
組換えヨウシュヤマゴボウ(pokeweed)抗ウイルスタンパク質、それに関連する組成物および方法
本発明は、新規修飾ヨウシュヤマゴボウ(pokeweed)抗ウイルスタンパク質、該タンパク質をコードする核酸、該タンパク質を取り込むコンジュゲート、および該タンパク質を作製しそして使用する方法を提供する。本発明はまた、特定の細胞に毒素を導く目的のため、該コンジュゲートを動物に投与する方法も提供する。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願
連邦政府が資金援助した研究に関する言及
[0002]なし
技術分野
[0003]本発明は、一般的に、分子生物学および生化学に関し、より詳細には、ヨウシュヤマゴボウ(pokeweed)植物由来の修飾リボソーム不活性化タンパク質に関する。ヨウシュヤマゴボウ植物はまた、Phytolacca americanaとしても知られ、そしてヨウシュヤマゴボウ・リボソーム不活性化タンパク質はまた、ヨウシュヤマゴボウ抗ウイルスタンパク質とも呼ばれ、しばしば「PAP」と略される。本発明はまた、獣医学的薬剤を含む薬剤にも関する。
【背景技術】
【0002】
[0004]化合物コンジュゲート化ヨウシュヤマゴボウ抗ウイルスタンパク質(PAP)、ならびにリシンおよびジフテリア毒素などの他の天然毒素のコンジュゲートは、長らく、療法的有効性の可能性を秘めてきた。理論的には、コンジュゲートの「化合物」部分としての天然リガンドの存在は、ターゲット細胞損傷を生じ、そして他の細胞損傷を生じない。実際には、部分的に、コンジュゲート化されていない毒素が、意図されない細胞損傷を生じるために、不正確なターゲティング毒性を生じる。PAPに関しては、1つの問題は、コンジュゲート化PAPおよび非コンジュゲート化PAPのサイズが非常に類似であるため、分離技術がこれらを区別不能であることである。
【0003】
[0005]自然(「天然」とも称される)PAPはヨウシュヤマゴボウ植物から単離され、そして天然PAPを化合物−毒素コンジュゲートにおいて利用しようとする試みがなされてきたが、こうした試みは信頼性があることを示していない。予期されるように、年ごとおよびバッチごとのアイソフォームの変動が、薬学的品質調節の背景では、厄介でそして実現不能であることがわかった。さらに、いくつかのアイソフォームはコンジュゲートを形成せず、そして異なるアイソフォームは互いに異なってコンジュゲートを形成した。
【0004】
[0006]理想的には、組換え発現は、これらの変動に対する調節を提供するであろう。しかし、PAPの組換え発現もまた、困難に遭遇してきた。大腸菌(E. coli)における以前の発現は、毒性および成長阻害、ならびに封入体における組換えヨウシュヤマゴボウ抗ウイルスタンパク質(rPAP)の集積を生じた。これに関連して、組換えPAPは、別個の可溶化工程およびそれに続くタンパク質の再フォールディングを必要とし、その結果、収量が劣り、そしてスケールアップが困難であった。大腸菌、S.セレビシエ(S. cerevisiae)、植物およびP.パストリス(P. pastoris)における他の試みは低収量を生じ、またはP.パストリスの場合、潜在的に炎症反応を誘導しうる配列が導入された。さらに、組換えPAP−化合物融合タンパク質は、毒素と結合するのに失敗するかまたはターゲット細胞に毒素を導くのに失敗するか、あるいは天然PAPに比較して、はるかに減少した活性を示すか、いずれかであった。
【発明の概要】
【発明が解決しようとする課題】
【0005】
[0007]したがって、未結合(free)システインを有するrPAP分子、該分子から作製されるコンジュゲート、およびrPAPを産生する方法、特に、高収量であり、容易にフォールディングし、そして精製されるrPAPを生じ、そして場合によってコンジュゲート化に化学的に利用可能なrPAPを提供する方法は、大きな貢献となる。
【課題を解決するための手段】
【0006】
[0008]一般論として、本発明は、未結合システイン、好ましくは末端システイン、より好ましくはN末端システインを有する組換えヨウシュヤマゴボウ抗ウイルスタンパク質を含む組成物を提供する。やはり提供するのは、PAPが全長rPAP、より好ましくは未結合システインを含む全長rPAP、最も好ましくは未結合システインおよびアミノ酸リンカーを含む全長rPAPである、rPAP分子である。好ましいのは、N末端Cysおよびアミノ酸リンカーを含むrPAP分子であり、最も好ましくは、Gly−Gly−Gly−Gly−Serの少なくとも1つの反復を有するものである。より好ましくは、Cys−Gly−Gly−Gly−Gly−Ser−全長rPAPおよびCys−Gly−Gly−Gly−Gly−Ser−Gly−Gly−Gly−Gly−Ser−全長rPAPである。
【0007】
[0009]本発明は、本発明の方法にしたがって発現された場合、宿主細胞を殺さないrPAPを提供する。本発明の組成物および方法で利用されるrPAPは、好ましくは、29.5ダルトン以上であり、より好ましくは30ダルトン以上であり、最も好ましくは30.5ダルトン以上である。しかし、31.5、31.75および32ダルトン以上の分子量を有する全長rPAPを利用する組成物および方法もまた、本発明の範囲内である。全長rPAPタンパク質(翻訳後修飾されていない天然PAPの分子量と一致するもの)が、本発明で用いるのに好ましい物質である。
【0008】
[00010]核酸、プラスミド、ならびに本発明の核酸およびタンパク質を含む細胞もまた提供し、好ましい細胞は大腸菌である。
[00011]構造:
X−Y−Z、
式中、Xは未結合システインを有する全長rPAPであり;Yは存在しないかまたは化学的リンカーであり、そしてZは化合物である
を有するコンジュゲートもまた提供する。
【0009】
[00012]細胞ターゲティング化タンパク質である化合物が好ましく、より好ましくは:抗体;ホルモン;修飾ホルモン放出因子;およびホルモン放出因子からなる群より選択されるものが好ましい。化学的リンカーが柔軟なリンカーである化合物が好ましく、ヘテロ二官能性リンカーを持つものがより好ましく、マレイミド基を有するリンカーを持つものが最も好ましい。リンカーがGMBS;EMCS;SMPH;SPDP;およびLC−SPDPからなる群より選択される、本明細書に記載するようなコンジュゲートが好ましい。前記リンカーがGMBSであり、そして前記タンパク質がd−lys6−ゴナドトロピン放出ホルモンであるコンジュゲートが最も好ましい。
【0010】
[00013]本明細書のrPAPを別の化合物とコンジュゲート化する方法であって、組換えヨウシュヤマゴボウ抗ウイルスタンパク質の前記の未結合システインおよび別の化合物間に、化学結合を誘導する工程を含む、前記方法もまた提供する。好ましい方法は、前記化学結合が、ヘテロ二官能性架橋剤を介して誘導されるものであり、より好ましくは、化学結合が、未結合システインおよび化合物上のマレイミド基間に誘導されるものである。ヘテロ二官能性架橋剤がGMBSであり、そして/または化合物がd−lys6−ゴナドトロピン放出ホルモンであるものが最も好ましい。
【0011】
[00014]GMBSリンカーをd−lys6−ゴナドトロピン放出ホルモンに結合させる方法であって、GMBSとd−lys6−ゴナドトロピン放出ホルモンを、非水性条件下でインキュベーションする工程を含む、前記方法もまた提供し、好ましくは、前記非水性条件は:GMBSをメタノール中に溶解させて、第一の非水性溶液を生成し;d−lys6−ゴナドトロピン放出ホルモンをメタノール中に溶解させて、第二の非水性溶液を生成し;前記の第一および第二の非水性溶液を、1.1:1のモル比で混合する工程を含む。
【0012】
[00015]大腸菌において、全長rPAPをコードする核酸を発現する工程を含む、rPAPを得る方法もまた提供する。
[00016]全長rPAPを含む核酸で形質転換された細胞をインキュベーションする工程を含み、rPAPがT7プロモーター系の調節下にあり、そして前記T7プロモーター系がアラビノース・プロモーターの調節下にあるRNAポリメラーゼを有する、細胞を増殖させる方法もまた提供する。前記細胞が大腸菌細胞である方法が好ましい。rPAPが未結合システインを含む方法が好ましく、rPAPが末端システインを含むものが最も好ましい。全長rPAPが:化学的に修飾されたrPAP、天然変異体rPAP、および遺伝子操作されたrPAPからなる群より選択される方法もまた好ましい。
【0013】
[00017]本明細書のPAP組成物を含むコンジュゲートもまた提供する。構造:
X−Y−Z、
式中、XはN末端Cys−Gly−Gly−Gly−Gly−Serを有する全長rPAPであり;Yは化学的リンカーであり、そしてZはタンパク質である
を有するものが特に好ましい。XがGMBSであり、そしてZがd−lys6−GnRHであるものが最も好ましい。
【0014】
定義
[00018]「未結合システイン」は、ジスルフヒドリル結合を介して別のシステインに結合しているもの以外の任意のシステインを意味する。これに関連して、「未結合システイン」には、システインがジスルフヒドリル結合を介して別のシステインに結合していない限り、別の残基または化合物に結合しているシステインが含まれる。
【0015】
[00019]「全長rPAP」は、毒素活性を有し、そして29,500ダルトン以上の分子量を有する、任意の組換えPAPを意味する。
[00020]本発明のこれらのおよび他の特徴および利点は、好ましい態様の詳細な説明から、当業者にはより明らかとなるであろう。詳細な説明に付随する図を以下に記載する:
【図面の簡単な説明】
【0016】
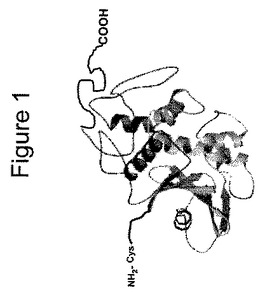
【図1】[00021]図1は、全長ヨウシュヤマゴボウ抗ウイルスタンパク質の構造の定型化された描写であり、翻訳後プロセシング中、植物において通常は切断されるC末端残基、およびさらなる非天然存在N末端残基を示す。
【図2】[00022]図2は:翻訳後プロセシング中、植物において通常は切断されるC末端;およびさらなる非天然存在N末端残基を示す、全長ヨウシュヤマゴボウ抗ウイルスタンパク質;N末端リンカー;ならびに例示的な化合物、修飾ゴナドトロピン放出ホルモン(GnRH)の構造の定型化された描写である。
【図3】[00023]図3は、組換え切断不能(「nc」と略され、そしてリンカーがジスルフィド切断部位を所持しないことを意味する)ヨウシュヤマゴボウ・コンジュゲートの構造の定型化された描写であり、リンカーおよび修飾ゴナドトロピン放出ホルモンを示すアミノ末端の詳細を伴う。
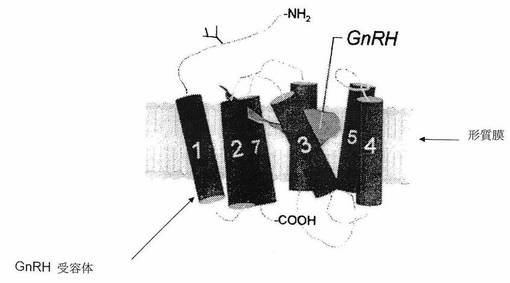
【図4】[00024]図4は、ゴナドトロピン放出ホルモンおよびゴナドトロピン放出ホルモン受容体の細胞外ドメインの相互作用の定型化された描写である。
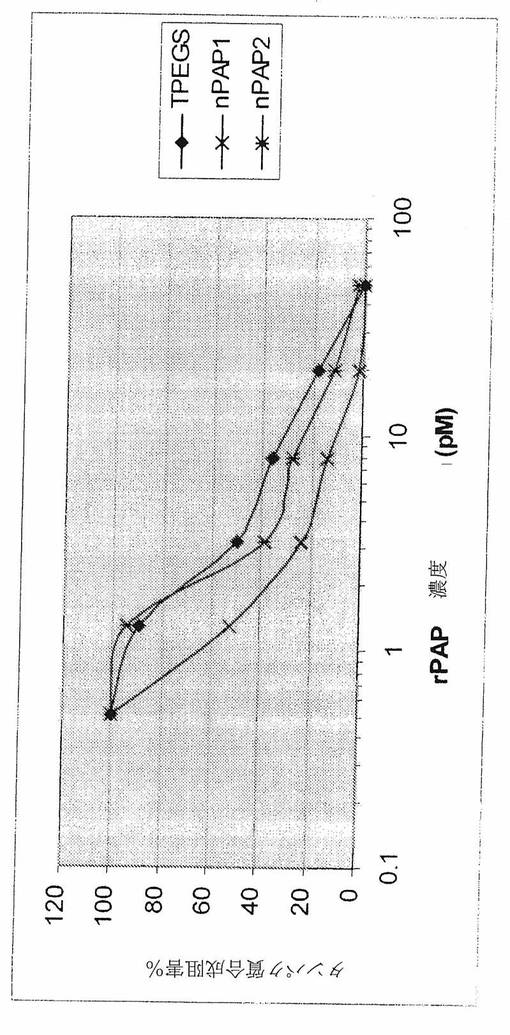
【図5】[00025]図5は、ウサギ網状赤血球溶解物アッセイにおいて、ルシフェラーゼmRNAのタンパク質合成阻害によって測定されるようなrPAPの生物学的活性を示すグラフである。
【図6】[00026]図6は、競合的ラジオイムノ受容体結合アッセイの結果を示すグラフである。X軸に最も近い曲線は、標準に関するデータを反映する。次の曲線は、PAPが植物部分から精製された(自然または天然PAPとしても知られる)d−lys6−GnRH−GMBS−PAPに関するデータを反映する。データ点が三角形で示される曲線は、d−lys6−GnRH−GMBS−rPAPに関するデータを反映する。データ点がXとして示される線は、非コンジュゲート化組換えPAP(d−lys6−GnRH−GMBSに結合していない)を反映する。
【図7】[00027]図7。本発明の核酸−rPAP DNA配列−配列番号__。
【図8】[00028]図8。本発明のアミノ酸配列−rPAPタンパク質配列−配列番号1。太字:ATGおよびリンカー配列;黒字:天然成熟PAP配列;太字および下線:植物において翻訳後切断されるタンパク質のC末端部分をコードする天然プレPAP配列。
【図9】[00029]図9。本発明のrPAP(上部)−配列番号2、および天然成熟(植物において翻訳後修飾されたもの)PAP(下部)発現DNA配列のアライメント。
【図10】[00030]図10。rPAP(上部)、および天然成熟PAP(下部)タンパク質配列のアライメント。
【図11】[00031]図11。大きな太字で示す内部ジスルフィド結合システインを含む、発現されたrPAPの配列:Cys−34はCys−258に結合し、そしてCys−84はCys−105に結合する;太字;N末端システインを伴う操作されたリンカー;VNTII....:天然PAP配列;太字および下線は天然プレPAP(植物において翻訳後修飾されたもの)のC末端である。
【発明を実施するための形態】
【0017】
[00032]本発明は、大腸菌において高収量で発現可能であり、そしていかなる他の組換えPAPよりも30〜40倍高い比活性(生物学的活性/単位質量)を有する、組換えヨウシュヤマゴボウ抗ウイルスタンパク質を提供する。さらに、本発明は、薬学的な量で、rPAPを産生するための方法を提供する。
【0018】
[00033]本発明のrPAP物質(タンパク質、核酸、構築物、細胞等)を用いて、rPAP、およびリンカーを介すかまたは直接のいずれかで、これらに結合されたターゲティング化合物を有する、rPAPコンジュゲートを産生してもよい。1つの態様において、rPAPは、別の化合物に対するリンカーを連結するのに場合によって使用するための、未結合システインを有する。1つのこうした態様において、本発明のrPAPタンパク質は、こうした目的のために都合のよいN末端システインを提供するが、本発明のrPAPの使用はN末端コンジュゲート化に限定されない。例えば、本発明のrPAPを、コンジュゲート化なしの毒素として用いてもよいし、あるいは、末端システインまたは内部システインの未結合システインを介して、コンジュゲート化してもよい。
【0019】
[00034]本明細書記載のrPAP分子は、これらにコンジュゲート化されたリンカーまたはターゲティング化合物を伴いまたは伴わず、ウサギ網状赤血球溶解物アッセイにおいて活性である。
【0020】
[00035]本発明には、組換えPAPを発現させ、再フォールディングさせ、コンジュゲート化し、そして精製する方法が含まれる。いくつかの障害を克服して、発現成功を達成した。非ヨウシュヤマゴボウ宿主細胞においてrPAPを発現するのに伴う基本的な問題は、これが毒素であり、そして宿主細胞を殺すことである。T7系を用いて、大腸菌において成熟(翻訳後切断された)PAPを発現する試みを行った。細胞はまったくではなくてもあまり増殖せず、そしてrPAPタンパク質誘導前に弱っているように見えた。続いて、大腸菌において、T7誘導性プロモーター系を用いて、全長rPAP(植物において通常は翻訳後に切断されるC末端部分を加えた成熟PAP)を発現する試みもまた失敗した。細胞はやはり、増殖期中およびrPAPの誘導前に、弱っているように見えた。最後に、2つの制御調節シグナル下の全長rPAPを、大腸菌において試みた。T7 RNAポリメラーゼをアラビノース(AraD)プロモーターの調節下に置き、T7プロモーターを全長rPAP配列の上流に置いた。アラビノース・プロモーターが厳重に抑制されていると、プラスミド上にrPAP遺伝子を宿する場合であっても、細胞は増殖可能であった。抑制の除去を介して誘導すると、薬学的に作業しうる収量のrPAPが生じた。
【0021】
[00036]本発明のrPAPを再フォールティングする多様な方法がある。最も成功した1つは、実施例2に記載するようなものである。別の方法は、スクロースの代わりに0.5M L−アルギニンを代用して、実施例2のプロトコルを用いるものである。さらに、実施例2のプロトコルにおいて、システアミンの代わりに、グルタチオンを用いてもよい。場合によって、8M尿素の代わりに6Mグアニジン−HClで封入体を可溶化してもよい。再フォールディングは、理想的には、塩基性pH範囲で行われる。
【0022】
[00037]すべて当該技術分野でよく記載される、イオン交換クロマトグラフィー、疎水性相互作用クロマトグラフィー、およびヒドロキシアパタイトクロマトグラフィーを含む多様な方法によって、タンパク質を精製してもよい。好ましい方法は、特に実施例5に記載するような陽イオン交換クロマトグラフィーである。
【0023】
[0038]さらに、この組換えタンパク質に対して行う実験に基づいて、本発明のrPAPの比活性(生物学的活性/単位質量)が、ウサギ網状赤血球溶解物におけるタンパク質翻訳の阻害において、別の報告されるrPAPよりも、30〜40x高い活性であることが決定された。非常に感受性であり、そしてナノモル未満の量のrPAPを検出可能なrPAP特異的放射免疫アッセイによって、rPAP濃度を決定した。
【0024】
[00039]理想的には毒素機能を保持するようにフォールティングされた組換えPAPタンパク質、好ましくは天然存在システインの天然ジスルフィド架橋を保持するもの、そして好ましくは少なくとも1つの未結合システイン(例えば天然存在配列には存在しないもの)、最も好ましくは、他の化合物に選択的に結合可能な末端未結合システインを有するものが、本明細書に提供される。当該技術分野の技術のように、任意のPAP配列が、本発明の出発物質として使用するのに適している。任意の既知のアイソタイプ、または明らかになる任意のものが、本発明を用意するのに有用であろう。
【0025】
[00040]全長PAPは、C末端に以下のアミノ酸配列を有する:YNQNAMFPQLIMSTYYNYVNLGDLFEGF−COOH。該配列は、通常、ヨウシュヤマゴボウ植物において、翻訳後に切断されるが、本発明の好ましい態様においては保持される。天然存在翻訳後切断PAPは、29,308.5ダルトンの分子量を有する。
【0026】
[00041]特に、配列番号1;未結合システインを含み、そしてデフォルト設定でBLASTソフトウェアバージョン2.2.21を用いて、配列番号1に対して少なくとも90%同一であるタンパク質;配列番号2にコードされるタンパク質;未結合システインを含み、そしてデフォルト設定でBLASTバージョン2.2.21ソフトウェアを用いて、配列番号2に対して少なくとも90%同一である核酸によってコードされるタンパク質からなる群より選択される、上述のようなrPAP組成物。しかし、配列同一性が:95%;96%;97%;98%;および99%からなる群より選択される、上述のような組成物もまた好ましい。
【0027】
[00042]配列番号2;デフォルト設定でBLASTソフトウェアバージョン2.2.21を用いて、配列番号2に対して少なくとも85%同一であり、そして未結合システインを有するタンパク質をコードする核酸;配列番号1をコードする核酸;および未結合システインを有するタンパク質をコードし、そしてデフォルト設定でBLASTソフトウェアバージョン2.2.21を用いて、配列番号1に対して少なくとも85%同一である核酸からなる群より選択される、核酸もまた提供する。しかし、配列同一性が:90%;95%;96%;97%;98%;および99%からなる群より選択される、上述のような組成物もまた好ましい。好ましい核酸は、本明細書のタンパク質をコードする核酸を含む。
【0028】
[00043]GMBSリンカーをd−lys6−ゴナドトロピン放出ホルモンに結合させる方法であって、GMBSとd−lys6−ゴナドトロピン放出ホルモンを、非水性条件下でインキュベーションする工程を含む、前記方法もまた提供する。この方法のより好ましい態様は、前記非水性条件が:GMBSをメタノール中に溶解させて、第一の非水性溶液を生成し;d−lys6−ゴナドトロピン放出ホルモンをメタノール中に溶解させて、第二の非水性溶液を生成し;前記の第一および第二の非水性溶液を、1.1:1のモル比で混合する工程を含むものである。
【0029】
[00044]特に、配列番号1に対して、少なくとも90%同一、好ましくは少なくとも95%同一、最も好ましくは少なくとも99%同一であるrPAPが、本発明の方法において有用である。未結合CYS残基もまた含むものが最も有用である。さらに、保存配列は変更されてはならず、そして非保存配列は場合によって変更可能である。PAPにおいて、天然存在システイン間のジスルフィド結合は、毒素機能に必要な三次構造を提供し、そして理想的には本発明の分子および方法において保存される。C末端ドメインにおける突然変異は、PAPの局在化プロセシングに影響を及ぼし、そして改変されたプロセシングが望ましい場合には、改変可能である。RNA結合ならびに脱プリン化に影響を及ぼす突然変異が知られる。例えば、最初の16アミノ酸の一部切除(truncation)によって、PAP細胞毒性およびリボソームを脱プリン化する能力が排除される。さらに、リボソーム脱プリン化は、アミノ酸がC末端から除去されるにつれて減少し、そしてGlu−244に停止コドンが導入されると排除される。さらに、特に毒性が高い化合物を得るために、既知の方法によって、過剰活性突然変異体をスクリーニングすることも可能である。本発明の機能を最適化するために、これらの突然変異効果を利用してもよい。さらに、これらの突然変異rPAPおよびこうした突然変異体を利用する組成物は、本発明の範囲内である。
【0030】
[00045]発現された際に、rPAPタンパク質を生じるヌクレオチドもまた本発明に含まれる。特に、配列番号2が好ましい。しかし、上記配列における特定の変化は、本発明の基本的な側面を改変しないであろうことを当業者は認識する。したがって、本発明には、本発明の核酸と、ストリンジェントな条件下のハイブリダイゼーションを用いて相同であるか、BLASTを用いて同一であるか、タンパク質配列を変化させない点突然変異、タンパク質配列を変化させないコドン変化などの、機能に影響を及ぼさない重要でない変化を有する核酸などが含まれる。
【0031】
[00046]本明細書のコンジュゲートおよび本明細書の化合物をコンジュゲート化する方法もまた提供する。コンジュゲートは、理想的には、細胞損傷が望ましい受容体に選択的に結合するよう設計される。一般的に、ターゲティング化合物を介して受容体に結合した後、コンジュゲートは、受容体が仲介するエンドサイトーシスによって取り込まれ、そしてコンジュゲートを細胞に送達する。取り込み後、コンジュゲートのrPAP部分は、大リボソームRNAの保存されたサルシン(sarcin)/リシンループを脱プリン化することによって、リボソームRNAに結合する。脱プリン化されたリボソームは、伸長因子2に結合不能であり、そしてしたがって、伸長サイクルの転位置工程が阻害され、タンパク質合成のシャットダウンが生じる。細胞は最終的に死ぬ。
【0032】
[00047]本明細書の特定のrPAPタンパク質に化合物をコンジュゲート化するための1つの特定の方法は、N末端システインおよび別の化合物間に化学結合を導入する工程を含む。化合物が抗体、ホルモン、修飾ホルモン放出因子、またはホルモン放出因子であるような、こうした方法が好ましい。特に、ホルモン放出因子がGnRHであるものがより好ましいが、d−lys6−修飾GnRHへのコンジュゲート化が最も好ましい。コンジュゲート化は、任意の既知の方法を介して実行可能であるが、好ましくは、リンカーまたは他の架橋化合物を介すかのいずれかで、ターゲティング化化合物およびrPAP間にスルフヒドリル結合を生成することを介する。言い換えると、未結合システインを利用して、rPAPにおける他のシステインへの結合を排除することが理想的であるが、当業者は、機能性を最適にするため、rPAPおよびそれにコンジュゲート化される化合物の両方を修飾する方法を知っている。
【0033】
[00048]本発明の好ましい態様において、修飾ゴナドトロピン放出ホルモン「d−lys6−GnRH」を全長rPAPにコンジュゲート化する。d−lys6−GnRHは、好ましくは、全長rPAP上の未結合システインへの結合を容易にするため、リンカーGMBSで活性化される。d−lys6−GnRHのGMBSでのこうした「活性化」は、通常試みられるような水性条件下で試みた場合には障害であることが証明された。水性条件下では、1つのd−lys6−GnRH分子は、2〜3分子のGMBSに結合し、これはd−lys6−GnRHあたり1つのrPAPを結合させるためには許容しえない。しかし、d−lys6−GnRHを非水性条件下(メタノール)で活性化すると、障害は克服され:1つのGMBSリンカー分子に対する1つのd−lys6−GnRHの比が達成された。したがって、rPAP対d−lys6−GnRHの1対1比もまた達成された。
【0034】
[00049]ヘテロ二官能性架橋剤、特にGMBSを利用する方法が好ましいが、リンカー上に位置するNHSエステル基を介してd−lys6−GnRHに結合するか、またはリンカー上に位置するマレイミド基を介してrPAP上の未結合スルフヒドリル基に付着するのを容易にするであろう任意のヘテロ二官能性架橋剤もまた好ましい。
【0035】
[00050]GnRH分子の両端が受容体への結合に必要である。GnRHおよびd−lys6−GnRH(交換可能に、「DK6」または「dK6」または「d−lys6」または「d−Lys6」とも称される)の唯一の相違は、6位のグリシンのD−リジンでの置換である。さらに、末端がブロッキングされる。C末端はエチルアミド基(ET−NH2)でブロッキングされ、それによって天然化合物の10位のグリシンが置換される。天然GnRH化合物は、NH2Glu−His−Trp−Ser−Tyr−Gly−Leu−Arg−Pro−GlyCOOHである。好ましい類似体は、dK6: Hp−Glu−His−Trp−Ser−Tyr−DLys−Leu−Arg−Pro−Et−NH2である。
【0036】
[00051]別の態様において、アミノ酸配列Cys−Gly−Gly−Gly−Gly−Serが全長rPAPに付加され、そしてターゲティング化合物に結合するのに用いられる。Cys−Gly−Gly−Gly−Gly−Serは、天然PAP配列の一部ではない。Val−Aspは、天然PAP配列の最初の2つのアミノ酸である。
【0037】
[00052]本発明の最も好ましいコンジュゲートは、以下の構造:
X−Y−Z
式中、Xはd−lys6−GnRHであり;YはGMBSであり;そしてZはN末端にCGGGGSを有する全長rPAPである
を有する。
【0038】
[0053]本明細書記載の方法、あるいは当該技術分野で知られるかまたは発展される任意の方法を介して、コンジュゲートを作製してもよい。さらに、当該技術分野に知られるように、任意の望ましい機能性を提供するため、コンジュゲートを修飾してもよい。実施例は好ましいコンジュゲート化法を記載する。
【0039】
[0054]本発明のコンジュゲートを投与するため、任意の塩、懸濁物、分散物等を用いてもよい。無菌であり、そして非発熱性である、0.7%〜10%、より好ましくは0.9%の塩化ナトリウム溶液が好ましく、より好ましくは、4.5〜7のpHでもある溶液が好ましい。さらに、コンジュゲートが適切な影響を提供するならば、任意の投与法が許容されうる。本発明の最も好ましい態様は、不妊処置の目的のために動物に注射するため、溶液中のrPAP−GNRH塩を用いるものである。不妊は完全である必要も、可逆的である必要もない;しかし、意図される最適な様式は、動物、特にイヌ、ネコ、ウマ、食物または他の製品のための家畜(畜牛、乳牛、ブタ、ヒツジ、ヤギ、バイソン、バイソン/畜牛種等)、労働する家畜、動物園の動物、および野生生物(特にシカ、エルク、および慢性消耗病に感受性である他の有蹄動物)で使用するための非可逆性rPAP−d−lys6−GnRH注射液である。
【0040】
[00055]前述の発明は、適切な法律基準にしたがって記載されてきており、したがって、説明は、事実上、限定するのではなく例示的である。開示する態様に対する変動および修飾が、当業者に明らかになることも可能であり、そしてこれらは本発明の範囲内である。
【実施例】
【0041】
実施例1. 大腸菌におけるrPAPの発現。
[00056]順方向プライマー、rPAP−F: 5’−CCCGGG CATATG TGC GGA GGC GGA GGC AGT GTG AAT ACA ATC ATC TAC AAT GTT GGA AGT ACC−3、および逆方向プライマー、rPAP−R: 5’−GCG CGC AAG CTT TCA GGA TTC TTC AAA TAG ATC ACC AAG ATT AAC Cを用いて、PCR増幅によって、全長配列(配列番号2)を得た。
【0042】
[00057]反応混合物は、以下の構成要素からなった: 600mM Tris−SO4 (pH 8.9)、180mM硫酸アンモニウム、0.2mM dATP、0.2mM dCTP、0.2mM dGTP、0.2mM dTTP、2mM MgSO4、0.2μM rPAP−Fプライマー、0.2μM rPAP−Rプライマー、1ngテンプレートDNA、1単位のPlatinumR Taq高忠実度ポリメラーゼ(Invitrogen社、カリフォルニア州カールスバッド)。PCR反応を以下の条件下で行った: 94℃x2分間(1サイクル)、94℃x30秒間、52℃x30秒間、68℃x1分間(15サイクル)、94℃x30秒間、55℃x30秒間、68℃xl分間(25サイクル)。
【0043】
[00058]製造者の説明書にしたがってNdeIおよびBamHI(New England Biolabs、マサチューセッツ州イプスウィッチ)を用いて、pET3a発現プラスミド中、T7プロモーターの下流に、rPAPをコードする全長配列を導入した(連結した)。
【0044】
[00059]rPAP配列含有プラスミドを用いて、One Shot(登録商標)TOP10化学的コンピテント大腸菌株(Invitrogen社、カリフォルニア州カールスバッド)を形質転換した。いくつかのコロニーを摘み取り、そして挿入物の存在に関して、DNA配列分析によってスクリーニングした。正しいrPAP配列を含有するプラスミドを宿することが示されたコロニー由来のプラスミドDNAを精製し、そして続いて、アンピシリン耐性選択可能マーカーとともに厳重に制御されるアラビノース・プロモーター(AraD)の調節下のT7 RNAポリメラーゼを所持する大腸菌BL21(AI)株を形質転換するのに、該プラスミドDNAを用いた。形質転換体を選択し、そしてrPAP発現を抑制するため、推定される形質転換体をLB選択培地およびグルコース上にプレーティングした。
【0045】
[00060]2つの単離体を研究用に選択し、そしてrPAP配列を含まない発現プラスミドを含有する対照を生成した。各単離体を別個に、ラクトースおよびアラビノースを含まず、そしてグルコースが存在する最少培地中で、振盪しながら37℃でおよそ12時間(一晩)増殖させた。対照を同じ条件下で増殖させた。アラビノース・プロモーター系の誘導を抑制する目的のために増殖培地を選択し、それによって、rPAP RNA発現/タンパク質翻訳を抑制した。
【0046】
[00061]結果は以下の通りである。
【0047】
【表1】

【0048】
[00062]少量の各一晩培養物を、アンピシリンを含有するLB培地に移し、そしてA600が0.4に到達した後、続いて、L−アラビノースを最終濃度0.2%に、そしてイソプロピルβ−D−1−チオガラクトピラノシドを1mMの濃度に添加することによって、rPAPを大腸菌細胞から誘導した。誘導をさらに3.5時間行った。
【0049】
実施例2. rPAPの再フォールディングおよび精製。
[00063]スナップ希釈によってrPAPを再フォールディングした。封入体の単離後、8M尿素、50mM Tris HCl、pH8.5中で封入体を可溶化した。DTTを最終濃度10mMまで添加し、そして混合物を室温で90分間攪拌した。次いで、可溶化されたタンパク質を室温で攪拌しながら、50mM Tris、pH8.5、0.4Mスクロース、0.05%ポリエチレングリコール−3550、0.9mM酸化システアミン(TPEGS)を含有する溶液内に一滴ずつ添加した。再フォールディング溶液中のrPAPの最終濃度は、10μg/mlおよび50μg/mlの間であった。可溶化されたrPAPを再フォールディング溶液に添加した後、混合物を4℃でさらに24時間攪拌した。24時間後、混合物を16000xgで15分間遠心分離し、上清をデカントし、そして再フォールディング後、50mM Tris、pH7.0、1mM EDTAを含有する緩衝液に対して、タンパク質溶液を透析した。緩衝液pHは、6.8〜8.5の範囲を有した。透析後、溶液を16000xgで15分間遠心分離し、そして上清を陽イオン交換樹脂上に置いた。次いで、カラムを50mM Tris−HCl、pH7.0、1.0mM EDTAで洗浄し、そして50mM Tris、pH7.0、1M NaClを含有する緩衝液でタンパク質を溶出させる。50mM NaPO4、pH7.2、100mM NaCl、1mM EDTAを含有するコンジュゲート化緩衝液に対して、溶出タンパク質を透析する。タンパク質濃度を、0.2mg/ml〜1.0mg/mlの濃度に調整する。
【0050】
実施例3. d−lys6修飾ゴナドトロピン放出ホルモン(GnRH)のマレイミドブチリルオキシ−スクシンイミドエステル(GMBS)リンカーでの活性化。
[00064]1224ダルトンの分子量を有するd−lys6−GnRHを、固相合成(Anaspec社、カリフォルニア州フレモント)によって調製した。6ミリグラムのd−lys6−GnRHを1.5ml脱イオン化メタノールと混合し、そしてジイソプロピルエタノールアミン(DIPEA)を用いて、pH7.0に調整した。
【0051】
[00065]Thermo Fisher Scientific(イリノイ州ロックフォード)からGMBSを購入した。1.25mgのGMBSを1.5ml脱イオン化メタノールと混合した。
【0052】
[00066]1.5mlのd−lys6−GnRH−メタノールおよび1.5mlのGMBS−メタノールを、キャップ付血清ボトル中で一緒に混合し、そしてDIPEAを用いてpH7.0に調整した。金属キャップを用いて血清ボトルを密封した。溶液を脱気し、そして窒素で4回パージした。血清ボトルをアルミニウムホイルで覆い、そして反応を攪拌しながら室温で90分間進行させた。
【0053】
[00067]生じたd−lys6−GnRH−GMBSは、およそ1421ダルトンの分子量を有し、1分子のGMBSが1分子のd−lys6−GnRHに結合したことが示された。これを質量分析によって確認した。
【0054】
実施例4. d−lys6−GnrH−GMBSへのrPAPのコンジュゲート化。
[00068]実施例3の溶液を遠心分離蒸発装置で蒸発させた。TCEP・HCl Tris(2−カルボキシエチル)ホスフィン塩酸塩を、コンジュゲート化緩衝液中に溶解され、再フォールディングされた組換えPAPに、最終濃度0.05mMまで添加した。混合物を室温で1〜2時間インキュベーションした。インキュベーション後、コンジュゲート化緩衝液中に溶解され、再フォールディングされたrPAPを、d−lys6−GnRH−GMBS対rPAPの比が20:1であるように、乾燥させたd−lys6−GnRH−GMBSに直接添加した。Tween 20を最終濃度0.25%まで添加した。必要であれば、10mMリン酸を用いて、pHを7.3に調整し、そして反応を暗所中、室温(70°F)でおよそ2〜3時間進行させた。
【0055】
実施例5. d−lys6−GnRH−GMBS−rPAPの精製
[00069]コンジュゲート化後、コンジュゲート化反応後に残った過剰のdK6を除去するために、10ml Bio−Rad Bio−Gel P10カラムを用いて、d−lys6−GnRH−GMBS−rPAPをサイズ排除クロマトグラフィーにさらに供した。50mM Tris、pH7.0、1mM EDTAを含有する緩衝液に対して、タンパク質溶液を透析した。緩衝液のpHは、6.8〜8.5の範囲を有した。透析後、溶液を16000xgで15分間遠心分離し、そして上清を陽イオン交換樹脂上に置いた。次いで、カラムを同じ緩衝液で洗浄し、そして50mM Tris、pH7.0、1M NaClを含有する緩衝液で、タンパク質を溶出させた。
【0056】
実施例6. 受容体結合アッセイ
[00070]実施例5の精製され、再フォールディングされたd−lys6−GnRH−GMBS−rPAPを、競合的ラジオイムノ受容体結合アッセイで用いた。ゴナドトロピン放出ホルモン受容体を有する精製下垂体膜をI125放射標識d−lys6−GnRHに浸した。続いて、異なる濃度のd−lys6−GnRH−GMBS−rPAPを膜に添加し、膜を1mM Tris−Cl pH7.4、1mM CaCl、1%BSAで洗浄した。反応を4時間インキュベーションし、同じ緩衝液で希釈した。希釈後、試験管を4℃、16000xgで15分間遠心分離し、試験管をデカントし、そして放射能の減少を測定した。d−lys6−GnRH−GMBS−植物由来成熟PAPに関して、同じ方法にしたがった。濃度を本実施例のついての表に記載する。
【0057】
[00071]図6は本研究の結果を示す。天然PAPに基づくコンジュゲートおよびrPAPに基づくコンジュゲートは70〜200nM範囲のIC50を有する。rPAP単独では結合せず、そしてしたがって、濃度依存応答を示さない。
【0058】
[00072]実施例6に関する表
【0059】
【表2】
【0060】
実施例7. ウサギ網状赤血球溶解物アッセイ
[00073]本実施例において、以下の材料を用いた: Promega Flexi(登録商標)ウサギ網状赤血球溶解物系: L4540; Promegaルシフェラーゼアッセイ試薬: L1483; Fischer Optizyme組換えRNアーゼ阻害剤: BP3222−5; ルミノメーター: Turner TD−20e。すべての緩衝液および溶液を、DEPC処理H2Oで調製した。試験しようとする毒素および/または毒素緩衝液用に、希釈緩衝液を調製した[0.5ml〜1mlの0.5Mストック(DEPC処理H2O、0.1M NaCl、希釈緩衝液(50mM NaCl 0.5%フラクションV BSA)]。
【0061】
[00074]プロトコルは以下の通りであった:
[00075]まず、0.5nMの毒素/コンジュゲート希釈物を調製した。次いで、0.5nM(500pM)ストックを用いて、100μL連続希釈(各希釈に関して1:2.5)の毒素/コンジュゲートを調製した。以下の希釈物を調製した: 200pM; 80pM; 32pM; 12.8pM; 5.12pM。
【0062】
[00076]アッセイをセットアップするため、500pMから開始して、上述の希釈各々に関して、2.5μLのDEPC処理H2Oおよび2.5μLの毒素/コンジュゲート希釈物を無菌0.65mlエッペンドルフ試験管に添加した。
【0063】
[00077]以下の対照反応もまた調製した:希釈緩衝液:RR溶解物に関する陽性対照; 0.5μM毒素/コンジュゲート:毒素/コンジュゲート活性に関する高濃度陽性対照。
【0064】
[00078]溶解物を氷上で融解し、そして17.5μLの試験希釈物または対照を各試験管に氷上で添加し、そしてピペットで穏やかに混合した。次いで、溶解物/試験または対照を氷上で15分間プレインキュベーションし、そして15分間のプレインキュベーション期間後、2.5μlの栄養プレミックスを添加した(アミノ酸(−lue); 4.2μL;アミノ酸(−met); 4.2μL; 2.5M KCl 11.76μL; RNAsin 8.4μL; DEPC H2O 10.92μL;ルシフェラーゼmRNA 2.52μL;総体積42μLまで)。15分間のプレインキュベーション期間中、mRNAをプレミックスに添加する。各反応試験管の総体積は25μLであった。
【0065】
[00079]各反応試験管の内容物をピペットで穏やかに混合し、そして30℃水槽中で90分間インキュベーションした。50μLの融解した室温のルシフェラーゼアッセイ試薬(LAR)のアリコットを、ルミノメーター試験管(3つ組)内に移し、そして1μLの反応試験管内容物をルミノメーター試験管に添加した。ルミノメーターで明度を計測した。各毒素/コンジュゲート希釈シリーズに関する最高カウントのパーセンテージに対して、濃度の対数をプロットした。各試料に関して、グラフからIC50を決定した。図5は、本実施例にしたがったデータから産生したグラフである。
【0066】
実施例8. 大腸菌に対する成熟rPAPの毒性
[00080]植物由来成熟PAPの成熟型と同じ構造を有するPAPの組換え型の生物学的活性を調べるため、4つの成熟PAPコード配列のうちの1つの上流にT7プロモーターを含有するpET3a発現プラスミド(各プラスミドは、植物由来PAPの翻訳後修飾型と同一のrPAPの成熟型をコードするDNA配列を含有する:クローン1−4.1、1−4.2、1−4.3、および1−4.4)を、アラビノース・プロモーター(AraD)の調節下のT7 RNAポリメラーゼを有する大腸菌BL21(AI)(Invitrogen社、カリフォルニア州カールスバッド)に形質転換した。細胞を振盪しながら、グルコースおよびアンピシリンを含有する最少培地中、37℃でおよそ12時間(一晩)増殖させた。朝、細胞をルリアブロスに移した。全長クローン(3.2)に関して、同じプロセスにしたがった。2時間増殖させた後、アラビノースを最終濃度0.2%に、そしてイソプロピルβ−D−1−チオガラクトピラノシドを1mMの濃度に添加することによって、プラスミドを宿する細胞を誘導した。その後、3時間に渡って1時間ごとにA600を測定した。結果を実施例8についての表に示す。
【0067】
[00081]実施例8についての表
【0068】
【表3】
【0069】
実施例9. 特定の発現条件下での全長rPAPの毒性
[00082]2つの異なる単離体およびrPAP挿入物を含まないプラスミドを宿する対照由来の単一コロニーを、各々、100μg/mlのアンピシリンを含有するルリアブロス培地内に接種した。次いで、3つの培養物を振盪しながら、37℃でおよそ18時間(一晩)増殖させた。各増殖培養物を、100μg/mlアンピシリンの存在下で、新鮮なルリアブロス培地内に1:25で希釈し、そして振盪しながら37℃で2時間増殖させた。
【0070】
[00083]結果は以下の通りであった:
【0071】
【表4】
【技術分野】
【0001】
関連出願
連邦政府が資金援助した研究に関する言及
[0002]なし
技術分野
[0003]本発明は、一般的に、分子生物学および生化学に関し、より詳細には、ヨウシュヤマゴボウ(pokeweed)植物由来の修飾リボソーム不活性化タンパク質に関する。ヨウシュヤマゴボウ植物はまた、Phytolacca americanaとしても知られ、そしてヨウシュヤマゴボウ・リボソーム不活性化タンパク質はまた、ヨウシュヤマゴボウ抗ウイルスタンパク質とも呼ばれ、しばしば「PAP」と略される。本発明はまた、獣医学的薬剤を含む薬剤にも関する。
【背景技術】
【0002】
[0004]化合物コンジュゲート化ヨウシュヤマゴボウ抗ウイルスタンパク質(PAP)、ならびにリシンおよびジフテリア毒素などの他の天然毒素のコンジュゲートは、長らく、療法的有効性の可能性を秘めてきた。理論的には、コンジュゲートの「化合物」部分としての天然リガンドの存在は、ターゲット細胞損傷を生じ、そして他の細胞損傷を生じない。実際には、部分的に、コンジュゲート化されていない毒素が、意図されない細胞損傷を生じるために、不正確なターゲティング毒性を生じる。PAPに関しては、1つの問題は、コンジュゲート化PAPおよび非コンジュゲート化PAPのサイズが非常に類似であるため、分離技術がこれらを区別不能であることである。
【0003】
[0005]自然(「天然」とも称される)PAPはヨウシュヤマゴボウ植物から単離され、そして天然PAPを化合物−毒素コンジュゲートにおいて利用しようとする試みがなされてきたが、こうした試みは信頼性があることを示していない。予期されるように、年ごとおよびバッチごとのアイソフォームの変動が、薬学的品質調節の背景では、厄介でそして実現不能であることがわかった。さらに、いくつかのアイソフォームはコンジュゲートを形成せず、そして異なるアイソフォームは互いに異なってコンジュゲートを形成した。
【0004】
[0006]理想的には、組換え発現は、これらの変動に対する調節を提供するであろう。しかし、PAPの組換え発現もまた、困難に遭遇してきた。大腸菌(E. coli)における以前の発現は、毒性および成長阻害、ならびに封入体における組換えヨウシュヤマゴボウ抗ウイルスタンパク質(rPAP)の集積を生じた。これに関連して、組換えPAPは、別個の可溶化工程およびそれに続くタンパク質の再フォールディングを必要とし、その結果、収量が劣り、そしてスケールアップが困難であった。大腸菌、S.セレビシエ(S. cerevisiae)、植物およびP.パストリス(P. pastoris)における他の試みは低収量を生じ、またはP.パストリスの場合、潜在的に炎症反応を誘導しうる配列が導入された。さらに、組換えPAP−化合物融合タンパク質は、毒素と結合するのに失敗するかまたはターゲット細胞に毒素を導くのに失敗するか、あるいは天然PAPに比較して、はるかに減少した活性を示すか、いずれかであった。
【発明の概要】
【発明が解決しようとする課題】
【0005】
[0007]したがって、未結合(free)システインを有するrPAP分子、該分子から作製されるコンジュゲート、およびrPAPを産生する方法、特に、高収量であり、容易にフォールディングし、そして精製されるrPAPを生じ、そして場合によってコンジュゲート化に化学的に利用可能なrPAPを提供する方法は、大きな貢献となる。
【課題を解決するための手段】
【0006】
[0008]一般論として、本発明は、未結合システイン、好ましくは末端システイン、より好ましくはN末端システインを有する組換えヨウシュヤマゴボウ抗ウイルスタンパク質を含む組成物を提供する。やはり提供するのは、PAPが全長rPAP、より好ましくは未結合システインを含む全長rPAP、最も好ましくは未結合システインおよびアミノ酸リンカーを含む全長rPAPである、rPAP分子である。好ましいのは、N末端Cysおよびアミノ酸リンカーを含むrPAP分子であり、最も好ましくは、Gly−Gly−Gly−Gly−Serの少なくとも1つの反復を有するものである。より好ましくは、Cys−Gly−Gly−Gly−Gly−Ser−全長rPAPおよびCys−Gly−Gly−Gly−Gly−Ser−Gly−Gly−Gly−Gly−Ser−全長rPAPである。
【0007】
[0009]本発明は、本発明の方法にしたがって発現された場合、宿主細胞を殺さないrPAPを提供する。本発明の組成物および方法で利用されるrPAPは、好ましくは、29.5ダルトン以上であり、より好ましくは30ダルトン以上であり、最も好ましくは30.5ダルトン以上である。しかし、31.5、31.75および32ダルトン以上の分子量を有する全長rPAPを利用する組成物および方法もまた、本発明の範囲内である。全長rPAPタンパク質(翻訳後修飾されていない天然PAPの分子量と一致するもの)が、本発明で用いるのに好ましい物質である。
【0008】
[00010]核酸、プラスミド、ならびに本発明の核酸およびタンパク質を含む細胞もまた提供し、好ましい細胞は大腸菌である。
[00011]構造:
X−Y−Z、
式中、Xは未結合システインを有する全長rPAPであり;Yは存在しないかまたは化学的リンカーであり、そしてZは化合物である
を有するコンジュゲートもまた提供する。
【0009】
[00012]細胞ターゲティング化タンパク質である化合物が好ましく、より好ましくは:抗体;ホルモン;修飾ホルモン放出因子;およびホルモン放出因子からなる群より選択されるものが好ましい。化学的リンカーが柔軟なリンカーである化合物が好ましく、ヘテロ二官能性リンカーを持つものがより好ましく、マレイミド基を有するリンカーを持つものが最も好ましい。リンカーがGMBS;EMCS;SMPH;SPDP;およびLC−SPDPからなる群より選択される、本明細書に記載するようなコンジュゲートが好ましい。前記リンカーがGMBSであり、そして前記タンパク質がd−lys6−ゴナドトロピン放出ホルモンであるコンジュゲートが最も好ましい。
【0010】
[00013]本明細書のrPAPを別の化合物とコンジュゲート化する方法であって、組換えヨウシュヤマゴボウ抗ウイルスタンパク質の前記の未結合システインおよび別の化合物間に、化学結合を誘導する工程を含む、前記方法もまた提供する。好ましい方法は、前記化学結合が、ヘテロ二官能性架橋剤を介して誘導されるものであり、より好ましくは、化学結合が、未結合システインおよび化合物上のマレイミド基間に誘導されるものである。ヘテロ二官能性架橋剤がGMBSであり、そして/または化合物がd−lys6−ゴナドトロピン放出ホルモンであるものが最も好ましい。
【0011】
[00014]GMBSリンカーをd−lys6−ゴナドトロピン放出ホルモンに結合させる方法であって、GMBSとd−lys6−ゴナドトロピン放出ホルモンを、非水性条件下でインキュベーションする工程を含む、前記方法もまた提供し、好ましくは、前記非水性条件は:GMBSをメタノール中に溶解させて、第一の非水性溶液を生成し;d−lys6−ゴナドトロピン放出ホルモンをメタノール中に溶解させて、第二の非水性溶液を生成し;前記の第一および第二の非水性溶液を、1.1:1のモル比で混合する工程を含む。
【0012】
[00015]大腸菌において、全長rPAPをコードする核酸を発現する工程を含む、rPAPを得る方法もまた提供する。
[00016]全長rPAPを含む核酸で形質転換された細胞をインキュベーションする工程を含み、rPAPがT7プロモーター系の調節下にあり、そして前記T7プロモーター系がアラビノース・プロモーターの調節下にあるRNAポリメラーゼを有する、細胞を増殖させる方法もまた提供する。前記細胞が大腸菌細胞である方法が好ましい。rPAPが未結合システインを含む方法が好ましく、rPAPが末端システインを含むものが最も好ましい。全長rPAPが:化学的に修飾されたrPAP、天然変異体rPAP、および遺伝子操作されたrPAPからなる群より選択される方法もまた好ましい。
【0013】
[00017]本明細書のPAP組成物を含むコンジュゲートもまた提供する。構造:
X−Y−Z、
式中、XはN末端Cys−Gly−Gly−Gly−Gly−Serを有する全長rPAPであり;Yは化学的リンカーであり、そしてZはタンパク質である
を有するものが特に好ましい。XがGMBSであり、そしてZがd−lys6−GnRHであるものが最も好ましい。
【0014】
定義
[00018]「未結合システイン」は、ジスルフヒドリル結合を介して別のシステインに結合しているもの以外の任意のシステインを意味する。これに関連して、「未結合システイン」には、システインがジスルフヒドリル結合を介して別のシステインに結合していない限り、別の残基または化合物に結合しているシステインが含まれる。
【0015】
[00019]「全長rPAP」は、毒素活性を有し、そして29,500ダルトン以上の分子量を有する、任意の組換えPAPを意味する。
[00020]本発明のこれらのおよび他の特徴および利点は、好ましい態様の詳細な説明から、当業者にはより明らかとなるであろう。詳細な説明に付随する図を以下に記載する:
【図面の簡単な説明】
【0016】
【図1】[00021]図1は、全長ヨウシュヤマゴボウ抗ウイルスタンパク質の構造の定型化された描写であり、翻訳後プロセシング中、植物において通常は切断されるC末端残基、およびさらなる非天然存在N末端残基を示す。
【図2】[00022]図2は:翻訳後プロセシング中、植物において通常は切断されるC末端;およびさらなる非天然存在N末端残基を示す、全長ヨウシュヤマゴボウ抗ウイルスタンパク質;N末端リンカー;ならびに例示的な化合物、修飾ゴナドトロピン放出ホルモン(GnRH)の構造の定型化された描写である。
【図3】[00023]図3は、組換え切断不能(「nc」と略され、そしてリンカーがジスルフィド切断部位を所持しないことを意味する)ヨウシュヤマゴボウ・コンジュゲートの構造の定型化された描写であり、リンカーおよび修飾ゴナドトロピン放出ホルモンを示すアミノ末端の詳細を伴う。
【図4】[00024]図4は、ゴナドトロピン放出ホルモンおよびゴナドトロピン放出ホルモン受容体の細胞外ドメインの相互作用の定型化された描写である。
【図5】[00025]図5は、ウサギ網状赤血球溶解物アッセイにおいて、ルシフェラーゼmRNAのタンパク質合成阻害によって測定されるようなrPAPの生物学的活性を示すグラフである。
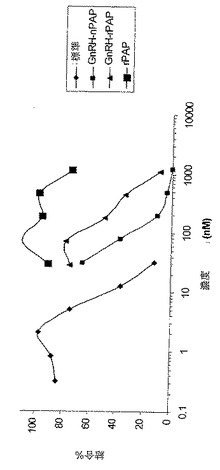
【図6】[00026]図6は、競合的ラジオイムノ受容体結合アッセイの結果を示すグラフである。X軸に最も近い曲線は、標準に関するデータを反映する。次の曲線は、PAPが植物部分から精製された(自然または天然PAPとしても知られる)d−lys6−GnRH−GMBS−PAPに関するデータを反映する。データ点が三角形で示される曲線は、d−lys6−GnRH−GMBS−rPAPに関するデータを反映する。データ点がXとして示される線は、非コンジュゲート化組換えPAP(d−lys6−GnRH−GMBSに結合していない)を反映する。
【図7】[00027]図7。本発明の核酸−rPAP DNA配列−配列番号__。
【図8】[00028]図8。本発明のアミノ酸配列−rPAPタンパク質配列−配列番号1。太字:ATGおよびリンカー配列;黒字:天然成熟PAP配列;太字および下線:植物において翻訳後切断されるタンパク質のC末端部分をコードする天然プレPAP配列。
【図9】[00029]図9。本発明のrPAP(上部)−配列番号2、および天然成熟(植物において翻訳後修飾されたもの)PAP(下部)発現DNA配列のアライメント。
【図10】[00030]図10。rPAP(上部)、および天然成熟PAP(下部)タンパク質配列のアライメント。
【図11】[00031]図11。大きな太字で示す内部ジスルフィド結合システインを含む、発現されたrPAPの配列:Cys−34はCys−258に結合し、そしてCys−84はCys−105に結合する;太字;N末端システインを伴う操作されたリンカー;VNTII....:天然PAP配列;太字および下線は天然プレPAP(植物において翻訳後修飾されたもの)のC末端である。
【発明を実施するための形態】
【0017】
[00032]本発明は、大腸菌において高収量で発現可能であり、そしていかなる他の組換えPAPよりも30〜40倍高い比活性(生物学的活性/単位質量)を有する、組換えヨウシュヤマゴボウ抗ウイルスタンパク質を提供する。さらに、本発明は、薬学的な量で、rPAPを産生するための方法を提供する。
【0018】
[00033]本発明のrPAP物質(タンパク質、核酸、構築物、細胞等)を用いて、rPAP、およびリンカーを介すかまたは直接のいずれかで、これらに結合されたターゲティング化合物を有する、rPAPコンジュゲートを産生してもよい。1つの態様において、rPAPは、別の化合物に対するリンカーを連結するのに場合によって使用するための、未結合システインを有する。1つのこうした態様において、本発明のrPAPタンパク質は、こうした目的のために都合のよいN末端システインを提供するが、本発明のrPAPの使用はN末端コンジュゲート化に限定されない。例えば、本発明のrPAPを、コンジュゲート化なしの毒素として用いてもよいし、あるいは、末端システインまたは内部システインの未結合システインを介して、コンジュゲート化してもよい。
【0019】
[00034]本明細書記載のrPAP分子は、これらにコンジュゲート化されたリンカーまたはターゲティング化合物を伴いまたは伴わず、ウサギ網状赤血球溶解物アッセイにおいて活性である。
【0020】
[00035]本発明には、組換えPAPを発現させ、再フォールディングさせ、コンジュゲート化し、そして精製する方法が含まれる。いくつかの障害を克服して、発現成功を達成した。非ヨウシュヤマゴボウ宿主細胞においてrPAPを発現するのに伴う基本的な問題は、これが毒素であり、そして宿主細胞を殺すことである。T7系を用いて、大腸菌において成熟(翻訳後切断された)PAPを発現する試みを行った。細胞はまったくではなくてもあまり増殖せず、そしてrPAPタンパク質誘導前に弱っているように見えた。続いて、大腸菌において、T7誘導性プロモーター系を用いて、全長rPAP(植物において通常は翻訳後に切断されるC末端部分を加えた成熟PAP)を発現する試みもまた失敗した。細胞はやはり、増殖期中およびrPAPの誘導前に、弱っているように見えた。最後に、2つの制御調節シグナル下の全長rPAPを、大腸菌において試みた。T7 RNAポリメラーゼをアラビノース(AraD)プロモーターの調節下に置き、T7プロモーターを全長rPAP配列の上流に置いた。アラビノース・プロモーターが厳重に抑制されていると、プラスミド上にrPAP遺伝子を宿する場合であっても、細胞は増殖可能であった。抑制の除去を介して誘導すると、薬学的に作業しうる収量のrPAPが生じた。
【0021】
[00036]本発明のrPAPを再フォールティングする多様な方法がある。最も成功した1つは、実施例2に記載するようなものである。別の方法は、スクロースの代わりに0.5M L−アルギニンを代用して、実施例2のプロトコルを用いるものである。さらに、実施例2のプロトコルにおいて、システアミンの代わりに、グルタチオンを用いてもよい。場合によって、8M尿素の代わりに6Mグアニジン−HClで封入体を可溶化してもよい。再フォールディングは、理想的には、塩基性pH範囲で行われる。
【0022】
[00037]すべて当該技術分野でよく記載される、イオン交換クロマトグラフィー、疎水性相互作用クロマトグラフィー、およびヒドロキシアパタイトクロマトグラフィーを含む多様な方法によって、タンパク質を精製してもよい。好ましい方法は、特に実施例5に記載するような陽イオン交換クロマトグラフィーである。
【0023】
[0038]さらに、この組換えタンパク質に対して行う実験に基づいて、本発明のrPAPの比活性(生物学的活性/単位質量)が、ウサギ網状赤血球溶解物におけるタンパク質翻訳の阻害において、別の報告されるrPAPよりも、30〜40x高い活性であることが決定された。非常に感受性であり、そしてナノモル未満の量のrPAPを検出可能なrPAP特異的放射免疫アッセイによって、rPAP濃度を決定した。
【0024】
[00039]理想的には毒素機能を保持するようにフォールティングされた組換えPAPタンパク質、好ましくは天然存在システインの天然ジスルフィド架橋を保持するもの、そして好ましくは少なくとも1つの未結合システイン(例えば天然存在配列には存在しないもの)、最も好ましくは、他の化合物に選択的に結合可能な末端未結合システインを有するものが、本明細書に提供される。当該技術分野の技術のように、任意のPAP配列が、本発明の出発物質として使用するのに適している。任意の既知のアイソタイプ、または明らかになる任意のものが、本発明を用意するのに有用であろう。
【0025】
[00040]全長PAPは、C末端に以下のアミノ酸配列を有する:YNQNAMFPQLIMSTYYNYVNLGDLFEGF−COOH。該配列は、通常、ヨウシュヤマゴボウ植物において、翻訳後に切断されるが、本発明の好ましい態様においては保持される。天然存在翻訳後切断PAPは、29,308.5ダルトンの分子量を有する。
【0026】
[00041]特に、配列番号1;未結合システインを含み、そしてデフォルト設定でBLASTソフトウェアバージョン2.2.21を用いて、配列番号1に対して少なくとも90%同一であるタンパク質;配列番号2にコードされるタンパク質;未結合システインを含み、そしてデフォルト設定でBLASTバージョン2.2.21ソフトウェアを用いて、配列番号2に対して少なくとも90%同一である核酸によってコードされるタンパク質からなる群より選択される、上述のようなrPAP組成物。しかし、配列同一性が:95%;96%;97%;98%;および99%からなる群より選択される、上述のような組成物もまた好ましい。
【0027】
[00042]配列番号2;デフォルト設定でBLASTソフトウェアバージョン2.2.21を用いて、配列番号2に対して少なくとも85%同一であり、そして未結合システインを有するタンパク質をコードする核酸;配列番号1をコードする核酸;および未結合システインを有するタンパク質をコードし、そしてデフォルト設定でBLASTソフトウェアバージョン2.2.21を用いて、配列番号1に対して少なくとも85%同一である核酸からなる群より選択される、核酸もまた提供する。しかし、配列同一性が:90%;95%;96%;97%;98%;および99%からなる群より選択される、上述のような組成物もまた好ましい。好ましい核酸は、本明細書のタンパク質をコードする核酸を含む。
【0028】
[00043]GMBSリンカーをd−lys6−ゴナドトロピン放出ホルモンに結合させる方法であって、GMBSとd−lys6−ゴナドトロピン放出ホルモンを、非水性条件下でインキュベーションする工程を含む、前記方法もまた提供する。この方法のより好ましい態様は、前記非水性条件が:GMBSをメタノール中に溶解させて、第一の非水性溶液を生成し;d−lys6−ゴナドトロピン放出ホルモンをメタノール中に溶解させて、第二の非水性溶液を生成し;前記の第一および第二の非水性溶液を、1.1:1のモル比で混合する工程を含むものである。
【0029】
[00044]特に、配列番号1に対して、少なくとも90%同一、好ましくは少なくとも95%同一、最も好ましくは少なくとも99%同一であるrPAPが、本発明の方法において有用である。未結合CYS残基もまた含むものが最も有用である。さらに、保存配列は変更されてはならず、そして非保存配列は場合によって変更可能である。PAPにおいて、天然存在システイン間のジスルフィド結合は、毒素機能に必要な三次構造を提供し、そして理想的には本発明の分子および方法において保存される。C末端ドメインにおける突然変異は、PAPの局在化プロセシングに影響を及ぼし、そして改変されたプロセシングが望ましい場合には、改変可能である。RNA結合ならびに脱プリン化に影響を及ぼす突然変異が知られる。例えば、最初の16アミノ酸の一部切除(truncation)によって、PAP細胞毒性およびリボソームを脱プリン化する能力が排除される。さらに、リボソーム脱プリン化は、アミノ酸がC末端から除去されるにつれて減少し、そしてGlu−244に停止コドンが導入されると排除される。さらに、特に毒性が高い化合物を得るために、既知の方法によって、過剰活性突然変異体をスクリーニングすることも可能である。本発明の機能を最適化するために、これらの突然変異効果を利用してもよい。さらに、これらの突然変異rPAPおよびこうした突然変異体を利用する組成物は、本発明の範囲内である。
【0030】
[00045]発現された際に、rPAPタンパク質を生じるヌクレオチドもまた本発明に含まれる。特に、配列番号2が好ましい。しかし、上記配列における特定の変化は、本発明の基本的な側面を改変しないであろうことを当業者は認識する。したがって、本発明には、本発明の核酸と、ストリンジェントな条件下のハイブリダイゼーションを用いて相同であるか、BLASTを用いて同一であるか、タンパク質配列を変化させない点突然変異、タンパク質配列を変化させないコドン変化などの、機能に影響を及ぼさない重要でない変化を有する核酸などが含まれる。
【0031】
[00046]本明細書のコンジュゲートおよび本明細書の化合物をコンジュゲート化する方法もまた提供する。コンジュゲートは、理想的には、細胞損傷が望ましい受容体に選択的に結合するよう設計される。一般的に、ターゲティング化合物を介して受容体に結合した後、コンジュゲートは、受容体が仲介するエンドサイトーシスによって取り込まれ、そしてコンジュゲートを細胞に送達する。取り込み後、コンジュゲートのrPAP部分は、大リボソームRNAの保存されたサルシン(sarcin)/リシンループを脱プリン化することによって、リボソームRNAに結合する。脱プリン化されたリボソームは、伸長因子2に結合不能であり、そしてしたがって、伸長サイクルの転位置工程が阻害され、タンパク質合成のシャットダウンが生じる。細胞は最終的に死ぬ。
【0032】
[00047]本明細書の特定のrPAPタンパク質に化合物をコンジュゲート化するための1つの特定の方法は、N末端システインおよび別の化合物間に化学結合を導入する工程を含む。化合物が抗体、ホルモン、修飾ホルモン放出因子、またはホルモン放出因子であるような、こうした方法が好ましい。特に、ホルモン放出因子がGnRHであるものがより好ましいが、d−lys6−修飾GnRHへのコンジュゲート化が最も好ましい。コンジュゲート化は、任意の既知の方法を介して実行可能であるが、好ましくは、リンカーまたは他の架橋化合物を介すかのいずれかで、ターゲティング化化合物およびrPAP間にスルフヒドリル結合を生成することを介する。言い換えると、未結合システインを利用して、rPAPにおける他のシステインへの結合を排除することが理想的であるが、当業者は、機能性を最適にするため、rPAPおよびそれにコンジュゲート化される化合物の両方を修飾する方法を知っている。
【0033】
[00048]本発明の好ましい態様において、修飾ゴナドトロピン放出ホルモン「d−lys6−GnRH」を全長rPAPにコンジュゲート化する。d−lys6−GnRHは、好ましくは、全長rPAP上の未結合システインへの結合を容易にするため、リンカーGMBSで活性化される。d−lys6−GnRHのGMBSでのこうした「活性化」は、通常試みられるような水性条件下で試みた場合には障害であることが証明された。水性条件下では、1つのd−lys6−GnRH分子は、2〜3分子のGMBSに結合し、これはd−lys6−GnRHあたり1つのrPAPを結合させるためには許容しえない。しかし、d−lys6−GnRHを非水性条件下(メタノール)で活性化すると、障害は克服され:1つのGMBSリンカー分子に対する1つのd−lys6−GnRHの比が達成された。したがって、rPAP対d−lys6−GnRHの1対1比もまた達成された。
【0034】
[00049]ヘテロ二官能性架橋剤、特にGMBSを利用する方法が好ましいが、リンカー上に位置するNHSエステル基を介してd−lys6−GnRHに結合するか、またはリンカー上に位置するマレイミド基を介してrPAP上の未結合スルフヒドリル基に付着するのを容易にするであろう任意のヘテロ二官能性架橋剤もまた好ましい。
【0035】
[00050]GnRH分子の両端が受容体への結合に必要である。GnRHおよびd−lys6−GnRH(交換可能に、「DK6」または「dK6」または「d−lys6」または「d−Lys6」とも称される)の唯一の相違は、6位のグリシンのD−リジンでの置換である。さらに、末端がブロッキングされる。C末端はエチルアミド基(ET−NH2)でブロッキングされ、それによって天然化合物の10位のグリシンが置換される。天然GnRH化合物は、NH2Glu−His−Trp−Ser−Tyr−Gly−Leu−Arg−Pro−GlyCOOHである。好ましい類似体は、dK6: Hp−Glu−His−Trp−Ser−Tyr−DLys−Leu−Arg−Pro−Et−NH2である。
【0036】
[00051]別の態様において、アミノ酸配列Cys−Gly−Gly−Gly−Gly−Serが全長rPAPに付加され、そしてターゲティング化合物に結合するのに用いられる。Cys−Gly−Gly−Gly−Gly−Serは、天然PAP配列の一部ではない。Val−Aspは、天然PAP配列の最初の2つのアミノ酸である。
【0037】
[00052]本発明の最も好ましいコンジュゲートは、以下の構造:
X−Y−Z
式中、Xはd−lys6−GnRHであり;YはGMBSであり;そしてZはN末端にCGGGGSを有する全長rPAPである
を有する。
【0038】
[0053]本明細書記載の方法、あるいは当該技術分野で知られるかまたは発展される任意の方法を介して、コンジュゲートを作製してもよい。さらに、当該技術分野に知られるように、任意の望ましい機能性を提供するため、コンジュゲートを修飾してもよい。実施例は好ましいコンジュゲート化法を記載する。
【0039】
[0054]本発明のコンジュゲートを投与するため、任意の塩、懸濁物、分散物等を用いてもよい。無菌であり、そして非発熱性である、0.7%〜10%、より好ましくは0.9%の塩化ナトリウム溶液が好ましく、より好ましくは、4.5〜7のpHでもある溶液が好ましい。さらに、コンジュゲートが適切な影響を提供するならば、任意の投与法が許容されうる。本発明の最も好ましい態様は、不妊処置の目的のために動物に注射するため、溶液中のrPAP−GNRH塩を用いるものである。不妊は完全である必要も、可逆的である必要もない;しかし、意図される最適な様式は、動物、特にイヌ、ネコ、ウマ、食物または他の製品のための家畜(畜牛、乳牛、ブタ、ヒツジ、ヤギ、バイソン、バイソン/畜牛種等)、労働する家畜、動物園の動物、および野生生物(特にシカ、エルク、および慢性消耗病に感受性である他の有蹄動物)で使用するための非可逆性rPAP−d−lys6−GnRH注射液である。
【0040】
[00055]前述の発明は、適切な法律基準にしたがって記載されてきており、したがって、説明は、事実上、限定するのではなく例示的である。開示する態様に対する変動および修飾が、当業者に明らかになることも可能であり、そしてこれらは本発明の範囲内である。
【実施例】
【0041】
実施例1. 大腸菌におけるrPAPの発現。
[00056]順方向プライマー、rPAP−F: 5’−CCCGGG CATATG TGC GGA GGC GGA GGC AGT GTG AAT ACA ATC ATC TAC AAT GTT GGA AGT ACC−3、および逆方向プライマー、rPAP−R: 5’−GCG CGC AAG CTT TCA GGA TTC TTC AAA TAG ATC ACC AAG ATT AAC Cを用いて、PCR増幅によって、全長配列(配列番号2)を得た。
【0042】
[00057]反応混合物は、以下の構成要素からなった: 600mM Tris−SO4 (pH 8.9)、180mM硫酸アンモニウム、0.2mM dATP、0.2mM dCTP、0.2mM dGTP、0.2mM dTTP、2mM MgSO4、0.2μM rPAP−Fプライマー、0.2μM rPAP−Rプライマー、1ngテンプレートDNA、1単位のPlatinumR Taq高忠実度ポリメラーゼ(Invitrogen社、カリフォルニア州カールスバッド)。PCR反応を以下の条件下で行った: 94℃x2分間(1サイクル)、94℃x30秒間、52℃x30秒間、68℃x1分間(15サイクル)、94℃x30秒間、55℃x30秒間、68℃xl分間(25サイクル)。
【0043】
[00058]製造者の説明書にしたがってNdeIおよびBamHI(New England Biolabs、マサチューセッツ州イプスウィッチ)を用いて、pET3a発現プラスミド中、T7プロモーターの下流に、rPAPをコードする全長配列を導入した(連結した)。
【0044】
[00059]rPAP配列含有プラスミドを用いて、One Shot(登録商標)TOP10化学的コンピテント大腸菌株(Invitrogen社、カリフォルニア州カールスバッド)を形質転換した。いくつかのコロニーを摘み取り、そして挿入物の存在に関して、DNA配列分析によってスクリーニングした。正しいrPAP配列を含有するプラスミドを宿することが示されたコロニー由来のプラスミドDNAを精製し、そして続いて、アンピシリン耐性選択可能マーカーとともに厳重に制御されるアラビノース・プロモーター(AraD)の調節下のT7 RNAポリメラーゼを所持する大腸菌BL21(AI)株を形質転換するのに、該プラスミドDNAを用いた。形質転換体を選択し、そしてrPAP発現を抑制するため、推定される形質転換体をLB選択培地およびグルコース上にプレーティングした。
【0045】
[00060]2つの単離体を研究用に選択し、そしてrPAP配列を含まない発現プラスミドを含有する対照を生成した。各単離体を別個に、ラクトースおよびアラビノースを含まず、そしてグルコースが存在する最少培地中で、振盪しながら37℃でおよそ12時間(一晩)増殖させた。対照を同じ条件下で増殖させた。アラビノース・プロモーター系の誘導を抑制する目的のために増殖培地を選択し、それによって、rPAP RNA発現/タンパク質翻訳を抑制した。
【0046】
[00061]結果は以下の通りである。
【0047】
【表1】
【0048】
[00062]少量の各一晩培養物を、アンピシリンを含有するLB培地に移し、そしてA600が0.4に到達した後、続いて、L−アラビノースを最終濃度0.2%に、そしてイソプロピルβ−D−1−チオガラクトピラノシドを1mMの濃度に添加することによって、rPAPを大腸菌細胞から誘導した。誘導をさらに3.5時間行った。
【0049】
実施例2. rPAPの再フォールディングおよび精製。
[00063]スナップ希釈によってrPAPを再フォールディングした。封入体の単離後、8M尿素、50mM Tris HCl、pH8.5中で封入体を可溶化した。DTTを最終濃度10mMまで添加し、そして混合物を室温で90分間攪拌した。次いで、可溶化されたタンパク質を室温で攪拌しながら、50mM Tris、pH8.5、0.4Mスクロース、0.05%ポリエチレングリコール−3550、0.9mM酸化システアミン(TPEGS)を含有する溶液内に一滴ずつ添加した。再フォールディング溶液中のrPAPの最終濃度は、10μg/mlおよび50μg/mlの間であった。可溶化されたrPAPを再フォールディング溶液に添加した後、混合物を4℃でさらに24時間攪拌した。24時間後、混合物を16000xgで15分間遠心分離し、上清をデカントし、そして再フォールディング後、50mM Tris、pH7.0、1mM EDTAを含有する緩衝液に対して、タンパク質溶液を透析した。緩衝液pHは、6.8〜8.5の範囲を有した。透析後、溶液を16000xgで15分間遠心分離し、そして上清を陽イオン交換樹脂上に置いた。次いで、カラムを50mM Tris−HCl、pH7.0、1.0mM EDTAで洗浄し、そして50mM Tris、pH7.0、1M NaClを含有する緩衝液でタンパク質を溶出させる。50mM NaPO4、pH7.2、100mM NaCl、1mM EDTAを含有するコンジュゲート化緩衝液に対して、溶出タンパク質を透析する。タンパク質濃度を、0.2mg/ml〜1.0mg/mlの濃度に調整する。
【0050】
実施例3. d−lys6修飾ゴナドトロピン放出ホルモン(GnRH)のマレイミドブチリルオキシ−スクシンイミドエステル(GMBS)リンカーでの活性化。
[00064]1224ダルトンの分子量を有するd−lys6−GnRHを、固相合成(Anaspec社、カリフォルニア州フレモント)によって調製した。6ミリグラムのd−lys6−GnRHを1.5ml脱イオン化メタノールと混合し、そしてジイソプロピルエタノールアミン(DIPEA)を用いて、pH7.0に調整した。
【0051】
[00065]Thermo Fisher Scientific(イリノイ州ロックフォード)からGMBSを購入した。1.25mgのGMBSを1.5ml脱イオン化メタノールと混合した。
【0052】
[00066]1.5mlのd−lys6−GnRH−メタノールおよび1.5mlのGMBS−メタノールを、キャップ付血清ボトル中で一緒に混合し、そしてDIPEAを用いてpH7.0に調整した。金属キャップを用いて血清ボトルを密封した。溶液を脱気し、そして窒素で4回パージした。血清ボトルをアルミニウムホイルで覆い、そして反応を攪拌しながら室温で90分間進行させた。
【0053】
[00067]生じたd−lys6−GnRH−GMBSは、およそ1421ダルトンの分子量を有し、1分子のGMBSが1分子のd−lys6−GnRHに結合したことが示された。これを質量分析によって確認した。
【0054】
実施例4. d−lys6−GnrH−GMBSへのrPAPのコンジュゲート化。
[00068]実施例3の溶液を遠心分離蒸発装置で蒸発させた。TCEP・HCl Tris(2−カルボキシエチル)ホスフィン塩酸塩を、コンジュゲート化緩衝液中に溶解され、再フォールディングされた組換えPAPに、最終濃度0.05mMまで添加した。混合物を室温で1〜2時間インキュベーションした。インキュベーション後、コンジュゲート化緩衝液中に溶解され、再フォールディングされたrPAPを、d−lys6−GnRH−GMBS対rPAPの比が20:1であるように、乾燥させたd−lys6−GnRH−GMBSに直接添加した。Tween 20を最終濃度0.25%まで添加した。必要であれば、10mMリン酸を用いて、pHを7.3に調整し、そして反応を暗所中、室温(70°F)でおよそ2〜3時間進行させた。
【0055】
実施例5. d−lys6−GnRH−GMBS−rPAPの精製
[00069]コンジュゲート化後、コンジュゲート化反応後に残った過剰のdK6を除去するために、10ml Bio−Rad Bio−Gel P10カラムを用いて、d−lys6−GnRH−GMBS−rPAPをサイズ排除クロマトグラフィーにさらに供した。50mM Tris、pH7.0、1mM EDTAを含有する緩衝液に対して、タンパク質溶液を透析した。緩衝液のpHは、6.8〜8.5の範囲を有した。透析後、溶液を16000xgで15分間遠心分離し、そして上清を陽イオン交換樹脂上に置いた。次いで、カラムを同じ緩衝液で洗浄し、そして50mM Tris、pH7.0、1M NaClを含有する緩衝液で、タンパク質を溶出させた。
【0056】
実施例6. 受容体結合アッセイ
[00070]実施例5の精製され、再フォールディングされたd−lys6−GnRH−GMBS−rPAPを、競合的ラジオイムノ受容体結合アッセイで用いた。ゴナドトロピン放出ホルモン受容体を有する精製下垂体膜をI125放射標識d−lys6−GnRHに浸した。続いて、異なる濃度のd−lys6−GnRH−GMBS−rPAPを膜に添加し、膜を1mM Tris−Cl pH7.4、1mM CaCl、1%BSAで洗浄した。反応を4時間インキュベーションし、同じ緩衝液で希釈した。希釈後、試験管を4℃、16000xgで15分間遠心分離し、試験管をデカントし、そして放射能の減少を測定した。d−lys6−GnRH−GMBS−植物由来成熟PAPに関して、同じ方法にしたがった。濃度を本実施例のついての表に記載する。
【0057】
[00071]図6は本研究の結果を示す。天然PAPに基づくコンジュゲートおよびrPAPに基づくコンジュゲートは70〜200nM範囲のIC50を有する。rPAP単独では結合せず、そしてしたがって、濃度依存応答を示さない。
【0058】
[00072]実施例6に関する表
【0059】
【表2】
【0060】
実施例7. ウサギ網状赤血球溶解物アッセイ
[00073]本実施例において、以下の材料を用いた: Promega Flexi(登録商標)ウサギ網状赤血球溶解物系: L4540; Promegaルシフェラーゼアッセイ試薬: L1483; Fischer Optizyme組換えRNアーゼ阻害剤: BP3222−5; ルミノメーター: Turner TD−20e。すべての緩衝液および溶液を、DEPC処理H2Oで調製した。試験しようとする毒素および/または毒素緩衝液用に、希釈緩衝液を調製した[0.5ml〜1mlの0.5Mストック(DEPC処理H2O、0.1M NaCl、希釈緩衝液(50mM NaCl 0.5%フラクションV BSA)]。
【0061】
[00074]プロトコルは以下の通りであった:
[00075]まず、0.5nMの毒素/コンジュゲート希釈物を調製した。次いで、0.5nM(500pM)ストックを用いて、100μL連続希釈(各希釈に関して1:2.5)の毒素/コンジュゲートを調製した。以下の希釈物を調製した: 200pM; 80pM; 32pM; 12.8pM; 5.12pM。
【0062】
[00076]アッセイをセットアップするため、500pMから開始して、上述の希釈各々に関して、2.5μLのDEPC処理H2Oおよび2.5μLの毒素/コンジュゲート希釈物を無菌0.65mlエッペンドルフ試験管に添加した。
【0063】
[00077]以下の対照反応もまた調製した:希釈緩衝液:RR溶解物に関する陽性対照; 0.5μM毒素/コンジュゲート:毒素/コンジュゲート活性に関する高濃度陽性対照。
【0064】
[00078]溶解物を氷上で融解し、そして17.5μLの試験希釈物または対照を各試験管に氷上で添加し、そしてピペットで穏やかに混合した。次いで、溶解物/試験または対照を氷上で15分間プレインキュベーションし、そして15分間のプレインキュベーション期間後、2.5μlの栄養プレミックスを添加した(アミノ酸(−lue); 4.2μL;アミノ酸(−met); 4.2μL; 2.5M KCl 11.76μL; RNAsin 8.4μL; DEPC H2O 10.92μL;ルシフェラーゼmRNA 2.52μL;総体積42μLまで)。15分間のプレインキュベーション期間中、mRNAをプレミックスに添加する。各反応試験管の総体積は25μLであった。
【0065】
[00079]各反応試験管の内容物をピペットで穏やかに混合し、そして30℃水槽中で90分間インキュベーションした。50μLの融解した室温のルシフェラーゼアッセイ試薬(LAR)のアリコットを、ルミノメーター試験管(3つ組)内に移し、そして1μLの反応試験管内容物をルミノメーター試験管に添加した。ルミノメーターで明度を計測した。各毒素/コンジュゲート希釈シリーズに関する最高カウントのパーセンテージに対して、濃度の対数をプロットした。各試料に関して、グラフからIC50を決定した。図5は、本実施例にしたがったデータから産生したグラフである。
【0066】
実施例8. 大腸菌に対する成熟rPAPの毒性
[00080]植物由来成熟PAPの成熟型と同じ構造を有するPAPの組換え型の生物学的活性を調べるため、4つの成熟PAPコード配列のうちの1つの上流にT7プロモーターを含有するpET3a発現プラスミド(各プラスミドは、植物由来PAPの翻訳後修飾型と同一のrPAPの成熟型をコードするDNA配列を含有する:クローン1−4.1、1−4.2、1−4.3、および1−4.4)を、アラビノース・プロモーター(AraD)の調節下のT7 RNAポリメラーゼを有する大腸菌BL21(AI)(Invitrogen社、カリフォルニア州カールスバッド)に形質転換した。細胞を振盪しながら、グルコースおよびアンピシリンを含有する最少培地中、37℃でおよそ12時間(一晩)増殖させた。朝、細胞をルリアブロスに移した。全長クローン(3.2)に関して、同じプロセスにしたがった。2時間増殖させた後、アラビノースを最終濃度0.2%に、そしてイソプロピルβ−D−1−チオガラクトピラノシドを1mMの濃度に添加することによって、プラスミドを宿する細胞を誘導した。その後、3時間に渡って1時間ごとにA600を測定した。結果を実施例8についての表に示す。
【0067】
[00081]実施例8についての表
【0068】
【表3】
【0069】
実施例9. 特定の発現条件下での全長rPAPの毒性
[00082]2つの異なる単離体およびrPAP挿入物を含まないプラスミドを宿する対照由来の単一コロニーを、各々、100μg/mlのアンピシリンを含有するルリアブロス培地内に接種した。次いで、3つの培養物を振盪しながら、37℃でおよそ18時間(一晩)増殖させた。各増殖培養物を、100μg/mlアンピシリンの存在下で、新鮮なルリアブロス培地内に1:25で希釈し、そして振盪しながら37℃で2時間増殖させた。
【0070】
[00083]結果は以下の通りであった:
【0071】
【表4】
【特許請求の範囲】
【請求項1】
未結合(free)システインを有する組換えヨウシュヤマゴボウ抗ウイルスタンパク質を含む物質の組成物。
【請求項2】
前記組換えヨウシュヤマゴボウ抗ウイルスタンパク質が全長PAPである、請求項1の組成物。
【請求項3】
前記未結合システインが末端システインである、請求項2の組成物。
【請求項4】
Cys−Gly−Gly−Gly−Gly−Ser−全長PAPを含む、請求項3の組成物。
【請求項5】
請求項1の組成物をコードする核酸を含む核酸。
【請求項6】
請求項5の核酸を含むプラスミド。
【請求項7】
請求項6の核酸を含む細胞。
【請求項8】
大腸菌である、請求項7の細胞。
【請求項9】
構造:
X−Y−Z、
式中、Xは未結合システインを有する全長rPAPであり;Yは存在しないかまたは化学的リンカーであり、そしてZは化合物である
を有するコンジュゲート。
【請求項10】
前記化合物が細胞ターゲティング・タンパク質である、請求項9のコンジュゲート。
【請求項11】
抗体;ホルモン;修飾ホルモン放出因子;およびホルモン放出因子からなる群より選択される、請求項10のコンジュゲート。
【請求項12】
前記リンカーが:GMBS;EMCS;SMPH;SPDP;およびLC−SPDPからなる群より選択される、請求項10のコンジュゲート。
【請求項13】
前記リンカーがGMBSであり、そして前記タンパク質がd−lys6−ゴナドトロピン放出ホルモンである、請求項10のコンジュゲート。
【請求項14】
請求項1の化合物を別の化合物とコンジュゲート化する方法であって、組換えヨウシュヤマゴボウ抗ウイルスタンパク質の前記未結合システインおよび別の化合物間に、化学結合を誘導する工程を含む、前記方法。
【請求項15】
前記化学結合が、ヘテロ二官能性架橋剤を介して誘導される、請求項14の方法。
【請求項16】
前記ヘテロ二官能性架橋剤がGMBSである、請求項15の方法。
【請求項17】
化合物がd−lys6−ゴナドトロピン放出ホルモンである、請求項15の方法。
【請求項18】
GMBSリンカーをd−lys6−ゴナドトロピン放出ホルモンに結合させる方法であって、GMBSとd−lys6−ゴナドトロピン放出ホルモンを、非水性条件下でインキュベーションする工程を含む、前記方法。
【請求項19】
前記非水性条件が:GMBSをメタノール中に溶解させて、第一の非水性溶液を生成し;d−lys6−ゴナドトロピン放出ホルモンをメタノール中に溶解させて、第二の非水性溶液を生成し;前記の第一および第二の非水性溶液を、1.1:1のモル比で混合する工程を含む、請求項19の方法。
【請求項20】
大腸菌(E. coli)において、全長rPAPをコードする核酸を発現する工程を含む、請求項1の組成物を作製する方法。
【請求項21】
細胞を増殖させる方法であって、T7プロモーター系の調節下にある全長rPAPを含む核酸で形質転換された細胞をインキュベーションする工程を含み、前記T7プロモーター系がアラビノース・プロモーターの調節下にあるRNAポリメラーゼを有し、前記インキュベーションが細胞増殖を生じる、前記方法。
【請求項22】
前記細胞が大腸菌細胞である、請求項21の方法。
【請求項23】
前記細胞増殖後に前記全長rPAPの発現を誘導する工程をさらに含む、請求項22の方法。
【請求項24】
前記の発現誘導後に、前記細胞から前記全長rPAPを単離する工程をさらに含む、請求項23の方法。
【請求項25】
前記全長rPAPが:化学的に修飾されたrPAP、天然変異体rPAP、および遺伝子操作されたrPAPからなる群より選択される、請求項22の方法。
【請求項1】
未結合(free)システインを有する組換えヨウシュヤマゴボウ抗ウイルスタンパク質を含む物質の組成物。
【請求項2】
前記組換えヨウシュヤマゴボウ抗ウイルスタンパク質が全長PAPである、請求項1の組成物。
【請求項3】
前記未結合システインが末端システインである、請求項2の組成物。
【請求項4】
Cys−Gly−Gly−Gly−Gly−Ser−全長PAPを含む、請求項3の組成物。
【請求項5】
請求項1の組成物をコードする核酸を含む核酸。
【請求項6】
請求項5の核酸を含むプラスミド。
【請求項7】
請求項6の核酸を含む細胞。
【請求項8】
大腸菌である、請求項7の細胞。
【請求項9】
構造:
X−Y−Z、
式中、Xは未結合システインを有する全長rPAPであり;Yは存在しないかまたは化学的リンカーであり、そしてZは化合物である
を有するコンジュゲート。
【請求項10】
前記化合物が細胞ターゲティング・タンパク質である、請求項9のコンジュゲート。
【請求項11】
抗体;ホルモン;修飾ホルモン放出因子;およびホルモン放出因子からなる群より選択される、請求項10のコンジュゲート。
【請求項12】
前記リンカーが:GMBS;EMCS;SMPH;SPDP;およびLC−SPDPからなる群より選択される、請求項10のコンジュゲート。
【請求項13】
前記リンカーがGMBSであり、そして前記タンパク質がd−lys6−ゴナドトロピン放出ホルモンである、請求項10のコンジュゲート。
【請求項14】
請求項1の化合物を別の化合物とコンジュゲート化する方法であって、組換えヨウシュヤマゴボウ抗ウイルスタンパク質の前記未結合システインおよび別の化合物間に、化学結合を誘導する工程を含む、前記方法。
【請求項15】
前記化学結合が、ヘテロ二官能性架橋剤を介して誘導される、請求項14の方法。
【請求項16】
前記ヘテロ二官能性架橋剤がGMBSである、請求項15の方法。
【請求項17】
化合物がd−lys6−ゴナドトロピン放出ホルモンである、請求項15の方法。
【請求項18】
GMBSリンカーをd−lys6−ゴナドトロピン放出ホルモンに結合させる方法であって、GMBSとd−lys6−ゴナドトロピン放出ホルモンを、非水性条件下でインキュベーションする工程を含む、前記方法。
【請求項19】
前記非水性条件が:GMBSをメタノール中に溶解させて、第一の非水性溶液を生成し;d−lys6−ゴナドトロピン放出ホルモンをメタノール中に溶解させて、第二の非水性溶液を生成し;前記の第一および第二の非水性溶液を、1.1:1のモル比で混合する工程を含む、請求項19の方法。
【請求項20】
大腸菌(E. coli)において、全長rPAPをコードする核酸を発現する工程を含む、請求項1の組成物を作製する方法。
【請求項21】
細胞を増殖させる方法であって、T7プロモーター系の調節下にある全長rPAPを含む核酸で形質転換された細胞をインキュベーションする工程を含み、前記T7プロモーター系がアラビノース・プロモーターの調節下にあるRNAポリメラーゼを有し、前記インキュベーションが細胞増殖を生じる、前記方法。
【請求項22】
前記細胞が大腸菌細胞である、請求項21の方法。
【請求項23】
前記細胞増殖後に前記全長rPAPの発現を誘導する工程をさらに含む、請求項22の方法。
【請求項24】
前記の発現誘導後に、前記細胞から前記全長rPAPを単離する工程をさらに含む、請求項23の方法。
【請求項25】
前記全長rPAPが:化学的に修飾されたrPAP、天然変異体rPAP、および遺伝子操作されたrPAPからなる群より選択される、請求項22の方法。
【図1】

【図2】
【図6】
【図7】

【図8】

【図9】

【図10】

【図11】

【図3】
【図4】
【図5】
【図2】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図3】
【図4】
【図5】
【公表番号】特表2011−528230(P2011−528230A)
【公表日】平成23年11月17日(2011.11.17)
【国際特許分類】
【出願番号】特願2011−518880(P2011−518880)
【出願日】平成21年7月15日(2009.7.15)
【国際出願番号】PCT/US2009/050685
【国際公開番号】WO2010/009227
【国際公開日】平成22年1月21日(2010.1.21)
【出願人】(511013522)セダス,インコーポレーテッド (1)
【出願人】(592246587)コロラド ステート ユニバーシティー リサーチ ファウンデーション (17)
【Fターム(参考)】
【公表日】平成23年11月17日(2011.11.17)
【国際特許分類】
【出願日】平成21年7月15日(2009.7.15)
【国際出願番号】PCT/US2009/050685
【国際公開番号】WO2010/009227
【国際公開日】平成22年1月21日(2010.1.21)
【出願人】(511013522)セダス,インコーポレーテッド (1)
【出願人】(592246587)コロラド ステート ユニバーシティー リサーチ ファウンデーション (17)
【Fターム(参考)】
[ Back to top ]