
組成物、その調製方法、およびその使用
組成物は、セッコク属からの多糖を含む。この組成物は、経口寛容を誘導し、先天性免疫を促進することにより自己免疫疾患および粘膜の疾患を治療するために使用される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、自己免疫疾患および粘膜障害を治療するための組成物および方法、より詳細には、経口寛容の誘導および先天性免疫の促進を介して自己免疫疾患および粘膜障害を治療するための、セッコク属(Dendrobium)に由来する多糖を含む組成物およびセッコク属に由来する多糖を投与することを含む方法に関する。
【背景技術】
【0002】
免疫系が、宿主自身に対して反応することなく外部の病原体から宿主を保護しうるよう、免疫系の基本的な特性は、自己/非自己の区別をもたらす免疫寛容である。免疫系が自身に対して反応すると、自己免疫疾患が起こる (Annu. Rev. Med. 48:341-351, 1997) 。
【0003】
ブドウ膜炎、コラーゲン誘導性関節炎、アジュバント関節炎、全身性エリテマトーデス、多発性硬化症、甲状腺炎、重症筋無力症、炎症性腸疾患、およびI型糖尿病を含む自己免疫疾患において経口寛容は治療上有益であることが証明された (Annals of the New York Academy of Sciences 778(1): 217-227, 1996) 。
【0004】
経口的に投与された抗原は、良く発達した免疫ネットワークである腸管関連リンパ組織(GALT)に出会う。GALTは、絨毛、固有層、上皮細胞間リンパ球、およびパイエル板から成り、このパイエル板は、絨毛の間に散在するリンパ小節から成る。腸管上に並ぶ上皮は、抗原および微生物を輸送するためのM細胞を含む。腸管内の抗原および病原体は、その下にある免疫細胞に抗原を直接送達することに特化したM細胞を通してのみバリアを突破しうる。次いで、抗原を認識する上皮細胞間リンパ球がパイエル板に移動し、免疫応答を引き起こす。パイエル板では、B細胞およびマクロファージがリンパ球に抗原を提示し、次いで、この抗原特異的リンパ球が、体の粘膜系(呼吸器官の粘膜表面等)の固有層に移動する (Science 277:949-952, 1997; Science 277: 910-911, 1997) 。
【0005】
腸管の免疫系は、経口投与を介して媒介された疾患を予防するための先天性免疫および適応免疫を含む。病原体に感染した組織部分は、マクロファージ、単球、顆粒球、およびナチュラルキラー細胞の非特異的先天性免疫応答を誘導するためのケモカインを分泌する。マクロファージおよび顆粒球は、食作用により病原体を直接捕らえることができる。ナチュラルキラー細胞は、ウイルス感染または他の病原体により感染した細胞に対して最初に応答する体内における免疫細胞である。ナチュラルキラー細胞は標的細胞を攻撃するだけでなく、IFN-γ、TGF-β、TNF-α、IL-5、およびIL-10等の免疫応答を調節するためのサイトカインを分泌する。マクロファージおよび樹状細胞等の他の抗原提示細胞は、免疫応答すなわち、T細胞およびB細胞によりそれぞれ実行された細胞性免疫および体液性免疫をさらに活性化させるために、細胞表面上のMHCクラスII分子および共刺激因子を介してT細胞に抗原を提示する。粘膜の免疫は、CD4+T細胞から分泌されたサイトカインにより主に調節される。2型ヘルパーT細胞から分泌されたIL-4、IL-5、IL-6、およびIL-10は、抗体産生のためにB細胞を刺激することに関与する。3型ヘルパーT細胞から分泌されたTGF-βはB細胞を誘導し、IgAを生成させる(Trends Immunol. 22:244-247, 2004; J. Clin. Invest. 106:935-937, 2000)。
【0006】
粘膜の免疫応答は腸管においてだけでなく全身の粘膜系においても誘導されることに留意されたい。腸内の誘導組織であるパイエル板において抗原特異的リンパ球が活性化されたら、このリンパ球は、リンパ排液路および血液を通って、直腸、生殖器官、および肺等を含むエフェクター部位に移動する。次いで、これらのエフェクター部位で抗体が産生され、検出される。このホーミング現象は、いわゆる一般的な粘膜の免疫系である。経口投与により誘導された抗体は一般的な粘膜の免疫応答により全身に配置される。経口投与を介して媒介された抗原が存在すると、組織的寛容またはIgA、すなわち経口寛容が誘導される。経口寛容は、組織的不応答および粘膜応答の両者を含み、前者は、自己免疫疾患から全身を予防し、後者は、特異抗原に対して能動的抑制を誘導する。したがって、粘膜の免疫系は、炎症性の免疫応答を抑制し、さらに、非炎症性の局所粘膜でのIgAの提示を増強させる傾向がある(Immunol. Today 18:335-343, 1997)。
【0007】
胃腸管の表面には単層円柱上皮が並んで、細菌、食餌抗原、および高分子の過剰吸収に対するバリアを形成し、さらに、低分子の輸送は、バリア中に形成された密着結合により制御される。結合組織に埋まっている陰窩は、腸管上皮細胞を再生するための幹細胞を含む。腸管上皮細胞の他に、この腸管幹細胞は、粘液を分泌するための杯細胞および腸内分泌細胞ならびに抗菌ペプチドを分泌するためのパネート細胞にも分化しうる。幹細胞領域にとどまるパネート細胞の他に、分化した細胞は絨毛の先端へ移動する (J. Clin. Invest. 105:1493-1499, 2000; Science 294:2115-2116, 2001)。
【0008】
腸管上皮細胞の特異的配置により、この上皮細胞が、腸管腔内の様々な抗原に対する免疫応答に影響を及ぼす重要な役割を果たすということは適切な意見である。腸管上皮細胞は、腸管粘膜内でのT細胞応答を調節するよう、抗原提示細胞(APC)として抗原提示分子を発現しうる (Immunol. Today 21:123-128, 2000)。腸管上皮細胞により発現された抗原提示分子は、MHCクラスI、MHCクラスII、およびCDldを含むことが調べられている (Gastroenterology 124:1420-1431, 2003)。MHCクラスIおよびMHCクラスIIは上皮細胞の側底膜上に発現し、MHCクラスIはCD8+細胞と結合する役割があり、MHCクラスIIはCD4+細胞と結合する役割がある。CDldは糖脂質を介してNK T細胞を活性化する。これらの証拠は、腸管上皮細胞が粘膜の免疫系で重要な役割を果たすことを示す。
【0009】
物理的バリアであることの他に、腸管上皮細胞は、胃腸管内の微生物に対する抗菌ペプチドを分泌し、他の細胞にシグナルを与える (Immunol. Today 21:123-128, 2000; J. Clin. Invest. 95:55-65, 1995)。特異的な抗菌ペプチドであるディフェンシンが存在することがわかっている。ディフェンシンは、3〜5kDのタンパク質であり、8つのペプチドを有するα-ディフェンシンファミリーおよびβ-ディフェンシンファミリーを含む。これらのディフェンシンは、口腔粘膜、肺、および胃腸管の上皮細胞中で器官特異的な発現パターンを有する (Eur. J. Gastroenterol. Hepatol. 13:771-776, 2001; Nat. Rev. Immunol. 3:710-720, 2003)。胃腸管が感染すると、腸管上皮細胞はディフェンシンを発現し、ディフェンシンはケモカインのように作用してNK細胞および樹状細胞を感染部位に誘導し、いわゆる先天性免疫を実行させる。先天性免疫の誘導の他に、T細胞の増殖を誘導するために、ディフェンシンはToll様受容体4を介して樹状細胞を誘導して共刺激因子(B7.2)を発現させる。したがって、ディフェンシンは、先天性免疫および適応免疫の間の関連に関与する。
【0010】
他の抗菌タンパク質、アンギオゲニンは癌細胞の血管形成に関与すると考えられていた。しかしながら、2003年、Hooperにより、アンギオゲニンは、正常な生理学的条件下で胃腸管におけるパネート細胞により産生され、細胞顆粒中に貯蔵されることが開示された。アンギオゲニンはリポ多糖刺激に反応して胃腸管内に分泌される。無菌マウスおよび腸内細菌を送り込まれたマウスを比較すると、腸内細菌が存在することによりアンギオゲニンが著しく増加することがわかる。ディフェンシンと同様に、アンギオゲニンは殺菌性で、感染部で遭遇した局所的な環境条件により調節される (Nat. Immunol. 4:269-273, 2003)。
【0011】
病原体の感染に対して抗菌ペプチドを分泌する他に、腸管上皮細胞は、胃腸管における免疫応答の調節のための多くのシグナルを産生する。侵入する微生物を破壊することにより宿主を保護するために、腸管上皮細胞は、直ちに、防衛機構を介して侵入する微生物に応答するであろう。この防衛機構は先天性免疫系として知られ、先天性免疫系は、グラム陰性菌の細胞壁中に存在するリポ多糖等の微生物病原体の巨大分子を認識するために生殖細胞系にコードされたパターン認識受容体を使用する。近年、パターン認識受容体がどのように細胞を活性化するかについて開示された。成体ショウジョウバエで、Tollが、感染の際に、抗真菌性ペプチドおよび抗菌性ペプチドを誘導することが示されている。MedzhitovおよびJanewaysらは、哺乳動物において、Toll様受容体(TLR)の活性化により、適応免疫応答の活性化に必要とされるサイトカインおよび共刺激分子の誘導がもたらされることを開示する (Cell 91:295-298, 1997)。TLRにより活性化された細胞内シグナル伝達経路は、保存されたTIR(Toll/IL-1R相同性)ドメインにより、IL-1Rシグナル伝達と共通する多くのものを共有する。エンドトキシンがTLRにより認識されると、NF-kBの移動による核移行が引き起こされ、免疫応答に関与する他の遺伝子の転写が調節される。続いて、炎症誘発性サイトカイン遺伝子が活性化され、抗菌ペプチドが分泌され、IL-8、MIP、およびMCP-1等のケモカインが分泌される。感染部に移動して感染細胞を破壊するために、マクロファージおよびNK細胞がこれらのケモカインにより誘導される(Cell 91:295-298,1997; Curr. Opin. Immunol 14:103-110, 2002)。よって、NF-kBの活性化により引き起こされた炎症誘発性サイトカインの分泌により、Toll様受容体は、先天性免疫系を誘導するだけでなく、間接的に適応免疫系も誘導する。
【0012】
ヒト細胞には、少なくとも10個の異なるTLRが存在し、TLR-2、TLR-4、およびTLR-9がTLRファミリーのうちでより特徴づけられたメンバーである。TLRは、細菌およびウイルスを含む多くのタイプの病原体の認識を媒介することが示された。TLR-4は、グラム陰性菌LPSの受容体であり、TLR-2は、グラム陽性菌ペプチドグリカンとリポペプチドの受容体であり、TLR-9は、非メチル化およびリン酸化されたシトシン-グアニンオリゴヌクレオチド、CpGの受容体である(Curr. Opin. Immunol. 14:103-110, 2002)。TLR3は、二本鎖RNAウイルスを認識することができ(Nature 433:887-892, 2005)、TLR7は、一本鎖RNAウイルスを認識しうる(Proc Natl Acad Sci USA. 101(15):5598-5603, 2004)。TLRシグナル経路が活性化されると、先天性免疫または抗ウイルス応答を宿主保護のために誘導させることができる。
【0013】
正常な腸管上皮細胞において、TLR-3およびTLR-5が恒常的に発現する一方、TLR2およびTLR4はほとんど検出することができない(Infection and Immunity 68:7010-7017, 2000)。IL-8は、細菌DNAの刺激に反応して上皮細胞により産生される(FASEB J 17:1319-1321, 2003)。腸管上皮細胞において、細菌のCpGオリゴヌクレオチドはTLR-9のリガンドである。さらに、腸管上皮細胞はすべての異物に反応して炎症を誘導するわけではなく、すなわち、腸管上皮細胞は共生する微生物相に耐性があり、一方、腸管上皮細胞は、APCにより媒介され、T細胞により形成された炎症を誘導するために、起こりうる病原性条件下または自己免疫疾患下で、APCに危険シグナルを与える(Cell 118:229-241, 2004)。
【0014】
通常の定常状態条件下でTLRにより認識された共生細菌は腸管の傷を予防するのに重大な役割を果たす(Cell 118:229-241, 2004)。しかし、ヒトの炎症性腸疾患(IBD)において、Th1媒介性の病的作用は微生物相に対する異常な粘膜免疫応答によるものと思われる(Gastroenterol. Clin. North Am. 31:41-62, 2002)。これらの発見は、TLRが、宿主-共生動物間の粘膜のホメオスタシスを制御することを明らかにする。
【0015】
中国の薬草抽出物の経口投与により免疫の発生を調節することが良く知られている。中国の薬草抽出物により腸管粘膜の免疫を介して免疫の発生を調節することは非常に可能性がある。セッコク属種は最も貴重な中国の薬草であると考えられる。セッコク属種はラン科に属し、茎が主な薬用部分である。茎はわずかに甘く、やや塩辛い味がする。いくつかの中国の医療用の薬局方には、セッコク属種は、粘膜障害、胃障害、および眼障害等のいくつかの疾病のための治療薬であることが開示される。我々の先の研究経験によれば、デンドロビーハーバ(Dendrobii Herba)が最も治癒力のある薬用種であると思われる。
【0016】
本発明は、セッコク属に由来する多糖を調製するための方法を提供し、さらに、セッコク属に由来する多糖の医薬としての使用を提供する。
【発明の開示】
【0017】
発明の概要
本発明の一態様として、植物からの多糖を調製するための方法を提供する。方法は、a)第1のアルコールで植物を抽出して第1の抽出物を得るステップ、b)溶媒で第1の抽出物を抽出して第2の抽出物を得るステップ、およびc) 多糖を得るために第2のアルコールで第2の抽出物を沈殿させるステップを含む。
【0018】
好ましくは、植物はセッコク属に属する。
【0019】
好ましくは、第1のアルコールはメタノールである。
【0020】
好ましくは、溶媒は水である。
【0021】
好ましくは、第2のアルコールはエタノールである。
【0022】
本発明において、前記方法は、ステップb)において溶媒で第1の抽出物を抽出した後、遠心分離し、濾過して第2の抽出物を得るステップをさらに含む。
【0023】
本発明の他の態様として、自己免疫疾患および粘膜障害を治療するための組成物を提供する。前記組成物は植物から調製した多糖を含み、該植物はセッコク属に属し、抗原は自己免疫疾患の誘導に関連する。
【0024】
好ましくは、多糖は本発明において提供される方法により調製される。
【0025】
本発明において、自己免疫疾患はブドウ膜炎であってもよい。
【0026】
本発明の他の態様として、哺乳動物の自己免疫疾患および粘膜障害を治療するための方法を提供する。前記方法は、哺乳動物に抗原および多糖を投与するステップを含み、該抗原は自己免疫疾患の誘導に関連し、該多糖はセッコク属に属する植物から調製される。
【0027】
本発明において、多糖の投与は経口投与の方法で実行されてもよい。
【0028】
本発明において、自己免疫疾患はブドウ膜炎であってもよい。
【0029】
本発明の他の態様として、哺乳動物の自己免疫疾患および粘膜障害を治療するための方法を提供する。前記方法は、哺乳動物に多糖を投与するステップを含み、該多糖がセッコク属に属する植物から調製される。
【0030】
本発明において、多糖の投与は経口投与の方法で実行される。
【0031】
本発明において、自己免疫疾患はブドウ膜炎であってもよい。
【0032】
以下の詳細な説明および添付の図面を検討すれば、上記態様および本発明の利点が当業者にとってより容易に明らかになろう。
【発明を実施するための最良の形態】
【0033】
本発明を、以下の実施形態を参照してより具体的に説明する。以下の本発明の最良の形態は、例証および説明のみを目的として本明細書に提示されるものであって、網羅的なものとすることを意図するものではない、または、開示した詳細な形態に限定されることを意図するものではないことに留意されたい。
【0034】
本発明は、植物からの多糖を調製するための方法であって、a)第1のアルコールで植物を抽出して第1の抽出物を得るステップ、b)溶媒で第1の抽出物を抽出して第2の抽出物を得るステップ、およびc) 多糖を得るために第2のアルコールで第2の抽出物を沈殿させるステップを含む方法を提供する。
【実施例1】
【0035】
セッコク属からの多糖の調製
10kgの新鮮なデンドロビーハーバを乾燥させて2.92kgの乾燥材料にし、次いで、乾燥材料を73.3Lメタノールに浸して抽出し、136.79グラムの粗抽出物と残渣を得た。粗抽出物を脱メタノール工程で処理し、次いで一晩水に浸した。反応溶液を、55℃〜60℃の範囲の温度の温水で30分間抽出し、5,000rpm、10℃で遠心分離機(ER-RC13 C-124、HITACH社)により遠心分離した。遠心分離後、上清を6mu m濾紙で濾過した。濾液をエタノールで沈殿させ、20.684gの多糖(以下DeCaPSと称する)を得た。
【実施例2】
【0036】
DeCaPSで処理したIEC-6細胞の免疫応答
a. IEC-6細胞の培養
正常ラット小腸から生じたIEC細胞を、5%CO2、37℃のインキュベータ中で、5%ウシ胎児血清、4.5g/Lグルコース、5mu g/mlウシインスリン、および2mM L-グルタミンを含むDMEM培地で培養した。
【0037】
b. DeCaPSによるIEC-6細胞の処理
IEC-6細胞を6ウェルプレートに流し、IEC細胞を、それぞれ、1mu g/ml、10mu g/ml、および100mu g/mlの濃度でDeCaPSにより6時間処理した。
【0038】
c. DeCaPSで処理したIEC細胞からの全RNAの単離
DeCaPSでの処理の後、IEC細胞を採取し、1ml Ultraspec(商標)RNA単離キット(Biotex laboratories Inc.(米国))中に懸濁させ、全RNAを、キットの標準プロトコールに従って得た。得られた全RNAを定量した。
【0039】
d. サイトカインおよびToll様受容体の逆転写およびPCR分析
26.5mu lの反応容量を有し、0.1gオリゴ-dT、5mu gの得られた全RNA、およびDEPCで処理した滅菌水を含む反応を70℃で10分間実行した。次いで、4mu lの10mM dNTP、0.5mu l rRNasin、1mu l AMV(鳥骨髄芽球症ウイルス(Avian Myeloblastosis virus))逆転写酵素(10ユニット)、および8mu l 5×RT緩衝液を反応に加え、全容量を40mu lとして42℃で60分間、次いで90℃で5分間インキュベートし、その結果として、cDNA生成物を逆転写反応により得た。その後、0.5muの10mM dNTP、0.5mu l 10×プロザイム緩衝液、プライマー(0.8mu lの5mu MセンスDNAおよび0.8mu lの5mu MアンチセンスDNA)、および滅菌水を2.5mu l cDNA生成物に加え、全容量25mu lとし、DNAサーマルサイクラー(Perkin-Elmer-Cetus社)中でインキュベートし、PCRを35サイクル実行した。標的遺伝子に対するセンスプライマーおよびアンチセンスプライマーを含むプライマー配列[配列番号:1〜32]ならびに対応するアニール温度を以下の表Iに記載する。
【表1】

【0040】
図1を参照されたい。図1は、DeCaPSで処理したIEC-6細胞中のサイトカインおよび抗菌ペプチドのmRNA発現を示す。1mu g/ml、10mu g/ml、および100mu g/mlのDeCaPSで6時間処理した後、IEC-6細胞中のTNF-α[配列番号:33]のmRNA発現は約29%、20%、および30%抑制されたが、IEC細胞中のIL-10[配列番号:34]のmRNA発現は9.89倍、6.55倍、および1.77倍までそれぞれ高められた。TH2/3経路を引き起こすために、免疫系の他の細胞に寛容シグナルを提供するよう、DeCaPSにより腸管上皮細胞を誘導しうることが示されている。さらに、1mu g/ml、10mu g/ml、および100mu g/mlのDeCaPSで6時間処理した後、a-ディフェンシン[配列番号:35]のmRNA発現は約42%、33%、および51%抑制されたが、アンギオゲニン[配列番号:36]のmRNA発現は1.96倍、1.42倍、および1.70倍までそれぞれ高められた。
【0041】
図2を参照されたい。図2は、DeCaPSで処理したIEC-6細胞中のToll様受容体のmRNA発現を示す。IEC-6細胞において、Toll様受容体であるTLR2[配列番号:37]、TNR4[配列番号:38]、TLR7[配列番号:39]、およびTLR9[配列番号:40]はDeCaPS処理なしで発現した。1mu g/ml、10mu g/ml、および100mu g/mlのDeCaPSで6時間処理した後、それぞれ、IEC-6細胞中のTLR4[配列番号:38]のmRNA発現は約6%、16%、および13%抑制され、IEC-6細胞中のTLR5[配列番号:41]のmRNA発現は約13%、37%、および35%抑制され、IEC-6細胞中のTLR7[配列番号:39]のmRNA発現は約11%、7%、および48%抑制された。しかしながら、TLR9[配列番号:40]のmRNA発現は2.21倍、1.09倍、および3.56倍までそれぞれ高められた。セッコク属に由来する多糖は、TNF-α[配列番号:33]の発現を抑制するだけでなく、TLR4[配列番号:38]、TLR5[配列番号:41]、TLR7[配列番号:39]、およびTLR9[配列番号:40]の発現もまた抑制するということが示されている。
【実施例3】
【0042】
DeCaPSの経口投与による動物モデルの免疫系に対する効果
C3Hマウス(13週令)に、実施例Iで調製したDeCaPSを、10mg/kg/日、50mg/kg/日、および250mg/kg/日の投与量で飲料水を介して5日間それぞれに与え、次いで、脾臓および骨髄を得るために屠殺した。
【0043】
a. C3HマウスからのコンカナバリンA刺激脾細胞に対するMTTアッセイ
DeCaPSで処理したC3Hマウスから得た脾細胞(4×105細胞/ウェル)を、10%ウシ胎児血清ならびに1mu g/mlおよび5mu g/mlのコンカナバリンA(ConA)を含むRPMI-1640培地で72時間培養し、次いで、1mg/ml MTTで3時間処理した。次いで、細胞培養物に、50%DMFおよび20%SDSを含む溶解緩衝液をそれぞれに加え、反応を16時間実行させた。反応における570nmでの吸収を分析すると、図3に示すような成長指数を示した。
【0044】
図3を参照されたい。図3は、DeCaPSで経口的に処理したC3HマウスからのConA刺激脾細胞の分裂応答に対する効果を示す。C3Hマウスを、DeCaPSにより、10mg/kg/日、50mg/kg/日、および250mg/kg/日の投与量で5日間経口的に処理した後、脾細胞の1mu g/ml ConA刺激に対する応答は、約28%、49%、および30%抑制された。
【0045】
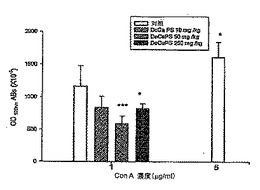
b. C3Hマウスから得たGM-CSF-刺激骨髄細胞に対するMTTアッセイ
DeCaPSで処理したC3Hマウスの脚から骨髄細胞を得た。2%FCSおよび4ng/ml GM-CSFを含むA-MEM培地で骨髄細胞を72時間培養し、骨髄細胞の陽性対照群を、2%FCSおよび20ng/ml GM-CSFを含むα-MEM培地で72時間培養した。次いで、骨髄細胞を3時間処理するために、培地を、1mg/ml MTTおよび2%FCSを含むRPMI-1640と置換し、細胞を、MTT溶解緩衝液(20%SDSおよび50%DMF、pH4.5)でさらに処理し、一晩インキュベートした。反応における570nmでの吸収を分析すると、図4に示すような成長指数を示した。
【0046】
図4を参照されたい。図4は、DeCaPSで経口的に処理したC3Hマウスから得たGM-CSF-刺激骨髄細胞の分裂応答に対する効果を示す。C3Hマウスを、DeCaPSにより、10mg/kg/日、50mg/kg/日、および250mg/kg/日の投与量で5日間それぞれ経口的に処理した後、骨髄細胞のGM-CSF刺激に対する分裂応答は、1.71倍、1.62倍、および1.76倍まで高められた。
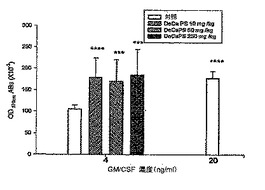
【0047】
c. C3Hマウスから得たパイエル板および脾細胞におけるサイトカインのmRNA発現
DeCaPSで処理したC3Hマウスからパイエル板および脾細胞を得た。パイエル板および脾細胞におけるサイトカイン特異的mRNAのPCR分析を実施例IIに記載する方法で実行し、PCR結果を図5に示した。
【0048】
図5を参照されたい。図5は、DeCaPSで経口的に処理したC3Hマウスから得たパイエル板および脾細胞におけるサイトカイン特異的mRNAのPCR結果を示すが、レーン1〜4は、DeCaPSにより、0mg/kg/日、10mg/kg/日、50mg/kg/日、および250mg/kg/日の投与量で経口的に処理したC3Hマウスをそれぞれ示す。
【0049】
C3HマウスをDeCaPSで経口的に5日間処理した後、パイエル板におけるIL-4[配列番号:42]、IL-6[配列番号:43]、IL-1β[配列番号:44]、IFN-γ[配列番号:45]、およびTGF-β [配列番号:46]のmRNA発現は図5に示すように増加した。C3Hマウスを、DeCaPSにより、10mg/kg/日および50mg/kg/日の投与量で5日間経口的に処理した後、IL-4[配列番号:42]のmRNA発現は2.64倍および2.46倍までそれぞれ高められた。C3Hマウスを、DeCaPSにより、10mg/kg/日、50mg/kg/日、および250mg/kg/日の投与量で5日間経口的に処理した後、IL-6[配列番号:43]のmRNA発現はそれぞれ2.61倍、3.99倍、および5.35倍まで高められ、IL-β[配列番号:44]のmRNA発現はそれぞれ1.83倍、2.00倍、および1.11倍まで高められ、IFN-γ[配列番号:45]のmRNA発現はそれぞれ3.47倍、5.47倍、および5.57倍まで高められ、TGF-β[配列番号:46]のmRNA発現はそれぞれ2.38倍、2.54倍、および1.89倍まで高められた。パイエル板は、抗原に対する免疫応答を測定するための主な組織であるので、Th2経路がIL-4[配列番号:42]およびIL-6[配列番号:43]の増加に反応して引き起こされ、Tヘルパー細胞がIL-1β[配列番号:44]の増加によりTGF-β[配列番号:46]等のサイトカインを分泌するよう活性化され、IgAクラススイッチB細胞は、ある種の免疫応答を抑制するために、TGF-β[配列番号:46]によりIgAを分泌するよう誘導される。
【0050】
C3HマウスをDeCaPSで経口的に5日間処理した後、脾細胞中のIL-1β[配列番号:44]、IL-4[配列番号:42]、およびTGF-β[配列番号:46]のmRNA発現は図5に示すように抑制された。C3Hマウスを、DeCaPSにより、10mg/kg/日、50mg/kg/日、および250mg/kg/日の投与量で5日間経口的に処理した後、IL-1β[配列番号:44]のmRNA発現は約32%、43%、および79%抑制され、IL-4[配列番号:42]のmRNA発現は約7%、67%、および14%抑制された。C3Hマウスを、DeCaPSにより、10mg/kg/日、50mg/kg/日、および250mg/kg/日の投与量で5日間経口的に処理した後、それぞれ、TGF-β[配列番号:46]のmRNA発現は約27%および44%抑制され、IFN-γ[配列番号:45]のmRNA発現は約33%および97%抑制された。しかしながら、C3Hマウスを、DeCaPSにより、10mg/kg/日、50mg/kg/日、および250mg/kg/日の投与量で5日間経口的に処理した後、IL-6[配列番号:43]のmRNA発現は1.84倍、1.4倍、および1.23倍まで高められた。DeCaPS処理に対する著しい細胞応答はなく、この現象はConA刺激に対する応答と一致することに留意されたい。
【実施例4】
【0051】
DeCaPSによる動物モデルの経口寛容の増強
自己免疫疾患用の動物モデルを確立するためにオボアルブミン(OVA)により動物を免疫できることは周知なので(The Journal of Pharmacology and Experimental Therapeutics 288:849-857, 1999)、本発明では、OVA誘導自己免疫疾患を有するマウスを確立し、OVA誘導自己免疫疾患を有するマウスにおけるDeCaPSによる経口寛容の増強を以下のとおり研究する。C57/BL6jマウス(65週令)を、4日目および5日目に、飲料水中0.5mg/mlオボアルブミン(OVA)で経口的に処理した。3〜7日目に、10mg/kg/日、40mg/kg/日、および160mg/kg/日の投与量でアジュバントとしてDeCaPSをC57/BL6jマウスに投与した。CFAをアジュバントとした50mu g OVAを抗原として8日目にi.p.注射によりマウスに投与した。血液採集物中でOVAに反応したIgGおよびIgMが22日目に検出され、50mu g OVAをi.p注射により24日目に抗原としてマウスに投与し、OVAに反応した抗体が30日目に検出され、腸管粘膜の腸管洗浄溶液および肺粘膜を得るために32日目にマウスを屠殺し、その後、OVAに反応したIgAが検出された。詳細な実験手順を以下の表IIに示す。
【表2】
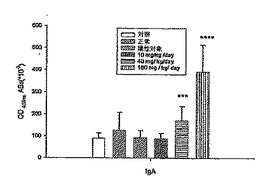
【0052】
腸管粘膜の腸管洗浄溶液を32日目にマウスから得て、図6に示すように、腸管洗浄溶液中のオボアルブミン特異的IgAの力価を測定した。図6に示すように、腸管粘膜から分泌されたIgAは、DeCaPS(40mg/kg/日)と共にOVAで処理したC57/BL6jマウスで1.9倍(p<0.01)まで高められ、腸管粘膜から分泌されたIgAは、DeCaPS(160mg/kg/日)と共にOVAで処理したC57/BL6jマウスで4.35倍(p<0.001)まで高められた。
【0053】
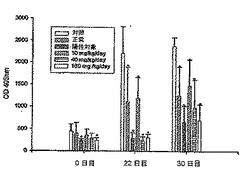
図7に示すように、22日目および30日目の血液採集物中のIgM抗体を測定した。22日目の対照群で発現したIgM抗体レベルと比較すると、正常群で発現したIgM抗体レベルは、約49%(p<0.05)抑制され、陽性対照群で発現したIgM抗体レベルは、約87%(p<0.05)抑制され、DC-1群で発現したIgM抗体レベルは、約46%(p<0.02)抑制され、DC-2群で発現したIgM抗体レベルは、約88%(p<0.001)抑制され、DC-3群で発現したIgM抗体レベルは、約86%(p<0.001)抑制された。さらに30日目の対照群で発現したIgM抗体レベルと比較すると、正常群で発現したIgM抗体レベルは、約47%(p<0.01)抑制され、陽性対照群で発現したIgM抗体レベルは、約72%(p<0.001)抑制され、DC-1群で発現したIgM抗体レベルは、約38%(p<0.01)抑制され、DC-2群で発現したIgM抗体レベルは、約58%(p<0.01)抑制され、DC-3群で発現したIgM抗体レベルは、約71%(p<0.001)抑制された。
【0054】
図8に示すように、22日目および30日目の血液採集物中のIgG抗体を測定した。22日目の対照群で発現したIgG抗体レベルと比較すると、陽性対照群で発現したIgG抗体レベルは、約94%(p<0.01)抑制され、DC-1群で発現したIgG抗体レベルは、約67%(p<0.01)抑制され、DC-2群で発現したIgG抗体レベルは、約76%(p<0.01)抑制され、DC-3群で発現したIgG抗体レベルは、約88%(p<0.001)抑制された。さらに30日目の対照群で発現したIgG抗体レベルと比較すると、陽性対照群で発現したIgG抗体レベルは、約75%(p<0.001)抑制され、DC-2群で発現したIgG抗体レベルは、約62%(p<0.001)抑制され、DC-3群で発現したIgG抗体レベルは、約63%(p<0.001)抑制された。
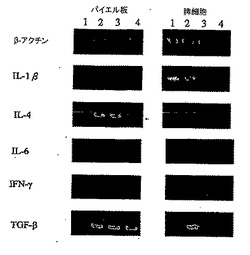
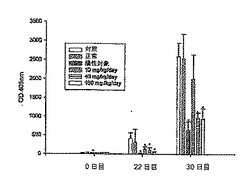
【0055】
さらに、パイエル板を32日目のマウスから得て、パイエル板におけるリンパ球のRNAを抽出し、サイトカインを分析した。図9に示すように、パイエル板におけるリンパ球のサイトカイン発現を測定したが、レーンAは正常群を示し、レーンBは対照群を示し、レーンCは陽性対照群を示し、レーンDはDC-1群を示し、レーンEはDC-2群を示し、レーンFはDC-3群を示す。PCR増幅を変性のために94℃で45秒間に設定し、アニーリングを61℃で45秒間実行し、伸長を72℃で1分間実行した。PCRプログラムは37サイクルとした。2%アガロースゲルの電気泳動によりPCR生成物を分離し、ゲルを臭化エチジウムで染色し、UV光下で可視化した。
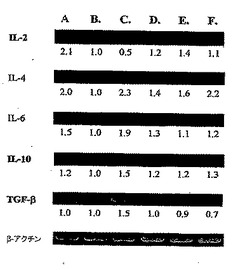
【0056】
対照群と比較すると、DC-3群で発現したTGF-β[配列番号:46]レベルは約30%抑制され、DC-3群で発現したIL-4[配列番号:42]レベルは2.2倍まで高められ、DC-3群で発現したIL-10[配列番号:34]レベルは1.3倍まで高められたということに留意されたい。対照群と比較すると、DC-2群で発現したIL-2[配列番号:47]レベルは1.4倍まで高められ、DC-2群で発現したIL-4[配列番号:42]レベルは1.6倍まで高められたことに留意されたい。さらに対照群と比較すると、DC-1群で発現したIL-4[配列番号:42]レベルは1.4倍まで高められ、DC-1群で発現したIL-6[配列番号:43]レベルは1.3倍まで高められた。
【0057】
IL-4[配列番号:42]およびIL-6[配列番号:43]のmRNA発現はパイエル板におけるリンパ球で高められた。パイエル板は、抗原に対する免疫応答を決定するための組織であるので、抗原に感作されたT細胞またはB細胞がホーミング機構により腸に戻ると、Th2経路が、IL-4[配列番号:42]およびIL-6[配列番号:43]の増加に反応して抗体を産生するよう活性化され、血清中のIgGおよびIgMの発現が、アレルゲン応答を抑制するように抑制され、経口寛容が達成されることに注目されたい。
【0058】
前述の説明を考慮して、脾臓中のT細胞の分裂応答はセッコク属の多糖により抑制されるが、骨髄中の顆粒球およびマクロファージはセッコク属の多糖により活性化される。言い換えれば、自己免疫疾患において活性化されたT細胞は、セッコク属の多糖の経口投与により抑制しうるが、顆粒球系細胞は、先天性免疫の促進により病原体の侵入を予防するためのセッコク属の多糖の経口投与により活性化される。
【0059】
腸管の免疫に関し、経口寛容を生じさせるために、IL-4[配列番号:42]、IL-6[配列番号:43]、IL-1β[配列番号:44]、IFN-γ [配列番号:45]、およびTGF-β[配列番号:46]のmRNA発現が高められて腸内のTh2/Th3経路が引き起こされる。セッコク属の多糖で処理したIEC-6細胞中のTNF-α[配列番号:33]、TLR2[配列番号:37]、TLR4[配列番号:38]、TLR5[配列番号:41]、およびTLR7[配列番号:39]の発現によれば、経口寛容関連シグナル、すなわちTh2/3経路を高め、自己免疫疾患および粘膜障害等の病原性状態における粘膜のホメオスタシスを維持するために、Toll様受容体により媒介されたシグナルを免疫抑制性のまたは抗炎症性の応答とするようセッコク属の多糖により調節しうることが示されている。興味深いことには、抗菌活性を有するアンギオゲニンの発現は、実施例2に記載されるように、セッコク属の多糖の投与により誘導しうる。したがって、セッコク属の多糖の経口投与は経口寛容を高めるだけでなく、病原体侵入を予防するための先天性免疫機構を引き起こす。これらの発見によれば、長引く炎症およびさらなる傷を免れるために、炎症性腸疾患(IBD)等の自己免疫疾患および粘膜障害にセッコク属の多糖を有利に使用してもよい。
【0060】
アンギオゲニンは、エンテロコッカスフェカリス(Enterococcus faecalis)等の腸内微生物だけでなくヒトにおいて組織的感染を引き起こすカンジダアルビカンス(Candida albicans)および肺炎連鎖球菌(Streptococcus pneumoniae)等の微生物に対しても活性を有することが開示されている (Nat. Immunol. 4:269-273, 2003) 。臨床研究では、女性の帯下は、多微生物性の表在性膣感染症とされる。この感染症は、G.バジナリス(G. vaginalis)、カンジダ種(Candida spp)、C.アルビカンス、T.バジナリス(T. vaginalis)、D群連鎖球菌(Streptococcus group D)、溶血性連鎖球菌b(Streptococcus b hemolytic)、大腸菌(E. coli)、およびクレブシエラ種(Klebsiella spp)等のような細菌により頻繁に引き起こされる (Salud publica Mex vol.45 suppl. 5, pS694-S697, 2003)。カンジダアルビカンス等の微生物により引き起こされた女性の帯下は、セッコク属の多糖の経口投与により誘導されたアンギオゲニンの活性により軽減することができたことは明らかである。
【実施例5】
【0061】
DeCaPSによる自己免疫疾患の治療
実験的自己免疫ブドウ膜炎は、いくつかの眼の自己免疫疾患用モデルとして役立つT細胞媒介自己免疫疾患である。実験的自己免疫ブドウ膜炎は、140kDのグリコリポプロテインである光受容体間レチノイド結合タンパク質(IRBP)での免疫化により誘導されることが確認されている。さらに、自己免疫ブドウ膜炎はIRBPペプチド1〜20(配列番号48:GPTHLFQPSLVLDMAKVLLD)、IRBPのアミノ酸1〜20により誘導しうることが確認されている。さらに、IRBPペプチド1〜20はマウスおよびヒトで保存されている。(Investigative Ophthalmology & Visual Science 41(1):127-131, 2000)よって、本発明において自己免疫疾患モデルを確立するためにIRBPペプチドをC57BL/6jマウスに投与した。
【0062】
a. IRBPペプチドの合成
IRBPペプチド(配列番号48:GPTHLFQPSLVLDMAKVLLD)をFmoc化学を使用してペプチド合成機(PS3)で合成し、Agilent社製HPLCで精製し、Brukeer社製esquire 2000 MSで特定した。
【0063】
b. IRBPおよびDeCaPSの投与
特定病原体フリー(SPF)マウスであるC57BL/6jマウスを、正常群、対照群、DC-10群、DC-40群、およびDC-160群に無作為に分類し、各群のマウスを6匹とした。0.2ml乳剤中のヒトIRBPペプチド[配列番号:48](150mu g/マウス)およびCFA(1:1、体積/体積)を群の各SPFマウスにi.p.注射で正常群以外に投与した。DC-10群のマウスに、DeCaPSを、10mg/kg/日の投与量で28日間与え、DC-40群のマウスに、DeCaPSを、40mg/kg/日の投与量で28日間与え、DC-160群のマウスに、DeCaPSを、飲料水を介して160mg/kg/日の投与量で28日間与えた。マウスの網膜の作用を網膜電図(ERG)分析により測定し、バイオアッセイおよび組織変化の測定用の眼を得るためにマウスを28日目に屠殺した。実験手順を以下の表IIIに示す。
【表3】
【0064】
c. 遅延型過敏症(DTH)試験
IRBPペプチドによる免疫化後21日目に、IFA(20mu l)中20mu g IRBPペプチド乳剤をマウスの左足蹠に皮下注射した。右足蹠にはIFAを注射した。48時間後、足蹠の厚さをキャリパで測定した。
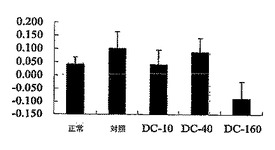
【0065】
図10を参照されたい。図10は、マウスのDTH応答を示す。DTH応答は、リン酸緩衝液生理食塩水(PBS)を注射したマウスの足蹠増加と水容量で比較した足蹠増加により示した。DeCaPS(160mg/kg/日)で共処理したマウスの足蹠増加が著しく低下したことに留意されたい。
【0066】
d. 網膜電図(ERG)分析
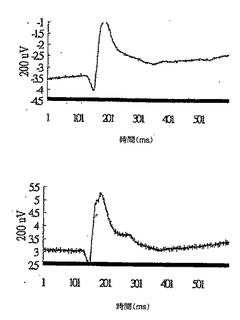
マウスを2時間暗順応させ、次いで、ペントバルビタールナトリウムで麻酔した。次いで、マウスの角膜に麻酔し、瞳孔を拡大させた。網膜を閃光で刺激し、閃光に対する網膜の応答を網膜電図として記録した。網膜電図は、光受容体の働きならびに双極細胞およびミュラー細胞等の網膜基部の作用を示す。全網膜の状態を反映させるために網膜電図を使用する。網膜電図では通常、陰性のa波に陽性のb波が続く。a波の立ち上がりにより、光受容体層内の錐体細胞および桿細胞の活性が直接測定され、b波は、内顆粒層内の双極細胞の働きを反映する。
【0067】
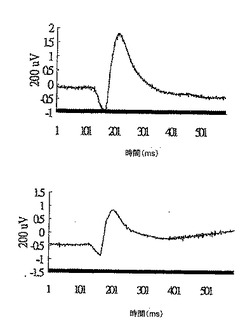
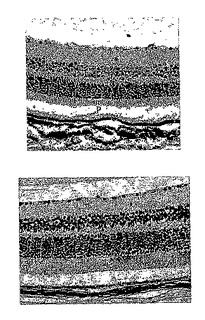
28日目に正常群から無作為に選択した2匹のマウスについて、網膜電図分析の結果を、図11Aの上方部および下方部にそれぞれ示したが、閃光開始は、100ミリ秒の時間のところであり、閃光持続期間は約600ミリ秒であった。28日目に正常群から無作為に選択した2匹のマウスについて、眼の組織変化を図11Bの上方部および下方部(倍率400倍)にそれぞれ示したが、光受容体層を「P」と示し、外顆粒層を「ONL」と示し、内顆粒層を「INL」と示した。
【0068】
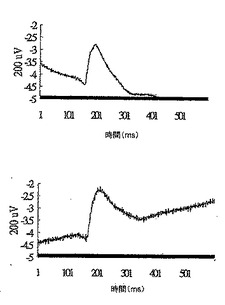
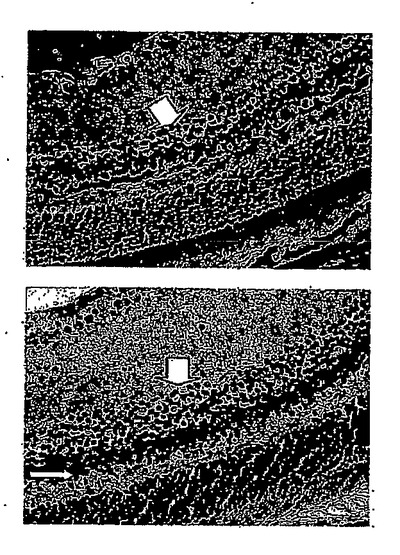
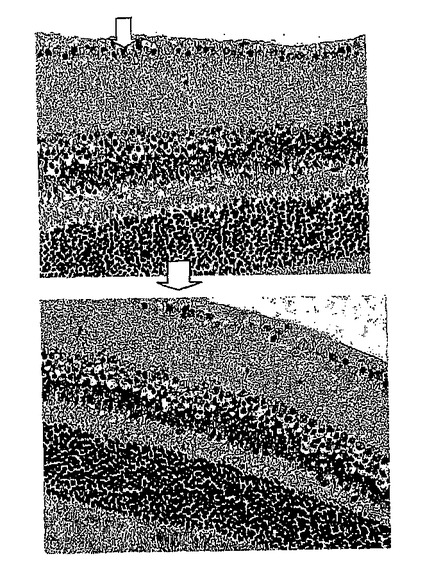
28日目に対照群から無作為に選択した2匹のマウスについて、網膜電図分析の結果を、図12Aの上方部および下方部にそれぞれ示したが、閃光開始は、100ミリ秒の時間のところであり、閃光持続期間は約600ミリ秒であった。28日目に対照群から無作為に選択した2匹のマウスについて、眼の組織変化を図12Bの上方部および下方部(倍率400倍)にそれぞれ示した。IRBPペプチドによる免疫化により、対照群のマウスの光受容体層(P)、外顆粒層(ONL)、および内顆粒層(INL)は激しく損傷を受けたことが示された。
【0069】
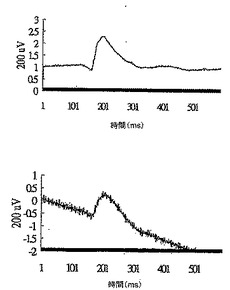
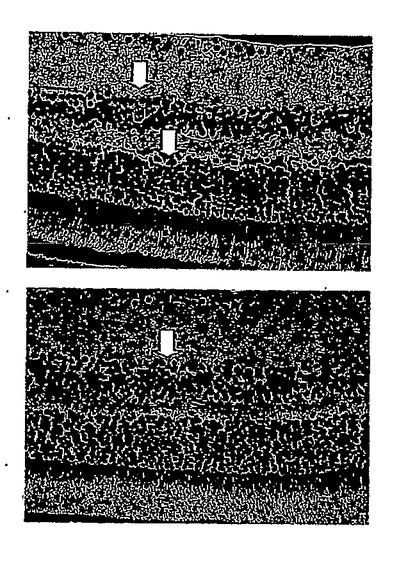
28日目にDC-10 群から無作為に選択した2匹のマウスについて、網膜電図分析の結果を、図13Aの上方部および下方部にそれぞれ示したが、閃光開始は、100ミリ秒の時間のところであり、閃光持続期間は約600ミリ秒であった。28日目にDC-10群から無作為に選択した2匹のマウスについて、眼の組織変化を図13Bの上方部および下方部(倍率400倍)にそれぞれ示した。IRBPペプチドによる免疫化およびDeCaPS(10mg/kg/日)の28日間の経口投与の後、DC-10群のマウスの光受容体層(P)、外顆粒層(ONL)、および内顆粒層(INL)は激しく損傷を受けたことが示された。
【0070】
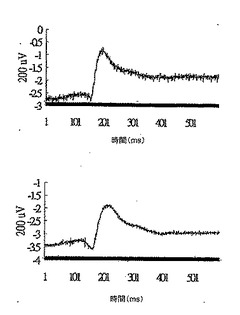
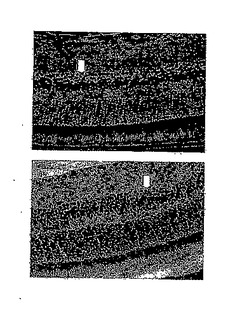
28日目にDC-40群から無作為に選択した2匹のマウスについて、網膜電図分析の結果を、図14Aの上方部および下方部にそれぞれ示したが、閃光開始は、100ミリ秒の時間のところであり、閃光持続期間は約600ミリ秒であった。28日目にDC-40群から無作為に選択した2匹のマウスについて、眼の組織変化を図14Bの上方部および下方部(倍率400倍)にそれぞれ示した。IRBPペプチドによる免疫化およびDeCaPS(40mg/kg/日)の28日間の経口投与の後、DC-40群のマウスの光受容体層(P)、外顆粒層(ONL)、および内顆粒層(INL)はわずかに損傷を受けたことが示された。
【0071】
28日目にDC-160群から無作為に選択した2匹のマウスについて、網膜電図分析の結果を、図15Aの上方部および下方部にそれぞれ示したが、閃光開始は、100ミリ秒の時間のところであり、閃光持続期間は約600ミリ秒であった。28日目にDC-160 群から無作為に選択した2匹のマウスについて、眼の組織変化を図15Bの上方部および下方部(倍率400倍)にそれぞれ示した。IRBPペプチドによる免疫化およびDeCaPS(160mg/kg/日)の28日間の経口投与の後、DC-160群のマウスの光受容体層(P)、外顆粒層(ONL)、および内顆粒層(INL)は損傷を受けなかったことが示された。
【0072】
その結果、IRBPペプチドでの免疫化の後、より多くの細胞死が内顆粒層内に観察されたが、死細胞は外顆粒層上に沈まなかった。内顆粒層内の細胞がIRBPペプチド処理により損傷を受けたが、細胞が内顆粒層から剥離することが160mg/kg/日の投与量でのDeCaPSによる共処理により著しく防止されたことがさらに観察された。
【0073】
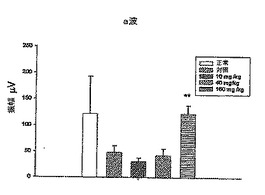
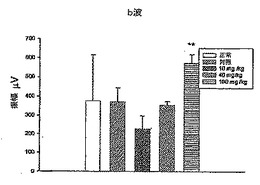
さらに、図11A、12A、13A、14A、および15Aにおいてそれぞれ示されるa波を図16Aと比較し(統計でp<0.02)、図11A、12A、13A、14A、および15Aにおいてそれぞれ示されるb波を図16Bと比較した(統計でp<0.02)。IRBPペプチドで免疫された対照群のマウスのa波は正常群のマウスのa波よりも非常に低いことに注目されたい。しかしながら、対照群と比較すると、DeCaPS(40mg/kg/日)で共処理したマウスの内顆粒層からの細胞の剥離が著しく予防される。さらに、DeCaPS(160mg/kg/日)で共処理したマウスのa波およびb波の振幅は正常群のマウスのものとほぼ等しい。セッコク属から調製した多糖(160/mg/kg/日)での共処理によりIRBPペプチドでの免疫化による損傷から救出しうることにさらに注目されたい。言い換えれば、セッコク属から調製した多糖の経口投与により、IRBPにより誘発された炎症を網膜において防止しうる。
【0074】
前述の実験によれば、本発明は、自己免疫疾患および粘膜障害を治療するための組成物であって、組成物が、セッコク属から調製した多糖および自己免疫疾患の誘導に関連した抗原を含んでおり、セッコク属から調製した多糖が経口寛容誘導および先天性免疫促進を高めることができる組成物を提供する。さらに、本発明は、セッコク属から調製した多糖が単独で経口寛容を誘導し、先天性免疫を促進しうるので、自己免疫疾患および粘膜障害を治療するためにセッコク属から調製した多糖のみを含む組成物を提供する。
【0075】
本発明は、現在最も実用的で好ましい実施形態であると考えられるものに関して記載するが、本発明は、開示された実施形態に限定されないことを理解されたい。それどころか、本発明は、変更および類似する構造すべてを包含するよう、最も広い解釈と一致する添付の請求項の精神および範囲に含まれる上記の様々な変更および類似する構成を包含することが意図される。
【図面の簡単な説明】
【0076】
【図1】DeCaPSで処理したIEC-6細胞中のサイトカインおよび抗菌ペプチドのmRNA発現を示すPCR分析である。
【図2】DeCaPSで処理したIEC-6細胞中のToll様受容体のmRNA発現を示すPCR分析である。
【図3】DeCaPSで経口的に処理したC3HマウスからのConA刺激脾細胞の分裂応答に対する効果を示す図である。
【図4】DeCaPSで経口的に処理したC3HマウスからのGM-CSF共刺激骨髄細胞の分裂応答に対する効果を示す図である。
【図5】DeCaPSで経口的に処理したC3Hマウスから得たパイエル板および脾細胞におけるサイトカイン特異的mRNAを示すPCR分析である。
【図6】DeCaPSの投与ありまたはなしでオボアルブミンにより経口的に免疫したC57BL/6jマウスからの腸管洗浄溶液中のオボアルブミン特異的IgAの力価を示す図である。
【図7】DeCaPSの投与ありまたはなしでオボアルブミンにより経口的に免疫したC57BL/6jマウスからの血清中のオボアルブミン特異的IgMの力価を示す図である。
【図8】DeCaPSの投与ありまたはなしでオボアルブミンにより経口的に免疫したC57BL/6jマウスからの血清中のオボアルブミン特異的IgGの力価を示す図である。
【図9】C57BL/6jマウスからのパイエル板におけるリンパ球のサイトカイン発現を示すPCR分析である。
【図10】C57BL/6jマウスのDTH応答を示す図である。
【図11A】図11Aは、正常群のマウスにおける一定の範囲の閃光強度に対する網膜電図を示す図である。
【図11B】図11Bは、正常群におけるマウスの眼の組織変化を示す図である。
【図12A】図12Aは、対照群のマウスにおける一定の範囲の閃光強度に対する網膜電図を示す図である。
【図12B】図12Bは、対照群におけるマウスの眼の組織変化を示す図である。
【図13A】図13Aは、DC-10群のマウスにおける一定の範囲の閃光強度に対する網膜電図を示す図である。
【図13B】図13Bは、DC-10群におけるマウスの眼の組織変化を示す図である。
【図14A】図14Aは、DC-40群のマウスにおける一定の範囲の閃光強度に対する網膜電図を示す図である。
【図14B】図14Bは、DC-40群におけるマウスの眼の組織変化を示す図である。
【図15A】図15Aは、DC-160群のマウスにおける一定の範囲の閃光強度に対する網膜電図を示す図である。
【図15B】図15Bは、DC-160群におけるマウスの眼の組織変化を示す図である。
【図16A】図16Aは、図11A、12A、13A、14A、および15Aにおけるa波の振幅を示す図である。
【図16B】図16Bは、図11A、12A、13A、14A、および15Aにおけるb波の振幅を示す図である。
【技術分野】
【0001】
本発明は、自己免疫疾患および粘膜障害を治療するための組成物および方法、より詳細には、経口寛容の誘導および先天性免疫の促進を介して自己免疫疾患および粘膜障害を治療するための、セッコク属(Dendrobium)に由来する多糖を含む組成物およびセッコク属に由来する多糖を投与することを含む方法に関する。
【背景技術】
【0002】
免疫系が、宿主自身に対して反応することなく外部の病原体から宿主を保護しうるよう、免疫系の基本的な特性は、自己/非自己の区別をもたらす免疫寛容である。免疫系が自身に対して反応すると、自己免疫疾患が起こる (Annu. Rev. Med. 48:341-351, 1997) 。
【0003】
ブドウ膜炎、コラーゲン誘導性関節炎、アジュバント関節炎、全身性エリテマトーデス、多発性硬化症、甲状腺炎、重症筋無力症、炎症性腸疾患、およびI型糖尿病を含む自己免疫疾患において経口寛容は治療上有益であることが証明された (Annals of the New York Academy of Sciences 778(1): 217-227, 1996) 。
【0004】
経口的に投与された抗原は、良く発達した免疫ネットワークである腸管関連リンパ組織(GALT)に出会う。GALTは、絨毛、固有層、上皮細胞間リンパ球、およびパイエル板から成り、このパイエル板は、絨毛の間に散在するリンパ小節から成る。腸管上に並ぶ上皮は、抗原および微生物を輸送するためのM細胞を含む。腸管内の抗原および病原体は、その下にある免疫細胞に抗原を直接送達することに特化したM細胞を通してのみバリアを突破しうる。次いで、抗原を認識する上皮細胞間リンパ球がパイエル板に移動し、免疫応答を引き起こす。パイエル板では、B細胞およびマクロファージがリンパ球に抗原を提示し、次いで、この抗原特異的リンパ球が、体の粘膜系(呼吸器官の粘膜表面等)の固有層に移動する (Science 277:949-952, 1997; Science 277: 910-911, 1997) 。
【0005】
腸管の免疫系は、経口投与を介して媒介された疾患を予防するための先天性免疫および適応免疫を含む。病原体に感染した組織部分は、マクロファージ、単球、顆粒球、およびナチュラルキラー細胞の非特異的先天性免疫応答を誘導するためのケモカインを分泌する。マクロファージおよび顆粒球は、食作用により病原体を直接捕らえることができる。ナチュラルキラー細胞は、ウイルス感染または他の病原体により感染した細胞に対して最初に応答する体内における免疫細胞である。ナチュラルキラー細胞は標的細胞を攻撃するだけでなく、IFN-γ、TGF-β、TNF-α、IL-5、およびIL-10等の免疫応答を調節するためのサイトカインを分泌する。マクロファージおよび樹状細胞等の他の抗原提示細胞は、免疫応答すなわち、T細胞およびB細胞によりそれぞれ実行された細胞性免疫および体液性免疫をさらに活性化させるために、細胞表面上のMHCクラスII分子および共刺激因子を介してT細胞に抗原を提示する。粘膜の免疫は、CD4+T細胞から分泌されたサイトカインにより主に調節される。2型ヘルパーT細胞から分泌されたIL-4、IL-5、IL-6、およびIL-10は、抗体産生のためにB細胞を刺激することに関与する。3型ヘルパーT細胞から分泌されたTGF-βはB細胞を誘導し、IgAを生成させる(Trends Immunol. 22:244-247, 2004; J. Clin. Invest. 106:935-937, 2000)。
【0006】
粘膜の免疫応答は腸管においてだけでなく全身の粘膜系においても誘導されることに留意されたい。腸内の誘導組織であるパイエル板において抗原特異的リンパ球が活性化されたら、このリンパ球は、リンパ排液路および血液を通って、直腸、生殖器官、および肺等を含むエフェクター部位に移動する。次いで、これらのエフェクター部位で抗体が産生され、検出される。このホーミング現象は、いわゆる一般的な粘膜の免疫系である。経口投与により誘導された抗体は一般的な粘膜の免疫応答により全身に配置される。経口投与を介して媒介された抗原が存在すると、組織的寛容またはIgA、すなわち経口寛容が誘導される。経口寛容は、組織的不応答および粘膜応答の両者を含み、前者は、自己免疫疾患から全身を予防し、後者は、特異抗原に対して能動的抑制を誘導する。したがって、粘膜の免疫系は、炎症性の免疫応答を抑制し、さらに、非炎症性の局所粘膜でのIgAの提示を増強させる傾向がある(Immunol. Today 18:335-343, 1997)。
【0007】
胃腸管の表面には単層円柱上皮が並んで、細菌、食餌抗原、および高分子の過剰吸収に対するバリアを形成し、さらに、低分子の輸送は、バリア中に形成された密着結合により制御される。結合組織に埋まっている陰窩は、腸管上皮細胞を再生するための幹細胞を含む。腸管上皮細胞の他に、この腸管幹細胞は、粘液を分泌するための杯細胞および腸内分泌細胞ならびに抗菌ペプチドを分泌するためのパネート細胞にも分化しうる。幹細胞領域にとどまるパネート細胞の他に、分化した細胞は絨毛の先端へ移動する (J. Clin. Invest. 105:1493-1499, 2000; Science 294:2115-2116, 2001)。
【0008】
腸管上皮細胞の特異的配置により、この上皮細胞が、腸管腔内の様々な抗原に対する免疫応答に影響を及ぼす重要な役割を果たすということは適切な意見である。腸管上皮細胞は、腸管粘膜内でのT細胞応答を調節するよう、抗原提示細胞(APC)として抗原提示分子を発現しうる (Immunol. Today 21:123-128, 2000)。腸管上皮細胞により発現された抗原提示分子は、MHCクラスI、MHCクラスII、およびCDldを含むことが調べられている (Gastroenterology 124:1420-1431, 2003)。MHCクラスIおよびMHCクラスIIは上皮細胞の側底膜上に発現し、MHCクラスIはCD8+細胞と結合する役割があり、MHCクラスIIはCD4+細胞と結合する役割がある。CDldは糖脂質を介してNK T細胞を活性化する。これらの証拠は、腸管上皮細胞が粘膜の免疫系で重要な役割を果たすことを示す。
【0009】
物理的バリアであることの他に、腸管上皮細胞は、胃腸管内の微生物に対する抗菌ペプチドを分泌し、他の細胞にシグナルを与える (Immunol. Today 21:123-128, 2000; J. Clin. Invest. 95:55-65, 1995)。特異的な抗菌ペプチドであるディフェンシンが存在することがわかっている。ディフェンシンは、3〜5kDのタンパク質であり、8つのペプチドを有するα-ディフェンシンファミリーおよびβ-ディフェンシンファミリーを含む。これらのディフェンシンは、口腔粘膜、肺、および胃腸管の上皮細胞中で器官特異的な発現パターンを有する (Eur. J. Gastroenterol. Hepatol. 13:771-776, 2001; Nat. Rev. Immunol. 3:710-720, 2003)。胃腸管が感染すると、腸管上皮細胞はディフェンシンを発現し、ディフェンシンはケモカインのように作用してNK細胞および樹状細胞を感染部位に誘導し、いわゆる先天性免疫を実行させる。先天性免疫の誘導の他に、T細胞の増殖を誘導するために、ディフェンシンはToll様受容体4を介して樹状細胞を誘導して共刺激因子(B7.2)を発現させる。したがって、ディフェンシンは、先天性免疫および適応免疫の間の関連に関与する。
【0010】
他の抗菌タンパク質、アンギオゲニンは癌細胞の血管形成に関与すると考えられていた。しかしながら、2003年、Hooperにより、アンギオゲニンは、正常な生理学的条件下で胃腸管におけるパネート細胞により産生され、細胞顆粒中に貯蔵されることが開示された。アンギオゲニンはリポ多糖刺激に反応して胃腸管内に分泌される。無菌マウスおよび腸内細菌を送り込まれたマウスを比較すると、腸内細菌が存在することによりアンギオゲニンが著しく増加することがわかる。ディフェンシンと同様に、アンギオゲニンは殺菌性で、感染部で遭遇した局所的な環境条件により調節される (Nat. Immunol. 4:269-273, 2003)。
【0011】
病原体の感染に対して抗菌ペプチドを分泌する他に、腸管上皮細胞は、胃腸管における免疫応答の調節のための多くのシグナルを産生する。侵入する微生物を破壊することにより宿主を保護するために、腸管上皮細胞は、直ちに、防衛機構を介して侵入する微生物に応答するであろう。この防衛機構は先天性免疫系として知られ、先天性免疫系は、グラム陰性菌の細胞壁中に存在するリポ多糖等の微生物病原体の巨大分子を認識するために生殖細胞系にコードされたパターン認識受容体を使用する。近年、パターン認識受容体がどのように細胞を活性化するかについて開示された。成体ショウジョウバエで、Tollが、感染の際に、抗真菌性ペプチドおよび抗菌性ペプチドを誘導することが示されている。MedzhitovおよびJanewaysらは、哺乳動物において、Toll様受容体(TLR)の活性化により、適応免疫応答の活性化に必要とされるサイトカインおよび共刺激分子の誘導がもたらされることを開示する (Cell 91:295-298, 1997)。TLRにより活性化された細胞内シグナル伝達経路は、保存されたTIR(Toll/IL-1R相同性)ドメインにより、IL-1Rシグナル伝達と共通する多くのものを共有する。エンドトキシンがTLRにより認識されると、NF-kBの移動による核移行が引き起こされ、免疫応答に関与する他の遺伝子の転写が調節される。続いて、炎症誘発性サイトカイン遺伝子が活性化され、抗菌ペプチドが分泌され、IL-8、MIP、およびMCP-1等のケモカインが分泌される。感染部に移動して感染細胞を破壊するために、マクロファージおよびNK細胞がこれらのケモカインにより誘導される(Cell 91:295-298,1997; Curr. Opin. Immunol 14:103-110, 2002)。よって、NF-kBの活性化により引き起こされた炎症誘発性サイトカインの分泌により、Toll様受容体は、先天性免疫系を誘導するだけでなく、間接的に適応免疫系も誘導する。
【0012】
ヒト細胞には、少なくとも10個の異なるTLRが存在し、TLR-2、TLR-4、およびTLR-9がTLRファミリーのうちでより特徴づけられたメンバーである。TLRは、細菌およびウイルスを含む多くのタイプの病原体の認識を媒介することが示された。TLR-4は、グラム陰性菌LPSの受容体であり、TLR-2は、グラム陽性菌ペプチドグリカンとリポペプチドの受容体であり、TLR-9は、非メチル化およびリン酸化されたシトシン-グアニンオリゴヌクレオチド、CpGの受容体である(Curr. Opin. Immunol. 14:103-110, 2002)。TLR3は、二本鎖RNAウイルスを認識することができ(Nature 433:887-892, 2005)、TLR7は、一本鎖RNAウイルスを認識しうる(Proc Natl Acad Sci USA. 101(15):5598-5603, 2004)。TLRシグナル経路が活性化されると、先天性免疫または抗ウイルス応答を宿主保護のために誘導させることができる。
【0013】
正常な腸管上皮細胞において、TLR-3およびTLR-5が恒常的に発現する一方、TLR2およびTLR4はほとんど検出することができない(Infection and Immunity 68:7010-7017, 2000)。IL-8は、細菌DNAの刺激に反応して上皮細胞により産生される(FASEB J 17:1319-1321, 2003)。腸管上皮細胞において、細菌のCpGオリゴヌクレオチドはTLR-9のリガンドである。さらに、腸管上皮細胞はすべての異物に反応して炎症を誘導するわけではなく、すなわち、腸管上皮細胞は共生する微生物相に耐性があり、一方、腸管上皮細胞は、APCにより媒介され、T細胞により形成された炎症を誘導するために、起こりうる病原性条件下または自己免疫疾患下で、APCに危険シグナルを与える(Cell 118:229-241, 2004)。
【0014】
通常の定常状態条件下でTLRにより認識された共生細菌は腸管の傷を予防するのに重大な役割を果たす(Cell 118:229-241, 2004)。しかし、ヒトの炎症性腸疾患(IBD)において、Th1媒介性の病的作用は微生物相に対する異常な粘膜免疫応答によるものと思われる(Gastroenterol. Clin. North Am. 31:41-62, 2002)。これらの発見は、TLRが、宿主-共生動物間の粘膜のホメオスタシスを制御することを明らかにする。
【0015】
中国の薬草抽出物の経口投与により免疫の発生を調節することが良く知られている。中国の薬草抽出物により腸管粘膜の免疫を介して免疫の発生を調節することは非常に可能性がある。セッコク属種は最も貴重な中国の薬草であると考えられる。セッコク属種はラン科に属し、茎が主な薬用部分である。茎はわずかに甘く、やや塩辛い味がする。いくつかの中国の医療用の薬局方には、セッコク属種は、粘膜障害、胃障害、および眼障害等のいくつかの疾病のための治療薬であることが開示される。我々の先の研究経験によれば、デンドロビーハーバ(Dendrobii Herba)が最も治癒力のある薬用種であると思われる。
【0016】
本発明は、セッコク属に由来する多糖を調製するための方法を提供し、さらに、セッコク属に由来する多糖の医薬としての使用を提供する。
【発明の開示】
【0017】
発明の概要
本発明の一態様として、植物からの多糖を調製するための方法を提供する。方法は、a)第1のアルコールで植物を抽出して第1の抽出物を得るステップ、b)溶媒で第1の抽出物を抽出して第2の抽出物を得るステップ、およびc) 多糖を得るために第2のアルコールで第2の抽出物を沈殿させるステップを含む。
【0018】
好ましくは、植物はセッコク属に属する。
【0019】
好ましくは、第1のアルコールはメタノールである。
【0020】
好ましくは、溶媒は水である。
【0021】
好ましくは、第2のアルコールはエタノールである。
【0022】
本発明において、前記方法は、ステップb)において溶媒で第1の抽出物を抽出した後、遠心分離し、濾過して第2の抽出物を得るステップをさらに含む。
【0023】
本発明の他の態様として、自己免疫疾患および粘膜障害を治療するための組成物を提供する。前記組成物は植物から調製した多糖を含み、該植物はセッコク属に属し、抗原は自己免疫疾患の誘導に関連する。
【0024】
好ましくは、多糖は本発明において提供される方法により調製される。
【0025】
本発明において、自己免疫疾患はブドウ膜炎であってもよい。
【0026】
本発明の他の態様として、哺乳動物の自己免疫疾患および粘膜障害を治療するための方法を提供する。前記方法は、哺乳動物に抗原および多糖を投与するステップを含み、該抗原は自己免疫疾患の誘導に関連し、該多糖はセッコク属に属する植物から調製される。
【0027】
本発明において、多糖の投与は経口投与の方法で実行されてもよい。
【0028】
本発明において、自己免疫疾患はブドウ膜炎であってもよい。
【0029】
本発明の他の態様として、哺乳動物の自己免疫疾患および粘膜障害を治療するための方法を提供する。前記方法は、哺乳動物に多糖を投与するステップを含み、該多糖がセッコク属に属する植物から調製される。
【0030】
本発明において、多糖の投与は経口投与の方法で実行される。
【0031】
本発明において、自己免疫疾患はブドウ膜炎であってもよい。
【0032】
以下の詳細な説明および添付の図面を検討すれば、上記態様および本発明の利点が当業者にとってより容易に明らかになろう。
【発明を実施するための最良の形態】
【0033】
本発明を、以下の実施形態を参照してより具体的に説明する。以下の本発明の最良の形態は、例証および説明のみを目的として本明細書に提示されるものであって、網羅的なものとすることを意図するものではない、または、開示した詳細な形態に限定されることを意図するものではないことに留意されたい。
【0034】
本発明は、植物からの多糖を調製するための方法であって、a)第1のアルコールで植物を抽出して第1の抽出物を得るステップ、b)溶媒で第1の抽出物を抽出して第2の抽出物を得るステップ、およびc) 多糖を得るために第2のアルコールで第2の抽出物を沈殿させるステップを含む方法を提供する。
【実施例1】
【0035】
セッコク属からの多糖の調製
10kgの新鮮なデンドロビーハーバを乾燥させて2.92kgの乾燥材料にし、次いで、乾燥材料を73.3Lメタノールに浸して抽出し、136.79グラムの粗抽出物と残渣を得た。粗抽出物を脱メタノール工程で処理し、次いで一晩水に浸した。反応溶液を、55℃〜60℃の範囲の温度の温水で30分間抽出し、5,000rpm、10℃で遠心分離機(ER-RC13 C-124、HITACH社)により遠心分離した。遠心分離後、上清を6mu m濾紙で濾過した。濾液をエタノールで沈殿させ、20.684gの多糖(以下DeCaPSと称する)を得た。
【実施例2】
【0036】
DeCaPSで処理したIEC-6細胞の免疫応答
a. IEC-6細胞の培養
正常ラット小腸から生じたIEC細胞を、5%CO2、37℃のインキュベータ中で、5%ウシ胎児血清、4.5g/Lグルコース、5mu g/mlウシインスリン、および2mM L-グルタミンを含むDMEM培地で培養した。
【0037】
b. DeCaPSによるIEC-6細胞の処理
IEC-6細胞を6ウェルプレートに流し、IEC細胞を、それぞれ、1mu g/ml、10mu g/ml、および100mu g/mlの濃度でDeCaPSにより6時間処理した。
【0038】
c. DeCaPSで処理したIEC細胞からの全RNAの単離
DeCaPSでの処理の後、IEC細胞を採取し、1ml Ultraspec(商標)RNA単離キット(Biotex laboratories Inc.(米国))中に懸濁させ、全RNAを、キットの標準プロトコールに従って得た。得られた全RNAを定量した。
【0039】
d. サイトカインおよびToll様受容体の逆転写およびPCR分析
26.5mu lの反応容量を有し、0.1gオリゴ-dT、5mu gの得られた全RNA、およびDEPCで処理した滅菌水を含む反応を70℃で10分間実行した。次いで、4mu lの10mM dNTP、0.5mu l rRNasin、1mu l AMV(鳥骨髄芽球症ウイルス(Avian Myeloblastosis virus))逆転写酵素(10ユニット)、および8mu l 5×RT緩衝液を反応に加え、全容量を40mu lとして42℃で60分間、次いで90℃で5分間インキュベートし、その結果として、cDNA生成物を逆転写反応により得た。その後、0.5muの10mM dNTP、0.5mu l 10×プロザイム緩衝液、プライマー(0.8mu lの5mu MセンスDNAおよび0.8mu lの5mu MアンチセンスDNA)、および滅菌水を2.5mu l cDNA生成物に加え、全容量25mu lとし、DNAサーマルサイクラー(Perkin-Elmer-Cetus社)中でインキュベートし、PCRを35サイクル実行した。標的遺伝子に対するセンスプライマーおよびアンチセンスプライマーを含むプライマー配列[配列番号:1〜32]ならびに対応するアニール温度を以下の表Iに記載する。
【表1】
【0040】
図1を参照されたい。図1は、DeCaPSで処理したIEC-6細胞中のサイトカインおよび抗菌ペプチドのmRNA発現を示す。1mu g/ml、10mu g/ml、および100mu g/mlのDeCaPSで6時間処理した後、IEC-6細胞中のTNF-α[配列番号:33]のmRNA発現は約29%、20%、および30%抑制されたが、IEC細胞中のIL-10[配列番号:34]のmRNA発現は9.89倍、6.55倍、および1.77倍までそれぞれ高められた。TH2/3経路を引き起こすために、免疫系の他の細胞に寛容シグナルを提供するよう、DeCaPSにより腸管上皮細胞を誘導しうることが示されている。さらに、1mu g/ml、10mu g/ml、および100mu g/mlのDeCaPSで6時間処理した後、a-ディフェンシン[配列番号:35]のmRNA発現は約42%、33%、および51%抑制されたが、アンギオゲニン[配列番号:36]のmRNA発現は1.96倍、1.42倍、および1.70倍までそれぞれ高められた。
【0041】
図2を参照されたい。図2は、DeCaPSで処理したIEC-6細胞中のToll様受容体のmRNA発現を示す。IEC-6細胞において、Toll様受容体であるTLR2[配列番号:37]、TNR4[配列番号:38]、TLR7[配列番号:39]、およびTLR9[配列番号:40]はDeCaPS処理なしで発現した。1mu g/ml、10mu g/ml、および100mu g/mlのDeCaPSで6時間処理した後、それぞれ、IEC-6細胞中のTLR4[配列番号:38]のmRNA発現は約6%、16%、および13%抑制され、IEC-6細胞中のTLR5[配列番号:41]のmRNA発現は約13%、37%、および35%抑制され、IEC-6細胞中のTLR7[配列番号:39]のmRNA発現は約11%、7%、および48%抑制された。しかしながら、TLR9[配列番号:40]のmRNA発現は2.21倍、1.09倍、および3.56倍までそれぞれ高められた。セッコク属に由来する多糖は、TNF-α[配列番号:33]の発現を抑制するだけでなく、TLR4[配列番号:38]、TLR5[配列番号:41]、TLR7[配列番号:39]、およびTLR9[配列番号:40]の発現もまた抑制するということが示されている。
【実施例3】
【0042】
DeCaPSの経口投与による動物モデルの免疫系に対する効果
C3Hマウス(13週令)に、実施例Iで調製したDeCaPSを、10mg/kg/日、50mg/kg/日、および250mg/kg/日の投与量で飲料水を介して5日間それぞれに与え、次いで、脾臓および骨髄を得るために屠殺した。
【0043】
a. C3HマウスからのコンカナバリンA刺激脾細胞に対するMTTアッセイ
DeCaPSで処理したC3Hマウスから得た脾細胞(4×105細胞/ウェル)を、10%ウシ胎児血清ならびに1mu g/mlおよび5mu g/mlのコンカナバリンA(ConA)を含むRPMI-1640培地で72時間培養し、次いで、1mg/ml MTTで3時間処理した。次いで、細胞培養物に、50%DMFおよび20%SDSを含む溶解緩衝液をそれぞれに加え、反応を16時間実行させた。反応における570nmでの吸収を分析すると、図3に示すような成長指数を示した。
【0044】
図3を参照されたい。図3は、DeCaPSで経口的に処理したC3HマウスからのConA刺激脾細胞の分裂応答に対する効果を示す。C3Hマウスを、DeCaPSにより、10mg/kg/日、50mg/kg/日、および250mg/kg/日の投与量で5日間経口的に処理した後、脾細胞の1mu g/ml ConA刺激に対する応答は、約28%、49%、および30%抑制された。
【0045】
b. C3Hマウスから得たGM-CSF-刺激骨髄細胞に対するMTTアッセイ
DeCaPSで処理したC3Hマウスの脚から骨髄細胞を得た。2%FCSおよび4ng/ml GM-CSFを含むA-MEM培地で骨髄細胞を72時間培養し、骨髄細胞の陽性対照群を、2%FCSおよび20ng/ml GM-CSFを含むα-MEM培地で72時間培養した。次いで、骨髄細胞を3時間処理するために、培地を、1mg/ml MTTおよび2%FCSを含むRPMI-1640と置換し、細胞を、MTT溶解緩衝液(20%SDSおよび50%DMF、pH4.5)でさらに処理し、一晩インキュベートした。反応における570nmでの吸収を分析すると、図4に示すような成長指数を示した。
【0046】
図4を参照されたい。図4は、DeCaPSで経口的に処理したC3Hマウスから得たGM-CSF-刺激骨髄細胞の分裂応答に対する効果を示す。C3Hマウスを、DeCaPSにより、10mg/kg/日、50mg/kg/日、および250mg/kg/日の投与量で5日間それぞれ経口的に処理した後、骨髄細胞のGM-CSF刺激に対する分裂応答は、1.71倍、1.62倍、および1.76倍まで高められた。
【0047】
c. C3Hマウスから得たパイエル板および脾細胞におけるサイトカインのmRNA発現
DeCaPSで処理したC3Hマウスからパイエル板および脾細胞を得た。パイエル板および脾細胞におけるサイトカイン特異的mRNAのPCR分析を実施例IIに記載する方法で実行し、PCR結果を図5に示した。
【0048】
図5を参照されたい。図5は、DeCaPSで経口的に処理したC3Hマウスから得たパイエル板および脾細胞におけるサイトカイン特異的mRNAのPCR結果を示すが、レーン1〜4は、DeCaPSにより、0mg/kg/日、10mg/kg/日、50mg/kg/日、および250mg/kg/日の投与量で経口的に処理したC3Hマウスをそれぞれ示す。
【0049】
C3HマウスをDeCaPSで経口的に5日間処理した後、パイエル板におけるIL-4[配列番号:42]、IL-6[配列番号:43]、IL-1β[配列番号:44]、IFN-γ[配列番号:45]、およびTGF-β [配列番号:46]のmRNA発現は図5に示すように増加した。C3Hマウスを、DeCaPSにより、10mg/kg/日および50mg/kg/日の投与量で5日間経口的に処理した後、IL-4[配列番号:42]のmRNA発現は2.64倍および2.46倍までそれぞれ高められた。C3Hマウスを、DeCaPSにより、10mg/kg/日、50mg/kg/日、および250mg/kg/日の投与量で5日間経口的に処理した後、IL-6[配列番号:43]のmRNA発現はそれぞれ2.61倍、3.99倍、および5.35倍まで高められ、IL-β[配列番号:44]のmRNA発現はそれぞれ1.83倍、2.00倍、および1.11倍まで高められ、IFN-γ[配列番号:45]のmRNA発現はそれぞれ3.47倍、5.47倍、および5.57倍まで高められ、TGF-β[配列番号:46]のmRNA発現はそれぞれ2.38倍、2.54倍、および1.89倍まで高められた。パイエル板は、抗原に対する免疫応答を測定するための主な組織であるので、Th2経路がIL-4[配列番号:42]およびIL-6[配列番号:43]の増加に反応して引き起こされ、Tヘルパー細胞がIL-1β[配列番号:44]の増加によりTGF-β[配列番号:46]等のサイトカインを分泌するよう活性化され、IgAクラススイッチB細胞は、ある種の免疫応答を抑制するために、TGF-β[配列番号:46]によりIgAを分泌するよう誘導される。
【0050】
C3HマウスをDeCaPSで経口的に5日間処理した後、脾細胞中のIL-1β[配列番号:44]、IL-4[配列番号:42]、およびTGF-β[配列番号:46]のmRNA発現は図5に示すように抑制された。C3Hマウスを、DeCaPSにより、10mg/kg/日、50mg/kg/日、および250mg/kg/日の投与量で5日間経口的に処理した後、IL-1β[配列番号:44]のmRNA発現は約32%、43%、および79%抑制され、IL-4[配列番号:42]のmRNA発現は約7%、67%、および14%抑制された。C3Hマウスを、DeCaPSにより、10mg/kg/日、50mg/kg/日、および250mg/kg/日の投与量で5日間経口的に処理した後、それぞれ、TGF-β[配列番号:46]のmRNA発現は約27%および44%抑制され、IFN-γ[配列番号:45]のmRNA発現は約33%および97%抑制された。しかしながら、C3Hマウスを、DeCaPSにより、10mg/kg/日、50mg/kg/日、および250mg/kg/日の投与量で5日間経口的に処理した後、IL-6[配列番号:43]のmRNA発現は1.84倍、1.4倍、および1.23倍まで高められた。DeCaPS処理に対する著しい細胞応答はなく、この現象はConA刺激に対する応答と一致することに留意されたい。
【実施例4】
【0051】
DeCaPSによる動物モデルの経口寛容の増強
自己免疫疾患用の動物モデルを確立するためにオボアルブミン(OVA)により動物を免疫できることは周知なので(The Journal of Pharmacology and Experimental Therapeutics 288:849-857, 1999)、本発明では、OVA誘導自己免疫疾患を有するマウスを確立し、OVA誘導自己免疫疾患を有するマウスにおけるDeCaPSによる経口寛容の増強を以下のとおり研究する。C57/BL6jマウス(65週令)を、4日目および5日目に、飲料水中0.5mg/mlオボアルブミン(OVA)で経口的に処理した。3〜7日目に、10mg/kg/日、40mg/kg/日、および160mg/kg/日の投与量でアジュバントとしてDeCaPSをC57/BL6jマウスに投与した。CFAをアジュバントとした50mu g OVAを抗原として8日目にi.p.注射によりマウスに投与した。血液採集物中でOVAに反応したIgGおよびIgMが22日目に検出され、50mu g OVAをi.p注射により24日目に抗原としてマウスに投与し、OVAに反応した抗体が30日目に検出され、腸管粘膜の腸管洗浄溶液および肺粘膜を得るために32日目にマウスを屠殺し、その後、OVAに反応したIgAが検出された。詳細な実験手順を以下の表IIに示す。
【表2】
【0052】
腸管粘膜の腸管洗浄溶液を32日目にマウスから得て、図6に示すように、腸管洗浄溶液中のオボアルブミン特異的IgAの力価を測定した。図6に示すように、腸管粘膜から分泌されたIgAは、DeCaPS(40mg/kg/日)と共にOVAで処理したC57/BL6jマウスで1.9倍(p<0.01)まで高められ、腸管粘膜から分泌されたIgAは、DeCaPS(160mg/kg/日)と共にOVAで処理したC57/BL6jマウスで4.35倍(p<0.001)まで高められた。
【0053】
図7に示すように、22日目および30日目の血液採集物中のIgM抗体を測定した。22日目の対照群で発現したIgM抗体レベルと比較すると、正常群で発現したIgM抗体レベルは、約49%(p<0.05)抑制され、陽性対照群で発現したIgM抗体レベルは、約87%(p<0.05)抑制され、DC-1群で発現したIgM抗体レベルは、約46%(p<0.02)抑制され、DC-2群で発現したIgM抗体レベルは、約88%(p<0.001)抑制され、DC-3群で発現したIgM抗体レベルは、約86%(p<0.001)抑制された。さらに30日目の対照群で発現したIgM抗体レベルと比較すると、正常群で発現したIgM抗体レベルは、約47%(p<0.01)抑制され、陽性対照群で発現したIgM抗体レベルは、約72%(p<0.001)抑制され、DC-1群で発現したIgM抗体レベルは、約38%(p<0.01)抑制され、DC-2群で発現したIgM抗体レベルは、約58%(p<0.01)抑制され、DC-3群で発現したIgM抗体レベルは、約71%(p<0.001)抑制された。
【0054】
図8に示すように、22日目および30日目の血液採集物中のIgG抗体を測定した。22日目の対照群で発現したIgG抗体レベルと比較すると、陽性対照群で発現したIgG抗体レベルは、約94%(p<0.01)抑制され、DC-1群で発現したIgG抗体レベルは、約67%(p<0.01)抑制され、DC-2群で発現したIgG抗体レベルは、約76%(p<0.01)抑制され、DC-3群で発現したIgG抗体レベルは、約88%(p<0.001)抑制された。さらに30日目の対照群で発現したIgG抗体レベルと比較すると、陽性対照群で発現したIgG抗体レベルは、約75%(p<0.001)抑制され、DC-2群で発現したIgG抗体レベルは、約62%(p<0.001)抑制され、DC-3群で発現したIgG抗体レベルは、約63%(p<0.001)抑制された。
【0055】
さらに、パイエル板を32日目のマウスから得て、パイエル板におけるリンパ球のRNAを抽出し、サイトカインを分析した。図9に示すように、パイエル板におけるリンパ球のサイトカイン発現を測定したが、レーンAは正常群を示し、レーンBは対照群を示し、レーンCは陽性対照群を示し、レーンDはDC-1群を示し、レーンEはDC-2群を示し、レーンFはDC-3群を示す。PCR増幅を変性のために94℃で45秒間に設定し、アニーリングを61℃で45秒間実行し、伸長を72℃で1分間実行した。PCRプログラムは37サイクルとした。2%アガロースゲルの電気泳動によりPCR生成物を分離し、ゲルを臭化エチジウムで染色し、UV光下で可視化した。
【0056】
対照群と比較すると、DC-3群で発現したTGF-β[配列番号:46]レベルは約30%抑制され、DC-3群で発現したIL-4[配列番号:42]レベルは2.2倍まで高められ、DC-3群で発現したIL-10[配列番号:34]レベルは1.3倍まで高められたということに留意されたい。対照群と比較すると、DC-2群で発現したIL-2[配列番号:47]レベルは1.4倍まで高められ、DC-2群で発現したIL-4[配列番号:42]レベルは1.6倍まで高められたことに留意されたい。さらに対照群と比較すると、DC-1群で発現したIL-4[配列番号:42]レベルは1.4倍まで高められ、DC-1群で発現したIL-6[配列番号:43]レベルは1.3倍まで高められた。
【0057】
IL-4[配列番号:42]およびIL-6[配列番号:43]のmRNA発現はパイエル板におけるリンパ球で高められた。パイエル板は、抗原に対する免疫応答を決定するための組織であるので、抗原に感作されたT細胞またはB細胞がホーミング機構により腸に戻ると、Th2経路が、IL-4[配列番号:42]およびIL-6[配列番号:43]の増加に反応して抗体を産生するよう活性化され、血清中のIgGおよびIgMの発現が、アレルゲン応答を抑制するように抑制され、経口寛容が達成されることに注目されたい。
【0058】
前述の説明を考慮して、脾臓中のT細胞の分裂応答はセッコク属の多糖により抑制されるが、骨髄中の顆粒球およびマクロファージはセッコク属の多糖により活性化される。言い換えれば、自己免疫疾患において活性化されたT細胞は、セッコク属の多糖の経口投与により抑制しうるが、顆粒球系細胞は、先天性免疫の促進により病原体の侵入を予防するためのセッコク属の多糖の経口投与により活性化される。
【0059】
腸管の免疫に関し、経口寛容を生じさせるために、IL-4[配列番号:42]、IL-6[配列番号:43]、IL-1β[配列番号:44]、IFN-γ [配列番号:45]、およびTGF-β[配列番号:46]のmRNA発現が高められて腸内のTh2/Th3経路が引き起こされる。セッコク属の多糖で処理したIEC-6細胞中のTNF-α[配列番号:33]、TLR2[配列番号:37]、TLR4[配列番号:38]、TLR5[配列番号:41]、およびTLR7[配列番号:39]の発現によれば、経口寛容関連シグナル、すなわちTh2/3経路を高め、自己免疫疾患および粘膜障害等の病原性状態における粘膜のホメオスタシスを維持するために、Toll様受容体により媒介されたシグナルを免疫抑制性のまたは抗炎症性の応答とするようセッコク属の多糖により調節しうることが示されている。興味深いことには、抗菌活性を有するアンギオゲニンの発現は、実施例2に記載されるように、セッコク属の多糖の投与により誘導しうる。したがって、セッコク属の多糖の経口投与は経口寛容を高めるだけでなく、病原体侵入を予防するための先天性免疫機構を引き起こす。これらの発見によれば、長引く炎症およびさらなる傷を免れるために、炎症性腸疾患(IBD)等の自己免疫疾患および粘膜障害にセッコク属の多糖を有利に使用してもよい。
【0060】
アンギオゲニンは、エンテロコッカスフェカリス(Enterococcus faecalis)等の腸内微生物だけでなくヒトにおいて組織的感染を引き起こすカンジダアルビカンス(Candida albicans)および肺炎連鎖球菌(Streptococcus pneumoniae)等の微生物に対しても活性を有することが開示されている (Nat. Immunol. 4:269-273, 2003) 。臨床研究では、女性の帯下は、多微生物性の表在性膣感染症とされる。この感染症は、G.バジナリス(G. vaginalis)、カンジダ種(Candida spp)、C.アルビカンス、T.バジナリス(T. vaginalis)、D群連鎖球菌(Streptococcus group D)、溶血性連鎖球菌b(Streptococcus b hemolytic)、大腸菌(E. coli)、およびクレブシエラ種(Klebsiella spp)等のような細菌により頻繁に引き起こされる (Salud publica Mex vol.45 suppl. 5, pS694-S697, 2003)。カンジダアルビカンス等の微生物により引き起こされた女性の帯下は、セッコク属の多糖の経口投与により誘導されたアンギオゲニンの活性により軽減することができたことは明らかである。
【実施例5】
【0061】
DeCaPSによる自己免疫疾患の治療
実験的自己免疫ブドウ膜炎は、いくつかの眼の自己免疫疾患用モデルとして役立つT細胞媒介自己免疫疾患である。実験的自己免疫ブドウ膜炎は、140kDのグリコリポプロテインである光受容体間レチノイド結合タンパク質(IRBP)での免疫化により誘導されることが確認されている。さらに、自己免疫ブドウ膜炎はIRBPペプチド1〜20(配列番号48:GPTHLFQPSLVLDMAKVLLD)、IRBPのアミノ酸1〜20により誘導しうることが確認されている。さらに、IRBPペプチド1〜20はマウスおよびヒトで保存されている。(Investigative Ophthalmology & Visual Science 41(1):127-131, 2000)よって、本発明において自己免疫疾患モデルを確立するためにIRBPペプチドをC57BL/6jマウスに投与した。
【0062】
a. IRBPペプチドの合成
IRBPペプチド(配列番号48:GPTHLFQPSLVLDMAKVLLD)をFmoc化学を使用してペプチド合成機(PS3)で合成し、Agilent社製HPLCで精製し、Brukeer社製esquire 2000 MSで特定した。
【0063】
b. IRBPおよびDeCaPSの投与
特定病原体フリー(SPF)マウスであるC57BL/6jマウスを、正常群、対照群、DC-10群、DC-40群、およびDC-160群に無作為に分類し、各群のマウスを6匹とした。0.2ml乳剤中のヒトIRBPペプチド[配列番号:48](150mu g/マウス)およびCFA(1:1、体積/体積)を群の各SPFマウスにi.p.注射で正常群以外に投与した。DC-10群のマウスに、DeCaPSを、10mg/kg/日の投与量で28日間与え、DC-40群のマウスに、DeCaPSを、40mg/kg/日の投与量で28日間与え、DC-160群のマウスに、DeCaPSを、飲料水を介して160mg/kg/日の投与量で28日間与えた。マウスの網膜の作用を網膜電図(ERG)分析により測定し、バイオアッセイおよび組織変化の測定用の眼を得るためにマウスを28日目に屠殺した。実験手順を以下の表IIIに示す。
【表3】
【0064】
c. 遅延型過敏症(DTH)試験
IRBPペプチドによる免疫化後21日目に、IFA(20mu l)中20mu g IRBPペプチド乳剤をマウスの左足蹠に皮下注射した。右足蹠にはIFAを注射した。48時間後、足蹠の厚さをキャリパで測定した。
【0065】
図10を参照されたい。図10は、マウスのDTH応答を示す。DTH応答は、リン酸緩衝液生理食塩水(PBS)を注射したマウスの足蹠増加と水容量で比較した足蹠増加により示した。DeCaPS(160mg/kg/日)で共処理したマウスの足蹠増加が著しく低下したことに留意されたい。
【0066】
d. 網膜電図(ERG)分析
マウスを2時間暗順応させ、次いで、ペントバルビタールナトリウムで麻酔した。次いで、マウスの角膜に麻酔し、瞳孔を拡大させた。網膜を閃光で刺激し、閃光に対する網膜の応答を網膜電図として記録した。網膜電図は、光受容体の働きならびに双極細胞およびミュラー細胞等の網膜基部の作用を示す。全網膜の状態を反映させるために網膜電図を使用する。網膜電図では通常、陰性のa波に陽性のb波が続く。a波の立ち上がりにより、光受容体層内の錐体細胞および桿細胞の活性が直接測定され、b波は、内顆粒層内の双極細胞の働きを反映する。
【0067】
28日目に正常群から無作為に選択した2匹のマウスについて、網膜電図分析の結果を、図11Aの上方部および下方部にそれぞれ示したが、閃光開始は、100ミリ秒の時間のところであり、閃光持続期間は約600ミリ秒であった。28日目に正常群から無作為に選択した2匹のマウスについて、眼の組織変化を図11Bの上方部および下方部(倍率400倍)にそれぞれ示したが、光受容体層を「P」と示し、外顆粒層を「ONL」と示し、内顆粒層を「INL」と示した。
【0068】
28日目に対照群から無作為に選択した2匹のマウスについて、網膜電図分析の結果を、図12Aの上方部および下方部にそれぞれ示したが、閃光開始は、100ミリ秒の時間のところであり、閃光持続期間は約600ミリ秒であった。28日目に対照群から無作為に選択した2匹のマウスについて、眼の組織変化を図12Bの上方部および下方部(倍率400倍)にそれぞれ示した。IRBPペプチドによる免疫化により、対照群のマウスの光受容体層(P)、外顆粒層(ONL)、および内顆粒層(INL)は激しく損傷を受けたことが示された。
【0069】
28日目にDC-10 群から無作為に選択した2匹のマウスについて、網膜電図分析の結果を、図13Aの上方部および下方部にそれぞれ示したが、閃光開始は、100ミリ秒の時間のところであり、閃光持続期間は約600ミリ秒であった。28日目にDC-10群から無作為に選択した2匹のマウスについて、眼の組織変化を図13Bの上方部および下方部(倍率400倍)にそれぞれ示した。IRBPペプチドによる免疫化およびDeCaPS(10mg/kg/日)の28日間の経口投与の後、DC-10群のマウスの光受容体層(P)、外顆粒層(ONL)、および内顆粒層(INL)は激しく損傷を受けたことが示された。
【0070】
28日目にDC-40群から無作為に選択した2匹のマウスについて、網膜電図分析の結果を、図14Aの上方部および下方部にそれぞれ示したが、閃光開始は、100ミリ秒の時間のところであり、閃光持続期間は約600ミリ秒であった。28日目にDC-40群から無作為に選択した2匹のマウスについて、眼の組織変化を図14Bの上方部および下方部(倍率400倍)にそれぞれ示した。IRBPペプチドによる免疫化およびDeCaPS(40mg/kg/日)の28日間の経口投与の後、DC-40群のマウスの光受容体層(P)、外顆粒層(ONL)、および内顆粒層(INL)はわずかに損傷を受けたことが示された。
【0071】
28日目にDC-160群から無作為に選択した2匹のマウスについて、網膜電図分析の結果を、図15Aの上方部および下方部にそれぞれ示したが、閃光開始は、100ミリ秒の時間のところであり、閃光持続期間は約600ミリ秒であった。28日目にDC-160 群から無作為に選択した2匹のマウスについて、眼の組織変化を図15Bの上方部および下方部(倍率400倍)にそれぞれ示した。IRBPペプチドによる免疫化およびDeCaPS(160mg/kg/日)の28日間の経口投与の後、DC-160群のマウスの光受容体層(P)、外顆粒層(ONL)、および内顆粒層(INL)は損傷を受けなかったことが示された。
【0072】
その結果、IRBPペプチドでの免疫化の後、より多くの細胞死が内顆粒層内に観察されたが、死細胞は外顆粒層上に沈まなかった。内顆粒層内の細胞がIRBPペプチド処理により損傷を受けたが、細胞が内顆粒層から剥離することが160mg/kg/日の投与量でのDeCaPSによる共処理により著しく防止されたことがさらに観察された。
【0073】
さらに、図11A、12A、13A、14A、および15Aにおいてそれぞれ示されるa波を図16Aと比較し(統計でp<0.02)、図11A、12A、13A、14A、および15Aにおいてそれぞれ示されるb波を図16Bと比較した(統計でp<0.02)。IRBPペプチドで免疫された対照群のマウスのa波は正常群のマウスのa波よりも非常に低いことに注目されたい。しかしながら、対照群と比較すると、DeCaPS(40mg/kg/日)で共処理したマウスの内顆粒層からの細胞の剥離が著しく予防される。さらに、DeCaPS(160mg/kg/日)で共処理したマウスのa波およびb波の振幅は正常群のマウスのものとほぼ等しい。セッコク属から調製した多糖(160/mg/kg/日)での共処理によりIRBPペプチドでの免疫化による損傷から救出しうることにさらに注目されたい。言い換えれば、セッコク属から調製した多糖の経口投与により、IRBPにより誘発された炎症を網膜において防止しうる。
【0074】
前述の実験によれば、本発明は、自己免疫疾患および粘膜障害を治療するための組成物であって、組成物が、セッコク属から調製した多糖および自己免疫疾患の誘導に関連した抗原を含んでおり、セッコク属から調製した多糖が経口寛容誘導および先天性免疫促進を高めることができる組成物を提供する。さらに、本発明は、セッコク属から調製した多糖が単独で経口寛容を誘導し、先天性免疫を促進しうるので、自己免疫疾患および粘膜障害を治療するためにセッコク属から調製した多糖のみを含む組成物を提供する。
【0075】
本発明は、現在最も実用的で好ましい実施形態であると考えられるものに関して記載するが、本発明は、開示された実施形態に限定されないことを理解されたい。それどころか、本発明は、変更および類似する構造すべてを包含するよう、最も広い解釈と一致する添付の請求項の精神および範囲に含まれる上記の様々な変更および類似する構成を包含することが意図される。
【図面の簡単な説明】
【0076】
【図1】DeCaPSで処理したIEC-6細胞中のサイトカインおよび抗菌ペプチドのmRNA発現を示すPCR分析である。
【図2】DeCaPSで処理したIEC-6細胞中のToll様受容体のmRNA発現を示すPCR分析である。
【図3】DeCaPSで経口的に処理したC3HマウスからのConA刺激脾細胞の分裂応答に対する効果を示す図である。
【図4】DeCaPSで経口的に処理したC3HマウスからのGM-CSF共刺激骨髄細胞の分裂応答に対する効果を示す図である。
【図5】DeCaPSで経口的に処理したC3Hマウスから得たパイエル板および脾細胞におけるサイトカイン特異的mRNAを示すPCR分析である。
【図6】DeCaPSの投与ありまたはなしでオボアルブミンにより経口的に免疫したC57BL/6jマウスからの腸管洗浄溶液中のオボアルブミン特異的IgAの力価を示す図である。
【図7】DeCaPSの投与ありまたはなしでオボアルブミンにより経口的に免疫したC57BL/6jマウスからの血清中のオボアルブミン特異的IgMの力価を示す図である。
【図8】DeCaPSの投与ありまたはなしでオボアルブミンにより経口的に免疫したC57BL/6jマウスからの血清中のオボアルブミン特異的IgGの力価を示す図である。
【図9】C57BL/6jマウスからのパイエル板におけるリンパ球のサイトカイン発現を示すPCR分析である。
【図10】C57BL/6jマウスのDTH応答を示す図である。
【図11A】図11Aは、正常群のマウスにおける一定の範囲の閃光強度に対する網膜電図を示す図である。
【図11B】図11Bは、正常群におけるマウスの眼の組織変化を示す図である。
【図12A】図12Aは、対照群のマウスにおける一定の範囲の閃光強度に対する網膜電図を示す図である。
【図12B】図12Bは、対照群におけるマウスの眼の組織変化を示す図である。
【図13A】図13Aは、DC-10群のマウスにおける一定の範囲の閃光強度に対する網膜電図を示す図である。
【図13B】図13Bは、DC-10群におけるマウスの眼の組織変化を示す図である。
【図14A】図14Aは、DC-40群のマウスにおける一定の範囲の閃光強度に対する網膜電図を示す図である。
【図14B】図14Bは、DC-40群におけるマウスの眼の組織変化を示す図である。
【図15A】図15Aは、DC-160群のマウスにおける一定の範囲の閃光強度に対する網膜電図を示す図である。
【図15B】図15Bは、DC-160群におけるマウスの眼の組織変化を示す図である。
【図16A】図16Aは、図11A、12A、13A、14A、および15Aにおけるa波の振幅を示す図である。
【図16B】図16Bは、図11A、12A、13A、14A、および15Aにおけるb波の振幅を示す図である。
【特許請求の範囲】
【請求項1】
自己免疫疾患および粘膜障害を治療するための組成物であって、セッコク属に属する植物から調製した多糖および免疫系における自己免疫疾患の誘導に関連する抗原を含む組成物。
【請求項2】
前記自己免疫疾患がブドウ膜炎である、請求項1に記載の組成物。
【請求項3】
哺乳動物の免疫系を調節しうる組成物の使用であって、前記哺乳動物に抗原および多糖を投与するステップを含み、前記抗原が免疫系において自己免疫疾患の誘導に関連し、前記多糖がセッコク属に属する植物から調製される使用。
【請求項4】
前記投与するステップが経口投与の方法で実行される、請求項3に記載の使用。
【請求項5】
前記自己免疫疾患がブドウ膜炎である、請求項3に記載の使用。
【請求項6】
セッコク属に属する植物から調製した多糖を含む、哺乳動物の免疫系を調節するための組成物。
【請求項7】
組成物により、哺乳動物の自己免疫疾患の発生を低下させることができることを特徴とする、請求項6に記載の組成物。
【請求項8】
前記自己免疫疾患がブドウ膜炎である、請求項7に記載の組成物。
【請求項9】
哺乳動物の粘膜細胞中で生体反応を調節するための組成物であって、
セッコク属に属する植物から調製した多糖および
前記粘膜の疾患の誘導に関連する抗原を含む組成物。
【請求項10】
哺乳動物の粘膜細胞中で生体反応を調節しうる組成物の使用であって、
前記哺乳動物に抗原および多糖を投与するステップを含み、
前記抗原が前記粘膜の疾患の誘導に関連し、前記多糖がセッコク属に属する植物から調製される使用。
【請求項11】
前記投与するステップが経口投与の方法で実行される、請求項10に記載の使用。
【請求項12】
哺乳動物の粘膜細胞中で生体反応を調節するための組成物であって、
セッコク属に属する植物から調製した多糖を含む組成物。
【請求項13】
哺乳動物の粘膜細胞中で免疫系および/または免疫反応を調節するための組成物であって、
セッコク属に属する植物から調製した多糖および
免疫系における自己免疫疾患の誘導および/または前記粘膜の疾患の誘導に関連する抗原を含む組成物。
【請求項14】
哺乳動物の自己免疫疾患および/または粘膜の疾患を治療するための組成物の使用であって、
前記哺乳動物に多糖を投与するステップを含み、
前記多糖がセッコク属に属する植物から調製される使用。
【請求項15】
前記投与するステップが経口投与の方法で実行される、請求項14に記載の使用。
【請求項16】
前記自己免疫疾患がブドウ膜炎である、請求項14に記載の使用。
【請求項17】
植物からの多糖を調製するための方法であって、
a)第1のアルコールで植物を抽出して第1の抽出物を得るステップ、
b)溶媒で第1の抽出物を抽出して第2の抽出物を得るステップ、および
c)第2のアルコールで第2の抽出物を沈殿させて多糖を得るステップ
を含む方法。
【請求項18】
植物がセッコク属に属する、請求項17に記載の方法。
【請求項19】
第1のアルコールが好ましくはメタノールである、請求項17に記載の方法。
【請求項20】
溶媒が好ましくは水である、請求項17に記載の方法。
【請求項21】
第2のアルコールが好ましくはエタノールである、請求項17に記載の方法。
【請求項22】
ステップb)において溶媒で第1の抽出物を抽出した後、遠心分離し、濾過して第2の抽出物を得るステップをさらに含む、請求項17に記載の方法。
【請求項1】
自己免疫疾患および粘膜障害を治療するための組成物であって、セッコク属に属する植物から調製した多糖および免疫系における自己免疫疾患の誘導に関連する抗原を含む組成物。
【請求項2】
前記自己免疫疾患がブドウ膜炎である、請求項1に記載の組成物。
【請求項3】
哺乳動物の免疫系を調節しうる組成物の使用であって、前記哺乳動物に抗原および多糖を投与するステップを含み、前記抗原が免疫系において自己免疫疾患の誘導に関連し、前記多糖がセッコク属に属する植物から調製される使用。
【請求項4】
前記投与するステップが経口投与の方法で実行される、請求項3に記載の使用。
【請求項5】
前記自己免疫疾患がブドウ膜炎である、請求項3に記載の使用。
【請求項6】
セッコク属に属する植物から調製した多糖を含む、哺乳動物の免疫系を調節するための組成物。
【請求項7】
組成物により、哺乳動物の自己免疫疾患の発生を低下させることができることを特徴とする、請求項6に記載の組成物。
【請求項8】
前記自己免疫疾患がブドウ膜炎である、請求項7に記載の組成物。
【請求項9】
哺乳動物の粘膜細胞中で生体反応を調節するための組成物であって、
セッコク属に属する植物から調製した多糖および
前記粘膜の疾患の誘導に関連する抗原を含む組成物。
【請求項10】
哺乳動物の粘膜細胞中で生体反応を調節しうる組成物の使用であって、
前記哺乳動物に抗原および多糖を投与するステップを含み、
前記抗原が前記粘膜の疾患の誘導に関連し、前記多糖がセッコク属に属する植物から調製される使用。
【請求項11】
前記投与するステップが経口投与の方法で実行される、請求項10に記載の使用。
【請求項12】
哺乳動物の粘膜細胞中で生体反応を調節するための組成物であって、
セッコク属に属する植物から調製した多糖を含む組成物。
【請求項13】
哺乳動物の粘膜細胞中で免疫系および/または免疫反応を調節するための組成物であって、
セッコク属に属する植物から調製した多糖および
免疫系における自己免疫疾患の誘導および/または前記粘膜の疾患の誘導に関連する抗原を含む組成物。
【請求項14】
哺乳動物の自己免疫疾患および/または粘膜の疾患を治療するための組成物の使用であって、
前記哺乳動物に多糖を投与するステップを含み、
前記多糖がセッコク属に属する植物から調製される使用。
【請求項15】
前記投与するステップが経口投与の方法で実行される、請求項14に記載の使用。
【請求項16】
前記自己免疫疾患がブドウ膜炎である、請求項14に記載の使用。
【請求項17】
植物からの多糖を調製するための方法であって、
a)第1のアルコールで植物を抽出して第1の抽出物を得るステップ、
b)溶媒で第1の抽出物を抽出して第2の抽出物を得るステップ、および
c)第2のアルコールで第2の抽出物を沈殿させて多糖を得るステップ
を含む方法。
【請求項18】
植物がセッコク属に属する、請求項17に記載の方法。
【請求項19】
第1のアルコールが好ましくはメタノールである、請求項17に記載の方法。
【請求項20】
溶媒が好ましくは水である、請求項17に記載の方法。
【請求項21】
第2のアルコールが好ましくはエタノールである、請求項17に記載の方法。
【請求項22】
ステップb)において溶媒で第1の抽出物を抽出した後、遠心分離し、濾過して第2の抽出物を得るステップをさらに含む、請求項17に記載の方法。
【図1】


【図2】

【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11A】
【図11B】
【図12A】
【図12B】
【図13A】
【図13B】
【図14A】
【図14B】
【図15A】
【図15B】
【図16A】
【図16B】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11A】
【図11B】
【図12A】
【図12B】
【図13A】
【図13B】
【図14A】
【図14B】
【図15A】
【図15B】
【図16A】
【図16B】
【公表番号】特表2008−540348(P2008−540348A)
【公表日】平成20年11月20日(2008.11.20)
【国際特許分類】
【出願番号】特願2008−509290(P2008−509290)
【出願日】平成18年4月30日(2006.4.30)
【国際出願番号】PCT/CN2006/000895
【国際公開番号】WO2006/116950
【国際公開日】平成18年11月9日(2006.11.9)
【出願人】(507366511)ナショナル ヤン−ミン ユニバーシティー (1)
【Fターム(参考)】
【公表日】平成20年11月20日(2008.11.20)
【国際特許分類】
【出願日】平成18年4月30日(2006.4.30)
【国際出願番号】PCT/CN2006/000895
【国際公開番号】WO2006/116950
【国際公開日】平成18年11月9日(2006.11.9)
【出願人】(507366511)ナショナル ヤン−ミン ユニバーシティー (1)
【Fターム(参考)】
[ Back to top ]