
組換え可溶性Fc受容体
【課題】生産するのが容易で医療または診断用途のために有利に使用できる受容体およびその用途の提供。
【解決手段】本発明による組換え可溶性Fc受容体は、膜貫通ドメイン、シグナルペプチドおよびグリコシル化の存在しない点に特徴がある。この種のFc受容体は、原核宿主細胞においてそれぞれの核酸を発現させ、得られた封入体を再生することにより容易に取得することができ、この方法からは非常に均質で純粋な産物が得られる。かかる産物は診断および医薬用途のために、さらに結晶構造データの生成のために使用することができる。このような結晶構造データは人工分子のモデリングに使用することができる。さらなる実施形態は、本発明によるFc受容体を、抗体の分離および/または濃縮に使用しうるクロマトグラフィー材料などの固相材料に結合させることを含む。
【解決手段】本発明による組換え可溶性Fc受容体は、膜貫通ドメイン、シグナルペプチドおよびグリコシル化の存在しない点に特徴がある。この種のFc受容体は、原核宿主細胞においてそれぞれの核酸を発現させ、得られた封入体を再生することにより容易に取得することができ、この方法からは非常に均質で純粋な産物が得られる。かかる産物は診断および医薬用途のために、さらに結晶構造データの生成のために使用することができる。このような結晶構造データは人工分子のモデリングに使用することができる。さらなる実施形態は、本発明によるFc受容体を、抗体の分離および/または濃縮に使用しうるクロマトグラフィー材料などの固相材料に結合させることを含む。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、組換え可溶性Fc受容体(FcR)、そのようなFc受容体をコードする組換え核酸、対応する核酸を含む宿主細胞、ならびに患者の血液、血漿または血清に含まれるある型の抗体の量を測定する方法、免疫系の慢性疾患を有する患者の免疫状態を調べる方法、および抗体のそれぞれの細胞性受容体による認識およ結合を阻害するインヒビターとして作用する能力に関して物質をスクリーニングする方法に関する。さらに、本発明は、組換え可溶性FcRを含有する医薬組成物、FcRおよびFcR/Ig複合体の結晶性調製物、特にFc受容体の結晶構造データを生成するための該結晶性調製物の使用、ならびにFcRインヒビターおよび該FcRインヒビターを含有する医薬組成物に関する。
【0002】
本発明の更なる主題は、固相(例えば、クロマトグラフィー担体材料)に結合された組換えFc受容体である。そのようなクロマトグラフィー材料(これは本発明のもう1つの主題である)の使用は、患者の体液からの、または免疫グロブリン産生細胞の培養上清からの免疫グロブリンの吸着にある。
【背景技術】
【0003】
Fc受容体(FcR)は感染からヒトを防御するうえで重要な役割を果たしている。病原体は、血液循環系に接近した後に、免疫グロブリン(Ig)によるオプソニン化を受ける。その結果生じる免疫複合体はその多価性のため高い結合力でFcR保有細胞に結合し、FcRのクラスター化へ至らせ、これがいくつかのエフェクター機能を開始させる(Metzger, H., 1992A)。こうした機能としては、発現されたFcRの型および関連タンパク質に応じて、病原体のその後の中和および抗原提示によるエンドサイトーシス、抗体依存性細胞傷害性(ADCC)、メディエーターの分泌または抗体産生の調節などがある(Fridmanら, 1992; van de Winkel and Capel, 1993)。
【0004】
全てのIgクラスに対して特異的なFcRが存在し、IgGに対するFcRは最も豊富で、多様性も広範囲にわたる。IgEの高親和性受容体(FcεRIa)と共に、FcγRI(CD64)、FcγRII(CD32)およびFcγRIIIa(CD16)はI型膜貫通タンパク質としてまたは可溶性形態(sFcR)で存在しているが、FcγRIIIのグリコシルホスファチジルイノシトールアンカー型(FcγRIIIb)も存在している。さらに、FcRはさまざまなイソ型(FcγRIa、b1、b2、c; FcγRIIa1-2、b1-3、c)および対立遺伝子(FcγRIIa1-HR、-LR; FcγRIIIb-NA1、-NA2)として存在している(van de Winkel and Capel, 1993)。全体的に相同性の細胞外部分とは対照的に、膜貫通ドメインおよび細胞質ドメインは相違している。それらは完全に欠失させることができ、また、8kDaの大きさのものでありうる。それらはFcγRIIaにおけるような26アミノ酸の免疫受容体チロシンベースの活性化モチーフ(ITAM)、またはシグナル伝達に関与するFcγRIIbにおけるような個々の13アミノ酸抑制性モチーフ(ITIM)のいずれかを含みうる(Amigorenaら, 1992)。
【0005】
保存されたシステイン間隔から判断すると、FcRの細胞外部分は3つ(FcγRI、CD64)または2つ(FcεRI、FcγRII、CD32およびFcγRIII、CD16)のIg様ドメイン(10kDa/ドメイン)からなり、それゆえに免疫グロブリンスーパーファミリーに属する。これらの高度にグリコシル化された受容体は相同体であり、FcγRとFcεRIa間のアミノ酸配列の全体的な同一性はそれらの細胞外領域では50%を上回る。それにもかかわらず、FcRのそのリガンドへの親和性はさまざまである。FcγRIのFcフラグメントに対する親和性は約108M-1と比較的高いが、これはその第3ドメインのせいであり、一方、2つのドメインをもつ他のFcγRのIgGに対する親和性は105〜107M-1の範囲で変化する。2つのドメインをもつFcεRIaのIgEに対する親和性は1010M-1と一定しており、これらの値をはるかに上回る(Metzger, H., 1992B)。上記のFcRとは対照的に、IgEに対する低親和性受容体であるFcεRIIはある型の膜貫通タンパク質であり、より低い相同性を示す。
【0006】
FcγRは免疫的に活動しているあらゆる細胞上に特定のパターンで発現されている。FcγRIは単球とマクロファージ上に構成的に発現され、好中球および好酸球上に誘導することができる。FcγRIの生理学的役割は、単球上での発現が必須ではないので、まだ不明である(Ceuppensら, 1988)。FcγRIIIのGPIアンカー型(FcγRIIIb)は顆粒球上に広く発現されている。その細胞質部分が欠失されているため、細胞へのシグナル伝達は、補体受容体3型(CR3)のような、少なくともFcγRIIIbと会合できる他の膜貫通タンパク質を介してのみ起こる(Zhouら, 1993; Pooら, 1995)。FcγRIIIaは主に単球とマクロファージ上に発現されるが、結合タンパク質(例えば、α鎖またはγ鎖)と一緒にしか発現されない。FcγRIIは免疫担当細胞上に最も広く分布している受容体であり、主として免疫複合体のエンドサイトーシスに関与している。
【0007】
FcγRIIaとFcγRIIbは、その細胞外領域のアミノ酸残基がたった7%しか違っていない。それにもかかわらず、両形態はヒトおよびマウスIgGサブクラスへのその結合特性(van de Winkel and Capel, 1993)およびヒトIgGへのその異なる親和性(Sondermannら, 1998A)によって区別され得る。こうした状態は、何人かの個体に由来するT細胞がマウスIgG1誘導有糸分裂に応答できることが判明した後に命名されたFcγRIIaの高応答性/低応答性(HR/LR)多型によってさらに一層複雑になってくる(Taxら, 1983)。その後、LR型とHR型の間のアミノ酸配列の2つの交換によりヒトIgG2への結合能が変化することが見いだされ、このことは、それらの少なくとも1つがIgG結合に関与することを示唆している(Hogarthら, 1992)。
【0008】
健康な個体においてFcRが果たす有益な役割とは対照的に、それらはまた、アレルギー(FcεRIa)または自己免疫疾患において免疫系の刺激を伝達する。さらに、一部のウイルス、例えばHIV(Homsyら, 1989)やデング熱ウイルス(Littauaら, 1990)は細胞に接近するためにFcγRを利用し、またエボラウイルス(Yangら, 1998)や麻疹ウイルス(Ravanelら, 1997)の場合にはFcγRをブロックすることにより免疫応答を遅らせる。
【発明の概要】
【発明が解決しようとする課題】
【0009】
それゆえに、本発明の基礎となる目的は、生産するのが容易で、医療または診断用途のために有利に使用できる受容体を提供することであった。さらに、本発明の目的は、ヒトの体内に存在する天然の受容体に類似した結合特異性および活性を示し、その上、構造決定に適する結晶を生成することを可能にする可溶性受容体を提供することであった。
【課題を解決するための手段】
【0010】
この目的は、Fc受容体の細胞外部分のみからなり、グリコシル化されていない組換え体の可溶性Fc受容体によって達成される。したがって、本発明による受容体は膜貫通ドメイン、シグナルペプチドおよびグリコシル化が存在しないことを特徴とする。
【図面の簡単な説明】
【0011】
【図1】図1は、sFcγRIIbの精製を示す15%還元SDS PAGEである。レーン1:分子量マーカー。レーン2:導入前の大腸菌溶解物。レーン3:導入1時間後の大腸菌溶解物。レーン4:導入4時間後の大腸菌溶解物。レーン5:精製したsFcγRIIbの封入体。レーン6:hIgGアフィニティーカラムの溶出液。レーン7:ゲル濾過カラムのプールした画分。
【図2】図2は、平衡化ゲル濾過である。1 μgのhFcを10 μlの平衡化バッファー(PBS1mlあたり10 μgのsFcγRIIbを含む)に溶解したものをサイズ排除クロマトグラフィーカラムにアプライし、溶出液の吸光度(280nm)を時間の関数として測定した。注入したFcフラグメントは、平衡化バッファー中でsFcγRIIbと複合体を形成する(t = 22分)。消費されたsFcγRIIbの負のピークは、t = 26分に観察される。
【図3】図3は、ヒトsFcγRIIbの全体構造である。sFcγRIIb構造の立体的リボン表示である。IgG結合に重要であると思われるループは、結合部位内のいくつかの残基とともに赤で示し、保存されたジスルフィド架橋は球および棒で示す。潜在的Nグリコシル化部位は緑の球で示す。末端に記を付け、β鎖においてはN末端ドメインには黒で、C末端ドメインには青で、連続的に番号を付した。この図は、プログラムMOLSCRIPT (Kraulis, 1991)およびRENDER (MerrittおよびMurphy, 1994)で作成した。
【図4】図4は、ドメイン間接触を表す。この図は、sFcγRIIbのドメイン間接触に関与する残基の拡大図を示す。N末端ドメインのアミノ酸残基を青色で示し、C末端ドメインの残基を黄色で示す。このモデルは、最終座標から得られた1σで等高線を付した2Fo-Fc電子密度によってカバーされる。ドメイン間の水素結合は白線で示す。この図は、プログラムMAIN(Turk、1992)を用いて作成した。
【図5A】図5aは、2つのFcγRIIbドメインとヒトIgG1のCH2ドメインとの重ね合わせを表す。FcγRIIbの双方のドメインとhIgG1のCH2ドメインを重ね合わせた。N末端ドメインは青色で示し、C末端ドメインは赤色で示し、hIgG1のCH2ドメインは緑色で示す。各末端には記を付け、保存されたジスルフィド架橋は細い線で示す。
【図5B】図5bは、sFcγRIIbドメインとFcRファミリーのその他のメンバーのドメインとの構造に基づく配列アライメントである。図の上部は、プログラムGBF-3D-FIT (Lessel & Schomburg, 1994)を用いて実施したFcγRIIbとhIgG1 Fcフラグメントドメインとの構造に基づく配列アライメントである。重ね合わせたドメインのうち、2.0Å未満のCα距離を有するアミノ酸残基は、Fcフラグメントドメイン間でマッチした残基についてはライラック色で記を付け、FcγRIIbドメイン内の残基には黄色で記を付け、4つのドメインすべてにおいて重ね合わせることができたときは緑色で記をつけている。β鎖はこのアライメントの部分の下に示し、図3と一致するように記をつけている。図の下部は、GCGパッケージ(Genetics Computer Group, 1994)からのルーチンを用い、図の上部に示されたプロファイルに従った、その他のFcγRのアミノ酸配列と相同体FcεRIaとのアミノ酸配列のアライメントを示す。上および下の列の数字は、FcγRIIbのN末端およびC末端ドメインに関する。保存されたシステインは、マゼンタ色でタイプし、潜在的グリコシル化部位は青色でタイプしている。第1ドメインで同一の残基には橙色で記を付け、第2ドメインで同一の残基にはピンク色で記をつけ、双方のドメインで残基が保存されている場合は緑色で記をつけている。あまり保存されていないFcγRIの第3ドメインは、第1および第2ドメイン間でアライメントを行った。赤色の矢印は第1ドメインと第2ドメイン間の側鎖の接触に関与する残基を指し、青色の矢印はIgG結合に関連する残基を示している。この図は、プログラムALSCRIPT(Barton, 1993)で作成した。
【図6】図6は、FcγRIIbの推定の結合部位を表す。GRASP (Nichollsら、1991)で作成したときのFcγRIIbの固体表面表示は、カラーコードは負(赤色)から正(青色)への相対的表面電位に従う。図6aは、図3で示した分子を垂直線を軸として反時計回りに約90°回転させたものを示す。図6bでは、該分子を同じ軸まわりで、時計回りに90°回転させている。両図は、C末端ドメイン(図6a)およびN末端ドメイン(図6b)上の推定結合領域を示す。本明細書中で考察したアミノ酸残基に記を付けている。
【図7】図7は、Fcγ受容体の重ね合わせ構造のCαトレースである。FcγRIIIは赤色、FcγRIIaは緑色、FcγRIIbは青色で表している。IgG結合に重要な残基は球と棒で表している。N末端およびC末端には記を付けている。
【図8】図8は、FcγRIII/Fcフラグメント結晶構造のリボン表示の概観である。Fcフラグメントに結合する糖残基を、球と棒で示している。FcγRIII(青色)は、Fcフラグメントの鎖B(赤色)および鎖A(緑色)間の下部のヒンジ領域に結合している。
【図9】図9は、FcγRIIIとFcフラグメントとの結合領域の拡大図である。着色は、図8と一致させ、複合体形成に重要な残基は、球と棒で表している。
【図10A】図10aの上部には、Fc受容体エクトドメイン(細胞外ドメイン)の構造に基づく配列アライメントを示す。保存された残基は黄色で陰を付け、同一の残基は橙色で陰を付けている。この図の下部は、ヒト抗体配列のアライメントの一部を示す。複合体結晶構造中においてFcフラグメントと接触しているヒトFcγRIIIの残基は、線でつないでいる(疎水性相互作用は黒色、塩橋(salt brigde)は赤色、水素橋は青色)。FcフラグメントのA鎖と接触しているFc受容体由来の残基は点線でつなぎ、FcフラグメントのB鎖と接触している残基は実線でつないでいる。赤色、青色および黒色の線はそれぞれ、荷電性の接触、極性の接触およびその他の接触を示す。
【図10B】図10bの上部は、Fc受容体エクトドメインの構造に基づく配列アライメントを示す。保存された残基には黄色で影を付け、同一の残基には橙色で影を付けている。あまり関連していないKirとFc受容体配列間の保存された残基は、青で影を付けている。図の下部は、ヒト抗体とマウスIgE(mIgE)配列とのアライメントの一部を示す。複合体結晶構造内でFcフラグメントと接触しているヒトFcγRIIIの残基は、線でつないでいる(疎水性相互作用は黒色、塩橋は赤色、水素結合は青色)。FcフラグメントのA鎖と接触しているFc受容体由来の残基は点線でつなぎ、FcフラグメントのB鎖と接触している残基は実線でつないでいる。赤色、青色および黒色の線はそれぞれ、荷電性の接触、極性の接触およびその他の接触を示す。
【図11】図11は生成したsFcγR、sFcεRIaおよびsFcεRIIの短い形態のもののアライメントを示す。
【図12】図12は生成したsFcγRとsFcεRIaのアライメントを示し、sFcεRIIは含ませていない。
【発明を実施するための形態】
【0012】
本発明にとって特に好ましいものはFcγまたはFcε受容体である。その理由は、IgGおよびIgE分子が多数の疾患および症状に特徴的であり、そのため、それらの定量およびそれらに影響を及ぼしうる方法が非常に関心を集めているからである。図11および12は、いくつかのFcγRとFcεRIとの細胞外部分のアミノ酸配列のアライメントを示す。本発明によるFcRは、これら全ての配列、または抗体への結合能および/または適切な結晶化を依然として保持するその一部を含むものである。
【0013】
本発明の特に好ましい実施形態において、組換え可溶性FcRはFcγRIIb受容体である。さらに、該受容体はヒト由来のものが特に好適である。特に好ましい実施形態において、それは配列番号1〜配列番号6のうちの1つに示したアミノ酸配列を含む。
【0014】
本発明によると、可溶性Fc受容体の調製は原核細胞において行なうことが好ましい。そのような発現の後には、組換えタンパク質を含む不溶性の封入体が原核細胞中に形成され、かくして封入体を他の細胞成分から分離することで精製が促進され、その後に封入体に含まれるタンパク質の再生(復元)が行なわれる。封入体に含まれる本発明によるFcRの再生は基本的には公知の方法に従って行なうことができる。原核細胞における調製の利点である、封入体およびこうして得られる組換え可溶性Fc受容体の生産は、非常に純度が高く、とりわけ非常に均質でもある、FcR調製物を得ることを可能にする。また、グリコシル化が存在しないため、得られた産物はきわめて均質なものである。
【0015】
これまでに組換え法で産生された可溶性Fc受容体は、とりわけ、相当に厄介な精製が必要であるという欠点を抱えていた。というのは、それらが真核細胞で発現され、真核細胞において常に均質であるとは限らないグリコシル化のため、これらの産物もそれほど均質でなかったからである。
【0016】
本発明による組換え可溶性Fc受容体は、本発明の別の実施形態の説明のところで後述するように、X線解析で使用するのに適した結晶を生成することを可能にする。本発明のFcRはさらに、in vivoで天然に存在する受容体と実質的に同じ活性および特異性を示す。
【0017】
本発明の更なる主題は、本発明による組換え可溶性Fc受容体をコードする配列を有する組換え核酸である。
【0018】
本発明による核酸は、コード配列のみを含むこともできるし、また、ベクター配列および/または特に組換えFcRをコードする配列に機能的に連結された発現制御配列(プロモーター、オペレーターなど)をさらに含むこともできる。
【0019】
特に好ましい実施形態において、本発明の核酸は配列番号7〜配列番号12のうちの1つに示した配列を含む。比較のために、配列番号13および配列番号14はそれぞれFcγRIIbおよびFcεRIaをコードする野性型配列を示す。配列番号15〜18はFcγRI、FcγRIIa、FcγRIIIおよびFcεRIIの野性型配列を示す。
【0020】
本発明の核酸がベクター配列を含む場合には、これらは好ましくは1種または数種の原核生物発現ベクターの配列であり、pETベクターの配列が好ましい。所望により、本発明の組換え核酸には発現ベクターの他の知られている機能または構成成分が含まれていてもよい。例えば、これらは形質転換された宿主細胞の効率的な選択を可能とする耐性遺伝子でありうる。
【0021】
本発明のさらに他の主題は、本発明による組換え核酸を含む宿主細胞である。上述したように、宿主細胞は原核宿主細胞であることが好ましく、特に大腸菌細胞が好ましい。
【0022】
本発明による組換え可溶性Fc受容体は、特に抗体と反応するので、多くの検査または用途に使用することができる。in vivoにおいては、可溶性Fc受容体は、上昇したレベルで存在する場合に、免疫系の著しい抑制(多くの部分的に知られた作用および部分的にまだ理解されていない作用へと至らせる)をもたらす強力な免疫調節因子となる。こうした作用に基づいて、本発明のFc受容体のいくつかの用途は本発明の更なる主題となる。
【0023】
そのような主題の1つは、患者の血液または血清中に含まれる、ある型の抗体の量を測定する方法であり、この方法は、イムノアッセイにおいて本発明の組換え可溶性FcRを使用し、FcR−抗体複合体の存在を測定することを特徴とする。かかるアッセイにより、患者の血液、血漿または血清中のある種の抗体の存在をスクリーニングすること、そしてまた、抗体の量を測定することが可能である。
【0024】
FcR−抗体複合体の存在を検出できるのであれば、どのようなタイプのイムノアッセイでも本発明に従って使用するのに基本的に適している。ELISA(固相酵素イムノアッセイ)、特にサンドイッチアッセイ、およびRIA(放射線イムノアッセイ)はどちらも適しているが、競合的試験法も好ましい。IgE抗体の存在および/量を調べる場合の本発明の好適な実施形態においては、本発明による組換え可溶性受容体としてFcεRを使用する。特に、この方法はアレルギーの素因または症状発現を判定するのに適している。
【0025】
さらに、可溶性FcRの存在を測定し、必要ならば定量する方法が好適である。そのような測定では、競合イムノアッセイ法を用いることが好ましく、その際、競合試薬として本発明の組換え可溶性受容体、最も好ましくは組換えFcγRを使用する。この試験によって、とりわけ、免疫系の慢性疾患をもつ患者の免疫状態を競合イムノアッセイで調べることができる。これらの過程における慢性疾患とは、例えば、AIDS、SLE(全身性エリテマトーデス)、MM(多発性骨髄腫)または慢性関節リウマチがあり、またFcεRIIの場合にはB-CLL (Gordonら, 1987)、高度IgE症候群(Sarfatiら, 1988)、またはHCL(Smallら, 1990)がある。
【0026】
本発明の組換え受容体のさらに有利な使用法は、抗体のそれぞれの細胞性受容体による認識および結合を阻害するインヒビターとして作用する能力に関して物質をスクリーニングすることにある。
【0027】
マルチウェルマイクロタイタープレートおよび自動ピペッティング装置と組み合わせたHTPS(ハイスループットスクリーニング)のような最近のスクリーニング技術を用いると、現在では、特定の性質について多数の物質を同時に試験することが可能である。本発明のFcRは、低コストで簡単に生産することができるので、そのような一連の試験法に使用することにより、阻害作用を示す物質を容易に同定することができる。
【0028】
特に、本発明のFc受容体を用いて、各抗体の特定の受容体による認識および結合を阻害する能力があるインヒビターを見つけるまたはスクリーニングするような使用法が好適である。
【0029】
本発明による物質の更なる応用分野は医薬分野にある。それゆえに、本発明の更なる主題は、本発明による組換え可溶性Fc受容体を活性薬剤として含有する医薬組成物である。本発明によれば、この医薬組成物はもちろん慣用の担体や補助物質を含んでいてもよい。そのような物質は当業者には公知であり、さらに投与方式も考慮する必要がある。本発明の医薬組成物は自己免疫疾患、アレルギーまたは腫瘍性疾患の治療や予防に有利に使用することができる。
【0030】
FcγRIIIなどのFc受容体の可溶性形態は、B細胞の増殖および免疫グロブリン産生のアイソタイプ特異的調節を媒介する。骨髄腫のマウスモデルにおいて、sFcRは腫瘍細胞の増殖および免疫グロブリン産生を抑制する(Mullerら, 1985; Romanら, 1988; Teillaudら, 1990)。さらに、sFcRはヒトIgGを分泌する骨髄腫細胞の培養物上の表面IgGと結合し、腫瘍細胞の増殖およびIgG分泌を抑制する。これらの細胞をsFcRに長期間暴露すると、腫瘍細胞の細胞溶解が生じる(Hooverら, 1995)。
【0031】
さらに、アレルギー反応における、または大量の抗原による、免疫系の過剰反応は、例えば、可溶性FcRの静脈内適用によって軽減される可能性がある(Lerinoら, 1993)。
【0032】
したがって、AIDS、慢性関節リウマチまたは多発性骨髄腫の治療に用いる本発明の好適な医薬組成物は、組換え可溶性Fcγ受容体、好ましくは配列番号1〜4に示したアミノ酸配列を有する受容体を含有する。
【0033】
また、Fc受容体および/またはFc受容体/Ig複合体の結晶構造データを得ることにも非常に関心がもてる。一方において、これらは免疫複合体認識の分子機構を理解するうえで鍵となるものであり、他方において、これらの構造データは、各種のFc受容体の構造に共通する特徴を見つけ出し、その構造の情報を用いてインヒビターを生成するために、または新たな人工抗体受容体を同定し生産するために使用することができる。
【0034】
さらに、天然の三次元分子としての免疫グロブリンのそれぞれの受容体に対する実際の結合部位に関する情報を得ることも非常に興味がもてる。それらから、抗体と受容体の相互作用に関する、さらに、その相互作用が如何にモジュレートされ得るかに関する、より一層正確な知見が得られる。これに関連して、モジュレーションとは、そのような相互作用の増強もしくは促進、または、例えば複合体の1以上の部分にある結合部位を塞ぐもしくは覆うことにより、阻害に至らせる相互作用の低下を意味する。
【0035】
そのような結晶構造データおよびコンフォメーション情報を得るためには、本発明による組換え可溶性Fc受容体の結晶性調製物が用いられる。本発明による組換え可溶性FcRは、驚いたことに、信頼できるX線構造解析データをもたらす結晶を生成するに足る純度で得られる。そのような結晶化は、たいていが均質性の欠如ゆえに、これまで生産された受容体分子では可能でなかった。
【0036】
したがって、本発明の別の実施形態は本発明によるFc受容体の結晶性調製物に関する。本発明のさらに別の実施形態は、関連した免疫グロブリンFc部分と一緒の本発明の可溶性Fc受容体からなる複合体の結晶性調製物である。特に好ましい実施形態は、適切な結晶構造データだけでなく実施例においても示される。この結晶性調製物の結晶構造解析により、Fc受容体/Ig複合体の、まさに結合を媒介するアミノ酸を検出することができた。これらのアミノ酸を図6aおよび6bに示してあり、複合体中の両分子の個々のアミノ酸間の結合のタイプも示してある。したがって、本発明の更なる実施形態は、Fc受容体の結晶構造データを生成するための組換え可溶性Fc受容体の結晶性調製物の使用である。この結晶構造データから、抗体の三次元構造および活性結合部位についての情報が得られる。特に、本発明の組換え可溶性Fc受容体と、対応する免疫グロブリン分子との複合体の結晶性調製物を使用して、該複合体の結晶構造データを生成することが好ましい。こうしたデータにより、2つの分子間で形成される実際の相互作用を調べることが可能となり、また、分子間の相互作用に関する正確な情報を入手することによって結合阻害または増強のための可能な部位についての情報を提供することが初めて可能となる。結晶構造データから得られた情報に基づいて、Fc受容体と免疫グロブリンとの相互作用をモジュレートするのに必要な知見が得られる。このモジュレーションは結合の増強から完全な阻害ないし一部の阻害までの範囲でありうる。
【0037】
上記の応用例は結晶構造データの好ましい実施形態にすぎず、他の多くの応用例も考えられる。
【0038】
好ましくは、インヒビターまたは新受容体をそれぞれ生成するおよび/または同定するための構造データはコンピュータ援用モデリングプログラムにおいて使用される。
【0039】
本発明にとっては、図面および実施例に示したFcRまたはFcR:Fcフラグメント複合体の構造が特に好適である。このような構造を用いてインヒビター、アンタゴニストおよび人工受容体分子を設計することができる。
【0040】
コンピュータ援用ドラッグデザインおよびスクリーニングに適するコンピュータプログラムは当業者に公知であり、一般に入手可能である。そのようなプログラムは、対応する構造データをコンピュータに入力したときに、多数の組成物を、ある特定の分子と結合するその能力に関してコンピュータで調べる可能性を提供する。この可能性の助けをかりて、多数の既知の化学組成物をその阻害作用つまりアンタゴニスト作用に関して調べることができる。当業者にとっては、本発明により提供された結晶構造データと市販のスクリーニングプログラム(Program Flexx: GMD-German Narional Research Center for Information Technology, Schloss Birlinghoven, D-53754 Sankt Augustin, Germany)が必要になるだけである。したがって、本発明の好ましい実施形態は、Fc受容体インヒビターを同定し生産するためのコンピュータ援用モデリングプログラムにおいて、本発明の組換え可溶性Fc受容体の得られた結晶構造データ、または本発明の組換え可溶性Fc受容体/対応する免疫グロブリン複合体の得られた結晶構造データを使用することである。
【0041】
同様に、本発明の更なる実施形態は、本発明の受容体または受容体/免疫グロブリン複合体の得られた結晶構造データを用いて、例えばアンタゴニストや競合物質として使用できる新Fc受容体を同定し調製することである。結晶構造データおよびそれから得られたFc受容体結合に関与するアミノ酸に関するデータは、例えば、インヒビターとしても使用できる変異型の免疫グロブリンを生成するのに役立つだろう。突然変異させたまたは化学的に改変したインヒビターが強固に受容体と結合して、受容体をブロックするということが考えられる。一方、免疫グロブリンの結合部位に関して得られたデータは、免疫グロブリン分子のインヒビターの同定および/または調製に使用することができる。本発明は受容体への結合部位を教示するので、比較的単純な分子を用いて結合部位のブロックを行なうことは容易である。それゆえに、本発明の更なる主題は、免疫グロブリンインヒビターを同定しかつ/また調製するためにFcR/Ig複合体の得られた結晶構造データを使用することである。
【0042】
したがって、本発明のさらに他の主題は、本発明の組換え可溶性FcRと相補的な三次元構造を有し、かつ抗体のFcRへの結合を阻害するFcRインヒビターである。
【0043】
本発明のさらに別の主題は、本発明の組換え可溶性Fc受容体の免疫グロブリン結合部位と相補的な三次元構造を有し、かつ免疫グロブリンのFc受容体への結合を阻害する免疫グロブリンインヒビターである。
【0044】
「相補的」という用語は、インヒビター分子が、Fc受容体と免疫グロブリンとの結合を少なくとも決定的に弱体化するほど多くの免疫グロブリン上またはFc受容体上の結合部位を塞ぐことができる物質でなければならない、とそのように本発明の明細書中では理解されるべきである。結合部位を塞ぐことは、いずれか一方の構成成分の複合体形成媒介アミノ酸に結合し、しかも少なくとも複合体形成をもはや可能にしないように結合することによって(立体的阻害)、または隣接アミノ酸に結合し、他方では、Fc受容体と免疫グロブリンとの複雑な結合に関与するアミノ酸を塞ぐことにより、行なうことができる。
【0045】
本発明に関連して、抗体と抗体受容体分子との結合に関与する、まさにその結合部位およびアミノ酸を決定することが初めて可能となった。いまや、特異的に結合する分子を設計し、コンピュータで候補組成物をスクリーニングすることができる。これにより、種々の候補組成物の中から、Fc受容体と免疫グロブリンとの複合体形成を十分に阻害しうる組成物を選択することが可能となる。
【0046】
本発明のインヒビターにとって重要なことは、その構造および特異性のために、それがFcRまたは免疫グロブリンと結合して、FcRと抗体の定常部との正常な結合を妨げる能力がある、という点である。
【0047】
そのようなFcRまたはIgGインヒビターは経口投与が簡単な小さい有機分子であることが好ましい。それらは自己免疫疾患および宿主/移植片拒絶反応の治療に用いられるコルチゾンの興味のもてる代替品となる。そのような分子はまた、いくつかのウイルス、例えば、デング熱ウイルス(抗体被覆ウイルスはFcγRIIb依存的インターナリゼーションを受ける; Littauaら, 1990)、HIV(CD4陽性T細胞上では、HIV感染の抗体増強がFcγRIIIにより媒介される; Homsyら, 1989)、またはエボラ(ウイルスにより分泌される糖タンパク質が、感染に対する宿主応答に影響を及ぼすsFcγRIIIをブロックすることにより初期の好中球活性化を抑制する; Yangら, 1998)による再感染率を低下させるだろう。
【0048】
インヒビターの開発はさらに、IgEのその受容体による認識を妨害する物質をもたらす。FcεRIのモデル化構造から、マスト細胞の脱顆粒をin vitroで抑制するペプチドがすでに開発されている。現在入手できる、相同受容体および受容体−抗体複合体の構造の原子レベルでの情報を用いると、合理的なドラッグデザインの新しい可能性が開けてくる。
【0049】
Fc受容体はいわゆる下部ヒンジ領域にあるFcフラグメントの2つのCH2ドメイン間に結合する(図8)。Fc受容体の結合領域は実施例1に記載される(IgGに対する接触境界面)。FcRと免疫グロブリンとの相互作用を促進する残基は図7、10aおよび10bに示してある。それにより、3つの相互作用領域が明らかになる(図5)。
【0050】
第1の領域: FcR(残基85-87および残基110)−Ig(A鎖残基326-328)
Igのプロリン328は、サンドイッチのようなやり方で残基Trp87および110により固定される。これらの残基はIgGおよびIgEだけでなくIgG受容体およびIgE受容体において保存されている。この突出した領域に結合するインヒビターは結合を強力に妨げるだろう。この領域はさらにインヒビターの設計にとっても魅力的である。なぜならば、受容体の残基Trp87、Ile85、Gly86を含む、露出された疎水性表面領域が更なる結合エネルギーを得るために利用される可能性があるからである。近傍にあるThr113、Glu18およびLys19側鎖の官能基は特に特異的インヒビター結合に寄与しうる。
【0051】
第2の領域: FcR(残基126-132および残基155-158)−Ig(A鎖およびB鎖残基234-239)
両Ig鎖のアミノ末端残基234-239はFcRによって異なって認識され、それにより2倍対称のFcフラグメントを破壊する。
【0052】
FcフラグメントA鎖のこの残基は受容体の残基Val155-Lys158と接触しており、FcフラグメントB鎖からの同残基は受容体の残基Gly126-His132と接触している。この領域は免疫グロブリンだけでなく受容体の配列アライメントにおいても最大の差異を示し、したがって特異性の発生に関与しているのであろう。Fcフラグメント鎖間のこの深い割れ目はインヒビターの設計によく適合し、特異性の問題が関係する場合のインヒビターを開発するための絶好の部位となろう。
【0053】
第3の領域: FcR(残基117、126および129-132)−Ig(B鎖残基264-265および残基296-297) この結合領域は、側鎖中に官能基をもつアミノ酸残基が集まっている点に特徴がある。これらは受容体およびIg接触面に対するインヒビターの設計のために様々なやり方で利用できる可能性がある。
【0054】
結合部位の情報を活用して明らかに設計またはスクリーニングされて、上記領域の1以上と相互作用する分子は、本発明によるインヒビターと見なされる。
【0055】
本発明の更なる主題は、上記のFcRインヒビターまたは免疫グロブリンインヒビターを活性薬剤として含有する医薬組成物である。そのような医薬組成物は例えば、免疫系の過剰反応または誤った反応に起因する疾患の治療または予防に、好ましくはアレルギー、自己免疫疾患またはアナフィラキシーショックの治療または予防に使用しうる。
【0056】
本発明の更なる主題は、固相に結合された本発明のsFcRである。このような固定化受容体は、イムノアッセイまたは固定化形態の受容体が有利に用いられる他の用途に使用することができる。
【0057】
本発明の好適な実施形態において、固相は、Fc受容体をその上に固定しうるクロマトグラフィー担体材料、例えば、セファロース、デキストラン硫酸などである。Fc受容体を結合させてあるそのようなクロマトグラフィー材料を用いて、患者の血液、血漿または血清から、または免疫グロブリン産生細胞の培養上清から免疫グロブリンを吸着させることができる(抗体の濃縮、富化および精製を意味する)。
【0058】
一方、クロマトグラフィー材料に結合された抗体を溶出して、例えば患者の免疫状態を調べることができる。他方、後続の試験を行なう前に、患者の血液由来の抗体を富化することもでき、これは本発明のさらに好ましい実施形態である。多くの場合、血液サンプルは同定すべき抗体をほんの少ししか含んでおらず、診断アッセイを血液サンプルから行なうことは難しい。本発明のFc受容体を結合させた特異的クロマトグラフカラムを用いた濃縮によって、関心のある抗体を簡単に濃縮し、試験を妨害する恐れのある他の多くの物質から分離することが可能である。
【0059】
基本的には、抗体の分離が決定的な役割を果たすような、ある種の疾患の場合には、血液洗浄用の体外灌流システムにおいて本発明のクロマトグラフィー材料を使用することも可能である。
【0060】
しかしながら、本発明の可溶性Fc受容体が結合する他の材料を固相として使用することも考えられ、例えば、壁面にFc受容体が直接または間接に結合する小型の反応容器、マイクロタイタープレートなどがある。そのような固相および容器は診断法には特に重要である。なんとなれば、それらは、例えば患者の血液または他の体液中のある種の免疫グロブリンの存在を検出するためのイムノアッセイを使用することにより、スクリーニングを可能にするからである。
【0061】
要約すると、本発明により提供された組換え可溶性Fc受容体、ならびに該受容体の結晶性調製物の、および受容体と免疫グロブリンとの結晶性複合体の、対応する構造解析は、合理的ドラッグデザインを行なうことを初めて可能にし、それらから、免疫グロブリンと細胞上のFc受容体または可溶性受容体との相互作用をモジュレートすることが可能である。かかるモジュレーションは好ましくは阻害であり、その場合に、IgGとFc受容体からの複合体形成の阻害は、塞ぐことによって、好ましくはFc受容体または免疫グロブリンへのインヒビター分子の結合によって行なわれる。このようなモジュレーター薬物、特にインヒビターの医療用途はいくつか存在しているが、本発明の明細書中ではほんの2、3の応用例を例示的に記載しているにすぎない。これは他の健康障害の治療または予防のためにここに開示した分子構造体またはFcR/Ig複合体についての知見に基づいて設計またはスクリーニングされた分子の利用可能性を決して排除するものではない。
【0062】
本発明は、以下の態様を含む。
【0063】
[1] 膜貫通ドメイン、シグナルペプチドおよびグリコシル化が存在しないことを特徴とする、組換え可溶性Fc受容体。
【0064】
[2] 前記受容体がFcγRまたはFcεRである、上記[1]に記載の組換えFc受容体。
【0065】
[3] 前記受容体がFcγRIIbである、上記[1]または[2]に記載の組換えFc受容体。
【0066】
[4] 前記受容体がヒト由来のものである、上記[1]〜[3]のいずれかに記載の組換えFc受容体。
【0067】
[5] 配列番号1〜6のうちの1つに示したアミノ酸配列を含む、上記[1]〜[4]のいずれかに記載の組換えFc受容体。
【0068】
[6] 上記[1]〜[5]のいずれかに記載の組換えFc受容体をコードする配列を含む組換え核酸。
【0069】
[7] 配列番号7〜12のうちの1つに示した配列を含む、上記[6]に記載の組換え核酸。
【0070】
[8] 組換えFc受容体をコードする配列に機能的に連結された発現制御配列をさらに含む、上記[6]または[7]に記載の組換え核酸。
【0071】
[9] 原核生物発現ベクター、好ましくはpETベクター上に保持されている、上記[6]〜[8]のいずれかに記載の組換え核酸。
【0072】
[10] 上記[6]〜[8]のいずれかに記載の組換え核酸が存在することを特徴とする宿主細胞。
【0073】
[11] 原核宿主細胞、好ましくは大腸菌細胞である、上記[10]に記載の宿主細胞。
【0074】
[12] 上記[1]〜[5]のいずれかに記載の組換え可溶性Fc受容体をイムノアッセイにおいて使用して、FcR−抗体複合体の存在を測定することを特徴とする、患者の血液、血漿または血清中に含まれるある型の抗体の量を測定する方法。
【0075】
[13] イムノアッセイがELISAであり、好ましくはサンドイッチアッセイである、上記[12]に記載の方法。
【0076】
[14] 測定すべき抗体がIgE抗体であり、組換え可溶性受容体がFcεRである、上記[12]または[13]に記載の方法。
【0077】
[15] アレルギーの素因または症状発現を判定するための上記[14]に記載の方法。
【0078】
[16] 測定すべき抗体がIgG抗体であり、組換え可溶性受容体がFcγRである、上記[12]または[13]に記載の方法。
【0079】
[17] 上記[1]〜[5]のいずれかに記載のFc受容体を競合イムノアッセイにおいて使用し、患者の血液、血漿または血清中に含まれる対応する可溶性Fc受容体の量を測定することを特徴とする、免疫系の慢性疾患を有する患者の免疫状態を調べる方法。
【0080】
[18] 慢性疾患がAIDS、SLE、MMまたは慢性関節リウマチである、上記[17]に記載の方法。
【0081】
[19] 抗体のそれぞれの細胞性受容体による認識および結合を阻害するインヒビターとして作用する能力に関して物質をスクリーニングするための、上記[1]〜[5]のいずれかに記載の組換え可溶性Fc受容体の使用。
【0082】
[20] 組換え可溶性FcγRを使用し、IgG抗体の認識および結合に関心がある、上記[19]に記載の使用。
【0083】
[21] 上記[1]〜[5]のいずれかに記載の組換え可溶性Fc受容体を活性薬剤として含有する医薬組成物。
【0084】
[22] 自己免疫疾患、アレルギーもしくは腫瘍性疾患の治療または予防に使用するための、上記[21]に記載の医薬組成物。
【0085】
[23] 組換え可溶性FcγR、好ましくは配列番号1に示したアミノ酸配列を有するもの、を含有する、AIDS、慢性関節リウマチまたは多発性骨髄腫の治療に使用するための上記[21]または[22]に記載の医薬組成物。
【0086】
[24] 上記[1]〜[5]のいずれかに記載の可溶性組換えFc受容体の結晶性調製物。
【0087】
[25] 可溶性組換えFc受容体/免疫グロブリン複合体の結晶性調製物。
【0088】
[26] Fc受容体の結晶構造データを得るための上記[1]〜[5]のいずれかに記載の組換え可溶性Fc受容体の結晶性調製物の使用。
【0089】
[27] 受容体/免疫グロブリン複合体およびそれらのそれぞれの結合部位の結晶構造データを生成するための、可溶性組換えFc受容体/免疫グロブリン複合体の結晶性調製物の使用。
【0090】
[28] Fc受容体インヒビターまたは免疫グロブリンインヒビターを同定および/または調製するための、上記[26]または[27]に記載の使用により得られた結晶構造データの使用。
【0091】
[29] 新しい抗体受容体を同定して調製するための、上記[26]または[27]に記載の使用により得られた結晶構造データの使用。
【0092】
[30] コンピュータ援用モデリングプログラムにおける上記[26]〜[29]のいずれかに記載の使用。
【0093】
[31] 上記[1]〜[5]のいずれか1項に記載の組換え可溶性FcRに相補的な三次元構造を有することを特徴とするFcRインヒビター。
【0094】
[32] 免疫グロブリンのFc受容体結合部位に相補的な三次元構造を有することを特徴とする免疫グロブリンインヒビター。
【0095】
[33] 上記[31]に記載のFcRインヒビターを活性薬剤として含有する医薬組成物。
【0096】
[34] 上記[32]に記載の免疫グロブリンインヒビターを活性薬剤として含有する医薬組成物。
【0097】
[35] 免疫系の過剰反応または誤った反応による疾患の治療または予防に使用するための、上記[33]または[34]に記載の医薬組成物。
【0098】
[36] アレルギー、自己免疫疾患またはアナフィラキシーショックを治療または予防するための、上記[33]、[34]または[35]に記載の医薬組成物。
【0099】
[37] Fc受容体と免疫グロブリンとの相互作用をモジュレートするための分子の使用であって、該分子が上記[24]または[25]に記載の結晶性調製物から得られた結晶構造データを用いて設計または同定されることを特徴とする、上記使用。
【0100】
[38] 前記モジュレーションがFc受容体と免疫グロブリンとの結合の部分的または完全な阻害である、上記[37]に記載の使用。
【0101】
[39] 固相に結合された、上記[1]〜[5]のいずれかに記載のFc受容体。
【0102】
[40] 固相がクロマトグラフィー担体材料である、上記[39]に記載のFc受容体。
【0103】
[41] 患者の血液、結晶もしくは血清から、または免疫グロブリン産生細胞の培養上清から免疫グロブリンを吸着させるための、上記[40]に記載のクロマトグラフィー担体材料の使用。
【0104】
[42] さらなる試験を行なうために患者の血液、結晶もしくは血清から、または免疫グロブリン産生細胞の培養上清から抗体を富化するための、上記[41]に記載の使用。
【実施例】
【0105】
以下の実施例は、図面と関連させて本発明をさらに説明するためのものである。
【0106】
実施例1
shFcγRIIb(可溶性ヒトFcγRIIb)
1.1 クローニングおよび発現
ヒトFcγRIIb2のcDNA (Engelhardtら、1990)を突然変異誘発(mutagenous)PCR (Dulauら、1989)を用いて改変した。すなわち、フォワードプライマーを用いて、新規の開始メチオニンを、NcoI部位内のシグナルペプチドの切断部位の後に導入した(5'-AAT AGA ATT CCA TGG GGA CAC CTG CAG CTC CC-3')。一方、リバースプライマー用いて、終止コドンを、SalI部位がその後に続く膜貫通領域と推定上の細胞外部分との間に導入した(5' CCC AGT GTC GAC AGC CTA AAT GAT CCC C-3')。PCR産物をNcoIおよびSalIで消化してpET11d発現ベクター(Novagen)中にクローン化し、予想される配列を確認した。最終的な構築物をBL21 (DE3)中で増殖させた(GrodbergおよびDunn, 1988)。FcγRIIbを過剰発現させるために、形質転換した細菌の単一のコロニーを、100μg/mlのアンピシリンを含有する5mlのLB培地(LB-Amp100)中に接種し、37℃で一晩インキュベートした。この培養物をLB-Amp100中で200倍に希釈し、OD600が0.7〜0.9に達するまでインキュベーションを続けた。タンパク質の過剰発現は、最終濃度1mMになるまでIPTGを添加することによって誘導した。4時間の増殖時間が経過した後、細胞を遠心(30分間, 4000 x g)により回収し、超音波処理バッファー(リン酸ナトリウム30 mM、塩化ナトリウム300 mM、アジ化ナトリウム 0.02%、pH 7.8)中に再懸濁した。懸濁液1mlあたり0.1mgのリゾチームを添加し、室温で30分間インキュベートした後、氷上で超音波処理を行った(Branson Sonifier、Danbury、CT;Macrotip、出力90%、インターバル80%、15分間)。この懸濁液を遠心(30分間、30,000 x g)にかけ、Dounceホモジナイザーを用いて、0.5%のLDAOを含有する超音波処理バッファー中に再懸濁した。この遠心工程およびLDAO含有バッファー中への再懸濁をもう一度繰り返し、その後、この工程をLDAOなしで2回繰り返した。精製した封入体を4℃で保存した。
【0107】
1.2 可溶性ヒトFcγRIIb (shFcγRIIb)の再生および精製
精製した封入体は、塩化グアニジン6 M、2-メルカプトエタノール100 mM中に、タンパク質濃度10mg/mlで溶解し、遠心により不溶性物質から分離した。再生は、急速な希釈によって達成された。そうして、1mlの該封入体溶液を、撹拌しながら、15時間以内に400mlの再生バッファー(0.1 M TRIS/HCl、アルギニン1.4 M、塩化ナトリウム150 mM、GSH 5 mM、GSSG 0.5 mM、PMSF 0.1 mM、アジ化ナトリウム 0.02%、pH 8.5、4℃)に滴下した。その後、この混合物を、空気酸化により、遊離のチオール基の濃度がEllman (Ellman, 1959)の方法で測定して1 mMに減じるまで、2〜3日間にわたって撹拌した。この溶液をPBSに対して透析して滅菌濾過し、その後、3kD MWCO限外濾過膜を備えた撹拌セル中で10倍に濃縮した。このタンパク質溶液を、hIgGセファロースカラム(セファロース4B 1mlあたりhIgG 50 mg)にアプライした。未結合のタンパク質は、50 mM TRIS pH 8.0で洗浄して取り除き、その後pHジャンプ(塩化ナトリウム150mM、グリシン100mM、アジ化ナトリウム 0.02%、pH 3.0)によって、FcγRIIbを溶出させた。溶出物は、1 M TRIS pH 8.0ですぐに中和させた。FcγRIIb含有溶液を濃縮し、結晶化バッファー(MOPS 2 mM、塩化ナトリウム150 mM、アジ化ナトリウム 0.02%、pH 7.0)で平衡化したSuperdex-75カラムを用いたゲル濾過に付した。FcγRIIbを含む画分をプールし、7 mg/mlまで濃縮して-20℃で保存した。
【0108】
1.3 平衡ゲル濾過実験
Superdex75カラムをFPLCに連結し、10μg/mlのshFcRIIbを含むPBSで平衡化させた。ヒトFcフラグメントを、1μg/10μlの濃度で、平衡化バッファー中に溶解して注入した。得られたクロマトグラムは、shFcγRIIbとFcフラグメントの複合体を含む正のピークを示した。一方、負のピークは複合体形成のためにランニングバッファーから消費された受容体の欠如を表す。
【0109】
1.4 結晶化およびデータ収集
96条件スパースマトリックススクリーニング(96 condition sparse matrix screen)(JancarikおよびKim, 1991)を使用した最初の結晶化試験は、蒸気拡散法を用い、20℃でのシッティングドロップ法で実施した。生成する結晶は、塩、沈殿剤および添加剤の濃度だけでなくpHを変化させることによっても向上した。適当な結晶の回折データを、RU200b回転アノード発生装置(50kV、100mAで操作;Rigaku)からのグラファイト単色CuKα放射線を用いて、イメージプレートシステム(MAR research)上に収集した。反射をプログラムMOSFLM (Leslie, 1997)で積分し、続いてそのデータは、CCP4プログラムセット(Collaborative Computational Project, 1994)の手順を用いて、基準化、減少および切り捨てを行い、構造因子の独立変数を得た。
【0110】
1.5 shFcγRIIbの発現、精製および再生の概要
FcγRIIbの細胞外部分は、T7 RNAポリメラーゼ陽性大腸菌株BL21/DE3 (Grodberg & Dunn, 1988)中、T7プロモーターの制御下で、高レベルで発現させた。該タンパク質は封入体中に集積していたが、これを最初の精製工程において使用した。封入体の単離は、リゾチームと超音波処理を組み合わせた強力な手法を用いて開始し、そうしなければ産物を汚染することになる細胞を実質的にすべて破裂させた。それに続く界面活性剤LDAO(この界面活性剤は、不純物を溶解させるが封入体自体は溶解しない点で優れた特性を有する)を用いた洗浄工程により、早くも90%より高い純度で産物が得られた(図1)。
【0111】
この産物は、さらに精製を行うことなく、再生試験に使用した。該封入体は高濃度の2-メルカプトエタノールおよびグアニジンに溶解し、共有結合凝集体および非共有結合凝集体を確実に単量体へシフトさせた。この溶液は、すぐに再生バッファーで希釈し、そうしなければ凝集体を形成してしまう折りたたまれていないタンパク質分子同士の接触を最小限に抑えた。再生バッファー中でアルギニンを用いることにより、尿素を用いた場合にしばしば認められるような側鎖の不可逆的改変を防ぐことができる。タンパク質を再生バッファーに添加した後、該溶液を、遊離のチオール基の濃度が1mMに減じるまで4℃で撹拌した。この工程は、初期の透析で不活性産物が生じたときに絶対的に必要であった。第2の精製工程において、透析して再生されたFcγRIIbを固定化したhIgGに結合させ、大腸菌タンパク質および不活性受容体のマイナー画分を取り除いた。該タンパクをpHジャンプによって溶出させ、すぐに中和した。このアフィニティークロマトグラフィー工程の後、shFcγRIIbは、繰り返し使用した後でさえもマトリックスから浸出してくる共溶出IgGによるわずかな汚染を除き、本質的に純粋である(図1)。還元SDS-PAGEで認識されないIgGならびに受容体多量体は、ゲル濾過によって容易に除去できた。この工程における汚染物質の除去と平行して、バッファーを定量的に交換した。この手法は、わずかな改変さえもが結晶化の再現不能を引き起こしたり、結晶形成を阻害するときでさえ、一定の組成のタンパク質溶液を確実にもたらす。大腸菌培養物1Lあたり全部で6 mgの純粋なタンパク質を得ることができた。これは、封入体のFcγRIIb含有量の約10%である。
【0112】
N末端タンパク質の配列決定により、検出可能な不純物を含まず、予想された配列H2N-GTPAAPを持つものであることが示された。ESI-MS分析により、結晶化試験で用いられた最終物質は、サイズに関して均一であることを示された。1次配列から、分子量は20434 Daであることが計算され、これは、質量分析で測定された20429 Daと一致している。誤差は、装置のエラー範囲にあり、リーディングメチオニンを含有する種についての追加のピークは、見られなかった。
【0113】
shFcγRIIbの結晶化は、蒸気拡散法を用い、シッティングドロップ法で実施した。スパースマトリックススクリーニング(Jancarik & Kim, 1991)を用いた最初の試験で、早くも、小さな針状結晶が得られた。これに続く、沈殿剤、塩、それらの濃度およびpHを変化させることによる予備的結晶化条件の最適化により、3つの異なる結晶形態が単離された。斜方晶結晶は、1.5 μlのリザーバー溶液(PEG2000 33%、酢酸ナトリウム 0.2 M、pH 5.4)と3 μlのタンパク質溶液の混合物から成長した。それらは3日以内に発生し、約80μm x 80μm x 500μmの最終サイズに1週間後に到達した。これらの結晶は1.7Åで回折した。結晶は、PEG8000 26%、酢酸ナトリウム0.2 M、pH 5.6、Zn(OAc)2 5 mM、塩化ナトリウム100 mMを含むリザーバー溶液から(六方晶形態)、およびPEG8000 26%、NaOAc 0.2 M、pH 5.6、1,4-ジオキサン 10% (v/v)、塩化ナトリウム100mMを含むリザーバー溶液から(正方晶形態)、2つの異なる空間群でも成長できた。これらの結晶はX線解析に適したサイズであるが、それぞれ正方晶形態で2.7Å、六方晶形態で3.8Åでしか回折しなかった(表1)。
【0114】
FcγRIIは大腸菌で発現させた。大腸菌は、比較的安価な製造コストおよび入手可能性に加え、IgG結合が炭水化物結合と独立に生じるFcγRIIの場合のように、哺乳動物細胞により起こるグリコシル化がタンパク質の機能に必要でないときに、特に、いくつかの利点を有する(Sondermannら、1998A)。大腸菌においては、均一な産物が再現可能に生成でき、バッチに依存した変動がしばしば観察される哺乳動物細胞における発現と対照的である。このような系においては、産物が数日間、30℃をこえる温度でプロテアーゼに暴露される。反対に、37℃で、強力なT7プロモーターの制御下、大腸菌内でタンパク質を発現させると、しばしばプロテアーゼが接近できない封入体が形成される。細菌内における発現のさらなる利点は、該物質が使用した仔牛血清または細胞系自体に由来しうる病原菌を含まないと考えられることである。哺乳動物細胞発現においては、標的タンパク質の精製の間、特別に注意を払わなければならない。なぜなら、存在しうる効果的なホルモンまたは成長因子が同時に精製される可能性があるからである。sFcγRの効果がTGFβ1汚染に起因する場合の1事例が、既に報告されている(Galonら、1995)。
【0115】
1.6 精製
精製手順は、簡便である。該手順は1日で容易に実施できる3つのステップからなる。該タンパク質は、純粋な形態でかつ高収率で得られ、その上、高価なIgGアフィニティーカラムを用いずに、かなりの品質で得ることができた。このプロトコルの成功は、封入体の注意深い調製に依るのだろう。なぜなら、ほとんどの不純物は、最初の精製ステップにおいて前もって除去できるからである。
【0116】
1.7 特性評価
精製FcγRIIbは、SDS-PAGEおよび等電点電気泳動ならびにN末端配列決定および質量分析によって特性評価した。従って、該物質は、その化学的組成に関して純粋かつ均一であると考えられる。しかし、受容体が正確に折りたたまれているかという興味深い疑問を考察する余地がある。Ellmanの試験では遊離のチオール基は検出されないことから、すべてのシステインは対になっている。該物質は単量体であり、サイズ排除クロマトグラフィーカラムから、対称形のピークとして、予想された保持時間に溶出する。さらに。FcγRIIbはIgGセファロースに結合し、大腸菌からの組換えFcγRIIbはIgGを特異的に結合することから活性である。
【0117】
1.8 結晶化
FcγRIIbの斜方晶結晶形態は、X線を、1.7Åの分解能で回折した。これは、以前に報告された昆虫細胞発現に由来する同一分子の結晶と比較して、劇的な改良である(Sondermannら、1998A)。これらの結晶は、2.9Åで回折し、空間群P3121である。従って、昆虫細胞由来の受容体のグリコシル化は、結晶化状態に影響を与えることになる。三方晶空間群のかわりに、3つの異なる結晶形態が見出される。その構造の解析が可能となった後は、これらの結晶形態は、結晶接触に起因するタンパク質の人工的コンフォメーションの同定を助けるであろう。
【0118】
FcγRは、その他のタンパク質と配列類似性を示さないが、保存されたシステイン配置から、それらは免疫グロブリンスーパーファミリーに属するとみなされる。従って、本発明者らはその構造を分子置換によって解析しようと試みたが、多様な分子からのIgGドメインを用いた広範囲にわたる試験は、失敗に終わった。従って、FcγRIIbの構造は、多重重原子同形置換法によって解析しなければならない。
【0119】
本発明者らは、初めて、FcγRIIbが大腸菌から活性な形態で得られることを示した。これは、結晶学的研究にとっての基礎であり、既に得られた異例の品質を有する結晶のため、すぐに、この重要な分子の構造解析をもたらすだろう。この構造は、IgG結合部位に関する情報を提供し、かつリガンドのその受容体による認識を阻害する薬物を知見に基づいて設計するための開始点を提供するだろう。さらに、FcγRIIbとFcεRIaを含むその他のFcRとの高度な相同性から、これらの分子は同様に作製することができると考えられ、進行中の研究に対して価値ある材料を提供するだろう。
【0120】
1.9 方法
タンパク質化学
組換え可溶性ヒトFcγRIIbは、大腸菌で発現させ、他で記載されたように、再生し、精製し、結晶化した(Sondermannら、1998B)。簡単に言うと、hFcγRIIb2の推定上の細胞外領域(Engelhardtら、1990)を、大腸菌内で過剰発現させた。 封入体は、細胞のリゾチーム処理およびこれに続く超音波処理によって精製した。得られた懸濁液を遠心(30分間、30,000 x g)にかけ、0.5% LDAOを含むバッファーで洗浄した。遠心工程およびLDAO含有バッファー中への再懸濁をもう一度繰り返し、その後、この手順をLDAOなしで2回繰り返した。封入体を6 M塩酸グアニジンに溶解し、タンパク質を上記のように再生した。透析し、濾過したタンパク質溶液をhIgGセファロースカラムにアプライし、pHジャンプによって溶出した。濃縮し、中和した画分を、Superdex-75カラム(26/60, Pharmacia)を用いるサイズ排除クロマトグラフィーにかけた。
【0121】
結晶化
結晶化は、蒸気拡散法を用い、シッティングドロップ法で20℃にて実施した。結晶化スクリーニングは、pH、塩、沈殿剤および添加剤を変化させることにより実施した。データ収集のために使用する最終的な結晶は、PEG2000 33%、酢酸ナトリウム0.2 M、pH 5.4中 (斜方晶形態)、PEG8000 26%、酢酸ナトリウム0.2 M、pH 5.6、1,4-ジオキサン 10% (v/v)、塩化ナトリウム100 mM中(正方晶形態)、およびPEG8000 26%、酢酸ナトリウム0.2 M、pH 5.6、ZN(OAc)2 5mM、塩化ナトリウム100mM中(六方晶形態)で成長させた。昆虫細胞由来のタンパク質は、PEG6000 32%、酢酸ナトリウム0.2 M、pH 5.3中で結晶化した。
【0122】
重原子誘導体の調製
重原子誘導体は、2 mMプラチナ(II)-(2,2'-6,2''テルピリジニウム)クロリドを含む結晶化バッファー中に24時間または塩化ウラニル10 mMを含む結晶化バッファー中に8日間、結晶を浸透することによって調製した。
【0123】
X線データ収集
回折データはRU200b回転アノード発生装置(50kV、100mAで操作;Rigaku)からのグラファイト単色CuKα放射を用い、イメージプレートシステム(MAR research)上に収集した。反射を、プログラムMOSFLM 5.50 (Leslie, 1997)で積分し、続いてそのデータは、CCP4プログラムセット(Collaborative Computational Project, 1994)の手順を用いて、基準化および切り捨てを行い、構造因子の独立変数を得た。
【0124】
構造決定
構造は、MIR法の標準的手法で解析した。様々な重原子成分を用いて実施した多数の浸透物から、2つの化合物のみが、解釈可能なパターソンマップをもたらした。各誘導体についての重原子位置は差パターソンマップから決定し、初期位相を算出した。2次元(Cross-phased)差フーリエ合成図を使用して、重原子位置を確認し、誘導体の共通の原点を確立した。異常分散データ(anomalous data)を含ませてエナンチオマー同士を区別した。重原子パラメーターは、CCP4パッケージのプログラムMLPHAREを用いてさらに精密化すると、表2の統計をもたらす。電子密度マップは、分解能2.1Åまで計算し、位相は、CCP4セットのプログラムDMを用いた溶媒平滑化(solvent flattening)およびヒストグラムマッチング(histogram matching)によりさらに改良した。得られた電子密度マップは、ほとんどのアミノ酸残基を構築するのに十分な品質であった。モデル構築は、Indigo2ワークステーション(Silicon Graphics Incorporation)上のO (Jonesら、1991)を用いて実施した。構造精密化はXPLOR (Brungerら、 1987)を利用し、EnghおよびHuber (Engh & Huber, 1991)のパラメーターセットを使用して分解能を1.7Åまで段階的に増加させることにより実施した。構造が、数ラウンドのモデル構築および個々の精密化B因子(Rfac = 29% / RFree = 36%)の後に完成したときは、3.5σで等高線を付けたFo-Fcマップが、1σで等高線を付けた十分に明確な電子密度の2Fo-Fcマップと一致するとき、150個の水分子を電子密度に組み込んだ。得られた精密化統計を表3に示す。
【0125】
1.10 構造決定
組換え可溶性ヒトFcγRIIbの結晶構造は、多重重原子同形置換法(MIR)によって1.7Åの分解能で解析した。なぜなら、ヒトIgG1から単離したFcフラグメントのドメインを用いた分子置換による構造解析(Huberら、 1976, PDB entry 1fc1; Deisenhofer, 1981)が失敗したからである。該受容体の推定上の細胞外部分(アミノ酸残基1-187、配列番号2に示す)を使用して、結晶化試験を行った(Sondermannら、1998B)。一方、モデルは、末端がフレキシブルで電子密度を調査できないときは、残基5-176を含むものである。さらに該モデルは150個の水分子を含み、精密化統計は表2にまとめている。該構造は位置11にcisプロリンを含有する。ラマチャンドラン(Ramachandran)プロットの非許容領域に位置する主鎖ねじれ角はなかった。完全に精密化したモデルを使用して、同一タンパク質の空間群P42212の結晶および昆虫細胞に由来するグリコシル化形態の空間群P3121の結晶の構造を解析した(表2)。
【0126】
FcγRIIbのポリペプチド鎖は、免疫グロブリンスーパーファミリーとの関係から予測されるように、2つのIg様ドメインに折りたたまれる。各ドメインは、向かい合っているシート上のB鎖とF鎖を連結する保存されたジスルフィド架橋によってサンドイッチ状に配置された2つのβシートから構成される(図3)。3つの逆平行のβ鎖(A1, B, E)は、5つのβ鎖(C', C, F, G, A2)のシートと向かいあっており、これにより、鎖A1は3本鎖のβシートを残し、5番目の短く平行な鎖A2を追加した4本鎖の逆平行シートに交差する。2次構造要素の配置ならびにその連結性はFcγRIIbの双方のドメインにおいて同一であり、一方のドメインのもう一方のドメイン上への強固な立体フィットは、67個のマッチしたCα原子の平方二乗平均で1.29Åの距離であった。
【0127】
このドメインは互いにほぼ垂直に配置され、その長軸間は70℃の角度を有し、ハート形の全体構造を形成している。この配置は、該ドメイン間に十分な接触領域をもたらす(図4)。鎖A2由来の残基およびN末端ドメインのA2とA1を連結するセグメント由来の残基は、C末端ドメインの鎖A1およびBの残基とかみ合っている。この領域は密にパッキングされ、相互作用はいくつかの水素結合によって強化され、強固な配置をもたらす。これは、3つの異なる空間群における構造の保存によって確認される。斜方晶、正方晶および六方晶(昆虫細胞由来)の結晶形態では、ドメイン間角度において2°未満の偏差が認められる。
【0128】
1.11 全体構造
大腸菌からの組換えヒトFcγRIIbの構造は、斜方晶結晶から、MIRによって1.7Åの分解能で解析した。実質的に同一の構造が、正方晶結晶および昆虫細胞由来タンパク質の六方晶結晶において見られる。これら3つの構造すべてにおいて、ポリペプチド鎖の最後の9残基は、無秩序であることが見出された。分子の構造中心と膜貫通部位間のC末端リンカー領域のフレキシビリティーは機能的に関連し、受容体のいくつかの再配向を可能にして、免疫複合体におけるFc部位の認識を強化しうる。
【0129】
1.12 相同受容体
タンパク質のIgスーパーファミリーに見られるIgドメインは、向かい合ったシートの2つの鎖を連結する保存されたジスルフィド架橋を有するβサンドイッチ構造によって特徴づけられる。FcγRIIb に見られるようなサンドイッチを形成する3および4個の逆平行β鎖の典型的な配置は、T細胞受容体、Fcフラグメント、CD4またはFabフラグメントにおいても生じる。これらの分子の個々のIgドメインとFcγRIIbの2つのドメインとの構造アライメントは、共通の、密接に関連した構造を示す。しかし、該ドメインの相対的配置は、これらの分子においては関連しておらず、広範なセクターに及ぶ。様々な分子に由来するIgドメイン間の構造類似性およびFcγRIIの2つのドメインが重なりあっている場合に生じるCα原子の著しく低い平均二乗平均偏差にもかからわず、有意な配列類似性は見られない(図5aおよび5b)。構造に基づく配列アライメントは、ドメインの配列にそって保存された疎水性パターンを示し、それとともにシステインの他には、同一のアミノ酸残基は少ししかなかった。本発明者らは、最初に、IgG1重鎖の2つのC末端ドメインとFcγRIIbとの構造に基づくアライメントを作成し、その他の関連するFcγRおよびFcεRIaドメインの配列を加えた。これにより、3つのドメインのFcγRIおよび2つのドメインの受容体の配列はIgドメインの疎水性パターンと一致し、いくつかの保存されたアミノ酸残基を表すことが示された。第1に、FcRの異なるドメインは、Igスーパーファミリーのその他の分子からのIgドメインに対するよりも、互いにより関連する。第2に、受容体のN末端ドメインは、第2ドメインと同様に互いに関連する。第3に、FcγRIの第3ドメインの配列は、ドメイングループの双方に由来する特徴を示す。まとめると、本発明者らは、FcRがIgスーパーファミリーに属することを確認し、すべてのFcRドメインは共通の先祖、遺伝子複製により第2ドメインを獲得した古代の1ドメインの受容体に源を有するものと考えられ、さらに、このような2ドメインの受容体のさらなる分枝発生は、第3ドメインを獲得したFcγRIを含む現在の多様性をもたらした。
【0130】
アライメントにおける、FcγRIIb内のドメイン間接触に寄与するこれらのアミノ酸残基の保存は、異なる受容体における類似のドメイン配置に対するヒントにる。表4では、その側鎖でドメイン間接触に寄与している残基(図4)を、図5bの構造に基づく配列アライメントに従って、FcγRIIbについて、その他の受容体内の対応するアミノ酸残基とともに編集している。FcR間で保存されていないAsn15を除き、含まれる残基は同一であるかまたは保存的に置換され、すべてのFcRにおける類似の構造およびドメイン配置を強力に支持している。
【0131】
1.13 IgGに対する接触境界面
FcRとそのリガンドとの相互作用に関する限定的な情報は、突然変異誘発研究(Hogarthら、1992; Hulettら、1994; Hulettら、1995)から入手可能である。FcγRIIa β鎖間のループをFcεRIaアミノ酸残基と系統的に交換することにより、C末端ドメインのB/C、C'/EおよびF/Gループは、リガンドの結合にとって重要であることが評価された(図3、図5b)。構造モデルにおいて、これらのループは隣接し、潜在的なリガンドに自由に接近可能である。さらに、これらのループ内のほとんどのアミノ酸残基を、単点突然変異によってアラニンに置換すると、FcγRIIaの2量体ヒトIgG1への親和性の劇的な変化が生じた。また、高応答性/低応答性多形におけるC末端ドメイン(C'/Eループ)内のArg131のHisへの単一アミノ酸置換は、FcγRIIaのマウスIgG1への親和性を変化させ、その領域を指す。従って、このエリア内のアミノ酸残基は、リガンド結合またはその領域の構造保全性のいずれにも重要である。ここで、該構造は、Tyr 157近傍の疎水性アミノ酸残基Pro 114、Leu 115およびVal 116のクラスター形成を示す。このパッチは中心構造からはみ出している正に荷電したアミノ酸残基Arg 131およびLys 117により、Leu 159、Phe 121およびPhe 129の領域から分離されている(図5b)。
【0132】
1.14 グリコシル化
FcγRIIbの配列中に、3つの潜在的Nグリコシル化部位が見出された。3つの部位はすべて、分子の表面にあり、接近可能である。それらは、両ドメインのE/Fループ内(N61およびN142)およびC末端ドメインの鎖E(N135)上に位置する(図3、図6)。構造解析に使用した材料は、大腸菌から得たものであるため、炭水化物を含まず、一方、哺乳動物細胞から単離したFcRは高度にグリコシル化されている。これらの3つの潜在的グリコシル化部位は、推定のIgG結合領域からかなり離れて位置し、グリコシル化されていないFcγRIIbはヒトIgGを結合することから、結合においてグリコシル化の役割が少ないことを示唆している。この事実は、昆虫細胞において産生されグリコシル化されているFcγRIIbの構造により確認された(Sondermannら、1998A)。おそらく異なる結晶接触によるものであるドメイン間角度の2°の変異を除き、グリコシル化タンパク質と非グリコシル化タンパク質の構造の間に違いは見られなかった。3つのグリコシル化部位は、物質が4つのバンドとして現れるSDS-PAGEで示されるように、場合により使用されるのみである。これらの糖についての追加の電子密度によっても、化学的および構造的な異質性の結果は見られなかった。
【0133】
実施例2
shFcγRIIa(可溶性ヒトFcγRIIa)
手順は、以下に示す変化を除き、実施例1に従って実施した。
【0134】
2.1 クローニングおよび発現
shFcγRIIaは、表5に挙げた突然変異(mutagenous)プライマーを用いた実施例1に従い、各野生型cDNAを突然変異させることによって産生し(Stengelinら、1988)、発現させた。タンパク質の発現のためには、pET22b+ベクターを選択した。
【0135】
2.2 再生および精製
shFcγRIIaは、実施例1に従って、表6に挙げた各再生バッファーを用いて、再生した。
【0136】
2.3 結晶化
shFcγRIIaは、表7に示した条件下で記載した通りに結晶化した。
【0137】
2.4 構造決定
構造は、調査モデルとしてshFcγRIIaを用い、同形置換法で解析した。
【0138】
実施例3
shFcγRIII(可溶性ヒトFcγRIII)
手順は、以下に示す変化を除き、実施例1に従って実施した。
【0139】
3.1 クローニングおよび発現
shFcγRIIIは、表5に挙げた突然変異(mutagenous)プライマーを用いた実施例1に従い、各野生型cDNAを突然変異させることによって産生し(Simmons&Seed、1988)、発現させた。タンパク質の発現のためには、pET22b+ベクターを選択した。
【0140】
3.2 再生および精製
shFcγRIIIは、実施例1に従って、表6に挙げた各再生バッファーを用いて再生した。
【0141】
3.3 結晶化
shFcγRIIIは、表7に示した条件下で記載した通りに結晶化した。
【0142】
3.4 構造決定
構造は、調査モデルとしてshFcγRIIbを用い、同形置換法で解析した。
【0143】
3.5 shFcγRIII:hFc1複合体の結晶化
骨髄腫患者の血清に由来するhIgG1を使用して、プラスミンを用いた消化によってFcフラグメント(hFc1)を調製した(Deisenhoferら、1976)。得られたFcフラグメントは、プロテインAクロマトグラフィーによって、Fabフラグメントから分離した。部分的に消化されたhIgGは、サイズ排除クロマトグラフィーにより、ランニングバッファーとしてMBS(MOPS 2mM、NaCl 150mM、アジ化ナトリウム 0.02%、pH 7.0)を用いて除去した。等モル量のhFc1およびshFcgRIIIを混合してMBSで希釈し、10mg/mlの濃度とした。複合体は、表5に示した条件下で記載の通りに結晶化した。
【0144】
実施例4
shFcεRII(可溶性ヒトFcεRII)
手順は、以下に示す変化を除き、実施例1に従って実施した。
【0145】
4.1 クローニングおよび発現
shFcεRIIは、表5に挙げた突然変異(mutagenous)プライマーを用いた実施例2に従い、各野生型cDNAを突然変異させることによって産生し(Kikutaniら、1986)、発現させた。タンパク質の発現のためには、pET23a+ベクターを選択した。
【0146】
4.2 再生および精製
shFcεRIIの再生は、急速な希釈の前に溶解した封入体を塩化グアニジン 6M、酢酸ナトリウム 20mM、pH 4.0に対して透析することを除き、実施例1に記載の通りに実施した。shFcεRIIは、実施例1に従って、表6に挙げた各再生バッファーを用いて再生した。再生の後、タンパク質溶液をPBSに対して透析し、100倍に濃縮し、Superdex 75を用いたゲル濾過クロマトグラフィーによって精製した。これにより、純粋なshFcεRIIが得られ、これをTRIS/HCL 2mM、NaCl 150mM、アジ化ナトリウム 0.02%、pH 8.0に対して透析し、10mg/mlまで濃縮し、4℃で保存した。
【0147】
実施例5
shFcγRI(可溶性ヒトFcγRI)
手順は、以下に示す変化を除き、実施例1に従って実施した。
【0148】
5.1 クローニングおよび発現
shFcγRIは、表5に挙げた突然変異(mutagenous)プライマーを用いた実施例1に従い、各野生型cDNAを突然変異させることによって産生し(Allen&Seed、1988)、発現させた。タンパク質の発現のためには、pET32a+ベクターを選択した。該ベクターは、N末端チオレドキシンの後にC末端トロンビン切断部位を有するヘキサヒスチジンタグを含み、その後に記載のタンパク質およびアミノ酸残基とインフレームのshFcγRIが続く。融合タンパク質の過剰発現のために、プラスミドpUBSおよびpLysS (Novagen)を含む大腸菌株BL21(DE3)を使用した。
【0149】
精製した封入体は、グアニジン-HCl 6M、β-メルカプトエタノール 10mM、Tris 50mM、pH 8.0に溶解し、Ni-NTAカラム(Qiagen)に結合させた。溶出は、0〜1 Mのイミダゾール勾配で行った。溶出したタンパク質は、NaCl 150mM、Tris 50mM、pH 8.0、GSH 2mM、GSSG 0.5 mMの1000倍量に対して、24時間4℃で透析した。タンパク質溶液を最初の容量の25%に濃縮した後、トロンビンを添加した。37℃で6時間のインキュベートした後、N末端チオレドキシンおよびHisタグをN末端配列決定により確認して完全に除去した。この消化の間、shFcgRIは溶液から定量的に沈殿析出した。
【0150】
5.2 再生および精製
shFcγRIは、実施例1に従って、表6に挙げた各再生バッファーを用いて再生した。酸化還元電位が1mMに減じた後、溶液をPBS pH 8.0に対して透析し、濃縮した。再生したタンパク質をサイズ排除クロマトグラフィーによって分析すると、予想された単量体受容体のピークが得られ、非還元SDS-PAGEは30 kDaに主要なバンドを示した。
【0151】
実施例6
shFcεRIa(可溶性ヒトFcεRIa)
手順は、以下に示す変化を除き、実施例1に従って実施した。
【0152】
6.1 クローニングおよび発現
shFcεRIは、表5に挙げた突然変異(mutagenous)プライマーを用いた実施例1に従い、各野生型cDNAを突然変異させることによって産生し(Kochanら、1988)、発現させた。タンパク質の発現のためには、pET23a+ベクターを選択した。
【0153】
【表1】

【0154】
【表2】
【0155】
【表3】
【0156】
【表4】
【0157】
【表5】
【0158】
【表6】
【0159】
【表7】
【0160】
参照文献
1. Ades, E.W., Phillips, D.J. , Shore, S.L., Gordon, D.S., LaVia, M.F., Black, C.M., Reimer, C.B. (1976),「Analysis of mononuclear cell surfaces with fluoresceinated Staphylococcal protein A complexed with IgG antibody or heat-aggregated γ-globulin(IgG抗体または熱凝集したγ-グロブリンと複合体形成する蛍光標識したブドウ球菌由来のプロテインAを有する単核細胞表面の解析)」,J. Immunol. 117, 2119.
2. Allen J.M., Seed B.;「Nucleotide sequence of three cDNAs for the human high affinity Fc receptor (FcRI)(ヒト高親和性Fc受容体(FcRI)に対する3種のcDNAのヌクレオチド配列)」; Nucleic Acids Res. 16:11824-11824(1988).
3. Amigorena, S., Bonnerot, C., Drake, J.R., Choquet, D., Hunziker, W., Guillet, J.G., Webster, P., Sautes, C., Mellman. I., Fridman, W.H. (1992),「Cytoplasmic domain heterogeneity and functions of IgG Fc receptors in B lymphocytes(Bリンパ球におけるlgG Fc 受容体の細胞質ドメイン異質性および機能)」, Science 256, 1808-1812.
4. Barton, G.C. (1993), ALSCRIPT:「tool to format multiple sequence alignments(多重配列アライメントを形式化する手段)」. Prot. Eng. 6, 37-40.
5. Bazil, V, and Strominger, J.L. (1994),「Metalloprotease and serine protease are involved in cleavage of CD43, CD44, and CD16 from stimulated human granulocytes(メタロプロテアーゼおよびセリンプロテアーゼは、刺激されたヒト顆粒球由来のCD43、CD44およびCD16の切断に関与している)」, J. Immunol. 152, 1314-1322.
6. Brunger. A.T., Kuriyan, J., Karplus. M. (1987),「Crystallographic R factor refinement by molecular dynamics(分子動力学による結晶学的なR因子精密化)」, Science 35. 458-460.
7. Burmeister, W.P., Huber, A.H., Bjorkman, P.J. (1994),「Crystal structureof the complex of rat neonatal Fc receptor with Fc(ラット新生児のFc受容体とFcとの複合体の結晶構造)」, Nature 372, 379-383.
8. Ceuppens, J.L., Baroja, M.L., van Vaeck, F., Anderson, C.L. (1988),「Defect in the membrane expression of high affinity 72kD Fcγ receptors on phagocytic cells in four healthy subjects(4人の健康な被験者由来の貪食細胞上における高親和性72kD Fcγ受容体の膜での発現欠陥)」, J. Clin. Invest. 82, 571-578.
9. Collaborative computational project, 第4号 (1994),「The CCP4 suite: Programs for protein crystallography(CCP4セット:タンパク質結晶学のプログラム)」, Acta crystallogr. D50, 760-763.
10. Deisenhofer, J., Jones, T.A., Huber, R., Sjodahl, J., Sjoquist. J. (1978),「Crystallization, crystal structure analysis and atomic model of the complex formed by a human Fc fragment and fragment B of protein A from Staphylococcus aureus(ヒトFcフラグメントと黄色ブドウ球菌由来プロテインAのフラグメントBにより形成される複合体の結晶化、結晶構造解析および原子モデル)」,Z. Phys. Chem. 359, 975-985.
11. Deisenhofer, J. (1981),「Crystallographic refinement and atomic models of a human Fc fragment and its complex with fragment B of protein A from Staphylococcus aureus at 2.9- and 2.8A resolution(2,9および2.8Å分解能における、ヒトFcフラグメントおよびその黄色ブドウ球菌由来プロテインAのフラグメントBとの複合体の結晶学的精密化および原子モデル)」, Biochemistry 20, 2361-2370.
12. Deisenhofer J., Colman PM., Huber R., Haupt H., Schwick G.;「Crystallographic structural studies of a human Fc-fragment. I. An electron-density map at 4Å resolution and a partial model(ヒトFcフラグメント.Iの結晶学的構造研究。4Å分解能における電子密度マップおよび部分モデル)」; Hoppe-Seyler's Z. Physiol. Chem. 357: 435-445 (1976).
13. Dulau, L., Cheyrou, A., Aigle, M. (1989),「Directed mutagenesis using PCR(PCRを使用した直接的突然変異誘発)」, Nucleic Acids Res. 17, 2873.
14. Ellman (1959),「Tissue sulfhydryl groups(組織スルフヒドリル基)」, Arch. Biochem. Biophys. 82, 79-77.
15. Engelhardt, W., Geerds, C., Frey, J. (1990), 「Distribution, inducibility and biological function of the cloned and expressed human βFc receptor II(クローン化し、発現させたヒトβFc受容体IIの分布、誘導性および生物学的機能)」, Eur. J. Immunol. 20, 1367-1377.
16. Engh, R.A. and Huber, R. (l991), 「Accurate bond and angle parameters for X-ray protein structure refinement(X線タンパク質構造精密化のための正確な結合および角度パラメーター)」, Acta crystallogr. A47, 392-400.
17. Fleit, H.B., Kobasiuk, C.D., Daly, C., Furie, R., Levy, P.C., Webster, R.O. (1992),「A soluble form of FcγRIII is present in human serum and other body fluids and is elevated at sites of inflammation(FcγRIIIの可溶性形態は、ヒト血清および他の体液に存在し、炎症部位で上昇する)」, Blood 79, 2721-2728.
18. Fridman, W.H., Bonnerot, C., Daeron, M., Amigorena, S., Teillaud, J.-L., Sautes, C. (1992),「Structural bases of Fcγ receptor functions(Fcγ受容体機能の構造的基盤)」, Immunol. Rev. 125, 49-76.
19. Fridman, W.H., Teillaud, J.-L., Bouchard, C., Teillaud, C., Astier, A., Tartour, E., Galon, J., Mathiot, C., Sautes, C. (1993),「Soluble Fcγreceptors(可溶性Fcγ受容体)」, J. Leukocyte Biol. 54, 504-512.
20. Gabb, H.A., Jackson, R.M., Sternberg, M.J.E. (1997),「Modelling protein docking using shape complementarity, electrostatics and biochemical information(形状相補性、静電気学および生化学的情報を用いたタンパク質連結のモデル化)」, J. Mol. Biol. 272, 106-120.
21. Galon, J., Bouchard, C., Fridman, W.H., Sautes, C. (1995),「Ligands and biological activities of soluble Fcγ receptors(可溶性Fcγ受容体のリガンドおよび生物学的活性)」, Immunol. Lett. 44, 175-181.
22. Genetics Computer Group(1994),「Program Manual for the Wisconsin Package Version 8(ウィスコンシンパッケージ第8版用のプログラムマニュアル)」,Madison, Wisconsin.
23. Gordon, J.ら, (1980),「The molecules controlling B lymphocytes(Bリンパ球を制御する分子群)」. Immunol. Today, 8: 339-344.
24. Grodberg, J. and Dunn, J.J. (1988), 「OmpT encodes the Escherichia coli outer membrane protease that cleaves T7 RNA polymerase during purification(OmpTは精製の間中、T7 RNA ポリメラーゼを切断する大腸菌外膜プロテアーゼをコード化する)」, J. Bacteriol. 170, 1245-1253.
25. Hogarth, P.M., Hulett, M.D., Ierino, F.L., Tate, B., Powell. M.S., Brinkworth, R.I. (1992),「Identification of the immunoglobulin binding regions (IBR) of FcγRII and FcεRI(FcγRIIおよびFcεRIの免疫グロブリン結合領域(IBR)の同定)」, Immunol. Rev. 125, 21-35.
26. Homsy, J., Meyer, M., Tateno, M., Clarkson, S., Levy, J.A. (1989),「The Fc and not CD4 receptor mediates antibody enhancement of HIV infection in human cells(CD4受容体ではなくFc受容体が、ヒト細胞でのHIV感染における抗体増強を媒介する)」, Science 244, 1357-1360.
27. Hoover, R.G., Lary, C., Page, R., Travis, P., Owens, R., Flick, J., Kornbluth, J., Barlogie, B. (1995), 「Autoregulatory circuits in myeloma: Tumor cell cytotoxity mediated by soluble CD16(骨髄腫における自己調節回路:可溶性CD16が媒介する腫瘍細胞細胞傷害)」, J. Clin. Invest. 95, 241-247.
28. Huber, R., Deisenhofer, J., Colman, P.M., Matsushima, M. and Palm, W. (1976), 「Crystallographic structure studies of an IgG molecule and an Fc fragment(IgG分子およびFcフラグメントの結晶学的構造研究)」, Nature 264, 415-420.
29. Hulett, M.D., Witort, E., Brinkworth, R.I., McKenzie, I.F.C., Hogarth, P.M. (1994), 「Identification of the IgG binding site of the human low affinity receptor for IgG FcγRII(IgG FcγRIIに対するヒト低親和性受容体のIgG結合部位の同定)」, J. Biol. Chem. 269, 15287-15293.
30. Hulett, M.D., Witort, E., Brinkworth, R.I., McKenzie, I.F.C., Hogarth, P.M. (1995), 「Multiple regions of human FcγRII (CD32) contribute to the binding of IgG (ヒトFcγRII(CD32)の多数の領域が、IgGの結合に寄与する)」, J. Biol. Chem. 270, 21188-21194.
31. lerino, F.L., Powell, M.S., McKenzie, I.F.C., Hogarth, P.M. (1993),「Recombinant soluble human FcγRII: Production, characterization, and inhibition of the arthus reaction(組換え可溶性ヒトFcγRII:産生、特性評価およびアルツス反応の阻害)」, J. Exp. Med. 178, 1617-1628.
32. Jancarik. J. and Kim, S.H. (1991),「Sparse matrix sampling: A screening method for crystallization of proteins(スパースマトリックスサンプリング:タンパク質結晶化のためのスクリーング方法)」, J. Appl. Crystallogr. 24, 409-411.
33. Jones, T.A., Zou, J.-Y., Cowan, S.W., Kjeldgaard, M. (l991),「Improved methods for building protein models in electron density maps and the location of errors in these models(改良した電子密度マップにおけるタンパク質モデルを構築する方法および該モデルの誤差位置)」, Acta crystallogr. A47, 110-119.
34. Kikutani H., Inui S., Sato R., Barsumian E.L., Owaki H., Yamasaki K., Kaisho T., Uchibayashi N., Hardy R.R., Hirano T., Tsunasawa S., Sakiyama F., Suemura M., Kishimoto T.;「Molecular structure of human lymphocyte receptor for immunoglobulin E(免疫グロブリンEのヒトリンパ球受容体の分子構造)」; Cell 47(5):657-665(1986).
35. Khayat, D., Soubrane, C., Andriew, J.M., Visonneau, S., Eme, D., Tourani, J.M., Beldjord, K., Weil, M., Fernandez, E., Jaquillat, C. (1990),「Changes of soluble CD16 levels in serum of HIV patients: Correlation with clinical and biological prognostic factors(HIV患者の血清中の可溶性CD16のレベル変化:臨床的および生物学的予後因子との相関関係)」, J. Infect. Dis. 161, 430-435.
36. Kochan J., Pettine L.F., Hakimi J., Kishi K., Kinet J.P.;「Isolation of the gene coding for the alpha subunit of the human high affinity IgE receptor(ヒト高親和性IgE受容体のαサブユニットをコードする遺伝子の単離)」; Nucleic Acids Res. 16: 3584-3584(1988).
37. Simmons D., Seed B.;「The Fc-gamma receptor of natural killer cells is a phospholipid-linked membrane protein(ナチュラルキラー細胞のFcγ受容体は、リン脂質連結膜タンパク質である)」; Nature 333: 568-570 (1988).
38. Kraulis, P.J. (1991) , MOLSCRIPT:「a program to produce both detailed and schematic plots of protein structures(タンパク質構造の詳細で図式的なプロットの作製用プログラム)」, J. Appl. Cryst. 24, 946-950.
39. Leslie, A.G.W. (1997),「Mosflm user guide, mosflm version 5.50(Mosflm使用者説明書, Mosflm 第5.50版)」, Laboratory of Molecular Biology, Cambridge. UK.
40. Lessel, U. and Schomburg, D. (1994),「Similarities between protein 3-D structures(タンパク質3次元構造間の類似性)」, Protein Eng. 7, 1175-1187.
41. Littaua, R., Kurane, I. and Ennis, F.A. (1990),「Human IgG Fc receptor II mediates antibody-dependent enhancement of dengue virus infection(ヒトIgG Fc受容体IIは、デング熱ウイルス感染の抗体依存性の増強を媒介する)」, J. Immunol. 144, 3183-3186.
42. Lynch, R.G., Hagen, M., Mueller, A., Sandor, M. (1995),「Potential role of FcγR in early development of murine lymphoid cells: Evidence for functional interaction between FcγR on pre-thymocytes and an alternative, non-Ig ligand on thymic stromal cells(マウスリンパ球の初期発生におけるFcγRの潜在的役割:前胸腺細胞上のFcγRと胸腺ストローマ細胞上の選択的な非Igのリガンド間の機能的相互作用の証拠)」, Immunol. Lett. 44, 105-109.
43. Mathiot, C., Teillaud, J.L., Elmalek, M., Mosseri, L., Euller-Ziegler, L., Daragon, A., Grosbois, B., Michaux, J.L., Facon, T., Bernard, J.F., Duclos, B., Monconduit. M., Fridman, W.H. (1993),「Correlation between serum soluble CD16 (sCD16) levels and disease stage in patients with multiple myeloma(血清に可溶性のCD16 (sCD16)レベルと多発性骨髄腫を有する患者の疾患の段階間の相関)」, J. Clin. Immunol. 13, 41-48.
44. Merritt, E.A. and Murphy, M.E.P. (1994),「Raster3D Version 2.0. A program for photorealistic molecular graphics(Raster3D 第2.0版:写真のように写実的な分子グラフィックス用プログラム)」, Acta Cryst. D50, 869-873.
45. Metzger, H, (1992A).「Transmembrane signaling: The joy of aggregation(膜貫通シグナル:凝集の価値)」, J. Immunol. 149, 1477-1487.
46. Metzger, H. (1992B). 「The receptor with high affinity for IgE(IgEと高親和性の受容体)」, Immunol. Rev. 125, 37-48.
47. Muller, S. and Hoover, R.G. (1985),「T cells with Fc receptors in myeloma; suppression of growth and secretion of MOPC-315 by T alpha cells(骨髄腫におけるFc受容体を有するT細胞;Tα細胞による増殖抑制およびMOPC-315の分泌)」, J. Immunol. 134, 644-7.
48. Nicholls, A., Sharp, K.A., Honig, B. (1991),「Protein folding and association: insights from the interfacial and thermodynamic properties of hydrocarbons(タンパク質の折たたみおよび会合:炭化水素の界面特性および熱力学的特性からの洞察)」, Proteins 11, 281-296.
49. Poo, H., Kraus, J.C., Mayo-Bond, L., Todd, R.F., Petty, H.R. (1995),「Interaction of Fcγ receptor IIIB with complement receptor type 3 in fibroblast transfectants: evidence from lateral diffusion and resonance energy transfer studies(繊維芽細胞トランスフェクト体におけるFcγ受容体IIIBと補体受容体3型との相互作用:側方拡散および共鳴エネルギー転移研究からの証拠)」, J. Mol. Biol. 247, 597‐603.
50. Rappaport, E.F., Cassel, D.L., Walterhouse, D.O., McKenzie, S.E., Surrey, S., Keller, M.A., Schreiber, A.D., Schwartz, E. (1993),「A soluble form of the human Fc receptor FcγRIIa: cloning, transcript analysis and detection.(ヒトFc受容体FcγRIIaの可溶性形態: クローニング、 転写産物解析および検出)」. Exp. Hematol. 21, 689-696.
51. Ravanel, K., Castelle, C., Defrance. T., Wild, T.F., Charron, D., Lotteau, V., Rabourdincombe, C. (1997), 「Measles virus nucleocapsid protein binds to FcγRII and inhibits human B cell antibody production(麻疹ウイルスのヌクレオキャプシドは、FcγRIIに結合し、ヒトB細胞の抗体産生を阻害する)」. J. Exp. Med. 186, 269-278.
52. Roman, S., Moore, J.S., Darby, C., Muller, S., Hoover, R.G. (l988),「Modulation of Ig gene expression by Ig binding factors. Suppression of alpha-H chain and lambda-2-L chain mRNA accumulation in MOPC-315 by IgA-binding factor(Ig結合因子によるIg遺伝子発現のモジュレーション。IgA結合因子による、MOPC-315におけるα-H鎖およびλ-2-L鎖mRNA蓄積の抑制)」, J. Immunology 140, 3622-30.
53. Sarfat, D.ら, (1988),「Elevation of IgE-binding factors of serum in patients with B-cell derived chronic lymphocytic leukemia(B細胞由来慢性リンパ性白血病を患う患者の血清におけるIgE結合因子の上昇)」. Blood, 71: 94-98.
54. Sauer-Eriksson, A.E., Kleywegt, G.J., Uhlen, M., Jones, T.A. (1995),「Crystal structure of the C2 fragment of streptococcal protein G in complex with the Fc domain of human IgG(ヒトIgGのFcドメインと複合体を形成するブドウ球菌プロテインGのC2フラグメントの結晶構造)」, Structure 3, 265-78.
55. Small, T.,ら, (1990),「B-cell differentiation following autologous, conventional or T-cell depleted bone marrow transplantation: a recapitulation of normal B-cell ontogeny(自己由来の骨髄の移植、通常の骨髄移植、またはT細胞を欠損した骨髄の移植後のB細胞分化:正常なB細胞発生における発生反復)」. Blood, 76: 1647-1656.
56. Sondermann, P., Huber, R., Jacob, U. (1998B),「Preparation and crystallization of active soluble human FcγRIIb derived from E.coli(大腸菌由来の活性型可溶性ヒトFcγRIIbの調製および結晶化)」, Protein Structure, (提出済み).
57. Sondermann, P., Kutscher, C., Jacob, U., Frey, J. (1998A),「Characterization and crystallization of soluble human Fcγ receptor 11 isoforms produced in insect cells(昆虫細胞中で産生した可溶性ヒトFcγ受容体の11個のアイソフォームの特性評価および結晶化)」, Biochemistry, (提出済み).
58. Sondermann, P., Kutscher, C., Jacob, U., Frey, J.,「Analysis of complexes of IgG and soluble human Fcγ-Receptor II produced in insect cells and its crystallization(昆虫細胞中で産生したIgGと可溶性ヒトFcγ受容体IIとの複合体の解析およびその結晶化)」, (提出済み).
59. Stengelin S., Stamenkovic I., Seed B.;「Isolation of cDNAs for two distinct human Fc receptors by ligand affinity cloning(リガンド親和性クローニングによる異なった2つのヒトFc 受容体のcDNAの単離)」: EMBO J. 7: 1053-1059 (l988).
60. Tax, W.J.M., Willems, H.W., Reekers, P.P.M., Capel, P.J.A., Koene, R.A.P. (1983),「Polymorphism in mitogenic effect of IgG1 monoclonal antibodies against T3 antigen on human T cells(ヒトT細胞上のT3抗原に対するIgG1モノクローナル抗体の有糸分裂促進効果における多形性)」, Nature 304, 445-447.
61. Teillaud, J.L., Brunati. S., Elmalek, M., Astier, A., Nicaise, P., Moncuit, J., Mathiot, C., Job-Deslandre, C., Fridman, W.H. (1990),「Involvement of FcR+ Tcells and of IgG-BF in the control of myeloma cells (骨髄腫細胞の制御におけるFcR+ T細胞およびIgG-BFの関与)」, Mol. Immunol. 27, 1209-17.
62. Turk, D. (1992),博士論文, TU Munchen, Germany.
63. Ulvestad, E., Matre, R., Tonder, O. (1988),「IgG Fc receptors in sera from patients with Rheumatoid Arthritis and Systemic Lupus Erythematosus(慢性関節リウマチおよび全身性エリテマトーデスを患う患者の血清中のIgG Fc受容体)」, Scand. J. Rheumatol., Suppl. 75, 203-208.
64. van de Winkel, J.G.J. and Capel, P.J.A. (1993),「Human IgG Fc receptor heterogeneity: Molecular aspects and clinical implications(ヒトIgG Fc 受容体異質性:分子面および臨床的関連性)」, Immunol. Today 14, 215-221.
65. Varin, N., Sautes, C., Galinha, A., Even, J., Hogarth, P.M., Fridman, W.H. (1989),「Recombinant soluble reseptors for the Fcγ portion inhibit antibody production in vitro(Fcγ部分に対する組換え可溶性受容体は、in vitroで抗体産生を阻害する)」, Eur. J. Immunol. 19, 2263-2268.
66. Yang, Z., Delgado, R., Xu, L., Todd, R.F., Nabel, E.G., Sanchez, A., Nabel, G.J. (1998),「Distinct cellular interactions of secreted and transmembrane Ebola virus glycoproteins(分泌型および膜貫通型エボラウイルス糖タンパク質の異なった細胞相互作用)」, Science 279, 983-984.
67. Zhou, M.-J., Todd, R.F., van de Winkel, J.G.J., Petty, H.R. (1993),「Cocapping of the leukoadhesin molecules complement receptor type 3 and lymphocyte function-associated antigen-1 with Fcγ receptor III on human neutrophils. Possible role of lectin-like interactions(ヒト好中球上の白血球粘着分子補体受容体3型およびリンパ球機能関連抗原-1とFcγ受容体IIIとの共キャップ形成。レクチン様相互作用の潜在的役割」, J. Immunol. 150, 3030-3041.
【0161】
配列情報
【技術分野】
【0001】
本発明は、組換え可溶性Fc受容体(FcR)、そのようなFc受容体をコードする組換え核酸、対応する核酸を含む宿主細胞、ならびに患者の血液、血漿または血清に含まれるある型の抗体の量を測定する方法、免疫系の慢性疾患を有する患者の免疫状態を調べる方法、および抗体のそれぞれの細胞性受容体による認識およ結合を阻害するインヒビターとして作用する能力に関して物質をスクリーニングする方法に関する。さらに、本発明は、組換え可溶性FcRを含有する医薬組成物、FcRおよびFcR/Ig複合体の結晶性調製物、特にFc受容体の結晶構造データを生成するための該結晶性調製物の使用、ならびにFcRインヒビターおよび該FcRインヒビターを含有する医薬組成物に関する。
【0002】
本発明の更なる主題は、固相(例えば、クロマトグラフィー担体材料)に結合された組換えFc受容体である。そのようなクロマトグラフィー材料(これは本発明のもう1つの主題である)の使用は、患者の体液からの、または免疫グロブリン産生細胞の培養上清からの免疫グロブリンの吸着にある。
【背景技術】
【0003】
Fc受容体(FcR)は感染からヒトを防御するうえで重要な役割を果たしている。病原体は、血液循環系に接近した後に、免疫グロブリン(Ig)によるオプソニン化を受ける。その結果生じる免疫複合体はその多価性のため高い結合力でFcR保有細胞に結合し、FcRのクラスター化へ至らせ、これがいくつかのエフェクター機能を開始させる(Metzger, H., 1992A)。こうした機能としては、発現されたFcRの型および関連タンパク質に応じて、病原体のその後の中和および抗原提示によるエンドサイトーシス、抗体依存性細胞傷害性(ADCC)、メディエーターの分泌または抗体産生の調節などがある(Fridmanら, 1992; van de Winkel and Capel, 1993)。
【0004】
全てのIgクラスに対して特異的なFcRが存在し、IgGに対するFcRは最も豊富で、多様性も広範囲にわたる。IgEの高親和性受容体(FcεRIa)と共に、FcγRI(CD64)、FcγRII(CD32)およびFcγRIIIa(CD16)はI型膜貫通タンパク質としてまたは可溶性形態(sFcR)で存在しているが、FcγRIIIのグリコシルホスファチジルイノシトールアンカー型(FcγRIIIb)も存在している。さらに、FcRはさまざまなイソ型(FcγRIa、b1、b2、c; FcγRIIa1-2、b1-3、c)および対立遺伝子(FcγRIIa1-HR、-LR; FcγRIIIb-NA1、-NA2)として存在している(van de Winkel and Capel, 1993)。全体的に相同性の細胞外部分とは対照的に、膜貫通ドメインおよび細胞質ドメインは相違している。それらは完全に欠失させることができ、また、8kDaの大きさのものでありうる。それらはFcγRIIaにおけるような26アミノ酸の免疫受容体チロシンベースの活性化モチーフ(ITAM)、またはシグナル伝達に関与するFcγRIIbにおけるような個々の13アミノ酸抑制性モチーフ(ITIM)のいずれかを含みうる(Amigorenaら, 1992)。
【0005】
保存されたシステイン間隔から判断すると、FcRの細胞外部分は3つ(FcγRI、CD64)または2つ(FcεRI、FcγRII、CD32およびFcγRIII、CD16)のIg様ドメイン(10kDa/ドメイン)からなり、それゆえに免疫グロブリンスーパーファミリーに属する。これらの高度にグリコシル化された受容体は相同体であり、FcγRとFcεRIa間のアミノ酸配列の全体的な同一性はそれらの細胞外領域では50%を上回る。それにもかかわらず、FcRのそのリガンドへの親和性はさまざまである。FcγRIのFcフラグメントに対する親和性は約108M-1と比較的高いが、これはその第3ドメインのせいであり、一方、2つのドメインをもつ他のFcγRのIgGに対する親和性は105〜107M-1の範囲で変化する。2つのドメインをもつFcεRIaのIgEに対する親和性は1010M-1と一定しており、これらの値をはるかに上回る(Metzger, H., 1992B)。上記のFcRとは対照的に、IgEに対する低親和性受容体であるFcεRIIはある型の膜貫通タンパク質であり、より低い相同性を示す。
【0006】
FcγRは免疫的に活動しているあらゆる細胞上に特定のパターンで発現されている。FcγRIは単球とマクロファージ上に構成的に発現され、好中球および好酸球上に誘導することができる。FcγRIの生理学的役割は、単球上での発現が必須ではないので、まだ不明である(Ceuppensら, 1988)。FcγRIIIのGPIアンカー型(FcγRIIIb)は顆粒球上に広く発現されている。その細胞質部分が欠失されているため、細胞へのシグナル伝達は、補体受容体3型(CR3)のような、少なくともFcγRIIIbと会合できる他の膜貫通タンパク質を介してのみ起こる(Zhouら, 1993; Pooら, 1995)。FcγRIIIaは主に単球とマクロファージ上に発現されるが、結合タンパク質(例えば、α鎖またはγ鎖)と一緒にしか発現されない。FcγRIIは免疫担当細胞上に最も広く分布している受容体であり、主として免疫複合体のエンドサイトーシスに関与している。
【0007】
FcγRIIaとFcγRIIbは、その細胞外領域のアミノ酸残基がたった7%しか違っていない。それにもかかわらず、両形態はヒトおよびマウスIgGサブクラスへのその結合特性(van de Winkel and Capel, 1993)およびヒトIgGへのその異なる親和性(Sondermannら, 1998A)によって区別され得る。こうした状態は、何人かの個体に由来するT細胞がマウスIgG1誘導有糸分裂に応答できることが判明した後に命名されたFcγRIIaの高応答性/低応答性(HR/LR)多型によってさらに一層複雑になってくる(Taxら, 1983)。その後、LR型とHR型の間のアミノ酸配列の2つの交換によりヒトIgG2への結合能が変化することが見いだされ、このことは、それらの少なくとも1つがIgG結合に関与することを示唆している(Hogarthら, 1992)。
【0008】
健康な個体においてFcRが果たす有益な役割とは対照的に、それらはまた、アレルギー(FcεRIa)または自己免疫疾患において免疫系の刺激を伝達する。さらに、一部のウイルス、例えばHIV(Homsyら, 1989)やデング熱ウイルス(Littauaら, 1990)は細胞に接近するためにFcγRを利用し、またエボラウイルス(Yangら, 1998)や麻疹ウイルス(Ravanelら, 1997)の場合にはFcγRをブロックすることにより免疫応答を遅らせる。
【発明の概要】
【発明が解決しようとする課題】
【0009】
それゆえに、本発明の基礎となる目的は、生産するのが容易で、医療または診断用途のために有利に使用できる受容体を提供することであった。さらに、本発明の目的は、ヒトの体内に存在する天然の受容体に類似した結合特異性および活性を示し、その上、構造決定に適する結晶を生成することを可能にする可溶性受容体を提供することであった。
【課題を解決するための手段】
【0010】
この目的は、Fc受容体の細胞外部分のみからなり、グリコシル化されていない組換え体の可溶性Fc受容体によって達成される。したがって、本発明による受容体は膜貫通ドメイン、シグナルペプチドおよびグリコシル化が存在しないことを特徴とする。
【図面の簡単な説明】
【0011】
【図1】図1は、sFcγRIIbの精製を示す15%還元SDS PAGEである。レーン1:分子量マーカー。レーン2:導入前の大腸菌溶解物。レーン3:導入1時間後の大腸菌溶解物。レーン4:導入4時間後の大腸菌溶解物。レーン5:精製したsFcγRIIbの封入体。レーン6:hIgGアフィニティーカラムの溶出液。レーン7:ゲル濾過カラムのプールした画分。
【図2】図2は、平衡化ゲル濾過である。1 μgのhFcを10 μlの平衡化バッファー(PBS1mlあたり10 μgのsFcγRIIbを含む)に溶解したものをサイズ排除クロマトグラフィーカラムにアプライし、溶出液の吸光度(280nm)を時間の関数として測定した。注入したFcフラグメントは、平衡化バッファー中でsFcγRIIbと複合体を形成する(t = 22分)。消費されたsFcγRIIbの負のピークは、t = 26分に観察される。
【図3】図3は、ヒトsFcγRIIbの全体構造である。sFcγRIIb構造の立体的リボン表示である。IgG結合に重要であると思われるループは、結合部位内のいくつかの残基とともに赤で示し、保存されたジスルフィド架橋は球および棒で示す。潜在的Nグリコシル化部位は緑の球で示す。末端に記を付け、β鎖においてはN末端ドメインには黒で、C末端ドメインには青で、連続的に番号を付した。この図は、プログラムMOLSCRIPT (Kraulis, 1991)およびRENDER (MerrittおよびMurphy, 1994)で作成した。
【図4】図4は、ドメイン間接触を表す。この図は、sFcγRIIbのドメイン間接触に関与する残基の拡大図を示す。N末端ドメインのアミノ酸残基を青色で示し、C末端ドメインの残基を黄色で示す。このモデルは、最終座標から得られた1σで等高線を付した2Fo-Fc電子密度によってカバーされる。ドメイン間の水素結合は白線で示す。この図は、プログラムMAIN(Turk、1992)を用いて作成した。
【図5A】図5aは、2つのFcγRIIbドメインとヒトIgG1のCH2ドメインとの重ね合わせを表す。FcγRIIbの双方のドメインとhIgG1のCH2ドメインを重ね合わせた。N末端ドメインは青色で示し、C末端ドメインは赤色で示し、hIgG1のCH2ドメインは緑色で示す。各末端には記を付け、保存されたジスルフィド架橋は細い線で示す。
【図5B】図5bは、sFcγRIIbドメインとFcRファミリーのその他のメンバーのドメインとの構造に基づく配列アライメントである。図の上部は、プログラムGBF-3D-FIT (Lessel & Schomburg, 1994)を用いて実施したFcγRIIbとhIgG1 Fcフラグメントドメインとの構造に基づく配列アライメントである。重ね合わせたドメインのうち、2.0Å未満のCα距離を有するアミノ酸残基は、Fcフラグメントドメイン間でマッチした残基についてはライラック色で記を付け、FcγRIIbドメイン内の残基には黄色で記を付け、4つのドメインすべてにおいて重ね合わせることができたときは緑色で記をつけている。β鎖はこのアライメントの部分の下に示し、図3と一致するように記をつけている。図の下部は、GCGパッケージ(Genetics Computer Group, 1994)からのルーチンを用い、図の上部に示されたプロファイルに従った、その他のFcγRのアミノ酸配列と相同体FcεRIaとのアミノ酸配列のアライメントを示す。上および下の列の数字は、FcγRIIbのN末端およびC末端ドメインに関する。保存されたシステインは、マゼンタ色でタイプし、潜在的グリコシル化部位は青色でタイプしている。第1ドメインで同一の残基には橙色で記を付け、第2ドメインで同一の残基にはピンク色で記をつけ、双方のドメインで残基が保存されている場合は緑色で記をつけている。あまり保存されていないFcγRIの第3ドメインは、第1および第2ドメイン間でアライメントを行った。赤色の矢印は第1ドメインと第2ドメイン間の側鎖の接触に関与する残基を指し、青色の矢印はIgG結合に関連する残基を示している。この図は、プログラムALSCRIPT(Barton, 1993)で作成した。
【図6】図6は、FcγRIIbの推定の結合部位を表す。GRASP (Nichollsら、1991)で作成したときのFcγRIIbの固体表面表示は、カラーコードは負(赤色)から正(青色)への相対的表面電位に従う。図6aは、図3で示した分子を垂直線を軸として反時計回りに約90°回転させたものを示す。図6bでは、該分子を同じ軸まわりで、時計回りに90°回転させている。両図は、C末端ドメイン(図6a)およびN末端ドメイン(図6b)上の推定結合領域を示す。本明細書中で考察したアミノ酸残基に記を付けている。
【図7】図7は、Fcγ受容体の重ね合わせ構造のCαトレースである。FcγRIIIは赤色、FcγRIIaは緑色、FcγRIIbは青色で表している。IgG結合に重要な残基は球と棒で表している。N末端およびC末端には記を付けている。
【図8】図8は、FcγRIII/Fcフラグメント結晶構造のリボン表示の概観である。Fcフラグメントに結合する糖残基を、球と棒で示している。FcγRIII(青色)は、Fcフラグメントの鎖B(赤色)および鎖A(緑色)間の下部のヒンジ領域に結合している。
【図9】図9は、FcγRIIIとFcフラグメントとの結合領域の拡大図である。着色は、図8と一致させ、複合体形成に重要な残基は、球と棒で表している。
【図10A】図10aの上部には、Fc受容体エクトドメイン(細胞外ドメイン)の構造に基づく配列アライメントを示す。保存された残基は黄色で陰を付け、同一の残基は橙色で陰を付けている。この図の下部は、ヒト抗体配列のアライメントの一部を示す。複合体結晶構造中においてFcフラグメントと接触しているヒトFcγRIIIの残基は、線でつないでいる(疎水性相互作用は黒色、塩橋(salt brigde)は赤色、水素橋は青色)。FcフラグメントのA鎖と接触しているFc受容体由来の残基は点線でつなぎ、FcフラグメントのB鎖と接触している残基は実線でつないでいる。赤色、青色および黒色の線はそれぞれ、荷電性の接触、極性の接触およびその他の接触を示す。
【図10B】図10bの上部は、Fc受容体エクトドメインの構造に基づく配列アライメントを示す。保存された残基には黄色で影を付け、同一の残基には橙色で影を付けている。あまり関連していないKirとFc受容体配列間の保存された残基は、青で影を付けている。図の下部は、ヒト抗体とマウスIgE(mIgE)配列とのアライメントの一部を示す。複合体結晶構造内でFcフラグメントと接触しているヒトFcγRIIIの残基は、線でつないでいる(疎水性相互作用は黒色、塩橋は赤色、水素結合は青色)。FcフラグメントのA鎖と接触しているFc受容体由来の残基は点線でつなぎ、FcフラグメントのB鎖と接触している残基は実線でつないでいる。赤色、青色および黒色の線はそれぞれ、荷電性の接触、極性の接触およびその他の接触を示す。
【図11】図11は生成したsFcγR、sFcεRIaおよびsFcεRIIの短い形態のもののアライメントを示す。
【図12】図12は生成したsFcγRとsFcεRIaのアライメントを示し、sFcεRIIは含ませていない。
【発明を実施するための形態】
【0012】
本発明にとって特に好ましいものはFcγまたはFcε受容体である。その理由は、IgGおよびIgE分子が多数の疾患および症状に特徴的であり、そのため、それらの定量およびそれらに影響を及ぼしうる方法が非常に関心を集めているからである。図11および12は、いくつかのFcγRとFcεRIとの細胞外部分のアミノ酸配列のアライメントを示す。本発明によるFcRは、これら全ての配列、または抗体への結合能および/または適切な結晶化を依然として保持するその一部を含むものである。
【0013】
本発明の特に好ましい実施形態において、組換え可溶性FcRはFcγRIIb受容体である。さらに、該受容体はヒト由来のものが特に好適である。特に好ましい実施形態において、それは配列番号1〜配列番号6のうちの1つに示したアミノ酸配列を含む。
【0014】
本発明によると、可溶性Fc受容体の調製は原核細胞において行なうことが好ましい。そのような発現の後には、組換えタンパク質を含む不溶性の封入体が原核細胞中に形成され、かくして封入体を他の細胞成分から分離することで精製が促進され、その後に封入体に含まれるタンパク質の再生(復元)が行なわれる。封入体に含まれる本発明によるFcRの再生は基本的には公知の方法に従って行なうことができる。原核細胞における調製の利点である、封入体およびこうして得られる組換え可溶性Fc受容体の生産は、非常に純度が高く、とりわけ非常に均質でもある、FcR調製物を得ることを可能にする。また、グリコシル化が存在しないため、得られた産物はきわめて均質なものである。
【0015】
これまでに組換え法で産生された可溶性Fc受容体は、とりわけ、相当に厄介な精製が必要であるという欠点を抱えていた。というのは、それらが真核細胞で発現され、真核細胞において常に均質であるとは限らないグリコシル化のため、これらの産物もそれほど均質でなかったからである。
【0016】
本発明による組換え可溶性Fc受容体は、本発明の別の実施形態の説明のところで後述するように、X線解析で使用するのに適した結晶を生成することを可能にする。本発明のFcRはさらに、in vivoで天然に存在する受容体と実質的に同じ活性および特異性を示す。
【0017】
本発明の更なる主題は、本発明による組換え可溶性Fc受容体をコードする配列を有する組換え核酸である。
【0018】
本発明による核酸は、コード配列のみを含むこともできるし、また、ベクター配列および/または特に組換えFcRをコードする配列に機能的に連結された発現制御配列(プロモーター、オペレーターなど)をさらに含むこともできる。
【0019】
特に好ましい実施形態において、本発明の核酸は配列番号7〜配列番号12のうちの1つに示した配列を含む。比較のために、配列番号13および配列番号14はそれぞれFcγRIIbおよびFcεRIaをコードする野性型配列を示す。配列番号15〜18はFcγRI、FcγRIIa、FcγRIIIおよびFcεRIIの野性型配列を示す。
【0020】
本発明の核酸がベクター配列を含む場合には、これらは好ましくは1種または数種の原核生物発現ベクターの配列であり、pETベクターの配列が好ましい。所望により、本発明の組換え核酸には発現ベクターの他の知られている機能または構成成分が含まれていてもよい。例えば、これらは形質転換された宿主細胞の効率的な選択を可能とする耐性遺伝子でありうる。
【0021】
本発明のさらに他の主題は、本発明による組換え核酸を含む宿主細胞である。上述したように、宿主細胞は原核宿主細胞であることが好ましく、特に大腸菌細胞が好ましい。
【0022】
本発明による組換え可溶性Fc受容体は、特に抗体と反応するので、多くの検査または用途に使用することができる。in vivoにおいては、可溶性Fc受容体は、上昇したレベルで存在する場合に、免疫系の著しい抑制(多くの部分的に知られた作用および部分的にまだ理解されていない作用へと至らせる)をもたらす強力な免疫調節因子となる。こうした作用に基づいて、本発明のFc受容体のいくつかの用途は本発明の更なる主題となる。
【0023】
そのような主題の1つは、患者の血液または血清中に含まれる、ある型の抗体の量を測定する方法であり、この方法は、イムノアッセイにおいて本発明の組換え可溶性FcRを使用し、FcR−抗体複合体の存在を測定することを特徴とする。かかるアッセイにより、患者の血液、血漿または血清中のある種の抗体の存在をスクリーニングすること、そしてまた、抗体の量を測定することが可能である。
【0024】
FcR−抗体複合体の存在を検出できるのであれば、どのようなタイプのイムノアッセイでも本発明に従って使用するのに基本的に適している。ELISA(固相酵素イムノアッセイ)、特にサンドイッチアッセイ、およびRIA(放射線イムノアッセイ)はどちらも適しているが、競合的試験法も好ましい。IgE抗体の存在および/量を調べる場合の本発明の好適な実施形態においては、本発明による組換え可溶性受容体としてFcεRを使用する。特に、この方法はアレルギーの素因または症状発現を判定するのに適している。
【0025】
さらに、可溶性FcRの存在を測定し、必要ならば定量する方法が好適である。そのような測定では、競合イムノアッセイ法を用いることが好ましく、その際、競合試薬として本発明の組換え可溶性受容体、最も好ましくは組換えFcγRを使用する。この試験によって、とりわけ、免疫系の慢性疾患をもつ患者の免疫状態を競合イムノアッセイで調べることができる。これらの過程における慢性疾患とは、例えば、AIDS、SLE(全身性エリテマトーデス)、MM(多発性骨髄腫)または慢性関節リウマチがあり、またFcεRIIの場合にはB-CLL (Gordonら, 1987)、高度IgE症候群(Sarfatiら, 1988)、またはHCL(Smallら, 1990)がある。
【0026】
本発明の組換え受容体のさらに有利な使用法は、抗体のそれぞれの細胞性受容体による認識および結合を阻害するインヒビターとして作用する能力に関して物質をスクリーニングすることにある。
【0027】
マルチウェルマイクロタイタープレートおよび自動ピペッティング装置と組み合わせたHTPS(ハイスループットスクリーニング)のような最近のスクリーニング技術を用いると、現在では、特定の性質について多数の物質を同時に試験することが可能である。本発明のFcRは、低コストで簡単に生産することができるので、そのような一連の試験法に使用することにより、阻害作用を示す物質を容易に同定することができる。
【0028】
特に、本発明のFc受容体を用いて、各抗体の特定の受容体による認識および結合を阻害する能力があるインヒビターを見つけるまたはスクリーニングするような使用法が好適である。
【0029】
本発明による物質の更なる応用分野は医薬分野にある。それゆえに、本発明の更なる主題は、本発明による組換え可溶性Fc受容体を活性薬剤として含有する医薬組成物である。本発明によれば、この医薬組成物はもちろん慣用の担体や補助物質を含んでいてもよい。そのような物質は当業者には公知であり、さらに投与方式も考慮する必要がある。本発明の医薬組成物は自己免疫疾患、アレルギーまたは腫瘍性疾患の治療や予防に有利に使用することができる。
【0030】
FcγRIIIなどのFc受容体の可溶性形態は、B細胞の増殖および免疫グロブリン産生のアイソタイプ特異的調節を媒介する。骨髄腫のマウスモデルにおいて、sFcRは腫瘍細胞の増殖および免疫グロブリン産生を抑制する(Mullerら, 1985; Romanら, 1988; Teillaudら, 1990)。さらに、sFcRはヒトIgGを分泌する骨髄腫細胞の培養物上の表面IgGと結合し、腫瘍細胞の増殖およびIgG分泌を抑制する。これらの細胞をsFcRに長期間暴露すると、腫瘍細胞の細胞溶解が生じる(Hooverら, 1995)。
【0031】
さらに、アレルギー反応における、または大量の抗原による、免疫系の過剰反応は、例えば、可溶性FcRの静脈内適用によって軽減される可能性がある(Lerinoら, 1993)。
【0032】
したがって、AIDS、慢性関節リウマチまたは多発性骨髄腫の治療に用いる本発明の好適な医薬組成物は、組換え可溶性Fcγ受容体、好ましくは配列番号1〜4に示したアミノ酸配列を有する受容体を含有する。
【0033】
また、Fc受容体および/またはFc受容体/Ig複合体の結晶構造データを得ることにも非常に関心がもてる。一方において、これらは免疫複合体認識の分子機構を理解するうえで鍵となるものであり、他方において、これらの構造データは、各種のFc受容体の構造に共通する特徴を見つけ出し、その構造の情報を用いてインヒビターを生成するために、または新たな人工抗体受容体を同定し生産するために使用することができる。
【0034】
さらに、天然の三次元分子としての免疫グロブリンのそれぞれの受容体に対する実際の結合部位に関する情報を得ることも非常に興味がもてる。それらから、抗体と受容体の相互作用に関する、さらに、その相互作用が如何にモジュレートされ得るかに関する、より一層正確な知見が得られる。これに関連して、モジュレーションとは、そのような相互作用の増強もしくは促進、または、例えば複合体の1以上の部分にある結合部位を塞ぐもしくは覆うことにより、阻害に至らせる相互作用の低下を意味する。
【0035】
そのような結晶構造データおよびコンフォメーション情報を得るためには、本発明による組換え可溶性Fc受容体の結晶性調製物が用いられる。本発明による組換え可溶性FcRは、驚いたことに、信頼できるX線構造解析データをもたらす結晶を生成するに足る純度で得られる。そのような結晶化は、たいていが均質性の欠如ゆえに、これまで生産された受容体分子では可能でなかった。
【0036】
したがって、本発明の別の実施形態は本発明によるFc受容体の結晶性調製物に関する。本発明のさらに別の実施形態は、関連した免疫グロブリンFc部分と一緒の本発明の可溶性Fc受容体からなる複合体の結晶性調製物である。特に好ましい実施形態は、適切な結晶構造データだけでなく実施例においても示される。この結晶性調製物の結晶構造解析により、Fc受容体/Ig複合体の、まさに結合を媒介するアミノ酸を検出することができた。これらのアミノ酸を図6aおよび6bに示してあり、複合体中の両分子の個々のアミノ酸間の結合のタイプも示してある。したがって、本発明の更なる実施形態は、Fc受容体の結晶構造データを生成するための組換え可溶性Fc受容体の結晶性調製物の使用である。この結晶構造データから、抗体の三次元構造および活性結合部位についての情報が得られる。特に、本発明の組換え可溶性Fc受容体と、対応する免疫グロブリン分子との複合体の結晶性調製物を使用して、該複合体の結晶構造データを生成することが好ましい。こうしたデータにより、2つの分子間で形成される実際の相互作用を調べることが可能となり、また、分子間の相互作用に関する正確な情報を入手することによって結合阻害または増強のための可能な部位についての情報を提供することが初めて可能となる。結晶構造データから得られた情報に基づいて、Fc受容体と免疫グロブリンとの相互作用をモジュレートするのに必要な知見が得られる。このモジュレーションは結合の増強から完全な阻害ないし一部の阻害までの範囲でありうる。
【0037】
上記の応用例は結晶構造データの好ましい実施形態にすぎず、他の多くの応用例も考えられる。
【0038】
好ましくは、インヒビターまたは新受容体をそれぞれ生成するおよび/または同定するための構造データはコンピュータ援用モデリングプログラムにおいて使用される。
【0039】
本発明にとっては、図面および実施例に示したFcRまたはFcR:Fcフラグメント複合体の構造が特に好適である。このような構造を用いてインヒビター、アンタゴニストおよび人工受容体分子を設計することができる。
【0040】
コンピュータ援用ドラッグデザインおよびスクリーニングに適するコンピュータプログラムは当業者に公知であり、一般に入手可能である。そのようなプログラムは、対応する構造データをコンピュータに入力したときに、多数の組成物を、ある特定の分子と結合するその能力に関してコンピュータで調べる可能性を提供する。この可能性の助けをかりて、多数の既知の化学組成物をその阻害作用つまりアンタゴニスト作用に関して調べることができる。当業者にとっては、本発明により提供された結晶構造データと市販のスクリーニングプログラム(Program Flexx: GMD-German Narional Research Center for Information Technology, Schloss Birlinghoven, D-53754 Sankt Augustin, Germany)が必要になるだけである。したがって、本発明の好ましい実施形態は、Fc受容体インヒビターを同定し生産するためのコンピュータ援用モデリングプログラムにおいて、本発明の組換え可溶性Fc受容体の得られた結晶構造データ、または本発明の組換え可溶性Fc受容体/対応する免疫グロブリン複合体の得られた結晶構造データを使用することである。
【0041】
同様に、本発明の更なる実施形態は、本発明の受容体または受容体/免疫グロブリン複合体の得られた結晶構造データを用いて、例えばアンタゴニストや競合物質として使用できる新Fc受容体を同定し調製することである。結晶構造データおよびそれから得られたFc受容体結合に関与するアミノ酸に関するデータは、例えば、インヒビターとしても使用できる変異型の免疫グロブリンを生成するのに役立つだろう。突然変異させたまたは化学的に改変したインヒビターが強固に受容体と結合して、受容体をブロックするということが考えられる。一方、免疫グロブリンの結合部位に関して得られたデータは、免疫グロブリン分子のインヒビターの同定および/または調製に使用することができる。本発明は受容体への結合部位を教示するので、比較的単純な分子を用いて結合部位のブロックを行なうことは容易である。それゆえに、本発明の更なる主題は、免疫グロブリンインヒビターを同定しかつ/また調製するためにFcR/Ig複合体の得られた結晶構造データを使用することである。
【0042】
したがって、本発明のさらに他の主題は、本発明の組換え可溶性FcRと相補的な三次元構造を有し、かつ抗体のFcRへの結合を阻害するFcRインヒビターである。
【0043】
本発明のさらに別の主題は、本発明の組換え可溶性Fc受容体の免疫グロブリン結合部位と相補的な三次元構造を有し、かつ免疫グロブリンのFc受容体への結合を阻害する免疫グロブリンインヒビターである。
【0044】
「相補的」という用語は、インヒビター分子が、Fc受容体と免疫グロブリンとの結合を少なくとも決定的に弱体化するほど多くの免疫グロブリン上またはFc受容体上の結合部位を塞ぐことができる物質でなければならない、とそのように本発明の明細書中では理解されるべきである。結合部位を塞ぐことは、いずれか一方の構成成分の複合体形成媒介アミノ酸に結合し、しかも少なくとも複合体形成をもはや可能にしないように結合することによって(立体的阻害)、または隣接アミノ酸に結合し、他方では、Fc受容体と免疫グロブリンとの複雑な結合に関与するアミノ酸を塞ぐことにより、行なうことができる。
【0045】
本発明に関連して、抗体と抗体受容体分子との結合に関与する、まさにその結合部位およびアミノ酸を決定することが初めて可能となった。いまや、特異的に結合する分子を設計し、コンピュータで候補組成物をスクリーニングすることができる。これにより、種々の候補組成物の中から、Fc受容体と免疫グロブリンとの複合体形成を十分に阻害しうる組成物を選択することが可能となる。
【0046】
本発明のインヒビターにとって重要なことは、その構造および特異性のために、それがFcRまたは免疫グロブリンと結合して、FcRと抗体の定常部との正常な結合を妨げる能力がある、という点である。
【0047】
そのようなFcRまたはIgGインヒビターは経口投与が簡単な小さい有機分子であることが好ましい。それらは自己免疫疾患および宿主/移植片拒絶反応の治療に用いられるコルチゾンの興味のもてる代替品となる。そのような分子はまた、いくつかのウイルス、例えば、デング熱ウイルス(抗体被覆ウイルスはFcγRIIb依存的インターナリゼーションを受ける; Littauaら, 1990)、HIV(CD4陽性T細胞上では、HIV感染の抗体増強がFcγRIIIにより媒介される; Homsyら, 1989)、またはエボラ(ウイルスにより分泌される糖タンパク質が、感染に対する宿主応答に影響を及ぼすsFcγRIIIをブロックすることにより初期の好中球活性化を抑制する; Yangら, 1998)による再感染率を低下させるだろう。
【0048】
インヒビターの開発はさらに、IgEのその受容体による認識を妨害する物質をもたらす。FcεRIのモデル化構造から、マスト細胞の脱顆粒をin vitroで抑制するペプチドがすでに開発されている。現在入手できる、相同受容体および受容体−抗体複合体の構造の原子レベルでの情報を用いると、合理的なドラッグデザインの新しい可能性が開けてくる。
【0049】
Fc受容体はいわゆる下部ヒンジ領域にあるFcフラグメントの2つのCH2ドメイン間に結合する(図8)。Fc受容体の結合領域は実施例1に記載される(IgGに対する接触境界面)。FcRと免疫グロブリンとの相互作用を促進する残基は図7、10aおよび10bに示してある。それにより、3つの相互作用領域が明らかになる(図5)。
【0050】
第1の領域: FcR(残基85-87および残基110)−Ig(A鎖残基326-328)
Igのプロリン328は、サンドイッチのようなやり方で残基Trp87および110により固定される。これらの残基はIgGおよびIgEだけでなくIgG受容体およびIgE受容体において保存されている。この突出した領域に結合するインヒビターは結合を強力に妨げるだろう。この領域はさらにインヒビターの設計にとっても魅力的である。なぜならば、受容体の残基Trp87、Ile85、Gly86を含む、露出された疎水性表面領域が更なる結合エネルギーを得るために利用される可能性があるからである。近傍にあるThr113、Glu18およびLys19側鎖の官能基は特に特異的インヒビター結合に寄与しうる。
【0051】
第2の領域: FcR(残基126-132および残基155-158)−Ig(A鎖およびB鎖残基234-239)
両Ig鎖のアミノ末端残基234-239はFcRによって異なって認識され、それにより2倍対称のFcフラグメントを破壊する。
【0052】
FcフラグメントA鎖のこの残基は受容体の残基Val155-Lys158と接触しており、FcフラグメントB鎖からの同残基は受容体の残基Gly126-His132と接触している。この領域は免疫グロブリンだけでなく受容体の配列アライメントにおいても最大の差異を示し、したがって特異性の発生に関与しているのであろう。Fcフラグメント鎖間のこの深い割れ目はインヒビターの設計によく適合し、特異性の問題が関係する場合のインヒビターを開発するための絶好の部位となろう。
【0053】
第3の領域: FcR(残基117、126および129-132)−Ig(B鎖残基264-265および残基296-297) この結合領域は、側鎖中に官能基をもつアミノ酸残基が集まっている点に特徴がある。これらは受容体およびIg接触面に対するインヒビターの設計のために様々なやり方で利用できる可能性がある。
【0054】
結合部位の情報を活用して明らかに設計またはスクリーニングされて、上記領域の1以上と相互作用する分子は、本発明によるインヒビターと見なされる。
【0055】
本発明の更なる主題は、上記のFcRインヒビターまたは免疫グロブリンインヒビターを活性薬剤として含有する医薬組成物である。そのような医薬組成物は例えば、免疫系の過剰反応または誤った反応に起因する疾患の治療または予防に、好ましくはアレルギー、自己免疫疾患またはアナフィラキシーショックの治療または予防に使用しうる。
【0056】
本発明の更なる主題は、固相に結合された本発明のsFcRである。このような固定化受容体は、イムノアッセイまたは固定化形態の受容体が有利に用いられる他の用途に使用することができる。
【0057】
本発明の好適な実施形態において、固相は、Fc受容体をその上に固定しうるクロマトグラフィー担体材料、例えば、セファロース、デキストラン硫酸などである。Fc受容体を結合させてあるそのようなクロマトグラフィー材料を用いて、患者の血液、血漿または血清から、または免疫グロブリン産生細胞の培養上清から免疫グロブリンを吸着させることができる(抗体の濃縮、富化および精製を意味する)。
【0058】
一方、クロマトグラフィー材料に結合された抗体を溶出して、例えば患者の免疫状態を調べることができる。他方、後続の試験を行なう前に、患者の血液由来の抗体を富化することもでき、これは本発明のさらに好ましい実施形態である。多くの場合、血液サンプルは同定すべき抗体をほんの少ししか含んでおらず、診断アッセイを血液サンプルから行なうことは難しい。本発明のFc受容体を結合させた特異的クロマトグラフカラムを用いた濃縮によって、関心のある抗体を簡単に濃縮し、試験を妨害する恐れのある他の多くの物質から分離することが可能である。
【0059】
基本的には、抗体の分離が決定的な役割を果たすような、ある種の疾患の場合には、血液洗浄用の体外灌流システムにおいて本発明のクロマトグラフィー材料を使用することも可能である。
【0060】
しかしながら、本発明の可溶性Fc受容体が結合する他の材料を固相として使用することも考えられ、例えば、壁面にFc受容体が直接または間接に結合する小型の反応容器、マイクロタイタープレートなどがある。そのような固相および容器は診断法には特に重要である。なんとなれば、それらは、例えば患者の血液または他の体液中のある種の免疫グロブリンの存在を検出するためのイムノアッセイを使用することにより、スクリーニングを可能にするからである。
【0061】
要約すると、本発明により提供された組換え可溶性Fc受容体、ならびに該受容体の結晶性調製物の、および受容体と免疫グロブリンとの結晶性複合体の、対応する構造解析は、合理的ドラッグデザインを行なうことを初めて可能にし、それらから、免疫グロブリンと細胞上のFc受容体または可溶性受容体との相互作用をモジュレートすることが可能である。かかるモジュレーションは好ましくは阻害であり、その場合に、IgGとFc受容体からの複合体形成の阻害は、塞ぐことによって、好ましくはFc受容体または免疫グロブリンへのインヒビター分子の結合によって行なわれる。このようなモジュレーター薬物、特にインヒビターの医療用途はいくつか存在しているが、本発明の明細書中ではほんの2、3の応用例を例示的に記載しているにすぎない。これは他の健康障害の治療または予防のためにここに開示した分子構造体またはFcR/Ig複合体についての知見に基づいて設計またはスクリーニングされた分子の利用可能性を決して排除するものではない。
【0062】
本発明は、以下の態様を含む。
【0063】
[1] 膜貫通ドメイン、シグナルペプチドおよびグリコシル化が存在しないことを特徴とする、組換え可溶性Fc受容体。
【0064】
[2] 前記受容体がFcγRまたはFcεRである、上記[1]に記載の組換えFc受容体。
【0065】
[3] 前記受容体がFcγRIIbである、上記[1]または[2]に記載の組換えFc受容体。
【0066】
[4] 前記受容体がヒト由来のものである、上記[1]〜[3]のいずれかに記載の組換えFc受容体。
【0067】
[5] 配列番号1〜6のうちの1つに示したアミノ酸配列を含む、上記[1]〜[4]のいずれかに記載の組換えFc受容体。
【0068】
[6] 上記[1]〜[5]のいずれかに記載の組換えFc受容体をコードする配列を含む組換え核酸。
【0069】
[7] 配列番号7〜12のうちの1つに示した配列を含む、上記[6]に記載の組換え核酸。
【0070】
[8] 組換えFc受容体をコードする配列に機能的に連結された発現制御配列をさらに含む、上記[6]または[7]に記載の組換え核酸。
【0071】
[9] 原核生物発現ベクター、好ましくはpETベクター上に保持されている、上記[6]〜[8]のいずれかに記載の組換え核酸。
【0072】
[10] 上記[6]〜[8]のいずれかに記載の組換え核酸が存在することを特徴とする宿主細胞。
【0073】
[11] 原核宿主細胞、好ましくは大腸菌細胞である、上記[10]に記載の宿主細胞。
【0074】
[12] 上記[1]〜[5]のいずれかに記載の組換え可溶性Fc受容体をイムノアッセイにおいて使用して、FcR−抗体複合体の存在を測定することを特徴とする、患者の血液、血漿または血清中に含まれるある型の抗体の量を測定する方法。
【0075】
[13] イムノアッセイがELISAであり、好ましくはサンドイッチアッセイである、上記[12]に記載の方法。
【0076】
[14] 測定すべき抗体がIgE抗体であり、組換え可溶性受容体がFcεRである、上記[12]または[13]に記載の方法。
【0077】
[15] アレルギーの素因または症状発現を判定するための上記[14]に記載の方法。
【0078】
[16] 測定すべき抗体がIgG抗体であり、組換え可溶性受容体がFcγRである、上記[12]または[13]に記載の方法。
【0079】
[17] 上記[1]〜[5]のいずれかに記載のFc受容体を競合イムノアッセイにおいて使用し、患者の血液、血漿または血清中に含まれる対応する可溶性Fc受容体の量を測定することを特徴とする、免疫系の慢性疾患を有する患者の免疫状態を調べる方法。
【0080】
[18] 慢性疾患がAIDS、SLE、MMまたは慢性関節リウマチである、上記[17]に記載の方法。
【0081】
[19] 抗体のそれぞれの細胞性受容体による認識および結合を阻害するインヒビターとして作用する能力に関して物質をスクリーニングするための、上記[1]〜[5]のいずれかに記載の組換え可溶性Fc受容体の使用。
【0082】
[20] 組換え可溶性FcγRを使用し、IgG抗体の認識および結合に関心がある、上記[19]に記載の使用。
【0083】
[21] 上記[1]〜[5]のいずれかに記載の組換え可溶性Fc受容体を活性薬剤として含有する医薬組成物。
【0084】
[22] 自己免疫疾患、アレルギーもしくは腫瘍性疾患の治療または予防に使用するための、上記[21]に記載の医薬組成物。
【0085】
[23] 組換え可溶性FcγR、好ましくは配列番号1に示したアミノ酸配列を有するもの、を含有する、AIDS、慢性関節リウマチまたは多発性骨髄腫の治療に使用するための上記[21]または[22]に記載の医薬組成物。
【0086】
[24] 上記[1]〜[5]のいずれかに記載の可溶性組換えFc受容体の結晶性調製物。
【0087】
[25] 可溶性組換えFc受容体/免疫グロブリン複合体の結晶性調製物。
【0088】
[26] Fc受容体の結晶構造データを得るための上記[1]〜[5]のいずれかに記載の組換え可溶性Fc受容体の結晶性調製物の使用。
【0089】
[27] 受容体/免疫グロブリン複合体およびそれらのそれぞれの結合部位の結晶構造データを生成するための、可溶性組換えFc受容体/免疫グロブリン複合体の結晶性調製物の使用。
【0090】
[28] Fc受容体インヒビターまたは免疫グロブリンインヒビターを同定および/または調製するための、上記[26]または[27]に記載の使用により得られた結晶構造データの使用。
【0091】
[29] 新しい抗体受容体を同定して調製するための、上記[26]または[27]に記載の使用により得られた結晶構造データの使用。
【0092】
[30] コンピュータ援用モデリングプログラムにおける上記[26]〜[29]のいずれかに記載の使用。
【0093】
[31] 上記[1]〜[5]のいずれか1項に記載の組換え可溶性FcRに相補的な三次元構造を有することを特徴とするFcRインヒビター。
【0094】
[32] 免疫グロブリンのFc受容体結合部位に相補的な三次元構造を有することを特徴とする免疫グロブリンインヒビター。
【0095】
[33] 上記[31]に記載のFcRインヒビターを活性薬剤として含有する医薬組成物。
【0096】
[34] 上記[32]に記載の免疫グロブリンインヒビターを活性薬剤として含有する医薬組成物。
【0097】
[35] 免疫系の過剰反応または誤った反応による疾患の治療または予防に使用するための、上記[33]または[34]に記載の医薬組成物。
【0098】
[36] アレルギー、自己免疫疾患またはアナフィラキシーショックを治療または予防するための、上記[33]、[34]または[35]に記載の医薬組成物。
【0099】
[37] Fc受容体と免疫グロブリンとの相互作用をモジュレートするための分子の使用であって、該分子が上記[24]または[25]に記載の結晶性調製物から得られた結晶構造データを用いて設計または同定されることを特徴とする、上記使用。
【0100】
[38] 前記モジュレーションがFc受容体と免疫グロブリンとの結合の部分的または完全な阻害である、上記[37]に記載の使用。
【0101】
[39] 固相に結合された、上記[1]〜[5]のいずれかに記載のFc受容体。
【0102】
[40] 固相がクロマトグラフィー担体材料である、上記[39]に記載のFc受容体。
【0103】
[41] 患者の血液、結晶もしくは血清から、または免疫グロブリン産生細胞の培養上清から免疫グロブリンを吸着させるための、上記[40]に記載のクロマトグラフィー担体材料の使用。
【0104】
[42] さらなる試験を行なうために患者の血液、結晶もしくは血清から、または免疫グロブリン産生細胞の培養上清から抗体を富化するための、上記[41]に記載の使用。
【実施例】
【0105】
以下の実施例は、図面と関連させて本発明をさらに説明するためのものである。
【0106】
実施例1
shFcγRIIb(可溶性ヒトFcγRIIb)
1.1 クローニングおよび発現
ヒトFcγRIIb2のcDNA (Engelhardtら、1990)を突然変異誘発(mutagenous)PCR (Dulauら、1989)を用いて改変した。すなわち、フォワードプライマーを用いて、新規の開始メチオニンを、NcoI部位内のシグナルペプチドの切断部位の後に導入した(5'-AAT AGA ATT CCA TGG GGA CAC CTG CAG CTC CC-3')。一方、リバースプライマー用いて、終止コドンを、SalI部位がその後に続く膜貫通領域と推定上の細胞外部分との間に導入した(5' CCC AGT GTC GAC AGC CTA AAT GAT CCC C-3')。PCR産物をNcoIおよびSalIで消化してpET11d発現ベクター(Novagen)中にクローン化し、予想される配列を確認した。最終的な構築物をBL21 (DE3)中で増殖させた(GrodbergおよびDunn, 1988)。FcγRIIbを過剰発現させるために、形質転換した細菌の単一のコロニーを、100μg/mlのアンピシリンを含有する5mlのLB培地(LB-Amp100)中に接種し、37℃で一晩インキュベートした。この培養物をLB-Amp100中で200倍に希釈し、OD600が0.7〜0.9に達するまでインキュベーションを続けた。タンパク質の過剰発現は、最終濃度1mMになるまでIPTGを添加することによって誘導した。4時間の増殖時間が経過した後、細胞を遠心(30分間, 4000 x g)により回収し、超音波処理バッファー(リン酸ナトリウム30 mM、塩化ナトリウム300 mM、アジ化ナトリウム 0.02%、pH 7.8)中に再懸濁した。懸濁液1mlあたり0.1mgのリゾチームを添加し、室温で30分間インキュベートした後、氷上で超音波処理を行った(Branson Sonifier、Danbury、CT;Macrotip、出力90%、インターバル80%、15分間)。この懸濁液を遠心(30分間、30,000 x g)にかけ、Dounceホモジナイザーを用いて、0.5%のLDAOを含有する超音波処理バッファー中に再懸濁した。この遠心工程およびLDAO含有バッファー中への再懸濁をもう一度繰り返し、その後、この工程をLDAOなしで2回繰り返した。精製した封入体を4℃で保存した。
【0107】
1.2 可溶性ヒトFcγRIIb (shFcγRIIb)の再生および精製
精製した封入体は、塩化グアニジン6 M、2-メルカプトエタノール100 mM中に、タンパク質濃度10mg/mlで溶解し、遠心により不溶性物質から分離した。再生は、急速な希釈によって達成された。そうして、1mlの該封入体溶液を、撹拌しながら、15時間以内に400mlの再生バッファー(0.1 M TRIS/HCl、アルギニン1.4 M、塩化ナトリウム150 mM、GSH 5 mM、GSSG 0.5 mM、PMSF 0.1 mM、アジ化ナトリウム 0.02%、pH 8.5、4℃)に滴下した。その後、この混合物を、空気酸化により、遊離のチオール基の濃度がEllman (Ellman, 1959)の方法で測定して1 mMに減じるまで、2〜3日間にわたって撹拌した。この溶液をPBSに対して透析して滅菌濾過し、その後、3kD MWCO限外濾過膜を備えた撹拌セル中で10倍に濃縮した。このタンパク質溶液を、hIgGセファロースカラム(セファロース4B 1mlあたりhIgG 50 mg)にアプライした。未結合のタンパク質は、50 mM TRIS pH 8.0で洗浄して取り除き、その後pHジャンプ(塩化ナトリウム150mM、グリシン100mM、アジ化ナトリウム 0.02%、pH 3.0)によって、FcγRIIbを溶出させた。溶出物は、1 M TRIS pH 8.0ですぐに中和させた。FcγRIIb含有溶液を濃縮し、結晶化バッファー(MOPS 2 mM、塩化ナトリウム150 mM、アジ化ナトリウム 0.02%、pH 7.0)で平衡化したSuperdex-75カラムを用いたゲル濾過に付した。FcγRIIbを含む画分をプールし、7 mg/mlまで濃縮して-20℃で保存した。
【0108】
1.3 平衡ゲル濾過実験
Superdex75カラムをFPLCに連結し、10μg/mlのshFcRIIbを含むPBSで平衡化させた。ヒトFcフラグメントを、1μg/10μlの濃度で、平衡化バッファー中に溶解して注入した。得られたクロマトグラムは、shFcγRIIbとFcフラグメントの複合体を含む正のピークを示した。一方、負のピークは複合体形成のためにランニングバッファーから消費された受容体の欠如を表す。
【0109】
1.4 結晶化およびデータ収集
96条件スパースマトリックススクリーニング(96 condition sparse matrix screen)(JancarikおよびKim, 1991)を使用した最初の結晶化試験は、蒸気拡散法を用い、20℃でのシッティングドロップ法で実施した。生成する結晶は、塩、沈殿剤および添加剤の濃度だけでなくpHを変化させることによっても向上した。適当な結晶の回折データを、RU200b回転アノード発生装置(50kV、100mAで操作;Rigaku)からのグラファイト単色CuKα放射線を用いて、イメージプレートシステム(MAR research)上に収集した。反射をプログラムMOSFLM (Leslie, 1997)で積分し、続いてそのデータは、CCP4プログラムセット(Collaborative Computational Project, 1994)の手順を用いて、基準化、減少および切り捨てを行い、構造因子の独立変数を得た。
【0110】
1.5 shFcγRIIbの発現、精製および再生の概要
FcγRIIbの細胞外部分は、T7 RNAポリメラーゼ陽性大腸菌株BL21/DE3 (Grodberg & Dunn, 1988)中、T7プロモーターの制御下で、高レベルで発現させた。該タンパク質は封入体中に集積していたが、これを最初の精製工程において使用した。封入体の単離は、リゾチームと超音波処理を組み合わせた強力な手法を用いて開始し、そうしなければ産物を汚染することになる細胞を実質的にすべて破裂させた。それに続く界面活性剤LDAO(この界面活性剤は、不純物を溶解させるが封入体自体は溶解しない点で優れた特性を有する)を用いた洗浄工程により、早くも90%より高い純度で産物が得られた(図1)。
【0111】
この産物は、さらに精製を行うことなく、再生試験に使用した。該封入体は高濃度の2-メルカプトエタノールおよびグアニジンに溶解し、共有結合凝集体および非共有結合凝集体を確実に単量体へシフトさせた。この溶液は、すぐに再生バッファーで希釈し、そうしなければ凝集体を形成してしまう折りたたまれていないタンパク質分子同士の接触を最小限に抑えた。再生バッファー中でアルギニンを用いることにより、尿素を用いた場合にしばしば認められるような側鎖の不可逆的改変を防ぐことができる。タンパク質を再生バッファーに添加した後、該溶液を、遊離のチオール基の濃度が1mMに減じるまで4℃で撹拌した。この工程は、初期の透析で不活性産物が生じたときに絶対的に必要であった。第2の精製工程において、透析して再生されたFcγRIIbを固定化したhIgGに結合させ、大腸菌タンパク質および不活性受容体のマイナー画分を取り除いた。該タンパクをpHジャンプによって溶出させ、すぐに中和した。このアフィニティークロマトグラフィー工程の後、shFcγRIIbは、繰り返し使用した後でさえもマトリックスから浸出してくる共溶出IgGによるわずかな汚染を除き、本質的に純粋である(図1)。還元SDS-PAGEで認識されないIgGならびに受容体多量体は、ゲル濾過によって容易に除去できた。この工程における汚染物質の除去と平行して、バッファーを定量的に交換した。この手法は、わずかな改変さえもが結晶化の再現不能を引き起こしたり、結晶形成を阻害するときでさえ、一定の組成のタンパク質溶液を確実にもたらす。大腸菌培養物1Lあたり全部で6 mgの純粋なタンパク質を得ることができた。これは、封入体のFcγRIIb含有量の約10%である。
【0112】
N末端タンパク質の配列決定により、検出可能な不純物を含まず、予想された配列H2N-GTPAAPを持つものであることが示された。ESI-MS分析により、結晶化試験で用いられた最終物質は、サイズに関して均一であることを示された。1次配列から、分子量は20434 Daであることが計算され、これは、質量分析で測定された20429 Daと一致している。誤差は、装置のエラー範囲にあり、リーディングメチオニンを含有する種についての追加のピークは、見られなかった。
【0113】
shFcγRIIbの結晶化は、蒸気拡散法を用い、シッティングドロップ法で実施した。スパースマトリックススクリーニング(Jancarik & Kim, 1991)を用いた最初の試験で、早くも、小さな針状結晶が得られた。これに続く、沈殿剤、塩、それらの濃度およびpHを変化させることによる予備的結晶化条件の最適化により、3つの異なる結晶形態が単離された。斜方晶結晶は、1.5 μlのリザーバー溶液(PEG2000 33%、酢酸ナトリウム 0.2 M、pH 5.4)と3 μlのタンパク質溶液の混合物から成長した。それらは3日以内に発生し、約80μm x 80μm x 500μmの最終サイズに1週間後に到達した。これらの結晶は1.7Åで回折した。結晶は、PEG8000 26%、酢酸ナトリウム0.2 M、pH 5.6、Zn(OAc)2 5 mM、塩化ナトリウム100 mMを含むリザーバー溶液から(六方晶形態)、およびPEG8000 26%、NaOAc 0.2 M、pH 5.6、1,4-ジオキサン 10% (v/v)、塩化ナトリウム100mMを含むリザーバー溶液から(正方晶形態)、2つの異なる空間群でも成長できた。これらの結晶はX線解析に適したサイズであるが、それぞれ正方晶形態で2.7Å、六方晶形態で3.8Åでしか回折しなかった(表1)。
【0114】
FcγRIIは大腸菌で発現させた。大腸菌は、比較的安価な製造コストおよび入手可能性に加え、IgG結合が炭水化物結合と独立に生じるFcγRIIの場合のように、哺乳動物細胞により起こるグリコシル化がタンパク質の機能に必要でないときに、特に、いくつかの利点を有する(Sondermannら、1998A)。大腸菌においては、均一な産物が再現可能に生成でき、バッチに依存した変動がしばしば観察される哺乳動物細胞における発現と対照的である。このような系においては、産物が数日間、30℃をこえる温度でプロテアーゼに暴露される。反対に、37℃で、強力なT7プロモーターの制御下、大腸菌内でタンパク質を発現させると、しばしばプロテアーゼが接近できない封入体が形成される。細菌内における発現のさらなる利点は、該物質が使用した仔牛血清または細胞系自体に由来しうる病原菌を含まないと考えられることである。哺乳動物細胞発現においては、標的タンパク質の精製の間、特別に注意を払わなければならない。なぜなら、存在しうる効果的なホルモンまたは成長因子が同時に精製される可能性があるからである。sFcγRの効果がTGFβ1汚染に起因する場合の1事例が、既に報告されている(Galonら、1995)。
【0115】
1.6 精製
精製手順は、簡便である。該手順は1日で容易に実施できる3つのステップからなる。該タンパク質は、純粋な形態でかつ高収率で得られ、その上、高価なIgGアフィニティーカラムを用いずに、かなりの品質で得ることができた。このプロトコルの成功は、封入体の注意深い調製に依るのだろう。なぜなら、ほとんどの不純物は、最初の精製ステップにおいて前もって除去できるからである。
【0116】
1.7 特性評価
精製FcγRIIbは、SDS-PAGEおよび等電点電気泳動ならびにN末端配列決定および質量分析によって特性評価した。従って、該物質は、その化学的組成に関して純粋かつ均一であると考えられる。しかし、受容体が正確に折りたたまれているかという興味深い疑問を考察する余地がある。Ellmanの試験では遊離のチオール基は検出されないことから、すべてのシステインは対になっている。該物質は単量体であり、サイズ排除クロマトグラフィーカラムから、対称形のピークとして、予想された保持時間に溶出する。さらに。FcγRIIbはIgGセファロースに結合し、大腸菌からの組換えFcγRIIbはIgGを特異的に結合することから活性である。
【0117】
1.8 結晶化
FcγRIIbの斜方晶結晶形態は、X線を、1.7Åの分解能で回折した。これは、以前に報告された昆虫細胞発現に由来する同一分子の結晶と比較して、劇的な改良である(Sondermannら、1998A)。これらの結晶は、2.9Åで回折し、空間群P3121である。従って、昆虫細胞由来の受容体のグリコシル化は、結晶化状態に影響を与えることになる。三方晶空間群のかわりに、3つの異なる結晶形態が見出される。その構造の解析が可能となった後は、これらの結晶形態は、結晶接触に起因するタンパク質の人工的コンフォメーションの同定を助けるであろう。
【0118】
FcγRは、その他のタンパク質と配列類似性を示さないが、保存されたシステイン配置から、それらは免疫グロブリンスーパーファミリーに属するとみなされる。従って、本発明者らはその構造を分子置換によって解析しようと試みたが、多様な分子からのIgGドメインを用いた広範囲にわたる試験は、失敗に終わった。従って、FcγRIIbの構造は、多重重原子同形置換法によって解析しなければならない。
【0119】
本発明者らは、初めて、FcγRIIbが大腸菌から活性な形態で得られることを示した。これは、結晶学的研究にとっての基礎であり、既に得られた異例の品質を有する結晶のため、すぐに、この重要な分子の構造解析をもたらすだろう。この構造は、IgG結合部位に関する情報を提供し、かつリガンドのその受容体による認識を阻害する薬物を知見に基づいて設計するための開始点を提供するだろう。さらに、FcγRIIbとFcεRIaを含むその他のFcRとの高度な相同性から、これらの分子は同様に作製することができると考えられ、進行中の研究に対して価値ある材料を提供するだろう。
【0120】
1.9 方法
タンパク質化学
組換え可溶性ヒトFcγRIIbは、大腸菌で発現させ、他で記載されたように、再生し、精製し、結晶化した(Sondermannら、1998B)。簡単に言うと、hFcγRIIb2の推定上の細胞外領域(Engelhardtら、1990)を、大腸菌内で過剰発現させた。 封入体は、細胞のリゾチーム処理およびこれに続く超音波処理によって精製した。得られた懸濁液を遠心(30分間、30,000 x g)にかけ、0.5% LDAOを含むバッファーで洗浄した。遠心工程およびLDAO含有バッファー中への再懸濁をもう一度繰り返し、その後、この手順をLDAOなしで2回繰り返した。封入体を6 M塩酸グアニジンに溶解し、タンパク質を上記のように再生した。透析し、濾過したタンパク質溶液をhIgGセファロースカラムにアプライし、pHジャンプによって溶出した。濃縮し、中和した画分を、Superdex-75カラム(26/60, Pharmacia)を用いるサイズ排除クロマトグラフィーにかけた。
【0121】
結晶化
結晶化は、蒸気拡散法を用い、シッティングドロップ法で20℃にて実施した。結晶化スクリーニングは、pH、塩、沈殿剤および添加剤を変化させることにより実施した。データ収集のために使用する最終的な結晶は、PEG2000 33%、酢酸ナトリウム0.2 M、pH 5.4中 (斜方晶形態)、PEG8000 26%、酢酸ナトリウム0.2 M、pH 5.6、1,4-ジオキサン 10% (v/v)、塩化ナトリウム100 mM中(正方晶形態)、およびPEG8000 26%、酢酸ナトリウム0.2 M、pH 5.6、ZN(OAc)2 5mM、塩化ナトリウム100mM中(六方晶形態)で成長させた。昆虫細胞由来のタンパク質は、PEG6000 32%、酢酸ナトリウム0.2 M、pH 5.3中で結晶化した。
【0122】
重原子誘導体の調製
重原子誘導体は、2 mMプラチナ(II)-(2,2'-6,2''テルピリジニウム)クロリドを含む結晶化バッファー中に24時間または塩化ウラニル10 mMを含む結晶化バッファー中に8日間、結晶を浸透することによって調製した。
【0123】
X線データ収集
回折データはRU200b回転アノード発生装置(50kV、100mAで操作;Rigaku)からのグラファイト単色CuKα放射を用い、イメージプレートシステム(MAR research)上に収集した。反射を、プログラムMOSFLM 5.50 (Leslie, 1997)で積分し、続いてそのデータは、CCP4プログラムセット(Collaborative Computational Project, 1994)の手順を用いて、基準化および切り捨てを行い、構造因子の独立変数を得た。
【0124】
構造決定
構造は、MIR法の標準的手法で解析した。様々な重原子成分を用いて実施した多数の浸透物から、2つの化合物のみが、解釈可能なパターソンマップをもたらした。各誘導体についての重原子位置は差パターソンマップから決定し、初期位相を算出した。2次元(Cross-phased)差フーリエ合成図を使用して、重原子位置を確認し、誘導体の共通の原点を確立した。異常分散データ(anomalous data)を含ませてエナンチオマー同士を区別した。重原子パラメーターは、CCP4パッケージのプログラムMLPHAREを用いてさらに精密化すると、表2の統計をもたらす。電子密度マップは、分解能2.1Åまで計算し、位相は、CCP4セットのプログラムDMを用いた溶媒平滑化(solvent flattening)およびヒストグラムマッチング(histogram matching)によりさらに改良した。得られた電子密度マップは、ほとんどのアミノ酸残基を構築するのに十分な品質であった。モデル構築は、Indigo2ワークステーション(Silicon Graphics Incorporation)上のO (Jonesら、1991)を用いて実施した。構造精密化はXPLOR (Brungerら、 1987)を利用し、EnghおよびHuber (Engh & Huber, 1991)のパラメーターセットを使用して分解能を1.7Åまで段階的に増加させることにより実施した。構造が、数ラウンドのモデル構築および個々の精密化B因子(Rfac = 29% / RFree = 36%)の後に完成したときは、3.5σで等高線を付けたFo-Fcマップが、1σで等高線を付けた十分に明確な電子密度の2Fo-Fcマップと一致するとき、150個の水分子を電子密度に組み込んだ。得られた精密化統計を表3に示す。
【0125】
1.10 構造決定
組換え可溶性ヒトFcγRIIbの結晶構造は、多重重原子同形置換法(MIR)によって1.7Åの分解能で解析した。なぜなら、ヒトIgG1から単離したFcフラグメントのドメインを用いた分子置換による構造解析(Huberら、 1976, PDB entry 1fc1; Deisenhofer, 1981)が失敗したからである。該受容体の推定上の細胞外部分(アミノ酸残基1-187、配列番号2に示す)を使用して、結晶化試験を行った(Sondermannら、1998B)。一方、モデルは、末端がフレキシブルで電子密度を調査できないときは、残基5-176を含むものである。さらに該モデルは150個の水分子を含み、精密化統計は表2にまとめている。該構造は位置11にcisプロリンを含有する。ラマチャンドラン(Ramachandran)プロットの非許容領域に位置する主鎖ねじれ角はなかった。完全に精密化したモデルを使用して、同一タンパク質の空間群P42212の結晶および昆虫細胞に由来するグリコシル化形態の空間群P3121の結晶の構造を解析した(表2)。
【0126】
FcγRIIbのポリペプチド鎖は、免疫グロブリンスーパーファミリーとの関係から予測されるように、2つのIg様ドメインに折りたたまれる。各ドメインは、向かい合っているシート上のB鎖とF鎖を連結する保存されたジスルフィド架橋によってサンドイッチ状に配置された2つのβシートから構成される(図3)。3つの逆平行のβ鎖(A1, B, E)は、5つのβ鎖(C', C, F, G, A2)のシートと向かいあっており、これにより、鎖A1は3本鎖のβシートを残し、5番目の短く平行な鎖A2を追加した4本鎖の逆平行シートに交差する。2次構造要素の配置ならびにその連結性はFcγRIIbの双方のドメインにおいて同一であり、一方のドメインのもう一方のドメイン上への強固な立体フィットは、67個のマッチしたCα原子の平方二乗平均で1.29Åの距離であった。
【0127】
このドメインは互いにほぼ垂直に配置され、その長軸間は70℃の角度を有し、ハート形の全体構造を形成している。この配置は、該ドメイン間に十分な接触領域をもたらす(図4)。鎖A2由来の残基およびN末端ドメインのA2とA1を連結するセグメント由来の残基は、C末端ドメインの鎖A1およびBの残基とかみ合っている。この領域は密にパッキングされ、相互作用はいくつかの水素結合によって強化され、強固な配置をもたらす。これは、3つの異なる空間群における構造の保存によって確認される。斜方晶、正方晶および六方晶(昆虫細胞由来)の結晶形態では、ドメイン間角度において2°未満の偏差が認められる。
【0128】
1.11 全体構造
大腸菌からの組換えヒトFcγRIIbの構造は、斜方晶結晶から、MIRによって1.7Åの分解能で解析した。実質的に同一の構造が、正方晶結晶および昆虫細胞由来タンパク質の六方晶結晶において見られる。これら3つの構造すべてにおいて、ポリペプチド鎖の最後の9残基は、無秩序であることが見出された。分子の構造中心と膜貫通部位間のC末端リンカー領域のフレキシビリティーは機能的に関連し、受容体のいくつかの再配向を可能にして、免疫複合体におけるFc部位の認識を強化しうる。
【0129】
1.12 相同受容体
タンパク質のIgスーパーファミリーに見られるIgドメインは、向かい合ったシートの2つの鎖を連結する保存されたジスルフィド架橋を有するβサンドイッチ構造によって特徴づけられる。FcγRIIb に見られるようなサンドイッチを形成する3および4個の逆平行β鎖の典型的な配置は、T細胞受容体、Fcフラグメント、CD4またはFabフラグメントにおいても生じる。これらの分子の個々のIgドメインとFcγRIIbの2つのドメインとの構造アライメントは、共通の、密接に関連した構造を示す。しかし、該ドメインの相対的配置は、これらの分子においては関連しておらず、広範なセクターに及ぶ。様々な分子に由来するIgドメイン間の構造類似性およびFcγRIIの2つのドメインが重なりあっている場合に生じるCα原子の著しく低い平均二乗平均偏差にもかからわず、有意な配列類似性は見られない(図5aおよび5b)。構造に基づく配列アライメントは、ドメインの配列にそって保存された疎水性パターンを示し、それとともにシステインの他には、同一のアミノ酸残基は少ししかなかった。本発明者らは、最初に、IgG1重鎖の2つのC末端ドメインとFcγRIIbとの構造に基づくアライメントを作成し、その他の関連するFcγRおよびFcεRIaドメインの配列を加えた。これにより、3つのドメインのFcγRIおよび2つのドメインの受容体の配列はIgドメインの疎水性パターンと一致し、いくつかの保存されたアミノ酸残基を表すことが示された。第1に、FcRの異なるドメインは、Igスーパーファミリーのその他の分子からのIgドメインに対するよりも、互いにより関連する。第2に、受容体のN末端ドメインは、第2ドメインと同様に互いに関連する。第3に、FcγRIの第3ドメインの配列は、ドメイングループの双方に由来する特徴を示す。まとめると、本発明者らは、FcRがIgスーパーファミリーに属することを確認し、すべてのFcRドメインは共通の先祖、遺伝子複製により第2ドメインを獲得した古代の1ドメインの受容体に源を有するものと考えられ、さらに、このような2ドメインの受容体のさらなる分枝発生は、第3ドメインを獲得したFcγRIを含む現在の多様性をもたらした。
【0130】
アライメントにおける、FcγRIIb内のドメイン間接触に寄与するこれらのアミノ酸残基の保存は、異なる受容体における類似のドメイン配置に対するヒントにる。表4では、その側鎖でドメイン間接触に寄与している残基(図4)を、図5bの構造に基づく配列アライメントに従って、FcγRIIbについて、その他の受容体内の対応するアミノ酸残基とともに編集している。FcR間で保存されていないAsn15を除き、含まれる残基は同一であるかまたは保存的に置換され、すべてのFcRにおける類似の構造およびドメイン配置を強力に支持している。
【0131】
1.13 IgGに対する接触境界面
FcRとそのリガンドとの相互作用に関する限定的な情報は、突然変異誘発研究(Hogarthら、1992; Hulettら、1994; Hulettら、1995)から入手可能である。FcγRIIa β鎖間のループをFcεRIaアミノ酸残基と系統的に交換することにより、C末端ドメインのB/C、C'/EおよびF/Gループは、リガンドの結合にとって重要であることが評価された(図3、図5b)。構造モデルにおいて、これらのループは隣接し、潜在的なリガンドに自由に接近可能である。さらに、これらのループ内のほとんどのアミノ酸残基を、単点突然変異によってアラニンに置換すると、FcγRIIaの2量体ヒトIgG1への親和性の劇的な変化が生じた。また、高応答性/低応答性多形におけるC末端ドメイン(C'/Eループ)内のArg131のHisへの単一アミノ酸置換は、FcγRIIaのマウスIgG1への親和性を変化させ、その領域を指す。従って、このエリア内のアミノ酸残基は、リガンド結合またはその領域の構造保全性のいずれにも重要である。ここで、該構造は、Tyr 157近傍の疎水性アミノ酸残基Pro 114、Leu 115およびVal 116のクラスター形成を示す。このパッチは中心構造からはみ出している正に荷電したアミノ酸残基Arg 131およびLys 117により、Leu 159、Phe 121およびPhe 129の領域から分離されている(図5b)。
【0132】
1.14 グリコシル化
FcγRIIbの配列中に、3つの潜在的Nグリコシル化部位が見出された。3つの部位はすべて、分子の表面にあり、接近可能である。それらは、両ドメインのE/Fループ内(N61およびN142)およびC末端ドメインの鎖E(N135)上に位置する(図3、図6)。構造解析に使用した材料は、大腸菌から得たものであるため、炭水化物を含まず、一方、哺乳動物細胞から単離したFcRは高度にグリコシル化されている。これらの3つの潜在的グリコシル化部位は、推定のIgG結合領域からかなり離れて位置し、グリコシル化されていないFcγRIIbはヒトIgGを結合することから、結合においてグリコシル化の役割が少ないことを示唆している。この事実は、昆虫細胞において産生されグリコシル化されているFcγRIIbの構造により確認された(Sondermannら、1998A)。おそらく異なる結晶接触によるものであるドメイン間角度の2°の変異を除き、グリコシル化タンパク質と非グリコシル化タンパク質の構造の間に違いは見られなかった。3つのグリコシル化部位は、物質が4つのバンドとして現れるSDS-PAGEで示されるように、場合により使用されるのみである。これらの糖についての追加の電子密度によっても、化学的および構造的な異質性の結果は見られなかった。
【0133】
実施例2
shFcγRIIa(可溶性ヒトFcγRIIa)
手順は、以下に示す変化を除き、実施例1に従って実施した。
【0134】
2.1 クローニングおよび発現
shFcγRIIaは、表5に挙げた突然変異(mutagenous)プライマーを用いた実施例1に従い、各野生型cDNAを突然変異させることによって産生し(Stengelinら、1988)、発現させた。タンパク質の発現のためには、pET22b+ベクターを選択した。
【0135】
2.2 再生および精製
shFcγRIIaは、実施例1に従って、表6に挙げた各再生バッファーを用いて、再生した。
【0136】
2.3 結晶化
shFcγRIIaは、表7に示した条件下で記載した通りに結晶化した。
【0137】
2.4 構造決定
構造は、調査モデルとしてshFcγRIIaを用い、同形置換法で解析した。
【0138】
実施例3
shFcγRIII(可溶性ヒトFcγRIII)
手順は、以下に示す変化を除き、実施例1に従って実施した。
【0139】
3.1 クローニングおよび発現
shFcγRIIIは、表5に挙げた突然変異(mutagenous)プライマーを用いた実施例1に従い、各野生型cDNAを突然変異させることによって産生し(Simmons&Seed、1988)、発現させた。タンパク質の発現のためには、pET22b+ベクターを選択した。
【0140】
3.2 再生および精製
shFcγRIIIは、実施例1に従って、表6に挙げた各再生バッファーを用いて再生した。
【0141】
3.3 結晶化
shFcγRIIIは、表7に示した条件下で記載した通りに結晶化した。
【0142】
3.4 構造決定
構造は、調査モデルとしてshFcγRIIbを用い、同形置換法で解析した。
【0143】
3.5 shFcγRIII:hFc1複合体の結晶化
骨髄腫患者の血清に由来するhIgG1を使用して、プラスミンを用いた消化によってFcフラグメント(hFc1)を調製した(Deisenhoferら、1976)。得られたFcフラグメントは、プロテインAクロマトグラフィーによって、Fabフラグメントから分離した。部分的に消化されたhIgGは、サイズ排除クロマトグラフィーにより、ランニングバッファーとしてMBS(MOPS 2mM、NaCl 150mM、アジ化ナトリウム 0.02%、pH 7.0)を用いて除去した。等モル量のhFc1およびshFcgRIIIを混合してMBSで希釈し、10mg/mlの濃度とした。複合体は、表5に示した条件下で記載の通りに結晶化した。
【0144】
実施例4
shFcεRII(可溶性ヒトFcεRII)
手順は、以下に示す変化を除き、実施例1に従って実施した。
【0145】
4.1 クローニングおよび発現
shFcεRIIは、表5に挙げた突然変異(mutagenous)プライマーを用いた実施例2に従い、各野生型cDNAを突然変異させることによって産生し(Kikutaniら、1986)、発現させた。タンパク質の発現のためには、pET23a+ベクターを選択した。
【0146】
4.2 再生および精製
shFcεRIIの再生は、急速な希釈の前に溶解した封入体を塩化グアニジン 6M、酢酸ナトリウム 20mM、pH 4.0に対して透析することを除き、実施例1に記載の通りに実施した。shFcεRIIは、実施例1に従って、表6に挙げた各再生バッファーを用いて再生した。再生の後、タンパク質溶液をPBSに対して透析し、100倍に濃縮し、Superdex 75を用いたゲル濾過クロマトグラフィーによって精製した。これにより、純粋なshFcεRIIが得られ、これをTRIS/HCL 2mM、NaCl 150mM、アジ化ナトリウム 0.02%、pH 8.0に対して透析し、10mg/mlまで濃縮し、4℃で保存した。
【0147】
実施例5
shFcγRI(可溶性ヒトFcγRI)
手順は、以下に示す変化を除き、実施例1に従って実施した。
【0148】
5.1 クローニングおよび発現
shFcγRIは、表5に挙げた突然変異(mutagenous)プライマーを用いた実施例1に従い、各野生型cDNAを突然変異させることによって産生し(Allen&Seed、1988)、発現させた。タンパク質の発現のためには、pET32a+ベクターを選択した。該ベクターは、N末端チオレドキシンの後にC末端トロンビン切断部位を有するヘキサヒスチジンタグを含み、その後に記載のタンパク質およびアミノ酸残基とインフレームのshFcγRIが続く。融合タンパク質の過剰発現のために、プラスミドpUBSおよびpLysS (Novagen)を含む大腸菌株BL21(DE3)を使用した。
【0149】
精製した封入体は、グアニジン-HCl 6M、β-メルカプトエタノール 10mM、Tris 50mM、pH 8.0に溶解し、Ni-NTAカラム(Qiagen)に結合させた。溶出は、0〜1 Mのイミダゾール勾配で行った。溶出したタンパク質は、NaCl 150mM、Tris 50mM、pH 8.0、GSH 2mM、GSSG 0.5 mMの1000倍量に対して、24時間4℃で透析した。タンパク質溶液を最初の容量の25%に濃縮した後、トロンビンを添加した。37℃で6時間のインキュベートした後、N末端チオレドキシンおよびHisタグをN末端配列決定により確認して完全に除去した。この消化の間、shFcgRIは溶液から定量的に沈殿析出した。
【0150】
5.2 再生および精製
shFcγRIは、実施例1に従って、表6に挙げた各再生バッファーを用いて再生した。酸化還元電位が1mMに減じた後、溶液をPBS pH 8.0に対して透析し、濃縮した。再生したタンパク質をサイズ排除クロマトグラフィーによって分析すると、予想された単量体受容体のピークが得られ、非還元SDS-PAGEは30 kDaに主要なバンドを示した。
【0151】
実施例6
shFcεRIa(可溶性ヒトFcεRIa)
手順は、以下に示す変化を除き、実施例1に従って実施した。
【0152】
6.1 クローニングおよび発現
shFcεRIは、表5に挙げた突然変異(mutagenous)プライマーを用いた実施例1に従い、各野生型cDNAを突然変異させることによって産生し(Kochanら、1988)、発現させた。タンパク質の発現のためには、pET23a+ベクターを選択した。
【0153】
【表1】
【0154】
【表2】
【0155】
【表3】
【0156】
【表4】
【0157】
【表5】
【0158】
【表6】
【0159】
【表7】
【0160】
参照文献
1. Ades, E.W., Phillips, D.J. , Shore, S.L., Gordon, D.S., LaVia, M.F., Black, C.M., Reimer, C.B. (1976),「Analysis of mononuclear cell surfaces with fluoresceinated Staphylococcal protein A complexed with IgG antibody or heat-aggregated γ-globulin(IgG抗体または熱凝集したγ-グロブリンと複合体形成する蛍光標識したブドウ球菌由来のプロテインAを有する単核細胞表面の解析)」,J. Immunol. 117, 2119.
2. Allen J.M., Seed B.;「Nucleotide sequence of three cDNAs for the human high affinity Fc receptor (FcRI)(ヒト高親和性Fc受容体(FcRI)に対する3種のcDNAのヌクレオチド配列)」; Nucleic Acids Res. 16:11824-11824(1988).
3. Amigorena, S., Bonnerot, C., Drake, J.R., Choquet, D., Hunziker, W., Guillet, J.G., Webster, P., Sautes, C., Mellman. I., Fridman, W.H. (1992),「Cytoplasmic domain heterogeneity and functions of IgG Fc receptors in B lymphocytes(Bリンパ球におけるlgG Fc 受容体の細胞質ドメイン異質性および機能)」, Science 256, 1808-1812.
4. Barton, G.C. (1993), ALSCRIPT:「tool to format multiple sequence alignments(多重配列アライメントを形式化する手段)」. Prot. Eng. 6, 37-40.
5. Bazil, V, and Strominger, J.L. (1994),「Metalloprotease and serine protease are involved in cleavage of CD43, CD44, and CD16 from stimulated human granulocytes(メタロプロテアーゼおよびセリンプロテアーゼは、刺激されたヒト顆粒球由来のCD43、CD44およびCD16の切断に関与している)」, J. Immunol. 152, 1314-1322.
6. Brunger. A.T., Kuriyan, J., Karplus. M. (1987),「Crystallographic R factor refinement by molecular dynamics(分子動力学による結晶学的なR因子精密化)」, Science 35. 458-460.
7. Burmeister, W.P., Huber, A.H., Bjorkman, P.J. (1994),「Crystal structureof the complex of rat neonatal Fc receptor with Fc(ラット新生児のFc受容体とFcとの複合体の結晶構造)」, Nature 372, 379-383.
8. Ceuppens, J.L., Baroja, M.L., van Vaeck, F., Anderson, C.L. (1988),「Defect in the membrane expression of high affinity 72kD Fcγ receptors on phagocytic cells in four healthy subjects(4人の健康な被験者由来の貪食細胞上における高親和性72kD Fcγ受容体の膜での発現欠陥)」, J. Clin. Invest. 82, 571-578.
9. Collaborative computational project, 第4号 (1994),「The CCP4 suite: Programs for protein crystallography(CCP4セット:タンパク質結晶学のプログラム)」, Acta crystallogr. D50, 760-763.
10. Deisenhofer, J., Jones, T.A., Huber, R., Sjodahl, J., Sjoquist. J. (1978),「Crystallization, crystal structure analysis and atomic model of the complex formed by a human Fc fragment and fragment B of protein A from Staphylococcus aureus(ヒトFcフラグメントと黄色ブドウ球菌由来プロテインAのフラグメントBにより形成される複合体の結晶化、結晶構造解析および原子モデル)」,Z. Phys. Chem. 359, 975-985.
11. Deisenhofer, J. (1981),「Crystallographic refinement and atomic models of a human Fc fragment and its complex with fragment B of protein A from Staphylococcus aureus at 2.9- and 2.8A resolution(2,9および2.8Å分解能における、ヒトFcフラグメントおよびその黄色ブドウ球菌由来プロテインAのフラグメントBとの複合体の結晶学的精密化および原子モデル)」, Biochemistry 20, 2361-2370.
12. Deisenhofer J., Colman PM., Huber R., Haupt H., Schwick G.;「Crystallographic structural studies of a human Fc-fragment. I. An electron-density map at 4Å resolution and a partial model(ヒトFcフラグメント.Iの結晶学的構造研究。4Å分解能における電子密度マップおよび部分モデル)」; Hoppe-Seyler's Z. Physiol. Chem. 357: 435-445 (1976).
13. Dulau, L., Cheyrou, A., Aigle, M. (1989),「Directed mutagenesis using PCR(PCRを使用した直接的突然変異誘発)」, Nucleic Acids Res. 17, 2873.
14. Ellman (1959),「Tissue sulfhydryl groups(組織スルフヒドリル基)」, Arch. Biochem. Biophys. 82, 79-77.
15. Engelhardt, W., Geerds, C., Frey, J. (1990), 「Distribution, inducibility and biological function of the cloned and expressed human βFc receptor II(クローン化し、発現させたヒトβFc受容体IIの分布、誘導性および生物学的機能)」, Eur. J. Immunol. 20, 1367-1377.
16. Engh, R.A. and Huber, R. (l991), 「Accurate bond and angle parameters for X-ray protein structure refinement(X線タンパク質構造精密化のための正確な結合および角度パラメーター)」, Acta crystallogr. A47, 392-400.
17. Fleit, H.B., Kobasiuk, C.D., Daly, C., Furie, R., Levy, P.C., Webster, R.O. (1992),「A soluble form of FcγRIII is present in human serum and other body fluids and is elevated at sites of inflammation(FcγRIIIの可溶性形態は、ヒト血清および他の体液に存在し、炎症部位で上昇する)」, Blood 79, 2721-2728.
18. Fridman, W.H., Bonnerot, C., Daeron, M., Amigorena, S., Teillaud, J.-L., Sautes, C. (1992),「Structural bases of Fcγ receptor functions(Fcγ受容体機能の構造的基盤)」, Immunol. Rev. 125, 49-76.
19. Fridman, W.H., Teillaud, J.-L., Bouchard, C., Teillaud, C., Astier, A., Tartour, E., Galon, J., Mathiot, C., Sautes, C. (1993),「Soluble Fcγreceptors(可溶性Fcγ受容体)」, J. Leukocyte Biol. 54, 504-512.
20. Gabb, H.A., Jackson, R.M., Sternberg, M.J.E. (1997),「Modelling protein docking using shape complementarity, electrostatics and biochemical information(形状相補性、静電気学および生化学的情報を用いたタンパク質連結のモデル化)」, J. Mol. Biol. 272, 106-120.
21. Galon, J., Bouchard, C., Fridman, W.H., Sautes, C. (1995),「Ligands and biological activities of soluble Fcγ receptors(可溶性Fcγ受容体のリガンドおよび生物学的活性)」, Immunol. Lett. 44, 175-181.
22. Genetics Computer Group(1994),「Program Manual for the Wisconsin Package Version 8(ウィスコンシンパッケージ第8版用のプログラムマニュアル)」,Madison, Wisconsin.
23. Gordon, J.ら, (1980),「The molecules controlling B lymphocytes(Bリンパ球を制御する分子群)」. Immunol. Today, 8: 339-344.
24. Grodberg, J. and Dunn, J.J. (1988), 「OmpT encodes the Escherichia coli outer membrane protease that cleaves T7 RNA polymerase during purification(OmpTは精製の間中、T7 RNA ポリメラーゼを切断する大腸菌外膜プロテアーゼをコード化する)」, J. Bacteriol. 170, 1245-1253.
25. Hogarth, P.M., Hulett, M.D., Ierino, F.L., Tate, B., Powell. M.S., Brinkworth, R.I. (1992),「Identification of the immunoglobulin binding regions (IBR) of FcγRII and FcεRI(FcγRIIおよびFcεRIの免疫グロブリン結合領域(IBR)の同定)」, Immunol. Rev. 125, 21-35.
26. Homsy, J., Meyer, M., Tateno, M., Clarkson, S., Levy, J.A. (1989),「The Fc and not CD4 receptor mediates antibody enhancement of HIV infection in human cells(CD4受容体ではなくFc受容体が、ヒト細胞でのHIV感染における抗体増強を媒介する)」, Science 244, 1357-1360.
27. Hoover, R.G., Lary, C., Page, R., Travis, P., Owens, R., Flick, J., Kornbluth, J., Barlogie, B. (1995), 「Autoregulatory circuits in myeloma: Tumor cell cytotoxity mediated by soluble CD16(骨髄腫における自己調節回路:可溶性CD16が媒介する腫瘍細胞細胞傷害)」, J. Clin. Invest. 95, 241-247.
28. Huber, R., Deisenhofer, J., Colman, P.M., Matsushima, M. and Palm, W. (1976), 「Crystallographic structure studies of an IgG molecule and an Fc fragment(IgG分子およびFcフラグメントの結晶学的構造研究)」, Nature 264, 415-420.
29. Hulett, M.D., Witort, E., Brinkworth, R.I., McKenzie, I.F.C., Hogarth, P.M. (1994), 「Identification of the IgG binding site of the human low affinity receptor for IgG FcγRII(IgG FcγRIIに対するヒト低親和性受容体のIgG結合部位の同定)」, J. Biol. Chem. 269, 15287-15293.
30. Hulett, M.D., Witort, E., Brinkworth, R.I., McKenzie, I.F.C., Hogarth, P.M. (1995), 「Multiple regions of human FcγRII (CD32) contribute to the binding of IgG (ヒトFcγRII(CD32)の多数の領域が、IgGの結合に寄与する)」, J. Biol. Chem. 270, 21188-21194.
31. lerino, F.L., Powell, M.S., McKenzie, I.F.C., Hogarth, P.M. (1993),「Recombinant soluble human FcγRII: Production, characterization, and inhibition of the arthus reaction(組換え可溶性ヒトFcγRII:産生、特性評価およびアルツス反応の阻害)」, J. Exp. Med. 178, 1617-1628.
32. Jancarik. J. and Kim, S.H. (1991),「Sparse matrix sampling: A screening method for crystallization of proteins(スパースマトリックスサンプリング:タンパク質結晶化のためのスクリーング方法)」, J. Appl. Crystallogr. 24, 409-411.
33. Jones, T.A., Zou, J.-Y., Cowan, S.W., Kjeldgaard, M. (l991),「Improved methods for building protein models in electron density maps and the location of errors in these models(改良した電子密度マップにおけるタンパク質モデルを構築する方法および該モデルの誤差位置)」, Acta crystallogr. A47, 110-119.
34. Kikutani H., Inui S., Sato R., Barsumian E.L., Owaki H., Yamasaki K., Kaisho T., Uchibayashi N., Hardy R.R., Hirano T., Tsunasawa S., Sakiyama F., Suemura M., Kishimoto T.;「Molecular structure of human lymphocyte receptor for immunoglobulin E(免疫グロブリンEのヒトリンパ球受容体の分子構造)」; Cell 47(5):657-665(1986).
35. Khayat, D., Soubrane, C., Andriew, J.M., Visonneau, S., Eme, D., Tourani, J.M., Beldjord, K., Weil, M., Fernandez, E., Jaquillat, C. (1990),「Changes of soluble CD16 levels in serum of HIV patients: Correlation with clinical and biological prognostic factors(HIV患者の血清中の可溶性CD16のレベル変化:臨床的および生物学的予後因子との相関関係)」, J. Infect. Dis. 161, 430-435.
36. Kochan J., Pettine L.F., Hakimi J., Kishi K., Kinet J.P.;「Isolation of the gene coding for the alpha subunit of the human high affinity IgE receptor(ヒト高親和性IgE受容体のαサブユニットをコードする遺伝子の単離)」; Nucleic Acids Res. 16: 3584-3584(1988).
37. Simmons D., Seed B.;「The Fc-gamma receptor of natural killer cells is a phospholipid-linked membrane protein(ナチュラルキラー細胞のFcγ受容体は、リン脂質連結膜タンパク質である)」; Nature 333: 568-570 (1988).
38. Kraulis, P.J. (1991) , MOLSCRIPT:「a program to produce both detailed and schematic plots of protein structures(タンパク質構造の詳細で図式的なプロットの作製用プログラム)」, J. Appl. Cryst. 24, 946-950.
39. Leslie, A.G.W. (1997),「Mosflm user guide, mosflm version 5.50(Mosflm使用者説明書, Mosflm 第5.50版)」, Laboratory of Molecular Biology, Cambridge. UK.
40. Lessel, U. and Schomburg, D. (1994),「Similarities between protein 3-D structures(タンパク質3次元構造間の類似性)」, Protein Eng. 7, 1175-1187.
41. Littaua, R., Kurane, I. and Ennis, F.A. (1990),「Human IgG Fc receptor II mediates antibody-dependent enhancement of dengue virus infection(ヒトIgG Fc受容体IIは、デング熱ウイルス感染の抗体依存性の増強を媒介する)」, J. Immunol. 144, 3183-3186.
42. Lynch, R.G., Hagen, M., Mueller, A., Sandor, M. (1995),「Potential role of FcγR in early development of murine lymphoid cells: Evidence for functional interaction between FcγR on pre-thymocytes and an alternative, non-Ig ligand on thymic stromal cells(マウスリンパ球の初期発生におけるFcγRの潜在的役割:前胸腺細胞上のFcγRと胸腺ストローマ細胞上の選択的な非Igのリガンド間の機能的相互作用の証拠)」, Immunol. Lett. 44, 105-109.
43. Mathiot, C., Teillaud, J.L., Elmalek, M., Mosseri, L., Euller-Ziegler, L., Daragon, A., Grosbois, B., Michaux, J.L., Facon, T., Bernard, J.F., Duclos, B., Monconduit. M., Fridman, W.H. (1993),「Correlation between serum soluble CD16 (sCD16) levels and disease stage in patients with multiple myeloma(血清に可溶性のCD16 (sCD16)レベルと多発性骨髄腫を有する患者の疾患の段階間の相関)」, J. Clin. Immunol. 13, 41-48.
44. Merritt, E.A. and Murphy, M.E.P. (1994),「Raster3D Version 2.0. A program for photorealistic molecular graphics(Raster3D 第2.0版:写真のように写実的な分子グラフィックス用プログラム)」, Acta Cryst. D50, 869-873.
45. Metzger, H, (1992A).「Transmembrane signaling: The joy of aggregation(膜貫通シグナル:凝集の価値)」, J. Immunol. 149, 1477-1487.
46. Metzger, H. (1992B). 「The receptor with high affinity for IgE(IgEと高親和性の受容体)」, Immunol. Rev. 125, 37-48.
47. Muller, S. and Hoover, R.G. (1985),「T cells with Fc receptors in myeloma; suppression of growth and secretion of MOPC-315 by T alpha cells(骨髄腫におけるFc受容体を有するT細胞;Tα細胞による増殖抑制およびMOPC-315の分泌)」, J. Immunol. 134, 644-7.
48. Nicholls, A., Sharp, K.A., Honig, B. (1991),「Protein folding and association: insights from the interfacial and thermodynamic properties of hydrocarbons(タンパク質の折たたみおよび会合:炭化水素の界面特性および熱力学的特性からの洞察)」, Proteins 11, 281-296.
49. Poo, H., Kraus, J.C., Mayo-Bond, L., Todd, R.F., Petty, H.R. (1995),「Interaction of Fcγ receptor IIIB with complement receptor type 3 in fibroblast transfectants: evidence from lateral diffusion and resonance energy transfer studies(繊維芽細胞トランスフェクト体におけるFcγ受容体IIIBと補体受容体3型との相互作用:側方拡散および共鳴エネルギー転移研究からの証拠)」, J. Mol. Biol. 247, 597‐603.
50. Rappaport, E.F., Cassel, D.L., Walterhouse, D.O., McKenzie, S.E., Surrey, S., Keller, M.A., Schreiber, A.D., Schwartz, E. (1993),「A soluble form of the human Fc receptor FcγRIIa: cloning, transcript analysis and detection.(ヒトFc受容体FcγRIIaの可溶性形態: クローニング、 転写産物解析および検出)」. Exp. Hematol. 21, 689-696.
51. Ravanel, K., Castelle, C., Defrance. T., Wild, T.F., Charron, D., Lotteau, V., Rabourdincombe, C. (1997), 「Measles virus nucleocapsid protein binds to FcγRII and inhibits human B cell antibody production(麻疹ウイルスのヌクレオキャプシドは、FcγRIIに結合し、ヒトB細胞の抗体産生を阻害する)」. J. Exp. Med. 186, 269-278.
52. Roman, S., Moore, J.S., Darby, C., Muller, S., Hoover, R.G. (l988),「Modulation of Ig gene expression by Ig binding factors. Suppression of alpha-H chain and lambda-2-L chain mRNA accumulation in MOPC-315 by IgA-binding factor(Ig結合因子によるIg遺伝子発現のモジュレーション。IgA結合因子による、MOPC-315におけるα-H鎖およびλ-2-L鎖mRNA蓄積の抑制)」, J. Immunology 140, 3622-30.
53. Sarfat, D.ら, (1988),「Elevation of IgE-binding factors of serum in patients with B-cell derived chronic lymphocytic leukemia(B細胞由来慢性リンパ性白血病を患う患者の血清におけるIgE結合因子の上昇)」. Blood, 71: 94-98.
54. Sauer-Eriksson, A.E., Kleywegt, G.J., Uhlen, M., Jones, T.A. (1995),「Crystal structure of the C2 fragment of streptococcal protein G in complex with the Fc domain of human IgG(ヒトIgGのFcドメインと複合体を形成するブドウ球菌プロテインGのC2フラグメントの結晶構造)」, Structure 3, 265-78.
55. Small, T.,ら, (1990),「B-cell differentiation following autologous, conventional or T-cell depleted bone marrow transplantation: a recapitulation of normal B-cell ontogeny(自己由来の骨髄の移植、通常の骨髄移植、またはT細胞を欠損した骨髄の移植後のB細胞分化:正常なB細胞発生における発生反復)」. Blood, 76: 1647-1656.
56. Sondermann, P., Huber, R., Jacob, U. (1998B),「Preparation and crystallization of active soluble human FcγRIIb derived from E.coli(大腸菌由来の活性型可溶性ヒトFcγRIIbの調製および結晶化)」, Protein Structure, (提出済み).
57. Sondermann, P., Kutscher, C., Jacob, U., Frey, J. (1998A),「Characterization and crystallization of soluble human Fcγ receptor 11 isoforms produced in insect cells(昆虫細胞中で産生した可溶性ヒトFcγ受容体の11個のアイソフォームの特性評価および結晶化)」, Biochemistry, (提出済み).
58. Sondermann, P., Kutscher, C., Jacob, U., Frey, J.,「Analysis of complexes of IgG and soluble human Fcγ-Receptor II produced in insect cells and its crystallization(昆虫細胞中で産生したIgGと可溶性ヒトFcγ受容体IIとの複合体の解析およびその結晶化)」, (提出済み).
59. Stengelin S., Stamenkovic I., Seed B.;「Isolation of cDNAs for two distinct human Fc receptors by ligand affinity cloning(リガンド親和性クローニングによる異なった2つのヒトFc 受容体のcDNAの単離)」: EMBO J. 7: 1053-1059 (l988).
60. Tax, W.J.M., Willems, H.W., Reekers, P.P.M., Capel, P.J.A., Koene, R.A.P. (1983),「Polymorphism in mitogenic effect of IgG1 monoclonal antibodies against T3 antigen on human T cells(ヒトT細胞上のT3抗原に対するIgG1モノクローナル抗体の有糸分裂促進効果における多形性)」, Nature 304, 445-447.
61. Teillaud, J.L., Brunati. S., Elmalek, M., Astier, A., Nicaise, P., Moncuit, J., Mathiot, C., Job-Deslandre, C., Fridman, W.H. (1990),「Involvement of FcR+ Tcells and of IgG-BF in the control of myeloma cells (骨髄腫細胞の制御におけるFcR+ T細胞およびIgG-BFの関与)」, Mol. Immunol. 27, 1209-17.
62. Turk, D. (1992),博士論文, TU Munchen, Germany.
63. Ulvestad, E., Matre, R., Tonder, O. (1988),「IgG Fc receptors in sera from patients with Rheumatoid Arthritis and Systemic Lupus Erythematosus(慢性関節リウマチおよび全身性エリテマトーデスを患う患者の血清中のIgG Fc受容体)」, Scand. J. Rheumatol., Suppl. 75, 203-208.
64. van de Winkel, J.G.J. and Capel, P.J.A. (1993),「Human IgG Fc receptor heterogeneity: Molecular aspects and clinical implications(ヒトIgG Fc 受容体異質性:分子面および臨床的関連性)」, Immunol. Today 14, 215-221.
65. Varin, N., Sautes, C., Galinha, A., Even, J., Hogarth, P.M., Fridman, W.H. (1989),「Recombinant soluble reseptors for the Fcγ portion inhibit antibody production in vitro(Fcγ部分に対する組換え可溶性受容体は、in vitroで抗体産生を阻害する)」, Eur. J. Immunol. 19, 2263-2268.
66. Yang, Z., Delgado, R., Xu, L., Todd, R.F., Nabel, E.G., Sanchez, A., Nabel, G.J. (1998),「Distinct cellular interactions of secreted and transmembrane Ebola virus glycoproteins(分泌型および膜貫通型エボラウイルス糖タンパク質の異なった細胞相互作用)」, Science 279, 983-984.
67. Zhou, M.-J., Todd, R.F., van de Winkel, J.G.J., Petty, H.R. (1993),「Cocapping of the leukoadhesin molecules complement receptor type 3 and lymphocyte function-associated antigen-1 with Fcγ receptor III on human neutrophils. Possible role of lectin-like interactions(ヒト好中球上の白血球粘着分子補体受容体3型およびリンパ球機能関連抗原-1とFcγ受容体IIIとの共キャップ形成。レクチン様相互作用の潜在的役割」, J. Immunol. 150, 3030-3041.
【0161】
配列情報
【特許請求の範囲】
【請求項1】
組換え可溶性受容体FcγRIIbもしくはFcγRIIIに相補的な受容体FcγRIIbもしくはFcγRIIIインヒビター、受容体FcγRIIbもしくはFcγRIII、および/または組換え可溶性受容体FcγRIIbもしくはFcγRIIIの免疫グロブリン結合部位に相補的な免疫グロブリンインヒビターを同定および/または生成するための、結晶性調製物を使用して得られる結晶構造データの、免疫グロブリンと受容体FcγRIIbまたはFcγRIIIとの間の相互作用のモジュレーションを生じさせる使用であって、
該結晶性調製物が組換え可溶性受容体FcγRIIbまたはFcγRIIIの結晶性調製物であり、
該受容体が、膜貫通ドメイン、シグナルペプチドおよびグリコシル化が存在しないこと、ならびに該受容体が、原核細胞において該可溶性受容体をコードする核酸を発現させて不溶性封入体の生成をもたらし、その封入体に含まれる受容体分子を再生させることにより得られることを特徴とする、使用。
【請求項2】
前記受容体がヒト由来のものである、請求項1に記載の使用。
【請求項3】
前記受容体が、配列番号3または4に示したアミノ酸配列を含む、請求項1または2に記載の使用。
【請求項4】
前記結晶性調製物が、組換え可溶性受容体FcγRIIbまたはFcγRIII/免疫グロブリン複合体の結晶性調製物である、請求項1〜3のいずれか1項に記載の使用。
【請求項5】
受容体FcγRIIbもしくはFcγRIIIまたは免疫グロブリンのインヒビター、受容体FcγRIIbもしくはFcγRIII、または免疫グロブリンを突然変異誘発により同定および/または調製もしくは生成する方法であって、
(i) 組換え可溶性受容体FcγRIIbまたはFcγRIIIの結晶性調製物を提供する工程であって、該受容体が、膜貫通ドメイン、シグナルペプチドおよびグリコシル化が存在しないことを特徴とし、該受容体が、原核細胞において該可溶性受容体をコードする核酸を発現させて不溶性封入体の生成をもたらし、その封入体に含まれる受容体分子を再生させることにより得られる、工程、
(ii) 三次元構造データの生成のために、工程(i)で得た結晶性調製物を使用する工程、ならびに
(iii) 工程(ii)で得た構造データに基づいて、以下を選択または設計する工程:
(a) 免疫グロブリンに対する受容体FcγRIIbまたはFcγRIIIの結合部位に相補的なインヒビター
(b) 受容体FcγRIIbまたはFcγRIIIに対する免疫グロブリンの結合部位に相補的なインヒビター
(c) アンタゴニストおよび競合物質として使用しうる受容体FcγRIIbまたはFcγRIII
を含むことを特徴とする、方法。
【請求項6】
組換え可溶性受容体FcγRIIbまたはFcγRIIIに相補的な三次元構造を有することを特徴とするFcγRIIbまたはFcγRIIIインヒビターであって、該受容体が膜貫通ドメイン、シグナルペプチドおよびグリコシル化が存在しないことを特徴とする、前記インヒビター。
【請求項7】
前記受容体が配列番号3および4に示したアミノ酸配列のいずれか1つを含む、請求項6に記載のインヒビター。
【請求項8】
前記受容体がヒト由来のものである、請求項6または7に記載のインヒビター。
【請求項9】
免疫グロブリンの受容体FcγRIIbまたはFcγRIIIへの結合部位に相補的な三次元構造を有することを特徴とする免疫グロブリンインヒビター。
【請求項10】
請求項6〜8のいずれか1項に記載のFcγRIIbまたはFcγRIIIインヒビターを活性薬剤として含有する医薬組成物。
【請求項11】
請求項9に記載の免疫グロブリンインヒビターを活性薬剤として含有する医薬組成物。
【請求項12】
免疫系の過剰反応または誤った反応による疾患の治療または予防に使用するための、請求項10または11に記載の医薬組成物。
【請求項13】
アレルギー、自己免疫疾患またはアナフィラキシーショックを治療または予防するための、請求項10、11または12に記載の医薬組成物。
【請求項14】
受容体FcγRIIbまたはFcγRIIIと免疫グロブリンとの相互作用をモジュレートするための、分子の使用であって、該分子が結晶性調製物から得られた結晶構造データを用いて設計または同定されることを特徴とし、ここで前記結晶性調製物が、膜貫通ドメイン、シグナルペプチドおよびグリコシル化が存在しないこと、ならびに該受容体が、原核細胞において該可溶性受容体をコードする核酸を発現させて不溶性封入体の生成をもたらし、その封入体に含まれる受容体分子を再生させることにより得られることを特徴とする組換え可溶性受容体FcγRIIbまたはFcγRIIIの結晶性調製物である、前記使用。
【請求項15】
前記結晶性調製物における前記受容体がヒト由来のものである、請求項14に記載の使用。
【請求項16】
前記結晶性調製物における前記受容体が、配列番号3または4に示したアミノ酸配列を含む、請求項14または15に記載の使用。
【請求項17】
前記モジュレーションが受容体FcγRIIbまたはFcγRIIIと免疫グロブリンとの結合の部分的または完全な阻害である、請求項14〜16のいずれか1項に記載の使用。
【請求項18】
固相に結合された、均質調製物由来のFc受容体であって、ここで該均質調製物が、組換え可溶性受容体FcγRIIbまたはFcγRIIIの均質調製物であり、該受容体が、膜貫通ドメイン、シグナルペプチドおよびグリコシル化が存在しないことを特徴とし、かつ、前記調製物が、原核細胞においてその可溶性受容体をコードする核酸を発現させて不溶性封入体の生成をもたらし、その封入体に含まれる受容体分子を再生させることにより得られることを特徴とする、前記受容体。
【請求項19】
前記受容体が配列番号3および4に示したアミノ酸配列のいずれか1つを含む、請求項18に記載の受容体。
【請求項20】
前記受容体がヒト由来のものである、請求項18または19に記載の受容体。
【請求項21】
固相がクロマトグラフィー担体材料である、請求項18〜20のいずれか1項に記載の受容体。
【請求項22】
患者の血液、血漿もしくは血清から、または免疫グロブリン産生細胞の培養上清から免疫グロブリンを吸着させるための、請求項21に記載のクロマトグラフィー担体材料の使用。
【請求項23】
さらなる試験を行なうために患者の血液、血漿もしくは血清から、または免疫グロブリン産生細胞の培養上清から抗体を富化するための、請求項22に記載の使用。
【請求項24】
受容体FcγRIIbまたはFcγRIIIと免疫グロブリンとの相互作用に対する阻害のための、またはFcγRIIbもしくはFcγRIIIインヒビターとしての、膜貫通ドメイン、シグナルペプチドおよびグリコシル化が存在しない組換え可溶性受容体FcγRIIbまたはFcγRIIIに相補的な三次元構造を有することを特徴とする物質の、使用。
【請求項25】
前記組換え可溶性受容体が配列番号3および4に示したアミノ酸配列のいずれか1つを含む、請求項24に記載の使用。
【請求項26】
前記組換え可溶性受容体がヒト由来のものである、請求項24または25に記載の使用。
【請求項27】
受容体FcγRIIbまたはFcγRIIIと免疫グロブリンとの相互作用に対する阻害のための、または免疫グロブリンインヒビターとしての、免疫グロブリンのFcγRIIbもしくはFcγRIII受容体結合部位に相補的な三次元構造を有することを特徴とする物質の、使用。
【請求項28】
膜貫通ドメイン、シグナルペプチドおよびグリコシル化が存在しない組換え可溶性FcγRIIbまたはFcγRIIIに相補的な三次元構造を有することを特徴とするFcγRIIbまたはFcγRIIIインヒビターの、医薬組成物を製造するための活性薬剤としての、使用。
【請求項29】
免疫グロブリンの受容体FcγRIIbまたはFcγRIIIへの結合部位に相補的な三次元構造を有することを特徴とする免疫グロブリンインヒビターの、医薬組成物を製造するための活性薬剤としての、使用。
【請求項30】
医薬組成物が、免疫系の過剰反応または誤った反応による疾患の治療または予防に使用するためのものである、請求項28または29に記載の使用。
【請求項31】
医薬組成物が、アレルギー、自己免疫疾患またはアナフィラキシーショックを治療または予防するためのものである、請求項28、29または30に記載の使用。
【請求項32】
組換え可溶性受容体FcγRIIbまたはFcγRIIIと免疫グロブリンとの複合体の結晶性調製物を使用して得られる結晶構造データを使用して、以下の(a)〜(c):
(a) 免疫グロブリンに対する受容体FcγRIIbまたはFcγRIIIの結合部位に相補的なインヒビター、
(b) 受容体FcγRIIbまたはFcγRIIIに対する免疫グロブリンの結合部位に相補的なインヒビター、及び
(c) アンタゴニストおよび競合物質として使用しうる受容体FcγRIIbまたはFcγRIII、
のいずれかを選択または設計することを特徴とする、
受容体FcγRIIbまたはFcγRIIIと免疫グロブリンの相互作用に対するインヒビターを同定および/または調製もしくは生成する方法であって、
ここで前記結晶性調製物が、組換え可溶性受容体FcγRIIbまたはFcγRIII/免疫グロブリン複合体の結晶性調製物であり、該受容体が、膜貫通ドメイン、シグナルペプチドおよびグリコシル化が存在しないこと、ならびに該受容体が、原核細胞において該可溶性受容体をコードする核酸を発現させて不溶性封入体の生成をもたらし、その封入体に含まれる受容体分子を再生させることにより得られることを特徴とする、方法。
【請求項1】
組換え可溶性受容体FcγRIIbもしくはFcγRIIIに相補的な受容体FcγRIIbもしくはFcγRIIIインヒビター、受容体FcγRIIbもしくはFcγRIII、および/または組換え可溶性受容体FcγRIIbもしくはFcγRIIIの免疫グロブリン結合部位に相補的な免疫グロブリンインヒビターを同定および/または生成するための、結晶性調製物を使用して得られる結晶構造データの、免疫グロブリンと受容体FcγRIIbまたはFcγRIIIとの間の相互作用のモジュレーションを生じさせる使用であって、
該結晶性調製物が組換え可溶性受容体FcγRIIbまたはFcγRIIIの結晶性調製物であり、
該受容体が、膜貫通ドメイン、シグナルペプチドおよびグリコシル化が存在しないこと、ならびに該受容体が、原核細胞において該可溶性受容体をコードする核酸を発現させて不溶性封入体の生成をもたらし、その封入体に含まれる受容体分子を再生させることにより得られることを特徴とする、使用。
【請求項2】
前記受容体がヒト由来のものである、請求項1に記載の使用。
【請求項3】
前記受容体が、配列番号3または4に示したアミノ酸配列を含む、請求項1または2に記載の使用。
【請求項4】
前記結晶性調製物が、組換え可溶性受容体FcγRIIbまたはFcγRIII/免疫グロブリン複合体の結晶性調製物である、請求項1〜3のいずれか1項に記載の使用。
【請求項5】
受容体FcγRIIbもしくはFcγRIIIまたは免疫グロブリンのインヒビター、受容体FcγRIIbもしくはFcγRIII、または免疫グロブリンを突然変異誘発により同定および/または調製もしくは生成する方法であって、
(i) 組換え可溶性受容体FcγRIIbまたはFcγRIIIの結晶性調製物を提供する工程であって、該受容体が、膜貫通ドメイン、シグナルペプチドおよびグリコシル化が存在しないことを特徴とし、該受容体が、原核細胞において該可溶性受容体をコードする核酸を発現させて不溶性封入体の生成をもたらし、その封入体に含まれる受容体分子を再生させることにより得られる、工程、
(ii) 三次元構造データの生成のために、工程(i)で得た結晶性調製物を使用する工程、ならびに
(iii) 工程(ii)で得た構造データに基づいて、以下を選択または設計する工程:
(a) 免疫グロブリンに対する受容体FcγRIIbまたはFcγRIIIの結合部位に相補的なインヒビター
(b) 受容体FcγRIIbまたはFcγRIIIに対する免疫グロブリンの結合部位に相補的なインヒビター
(c) アンタゴニストおよび競合物質として使用しうる受容体FcγRIIbまたはFcγRIII
を含むことを特徴とする、方法。
【請求項6】
組換え可溶性受容体FcγRIIbまたはFcγRIIIに相補的な三次元構造を有することを特徴とするFcγRIIbまたはFcγRIIIインヒビターであって、該受容体が膜貫通ドメイン、シグナルペプチドおよびグリコシル化が存在しないことを特徴とする、前記インヒビター。
【請求項7】
前記受容体が配列番号3および4に示したアミノ酸配列のいずれか1つを含む、請求項6に記載のインヒビター。
【請求項8】
前記受容体がヒト由来のものである、請求項6または7に記載のインヒビター。
【請求項9】
免疫グロブリンの受容体FcγRIIbまたはFcγRIIIへの結合部位に相補的な三次元構造を有することを特徴とする免疫グロブリンインヒビター。
【請求項10】
請求項6〜8のいずれか1項に記載のFcγRIIbまたはFcγRIIIインヒビターを活性薬剤として含有する医薬組成物。
【請求項11】
請求項9に記載の免疫グロブリンインヒビターを活性薬剤として含有する医薬組成物。
【請求項12】
免疫系の過剰反応または誤った反応による疾患の治療または予防に使用するための、請求項10または11に記載の医薬組成物。
【請求項13】
アレルギー、自己免疫疾患またはアナフィラキシーショックを治療または予防するための、請求項10、11または12に記載の医薬組成物。
【請求項14】
受容体FcγRIIbまたはFcγRIIIと免疫グロブリンとの相互作用をモジュレートするための、分子の使用であって、該分子が結晶性調製物から得られた結晶構造データを用いて設計または同定されることを特徴とし、ここで前記結晶性調製物が、膜貫通ドメイン、シグナルペプチドおよびグリコシル化が存在しないこと、ならびに該受容体が、原核細胞において該可溶性受容体をコードする核酸を発現させて不溶性封入体の生成をもたらし、その封入体に含まれる受容体分子を再生させることにより得られることを特徴とする組換え可溶性受容体FcγRIIbまたはFcγRIIIの結晶性調製物である、前記使用。
【請求項15】
前記結晶性調製物における前記受容体がヒト由来のものである、請求項14に記載の使用。
【請求項16】
前記結晶性調製物における前記受容体が、配列番号3または4に示したアミノ酸配列を含む、請求項14または15に記載の使用。
【請求項17】
前記モジュレーションが受容体FcγRIIbまたはFcγRIIIと免疫グロブリンとの結合の部分的または完全な阻害である、請求項14〜16のいずれか1項に記載の使用。
【請求項18】
固相に結合された、均質調製物由来のFc受容体であって、ここで該均質調製物が、組換え可溶性受容体FcγRIIbまたはFcγRIIIの均質調製物であり、該受容体が、膜貫通ドメイン、シグナルペプチドおよびグリコシル化が存在しないことを特徴とし、かつ、前記調製物が、原核細胞においてその可溶性受容体をコードする核酸を発現させて不溶性封入体の生成をもたらし、その封入体に含まれる受容体分子を再生させることにより得られることを特徴とする、前記受容体。
【請求項19】
前記受容体が配列番号3および4に示したアミノ酸配列のいずれか1つを含む、請求項18に記載の受容体。
【請求項20】
前記受容体がヒト由来のものである、請求項18または19に記載の受容体。
【請求項21】
固相がクロマトグラフィー担体材料である、請求項18〜20のいずれか1項に記載の受容体。
【請求項22】
患者の血液、血漿もしくは血清から、または免疫グロブリン産生細胞の培養上清から免疫グロブリンを吸着させるための、請求項21に記載のクロマトグラフィー担体材料の使用。
【請求項23】
さらなる試験を行なうために患者の血液、血漿もしくは血清から、または免疫グロブリン産生細胞の培養上清から抗体を富化するための、請求項22に記載の使用。
【請求項24】
受容体FcγRIIbまたはFcγRIIIと免疫グロブリンとの相互作用に対する阻害のための、またはFcγRIIbもしくはFcγRIIIインヒビターとしての、膜貫通ドメイン、シグナルペプチドおよびグリコシル化が存在しない組換え可溶性受容体FcγRIIbまたはFcγRIIIに相補的な三次元構造を有することを特徴とする物質の、使用。
【請求項25】
前記組換え可溶性受容体が配列番号3および4に示したアミノ酸配列のいずれか1つを含む、請求項24に記載の使用。
【請求項26】
前記組換え可溶性受容体がヒト由来のものである、請求項24または25に記載の使用。
【請求項27】
受容体FcγRIIbまたはFcγRIIIと免疫グロブリンとの相互作用に対する阻害のための、または免疫グロブリンインヒビターとしての、免疫グロブリンのFcγRIIbもしくはFcγRIII受容体結合部位に相補的な三次元構造を有することを特徴とする物質の、使用。
【請求項28】
膜貫通ドメイン、シグナルペプチドおよびグリコシル化が存在しない組換え可溶性FcγRIIbまたはFcγRIIIに相補的な三次元構造を有することを特徴とするFcγRIIbまたはFcγRIIIインヒビターの、医薬組成物を製造するための活性薬剤としての、使用。
【請求項29】
免疫グロブリンの受容体FcγRIIbまたはFcγRIIIへの結合部位に相補的な三次元構造を有することを特徴とする免疫グロブリンインヒビターの、医薬組成物を製造するための活性薬剤としての、使用。
【請求項30】
医薬組成物が、免疫系の過剰反応または誤った反応による疾患の治療または予防に使用するためのものである、請求項28または29に記載の使用。
【請求項31】
医薬組成物が、アレルギー、自己免疫疾患またはアナフィラキシーショックを治療または予防するためのものである、請求項28、29または30に記載の使用。
【請求項32】
組換え可溶性受容体FcγRIIbまたはFcγRIIIと免疫グロブリンとの複合体の結晶性調製物を使用して得られる結晶構造データを使用して、以下の(a)〜(c):
(a) 免疫グロブリンに対する受容体FcγRIIbまたはFcγRIIIの結合部位に相補的なインヒビター、
(b) 受容体FcγRIIbまたはFcγRIIIに対する免疫グロブリンの結合部位に相補的なインヒビター、及び
(c) アンタゴニストおよび競合物質として使用しうる受容体FcγRIIbまたはFcγRIII、
のいずれかを選択または設計することを特徴とする、
受容体FcγRIIbまたはFcγRIIIと免疫グロブリンの相互作用に対するインヒビターを同定および/または調製もしくは生成する方法であって、
ここで前記結晶性調製物が、組換え可溶性受容体FcγRIIbまたはFcγRIII/免疫グロブリン複合体の結晶性調製物であり、該受容体が、膜貫通ドメイン、シグナルペプチドおよびグリコシル化が存在しないこと、ならびに該受容体が、原核細胞において該可溶性受容体をコードする核酸を発現させて不溶性封入体の生成をもたらし、その封入体に含まれる受容体分子を再生させることにより得られることを特徴とする、方法。
【図1】

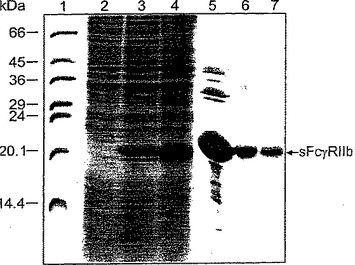
【図2】
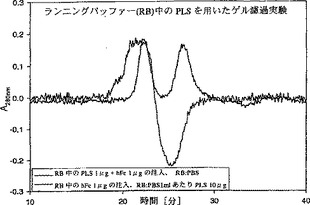
【図3】

【図4】
【図5A】
【図5B】
【図6】
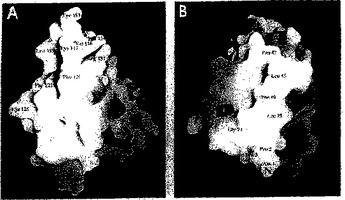
【図7】
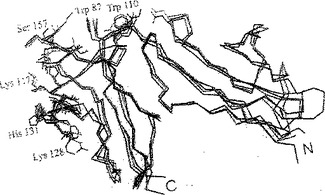
【図8】

【図9】
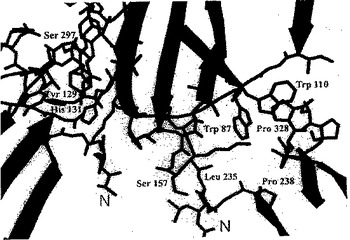
【図10A】
【図10B】

【図11】
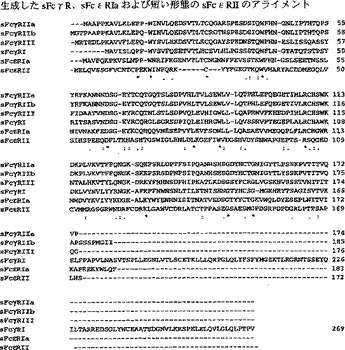
【図12】

【図2】
【図3】
【図4】
【図5A】
【図5B】
【図6】
【図7】
【図8】
【図9】
【図10A】
【図10B】
【図11】
【図12】
【公開番号】特開2011−105716(P2011−105716A)
【公開日】平成23年6月2日(2011.6.2)
【国際特許分類】
【出願番号】特願2010−243504(P2010−243504)
【出願日】平成22年10月29日(2010.10.29)
【分割の表示】特願2000−585398(P2000−585398)の分割
【原出願日】平成11年12月3日(1999.12.3)
【出願人】(301033396)マックス−プランク−ゲゼルシャフト ツール フォーデルング デル ヴィッセンシャフテン エー.ヴェー. (3)
【Fターム(参考)】
【公開日】平成23年6月2日(2011.6.2)
【国際特許分類】
【出願日】平成22年10月29日(2010.10.29)
【分割の表示】特願2000−585398(P2000−585398)の分割
【原出願日】平成11年12月3日(1999.12.3)
【出願人】(301033396)マックス−プランク−ゲゼルシャフト ツール フォーデルング デル ヴィッセンシャフテン エー.ヴェー. (3)
【Fターム(参考)】
[ Back to top ]