッドする、アルドース還元酵素をコードするヌクレオチド配列によって定義されてもよい。
【0099】
[配列同一性]
本明細書で配列同一性(または配列類似性)とは、2つ以上のアミノ酸(ポリペプチドまたはタンパク質)配列または2つ以上の核酸(ポリヌクレオチド)配列を比較して判定される、配列間の関係と定義される。通常、配列アイデンティティまたは類似性が比較され、典型的に全長にわたって配列が比較される。しかし配列は、より短い比較ウィンドウにわたって比較されてもよい。当該技術分野で「アイデンティティ」はまた、場合によってはそのような配列のストリング間のマッチによって判定される。アミノ酸または核酸配列間の配列関連性の程度を意味する。
【0100】
アイデンティティを判定する好ましい方法は、試験される配列間に最大のマッチを与えるようにデザインされる。アイデンティティおよび類似性を判定する方法は、公的に入手可能なコンピュータプログラムにおいて体系化される。2つの配列間のアイデンティティおよび類似性を判定する好ましいコンピュータプログラム法としては、例えばBestFit、BLASTP、BLASTN、およびFASTA(Altschul,S.F.et al.,J.Mol.Biol.215:403−410(1990)、NCBIおよびその他の情報源から公的に入手可能(BLAST Manual,Altschul,S.,et al.,NCBI NLM NIH Bethesda,MD 20894)が挙げられる。BLASTPを使用したアミノ酸配列比較の好ましいパラメーターは、gap open 11.0、gap extend 1、Blosum62 matrixである。BLASTPを使用した核酸配列比較の好ましいパラメーターは、gap open 11.0、gap extend 1、DNA full matrix(DNA identity matrix)である。
【0101】
任意選択的に、アミノ酸の類似度を判定する上で、当業者はまた、当業者には明らかであるように、いわゆる「保存的」アミノ酸置換を考慮してもよい。
【0102】
保存的アミノ酸置換とは、同様の側鎖を有する残基の互換性を指す。例えば脂肪族側鎖を有する一群のアミノ酸はグリシン、アラニン、バリン、ロイシン、およびイソロイシンであり;脂肪族ヒドロキシル側鎖を有する一群のアミノ酸はセリンおよびスレオニンであり;アミド含有側鎖を有する一群のアミノ酸はアスパラギンおよびグルタミンであり;芳香族側鎖を有する一群のアミノ酸はフェニルアラニン、チロシン、およびトリプトファンであり;塩基性側鎖を有する一群のアミノ酸はリジン、アルギニン、およびヒスチジンであり;イオウ含有側鎖を有する一群のアミノ酸はシステインおよびメチオニンである。
【0103】
好ましい保存的アミノ酸置換基は、バリン−ロイシン−イソロイシン、フェニルアラニン−チロシン、リジン−アルギニン、アラニン−バリン、およびアスパラギン−グルタミンである。本明細書で開示される置換変異型アミノ酸配列とは、その中で開示される配列の少なくとも1つの残基が除去され、その場所に異なる残基が挿入されるものである。好ましくは、アミノ酸変化は保存的である。天然アミノ酸のそれぞれについて、好ましい保存的置換を下に示す。Alaからser;Argからlys;Asnからglnまたはhis;Aspからglu;Cysからserまたはala;Glnからasn;Gluからasp;Glyからpro;Hisからasnまたはgln;Heからleuまたはval;Leuからileまたはval;Lysからarg;glnまたはglu;Metからleuまたはile;Pheからmet、leuまたはtyr;Serからthr;Thrからser;Trpからtyr;Tyrからtrpまたはphe;およびValからileまたはleu。
【0104】
ストリンジェントなハイブリダイゼーション条件は、本明細書で、少なくとも約25個、好ましくは約50個のヌクレオチド、75個または100個および最も好ましくは約200個以上のヌクレオチドの核酸配列を約1Mの塩、好ましくは6×SSC(塩化ナトリウム、クエン酸ナトリウム)を含んでなる溶液中、または比較できるイオン強度を有するあらゆるその他の溶液中において、約65℃の温度でハイブリッド形成させ、約0.1M以下の塩、好ましくは0.2×SSCを含んでなる溶液、または比較できるイオン強度を有するあらゆるその他の溶液中において、65℃で洗浄する条件と定義される。好ましくはハイブリダイゼーションは、一晩、すなわち少なくとも10時間にわたり実施され、好ましくは洗浄は、洗浄溶液を少なくとも2回取り替えて少なくとも1時間にわたり実施される。これらの条件は、通常、約90%以上の配列同一性を有する配列が特異的にハイブリダイゼーションできるようにする。
【0105】
本明細書で中程度の条件は、少なくとも50個のヌクレオチド、好ましくは約200個以上のヌクレオチドの核酸配列を約1Mの塩、好ましくは6×SSCを含んでなる溶液中、または比較できるイオン強度を有するあらゆるその他の溶液中において、約45℃の温度でハイブリッド形成させ、約1M塩、好ましくは6×SSCを含んでなる溶液、または比較できるイオン強度を有するあらゆるその他の溶液中において、室温で洗浄する条件と定義される。好ましくはハイブリダイゼーションは、一晩、すなわち少なくとも10時間にわたり実施され、好ましくは洗浄は、洗浄溶液を少なくとも2回取り替えて少なくとも1時間にわたり実施される。これらの条件は、通常、50%以下の配列同一性を有する配列が特異的にハイブリダイゼーションできるようにする。当業者は、アイデンティティが50%〜90%で変動する配列を特異的に同定するために、これらのハイブリダイゼーション条件を変更できる。
【0106】
導入された酵素が本発明の細胞中において活性形態で発現する可能性を増大させるために、対応するコードディングヌクレオチド配列を適応させて、そのコドン使用頻度を選択された酵母細胞にとって最適化させてもよい。いくつかのコドン最適化法が、当該技術分野で知られている。酵母のヌクレオチド配列のコドン使用頻度を最適化する好ましい方法は、国際公開第2006/077258号パンフレットおよび/または国際公開第2008/000632号パンフレットで開示されるようなコドンペア最適化技術である。国際公開第2008/000632号パンフレットは、コドンペア最適化に対処する。コドンペア最適化は、ポリペプチドコードするヌクレオチド配列をそれらのコドン、特に使用されるコドンペアの使用頻度について改変し、ポリペプチドをコードするヌクレオチド配列の改善された発現および/またはコードされたポリペプチドの改善された生成を得る方法である。コドンペアは、コード配中の2個の引き続く三塩基(コドン)の組と定義される。
【0107】
本明細書では、遺伝子発現および翻訳効率の簡単な尺度として、Xuhua Xia,Evolutionary Bioinformatics 2007,:3 53−58で記載されるコドン適応指数(CAI)を使用する。指数は、高度に発現される種からの遺伝子の参照集合を使用して各コドンの優劣を評価し、遺伝子のスコアをその遺伝子中の全コドンの使用頻度から計算する。指数は、コドン使用頻度パターンを形成する上で選択が効果的である程度を評価する。その点において、それらの宿主に対するウィルス性遺伝子の適応を評価するために、異なる生物中のコドン使用頻度の比較を行うために、遺伝子発現レベルを予測することが有用である。指数はまた、異種遺伝子発現の成功可能性のおよその指標も与えてもよい。本発明に従ったコドンペア最適化された遺伝子中では、CAIは0.6以上、0.7以上、0.8以上、0.85以上、0.87以上、0.90以上、0.95以上、または約1.0である。
【0108】
したがって本発明の細胞は、上で定義されるaraA、araB、およびaraD遺伝子をコードするヌクレオチド配列を含んでなる核酸コンストラクトを含んでなる、すなわちそれで形質転換された細胞である。araAコード配列を含んでなる核酸コンストラクトは、好ましくは宿主細胞中でaraA遺伝子を発現できる。
【0109】
好ましくは遺伝子は、細胞質ゾル中で発現される。細胞質性発現は、ミトコンドリアまたはペルオキシソーム標的シグナルの欠失または修飾によって達成されてもよい。
【0110】
[バイオ製品生産]
長年にわたって、糖科作物からバイオエタノールを生産するために、様々な生物の導入が提案されてきた。しかし実際には、全ての主要なバイオエタノール製造工程は、エタノール生産菌としてサッカロミセス(Saccharomyces)属の酵母を使用し続けている。これは工業的方法のための、サッカロミセス(Saccharomyces)種の数多くの魅力的な特徴、すなわち高い酸、エタノール、および浸透圧耐性、嫌気的成長能力、そしてもちろんその高いアルコール発酵能力のためである。宿主細胞として好ましい酵母種としては、S.セレヴィシエ(S.cerevisiae)、S.ブルデリ(S.bulderi)、S.バルネッチ(S.barnetti)、S.エクシグウス(S.exiguus)、S.ウバラム(S.uvarum)、S.ディアスタティカス(S.diastaticus)、K.ラクチス(K.lactis)、K.マルキシアヌス(K.marxianus)、またはK.フラギリス(K.fragilis)が挙げられる。
【0111】
本発明の細胞は、植物バイオマス、セルロース、ヘミセルロース、ペクチン、ラムノース、ガラクトース、果糖、マルトース、マルトデキストリン、リボース、リブロース、またはデンプン、デンプン誘導体、スクロース、乳糖およびグリセロールを例えば発酵性糖に転換できてもよい。したがって本発明の細胞は、セルロースのグルコースモノマーへの、そしてヘミセルロースのキシロースおよびアラビノースモノマーへの転換に必要なセルラーゼ(エンドセルラーゼまたはエキソセルラーゼ)、ヘミセルラーゼ(エンドまたはエキソキシラナーゼまたはアラビナーゼ);ペクチンをグルクロン酸およびガラクツロン酸に転換できるペクチナーゼ;またはデンプンをグルコースモノマーに転換するアミラーゼなどの1つ以上の酵素を発現してもよい。
【0112】
本細胞は、さらに好ましくは、ピルビン酸から、エタノール、ブタノール、乳酸、3−ヒドロキシ−プロピオン酸、アクリル酸、酢酸、コハク酸、クエン酸、フマル酸、リンゴ酸、イタコン酸、アミノ酸、1,3−プロパン−ジオール、エチレン、グリセロール、β−ラクタム抗生物質またはセファロスポリンなどの所望の発酵産物への転換に必要とされる酵素活性をさらに含んでなる。
【0113】
好ましい本発明の細胞は、天然でアルコール発酵、好ましくは嫌気性アルコール発酵できる細胞である。本発明の細胞は好ましくは、高いエタノール耐性、高い低pH耐性(すなわち約5、約4、約3、または約2.5よりも低いpHで成長できる)、および乳酸、酢酸またはギ酸のような有機酸、および/またはフルフラールおよびヒドロキシメチルフルフラールなどの糖分解生成物に対する高い耐性、および/または高い高温耐性を有する。
【0114】
上記のいずれかの本発明の細胞の特徴または活性は、細胞内に天然に存在してもよく、または遺伝子改変によって導入または改変されてもよい。
【0115】
本発明の細胞は、エタノール生産に適した細胞であってもよい。しかし本発明の細胞は、エタノール以外の発酵産物生産に適していてもよい。このような非エタノール発酵産物としては、原則的には、酵母または糸状菌などの真核微生物が生成可能なあらゆるバルクまたはファインケミカルが挙げられる。
【0116】
このような発酵産物は、例えばブタノール、乳酸、3−ヒドロキシ−プロピオン酸、アクリル酸、酢酸、コハク酸、クエン酸、リンゴ酸、フマル酸、イタコン酸、アミノ酸、1,3−プロパン−ジオール、エチレン、グリセロール、β−ラクタム抗生物質またはセファロスポリンであってもよい。非エタノール発酵産物のための好ましい本発明の細胞は、アルコールデヒドロゲナーゼ活性の低下をもたらす遺伝子改変を含有する宿主細胞である。
【0117】
さらなる態様では、本発明は、その中で本発明細胞が、キシロースなどのキシロース源を含んでなる炭素源発酵のために使用される発酵工程に関する。キシロース源に加えて発酵培地中の炭素源は、グルコース源もまた含んでなってもよい。キシロースまたはグルコース源は、キシロースまたはグルコースそれ自体であってもよく、または例えばリグノセルロース、キシラン、セルロース、デンプンなどのキシロースまたはグルコース単位を含んでなる、あらゆる炭水化物オリゴマーまたはポリマーであってもよい。このような炭水化物からのキシロースまたはグルコース単位の放出のために、適切なカルボヒドラーゼ(キシラナーゼ、グルカナーゼ、アミラーゼなど)が発酵培地に添加されてもよく、または細胞によって生成されてもよい。後者の場合、細胞は遺伝子操作されて、このようなカルボヒドラーゼを生成し分泌してもよい。オリゴまたはポリマーグルコース源を使用することの追加的利点は、例えば律速量のカルボヒドラーゼを使用することにより、それが発酵中に(より)低い濃度の遊離グルコースを維持できるようにすることである。これは、次に、キシロースなどの非グルコース糖の代謝および輸送に必要とされるシステムの抑制を防止する。
【0118】
好ましい方法では、細胞はキシロースおよびグルコースの双方を好ましくは同時に発酵し、その場合、好ましくはジオーキシー成長を妨げるグルコース抑制に非感受性の細胞が使用される。炭素源としてのキシロース(およびグルコース)源に加えて、発酵培地は、細胞成長に必要とされる適切な成分をさらに含んでなる。酵母などの微生物の成長のための発酵培地組成物は、当該技術分野で良く知られている。発酵過程は、例えばエタノール、ブタノール、乳酸、3−ヒドロキシ−プロピオン酸、アクリル酸、酢酸、コハク酸、クエン酸、リンゴ酸、フマル酸、イタコン酸、アミノ酸、1,3−プロパン−ジオール、エチレン、グリセロール、ペニシリンGまたはペニシリンVとその発酵誘導体などのβ−ラクタム抗生物質、およびセファロスポリンなどの発酵産物生産のための過程である。
【0119】
[リグノセルロース]
潜在的な再生可能原材料とみなされてもよいリグノセルロースは、一般に多糖類セルロース(グルカン)およびヘミセルロース(キシラン、ヘテロキシラン、およびキシログルカン)を含んでなる。これに加えて例えば木材由来原材料の中に、ある種のヘミセルロースがグルコマンナンとして存在してもよい。例えばこれらの多糖類のグルコース、セロビオース、キシロース、アラビノース、ガラクトース、果糖、マンノース、ラムノース、リボース、ガラクツロン酸、グルクロン酸、およびその他のヘキソースおよび五炭糖などのモノマーおよび多量体の双方をはじめとする可溶性糖への酵素加水分解は、協調して作用する異なる酵素作用の下で起きる。
【0120】
さらにペクチン、およびアラビナンなどのその他のペクチン質が、典型的な非木本組織からの細胞壁の乾燥質量のかなりの部分を構成してもよい(乾燥質量の約4分の1から2分の1がペクチンであってもよい)。
【0121】
[前処理]
(ヘミセルロース系をはじめとする)リグノセルロース系材料から、本発明に従って発酵させてもよい糖を放出させるために前処理が望ましいこともあり得る。このステップは、例えば以下のような従来の方法で実施してもよい。
【0122】
[酵素加水分解]
酵素加水分解は、従来の方法で実施してもよい。
【0123】
[発酵]
発酵工程は、好気的または嫌気的発酵工程であってもよい。嫌気的発酵工程は、本明細書で、酸素不在下で進行する発酵工程、またはその中で実質的に酸素が消費されず、好ましくは約5、約2.5または約1mmol/L/h未満、より好ましくは0mmol/L/h未満が消費され(すなわち酸素消費が検出可能でない)、その中で有機分子が電子供与体および電子受容体の双方の役割を果たす発酵工程と定義される。酸素の不在下で、解糖およびバイオマス形成中に生じるNADHは、酸化的リン酸化によって酸化され得ない。この問題を解決するために、多数の微生物は、電子および水素受容体としてピルビン酸またはその誘導体の1つを使用し、それによってNAD+を再生する。
【0124】
したがって好ましい嫌気的発酵工程では、ピルビン酸が電子(および水素受容体)として使用され、還元されて、エタノール、ブタノール、乳酸、3−ヒドロキシ−プロピオン酸、アクリル酸、酢酸、コハク酸、クエン酸、リンゴ酸、フマル酸、アミノ酸、1,3−プロパン−ジオール、エチレン、グリセロール、β−ラクタム抗生物質、およびセファロスポリンなどの発酵産物になる。
【0125】
発酵工程は、好ましくは細胞にとって最適の温度で実施される。したがってほとんどの酵母または真菌宿主細胞では、発酵工程は、約42℃未満、好ましくは約38℃未満の温度で実施される。酵母または糸状菌宿主細胞では、発酵工程は好ましくは約35、約33、約30または約28℃よりも低い温度、および約20、約22、または約25℃よりも高い温度で実施される。
【0126】
工程におけるキシロースおよび/またはグルコース上のエタノール収率は、好ましくは少なくとも約50、約60、約70、約80、約90、約95または約98%である。エタノール収率は、本明細書で理論的最大収率の百分率と定義される。
【0127】
本発明はまた、発酵産物を製造する方法にも関する。
【0128】
発酵工程は、バッチ、流加または連続様式で実施してもよい。別個の加水分解および発酵(SHF)工程、または同時の糖化および発酵(SSF)工程もまた適用してもよい。最適生産性のために、これらの発酵工程様式の併用もまた可能であってもよい。
【0129】
本発明に従った発酵工程は、好気的および嫌気的条件下で進行してもよい。好ましくは工程は、微好気的または酸素限定条件下で実施される。
【0130】
嫌気的発酵工程は、本明細書では、酸素不在下で進行し、またはその中で実質的に酸素が消費されず、好ましくは約5、約2.5または約1mmol/L/h未満が消費され、有機分子が電子供与体および電子受容体の双方の役割を果たす発酵工程、と定義される。
【0131】
酸素限定発酵工程は、気体から液体への酸素移動によって、酸素消費が制限される工程である。酸素制限の程度は、流入ガス流量および組成、ならびに使用される発酵装置の実際の混合/質量移動特性によって定まる。好ましくは、酸素限定条件下の工程では、酸素消費速度は、少なくとも約5.5、より好ましくは少なくとも約6、少なくとも7mmol/L/hなどである。本発明の方法は、発酵産物の回収を含んでなる。
【0132】
好ましい方法では、細胞はキシロースおよびグルコースの双方を好ましくは同時に発酵し、その場合、好ましくはジオーキシー成長を妨げるグルコース抑制に非感受性の細胞が使用される。炭素源としてのキシロース(およびグルコース)源に加えて、発酵培地は、細胞成長に必要とされる適切な成分をさらに含んでなる。酵母などの微生物の成長のための発酵培地組成物は、当該技術分野で周知である。
【0133】
発酵工程は、バッチ、流加または連続様式で実施してもよい。別個の加水分解および発酵(SHF)工程、または同時の糖化および発酵(SSF)工程もまた応用してもよい。最適生産性のために、これらの発酵工程様式の併用もまた可能であり得る。これらの工程については、以下でより詳細に説明する。
【0134】
[SSF様式]
同時糖化および発酵(SSF)様式では、液化/加水分解または前糖化工程のための反応時間は、所望の収率、すなわちセルロースからグルコースへの収率を実現する時間に左右される。このような収率は、好ましくは可能な限り高く、好ましくは60%以上、65%以上、70%以上、75%以上80%以上、85%以上、90%以上、95%以上、96%以上、97%以上、98%以上、99%以上、99.5%以上または99.9%以上でさえある。
【0135】
本発明に従って、SHF様式における非常に高い糖濃度力、およびSSF様式における非常に高い生成物(例えばエタノール)濃度が実現する。SHF操作では、グルコース濃度は、25g/L以上、30g/L以上、35g/L以上、40g/L以上、45g/L以上、50g/L以上、55g/L以上、60g/L以上、65g/L以上、70g/L以上、75g/L以上、80g/L以上、85g/L以上、90g/L以上、95g/L以上、100g/L以上、110g/L以上、120g/L以上であり、または例えば25g/L〜250g/L、30gl/L〜200g/L、40g/L〜200g/L、50g/L〜200g/L、60g/L〜200g/L、70g/L〜200g/L、80g/L〜200g/L、90g/L、80g/L〜200g/Lであってもよい。
【0136】
[SSF様式における生成物濃度]
SSF操作では、生成物濃度(g/L)は生成されるグルコース量に左右されるが、糖はSSF中で生成物に転換され、生成物濃度は、理論的な最小収率の増大により根底にあるグルコース濃度に関連し得る(Yps maxは、グルコース1グラム当たりの生成物グラムである)ので、これは目に見えない。
【0137】
発酵産物の理論的最大収率(Yps maxは、グルコース1グラム当たりの生成物グラムである)は、教科書通りの生化学から導かれ得る。エタノールでは、1モルのグルコース(180g)は、酵母中の通常解糖発酵経路に従って、2モルのエタノール(=2×46=92グラムのエタノール)を生じる。したがってグルコース上のエタノールの理論的最大収率は、92/180=0.511gのエタノール/グルコース1グラムである。
【0138】
ブタノール(MW74g/モル)またはイソブタノールでは、理論的最大収率はグルコース1グラム当たり1モルのブタノールである。したがって(イソ−)ブタノールのYps max=74/180=0.411g(イソ−)ブタノール/グルコース1グラムである。
【0139】
乳酸では、ホモ乳酸発酵の発酵収率は、1モルのグルコース当たり2モルの乳酸(MW=90g/モル)である。この化学量論によれば、Yps max=乳酸1g/グルコース1gである。
【0140】
その他の発酵産物についても、同様の計算を実施してもよい。
【0141】
[SSF様式]
SSF操作では、生成物濃度は、25g*Yps g/L/L以上、30*Yps g/L以上、35g*Yps/L以上、40*Yps g/L以上、45*Yps g/L以上、50*Yps g/L以上、55*Yps g/L以上、60*Yps g/L以上、65*Yps g/L以上、70*Yps g/L以上、75*Yps g/L以上、80*Yps g/L以上、85*Yps g/L以上、90*Yps g/L以上、95*Yps g/L以上、100*Yps g/L以上、110*Yps g/L以上、120g/L*Yps以上であり、または例えば25*Yps g/L〜250*Yps g/L、30*Yps gl/L〜200*Yps g/L、40*Yps g/L〜200*Yps g/L、50*Yps g/L〜200*Yps g/L、60*Yps g/L〜200*Yps g/L、70*Yps g/L〜200*Yps g/L、80*Yps g/L〜200*Yps g/L、90*Yps g/L、80*Yps g/L〜200*Yps g/Lであってもよい。
【0142】
したがって本発明は発酵産物を調製する方法を提供し、方法は、
a.本明細書に記載される方法を使用してリグノセルロースを分解するステップと;
b.得られた物質を発酵し、
それによって発酵産物を調製するステップ
を含んでなる。
【0143】
[発酵産物]
本発明の発酵産物は、いかなる有用な生成物であってもよい。一実施態様では、これは、エタノール;n−ブタノール;イソブタノール;乳酸;3−ヒドロキシ−プロピオン酸;アクリル酸;酢酸;コハク酸;フマル酸;リンゴ酸;イタコン酸;マレイン酸;クエン酸;アジピン酸;リジン、メチオニン、トリプトファン、スレオニン、およびアスパラギン酸などのアミノ酸;1,3−プロパン−ジオール;エチレン;グリセロール;β−ラクタム抗生物質およびセファロスポリン;ビタミン;医薬品;動物飼料補給剤;特殊化学薬品;化学原料;プラスチック;溶剤;生物燃料およびバイオガスまたは有機ポリマーをはじめとする燃料;およびプロテアーゼ、セルラーゼ、アミラーゼ、グルカナーゼ、ラクターゼ、リパーゼ、リアーゼ、オキシド還元酵素、トランスフェラーゼまたはキシラナーゼなどの工業用酵素からなる群から選択される生成物である。
【0144】
[発酵産物の回収]
発酵生成物の回収のためには、既存の技術が使用される。異なる発酵産物では、異なる回収工程が適切である。水性混合物からエタノールを回収する既存の方法は、一般に分留および吸着技術を使用する。例えばビール蒸留缶を使用して、エタノールを水性混合物中に含有する発酵産物を処理して濃縮エタノール含有混合物を生成し、次に分留(例えば分別蒸留またはその他の類似技術)を実施し得る。次に最高濃度のエタノールを含有する留分を吸収材に通過させ、エタノールから残りの水の全部ではないが大部分を除去し得る。
【0145】
以下の実施例は、本発明を例証する。
【0146】
[実施例]
特に断りのない限り、使用された方法は標準生化学的技術である。適切な一般方法の教科書は例えば、Sambrook et al.,Molecular Cloning,a Laboratory Manual(1989)and Ausubel et al.,Current Protocols in Molecular Biology(1995),John Wiley & Sons,Inc.を含む。
【0147】
[S.セレヴィシエ(S.cerevisiae)の形質転換]
S.セレヴィシエ(S.cerevisiae)の形質転換は、Gietz and Woods(2002;Transformation of the yeast by the LiAc/SS carrier DNA/PEG method.Methods in Enzymology 350:87−96)に記載されるようにして実施した。
【0148】
[コロニーPCR]
単一コロニー分離株をプラスチック爪楊枝で拾って50μlミリQ水に再懸濁した。サンプルを99℃で10分間インキュベートした。5μlのインキュベートしたサンプルをPhusion(登録商標)DNAポリメラーゼ(Finnzymes)を使用したPCR反応のためのテンプレートとして、供給元によって提供される使用説明書に従って使用した。
【0149】
【表2】
【0150】
[培地組成]
成長実験:サッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)株を以下の組成を有する培地上で成長させた。0.67%(w/v)酵母窒素ベースまたは合成培地(Verduyn et al.,Yeast 8:501−517,1992)およびグルコース、アラビノース、ガラクトースまたはキシロースのいずれか、またはこれらの基質の組み合わせ(下記参照)。寒天プレートでは、培地に2%(w/v)細菌用寒天を添加した。
【0151】
[エタノール生成]
培養は、30℃でBAM(Biological Activity Monitor;オランダ国(The Netherlands)のHalotec)内において、100mlの合成モデル培地(5%グルコース、5%キシロース、3.5%アラビノースおよび1〜1.5%ガラクトースを添加したVerduyn培地(Verduyn et al.,Yeast 8:501−517,1992))中で実施した。滅菌に先だって、培地のpHを2MのNaOH/H2SO4で4.2に調節した。嫌気的培養のための合成培地に、エタノールに溶解させた0.01g/Lのエルゴステロールおよび0.42g/LのTween80を添加した(Andreasen and Stier.J.Cell Physiol.41:23−36,1953;およびAndreasen and Stier.J.Cell Physiol.43:271−281,1954)。培養物を磁気撹拌機によって撹拌した。培養物に通気しなかったために、嫌気的条件は発酵中に迅速に発生した。CO2の生成を絶えずモニターした。糖転換および生成物形成をNMRによって分析した。培養物の光学濃度をLKB Ultrospec K分光光度計上で600nmで追跡することにより、成長をモニターした。
【0152】
100ml振盪フラスコ内の2%グルコースを添加した25mlのVerduyn培地(Verduyn et al.,Yeast 8:501−517,1992)に、冷凍保存培養または寒天プレートからの単一コロニーを接種して、前培養を調製した。軌道振盪機(200rpm)内における30℃でおよそ24時間のインキュベーション後にこの培養物を収集し、およそ2のOD600でBAM接種のために使用した。
【0153】
[実施例1]
「S.セレヴィシエ(S.cerevisiae)ゲノムへの遺伝子araA、araB、およびaraDの導入]
[1.1 アラビノース経路の遺伝子を含有する発現ベクターの構築]
図2に記載されるプラスミドpPWT018を次のようにして構築した。ベクターpPWT006(図1、SIT2遺伝子座からなる(Gottlin−Ninfa and Kaback(1986)Molecular and Cell Biology vol.6,no.6,2185−2197))と、抗生物質G418上およびアセトアミド上で成長する能力(上記参照)により形質転換体を選択できるようにするマーカーとを制限酵素BsiWIおよびMluIで消化した。G418耐性をもたらすkanMX−マーカーは、p427TEF(Dualsystems Biotech)から単離され、amdSマーカーを含有する断片については文献で記載されている(Swinkels,B.W.,Noordermeer,A.C.M.and Renniers,A.C.H.M(1995)The use of the amdS cDNA of Aspergillus nidulans as a dominant,bidirectional selectable marker for yeast transformation.Yeast Volume 11,Issue 1995A,page S579;および米国特許第6051431号明細書)。国際公開第2008/041840号パンフレットで開示されるような、ラクトバチルス・プランタルム(Lactobacillus plantarum)からのアラビノースイソメラーゼ(araA)、L−リブロキナーゼ(araB)、およびL−リブロース−5−リン酸−4−エピメラーゼ(araD)をコードする遺伝子は、のBaseClear(オランダ国ライデン(Leiden,The Netherlands))によって合成された。S.セレヴィシエ(S.cerevisiae)からの強力なプロモーター、すなわちaraA遺伝子の発現を制御するTDH3プロモーター、araB遺伝子を制御するENO1プロモーター、およびaraD遺伝子を制御するPGI1プロモーターの制御下にある(またはそれらと作動可能に結合している)上述の3つのアラビノース遺伝子を含む、1個の大きな断片が合成された。この断片は、ユニークな制限酵素Acc65lおよびMluIで取り囲まれた。この断片のpPWT006へのクローニング、BsiWIおよびMluIによる消化は、プラスミドpPWT018をもたらした(図2)。プラスミドpPWT018の配列を配列番号17で記載する。
【0154】
[1.2 酵母形質転換]
供給元によって提供される使用説明書に従って、あらかじめSfiI(New England Biolabs)で直線化したプラスミドpPWT018でCEN.PK113−7D[MATa URA3 HIS3 LEU2 TRP1 MAL2−8 SUC2)を形質転換した。合成SfiI部位をSIT2遺伝子の5’−flank中にデザインした(図2参照)。形質転換混合物を1ml当たり100μgのG418(シグマアルドリッチ(Sigma Aldrich))を含有するYPD−寒天(1リットル当たり10グラムの酵母抽出物、1リットル当たり20グラムのペプトン、1リットル当たり20グラムのデキストロース、20グラムの寒天)上に播種した。2〜4日後にはコロニーがプレート上に出現したが、陰性対照(すなわち形質転換実験中でDNA添加なし)はブランクのYPD/G418プレートをもたらした。プラスミドpPWT018の組み込みは、SIT2遺伝子座を対象とする。質転換体は、PCRおよびサザンブロット技術を使用して特性決定した。
【0155】
配列番号18および15と、15および14で示されるプライマーを用いて、プラスミドpPWT018の1コピーの正しい組み込みの指標となるPCR反応を実施した(図4参照)。配列番号18および15のプライマー対によって、SIT2遺伝子座での正しい組み込みをチェックした。プラスミドpPWT018が複数コピー組み込まれれば(頭尾組み込み)、配列番号15および14のプライマー対はPCR生成物を与える。後者のPCR産物が不在であれば、これはpPWT018の1コピー組み込みの徴候である。プラスミドpPWT018の1コピーがSIT2遺伝子座中に組み込まれた株をBIE104R2と命名した。
【0156】
[1.3 マーカーレスキュー]
同一選択マーカーを使用して、酵母株をその他のコンストラクトで形質転換できるようにするためには、選択可能なマーカーを除去することが必要である。プラスミドpPWT018のデザインによって、染色体へのpPWT018組み込みに際し、相同的な配列が互いにごく接近するようになる。このデザインは、これらの相同領域の突発性分子内組み換えによって、選択可能なマーカーが喪失できるようにする。
【0157】
栄養成長に際して、低頻度ではあるが分子内組み換えが起きる。この遺伝子組み換え頻度は、相同性の長さおよびゲノム中の遺伝子座(結果未公開)に左右される。培養物副画分の新鮮培地への連続的移動に際して、分子内組み換えは経時的に蓄積する。
【0158】
この目的を達成するために、単一コロニー分離株から開始して、YPD培地(1リットル当たり10グラムの酵母抽出物、1リットル当たり20グラムのペプトン、1リットル当たり20グラムのデキストロース)中でBIE104R2株を培養した。25μlの一晩培養物を使用して、新鮮なYPD培地に接種した。少なくとも5回のこのような連続的移動後、培養物の光学濃度を測定し、1mlあたりおよそ5000個の濃度に細胞を希釈した。100μlの細胞懸濁液を30mMのKPi(pH6.8)、0.1%(NH4)2SO4、40mMのフルオロ−アセトアミド(アマシャム(Amersham))、および1.8%の寒天(ディフコ(Difco))を含有する酵母炭素ベース培地(ディフコ)に播種した。BIE104R2株の細胞と同一である、すなわち細胞内組み換えのない細胞は、依然としてamdS遺伝子を含有した。これらの細胞にとって、フルオロ−アセトアミドは有毒である。これらの細胞はフルオロアセトアミド含有培地上で成長できず、コロニーを形成しない。しかし分子内組み換えが起きた場合、選択可能なマーカーを失ったBIE104R2変異株はフルオロ−アセトアミドを成長阻害化合物に転換できないので、フルオロアセトアミド培地上で成長できる。これらの細胞はこの寒天培地上でコロニーを形成する。
【0159】
このようにして得られたフルオロ−アセトアミド抵抗性コロニーに、配列番号18および15と、14および19のプライマーを使用したPCR分析を実施した。配列番号18および5のプライマーは、選択可能なマーカーの組み換えが意図したように起きていればバンドを与える。その結果、強力酵母プロモーター制御下の遺伝子araA、araB、およびaraDのあるカセットが、宿主株のゲノムSIT2遺伝子座に組み込まれた。この場合、プライマー14は組み換えのために失われたはずの領域内でプライムするので、配列番号14および19のプライマーを使用したPCR反応は、PCR産物をもたらさないはずである。後者のプライマーによってバンドが得られた場合、これはゲノム中の完全なプラスミドpPWT018の存在の徴候であり、したがって組み換えは起きていない。
【0160】
配列番号18および15のプライマーがPCR産物をもたらさなければ、組み換えは起きているが、完全なプラスミドpPWT018は組み換えによってゲノムの外にある。選択可能なマーカーだけでなく、アラビノース遺伝子もまた失われた。実際、野生型酵母が回復されていた。
【0161】
pPWT018の1コピーの組み込みに従ってPCR結果を示す分離株に、サザンブロット分析を実施した。CEN.PK113−7D株の染色体DNAと正確な組み換体えをEcoRIおよびHindIIIで消化した(二重消化)。テンプレートとしてCEN.PK113−7Dの染色体DNAを使用して、配列番号20および21プライマーによりSIT2プローブを調製した。ハイブリダイゼーション実験の結果を図3に示す。予期されるハイブリダイゼーションパターンは、図4(パネルaおよびb)に示すような物理的地図から推定されてもよい。
【0162】
野生型株では、2.35kbのバンドが観察され、それは野性型遺伝子(図4、パネルa)の予期されるサイズに一致した。プラスミドpPWT018の組み換えによる組み込みおよび部分的損失に際して、1.06kbのバンドが予期された(図4、パネルb)。実際、このバンドは図3(レーン2)に示すように観察された。
【0163】
サザンブロット法でバンドの正確なパターン(図3から推定し得るような)を示した株の1つは、BIE104A2と命名された株である。
【0164】
[1.4 非酸化的ペントースリン酸経路の4つの構成的に発現される遺伝子の導入]
遺伝子araA、araB、およびaraDを構成的に発現するサッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)BIE104A2をプラスミドpPWT080で形質転換した(図5)。プラスミドpPWT080の配列は、配列番号4に記載される。1コピーの形質転換体を選択した後の形質転換および選択手順は、1.1、1.2、および1.3節で上述したのと同じであった。手短に言えば、BIE104A2をSfiI消化pPWT080で形質転換した。形質転換混合物を1mlあたり100μgのG418(シグマアルドリッチ)を含有するYPD寒天(1リットル当たり10グラムの酵母抽出物、1リットル当たり20グラムのペプトン、1リットル当たり20グラムのデキストロース、20グラムの寒天)上に播種した。
【0165】
2〜4日後にはコロニーがプレート上に出現したが、陰性対照(すなわち形質転換実験中でDNA添加なし)はブランクのYPD/G418プレートをもたらした。
【0166】
プラスミドpPWT080の組み込みは、GRE3遺伝子座を対象とする。形質転換体は、PCRおよびサザンブロット技術を使用して特性決定した。
【0167】
予期されるハイブリダイゼーションパターンに従って、プラスミドpPWT080の1コピーの正しい組み込みを示す形質転換体をBIE104A2F1と命名した。
【0168】
キシロースイソメラーゼおよびキシルロキナーゼをコードする遺伝子を導入できるように(実施例5)、プラスミドpPWT080の組み込みによって導入された選択マーカーを除去することが必要である。この目的を達成するために、コロニー分離株から開始して、BIE104A2F1株をYPD培地中で培養した。25μlの一晩培養物を使用して新鮮なYPD培地に接種した。5回の連続的移動後、培養物の光学濃度を測定し、1ml当たりおよそ5000個の濃度に細胞を希釈した。100μlの細胞懸濁液を30mMのKPi(pH6.8)、0.1%のH2SO4、40mMのフルオロ−アセトアミド(アマシャム)、および1.8%の寒天(ディフコ)を含有する酵母炭素ベース培地(ディフコ)に播種した。フルオロアセトアミド抵抗性コロニーにPCR分析を実施して、正確なPCRプロフィールの場合は、サザンブロット分析を実施した(実施例1の1.3節)。サザンブロット法で正確なバンドパターンを示す株の1つは、BIE104A2P1と命名された株である。
【0169】
[実施例2]
[適応進化]
[2.1 適応進化(好気的)]
BIE104A2P1株の単一コロニー分離株を使用して、2%ガラクトースを添加したYNB培地(ディフコ)に接種した。前培養を30℃および280rpmでおよそ24時間インキュベートした。細胞を収集して、0.2の開始OD600で1%ガラクトースおよび1%アラビノースを含有するYNB培地に接種した(図8)。細胞を30℃および280rpmで成長させた。600nmでの光学濃度を定期的にモニターした。
【0170】
光学濃度の値が5に達した際、培養物のアリコートを同一培地を含有する新鮮なYNB培地に移した。添加された細胞量は、培養物の開始OD600が0.2になるような量であった。OD600が再度5に達したら、培養物のアリコートを唯一の炭素源として2%アラビノースを含有するYNB培地に移した(図8の(1)で示される事象)。
【0171】
唯一の炭素源として2%アラビノース添加YNBに移すと、およそ2週間後に成長が観察できた。600nmでの光学濃度の値が少なくとも1に達した際、細胞を0.2の開始OD600で、2%アラビノースを添加した新鮮なYNB培地を入れた振盪フラスコに移した(図8)。
【0172】
図8に示すように連続的移動を3回繰り返した。得られたアラビノースで迅速に成長できる株をBIE104A2P1cと命名した。
【0173】
[2.2適応進化(嫌気的)]
好気的条件下におけるアラビノース上での成長適応後、BIE104A2P1c株からの単一コロニーを2%グルコースを添加したYNB培地上に接種した。前培養を30℃および280rpmでおよそ24時間インキュベートした。
【0174】
細胞を収集して2%アラビノースを含有するYNB培地中に、0.2の光学濃度OD600で接種した。フラスコをウォーターロックで閉鎖して、培地およびヘッドスペースから酸素が枯渇した後の嫌気的成長条件を確実にした。OD600が最低3に達したら、培養物のアリコートを2%アラビノースを含有する新鮮なYNB培地にそれぞれ0.2の光学濃度OD600で移した(図9)。
【0175】
数回の移動後、得られた株をBIE104A2P1d(=BIE201)と命名した。
【0176】
[実施例3]
[発酵能力測定]
BIE104、BIE104A2P1c、およびBIE201株の単一コロニー分離株を使用して、2%グルコースを添加したYNB培地(ディフコ)に接種した。前培養を30℃および280rpmでおよそ24時間インキュベートした。細胞を収集し、BAM内で合成モデル培地(Verduyn et al.,Yeast 8:501−517、1992;5%グルコース、5%キシロース、3.5%アラビノース、1%ガラクトース)におよそ2の開始OD600で接種した。CO2生成を絶えずモニターした。糖転換および生成物形成をNMRによって分析した。データは指定の糖の残留量(1リットルあたりのグルコース、アラビノース、ガラクトースおよびキシロースのグラム数)および(副)生成物形成(エタノール、グリセロール)を表す。600nmで培養物の光学濃度を追跡して成長をモニターした(図10、11、12)。実験はおよそ140時間実施した。
【0177】
実験は参照株BIE104がグルコースを迅速に転換したが、アラビノースまたはガラクトースのどちらも140時間以内に転換できなかったことを明らかに示した(図10)。しかしBIE104A2P1cおよびBIE201株は、アラビノースおよびガラクトースを転換できた(それぞれ図11および12)。ガラクトースおよびアラビノース利用は、20時間以内にグルコース枯渇の直後に始まった。どちらの糖も同時に転換された。しかし嫌気的条件下におけるアラビノース成長のために改善されたBIE201株は、双方の糖をより迅速に消費した(図12)。全ての発酵で、副産物としてはグリセロールのみが生じた。BIE201の発酵のデータは、本明細書で表2に示す。
【0178】
【表3】
【0179】
以下からの最大エタノール濃度(g/l)
グルコース 21.8
アラビノース 16.1
ガラクトース 6.6
合計 44.5
実験的エタノール収率0.45gエタノール/g糖
【0180】
この計算から、糖グルコース、ガラクトース、およびアラビノースが、それぞれエタノールに変換されることが明らかである。
【0181】
[実施例4]
[糖転換に対するPPP遺伝子の影響]
PPP遺伝子が糖転換におよぼす影響を試験するために、BIE104A2およびBIE105A2株からの単一コロニーを2%グルコースを添加したYNB培地(ディフコ)に接種した。どちらの株もアラビノース遺伝子を含有し、アラビノース上での成長のために進化させた(実施例2、2.1節に記載されるように)。BIE105A2株は工業株のバックグラウンドを有する。しかしこれは前述したのと同じ方法およびコンストラクトで形質転換された(実施例1、1.2節)。
【0182】
前培養を収集し、BAM内で合成トウモロコシ繊維モデル培地(Verduyn et al.,Yeast 8:501−517、1992;5%グルコース、5%キシロース、3.5%アラビノース、1.5%ガラクトース)におよそ2の開始OD600で接種した。CO2生成を絶えずモニターした。糖転換および生成物形成をNMRによって分析した。データは指定の糖の残留量(1リットルあたりのグルコース、アラビノース、ガラクトースおよびキシロースのグラム数)および(副)生成物形成(エタノール、グリセロール)を表す。600nmで培養物の光学濃度を追跡して成長をモニターした。実験はおよそ160時間実施した。
【0183】
実験はどちらの株もグルコース枯渇直後に、PPP遺伝子を過剰発現することなく、アラビノースおよびガラクトースを転換できることを示す(図13および14)。
【0184】
[実施例5]
[キシロースイソメラーゼおよびキシルロキナーゼをコードする構成的に発現される遺伝子の導入]
[5.1 酵母形質転換]
BIE104A2P1株(MATa URA3 HIS3 LEU2 TRP1 MAL2−8 SUC2 SIT2::[TDH3−araA、ENO1−araB、PGI1−araD] ΔGRE3::[TPI1p−TAL1、ADH1p−TKL1、PGI1p−RPE1、ENO1 P−RKI1])をプラスミドpPWT042で形質転換した(図16)。プラスミドpPWT042は、ベクターpPWT007に由来する(図15)。それはPCT欧州特許第2009/52623号明細書で開示されるように、S.セレヴィシエ(S.cerevisiae)からのコドンペア最適化されたキシルロキナーゼ、およびバクテロイデス・ユニフォルミス(Bacteroides uniformis)からのコドンペア最適化されたキシロースイソメラーゼ(配列番号Q2)を含有する。BIE104A2P1の形質転換に先だって、供給元が提供する使用説明書に従って、制限酵素SfiIを使用してpPWT042を直線化した。形質転換混合物を1mlあたり100μgのG418(シグマアルドリッチ)を含有するYPD寒天(1リットル当たり10グラムの酵母抽出物、1リットル当たり20グラムのペプトン、1リットル当たり20グラムのデキストロース、20グラムの寒天)上に播種した。
【0185】
2〜4日後にはコロニーがプレート上に出現したが、陰性対照(すなわち形質転換実験中でDNA添加なし)はブランクのYPD/G418プレートをもたらした。
【0186】
プラスミドをSfiIでpPWT042消化すると、その組み込みはゲノム中のSIT4遺伝子座を対象とする(Gottlin−Ninfa and Kaback(1986)Molecular and Cellular Biology Vol.6,No.6,2185−2197)(図17)。形質転換体は、実施例1(1.2節)に記載されるようにして、PCRおよびサザンブロティング技術を使用して特性決定した。
【0187】
1コピーのpPWT042プラスミドがゲノムに組み込まれた株をBIE104A2P1Y9と命名した。
【0188】
[5.2 成長実験]
BIE104A2P1Y9株の単一コロニー分離株を使用して、2%グルコースまたは2%ガラクトースを添加したYNB培地(ディフコ)に接種した。接種されたフラスコは、600nmでの光学濃度が少なくとも2.0の値に達するまで、30℃および280rpmでインキュベートした。
【0189】
1%アラビノースおよび1%キシロースを添加したYNB培地に、0.2の開始OD600で一晩培養物を接種した。細胞を30℃および280rpmで成長させた。600nmの光学濃度を定期的にモニターした。光学濃度が2.0を超える値に達した際、培養物のアリコートを2%キシロースおよび0.2%アラビノースを含有する新鮮なYNB培地に移した。添加された細胞量は、培養物の開始OD600が0.2になるような量であった。
【0190】
光学濃度を定期的にモニターした。結果を図18のパネルa(ガラクトース上の前培養)およびパネルb(グルコース上の前培養)に示す。
【0191】
結果は株が、グルコース、ガラクトース、アラビノース、およびキシロースを利用できることを明らかに示す。
【0192】
[5.3 マーカーレスキュー]
プラスミドpPWT042の組み込みによって導入された選択マーカー除去するために、コロニー分離株から開始して、BIE104A2P1Y9株をYPD培地中で培養した。25μlの一晩培養物を使用して新鮮なYPD培地に接種した。連続的移動後、培養物の光学濃度を測定し、1ml当たりおよそ5000個の濃度に細胞を希釈した。100μlの細胞懸濁液を30mMのKPi(pH6.8)、0.1%の(NH4)2SO4、40mMのフルオロ−アセトアミド(アマシャム)、および1.8%の寒天(ディフコ)を含有する酵母炭素ベース培地(ディフコ)に播種した。フルオロアセトアミド抵抗性コロニーにPCR分析を実施して、正確なPCRプロフィールの場合は、サザンブロット分析を実施した(実施例1、1.3節)。サザンブロット法で正確なバンドパターンを示す株の1つは、BIE104A2P1X9と命名された株である。
【0193】
[5.4 成長実験]
BIE104A2P1X9(BIE104A2P1X9a1およびBIE104A2P1X9a2)株の単一コロニー分離株を使用して、2%グルコースを添加したVerduyn培地(ディフコ)に接種した。接種したフラスコを30℃および280rpmでおよそ24時間インキュベートした。
【0194】
2%キシロースを添加したVerduyn培地に一晩培養物を0.2の開始OD600で接種した。細胞を30℃および280rpmで成長させた。600nmでの光学濃度を定期的にモニターした。結果を図19に示す。結果は、BIE104A2P1X9株の独立したコロニーの双方が、マーカーレスキュー後に依然としてキシロースを利用できることを明らかに示す。実施例3で既に示されたように、株はグルコース、アラビノース、およびガラクトースを利用できる(図11および図12)。
【0195】
[実施例6]
[アラビノースおよびガラクトース上のコハク酸生成のためのS.セレヴィシエ(S.cerevisiae)形質転換]
[6.1. 発現コンストラクト]
アクチノバチルス・サクシノジェネス(Actinobacillus succinogenes)からのホスホエノールピルビン酸カルボキシキナーゼPCKa(E.C.4.1.1.49)と、トリパノソーマ・ブルセイ(Trypanosoma brucei)からのグリコソームフマル酸還元酵素FRDg(E.C.1.3.1.6)とを含んでなる発現コンストラクトpGBS414PPK−3、およびリゾプス・オリゼー(Rhizopus oryzae)からのフマラーゼ(E.C.4.2.1.2.)と、ペルオキシソームリンゴ酸デヒドロゲナーゼMDH3(E.C.1.1.1.37)を含んでなる発現コンストラクトpGBS415FUM3を本明細書に図および配列表を含めて参照として組み込んだ国際公開第2009/065778号パンフレットの19〜20および22〜30頁で既述されたようにして作成する。
【0196】
プラスミドpPWT018からのaraABD発現カセットを含んでなるPCR産物をプラスミドpRS416にクローニングすることで、ラクトバチルス・プランタルム(Lactobacillus plantarum)に由来する遺伝子araA、araB、およびaraDを含んでなる発現コンストラクトpGBS416ARAABDを構築する。Phusion(登録商標)DNAポリメラーゼ(Finnzymes)と、本明細書で配列番号22および配列番号23で定義されるPCRプライマーとを使用してPCR断片を作成する。PCR産物を制限酵素SalIおよびNotIで切断し、プラスミドpRS416も同様にする。ライゲーションおよび大腸菌(E.coli)TOP10への形質転換後、制限酵素分析に基づいて正しい組み換え体を選択する。プラスミドpGBS416ARAABDの物理的地図を図20に記載する。
【0197】
[6.2. S.セレヴィシエ(S.cerevisiae)株]
プラスミドpGBS414PPK−3、pGBS415−FUM−3をS.セレヴィシエ(S.cerevisiae)株CEN.PK113−6B[MATA ura3−52 Ieu2−112 trp1−289)に形質転換する。さらにプラスミドpGBS416ARAABDをこの酵母に形質転換して、原栄養酵母株を作り出す。発現ベクターを電気穿孔によって酵母に形質転換する。形質転換混合物を酵母窒素ベース(YNB)w/o AA(ディフコ)+2%グルコース上に播種する。
【0198】
株に、唯一の炭素源としてのアラビノース上での成長のための適応進化(2節参照)を実施する。
【0199】
[6.3. 成長実験およびコハク酸生成]
形質転換体を2%ガラクトース(w/v)を含んでなるVerduyn培地(Verduyn et al.,1992,Yeast.Jul;8(7):501−17)からなる20mlの前培養液に接種して、好気的条件下で振盪恒温器内の100ml振盪フラスコ内で30℃および250rpmで成長させる。およそ24時間後、細胞を2%グルコース、2%ガラクトースまたは2%アラビノース、またはそれらの混合物のいずれかを含有する新鮮なVerduyn培地に4連で移す。2本のフラスコは好気的条件下でインキュベートし、2本のフラスコは、例えばウォーターロックを使用してフラスコを閉鎖して、または嫌気性軌道振盪機内でのインキュベーションによって、嫌気的条件下でインキュベートする。周期的に培養サンプルを採取する。サンプルを4750rpmで5分間遠心分離する。1mlの上清を使用し、6.4.節に記載されるようにしてHPLCによりコハク酸レベルを測定する。
【0200】
[6.4. HPLC分析]
有機酸および糖の定量のためにHPLCを実施する。Phenomenex Rezex−RHM−単糖類カラムの分離の原理は、逆相機序を使用した、サイズ排除、イオン排除、およびイオン交換に基づく。検出を示差屈折率および紫外線検出器によって実施する。
【0201】
[文献]
文献番号 出典
(1)Bioresource Technology 1994 Vol.47 283−284頁
(2)Micard,Enzyme Microbiol Technology 1996 Vol 19 163−170頁
(3)DOE Radke,Idaho wheat straw composition
(4)Grohman and Botast Process Biochemistry 1997 Vol.32 No 5 405−415
(5)Saska B & B 1995 517−523
(6)PCT/欧州特許第2009/52623号明細書
(7)Zheng Appl.Biochem.Microbiol.2007,Vol.136−140 423−436頁
(8)Bradshaw Appl Biochem.Microbiol.2007 Vol 136−140 395−406頁
(9)Cara Appl Biochem.Microbiol.2007 Vol 136−140 379−394頁
【特許請求の範囲】
【請求項1】
a)サッカロミセス(Saccharomyces)、クリヴェロミセス(Kluyveromyces)、カンジダ(Candida)、ピチア(Pichia)、分裂酵母(SchizoSaccharomyces)、ハンゼヌラ(Hansenula)、クロエケラ(Kloeckera)、シュワニオミセス(Schwanniomyces)またはヤロウィア(Yarrowia)属に属する酵母の存在下で糖組成物を発酵させるステップと、
b)発酵産物を回収するステップを含んでなり、
前記酵母が遺伝子araA、araB、およびaraDを含んでなり、前記糖組成物がグルコース、ガラクトース、およびアラビノースを含んでなる、
糖組成物から1つ以上の発酵産物を生成する方法。
【請求項2】
前記グルコース、ガラクトース、およびアラビノース糖が発酵産物に変換される、請求項1に記載の方法。
【請求項3】
前記1つ以上の発酵産物がエタノールである、請求項1または2に記載の方法。
【請求項4】
a)1つ以上のリグノセルロース系材料を前処理して前処理されたリグノセルロース系材料を生成するステップと;
b)前記前処理されたリグノセルロース系材料を酵素処理して前記糖組成物を生成するステップ
によって、前記糖組成物がリグノセルロース系材料から生成される、請求項の1〜3のいずれか一項に記載の方法。
【請求項5】
混合糖細胞がサッカロミセス(Saccharomyces)属のものである、請求項1〜4のいずれか一項に記載の方法。
【請求項6】
前記混合糖細胞がサッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)である、請求項5に記載の方法。
【請求項7】
前記混合糖細胞がアルドース還元酵素遺伝子の欠失を含んでなる、請求項1〜6のいずれか一項に記載の方法。
【請求項8】
前記発酵が嫌気的または酸素限定条件下で実施される、請求項1〜7のいずれか一項に記載の方法。
【請求項9】
前記混合糖細胞が、過剰発現されたPPP遺伝子TAL1、TKL1、RPE1、およびRKI1を含んでなる、請求項1〜8のいずれか一項に記載の方法。
【請求項10】
前記混合糖細胞がxylA遺伝子および/またはXKS1遺伝子を含んでなる、請求項1〜9のいずれか一項に記載の方法。
【請求項11】
前記遺伝子が、
a)強力なプロモーター制御下のPPP遺伝子TAL1、TKL1、RPE1およびRKI1からなるクラスター;
b)どちらも構成的プロモーター制御下のxylA遺伝子およびXKS1遺伝子からなるクラスター;
c)遺伝子araA、araB、およびaraDからなるクラスターおよび/またはxylA遺伝子およびXKS1遺伝子のクラスター;
d)アルドース還元酵素遺伝子の欠失
の宿主細胞への導入と、前記混合糖細胞を生じる混合糖コンストラクトの適応進化によって、前記混合糖細胞に導入される、請求項1〜10のいずれか一項に記載の方法。
【請求項12】
前記宿主細胞が阻害物質抵抗性細胞である、請求項11に記載の方法。
【請求項13】
前記宿主細胞が工業株である、請求項11または12に記載の方法。
【請求項14】
前記発酵産物が、エタノール;n−ブタノール;イソブタノール;乳酸;3−ヒドロキシ−プロピオン酸;アクリル酸;酢酸;コハク酸;フマル酸;リンゴ酸;イタコン酸;マレイン酸;クエン酸;アジピン酸;リジン、メチオニン、トリプトファン、スレオニン、およびアスパラギン酸などのアミノ酸;1,3−プロパン−ジオール;エチレン;グリセロール;β−ラクタム抗生物質およびセファロスポリン;ビタミン;医薬品;動物飼料補給剤;特殊化学薬品;化学原料;プラスチック;溶剤;生物燃料およびバイオガスまたは有機ポリマーを含む燃料;およびプロテアーゼ、セルラーゼ、アミラーゼ、グルカナーゼ、ラクターゼ、リパーゼ、リアーゼ、オキシド還元酵素、トランスフェラーゼまたはキシラナーゼなどの工業用酵素からなる群から選択される、請求項1〜13のいずれか一項に記載の方法。
【請求項15】
発現を通じて、サッカロミセス(Saccharomyces)、クリヴェロミセス(Kluyveromyces)、カンジダ(Candida)、ピチア(Pichia)、分裂酵母(SchizoSaccharomyces)、ハンゼヌラ(Hansenula)、クロエケラ(Kloeckera)、シュワニオミセス(Schwanniomyces)またはヤロウィア(Yarrowia)属に属するグルコース発酵酵母に、グルコースの枯渇後に、アラビノースの存在下でガラクトースを嫌気的に発酵する能力をもたらす、遺伝子araA、araB、およびaraDの使用。
【請求項1】
a)サッカロミセス(Saccharomyces)、クリヴェロミセス(Kluyveromyces)、カンジダ(Candida)、ピチア(Pichia)、分裂酵母(SchizoSaccharomyces)、ハンゼヌラ(Hansenula)、クロエケラ(Kloeckera)、シュワニオミセス(Schwanniomyces)またはヤロウィア(Yarrowia)属に属する酵母の存在下で糖組成物を発酵させるステップと、
b)発酵産物を回収するステップを含んでなり、
前記酵母が遺伝子araA、araB、およびaraDを含んでなり、前記糖組成物がグルコース、ガラクトース、およびアラビノースを含んでなる、
糖組成物から1つ以上の発酵産物を生成する方法。
【請求項2】
前記グルコース、ガラクトース、およびアラビノース糖が発酵産物に変換される、請求項1に記載の方法。
【請求項3】
前記1つ以上の発酵産物がエタノールである、請求項1または2に記載の方法。
【請求項4】
a)1つ以上のリグノセルロース系材料を前処理して前処理されたリグノセルロース系材料を生成するステップと;
b)前記前処理されたリグノセルロース系材料を酵素処理して前記糖組成物を生成するステップ
によって、前記糖組成物がリグノセルロース系材料から生成される、請求項の1〜3のいずれか一項に記載の方法。
【請求項5】
混合糖細胞がサッカロミセス(Saccharomyces)属のものである、請求項1〜4のいずれか一項に記載の方法。
【請求項6】
前記混合糖細胞がサッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)である、請求項5に記載の方法。
【請求項7】
前記混合糖細胞がアルドース還元酵素遺伝子の欠失を含んでなる、請求項1〜6のいずれか一項に記載の方法。
【請求項8】
前記発酵が嫌気的または酸素限定条件下で実施される、請求項1〜7のいずれか一項に記載の方法。
【請求項9】
前記混合糖細胞が、過剰発現されたPPP遺伝子TAL1、TKL1、RPE1、およびRKI1を含んでなる、請求項1〜8のいずれか一項に記載の方法。
【請求項10】
前記混合糖細胞がxylA遺伝子および/またはXKS1遺伝子を含んでなる、請求項1〜9のいずれか一項に記載の方法。
【請求項11】
前記遺伝子が、
a)強力なプロモーター制御下のPPP遺伝子TAL1、TKL1、RPE1およびRKI1からなるクラスター;
b)どちらも構成的プロモーター制御下のxylA遺伝子およびXKS1遺伝子からなるクラスター;
c)遺伝子araA、araB、およびaraDからなるクラスターおよび/またはxylA遺伝子およびXKS1遺伝子のクラスター;
d)アルドース還元酵素遺伝子の欠失
の宿主細胞への導入と、前記混合糖細胞を生じる混合糖コンストラクトの適応進化によって、前記混合糖細胞に導入される、請求項1〜10のいずれか一項に記載の方法。
【請求項12】
前記宿主細胞が阻害物質抵抗性細胞である、請求項11に記載の方法。
【請求項13】
前記宿主細胞が工業株である、請求項11または12に記載の方法。
【請求項14】
前記発酵産物が、エタノール;n−ブタノール;イソブタノール;乳酸;3−ヒドロキシ−プロピオン酸;アクリル酸;酢酸;コハク酸;フマル酸;リンゴ酸;イタコン酸;マレイン酸;クエン酸;アジピン酸;リジン、メチオニン、トリプトファン、スレオニン、およびアスパラギン酸などのアミノ酸;1,3−プロパン−ジオール;エチレン;グリセロール;β−ラクタム抗生物質およびセファロスポリン;ビタミン;医薬品;動物飼料補給剤;特殊化学薬品;化学原料;プラスチック;溶剤;生物燃料およびバイオガスまたは有機ポリマーを含む燃料;およびプロテアーゼ、セルラーゼ、アミラーゼ、グルカナーゼ、ラクターゼ、リパーゼ、リアーゼ、オキシド還元酵素、トランスフェラーゼまたはキシラナーゼなどの工業用酵素からなる群から選択される、請求項1〜13のいずれか一項に記載の方法。
【請求項15】
発現を通じて、サッカロミセス(Saccharomyces)、クリヴェロミセス(Kluyveromyces)、カンジダ(Candida)、ピチア(Pichia)、分裂酵母(SchizoSaccharomyces)、ハンゼヌラ(Hansenula)、クロエケラ(Kloeckera)、シュワニオミセス(Schwanniomyces)またはヤロウィア(Yarrowia)属に属するグルコース発酵酵母に、グルコースの枯渇後に、アラビノースの存在下でガラクトースを嫌気的に発酵する能力をもたらす、遺伝子araA、araB、およびaraDの使用。
【図1】

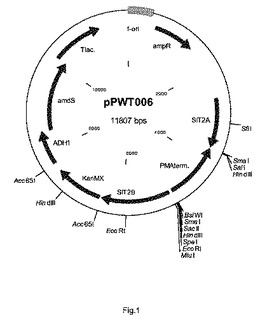
【図2】
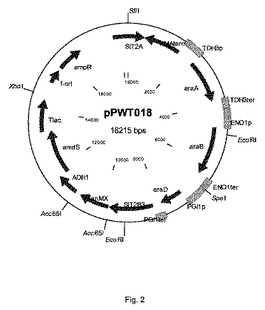
【図3】

【図4】
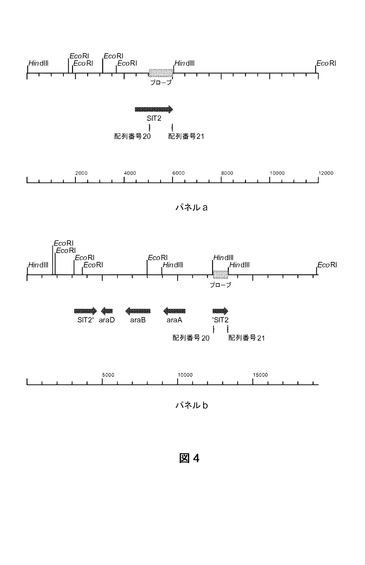
【図5】
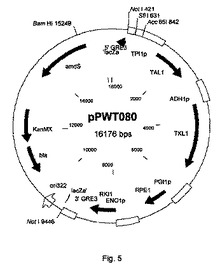
【図6−1】

【図6−2】

【図7】

【図8】
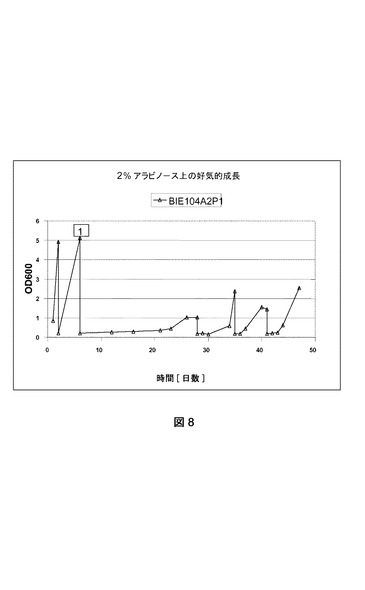
【図9】
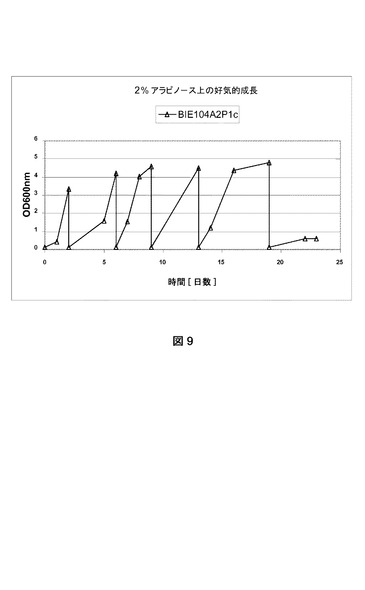
【図10】
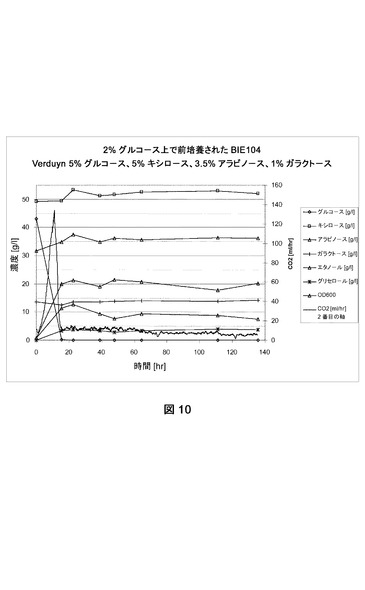
【図11】
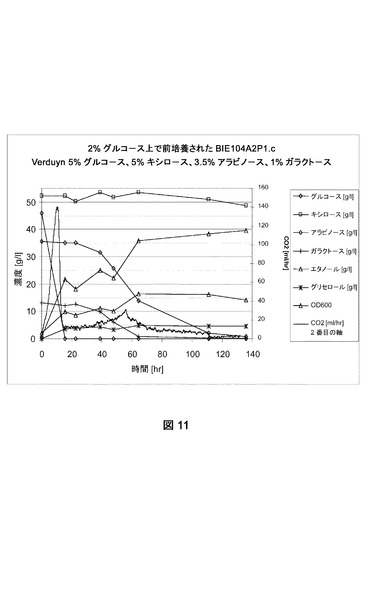
【図12】
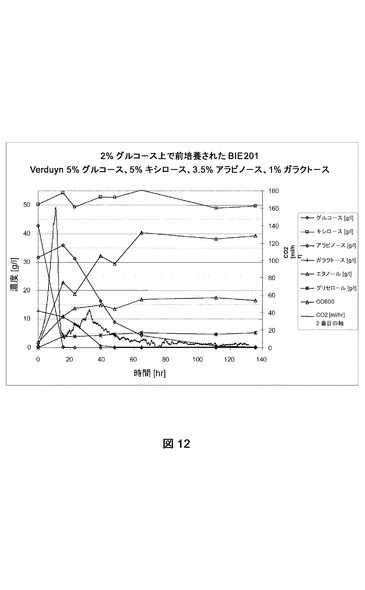
【図13】
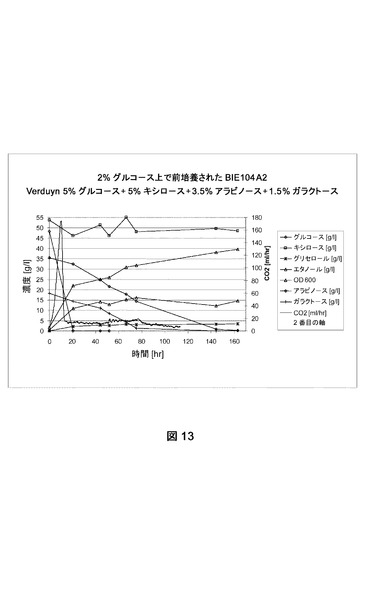
【図14】
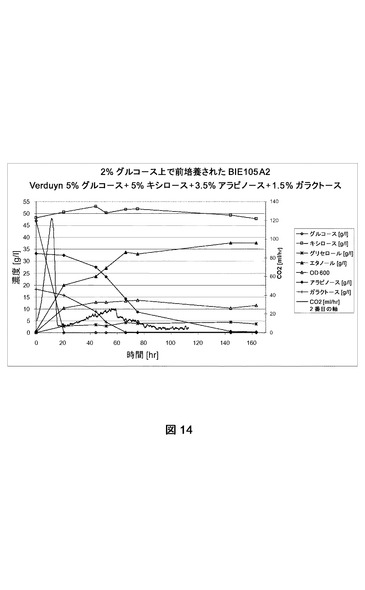
【図15】
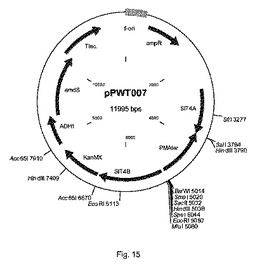
【図16】
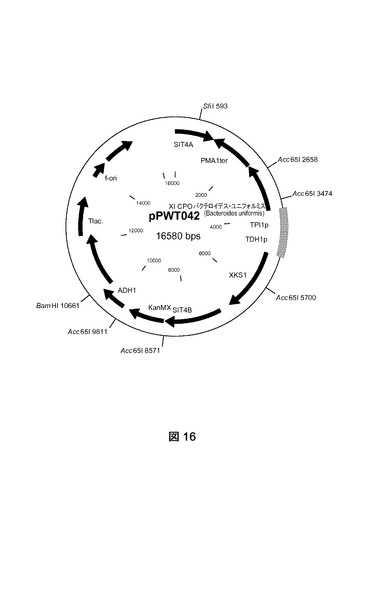
【図17】

【図18−1】
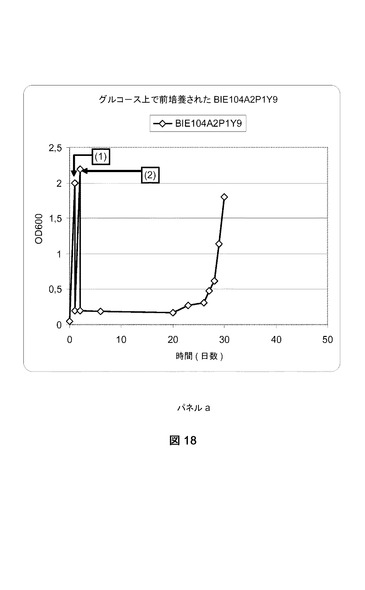
【図18−2】
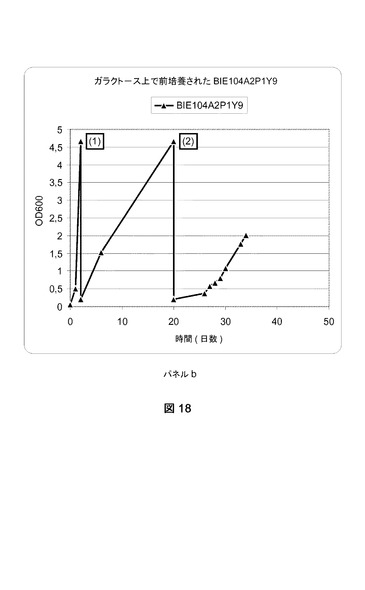
【図19】
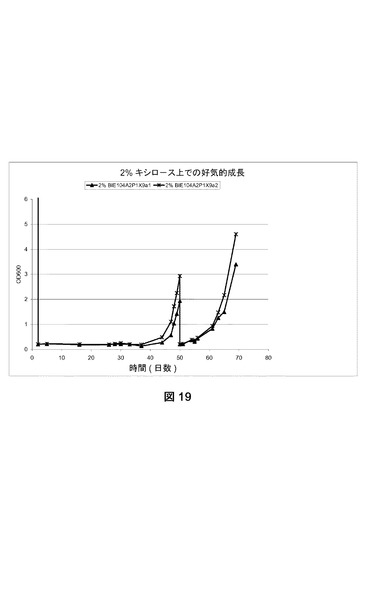
【図20】
【図2】
【図3】
【図4】
【図5】
【図6−1】
【図6−2】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18−1】
【図18−2】
【図19】
【図20】
【公表番号】特表2012−532591(P2012−532591A)
【公表日】平成24年12月20日(2012.12.20)
【国際特許分類】
【出願番号】特願2012−518961(P2012−518961)
【出願日】平成22年7月6日(2010.7.6)
【国際出願番号】PCT/EP2010/059618
【国際公開番号】WO2011/003893
【国際公開日】平成23年1月13日(2011.1.13)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
【公表日】平成24年12月20日(2012.12.20)
【国際特許分類】
【出願日】平成22年7月6日(2010.7.6)
【国際出願番号】PCT/EP2010/059618
【国際公開番号】WO2011/003893
【国際公開日】平成23年1月13日(2011.1.13)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
[ Back to top ]