
組換え微生物
【課題】タンパク質又はポリペプチドの生産性向上を可能とする宿主微生物、及び当該微生物を用いるタンパク質又はポリペプチドの製造法の提供。
【解決手段】枯草菌の遺伝子yopO、treR、yvbA、cspB、yvaN、licR、sigL、glcT、yvdE、slr、rocR、yycH及びyacPのいずれか1以上の遺伝子が削除又は不活性化された枯草菌又はその他のバチルス属細菌株に、転写開始制御領域、翻訳開始制御領域及び分泌用シグナル領域が上流に結合した異種のタンパク質又はポリペプチドをコードする遺伝子を導入した組換え微生物であって、当該転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域が、配列番号1で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜659の塩基配列、配列番号3で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜696の塩基配列又は当該塩基配列のいずれかと90%以上の同一性を有する塩基配列からなり、且つ転写開始制御機能、翻訳開始制御機能及び分泌用シグナル機能を有するDNA断片である、組換え微生物。
【解決手段】枯草菌の遺伝子yopO、treR、yvbA、cspB、yvaN、licR、sigL、glcT、yvdE、slr、rocR、yycH及びyacPのいずれか1以上の遺伝子が削除又は不活性化された枯草菌又はその他のバチルス属細菌株に、転写開始制御領域、翻訳開始制御領域及び分泌用シグナル領域が上流に結合した異種のタンパク質又はポリペプチドをコードする遺伝子を導入した組換え微生物であって、当該転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域が、配列番号1で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜659の塩基配列、配列番号3で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜696の塩基配列又は当該塩基配列のいずれかと90%以上の同一性を有する塩基配列からなり、且つ転写開始制御機能、翻訳開始制御機能及び分泌用シグナル機能を有するDNA断片である、組換え微生物。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、有用なタンパク質又はポリペプチドの生産に用いる組換え微生物、及びタンパク質又はポリペプチドの生産方法に関する。
【背景技術】
【0002】
微生物による有用物質の工業的生産は、アルコール飲料や味噌、醤油等の食品類をはじめとし、アミノ酸、有機酸、核酸関連物質、抗生物質、糖質、脂質、タンパク質等、その種類は多岐に渡っており、またその用途についても食品、医薬や、洗剤、化粧品等の日用品、或いは各種化成品原料に至るまで幅広い分野に広がっている。
【0003】
こうした微生物による有用物質の工業生産においては、その生産性の向上が重要な課題の一つであり、その手法として、突然変異等の遺伝学的手法による生産菌の育種が行われてきた。特に最近では、微生物遺伝学、バイオテクノロジーの発展により、遺伝子組換え技術等を用いたより効率的な生産菌の育種が行われるようになっており、遺伝子組換えのための宿主微生物の開発が進められている。例えば、枯草菌(Bacillus subtilis) Marburg No.168系統株の様に宿主微生物として安全かつ優良と認められた微生物菌株に更に改良を加えた菌株が開発されている。
【0004】
しかしながら、微生物は元来、自然界における環境変化に対応するための多種多様な遺伝子群を有しており、限定された生産培地が使用されるタンパク質等の工業的生産においては、必ずしも生産効率が高いとは言えない状況であった。
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明は、タンパク質又はポリペプチドの生産性向上を可能とする宿主微生物にタンパク質又はポリペプチドをコードする遺伝子を導入して得られる組換え微生物、更に、当該組換え微生物を用いるタンパク質又はポリペプチドの製造法を提供することを目的とする。
【課題を解決するための手段】
【0006】
本発明者らは、微生物ゲノム上にコードされる各種遺伝子において、有用なタンパク質又はポリペプチドの生産にとって不要或いは有害な働きをする遺伝子群を鋭意探索したところ、枯草菌等の微生物の特定の遺伝子をゲノム上から削除又は不活性化した後、目的のタンパク質又はポリペプチドをコードする遺伝子を導入することにより、目的のタンパク質又はポリペプチドの生産性が、削除又は不活性化前と比較して向上することを見出した。
【0007】
すなわち本発明は、枯草菌の遺伝子comA、yopO、treR、yvbA、cspB、yvaN、yttP、yurK、yozA、licR、sigL、mntR、glcT、yvdE、ykvE、slr、rocR、ccpA、yaaT、yyaA、yycH、yacP、hprK、rsiX、yhdK及びylbOのいずれか、又は当該遺伝子に相当する遺伝子のいずれか1以上の遺伝子が削除又は不活性化された微生物変異株に、異種のタンパク質又はポリペプチドをコードする遺伝子を導入した組換え微生物、特に異種のタンパク質又はポリペプチドをコードする遺伝子の上流に転写開始制御領域、翻訳開始制御領域、又は分泌用シグナル領域を結合した当該組換え微生物、また当該組換え微生物を用いたタンパク質又はポリペプチドの製造方法を提供するものである。
【0008】
また本発明は、枯草菌の遺伝子yopO、treR、yvbA、cspB、yvaN、licR、sigL、glcT、yvdE、slr、rocR、yycH及びyacPのいずれか1以上の遺伝子が削除又は不活性化された枯草菌又はその他のバチルス属細菌株に、転写開始制御領域、翻訳開始制御領域及び分泌用シグナル領域が上流に結合した異種のタンパク質又はポリペプチドをコードする遺伝子を導入した組換え微生物であって、当該転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域が、配列番号1で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜659の塩基配列、配列番号3で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜696の塩基配列又は当該塩基配列のいずれかと90%以上の同一性を有する塩基配列からなり、且つ転写開始制御機能、翻訳開始制御機能及び分泌用シグナル機能を有するDNA断片である、組換え微生物、また当該組換え微生物を用いた異種タンパク質又はポリペプチドの製造方法を提供するものである。
一態様において、前記異種のタンパク質又はポリペプチドは、バチルス属細菌由来のアルカリセルラーゼ又はアルカリアミラーゼである。
【発明の効果】
【0009】
本発明の微生物は、目的タンパク質又はポリペプチドの生産にとって不要、或いは有害な遺伝子が削除、又は不活性化されているため、エネルギーロス、副産物の生産や比生産速度の低下等、培地の浪費が大幅に減少でき、また、タンパク質又はポリペプチドの生産期間が長期化することによって効率よく目的生産物を生産することができる。
【図面の簡単な説明】
【0010】
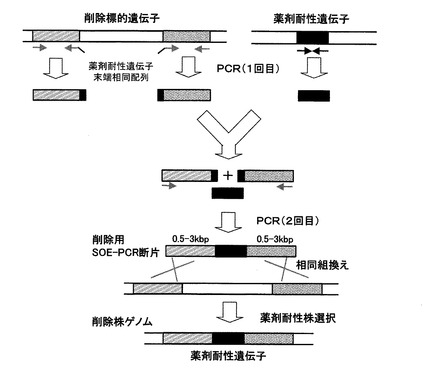
【図1】図1は、SOE−PCRによる遺伝子削除用DNA断片の調製、及び当該DNA断片を用いて標的遺伝子を削除(薬剤耐性遺伝子と置換)する方法を模式的に示したものである。
【発明を実施するための形態】
【0011】
本発明においてアミノ酸配列および塩基配列の同一性はLipman-Pearson法 (Science, 227, 1435, (1985))によって計算される。具体的には、遺伝情報処理ソフトウェアGenetyx-Win(ソフトウェア開発)のホモロジー解析(Search homology)プログラムを用いて、Unit size to compare(ktup)を2として解析を行うことにより算出される。
【0012】
本発明の微生物を構築するための親微生物としては、有用なタンパク質又はポリペプチドの生産にとって不要な遺伝子、具体的には表1に示す枯草菌の遺伝子又は当該遺伝子に相当する遺伝子を有するものであればよく、野生型のものでも変異を施したものでものよい。具体的には、枯草菌などのバチルス(Bacillus)属細菌や、クロストリジウム(Clostridium)属細菌、或いは酵母等が挙げられ、中でもバチルス(Bacillus)属細菌が好ましい。更に、全ゲノム情報が明らかにされ、遺伝子工学、ゲノム工学技術が確立されている点、またタンパク質と菌体外に分泌生産させる能力を有する点から特に枯草菌が好ましい。
【0013】
本発明の微生物を用いて生産する目的タンパク質又はポリペプチドとしては、例えば食品用、医薬品用、化粧品用、洗浄剤用、繊維処理用、医療検査薬用等として有用な酵素や生理活性因子等のタンパク質やポリペプチドが挙げられる。
【0014】
例えば、枯草菌にはゲノム上に4106個の遺伝子が存在することが知られているが、本発明において削除、又は不活性の対象となる遺伝子群は、目的タンパク質又はポリペプチドの生産にとって不要或いは有害な働きをする遺伝子群であり、表1に示される枯草菌の遺伝子のいずれか、又は当該遺伝子に相当する遺伝子群の中から選択されるものである。斯かる遺伝子群は、目的タンパク質又はポリペプチドの生産には直接関与しておらず、また、通常の工業的生産培地における微生物の生育にも不要であることが本発明者らによって見出された。
【0015】
尚、表中の各遺伝子の名称、番号及び機能等は、Nature, 390, 249-256, (1997) で報告され、JAFAN: Japan Functional Analysis Network for Bacillussubtilis (BSORF DB)でインターネット公開(http://bacillus.genome.ad.jp/、2003年6月17日更新)された枯草菌ゲノムデーターに基づいて記載している。
【0016】
【表1】

【0017】
また、表1に示される枯草菌の各遺伝子と同じ機能を有する、または、表1の各遺伝子と塩基配列において70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有する、他の微生物由来、好ましくはバチルス属細菌由来の遺伝子は、表1に記載の遺伝子に相当する遺伝子と考えられ、本発明において削除、不活性化すべき遺伝子に含まれる。尚、塩基配列の同一性はLipman-Pearson法 (Science, 227, 1435, (1985))によって計算される。
【0018】
表1に示される枯草菌の遺伝子群の中には、各種遺伝子発現の活性化や抑制に関わる制御遺伝子、或いは構造の類似性から制御遺伝子であろうと推定されるものが多く存在していることから、タンパク質又はポリペプチドの生産にとって不要或いは有害な遺伝子制御の存在が本発明によって明らかになった。
【0019】
特に、グルコースのPTS取り込み系オペロンのアンチターミネーターであるglcT遺伝子、リケナン分解系オペロンのアンチターミネーターであるlicT遺伝子、トレハロース取り込み、代謝系オペロンのリプレッサーであるtreR遺伝子、更には、グルコースカタボライトリプレッションに関わるccpA遺伝子やhprK遺伝子など、糖の取り込み、代謝に関わる制御遺伝子が多く存在しており、注目に値する。
【0020】
その他、糖の取り込み、代謝に関わる制御遺伝子以外の制御遺伝子として、アルギニン資化の活性化に関与するrocR遺伝子やコンピテンス関連の制御遺伝子comA、slr等を削除又は不活性化することによりタンパク質又はポリペプチドの生産性が向上する。
【0021】
また、表1に示される遺伝子群の中には、ECFシグマ因子のひとつであるシグマXの発現を抑制するアンチECFシグマ因子をコードするrsiX遺伝子、及び同じくシグマMの発現抑制に関与するとの報告例があるyhdK遺伝子(Mol Microbiol., 32, 41, (1999))などが存在し、また逆にシグマLをコードするsigL遺伝子が含まれており、シグマXやシグマMの制御下にある遺伝子の発現がタンパク質生産にとって好ましく、逆にシグマL制御下の何らかの遺伝子発現がタンパク質生産にとって好ましくないことが示唆されている。
【0022】
斯かる遺伝子群の中から選ばれる1又は複数の遺伝子を削除又は不活性化することにより、タンパク質又はポリペプチドの生産にとって不要な、或いは有害な遺伝子の発現が生じないため、当該タンパク質又はポリペプチドの生産において、その生産性の向上が達成される。
【0023】
尚、削除又は不活性化する遺伝子は1以上であればよく、複数、特に3以上が好ましく、更には5以上であることが好ましい。更に本発明の微生物の構築には、上記以外の遺伝子群の削除又は不活性化を組み合わせることも可能であり、生産性向上に対してより大きな効果が期待される。また、本発明は目的遺伝子中に他のDNA断片を挿入する、あるいは、当該遺伝子の転写・翻訳開始領域に変異を与える等の方法によって目的遺伝子を不活性化することによっても達成できるが、好適には、標的遺伝子を物理的に削除する方がより望ましい。
【0024】
遺伝子群の削除又は不活性化手順としては、表1に示した標的遺伝子を計画的に削除又は不活性化する方法のほか、ランダムな遺伝子の削除又は不活性化変異を与えた後、適当な方法によりタンパク質生産性の評価及び遺伝子解析を行う方法が挙げられる。
【0025】
標的とする遺伝子を削除又は不活性化するには、例えば相同組換えによる方法を用いればよい。すなわち、標的遺伝子の一部を含むDNA断片を適当なプラスミドベクターにクローニングして得られる環状の組換えプラスミドを親微生物細胞内に取り込ませ、標的遺伝子の一部領域に於ける相同組換えによって親微生物ゲノム上の標的遺伝子を分断して不活性化することが可能である。或いは、塩基置換や塩基挿入等によって不活性化した標的遺伝子、又は標的遺伝子の外側領域を含むが標的遺伝子を含まない直鎖状のDNA断片等をPCR等の方法によって構築し、これを親微生物細胞内に取り込ませて親微生物ゲノムの標的遺伝子内の変異箇所の外側の2ヶ所、又は標的遺伝子外側の2ヶ所の領域で2回交差の相同組換えを起こさせることにより、ゲノム上の標的遺伝子を削除或いは不活性化した遺伝子断片と置換することが可能である。
【0026】
特に、本発明微生物を構築するための親微生物として枯草菌を用いる場合、相同組換えにより標的遺伝子を削除又は不活性化する方法については、既にいくつかの報告例があり(Mol. Gen. Genet., 223, 268 (1990)等)、こうした方法を繰り返すことによって、本発明の宿主微生物を得ることができる。
【0027】
また、ランダムな遺伝子の削除又は不活性化についてもランダムにクローニングしたDNA断片を用いて上述の方法と同様な相同組換えを起こさせる方法や、親微生物にγ線等を照射すること等によっても実施可能である。
【0028】
以下、より具体的にSOE(splicing by overlap extension)−PCR法(Gene, 77, 61, (1989))によって調製される削除用DNA断片を用いた二重交差法による削除方法について説明するが、本発明に於ける遺伝子削除方法は下記に限定されるものではない。
【0029】
本方法で用いる削除用DNA断片は、削除対象遺伝子の上流に隣接する約0.5〜3kb断片と、同じく下流に隣接する約0.5〜3kb断片の間に、薬剤耐性マーカー遺伝子断片を挿入した断片である。まず、1回目のPCRによって、削除対象遺伝子の上流断片及び下流断片、並びに薬剤耐性マーカー遺伝子断片の3断片を調製するが、この際、例えば、上流断片の下流末端に薬剤耐性マーカー遺伝子の上流側10〜30塩基対配列、逆に下流断片の上流末端には薬剤耐性マーカー遺伝子の下流側10〜30塩基対配列が付加される様にデザインしたプライマーを用いる(図1)。
【0030】
次いで、1回目に調製した3種類のPCR断片を鋳型とし、上流断片の上流側プライマーと下流断片の下流側プライマーを用いて2回目のPCRを行うことによって、上流断片の下流末端及び下流断片の上流末端に付加した薬剤耐性マーカー遺伝子配列に於いて、薬剤耐性マーカー遺伝子断片とのアニールが生じ、PCR増幅の結果、上流側断片と下流側断片の間に、薬剤耐性マーカー遺伝子が挿入したDNA断片を得ることができる(図1)。
【0031】
薬剤耐性マーカー遺伝子として、クロラムフェニコール耐性遺伝子を用いる場合、例えば表2に示したプライマーセットを用い、Pyrobest DNAポリメーラーゼ(宝酒造)などの一般のPCR用酵素キット等を用いて、成書(PCR Protocols. Current Methods and Applications, Edited by B.A.White, Humana Press, pp251 (1993)、Gene, 77, 61, (1989)等)に示される通常の条件によりSOE−PCRを行うことによって、各遺伝子の削除用DNA断片が得られる。
【0032】
かくして得られた削除用DNA断片を、コンピテント法等によって細胞内に導入すると、同一性のある削除対象遺伝子の上流及び下流の相同領域おいて、細胞内での遺伝子組換えが生じ、目標遺伝子が薬剤耐性遺伝子と置換した細胞を薬剤耐性マーカーによる選択によって分離することができる(図1)。即ち、表2に示したプライマーセットを用いて調製した削除用DNA断片を導入した場合、クロラムフェニコールを含む寒天培地上に生育するコロニーを分離し、目的の遺伝子が削除されてクロラムフェニコール耐性遺伝子と置換していることを、ゲノムを鋳型としたPCR法などによって確認すれば良い。
【0033】
次に、表1に示される枯草菌の遺伝子のいずれか、又は当該遺伝子に相当する遺伝子から選ばれた1以上の遺伝子が削除又は不活性化された宿主微生物変異株に、目的とするタンパク質又はポリペプチドをコードする遺伝子を導入することによって、本発明の組換え微生物を得ることができる。
【0034】
目的タンパク質又はポリペプチド遺伝子は特に限定されず、洗剤、食品、繊維、飼料、化学品、医療、診断など各種産業用酵素や、生理活性ペプチドなどが含まれる。また、産業用酵素の機能別には、酸化還元酵素 (Oxidoreductase) 、転移酵素 (Transferase) 、加水分解酵素 (Hydrolase) 、脱離酵素 (Lyase)、異性化酵素 (Isomerase) 、合成酵素 (Ligase/Synthetase) 等が含まれるが、好適にはセルラーゼ、α-アミラーゼ、プロテアーゼ等の加水分解酵素の遺伝子が挙げられる。具体的には、多糖加水分解酵素の分類(Biochem. J., 280, 309 (1991))中でファミリー5に属するセルラーゼが挙げられ、中でも微生物由来、特にバチルス属細菌由来のセルラーゼが挙げられる。より具体的な例として、配列番号2又は4で示されるアミノ酸配列からなるバチルス属細菌由来のアルカリセルラーゼや、当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列からなるセルラーゼが挙げられる。
【0035】
また、α−アミラーゼの具体例としては、微生物由来のα−アミラーゼが挙げられ、特にバチルス属細菌由来の液化型アミラーゼが好ましい。より具体的な例として、配列番号6で示されるアミノ酸配列からなるバチルス属細菌由来のアルカリアミラーゼや、当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列からなるアミラーゼが挙げられる。尚、アミノ酸配列の同一性はLipman-Pearson法 (Science, 227, 1435, (1985))によって計算される。また、プロテアーゼの具体例としては、微生物由来、特にバチルス属細菌由来のセリンプロテアーゼや金属プロテアーゼ等が挙げられる。
【0036】
また、目的タンパク質又はポリペプチド遺伝子は、その上流に当該遺伝子の転写、翻訳、分泌に関わる制御領域、即ち、プロモーターおよび転写開始点を含む転写開始制御領域、リボソーム結合部位および開始コドンを含む翻訳開始領域、又、分泌用シグナルペプチド領域が適正な形で結合されていることが望ましい。例えば、特開2000-210081号公報や特開平4-190793号公報等に記載されているバチルス属細菌、すなわちKSM−S237株(FERM BP-7875)、KSM−64株(FERM BP-2886)由来のセルラーゼ遺伝子と当該セルラーゼ遺伝子の転写開始制御領域、翻訳開始領域、分泌用シグナルペプチド領域、より具体的には配列番号1で示される塩基配列の塩基番号1〜659の塩基配列、配列番号3で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜696の塩基配列、また当該塩基配列に対して70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有する塩基配列からなるDNA断片、あるいは上記いずれかの塩基配列の一部が欠失した塩基配列からなるDNA断片が、目的タンパク質又はポリペプチドの構造遺伝子と適正に結合されていることが望ましい。
【0037】
上記の目的タンパク質又はポリペプチド遺伝子を含むDNA断片と適当なプラスミドベクターを結合させた組換えプラスミドを、一般的な形質転換法によって宿主微生物細胞に取り込ませることによって、本発明の組換え微生物を得ることができる。また、当該DNA断片に宿主微生物ゲノムとの適当な相同領域を結合したDNA断片を用い、宿主微生物ゲノムに直接組み込むことによっても本発明の組換え微生物を得ることができる。
【0038】
本発明の組換え微生物を用いた目的タンパク質又はポリペプチドの生産は、当該菌株を同化性の炭素源、窒素源、その他の必須成分を含む培地に接種し、通常の微生物培養法にて培養し、培養終了後、タンパク質又はポリペプチドを採取・精製することにより行えばよい。
【0039】
以上より、表1に示される枯草菌の遺伝子のいずれか、又は当該遺伝子に相当する遺伝子から選ばれた1以上の遺伝子が削除又は不活性化された宿主微生物変異株、及び当該変異株を用いて組換え微生物を構築することができ、これを用いれば有用なタンパク質又はポリペプチドを効率的に生産することができる。
【0040】
以下に、枯草菌のccpA遺伝子(BG10376)を削除した組換え枯草菌株構築の実施例を中心に、当該発明の組換え微生物の構築方法と、当該組換え微生物を用いたセルラーゼ及びα―アミラーゼの生産方法について具体的に説明する。
【実施例】
【0041】
実施例1
枯草菌168株から抽出したゲノムDNAを鋳型とし、表2に示したccpA-AFとccpA-A/CmR、及びccpA-B/CmFとccpA-BRの各プライマーセットを用いて、ゲノム上のccpA遺伝子の上流に隣接する0.6kb断片(A)、及び下流に隣接する0.6kb断片(B)をそれぞれ調製した。一方、プラスミドpC194(J. Bacteriol. 150 (2), 815 (1982))のクロラムフェニコール耐性遺伝子をプラスミドpUC18のXbaI−BamHI切断点に挿入した組換えプラスミドpCBB31を鋳型とし、表2に示したCmFとCmRプライマーセットを用いて、クロラムフェニコール耐性遺伝子を含む1kb断片(C)を調製した。次に、得られた(A)(B)(C)3断片を混合して鋳型とし、表2のプライマーccpA-AFとccpA-BRを用いたSOE−PCRを行うことによって、3断片を(A)(C)(B)の順になる様に結合させ、2.2kbのDNA断片を得た(図1参照)。このDNA断片を用いてコンピテント法により枯草菌168株の形質転換を行い、クロラムフェニコールを含むLB寒天培地上に生育したコロニーを形質転換体として分離した。得られた形質転換体のゲノムを抽出し、PCRによってccpA遺伝子が削除され、クロラムフェニコール耐性遺伝子に置換していることを確認した。
【0042】
【表2−1】
【0043】
【表2−2】
【0044】
【表2−3】
【0045】
【表2−4】
【0046】
実施例2
一方、実施例1と同様に、表2に示した各遺伝子-AF、各遺伝子-A/CmR、各遺伝子-B/CmF、各遺伝子-BR、CmF、CmRのプライマーセットにより調製した削除用DNA断片を用いて、ゲノム上のcomA、yopO、treR、yvbA、yvaN、yttP、yurK、yozA、licR、sigL、mntR、glcT、ykvE、slr、rocR、yyaA、及びrsiX、遺伝子が削除され、クロラムフェニコール耐性
遺伝子に置換した胞子形成遺伝子削除株をそれぞれ分離した。
【0047】
実施例3
また、表2に示した各遺伝子-AF、各遺伝子-A/Cm2R、各遺伝子-B/Cm2F、各遺伝子-BR、Cm2F、Cm2Rのプライマーセットにより、実施例2と同様に調製した削除用DNA断片を用いて、ゲノム上のcspB、yvdE、yaaT、yycH、及びylbO、各遺伝子が削除され、クロラムフェニコール耐性遺伝子に置換した胞子形成遺伝子削除株をそれぞれ分離した。
【0048】
実施例4
更に、表2に示した各遺伝子-AF、各遺伝子-A/Cm4R、各遺伝子-B/Cm4F、各遺伝子-BR、各遺伝子-A/Cm4F、各遺伝子-B/Cm4Rのプライマーセットにより、実施例2と同様に調製した削除用DNA断片を用いて、ゲノム上のyacP、hprK、及びyhdK、各遺伝子が削除され、クロラムフェニコール耐性遺伝子に置換した胞子形成遺伝子削除株をそれぞれ分離した。
【0049】
実施例5
実施例1〜4にて得られた各遺伝子削除株、及び対照として枯草菌168株に、バチルス エスピー(Bacillus sp.)KSM−S237株由来のアルカリセルラーゼ遺伝子(特開2000-210081号公報)をコードするDNA断片(3.1kb)がシャトルベ
クターpHY300PLKのBamHI制限酵素切断点に挿入された組換えプラスミドpHY−S237を、プロトプラスト形質転換法によって導入した。これによって得られた菌株を5mLのLB培地で一夜30℃で振盪培養を行い、更にこの培養液0.03mLを30mLの2×L−マルトース培地(2%トリプトン、1%酵母エキス、1%NaCl、7.5%マルトース、7.5ppm硫酸マンガン4−5水和物、15ppmテトラサイクリン)に接種し、30℃で3日間、振盪培養を行った。培養後、遠心分離によって菌体を除いた培養液上清のアルカリセルラーゼ活性を測定し、培養によって菌体外に分泌生産されたアルカリセルラーゼの量を求めた。この結果、表3に示した様に、宿主として各遺伝子削除株を用いた場合、対照の168株(野生型)の場合と比較して高いアルカリセルラーゼの分泌生産が認められた。
【0050】
【表3】
【0051】
実施例6
実施例1〜4にて得られた各遺伝子削除株、及び対照として枯草菌168株に、バチルス エスピー(Bacillus sp.)KSM−K38株由来のアルカリアミラーゼ遺伝子(特開2000−184882号公報、Eur. J. Biochem., 268, 2974 (2001))の成熟酵素領域(Asp1−Gln480)をコードするDNA断片(1.5kb)の上流に配列番号3に示されるアルカリセルラーゼ遺伝子のプロモーター領域とシグナル配列領域の一部を含む上流側0.6kb断片を結合して成る2.1kb断片(配列番号5)をシャトルベクターpHY300PLKのBglII−XbaI制限酵素切断部位に挿入された組換えプラスミドpHSP−K38を、プロトプラスト形質転換法によって導入した。これによって得られた菌株を5mLのLB培地で一夜30℃で振盪培養を行い、更にこの培養液0.6mLを30mLの2×L−マルトース培地(2%トリプトン、1%酵母エキス、1%NaCl、7.5%マルトース、7.5ppm硫酸マンガン4−5水和物、15ppmテトラサイクリン)に接種し、30℃で3〜6日間、振盪培養を行った。培養後、遠心分離によって菌体を除いた培養液上清のアルカリアミラーゼ活性を測定し、培養によって菌体外に分泌生産されたアルカリアミラーゼの量を求めた。この結果、表4に示した様に、各遺伝子削除株を宿主として用いた場合、対照の168株(野生型)の場合と比較して高いアルカリアミラーゼの分泌生産が認められた。
【0052】
【表4】
【技術分野】
【0001】
本発明は、有用なタンパク質又はポリペプチドの生産に用いる組換え微生物、及びタンパク質又はポリペプチドの生産方法に関する。
【背景技術】
【0002】
微生物による有用物質の工業的生産は、アルコール飲料や味噌、醤油等の食品類をはじめとし、アミノ酸、有機酸、核酸関連物質、抗生物質、糖質、脂質、タンパク質等、その種類は多岐に渡っており、またその用途についても食品、医薬や、洗剤、化粧品等の日用品、或いは各種化成品原料に至るまで幅広い分野に広がっている。
【0003】
こうした微生物による有用物質の工業生産においては、その生産性の向上が重要な課題の一つであり、その手法として、突然変異等の遺伝学的手法による生産菌の育種が行われてきた。特に最近では、微生物遺伝学、バイオテクノロジーの発展により、遺伝子組換え技術等を用いたより効率的な生産菌の育種が行われるようになっており、遺伝子組換えのための宿主微生物の開発が進められている。例えば、枯草菌(Bacillus subtilis) Marburg No.168系統株の様に宿主微生物として安全かつ優良と認められた微生物菌株に更に改良を加えた菌株が開発されている。
【0004】
しかしながら、微生物は元来、自然界における環境変化に対応するための多種多様な遺伝子群を有しており、限定された生産培地が使用されるタンパク質等の工業的生産においては、必ずしも生産効率が高いとは言えない状況であった。
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明は、タンパク質又はポリペプチドの生産性向上を可能とする宿主微生物にタンパク質又はポリペプチドをコードする遺伝子を導入して得られる組換え微生物、更に、当該組換え微生物を用いるタンパク質又はポリペプチドの製造法を提供することを目的とする。
【課題を解決するための手段】
【0006】
本発明者らは、微生物ゲノム上にコードされる各種遺伝子において、有用なタンパク質又はポリペプチドの生産にとって不要或いは有害な働きをする遺伝子群を鋭意探索したところ、枯草菌等の微生物の特定の遺伝子をゲノム上から削除又は不活性化した後、目的のタンパク質又はポリペプチドをコードする遺伝子を導入することにより、目的のタンパク質又はポリペプチドの生産性が、削除又は不活性化前と比較して向上することを見出した。
【0007】
すなわち本発明は、枯草菌の遺伝子comA、yopO、treR、yvbA、cspB、yvaN、yttP、yurK、yozA、licR、sigL、mntR、glcT、yvdE、ykvE、slr、rocR、ccpA、yaaT、yyaA、yycH、yacP、hprK、rsiX、yhdK及びylbOのいずれか、又は当該遺伝子に相当する遺伝子のいずれか1以上の遺伝子が削除又は不活性化された微生物変異株に、異種のタンパク質又はポリペプチドをコードする遺伝子を導入した組換え微生物、特に異種のタンパク質又はポリペプチドをコードする遺伝子の上流に転写開始制御領域、翻訳開始制御領域、又は分泌用シグナル領域を結合した当該組換え微生物、また当該組換え微生物を用いたタンパク質又はポリペプチドの製造方法を提供するものである。
【0008】
また本発明は、枯草菌の遺伝子yopO、treR、yvbA、cspB、yvaN、licR、sigL、glcT、yvdE、slr、rocR、yycH及びyacPのいずれか1以上の遺伝子が削除又は不活性化された枯草菌又はその他のバチルス属細菌株に、転写開始制御領域、翻訳開始制御領域及び分泌用シグナル領域が上流に結合した異種のタンパク質又はポリペプチドをコードする遺伝子を導入した組換え微生物であって、当該転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域が、配列番号1で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜659の塩基配列、配列番号3で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜696の塩基配列又は当該塩基配列のいずれかと90%以上の同一性を有する塩基配列からなり、且つ転写開始制御機能、翻訳開始制御機能及び分泌用シグナル機能を有するDNA断片である、組換え微生物、また当該組換え微生物を用いた異種タンパク質又はポリペプチドの製造方法を提供するものである。
一態様において、前記異種のタンパク質又はポリペプチドは、バチルス属細菌由来のアルカリセルラーゼ又はアルカリアミラーゼである。
【発明の効果】
【0009】
本発明の微生物は、目的タンパク質又はポリペプチドの生産にとって不要、或いは有害な遺伝子が削除、又は不活性化されているため、エネルギーロス、副産物の生産や比生産速度の低下等、培地の浪費が大幅に減少でき、また、タンパク質又はポリペプチドの生産期間が長期化することによって効率よく目的生産物を生産することができる。
【図面の簡単な説明】
【0010】
【図1】図1は、SOE−PCRによる遺伝子削除用DNA断片の調製、及び当該DNA断片を用いて標的遺伝子を削除(薬剤耐性遺伝子と置換)する方法を模式的に示したものである。
【発明を実施するための形態】
【0011】
本発明においてアミノ酸配列および塩基配列の同一性はLipman-Pearson法 (Science, 227, 1435, (1985))によって計算される。具体的には、遺伝情報処理ソフトウェアGenetyx-Win(ソフトウェア開発)のホモロジー解析(Search homology)プログラムを用いて、Unit size to compare(ktup)を2として解析を行うことにより算出される。
【0012】
本発明の微生物を構築するための親微生物としては、有用なタンパク質又はポリペプチドの生産にとって不要な遺伝子、具体的には表1に示す枯草菌の遺伝子又は当該遺伝子に相当する遺伝子を有するものであればよく、野生型のものでも変異を施したものでものよい。具体的には、枯草菌などのバチルス(Bacillus)属細菌や、クロストリジウム(Clostridium)属細菌、或いは酵母等が挙げられ、中でもバチルス(Bacillus)属細菌が好ましい。更に、全ゲノム情報が明らかにされ、遺伝子工学、ゲノム工学技術が確立されている点、またタンパク質と菌体外に分泌生産させる能力を有する点から特に枯草菌が好ましい。
【0013】
本発明の微生物を用いて生産する目的タンパク質又はポリペプチドとしては、例えば食品用、医薬品用、化粧品用、洗浄剤用、繊維処理用、医療検査薬用等として有用な酵素や生理活性因子等のタンパク質やポリペプチドが挙げられる。
【0014】
例えば、枯草菌にはゲノム上に4106個の遺伝子が存在することが知られているが、本発明において削除、又は不活性の対象となる遺伝子群は、目的タンパク質又はポリペプチドの生産にとって不要或いは有害な働きをする遺伝子群であり、表1に示される枯草菌の遺伝子のいずれか、又は当該遺伝子に相当する遺伝子群の中から選択されるものである。斯かる遺伝子群は、目的タンパク質又はポリペプチドの生産には直接関与しておらず、また、通常の工業的生産培地における微生物の生育にも不要であることが本発明者らによって見出された。
【0015】
尚、表中の各遺伝子の名称、番号及び機能等は、Nature, 390, 249-256, (1997) で報告され、JAFAN: Japan Functional Analysis Network for Bacillussubtilis (BSORF DB)でインターネット公開(http://bacillus.genome.ad.jp/、2003年6月17日更新)された枯草菌ゲノムデーターに基づいて記載している。
【0016】
【表1】
【0017】
また、表1に示される枯草菌の各遺伝子と同じ機能を有する、または、表1の各遺伝子と塩基配列において70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有する、他の微生物由来、好ましくはバチルス属細菌由来の遺伝子は、表1に記載の遺伝子に相当する遺伝子と考えられ、本発明において削除、不活性化すべき遺伝子に含まれる。尚、塩基配列の同一性はLipman-Pearson法 (Science, 227, 1435, (1985))によって計算される。
【0018】
表1に示される枯草菌の遺伝子群の中には、各種遺伝子発現の活性化や抑制に関わる制御遺伝子、或いは構造の類似性から制御遺伝子であろうと推定されるものが多く存在していることから、タンパク質又はポリペプチドの生産にとって不要或いは有害な遺伝子制御の存在が本発明によって明らかになった。
【0019】
特に、グルコースのPTS取り込み系オペロンのアンチターミネーターであるglcT遺伝子、リケナン分解系オペロンのアンチターミネーターであるlicT遺伝子、トレハロース取り込み、代謝系オペロンのリプレッサーであるtreR遺伝子、更には、グルコースカタボライトリプレッションに関わるccpA遺伝子やhprK遺伝子など、糖の取り込み、代謝に関わる制御遺伝子が多く存在しており、注目に値する。
【0020】
その他、糖の取り込み、代謝に関わる制御遺伝子以外の制御遺伝子として、アルギニン資化の活性化に関与するrocR遺伝子やコンピテンス関連の制御遺伝子comA、slr等を削除又は不活性化することによりタンパク質又はポリペプチドの生産性が向上する。
【0021】
また、表1に示される遺伝子群の中には、ECFシグマ因子のひとつであるシグマXの発現を抑制するアンチECFシグマ因子をコードするrsiX遺伝子、及び同じくシグマMの発現抑制に関与するとの報告例があるyhdK遺伝子(Mol Microbiol., 32, 41, (1999))などが存在し、また逆にシグマLをコードするsigL遺伝子が含まれており、シグマXやシグマMの制御下にある遺伝子の発現がタンパク質生産にとって好ましく、逆にシグマL制御下の何らかの遺伝子発現がタンパク質生産にとって好ましくないことが示唆されている。
【0022】
斯かる遺伝子群の中から選ばれる1又は複数の遺伝子を削除又は不活性化することにより、タンパク質又はポリペプチドの生産にとって不要な、或いは有害な遺伝子の発現が生じないため、当該タンパク質又はポリペプチドの生産において、その生産性の向上が達成される。
【0023】
尚、削除又は不活性化する遺伝子は1以上であればよく、複数、特に3以上が好ましく、更には5以上であることが好ましい。更に本発明の微生物の構築には、上記以外の遺伝子群の削除又は不活性化を組み合わせることも可能であり、生産性向上に対してより大きな効果が期待される。また、本発明は目的遺伝子中に他のDNA断片を挿入する、あるいは、当該遺伝子の転写・翻訳開始領域に変異を与える等の方法によって目的遺伝子を不活性化することによっても達成できるが、好適には、標的遺伝子を物理的に削除する方がより望ましい。
【0024】
遺伝子群の削除又は不活性化手順としては、表1に示した標的遺伝子を計画的に削除又は不活性化する方法のほか、ランダムな遺伝子の削除又は不活性化変異を与えた後、適当な方法によりタンパク質生産性の評価及び遺伝子解析を行う方法が挙げられる。
【0025】
標的とする遺伝子を削除又は不活性化するには、例えば相同組換えによる方法を用いればよい。すなわち、標的遺伝子の一部を含むDNA断片を適当なプラスミドベクターにクローニングして得られる環状の組換えプラスミドを親微生物細胞内に取り込ませ、標的遺伝子の一部領域に於ける相同組換えによって親微生物ゲノム上の標的遺伝子を分断して不活性化することが可能である。或いは、塩基置換や塩基挿入等によって不活性化した標的遺伝子、又は標的遺伝子の外側領域を含むが標的遺伝子を含まない直鎖状のDNA断片等をPCR等の方法によって構築し、これを親微生物細胞内に取り込ませて親微生物ゲノムの標的遺伝子内の変異箇所の外側の2ヶ所、又は標的遺伝子外側の2ヶ所の領域で2回交差の相同組換えを起こさせることにより、ゲノム上の標的遺伝子を削除或いは不活性化した遺伝子断片と置換することが可能である。
【0026】
特に、本発明微生物を構築するための親微生物として枯草菌を用いる場合、相同組換えにより標的遺伝子を削除又は不活性化する方法については、既にいくつかの報告例があり(Mol. Gen. Genet., 223, 268 (1990)等)、こうした方法を繰り返すことによって、本発明の宿主微生物を得ることができる。
【0027】
また、ランダムな遺伝子の削除又は不活性化についてもランダムにクローニングしたDNA断片を用いて上述の方法と同様な相同組換えを起こさせる方法や、親微生物にγ線等を照射すること等によっても実施可能である。
【0028】
以下、より具体的にSOE(splicing by overlap extension)−PCR法(Gene, 77, 61, (1989))によって調製される削除用DNA断片を用いた二重交差法による削除方法について説明するが、本発明に於ける遺伝子削除方法は下記に限定されるものではない。
【0029】
本方法で用いる削除用DNA断片は、削除対象遺伝子の上流に隣接する約0.5〜3kb断片と、同じく下流に隣接する約0.5〜3kb断片の間に、薬剤耐性マーカー遺伝子断片を挿入した断片である。まず、1回目のPCRによって、削除対象遺伝子の上流断片及び下流断片、並びに薬剤耐性マーカー遺伝子断片の3断片を調製するが、この際、例えば、上流断片の下流末端に薬剤耐性マーカー遺伝子の上流側10〜30塩基対配列、逆に下流断片の上流末端には薬剤耐性マーカー遺伝子の下流側10〜30塩基対配列が付加される様にデザインしたプライマーを用いる(図1)。
【0030】
次いで、1回目に調製した3種類のPCR断片を鋳型とし、上流断片の上流側プライマーと下流断片の下流側プライマーを用いて2回目のPCRを行うことによって、上流断片の下流末端及び下流断片の上流末端に付加した薬剤耐性マーカー遺伝子配列に於いて、薬剤耐性マーカー遺伝子断片とのアニールが生じ、PCR増幅の結果、上流側断片と下流側断片の間に、薬剤耐性マーカー遺伝子が挿入したDNA断片を得ることができる(図1)。
【0031】
薬剤耐性マーカー遺伝子として、クロラムフェニコール耐性遺伝子を用いる場合、例えば表2に示したプライマーセットを用い、Pyrobest DNAポリメーラーゼ(宝酒造)などの一般のPCR用酵素キット等を用いて、成書(PCR Protocols. Current Methods and Applications, Edited by B.A.White, Humana Press, pp251 (1993)、Gene, 77, 61, (1989)等)に示される通常の条件によりSOE−PCRを行うことによって、各遺伝子の削除用DNA断片が得られる。
【0032】
かくして得られた削除用DNA断片を、コンピテント法等によって細胞内に導入すると、同一性のある削除対象遺伝子の上流及び下流の相同領域おいて、細胞内での遺伝子組換えが生じ、目標遺伝子が薬剤耐性遺伝子と置換した細胞を薬剤耐性マーカーによる選択によって分離することができる(図1)。即ち、表2に示したプライマーセットを用いて調製した削除用DNA断片を導入した場合、クロラムフェニコールを含む寒天培地上に生育するコロニーを分離し、目的の遺伝子が削除されてクロラムフェニコール耐性遺伝子と置換していることを、ゲノムを鋳型としたPCR法などによって確認すれば良い。
【0033】
次に、表1に示される枯草菌の遺伝子のいずれか、又は当該遺伝子に相当する遺伝子から選ばれた1以上の遺伝子が削除又は不活性化された宿主微生物変異株に、目的とするタンパク質又はポリペプチドをコードする遺伝子を導入することによって、本発明の組換え微生物を得ることができる。
【0034】
目的タンパク質又はポリペプチド遺伝子は特に限定されず、洗剤、食品、繊維、飼料、化学品、医療、診断など各種産業用酵素や、生理活性ペプチドなどが含まれる。また、産業用酵素の機能別には、酸化還元酵素 (Oxidoreductase) 、転移酵素 (Transferase) 、加水分解酵素 (Hydrolase) 、脱離酵素 (Lyase)、異性化酵素 (Isomerase) 、合成酵素 (Ligase/Synthetase) 等が含まれるが、好適にはセルラーゼ、α-アミラーゼ、プロテアーゼ等の加水分解酵素の遺伝子が挙げられる。具体的には、多糖加水分解酵素の分類(Biochem. J., 280, 309 (1991))中でファミリー5に属するセルラーゼが挙げられ、中でも微生物由来、特にバチルス属細菌由来のセルラーゼが挙げられる。より具体的な例として、配列番号2又は4で示されるアミノ酸配列からなるバチルス属細菌由来のアルカリセルラーゼや、当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列からなるセルラーゼが挙げられる。
【0035】
また、α−アミラーゼの具体例としては、微生物由来のα−アミラーゼが挙げられ、特にバチルス属細菌由来の液化型アミラーゼが好ましい。より具体的な例として、配列番号6で示されるアミノ酸配列からなるバチルス属細菌由来のアルカリアミラーゼや、当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列からなるアミラーゼが挙げられる。尚、アミノ酸配列の同一性はLipman-Pearson法 (Science, 227, 1435, (1985))によって計算される。また、プロテアーゼの具体例としては、微生物由来、特にバチルス属細菌由来のセリンプロテアーゼや金属プロテアーゼ等が挙げられる。
【0036】
また、目的タンパク質又はポリペプチド遺伝子は、その上流に当該遺伝子の転写、翻訳、分泌に関わる制御領域、即ち、プロモーターおよび転写開始点を含む転写開始制御領域、リボソーム結合部位および開始コドンを含む翻訳開始領域、又、分泌用シグナルペプチド領域が適正な形で結合されていることが望ましい。例えば、特開2000-210081号公報や特開平4-190793号公報等に記載されているバチルス属細菌、すなわちKSM−S237株(FERM BP-7875)、KSM−64株(FERM BP-2886)由来のセルラーゼ遺伝子と当該セルラーゼ遺伝子の転写開始制御領域、翻訳開始領域、分泌用シグナルペプチド領域、より具体的には配列番号1で示される塩基配列の塩基番号1〜659の塩基配列、配列番号3で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜696の塩基配列、また当該塩基配列に対して70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有する塩基配列からなるDNA断片、あるいは上記いずれかの塩基配列の一部が欠失した塩基配列からなるDNA断片が、目的タンパク質又はポリペプチドの構造遺伝子と適正に結合されていることが望ましい。
【0037】
上記の目的タンパク質又はポリペプチド遺伝子を含むDNA断片と適当なプラスミドベクターを結合させた組換えプラスミドを、一般的な形質転換法によって宿主微生物細胞に取り込ませることによって、本発明の組換え微生物を得ることができる。また、当該DNA断片に宿主微生物ゲノムとの適当な相同領域を結合したDNA断片を用い、宿主微生物ゲノムに直接組み込むことによっても本発明の組換え微生物を得ることができる。
【0038】
本発明の組換え微生物を用いた目的タンパク質又はポリペプチドの生産は、当該菌株を同化性の炭素源、窒素源、その他の必須成分を含む培地に接種し、通常の微生物培養法にて培養し、培養終了後、タンパク質又はポリペプチドを採取・精製することにより行えばよい。
【0039】
以上より、表1に示される枯草菌の遺伝子のいずれか、又は当該遺伝子に相当する遺伝子から選ばれた1以上の遺伝子が削除又は不活性化された宿主微生物変異株、及び当該変異株を用いて組換え微生物を構築することができ、これを用いれば有用なタンパク質又はポリペプチドを効率的に生産することができる。
【0040】
以下に、枯草菌のccpA遺伝子(BG10376)を削除した組換え枯草菌株構築の実施例を中心に、当該発明の組換え微生物の構築方法と、当該組換え微生物を用いたセルラーゼ及びα―アミラーゼの生産方法について具体的に説明する。
【実施例】
【0041】
実施例1
枯草菌168株から抽出したゲノムDNAを鋳型とし、表2に示したccpA-AFとccpA-A/CmR、及びccpA-B/CmFとccpA-BRの各プライマーセットを用いて、ゲノム上のccpA遺伝子の上流に隣接する0.6kb断片(A)、及び下流に隣接する0.6kb断片(B)をそれぞれ調製した。一方、プラスミドpC194(J. Bacteriol. 150 (2), 815 (1982))のクロラムフェニコール耐性遺伝子をプラスミドpUC18のXbaI−BamHI切断点に挿入した組換えプラスミドpCBB31を鋳型とし、表2に示したCmFとCmRプライマーセットを用いて、クロラムフェニコール耐性遺伝子を含む1kb断片(C)を調製した。次に、得られた(A)(B)(C)3断片を混合して鋳型とし、表2のプライマーccpA-AFとccpA-BRを用いたSOE−PCRを行うことによって、3断片を(A)(C)(B)の順になる様に結合させ、2.2kbのDNA断片を得た(図1参照)。このDNA断片を用いてコンピテント法により枯草菌168株の形質転換を行い、クロラムフェニコールを含むLB寒天培地上に生育したコロニーを形質転換体として分離した。得られた形質転換体のゲノムを抽出し、PCRによってccpA遺伝子が削除され、クロラムフェニコール耐性遺伝子に置換していることを確認した。
【0042】
【表2−1】
【0043】
【表2−2】
【0044】
【表2−3】
【0045】
【表2−4】
【0046】
実施例2
一方、実施例1と同様に、表2に示した各遺伝子-AF、各遺伝子-A/CmR、各遺伝子-B/CmF、各遺伝子-BR、CmF、CmRのプライマーセットにより調製した削除用DNA断片を用いて、ゲノム上のcomA、yopO、treR、yvbA、yvaN、yttP、yurK、yozA、licR、sigL、mntR、glcT、ykvE、slr、rocR、yyaA、及びrsiX、遺伝子が削除され、クロラムフェニコール耐性
遺伝子に置換した胞子形成遺伝子削除株をそれぞれ分離した。
【0047】
実施例3
また、表2に示した各遺伝子-AF、各遺伝子-A/Cm2R、各遺伝子-B/Cm2F、各遺伝子-BR、Cm2F、Cm2Rのプライマーセットにより、実施例2と同様に調製した削除用DNA断片を用いて、ゲノム上のcspB、yvdE、yaaT、yycH、及びylbO、各遺伝子が削除され、クロラムフェニコール耐性遺伝子に置換した胞子形成遺伝子削除株をそれぞれ分離した。
【0048】
実施例4
更に、表2に示した各遺伝子-AF、各遺伝子-A/Cm4R、各遺伝子-B/Cm4F、各遺伝子-BR、各遺伝子-A/Cm4F、各遺伝子-B/Cm4Rのプライマーセットにより、実施例2と同様に調製した削除用DNA断片を用いて、ゲノム上のyacP、hprK、及びyhdK、各遺伝子が削除され、クロラムフェニコール耐性遺伝子に置換した胞子形成遺伝子削除株をそれぞれ分離した。
【0049】
実施例5
実施例1〜4にて得られた各遺伝子削除株、及び対照として枯草菌168株に、バチルス エスピー(Bacillus sp.)KSM−S237株由来のアルカリセルラーゼ遺伝子(特開2000-210081号公報)をコードするDNA断片(3.1kb)がシャトルベ
クターpHY300PLKのBamHI制限酵素切断点に挿入された組換えプラスミドpHY−S237を、プロトプラスト形質転換法によって導入した。これによって得られた菌株を5mLのLB培地で一夜30℃で振盪培養を行い、更にこの培養液0.03mLを30mLの2×L−マルトース培地(2%トリプトン、1%酵母エキス、1%NaCl、7.5%マルトース、7.5ppm硫酸マンガン4−5水和物、15ppmテトラサイクリン)に接種し、30℃で3日間、振盪培養を行った。培養後、遠心分離によって菌体を除いた培養液上清のアルカリセルラーゼ活性を測定し、培養によって菌体外に分泌生産されたアルカリセルラーゼの量を求めた。この結果、表3に示した様に、宿主として各遺伝子削除株を用いた場合、対照の168株(野生型)の場合と比較して高いアルカリセルラーゼの分泌生産が認められた。
【0050】
【表3】
【0051】
実施例6
実施例1〜4にて得られた各遺伝子削除株、及び対照として枯草菌168株に、バチルス エスピー(Bacillus sp.)KSM−K38株由来のアルカリアミラーゼ遺伝子(特開2000−184882号公報、Eur. J. Biochem., 268, 2974 (2001))の成熟酵素領域(Asp1−Gln480)をコードするDNA断片(1.5kb)の上流に配列番号3に示されるアルカリセルラーゼ遺伝子のプロモーター領域とシグナル配列領域の一部を含む上流側0.6kb断片を結合して成る2.1kb断片(配列番号5)をシャトルベクターpHY300PLKのBglII−XbaI制限酵素切断部位に挿入された組換えプラスミドpHSP−K38を、プロトプラスト形質転換法によって導入した。これによって得られた菌株を5mLのLB培地で一夜30℃で振盪培養を行い、更にこの培養液0.6mLを30mLの2×L−マルトース培地(2%トリプトン、1%酵母エキス、1%NaCl、7.5%マルトース、7.5ppm硫酸マンガン4−5水和物、15ppmテトラサイクリン)に接種し、30℃で3〜6日間、振盪培養を行った。培養後、遠心分離によって菌体を除いた培養液上清のアルカリアミラーゼ活性を測定し、培養によって菌体外に分泌生産されたアルカリアミラーゼの量を求めた。この結果、表4に示した様に、各遺伝子削除株を宿主として用いた場合、対照の168株(野生型)の場合と比較して高いアルカリアミラーゼの分泌生産が認められた。
【0052】
【表4】
【特許請求の範囲】
【請求項1】
枯草菌の遺伝子yopO、treR、yvbA、cspB、yvaN、licR、sigL、glcT、yvdE、slr、rocR、yycH及びyacPのいずれか1以上の遺伝子が削除又は不活性化された枯草菌又はその他のバチルス属細菌株に、転写開始制御領域、翻訳開始制御領域及び分泌用シグナル領域が上流に結合した異種のタンパク質又はポリペプチドをコードする遺伝子を導入した組換え微生物であって、当該転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域が、配列番号1で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜659の塩基配列、配列番号3で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜696の塩基配列又は当該塩基配列のいずれかと90%以上の同一性を有する塩基配列からなり、且つ転写開始制御機能、翻訳開始制御機能及び分泌用シグナル機能を有するDNA断片である、組換え微生物。
【請求項2】
異種のタンパク質又はポリペプチドがバチルス属細菌由来のアルカリセルラーゼ又はアルカリアミラーゼである、請求項1記載の組換え微生物。
【請求項3】
請求項1又は2記載の組換え微生物を用いる異種のタンパク質又はポリペプチドの製造方法。
【請求項1】
枯草菌の遺伝子yopO、treR、yvbA、cspB、yvaN、licR、sigL、glcT、yvdE、slr、rocR、yycH及びyacPのいずれか1以上の遺伝子が削除又は不活性化された枯草菌又はその他のバチルス属細菌株に、転写開始制御領域、翻訳開始制御領域及び分泌用シグナル領域が上流に結合した異種のタンパク質又はポリペプチドをコードする遺伝子を導入した組換え微生物であって、当該転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域が、配列番号1で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜659の塩基配列、配列番号3で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜696の塩基配列又は当該塩基配列のいずれかと90%以上の同一性を有する塩基配列からなり、且つ転写開始制御機能、翻訳開始制御機能及び分泌用シグナル機能を有するDNA断片である、組換え微生物。
【請求項2】
異種のタンパク質又はポリペプチドがバチルス属細菌由来のアルカリセルラーゼ又はアルカリアミラーゼである、請求項1記載の組換え微生物。
【請求項3】
請求項1又は2記載の組換え微生物を用いる異種のタンパク質又はポリペプチドの製造方法。
【図1】

【公開番号】特開2009−165487(P2009−165487A)
【公開日】平成21年7月30日(2009.7.30)
【国際特許分類】
【出願番号】特願2009−95379(P2009−95379)
【出願日】平成21年4月10日(2009.4.10)
【分割の表示】特願2003−379167(P2003−379167)の分割
【原出願日】平成15年11月7日(2003.11.7)
【国等の委託研究の成果に係る記載事項】(出願人による申告)国等の委託研究の成果に係る特許出願(平成14年度、新エネルギー・産業技術総合開発機構、生物機能を活用した生産プロセスの基盤技術開発、産業活力再生特別措置法第30条の適用を受けるもの)
【出願人】(000000918)花王株式会社 (8,290)
【Fターム(参考)】
【公開日】平成21年7月30日(2009.7.30)
【国際特許分類】
【出願日】平成21年4月10日(2009.4.10)
【分割の表示】特願2003−379167(P2003−379167)の分割
【原出願日】平成15年11月7日(2003.11.7)
【国等の委託研究の成果に係る記載事項】(出願人による申告)国等の委託研究の成果に係る特許出願(平成14年度、新エネルギー・産業技術総合開発機構、生物機能を活用した生産プロセスの基盤技術開発、産業活力再生特別措置法第30条の適用を受けるもの)
【出願人】(000000918)花王株式会社 (8,290)
【Fターム(参考)】
[ Back to top ]