
組換え微生物
【課題】タンパク質又はポリペプチドの生産性向上させた組換え微生物及び当該組換え微生物を用いるタンパク質又はポリペプチドの製造方法を提供する。
【解決手段】枯草菌の遺伝子yclJ、ycnC、yhcF、yhgD、ykvZ、yotL、yybE及びylnFのいずれか、又は当該遺伝子に相当する遺伝子のいずれか1以上の遺伝子が欠失又は不活性化された微生物株に、目的のタンパク質又はポリペプチドをコードする遺伝子を導入した組換え微生物。
【解決手段】枯草菌の遺伝子yclJ、ycnC、yhcF、yhgD、ykvZ、yotL、yybE及びylnFのいずれか、又は当該遺伝子に相当する遺伝子のいずれか1以上の遺伝子が欠失又は不活性化された微生物株に、目的のタンパク質又はポリペプチドをコードする遺伝子を導入した組換え微生物。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、有用なタンパク質又はポリペプチドの生産に用いる組換え微生物、及びタンパク質又はポリペプチドの生産方法に関する。
【背景技術】
【0002】
微生物による有用物質の工業的生産は、アルコール飲料や味噌、醤油等の食品類をはじめとし、アミノ酸、有機酸、核酸関連物質、抗生物質、糖質、脂質、タンパク質等、その種類は多岐に渡っており、またその用途についても食品、医薬や、洗剤、化粧品等の日用品、或いは各種化成品原料に至るまで幅広い分野に広がっている。
【0003】
こうした微生物による有用物質の工業生産においては、その生産性の向上が重要な課題の一つであり、その手法として、突然変異等の遺伝学的手法による生産菌の育種が行われてきた。特に最近では、微生物遺伝学、バイオテクノロジーの発展により、遺伝子組換え技術等を用いたより効率的な生産菌の育種が行われるようになっている。さらに、近年のゲノム解析技術の急速な発展を受けて、対象とする微生物のゲノム情報を解読し、これらを積極的に産業に応用しようとする試みもなされている。ゲノム情報の公開されている産業的に有用な宿主微生物としては、枯草菌Bacillus subtilis Marburg No.168(非特許文献1)、大腸菌Escherichia coli K-12 MG1655(非特許文献2)、コリネバクテリウムCorynebacterium glutamicum ATCC132032などが挙げられ、これらのゲノム情報を利用し、改良を加えた菌株が開発されている。しかしながら、上記のような取り組みにも関わらず、生産効率は必ずしも満足できるものではなかった。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】Nature,390,249,1997
【非特許文献2】Science,277,1453,1997
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明は、タンパク質又はポリペプチドの生産性がより向上された微生物、及び当該微生物を利用して、目的のタンパク質又はポリペプチドを製造する方法を提供することに関する。
【課題を解決するための手段】
【0006】
本発明者らは、微生物ゲノム上にコードされる各種遺伝子において、有用なタンパク質又はポリペプチドの生産に影響を及ぼす遺伝子を探索したところ、特定の遺伝子をゲノム上から欠失又は不活性化した後、目的のタンパク質又はポリペプチドをコードする遺伝子を導入した場合に、目的のタンパク質又はポリペプチドの生産性が、当該遺伝子の欠失又は不活性化前と比較して向上することを見出した。
【0007】
すなわち本発明は、枯草菌の遺伝子yclJ、ycnC、yhcF、yhgD、ykvZ、yotL、yybE及びylnFのいずれか、又は当該遺伝子に相当する遺伝子のいずれか1以上の遺伝子が欠失又は不活性化された微生物株に、目的のタンパク質又はポリペプチドをコードする遺伝子を導入した組換え微生物に関する。
【0008】
また本発明は、当該組換え微生物を用いた目的のタンパク質又はポリペプチドの製造方法に関する。
【発明の効果】
【0009】
本発明の組換え微生物を用いることにより、目的のタンパク質又はポリペプチドの生産性向上を図ることができる。
【図面の簡単な説明】
【0010】
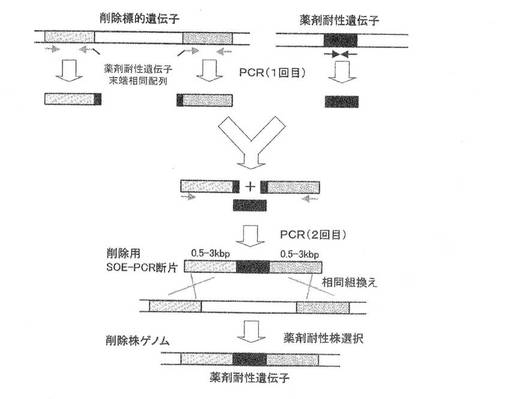
【図1】SOE−PCRによる遺伝子欠失用DNA断片の調製、及び当該DNA断片を用いて標的遺伝子を欠失する(薬剤耐性遺伝子と置換)方法を模式的に示したものである。
【発明を実施するための形態】
【0011】
本発明においてアミノ酸配列および塩基配列の同一性はLipman-Pearson法 (Science, 227, 1435, (1985))によって計算される。具体的には、遺伝情報処理ソフトウェアGenetyx-Win(ソフトウェア開発)のホモロジー解析(Search homology)プログラムを用いて、Unit size to compare(ktup)を2として解析を行うことにより算出される。
【0012】
本発明の微生物を構築するための親微生物としては、枯草菌の遺伝子yclJ、ycnC、yhcF、yhgD、ykvZ、yotL、yybE及びylnFのいずれか、又は当該遺伝子に相当する遺伝子を有するものであればよく、これらは野生型のものでも変異を施したものでもよい。具体的には、バチルス(Bacillus)属細菌や、クロストリジウム(Clostridium)属細菌、或いは酵母等が挙げられ、中でもバチルス(Bacillus)属細菌が好ましい。更に、全ゲノム情報が明らかにされ、遺伝子工学、ゲノム工学技術が確立されている点、またタンパク質を菌体外に分泌生産させる能力を有する点から特に枯草菌(Bacillus subtilis)が好ましい。
【0013】
本発明において欠失又は不活性化の対象となる枯草菌の遺伝子は、yclJ、ycnC、yhcF、yhgD、ykvZ、yotL、yybE及びylnFのいずれか、又は当該遺伝子に相当する遺伝子のいずれか1以上の遺伝子である。
yclJ、ycnC、yhcF、yhgD、ykvZ、yotL、yybE及びylnFは全て、現在機能未知の遺伝子である。以下の表1に、これらの遺伝子の名称及び番号を示す。これらは、Kunstらによって報告され(Nature,390,249-256,1997)、JAFAN: Japan Functional Analysis Network for Bacillus subtilis (BSORF DB)でインターネット公開(http://bacillus.genome.ad.jp/、2004年3月10日更新)された枯草菌ゲノムデーターに基づいて記載している。
【0014】
【表1】

【0015】
表1に示される枯草菌の各遺伝子と塩基配列において70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有する、他の微生物由来、好ましくはBacillus属細菌由来の遺伝子は、表1に記載の遺伝子に相当する遺伝子と考えられ、本発明において欠失、不活性化すべき遺伝子に含まれる。
【0016】
欠失又は不活性化する遺伝子は1以上であればよく、上記以外の遺伝子群の欠失又は不活性化を組み合わせることも可能である。更には上記遺伝子群の欠失の他に、それ以外の遺伝子群(例えば、prsA遺伝子等)の発現強化及び機能強化を組み合わせることも可能であり、生産性向上に対してより大きな効果が期待される。
また、上記遺伝子の不活性化は、当該遺伝子中に他のDNA断片を挿入する、或いは当該遺伝子の転写・翻訳開始領域に変異を与える等の方法によって行うことができるが、好適には、標的遺伝子を物理的に欠失させる方がより望ましい。
【0017】
本発明遺伝子の欠失又は不活性化の手順としては、yclJ、ycnC、yhcF、yhgD、ykvZ、yotL、yybE及びylnFのいずれか、又は当該遺伝子に相当する遺伝子(標的遺伝子)を計画的に欠失又は不活性化する方法のほか、ランダムな遺伝子の欠失又は不活性化変異を与えた後、適当な方法によりタンパク質生産性の評価及び遺伝子解析を行う方法が挙げられる。
【0018】
標的遺伝子を欠失又は不活性化するには、例えば相同組換えによる方法を用いればよい。すなわち、標的遺伝子の一部を含むDNA断片を適当なプラスミドベクターにクローニングして得られる環状の組換えプラスミドを親微生物細胞内に取り込ませ、標的遺伝子の一部領域に於ける相同組換えによって親微生物ゲノム上の標的遺伝子を分断して不活性化することが可能である。或いは、塩基置換や塩基挿入等の変異によって不活性化した標的遺伝子、又は図1のように標的遺伝子の上流、下流領域を含むが標的遺伝子を含まない直鎖状のDNA断片等をPCR等の方法によって構築し、これを親微生物細胞内に取り込ませて親微生物ゲノムの標的遺伝子内の変異箇所の外側の2ヶ所、又は標的遺伝子上流側、下流側で2回交差の相同組換えを起こさせることにより、ゲノム上の標的遺伝子を欠失或いは不活性化した遺伝子断片と置換することが可能である。
【0019】
特に、本発明微生物を構築するための親微生物として枯草菌を用いる場合、相同組換えにより標的遺伝子を欠失又は不活性化する方法については、既にいくつかの報告例があり(Mol.Gen.Genet.,223,268,1990等)、こうした方法を繰り返すことによって、本発明の宿主微生物を得ることができる。
【0020】
また、ランダムな遺伝子の欠失又は不活性化についてもランダムにクローニングしたDNA断片を用いて上述の方法と同様な相同組換えを起こさせる方法や、親微生物にγ線等を照射すること等によっても実施可能である。
【0021】
以下に、より具体的にSOE(splicing by overlap extension)−PCR法(Gene,77,61,1989)によって調製される欠失用DNA断片を用いた二重交差法による欠失方法について説明するが、本発明に於ける遺伝子欠失方法は下記に限定されるものではない。
【0022】
本方法で用いる欠失用DNA断片は、欠失対象遺伝子の上流に隣接する約1.0kb断片と、同じく下流に隣接する約1.0kb断片の間に、薬剤耐性マーカー遺伝子断片を挿入した断片である。まず、1回目のPCRによって、欠失対象遺伝子の上流断片及び下流断片、並びに薬剤耐性マーカー遺伝子断片の3断片を調製するが、この際、例えば、上流断片の下流末端に薬剤耐性マーカー遺伝子の上流側10〜30塩基対配列、逆に下流断片の上流末端には薬剤耐性マーカー遺伝子の下流側10〜30塩基対配列が付加される様にデザインしたプライマーを用いる(図1)。
【0023】
次いで、1回目に調製した3種類のPCR断片を鋳型とし、上流断片の上流側プライマーと下流断片の下流側プライマーを用いて2回目のPCRを行うことによって、上流断片の下流末端及び下流断片の上流末端に付加した薬剤耐性マーカー遺伝子配列に於いて、薬剤耐性マーカー遺伝子断片とのアニールが生じ、PCR増幅の結果、上流側断片と下流側断片の間に、薬剤耐性マーカー遺伝子を挿入したDNA断片を得ることができる(図1)。
【0024】
薬剤耐性マーカー遺伝子として、クロラムフェニコール耐性遺伝子を用いる場合、例えば表2に示したプライマーセットを用い、Pyrobest DNAポリメーラーゼ(宝酒造)などの一般のPCR用酵素キット等を用いて、成書(PCR Protocols. Current Methods and Applications, Edited by B.A.White, Humana Press pp251 ,1993、Gene,77,61,1989)等に示される通常の条件によりSOE−PCRを行うことによって、各遺伝子の欠失用DNA断片が得られる。
【0025】
かくして得られた遺伝子欠失用DNA断片を、コンピテント法等によって細胞内に導入すると、同一性のある欠失対象遺伝子の上流及び下流の相同領域において、細胞内での遺伝子組換えが生じ、標的遺伝子が薬剤耐性遺伝子と置換した細胞、或いは標的遺伝子内に薬剤耐性遺伝子が挿入された細胞が薬剤耐性マーカーによる選択によって分離できる(図1)。即ち、後記表2に示したプライマーセットを用いて調製した欠失用DNA断片を導入した場合、クロラムフェニコールを含む寒天培地上に生育するコロニーを分離し、ゲノムを鋳型としたPCR法などによってゲノム上の目的遺伝子がクロラムフェニコール耐性遺伝子と置換されていることを確認すれば良い。
【0026】
本発明の組換え微生物を用いて生産される目的タンパク質又は目的ポリペプチドは、特に限定されず、例えば洗剤、食品、繊維、飼料、化学品、医療、診断など各種産業用酵素や、生理活性ペプチドなどが挙げられるが、産業用酵素が好ましい。また、産業用酵素は、機能別に、酸化還元酵素(Oxidoreductase)、転移酵素(Transferase)、加水分解酵素(Hydrolase)、脱離酵素(Lyase)、異性化酵素(Isomerase)、合成酵素(Ligase/Synthetase)等が含まれる。好適には、セルラーゼ、α-アミラーゼ、プロテアーゼ等の加水分解酵素やクロラムフェニコールアセチルトランスフェラーゼ(CAT)等の転移酵素が挙げられ、より好適には、α-アミラーゼが挙げられる。
【0027】
セルラーゼとしては、例えば、多糖加水分解酵素の分類(Biochem.J.,280,309,1991)中でファミリー5に属するセルラーゼが挙げられ、中でも微生物由来、特にBacillus属細菌由来のセルラーゼが挙げられる。より具体的な例として、配列番号2で示されるアミノ酸配列からなるBacillus属細菌KSM-S237株(FERM BP-7875)由来のアルカリセルラーゼ、または、配列番号4で示されるアミノ酸配列からなるBacillus属細菌KSM-64株(FERM BP-2886)由来のアルカリセルラーゼ、或いは、当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列からなるセルラーゼが挙げられる。
【0028】
α−アミラーゼの具体例としては、微生物由来のα−アミラーゼが挙げられ、特にBacillus属細菌由来の液化型アミラーゼが好ましい。より具体的な例として、配列番号6で示されるアミノ酸配列からなるBacillus属細菌KSM-K38株(FERM BP-6946)由来のアルカリアミラーゼや、当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列からなるアミラーゼが挙げられる。
【0029】
プロテアーゼの具体例としては、微生物由来、特にBacillus属細菌由来のセリンプロテアーゼや金属プロテアーゼ等が挙げられる。より具体的な例として、配列番号8で示されるアミノ酸配列からなるバチルス クラウジ(Bacillus clausii)KSM-K16株(FERM BP-3376)由来のアルカリプロテアーゼや、当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列からなるプロテアーゼが挙げられる。
【0030】
クロラムフェニコールアセチルトランスフェラーゼ(CAT)は、アセチル-CoAのアセチル基をクロラムフェニコールの3位に転移する酵素である。具体的には、配列番号10で示されるアミノ酸配列からなるStaphylococcus aureus由来のCATや、当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有し、クロラムフェニコールアセチルトランスフェラーゼ活性を有するタンパク質が挙げられる。
【0031】
本発明の微生物に導入される目的タンパク質又はポリペプチドの遺伝子は、その上流に当該遺伝子の転写、翻訳、分泌に関わる制御領域、即ち、プロモーター及び転写開始点を含む転写開始制御領域、リボソーム結合部位及び開始コドンを含む翻訳開始領域並びに分泌シグナルペプチド領域から選ばれる1以上の領域が適正な形で結合されていることが望ましい。特に、転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が結合されていることが好ましく、更に分泌シグナルペプチド領域がバチルス(Bacillus)属細菌のセルラーゼ遺伝子由来のものであり、転写開始領域及び翻訳開始領域が当該セルラーゼ遺伝子の上流0.6〜1 kb領域であるものが、目的のタンパク質又はポリペプチド遺伝子と適正な形で結合されていることが望ましい。例えば、特開2000-210081号公報や特開平4-190793号公報等に記載されているバチルス(Bacillus)属細菌、すなわちKSM-S237株(FERM BP-7875)、KSM-64株(FERM BP-2886)由来のセルラーゼ遺伝子の転写開始制御領域、翻訳開始領域及び分泌シグナルペプチド領域が目的のタンパク質又はポリペプチドの構造遺伝子と適正に結合されていることが望ましい。より具体的には配列番号1で示される塩基配列の塩基番号1〜659の塩基配列、配列番号3で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜696の塩基配列、また当該塩基配列に対して70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有する塩基配列からなるDNA断片、或いは上記いずれかの塩基配列の一部が欠失した塩基配列からなるDNA断片が、目的のタンパク質又はポリペプチドの構造遺伝子と適正に結合されていることが望ましい。尚、ここで、上記塩基配列の一部が欠失した塩基配列からなるDNA断片とは、上記塩基配列の一部を欠失しているが、遺伝子の転写、翻訳、分泌に関わる機能を保持しているDNA断片を意味する。
【0032】
上記の目的のタンパク質又はポリペプチド遺伝子を含むDNA断片と適当なプラスミドベクターを結合させた組換えプラスミドを、一般的な形質転換法を用いて宿主微生物細胞に取り込ませることによって、本発明の組換え微生物を得ることができる。また、当該DNA断片に宿主微生物ゲノムとの適当な相同領域を結合したDNA断片を用い、宿主微生物ゲノムに直接組み込むことによっても本発明の組換え微生物を得ることができる。
【0033】
本発明の組換え微生物を用いた目的のタンパク質又はポリペプチドの生産は、当該菌株を同化性の炭素源、窒素源、その他の必須成分を含む培地に接種し、通常の微生物培養法にて培養し、培養終了後、タンパク質又はポリペプチドを採取・精製することにより行えばよい。そして、後記実施例に示すように、目的のタンパク質又はポリペプチドの生産性は、本発明の遺伝子を欠失又は不活性化していない微生物を用いた場合と比較して、その向上が達成されている。
【0034】
以下に、本発明の組換え微生物の構築方法及び当該組換え微生物を用いたアルカリアミラーゼの生産方法について具体的に説明する。
【実施例】
【0035】
実施例1
枯草菌168株から抽出したゲノムDNAを鋳型とし、表2に示したyclJ-AFとyclJ-A/CmR、及びyclJ-B/CmFとyclJ-BRの各プライマーセットを用いて、ゲノム上のyclJ遺伝子の上流に隣接する0.6kb断片(A)、及び下流に隣接する0.6kb断片(B)をそれぞれ調製した。一方、プラスミドpC194(J. Bacteriol. 150 (2), 815 (1982))のクロラムフェニコール耐性遺伝子をプラスミドpUC18のXbaI−BamHI切断点に挿入した組換えプラスミドpCBB31を鋳型とし、表2に示したCmFとCmRプライマーセットを用いて、クロラムフェニコール耐性遺伝子を含む1kb断片(C)を調製した。次に、得られた(A)(B)(C)3断片を混合して鋳型とし、表2に記載のプライマーyclJ-AFとyclJ-BRを用いたSOE−PCRを行うことによって、3断片を(A)(C)(B)の順になる様に結合させ、2.2kbのDNA断片を得た(図1参照)。このDNA断片を用いてコンピテント法により枯草菌168株の形質転換を行い、クロラムフェニコールを含むLB寒天培地上に生育したコロニーを形質転換体として分離した。得られた形質転換体のゲノムを抽出し、PCRによってyclJ遺伝子が欠失され、クロラムフェニコール耐性遺伝子に置換していることを確認した。
【0036】
【表2】
【0037】
実施例2
一方、実施例1と同様に、表2に示した各遺伝子-AF、各遺伝子-A/CmR、各遺伝子-B/CmF、各遺伝子-BR、CmF、CmRのプライマーセットにより調製した欠失用DNA断片を用いて、ゲノム上のycnC、yhcF、yhgD、ykvZ、yotL及びyybEが欠失され、クロラムフェニコール耐性遺伝子に置換した遺伝子欠失株をそれぞれ分離した。
【0038】
実施例3
枯草菌168株から抽出したゲノムDNAを鋳型とし、表2に示したylnF-AFとylnF-A/Cm2R、及びylnF-B/Cm2FとylnF-BRの各プライマーセットを用いて、ゲノム上のylnF遺伝子の上流に隣接する0.5kb断片(A)、及び下流に隣接する0.5kb断片(B)をそれぞれ調製した。一方、プラスミドpC194(J. Bacteriol. 150 (2), 815 (1982))のクロラムフェニコール耐性遺伝子をプラスミドpUC18のXbaI−BamHI切断点に挿入した組換えプラスミドpCBB31を鋳型とし、表2に示したCm2FとCm2Rプライマーセットを用いて、クロラムフェニコール耐性遺伝子を含む1kb断片(C)を調製した。次に、得られた(A)(B)(C)3断片を混合して鋳型とし、表2に記載のプライマーylnF-AFとylnF-BRを用いたSOE−PCRを行うことによって、3断片を(A)(C)(B)の順になる様に結合させ、2.0kbのDNA断片を得た(図1参照)。このDNA断片を用いてコンピテント法により枯草菌168株の形質転換を行い、クロラムフェニコールを含むLB寒天培地上に生育したコロニーを形質転換体として分離した。得られた形質転換体のゲノムを抽出し、PCRによってylnF遺伝子が欠失され、クロラムフェニコール耐性遺伝子に置換していることを確認した。
【0039】
実施例4
バチルス エスピー(Bacillus sp.)KSM-K38株(FERM BP-6946)より抽出したゲノムDNAを鋳型として、表3に示されるK38matu-F2(ALAA)とSP64K38-R(XbaI)のプライマーセットを用いてPCRを行い、アルカリアミラーゼ(特開2000-184882号公報、Eur.J.Biochem.,268,2974,2001)をコードする配列番号5で示される塩基配列のうちアルカリアミラーゼ成熟酵素領域1.6kbのDNA断片(D)を増幅した。またバチルス エスピー(Bacillus sp.)KSM-S237株(FERM BP-7875)より抽出したゲノムDNAを鋳型として、表3に示されるS237ppp-F2(BamHI)とS237ppp-R2(ALAA)のプライマーセットを用いてPCRを行い、配列番号1で示されるアルカリセルラーゼ遺伝子(特開2000-210081号公報)のうち転写開始制御領域、翻訳開始制御プロモーター領域と、及び分泌シグナル配列をコードする領域を含む0.6kbのDNA断片(E)を増幅した。次いで、得られた(D)(E)の2断片を混合して鋳型とし、表3に示されるS237ppp-F2(BamHI)とSP64K38-R(XbaI)のプライマーセットを用いたSOE-PCRを行うことによって、アルカリセルラーゼ遺伝子の転写開始制御領域、翻訳開始制御プロモーター領域と、及び分泌シグナル配列をコードする領域の下流にアルカリアミラーゼ遺伝子が連結した2.2 kbのDNA断片(F)を得た。得られた2.2kbのDNA断片(F)をシャトルベクターpHY300PLK(ヤクルト)のBamHI-XbaI制限酵素切断点に挿入し、アルカリアミラーゼ生産性評価用プラスミドpHYK38(S237ps)を構築した。
【0040】
【表3】
【0041】
実施例1〜3にて得られた各遺伝子欠失株、及び対照として枯草菌168株に、構築したアルカリアミラーゼ生産性評価用プラスミドpHYK38(S237ps)をプロトプラスト形質転換法によって導入した。これによって得られた菌株を10mLのLB培地で一夜30℃で振盪培養を行い、更にこの培養液0.05mLを50mLの2×L−マルトース培地(2%トリプトン、1%酵母エキス、1%NaCl、7.5%マルトース、7.5ppm硫酸マンガン4−5水和物、15ppmテトラサイクリン)に接種し、30℃で7日間、振盪培養を行った。培養後、遠心分離によって菌体を除いた培養液上清のアルカリアミラーゼ活性を測定し、培養によって菌体外に分泌生産されたアルカリアミラーゼの量を求めた。この結果、表4に示した様に、各遺伝子欠失株を宿主として用いた場合、対照の168株(野生型)の場合と比較して高いアルカリアミラーゼの分泌生産が認められた。
【0042】
【表4】
【技術分野】
【0001】
本発明は、有用なタンパク質又はポリペプチドの生産に用いる組換え微生物、及びタンパク質又はポリペプチドの生産方法に関する。
【背景技術】
【0002】
微生物による有用物質の工業的生産は、アルコール飲料や味噌、醤油等の食品類をはじめとし、アミノ酸、有機酸、核酸関連物質、抗生物質、糖質、脂質、タンパク質等、その種類は多岐に渡っており、またその用途についても食品、医薬や、洗剤、化粧品等の日用品、或いは各種化成品原料に至るまで幅広い分野に広がっている。
【0003】
こうした微生物による有用物質の工業生産においては、その生産性の向上が重要な課題の一つであり、その手法として、突然変異等の遺伝学的手法による生産菌の育種が行われてきた。特に最近では、微生物遺伝学、バイオテクノロジーの発展により、遺伝子組換え技術等を用いたより効率的な生産菌の育種が行われるようになっている。さらに、近年のゲノム解析技術の急速な発展を受けて、対象とする微生物のゲノム情報を解読し、これらを積極的に産業に応用しようとする試みもなされている。ゲノム情報の公開されている産業的に有用な宿主微生物としては、枯草菌Bacillus subtilis Marburg No.168(非特許文献1)、大腸菌Escherichia coli K-12 MG1655(非特許文献2)、コリネバクテリウムCorynebacterium glutamicum ATCC132032などが挙げられ、これらのゲノム情報を利用し、改良を加えた菌株が開発されている。しかしながら、上記のような取り組みにも関わらず、生産効率は必ずしも満足できるものではなかった。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】Nature,390,249,1997
【非特許文献2】Science,277,1453,1997
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明は、タンパク質又はポリペプチドの生産性がより向上された微生物、及び当該微生物を利用して、目的のタンパク質又はポリペプチドを製造する方法を提供することに関する。
【課題を解決するための手段】
【0006】
本発明者らは、微生物ゲノム上にコードされる各種遺伝子において、有用なタンパク質又はポリペプチドの生産に影響を及ぼす遺伝子を探索したところ、特定の遺伝子をゲノム上から欠失又は不活性化した後、目的のタンパク質又はポリペプチドをコードする遺伝子を導入した場合に、目的のタンパク質又はポリペプチドの生産性が、当該遺伝子の欠失又は不活性化前と比較して向上することを見出した。
【0007】
すなわち本発明は、枯草菌の遺伝子yclJ、ycnC、yhcF、yhgD、ykvZ、yotL、yybE及びylnFのいずれか、又は当該遺伝子に相当する遺伝子のいずれか1以上の遺伝子が欠失又は不活性化された微生物株に、目的のタンパク質又はポリペプチドをコードする遺伝子を導入した組換え微生物に関する。
【0008】
また本発明は、当該組換え微生物を用いた目的のタンパク質又はポリペプチドの製造方法に関する。
【発明の効果】
【0009】
本発明の組換え微生物を用いることにより、目的のタンパク質又はポリペプチドの生産性向上を図ることができる。
【図面の簡単な説明】
【0010】
【図1】SOE−PCRによる遺伝子欠失用DNA断片の調製、及び当該DNA断片を用いて標的遺伝子を欠失する(薬剤耐性遺伝子と置換)方法を模式的に示したものである。
【発明を実施するための形態】
【0011】
本発明においてアミノ酸配列および塩基配列の同一性はLipman-Pearson法 (Science, 227, 1435, (1985))によって計算される。具体的には、遺伝情報処理ソフトウェアGenetyx-Win(ソフトウェア開発)のホモロジー解析(Search homology)プログラムを用いて、Unit size to compare(ktup)を2として解析を行うことにより算出される。
【0012】
本発明の微生物を構築するための親微生物としては、枯草菌の遺伝子yclJ、ycnC、yhcF、yhgD、ykvZ、yotL、yybE及びylnFのいずれか、又は当該遺伝子に相当する遺伝子を有するものであればよく、これらは野生型のものでも変異を施したものでもよい。具体的には、バチルス(Bacillus)属細菌や、クロストリジウム(Clostridium)属細菌、或いは酵母等が挙げられ、中でもバチルス(Bacillus)属細菌が好ましい。更に、全ゲノム情報が明らかにされ、遺伝子工学、ゲノム工学技術が確立されている点、またタンパク質を菌体外に分泌生産させる能力を有する点から特に枯草菌(Bacillus subtilis)が好ましい。
【0013】
本発明において欠失又は不活性化の対象となる枯草菌の遺伝子は、yclJ、ycnC、yhcF、yhgD、ykvZ、yotL、yybE及びylnFのいずれか、又は当該遺伝子に相当する遺伝子のいずれか1以上の遺伝子である。
yclJ、ycnC、yhcF、yhgD、ykvZ、yotL、yybE及びylnFは全て、現在機能未知の遺伝子である。以下の表1に、これらの遺伝子の名称及び番号を示す。これらは、Kunstらによって報告され(Nature,390,249-256,1997)、JAFAN: Japan Functional Analysis Network for Bacillus subtilis (BSORF DB)でインターネット公開(http://bacillus.genome.ad.jp/、2004年3月10日更新)された枯草菌ゲノムデーターに基づいて記載している。
【0014】
【表1】
【0015】
表1に示される枯草菌の各遺伝子と塩基配列において70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有する、他の微生物由来、好ましくはBacillus属細菌由来の遺伝子は、表1に記載の遺伝子に相当する遺伝子と考えられ、本発明において欠失、不活性化すべき遺伝子に含まれる。
【0016】
欠失又は不活性化する遺伝子は1以上であればよく、上記以外の遺伝子群の欠失又は不活性化を組み合わせることも可能である。更には上記遺伝子群の欠失の他に、それ以外の遺伝子群(例えば、prsA遺伝子等)の発現強化及び機能強化を組み合わせることも可能であり、生産性向上に対してより大きな効果が期待される。
また、上記遺伝子の不活性化は、当該遺伝子中に他のDNA断片を挿入する、或いは当該遺伝子の転写・翻訳開始領域に変異を与える等の方法によって行うことができるが、好適には、標的遺伝子を物理的に欠失させる方がより望ましい。
【0017】
本発明遺伝子の欠失又は不活性化の手順としては、yclJ、ycnC、yhcF、yhgD、ykvZ、yotL、yybE及びylnFのいずれか、又は当該遺伝子に相当する遺伝子(標的遺伝子)を計画的に欠失又は不活性化する方法のほか、ランダムな遺伝子の欠失又は不活性化変異を与えた後、適当な方法によりタンパク質生産性の評価及び遺伝子解析を行う方法が挙げられる。
【0018】
標的遺伝子を欠失又は不活性化するには、例えば相同組換えによる方法を用いればよい。すなわち、標的遺伝子の一部を含むDNA断片を適当なプラスミドベクターにクローニングして得られる環状の組換えプラスミドを親微生物細胞内に取り込ませ、標的遺伝子の一部領域に於ける相同組換えによって親微生物ゲノム上の標的遺伝子を分断して不活性化することが可能である。或いは、塩基置換や塩基挿入等の変異によって不活性化した標的遺伝子、又は図1のように標的遺伝子の上流、下流領域を含むが標的遺伝子を含まない直鎖状のDNA断片等をPCR等の方法によって構築し、これを親微生物細胞内に取り込ませて親微生物ゲノムの標的遺伝子内の変異箇所の外側の2ヶ所、又は標的遺伝子上流側、下流側で2回交差の相同組換えを起こさせることにより、ゲノム上の標的遺伝子を欠失或いは不活性化した遺伝子断片と置換することが可能である。
【0019】
特に、本発明微生物を構築するための親微生物として枯草菌を用いる場合、相同組換えにより標的遺伝子を欠失又は不活性化する方法については、既にいくつかの報告例があり(Mol.Gen.Genet.,223,268,1990等)、こうした方法を繰り返すことによって、本発明の宿主微生物を得ることができる。
【0020】
また、ランダムな遺伝子の欠失又は不活性化についてもランダムにクローニングしたDNA断片を用いて上述の方法と同様な相同組換えを起こさせる方法や、親微生物にγ線等を照射すること等によっても実施可能である。
【0021】
以下に、より具体的にSOE(splicing by overlap extension)−PCR法(Gene,77,61,1989)によって調製される欠失用DNA断片を用いた二重交差法による欠失方法について説明するが、本発明に於ける遺伝子欠失方法は下記に限定されるものではない。
【0022】
本方法で用いる欠失用DNA断片は、欠失対象遺伝子の上流に隣接する約1.0kb断片と、同じく下流に隣接する約1.0kb断片の間に、薬剤耐性マーカー遺伝子断片を挿入した断片である。まず、1回目のPCRによって、欠失対象遺伝子の上流断片及び下流断片、並びに薬剤耐性マーカー遺伝子断片の3断片を調製するが、この際、例えば、上流断片の下流末端に薬剤耐性マーカー遺伝子の上流側10〜30塩基対配列、逆に下流断片の上流末端には薬剤耐性マーカー遺伝子の下流側10〜30塩基対配列が付加される様にデザインしたプライマーを用いる(図1)。
【0023】
次いで、1回目に調製した3種類のPCR断片を鋳型とし、上流断片の上流側プライマーと下流断片の下流側プライマーを用いて2回目のPCRを行うことによって、上流断片の下流末端及び下流断片の上流末端に付加した薬剤耐性マーカー遺伝子配列に於いて、薬剤耐性マーカー遺伝子断片とのアニールが生じ、PCR増幅の結果、上流側断片と下流側断片の間に、薬剤耐性マーカー遺伝子を挿入したDNA断片を得ることができる(図1)。
【0024】
薬剤耐性マーカー遺伝子として、クロラムフェニコール耐性遺伝子を用いる場合、例えば表2に示したプライマーセットを用い、Pyrobest DNAポリメーラーゼ(宝酒造)などの一般のPCR用酵素キット等を用いて、成書(PCR Protocols. Current Methods and Applications, Edited by B.A.White, Humana Press pp251 ,1993、Gene,77,61,1989)等に示される通常の条件によりSOE−PCRを行うことによって、各遺伝子の欠失用DNA断片が得られる。
【0025】
かくして得られた遺伝子欠失用DNA断片を、コンピテント法等によって細胞内に導入すると、同一性のある欠失対象遺伝子の上流及び下流の相同領域において、細胞内での遺伝子組換えが生じ、標的遺伝子が薬剤耐性遺伝子と置換した細胞、或いは標的遺伝子内に薬剤耐性遺伝子が挿入された細胞が薬剤耐性マーカーによる選択によって分離できる(図1)。即ち、後記表2に示したプライマーセットを用いて調製した欠失用DNA断片を導入した場合、クロラムフェニコールを含む寒天培地上に生育するコロニーを分離し、ゲノムを鋳型としたPCR法などによってゲノム上の目的遺伝子がクロラムフェニコール耐性遺伝子と置換されていることを確認すれば良い。
【0026】
本発明の組換え微生物を用いて生産される目的タンパク質又は目的ポリペプチドは、特に限定されず、例えば洗剤、食品、繊維、飼料、化学品、医療、診断など各種産業用酵素や、生理活性ペプチドなどが挙げられるが、産業用酵素が好ましい。また、産業用酵素は、機能別に、酸化還元酵素(Oxidoreductase)、転移酵素(Transferase)、加水分解酵素(Hydrolase)、脱離酵素(Lyase)、異性化酵素(Isomerase)、合成酵素(Ligase/Synthetase)等が含まれる。好適には、セルラーゼ、α-アミラーゼ、プロテアーゼ等の加水分解酵素やクロラムフェニコールアセチルトランスフェラーゼ(CAT)等の転移酵素が挙げられ、より好適には、α-アミラーゼが挙げられる。
【0027】
セルラーゼとしては、例えば、多糖加水分解酵素の分類(Biochem.J.,280,309,1991)中でファミリー5に属するセルラーゼが挙げられ、中でも微生物由来、特にBacillus属細菌由来のセルラーゼが挙げられる。より具体的な例として、配列番号2で示されるアミノ酸配列からなるBacillus属細菌KSM-S237株(FERM BP-7875)由来のアルカリセルラーゼ、または、配列番号4で示されるアミノ酸配列からなるBacillus属細菌KSM-64株(FERM BP-2886)由来のアルカリセルラーゼ、或いは、当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列からなるセルラーゼが挙げられる。
【0028】
α−アミラーゼの具体例としては、微生物由来のα−アミラーゼが挙げられ、特にBacillus属細菌由来の液化型アミラーゼが好ましい。より具体的な例として、配列番号6で示されるアミノ酸配列からなるBacillus属細菌KSM-K38株(FERM BP-6946)由来のアルカリアミラーゼや、当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列からなるアミラーゼが挙げられる。
【0029】
プロテアーゼの具体例としては、微生物由来、特にBacillus属細菌由来のセリンプロテアーゼや金属プロテアーゼ等が挙げられる。より具体的な例として、配列番号8で示されるアミノ酸配列からなるバチルス クラウジ(Bacillus clausii)KSM-K16株(FERM BP-3376)由来のアルカリプロテアーゼや、当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列からなるプロテアーゼが挙げられる。
【0030】
クロラムフェニコールアセチルトランスフェラーゼ(CAT)は、アセチル-CoAのアセチル基をクロラムフェニコールの3位に転移する酵素である。具体的には、配列番号10で示されるアミノ酸配列からなるStaphylococcus aureus由来のCATや、当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有し、クロラムフェニコールアセチルトランスフェラーゼ活性を有するタンパク質が挙げられる。
【0031】
本発明の微生物に導入される目的タンパク質又はポリペプチドの遺伝子は、その上流に当該遺伝子の転写、翻訳、分泌に関わる制御領域、即ち、プロモーター及び転写開始点を含む転写開始制御領域、リボソーム結合部位及び開始コドンを含む翻訳開始領域並びに分泌シグナルペプチド領域から選ばれる1以上の領域が適正な形で結合されていることが望ましい。特に、転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が結合されていることが好ましく、更に分泌シグナルペプチド領域がバチルス(Bacillus)属細菌のセルラーゼ遺伝子由来のものであり、転写開始領域及び翻訳開始領域が当該セルラーゼ遺伝子の上流0.6〜1 kb領域であるものが、目的のタンパク質又はポリペプチド遺伝子と適正な形で結合されていることが望ましい。例えば、特開2000-210081号公報や特開平4-190793号公報等に記載されているバチルス(Bacillus)属細菌、すなわちKSM-S237株(FERM BP-7875)、KSM-64株(FERM BP-2886)由来のセルラーゼ遺伝子の転写開始制御領域、翻訳開始領域及び分泌シグナルペプチド領域が目的のタンパク質又はポリペプチドの構造遺伝子と適正に結合されていることが望ましい。より具体的には配列番号1で示される塩基配列の塩基番号1〜659の塩基配列、配列番号3で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜696の塩基配列、また当該塩基配列に対して70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有する塩基配列からなるDNA断片、或いは上記いずれかの塩基配列の一部が欠失した塩基配列からなるDNA断片が、目的のタンパク質又はポリペプチドの構造遺伝子と適正に結合されていることが望ましい。尚、ここで、上記塩基配列の一部が欠失した塩基配列からなるDNA断片とは、上記塩基配列の一部を欠失しているが、遺伝子の転写、翻訳、分泌に関わる機能を保持しているDNA断片を意味する。
【0032】
上記の目的のタンパク質又はポリペプチド遺伝子を含むDNA断片と適当なプラスミドベクターを結合させた組換えプラスミドを、一般的な形質転換法を用いて宿主微生物細胞に取り込ませることによって、本発明の組換え微生物を得ることができる。また、当該DNA断片に宿主微生物ゲノムとの適当な相同領域を結合したDNA断片を用い、宿主微生物ゲノムに直接組み込むことによっても本発明の組換え微生物を得ることができる。
【0033】
本発明の組換え微生物を用いた目的のタンパク質又はポリペプチドの生産は、当該菌株を同化性の炭素源、窒素源、その他の必須成分を含む培地に接種し、通常の微生物培養法にて培養し、培養終了後、タンパク質又はポリペプチドを採取・精製することにより行えばよい。そして、後記実施例に示すように、目的のタンパク質又はポリペプチドの生産性は、本発明の遺伝子を欠失又は不活性化していない微生物を用いた場合と比較して、その向上が達成されている。
【0034】
以下に、本発明の組換え微生物の構築方法及び当該組換え微生物を用いたアルカリアミラーゼの生産方法について具体的に説明する。
【実施例】
【0035】
実施例1
枯草菌168株から抽出したゲノムDNAを鋳型とし、表2に示したyclJ-AFとyclJ-A/CmR、及びyclJ-B/CmFとyclJ-BRの各プライマーセットを用いて、ゲノム上のyclJ遺伝子の上流に隣接する0.6kb断片(A)、及び下流に隣接する0.6kb断片(B)をそれぞれ調製した。一方、プラスミドpC194(J. Bacteriol. 150 (2), 815 (1982))のクロラムフェニコール耐性遺伝子をプラスミドpUC18のXbaI−BamHI切断点に挿入した組換えプラスミドpCBB31を鋳型とし、表2に示したCmFとCmRプライマーセットを用いて、クロラムフェニコール耐性遺伝子を含む1kb断片(C)を調製した。次に、得られた(A)(B)(C)3断片を混合して鋳型とし、表2に記載のプライマーyclJ-AFとyclJ-BRを用いたSOE−PCRを行うことによって、3断片を(A)(C)(B)の順になる様に結合させ、2.2kbのDNA断片を得た(図1参照)。このDNA断片を用いてコンピテント法により枯草菌168株の形質転換を行い、クロラムフェニコールを含むLB寒天培地上に生育したコロニーを形質転換体として分離した。得られた形質転換体のゲノムを抽出し、PCRによってyclJ遺伝子が欠失され、クロラムフェニコール耐性遺伝子に置換していることを確認した。
【0036】
【表2】
【0037】
実施例2
一方、実施例1と同様に、表2に示した各遺伝子-AF、各遺伝子-A/CmR、各遺伝子-B/CmF、各遺伝子-BR、CmF、CmRのプライマーセットにより調製した欠失用DNA断片を用いて、ゲノム上のycnC、yhcF、yhgD、ykvZ、yotL及びyybEが欠失され、クロラムフェニコール耐性遺伝子に置換した遺伝子欠失株をそれぞれ分離した。
【0038】
実施例3
枯草菌168株から抽出したゲノムDNAを鋳型とし、表2に示したylnF-AFとylnF-A/Cm2R、及びylnF-B/Cm2FとylnF-BRの各プライマーセットを用いて、ゲノム上のylnF遺伝子の上流に隣接する0.5kb断片(A)、及び下流に隣接する0.5kb断片(B)をそれぞれ調製した。一方、プラスミドpC194(J. Bacteriol. 150 (2), 815 (1982))のクロラムフェニコール耐性遺伝子をプラスミドpUC18のXbaI−BamHI切断点に挿入した組換えプラスミドpCBB31を鋳型とし、表2に示したCm2FとCm2Rプライマーセットを用いて、クロラムフェニコール耐性遺伝子を含む1kb断片(C)を調製した。次に、得られた(A)(B)(C)3断片を混合して鋳型とし、表2に記載のプライマーylnF-AFとylnF-BRを用いたSOE−PCRを行うことによって、3断片を(A)(C)(B)の順になる様に結合させ、2.0kbのDNA断片を得た(図1参照)。このDNA断片を用いてコンピテント法により枯草菌168株の形質転換を行い、クロラムフェニコールを含むLB寒天培地上に生育したコロニーを形質転換体として分離した。得られた形質転換体のゲノムを抽出し、PCRによってylnF遺伝子が欠失され、クロラムフェニコール耐性遺伝子に置換していることを確認した。
【0039】
実施例4
バチルス エスピー(Bacillus sp.)KSM-K38株(FERM BP-6946)より抽出したゲノムDNAを鋳型として、表3に示されるK38matu-F2(ALAA)とSP64K38-R(XbaI)のプライマーセットを用いてPCRを行い、アルカリアミラーゼ(特開2000-184882号公報、Eur.J.Biochem.,268,2974,2001)をコードする配列番号5で示される塩基配列のうちアルカリアミラーゼ成熟酵素領域1.6kbのDNA断片(D)を増幅した。またバチルス エスピー(Bacillus sp.)KSM-S237株(FERM BP-7875)より抽出したゲノムDNAを鋳型として、表3に示されるS237ppp-F2(BamHI)とS237ppp-R2(ALAA)のプライマーセットを用いてPCRを行い、配列番号1で示されるアルカリセルラーゼ遺伝子(特開2000-210081号公報)のうち転写開始制御領域、翻訳開始制御プロモーター領域と、及び分泌シグナル配列をコードする領域を含む0.6kbのDNA断片(E)を増幅した。次いで、得られた(D)(E)の2断片を混合して鋳型とし、表3に示されるS237ppp-F2(BamHI)とSP64K38-R(XbaI)のプライマーセットを用いたSOE-PCRを行うことによって、アルカリセルラーゼ遺伝子の転写開始制御領域、翻訳開始制御プロモーター領域と、及び分泌シグナル配列をコードする領域の下流にアルカリアミラーゼ遺伝子が連結した2.2 kbのDNA断片(F)を得た。得られた2.2kbのDNA断片(F)をシャトルベクターpHY300PLK(ヤクルト)のBamHI-XbaI制限酵素切断点に挿入し、アルカリアミラーゼ生産性評価用プラスミドpHYK38(S237ps)を構築した。
【0040】
【表3】
【0041】
実施例1〜3にて得られた各遺伝子欠失株、及び対照として枯草菌168株に、構築したアルカリアミラーゼ生産性評価用プラスミドpHYK38(S237ps)をプロトプラスト形質転換法によって導入した。これによって得られた菌株を10mLのLB培地で一夜30℃で振盪培養を行い、更にこの培養液0.05mLを50mLの2×L−マルトース培地(2%トリプトン、1%酵母エキス、1%NaCl、7.5%マルトース、7.5ppm硫酸マンガン4−5水和物、15ppmテトラサイクリン)に接種し、30℃で7日間、振盪培養を行った。培養後、遠心分離によって菌体を除いた培養液上清のアルカリアミラーゼ活性を測定し、培養によって菌体外に分泌生産されたアルカリアミラーゼの量を求めた。この結果、表4に示した様に、各遺伝子欠失株を宿主として用いた場合、対照の168株(野生型)の場合と比較して高いアルカリアミラーゼの分泌生産が認められた。
【0042】
【表4】
【特許請求の範囲】
【請求項1】
枯草菌の遺伝子yybE及びylnFのいずれか、又は当該遺伝子に相当する遺伝子のいずれか1以上の遺伝子が欠失又は不活性化された微生物株に、目的のタンパク質又はポリペプチドをコードする遺伝子を導入した組換え微生物。
【請求項2】
微生物がバチルス属(Bacillus)細菌である請求項1記載の組換え微生物。
【請求項3】
バチルス(Bacillus)属細菌が枯草菌(Bacillus subtilis)である請求項2記載の組換え微生物。
【請求項4】
目的のタンパク質又はポリペプチドをコードする遺伝子の上流に転写開始制御領域、翻訳開始制御領域又は分泌シグナル領域のいずれか1以上の領域を結合した請求項1〜3のいずれか1項記載の組換え微生物。
【請求項5】
転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域を結合した請求項4記載の組換え微生物。
【請求項6】
分泌シグナル領域がバチルス(Bacillus)属細菌のセルラーゼ遺伝子由来のものであり、転写開始制御領域及び翻訳開始制御領域が当該セルラーゼ遺伝子の上流0.6〜1kb領域由来のものである請求項4又は5記載の組換え微生物。
【請求項7】
転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が、配列番号1で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜659の塩基配列、配列番号3で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜696の塩基配列又は当該塩基配列のいずれかと70%以上の同一性を有する塩基配列からなるDNA断片、又は当該塩基配列の一部が欠失した塩基配列からなるDNA断片である請求項4〜6のいずれか1項記載の組換え微生物。
【請求項8】
請求項1〜7のいずれか1項記載の組換え微生物を用いる目的のタンパク質又はポリペプチドの製造方法。
【請求項1】
枯草菌の遺伝子yybE及びylnFのいずれか、又は当該遺伝子に相当する遺伝子のいずれか1以上の遺伝子が欠失又は不活性化された微生物株に、目的のタンパク質又はポリペプチドをコードする遺伝子を導入した組換え微生物。
【請求項2】
微生物がバチルス属(Bacillus)細菌である請求項1記載の組換え微生物。
【請求項3】
バチルス(Bacillus)属細菌が枯草菌(Bacillus subtilis)である請求項2記載の組換え微生物。
【請求項4】
目的のタンパク質又はポリペプチドをコードする遺伝子の上流に転写開始制御領域、翻訳開始制御領域又は分泌シグナル領域のいずれか1以上の領域を結合した請求項1〜3のいずれか1項記載の組換え微生物。
【請求項5】
転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域を結合した請求項4記載の組換え微生物。
【請求項6】
分泌シグナル領域がバチルス(Bacillus)属細菌のセルラーゼ遺伝子由来のものであり、転写開始制御領域及び翻訳開始制御領域が当該セルラーゼ遺伝子の上流0.6〜1kb領域由来のものである請求項4又は5記載の組換え微生物。
【請求項7】
転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が、配列番号1で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜659の塩基配列、配列番号3で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜696の塩基配列又は当該塩基配列のいずれかと70%以上の同一性を有する塩基配列からなるDNA断片、又は当該塩基配列の一部が欠失した塩基配列からなるDNA断片である請求項4〜6のいずれか1項記載の組換え微生物。
【請求項8】
請求項1〜7のいずれか1項記載の組換え微生物を用いる目的のタンパク質又はポリペプチドの製造方法。
【図1】

【公開番号】特開2013−9688(P2013−9688A)
【公開日】平成25年1月17日(2013.1.17)
【国際特許分類】
【出願番号】特願2012−228015(P2012−228015)
【出願日】平成24年10月15日(2012.10.15)
【分割の表示】特願2007−203046(P2007−203046)の分割
【原出願日】平成19年8月3日(2007.8.3)
【出願人】(000000918)花王株式会社 (8,290)
【Fターム(参考)】
【公開日】平成25年1月17日(2013.1.17)
【国際特許分類】
【出願日】平成24年10月15日(2012.10.15)
【分割の表示】特願2007−203046(P2007−203046)の分割
【原出願日】平成19年8月3日(2007.8.3)
【出願人】(000000918)花王株式会社 (8,290)
【Fターム(参考)】
[ Back to top ]