
組換えポリクローナルタンパク質またはポリクローナル細胞株を構造上特徴付ける方法
本発明は、産生中のポリクローナル細胞株の安定性、ならびに最終ポリクローナル産物のバッチとバッチとの間の一貫性を評価するために用いられ得る構造的特徴決定基盤を提供する。構造上の特徴決定基盤は、単独または組み合わせて、ポリクローナル細胞株および最終産物を特徴決定するのに必須の情報を提供する遺伝子分析ならびにタンパク質の特徴決定技術に基づくものである。基盤技術を用いて分析されるべき異なる相同タンパク質の収集物は、例えば、組換えポリクローナル抗体またはモノクローナル抗体の混合物である。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、最終生産物のバッチとバッチとの間の一貫性、ならびに1回の生産工程における組成物の安定性を証明するために、組換えポリクローナルタンパク質またはかかるタンパク質を産生するポリクローナル細胞株を構造上特徴付ける方法に関するものである。特に、本発明は、組換えポリクローナル抗体を特徴付ける方法に関するものである。
【0002】
発明の背景
抗体の予防上または治療上の投与(いわゆる受動免疫)は、生体の免疫系の能力を増強して、感染性因子を排除し得ると長い間考えられてきた。歴史的に、かかる治療用抗体は、ヒト血漿から得られてきた(そのため、抗体組成物は免疫グロブリンまたはγ−グロブリンと称される)。これらの抗体を得るために、免疫ヒトドナーから血液プールが集められ、そして免疫グロブリンフラクションが抽出され、精製される。免疫グロブリンのフラクションのみが、特定の抗原に特異的となる。ドナーの数が限られていること、製造に費用がかかること、ドナー由来の感染性汚染物質のリスクがあること、バッチとバッチとの間の変動が不可避であること、ならびに投与計画が複雑であることといったいくつかの制限が原因で、免疫グロブリンの治療上の使用は複雑である。
【0003】
最近、組換えモノクローナル抗体が免疫グロブリン製品に取って代わりつつある。しかしながら、それらは1つの標的に対してのみ方向付けられており、それゆえ、感染性因子のような複雑または動的な標的に対しては有効でないかもしれない。この問題を解決するために、モノクローナル抗体を混合する例がいくつか存在する(例えば、Nowakowski, A. et al. 2002. Proc Natl Acad Sci U S A 99, 11346-11350および米国特許第5,126,130号)。
【0004】
最近、予防または治療投与に適した高度に特異的なポリクローナル抗体の組換え生産技術が開発された(WO2004/061104)。組換えポリクローナルタンパク質を構成する個々のメンバーを別々に取り扱うこと、製造すること、精製すること、または特徴付けることなく、組換えポリクローナル抗体(rpAb)は、生産物バイオリアクターから1つの調製物として精製され得る。しかしながら、かかる産生ストラテジーには、同一性を証明し、抗体分子の複雑な混合物を長期にわたり一貫して産生することを明らかにする方法が必要である。
【0005】
さらに、国家的かつ超国家的監督機関から研究薬または治療薬としての認可を得るために、組換え技術を用いて産業上生産されるポリクローナル抗体はある程度特徴付けられなければならないだろう。組換えポリクローナル抗体アプローチは完全に新しいコンセプトであるので、試料における個々のタンパク質の相対的比率について、複数の、異なるが高度に相同なタンパク質を含む試料を特徴付けるという問題がこれまでに取り扱われたことはない。従って、一般に、非臨床上および臨床上の有効性データ、およびしばしば歴史上安全性のデータ、ならびに粗化学反応、製造、および純度、結合タイター、および偶発因子の不存のようなコントロール(CMC)パラメーターに基づいて、血液由来の免疫グロブリンは認可される。もちろん、かかる単純化したアプローチは、組換え産生されたタンパク質には容認されない。ゆえに、2、3種のモノクローナル抗体の混合物について、規制ガイドラインには、かかる混合物が、生物学的アッセイと一体となった包括的なタンパク質化学的特徴決定技術を用いてそれぞれの構成タンパク質について個々に特徴決定されなければならないことが述べられている。しかしながら、これは、技術上実現可能なアプローチでもなく、また10、20、ましてやそれより多くの異なる抗体に基づいた真のポリクローナル組成物に適当でもない。
【0006】
貢献の開示
本発明は、ポリクローナル細胞株からの組換えポリクローナルタンパク質、特に、組換えポリクローナル抗体のような異なる相同タンパク質の混合物の一貫した生産を示すための構造上の特徴決定基盤を提供する。
【0007】
発明の記載
予防上および治療上使用する組換えポリクローナルタンパク質の産業上の生産に前もって必要なものは、発現中のクローンの多様性の維持である。それゆえ、ポリクローナル抗体を産生するポリクローナル細胞株のクローンの多様性、ならびに任意の所望の時間ポイントでのポリクローナルタンパク質、および任意の関連試料における個々のタンパク質の相対的提示をモニターし、測定できることが重要であり、これにより1回の実施における発現系の安定性、ならびに最終生産物のバッチとバッチとの間の変動の分析が可能になる。
【0008】
組換えポリクローナル抗体または組換えポリクローナルT細胞受容体(TcR)のような相同タンパク質の組成物は、非常に類似した物理化学的特性を有する変異体タンパク質から成る。このことは、組換え生産されたポリクローナルタンパク質を精製する場合に利点がある。それが1つのタンパク質であるかのうように、工程において多様性を喪失することなく、精製が行われ得るからである。しかしながら、この類似性は、ポリクローナルタンパク質の個々のメンバーの相対的分布を特徴付ける場合の難問をもたらす。なぜなら、物理化学的特性における類似性は、メンバー1つ1つを互いに区別することを難しくするからである。
【0009】
最も一般的には、組換えポリクローナルタンパク質を産生する場合、オリジナルの組成物は既知である。なぜなら、組換えポリクローナルタンパク質産生用のポリクローナル製造細胞株の創出に先立ち、ポリクローナルタンパク質をコードする配列が単離され、スクリーニングされ、そして配列決定されているからである。かかる細胞株の創出については、WO2004/061104(本明細書において引用により取り込まれる)を参照されたい。これに対する希な例外は、例えば、回復期の患者由来のスクリーニングまたはセレクションされていないライブラリーを直接用いて、組換えポリクローナル抗体を創出することであり得る。
【0010】
アウトプット(組換えポリクローナルタンパク質)の多様性が、培養および精製後にインプット(コード配列のライブラリー)の多様性と類似することを確実にするために、ポリクローナルタンパク質の個々のメンバーおよび/またはポリクローナル製造細胞株内のそれらのコード配列の相対的比率についての情報を得ることが必要だろう。本発明は、ポリクローナル細胞株およびポリクローナルタンパク質両方の多様性についての情報をもたらす能力のある、遺伝子分析ならびにタンパク質の特徴決定技術に基づく、構造上の特徴決定基盤を提供する。
【0011】
定義
用語「抗イディオタイプ抗体」は、ポリクローナルタンパク質の個々のメンバーの変異体部分に特異的に結合する全長抗体またはそのフラグメント(例えば、Fv、scFv、Fab、Fab’、またはF(ab)2)を意味する。好ましくは、本発明の抗イディオタイプ抗体は、ポリクローナル抗体またはポリクローナルTcRの個々のメンバーの変異体部分に特異的に結合する。好ましくは、抗イディオタイプ抗体の特異性は、ポリクローナル抗体またはポリクローナルT細胞受容体の個々のメンバーの抗原特異的部分、いわゆるV領域に対して方向付けられる。しかしながら、それは、個々のメンバーの所定のサブ集団、例えば、混合物において表された特異的VH遺伝子ファミリーに対する特異性も示し得る。
【0012】
用語「抗イディオタイプペプチド」は、相同タンパク質の混合物内の個々のタンパク質メンバーを特異的に関連付け、これにより同定する能力のある特異的なペプチドリガンドを意味する。好ましくは、本発明の抗イディオタイプペプチドは、ポリクローナル抗体またはポリクローナルTcRの個々のメンバーに特異的に結合する。好ましくは、本発明の抗イディオタイプペプチドは、個々の抗体または個々のT細胞受容体の配列の抗原特異的部分に対して方向付けられる。しかしながら、抗イディオタイプペプチドは、個々のメンバーの所定のサブ集団に対する特異性も示し得る。
【0013】
用語「「大量」N−末端配列決定」は、多数の変異体相同タンパク質分子、例えば、ポリクローナルタンパク質を含む試料のN−末端のタンパク質の配列決定を意味する。この大量配列決定により、試料内に存在する異なるタンパク質全ての配列情報が一度に提供される。アミノ酸が試料中の個々のメンバー間で変動する位置において、これらは定量され、可変位置での個々のアミノ酸の異なる量により、特定の変動を含有するタンパク質サブ集団に関する情報がもたらされるだろう。N−末端を配列決定すべきタンパク質が1個より多くのサブユニットを含有する場合、好ましくは、これらが分けられて、複雑性が低減され、配列決定される(例えば、試料がポリクローナル抗体の場合、重鎖を軽鎖が分けられて、配列決定される)。
【0014】
用語「クローンの多様性」または「ポリクローナル性(polyclonality)」は、ポリクローナルタンパク質、それをコードする核酸配列、またはそれを産生するポリクローナル細胞株の変動性または多様性を意味する。変動性は、ポリクローナルタンパク質またはコード配列ライブラリーの個々のメンバー間でのアミノ酸配列または核酸配列の相違により特徴付けられる。ポリクローナル細胞株について、クローンの多様性は、例えば、個々の細胞のゲノムへの一部位の組込みとして細胞株において表される核酸配列の変動性により評価され得る。しかしながら、それは、細胞株において細胞表面上に表されるアミノ酸配列の変動性としても評価され得る。
【0015】
用語「エピトープ」は、T細胞受容体または抗体が結合する抗原分子の部分を意味する。一般に、抗原または抗原分子は、いくつか、あるいは多数のエピトープを同時に提示するだろう。
【0016】
通常、用語「免疫グロブリン」は、血液または血清において見出される抗体の混合物の総称として用いられる。ゆえに、血清由来のポリクローナル抗体は、しばしば、免疫グロブリンまたはγ−グロブリンと称される。しかしながら、免疫グロブリンは、他の供給源からもたらされる抗体の混合物、例えば、組換え免疫グロブリンを称するためにも用いられ得る。
【0017】
本明細書において用いられる用語「個々のクローン」は、特定のタンパク質、例えば、モノクローナル抗体を発現する細胞のアイソジェニックな集団を示す。かかる個々のクローンは、例えば、宿主細胞を所望の核酸でトランスフェクションし、次に、陽性のトランスフェクタントを選択することにより得られ、単一クローンが拡大されるか、あるいは多数の単一クローンがプールされ、次に拡大され得る。ポリクローナル細胞株は、ポリクローナルタンパク質の異なる個々のメンバーを発現する個々のクローンを混合することにより生成され得る。
【0018】
用語「個々のメンバー」または「別個のメンバー」は、ポリクローナルタンパク質のような異なるが相同なタンパク質を含むタンパク質組成物のタンパク質分子を示し、個々のタンパク質分子は、組成物の他の分子と相同だが、ポリペプチド配列の1個以上のストレッチ(stretch)を含有し、そしてそれは、可変領域とも称される、ポリクローナルタンパク質の個々のメンバー間のアミノ酸配列の相違により、特徴付けられる。例えば、Ab1からAb50からなるポリクローナル抗体において、Ab1配列を有するタンパク質の全てが、ポリクローナル抗体の個々のメンバーと考えられ、Ab1は、例えば、CDR3領域においてAb2タンパク質と異なっていてもよい。個々のメンバーのサブ集団は、例えば、Ab1、Ab12、およびAb33に属する抗体から構成され得る。
【0019】
用語「ポリクローナル抗体」は、同じ抗原または異なる抗原上のいくつかの異なる特異的抗原決定基と結合するか、あるいは反応する能力を有する異なる抗体分子の組成物を表す。ポリクローナル抗体の変動性は、ポリクローナル抗体を構成する個々の抗体のいわゆる可変領域、具体的には、相補性決定領域(CDR)1、CDR2、およびCDR3領域に位置する。
【0020】
用語「ポリクローナル製造細胞株」、「ポリクローナル細胞株」、「ポリクローナルマスターセルバンク(pMCB)」、および「ポリクローナルワーキングセルバンク(pWBC)」は、互換使用され、目的の変異体核酸配列ライブラリーでトランスフェクションされたタンパク質発現細胞の集団を意味する。好ましくは、個々の細胞は、一体となって組換えポリクローナル製造細胞株を構成し、目的の組換えポリクローナルタンパク質の1メンバーをコードする目的の異なる核酸配列を1コピー有し、そしてそれぞれのコピーは、各細胞のゲノムの同じ部位に組み込まれる。かかる製造細胞株を構成し得る細胞は、例えば、細菌、真菌、真核細胞、例えば、酵母、昆虫細胞、または哺乳類細胞、特に、CHO細胞、COS細胞、BHK細胞、ミエローマ細胞(例えば、Sp2/0細胞、NS0)、NIH3T3、YB2/0のような不死の哺乳類細胞株、およびHeLa細胞、HEK293細胞、またはPER C6細胞のような不死化ヒト細胞であり得る。
【0021】
本明細書で用いられる用語「ポリクローナルタンパク質」は、異なるが、相同なタンパク質分子を含むタンパク質組成物を意味し、好ましくは、それは、免疫グロブリンスーパーファミリーから選択される。なおより好ましくは、相同なタンパク質分子は、抗体またはT細胞受容体(TcR)である。従って、それぞれのタンパク質分子は、組成物の他の分子と相同だが、変動可能なポリペプチド配列の1個以上のストレッチも含有し、そしてそれは、ポリクローナルタンパク質の別の変異体メンバーとも称される個々のメンバー間でのアミノ酸配列の相違により特徴付けられる。かかるポリクローナルタンパク質の既知の例は、抗体、T細胞受容体、およびB細胞受容体を含む。ポリクローナルタンパク質は、タンパク質分子の所定のサブセットからなり得、そしてそれは、例えば、所望の標的抗原に対するポリクローナル抗体の場合、所望の標的に対する共有結合活性のような共通の特性により定義されてきた。一般に、組換えポリクローナルタンパク質は、かかる分子の所定のサブセットからなり、ここで、それぞれのメンバーの配列は既知である。希にだが、組換えポリクローナルタンパク質は、非標的特異的タンパク質の重要な部分も含有するという意味で、血清由来の免疫グロブリンと類似し得る。
【0022】
用語「ポリクローナルT細胞受容体(TcR)」は、同じ抗原または異なる抗原由来のいくつかの異なる特異的な抗原決定基に結合するか、あるいはそれと反応する能力を有する、異なるTcR分子の組成物を表す。ポリクローナルTcRの変動性は、ポリクローナルTcRを構成する個々のTcR分子のいわゆる可変領域、具体的には、CDR1、CDR2、CDR3、またはCDR4領域に位置する。本発明のTcR分子は、α−β鎖またはγ−δ鎖の改変可溶性ダイマーである。かかる改変TcRは、例えば、(Willcox, B.E. et al. 1999. Protein Sci 8, 2418-2423)に記載されている。
【0023】
用語「タンパク質」は、長さまたは翻訳後修飾の如何に関わらず、アミノ酸の任意の鎖を意味する。タンパク質は、2個以上のアセンブリされたポリペプチド鎖、タンパク質のフラグメント、ポリペプチド、オリゴペプチド、またはペプチドを含む単量体または多量体として存在し得る。
【0024】
用語「センチネルタンパク質」は、ポリクローナルタンパク質の個々のメンバーを表し、そしてポリクローナルタンパク質の産生中、または異なるバッチにおいてその存在についてモニターされ得る。一連の関連試料におけるセンチネルタンパク質の存在の一貫性は、バッチ間あるいは1回の産生での長時間のポリクローナルタンパク質の発現の安定性を反映するだろう。さらにそれは、組換え産生されたポリクローナルタンパク質の精製のような下流処理における多様性の維持を反映するだろう。
【0025】
用語「固有のマーカーペプチド」は、ポリクローナルタンパク質の個々のメンバーの可変領域を起源とする多数のペプチドを表す。好ましくは、ペプチドは、タンパク質の断片化であるプロテアーゼ処理または他の手段により生成され、ポリクローナルタンパク質の単一の個々のメンバーに明確に割り当てられ得るペプチドが、固有のマーカーペプチドと称される。
【0026】
発明の詳細な説明
本発明の態様は、(i)異なる可変領域を有する異なる相同タンパク質、または(ii)かかるタンパク質を産生する細胞株を含む試料における個々のメンバーの相対的比率に関する情報を得るための構造上の特徴決定用基盤を提供することである。特徴決定基盤を用いて、異なる相同タンパク質を含む組成物の産生または精製過程、あるいは長期保存中に異なる態様が評価され得る。好ましくは、本発明の特徴決定基盤は、次の目的:i)1つの試料内で互いに関連する個々のメンバーまたは個々のメンバーのいくつかの相対的出現量を決定すること、ii)バッチとバッチとの間の一貫性の決定のため、異なる試料における1個以上の個々のメンバーの相対的比率を評価すること、およびiii)1個以上の個々のメンバーの実際の比率を評価すること、のうちの1つのために用いられる。必要に応じて、これは、ポリクローナル製造細胞株を生成するためにもともと用いられたベクターライブラリーと比較されてもよい。特徴決定基盤は、ポリクローナル細胞株のクローンの多様性および/または細胞株により産生されたポリクローナルタンパク質における個々のタンパク質の出現量をモニタリングするのに特に有用である。個々の生産工程における組成物の安定性およびバッチとバッチとの間の一貫性の両方がモニターされ得る。別法として、基盤手段はまた、ポリクローナルタンパク質またはモノクローナル抗体の混合物を含む異なる相同タンパク質の混合物の精製組成物に適用され、例えば、かかる組成物における個々のメンバーの長期安定性が評価され得る。
【0027】
本発明の1つの実施態様は、タンパク質またはそれらのコード配列の個々のメンバーの相対的比率または存在に関する情報が得られるよう、異なる可変領域を有する異なる相同タンパク質またはかかるタンパク質を産生する細胞を含む試料を特徴付ける方法であって、該方法は、1個以上のタンパク質特徴決定技術により、および/または1個以上のタンパク質コード配列の遺伝子分析により、該試料のアリコートを分析することを含む。
【0028】
本発明のさらなる実施態様において、構造上の特徴決定基盤は、タンパク質の特徴決定技術ならびに遺伝子分析から選択される多数の分析技術からなる。従って、構造上の特徴決定基盤は、任意の数の個々の実施態様(次のセクションに記載)からなる。実施態様に記載の分析技術の1つからのみ、試料について情報を得れば足り得る。しかしながら、好ましくは、これらの分析技術の少なくとも2、3、4、5、6、7、8、9、または10個から情報が得られ、それにより、以下に詳述される個々の実施態様が組み合わされ、特徴決定基盤が生成される。いくつかの分析技術の組み合わせにより、ポリクローナル混合物の相対的または絶対的組成に関するより記述的なデータセットの創出が可能となる。これらの技術から得られる情報は、定量的性質ならびに定性的性質のものであり、一体となって蓄積された場合、分析された試料の全体的な特徴決定を提供する。
【0029】
本発明の好ましい実施態様において、1つの分析技術は、タンパク質の特徴決定技術であり、別の分析技術は遺伝子分析である。
【0030】
遺伝子分析は、制限断片長多型(RFLP)分析、末端RFLP(T−RFLP)、マイクロアレイ分析、リアルタイムPCRのような定量的PCR、および核酸配列決定のような技術を意味する。
【0031】
タンパク質の特徴決定技術は、i)物理化学的特性によりタンパク質を分けるクロマトグラフィー分析、ii)相同タンパク質のタンパク分解性消化物の分析、iii)「大量」N−末端配列決定、およびiv)相同タンパク質に特異的な検出分子を用いた分析のような未知のタンパク質を特徴付けるため、プロテオミクス分野で一般に用いられる技術を意味する。
【0032】
本発明のさらなるコンセプトは、相同タンパク質の複雑なプールを特徴付けるため、上述の分析技術と組み合わせて適用でき、本発明と関連して開発された。該コンセプトは、相同タンパク質のプール(例えば、ポリクローナル抗体またはポリクローナルTcR)、または相同タンパク質を産生する細胞のプール(例えば、ポリクローナル製造細胞株)の表面に存在する多数のセンチネルタンパク質の選択に基づくものである。センチネルタンパク質は、定量的および定性的に特徴付けられ、タンパク質のこのサブ集団が、ポリクローナル細胞培養物の上清中または産生中の細胞表面上のいずれかで、一貫した方法で存在することが証明される。センチネルタンパク質は、例えば、抗イディオタイプ分子のような相同タンパク質の個々のメンバーに特異的な検出分子を用いて分析され得る。センチネルタンパク質のコンセプトはさらに、異なる細胞培養物のバッチ間の一貫性を評価するために適用され得る。センチネルタンパク質のコンセプトは、ポリクローナルタンパク質がポリクローナル抗体またはTcRである場合、好ましくは、センチネルペプチドがCDRの一部を含有するよう、プロテアーゼ処理によりポリクローナルタンパク質からもたらされた固有のペプチドに拡大され得る。遺伝子レベルで行われる分析はまた、センチネルの原理を適用し、これは、ポリクローナルタンパク質をコードするライブラリーの個々のメンバー由来の固有の核酸配列に基づくものである。具体的には、抗体またはTcRのCDR領域に対応する核酸配列は、センチネル核酸配列として選択される。最も好ましくは、CDR3領域である。センチネルタンパク質、ペプチド、または核酸配列は、ポリクローナルタンパク質のメンバー、またはそれをコードする核酸配列に依存して、個々の分析技術について変動し得、それは、選択された分析技術を用いて区別され得る。
【0033】
ポリクローナル製造細胞株のクローンの多様性の遺伝子分析
本発明のある実施態様において、ポリクローナルタンパク質を産生するための発現系におけるポリクローナル性は、ポリクローナルタンパク質の特定のメンバーをコードする細胞の量および/またはポリクローナルタンパク質の個々のメンバーをコードするmRNAレベルを評価することによりモニターされる。これは、例えば、RFLPまたはT−RFLP分析、オリゴヌクレオチドマイクロアレイ分析、リアルタイムPCRのような定量的PCR、および製造細胞株から得られた遺伝子配列の可変領域の核酸配列決定を用いて、mRNAレベルまたはゲノムレベルでモニターされ得る。別法として、同じ技術を定性的に用いて、ポリクローナル細胞株の多様性が示され得る。ポリクローナルタンパク質をコードする核酸配列は、培養中の異なる時間ポイントで単一のポリクローナル細胞培養物から得られた試料においてモニターされ、それにより、生産工程を通じて個々のコード配列の相対的比率をモニターし、その組成物の安定性が評価され得る。別法として、ポリクローナルタンパク質をコードする核酸配列は、特定の時間ポイントで異なるポリクローナル細胞培養物から得られる試料においてモニターされ、それにより、異なるバッチにおける個々のコード配列の相対的比率をモニターし、バッチとバッチとの間の変動が評価され得る。好ましくは、遺伝子分析において用いられる試料は、例えば、沈殿により、培養物の細胞について濃縮された細胞培養物フラクションである。一般に、試料は、所望の時間ポイントで細胞培養物のフラクションを回収し、次に、例えば、遠心分離により培地を除去して得られる。バッチとバッチとの間の一貫性の比較のための試料は、好ましくは、産生についてのインビトロの細胞継代数(age)の限界で、細胞から得られる。
【0034】
RFLP/T−RFLP
RFLPおよびT−RFLP分析は、ゲノムレベルまたはmRNAレベルで行われ得る。各細胞が目的の配列を1コピーのみ含有するように、ポリクローナル製造細胞株が生成される場合、ゲノムレベルでの分析は、ポリクローナルタンパク質の個々のメンバーを産生する製造細胞株における細胞の相対的比率に関する情報をもたらすだろう。一方、mRNAレベルでの分析は、ポリクローナルタンパク質の個々のメンバーの可能性のある発現レベルに関する情報をもたらすだろう。一般に、mRNAレベルでの分析は、mRNAをcDNAに逆転写し、次に制限酵素分析することにより、行われる。しかしながら、mRNAにおいて分析を直接行うことも可能である。
【0035】
末端RFLP分析において、PCRまたはRT−PCRのために用いられるフォワードおよび/またはリバースプライマーは標識され、これにより、PCRフラグメントの末端標識がもたらされる。適当な制限酵素を用いて消化した後、異なる大きさのフラグメントが生成され、電気泳動、好ましくは、キャピラリー電気泳動により分けられ、そしてフラグメントが標識単位複製配列により検出され得る(Liu et al. 1997, Applied and Environmental Microbiology 63, 4516-4522)。適当な標識は、蛍光、放射能、比色、X線回析または吸収、磁性、または酵素活性により検出可能なシグナルをもたらし、そして、例えば、フルオロフォア、発色団、放射性同位元素(特に、32P、33P、35S、および125I)、高電子密度試薬、酵素、および特異的結合パートナーを有するリガンドを含む。好ましくは、フルオロフォアが標識として用いられる。
【0036】
巨大な多様性を有するポリクローナル製造細胞株において、それぞれ個々のコード配列について固有の制限酵素フラグメントを得ることはできないかもしれない。かかる状況が生じる場合、センチネル核酸配列は、ポリクローナル製造細胞株のクローンの多様性をモニターするために選択され得る。別法として、大きさにより分けられないフラグメントが、個々のコード配列全ての分布を評価するために配列決定され得る。
【0037】
オリゴヌクレオチドマイクロアレイ分析
DNAチップのようなオリゴヌクレオチドマイクロアレイを用いて、細胞株から生成された標識DNAのハイブリダイゼーションを測定することにより、ポリクローナル細胞株におけるゲノムDNAレベルまたはmRNAレベルが測定され得る(Guo, Z. et al 1994. Nucleic Acids Res. 22, 5456-5465)。
【0038】
プローブは、ポリクローナル細胞株に存在すると予測される配列の2本鎖cDNA配列の代表(ポリクローナル細胞株自体、またはポリクローナル細胞株を含む宿主細胞をトランスフェクションするために用いられるDNAライブラリーのいずれかからもたらされる)、またはセンスオリゴヌクレオチド(長さ20〜90nt)のいずれかであり得る。プローブは、ガラス、プラスチック、またはゲルマトリックスのような固体表面に付着し、そして2本鎖プローブが用いられる場合、それは変性されて、アッセイが行われる。相同タンパク質、例えば、ポリクローナル抗体またはポリクローナルTcRを発現するポリクローナル細胞株を分析するとき、好ましくは、プローブとポリクローナル細胞株からもたらされる標識cDNAとの間でのクロスハイブリダイゼーションを防ぐよう、注意深く設計されたオリゴヌクレオチドプローブが用いられる。かかるプローブは、ポリクローナル産物の個々のメンバーそれぞれに特異的なプローブを設計するためにポリクローナル製造細胞株を生成するために用いられた可変領域コード配列のアライメントに基づき設計される。抗体について、コード配列はCDR領域において主として異なり、CDR3領域における変動性の程度が最も高いであろう。好ましくは、最大の変動性を有する領域が、センスオリゴヌクレオチドの設計のため用いられる。好ましくは、プローブは、ポリクローナル細胞株において含まれる個々のメンバーに対して配列が相補であり、他のメンバーに対して出来るだけ高度な相違を有する。それぞれの可変領域に特異的な1個以上のプローブが用いられ得る。標準的な目的のため、定常領域の配列とハイブリダイズするプローブが用いられ得る。プローブは、ハイブリダイゼーションに用いられる表面に直接スポットされるか、あるいは表面上にインサイツで合成される(Pease et al. 1994. PNAS 91: 5022-5026, Singh-Gasson et al. 1999. Nature Biotech. 17: 974-978)。
【0039】
分析されるべき標識DNAは、ポリクローナル細胞集団を回収し、細胞からゲノムDNA、トータルRNA、またはmRNAを調製することにより、生成される。ゲノムDNAを用いる場合、標識は、関連コード配列のPCR増幅において適当に標識されたプライマーまたは標識ヌクレオチドのいずれかを用いて得られる。トータルRNAまたはmRNAを用いる場合、逆転写のみ、または標識プライマーまたはヌクレオチドを用いたPCR工程と組み合わせて、標識cDNAを得ることが可能である。適当な標識は、蛍光、放射能、比色、X線回析または吸収、磁性、または酵素活性により検出可能なシグナルをもたらし得、そして例えば、フルオロフォア、発色団、放射性同位元素(特に、32P、33P、35S、および125I)、高電子密度試薬、酵素、および特異的結合パートナーを有するリガンドを含む。好ましくは、フルオロフォアが標識として用いられる。分析されるべきコード配列が抗体重鎖および軽鎖である場合、1本鎖cDNAは、可変領域の3’側の定常領域に位置するアンチセンスプライマーを用いたプライミングによる逆転写により調製される。さらにPCRが行われない場合、好ましくは、合成は、標識ヌクレオチドを用いて行われる。逆転写の後PCRが行われ、一連のセンスプライマーが適用され、確実に可変領域の全ファミーが増幅される。別法として、全てのmRNAにおいて一致する領域(例えば、5’非翻訳領域またはシグナルペプチドコード配列)にハイブリダイズするセンスプライマーが用いられ得る。センス、および/またはアンチセンスプライマーは蛍光標識されるか、あるいは標識ヌクレオチドがこのアプローチにおいて用いられ得る。
【0040】
プローブおよび標識DNAが調製された場合、マイクロアレイアッセイは、低ノイズかつ高度に特異的なシグナルについて最適化された条件下で、変性させた標識DNAを固定オリゴヌクレオチドとハイブリダイズさせることにより、行われる。洗浄後、ハイブリダイズしたプローブそれぞれが測定され、特異的なメッセージの量が計算される。
【0041】
定量的PCR
PCR法は、試料中の核酸配列の検出および定量の両方をもたらすよう既に適合されている。例えば、Higuchi, R. et al. 1993. Kinetic Biotechnology 11, 1026-1030;Holland, P.M. et al. 1991. PNAS 88, 7276-7280;Livak, K.J. et al. 1995 PCR Methods Appl. 4, 357-362を参照されたい。これらの方法は、標準的PCRなどでのフォワードおよびリバースプライマー、および増幅される核酸とハイブリダイズする1個以上のさらなる核酸配列を利用する。このさらなる核酸配列は、「プローブ」と称され、2つのプライマーにハイブリダイズする部分の間で増幅されるべき核酸の一部にハイブリダイズし、各連続的PCRサイクルがプローブまたはその標識の変化を生じるような方法で標識される。プローブまたはその標識のこの変化により、各PCRサイクルにおいて増幅された核酸の追加コピー数に関連する程度まで、標識は活性化されるか、あるいは強調される。一般に、かかる方法は、「リアルタイム」PCRと称され、熱サイクルを標識の検出と組み合わせることにより、PCR産物の増大をサイクル毎に検出する。リアルタイムPCRの特定のバージョンにおいて、プローブまたはその標識の変化は、ポリメラーゼ、例えば、Taqポリメラーゼのエキソヌクレアーゼ活性により引き起こされ、それゆえ、この技術は、一般にTaqまたはTaqManリアルタイムPCRと称される(例えば、Holland, P.M. et al. 1991. PNAS 88, 7276-7280)。
【0042】
適当な標識は、蛍光、放射能、比色、X線回析または吸収、磁性、または酵素活性により検出可能なシグナルをもたらし、そして例えば、フルオロフォア、発色団、放射性同位元素(特に、32P、33P、35S、および125I)、高電子密度試薬、酵素、および特異的結合パートナーを有するリガンドを含む。最も一般的には、プローブの標識は蛍光標識であり、これは蛍光出力アウトプットシグナルをもたらす。これは、一方の端、典型的には、5’末端で蛍光レポーターダイ、そしてもう一方の端、3’末端でクエンチャーダイで2重標識されたプローブを用意することにより、達成され得る(例えば、Livak, K.J. et al. 1995 PCR Methods Appl. 4, 357-362)。プローブがインタクトであるとき、クエンチャーダイがレポーターダイに近づくと、レポーターダイの蛍光が抑制される。適当なダイは、Wilhelm, J. and Pingoud, A., 2003. Chembiochem. 4, 1120-1128において概説されている。それぞれのPCRサイクルにおいて、DNAポリメラーゼの5’→3’エキソヌクレアーゼ活性によりプローブが開裂され、そしてそれにより、レポーターダイをクエンチャーダイと分離させる。この分離により、レポーターダイの蛍光の増大が生じる。
【0043】
PCRにおいて、目的の標的が試料に存在する場合、プローブは、フォワードとリバースPCRプライマー部位の間で特異的にアニーリングする。プローブが標的分子とハイブリダイズする場合のみ、DNAポリメラーゼのエキソヌクレアーゼ活性により、レポーターとクエンチャーダイの間でプローブが開裂される。これらのプローブは、しばしばTaqManプローブと称される。標的配列がプローブと相補的であり、かつPCRにおいて増幅される場合のみ、蛍光の増大が検出される。これらの要件のため、非特異的な増幅は検出されない。プローブに相補的な配列を含有する増幅産物のみが、蛍光シグナルの存在により認識され、それにより、擬陽性の分析と関連するある種の要素が排除される。加えて、1種類以上の他の酵素を用いて、キャリーオーバーの転写産物の増幅の制限が手助けされてもよい。
【0044】
この種の定量的PCRは、ピペット誤差および用量変化の校正を許可し、そしてそれは、各反応に含有されるレポーターの蛍光をパッシブリファレンスで割ることにより行われ、個々の反応それぞれについて校正されたレポーターシグナルが決定され得る。絶対的な定量化のため出発コピー数を決定するか、あるいは相対的な量の比較のため他の未知試料に対して比較するために、ソフトウエアを用いて、サイクル毎の蛍光強度の増大が分析され、次に、これらのデータが標準と比較され得る。
【0045】
具体的には、本発明においてTaqManリアルタイムPCRが、ポリクローナル細胞株の特徴決定に適していることが見出された。従って、ポリクローナル抗体発現細胞株に適用される場合、固有のTaqManプローブはポリクローナル細胞株において表されメンバーそれぞれの重鎖および/または軽鎖について設計され得るので、該技術により、個々の抗体のコード配列の相対的比率が定量される。好ましくは、CDR領域の1つであるCDR1、CDR2、またはCDR3が、TaqManプローブを設計するために選択される。最も好ましくは、CDR3領域がTaqManプローブを設計するために選択される。かかる可変重鎖CDR3のTaqManプローブの例は、Rasmussen, T. et al. 2000. Exp. Hematol. 28, 1039-1045(本明細書において引用により取り込まれる)において見られる。
【0046】
核酸の配列決定
核酸の配列決定はよく知られた技術であり、本発明と共に利用され、ポリクローナル製造細胞株の多様性に関する定性的情報をもたらし得る。配列決定は、単一細胞クローニングによりポリクローナル細胞株からもたらされる単一細胞、またはポリクローナル製造細胞株から得られる変異体細胞の未処理試料のいずれかにおいて行われ得る。
【0047】
単一細胞レベルでの配列決定は、ポリクローナルタンパク質の個々のメンバーを産生する、製造細胞株における細胞の相対的比率に関する情報をもたらすだろう。この方法において、ポリクローナル製造細胞株由来試料は、所望の時間ポイントで得られ、特徴付けられるべきポリクローナルタンパク質をコードする細胞は、例えば、限界希釈またはFACS Ariaのようなセルソーターにより、単一細胞クローニングされる。ポリクローナル細胞株試料から得られる単一細胞の数は、細胞株において表されると予測される配列の多様性に依存する。好ましくは、少なくとも3回、細胞株の生成におけるインプットを形成する個々のコード配列の数がシングルセルソートされ、試験試料中においてそれら全てを再び見出す見込みは95%となるだろう。従って、25種類の異なるコード配列のライブラリーを用いて、細胞株が創出される場合、少なくとも75種類の単一細胞クローンが、配列決定用試料から得られ、これにより、等しい量において表される異なる配列が25種類より多くなる。これにより、ポリクローナル細胞株において表される個々のコード配列の大部分が、製造過程において喪失されなかったなら、単一細胞クローン間で表されることが確実になるだろう。単一細胞は別々のウェルにおいてコンフレントまで成長され、それぞれのウェル由来のアリコートが、核酸の配列決定反応における鋳型として用いられる。配列決定は、配列決定に先立ちRT−PCRまたはPCR増幅工程をそれぞれ用いて、mRNAレベルまたはゲノムレベルで行われ得る。mRNAまたはゲノムレベルのいずれかで得られた配列情報により、個々の抗体成分のそれぞれをコードする細胞の割合が決定され得る。さらに、mRNAレベルで得られた配列情報を用いて、ポリクローナル組成物における個々の抗体それぞれの発現レベルが評価され得る。配列決定に加えて、mRNAレベルでTaqManリアルタイムPCRを行い、単一細胞クローンの可能性のある発現レベルと関連する情報を得ることが可能である。上述のRFLPまたはT−RFLP分析は、同様に、単一細胞レベルで行われ得る。
【0048】
ポリクローナル製造細胞株から得られる変異体細胞の未処理試料における配列決定はまた、細胞株から産生されるポリクローナルタンパク質の個々のメンバーの可能性のある発現レベルに関する情報をもたらすことができ、そしてそれは、ポリクローナルタンパク質の個々のメンバーのコード配列の相対的mRNAレベルに基づくものである。この方法において、ポリクローナル製造細胞株由来試料は、所望の時間ポイントで得られる。RT−PCRは、試料中の溶解細胞において直接行われる。RT−PCR反応に適用されるプライマーセットは、センスおよびアンチセンスプライマーが全てのmRNAにおいて一致する領域にハイブリダイズする場合、同じ有効性を有するコード配列全てを増幅すると予測されるような方法で、設計される(例えば、5’非翻訳領域またはシグナルペプチドコード配列中のセンスプライマー、および定常領域配列中ののアンチセンスプライマーが用いられ得る)。増幅されたPCRフラグメントは、配列決定ベクターにクローン化され、宿主細胞、好ましくは、エスケリキア・コリ(E.coli)にトランスフェクションされる。ポリクローナル製造細胞株由来の個々のコード配列を表す単一クローン由来のプラスミドDNAが配列決定され、得られた個々のコード配列の比率は、ポリクローナル細胞株における個々のコード配列それぞれのmRNAレベル、ならびに個々のタンパク質メンバーの可能性のある発現レベルを反映するだろう。
【0049】
本発明のさらなる実施態様において、上述の遺伝子分析が別の分析として適用される。好ましくは、1種類以上の分析が、細胞株のクローンの多様性についての情報をできるだけ多く得るために、同じ試料由来のアリコートにおいて行われる。別法として、遺伝子分析が、多次元様式で組み合わされ得る(例えば、マイクロアレイ分析が、この分析後、RFLPまたはT−RFLPフラグメントにおいて行われるか、あるいはRFLPフラグメントが、RFLP分析後に配列決定される)。特に、1種類より多くの個々の成分を表し、かつ制限酵素フラグメントのサイズが一致するために分けられないRFLPフラグメントでの配列決定の実施に有利である。
【0050】
ポリクローナル性を評価するためのタンパク質の特徴決定技術
本発明の実施態様において、相同タンパク質のプールまたは相同タンパク質を産生するための発現系のポリクローナル性は、1種類以上のタンパク質の特徴決定技術によりモニターされる。タンパク質の特徴決定技術は、単独または他の技術と組み合わされて、溶液中またはポリクローナル細胞株に存在する細胞の表面上のモノクローナルタンパク質の混合物または組換えポリクローナルタンパク質の個々のメンバーの存在および相対的比率に関する情報をもたらす能力のある任意の技術を意味する。組換えポリクローナルタンパク質の複雑性に依存して、次の技術のうちの1種類以上が用いられ得る:i)クロマトグラフ分離技術、ii)ポリクローナルタンパク質の個々のメンバーを表す固有のマーカーペプチドの同定のためのポリクローナルタンパク質のタンパク分解消化産物の分析、iii)「大量」N−末端配列決定、およびiv)例えば、ポリクローナルタンパク質のセンチネルタンパク質メンバーの特徴決定のための特異的検出分子を用いた分析。
【0051】
異なる相同タンパク質を含有する試料は、精製モノクローナルタンパク質の混合物、またはポリクローナルタンパク質であり得る。ポリクローナルタンパク質は、例えば、遠心分離により細胞から分離されただけの「未処理」の上清、または、例えば、タンパク質Aアフィニティー精製、免疫沈降、またはゲル濾過により精製された上清の形のポリクローナル細胞培養物から得られた細胞培養上清であり得る。しかしながら、これらの前精製工程は、組成物中の異なる相同タンパク質の分離を全くもたらさないので、組換えポリクローナルタンパク質の特徴決定の一部ではない。好ましくは、本発明の特徴決定過程の対象となる試料は、精製工程を少なくとも1回受けている。最も好ましくは、試料は、90%、95%、または99%純粋な相同タンパク質を含む。
【0052】
ポリクローナルタンパク質を構成する異なる相同タンパク質は、培養中の異なる時間ポイントで単一のポリクローナル細胞培養物から得られた試料においてモニターされ、それにより、個々のポリクローナルタンパク質の相対的比率が生産工程を通じてモニターされ、その組成の安定性が評価され得る。別法として、ポリクローナルタンパク質を構成する異なる相同タンパク質が、特定の時間ポイントで異なるポリクローナル細胞培養物から得られた試料においてモニターされ、それにより、異なるバッチにおける個々のコード配列の相対的比率がモニターされ、バッチとバッチとの間の一貫性が評価され得る。
【0053】
クロマトグラフ分離技術
ポリクローナルタンパク質の個々のメンバーのクロマトグラフ分離は、i)正味電荷(例えば、イオン交換クロマトグラフィー(IEX))、ii)疎水性(例えば、逆相クロマトグラフィー(RP−HPLC)、および塩濃度による疎水性相互作用クロマトグラフィー(HIC))、iii)等電点(pI値)(例えば、クロマトフォーカシング)、またはiv)アフィニティー(例えば、抗イディオタイプペプチド/抗体を用いたアフィニティークロマトグラフィー、またはκおよびλ抗体軽鎖の分離のためのタンパク質Lクロマトグラフィー)のような物理化学的特性の相違に基づくものであり得る。5種類のよく知られたクロマトグラフ技術は、次の物理化学的特性:大きさに基づくものである。しかしながら、全てのメンバーは本質的に同じ大きさのものであるので、これは、ポリクローナル抗体またはポリクローナルTcRのような相同タンパク質の特徴決定に特に適した技術ではない。大きさによる分離は、特徴決定基盤から完全に省かれる。上述のこれらのクロマトグラフ技術のいくつかは、IgA、IgG、およびIgMのような免疫グロブリンのクラス(Gallo, P. et al. 1987. J. Chromatogr. 416, 53-62)、またはIgG1、IgG2、IgG3のようなサブクラス(Scharf, O et al. 2001. J. Virol. 75, 6558-6565)をヒト血清から分離する際に利用されてきた。しかしながら、血清由来の免疫グロブリンまたは組換えポリクローナル抗体における個々の抗体の多様性に関連する分離は、これまで行われていない。
【0054】
a)イオン交換クロマトグラフィー
本発明の実施態様において、イオン交換クロマトグラフィーを用いて、組換えポリクローナルタンパク質の個々のメンバー、またはポリクローナルタンパク質の個々のメンバーのサブ集団が分離される。イオン交換クロマトグラフィーによる分離は、分離されるべき組成物中の個々のタンパク質の正味電荷に基づくものである。組換えポリクローナルタンパク質のpI値、選択されたカラムバッファーのpH値および塩濃度に依存し、組換えポリクローナルタンパク質の個々のメンバーは、陰イオンまたは陽イオン交換クロマトグラフィーのいずれかを用いて、少なくともある程度まで分離され得る。例えば、pHが、組換えポリクローナルタンパク質組成物の個々のメンバーの最も低いpI値より十分に低いものである限り、通常、組換えポリクローナルタンパク質の個々のメンバーの全てが、負に荷電した陽イオン交換媒体に結合するだろう。続いて、結合した組換えポリクローナルタンパク質の個々のメンバーは、典型的には漸増勾配の塩(例えば、塩化ナトリウム)または漸増pH値を用いて、個々のタンパク質の正味電荷に依存して、カラムから溶出され得る。いくつかのフラクションが溶出中に得られるだろう。好ましくは、単一のフラクションは、ポリクローナルタンパク質の個々のメンバーを含有するが、ポリクローナルタンパク質の2、3、4、5、6、7、8、9、10、15、20、またはそれ以上の別のメンバーも含有し得る。陽イオンおよび陰イオン交換の一般的な原理は、当該技術分野においてよく知られており、イオン交換クロマトグラフィー用のカラムは市販されている。
b)クロマトフォーカシング
本発明のさらなる実施態様において、クロマトフォーカシングを用いて、組換えポリクローナルタンパク質の個々のメンバー、またはポリクローナルタンパク質の個々のメンバーのサブ集団が分離される。クロマトフォーカシングによる分離は、個々のタンパク質のpI値の相違に基づくものであり、組換えポリクローナルタンパク質のpI値より上のpH値を有するカラムバッファーを用いて行われる。個々のメンバーが比較的低いpI値を有する組換えポリクローナルタンパク質は、正に荷電した弱陽イオン交換媒体に結合するだろう。続いて、結合した組換えポリクローナルタンパク質の個々のメンバーは、個々のメンバーのpI値のpH範囲をカバーするよう設計されたポリバッファーを用いてカラム内の漸減pH勾配を創出することにより、個々のメンバーのpI値に依存して、カラムから溶出され得る。いくつかのフラクションが溶出中に得られるだろう。好ましくは、単一のフラクションは、ポリクローナルタンパク質の個々のメンバーを含有するが、ポリクローナルタンパク質の2、3、4、5、6、7、8、9、10、15、20、またはそれ以上の別のメンバーも含有し得る。陽イオン交換体を用いたクロマトフォーカシングの一般的な原理は、当該技術分野においてよく知られており、陽イオンカラムは市販されている。陽イオン交換体を用いたクロマトフォーカシングも当該技術分野において知られている(Kang, X. and Frey, D.D., 2003. J. Chromatogr. 991, 117-128(引用により、本明細書に取り込まれる))。
【0055】
c)疎水性相互作用クロマトグラフィー
本発明のさらなる実施態様において、疎水性相互作用クロマトグラフィーを用いて、組換えポリクローナルタンパク質の個々のメンバー、またはポリクローナルタンパク質の個々のメンバーのサブ集団が分離される。疎水性相互作用クロマトグラフィーによる分離は、分離されるべき組成物における個々のタンパク質の疎水性の相違に基づくものである。組換え産生されたポリクローナルタンパク質は、疎水性相互作用に有利に働くバッファー中の疎水性リガンドで修飾されたクロマトグラフィー媒体に結合する。一般に、これは、有機溶媒を低い割合で含有するバッファー(RP−HPLC)、または選択された塩を非常に高濃度含有するバッファー(HIC)において達成される。続いて、結合した組換えポリクローナルタンパク質の個々のメンバーが、典型的には、漸増勾配の有機溶媒(RP−HPLC)または漸減勾配の選択された塩(HIC)を用いて、個々のメンバーの疎水性に依存して、カラムから溶出されるだろう。いくつかのフラクションが溶出中に得られるだろう。好ましくは、単一のフラクションは、ポリクローナルタンパク質の個々のメンバーを含有するが、ポリクローナルタンパク質の2、3、4、5、6、7、8、9、10、15、20、またはそれ以上のメンバーも含有し得る。疎水性相互作用クロマトグラフィーの一般的な原理は、当該技術分野においてよく知られており、RP−HPLCならびにHIC用カラムは市販されている。
【0056】
d)アフィニティークロマトグラフィー
本発明のさらなる実施態様において、アフィニティークロマトグラフィーを用いて、ポリクローナルタンパク質の個々のメンバー、またはポリクローナルタンパク質の個々のメンバーのサブ集団が分離される。アフィニティークロマトグラフィーによる分離は、特異的検出分子、リガンド、またはタンパク質に対するアフィニティーの相違に基づくものである。検出分子、リガンド、またはタンパク質、またはこれらのうちの複数のもの(これらの異なるオプションを、以下、ただリガンドと称する)は、クロマトグラフィー媒体に固定され、組換えポリクローナルタンパク質は、個々のメンバーと固定されたリガンドとの間の相互作用に有利に働く条件下でアフィニティーカラムにアプライされる。固定されたリガンドに対するアフィニティーを示さないタンパク質は、カラムフロースロー(flow-through)に集められ、次に、固定されたリガンドに対するアフィニティーを示すタンパク質は、結合に不都合に働く条件(例えば、低pH、高塩濃度、または高いリガンド濃度)下でカラムから溶出される。いくつかのフラクションが溶出中に得られ得る。好ましくは、単一のフラクションは、ポリクローナルタンパク質の個々のメンバーを含有するが、ポリクローナルタンパク質の2、3、4、5、6、7、8、9、10、15、20、またはそれ以上の別メンバーも含有し得る。組換えポリクローナルタンパク質を特徴付けるために用いられ得るリガンドは、例えば、標的抗原、抗イディオタイプ分子、またはκまたはλ軽鎖を有する抗体の分離のためのタンパク質Lである。
【0057】
標的抗原を用いたアフィニティークロマトグラフィーは、組換えポリクローナルタンパク質が1種類以上のエピトープに対するアフィニティーを含む場合、特に適切だろう。標的は、例えば、癌細胞もしくはウイルス、または標的の組合せであってもよく、そしてそれは、多くのエピトープを含有する。これらのエピトープは合成され、クロマトグラフ媒体上に固定される。1個のカラム当たり1個のエピトープ、または1個のカラム当たりいくつかの異なるエピトープを用いて、アッセイは設計され、それにより、特定のエピトープに対する個々のメンバーの分布に関して、組換えポリクローナルタンパク質混合物を特徴付けることが可能になる。別法として、完全抗原または標的分子は、クロマトグラフ媒体上に固定され得る。
【0058】
ポリクローナルタンパク質の個々のメンバー、またはかかる個々のメンバーのサブ集団に特異的に結合する抗イディオタイプ分子(例えば、抗イディオタイプペプチドまたは抗イディオタイプ抗体)を用いたアフィニティークロマトグラフィーを行い、組換えポリクローナルタンパク質の選択されたメンバー(センチネルタンパク質とも称される)、または個々のメンバーのサブ集団の相対的比率に関する情報が得られ得る。理想的には、個々の抗イディオタイプ分子は、1種類の個々のメンバーのみに特異的に結合し、組換えポリクローナルタンパク質の他のメンバーには結合しないものであるが、メンバーの所定のサブセットと結合する抗イディオタイプ分子も本発明において適用可能である。好ましくは、完全なポリクローナル組成物が特徴付けられ得るように、抗イディオタイプ分子は個々のメンバー全てに対して生成される。組換えポリクローナルタンパク質がポリクローナル抗体またはTcRである場合、抗イディオタイプ分子は、抗体またはT細胞受容体の配列の抗原特異的部分に対して方向付けられる。1個のカラムが1種類の抗イディオタイプ分子を含有するように、抗イディオタイプ分子は個々にクロマトグラフ媒体に固定され、それにより、特定のタンパク質メンバーまたはタンパク質のサブ集団についての情報がえられる。次に、フロースローが、第2の固定化抗イディオタイプ分子などを有する第2のカラムにアプライされる。別法として、いくつかの異なる抗イディオタイプ分子が、同じカラムに適用されたのと同じクロマトグラフ媒体上に固定される。次に、例えば、漸増量の遊離イディオタイプ分子をカラムに添加することにより、あるいはpHまたは塩勾配を用いることにより、個々のタンパク質が異なるフラクションに溶出されることを可能にする条件下で、溶出が行われる。このアプローチにより、1次元分析を用いて、ポリクローナルタンパク質のいくつかのメンバーの比率についての情報を得ることが可能だろう。
【0059】
組換えポリクローナルタンパク質がポリクローナル抗体である場合、これは、κ軽鎖またはλ軽鎖のいずれかを含有する個々のメンバーからなり得る。かかるポリクローナル抗体において、λ軽鎖の抗体がタンパク質Lに対するアフィニティーを欠いていることを用いて、λ軽鎖を有する抗体がκ軽鎖を有する抗体から分離され得る。従って、λ軽鎖を含有する抗体メンバーのサブセットは、タンパク質Lアフィニティークロマトグラフィーを用いて、κ軽鎖を含有する抗体メンバーのサブセットから分離され得る。続いて、κおよびλ抗体サブセットが、例えば、上述の個々の抗体の定量のための別のクロマトグラフ技術をさらに用いて、特徴付けられ得る。
【0060】
多次元クロマトグラフィー
分析されるべき試料中の変異体相同タンパク質、例えば、組換えポリクローナルタンパク質の複雑性に依存して、上記のa)からd)のクロマトラフ技術のうちの2種類以上を2次元、3次元、または多次元フォーマットで組み合わせることが所望され得る。好ましくは、2次元ゲル電気泳動の代わりに全次元において液体クロマトグラフィーが用いられる。しかしながら、これは、組換えポリクローナルタンパク質を特徴付けるための1以上の次元でのゲル電気泳動または沈殿技術の使用を排除するものではない。
【0061】
液体2次元クロマトグラフィーは、例えば、Lubman, D.M. et al. 2002. J.Chromatogr.B Analyt.Technol.Biomed.Life Sci. 782, 183-196;WO01/58925およびWO01/58926に記載されている。該方法を用いて、健常な細胞と癌細胞のタンパク質の発現が比較され、それにより、タンパク質レベルでの異なるディスプレイが創出されてきた。さらに、3次元が大きさによる分離である3次元クロマトグラフィーがWO03/102539に記載されており、例えば、細胞抽出物中のタンパク質が分離され、それにより、同様に、差次的ディスプレイが創出される。
【0062】
本発明のさらなる実施態様において、多次元クロマトグラフィーを用いて、試料中の異なる相同タンパク質が特徴付けられる。具体的には、抗体およびT細胞受容体のような異なる可変領域を有する相同タンパク質試料が、多次元クロマトグラフィーを用いて特徴付けられる。好ましくは、さらなる次元が、前の次元における溶出中に得られたフラクションにおいて行われる。しかしながら、フロースルーもさらなる次元の分析のために用いられ得る。これは、前の次元がアフィニティークロマトグラフィーである場合、特に適切であり得る。
【0063】
本発明の1つの実施態様において、多次元クロマトグラフィーは、ポリクローナル抗体(血清からもたらされた免疫グロブリン、または組換え体)、またはモノクローナル抗体の混合物のいずれかから、個々の抗体分子をそれらの多様性に関して分離させる際に用いられる。好ましくは、多次元クロマトグラフィーは液体クロマトグラフィーである。
【0064】
本発明の別の実施態様において、多次元クロマトグラフィーは、ポリクローナルT細胞受容体またはモノクローナルT細胞受容体の混合物のいずれかから、個々のT細胞分子をそれらの多様性に関して分離させる際に用いられる。好ましくは、多次元クロマトグラフィーは液体クロマトグラフィーである。
【0065】
一般に、多次元クロマトグラフィーにおける異なる次元の異なる物理化学的特性に基づくクロマトグラフ技術、例えば、1次元の電荷による分離、2次元の疎水性による分離、そして3次元のアフィニティーによる分離を用いることが試みられる。しかしながら、クロマトグラフ技術のなかには、次の次元(たとえそれらが同様の物理化学的特性を利用するものであっても)で用いられたときに、さらなる分離をもたらし得るものがある。例えば、クロマトフォーカシング、そしてイオン交換クロマトグラフィーまたはアフィニティークロマトグラフィー(異なるリガンドを用い、互いに成功)が続く場合、さらなる分離が得られ得る。
【0066】
表1には5つの次元が挙げられ、そこでは、クロマトグラフ技術が本発明の特徴決定基盤の一部として利用され得る。しかしながら、これは必須の次元数と考えられるべきではない。組換えポリクローナルタンパク質を特徴決定するのに十分な分離が、1、2、3、または4次元の後得られたなら、残りの次元は省れ得る。ゆえに、十分な分離が、イオン交換クロマトグラフィー(IEX)を用いて得られたなら、クロマトフォーカシング、RP−HPLCなどを行う必要はない。他方、5つの次元が不十分な分離をもたらしたなら、さらなる次元が加えられ得る。さらに、表1は、クロマトグラフ技術の可能性のある組合せの包括的リスト、または技術自体の包括的リストであると考えられるべきではない。
【0067】
表1
【表1】

【0068】
【表2】
【0069】
本発明の好ましい実施態様において、多次元液体クロマトグラフィー(LC)は、表1に示される最初の2つの次元から選択される2次元LC技術である。
【0070】
本発明のさらに好ましい実施態様において、多次元LCは、表1に示される最初の3つの次元から選択される3次元LC技術である。
【0071】
多次元LC技術の代替として、ゲル電気泳動またはキャピラリー電気泳動のような適当な電気泳動技術と組み合わされた免疫沈降、そして続いて抗原の定量化を用いて、組換えポリクローナルタンパク質が特徴付けられ得る。この技術は、複雑な抗原に対して標的化された組換えポリクローナル抗体を特徴付けるのに特に有用だろう。例えば、複雑なウイルス抗原に対して標的化された組換えポリクローナル抗体は、標識抗原混合物およびタンパク質Aビーズを用いて、免疫沈降され得る。続いて、抗原は、等電点電気泳動法または2D PAGEを用いて分離され、次に、個々の抗原が定量される。これは、特異的抗原に対して標的化された組換えポリクローナル抗体における抗体量を反映する。
【0072】
組換えタンパク質のN−末端荷電の不均一性の排除
上述のタンパク質の特徴決定技術において、単一のタンパク質は、例えば、IEXプロファイルにおいていくつかのピークを生じうるので、相同タンパク質プールにおける個々のタンパク質の不均一性は、特徴決定をなおさら複雑にし得る。不均一性は、抗体および他の組換えタンパク質において共通する現象であり、酵素性または非酵素性翻訳後修飾によるものである。共通する翻訳後修飾は、N−グリコシル化、メチオニン酸化、タンパク分解性断片化、およびアミド分解を含む。不均一性は、トランスフェクションにおいて誘発される変異(Harris, J.R, et al. 1993. Biotechnology 11,1293-7)、および転写中の重鎖と軽鎖の可変遺伝子間でのクロスオーバー現象(Wan, M. et al. 1999. Biotechnol Bioeng. 62,485-8)のような、遺伝子レベルでの修飾からも生じ得る。これらの修飾は、エピジェネティックであり、従って、構築物の遺伝子構造単独から予測することはできない。
【0073】
不均一性を生じ得るこれらの翻訳後修飾のいくつかは、特徴決定に先立ち処理され得る。C−末端のリジンの酵素的除去から生じる電荷のバリエーションは、特異的なカルボキシペプチダーゼインヒビターの使用、または全体的パターンを単純化するために抗体をカルボキシペプチダーゼで処理することにより、解決され得る(Perkins, M. et al. 2000. Pharm Res. 17,1110-7)。グリコシル化パターンの相違から生じる大きさのバリエーションは、例えば、PNGase F、Endo H、O−グリコシダーゼ、またはノイラミニダーゼを用いた酵素による脱グリコシル化によっても処理され得る。
【0074】
アミド分解のようなタンパク質の化学的分解は、産生および保存中の重大な問題であり、電荷の不均一性を生じることが示されてきた。AsnからAspへのアミド分解、およびisoAsp(イソアスパルチルペプチド結合)の形成は、穏和な条件下で生じる(Aswad, D.W. et al. 2000. J Pharm Biomed Anal. 21, 1129-36)。これらの転位は、部分的なポリペプチド鎖の柔軟性が高いAsn−Gly、Asn−Ser、およびAsp−Gly配列にて最も容易に生じる。
【0075】
電荷の不均一性の別の要因は、N−末端グルタミン残基(アミド分解)の環化から生じるピログルタミン酸(PyroGlu)によるN−末端の封鎖から生じ得る。かかる翻訳後修飾は、IgGならびに他のタンパク質について述べられてきた。特に、HCおよびLCの両方が含まれると、抗体のN−末端の部分的環化は電荷の不均一性を生じ、これにより、複雑なIEXパターンが与えられるだろう。抗体の1個以上のVH鎖およびVL鎖の上でのN−末端PyroGluの形成に起因する可能性のあるIEXパターンが、図16に示される。組換えポリクローナル抗体のような、異なる可変領域を有する異なる相同タンパク質を含む試料が、正味電荷に基づく技術により特徴決定されるべきでものであるなら、たとえ2、3種の試料成分が図16BおよびCに示されるIEXパターンを有するとしても、これは、例えば、ポリクローナル抗体組成物のIEXパターンにおけるクローンの多様性を隠すであろうから、かかる分析が複雑になることは明らかである。この問題は、特定の酵素であるピログルタミン酸アミノペプチダーゼの使用によっては解決されない。なぜなら、まず第一に、脱閉鎖抗体を高収率で得るために、脱閉鎖が還元またはアルキル化抗体において行われなければならないからであり(Mozdzanowski, J. et al. 1998Anal Biochem. 260,183-7)、そして第二に、全ての抗体について100%の開裂を得ることは可能でないだろうからである。
【0076】
それゆえ、本発明のさらなる態様は、N−末端グルタミン残基の環化により引き起こされる電荷の不均一性の排除に関するものである。本発明のこの態様は、物理化学的特性である正味電荷に基づく上述の特徴決定ツール、例えば、IEXクロマトグラフィーおよびクロマトフォーカシングのいずれかと組み合わされると、特に有用である。N−末端のPyroGlu残基の形成は、例えば、N−末端グルタミン残基を別のアミノ酸に変えることにより、ポリペプチド鎖の全てがN−末端グルタミンを含有しないことを確実にすることにより、排除される。タンパク質が異なるサブユニットを含むヘテロマーのタンパク質であるなら、好ましくは、N−末端Gln残基の全てが他の残基と交換される。抗体については、重鎖および/または軽鎖のN−末端のGln残基が変換される。これは、N−末端にグルタミンを有するポリペプチドをコードする核酸配列の部位指定の変異誘発により行われる。好ましくは、N−末端グルタミン残基は、グルタミン酸残基によって置き換えられる。なぜなら、これはグルタミンの非荷電誘導体だからである。組換えポリクローナルタンパク質において、メンバーをコードする個々の配列は、荷電タンパク質を発現する新規細胞株を創出するために、変更され、発現ベクターに再挿入されなければならない。次に、この細胞株は、ポリクローナルタンパク質を産生する細胞の収集物内に含まれ得る。
【0077】
相同タンパク質の可変領域のタンパク分解性消化産物の分析
異なる可変領域を有する異なる相同タンパク質を含むタンパク質試料は、上述の通り、広範なクロマトグラフ技術を用いて、インタクトなタンパク質の物理化学的特性に基づき特徴付けられ得る。上述のこれらの分析から得られた情報は、相同タンパク質のタンパク分解性消化産物の分析から得られた情報でさらに補足され得る。好ましくは、タンパク分解性消化は、インタクトなタンパク質のクロマトグラフ分析が行われるのと同じ試料のアリコート上で行われる。
【0078】
本発明のさらなる実施態様において、相同タンパク質の組成物の個々のメンバーの可変領域を起源とする固有のマーカーペプチドの同定を用いて、タンパク質組成物が個々のメンバーの存在について定性的方法で特徴付けられる。これらの固有のマーカーペプチドは、異なる相同タンパク質を含むタンパク質組成物(試料)のタンパク分解性消化により生成される。
【0079】
相同タンパク質の混合物の可変領域のペプチドマッピングを行うために、混合物中の個々のメンバーを他のメンバーと区別する可変領域の一部または複数の部分が、タンパク分解性消化後もインタクトなままであることが重要である。ゆえに、少なくとも1種類の固有の配列(マーカーペプチドとも称される)が組換えポリクローナルタンパク質の個々のメンバーそれぞれについて得られ得るように、1種類以上のプロテアーゼが選択されるだろう。相同タンパク質の混合物が、組換えポリクローナル抗体または組換えポリクローナルTcRである場合、通常、個々のメンバーを互いに区別する配列が、CDRにより特徴付けられる。固有のマーカーペプチドを生成するために用いられるプロテアーゼ、複数のプロテアーゼ、または化合物の選択は、相同タンパク質試料を構成するタンパク質配列の分析に基づくものである。一般に、プロテアーゼまたは化合物は、高い特異性でもってタンパク質の所定の部位で開裂させるだろう。かかる開裂特異的なプロテアーゼは、当該技術分野においてよく知られており、例えば、トリプシン、エンドGlu−C、リジルエンドペプチダーゼ、エンドArg−C、エンドAsp−N、またはエンドAsn−Cであり得る。これらはただの例であり、本実施態様に対する限定と考えられるべきではない。
【0080】
ポリクローナルタンパク質試料は、1種類以上の選択されたプロテアーゼで消化される場合、全ての個々のメンバーに由来する定常領域および可変領域の両方を起源とするペプチドのプールが生成されるだろう。固有のマーカーペプチドの比率は、定常領域を起源とするペプチドの主な集団と比較して、それらの物理化学的特性の相違を示すだろう。それゆえ、固有のマーカーペプチドは、上述のクロマトグラフ技術のうち1種類を用いて、単離され得る。好ましくは、ペプチド分離のために特別に設計されたイオン交換クロマトグラフィーまたはRP−HPLCを用いて、固有のペプチドが定常領域ペプチドの主要フラクションと区別される。同様に、上述の多次元クロマトグラフ技術を適用して、固有のマーカーペプチドが分離され得る。1以上の次元の分離に続き、質量分析(MS)が、異なるペプチドの同定のために用いられ得る。プロテオミクス分野の技術者によく知られたMS技術が、ペプチドの同定のために用いられ得る。好ましいMS技術は、マトリックス支援レーザー脱離イオン化法(MALDI)、飛行時間(TOF)質量分析法、およびエレクトロスプレーイオン化飛行時間(ESI−TOF)質量分析法である。
【0081】
別法として、タンパク分解性消化は、上のa)からd)および「多次元液体クロマトグラフィー」のセクションに記載の1次元または多次元クロマトグラフィーにより分離されたインタクトなタンパク質において行われ、次に、分離されたタンパク質フラクションがタンパク分解性開裂され得る。次に、これらの消化産物がMSにより分析され得る(Kachman, M.T. et al. 2002. Anal.Chem. 74, 1779-1791)。このアプローチは、これらをさらに特徴付けるためにインタクトなタンパク質の1次元または多次元分析により得られたフラクションの比率に選択的に適用され得るので、非常に複雑なポリクローナルタンパク質の特徴決定の有利であり得る。
【0082】
タンパク分解性消化は、N−末端マーカーペプチドが固有の可変領域を含有するなら、これらを単離するためにさらに行われ得る。N−末端ペプチドは、本質的にはGevaert, K. et al., 2003. Nat.Biotechnol. 21, 566-569(引用により本明細書に取り込まれる)に記載の通り、単離され得る。簡単にいうと、組換えポリクローナルタンパク質の遊離アミノ基は、例えば、アセチル化によりブロックされ、次に、タンパク質混合物が適当なプロテアーゼを用いて消化される。消化により、内部ペプチド上に遊離N−末端アミノ基が創出され、続いて、これが化合物でブロックされ、これにより、内部ペプチドをN−末端ペプチドと分離することが可能になる。かかる化合物は、i)疎水性相互作用による単離を可能にする、Gevaertにより記載されている2,4,6−トリニトロベンゼンスルホン酸(TNBS)、ii)ビオチン(これは、固定化ストレプトアビジンに結合後、除去される)、またはiii)予め活性化された物質(例えば、NHS−活性化、CNBr−活性化、ECH セファロース物質、またはアズラクトン基を有するUltraLink ビス−アクリルアミド支持体)であり得、続いて、遠心により、結合した内部ペプチドが、アセチル化N−末端ペプチドから分離される(それは上清に存在する)。次に、単離アセチル化N−末端ペプチドが、上述のMS分析と組み合わされた1次元または多次元液体クロマトグラフィーにより分析され得る。別法として、インタクトなタンパク質のN−末端が、特異的な分離を可能にする化合物でブロックされ、開裂後に生成された内部ペプチドが、第2の化合物でアセチル化またはブロックされる。
【0083】
加えて、タンパク分解性消化後の固有マーカーペプチドの同定が、特徴的なアミノ酸側鎖の機能性を用いて行われ得る。関連するアミノ酸側鎖修飾を有する特異的なアミノ酸残基を含有するペプチド、およびそれを有さない特異的なアミノ酸残基を含有するペプチドを捕獲する、異なるアフィニティー技術の1つまたは組合せが用いられ得る。例えば、システイン、メチオニン、トリプトファン、ヒスチジン、およびチロシンを含有するペプチドは、これらのアミノ酸残基を含有するペプチドを捕獲する特異的なアフィニティータグで固定されたカラム材料またはビーズを用いて精製され得る(Bernhard, O.K. et al. 2003. Proteomics 3, 139-146;Chelius, D. and Shaler, T.A., 2003. Bioconjug. Chem. 14, 205-211;Gevaert, K. et al. 2002. Mol.Cell Proteomics 1, 896-903;Gygi, S.P. et al., 1999. Nat.Biotechnol 17, 994-999)。システインおよびチロシンを含有する固有の可変領域ペプチドは、例えば、システイン残基をビオチン化した後、ストレプトアビジンカラム上で捕獲され、続いて、ペプチドを含有するシステインが溶出され、これらが、チロシン残基と特異的に結合するカラムまたはビーズのいずれかにアプライされ得る。特異的なアミノ酸残基に対するアフィニティーに基づくこのペプチドの捕獲は、RP−HPLCおよびイオン交換クロマトグラフィーのような上述のクロマトグラフ技術に対するさらなる次元として行われ得る。まず、クロマトグラフ技術が、1以上の次元の組換えポリクローナルタンパク質のタンパク分解性消化に適用され、次に、アミノ酸に特異的な捕獲が、最終次元の1個以上のフラクションにおいて行われ、続いて、MSによる分析される。側鎖の機能性に基づくペプチドの単離は、さらに、N−末端ペプチド単離技術と組合せて行われ得る。
【0084】
タンパク分解性消化、続いてペプチドの単離により分析されるべき組換えポリクローナルタンパク質が、多量体タンパク質である場合、好ましくは、サブユニットの分離が、タンパク分解性消化産物の「フィンガープリンティング」を単純化するために行われ、その後、タンパク分解される。これは、例えば、遊離システイン残基の還元およびアルキル化、次に、ゲルろ過により行われ、サブユニットが分離(例えば、ポリクローナルタンパク質が抗体またはTcRである場合、それぞれ、重鎖の軽鎖との分離、またはα鎖のβ鎖との分離)され得る。別法として、タンパク分解性消化が天然条件下で行われ得る。特に、抗体にとってこれは適当な代替であり得る。抗体の4要素からなる構造により、定常領域内でタンパク分解性開裂に対して高度に耐性となるからである。従って、インタクトな非還元ポリクローナル抗体のタンパク分解性開裂は、主として可変領域からペプチドを生成するようである。
【0085】
上述のタンパク分解性消化技術はまた、センチネルの概念に従い、タンパク分解性消化物において特徴付けられ得るセンチネルペプチドを選択することにより、応用され得る。
【0086】
「大量」N−末端配列決定
上述のN−末端配列は、単離され、ポリクローナルタンパク質のタンパク分解性消化産物をフィンガープリンティングするために用いられ得る。別法として、N−末端配列が、インタクトなタンパク質から直接配列決定され、それにより、タンパク分解工程が省かれ得る。異なる可変領域を有する異なる相同タンパク質を含むタンパク質試料の「大量」N−末端配列分析を用いて、例えば、組換えポリクローナルタンパク質の精製バッチ産物が比較され得る。ポリクローナルタンパク質が組換えポリクローナル抗体またはTcRである場合、好ましくは、「大量」N−末端配列決定は、それぞれ、分離された重鎖および軽鎖、または分離されたα鎖およびβ鎖のプールにおいて行われる。例えば、相同な重鎖のプールにおいて、アミノ酸位置のいくつかは完全に保存され得るのに対し、他の位置は変動し得、これはアミノ酸配列のアライメントにより評価され得る。従って、いくつかの異なるアミノ酸が、特定ラウンドの配列決定において得られ得る。例えば、4位が、相同配列のアライメントにより予め決定された、ポリクローナル試料中の5種類の異なるアミノ酸により表され得る。「大量」N−末端配列分析において、アミノ酸が変動するこれらのものが定量され、例えば、組換えポリクローナル抗体の4位を表す個々のアミノ酸の異なる量を用いて、関連する組成物である異なる試料が比較され得る。
【0087】
特異的検出分子を用いた複雑な相同タンパク質混合物の特徴決定
本発明の特徴決定基盤はさらに、特異的検出分子を利用し、それぞれの特異的検出分子は、相同タンパク質の複雑な混合物内の個々のタンパク質メンバーを同定する能力を有し、それにより、試料中の特定のメンバーの存在のモニタリングを助ける。特異的な検出分子は、例えば、ポリクローナルタンパク質の個々のメンバーに対する特異性を有する小有機分子、ペプチド、またはタンパク質のような特異的リガンドであり得る。特に、抗イディオタイプペプチドのようなリガンドペプチドまたはタンパク質、あるいは抗イディオタイプ抗体が、本発明の好ましい実施態様である。相同タンパク質の複雑な混合物の所定のサブセットに結合する検出分子も、本発明において適用可能である。
【0088】
i)相同タンパク質の複雑な混合物を含む試料中の1個以上の個々のタンパク質の濃度または相対的比率の決定を可能にすること、ii)クロマトグラフ分析のさらなる次元として働くこと、iii)相同タンパク質の複雑な混合物の発酵中に得られる試料における個々のタンパク質の濃度の決定を可能にすること、およびiv)相同タンパク質の混合物を発現するワーキングセルバンクまたはバイオリアクター細胞試料のようなポリクローナル細胞株における個々のタンパク質産生細胞の決定を可能にすることにより、特異的な検出分子を用いて、相同タンパク質の複雑な混合物が特徴付けられ得る。工程iv)は、ポリクローナル細胞株、またはポリクローナル細胞株由来の単一チューブ内に分布された単一細胞のいずれかにおいて行われ、その後、連続的培養が行われ得る。
【0089】
相同タンパク質の複雑な混合物中の個々のタンパク質メンバーを同定する能力を有する特異的ペプチド−リガンドを生成するため、ビリオン表面上に外来オリゴマーペプチドをディスプレイする発酵ファージ発現ベクターの巨大ライブラリーが、アフィニティーによりスクリーニングされ、次に、抗体、TcR、または別の所望の個々のタンパク質メンバーに結合する外来ペプチドをディスプレイするファージが精製され得る(Scott and Smith 1990. Science 249, 386-90)。EP 1 106 625には、特に、免疫目的のための抗RhD抗体に結合する能力を有するペプチドの生成が記載されている。ディスプレイされたペプチドライブラリーは、約5から50の間のアミノ酸長、好ましくは、7から20の間のアミノ酸長、なおより好ましくは、8から15の間のアミノ酸長、そして最も好ましくは、9から12の間のアミノ酸長である。関連ペプチドが同定されたなら、それらが合成され得る。
【0090】
抗イディオタイプ抗体の生成は当該技術分野において一般的に知られている。簡単に言うと、所望の抗イディオタイプ抗体に対する抗体を用いて、マウスが免疫される。モノクローナル抗体は免疫マウスから生成され、そしてそれは、例えば、ハイブリドーマ技術またはファージディスプレイを用いて所望の特異性を有する抗イディオタイプ抗体の産生についてスクリーニングされる。抗イディオタイプペプチドまたは抗イディオタイプ抗体は、特異性および可能性のある交差反応性に関して特徴付けられるだろう。この分析により、抗イディオタイプペプチドまたは抗イディオタイプ抗体が、特異的メンバーを認識するか、あるいは別法として、ポリクローナルタンパク質中の密接に関連するメンバーのサブセット(抗体について、関連するメンバーは、例えば、特異的VH遺伝子ファミリーであり得る)を認識するかどうかが証明されるだろう。
【0091】
抗イディオタイプペプチド/抗体は、個々のメンバータンパク質(例えば、特異的抗体または特異的TcR)の直接的定量化のためのELISA、FLISA、またはRIAのような免疫検出アッセイにおいて適用され得る。別法として、抗イディオタイプペプチド/抗体は、単独、あるいは1次元または上述の他のクロマトグラフ分離後のさらなる次元として、アフィニティークロマトグラフィーにおいて適用され得る。免疫沈降は、検出分子を用いて、ポリクローナルタンパク質の個々のメンバーが分離され、特徴付けられ得る、さらなる手段である。さらに、抗イディオタイプペプチドまたは抗イディオタイプ抗体は、ポリクローナル細胞株における個々のタンパク質産生細胞の単離および/または決定のために用いられ得る。該技術は、Borth, N. et al. 2000-2001に記載されている。Biotechnol Bioeng. 71, 266-273およびBrezinsky, S.C. et al. 2003. J. Immunol. Methods 277, 141-155の両方が、個々のタンパク質産生細胞の細胞培養物からの単離に適用可能である。
【0092】
可能性として、完全な特徴決定を得るために、ポリクローナルタンパク質におけるそれぞれの個々のメンバーおよび全ての個々のメンバーに特異的な検出分子を生成することが可能である。しかしながら、発現の安定性、またはバッチとバッチとの間の一貫性をモニターするためには、それは、本発明をふまえて、組換えポリクローナルタンパク質内の多数の個々のメンバー、いわゆるセンチネルタンパク質を同定するのに十分であり、そしてその同定は、個々のタンパク質メンバーのこの収集物が、一貫して発現され、組換え産生ポリクローナルタンパク質の異なるバッチにおいて精製されることを確かにする定量的および/または定性的特徴決定についてのものである。特に、このアプローチを用いて、タンパク質分子の複雑なプールの特徴決定が単純化され得る。組換えポリクローナルタンパク質の代表としてのセンチネルタンパク質のコンセプトは、特異的検出分子に適用されるばかりでなく、事実上、上述の特徴決定技術のいずれかまたはこれらの組合せが、センチネルタンパク質またはペプチドのコンセプトを適用し得る。さらに、センチネルタンパク質は、技術から技術で変わり得る。ポリクローナルタンパク質の特異的メンバーのうち、それらの物理化学的特性の相違に基づき、特に十分に分離され得るものもあるが、高アフィニティーを有する抗イディオタイプペプチドは、同一の物理化学的特性を有するタンパク質の分離に特に有用である。
【0093】
本発明の実施態様において、1種類以上の特異的検出分子を用いて、異なる可変領域を有する異なる相同タンパク質を含む試料中の1種類以上のセンチネルタンパク質の相対的比率がモニターされる。一連の関連試料中の1種類以上のセンチネルタンパク質の比率の一貫性は、バッチ間、ならびに1回の生産工程での長時間にわたるポリクローナルタンパク質の発現における組成物の安定性を反映するだろう。さらにそれは、組換えポリクローナルタンパク質またはモノクローナルタンパク質の混合物の長期保存中の組成物の安定性を評価することができる。
【0094】
本発明の好ましい実施態様において、組換えポリクローナルタンパク質のセンチネルタンパク質は、次の技術、i)抗イディオタイプペプチド/抗体アフィニティークロマトグラフィー、ii)抗イディオタイプペプチド/抗体での免疫的検出、iii)それらの特徴的物理化学的特性に関するインタクトなメンバーの多次元クロマトグラフ単離、およびiv)クロマトグラフィーおよびMSを用いたタンパク分解性ペプチドマッピングのうちの1個以上により特徴決定される。
【0095】
特徴付けられるべき異なる相同タンパク質混合物の複雑性
本発明の基盤により特徴付けられるべき試料は、異なる可変領域タンパク質を有する異なる相同タンパク質の所定のサブセット、例えば、異なるCDR領域を有するポリクローナルタンパク質または抗体(例えば、ポリクローナル抗体またはモノクローナル抗体の混合物)、または異なるCDR領域を有するT細胞受容体(例えば、ポリクローナルTcRまたはモノクローナルTcRの混合物)を含む。好ましくは、異なる可変領域を有する異なる相同タンパク質は組換えタンパク質である。加えて、好ましくは、ポリクローナルタンパク質またはモノクローナルタンパク質の混合物の個々のメンバーは、例えば、所望の標的抗原に対する抗体またはTcRの場合、所望の標的に対する共有結合活性のような共通する特徴により定義されてきた。典型的には、本発明の特徴決定基盤により分析されるべきポリクローナルタンパク質組成物は、少なくとも3、4、5、10、20、50、100、1000、104、105、または106の別の変異体メンバーを含む。通常、ポリクローナルタンパク質組成物中の個々のメンバーの総数の75%より多くを構成する単一の変異体メンバーは存在しない。好ましくは、最終ポリクローナル組成物中の個々のメンバーの総数の50%、より好ましくは、25%、最も好ましくは、10%を超える個々のメンバーは存在しない。
【0096】
抗体の場合、標的とされる抗原の複雑さは、本発明の基盤を利用して特徴付けられるべきポリクローナルタンパク質組成物中の別の変異体メンバーの数に影響するだろう。小さな標的または全く複雑でない標的、例えば、小さな標的タンパク質を用いて、3から100種類の間の別の変異体メンバーを含むポリクローナルタンパク質組成物が、特徴決定のために確立されるだろう。そして、好ましくは、変異体の数は、90、または80もしくは70を超えない。多くの場合、異なる変異体の数は、60または50を超えず、好ましくは、変異体の数は、5と40の間、例えば、5と30の間の範囲にある。一方、より複雑な標的、例えば、複雑な表面タンパク質または互換可能な表面タンパク質を有するか、あるいはいくつかのウイルスサブタイプを包含するウイルスについては、20から500種類の間の異なる変異体メンバーを含むポリクローナルタンパク質組成物が、特徴決定のために確立されるだろう。抗原が多数の異なる分子を含む非常に複雑な標的については、50から10,000種類の間の異なる変異体メンバーを含むポリクローナルタンパク質組成物が本発明により特徴付けられるのに必要であり得る。
【0097】
本発明の1つの実施態様において、異なる可変領域を有する異なる相同タンパク質を含む試料は、ポリクローナル抗体である。ポリクローナル抗体は、ヒトアイソタイプIgG1、IgG2、IgG3、IgG4、IgA1、およびIgA2、またはマウスアイソタイプIgG1、IgG2a、IgG2b、IgG3、およびIgAのような1種類以上の異なる抗体アイソタイプからなり得る。
【0098】
本発明の1つの実施態様において、異なる可変領域を有する異なる相同タンパク質を含む試料はポリクローナルTcRである。
【0099】
実施例
以下の実施例において、異なる個々の抗RhD抗体メンバー、または抗RhD rpAbを産生する細胞株からなる種々の抗RhD組換えポリクローナル抗体(抗RhD rpAb)組成物を用いて、本発明の構造上の特徴決定基盤を説明する。個々の抗RhD特異的抗体およびそれらを産生する細胞株は、2004年7月20日に出願した譲受人のデンマーク特許出願PA 2004 01133に記載に対応するものであった。簡単にいうと、重鎖可変領域およびκ/λ軽鎖のコンビナトリアルファージディスプレイライブラリーを、RhD陽性赤血球で免疫したrhesus D陰性ドナーから調製した。ライブラリーを、抗RhD特異的抗体産生クローンについてピックアップした。抗原特異的ファージ由来の可変重鎖および軽鎖遺伝子対を哺乳類発現ベクターに導入した。哺乳類ベクターを、Flp−FRT組換え系を用いた部位特異的方法でCHO Flp−In細胞株(Invitrogen, CA)に個々にトランスフェクションした。完全な軽鎖(LC)ならびに重鎖(VH)の可変領域の核酸(nuc.)ならびにタンパク質(a.a.)配列を、表2に示す配列同一性ナンバー(配列番号)により同定する。これらのナンバーは、2005年7月18日に出願した譲受人の国際特許出願PCT/DK2005/000501(発明の名称「抗RHESUS D組換えポリクローナル抗体、および製造方法」)における配列番号に対応する。表2の配列番号を本発明の出願の配列番号と区別しなければならない。これらは同一でないからである。重鎖の定常領域は、ヒトIgG1に対応する。
【0100】
表2 以下の実施例において用いられる個々の抗RhD抗体/細胞クローンのリスト
【表3】
【0101】
実施例1
本実施例は、ポリクローナル製造細胞株の生成、および1次元のクロマトグラフ技術を用いたタンパク質レベル、およびRFLP分析を用いた遺伝子レベルでバッチとバッチとの間のバリエーションの特徴決定を説明するものである。
【0102】
抗Rhesus D組換えポリクローナル抗体産生のための製造細胞株の樹立
ゲノム上の特定の部位と異なる組換え抗Rhesus D抗体をそれぞれ発現する10種類の細胞株(RhD157.119D11、RhD158.119B06、RhD159.119B09、RhD161.119E09、RhD163.119A02、RhD190.119F05、RhD191.119E08、RhD192.119G06、RhD197.127A08、およびRhD204.128A03)を選択し、混合して、組換えポリクローナル製造細胞株を構築した。RhD197およびRhD204はλクローンであり、残りはκクローンであった。
【0103】
個々の抗Rhesus抗体を発現する細胞培養物を、シェーカーフラスコでの無血清懸濁培養(等しい細胞数で混合する)に完全に適合させ、それにより、ポリクローナルCHO−Flp−In(019)細胞株を生成した。混合した細胞培養物を遠心し、アリコート10×106細胞/チューブにて凍結した。
【0104】
2本のチューブ(3948 FCW065および3949 FCW065)を解凍し、ネオマイシンを含むExcell 302無血清培地100mlを含有する1000mlのシェーカーフラスコにて11週間個々に培養した。
上清を回収し、ろ過後、抗RhD rpAbを精製した。
【0105】
クローン多様性
タンパク質レベルならびにmRNAレベルの両方でクローン多様性をアッセイした。9週間培養後に抗体組成物の分析のために用いる上清試料を採取し、11週間培養後にmRNA組成物を分析するために用いる細胞試料を採取した。
【0106】
抗体組成物:
ポリクローナルCHO−Flp−In(019)細胞株から発現した抗RhD rpAbは、IgG1アイソタイプ抗体である。タンパク質Aを固定化したカラムを用いて、両方のアリコート(3948および3949)から抗RhD rpAbを精製した。個々の抗体は、固定化したタンパク質AとpH7.4で相互作用した。一方、コンタミしているタンパク質をカラムから洗い流した。次に、結合抗体を低pH値(pH2.7)にてカラムから溶出した。280nmでの吸光度測定により決定した抗体含有フラクションをプールし、5mM 酢酸ナトリウム(pH5)に対して透析した。
【0107】
9週間培養後にアリコート3948および3949(FCW065)から得られた抗RhD rpAb組成物を、陽イオン交換クロマトグラフィーを用いて分析した。タンパク質Aにより精製した抗RhD rpAbを、PolyCatAカラム(4.6×100mm)に25mM 酢酸ナトリウム、150mM 塩化ナトリウム、pH5.0中、流速60ml/時間、室温にてアプライした。次に、25mM 酢酸ナトリウム中の150〜350mMの直線的勾配の塩化ナトリウム、pH5.0を用いて、流速60ml/時間にて抗体成分を溶出した。分光光度法により280nmにて抗体成分を検出した。次に、クロマトグラム(図1)を合計し、個々のピークA−Jの面積を用いて、抗体成分を定量した(表3)。ピークの全面積を100%と設定した。2つのアリコートのクロマトグラムは、同一のピーク分布、ならびに各ピークにおける類似の成分濃度を示した。これらの結果から、同一条件下で成長させた同じポリクローナル細胞株のアリコートは、個々の抗体メンバーからなる類似の分布を有する抗RhD rpAbを産生すると結論付けられる。
【0108】
抗RhD rpAbの個々のメンバーを、1種類以上の特定のピークに割り当てた(表3にて概説)。この割り当ては、同一条件下で分析した個々の抗体について得られたクロマトグラムに基づくものである。RhD158Abについて、個々のクロマトグラムを得なかったので、このクローンをピークのいずれにも割り当てなかった。しかしながら、ピークDはRhD158を構成し、この抗体は不均一性に起因して他のピークのいくつかにも現れ得るように思われる。特に、クローンRhD197由来の抗体産物は、IEXプロファイルにおいて高程度の不均一性を示す。RhD190 Abは、保持時間15.3分にて視認できなければならい。しかしながら、それは検出可能ではなかった。このことは、このクローンが失われたか、あるいは組換えポリクローナル製造細胞株における検出限界より下の量で産生されたことを示している。クローンRhD190の喪失は、多様性の10%の低減に該当し、それは、最終抗RhD rpAB組成物の多様性に関して受容されると考えられる。
【0109】
表3
【表4】
【0110】
mRNA組成物:
11週間培養後のポリクローナルCHO−Flp−In(019)細胞株内のクローン多様性をRT−PCR−RFLP分析により評価した。簡単にいうと、200個の細胞に対応する細胞懸濁液を凍結融解の対象とし、これらの溶解物を、軽鎖を増幅するプライマーを含むOne−STEP RT−PCRキット(Qiagen)を用いたRT−PCRにおける鋳型として用いた。プライマー配列は次の通りであった:
フォワードプライマー:5’−TCTCTTCCGCATCGCTGTCT(配列番号1)
リバースプライマー:5’−AGGAAAGGACAGTGGGAGTGGCAC(配列番号2)。
【0111】
RT−PCR産物をHinfIで消化し、制限酵素消化産物をアガロースゲル電気泳動により分析した(エチジウムブロマイド染色を用いて視覚化)(図2)。
【0112】
それぞれの個々のクローンについて、RT−PCRにより増幅した軽鎖のHinfI消化により得られた制限酵素フラグメントの予測サイズを表4に示す。ポリクローナル抗Rhesus D抗体コード遺伝子の個々のメンバーに割り当てられ得るゲル上の6種類の固有のフラグメントサイズを太字で示す。全ての固有のフラグメントをゲル上で同定し得るとは限らず、これらをイタリック体で示す。しかしながら、これは、これらのクローンが培養物中に実際に現れることを必ずしも排除しない。なぜなら、フラグメントが同定可能なように他のフラグメントと十分に分離されていないか、あるいは濃度がより強く出現したバンドと比較してあまりに弱かったからである。このことは、短いフラグメントについてほど断言され得る。なぜなら、それらはより少量のエチジウムブロマイド分子と結合し、それゆえ、ほとんど視認できないからである。
【0113】
表4
【表5】
【0114】
同じポリクローナル細胞株の2つのアリコート(3948および3949)は、ゲルにおいて類似の発現パターンを示したが、バンドの強度は完全に同一ではなかった。これは、同一条件下で成長させた同じポリクローナル細胞株のアリコートは、類似のクローン多様性を有する抗RhD rpAbを産生することを示す。
【0115】
要約
それぞれモノクローナル抗RhD抗体を発現する10種類の細胞株を混合して、抗RhD rpAb製造細胞株を生成した。9週間培養後、それはなお、最初の多様性の90%を維持していた。11週間培養後、6種類の異なるクローン由来のmRNAを明確に同定することができ、そして他のいくつかのクローンは、約500bpのバンド中に現われているようであった。
【0116】
ポリクローナルCHO−Flp−In(019)細胞株の2つのアリコートがクローン多様性に関して類似の結果を示したという事実は、再現可能な結果が異なるバッチ間で得られ得ることを説明するものである。
【0117】
実施例2
本実施例は、8種類のメンバーを有するポリクローナル細胞培養物の長期にわたる特徴決定を説明するものである。RFLP分析を用いて遺伝子レベルで、および1次元クロマトグラフ技術を用いてタンパク質レベルで培養物のクローン多様性を評価した。
【0118】
ポリクローナル細胞培養物におけるクローン多様性を評価するためのRFLP分析
8種の異なる抗Rhesus D抗体を発現するポリクローナル細胞培養物における個々のクローンの分布を、ポリクローナル細胞株由来のRT−PCR産物の末端RFLP(T−RFLP)により評価した。T−RFLP方法において、フォワードおよび/またはリバースプライマーを蛍光標識した。それゆえ、単位複製配列から生じた制限酵素消化フラグメントの部分は標識を含有するだろう。次に、標識フラグメントを、キャピラリー電気泳動により分離させ、蛍光により検出することができる。適用したプライマーに依存して、軽鎖および重鎖の可変領域をコードする配列両方において分析を行うことができる。
【0119】
簡単にいうと、200個の細胞に該当する細胞懸濁液をPBSにて1回洗浄し、凍結融解手段の対象として、溶解物を生成し、これをOne−Step RT−PCRキット(Qiagen)および適当なプライマーを用いたRT−PCR増幅における鋳型として用いた。標準的サーマルサイクラーにて次の条件を用いてRT−PCRを行った。
【表6】
【0120】
軽鎖の分析のため、次のプライマーを用いてRT−PCR増幅した。リバースプライマーを6−カルボキシフルオレッセイン(FAM)標識し、プライマー配列は次の通りであった:
VLフォワードプライマー:5’−TCTCTTCCGCATCGCTGTCT(配列番号1)
CLリバースプライマー:5’−FAM−AGGAAAGGACAGTGGGAGTGGCAC(配列番号2)。
【0121】
RT−PCR産物20μlを、NheI 1U、PstI 1U、およびHinfI 1U(全てNew England Biolabs)を用いてNEB1中にて2時間消化した。
【0122】
Statens Serum Institute, Copenhagen, DKのABI3700(Applied Biosystems)上での蛍光キャピラリー電気泳動により、標識フラグメントを検出した。
【0123】
抗RhD抗体産生細胞クローンそれぞれについての予測フラグメントを表5に示し、そしてFAM標識フラグメントを太字で示す。
【0124】
表5
【表7】
【0125】
T−RFLPパターンを図3に示し、8種類の抗Rhesus D抗体産生クローン全てを特定のピークに割り当てた。RT−PCR中に鋳型/プライマーの競合がないという想定下で、相対的なピーク面積は、ポリクローナル細胞株に表されるそれぞれの抗体の軽鎖遺伝子から転写されたmRNAの相対量に該当する。
【0126】
同じポリクローナル細胞株内の重鎖可変領域の分析のため、VH特異的プライマーを用いてRT−PCR増幅を行った。プライマー配列は次の通りであった:
VHフォワードプライマー:5’−FAM CGTAGCTCTTTTAAGAGGTG(配列番号3)
VHリバースプライマー:5’−HEX−ACCGATGGGCCCTTGGTGGA(配列番号4)。
【0127】
RT−PCR産物20μlを、RsaI 1U、およびNdeI 1U(全てNew England Biolabs)を用いて、NEB2にて、2時間消化した。
【0128】
ABI3700上の蛍光キャピラリー電気泳動により、標識フラグメントを検出した。Statens Serum Institute, Copenhagen, DKにより分析を行った。
【0129】
予測T−RFLPパターンを表6に示す。表中、FAM標識フラグメントを太字で示し、そしてHEX(6−カルボキシ−2’,4,4’,5,7,7’−ヘキサクロロフルオレッセインスクシンイミジルエステル)標識フラグメントに下線を付す。
【0130】
表6
【表8】
【0131】
ポリクローナル細胞株を5週間培養し、1週間に1回T−RFLP分析のために試料を採取した。可変重鎖において分析を行ったが、必要に応じて軽鎖についても行うことができた。
【0132】
制限酵素消化フラグメントをキャピラリー電気泳動した後、相対的ピーク面積を合計し、これを用いて、ポリクローナル細胞培養物のクローン多様性を評価した。経時的な相対量を図5に示す。
【0133】
これらの結果から、時間と共にRhD196は増大するのに対して、RhD203は低減するようである。他のクローンの量は培養期間中非常に安定し、5週間培養後、8種類のcDNA全てを検出することができた。
【0134】
軽鎖および重鎖の両方、ならびにmRNAおよびDNAの両方においてT−RFLPを行うことにより、例えば、インビトロの細胞年齢の限界にある細胞、または培養中の任意の所定の時間ポイントの細胞における、ポリクローナル細胞培養物内のクローン多様性の正確なフィンガープリントを得ることが可能となる。
【0135】
それゆえ、この技術を用いて、長時間にわたり細胞培養物におけるクローン多様性の安定性を抗体産生中にモニターすることができる。この技術を応用して、例えば、同じポリクローナルワーキングセルバンク(pWCB)から凍結された異なるアンプル、あるいは2回以上製造工程を行った後回収された細胞における、バッチとバッチとの間の一貫性をモニターすることもできる。
【0136】
ポリクローナル細胞培養物におけるクローン多様性を評価するための陽イオンクロマトグラフィー分析
上述のT−RFLP分析において用いたのと同じポリクローナル細胞培養物から産生された抗RhD rpAbを、陽イオン交換クロマトグラフィーを用いて分析した。タンパク質Aにより精製した組換え産生ポリクローナル抗体を、25mM 酢酸ナトリウム、150mM 塩化ナトリウム,pH5.0中、流速60ml/時間、室温にてPolyCatAカラム(4.6×100mm)にアプライした。次に、25mM 酢酸ナトリウム、pH5.0中の150〜350mMの直線的勾配の塩化ナトリウムを用いて、流速60ml/時間にて抗体成分を溶出した。抗体成分を分光光度法により280nmにて検出し、次に、クロマトグラムを統合し、個々のピーク面積を用いて、抗体成分を定量した。経時的な相対量を図6に示す。
【0137】
要約
RFLP分析により遺伝子レベルで得られた結果と陽イオン交換クロマトグラフィーによりタンパク質レベルで得られた結果は比較可能である。図5および6は、ポリクローナル細胞株における個々のクローンの大部分、ならびに細胞株から発現されたポリクローナル抗体の個々の抗体が、5週間培養中、同じ傾向をたどることを明瞭に説明する。従って、遺伝子レベルならびにタンパク質レベルでの分析は、遺伝子レベルでの細胞株の組成の多様性、および細胞株から産生された組換えポリクローナルタンパク質の組成の多様性の評価について十分に同等なものである。
【0138】
実施例3
本実施例は、25種類のメンバーを有するポリクローナル細胞培養物の長期にわたる特徴決定を説明するものである。T−RFLP分析を用いて遺伝子レベルで、および1次元クロマトグラフ技術を用いてタンパク質レベルで、培養物のクローン多様性を評価した。
【0139】
培養期間5週間にわたる、25種類の異なる抗Rhesus D抗体発現ポリクローナル細胞培養物からもたらされた重鎖遺伝子の可変部分のT−RFLP分析
本実施例において調べたポリクローナル細胞培養物は、25種類の異なる抗Rhesus D抗体発現細胞培養物の混合物(実施例1に記載のとおり生成したもの)から構成されるものであった。ポリクローナル細胞培養物を5週間培養し、1週間に1回、T−RFLP分析のために試料を採取した。
【0140】
実施例2に記載のVH特異的プライマーを用いてRT−PCRを行い、同様に、制限酵素断片化を行った。
【0141】
全ての遺伝子型が存在するなら、25種類の異なる抗Rhesus Dコード配列のT−RFLPは、17種類の異なるFAM標識フラグメントを生じるだろう。フラグメントには、3種類までの異なる遺伝子型を表すものもあれば、単一の遺伝子型を表すものもあるだろう。FAM標識フラグメントの予測サイズを、経時的な異なるFAM標識フラグメントの相対量と共に表7に示す。さらに、T−RFLPプロファイルの1例を図4に示す。
【0142】
表7
【表9】
【0143】
細胞株を構成する25種類のクローンのうち12種類の個々のクローンについて情報を得ることを可能にする程度まで、消化フラグメントを分離させることが可能であった。残りのフラクションを配列決定の対象とし、残りのクローンについてさらなる情報を得られる可能性があった。
【0144】
25種類の異なる抗Rhesus D抗体発現ポリクローナル細胞培養物におけるクローン多様性を評価するための陽イオンクロマトグラフィー分析
上述のT−RFLP分析において用いたのと同じポリクローナル細胞培養物から産生された抗RhD rpAbを、陽イオン交換クロマトグラフィーを用いて分析した。タンパク質Aにより精製した組換え産生ポリクローナル抗体を、25mM 酢酸ナトリウム、150mM 塩化ナトリウム、pH5.0中、流速60ml/時間、室温にてPolyCatAカラム(4.6×100mm)にアプライした。次に、25mM 酢酸ナトリウム、pH5.0中の150〜350mMの直線的勾配の塩化ナトリウムを用いて、流速60ml/時間にて抗体成分を溶出した。280nmにて分光光度法により抗体成分を検出し、次に、クロマトグラムを統合して、個々のピーク面積を用いて、異なる抗体成分を定量した。図7は、4週目に得られた試料から生じたクロマトグラムを示し、抗体含有ピークに1から25までの番号を付けている。クロマトグラムが、分析したポリクローナル抗体における個々の抗体の数と同一数のピークを含有することは、正真正銘一致する。表8は、トータルの抗体成分(AC1から25)のパーセントにおける相対含有量、ならびに各抗体成分(ピーク)における個々の抗体の出現量を示す。合計したクロマトグラムのピークへの個々の抗体の割り当ては、陽イオン交換クロマトグラフィーを用いて同一条件下で分析したモノクローナル抗体から得られた保持時間およびピークパターンに基づくものであった。
【0145】
表8
【表10】
【0146】
陽イオン交換クロマトグラフィーは、個々のメンバー間の正味電荷の相違に基づき、ポリクローナル抗体から個々の抗体メンバーを分離させ、加えて、電荷の不均一性を明らかにする個々の抗体の形態を分離させる。それゆえ、いくつかの抗体は単一ピーク中に表された。例えば、AC1が含有するRhD293およびRhD319(表8参照)、ならびにいくつかの個々の抗体はさらに、いくつかのクロマトグラムピーク中に表される(例えば、AC1および5の両方に存在するRhD319(表8参照))。
【0147】
1種類より多くの個々の抗体を含有するピークを、抗イディオタイプペプチドを用いた定量的分析、タンパク分解性ペプチドマッピング、N−末端配列決定、または2次元クロマトグラフィーのようなさらなるタンパク質化学的特徴決定技術の対象とすることができた。
【0148】
要約
本実施例は、培養期間にわたる1次転写物の分布および抗体成分の分布をそれぞれ評価するためのT−RFLP分析と陽イオン交換クロマトグラフィーを組み合わせて使用することを説明するものである。T−RFLP分析により、ポリクローナル細胞株において発現した25種類のクローンのうち12種類の個々のクローンを固有に同定することが可能となり、そして本実施例において、4週間の培養中T−RFLP分析を用いて、これらの12種類のクローンを検出できることを説明する。可能性としては、1種類より多くのクローンを表すフラグメントの配列分析により、より多くのクローンを同定できる。陽イオン交換クロマトグラフィーを用いて、抗体成分の分布を分析し、そして本実施例において、25種類の分析した成分の分布が培養中比較的安定していることを見出した。個々の抗体全てを固有に同定することは、発現抗体の本来の電荷の不均一性質に起因して困難であるが、本実施例において、RhD160抗体を表す抗体成分8が培養中最も高い抗体レベルを示し、そしてこれはRhD160、293、および196クローンを表す群13について得られた高いT−RFLP値とも一致することを示した。さらに、T−RFLP、ならびに陽イオン交換クロマトグラフィーにより固有に同定することのできるRhD 207成分は、T−RFLPレベル10〜11%を示し、抗体レベルで得られたものは若干低い5.5〜10%を示した。とりわけ、2つの技術は一体となって、培養中のmRNAおよび抗体レベルでの比較的安定な産生を示す;しかしながら、2つの技術の間の可能性のある矛盾も見出され、これは、抗体レベルで得られた結果と対照的な、培養5週間でのいくつかのクローンの転写の明らかな喪失により説明される。従って、本実施例は、両方の技術を相補的に用いて、複雑なポリクローナルタンパク質の安定的産生を得られる培養間隔を定義することを十分に示すものである。
【0149】
実施例4
本実施例は、ポリクローナル細胞培養物からもたらされた10種類の個々のメンバーを有するポリクローナル抗RhD抗体の組成の分析を説明するものである。1次元陽イオン交換および2次元逆相(RP)−HPLCをそれぞれ用いて、正味電荷および疎水性における相違に基づき抗体を分離する2次元液体クロマトグラフィーを用いて、ポリクローナル抗体試料の多様性を評価した。
【0150】
10種類の個々のメンバーを有するポリクローナル抗RhD抗体試料は、ポリクローナル細胞培養物からもたらされたものであった。タンパク質Aカラム(HiTrap[登録商標]タンパク質Aカラム、Amersham Biosciences GE Healthcare, England)を用いて、上清から抗RhD rpAbを精製した。
【0151】
25mM 酢酸ナトリウム、150mM 塩化ナトリウム、pH5.0中、流速60ml/時間、室温、Ettan LCシステム(Amersham Biosystems, GE Healthcare, England)上にて、ProPac WCE10カラム(4×250mm)に精製ポリクローナル抗体をアプライすることにより、1次元を実行した。次に、25mM 酢酸ナトリウム、pH5.0中の150から350mMの直線的勾配のNaClを用いて、流速60ml/時間にて、抗体成分を溶出した。抗体成分を280nmにて分光光度法により検出し、特定のピークに対応するフラクションを収集し、さらに濃縮して、RP−HPLCにより分析した。
【0152】
図8に示すフラクションを、RP−HPLCを用いて2次元目においてさらに分離した。Summit HPLCシステム(Dionex, CA)にて、Zorbax Poroshell 300SB−C8カラム(2.1×75mm(5μm))を用いて2次元目を行った。Poroshellカラム(Agilent Technologies, CA)についての指示書において推奨された通り、HPLCシステムを設定した。陽イオン交換クロマトグラフィーから収集した抗体成分を、10% CH3CN、0.1% TFA、0.3% PEG中、流速120ml/時間にてカラム(5μl)にアプライし、次に、直線的勾配の90% CH3CN、0.08% TFA、0.3% PEGにより溶出した。カラムを70℃にて操作した。全ての抗体成分試料が、1または2個の細いピークを有するクロマトグラムとなった。抗体成分B5のRP−HPLCプロファイルを図9に示す。
【0153】
要約
1次元の陽イオン交換クロマトグラフィーにより、正味電荷の異なる個々の抗体、ならびに電荷不均一性を示す個々の抗体が分離されるので、いくつかの抗体は単一ピークに表され得る。
【0154】
いくつかの抗体成分を分離し、これにより、図8に示すようなやや複雑なプロファイルを得た。実施例2および3において説明した通り、同一条件下で分析したモノクローナル抗体を用いた比較分析により、それぞれのピーク中の個々の成分を同定することが可能である。しかしながら、本実施例においてこれを行わなかった。rpAb中の各モノクローナル抗体を割り当てなくても、互いに試料を比較するためのフィンガープリントを提供することが目的だったからである。従って、複雑なrpABの個々のメンバーを特徴付けるためにモノクローナル抗体を分析することを必要とすることなく、バッチとバッチとの間の一貫性を評価するために、陽イオン交換、次に、RP−HPLCの組合せにより、2つの次元からデータを創出し、図10に説明する詳細に色分けしたタンパク質マップ(ProteoVue software, Eprogen, USA)を構築することができる。
【0155】
実施例5
本実施例は、ポリクローナル細胞培養物からもたらされた8種類の個々のメンバーを有するポリクローナル抗RhD抗体の特徴決定を説明するものである。「大量」N−末端配列分析を用いて、ポリクローナル抗体の多様性を評価した。
【0156】
本実施例で分析したポリクローナル抗RhD抗体試料中に存在する個々のメンバーのN−末端配列を以下の表9に示す。λ軽鎖配列をイタリック体で示す。
【0157】
表9
【表11】
【0158】
タンパク質Aにより精製した抗RhD rpAbを、還元SDS−PAGE(NuPAGE 4〜12%)により分析した。ポリペプチドをPVDFメンブレンに電気的に移動させ、次に、製造元の指示に従いクーマシーブルーで染色した。
【0159】
重鎖(HC)に対応する約53kDaの1本のバンド、およびκおよびλ+κ軽鎖にそれぞれ対応する約25および30kDaの2本のバンドを、クーマシーブルー染色したPVDFメンブレン上にてはっきりと認めた。これらのバンドを切り出し、ABI Prociseタンパク質シークエンサー(Applied Biosystems, CA)および標準的プログラムを用いたN−末端配列分析の対象とした。配列決定の結果を以下の表10に概説する。
【0160】
表10
【表12】
ND=決定していない
【0161】
HCの配列は第1の残基以外一致し、一方κLCは残基2、5、6、および7について保存され、λLCは、残基1、5、および6について保存されている(表9参照)。
【0162】
HCの配列決定から得られた結果は、表10に示す予測配列と一致する。〜25kDaのκLC由来の配列データは、RhD191、324、201、および306に対応するN−末端配列EIVLTQS(配列番号7)を有する抗体、およびRhD244および196に対応するN−末端配列DIQMTQS(配列番号8)を有する抗体の存在を示した。しかしながら、本技術を用いて、個々のメンバー全てがポリクローナル抗体試料に存在するかどうかを評価することは不可能である。〜30kDaのLCバンドの配列決定は、RhD319抗体の存在を示し、これは3および4サイクルにおけるValの存在により判断した。RhD203抗体の存在についての証拠は全く得られなかった(2および3サイクルそれぞれにおいてSもAもないこと)。しかしながら、この組換えモノクローナル抗体のイオン交換クロマトグラフィーおよびN−末端配列分析は、RhD203のLCが、部分的にブロックされたN−末端を有することを強く示唆している。従って、この抗体が分析混合物中に存在するか否か、N−末端配列分析により結論付けることはできない。加えて、サイクル1においてEおよびD残基が存在し、サイクル4においてM残基が存在するので、30kDaのバンドはκLCをいくつか含有するようである。
【0163】
簡単にいうと、HCまたはKCのいずれかにおいて配列が異なり、かつN−末端においてブロックされていない場合、大量N−末端配列分析を用いて、個々の抗体の存在を同定することができる。この方法は、個々のポリペプチドのN−末端が部分的にブロックされない限り、定量的である。
【0164】
実施例6
本実施例は、ポリクローナル細胞培養物からもたらされた8種類の個々のメンバーを有するポリクローナル抗RhD抗体の特徴決定を説明するものである。ペプチドを分離するためRP−HPLCまたはイオン交換クロマトグラフィー(IEX)のいずれかを用いて、可変領域を起源とする固有マーカーペプチドを単離することにより、抗体の多様性を分析した。
【0165】
単離した重鎖および軽鎖の消化によるペプチドの生成
8種類の個々のメンバーを有するポリクローナル抗RhD抗体試料を、HiTraprタンパク質Aカラムを用いたアフィニティークロマトグラフィーによりポリクローナル細胞培養物の上清から精製した。凍結乾燥した材料を6M 塩酸グアニジウム、0.5M EDTA、0.2M Tris HCl、pH8.4に溶解し、還元(DTT)し、カルボキシメチル化(ヨード酢酸)した。Ettan LCシステム(Amersham Biosciences, GE Healthcare, England)において、6M グアニジウムHCl、50mM リン酸ナトリウム、pH8.4中、Superose 12カラム(10/300 Amersham Biosciences, GE Healthcare)上でのゲルろ過により、重鎖および軽鎖を分離した。分離したHC(〜3.5mg/ml)およびLC(6.5mg/ml)を、リン酸ナトリウム、pH8中の酵素対基質濃度1:500にて、Endoproteinase Asp−N(Roche, 1 054 589)で消化した。
【0166】
RP−HPLCによる固有ペプチドの単離
ポリクローナル抗体試料から得られた単離HCおよびLCの個々のAsp−N消化物のアリコートを、流速0.2ml/分を用いて0.1% TFAにて平衡化したガードカラム(Zorbax 300SB−C8、2.1×12.5mm、5μm)と結合したZorbax 300SB−C18(2.1×150mm)5μmカラムを装備したAgilent 1100 LC/MSD SLシステムに適用した。直線的勾配の0.08% TFA、70% アセトニトリルを用いてペプチドを溶出した。ペプチドを220nmにおいて分光光度法により検出し、オンラインのMS(大気圧イオン化(API)エレクトロスプレー)により分析した。75% プロピオン酸/25% イソプロパノールの混合物を移動相にポストカラム添加し、シグナルを増大させた。得られた質量スペクトルをChemstationソフトウェア(Agilent Technologies, CA)およびBioLynxソフトウェア(Micromass, Waters Corporation, MA)を用いて分析した。
【0167】
HCおよびLCのAsp−N消化物のMS分析の結果を、表11および12にそれぞれ概説する。両方の表が、平均質量として与えられる理論上の質量および検出質量を示す。
【0168】
表11:重鎖についての結果
【表13】
a同じ質量を有する異なるペプチド。bほぼ同じ質量を有するペプチド(4個が一致し、1個が異なる)。*N−末端環化Gln(PyroGlu)を示す。
【0169】
表12:軽鎖についての結果
【表14】
【0170】
表11において見られるように、可変HC由来の13種類のペプチド、および可変LC由来の16種類のペプチドを固有のマーカーペプチドとして同定することができ、HCの可変領域由来のいくつかのペプチド(例えば、aにより示されるRhD196、RhD244、およびRhD306由来のD1)は同じ質量を有し、従って、これらの質量を明確に割り当てることができない。しかしながら、全ての場合で他の質量を固有のペプチドに明確に割り当てることができるので、8種類の抗体全てを明確に同定した。LCについて、8種類の抗体の中から7種類について、固有ペプチドを割り当てた(表12)。LCについての情報がない抗体はRhD324であった。従って、HCおよびLC由来の統合MSデータは、各抗体由来の固有ペプチドの検出に基づき、抗RhD rpAb試料において、8種類の抗体全てを同定できたことを示している。
【0171】
陽イオン交換クロマトグラフィーによる固有ペプチドの単離
HCおよびLCのAsp−N消化物を強陽イオン交換クロマトグラフィーにより次の通り分離した:上述のポリクローナル抗体から得られた単離HCおよびLCの個々のAsp−N消化物のアリコートを、Ettan LCシステム(Amersham Biosciences, GE Healthcare, England)において、10mM リン酸カリウム、20%(v/v) アセトニトリル、pH3.0にて平衡化したPolySulfoethyl Aカラム(2.1×100mm)に、流速0.2ml/分、室温にてアプライした。次に、10mM リン酸カリウム、20%または30%(v/v) アセトニトリル、pH3.0中の0〜500mMの直線的勾配の塩化カリウムを用いて、ペプチドを溶出した。溶出したペプチドを215nmにて分光光度法により検出し、時間フラクションに基づきフラクションを収集した。フラクションのアリコート(1μl)を、70% アセトニトリル/30%、0.1% TFA中のα−シアノ−4−ヒドロキシ桂皮酸(20mg/mL)溶液1μlと混合し、MS標的上にアプライし、次に、0.1% TFAで洗浄した。試料を、Autoflex TOF(Bruker Daltronics, Bremen, Germany)上のMALDI−TOFにより分析し、Bruker Daltronics(Bremen, Germany)の校正用混合物を用いて、質量の外部校正を行った。GPMAW 6.1ソフトウェア(Lighthouse data, Odense, Denmark)を用いて、MALDIスペクトルを分析した(質量検索および内部校正)。
【0172】
LCのAsp−N消化物由来のいくつかのピークを含有する代表的なクロマトグラムを、図11に示す。LCおよびHCのAsp−N消化物のフラクションのMALDI−TOF分析の結果を、それぞれ表13および14に示す。理論上の質量および見出した質量を、質量<3500Daについてモノアイソトピック質量として、質量>3500Daについて平均質量として示す。
【0173】
表13:軽鎖の結果
【表15】
a同じペプチド b同じペプチド c同じペプチド
【0174】
表14:重鎖の結果
【表16】
a同じペプチド b酸化Metを有する同じペプチドの同定により証明した
【0175】
表13および14において見られるように、可変LC由来の15種類のペプチド、および可変HC由来の9種類のペプチドを、固有のマーカーペプチドとして同定することができる。HCの可変領域ならびにLC由来のいくつかのペプチドは同じ質量を有し、これを明確に割り当てることができない。従って、強陽イオン交換クロマトグラフィーを用いて、HC RhD201、203、および324、ならびにLC RhD201および319について固有ペプチドを割り当てることはできなかった。
【0176】
要約
2種類の異なるマーカーペプチド分析から得られた結果は、HCおよびLCのMS分析から得られた統合データにより、RP−HPLCを用いて抗RhD rpABを構成する8種類の抗体全てから可変領域由来の固有ペプチドの同定を可能とすることを実証するのに十分である。強陽イオン交換クロマトグラフィーを用いることにより、抗RhD rpAb組成物中の8種類の個々のメンバーの中から6種類を同定することができた。完全に詳細までではないが、表11から14に示す程度まで、MS分析の結果を分析した。
【0177】
実施例6A
本実施例は、ポリクローナル細胞培養物(バイオリアクターラン)からもたらされた25種類の個々のメンバーを有する組換えポリクローナル抗RhD抗体の特徴決定を説明するものである。ペプチドの同定のための質量分析と組み合わされたRP−HPLCを用いてLCまたはHC由来の可変領域を起源とする固有マーカーペプチドを単離することにより、抗体の多様性を分析した。
【0178】
単離した重鎖および軽鎖の消化によるペプチドの生成
25種類の個々のメンバーを有するポリクローナル抗RhD抗体試料を、バイオリアクターラン由来のポリクローナル細胞培養物の上清から精製した。MabSelect(Amersham Biosciences, GE Healthcare)カラムを用いたアフィニティークロマトグラフィーにより、精製を行い、G25カラム(Amersham Biosciences, GE Healthcare)上で脱塩した。凍結乾燥した材料を6M 塩酸グアニジウム、0.2M Tris HCl、pH8.4に溶解し、還元(DTT)し、次に、カルボキシメチル化(ヨード酢酸)した。6M グアニジウムHCl、50mM リン酸ナトリウム、pH8.4中Superose 12カラム(10/300 GL Amersham Biosciences, GE Healthcare)上でのゲルろ過により、重鎖および軽鎖を分離した。分離したHCおよびLCを、1M 尿素、50mM リン酸ナトリウム、pH8中酵素対基質濃度1:200、37℃にて一晩Endoproteinase Asp−N(Roche, 1 054 589)で消化した。
【0179】
LC−MSによる固有ペプチドの単離
ポリクローナル抗体試料から得られた単離HCおよびLCの個々のAsp−N消化物のアリコートを、0.1% TFA、14% ACN中、流速0.2ml/分を用いて平衡化したガードカラム(Zorbax 300SB−C8、2.1×12.5mm、5μm)と結合したZorbax 300SB−C18(2.1×150mm)5μmカラムを装備したAgilent 1100 LC/MSD SLシステムに適用した。直線的勾配の0.08% TFA、70% アセトニトリルを用いて、ペプチドを溶出した。ペプチドを220nmにて分光光度法により検出し、オンラインのMS(大気圧イオン化(API)エレクトロスプレー)により分析した。75% プロピオン酸/25% イソプロパノールの混合物を、移動相にポストカラム添加し、シグナルを増大させた。得られた質量スペクトルをChemstationソフトウェア(Agilent Technologies, CA)およびGPMAW 6.2ソフトウェア(Lighthouse data, Odense, Denmark)を用いて分析した。
【0180】
HCおよびLCのAsp−N消化物のMS分析の結果を表14に概説する。表中、理論上の質量および検出した質量を平均分子量として示す。
【0181】
表14A:Asp−N消化およびLC−MS分析を利用した抗RhD rpAb中の25種類の抗体由来の固有疎水性ペプチドの同定
【表17】
aDおよびdは、それぞれAsp−Nにより生じたHCおよびLC由来ペプチドを示し、予測配列のN−末端からC−末端までペプチドに番号を付けた。ゆえに、d4は、LCペプチドの3番目と4番目のAsp−N部位での開裂により生じたペプチドを示す。bこのペプチドは、見逃された開裂部位を含有する。*N−末端環化Gln(pyroGlu)を示す。
【0182】
表14Aにおいて見られる通り、LCの可変部分由来の22種類のペプチド、およびHCの可変部分由来の3種類のペプチドを固有マーカーペプチドと同定することができる。従って、HCおよびLCのMSデータは、各抗体由来の固有ペプチドの検出に基づき、抗RhD rpAb試料において25種類の抗体全てを同定したことを明確に示している。
【0183】
実施例7
本実施例は、組換えポリクローナル抗RhD抗体の個々のメンバーに対する特異性を有する抗イディオタイプペプチドの生成、ならびに組換えポリクローナル抗体における1種類の個々のメンバーの濃度の評価を説明するものである。
【0184】
抗RhD抗体特異的ペプチドリガンドの生成
PIII(New England Biolab)のN−末端でランダムな配列順序で7種類のアミノ酸を提示するファージライブラリーを、個々の抗RhD抗体に対するペプチド結合体のアフィニティー選択のために用いた。ペプチドライブラリーの線形および制約バージョンの両方を選択のために用いた。4℃にて12〜16時間、10μg/ml(1ウェル当たり100μlを用いる)精製モノクローナル抗RhD抗体でマイクロタイタープレート(Maxisorb, NUNC)をコートした。組換えポリクローナル抗RhD抗体に含有される25種類の個々の抗体全てを用いて、抗イディオタイプペプチドについてスクリーニングした。しかしながら、組換えポリクローナル抗体が多数の個々のメンバー(例えば、50より多い)を含有する状況下では、センチネル抗体をスクリーニングのために選択してもよい。好ましくは、組換えポリクローナルタンパク質を構成する抗体の総数の少なくとも4%に該当する多数のセンチネル抗体を選択し、なおより好ましくは、組換えポリクローナルタンパク質を構成する抗体の総数の少なくとも8%、12%、16%、20%、30%、または50%で構成されるセンチネル抗体が選択される。次に、コートしたプレートをPBS、0.05% Tween−20で洗浄し、2% スキムミルク/PBSを用いてブロックした。〜1011pfu/100μlのバクテリオファージを、各パニングラウンドに用いた。制約ライブラリーおよび線形ライブラリーを混合し、2% スキムミルク/PBS中の混合物として一緒にパニングした。室温にて1時間インキュベーションした後、結合ファージを、グリシン/HCl、pH=2.2で10分間溶出し、次に、Tris−HCl、pH=9.0を用いて中和した。パニング3から4ラウンド後、単一クローンを単離し、DNAを抽出し、次に、ランダムなペプチド領域に対応する領域において配列決定した。以下の表15は、単一クローン由来の推定アミノ酸配列のアラインメントを示す。
【0185】
表15
【表18】
【0186】
それぞれ抗RhD162、202、または305のいずれかに対して特異的アフィニティーを有する3種類の合成ペプチドを、関連配列群の推定一致アミノ酸配列に従い合成した。各合成ペプチドのC−末端にビオチンを結合させた。
【0187】
各ペプチドの特異性をELISAにより試験した。簡単にいうと、ELISAプレートを、5μg/ml(1ウェル当たり100μlを用いた)Streptavidinを用いて、4℃にて12〜16時間コートした。次に、PBS中〜10μg/mlまで希釈したペプチドを添加し、1時間インキュベーションし、洗浄により過剰なペプチドを除去した。次に、プレートを、2% スキムミルク/PBSにてブロックし、PBSにて3回洗浄した。種々の希釈(10μg/mlにて出発)にて個々の抗RhD抗体それぞれを添加した。抗ヒトIgG−結合体(カタログ番号H10307)を用いて、結合抗体を検出した。プレートを5回洗浄し、色素原(TMB, Kem-En-Tech)25μlを添加して、検出を行った。1M H2SO4 25μlを添加することにより、15〜25分後に反応を終結させた。吸光度を450nmにて測定した。モノクローナル抗RhD抗体のパネルに対する各ペプチドの試験は、反応性が適当な個々のメンバータンパク質に特異的であることを示した。ゆえに、10より高いシグナル対ノイズ比で、PEP162のみが抗RhD162抗体に結合し、PEP202のみが抗RhD202に結合し、そしてPRP305のみが抗RhD305に結合した。
【0188】
組換えポリクローナル抗RhD抗体における抗RhD305抗体量の決定
適切な希釈の精製抗RhD305モノクローナル抗体を参照標準として用いて、組換えポリクローナル抗RhD抗体混合物中の抗体総量に対する抗RhD305抗体量を決定することが可能であった。簡単にいうと、ELISAプレートをストレプトアビジンでコートし、PBS中〜10μg/mlまで希釈したPEP305と1時間インキュベーションした。インキュベーション後、過剰なペプチドを洗浄して除去した。次に、プレートを2% スキムミルク/PBSにおいてブロッキングし、3回洗浄した。25種類の個々の抗RhD抗体(試料)からなる組換えポリクローナル抗RhD抗体を、1から16384倍の範囲の希釈率で添加した。試料を4重に分析した。同じプレート上の別々のウェルにおいて、標準曲線を作成するために、参照試料としてモノクローナル抗RhD305抗体の連続希釈液(3重)(10μg/mlにて開始)を添加した。抗ヒトIgG結合体(カタログ番号H10307)を用いて、結合抗体を検出した。プレートを5回洗浄し、色素原(TMB, Kem-En-Tech)25μlを添加することにより、検出を行った。1M H2SO425μlを添加することにより、15〜25分後に反応を終結させた。吸光度を450nmにて測定した。
【0189】
標準曲線は次の範囲の濃度に直線的に比例した:
【表19】
【0190】
これらのデータは、方程式y=0.1161x−0.0256、およびR2=0.9812を有する標準曲線となった。
【0191】
標準曲線について決定した方程式、ならびに試料の希釈因子を用いて、組換えポリクローナル抗RhD抗体試料中の抗RhD305抗体濃度を計算した。
【0192】
32倍希釈において、試料について測定した平均OD450は1.24±0.14であり、これは、ポリクローナル抗RhD抗体中の抗RhD305抗体濃度3.8±0.5μg/mlに対応するものであった。組換えポリクローナル抗RhD抗体試料中のトータルの抗体濃度は、100μg/mlであった。従って、抗RhD305抗体はポリクローナル抗体試料の3.8%を表す。
【0193】
実施例8
本実施例は、組換えポリクローナル抗RhD抗体の液体イオン交換クロマトグラフィー分析による1次元の分離後、特定のフラクション/ピークにおけるセンチネル抗体を同定するための抗イディオタイプペプチドの使用を説明するものである。
【0194】
25種類の個々の抗RhD抗体からなる組換えポリクローナル抗RhD抗体を陽イオン交換クロマトグラフィーにより分離し、フラクションを収集した。3種類の抗イディオタイプペプチドを用いたELISA(実施例7に記載の通り)により、各フラクションを調べ、各フラクションにおける特定の抗RhD抗体の存在を検出した。各フラクションにおいて行われたELISAデータとのクロマトグラムのオーバーレイは、この方法を用いて、特定のフラクション中の個々の抗体を同定できることを示した(図12)。従って、特定のピークにおける吸光度をELISAデータと比較することにより、相同タンパク質の複雑な混合物の組成物を半定量的に評価することが可能となる。
【0195】
実施例9
医薬上使用するためにポリクローナルタンパク質を製造する場合、組成物の安定性の確保は重要な問題である。相同タンパク質の複雑な混合物中の個々のタンパク質メンバーを同定する能力のある特異的なペプチド−リガンドを用いて、発酵中、異なる生成時間ポイントで培地試料を抽出し、実施例7に記載のELISA法のような定量的検出法を適用することにより、製造中のポリクローナルタンパク質の組成物の安定性をモニターすることができる。
【0196】
本実施例において、3種類のセンチネルタンパク質、抗RhD162、202、および305抗体の実際の量を、25種類の固有の抗RhD抗体からなる組換えポリクローナル抗RhD抗体産生pWCBのかん流発酵過程において評価した。図13は、バイオリアクターの播種の8日後に対応するG8を用いて、発酵中の異なる培養時間ポイントでの3種類のセンチネル抗体(抗RhD162、202、および305)の分布を説明するものである。
【0197】
実施例10
本実施例は、細胞培養混合物中の特定の抗RhD抗体産生細胞の同定方法を説明するものである。実施例において、抗イディオタイプペプチドおよび検出用フローサイトメトリーを用いて、2種類の異なる抗体産生細胞株を分析した。
【0198】
2種類の個々の抗RhD抗体産生細胞株、RhD162およびRhD202を所定の比率で混合した。クローンRhD202の割合を表16に示す。
【0199】
ビオチン化ペプチド202(実施例7において調製したPEP202)を、フィコエリトリン結合ストレプトアビジン(SA−PE)とインキュベーションし、ペプチドテトラマーを形成させた。テトラマーを細胞株混合物と室温にて20分間インキュベーションし、FACS CAliburフローサイトメーター(Becton Dickinson)に細胞を通した。図14のR1によって示す通り、テトラマー陽性細胞にゲートをかけた。個々の非混合細胞株を同様に測定した(図14AおよびB)。
【0200】
抗RhD抗体産生細胞株について見られる特徴は、細胞株中の抗体発現細胞および非抗体発現細胞両方の存在である。混合物中のPEP202陽性細胞のシェアを評価するために、発現細胞の総数を決定することが必要であった。この実施例において、RhD162とRhD202の混合物中の発現細胞のシェアは、RhD202単独におけるものと同じであると仮定した。これは、Pep202および抗IgG抗体で細胞を2重染色することにより、評価することができた(行ってはいない)。しかしながら、結果は、仮定が正確であることを示すものであった。図14に示す通り、ゲート6(R6)における細胞の割合から、混合物中のPEP202テトラマーによって結合したRhD202細胞の割合を計算した。
【0201】
mixa)におけるRhD202細胞株の割合を、次の方程式(mix aについての測定値を用いて例示)により計算した。
【数1】
【0202】
表16
【表20】
【0203】
実施例11
本実施例は、個々のクローンの分布、またはポリクローナル細胞培養物中のかかるクローンのセンチネルセレクションを評価するためのリアルタイムPCRの使用を説明するものである。
【0204】
技術は、個々の抗体コード核酸配列間の配列の相違に基づくものである。個々の抗体コード配列の多様性により、ポリクローナル細胞株に表される各メンバーの重鎖および/または軽鎖について、固有のTaqManプローブを設計することができる。好ましくは、CDR領域であるCDR1、CDR2、またはCDR3の1つを、TaqManプローブの設計のために選択する。最も好ましくは、CDR3領域をTaqManプローブの設計のため選択する。
【0205】
オリゴヌクレオチドの設計
好ましくは、70〜150ヌクレオチドの単位複製配列を得るように、プライマーを設計する。いくつかの可能性のあるプライマーの設計は、重鎖または軽鎖可変領域のFR3領域においてアニーリングする一致フォワードプライマー、および定常領域においてアニーリングするリバースプライマーである。試料の個々のメンバー間で異なるCDR領域の一部、好ましくは、CDR3領域に特異的なTaqManプローブを、目的の各クローンについて設計する。
【0206】
次の8種類の抗RhD抗体発現ポリクローナル細胞株の分析のためのプライマーおよびプローブの可能性のあるセットを、以下に示す通り設計し得る。
【0207】
全てのクローンについてフォワードおよびリバースプライマー:
Fwプライマー:CAC GGC TGA GTA TTA CTG TGC(配列番号24)
Rwプライマー:TTG GTG GAG CCA CTC GA(配列番号25)
【0208】
個々のクローン全てについてのTaqManプローブを表17に示す。
表17
【表21】
【0209】
重鎖コード配列についてのプライマー設計の別法は、VH−DH結合においてアニーリングするフォワードプライマーおよび定常領域におけるリバースプライマー、ならびにJH−C結合におけるTaqManプローブ(Rasmussen, T. et al. 2000. Exp.Hematol. 28, 1039-1045に記載)を構成する。
【0210】
リアルタイム定量的PCR
mRNAまたはゲノムDNAをペレット化細胞から抽出する。mRNAを鋳型として用いる場合、リアルタイムPCRに先立ち、逆転写してcDNAを生成する。分析すべきクローン数に対応する数のリアルタイムPCR反応をセットアップする。
【0211】
プライマー濃度およびTaqManプローブ濃度について、リアルタイムアッセイを最適化する。光学的接着性カバーでシールした96ウェルプレートにおいて、反応をトリプリケートで行う。市販のPCRマスターミックスにおいてPCR反応を行い、次に、ABIプリズム7000(Applied Biosystems)において、ABIプリズム7000 SDSソフトウェアを用いて分析を行った。
【0212】
多様性の分析
異なるクローンのCT値を互いに比較し、ポリクローナル細胞株における各クローンの分布を計算する。方法を応用して、バッチとバッチとの間の変動ならびに個々の生産工程における経時的なクローン安定性を評価し得る。
【0213】
実施例12
本実施例は、ポリクローナル細胞株からもたらされた単一細胞クローン由来の重鎖および/または軽鎖抗体遺伝子の可変領域のDNA配列決定により、組換えポリクローナル抗体を産生する能力のあるポリクローナル細胞株(例えば、pWCB)のポリクローナル性質を評価し、明らかにする方法を説明するものである。
【0214】
シングルセルクローニング
pWCBのアンプルを解凍し、完全培地にて2、3日間培養し、良好な細胞バイアビリティーを再構築した。続いて、96ウェル細胞培養プレート中、完全培地中1細胞/ウェルの密度にて細胞を播種する限界希釈により、シングルセルクローンを得る。37℃、5% CO2にて、10〜20日間細胞を培養し、次に、顕微鏡下でシングルコロニーを有するウェルについて、プレートを視覚的にスコア化する。別法として、FACS細胞ソーターを用いて、pWCB由来のシングルセルクローンを得る。生存可能なpWCB細胞にゲートをかけ、予め条件完全培地100μlで満たした96ウェルプレートに、1細胞/ウェルにて播種する。細胞をインキュベーションし、上述の通り、シングルコロニーについてスコア化する。
【0215】
核酸配列決定
ウェル中のシングルセルコロニーをコンフレントまで成長させるとき、所望数(例えば、100)のウェルのそれぞれからのアリコート(10〜20μl)を新しい96ウェルプレートに移し、DNA配列決定反応の鋳型として用いる。1から100、または1から1000個の細胞をそれぞれ用いてmRNAレベルまたはゲノムレベルのいずれかで、配列決定を行う。前者の場合、pWCBに存在する異なる抗体重鎖および軽鎖遺伝子を区別するのに十分な可変領域(典型的には、少なくともCDR3領域)をカバーするPCRフラグメントを、例えば、市販のQiagen one−step RT−PCRキットを用いて、製造元の指示に従い、標準的RT−PCR技術により生成する。PCR反応に先立ち、細胞を溶解させる。得られたPCRフラグメントを、例えば、Qiagen Qiaquick Gel Extractionキットを用いてゲル精製し、標準的配列決定反応における鋳型として用い、次に、ABI Prism[登録商標]3100 Genetic Analyzer(Applied Biosystems)のような自動DNA配列決定マシーンにおいて分析する。別法として、DNA配列決定を、逆転写工程を省略する以外上述の通り、ゲノムDNAにおいて行う。
【0216】
抗RhD組換えポリクローナル抗体の特徴決定のため、次のプライマーを用いた:
VH増幅用PCRプライマー:
RhD番号001:5’TCTCTTCCGCATCGCTGTCT(配列番号34)
RhD番号007:5’AGGAAAGGACAGTGGGAGTGGCAC(配列番号35)
VL増幅用PCRプライマー:
RhD番号005:5’CGTTCTTTTTCGCAACGGGTTTG(配列番号36)
RhD番号008:5’AAGACCGATGGGCCCTTGGTGGA(配列番号37)
配列決定プライマー:
VH:5’AACGGGTTTGCCGCCAGAACA(配列番号38)
VL:5’CCGAGGGACCTGAGCGAGT(配列番号39)。
【0217】
抗イディオタイプペプチドを用いたシングルセルにおけるELISA
核酸配列決定に対する手助けとして、ポリクローナルワーキングセルバンクのような抗体産生細胞の混合物のクローナルな組成物を、抗イディオタイプペプチドELISAを用いて評価することができる。
【0218】
ソートしたシングルセルを約14日間培養し、それにより、シングルクローン由来のアイソジェニックな細胞培養物を生成した。実施例7に記載の通り、ELISAアッセイにおいて抗イディオタイプペプチドを用いて、特異的抗RhD抗体の存在について、これらの培養物由来の上清を分析することができる。これにより、特定の個々のメンバーを産生するクローンの数に関する情報がもたらされるだろう。個々のメンバーの量を抗体産生細胞の総量と比較する(例えば、全てのアイソジェニックな細胞培養物においてIgGを測定することによる)場合、ポリクローナル細胞培養物における個々の抗RhD抗体産生細胞のフラクションについての定量的測定値を得ることができる。
【0219】
実施例13
本実施例は、組換えポリクローナル抗体の下流プロセッシング(DSP)中のクローン多様性を評価するための陽イオンクロマトグラフィー分析の使用を説明するものである。
【0220】
下流プロセッシング
25種類の個々のメンバーを含有する抗RhD rpAb試料を、発展的バイオリアクターランから、次のDSP工程を用いて精製した:
1.MAbSelectカラムを用いた抗体の捕獲
2.pH3でのウイルスの不活化
3.sephadex G−25カラムを用いたバッファー交換
4.DEAE−セファロースカラムを用いたイオン交換クロマトグラフィー
5.Planova 15Nフィルターを用いたウイルスフィルトレーション、および
6.MEP hypercelカラムを用いた疎水性電荷導入クロマトグラフィー
7.milipore biomaxフィルターを用いた限外ろ過/ダイアフィルトレーション。
【0221】
個々のDSP工程後のクローン多様性の分析
陽イオン交換クロマトグラフィーを用いて、組換えポリクローナル抗体組成物のDSP中のクローン多様性を分析した。抗RhD rpAbのDSP中、工程1、3、4、および6の後に採取した試料を、25mM 酢酸ナトリウム、150mM 塩化ナトリウム、pH5.0中、流速60ml/時間、室温にてPolyCatAカラム(4.6×100mm)にアプライした。次に、25mM 酢酸ナトリウム、pH5.0中150〜500mMの直線的勾配の塩化ナトリウムを用いて、流速60ml/時間にて抗体成分を溶出した。抗体成分を280nmにて分光光度法により検出し、クロマトグラム(図15)を比較して、DSP中のクローン多様性の可能性のある損失を検出した。本実施例において、陽イオン交換クロマトグラフィーを用いて、クローン多様性が組換えポリクローナル抗体のDSP中に本質的に変化しないことを明らかにした。
【0222】
実施例14
RhDに対する40種類より多くの抗体のIEX分析により、相当数の個々の抗体が図16Bに示す通り「3ピークパターン」を表すことを明らかにした。カルボキシペプチダーゼB処理、ならびに炭水化物分析は、この電荷不均一性が、C−末端リジンクリッピングまたはシアル酸の存在により引き起こされることを示した(データは示していない)。
【0223】
本実施例は、電荷不均一性がPyroGlu形成に起因するものであり、部位指定変異誘発を用いて、均一なIEXパターンを得ることができることを明らかにするものである。
【0224】
抗体の発現および精製
ゲノム上の特定の部位と異なる組換え抗Rhesus Dモノクローナル抗体をそれぞれ発現する安定な細胞株(2004年7月20日に出願されたデンマーク特許出願PA 2004 01133に記載の通り、得たもの)を、4mM L−グルタミン(Invitrogen)および1:250希釈した抗凝集剤(Invitrogen)を添加した無血清Excell 302培地(JRH Biosciences, Andover, UK)における懸濁培養に適用し、拡大させ、次に、通常の凍結手段を用いて−150℃にて保存した。
【0225】
保存前に、細胞培養物から上清を収集し、上清をろ過して、本質的に実施例1に記載の通り、アフィニティークロマトグラフィー(タンパク質A)を用いて抗RhDモノクローナル抗体を精製した。
【0226】
強陽イオン交換クロマトグラフィー
上述の工程において精製したモノクローナル抗体を、本質的に実施例1に記載の強IEXクロマトグラフィーの対象とした。表18のIEX列は、選択した抗体のIEXプロファイルに存在するピーク数を概説するものである。かかるIEXプロファイルを図16においても示す。
【0227】
N−末端配列分析
2種類の選択した抗体(RhD198およびRhD307)のIEX分析から分離したピークのN−末端配列分析を、Procise 494 Sequencer(Applied Biosystems, CA)(製造元によより記載された通り操作)を用いて、Edman配列決定により、溶液中で行った。配列分析により、電荷不均一性が、HCのN−末端Glnの部分的環化に起因するものであることが明らかになった(表18参照)。従って、第1のピークは、HCの全体的にブロックされたN−末端(HCのN−末端が0の電荷を有する)を有する抗体を含有し;第2のピークは、HCのN−末端の1つがブロックされている(HCのN−末端が+1の電荷を有する)抗体に対応し、そして第3のピークは、HCのN−末端Glnが非修飾されている(HCのN−末端が+2の電荷を有する)抗体を表す可能性が最も高かった。N−末端グルタミン残基のPyroGluへの環化は、それをEdman配列決定に不応性にする。
【0228】
かかる「3ピークパターン」または「1ピークパターン」を同様に示す多数の他の抗RhD抗体を、SDS−PAGEの対象にし、これをPVDFメンブレンに電気的にブロットすることによってN−末端配列分析により、分析した。これらのブロット上のHCおよびLCバンドを、Edman配列決定の対象とした。
【0229】
HCにおいてN−末端Glnを有する2、3種類の抗体(RhD162、RhD240)は、それらのIEXプロファイル(「1ピークパターン」)をふまえて、全体的にブロックされていることを示し、一方「3ピークパターン」を有する抗体(RhD196、RhD305、およびRhD306)は、予測通り部分的にブロックされていることを見出した(表18参照)。説明は、配列の収量、ならびにIEXプロファイルにおける異なる電荷変異体(0、+1、および2+)の相対的割合に基づくものである。
【0230】
表18
【表22】
a予測および得られたN−末端配列を、それぞれ通常および太字のフォント形式で示す。bブロットから得られたデータ。cIEX分析(溶液において分析)から単離したフラクション(ピーク1&2)から得られたデータ。dN−末端グルタミン残基のPyroGluへの環化はそれをEdman配列決定に不応性にする。n.d.;決定していない。
【0231】
部位指定変異誘発
部位指定変異誘発を用いて、N−末端GlnをGluに変えることにより、選択した抗体から電荷不均一性を除いた。重鎖中にN−末端Gln、および軽鎖中にN−末端Gluを有する全長抗体をコードする発現プラスミドRhD189を用いた。このプラスミドのVH領域には、シグナルペプチドコード領域の3’末端のAscI部位、およびJ領域のサイレントXhoI部位がフランクする。
【0232】
変異誘発を次のプライマー:
を用いて行った。
【0233】
フォワードプライマー中のAscI部位およびリバースプライマー中のXhoI部位に下線を付し、VH領域のN−末端のGluコドン(GAG)を太字で示す。上述のプライマーを用いたPCR反応における鋳型として、RhD189プラスミドを用いた。製造元の指示に従い、Phusion DNAポリメラーゼ(Finnzymes, Finland)を用いて25サイクルPCR反応を行った。約400bpのVHバンドを1% アガロースゲル上で精製し、BioTaq DNAポリメラーゼとインキュベーションし、アガロースゲル上で再び精製し、製造元の指示に従い、pCR2.1TOPOベクター(Invitrogen, CA)にクローニングした。VHインサートを含有するクローンを配列決定により確かめた。オリジナルのVHフラグメントをAscIおよびXhoIを用いてプラスミドRhD189から切り出し、pCR2.1TOPOプラスミド由来の変異誘発フラグメントを代わりに挿入した。エンドトキシンフリーのプラスミドミディプレップ(Macherey-Nagel, Germany)をクローニングの陽性コロニーから調製し、配列決定して、正しいフラグメントの存在を確かめた。
【0234】
抗体をSDS−PAGE分析の対象とし、電気的にブロットして、HCバンドのN−末端配列決定を行い、GlnからGluへの置換を確かめた(データは示していない)。「3ピーク」IEXプロファイルを示す抗体RhD189のHCのN−末端のGlnからGluへの置換は、1ピークのみを有する有意に異なるプロファイルとなった(図17)。従って、N−末端Gln残基からGlu残基に置換することにより、電荷の不均一性を首尾よく排除できた。
【0235】
結合アッセイ
N−末端の変異が抗体の機能に影響するかどうかを評価するために、天然の抗体であるRhD189、ならびにその変異誘発Glu対応物であるRhD189Eを、RhD陽性赤血球に対する結合についてアッセイした。
【0236】
Blood Bank, Aalborg Hospital, DKにおいてインフォームドコンセントの後健常ドナーから得た全血から、1% ウシ血清アルブミン(BSA、Sigma-Aldrich, Germany)を含有するPBS(Gibco, Invitrogen, United Kingdom)において3回洗浄することにより、赤血球を調製した。赤血球を再懸濁し、ID−Cellstab((DiaMed, Switzerland)中の10%溶液として4℃にて保存した。
【0237】
PBS、1% BSA中5×104細胞/μlのRhD陽性赤血球を用いて、抗体の結合能を測定した。PBS、1% BSAにて、96ウェルプレート(Becton Dickinson Labware, NJ, USA)中トリプリケートにて抗体希釈を行った。抗体溶液50μlを赤血球50μlと混合し、37℃にて40分間インキュベーションした。細胞をPBS、1% BSAにて2回(300g、2分間)洗浄した。PBS、1% BSAに1:20希釈したフィコエリトリン結合ヤギ抗ヒトIgG(Beckman Coulter, CA, USA)80μlを各試料に添加し、4℃にて30分間置いた。試料をPBS、1% BSA、次にFacsFlow(Becton Dickinson, Belgium)(300g、2分間)にて洗浄し、FACSFlow200μlに再懸濁した。FACSCalibur(Becton Dickinson, CA, USA)に試料を通し、CellQuest ProおよびExcelを用いてデータ分析を行った。
【0238】
図18において示す通り、RhD陽性赤血球に対する結合能における有意差を、Glu変異体とその天然の対応物の間で観察しなかった。
【0239】
要約
多くの抗RhD抗体のIEXプロファイルにおいて観察される不均一性は、これらの抗体におけるN−末端Gln残基の部分的環化に起因するものであった。抗RhD抗体におけるHCのN−末端GlnをGlu残基に置換することにより、おそらくRhD陽性赤血球に対する結合力に影響することなく、生来のN−末端の電荷不均一性を排除する。
【図面の簡単な説明】
【0240】
【図1】図1:9週間培養後のアリコート3948および3949由来の抗RhD組換えポリクローナル抗体(抗RhD rpAb)組成物の陽イオンクロマトグラム。下側の線図がアリコート3949に対応し、上側の線図がアリコート3948に対応する。線図の頂点のY軸を、下側の線図と区別するためにはずした。ピークのA〜Jは、正味電荷の異なる抗体、および電荷不均一性を現す個々の抗体を含む。
【図2】図2:11週間培養後の抗RhD rpAb産生ポリクローナル細胞株のアリコート3948+および3949+(FCW065)からもたらされたRT−PCR産物におけるHinfI RFLP分析を示すゲルの写真。特定のクローンに割り当てられるバンドを同定する。
【図3】図3:8種類の異なる抗Rhesus D抗体を有する抗RhD rpAb発現ポリクローナル細胞培養物由来の抗Rhesus D抗体軽鎖のT−RFLPパターン。8種類の異なる抗Rhesus Dクローンを、矢印により示されるピークに割り当てた。
【図4】図4:所定の時間ポイントで25種類の異なる抗Rhesus D抗体を有する抗RhD rpAbを発現するポリクローナル細胞培養物由来の抗Rhesus D抗体重鎖可変領域のT−RFLPパターン。25種類の異なる抗Rhesus Dクローンを、矢印により示されるピークに割り当てた。
【図5】図5:5週間培養したポリクローナル細胞培養物由来の8種類の異なる抗Rhesus D重鎖コード配列のT−RFLPにより評価したcDNA分布。
【図6】図6:陽イオン交換クロマトグラフィーを用いて分析した8種類の異なる抗体を有する抗RhD rpAbの相対的含有量(%)を示す。統合したクロマトグラムピークを、保持時間、および同一条件下で陽イオン交換クロマトグラフィーを用いて個々に分析した単一抗体から得られたピークパターンから個々の抗体に割り当てた。
【図7】図7:4週間培養後に得られた試料由来の25種類の個々のメンバーを有する抗RhD rpAbの陽イオンクロマトグラム。ピークAC1から25は、正味電荷の異なる抗体、および電荷不均一性を現す個々の抗体を含む。
【図8】図8:10種類の個々のメンバーを有する組換えポリクローナル抗RhD抗体の陽イオン交換クロマトグラフィー由来の溶出プロファイル。文字は、2次元のRP−HPLCの対象としたピークを示す。
【図9】図9:図8のフラクションB5のRP−HPLC由来の溶出プロファイルを示す。
【図10】図10:色分けしたタンパク質マップ(グレースケールで表す)により視覚化した10種類の個々のメンバーを有する組換えポリクローナル抗RhD抗体の2D LC組成分析を示す。
【図11】図11:8種類の個々のメンバーを有する組換えポリクローナル抗RhD抗体由来の精製LCのAsp−N消化物の強陽イオン交換クロマトグラフィー由来の溶出プロファイルを示す。太い横線は、マーカーペプチドの同定のためMALDI−TOF分析の対象としたフラクションを示す。
【図12】図12:抗イディオタイプペプチドPEP162、PEP202、およびPEP305を用いた3種類の個々のELISA分析から得られたELISAデータを用いて、25種類の個々のメンバーをを有する抗RhD rpAbにおいて得られたOD280IEXクロマトグラムのオーバーレイを示す。ELISA分析を、イオン交換クロマトグラフィーにより得られた各フラクションにおいて行った。ELISAデータを、それぞれ互いに比較可能なPEP162、PEP202、およびPEP305を用いてELISA分析を3回行うために、トータルのOD%に対して校正する。
【図13】図13:発酵中の異なる培養時間ポイントで3種類のセンチネル抗体である抗RhD162、202、および305の分布を示す。G8は、バイオリアクターの播種の8日後に該当する。
【図14】図14:PEP202テトラマーを用いて染色した3種類の細胞株のFACSデータ。(A)は、PEP202陰性細胞株RhD162を示す。(B)は、細胞株RhD202を示し、そして(C)は、細胞株RhD162とRhd202の50%混合物(実験のmix aに対応する)を示す。A、B、およびCの第1パネルは、FSC−SSCドットプロットを示し、R1は、サイズ(FSC)および粒度(SSC)に基づき、生存かつ健常な細胞についてのゲートである。中央のパネルのヒストグラムは、細胞の蛍光強度を表す。R6ゲートは、テトラマー染色細胞を囲うものである。最後のパネルは、計算で用いたR6の細胞の割合を示す。
【図15】図15:捕獲溶出(A)、Sephadex G−25(B)、DEAE−Sepharose(C)、およびMEP Hypercel(D)後に収集した材料により表される25種類の個々のメンバーを含有する抗RhD rpAb試料の下流処理中の異なるステージで採取した試料を示す陽イオン交換クロマトグラフィープロファイル。
【図16】図16:異なる電荷パターンを示す3個の代表的なモノクローナル抗RhD抗体のIEXプロファイル。(A)均一性、(B)「3ピーク」パターン、(C)複雑なパターン。
【図17】図17:RhD189、および変異誘発Glu変異体RhD189EのIEX分析。
【図18】図18:Glu変異体RhD189E、およびその天然対応物RhD189の結合活性。RhD陽性赤血球に対する抗体の結合をFACSにより測定し、平均蛍光強度(MFI)を抗体濃度のフラクションとして示す。
【技術分野】
【0001】
発明の分野
本発明は、最終生産物のバッチとバッチとの間の一貫性、ならびに1回の生産工程における組成物の安定性を証明するために、組換えポリクローナルタンパク質またはかかるタンパク質を産生するポリクローナル細胞株を構造上特徴付ける方法に関するものである。特に、本発明は、組換えポリクローナル抗体を特徴付ける方法に関するものである。
【0002】
発明の背景
抗体の予防上または治療上の投与(いわゆる受動免疫)は、生体の免疫系の能力を増強して、感染性因子を排除し得ると長い間考えられてきた。歴史的に、かかる治療用抗体は、ヒト血漿から得られてきた(そのため、抗体組成物は免疫グロブリンまたはγ−グロブリンと称される)。これらの抗体を得るために、免疫ヒトドナーから血液プールが集められ、そして免疫グロブリンフラクションが抽出され、精製される。免疫グロブリンのフラクションのみが、特定の抗原に特異的となる。ドナーの数が限られていること、製造に費用がかかること、ドナー由来の感染性汚染物質のリスクがあること、バッチとバッチとの間の変動が不可避であること、ならびに投与計画が複雑であることといったいくつかの制限が原因で、免疫グロブリンの治療上の使用は複雑である。
【0003】
最近、組換えモノクローナル抗体が免疫グロブリン製品に取って代わりつつある。しかしながら、それらは1つの標的に対してのみ方向付けられており、それゆえ、感染性因子のような複雑または動的な標的に対しては有効でないかもしれない。この問題を解決するために、モノクローナル抗体を混合する例がいくつか存在する(例えば、Nowakowski, A. et al. 2002. Proc Natl Acad Sci U S A 99, 11346-11350および米国特許第5,126,130号)。
【0004】
最近、予防または治療投与に適した高度に特異的なポリクローナル抗体の組換え生産技術が開発された(WO2004/061104)。組換えポリクローナルタンパク質を構成する個々のメンバーを別々に取り扱うこと、製造すること、精製すること、または特徴付けることなく、組換えポリクローナル抗体(rpAb)は、生産物バイオリアクターから1つの調製物として精製され得る。しかしながら、かかる産生ストラテジーには、同一性を証明し、抗体分子の複雑な混合物を長期にわたり一貫して産生することを明らかにする方法が必要である。
【0005】
さらに、国家的かつ超国家的監督機関から研究薬または治療薬としての認可を得るために、組換え技術を用いて産業上生産されるポリクローナル抗体はある程度特徴付けられなければならないだろう。組換えポリクローナル抗体アプローチは完全に新しいコンセプトであるので、試料における個々のタンパク質の相対的比率について、複数の、異なるが高度に相同なタンパク質を含む試料を特徴付けるという問題がこれまでに取り扱われたことはない。従って、一般に、非臨床上および臨床上の有効性データ、およびしばしば歴史上安全性のデータ、ならびに粗化学反応、製造、および純度、結合タイター、および偶発因子の不存のようなコントロール(CMC)パラメーターに基づいて、血液由来の免疫グロブリンは認可される。もちろん、かかる単純化したアプローチは、組換え産生されたタンパク質には容認されない。ゆえに、2、3種のモノクローナル抗体の混合物について、規制ガイドラインには、かかる混合物が、生物学的アッセイと一体となった包括的なタンパク質化学的特徴決定技術を用いてそれぞれの構成タンパク質について個々に特徴決定されなければならないことが述べられている。しかしながら、これは、技術上実現可能なアプローチでもなく、また10、20、ましてやそれより多くの異なる抗体に基づいた真のポリクローナル組成物に適当でもない。
【0006】
貢献の開示
本発明は、ポリクローナル細胞株からの組換えポリクローナルタンパク質、特に、組換えポリクローナル抗体のような異なる相同タンパク質の混合物の一貫した生産を示すための構造上の特徴決定基盤を提供する。
【0007】
発明の記載
予防上および治療上使用する組換えポリクローナルタンパク質の産業上の生産に前もって必要なものは、発現中のクローンの多様性の維持である。それゆえ、ポリクローナル抗体を産生するポリクローナル細胞株のクローンの多様性、ならびに任意の所望の時間ポイントでのポリクローナルタンパク質、および任意の関連試料における個々のタンパク質の相対的提示をモニターし、測定できることが重要であり、これにより1回の実施における発現系の安定性、ならびに最終生産物のバッチとバッチとの間の変動の分析が可能になる。
【0008】
組換えポリクローナル抗体または組換えポリクローナルT細胞受容体(TcR)のような相同タンパク質の組成物は、非常に類似した物理化学的特性を有する変異体タンパク質から成る。このことは、組換え生産されたポリクローナルタンパク質を精製する場合に利点がある。それが1つのタンパク質であるかのうように、工程において多様性を喪失することなく、精製が行われ得るからである。しかしながら、この類似性は、ポリクローナルタンパク質の個々のメンバーの相対的分布を特徴付ける場合の難問をもたらす。なぜなら、物理化学的特性における類似性は、メンバー1つ1つを互いに区別することを難しくするからである。
【0009】
最も一般的には、組換えポリクローナルタンパク質を産生する場合、オリジナルの組成物は既知である。なぜなら、組換えポリクローナルタンパク質産生用のポリクローナル製造細胞株の創出に先立ち、ポリクローナルタンパク質をコードする配列が単離され、スクリーニングされ、そして配列決定されているからである。かかる細胞株の創出については、WO2004/061104(本明細書において引用により取り込まれる)を参照されたい。これに対する希な例外は、例えば、回復期の患者由来のスクリーニングまたはセレクションされていないライブラリーを直接用いて、組換えポリクローナル抗体を創出することであり得る。
【0010】
アウトプット(組換えポリクローナルタンパク質)の多様性が、培養および精製後にインプット(コード配列のライブラリー)の多様性と類似することを確実にするために、ポリクローナルタンパク質の個々のメンバーおよび/またはポリクローナル製造細胞株内のそれらのコード配列の相対的比率についての情報を得ることが必要だろう。本発明は、ポリクローナル細胞株およびポリクローナルタンパク質両方の多様性についての情報をもたらす能力のある、遺伝子分析ならびにタンパク質の特徴決定技術に基づく、構造上の特徴決定基盤を提供する。
【0011】
定義
用語「抗イディオタイプ抗体」は、ポリクローナルタンパク質の個々のメンバーの変異体部分に特異的に結合する全長抗体またはそのフラグメント(例えば、Fv、scFv、Fab、Fab’、またはF(ab)2)を意味する。好ましくは、本発明の抗イディオタイプ抗体は、ポリクローナル抗体またはポリクローナルTcRの個々のメンバーの変異体部分に特異的に結合する。好ましくは、抗イディオタイプ抗体の特異性は、ポリクローナル抗体またはポリクローナルT細胞受容体の個々のメンバーの抗原特異的部分、いわゆるV領域に対して方向付けられる。しかしながら、それは、個々のメンバーの所定のサブ集団、例えば、混合物において表された特異的VH遺伝子ファミリーに対する特異性も示し得る。
【0012】
用語「抗イディオタイプペプチド」は、相同タンパク質の混合物内の個々のタンパク質メンバーを特異的に関連付け、これにより同定する能力のある特異的なペプチドリガンドを意味する。好ましくは、本発明の抗イディオタイプペプチドは、ポリクローナル抗体またはポリクローナルTcRの個々のメンバーに特異的に結合する。好ましくは、本発明の抗イディオタイプペプチドは、個々の抗体または個々のT細胞受容体の配列の抗原特異的部分に対して方向付けられる。しかしながら、抗イディオタイプペプチドは、個々のメンバーの所定のサブ集団に対する特異性も示し得る。
【0013】
用語「「大量」N−末端配列決定」は、多数の変異体相同タンパク質分子、例えば、ポリクローナルタンパク質を含む試料のN−末端のタンパク質の配列決定を意味する。この大量配列決定により、試料内に存在する異なるタンパク質全ての配列情報が一度に提供される。アミノ酸が試料中の個々のメンバー間で変動する位置において、これらは定量され、可変位置での個々のアミノ酸の異なる量により、特定の変動を含有するタンパク質サブ集団に関する情報がもたらされるだろう。N−末端を配列決定すべきタンパク質が1個より多くのサブユニットを含有する場合、好ましくは、これらが分けられて、複雑性が低減され、配列決定される(例えば、試料がポリクローナル抗体の場合、重鎖を軽鎖が分けられて、配列決定される)。
【0014】
用語「クローンの多様性」または「ポリクローナル性(polyclonality)」は、ポリクローナルタンパク質、それをコードする核酸配列、またはそれを産生するポリクローナル細胞株の変動性または多様性を意味する。変動性は、ポリクローナルタンパク質またはコード配列ライブラリーの個々のメンバー間でのアミノ酸配列または核酸配列の相違により特徴付けられる。ポリクローナル細胞株について、クローンの多様性は、例えば、個々の細胞のゲノムへの一部位の組込みとして細胞株において表される核酸配列の変動性により評価され得る。しかしながら、それは、細胞株において細胞表面上に表されるアミノ酸配列の変動性としても評価され得る。
【0015】
用語「エピトープ」は、T細胞受容体または抗体が結合する抗原分子の部分を意味する。一般に、抗原または抗原分子は、いくつか、あるいは多数のエピトープを同時に提示するだろう。
【0016】
通常、用語「免疫グロブリン」は、血液または血清において見出される抗体の混合物の総称として用いられる。ゆえに、血清由来のポリクローナル抗体は、しばしば、免疫グロブリンまたはγ−グロブリンと称される。しかしながら、免疫グロブリンは、他の供給源からもたらされる抗体の混合物、例えば、組換え免疫グロブリンを称するためにも用いられ得る。
【0017】
本明細書において用いられる用語「個々のクローン」は、特定のタンパク質、例えば、モノクローナル抗体を発現する細胞のアイソジェニックな集団を示す。かかる個々のクローンは、例えば、宿主細胞を所望の核酸でトランスフェクションし、次に、陽性のトランスフェクタントを選択することにより得られ、単一クローンが拡大されるか、あるいは多数の単一クローンがプールされ、次に拡大され得る。ポリクローナル細胞株は、ポリクローナルタンパク質の異なる個々のメンバーを発現する個々のクローンを混合することにより生成され得る。
【0018】
用語「個々のメンバー」または「別個のメンバー」は、ポリクローナルタンパク質のような異なるが相同なタンパク質を含むタンパク質組成物のタンパク質分子を示し、個々のタンパク質分子は、組成物の他の分子と相同だが、ポリペプチド配列の1個以上のストレッチ(stretch)を含有し、そしてそれは、可変領域とも称される、ポリクローナルタンパク質の個々のメンバー間のアミノ酸配列の相違により、特徴付けられる。例えば、Ab1からAb50からなるポリクローナル抗体において、Ab1配列を有するタンパク質の全てが、ポリクローナル抗体の個々のメンバーと考えられ、Ab1は、例えば、CDR3領域においてAb2タンパク質と異なっていてもよい。個々のメンバーのサブ集団は、例えば、Ab1、Ab12、およびAb33に属する抗体から構成され得る。
【0019】
用語「ポリクローナル抗体」は、同じ抗原または異なる抗原上のいくつかの異なる特異的抗原決定基と結合するか、あるいは反応する能力を有する異なる抗体分子の組成物を表す。ポリクローナル抗体の変動性は、ポリクローナル抗体を構成する個々の抗体のいわゆる可変領域、具体的には、相補性決定領域(CDR)1、CDR2、およびCDR3領域に位置する。
【0020】
用語「ポリクローナル製造細胞株」、「ポリクローナル細胞株」、「ポリクローナルマスターセルバンク(pMCB)」、および「ポリクローナルワーキングセルバンク(pWBC)」は、互換使用され、目的の変異体核酸配列ライブラリーでトランスフェクションされたタンパク質発現細胞の集団を意味する。好ましくは、個々の細胞は、一体となって組換えポリクローナル製造細胞株を構成し、目的の組換えポリクローナルタンパク質の1メンバーをコードする目的の異なる核酸配列を1コピー有し、そしてそれぞれのコピーは、各細胞のゲノムの同じ部位に組み込まれる。かかる製造細胞株を構成し得る細胞は、例えば、細菌、真菌、真核細胞、例えば、酵母、昆虫細胞、または哺乳類細胞、特に、CHO細胞、COS細胞、BHK細胞、ミエローマ細胞(例えば、Sp2/0細胞、NS0)、NIH3T3、YB2/0のような不死の哺乳類細胞株、およびHeLa細胞、HEK293細胞、またはPER C6細胞のような不死化ヒト細胞であり得る。
【0021】
本明細書で用いられる用語「ポリクローナルタンパク質」は、異なるが、相同なタンパク質分子を含むタンパク質組成物を意味し、好ましくは、それは、免疫グロブリンスーパーファミリーから選択される。なおより好ましくは、相同なタンパク質分子は、抗体またはT細胞受容体(TcR)である。従って、それぞれのタンパク質分子は、組成物の他の分子と相同だが、変動可能なポリペプチド配列の1個以上のストレッチも含有し、そしてそれは、ポリクローナルタンパク質の別の変異体メンバーとも称される個々のメンバー間でのアミノ酸配列の相違により特徴付けられる。かかるポリクローナルタンパク質の既知の例は、抗体、T細胞受容体、およびB細胞受容体を含む。ポリクローナルタンパク質は、タンパク質分子の所定のサブセットからなり得、そしてそれは、例えば、所望の標的抗原に対するポリクローナル抗体の場合、所望の標的に対する共有結合活性のような共通の特性により定義されてきた。一般に、組換えポリクローナルタンパク質は、かかる分子の所定のサブセットからなり、ここで、それぞれのメンバーの配列は既知である。希にだが、組換えポリクローナルタンパク質は、非標的特異的タンパク質の重要な部分も含有するという意味で、血清由来の免疫グロブリンと類似し得る。
【0022】
用語「ポリクローナルT細胞受容体(TcR)」は、同じ抗原または異なる抗原由来のいくつかの異なる特異的な抗原決定基に結合するか、あるいはそれと反応する能力を有する、異なるTcR分子の組成物を表す。ポリクローナルTcRの変動性は、ポリクローナルTcRを構成する個々のTcR分子のいわゆる可変領域、具体的には、CDR1、CDR2、CDR3、またはCDR4領域に位置する。本発明のTcR分子は、α−β鎖またはγ−δ鎖の改変可溶性ダイマーである。かかる改変TcRは、例えば、(Willcox, B.E. et al. 1999. Protein Sci 8, 2418-2423)に記載されている。
【0023】
用語「タンパク質」は、長さまたは翻訳後修飾の如何に関わらず、アミノ酸の任意の鎖を意味する。タンパク質は、2個以上のアセンブリされたポリペプチド鎖、タンパク質のフラグメント、ポリペプチド、オリゴペプチド、またはペプチドを含む単量体または多量体として存在し得る。
【0024】
用語「センチネルタンパク質」は、ポリクローナルタンパク質の個々のメンバーを表し、そしてポリクローナルタンパク質の産生中、または異なるバッチにおいてその存在についてモニターされ得る。一連の関連試料におけるセンチネルタンパク質の存在の一貫性は、バッチ間あるいは1回の産生での長時間のポリクローナルタンパク質の発現の安定性を反映するだろう。さらにそれは、組換え産生されたポリクローナルタンパク質の精製のような下流処理における多様性の維持を反映するだろう。
【0025】
用語「固有のマーカーペプチド」は、ポリクローナルタンパク質の個々のメンバーの可変領域を起源とする多数のペプチドを表す。好ましくは、ペプチドは、タンパク質の断片化であるプロテアーゼ処理または他の手段により生成され、ポリクローナルタンパク質の単一の個々のメンバーに明確に割り当てられ得るペプチドが、固有のマーカーペプチドと称される。
【0026】
発明の詳細な説明
本発明の態様は、(i)異なる可変領域を有する異なる相同タンパク質、または(ii)かかるタンパク質を産生する細胞株を含む試料における個々のメンバーの相対的比率に関する情報を得るための構造上の特徴決定用基盤を提供することである。特徴決定基盤を用いて、異なる相同タンパク質を含む組成物の産生または精製過程、あるいは長期保存中に異なる態様が評価され得る。好ましくは、本発明の特徴決定基盤は、次の目的:i)1つの試料内で互いに関連する個々のメンバーまたは個々のメンバーのいくつかの相対的出現量を決定すること、ii)バッチとバッチとの間の一貫性の決定のため、異なる試料における1個以上の個々のメンバーの相対的比率を評価すること、およびiii)1個以上の個々のメンバーの実際の比率を評価すること、のうちの1つのために用いられる。必要に応じて、これは、ポリクローナル製造細胞株を生成するためにもともと用いられたベクターライブラリーと比較されてもよい。特徴決定基盤は、ポリクローナル細胞株のクローンの多様性および/または細胞株により産生されたポリクローナルタンパク質における個々のタンパク質の出現量をモニタリングするのに特に有用である。個々の生産工程における組成物の安定性およびバッチとバッチとの間の一貫性の両方がモニターされ得る。別法として、基盤手段はまた、ポリクローナルタンパク質またはモノクローナル抗体の混合物を含む異なる相同タンパク質の混合物の精製組成物に適用され、例えば、かかる組成物における個々のメンバーの長期安定性が評価され得る。
【0027】
本発明の1つの実施態様は、タンパク質またはそれらのコード配列の個々のメンバーの相対的比率または存在に関する情報が得られるよう、異なる可変領域を有する異なる相同タンパク質またはかかるタンパク質を産生する細胞を含む試料を特徴付ける方法であって、該方法は、1個以上のタンパク質特徴決定技術により、および/または1個以上のタンパク質コード配列の遺伝子分析により、該試料のアリコートを分析することを含む。
【0028】
本発明のさらなる実施態様において、構造上の特徴決定基盤は、タンパク質の特徴決定技術ならびに遺伝子分析から選択される多数の分析技術からなる。従って、構造上の特徴決定基盤は、任意の数の個々の実施態様(次のセクションに記載)からなる。実施態様に記載の分析技術の1つからのみ、試料について情報を得れば足り得る。しかしながら、好ましくは、これらの分析技術の少なくとも2、3、4、5、6、7、8、9、または10個から情報が得られ、それにより、以下に詳述される個々の実施態様が組み合わされ、特徴決定基盤が生成される。いくつかの分析技術の組み合わせにより、ポリクローナル混合物の相対的または絶対的組成に関するより記述的なデータセットの創出が可能となる。これらの技術から得られる情報は、定量的性質ならびに定性的性質のものであり、一体となって蓄積された場合、分析された試料の全体的な特徴決定を提供する。
【0029】
本発明の好ましい実施態様において、1つの分析技術は、タンパク質の特徴決定技術であり、別の分析技術は遺伝子分析である。
【0030】
遺伝子分析は、制限断片長多型(RFLP)分析、末端RFLP(T−RFLP)、マイクロアレイ分析、リアルタイムPCRのような定量的PCR、および核酸配列決定のような技術を意味する。
【0031】
タンパク質の特徴決定技術は、i)物理化学的特性によりタンパク質を分けるクロマトグラフィー分析、ii)相同タンパク質のタンパク分解性消化物の分析、iii)「大量」N−末端配列決定、およびiv)相同タンパク質に特異的な検出分子を用いた分析のような未知のタンパク質を特徴付けるため、プロテオミクス分野で一般に用いられる技術を意味する。
【0032】
本発明のさらなるコンセプトは、相同タンパク質の複雑なプールを特徴付けるため、上述の分析技術と組み合わせて適用でき、本発明と関連して開発された。該コンセプトは、相同タンパク質のプール(例えば、ポリクローナル抗体またはポリクローナルTcR)、または相同タンパク質を産生する細胞のプール(例えば、ポリクローナル製造細胞株)の表面に存在する多数のセンチネルタンパク質の選択に基づくものである。センチネルタンパク質は、定量的および定性的に特徴付けられ、タンパク質のこのサブ集団が、ポリクローナル細胞培養物の上清中または産生中の細胞表面上のいずれかで、一貫した方法で存在することが証明される。センチネルタンパク質は、例えば、抗イディオタイプ分子のような相同タンパク質の個々のメンバーに特異的な検出分子を用いて分析され得る。センチネルタンパク質のコンセプトはさらに、異なる細胞培養物のバッチ間の一貫性を評価するために適用され得る。センチネルタンパク質のコンセプトは、ポリクローナルタンパク質がポリクローナル抗体またはTcRである場合、好ましくは、センチネルペプチドがCDRの一部を含有するよう、プロテアーゼ処理によりポリクローナルタンパク質からもたらされた固有のペプチドに拡大され得る。遺伝子レベルで行われる分析はまた、センチネルの原理を適用し、これは、ポリクローナルタンパク質をコードするライブラリーの個々のメンバー由来の固有の核酸配列に基づくものである。具体的には、抗体またはTcRのCDR領域に対応する核酸配列は、センチネル核酸配列として選択される。最も好ましくは、CDR3領域である。センチネルタンパク質、ペプチド、または核酸配列は、ポリクローナルタンパク質のメンバー、またはそれをコードする核酸配列に依存して、個々の分析技術について変動し得、それは、選択された分析技術を用いて区別され得る。
【0033】
ポリクローナル製造細胞株のクローンの多様性の遺伝子分析
本発明のある実施態様において、ポリクローナルタンパク質を産生するための発現系におけるポリクローナル性は、ポリクローナルタンパク質の特定のメンバーをコードする細胞の量および/またはポリクローナルタンパク質の個々のメンバーをコードするmRNAレベルを評価することによりモニターされる。これは、例えば、RFLPまたはT−RFLP分析、オリゴヌクレオチドマイクロアレイ分析、リアルタイムPCRのような定量的PCR、および製造細胞株から得られた遺伝子配列の可変領域の核酸配列決定を用いて、mRNAレベルまたはゲノムレベルでモニターされ得る。別法として、同じ技術を定性的に用いて、ポリクローナル細胞株の多様性が示され得る。ポリクローナルタンパク質をコードする核酸配列は、培養中の異なる時間ポイントで単一のポリクローナル細胞培養物から得られた試料においてモニターされ、それにより、生産工程を通じて個々のコード配列の相対的比率をモニターし、その組成物の安定性が評価され得る。別法として、ポリクローナルタンパク質をコードする核酸配列は、特定の時間ポイントで異なるポリクローナル細胞培養物から得られる試料においてモニターされ、それにより、異なるバッチにおける個々のコード配列の相対的比率をモニターし、バッチとバッチとの間の変動が評価され得る。好ましくは、遺伝子分析において用いられる試料は、例えば、沈殿により、培養物の細胞について濃縮された細胞培養物フラクションである。一般に、試料は、所望の時間ポイントで細胞培養物のフラクションを回収し、次に、例えば、遠心分離により培地を除去して得られる。バッチとバッチとの間の一貫性の比較のための試料は、好ましくは、産生についてのインビトロの細胞継代数(age)の限界で、細胞から得られる。
【0034】
RFLP/T−RFLP
RFLPおよびT−RFLP分析は、ゲノムレベルまたはmRNAレベルで行われ得る。各細胞が目的の配列を1コピーのみ含有するように、ポリクローナル製造細胞株が生成される場合、ゲノムレベルでの分析は、ポリクローナルタンパク質の個々のメンバーを産生する製造細胞株における細胞の相対的比率に関する情報をもたらすだろう。一方、mRNAレベルでの分析は、ポリクローナルタンパク質の個々のメンバーの可能性のある発現レベルに関する情報をもたらすだろう。一般に、mRNAレベルでの分析は、mRNAをcDNAに逆転写し、次に制限酵素分析することにより、行われる。しかしながら、mRNAにおいて分析を直接行うことも可能である。
【0035】
末端RFLP分析において、PCRまたはRT−PCRのために用いられるフォワードおよび/またはリバースプライマーは標識され、これにより、PCRフラグメントの末端標識がもたらされる。適当な制限酵素を用いて消化した後、異なる大きさのフラグメントが生成され、電気泳動、好ましくは、キャピラリー電気泳動により分けられ、そしてフラグメントが標識単位複製配列により検出され得る(Liu et al. 1997, Applied and Environmental Microbiology 63, 4516-4522)。適当な標識は、蛍光、放射能、比色、X線回析または吸収、磁性、または酵素活性により検出可能なシグナルをもたらし、そして、例えば、フルオロフォア、発色団、放射性同位元素(特に、32P、33P、35S、および125I)、高電子密度試薬、酵素、および特異的結合パートナーを有するリガンドを含む。好ましくは、フルオロフォアが標識として用いられる。
【0036】
巨大な多様性を有するポリクローナル製造細胞株において、それぞれ個々のコード配列について固有の制限酵素フラグメントを得ることはできないかもしれない。かかる状況が生じる場合、センチネル核酸配列は、ポリクローナル製造細胞株のクローンの多様性をモニターするために選択され得る。別法として、大きさにより分けられないフラグメントが、個々のコード配列全ての分布を評価するために配列決定され得る。
【0037】
オリゴヌクレオチドマイクロアレイ分析
DNAチップのようなオリゴヌクレオチドマイクロアレイを用いて、細胞株から生成された標識DNAのハイブリダイゼーションを測定することにより、ポリクローナル細胞株におけるゲノムDNAレベルまたはmRNAレベルが測定され得る(Guo, Z. et al 1994. Nucleic Acids Res. 22, 5456-5465)。
【0038】
プローブは、ポリクローナル細胞株に存在すると予測される配列の2本鎖cDNA配列の代表(ポリクローナル細胞株自体、またはポリクローナル細胞株を含む宿主細胞をトランスフェクションするために用いられるDNAライブラリーのいずれかからもたらされる)、またはセンスオリゴヌクレオチド(長さ20〜90nt)のいずれかであり得る。プローブは、ガラス、プラスチック、またはゲルマトリックスのような固体表面に付着し、そして2本鎖プローブが用いられる場合、それは変性されて、アッセイが行われる。相同タンパク質、例えば、ポリクローナル抗体またはポリクローナルTcRを発現するポリクローナル細胞株を分析するとき、好ましくは、プローブとポリクローナル細胞株からもたらされる標識cDNAとの間でのクロスハイブリダイゼーションを防ぐよう、注意深く設計されたオリゴヌクレオチドプローブが用いられる。かかるプローブは、ポリクローナル産物の個々のメンバーそれぞれに特異的なプローブを設計するためにポリクローナル製造細胞株を生成するために用いられた可変領域コード配列のアライメントに基づき設計される。抗体について、コード配列はCDR領域において主として異なり、CDR3領域における変動性の程度が最も高いであろう。好ましくは、最大の変動性を有する領域が、センスオリゴヌクレオチドの設計のため用いられる。好ましくは、プローブは、ポリクローナル細胞株において含まれる個々のメンバーに対して配列が相補であり、他のメンバーに対して出来るだけ高度な相違を有する。それぞれの可変領域に特異的な1個以上のプローブが用いられ得る。標準的な目的のため、定常領域の配列とハイブリダイズするプローブが用いられ得る。プローブは、ハイブリダイゼーションに用いられる表面に直接スポットされるか、あるいは表面上にインサイツで合成される(Pease et al. 1994. PNAS 91: 5022-5026, Singh-Gasson et al. 1999. Nature Biotech. 17: 974-978)。
【0039】
分析されるべき標識DNAは、ポリクローナル細胞集団を回収し、細胞からゲノムDNA、トータルRNA、またはmRNAを調製することにより、生成される。ゲノムDNAを用いる場合、標識は、関連コード配列のPCR増幅において適当に標識されたプライマーまたは標識ヌクレオチドのいずれかを用いて得られる。トータルRNAまたはmRNAを用いる場合、逆転写のみ、または標識プライマーまたはヌクレオチドを用いたPCR工程と組み合わせて、標識cDNAを得ることが可能である。適当な標識は、蛍光、放射能、比色、X線回析または吸収、磁性、または酵素活性により検出可能なシグナルをもたらし得、そして例えば、フルオロフォア、発色団、放射性同位元素(特に、32P、33P、35S、および125I)、高電子密度試薬、酵素、および特異的結合パートナーを有するリガンドを含む。好ましくは、フルオロフォアが標識として用いられる。分析されるべきコード配列が抗体重鎖および軽鎖である場合、1本鎖cDNAは、可変領域の3’側の定常領域に位置するアンチセンスプライマーを用いたプライミングによる逆転写により調製される。さらにPCRが行われない場合、好ましくは、合成は、標識ヌクレオチドを用いて行われる。逆転写の後PCRが行われ、一連のセンスプライマーが適用され、確実に可変領域の全ファミーが増幅される。別法として、全てのmRNAにおいて一致する領域(例えば、5’非翻訳領域またはシグナルペプチドコード配列)にハイブリダイズするセンスプライマーが用いられ得る。センス、および/またはアンチセンスプライマーは蛍光標識されるか、あるいは標識ヌクレオチドがこのアプローチにおいて用いられ得る。
【0040】
プローブおよび標識DNAが調製された場合、マイクロアレイアッセイは、低ノイズかつ高度に特異的なシグナルについて最適化された条件下で、変性させた標識DNAを固定オリゴヌクレオチドとハイブリダイズさせることにより、行われる。洗浄後、ハイブリダイズしたプローブそれぞれが測定され、特異的なメッセージの量が計算される。
【0041】
定量的PCR
PCR法は、試料中の核酸配列の検出および定量の両方をもたらすよう既に適合されている。例えば、Higuchi, R. et al. 1993. Kinetic Biotechnology 11, 1026-1030;Holland, P.M. et al. 1991. PNAS 88, 7276-7280;Livak, K.J. et al. 1995 PCR Methods Appl. 4, 357-362を参照されたい。これらの方法は、標準的PCRなどでのフォワードおよびリバースプライマー、および増幅される核酸とハイブリダイズする1個以上のさらなる核酸配列を利用する。このさらなる核酸配列は、「プローブ」と称され、2つのプライマーにハイブリダイズする部分の間で増幅されるべき核酸の一部にハイブリダイズし、各連続的PCRサイクルがプローブまたはその標識の変化を生じるような方法で標識される。プローブまたはその標識のこの変化により、各PCRサイクルにおいて増幅された核酸の追加コピー数に関連する程度まで、標識は活性化されるか、あるいは強調される。一般に、かかる方法は、「リアルタイム」PCRと称され、熱サイクルを標識の検出と組み合わせることにより、PCR産物の増大をサイクル毎に検出する。リアルタイムPCRの特定のバージョンにおいて、プローブまたはその標識の変化は、ポリメラーゼ、例えば、Taqポリメラーゼのエキソヌクレアーゼ活性により引き起こされ、それゆえ、この技術は、一般にTaqまたはTaqManリアルタイムPCRと称される(例えば、Holland, P.M. et al. 1991. PNAS 88, 7276-7280)。
【0042】
適当な標識は、蛍光、放射能、比色、X線回析または吸収、磁性、または酵素活性により検出可能なシグナルをもたらし、そして例えば、フルオロフォア、発色団、放射性同位元素(特に、32P、33P、35S、および125I)、高電子密度試薬、酵素、および特異的結合パートナーを有するリガンドを含む。最も一般的には、プローブの標識は蛍光標識であり、これは蛍光出力アウトプットシグナルをもたらす。これは、一方の端、典型的には、5’末端で蛍光レポーターダイ、そしてもう一方の端、3’末端でクエンチャーダイで2重標識されたプローブを用意することにより、達成され得る(例えば、Livak, K.J. et al. 1995 PCR Methods Appl. 4, 357-362)。プローブがインタクトであるとき、クエンチャーダイがレポーターダイに近づくと、レポーターダイの蛍光が抑制される。適当なダイは、Wilhelm, J. and Pingoud, A., 2003. Chembiochem. 4, 1120-1128において概説されている。それぞれのPCRサイクルにおいて、DNAポリメラーゼの5’→3’エキソヌクレアーゼ活性によりプローブが開裂され、そしてそれにより、レポーターダイをクエンチャーダイと分離させる。この分離により、レポーターダイの蛍光の増大が生じる。
【0043】
PCRにおいて、目的の標的が試料に存在する場合、プローブは、フォワードとリバースPCRプライマー部位の間で特異的にアニーリングする。プローブが標的分子とハイブリダイズする場合のみ、DNAポリメラーゼのエキソヌクレアーゼ活性により、レポーターとクエンチャーダイの間でプローブが開裂される。これらのプローブは、しばしばTaqManプローブと称される。標的配列がプローブと相補的であり、かつPCRにおいて増幅される場合のみ、蛍光の増大が検出される。これらの要件のため、非特異的な増幅は検出されない。プローブに相補的な配列を含有する増幅産物のみが、蛍光シグナルの存在により認識され、それにより、擬陽性の分析と関連するある種の要素が排除される。加えて、1種類以上の他の酵素を用いて、キャリーオーバーの転写産物の増幅の制限が手助けされてもよい。
【0044】
この種の定量的PCRは、ピペット誤差および用量変化の校正を許可し、そしてそれは、各反応に含有されるレポーターの蛍光をパッシブリファレンスで割ることにより行われ、個々の反応それぞれについて校正されたレポーターシグナルが決定され得る。絶対的な定量化のため出発コピー数を決定するか、あるいは相対的な量の比較のため他の未知試料に対して比較するために、ソフトウエアを用いて、サイクル毎の蛍光強度の増大が分析され、次に、これらのデータが標準と比較され得る。
【0045】
具体的には、本発明においてTaqManリアルタイムPCRが、ポリクローナル細胞株の特徴決定に適していることが見出された。従って、ポリクローナル抗体発現細胞株に適用される場合、固有のTaqManプローブはポリクローナル細胞株において表されメンバーそれぞれの重鎖および/または軽鎖について設計され得るので、該技術により、個々の抗体のコード配列の相対的比率が定量される。好ましくは、CDR領域の1つであるCDR1、CDR2、またはCDR3が、TaqManプローブを設計するために選択される。最も好ましくは、CDR3領域がTaqManプローブを設計するために選択される。かかる可変重鎖CDR3のTaqManプローブの例は、Rasmussen, T. et al. 2000. Exp. Hematol. 28, 1039-1045(本明細書において引用により取り込まれる)において見られる。
【0046】
核酸の配列決定
核酸の配列決定はよく知られた技術であり、本発明と共に利用され、ポリクローナル製造細胞株の多様性に関する定性的情報をもたらし得る。配列決定は、単一細胞クローニングによりポリクローナル細胞株からもたらされる単一細胞、またはポリクローナル製造細胞株から得られる変異体細胞の未処理試料のいずれかにおいて行われ得る。
【0047】
単一細胞レベルでの配列決定は、ポリクローナルタンパク質の個々のメンバーを産生する、製造細胞株における細胞の相対的比率に関する情報をもたらすだろう。この方法において、ポリクローナル製造細胞株由来試料は、所望の時間ポイントで得られ、特徴付けられるべきポリクローナルタンパク質をコードする細胞は、例えば、限界希釈またはFACS Ariaのようなセルソーターにより、単一細胞クローニングされる。ポリクローナル細胞株試料から得られる単一細胞の数は、細胞株において表されると予測される配列の多様性に依存する。好ましくは、少なくとも3回、細胞株の生成におけるインプットを形成する個々のコード配列の数がシングルセルソートされ、試験試料中においてそれら全てを再び見出す見込みは95%となるだろう。従って、25種類の異なるコード配列のライブラリーを用いて、細胞株が創出される場合、少なくとも75種類の単一細胞クローンが、配列決定用試料から得られ、これにより、等しい量において表される異なる配列が25種類より多くなる。これにより、ポリクローナル細胞株において表される個々のコード配列の大部分が、製造過程において喪失されなかったなら、単一細胞クローン間で表されることが確実になるだろう。単一細胞は別々のウェルにおいてコンフレントまで成長され、それぞれのウェル由来のアリコートが、核酸の配列決定反応における鋳型として用いられる。配列決定は、配列決定に先立ちRT−PCRまたはPCR増幅工程をそれぞれ用いて、mRNAレベルまたはゲノムレベルで行われ得る。mRNAまたはゲノムレベルのいずれかで得られた配列情報により、個々の抗体成分のそれぞれをコードする細胞の割合が決定され得る。さらに、mRNAレベルで得られた配列情報を用いて、ポリクローナル組成物における個々の抗体それぞれの発現レベルが評価され得る。配列決定に加えて、mRNAレベルでTaqManリアルタイムPCRを行い、単一細胞クローンの可能性のある発現レベルと関連する情報を得ることが可能である。上述のRFLPまたはT−RFLP分析は、同様に、単一細胞レベルで行われ得る。
【0048】
ポリクローナル製造細胞株から得られる変異体細胞の未処理試料における配列決定はまた、細胞株から産生されるポリクローナルタンパク質の個々のメンバーの可能性のある発現レベルに関する情報をもたらすことができ、そしてそれは、ポリクローナルタンパク質の個々のメンバーのコード配列の相対的mRNAレベルに基づくものである。この方法において、ポリクローナル製造細胞株由来試料は、所望の時間ポイントで得られる。RT−PCRは、試料中の溶解細胞において直接行われる。RT−PCR反応に適用されるプライマーセットは、センスおよびアンチセンスプライマーが全てのmRNAにおいて一致する領域にハイブリダイズする場合、同じ有効性を有するコード配列全てを増幅すると予測されるような方法で、設計される(例えば、5’非翻訳領域またはシグナルペプチドコード配列中のセンスプライマー、および定常領域配列中ののアンチセンスプライマーが用いられ得る)。増幅されたPCRフラグメントは、配列決定ベクターにクローン化され、宿主細胞、好ましくは、エスケリキア・コリ(E.coli)にトランスフェクションされる。ポリクローナル製造細胞株由来の個々のコード配列を表す単一クローン由来のプラスミドDNAが配列決定され、得られた個々のコード配列の比率は、ポリクローナル細胞株における個々のコード配列それぞれのmRNAレベル、ならびに個々のタンパク質メンバーの可能性のある発現レベルを反映するだろう。
【0049】
本発明のさらなる実施態様において、上述の遺伝子分析が別の分析として適用される。好ましくは、1種類以上の分析が、細胞株のクローンの多様性についての情報をできるだけ多く得るために、同じ試料由来のアリコートにおいて行われる。別法として、遺伝子分析が、多次元様式で組み合わされ得る(例えば、マイクロアレイ分析が、この分析後、RFLPまたはT−RFLPフラグメントにおいて行われるか、あるいはRFLPフラグメントが、RFLP分析後に配列決定される)。特に、1種類より多くの個々の成分を表し、かつ制限酵素フラグメントのサイズが一致するために分けられないRFLPフラグメントでの配列決定の実施に有利である。
【0050】
ポリクローナル性を評価するためのタンパク質の特徴決定技術
本発明の実施態様において、相同タンパク質のプールまたは相同タンパク質を産生するための発現系のポリクローナル性は、1種類以上のタンパク質の特徴決定技術によりモニターされる。タンパク質の特徴決定技術は、単独または他の技術と組み合わされて、溶液中またはポリクローナル細胞株に存在する細胞の表面上のモノクローナルタンパク質の混合物または組換えポリクローナルタンパク質の個々のメンバーの存在および相対的比率に関する情報をもたらす能力のある任意の技術を意味する。組換えポリクローナルタンパク質の複雑性に依存して、次の技術のうちの1種類以上が用いられ得る:i)クロマトグラフ分離技術、ii)ポリクローナルタンパク質の個々のメンバーを表す固有のマーカーペプチドの同定のためのポリクローナルタンパク質のタンパク分解消化産物の分析、iii)「大量」N−末端配列決定、およびiv)例えば、ポリクローナルタンパク質のセンチネルタンパク質メンバーの特徴決定のための特異的検出分子を用いた分析。
【0051】
異なる相同タンパク質を含有する試料は、精製モノクローナルタンパク質の混合物、またはポリクローナルタンパク質であり得る。ポリクローナルタンパク質は、例えば、遠心分離により細胞から分離されただけの「未処理」の上清、または、例えば、タンパク質Aアフィニティー精製、免疫沈降、またはゲル濾過により精製された上清の形のポリクローナル細胞培養物から得られた細胞培養上清であり得る。しかしながら、これらの前精製工程は、組成物中の異なる相同タンパク質の分離を全くもたらさないので、組換えポリクローナルタンパク質の特徴決定の一部ではない。好ましくは、本発明の特徴決定過程の対象となる試料は、精製工程を少なくとも1回受けている。最も好ましくは、試料は、90%、95%、または99%純粋な相同タンパク質を含む。
【0052】
ポリクローナルタンパク質を構成する異なる相同タンパク質は、培養中の異なる時間ポイントで単一のポリクローナル細胞培養物から得られた試料においてモニターされ、それにより、個々のポリクローナルタンパク質の相対的比率が生産工程を通じてモニターされ、その組成の安定性が評価され得る。別法として、ポリクローナルタンパク質を構成する異なる相同タンパク質が、特定の時間ポイントで異なるポリクローナル細胞培養物から得られた試料においてモニターされ、それにより、異なるバッチにおける個々のコード配列の相対的比率がモニターされ、バッチとバッチとの間の一貫性が評価され得る。
【0053】
クロマトグラフ分離技術
ポリクローナルタンパク質の個々のメンバーのクロマトグラフ分離は、i)正味電荷(例えば、イオン交換クロマトグラフィー(IEX))、ii)疎水性(例えば、逆相クロマトグラフィー(RP−HPLC)、および塩濃度による疎水性相互作用クロマトグラフィー(HIC))、iii)等電点(pI値)(例えば、クロマトフォーカシング)、またはiv)アフィニティー(例えば、抗イディオタイプペプチド/抗体を用いたアフィニティークロマトグラフィー、またはκおよびλ抗体軽鎖の分離のためのタンパク質Lクロマトグラフィー)のような物理化学的特性の相違に基づくものであり得る。5種類のよく知られたクロマトグラフ技術は、次の物理化学的特性:大きさに基づくものである。しかしながら、全てのメンバーは本質的に同じ大きさのものであるので、これは、ポリクローナル抗体またはポリクローナルTcRのような相同タンパク質の特徴決定に特に適した技術ではない。大きさによる分離は、特徴決定基盤から完全に省かれる。上述のこれらのクロマトグラフ技術のいくつかは、IgA、IgG、およびIgMのような免疫グロブリンのクラス(Gallo, P. et al. 1987. J. Chromatogr. 416, 53-62)、またはIgG1、IgG2、IgG3のようなサブクラス(Scharf, O et al. 2001. J. Virol. 75, 6558-6565)をヒト血清から分離する際に利用されてきた。しかしながら、血清由来の免疫グロブリンまたは組換えポリクローナル抗体における個々の抗体の多様性に関連する分離は、これまで行われていない。
【0054】
a)イオン交換クロマトグラフィー
本発明の実施態様において、イオン交換クロマトグラフィーを用いて、組換えポリクローナルタンパク質の個々のメンバー、またはポリクローナルタンパク質の個々のメンバーのサブ集団が分離される。イオン交換クロマトグラフィーによる分離は、分離されるべき組成物中の個々のタンパク質の正味電荷に基づくものである。組換えポリクローナルタンパク質のpI値、選択されたカラムバッファーのpH値および塩濃度に依存し、組換えポリクローナルタンパク質の個々のメンバーは、陰イオンまたは陽イオン交換クロマトグラフィーのいずれかを用いて、少なくともある程度まで分離され得る。例えば、pHが、組換えポリクローナルタンパク質組成物の個々のメンバーの最も低いpI値より十分に低いものである限り、通常、組換えポリクローナルタンパク質の個々のメンバーの全てが、負に荷電した陽イオン交換媒体に結合するだろう。続いて、結合した組換えポリクローナルタンパク質の個々のメンバーは、典型的には漸増勾配の塩(例えば、塩化ナトリウム)または漸増pH値を用いて、個々のタンパク質の正味電荷に依存して、カラムから溶出され得る。いくつかのフラクションが溶出中に得られるだろう。好ましくは、単一のフラクションは、ポリクローナルタンパク質の個々のメンバーを含有するが、ポリクローナルタンパク質の2、3、4、5、6、7、8、9、10、15、20、またはそれ以上の別のメンバーも含有し得る。陽イオンおよび陰イオン交換の一般的な原理は、当該技術分野においてよく知られており、イオン交換クロマトグラフィー用のカラムは市販されている。
b)クロマトフォーカシング
本発明のさらなる実施態様において、クロマトフォーカシングを用いて、組換えポリクローナルタンパク質の個々のメンバー、またはポリクローナルタンパク質の個々のメンバーのサブ集団が分離される。クロマトフォーカシングによる分離は、個々のタンパク質のpI値の相違に基づくものであり、組換えポリクローナルタンパク質のpI値より上のpH値を有するカラムバッファーを用いて行われる。個々のメンバーが比較的低いpI値を有する組換えポリクローナルタンパク質は、正に荷電した弱陽イオン交換媒体に結合するだろう。続いて、結合した組換えポリクローナルタンパク質の個々のメンバーは、個々のメンバーのpI値のpH範囲をカバーするよう設計されたポリバッファーを用いてカラム内の漸減pH勾配を創出することにより、個々のメンバーのpI値に依存して、カラムから溶出され得る。いくつかのフラクションが溶出中に得られるだろう。好ましくは、単一のフラクションは、ポリクローナルタンパク質の個々のメンバーを含有するが、ポリクローナルタンパク質の2、3、4、5、6、7、8、9、10、15、20、またはそれ以上の別のメンバーも含有し得る。陽イオン交換体を用いたクロマトフォーカシングの一般的な原理は、当該技術分野においてよく知られており、陽イオンカラムは市販されている。陽イオン交換体を用いたクロマトフォーカシングも当該技術分野において知られている(Kang, X. and Frey, D.D., 2003. J. Chromatogr. 991, 117-128(引用により、本明細書に取り込まれる))。
【0055】
c)疎水性相互作用クロマトグラフィー
本発明のさらなる実施態様において、疎水性相互作用クロマトグラフィーを用いて、組換えポリクローナルタンパク質の個々のメンバー、またはポリクローナルタンパク質の個々のメンバーのサブ集団が分離される。疎水性相互作用クロマトグラフィーによる分離は、分離されるべき組成物における個々のタンパク質の疎水性の相違に基づくものである。組換え産生されたポリクローナルタンパク質は、疎水性相互作用に有利に働くバッファー中の疎水性リガンドで修飾されたクロマトグラフィー媒体に結合する。一般に、これは、有機溶媒を低い割合で含有するバッファー(RP−HPLC)、または選択された塩を非常に高濃度含有するバッファー(HIC)において達成される。続いて、結合した組換えポリクローナルタンパク質の個々のメンバーが、典型的には、漸増勾配の有機溶媒(RP−HPLC)または漸減勾配の選択された塩(HIC)を用いて、個々のメンバーの疎水性に依存して、カラムから溶出されるだろう。いくつかのフラクションが溶出中に得られるだろう。好ましくは、単一のフラクションは、ポリクローナルタンパク質の個々のメンバーを含有するが、ポリクローナルタンパク質の2、3、4、5、6、7、8、9、10、15、20、またはそれ以上のメンバーも含有し得る。疎水性相互作用クロマトグラフィーの一般的な原理は、当該技術分野においてよく知られており、RP−HPLCならびにHIC用カラムは市販されている。
【0056】
d)アフィニティークロマトグラフィー
本発明のさらなる実施態様において、アフィニティークロマトグラフィーを用いて、ポリクローナルタンパク質の個々のメンバー、またはポリクローナルタンパク質の個々のメンバーのサブ集団が分離される。アフィニティークロマトグラフィーによる分離は、特異的検出分子、リガンド、またはタンパク質に対するアフィニティーの相違に基づくものである。検出分子、リガンド、またはタンパク質、またはこれらのうちの複数のもの(これらの異なるオプションを、以下、ただリガンドと称する)は、クロマトグラフィー媒体に固定され、組換えポリクローナルタンパク質は、個々のメンバーと固定されたリガンドとの間の相互作用に有利に働く条件下でアフィニティーカラムにアプライされる。固定されたリガンドに対するアフィニティーを示さないタンパク質は、カラムフロースロー(flow-through)に集められ、次に、固定されたリガンドに対するアフィニティーを示すタンパク質は、結合に不都合に働く条件(例えば、低pH、高塩濃度、または高いリガンド濃度)下でカラムから溶出される。いくつかのフラクションが溶出中に得られ得る。好ましくは、単一のフラクションは、ポリクローナルタンパク質の個々のメンバーを含有するが、ポリクローナルタンパク質の2、3、4、5、6、7、8、9、10、15、20、またはそれ以上の別メンバーも含有し得る。組換えポリクローナルタンパク質を特徴付けるために用いられ得るリガンドは、例えば、標的抗原、抗イディオタイプ分子、またはκまたはλ軽鎖を有する抗体の分離のためのタンパク質Lである。
【0057】
標的抗原を用いたアフィニティークロマトグラフィーは、組換えポリクローナルタンパク質が1種類以上のエピトープに対するアフィニティーを含む場合、特に適切だろう。標的は、例えば、癌細胞もしくはウイルス、または標的の組合せであってもよく、そしてそれは、多くのエピトープを含有する。これらのエピトープは合成され、クロマトグラフ媒体上に固定される。1個のカラム当たり1個のエピトープ、または1個のカラム当たりいくつかの異なるエピトープを用いて、アッセイは設計され、それにより、特定のエピトープに対する個々のメンバーの分布に関して、組換えポリクローナルタンパク質混合物を特徴付けることが可能になる。別法として、完全抗原または標的分子は、クロマトグラフ媒体上に固定され得る。
【0058】
ポリクローナルタンパク質の個々のメンバー、またはかかる個々のメンバーのサブ集団に特異的に結合する抗イディオタイプ分子(例えば、抗イディオタイプペプチドまたは抗イディオタイプ抗体)を用いたアフィニティークロマトグラフィーを行い、組換えポリクローナルタンパク質の選択されたメンバー(センチネルタンパク質とも称される)、または個々のメンバーのサブ集団の相対的比率に関する情報が得られ得る。理想的には、個々の抗イディオタイプ分子は、1種類の個々のメンバーのみに特異的に結合し、組換えポリクローナルタンパク質の他のメンバーには結合しないものであるが、メンバーの所定のサブセットと結合する抗イディオタイプ分子も本発明において適用可能である。好ましくは、完全なポリクローナル組成物が特徴付けられ得るように、抗イディオタイプ分子は個々のメンバー全てに対して生成される。組換えポリクローナルタンパク質がポリクローナル抗体またはTcRである場合、抗イディオタイプ分子は、抗体またはT細胞受容体の配列の抗原特異的部分に対して方向付けられる。1個のカラムが1種類の抗イディオタイプ分子を含有するように、抗イディオタイプ分子は個々にクロマトグラフ媒体に固定され、それにより、特定のタンパク質メンバーまたはタンパク質のサブ集団についての情報がえられる。次に、フロースローが、第2の固定化抗イディオタイプ分子などを有する第2のカラムにアプライされる。別法として、いくつかの異なる抗イディオタイプ分子が、同じカラムに適用されたのと同じクロマトグラフ媒体上に固定される。次に、例えば、漸増量の遊離イディオタイプ分子をカラムに添加することにより、あるいはpHまたは塩勾配を用いることにより、個々のタンパク質が異なるフラクションに溶出されることを可能にする条件下で、溶出が行われる。このアプローチにより、1次元分析を用いて、ポリクローナルタンパク質のいくつかのメンバーの比率についての情報を得ることが可能だろう。
【0059】
組換えポリクローナルタンパク質がポリクローナル抗体である場合、これは、κ軽鎖またはλ軽鎖のいずれかを含有する個々のメンバーからなり得る。かかるポリクローナル抗体において、λ軽鎖の抗体がタンパク質Lに対するアフィニティーを欠いていることを用いて、λ軽鎖を有する抗体がκ軽鎖を有する抗体から分離され得る。従って、λ軽鎖を含有する抗体メンバーのサブセットは、タンパク質Lアフィニティークロマトグラフィーを用いて、κ軽鎖を含有する抗体メンバーのサブセットから分離され得る。続いて、κおよびλ抗体サブセットが、例えば、上述の個々の抗体の定量のための別のクロマトグラフ技術をさらに用いて、特徴付けられ得る。
【0060】
多次元クロマトグラフィー
分析されるべき試料中の変異体相同タンパク質、例えば、組換えポリクローナルタンパク質の複雑性に依存して、上記のa)からd)のクロマトラフ技術のうちの2種類以上を2次元、3次元、または多次元フォーマットで組み合わせることが所望され得る。好ましくは、2次元ゲル電気泳動の代わりに全次元において液体クロマトグラフィーが用いられる。しかしながら、これは、組換えポリクローナルタンパク質を特徴付けるための1以上の次元でのゲル電気泳動または沈殿技術の使用を排除するものではない。
【0061】
液体2次元クロマトグラフィーは、例えば、Lubman, D.M. et al. 2002. J.Chromatogr.B Analyt.Technol.Biomed.Life Sci. 782, 183-196;WO01/58925およびWO01/58926に記載されている。該方法を用いて、健常な細胞と癌細胞のタンパク質の発現が比較され、それにより、タンパク質レベルでの異なるディスプレイが創出されてきた。さらに、3次元が大きさによる分離である3次元クロマトグラフィーがWO03/102539に記載されており、例えば、細胞抽出物中のタンパク質が分離され、それにより、同様に、差次的ディスプレイが創出される。
【0062】
本発明のさらなる実施態様において、多次元クロマトグラフィーを用いて、試料中の異なる相同タンパク質が特徴付けられる。具体的には、抗体およびT細胞受容体のような異なる可変領域を有する相同タンパク質試料が、多次元クロマトグラフィーを用いて特徴付けられる。好ましくは、さらなる次元が、前の次元における溶出中に得られたフラクションにおいて行われる。しかしながら、フロースルーもさらなる次元の分析のために用いられ得る。これは、前の次元がアフィニティークロマトグラフィーである場合、特に適切であり得る。
【0063】
本発明の1つの実施態様において、多次元クロマトグラフィーは、ポリクローナル抗体(血清からもたらされた免疫グロブリン、または組換え体)、またはモノクローナル抗体の混合物のいずれかから、個々の抗体分子をそれらの多様性に関して分離させる際に用いられる。好ましくは、多次元クロマトグラフィーは液体クロマトグラフィーである。
【0064】
本発明の別の実施態様において、多次元クロマトグラフィーは、ポリクローナルT細胞受容体またはモノクローナルT細胞受容体の混合物のいずれかから、個々のT細胞分子をそれらの多様性に関して分離させる際に用いられる。好ましくは、多次元クロマトグラフィーは液体クロマトグラフィーである。
【0065】
一般に、多次元クロマトグラフィーにおける異なる次元の異なる物理化学的特性に基づくクロマトグラフ技術、例えば、1次元の電荷による分離、2次元の疎水性による分離、そして3次元のアフィニティーによる分離を用いることが試みられる。しかしながら、クロマトグラフ技術のなかには、次の次元(たとえそれらが同様の物理化学的特性を利用するものであっても)で用いられたときに、さらなる分離をもたらし得るものがある。例えば、クロマトフォーカシング、そしてイオン交換クロマトグラフィーまたはアフィニティークロマトグラフィー(異なるリガンドを用い、互いに成功)が続く場合、さらなる分離が得られ得る。
【0066】
表1には5つの次元が挙げられ、そこでは、クロマトグラフ技術が本発明の特徴決定基盤の一部として利用され得る。しかしながら、これは必須の次元数と考えられるべきではない。組換えポリクローナルタンパク質を特徴決定するのに十分な分離が、1、2、3、または4次元の後得られたなら、残りの次元は省れ得る。ゆえに、十分な分離が、イオン交換クロマトグラフィー(IEX)を用いて得られたなら、クロマトフォーカシング、RP−HPLCなどを行う必要はない。他方、5つの次元が不十分な分離をもたらしたなら、さらなる次元が加えられ得る。さらに、表1は、クロマトグラフ技術の可能性のある組合せの包括的リスト、または技術自体の包括的リストであると考えられるべきではない。
【0067】
表1
【表1】
【0068】
【表2】
【0069】
本発明の好ましい実施態様において、多次元液体クロマトグラフィー(LC)は、表1に示される最初の2つの次元から選択される2次元LC技術である。
【0070】
本発明のさらに好ましい実施態様において、多次元LCは、表1に示される最初の3つの次元から選択される3次元LC技術である。
【0071】
多次元LC技術の代替として、ゲル電気泳動またはキャピラリー電気泳動のような適当な電気泳動技術と組み合わされた免疫沈降、そして続いて抗原の定量化を用いて、組換えポリクローナルタンパク質が特徴付けられ得る。この技術は、複雑な抗原に対して標的化された組換えポリクローナル抗体を特徴付けるのに特に有用だろう。例えば、複雑なウイルス抗原に対して標的化された組換えポリクローナル抗体は、標識抗原混合物およびタンパク質Aビーズを用いて、免疫沈降され得る。続いて、抗原は、等電点電気泳動法または2D PAGEを用いて分離され、次に、個々の抗原が定量される。これは、特異的抗原に対して標的化された組換えポリクローナル抗体における抗体量を反映する。
【0072】
組換えタンパク質のN−末端荷電の不均一性の排除
上述のタンパク質の特徴決定技術において、単一のタンパク質は、例えば、IEXプロファイルにおいていくつかのピークを生じうるので、相同タンパク質プールにおける個々のタンパク質の不均一性は、特徴決定をなおさら複雑にし得る。不均一性は、抗体および他の組換えタンパク質において共通する現象であり、酵素性または非酵素性翻訳後修飾によるものである。共通する翻訳後修飾は、N−グリコシル化、メチオニン酸化、タンパク分解性断片化、およびアミド分解を含む。不均一性は、トランスフェクションにおいて誘発される変異(Harris, J.R, et al. 1993. Biotechnology 11,1293-7)、および転写中の重鎖と軽鎖の可変遺伝子間でのクロスオーバー現象(Wan, M. et al. 1999. Biotechnol Bioeng. 62,485-8)のような、遺伝子レベルでの修飾からも生じ得る。これらの修飾は、エピジェネティックであり、従って、構築物の遺伝子構造単独から予測することはできない。
【0073】
不均一性を生じ得るこれらの翻訳後修飾のいくつかは、特徴決定に先立ち処理され得る。C−末端のリジンの酵素的除去から生じる電荷のバリエーションは、特異的なカルボキシペプチダーゼインヒビターの使用、または全体的パターンを単純化するために抗体をカルボキシペプチダーゼで処理することにより、解決され得る(Perkins, M. et al. 2000. Pharm Res. 17,1110-7)。グリコシル化パターンの相違から生じる大きさのバリエーションは、例えば、PNGase F、Endo H、O−グリコシダーゼ、またはノイラミニダーゼを用いた酵素による脱グリコシル化によっても処理され得る。
【0074】
アミド分解のようなタンパク質の化学的分解は、産生および保存中の重大な問題であり、電荷の不均一性を生じることが示されてきた。AsnからAspへのアミド分解、およびisoAsp(イソアスパルチルペプチド結合)の形成は、穏和な条件下で生じる(Aswad, D.W. et al. 2000. J Pharm Biomed Anal. 21, 1129-36)。これらの転位は、部分的なポリペプチド鎖の柔軟性が高いAsn−Gly、Asn−Ser、およびAsp−Gly配列にて最も容易に生じる。
【0075】
電荷の不均一性の別の要因は、N−末端グルタミン残基(アミド分解)の環化から生じるピログルタミン酸(PyroGlu)によるN−末端の封鎖から生じ得る。かかる翻訳後修飾は、IgGならびに他のタンパク質について述べられてきた。特に、HCおよびLCの両方が含まれると、抗体のN−末端の部分的環化は電荷の不均一性を生じ、これにより、複雑なIEXパターンが与えられるだろう。抗体の1個以上のVH鎖およびVL鎖の上でのN−末端PyroGluの形成に起因する可能性のあるIEXパターンが、図16に示される。組換えポリクローナル抗体のような、異なる可変領域を有する異なる相同タンパク質を含む試料が、正味電荷に基づく技術により特徴決定されるべきでものであるなら、たとえ2、3種の試料成分が図16BおよびCに示されるIEXパターンを有するとしても、これは、例えば、ポリクローナル抗体組成物のIEXパターンにおけるクローンの多様性を隠すであろうから、かかる分析が複雑になることは明らかである。この問題は、特定の酵素であるピログルタミン酸アミノペプチダーゼの使用によっては解決されない。なぜなら、まず第一に、脱閉鎖抗体を高収率で得るために、脱閉鎖が還元またはアルキル化抗体において行われなければならないからであり(Mozdzanowski, J. et al. 1998Anal Biochem. 260,183-7)、そして第二に、全ての抗体について100%の開裂を得ることは可能でないだろうからである。
【0076】
それゆえ、本発明のさらなる態様は、N−末端グルタミン残基の環化により引き起こされる電荷の不均一性の排除に関するものである。本発明のこの態様は、物理化学的特性である正味電荷に基づく上述の特徴決定ツール、例えば、IEXクロマトグラフィーおよびクロマトフォーカシングのいずれかと組み合わされると、特に有用である。N−末端のPyroGlu残基の形成は、例えば、N−末端グルタミン残基を別のアミノ酸に変えることにより、ポリペプチド鎖の全てがN−末端グルタミンを含有しないことを確実にすることにより、排除される。タンパク質が異なるサブユニットを含むヘテロマーのタンパク質であるなら、好ましくは、N−末端Gln残基の全てが他の残基と交換される。抗体については、重鎖および/または軽鎖のN−末端のGln残基が変換される。これは、N−末端にグルタミンを有するポリペプチドをコードする核酸配列の部位指定の変異誘発により行われる。好ましくは、N−末端グルタミン残基は、グルタミン酸残基によって置き換えられる。なぜなら、これはグルタミンの非荷電誘導体だからである。組換えポリクローナルタンパク質において、メンバーをコードする個々の配列は、荷電タンパク質を発現する新規細胞株を創出するために、変更され、発現ベクターに再挿入されなければならない。次に、この細胞株は、ポリクローナルタンパク質を産生する細胞の収集物内に含まれ得る。
【0077】
相同タンパク質の可変領域のタンパク分解性消化産物の分析
異なる可変領域を有する異なる相同タンパク質を含むタンパク質試料は、上述の通り、広範なクロマトグラフ技術を用いて、インタクトなタンパク質の物理化学的特性に基づき特徴付けられ得る。上述のこれらの分析から得られた情報は、相同タンパク質のタンパク分解性消化産物の分析から得られた情報でさらに補足され得る。好ましくは、タンパク分解性消化は、インタクトなタンパク質のクロマトグラフ分析が行われるのと同じ試料のアリコート上で行われる。
【0078】
本発明のさらなる実施態様において、相同タンパク質の組成物の個々のメンバーの可変領域を起源とする固有のマーカーペプチドの同定を用いて、タンパク質組成物が個々のメンバーの存在について定性的方法で特徴付けられる。これらの固有のマーカーペプチドは、異なる相同タンパク質を含むタンパク質組成物(試料)のタンパク分解性消化により生成される。
【0079】
相同タンパク質の混合物の可変領域のペプチドマッピングを行うために、混合物中の個々のメンバーを他のメンバーと区別する可変領域の一部または複数の部分が、タンパク分解性消化後もインタクトなままであることが重要である。ゆえに、少なくとも1種類の固有の配列(マーカーペプチドとも称される)が組換えポリクローナルタンパク質の個々のメンバーそれぞれについて得られ得るように、1種類以上のプロテアーゼが選択されるだろう。相同タンパク質の混合物が、組換えポリクローナル抗体または組換えポリクローナルTcRである場合、通常、個々のメンバーを互いに区別する配列が、CDRにより特徴付けられる。固有のマーカーペプチドを生成するために用いられるプロテアーゼ、複数のプロテアーゼ、または化合物の選択は、相同タンパク質試料を構成するタンパク質配列の分析に基づくものである。一般に、プロテアーゼまたは化合物は、高い特異性でもってタンパク質の所定の部位で開裂させるだろう。かかる開裂特異的なプロテアーゼは、当該技術分野においてよく知られており、例えば、トリプシン、エンドGlu−C、リジルエンドペプチダーゼ、エンドArg−C、エンドAsp−N、またはエンドAsn−Cであり得る。これらはただの例であり、本実施態様に対する限定と考えられるべきではない。
【0080】
ポリクローナルタンパク質試料は、1種類以上の選択されたプロテアーゼで消化される場合、全ての個々のメンバーに由来する定常領域および可変領域の両方を起源とするペプチドのプールが生成されるだろう。固有のマーカーペプチドの比率は、定常領域を起源とするペプチドの主な集団と比較して、それらの物理化学的特性の相違を示すだろう。それゆえ、固有のマーカーペプチドは、上述のクロマトグラフ技術のうち1種類を用いて、単離され得る。好ましくは、ペプチド分離のために特別に設計されたイオン交換クロマトグラフィーまたはRP−HPLCを用いて、固有のペプチドが定常領域ペプチドの主要フラクションと区別される。同様に、上述の多次元クロマトグラフ技術を適用して、固有のマーカーペプチドが分離され得る。1以上の次元の分離に続き、質量分析(MS)が、異なるペプチドの同定のために用いられ得る。プロテオミクス分野の技術者によく知られたMS技術が、ペプチドの同定のために用いられ得る。好ましいMS技術は、マトリックス支援レーザー脱離イオン化法(MALDI)、飛行時間(TOF)質量分析法、およびエレクトロスプレーイオン化飛行時間(ESI−TOF)質量分析法である。
【0081】
別法として、タンパク分解性消化は、上のa)からd)および「多次元液体クロマトグラフィー」のセクションに記載の1次元または多次元クロマトグラフィーにより分離されたインタクトなタンパク質において行われ、次に、分離されたタンパク質フラクションがタンパク分解性開裂され得る。次に、これらの消化産物がMSにより分析され得る(Kachman, M.T. et al. 2002. Anal.Chem. 74, 1779-1791)。このアプローチは、これらをさらに特徴付けるためにインタクトなタンパク質の1次元または多次元分析により得られたフラクションの比率に選択的に適用され得るので、非常に複雑なポリクローナルタンパク質の特徴決定の有利であり得る。
【0082】
タンパク分解性消化は、N−末端マーカーペプチドが固有の可変領域を含有するなら、これらを単離するためにさらに行われ得る。N−末端ペプチドは、本質的にはGevaert, K. et al., 2003. Nat.Biotechnol. 21, 566-569(引用により本明細書に取り込まれる)に記載の通り、単離され得る。簡単にいうと、組換えポリクローナルタンパク質の遊離アミノ基は、例えば、アセチル化によりブロックされ、次に、タンパク質混合物が適当なプロテアーゼを用いて消化される。消化により、内部ペプチド上に遊離N−末端アミノ基が創出され、続いて、これが化合物でブロックされ、これにより、内部ペプチドをN−末端ペプチドと分離することが可能になる。かかる化合物は、i)疎水性相互作用による単離を可能にする、Gevaertにより記載されている2,4,6−トリニトロベンゼンスルホン酸(TNBS)、ii)ビオチン(これは、固定化ストレプトアビジンに結合後、除去される)、またはiii)予め活性化された物質(例えば、NHS−活性化、CNBr−活性化、ECH セファロース物質、またはアズラクトン基を有するUltraLink ビス−アクリルアミド支持体)であり得、続いて、遠心により、結合した内部ペプチドが、アセチル化N−末端ペプチドから分離される(それは上清に存在する)。次に、単離アセチル化N−末端ペプチドが、上述のMS分析と組み合わされた1次元または多次元液体クロマトグラフィーにより分析され得る。別法として、インタクトなタンパク質のN−末端が、特異的な分離を可能にする化合物でブロックされ、開裂後に生成された内部ペプチドが、第2の化合物でアセチル化またはブロックされる。
【0083】
加えて、タンパク分解性消化後の固有マーカーペプチドの同定が、特徴的なアミノ酸側鎖の機能性を用いて行われ得る。関連するアミノ酸側鎖修飾を有する特異的なアミノ酸残基を含有するペプチド、およびそれを有さない特異的なアミノ酸残基を含有するペプチドを捕獲する、異なるアフィニティー技術の1つまたは組合せが用いられ得る。例えば、システイン、メチオニン、トリプトファン、ヒスチジン、およびチロシンを含有するペプチドは、これらのアミノ酸残基を含有するペプチドを捕獲する特異的なアフィニティータグで固定されたカラム材料またはビーズを用いて精製され得る(Bernhard, O.K. et al. 2003. Proteomics 3, 139-146;Chelius, D. and Shaler, T.A., 2003. Bioconjug. Chem. 14, 205-211;Gevaert, K. et al. 2002. Mol.Cell Proteomics 1, 896-903;Gygi, S.P. et al., 1999. Nat.Biotechnol 17, 994-999)。システインおよびチロシンを含有する固有の可変領域ペプチドは、例えば、システイン残基をビオチン化した後、ストレプトアビジンカラム上で捕獲され、続いて、ペプチドを含有するシステインが溶出され、これらが、チロシン残基と特異的に結合するカラムまたはビーズのいずれかにアプライされ得る。特異的なアミノ酸残基に対するアフィニティーに基づくこのペプチドの捕獲は、RP−HPLCおよびイオン交換クロマトグラフィーのような上述のクロマトグラフ技術に対するさらなる次元として行われ得る。まず、クロマトグラフ技術が、1以上の次元の組換えポリクローナルタンパク質のタンパク分解性消化に適用され、次に、アミノ酸に特異的な捕獲が、最終次元の1個以上のフラクションにおいて行われ、続いて、MSによる分析される。側鎖の機能性に基づくペプチドの単離は、さらに、N−末端ペプチド単離技術と組合せて行われ得る。
【0084】
タンパク分解性消化、続いてペプチドの単離により分析されるべき組換えポリクローナルタンパク質が、多量体タンパク質である場合、好ましくは、サブユニットの分離が、タンパク分解性消化産物の「フィンガープリンティング」を単純化するために行われ、その後、タンパク分解される。これは、例えば、遊離システイン残基の還元およびアルキル化、次に、ゲルろ過により行われ、サブユニットが分離(例えば、ポリクローナルタンパク質が抗体またはTcRである場合、それぞれ、重鎖の軽鎖との分離、またはα鎖のβ鎖との分離)され得る。別法として、タンパク分解性消化が天然条件下で行われ得る。特に、抗体にとってこれは適当な代替であり得る。抗体の4要素からなる構造により、定常領域内でタンパク分解性開裂に対して高度に耐性となるからである。従って、インタクトな非還元ポリクローナル抗体のタンパク分解性開裂は、主として可変領域からペプチドを生成するようである。
【0085】
上述のタンパク分解性消化技術はまた、センチネルの概念に従い、タンパク分解性消化物において特徴付けられ得るセンチネルペプチドを選択することにより、応用され得る。
【0086】
「大量」N−末端配列決定
上述のN−末端配列は、単離され、ポリクローナルタンパク質のタンパク分解性消化産物をフィンガープリンティングするために用いられ得る。別法として、N−末端配列が、インタクトなタンパク質から直接配列決定され、それにより、タンパク分解工程が省かれ得る。異なる可変領域を有する異なる相同タンパク質を含むタンパク質試料の「大量」N−末端配列分析を用いて、例えば、組換えポリクローナルタンパク質の精製バッチ産物が比較され得る。ポリクローナルタンパク質が組換えポリクローナル抗体またはTcRである場合、好ましくは、「大量」N−末端配列決定は、それぞれ、分離された重鎖および軽鎖、または分離されたα鎖およびβ鎖のプールにおいて行われる。例えば、相同な重鎖のプールにおいて、アミノ酸位置のいくつかは完全に保存され得るのに対し、他の位置は変動し得、これはアミノ酸配列のアライメントにより評価され得る。従って、いくつかの異なるアミノ酸が、特定ラウンドの配列決定において得られ得る。例えば、4位が、相同配列のアライメントにより予め決定された、ポリクローナル試料中の5種類の異なるアミノ酸により表され得る。「大量」N−末端配列分析において、アミノ酸が変動するこれらのものが定量され、例えば、組換えポリクローナル抗体の4位を表す個々のアミノ酸の異なる量を用いて、関連する組成物である異なる試料が比較され得る。
【0087】
特異的検出分子を用いた複雑な相同タンパク質混合物の特徴決定
本発明の特徴決定基盤はさらに、特異的検出分子を利用し、それぞれの特異的検出分子は、相同タンパク質の複雑な混合物内の個々のタンパク質メンバーを同定する能力を有し、それにより、試料中の特定のメンバーの存在のモニタリングを助ける。特異的な検出分子は、例えば、ポリクローナルタンパク質の個々のメンバーに対する特異性を有する小有機分子、ペプチド、またはタンパク質のような特異的リガンドであり得る。特に、抗イディオタイプペプチドのようなリガンドペプチドまたはタンパク質、あるいは抗イディオタイプ抗体が、本発明の好ましい実施態様である。相同タンパク質の複雑な混合物の所定のサブセットに結合する検出分子も、本発明において適用可能である。
【0088】
i)相同タンパク質の複雑な混合物を含む試料中の1個以上の個々のタンパク質の濃度または相対的比率の決定を可能にすること、ii)クロマトグラフ分析のさらなる次元として働くこと、iii)相同タンパク質の複雑な混合物の発酵中に得られる試料における個々のタンパク質の濃度の決定を可能にすること、およびiv)相同タンパク質の混合物を発現するワーキングセルバンクまたはバイオリアクター細胞試料のようなポリクローナル細胞株における個々のタンパク質産生細胞の決定を可能にすることにより、特異的な検出分子を用いて、相同タンパク質の複雑な混合物が特徴付けられ得る。工程iv)は、ポリクローナル細胞株、またはポリクローナル細胞株由来の単一チューブ内に分布された単一細胞のいずれかにおいて行われ、その後、連続的培養が行われ得る。
【0089】
相同タンパク質の複雑な混合物中の個々のタンパク質メンバーを同定する能力を有する特異的ペプチド−リガンドを生成するため、ビリオン表面上に外来オリゴマーペプチドをディスプレイする発酵ファージ発現ベクターの巨大ライブラリーが、アフィニティーによりスクリーニングされ、次に、抗体、TcR、または別の所望の個々のタンパク質メンバーに結合する外来ペプチドをディスプレイするファージが精製され得る(Scott and Smith 1990. Science 249, 386-90)。EP 1 106 625には、特に、免疫目的のための抗RhD抗体に結合する能力を有するペプチドの生成が記載されている。ディスプレイされたペプチドライブラリーは、約5から50の間のアミノ酸長、好ましくは、7から20の間のアミノ酸長、なおより好ましくは、8から15の間のアミノ酸長、そして最も好ましくは、9から12の間のアミノ酸長である。関連ペプチドが同定されたなら、それらが合成され得る。
【0090】
抗イディオタイプ抗体の生成は当該技術分野において一般的に知られている。簡単に言うと、所望の抗イディオタイプ抗体に対する抗体を用いて、マウスが免疫される。モノクローナル抗体は免疫マウスから生成され、そしてそれは、例えば、ハイブリドーマ技術またはファージディスプレイを用いて所望の特異性を有する抗イディオタイプ抗体の産生についてスクリーニングされる。抗イディオタイプペプチドまたは抗イディオタイプ抗体は、特異性および可能性のある交差反応性に関して特徴付けられるだろう。この分析により、抗イディオタイプペプチドまたは抗イディオタイプ抗体が、特異的メンバーを認識するか、あるいは別法として、ポリクローナルタンパク質中の密接に関連するメンバーのサブセット(抗体について、関連するメンバーは、例えば、特異的VH遺伝子ファミリーであり得る)を認識するかどうかが証明されるだろう。
【0091】
抗イディオタイプペプチド/抗体は、個々のメンバータンパク質(例えば、特異的抗体または特異的TcR)の直接的定量化のためのELISA、FLISA、またはRIAのような免疫検出アッセイにおいて適用され得る。別法として、抗イディオタイプペプチド/抗体は、単独、あるいは1次元または上述の他のクロマトグラフ分離後のさらなる次元として、アフィニティークロマトグラフィーにおいて適用され得る。免疫沈降は、検出分子を用いて、ポリクローナルタンパク質の個々のメンバーが分離され、特徴付けられ得る、さらなる手段である。さらに、抗イディオタイプペプチドまたは抗イディオタイプ抗体は、ポリクローナル細胞株における個々のタンパク質産生細胞の単離および/または決定のために用いられ得る。該技術は、Borth, N. et al. 2000-2001に記載されている。Biotechnol Bioeng. 71, 266-273およびBrezinsky, S.C. et al. 2003. J. Immunol. Methods 277, 141-155の両方が、個々のタンパク質産生細胞の細胞培養物からの単離に適用可能である。
【0092】
可能性として、完全な特徴決定を得るために、ポリクローナルタンパク質におけるそれぞれの個々のメンバーおよび全ての個々のメンバーに特異的な検出分子を生成することが可能である。しかしながら、発現の安定性、またはバッチとバッチとの間の一貫性をモニターするためには、それは、本発明をふまえて、組換えポリクローナルタンパク質内の多数の個々のメンバー、いわゆるセンチネルタンパク質を同定するのに十分であり、そしてその同定は、個々のタンパク質メンバーのこの収集物が、一貫して発現され、組換え産生ポリクローナルタンパク質の異なるバッチにおいて精製されることを確かにする定量的および/または定性的特徴決定についてのものである。特に、このアプローチを用いて、タンパク質分子の複雑なプールの特徴決定が単純化され得る。組換えポリクローナルタンパク質の代表としてのセンチネルタンパク質のコンセプトは、特異的検出分子に適用されるばかりでなく、事実上、上述の特徴決定技術のいずれかまたはこれらの組合せが、センチネルタンパク質またはペプチドのコンセプトを適用し得る。さらに、センチネルタンパク質は、技術から技術で変わり得る。ポリクローナルタンパク質の特異的メンバーのうち、それらの物理化学的特性の相違に基づき、特に十分に分離され得るものもあるが、高アフィニティーを有する抗イディオタイプペプチドは、同一の物理化学的特性を有するタンパク質の分離に特に有用である。
【0093】
本発明の実施態様において、1種類以上の特異的検出分子を用いて、異なる可変領域を有する異なる相同タンパク質を含む試料中の1種類以上のセンチネルタンパク質の相対的比率がモニターされる。一連の関連試料中の1種類以上のセンチネルタンパク質の比率の一貫性は、バッチ間、ならびに1回の生産工程での長時間にわたるポリクローナルタンパク質の発現における組成物の安定性を反映するだろう。さらにそれは、組換えポリクローナルタンパク質またはモノクローナルタンパク質の混合物の長期保存中の組成物の安定性を評価することができる。
【0094】
本発明の好ましい実施態様において、組換えポリクローナルタンパク質のセンチネルタンパク質は、次の技術、i)抗イディオタイプペプチド/抗体アフィニティークロマトグラフィー、ii)抗イディオタイプペプチド/抗体での免疫的検出、iii)それらの特徴的物理化学的特性に関するインタクトなメンバーの多次元クロマトグラフ単離、およびiv)クロマトグラフィーおよびMSを用いたタンパク分解性ペプチドマッピングのうちの1個以上により特徴決定される。
【0095】
特徴付けられるべき異なる相同タンパク質混合物の複雑性
本発明の基盤により特徴付けられるべき試料は、異なる可変領域タンパク質を有する異なる相同タンパク質の所定のサブセット、例えば、異なるCDR領域を有するポリクローナルタンパク質または抗体(例えば、ポリクローナル抗体またはモノクローナル抗体の混合物)、または異なるCDR領域を有するT細胞受容体(例えば、ポリクローナルTcRまたはモノクローナルTcRの混合物)を含む。好ましくは、異なる可変領域を有する異なる相同タンパク質は組換えタンパク質である。加えて、好ましくは、ポリクローナルタンパク質またはモノクローナルタンパク質の混合物の個々のメンバーは、例えば、所望の標的抗原に対する抗体またはTcRの場合、所望の標的に対する共有結合活性のような共通する特徴により定義されてきた。典型的には、本発明の特徴決定基盤により分析されるべきポリクローナルタンパク質組成物は、少なくとも3、4、5、10、20、50、100、1000、104、105、または106の別の変異体メンバーを含む。通常、ポリクローナルタンパク質組成物中の個々のメンバーの総数の75%より多くを構成する単一の変異体メンバーは存在しない。好ましくは、最終ポリクローナル組成物中の個々のメンバーの総数の50%、より好ましくは、25%、最も好ましくは、10%を超える個々のメンバーは存在しない。
【0096】
抗体の場合、標的とされる抗原の複雑さは、本発明の基盤を利用して特徴付けられるべきポリクローナルタンパク質組成物中の別の変異体メンバーの数に影響するだろう。小さな標的または全く複雑でない標的、例えば、小さな標的タンパク質を用いて、3から100種類の間の別の変異体メンバーを含むポリクローナルタンパク質組成物が、特徴決定のために確立されるだろう。そして、好ましくは、変異体の数は、90、または80もしくは70を超えない。多くの場合、異なる変異体の数は、60または50を超えず、好ましくは、変異体の数は、5と40の間、例えば、5と30の間の範囲にある。一方、より複雑な標的、例えば、複雑な表面タンパク質または互換可能な表面タンパク質を有するか、あるいはいくつかのウイルスサブタイプを包含するウイルスについては、20から500種類の間の異なる変異体メンバーを含むポリクローナルタンパク質組成物が、特徴決定のために確立されるだろう。抗原が多数の異なる分子を含む非常に複雑な標的については、50から10,000種類の間の異なる変異体メンバーを含むポリクローナルタンパク質組成物が本発明により特徴付けられるのに必要であり得る。
【0097】
本発明の1つの実施態様において、異なる可変領域を有する異なる相同タンパク質を含む試料は、ポリクローナル抗体である。ポリクローナル抗体は、ヒトアイソタイプIgG1、IgG2、IgG3、IgG4、IgA1、およびIgA2、またはマウスアイソタイプIgG1、IgG2a、IgG2b、IgG3、およびIgAのような1種類以上の異なる抗体アイソタイプからなり得る。
【0098】
本発明の1つの実施態様において、異なる可変領域を有する異なる相同タンパク質を含む試料はポリクローナルTcRである。
【0099】
実施例
以下の実施例において、異なる個々の抗RhD抗体メンバー、または抗RhD rpAbを産生する細胞株からなる種々の抗RhD組換えポリクローナル抗体(抗RhD rpAb)組成物を用いて、本発明の構造上の特徴決定基盤を説明する。個々の抗RhD特異的抗体およびそれらを産生する細胞株は、2004年7月20日に出願した譲受人のデンマーク特許出願PA 2004 01133に記載に対応するものであった。簡単にいうと、重鎖可変領域およびκ/λ軽鎖のコンビナトリアルファージディスプレイライブラリーを、RhD陽性赤血球で免疫したrhesus D陰性ドナーから調製した。ライブラリーを、抗RhD特異的抗体産生クローンについてピックアップした。抗原特異的ファージ由来の可変重鎖および軽鎖遺伝子対を哺乳類発現ベクターに導入した。哺乳類ベクターを、Flp−FRT組換え系を用いた部位特異的方法でCHO Flp−In細胞株(Invitrogen, CA)に個々にトランスフェクションした。完全な軽鎖(LC)ならびに重鎖(VH)の可変領域の核酸(nuc.)ならびにタンパク質(a.a.)配列を、表2に示す配列同一性ナンバー(配列番号)により同定する。これらのナンバーは、2005年7月18日に出願した譲受人の国際特許出願PCT/DK2005/000501(発明の名称「抗RHESUS D組換えポリクローナル抗体、および製造方法」)における配列番号に対応する。表2の配列番号を本発明の出願の配列番号と区別しなければならない。これらは同一でないからである。重鎖の定常領域は、ヒトIgG1に対応する。
【0100】
表2 以下の実施例において用いられる個々の抗RhD抗体/細胞クローンのリスト
【表3】
【0101】
実施例1
本実施例は、ポリクローナル製造細胞株の生成、および1次元のクロマトグラフ技術を用いたタンパク質レベル、およびRFLP分析を用いた遺伝子レベルでバッチとバッチとの間のバリエーションの特徴決定を説明するものである。
【0102】
抗Rhesus D組換えポリクローナル抗体産生のための製造細胞株の樹立
ゲノム上の特定の部位と異なる組換え抗Rhesus D抗体をそれぞれ発現する10種類の細胞株(RhD157.119D11、RhD158.119B06、RhD159.119B09、RhD161.119E09、RhD163.119A02、RhD190.119F05、RhD191.119E08、RhD192.119G06、RhD197.127A08、およびRhD204.128A03)を選択し、混合して、組換えポリクローナル製造細胞株を構築した。RhD197およびRhD204はλクローンであり、残りはκクローンであった。
【0103】
個々の抗Rhesus抗体を発現する細胞培養物を、シェーカーフラスコでの無血清懸濁培養(等しい細胞数で混合する)に完全に適合させ、それにより、ポリクローナルCHO−Flp−In(019)細胞株を生成した。混合した細胞培養物を遠心し、アリコート10×106細胞/チューブにて凍結した。
【0104】
2本のチューブ(3948 FCW065および3949 FCW065)を解凍し、ネオマイシンを含むExcell 302無血清培地100mlを含有する1000mlのシェーカーフラスコにて11週間個々に培養した。
上清を回収し、ろ過後、抗RhD rpAbを精製した。
【0105】
クローン多様性
タンパク質レベルならびにmRNAレベルの両方でクローン多様性をアッセイした。9週間培養後に抗体組成物の分析のために用いる上清試料を採取し、11週間培養後にmRNA組成物を分析するために用いる細胞試料を採取した。
【0106】
抗体組成物:
ポリクローナルCHO−Flp−In(019)細胞株から発現した抗RhD rpAbは、IgG1アイソタイプ抗体である。タンパク質Aを固定化したカラムを用いて、両方のアリコート(3948および3949)から抗RhD rpAbを精製した。個々の抗体は、固定化したタンパク質AとpH7.4で相互作用した。一方、コンタミしているタンパク質をカラムから洗い流した。次に、結合抗体を低pH値(pH2.7)にてカラムから溶出した。280nmでの吸光度測定により決定した抗体含有フラクションをプールし、5mM 酢酸ナトリウム(pH5)に対して透析した。
【0107】
9週間培養後にアリコート3948および3949(FCW065)から得られた抗RhD rpAb組成物を、陽イオン交換クロマトグラフィーを用いて分析した。タンパク質Aにより精製した抗RhD rpAbを、PolyCatAカラム(4.6×100mm)に25mM 酢酸ナトリウム、150mM 塩化ナトリウム、pH5.0中、流速60ml/時間、室温にてアプライした。次に、25mM 酢酸ナトリウム中の150〜350mMの直線的勾配の塩化ナトリウム、pH5.0を用いて、流速60ml/時間にて抗体成分を溶出した。分光光度法により280nmにて抗体成分を検出した。次に、クロマトグラム(図1)を合計し、個々のピークA−Jの面積を用いて、抗体成分を定量した(表3)。ピークの全面積を100%と設定した。2つのアリコートのクロマトグラムは、同一のピーク分布、ならびに各ピークにおける類似の成分濃度を示した。これらの結果から、同一条件下で成長させた同じポリクローナル細胞株のアリコートは、個々の抗体メンバーからなる類似の分布を有する抗RhD rpAbを産生すると結論付けられる。
【0108】
抗RhD rpAbの個々のメンバーを、1種類以上の特定のピークに割り当てた(表3にて概説)。この割り当ては、同一条件下で分析した個々の抗体について得られたクロマトグラムに基づくものである。RhD158Abについて、個々のクロマトグラムを得なかったので、このクローンをピークのいずれにも割り当てなかった。しかしながら、ピークDはRhD158を構成し、この抗体は不均一性に起因して他のピークのいくつかにも現れ得るように思われる。特に、クローンRhD197由来の抗体産物は、IEXプロファイルにおいて高程度の不均一性を示す。RhD190 Abは、保持時間15.3分にて視認できなければならい。しかしながら、それは検出可能ではなかった。このことは、このクローンが失われたか、あるいは組換えポリクローナル製造細胞株における検出限界より下の量で産生されたことを示している。クローンRhD190の喪失は、多様性の10%の低減に該当し、それは、最終抗RhD rpAB組成物の多様性に関して受容されると考えられる。
【0109】
表3
【表4】
【0110】
mRNA組成物:
11週間培養後のポリクローナルCHO−Flp−In(019)細胞株内のクローン多様性をRT−PCR−RFLP分析により評価した。簡単にいうと、200個の細胞に対応する細胞懸濁液を凍結融解の対象とし、これらの溶解物を、軽鎖を増幅するプライマーを含むOne−STEP RT−PCRキット(Qiagen)を用いたRT−PCRにおける鋳型として用いた。プライマー配列は次の通りであった:
フォワードプライマー:5’−TCTCTTCCGCATCGCTGTCT(配列番号1)
リバースプライマー:5’−AGGAAAGGACAGTGGGAGTGGCAC(配列番号2)。
【0111】
RT−PCR産物をHinfIで消化し、制限酵素消化産物をアガロースゲル電気泳動により分析した(エチジウムブロマイド染色を用いて視覚化)(図2)。
【0112】
それぞれの個々のクローンについて、RT−PCRにより増幅した軽鎖のHinfI消化により得られた制限酵素フラグメントの予測サイズを表4に示す。ポリクローナル抗Rhesus D抗体コード遺伝子の個々のメンバーに割り当てられ得るゲル上の6種類の固有のフラグメントサイズを太字で示す。全ての固有のフラグメントをゲル上で同定し得るとは限らず、これらをイタリック体で示す。しかしながら、これは、これらのクローンが培養物中に実際に現れることを必ずしも排除しない。なぜなら、フラグメントが同定可能なように他のフラグメントと十分に分離されていないか、あるいは濃度がより強く出現したバンドと比較してあまりに弱かったからである。このことは、短いフラグメントについてほど断言され得る。なぜなら、それらはより少量のエチジウムブロマイド分子と結合し、それゆえ、ほとんど視認できないからである。
【0113】
表4
【表5】
【0114】
同じポリクローナル細胞株の2つのアリコート(3948および3949)は、ゲルにおいて類似の発現パターンを示したが、バンドの強度は完全に同一ではなかった。これは、同一条件下で成長させた同じポリクローナル細胞株のアリコートは、類似のクローン多様性を有する抗RhD rpAbを産生することを示す。
【0115】
要約
それぞれモノクローナル抗RhD抗体を発現する10種類の細胞株を混合して、抗RhD rpAb製造細胞株を生成した。9週間培養後、それはなお、最初の多様性の90%を維持していた。11週間培養後、6種類の異なるクローン由来のmRNAを明確に同定することができ、そして他のいくつかのクローンは、約500bpのバンド中に現われているようであった。
【0116】
ポリクローナルCHO−Flp−In(019)細胞株の2つのアリコートがクローン多様性に関して類似の結果を示したという事実は、再現可能な結果が異なるバッチ間で得られ得ることを説明するものである。
【0117】
実施例2
本実施例は、8種類のメンバーを有するポリクローナル細胞培養物の長期にわたる特徴決定を説明するものである。RFLP分析を用いて遺伝子レベルで、および1次元クロマトグラフ技術を用いてタンパク質レベルで培養物のクローン多様性を評価した。
【0118】
ポリクローナル細胞培養物におけるクローン多様性を評価するためのRFLP分析
8種の異なる抗Rhesus D抗体を発現するポリクローナル細胞培養物における個々のクローンの分布を、ポリクローナル細胞株由来のRT−PCR産物の末端RFLP(T−RFLP)により評価した。T−RFLP方法において、フォワードおよび/またはリバースプライマーを蛍光標識した。それゆえ、単位複製配列から生じた制限酵素消化フラグメントの部分は標識を含有するだろう。次に、標識フラグメントを、キャピラリー電気泳動により分離させ、蛍光により検出することができる。適用したプライマーに依存して、軽鎖および重鎖の可変領域をコードする配列両方において分析を行うことができる。
【0119】
簡単にいうと、200個の細胞に該当する細胞懸濁液をPBSにて1回洗浄し、凍結融解手段の対象として、溶解物を生成し、これをOne−Step RT−PCRキット(Qiagen)および適当なプライマーを用いたRT−PCR増幅における鋳型として用いた。標準的サーマルサイクラーにて次の条件を用いてRT−PCRを行った。
【表6】
【0120】
軽鎖の分析のため、次のプライマーを用いてRT−PCR増幅した。リバースプライマーを6−カルボキシフルオレッセイン(FAM)標識し、プライマー配列は次の通りであった:
VLフォワードプライマー:5’−TCTCTTCCGCATCGCTGTCT(配列番号1)
CLリバースプライマー:5’−FAM−AGGAAAGGACAGTGGGAGTGGCAC(配列番号2)。
【0121】
RT−PCR産物20μlを、NheI 1U、PstI 1U、およびHinfI 1U(全てNew England Biolabs)を用いてNEB1中にて2時間消化した。
【0122】
Statens Serum Institute, Copenhagen, DKのABI3700(Applied Biosystems)上での蛍光キャピラリー電気泳動により、標識フラグメントを検出した。
【0123】
抗RhD抗体産生細胞クローンそれぞれについての予測フラグメントを表5に示し、そしてFAM標識フラグメントを太字で示す。
【0124】
表5
【表7】
【0125】
T−RFLPパターンを図3に示し、8種類の抗Rhesus D抗体産生クローン全てを特定のピークに割り当てた。RT−PCR中に鋳型/プライマーの競合がないという想定下で、相対的なピーク面積は、ポリクローナル細胞株に表されるそれぞれの抗体の軽鎖遺伝子から転写されたmRNAの相対量に該当する。
【0126】
同じポリクローナル細胞株内の重鎖可変領域の分析のため、VH特異的プライマーを用いてRT−PCR増幅を行った。プライマー配列は次の通りであった:
VHフォワードプライマー:5’−FAM CGTAGCTCTTTTAAGAGGTG(配列番号3)
VHリバースプライマー:5’−HEX−ACCGATGGGCCCTTGGTGGA(配列番号4)。
【0127】
RT−PCR産物20μlを、RsaI 1U、およびNdeI 1U(全てNew England Biolabs)を用いて、NEB2にて、2時間消化した。
【0128】
ABI3700上の蛍光キャピラリー電気泳動により、標識フラグメントを検出した。Statens Serum Institute, Copenhagen, DKにより分析を行った。
【0129】
予測T−RFLPパターンを表6に示す。表中、FAM標識フラグメントを太字で示し、そしてHEX(6−カルボキシ−2’,4,4’,5,7,7’−ヘキサクロロフルオレッセインスクシンイミジルエステル)標識フラグメントに下線を付す。
【0130】
表6
【表8】
【0131】
ポリクローナル細胞株を5週間培養し、1週間に1回T−RFLP分析のために試料を採取した。可変重鎖において分析を行ったが、必要に応じて軽鎖についても行うことができた。
【0132】
制限酵素消化フラグメントをキャピラリー電気泳動した後、相対的ピーク面積を合計し、これを用いて、ポリクローナル細胞培養物のクローン多様性を評価した。経時的な相対量を図5に示す。
【0133】
これらの結果から、時間と共にRhD196は増大するのに対して、RhD203は低減するようである。他のクローンの量は培養期間中非常に安定し、5週間培養後、8種類のcDNA全てを検出することができた。
【0134】
軽鎖および重鎖の両方、ならびにmRNAおよびDNAの両方においてT−RFLPを行うことにより、例えば、インビトロの細胞年齢の限界にある細胞、または培養中の任意の所定の時間ポイントの細胞における、ポリクローナル細胞培養物内のクローン多様性の正確なフィンガープリントを得ることが可能となる。
【0135】
それゆえ、この技術を用いて、長時間にわたり細胞培養物におけるクローン多様性の安定性を抗体産生中にモニターすることができる。この技術を応用して、例えば、同じポリクローナルワーキングセルバンク(pWCB)から凍結された異なるアンプル、あるいは2回以上製造工程を行った後回収された細胞における、バッチとバッチとの間の一貫性をモニターすることもできる。
【0136】
ポリクローナル細胞培養物におけるクローン多様性を評価するための陽イオンクロマトグラフィー分析
上述のT−RFLP分析において用いたのと同じポリクローナル細胞培養物から産生された抗RhD rpAbを、陽イオン交換クロマトグラフィーを用いて分析した。タンパク質Aにより精製した組換え産生ポリクローナル抗体を、25mM 酢酸ナトリウム、150mM 塩化ナトリウム,pH5.0中、流速60ml/時間、室温にてPolyCatAカラム(4.6×100mm)にアプライした。次に、25mM 酢酸ナトリウム、pH5.0中の150〜350mMの直線的勾配の塩化ナトリウムを用いて、流速60ml/時間にて抗体成分を溶出した。抗体成分を分光光度法により280nmにて検出し、次に、クロマトグラムを統合し、個々のピーク面積を用いて、抗体成分を定量した。経時的な相対量を図6に示す。
【0137】
要約
RFLP分析により遺伝子レベルで得られた結果と陽イオン交換クロマトグラフィーによりタンパク質レベルで得られた結果は比較可能である。図5および6は、ポリクローナル細胞株における個々のクローンの大部分、ならびに細胞株から発現されたポリクローナル抗体の個々の抗体が、5週間培養中、同じ傾向をたどることを明瞭に説明する。従って、遺伝子レベルならびにタンパク質レベルでの分析は、遺伝子レベルでの細胞株の組成の多様性、および細胞株から産生された組換えポリクローナルタンパク質の組成の多様性の評価について十分に同等なものである。
【0138】
実施例3
本実施例は、25種類のメンバーを有するポリクローナル細胞培養物の長期にわたる特徴決定を説明するものである。T−RFLP分析を用いて遺伝子レベルで、および1次元クロマトグラフ技術を用いてタンパク質レベルで、培養物のクローン多様性を評価した。
【0139】
培養期間5週間にわたる、25種類の異なる抗Rhesus D抗体発現ポリクローナル細胞培養物からもたらされた重鎖遺伝子の可変部分のT−RFLP分析
本実施例において調べたポリクローナル細胞培養物は、25種類の異なる抗Rhesus D抗体発現細胞培養物の混合物(実施例1に記載のとおり生成したもの)から構成されるものであった。ポリクローナル細胞培養物を5週間培養し、1週間に1回、T−RFLP分析のために試料を採取した。
【0140】
実施例2に記載のVH特異的プライマーを用いてRT−PCRを行い、同様に、制限酵素断片化を行った。
【0141】
全ての遺伝子型が存在するなら、25種類の異なる抗Rhesus Dコード配列のT−RFLPは、17種類の異なるFAM標識フラグメントを生じるだろう。フラグメントには、3種類までの異なる遺伝子型を表すものもあれば、単一の遺伝子型を表すものもあるだろう。FAM標識フラグメントの予測サイズを、経時的な異なるFAM標識フラグメントの相対量と共に表7に示す。さらに、T−RFLPプロファイルの1例を図4に示す。
【0142】
表7
【表9】
【0143】
細胞株を構成する25種類のクローンのうち12種類の個々のクローンについて情報を得ることを可能にする程度まで、消化フラグメントを分離させることが可能であった。残りのフラクションを配列決定の対象とし、残りのクローンについてさらなる情報を得られる可能性があった。
【0144】
25種類の異なる抗Rhesus D抗体発現ポリクローナル細胞培養物におけるクローン多様性を評価するための陽イオンクロマトグラフィー分析
上述のT−RFLP分析において用いたのと同じポリクローナル細胞培養物から産生された抗RhD rpAbを、陽イオン交換クロマトグラフィーを用いて分析した。タンパク質Aにより精製した組換え産生ポリクローナル抗体を、25mM 酢酸ナトリウム、150mM 塩化ナトリウム、pH5.0中、流速60ml/時間、室温にてPolyCatAカラム(4.6×100mm)にアプライした。次に、25mM 酢酸ナトリウム、pH5.0中の150〜350mMの直線的勾配の塩化ナトリウムを用いて、流速60ml/時間にて抗体成分を溶出した。280nmにて分光光度法により抗体成分を検出し、次に、クロマトグラムを統合して、個々のピーク面積を用いて、異なる抗体成分を定量した。図7は、4週目に得られた試料から生じたクロマトグラムを示し、抗体含有ピークに1から25までの番号を付けている。クロマトグラムが、分析したポリクローナル抗体における個々の抗体の数と同一数のピークを含有することは、正真正銘一致する。表8は、トータルの抗体成分(AC1から25)のパーセントにおける相対含有量、ならびに各抗体成分(ピーク)における個々の抗体の出現量を示す。合計したクロマトグラムのピークへの個々の抗体の割り当ては、陽イオン交換クロマトグラフィーを用いて同一条件下で分析したモノクローナル抗体から得られた保持時間およびピークパターンに基づくものであった。
【0145】
表8
【表10】
【0146】
陽イオン交換クロマトグラフィーは、個々のメンバー間の正味電荷の相違に基づき、ポリクローナル抗体から個々の抗体メンバーを分離させ、加えて、電荷の不均一性を明らかにする個々の抗体の形態を分離させる。それゆえ、いくつかの抗体は単一ピーク中に表された。例えば、AC1が含有するRhD293およびRhD319(表8参照)、ならびにいくつかの個々の抗体はさらに、いくつかのクロマトグラムピーク中に表される(例えば、AC1および5の両方に存在するRhD319(表8参照))。
【0147】
1種類より多くの個々の抗体を含有するピークを、抗イディオタイプペプチドを用いた定量的分析、タンパク分解性ペプチドマッピング、N−末端配列決定、または2次元クロマトグラフィーのようなさらなるタンパク質化学的特徴決定技術の対象とすることができた。
【0148】
要約
本実施例は、培養期間にわたる1次転写物の分布および抗体成分の分布をそれぞれ評価するためのT−RFLP分析と陽イオン交換クロマトグラフィーを組み合わせて使用することを説明するものである。T−RFLP分析により、ポリクローナル細胞株において発現した25種類のクローンのうち12種類の個々のクローンを固有に同定することが可能となり、そして本実施例において、4週間の培養中T−RFLP分析を用いて、これらの12種類のクローンを検出できることを説明する。可能性としては、1種類より多くのクローンを表すフラグメントの配列分析により、より多くのクローンを同定できる。陽イオン交換クロマトグラフィーを用いて、抗体成分の分布を分析し、そして本実施例において、25種類の分析した成分の分布が培養中比較的安定していることを見出した。個々の抗体全てを固有に同定することは、発現抗体の本来の電荷の不均一性質に起因して困難であるが、本実施例において、RhD160抗体を表す抗体成分8が培養中最も高い抗体レベルを示し、そしてこれはRhD160、293、および196クローンを表す群13について得られた高いT−RFLP値とも一致することを示した。さらに、T−RFLP、ならびに陽イオン交換クロマトグラフィーにより固有に同定することのできるRhD 207成分は、T−RFLPレベル10〜11%を示し、抗体レベルで得られたものは若干低い5.5〜10%を示した。とりわけ、2つの技術は一体となって、培養中のmRNAおよび抗体レベルでの比較的安定な産生を示す;しかしながら、2つの技術の間の可能性のある矛盾も見出され、これは、抗体レベルで得られた結果と対照的な、培養5週間でのいくつかのクローンの転写の明らかな喪失により説明される。従って、本実施例は、両方の技術を相補的に用いて、複雑なポリクローナルタンパク質の安定的産生を得られる培養間隔を定義することを十分に示すものである。
【0149】
実施例4
本実施例は、ポリクローナル細胞培養物からもたらされた10種類の個々のメンバーを有するポリクローナル抗RhD抗体の組成の分析を説明するものである。1次元陽イオン交換および2次元逆相(RP)−HPLCをそれぞれ用いて、正味電荷および疎水性における相違に基づき抗体を分離する2次元液体クロマトグラフィーを用いて、ポリクローナル抗体試料の多様性を評価した。
【0150】
10種類の個々のメンバーを有するポリクローナル抗RhD抗体試料は、ポリクローナル細胞培養物からもたらされたものであった。タンパク質Aカラム(HiTrap[登録商標]タンパク質Aカラム、Amersham Biosciences GE Healthcare, England)を用いて、上清から抗RhD rpAbを精製した。
【0151】
25mM 酢酸ナトリウム、150mM 塩化ナトリウム、pH5.0中、流速60ml/時間、室温、Ettan LCシステム(Amersham Biosystems, GE Healthcare, England)上にて、ProPac WCE10カラム(4×250mm)に精製ポリクローナル抗体をアプライすることにより、1次元を実行した。次に、25mM 酢酸ナトリウム、pH5.0中の150から350mMの直線的勾配のNaClを用いて、流速60ml/時間にて、抗体成分を溶出した。抗体成分を280nmにて分光光度法により検出し、特定のピークに対応するフラクションを収集し、さらに濃縮して、RP−HPLCにより分析した。
【0152】
図8に示すフラクションを、RP−HPLCを用いて2次元目においてさらに分離した。Summit HPLCシステム(Dionex, CA)にて、Zorbax Poroshell 300SB−C8カラム(2.1×75mm(5μm))を用いて2次元目を行った。Poroshellカラム(Agilent Technologies, CA)についての指示書において推奨された通り、HPLCシステムを設定した。陽イオン交換クロマトグラフィーから収集した抗体成分を、10% CH3CN、0.1% TFA、0.3% PEG中、流速120ml/時間にてカラム(5μl)にアプライし、次に、直線的勾配の90% CH3CN、0.08% TFA、0.3% PEGにより溶出した。カラムを70℃にて操作した。全ての抗体成分試料が、1または2個の細いピークを有するクロマトグラムとなった。抗体成分B5のRP−HPLCプロファイルを図9に示す。
【0153】
要約
1次元の陽イオン交換クロマトグラフィーにより、正味電荷の異なる個々の抗体、ならびに電荷不均一性を示す個々の抗体が分離されるので、いくつかの抗体は単一ピークに表され得る。
【0154】
いくつかの抗体成分を分離し、これにより、図8に示すようなやや複雑なプロファイルを得た。実施例2および3において説明した通り、同一条件下で分析したモノクローナル抗体を用いた比較分析により、それぞれのピーク中の個々の成分を同定することが可能である。しかしながら、本実施例においてこれを行わなかった。rpAb中の各モノクローナル抗体を割り当てなくても、互いに試料を比較するためのフィンガープリントを提供することが目的だったからである。従って、複雑なrpABの個々のメンバーを特徴付けるためにモノクローナル抗体を分析することを必要とすることなく、バッチとバッチとの間の一貫性を評価するために、陽イオン交換、次に、RP−HPLCの組合せにより、2つの次元からデータを創出し、図10に説明する詳細に色分けしたタンパク質マップ(ProteoVue software, Eprogen, USA)を構築することができる。
【0155】
実施例5
本実施例は、ポリクローナル細胞培養物からもたらされた8種類の個々のメンバーを有するポリクローナル抗RhD抗体の特徴決定を説明するものである。「大量」N−末端配列分析を用いて、ポリクローナル抗体の多様性を評価した。
【0156】
本実施例で分析したポリクローナル抗RhD抗体試料中に存在する個々のメンバーのN−末端配列を以下の表9に示す。λ軽鎖配列をイタリック体で示す。
【0157】
表9
【表11】
【0158】
タンパク質Aにより精製した抗RhD rpAbを、還元SDS−PAGE(NuPAGE 4〜12%)により分析した。ポリペプチドをPVDFメンブレンに電気的に移動させ、次に、製造元の指示に従いクーマシーブルーで染色した。
【0159】
重鎖(HC)に対応する約53kDaの1本のバンド、およびκおよびλ+κ軽鎖にそれぞれ対応する約25および30kDaの2本のバンドを、クーマシーブルー染色したPVDFメンブレン上にてはっきりと認めた。これらのバンドを切り出し、ABI Prociseタンパク質シークエンサー(Applied Biosystems, CA)および標準的プログラムを用いたN−末端配列分析の対象とした。配列決定の結果を以下の表10に概説する。
【0160】
表10
【表12】
ND=決定していない
【0161】
HCの配列は第1の残基以外一致し、一方κLCは残基2、5、6、および7について保存され、λLCは、残基1、5、および6について保存されている(表9参照)。
【0162】
HCの配列決定から得られた結果は、表10に示す予測配列と一致する。〜25kDaのκLC由来の配列データは、RhD191、324、201、および306に対応するN−末端配列EIVLTQS(配列番号7)を有する抗体、およびRhD244および196に対応するN−末端配列DIQMTQS(配列番号8)を有する抗体の存在を示した。しかしながら、本技術を用いて、個々のメンバー全てがポリクローナル抗体試料に存在するかどうかを評価することは不可能である。〜30kDaのLCバンドの配列決定は、RhD319抗体の存在を示し、これは3および4サイクルにおけるValの存在により判断した。RhD203抗体の存在についての証拠は全く得られなかった(2および3サイクルそれぞれにおいてSもAもないこと)。しかしながら、この組換えモノクローナル抗体のイオン交換クロマトグラフィーおよびN−末端配列分析は、RhD203のLCが、部分的にブロックされたN−末端を有することを強く示唆している。従って、この抗体が分析混合物中に存在するか否か、N−末端配列分析により結論付けることはできない。加えて、サイクル1においてEおよびD残基が存在し、サイクル4においてM残基が存在するので、30kDaのバンドはκLCをいくつか含有するようである。
【0163】
簡単にいうと、HCまたはKCのいずれかにおいて配列が異なり、かつN−末端においてブロックされていない場合、大量N−末端配列分析を用いて、個々の抗体の存在を同定することができる。この方法は、個々のポリペプチドのN−末端が部分的にブロックされない限り、定量的である。
【0164】
実施例6
本実施例は、ポリクローナル細胞培養物からもたらされた8種類の個々のメンバーを有するポリクローナル抗RhD抗体の特徴決定を説明するものである。ペプチドを分離するためRP−HPLCまたはイオン交換クロマトグラフィー(IEX)のいずれかを用いて、可変領域を起源とする固有マーカーペプチドを単離することにより、抗体の多様性を分析した。
【0165】
単離した重鎖および軽鎖の消化によるペプチドの生成
8種類の個々のメンバーを有するポリクローナル抗RhD抗体試料を、HiTraprタンパク質Aカラムを用いたアフィニティークロマトグラフィーによりポリクローナル細胞培養物の上清から精製した。凍結乾燥した材料を6M 塩酸グアニジウム、0.5M EDTA、0.2M Tris HCl、pH8.4に溶解し、還元(DTT)し、カルボキシメチル化(ヨード酢酸)した。Ettan LCシステム(Amersham Biosciences, GE Healthcare, England)において、6M グアニジウムHCl、50mM リン酸ナトリウム、pH8.4中、Superose 12カラム(10/300 Amersham Biosciences, GE Healthcare)上でのゲルろ過により、重鎖および軽鎖を分離した。分離したHC(〜3.5mg/ml)およびLC(6.5mg/ml)を、リン酸ナトリウム、pH8中の酵素対基質濃度1:500にて、Endoproteinase Asp−N(Roche, 1 054 589)で消化した。
【0166】
RP−HPLCによる固有ペプチドの単離
ポリクローナル抗体試料から得られた単離HCおよびLCの個々のAsp−N消化物のアリコートを、流速0.2ml/分を用いて0.1% TFAにて平衡化したガードカラム(Zorbax 300SB−C8、2.1×12.5mm、5μm)と結合したZorbax 300SB−C18(2.1×150mm)5μmカラムを装備したAgilent 1100 LC/MSD SLシステムに適用した。直線的勾配の0.08% TFA、70% アセトニトリルを用いてペプチドを溶出した。ペプチドを220nmにおいて分光光度法により検出し、オンラインのMS(大気圧イオン化(API)エレクトロスプレー)により分析した。75% プロピオン酸/25% イソプロパノールの混合物を移動相にポストカラム添加し、シグナルを増大させた。得られた質量スペクトルをChemstationソフトウェア(Agilent Technologies, CA)およびBioLynxソフトウェア(Micromass, Waters Corporation, MA)を用いて分析した。
【0167】
HCおよびLCのAsp−N消化物のMS分析の結果を、表11および12にそれぞれ概説する。両方の表が、平均質量として与えられる理論上の質量および検出質量を示す。
【0168】
表11:重鎖についての結果
【表13】
a同じ質量を有する異なるペプチド。bほぼ同じ質量を有するペプチド(4個が一致し、1個が異なる)。*N−末端環化Gln(PyroGlu)を示す。
【0169】
表12:軽鎖についての結果
【表14】
【0170】
表11において見られるように、可変HC由来の13種類のペプチド、および可変LC由来の16種類のペプチドを固有のマーカーペプチドとして同定することができ、HCの可変領域由来のいくつかのペプチド(例えば、aにより示されるRhD196、RhD244、およびRhD306由来のD1)は同じ質量を有し、従って、これらの質量を明確に割り当てることができない。しかしながら、全ての場合で他の質量を固有のペプチドに明確に割り当てることができるので、8種類の抗体全てを明確に同定した。LCについて、8種類の抗体の中から7種類について、固有ペプチドを割り当てた(表12)。LCについての情報がない抗体はRhD324であった。従って、HCおよびLC由来の統合MSデータは、各抗体由来の固有ペプチドの検出に基づき、抗RhD rpAb試料において、8種類の抗体全てを同定できたことを示している。
【0171】
陽イオン交換クロマトグラフィーによる固有ペプチドの単離
HCおよびLCのAsp−N消化物を強陽イオン交換クロマトグラフィーにより次の通り分離した:上述のポリクローナル抗体から得られた単離HCおよびLCの個々のAsp−N消化物のアリコートを、Ettan LCシステム(Amersham Biosciences, GE Healthcare, England)において、10mM リン酸カリウム、20%(v/v) アセトニトリル、pH3.0にて平衡化したPolySulfoethyl Aカラム(2.1×100mm)に、流速0.2ml/分、室温にてアプライした。次に、10mM リン酸カリウム、20%または30%(v/v) アセトニトリル、pH3.0中の0〜500mMの直線的勾配の塩化カリウムを用いて、ペプチドを溶出した。溶出したペプチドを215nmにて分光光度法により検出し、時間フラクションに基づきフラクションを収集した。フラクションのアリコート(1μl)を、70% アセトニトリル/30%、0.1% TFA中のα−シアノ−4−ヒドロキシ桂皮酸(20mg/mL)溶液1μlと混合し、MS標的上にアプライし、次に、0.1% TFAで洗浄した。試料を、Autoflex TOF(Bruker Daltronics, Bremen, Germany)上のMALDI−TOFにより分析し、Bruker Daltronics(Bremen, Germany)の校正用混合物を用いて、質量の外部校正を行った。GPMAW 6.1ソフトウェア(Lighthouse data, Odense, Denmark)を用いて、MALDIスペクトルを分析した(質量検索および内部校正)。
【0172】
LCのAsp−N消化物由来のいくつかのピークを含有する代表的なクロマトグラムを、図11に示す。LCおよびHCのAsp−N消化物のフラクションのMALDI−TOF分析の結果を、それぞれ表13および14に示す。理論上の質量および見出した質量を、質量<3500Daについてモノアイソトピック質量として、質量>3500Daについて平均質量として示す。
【0173】
表13:軽鎖の結果
【表15】
a同じペプチド b同じペプチド c同じペプチド
【0174】
表14:重鎖の結果
【表16】
a同じペプチド b酸化Metを有する同じペプチドの同定により証明した
【0175】
表13および14において見られるように、可変LC由来の15種類のペプチド、および可変HC由来の9種類のペプチドを、固有のマーカーペプチドとして同定することができる。HCの可変領域ならびにLC由来のいくつかのペプチドは同じ質量を有し、これを明確に割り当てることができない。従って、強陽イオン交換クロマトグラフィーを用いて、HC RhD201、203、および324、ならびにLC RhD201および319について固有ペプチドを割り当てることはできなかった。
【0176】
要約
2種類の異なるマーカーペプチド分析から得られた結果は、HCおよびLCのMS分析から得られた統合データにより、RP−HPLCを用いて抗RhD rpABを構成する8種類の抗体全てから可変領域由来の固有ペプチドの同定を可能とすることを実証するのに十分である。強陽イオン交換クロマトグラフィーを用いることにより、抗RhD rpAb組成物中の8種類の個々のメンバーの中から6種類を同定することができた。完全に詳細までではないが、表11から14に示す程度まで、MS分析の結果を分析した。
【0177】
実施例6A
本実施例は、ポリクローナル細胞培養物(バイオリアクターラン)からもたらされた25種類の個々のメンバーを有する組換えポリクローナル抗RhD抗体の特徴決定を説明するものである。ペプチドの同定のための質量分析と組み合わされたRP−HPLCを用いてLCまたはHC由来の可変領域を起源とする固有マーカーペプチドを単離することにより、抗体の多様性を分析した。
【0178】
単離した重鎖および軽鎖の消化によるペプチドの生成
25種類の個々のメンバーを有するポリクローナル抗RhD抗体試料を、バイオリアクターラン由来のポリクローナル細胞培養物の上清から精製した。MabSelect(Amersham Biosciences, GE Healthcare)カラムを用いたアフィニティークロマトグラフィーにより、精製を行い、G25カラム(Amersham Biosciences, GE Healthcare)上で脱塩した。凍結乾燥した材料を6M 塩酸グアニジウム、0.2M Tris HCl、pH8.4に溶解し、還元(DTT)し、次に、カルボキシメチル化(ヨード酢酸)した。6M グアニジウムHCl、50mM リン酸ナトリウム、pH8.4中Superose 12カラム(10/300 GL Amersham Biosciences, GE Healthcare)上でのゲルろ過により、重鎖および軽鎖を分離した。分離したHCおよびLCを、1M 尿素、50mM リン酸ナトリウム、pH8中酵素対基質濃度1:200、37℃にて一晩Endoproteinase Asp−N(Roche, 1 054 589)で消化した。
【0179】
LC−MSによる固有ペプチドの単離
ポリクローナル抗体試料から得られた単離HCおよびLCの個々のAsp−N消化物のアリコートを、0.1% TFA、14% ACN中、流速0.2ml/分を用いて平衡化したガードカラム(Zorbax 300SB−C8、2.1×12.5mm、5μm)と結合したZorbax 300SB−C18(2.1×150mm)5μmカラムを装備したAgilent 1100 LC/MSD SLシステムに適用した。直線的勾配の0.08% TFA、70% アセトニトリルを用いて、ペプチドを溶出した。ペプチドを220nmにて分光光度法により検出し、オンラインのMS(大気圧イオン化(API)エレクトロスプレー)により分析した。75% プロピオン酸/25% イソプロパノールの混合物を、移動相にポストカラム添加し、シグナルを増大させた。得られた質量スペクトルをChemstationソフトウェア(Agilent Technologies, CA)およびGPMAW 6.2ソフトウェア(Lighthouse data, Odense, Denmark)を用いて分析した。
【0180】
HCおよびLCのAsp−N消化物のMS分析の結果を表14に概説する。表中、理論上の質量および検出した質量を平均分子量として示す。
【0181】
表14A:Asp−N消化およびLC−MS分析を利用した抗RhD rpAb中の25種類の抗体由来の固有疎水性ペプチドの同定
【表17】
aDおよびdは、それぞれAsp−Nにより生じたHCおよびLC由来ペプチドを示し、予測配列のN−末端からC−末端までペプチドに番号を付けた。ゆえに、d4は、LCペプチドの3番目と4番目のAsp−N部位での開裂により生じたペプチドを示す。bこのペプチドは、見逃された開裂部位を含有する。*N−末端環化Gln(pyroGlu)を示す。
【0182】
表14Aにおいて見られる通り、LCの可変部分由来の22種類のペプチド、およびHCの可変部分由来の3種類のペプチドを固有マーカーペプチドと同定することができる。従って、HCおよびLCのMSデータは、各抗体由来の固有ペプチドの検出に基づき、抗RhD rpAb試料において25種類の抗体全てを同定したことを明確に示している。
【0183】
実施例7
本実施例は、組換えポリクローナル抗RhD抗体の個々のメンバーに対する特異性を有する抗イディオタイプペプチドの生成、ならびに組換えポリクローナル抗体における1種類の個々のメンバーの濃度の評価を説明するものである。
【0184】
抗RhD抗体特異的ペプチドリガンドの生成
PIII(New England Biolab)のN−末端でランダムな配列順序で7種類のアミノ酸を提示するファージライブラリーを、個々の抗RhD抗体に対するペプチド結合体のアフィニティー選択のために用いた。ペプチドライブラリーの線形および制約バージョンの両方を選択のために用いた。4℃にて12〜16時間、10μg/ml(1ウェル当たり100μlを用いる)精製モノクローナル抗RhD抗体でマイクロタイタープレート(Maxisorb, NUNC)をコートした。組換えポリクローナル抗RhD抗体に含有される25種類の個々の抗体全てを用いて、抗イディオタイプペプチドについてスクリーニングした。しかしながら、組換えポリクローナル抗体が多数の個々のメンバー(例えば、50より多い)を含有する状況下では、センチネル抗体をスクリーニングのために選択してもよい。好ましくは、組換えポリクローナルタンパク質を構成する抗体の総数の少なくとも4%に該当する多数のセンチネル抗体を選択し、なおより好ましくは、組換えポリクローナルタンパク質を構成する抗体の総数の少なくとも8%、12%、16%、20%、30%、または50%で構成されるセンチネル抗体が選択される。次に、コートしたプレートをPBS、0.05% Tween−20で洗浄し、2% スキムミルク/PBSを用いてブロックした。〜1011pfu/100μlのバクテリオファージを、各パニングラウンドに用いた。制約ライブラリーおよび線形ライブラリーを混合し、2% スキムミルク/PBS中の混合物として一緒にパニングした。室温にて1時間インキュベーションした後、結合ファージを、グリシン/HCl、pH=2.2で10分間溶出し、次に、Tris−HCl、pH=9.0を用いて中和した。パニング3から4ラウンド後、単一クローンを単離し、DNAを抽出し、次に、ランダムなペプチド領域に対応する領域において配列決定した。以下の表15は、単一クローン由来の推定アミノ酸配列のアラインメントを示す。
【0185】
表15
【表18】
【0186】
それぞれ抗RhD162、202、または305のいずれかに対して特異的アフィニティーを有する3種類の合成ペプチドを、関連配列群の推定一致アミノ酸配列に従い合成した。各合成ペプチドのC−末端にビオチンを結合させた。
【0187】
各ペプチドの特異性をELISAにより試験した。簡単にいうと、ELISAプレートを、5μg/ml(1ウェル当たり100μlを用いた)Streptavidinを用いて、4℃にて12〜16時間コートした。次に、PBS中〜10μg/mlまで希釈したペプチドを添加し、1時間インキュベーションし、洗浄により過剰なペプチドを除去した。次に、プレートを、2% スキムミルク/PBSにてブロックし、PBSにて3回洗浄した。種々の希釈(10μg/mlにて出発)にて個々の抗RhD抗体それぞれを添加した。抗ヒトIgG−結合体(カタログ番号H10307)を用いて、結合抗体を検出した。プレートを5回洗浄し、色素原(TMB, Kem-En-Tech)25μlを添加して、検出を行った。1M H2SO4 25μlを添加することにより、15〜25分後に反応を終結させた。吸光度を450nmにて測定した。モノクローナル抗RhD抗体のパネルに対する各ペプチドの試験は、反応性が適当な個々のメンバータンパク質に特異的であることを示した。ゆえに、10より高いシグナル対ノイズ比で、PEP162のみが抗RhD162抗体に結合し、PEP202のみが抗RhD202に結合し、そしてPRP305のみが抗RhD305に結合した。
【0188】
組換えポリクローナル抗RhD抗体における抗RhD305抗体量の決定
適切な希釈の精製抗RhD305モノクローナル抗体を参照標準として用いて、組換えポリクローナル抗RhD抗体混合物中の抗体総量に対する抗RhD305抗体量を決定することが可能であった。簡単にいうと、ELISAプレートをストレプトアビジンでコートし、PBS中〜10μg/mlまで希釈したPEP305と1時間インキュベーションした。インキュベーション後、過剰なペプチドを洗浄して除去した。次に、プレートを2% スキムミルク/PBSにおいてブロッキングし、3回洗浄した。25種類の個々の抗RhD抗体(試料)からなる組換えポリクローナル抗RhD抗体を、1から16384倍の範囲の希釈率で添加した。試料を4重に分析した。同じプレート上の別々のウェルにおいて、標準曲線を作成するために、参照試料としてモノクローナル抗RhD305抗体の連続希釈液(3重)(10μg/mlにて開始)を添加した。抗ヒトIgG結合体(カタログ番号H10307)を用いて、結合抗体を検出した。プレートを5回洗浄し、色素原(TMB, Kem-En-Tech)25μlを添加することにより、検出を行った。1M H2SO425μlを添加することにより、15〜25分後に反応を終結させた。吸光度を450nmにて測定した。
【0189】
標準曲線は次の範囲の濃度に直線的に比例した:
【表19】
【0190】
これらのデータは、方程式y=0.1161x−0.0256、およびR2=0.9812を有する標準曲線となった。
【0191】
標準曲線について決定した方程式、ならびに試料の希釈因子を用いて、組換えポリクローナル抗RhD抗体試料中の抗RhD305抗体濃度を計算した。
【0192】
32倍希釈において、試料について測定した平均OD450は1.24±0.14であり、これは、ポリクローナル抗RhD抗体中の抗RhD305抗体濃度3.8±0.5μg/mlに対応するものであった。組換えポリクローナル抗RhD抗体試料中のトータルの抗体濃度は、100μg/mlであった。従って、抗RhD305抗体はポリクローナル抗体試料の3.8%を表す。
【0193】
実施例8
本実施例は、組換えポリクローナル抗RhD抗体の液体イオン交換クロマトグラフィー分析による1次元の分離後、特定のフラクション/ピークにおけるセンチネル抗体を同定するための抗イディオタイプペプチドの使用を説明するものである。
【0194】
25種類の個々の抗RhD抗体からなる組換えポリクローナル抗RhD抗体を陽イオン交換クロマトグラフィーにより分離し、フラクションを収集した。3種類の抗イディオタイプペプチドを用いたELISA(実施例7に記載の通り)により、各フラクションを調べ、各フラクションにおける特定の抗RhD抗体の存在を検出した。各フラクションにおいて行われたELISAデータとのクロマトグラムのオーバーレイは、この方法を用いて、特定のフラクション中の個々の抗体を同定できることを示した(図12)。従って、特定のピークにおける吸光度をELISAデータと比較することにより、相同タンパク質の複雑な混合物の組成物を半定量的に評価することが可能となる。
【0195】
実施例9
医薬上使用するためにポリクローナルタンパク質を製造する場合、組成物の安定性の確保は重要な問題である。相同タンパク質の複雑な混合物中の個々のタンパク質メンバーを同定する能力のある特異的なペプチド−リガンドを用いて、発酵中、異なる生成時間ポイントで培地試料を抽出し、実施例7に記載のELISA法のような定量的検出法を適用することにより、製造中のポリクローナルタンパク質の組成物の安定性をモニターすることができる。
【0196】
本実施例において、3種類のセンチネルタンパク質、抗RhD162、202、および305抗体の実際の量を、25種類の固有の抗RhD抗体からなる組換えポリクローナル抗RhD抗体産生pWCBのかん流発酵過程において評価した。図13は、バイオリアクターの播種の8日後に対応するG8を用いて、発酵中の異なる培養時間ポイントでの3種類のセンチネル抗体(抗RhD162、202、および305)の分布を説明するものである。
【0197】
実施例10
本実施例は、細胞培養混合物中の特定の抗RhD抗体産生細胞の同定方法を説明するものである。実施例において、抗イディオタイプペプチドおよび検出用フローサイトメトリーを用いて、2種類の異なる抗体産生細胞株を分析した。
【0198】
2種類の個々の抗RhD抗体産生細胞株、RhD162およびRhD202を所定の比率で混合した。クローンRhD202の割合を表16に示す。
【0199】
ビオチン化ペプチド202(実施例7において調製したPEP202)を、フィコエリトリン結合ストレプトアビジン(SA−PE)とインキュベーションし、ペプチドテトラマーを形成させた。テトラマーを細胞株混合物と室温にて20分間インキュベーションし、FACS CAliburフローサイトメーター(Becton Dickinson)に細胞を通した。図14のR1によって示す通り、テトラマー陽性細胞にゲートをかけた。個々の非混合細胞株を同様に測定した(図14AおよびB)。
【0200】
抗RhD抗体産生細胞株について見られる特徴は、細胞株中の抗体発現細胞および非抗体発現細胞両方の存在である。混合物中のPEP202陽性細胞のシェアを評価するために、発現細胞の総数を決定することが必要であった。この実施例において、RhD162とRhD202の混合物中の発現細胞のシェアは、RhD202単独におけるものと同じであると仮定した。これは、Pep202および抗IgG抗体で細胞を2重染色することにより、評価することができた(行ってはいない)。しかしながら、結果は、仮定が正確であることを示すものであった。図14に示す通り、ゲート6(R6)における細胞の割合から、混合物中のPEP202テトラマーによって結合したRhD202細胞の割合を計算した。
【0201】
mixa)におけるRhD202細胞株の割合を、次の方程式(mix aについての測定値を用いて例示)により計算した。
【数1】
【0202】
表16
【表20】
【0203】
実施例11
本実施例は、個々のクローンの分布、またはポリクローナル細胞培養物中のかかるクローンのセンチネルセレクションを評価するためのリアルタイムPCRの使用を説明するものである。
【0204】
技術は、個々の抗体コード核酸配列間の配列の相違に基づくものである。個々の抗体コード配列の多様性により、ポリクローナル細胞株に表される各メンバーの重鎖および/または軽鎖について、固有のTaqManプローブを設計することができる。好ましくは、CDR領域であるCDR1、CDR2、またはCDR3の1つを、TaqManプローブの設計のために選択する。最も好ましくは、CDR3領域をTaqManプローブの設計のため選択する。
【0205】
オリゴヌクレオチドの設計
好ましくは、70〜150ヌクレオチドの単位複製配列を得るように、プライマーを設計する。いくつかの可能性のあるプライマーの設計は、重鎖または軽鎖可変領域のFR3領域においてアニーリングする一致フォワードプライマー、および定常領域においてアニーリングするリバースプライマーである。試料の個々のメンバー間で異なるCDR領域の一部、好ましくは、CDR3領域に特異的なTaqManプローブを、目的の各クローンについて設計する。
【0206】
次の8種類の抗RhD抗体発現ポリクローナル細胞株の分析のためのプライマーおよびプローブの可能性のあるセットを、以下に示す通り設計し得る。
【0207】
全てのクローンについてフォワードおよびリバースプライマー:
Fwプライマー:CAC GGC TGA GTA TTA CTG TGC(配列番号24)
Rwプライマー:TTG GTG GAG CCA CTC GA(配列番号25)
【0208】
個々のクローン全てについてのTaqManプローブを表17に示す。
表17
【表21】
【0209】
重鎖コード配列についてのプライマー設計の別法は、VH−DH結合においてアニーリングするフォワードプライマーおよび定常領域におけるリバースプライマー、ならびにJH−C結合におけるTaqManプローブ(Rasmussen, T. et al. 2000. Exp.Hematol. 28, 1039-1045に記載)を構成する。
【0210】
リアルタイム定量的PCR
mRNAまたはゲノムDNAをペレット化細胞から抽出する。mRNAを鋳型として用いる場合、リアルタイムPCRに先立ち、逆転写してcDNAを生成する。分析すべきクローン数に対応する数のリアルタイムPCR反応をセットアップする。
【0211】
プライマー濃度およびTaqManプローブ濃度について、リアルタイムアッセイを最適化する。光学的接着性カバーでシールした96ウェルプレートにおいて、反応をトリプリケートで行う。市販のPCRマスターミックスにおいてPCR反応を行い、次に、ABIプリズム7000(Applied Biosystems)において、ABIプリズム7000 SDSソフトウェアを用いて分析を行った。
【0212】
多様性の分析
異なるクローンのCT値を互いに比較し、ポリクローナル細胞株における各クローンの分布を計算する。方法を応用して、バッチとバッチとの間の変動ならびに個々の生産工程における経時的なクローン安定性を評価し得る。
【0213】
実施例12
本実施例は、ポリクローナル細胞株からもたらされた単一細胞クローン由来の重鎖および/または軽鎖抗体遺伝子の可変領域のDNA配列決定により、組換えポリクローナル抗体を産生する能力のあるポリクローナル細胞株(例えば、pWCB)のポリクローナル性質を評価し、明らかにする方法を説明するものである。
【0214】
シングルセルクローニング
pWCBのアンプルを解凍し、完全培地にて2、3日間培養し、良好な細胞バイアビリティーを再構築した。続いて、96ウェル細胞培養プレート中、完全培地中1細胞/ウェルの密度にて細胞を播種する限界希釈により、シングルセルクローンを得る。37℃、5% CO2にて、10〜20日間細胞を培養し、次に、顕微鏡下でシングルコロニーを有するウェルについて、プレートを視覚的にスコア化する。別法として、FACS細胞ソーターを用いて、pWCB由来のシングルセルクローンを得る。生存可能なpWCB細胞にゲートをかけ、予め条件完全培地100μlで満たした96ウェルプレートに、1細胞/ウェルにて播種する。細胞をインキュベーションし、上述の通り、シングルコロニーについてスコア化する。
【0215】
核酸配列決定
ウェル中のシングルセルコロニーをコンフレントまで成長させるとき、所望数(例えば、100)のウェルのそれぞれからのアリコート(10〜20μl)を新しい96ウェルプレートに移し、DNA配列決定反応の鋳型として用いる。1から100、または1から1000個の細胞をそれぞれ用いてmRNAレベルまたはゲノムレベルのいずれかで、配列決定を行う。前者の場合、pWCBに存在する異なる抗体重鎖および軽鎖遺伝子を区別するのに十分な可変領域(典型的には、少なくともCDR3領域)をカバーするPCRフラグメントを、例えば、市販のQiagen one−step RT−PCRキットを用いて、製造元の指示に従い、標準的RT−PCR技術により生成する。PCR反応に先立ち、細胞を溶解させる。得られたPCRフラグメントを、例えば、Qiagen Qiaquick Gel Extractionキットを用いてゲル精製し、標準的配列決定反応における鋳型として用い、次に、ABI Prism[登録商標]3100 Genetic Analyzer(Applied Biosystems)のような自動DNA配列決定マシーンにおいて分析する。別法として、DNA配列決定を、逆転写工程を省略する以外上述の通り、ゲノムDNAにおいて行う。
【0216】
抗RhD組換えポリクローナル抗体の特徴決定のため、次のプライマーを用いた:
VH増幅用PCRプライマー:
RhD番号001:5’TCTCTTCCGCATCGCTGTCT(配列番号34)
RhD番号007:5’AGGAAAGGACAGTGGGAGTGGCAC(配列番号35)
VL増幅用PCRプライマー:
RhD番号005:5’CGTTCTTTTTCGCAACGGGTTTG(配列番号36)
RhD番号008:5’AAGACCGATGGGCCCTTGGTGGA(配列番号37)
配列決定プライマー:
VH:5’AACGGGTTTGCCGCCAGAACA(配列番号38)
VL:5’CCGAGGGACCTGAGCGAGT(配列番号39)。
【0217】
抗イディオタイプペプチドを用いたシングルセルにおけるELISA
核酸配列決定に対する手助けとして、ポリクローナルワーキングセルバンクのような抗体産生細胞の混合物のクローナルな組成物を、抗イディオタイプペプチドELISAを用いて評価することができる。
【0218】
ソートしたシングルセルを約14日間培養し、それにより、シングルクローン由来のアイソジェニックな細胞培養物を生成した。実施例7に記載の通り、ELISAアッセイにおいて抗イディオタイプペプチドを用いて、特異的抗RhD抗体の存在について、これらの培養物由来の上清を分析することができる。これにより、特定の個々のメンバーを産生するクローンの数に関する情報がもたらされるだろう。個々のメンバーの量を抗体産生細胞の総量と比較する(例えば、全てのアイソジェニックな細胞培養物においてIgGを測定することによる)場合、ポリクローナル細胞培養物における個々の抗RhD抗体産生細胞のフラクションについての定量的測定値を得ることができる。
【0219】
実施例13
本実施例は、組換えポリクローナル抗体の下流プロセッシング(DSP)中のクローン多様性を評価するための陽イオンクロマトグラフィー分析の使用を説明するものである。
【0220】
下流プロセッシング
25種類の個々のメンバーを含有する抗RhD rpAb試料を、発展的バイオリアクターランから、次のDSP工程を用いて精製した:
1.MAbSelectカラムを用いた抗体の捕獲
2.pH3でのウイルスの不活化
3.sephadex G−25カラムを用いたバッファー交換
4.DEAE−セファロースカラムを用いたイオン交換クロマトグラフィー
5.Planova 15Nフィルターを用いたウイルスフィルトレーション、および
6.MEP hypercelカラムを用いた疎水性電荷導入クロマトグラフィー
7.milipore biomaxフィルターを用いた限外ろ過/ダイアフィルトレーション。
【0221】
個々のDSP工程後のクローン多様性の分析
陽イオン交換クロマトグラフィーを用いて、組換えポリクローナル抗体組成物のDSP中のクローン多様性を分析した。抗RhD rpAbのDSP中、工程1、3、4、および6の後に採取した試料を、25mM 酢酸ナトリウム、150mM 塩化ナトリウム、pH5.0中、流速60ml/時間、室温にてPolyCatAカラム(4.6×100mm)にアプライした。次に、25mM 酢酸ナトリウム、pH5.0中150〜500mMの直線的勾配の塩化ナトリウムを用いて、流速60ml/時間にて抗体成分を溶出した。抗体成分を280nmにて分光光度法により検出し、クロマトグラム(図15)を比較して、DSP中のクローン多様性の可能性のある損失を検出した。本実施例において、陽イオン交換クロマトグラフィーを用いて、クローン多様性が組換えポリクローナル抗体のDSP中に本質的に変化しないことを明らかにした。
【0222】
実施例14
RhDに対する40種類より多くの抗体のIEX分析により、相当数の個々の抗体が図16Bに示す通り「3ピークパターン」を表すことを明らかにした。カルボキシペプチダーゼB処理、ならびに炭水化物分析は、この電荷不均一性が、C−末端リジンクリッピングまたはシアル酸の存在により引き起こされることを示した(データは示していない)。
【0223】
本実施例は、電荷不均一性がPyroGlu形成に起因するものであり、部位指定変異誘発を用いて、均一なIEXパターンを得ることができることを明らかにするものである。
【0224】
抗体の発現および精製
ゲノム上の特定の部位と異なる組換え抗Rhesus Dモノクローナル抗体をそれぞれ発現する安定な細胞株(2004年7月20日に出願されたデンマーク特許出願PA 2004 01133に記載の通り、得たもの)を、4mM L−グルタミン(Invitrogen)および1:250希釈した抗凝集剤(Invitrogen)を添加した無血清Excell 302培地(JRH Biosciences, Andover, UK)における懸濁培養に適用し、拡大させ、次に、通常の凍結手段を用いて−150℃にて保存した。
【0225】
保存前に、細胞培養物から上清を収集し、上清をろ過して、本質的に実施例1に記載の通り、アフィニティークロマトグラフィー(タンパク質A)を用いて抗RhDモノクローナル抗体を精製した。
【0226】
強陽イオン交換クロマトグラフィー
上述の工程において精製したモノクローナル抗体を、本質的に実施例1に記載の強IEXクロマトグラフィーの対象とした。表18のIEX列は、選択した抗体のIEXプロファイルに存在するピーク数を概説するものである。かかるIEXプロファイルを図16においても示す。
【0227】
N−末端配列分析
2種類の選択した抗体(RhD198およびRhD307)のIEX分析から分離したピークのN−末端配列分析を、Procise 494 Sequencer(Applied Biosystems, CA)(製造元によより記載された通り操作)を用いて、Edman配列決定により、溶液中で行った。配列分析により、電荷不均一性が、HCのN−末端Glnの部分的環化に起因するものであることが明らかになった(表18参照)。従って、第1のピークは、HCの全体的にブロックされたN−末端(HCのN−末端が0の電荷を有する)を有する抗体を含有し;第2のピークは、HCのN−末端の1つがブロックされている(HCのN−末端が+1の電荷を有する)抗体に対応し、そして第3のピークは、HCのN−末端Glnが非修飾されている(HCのN−末端が+2の電荷を有する)抗体を表す可能性が最も高かった。N−末端グルタミン残基のPyroGluへの環化は、それをEdman配列決定に不応性にする。
【0228】
かかる「3ピークパターン」または「1ピークパターン」を同様に示す多数の他の抗RhD抗体を、SDS−PAGEの対象にし、これをPVDFメンブレンに電気的にブロットすることによってN−末端配列分析により、分析した。これらのブロット上のHCおよびLCバンドを、Edman配列決定の対象とした。
【0229】
HCにおいてN−末端Glnを有する2、3種類の抗体(RhD162、RhD240)は、それらのIEXプロファイル(「1ピークパターン」)をふまえて、全体的にブロックされていることを示し、一方「3ピークパターン」を有する抗体(RhD196、RhD305、およびRhD306)は、予測通り部分的にブロックされていることを見出した(表18参照)。説明は、配列の収量、ならびにIEXプロファイルにおける異なる電荷変異体(0、+1、および2+)の相対的割合に基づくものである。
【0230】
表18
【表22】
a予測および得られたN−末端配列を、それぞれ通常および太字のフォント形式で示す。bブロットから得られたデータ。cIEX分析(溶液において分析)から単離したフラクション(ピーク1&2)から得られたデータ。dN−末端グルタミン残基のPyroGluへの環化はそれをEdman配列決定に不応性にする。n.d.;決定していない。
【0231】
部位指定変異誘発
部位指定変異誘発を用いて、N−末端GlnをGluに変えることにより、選択した抗体から電荷不均一性を除いた。重鎖中にN−末端Gln、および軽鎖中にN−末端Gluを有する全長抗体をコードする発現プラスミドRhD189を用いた。このプラスミドのVH領域には、シグナルペプチドコード領域の3’末端のAscI部位、およびJ領域のサイレントXhoI部位がフランクする。
【0232】
変異誘発を次のプライマー:
を用いて行った。
【0233】
フォワードプライマー中のAscI部位およびリバースプライマー中のXhoI部位に下線を付し、VH領域のN−末端のGluコドン(GAG)を太字で示す。上述のプライマーを用いたPCR反応における鋳型として、RhD189プラスミドを用いた。製造元の指示に従い、Phusion DNAポリメラーゼ(Finnzymes, Finland)を用いて25サイクルPCR反応を行った。約400bpのVHバンドを1% アガロースゲル上で精製し、BioTaq DNAポリメラーゼとインキュベーションし、アガロースゲル上で再び精製し、製造元の指示に従い、pCR2.1TOPOベクター(Invitrogen, CA)にクローニングした。VHインサートを含有するクローンを配列決定により確かめた。オリジナルのVHフラグメントをAscIおよびXhoIを用いてプラスミドRhD189から切り出し、pCR2.1TOPOプラスミド由来の変異誘発フラグメントを代わりに挿入した。エンドトキシンフリーのプラスミドミディプレップ(Macherey-Nagel, Germany)をクローニングの陽性コロニーから調製し、配列決定して、正しいフラグメントの存在を確かめた。
【0234】
抗体をSDS−PAGE分析の対象とし、電気的にブロットして、HCバンドのN−末端配列決定を行い、GlnからGluへの置換を確かめた(データは示していない)。「3ピーク」IEXプロファイルを示す抗体RhD189のHCのN−末端のGlnからGluへの置換は、1ピークのみを有する有意に異なるプロファイルとなった(図17)。従って、N−末端Gln残基からGlu残基に置換することにより、電荷の不均一性を首尾よく排除できた。
【0235】
結合アッセイ
N−末端の変異が抗体の機能に影響するかどうかを評価するために、天然の抗体であるRhD189、ならびにその変異誘発Glu対応物であるRhD189Eを、RhD陽性赤血球に対する結合についてアッセイした。
【0236】
Blood Bank, Aalborg Hospital, DKにおいてインフォームドコンセントの後健常ドナーから得た全血から、1% ウシ血清アルブミン(BSA、Sigma-Aldrich, Germany)を含有するPBS(Gibco, Invitrogen, United Kingdom)において3回洗浄することにより、赤血球を調製した。赤血球を再懸濁し、ID−Cellstab((DiaMed, Switzerland)中の10%溶液として4℃にて保存した。
【0237】
PBS、1% BSA中5×104細胞/μlのRhD陽性赤血球を用いて、抗体の結合能を測定した。PBS、1% BSAにて、96ウェルプレート(Becton Dickinson Labware, NJ, USA)中トリプリケートにて抗体希釈を行った。抗体溶液50μlを赤血球50μlと混合し、37℃にて40分間インキュベーションした。細胞をPBS、1% BSAにて2回(300g、2分間)洗浄した。PBS、1% BSAに1:20希釈したフィコエリトリン結合ヤギ抗ヒトIgG(Beckman Coulter, CA, USA)80μlを各試料に添加し、4℃にて30分間置いた。試料をPBS、1% BSA、次にFacsFlow(Becton Dickinson, Belgium)(300g、2分間)にて洗浄し、FACSFlow200μlに再懸濁した。FACSCalibur(Becton Dickinson, CA, USA)に試料を通し、CellQuest ProおよびExcelを用いてデータ分析を行った。
【0238】
図18において示す通り、RhD陽性赤血球に対する結合能における有意差を、Glu変異体とその天然の対応物の間で観察しなかった。
【0239】
要約
多くの抗RhD抗体のIEXプロファイルにおいて観察される不均一性は、これらの抗体におけるN−末端Gln残基の部分的環化に起因するものであった。抗RhD抗体におけるHCのN−末端GlnをGlu残基に置換することにより、おそらくRhD陽性赤血球に対する結合力に影響することなく、生来のN−末端の電荷不均一性を排除する。
【図面の簡単な説明】
【0240】
【図1】図1:9週間培養後のアリコート3948および3949由来の抗RhD組換えポリクローナル抗体(抗RhD rpAb)組成物の陽イオンクロマトグラム。下側の線図がアリコート3949に対応し、上側の線図がアリコート3948に対応する。線図の頂点のY軸を、下側の線図と区別するためにはずした。ピークのA〜Jは、正味電荷の異なる抗体、および電荷不均一性を現す個々の抗体を含む。
【図2】図2:11週間培養後の抗RhD rpAb産生ポリクローナル細胞株のアリコート3948+および3949+(FCW065)からもたらされたRT−PCR産物におけるHinfI RFLP分析を示すゲルの写真。特定のクローンに割り当てられるバンドを同定する。
【図3】図3:8種類の異なる抗Rhesus D抗体を有する抗RhD rpAb発現ポリクローナル細胞培養物由来の抗Rhesus D抗体軽鎖のT−RFLPパターン。8種類の異なる抗Rhesus Dクローンを、矢印により示されるピークに割り当てた。
【図4】図4:所定の時間ポイントで25種類の異なる抗Rhesus D抗体を有する抗RhD rpAbを発現するポリクローナル細胞培養物由来の抗Rhesus D抗体重鎖可変領域のT−RFLPパターン。25種類の異なる抗Rhesus Dクローンを、矢印により示されるピークに割り当てた。
【図5】図5:5週間培養したポリクローナル細胞培養物由来の8種類の異なる抗Rhesus D重鎖コード配列のT−RFLPにより評価したcDNA分布。
【図6】図6:陽イオン交換クロマトグラフィーを用いて分析した8種類の異なる抗体を有する抗RhD rpAbの相対的含有量(%)を示す。統合したクロマトグラムピークを、保持時間、および同一条件下で陽イオン交換クロマトグラフィーを用いて個々に分析した単一抗体から得られたピークパターンから個々の抗体に割り当てた。
【図7】図7:4週間培養後に得られた試料由来の25種類の個々のメンバーを有する抗RhD rpAbの陽イオンクロマトグラム。ピークAC1から25は、正味電荷の異なる抗体、および電荷不均一性を現す個々の抗体を含む。
【図8】図8:10種類の個々のメンバーを有する組換えポリクローナル抗RhD抗体の陽イオン交換クロマトグラフィー由来の溶出プロファイル。文字は、2次元のRP−HPLCの対象としたピークを示す。
【図9】図9:図8のフラクションB5のRP−HPLC由来の溶出プロファイルを示す。
【図10】図10:色分けしたタンパク質マップ(グレースケールで表す)により視覚化した10種類の個々のメンバーを有する組換えポリクローナル抗RhD抗体の2D LC組成分析を示す。
【図11】図11:8種類の個々のメンバーを有する組換えポリクローナル抗RhD抗体由来の精製LCのAsp−N消化物の強陽イオン交換クロマトグラフィー由来の溶出プロファイルを示す。太い横線は、マーカーペプチドの同定のためMALDI−TOF分析の対象としたフラクションを示す。
【図12】図12:抗イディオタイプペプチドPEP162、PEP202、およびPEP305を用いた3種類の個々のELISA分析から得られたELISAデータを用いて、25種類の個々のメンバーをを有する抗RhD rpAbにおいて得られたOD280IEXクロマトグラムのオーバーレイを示す。ELISA分析を、イオン交換クロマトグラフィーにより得られた各フラクションにおいて行った。ELISAデータを、それぞれ互いに比較可能なPEP162、PEP202、およびPEP305を用いてELISA分析を3回行うために、トータルのOD%に対して校正する。
【図13】図13:発酵中の異なる培養時間ポイントで3種類のセンチネル抗体である抗RhD162、202、および305の分布を示す。G8は、バイオリアクターの播種の8日後に該当する。
【図14】図14:PEP202テトラマーを用いて染色した3種類の細胞株のFACSデータ。(A)は、PEP202陰性細胞株RhD162を示す。(B)は、細胞株RhD202を示し、そして(C)は、細胞株RhD162とRhd202の50%混合物(実験のmix aに対応する)を示す。A、B、およびCの第1パネルは、FSC−SSCドットプロットを示し、R1は、サイズ(FSC)および粒度(SSC)に基づき、生存かつ健常な細胞についてのゲートである。中央のパネルのヒストグラムは、細胞の蛍光強度を表す。R6ゲートは、テトラマー染色細胞を囲うものである。最後のパネルは、計算で用いたR6の細胞の割合を示す。
【図15】図15:捕獲溶出(A)、Sephadex G−25(B)、DEAE−Sepharose(C)、およびMEP Hypercel(D)後に収集した材料により表される25種類の個々のメンバーを含有する抗RhD rpAb試料の下流処理中の異なるステージで採取した試料を示す陽イオン交換クロマトグラフィープロファイル。
【図16】図16:異なる電荷パターンを示す3個の代表的なモノクローナル抗RhD抗体のIEXプロファイル。(A)均一性、(B)「3ピーク」パターン、(C)複雑なパターン。
【図17】図17:RhD189、および変異誘発Glu変異体RhD189EのIEX分析。
【図18】図18:Glu変異体RhD189E、およびその天然対応物RhD189の結合活性。RhD陽性赤血球に対する抗体の結合をFACSにより測定し、平均蛍光強度(MFI)を抗体濃度のフラクションとして示す。
【特許請求の範囲】
【請求項1】
異なる可変領域を有する異なる相同タンパク質またはかかるタンパク質を産生する細胞を含む試料を、該タンパク質の個々のメンバーまたはそれらのコード配列の相対的比率または存在に関する情報が得られるように特徴付ける方法であって、1種類以上のタンパク質特徴決定技術により、および/またはタンパク質コード配列の1種類以上の遺伝子分析により該試料のアリコートを分析することを含む、方法。
【請求項2】
該遺伝子分析が、RFLP、T−RFLP、マイクロアレイ分析、定量的PCR、および核酸配列決定から選択されるものである、請求項1記載の方法。
【請求項3】
試料が該培養物の細胞を含む細胞培養物フラクションである、請求項1または2記載の方法。
【請求項4】
該タンパク質特徴決定技術が、i)物理化学的特性によりタンパク質を分離するクロマトグラフ分析、ii)相同タンパク質のタンパク分解消化産物の分析、iii)「大量」N−末端配列決定、およびiv)相同タンパク質に特異的な検出分子を用いた分析から選択されるものである、請求項1記載の方法。
【請求項5】
該クロマトグラフ分析がサイズよりむしろ別の物理化学的特性に基づくものである、請求項4記載の方法。
【請求項6】
個々のクロマトグラフ分析が、i)正味電荷、ii)疎水性、iii)等電点、およびiv)アフィニティーから選択される物理化学的特性に基づくものである、請求項4記載の方法。
【請求項7】
クロマトグラフ分析が多次元クロマトグラフィーとして行われる、請求項4から6のいずれか1項記載の方法。
【請求項8】
相同タンパク質のタンパク分解性消化産物の分析が、特徴的なアミノ酸側鎖官能基を有するN−末端マーカーペプチドまたはペプチドを単離するために行われる、請求項4記載の方法。
【請求項9】
該特異的検出分子が、抗イディオタイプペプチドまたは抗イディオタイプ抗体である、請求項4記載の方法。
【請求項10】
該抗イディオタイプペプチドまたは抗イディオタイプ抗体が、ポリクローナル細胞株における個々のタンパク質産生細胞の決定において利用される、請求項9記載の方法。
【請求項11】
該試料が細胞培養上清からもたらされるものである、請求項1または4から9のいずれか1項記載の方法。
【請求項12】
該特徴決定が該試料に存在する1種類以上のセンチネルタンパク質またはセンチネル核酸配列の分析に基づくものである、請求項1から11のいずれか1項記載の方法。
【請求項13】
少なくとも2種類の分析技術が該試料を分析すのに適用される、請求項1から12のいずれか1項記載の方法。
【請求項14】
一方の分析技術が請求項4から9のいずれか1項記載のタンパク質特徴決定技術であり、そしてもう一方の分析技術が請求項2記載の遺伝子分析である、請求項13記載の方法。
【請求項15】
該試料が培養中の異なる時間ポイントで単一のポリクローナル細胞培養物から得られものであり、そして該個々の相同タンパク質および/またはコード配列の相対的比率が比較される、請求項1から14のいずれか1項記載の方法。
【請求項16】
該試料が特定の時間ポイントで異なるポリクローナル細胞培養物から得られものであり、そして該個々の相同タンパク質および/またはコード配列の相対的比率が比較される、請求項1から14のいずれか1項記載の方法。
【請求項17】
異なる可変領域を有する異なる相同タンパク質が組換えタンパク質である、請求項1から16のいずれか1項記載の方法。
【請求項18】
組換えタンパク質が組換えポリクローナルタンパク質を構成するものである、請求項17記載の方法。
【請求項19】
異なる可変領域を有する異なる相同タンパク質が異なるCDR領域を有する抗体である、請求項1から18のいずれか1項記載の方法。
【請求項20】
異なる可変領域を有する異なる相同タンパク質が異なるCDR領域を有するT細胞受容体である、請求項1から18のいずれか1項記載の方法。
【請求項21】
ポリクローナル細胞株における個々のタンパク質産生細胞の単離および/または決定のための、抗イディオタイプペプチドまたは抗イディオタイプ抗体の使用。
【請求項22】
ポリクローナル抗体またはモノクローナル抗体の混合物からの多様性に関する個々の抗体分子の分離における、多次元クロマトグラフィーの使用。
【請求項23】
ポリクローナルT細胞受容体またはモノクローナルT細胞受容体の混合物からの多様性に関する個々のT細胞受容体分子の分離における、多次元クロマトグラフィーの使用。
【請求項24】
組換えタンパク質中のN−末端グルタミン残基の環化により引き起こされるN−末端の電荷不均一性を解消する方法であって、該N−末端グルタミン残基を別のアミノ酸に置換することを含む、方法。
【請求項25】
該置換が該タンパク質をコードする核酸配列の部位指定変異誘発により行われる、請求項24記載の方法。
【請求項26】
該置換がポリクローナルタンパク質の1種類以上の個々のメンバーにおいて行われる、請求項24または25記載の方法。
【請求項27】
該タンパク質が抗体またはT細胞受容体である、請求項24から26のいずれか1項記載の方法。
【請求項28】
正味電荷に基づくクロマトグラフ分析を適用する方法であって、N−末端グルタミン残基の環化により引き起こされる電荷不均一性を示すタンパク質が請求項24から27記載の方法の対象とされる、請求項1またはその従属請求項に記載の方法。
【請求項1】
異なる可変領域を有する異なる相同タンパク質またはかかるタンパク質を産生する細胞を含む試料を、該タンパク質の個々のメンバーまたはそれらのコード配列の相対的比率または存在に関する情報が得られるように特徴付ける方法であって、1種類以上のタンパク質特徴決定技術により、および/またはタンパク質コード配列の1種類以上の遺伝子分析により該試料のアリコートを分析することを含む、方法。
【請求項2】
該遺伝子分析が、RFLP、T−RFLP、マイクロアレイ分析、定量的PCR、および核酸配列決定から選択されるものである、請求項1記載の方法。
【請求項3】
試料が該培養物の細胞を含む細胞培養物フラクションである、請求項1または2記載の方法。
【請求項4】
該タンパク質特徴決定技術が、i)物理化学的特性によりタンパク質を分離するクロマトグラフ分析、ii)相同タンパク質のタンパク分解消化産物の分析、iii)「大量」N−末端配列決定、およびiv)相同タンパク質に特異的な検出分子を用いた分析から選択されるものである、請求項1記載の方法。
【請求項5】
該クロマトグラフ分析がサイズよりむしろ別の物理化学的特性に基づくものである、請求項4記載の方法。
【請求項6】
個々のクロマトグラフ分析が、i)正味電荷、ii)疎水性、iii)等電点、およびiv)アフィニティーから選択される物理化学的特性に基づくものである、請求項4記載の方法。
【請求項7】
クロマトグラフ分析が多次元クロマトグラフィーとして行われる、請求項4から6のいずれか1項記載の方法。
【請求項8】
相同タンパク質のタンパク分解性消化産物の分析が、特徴的なアミノ酸側鎖官能基を有するN−末端マーカーペプチドまたはペプチドを単離するために行われる、請求項4記載の方法。
【請求項9】
該特異的検出分子が、抗イディオタイプペプチドまたは抗イディオタイプ抗体である、請求項4記載の方法。
【請求項10】
該抗イディオタイプペプチドまたは抗イディオタイプ抗体が、ポリクローナル細胞株における個々のタンパク質産生細胞の決定において利用される、請求項9記載の方法。
【請求項11】
該試料が細胞培養上清からもたらされるものである、請求項1または4から9のいずれか1項記載の方法。
【請求項12】
該特徴決定が該試料に存在する1種類以上のセンチネルタンパク質またはセンチネル核酸配列の分析に基づくものである、請求項1から11のいずれか1項記載の方法。
【請求項13】
少なくとも2種類の分析技術が該試料を分析すのに適用される、請求項1から12のいずれか1項記載の方法。
【請求項14】
一方の分析技術が請求項4から9のいずれか1項記載のタンパク質特徴決定技術であり、そしてもう一方の分析技術が請求項2記載の遺伝子分析である、請求項13記載の方法。
【請求項15】
該試料が培養中の異なる時間ポイントで単一のポリクローナル細胞培養物から得られものであり、そして該個々の相同タンパク質および/またはコード配列の相対的比率が比較される、請求項1から14のいずれか1項記載の方法。
【請求項16】
該試料が特定の時間ポイントで異なるポリクローナル細胞培養物から得られものであり、そして該個々の相同タンパク質および/またはコード配列の相対的比率が比較される、請求項1から14のいずれか1項記載の方法。
【請求項17】
異なる可変領域を有する異なる相同タンパク質が組換えタンパク質である、請求項1から16のいずれか1項記載の方法。
【請求項18】
組換えタンパク質が組換えポリクローナルタンパク質を構成するものである、請求項17記載の方法。
【請求項19】
異なる可変領域を有する異なる相同タンパク質が異なるCDR領域を有する抗体である、請求項1から18のいずれか1項記載の方法。
【請求項20】
異なる可変領域を有する異なる相同タンパク質が異なるCDR領域を有するT細胞受容体である、請求項1から18のいずれか1項記載の方法。
【請求項21】
ポリクローナル細胞株における個々のタンパク質産生細胞の単離および/または決定のための、抗イディオタイプペプチドまたは抗イディオタイプ抗体の使用。
【請求項22】
ポリクローナル抗体またはモノクローナル抗体の混合物からの多様性に関する個々の抗体分子の分離における、多次元クロマトグラフィーの使用。
【請求項23】
ポリクローナルT細胞受容体またはモノクローナルT細胞受容体の混合物からの多様性に関する個々のT細胞受容体分子の分離における、多次元クロマトグラフィーの使用。
【請求項24】
組換えタンパク質中のN−末端グルタミン残基の環化により引き起こされるN−末端の電荷不均一性を解消する方法であって、該N−末端グルタミン残基を別のアミノ酸に置換することを含む、方法。
【請求項25】
該置換が該タンパク質をコードする核酸配列の部位指定変異誘発により行われる、請求項24記載の方法。
【請求項26】
該置換がポリクローナルタンパク質の1種類以上の個々のメンバーにおいて行われる、請求項24または25記載の方法。
【請求項27】
該タンパク質が抗体またはT細胞受容体である、請求項24から26のいずれか1項記載の方法。
【請求項28】
正味電荷に基づくクロマトグラフ分析を適用する方法であって、N−末端グルタミン残基の環化により引き起こされる電荷不均一性を示すタンパク質が請求項24から27記載の方法の対象とされる、請求項1またはその従属請求項に記載の方法。
【図1】

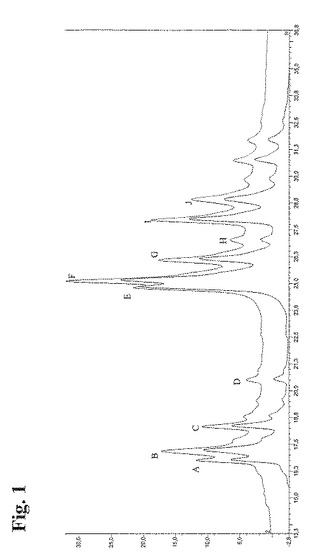
【図2】
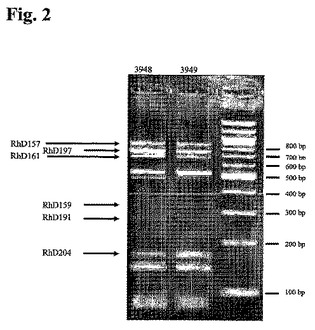
【図3】
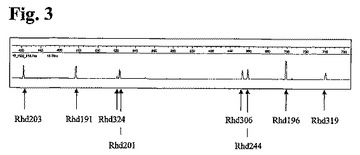
【図4】
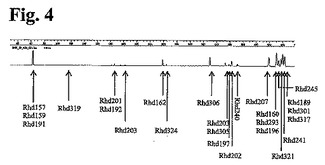
【図5】
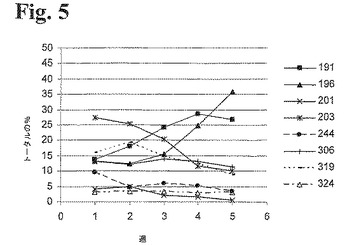
【図6】
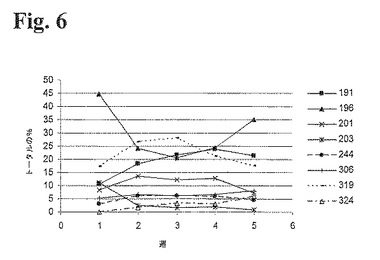
【図7】
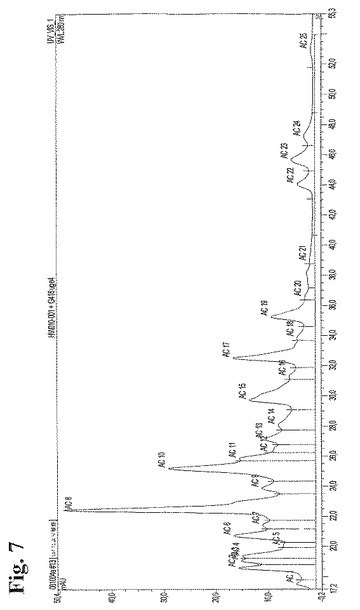
【図8】
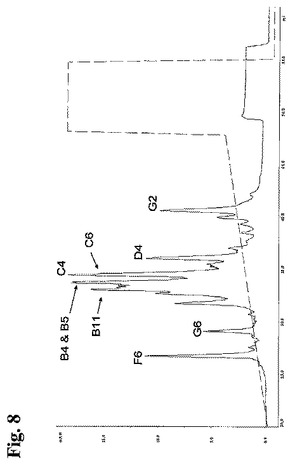
【図9】
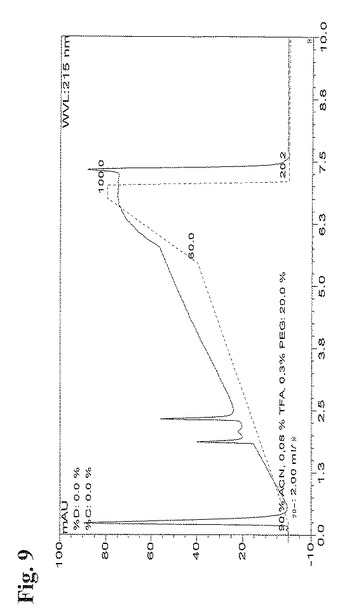
【図10】
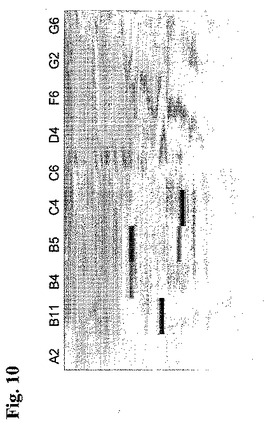
【図11】
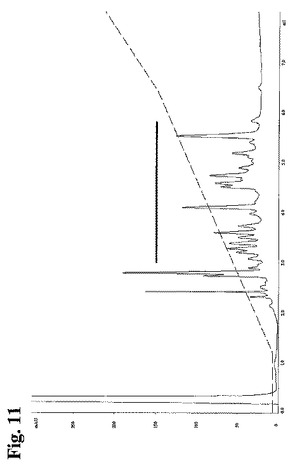
【図12】
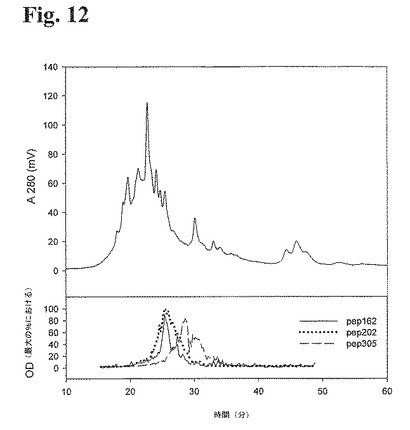
【図13】
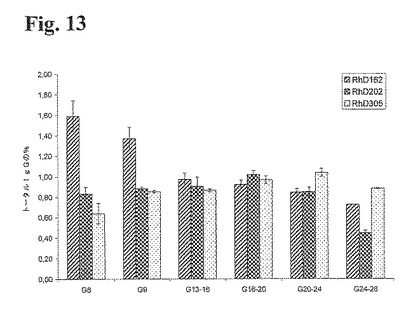
【図14】
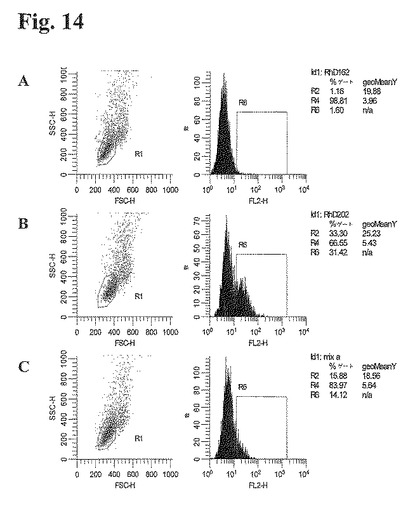
【図15】
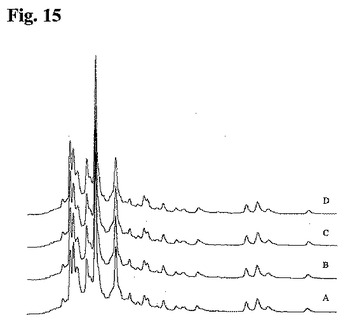
【図16】
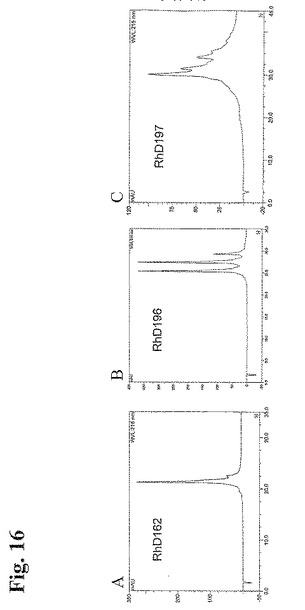
【図17】
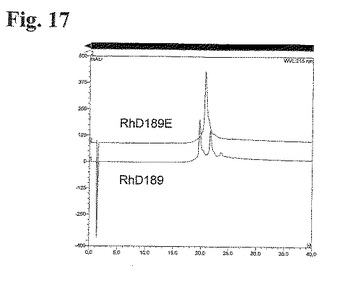
【図18】
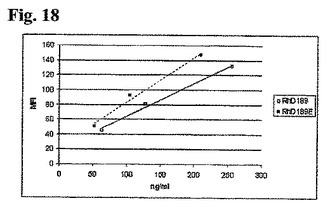
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【公表番号】特表2008−506960(P2008−506960A)
【公表日】平成20年3月6日(2008.3.6)
【国際特許分類】
【出願番号】特願2007−521795(P2007−521795)
【出願日】平成17年7月20日(2005.7.20)
【国際出願番号】PCT/DK2005/000504
【国際公開番号】WO2006/007853
【国際公開日】平成18年1月26日(2006.1.26)
【出願人】(505257682)シムフォゲン・アクティーゼルスカブ (24)
【氏名又は名称原語表記】SYMPHOGEN A/S
【Fターム(参考)】
【公表日】平成20年3月6日(2008.3.6)
【国際特許分類】
【出願日】平成17年7月20日(2005.7.20)
【国際出願番号】PCT/DK2005/000504
【国際公開番号】WO2006/007853
【国際公開日】平成18年1月26日(2006.1.26)
【出願人】(505257682)シムフォゲン・アクティーゼルスカブ (24)
【氏名又は名称原語表記】SYMPHOGEN A/S
【Fターム(参考)】
[ Back to top ]