
組換え枯草菌
【課題】タンパク質又はポリペプチドの生産性向上を可能とする宿主枯草菌を用い、これにタンパク質又はポリペプチドをコードする遺伝子を導入して得られる組換え枯草菌、更に当該組換え枯草菌を用いるタンパク質又はポリペプチドの製造法を提供する。
【解決手段】prophage6領域、prophage1領域、prophage4領域、PBSX領域、prophage5領域、prophage3領域、spb領域、pks領域、skin領域、pps領域、prophage2領域、ydcL−ydeK−ydhU領域、yisB−yitD領域、yunA−yurT領域、cgeE−ypmQ領域、yeeK−yesX領域、pdp−rocR領域、ycxB−sipU領域、SKIN−Pro7領域、sbo−ywhH領域、yybP−yyaJ領域及びyncM−fosB領域が欠失した枯草菌変異株の、malA遺伝子を削除又は不活性化させ、更に異種タンパク質又は異種ポリペプチドをコードする遺伝子を導入して作製した組換え枯草菌。
【解決手段】prophage6領域、prophage1領域、prophage4領域、PBSX領域、prophage5領域、prophage3領域、spb領域、pks領域、skin領域、pps領域、prophage2領域、ydcL−ydeK−ydhU領域、yisB−yitD領域、yunA−yurT領域、cgeE−ypmQ領域、yeeK−yesX領域、pdp−rocR領域、ycxB−sipU領域、SKIN−Pro7領域、sbo−ywhH領域、yybP−yyaJ領域及びyncM−fosB領域が欠失した枯草菌変異株の、malA遺伝子を削除又は不活性化させ、更に異種タンパク質又は異種ポリペプチドをコードする遺伝子を導入して作製した組換え枯草菌。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、有用なタンパク質又はポリペプチドの生産に用いる組換え枯草菌、及びそれを用いたタンパク質又はポリペプチドの生産方法に関する。
【背景技術】
【0002】
微生物による有用物質の工業的生産は、アルコール飲料や味噌、醤油等の食品の醸造をはじめとして、アミノ酸、有機酸、核酸関連物質、抗生物質、糖質、脂質、タンパク質等の工業的生産など、多岐に渡って実施されている。またその用途も、食品をはじめとして、医薬、洗剤、化粧品等の日用品、あるいは各種化成品原料に至るまで、幅広い分野に広がっている。
【0003】
微生物を用いた有用物質の工業的生産における一つの重要課題として、当該有用物質の生産性向上が挙げられる。従来、当該課題を解決するため、突然変異等の遺伝学的手法による高生産菌の育種が行われてきた。特に最近では、微生物遺伝学、バイオテクノロジーの発展により、遺伝子組換え技術等を用いた、より効率的な高生産菌の育種が行われている。更に、近年のゲノム解析技術の急速な発展を受け、対象とする微生物のゲノム情報を解読し、これらを積極的に産業に応用する試みもなされている。例えば、枯草菌(Bacillus subtilis)Marburg No.168系統株など、宿主微生物として安全かつ優良と認められた微生物菌株に更に改良を加えた菌株が開発されている。
【0004】
枯草菌(Bacillus subtilis)において、マルトースは、フォスフォエノールピルビン酸依存性の糖トランスポーター(PTS:Phosphoenol pyruvate−dependent phosphotransferase system)の一つであるMalPにより細胞内に取り込まれた後(非特許文献1)リン酸化され、マルトース6リン酸に変換される。更に、マルトース6リン酸は、6−ホスホ−α−グルコシダーゼ活性を有するMalAによりグルコース及びグルコース6リン酸へと分解され、解糖系で代謝される。MalAをコードする遺伝子malAはmalA−glvR−malPオペロンを構成することが知られている(非特許文献2)。
【0005】
本出願人は、過去の特許出願(特許文献1)において、枯草菌168株(野生株)を親株としてmalP(当時の名称はglvC)及びglvRを欠失させた株で酵素セルラーゼ及びα−アミラーゼの生産量が向上することを報告している。しかしながら、malA遺伝子の欠失による酵素生産量への影響に関しては明らかにされていない。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】特開2005−151983号公報
【特許文献2】特開2007−130013号公報
【非特許文献】
【0007】
【非特許文献1】J.bacteriol,188,11,p.3911−3922,2006
【非特許文献2】J.bacteriol,183,17,p.5110−5121,2001
【発明の開示】
【発明が解決しようとする課題】
【0008】
本発明は、タンパク質又はポリペプチドの生産性向上を可能とする組換え枯草菌、更には当該組換え枯草菌を用いたタンパク質又はポリペプチドの製造方法の提供に関する。
【課題を解決するための手段】
【0009】
本発明者らは、有用なタンパク質又はポリペプチドの生産にとって不要あるいは有害な働きをする遺伝子群をゲノム上から削除した枯草菌を宿主として用い、その宿主のmalA遺伝子を削除又は不活性化し、更に目的の異種タンパク質又は異種ポリペプチドをコードする遺伝子を導入することにより、当該異種タンパク質又はポリペプチドの生産が顕著に向上することを見出し、本発明を完成させるに至った。
【0010】
すなわち本発明は、以下の(1)から(9)に係るものである。
(1) prophage6領域、prophage1領域、prophage4領域、PBSX領域、prophage5領域、prophage3領域、spb領域、pks領域、skin領域、pps領域、prophage2領域、ydcL−ydeK−ydhU領域、yisB−yitD領域、yunA−yurT領域、cgeE−ypmQ領域、yeeK−yesX領域、pdp−rocR領域、ycxB−sipU領域、SKIN−Pro7領域、sbo−ywhH領域、yybP−yyaJ領域及びyncM−fosB領域が欠失したゲノム構造を有する枯草菌変異株に、異種タンパク質又は異種ポリペプチドをコードする遺伝子を導入して作製される組換え枯草菌であって、malA遺伝子が削除又は不活性化された組換え枯草菌。
(2) 前記枯草菌変異株が、更に以下の領域のうちの1つ又は2つ以上が欠失したゲノム構造を有する、上記(1)記載の組換え枯草菌:
ybbU−ybfI領域、ydjM−cotA領域、yefA−yesX領域、yfiB−yfiX領域、yhcE−yhcU領域、yhaU−yhaL領域、yjbX−yjlB領域、xkdA−ykcC領域、bpr−ylmA領域、flgB−cheD領域、ynfF−ppsA領域、yoxC−yobO領域、spoVAF−spoIIAA領域、spoIIIAH−yqhV領域、ytvB−ytqB領域、yteA−ytaB領域、yuaJ−yugO領域、yusJ−mrgA領域、gerAA−yvrI領域、yvaM−yvbK領域、araE−yveK領域、yvdE−yvcP領域、gerBA−ywsC領域、ywrK−ywqM領域、spoIIID−ywoB領域、slp−ylaF領域、licH−sigY領域、yqeF−yrhK領域、yuzE−yukJ領域及びyncM−yndN領域。
(3) 異種タンパク質又は異種ポリペプチドをコードする遺伝子の上流に、転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域から選ばれる1以上の領域が作動可能に連結されている、上記(1)又は(2)に記載の組換え枯草菌。
(4) 異種タンパク質又は異種ポリペプチドをコードする遺伝子の上流に、転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が作動可能に連結されている、上記(1)から(3)のいずれか1つに記載の組換え枯草菌。
(5) 前記分泌シグナル領域が、バチルス属細菌のセルラーゼ遺伝子由来のものであり、前記転写開始制御領域及び前記翻訳開始制御領域が、当該セルラーゼ遺伝子の上流0.6〜1kb領域由来のものである、上記(3)又は(4)記載の組換え枯草菌。
(6) 前記転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が、配列番号3で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜659の塩基配列、又は当該塩基配列のいずれかと70%以上の同一性を有する塩基配列からなるDNA断片、又は当該塩基配列の一部が欠失した塩基配列からなるDNA断片である、上記(4)記載の組換え枯草菌。
(7) 前記異種タンパク質又は異種ポリペプチドをコードする遺伝子が、バチルス属細菌のアルカリセルラーゼ遺伝子である、上記(1)から(6)のいずれか1つに記載の組換え枯草菌。
(8) 前記アルカリセルラーゼ遺伝子が、配列番号3で示される塩基配列からなるセルラーゼ遺伝子、又は当該塩基配列と70%以上の同一性を有する塩基配列からなるDNA断片、又は当該塩基配列の一部が欠失、置換若しくは付加された塩基配列からなるDNA断片である、上記(7)記載の組換え枯草菌。
(9) 上記(1)から(8)のいずれか1つに記載の組換え枯草菌を用いる異種タンパク質又は異種ポリペプチドの製造方法。
【発明の効果】
【0011】
本願発明の組換え枯草菌を用いれば、目的タンパク質又はポリペプチドを効率よく大量生産することが可能となる。
【図面の簡単な説明】
【0012】
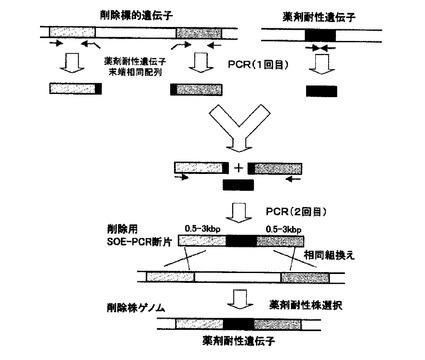
【図1】SOE−PCRによる遺伝子欠失用DNA断片の調製、及び当該DNA断片を用いて標的遺伝子を欠失する(薬剤耐性遺伝子と置換)方法を模式的に示した図。
【発明を実施するための形態】
【0013】
本願発明の組換え枯草菌は、そのゲノムの一部が欠失した枯草菌変異株を親株として用い、malA遺伝子を削除又は不活性化させ、更に異種タンパク質又は異種ポリペプチドをコードする遺伝子を導入して作製される。
【0014】
枯草菌(Bacillus subtilis)とは、好気性のグラム陽性桿菌で、芽胞を形成する真正細菌の一種である。枯草菌は、全ゲノム情報が明らかにされ、遺伝子工学、ゲノム工学技術が確立されており、またタンパク質と菌体外に分泌生産させる能力を有するため、本願発明にとり有用な微生物といえる。
【0015】
本願発明において用いる枯草菌としては、そのゲノムの大領域、すなわちprophage6(yoaV−yobO)領域、prophage1(ybbU−ybdE)領域、prophage4(yjcM−yjdJ)領域、PBSX(ykdA−xlyA)領域、prophage5(ynxB−dut)領域、prophage3(ydiM−ydjC)領域、spb(yodU−ypqP)領域、pks(pksA−ymaC)領域、skin(spoIVCB−spoIIIC)領域、pps(ppsE−ppsA)領域、prophage2(ydcL−ydeJ)領域、ydcL−ydeK−ydhU領域、yisB−yitD領域、yunA−yurT領域、cgeE−ypmQ領域、yeeK−yesX領域、pdp−rocR領域、ycxB−sipU領域、SKIN−Pro7(spoIVCB−yraK)領域、sbo−ywhH領域、yybP−yyaJ領域及びyncM−fosB領域を欠失させて構築した株が用いられ、具体的には、枯草菌Bacillus subtilis Marburg No.168(枯草菌168株)を野生株とし、当該大領域を欠失させて構築した枯草菌MGB874株が挙げられる(特許文献2)。これらの欠失領域はまた、表1に示す一対のオリゴヌクレオチドセットにより挟み込まれる領域として表すことができる。
【0016】
【表1】

【0017】
枯草菌168株の全塩基配列及び遺伝子は既に報告されており、またインターネット公開されている(Nature,390,249−256,1997及びBSORF Bacillus subtilis Genome Database[http://bacillus.genome.jp/];GenBank:AL009126.2[http://www.ncbi.nlm.nih.gov/nuccore/38680335])。当業者は、これらの情報源から得た枯草菌168株のゲノム情報、例えばGenBank:AL009126.2[http://www.ncbi.nlm.nih.gov/nuccore/38680335]に基づいて、MGB874株のゲノムから欠失させるべき上記表2に示した遺伝子領域を見出すことができる。ここで、欠失させるべき遺伝子領域は、公開されている枯草菌168株の遺伝子の塩基配列に対して1又は複数個の塩基における天然又は人工的に引き起こされた欠失、置換、挿入、付加等の変異を含み得る塩基配列を有し得る。変異し得る塩基の個数の範囲は、枯草菌168株のゲノムとの対応関係が認識できる限り特に限定されないが、例えば、枯草菌168株のゲノム領域の塩基配列に対して50%以上の同一性、好ましくは60%以上の同一性、より好ましくは70%以上の同一性、更に好ましくは80%以上の同一性、更により好ましくは90%以上の同一性、なお好ましくは95%以上の同一性を有する範囲が挙げられる。上記「付加」には、塩基配列の一末端及び両末端への塩基の付加が含まれる。
【0018】
本発明に係る枯草菌変異株は、上記枯草菌変異株MGB874株のゲノム領域から、更にゲノム領域を欠失させたゲノム構造を有してもよい。具体的には、当該枯草菌変異株は、枯草菌168株のゲノム上における表2に示す遺伝子領域のうちの少なくとも1つを、枯草菌変異株MGB874株のゲノム領域から更に欠失させたゲノム構造を有してもよい。当該欠失領域としては、表2に示される遺伝子領域及び当該領域に相当する領域が挙げられる。
【0019】
【表2】
【0020】
本発明において削除すべきmalA遺伝子(BSORF Gene Entry No.:BG11839、EC3.2.1.122)の遺伝子産物であるMalAタンパク質は、α−グルコシダーゼ活性を有し、マルトースを分解してグルコースに変換する。malA遺伝子はmalP遺伝子及びglvR遺伝子と共にmalA−glvR−malPオペロンを構成し、マルトースの代謝活動に関与する。
【0021】
これらmalA遺伝子以外の遺伝子の欠失又は不活化との組み合わせにより、異種タンパク質又は異種ポリペプチドの生産性向上に対するより大きな効果が期待される。
【0022】
本発明に係るmalA遺伝子が削除又は不活性化された組換え枯草菌は、例えば相同組換えにより標的遺伝子を削除又は不活性化する方法など、既に報告された方法(Mol.Gen.Genet.,223,268(1990)等)を繰り返すことによって構築される。
【0023】
また、ランダムな遺伝子の削除又は不活性化についてもランダムにクローニングしたDNA断片を用いて上述の方法と同様な相同組換えを起こさせる方法や、親微生物にγ線等を照射すること等によっても実施可能である。
【0024】
本発明では、上記のmalA遺伝子内に他のDNA断片を挿入する方法、あるいは当該遺伝子の転写・翻訳開始領域を変異させる方法等によって目的遺伝子を不活性化しても本発明の目的を達成できるが、標的遺伝子を物理的に削除する方がより好ましい。また、削除又は不活性化する遺伝子は1つでもよいが、2つ以上の遺伝子の削除を組み合わせてもよい。更に本発明の組換え枯草菌の構築には、マルトースの膜透過関与以外の遺伝子群の削除又は不活性化を組み合わせることも可能であり、生産性向上に対してより大きな効果が期待される。
【0025】
本発明における、malA遺伝子の削除又は不活性化の具体例として、SOE(splicing by overlap extension)−PCR法(Gene,77,61,(1989))によって調製される削除用DNA断片を用いた二重交差法による削除方法について説明するが、本発明における遺伝子削除方法はこの方法に限定されるものではない。
【0026】
本方法で用いる削除用DNA断片は、削除対象遺伝子の上流に隣接する約0.2〜3kb断片と、同じく下流に隣接する約0.2〜3kb断片の間に、薬剤耐性マーカー遺伝子断片を挿入して構築した断片である。まず、1回目のPCRによって、削除対象遺伝子の上流断片及び下流断片、並びに薬剤耐性マーカー遺伝子断片の3断片を調製するが、この際、例えば、上流断片の下流末端に薬剤耐性マーカー遺伝子の上流側10〜30塩基対配列、逆に下流断片の上流末端には薬剤耐性マーカー遺伝子の下流側10〜30塩基対配列が付加されるようにデザインされたプライマーを用いる(図1)。
【0027】
次いで、1回目に調製した3種類のPCR断片を鋳型とし、上流断片の上流側プライマーと下流断片の下流側プライマーを用いて2回目のPCRを行うことによって、上流断片の下流末端及び下流断片の上流末端に付加した薬剤耐性マーカー遺伝子配列において、薬剤耐性マーカー遺伝子断片とのアニーリングが生じ、PCR増幅の結果、上流側断片と下流側断片の間に、薬剤耐性マーカー遺伝子が挿入されたDNA断片が得られる(図1)。
【0028】
表3に示したプライマーセットと適当な鋳型DNAを用い、Pyrobest DNAポリメーラーゼ(宝酒造)などの一般のPCR用酵素キット等を用いて、“PCR Protocols.Current Methods and Applications”,Edited by B.A.White,Humana Press,pp251(1993)、Gene,77,61,(1989)等に示される通常の条件によりSOE−PCRを行うことによって、各遺伝子の削除用DNA断片が得られる。
【0029】
かくして得られた削除用DNA断片を、コンピテント法等によって細胞内に導入すると、削除対象遺伝子の上流及び下流の、上記削除用DNA断片との相同領域おいて、細胞内での遺伝子組換えが生じ、削除対象遺伝子が薬剤耐性遺伝子で置換された細胞を、薬剤耐性マーカーによる選抜により単離できる(図1)。即ち、表3に示したプライマーセットを用いて調製した削除用DNA断片を導入した場合、薬剤を含む寒天培地上に生育するコロニーを単離し、目的の遺伝子が削除されて薬剤耐性遺伝子と置換していることを、ゲノムを鋳型としたPCR法などによって確認すればよい。
【0030】
次に、かくして得られた本発明に係る枯草菌変異株に、目的とする異種タンパク質又はポリペプチドをコードする遺伝子を導入することによって、本発明の組換え枯草菌を得ることができる。
【0031】
当該組換え枯草菌を用いて異種タンパク質又はポリペプチドを製造する場合、上記目的タンパク質又はポリペプチドの遺伝子の上流に、当該遺伝子の転写、翻訳、分泌を制御する制御領域を、適切な形で結合させるのが望ましい。かかる制御領域としては、プロモーター及び転写開始点を含む転写開始制御領域、リボソーム結合部位及び開始コドンを含む翻訳開始領域並びに分泌シグナルペプチド領域から選ばれる1以上の領域などが挙げられる。特に転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が結合されていることが好ましく、更に分泌シグナルペプチド領域がバチルス(Bacillus)属細菌のセルラーゼ遺伝子由来のものであり、転写開始制御領域及び翻訳開始制御領域が当該セルラーゼ遺伝子の開始コドンから始まる長さ0.6〜1kbの上流領域であるものが、異種タンパク質又はポリペプチド遺伝子と作動可能に連結されていることが望ましい。例えば、特開2000−210081号公報や特開平4−190793号公報等に記載されているバチルス(Bacillus)属細菌、すなわちKSM−S237株(FERM BP−7875)、KSM−64株(FERM BP−2886)由来のセルラーゼ遺伝子の転写開始制御領域、翻訳開始領域及び分泌シグナルペプチド領域が目的のタンパク質又はポリペプチドの構造遺伝子と作動可能に連結されていることが望ましい。より具体的には、前記転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が、配列番号3で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜659の塩基配列、又は当該塩基配列のいずれかと70%以上、好ましくは80%以上、さらに好ましくは90%以上、特に好ましくは95%以上の同一性を有し、且つ遺伝子の転写、翻訳、分泌に関わる機能を有する塩基配列からなるDNA断片、又は当該塩基配列の一部が欠失した塩基配列からなるDNA断片が、上記の異種タンパク質又はポリペプチド遺伝子と作動可能に連結していることが望ましい。ここで、上記塩基配列の一部が欠失した塩基配列からなるDNA断片とは、上記塩基配列の一部を欠失しているが、遺伝子の転写、翻訳、分泌に関わる機能を保持しているDNA断片を意味する。また、ここで、作動可能に連結されている(operably linked)とは、上記の制御配列が機能してコード配列によりコードされた目的タンパク質の発現が制御されうる位置関係で、当該制御配列と当該コード配列とが配置していることを指す。具体例としては、プロモーターとそれに隣接する目的遺伝子とが、当該プロモーターの方向に沿って、当該遺伝子が発現しうる状態で配置していることを指す。また、ここで、ストリンジェントな条件とは、例えば[Molecular cloning−a Laboratory manual 2nd edition(Sambrookら、1989)]に記載の条件等が挙げられる。例えば、6×SSC(1×SSCの組成:0.15M 塩化ナトリウム、0.015M クエン酸ナトリウム、pH7.0)、0.5% SDS、5×デンハート液及び100mg/mL ニシン精子DNAを含む溶液中で、プローブと共に65℃で8〜16時間インキュベートし、ハイブリダイズさせる条件が挙げられる。
【0032】
異種タンパク質又はポリペプチドをコードする遺伝子を細胞内で発現させるためには、当該遺伝子を含むDNA断片を、適切なベクターに挿入した発現プラスミドを構築し、当該発現プラスミドを宿主に導入して形質転換する必要がある。当該発現ベクターとしては、枯草菌体内で自立複製可能なベクターが好適であり、例えばシャトルベクターpHY300PLK等が挙げられるが、特に限定されない。
【0033】
ポリペプチドとは一般に、直鎖状に連結したアミノ酸のポリマーを指し、タンパク質とは一般に、50以上のアミノ酸からなる1つ以上のポリペプチドのことを指すが、本願においてはこれらの用語は交換可能に用いられるものとする。
【0034】
本発明の組換え枯草菌を用いて生産する異種タンパク質又はポリペプチドとしては、例えば食品用、医薬用、化粧用、洗浄用、繊維処理用、検査用に用いられる各種産業用酵素や、生理活性ペプチドなどが挙げられる。また、産業用酵素の機能別には、酸化還元酵素(オキシドレダクターゼ)、転移酵素(トランスフェラーゼ)、加水分解酵素(ヒドロラーゼ)、脱離酵素(リアーゼ)、異性化酵素(イソメラーゼ)、合成酵素(リガーゼ/シンセターゼ)等が含まれるが、好適にはセルラーゼ、α−アミラーゼ、プロテアーゼ等の加水分解酵素が挙げられ、より好ましくはセルラーゼが挙げられる。
【0035】
より具体的な例として、配列番号3で示される塩基配列によりコードされるバチルス属細菌KSM−S237(FERM BP−7875)由来のアルカリセルラーゼや、配列番号119で示されるアミノ酸配列からなるバチルス属細菌KSM−64株(FERM BP−2886)由来のアルカリセルラーゼ、配列番号4又は119で示されるアミノ酸配列の1個もしくは数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列を有するアルカリセルラーゼが挙げられ、更には、当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、更に好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列からなるセルラーゼが挙げられる。
【0036】
また、α−アミラーゼの具体例としては、微生物由来のα−アミラーゼが挙げられ、特にバチルス属細菌由来の液化型アミラーゼや、配列番号120で示されるアミノ酸配列からなるバチルス属細菌KSM−K38株(FERM BP−6946)由来のアルカリアミラーゼ、当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、更に好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列からなるアミラーゼ、配列番号120で示されるアミノ酸配列において1〜数個のアミノ酸が置換、欠失若しくは付加されたアミノ酸配列からなりアミラーゼ活性を有するタンパク質が挙げられる。また、プロテアーゼの具体例としては、微生物由来、特にバチルス属細菌由来のセリンプロテアーゼや金属プロテアーゼや、配列番号121で示されるアミノ酸配列からなるバチルス クラウジ(Bacillus clausii)KSM−K16株(FERM BP−3376)由来のアルカリプロテアーゼ、あるいは当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、さらにより好ましくは98%以上の同一性を有するアミノ酸配列からなりプロテアーゼ活性を有するタンパク質、配列番号121で示されるアミノ酸配列において1〜数個のアミノ酸が置換、欠失若しくは付加されたアミノ酸配列からなりプロテアーゼ活性を有するタンパク質が挙げられる。
【0037】
本発明において、アミノ酸配列及び塩基配列の同一性は、Lipman−Pearson法(Science,227,1435,(1985))によって計算される。より具体的には、遺伝情報処理ソフトウェアGenetyx−Win(ソフトウェア開発)のホモロジー解析(Search homology)プログラムを用いて、Unit size to compare(ktup)を2として解析を行うことにより算出される。
【0038】
本願発明の別の態様は、上述した組換え枯草菌を用いる異種タンパク質又は異種ポリペプチドの製造方法に関する。
【0039】
本発明の組換え枯草菌は、上記の異種タンパク質又はポリペプチド遺伝子を含むDNA断片と適当なプラスミドベクターを結合させた組換えプラスミドを、一般的な形質転換法によって宿主となる枯草菌変異株に取り込ませることによって得られる。また、当該DNA断片に宿主ゲノムとの適当な相同領域を結合したDNA断片を用い、宿主ゲノムに直接組み込むことによって本発明の組換え枯草菌を得てもよい。
【0040】
上記の組換え枯草菌を用いた異種タンパク質又はポリペプチドの製造は、上記異種タンパク質又はポリペプチドの遺伝子を上記のとおり宿主となる枯草菌変異株に導入して得られる菌株を、例えば同化性の炭素源、窒素源、その他の必須成分を含む培地に接種して培養して行うことができる。培養方法は、原則的には一般的な微生物の培養方法であってもよく、通常、液体培養による振盪培養、通気撹拌培養等の好気的条件下で実施するのが好ましい。培養終了後、培養液を遠心分離し、得られる上清又は菌体から、目的の異種タンパク質又はポリペプチドを、硫安沈殿やクロマトグラフィなどを適宜組み合わせ、常法に従い抽出・精製することにより得ることができる。
【0041】
以下の実施例において、本発明の組換え枯草菌の構築方法及び当該組換え枯草菌を用いたタンパク質の製造方法について説明するが、本願発明は以下の実施例に限定されるものではない。
【実施例】
【0042】
枯草菌MGB874株のmalA遺伝子を削除した組換え枯草菌株の構築、並びに当該組換え枯草菌を用いたセルラーゼの生産方法について、以下に具体的に説明する。
【0043】
以下の実施例におけるDNA断片増幅のためのポリメラーゼ連鎖反応(PCR)は、Pyrobest DNAポリメラーゼ(タカラバイオ社製)と付属の試薬類を用いた、GeneAmp PCR System(アプライドバイオシステムズ社製)によるDNA増幅により行った。PCR反応液は、適宜希釈した鋳型DNAを1μL、センス及びアンチセンスプライマーを各々20pmol、及びPyrobest DNAポリメラーゼを2.5U添加し、更に総反応液量を50μLとすることにより調製した。PCR反応条件は、98℃で10秒間、55℃で30秒間及び72℃で1〜5分間(目的増幅産物に応じて調整。目安は1kbあたり1分間)の3段階の温度変化を30サイクル繰り返した後、72℃で5分間反応、とした。
【0044】
また、以下の実施例において、遺伝子の上流及び下流とは、複製開始点からの位置を指すのではなく、上流とは各操作・工程において対象となる遺伝子の開始コドンの5’側に続く領域を指し、一方下流とは各操作・工程において対象となる遺伝子の終止コドンの3’側に続く領域を指すものとする。
【0045】
更に、以下の実施例における各遺伝子及び遺伝子領域の名称は、Nature,390,249−256,(1997)で報告され、JAFAN:Japan Functional Analysis Network for Bacillus subtilis(BSORF DB)としてインターネットで公開された枯草菌ゲノムデータ(http://bacillus.genome.ad.jp/2004年3月10日更新)に基づき記載する。
【0046】
枯草菌の形質転換は、コンピテントセル法(J.Bacteriol.93,1925(1967))により行った。すなわち、枯草菌株をSPI培地(0.20% 硫酸アンモニウム、1.40% リン酸水素二カリウム、0.60% リン酸二水素カリウム、0.10% クエン酸三ナトリウム二水和物、0.50% グルコース、0.02% カザミノ酸(Difco社製)、5mM 硫酸マグネシウム、0.25μM 塩化マンガン、50μg/mL トリプトファン)中で、37℃で、生育度(OD600)の値が約1となるまで振盪培養し、振盪培養後、培養液の一部を9倍量のSPII培地(0.20% 硫酸アンモニウム、1.40% リン酸水素二カリウム、0.60% リン酸二水素カリウム、0.10% クエン酸三ナトリウム二水和物、0.50% グルコース、0.01% カザミノ酸(Difco社製)、5mM 硫酸マグネシウム、0.40μM 塩化マンガン、5μg/mL トリプトファン)に接種し、更に生育度(OD600)の値が約0.4となるまで振盪培養することにより、枯草菌株のコンピテントセルを調製した。
【0047】
次いで調製したコンピテントセル懸濁液(SPII培地における培養液)100μLに、各種DNA断片を含む溶液(SOE−PCRの反応液等)を5μL添加し、37℃で1時間振盪培養後、適切な薬剤を含むLB寒天培地(1% トリプトン、0.5% 酵母エキス、1% NaCl、1.5% 寒天)に全量を塗沫した。37℃での静置培養の後、生育したコロニーを形質転換体として分離した。得られた形質転換体のゲノムを抽出し、これを鋳型とするPCRを行い、目的とするゲノム構造の改変がなされたことを確認した。
【0048】
目的のタンパク質を発現するプラスミドの宿主微生物への導入は、プロトプラスト形質転換法(Mol.Gen.Genet.168,111(1979))により行った。組換え枯草菌によるタンパク質生産用の培養として、LB培地(1% トリプトン、0.5% 酵母エキス、1% NaCl)、及び2×L−マルトース培地(2% トリプトン、1% 酵母エキス、1% NaCl、7.5% マルトース、7.5ppm 硫酸マンガン4〜5水和物)を用いた。
【0049】
(実施例1:malA遺伝子欠失株の構築)
malA遺伝子(配列番号1)はmalA−glvR−malPオペロンの先頭に位置する遺伝子であり、本遺伝子を欠失する際には、malAの下流遺伝子であるglvR及びmalPの発現に影響を与えない形で欠失株の構築を行わなければならない。そこで、本発明においては、malAの構造遺伝子をネオマイシン耐性遺伝子のORF(open reading frame)と置換する方法により、malA遺伝子欠失株を構築した。枯草菌MGB874株から抽出したゲノムDNAを鋳型とし、表3に示したmalAFW1とmalA/neof2.R、及びmalA/neor2.FとmalA RVの各プライマーセットを用いて、ゲノム上のmalA遺伝子の上流に隣接する1.0kbp断片(A)、及び下流に隣接する1.0kbp断片(B)をそれぞれ調製した。また、ネオマイシン耐性遺伝子を有するプラスミドpUB110(Plasmid 15:93−103.(1986))を鋳型として、表3に示したneof2及びneor2のプライマーセットを用いて、ネオマイシン耐性遺伝子(C)を調製した。次に、得られた(A)、(B)及び(C)の3断片を混合して鋳型とし、malA FW2とmalA RV2のプライマーを用いてPCRを行ない、3断片を(A)−(C)−(B)の順になるように結合させ、遺伝子欠失用のDNA断片を得た(図1参照)。このDNA断片を用いて、上述したコンピテントセル法により、枯草菌168株及びMGB874株を形質転換し、ネオマイシン(10mg/L)及びマルトース(1g/L)を含むLB寒天培地上で生育したコロニーを形質転換体として単離した。マルトース存在下でmalA−glvR−malPオペロンの発現が誘導されるため(J.bacteriol.,183,17,5110,(2001))、形質転換体ではネオマイシン耐性遺伝子が発現し、ネオマイシンを含むLB寒天培地上でも生育可能となる。
【0050】
得られた形質転換体からゲノムDNAを抽出し、これを鋳型としてPCRを行った結果、malA遺伝子がネオマイシン耐性遺伝子と置換され、malA遺伝子欠失株となっていることが確認された。以上のようにして得られたmalA欠失株のうち、枯草菌168株を親株として構築した株を168ΔmalA株と、枯草菌MGB874株を親株として構築した株を874ΔmalA株とした。
【0051】
【表3】
【0052】
(実施例2:セルラーゼ生産性評価)
実施例1にて得られた168ΔmalA株、874ΔmalA株及び対照として、親株である枯草菌168株、MGB874株に、アルカリセルラーゼ遺伝子を導入した。具体的には、バチルス属細菌 KSM−S237株(FERM BP−7875)由来のS237セルラーゼ遺伝子(特開2000−210081号公報参照)(配列番号3)をコードするDNA断片(3.1kb;配列番号3の塩基番号13〜3124)を鋳型として、表4に示されるプライマーEgl−S237.F及びプライマーEgl−S237.Rのプライマーセットを用いてPCRを行い、シャトルベクターpHY300PLKのBamHI制限酵素切断点に挿入された組換えプラスミドpHY−S237を構築し、プロトプラスト形質転換法によって各菌株に導入した。
【0053】
【表4】
【0054】
得られた菌株を、5mLのLB培地で、30℃で15時間振盪培養し、更にこの培養液0.6mLを30mLの2×L−マルトース培地(2% トリプトン、1% 酵母エキス、1% 塩化ナトリウム、7.5% マルトース、7.5ppm 硫酸マンガン4−5水和物、15ppm テトラサイクリン)に接種し、30℃で3日間振盪培養した。この際、測定誤差を算出する目的で、培養を3回行った。培養後、遠心分離によって菌体を除いた培養上清のアルカリセルラーゼ活性を測定し、菌体外に分泌生産されたアルカリセルラーゼの量を算出した。セルラーゼ活性測定は、1/7.5M リン酸緩衝液(pH7.4 和光純薬工業社製)で適宜希釈したサンプル溶液50μLに0.4mM p−ニトロフェニル−β−D−セロトリオシド(生化学工業社製)を50μL添加して混合し、30℃にて反応させた際に遊離するp−ニトロフェノールの量を、420nmにおける吸光度(OD420nm)変化により定量することにより行った。セルラーゼ活性は、1分間に1μmolのp−ニトロフェノールを遊離させる酵素量を1Uとして定義した。生産性の比較は、親株の生産性を100%とする相対値により行った(表5:表中の値は平均値±標準偏差(N=3)を示す)。この結果、親株としてMGB874株を用いた場合、親株に168株を用いた場合と比較し、malA欠失により高いアルカリセルラーゼの分泌生産量向上効果が確認された。
【0055】
【表5】
【技術分野】
【0001】
本発明は、有用なタンパク質又はポリペプチドの生産に用いる組換え枯草菌、及びそれを用いたタンパク質又はポリペプチドの生産方法に関する。
【背景技術】
【0002】
微生物による有用物質の工業的生産は、アルコール飲料や味噌、醤油等の食品の醸造をはじめとして、アミノ酸、有機酸、核酸関連物質、抗生物質、糖質、脂質、タンパク質等の工業的生産など、多岐に渡って実施されている。またその用途も、食品をはじめとして、医薬、洗剤、化粧品等の日用品、あるいは各種化成品原料に至るまで、幅広い分野に広がっている。
【0003】
微生物を用いた有用物質の工業的生産における一つの重要課題として、当該有用物質の生産性向上が挙げられる。従来、当該課題を解決するため、突然変異等の遺伝学的手法による高生産菌の育種が行われてきた。特に最近では、微生物遺伝学、バイオテクノロジーの発展により、遺伝子組換え技術等を用いた、より効率的な高生産菌の育種が行われている。更に、近年のゲノム解析技術の急速な発展を受け、対象とする微生物のゲノム情報を解読し、これらを積極的に産業に応用する試みもなされている。例えば、枯草菌(Bacillus subtilis)Marburg No.168系統株など、宿主微生物として安全かつ優良と認められた微生物菌株に更に改良を加えた菌株が開発されている。
【0004】
枯草菌(Bacillus subtilis)において、マルトースは、フォスフォエノールピルビン酸依存性の糖トランスポーター(PTS:Phosphoenol pyruvate−dependent phosphotransferase system)の一つであるMalPにより細胞内に取り込まれた後(非特許文献1)リン酸化され、マルトース6リン酸に変換される。更に、マルトース6リン酸は、6−ホスホ−α−グルコシダーゼ活性を有するMalAによりグルコース及びグルコース6リン酸へと分解され、解糖系で代謝される。MalAをコードする遺伝子malAはmalA−glvR−malPオペロンを構成することが知られている(非特許文献2)。
【0005】
本出願人は、過去の特許出願(特許文献1)において、枯草菌168株(野生株)を親株としてmalP(当時の名称はglvC)及びglvRを欠失させた株で酵素セルラーゼ及びα−アミラーゼの生産量が向上することを報告している。しかしながら、malA遺伝子の欠失による酵素生産量への影響に関しては明らかにされていない。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】特開2005−151983号公報
【特許文献2】特開2007−130013号公報
【非特許文献】
【0007】
【非特許文献1】J.bacteriol,188,11,p.3911−3922,2006
【非特許文献2】J.bacteriol,183,17,p.5110−5121,2001
【発明の開示】
【発明が解決しようとする課題】
【0008】
本発明は、タンパク質又はポリペプチドの生産性向上を可能とする組換え枯草菌、更には当該組換え枯草菌を用いたタンパク質又はポリペプチドの製造方法の提供に関する。
【課題を解決するための手段】
【0009】
本発明者らは、有用なタンパク質又はポリペプチドの生産にとって不要あるいは有害な働きをする遺伝子群をゲノム上から削除した枯草菌を宿主として用い、その宿主のmalA遺伝子を削除又は不活性化し、更に目的の異種タンパク質又は異種ポリペプチドをコードする遺伝子を導入することにより、当該異種タンパク質又はポリペプチドの生産が顕著に向上することを見出し、本発明を完成させるに至った。
【0010】
すなわち本発明は、以下の(1)から(9)に係るものである。
(1) prophage6領域、prophage1領域、prophage4領域、PBSX領域、prophage5領域、prophage3領域、spb領域、pks領域、skin領域、pps領域、prophage2領域、ydcL−ydeK−ydhU領域、yisB−yitD領域、yunA−yurT領域、cgeE−ypmQ領域、yeeK−yesX領域、pdp−rocR領域、ycxB−sipU領域、SKIN−Pro7領域、sbo−ywhH領域、yybP−yyaJ領域及びyncM−fosB領域が欠失したゲノム構造を有する枯草菌変異株に、異種タンパク質又は異種ポリペプチドをコードする遺伝子を導入して作製される組換え枯草菌であって、malA遺伝子が削除又は不活性化された組換え枯草菌。
(2) 前記枯草菌変異株が、更に以下の領域のうちの1つ又は2つ以上が欠失したゲノム構造を有する、上記(1)記載の組換え枯草菌:
ybbU−ybfI領域、ydjM−cotA領域、yefA−yesX領域、yfiB−yfiX領域、yhcE−yhcU領域、yhaU−yhaL領域、yjbX−yjlB領域、xkdA−ykcC領域、bpr−ylmA領域、flgB−cheD領域、ynfF−ppsA領域、yoxC−yobO領域、spoVAF−spoIIAA領域、spoIIIAH−yqhV領域、ytvB−ytqB領域、yteA−ytaB領域、yuaJ−yugO領域、yusJ−mrgA領域、gerAA−yvrI領域、yvaM−yvbK領域、araE−yveK領域、yvdE−yvcP領域、gerBA−ywsC領域、ywrK−ywqM領域、spoIIID−ywoB領域、slp−ylaF領域、licH−sigY領域、yqeF−yrhK領域、yuzE−yukJ領域及びyncM−yndN領域。
(3) 異種タンパク質又は異種ポリペプチドをコードする遺伝子の上流に、転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域から選ばれる1以上の領域が作動可能に連結されている、上記(1)又は(2)に記載の組換え枯草菌。
(4) 異種タンパク質又は異種ポリペプチドをコードする遺伝子の上流に、転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が作動可能に連結されている、上記(1)から(3)のいずれか1つに記載の組換え枯草菌。
(5) 前記分泌シグナル領域が、バチルス属細菌のセルラーゼ遺伝子由来のものであり、前記転写開始制御領域及び前記翻訳開始制御領域が、当該セルラーゼ遺伝子の上流0.6〜1kb領域由来のものである、上記(3)又は(4)記載の組換え枯草菌。
(6) 前記転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が、配列番号3で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜659の塩基配列、又は当該塩基配列のいずれかと70%以上の同一性を有する塩基配列からなるDNA断片、又は当該塩基配列の一部が欠失した塩基配列からなるDNA断片である、上記(4)記載の組換え枯草菌。
(7) 前記異種タンパク質又は異種ポリペプチドをコードする遺伝子が、バチルス属細菌のアルカリセルラーゼ遺伝子である、上記(1)から(6)のいずれか1つに記載の組換え枯草菌。
(8) 前記アルカリセルラーゼ遺伝子が、配列番号3で示される塩基配列からなるセルラーゼ遺伝子、又は当該塩基配列と70%以上の同一性を有する塩基配列からなるDNA断片、又は当該塩基配列の一部が欠失、置換若しくは付加された塩基配列からなるDNA断片である、上記(7)記載の組換え枯草菌。
(9) 上記(1)から(8)のいずれか1つに記載の組換え枯草菌を用いる異種タンパク質又は異種ポリペプチドの製造方法。
【発明の効果】
【0011】
本願発明の組換え枯草菌を用いれば、目的タンパク質又はポリペプチドを効率よく大量生産することが可能となる。
【図面の簡単な説明】
【0012】
【図1】SOE−PCRによる遺伝子欠失用DNA断片の調製、及び当該DNA断片を用いて標的遺伝子を欠失する(薬剤耐性遺伝子と置換)方法を模式的に示した図。
【発明を実施するための形態】
【0013】
本願発明の組換え枯草菌は、そのゲノムの一部が欠失した枯草菌変異株を親株として用い、malA遺伝子を削除又は不活性化させ、更に異種タンパク質又は異種ポリペプチドをコードする遺伝子を導入して作製される。
【0014】
枯草菌(Bacillus subtilis)とは、好気性のグラム陽性桿菌で、芽胞を形成する真正細菌の一種である。枯草菌は、全ゲノム情報が明らかにされ、遺伝子工学、ゲノム工学技術が確立されており、またタンパク質と菌体外に分泌生産させる能力を有するため、本願発明にとり有用な微生物といえる。
【0015】
本願発明において用いる枯草菌としては、そのゲノムの大領域、すなわちprophage6(yoaV−yobO)領域、prophage1(ybbU−ybdE)領域、prophage4(yjcM−yjdJ)領域、PBSX(ykdA−xlyA)領域、prophage5(ynxB−dut)領域、prophage3(ydiM−ydjC)領域、spb(yodU−ypqP)領域、pks(pksA−ymaC)領域、skin(spoIVCB−spoIIIC)領域、pps(ppsE−ppsA)領域、prophage2(ydcL−ydeJ)領域、ydcL−ydeK−ydhU領域、yisB−yitD領域、yunA−yurT領域、cgeE−ypmQ領域、yeeK−yesX領域、pdp−rocR領域、ycxB−sipU領域、SKIN−Pro7(spoIVCB−yraK)領域、sbo−ywhH領域、yybP−yyaJ領域及びyncM−fosB領域を欠失させて構築した株が用いられ、具体的には、枯草菌Bacillus subtilis Marburg No.168(枯草菌168株)を野生株とし、当該大領域を欠失させて構築した枯草菌MGB874株が挙げられる(特許文献2)。これらの欠失領域はまた、表1に示す一対のオリゴヌクレオチドセットにより挟み込まれる領域として表すことができる。
【0016】
【表1】
【0017】
枯草菌168株の全塩基配列及び遺伝子は既に報告されており、またインターネット公開されている(Nature,390,249−256,1997及びBSORF Bacillus subtilis Genome Database[http://bacillus.genome.jp/];GenBank:AL009126.2[http://www.ncbi.nlm.nih.gov/nuccore/38680335])。当業者は、これらの情報源から得た枯草菌168株のゲノム情報、例えばGenBank:AL009126.2[http://www.ncbi.nlm.nih.gov/nuccore/38680335]に基づいて、MGB874株のゲノムから欠失させるべき上記表2に示した遺伝子領域を見出すことができる。ここで、欠失させるべき遺伝子領域は、公開されている枯草菌168株の遺伝子の塩基配列に対して1又は複数個の塩基における天然又は人工的に引き起こされた欠失、置換、挿入、付加等の変異を含み得る塩基配列を有し得る。変異し得る塩基の個数の範囲は、枯草菌168株のゲノムとの対応関係が認識できる限り特に限定されないが、例えば、枯草菌168株のゲノム領域の塩基配列に対して50%以上の同一性、好ましくは60%以上の同一性、より好ましくは70%以上の同一性、更に好ましくは80%以上の同一性、更により好ましくは90%以上の同一性、なお好ましくは95%以上の同一性を有する範囲が挙げられる。上記「付加」には、塩基配列の一末端及び両末端への塩基の付加が含まれる。
【0018】
本発明に係る枯草菌変異株は、上記枯草菌変異株MGB874株のゲノム領域から、更にゲノム領域を欠失させたゲノム構造を有してもよい。具体的には、当該枯草菌変異株は、枯草菌168株のゲノム上における表2に示す遺伝子領域のうちの少なくとも1つを、枯草菌変異株MGB874株のゲノム領域から更に欠失させたゲノム構造を有してもよい。当該欠失領域としては、表2に示される遺伝子領域及び当該領域に相当する領域が挙げられる。
【0019】
【表2】
【0020】
本発明において削除すべきmalA遺伝子(BSORF Gene Entry No.:BG11839、EC3.2.1.122)の遺伝子産物であるMalAタンパク質は、α−グルコシダーゼ活性を有し、マルトースを分解してグルコースに変換する。malA遺伝子はmalP遺伝子及びglvR遺伝子と共にmalA−glvR−malPオペロンを構成し、マルトースの代謝活動に関与する。
【0021】
これらmalA遺伝子以外の遺伝子の欠失又は不活化との組み合わせにより、異種タンパク質又は異種ポリペプチドの生産性向上に対するより大きな効果が期待される。
【0022】
本発明に係るmalA遺伝子が削除又は不活性化された組換え枯草菌は、例えば相同組換えにより標的遺伝子を削除又は不活性化する方法など、既に報告された方法(Mol.Gen.Genet.,223,268(1990)等)を繰り返すことによって構築される。
【0023】
また、ランダムな遺伝子の削除又は不活性化についてもランダムにクローニングしたDNA断片を用いて上述の方法と同様な相同組換えを起こさせる方法や、親微生物にγ線等を照射すること等によっても実施可能である。
【0024】
本発明では、上記のmalA遺伝子内に他のDNA断片を挿入する方法、あるいは当該遺伝子の転写・翻訳開始領域を変異させる方法等によって目的遺伝子を不活性化しても本発明の目的を達成できるが、標的遺伝子を物理的に削除する方がより好ましい。また、削除又は不活性化する遺伝子は1つでもよいが、2つ以上の遺伝子の削除を組み合わせてもよい。更に本発明の組換え枯草菌の構築には、マルトースの膜透過関与以外の遺伝子群の削除又は不活性化を組み合わせることも可能であり、生産性向上に対してより大きな効果が期待される。
【0025】
本発明における、malA遺伝子の削除又は不活性化の具体例として、SOE(splicing by overlap extension)−PCR法(Gene,77,61,(1989))によって調製される削除用DNA断片を用いた二重交差法による削除方法について説明するが、本発明における遺伝子削除方法はこの方法に限定されるものではない。
【0026】
本方法で用いる削除用DNA断片は、削除対象遺伝子の上流に隣接する約0.2〜3kb断片と、同じく下流に隣接する約0.2〜3kb断片の間に、薬剤耐性マーカー遺伝子断片を挿入して構築した断片である。まず、1回目のPCRによって、削除対象遺伝子の上流断片及び下流断片、並びに薬剤耐性マーカー遺伝子断片の3断片を調製するが、この際、例えば、上流断片の下流末端に薬剤耐性マーカー遺伝子の上流側10〜30塩基対配列、逆に下流断片の上流末端には薬剤耐性マーカー遺伝子の下流側10〜30塩基対配列が付加されるようにデザインされたプライマーを用いる(図1)。
【0027】
次いで、1回目に調製した3種類のPCR断片を鋳型とし、上流断片の上流側プライマーと下流断片の下流側プライマーを用いて2回目のPCRを行うことによって、上流断片の下流末端及び下流断片の上流末端に付加した薬剤耐性マーカー遺伝子配列において、薬剤耐性マーカー遺伝子断片とのアニーリングが生じ、PCR増幅の結果、上流側断片と下流側断片の間に、薬剤耐性マーカー遺伝子が挿入されたDNA断片が得られる(図1)。
【0028】
表3に示したプライマーセットと適当な鋳型DNAを用い、Pyrobest DNAポリメーラーゼ(宝酒造)などの一般のPCR用酵素キット等を用いて、“PCR Protocols.Current Methods and Applications”,Edited by B.A.White,Humana Press,pp251(1993)、Gene,77,61,(1989)等に示される通常の条件によりSOE−PCRを行うことによって、各遺伝子の削除用DNA断片が得られる。
【0029】
かくして得られた削除用DNA断片を、コンピテント法等によって細胞内に導入すると、削除対象遺伝子の上流及び下流の、上記削除用DNA断片との相同領域おいて、細胞内での遺伝子組換えが生じ、削除対象遺伝子が薬剤耐性遺伝子で置換された細胞を、薬剤耐性マーカーによる選抜により単離できる(図1)。即ち、表3に示したプライマーセットを用いて調製した削除用DNA断片を導入した場合、薬剤を含む寒天培地上に生育するコロニーを単離し、目的の遺伝子が削除されて薬剤耐性遺伝子と置換していることを、ゲノムを鋳型としたPCR法などによって確認すればよい。
【0030】
次に、かくして得られた本発明に係る枯草菌変異株に、目的とする異種タンパク質又はポリペプチドをコードする遺伝子を導入することによって、本発明の組換え枯草菌を得ることができる。
【0031】
当該組換え枯草菌を用いて異種タンパク質又はポリペプチドを製造する場合、上記目的タンパク質又はポリペプチドの遺伝子の上流に、当該遺伝子の転写、翻訳、分泌を制御する制御領域を、適切な形で結合させるのが望ましい。かかる制御領域としては、プロモーター及び転写開始点を含む転写開始制御領域、リボソーム結合部位及び開始コドンを含む翻訳開始領域並びに分泌シグナルペプチド領域から選ばれる1以上の領域などが挙げられる。特に転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が結合されていることが好ましく、更に分泌シグナルペプチド領域がバチルス(Bacillus)属細菌のセルラーゼ遺伝子由来のものであり、転写開始制御領域及び翻訳開始制御領域が当該セルラーゼ遺伝子の開始コドンから始まる長さ0.6〜1kbの上流領域であるものが、異種タンパク質又はポリペプチド遺伝子と作動可能に連結されていることが望ましい。例えば、特開2000−210081号公報や特開平4−190793号公報等に記載されているバチルス(Bacillus)属細菌、すなわちKSM−S237株(FERM BP−7875)、KSM−64株(FERM BP−2886)由来のセルラーゼ遺伝子の転写開始制御領域、翻訳開始領域及び分泌シグナルペプチド領域が目的のタンパク質又はポリペプチドの構造遺伝子と作動可能に連結されていることが望ましい。より具体的には、前記転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が、配列番号3で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜659の塩基配列、又は当該塩基配列のいずれかと70%以上、好ましくは80%以上、さらに好ましくは90%以上、特に好ましくは95%以上の同一性を有し、且つ遺伝子の転写、翻訳、分泌に関わる機能を有する塩基配列からなるDNA断片、又は当該塩基配列の一部が欠失した塩基配列からなるDNA断片が、上記の異種タンパク質又はポリペプチド遺伝子と作動可能に連結していることが望ましい。ここで、上記塩基配列の一部が欠失した塩基配列からなるDNA断片とは、上記塩基配列の一部を欠失しているが、遺伝子の転写、翻訳、分泌に関わる機能を保持しているDNA断片を意味する。また、ここで、作動可能に連結されている(operably linked)とは、上記の制御配列が機能してコード配列によりコードされた目的タンパク質の発現が制御されうる位置関係で、当該制御配列と当該コード配列とが配置していることを指す。具体例としては、プロモーターとそれに隣接する目的遺伝子とが、当該プロモーターの方向に沿って、当該遺伝子が発現しうる状態で配置していることを指す。また、ここで、ストリンジェントな条件とは、例えば[Molecular cloning−a Laboratory manual 2nd edition(Sambrookら、1989)]に記載の条件等が挙げられる。例えば、6×SSC(1×SSCの組成:0.15M 塩化ナトリウム、0.015M クエン酸ナトリウム、pH7.0)、0.5% SDS、5×デンハート液及び100mg/mL ニシン精子DNAを含む溶液中で、プローブと共に65℃で8〜16時間インキュベートし、ハイブリダイズさせる条件が挙げられる。
【0032】
異種タンパク質又はポリペプチドをコードする遺伝子を細胞内で発現させるためには、当該遺伝子を含むDNA断片を、適切なベクターに挿入した発現プラスミドを構築し、当該発現プラスミドを宿主に導入して形質転換する必要がある。当該発現ベクターとしては、枯草菌体内で自立複製可能なベクターが好適であり、例えばシャトルベクターpHY300PLK等が挙げられるが、特に限定されない。
【0033】
ポリペプチドとは一般に、直鎖状に連結したアミノ酸のポリマーを指し、タンパク質とは一般に、50以上のアミノ酸からなる1つ以上のポリペプチドのことを指すが、本願においてはこれらの用語は交換可能に用いられるものとする。
【0034】
本発明の組換え枯草菌を用いて生産する異種タンパク質又はポリペプチドとしては、例えば食品用、医薬用、化粧用、洗浄用、繊維処理用、検査用に用いられる各種産業用酵素や、生理活性ペプチドなどが挙げられる。また、産業用酵素の機能別には、酸化還元酵素(オキシドレダクターゼ)、転移酵素(トランスフェラーゼ)、加水分解酵素(ヒドロラーゼ)、脱離酵素(リアーゼ)、異性化酵素(イソメラーゼ)、合成酵素(リガーゼ/シンセターゼ)等が含まれるが、好適にはセルラーゼ、α−アミラーゼ、プロテアーゼ等の加水分解酵素が挙げられ、より好ましくはセルラーゼが挙げられる。
【0035】
より具体的な例として、配列番号3で示される塩基配列によりコードされるバチルス属細菌KSM−S237(FERM BP−7875)由来のアルカリセルラーゼや、配列番号119で示されるアミノ酸配列からなるバチルス属細菌KSM−64株(FERM BP−2886)由来のアルカリセルラーゼ、配列番号4又は119で示されるアミノ酸配列の1個もしくは数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列を有するアルカリセルラーゼが挙げられ、更には、当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、更に好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列からなるセルラーゼが挙げられる。
【0036】
また、α−アミラーゼの具体例としては、微生物由来のα−アミラーゼが挙げられ、特にバチルス属細菌由来の液化型アミラーゼや、配列番号120で示されるアミノ酸配列からなるバチルス属細菌KSM−K38株(FERM BP−6946)由来のアルカリアミラーゼ、当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、更に好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列からなるアミラーゼ、配列番号120で示されるアミノ酸配列において1〜数個のアミノ酸が置換、欠失若しくは付加されたアミノ酸配列からなりアミラーゼ活性を有するタンパク質が挙げられる。また、プロテアーゼの具体例としては、微生物由来、特にバチルス属細菌由来のセリンプロテアーゼや金属プロテアーゼや、配列番号121で示されるアミノ酸配列からなるバチルス クラウジ(Bacillus clausii)KSM−K16株(FERM BP−3376)由来のアルカリプロテアーゼ、あるいは当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、さらにより好ましくは98%以上の同一性を有するアミノ酸配列からなりプロテアーゼ活性を有するタンパク質、配列番号121で示されるアミノ酸配列において1〜数個のアミノ酸が置換、欠失若しくは付加されたアミノ酸配列からなりプロテアーゼ活性を有するタンパク質が挙げられる。
【0037】
本発明において、アミノ酸配列及び塩基配列の同一性は、Lipman−Pearson法(Science,227,1435,(1985))によって計算される。より具体的には、遺伝情報処理ソフトウェアGenetyx−Win(ソフトウェア開発)のホモロジー解析(Search homology)プログラムを用いて、Unit size to compare(ktup)を2として解析を行うことにより算出される。
【0038】
本願発明の別の態様は、上述した組換え枯草菌を用いる異種タンパク質又は異種ポリペプチドの製造方法に関する。
【0039】
本発明の組換え枯草菌は、上記の異種タンパク質又はポリペプチド遺伝子を含むDNA断片と適当なプラスミドベクターを結合させた組換えプラスミドを、一般的な形質転換法によって宿主となる枯草菌変異株に取り込ませることによって得られる。また、当該DNA断片に宿主ゲノムとの適当な相同領域を結合したDNA断片を用い、宿主ゲノムに直接組み込むことによって本発明の組換え枯草菌を得てもよい。
【0040】
上記の組換え枯草菌を用いた異種タンパク質又はポリペプチドの製造は、上記異種タンパク質又はポリペプチドの遺伝子を上記のとおり宿主となる枯草菌変異株に導入して得られる菌株を、例えば同化性の炭素源、窒素源、その他の必須成分を含む培地に接種して培養して行うことができる。培養方法は、原則的には一般的な微生物の培養方法であってもよく、通常、液体培養による振盪培養、通気撹拌培養等の好気的条件下で実施するのが好ましい。培養終了後、培養液を遠心分離し、得られる上清又は菌体から、目的の異種タンパク質又はポリペプチドを、硫安沈殿やクロマトグラフィなどを適宜組み合わせ、常法に従い抽出・精製することにより得ることができる。
【0041】
以下の実施例において、本発明の組換え枯草菌の構築方法及び当該組換え枯草菌を用いたタンパク質の製造方法について説明するが、本願発明は以下の実施例に限定されるものではない。
【実施例】
【0042】
枯草菌MGB874株のmalA遺伝子を削除した組換え枯草菌株の構築、並びに当該組換え枯草菌を用いたセルラーゼの生産方法について、以下に具体的に説明する。
【0043】
以下の実施例におけるDNA断片増幅のためのポリメラーゼ連鎖反応(PCR)は、Pyrobest DNAポリメラーゼ(タカラバイオ社製)と付属の試薬類を用いた、GeneAmp PCR System(アプライドバイオシステムズ社製)によるDNA増幅により行った。PCR反応液は、適宜希釈した鋳型DNAを1μL、センス及びアンチセンスプライマーを各々20pmol、及びPyrobest DNAポリメラーゼを2.5U添加し、更に総反応液量を50μLとすることにより調製した。PCR反応条件は、98℃で10秒間、55℃で30秒間及び72℃で1〜5分間(目的増幅産物に応じて調整。目安は1kbあたり1分間)の3段階の温度変化を30サイクル繰り返した後、72℃で5分間反応、とした。
【0044】
また、以下の実施例において、遺伝子の上流及び下流とは、複製開始点からの位置を指すのではなく、上流とは各操作・工程において対象となる遺伝子の開始コドンの5’側に続く領域を指し、一方下流とは各操作・工程において対象となる遺伝子の終止コドンの3’側に続く領域を指すものとする。
【0045】
更に、以下の実施例における各遺伝子及び遺伝子領域の名称は、Nature,390,249−256,(1997)で報告され、JAFAN:Japan Functional Analysis Network for Bacillus subtilis(BSORF DB)としてインターネットで公開された枯草菌ゲノムデータ(http://bacillus.genome.ad.jp/2004年3月10日更新)に基づき記載する。
【0046】
枯草菌の形質転換は、コンピテントセル法(J.Bacteriol.93,1925(1967))により行った。すなわち、枯草菌株をSPI培地(0.20% 硫酸アンモニウム、1.40% リン酸水素二カリウム、0.60% リン酸二水素カリウム、0.10% クエン酸三ナトリウム二水和物、0.50% グルコース、0.02% カザミノ酸(Difco社製)、5mM 硫酸マグネシウム、0.25μM 塩化マンガン、50μg/mL トリプトファン)中で、37℃で、生育度(OD600)の値が約1となるまで振盪培養し、振盪培養後、培養液の一部を9倍量のSPII培地(0.20% 硫酸アンモニウム、1.40% リン酸水素二カリウム、0.60% リン酸二水素カリウム、0.10% クエン酸三ナトリウム二水和物、0.50% グルコース、0.01% カザミノ酸(Difco社製)、5mM 硫酸マグネシウム、0.40μM 塩化マンガン、5μg/mL トリプトファン)に接種し、更に生育度(OD600)の値が約0.4となるまで振盪培養することにより、枯草菌株のコンピテントセルを調製した。
【0047】
次いで調製したコンピテントセル懸濁液(SPII培地における培養液)100μLに、各種DNA断片を含む溶液(SOE−PCRの反応液等)を5μL添加し、37℃で1時間振盪培養後、適切な薬剤を含むLB寒天培地(1% トリプトン、0.5% 酵母エキス、1% NaCl、1.5% 寒天)に全量を塗沫した。37℃での静置培養の後、生育したコロニーを形質転換体として分離した。得られた形質転換体のゲノムを抽出し、これを鋳型とするPCRを行い、目的とするゲノム構造の改変がなされたことを確認した。
【0048】
目的のタンパク質を発現するプラスミドの宿主微生物への導入は、プロトプラスト形質転換法(Mol.Gen.Genet.168,111(1979))により行った。組換え枯草菌によるタンパク質生産用の培養として、LB培地(1% トリプトン、0.5% 酵母エキス、1% NaCl)、及び2×L−マルトース培地(2% トリプトン、1% 酵母エキス、1% NaCl、7.5% マルトース、7.5ppm 硫酸マンガン4〜5水和物)を用いた。
【0049】
(実施例1:malA遺伝子欠失株の構築)
malA遺伝子(配列番号1)はmalA−glvR−malPオペロンの先頭に位置する遺伝子であり、本遺伝子を欠失する際には、malAの下流遺伝子であるglvR及びmalPの発現に影響を与えない形で欠失株の構築を行わなければならない。そこで、本発明においては、malAの構造遺伝子をネオマイシン耐性遺伝子のORF(open reading frame)と置換する方法により、malA遺伝子欠失株を構築した。枯草菌MGB874株から抽出したゲノムDNAを鋳型とし、表3に示したmalAFW1とmalA/neof2.R、及びmalA/neor2.FとmalA RVの各プライマーセットを用いて、ゲノム上のmalA遺伝子の上流に隣接する1.0kbp断片(A)、及び下流に隣接する1.0kbp断片(B)をそれぞれ調製した。また、ネオマイシン耐性遺伝子を有するプラスミドpUB110(Plasmid 15:93−103.(1986))を鋳型として、表3に示したneof2及びneor2のプライマーセットを用いて、ネオマイシン耐性遺伝子(C)を調製した。次に、得られた(A)、(B)及び(C)の3断片を混合して鋳型とし、malA FW2とmalA RV2のプライマーを用いてPCRを行ない、3断片を(A)−(C)−(B)の順になるように結合させ、遺伝子欠失用のDNA断片を得た(図1参照)。このDNA断片を用いて、上述したコンピテントセル法により、枯草菌168株及びMGB874株を形質転換し、ネオマイシン(10mg/L)及びマルトース(1g/L)を含むLB寒天培地上で生育したコロニーを形質転換体として単離した。マルトース存在下でmalA−glvR−malPオペロンの発現が誘導されるため(J.bacteriol.,183,17,5110,(2001))、形質転換体ではネオマイシン耐性遺伝子が発現し、ネオマイシンを含むLB寒天培地上でも生育可能となる。
【0050】
得られた形質転換体からゲノムDNAを抽出し、これを鋳型としてPCRを行った結果、malA遺伝子がネオマイシン耐性遺伝子と置換され、malA遺伝子欠失株となっていることが確認された。以上のようにして得られたmalA欠失株のうち、枯草菌168株を親株として構築した株を168ΔmalA株と、枯草菌MGB874株を親株として構築した株を874ΔmalA株とした。
【0051】
【表3】
【0052】
(実施例2:セルラーゼ生産性評価)
実施例1にて得られた168ΔmalA株、874ΔmalA株及び対照として、親株である枯草菌168株、MGB874株に、アルカリセルラーゼ遺伝子を導入した。具体的には、バチルス属細菌 KSM−S237株(FERM BP−7875)由来のS237セルラーゼ遺伝子(特開2000−210081号公報参照)(配列番号3)をコードするDNA断片(3.1kb;配列番号3の塩基番号13〜3124)を鋳型として、表4に示されるプライマーEgl−S237.F及びプライマーEgl−S237.Rのプライマーセットを用いてPCRを行い、シャトルベクターpHY300PLKのBamHI制限酵素切断点に挿入された組換えプラスミドpHY−S237を構築し、プロトプラスト形質転換法によって各菌株に導入した。
【0053】
【表4】
【0054】
得られた菌株を、5mLのLB培地で、30℃で15時間振盪培養し、更にこの培養液0.6mLを30mLの2×L−マルトース培地(2% トリプトン、1% 酵母エキス、1% 塩化ナトリウム、7.5% マルトース、7.5ppm 硫酸マンガン4−5水和物、15ppm テトラサイクリン)に接種し、30℃で3日間振盪培養した。この際、測定誤差を算出する目的で、培養を3回行った。培養後、遠心分離によって菌体を除いた培養上清のアルカリセルラーゼ活性を測定し、菌体外に分泌生産されたアルカリセルラーゼの量を算出した。セルラーゼ活性測定は、1/7.5M リン酸緩衝液(pH7.4 和光純薬工業社製)で適宜希釈したサンプル溶液50μLに0.4mM p−ニトロフェニル−β−D−セロトリオシド(生化学工業社製)を50μL添加して混合し、30℃にて反応させた際に遊離するp−ニトロフェノールの量を、420nmにおける吸光度(OD420nm)変化により定量することにより行った。セルラーゼ活性は、1分間に1μmolのp−ニトロフェノールを遊離させる酵素量を1Uとして定義した。生産性の比較は、親株の生産性を100%とする相対値により行った(表5:表中の値は平均値±標準偏差(N=3)を示す)。この結果、親株としてMGB874株を用いた場合、親株に168株を用いた場合と比較し、malA欠失により高いアルカリセルラーゼの分泌生産量向上効果が確認された。
【0055】
【表5】
【特許請求の範囲】
【請求項1】
prophage6領域、prophage1領域、prophage4領域、PBSX領域、prophage5領域、prophage3領域、spb領域、pks領域、skin領域、pps領域、prophage2領域、ydcL−ydeK−ydhU領域、yisB−yitD領域、yunA−yurT領域、cgeE−ypmQ領域、yeeK−yesX領域、pdp−rocR領域、ycxB−sipU領域、SKIN−Pro7領域、sbo−ywhH領域、yybP−yyaJ領域及びyncM−fosB領域が欠失したゲノム構造を有する枯草菌変異株に、異種タンパク質又は異種ポリペプチドをコードする遺伝子を導入して作製される組換え枯草菌であって、malA遺伝子が削除又は不活性化された組換え枯草菌。
【請求項2】
前記枯草菌変異株が、更に以下の領域のうちの1つ又は2つ以上が欠失したゲノム構造を有する、請求項1記載の組換え枯草菌:
ybbU−ybfI領域、ydjM−cotA領域、yefA−yesX領域、yfiB−yfiX領域、yhcE−yhcU領域、yhaU−yhaL領域、yjbX−yjlB領域、xkdA−ykcC領域、bpr−ylmA領域、flgB−cheD領域、ynfF−ppsA領域、yoxC−yobO領域、spoVAF−spoIIAA領域、spoIIIAH−yqhV領域、ytvB−ytqB領域、yteA−ytaB領域、yuaJ−yugO領域、yusJ−mrgA領域、gerAA−yvrI領域、yvaM−yvbK領域、araE−yveK領域、yvdE−yvcP領域、gerBA−ywsC領域、ywrK−ywqM領域、spoIIID−ywoB領域、slp−ylaF領域、licH−sigY領域、yqeF−yrhK領域、yuzE−yukJ領域及びyncM−yndN領域。
【請求項3】
異種タンパク質又は異種ポリペプチドをコードする遺伝子の上流に、転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域から選ばれる1以上の領域が作動可能に連結されている、請求項1又は2記載の組換え枯草菌。
【請求項4】
異種タンパク質又は異種ポリペプチドをコードする遺伝子の上流に、転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が作動可能に連結されている、請求項1から3のいずれか1項記載の組換え枯草菌。
【請求項5】
前記分泌シグナル領域が、バチルス属細菌のセルラーゼ遺伝子由来のものであり、前記転写開始制御領域及び前記翻訳開始制御領域が、当該セルラーゼ遺伝子の上流0.6〜1kb領域由来のものである、請求項3又は4記載の組換え枯草菌。
【請求項6】
前記転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が、配列番号3で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜659の塩基配列、又は当該塩基配列のいずれかと70%以上の同一性を有する塩基配列からなるDNA断片、又は当該塩基配列の一部が欠失した塩基配列からなるDNA断片である、請求項5記載の組換え枯草菌。
【請求項7】
前記異種タンパク質又は異種ポリペプチドをコードする遺伝子が、バチルス属細菌のアルカリセルラーゼ遺伝子である、請求項1から6のいずれか1項記載の組換え枯草菌。
【請求項8】
前記アルカリセルラーゼ遺伝子が、配列番号3で示される塩基配列からなるセルラーゼ遺伝子、又は当該塩基配列と70%以上の同一性を有する塩基配列からなるDNA断片、又は当該塩基配列の一部が欠失、置換若しくは付加された塩基配列からなるDNA断片である、請求項7記載の組換え枯草菌。
【請求項9】
請求項1から8のいずれか1項記載の組換え枯草菌を用いる異種タンパク質又は異種ポリペプチドの製造方法。
【請求項1】
prophage6領域、prophage1領域、prophage4領域、PBSX領域、prophage5領域、prophage3領域、spb領域、pks領域、skin領域、pps領域、prophage2領域、ydcL−ydeK−ydhU領域、yisB−yitD領域、yunA−yurT領域、cgeE−ypmQ領域、yeeK−yesX領域、pdp−rocR領域、ycxB−sipU領域、SKIN−Pro7領域、sbo−ywhH領域、yybP−yyaJ領域及びyncM−fosB領域が欠失したゲノム構造を有する枯草菌変異株に、異種タンパク質又は異種ポリペプチドをコードする遺伝子を導入して作製される組換え枯草菌であって、malA遺伝子が削除又は不活性化された組換え枯草菌。
【請求項2】
前記枯草菌変異株が、更に以下の領域のうちの1つ又は2つ以上が欠失したゲノム構造を有する、請求項1記載の組換え枯草菌:
ybbU−ybfI領域、ydjM−cotA領域、yefA−yesX領域、yfiB−yfiX領域、yhcE−yhcU領域、yhaU−yhaL領域、yjbX−yjlB領域、xkdA−ykcC領域、bpr−ylmA領域、flgB−cheD領域、ynfF−ppsA領域、yoxC−yobO領域、spoVAF−spoIIAA領域、spoIIIAH−yqhV領域、ytvB−ytqB領域、yteA−ytaB領域、yuaJ−yugO領域、yusJ−mrgA領域、gerAA−yvrI領域、yvaM−yvbK領域、araE−yveK領域、yvdE−yvcP領域、gerBA−ywsC領域、ywrK−ywqM領域、spoIIID−ywoB領域、slp−ylaF領域、licH−sigY領域、yqeF−yrhK領域、yuzE−yukJ領域及びyncM−yndN領域。
【請求項3】
異種タンパク質又は異種ポリペプチドをコードする遺伝子の上流に、転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域から選ばれる1以上の領域が作動可能に連結されている、請求項1又は2記載の組換え枯草菌。
【請求項4】
異種タンパク質又は異種ポリペプチドをコードする遺伝子の上流に、転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が作動可能に連結されている、請求項1から3のいずれか1項記載の組換え枯草菌。
【請求項5】
前記分泌シグナル領域が、バチルス属細菌のセルラーゼ遺伝子由来のものであり、前記転写開始制御領域及び前記翻訳開始制御領域が、当該セルラーゼ遺伝子の上流0.6〜1kb領域由来のものである、請求項3又は4記載の組換え枯草菌。
【請求項6】
前記転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が、配列番号3で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜659の塩基配列、又は当該塩基配列のいずれかと70%以上の同一性を有する塩基配列からなるDNA断片、又は当該塩基配列の一部が欠失した塩基配列からなるDNA断片である、請求項5記載の組換え枯草菌。
【請求項7】
前記異種タンパク質又は異種ポリペプチドをコードする遺伝子が、バチルス属細菌のアルカリセルラーゼ遺伝子である、請求項1から6のいずれか1項記載の組換え枯草菌。
【請求項8】
前記アルカリセルラーゼ遺伝子が、配列番号3で示される塩基配列からなるセルラーゼ遺伝子、又は当該塩基配列と70%以上の同一性を有する塩基配列からなるDNA断片、又は当該塩基配列の一部が欠失、置換若しくは付加された塩基配列からなるDNA断片である、請求項7記載の組換え枯草菌。
【請求項9】
請求項1から8のいずれか1項記載の組換え枯草菌を用いる異種タンパク質又は異種ポリペプチドの製造方法。
【図1】

【公開番号】特開2012−95604(P2012−95604A)
【公開日】平成24年5月24日(2012.5.24)
【国際特許分類】
【出願番号】特願2010−246270(P2010−246270)
【出願日】平成22年11月2日(2010.11.2)
【出願人】(000000918)花王株式会社 (8,290)
【Fターム(参考)】
【公開日】平成24年5月24日(2012.5.24)
【国際特許分類】
【出願日】平成22年11月2日(2010.11.2)
【出願人】(000000918)花王株式会社 (8,290)
【Fターム(参考)】
[ Back to top ]