
組換え型Yarrowiasp.を用いた発酵による超長鎖脂肪酸(VLCFA)の生産方法
本発明は、発酵による超長鎖脂肪酸(VLCFA)の生産方法に関するものであって、該方法は、Yarrowia sp.における異種遺伝子の発現を可能にする調節エレメントの制御下でヒドロキシアシル−CoAデヒドラターゼをコードする前記異種遺伝子を含むYarrowia sp.の組換え株を培養するステップを含む。また、本発明は、この組換え型Yarrowia sp.にも関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、発酵による超長鎖脂肪酸(VLCFA)の生産方法に関するものであり、該方法は、Yarrowia sp.における異種遺伝子の発現を可能にする調節エレメントの制御下でヒドロキシアシル−CoAデヒドラターゼをコードする前記異種遺伝子を含むYarrowia sp.の組換え株を培養するステップを含む。
【0002】
本発明は、この組換え型Yarrowia sp.にも関するものである。
【背景技術】
【0003】
生命体は、複合脂質内に取り込まれるおびただしい種類の異なる脂肪酸を合成する。これらの複合脂質は、主要な構造的構成要素膜であると同時に、植物および動物の両方における主要な貯蔵物質である。
【0004】
超長鎖脂肪酸(VLCFA)は真核細胞の構成要素であり、炭素20個以上の長さで構成されている(すなわち >C18)。VLCFAは、異なる生物内で多くの異なる生理学的機能に関与している。これらは、脳(ミエリン)または植物種子(貯蔵トリアシルグリセロール、TAG)などの一部の組織の豊富な構成成分である。VLCFAは、皮膚の脂質バリアおよび植物のクチクラワックスの構成要素である。一部のVLCFAの長いアシル鎖は、例えば核孔内に見られる高い膜湾曲に必要なものである。同様に、VLCFAは、タンパク質輸送およびGPI脂質アンカー合成のための分泌経路にも関与する。最後に、VLCFAは、膜構成成分であると同時にシグナル伝達分子でもあるスフィンゴ脂質の構成要素である。
【0005】
VLCFAは、C18より長いアシル鎖を有する脂肪酸である。多価不飽和VLCFAは、主としてエイコサペンタエン酸(EPA)またはドコサヘキサエン酸(DHA)のように人間の食生活に重要な栄養成分とみなされている。国際公開第2005/118814号パンフレットは、Saccharomyces cerevisiaeにおける多価不飽和脂肪酸の産生を改善する方法を開示している。不飽和VLCFAは、洗浄剤または潤滑剤としても作用することから、工業的にも有益である。
【0006】
VLCFAは、四つの相異なる酵素(KCS、KCR、HCDおよびECR)を含む膜結合酵素複合体であるエロンガーゼ複合体という名称のタンパク質複合体の内部で小胞体内に集められた四つの連続する酵素反応を通して、二個の炭素の逐次的付加によって合成される。脂肪伸長の第一のステップは、3−ケトアシル−CoAシンターゼ(KCSまたは縮合酵素)による、マロニル−CoAを用いた長鎖アシル−CoAの縮合である。結果として得られた3−ケトアシル−CoAは、次に3−ケトアシル−CoAレダクターゼ(KCR)により還元されて、3−ヒドロキシアシル−CoAを生成する。第三のステップは3−ヒドロキシアシル−CoAデヒドラターゼ(HCD)により3−ヒドロキシアシル−CoAをトランス−2,3−エノイル−CoAへと脱水するステップであり、このトランス−2,3−エノイル−CoAは最終的にトランス−2,3−エノイル−CoAレダクターゼ(ECR)により還元されて二炭素伸長されたアシル−CoAを生成する。最後の三つの酵素は、アシル−CoAの特異性に関与しないことから、コア酵素と呼ばれる。アシル−CoAがひとたびエロンガーゼ複合体から伸長されると、これらは、リン脂質、トリアシルグリセロール、スフィンゴ脂質、および植物エピクチクラワックスなどの特異的脂質といった異なる脂質クラスの中に取り込まれ得る。
【0007】
Yarrowia lipolyticaは、この酵母がその乾燥重量の50%超を脂質として蓄積できるだけでなく、炭素源として脂質を効率良く使用することもできることから、油性酵母とみなされている(Beopoulosら、2009)。そのゲノムの完全な配列決定ならびにこの酵母についての分子遺伝学的手段の開発によって、この生物は脂質蓄積の機序の研究用モデルとなったばかりでなく、油脂化学バイオテクノロジーのための細胞工場にもなった。近年になって、β−酸化に関与するPOX1〜6遺伝子とグルコース−3−ホスフェートデヒトロゲナーゼGUT2の組合せ欠失が、脂質、主として遊離脂肪酸のきわめて高い蓄積を導くことが示された(Beopoulosら、2008、FR0854786;11 2008年7月11日)。この肥満菌株は、それぞれグルコースまたはオレイン酸上で成長させた場合に野生型に比べて二倍および三倍多い脂肪酸を蓄積した。興味深いことに、これらの脂質は、単一の大きな脂肪体に蓄積した。Yarrowia lipolyticaは、主として長鎖脂肪酸c18:2、c18:1(n−9)、c16:1(n−7)およびc16:0を蓄積する。しかしながら、Y.lipolyticaにおける超長鎖脂肪酸(VLCFA)については、わずかな情報しかない。
【0008】
現在、Yarrowia sp.、特にYarrowia lipolyticaにおいてヒドロキシアシル−CoAデヒドラターゼをコードする異種遺伝子を発現させることで、菌株による脂肪酸およびVLCFAの産生に対して、品質および/または量の観点において直接的影響が及ぼされることが発見されている。
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は、Yarrowia sp.におけるヒドロキシアシル−CoAデヒドラターゼをコードする異種遺伝子の発現を可能にする調節エレメントの制御下で、前記異種遺伝子を含む、Yarrowia sp.の組換え株に関するものである。
【0010】
ヒドロキシアシル−CoAデヒドラターゼをコードする遺伝子は、詳細には、ヒドロキシアシル−CoAデヒドラターゼをコードする植物種の遺伝子、その機能的相同体およびフラグメントからなる群から選択される。植物種の遺伝子は、有利には、Arabidopsis thaliana、Vitis vinifera、Oryza sativa、Brassica rapa、Hyacinthus orientalis、Ostreacoccus lucimarinus、Chlamydomonas reinhardtii、Brassica napus、Raphanus sativus、およびBrassica oleracea由来のヒドロキシアシル−CoAデヒドラターゼをコードする遺伝子から選択され、より詳細には、Arabidopsis thaliana由来の遺伝子PAS2である。
【0011】
本発明は同様に、発酵による超長鎖脂肪酸(VLCFA)の生産方法にも関するものであり、該方法は、適切な培養培地内で本発明の組換え株を培養するステップと、菌株および/または培地からVCLFAを回収するステップとを含む。
【0012】
前記方法により生産される脂肪酸も、同じく本発明の一部である。
【課題を解決するための手段】
【0013】
本発明は、Yarrowia sp.の組換え株に関するものであり、該組換え株は、Yarrowia sp.におけるヒドロキシアシル−CoAデヒドラターゼをコードする異種遺伝子の発現を可能にする調節エレメントの制御下にある、前記異種遺伝子を含む。
【0014】
組換え株
本発明によると、菌株は、それが遺伝子置換またはプラスミド導入などの細胞生物学的手段によって遺伝的に修飾されている場合に、組換え型となる。これは、遺伝子内に新しい遺伝子もしくは変異もしくは新しい調節エレメントを導入するため、または内因性遺伝子を欠失させるための定方向突然変異誘発により得られるであろう。組換え型微生物は、ランダム突然変異誘発の唯一の結果ではない。
【0015】
新しい遺伝子が菌株内に導入される場合、それは、発現プラスミドと共に導入されてよく、あるいは、その菌株のゲノム内に組込まれてもよい。
【0016】
菌株のゲノム内に組込まれる場合、遺伝子は、相同的組換え技術などの公知の遺伝子置換方法によって特異的部位にまたは無作為に組込まれてよい。
【0017】
異種遺伝子は、導入される場合、Yarrowia sp.におけるこの異種遺伝子の発現を可能にする調節エレメントの制御下のコード配列を含むことができる。代替的には、異種遺伝子は、既存の内因性調節エレメントの制御下で微生物のゲノム内に導入されて、欠失させられる対応する内因性コード配列から置き換わる、コード配列を含むことができる。
【0018】
特に新しい遺伝子を導入するかまたは遺伝子を欠失させるためのYarrowia sp.の修飾方法は、当該技術分野において公知であり、BarthおよびGaillardin(1996)ならびにFickersら、(2003)がある。
【0019】
ヒドロキシアシル−CoAデヒドラターゼをコードする異種遺伝子
「ヒドロキシアシル−CoAデヒドラターゼ」(HCD)という用語は、3−ヒドロキシアシル−CoAをトランス−2,3−エノイル−CoAに脱水する反応を触媒する酵素を意味する。それは、ヒドロリアーゼのファミリーに属する。この酵素は、エロンガーゼ複合体の一部を成し、植物内のVLCFAの合成のみに関与し、その分解には関与しない。
【0020】
ヒドロキシアシル−CoAデヒドラターゼをコードする遺伝子は、異種遺伝子である。本発明によると、遺伝子は、天然の菌株中で同一のものが発見されない場合、異種である。それは、異種調節エレメントの制御下の天然コード配列であるかまたは天然調節エレメントの制御下の異種コード配列であり得る。それは同様に、修飾されるべき菌株中には見られるものの未修飾菌株中に同じ遺伝子が発見されないプラスミド上またはゲノム内遺伝子座には見られない天然の構成要素を有する遺伝子でもあり得る。
【0021】
ヒドロキシアシル−CoAデヒドラターゼをコードする異種遺伝子は、特に、ヒドロキシアシル−CoAデヒドラターゼをコードする植物種の遺伝子由来のコード配列を含む遺伝子、その機能的相同体およびフラグメントからなる群から選択される。
【0022】
ヒドロキシアシル−CoAデヒドラターゼをコードする植物種の遺伝子は、当該技術分野において公知であり、Vitis vinifera(CAN64341.1仮想タンパク質をコードする)、Oryza sativa(CAD39891.2、EAY72548.1仮想タンパク質OsI_000395、EAZ30025.1仮想タンパク質OsJ_013508およびBAD61107.1チロシンホスファターゼ様)、Brassica rapa(AAZ66946.1)、Hyacinthus orientalis(AAT08740.1タンパク質チロシンホスファターゼ)、Ostreacoccus lucimarinus(XP_001420997.1推定タンパク質およびXP_001422898.1推定タンパク質)、Chlamydomonas reinhardtii(EDP01055.1推定タンパク質)、および同様にBrassica napus、Raphanus sativus、Brassica oleracea由来の遺伝子の中から特に選択されたものを含む。
【0023】
本発明の好ましい実施形態において、異種遺伝子は、UniGeneデータバンクにNP_196610.2の番号で登録済であり、F12B17.170、F12B17_170、PASTICCINO 2、PEP、およびPEPINOとしても同様に知られている、Arabidopsis thaliana由来の遺伝子PAS2(Bachら、2008)である。
【0024】
本発明の別の具体的実施形態において、異種遺伝子は、遺伝子データバンクにNP_012438.1の番号で登録済である、Saccharomyces cerevisiae由来のPHS1遺伝子(Denicら、2007)、その機能的相同体およびフラグメントである。
【0025】
異種遺伝子のコード配列が別のものに由来する場合、このコード配列は実際、Yarrowia sp.について公知の好ましいコドン使用頻度でこれを再度コードすることができる。当業者であれば、Yarrowia sp.において使用される好ましいコドンおよびこのような再コードされたコード配列の調製方法を知っている。
【0026】
本発明によると、「機能的相同体」とは、ヒドロキシアシル−CoAデヒドラターゼをコードする異種遺伝子と相同性を共有する遺伝子、または、ヒドロキシアシル−CoAデヒドラターゼをコードする異種遺伝子によりコードされるタンパク質と相同性を共有するタンパク質をコードする遺伝子である。
【0027】
ヒドロキシアシル−CoAデヒドラターゼをコードする遺伝子によりコードされるタンパク質と相同性を共有するタンパク質は、植物から得られるか、または、植物に由来する天然タンパク質のバリアントもしくは機能的フラグメントであってよい。
【0028】
「天然タンパク質のバリアントまたは機能的フラグメント」という用語は、ポリペプチドのアミノ酸配列が、天然に観察される配列に厳密に限定されなくてよく、追加のアミノ酸を含んでいてよい、ということを意味する。「フラグメント」という用語は、ポリペプチドの配列が、もとの配列よりもアミノ酸を少なく含むがヒドロキシアシル−CoAデヒドラターゼ活性を付与するのに充分なアミノ酸を依然として含んでいてよいことを意味する。ポリペプチドが、その酵素活性を保持しながら、一つ以上のアミノ酸の置換、挿入、欠失および/または付加により修飾可能であることは、当該技術分野において周知である。例えば、タンパク質の機能的特性に影響を及ぼさない、化学的に等価のアミノ酸による所与の位置にある一つのアミノ酸の置換が一般的である。本発明に関しては、置換は、以下の群のうちの一つの中での交換として定義される:
・小さい脂肪族の非極性またはわずかに極性の残基:Ala、Ser、Thr、Pro、Gly
・極性の、負荷電残基およびそのアミド:Asp、Asn、Glu、Gln
・極性の、正荷電残基:His、Arg、Lys
・大きい脂肪族の非極性残基:Met、Leu、Ile、Val、Cys
・大きい芳香族残基:Phe、Tyr、Trp。
【0029】
したがって、一つの負荷電残基での別の残基の置換(例えばグルタミン酸でアスパラギン酸を置換)または一つの正荷電残基での別の残基の置換(例えばリジンでアルギニンを置換)を結果としてもたらす変化が、機能的に等価の生成物を生成すると予想することができる。
【0030】
アミノ酸配列内の、アミノ酸が修飾される位置および修飾を受けるアミノ酸の数は、特に限定されない。当業者であれば、タンパク質の活性に影響を及ぼすことなく導入可能な修飾を認識することができる。例えば、タンパク質のNまたはC末端部分での修飾は、一部の状況下でタンパク質の活性を改変しないものと予想され得る。
【0031】
「バリアント」という用語は、もとの酵素活性をなおも保持しながら、以上で定義したような修飾に付されたポリペプチドを意味する。
【0032】
本発明によると、ヒドロアシル−CoAデヒドラターゼ酵素活性を有するポリペプチドは、PAS2の配列と少なくとも30%の相同性、好ましくは少なくとも50%の相同性、そしてより好ましくは少なくとも70%の相同性を有する配列を含んでいてよい。
【0033】
二つのタンパク質配列間の相同性百分率の決定方法は、当業者にとって公知である。例えば、これは、http://www.ebi.ac.uk/clustalw/のウェブサイト上で入手可能なソフトウェアCLUSATALWを、このウェブサイト上に記されたデフォルトパラメータと共に使用することによって、配列のアラインメントの後に行なうことができる。残基の総数と比較した同じ位置における同一の残基の数を記録することにより、アラインメントから同一性百分率を容易に計算することができる。代替的には、例えば、http://www.ncbi.nlm.nih.gov/BLAST/のウェブサイト上で入手可能なBLASTプログラムをそのウェブサイト上に記されたデフォルトパラメータと共に使用することによって、自動計算を行なうことも可能である。
【0034】
Yarrowia sp.における異種遺伝子の発現を可能にする調節エレメント
このような調節エレメントは、当該技術分野において周知であり、アシル−CoAオキシダーゼ2由来のPOX2プロモーター、イソシトレートデヒドロゲナーゼ由来のICLプロモーター、プロモーターHp4d、プロモーターGPDおよびGPM、プロモーターFBP、ならびにプロモーターXPR2を含む。前記複数のプロモーターは当該技術分野において公知であり、なかでもJuretzekら(2000)、Madzakら(2004)、Madzakら(2000)、米国特許第7259255号明細書、米国特許第7202356号明細書およびBlanchin−Rolandら(1994)中で開示されている。
【0035】
Yarrowia sp.
本発明によると、Yarrowia sp.の任意の菌株を形質転換し、本発明の方法において使用してよい。好ましくは、Yarrowia sp.の菌株はYarrowia lipolytica属に属する。
【0036】
Yarrowia lipolytica属の菌株は、この菌株の形質転換方法と同様、当該技術分野において周知である。対象となるコード領域を含む構築物を、任意の標準的技術により宿主細胞中に導入してよい。これらの技術には、形質転換(例えば酢酸リチウム形質転換(Methods in Enzymology,194:186−187(1991)))、原形質融合、遺伝子銃衝撃、電気穿孔法、マイクロインジェクション、または対象となる遺伝子を宿主細胞内に導入する任意の他の方法が含まれる。油性酵母(すなわちYarrowia lipolytica)に適用可能なより具体的な教示としては、米国特許第4880741号明細書および米国特許第5071764号明細書が含まれる。
【0037】
参照として本明細書に援用されている、脂質、主として遊離脂肪酸の非常に高い蓄積を有する菌株(仏国特許出願第08/54786号明細書;2008年7月11日)のように、改良された脂肪酸生産のために修飾された菌株も同様に開示されている。このような菌株は、ヒドロキシアシル−CoAデヒドラターゼをコードする異種遺伝子を用いて本発明にしたがってさらに修飾されてよい。
【0038】
また、本発明の組換え株は、脂肪酸のβ−酸化に関与する少なくとも一つの遺伝子の欠失、特にアシルCoAオキシダーゼをコードする遺伝子POX1〜POX6のうちの一つの欠失、特に六つのアシルCoAオキシダーゼをコードする六つの遺伝子POX1〜6ならびに/または脂肪酸およびTAG合成の経路に関与する一つの遺伝子の欠失、特にグリセロール3−ホスフェートデヒドロゲナーゼをコードする遺伝子の欠失も含み得る。
【0039】
組換え株の培養
脂肪酸、特にVLCFAは、適切な培養培地内で発酵によって組換え株を培養する場合に生産される。
【0040】
発酵による培養とは、微生物が培養培地で発育させられ、培養培地の炭素源を形質転換することによって、この培養ステップ中にVLCFAを産生することを意味する。VLCFAは、バイオマスと共に、細胞内および/または培地内に蓄積される。
【0041】
発酵は、酵素変換プロセスでさらに使用される、酵素を生産するために培養を用いる生物変換反応とは全く異なるものである。
【0042】
適切な培養培地
Yarrowia sp.のための培養培地は、BarthおよびGaillardin(1996)、Nicaudら(2002)およびMauersbergerおよびNicaud(2002)を含め、当該技術分野において周知である。発酵に関して規定された培地は、Leblondら(2009)および韓国特許第2009/0029808号公報中で詳細に開示されている。
【0043】
スクロース培地は、Nicaudら(1989)において詳細に開示されている。
【0044】
適切な培養培地は、Yarrowia sp.が成長でき、かつ菌株の成長およびVLCFAの産生を可能にする全ての栄養素、特に炭素源を含む培地である。
【0045】
炭素源は、スクロースまたは他の炭水化物などの、任意の炭素源であってよい。
【0046】
菌株および/または培地からのVCLFAの回収
VCLFAは、バイオマスと共に菌株中および/または培養培地内に蓄積される。VCFLAの回収には、一般に、細胞溶解、濾過および培地からの回収というステップが含まれる。脂肪酸バイオプロダクションの当業者は、本発明の方法に、バイオマスからの脂肪酸の通常の回収方法をいかに適応させるべきかを知っている。
【0047】
本発明の方法を用いて生産されたVCFLAは、そのままの状態で、本発明の菌株により産生された脂肪酸の混合物中で使用されてよい。これらをさらに精製し単離することもできる。
【図面の簡単な説明】
【0048】
【図1】Yarrowia lipolytica発現のために最適化された合成PAS2を表わす。(A)PAS2Ylの遺伝子およびタンパク質の配列。(B)Arabidopsis PAS2AtとPAS2Ylのアラインメント。
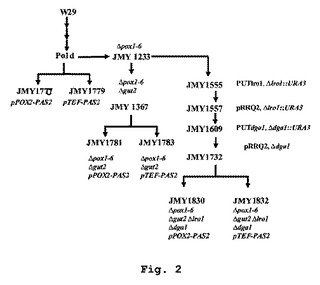
【図2】使用または創出された、異なる菌株の概略図と共に、YarrowiaにおけるPAS2発現を表わす。
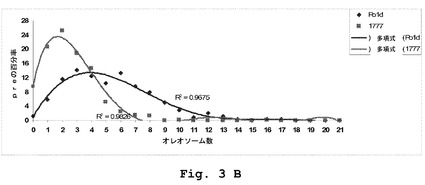
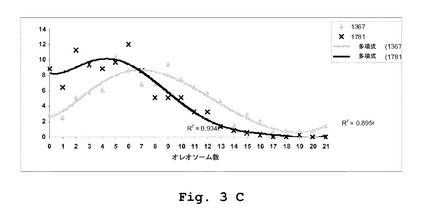
【図3】YarrowiaにおけるPAS2発現の効果を表わす。(A)JM1367およびJM1781菌株中でのナイルレッドを用いた脂肪体の染色。(B)Po1dおよびJM1777における細胞一個あたりの脂肪体の数。分布を比較するために、多項回帰を適用した。(C)JM1367およびJM1381における細胞一個あたりの脂肪体の数。分布を比較するために、多項回帰を適用した。
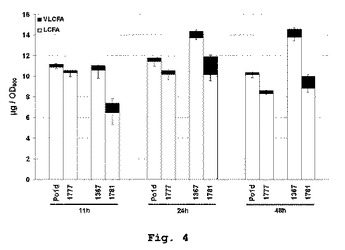
【図4】YarrowiaにおけるPAS2発現を伴う、生産されたLCFA/VLCFAの比を示す。
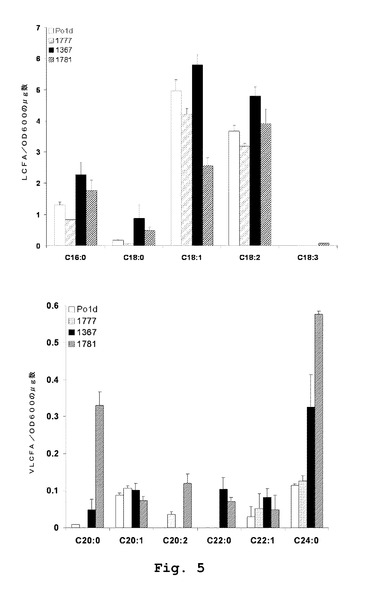
【図5】YarrowiaにおけるPAS2発現を伴う、修飾されたLCFAおよびVLCFAの含有量を示す。(A)LCFA含有量。(B)VLCFA含有量。
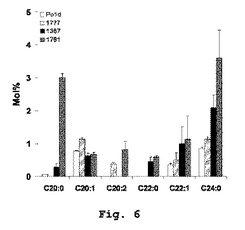
【図6】YarrowiaにおけるPAS2発現を伴う、修飾されたVLCFAのプロファイルを示す。
【実施例】
【0049】
材料と方法
Yarrowia lipolyticaコドン使用頻度にしたがって、合成PAS2遺伝子(PAS2Yl)を合成して、プラスミドJME1107を得た。PASYlをプラスミドJMP62−POX2−URA3ex(JME803)内そしてプラスミドJMP62−TEF−URA3ex(JME1012)内に以下の通りにクローニングした:プラスミドJME1107をBamHI−AvrIIにより消化し、PAS2遺伝子を担持する対応するフラグメントを、プラスミドJME1012およびJME1107の対応する部位でクローニングして、それぞれプラスミドJME1108(POX2−PAS2)およびJME1110(TEF−PAS2)を得た。プラスミドをNotIで消化し、発現カセットを担持するフラグメントを、酢酸リチウム法によるYarrowia lipolyticaの形質転換(G.BarthおよびGaillardinの概説:すなわち「Nonconventional Yeasts in Biotechnology A Handbook」(Wolf,K.、Ed.)、Vol.1、1996、pp.313−388.Springer−Verlag中のYarrowia lipolyticaに記載されている)のために使用した。形質転換体をYNBcasa上で選択した。典型的には、フラグメント1μgあたり約5×103個の形質転換体が得られた。それぞれPOX2−PAS2およびTEF−PAS2を含むクローンのためのプライマー対61start/61stopおよびTEFstart/61stopを用いたPCRにより、4〜8個の形質転換体を分析した。PCR産物をさらに、PAS2遺伝子内のAvaI固有の制限部位により消化した。
【0050】
Yarrowia lipolytica中のPAS2の安定した発現
Arabidopsis PAS2遺伝子のオープンリーディングフレームを再コードして、Yarrowiaコドン使用頻度でのその発現を改善した(図1A〜B)。クローニングを促すため、PAS2 ORFの5’および3’末端に二つの制限部位BamHIおよびAvrIIをそれぞれ付加した。PAS2Ylと命名し直した、新規の配列を化学的に合成し(GeneArt inc.)、二つの発現ベクターJME1110およびJME1108内にクローニングした。その二つのベクターは、構成的プロモーター(pTEF、JME1110)またはオレイン酸誘発性プロモーター(pPOX2、JME1108)の下でのPAS2の発現を可能にする。
【0051】
野生型菌株Po1d(JMY195)およびΔgut2、Δpox1−6肥満菌株(JMY1367)を形質転換するために、両方の構築物を使用した。形質転換体をウラシル上で選択し、発現カセットの組込みをPCRで確認した。さらなる分析のために、複数のクローンを選択し使用した。しかしながら、POX2プロモーターはインデューサーの不在下でも強い発現を可能にすることから、我々は主として、JME1108構築物を用いて形質転換体を特徴づけした。菌株JMY1777およびJMY1778は、pPOX2−PAS2で形質転換されたPo1dの二つの独立したクローンである。同様に、JMY1781およびJMY1782は、pPOX2−PAS2で形質転換されたJMY1367(Δgut Δpox1−6)の二つの独立したクローンである。
【0052】
PAS2発現は細胞の成長を改善し、脂肪体の断片化を導く
複数の異なるPSA2発現菌株の成長を、グルコースを補足した培地上でそれらの未形質転換相対物と比較した。全ての菌株を、30mlのYPD培地中OD600=0.6で植菌した。全ての菌株は、植菌後、40時間の培養後に静止期の開始に達するまでの間に、約4〜5時間の遅滞期、および9〜12時間の平担域を伴う二峰性曲線を示した。
【0053】
Yarrowia lipolyticaは、脂肪体内で脂質を蓄積するものとして知られている。肥満菌株JMY1367はより大きい構造まで脂肪体を融合することを特徴とすることが報告された。こうして、脂肪体の構造に対するPAS2発現の効果を、ナイルレッドによる複数の異なる菌株の染色によって調べた(図3A)。静止段階は、脂質の高い蓄積度を特徴とすることから、48時間後に細胞を収集し、細胞一個あたりの脂肪体の総数を定量した(図3B〜C)。
【0054】
Po1d内ならびに肥満菌株JM1367内の両方におけるPAS2の発現は、脂肪体数の減少を導く(図3B〜C)。
【0055】
PAS2の発現がVLCFAレベルを増強する
成長曲線の三つの異なる時点、すなわち第一成長期の終り(11時間)、第二成長期の終り(24時間)および静止期中(48時間)に、四つの菌株中で、脂肪酸アシルメチルエステル(FAMES)の脂質総含有量をガスクロマトグラフィにより分析した。予想通り、肥満菌株JMY1367は、24時間後と48時間後にそれぞれ22%および40%という、野生型Po1dに比べて高い脂肪酸含有量を有する(図4)。PAS2の発現は、全ての時点において脂肪酸総含有量を減少させた。最も強い減少は、48時間後に観察され、Po1dバックグラウンドで20%、JMY1367バックグラウンドで46%であった。
【0056】
Yarrowia lipolyticaの最も豊富な脂肪酸に相当する長鎖脂肪酸(LCFA)の全量は、PAS2発現菌株において18%および36%減少した。LCFAの分析から、最も豊富なLCFAの一つであるc18:1が最も影響を受け、肥満菌株JMY1367バックグラウンドにおいて48時間後に例えば126%の減少であったことを除いて、異なるクラス全てがPAS2の発現時点で減少したレベルを示していることがわかった(図5)。合計LCFAの減少は、成長曲線の始め(11時間)においてさえ有効であった。
【0057】
VLCFAは、Yarrowia lipolyticaにおけるわずかな脂質種に相当するにすぎない(総脂肪酸の2.2〜3.2%)(図4)。三つの主要な種、すなわち、それぞれ48時間の培養時点で総脂肪酸の0.86、0.77および0.34%(モル%)に相当する24:0、20:1および22:1が有意に蓄積された(図6)。Δgut Δpox1−6は、24時間の培養において倍増した(Po1dにおいて、3.23に比べ6.53μg/10OD)ことから、VLCFAレベルに対し明らかな効果を有していた。関与した主要なVLCFAはc24:0およびc22:1であり、その含有量は総脂肪酸のそれぞれ2.09%と0.99%(モル%)に達した。新しいVLCFAを、0.45%に達するc22:0として検出することができた。Po1dバックグラウンドでのPAS2の発現は、蓄積したVLCFAの量または性質をさほど変化させなかった。しかしながら、肥満菌株JMY1367バックグラウンドでのPAS2の発現は、VLCFAの含有量を極めて有意に増大させた。24時間の培養後、JMY1781は、17.35μg/10ODを蓄積し、これは肥満菌株および野生型Po1d株よりそれぞれ2.65倍および5.3倍多いものであった。蓄積した主要なVLCFAは、c20:0およびc24:0であり、総VLCFAの半分超を占めていた。JMY1781では、エルカ酸c22:1、c22:0、c20:2も同様に有意に蓄積した。
【0058】
PAS2発現は、新しいモノメチル分岐脂肪酸の蓄積を誘発する
FAMESの詳細な分析から、PAS2発現菌株JMY1781が新しい脂肪酸を蓄積させることが明らかになった。質量分析は、脂質が偶数または奇数のアシル鎖を有するモノメチル分岐脂肪酸であることを判定した。JMY1781は、詳細には、cl4:0(Me)、cl5:0(Me)、cl6:0(Me)、cl7:0(Me)、cl8:0(Me)ならびにcl9:0(Me)の存在を示した。化合物は、野生型Po1d菌株だけでなく肥満菌株JMY1367においてもほとんど検出不可能であった。テトラデカン酸、12メチル、メチルエステル、14:0(Me)は、高レベルで蓄積するように思われた(少なくともオクタデカン酸、メチルエステル、c18:0のレベルまで)。10.7分、14.4分、18.4分および23分でのピークのように、JMY1781菌株中に複数の他の生成物が蓄積した。
【0059】
考察
Yarrowia lipolyticaにおけるArabidopsis由来の3−ヒドロキシアシル−CoAデヒドラターゼPASTICCINOの発現は、油脂化学バイオテクノロジーのための細胞工場としてのこの酵母の使用に関するいくつかの革新的形質を導いた。
【0060】
1− PAS2Ylの発現は、二つの異なるYarrowia菌株すなわち野生型Po1dならびに細胞内部における脂肪酸の高い蓄積を特徴とするΔgutΔpox1−6における油体数を変化させる。脂肪体数の減少によってVLCFAの蓄積が損なわれることはない。脂肪体数の減少は、プレス処理を介した油の抽出を改善するかもしれない。可能性としては、PAS2は、脂質分泌を変化させ得る。
2− PAS2Ylの発現は、VLCFA蓄積の非常に有意な増大をひき起こす。工業的生産に直接用いることのできるVLCFAのレベルは、当業者により公知のものなどのエロンガーゼ複合体の他の遺伝子をYarrowia sp.中で同時発現させることによって得られるはずである。Arabidopsis遺伝子の発現は、YarrowiaにおけるVLCFAホメオスタシスを変更するために効果的であることから、我々は、植物由来の他のエロンガーゼ遺伝子を使用することを提案する。
【0061】
【表1】

【0062】
【表2】
【0063】
参考文献
・Bach L、Michaelson LV、Haslam R、Bellec Y、Gissot L、Marion J、Da Costa M、Boutin JP、Miquel M、Tellier F、Domergue F、Markham JE、Beaudoin F、Napier JA、Faure JD.(2008):「The very−long−chain hydroxy fatty acyl−CoA dehydratase PASTICCINO2 is essential and limiting for plant development」、Proc Nati Acad Sci USA.2008 Sep23;105(38):14727−31
・BarthおよびGaillardin:「Yarrowia lipolytica」、「Nonconventional Yeasts in Biotechnology」、A Handbook(Wolf,K.、Ed.)、Vol.1、1996、pp.313−388.Springer−Verlag)中。
・Beopoulos,A.、Mrozova,Z.、Thevenieau,F.、Le Dall,M.T.、Hapala,I.、Papanikolaou,S.、Chardot,T.、およびNicaud,J.M.(2008)。「Control of lipid accumulation in the yeast Yarrowia lipolytica」、Appl Environ Microbiol 74、7779−7789。
・Beopoulos,A.、Cescut,J.、Haddouche,R.、Uribelarrea,J.L.、Molina−Jouve,C.、およびNicaud,J.M.(2009)。「Yarrowia lipolytica as a model for bio−oil production」、Prog Lipid Res 48、375−387。
・Blanchin−Rolandら、「Two Upstream Activation Sequences Control the Expression of the XPR2 Gene in the Yeast Yarrowia lipolytica」、Molecular and Cellular Biology、vol.14(l):327−338、1994。
・Denic V、Weissman JS.(2007):「A molecular caliper mechanism for determining very long−chain fatty acid length」、Cell.2007 Aug 24; 130(4):663−77。
・Fickers P.、Le Dall M.T.、Gaillardin C.Thonart P.Nicaud J−M.(2003)「New disruption cassettes for rapid gene disruption and marker rescue in the yeast Yarrowia lipolytica」、J.Microbiol.Methods 55/3:727−737。
・Juretzek T、Wang H.、Nicaud,J−M.、Mauersberger S、Barth G.(2000)、「Comparison of promoters suitable for regulated overexpression of β−galactosidase in the alkane− utilizing yeast Yarrowia lipolytica」、Biotechnol.Bioprocess Eng.5:320−326。
・Leblond,Y.、A.Marty、N.Mouz、およびJ.L.Uribelarrea.2009、「Method for producing lipase、transformed Yarrowia lipolytica cell capable of producing said lipase and their uses」。
・Madzak,C、Gaillardin,C.およびBeckerich,J.M.(2004)。J.Biotechnol.109:63−81。(概説)。
・Madzak,C、Treton,B.およびBlanchin−Roland,S.(2000)。J.Mol.Microbiol.Biotechnol.2:207−216。
・Mauersberger,S.、Nicaud,J−M(2002)、Chapter56、「Tagging of genes by insertional mutagenesis in the yeast Yarrowia lipolytica」、「Laboratory Manual on Non−conventional Yeasts in Genetics」Biochemistry and Biotechnology中。
・Nicaud J−M.、Fabre E.、Gaillardin C.(1989)、「Expression of invertase activity in Yarrowia lipolytica and its use as a selective marker」、Cur.Genet.16:253−260。
・Nicaud,J−M、Madzak,C.、van den Broek,P.、Gysler,C.、Duboc,P.、Niederberger,P.、Gaillardin,C.(2002)、「Protein expression and secretion in the yeast Yarrowia lipolytica」、FEMS Yeast Research 2/3:371−379。
・Wolf K、Breunig K、Barth G(編)、Springer、Berlin Heidelberg−New York、pp343−356。(Buchbeitrag)。
【先行技術文献】
【特許文献】
【0064】
【特許文献1】仏国特許出願第08/54786号明細書
【図1(A)−1】
【図1(A)−2】
【図1(B)】
【技術分野】
【0001】
本発明は、発酵による超長鎖脂肪酸(VLCFA)の生産方法に関するものであり、該方法は、Yarrowia sp.における異種遺伝子の発現を可能にする調節エレメントの制御下でヒドロキシアシル−CoAデヒドラターゼをコードする前記異種遺伝子を含むYarrowia sp.の組換え株を培養するステップを含む。
【0002】
本発明は、この組換え型Yarrowia sp.にも関するものである。
【背景技術】
【0003】
生命体は、複合脂質内に取り込まれるおびただしい種類の異なる脂肪酸を合成する。これらの複合脂質は、主要な構造的構成要素膜であると同時に、植物および動物の両方における主要な貯蔵物質である。
【0004】
超長鎖脂肪酸(VLCFA)は真核細胞の構成要素であり、炭素20個以上の長さで構成されている(すなわち >C18)。VLCFAは、異なる生物内で多くの異なる生理学的機能に関与している。これらは、脳(ミエリン)または植物種子(貯蔵トリアシルグリセロール、TAG)などの一部の組織の豊富な構成成分である。VLCFAは、皮膚の脂質バリアおよび植物のクチクラワックスの構成要素である。一部のVLCFAの長いアシル鎖は、例えば核孔内に見られる高い膜湾曲に必要なものである。同様に、VLCFAは、タンパク質輸送およびGPI脂質アンカー合成のための分泌経路にも関与する。最後に、VLCFAは、膜構成成分であると同時にシグナル伝達分子でもあるスフィンゴ脂質の構成要素である。
【0005】
VLCFAは、C18より長いアシル鎖を有する脂肪酸である。多価不飽和VLCFAは、主としてエイコサペンタエン酸(EPA)またはドコサヘキサエン酸(DHA)のように人間の食生活に重要な栄養成分とみなされている。国際公開第2005/118814号パンフレットは、Saccharomyces cerevisiaeにおける多価不飽和脂肪酸の産生を改善する方法を開示している。不飽和VLCFAは、洗浄剤または潤滑剤としても作用することから、工業的にも有益である。
【0006】
VLCFAは、四つの相異なる酵素(KCS、KCR、HCDおよびECR)を含む膜結合酵素複合体であるエロンガーゼ複合体という名称のタンパク質複合体の内部で小胞体内に集められた四つの連続する酵素反応を通して、二個の炭素の逐次的付加によって合成される。脂肪伸長の第一のステップは、3−ケトアシル−CoAシンターゼ(KCSまたは縮合酵素)による、マロニル−CoAを用いた長鎖アシル−CoAの縮合である。結果として得られた3−ケトアシル−CoAは、次に3−ケトアシル−CoAレダクターゼ(KCR)により還元されて、3−ヒドロキシアシル−CoAを生成する。第三のステップは3−ヒドロキシアシル−CoAデヒドラターゼ(HCD)により3−ヒドロキシアシル−CoAをトランス−2,3−エノイル−CoAへと脱水するステップであり、このトランス−2,3−エノイル−CoAは最終的にトランス−2,3−エノイル−CoAレダクターゼ(ECR)により還元されて二炭素伸長されたアシル−CoAを生成する。最後の三つの酵素は、アシル−CoAの特異性に関与しないことから、コア酵素と呼ばれる。アシル−CoAがひとたびエロンガーゼ複合体から伸長されると、これらは、リン脂質、トリアシルグリセロール、スフィンゴ脂質、および植物エピクチクラワックスなどの特異的脂質といった異なる脂質クラスの中に取り込まれ得る。
【0007】
Yarrowia lipolyticaは、この酵母がその乾燥重量の50%超を脂質として蓄積できるだけでなく、炭素源として脂質を効率良く使用することもできることから、油性酵母とみなされている(Beopoulosら、2009)。そのゲノムの完全な配列決定ならびにこの酵母についての分子遺伝学的手段の開発によって、この生物は脂質蓄積の機序の研究用モデルとなったばかりでなく、油脂化学バイオテクノロジーのための細胞工場にもなった。近年になって、β−酸化に関与するPOX1〜6遺伝子とグルコース−3−ホスフェートデヒトロゲナーゼGUT2の組合せ欠失が、脂質、主として遊離脂肪酸のきわめて高い蓄積を導くことが示された(Beopoulosら、2008、FR0854786;11 2008年7月11日)。この肥満菌株は、それぞれグルコースまたはオレイン酸上で成長させた場合に野生型に比べて二倍および三倍多い脂肪酸を蓄積した。興味深いことに、これらの脂質は、単一の大きな脂肪体に蓄積した。Yarrowia lipolyticaは、主として長鎖脂肪酸c18:2、c18:1(n−9)、c16:1(n−7)およびc16:0を蓄積する。しかしながら、Y.lipolyticaにおける超長鎖脂肪酸(VLCFA)については、わずかな情報しかない。
【0008】
現在、Yarrowia sp.、特にYarrowia lipolyticaにおいてヒドロキシアシル−CoAデヒドラターゼをコードする異種遺伝子を発現させることで、菌株による脂肪酸およびVLCFAの産生に対して、品質および/または量の観点において直接的影響が及ぼされることが発見されている。
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は、Yarrowia sp.におけるヒドロキシアシル−CoAデヒドラターゼをコードする異種遺伝子の発現を可能にする調節エレメントの制御下で、前記異種遺伝子を含む、Yarrowia sp.の組換え株に関するものである。
【0010】
ヒドロキシアシル−CoAデヒドラターゼをコードする遺伝子は、詳細には、ヒドロキシアシル−CoAデヒドラターゼをコードする植物種の遺伝子、その機能的相同体およびフラグメントからなる群から選択される。植物種の遺伝子は、有利には、Arabidopsis thaliana、Vitis vinifera、Oryza sativa、Brassica rapa、Hyacinthus orientalis、Ostreacoccus lucimarinus、Chlamydomonas reinhardtii、Brassica napus、Raphanus sativus、およびBrassica oleracea由来のヒドロキシアシル−CoAデヒドラターゼをコードする遺伝子から選択され、より詳細には、Arabidopsis thaliana由来の遺伝子PAS2である。
【0011】
本発明は同様に、発酵による超長鎖脂肪酸(VLCFA)の生産方法にも関するものであり、該方法は、適切な培養培地内で本発明の組換え株を培養するステップと、菌株および/または培地からVCLFAを回収するステップとを含む。
【0012】
前記方法により生産される脂肪酸も、同じく本発明の一部である。
【課題を解決するための手段】
【0013】
本発明は、Yarrowia sp.の組換え株に関するものであり、該組換え株は、Yarrowia sp.におけるヒドロキシアシル−CoAデヒドラターゼをコードする異種遺伝子の発現を可能にする調節エレメントの制御下にある、前記異種遺伝子を含む。
【0014】
組換え株
本発明によると、菌株は、それが遺伝子置換またはプラスミド導入などの細胞生物学的手段によって遺伝的に修飾されている場合に、組換え型となる。これは、遺伝子内に新しい遺伝子もしくは変異もしくは新しい調節エレメントを導入するため、または内因性遺伝子を欠失させるための定方向突然変異誘発により得られるであろう。組換え型微生物は、ランダム突然変異誘発の唯一の結果ではない。
【0015】
新しい遺伝子が菌株内に導入される場合、それは、発現プラスミドと共に導入されてよく、あるいは、その菌株のゲノム内に組込まれてもよい。
【0016】
菌株のゲノム内に組込まれる場合、遺伝子は、相同的組換え技術などの公知の遺伝子置換方法によって特異的部位にまたは無作為に組込まれてよい。
【0017】
異種遺伝子は、導入される場合、Yarrowia sp.におけるこの異種遺伝子の発現を可能にする調節エレメントの制御下のコード配列を含むことができる。代替的には、異種遺伝子は、既存の内因性調節エレメントの制御下で微生物のゲノム内に導入されて、欠失させられる対応する内因性コード配列から置き換わる、コード配列を含むことができる。
【0018】
特に新しい遺伝子を導入するかまたは遺伝子を欠失させるためのYarrowia sp.の修飾方法は、当該技術分野において公知であり、BarthおよびGaillardin(1996)ならびにFickersら、(2003)がある。
【0019】
ヒドロキシアシル−CoAデヒドラターゼをコードする異種遺伝子
「ヒドロキシアシル−CoAデヒドラターゼ」(HCD)という用語は、3−ヒドロキシアシル−CoAをトランス−2,3−エノイル−CoAに脱水する反応を触媒する酵素を意味する。それは、ヒドロリアーゼのファミリーに属する。この酵素は、エロンガーゼ複合体の一部を成し、植物内のVLCFAの合成のみに関与し、その分解には関与しない。
【0020】
ヒドロキシアシル−CoAデヒドラターゼをコードする遺伝子は、異種遺伝子である。本発明によると、遺伝子は、天然の菌株中で同一のものが発見されない場合、異種である。それは、異種調節エレメントの制御下の天然コード配列であるかまたは天然調節エレメントの制御下の異種コード配列であり得る。それは同様に、修飾されるべき菌株中には見られるものの未修飾菌株中に同じ遺伝子が発見されないプラスミド上またはゲノム内遺伝子座には見られない天然の構成要素を有する遺伝子でもあり得る。
【0021】
ヒドロキシアシル−CoAデヒドラターゼをコードする異種遺伝子は、特に、ヒドロキシアシル−CoAデヒドラターゼをコードする植物種の遺伝子由来のコード配列を含む遺伝子、その機能的相同体およびフラグメントからなる群から選択される。
【0022】
ヒドロキシアシル−CoAデヒドラターゼをコードする植物種の遺伝子は、当該技術分野において公知であり、Vitis vinifera(CAN64341.1仮想タンパク質をコードする)、Oryza sativa(CAD39891.2、EAY72548.1仮想タンパク質OsI_000395、EAZ30025.1仮想タンパク質OsJ_013508およびBAD61107.1チロシンホスファターゼ様)、Brassica rapa(AAZ66946.1)、Hyacinthus orientalis(AAT08740.1タンパク質チロシンホスファターゼ)、Ostreacoccus lucimarinus(XP_001420997.1推定タンパク質およびXP_001422898.1推定タンパク質)、Chlamydomonas reinhardtii(EDP01055.1推定タンパク質)、および同様にBrassica napus、Raphanus sativus、Brassica oleracea由来の遺伝子の中から特に選択されたものを含む。
【0023】
本発明の好ましい実施形態において、異種遺伝子は、UniGeneデータバンクにNP_196610.2の番号で登録済であり、F12B17.170、F12B17_170、PASTICCINO 2、PEP、およびPEPINOとしても同様に知られている、Arabidopsis thaliana由来の遺伝子PAS2(Bachら、2008)である。
【0024】
本発明の別の具体的実施形態において、異種遺伝子は、遺伝子データバンクにNP_012438.1の番号で登録済である、Saccharomyces cerevisiae由来のPHS1遺伝子(Denicら、2007)、その機能的相同体およびフラグメントである。
【0025】
異種遺伝子のコード配列が別のものに由来する場合、このコード配列は実際、Yarrowia sp.について公知の好ましいコドン使用頻度でこれを再度コードすることができる。当業者であれば、Yarrowia sp.において使用される好ましいコドンおよびこのような再コードされたコード配列の調製方法を知っている。
【0026】
本発明によると、「機能的相同体」とは、ヒドロキシアシル−CoAデヒドラターゼをコードする異種遺伝子と相同性を共有する遺伝子、または、ヒドロキシアシル−CoAデヒドラターゼをコードする異種遺伝子によりコードされるタンパク質と相同性を共有するタンパク質をコードする遺伝子である。
【0027】
ヒドロキシアシル−CoAデヒドラターゼをコードする遺伝子によりコードされるタンパク質と相同性を共有するタンパク質は、植物から得られるか、または、植物に由来する天然タンパク質のバリアントもしくは機能的フラグメントであってよい。
【0028】
「天然タンパク質のバリアントまたは機能的フラグメント」という用語は、ポリペプチドのアミノ酸配列が、天然に観察される配列に厳密に限定されなくてよく、追加のアミノ酸を含んでいてよい、ということを意味する。「フラグメント」という用語は、ポリペプチドの配列が、もとの配列よりもアミノ酸を少なく含むがヒドロキシアシル−CoAデヒドラターゼ活性を付与するのに充分なアミノ酸を依然として含んでいてよいことを意味する。ポリペプチドが、その酵素活性を保持しながら、一つ以上のアミノ酸の置換、挿入、欠失および/または付加により修飾可能であることは、当該技術分野において周知である。例えば、タンパク質の機能的特性に影響を及ぼさない、化学的に等価のアミノ酸による所与の位置にある一つのアミノ酸の置換が一般的である。本発明に関しては、置換は、以下の群のうちの一つの中での交換として定義される:
・小さい脂肪族の非極性またはわずかに極性の残基:Ala、Ser、Thr、Pro、Gly
・極性の、負荷電残基およびそのアミド:Asp、Asn、Glu、Gln
・極性の、正荷電残基:His、Arg、Lys
・大きい脂肪族の非極性残基:Met、Leu、Ile、Val、Cys
・大きい芳香族残基:Phe、Tyr、Trp。
【0029】
したがって、一つの負荷電残基での別の残基の置換(例えばグルタミン酸でアスパラギン酸を置換)または一つの正荷電残基での別の残基の置換(例えばリジンでアルギニンを置換)を結果としてもたらす変化が、機能的に等価の生成物を生成すると予想することができる。
【0030】
アミノ酸配列内の、アミノ酸が修飾される位置および修飾を受けるアミノ酸の数は、特に限定されない。当業者であれば、タンパク質の活性に影響を及ぼすことなく導入可能な修飾を認識することができる。例えば、タンパク質のNまたはC末端部分での修飾は、一部の状況下でタンパク質の活性を改変しないものと予想され得る。
【0031】
「バリアント」という用語は、もとの酵素活性をなおも保持しながら、以上で定義したような修飾に付されたポリペプチドを意味する。
【0032】
本発明によると、ヒドロアシル−CoAデヒドラターゼ酵素活性を有するポリペプチドは、PAS2の配列と少なくとも30%の相同性、好ましくは少なくとも50%の相同性、そしてより好ましくは少なくとも70%の相同性を有する配列を含んでいてよい。
【0033】
二つのタンパク質配列間の相同性百分率の決定方法は、当業者にとって公知である。例えば、これは、http://www.ebi.ac.uk/clustalw/のウェブサイト上で入手可能なソフトウェアCLUSATALWを、このウェブサイト上に記されたデフォルトパラメータと共に使用することによって、配列のアラインメントの後に行なうことができる。残基の総数と比較した同じ位置における同一の残基の数を記録することにより、アラインメントから同一性百分率を容易に計算することができる。代替的には、例えば、http://www.ncbi.nlm.nih.gov/BLAST/のウェブサイト上で入手可能なBLASTプログラムをそのウェブサイト上に記されたデフォルトパラメータと共に使用することによって、自動計算を行なうことも可能である。
【0034】
Yarrowia sp.における異種遺伝子の発現を可能にする調節エレメント
このような調節エレメントは、当該技術分野において周知であり、アシル−CoAオキシダーゼ2由来のPOX2プロモーター、イソシトレートデヒドロゲナーゼ由来のICLプロモーター、プロモーターHp4d、プロモーターGPDおよびGPM、プロモーターFBP、ならびにプロモーターXPR2を含む。前記複数のプロモーターは当該技術分野において公知であり、なかでもJuretzekら(2000)、Madzakら(2004)、Madzakら(2000)、米国特許第7259255号明細書、米国特許第7202356号明細書およびBlanchin−Rolandら(1994)中で開示されている。
【0035】
Yarrowia sp.
本発明によると、Yarrowia sp.の任意の菌株を形質転換し、本発明の方法において使用してよい。好ましくは、Yarrowia sp.の菌株はYarrowia lipolytica属に属する。
【0036】
Yarrowia lipolytica属の菌株は、この菌株の形質転換方法と同様、当該技術分野において周知である。対象となるコード領域を含む構築物を、任意の標準的技術により宿主細胞中に導入してよい。これらの技術には、形質転換(例えば酢酸リチウム形質転換(Methods in Enzymology,194:186−187(1991)))、原形質融合、遺伝子銃衝撃、電気穿孔法、マイクロインジェクション、または対象となる遺伝子を宿主細胞内に導入する任意の他の方法が含まれる。油性酵母(すなわちYarrowia lipolytica)に適用可能なより具体的な教示としては、米国特許第4880741号明細書および米国特許第5071764号明細書が含まれる。
【0037】
参照として本明細書に援用されている、脂質、主として遊離脂肪酸の非常に高い蓄積を有する菌株(仏国特許出願第08/54786号明細書;2008年7月11日)のように、改良された脂肪酸生産のために修飾された菌株も同様に開示されている。このような菌株は、ヒドロキシアシル−CoAデヒドラターゼをコードする異種遺伝子を用いて本発明にしたがってさらに修飾されてよい。
【0038】
また、本発明の組換え株は、脂肪酸のβ−酸化に関与する少なくとも一つの遺伝子の欠失、特にアシルCoAオキシダーゼをコードする遺伝子POX1〜POX6のうちの一つの欠失、特に六つのアシルCoAオキシダーゼをコードする六つの遺伝子POX1〜6ならびに/または脂肪酸およびTAG合成の経路に関与する一つの遺伝子の欠失、特にグリセロール3−ホスフェートデヒドロゲナーゼをコードする遺伝子の欠失も含み得る。
【0039】
組換え株の培養
脂肪酸、特にVLCFAは、適切な培養培地内で発酵によって組換え株を培養する場合に生産される。
【0040】
発酵による培養とは、微生物が培養培地で発育させられ、培養培地の炭素源を形質転換することによって、この培養ステップ中にVLCFAを産生することを意味する。VLCFAは、バイオマスと共に、細胞内および/または培地内に蓄積される。
【0041】
発酵は、酵素変換プロセスでさらに使用される、酵素を生産するために培養を用いる生物変換反応とは全く異なるものである。
【0042】
適切な培養培地
Yarrowia sp.のための培養培地は、BarthおよびGaillardin(1996)、Nicaudら(2002)およびMauersbergerおよびNicaud(2002)を含め、当該技術分野において周知である。発酵に関して規定された培地は、Leblondら(2009)および韓国特許第2009/0029808号公報中で詳細に開示されている。
【0043】
スクロース培地は、Nicaudら(1989)において詳細に開示されている。
【0044】
適切な培養培地は、Yarrowia sp.が成長でき、かつ菌株の成長およびVLCFAの産生を可能にする全ての栄養素、特に炭素源を含む培地である。
【0045】
炭素源は、スクロースまたは他の炭水化物などの、任意の炭素源であってよい。
【0046】
菌株および/または培地からのVCLFAの回収
VCLFAは、バイオマスと共に菌株中および/または培養培地内に蓄積される。VCFLAの回収には、一般に、細胞溶解、濾過および培地からの回収というステップが含まれる。脂肪酸バイオプロダクションの当業者は、本発明の方法に、バイオマスからの脂肪酸の通常の回収方法をいかに適応させるべきかを知っている。
【0047】
本発明の方法を用いて生産されたVCFLAは、そのままの状態で、本発明の菌株により産生された脂肪酸の混合物中で使用されてよい。これらをさらに精製し単離することもできる。
【図面の簡単な説明】
【0048】
【図1】Yarrowia lipolytica発現のために最適化された合成PAS2を表わす。(A)PAS2Ylの遺伝子およびタンパク質の配列。(B)Arabidopsis PAS2AtとPAS2Ylのアラインメント。
【図2】使用または創出された、異なる菌株の概略図と共に、YarrowiaにおけるPAS2発現を表わす。
【図3】YarrowiaにおけるPAS2発現の効果を表わす。(A)JM1367およびJM1781菌株中でのナイルレッドを用いた脂肪体の染色。(B)Po1dおよびJM1777における細胞一個あたりの脂肪体の数。分布を比較するために、多項回帰を適用した。(C)JM1367およびJM1381における細胞一個あたりの脂肪体の数。分布を比較するために、多項回帰を適用した。
【図4】YarrowiaにおけるPAS2発現を伴う、生産されたLCFA/VLCFAの比を示す。
【図5】YarrowiaにおけるPAS2発現を伴う、修飾されたLCFAおよびVLCFAの含有量を示す。(A)LCFA含有量。(B)VLCFA含有量。
【図6】YarrowiaにおけるPAS2発現を伴う、修飾されたVLCFAのプロファイルを示す。
【実施例】
【0049】
材料と方法
Yarrowia lipolyticaコドン使用頻度にしたがって、合成PAS2遺伝子(PAS2Yl)を合成して、プラスミドJME1107を得た。PASYlをプラスミドJMP62−POX2−URA3ex(JME803)内そしてプラスミドJMP62−TEF−URA3ex(JME1012)内に以下の通りにクローニングした:プラスミドJME1107をBamHI−AvrIIにより消化し、PAS2遺伝子を担持する対応するフラグメントを、プラスミドJME1012およびJME1107の対応する部位でクローニングして、それぞれプラスミドJME1108(POX2−PAS2)およびJME1110(TEF−PAS2)を得た。プラスミドをNotIで消化し、発現カセットを担持するフラグメントを、酢酸リチウム法によるYarrowia lipolyticaの形質転換(G.BarthおよびGaillardinの概説:すなわち「Nonconventional Yeasts in Biotechnology A Handbook」(Wolf,K.、Ed.)、Vol.1、1996、pp.313−388.Springer−Verlag中のYarrowia lipolyticaに記載されている)のために使用した。形質転換体をYNBcasa上で選択した。典型的には、フラグメント1μgあたり約5×103個の形質転換体が得られた。それぞれPOX2−PAS2およびTEF−PAS2を含むクローンのためのプライマー対61start/61stopおよびTEFstart/61stopを用いたPCRにより、4〜8個の形質転換体を分析した。PCR産物をさらに、PAS2遺伝子内のAvaI固有の制限部位により消化した。
【0050】
Yarrowia lipolytica中のPAS2の安定した発現
Arabidopsis PAS2遺伝子のオープンリーディングフレームを再コードして、Yarrowiaコドン使用頻度でのその発現を改善した(図1A〜B)。クローニングを促すため、PAS2 ORFの5’および3’末端に二つの制限部位BamHIおよびAvrIIをそれぞれ付加した。PAS2Ylと命名し直した、新規の配列を化学的に合成し(GeneArt inc.)、二つの発現ベクターJME1110およびJME1108内にクローニングした。その二つのベクターは、構成的プロモーター(pTEF、JME1110)またはオレイン酸誘発性プロモーター(pPOX2、JME1108)の下でのPAS2の発現を可能にする。
【0051】
野生型菌株Po1d(JMY195)およびΔgut2、Δpox1−6肥満菌株(JMY1367)を形質転換するために、両方の構築物を使用した。形質転換体をウラシル上で選択し、発現カセットの組込みをPCRで確認した。さらなる分析のために、複数のクローンを選択し使用した。しかしながら、POX2プロモーターはインデューサーの不在下でも強い発現を可能にすることから、我々は主として、JME1108構築物を用いて形質転換体を特徴づけした。菌株JMY1777およびJMY1778は、pPOX2−PAS2で形質転換されたPo1dの二つの独立したクローンである。同様に、JMY1781およびJMY1782は、pPOX2−PAS2で形質転換されたJMY1367(Δgut Δpox1−6)の二つの独立したクローンである。
【0052】
PAS2発現は細胞の成長を改善し、脂肪体の断片化を導く
複数の異なるPSA2発現菌株の成長を、グルコースを補足した培地上でそれらの未形質転換相対物と比較した。全ての菌株を、30mlのYPD培地中OD600=0.6で植菌した。全ての菌株は、植菌後、40時間の培養後に静止期の開始に達するまでの間に、約4〜5時間の遅滞期、および9〜12時間の平担域を伴う二峰性曲線を示した。
【0053】
Yarrowia lipolyticaは、脂肪体内で脂質を蓄積するものとして知られている。肥満菌株JMY1367はより大きい構造まで脂肪体を融合することを特徴とすることが報告された。こうして、脂肪体の構造に対するPAS2発現の効果を、ナイルレッドによる複数の異なる菌株の染色によって調べた(図3A)。静止段階は、脂質の高い蓄積度を特徴とすることから、48時間後に細胞を収集し、細胞一個あたりの脂肪体の総数を定量した(図3B〜C)。
【0054】
Po1d内ならびに肥満菌株JM1367内の両方におけるPAS2の発現は、脂肪体数の減少を導く(図3B〜C)。
【0055】
PAS2の発現がVLCFAレベルを増強する
成長曲線の三つの異なる時点、すなわち第一成長期の終り(11時間)、第二成長期の終り(24時間)および静止期中(48時間)に、四つの菌株中で、脂肪酸アシルメチルエステル(FAMES)の脂質総含有量をガスクロマトグラフィにより分析した。予想通り、肥満菌株JMY1367は、24時間後と48時間後にそれぞれ22%および40%という、野生型Po1dに比べて高い脂肪酸含有量を有する(図4)。PAS2の発現は、全ての時点において脂肪酸総含有量を減少させた。最も強い減少は、48時間後に観察され、Po1dバックグラウンドで20%、JMY1367バックグラウンドで46%であった。
【0056】
Yarrowia lipolyticaの最も豊富な脂肪酸に相当する長鎖脂肪酸(LCFA)の全量は、PAS2発現菌株において18%および36%減少した。LCFAの分析から、最も豊富なLCFAの一つであるc18:1が最も影響を受け、肥満菌株JMY1367バックグラウンドにおいて48時間後に例えば126%の減少であったことを除いて、異なるクラス全てがPAS2の発現時点で減少したレベルを示していることがわかった(図5)。合計LCFAの減少は、成長曲線の始め(11時間)においてさえ有効であった。
【0057】
VLCFAは、Yarrowia lipolyticaにおけるわずかな脂質種に相当するにすぎない(総脂肪酸の2.2〜3.2%)(図4)。三つの主要な種、すなわち、それぞれ48時間の培養時点で総脂肪酸の0.86、0.77および0.34%(モル%)に相当する24:0、20:1および22:1が有意に蓄積された(図6)。Δgut Δpox1−6は、24時間の培養において倍増した(Po1dにおいて、3.23に比べ6.53μg/10OD)ことから、VLCFAレベルに対し明らかな効果を有していた。関与した主要なVLCFAはc24:0およびc22:1であり、その含有量は総脂肪酸のそれぞれ2.09%と0.99%(モル%)に達した。新しいVLCFAを、0.45%に達するc22:0として検出することができた。Po1dバックグラウンドでのPAS2の発現は、蓄積したVLCFAの量または性質をさほど変化させなかった。しかしながら、肥満菌株JMY1367バックグラウンドでのPAS2の発現は、VLCFAの含有量を極めて有意に増大させた。24時間の培養後、JMY1781は、17.35μg/10ODを蓄積し、これは肥満菌株および野生型Po1d株よりそれぞれ2.65倍および5.3倍多いものであった。蓄積した主要なVLCFAは、c20:0およびc24:0であり、総VLCFAの半分超を占めていた。JMY1781では、エルカ酸c22:1、c22:0、c20:2も同様に有意に蓄積した。
【0058】
PAS2発現は、新しいモノメチル分岐脂肪酸の蓄積を誘発する
FAMESの詳細な分析から、PAS2発現菌株JMY1781が新しい脂肪酸を蓄積させることが明らかになった。質量分析は、脂質が偶数または奇数のアシル鎖を有するモノメチル分岐脂肪酸であることを判定した。JMY1781は、詳細には、cl4:0(Me)、cl5:0(Me)、cl6:0(Me)、cl7:0(Me)、cl8:0(Me)ならびにcl9:0(Me)の存在を示した。化合物は、野生型Po1d菌株だけでなく肥満菌株JMY1367においてもほとんど検出不可能であった。テトラデカン酸、12メチル、メチルエステル、14:0(Me)は、高レベルで蓄積するように思われた(少なくともオクタデカン酸、メチルエステル、c18:0のレベルまで)。10.7分、14.4分、18.4分および23分でのピークのように、JMY1781菌株中に複数の他の生成物が蓄積した。
【0059】
考察
Yarrowia lipolyticaにおけるArabidopsis由来の3−ヒドロキシアシル−CoAデヒドラターゼPASTICCINOの発現は、油脂化学バイオテクノロジーのための細胞工場としてのこの酵母の使用に関するいくつかの革新的形質を導いた。
【0060】
1− PAS2Ylの発現は、二つの異なるYarrowia菌株すなわち野生型Po1dならびに細胞内部における脂肪酸の高い蓄積を特徴とするΔgutΔpox1−6における油体数を変化させる。脂肪体数の減少によってVLCFAの蓄積が損なわれることはない。脂肪体数の減少は、プレス処理を介した油の抽出を改善するかもしれない。可能性としては、PAS2は、脂質分泌を変化させ得る。
2− PAS2Ylの発現は、VLCFA蓄積の非常に有意な増大をひき起こす。工業的生産に直接用いることのできるVLCFAのレベルは、当業者により公知のものなどのエロンガーゼ複合体の他の遺伝子をYarrowia sp.中で同時発現させることによって得られるはずである。Arabidopsis遺伝子の発現は、YarrowiaにおけるVLCFAホメオスタシスを変更するために効果的であることから、我々は、植物由来の他のエロンガーゼ遺伝子を使用することを提案する。
【0061】
【表1】
【0062】
【表2】
【0063】
参考文献
・Bach L、Michaelson LV、Haslam R、Bellec Y、Gissot L、Marion J、Da Costa M、Boutin JP、Miquel M、Tellier F、Domergue F、Markham JE、Beaudoin F、Napier JA、Faure JD.(2008):「The very−long−chain hydroxy fatty acyl−CoA dehydratase PASTICCINO2 is essential and limiting for plant development」、Proc Nati Acad Sci USA.2008 Sep23;105(38):14727−31
・BarthおよびGaillardin:「Yarrowia lipolytica」、「Nonconventional Yeasts in Biotechnology」、A Handbook(Wolf,K.、Ed.)、Vol.1、1996、pp.313−388.Springer−Verlag)中。
・Beopoulos,A.、Mrozova,Z.、Thevenieau,F.、Le Dall,M.T.、Hapala,I.、Papanikolaou,S.、Chardot,T.、およびNicaud,J.M.(2008)。「Control of lipid accumulation in the yeast Yarrowia lipolytica」、Appl Environ Microbiol 74、7779−7789。
・Beopoulos,A.、Cescut,J.、Haddouche,R.、Uribelarrea,J.L.、Molina−Jouve,C.、およびNicaud,J.M.(2009)。「Yarrowia lipolytica as a model for bio−oil production」、Prog Lipid Res 48、375−387。
・Blanchin−Rolandら、「Two Upstream Activation Sequences Control the Expression of the XPR2 Gene in the Yeast Yarrowia lipolytica」、Molecular and Cellular Biology、vol.14(l):327−338、1994。
・Denic V、Weissman JS.(2007):「A molecular caliper mechanism for determining very long−chain fatty acid length」、Cell.2007 Aug 24; 130(4):663−77。
・Fickers P.、Le Dall M.T.、Gaillardin C.Thonart P.Nicaud J−M.(2003)「New disruption cassettes for rapid gene disruption and marker rescue in the yeast Yarrowia lipolytica」、J.Microbiol.Methods 55/3:727−737。
・Juretzek T、Wang H.、Nicaud,J−M.、Mauersberger S、Barth G.(2000)、「Comparison of promoters suitable for regulated overexpression of β−galactosidase in the alkane− utilizing yeast Yarrowia lipolytica」、Biotechnol.Bioprocess Eng.5:320−326。
・Leblond,Y.、A.Marty、N.Mouz、およびJ.L.Uribelarrea.2009、「Method for producing lipase、transformed Yarrowia lipolytica cell capable of producing said lipase and their uses」。
・Madzak,C、Gaillardin,C.およびBeckerich,J.M.(2004)。J.Biotechnol.109:63−81。(概説)。
・Madzak,C、Treton,B.およびBlanchin−Roland,S.(2000)。J.Mol.Microbiol.Biotechnol.2:207−216。
・Mauersberger,S.、Nicaud,J−M(2002)、Chapter56、「Tagging of genes by insertional mutagenesis in the yeast Yarrowia lipolytica」、「Laboratory Manual on Non−conventional Yeasts in Genetics」Biochemistry and Biotechnology中。
・Nicaud J−M.、Fabre E.、Gaillardin C.(1989)、「Expression of invertase activity in Yarrowia lipolytica and its use as a selective marker」、Cur.Genet.16:253−260。
・Nicaud,J−M、Madzak,C.、van den Broek,P.、Gysler,C.、Duboc,P.、Niederberger,P.、Gaillardin,C.(2002)、「Protein expression and secretion in the yeast Yarrowia lipolytica」、FEMS Yeast Research 2/3:371−379。
・Wolf K、Breunig K、Barth G(編)、Springer、Berlin Heidelberg−New York、pp343−356。(Buchbeitrag)。
【先行技術文献】
【特許文献】
【0064】
【特許文献1】仏国特許出願第08/54786号明細書
【図1(A)−1】
【図1(A)−2】
【図1(B)】
【特許請求の範囲】
【請求項1】
Yarrowia sp.の組換え株であって、Yarrowia sp.における異種遺伝子の発現を可能にする調節エレメントの制御下でヒドロキシアシル−CoAデヒドラターゼをコードする前記異種遺伝子を含む組換え株。
【請求項2】
ヒドロキシアシル−CoAデヒドラターゼをコードする遺伝子が、ヒドロキシアシル−CoAデヒドラターゼをコードする植物種の遺伝子、その機能的相同体およびフラグメントからなる群から選択される、請求項1に記載の組換え株。
【請求項3】
植物種の遺伝子が、Arabidopsis thaliana、Vitis vinifera、Oryza sativa、Brassica rapa、Hyacinthus orientalis、Ostreacoccus lucimarinus、Chlamydomonas reinhardtii、Brassica napus、Raphanus sativus、およびBrassica oleracea由来のヒドロキシアシル−CoAデヒドラターゼをコードする遺伝子から選択される、請求項2に記載の組換え株。
【請求項4】
植物種の遺伝子がArabidopsis thaliana由来の遺伝子PAS2である、請求項3に記載の組換え株。
【請求項5】
Yarrowia sp.の菌株がYarrowia lipolytica属に属する、請求項1〜3のいずれか一つに記載の組換え株。
【請求項6】
菌株が、脂肪酸のβ酸化に関与する少なくとも一つの遺伝子の欠失を含む、請求項1〜5のいずれか一つに記載の組換え株。
【請求項7】
グルコース−3−ホスフェートデヒドロゲナーゼをコードする遺伝子および/または遺伝子POX1〜6の欠失を含む、請求項6に記載の組換え株。
【請求項8】
発酵による超長鎖脂肪酸(VLCFA)の生産方法であって、適切な培養培地内で請求項1〜7のいずれか一つに記載の組換え株を培養するステップと、菌株および/または培地からVCLAを回収するステップとを含む生産方法。
【請求項9】
請求項8に記載の方法により得られる、VLCFA組成物。
【請求項1】
Yarrowia sp.の組換え株であって、Yarrowia sp.における異種遺伝子の発現を可能にする調節エレメントの制御下でヒドロキシアシル−CoAデヒドラターゼをコードする前記異種遺伝子を含む組換え株。
【請求項2】
ヒドロキシアシル−CoAデヒドラターゼをコードする遺伝子が、ヒドロキシアシル−CoAデヒドラターゼをコードする植物種の遺伝子、その機能的相同体およびフラグメントからなる群から選択される、請求項1に記載の組換え株。
【請求項3】
植物種の遺伝子が、Arabidopsis thaliana、Vitis vinifera、Oryza sativa、Brassica rapa、Hyacinthus orientalis、Ostreacoccus lucimarinus、Chlamydomonas reinhardtii、Brassica napus、Raphanus sativus、およびBrassica oleracea由来のヒドロキシアシル−CoAデヒドラターゼをコードする遺伝子から選択される、請求項2に記載の組換え株。
【請求項4】
植物種の遺伝子がArabidopsis thaliana由来の遺伝子PAS2である、請求項3に記載の組換え株。
【請求項5】
Yarrowia sp.の菌株がYarrowia lipolytica属に属する、請求項1〜3のいずれか一つに記載の組換え株。
【請求項6】
菌株が、脂肪酸のβ酸化に関与する少なくとも一つの遺伝子の欠失を含む、請求項1〜5のいずれか一つに記載の組換え株。
【請求項7】
グルコース−3−ホスフェートデヒドロゲナーゼをコードする遺伝子および/または遺伝子POX1〜6の欠失を含む、請求項6に記載の組換え株。
【請求項8】
発酵による超長鎖脂肪酸(VLCFA)の生産方法であって、適切な培養培地内で請求項1〜7のいずれか一つに記載の組換え株を培養するステップと、菌株および/または培地からVCLAを回収するステップとを含む生産方法。
【請求項9】
請求項8に記載の方法により得られる、VLCFA組成物。
【図2】

【図3A】

【図3B】
【図3C】
【図4】
【図5】
【図6】
【図3A】
【図3B】
【図3C】
【図4】
【図5】
【図6】
【公表番号】特表2013−511972(P2013−511972A)
【公表日】平成25年4月11日(2013.4.11)
【国際特許分類】
【出願番号】特願2012−540461(P2012−540461)
【出願日】平成22年11月30日(2010.11.30)
【国際出願番号】PCT/EP2010/068541
【国際公開番号】WO2011/064393
【国際公開日】平成23年6月3日(2011.6.3)
【出願人】(505129079)アンスティテュ ナシオナル ドゥ ラ ルシェルシュ アグロノミック (15)
【氏名又は名称原語表記】INSTITUT NATIONAL DE LA RECHERCHE AGRONOMIQUE
【Fターム(参考)】
【公表日】平成25年4月11日(2013.4.11)
【国際特許分類】
【出願日】平成22年11月30日(2010.11.30)
【国際出願番号】PCT/EP2010/068541
【国際公開番号】WO2011/064393
【国際公開日】平成23年6月3日(2011.6.3)
【出願人】(505129079)アンスティテュ ナシオナル ドゥ ラ ルシェルシュ アグロノミック (15)
【氏名又は名称原語表記】INSTITUT NATIONAL DE LA RECHERCHE AGRONOMIQUE
【Fターム(参考)】
[ Back to top ]