
組換えインフルエンザ菌アドヘシンタンパク質
【技術課題】組換えアドヘシン(rHia)を提供すること。
【解決手段】完全長でN末端が切断されている形の、インフルエンザ菌の分類不能型菌株のHiaタンパク質の組み換え生成物、分類不能型およびc型インフルエンザ菌のさまざまな菌株のHia遺伝子の核酸およびアミノ酸配列を提供する。
【解決手段】完全長でN末端が切断されている形の、インフルエンザ菌の分類不能型菌株のHiaタンパク質の組み換え生成物、分類不能型およびc型インフルエンザ菌のさまざまな菌株のHia遺伝子の核酸およびアミノ酸配列を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
(関連出願の参照)
本出願は、同時係続の米国特許出願第09/268,347号の一部継続出願である。
【0002】
(発明の分野)
本発明は分子遺伝学の分野に関し、詳細には組換えインフルエンザ菌(ヘモフィルス・インフルエンジー(Haemophilus influenzae))アドヘシン(Hia)タンパク質の産生に関する。
【背景技術】
【0003】
(発明の背景)
インフルエンザ菌は、髄膜炎、喉頭蓋炎、敗血症および中耳炎などのいくつかの重大なヒト疾患の原因である。インフルエンザ菌には、aからfで表され、それらの莢膜多糖によって識別される6種類の血清型がある。b型インフルエンザ菌(Hib)は、1980年代にいくつかのHib結合ワクチンが導入されるまでは細菌性髄膜炎の主な原因であった(ref.1.本出願全体にわたって、さまざまな参考文献を括弧内に引用し、本発明が関係する分野の現状をより完全に説明する。それぞれの引用に関する完全な文献情報は、明細書の最後、特許請求の範囲の直前に見出される。これらの参考文献の開示を、参照により本開示に組み込む)。ジフテリアトキソイド(ref.2)、破傷風トキソイド(ref.3および米国特許第4,496,538号)または髄膜炎菌外膜タンパク質(ref.4)に結合したb型インフルエンザ菌莢膜多糖をベースとするワクチンは、b型インフルエンザ菌誘発髄膜炎を減らすのに有効とされてきた。他の血清型インフルエンザ菌が侵襲性疾患に関連する頻度は低いが、Hib疾患の発生率が減少するにつれて、これらの菌株に起因する疾患の発生率が増加するようである(ref.5、6)。非被包性すなわち分類不能型インフルエンザ菌(NTHi)もまた中耳炎、喉頭蓋炎、肺炎および気管気管支炎を含む広範囲のヒト疾患の原因となる。NTHi誘発疾患の発生率は、Hibワクチンの導入によって影響を受けなかった(ref.7)。
【0004】
中耳炎は幼児期に最もよく見られる病気であり、2歳未満の子供全体の60から70%が1回から3回の耳感染症を経験する(ref.8)。子供の慢性中耳炎は聴力、発話および認知障害の原因となる。インフルエンザ菌感染症は、急性中耳炎の症例の約30%、慢性中耳炎の約60%を占める。米国だけでも中耳炎の治療には、抗生物質ならびに扁桃摘出術、咽頭扁桃切除術および中耳腔換気用チューブの挿入などの外科的処置に年間10億ドルから20億ドルが費やされている。言語療法および特殊教育授業などの付加的治療に対し、年間さらに300億ドルが費やされていると推定される。さらに、中耳炎の原因となる生物の多くは、抗生物質治療に対して耐性になりつつある。したがって、中耳炎に対する有効な予防ワクチンが望まれている。NTHiによる自然感染では、抗体の応答を刺激する露出した外膜タンパク質が殺菌および/または防御的な抗体として潜在的に重要な標的であり、したがって潜在的なワクチンの候補である。NTHiの上皮細胞への付着の際に重要である高分子量タンパク質(HMW1およびHMW2)のファミリーが、約70から75%のNTHi菌株において同定されている(ref.9、ref.10)。これらの高分子量アドヘシン(adhesin)は、中耳炎のチンチラモデルにおいていくらかの防御を可能にすることが分かっている(ref.11)。高分子量アドヘシンタンパク質の第2のファミリーが、約25%のNTHiおよび被包性インフルエンザ菌株において同定されている(ref.12、ref.13、ref.14)。この第2のファミリーのNTHiメンバーは、インフルエンザ菌アドヘシンすなわちHiaと呼ばれ、被包性菌株において発見される相同タンパク質はインフルエンザ菌表面原繊維タンパク質すなわちHsfと呼ばれる。hia遺伝子は、中耳炎患者の回復期血清を使用して発現ライブラリーから最初にクローニングされ、このことはhia遺伝子が疾患における重要な免疫原であることを示す。プロトタイプのHiaおよびHsfタンパク質は約82%の配列類似性を示すが、Hsfタンパク質はかなり高い。これらのタンパク質には保存されたアミノ酸およびカルボキシ末端およびいくつかの繰り返しモチーフが含まれ、HsfはHiaと比較してより多くの繰り返し配列を含む。高分子量タンパク質(200kDa)も、HsfおよびHiaタンパク質といくらかの配列相同性があるMoraxella catarrhalisから同定されている(米国特許第5,808,024号)。
【0005】
HiaおよびHsfはインフルエンザ菌の被包性菌株および約20〜25%の非被包性菌株中で保存され、アドヘシンであることが示されているので、HiaまたはHiaと特異的に反応する抗体を産生する能力のあるタンパク質を産生する、インフルエンザ菌または他の細菌性病原体によって引き起こされる疾患に対する診断および予防接種において、このタンパク質は有用性がある。
【0006】
診断において有用であり予防接種における免疫原としてのアンチHia抗体を産生するために、診断において抗原としてHiaを使用することの欠点は、インフルエンザ菌種からの天然タンパク質の回収があまりできないことである。
【0007】
他の免疫原用のワクチン、担体を含めた免疫調製、および診断試薬の生成において、抗原として使用するための組換えHiaタンパク質を提供することが有利であろう。
【発明の開示】
【発明が解決しようとする課題】
【0008】
(発明の概要)
本発明は組換えインフルエンザ菌アドヘシン(rHia)の提供を対象とする。
【課題を解決するための手段】
【0009】
このような組換えタンパク質の提供に関して、本発明は単離され精製されたある種の核酸分子を提供する。したがってその一態様において、本発明は単離され精製された核酸分子を提供し、この核酸分子はインフルエンザ菌株のインフルエンザ菌アドヘシン(Hia)タンパク質をエンコードしており、以下のものを含む。(a)図18、19、20、21、22、23、24および25に示される配列(配列番号23、25、27、29、31、33、35、37)からなる群から選択されるDNA配列、または(b)図18、19、20、21、22、23、24および25に示される配列(配列番号24、26、28、30、32、34、36、38)からなる群から選択されるアミノ酸配列を有するインフルエンザ菌アドヘシン(rHia)タンパク質をエンコードしているDNA配列。
【0010】
このような核酸は、プラスミドベクターである可能性のあるベクター中に含まれていてよい。詳細には、分類不能型ヘモフィルスの菌株11または33からのHiaタンパク質を、核酸分子はエンコードしていてよい。
【0011】
本発明の他の態様では、インフルエンザ菌の菌株のN末端切断型インフルエンザ菌アドヘシン(Hia)タンパク質をエンコードしており、1対のヌクレオチドによって増幅可能な単離され精製された核酸分子が提供される。ヌクレオチドは配列番号7と配列番号15、配列番号9と配列番号15、配列番号11と配列番号15、配列番号13、配列番号15、配列番号49、および配列番号51からなる群から選択される。
【0012】
このような核酸は、プラスミドベクターである可能性のあるベクター中に含まれていてよい。詳細には、分類不能型ヘモフィルスの菌株11または33からのコドンV38またはS44で始まるN末端切断型Hiaタンパク質を、核酸分子はエンコードしていてよい。
【0013】
本発明のこれらの態様に従って提供される単離され精製された核酸を組み込んでいるプラスミドベクターは、
図1A中に示されるDS-2008-2-3
図5A中に示されるDS-2186-1-1
図5A中に示されるDS-2201-1
図5A中に示されるDS-2186-2-1
図5A中に示されるDS-2168-2-6
図32中に示される1A-191-3-1
からなる群から選択されるプラスミドの同定特性を有する可能性がある。
【0014】
本明細書で提供されるベクターは、大腸菌からのcer遺伝子を含んでよい。したがって、本発明の他の態様では、完全長またはN末端切断型インフルエンザ菌アドヘシン(Hia)タンパク質をエンコードしている核酸分子、前記完全長または切断型Hiaタンパク質を発現するためのプロモーター、任意選択で大腸菌のcer遺伝子含む、宿主を形質転換するためのベクターが提供される。ベクターはプラスミドベクターまたは他の非複製ベクターであってよく、
図6A中に示されるBK-96-2-11
図7A中に示されるDS-2242-1
図7A中に示されるDS-2242-2
図8A中に示されるDS-2340-2-3
図9A中に示されるDS-2247-2
図9B中に示されるDS-2448-17
図32中に示されるJB-2930-3
からなる群から選択されるプラスミドベクターの同定特性を有する可能性がある。
【0015】
本明細書で提供されるベクターは、サルモネラ、BCG、アデノウイルス、ポックスウイルス、ワクシニアまたはポリオウイルスからのベクターを含めた複製ベクターを含んでよい。
【0016】
本明細書で提供される任意のベクターを使用して、完全長または切断型である可能性のある、ヘモフィルスの分類不能型菌株の防御性インフルエンザ菌アドヘシン(Hia)タンパク質を発現するのに適切な宿主細胞を形質転換することができる。このような宿主は、好都合には大腸菌であってよい。このような発現はT7プロモーターの制御下で行うことができ、形質転換された宿主からの組換えHiaの発現はラクトースまたは他の好都合な誘導剤の誘導用濃縮液中で培養することによって実施することができる。
【0017】
本発明は他の態様において、形質転換された宿主、特に本明細書で提供される大腸菌によって産生される、分類不能型ヘモフィルス菌株の組換え防御性インフルエンザ菌アドヘシン(Hia)タンパク質をさらに含む。このようなHiaタンパク質を組換えタンパク質の免疫原性断片またはアドヘシン官能性類縁体の形で提供することができる。
【0018】
完全長またはN末端切断型である本明細書で提供される組換えHiaタンパク質は、免疫原性組成物中の抗原、他の免疫原用の担体、診断用薬としておよび診断用薬の生成において有用である。完全長またはN末端切断型であるHiaタンパク質をエンコードしている核酸分子も、診断のための使用用、および免疫原性組成物中においてもプローブとして有用である。
【0019】
追加的な態様において本発明は、本明細書で提供されるのと同様な単離され精製された核酸分子、本明細書で提供されるのと同様なヘモフィルスの菌株の完全長またはN末端切断型である組換え防御性Hiaタンパク質からなる群から選択される少なくとも1つの免疫学的に活性のある成分、および薬学的に許容可能なこれらの担体を含む、免疫原性組成物を提供する。
【0020】
本明細書で提供される免疫原性組成物を、宿主へのin vitro投与用ワクチンとして製剤化して、インフルエンザ菌に起因する疾患に対する防御を提供することができる。このような目的のために、組成物をミクロ粒子、カプセル、ISCOMまたはリポソーム調製物として製剤化することができる。免疫原性組成物を免疫系の特定の細胞または粘膜表面への送達用の標的分子と組み合わせて提供することができる。
【0021】
本発明の免疫原性組成物(ワクチンを含めて)は少なくとも1種の他の免疫原性または免疫刺激性材料をさらに含んでよく、この免疫刺激性材料少なくとも1種のアジュバントまたは少なくとも1種のサイトカインであってよい。本発明で使用するのに適切なアジュバントには(これらに限らないが)リン酸アルミニウム、水酸化アルミニウム、QS21、Quil A、その誘導体および成分、ISCOMマトリックス、リン酸カルシウム、水酸化カルシウム、水酸化亜鉛、糖脂質類縁体、アミノ酸のオクタデシルエステル、ムラミルジペプチド、ポリホスファゼン、ISCOPREP、DC-chol、DDBAおよびリポタンパク質ならびに他のアジュバントがある。
【0022】
アジュバントの有利な組み合わせは、本出願人と同じ譲受人に譲渡され、その開示を参照(1995年11月21日発行のWO 95/34308)により本明細書に組み込む1994年6月16日出願の同時係続の米国特許第08/261,194号、および1995年6月7日出願の第08/483,856号に記載されている。
【0023】
本発明の他の態様に従って、感受性のある宿主に有効量の前述の免疫原性組成物を投与するステップを含む、宿主中で免疫応答を生み出すための方法が提供される。免疫応答は体液の免疫応答であるか、または細胞を介する免疫応答であってよい。その内部で疾患に対する防御を与えることができる宿主には、ヒトを含めた霊長類がある。
【0024】
本発明の他の態様に従って、薬剤として使用されるとき本明細書で提供される免疫原性組成物、および疫原性組成物の製造における免疫原性組成物のこれらの成分の使用法が提供される。
【0025】
他の追加的な態様において本発明は、インフルエンザ菌の分類不能型菌株の防御性インフルエンザ菌アドヘシン(Hia)タンパク質の産生のための方法を提供し、この方法は
本明細書で提供されるのと同様な菌株のN末端切断型のインフルエンザ菌アドヘシンタンパク質をエンコードしている核酸分子を含むベクターによって、大腸菌などの宿主を形質転換すること、
宿主を成長させてエンコードされている切断型Hiaを発現させること、
発現したHiaタンパク質を単離および精製することを含む。
【0026】
エンコードされている切断型Hiaを封入体中で発現させることができる。成長し形質転換された細胞を破壊して上澄みおよびHiaを含む封入体を生成し、上澄みからの分離の後に封入体を可溶化して組換えHiaの溶液を生成し、クロマトグラフィによって細胞の残骸のない組換えHiaの溶液を精製し、精製された組換えHiaタンパク質を単離することによって、単離および精製ステップを実施することができる。
【0027】
大腸菌などの宿主細胞を形質転換するベクターはT7プロモーターを含んでよく、大腸菌または他の宿主細胞を誘導量のラクトースまたは他の好都合な誘導剤の存在下で培養することができる。
【0028】
本明細書のインフルエンザ菌の菌株は、菌株11、33、32、29、M4071、K9、K22および12からなる分類不能型菌株の群から選択することができる。このような菌株からの代表的なHiaタンパク質をエンコードしている遺伝子に特異的な核酸配列が、本明細書で提供され以下に記載される。
【0029】
本明細書で提供される核酸配列は、診断に関する適用例において有用である。したがって本発明の他の態様では、サンプル中でインフルエンザ菌アドヘシンタンパク質をエンコードしている核酸の存在を判定するするための方法が提供され、この方法は、
a)サンプルを本明細書で提供されるのと同様な核酸分子と接触させて、本明細書で提供される核酸配列、およびサンプル中に存在し特異的にハイブリダイズすることができるヘモフィルスの菌株のHiaタンパク質をエンコードしている核酸を含むデュプレックス(duplex)を生成するステップと、
b)デュプレックスの生成を判定するステップを含む。
【0030】
さらに本発明は、サンプル中でインフルエンザ菌アドヘシンタンパク質をエンコードしている核酸の存在を判定するための診断キットを提供し、このキットは、
a)本明細書で提供されるのと同様な核酸分子と、
b)核酸分子とサンプルを接触させて、この核酸分子および任意のこのような核酸分子を含むデュプレックスを生成するための手段と、
c)デュプレックスの生成を判定するための手段をを含む。
【0031】
本明細書で提供される組換えによって産生された切断型Hiaタンパク質も、診断に関する適用例において有用である。したがって本発明の他の態様では、サンプル中のHiaタンパク質と特異的に反応する抗体の存在を判定するための方法が提供され、この方法は、
(a)サンプルと本明細書で提供される組換えHiaタンパク質を接触させて、この組換えHiaタンパク質とサンプル中に存在しタンパク質と特異的に反応する任意のこのような抗体の複合体を提供するステップと、
(b)複合体の生成を判定するステップを含む。
【0032】
本発明の利点には以下のものがある。
【0033】
インフルエンザ菌アドヘシンタンパク質またはこのHiaタンパク質の断片または類縁体をエンコードしている単離され精製された核酸分子、
他のいかなるヘモフィルスのタンパク質も含まない組換えによって産生されたHiaタンパク質、
ヘモフィルスの特異的な同定用の診断キットおよび免疫試薬。
【発明を実施するための最良の形態】
【0034】
本発明は、図面を参照しながら以下の記述からさらに理解されるはずである。
【0035】
(発明の全般的な説明)
H.influenzae株は、少量のHiaおよびHsfタンパク質を発現するので、NTHi株由来のhia遺伝子を、E.Coli中で組み換えタンパク質を過剰発現させるための発現ベクター中にクローン化した。全長組み換えHia(rHia)タンパク質が発現する際、それは比較的少量であった。組み換えタンパク質が発現されているのを確認するために、本出願と同じ譲受人に譲受され、その開示を参照により本明細書に組み込む米国特許第5,808,024号で200kDaであると同定されたMoraxella catarrhalis高分子量アドヘシンタンパク質に対する抗体を用いてイムノブロットを実施した。ゲル精製した未変性200kDaタンパク質に対する抗体は、rHiaタンパク質サンプル中に特異的に誘導されたバンドを認識した。rHiaの収量はT7hia遺伝子カセットの遺伝子コピー数を増加させることによっても顕著には増加しない。
【0036】
E.Coliのcer遺伝子は、大きな挿入部を含むプラスミドを安定化するが(参考文献15)、E.Coliのcer遺伝子を発現ベクターに添加しても、rHiaの収量は顕著には増加しない。しかし、E.Coli細胞は培養中に凝集し、このことは、Hiaアドヘシンタンパク質の表面発現が起きていることを示唆している。rHiaタンパク質が封入体として生成されればrHiaタンパク質の強い毒性は克服されるので、推定シグナル配列を欠失させるために遺伝子の5’-末端を切断した。この変性の結果、V38位置から出発している切断型rHiaの生産および回収が良好となった。
【0037】
全長およびV-38切断型rHiaタンパク質は免疫原性であり、得られる抗-rHia抗体は、H.influrnzaeのタイプaまたはタイプb株による菌血症の受動的幼ラットモデルを防護する。さらに、切断型V38rHiaタンパク質は、チンチラの能動的抗原投与モデルにおける鼻咽頭コロニー形成に対して特に防護に働くことが見出された。NTHi株からのrHiaによってもたらされる防護が、NTHiおよび被包性タイプaまたはタイプb株が原因となって起こる疾患に対するものであることは、一般的な防護エピトープが存在することを意味している。付加的なhia遺伝子のクローニングおよび配列分析は、保存領域の同定の助けとなる。全長またはN-末端切断型rHiaタンパク質を、Haemophilus influenzae疾患に対する防護のためのワクチン成分として使用できる。
【0038】
hia遺伝子を有する任意のHaemophilus株は、少なくとも本発明の態様で特徴付けるHiaタンパク質をコードする部分を含有する核酸分子(おそらくDNA分子の形)の精製および単離に利用できる。このような株は一般的な臨床所およびバクテリア培養収集所、例えばAmerican Type Culture Collectionから入手される。Haemophilus influenzaeの適切な株:
分類不能型 Haemophilus 株11;
分類不能型 Haemophilus 株33;
分類不能型 Haemophilus 株32;
分類不能型 Haemophilus 株29;
分類不能型 Haemophilus 株 M4071;
分類不能型 Haemophilus 株 K9;
分類不能型 Haemophilus 株 K22;
分類不能型 Haemophilus 株 12;
タイプC Haemophilus 株 API。
【0039】
本明細書において、“Hia”タンパク質という用語は、Hiaタンパク質のファミリーを定義するために使用され、Hiaタンパク質には種々のHaemophilus株中で見出されるアミノ酸配列を有する天然の変異体が含まれる。
【0040】
図1Aは、T7プロモーターの影響下での、分類不能型 Haemophilus influenzae株11由来全長hia遺伝子を含むプラスミドDS-2008-2-3の制限地図を示す図である。株11由来のhia遺伝子の核酸(配列番号:43)および推測アミノ酸配列(配列番号:44)は、前記米国特許第5,646,259に記載されている(該明細書中では“HA1”と定義される)。hia遺伝子を株11の遺伝子のATG開始コドンからPCR増幅するために使用したオリゴヌクレオチドを、図1Bに示す。
【0041】
図2は、抗-未変性Moraxella catarrhalis高分子量アドヘシン抗体によりrHia(11)タンパク質を確認するためのイムノブロットを示す図である。前記米国特許第5,808,024に記載される、M.catarrhalis高分子量アドヘシンタンパク質または200kDaタンパク質は、HiaおよびHsfタンパク質と、特にカルボキシ末端部に、何らかの配列相同性を有する(図28)。
【0042】
図3は、NTHi株11由来の全長hia遺伝子から成るT7hia遺伝子カセットのタンデムコピーを含むプラスミドDS-2092-1およびDS-2092-40の構築スキームを示す図である。増加した遺伝子コピー数を示すこのようなプラスミドは、しばしば組み換えタンパク質の生産レベルを上昇させる。しかし、以下に示すように、組み換えHiaの低い収率は、遺伝子コピー数の増加によって顕著には増加しない。
【0043】
図4は、NTHi株11タンパク質のN-末端配列および経時的にN-末端切断されたrHiaタンパク質の位置を示す図である。E21部分までのN-末端切断型は、Hiaのシグナル配列の構成部分であると考えられる長い疎水性領域が欠失している。T33部分までの欠失は、長い疎水性領域を含み、潜在的なAla-X-Alaシグナル切断部位を有する。V38部分までの欠失は、長い疎水性領域を含み、潜在的なAla-X-Alaシグナル切断部位を有する。S44部分から始まる組み換えHiaタンパク質は、長い疎水性領域を含み、潜在的なAla-X-Alaシグナル切断部位を有する。N52部分から始まる組み換えHiaタンパク質は、Moraxella catarrhalis由来の高分子量(200kDa)アドヘシンの近接した開始点を模倣しており、アドヘシンは米国特許第5,808,024に記載され、該組み換えタンパク質は、V56から開始するようにN-末端切断型されている場合に過剰発現する。
【0044】
図5Aは、4種のN―末端切断型rHiaを製造するための、プラスミドDS-2186-1-1、DS-2201-1、DS-2186-2-1、およびDS-2168-2-6を作製するための構築スキームを示す図である。5’-末端フラグメントをPCR増幅するために使用されるこのオリゴヌクレオチドを図5Bに示す。図30は、S44欠失を製造する、プラスミドJB2930-3を作製するための構築スキームを示す図である。5’-フラグメントをPCR増幅するために使用するオリゴヌクレオチドを図29に示す。
【0045】
図6Aは、NTHi株11由来のV38hia遺伝子ならびにプラスミドを安定化するE.Coli cer遺伝子を含むプラスミドBK-96-2-11を作製するための構築スキームを示す図である。毒性のタンパク質を産生するプラスミドへのcer遺伝子の導入により、タンパク質産生の強化が予測された。cer遺伝子の存在下で全長rHia産生E.Coli細胞の形態に変化が生じ、すなわち細胞が凝集した。このことは、凝集を起こす細胞表面のアドヘシンの発現が強化されたことを示唆する。発現プラスミドBK-96-2-11はさらに、遺伝子安定性を高めると考えられているT7V38hia遺伝子カセットの上流および下流に転写ターミネーターを含んでいる。マルチクローニング部位および転写ターミネーターを作製するために使用されるオリゴヌクレオチドを図6Bに示す。
【0046】
図7Aは、T7プロモーターの影響下に、分類不能型Haemophilus influenzae株33由来の全長hia遺伝子を含むプラスミドDS-2242-1およびDS-2242-2のための構築スキームを示す図である。発現プラスミドは、T7hia(33)遺伝子カセットの上流および下流にE.Coli cer遺伝子および形質転換ターミネーターをも含有する。DS-2242-1は、T7hia(33)遺伝子と同一の鎖上にコードされたターミネーターを有する。しかし、2つのプラスミドに由来するrHiaの発現には違いが見られなかった。NTHi株33の真性5’末端の構築に使用されるオリゴヌクレオチドを図7Bに示す。
【0047】
図8Aは、NTHi株33由来のV38Hia遺伝子ならびにE.Coli cer遺伝子を含むプラスミドDS-2340-2-3のための構築スキームを示す図である。T7V38hia遺伝子カセットの上流および下流の同一の鎖上に転写ターミネーターも存在する。NTHi株33hia遺伝子をV38コドンからPCR増幅するために使用されるオリゴヌクレオチドを図8Bに示す。
【0048】
図9は、T7V38hia(11)またはT7V38hia(33)遺伝子カセットのタンデムコピーをそれぞれ含有するプラスミドDS-2447-2およびDS-2448-17の構築を示す図である。
【0049】
図10、パネルAは、NTHi株11由来の全長または切断型hia遺伝子をコードするプラスミドからのrHiaタンパク質の産生を示す図である。全長rHia(11)タンパク質の産生は非常に低かった。E21およびT33切断型rHiaタンパク質の場合も発現は低かった。しかし、V38およびN52切断型rHiaタンパク質は顕著に改善された発現レベルを示す。図10、パネルBから明らかなように、V38rHia(11)の産生は、E.Coli cer遺伝子を発現プラスミドへ添加した場合に明らかに増加している。
【0050】
図11は、封入体として製造されるrHiaタンパク質の精製スキームを示す図である。細胞を、超音波破砕により溶解し、封入体を連続した抽出により精製した。封入体を、グアニジニウムクロライド中に溶解し、ポリエチレングリコール(PEG)を添加して不純物を沈殿させた。(NH4)2SO4の添加によりrHiaが沈殿し、粗rHiaをさらにゲル濾過により精製した。
【0051】
図12は、株11および33由来の精製されたV38rHiaタンパク質を示す図である。封入体はレーン3に示され、最終精製タンパク質はレーン4に示される。精製されたタンパク質の推定純度は、SDS-PAGEデンシトメトリーにより測定して、約90%を上回る。
【0052】
図13は、ここに記載されるようにして製造されたrHiaタンパク質の、グリ
セロールを添加した場合としない場合でかつ4℃で、およびグリセロールを添加
して−20℃で8週間貯蔵する際の安定性に関する、SDS-PAGE分析を示
す図である。タンパク質はいずれの条件でも安定である。
【0053】
図14は、CD-1マウス中の株11および33由来V38rHiaタンパク質の免疫原性を示す図である。0.3〜10μgの用量では、いずれのタンパク質でも、1ないし2回投与した後に強い免疫応答を示す。これらのレベルでは顕著な用量応答は認められない。同様の結果が、BALB/cマウス(図15A)およびモルモット(図15B)でも見られ、このことはrHiaが投与量あたり0.3μgであっても強い免疫原性を有することを示唆している。
【0054】
図16は、V38rHia(33)によってもたらされる、NTHi株33が引き起こすコロニー形成に対する防護を示す図である。Yang 等(参考文献20)が記載しているように、チンチラ鼻咽頭コロニー形成モデルは、この疾患の初期の段階での防護を評価するために開発された。このモデルは、最初に、hmw遺伝子を発現するNTHi株のために確立され、hia遺伝子を発現するNTHi株に適応させなければならなかった。プロトタイプhmw-発現株(NTHi12)における感染の確立には102〜108cfuを使用するが、NTHi33株の場合には5×108cfuが必要であり、このような高レベルであってもプロトタイプhia-発現株11では感染が確立されない。100μg量で免疫化コホートは明らかに部分的に防護されるが、50μg量では防護されない。疾患の初期の段階でのこのような防護は、ワクチン抗原としてのrHiaアドヘシンの利用性を示している。
【0055】
図17は、別のHaemophilus influenzae hia遺伝子をPCR増幅するために使用されるオリゴヌクレオチドを示す図である。配列は、HiaおよびHsfタンパク質の保存アミノ酸およびカルボキシ末端配列に基づいている。
【0056】
図18は、NTHi株33hia遺伝子の完全なヌクレオチド配列および推定アミノ酸配列を示す図である。図19は、NTHi32hia遺伝子の完全なヌクレオチド配列および推定アミノ酸配列を示す図である。図20は、NTHi株29hia遺伝子の完全なヌクレオチド配列および推定アミノ酸配列を示す図である。図21は、NTHi株 M4071hia遺伝子の完全なヌクレオチド配列および推定アミノ酸配列を示す図である。図22は、NTHi株 K9hia遺伝子の完全なヌクレオチド配列および推定アミノ酸配列を示す図である。図23は、NTHi株 K22hia遺伝子の完全なヌクレオチド配列および推定アミノ酸配列を示す図である。図24は、Haemophilus influenzae タイプc株APIhia遺伝子の完全なヌクレオチド配列および推定アミノ酸配列を示す図である。図25は、NTHi株12由来のhia座の完全なヌクレオチド配列および推定アミノ酸配列を示す図である。PCR増幅されたフラグメントは、hia遺伝子の3’-末端へ連結したHaemophilus influenzaeタイプd株RdゲノムのHI1733遺伝子に関連する遺伝子の3’-末端を含有する。HI1733タンパク質の上流ORFのアラインメントを図27に示す。
【0057】
図26は、前記米国特許5,646,259に記載されるような、NTHi株11由来のHia遺伝子の完全なヌクレオチド配列および推定アミノ酸配列を示す図である。
【0058】
図28は、Hsf、Hiaから推定されるタンパク質の配列のアラインメント、およびM.catarrhalis200kDaタンパク質の部分配列を示す図である。
【0059】
本発明の種々の実施態様を、ワクチン摂取、診療、Haemophilus感染の治療、免疫剤の製造の分野の応用に利用できることは、当業者に明らかである。このような使用について以下に述べるが、これに限定するものではない。
【0060】
ワクチンの製造および使用
ワクチンとして使用するのに好適な免疫原性組成物は、ここに記載されるような、分類不能型Haemophilus株の免疫原性組み換えHaemophilus influenzaeアドヘシン(rHia)タンパク質、その免疫原性類似体およびフラグメントおよび/または免疫原性ペプチドから製造できる。ワクチンは免疫応答を誘起し、これにより抗-rHia抗体およびオプソニン化または死菌化された抗体を含む抗体が産生される。
【0061】
ワクチンを含む免疫原性組成物は、液体溶液またはエマルジョンのような注入可能な形で製造できる。rHiaタンパク質、その免疫原性類似体およびフラグメントおよび/または免疫原性ペプチドを、製薬学的に認容性の添加剤と混合してよく、該添加剤はrHiaタンパク質、免疫原性フラグメント類似体または免疫原性ペプチドと混和性である。このような添加物には、水、生理食塩水、デキストロース、グリセロール、エタノールおよびそれらの組合せが含まれる。
【0062】
免疫原性組成物およびワクチンはさらに、補助物質、例えば湿潤剤または乳化剤、pH緩衝剤、またはワクチンの効果を増強するためのアジュバントを含有してよい。
【0063】
免疫原性組成物およびワクチンは、皮下注射または筋肉内注射のような非経口投与が可能である。また、本発明で製造される免疫原性組成物を処方し、粘膜表面で免疫応答を喚起する方法で輸送する。従って、免疫原性組成物は例えば鼻または口(胃)のような経路で粘膜表面に適用してよい。
【0064】
免疫原性組成物は免疫系の特異的な細胞または粘膜表面へ輸送するために、標的分子と組み合わせてよい。標的分子には、WO92/17167に記載されるような(Biotech Australia Pty. Ltd.)、ビタミンB12およびバクテリア毒素、および米国特許第5,194,254(Barber等)に記載されるようなモノクローナル抗体が含まれる。
【0065】
また、座剤および経口処方を含む別の投与形態が望ましい。座剤には、結合剤および担体が含まれていてよく、例えばポリアルカレングリコールまたはトリグリセリドである。経口処方には、一般的に使用される添加剤、例えば製薬学的な量のサッカリン、セルロースおよび炭酸マグネシウムが含まれてよい。これらの組成物は、溶剤、懸濁剤、錠剤、丸剤、カプセル剤、持続性製剤または粉剤の形で投与され、rHiaタンパク質、フラグメント類似体および/またはペプチドを約1〜95%含有する。
【0066】
ワクチンは、投与処方に適合する方法で、治療効果、防護および免疫原性として機能する量で投与される。投与量は治療対象、例えば個人の免疫系の抗体合成能力および必要であれば細胞-仲介免疫応答の発現能力により変化する。投与に必要とされる活性成分の適切な量は、専門医の評価に依存する。しかし、好適な投与範囲は当業者により容易に決定可能であり、rHia、その類似体およびフラグメントおよび/またはペプチドはマイクログラム程度でよい。初期投与およびブースター投与のための好適な治療法も効果的であるが、初期投与には連続投与も含まれる。ワクチンの投与量は、投与経路に依存し、宿主の大きさによっても変化する。
【0067】
分類不能型HaemophilusのrHiaタンパク質をコードする核酸分子を免疫化に直接使用してよく、その際、DNAを直接投与、例えば遺伝子的免疫化として注入するかまたは生ベクター、例えば核酸分子を含むサルモネラ、BCG、アデノウイルス、ポックスウイルス、ワクシニアまたはポリオウイルスを構築する。異種抗体を免疫系へ運ぶために使用される数種の生ベクターについては、例えばO,Hagan(1992)(参考文献16)に議論されている。遺伝子的免疫化のための、DNAの被検対象への直接的な注入方法は、例えばUlmer等、1993(参考文献17)に記載されている。
【0068】
抗原と一般的にリン酸バッファー生理食塩水中の0.05〜1.0%溶液として使用されるアジュバントとを同時に投与することにより、免疫原性は著しく向上する。アジュバントは抗原の免疫原性を強化するが、それ自体は免疫原性を必ずしも有さない。アジュバントは、抗原を投与部位の近くに局所的に維持することにより、抗原の免疫系の細胞へのゆっくりとした継続的な放出を助ける、蓄積効果を生み出す。アジュバントはまた、免疫系細胞を蓄積した抗原へと誘引し、そのような細胞を刺激して免疫応答を促す。
【0069】
免疫促進剤またはアジュバントは、宿主の免疫応答を向上させるために長年使用されてきており、例えばワクチンが挙げられる。内因性アジュバント、例えばリポポリサッカライドは、一般的にワクチンとして使用される死菌または弱毒化バクテリアの成分である。外因性アジュバントは、典型的に、抗原と非-共役結合した免疫調節物質であり、宿主の免疫応答を強化するために処方される。従って、アジュバントは非経口的に導入された抗体への免疫応答を強化するものである。これらのアジュバントのいくつかは毒性であり、不所望の副作用を起こすので、ヒトおよび多くの動物に使用するのは不適当である。実際、水酸化アルミニウムおよびリン酸アルミニウム(まとめて一般的にアルムと称される)のみが、ヒトおよび家畜用のワクチン中にアジュバントとして慣用される。アルムによるジフテリアおよび破傷風トキソイドに対する抗体応答の増強は、よく確立されている。
【0070】
外因性アジュバントの多くは抗原に対する潜在的な免疫応答を呼び起こす。これらには、前記した特異的アジュバントならびに膜タンパク質抗原と複合するサポニン(免疫刺激複合体)、鉱油を含む複合ポリマー、死んだ微生物および鉱油、フロイドの完全アジュバント、微生物の生成物、例えばムラミルジペプチド(MDP)およびリポポリサッカライド(LPS)ならびにリピドAおよびリポソームが含まれる。
【0071】
効果的に体液性免疫応答(HIR)および細胞-仲介免疫(CMI)を誘導するには、免疫原をアジュバント中に乳化させる。多くのアジュバントは毒性であり、肉芽腫、急性および慢性の炎症(フロイドの完全アジュバント、FCA)、細胞溶解(サポニンおよび複合ポリマー)および発熱、関節炎および前ブドウ膜炎(LPSおよびMDP)を引き起こす。FCAは優れたアジュバントであり研究に汎用されるが、その毒性のために、ヒトまたは家畜用ワクチン中に使用することを許可されていない。
【0072】
理想的なアジュバントの所望の特徴は:
(1)毒性の欠如;
(2)長期にわたる免疫応答刺激能力;
(3)製造の容易性および長期貯蔵における安定性;
(4)種々の経路で投与された抗原に対して、CMIおよびHIRの両方を誘起する能力;所望であれば、
(5)他のアジュバントとの相乗作用;
(6)抗原提示細胞(APC)群と選択的に相互作用する能力;
(7)適切なTH1またはTH2細胞-特異的免疫応答を特異的に誘起する能力;
および
(8)抗原に対する適切な抗体イソタイプレベル(例えば、IgA)を選択的に増加させる能力である。
【0073】
参照することでここに組み込んだものとする、Lockhoff等の1989年8月8日付けの米国特許第4,855,283には、免疫調節剤またはアジュバントとして、N-グリコシルアミド、N-グリコシルウレアおよびN-グリコシルカルバメートを含む糖脂質類似体が記載され、それぞれ糖残基をアミノ酸で置換されていてよい。また、Lockhoff等、1991(参考文献18)は、天然の糖脂質と構造的類似性を有するN-糖脂質類似体、例えばグリコスフィンゴリピドおよびグリコグリセロリピドが、ヘルペス単純ウイルスワクチンおよび偽性狂犬病ウイルスワクチンの両方で強い免疫応答を誘起することを報告している。ある種の糖脂質は長鎖-アルキルアミンと脂肪酸とから合成され、芳香族炭素を介して糖と直接結合し、天然の脂質残基の機能を模倣する。
【0074】
本出願と同じ譲受人に譲渡され、参照により本明細書に組み込む、Molony等による米国特許第4,258,029には、破傷風トキソイドおよびホルマリン不活化タイプI、IIおよびIIIポリオウイルスワクチンと複合させた場合に、オクタデシルチロシンヒドロクロライド(OTH)がアジュバントとして機能することが記載されている。またNixon-George等、1990(参考文献19)は、組み換えB型肝炎表面抗原と組み合わせた、芳香族アミノ酸のオクタデシルエステルが、B型肝炎ウイルスに対する宿主の免疫応答を強化することを報告している。
【0075】
イムノアッセイ
本発明により製造されるHaemophilusの分類不能型株のrHiaタンパク質、その類似体およびフラグメントは、免疫原性として、酵素結合性免疫吸着剤検定(ELISA)、RIAおよび他の非-酵素結合性抗体結合アッセイまたは抗菌、Haemophilus、および/またはHia抗体を検出するための従来公知の方法を含むイムノアッセイ中の抗原として、有用である。ELISAアッセイでは、Hiaタンパク質、類似体およびフラグメントを、選択された表面、例えばポリスチレンマイクロタイタープレートの壁面のようなタンパク質またはペプチドが結合可能な表面上へ固定する。不完全に吸着されているHiaタンパク質、類似体および/またはフラグメントを除くために洗浄し、その後、非特異的タンパク質、例えば試験サンプルに対する抗原性が中性であることを知られているウシ血清アルブミン(BSA)またはカゼイン溶液を選択表面へ結合させてよい。これにより、固定表面上の非特異的吸着部位が遮断され、結果的に、表面上への抗血清の非特異的結合によって起こるバックグラウンドが減少する。
【0076】
次いで固定表面を、臨床または生化学的材料のようなサンプルと接触させ、免疫複合体(抗原/抗体)の形成が起こる方法で試験する。この際、BSA、ウシガンマグロブリン(BGG)および/またはリン酸バッファー生理食塩水(PBS)/Tweenのような希釈剤でサンプルを希釈してよい。サンプルを次に約25℃〜37℃程度の温度で約2〜約4時間インキュベートする。インキュベートに続き、サンプル-接触表面を洗浄して、非-免疫複合材料を除去する。洗浄方法には、PBS/Tween、またはホウ酸バッファーのような溶液による洗浄が含まれてよい。
【0077】
試験サンプルと結合Hiaタンパク質、類似体および/またはフラグメントとの間に特異的な免疫複合体が形成された後、次いで洗浄し、複合体形成の出現およびその量を、一次抗体に特異的な二次抗体を免疫複合体へ添加することにより測定する。試験サンプルがヒト起源である場合、二次抗体はヒトイムノグロブリンに特異的な抗体であり、一般的にIgGである。検出可能にするために、二次抗体は、例えば適当な発色性物質とインキュベートすることにより呈色が見られるような酵素活性等の関連活性を有する。定量化は、例えば可視スペクトル分光光度計を用いて呈色の度合いを測定することにより実施される。
【0078】
ハイブリダイゼーションプローブとしての配列の使用
本発明のヌクレオチド配列は、新規に単離および特徴付けされたhia遺伝子配列を含有し、他のHaemophilus分類不能型株から、hia遺伝子を同定およびクローニングすることができる。
【0079】
本発明のhia遺伝子配列を含有するヌクレオチド配列は、その能力のために、他のhia遺伝子の補足的部分と倍体分子を選択的に形成するのが有用である。適用に応じて、種々のハイブリダイゼーション条件を利用し、他の分類不能型Haemophilus株中の別のhia遺伝子のための、プローブの選択性の度合いを変化させる。高い選択性のためには、比較的厳しい条件を利用して倍体を形成し、例えば低塩および/または高温条件、例えばNaCl 0.02M〜0.15Mでありかつ約50〜70℃の温度という条件が挙げられる。ある種の適用では、それほど厳しいハイブリダイゼーション条件は必要とされず、例えば0.15〜0.9Mの塩で20〜55℃の温度範囲でよい。ハイブリダイゼーション条件は、ホルムアミドの添加量が増加すると、ハイブリッドした倍体が不安定になるので、より厳しい条件が必要とされる。従って、特定のハイブリダイゼーション条件は容易に操作でき、所望の結果に応じて方法を選択するのが一般的である。一般的に、便利なハイブリダイゼーション温度は、50%ホルムアミドの存在下かつNaCl 0.15Mで:hia遺伝子が標的核酸フラグメントと95〜100%の相同性を有する場合に42℃、90〜95%の相同性で37℃および85〜90%の相同性で32℃である。
【0080】
臨床診断の態様において、本発明のhia遺伝子の核酸配列は、適当な手段、例えばハイブリダイゼーションを測定するための標識、と組み合わせて利用してよい。種々の適当な指示手段が知られており、放射活性、酵素または他のリガンド、例えばアビジン/ビオチンが含まれ、検出可能なシグナルの提供を可能にする。ある診断の態様において、放射活性タグの代わりに、酵素タグ、例えばウレアーゼ、アルカリンホスファターゼまたはペルオキシダーゼを使用してもよい。酵素タグの場合、肉眼または分光光度計で視覚的に捉えられ、Hia遺伝子配列を含むサンプルとの特異的ハイブリダイゼーションを同定できる手段として使用されるものとして、発色指示物質が公知である。
【0081】
本発明のHia遺伝子核酸配列は、溶液ハイブリダイゼーション中のハイブリダイゼーションプローブとして、および液相法で使用される態様として有用である。固相法に関する態様において、滲出液、体液(例えば血清、羊水、中耳滲出液、唾液、気管支肺胞洗浄液)または組織のような臨床サンプルを含むサンプルの試験DNA(またはRNA)を、選択したマトリクスまたは表面へ吸着あるいは固定させる。固定された一本鎖核酸を、次いで、所望の条件下に、本発明のhia遺伝子またはそのフラグメントの核酸配列を含む選択プローブと特異的にハイブリダイゼーションさせる。選択する条件は、例えばG+C含量、標的核酸のタイプ、核酸源、ハイブリダイゼーションプローブの大きさ等に応じて必要とされる特定の基準に基づく特定の状況に応じて変化する。続いてハイブリダイゼーション表面を洗浄して非特異的に結合したプローブ分子を除去し、標識の手段により、特異的ハイブリダイゼーションを検出または定量化する。Haemophilus種で保存されている核酸配列部分を選択するのが好ましい。選択されたプローブは、少なくとも長さ18bpであり、多くの場合30〜90bp長さである。
【0082】
Haemophilus influenzaeアドヘシン遺伝子の発現
宿主細胞に適合性の種に由来するレプリコンおよび制御配列を、発現系中のhia遺伝子の発現に利用してよい。ベクターは、通常、複製部位ならびに形質転換細胞中の表現型の選択を行うマーカー配列を有する。例えば、E.Coliは、アンピシリンおよびテトラサイクリン耐性遺伝子を含むpBR322を使用して形質転換でき、従って、形質転換細胞を同定するための簡易な方法を提供する。pBR322プラスミド、または他の微生物プラスミドまたはファージは、宿主細胞が自身のタンパク質を発現させるために使用できるプロモーターも含有するか、または含有するように変性する。
【0083】
さらに、宿主細胞に適合性のレプリコンおよび制御配列を含有するファージベクターは、これらの宿主と一緒に形質転換ベクターとして使用できる。例えば、ラムダGEMTM-11中のファージを、E.ColiLE392のような宿主細胞を形質転換させるために使用できる組み換えファージベクターを製造するために使用してよい。
【0084】
組み換えDNA構築体中で一般的に使用されるプロモーターには、β-ラクタマーゼ(ペニシリナーゼ)およびラクトースプロモーターシステムおよび他の微生物プロモーター、例えばここで好ましい態様として使用されるT7プロモーターシステム(米国特許第4,952,496)が含まれる。プロモーターのヌクレオチド配列の詳細は公知であり、当業者にはそれらを遺伝子を用いて機能的に結合することが可能である。使用される特定のプロモーターは、一般的に、所望の結果に応じて選択されたプロモーターである。Hiaタンパク質および免疫学的フラグメントまたはその類似体の発現に適当な宿主には、E.Coliが含まれ、Bordetella種、Bacillus種、Haemophilusu種、菌体、酵母またはバクロウイルスシステムを使用してもよい。E.Coliがここで使用される好ましい宿主である。
【0085】
本発明において、特にHaemophilus種を培養して精製した天然Hiaタンパク質が微量の毒性物質または他の汚染物質を含む場合に、Hiaタンパク質の製造は、組み換え法によるのが好ましい。この問題点は、組み換えにより製造されたHiaタンパク質を異種システム中で使用することにより解決され、この際、Hiaタンパク質は、特に記載された構築体を使用して、精製物質中の汚染物質を最小化する方法で宿主から単離される。
【0086】
微生物寄託
ここに記載し参照した分類不能型Haemophilus株の高分子量タンパク質をコードする核酸を含むベクターを、本明細書の出願に先立って、ブタペスト条約に従い、10801 University Boulevard、Manassas、Virfinia 20110-2209、米国に所在するAmerica Type Culture Collection(ATCC)に寄託した。寄託されたベクターのサンプルは、公共に利用可能であり、課される全ての制限または寄託物を利用する権利は、この米国特許出願に基づく特許の許可を受けることとする。さらに、受託者が生存可能なサンプルを分配できない場合には、寄託物を受託者に戻すこととする。寄託された実施形態は本発明を例示するためだけのものであるので、本明細書に記載および請求された発明が、寄託された生物材料によって範囲を限定されることはない。本出願の記載と等価または同等な抗原をコードする核酸を含むいずれの等価または類似のベクターも本発明の範疇に入るものとする。
寄託概要
プラスミド ATCC 寄託年月日
BK-96-2-11 203771 1999年2月11日
【実施例】
【0087】
上記の開示は概して本発明を説明している。以下の特定した実施例を参照するとさらに完全な理解が得られる。これらの実施例はただ例示の目的で説明されるものであり、本発明の範囲を限定しようとするものではない。状況によって形状の変化および同等品への置き換えを図ることで好都合なことを示唆できるかまたは提供できる。ここでは特殊な用語を使用したが、そのような用語は説明的感覚で使用するつもりであって限定を目的とするものではない。
【0088】
本開示およびこれらの実施例には明記していないが、使用した分子遺伝学、蛋白質生化学、免疫学および発酵技術による方法は科学文献で広く報告されており、当業者の能力に充分に保有されたものである。
【0089】
実施例1
この実施例はNTHi菌株11由来の全長のそろったrHia蛋白質を発現するプラスミドDS-2008-2-3の構造を説明するものである。
【0090】
NTHi菌株11から染色体DNAを精製し、図1Bに記載したオリゴヌクレオチド(5038.SLおよび5039.SL)を使用して全長のそろったhia遺伝子をPCR増幅した。pT7-7発現ベクター(ref.21)にクローン化するためにNde I部位をこの遺伝子の5’末端で設計し、BamH I部位を3’末端で設計した。増幅した断片をNde I/BamH Iで切断し、同じ酵素で切断したpT7-7にクローン化した。プラスミドDS-2008-2-3はT7プロモーターの下流に3.4kbの11株hia遺伝子を含む(図1A)。このプラスミドは組換えHiaを発現するために使用した(下記の実施例9)。
【0091】
実施例2
この実施例は抗Moraxella catarrhalis野生株高分子量アドヘシン抗体によるrHiaの識別を例示するものである。
【0092】
インフルエンザ菌のHia蛋白質と前述した米国特許第5,808,024号の中でM.catarrhalis200kDa蛋白質として分類されるMoraxella catarrhalis高分子量アドヘシン(図28)との間にはいくらかの配列不変性が認められる。米国特許第5,808,024号の記載では、M.catarrhalis野生株200kDa蛋白質がゲル精製され、テンジクネズミの抗野生株200kDa抗体が生成された。T7 hia遺伝子はプラスミドDS-2008-2-3から発現され、rHia蛋白質を含む細胞培養物がニトロセルロース膜に電気泳動的に転写された。抗野生株200kDa抗体を使用した免疫ブロット分析から、この抗体が図2に見られるように、rHia蛋白質を認識することが示された。
【0093】
実施例3
この実施例はT7 hia(11)遺伝子カセットのタンデムコピーを含むプラスミドDS-2092-1およびDS-2092-40の構造を説明するものである。
【0094】
全長のそろった組換えHia蛋白質の生成を向上させるために、菌株11のhia遺伝子(実施例1)を含むT7 hia遺伝子カセットのタンデムコピーを単一のベクターに挿入した。プラスミドDS-2008-2-3をBgl IIで直線化させ、脱リン酸化した。プラスミドDS-2008-2-3はまた、T7 hia遺伝子カセットを切り取るためにBgl IIおよびBamH Iによっても切断された。このT7 hia断片を直線化したベクターに連結し、反時計回り方向に配向して2つのT7 hia遺伝子コピーを含むプラスミドDS-2092-1(a,a)および反対方向に配向してタンデムコピーを含むプラスミドDS-2092-40(a,c)(図3)を生成した。どちらの作製試料からもrHia発現における顕著な向上は認められなかった(下記の実施例9参照)。
【0095】
実施例4
この実施例は末端欠失した菌株11のhia遺伝子を発現するプラスミドの作製を説明するものである。
【0096】
T7 hia遺伝子カセットの単一またはタンデムのコピーからのrHia蛋白質生成量は非常に低く、この蛋白質は(下記の実施例9で説明するように)E.coliに対する毒性を有するようであった。インフルエンザ菌のHiaは表面露出したアドヘシン分子であるので、分泌をするためにはシグナル配列または補助蛋白質のいずれかを使用しなければならないが、しかし関連の補助遺伝子は判明していない。もしもE.coli中での組換え蛋白質発現のためにシグナル配列が除去された場合、rHiaは封入体として発現され、毒性効果が低減される可能性がある。推定されるシグナル配列および切断部位を分類し、N末端で末端欠失したrHia蛋白質を発現する4種類の作製試料を設計した(図4)。菌株11のhia遺伝子では開始コドンから約500bpに独特のSty I切断部位が存在する。プラスミドDS-2008-2-3をNde IおよびSty Iで切断し、5.7kbのベクター断片を精製した(図5A)。末端欠失部位からSty I切断部位までを増幅するためにPCRプライマーを設計し、末端欠失したクローンを選別するためにアンチセンスプライマーに独特のNhe I切断部位を導入した(図5B)。増幅した断片は、より操作を容易にするためにpCRII中でサブクローン化し、プラスミドDS-2153R-1-2(E21)、DS-2165-4-8(T33)、DS-2153-3-5(V38)、およびDS-2153-4-4(N52)を生成した。このpCRII hiaプラスミドをNde IおよびSty Iで切断し、断片をDS-2008-2-3から得たベクター片と連結した。プラスミドDS-2186-1-1(E21)、DS-2201-1(T33)、DS-2186-2-1(V38)、およびDS-2168-2-6(N52)を、カッコ内に示したT7プロモーターおよび末端欠失したhia遺伝子を含ませて生成した。これらのプラスミドは組換えHiaを発現させるために使用した(下記の実施例9参照)。
【0097】
実施例5
この実施例はT7 V38 hia(11)カセット、E.coli cer遺伝子、およびカナマイシン耐性遺伝子を含むプラスミドBK-96-2-11の作製を説明するものである。
【0098】
プラスミドDS-1843-2は多数のクローン化部位および2つの転写終結部がEcoR IおよびPst I部位の間でオリゴヌクレオチド上に導入され、これによってクロラムフェニコールおよびアンピシリン耐性遺伝子の両方が破壊されたpBR328を基にしたプラスミドである(図6B)。pUC-4Kから得たカナマイシン耐性遺伝子をSal I部位に挿入し、カナマイシン耐性でかつテトラサイクリン感受性のプラスミドDS-2147-1を生成した。プラスミドDS-2224-1-4はオリゴヌクレオチドから作製してBamH I切断部位に接する合成のE.coli cer遺伝子(ref.15)を含むpUCプラスミドである。このcer遺伝子の290bpのBamH I断片をDS-2147-1のBamH I部位に挿入してプラスミドBK-2-1-2を作製した。その結果、このpBRを基にしたプラスミドは多数のクローン化部位、カナマイシン耐性遺伝子およびcer遺伝子を含む。プラスミドBK-2-1-2はBgl IIで直線化し、脱リン酸化した。プラスミドDS-2186-2-1をBgl IIおよびBamH Iで切断し、3.6kbのT7 V38 hia断片をBK-2-1-2に挿入してプラスミドBK-96-2-11を作製した(図6A)。
【0099】
実施例6
この実施例はE.coli cer遺伝子の存在下で全長のそろったNTHi菌株33のhia遺伝子を発現するプラスミドDS-2242-1およびDS-2242-2の作製を説明するものである。
【0100】
NTHi菌株33から染色体DNAを精製し、オリゴヌクレオチド5039.SLおよび5040.SL(図17)を使用してPCR増幅を実施した。センスプライマー(5040.SL)は菌株11のhiaの5’隣接配列を基にして設計したものであって、NTHi HiaおよびHib Hsf蛋白質のアミノ末端配列を保存していた。アンチセンスプライマー(5039.SL)は実施例1で述べたものと同じであって、HiaおよびHsf蛋白質のカルボキシ末端配列の保存を基本とした。菌株33の3kbのhia PCR断片をpCR II中でクローン化し、プラスミドDS-1917-3-8を作成した。
【0101】
全長のそろった菌株33 hia遺伝子を発現させるために、この遺伝子の5’末端の、開始コドンからAlwN I切断部位まで(図7B)の、およそ106bpをオリゴヌクレオチドから合成した。プラスミドDS-1917-3-8をAlwN IおよびBamH Iで切断し、hia遺伝子を含む約2.9kbの断片を精製した。プラスミドpT7-7をNde IおよびBamH Iで切断し、Nde I-AlwN IオリゴヌクレオチドおよびAlwN I-BamH I hia断片をこのpT7-7ベクターに連結してプラスミドDS-2103-4を作製した。
【0102】
E.coli cer遺伝子を含ませ、カナマイシンによる選択を使用するために、T7 hia(33)遺伝子カセットを含むBgl II-BamH I断片をDS-2103-4から切り取り、Bgl IIで切断されて脱リン酸化されたBK-2-1-1中でクローン化した。プラスミドDS-2242-1およびDS-2242-2は反対方向に配向したT7 hia(33)遺伝子カセットの単一コピー、E.coli cer遺伝子、およびカナマイシン耐性遺伝子を含む(図7A)。
【0103】
実施例7
この実施例は末端欠失したV38菌株33のhia遺伝子をもつT7 hia遺伝子カセット、E.coli cer遺伝子、およびカナマイシン耐性遺伝子を含むプラスミドDS-2340-2-3の作製を説明するものである。
【0104】
NTHi菌株33 hia遺伝子の、V38開始コドンから内側のSnaB I切断部位に至るまでの、5’末端の250bp断片を増幅するためにPCRプライマーを設計した。クローン化を目的として5’末端にNde I切断部位を追加し、鋳型としてプラスミドDS-2242-1を使用してこの断片を増幅した。この構成図式は図8Aに示されており、PCRプライマーは図8Bに示されている。この断片をpCR IIにクローン化し、プラスミドDS-2328-1-1を作製した。DS-2242-1をNde IおよびSnaB Iで切断し、8.5kbのベクター断片を精製した。DS-2328-1-1をNde IおよびSnaB Iで切断し、その0.25kbの5’hia断片をDS-2242-1から得た8.5kbのベクター断片と連結してプラスミドDS-2340-2-3を作製した。
【0105】
実施例8
この実施例はT7 V38 hia(11)またはT7 V38 hia(33)遺伝子カセットのタンデムコピーをそれぞれと、E.coli cer遺伝子、およびカナマイシン耐性遺伝子を含むプラスミドDS-2447-2およびDS-2448-17の作製を説明するものである。
【0106】
T7 V38 hia(11)遺伝子カセットを含むプラスミドBK-96-2-11をBgl IIで直線化し、脱リン酸化した。DS-2186-2-1から得たBgl II-BamH I T7 V38 hia(11)遺伝子カセットをBK-96-2-11に連結し、同じ配向でT7 V38 hia(11)遺伝子のタンデムコピーを含むプラスミドDS-2447-2を作製した(図9A)。
【0107】
プラスミドDS-2340-2-3をEcoR Iで切断し、そのT7 V38 hia(33)遺伝子カセットを、EcoR Iで切断して脱リン酸化したpUC-BgXbにサブクローン化した。その結果得られたプラスミドDS-2440-2をBgl IIおよびBamH Iで切断してT7 V38 hia(33)カセットを放出させ、Bgl IIで直線化して脱リン酸化したDS-2340-2-3とそれを連結した。プラスミドDS-2448-17は同じ配向でタンデムのT7 V38 hia(33)遺伝子を含む(図9B)。
【0108】
実施例9
この実施例は全長のそろったおよび末端欠失した組換えhia遺伝子の発現を説明するものである。
【0109】
先行の実施例で説明したようにして調製した発現プラスミドのDNAを、BioRad社のエレクトロポレータを使用して電子的に受容可能となったE.coli BL21(DE3)細胞に導入した。細胞は適切な抗生物質選別を使用して、ラクトースを1.0%になるまで加える前に、A578が0.3となるまでNZCYM培地中で37℃にて4時間成育させた。サンプルはSDS-PAGE分解液+負荷バッファで0.2OD/μlとなるよう調整し、それぞれの蛋白質サンプルを同じ量でSDS-PAGEゲル(ref.22)に流した。図10は種々の作製試料からのrHia(11)蛋白質相対生成量を図解したものである。パネルAに見られるように、rHiaのサイズが低下するにつれて生成量の増加が認められる。V38-(レーン5)およびN52-末端欠失rHia(レーン6)はそれらのさらに長い相当物質(レーン2、3、4)よりも有意差をもって高い発現レベルを有する。加えて、パネルBはV38 rHiaの生成がcer遺伝子の存在下で明らかに増加することを示している。
【0110】
実施例10
この実施例はrHia蛋白質の精製について説明するものである。
【0111】
すべての組換えHia蛋白質はE.coli中で封入体として発現され、同じ手法によって精製された(図11)。500mlの培養物から得たE.coli細胞ペレットを0.1MのNaClを含むpH8.0の50mMトリス塩酸50ml中に再懸濁し、超音波破砕した。この抽出物を20,000×gで30分間遠心分離し、その結果生じた上清を廃棄した。このペレット(PPT1)をさらに0.5%Triton X-100および10mM EDTAを含む50mlの50mMトリス塩酸、pH8.0で抽出処理し、それから20,000×gで30分間遠心分離して上清を廃棄した。このペレット(PPT2)をさらに1%のオクチルグルコシドを含む50mlの50mMトリス塩酸、pH8.0で抽出し、それから20,000×gで30分間遠心分離して上清を廃棄した。
【0112】
以上の抽出処理後に結果として得られたペレット(PPT3)は封入体を含んでいる。このペレットを6Mのグアニジンおよび5mMのDTTを含む6mlの50mMトリス塩酸、pH8.0に溶解した。この溶液に12mlの50mMトリス塩酸、pH8.0を加え、混合液を20,000×gで30分間遠心分離した。この上清(SUP4)を終濃度7%のポリエチレングリコール(PEG)4000で沈降させた。その結果生じたペレット(PPT5)を20,000×gで30分間遠心分離して除去し、上清を50%飽和の硫酸アンモニウムで沈降させた。硫酸アンモニウムによる沈降物は20,000×gで30分間遠心分離して採取した。結果として得られたペレット(PPT6)を6Mのグアニジン塩酸および5mMのDTTを含む2mlの50mMトリス塩酸、pH8.0に溶解し、この透明な液体を、2Mのグアニジン塩酸を含む50mMトリス塩酸、pH8.0で平衡化したSuperdex200ゲル濾過カラムで精製した。画分はSDS-PAGEによって分析し、精製されたrHiaを含む画分をプールし、4℃でPBSに対して一晩透析し、その後、20,000×gで30分間遠心分離した。これらの条件では蛋白質は可溶性を維持しており、−20℃で保存するためにこのrHia調製液にグリセロールを終濃度20%で加えた。精製されたV38 rHia(11)およびV38 rHia(33)のSDS-PAGE分析は図12に図解されている。精製V38 rHia蛋白質の平均収率は約10mg/L培地である。
【0113】
rHiaの安定性を調べるために、精製したV38 rHia(11)蛋白質は4℃でグリセロールなしかまたは入れて保存するか、あるいはグリセロールを入れて-20℃で保存した。この蛋白質は3つの条件すべてにおいて安定であることが分かり、凍結と解凍を繰り返して少なくとも8週間は完全さを維持した(図13)。
【0114】
実施例11
この実施例はV38 rHia(11)およびV38 rHia(33)蛋白質の免疫原性を説明するものである。
【0115】
日程1において完全フロイントアジュバント(CFA、Difco)で乳化した5μgの抗原を筋肉内(i.m.)注入して二匹のテンジクネズミ(Charles River)を免疫感作させることによってrHia蛋白質に対する高度免疫抗血清を生成させた。日程14および28において不完全フロイントアジュバント(IFA)に加えた蛋白質5μgを投与して動物を追加免疫応答させ、日程42において血清を集めた。免疫原として熱で不活化したバクテリア調製物(1回の投与量1×108cfu)を使用した以外は同じプロトコルを使用して、抗Hib菌株MinnAおよび抗ヘモフィルス属菌株ATCC9006抗血清を作製した。
【0116】
V38 rHia蛋白質の免疫原性を調べるために、5匹のCD-1マウス(Charles River,Quebec)の複数グループを日程1および28において、リン酸アルミニウム(alum)の存在下(1回の投与量1.5mg)で0.3、1、3、および10μgの抗原によりs.c.免疫感作させた。血液サンプルを日程1、28および42において集めた。顕著に抗V38 rHia抗体を生成したマウスは抗体0.3μgを1回注入されても応答を示し(図14、パネルA)、両方の蛋白質ともに封入体抽出および可溶化処理の後でも免疫原性を維持することを示唆している。菌株11または33に由来するV38 rHia蛋白質によって誘導される抗体力価において統計学的に顕著な差異は認められなかった。
【0117】
BALB/cマウスにおいてV38 rHia(11)蛋白質の免疫原性を調べるために、5匹の動物(Charles River,Quebec)の複数グループを日程1、28および42において、リン酸アルミニウムの存在下(1回の投与量1.5mg)で抗原0.3、1、3、および10μgによりs.c.免疫感作させた。血液サンプルは日程1、14、28、42および56において集めた。すべてのグループにおいて高い抗体力価が認められ、この蛋白質が1回の投与量0.3μgにおいてさえ強度の免疫原性であることを示した(図15、パネルA)。
【0118】
テンジクネズミにおいてV38 rHia(11)蛋白質の免疫原性を調べるために、5匹の動物(Charles River,Quebec)の複数グループを日程1、28および42において、リン酸アルミニウムの存在下(1回の投与量1.5mg)で抗原0.3、1、3、および10μgによりs.c.免疫感作させた。血液サンプルは日程1、14、28、42および56において集めた。すべてのグループにおいて高い抗体力価が認められ、この蛋白質がテンジクネズミにおいてもまた強度の免疫原性であることを示した(図15、パネルB)。
【0119】
実施例12
この実施例は菌血症の無抵抗な幼齢ラットのモデルにおいて抗rHia抗体によって生じる防御の分析を説明するものである。
【0120】
妊娠中のWistarラットをCharles Riverから入手した。インフルエンザ菌b型の菌血症モデルでは、6〜10匹の5日齢の幼齢ラットのグループを、テンジクネズミの抗rHiaまたは抗菌株MinnA抗血清0.1ml、背面 でs.c.注入した。比較対照の動物には免疫感作前の血清のみを注入した。20時間後、元気に生育したHib菌株MinnAを200〜240コロニー形成ユニット(cfu)で(0.1ml)腹腔内(i.p.)抗原投与した。血液サンプルを抗原投与後20時間に、イソフルラン麻酔して心臓に針を刺して集め、チョコレート寒天培地にまいた。1日後、コロニーをカウントしてその結果をフィッシャー検定(Fisher’s Exact test)によって統計学的に分析した。
【0121】
インフルエンザ菌a型菌血症モデル(ref.23)では、9〜10匹の5日齢の幼齢ラットのグループを、テンジクネズミの抗rHiaまたは抗菌株ATCC9006抗血清0.1ml、背面でs.c.注入した。比較対照グループの動物にはテンジクネズミの免疫感作前の血清を注入した。20時間後、元気に生育したインフルエンザ菌a型菌株ATCC9006を(0.1ml)100,000cfuで動物にi.p.抗原投与した。血液サンプルを抗原投与の20時間後に集めて上述のように分析した。
【0122】
後述の表1および2に示したように、テンジクネズミの抗rHia(11)または抗V38 rHia(11)抗血清のいずれかで受動免疫感作された幼齢ラットは、菌血症を引き起こしたインフルエンザ菌a型またはb型に対してすべてが顕著に防御された。これらの結果は、インフルエンザ菌a型またはb型によって引き起こされる菌血症に対して動物を防御することにおいて、わずかに末端欠失したHia蛋白質(V38 rHia)に対応して生成した抗体が全長のそろった蛋白質に対応して生成したものと同程度に効果的であることを示している。NTHi由来の組換え蛋白質によって生じる、被包性のバクテリアによって引き起こされる侵襲性の疾病に対するそのような防御はrHia蛋白質のワクチン抗原としての有用性を説明するものである。
【0123】
実施例13
この実施例は鼻咽頭部でのコロニー形成のチンチラのモデルにおいて、V38 rHia蛋白質の免疫感作によって生じる防御を説明するものである。
【0124】
鼻咽頭部のコロニー形成モデルはYangらによって報告された(ref.20)。このモデルは、HMWアドヘシンを産生するNTHi菌株については適切であるが、しかしHiaを産生する菌株では同じ条件で再現性のあるコロニー形成は達成できなかった。原型のHia産生NTHi菌株11で繰り返したコロニー形成の試みは不成功であった。原型のHMW産生NTHi菌株12ではわずか108cfuしか必要としなかったのに比べて、NTHi菌株33では1回の植菌当たり5×108cfuでコロニー形成が達成された。これらの条件下では、100μgのV38 rHia(33)で免疫感作され、同様のNTHi菌株33で抗原投与された動物において部分的な防御が認められた。
【0125】
実施例14
この実施例はインフルエンザ菌株に由来する追加のhia遺伝子のクローン化および配列分析を説明するものである。
【0126】
保存されたプロモーター、hiaおよびhsf遺伝子および蛋白質のN末端およびC末端配列に基づいて、PCR増幅のためのオリゴヌクレオチド(5040.SLおよび5039.SL)を設計した(図17)。PCR増幅のための選択菌株は抗rHia(11)抗血清との反応性に基づいて選んだ。
【0127】
NTHi菌株12、29、32、M4071、K9および、K22およびヘモフィルスc型菌株APIから染色体DNAを調製した。PCR増幅は以下のようにして実施した。それぞれの反応混合液には5〜100ngのDNA、それぞれ1μgのプライマー、5ユニットのtaq+またはtsg+(Sangon)またはtaqプラスロング(Stratagene)、2mMのdNTPs、20mMのトリス塩酸(pH8.8)、10mMの塩化カリウム、10mMの硫酸アンモニウム、2mMの硫酸マグネシウム、0.1%のTriton X-100、BSAを含ませた。サイクル条件は、95℃で1分間、その後、95℃で30秒、45℃で1分間、72℃で2分間のサイクルを25サイクル行い、その後、72℃で10分間とした。
【0128】
菌株33のhia遺伝子のヌクレオチドおよびそれに由来するアミノ酸の配列が図18に示されている。予測された菌株33由来のHia蛋白質は103.6kDaの分子量を有し、pIが9.47であった。菌株32のhia遺伝子のヌクレオチドおよびそれに由来するアミノ酸の配列は図19に示されている。予測された菌株32由来のHia蛋白質は70.4kDaの分子量を有し、pIが5.67であった。残基493と496との間にはKDEL配列が存在する。そのような配列は小胞体への固定蛋白質に付随するものであった。由来する菌株32のHia蛋白質は顕著により小さくかつ顕著に異なるpIを有するが、しかしながら他のHia蛋白質に存在する特色の多くを含んでいる。
【0129】
菌株29のhia遺伝子のヌクレオチドおよびそれに由来するアミノ酸の配列が図20に示されている。予測された菌株29のHia蛋白質は114.4kDaの分子量および7.58のpIを有する。菌株K22のhia遺伝子のヌクレオチドおよびそれに由来するアミノ酸の配列が図23に示されている。予測された菌株K22のHia蛋白質は114.4kDaの分子量および7.58のpIを有する。NTHi菌株29およびK22に由来するHia配列は同一であることが判明した。菌株29はオハイオ州クリーブランドの中耳炎の生後7カ月の乳児から単離され、それに対して菌株K22はオーストラリアのキンバリー近郊のアボリジニから単離された。
【0130】
菌株4071のhia遺伝子のヌクレオチドおよびそれに由来するアミノ酸の配列が図21に示されている。予測されたM4071のHia蛋白質は103.4kDaの分子量および9.49のpIを有する。残基534と537との間にはKDEL配列が存在する。
【0131】
菌株K9のhia遺伝子のヌクレオチドおよびそれに由来するアミノ酸の配列が図22に示されている。予測された菌株K9のHia蛋白質は113.8kDaの分子量および6.45のpIを有する。
【0132】
ヘモフィルスc型菌株APIのhia遺伝子のヌクレオチドおよびそれに由来するアミノ酸の配列が図24に示されている。予測されたAPIのHia蛋白質は249.4kDaの分子量および5.34のpIを有する。c型菌株APIの由来するHia/Hsf配列は60残基の挿入部があることを除いて公表されているb型のHsf配列と殆ど同一である。ここに提示したNTHiに基づくHia蛋白質はa型およびb型感染の受動モデルにおいて防御をするので、b型とc型蛋白質との間の配列類似性に起因するc型の疾病に対してもまた防御をすることはありそうである。
【0133】
菌株12のhia遺伝子座のヌクレオチドおよびそれに由来するアミノ酸の配列が図25に示されている。NTHi菌株12はHiaを生成しない。しかしながら、hia遺伝子部分はPCR増幅可能であり、SB12細胞溶解物と抗rHia抗体とのつじつまの合わない陽性の反応性が存在し、サザンブロットでは菌株11のhia遺伝子の3’末端に由来するDNAプローブとの反応性がある。PCR増幅したDNAの分析から、上流HI1732関連遺伝子の3’末端の1kbおよびhia遺伝子の3’末端の0.8kbを含む1.8kbの断片が明らかになった。
【0134】
菌株12中のこれら2つの遺伝子の推定される接合部を交差して増幅するであろうプライマーを使用してPCR増幅したところ、この遺伝子座の遺伝子組成が確認された。したがって、菌株12はhia遺伝子の5’末端の欠失を生じたためにHiaを生成しないようであった。図27は菌株12の上流ORFとRdゲノム由来HI1733蛋白質との配列比較を示すものである。相同の領域によって、これら2つの蛋白質は95%が同一である。
【0135】
NTHi菌株33、32、29、K22、M4071、11、9およびc型菌株APIからの由来Hia配列のアラインメントが、インフルエンザ菌b型Hsf、RdゲノムのaidA類似(Hsf/Hia)HI1732遺伝子、と菌株4223およびLES-1のM.catarrhalis 200kDa蛋白質と比較して図28に示されている。HI1732蛋白質の早期末端欠失の結果生じる、Rdゲノム配列内のフレーム移動が認められる。hiaに関連する付加的な下流配列はここに含まれる。配列の下にある星印は保存された残基を示している。N末端(約50残基)およびC末端配列(約150残基)はヘモフィルス菌株の中で高度に保存されており、M.catarrhalisの相応部分とのある程度の類似性は明白である。配列分析から、Hia蛋白質について2つの可能性のある遺伝子集団が存在し、1つは原型の菌株11に関係し、他方はさらに密接に菌株33に関係することは明らかである。菌株11およびK9の蛋白質は、より一層ヘモフィルス菌株のb型、c型またはd型由来のHsf蛋白質に類似しているようであるが、一方で、菌株33、32、29、K22およびM4071の蛋白質は第2の集団を構成するようである。
【0136】
実施例15
この実施例は末端欠失したS44 菌株11 hia遺伝子をもつT7 hia遺伝子カセット、E.coli cer遺伝子、およびカナマイシン抗生物質耐性遺伝子を含むプラスミドJB-2930-3の作製、およびS44 Hia蛋白質の発現を説明するものである。
【0137】
S44アミノ酸から内側のSty I切断部位までのNTHi菌株11 hia遺伝子のS44 Hia N末端(図29)を増幅するためにPCRプライマーを設計した。クローン化を目的として5’末端にNde I切断部位を追加し、鋳型としてプラスミドDS-2242-1を使用してこの断片を増幅した。この断片をpCR IIにクローン化してプラスミドJB-2910-1-1を作製した。この構成図式を図30に示す。プラスミドJB-2910-1-1をNde IおよびSty Iで切断し、5’PCR hia断片を単離した。V38 hia遺伝子を含むプラスミドIA-46-5をNde IおよびSty Iで切断し、大きい方の約8.5kbの断片を精製した。これら2つの精製断片を連結してプラスミドJB-2917-1を作製した。その後、このプラスミドをNde Iで切断し、ウシ腸のホスファターゼ(CAP)で処理し、プラスミドIA-46-5から得たT7プロモーターをその中でクローン化した。このプロモーターはIA-46-5のNde I切断により切り取った。結果として得られたプラスミドJB-2925-3をBgl IIおよびBamH Iで切断してhia遺伝子を単離した。Bgl II/CAP処理したプラスミドBK-2-1-2にこの断片を連結し、プラスミドJB-2930-3を作製した。このプラスミドはT7プロモーターS44 hia遺伝子およびE.coli cer遺伝子を含み、カナマイシン耐性である。
【0138】
発現を調べるため、組換えS44 hiaベクターをE.coli BL21(DE3)に転換した。E.coliにおける発現のための手法は実施例9に記載したりである。図31のSDS-PAGE分析は、2つの異なるベクター、(上述の)JB-2930-3およびpETベクターのIA-191-3-1からの組換えS44 hiaの発現を示している。プラスミドIA-191-3-1は、それがlacIqリプレッサーを含むpETベクターであり、したがって生成するS44 Hiaの量がJB-2930-3からのT7 S44よりも少ないということを除くとJB-2930-3と同じである。このプラスミドは、プラスミドJB-2930-3とともに、図32に示されている。図31は二重のバンド(レーン3)としておよそ116kDaのS44 Hiaを示している。JB-2930-3から精製したS44 hiaを使用した追加の分析によると、この二重バンドの下側のバンドはC末端の94アミノ酸の末端欠失を有するがN末端と考えられるものはそのまま存在することが判明した。末端欠失したHiaの単離に使用した精製方法は実施例10に記載した通りであった。
【0139】
(開示の概要)
本開示の概要では、本発明は新たに単離精製した、ヘモフィルスから得たインフルエンザ菌アドヘシン(Hia)蛋白質の全長を有するものおよびN末端欠失したものをコードし、防御力のあるHia蛋白質を遺伝子組換えにより生成することを可能にする核酸分子を提供する。本発明の範囲内での変更態様は可能である。
【0140】
【表1】

【0141】
【表2】
【0142】
参考文献
【0143】
【表3】
【0144】
【表4】
【0145】
【表5】
【図面の簡単な説明】
【0146】
【図1A】T7プロモーターおよび完全長NTHi菌株11のhia遺伝子を含むプラスミドDS-2008-2-3の制限地図を示す図である。
【図1B】菌株11のhia遺伝子をPCR増幅するために使用されるオリゴヌクレオチドを示す図である。センス鎖(5038.SL):配列番号1、エンコードされているアミノ酸配列番号2;アンチセンス鎖(5039.SL):配列番号3、相補配列番号4、エンコードされているアミノ酸配列番号5。制限酵素部位はB、BamH I;Bg、Bgl II;H、Hind III;N、Nde I;Ps、Pst I;Sty、Sty Iである。他の略語はT7p、T7プロモーター;ApR、アンピシリン耐性である。
【図2】抗ネイティブMoraxella catarrhalisの高分子量アドヘシン抗体による、完全長rHiaタンパク質の認識のイムノブロットを示す図である。レーン1、非誘導型DS-2043-1;レーン2、4時間かけて誘導されたDS-2043-1;レーン3、非誘導型DS-2043-2;レーン4、4時間かけて誘導されたDS-2043-2;レーン5、分子量マーカー。DS-2043-1およびDS-2043-2は、BL21(DE3)内のpT7hia(11)の独立したクローンである。
【図3】菌株11のhia遺伝子用のT7hia遺伝子カセットのタンデムコピーを含むプラスミドDS-2092-1およびDS-2092-40の構築を示す図である。制限酵素部位はB、BamH I;Bg、Bgl II;H、Hind III;Ps、Pst I;Xb、Xba Iである。他の略語はCAP、子牛のアルカリホスファターゼ;T7p、T7プロモーター;ApR、アンピシリン耐性である。
【図4】菌株11のHiaタンパク質の切断部位(配列番号6)を示す図である。
【図5A】菌株11からの切断型hia遺伝子を発現するプラスミドの構築を示す図である。制限酵素部位はB、BamH I;Bg、Bgl II;H、Hind III;N、Nde I;Nhe、Nhe I;Ps、Pst I;R、EcoR I;Sty、Sty I;Xb、Xba Iである。他の略語はT7p、T7プロモーター;ApR、アンピシリン耐性;KanR、カナマイシン耐性である。
【図5B】切断型遺伝子用に5’断片をPCR増幅するために使用されるオリゴヌクレオチドを示す図である。E21切断部:センス(5524.SL):配列番号7、エンコードされているアミノ酸配列番号8;T33切断部:センス(5525.SL):配列番号9、エンコードされているアミノ酸配列番号10;V38切断部:センス(5526.SL):配列番号11、エンコードされているアミノ酸配列番号12;N52切断部:センス(5527.SL):配列番号13、エンコードされているアミノ酸配列番号14;アンチセンス(5528.SL):配列番号15、相補配列番号16、エンコードされているアミノ酸配列番号17。
【図5B1】切断型遺伝子用に5’断片をPCR増幅するために使用されるオリゴヌクレオチドを示す図である。E21切断部:センス(5524.SL):配列番号7、エンコードされているアミノ酸配列番号8;T33切断部:センス(5525.SL):配列番号9、エンコードされているアミノ酸配列番号10;V38切断部:センス(5526.SL):配列番号11、エンコードされているアミノ酸配列番号12;N52切断部:センス(5527.SL):配列番号13、エンコードされているアミノ酸配列番号14;アンチセンス(5528.SL):配列番号15、相補配列番号16、エンコードされているアミノ酸配列番号17。
【図6A】NTHi菌株11および大腸菌のcer遺伝子からのV38hia遺伝子を含むプラスミドBK-96-2-11の構築を示す図である。制限酵素部位はB、BamH I;Bg、Bgl II;K、Kpn I;N、Nde I;P、Pst I;R、EcoR I;S、Sal I;Sm、Sma I;Sty、Sty I;Xb、Xba I;Xho、Xho Iである。他の略語はT7p、T7プロモーター;ApR、アンピシリン耐性;KanR、カナマイシン耐性;CAP、子牛のアルカリホスファターゼ;tt1、trpAからの転写ターミネーター1;tt2、T7遺伝子10からの転写ターミネーター2である。
【図6B】複数のクローン部位および転写ターミネーターを構築するために使用されるオリゴヌクレオチドを示す図である。「R」および「Ps」はEcoR IまたはPst Iの端部とオーバーラップする末端を示すが、これらの部位を再び生成するわけではない。上側鎖(配列番号50)、下側(配列番号51)。
【図7A】T7プロモーターおよび完全長のNTHi菌株33のhia遺伝子、大腸菌のcer遺伝子およびカナマイシン耐性遺伝子を含むプラスミドDS-2241-1およびDS-2242-2の構築を示す図である。制限酵素部位はA、AlwN I;B、BamH I;Bg、Bgl II;H、Hind III;K、Kpn I;N、Nde I;Ps、Pst I;R、EcoR I;S、Sal I;Sm、Sma I;;Xb、Xba I;Xho、Xho Iである。他の略語はT7p、T7プロモーター;ApR、アンピシリン耐性;KanR、カナマイシン耐性;tt1、trpAからの転写ターミネーター1;tt2、T7遺伝子10からの転写ターミネーター2である。
【図7B】鎖(配列番号52)、相補鎖(配列番号53)、およびエンコードされているアミノ酸配列(配列番号54)をコードしている菌株33のhia遺伝子の5’末端を生成するために使用されるオリゴヌクレオチドを示す図である。
【図8A】T7プロモーターおよび菌株33からのV38遺伝子、大腸菌のcer遺伝子およびカナマイシン耐性遺伝子を含むプラスミドDS-2340-2-3の構築を示す図である。制限酵素部位はB、BamH I;Bg、Bgl II;H、Hind III;N、Nde I;Ps、Pst I;R、EcoR I;S、Sal I;Sn、SnaB I;;Xb、Xba Iである。他の略語はT7p、T7プロモーター;ApR、アンピシリン耐性;KanR、カナマイシン耐性;tt1、trpAからの転写ターミネーター1;tt2、T7遺伝子10からの転写ターミネーター2である。
【図8B】切断型遺伝子用の5’端をPCR増幅するために使用されるオリゴヌクレオチドを示す図である。センス(6286.SL):配列番号16、エンコードされているアミノ酸配列番号17;アンチセンス(6287.SL):配列番号18、相補配列番号19、エンコードされているアミノ酸配列番号20。
【図9A】T7のV38hia(11)およびT7のV38hia(33)遺伝子のタンデムコピーをそれぞれ含むプラスミドDS-2447-2およびDS-2448-17の構築を示す図である。制限酵素部位はB、BamH I;Bg、Bgl II;H、Hind III;Ps、Pst I;R、EcoR I;S、Sal I;Xb、Xba Iである。他の略語はT7p、T7プロモーター;ApR、アンピシリン耐性;KanR、カナマイシン耐性;CAP、子牛のアルカリホスファターゼ;tt1、trpAからの転写ターミネーター1;tt2、T7遺伝子10からの転写ターミネーター2である。
【図9B】T7のV38hia(11)およびT7のV38hia(33)遺伝子のタンデムコピーをそれぞれ含むプラスミドDS-2447-2およびDS-2448-17の構築を示す図である。制限酵素部位はB、BamH I;Bg、Bgl II;H、Hind III;Ps、Pst I;R、EcoR I;S、Sal I;Xb、Xba Iである。他の略語はT7p、T7プロモーター;ApR、アンピシリン耐性;KanR、カナマイシン耐性;CAP、子牛のアルカリホスファターゼ;tt1、trpAからの転写ターミネーター1;tt2、T7遺伝子10からの転写ターミネーター2である。
【図10】rHiaの発現を示す図である。パネルA:レーン1、誘導されていない完全長rHia(11);レーン2、完全長rHia(11);レーン3、E21rHia(11);レーン4、T33rHia(11);レーン5、V38rHia(11);レーン6、N52rHia(11)。パネルB:レーン1、誘導されていないV38rHia(11);レーン2、V38rHia(11);レーン3、V38rHia(11)/cer。
【図11】rHiaの精製スキームを示す図である。略語はSP、上澄み;PPT、沈殿;DTT、ジチオスレイトール;OG、オクチルグルコシドであり、(x)は捨てられたことを意味する。
【図12】パネルAおよびBを有し、精製されたrHiaのSDS-PAGE分析を示す図である。パネルAは菌株11からの精製されたV38rHiaタンパク質を示し、パネルBは菌株33からのV38rHiaタンパク質を示す。レーン1、分子量マーカー;レーン2、全細胞の溶解物;レーン3、未精製抽出物;レーン4、精製されたrHiaタンパク質。
【図13A】パネルA、BおよびCを有し、V38rHia(11)の安定性を示す図である。パネルAは、グリセロールなしで4℃で保存されたサンプルを示す。パネルBは、20%グリセロールの存在下で4℃で保存されたサンプルを示す。パネルCは、20%グリセロールの存在下で-20℃で保存されたサンプルを示す。レーン0はt0を示し、レーン1〜8は1〜8週間保存されたサンプルを示す。
【図13B】パネルA、BおよびCを有し、V38rHia(11)の安定性を示す図である。パネルAは、グリセロールなしで4℃で保存されたサンプルを示す。パネルBは、20%グリセロールの存在下で4℃で保存されたサンプルを示す。パネルCは、20%グリセロールの存在下で-20℃で保存されたサンプルを示す。レーン0はt0を示し、レーン1〜8は1〜8週間保存されたサンプルを示す。
【図13C】パネルA、BおよびCを有し、V38rHia(11)の安定性を示す図である。パネルAは、グリセロールなしで4℃で保存されたサンプルを示す。パネルBは、20%グリセロールの存在下で4℃で保存されたサンプルを示す。パネルCは、20%グリセロールの存在下で-20℃で保存されたサンプルを示す。レーン0はt0を示し、レーン1〜8は1〜8週間保存されたサンプルを示す。
【図14A】パネルAおよびBを有し、CD-1マウスのV38rHia(11)またはV38rHia(33)の免疫原性を示す図である。パネルAは単一の免疫化後の応答を示し、パネルBは最初の/追加の免疫化の応答を示す。
【図14B】パネルAおよびBを有し、CD-1マウスのV38rHia(11)またはV38rHia(33)の免疫原性を示す図である。パネルAは単一の免疫化後の応答を示し、パネルBは最初の/追加の免疫化の応答を示す。
【図15A】BALB/cマウスおよびモルモットのV38rHia(11)の免疫原性を示す図である。図15Aはマウスにおける抗体の応答を示し、図15Bはモルモットにおける応答を示す。
【図15B】BALB/cマウスおよびモルモットのV38rHia(11)の免疫原性を示す図である。図15Aはマウスにおける抗体の応答を示し、図15Bはモルモットにおける応答を示す。
【図16】チンチラモデルにおける鼻咽頭コロニー形成に対するV38rHia(33)の防御能力を示す図である。
【図17】追加のhia遺伝子をPCR増幅するために使用されるオリゴヌクレオチドを示す図である。センス(5040.SL):配列番号21、エンコードされているアミノ酸配列番号22;アンチセンス(5039.SL):配列番号3、相補配列番号4、エンコードされているアミノ酸配列番号5。
【図18A】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18B】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18C】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18D】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18E】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18F】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18G】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18H】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18I】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18J】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18K】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18L】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18M】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18N】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18O】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図19A】NTHi菌株32からのhia遺伝子のヌクレオチド配列(配列番号25)および演繹アミノ酸配列(配列番号26)を示す図である。
【図19B】NTHi菌株32からのhia遺伝子のヌクレオチド配列(配列番号25)および演繹アミノ酸配列(配列番号26)を示す図である。
【図19C】NTHi菌株32からのhia遺伝子のヌクレオチド配列(配列番号25)および演繹アミノ酸配列(配列番号26)を示す図である。
【図19D】NTHi菌株32からのhia遺伝子のヌクレオチド配列(配列番号25)および演繹アミノ酸配列(配列番号26)を示す図である。
【図19E】NTHi菌株32からのhia遺伝子のヌクレオチド配列(配列番号25)および演繹アミノ酸配列(配列番号26)を示す図である。
【図19F】NTHi菌株32からのhia遺伝子のヌクレオチド配列(配列番号25)および演繹アミノ酸配列(配列番号26)を示す図である。
【図19G】NTHi菌株32からのhia遺伝子のヌクレオチド配列(配列番号25)および演繹アミノ酸配列(配列番号26)を示す図である。
【図19H】NTHi菌株32からのhia遺伝子のヌクレオチド配列(配列番号25)および演繹アミノ酸配列(配列番号26)を示す図である。
【図19I】NTHi菌株32からのhia遺伝子のヌクレオチド配列(配列番号25)および演繹アミノ酸配列(配列番号26)を示す図である。
【図19J】NTHi菌株32からのhia遺伝子のヌクレオチド配列(配列番号25)および演繹アミノ酸配列(配列番号26)を示す図である。
【図20A】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20B】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20C】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20D】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20E】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20F】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20G】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20H】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20I】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20J】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20K】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20L】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20M】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20N】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20O】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20P】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図21A】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21B】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21C】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21D】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21E】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21F】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21G】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21H】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21I】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21J】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21K】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21L】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21M】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21N】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21O】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図22A】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22B】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22C】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22D】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22E】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22F】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22G】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22H】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22I】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22J】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22K】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22L】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22M】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22N】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22O】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22P】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図23A】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23B】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23C】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23D】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23E】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23F】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23G】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23H】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23I】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23J】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23K】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23L】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23M】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23N】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23O】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23P】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図24A】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24B】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24C】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24D】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24E】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24F】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24G】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24H】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24I】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24J】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24K】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24L】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24M】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24N】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24O】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24P】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24Q】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24R】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24S】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24T】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24U】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24V】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24W】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24X】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24Y】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24Z】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24A1】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24B1】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24C1】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24D1】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24E1】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24F1】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24G1】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24H1】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24I1】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図25A】NTHi菌株12からのhia遺伝子座のヌクレオチド配列(配列番号37)および演繹アミノ酸配列(配列番号38)を示す図である。上線または下線を引かれた配列は、2つのorfの接合部を越えてPCR増幅するために使用されるオリゴヌクレオチドを示す。センス(6431.SL):配列番号39、(6432.SL)配列番号40;アンチセンス(6295.SL)配列番号41、(6271.SL)配列番号42。
【図25B】NTHi菌株12からのhia遺伝子座のヌクレオチド配列(配列番号37)および演繹アミノ酸配列(配列番号38)を示す図である。上線または下線を引かれた配列は、2つのorfの接合部を越えてPCR増幅するために使用されるオリゴヌクレオチドを示す。センス(6431.SL):配列番号39、(6432.SL)配列番号40;アンチセンス(6295.SL)配列番号41、(6271.SL)配列番号42。
【図25C】NTHi菌株12からのhia遺伝子座のヌクレオチド配列(配列番号37)および演繹アミノ酸配列(配列番号38)を示す図である。上線または下線を引かれた配列は、2つのorfの接合部を越えてPCR増幅するために使用されるオリゴヌクレオチドを示す。センス(6431.SL):配列番号39、(6432.SL)配列番号40;アンチセンス(6295.SL)配列番号41、(6271.SL)配列番号42。
【図25D】NTHi菌株12からのhia遺伝子座のヌクレオチド配列(配列番号37)および演繹アミノ酸配列(配列番号38)を示す図である。上線または下線を引かれた配列は、2つのorfの接合部を越えてPCR増幅するために使用されるオリゴヌクレオチドを示す。センス(6431.SL):配列番号39、(6432.SL)配列番号40;アンチセンス(6295.SL)配列番号41、(6271.SL)配列番号42。
【図25E】NTHi菌株12からのhia遺伝子座のヌクレオチド配列(配列番号37)および演繹アミノ酸配列(配列番号38)を示す図である。上線または下線を引かれた配列は、2つのorfの接合部を越えてPCR増幅するために使用されるオリゴヌクレオチドを示す。センス(6431.SL):配列番号39、(6432.SL)配列番号40;アンチセンス(6295.SL)配列番号41、(6271.SL)配列番号42。
【図25F】NTHi菌株12からのhia遺伝子座のヌクレオチド配列(配列番号37)および演繹アミノ酸配列(配列番号38)を示す図である。上線または下線を引かれた配列は、2つのorfの接合部を越えてPCR増幅するために使用されるオリゴヌクレオチドを示す。センス(6431.SL):配列番号39、(6432.SL)配列番号40;アンチセンス(6295.SL)配列番号41、(6271.SL)配列番号42。
【図25G】NTHi菌株12からのhia遺伝子座のヌクレオチド配列(配列番号37)および演繹アミノ酸配列(配列番号38)を示す図である。上線または下線を引かれた配列は、2つのorfの接合部を越えてPCR増幅するために使用されるオリゴヌクレオチドを示す。センス(6431.SL):配列番号39、(6432.SL)配列番号40;アンチセンス(6295.SL)配列番号41、(6271.SL)配列番号42。
【図25H】NTHi菌株12からのhia遺伝子座のヌクレオチド配列(配列番号37)および演繹アミノ酸配列(配列番号38)を示す図である。上線または下線を引かれた配列は、2つのorfの接合部を越えてPCR増幅するために使用されるオリゴヌクレオチドを示す。センス(6431.SL):配列番号39、(6432.SL)配列番号40;アンチセンス(6295.SL)配列番号41、(6271.SL)配列番号42。
【図25I】NTHi菌株12からのhia遺伝子座のヌクレオチド配列(配列番号37)および演繹アミノ酸配列(配列番号38)を示す図である。上線または下線を引かれた配列は、2つのorfの接合部を越えてPCR増幅するために使用されるオリゴヌクレオチドを示す。センス(6431.SL):配列番号39、(6432.SL)配列番号40;アンチセンス(6295.SL)配列番号41、(6271.SL)配列番号42。
【図26A】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図26B】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図26C】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図26D】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図26E】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図26F】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図26G】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図26H】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図26I】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図26J】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図26K】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図26L】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図27A】菌株12のhia遺伝子座からの上流ORF(配列番号45)の配列およびb型インフルエンザ菌株RdからのHI1732タンパク質(配列番号46)の一部の整列を示す図である。
【図27B】菌株12のhia遺伝子座からの上流ORF(配列番号45)の配列およびb型インフルエンザ菌株RdからのHI1732タンパク質(配列番号46)の一部の整列を示す図である。
【図28A】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28B】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28C】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28D】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28E】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28F】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28G】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28H】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28I】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28J】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28K】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28L】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28M】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28N】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28O】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28P】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28Q】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28R】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28S】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28T】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28U】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28V】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28W】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28X】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28Y】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28Z】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28A1】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28B1】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28C1】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28D1】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図29】hia遺伝子の5’端をS44切断位置でPCR増幅するために使用されるオリゴヌクレオチドを示す図である。センス(6817.SL)配列番号49、エンコードされているアミノ酸配列番号50;アンチセンス(6818.SL)配列番号51、相補配列番号52、エンコードされているアミノ酸配列番号53。
【図30A】NTHi菌株11からのS44hia遺伝子および大腸菌のcer遺伝子およびT7プロモーターを含む、プラスミドJB-2930-3の構築を示す図である。制限酵素部位はB、BamH I;Bg、Bgl II;K、Kpn I;N、Nde I;P、Pst I;R、EcoR I;S、Sal I;Sm、Sma I;Sty、Sty I;Xb、Xba I;Xho、Xho Iである。他の略語はT7p、T7プロモーター;ApR、アンピシリン耐性;KanR、カナマイシン耐性;CAP、子牛のアルカリホスファターゼ;tt1、trpAからの転写ターミネーター1;tt2、T7遺伝子10からの転写ターミネーター2である。
【図30B】NTHi菌株11からのS44hia遺伝子および大腸菌のcer遺伝子およびT7プロモーターを含む、プラスミドJB-2930-3の構築を示す図である。制限酵素部位はB、BamH I;Bg、Bgl II;K、Kpn I;N、Nde I;P、Pst I;R、EcoR I;S、Sal I;Sm、Sma I;Sty、Sty I;Xb、Xba I;Xho、Xho Iである。他の略語はT7p、T7プロモーター;ApR、アンピシリン耐性;KanR、カナマイシン耐性;CAP、子牛のアルカリホスファターゼ;tt1、trpAからの転写ターミネーター1;tt2、T7遺伝子10からの転写ターミネーター2である。
【図31】S44からのrHiaの発現のSDS-PAGE分析を示す図である。レーン1、時間0での(誘導なしで)pETS44ベクターからの発現体;レーン2、4時間の誘導後のpETS44ベクターからの発現体;レーン3、4時間の誘導後のJB-2930-3からの発現体。
【図32】発現試験用に使用された2つのベクター、S44切断型rHiaのJB-2930-3およびIA-191-3-1の概略を示す図である。
【技術分野】
【0001】
(関連出願の参照)
本出願は、同時係続の米国特許出願第09/268,347号の一部継続出願である。
【0002】
(発明の分野)
本発明は分子遺伝学の分野に関し、詳細には組換えインフルエンザ菌(ヘモフィルス・インフルエンジー(Haemophilus influenzae))アドヘシン(Hia)タンパク質の産生に関する。
【背景技術】
【0003】
(発明の背景)
インフルエンザ菌は、髄膜炎、喉頭蓋炎、敗血症および中耳炎などのいくつかの重大なヒト疾患の原因である。インフルエンザ菌には、aからfで表され、それらの莢膜多糖によって識別される6種類の血清型がある。b型インフルエンザ菌(Hib)は、1980年代にいくつかのHib結合ワクチンが導入されるまでは細菌性髄膜炎の主な原因であった(ref.1.本出願全体にわたって、さまざまな参考文献を括弧内に引用し、本発明が関係する分野の現状をより完全に説明する。それぞれの引用に関する完全な文献情報は、明細書の最後、特許請求の範囲の直前に見出される。これらの参考文献の開示を、参照により本開示に組み込む)。ジフテリアトキソイド(ref.2)、破傷風トキソイド(ref.3および米国特許第4,496,538号)または髄膜炎菌外膜タンパク質(ref.4)に結合したb型インフルエンザ菌莢膜多糖をベースとするワクチンは、b型インフルエンザ菌誘発髄膜炎を減らすのに有効とされてきた。他の血清型インフルエンザ菌が侵襲性疾患に関連する頻度は低いが、Hib疾患の発生率が減少するにつれて、これらの菌株に起因する疾患の発生率が増加するようである(ref.5、6)。非被包性すなわち分類不能型インフルエンザ菌(NTHi)もまた中耳炎、喉頭蓋炎、肺炎および気管気管支炎を含む広範囲のヒト疾患の原因となる。NTHi誘発疾患の発生率は、Hibワクチンの導入によって影響を受けなかった(ref.7)。
【0004】
中耳炎は幼児期に最もよく見られる病気であり、2歳未満の子供全体の60から70%が1回から3回の耳感染症を経験する(ref.8)。子供の慢性中耳炎は聴力、発話および認知障害の原因となる。インフルエンザ菌感染症は、急性中耳炎の症例の約30%、慢性中耳炎の約60%を占める。米国だけでも中耳炎の治療には、抗生物質ならびに扁桃摘出術、咽頭扁桃切除術および中耳腔換気用チューブの挿入などの外科的処置に年間10億ドルから20億ドルが費やされている。言語療法および特殊教育授業などの付加的治療に対し、年間さらに300億ドルが費やされていると推定される。さらに、中耳炎の原因となる生物の多くは、抗生物質治療に対して耐性になりつつある。したがって、中耳炎に対する有効な予防ワクチンが望まれている。NTHiによる自然感染では、抗体の応答を刺激する露出した外膜タンパク質が殺菌および/または防御的な抗体として潜在的に重要な標的であり、したがって潜在的なワクチンの候補である。NTHiの上皮細胞への付着の際に重要である高分子量タンパク質(HMW1およびHMW2)のファミリーが、約70から75%のNTHi菌株において同定されている(ref.9、ref.10)。これらの高分子量アドヘシン(adhesin)は、中耳炎のチンチラモデルにおいていくらかの防御を可能にすることが分かっている(ref.11)。高分子量アドヘシンタンパク質の第2のファミリーが、約25%のNTHiおよび被包性インフルエンザ菌株において同定されている(ref.12、ref.13、ref.14)。この第2のファミリーのNTHiメンバーは、インフルエンザ菌アドヘシンすなわちHiaと呼ばれ、被包性菌株において発見される相同タンパク質はインフルエンザ菌表面原繊維タンパク質すなわちHsfと呼ばれる。hia遺伝子は、中耳炎患者の回復期血清を使用して発現ライブラリーから最初にクローニングされ、このことはhia遺伝子が疾患における重要な免疫原であることを示す。プロトタイプのHiaおよびHsfタンパク質は約82%の配列類似性を示すが、Hsfタンパク質はかなり高い。これらのタンパク質には保存されたアミノ酸およびカルボキシ末端およびいくつかの繰り返しモチーフが含まれ、HsfはHiaと比較してより多くの繰り返し配列を含む。高分子量タンパク質(200kDa)も、HsfおよびHiaタンパク質といくらかの配列相同性があるMoraxella catarrhalisから同定されている(米国特許第5,808,024号)。
【0005】
HiaおよびHsfはインフルエンザ菌の被包性菌株および約20〜25%の非被包性菌株中で保存され、アドヘシンであることが示されているので、HiaまたはHiaと特異的に反応する抗体を産生する能力のあるタンパク質を産生する、インフルエンザ菌または他の細菌性病原体によって引き起こされる疾患に対する診断および予防接種において、このタンパク質は有用性がある。
【0006】
診断において有用であり予防接種における免疫原としてのアンチHia抗体を産生するために、診断において抗原としてHiaを使用することの欠点は、インフルエンザ菌種からの天然タンパク質の回収があまりできないことである。
【0007】
他の免疫原用のワクチン、担体を含めた免疫調製、および診断試薬の生成において、抗原として使用するための組換えHiaタンパク質を提供することが有利であろう。
【発明の開示】
【発明が解決しようとする課題】
【0008】
(発明の概要)
本発明は組換えインフルエンザ菌アドヘシン(rHia)の提供を対象とする。
【課題を解決するための手段】
【0009】
このような組換えタンパク質の提供に関して、本発明は単離され精製されたある種の核酸分子を提供する。したがってその一態様において、本発明は単離され精製された核酸分子を提供し、この核酸分子はインフルエンザ菌株のインフルエンザ菌アドヘシン(Hia)タンパク質をエンコードしており、以下のものを含む。(a)図18、19、20、21、22、23、24および25に示される配列(配列番号23、25、27、29、31、33、35、37)からなる群から選択されるDNA配列、または(b)図18、19、20、21、22、23、24および25に示される配列(配列番号24、26、28、30、32、34、36、38)からなる群から選択されるアミノ酸配列を有するインフルエンザ菌アドヘシン(rHia)タンパク質をエンコードしているDNA配列。
【0010】
このような核酸は、プラスミドベクターである可能性のあるベクター中に含まれていてよい。詳細には、分類不能型ヘモフィルスの菌株11または33からのHiaタンパク質を、核酸分子はエンコードしていてよい。
【0011】
本発明の他の態様では、インフルエンザ菌の菌株のN末端切断型インフルエンザ菌アドヘシン(Hia)タンパク質をエンコードしており、1対のヌクレオチドによって増幅可能な単離され精製された核酸分子が提供される。ヌクレオチドは配列番号7と配列番号15、配列番号9と配列番号15、配列番号11と配列番号15、配列番号13、配列番号15、配列番号49、および配列番号51からなる群から選択される。
【0012】
このような核酸は、プラスミドベクターである可能性のあるベクター中に含まれていてよい。詳細には、分類不能型ヘモフィルスの菌株11または33からのコドンV38またはS44で始まるN末端切断型Hiaタンパク質を、核酸分子はエンコードしていてよい。
【0013】
本発明のこれらの態様に従って提供される単離され精製された核酸を組み込んでいるプラスミドベクターは、
図1A中に示されるDS-2008-2-3
図5A中に示されるDS-2186-1-1
図5A中に示されるDS-2201-1
図5A中に示されるDS-2186-2-1
図5A中に示されるDS-2168-2-6
図32中に示される1A-191-3-1
からなる群から選択されるプラスミドの同定特性を有する可能性がある。
【0014】
本明細書で提供されるベクターは、大腸菌からのcer遺伝子を含んでよい。したがって、本発明の他の態様では、完全長またはN末端切断型インフルエンザ菌アドヘシン(Hia)タンパク質をエンコードしている核酸分子、前記完全長または切断型Hiaタンパク質を発現するためのプロモーター、任意選択で大腸菌のcer遺伝子含む、宿主を形質転換するためのベクターが提供される。ベクターはプラスミドベクターまたは他の非複製ベクターであってよく、
図6A中に示されるBK-96-2-11
図7A中に示されるDS-2242-1
図7A中に示されるDS-2242-2
図8A中に示されるDS-2340-2-3
図9A中に示されるDS-2247-2
図9B中に示されるDS-2448-17
図32中に示されるJB-2930-3
からなる群から選択されるプラスミドベクターの同定特性を有する可能性がある。
【0015】
本明細書で提供されるベクターは、サルモネラ、BCG、アデノウイルス、ポックスウイルス、ワクシニアまたはポリオウイルスからのベクターを含めた複製ベクターを含んでよい。
【0016】
本明細書で提供される任意のベクターを使用して、完全長または切断型である可能性のある、ヘモフィルスの分類不能型菌株の防御性インフルエンザ菌アドヘシン(Hia)タンパク質を発現するのに適切な宿主細胞を形質転換することができる。このような宿主は、好都合には大腸菌であってよい。このような発現はT7プロモーターの制御下で行うことができ、形質転換された宿主からの組換えHiaの発現はラクトースまたは他の好都合な誘導剤の誘導用濃縮液中で培養することによって実施することができる。
【0017】
本発明は他の態様において、形質転換された宿主、特に本明細書で提供される大腸菌によって産生される、分類不能型ヘモフィルス菌株の組換え防御性インフルエンザ菌アドヘシン(Hia)タンパク質をさらに含む。このようなHiaタンパク質を組換えタンパク質の免疫原性断片またはアドヘシン官能性類縁体の形で提供することができる。
【0018】
完全長またはN末端切断型である本明細書で提供される組換えHiaタンパク質は、免疫原性組成物中の抗原、他の免疫原用の担体、診断用薬としておよび診断用薬の生成において有用である。完全長またはN末端切断型であるHiaタンパク質をエンコードしている核酸分子も、診断のための使用用、および免疫原性組成物中においてもプローブとして有用である。
【0019】
追加的な態様において本発明は、本明細書で提供されるのと同様な単離され精製された核酸分子、本明細書で提供されるのと同様なヘモフィルスの菌株の完全長またはN末端切断型である組換え防御性Hiaタンパク質からなる群から選択される少なくとも1つの免疫学的に活性のある成分、および薬学的に許容可能なこれらの担体を含む、免疫原性組成物を提供する。
【0020】
本明細書で提供される免疫原性組成物を、宿主へのin vitro投与用ワクチンとして製剤化して、インフルエンザ菌に起因する疾患に対する防御を提供することができる。このような目的のために、組成物をミクロ粒子、カプセル、ISCOMまたはリポソーム調製物として製剤化することができる。免疫原性組成物を免疫系の特定の細胞または粘膜表面への送達用の標的分子と組み合わせて提供することができる。
【0021】
本発明の免疫原性組成物(ワクチンを含めて)は少なくとも1種の他の免疫原性または免疫刺激性材料をさらに含んでよく、この免疫刺激性材料少なくとも1種のアジュバントまたは少なくとも1種のサイトカインであってよい。本発明で使用するのに適切なアジュバントには(これらに限らないが)リン酸アルミニウム、水酸化アルミニウム、QS21、Quil A、その誘導体および成分、ISCOMマトリックス、リン酸カルシウム、水酸化カルシウム、水酸化亜鉛、糖脂質類縁体、アミノ酸のオクタデシルエステル、ムラミルジペプチド、ポリホスファゼン、ISCOPREP、DC-chol、DDBAおよびリポタンパク質ならびに他のアジュバントがある。
【0022】
アジュバントの有利な組み合わせは、本出願人と同じ譲受人に譲渡され、その開示を参照(1995年11月21日発行のWO 95/34308)により本明細書に組み込む1994年6月16日出願の同時係続の米国特許第08/261,194号、および1995年6月7日出願の第08/483,856号に記載されている。
【0023】
本発明の他の態様に従って、感受性のある宿主に有効量の前述の免疫原性組成物を投与するステップを含む、宿主中で免疫応答を生み出すための方法が提供される。免疫応答は体液の免疫応答であるか、または細胞を介する免疫応答であってよい。その内部で疾患に対する防御を与えることができる宿主には、ヒトを含めた霊長類がある。
【0024】
本発明の他の態様に従って、薬剤として使用されるとき本明細書で提供される免疫原性組成物、および疫原性組成物の製造における免疫原性組成物のこれらの成分の使用法が提供される。
【0025】
他の追加的な態様において本発明は、インフルエンザ菌の分類不能型菌株の防御性インフルエンザ菌アドヘシン(Hia)タンパク質の産生のための方法を提供し、この方法は
本明細書で提供されるのと同様な菌株のN末端切断型のインフルエンザ菌アドヘシンタンパク質をエンコードしている核酸分子を含むベクターによって、大腸菌などの宿主を形質転換すること、
宿主を成長させてエンコードされている切断型Hiaを発現させること、
発現したHiaタンパク質を単離および精製することを含む。
【0026】
エンコードされている切断型Hiaを封入体中で発現させることができる。成長し形質転換された細胞を破壊して上澄みおよびHiaを含む封入体を生成し、上澄みからの分離の後に封入体を可溶化して組換えHiaの溶液を生成し、クロマトグラフィによって細胞の残骸のない組換えHiaの溶液を精製し、精製された組換えHiaタンパク質を単離することによって、単離および精製ステップを実施することができる。
【0027】
大腸菌などの宿主細胞を形質転換するベクターはT7プロモーターを含んでよく、大腸菌または他の宿主細胞を誘導量のラクトースまたは他の好都合な誘導剤の存在下で培養することができる。
【0028】
本明細書のインフルエンザ菌の菌株は、菌株11、33、32、29、M4071、K9、K22および12からなる分類不能型菌株の群から選択することができる。このような菌株からの代表的なHiaタンパク質をエンコードしている遺伝子に特異的な核酸配列が、本明細書で提供され以下に記載される。
【0029】
本明細書で提供される核酸配列は、診断に関する適用例において有用である。したがって本発明の他の態様では、サンプル中でインフルエンザ菌アドヘシンタンパク質をエンコードしている核酸の存在を判定するするための方法が提供され、この方法は、
a)サンプルを本明細書で提供されるのと同様な核酸分子と接触させて、本明細書で提供される核酸配列、およびサンプル中に存在し特異的にハイブリダイズすることができるヘモフィルスの菌株のHiaタンパク質をエンコードしている核酸を含むデュプレックス(duplex)を生成するステップと、
b)デュプレックスの生成を判定するステップを含む。
【0030】
さらに本発明は、サンプル中でインフルエンザ菌アドヘシンタンパク質をエンコードしている核酸の存在を判定するための診断キットを提供し、このキットは、
a)本明細書で提供されるのと同様な核酸分子と、
b)核酸分子とサンプルを接触させて、この核酸分子および任意のこのような核酸分子を含むデュプレックスを生成するための手段と、
c)デュプレックスの生成を判定するための手段をを含む。
【0031】
本明細書で提供される組換えによって産生された切断型Hiaタンパク質も、診断に関する適用例において有用である。したがって本発明の他の態様では、サンプル中のHiaタンパク質と特異的に反応する抗体の存在を判定するための方法が提供され、この方法は、
(a)サンプルと本明細書で提供される組換えHiaタンパク質を接触させて、この組換えHiaタンパク質とサンプル中に存在しタンパク質と特異的に反応する任意のこのような抗体の複合体を提供するステップと、
(b)複合体の生成を判定するステップを含む。
【0032】
本発明の利点には以下のものがある。
【0033】
インフルエンザ菌アドヘシンタンパク質またはこのHiaタンパク質の断片または類縁体をエンコードしている単離され精製された核酸分子、
他のいかなるヘモフィルスのタンパク質も含まない組換えによって産生されたHiaタンパク質、
ヘモフィルスの特異的な同定用の診断キットおよび免疫試薬。
【発明を実施するための最良の形態】
【0034】
本発明は、図面を参照しながら以下の記述からさらに理解されるはずである。
【0035】
(発明の全般的な説明)
H.influenzae株は、少量のHiaおよびHsfタンパク質を発現するので、NTHi株由来のhia遺伝子を、E.Coli中で組み換えタンパク質を過剰発現させるための発現ベクター中にクローン化した。全長組み換えHia(rHia)タンパク質が発現する際、それは比較的少量であった。組み換えタンパク質が発現されているのを確認するために、本出願と同じ譲受人に譲受され、その開示を参照により本明細書に組み込む米国特許第5,808,024号で200kDaであると同定されたMoraxella catarrhalis高分子量アドヘシンタンパク質に対する抗体を用いてイムノブロットを実施した。ゲル精製した未変性200kDaタンパク質に対する抗体は、rHiaタンパク質サンプル中に特異的に誘導されたバンドを認識した。rHiaの収量はT7hia遺伝子カセットの遺伝子コピー数を増加させることによっても顕著には増加しない。
【0036】
E.Coliのcer遺伝子は、大きな挿入部を含むプラスミドを安定化するが(参考文献15)、E.Coliのcer遺伝子を発現ベクターに添加しても、rHiaの収量は顕著には増加しない。しかし、E.Coli細胞は培養中に凝集し、このことは、Hiaアドヘシンタンパク質の表面発現が起きていることを示唆している。rHiaタンパク質が封入体として生成されればrHiaタンパク質の強い毒性は克服されるので、推定シグナル配列を欠失させるために遺伝子の5’-末端を切断した。この変性の結果、V38位置から出発している切断型rHiaの生産および回収が良好となった。
【0037】
全長およびV-38切断型rHiaタンパク質は免疫原性であり、得られる抗-rHia抗体は、H.influrnzaeのタイプaまたはタイプb株による菌血症の受動的幼ラットモデルを防護する。さらに、切断型V38rHiaタンパク質は、チンチラの能動的抗原投与モデルにおける鼻咽頭コロニー形成に対して特に防護に働くことが見出された。NTHi株からのrHiaによってもたらされる防護が、NTHiおよび被包性タイプaまたはタイプb株が原因となって起こる疾患に対するものであることは、一般的な防護エピトープが存在することを意味している。付加的なhia遺伝子のクローニングおよび配列分析は、保存領域の同定の助けとなる。全長またはN-末端切断型rHiaタンパク質を、Haemophilus influenzae疾患に対する防護のためのワクチン成分として使用できる。
【0038】
hia遺伝子を有する任意のHaemophilus株は、少なくとも本発明の態様で特徴付けるHiaタンパク質をコードする部分を含有する核酸分子(おそらくDNA分子の形)の精製および単離に利用できる。このような株は一般的な臨床所およびバクテリア培養収集所、例えばAmerican Type Culture Collectionから入手される。Haemophilus influenzaeの適切な株:
分類不能型 Haemophilus 株11;
分類不能型 Haemophilus 株33;
分類不能型 Haemophilus 株32;
分類不能型 Haemophilus 株29;
分類不能型 Haemophilus 株 M4071;
分類不能型 Haemophilus 株 K9;
分類不能型 Haemophilus 株 K22;
分類不能型 Haemophilus 株 12;
タイプC Haemophilus 株 API。
【0039】
本明細書において、“Hia”タンパク質という用語は、Hiaタンパク質のファミリーを定義するために使用され、Hiaタンパク質には種々のHaemophilus株中で見出されるアミノ酸配列を有する天然の変異体が含まれる。
【0040】
図1Aは、T7プロモーターの影響下での、分類不能型 Haemophilus influenzae株11由来全長hia遺伝子を含むプラスミドDS-2008-2-3の制限地図を示す図である。株11由来のhia遺伝子の核酸(配列番号:43)および推測アミノ酸配列(配列番号:44)は、前記米国特許第5,646,259に記載されている(該明細書中では“HA1”と定義される)。hia遺伝子を株11の遺伝子のATG開始コドンからPCR増幅するために使用したオリゴヌクレオチドを、図1Bに示す。
【0041】
図2は、抗-未変性Moraxella catarrhalis高分子量アドヘシン抗体によりrHia(11)タンパク質を確認するためのイムノブロットを示す図である。前記米国特許第5,808,024に記載される、M.catarrhalis高分子量アドヘシンタンパク質または200kDaタンパク質は、HiaおよびHsfタンパク質と、特にカルボキシ末端部に、何らかの配列相同性を有する(図28)。
【0042】
図3は、NTHi株11由来の全長hia遺伝子から成るT7hia遺伝子カセットのタンデムコピーを含むプラスミドDS-2092-1およびDS-2092-40の構築スキームを示す図である。増加した遺伝子コピー数を示すこのようなプラスミドは、しばしば組み換えタンパク質の生産レベルを上昇させる。しかし、以下に示すように、組み換えHiaの低い収率は、遺伝子コピー数の増加によって顕著には増加しない。
【0043】
図4は、NTHi株11タンパク質のN-末端配列および経時的にN-末端切断されたrHiaタンパク質の位置を示す図である。E21部分までのN-末端切断型は、Hiaのシグナル配列の構成部分であると考えられる長い疎水性領域が欠失している。T33部分までの欠失は、長い疎水性領域を含み、潜在的なAla-X-Alaシグナル切断部位を有する。V38部分までの欠失は、長い疎水性領域を含み、潜在的なAla-X-Alaシグナル切断部位を有する。S44部分から始まる組み換えHiaタンパク質は、長い疎水性領域を含み、潜在的なAla-X-Alaシグナル切断部位を有する。N52部分から始まる組み換えHiaタンパク質は、Moraxella catarrhalis由来の高分子量(200kDa)アドヘシンの近接した開始点を模倣しており、アドヘシンは米国特許第5,808,024に記載され、該組み換えタンパク質は、V56から開始するようにN-末端切断型されている場合に過剰発現する。
【0044】
図5Aは、4種のN―末端切断型rHiaを製造するための、プラスミドDS-2186-1-1、DS-2201-1、DS-2186-2-1、およびDS-2168-2-6を作製するための構築スキームを示す図である。5’-末端フラグメントをPCR増幅するために使用されるこのオリゴヌクレオチドを図5Bに示す。図30は、S44欠失を製造する、プラスミドJB2930-3を作製するための構築スキームを示す図である。5’-フラグメントをPCR増幅するために使用するオリゴヌクレオチドを図29に示す。
【0045】
図6Aは、NTHi株11由来のV38hia遺伝子ならびにプラスミドを安定化するE.Coli cer遺伝子を含むプラスミドBK-96-2-11を作製するための構築スキームを示す図である。毒性のタンパク質を産生するプラスミドへのcer遺伝子の導入により、タンパク質産生の強化が予測された。cer遺伝子の存在下で全長rHia産生E.Coli細胞の形態に変化が生じ、すなわち細胞が凝集した。このことは、凝集を起こす細胞表面のアドヘシンの発現が強化されたことを示唆する。発現プラスミドBK-96-2-11はさらに、遺伝子安定性を高めると考えられているT7V38hia遺伝子カセットの上流および下流に転写ターミネーターを含んでいる。マルチクローニング部位および転写ターミネーターを作製するために使用されるオリゴヌクレオチドを図6Bに示す。
【0046】
図7Aは、T7プロモーターの影響下に、分類不能型Haemophilus influenzae株33由来の全長hia遺伝子を含むプラスミドDS-2242-1およびDS-2242-2のための構築スキームを示す図である。発現プラスミドは、T7hia(33)遺伝子カセットの上流および下流にE.Coli cer遺伝子および形質転換ターミネーターをも含有する。DS-2242-1は、T7hia(33)遺伝子と同一の鎖上にコードされたターミネーターを有する。しかし、2つのプラスミドに由来するrHiaの発現には違いが見られなかった。NTHi株33の真性5’末端の構築に使用されるオリゴヌクレオチドを図7Bに示す。
【0047】
図8Aは、NTHi株33由来のV38Hia遺伝子ならびにE.Coli cer遺伝子を含むプラスミドDS-2340-2-3のための構築スキームを示す図である。T7V38hia遺伝子カセットの上流および下流の同一の鎖上に転写ターミネーターも存在する。NTHi株33hia遺伝子をV38コドンからPCR増幅するために使用されるオリゴヌクレオチドを図8Bに示す。
【0048】
図9は、T7V38hia(11)またはT7V38hia(33)遺伝子カセットのタンデムコピーをそれぞれ含有するプラスミドDS-2447-2およびDS-2448-17の構築を示す図である。
【0049】
図10、パネルAは、NTHi株11由来の全長または切断型hia遺伝子をコードするプラスミドからのrHiaタンパク質の産生を示す図である。全長rHia(11)タンパク質の産生は非常に低かった。E21およびT33切断型rHiaタンパク質の場合も発現は低かった。しかし、V38およびN52切断型rHiaタンパク質は顕著に改善された発現レベルを示す。図10、パネルBから明らかなように、V38rHia(11)の産生は、E.Coli cer遺伝子を発現プラスミドへ添加した場合に明らかに増加している。
【0050】
図11は、封入体として製造されるrHiaタンパク質の精製スキームを示す図である。細胞を、超音波破砕により溶解し、封入体を連続した抽出により精製した。封入体を、グアニジニウムクロライド中に溶解し、ポリエチレングリコール(PEG)を添加して不純物を沈殿させた。(NH4)2SO4の添加によりrHiaが沈殿し、粗rHiaをさらにゲル濾過により精製した。
【0051】
図12は、株11および33由来の精製されたV38rHiaタンパク質を示す図である。封入体はレーン3に示され、最終精製タンパク質はレーン4に示される。精製されたタンパク質の推定純度は、SDS-PAGEデンシトメトリーにより測定して、約90%を上回る。
【0052】
図13は、ここに記載されるようにして製造されたrHiaタンパク質の、グリ
セロールを添加した場合としない場合でかつ4℃で、およびグリセロールを添加
して−20℃で8週間貯蔵する際の安定性に関する、SDS-PAGE分析を示
す図である。タンパク質はいずれの条件でも安定である。
【0053】
図14は、CD-1マウス中の株11および33由来V38rHiaタンパク質の免疫原性を示す図である。0.3〜10μgの用量では、いずれのタンパク質でも、1ないし2回投与した後に強い免疫応答を示す。これらのレベルでは顕著な用量応答は認められない。同様の結果が、BALB/cマウス(図15A)およびモルモット(図15B)でも見られ、このことはrHiaが投与量あたり0.3μgであっても強い免疫原性を有することを示唆している。
【0054】
図16は、V38rHia(33)によってもたらされる、NTHi株33が引き起こすコロニー形成に対する防護を示す図である。Yang 等(参考文献20)が記載しているように、チンチラ鼻咽頭コロニー形成モデルは、この疾患の初期の段階での防護を評価するために開発された。このモデルは、最初に、hmw遺伝子を発現するNTHi株のために確立され、hia遺伝子を発現するNTHi株に適応させなければならなかった。プロトタイプhmw-発現株(NTHi12)における感染の確立には102〜108cfuを使用するが、NTHi33株の場合には5×108cfuが必要であり、このような高レベルであってもプロトタイプhia-発現株11では感染が確立されない。100μg量で免疫化コホートは明らかに部分的に防護されるが、50μg量では防護されない。疾患の初期の段階でのこのような防護は、ワクチン抗原としてのrHiaアドヘシンの利用性を示している。
【0055】
図17は、別のHaemophilus influenzae hia遺伝子をPCR増幅するために使用されるオリゴヌクレオチドを示す図である。配列は、HiaおよびHsfタンパク質の保存アミノ酸およびカルボキシ末端配列に基づいている。
【0056】
図18は、NTHi株33hia遺伝子の完全なヌクレオチド配列および推定アミノ酸配列を示す図である。図19は、NTHi32hia遺伝子の完全なヌクレオチド配列および推定アミノ酸配列を示す図である。図20は、NTHi株29hia遺伝子の完全なヌクレオチド配列および推定アミノ酸配列を示す図である。図21は、NTHi株 M4071hia遺伝子の完全なヌクレオチド配列および推定アミノ酸配列を示す図である。図22は、NTHi株 K9hia遺伝子の完全なヌクレオチド配列および推定アミノ酸配列を示す図である。図23は、NTHi株 K22hia遺伝子の完全なヌクレオチド配列および推定アミノ酸配列を示す図である。図24は、Haemophilus influenzae タイプc株APIhia遺伝子の完全なヌクレオチド配列および推定アミノ酸配列を示す図である。図25は、NTHi株12由来のhia座の完全なヌクレオチド配列および推定アミノ酸配列を示す図である。PCR増幅されたフラグメントは、hia遺伝子の3’-末端へ連結したHaemophilus influenzaeタイプd株RdゲノムのHI1733遺伝子に関連する遺伝子の3’-末端を含有する。HI1733タンパク質の上流ORFのアラインメントを図27に示す。
【0057】
図26は、前記米国特許5,646,259に記載されるような、NTHi株11由来のHia遺伝子の完全なヌクレオチド配列および推定アミノ酸配列を示す図である。
【0058】
図28は、Hsf、Hiaから推定されるタンパク質の配列のアラインメント、およびM.catarrhalis200kDaタンパク質の部分配列を示す図である。
【0059】
本発明の種々の実施態様を、ワクチン摂取、診療、Haemophilus感染の治療、免疫剤の製造の分野の応用に利用できることは、当業者に明らかである。このような使用について以下に述べるが、これに限定するものではない。
【0060】
ワクチンの製造および使用
ワクチンとして使用するのに好適な免疫原性組成物は、ここに記載されるような、分類不能型Haemophilus株の免疫原性組み換えHaemophilus influenzaeアドヘシン(rHia)タンパク質、その免疫原性類似体およびフラグメントおよび/または免疫原性ペプチドから製造できる。ワクチンは免疫応答を誘起し、これにより抗-rHia抗体およびオプソニン化または死菌化された抗体を含む抗体が産生される。
【0061】
ワクチンを含む免疫原性組成物は、液体溶液またはエマルジョンのような注入可能な形で製造できる。rHiaタンパク質、その免疫原性類似体およびフラグメントおよび/または免疫原性ペプチドを、製薬学的に認容性の添加剤と混合してよく、該添加剤はrHiaタンパク質、免疫原性フラグメント類似体または免疫原性ペプチドと混和性である。このような添加物には、水、生理食塩水、デキストロース、グリセロール、エタノールおよびそれらの組合せが含まれる。
【0062】
免疫原性組成物およびワクチンはさらに、補助物質、例えば湿潤剤または乳化剤、pH緩衝剤、またはワクチンの効果を増強するためのアジュバントを含有してよい。
【0063】
免疫原性組成物およびワクチンは、皮下注射または筋肉内注射のような非経口投与が可能である。また、本発明で製造される免疫原性組成物を処方し、粘膜表面で免疫応答を喚起する方法で輸送する。従って、免疫原性組成物は例えば鼻または口(胃)のような経路で粘膜表面に適用してよい。
【0064】
免疫原性組成物は免疫系の特異的な細胞または粘膜表面へ輸送するために、標的分子と組み合わせてよい。標的分子には、WO92/17167に記載されるような(Biotech Australia Pty. Ltd.)、ビタミンB12およびバクテリア毒素、および米国特許第5,194,254(Barber等)に記載されるようなモノクローナル抗体が含まれる。
【0065】
また、座剤および経口処方を含む別の投与形態が望ましい。座剤には、結合剤および担体が含まれていてよく、例えばポリアルカレングリコールまたはトリグリセリドである。経口処方には、一般的に使用される添加剤、例えば製薬学的な量のサッカリン、セルロースおよび炭酸マグネシウムが含まれてよい。これらの組成物は、溶剤、懸濁剤、錠剤、丸剤、カプセル剤、持続性製剤または粉剤の形で投与され、rHiaタンパク質、フラグメント類似体および/またはペプチドを約1〜95%含有する。
【0066】
ワクチンは、投与処方に適合する方法で、治療効果、防護および免疫原性として機能する量で投与される。投与量は治療対象、例えば個人の免疫系の抗体合成能力および必要であれば細胞-仲介免疫応答の発現能力により変化する。投与に必要とされる活性成分の適切な量は、専門医の評価に依存する。しかし、好適な投与範囲は当業者により容易に決定可能であり、rHia、その類似体およびフラグメントおよび/またはペプチドはマイクログラム程度でよい。初期投与およびブースター投与のための好適な治療法も効果的であるが、初期投与には連続投与も含まれる。ワクチンの投与量は、投与経路に依存し、宿主の大きさによっても変化する。
【0067】
分類不能型HaemophilusのrHiaタンパク質をコードする核酸分子を免疫化に直接使用してよく、その際、DNAを直接投与、例えば遺伝子的免疫化として注入するかまたは生ベクター、例えば核酸分子を含むサルモネラ、BCG、アデノウイルス、ポックスウイルス、ワクシニアまたはポリオウイルスを構築する。異種抗体を免疫系へ運ぶために使用される数種の生ベクターについては、例えばO,Hagan(1992)(参考文献16)に議論されている。遺伝子的免疫化のための、DNAの被検対象への直接的な注入方法は、例えばUlmer等、1993(参考文献17)に記載されている。
【0068】
抗原と一般的にリン酸バッファー生理食塩水中の0.05〜1.0%溶液として使用されるアジュバントとを同時に投与することにより、免疫原性は著しく向上する。アジュバントは抗原の免疫原性を強化するが、それ自体は免疫原性を必ずしも有さない。アジュバントは、抗原を投与部位の近くに局所的に維持することにより、抗原の免疫系の細胞へのゆっくりとした継続的な放出を助ける、蓄積効果を生み出す。アジュバントはまた、免疫系細胞を蓄積した抗原へと誘引し、そのような細胞を刺激して免疫応答を促す。
【0069】
免疫促進剤またはアジュバントは、宿主の免疫応答を向上させるために長年使用されてきており、例えばワクチンが挙げられる。内因性アジュバント、例えばリポポリサッカライドは、一般的にワクチンとして使用される死菌または弱毒化バクテリアの成分である。外因性アジュバントは、典型的に、抗原と非-共役結合した免疫調節物質であり、宿主の免疫応答を強化するために処方される。従って、アジュバントは非経口的に導入された抗体への免疫応答を強化するものである。これらのアジュバントのいくつかは毒性であり、不所望の副作用を起こすので、ヒトおよび多くの動物に使用するのは不適当である。実際、水酸化アルミニウムおよびリン酸アルミニウム(まとめて一般的にアルムと称される)のみが、ヒトおよび家畜用のワクチン中にアジュバントとして慣用される。アルムによるジフテリアおよび破傷風トキソイドに対する抗体応答の増強は、よく確立されている。
【0070】
外因性アジュバントの多くは抗原に対する潜在的な免疫応答を呼び起こす。これらには、前記した特異的アジュバントならびに膜タンパク質抗原と複合するサポニン(免疫刺激複合体)、鉱油を含む複合ポリマー、死んだ微生物および鉱油、フロイドの完全アジュバント、微生物の生成物、例えばムラミルジペプチド(MDP)およびリポポリサッカライド(LPS)ならびにリピドAおよびリポソームが含まれる。
【0071】
効果的に体液性免疫応答(HIR)および細胞-仲介免疫(CMI)を誘導するには、免疫原をアジュバント中に乳化させる。多くのアジュバントは毒性であり、肉芽腫、急性および慢性の炎症(フロイドの完全アジュバント、FCA)、細胞溶解(サポニンおよび複合ポリマー)および発熱、関節炎および前ブドウ膜炎(LPSおよびMDP)を引き起こす。FCAは優れたアジュバントであり研究に汎用されるが、その毒性のために、ヒトまたは家畜用ワクチン中に使用することを許可されていない。
【0072】
理想的なアジュバントの所望の特徴は:
(1)毒性の欠如;
(2)長期にわたる免疫応答刺激能力;
(3)製造の容易性および長期貯蔵における安定性;
(4)種々の経路で投与された抗原に対して、CMIおよびHIRの両方を誘起する能力;所望であれば、
(5)他のアジュバントとの相乗作用;
(6)抗原提示細胞(APC)群と選択的に相互作用する能力;
(7)適切なTH1またはTH2細胞-特異的免疫応答を特異的に誘起する能力;
および
(8)抗原に対する適切な抗体イソタイプレベル(例えば、IgA)を選択的に増加させる能力である。
【0073】
参照することでここに組み込んだものとする、Lockhoff等の1989年8月8日付けの米国特許第4,855,283には、免疫調節剤またはアジュバントとして、N-グリコシルアミド、N-グリコシルウレアおよびN-グリコシルカルバメートを含む糖脂質類似体が記載され、それぞれ糖残基をアミノ酸で置換されていてよい。また、Lockhoff等、1991(参考文献18)は、天然の糖脂質と構造的類似性を有するN-糖脂質類似体、例えばグリコスフィンゴリピドおよびグリコグリセロリピドが、ヘルペス単純ウイルスワクチンおよび偽性狂犬病ウイルスワクチンの両方で強い免疫応答を誘起することを報告している。ある種の糖脂質は長鎖-アルキルアミンと脂肪酸とから合成され、芳香族炭素を介して糖と直接結合し、天然の脂質残基の機能を模倣する。
【0074】
本出願と同じ譲受人に譲渡され、参照により本明細書に組み込む、Molony等による米国特許第4,258,029には、破傷風トキソイドおよびホルマリン不活化タイプI、IIおよびIIIポリオウイルスワクチンと複合させた場合に、オクタデシルチロシンヒドロクロライド(OTH)がアジュバントとして機能することが記載されている。またNixon-George等、1990(参考文献19)は、組み換えB型肝炎表面抗原と組み合わせた、芳香族アミノ酸のオクタデシルエステルが、B型肝炎ウイルスに対する宿主の免疫応答を強化することを報告している。
【0075】
イムノアッセイ
本発明により製造されるHaemophilusの分類不能型株のrHiaタンパク質、その類似体およびフラグメントは、免疫原性として、酵素結合性免疫吸着剤検定(ELISA)、RIAおよび他の非-酵素結合性抗体結合アッセイまたは抗菌、Haemophilus、および/またはHia抗体を検出するための従来公知の方法を含むイムノアッセイ中の抗原として、有用である。ELISAアッセイでは、Hiaタンパク質、類似体およびフラグメントを、選択された表面、例えばポリスチレンマイクロタイタープレートの壁面のようなタンパク質またはペプチドが結合可能な表面上へ固定する。不完全に吸着されているHiaタンパク質、類似体および/またはフラグメントを除くために洗浄し、その後、非特異的タンパク質、例えば試験サンプルに対する抗原性が中性であることを知られているウシ血清アルブミン(BSA)またはカゼイン溶液を選択表面へ結合させてよい。これにより、固定表面上の非特異的吸着部位が遮断され、結果的に、表面上への抗血清の非特異的結合によって起こるバックグラウンドが減少する。
【0076】
次いで固定表面を、臨床または生化学的材料のようなサンプルと接触させ、免疫複合体(抗原/抗体)の形成が起こる方法で試験する。この際、BSA、ウシガンマグロブリン(BGG)および/またはリン酸バッファー生理食塩水(PBS)/Tweenのような希釈剤でサンプルを希釈してよい。サンプルを次に約25℃〜37℃程度の温度で約2〜約4時間インキュベートする。インキュベートに続き、サンプル-接触表面を洗浄して、非-免疫複合材料を除去する。洗浄方法には、PBS/Tween、またはホウ酸バッファーのような溶液による洗浄が含まれてよい。
【0077】
試験サンプルと結合Hiaタンパク質、類似体および/またはフラグメントとの間に特異的な免疫複合体が形成された後、次いで洗浄し、複合体形成の出現およびその量を、一次抗体に特異的な二次抗体を免疫複合体へ添加することにより測定する。試験サンプルがヒト起源である場合、二次抗体はヒトイムノグロブリンに特異的な抗体であり、一般的にIgGである。検出可能にするために、二次抗体は、例えば適当な発色性物質とインキュベートすることにより呈色が見られるような酵素活性等の関連活性を有する。定量化は、例えば可視スペクトル分光光度計を用いて呈色の度合いを測定することにより実施される。
【0078】
ハイブリダイゼーションプローブとしての配列の使用
本発明のヌクレオチド配列は、新規に単離および特徴付けされたhia遺伝子配列を含有し、他のHaemophilus分類不能型株から、hia遺伝子を同定およびクローニングすることができる。
【0079】
本発明のhia遺伝子配列を含有するヌクレオチド配列は、その能力のために、他のhia遺伝子の補足的部分と倍体分子を選択的に形成するのが有用である。適用に応じて、種々のハイブリダイゼーション条件を利用し、他の分類不能型Haemophilus株中の別のhia遺伝子のための、プローブの選択性の度合いを変化させる。高い選択性のためには、比較的厳しい条件を利用して倍体を形成し、例えば低塩および/または高温条件、例えばNaCl 0.02M〜0.15Mでありかつ約50〜70℃の温度という条件が挙げられる。ある種の適用では、それほど厳しいハイブリダイゼーション条件は必要とされず、例えば0.15〜0.9Mの塩で20〜55℃の温度範囲でよい。ハイブリダイゼーション条件は、ホルムアミドの添加量が増加すると、ハイブリッドした倍体が不安定になるので、より厳しい条件が必要とされる。従って、特定のハイブリダイゼーション条件は容易に操作でき、所望の結果に応じて方法を選択するのが一般的である。一般的に、便利なハイブリダイゼーション温度は、50%ホルムアミドの存在下かつNaCl 0.15Mで:hia遺伝子が標的核酸フラグメントと95〜100%の相同性を有する場合に42℃、90〜95%の相同性で37℃および85〜90%の相同性で32℃である。
【0080】
臨床診断の態様において、本発明のhia遺伝子の核酸配列は、適当な手段、例えばハイブリダイゼーションを測定するための標識、と組み合わせて利用してよい。種々の適当な指示手段が知られており、放射活性、酵素または他のリガンド、例えばアビジン/ビオチンが含まれ、検出可能なシグナルの提供を可能にする。ある診断の態様において、放射活性タグの代わりに、酵素タグ、例えばウレアーゼ、アルカリンホスファターゼまたはペルオキシダーゼを使用してもよい。酵素タグの場合、肉眼または分光光度計で視覚的に捉えられ、Hia遺伝子配列を含むサンプルとの特異的ハイブリダイゼーションを同定できる手段として使用されるものとして、発色指示物質が公知である。
【0081】
本発明のHia遺伝子核酸配列は、溶液ハイブリダイゼーション中のハイブリダイゼーションプローブとして、および液相法で使用される態様として有用である。固相法に関する態様において、滲出液、体液(例えば血清、羊水、中耳滲出液、唾液、気管支肺胞洗浄液)または組織のような臨床サンプルを含むサンプルの試験DNA(またはRNA)を、選択したマトリクスまたは表面へ吸着あるいは固定させる。固定された一本鎖核酸を、次いで、所望の条件下に、本発明のhia遺伝子またはそのフラグメントの核酸配列を含む選択プローブと特異的にハイブリダイゼーションさせる。選択する条件は、例えばG+C含量、標的核酸のタイプ、核酸源、ハイブリダイゼーションプローブの大きさ等に応じて必要とされる特定の基準に基づく特定の状況に応じて変化する。続いてハイブリダイゼーション表面を洗浄して非特異的に結合したプローブ分子を除去し、標識の手段により、特異的ハイブリダイゼーションを検出または定量化する。Haemophilus種で保存されている核酸配列部分を選択するのが好ましい。選択されたプローブは、少なくとも長さ18bpであり、多くの場合30〜90bp長さである。
【0082】
Haemophilus influenzaeアドヘシン遺伝子の発現
宿主細胞に適合性の種に由来するレプリコンおよび制御配列を、発現系中のhia遺伝子の発現に利用してよい。ベクターは、通常、複製部位ならびに形質転換細胞中の表現型の選択を行うマーカー配列を有する。例えば、E.Coliは、アンピシリンおよびテトラサイクリン耐性遺伝子を含むpBR322を使用して形質転換でき、従って、形質転換細胞を同定するための簡易な方法を提供する。pBR322プラスミド、または他の微生物プラスミドまたはファージは、宿主細胞が自身のタンパク質を発現させるために使用できるプロモーターも含有するか、または含有するように変性する。
【0083】
さらに、宿主細胞に適合性のレプリコンおよび制御配列を含有するファージベクターは、これらの宿主と一緒に形質転換ベクターとして使用できる。例えば、ラムダGEMTM-11中のファージを、E.ColiLE392のような宿主細胞を形質転換させるために使用できる組み換えファージベクターを製造するために使用してよい。
【0084】
組み換えDNA構築体中で一般的に使用されるプロモーターには、β-ラクタマーゼ(ペニシリナーゼ)およびラクトースプロモーターシステムおよび他の微生物プロモーター、例えばここで好ましい態様として使用されるT7プロモーターシステム(米国特許第4,952,496)が含まれる。プロモーターのヌクレオチド配列の詳細は公知であり、当業者にはそれらを遺伝子を用いて機能的に結合することが可能である。使用される特定のプロモーターは、一般的に、所望の結果に応じて選択されたプロモーターである。Hiaタンパク質および免疫学的フラグメントまたはその類似体の発現に適当な宿主には、E.Coliが含まれ、Bordetella種、Bacillus種、Haemophilusu種、菌体、酵母またはバクロウイルスシステムを使用してもよい。E.Coliがここで使用される好ましい宿主である。
【0085】
本発明において、特にHaemophilus種を培養して精製した天然Hiaタンパク質が微量の毒性物質または他の汚染物質を含む場合に、Hiaタンパク質の製造は、組み換え法によるのが好ましい。この問題点は、組み換えにより製造されたHiaタンパク質を異種システム中で使用することにより解決され、この際、Hiaタンパク質は、特に記載された構築体を使用して、精製物質中の汚染物質を最小化する方法で宿主から単離される。
【0086】
微生物寄託
ここに記載し参照した分類不能型Haemophilus株の高分子量タンパク質をコードする核酸を含むベクターを、本明細書の出願に先立って、ブタペスト条約に従い、10801 University Boulevard、Manassas、Virfinia 20110-2209、米国に所在するAmerica Type Culture Collection(ATCC)に寄託した。寄託されたベクターのサンプルは、公共に利用可能であり、課される全ての制限または寄託物を利用する権利は、この米国特許出願に基づく特許の許可を受けることとする。さらに、受託者が生存可能なサンプルを分配できない場合には、寄託物を受託者に戻すこととする。寄託された実施形態は本発明を例示するためだけのものであるので、本明細書に記載および請求された発明が、寄託された生物材料によって範囲を限定されることはない。本出願の記載と等価または同等な抗原をコードする核酸を含むいずれの等価または類似のベクターも本発明の範疇に入るものとする。
寄託概要
プラスミド ATCC 寄託年月日
BK-96-2-11 203771 1999年2月11日
【実施例】
【0087】
上記の開示は概して本発明を説明している。以下の特定した実施例を参照するとさらに完全な理解が得られる。これらの実施例はただ例示の目的で説明されるものであり、本発明の範囲を限定しようとするものではない。状況によって形状の変化および同等品への置き換えを図ることで好都合なことを示唆できるかまたは提供できる。ここでは特殊な用語を使用したが、そのような用語は説明的感覚で使用するつもりであって限定を目的とするものではない。
【0088】
本開示およびこれらの実施例には明記していないが、使用した分子遺伝学、蛋白質生化学、免疫学および発酵技術による方法は科学文献で広く報告されており、当業者の能力に充分に保有されたものである。
【0089】
実施例1
この実施例はNTHi菌株11由来の全長のそろったrHia蛋白質を発現するプラスミドDS-2008-2-3の構造を説明するものである。
【0090】
NTHi菌株11から染色体DNAを精製し、図1Bに記載したオリゴヌクレオチド(5038.SLおよび5039.SL)を使用して全長のそろったhia遺伝子をPCR増幅した。pT7-7発現ベクター(ref.21)にクローン化するためにNde I部位をこの遺伝子の5’末端で設計し、BamH I部位を3’末端で設計した。増幅した断片をNde I/BamH Iで切断し、同じ酵素で切断したpT7-7にクローン化した。プラスミドDS-2008-2-3はT7プロモーターの下流に3.4kbの11株hia遺伝子を含む(図1A)。このプラスミドは組換えHiaを発現するために使用した(下記の実施例9)。
【0091】
実施例2
この実施例は抗Moraxella catarrhalis野生株高分子量アドヘシン抗体によるrHiaの識別を例示するものである。
【0092】
インフルエンザ菌のHia蛋白質と前述した米国特許第5,808,024号の中でM.catarrhalis200kDa蛋白質として分類されるMoraxella catarrhalis高分子量アドヘシン(図28)との間にはいくらかの配列不変性が認められる。米国特許第5,808,024号の記載では、M.catarrhalis野生株200kDa蛋白質がゲル精製され、テンジクネズミの抗野生株200kDa抗体が生成された。T7 hia遺伝子はプラスミドDS-2008-2-3から発現され、rHia蛋白質を含む細胞培養物がニトロセルロース膜に電気泳動的に転写された。抗野生株200kDa抗体を使用した免疫ブロット分析から、この抗体が図2に見られるように、rHia蛋白質を認識することが示された。
【0093】
実施例3
この実施例はT7 hia(11)遺伝子カセットのタンデムコピーを含むプラスミドDS-2092-1およびDS-2092-40の構造を説明するものである。
【0094】
全長のそろった組換えHia蛋白質の生成を向上させるために、菌株11のhia遺伝子(実施例1)を含むT7 hia遺伝子カセットのタンデムコピーを単一のベクターに挿入した。プラスミドDS-2008-2-3をBgl IIで直線化させ、脱リン酸化した。プラスミドDS-2008-2-3はまた、T7 hia遺伝子カセットを切り取るためにBgl IIおよびBamH Iによっても切断された。このT7 hia断片を直線化したベクターに連結し、反時計回り方向に配向して2つのT7 hia遺伝子コピーを含むプラスミドDS-2092-1(a,a)および反対方向に配向してタンデムコピーを含むプラスミドDS-2092-40(a,c)(図3)を生成した。どちらの作製試料からもrHia発現における顕著な向上は認められなかった(下記の実施例9参照)。
【0095】
実施例4
この実施例は末端欠失した菌株11のhia遺伝子を発現するプラスミドの作製を説明するものである。
【0096】
T7 hia遺伝子カセットの単一またはタンデムのコピーからのrHia蛋白質生成量は非常に低く、この蛋白質は(下記の実施例9で説明するように)E.coliに対する毒性を有するようであった。インフルエンザ菌のHiaは表面露出したアドヘシン分子であるので、分泌をするためにはシグナル配列または補助蛋白質のいずれかを使用しなければならないが、しかし関連の補助遺伝子は判明していない。もしもE.coli中での組換え蛋白質発現のためにシグナル配列が除去された場合、rHiaは封入体として発現され、毒性効果が低減される可能性がある。推定されるシグナル配列および切断部位を分類し、N末端で末端欠失したrHia蛋白質を発現する4種類の作製試料を設計した(図4)。菌株11のhia遺伝子では開始コドンから約500bpに独特のSty I切断部位が存在する。プラスミドDS-2008-2-3をNde IおよびSty Iで切断し、5.7kbのベクター断片を精製した(図5A)。末端欠失部位からSty I切断部位までを増幅するためにPCRプライマーを設計し、末端欠失したクローンを選別するためにアンチセンスプライマーに独特のNhe I切断部位を導入した(図5B)。増幅した断片は、より操作を容易にするためにpCRII中でサブクローン化し、プラスミドDS-2153R-1-2(E21)、DS-2165-4-8(T33)、DS-2153-3-5(V38)、およびDS-2153-4-4(N52)を生成した。このpCRII hiaプラスミドをNde IおよびSty Iで切断し、断片をDS-2008-2-3から得たベクター片と連結した。プラスミドDS-2186-1-1(E21)、DS-2201-1(T33)、DS-2186-2-1(V38)、およびDS-2168-2-6(N52)を、カッコ内に示したT7プロモーターおよび末端欠失したhia遺伝子を含ませて生成した。これらのプラスミドは組換えHiaを発現させるために使用した(下記の実施例9参照)。
【0097】
実施例5
この実施例はT7 V38 hia(11)カセット、E.coli cer遺伝子、およびカナマイシン耐性遺伝子を含むプラスミドBK-96-2-11の作製を説明するものである。
【0098】
プラスミドDS-1843-2は多数のクローン化部位および2つの転写終結部がEcoR IおよびPst I部位の間でオリゴヌクレオチド上に導入され、これによってクロラムフェニコールおよびアンピシリン耐性遺伝子の両方が破壊されたpBR328を基にしたプラスミドである(図6B)。pUC-4Kから得たカナマイシン耐性遺伝子をSal I部位に挿入し、カナマイシン耐性でかつテトラサイクリン感受性のプラスミドDS-2147-1を生成した。プラスミドDS-2224-1-4はオリゴヌクレオチドから作製してBamH I切断部位に接する合成のE.coli cer遺伝子(ref.15)を含むpUCプラスミドである。このcer遺伝子の290bpのBamH I断片をDS-2147-1のBamH I部位に挿入してプラスミドBK-2-1-2を作製した。その結果、このpBRを基にしたプラスミドは多数のクローン化部位、カナマイシン耐性遺伝子およびcer遺伝子を含む。プラスミドBK-2-1-2はBgl IIで直線化し、脱リン酸化した。プラスミドDS-2186-2-1をBgl IIおよびBamH Iで切断し、3.6kbのT7 V38 hia断片をBK-2-1-2に挿入してプラスミドBK-96-2-11を作製した(図6A)。
【0099】
実施例6
この実施例はE.coli cer遺伝子の存在下で全長のそろったNTHi菌株33のhia遺伝子を発現するプラスミドDS-2242-1およびDS-2242-2の作製を説明するものである。
【0100】
NTHi菌株33から染色体DNAを精製し、オリゴヌクレオチド5039.SLおよび5040.SL(図17)を使用してPCR増幅を実施した。センスプライマー(5040.SL)は菌株11のhiaの5’隣接配列を基にして設計したものであって、NTHi HiaおよびHib Hsf蛋白質のアミノ末端配列を保存していた。アンチセンスプライマー(5039.SL)は実施例1で述べたものと同じであって、HiaおよびHsf蛋白質のカルボキシ末端配列の保存を基本とした。菌株33の3kbのhia PCR断片をpCR II中でクローン化し、プラスミドDS-1917-3-8を作成した。
【0101】
全長のそろった菌株33 hia遺伝子を発現させるために、この遺伝子の5’末端の、開始コドンからAlwN I切断部位まで(図7B)の、およそ106bpをオリゴヌクレオチドから合成した。プラスミドDS-1917-3-8をAlwN IおよびBamH Iで切断し、hia遺伝子を含む約2.9kbの断片を精製した。プラスミドpT7-7をNde IおよびBamH Iで切断し、Nde I-AlwN IオリゴヌクレオチドおよびAlwN I-BamH I hia断片をこのpT7-7ベクターに連結してプラスミドDS-2103-4を作製した。
【0102】
E.coli cer遺伝子を含ませ、カナマイシンによる選択を使用するために、T7 hia(33)遺伝子カセットを含むBgl II-BamH I断片をDS-2103-4から切り取り、Bgl IIで切断されて脱リン酸化されたBK-2-1-1中でクローン化した。プラスミドDS-2242-1およびDS-2242-2は反対方向に配向したT7 hia(33)遺伝子カセットの単一コピー、E.coli cer遺伝子、およびカナマイシン耐性遺伝子を含む(図7A)。
【0103】
実施例7
この実施例は末端欠失したV38菌株33のhia遺伝子をもつT7 hia遺伝子カセット、E.coli cer遺伝子、およびカナマイシン耐性遺伝子を含むプラスミドDS-2340-2-3の作製を説明するものである。
【0104】
NTHi菌株33 hia遺伝子の、V38開始コドンから内側のSnaB I切断部位に至るまでの、5’末端の250bp断片を増幅するためにPCRプライマーを設計した。クローン化を目的として5’末端にNde I切断部位を追加し、鋳型としてプラスミドDS-2242-1を使用してこの断片を増幅した。この構成図式は図8Aに示されており、PCRプライマーは図8Bに示されている。この断片をpCR IIにクローン化し、プラスミドDS-2328-1-1を作製した。DS-2242-1をNde IおよびSnaB Iで切断し、8.5kbのベクター断片を精製した。DS-2328-1-1をNde IおよびSnaB Iで切断し、その0.25kbの5’hia断片をDS-2242-1から得た8.5kbのベクター断片と連結してプラスミドDS-2340-2-3を作製した。
【0105】
実施例8
この実施例はT7 V38 hia(11)またはT7 V38 hia(33)遺伝子カセットのタンデムコピーをそれぞれと、E.coli cer遺伝子、およびカナマイシン耐性遺伝子を含むプラスミドDS-2447-2およびDS-2448-17の作製を説明するものである。
【0106】
T7 V38 hia(11)遺伝子カセットを含むプラスミドBK-96-2-11をBgl IIで直線化し、脱リン酸化した。DS-2186-2-1から得たBgl II-BamH I T7 V38 hia(11)遺伝子カセットをBK-96-2-11に連結し、同じ配向でT7 V38 hia(11)遺伝子のタンデムコピーを含むプラスミドDS-2447-2を作製した(図9A)。
【0107】
プラスミドDS-2340-2-3をEcoR Iで切断し、そのT7 V38 hia(33)遺伝子カセットを、EcoR Iで切断して脱リン酸化したpUC-BgXbにサブクローン化した。その結果得られたプラスミドDS-2440-2をBgl IIおよびBamH Iで切断してT7 V38 hia(33)カセットを放出させ、Bgl IIで直線化して脱リン酸化したDS-2340-2-3とそれを連結した。プラスミドDS-2448-17は同じ配向でタンデムのT7 V38 hia(33)遺伝子を含む(図9B)。
【0108】
実施例9
この実施例は全長のそろったおよび末端欠失した組換えhia遺伝子の発現を説明するものである。
【0109】
先行の実施例で説明したようにして調製した発現プラスミドのDNAを、BioRad社のエレクトロポレータを使用して電子的に受容可能となったE.coli BL21(DE3)細胞に導入した。細胞は適切な抗生物質選別を使用して、ラクトースを1.0%になるまで加える前に、A578が0.3となるまでNZCYM培地中で37℃にて4時間成育させた。サンプルはSDS-PAGE分解液+負荷バッファで0.2OD/μlとなるよう調整し、それぞれの蛋白質サンプルを同じ量でSDS-PAGEゲル(ref.22)に流した。図10は種々の作製試料からのrHia(11)蛋白質相対生成量を図解したものである。パネルAに見られるように、rHiaのサイズが低下するにつれて生成量の増加が認められる。V38-(レーン5)およびN52-末端欠失rHia(レーン6)はそれらのさらに長い相当物質(レーン2、3、4)よりも有意差をもって高い発現レベルを有する。加えて、パネルBはV38 rHiaの生成がcer遺伝子の存在下で明らかに増加することを示している。
【0110】
実施例10
この実施例はrHia蛋白質の精製について説明するものである。
【0111】
すべての組換えHia蛋白質はE.coli中で封入体として発現され、同じ手法によって精製された(図11)。500mlの培養物から得たE.coli細胞ペレットを0.1MのNaClを含むpH8.0の50mMトリス塩酸50ml中に再懸濁し、超音波破砕した。この抽出物を20,000×gで30分間遠心分離し、その結果生じた上清を廃棄した。このペレット(PPT1)をさらに0.5%Triton X-100および10mM EDTAを含む50mlの50mMトリス塩酸、pH8.0で抽出処理し、それから20,000×gで30分間遠心分離して上清を廃棄した。このペレット(PPT2)をさらに1%のオクチルグルコシドを含む50mlの50mMトリス塩酸、pH8.0で抽出し、それから20,000×gで30分間遠心分離して上清を廃棄した。
【0112】
以上の抽出処理後に結果として得られたペレット(PPT3)は封入体を含んでいる。このペレットを6Mのグアニジンおよび5mMのDTTを含む6mlの50mMトリス塩酸、pH8.0に溶解した。この溶液に12mlの50mMトリス塩酸、pH8.0を加え、混合液を20,000×gで30分間遠心分離した。この上清(SUP4)を終濃度7%のポリエチレングリコール(PEG)4000で沈降させた。その結果生じたペレット(PPT5)を20,000×gで30分間遠心分離して除去し、上清を50%飽和の硫酸アンモニウムで沈降させた。硫酸アンモニウムによる沈降物は20,000×gで30分間遠心分離して採取した。結果として得られたペレット(PPT6)を6Mのグアニジン塩酸および5mMのDTTを含む2mlの50mMトリス塩酸、pH8.0に溶解し、この透明な液体を、2Mのグアニジン塩酸を含む50mMトリス塩酸、pH8.0で平衡化したSuperdex200ゲル濾過カラムで精製した。画分はSDS-PAGEによって分析し、精製されたrHiaを含む画分をプールし、4℃でPBSに対して一晩透析し、その後、20,000×gで30分間遠心分離した。これらの条件では蛋白質は可溶性を維持しており、−20℃で保存するためにこのrHia調製液にグリセロールを終濃度20%で加えた。精製されたV38 rHia(11)およびV38 rHia(33)のSDS-PAGE分析は図12に図解されている。精製V38 rHia蛋白質の平均収率は約10mg/L培地である。
【0113】
rHiaの安定性を調べるために、精製したV38 rHia(11)蛋白質は4℃でグリセロールなしかまたは入れて保存するか、あるいはグリセロールを入れて-20℃で保存した。この蛋白質は3つの条件すべてにおいて安定であることが分かり、凍結と解凍を繰り返して少なくとも8週間は完全さを維持した(図13)。
【0114】
実施例11
この実施例はV38 rHia(11)およびV38 rHia(33)蛋白質の免疫原性を説明するものである。
【0115】
日程1において完全フロイントアジュバント(CFA、Difco)で乳化した5μgの抗原を筋肉内(i.m.)注入して二匹のテンジクネズミ(Charles River)を免疫感作させることによってrHia蛋白質に対する高度免疫抗血清を生成させた。日程14および28において不完全フロイントアジュバント(IFA)に加えた蛋白質5μgを投与して動物を追加免疫応答させ、日程42において血清を集めた。免疫原として熱で不活化したバクテリア調製物(1回の投与量1×108cfu)を使用した以外は同じプロトコルを使用して、抗Hib菌株MinnAおよび抗ヘモフィルス属菌株ATCC9006抗血清を作製した。
【0116】
V38 rHia蛋白質の免疫原性を調べるために、5匹のCD-1マウス(Charles River,Quebec)の複数グループを日程1および28において、リン酸アルミニウム(alum)の存在下(1回の投与量1.5mg)で0.3、1、3、および10μgの抗原によりs.c.免疫感作させた。血液サンプルを日程1、28および42において集めた。顕著に抗V38 rHia抗体を生成したマウスは抗体0.3μgを1回注入されても応答を示し(図14、パネルA)、両方の蛋白質ともに封入体抽出および可溶化処理の後でも免疫原性を維持することを示唆している。菌株11または33に由来するV38 rHia蛋白質によって誘導される抗体力価において統計学的に顕著な差異は認められなかった。
【0117】
BALB/cマウスにおいてV38 rHia(11)蛋白質の免疫原性を調べるために、5匹の動物(Charles River,Quebec)の複数グループを日程1、28および42において、リン酸アルミニウムの存在下(1回の投与量1.5mg)で抗原0.3、1、3、および10μgによりs.c.免疫感作させた。血液サンプルは日程1、14、28、42および56において集めた。すべてのグループにおいて高い抗体力価が認められ、この蛋白質が1回の投与量0.3μgにおいてさえ強度の免疫原性であることを示した(図15、パネルA)。
【0118】
テンジクネズミにおいてV38 rHia(11)蛋白質の免疫原性を調べるために、5匹の動物(Charles River,Quebec)の複数グループを日程1、28および42において、リン酸アルミニウムの存在下(1回の投与量1.5mg)で抗原0.3、1、3、および10μgによりs.c.免疫感作させた。血液サンプルは日程1、14、28、42および56において集めた。すべてのグループにおいて高い抗体力価が認められ、この蛋白質がテンジクネズミにおいてもまた強度の免疫原性であることを示した(図15、パネルB)。
【0119】
実施例12
この実施例は菌血症の無抵抗な幼齢ラットのモデルにおいて抗rHia抗体によって生じる防御の分析を説明するものである。
【0120】
妊娠中のWistarラットをCharles Riverから入手した。インフルエンザ菌b型の菌血症モデルでは、6〜10匹の5日齢の幼齢ラットのグループを、テンジクネズミの抗rHiaまたは抗菌株MinnA抗血清0.1ml、背面 でs.c.注入した。比較対照の動物には免疫感作前の血清のみを注入した。20時間後、元気に生育したHib菌株MinnAを200〜240コロニー形成ユニット(cfu)で(0.1ml)腹腔内(i.p.)抗原投与した。血液サンプルを抗原投与後20時間に、イソフルラン麻酔して心臓に針を刺して集め、チョコレート寒天培地にまいた。1日後、コロニーをカウントしてその結果をフィッシャー検定(Fisher’s Exact test)によって統計学的に分析した。
【0121】
インフルエンザ菌a型菌血症モデル(ref.23)では、9〜10匹の5日齢の幼齢ラットのグループを、テンジクネズミの抗rHiaまたは抗菌株ATCC9006抗血清0.1ml、背面でs.c.注入した。比較対照グループの動物にはテンジクネズミの免疫感作前の血清を注入した。20時間後、元気に生育したインフルエンザ菌a型菌株ATCC9006を(0.1ml)100,000cfuで動物にi.p.抗原投与した。血液サンプルを抗原投与の20時間後に集めて上述のように分析した。
【0122】
後述の表1および2に示したように、テンジクネズミの抗rHia(11)または抗V38 rHia(11)抗血清のいずれかで受動免疫感作された幼齢ラットは、菌血症を引き起こしたインフルエンザ菌a型またはb型に対してすべてが顕著に防御された。これらの結果は、インフルエンザ菌a型またはb型によって引き起こされる菌血症に対して動物を防御することにおいて、わずかに末端欠失したHia蛋白質(V38 rHia)に対応して生成した抗体が全長のそろった蛋白質に対応して生成したものと同程度に効果的であることを示している。NTHi由来の組換え蛋白質によって生じる、被包性のバクテリアによって引き起こされる侵襲性の疾病に対するそのような防御はrHia蛋白質のワクチン抗原としての有用性を説明するものである。
【0123】
実施例13
この実施例は鼻咽頭部でのコロニー形成のチンチラのモデルにおいて、V38 rHia蛋白質の免疫感作によって生じる防御を説明するものである。
【0124】
鼻咽頭部のコロニー形成モデルはYangらによって報告された(ref.20)。このモデルは、HMWアドヘシンを産生するNTHi菌株については適切であるが、しかしHiaを産生する菌株では同じ条件で再現性のあるコロニー形成は達成できなかった。原型のHia産生NTHi菌株11で繰り返したコロニー形成の試みは不成功であった。原型のHMW産生NTHi菌株12ではわずか108cfuしか必要としなかったのに比べて、NTHi菌株33では1回の植菌当たり5×108cfuでコロニー形成が達成された。これらの条件下では、100μgのV38 rHia(33)で免疫感作され、同様のNTHi菌株33で抗原投与された動物において部分的な防御が認められた。
【0125】
実施例14
この実施例はインフルエンザ菌株に由来する追加のhia遺伝子のクローン化および配列分析を説明するものである。
【0126】
保存されたプロモーター、hiaおよびhsf遺伝子および蛋白質のN末端およびC末端配列に基づいて、PCR増幅のためのオリゴヌクレオチド(5040.SLおよび5039.SL)を設計した(図17)。PCR増幅のための選択菌株は抗rHia(11)抗血清との反応性に基づいて選んだ。
【0127】
NTHi菌株12、29、32、M4071、K9および、K22およびヘモフィルスc型菌株APIから染色体DNAを調製した。PCR増幅は以下のようにして実施した。それぞれの反応混合液には5〜100ngのDNA、それぞれ1μgのプライマー、5ユニットのtaq+またはtsg+(Sangon)またはtaqプラスロング(Stratagene)、2mMのdNTPs、20mMのトリス塩酸(pH8.8)、10mMの塩化カリウム、10mMの硫酸アンモニウム、2mMの硫酸マグネシウム、0.1%のTriton X-100、BSAを含ませた。サイクル条件は、95℃で1分間、その後、95℃で30秒、45℃で1分間、72℃で2分間のサイクルを25サイクル行い、その後、72℃で10分間とした。
【0128】
菌株33のhia遺伝子のヌクレオチドおよびそれに由来するアミノ酸の配列が図18に示されている。予測された菌株33由来のHia蛋白質は103.6kDaの分子量を有し、pIが9.47であった。菌株32のhia遺伝子のヌクレオチドおよびそれに由来するアミノ酸の配列は図19に示されている。予測された菌株32由来のHia蛋白質は70.4kDaの分子量を有し、pIが5.67であった。残基493と496との間にはKDEL配列が存在する。そのような配列は小胞体への固定蛋白質に付随するものであった。由来する菌株32のHia蛋白質は顕著により小さくかつ顕著に異なるpIを有するが、しかしながら他のHia蛋白質に存在する特色の多くを含んでいる。
【0129】
菌株29のhia遺伝子のヌクレオチドおよびそれに由来するアミノ酸の配列が図20に示されている。予測された菌株29のHia蛋白質は114.4kDaの分子量および7.58のpIを有する。菌株K22のhia遺伝子のヌクレオチドおよびそれに由来するアミノ酸の配列が図23に示されている。予測された菌株K22のHia蛋白質は114.4kDaの分子量および7.58のpIを有する。NTHi菌株29およびK22に由来するHia配列は同一であることが判明した。菌株29はオハイオ州クリーブランドの中耳炎の生後7カ月の乳児から単離され、それに対して菌株K22はオーストラリアのキンバリー近郊のアボリジニから単離された。
【0130】
菌株4071のhia遺伝子のヌクレオチドおよびそれに由来するアミノ酸の配列が図21に示されている。予測されたM4071のHia蛋白質は103.4kDaの分子量および9.49のpIを有する。残基534と537との間にはKDEL配列が存在する。
【0131】
菌株K9のhia遺伝子のヌクレオチドおよびそれに由来するアミノ酸の配列が図22に示されている。予測された菌株K9のHia蛋白質は113.8kDaの分子量および6.45のpIを有する。
【0132】
ヘモフィルスc型菌株APIのhia遺伝子のヌクレオチドおよびそれに由来するアミノ酸の配列が図24に示されている。予測されたAPIのHia蛋白質は249.4kDaの分子量および5.34のpIを有する。c型菌株APIの由来するHia/Hsf配列は60残基の挿入部があることを除いて公表されているb型のHsf配列と殆ど同一である。ここに提示したNTHiに基づくHia蛋白質はa型およびb型感染の受動モデルにおいて防御をするので、b型とc型蛋白質との間の配列類似性に起因するc型の疾病に対してもまた防御をすることはありそうである。
【0133】
菌株12のhia遺伝子座のヌクレオチドおよびそれに由来するアミノ酸の配列が図25に示されている。NTHi菌株12はHiaを生成しない。しかしながら、hia遺伝子部分はPCR増幅可能であり、SB12細胞溶解物と抗rHia抗体とのつじつまの合わない陽性の反応性が存在し、サザンブロットでは菌株11のhia遺伝子の3’末端に由来するDNAプローブとの反応性がある。PCR増幅したDNAの分析から、上流HI1732関連遺伝子の3’末端の1kbおよびhia遺伝子の3’末端の0.8kbを含む1.8kbの断片が明らかになった。
【0134】
菌株12中のこれら2つの遺伝子の推定される接合部を交差して増幅するであろうプライマーを使用してPCR増幅したところ、この遺伝子座の遺伝子組成が確認された。したがって、菌株12はhia遺伝子の5’末端の欠失を生じたためにHiaを生成しないようであった。図27は菌株12の上流ORFとRdゲノム由来HI1733蛋白質との配列比較を示すものである。相同の領域によって、これら2つの蛋白質は95%が同一である。
【0135】
NTHi菌株33、32、29、K22、M4071、11、9およびc型菌株APIからの由来Hia配列のアラインメントが、インフルエンザ菌b型Hsf、RdゲノムのaidA類似(Hsf/Hia)HI1732遺伝子、と菌株4223およびLES-1のM.catarrhalis 200kDa蛋白質と比較して図28に示されている。HI1732蛋白質の早期末端欠失の結果生じる、Rdゲノム配列内のフレーム移動が認められる。hiaに関連する付加的な下流配列はここに含まれる。配列の下にある星印は保存された残基を示している。N末端(約50残基)およびC末端配列(約150残基)はヘモフィルス菌株の中で高度に保存されており、M.catarrhalisの相応部分とのある程度の類似性は明白である。配列分析から、Hia蛋白質について2つの可能性のある遺伝子集団が存在し、1つは原型の菌株11に関係し、他方はさらに密接に菌株33に関係することは明らかである。菌株11およびK9の蛋白質は、より一層ヘモフィルス菌株のb型、c型またはd型由来のHsf蛋白質に類似しているようであるが、一方で、菌株33、32、29、K22およびM4071の蛋白質は第2の集団を構成するようである。
【0136】
実施例15
この実施例は末端欠失したS44 菌株11 hia遺伝子をもつT7 hia遺伝子カセット、E.coli cer遺伝子、およびカナマイシン抗生物質耐性遺伝子を含むプラスミドJB-2930-3の作製、およびS44 Hia蛋白質の発現を説明するものである。
【0137】
S44アミノ酸から内側のSty I切断部位までのNTHi菌株11 hia遺伝子のS44 Hia N末端(図29)を増幅するためにPCRプライマーを設計した。クローン化を目的として5’末端にNde I切断部位を追加し、鋳型としてプラスミドDS-2242-1を使用してこの断片を増幅した。この断片をpCR IIにクローン化してプラスミドJB-2910-1-1を作製した。この構成図式を図30に示す。プラスミドJB-2910-1-1をNde IおよびSty Iで切断し、5’PCR hia断片を単離した。V38 hia遺伝子を含むプラスミドIA-46-5をNde IおよびSty Iで切断し、大きい方の約8.5kbの断片を精製した。これら2つの精製断片を連結してプラスミドJB-2917-1を作製した。その後、このプラスミドをNde Iで切断し、ウシ腸のホスファターゼ(CAP)で処理し、プラスミドIA-46-5から得たT7プロモーターをその中でクローン化した。このプロモーターはIA-46-5のNde I切断により切り取った。結果として得られたプラスミドJB-2925-3をBgl IIおよびBamH Iで切断してhia遺伝子を単離した。Bgl II/CAP処理したプラスミドBK-2-1-2にこの断片を連結し、プラスミドJB-2930-3を作製した。このプラスミドはT7プロモーターS44 hia遺伝子およびE.coli cer遺伝子を含み、カナマイシン耐性である。
【0138】
発現を調べるため、組換えS44 hiaベクターをE.coli BL21(DE3)に転換した。E.coliにおける発現のための手法は実施例9に記載したりである。図31のSDS-PAGE分析は、2つの異なるベクター、(上述の)JB-2930-3およびpETベクターのIA-191-3-1からの組換えS44 hiaの発現を示している。プラスミドIA-191-3-1は、それがlacIqリプレッサーを含むpETベクターであり、したがって生成するS44 Hiaの量がJB-2930-3からのT7 S44よりも少ないということを除くとJB-2930-3と同じである。このプラスミドは、プラスミドJB-2930-3とともに、図32に示されている。図31は二重のバンド(レーン3)としておよそ116kDaのS44 Hiaを示している。JB-2930-3から精製したS44 hiaを使用した追加の分析によると、この二重バンドの下側のバンドはC末端の94アミノ酸の末端欠失を有するがN末端と考えられるものはそのまま存在することが判明した。末端欠失したHiaの単離に使用した精製方法は実施例10に記載した通りであった。
【0139】
(開示の概要)
本開示の概要では、本発明は新たに単離精製した、ヘモフィルスから得たインフルエンザ菌アドヘシン(Hia)蛋白質の全長を有するものおよびN末端欠失したものをコードし、防御力のあるHia蛋白質を遺伝子組換えにより生成することを可能にする核酸分子を提供する。本発明の範囲内での変更態様は可能である。
【0140】
【表1】
【0141】
【表2】
【0142】
参考文献
【0143】
【表3】
【0144】
【表4】
【0145】
【表5】
【図面の簡単な説明】
【0146】
【図1A】T7プロモーターおよび完全長NTHi菌株11のhia遺伝子を含むプラスミドDS-2008-2-3の制限地図を示す図である。
【図1B】菌株11のhia遺伝子をPCR増幅するために使用されるオリゴヌクレオチドを示す図である。センス鎖(5038.SL):配列番号1、エンコードされているアミノ酸配列番号2;アンチセンス鎖(5039.SL):配列番号3、相補配列番号4、エンコードされているアミノ酸配列番号5。制限酵素部位はB、BamH I;Bg、Bgl II;H、Hind III;N、Nde I;Ps、Pst I;Sty、Sty Iである。他の略語はT7p、T7プロモーター;ApR、アンピシリン耐性である。
【図2】抗ネイティブMoraxella catarrhalisの高分子量アドヘシン抗体による、完全長rHiaタンパク質の認識のイムノブロットを示す図である。レーン1、非誘導型DS-2043-1;レーン2、4時間かけて誘導されたDS-2043-1;レーン3、非誘導型DS-2043-2;レーン4、4時間かけて誘導されたDS-2043-2;レーン5、分子量マーカー。DS-2043-1およびDS-2043-2は、BL21(DE3)内のpT7hia(11)の独立したクローンである。
【図3】菌株11のhia遺伝子用のT7hia遺伝子カセットのタンデムコピーを含むプラスミドDS-2092-1およびDS-2092-40の構築を示す図である。制限酵素部位はB、BamH I;Bg、Bgl II;H、Hind III;Ps、Pst I;Xb、Xba Iである。他の略語はCAP、子牛のアルカリホスファターゼ;T7p、T7プロモーター;ApR、アンピシリン耐性である。
【図4】菌株11のHiaタンパク質の切断部位(配列番号6)を示す図である。
【図5A】菌株11からの切断型hia遺伝子を発現するプラスミドの構築を示す図である。制限酵素部位はB、BamH I;Bg、Bgl II;H、Hind III;N、Nde I;Nhe、Nhe I;Ps、Pst I;R、EcoR I;Sty、Sty I;Xb、Xba Iである。他の略語はT7p、T7プロモーター;ApR、アンピシリン耐性;KanR、カナマイシン耐性である。
【図5B】切断型遺伝子用に5’断片をPCR増幅するために使用されるオリゴヌクレオチドを示す図である。E21切断部:センス(5524.SL):配列番号7、エンコードされているアミノ酸配列番号8;T33切断部:センス(5525.SL):配列番号9、エンコードされているアミノ酸配列番号10;V38切断部:センス(5526.SL):配列番号11、エンコードされているアミノ酸配列番号12;N52切断部:センス(5527.SL):配列番号13、エンコードされているアミノ酸配列番号14;アンチセンス(5528.SL):配列番号15、相補配列番号16、エンコードされているアミノ酸配列番号17。
【図5B1】切断型遺伝子用に5’断片をPCR増幅するために使用されるオリゴヌクレオチドを示す図である。E21切断部:センス(5524.SL):配列番号7、エンコードされているアミノ酸配列番号8;T33切断部:センス(5525.SL):配列番号9、エンコードされているアミノ酸配列番号10;V38切断部:センス(5526.SL):配列番号11、エンコードされているアミノ酸配列番号12;N52切断部:センス(5527.SL):配列番号13、エンコードされているアミノ酸配列番号14;アンチセンス(5528.SL):配列番号15、相補配列番号16、エンコードされているアミノ酸配列番号17。
【図6A】NTHi菌株11および大腸菌のcer遺伝子からのV38hia遺伝子を含むプラスミドBK-96-2-11の構築を示す図である。制限酵素部位はB、BamH I;Bg、Bgl II;K、Kpn I;N、Nde I;P、Pst I;R、EcoR I;S、Sal I;Sm、Sma I;Sty、Sty I;Xb、Xba I;Xho、Xho Iである。他の略語はT7p、T7プロモーター;ApR、アンピシリン耐性;KanR、カナマイシン耐性;CAP、子牛のアルカリホスファターゼ;tt1、trpAからの転写ターミネーター1;tt2、T7遺伝子10からの転写ターミネーター2である。
【図6B】複数のクローン部位および転写ターミネーターを構築するために使用されるオリゴヌクレオチドを示す図である。「R」および「Ps」はEcoR IまたはPst Iの端部とオーバーラップする末端を示すが、これらの部位を再び生成するわけではない。上側鎖(配列番号50)、下側(配列番号51)。
【図7A】T7プロモーターおよび完全長のNTHi菌株33のhia遺伝子、大腸菌のcer遺伝子およびカナマイシン耐性遺伝子を含むプラスミドDS-2241-1およびDS-2242-2の構築を示す図である。制限酵素部位はA、AlwN I;B、BamH I;Bg、Bgl II;H、Hind III;K、Kpn I;N、Nde I;Ps、Pst I;R、EcoR I;S、Sal I;Sm、Sma I;;Xb、Xba I;Xho、Xho Iである。他の略語はT7p、T7プロモーター;ApR、アンピシリン耐性;KanR、カナマイシン耐性;tt1、trpAからの転写ターミネーター1;tt2、T7遺伝子10からの転写ターミネーター2である。
【図7B】鎖(配列番号52)、相補鎖(配列番号53)、およびエンコードされているアミノ酸配列(配列番号54)をコードしている菌株33のhia遺伝子の5’末端を生成するために使用されるオリゴヌクレオチドを示す図である。
【図8A】T7プロモーターおよび菌株33からのV38遺伝子、大腸菌のcer遺伝子およびカナマイシン耐性遺伝子を含むプラスミドDS-2340-2-3の構築を示す図である。制限酵素部位はB、BamH I;Bg、Bgl II;H、Hind III;N、Nde I;Ps、Pst I;R、EcoR I;S、Sal I;Sn、SnaB I;;Xb、Xba Iである。他の略語はT7p、T7プロモーター;ApR、アンピシリン耐性;KanR、カナマイシン耐性;tt1、trpAからの転写ターミネーター1;tt2、T7遺伝子10からの転写ターミネーター2である。
【図8B】切断型遺伝子用の5’端をPCR増幅するために使用されるオリゴヌクレオチドを示す図である。センス(6286.SL):配列番号16、エンコードされているアミノ酸配列番号17;アンチセンス(6287.SL):配列番号18、相補配列番号19、エンコードされているアミノ酸配列番号20。
【図9A】T7のV38hia(11)およびT7のV38hia(33)遺伝子のタンデムコピーをそれぞれ含むプラスミドDS-2447-2およびDS-2448-17の構築を示す図である。制限酵素部位はB、BamH I;Bg、Bgl II;H、Hind III;Ps、Pst I;R、EcoR I;S、Sal I;Xb、Xba Iである。他の略語はT7p、T7プロモーター;ApR、アンピシリン耐性;KanR、カナマイシン耐性;CAP、子牛のアルカリホスファターゼ;tt1、trpAからの転写ターミネーター1;tt2、T7遺伝子10からの転写ターミネーター2である。
【図9B】T7のV38hia(11)およびT7のV38hia(33)遺伝子のタンデムコピーをそれぞれ含むプラスミドDS-2447-2およびDS-2448-17の構築を示す図である。制限酵素部位はB、BamH I;Bg、Bgl II;H、Hind III;Ps、Pst I;R、EcoR I;S、Sal I;Xb、Xba Iである。他の略語はT7p、T7プロモーター;ApR、アンピシリン耐性;KanR、カナマイシン耐性;CAP、子牛のアルカリホスファターゼ;tt1、trpAからの転写ターミネーター1;tt2、T7遺伝子10からの転写ターミネーター2である。
【図10】rHiaの発現を示す図である。パネルA:レーン1、誘導されていない完全長rHia(11);レーン2、完全長rHia(11);レーン3、E21rHia(11);レーン4、T33rHia(11);レーン5、V38rHia(11);レーン6、N52rHia(11)。パネルB:レーン1、誘導されていないV38rHia(11);レーン2、V38rHia(11);レーン3、V38rHia(11)/cer。
【図11】rHiaの精製スキームを示す図である。略語はSP、上澄み;PPT、沈殿;DTT、ジチオスレイトール;OG、オクチルグルコシドであり、(x)は捨てられたことを意味する。
【図12】パネルAおよびBを有し、精製されたrHiaのSDS-PAGE分析を示す図である。パネルAは菌株11からの精製されたV38rHiaタンパク質を示し、パネルBは菌株33からのV38rHiaタンパク質を示す。レーン1、分子量マーカー;レーン2、全細胞の溶解物;レーン3、未精製抽出物;レーン4、精製されたrHiaタンパク質。
【図13A】パネルA、BおよびCを有し、V38rHia(11)の安定性を示す図である。パネルAは、グリセロールなしで4℃で保存されたサンプルを示す。パネルBは、20%グリセロールの存在下で4℃で保存されたサンプルを示す。パネルCは、20%グリセロールの存在下で-20℃で保存されたサンプルを示す。レーン0はt0を示し、レーン1〜8は1〜8週間保存されたサンプルを示す。
【図13B】パネルA、BおよびCを有し、V38rHia(11)の安定性を示す図である。パネルAは、グリセロールなしで4℃で保存されたサンプルを示す。パネルBは、20%グリセロールの存在下で4℃で保存されたサンプルを示す。パネルCは、20%グリセロールの存在下で-20℃で保存されたサンプルを示す。レーン0はt0を示し、レーン1〜8は1〜8週間保存されたサンプルを示す。
【図13C】パネルA、BおよびCを有し、V38rHia(11)の安定性を示す図である。パネルAは、グリセロールなしで4℃で保存されたサンプルを示す。パネルBは、20%グリセロールの存在下で4℃で保存されたサンプルを示す。パネルCは、20%グリセロールの存在下で-20℃で保存されたサンプルを示す。レーン0はt0を示し、レーン1〜8は1〜8週間保存されたサンプルを示す。
【図14A】パネルAおよびBを有し、CD-1マウスのV38rHia(11)またはV38rHia(33)の免疫原性を示す図である。パネルAは単一の免疫化後の応答を示し、パネルBは最初の/追加の免疫化の応答を示す。
【図14B】パネルAおよびBを有し、CD-1マウスのV38rHia(11)またはV38rHia(33)の免疫原性を示す図である。パネルAは単一の免疫化後の応答を示し、パネルBは最初の/追加の免疫化の応答を示す。
【図15A】BALB/cマウスおよびモルモットのV38rHia(11)の免疫原性を示す図である。図15Aはマウスにおける抗体の応答を示し、図15Bはモルモットにおける応答を示す。
【図15B】BALB/cマウスおよびモルモットのV38rHia(11)の免疫原性を示す図である。図15Aはマウスにおける抗体の応答を示し、図15Bはモルモットにおける応答を示す。
【図16】チンチラモデルにおける鼻咽頭コロニー形成に対するV38rHia(33)の防御能力を示す図である。
【図17】追加のhia遺伝子をPCR増幅するために使用されるオリゴヌクレオチドを示す図である。センス(5040.SL):配列番号21、エンコードされているアミノ酸配列番号22;アンチセンス(5039.SL):配列番号3、相補配列番号4、エンコードされているアミノ酸配列番号5。
【図18A】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18B】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18C】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18D】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18E】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18F】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18G】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18H】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18I】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18J】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18K】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18L】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18M】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18N】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図18O】NTHi菌株33からのhia遺伝子のヌクレオチド配列(配列番号23)および演繹アミノ酸配列(配列番号24)を示す図である。
【図19A】NTHi菌株32からのhia遺伝子のヌクレオチド配列(配列番号25)および演繹アミノ酸配列(配列番号26)を示す図である。
【図19B】NTHi菌株32からのhia遺伝子のヌクレオチド配列(配列番号25)および演繹アミノ酸配列(配列番号26)を示す図である。
【図19C】NTHi菌株32からのhia遺伝子のヌクレオチド配列(配列番号25)および演繹アミノ酸配列(配列番号26)を示す図である。
【図19D】NTHi菌株32からのhia遺伝子のヌクレオチド配列(配列番号25)および演繹アミノ酸配列(配列番号26)を示す図である。
【図19E】NTHi菌株32からのhia遺伝子のヌクレオチド配列(配列番号25)および演繹アミノ酸配列(配列番号26)を示す図である。
【図19F】NTHi菌株32からのhia遺伝子のヌクレオチド配列(配列番号25)および演繹アミノ酸配列(配列番号26)を示す図である。
【図19G】NTHi菌株32からのhia遺伝子のヌクレオチド配列(配列番号25)および演繹アミノ酸配列(配列番号26)を示す図である。
【図19H】NTHi菌株32からのhia遺伝子のヌクレオチド配列(配列番号25)および演繹アミノ酸配列(配列番号26)を示す図である。
【図19I】NTHi菌株32からのhia遺伝子のヌクレオチド配列(配列番号25)および演繹アミノ酸配列(配列番号26)を示す図である。
【図19J】NTHi菌株32からのhia遺伝子のヌクレオチド配列(配列番号25)および演繹アミノ酸配列(配列番号26)を示す図である。
【図20A】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20B】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20C】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20D】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20E】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20F】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20G】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20H】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20I】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20J】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20K】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20L】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20M】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20N】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20O】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図20P】NTHi菌株29からのhia遺伝子のヌクレオチド配列(配列番号27)および演繹アミノ酸配列(配列番号28)を示す図である。
【図21A】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21B】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21C】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21D】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21E】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21F】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21G】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21H】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21I】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21J】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21K】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21L】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21M】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21N】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図21O】NTHi菌株M4071からのhia遺伝子のヌクレオチド配列(配列番号29)および演繹アミノ酸配列(配列番号30)を示す図である。
【図22A】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22B】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22C】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22D】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22E】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22F】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22G】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22H】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22I】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22J】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22K】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22L】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22M】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22N】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22O】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図22P】NTHi菌株K9からのhia遺伝子のヌクレオチド配列(配列番号31)および演繹アミノ酸配列(配列番号32)を示す図である。
【図23A】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23B】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23C】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23D】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23E】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23F】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23G】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23H】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23I】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23J】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23K】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23L】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23M】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23N】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23O】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図23P】NTHi菌株K22からのhia遺伝子のヌクレオチド配列(配列番号33)および演繹アミノ酸配列(配列番号34)を示す図である。
【図24A】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24B】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24C】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24D】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24E】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24F】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24G】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24H】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24I】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24J】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24K】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24L】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24M】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24N】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24O】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24P】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24Q】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24R】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24S】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24T】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24U】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24V】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24W】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24X】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24Y】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24Z】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24A1】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24B1】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24C1】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24D1】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24E1】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24F1】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24G1】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24H1】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図24I1】c型菌株APIからのhia遺伝子のヌクレオチド配列(配列番号35)および演繹アミノ酸配列(配列番号36)を示す図である。
【図25A】NTHi菌株12からのhia遺伝子座のヌクレオチド配列(配列番号37)および演繹アミノ酸配列(配列番号38)を示す図である。上線または下線を引かれた配列は、2つのorfの接合部を越えてPCR増幅するために使用されるオリゴヌクレオチドを示す。センス(6431.SL):配列番号39、(6432.SL)配列番号40;アンチセンス(6295.SL)配列番号41、(6271.SL)配列番号42。
【図25B】NTHi菌株12からのhia遺伝子座のヌクレオチド配列(配列番号37)および演繹アミノ酸配列(配列番号38)を示す図である。上線または下線を引かれた配列は、2つのorfの接合部を越えてPCR増幅するために使用されるオリゴヌクレオチドを示す。センス(6431.SL):配列番号39、(6432.SL)配列番号40;アンチセンス(6295.SL)配列番号41、(6271.SL)配列番号42。
【図25C】NTHi菌株12からのhia遺伝子座のヌクレオチド配列(配列番号37)および演繹アミノ酸配列(配列番号38)を示す図である。上線または下線を引かれた配列は、2つのorfの接合部を越えてPCR増幅するために使用されるオリゴヌクレオチドを示す。センス(6431.SL):配列番号39、(6432.SL)配列番号40;アンチセンス(6295.SL)配列番号41、(6271.SL)配列番号42。
【図25D】NTHi菌株12からのhia遺伝子座のヌクレオチド配列(配列番号37)および演繹アミノ酸配列(配列番号38)を示す図である。上線または下線を引かれた配列は、2つのorfの接合部を越えてPCR増幅するために使用されるオリゴヌクレオチドを示す。センス(6431.SL):配列番号39、(6432.SL)配列番号40;アンチセンス(6295.SL)配列番号41、(6271.SL)配列番号42。
【図25E】NTHi菌株12からのhia遺伝子座のヌクレオチド配列(配列番号37)および演繹アミノ酸配列(配列番号38)を示す図である。上線または下線を引かれた配列は、2つのorfの接合部を越えてPCR増幅するために使用されるオリゴヌクレオチドを示す。センス(6431.SL):配列番号39、(6432.SL)配列番号40;アンチセンス(6295.SL)配列番号41、(6271.SL)配列番号42。
【図25F】NTHi菌株12からのhia遺伝子座のヌクレオチド配列(配列番号37)および演繹アミノ酸配列(配列番号38)を示す図である。上線または下線を引かれた配列は、2つのorfの接合部を越えてPCR増幅するために使用されるオリゴヌクレオチドを示す。センス(6431.SL):配列番号39、(6432.SL)配列番号40;アンチセンス(6295.SL)配列番号41、(6271.SL)配列番号42。
【図25G】NTHi菌株12からのhia遺伝子座のヌクレオチド配列(配列番号37)および演繹アミノ酸配列(配列番号38)を示す図である。上線または下線を引かれた配列は、2つのorfの接合部を越えてPCR増幅するために使用されるオリゴヌクレオチドを示す。センス(6431.SL):配列番号39、(6432.SL)配列番号40;アンチセンス(6295.SL)配列番号41、(6271.SL)配列番号42。
【図25H】NTHi菌株12からのhia遺伝子座のヌクレオチド配列(配列番号37)および演繹アミノ酸配列(配列番号38)を示す図である。上線または下線を引かれた配列は、2つのorfの接合部を越えてPCR増幅するために使用されるオリゴヌクレオチドを示す。センス(6431.SL):配列番号39、(6432.SL)配列番号40;アンチセンス(6295.SL)配列番号41、(6271.SL)配列番号42。
【図25I】NTHi菌株12からのhia遺伝子座のヌクレオチド配列(配列番号37)および演繹アミノ酸配列(配列番号38)を示す図である。上線または下線を引かれた配列は、2つのorfの接合部を越えてPCR増幅するために使用されるオリゴヌクレオチドを示す。センス(6431.SL):配列番号39、(6432.SL)配列番号40;アンチセンス(6295.SL)配列番号41、(6271.SL)配列番号42。
【図26A】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図26B】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図26C】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図26D】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図26E】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図26F】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図26G】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図26H】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図26I】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図26J】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図26K】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図26L】米国特許第5,646,259号において公開されている、NTHi菌株11からのhia遺伝子座のヌクレオチド配列(配列番号43)および演繹アミノ酸配列(配列番号44)を示す図である。
【図27A】菌株12のhia遺伝子座からの上流ORF(配列番号45)の配列およびb型インフルエンザ菌株RdからのHI1732タンパク質(配列番号46)の一部の整列を示す図である。
【図27B】菌株12のhia遺伝子座からの上流ORF(配列番号45)の配列およびb型インフルエンザ菌株RdからのHI1732タンパク質(配列番号46)の一部の整列を示す図である。
【図28A】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28B】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28C】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28D】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28E】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28F】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28G】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28H】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28I】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28J】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28K】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28L】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28M】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28N】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28O】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28P】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28Q】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28R】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28S】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28T】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28U】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28V】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28W】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28X】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28Y】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28Z】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28A1】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28B1】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28C1】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図28D1】Hia(配列番号24、26、28、34、30、44、32)、Hsf(配列番号47)、および菌株4223およびLES-1(配列番号48、49)からのMoraxella catarrhalisの高分子量タンパク質(200kDa)からの一部の配列からのアミノ酸配列の整列を示す図である。配列内の星印はストップコドンを示すが、配列の下部では配列の相同性が示された。ドットは同一の残基を示す。配列の整列は、それぞれのタンパク質のアミノ酸配列を直接比較することによって作製された。
【図29】hia遺伝子の5’端をS44切断位置でPCR増幅するために使用されるオリゴヌクレオチドを示す図である。センス(6817.SL)配列番号49、エンコードされているアミノ酸配列番号50;アンチセンス(6818.SL)配列番号51、相補配列番号52、エンコードされているアミノ酸配列番号53。
【図30A】NTHi菌株11からのS44hia遺伝子および大腸菌のcer遺伝子およびT7プロモーターを含む、プラスミドJB-2930-3の構築を示す図である。制限酵素部位はB、BamH I;Bg、Bgl II;K、Kpn I;N、Nde I;P、Pst I;R、EcoR I;S、Sal I;Sm、Sma I;Sty、Sty I;Xb、Xba I;Xho、Xho Iである。他の略語はT7p、T7プロモーター;ApR、アンピシリン耐性;KanR、カナマイシン耐性;CAP、子牛のアルカリホスファターゼ;tt1、trpAからの転写ターミネーター1;tt2、T7遺伝子10からの転写ターミネーター2である。
【図30B】NTHi菌株11からのS44hia遺伝子および大腸菌のcer遺伝子およびT7プロモーターを含む、プラスミドJB-2930-3の構築を示す図である。制限酵素部位はB、BamH I;Bg、Bgl II;K、Kpn I;N、Nde I;P、Pst I;R、EcoR I;S、Sal I;Sm、Sma I;Sty、Sty I;Xb、Xba I;Xho、Xho Iである。他の略語はT7p、T7プロモーター;ApR、アンピシリン耐性;KanR、カナマイシン耐性;CAP、子牛のアルカリホスファターゼ;tt1、trpAからの転写ターミネーター1;tt2、T7遺伝子10からの転写ターミネーター2である。
【図31】S44からのrHiaの発現のSDS-PAGE分析を示す図である。レーン1、時間0での(誘導なしで)pETS44ベクターからの発現体;レーン2、4時間の誘導後のpETS44ベクターからの発現体;レーン3、4時間の誘導後のJB-2930-3からの発現体。
【図32】発現試験用に使用された2つのベクター、S44切断型rHiaのJB-2930-3およびIA-191-3-1の概略を示す図である。
【特許請求の範囲】
【請求項1】
インフルエンザ菌(ヘモフィルス・インフルエンジー(Haemophilus influenzae))の菌株のインフルエンザ菌アドヘシン(Hia)タンパク質をエンコードしている単離され精製された核酸分子であって、
(a)図18、19、20、21、22、23、24および25に示される配列(配列番号23、25、27、29、31、33、35、37)からなる群から選択されるDNA配列、または
(b)図18、19、20、21、22、23、24および25に示される配列(配列番号24、26、28、30、32、34、36、38)からなる群から選択されるアミノ酸配列を有するインフルエンザ菌アドヘシン(rHia)タンパク質をエンコードしているDNA配列を有する単離され精製された核酸分子。
【請求項2】
インフルエンザ菌の菌株のN末端切断型インフルエンザ菌アドヘシン(Hia)タンパク質をエンコードしており、
配列番号7と配列番号15
配列番号9と配列番号15
配列番号11と配列番号15
配列番号13と配列番号15
配列番号49と配列番号51
からなる群から選択されるヌクレオチドの1対によって増幅可能な、単離され精製された核酸分子。
【請求項3】
封入体として発現可能なインフルエンザ菌の菌株の切断型インフルエンザ菌アドヘシン(Hia)タンパク質をエンコードしている、単離され精製された核酸。
【請求項4】
請求項1に記載の前記核酸分子を含む宿主を形質転換するためのベクター。
【請求項5】
請求項2または3に記載の前記核酸分子を含む宿主を形質転換するためのベクター。
【請求項6】
プラスミドベクターである、請求項4または5に記載のベクター。
【請求項7】
前記プラスミドベクターが、
図1A中に示されるDS-2008-2-3
図5A中に示されるDS-2186-1-1
図5A中に示されるDS-2201-1
図5A中に示されるDS-2186-2-1
図5A中に示されるDS-2168-2-6
図32中に示されるIA-191-3-1
からなる群から選択されるプラスミドの同定特性を有する、請求項6に記載のベクター。
【請求項8】
完全長またはN末端切断型インフルエンザ菌アドヘシン(Hia)タンパク質および前記完全長または切断型Hiaタンパク質を発現するためのプロモーターを含む、宿主を形質転換するためのベクター。
【請求項9】
大腸菌の遺伝子をさらに含む、請求項8に記載のベクター。
【請求項10】
プラスミドベクターである、請求項8に記載のベクター。
【請求項11】
前記プラスミドベクターが、
図6A中に示されるBK-96-2-11
図7A中に示されるDS-2242-1
図7A中に示されるDS-2242-2
図8A中に示されるDS-2340-2-3
図9A中に示されるDS-2447-2
図9B中に示されるDS-2448-17
図32中に示されるJB-2930-3
からなる群から選択されるプラスミドベクターの同定特性を有する請求項10に記載のベクター。
【請求項12】
請求項4、5または8に記載のベクターによって形質転換され、ヘモフィルスの分類不能型菌株の防御性インフルエンザ菌アドヘシン(Hia)タンパク質を発現する宿主細胞。
【請求項13】
大腸菌の菌株である、請求項12に記載の宿主細胞。
【請求項14】
請求項13に記載の形質転換された大腸菌またはその免疫原性断片またはアドヘシン官能性類縁体によって産生可能な、インフルエンザ菌の菌株の組換え防御性インフルエンザ菌アドヘシン(Hia)タンパク質。
【請求項15】
(A)インフルエンザ菌の菌株のインフルエンザ菌アドヘシン(Hia)タンパク質をエンコードしている単離され精製された核酸分子であって、
(a)図18、19、20、21、22、23、24および25に示される配列(配列番号23、25、27、29、31、33、35、37)からなる群から選択されるDNA配列、または
(b)図18、19、20、21、22、23、24および25に示される配列(配列番号24、26、28、30、32、34、36、38)からなる群から選択されるアミノ酸配列を有するインフルエンザ菌アドヘシン(rHia)タンパク質をエンコードしているDNA配列を有する単離され精製された核酸分子、
(B)インフルエンザ菌の菌株のN末端切断型インフルエンザ菌アドヘシン(Hia)タンパク質をエンコードしており、
配列番号7と配列番号15
配列番号9と配列番号15
配列番号11と配列番号15
配列番号13と配列番号15
配列番号49と配列番号51
からなる群から選択されるヌクレオチドの1対によって増幅可能な、単離され精製された核酸分子、
(C)封入体として発現可能なインフルエンザ菌の菌株の切断型インフルエンザ菌アドヘシン(Hia)タンパク質をエンコードしている、単離され精製された核酸分子、
(D)請求項4、5または8に記載の発現ベクターによって形質転換された大腸菌によって産生可能な、インフルエンザ菌の菌株の組換え防御性インフルエンザ菌アドヘシン(Hia)タンパク質からなる群から選択される少なくとも1つの免疫学的に活性のある成分、および薬学的に許容可能なこれらの担体を含む免疫原性組成物。
【請求項16】
ヘモフィルスに起因する疾患に対して防御するために宿主へのin vitro投与用ワクチンとして製剤化される、請求項15に記載の免疫原性組成物。
【請求項17】
免疫系の特定の細胞または粘膜表面への送達用の標的分子と組み合わせた、請求項15に記載の免疫原性組成物。
【請求項18】
ミクロ粒子、カプセル、またはリポソーム調製物として製剤化される、請求項15に記載の免疫原性組成物。
【請求項19】
アジュバントをさらに含む、請求項15に記載の免疫原性組成物。
【請求項20】
感受性のある宿主に請求項15に記載の免疫原性組成物を有効量投与すること含む、ヘモフィルスに起因する疾患に対する防御を誘導するための方法。
【請求項21】
前記感受性のある宿主がヒトである、請求項20に記載の方法。
【請求項22】
インフルエンザ菌の分類不能型菌株の防御性インフルエンザ菌アドヘシン(Hia)タンパク質の産生のための方法であって、
請求項5と同様にベクターによって宿主を形質転換すること、
宿主を成長させてエンコードされている切断型Hiaを発現させること、
前記発現されたHiaタンパク質を単離および精製することを含む方法。
【請求項23】
前記宿主細胞が大腸菌である、請求項22に記載の方法。
【請求項24】
前記エンコードされている切断型Hiaが封入体内で発現される、請求項22に記載の方法。
【請求項25】
前記発現されたHiaの前記単離および精製が、
成長し形質転換された細胞を破壊して上澄みおよび前記封入体を生成すること、
前記封入体を可溶化して組換えHiaの溶液を生成すること、
クロマトグラフィによって細胞の残骸のない組換えHiaの前記溶液を精製すること、および
前記精製された組換えHiaタンパク質を単離することによって実施される、請求項24に記載の方法。
【請求項26】
前記ヘモフィルスの分類不能型菌株が菌株11、33、32、29、M4071、K9、K22および12からなる群から選択される、請求項22に記載の方法。
【請求項27】
前記ベクターがT7プロモーターを含み、前記大腸菌が誘導量のラクトースの存在下で培養される、請求項22に記載の方法。
【請求項1】
インフルエンザ菌(ヘモフィルス・インフルエンジー(Haemophilus influenzae))の菌株のインフルエンザ菌アドヘシン(Hia)タンパク質をエンコードしている単離され精製された核酸分子であって、
(a)図18、19、20、21、22、23、24および25に示される配列(配列番号23、25、27、29、31、33、35、37)からなる群から選択されるDNA配列、または
(b)図18、19、20、21、22、23、24および25に示される配列(配列番号24、26、28、30、32、34、36、38)からなる群から選択されるアミノ酸配列を有するインフルエンザ菌アドヘシン(rHia)タンパク質をエンコードしているDNA配列を有する単離され精製された核酸分子。
【請求項2】
インフルエンザ菌の菌株のN末端切断型インフルエンザ菌アドヘシン(Hia)タンパク質をエンコードしており、
配列番号7と配列番号15
配列番号9と配列番号15
配列番号11と配列番号15
配列番号13と配列番号15
配列番号49と配列番号51
からなる群から選択されるヌクレオチドの1対によって増幅可能な、単離され精製された核酸分子。
【請求項3】
封入体として発現可能なインフルエンザ菌の菌株の切断型インフルエンザ菌アドヘシン(Hia)タンパク質をエンコードしている、単離され精製された核酸。
【請求項4】
請求項1に記載の前記核酸分子を含む宿主を形質転換するためのベクター。
【請求項5】
請求項2または3に記載の前記核酸分子を含む宿主を形質転換するためのベクター。
【請求項6】
プラスミドベクターである、請求項4または5に記載のベクター。
【請求項7】
前記プラスミドベクターが、
図1A中に示されるDS-2008-2-3
図5A中に示されるDS-2186-1-1
図5A中に示されるDS-2201-1
図5A中に示されるDS-2186-2-1
図5A中に示されるDS-2168-2-6
図32中に示されるIA-191-3-1
からなる群から選択されるプラスミドの同定特性を有する、請求項6に記載のベクター。
【請求項8】
完全長またはN末端切断型インフルエンザ菌アドヘシン(Hia)タンパク質および前記完全長または切断型Hiaタンパク質を発現するためのプロモーターを含む、宿主を形質転換するためのベクター。
【請求項9】
大腸菌の遺伝子をさらに含む、請求項8に記載のベクター。
【請求項10】
プラスミドベクターである、請求項8に記載のベクター。
【請求項11】
前記プラスミドベクターが、
図6A中に示されるBK-96-2-11
図7A中に示されるDS-2242-1
図7A中に示されるDS-2242-2
図8A中に示されるDS-2340-2-3
図9A中に示されるDS-2447-2
図9B中に示されるDS-2448-17
図32中に示されるJB-2930-3
からなる群から選択されるプラスミドベクターの同定特性を有する請求項10に記載のベクター。
【請求項12】
請求項4、5または8に記載のベクターによって形質転換され、ヘモフィルスの分類不能型菌株の防御性インフルエンザ菌アドヘシン(Hia)タンパク質を発現する宿主細胞。
【請求項13】
大腸菌の菌株である、請求項12に記載の宿主細胞。
【請求項14】
請求項13に記載の形質転換された大腸菌またはその免疫原性断片またはアドヘシン官能性類縁体によって産生可能な、インフルエンザ菌の菌株の組換え防御性インフルエンザ菌アドヘシン(Hia)タンパク質。
【請求項15】
(A)インフルエンザ菌の菌株のインフルエンザ菌アドヘシン(Hia)タンパク質をエンコードしている単離され精製された核酸分子であって、
(a)図18、19、20、21、22、23、24および25に示される配列(配列番号23、25、27、29、31、33、35、37)からなる群から選択されるDNA配列、または
(b)図18、19、20、21、22、23、24および25に示される配列(配列番号24、26、28、30、32、34、36、38)からなる群から選択されるアミノ酸配列を有するインフルエンザ菌アドヘシン(rHia)タンパク質をエンコードしているDNA配列を有する単離され精製された核酸分子、
(B)インフルエンザ菌の菌株のN末端切断型インフルエンザ菌アドヘシン(Hia)タンパク質をエンコードしており、
配列番号7と配列番号15
配列番号9と配列番号15
配列番号11と配列番号15
配列番号13と配列番号15
配列番号49と配列番号51
からなる群から選択されるヌクレオチドの1対によって増幅可能な、単離され精製された核酸分子、
(C)封入体として発現可能なインフルエンザ菌の菌株の切断型インフルエンザ菌アドヘシン(Hia)タンパク質をエンコードしている、単離され精製された核酸分子、
(D)請求項4、5または8に記載の発現ベクターによって形質転換された大腸菌によって産生可能な、インフルエンザ菌の菌株の組換え防御性インフルエンザ菌アドヘシン(Hia)タンパク質からなる群から選択される少なくとも1つの免疫学的に活性のある成分、および薬学的に許容可能なこれらの担体を含む免疫原性組成物。
【請求項16】
ヘモフィルスに起因する疾患に対して防御するために宿主へのin vitro投与用ワクチンとして製剤化される、請求項15に記載の免疫原性組成物。
【請求項17】
免疫系の特定の細胞または粘膜表面への送達用の標的分子と組み合わせた、請求項15に記載の免疫原性組成物。
【請求項18】
ミクロ粒子、カプセル、またはリポソーム調製物として製剤化される、請求項15に記載の免疫原性組成物。
【請求項19】
アジュバントをさらに含む、請求項15に記載の免疫原性組成物。
【請求項20】
感受性のある宿主に請求項15に記載の免疫原性組成物を有効量投与すること含む、ヘモフィルスに起因する疾患に対する防御を誘導するための方法。
【請求項21】
前記感受性のある宿主がヒトである、請求項20に記載の方法。
【請求項22】
インフルエンザ菌の分類不能型菌株の防御性インフルエンザ菌アドヘシン(Hia)タンパク質の産生のための方法であって、
請求項5と同様にベクターによって宿主を形質転換すること、
宿主を成長させてエンコードされている切断型Hiaを発現させること、
前記発現されたHiaタンパク質を単離および精製することを含む方法。
【請求項23】
前記宿主細胞が大腸菌である、請求項22に記載の方法。
【請求項24】
前記エンコードされている切断型Hiaが封入体内で発現される、請求項22に記載の方法。
【請求項25】
前記発現されたHiaの前記単離および精製が、
成長し形質転換された細胞を破壊して上澄みおよび前記封入体を生成すること、
前記封入体を可溶化して組換えHiaの溶液を生成すること、
クロマトグラフィによって細胞の残骸のない組換えHiaの前記溶液を精製すること、および
前記精製された組換えHiaタンパク質を単離することによって実施される、請求項24に記載の方法。
【請求項26】
前記ヘモフィルスの分類不能型菌株が菌株11、33、32、29、M4071、K9、K22および12からなる群から選択される、請求項22に記載の方法。
【請求項27】
前記ベクターがT7プロモーターを含み、前記大腸菌が誘導量のラクトースの存在下で培養される、請求項22に記載の方法。
【図1A】

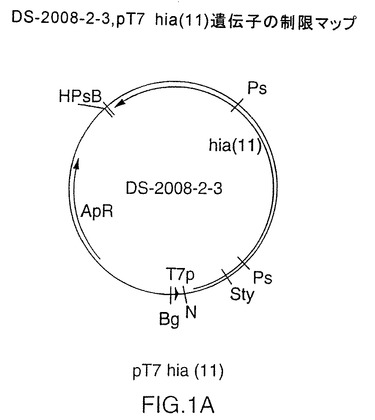
【図1B】

【図2】
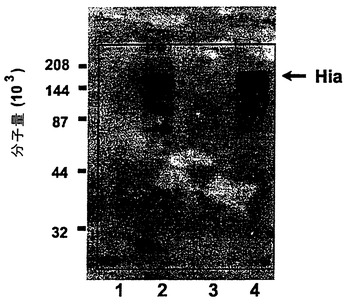
【図3】
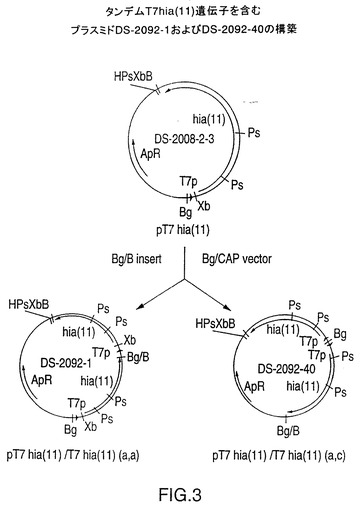
【図4】
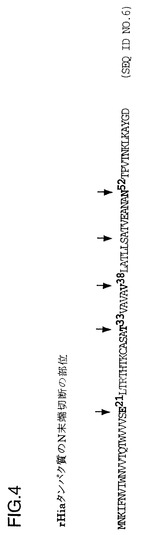
【図5A】
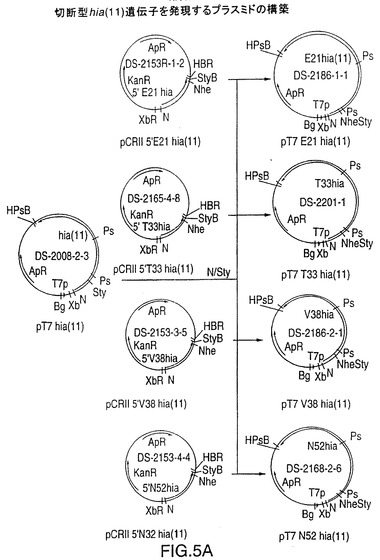
【図5B】

【図5B1】
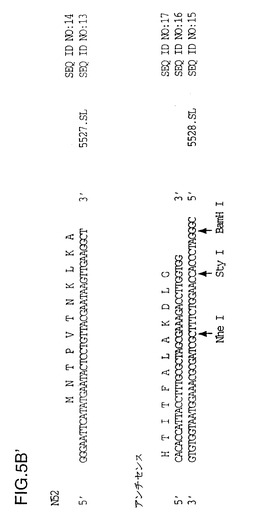
【図6A】
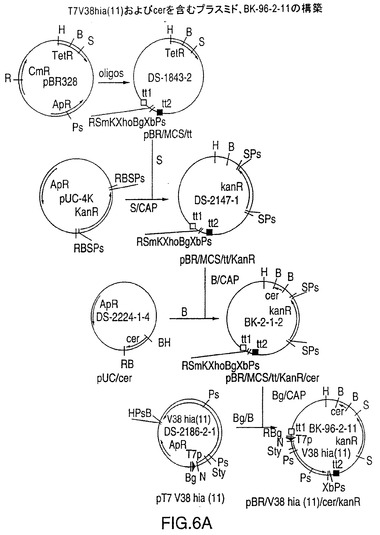
【図6B】
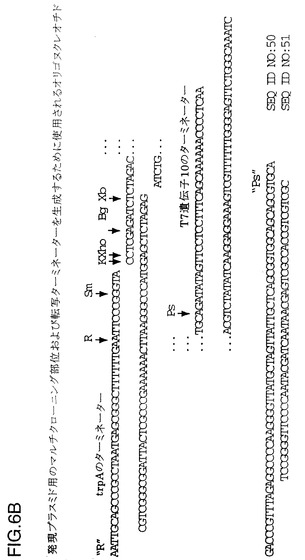
【図7A】
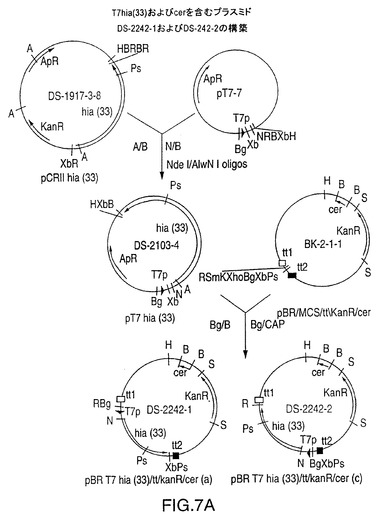
【図7B】

【図8A】
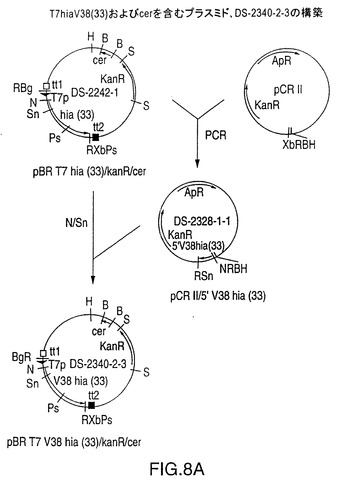
【図8B】

【図9A】
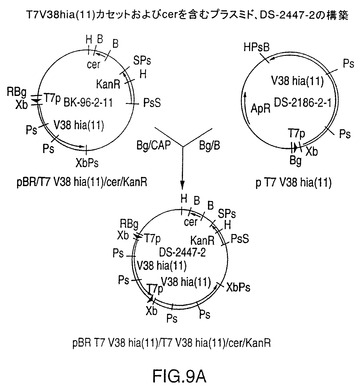
【図9B】
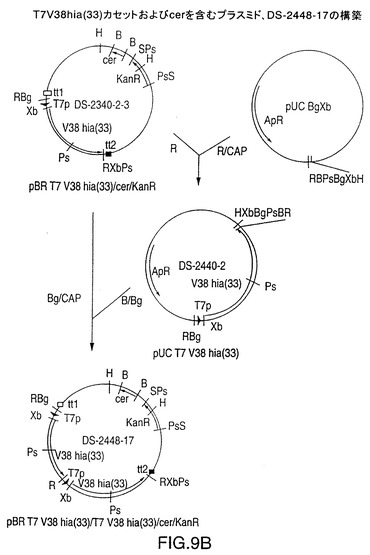
【図10】
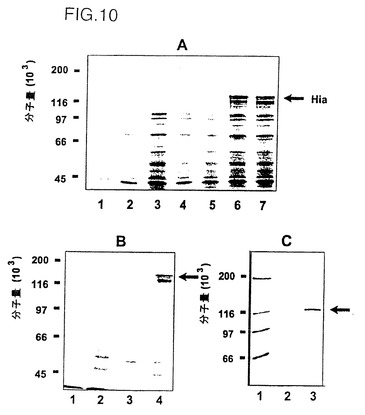
【図11】
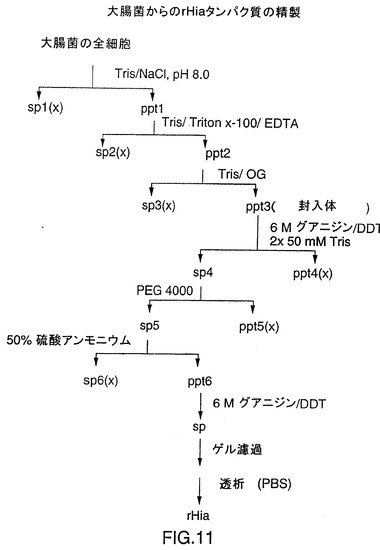
【図12】
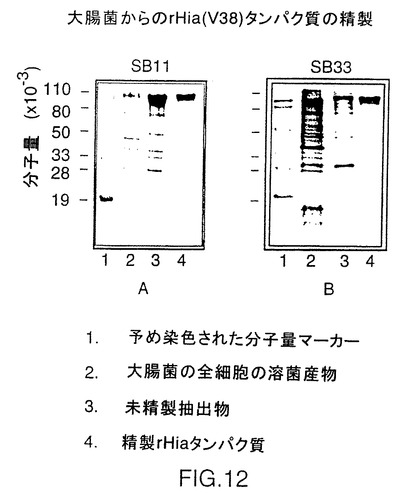
【図13A】
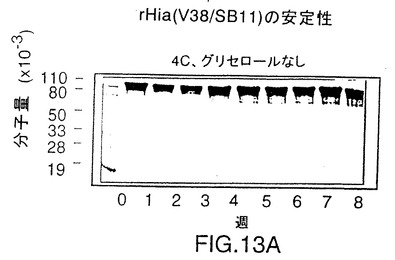
【図13B】
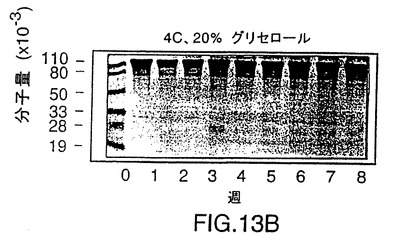
【図13C】
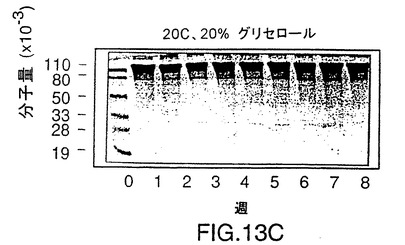
【図14A】
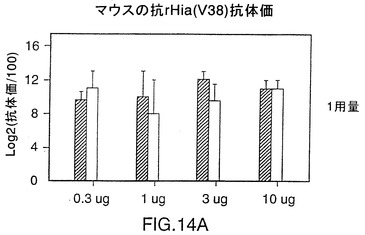
【図14B】
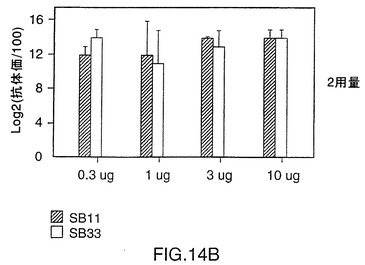
【図15A】
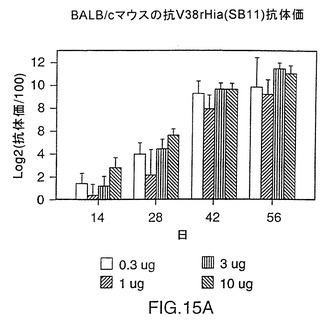
【図15B】
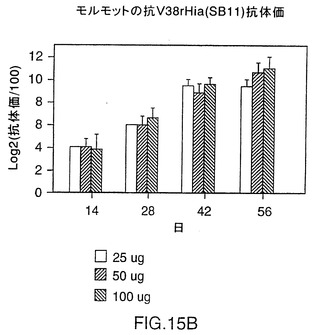
【図16】

【図17】

【図18A】

【図18B】

【図18C】

【図18D】

【図18E】

【図18F】

【図18G】

【図18H】

【図18I】

【図18J】

【図18K】

【図18L】

【図18M】

【図18N】

【図18O】

【図19A】

【図19B】

【図19C】

【図19D】

【図19E】

【図19F】

【図19G】

【図19H】

【図19I】

【図19J】

【図20A】

【図20B】

【図20C】

【図20D】

【図20E】

【図20F】

【図20G】

【図20H】

【図20I】

【図20J】

【図20K】

【図20L】

【図20M】

【図20N】

【図20O】

【図20P】

【図21A】

【図21B】

【図21C】

【図21D】

【図21E】

【図21F】

【図21G】

【図21H】

【図21I】

【図21J】

【図21K】

【図21L】

【図21M】

【図21N】

【図21O】

【図22A】

【図22B】

【図22C】

【図22D】

【図22E】

【図22F】

【図22G】

【図22H】

【図22I】

【図22J】

【図22K】

【図22L】

【図22M】

【図22N】

【図22O】

【図22P】

【図23A】

【図23B】

【図23C】

【図23D】

【図23E】

【図23F】

【図23G】

【図23H】

【図23I】

【図23J】

【図23K】

【図23L】

【図23M】

【図23N】

【図23O】

【図23P】

【図24A】

【図24B】

【図24C】

【図24D】

【図24E】

【図24F】

【図24G】

【図24H】

【図24I】

【図24J】

【図24K】

【図24L】

【図24M】

【図24N】

【図24O】

【図24P】

【図24Q】

【図24R】

【図24S】

【図24T】

【図24U】

【図24V】

【図24W】

【図24X】

【図24Y】

【図24Z】

【図24A1】

【図24B1】

【図24C1】

【図24D1】

【図24E1】

【図24F1】

【図24G1】

【図24H1】

【図24I1】

【図25A】

【図25B】

【図25C】

【図25D】

【図25E】

【図25F】

【図25G】

【図25H】

【図25I】

【図26A】

【図26B】

【図26C】

【図26D】

【図26E】

【図26F】

【図26G】

【図26H】

【図26I】

【図26J】

【図26K】

【図26L】

【図27A】

【図27B】
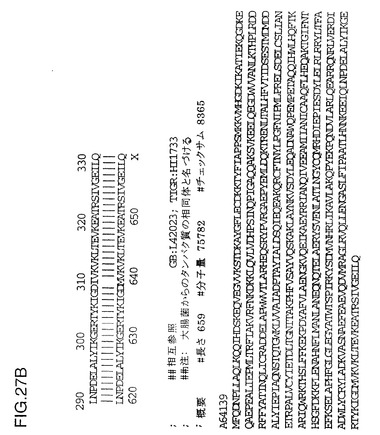
【図28A】
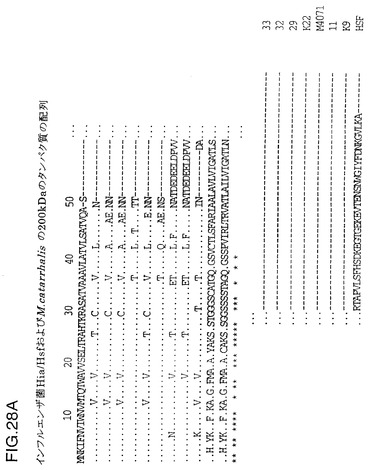
【図28B】

【図28C】

【図28D】

【図28E】

【図28F】

【図28G】
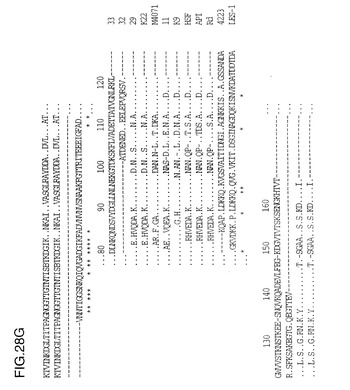
【図28H】

【図28I】

【図28J】

【図28K】

【図28L】

【図28M】

【図28N】

【図28O】

【図28P】

【図28Q】

【図28R】

【図28S】

【図28T】

【図28U】

【図28V】

【図28W】

【図28X】

【図28Y】

【図28Z】

【図28A1】

【図28B1】
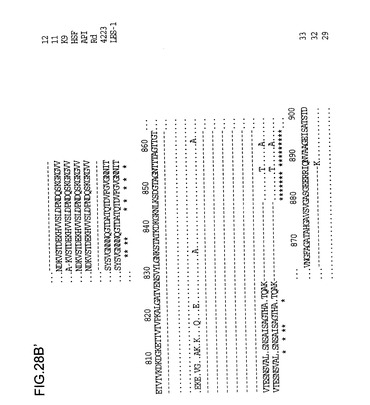
【図28C1】

【図29】

【図30A】
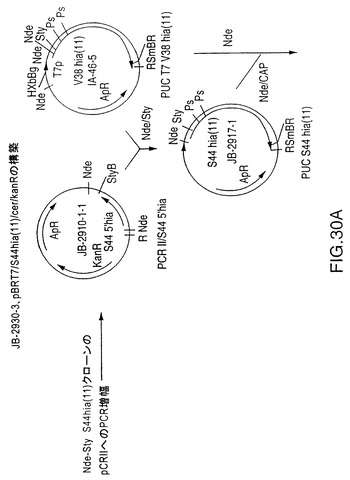
【図30B】
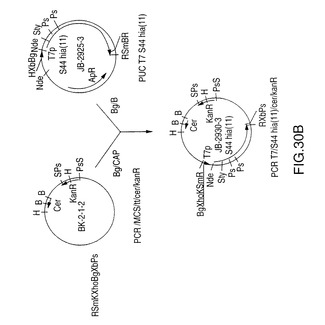
【図31】
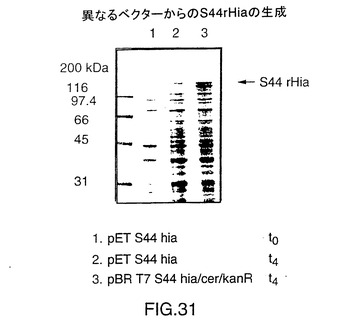
【図28D1】
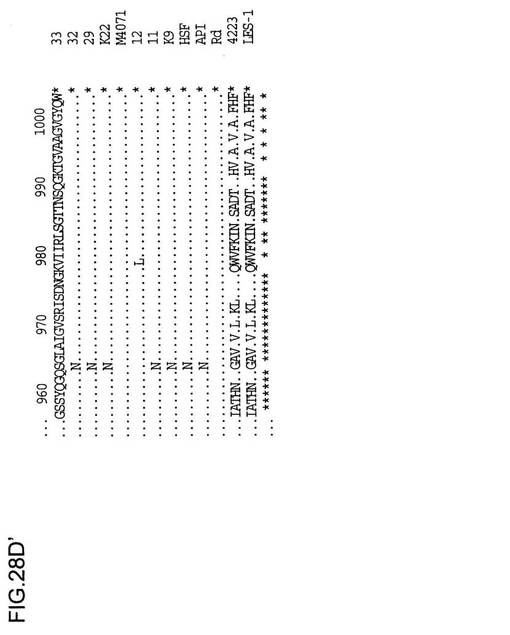
【図32】
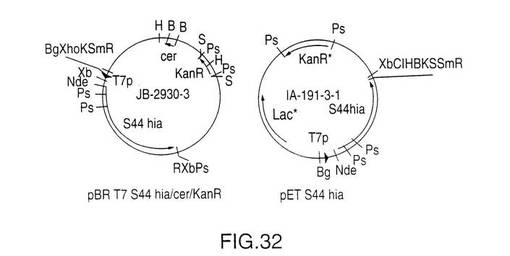
【図1B】
【図2】
【図3】
【図4】
【図5A】
【図5B】
【図5B1】
【図6A】
【図6B】
【図7A】
【図7B】
【図8A】
【図8B】
【図9A】
【図9B】
【図10】
【図11】
【図12】
【図13A】
【図13B】
【図13C】
【図14A】
【図14B】
【図15A】
【図15B】
【図16】
【図17】
【図18A】
【図18B】
【図18C】
【図18D】
【図18E】
【図18F】
【図18G】
【図18H】
【図18I】
【図18J】
【図18K】
【図18L】
【図18M】
【図18N】
【図18O】
【図19A】
【図19B】
【図19C】
【図19D】
【図19E】
【図19F】
【図19G】
【図19H】
【図19I】
【図19J】
【図20A】
【図20B】
【図20C】
【図20D】
【図20E】
【図20F】
【図20G】
【図20H】
【図20I】
【図20J】
【図20K】
【図20L】
【図20M】
【図20N】
【図20O】
【図20P】
【図21A】
【図21B】
【図21C】
【図21D】
【図21E】
【図21F】
【図21G】
【図21H】
【図21I】
【図21J】
【図21K】
【図21L】
【図21M】
【図21N】
【図21O】
【図22A】
【図22B】
【図22C】
【図22D】
【図22E】
【図22F】
【図22G】
【図22H】
【図22I】
【図22J】
【図22K】
【図22L】
【図22M】
【図22N】
【図22O】
【図22P】
【図23A】
【図23B】
【図23C】
【図23D】
【図23E】
【図23F】
【図23G】
【図23H】
【図23I】
【図23J】
【図23K】
【図23L】
【図23M】
【図23N】
【図23O】
【図23P】
【図24A】
【図24B】
【図24C】
【図24D】
【図24E】
【図24F】
【図24G】
【図24H】
【図24I】
【図24J】
【図24K】
【図24L】
【図24M】
【図24N】
【図24O】
【図24P】
【図24Q】
【図24R】
【図24S】
【図24T】
【図24U】
【図24V】
【図24W】
【図24X】
【図24Y】
【図24Z】
【図24A1】
【図24B1】
【図24C1】
【図24D1】
【図24E1】
【図24F1】
【図24G1】
【図24H1】
【図24I1】
【図25A】
【図25B】
【図25C】
【図25D】
【図25E】
【図25F】
【図25G】
【図25H】
【図25I】
【図26A】
【図26B】
【図26C】
【図26D】
【図26E】
【図26F】
【図26G】
【図26H】
【図26I】
【図26J】
【図26K】
【図26L】
【図27A】
【図27B】
【図28A】
【図28B】
【図28C】
【図28D】
【図28E】
【図28F】
【図28G】
【図28H】
【図28I】
【図28J】
【図28K】
【図28L】
【図28M】
【図28N】
【図28O】
【図28P】
【図28Q】
【図28R】
【図28S】
【図28T】
【図28U】
【図28V】
【図28W】
【図28X】
【図28Y】
【図28Z】
【図28A1】
【図28B1】
【図28C1】
【図29】
【図30A】
【図30B】
【図31】
【図28D1】
【図32】
【公開番号】特開2008−188018(P2008−188018A)
【公開日】平成20年8月21日(2008.8.21)
【国際特許分類】
【出願番号】特願2008−84499(P2008−84499)
【出願日】平成20年3月27日(2008.3.27)
【分割の表示】特願2000−605617(P2000−605617)の分割
【原出願日】平成12年3月16日(2000.3.16)
【出願人】(500096994)サノフィ パスツ−ル リミテッド (12)
【氏名又は名称原語表記】SANOFI PASTEUR LIMITED
【Fターム(参考)】
【公開日】平成20年8月21日(2008.8.21)
【国際特許分類】
【出願日】平成20年3月27日(2008.3.27)
【分割の表示】特願2000−605617(P2000−605617)の分割
【原出願日】平成12年3月16日(2000.3.16)
【出願人】(500096994)サノフィ パスツ−ル リミテッド (12)
【氏名又は名称原語表記】SANOFI PASTEUR LIMITED
【Fターム(参考)】
[ Back to top ]