
組換えタンパク質イムノテタヌストキシン
【課題】標的細胞の機能を一過的に抑制することができるような新たなイムノトキシン細胞標的法及びその為の手段を提供すること。
【解決手段】N末側から順に、ヒトインターロイキン‐2受容体α鎖(IL−2Rα)を選択的に認識する抗体の可変領域ポリペプチド、緑濃菌毒素PIIドメインポリペプチド、破傷風菌毒素L鎖ポリペプチド、標識タグ及びKDEL配列をこの順に含む、組換えタンパク質、及び、該組換えタンパク質をヒトインターロイキン‐2受容体α鎖を発現する標的神経細胞に接触させ、該可変領域ポリペプチドとヒトインターロイキン‐2受容体α鎖との特異的結合を介して、該組換えタンパク質を該標的神経細胞内に取り込ませ、破傷風菌毒素L鎖ポリペプチドの作用により該標的神経細胞内のVAMP−2を切断する方法、又は、該標的神経細胞における神経伝達物質の放出を一過的に阻害する方法。
【解決手段】N末側から順に、ヒトインターロイキン‐2受容体α鎖(IL−2Rα)を選択的に認識する抗体の可変領域ポリペプチド、緑濃菌毒素PIIドメインポリペプチド、破傷風菌毒素L鎖ポリペプチド、標識タグ及びKDEL配列をこの順に含む、組換えタンパク質、及び、該組換えタンパク質をヒトインターロイキン‐2受容体α鎖を発現する標的神経細胞に接触させ、該可変領域ポリペプチドとヒトインターロイキン‐2受容体α鎖との特異的結合を介して、該組換えタンパク質を該標的神経細胞内に取り込ませ、破傷風菌毒素L鎖ポリペプチドの作用により該標的神経細胞内のVAMP−2を切断する方法、又は、該標的神経細胞における神経伝達物質の放出を一過的に阻害する方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、組換えタンパク質イムノテタヌストキシン及び該組換えタンパク質を使用した、標的神経細胞を破壊せずに該細胞の特定の機能を一過的に抑制する方法等に関するものである。
【背景技術】
【0002】
さまざまな脳機能は、複雑な神経回路における情報の処理とその調節の機構に基づいている。脳機能を媒介する神経機構を理解するためには、神経回路を構成する特定のニューロンの行動生理学的な役割の解明が必須である。また、ある種の神経疾患は、特定の脳領域の活動異常と関係することが良く知られている。特定のニューロンタイプの機能を抑制する技術の開発は、脳機能の機構の解明や神経疾患の治療法の開発にとって重要なアプローチを提供する。
【0003】
本発明者の研究グループは、以前に、遺伝子発現の特異性に基づいて特定のニューロンの除去を誘導する遺伝子改変技術(イムノトキシン細胞標的法)を開発した(特許文献1、非特許文献1)。この技術では、細胞タイプに特異的なプロモーターを用いてヒトインターロイキン-2受容体αサブユニット(IL-2Rα) を発現するトランスジェニックマウスを作製し、このマウスに組換え体イムノトキシン(ヒトIL-2Rαに対するモノクローナル抗体可変部位と緑膿菌毒素の融合タンパク質)を投与することによって目的の細胞タイプの除去を誘導する。この方法は脳神経回路を構成する特定ニューロンの選択的除去を行い、その行動生理学的な役割を明らかにする研究に応用された(非特許文献2、非特許文献3)。
【特許文献1】特開平8−322427号公報
【非特許文献1】Kobayashi et al., Proc. Natl. Acad. Sci. USA, Vol.92, pp.1132-1136,1995
【非特許文献2】Sano et al., J. Neurosci., Vol.23, pp. 9078-9088, 2003
【非特許文献3】Yasoshima et al., J. Neurosci., Vol.25, pp. 7743-7753, 2005
【発明の開示】
【発明が解決しようとする課題】
【0004】
しかしながら、従来のイムノトキシン細胞標的法においては、標的細胞は完全に破壊又は損傷されてしまい、その後、必要に応じてその標的細胞が関与する機能を回復させることによって、神経回路メカニズムを解明することが不可能であった。そこで、このような解明に有効である、標的細胞の機能を一過的に抑制することができるようなイムノトキシン細胞標的法の開発が望まれている。従って、本発明の目的は、この課題を解決する手段を提供することにある。
【課題を解決するための手段】
【0005】
即ち、本発明は以下の態様に係るものである。
[1]ヒトインターロイキン‐2受容体α鎖(IL−2Rα)を選択的に認識する抗体の可変領域ポリペプチド及び破傷風菌毒素L鎖ポリペプチドを含む組換えタンパク質(イムノテタヌストキシン)。
[2]本発明の組換えタンパク質をコードする核酸分子。
[3]本発明の核酸分子を含む発現ベクター。
[4]本発明の組換えタンパク質の製造方法。
[5]本発明の組換えタンパク質の破傷風菌毒素L鎖ポリペプチドの作用により、標的神経細胞内のVAMP−2を切断する方法。
[6]本発明の組換えタンパク質の破傷風菌毒素L鎖ポリペプチドの作用により、標的神経細胞における神経伝達物質の放出を一過的に阻害する方法。
[7]本発明の方法により、標的神経細胞の損傷を誘導せずに、該細胞の特定の機能を一過的に抑制する方法。
[8]本発明の方法により、動物に一過性の行動障害を誘発させる方法。
【発明の効果】
【0006】
本発明の組換えタンパク質であるイムノテタヌストキシンは、ヒトIL-2Rαを発現する特定の細胞タイプに作用し、標的細胞タイプの損傷を誘導せずに、該細胞の機能を一過性に抑制することができる。
【発明を実施するための最良の形態】
【0007】
以下、本発明を実施する形態について、詳細に説明する。
【0008】
本発明の組換えタンパク質は、標的細胞で特異的に発現(細胞表面上に提示されている等)されているヒトインターロイキン‐2受容体α鎖(IL−2Rα)を選択的(特異的)に認識する抗体の可変領域ポリペプチド及び破傷風菌毒素(テタヌストキシン)L鎖ポリペプチトを構成要素として含む、「イムノトキシン(Immunotoxin)」の誘導体であるイムノテタヌストキシン(以下、「ITet」とも略す)である。
【0009】
テタヌストキシンL鎖は、神経伝達物質の放出に必須の役割を持つvesicle-associated membrane protein-2 (VAMP-2) を切断するプロテアーゼである。尚、テタヌストキシンL鎖のアミノ酸配列は、例えば、Eisel et al., EMBO J., Vol.12, pp. 3365-3372,1993に記載されている (GenBank accession number L19522)。
【0010】
ここで、該可変領域ポリペプチドはIL−2Rαを選択的に認識する任意の抗体に由来するものであり得、その好適例として、ヒトIL−2Rαに対するモノクローナル抗体である抗Tac抗体(Uchiyama, T., et al., J Immunol(1981)126:1393-1397)に由来するものを挙げることが出来る。より具体的には、抗Tac抗体のH鎖及びL鎖の可変領域がペプチドリンカーを介して結合して成る一本鎖Fvフラグメントを挙げることが出来る(Chaudhary,V.K., et al., Nature (1989) 339:394-397; Batra, J.K., et al., J Biol Chem (1990) 265:15198-15202)。
【0011】
更に、該可変領域ポリペプチド及び破傷風菌毒素L鎖ポリペプチドの間に、エンドゾームから細胞質への輸送に関わるポリペプチド、例えば、緑濃菌毒素PIIドメインポリペプチドを有することが好ましい。従って、本発明の組換えタンパク質の好適一例として、N末側から順に、ヒトIL−2Rαを選択的に認識する抗体の可変領域ポリペプチド、緑濃菌毒素PIIドメインポリペプチド、破傷風菌毒素L鎖ポリペプチド、標識タグ配列及びKDEL配列を含む、組換えタンパク質を挙げることが出来る。
尚、毒素PIIドメインポリペプチドのアミノ酸配列及びそれをコードする塩基配列はGray et al., Proc. Natl. Acad. Sci. USA Vol. 81, pp. 2645-2649 (1984)に記載されている。
【0012】
ここで、標識タグ配列の例として、当業者に公知の、例えば、ポリヒスチジン配列、MBP融合タンパク質、チオレドキシン融合タンパク質、及びFLAG融合タンパク質(FLAG配列)を挙げることが出来る。これら標識タグを有していると、各種のアフィニティクロマトグラフィーを用いて本発明の組換えタンパク質を容易に精製することが可能となる。又、KDEL配列はゴルジ膜に存在するKDEL受容体に特異的に結合する配列である。
【0013】
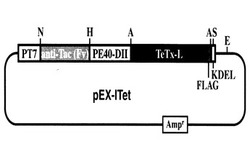
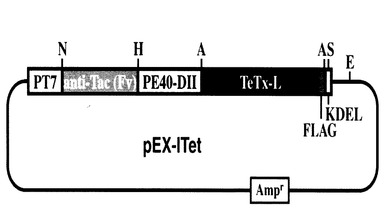
本発明の発現ベクターは、以上の各要素(ポリペプチド)をコードする核酸分子(DNA配列)を含むことを特徴とし、例えば、上記引用文献に記載のような、当業者に公知の任意の方法で調製することができる。該発現ベクターにおいて、上記ポリペプチドをコードする核酸分子は発現調節配列の発現制御下に結合されている。本発明の好適一具体例である発現ベクター (pEX-ITet)の構成を図1に示す。
【0014】
「発現制御下」とは、所定のアミノ酸配列をコードしたDNAが、所定の条件下で、そのアミノ酸配列を有するタンパク質を発現させる能力を有するという意味である。所定のアミノ酸配列をコードしたDNAが発現調節配列の発現制御下に結合されていると、そのDNAは、所定の条件下で、所定のタンパク質を発現するということになる。ここで、「発現調節配列」とは、他の核酸配列の発現を調節する核酸配列のことを意味しており、他の核酸配列の転写、及び、好ましくは翻訳をも制御及び調節する。発現調節配列には、適当なプロモーター、エンハンサー、転写ターミネーター、タンパク質をコードする遺伝子における開始コドン(すなわちATG)、イントロンのためのスプライシングシグナル、ポリアデニル化部位、及びストップコドンが含まれる。
【0015】
「プロモーター」とは、転写を行うために必要最小限な配列のことを意味している。プロモーターには、細胞タイプ特異的、組織特異的、又は外部からの信号や調節剤によってプロモーター依存的に遺伝子の発現を制御するプロモーター要素も含まれる。プロモーター要素は、発現されるDNAの5'領域、又は3'領域のいずれかに結合される。また、プロモーターには、構成的なもの、又は誘導的なもののいずれも含まれる。例えば、バクテリアにおいて発現ベクターを作製する場合には、誘導的なプロモーターとして、T7RNAポリメラーゼプロモーター、バクテリオファージγのpL、plac、ptrp、ptac(ptrp−lacのハイブリッドプロモーター)などのプロモーターが使用できる。
【0016】
このような発現ベクターで、例えば、上記引用文献に記載の方法に準じて、大腸菌等の当業者に公知の適当な宿主を形質転換し、得られた形質転換体を培養し、その細胞懸濁液から本発明の組換えタンパク質を精製することが出来る。
【0017】
「形質転換」とは、ある細胞にとって新しい(外来性の)DNAを導入することによって、その細胞が永久的な遺伝子変化を起こすことを意味している。細胞が哺乳動物細胞である場合には、永久的な遺伝子変化は、その細胞のゲノム中にDNAが導入されることによって達成される。
【0018】
「形質転換体」とは、その細胞、あるいはその細胞の祖先の細胞が、組換えDNA技術によって、本発明の組換えタンパク質をコードしているDNA(を含むベクター)が導入された細胞であることを意味している。組換えDNAで宿主細胞を形質転換することは、当業者にとって周知な方法によって行うことができる。宿主が原核細胞(例えば大腸菌)であるときには、DNAの取り込み能力を有するコンピテント細胞は、当業者に周知の手順によって、塩化カルシウム法で処理することで準備できる。トランスフォーメーションは、エレクトロポレーション等の代替方法によっても実行することができる。
【0019】
本発明の組換えタンパク質は、原核細胞由来の形質転換体においてそのタンパク質をコードしている核酸を発現させることによって得ることができる。原核細胞としては、本発明のタンパク質をコードする核酸配列を含むバクテリオファージ・プラスミドDNA・コスミドDNA発現用組換えベクターによって形質転換された微生物(例えばバクテリア)が例示できるが、本発明の技術的範囲は、これによって限定されるものではない。また、本発明の組換えタンパク質は、インビトロ試験系において、大腸菌を大規模スケールで培養することによって、発現させることが可能である。
【0020】
本発明の組換えタンパク質が発現された後の単離又は精製の技術については、当業者に公知の任意の手段、例えば、イオン交換、アフィニティ及びゲルろ過等の各種クロマトグラフィー、硫安沈殿、並びに、抗原又は抗体(モノクローナル抗体、ポリクローナル抗体)を利用した免疫学的な分離方法によって行うことができる。
【0021】
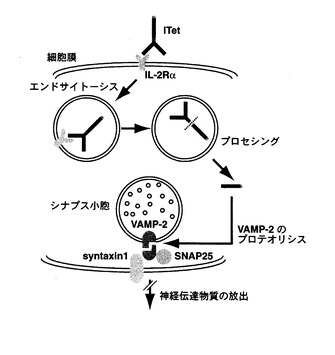
本発明の組換えタンパク質がヒトインターロイキン‐2受容体α鎖を発現する標的神経細胞に接触することによって、該可変領域ポリペプチドとヒトインターロイキン‐2受容体α鎖との特異的結合を介して、該組換えタンパク質を該標的神経細胞内に取り込まれ、破傷風菌毒素L鎖ポリペプチドの作用によりVAMP−2(vesicle-associated membrane protein-2)が切断され、その結果、神経伝達物質の放出が阻害される(図2)。これによって、例えば、神経伝達等の該細胞の特定機能を抑制することができる。この抑制は一過性であり標的神経細胞の細胞死を誘導しない。
【0022】
本発明の組換えタンパク質をヒトインターロイキン‐2受容体α鎖を発現する標的神経細胞に接触させるには、例えば、動物は標的細胞又は組織に該組換えタンパク質を当業者に公知の適当な方法で投与する。標的神経細胞(組織)が血流の影響を受け易い(に近い)場合には、簡単な注入又は他の組織的投与が用いられる。しかしながら、脳内の神経細胞が標的細胞である場合には、該組換えタンパク質は大脳脳室内の適当な区画(例えば、線条体)に直接投与(注入)されることが好ましい。本発明の組換えタンパク質は適当な製剤の形態で投与されることが好ましい。このような製剤は、投与方法及び該タンパク質自体の性質等を考慮して、本発明の組換えタンパク質以外の溶媒及び各種の補助成分を適宜含み、当該技術分野で公知の方法を用いて液体等の任意の形態に調製することが出来る。例えば、適当な調製方法についてはレミントンのファルマスーティカル・サイエンス(Remington's Pharmaceutical Science)最新版、マックパブリッシングカンパニー、イーストン、PA(Mack Publishing Company, Easton, PA)に記載されている。
【0023】
本発明の組換えタンパク質は、標的神経細胞又は組織における神経伝達物質の放出を選択的に且つ有効に一過的に阻害するように、適当な量で使用される。
【0024】
動物の例として、線条体ニューロンのような標的神経細胞においてヒトインターロイキン‐2受容体α鎖がドーパミンD2受容体遺伝子プロモーターの発現制御下に発現するトランスジェニック動物(例えば、マウス及びラット等の実験用齧歯類を含む哺乳動物、但し、ヒトは除く)を挙げることが出来る。このようなトランスジェニック動物の例として、本発明者らが作製したトランスジェニックマウス(非特許文献2)がある。
【実施例】
【0025】
以下、実施例および試験例により本発明をより詳細に説明する。これらの例は、本発明の一部を示すものであり、本発明の技術的範囲は、これらの実施例によって何ら限定されるものではない。又、特に記載のない場合には、各操作における実験条件等は当該技術分野における標準的な方法に従った。
【0026】
実施例1:ITetの精製と活性
A)ITetの作製及び精製:本発明の発現ベクターである pEX-ITetは、当業者に周知の遺伝子工学的手法を用いて、引用文献(Chaudhary,V.K., et al., Nature (1989) 339:394-397; Batra, J.K., et al., J Biol Chem (1990) 265:15198-15202)に記載されている抗Tac(Fv)-PE40(抗Tac抗体由来の一本鎖Fvフラグメントに緑濃菌外毒素のトランケート型PE40が融合した組換えタンパク質)をコードする発現ベクターに基づき、該PE40をコードする配列の一部(触媒部位)を上記の引用文献の記載等に基づき調製した、破傷風菌毒素L鎖ポリペプチドをコードする配列に換えることによって作製した。
【0027】
こうして作製したpEX-ITetを大腸菌に形質転換し、IPTGで誘導後、細胞懸濁液を作成した。その可溶性分画からイオン交換クロマトグラフィー、アフィニティクロマトグラフィー、ゲルろ過クロマトグラフィーを組み合わせて、SDS電気泳動上で単一バンドを示すまで精製した(図3A)。図中、(1)細胞懸濁液、(2)DEAEイオン交換クロマトグラフィー、(3)FLAGアフィニティクロマトグラフィー、(4)Superdex G-200ゲルろ過クロマトグラフィーから得られたピーク画分をSDS電気泳動により分析した結果を示す。
【0028】
B)IL-2Rαへの結合活性:精製ヒトIL-2Rα(Nikaido et al. Nature Vol.311, pp.631-635 (1984))を96ウェルにコーティングし、種々の濃度の精製ITetとインキュベーションした。さらに、PIIドメインに対する抗体(Prior TI, FitzGerald DJ, Pastan I.: Cell 64, 1017-1023 (1991))、HRP結合型の2次抗体(Jackson laboratory)とインキュベーションした。結合タンパク質を発色法によって検出し、450 nmの吸収を測定した。その結果、精製ITetがヒトIL-2Rαへの結合活性を有することがELISAによる結合試験で明らかになった(図3B)。
【0029】
C)In vitroプロテアーゼ活性:シナプトソームに種々の量のITetを添加し、3時間反応させ、一定量のタンパク質をSDS電気泳動により分離した。タンパク質をニトロセルロース膜に転写し、VAMP-2に対する抗体(和光純薬)又はSyntaxin-1Aに対する抗体(和光純薬)を用いてウェスタンブロッティングを行った。バンドを定量し、ITet無添加のサンプルに対する比率を計算し、プロットした( n = 6、*p < 0.05, **p < 0.01)。その結果、ITetがVAMP-2を切断するプロテアーゼ活性を有することが確認された(図3C)。
【0030】
実施例2:ITet注入による一過性の行動障害
野生型とトランスジェニックマウス(非特許文献2に記載のもの)の片側線条体(4箇所)へITetを脳定位固定装置により注入し、2日おきに回転運動を測定して脳組織への影響を調べた。図4に示した結果によれば、ITetの注入によって、トランスジェニックマウスにおいて、処理と反対側への回転運動を誘導し、この回転運動は注入4日後に顕著になった (n = 5-6, *p < 0.05)。その後、回転運動は注入6日後までに減弱し、野生型と同程度になった。この結果は、ITetがヒトIL-2Rαを発現する神経細胞に作用し、この細胞機能を一過性に抑制している可能性を示した。尚、ニッスル染色、in situ hybridizationの解析の結果、ITet注入による線条体ニューロンの損傷は観察されなかった。また、線条体に入力する中脳ドーパミン細胞の神経終末や細胞体への影響も認められなかった。従って、ITet投与は細胞損傷を誘導しないことが明らかとなった。
【0031】
実施例3: ITet注入による一過性の線条体VAMP-2レベルの変化
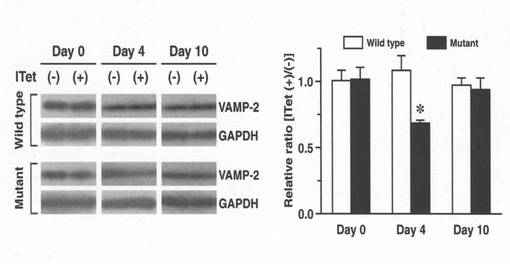
野生型とトランスジェニックマウスの片側線条体(4箇所)へITetを脳定位固定装置により注入した。処理前(day 0)、処理後4日 (day 4)、10日 (day 10) に、未処理側(−)と処理側(+)の線条体を摘出し、組織懸濁液を作製した後、抗VAMP-2抗体又は抗GAPDH (glyceraldehyde 3-phosphate dehydrogenase)抗体(Chemicon)を用いたウェスタンブロッティングを行った。それぞれの線条体について、GAPDHバンドの強度に対するVAMP-2バンドの強度の比率を求めた。2種類の遺伝子型についてITet未処理側に対する処理側の相対値を計算し、比較した (n = 5-6, *p < 0.05)。その結果(図5)によれば、注入4日後のトランスジェニックマウスにおいて、VAMP-2タンパク質レベルの顕著な低下が観察され、注入10日後においてトランスジェニックマウスマウスの線条体VAMP-2レベルは野生型と同程度にまで回復した。この結果は、生体内でもITetはVAMP-2を切断し、特定の細胞機能を一過性に抑制している可能性を支持した。
【産業上の利用可能性】
【0032】
本発明は、従来のイムノトキシン細胞標的法に加えて、行動の制御を媒介する神経回路メカニズムの解明に極めて有効な手法となる。従来のイムノトキシン細胞標的法では特定細胞を除去したが、本発明のイムノトキシン細胞標的法では、同一のマウス系統を用いてその細胞を破壊することなく活動を一過的に抑制できる。本発明は、ウイルスベクター系など遺伝子導入法を工夫することにより、齧歯類ばかりでなくサルも含めた様々な実験動物に適用することが可能であり、脳機能や行動の研究において有力な技術になることが期待される。さらに、ある種の神経疾患で特定の神経細胞の過剰な活動が病態に関係することが知られており、本発明を応用することによって、疾患の治療に結びつく可能性がある。
【0033】
本明細書中に引用される文献に記載された内容は、本明細書の一部として本明細書の開示内容を構成するものである。
【図面の簡単な説明】
【0034】
【図1】本発明の発現ベクター (pEX-ITet) の構成を模式的に示す。T7 RNAポリメラーゼのプロモーターの下流に本発明の組換えタンパク質(ITet)をコードする配列を持つ。ITetはヒトIL-2Rα モノクローナル抗体可変部位、緑膿菌毒素PIIドメイン、テタヌストキシンL鎖、FLAG配列、及びKDEL配列から構成される。
【図2】組換えタンパク質(ITet)の作用機序を示す。ITetはヒトIL-2Rαを発現する神経細胞に結合し、エンドサイトーシスを介して取り込まれる。小泡内でプロセッシングを受け、テタヌストキシンL鎖が細胞質に移行する。テタヌストキシンL鎖はVAMP-2を切断し、神経伝達物質の放出を阻害する。
【図3】A:各種クロマトグラフィーのピーク画分をSDS電気泳動により分析した結果を示す電気泳動の写真。B:精製ITetがヒトIL-2Rαへの結合活性を有することを示すELISAによる結合試験の結果。C: ITetがVAMP-2を切断するプロテアーゼ活性を有することを示すウェスタンブロットの結果。
【図4】ITetが注入されたマウスにおける一過性の行動障害を示す。
【図5】ITetが注入されたマウスにおける一過性の線条体VAMP-2レベルの変化を示す。
【技術分野】
【0001】
本発明は、組換えタンパク質イムノテタヌストキシン及び該組換えタンパク質を使用した、標的神経細胞を破壊せずに該細胞の特定の機能を一過的に抑制する方法等に関するものである。
【背景技術】
【0002】
さまざまな脳機能は、複雑な神経回路における情報の処理とその調節の機構に基づいている。脳機能を媒介する神経機構を理解するためには、神経回路を構成する特定のニューロンの行動生理学的な役割の解明が必須である。また、ある種の神経疾患は、特定の脳領域の活動異常と関係することが良く知られている。特定のニューロンタイプの機能を抑制する技術の開発は、脳機能の機構の解明や神経疾患の治療法の開発にとって重要なアプローチを提供する。
【0003】
本発明者の研究グループは、以前に、遺伝子発現の特異性に基づいて特定のニューロンの除去を誘導する遺伝子改変技術(イムノトキシン細胞標的法)を開発した(特許文献1、非特許文献1)。この技術では、細胞タイプに特異的なプロモーターを用いてヒトインターロイキン-2受容体αサブユニット(IL-2Rα) を発現するトランスジェニックマウスを作製し、このマウスに組換え体イムノトキシン(ヒトIL-2Rαに対するモノクローナル抗体可変部位と緑膿菌毒素の融合タンパク質)を投与することによって目的の細胞タイプの除去を誘導する。この方法は脳神経回路を構成する特定ニューロンの選択的除去を行い、その行動生理学的な役割を明らかにする研究に応用された(非特許文献2、非特許文献3)。
【特許文献1】特開平8−322427号公報
【非特許文献1】Kobayashi et al., Proc. Natl. Acad. Sci. USA, Vol.92, pp.1132-1136,1995
【非特許文献2】Sano et al., J. Neurosci., Vol.23, pp. 9078-9088, 2003
【非特許文献3】Yasoshima et al., J. Neurosci., Vol.25, pp. 7743-7753, 2005
【発明の開示】
【発明が解決しようとする課題】
【0004】
しかしながら、従来のイムノトキシン細胞標的法においては、標的細胞は完全に破壊又は損傷されてしまい、その後、必要に応じてその標的細胞が関与する機能を回復させることによって、神経回路メカニズムを解明することが不可能であった。そこで、このような解明に有効である、標的細胞の機能を一過的に抑制することができるようなイムノトキシン細胞標的法の開発が望まれている。従って、本発明の目的は、この課題を解決する手段を提供することにある。
【課題を解決するための手段】
【0005】
即ち、本発明は以下の態様に係るものである。
[1]ヒトインターロイキン‐2受容体α鎖(IL−2Rα)を選択的に認識する抗体の可変領域ポリペプチド及び破傷風菌毒素L鎖ポリペプチドを含む組換えタンパク質(イムノテタヌストキシン)。
[2]本発明の組換えタンパク質をコードする核酸分子。
[3]本発明の核酸分子を含む発現ベクター。
[4]本発明の組換えタンパク質の製造方法。
[5]本発明の組換えタンパク質の破傷風菌毒素L鎖ポリペプチドの作用により、標的神経細胞内のVAMP−2を切断する方法。
[6]本発明の組換えタンパク質の破傷風菌毒素L鎖ポリペプチドの作用により、標的神経細胞における神経伝達物質の放出を一過的に阻害する方法。
[7]本発明の方法により、標的神経細胞の損傷を誘導せずに、該細胞の特定の機能を一過的に抑制する方法。
[8]本発明の方法により、動物に一過性の行動障害を誘発させる方法。
【発明の効果】
【0006】
本発明の組換えタンパク質であるイムノテタヌストキシンは、ヒトIL-2Rαを発現する特定の細胞タイプに作用し、標的細胞タイプの損傷を誘導せずに、該細胞の機能を一過性に抑制することができる。
【発明を実施するための最良の形態】
【0007】
以下、本発明を実施する形態について、詳細に説明する。
【0008】
本発明の組換えタンパク質は、標的細胞で特異的に発現(細胞表面上に提示されている等)されているヒトインターロイキン‐2受容体α鎖(IL−2Rα)を選択的(特異的)に認識する抗体の可変領域ポリペプチド及び破傷風菌毒素(テタヌストキシン)L鎖ポリペプチトを構成要素として含む、「イムノトキシン(Immunotoxin)」の誘導体であるイムノテタヌストキシン(以下、「ITet」とも略す)である。
【0009】
テタヌストキシンL鎖は、神経伝達物質の放出に必須の役割を持つvesicle-associated membrane protein-2 (VAMP-2) を切断するプロテアーゼである。尚、テタヌストキシンL鎖のアミノ酸配列は、例えば、Eisel et al., EMBO J., Vol.12, pp. 3365-3372,1993に記載されている (GenBank accession number L19522)。
【0010】
ここで、該可変領域ポリペプチドはIL−2Rαを選択的に認識する任意の抗体に由来するものであり得、その好適例として、ヒトIL−2Rαに対するモノクローナル抗体である抗Tac抗体(Uchiyama, T., et al., J Immunol(1981)126:1393-1397)に由来するものを挙げることが出来る。より具体的には、抗Tac抗体のH鎖及びL鎖の可変領域がペプチドリンカーを介して結合して成る一本鎖Fvフラグメントを挙げることが出来る(Chaudhary,V.K., et al., Nature (1989) 339:394-397; Batra, J.K., et al., J Biol Chem (1990) 265:15198-15202)。
【0011】
更に、該可変領域ポリペプチド及び破傷風菌毒素L鎖ポリペプチドの間に、エンドゾームから細胞質への輸送に関わるポリペプチド、例えば、緑濃菌毒素PIIドメインポリペプチドを有することが好ましい。従って、本発明の組換えタンパク質の好適一例として、N末側から順に、ヒトIL−2Rαを選択的に認識する抗体の可変領域ポリペプチド、緑濃菌毒素PIIドメインポリペプチド、破傷風菌毒素L鎖ポリペプチド、標識タグ配列及びKDEL配列を含む、組換えタンパク質を挙げることが出来る。
尚、毒素PIIドメインポリペプチドのアミノ酸配列及びそれをコードする塩基配列はGray et al., Proc. Natl. Acad. Sci. USA Vol. 81, pp. 2645-2649 (1984)に記載されている。
【0012】
ここで、標識タグ配列の例として、当業者に公知の、例えば、ポリヒスチジン配列、MBP融合タンパク質、チオレドキシン融合タンパク質、及びFLAG融合タンパク質(FLAG配列)を挙げることが出来る。これら標識タグを有していると、各種のアフィニティクロマトグラフィーを用いて本発明の組換えタンパク質を容易に精製することが可能となる。又、KDEL配列はゴルジ膜に存在するKDEL受容体に特異的に結合する配列である。
【0013】
本発明の発現ベクターは、以上の各要素(ポリペプチド)をコードする核酸分子(DNA配列)を含むことを特徴とし、例えば、上記引用文献に記載のような、当業者に公知の任意の方法で調製することができる。該発現ベクターにおいて、上記ポリペプチドをコードする核酸分子は発現調節配列の発現制御下に結合されている。本発明の好適一具体例である発現ベクター (pEX-ITet)の構成を図1に示す。
【0014】
「発現制御下」とは、所定のアミノ酸配列をコードしたDNAが、所定の条件下で、そのアミノ酸配列を有するタンパク質を発現させる能力を有するという意味である。所定のアミノ酸配列をコードしたDNAが発現調節配列の発現制御下に結合されていると、そのDNAは、所定の条件下で、所定のタンパク質を発現するということになる。ここで、「発現調節配列」とは、他の核酸配列の発現を調節する核酸配列のことを意味しており、他の核酸配列の転写、及び、好ましくは翻訳をも制御及び調節する。発現調節配列には、適当なプロモーター、エンハンサー、転写ターミネーター、タンパク質をコードする遺伝子における開始コドン(すなわちATG)、イントロンのためのスプライシングシグナル、ポリアデニル化部位、及びストップコドンが含まれる。
【0015】
「プロモーター」とは、転写を行うために必要最小限な配列のことを意味している。プロモーターには、細胞タイプ特異的、組織特異的、又は外部からの信号や調節剤によってプロモーター依存的に遺伝子の発現を制御するプロモーター要素も含まれる。プロモーター要素は、発現されるDNAの5'領域、又は3'領域のいずれかに結合される。また、プロモーターには、構成的なもの、又は誘導的なもののいずれも含まれる。例えば、バクテリアにおいて発現ベクターを作製する場合には、誘導的なプロモーターとして、T7RNAポリメラーゼプロモーター、バクテリオファージγのpL、plac、ptrp、ptac(ptrp−lacのハイブリッドプロモーター)などのプロモーターが使用できる。
【0016】
このような発現ベクターで、例えば、上記引用文献に記載の方法に準じて、大腸菌等の当業者に公知の適当な宿主を形質転換し、得られた形質転換体を培養し、その細胞懸濁液から本発明の組換えタンパク質を精製することが出来る。
【0017】
「形質転換」とは、ある細胞にとって新しい(外来性の)DNAを導入することによって、その細胞が永久的な遺伝子変化を起こすことを意味している。細胞が哺乳動物細胞である場合には、永久的な遺伝子変化は、その細胞のゲノム中にDNAが導入されることによって達成される。
【0018】
「形質転換体」とは、その細胞、あるいはその細胞の祖先の細胞が、組換えDNA技術によって、本発明の組換えタンパク質をコードしているDNA(を含むベクター)が導入された細胞であることを意味している。組換えDNAで宿主細胞を形質転換することは、当業者にとって周知な方法によって行うことができる。宿主が原核細胞(例えば大腸菌)であるときには、DNAの取り込み能力を有するコンピテント細胞は、当業者に周知の手順によって、塩化カルシウム法で処理することで準備できる。トランスフォーメーションは、エレクトロポレーション等の代替方法によっても実行することができる。
【0019】
本発明の組換えタンパク質は、原核細胞由来の形質転換体においてそのタンパク質をコードしている核酸を発現させることによって得ることができる。原核細胞としては、本発明のタンパク質をコードする核酸配列を含むバクテリオファージ・プラスミドDNA・コスミドDNA発現用組換えベクターによって形質転換された微生物(例えばバクテリア)が例示できるが、本発明の技術的範囲は、これによって限定されるものではない。また、本発明の組換えタンパク質は、インビトロ試験系において、大腸菌を大規模スケールで培養することによって、発現させることが可能である。
【0020】
本発明の組換えタンパク質が発現された後の単離又は精製の技術については、当業者に公知の任意の手段、例えば、イオン交換、アフィニティ及びゲルろ過等の各種クロマトグラフィー、硫安沈殿、並びに、抗原又は抗体(モノクローナル抗体、ポリクローナル抗体)を利用した免疫学的な分離方法によって行うことができる。
【0021】
本発明の組換えタンパク質がヒトインターロイキン‐2受容体α鎖を発現する標的神経細胞に接触することによって、該可変領域ポリペプチドとヒトインターロイキン‐2受容体α鎖との特異的結合を介して、該組換えタンパク質を該標的神経細胞内に取り込まれ、破傷風菌毒素L鎖ポリペプチドの作用によりVAMP−2(vesicle-associated membrane protein-2)が切断され、その結果、神経伝達物質の放出が阻害される(図2)。これによって、例えば、神経伝達等の該細胞の特定機能を抑制することができる。この抑制は一過性であり標的神経細胞の細胞死を誘導しない。
【0022】
本発明の組換えタンパク質をヒトインターロイキン‐2受容体α鎖を発現する標的神経細胞に接触させるには、例えば、動物は標的細胞又は組織に該組換えタンパク質を当業者に公知の適当な方法で投与する。標的神経細胞(組織)が血流の影響を受け易い(に近い)場合には、簡単な注入又は他の組織的投与が用いられる。しかしながら、脳内の神経細胞が標的細胞である場合には、該組換えタンパク質は大脳脳室内の適当な区画(例えば、線条体)に直接投与(注入)されることが好ましい。本発明の組換えタンパク質は適当な製剤の形態で投与されることが好ましい。このような製剤は、投与方法及び該タンパク質自体の性質等を考慮して、本発明の組換えタンパク質以外の溶媒及び各種の補助成分を適宜含み、当該技術分野で公知の方法を用いて液体等の任意の形態に調製することが出来る。例えば、適当な調製方法についてはレミントンのファルマスーティカル・サイエンス(Remington's Pharmaceutical Science)最新版、マックパブリッシングカンパニー、イーストン、PA(Mack Publishing Company, Easton, PA)に記載されている。
【0023】
本発明の組換えタンパク質は、標的神経細胞又は組織における神経伝達物質の放出を選択的に且つ有効に一過的に阻害するように、適当な量で使用される。
【0024】
動物の例として、線条体ニューロンのような標的神経細胞においてヒトインターロイキン‐2受容体α鎖がドーパミンD2受容体遺伝子プロモーターの発現制御下に発現するトランスジェニック動物(例えば、マウス及びラット等の実験用齧歯類を含む哺乳動物、但し、ヒトは除く)を挙げることが出来る。このようなトランスジェニック動物の例として、本発明者らが作製したトランスジェニックマウス(非特許文献2)がある。
【実施例】
【0025】
以下、実施例および試験例により本発明をより詳細に説明する。これらの例は、本発明の一部を示すものであり、本発明の技術的範囲は、これらの実施例によって何ら限定されるものではない。又、特に記載のない場合には、各操作における実験条件等は当該技術分野における標準的な方法に従った。
【0026】
実施例1:ITetの精製と活性
A)ITetの作製及び精製:本発明の発現ベクターである pEX-ITetは、当業者に周知の遺伝子工学的手法を用いて、引用文献(Chaudhary,V.K., et al., Nature (1989) 339:394-397; Batra, J.K., et al., J Biol Chem (1990) 265:15198-15202)に記載されている抗Tac(Fv)-PE40(抗Tac抗体由来の一本鎖Fvフラグメントに緑濃菌外毒素のトランケート型PE40が融合した組換えタンパク質)をコードする発現ベクターに基づき、該PE40をコードする配列の一部(触媒部位)を上記の引用文献の記載等に基づき調製した、破傷風菌毒素L鎖ポリペプチドをコードする配列に換えることによって作製した。
【0027】
こうして作製したpEX-ITetを大腸菌に形質転換し、IPTGで誘導後、細胞懸濁液を作成した。その可溶性分画からイオン交換クロマトグラフィー、アフィニティクロマトグラフィー、ゲルろ過クロマトグラフィーを組み合わせて、SDS電気泳動上で単一バンドを示すまで精製した(図3A)。図中、(1)細胞懸濁液、(2)DEAEイオン交換クロマトグラフィー、(3)FLAGアフィニティクロマトグラフィー、(4)Superdex G-200ゲルろ過クロマトグラフィーから得られたピーク画分をSDS電気泳動により分析した結果を示す。
【0028】
B)IL-2Rαへの結合活性:精製ヒトIL-2Rα(Nikaido et al. Nature Vol.311, pp.631-635 (1984))を96ウェルにコーティングし、種々の濃度の精製ITetとインキュベーションした。さらに、PIIドメインに対する抗体(Prior TI, FitzGerald DJ, Pastan I.: Cell 64, 1017-1023 (1991))、HRP結合型の2次抗体(Jackson laboratory)とインキュベーションした。結合タンパク質を発色法によって検出し、450 nmの吸収を測定した。その結果、精製ITetがヒトIL-2Rαへの結合活性を有することがELISAによる結合試験で明らかになった(図3B)。
【0029】
C)In vitroプロテアーゼ活性:シナプトソームに種々の量のITetを添加し、3時間反応させ、一定量のタンパク質をSDS電気泳動により分離した。タンパク質をニトロセルロース膜に転写し、VAMP-2に対する抗体(和光純薬)又はSyntaxin-1Aに対する抗体(和光純薬)を用いてウェスタンブロッティングを行った。バンドを定量し、ITet無添加のサンプルに対する比率を計算し、プロットした( n = 6、*p < 0.05, **p < 0.01)。その結果、ITetがVAMP-2を切断するプロテアーゼ活性を有することが確認された(図3C)。
【0030】
実施例2:ITet注入による一過性の行動障害
野生型とトランスジェニックマウス(非特許文献2に記載のもの)の片側線条体(4箇所)へITetを脳定位固定装置により注入し、2日おきに回転運動を測定して脳組織への影響を調べた。図4に示した結果によれば、ITetの注入によって、トランスジェニックマウスにおいて、処理と反対側への回転運動を誘導し、この回転運動は注入4日後に顕著になった (n = 5-6, *p < 0.05)。その後、回転運動は注入6日後までに減弱し、野生型と同程度になった。この結果は、ITetがヒトIL-2Rαを発現する神経細胞に作用し、この細胞機能を一過性に抑制している可能性を示した。尚、ニッスル染色、in situ hybridizationの解析の結果、ITet注入による線条体ニューロンの損傷は観察されなかった。また、線条体に入力する中脳ドーパミン細胞の神経終末や細胞体への影響も認められなかった。従って、ITet投与は細胞損傷を誘導しないことが明らかとなった。
【0031】
実施例3: ITet注入による一過性の線条体VAMP-2レベルの変化
野生型とトランスジェニックマウスの片側線条体(4箇所)へITetを脳定位固定装置により注入した。処理前(day 0)、処理後4日 (day 4)、10日 (day 10) に、未処理側(−)と処理側(+)の線条体を摘出し、組織懸濁液を作製した後、抗VAMP-2抗体又は抗GAPDH (glyceraldehyde 3-phosphate dehydrogenase)抗体(Chemicon)を用いたウェスタンブロッティングを行った。それぞれの線条体について、GAPDHバンドの強度に対するVAMP-2バンドの強度の比率を求めた。2種類の遺伝子型についてITet未処理側に対する処理側の相対値を計算し、比較した (n = 5-6, *p < 0.05)。その結果(図5)によれば、注入4日後のトランスジェニックマウスにおいて、VAMP-2タンパク質レベルの顕著な低下が観察され、注入10日後においてトランスジェニックマウスマウスの線条体VAMP-2レベルは野生型と同程度にまで回復した。この結果は、生体内でもITetはVAMP-2を切断し、特定の細胞機能を一過性に抑制している可能性を支持した。
【産業上の利用可能性】
【0032】
本発明は、従来のイムノトキシン細胞標的法に加えて、行動の制御を媒介する神経回路メカニズムの解明に極めて有効な手法となる。従来のイムノトキシン細胞標的法では特定細胞を除去したが、本発明のイムノトキシン細胞標的法では、同一のマウス系統を用いてその細胞を破壊することなく活動を一過的に抑制できる。本発明は、ウイルスベクター系など遺伝子導入法を工夫することにより、齧歯類ばかりでなくサルも含めた様々な実験動物に適用することが可能であり、脳機能や行動の研究において有力な技術になることが期待される。さらに、ある種の神経疾患で特定の神経細胞の過剰な活動が病態に関係することが知られており、本発明を応用することによって、疾患の治療に結びつく可能性がある。
【0033】
本明細書中に引用される文献に記載された内容は、本明細書の一部として本明細書の開示内容を構成するものである。
【図面の簡単な説明】
【0034】
【図1】本発明の発現ベクター (pEX-ITet) の構成を模式的に示す。T7 RNAポリメラーゼのプロモーターの下流に本発明の組換えタンパク質(ITet)をコードする配列を持つ。ITetはヒトIL-2Rα モノクローナル抗体可変部位、緑膿菌毒素PIIドメイン、テタヌストキシンL鎖、FLAG配列、及びKDEL配列から構成される。
【図2】組換えタンパク質(ITet)の作用機序を示す。ITetはヒトIL-2Rαを発現する神経細胞に結合し、エンドサイトーシスを介して取り込まれる。小泡内でプロセッシングを受け、テタヌストキシンL鎖が細胞質に移行する。テタヌストキシンL鎖はVAMP-2を切断し、神経伝達物質の放出を阻害する。
【図3】A:各種クロマトグラフィーのピーク画分をSDS電気泳動により分析した結果を示す電気泳動の写真。B:精製ITetがヒトIL-2Rαへの結合活性を有することを示すELISAによる結合試験の結果。C: ITetがVAMP-2を切断するプロテアーゼ活性を有することを示すウェスタンブロットの結果。
【図4】ITetが注入されたマウスにおける一過性の行動障害を示す。
【図5】ITetが注入されたマウスにおける一過性の線条体VAMP-2レベルの変化を示す。
【特許請求の範囲】
【請求項1】
ヒトインターロイキン‐2受容体α鎖(IL−2Rα)を選択的に認識する抗体の可変領域ポリペプチド及び破傷風菌毒素L鎖ポリペプチドを含む組換えタンパク質。
【請求項2】
該可変領域ポリペプチドが、抗Tac抗体のH鎖及びL鎖の可変領域がペプチドリンカーを介して結合して成る一本鎖Fvフラグメントである、請求項1記載の組換えタンパク質。
【請求項3】
該可変領域ポリペプチド及び破傷風菌毒素L鎖ポリペプチドの間に緑濃菌毒素PIIドメインポリペプチドを有する、請求項1又は2記載の組換えタンパク質。
【請求項4】
N末側から順に、ヒトインターロイキン‐2受容体α鎖(IL−2Rα)を選択的に認識する抗体の可変領域ポリペプチド、緑濃菌毒素PIIドメインポリペプチド、破傷風菌毒素L鎖ポリペプチド、標識タグ及びKDEL配列を含む、請求項1〜3のいずれか一項に記載の組換えタンパク質。
【請求項5】
標識タグがFLAG配列である、請求項4記載の組換えタンパク質。
【請求項6】
請求項1〜5のいずれか一項に記載の組換えタンパク質をコードする核酸分子。
【請求項7】
請求項6記載の核酸分子を含む発現ベクター。
【請求項8】
請求項6記載の核酸分子が発現調節配列の発現制御下に結合されている、請求項6記載の発現ベクター。
【請求項9】
発現調節配列がT7RNAポリメラーゼプロモーターである、請求項8記載の発現ベクター。
【請求項10】
pEX−ITetである、請求項9記載の発現ベクター。
【請求項11】
請求項7〜9のいずれか一項に記載の発現ベクターで宿主を形質転換し、得られた形質転換体を培養し、その細胞懸濁液から請求項1〜4のいずれか一項に記載の組換えタンパク質を精製することから成る、該組換えタンパク質の製造方法。
【請求項12】
宿主が大腸菌である、請求項11記載の製造方法。
【請求項13】
請求項1〜5のいずれか一項に記載の組換えタンパク質をヒトインターロイキン‐2受容体α鎖を発現する標的神経細胞に接触させ、該可変領域ポリペプチドとヒトインターロイキン‐2受容体α鎖との特異的結合を介して、該組換えタンパク質を該標的神経細胞内に取り込ませ、破傷風菌毒素L鎖ポリペプチドの作用により該標的神経細胞内のVAMP−2を切断する方法。
【請求項14】
更に、請求項13記載の方法により、該標的神経細胞における神経伝達物質の放出を一過的に阻害する方法。
【請求項15】
請求項13記載の方法により、標的神経細胞の損傷を誘導せずに該細胞の特定の機能を一過的に抑制する方法。
【請求項16】
請求項1〜5のいずれか一項に記載の組換えタンパク質を動物体内に投与することにより、該組換えタンパク質をヒトインターロイキン‐2受容体α鎖を発現する標的神経細胞に接触させる、請求項13〜15のいずれか一項に記載の方法。
【請求項17】
動物が、標的神経細胞においてヒトインターロイキン‐2受容体α鎖がドーパミンD2受容体遺伝子プロモーターの発現制御下に発現するトランスジェニック動物である、請求項16記載の方法。
【請求項18】
標的神経細胞が線条体ニューロンである、請求項16または17記載の方法。
【請求項19】
請求項1〜5のいずれか一項に記載の組換えタンパク質を動物の片側線条体へ注入することにより動物体内に投与する、請求項16〜18のいずれか一項に記載の方法。
【請求項20】
請求項16〜19のいずれか一項に記載の方法により、動物に一過性の行動障害を誘発させる方法。
【請求項1】
ヒトインターロイキン‐2受容体α鎖(IL−2Rα)を選択的に認識する抗体の可変領域ポリペプチド及び破傷風菌毒素L鎖ポリペプチドを含む組換えタンパク質。
【請求項2】
該可変領域ポリペプチドが、抗Tac抗体のH鎖及びL鎖の可変領域がペプチドリンカーを介して結合して成る一本鎖Fvフラグメントである、請求項1記載の組換えタンパク質。
【請求項3】
該可変領域ポリペプチド及び破傷風菌毒素L鎖ポリペプチドの間に緑濃菌毒素PIIドメインポリペプチドを有する、請求項1又は2記載の組換えタンパク質。
【請求項4】
N末側から順に、ヒトインターロイキン‐2受容体α鎖(IL−2Rα)を選択的に認識する抗体の可変領域ポリペプチド、緑濃菌毒素PIIドメインポリペプチド、破傷風菌毒素L鎖ポリペプチド、標識タグ及びKDEL配列を含む、請求項1〜3のいずれか一項に記載の組換えタンパク質。
【請求項5】
標識タグがFLAG配列である、請求項4記載の組換えタンパク質。
【請求項6】
請求項1〜5のいずれか一項に記載の組換えタンパク質をコードする核酸分子。
【請求項7】
請求項6記載の核酸分子を含む発現ベクター。
【請求項8】
請求項6記載の核酸分子が発現調節配列の発現制御下に結合されている、請求項6記載の発現ベクター。
【請求項9】
発現調節配列がT7RNAポリメラーゼプロモーターである、請求項8記載の発現ベクター。
【請求項10】
pEX−ITetである、請求項9記載の発現ベクター。
【請求項11】
請求項7〜9のいずれか一項に記載の発現ベクターで宿主を形質転換し、得られた形質転換体を培養し、その細胞懸濁液から請求項1〜4のいずれか一項に記載の組換えタンパク質を精製することから成る、該組換えタンパク質の製造方法。
【請求項12】
宿主が大腸菌である、請求項11記載の製造方法。
【請求項13】
請求項1〜5のいずれか一項に記載の組換えタンパク質をヒトインターロイキン‐2受容体α鎖を発現する標的神経細胞に接触させ、該可変領域ポリペプチドとヒトインターロイキン‐2受容体α鎖との特異的結合を介して、該組換えタンパク質を該標的神経細胞内に取り込ませ、破傷風菌毒素L鎖ポリペプチドの作用により該標的神経細胞内のVAMP−2を切断する方法。
【請求項14】
更に、請求項13記載の方法により、該標的神経細胞における神経伝達物質の放出を一過的に阻害する方法。
【請求項15】
請求項13記載の方法により、標的神経細胞の損傷を誘導せずに該細胞の特定の機能を一過的に抑制する方法。
【請求項16】
請求項1〜5のいずれか一項に記載の組換えタンパク質を動物体内に投与することにより、該組換えタンパク質をヒトインターロイキン‐2受容体α鎖を発現する標的神経細胞に接触させる、請求項13〜15のいずれか一項に記載の方法。
【請求項17】
動物が、標的神経細胞においてヒトインターロイキン‐2受容体α鎖がドーパミンD2受容体遺伝子プロモーターの発現制御下に発現するトランスジェニック動物である、請求項16記載の方法。
【請求項18】
標的神経細胞が線条体ニューロンである、請求項16または17記載の方法。
【請求項19】
請求項1〜5のいずれか一項に記載の組換えタンパク質を動物の片側線条体へ注入することにより動物体内に投与する、請求項16〜18のいずれか一項に記載の方法。
【請求項20】
請求項16〜19のいずれか一項に記載の方法により、動物に一過性の行動障害を誘発させる方法。
【図1】


【図2】
【図4】
【図3】
【図5】
【図2】
【図4】
【図3】
【図5】
【公開番号】特開2008−228701(P2008−228701A)
【公開日】平成20年10月2日(2008.10.2)
【国際特許分類】
【出願番号】特願2007−76881(P2007−76881)
【出願日】平成19年3月23日(2007.3.23)
【出願人】(503360115)独立行政法人科学技術振興機構 (1,734)
【Fターム(参考)】
【公開日】平成20年10月2日(2008.10.2)
【国際特許分類】
【出願日】平成19年3月23日(2007.3.23)
【出願人】(503360115)独立行政法人科学技術振興機構 (1,734)
【Fターム(参考)】
[ Back to top ]