
組換え熱帯熱マラリア原虫(Plasmodiumfalciparum)メロゾイト表面タンパク質4および5およびこれらの使用
【課題】組換え熱帯熱マラリア原虫(Plasmodium falciparum)メロゾイト表面タンパク質4および5およびこれらの使用方法を提供する。
【解決手段】熱帯熱マラリア原虫(Plasmodium falciparum)MSP4およびMSP5、並びに生じるポリペプチドをコードする核酸が修飾された構築物。または、バキュロウイルス-昆虫細胞発現系において可溶性の、分泌されたポリペプチドとして発現される組換えMSP4およびMSP5ポリペプチド。前記ポリペプチドに対して産生される抗体および診断法、キット、ワクチン、または抗寄生虫療法におけるこのようなポリペプチドおよび抗体の使用
【解決手段】熱帯熱マラリア原虫(Plasmodium falciparum)MSP4およびMSP5、並びに生じるポリペプチドをコードする核酸が修飾された構築物。または、バキュロウイルス-昆虫細胞発現系において可溶性の、分泌されたポリペプチドとして発現される組換えMSP4およびMSP5ポリペプチド。前記ポリペプチドに対して産生される抗体および診断法、キット、ワクチン、または抗寄生虫療法におけるこのようなポリペプチドおよび抗体の使用
【発明の詳細な説明】
【技術分野】
【0001】
本出願は、組換え熱帯熱マラリア原虫(Plasmodium falciparum)メロゾイト表面タンパク質4および5およびこれらの使用に関する。
【発明の分野】
【0002】
本発明は、単離された組換えポリペプチド、このようなポリペプチドをコードする核酸、このようなポリペプチドの組換え形態の産生のための方法、これらのポリペプチドに対して産生される抗体および診断法、キット、ワクチンまたは抗寄生虫療法におけるこのようなポリペプチドおよび抗体の使用に向けられる。
【発明の背景】
【0003】
マラリアは、世界中で、大部分は5歳以下のアフリカの子供の、1年あたりおよそ2,000,000人の死亡の原因であり、多くの世界の最も乏しい国に対して莫大な公衆衛生負担を負わせている。この負担は、寄生虫およびその蚊媒介伝播における薬物耐性のため、驚くべき速度で増加しており、有効なワクチンに対する緊急の需要を募らせている。
【0004】
今までに調べられた最も有望な血液段階ワクチン候補は、メロゾイト表面タンパク質1(MSP1)および頂端膜抗原(AMA1)である。これらの表面抗原をターゲットする体液性免疫応答は、疾患発病率の減少と相関することが見いだされており、インビトロにおいて、このような抗体は、赤血球(RBC)の寄生虫再侵入を阻害することができる[1-3]。しかし、これらの抗原遺伝子は、一般に免疫エフェクターにアクセス可能ではないタンパク質をコードする遺伝子と比較して、不釣合に高い数の非同義一塩基多型(nsSNPs)を示し[4-6]、これらのnsSNPsのいくつかは、宿主免疫系に最もアクセス可能なタンパク質の領域内にクラスター形成したラジカルアミノ酸置換をコードする[7]。このようなアミノ酸多型は、BおよびT細胞エピトープの両方を変化させることによって免疫回避において機能し得る。現在、いずれの機能的なマラリアワクチンも、多対立遺伝子反応を誘導することを願って、それぞれの標的抗原のいくつかの対立遺伝子型および/またはいくつかの標的抗原の保存領域で構成されることを必要とするであろうことが一般に認められている。
【0005】
より詳細には、当該技術分野において、マラリアの、特に熱帯熱マラリア原虫(Plasmodium falciparum)マラリアおよび三日熱マラリア原虫(Plasmodium vivax)マラリアの診断、並びに治療に使用することができる抗原に対する需要が存在する。特に、風土病領域における疾患から保護する特異的免疫応答と関連した保存された抗原および多変異体(multi-valiant)マラリアワクチンの成分としてのこれらの適合性評価に対する需要がある。
【発明の概要】
【0006】
本発明は、当該技術分野におけるこれらの需要を果たすことを助ける。本発明は、配列番号:1、2、3、4、5、6、7、または8のDNA配列を含む精製され、または組換えもしくは合成の核酸分子および配列番号:9、10、11、12、13、14、15、16、17、18、19、20、21または22のアミノ酸配列をコードする精製され、または組換えもしくは合成の核酸分子を提供する。また、本発明は、これらの配列に相補的な精製された核酸分子も包含する。
【0007】
また、本発明は、配列によって予測される、配列番号:1〜8のDNA配列を含む精製されたか、または組換えの核酸分子によってコードされる精製されたポリペプチドを包含する。
【0008】
配列番号:1〜8のDNA配列または配列番号9〜22のアミノ酸配列に対する言及は、これらの配列のいずれかおよび各々に対する言及である。
【0009】
本発明は、配列番号:1〜8のDNA配列を含む精製された二本鎖核酸分子および配列番号:9〜22のアミノ酸配列をコードする精製された二本鎖核酸分子を含む。一本鎖および二本鎖の両方のRNAおよびDNA核酸分子が、本発明によって包含される。これらの分子は、本発明に包含されるポリペプチドをコードする一本鎖および二本鎖のRNA並びにDNA変異体を検出するために使用することができる。二本鎖DNAプローブにより、核酸分子のいずれの鎖にも相当する核酸分子の検出ができる。
【0010】
42℃にて50%のホルムアミドおよび6×SSCにおける適度なストリンジェンシーの条件下において、60℃、0.5×SSC、0.1%のSDSの洗浄条件で、配列番号:1〜8および配列番号:9〜22のアミノ酸配列をコードするDNA配列を含む変性された、二本鎖DNAにハイブリダイズする精製された核酸分子が本発明に包含される。
【0011】
本発明は、インビトロでの突然変異誘発によって配列番号:1〜8に由来する精製された核酸分子をさらに包含する。インビトロでの突然変異誘発は、部位特異的変異誘発、ランダムな突然変異誘発およびインビトロでの核酸合成を含むが、限定されない当該技術分野において公知の多数の技術を含む。引用した配列からインビトロでの突然変異誘発によって誘導されるこのような核酸分子は、詳細な態様に従って、本来のものと同じ長さのまたはより短い配列を有する。
【0012】
DNAおよびRNAを含む本発明の核酸分子は、本明細書において「組換えMSP4およびMSP5核酸」または「組換えMSP4およびMSP5 DNA」と称され、これらの分子によってコードされるアミノ酸は、本明細書において「組換えMSP4およびMSP5ポリペプチド」または「本発明のポリペプチド」と称される。
【0013】
また、本発明は、遺伝暗号の結果として、配列番号:1〜8から縮重する精製された核酸分子、組換えMSP4およびMSP5核酸の対立遺伝子変異体である精製された核酸分子または組換えMSP4およびMSP5核酸の種相同体を包含する。また、本発明は、これらの核酸分子の発現を指揮する組換えベクターおよびこれらのベクターで形質転換され、トランスフェクトされ、または感染させた宿主細胞を包含する。組換えMSP4またはMSP5ポリペプチドと結合する精製されたポリクローナルまたはモノクローナル抗体は、本発明に包含され、本明細書において「本発明の抗体」と称する。
【0014】
本発明は、組換えMSP4およびMSP5ポリペプチドの産生のための方法であって、発現を促進する条件下で宿主細胞を培養すること、および培地または細胞のペレットからポリペプチドを回収することを含む方法をさらに包含する。特に、バキュロウイルス昆虫発現系における組換えMSP4およびMSP5ポリペプチドの発現も、本発明に包含される。
【0015】
また、本発明は、標識された組換えMSP4およびMSP5ポリペプチドを提供する。好ましくは、標識されたポリペプチドは、精製された形態である。また、非標識または標識されたポリペプチドは、マラリアに対する抗体を含む体液によって免疫学的に認識することができることが好ましい。ポリペプチドは、たとえば放射標識、酵素標識、蛍光標識、化学発光標識および発色団からなる群より選択される免疫アッセイ標識で標識することができる。
【0016】
また、本発明の組換えMSP4またはMSP5ポリペプチドとこれらを認識する抗体との間の免疫学的複合体が提供される。免疫学的複合体は、放射標識、酵素標識、蛍光標識、化学発光標識および発色団からなる群より選択される免疫アッセイ標識で標識することができる。
【0017】
さらにまた、本発明は、プラスモディウム属寄生虫のMSP4および/またはMSP5ポリペプチドを検出するためのインビトロにおける方法を提供する。本方法には、プラスモディウム属寄生虫(マラリア原虫)のMSP4および/またはMSP5ポリペプチドを含むことが疑われる生物材料を含む組成物を提供することと、およびプラスモディウム属寄生虫のMSP4および/またはMSP5ポリペプチドの存在についてアッセイすることとを含む。プラスモディウム属寄生虫のMSP4およびMSP5ポリペプチドは、典型的には電気泳動法によって、または本発明の抗体での免疫アッセイ法によってアッセイされる。本方法は、生体試料におけるプラスモディウム属寄生虫の検出のために、および好ましい態様において熱帯熱マラリア原虫(Plasmodium falciparum)および三日熱マラリア原虫(Plasmodium vivax)寄生虫の検出のために使用することができる。
【0018】
また、本発明は、組換え、または精製された本発明のMSP4もしくはMSP5ポリペプチドまたはこれらの混合物を含む抗原と結合する抗体の有無の検出のためのインビトロでの診断法を提供する。本方法には、抗原を体液と、生物学的体液中の抗原および抗体が抗原抗体複合体を形成するために十分な時間の間および条件下で接触させることと、および複合体の形成を検出することとを含む。検出工程は、抗原抗体複合体の形成を測定することをさらに含むことができる。抗原抗体複合体の形成は、好ましくはウエスタンブロット法、ELISA(酵素結合免疫吸着検定法)、間接的免疫蛍光アッセイ法または免疫沈降アッセイ法に基づいた免疫アッセイ法によって測定される。本方法は、感染した動物またはヒト患者マラリア由来の体液中のプラスモディウム属寄生虫に対する免疫応答の検出のために使用することができる。好ましい態様において、本方法は、熱帯熱マラリア原虫(Plasmodium falciparum)感染または三日熱マラリア原虫(Plasmodium vivax)感染に対する免疫応答の検出のために使用することができる。
【0019】
本発明の組換えMSP4もしくはMSP5ポリペプチドまたはこれらの混合物と結合する抗体の有無の検出のための診断キットは、組換えMSP4および/またはMSP5ポリペプチドまたはこれらの混合物を含む抗原と、抗原と抗体との間の免疫複合体の形成を検出するための手段とを含む。抗原および手段は、検出を行うために十分な量で存在する。
【0020】
プラスモディウム属寄生虫のMSP4および/またはMSP5ポリペプチドの有無の検出のための診断キットには、本発明の抗体と抗原と抗体との間の免疫複合体の形成を検出するための手段が含まれる。抗体および手段は、検出を行うために十分な量で存在する。
【0021】
また、本発明は、インビボにおいて免疫原性反応または保護反応を誘導するために十分な量で、本発明の組換えMSP4もしくはMSP5ポリペプチドまたはその混合物を、薬学的に許容される免疫賦活剤に付随して含む免疫原性組成物を提供する。本発明のワクチン組成物は、十分な量の組換えMSP4および/またはMSP5ポリペプチドと、中和抗体を誘導する薬学的に許容される免疫賦活剤とを含む。
【0022】
したがって、本発明のMSP4およびMSP5ポリペプチドは、マラリアに関連した抗原タンパク質に対する抗体の存在を検出するための診断の組成物の一部として有用である。
【0023】
加えて、組換えMSP4およびMSP5ポリペプチドは、マラリアに関連した抗原タンパク質の存在を検出するための抗体を生じさせるために使用することができる。
【0024】
また、本発明のポリペプチドは、寄生虫を不活性化し、インビボにおける寄生虫の生存度を減少させ、または寄生虫複製を阻害し、もしくは予防する、いずれかの中和抗体を生じさせるためにも使用することができる。寄生虫中和抗体を誘発する能力は、本発明のポリペプチドが免疫組成物またはワクチン組成物に使用されるときに、特に重要である。
【0025】
以下は、(A)天然のシグナル配列、(B)C末端Hisタグおよび(C)本分野(MSP4p20)における感染防御抗体反応と最も強く相関される熱帯熱マラリア原虫(P. falciparum)抗原のものに似ている酸反復領域を含むワクチンである。
【0026】
MKVAYFLSVLDLLIIFSLYFDGRRSAFAGIAACIRHGRILGEGGE/QNSTPGSGGQTGDHSAEAENGDYNEQGDDHGDDHGDDHGDDHGDEQDGEDYDDAEDDDLYELSEVDENANLCLDNNGGCGDDKICENLGKGIVKCLCKPGYKLVGTECVEHHHHHH[配列番号:30]
本発明は、図面を参照して記述する
【図面の簡単な説明】
【0027】
【図1】プラスモディウム属のいくつかの異なる種におけるmsp4およびmsp5遺伝子座位のゲノム構成を示す。これらの遺伝子の類縁種保存は、基本的な機能を示している。
【図2】SALSAの配列は、熱帯熱マラリア原虫(Plasmodium falciparum)のMSP-4(PfMSP4)と92%一致であることを示す。SALSA配列は、BおよびT細胞エピトープの両方を保持することが公知である。PfMSP4配列は、黒色で示してあり、SALSA配列はグレーで示してある。
【図3−1】MF54および合成遺伝子のMSP4コドン選択性を示す。
【図3−2】PfMSP4の完全合成遺伝子配列を示す。この図に示したすべての配列は、オリゴヌクレオチドで示した。PfMSP4の遺伝子配列は、番号付けした重複するオリゴ配列(1-38)によってコードされ、太字および通常のテキストを交互にすることによって強調してある。制限酵素部位は、小文字のテキストで記される。
【図4】合成遺伝子のPCR製作を示す。「遺伝子構築反応」(レーン1および2それぞれ5または10μL)および「遺伝子増幅反応」(レーン3および4それぞれ5またはμL)の試料は、DNAサイズ標準に隣接し、かつ離した1%のアガロースゲルで分離した。
【図5】バキュロウイルス系における組換えタンパク質発現のために産生した4つの異なるMSP4構築物の整列を示す。推定上のシグナル配列(現在利用できるアルゴリズムによって予測される)は、グレーで強調してあり、共通の多形部位は、太字で強調してあり、最も平衡選択下にありそうで、したがって免疫回避に関与する部位は、*で示してある。
【図6−1】3D7および合成遺伝子のMSP5コドンの選択性を示す。
【図6−2】熱帯熱マラリア原虫(Plasmodium falciparum)のMSP5(PfMSP5)の完全合成遺伝子配列を示す。この図に示したすべての配列は、オリゴヌクレオチドで示した。PfMSP5の遺伝子配列は、番号付けした重複するオリゴ配列(1-38)によってコードされ、太字および通常のテキストを交互にすることによって強調してある。制限酵素部位は、小文字のテキストで記してある。
【図7】合成遺伝子のPCR製作を示す。「遺伝子増幅反応」の試料(5μL)は、DNAサイズ標準に隣接し、かつ離した1%のアガロースゲルで分離した。
【図8】2つの異なる昆虫株化細胞におけるある期間にわたったPfMSP4の発現を示す。細胞死の割合は、細胞懸濁液を4%のトリパンブルーと1:1混合し、カバーガラス下の格子に対して総細胞および青い細胞を計数することにより算出した。タンパク質を、感染後24、30、36、42、48、54、60および72時間(それぞれレーン1〜8)にて感染された培養SNから精製し、20μLの溶出タンパク質を、4〜12%のBis-Trisゲル(Invitrogen)において分離して、SimplyBlue safe stain(Invitrogen)で染色した。これらのタンパク質種は、非感染性の細胞培養(レーン9)において見られなかった。
【図9】それぞれのMSP4産物p40、p30およびp20のN末端の配列を強調している整列配列である。この研究において同定されたシグナル配列は、グレーで強調してある。それぞれの産物のN末端の配列は、青い太字に示してある。
【図10】3つの公知のマラリア抗原について報告されたシグナル切断部位付近に集中するおよそ80アミノ酸残基の疎水性プロフィールを示す。成熟したタンパク質の最初の残基は、黒色ダイヤモンドによって示してある。S抗原シグナル配列切断のプロフィールは、バキュロウイルス系におけるMSP4について見られるものと同等である。
【図11】2つの異なる昆虫株化細胞におけるある期間にわたったPfMSP5の発現を示す。細胞死は、細胞懸濁液を4%のトリパンブルーと1:1に混合し、カバーガラス下の格子に対して総細胞および青い細胞を計数することにより算出した。タンパク質を、感染後24、30、36、42、48、54、60および72時間(それぞれレーン1〜8)にて培養SNから精製し、20μLの溶出タンパク質を、4〜12%のBis-Trisゲル(Invitrogen)において分離して、SimplyBlue safe stain(Invitrogen)で染色した。これらのタンパク質種は、非感染性の細胞培養(レーン9)において見られなかった。
【図12】ウサギで生じたポリクローナル抗血清に関する。(A)全長MSP4p40(R6-7)および(B)MSP4分解生成物p20(R8-9)に対して生じた血清のELISA指標力価を、コーティング抗原として天然(NR)または不可逆的に還元した(R)MSP4-p40を使用して解析した。血清R8は、MSP4抗原が還元されると、50%以上の結合活性を失う。免疫前血清は、NEGとしてマークしてある。パネル(C)は、コーティング抗原としてMSP5-p45およびp35を使用して解析された抗MSP5血清(56-57)の力価を示す。
【図13】バキュロウイルスで発現した抗原に対して生じたポリクローナル血清、およびDielmo由来のヒト血清のプールまたは抗MSP4p40ウサギ血清のプールのいずれか由来のMSP4p40アフィニティー精製血清で寄生虫材料をプローブすることによって得られた結果を示す。非同期的に発達する寄生虫をiRBCから抽出して、Wangら(2003)のプロトコルを使用して粗雑に分画した。各々の免疫ブロットのそれぞれのレーン1には、非還元Triton X100可溶性寄生虫画分、レーン2には、非還元膜画分、レーン3には、還元Triton X100可溶性寄生虫画分、レーン4には、還元膜画分を充填した(パネルA,BおよびD)。パネルCは、風乾した熱帯熱マラリア原虫(P. falciparum)を感染したRBCで得られたIFAデータを示す。
【図14】1990年に雨季のピーク時に集めたDielmoからの9人の免疫成人(18〜49歳)由来の血清のELISA解析を示す。プレートをMSP4p40(A)またはMSP5p45およびp35(B)でコートした。
【図15】NO.3および9の個体由来のヒト免疫血清を使用する、組換えMSP4 p40、p30およびp20(それぞれレーン3、2および1)の免疫ブロットである(A)。(B)NO.8の個体由来のヒト免疫血清を使用する組換えMSP5の免疫ブロット解析。
【図16】それぞれの血清について産生されるELISAプロフィールは、図16Aおよび16Bの上のそれぞれのブロットに示してあり、これは、PfMSP4、p40およびp20のエピトープを特異的に認識する2つのモノクローナル抗体mAb L11-16およびmAb F12-7についてのウエスタンブロットを示す。レジェンド:nR=非還元、R=invitrogenの市販の緩衝液により還元、iR=DTTおよびアクリルアミドによって不可逆的に還元。
【図17】図17Aおよび17Bは、それぞれ、図16Aおよび16BにおけるmAbsについてのODまたは吸光度(y軸)の関数として希釈比(x軸)を示す。
【図18】図18Aおよび18Bは、PfMSP5、p45およびp35のエピトープを認識する2つのモノクローナル抗体mAb G21-2およびmAb J18-4についてのウエスタンブロットを示す。レジェンド:nR=非還元、R=invitrogenの市販の緩衝液により還元、iR=DTTおよびアクリルアミドによって不可逆的に還元。
【図19】図19Aおよび19Bは、それぞれ図18Aおよび18BにおけるmAbsについての希釈比(x軸)およびODまたは吸光度(y軸)の関数を示す。
【図20】図20A-Bは、タンパク質をコーティングする濃度を最適化するために行ったELISA解析を示す。プレートは、0.5または1μg/mLのタンパク質のPBS溶液をウェルあたり100μL添加して一晩コートした。パネル(A)は、3つの異なるMSP4由来の抗原、MSP4p40、MSP4p30(本明細書において、MSP4p40/2と命名した)およびMSP4p20で得られたデータを示す。パネル(B)は、MSP5で得られたデータを示し、反応の異なる強度を証明するためにパネル(A)からのMSP4p20のデータをプロットした。観察される光学濃度は、いずれの濃度を使用してもほとんど同一だったので、より低いコーティング濃度(0.5g/mL)をすべてのその後の解析において使用した。
【図21】図21A-Dは、ナイーブ個体(naif)、高免疫成人(shi)および2005年のDielmo集団からの個体(60605)からの対照血清のELISA解析を示す。プレートを0.5μg/mLのMSP5(A)、MSP4p40(B)、MSP4p20(C)およびMSP4p30(D)でコートし、抗血清の希釈を変化させて反応を実施した(3倍段階希釈)。すべての抗原は、免疫血清によって認識され、マラリアナイーブ対照によって認識されなかった。
【図22】図22A-Dは、15歳以上および以下の年齢によって層別化される2000年のNdiop集団からの個体の抗体反応を示す。パネル(A)は、抗原(左から右に)MSP4p20、MSP4p40、MSP4p30およびMSP5についての光学濃度の形態のELISAデータを示す。パネル(C)は、OD比率の形態での同一データを示す。パネル(B)のMSP1p19に対する個々の抗体反応は、Ndiop 2000年集団についての年齢の関数としてのOD比率としてプロットしてあり、その一方で、パネル(D)は、同じ血清でMSP5に対して得られたELISA OD比率を示す。
【図23】Ndiop 2000年集団からの血清によるMSP4およびその誘導体の認識の変動を示す。左のパネルは、MSP4p20、MSP4p40およびMSP4p30(MSP4md2に等しい)についてのOD比率としてプロットした平均反応を示す。右のパネルは、年齢の関数としてそれぞれのMSP4抗原に対する抗体の反応について、上下の四分位値およびデータ分布を示す。
【図24】特異的血清の存在下において誘導される酸化的破裂として測定された、抗体依存的な単球媒介性細胞障害を示す。血清は、集団の中央値の反応を上回るまたは下回るOD比率を与えるものに分けられた。パネル(A)は、MSP5反応性の関数(2.4のOD比を上回るまたは下回る)として分けられた血清からのデータを示し、パネル(B)は、MSP4p40反応性の関数(20のOD比を上回るまたは下回る)として分けられた血清からのデータを示す。これらがPBMC(末梢血単核細胞)および熱帯熱マラリア原虫(P. falciparum)メロゾイトの存在下において酸化的破裂を誘導する能力は、化学発光を使用して解析した。両群において、酸化的破裂における増大は、高OD比を示す個体で見られた。
【図25】IgG特異的二次抗体試薬を使用して解析したそれぞれの抗原についての抗体アイソタイププロフィールを示す。アイソタイププロフィールは、MSP4p40(パネルA)およびMSP5(パネルB)について示してある。データは、年齢(15齢を上回るおよび下回る)の関数としてプロットしてある。
【図26】PfMSP4およびPvMSP4のアミノ酸配列の整列である。Pf配列のいくつかの特色が強調され、(i)分泌性およびGPI付着シグナルは、緑に示してあり、(ii)SALSAとして公知の配列は、下線を引いてある、(iii)すべての負に荷電した残基(DおよびE)は、赤において示してあり、および(iv)PfMSP4p20のN末端配列および重要なシステイン残基は、青で強調してある。
【図27】バキュロウイルスPfMSP5を示す。(A)精製された組換えPfMSP5をSDS-PAGEゲル上で移動させ、ニトロセルロースへ転写し、ヒト免疫血清でプローブした。(B)PfMSP5をトリチウム化したミリスチン酸の存在下において発現させた。精製したタンパク質をSDS-PAGEゲル上で移動させ、ブルー染色およびオートラジオグラフィーを7週間行った。
【図28】RBC期におけるある時間にわたったPfMSP4の発現を示す。(A)アセトン固定したリング期の寄生虫および風乾した後期のおよび自由なメロゾイトを、PfMSP1p19または組換えPfMSP4p20(Rp20)に対してアフィニティー精製されたポリクローナルウサギ血清に特異的であるモノクローナル抗体G17.12でプローブした。抗体染色は、Alexa Fluor(登録商標)488のヤギ抗マウスまたは抗ウサギ結合抗体で明らかにされ、寄生虫DNAは、ヘキスト33342で染色した。(B)アフィニティー精製したウサギ(Rp20)およびヒト血清(NdiopおよびDielmoの1:1のプール:Hp20)を使用する、成熟シゾント(S)、自由なメロゾイト(M)、およびリング(R)からの寄生虫抽出物のウエスタンブロット分析。
【図29】組換えPfMSP4発現のために使用する構築物によってコードされるタンパク質の配列を示す。PfMSP4p20の直接の発現を促進するようにデザインされた構築物を示す。構築物の名前を配列テキストの左に収載してあり、それぞれの分泌タンパク質のN末端の配列をテキスト内で太字体に強調してあり、および組換えタンパク質の名前を右側に収載してある。
【図30】ある時間にわたったPfMSP4p20の直接発現を示す。PfMSP4p40、PfMSP4p21、PfMSP4p21ss1またはPfMSP4p21ss2バキュロウイルスが感染昆虫細胞の培養液上清の試料を、感染後24〜66時間の間6時間間隔にて収集し、透析し、TALON樹脂上でバッチ精製した。精製したタンパク質試料をNuPAGE 4〜12%の勾配ゲルで分離し、ブルー染色し、タンパク質のサイズを右に示す。おおよそのタンパク質収率を感染後66時間にて残存している培養液上清の最後の130 mLを使用して算出した。タンパク質をIMACおよびHPLCによって精製し、タンパク質収率をBCAタンパク質量キット(PIERCE)を使用して算出する。
【図31】PfMSP4p40およびPvMSP4/Hisのタンパク質配列の整列を示す。バキュロウイルスP. vivax MSP4およびP. falciparum MSP4発現構築物のアミノ酸(一文字表記)配列は、clustalxを使用して整列させた。アミノ酸同一性を(*)、保存的置換を(:)、半保存的な置換を(.)およびラジカル変化を空白で示してある。同定されたN末端の配列には、下線を引いてあり、かつ太字体である。異なるタンパク質産物についてのN末端の配列には、下線を引いてあり、かつ太字体である。
【図32】ある時間にわたったPvMSP4/His発現を示す。簡潔には、8 mLの撹拌培養のSN試料を感染後24および66時間の間6時間の間隔で収集し、透析し、TALON樹脂上でバッチ精製する。精製したタンパク質試料をNuPAGE 4-12%の勾配ゲルで分離し、SimplyBlue SafeStainで染色し、タンパク質サイズを左右に示す。
【図33】PvMSP4/Hisに対するヒト免疫血清の反応性を示す。(A)ELISAプレートには、バキュロウイルス発現系で発現したPvMSP4/Hisをコートした。三倍段階希釈またはすべての24血清を試験した。(B)平行して、それぞれの希釈系列を不可逆的に還元したPvMSP4に対して試験した。このグラフは、天然抗原に対する負の対照を上回るODを示すそれぞれの血清の下方パネルにおける、1/2700希釈でのODを示す。上方パネルは、1/2700の血清希釈での抗原減少において見られるODの減少を示す。
【図34】図34は、PMNによるメロゾイトの抗体依存的食作用(APDm)に基づいた機能的アッセイ法における血清(非動化、IgG除去、または精製IgG)の処理の結果を示す。アッセイ法は、実施例19に記してある。
【図35】図35は、PMNによるメロゾイトの抗体依存的食作用(APDm)に基づいた機能的アッセイ法における血清(非動化、IgG除去、または精製IgG)の処理の結果を示す。アッセイ法は、実施例19に記してある。
【発明の詳細な説明】
【0028】
2つの最近同定されたメロゾイト表面抗原の熱帯熱マラリア原虫(Plasmodium falciparum)メロゾイト表面タンパク質4および5(PfMSP4およびPfMSP5)は、潜在性多成分抗マラリアワクチンのタンパク質成分になる見込みがある。msp4およびmsp5遺伝子は、両方とも272残基タンパク質をコードする、それぞれの単一のC末端EGF様ドメインおよびGPI付着モチーフをもち、染色体2上でmsp2のすぐ上流に直列に位置する。メロゾイト表面での膜会合は、両タンパク質について証明されており、ヒト免疫血清は、大腸菌(Escherichia coli)において発現させた組換えMSP4と反応することが示された[12,13]。
【0029】
プラスモディウム属の3つのマウス種、P. ヨエリ(P. yoelii)、P. チャバウジ(P. chabaudi)およびP. ベルゲイ(P. berghei)において、MSP4およびMSP5座位(MSP4/5)に単一遺伝子のみがあり、これらは、それぞれに対してある程度の相同性を示す[14-16]。この遺伝子は、MSP4/5を示し、P. ヨエリ(P. yoelii)致死チャレンジモデルにおいて感染防御免疫を調査するために使用されてきた[17]。
【0030】
MSP4/5は、種々の免疫化ストラテジーを使用して保護を与えることが示されており、MSP1p19と組み合わせて送達したときに、有効性が最大化される[17-20]。加えて、MSP4/5タンパク質によって誘導されるマウス免疫応答における系統特異性はないと思われる[21]。熱帯熱マラリア原虫(P. falciparum)では、msp4およびmsp5は、それぞれ相同的な位置に単一のイントロンを有する[22]。
【0031】
熱帯熱マラリア原虫(Plasmodium falciparum)メロゾイト表面タンパク質4(PfMSP4)タンパク質配列は、分泌シグナル配列、C末端のEGF様ドメインおよびGPI-付着シグナルを含む[10]。
【0032】
Pfmsp4遺伝子配列は、960bpの長さであり、144bpの1つのイントロンを含み、272アミノ酸残基をコードする。Pfmsp5遺伝子配列は、955bpの長さであり、136 bpの1つのイントロンを含み、272アミノ酸残基をコードする。PfMSP4と同様に、PfMSP5タンパク質配列は、分泌シグナル配列、C末端のEGF様ドメインおよびGPI-付着シグナルからなる[11,22]。MSP遺伝子のクラスターのこの下流には、高度に保存されたアデニロコハク酸ライセース(lysase)(ASL)遺伝子があり、これは、その他の種におけるこの座位の同定を促進する有用なハンドルであることが判明した(図1)。
【0033】
多くの現在のデータは、PfMSP4およびPfMSP5が優れたワクチン候補であるという考えを支持する。それにもかかわらず、いくつかの公開された知見は、2つの異なる発現系(大腸菌(E.coli)および酵母)において産生されたPfMSP4の組換え類似体は抗原性が異なり、かつヒト免疫血清では見られない現象である、タンパク質のEGF様ドメインに対して高次構造非依存的な反応を誘導することを示している[13]。タンパク質のこの領域における高次構造上のエピトープの認識が、保護に重要である場合、MSP1-19の例のように、すべてのエピトープを忠実に再生する産物を産生することは避けられない。
【0034】
従って、本発明は、熱帯熱マラリア原虫(Plasmodium falciparum)MSP4およびMSP5をコードする核酸、並びに生じるポリペプチドが、昆虫細胞における最適な発現を達成するように修飾された構築物を提供する。より詳細には、本発明は、バキュロウイルス-昆虫細胞発現系において可溶性の分泌されるポリペプチドとして発現される組換えMSP4ポリペプチドをコードする構築物を提供する。組換えポリペプチドは、C末端にEGF様ドメインを含み、これは適切に折り畳まれるようである。これは、タンパク質をELISAプレート上で不可逆的に減少させるときに、MSP4のポリクローナルウサギ血清認識において顕著な減少(50〜60%)があることによって示される(図12を参照されたい)。
【0035】
本発明の組換えMSP4ポリペプチドの1つは、N末端の近くの多形領域からの30アミノ酸の欠失を伴うMSP4外抗原(GPI付着点のC末端疎水性残基がなく、したがって、タンパク質分泌が可能)である。本発明のこのポリペプチドは、MSP4p30と呼ばれる。
【0036】
本発明の組換えMSP4ポリペプチドのもう1つのものは、N末端の近くの多形領域からの30アミノ酸の欠失を伴わないMSP4外抗原(GPI付着点のC末端の疎水性残基がなく、したがってタンパク質分泌が可能)である。本発明のこのポリペプチドは、MSP4p40と呼ばれ、全長遺伝子産物である。
【0037】
本発明のもう一つの組換えMSP4ポリペプチドは、配列KSPKEモチーフ周辺で始まり、EGFドメインを含む、およそMSP4のC末端の半分に対応する20kDaのポリペプチドである。上流の補充アミノ酸残基、特にKSPKEモチーフ上流に局在化するMSP4p40配列の残基を含めることができる。本発明のこの組換えポリペプチドは、MSP4p20と呼ばれる。
【0038】
加えて、本発明は、同時に産生されるp35およびp45形態を含む組換えMSP5ポリペプチドを提供する。両方とも、翻訳後修飾を含み、免疫原性を押し上げることができる脂肪酸残基(ミリストイル化)の共有結合を含む可能性が高い。本発明のこれらの組換えポリペプチドは、MSP5p45およびMSP5p35と呼ばれる。
【0039】
それにもかかわらず、本発明の組換えMSP4およびMSP5ポリペプチドは、また、これらの天然のGPIシグナル配列またはGPI修飾のための別のGPIアンカータンパク質配列シグナリングからのもののいずれかを使用して、C末端GPIアンカー実体として発現することができる。このようなGPI修飾された実体は、任意の免疫のアジュバントの非存在下において、組換えMSP4およびMSP5ポリペプチドの免疫原性を実質的に増強することが予想されるであろう。
【0040】
本発明の組換えポリペプチドは、以下の通りのこれらの対応する配列番号に関して、より詳細に記述してある:
核酸
MSP4p20 配列番号:1
MSP4p30 配列番号:2
MSP4p40 配列番号:3
MSP5 配列番号:4
MSP5p10 配列番号:5
MSP4p21 配列番号:6
MSP4p21 ss1 配列番号:7および配列番号:28
MSP4p21 ss2 配列番号:8および配列番号:29
ポリペプチド
MSP4p20(分解生成物) 配列番号:9
MSP4p30(構築物に存在するORF=構築物によってコードされるポリペプチド) 配列番号:10
MSP4p30(発現の最終生成物=昆虫細胞によって産生されるポリペプチド) 配列番号:11
MSP4p40(構築物に存在するORF) 配列番号:12
MSP4p40(発現の最終生成物) 配列番号:13
MSP5(構築物に存在するORF) 配列番号:14
MSP5p10 配列番号:15
PvMSP4/His 配列番号:16
MSP4p21(構築物に存在するORF) 配列番号:17
MSP4p21(発現の最終生成物) 配列番号:18
MSP4p21 ss1(構築物に存在するORF) 配列番号:19
MSP4p21 ss1(発現の最終生成物) 配列番号:20
MSP4p21 ss2(構築物に存在するORF) 配列番号:21
MSP4p21 ss2(発現の最終生成物) 配列番号:22
PvMSP4p20(「PfMSP4およびPvMSP4のアミノ酸配列の整列に示したように、MSP4の三日熱マラリア原虫(Plasmodium vivax)配列に基づいて提唱されたワクチン構築物の配列) 配列番号:30。
【0041】
これらのポリペプチドは、本明細書において、個々におよびひとまとめにして、本発明の「組換えMSP4およびMSP5ポリペプチド」と称される。同様に、これらのポリペプチドをコードする核酸は、本発明の「組換えMSP4およびMSP5核酸」と呼ばれる。
【0042】
本発明に対する意味は、広範囲にわたる。組換えMSP4およびMSP5ポリペプチドのこの発見により、本発明の組換えMSP4およびMSP5ポリペプチドをコードする核酸配列を含む発現ベクターの構築;発現ベクターをトランスフェクトさせ、または形質転換した宿主細胞;生物学的に活性な組換えMSP4およびMSP5ポリペプチド、並びに単離され、または精製されたタンパク質としての組換えMSP4およびMSP5ポリペプチド;組換えMSP4およびMSP5ポリペプチドと免疫反応性の抗体、組換えMSP4およびMSP5ポリペプチド並びにプラスモディウム属寄生虫およびマラリア感染の検出における組換えMSP4およびMSP5ポリペプチドに対して向けられた抗体の診断使用、並びにプラスモディウム属感染から守るための組換えMSP4およびMSP5ポリペプチドのワクチン使用が可能になる。
【0043】
また、本明細書に使用される「組換えMSP4およびMSP5ポリペプチド」という用語は、配列番号:9〜22のアミノ酸配列を有するタンパク質をさらに包含するポリペプチドの属、並びにこのようなアミノ酸配列と高度な類似性(少なくとも90%の相同性)を有し、かつ該タンパク質およびポリペプチドが免疫反応性であるタンパク質およびポリペプチドをいう。加えて、組換えMSP4およびMSP5ポリペプチドは、配列番号:1〜8のヌクレオチドの遺伝子産物をいう。
【0044】
本明細書に使用される「精製された」という用語は、たとえば組換え宿主細胞培養の精製産物として、または非組換え供与源から精製された産物として、組換えMSP4およびMSP5ポリペプチドがその他のタンパク質またはポリペプチドを本質的に付随しないことを意味する。本明細書に使用される「実質的に精製された」という用語は、組換えMSP4およびMSP5ポリペプチドを含み、かつ特異抗体を使用して除去することができる公知のタンパク質の存在を除けば、その他のタンパク質またはポリペプチドを本質的に付随しない混合物をいい、該実質的に精製された組換えMSP4およびMSP5ポリペプチドは、抗原として使用することができる。
【0045】
本明細書で言及される組換えMSP4およびMSP5ポリペプチド「変異体」とは、1つまたは複数の欠失、挿入または置換のために、組換えMSP4およびMSP5ポリペプチドのものとは異なるアミノ酸配列を有する、組換えMSP4およびMSP5ポリペプチドに対して実質的に相同的なポリペプチドを意味する。変異体アミノ酸配列は、好ましくは組換えMSP4およびMSP5ポリペプチドアミノ酸配列と少なくとも80%同一で、最も好ましくは少なくとも90%同一である。パーセント同一性は、たとえばDevereuxらによって記述され、およびウィスコンシン大学Genetics Computer Group(UWGCG)から入手可能なGAPコンピュータプログラム、バージョン6.0(Nucl. Acids Res. 12:387、1984)を使用して配列情報を比較することにより、決定することができる。GAPプログラムは、SmithおよびWaterman (Adv. Appl. Math 2:482, 1981)によって修正されたNeedlemanおよびWunschの整列方法を利用する(J. Mol. Biol. 48:443, 1970)。GAPプログラムのための好ましいデフォルトパラメーターには、以下が含まれる:(1)Schwartz and Dayhoff, eds., Atlas of Protein Sequence and Structure, National Biomedical Research Foundation, pp. 353-358, 1979によって記述されたような、Gribskov and Burgess, Nucl. Acids Res. 14:6745, 1986の、ヌクレオチドのための単一比較マトリックス(同一性について1および非同一性について0の値を含む);(2)それぞれのギャップに対する3.0のペナルティーおよびそれぞれのギャップにおけるそれぞれのシンボルに対するさらなる0.10のペナルティー;および(3)末端ギャップに対するペナルティーなし。
【0046】
変異体には、所与のアミノ酸残基が同様の生理化学的特徴を有する残基によって置換されたことを意味する、保存的に置換された配列を含むことができる。保存的置換の例には、Ile、Val、LeuもしくはAlaを互いになどの1つの脂肪族残基の別ものへの置換またはLysとArg;GluとAsp;もしくはGlnとAsnとの間などの1つの極性残基の別のものへの置換を含む。その他のこのような保存的置換、例えば同様の疎水性特徴を有する全ての領域の置換も周知である。本発明のMSP4およびMSP5ポリペプチドの変異体の例は、選択的mRNAスプライシングイベントにより、、または組換えMSP4およびMSP5ポリペプチドのタンパク分解切断により生じるタンパク質である。タンパク質分解に起因する変異には、たとえば組換えMSP4およびMSP5ポリペプチドからの1つまたは複数の末端アミノ酸のタンパク分解性の除去による、宿主細胞の異なるタイプにおける発現による末端の相違が含まれる。フレームシフトに起因する変異には、たとえば宿主細胞の異なるタイプにおける発現による末端の相違が含まれる。
【0047】
前述の通り、本発明は、単離され、および精製された、または同種の組換えMSP4およびMSP5ポリペプチドを提供する。抗原として使用することができる組換えMSP4およびMSP5ポリペプチドの変異体および誘導体は、組換えMSP4およびMSP5ポリペプチドをコードするヌクレオチド配列の突然変異によって得ることができる。アミノ酸配列の変化は、任意の多数の従来法によって達成することができる。突然変異は、天然の配列の断片に対するライゲーションを可能にする制限部位に隣接した、突然変異体配列を含むオリゴヌクレオチドを合成することにより、特定の座位に導入することができる。ライゲーションに続いて、生じる再構築配列は、所望のアミノ酸挿入、置換または欠失を有する類似体をコードする。
【0048】
あるいは、オリゴヌクレオチド特異的、部位特異的突然変異誘発法を使用して、予め定められたコドンを置換、欠失または挿入によって変化させることができる変化させられた遺伝子を提供することができる。上記の変化を作製する例示的方法は、Walderら(Gene 42:133, 1986); Bauerら(Gene 37:73, 1985); Craik (BioTechniques, January 1985, 12-19); Smithら(Genetic Engineering: Principles and Methods, Plenum Press, 1981); Kunkel(Proc. Natl. Acad. Sci. USA 82:488, 1985); Kunkelら(Methods in Enzymol. 154:367, 1987);並びに米国特許第4,518,584号および第4,737,462号によって開示されており、これらの全ては、参照により援用される。
【0049】
本発明の側面内において、組換えMSP4およびMSP5ポリペプチドは、組換えMSP4およびMSP5ポリペプチドと特異的に結合する抗体を調製するために利用することができる。「抗体」という用語は、ポリクローナル抗体、モノクローナル抗体、F(ab')2およびFab断片などのこれらの断片、並びに任意の組換えで産生される結合パートナーを含むことが意味される。抗体は、これらが約107 M-1以上のKaで組換えMSP4およびMSP5ポリペプチドに結合する場合、特異的に結合すると定義される。たとえば、結合パートナーまたは抗体の親和性は、従来技術、たとえばScatchardrら、Ann. N.Y Acad. Sci., 51:660 (1949)によって記述されたものを使用して容易に決定することができる。ポリクローナル抗体は、当該技術分野において周知手順を使用して種々の供与源、例えばウマ、ウシ、ヤギ、ヒツジ、イヌ、ニワトリ、ウサギ、マウスまたはラットから容易に産生することができる。
【0050】
本発明は、配列番号:1、2、3、4、5、6、7および8のヌクレオチド配列に由来する単離された断片およびオリゴヌクレオチドをさらに包含する。また、本発明は、これらの断片およびオリゴヌクレオチドによってコードされるポリペプチドを包含する。
【0051】
本発明の範囲内の核酸配列には、適度なまたは厳密なストリンジェンシー条件下で、本明細書に開示した天然の組換えMSP4およびMSP5核酸にハイブリダイズし、かつ組換えMSP4およびMSP5ポリペプチドをコードする単離されたDNAおよびRNA配列が含まれる。本明細書に使用される適度なストリンジェンシーの条件は、当業者に公知のとおり、およびSambrookら、Molecular Cloning: A Laboratory Manual, 2 ed. Vol. 1, pp. 1.101-104, Cold Spring Harbor Laboratory Press, (1989)によって定義されるとおり、ニトロセルロースフィルターのための予洗溶液5×SSC、0.5% SDS、1.0mM EDTA(pH 8.0)、50% ホルムアミド、6×SSC、42℃におけるハイブリダイゼーション条件(または42℃にて50%ホルムアミドにおけるStark溶液などのその他の同様のハイブリダイゼーション溶液)、並びにおよび約60℃、0.5×SSC、0.1% SDSの洗浄条件の使用を含む。高ストリンジェンシーの条件は、上記のハイブリダイゼーション条件と同様に定義され、68℃、0.2×SSC、0.1% SDSにて洗浄する。当業者であれば、温度および洗浄液溶液塩濃度をプローブの長さなどの要因に従って必要に応じて調整することができることを認識するであろう。
【0052】
複数のコドンが同じアミノ酸をコードする遺伝暗号の公知の縮重のため、DNA配列は、配列番号:1〜8に示され、かつ配列番号:9〜22のアミノ酸配列を有する組換えMSP4およびMSP5ポリペプチドをコードするたものから変化することができる。このような変異体DNA配列は、サイレント突然変異(たとえば、PCR増幅の間に生じる)により生じ得るか、または天然の配列の慎重な突然変異誘発の産物であることができる。
【0053】
したがって、本発明は、(a);(a)配列番号:1〜8のヌクレオチド配列を含むDNA、(b)適度なストリンジェンシー条件下で(a)のDNAに対してハイブリダイゼーションすることができ、かつ組換えMSP4およびMSP5ポリペプチドをコードするDNA;並びに(c)(a)または(b)で定義したDNAに対する遺伝暗号の結果としての縮重であり、かつ組換えMSP4およびMSP5ポリペプチドをコードするDNAから選択される、組換えMSP4およびMSP5ポリペプチドをコードする対応する単離されたDNA配列を提供する。このようなDNA均等物配列によってコードされるポリペプチドは、本発明によって包含される。
【0054】
配列番号:1〜8のDNA配列に対応するDNAは、適度にストリンジェント条件下で、配列番号:9〜22のアミノ酸配列を含むポリペプチドをコードするDNA配列にハイブリダイズする。このようなDNAによってコードされる組換えMSP4およびMSP5ポリペプチドの例は、組換えMSP4およびMSP5ポリペプチド断片、並びに上記のように、不活性化されたN-グリコシル化部位、不活性化されたプロテアーゼプロセシング部位または保存的アミノ酸置換を含む組換えMSP4およびMSP5ポリペプチドを含むが、限定されない。DNAが配列番号:1〜8のDNAの相補物にハイブリダイズするプラスモディウム属のその他の種に由来するDNAによってコードされるポリペプチドも包含される。
【0055】
組換えMSP4およびMSP5ポリペプチドをコードする核酸配列を含む組換え発現ベクターは、周知の方法を使用して調製することができる。発現ベクターには、哺乳動物、微生物、ウイルスまたは昆虫遺伝子に由来するものなどの、適切な転写または翻訳制御ヌクレオチド配列に作動可能に連結された組換えMSP4およびMSP5 DNA配列を含む。制御配列の例には、転写プロモーター、オペレーターまたはエンハンサー、mRNAリボソーム結合部位および転写および翻訳の開始と終了を制御する適切な配列が含まれる。ヌクレオチド配列は、制御配列が組換えMSP4およびMSP5 DNA配列に機能的に関連するときに、「作動可能に連結される」。したがって、プロモーターヌクレオチド配列は、プロモーターヌクレオチド配列が、組換えMSP4およびMSP5 DNA配列の転写を制御する場合に、組換えMSP4およびMSP5 DNA配列に作動可能に連結されている。所望の宿主細胞において複製する能力は、通常複製開始点によって与えられ、形質転換体を同定することにより選択される遺伝子を、加えて発現ベクターに組み込むことができる。
【0056】
加えて、天然では組換えMSP4およびMSP5ポリペプチドと付随していない適切なシグナルペプチドをコードする配列を発現ベクターに組み込むことができる。たとえば、組換えMSP4およびMSP5ポリペプチドが、最初にシグナルペプチドを含む融合タンパク質として翻訳されるように、シグナルペプチド(分泌性のリーダー)のためのDNA配列を組換えMSP4およびMSP5ヌクレオチド配列に対してインフレームで融合させることができる。意図した宿主細胞において機能的であるシグナルペプチドは、組換えMSP4およびMSP5ポリペプチドの細胞外分泌を増強する。シグナルペプチドは、細胞からの組換えMSP4およびMSP5ポリペプチドの分泌により、組換えMSP4およびMSP5ポリペプチドから切断することができる。
【0057】
原核生物の宿主細胞に使用するための発現ベクターは、一般に1つまたは複数の表現型の選択可能なマーカー遺伝子を含む。表現型の選択可能なマーカー遺伝子は、たとえば抗生物質抵抗性を与えるか、または独立栄養性の要求を供給するタンパク質をコードする遺伝子である。原核生物の宿主細胞のための有用な発現ベクターの例には、市販のプラスミドに由来するものが含まれる。市販のベクターには、タンパク質の発現のために特異的にデザインされているものを含む。これらには、マルトース結合タンパク質に融合されるタンパク質の発現のために使用されるpMAL-p2およびpMAL-c2ベクターが含まれる(New England Biolabs Beverly、MA、USA)。
【0058】
組換え原核生物の宿主細胞発現ベクターのために一般に使用されるプロモーター配列は、β-ラクタマーゼ(ペニシリナーゼ)、ラクトースプロモータ系(Chang et al., Nature 275:615, 1978;およびGoeddel et al., Nature 281:544, 1979)、トリプトファン(trp)プロモーター系(Goeddelら、Nucl. Acids Res. 8:4057, 1980;およびEP-A-36776)およびtacプロモーター、(Maniatis, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, p. 412, 1982)を含む。
【0059】
組換えMSP4およびMSP5ポリペプチドの発現のための適した宿主細胞には、原核生物、酵母または高等真核細胞が含まれる。昆虫細胞が好ましい。細菌、真菌、酵母および哺乳動物宿主細胞で使用するために適したクローニングおよび発現ベクターは、たとえばPouwels ら、Cloning Vectors: A Laboratory Manual, Elsevier, New York, (1985)に記述されている。無細胞翻訳系も、本明細書に開示したDNA構築物に由来するRNAを使用して組換えMSP4およびMSP5ポリペプチドを産生するために使用することができるであろう。
【0060】
本発明は、本明細書に記述した技術またはその他の方法を使用して得られるかどうかにかかわらず、単離され、もしくは精製された形態の前述したタンパク質またはポリペプチドを包含することが意図されることが理解されよう。本発明の好ましい態様において、組換えMSP4およびMSP5ポリペプチドは、人体組織および人体組織成分、核酸、外来タンパク質および脂質、並びに細菌およびウイルスなどの外来微生物を実質的に含まない。また、本発明は、実質的に同じ生物学的および免疫原特性を有する同等のタンパク質を包含することが理解される。
【0061】
本発明の組換えMSP4およびMSP5ポリペプチドを作製するための使用に応じて、これらを標識することが望ましいであろう。適切な標識の例は、放射性標識、酵素標識、蛍光標識、化学発光標識および発色団である。本発明のタンパク質を標識するための方法は、標識化免疫グロブリンのために広く使用されているものと本質的に異ならない。標識する必要は、間接的マーカーとして本発明の抗原に対する標識抗体または抗原に対する抗体に対する抗免疫グロブリンを使用することによって回避してもよい。
【0062】
一旦本発明の組換えMSP4およびMSP5ポリペプチドが得られたら、これらを使用してそれと反応性のポリクローナル抗体およびモノクローナル抗体を産生した。したがって、本発明のタンパク質またはポリペプチドは、当該技術分野において公知の技術によって動物宿主を免疫するために使用することができる。このような技術には、通常接種が含まれるが、これらには、その他の投与様式が含まれていてもよい。動物宿主において免疫原性反応を生じさせるために、ポリペプチドの十分な量が投与される。本発明の抗原に対する抗体を産生するために任意の宿主を使用することができる。一旦動物を免疫して、これが抗原に対する抗体を産生し始めるのに十分な時間が過ぎたら、ポリクローナル抗体を回収することができる。一般的な方法には、動物から血液を除去すること、および血液から血清を分離することを含む。抗原に対する抗体を含む血清は、抗原に対する抗血清として使用することができる。あるいは、抗体を血清から回収することができる。親和性精製は、精製された、抗原に対するポリクローナル抗体を血清から回収するための好ましい技術である。
【0063】
また、本発明の抗原に対するモノクローナル抗体を調製することができる。抗原と反応性のモノクローナル抗体を産生するための1つの方法は、宿主を抗原で免疫すること;宿主の脾臓から抗体産生細胞を回収すること;抗体産生細胞を酵素ヒポキサンチン-グアニンホスホリボシルトランスフェラーゼを欠損した骨髄腫細胞と融合して、ハイブリドーマを形成すること;ヒポキサンチン、アミノプテリンおよびチミジンを含む培地における培養によって少なくともハイブリドーマの1つを選択すること;抗原に対する抗体を産生する少なくとも1つのハイブリドーマを同定すること;同定されたハイブリドーマを培養して、回収可能な量の抗体を産生すること;および培養されたハイブリドーマによって産生される抗体を回収することを含む。
【0064】
これらのポリクローナル抗体またはモノクローナル抗体は、種々の適用に使用することができる。対応するタンパク質の中和は、これらの一つである。また、これらは、生物学的製剤におけるプラスモディウム属寄生虫抗原を検出するために、またはたとえば親和性クロマトグラフィーカラムに使用したときに、対応するタンパク質、糖タンパク質もしくはこれらの混合物を精製する際に、使用することができる。
【0065】
組換えMSP4およびMSP5ポリペプチドは、生体試料におけるマラリア原虫に対して特異的な抗体の存在を検出するための、および現在または以前の感染に関するシグナルを構成する、これらの生体試料におけるこのような抗体のレベルを評価するための抗原として使用することができる。このような生体試料には、もちろんヒト組織およびヒト細胞、並びにヒト血清を含むヒト体液などの生物学的液体を含む。マラリアに対する抗体の存在または濃度を決定するための免疫アッセイ法の試薬として使用されるときに、本発明の抗原は、便利で、迅速で、感受性で、および特異的なアッセイ法を提供する。
【0066】
より詳細には、本発明の抗原は、液体中の体液性成分を検出し、または定量化する際に使用するための周知の免疫アッセイ法によるマラリアの発見のために使用することができる。したがって、抗原-抗体相互作用は、沈澱または凝集などの二次反応によって、直接観察すること、または決定することができる。加えて、免疫電気泳動技術も使用することができる。たとえば、寒天における電気泳動法、続く抗血清との反応の古典的な組み合わせ、並びに二次元電気泳動、ロケット電気泳動およびポリアクリルアミドゲルパターンの免疫標識を利用して、(ウエスタンブロットまたは免疫ブロット)を利用することができる。本発明の抗原が使用することができるその他の免疫アッセイ法には、放射免疫アッセイ法、競合的免疫沈降アッセイ法、酵素免疫測定法および免疫蛍光アッセイ法を含むが、限定されない。比濁技術、比色技術および比濁分析技術を使用することもできることが理解されるであろう。ウエスタンブロット技術に基づいた免疫アッセイ法が好ましい。
【0067】
免疫アッセイ法は、本発明の抗原または本発明の抗原に対する抗体のいずれかの免疫試薬の1つを、試薬の免疫反応性を保持すると共に、担体表面上に固定することによって実施することができる。相互免疫試薬は、免疫反応性も保持されるような方法で、非標識であること、または標識することができる。これらの技術は、特に酵素結合免疫吸着検定法(ELISA)および競合阻害酵素免疫測定法(CIEIA)などの酵素免疫測定法に使用するための適している。
【0068】
本発明の抗原または抗原に対する抗体が固体支持体に付着されるときに、支持体は、通常ガラスまたはプラスチック材料である。プレート、チューブ、ビーズまたは円板の形態で成形されるプラスチック材料が好ましい。適切なプラスチック材料の例は、ポリスチレンおよび塩化ビニルである。免疫試薬が固体支持体と容易に結合しない場合、担体材料は、試薬と支持体との間に配置することができる。適切な担体材料の例は、ウシ血清アルブミンなどのタンパク質またはグルタルアルデヒドもしくは尿素などの化学試薬である。固相のコーティングは、従来の技術使用することにより実施することができる。
【0069】
本発明は、免疫原性の組換えMSP4およびMSP5ポリペプチドおよびより詳しくは、マラリアに対するワクチン組成物の調製に使用するための保護薬ポリペプチドを提供する。したがって、これらのポリペプチドは、マラリア感染に感受性の哺乳類に対してポリペプチドを投与することによるワクチンとして使用することができる。従来の投与様式を使用することができる。たとえば、投与は、経口、舌下、呼吸器または非経口的経路によって実施することができる。ワクチンが非経口的に投与されるときは、皮内、皮下、筋肉内および静脈内の投与経路が好ましい。
【0070】
マラリアに感染した哺乳類における免疫応答の主目的は、マラリア原虫を不活性化すること、並びにマラリア原虫死滅および寄生虫が感染した赤血球のクリアランスを促進することである。免疫応答のB細胞腕は、血液段階でマラリア原虫を不活性化するための主要な役割を担う。これが達成される主要な様式は、感染力の中和(赤血球浸潤の阻害)および抗体依存性細胞障害作用(ADCC)による。標的抗原は、その他の寄生虫株による再感染に対して有効で、かつTヘルパー細胞活性(CD4+)を誘導して長期記憶反応を生じさせることができるように保存されなければならない。寄生虫感染細胞の破壊についてのT細胞を媒介したメカニズムは、細胞障害性(CD8+)Tリンパ球(CTL)によって提供され、これが感染のプレ赤血球相の肝細胞表面にてクラスI組織適合抗原と組み合わせて発現された組換えMSP4およびMSP5抗原を認識することができるであろう。
【0071】
以下は、(A)天然のシグナル配列、(B)C末端Hisタグおよび(C)本分野における感染防御抗体反応と最も強く相関される熱帯熱マラリア原虫(P. falciparum)抗原のものに似ている酸反復領域(MSP4p20)を含むワクチンである。
【0072】
MKVAYFLSVLDLLIIFSLYFDGRRSAFAGIAACIRHGRILGEGGE/QNSTPGSGGQTGDHSAEAENGDYNEQGDDHGDDHGDDHGDDHGDEQDGEDYDDAEDDDLYELSEVDENANLCLDNNGGCGDDKICENLGKGIVKCLCKPGYKLVGTECVEHHHHHH[配列番号:30]
本発明の組換えMSP4およびMSP5ポリペプチド、並びにワクチンが宿主において保護薬レベルの中和抗体(すなわち、病原体が赤血球に感染する能力を直接遮断する、抗原に対する体液性応答の間に誘発される抗体)を誘導する能力は、アジュバント(免疫賦活剤)との乳化、リポソームに組み込むこと、適切な担体に対して結合することにより、またはこれらの技術の組み合わせにより増強することができる。たとえば、本発明の組換えMSP4およびMSP5ポリペプチドは、リン酸アルミニウムおよび水酸化アルミニウムゲルなどの従来のアジュバントと共に、宿主において体液性免疫応答または細胞媒介免疫応答を増強するために十分な量で投与することができる。
【0073】
免疫化スケジュールは、感染に対する宿主の感受性および宿主の年齢などのいくつかの要因に依存する。本発明のワクチンの単一用量を宿主に投与することができ、または時間間隔をおいて数回の用量が投与される免疫化の一次経過を行うことができる。ブースターとして使用するその後の用量を、一次経過後に必要に応じて、投与することができる。
【0074】
本発明の組換えMSP4およびMSP5タンパク質、ポリペプチドおよびワクチンは、宿主における寄生虫負担を減少させ、臨床症状を減弱させるために、寄生虫感染およびインビボにおける複製を予防し、または阻害する免疫応答を誘導するために十分な量で宿主に投与することができる。免疫原性反応は、宿主に対して1用量あたり10〜500マイクログラム、好ましくは1用量あたり約50〜100マイクログラムの範囲である量で本発明のポリペプチドを投与することによって得ることができる。本発明のタンパク質およびワクチンは、生理的に許容される担体と共に投与することができる。たとえば、水または食塩水などの希釈剤を使用することができる。
【0075】
本発明のもう一つの側面は、DNAワクチン接種の方法を提供する。また、本方法には、組換えMSP4およびMSP5ポリペプチド、タンパク質およびポリペプチドをコードする核酸の任意の組み合わせを、それ自体、キャリア分子の有無において、個体に対して投与することを含む。態様において、個体は、動物であり、および好ましくは哺乳類である。より好ましくは、哺乳類は、ヒト、イヌ、ネコ、ウシ、ブタ、およびウマからなる群から選択される。特に好ましい態様において、哺乳類は、ヒトである。
【0076】
当業者であれば、核酸ワクチン(たとえば、DNAワクチン)および核酸ワクチン技術、並びにタンパク質およびポリペプチドに基づいた技術の概念、適用および有効性を認識するであろう。核酸に基づいた技術により、裸の、またはカプセル化された組換えMSP4およびMSP5ポリペプチドをコードする核酸を、投与前にコードされたタンパク質を産生する必要なく、組織および細胞に直接投与することができる。本技術は、これらの核酸が、レシピエント生物体の細胞によって取り込まれれ、発現してレシピエントの免疫系が反応する免疫原性決定因子を産生する能力に基づく。典型的には、発現された抗原は、核酸を取り込んで発現した細胞表面上に示されが、コードされた抗原の発現およびレシピエント個体の循環系への搬出も本発明の範囲内である。このような核酸ワクチン技術は、裸のDNAおよびRNAの送達および組換えMSP4およびMSP5ポリペプチドをコードする発現ベクターの送達を含むが、限定されない。本技術は、「ワクチン」と称されるが、これは保護反応を生じない免疫原性組成物にも同様に適用できる。このような非保護を誘導する組成物および方法は、本発明に包含される。
【0077】
裸の核酸として組換えMSP4およびMSP5ポリペプチドをコードする核酸および担体分子を送達することも本発明の範囲内であるが、本発明は、より大きな、またはより多くの複合体組成物の一部としての核酸の送達も包含する。これらの送達系に含まれるものには、組換えMSP4およびMSP5ポリペプチドをコードする核酸を含むウイルス、ウイルス様粒子または細菌である。また、リポソームなどの細胞透過化化合物を伴う本発明の核酸の複合体および担体分子も、本発明の範囲内に含まれる。分子ベクター(EP 696,191、Samainらの論文)などの核酸ワクチンのためのその他の化合物および送達系は、当業者に公知であり、たとえば、WO93 06223およびWO90 11092、U.S. 5,580,859およびU.S.5,589,466(Vicalの特許)において例証されており、これらは本明細書に引用として組み込まれ、過度または過剰の実験なしで製作して使用することができる。
【0078】
本発明は、以下の具体的態様において更に詳細に後述する。
【0079】
バキュロウイルス発現系を使用して産生した抗原は、システイン結合の形成を含む複合体構造を忠実に含み、結晶化研究のために適していることが、長い間公知であった。これらの抗原は、その他の系において産生された抗原が点検される「金本位(the gold standard)」として一般に保持される[24,25]。PfMSP4およびPfMSP5の両者は、ジスルフィド結合によって形成されたEGF様ドメインを含むので、これらは、バキュロウイルス-系において産生させることによる利益を受けるであろう。しかし、すべての一般に使用される発現系と同様に、タンパク質収量は、熱帯熱マラリア原虫(P. falciparum)の異常なコドンの選択性による悪影響を受ける[26,27]。加えて、これらの高度にA+Tリッチな配列は、大腸菌(E. coli)において頻繁に変異し[28]、また独特の制限酵素部位をまれに含む。これらの理由により、合成遺伝子は、Withers-Martinez(1999)によって記述された方法を使用してデザインして、構築した。
【0080】
合成遺伝子のデザイン
その時にGenBankから利用可能な配列データに基づいて、およびヒト検証モデルを含む将来の適用を念頭において、共通アミノ酸配列のPfMSP4 NF54配列(Ac番号AF295318)およびPfMSP5 3D7配列(Ac番号AF106476)を修飾のために選択した。平行して、配列データセットを濃縮し、および集団間および集団内解析および種間解析を行うために、以下の通りに風土が異なる場所から採取した試料を使用して多型研究を行った。
【0081】
msp4およびmsp5遺伝子多型の概要
抗原多型は、熱帯熱マラリア原虫(P. falciparum)のためのワクチンを開発するときにの、重要な検討事項である。今までに調べた最も有望な血液段階ワクチン候補は、メロゾイト表面タンパク質1(MSP1)および頂端膜抗原、(AMA1)である[1-3]。しかし、これらおよびその他の表面抗原遺伝子は、免疫エフェクター-メカニズムにアクセス可能でない抗原をコードする遺伝子と比較したときに、不相応に高頻度に同義一塩基多型(nsSNPs)を示す[4-6]。複雑にする要因として、これらのnsSNPsは、最も宿主免疫系にアクセス可能なペプチドの領域内に主にクラスター形成しているラジカルアミノ酸置換を引き起こすことが多い。生じるアミノ酸置換は、重要なBおよびT細胞エピトープ[34,38]を変化させることによって、免疫回避において機能すると考えられる。したがって、このような抗原の統合された組換えバージョンによるワクチン接種は、株特異的な保護を生じる可能性が高い[4,8]。世界的に有効なワクチンには、多段形標的に由来するいくつかの表面タンパク質の保存された部分を含むと広く考えられている[30]。
【0082】
現在まで、限られた量の多型データのみが、PfMSP4およびPfMSP5につて利用できる[11,31,32]。高度に固有な場所のPfMSP4およびPfMSP5の両方の多型を完全に探索するために、配列分析研究が行われ、その結果がPolsonら、2005に示されている[33]。要約すると、Pfmsp5遺伝子配列は、高度に保存されており、潜在的に純化選択のものとにあることが見いだされた。Pfmsp4遺伝子配列は、熱帯熱マラリア原虫(P. falciparum)表面抗原に関して、相対的に保存されていること[5]、および潜在的に平衡選択(N52およびG74)の下に2つの部位を含む多形部位のN末端のクラスターを含むこと(残基45〜81)が見いだされた。これは、フランキング配列内の明らかに中立な(または、ヒッチハイク)多型のクラスター形成が観察されたことを説明するだろうし、BまたはT細胞エピトープ変異に連関させることができる。平衡選択は、古典的に分子の免疫認識に対する影響を有する「相違」を内部に持つ2つの対立遺伝子遺伝子配列の存在により生じる。対立遺伝子Aが、集団において最も一般的なときは、対立遺伝子Bは、この対立遺伝子が免疫系、並びに優性対立遺伝子よって認識されないので、有利である。したがって、対立遺伝子Bは、それが最も多くの型になるまで、より一般的であり、免疫系は、対立遺伝子Bに対してコンピテントとなり、この時点で、対立遺伝子Aであることが有利になる。最初に、平衡選択の存在は、MSP4タンパク質が有効な免疫メカニズムの標的であることを示唆する。第2に、これは、貴方が両方の対立遺伝子でワクチン接種をすること、またはタンパク質のこの領域を除外することを必要とするであろう分野の人々を保護するために、保存された領域に対して免疫系を作用させることを示唆する。肝細胞に特異的に結合することができる2つのPfMSP4由来ペプチドの最近の同定に関して[34]、第1のもの(それは、MSP4p40の残基76〜92を表す)は、1つの準保存的多型(A81)をもち、第2のもの(それは、MSP4p40の残基113〜135を表す)は、1つの欠失(111〜119)および/または1つの準保存的多型(G119)を含む。既存のPfMSP4データセットにおいて、これらの配列内の相違は、相対的に低い頻度で存在し、これらが機能的に重要な部位であるというアイデアをサポートする、(注;これらの配列は、SALSAによっても表される(図2))。最後に、潜在的に平衡選択の下にもう一つの部位V190があり、これはEGF様ドメインの最初のシステインの17残基上流に位置し、再びこれも、BまたはT細胞エピトープ修飾に関与し得る。
【0083】
合成遺伝子;PfMSP4の構築
最初に、タンパク質分泌を促進するために、EGFドメインの3残基下流から始まるすべての配列を除去した。バキュロウイルス系は、このような配列を使用して昆虫細胞GPI部分を組み込んで、およびタンパク質の細胞表面局在化を生じることができる(Bonnet et al. 2006)ことが公知であるので、これには、GPI付着シグナルを含んだ。加えて、バキュロウイルス発現系は、組換え類似体の天然のMSP1シグナル配列を正確に切断するが、天然のMSP4の切断されたN末端は定義されされていないことが公知であるので、天然のシグナルMSP4配列を保持した。プラスモディウム属において生じることは知られていないが、バキュロウイルスにおいてPfMSP4配列の2つの潜在性部位にて生じる)N-グリコシル化を防止するために、セリン残基S65およびS73をアラニンに置換した。両方の残基は、小さな側鎖(Ser = HO-CH2-、Ala = CH3-)を有し、これらの部位は、天然に多形であることは報告されていなかったが、これらの変化が、タンパク質の局部的またはマクロ構造に対して最小限の効果を有することが考えられる。一旦、C末端の6-his-タグおよび終止コドンを含む残りの特色を付加したら、CODOPプログラムセットを使用して配列をキンウワバTrichoplusia ni (High Five)細胞コドンの選択性に翻訳し戻した(表1)。
【0084】
NF54および合成遺伝子のMSP4コドン選択性
【表1】

【0085】
CODOPは、HaleおよびThompsonによって提唱されたように、コドン最適化を実行するUnix(登録商標) perlスクリプトであり[35]、他に詳細に記述されている[29]。再コドンヌクレオチド配列は、その後に手動処理によって最適化し、60〜65℃のTmで38×40mersを生じさせ、およびサブクローニングを促進するためにいくつかの独特の制限部位を含めた(図3)。
【0086】
遺伝子の全体のGC含量を15%まで増加し、サブクローニングの容易さおよび効率を上昇させ、かつ構築物操作の間に大腸菌(E. coli)によってなされる複製エラーの頻度を非常に減少させた。38個のオーバーラップオリゴヌクレオチド(それぞれ40塩基の長さ)は、Eurogentecから得た(標準的精製のみで)。遺伝子構築および増幅は、前述したように達成した。
【0087】
簡潔には、遺伝子オリゴヌクレオチドを等モルの量に混合し(それぞれ25μM)、1μl pfu DNAポリメラーゼ(Stratagene)、5μL、10×産物緩衝液、200μM dNTPおよび2μLの25mM MgSO4を含む50μL PCR反応に10倍希釈した。PCRプログラムは、94℃1分間の1回の変性工程、続く94℃30s間、52℃30s間および72℃2分間の25サイクルから成った。全長遺伝子産物を増幅するためには、5μLの遺伝子構築反応を1μl pfu DNAポリメラーゼ(Stratagene)、5μL、10×産物緩衝液、200μM dNTP、2μLの25mM MgSO4およびそれぞれ500nMの外側遺伝子構築オリゴヌクレオチド番号1および20を含む50μL PCR反応に10倍希釈した。PCRプログラムは、94℃1分間の1サイクル、94℃45s間、68℃45s間および72℃2分間の25のサイクル、並びに72℃10分間の最終伸長工程から成った。図4に示した主要な、および正確な大きさの断片をゲル抽出して、シーケンシングのためにpMOSBlueにクローニングし、Baculoviralゲノム(Baculogold、PharMingen)に組込みのためにベクターpVL1393に最後に移した。この発現構築物をMSP4p40/Hisと呼ぶ。
【0088】
修飾されたPfMSP4構築物
また、2つのさらなるMSP4構築物を構築し、MSP4-EGF/HisおよびMSP4p30/Hisと命名した(図5)。主に結晶化研究の目的で、構築物MSP4-EFG/Hisは、MSP4 EGF様ドメインのみを発現するようにデザインした。PlasmoDBによって記述された予測されるシグナル配列(残基1-20)および2つの下流の残基を含むPCR断片は、高忠実度PCRによって産生した。反応には、遺伝子構築オリゴヌクレオチド1およびBgl II制限部位を含むリバースプライマーMSP4modI(5'.-TAT-AGC-AGA-TCT-TTG-TCG-AAG-TTG-ATG-GTG-CA-3')[配列番号:23]を含んだ。合成遺伝子クローンpMosMSP4を鋳型として使用した。生じる86bp PCR産物を制限酵素Bam HIおよびBgl IIで切断し、ゲル抽出した。次いで、以前に調製し、またBam HIおよびBgl IIで消化してEGF様ドメインの上流のすべての配列を除去して残基D205〜H252を残したpMosMSP4ベクターに産物を結合させた(T4DNAリガーゼ;NEB)。
【0089】
MSP4p30/His構築物は、大部分の報告された多型が属する(図5において強調した)MSP4のC末端から30残基(残基45〜74)を除去するようにデザインした。残基1〜44をコードするPCR断片は、遺伝子構築オリゴヌクレオチド1、リバースプライマーMSP4modII(5'-ATA TGG CTG CAG CCA AGA TCC TCA TGT TAA GCA T-3')[配列番号:24] および鋳型としてのpMosMSP4ベクターを使用して産生した。断片は、154bpのサイズであり、3'および5'最末端にてそれぞれ制限部位Bam HIおよびPst Iを有した。残基A75の顆粒のすべての配列をコードするDNAの断片を、制限酵素Pst IおよびSpe I(これは、pMOSBlueベクター配列のMSP4 ORFの下流にある)を使用して、ベクターpMosMSP4から切断した。2つのDNA断片をT4DNAリガーゼ(NEB)反応中で等モルの量で合わせて、3日にわたって4℃にてインキュベートした。臭化エチジウムで染色した1%のアガロースゲルにおける2μLリガーゼ反応の分離により、724bpの長さの必要とされる産物を含むいくつかの産物の存在が明らかになった。この産物を、遺伝子構築オリゴヌクレオチド1および20と鋳型としての1μL結合反応液とを使用する高忠実度PCRによって増幅し、ゲル抽出し、pMOSBlueへクローニングし戻した。合成遺伝子と同様に、MSP4-EGF/HisおよびMSP4p30/His遺伝子配列は、発現ベクターpVL1393にクローニングされる前に確認した。
【0090】
更なる発現構築物は、上記の合成MSP4遺伝子からデザインして、構築した。この発現カセットは、非多形、プロテアーゼ耐性およびタンパク質の構造的に無処置のC末端の領域(p20と呼ばれ、後の節に記述した)の発現をできるようにデザインした。この構築物は、MSP4p40/HisのxbaIクローニング部位の下流のすべてのヌクレオチド配列を含み、したがって、Leu 130(含まれない)の下流のすべての残基をコードする。タンパク質分泌を促進するために、MSP4シグナル配列さらに下流の9残基(Met1〜Pro49)をコードするPCR断片を、遺伝子構築オリゴ1およびリバースプライマーMSP4p21rev(ATT AAT CTA GAG GCT TTT CTT CAC CCA AGA TCC TCA TG)配列番号:25を使用して産生し、Bam HI/Xba I二重消化したpMosMSP4p40/Hisに結合した。この構築物は、MSP4p21/Hisと命名し、配列をバクロウイルスゲノムに組込むためにpVL1319に結合される前に確認した。2つの補助的な同様の発現構築物は、上記の合成のMSP4遺伝子からデザインして、構築した。これらの構築物は、MSP4p21 ss1およびMSP4p21 ss2と命名し、両方ともLeu130の下流のすべての残基をコードする。タンパク質分泌を促進するために、MSP4p21/Hisと同様に、MSP4シグナル配列(Met1-Pro49)をコードする同じ配列を付加した。さらに、MSP4p21 ss1構築物の配列(Asn50-Ser57およびLeu130)をコードし、およびMSP4p21 ss2のアミノ酸残基Leu130をコードする、MSP4p40/His構築物のいくつかのコドンを付加した。
【0091】
PfMSP5合成遺伝子の構築
Pfmsp4について記述したのと同じ手順を使用して、Pfmsp5合成遺伝子をデザインした。3D7株のGenBankタンパク質配列で開始して、GPI付着シグナルの上流の3残基で始まるすべての配列を除去し、6-His-タグと置換して、3つのグリコシル化部位をセリンからアラニンへの突然変異(S83、S102およびS126)によって崩壊させ、N末端のシグナル配列を無処置のままにした。CODOPによって実行したコドン修飾を表2において考証してあり、全ての合成遺伝子配列を図6に示してある。
【0092】
3D7および合成遺伝子のMSP5コドンの選択性
【表2】
【0093】
PfMSP5合成遺伝子のPCR構築は、前述したように行い、配列内にリピートモチーフが存在しないことにより、高度に純粋な最終産物が産生された(図7)。PCR産物をゲル抽出して、pMOSBlueにクローニングし、Baculoviralゲノム(Baculo-gold、Stratagene)に組込みのためにpVL1393ベクターに移す前にシーケンスした。
【0094】
こうして調製された構築物を評価し、結果を以下の実施例に報告してある。
【実施例】
【0095】
実施例1
昆虫細胞におけるPfMSP4の発現
標準的方法[36]を使用してウイルスを産出した後で、試験感染を、150cm2培養瓶(Corning)において行った。感染は、1時間、10のウイルス多重度にて行った。3日後に上清を収集し、talon樹脂上でバッチ精製した。予測されるサイズのタンパク質産物は、構築物MSP4p40/His(40kDa)およびMSP4p30/His(30kDa)で見られたが、構築物MSP4-EGF/Hisでは、タンパク質が見られなかった(さらに下記で論議した)。MSP4p21/Hisの発現は、いまだ解析の予備的段階である。
【0096】
MSP4p40およびMSP4p30の両方の産生をスケールアップして、箱からスピナフラスコ培養へ移した。タンパク質収量を最適化し、タンパク質分解(スピナのより激しい条件下で頻繁に見られる)を制限するために、時間経過実験を行った。MSP4p40/HisまたはMSP4p30/Hisが感染したスピナ培養(210mL)をスポドプトレア・フルギペルダ(Spodoptrea frugiperda)(SF9)昆虫細胞およびHigh Five昆虫細胞の両方のために準備した。試料(8 mL)を、24時間、30時間、36時間、42時間、48時間、54時間、60時間および72時間にて収集し、細胞死統計のために顕微鏡によって解析し、次いで、20 mM Tris HCl(pH8);500 mM NaClに対して透析した。タンパク質を、試験管プロトコルを使用してtalon樹脂上で精製して、100μLの100 mMイミダゾール;20 mM Tris HCl(pH8);500 mM NaClで溶出した。ある時間にわたった細胞死の割合をそれぞれの感染についてプロットし、それぞれの溶出の20μLの試料を構築物および細胞型の両方について並行して解析した(図8)。
【0097】
予想通りに、High five細胞は、SF9細胞よりはるかに高いタンパク質収率を生じた。しかし、タンパク質分解が始まる長い発現期間(48時間を超える)の後で、MSP4タンパク質は完全に分解せず、むしろより小さな安定な、約20 kDaの産物(MSP4p20)へと分解した。この産物は、MSP4p30感染の後期の段階においてもまた見られた。すべてのタンパク質は、C末端のヒスチジンタグを介して精製したので、この産物は、EGF様のドメインおよびプロテアーゼに耐性な堅い構造を形成するいくつかの上流配列を含むにちがいない。これは、EGF様のドメインの上流領域が、還元感受性のB細胞エピトープを有しているというWangら(2001)[13]の発見と一致する。
【0098】
実施例2
N末端シーケンシングおよび質量分析
時間経過実験から得られる情報を使用して、3つの異なる純粋なMSP4タンパク質標品を、異なる時点にて培養SNを収集することによって産出した。第1に、MSP4p40と表した全長MSP4、第2に、全長の修飾産物であるMSP4p30および第3に、分解産物であるMSP4p20。標準的なHigh Fiveスピナ培養におけるそれぞれの産物についての平均タンパク質収量は、それぞれ8 mg/L、6 mg/Lおよび8 mg/Lであり、収集の時間を反映しており、より早い時点での収集産物ほど、より少ないタンパク質を生じる。市販のバキュロウイルスコンピテンス(Henogen)においてなされた最近の進展を考慮すると、これらの収率は、最小限の見積もりであると考えられる。
【0099】
それぞれの産物のN末端のシーケンシングは、強力で明らかなシグナルを産出し(p40:41残基にて始まるMRILG、p30:41残基にて始まるMRILA、p20:132残基にて始まるKSPKE)、in vivoの熱帯熱マラリア原虫(P. falciparum)シグナル配列は、実際、PlasmoDB(http://plasmodb.org/plasmodb/servlet/sv?page= gene& source_id=PFB0310c)によって予測されるような長さの2倍(20に対しての40残基)であってもよい。MSP4p40およびMSP4p20の同一性は質量分析によってまた確認して、タンパク質は、実際にそれぞれ23.554 kDaおよび13.714 kDaの分子質量を有している。
【0100】
バキュロウイルスで発現したMSP4のN末端の配列は、寄生虫によって使用されるシグナル配列を同定する可能性が高い。第1に、MSP4-EGF/His構築物は、PlasmoDB予測されたシグナル配列に加えてさらに3つの下流の残基を含み、培養SNにおいて決して検出されなかった。第2に、Pfmsp4遺伝子内で同定される多形の部位のN末端のクラスターは、45番目のコドンにて始まる。Polsonら2005[33]で記録された解析によって示唆されるように、コドン45の下流の遺伝子の領域が、集団内の対立遺伝子型のバランスをとっることによって回避メカニズムを展開すると思われる効果的な免疫反応の標的である(52および74残基が関与する。図5を参照されたい)。したがって、シグナル配列切断部位(MRIL)に寄与する4つの保存された残基からなるPfMSP4のプロセスされたN末端は、このタンパク質のよりアクセスしやすい領域に優先的に方向付けられる免疫反応の有効性を減少させることができる多形部位のクラスターに直接続く可能性が高いようである。
【0101】
実施例3
シグナル配列の疎水性度
バキュロウイルス系によるMSP4構築物のために使用した切断部位は、天然の切断を再生することをさらに確認するために、バイオインフォマティクス研究を、以前に報告された天然の熱帯熱マラリア原虫(P. falciparum)シグナル配列に注目して行った。2001年において、公知の熱帯熱マラリア原虫(P. falciparum)シグナル配列切断部位を比較する研究が公表された[37]。このようなデータを得る際に関与する技術的困難性のため、わずか10個のこのような配列が存在するにすぎない。この研究の目的のために、それぞれのタンパク質の、報告されたシグナル配列切断部位に集中する、80残基のタンパク質配列を選択して、局部的な疎水性度の値を算出するために提出した(パスツールサーバーを介して)。Nacer 2001において報告された3つのタンパク質切断部位の疎水性プロフィールおよびバキュロウイルス系におけるMSP4p40のものを図10に示してある。上部パネルにおいて、KAHRPおよびMSP1シグナル配列の両方は、古典的な疎水性プロフィールを表し、疎水性度の値が中性のラインを(ポジティブからネガティブに)横切る直後に切断が生じている。PfMSP1シグナルの切断は、バキュロウイルス系によって忠実に行われることが知られている(Bonnet et.al. 2005)。局部的な疎水性度の同様の顕著な変化が、予測された切断部位でのMSP4p40タンパク質配列のN末端内に存在していない場合、疎水性度における最大の局部的変化(ネガティブからポジティブへ)の後のさらに下流で切断が実際に生じる。このプロフィールは、S抗原で見られたものにマッチし、したがって、独特ではないが、MSP4p40シグナル配列が40残基の長さであるという知見に重きが置かれる。
【0102】
実施例4
昆虫細胞におけるPfMSP5の発現
MSP5/Hisウイルスのストックを以前に記述したように産生し、試験感染および時間経過研究を、MSP4p40/Hisについて記述したように行った。図11に示したように、3つの異なるPfMSP5に由来する産物は、SDS-PAGEゲル電気泳動下でおよそ45、35および10 kDaにて移動して検出された。興味深いことに、2つのより大きな産物は、より大きなものが産生されて、その後に分解してより小さなものが形成するのではなく、時間経過実験の初期の間に同時に存在するように見えた。PfMSP4で見られた状況に対するもう一つの著明な相違は、研究した2つの細胞株の逆のタンパク質産生能力であり、SF9は、桁違いに早い増殖を行い、High Five細胞はより脆弱であった。これは、本来、PfMSP5がPfMSP4とは非常に異なる特性を有することを示唆する証拠の最初の部分であった。第2は、N末端のシーケンシングデータに由来する。
【0103】
実施例5
N末端シーケンシングおよび質量分析
MSP5発現の2つの最も大きな産物のN末端のためのシグナルを得ることはできなかったが、これらの両方がN末端の修飾を有し、これがN末端のシーケンシングにおける必須なイベントであるペプチド主鎖の破壊を遮断することを示唆している。より小さな10 kDaの産物のシーケンシングは良好であり、この産物のN末端が189残基にて始まると定義した:YNKVE[配列番号:26]。この産物の同定は、質量分析によっても確認され、これは、7.746 kDaの実際の分子量を有する。加えて、MSP5ウイルスゲノムの配列も確認され、YNKVE [配列番号:27]の残基は、正確なORFにおいてのみ存在し、したがって、これまでに未決定の産物としてこれらは、PfMSP5にちがいない。
【0104】
2つの最も大きなMSP5産物についてのMALDI-MSによって得られた質量値は、それぞれ、24.679および20.551 kDaであり、MSP5タンパク質配列のみに由来する任意のポリペプチドにマッチせず、これらのタンパク質が転写後修飾を有するという考えを支持した。起こりうるN末端遮断修飾を調べるために、MSP5配列をExPASYホームページ経由でprositeに提出した。これにより、C14脂肪酸の同時翻訳付加を指揮することができる2つのN-ミリストイル化部位の存在が明らかになった(42残基目:GGFTSK、および66残基目:GSLPTK)[38]。しかし、これらの修飾は、N末端に限定的であり、N末端のシーケンシングを遮断することができる一方、これらは、適切な質量値(それぞれ、24,015および21,301)を生じない。しかし、グリシン残基のN末端修飾を導くモチーフは、真核細胞によって行われるミリストイル化の唯一の形式ではない。また、これらは、ミリストイル基で、任意のN末端のグリシンおよび任意のリジンを転写後修飾することができる(下記に概説する)。これらの可能性と共に考えると、推定上の位置は、現在未決定であるが、それぞれのMSP5産物(p45およびp35)が2つの脂肪酸修飾を有すると仮定することによって、24.673kDaおよび20.553kDaに近接した一致が得られる。
【0105】
実施例6
脂肪酸修飾およびプラスモディウム属
グリコシル化、パルミトイル化、N-ミリストイル化およびミリストイル化を含む、真核細胞によって使用される、いくつかの異なるタイプの特別な翻訳修飾がある。これらのうちで、第1のものは、ここで発現されるMSP5タンパク質には存在しない。この種の修飾が熱帯熱マラリア原虫(P. falciparum)によって行われないことが知られているので、すべてのグリコシル化部位を、合成遺伝子の構築の間にタンパク質配列から除去した。第2の修飾のタイプは、全体として真核生物においてあまりよく理解されておらず、パルミトイル化が酵素依存的および酵素非依存的な両方の経路を介して生じうると考えられる[39]。現在まで、任意のパルミトイル化された熱帯熱マラリア原虫(P. falciparum)タンパク質の報告はなかった。しかし、タンパク質のN-ミリストイル化は、プラスモディウム属において重要であると示された。Pf N-ミリストイルトランスフェラーゼ遺伝子相同体を同定し(PlasmoDB PF14_10127)、ヒトおよび酵母の遺伝子オルソログに高度に相同的であることが見いだされた[40, 41]。もしこの酵素がブルーストリパノソーマ(Trypanosoma brucei)および森林型熱帯リーシュマニア(Leishmania major)などのその他の寄生性原虫に対するのと同様に、プラスモディウム属に対して重要であるなら、複数のタンパク質のN-ミリストイル化が寄生虫成長のために必須であると予想することができる。実際、PfADF(リボソームの因子)のN末端のミリストイル化は、GTPの結合を媒介し、酵素活性を促進するために重要であることが示された[42]。この脂肪酸修飾(N末端のグリシンおよび内部のリジンの両方での)は、膜へのタンパク質の局在化およびタンパク質-タンパク相互作用の安定化を含む、より高度な真核生物における種々の役割を演ずることが示された[39, 43-45]。
【0106】
実施例7
バキュロ系において
昆虫細胞のバキュロウイルス感染は高次元の真核細胞系であるので、多くのタイプのより進化したタンパク質修飾を行うことができる。実際、この系は、いくつかのNミリストイル化されたタンパク質を産出するために特異的に使用されており、NAP-22が最も記述されている[46]。
【0107】
実施例8
抗体産生
ウサギポリクローナル抗血清は、組換えMSP4p40、MSP4p20およびMSP5(p45およびp35)にたいして、標準的な免疫化手順を使用して、EUROGENTECによって産出した。すべてのウサギは、免疫化(抗原あたり2羽のウサギ)前にMSP4またはMSP5について陰性であり、3または4回の免疫化の後のELISAによって決定されるように高力価の特異的な血清抗体を産生した。還元および非還元組換えタンパク質に対する最終時点の力価を図12に示してある。パネルBに示したように、ウサギNo.8からの抗体によるMSP4p20の認識は、抗原が還元される時に50%以上減少し、MSP4p20タンパク質のエピトープの多くが高次構造的であることを示している。還元は、主にジスルフィド結合構造に影響を及ぼすので、このデータは、MSP4p20のEGF様ドメインが無傷であり、少なくとも1つのB細胞エピトープを形成することに関与することを示す。
【0108】
実施例9
寄生虫由来MSP4
予備的な免疫ブロットデータは、前述したポリクローナル免疫血清を使用して作製されており、図13に示してあり。MSP4についてのデータは、40kDaのタンパク質が専ら後期寄生虫の調製物のTriton X100可溶性画分において検出されるというR. Coppelのグループにより公開された結果[9]と一致する(図13A)。MSP4、MSP4p20のC末端の領域の特性を調べるために、プールしたウサギ血清(R6-9)および2002年においてSenegal のDielmoで収集したヒト血清を、固定したMSP4p20に対してアフィニティー精製した(Amersham: NHS-activated Sepharose 4 fast flow)。非同調性寄生虫材料(SDS-PAGEによって分離し、ニトロセルロース膜へ転写した)をこれらの血清でプローブしたときに、以前に観察された40 kDaでのバンドに加えて、およそ18 kDaのバンドも見られ(図13B)、自然感染において、MSP4抗原がMSP1p42に類似のタンパク質分解のプロセシングを受け得るか、またはバキュロウイルス感染の後期において得られるような自己制御様式でのプロテアーゼによって分解され得ることを示唆している。熱帯熱マラリア原虫(P. falciparum)メロゾイトのMSP4p20エピトープの天然の存在をさらに確認するために、間接的な免疫蛍光アッセイ法(IFA)を、風乾した非同期成長の寄生虫培養で行った。メロゾイト特異的な染色は、ヒト抗MSP4p20で見られ、二次抗体単独またはマラリア未感染血清で何も染色されなかった(図13C)。
【0109】
実施例10
寄生虫由来MSP5
バキュロウイルスが発現したMSP5に対して生じたポリクローナル血清を使用して得られた免疫ブロットデータは、その他のグループによって公開されたもの[9]とは非常に異なるプロフィールを示し、彼らは、MSP5の検出に明らかな問題を抱えており、それが低レベルで発現されていることを示唆する。本発明の免疫ブロットデータは、抗MSP5特異的な血清で強力なシグナルを示す(図13D)。MSP4とは異なり、寄生虫由来MSP5は、また、粗雑に調製した寄生虫抽出物の膜画分にも存在しており、これはGPIアンカーを有するだけのタンパク質と変わらぬ様式で分画しないことを示唆する。実際に、還元した寄生虫材料に注目した場合、推定のNミリストイル化MSP5に対して調製したポリクローナル血清は、その他のNミリストイル化されたタンパク質とも交差反応し得ると結論づけることができ、さらに、これが熱帯熱マラリア原虫(P. falciparum)における重要かつ頻繁に使用される翻訳後修飾であるという考えを支持する。
【0110】
実施例11
ヒト免疫血清の解析
本発明のタンパク質産物がマラリアに感染した個体の抗体によって認識されることを確実にするために、Senegal(Dielmo)における風土病村で生活している9人のランダムに選択された準免疫成人の血清を、ELISAによって解析した(図14)。すべての個体は、すべてのタンパク質産物について陽性であった(標準偏差3倍に加えて対照血清より大きいOD値を有する)。相対力価は、個体間でおよび産物間で変化したが、全体の最終時点の力価は、MSP4について1x105およびMSP5について1x104であった。ウエスタンブロット解析(図15)は、MSP4について高度に陽性の2人の個体(個体3および9)並びにMSP5について高度に陽性の1人の個体(個体8)からの血清を使用して行った。以前に抗MSP1p19で見られたように、EGF含有メロゾイト表面タンパク質に対するヒト抗体は、凝集した材料に対して特に親和性を有していた(図15)。
【0111】
実施例12
Senegalにおける抗MSP4抗体;保護における役割
自然感染における抗MSP4抗体のための推定上の役割をさらに調べるために、疫学調査を行った。MSP4の3つの異なる精製された形態であるp40、p30およびp20は、他で考証された大規模な先見の明ある研究で評価されたており(参照[47]を参照されたい)、実施例16において下記に詳述してある。
【0112】
要約すると、組換えMSP4タンパク質のそれぞれの形態は、90%を超える血清有病率で強く認識されたが、有意に異なる平均のIgG OD比(p40 = 20±10、p30 = 12.3±7.3およびp20 17.3±10)を示した。すべてのマラリア抗原で見られるように、抗体力価は、年齢と共に増大し、保護との相関は、年齢で調整した進行モデルを使用して対処しなければならない。この形態の解析を使用して、MSP4p40抗体ではなく抗MSP4-p30および抗MSP4-p20は、臨床的なマラリア発症の減少した発病率に正に相関した(それぞれP = 0.036、P = 0.018+ 0.067)。これらの知見は、MSP4タンパク質のN末端領域内の潜在的にバランスをとっている多型の同定を強化し、MSP4に基づいたワクチン構築物においてタンパク質のこの領域を欠失させるという考えを特に支持する。
【0113】
実施例13
Senegalにおける抗MSP5抗体;保護における役割
自然感染における抗MSP5抗体の役割をさらに調べるために、疫学調査を行った。MSP5の2つの高分子量型を含む標品は、他に記述されたように解析した(参照[47]を参照されたい)。血清有病率は、MSP4に関するよりもMSP5に関してより低い(59%)ものの、臨床的なマラリア発症の減少数との高度に有意な統計的相関が、年齢を調整した進行モデルを使用して観察された(P = 0.0028)。以前に考察したように、この抗原は、GPI修飾について見られるように、脂質が誘導する免疫原性および抗感染免疫における免疫反応にとって潜在的な重要性を持つ脂肪酸修飾を有し得る[48];Bonnetら、2005(作成中の刊行物)。
【0114】
要約すると、PfMSP4は、C末端にEGF様ドメインを有し、GPIアンカーによってメロゾイト表面に結合する。その機能は、未知であるが、生存可能なPfMSP4「ノックアウト」変異体は観察されなかったため、寄生虫生存のために必須と思われる。3つの組換えPfMSP4構築物は、EGF様ドメインの適切な折りたたみを含む天然の抗原の複製を最適化するために、バキュロウイルス昆虫細胞発現系において、可溶性な分泌タンパク質として発現される。2つの構築物は、それぞれN末端の近くの多形領域からの30アミノ酸の欠失を伴い、または伴わないMSP4外抗原(分泌を可能にするC末端の疎水性残基なし)のMSP4p30およびMSP4p40に対応する。第3の20 kDaのタンパク質は、EGFドメインを含むMSP4のC末端半分にほぼ対応する(MSP4p20)。より直接的様式でのこの抗原の発現は、前述した構築物MSP4p21/Hisを使用して評価されている。3つの異なる抗原に反応する抗MSP4抗体は、マラリアに曝露されたヒトにおける自然獲得免疫に関して、伝播が季節的である中間的な風土病の村で生活している205個体の断面調査で比較した。
【0115】
伝播期以前には、3つの構築物を強く認識した:血清有病率は90%を超えないが、MSP4p40、MSP4p30およびMSP4p20についてそれぞれ20±10、12.3±7.3および17.3±10の有意に異なる平均IgG OD比であった(約5×10-3力価)。MSP4構築物に反応する抗体は、成人よりも有意に低いIgGレベルを有する15歳より下の個体と年齢相関した(P<0.001、Rho 0.25-0.33)。伝播期を含むその後の6月の期間の間の臨床的発病の解析は、年齢補正モデルにおいて、両者とも多形性の領域を欠いているMSP4p30およびMSP4p20の両方に反応するIgGの存在は、マラリアの臨床的発症の減少した発病率と有意に関連したが、多形領域を有するMSP4p40は、関連していなかった(P<0.05、比# 0.75)ことを示した。まとめると、これらの結果は:(i)バキュロウイルス組換え抗原を使用してモニターすると、PfMSP4に反応する強力な自然に獲得したIgGが存在する;(ii)抗MSP4 IgGは、臨床的なマラリアに対する保護と相関する、および;(iii)可変領域に反応するIgGは、別の保護に関連する抗MSP4 IgGを妨げるかもしれない。
【0116】
実施例14
PfMSP4モノクローナル抗体
PfMSP4に特異的な4種のモノクローナル抗体(mAb)は、3つの異なるエピトープを認識する。
【0117】
モノクローナル抗体L11-16は、p40(完全なPfMSP4の多形性外部ドメイン)によって誘導される2つのその他の代表である。還元非感受性のそのエピトープ(17B)は、PfMSP4のN末端の半分に位置する(non-p20;16A)。これは、特にPfMSP4の不均一な凝集体と反応し、その形成は、還元感受性であり(図16A)、非還元(nR)および部分的還元(R)SDS-PAGEにおける高分子量物質が、実際にPfMSP4であることを証明するために役立った(バキュロウイルスPfMSP1p19について以前に見られたとおり)。
【0118】
モノクローナル抗体F12-7は、PfMSP4 p30(多形性領域を欠く)によって誘導され、PfMSP4のp20のC末端半分における還元に反応しないエピトープを認識する(図16Bおよび17B)。L11-16と比べて、その明らかな減少した凝集体との反応性は、そのかなりより低い親和性のためであろう:
クローン アイソタイプ KD-MSP5 エピトープ
L11-19 IgG1 3.2 10-9 A
F12-7 IgG1 1.0 10-7 C
【0119】
実施例15
PfMSP5モノクローナル抗体
PfMSP5(p45+p35)に特異的な7種のモノクローナル抗体(mAb)は、5つの異なるエピトープを認識する。
【0120】
モノクローナル抗体G21-2は、p45およびp35のPfMSP5産物を認識し(図18A)、そのエピトープは、還元感受性でない(図18Aおよび19A)。別々のより高分子量の凝集体と反応し(図18A、nR)、その形成は、還元感受性である(図18A、R)。
【0121】
モノクローナル抗体J18-14は、p45のみを認識し、そのエピトープは、還元感受性である(図19B)。また、これは、凝集体と反応する。
【0122】
これらの抗原は、C末端のヘキサヒスチジンタグを介して精製したので、p45およびp35がN末端で異なることは考えられる。これらの特徴は、J18-14エピトープがC末端のEGFドメインおよびN末端の両方を含む高次構造に対応し得ることを示唆する。両方のモノクローナル抗体は、同様の親和性を有する:
クローン アイソタイプ KD-MSP5 エピトープ
G21-2 IgG1 2.8 10-8 B
J18-14 IgG1 4.2 10-8 D
これらの2つのモノクローナル抗体の反応性は、PfMSP5産物を定義するために使用することができる。
【0123】
実施例16
曝露された個体の免疫血清および集団
以下の結果は、(A)風土病の地域で生活している病気の症候が明らかでない個体から採った過免疫血清の対照プール、(B)2005年6月に収集したDielmo(完全な風土病の伝播)の村からの個体の血清および(C)伝播シーズンが始まる前の2000年7月および8月にNdiop(中間的な風土病の伝播)の村において収集した205血清の集団を使用して得られた。その集団は、3および75歳の年齢の間に、108人の男性および97人の女性を含んだ。臨床的発作を以降5月の間に活発に記録し、合計278の臨床的発症を治療した(Perraut et al, JID 2005 191 264-271)。
【0124】
解析プロトコル
(1)抗原コーティングの最適化、(2)試験される標準的な血清希釈の選択、(3)血清の集団の系統分析法、(4)中央OD比値の算出、(5)年齢による結果の層別化、(6)浸潤および増殖阻害についての選択した血清の機能解析、(7)選択した血清グループについてのアイソタイピング、(8)臨床発症データに対する中央OD比の関数として層別化される結果の統計分析。
【0125】
(1)較正:抗原をコーティングする2つの濃度の1および0.5μg/mlを使用して、「血清過剰免役」(SHI)およびDielmo(2005)の村からの選択された個体の希釈系列を試験する。
【0126】
結果:0.5μg/mlでのコーティングは、結果が村およびSHIからの両者の個体について、1μg/ml の抗原で見られたのと同じであったので、至適であることが分かった。選択した個体の反応は、NdiopよりもDielmoにおいて強力であり、MSP5は、MSP4より低い程度の抗体反応を示した。図20Aおよび20Bを参照されたい。
【0127】
(2)較正:SHIおよびDielmoからの陽性血清の滴定
結果:図21A、21B、21Cおよび21Dを参照すると、Dielmoからの血清(60605)を使用したときに、抗MSP4反応は、高度に陽性であった(>SHI)。しかし、ODは、2000年のNdiop集団で後に得られたODに比べて比較的低く、それにもかかわらず、SHIの力価は、同程度であった。一見すると、検出された抗体は、鋭角の曲線で示されていたので、以前の研究からのMSP1に対するものと比較して、相対的に低い親和性を有するように見えた。
【0128】
SHI力価に対して MSP5=1/1600
MSP4-40 MSP4-30 MSP4-20=1/3200。
【0129】
(3)Ndiop集団2000年の解析:一次結果
SHIは、それぞれのアッセイ法において系統的に滴定した。OD比を、それぞれのプレート上の一連のSHI対照に関して算出した。この集団については、1/200希釈でのOD比は、すべて高かったが、実際の力価は、平均的であった(4〜6の間のOD比値は、約1/2000の最終時点の力価を与える)。
【0130】
(下記の結果)
力価の結果/OD比
【表3】
【0131】
(4)反応者の発病率
以下の表には、2000年のNdiopからの集団の個体からの抗体反応の特徴を示してある。陽性反応者は、2を超えるOD比をもつとみなした。以下に示すように、MSP4は、ほとんどすべての個体(>90%)によって認められ、MSP5認識は、実質的により低かった(約60%)。
【表4】
【0132】
(5)年齢およびその他の抗MSP反応との相関
図22A、22B、22Cおよび22Dを参照すると、その他の抗MSP反応との有意な相関があった。
【0133】
- MSP1で、有意なRho # 0.27-0.36
-種々のMSP4抗原との間で、Rho >0.9
-MSP4およびMSP5との間で、Rho # 0.44-0.47
Ndiop集団において、個体の年齢との有意な相関があったが(P<0.001、Rho0.33〜0.22)、MSP1とのものよりもわずかに目立たなかった。抗MSP5抗体反応は、年齢と最も相関しなかったが、<15歳の個体は、有意に低いレベルの抗体を有した。
【0134】
MSP4に対する着目:MSP4抗原の相違
図23を参照すると、MSP4の異なる断片の認識のレベルには、高度に有意な相違がある(P<0.001)。逆説的に、最も小さなおよび最も大きな抗原(p20およびp40)は、抗原MSP4-30(またはMSP4md2)よりも優れて認識され、それ自体は、全長MSP4(MSP4-40)よりも小さい30残基だけである。この結果は、それが免疫回避ストラテジーの存在と一致するようであるので、興味深く、この場合、抗体反応は、可変ドメインに対して集中する。これにより、IgG反応と浸潤阻害との間の相関が無いことを説明することができるであろう(下記を参照されたい)。
【0135】
(6)機能試験との関係
図24Aおよび24Bを参照すると、MSP4p40、MSP4p20、MSP4_mod(p30)(<20>)またはMSP5(<2.4>)についての中央OD比値の間の二比較に基づいて、成長または浸潤阻害における有意差は無い。(この特性は、この集団における抗MSP1p19抗体について統計学的に有意であるだけのように見えた)。しかし、メロゾイト食作用に関しては、有意差がある。メロゾイト食作用とのこの機能的相関は、この現象が特異的なAb力価(したがって、OD比率)の全体の増大と関連し、これは、風土病の地域における年齢の関数として増大することが公知であることから、予想された。
【0136】
下記の図を参照されたい:
(7)アイソタイピング
解析は、MSP4-40およびMSP5についての限られた数の血清に限定された。結果は、図25Aおよび25Bに示してある。MSP4-p40については、アイソタイププロフィールは、IgG1+、IgG3+であり、ごくわずかな量のIgG4であった:
-力価と最近の循環寄生虫血症との間に相関はない。
【0137】
-IgG3レベルは、15歳>個体についてより高く(P=0.006)、最近マラリア発症にかかった人々と同じであり(P=0.03)、これは、発症が風土病の地域において年齢と関連した事象であるので、予想できる結果である。
【0138】
-IgG1およびIgG3は、IgG反応の有意な成分であり、それぞれIgGの40%および30%を表した。
【0139】
-強力な反応者については、IgG反応と浸潤阻害との間に有意な関係が存在する(IgGサブクラスに非依存的)。
【0140】
MSP-5については、アイソタイププロフィールは、主にIgG1である。
【0141】
-最近の循環寄生虫血症(有意性P=0.06近く)または年齢との関係において相違がない。
【0142】
-IgG1は、IgG反応の有意な主要構成要素であり、反応の75%の合計となった。
【0143】
-IgG反応と浸潤または増殖阻害との間に相関はなかった(特定のアイソタイプは関連しなかった)。
【0144】
(8)マラリア発症との相関
年齢補正したポアソン回帰モデルを使用して、追跡調査期間の間にAg構築物およびマラリア発症の発病率に対して抗体反応性との間の関係を解析した。これらの抗原は、2000年のNdiopの村からの205個体の先見の明ある集団研究において、臨床的発症に対する保護と有意な相関を示した。
【0145】
・MSP4に関して
MSP4-p40およびMSP4-p30に対するIgG力価の間に有意差が見られたという前述の観察と一致して、全長抗原(MSP4-p40)に対する抗体反応は、保護と相関しない。しかし、抗原の30残基の可変ドメインの無い構築物(MSP4-p30)および最も小さな抗原(MSP4p20)構築物に対して向けられた抗体の反応性は、保護と有意に関連する(P<0.05)。これらの知見は、タンパク質のN末端での残基多型が免疫回避に関与すること、およびタンパク質のこの領域に向けられた抗体は、(A)限られた親和性を有し(B)株の特異性を示し、したがって大多数の株に限定された効果を有し、または/および(C)APLの存在のために、親和性成熟を達成できないか、もしくは必要とされるT細胞補助を欠くという考えを補強する。関与するメカニズムに関係なく、このデータは、保存されたタンパク質サブユニットが、免疫回避において機能すると考えられる多型の領域を含むことが多い完全な実体よりも広範な特異性免疫を誘導するより良い機会を有するという考えを支持する。
【表5】
【表6】
【表7】
【0146】
高力価と疾患発症の減少との間に高度に有意な相関を見いだした。抗原によって保持されると予測される脂肪酸修飾からみて、これらの結果は、重症度および抗脂質反応に関して興味深い意味を有する。
【表8】
【0147】
上に示されたデータから描区ことができる主要な結論は、(1)バキュロウイルスで発現したMSP4 およびMSP5は、MSP1p19について見られたものと同様の活性を有するワクチンとして有用であり、(2)これらの抗原は、個体を免疫することによって強く認識され、(3)MSP4の保存されたドメイン(p30およびより相応しいp20)のIgG認識の中央値および高度に保存されたMSP5抗原のものよりも高いことは、臨床的なマラリアに対する天然の保護と相関する。
【0148】
実施例17
MSP4p20発現構築物
MSP4p20の発現のために、熱帯熱マラリア原虫(Plasmodium falciparum)MSP4合成の遺伝子配列から2つの構築物をデザインした。1つの構築物は、PfMSP4p21ss1と他に名付けられたMSP4p21ss1として同定される[配列番号:28]。構築物MPS4p21ss1によってコードされるオープンリーディングフレーム[配列番号:19]および予想アミノ酸配列[配列番号:20]を以下に記述してある:
構築butMPS4p21ss1のヌクレオチド配列
>MSP4p21 ss1 ATGTGGATCGTAAAGTTCTTGATTGTGGTCCACTTCTTCATCATATGCACCATCAACTTCGACAAGCTCTACATTAGTTACTCTTACAACATCGTCCCTGAAAACGGACGTATGCTTAACATGAGGATCTTGGGTGAAGAAAAGCCTAACGTTGACGGTGTGTCAACATCTctaGAAAAGAGTCCCAAGGAGAGTCAAATGGTCGACGACAAGAAGAAGACCGAGGCCATTCCAAAGAAAGTCGTGCAGCCAAGCTCGAGCAACTCTGGAGGTCACGTCGGTGAAGAAGAAGACCACAACGAAGGAGAGGGAGAGCACGAAGAGGAGGAAGAACACGAAGAAGACGATGACGACGAGGACGACGACACATACAACAAAGACGACTTGGAGGACGAAGATCTTTGCAAGCACAACAACGGAGGATGTGGAGATGACAAGCTCTGCGAGTACGTTGGAAACCGTCGCGTAAAATGTAAATGTAAGGAAGGATACAAGTTGGAAGGAATTGAGTGCGTTGAACACCACCACCACCATCACTAA[配列番号:28]
構築物MSP4p21 ss1によってコードされるオープンリーディングフレーム
>MSP4p21 ss1 MWIVKFLIVVHFFIICTINFDKLYISYSYNIVPENGRMLNMRILGEEKPNVDGVSTSLEKSPKESQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEEDHNEGEGEHEEEEEHEEDDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLCEYVGNRRVKCKCKEGYKLEGIECVEHHHHHH[配列番号:19]
明るい灰色(GEEKPNVDGVSTS): MSP4p40の配列における残基MRIL直後の付加されたアミノ酸
濃い灰色(L):前記明るい灰色で表示されたアミノ酸配列直後の、MSP4p40の配列におけるMSP4p20配列の開始に直接先行する付加されたアミノ酸L
予想されるタンパク質産物MSP4p21 ss1
MRILGEEKPNVDGVSTSLEKSPKESQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEEDHNEGEGEHEEEEEHEEDDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLCEYVGNRRVKCKCKEGYKLEGIECVEHHHHHH[配列番号:20]
その他の構築物は、PfMSP4p21ss2と他に名付けられていたMSP4p21ss2として同定される[配列番号:29]。構築物MPS4p21ss2によってコードされるオープンリーディングフレーム[配列番号:21]および予想アミノ酸配列[配列番号:22]を本明細書に記述してある。
【0149】
構築物MSP4p21 ss2のヌクレオチド配列
>MSP4p21 ss2 ATGTGGATCGTAAAGTTCTTGATTGTGGTCCACTTCTTCATCATATGCACCATCAACTTCGACAAGCTCTACATTAGTTACTCTTACAACATCGTCCCTGAAAACGGACGTATGCTTAACATGAGGATCTTGGGTGAAGAAAAGCCTCTAGAAAAGAGTCCCAAGGAGAGTCAAATGGTCGACGACAAGAAGAAGACCGAGGCCATTCCAAAGAAAGTCGTGCAGCCAAGCTCGAGCAACTCTGGAGGTCACGTCGGTGAAGAAGAAGACCACAACGAAGGAGAGGGAGAGCACGAAGAGGAGGAAGAACACGAAGAAGACGATGACGACGAGGACGACGACACATACAACAAAGACGACTTGGAGGACGAAGATCTTTGCAAGCACAACAACGGAGGATGTGGAGATGACAAGCTCTGCGAGTACGTTGGAAACCGTCGCGTAAAATGTAAATGTAAGGAAGGATACAAGTTGGAAGGAATTGAGTGCGTTGAACACCACCACCACCATCACTAA[配列番号:29]
構築物MSP4p21 ss2によってコードされるORF
>MSP4p21 ss2 MWIVKFLIVVHFFIICTINFDKLYISYSYNIVPENGRMLNMRILGEEKPLEKSPKESQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEEDHNEGEGEHEEEEEHEEDDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLCEYVGNRRVKCKCKEGYKLEGIECVEHHHHHH[配列番号:21]
明るい灰色(GEEKP): MSP4p40の配列の切断部位MRILの直後に付加されたアミノ酸
濃い灰色(L):前記明るい灰色で表示されたアミノ酸配列直後の、MSP4p40の配列におけるMSP4p20配列の開始に直接先行する付加されたアミノ酸L
予想されるタンパク質産物MSP4p21 ss2
MRILGEEKPLEKSPKESQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEEDHNEGEGEHEEEEEHEEDDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLCEYVGNRRVKCKCKEGYKLEGIECVEHHHHHH[配列番号:22]
【0150】
実施例18
MSP4特色の進化的保存
熱帯熱マラリア原虫(P. falciparum)および三日熱マラリア原虫(P. vivax)由来のMSP4のアミノ酸配列を、一般の整列プログラム(ClustalX)を使用して整列させられると、いくつかの著明な観察がなされ得る。図26を参照されたい。第1に、熱帯熱マラリア原虫(P. falciparum)のシグナル配列切断部位の下流の残基は(本明細書において定義したとおり)、その種の間でほとんど100%保存される(Pf MRILGEおよびPv GRILGE)。この知見は、成熟したPfMSP4タンパク質が41残基にて始まり、直前の残基は、保存された切断部位の一部分であるという考えをもう一度強く支持する。第2に、SALSAとして公知の配列エレメント(下線を引いてある)は、三日熱マラリア原虫(P. vivax)タンパク質に存在しない。これは、肝細胞結合(インビボではPfMSP4によって媒介される)は、このタンパク質の保存されておらず、したがって重要な機能ではないことを示唆するであろう。その代わりに、三日熱マラリア原虫(P. vivax)タンパク質のこの領域は、熱帯熱マラリア原虫(P. falciparum)のものとは完全に異なり、非常に低い複雑さの配列を含む。明らかに熱帯熱マラリア原虫(P. falciparum)の配列と同等ではないが、免疫系における効果が同様であると想像することができる。熱帯熱マラリア原虫(P. falciparum)には、タンパク質の最N末端に、N末端の高頻度可変領域がある。多型頻度の統計分析は、免疫回避に関与し得るこの領域内の2つの部位を示す。これは、変異体配列に曝露されたときに、株特異的反応および/または免疫寛容を引き起こすBおよびT細胞エピトープの修飾または切除を介して達成されるであろう。また、このタイプの免疫調整は、反復配列の特性である。(A)これらは、それ自体変化したペプチドリガンドを含み、(B)これらは、T細胞非依存的なB細胞活性化を誘導することができ、これが古典的に短命で低親和性の反応の産生を引き起こす。整列した配列からなされ得る第3の観察は、MSP4p20の一次配列エレメントが両方の種に存在していることである。驚くことではないが、コアEFG-ドメインおよびGPI-付着シグナルの両方が保存されており、より面白いことに、酸性リピートは、同等の様式で配置されるようにである。これにより、自然感染において、MSP4は、メロゾイトの表面上の膜結合の小構造実体を残す様に分解または切断されるという考えに重きが置かれる。この実体は、MSP1p19で見られるように、新たに侵入したRBCに入るかどうかはいまだ確認されないが、本モデルは適合するようである。この比較に基づいて、以下のタンパク質配列は、抗三日熱マラリア原虫(P. vivax)マラリアワクチンの候補として提唱することができよう。
【0151】
実施例19
多形核好中球によるメロゾイトの抗体依存的食作用におけるMSP4p20に対する抗体の役割
好中球は、病原体を内部に取り込み、活性酸素種(ROS)および顆粒加水分解性タンパク質を使用してこれらを破壊する。活性化された好中球は、NADPHオキシダーゼを介して作動する呼吸バーストとして公知のプロセスにより、O2・-、H2O2...のようなROSを産出する際に非常に効果的である。ROSは、化学発光色素によって検出され、抗体が誘導する食細胞活性の測定として使用される。化学発光色素、はルミノールであり、ROSの存在下において、これが光を放射し、コンピュータに連結したルミノメーター(Microlumat+)で定量化する。
【0152】
多形核好中球(PMN)によるメロゾイトの抗体依存的食作用(ADPm)に基づいた新たな機能的アッセイ法を使用して、自然に獲得され、およびワクチン接種誘導された抗体によって誘導される熱帯熱マラリア原虫(Plasmodium falciparum)メロゾイト食作用を測定した。マラリア風土病地域における個体由来の免疫血清および凍結したメロゾイト標品を使用して、この試験では、ごくわずかな非免疫性バックグラウンドシグナルおよび優れたアッセイ内再現性を有し、高度に特異的であることが示された。アッセイ間の比較は、標準的な正の対照を使用することによって行う。
【0153】
季節的な伝播の地域に生活している個体からのデータは、ADPm活性が、年齢、並びにELISAによって測定されるメロゾイトおよびMSP組換え抗原に対するIgG抗体の反応性と有意に相関したことを示した。SenegalにおけるDielmoおよびNdiop村の居住者の風土病の免疫血清から、バキュロウイルス組換えMSP1p19およびMSP4p20に特異的な抗体を取り除くことは、これらの抗原に対する抗体がADPm活性の実質的な成分であることを示した。
【0154】
より詳細には、熱帯熱マラリア原虫(P. falciparum)メロゾイトを培養液上清の遠心分離によって収集し、凍結した一定量として貯蔵した。新鮮な献血から得られた多形核好中球を単純なFicoll-Hystopaque 1077勾配で単離して、すぐにADPmアッセイ法に使用した。風土病免疫血清は、Ndiop(中間的風土病)およびDielmo(完全風土病)の住民から得た。C末端のヘキサヒスチジンタグを有する組換えMSP1p19およびMSP4p20抗原をバキュロウイルス発現系において産生して、固定化金属-アフィニティークロマトグラフィによって精製した。対応する組換え抗原を荷電したTALON金属親和性樹脂を使用して、血清の抗MSP1p19または/および抗MSP4p20抗体を除去した。反応を開始するためのルミノールおよびPMN(ウェルあたり5.106細胞)の添加の前に、メロゾイトを96ウェルプレートにおいて血清と共にインキュベートした。発光の出力は、Berthold MicroLumat Plus 96ウェルを使用して1時間測定した。標準的なヒト免疫血清の陽性対照(HIS)を、反応を定量化するために使用した:
メロゾイトの抗原特異的のIgGオプソニン作用(結合)(補体の有無にかかわらず)は、非働化血清(56℃にて30分間の処理)またはプロテインGで精製された総IgGが初期の血清に対する同様の反応を誘導し、総IgGを除去した血清がもはや反応を産生しない(図34)ため、このアッセイ法における陽性の読み出しのために必要であった。しかし、特異的IgGによって産出される化学発光効果の大きさは変動する。図35は、バキュロウイルスMSP4p20およびMSP1p19に特異的な抗体は、このアッセイ法において両方とも、同じ程度ではないが、機能的であることを示す。MSP1p19抗体の枯渇による34%の減少と比較して、MSP4p20抗体の枯渇の後では、化学発光シグナルは、67%減少する。MSP4p20およびMSP1p19の両方に特異的な抗体の枯渇は、化学発光シグナルを78%にまで減少させた。
【0155】
これらの結果は、バキュロウイルスMSP4p20およびMSP1p19の組換えタンパク質を認識する自然抗体は、PMN免疫エフェクターによるメロゾイト食作用および破壊を媒介する重要な成分であることを示し、ワクチンとしてのこれらの抗原の使用を強く支持する。
【0156】
実施例20
PfMSP5修飾
バキュロウイルスで発現したPfMSP5に存在するように見えるN末端遮断の修飾の性質に注目するために、放射性標識実験を行った。質量分析は、その実体が共有結合で相互作用したミリストイル基であることを示唆したので、ミリストイル基の組み込みをず最初に調べた。実施例1において前述した様に、昆虫細胞感染を、SF9細胞を使用してT25細胞培養フラスコにおいて行った。感染後24時間にて、BSAを含まない脂肪酸との複合体の200μCiの[9,10(n)-3H]ミリスチン酸(Amersham)を培養液上清に添加した。感染後70時間にて、培養上清を収集し、20 mM Tris pH8;500 mM NaClに対して透析し、Talon(登録商標)樹脂上でバッチ精製した。ブルー染色したSDS-PAGEゲルにおいて、タンパク質発現は、非ラベル対照および放射標識の存在下において行った培養と同一であることが分かった。オートラジオグラフィーによって解析したときに、両方のPfMSP5バキュロウイルス発現タンパク質(35および45kDa)は、トリチウム化されたミリストイルを組み込んでいた(図27)。バックグランド活性は、非標識タンパク質では見られず、2つのPfMSP5タンパク質バンドは、ウイルス感染無しでは見られなかった。
【0157】
これらの結果は、バキュロウイルスに発現される組換えPfMSP5タンパク質は、ミリストイル化されることを示す。これらのデータは、昆虫細胞におけるPfMSP5の発現に関連した、実施例4〜7における以前のデータを確認する。
【0158】
実施例21
PfMSP4の運命
PfMSP4の自然の運命を追跡するために、いくつかの異なる精製された血清を産生した。最初に、4羽のMSP4ワクチン接種したウサギから収集した抗血清のプールを、バキュロウイルスで発現したPfMSP4p20を固定したNHS活性化セファロースに対してアフィニティー精製した。この仮定をNdiopまたはDielmoからのPfMSP4について陽性であることが知られている5つのヒト免疫血清の2つの異なるプールを使用して繰り返した。これらの抗体試薬を使用して、PfMSP4の運命を、3D7株を適応した培養において、IFAおよびウエスタンブロットによって追跡し、その知見を、PfMSP1p19に特異的モノクローナル抗体(G17.12)で見られたものと比較した。図28Aに示したように、PfMSP4は、後期寄生虫および自由なメロゾイトの表面にて検出されたが、PfMSP1p19を同定することができたアセトン固定リング期の寄生虫では検出されなかった。加えて、ウエスタンブロットによる寄生虫抽出物の同等量の平行分析に注目すると(図28B)、PfMSP4レベルは、寄生虫培養上清をから収集した自由なメロゾイトにおいて、成熟シゾントにおけるよりもはるかにより少ないよう見え、該タンパク質は、リング期の寄生虫抽出物において検出されない。このデータは、MSP1p19についてみられたように、PfMSP4がタンパク質分解的にプロセスされたというよりむしろ、Apical Membrane Antigen-1と同様の様式でメロゾイト表面から分断され、新たに侵入されたRBCに運ばれるという考えに一致する。
【0159】
面白いことに、両方のアフィニティー精製した血清(ウサギおよびヒト)は、成熟したシゾント抽出物に限定的な複数の高分子量タンパク質バンドを認識する。これは、以前にMSP3およびMSP6で見いだされたような交差反応性エピトープを含む成熟したシゾントに存在する抗原であり得るか、または寄生虫抽出物が還元されていないために、これが天然のPfMSP4タンパク質三量体であり得る。
【0160】
肝臓段階前の寄生虫におけるPfMSP4発現を調べるために、抗PfMSP4p20のアフィニティー精製したウサギ血清を使用して、熱帯熱マラリア原虫(P. falciparum)スポロゾイトに対してIFAを行った。陽性対照の抗CSPモノクローナル抗体は、強力で明らかな表面染色を与えたが、抗PfMSP4p20抗体では、染色は達成されなかった。
【0161】
これらの結果は、天然MSP4タンパク質がメロゾイトの表面上に局在化しており、寄生虫による血液細胞の再侵略において、もはや存在していないことを示す(リング期に感染した血液細胞のIFAを参照されたい(図28A)。図28Bのウエスタンブロットは、天然のMSP4に対して向けられた抗体(ヒト血清)および組換えMSP4に対して向けられた抗体(ウサギ血清)で一致したパターンを示す。そして、バキュロウイルス組換え抗原に対して向けられた抗体は、天然の形態の抗原を認識することができることがこれから推論することができる。
【0162】
実施例22
p40と独立したp20サブドメインの発現
p20サブドメインの直接の発現を促進するために、一連の新たな発現構築物を構築し(図29および実施例17)、試験した。図30に示したように、最小のシグナル配列をコードする構築物(PfMSP4p21)は、タンパク質分泌に至らなかったが、高レベルの細胞内タンパク質が検出された(データ示さず)。より優れたレベルの発現は、図30に示すように構築物PfMSP4p21ss1およびss2で達成され、N末端のシーケンシング(図29参照されたい)によって明らかにされるように、両タンパク質は、迅速に残りのN末端の残基を失ってバキュロウイルスPfMSP4p40で見られるのと同じ安定なp20実体を産生する。しかし、感染後66時間でのタンパク質発現を、バキュロウイルスPfMSP4p40およびPfMSP4p21ss1を使用して行った並行感染と比較すると、オリジナルの構築物は、はるかに優れたタンパク質収率(2倍)を引き起こす。これらの結果は、上記の実施例17を補完する。
【0163】
実施例23
バキュロウイルス発現系におけるPvMSP4の発現
PvMSP4オープンリーディングフレーム(ORF)を得るために、ネスティッドPCRを、寄生虫株BelemからのゲノムDNAに対して行った。三日熱マラリア原虫(P. vivax)ゲノムDNAは、約50%がGCリッチであるので、合成遺伝子を作る必要はなかった。全てのORFは、pMosBLUEにクローニングして、シーケンスした。次に、両方のエキソンをPCRによって増幅し、エキソン2の3'末端のGPI付着シグナルをヘキサhisタグによって置換した。加えて、適切な制限部位をエキソン1および2の3'または5'末端にそれぞれ導入し、これらをpMosBLUEベクターに連続してクローニングし、his-タグにおける連続したORF終末を形成した(図31)。
【0164】
バキュロイルスをコードするPvMSP4は、標準的なプロトコルを使用して産生し、ある時間にわたったタンパク質発現を、前述したようにHigh Five昆虫細胞を使用して評価した。図32に示したように、タンパク質発現は、感染後約30時間で検出可能であった。
【0165】
このタンパク質バンドについて行ったN末端の配列解析により、GIAACのN末端が明らかになった。タンパク質発現は、感染後54時間ごろにピークに達し、この時点にて、いくつかのタンパク質分解が明白であった。しかし、PvMSP4は、PfMSP4と同様の様式では分解しない。最終産物のN末端のシーケンシングでは、該タンパク質が13または30〜32アミノ酸にまで分解されて、EGGEQのN末端の配列、並びにGDSSG、DSSGGおよびSSGGLを示すの混合シグナルを示した。検出される最も小さなタンパク質産物は、サイズで約8 kDaであり、N末端の配列LDNNGを保持している。
【0166】
実施例24
ヒト免疫血清のPvMSP4の認識
時間経過解析の最後に(図32で示す66時間)、130 mLの培養液上清をTalon樹脂上でのHPLCによって精製した。精製した産物は、図32において見られる30、25 kDaおよび8 kDaのバンドからなる。
【0167】
ヒト血清認識に注目するために、ELISAプレートをバキュロイルス発現したPvMSP4、0.5 g/mLでコートした。
【0168】
合計24の血清試料を、スリランカにおいて三日熱マラリア原虫(P. vivax)マラリアの少なくとも1回の発症を経験した個体から収集して、平行して試験した。これらの試料のうちの17は、PvMSP4に対して陽性の血清で、1/25,000〜1/200,000の間のエンドポイント力価であった(図33A)。これらの17の陽性血清を、DTTおよびヨードアセトアミドでの処理によって不可逆的に還元したバキュロウイルスPvMSP4をコートしたELISAプレート上で、1/2700希釈にて試験した。17の陽性血清のうち11は、1/2700の固定された希釈にて、またはそれぞれの曲線(図33B)の中点値に当てはまる希釈を使用して算出した場合に、12〜80%からの範囲で、ある程度の感受性の減少を示した。
【0169】
三日熱マラリア原虫(Plasmodium vivax)MSP4(PvMSP4)に関する実施例23および24は、実施例18を補完する。図33Bに示したように、血清番号2、3、4、6、7、8、9、10、11、14および17は、、DTTで還元した抗原に対して非還元抗原に対してよりも少ない親和性を有する一方、血清番号12および15は、還元した抗原に対して、非還元のものに対してよりも優れた親和性を有する。抗原の高次構造は、抗体-抗原結合に影響を及ぼすと結論することができるが、プラスモディウム属感染に対して試験した血清のそれぞれの保護能力が確立されていないので、還元または非還元の抗原に対する抗体親和性は、2つのタイプの抗体の保護値に相関させることができない。
【0170】
以下のPfMSP5/His、PfMSP4p30/HisおよびPfMSP4p40/Hisバキュロウイルスは、2005年11月10日に、Institut PasteurのCollection Nationale de Cultures de Microorganismes(C.N.C.M.)、28, rue du Docteur Roux, 75724 Paris, Cedex 15, FRANCEに寄託し、以下の受託番号(Accession number)が割り当てられた。
【0171】
材料 受託番号
PfMSP5/His I-3512
PfMSP4p30/His I-3513
PfMSP4p40/His I-3514。
【0172】
以下のF12-7、G21-2、J18-14、およびL11-6ハイブリドーマは、2005年11月16日に、Institut PasteurのCollection Nationale de Cultures de Microorganismes(C.N.C.M.)、28, rue du Docteur Roux, 75724 Paris, Cedex 15, FRANCEに寄託し、以下の受託番号が割り当てられた。
【0173】
材料 受託番号
F12-7 I-3517
G21-2 I-3518
J18-147 I-3519
L11-6 I-3520。
【0174】
以下のPfMSP4p21ss1/His、PfMSP4p21ss2/HisおよびPvMSP4/Hisは、2006年11月21日に、Institut PasteurのCollection Nationale de Cultures de Microorganismes(C.N.C.M.)、28, rue du Docteur Roux, 75724 Paris, Cedex 15, FRANCEに寄託し、以下の受託番号が割り当てられた。
【0175】
材料 受託番号
PfMSP4p21ss1/His I-3695
PfMSP4p21ss2/His I-3696
PvMSP4/His I-3694。
【0176】
以下は、上記に記述したペプチドおよびポリヌクレオチドのアミノ酸および核酸配列である:
MSP5p10(MSP5の分解生成物、186-253残基)YNKVEKNVTDEMLLYNMMSDQNRKSCAINNGGCSDDQICININNIGVKCICKDGYLLGTKCIHHHHHH[配列番号:16] 。
【0177】
MSP5p10をコードする核酸配列
>readseq-48009_tmp_1 207 bp TACAACAAGGTCGAGAAGAACGTAACCGACGAGATGCTCTTGTACAACATGATGTCCGACCAAAACCGCAAAAGCTGTGCTATCAACAACGGTGGCTGCAGTGACGACCAGATCTGCATCAACATCAACAACATCGGTGTGAAGTGCATTTGTAAGGATGGATACCTACTTGGTACCAAGTGCATTCACCACCACCACCACCACTGA[配列番号:5]。
【0178】
配列番号:1 =配列番号:9のMSP4p20の分解生成物をコードする核酸(132-251残基)
>readseq-41112_tmp_1 363 bp AAGAGTCCCAAGGAGAGTCAAATGGTCGACGACAAGAAGAAGACCGAGGCCATTCCAAAGAAAGTCGTGCAGCCAAGCTCGAGCAACTCTGGAGGTCACGTCGGTGAAGAAGAAGACCACAACGAAGGAGAGGGAGAGCACGAAGAGGAGGAAGAACACGAAGAAGACGATGACGACGAGGACGACGACACATACAACAAAGACGACTTGGAGGACGAAGATCTTTGCAAGCACAACAACGGAGGATGTGGAGATGACAAGCTCTGCGAGTACGTTGGAAACCGTCGCGTAAAATGTAAATGTAAGGAAGGATACAAGTTGGAAGGAATTGAGTGCGTTGAACACCACCACCACCATCACTAA[配列番号:1]。
【0179】
MSP5
>readseq.input(1)762 bases, 6761DE71 checksum.
ATGAACATTCTCTGTATTCTCAGCTACATTTACTTCTTCGTCATCTTCTA CAGTTTAAACCTCAACAACAAAAACGAGAACTTCTTGGTGGTCCGCAGAC TCATGAACGACGAAAAGGGAGAAGGTGGCTTCACTAGTAAGAACAAGGAA AACGGAAACAACAACAGGAACAACGAGAACGAACTCAAAGAAGAAGGATC TTTGCCCACTAAGATGAACGAGAAAAACAGTAACTCCGCGGATAAGCAAC CAAACGACATCTCCCACGACGAAAGCAAGAGCAACAGTAACAACGCCCAA AACATCCAAAAGGAACCTGAAGAGAAGGAAAACTCAAACCCCAACCTCGA CTCGAGTGAAAACTCCGCTGAAAGTGCTACTAGAAGCGTCGACATCAGTG AACACAACTCAAACAACCCCGAAACTAAAGAAGAAAACGGAGAAGAACCT CTAGACCTGGAAATTAACGAAAACGCAGAAATCGGCCAGGAACCTCCAAA CCGTCTTCACTTCGACAACGTTGACGACGAAGTACCACATTACTCAGCCC TGAGGTACAACAAGGTCGAGAAGAACGTAACCGACGAGATGCTCTTGTAC AACATGATGTCCGACCAAAACCGCAAAAGCTGTGCTATCAACAACGGTGG CTGCAGTGACGACCAGATCTGCATCAACATCAACAACATCGGTGTGAAGT GCATTTGTAAGGATGGATACCTACTTGGTACCAAGTGCATTCACCACCAC CACCACCACTGA[配列番号:4]。
【0180】
ORFによってコードされるタンパク質
>readseq.input(1)253 bases, 7870B947 checksum.
MNILCILSYIYFFVIFYSLNLNNKNENFLVVRRLMNDEKGEGGFTSKNKE NGNNNRNNENELKEEGSLPTKMNEKNSNSADKQPNDISHDESKSNSNNAQ NIQKEPEEKENSNPNLDSSENSAESATRSVDISEHNSNNPETKEENGEEP LDLEINENAEIGQEPPNRLHFDNVDDEVPHYSALRYNKVEKNVTDEMLLY NMMSDQNRKSCAINNGGCSDDQICININNIGVKCICKDGYLLGTKCIHHH HHH[配列番号:14]。
【0181】
MSP4p40
>readseq.input(1)756 bases, 733F5E48 checksum.
ATGTGGATCGTAAAGTTCTTGATTGTGGTCCACTTCTTCATCATATGCAC CATCAACTTCGACAAGCTCTACATTAGTTACTCTTACAACATCGTCCCTG AAAACGGACGTATGCTTAACATGAGGATCTTGGGTGAAGAAAAGCCTAAC GTTGACGGTGTGTCAACATCTAACACACCTGGCGGAAACGAGGCATCTAG TGCTTCTCCTAACCTTGCTGACGCTGCAGAAAAGAAGGACGAAAAGGAAG CAAGCGAGCAAGGCGAAGAATCCCACAAGAAGGAAAACTCTCAGGAATCT GCAAACGGAAAAGACGACGTTAAGGAGGAGAAGAAGACCAACGAGAAGAA GGACGACGGAAAGACTGACAAGGTACAAGAAAAGGTTCTAGAAAAGAGTC CCAAGGAGAGTCAAATGGTCGACGACAAGAAGAAGACCGAGGCCATTCCA AAGAAAGTCGTGCAGCCAAGCTCGAGCAACTCTGGAGGTCACGTCGGTGA AGAAGAAGACCACAACGAAGGAGAGGGAGAGCACGAAGAGGAGGAAGAAC ACGAAGAAGACGATGACGACGAGGACGACGACACATACAACAAAGACGAC TTGGAGGACGAAGATCTTTGCAAGCACAACAACGGAGGATGTGGAGATGA CAAGCTCTGCGAGTACGTTGGAAACCGTCGCGTAAAATGTAAATGTAAGG AAGGATACAAGTTGGAAGGAATTGAGTGCGTTGAACACCACCACCACCAT CACTAA[配列番号:3]。
【0182】
ORFによってコードされるタンパク質
>readseq.input(1)251 bases, 1EFAEF35 checksum.
MWIVKFLIVVHFFIICTINFDKLYISYSYNIVPENGRMLNMRILGEEKPN VDGVSTSNTPGGNEASSASPNLADAAEKKDEKEASEQGEESHKKENSQES ANGKDDVKEEKKTNEKKDDGKTDKVQEKVLEKSPKESQMVDDKKKTEAIP KKVVQPSSSNSGGHVGEEEDHNEGEGEHEEEEEHEEDDDDEDDDTYNKDD LEDEDLCKHNNGGCGDDKLCEYVGNRRVKCKCKEGYKLEGIECVEHHHHH H[配列番号:12]。
【0183】
最終的に産生されたタンパク質
>readseq.input(1)211 bases, 67EAFE8C checksum.
MRILGEEKPNVDGVSTSNTPGGNEASSASPNLADAAEKKDEKEASEQGEE SHKKENSQESANGKDDVKEEKKTNEKKDDGKTDKVQEKVLEKSPKESQMV DDKKKTEAIPKKVVQPSSSNSGGHVGEEEDHNEGEGEHEEEEEHEEDDDD EDDDTYNKDDLEDEDLCKHNNGGCGDDKLCEYVGNRRVKCKCKEGYKLEG IECVEHHHHHH[配列番号:13]。
【0184】
最終的に産生されたタンパク質で分泌されるP20タンパク質
>readseq.input(1)120 bases, 9358BD28 checksum.
KSPKESQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEEDHNEGEGEHEEE EEHEEDDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLCEYVGNRRVKCK CKEGYKLEGIECVEHHHHHH[配列番号:9]。
【0185】
MSP4p30
>readseq.input(1)666 bases, A9B2DCBC checksum.
ATGTGGATCGTAAAGTTCTTGATTGTGGTCCACTTCTTCATCATATGCAC CATCAACTTCGACAAGCTCTACATTAGTTACTCTTACAACATCGTCCCTG AAAACGGACGTATGCTTAACATGAGGATCTTGGCTGCAGAAAAGAAGGAC GAAAAGGAAGCAAGCGAGCAAGGCGAAGAATCCCACAAGAAGGAAAACTC TCAGGAATCTGCAAACGGAAAAGACGACGTTAAGGAGGAGAAGAAGACCA ACGAGAAGAAGGACGACGGAAAGACTGACAAGGTACAAGAAAAGGTTCTA GAAAAGAGTCCCAAGGAGAGTCAAATGGTCGACGACAAGAAGAAGACCGA GGCCATTCCAAAGAAAGTCGTGCAGCCAAGCTCGAGCAACTCTGGAGGTC ACGTCGGTGAAGAAGAAGACCACAACGAAGGAGAGGGAGAGCACGAAGAG GAGGAAGAACACGAAGAAGACGATGACGACGAGGACGACGACACATACAA CAAAGACGACTTGGAGGACGAAGATCTTTGCAAGCACAACAACGGAGGAT GTGGAGATGACAAGCTCTGCGAGTACGTTGGAAACCGTCGCGTAAAATGT AAATGTAAGGAAGGATACAAGTTGGAAGGAATTGAGTGCGTTGAACACCA CCACCACCATCACTAA[配列番号:2]。
【0186】
ORFによってコードされるタンパク質
>readseq.input(1)221 bases, F982D7F0 checksum.
MWIVKFLIVVHFFIICTINFDKLYISYSYNIVPENGRMLNMRILAAEKKD EKEASEQGEESHKKENSQESANGKDDVKEEKKTNEKKDDGKTDKVQEKVL EKSPKESQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEEDHNEGEGEHEE EEEHEEDDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLCEYVGNRRVKC KCKEGYKLEGIECVEHHHHHH[配列番号:10]。
【0187】
最終的に産生されたタンパク質
>readseq.input(1)181 bases, FB2C49DD checksum.
MRILAAEKKDEKEASEQGEESHKKENSQESANGKDDVKEEKKTNEKKDDG KTDKVQEKVLEKSPKESQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEED HNEGEGEHEEEEEHEEDDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLC EYVGNRRVKCKCKEGYKLEGIECVEHHHHHH[配列番号:11]。
【0188】
MSP4p21
>readseq.input(1)498 bases, C2453A47 checksum.
ATGTGGATCGTAAAGTTCTTGATTGTGGTCCACTTCTTCATCATATGCAC CATCAACTTCGACAAGCTCTACATTAGTTACTCTTACAACATCGTCCCTG AAAACGGACGTATGCTTAACATGAGGATTCTAGAAAAGAGTCCCAAGGAG AGTCAAATGGTCGACGACAAGAAGAAGACCGAGGCCATTCCAAAGAAAGT CGTGCAGCCAAGCTCGAGCAACTCTGGAGGTCACGTCGGTGAAGAAGAAG ACCACAACGAAGGAGAGGGAGAGCACGAAGAGGAGGAAGAACACGAAGAA GACGATGACGACGAGGACGACGACACATACAACAAAGACGACTTGGAGGA CGAAGATCTTTGCAAGCACAACAACGGAGGATGTGGAGATGACAAGCTCT GCGAGTACGTTGGAAACCGTCGCGTAAAATGTAAATGTAAGGAAGGATAC AAGTTGGAAGGAATTGAGTGCGTTGAACACCACCACCACCATCACTAA[配列番号:6]。
【0189】
ORFによってコードされるタンパク質
>readseq.input(1)165 bases, 27BB4F2E checksum.
MWIVKFLIVVHFFIICTINFDKLYISYSYNIVPENGRMLNMRILEKSPKE SQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEEDHNEGEGEHEEEEEHEE DDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLCEYVGNRRVKCKCKEGY KLEGIECVEHHHHHH[配列番号:17]。
【0190】
最終的に産生されたタンパク質
readseq.input(1)125 bases, 50D64DD2 checksum.
MRILEKSPKESQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEEDHNEGEG EHEEEEEHEEDDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLCEYVGNR RVKCKCKEGYKLEGIECVEHHHHHH[配列番号:18]。
【0191】
MSP4p21 ss1
>readseq.input(1)540 bases, 96DADE5 checksum.
ATGTGGATCGTAAAGTTCTTGATTGTGGTCCACTTCTTCATCATATGCAC CATCAACTTCGACAAGCTCTACATTAGTTACTCTTACAACATCGTCCCTG AAAACGGACGTATGCTTAACATGAGGATCTTGGGTGAAGAAAAGCCTAAC GTTGACGGTGTGTCAACATCTctaGAAAAGAGTCCCAAGGAGAGTCAAAT GGTCGACGACAAGAAGAAGACCGAGGCCATTCCAAAGAAAGTCGTGCAGC CAAGCTCGAGCAACTCTGGAGGTCACGTCGGTGAAGAAGAAGACCACAAC GAAGGAGAGGGAGAGCACGAAGAGGAGGAAGAACACGAAGAAGACGATGA CGACGAGGACGACGACACATACAACAAAGACGACTTGGAGGACGAAGATC TTTGCAAGCACAACAACGGAGGATGTGGAGATGACAAGCTCTGCGAGTAC GTTGGAAACCGTCGCGTAAAATGTAAATGTAAGGAAGGATACAAGTTGGA AGGAATTGAGTGCGTTGAACACCACCACCACCATCACTAA[配列番号:7]。
【0192】
ORFによってコードされるタンパク質
>readseq.input(1)179 bases, F0D6C58 checksum.
MWIVKFLIVVHFFIICTINFDKLYISYSYNIVPENGRMLNMRILGEEKPN VDGVSTSLEKSPKESQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEEDHN EGEGEHEEEEEHEEDDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLCEY VGNRRVKCKCKEGYKLEGIECVEHHHHHH[配列番号:19]。
【0193】
最終的に産生されたタンパク質
>readseq.input(1)139 bases, 4D5C3E0F checksum.
MRILGEEKPNVDGVSTSLEKSPKESQMVDDKKKTEAIPKKVVQPSSSNSG GHVGEEEDHNEGEGEHEEEEEHEEDDDDEDDDTYNKDDLEDEDLCKHNNG GCGDDKLCEYVGNRRVKCKCKEGYKLEGIECVEHHHHHH[配列番号:20]。
【0194】
MSP4p21 ss2
>readseq.input(1)516 bases, 1B3CABBA checksum.
ATGTGGATCGTAAAGTTCTTGATTGTGGTCCACTTCTTCATCATATGCAC CATCAACTTCGACAAGCTCTACATTAGTTACTCTTACAACATCGTCCCTG AAAACGGACGTATGCTTAACATGAGGATCTTGGGTGAAGAAAAGCCTCTA GAAAAGAGTCCCAAGGAGAGTCAAATGGTCGACGACAAGAAGAAGACCGA GGCCATTCCAAAGAAAGTCGTGCAGCCAAGCTCGAGCAACTCTGGAGGTC ACGTCGGTGAAGAAGAAGACCACAACGAAGGAGAGGGAGAGCACGAAGAG GAGGAAGAACACGAAGAAGACGATGACGACGAGGACGACGACACATACAA CAAAGACGACTTGGAGGACGAAGATCTTTGCAAGCACAACAACGGAGGAT GTGGAGATGACAAGCTCTGCGAGTACGTTGGAAACCGTCGCGTAAAATGT AAATGTAAGGAAGGATACAAGTTGGAAGGAATTGAGTGCGTTGAACACCA CCACCACCATCACTAA[配列番号:8]。
【0195】
ORFによってコードされる予想タンパク質
>readseq.input(1)171 bases, EB09D214 checksum.
MWIVKFLIVVHFFIICTINFDKLYISYSYNIVPENGRMLNMRILGEEKPL EKSPKESQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEEDHNEGEGEHEE EEEHEEDDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLCEYVGNRRVKC KCKEGYKLEGIECVEHHHHHH[配列番号:21]。
【0196】
最終的に産生されたタンパク質
>readseq.input(1)131 bases, 6B6B091D checksum.
MRILGEEKPLEKSPKESQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEED HNEGEGEHEEEEEHEEDDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLC EYVGNRRVKCKCKEGYKLEGIECVEHHHHHH[配列番号:22]。
【0197】
PvMSP4/Hisタンパク質
MKVAYFLSVLDLLIIFSLYFDGRRSAFAGIAACIRHGRILGEGGEQSGGASGGSSGGSSGDSSGGLSGGSSGGPSPPAGSSGSGGSDPANSATGPQNSTPGSGGQTGDHSAEAENGDYNEQGDDHGDDHGDDHGDDHGDEQDGEDYDDAEDDDLYELSEVDENANLCLDNNGGCGDDKICENLGKGIVKCLCKPGYKLVGTECVESHHHHHH[配列番号:16]。
【0198】
PfMSP5/His、PfMSP4p30/His、PfMSP4p40/His、PfMSP4p21ss1/His、PfMSP4p21ss2/HisおよびPvMSP4/Hisとして開示した生物材料は、本出願において、特に実施例において、記述してある。それは、以下の要求に従う:
・ウイルスを繁殖させるための必要物:
-細胞型:Spodoptera frugiperda(Sf9)
-培地:
L-グルタミンを含むSF-900 II培地(GIBCO, Invitrogen Corporation)Cat番号10902-088
L-グルタミン200 mM(GIBCO, Invitrogen Corporation):2 mM終濃度Cat番号25030-024
ゲンタマイシン(50 mg/ml;GIBCO, Invitrogen Corporation)):50μg/ml終濃度Cat番号15750-037
-血清:
牛胎児血清5%
-pH 6.2
-温度 27-28℃
-気体相:大気(CO2濃縮のない)
・ウイルス懸濁液を得るために使用した方法:
-増殖についての詳細:新鮮な完全培地(5%FCSと共に)にSf9細胞の単層を有する容器(T-25、T-75またはT-150)における培養を、少量のウイルスの懸濁液(またはプラークもしくは限界希釈によるクローニング手順から得られるマイクロタイトレーションプレートのウェル)で播種する。培養は、すべての細胞の溶解まで、5〜6日ごろ、27〜28℃にてインキュベートする。
【0199】
-感染の証拠のための基準:伸長した核を有する細胞、続いて数日後に顆粒状の様相および細胞溶解となる。
【0200】
-ウイルス懸濁液を調製するための詳細
-細胞上清を、4000gで10分遠心分離し、細胞破片を除去する。
【0201】
・滴定のために使用した方法
限界希釈(1つのウェルには、96ウェルの陽性マイクロタイタープレートの約1/10を生じる希釈から開始して感染させる)。
結果を6日後に読み込み、予想される力価は、107〜108 pfu/mlであった。
【0202】
・貯蔵のための条件
長期の維持は、-80℃での凍結により可能となる。
懸濁液体は、標準的な培地である。
【0203】
上記のハイブリドーマF12-7、G21-2、J18-14およびL11-16は、マウス骨髄腫細胞(plasmocytome Ag8 X 63 6.5.3)と特定の抗原で免疫した動物の脾臓から得られるマウス免疫細胞との間の細胞融合の結果として得られた。
【0204】
・細胞の特徴付けおよび産生
-骨髄腫細胞の名称:X63-Ag8-6.5.3
-抗原特異性: F 12-7についてはMSP4 II、G21-2および518-14についてはMSP5、L11-16についてはMSP4。
【0205】
-抗体サブクラス:IgG1k
-分泌の安定性:良および限定的
・培養の条件:
-培地
Eagle MEM-Eurobio
ピルビン酸ナトリウム−Gibco BRL
グルタミン−Gibco BRL
抗生物質:ペニ-ストレプトマイシン−Gibco BRL
ウマ血清:10%
-血清炭酸水素塩2.2 g/l
血清:ウマ10%
pH7,4
至適温度:37℃
気体相:7% CO2
解凍のために講じた警戒:迅速に37℃にて非凍結し、培地+血清で即座に洗浄する。1 mlの24ウェルを有するプレート上で培養する。
【0206】
・培養のための条件:細胞懸濁液
集団倍加時間:15時間
最適な分割比:3日
予想される細胞密度:5.106
限られた寿命
ルーチンのサブカルチャー技術:分裂
・貯蔵のための条件
懸濁液:95% ウマ血清- 5% DMSO
細胞収集の技術:細胞懸濁液を取り、遠心分離し、ペレットを凍結培地に取り込む。
【0207】
凍結するための技術:1 mlの凍結培地における5.106の細胞を-20℃にて2時間、-80℃にて3-4日、そして液体窒素へ。懸濁液は、標準的な培地である。
【0208】
参照文献
以下の刊行物のそれぞれの全ての開示は、本明細書に参照に依拠し、および組み込まれる。
【0209】
1. Genton, B., Al-Yaman, F., Betuela, I. et al. Safety and immunogenicity of a three-component blood-stage malaria vaccine (MSP1, MSP2, RESA) against Plasmodium falciparum in Papua New Guinean children. Vaccine 2003, 22(1), 30-41.
2. Stowers, A.W., Kennedy, M.C., Keegan, B.P., Saul, A., Long, C.A. & Miller, L.H. Vaccination of monkeys with recombinant Plasmodium falciparum apical membrane antigen 1 confers protection against blood-stage malaria. Infect Immun 2002, 70(12), 6961-6967.
3. O'Donnell, R.A., de Koning-Ward, T.F., Burt, R.A. et al. Antibodies against merozoite surface protein (MSP)-1(19) are a major component of the invasion-inhibitory response in individuals immune to malaria. J Exp Med 2001, 193(12), 1403-1412.
4. Hughes, M.K. & Hughes, A.L. Natural selection on Plasmodium surface proteins. Mol Biochem Parasitol 1995, 71(1), 99-113.
5. Escalante, A.A., Lal, A.A. & Ayala, F.J. Genetic polymorphism and natural selection in the malaria parasite Plasmodium falciparum. Genetics 1998, 149(1), 189-202.
6. Volkman, S.K., Hartl, D.L., Wirth, D.F. et al. Excess polymorphisms in genes for membrane proteins in Plasmodium falciparum. Science 2002, 298(5591), 216-218.
7. Rayner, J.C., Corredor, V., Feldman, D. et al. Extensive polymorphism in the plasmodium vivax merozoite surface coat protein MSP-3alpha is limited to specific domains. Parasitology 2002, 125(Pt 5), 393-405.
8. Taylor, R.R., Smith, D.B., Robinson, V.J., McBride, J.S. & Riley, E.M. Human antibody response to Plasmodium falciparum merozoite surface protein 2 is serogroup specific and predominantly of the immunoglobulin G3 subclass. Infect Immun 1995, 63(11), 4382-4388.
9. Wang, L., Mohandas, N., Thomas, A. & Coppel, R.L. Detection of detergent-resistant membranes in asexual blood-stage parasites of Plasmodium falciparum. Mol Biochem Parasitol 2003, 130(2), 149-153.
10. Marshall, V.M., Silva, A., Foley, M. et al. A second merozoite surface protein (MSP-4) of Plasmodium falciparum that contains an epidermal growth factor-like domain. Infect Immun 1997, 65(11), 4460-4467.
11. Wu, T., Black, C.G., Wang, L., Hibbs, A.R. & Coppel, R.L. Lack of sequence diversity in the gene encoding merozoite surface protein 5 of Plasmodium falciparum. Mol Biochem Parasitol 1999, 103(2), 243-250.
12. Wang, L., Black, C.G., Marshall, V.M. & Coppel, R.L. Structural and antigenic properties of merozoite surface protein 4 of Plasmodium falciparum. Infect Immun 1999, 67(5), 2193-2200.
13. Wang, L., Richie, T.L., Stowers, A., Nhan, D.H. & Coppel, R.L. Naturally acquired antibody responses to Plasmodium falciparum merozoite surface protein 4 in a population living in an area of endemicity in Vietnam. Infect Immun 2001, 69(7), 4390-4397.
14. Black, C.G., Wang, L., Hibbs, A.R., Werner, E. & Coppel, R.L. Identification of the Plasmodium chabaudi homologue of merozoite surface proteins 4 and 5 of Plasmodium falciparum. Infect Immun 1999, 67(5), 2075-2081.
15. Black, C.G. & Coppel, R.L. Synonymous and non-synonymous mutations in a region of the Plasmodium chabaudi genome and evidence for selection acting on a malaria vaccine candidate. Mol Biochem Parasitol 2000, 111(2), 447-451.
16. Kedzierski, L., Black, C.G. & Coppel, R.L. Characterization of the merozoite surface protein 4/5 gene of Plasmodium berghei and Plasmodium yoelii. Mol Biochem Parasitol 2000, 105(1), 137-147.
17. Kedzierski, L., Black, C.G., Goschnick, M.W., Stowers, A.W. & Coppel, R.L. Immunization with a Combination of Merozoite Surface Proteins 4/5 and 1 Enhances Protection against Lethal Challenge with Plasmodium yoelii. Infect Immun 2002, 70(12), 6606-6613.
18. Kedzierski, L., Black, C.G. & Coppel, R.L. Immunization with recombinant Plasmodium yoelii merozoite surface protein 4/5 protects mice against lethal challenge. Infect Immun 2000, 68(10), 6034-6037.
19. Kedzierski, L., Black, C.G., Stowers, A.W., Goschnick, M.W., Kaslow, D.C. & Coppel, R.L. Comparison of the protective efficacy of yeast-derived and Escherichia coli-derived recombinant merozoite surface protein 4/5 against lethal challenge by Plasmodium yoelii. Vaccine 2001, 19(32), 4661-4668.
20. Wang, L., Goschnick, M.W. & Coppel, R.L. Oral Immunization with a Combination of Plasmodium yoelii Merozoite Surface Proteins 1 and 4/5 Enhances Protection against Lethal Malaria Challenge. Infect Immun 2004, 72(10), 6172-6175.
21. Goschnick, M.W., Black, C.G., Kedzierski, L., Holder, A.A. & Coppel, R.L. Merozoite Surface Protein 4/5 Provides Protection against Lethal Challenge with a Heterologous Malaria Parasite Strain. Infect Immun 2004, 72(10), 5840-5849.
22. Marshall, V.M., Tieqiao, W. & Coppel, R.L. Close linkage of three merozoite surface protein genes on chromosome 2 of Plasmodium falciparum. Mol Biochem Parasitol 1998, 94(1), 13-25.
23. Nishimura, Y., Chen, Y.Z., Uemura, Y. et al. Degenerate recognition and response of human CD4+ Th cell clones: implications for basic and applied immunology. Mol Immunol 2004, 40(14-15), 1089-1094.
24. Brady, C.P., Shimp, R.L., Miles, A.P., Whitmore, M. & Stowers, A.W. High-level production and purification of P30P2MSP1(19), an important vaccine antigen for malaria, expressed in the methylotropic yeast Pichia pastoris. Protein Expr Purif 2001, 23(3), 468-475.
25. Epp, C., Kauth, C.W., Bujard, H. & Lutz, R. Expression and purification of Plasmodium falciparum MSP-1(42): A malaria vaccine candidate. J Chromatogr B Analyt Technol Biomed Life Sci 2003, 786(1-2), 61-72.
26. Withers-Martinez, C., Saldanha, J.W., Ely, B. et al. Expression of recombinant Plasmodium falciparum subtilisin-like protease-1 in insect cells. Characterization, comparison with the parasite protease, and homology modeling PCR-based gene synthesis as an efficient approach for expression of the A+T-rich malaria genome. J Biol Chem 2002, 277(33), 29698-29709. Epub 22002 Jun 29696.
27. Zhou, Z., Schnake, P., Xiao, L. & Lal, A.A. Enhanced expression of a recombinant malaria candidate vaccine in Escherichia coli by codon optimization. Protein Expr Purif 2004, 34(1), 87-94.
28. Weber, J.L. Molecular Biology of Malaria Parasites. Exp. Parasitol. 1988, 66, 143-170.
29. Withers-Martinez, C., Carpenter, E.P., Hackett, F. et al. PCR-based gene synthesis as an efficient approach for expression of the A+T-rich malaria genome. Protein Eng 1999, 12(12), 1113-1120.
30. Ballou, W.R., Arevalo-Herrera, M., Carucci, D. et al. Update on the clinical development of candidate malaria vaccines. Am J Trop Med Hyg 2004, 71(2 Suppl), 239-247.
31. Wang, L., Marshall, V.M. & Coppel, R.L. Limited polymorphism of the vaccine candidate merozoite surface protein 4 of Plasmodium falciparum. Mol Biochem Parasitol 2002, 120(2), 301-303.
32. Jongwutiwes, S., Putaporntip, C., Friedman, R. & Hughes, A.L. The Extent of Nucleotide Polymorphism is Highly Variable Across a 3-kb Region on Plasmodium falciparum Chromosome 2. Mol Biol Evol 2002, 19(9), 1585-1590.
33. Polson, H.E., Conway, D.J., Fandeur, T., Mercereau-Puijalon, O. & Longacre, S. Gene polymorphism of Plasmodium falciparum merozoite surface proteins 4 and 5. Mol Biochem Parasitol 2005, 142(1), 110-115.
34. Puentes, A.G., J. Vera, R. Oopez, R. Suarez, J. Rodriguez, L. Curtidor, H. Sporozoite and Liver Stage Antigen Plasmodium falciparum peptides bind specifically to human hepatocytes. Vaccine 2004, 22, 1150-1156.
35. Hale, R.S. & Thompson, G. Codon optimization of the gene encoding a domain from human type 1 neurofibromin protein results in a threefold improvement in expression level in Escherichia coli. Protein Expr Purif 1998, 12(2), 185-188.
36. Longacre, S., Mendis, K.N. & David, P.H. Plasmodium vivax merozoite surface protein 1 C-terminal recombinant proteins in baculovirus. Mol Biochem Parasitol 1994, 64(2), 191-205.
37. Nacer, A., Berry, L., Slomianny, C. & Mattei, D. Plasmodium falciparum signal sequences: simply sequences or special signals? Int J Parasitol 2001, 31(12), 1371-1379.
38. Utsumi, T., Nakano, K., Funakoshi, T. et al. Vertical-scanning mutagenesis of amino acids in a model N-myristoylation motif reveals the major amino-terminal sequence requirements for protein N-myristoylation. Eur J Biochem 2004, 271(4), 863-874.
39. Resh, M.D. Fatty acylation of proteins: new insights into membrane targeting of myristoylated and palmitoylated proteins. Biochim Biophys Acta 1999, 1451(1), 1-16.
40. Gunaratne, R.S., Sajid, M., Ling, I.T., Tripathi, R., Pachebat, J.A. & Holder, A.A. Characterization of N-myristoyltransferase from Plasmodium falciparum. Biochem J 2000, 348 Pt 2, 459-463.
41. Price, H.P., Menon, M.R., Panethymitaki, C., Goulding, D., McKean, P.G. & Smith, D.F. Myristoyl-CoA:protein N-myristoyltransferase, an essential enzyme and potential drug target in kinetoplastid parasites. J Biol Chem 2003, 278(9), 7206-7214.
42. Stafford, W.H., Stockley, R.W., Ludbrook, S.B. & Holder, A.A. Isolation, expression and characterization of the gene for an ADP-ribosylation factor from the human malaria parasite, Plasmodium falciparum. Eur J Biochem 1996, 242(1), 104-113.
43. Stevenson, F.T., Bursten, S.L., Locksley, R.M. & Lovett, D.H. Myristyl acylation of the tumor necrosis factor alpha precursor on specific lysine residues. J Exp Med 1992, 176(4), 1053-1062.
44. Farazi, T.A., Waksman, G. & Gordon, J.I. The biology and enzymology of protein N-myristoylation. J Biol Chem 2001, 276(43), 39501-39504.
45. Hayashi, N., Nakagawa, C., Ito, Y. et al. Myristoylation-regulated direct interaction between calcium-bound calmodulin and N-terminal region of pp60v-src. J Mol Biol 2004, 338(1), 169-180.
46. Maekawa, S., Matsuura, Y. & Nakamura, S. Expression and myristoylation of NAP-22 using a baculovirus transfer vector system. Biochim Biophys Acta 1994, 1218(1), 119-122.
47. Perraut, R., Marrama, L., Diouf, B. et al. Distinct surrogate markers for protection against Plasmodium falciparum infection and clinical malaria identified in a Senegalese community after radical drug cure. J Infect Dis 2003, 188(12), 1940-1950.
48. Schofield, L., Hewitt, M.C., Evans, K., Siomos, M.A. & Seeberger, P.H. Synthetic GPI as a candidate anti-toxic vaccine in a model of malaria. Nature 2002, 418(6899), 785-789.
49. Bonnet S. et al, Vaccine 2006, vol. 24, p. 5997-6008, soluble and glyco-lipid modified Baculovirus Plasmodium falciparum C-terminal Merozoite Surface Protein 1, two forms of leading malaria vaccine candidate.
【技術分野】
【0001】
本出願は、組換え熱帯熱マラリア原虫(Plasmodium falciparum)メロゾイト表面タンパク質4および5およびこれらの使用に関する。
【発明の分野】
【0002】
本発明は、単離された組換えポリペプチド、このようなポリペプチドをコードする核酸、このようなポリペプチドの組換え形態の産生のための方法、これらのポリペプチドに対して産生される抗体および診断法、キット、ワクチンまたは抗寄生虫療法におけるこのようなポリペプチドおよび抗体の使用に向けられる。
【発明の背景】
【0003】
マラリアは、世界中で、大部分は5歳以下のアフリカの子供の、1年あたりおよそ2,000,000人の死亡の原因であり、多くの世界の最も乏しい国に対して莫大な公衆衛生負担を負わせている。この負担は、寄生虫およびその蚊媒介伝播における薬物耐性のため、驚くべき速度で増加しており、有効なワクチンに対する緊急の需要を募らせている。
【0004】
今までに調べられた最も有望な血液段階ワクチン候補は、メロゾイト表面タンパク質1(MSP1)および頂端膜抗原(AMA1)である。これらの表面抗原をターゲットする体液性免疫応答は、疾患発病率の減少と相関することが見いだされており、インビトロにおいて、このような抗体は、赤血球(RBC)の寄生虫再侵入を阻害することができる[1-3]。しかし、これらの抗原遺伝子は、一般に免疫エフェクターにアクセス可能ではないタンパク質をコードする遺伝子と比較して、不釣合に高い数の非同義一塩基多型(nsSNPs)を示し[4-6]、これらのnsSNPsのいくつかは、宿主免疫系に最もアクセス可能なタンパク質の領域内にクラスター形成したラジカルアミノ酸置換をコードする[7]。このようなアミノ酸多型は、BおよびT細胞エピトープの両方を変化させることによって免疫回避において機能し得る。現在、いずれの機能的なマラリアワクチンも、多対立遺伝子反応を誘導することを願って、それぞれの標的抗原のいくつかの対立遺伝子型および/またはいくつかの標的抗原の保存領域で構成されることを必要とするであろうことが一般に認められている。
【0005】
より詳細には、当該技術分野において、マラリアの、特に熱帯熱マラリア原虫(Plasmodium falciparum)マラリアおよび三日熱マラリア原虫(Plasmodium vivax)マラリアの診断、並びに治療に使用することができる抗原に対する需要が存在する。特に、風土病領域における疾患から保護する特異的免疫応答と関連した保存された抗原および多変異体(multi-valiant)マラリアワクチンの成分としてのこれらの適合性評価に対する需要がある。
【発明の概要】
【0006】
本発明は、当該技術分野におけるこれらの需要を果たすことを助ける。本発明は、配列番号:1、2、3、4、5、6、7、または8のDNA配列を含む精製され、または組換えもしくは合成の核酸分子および配列番号:9、10、11、12、13、14、15、16、17、18、19、20、21または22のアミノ酸配列をコードする精製され、または組換えもしくは合成の核酸分子を提供する。また、本発明は、これらの配列に相補的な精製された核酸分子も包含する。
【0007】
また、本発明は、配列によって予測される、配列番号:1〜8のDNA配列を含む精製されたか、または組換えの核酸分子によってコードされる精製されたポリペプチドを包含する。
【0008】
配列番号:1〜8のDNA配列または配列番号9〜22のアミノ酸配列に対する言及は、これらの配列のいずれかおよび各々に対する言及である。
【0009】
本発明は、配列番号:1〜8のDNA配列を含む精製された二本鎖核酸分子および配列番号:9〜22のアミノ酸配列をコードする精製された二本鎖核酸分子を含む。一本鎖および二本鎖の両方のRNAおよびDNA核酸分子が、本発明によって包含される。これらの分子は、本発明に包含されるポリペプチドをコードする一本鎖および二本鎖のRNA並びにDNA変異体を検出するために使用することができる。二本鎖DNAプローブにより、核酸分子のいずれの鎖にも相当する核酸分子の検出ができる。
【0010】
42℃にて50%のホルムアミドおよび6×SSCにおける適度なストリンジェンシーの条件下において、60℃、0.5×SSC、0.1%のSDSの洗浄条件で、配列番号:1〜8および配列番号:9〜22のアミノ酸配列をコードするDNA配列を含む変性された、二本鎖DNAにハイブリダイズする精製された核酸分子が本発明に包含される。
【0011】
本発明は、インビトロでの突然変異誘発によって配列番号:1〜8に由来する精製された核酸分子をさらに包含する。インビトロでの突然変異誘発は、部位特異的変異誘発、ランダムな突然変異誘発およびインビトロでの核酸合成を含むが、限定されない当該技術分野において公知の多数の技術を含む。引用した配列からインビトロでの突然変異誘発によって誘導されるこのような核酸分子は、詳細な態様に従って、本来のものと同じ長さのまたはより短い配列を有する。
【0012】
DNAおよびRNAを含む本発明の核酸分子は、本明細書において「組換えMSP4およびMSP5核酸」または「組換えMSP4およびMSP5 DNA」と称され、これらの分子によってコードされるアミノ酸は、本明細書において「組換えMSP4およびMSP5ポリペプチド」または「本発明のポリペプチド」と称される。
【0013】
また、本発明は、遺伝暗号の結果として、配列番号:1〜8から縮重する精製された核酸分子、組換えMSP4およびMSP5核酸の対立遺伝子変異体である精製された核酸分子または組換えMSP4およびMSP5核酸の種相同体を包含する。また、本発明は、これらの核酸分子の発現を指揮する組換えベクターおよびこれらのベクターで形質転換され、トランスフェクトされ、または感染させた宿主細胞を包含する。組換えMSP4またはMSP5ポリペプチドと結合する精製されたポリクローナルまたはモノクローナル抗体は、本発明に包含され、本明細書において「本発明の抗体」と称する。
【0014】
本発明は、組換えMSP4およびMSP5ポリペプチドの産生のための方法であって、発現を促進する条件下で宿主細胞を培養すること、および培地または細胞のペレットからポリペプチドを回収することを含む方法をさらに包含する。特に、バキュロウイルス昆虫発現系における組換えMSP4およびMSP5ポリペプチドの発現も、本発明に包含される。
【0015】
また、本発明は、標識された組換えMSP4およびMSP5ポリペプチドを提供する。好ましくは、標識されたポリペプチドは、精製された形態である。また、非標識または標識されたポリペプチドは、マラリアに対する抗体を含む体液によって免疫学的に認識することができることが好ましい。ポリペプチドは、たとえば放射標識、酵素標識、蛍光標識、化学発光標識および発色団からなる群より選択される免疫アッセイ標識で標識することができる。
【0016】
また、本発明の組換えMSP4またはMSP5ポリペプチドとこれらを認識する抗体との間の免疫学的複合体が提供される。免疫学的複合体は、放射標識、酵素標識、蛍光標識、化学発光標識および発色団からなる群より選択される免疫アッセイ標識で標識することができる。
【0017】
さらにまた、本発明は、プラスモディウム属寄生虫のMSP4および/またはMSP5ポリペプチドを検出するためのインビトロにおける方法を提供する。本方法には、プラスモディウム属寄生虫(マラリア原虫)のMSP4および/またはMSP5ポリペプチドを含むことが疑われる生物材料を含む組成物を提供することと、およびプラスモディウム属寄生虫のMSP4および/またはMSP5ポリペプチドの存在についてアッセイすることとを含む。プラスモディウム属寄生虫のMSP4およびMSP5ポリペプチドは、典型的には電気泳動法によって、または本発明の抗体での免疫アッセイ法によってアッセイされる。本方法は、生体試料におけるプラスモディウム属寄生虫の検出のために、および好ましい態様において熱帯熱マラリア原虫(Plasmodium falciparum)および三日熱マラリア原虫(Plasmodium vivax)寄生虫の検出のために使用することができる。
【0018】
また、本発明は、組換え、または精製された本発明のMSP4もしくはMSP5ポリペプチドまたはこれらの混合物を含む抗原と結合する抗体の有無の検出のためのインビトロでの診断法を提供する。本方法には、抗原を体液と、生物学的体液中の抗原および抗体が抗原抗体複合体を形成するために十分な時間の間および条件下で接触させることと、および複合体の形成を検出することとを含む。検出工程は、抗原抗体複合体の形成を測定することをさらに含むことができる。抗原抗体複合体の形成は、好ましくはウエスタンブロット法、ELISA(酵素結合免疫吸着検定法)、間接的免疫蛍光アッセイ法または免疫沈降アッセイ法に基づいた免疫アッセイ法によって測定される。本方法は、感染した動物またはヒト患者マラリア由来の体液中のプラスモディウム属寄生虫に対する免疫応答の検出のために使用することができる。好ましい態様において、本方法は、熱帯熱マラリア原虫(Plasmodium falciparum)感染または三日熱マラリア原虫(Plasmodium vivax)感染に対する免疫応答の検出のために使用することができる。
【0019】
本発明の組換えMSP4もしくはMSP5ポリペプチドまたはこれらの混合物と結合する抗体の有無の検出のための診断キットは、組換えMSP4および/またはMSP5ポリペプチドまたはこれらの混合物を含む抗原と、抗原と抗体との間の免疫複合体の形成を検出するための手段とを含む。抗原および手段は、検出を行うために十分な量で存在する。
【0020】
プラスモディウム属寄生虫のMSP4および/またはMSP5ポリペプチドの有無の検出のための診断キットには、本発明の抗体と抗原と抗体との間の免疫複合体の形成を検出するための手段が含まれる。抗体および手段は、検出を行うために十分な量で存在する。
【0021】
また、本発明は、インビボにおいて免疫原性反応または保護反応を誘導するために十分な量で、本発明の組換えMSP4もしくはMSP5ポリペプチドまたはその混合物を、薬学的に許容される免疫賦活剤に付随して含む免疫原性組成物を提供する。本発明のワクチン組成物は、十分な量の組換えMSP4および/またはMSP5ポリペプチドと、中和抗体を誘導する薬学的に許容される免疫賦活剤とを含む。
【0022】
したがって、本発明のMSP4およびMSP5ポリペプチドは、マラリアに関連した抗原タンパク質に対する抗体の存在を検出するための診断の組成物の一部として有用である。
【0023】
加えて、組換えMSP4およびMSP5ポリペプチドは、マラリアに関連した抗原タンパク質の存在を検出するための抗体を生じさせるために使用することができる。
【0024】
また、本発明のポリペプチドは、寄生虫を不活性化し、インビボにおける寄生虫の生存度を減少させ、または寄生虫複製を阻害し、もしくは予防する、いずれかの中和抗体を生じさせるためにも使用することができる。寄生虫中和抗体を誘発する能力は、本発明のポリペプチドが免疫組成物またはワクチン組成物に使用されるときに、特に重要である。
【0025】
以下は、(A)天然のシグナル配列、(B)C末端Hisタグおよび(C)本分野(MSP4p20)における感染防御抗体反応と最も強く相関される熱帯熱マラリア原虫(P. falciparum)抗原のものに似ている酸反復領域を含むワクチンである。
【0026】
MKVAYFLSVLDLLIIFSLYFDGRRSAFAGIAACIRHGRILGEGGE/QNSTPGSGGQTGDHSAEAENGDYNEQGDDHGDDHGDDHGDDHGDEQDGEDYDDAEDDDLYELSEVDENANLCLDNNGGCGDDKICENLGKGIVKCLCKPGYKLVGTECVEHHHHHH[配列番号:30]
本発明は、図面を参照して記述する
【図面の簡単な説明】
【0027】
【図1】プラスモディウム属のいくつかの異なる種におけるmsp4およびmsp5遺伝子座位のゲノム構成を示す。これらの遺伝子の類縁種保存は、基本的な機能を示している。
【図2】SALSAの配列は、熱帯熱マラリア原虫(Plasmodium falciparum)のMSP-4(PfMSP4)と92%一致であることを示す。SALSA配列は、BおよびT細胞エピトープの両方を保持することが公知である。PfMSP4配列は、黒色で示してあり、SALSA配列はグレーで示してある。
【図3−1】MF54および合成遺伝子のMSP4コドン選択性を示す。
【図3−2】PfMSP4の完全合成遺伝子配列を示す。この図に示したすべての配列は、オリゴヌクレオチドで示した。PfMSP4の遺伝子配列は、番号付けした重複するオリゴ配列(1-38)によってコードされ、太字および通常のテキストを交互にすることによって強調してある。制限酵素部位は、小文字のテキストで記される。
【図4】合成遺伝子のPCR製作を示す。「遺伝子構築反応」(レーン1および2それぞれ5または10μL)および「遺伝子増幅反応」(レーン3および4それぞれ5またはμL)の試料は、DNAサイズ標準に隣接し、かつ離した1%のアガロースゲルで分離した。
【図5】バキュロウイルス系における組換えタンパク質発現のために産生した4つの異なるMSP4構築物の整列を示す。推定上のシグナル配列(現在利用できるアルゴリズムによって予測される)は、グレーで強調してあり、共通の多形部位は、太字で強調してあり、最も平衡選択下にありそうで、したがって免疫回避に関与する部位は、*で示してある。
【図6−1】3D7および合成遺伝子のMSP5コドンの選択性を示す。
【図6−2】熱帯熱マラリア原虫(Plasmodium falciparum)のMSP5(PfMSP5)の完全合成遺伝子配列を示す。この図に示したすべての配列は、オリゴヌクレオチドで示した。PfMSP5の遺伝子配列は、番号付けした重複するオリゴ配列(1-38)によってコードされ、太字および通常のテキストを交互にすることによって強調してある。制限酵素部位は、小文字のテキストで記してある。
【図7】合成遺伝子のPCR製作を示す。「遺伝子増幅反応」の試料(5μL)は、DNAサイズ標準に隣接し、かつ離した1%のアガロースゲルで分離した。
【図8】2つの異なる昆虫株化細胞におけるある期間にわたったPfMSP4の発現を示す。細胞死の割合は、細胞懸濁液を4%のトリパンブルーと1:1混合し、カバーガラス下の格子に対して総細胞および青い細胞を計数することにより算出した。タンパク質を、感染後24、30、36、42、48、54、60および72時間(それぞれレーン1〜8)にて感染された培養SNから精製し、20μLの溶出タンパク質を、4〜12%のBis-Trisゲル(Invitrogen)において分離して、SimplyBlue safe stain(Invitrogen)で染色した。これらのタンパク質種は、非感染性の細胞培養(レーン9)において見られなかった。
【図9】それぞれのMSP4産物p40、p30およびp20のN末端の配列を強調している整列配列である。この研究において同定されたシグナル配列は、グレーで強調してある。それぞれの産物のN末端の配列は、青い太字に示してある。
【図10】3つの公知のマラリア抗原について報告されたシグナル切断部位付近に集中するおよそ80アミノ酸残基の疎水性プロフィールを示す。成熟したタンパク質の最初の残基は、黒色ダイヤモンドによって示してある。S抗原シグナル配列切断のプロフィールは、バキュロウイルス系におけるMSP4について見られるものと同等である。
【図11】2つの異なる昆虫株化細胞におけるある期間にわたったPfMSP5の発現を示す。細胞死は、細胞懸濁液を4%のトリパンブルーと1:1に混合し、カバーガラス下の格子に対して総細胞および青い細胞を計数することにより算出した。タンパク質を、感染後24、30、36、42、48、54、60および72時間(それぞれレーン1〜8)にて培養SNから精製し、20μLの溶出タンパク質を、4〜12%のBis-Trisゲル(Invitrogen)において分離して、SimplyBlue safe stain(Invitrogen)で染色した。これらのタンパク質種は、非感染性の細胞培養(レーン9)において見られなかった。
【図12】ウサギで生じたポリクローナル抗血清に関する。(A)全長MSP4p40(R6-7)および(B)MSP4分解生成物p20(R8-9)に対して生じた血清のELISA指標力価を、コーティング抗原として天然(NR)または不可逆的に還元した(R)MSP4-p40を使用して解析した。血清R8は、MSP4抗原が還元されると、50%以上の結合活性を失う。免疫前血清は、NEGとしてマークしてある。パネル(C)は、コーティング抗原としてMSP5-p45およびp35を使用して解析された抗MSP5血清(56-57)の力価を示す。
【図13】バキュロウイルスで発現した抗原に対して生じたポリクローナル血清、およびDielmo由来のヒト血清のプールまたは抗MSP4p40ウサギ血清のプールのいずれか由来のMSP4p40アフィニティー精製血清で寄生虫材料をプローブすることによって得られた結果を示す。非同期的に発達する寄生虫をiRBCから抽出して、Wangら(2003)のプロトコルを使用して粗雑に分画した。各々の免疫ブロットのそれぞれのレーン1には、非還元Triton X100可溶性寄生虫画分、レーン2には、非還元膜画分、レーン3には、還元Triton X100可溶性寄生虫画分、レーン4には、還元膜画分を充填した(パネルA,BおよびD)。パネルCは、風乾した熱帯熱マラリア原虫(P. falciparum)を感染したRBCで得られたIFAデータを示す。
【図14】1990年に雨季のピーク時に集めたDielmoからの9人の免疫成人(18〜49歳)由来の血清のELISA解析を示す。プレートをMSP4p40(A)またはMSP5p45およびp35(B)でコートした。
【図15】NO.3および9の個体由来のヒト免疫血清を使用する、組換えMSP4 p40、p30およびp20(それぞれレーン3、2および1)の免疫ブロットである(A)。(B)NO.8の個体由来のヒト免疫血清を使用する組換えMSP5の免疫ブロット解析。
【図16】それぞれの血清について産生されるELISAプロフィールは、図16Aおよび16Bの上のそれぞれのブロットに示してあり、これは、PfMSP4、p40およびp20のエピトープを特異的に認識する2つのモノクローナル抗体mAb L11-16およびmAb F12-7についてのウエスタンブロットを示す。レジェンド:nR=非還元、R=invitrogenの市販の緩衝液により還元、iR=DTTおよびアクリルアミドによって不可逆的に還元。
【図17】図17Aおよび17Bは、それぞれ、図16Aおよび16BにおけるmAbsについてのODまたは吸光度(y軸)の関数として希釈比(x軸)を示す。
【図18】図18Aおよび18Bは、PfMSP5、p45およびp35のエピトープを認識する2つのモノクローナル抗体mAb G21-2およびmAb J18-4についてのウエスタンブロットを示す。レジェンド:nR=非還元、R=invitrogenの市販の緩衝液により還元、iR=DTTおよびアクリルアミドによって不可逆的に還元。
【図19】図19Aおよび19Bは、それぞれ図18Aおよび18BにおけるmAbsについての希釈比(x軸)およびODまたは吸光度(y軸)の関数を示す。
【図20】図20A-Bは、タンパク質をコーティングする濃度を最適化するために行ったELISA解析を示す。プレートは、0.5または1μg/mLのタンパク質のPBS溶液をウェルあたり100μL添加して一晩コートした。パネル(A)は、3つの異なるMSP4由来の抗原、MSP4p40、MSP4p30(本明細書において、MSP4p40/2と命名した)およびMSP4p20で得られたデータを示す。パネル(B)は、MSP5で得られたデータを示し、反応の異なる強度を証明するためにパネル(A)からのMSP4p20のデータをプロットした。観察される光学濃度は、いずれの濃度を使用してもほとんど同一だったので、より低いコーティング濃度(0.5g/mL)をすべてのその後の解析において使用した。
【図21】図21A-Dは、ナイーブ個体(naif)、高免疫成人(shi)および2005年のDielmo集団からの個体(60605)からの対照血清のELISA解析を示す。プレートを0.5μg/mLのMSP5(A)、MSP4p40(B)、MSP4p20(C)およびMSP4p30(D)でコートし、抗血清の希釈を変化させて反応を実施した(3倍段階希釈)。すべての抗原は、免疫血清によって認識され、マラリアナイーブ対照によって認識されなかった。
【図22】図22A-Dは、15歳以上および以下の年齢によって層別化される2000年のNdiop集団からの個体の抗体反応を示す。パネル(A)は、抗原(左から右に)MSP4p20、MSP4p40、MSP4p30およびMSP5についての光学濃度の形態のELISAデータを示す。パネル(C)は、OD比率の形態での同一データを示す。パネル(B)のMSP1p19に対する個々の抗体反応は、Ndiop 2000年集団についての年齢の関数としてのOD比率としてプロットしてあり、その一方で、パネル(D)は、同じ血清でMSP5に対して得られたELISA OD比率を示す。
【図23】Ndiop 2000年集団からの血清によるMSP4およびその誘導体の認識の変動を示す。左のパネルは、MSP4p20、MSP4p40およびMSP4p30(MSP4md2に等しい)についてのOD比率としてプロットした平均反応を示す。右のパネルは、年齢の関数としてそれぞれのMSP4抗原に対する抗体の反応について、上下の四分位値およびデータ分布を示す。
【図24】特異的血清の存在下において誘導される酸化的破裂として測定された、抗体依存的な単球媒介性細胞障害を示す。血清は、集団の中央値の反応を上回るまたは下回るOD比率を与えるものに分けられた。パネル(A)は、MSP5反応性の関数(2.4のOD比を上回るまたは下回る)として分けられた血清からのデータを示し、パネル(B)は、MSP4p40反応性の関数(20のOD比を上回るまたは下回る)として分けられた血清からのデータを示す。これらがPBMC(末梢血単核細胞)および熱帯熱マラリア原虫(P. falciparum)メロゾイトの存在下において酸化的破裂を誘導する能力は、化学発光を使用して解析した。両群において、酸化的破裂における増大は、高OD比を示す個体で見られた。
【図25】IgG特異的二次抗体試薬を使用して解析したそれぞれの抗原についての抗体アイソタイププロフィールを示す。アイソタイププロフィールは、MSP4p40(パネルA)およびMSP5(パネルB)について示してある。データは、年齢(15齢を上回るおよび下回る)の関数としてプロットしてある。
【図26】PfMSP4およびPvMSP4のアミノ酸配列の整列である。Pf配列のいくつかの特色が強調され、(i)分泌性およびGPI付着シグナルは、緑に示してあり、(ii)SALSAとして公知の配列は、下線を引いてある、(iii)すべての負に荷電した残基(DおよびE)は、赤において示してあり、および(iv)PfMSP4p20のN末端配列および重要なシステイン残基は、青で強調してある。
【図27】バキュロウイルスPfMSP5を示す。(A)精製された組換えPfMSP5をSDS-PAGEゲル上で移動させ、ニトロセルロースへ転写し、ヒト免疫血清でプローブした。(B)PfMSP5をトリチウム化したミリスチン酸の存在下において発現させた。精製したタンパク質をSDS-PAGEゲル上で移動させ、ブルー染色およびオートラジオグラフィーを7週間行った。
【図28】RBC期におけるある時間にわたったPfMSP4の発現を示す。(A)アセトン固定したリング期の寄生虫および風乾した後期のおよび自由なメロゾイトを、PfMSP1p19または組換えPfMSP4p20(Rp20)に対してアフィニティー精製されたポリクローナルウサギ血清に特異的であるモノクローナル抗体G17.12でプローブした。抗体染色は、Alexa Fluor(登録商標)488のヤギ抗マウスまたは抗ウサギ結合抗体で明らかにされ、寄生虫DNAは、ヘキスト33342で染色した。(B)アフィニティー精製したウサギ(Rp20)およびヒト血清(NdiopおよびDielmoの1:1のプール:Hp20)を使用する、成熟シゾント(S)、自由なメロゾイト(M)、およびリング(R)からの寄生虫抽出物のウエスタンブロット分析。
【図29】組換えPfMSP4発現のために使用する構築物によってコードされるタンパク質の配列を示す。PfMSP4p20の直接の発現を促進するようにデザインされた構築物を示す。構築物の名前を配列テキストの左に収載してあり、それぞれの分泌タンパク質のN末端の配列をテキスト内で太字体に強調してあり、および組換えタンパク質の名前を右側に収載してある。
【図30】ある時間にわたったPfMSP4p20の直接発現を示す。PfMSP4p40、PfMSP4p21、PfMSP4p21ss1またはPfMSP4p21ss2バキュロウイルスが感染昆虫細胞の培養液上清の試料を、感染後24〜66時間の間6時間間隔にて収集し、透析し、TALON樹脂上でバッチ精製した。精製したタンパク質試料をNuPAGE 4〜12%の勾配ゲルで分離し、ブルー染色し、タンパク質のサイズを右に示す。おおよそのタンパク質収率を感染後66時間にて残存している培養液上清の最後の130 mLを使用して算出した。タンパク質をIMACおよびHPLCによって精製し、タンパク質収率をBCAタンパク質量キット(PIERCE)を使用して算出する。
【図31】PfMSP4p40およびPvMSP4/Hisのタンパク質配列の整列を示す。バキュロウイルスP. vivax MSP4およびP. falciparum MSP4発現構築物のアミノ酸(一文字表記)配列は、clustalxを使用して整列させた。アミノ酸同一性を(*)、保存的置換を(:)、半保存的な置換を(.)およびラジカル変化を空白で示してある。同定されたN末端の配列には、下線を引いてあり、かつ太字体である。異なるタンパク質産物についてのN末端の配列には、下線を引いてあり、かつ太字体である。
【図32】ある時間にわたったPvMSP4/His発現を示す。簡潔には、8 mLの撹拌培養のSN試料を感染後24および66時間の間6時間の間隔で収集し、透析し、TALON樹脂上でバッチ精製する。精製したタンパク質試料をNuPAGE 4-12%の勾配ゲルで分離し、SimplyBlue SafeStainで染色し、タンパク質サイズを左右に示す。
【図33】PvMSP4/Hisに対するヒト免疫血清の反応性を示す。(A)ELISAプレートには、バキュロウイルス発現系で発現したPvMSP4/Hisをコートした。三倍段階希釈またはすべての24血清を試験した。(B)平行して、それぞれの希釈系列を不可逆的に還元したPvMSP4に対して試験した。このグラフは、天然抗原に対する負の対照を上回るODを示すそれぞれの血清の下方パネルにおける、1/2700希釈でのODを示す。上方パネルは、1/2700の血清希釈での抗原減少において見られるODの減少を示す。
【図34】図34は、PMNによるメロゾイトの抗体依存的食作用(APDm)に基づいた機能的アッセイ法における血清(非動化、IgG除去、または精製IgG)の処理の結果を示す。アッセイ法は、実施例19に記してある。
【図35】図35は、PMNによるメロゾイトの抗体依存的食作用(APDm)に基づいた機能的アッセイ法における血清(非動化、IgG除去、または精製IgG)の処理の結果を示す。アッセイ法は、実施例19に記してある。
【発明の詳細な説明】
【0028】
2つの最近同定されたメロゾイト表面抗原の熱帯熱マラリア原虫(Plasmodium falciparum)メロゾイト表面タンパク質4および5(PfMSP4およびPfMSP5)は、潜在性多成分抗マラリアワクチンのタンパク質成分になる見込みがある。msp4およびmsp5遺伝子は、両方とも272残基タンパク質をコードする、それぞれの単一のC末端EGF様ドメインおよびGPI付着モチーフをもち、染色体2上でmsp2のすぐ上流に直列に位置する。メロゾイト表面での膜会合は、両タンパク質について証明されており、ヒト免疫血清は、大腸菌(Escherichia coli)において発現させた組換えMSP4と反応することが示された[12,13]。
【0029】
プラスモディウム属の3つのマウス種、P. ヨエリ(P. yoelii)、P. チャバウジ(P. chabaudi)およびP. ベルゲイ(P. berghei)において、MSP4およびMSP5座位(MSP4/5)に単一遺伝子のみがあり、これらは、それぞれに対してある程度の相同性を示す[14-16]。この遺伝子は、MSP4/5を示し、P. ヨエリ(P. yoelii)致死チャレンジモデルにおいて感染防御免疫を調査するために使用されてきた[17]。
【0030】
MSP4/5は、種々の免疫化ストラテジーを使用して保護を与えることが示されており、MSP1p19と組み合わせて送達したときに、有効性が最大化される[17-20]。加えて、MSP4/5タンパク質によって誘導されるマウス免疫応答における系統特異性はないと思われる[21]。熱帯熱マラリア原虫(P. falciparum)では、msp4およびmsp5は、それぞれ相同的な位置に単一のイントロンを有する[22]。
【0031】
熱帯熱マラリア原虫(Plasmodium falciparum)メロゾイト表面タンパク質4(PfMSP4)タンパク質配列は、分泌シグナル配列、C末端のEGF様ドメインおよびGPI-付着シグナルを含む[10]。
【0032】
Pfmsp4遺伝子配列は、960bpの長さであり、144bpの1つのイントロンを含み、272アミノ酸残基をコードする。Pfmsp5遺伝子配列は、955bpの長さであり、136 bpの1つのイントロンを含み、272アミノ酸残基をコードする。PfMSP4と同様に、PfMSP5タンパク質配列は、分泌シグナル配列、C末端のEGF様ドメインおよびGPI-付着シグナルからなる[11,22]。MSP遺伝子のクラスターのこの下流には、高度に保存されたアデニロコハク酸ライセース(lysase)(ASL)遺伝子があり、これは、その他の種におけるこの座位の同定を促進する有用なハンドルであることが判明した(図1)。
【0033】
多くの現在のデータは、PfMSP4およびPfMSP5が優れたワクチン候補であるという考えを支持する。それにもかかわらず、いくつかの公開された知見は、2つの異なる発現系(大腸菌(E.coli)および酵母)において産生されたPfMSP4の組換え類似体は抗原性が異なり、かつヒト免疫血清では見られない現象である、タンパク質のEGF様ドメインに対して高次構造非依存的な反応を誘導することを示している[13]。タンパク質のこの領域における高次構造上のエピトープの認識が、保護に重要である場合、MSP1-19の例のように、すべてのエピトープを忠実に再生する産物を産生することは避けられない。
【0034】
従って、本発明は、熱帯熱マラリア原虫(Plasmodium falciparum)MSP4およびMSP5をコードする核酸、並びに生じるポリペプチドが、昆虫細胞における最適な発現を達成するように修飾された構築物を提供する。より詳細には、本発明は、バキュロウイルス-昆虫細胞発現系において可溶性の分泌されるポリペプチドとして発現される組換えMSP4ポリペプチドをコードする構築物を提供する。組換えポリペプチドは、C末端にEGF様ドメインを含み、これは適切に折り畳まれるようである。これは、タンパク質をELISAプレート上で不可逆的に減少させるときに、MSP4のポリクローナルウサギ血清認識において顕著な減少(50〜60%)があることによって示される(図12を参照されたい)。
【0035】
本発明の組換えMSP4ポリペプチドの1つは、N末端の近くの多形領域からの30アミノ酸の欠失を伴うMSP4外抗原(GPI付着点のC末端疎水性残基がなく、したがって、タンパク質分泌が可能)である。本発明のこのポリペプチドは、MSP4p30と呼ばれる。
【0036】
本発明の組換えMSP4ポリペプチドのもう1つのものは、N末端の近くの多形領域からの30アミノ酸の欠失を伴わないMSP4外抗原(GPI付着点のC末端の疎水性残基がなく、したがってタンパク質分泌が可能)である。本発明のこのポリペプチドは、MSP4p40と呼ばれ、全長遺伝子産物である。
【0037】
本発明のもう一つの組換えMSP4ポリペプチドは、配列KSPKEモチーフ周辺で始まり、EGFドメインを含む、およそMSP4のC末端の半分に対応する20kDaのポリペプチドである。上流の補充アミノ酸残基、特にKSPKEモチーフ上流に局在化するMSP4p40配列の残基を含めることができる。本発明のこの組換えポリペプチドは、MSP4p20と呼ばれる。
【0038】
加えて、本発明は、同時に産生されるp35およびp45形態を含む組換えMSP5ポリペプチドを提供する。両方とも、翻訳後修飾を含み、免疫原性を押し上げることができる脂肪酸残基(ミリストイル化)の共有結合を含む可能性が高い。本発明のこれらの組換えポリペプチドは、MSP5p45およびMSP5p35と呼ばれる。
【0039】
それにもかかわらず、本発明の組換えMSP4およびMSP5ポリペプチドは、また、これらの天然のGPIシグナル配列またはGPI修飾のための別のGPIアンカータンパク質配列シグナリングからのもののいずれかを使用して、C末端GPIアンカー実体として発現することができる。このようなGPI修飾された実体は、任意の免疫のアジュバントの非存在下において、組換えMSP4およびMSP5ポリペプチドの免疫原性を実質的に増強することが予想されるであろう。
【0040】
本発明の組換えポリペプチドは、以下の通りのこれらの対応する配列番号に関して、より詳細に記述してある:
核酸
MSP4p20 配列番号:1
MSP4p30 配列番号:2
MSP4p40 配列番号:3
MSP5 配列番号:4
MSP5p10 配列番号:5
MSP4p21 配列番号:6
MSP4p21 ss1 配列番号:7および配列番号:28
MSP4p21 ss2 配列番号:8および配列番号:29
ポリペプチド
MSP4p20(分解生成物) 配列番号:9
MSP4p30(構築物に存在するORF=構築物によってコードされるポリペプチド) 配列番号:10
MSP4p30(発現の最終生成物=昆虫細胞によって産生されるポリペプチド) 配列番号:11
MSP4p40(構築物に存在するORF) 配列番号:12
MSP4p40(発現の最終生成物) 配列番号:13
MSP5(構築物に存在するORF) 配列番号:14
MSP5p10 配列番号:15
PvMSP4/His 配列番号:16
MSP4p21(構築物に存在するORF) 配列番号:17
MSP4p21(発現の最終生成物) 配列番号:18
MSP4p21 ss1(構築物に存在するORF) 配列番号:19
MSP4p21 ss1(発現の最終生成物) 配列番号:20
MSP4p21 ss2(構築物に存在するORF) 配列番号:21
MSP4p21 ss2(発現の最終生成物) 配列番号:22
PvMSP4p20(「PfMSP4およびPvMSP4のアミノ酸配列の整列に示したように、MSP4の三日熱マラリア原虫(Plasmodium vivax)配列に基づいて提唱されたワクチン構築物の配列) 配列番号:30。
【0041】
これらのポリペプチドは、本明細書において、個々におよびひとまとめにして、本発明の「組換えMSP4およびMSP5ポリペプチド」と称される。同様に、これらのポリペプチドをコードする核酸は、本発明の「組換えMSP4およびMSP5核酸」と呼ばれる。
【0042】
本発明に対する意味は、広範囲にわたる。組換えMSP4およびMSP5ポリペプチドのこの発見により、本発明の組換えMSP4およびMSP5ポリペプチドをコードする核酸配列を含む発現ベクターの構築;発現ベクターをトランスフェクトさせ、または形質転換した宿主細胞;生物学的に活性な組換えMSP4およびMSP5ポリペプチド、並びに単離され、または精製されたタンパク質としての組換えMSP4およびMSP5ポリペプチド;組換えMSP4およびMSP5ポリペプチドと免疫反応性の抗体、組換えMSP4およびMSP5ポリペプチド並びにプラスモディウム属寄生虫およびマラリア感染の検出における組換えMSP4およびMSP5ポリペプチドに対して向けられた抗体の診断使用、並びにプラスモディウム属感染から守るための組換えMSP4およびMSP5ポリペプチドのワクチン使用が可能になる。
【0043】
また、本明細書に使用される「組換えMSP4およびMSP5ポリペプチド」という用語は、配列番号:9〜22のアミノ酸配列を有するタンパク質をさらに包含するポリペプチドの属、並びにこのようなアミノ酸配列と高度な類似性(少なくとも90%の相同性)を有し、かつ該タンパク質およびポリペプチドが免疫反応性であるタンパク質およびポリペプチドをいう。加えて、組換えMSP4およびMSP5ポリペプチドは、配列番号:1〜8のヌクレオチドの遺伝子産物をいう。
【0044】
本明細書に使用される「精製された」という用語は、たとえば組換え宿主細胞培養の精製産物として、または非組換え供与源から精製された産物として、組換えMSP4およびMSP5ポリペプチドがその他のタンパク質またはポリペプチドを本質的に付随しないことを意味する。本明細書に使用される「実質的に精製された」という用語は、組換えMSP4およびMSP5ポリペプチドを含み、かつ特異抗体を使用して除去することができる公知のタンパク質の存在を除けば、その他のタンパク質またはポリペプチドを本質的に付随しない混合物をいい、該実質的に精製された組換えMSP4およびMSP5ポリペプチドは、抗原として使用することができる。
【0045】
本明細書で言及される組換えMSP4およびMSP5ポリペプチド「変異体」とは、1つまたは複数の欠失、挿入または置換のために、組換えMSP4およびMSP5ポリペプチドのものとは異なるアミノ酸配列を有する、組換えMSP4およびMSP5ポリペプチドに対して実質的に相同的なポリペプチドを意味する。変異体アミノ酸配列は、好ましくは組換えMSP4およびMSP5ポリペプチドアミノ酸配列と少なくとも80%同一で、最も好ましくは少なくとも90%同一である。パーセント同一性は、たとえばDevereuxらによって記述され、およびウィスコンシン大学Genetics Computer Group(UWGCG)から入手可能なGAPコンピュータプログラム、バージョン6.0(Nucl. Acids Res. 12:387、1984)を使用して配列情報を比較することにより、決定することができる。GAPプログラムは、SmithおよびWaterman (Adv. Appl. Math 2:482, 1981)によって修正されたNeedlemanおよびWunschの整列方法を利用する(J. Mol. Biol. 48:443, 1970)。GAPプログラムのための好ましいデフォルトパラメーターには、以下が含まれる:(1)Schwartz and Dayhoff, eds., Atlas of Protein Sequence and Structure, National Biomedical Research Foundation, pp. 353-358, 1979によって記述されたような、Gribskov and Burgess, Nucl. Acids Res. 14:6745, 1986の、ヌクレオチドのための単一比較マトリックス(同一性について1および非同一性について0の値を含む);(2)それぞれのギャップに対する3.0のペナルティーおよびそれぞれのギャップにおけるそれぞれのシンボルに対するさらなる0.10のペナルティー;および(3)末端ギャップに対するペナルティーなし。
【0046】
変異体には、所与のアミノ酸残基が同様の生理化学的特徴を有する残基によって置換されたことを意味する、保存的に置換された配列を含むことができる。保存的置換の例には、Ile、Val、LeuもしくはAlaを互いになどの1つの脂肪族残基の別ものへの置換またはLysとArg;GluとAsp;もしくはGlnとAsnとの間などの1つの極性残基の別のものへの置換を含む。その他のこのような保存的置換、例えば同様の疎水性特徴を有する全ての領域の置換も周知である。本発明のMSP4およびMSP5ポリペプチドの変異体の例は、選択的mRNAスプライシングイベントにより、、または組換えMSP4およびMSP5ポリペプチドのタンパク分解切断により生じるタンパク質である。タンパク質分解に起因する変異には、たとえば組換えMSP4およびMSP5ポリペプチドからの1つまたは複数の末端アミノ酸のタンパク分解性の除去による、宿主細胞の異なるタイプにおける発現による末端の相違が含まれる。フレームシフトに起因する変異には、たとえば宿主細胞の異なるタイプにおける発現による末端の相違が含まれる。
【0047】
前述の通り、本発明は、単離され、および精製された、または同種の組換えMSP4およびMSP5ポリペプチドを提供する。抗原として使用することができる組換えMSP4およびMSP5ポリペプチドの変異体および誘導体は、組換えMSP4およびMSP5ポリペプチドをコードするヌクレオチド配列の突然変異によって得ることができる。アミノ酸配列の変化は、任意の多数の従来法によって達成することができる。突然変異は、天然の配列の断片に対するライゲーションを可能にする制限部位に隣接した、突然変異体配列を含むオリゴヌクレオチドを合成することにより、特定の座位に導入することができる。ライゲーションに続いて、生じる再構築配列は、所望のアミノ酸挿入、置換または欠失を有する類似体をコードする。
【0048】
あるいは、オリゴヌクレオチド特異的、部位特異的突然変異誘発法を使用して、予め定められたコドンを置換、欠失または挿入によって変化させることができる変化させられた遺伝子を提供することができる。上記の変化を作製する例示的方法は、Walderら(Gene 42:133, 1986); Bauerら(Gene 37:73, 1985); Craik (BioTechniques, January 1985, 12-19); Smithら(Genetic Engineering: Principles and Methods, Plenum Press, 1981); Kunkel(Proc. Natl. Acad. Sci. USA 82:488, 1985); Kunkelら(Methods in Enzymol. 154:367, 1987);並びに米国特許第4,518,584号および第4,737,462号によって開示されており、これらの全ては、参照により援用される。
【0049】
本発明の側面内において、組換えMSP4およびMSP5ポリペプチドは、組換えMSP4およびMSP5ポリペプチドと特異的に結合する抗体を調製するために利用することができる。「抗体」という用語は、ポリクローナル抗体、モノクローナル抗体、F(ab')2およびFab断片などのこれらの断片、並びに任意の組換えで産生される結合パートナーを含むことが意味される。抗体は、これらが約107 M-1以上のKaで組換えMSP4およびMSP5ポリペプチドに結合する場合、特異的に結合すると定義される。たとえば、結合パートナーまたは抗体の親和性は、従来技術、たとえばScatchardrら、Ann. N.Y Acad. Sci., 51:660 (1949)によって記述されたものを使用して容易に決定することができる。ポリクローナル抗体は、当該技術分野において周知手順を使用して種々の供与源、例えばウマ、ウシ、ヤギ、ヒツジ、イヌ、ニワトリ、ウサギ、マウスまたはラットから容易に産生することができる。
【0050】
本発明は、配列番号:1、2、3、4、5、6、7および8のヌクレオチド配列に由来する単離された断片およびオリゴヌクレオチドをさらに包含する。また、本発明は、これらの断片およびオリゴヌクレオチドによってコードされるポリペプチドを包含する。
【0051】
本発明の範囲内の核酸配列には、適度なまたは厳密なストリンジェンシー条件下で、本明細書に開示した天然の組換えMSP4およびMSP5核酸にハイブリダイズし、かつ組換えMSP4およびMSP5ポリペプチドをコードする単離されたDNAおよびRNA配列が含まれる。本明細書に使用される適度なストリンジェンシーの条件は、当業者に公知のとおり、およびSambrookら、Molecular Cloning: A Laboratory Manual, 2 ed. Vol. 1, pp. 1.101-104, Cold Spring Harbor Laboratory Press, (1989)によって定義されるとおり、ニトロセルロースフィルターのための予洗溶液5×SSC、0.5% SDS、1.0mM EDTA(pH 8.0)、50% ホルムアミド、6×SSC、42℃におけるハイブリダイゼーション条件(または42℃にて50%ホルムアミドにおけるStark溶液などのその他の同様のハイブリダイゼーション溶液)、並びにおよび約60℃、0.5×SSC、0.1% SDSの洗浄条件の使用を含む。高ストリンジェンシーの条件は、上記のハイブリダイゼーション条件と同様に定義され、68℃、0.2×SSC、0.1% SDSにて洗浄する。当業者であれば、温度および洗浄液溶液塩濃度をプローブの長さなどの要因に従って必要に応じて調整することができることを認識するであろう。
【0052】
複数のコドンが同じアミノ酸をコードする遺伝暗号の公知の縮重のため、DNA配列は、配列番号:1〜8に示され、かつ配列番号:9〜22のアミノ酸配列を有する組換えMSP4およびMSP5ポリペプチドをコードするたものから変化することができる。このような変異体DNA配列は、サイレント突然変異(たとえば、PCR増幅の間に生じる)により生じ得るか、または天然の配列の慎重な突然変異誘発の産物であることができる。
【0053】
したがって、本発明は、(a);(a)配列番号:1〜8のヌクレオチド配列を含むDNA、(b)適度なストリンジェンシー条件下で(a)のDNAに対してハイブリダイゼーションすることができ、かつ組換えMSP4およびMSP5ポリペプチドをコードするDNA;並びに(c)(a)または(b)で定義したDNAに対する遺伝暗号の結果としての縮重であり、かつ組換えMSP4およびMSP5ポリペプチドをコードするDNAから選択される、組換えMSP4およびMSP5ポリペプチドをコードする対応する単離されたDNA配列を提供する。このようなDNA均等物配列によってコードされるポリペプチドは、本発明によって包含される。
【0054】
配列番号:1〜8のDNA配列に対応するDNAは、適度にストリンジェント条件下で、配列番号:9〜22のアミノ酸配列を含むポリペプチドをコードするDNA配列にハイブリダイズする。このようなDNAによってコードされる組換えMSP4およびMSP5ポリペプチドの例は、組換えMSP4およびMSP5ポリペプチド断片、並びに上記のように、不活性化されたN-グリコシル化部位、不活性化されたプロテアーゼプロセシング部位または保存的アミノ酸置換を含む組換えMSP4およびMSP5ポリペプチドを含むが、限定されない。DNAが配列番号:1〜8のDNAの相補物にハイブリダイズするプラスモディウム属のその他の種に由来するDNAによってコードされるポリペプチドも包含される。
【0055】
組換えMSP4およびMSP5ポリペプチドをコードする核酸配列を含む組換え発現ベクターは、周知の方法を使用して調製することができる。発現ベクターには、哺乳動物、微生物、ウイルスまたは昆虫遺伝子に由来するものなどの、適切な転写または翻訳制御ヌクレオチド配列に作動可能に連結された組換えMSP4およびMSP5 DNA配列を含む。制御配列の例には、転写プロモーター、オペレーターまたはエンハンサー、mRNAリボソーム結合部位および転写および翻訳の開始と終了を制御する適切な配列が含まれる。ヌクレオチド配列は、制御配列が組換えMSP4およびMSP5 DNA配列に機能的に関連するときに、「作動可能に連結される」。したがって、プロモーターヌクレオチド配列は、プロモーターヌクレオチド配列が、組換えMSP4およびMSP5 DNA配列の転写を制御する場合に、組換えMSP4およびMSP5 DNA配列に作動可能に連結されている。所望の宿主細胞において複製する能力は、通常複製開始点によって与えられ、形質転換体を同定することにより選択される遺伝子を、加えて発現ベクターに組み込むことができる。
【0056】
加えて、天然では組換えMSP4およびMSP5ポリペプチドと付随していない適切なシグナルペプチドをコードする配列を発現ベクターに組み込むことができる。たとえば、組換えMSP4およびMSP5ポリペプチドが、最初にシグナルペプチドを含む融合タンパク質として翻訳されるように、シグナルペプチド(分泌性のリーダー)のためのDNA配列を組換えMSP4およびMSP5ヌクレオチド配列に対してインフレームで融合させることができる。意図した宿主細胞において機能的であるシグナルペプチドは、組換えMSP4およびMSP5ポリペプチドの細胞外分泌を増強する。シグナルペプチドは、細胞からの組換えMSP4およびMSP5ポリペプチドの分泌により、組換えMSP4およびMSP5ポリペプチドから切断することができる。
【0057】
原核生物の宿主細胞に使用するための発現ベクターは、一般に1つまたは複数の表現型の選択可能なマーカー遺伝子を含む。表現型の選択可能なマーカー遺伝子は、たとえば抗生物質抵抗性を与えるか、または独立栄養性の要求を供給するタンパク質をコードする遺伝子である。原核生物の宿主細胞のための有用な発現ベクターの例には、市販のプラスミドに由来するものが含まれる。市販のベクターには、タンパク質の発現のために特異的にデザインされているものを含む。これらには、マルトース結合タンパク質に融合されるタンパク質の発現のために使用されるpMAL-p2およびpMAL-c2ベクターが含まれる(New England Biolabs Beverly、MA、USA)。
【0058】
組換え原核生物の宿主細胞発現ベクターのために一般に使用されるプロモーター配列は、β-ラクタマーゼ(ペニシリナーゼ)、ラクトースプロモータ系(Chang et al., Nature 275:615, 1978;およびGoeddel et al., Nature 281:544, 1979)、トリプトファン(trp)プロモーター系(Goeddelら、Nucl. Acids Res. 8:4057, 1980;およびEP-A-36776)およびtacプロモーター、(Maniatis, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, p. 412, 1982)を含む。
【0059】
組換えMSP4およびMSP5ポリペプチドの発現のための適した宿主細胞には、原核生物、酵母または高等真核細胞が含まれる。昆虫細胞が好ましい。細菌、真菌、酵母および哺乳動物宿主細胞で使用するために適したクローニングおよび発現ベクターは、たとえばPouwels ら、Cloning Vectors: A Laboratory Manual, Elsevier, New York, (1985)に記述されている。無細胞翻訳系も、本明細書に開示したDNA構築物に由来するRNAを使用して組換えMSP4およびMSP5ポリペプチドを産生するために使用することができるであろう。
【0060】
本発明は、本明細書に記述した技術またはその他の方法を使用して得られるかどうかにかかわらず、単離され、もしくは精製された形態の前述したタンパク質またはポリペプチドを包含することが意図されることが理解されよう。本発明の好ましい態様において、組換えMSP4およびMSP5ポリペプチドは、人体組織および人体組織成分、核酸、外来タンパク質および脂質、並びに細菌およびウイルスなどの外来微生物を実質的に含まない。また、本発明は、実質的に同じ生物学的および免疫原特性を有する同等のタンパク質を包含することが理解される。
【0061】
本発明の組換えMSP4およびMSP5ポリペプチドを作製するための使用に応じて、これらを標識することが望ましいであろう。適切な標識の例は、放射性標識、酵素標識、蛍光標識、化学発光標識および発色団である。本発明のタンパク質を標識するための方法は、標識化免疫グロブリンのために広く使用されているものと本質的に異ならない。標識する必要は、間接的マーカーとして本発明の抗原に対する標識抗体または抗原に対する抗体に対する抗免疫グロブリンを使用することによって回避してもよい。
【0062】
一旦本発明の組換えMSP4およびMSP5ポリペプチドが得られたら、これらを使用してそれと反応性のポリクローナル抗体およびモノクローナル抗体を産生した。したがって、本発明のタンパク質またはポリペプチドは、当該技術分野において公知の技術によって動物宿主を免疫するために使用することができる。このような技術には、通常接種が含まれるが、これらには、その他の投与様式が含まれていてもよい。動物宿主において免疫原性反応を生じさせるために、ポリペプチドの十分な量が投与される。本発明の抗原に対する抗体を産生するために任意の宿主を使用することができる。一旦動物を免疫して、これが抗原に対する抗体を産生し始めるのに十分な時間が過ぎたら、ポリクローナル抗体を回収することができる。一般的な方法には、動物から血液を除去すること、および血液から血清を分離することを含む。抗原に対する抗体を含む血清は、抗原に対する抗血清として使用することができる。あるいは、抗体を血清から回収することができる。親和性精製は、精製された、抗原に対するポリクローナル抗体を血清から回収するための好ましい技術である。
【0063】
また、本発明の抗原に対するモノクローナル抗体を調製することができる。抗原と反応性のモノクローナル抗体を産生するための1つの方法は、宿主を抗原で免疫すること;宿主の脾臓から抗体産生細胞を回収すること;抗体産生細胞を酵素ヒポキサンチン-グアニンホスホリボシルトランスフェラーゼを欠損した骨髄腫細胞と融合して、ハイブリドーマを形成すること;ヒポキサンチン、アミノプテリンおよびチミジンを含む培地における培養によって少なくともハイブリドーマの1つを選択すること;抗原に対する抗体を産生する少なくとも1つのハイブリドーマを同定すること;同定されたハイブリドーマを培養して、回収可能な量の抗体を産生すること;および培養されたハイブリドーマによって産生される抗体を回収することを含む。
【0064】
これらのポリクローナル抗体またはモノクローナル抗体は、種々の適用に使用することができる。対応するタンパク質の中和は、これらの一つである。また、これらは、生物学的製剤におけるプラスモディウム属寄生虫抗原を検出するために、またはたとえば親和性クロマトグラフィーカラムに使用したときに、対応するタンパク質、糖タンパク質もしくはこれらの混合物を精製する際に、使用することができる。
【0065】
組換えMSP4およびMSP5ポリペプチドは、生体試料におけるマラリア原虫に対して特異的な抗体の存在を検出するための、および現在または以前の感染に関するシグナルを構成する、これらの生体試料におけるこのような抗体のレベルを評価するための抗原として使用することができる。このような生体試料には、もちろんヒト組織およびヒト細胞、並びにヒト血清を含むヒト体液などの生物学的液体を含む。マラリアに対する抗体の存在または濃度を決定するための免疫アッセイ法の試薬として使用されるときに、本発明の抗原は、便利で、迅速で、感受性で、および特異的なアッセイ法を提供する。
【0066】
より詳細には、本発明の抗原は、液体中の体液性成分を検出し、または定量化する際に使用するための周知の免疫アッセイ法によるマラリアの発見のために使用することができる。したがって、抗原-抗体相互作用は、沈澱または凝集などの二次反応によって、直接観察すること、または決定することができる。加えて、免疫電気泳動技術も使用することができる。たとえば、寒天における電気泳動法、続く抗血清との反応の古典的な組み合わせ、並びに二次元電気泳動、ロケット電気泳動およびポリアクリルアミドゲルパターンの免疫標識を利用して、(ウエスタンブロットまたは免疫ブロット)を利用することができる。本発明の抗原が使用することができるその他の免疫アッセイ法には、放射免疫アッセイ法、競合的免疫沈降アッセイ法、酵素免疫測定法および免疫蛍光アッセイ法を含むが、限定されない。比濁技術、比色技術および比濁分析技術を使用することもできることが理解されるであろう。ウエスタンブロット技術に基づいた免疫アッセイ法が好ましい。
【0067】
免疫アッセイ法は、本発明の抗原または本発明の抗原に対する抗体のいずれかの免疫試薬の1つを、試薬の免疫反応性を保持すると共に、担体表面上に固定することによって実施することができる。相互免疫試薬は、免疫反応性も保持されるような方法で、非標識であること、または標識することができる。これらの技術は、特に酵素結合免疫吸着検定法(ELISA)および競合阻害酵素免疫測定法(CIEIA)などの酵素免疫測定法に使用するための適している。
【0068】
本発明の抗原または抗原に対する抗体が固体支持体に付着されるときに、支持体は、通常ガラスまたはプラスチック材料である。プレート、チューブ、ビーズまたは円板の形態で成形されるプラスチック材料が好ましい。適切なプラスチック材料の例は、ポリスチレンおよび塩化ビニルである。免疫試薬が固体支持体と容易に結合しない場合、担体材料は、試薬と支持体との間に配置することができる。適切な担体材料の例は、ウシ血清アルブミンなどのタンパク質またはグルタルアルデヒドもしくは尿素などの化学試薬である。固相のコーティングは、従来の技術使用することにより実施することができる。
【0069】
本発明は、免疫原性の組換えMSP4およびMSP5ポリペプチドおよびより詳しくは、マラリアに対するワクチン組成物の調製に使用するための保護薬ポリペプチドを提供する。したがって、これらのポリペプチドは、マラリア感染に感受性の哺乳類に対してポリペプチドを投与することによるワクチンとして使用することができる。従来の投与様式を使用することができる。たとえば、投与は、経口、舌下、呼吸器または非経口的経路によって実施することができる。ワクチンが非経口的に投与されるときは、皮内、皮下、筋肉内および静脈内の投与経路が好ましい。
【0070】
マラリアに感染した哺乳類における免疫応答の主目的は、マラリア原虫を不活性化すること、並びにマラリア原虫死滅および寄生虫が感染した赤血球のクリアランスを促進することである。免疫応答のB細胞腕は、血液段階でマラリア原虫を不活性化するための主要な役割を担う。これが達成される主要な様式は、感染力の中和(赤血球浸潤の阻害)および抗体依存性細胞障害作用(ADCC)による。標的抗原は、その他の寄生虫株による再感染に対して有効で、かつTヘルパー細胞活性(CD4+)を誘導して長期記憶反応を生じさせることができるように保存されなければならない。寄生虫感染細胞の破壊についてのT細胞を媒介したメカニズムは、細胞障害性(CD8+)Tリンパ球(CTL)によって提供され、これが感染のプレ赤血球相の肝細胞表面にてクラスI組織適合抗原と組み合わせて発現された組換えMSP4およびMSP5抗原を認識することができるであろう。
【0071】
以下は、(A)天然のシグナル配列、(B)C末端Hisタグおよび(C)本分野における感染防御抗体反応と最も強く相関される熱帯熱マラリア原虫(P. falciparum)抗原のものに似ている酸反復領域(MSP4p20)を含むワクチンである。
【0072】
MKVAYFLSVLDLLIIFSLYFDGRRSAFAGIAACIRHGRILGEGGE/QNSTPGSGGQTGDHSAEAENGDYNEQGDDHGDDHGDDHGDDHGDEQDGEDYDDAEDDDLYELSEVDENANLCLDNNGGCGDDKICENLGKGIVKCLCKPGYKLVGTECVEHHHHHH[配列番号:30]
本発明の組換えMSP4およびMSP5ポリペプチド、並びにワクチンが宿主において保護薬レベルの中和抗体(すなわち、病原体が赤血球に感染する能力を直接遮断する、抗原に対する体液性応答の間に誘発される抗体)を誘導する能力は、アジュバント(免疫賦活剤)との乳化、リポソームに組み込むこと、適切な担体に対して結合することにより、またはこれらの技術の組み合わせにより増強することができる。たとえば、本発明の組換えMSP4およびMSP5ポリペプチドは、リン酸アルミニウムおよび水酸化アルミニウムゲルなどの従来のアジュバントと共に、宿主において体液性免疫応答または細胞媒介免疫応答を増強するために十分な量で投与することができる。
【0073】
免疫化スケジュールは、感染に対する宿主の感受性および宿主の年齢などのいくつかの要因に依存する。本発明のワクチンの単一用量を宿主に投与することができ、または時間間隔をおいて数回の用量が投与される免疫化の一次経過を行うことができる。ブースターとして使用するその後の用量を、一次経過後に必要に応じて、投与することができる。
【0074】
本発明の組換えMSP4およびMSP5タンパク質、ポリペプチドおよびワクチンは、宿主における寄生虫負担を減少させ、臨床症状を減弱させるために、寄生虫感染およびインビボにおける複製を予防し、または阻害する免疫応答を誘導するために十分な量で宿主に投与することができる。免疫原性反応は、宿主に対して1用量あたり10〜500マイクログラム、好ましくは1用量あたり約50〜100マイクログラムの範囲である量で本発明のポリペプチドを投与することによって得ることができる。本発明のタンパク質およびワクチンは、生理的に許容される担体と共に投与することができる。たとえば、水または食塩水などの希釈剤を使用することができる。
【0075】
本発明のもう一つの側面は、DNAワクチン接種の方法を提供する。また、本方法には、組換えMSP4およびMSP5ポリペプチド、タンパク質およびポリペプチドをコードする核酸の任意の組み合わせを、それ自体、キャリア分子の有無において、個体に対して投与することを含む。態様において、個体は、動物であり、および好ましくは哺乳類である。より好ましくは、哺乳類は、ヒト、イヌ、ネコ、ウシ、ブタ、およびウマからなる群から選択される。特に好ましい態様において、哺乳類は、ヒトである。
【0076】
当業者であれば、核酸ワクチン(たとえば、DNAワクチン)および核酸ワクチン技術、並びにタンパク質およびポリペプチドに基づいた技術の概念、適用および有効性を認識するであろう。核酸に基づいた技術により、裸の、またはカプセル化された組換えMSP4およびMSP5ポリペプチドをコードする核酸を、投与前にコードされたタンパク質を産生する必要なく、組織および細胞に直接投与することができる。本技術は、これらの核酸が、レシピエント生物体の細胞によって取り込まれれ、発現してレシピエントの免疫系が反応する免疫原性決定因子を産生する能力に基づく。典型的には、発現された抗原は、核酸を取り込んで発現した細胞表面上に示されが、コードされた抗原の発現およびレシピエント個体の循環系への搬出も本発明の範囲内である。このような核酸ワクチン技術は、裸のDNAおよびRNAの送達および組換えMSP4およびMSP5ポリペプチドをコードする発現ベクターの送達を含むが、限定されない。本技術は、「ワクチン」と称されるが、これは保護反応を生じない免疫原性組成物にも同様に適用できる。このような非保護を誘導する組成物および方法は、本発明に包含される。
【0077】
裸の核酸として組換えMSP4およびMSP5ポリペプチドをコードする核酸および担体分子を送達することも本発明の範囲内であるが、本発明は、より大きな、またはより多くの複合体組成物の一部としての核酸の送達も包含する。これらの送達系に含まれるものには、組換えMSP4およびMSP5ポリペプチドをコードする核酸を含むウイルス、ウイルス様粒子または細菌である。また、リポソームなどの細胞透過化化合物を伴う本発明の核酸の複合体および担体分子も、本発明の範囲内に含まれる。分子ベクター(EP 696,191、Samainらの論文)などの核酸ワクチンのためのその他の化合物および送達系は、当業者に公知であり、たとえば、WO93 06223およびWO90 11092、U.S. 5,580,859およびU.S.5,589,466(Vicalの特許)において例証されており、これらは本明細書に引用として組み込まれ、過度または過剰の実験なしで製作して使用することができる。
【0078】
本発明は、以下の具体的態様において更に詳細に後述する。
【0079】
バキュロウイルス発現系を使用して産生した抗原は、システイン結合の形成を含む複合体構造を忠実に含み、結晶化研究のために適していることが、長い間公知であった。これらの抗原は、その他の系において産生された抗原が点検される「金本位(the gold standard)」として一般に保持される[24,25]。PfMSP4およびPfMSP5の両者は、ジスルフィド結合によって形成されたEGF様ドメインを含むので、これらは、バキュロウイルス-系において産生させることによる利益を受けるであろう。しかし、すべての一般に使用される発現系と同様に、タンパク質収量は、熱帯熱マラリア原虫(P. falciparum)の異常なコドンの選択性による悪影響を受ける[26,27]。加えて、これらの高度にA+Tリッチな配列は、大腸菌(E. coli)において頻繁に変異し[28]、また独特の制限酵素部位をまれに含む。これらの理由により、合成遺伝子は、Withers-Martinez(1999)によって記述された方法を使用してデザインして、構築した。
【0080】
合成遺伝子のデザイン
その時にGenBankから利用可能な配列データに基づいて、およびヒト検証モデルを含む将来の適用を念頭において、共通アミノ酸配列のPfMSP4 NF54配列(Ac番号AF295318)およびPfMSP5 3D7配列(Ac番号AF106476)を修飾のために選択した。平行して、配列データセットを濃縮し、および集団間および集団内解析および種間解析を行うために、以下の通りに風土が異なる場所から採取した試料を使用して多型研究を行った。
【0081】
msp4およびmsp5遺伝子多型の概要
抗原多型は、熱帯熱マラリア原虫(P. falciparum)のためのワクチンを開発するときにの、重要な検討事項である。今までに調べた最も有望な血液段階ワクチン候補は、メロゾイト表面タンパク質1(MSP1)および頂端膜抗原、(AMA1)である[1-3]。しかし、これらおよびその他の表面抗原遺伝子は、免疫エフェクター-メカニズムにアクセス可能でない抗原をコードする遺伝子と比較したときに、不相応に高頻度に同義一塩基多型(nsSNPs)を示す[4-6]。複雑にする要因として、これらのnsSNPsは、最も宿主免疫系にアクセス可能なペプチドの領域内に主にクラスター形成しているラジカルアミノ酸置換を引き起こすことが多い。生じるアミノ酸置換は、重要なBおよびT細胞エピトープ[34,38]を変化させることによって、免疫回避において機能すると考えられる。したがって、このような抗原の統合された組換えバージョンによるワクチン接種は、株特異的な保護を生じる可能性が高い[4,8]。世界的に有効なワクチンには、多段形標的に由来するいくつかの表面タンパク質の保存された部分を含むと広く考えられている[30]。
【0082】
現在まで、限られた量の多型データのみが、PfMSP4およびPfMSP5につて利用できる[11,31,32]。高度に固有な場所のPfMSP4およびPfMSP5の両方の多型を完全に探索するために、配列分析研究が行われ、その結果がPolsonら、2005に示されている[33]。要約すると、Pfmsp5遺伝子配列は、高度に保存されており、潜在的に純化選択のものとにあることが見いだされた。Pfmsp4遺伝子配列は、熱帯熱マラリア原虫(P. falciparum)表面抗原に関して、相対的に保存されていること[5]、および潜在的に平衡選択(N52およびG74)の下に2つの部位を含む多形部位のN末端のクラスターを含むこと(残基45〜81)が見いだされた。これは、フランキング配列内の明らかに中立な(または、ヒッチハイク)多型のクラスター形成が観察されたことを説明するだろうし、BまたはT細胞エピトープ変異に連関させることができる。平衡選択は、古典的に分子の免疫認識に対する影響を有する「相違」を内部に持つ2つの対立遺伝子遺伝子配列の存在により生じる。対立遺伝子Aが、集団において最も一般的なときは、対立遺伝子Bは、この対立遺伝子が免疫系、並びに優性対立遺伝子よって認識されないので、有利である。したがって、対立遺伝子Bは、それが最も多くの型になるまで、より一般的であり、免疫系は、対立遺伝子Bに対してコンピテントとなり、この時点で、対立遺伝子Aであることが有利になる。最初に、平衡選択の存在は、MSP4タンパク質が有効な免疫メカニズムの標的であることを示唆する。第2に、これは、貴方が両方の対立遺伝子でワクチン接種をすること、またはタンパク質のこの領域を除外することを必要とするであろう分野の人々を保護するために、保存された領域に対して免疫系を作用させることを示唆する。肝細胞に特異的に結合することができる2つのPfMSP4由来ペプチドの最近の同定に関して[34]、第1のもの(それは、MSP4p40の残基76〜92を表す)は、1つの準保存的多型(A81)をもち、第2のもの(それは、MSP4p40の残基113〜135を表す)は、1つの欠失(111〜119)および/または1つの準保存的多型(G119)を含む。既存のPfMSP4データセットにおいて、これらの配列内の相違は、相対的に低い頻度で存在し、これらが機能的に重要な部位であるというアイデアをサポートする、(注;これらの配列は、SALSAによっても表される(図2))。最後に、潜在的に平衡選択の下にもう一つの部位V190があり、これはEGF様ドメインの最初のシステインの17残基上流に位置し、再びこれも、BまたはT細胞エピトープ修飾に関与し得る。
【0083】
合成遺伝子;PfMSP4の構築
最初に、タンパク質分泌を促進するために、EGFドメインの3残基下流から始まるすべての配列を除去した。バキュロウイルス系は、このような配列を使用して昆虫細胞GPI部分を組み込んで、およびタンパク質の細胞表面局在化を生じることができる(Bonnet et al. 2006)ことが公知であるので、これには、GPI付着シグナルを含んだ。加えて、バキュロウイルス発現系は、組換え類似体の天然のMSP1シグナル配列を正確に切断するが、天然のMSP4の切断されたN末端は定義されされていないことが公知であるので、天然のシグナルMSP4配列を保持した。プラスモディウム属において生じることは知られていないが、バキュロウイルスにおいてPfMSP4配列の2つの潜在性部位にて生じる)N-グリコシル化を防止するために、セリン残基S65およびS73をアラニンに置換した。両方の残基は、小さな側鎖(Ser = HO-CH2-、Ala = CH3-)を有し、これらの部位は、天然に多形であることは報告されていなかったが、これらの変化が、タンパク質の局部的またはマクロ構造に対して最小限の効果を有することが考えられる。一旦、C末端の6-his-タグおよび終止コドンを含む残りの特色を付加したら、CODOPプログラムセットを使用して配列をキンウワバTrichoplusia ni (High Five)細胞コドンの選択性に翻訳し戻した(表1)。
【0084】
NF54および合成遺伝子のMSP4コドン選択性
【表1】
【0085】
CODOPは、HaleおよびThompsonによって提唱されたように、コドン最適化を実行するUnix(登録商標) perlスクリプトであり[35]、他に詳細に記述されている[29]。再コドンヌクレオチド配列は、その後に手動処理によって最適化し、60〜65℃のTmで38×40mersを生じさせ、およびサブクローニングを促進するためにいくつかの独特の制限部位を含めた(図3)。
【0086】
遺伝子の全体のGC含量を15%まで増加し、サブクローニングの容易さおよび効率を上昇させ、かつ構築物操作の間に大腸菌(E. coli)によってなされる複製エラーの頻度を非常に減少させた。38個のオーバーラップオリゴヌクレオチド(それぞれ40塩基の長さ)は、Eurogentecから得た(標準的精製のみで)。遺伝子構築および増幅は、前述したように達成した。
【0087】
簡潔には、遺伝子オリゴヌクレオチドを等モルの量に混合し(それぞれ25μM)、1μl pfu DNAポリメラーゼ(Stratagene)、5μL、10×産物緩衝液、200μM dNTPおよび2μLの25mM MgSO4を含む50μL PCR反応に10倍希釈した。PCRプログラムは、94℃1分間の1回の変性工程、続く94℃30s間、52℃30s間および72℃2分間の25サイクルから成った。全長遺伝子産物を増幅するためには、5μLの遺伝子構築反応を1μl pfu DNAポリメラーゼ(Stratagene)、5μL、10×産物緩衝液、200μM dNTP、2μLの25mM MgSO4およびそれぞれ500nMの外側遺伝子構築オリゴヌクレオチド番号1および20を含む50μL PCR反応に10倍希釈した。PCRプログラムは、94℃1分間の1サイクル、94℃45s間、68℃45s間および72℃2分間の25のサイクル、並びに72℃10分間の最終伸長工程から成った。図4に示した主要な、および正確な大きさの断片をゲル抽出して、シーケンシングのためにpMOSBlueにクローニングし、Baculoviralゲノム(Baculogold、PharMingen)に組込みのためにベクターpVL1393に最後に移した。この発現構築物をMSP4p40/Hisと呼ぶ。
【0088】
修飾されたPfMSP4構築物
また、2つのさらなるMSP4構築物を構築し、MSP4-EGF/HisおよびMSP4p30/Hisと命名した(図5)。主に結晶化研究の目的で、構築物MSP4-EFG/Hisは、MSP4 EGF様ドメインのみを発現するようにデザインした。PlasmoDBによって記述された予測されるシグナル配列(残基1-20)および2つの下流の残基を含むPCR断片は、高忠実度PCRによって産生した。反応には、遺伝子構築オリゴヌクレオチド1およびBgl II制限部位を含むリバースプライマーMSP4modI(5'.-TAT-AGC-AGA-TCT-TTG-TCG-AAG-TTG-ATG-GTG-CA-3')[配列番号:23]を含んだ。合成遺伝子クローンpMosMSP4を鋳型として使用した。生じる86bp PCR産物を制限酵素Bam HIおよびBgl IIで切断し、ゲル抽出した。次いで、以前に調製し、またBam HIおよびBgl IIで消化してEGF様ドメインの上流のすべての配列を除去して残基D205〜H252を残したpMosMSP4ベクターに産物を結合させた(T4DNAリガーゼ;NEB)。
【0089】
MSP4p30/His構築物は、大部分の報告された多型が属する(図5において強調した)MSP4のC末端から30残基(残基45〜74)を除去するようにデザインした。残基1〜44をコードするPCR断片は、遺伝子構築オリゴヌクレオチド1、リバースプライマーMSP4modII(5'-ATA TGG CTG CAG CCA AGA TCC TCA TGT TAA GCA T-3')[配列番号:24] および鋳型としてのpMosMSP4ベクターを使用して産生した。断片は、154bpのサイズであり、3'および5'最末端にてそれぞれ制限部位Bam HIおよびPst Iを有した。残基A75の顆粒のすべての配列をコードするDNAの断片を、制限酵素Pst IおよびSpe I(これは、pMOSBlueベクター配列のMSP4 ORFの下流にある)を使用して、ベクターpMosMSP4から切断した。2つのDNA断片をT4DNAリガーゼ(NEB)反応中で等モルの量で合わせて、3日にわたって4℃にてインキュベートした。臭化エチジウムで染色した1%のアガロースゲルにおける2μLリガーゼ反応の分離により、724bpの長さの必要とされる産物を含むいくつかの産物の存在が明らかになった。この産物を、遺伝子構築オリゴヌクレオチド1および20と鋳型としての1μL結合反応液とを使用する高忠実度PCRによって増幅し、ゲル抽出し、pMOSBlueへクローニングし戻した。合成遺伝子と同様に、MSP4-EGF/HisおよびMSP4p30/His遺伝子配列は、発現ベクターpVL1393にクローニングされる前に確認した。
【0090】
更なる発現構築物は、上記の合成MSP4遺伝子からデザインして、構築した。この発現カセットは、非多形、プロテアーゼ耐性およびタンパク質の構造的に無処置のC末端の領域(p20と呼ばれ、後の節に記述した)の発現をできるようにデザインした。この構築物は、MSP4p40/HisのxbaIクローニング部位の下流のすべてのヌクレオチド配列を含み、したがって、Leu 130(含まれない)の下流のすべての残基をコードする。タンパク質分泌を促進するために、MSP4シグナル配列さらに下流の9残基(Met1〜Pro49)をコードするPCR断片を、遺伝子構築オリゴ1およびリバースプライマーMSP4p21rev(ATT AAT CTA GAG GCT TTT CTT CAC CCA AGA TCC TCA TG)配列番号:25を使用して産生し、Bam HI/Xba I二重消化したpMosMSP4p40/Hisに結合した。この構築物は、MSP4p21/Hisと命名し、配列をバクロウイルスゲノムに組込むためにpVL1319に結合される前に確認した。2つの補助的な同様の発現構築物は、上記の合成のMSP4遺伝子からデザインして、構築した。これらの構築物は、MSP4p21 ss1およびMSP4p21 ss2と命名し、両方ともLeu130の下流のすべての残基をコードする。タンパク質分泌を促進するために、MSP4p21/Hisと同様に、MSP4シグナル配列(Met1-Pro49)をコードする同じ配列を付加した。さらに、MSP4p21 ss1構築物の配列(Asn50-Ser57およびLeu130)をコードし、およびMSP4p21 ss2のアミノ酸残基Leu130をコードする、MSP4p40/His構築物のいくつかのコドンを付加した。
【0091】
PfMSP5合成遺伝子の構築
Pfmsp4について記述したのと同じ手順を使用して、Pfmsp5合成遺伝子をデザインした。3D7株のGenBankタンパク質配列で開始して、GPI付着シグナルの上流の3残基で始まるすべての配列を除去し、6-His-タグと置換して、3つのグリコシル化部位をセリンからアラニンへの突然変異(S83、S102およびS126)によって崩壊させ、N末端のシグナル配列を無処置のままにした。CODOPによって実行したコドン修飾を表2において考証してあり、全ての合成遺伝子配列を図6に示してある。
【0092】
3D7および合成遺伝子のMSP5コドンの選択性
【表2】
【0093】
PfMSP5合成遺伝子のPCR構築は、前述したように行い、配列内にリピートモチーフが存在しないことにより、高度に純粋な最終産物が産生された(図7)。PCR産物をゲル抽出して、pMOSBlueにクローニングし、Baculoviralゲノム(Baculo-gold、Stratagene)に組込みのためにpVL1393ベクターに移す前にシーケンスした。
【0094】
こうして調製された構築物を評価し、結果を以下の実施例に報告してある。
【実施例】
【0095】
実施例1
昆虫細胞におけるPfMSP4の発現
標準的方法[36]を使用してウイルスを産出した後で、試験感染を、150cm2培養瓶(Corning)において行った。感染は、1時間、10のウイルス多重度にて行った。3日後に上清を収集し、talon樹脂上でバッチ精製した。予測されるサイズのタンパク質産物は、構築物MSP4p40/His(40kDa)およびMSP4p30/His(30kDa)で見られたが、構築物MSP4-EGF/Hisでは、タンパク質が見られなかった(さらに下記で論議した)。MSP4p21/Hisの発現は、いまだ解析の予備的段階である。
【0096】
MSP4p40およびMSP4p30の両方の産生をスケールアップして、箱からスピナフラスコ培養へ移した。タンパク質収量を最適化し、タンパク質分解(スピナのより激しい条件下で頻繁に見られる)を制限するために、時間経過実験を行った。MSP4p40/HisまたはMSP4p30/Hisが感染したスピナ培養(210mL)をスポドプトレア・フルギペルダ(Spodoptrea frugiperda)(SF9)昆虫細胞およびHigh Five昆虫細胞の両方のために準備した。試料(8 mL)を、24時間、30時間、36時間、42時間、48時間、54時間、60時間および72時間にて収集し、細胞死統計のために顕微鏡によって解析し、次いで、20 mM Tris HCl(pH8);500 mM NaClに対して透析した。タンパク質を、試験管プロトコルを使用してtalon樹脂上で精製して、100μLの100 mMイミダゾール;20 mM Tris HCl(pH8);500 mM NaClで溶出した。ある時間にわたった細胞死の割合をそれぞれの感染についてプロットし、それぞれの溶出の20μLの試料を構築物および細胞型の両方について並行して解析した(図8)。
【0097】
予想通りに、High five細胞は、SF9細胞よりはるかに高いタンパク質収率を生じた。しかし、タンパク質分解が始まる長い発現期間(48時間を超える)の後で、MSP4タンパク質は完全に分解せず、むしろより小さな安定な、約20 kDaの産物(MSP4p20)へと分解した。この産物は、MSP4p30感染の後期の段階においてもまた見られた。すべてのタンパク質は、C末端のヒスチジンタグを介して精製したので、この産物は、EGF様のドメインおよびプロテアーゼに耐性な堅い構造を形成するいくつかの上流配列を含むにちがいない。これは、EGF様のドメインの上流領域が、還元感受性のB細胞エピトープを有しているというWangら(2001)[13]の発見と一致する。
【0098】
実施例2
N末端シーケンシングおよび質量分析
時間経過実験から得られる情報を使用して、3つの異なる純粋なMSP4タンパク質標品を、異なる時点にて培養SNを収集することによって産出した。第1に、MSP4p40と表した全長MSP4、第2に、全長の修飾産物であるMSP4p30および第3に、分解産物であるMSP4p20。標準的なHigh Fiveスピナ培養におけるそれぞれの産物についての平均タンパク質収量は、それぞれ8 mg/L、6 mg/Lおよび8 mg/Lであり、収集の時間を反映しており、より早い時点での収集産物ほど、より少ないタンパク質を生じる。市販のバキュロウイルスコンピテンス(Henogen)においてなされた最近の進展を考慮すると、これらの収率は、最小限の見積もりであると考えられる。
【0099】
それぞれの産物のN末端のシーケンシングは、強力で明らかなシグナルを産出し(p40:41残基にて始まるMRILG、p30:41残基にて始まるMRILA、p20:132残基にて始まるKSPKE)、in vivoの熱帯熱マラリア原虫(P. falciparum)シグナル配列は、実際、PlasmoDB(http://plasmodb.org/plasmodb/servlet/sv?page= gene& source_id=PFB0310c)によって予測されるような長さの2倍(20に対しての40残基)であってもよい。MSP4p40およびMSP4p20の同一性は質量分析によってまた確認して、タンパク質は、実際にそれぞれ23.554 kDaおよび13.714 kDaの分子質量を有している。
【0100】
バキュロウイルスで発現したMSP4のN末端の配列は、寄生虫によって使用されるシグナル配列を同定する可能性が高い。第1に、MSP4-EGF/His構築物は、PlasmoDB予測されたシグナル配列に加えてさらに3つの下流の残基を含み、培養SNにおいて決して検出されなかった。第2に、Pfmsp4遺伝子内で同定される多形の部位のN末端のクラスターは、45番目のコドンにて始まる。Polsonら2005[33]で記録された解析によって示唆されるように、コドン45の下流の遺伝子の領域が、集団内の対立遺伝子型のバランスをとっることによって回避メカニズムを展開すると思われる効果的な免疫反応の標的である(52および74残基が関与する。図5を参照されたい)。したがって、シグナル配列切断部位(MRIL)に寄与する4つの保存された残基からなるPfMSP4のプロセスされたN末端は、このタンパク質のよりアクセスしやすい領域に優先的に方向付けられる免疫反応の有効性を減少させることができる多形部位のクラスターに直接続く可能性が高いようである。
【0101】
実施例3
シグナル配列の疎水性度
バキュロウイルス系によるMSP4構築物のために使用した切断部位は、天然の切断を再生することをさらに確認するために、バイオインフォマティクス研究を、以前に報告された天然の熱帯熱マラリア原虫(P. falciparum)シグナル配列に注目して行った。2001年において、公知の熱帯熱マラリア原虫(P. falciparum)シグナル配列切断部位を比較する研究が公表された[37]。このようなデータを得る際に関与する技術的困難性のため、わずか10個のこのような配列が存在するにすぎない。この研究の目的のために、それぞれのタンパク質の、報告されたシグナル配列切断部位に集中する、80残基のタンパク質配列を選択して、局部的な疎水性度の値を算出するために提出した(パスツールサーバーを介して)。Nacer 2001において報告された3つのタンパク質切断部位の疎水性プロフィールおよびバキュロウイルス系におけるMSP4p40のものを図10に示してある。上部パネルにおいて、KAHRPおよびMSP1シグナル配列の両方は、古典的な疎水性プロフィールを表し、疎水性度の値が中性のラインを(ポジティブからネガティブに)横切る直後に切断が生じている。PfMSP1シグナルの切断は、バキュロウイルス系によって忠実に行われることが知られている(Bonnet et.al. 2005)。局部的な疎水性度の同様の顕著な変化が、予測された切断部位でのMSP4p40タンパク質配列のN末端内に存在していない場合、疎水性度における最大の局部的変化(ネガティブからポジティブへ)の後のさらに下流で切断が実際に生じる。このプロフィールは、S抗原で見られたものにマッチし、したがって、独特ではないが、MSP4p40シグナル配列が40残基の長さであるという知見に重きが置かれる。
【0102】
実施例4
昆虫細胞におけるPfMSP5の発現
MSP5/Hisウイルスのストックを以前に記述したように産生し、試験感染および時間経過研究を、MSP4p40/Hisについて記述したように行った。図11に示したように、3つの異なるPfMSP5に由来する産物は、SDS-PAGEゲル電気泳動下でおよそ45、35および10 kDaにて移動して検出された。興味深いことに、2つのより大きな産物は、より大きなものが産生されて、その後に分解してより小さなものが形成するのではなく、時間経過実験の初期の間に同時に存在するように見えた。PfMSP4で見られた状況に対するもう一つの著明な相違は、研究した2つの細胞株の逆のタンパク質産生能力であり、SF9は、桁違いに早い増殖を行い、High Five細胞はより脆弱であった。これは、本来、PfMSP5がPfMSP4とは非常に異なる特性を有することを示唆する証拠の最初の部分であった。第2は、N末端のシーケンシングデータに由来する。
【0103】
実施例5
N末端シーケンシングおよび質量分析
MSP5発現の2つの最も大きな産物のN末端のためのシグナルを得ることはできなかったが、これらの両方がN末端の修飾を有し、これがN末端のシーケンシングにおける必須なイベントであるペプチド主鎖の破壊を遮断することを示唆している。より小さな10 kDaの産物のシーケンシングは良好であり、この産物のN末端が189残基にて始まると定義した:YNKVE[配列番号:26]。この産物の同定は、質量分析によっても確認され、これは、7.746 kDaの実際の分子量を有する。加えて、MSP5ウイルスゲノムの配列も確認され、YNKVE [配列番号:27]の残基は、正確なORFにおいてのみ存在し、したがって、これまでに未決定の産物としてこれらは、PfMSP5にちがいない。
【0104】
2つの最も大きなMSP5産物についてのMALDI-MSによって得られた質量値は、それぞれ、24.679および20.551 kDaであり、MSP5タンパク質配列のみに由来する任意のポリペプチドにマッチせず、これらのタンパク質が転写後修飾を有するという考えを支持した。起こりうるN末端遮断修飾を調べるために、MSP5配列をExPASYホームページ経由でprositeに提出した。これにより、C14脂肪酸の同時翻訳付加を指揮することができる2つのN-ミリストイル化部位の存在が明らかになった(42残基目:GGFTSK、および66残基目:GSLPTK)[38]。しかし、これらの修飾は、N末端に限定的であり、N末端のシーケンシングを遮断することができる一方、これらは、適切な質量値(それぞれ、24,015および21,301)を生じない。しかし、グリシン残基のN末端修飾を導くモチーフは、真核細胞によって行われるミリストイル化の唯一の形式ではない。また、これらは、ミリストイル基で、任意のN末端のグリシンおよび任意のリジンを転写後修飾することができる(下記に概説する)。これらの可能性と共に考えると、推定上の位置は、現在未決定であるが、それぞれのMSP5産物(p45およびp35)が2つの脂肪酸修飾を有すると仮定することによって、24.673kDaおよび20.553kDaに近接した一致が得られる。
【0105】
実施例6
脂肪酸修飾およびプラスモディウム属
グリコシル化、パルミトイル化、N-ミリストイル化およびミリストイル化を含む、真核細胞によって使用される、いくつかの異なるタイプの特別な翻訳修飾がある。これらのうちで、第1のものは、ここで発現されるMSP5タンパク質には存在しない。この種の修飾が熱帯熱マラリア原虫(P. falciparum)によって行われないことが知られているので、すべてのグリコシル化部位を、合成遺伝子の構築の間にタンパク質配列から除去した。第2の修飾のタイプは、全体として真核生物においてあまりよく理解されておらず、パルミトイル化が酵素依存的および酵素非依存的な両方の経路を介して生じうると考えられる[39]。現在まで、任意のパルミトイル化された熱帯熱マラリア原虫(P. falciparum)タンパク質の報告はなかった。しかし、タンパク質のN-ミリストイル化は、プラスモディウム属において重要であると示された。Pf N-ミリストイルトランスフェラーゼ遺伝子相同体を同定し(PlasmoDB PF14_10127)、ヒトおよび酵母の遺伝子オルソログに高度に相同的であることが見いだされた[40, 41]。もしこの酵素がブルーストリパノソーマ(Trypanosoma brucei)および森林型熱帯リーシュマニア(Leishmania major)などのその他の寄生性原虫に対するのと同様に、プラスモディウム属に対して重要であるなら、複数のタンパク質のN-ミリストイル化が寄生虫成長のために必須であると予想することができる。実際、PfADF(リボソームの因子)のN末端のミリストイル化は、GTPの結合を媒介し、酵素活性を促進するために重要であることが示された[42]。この脂肪酸修飾(N末端のグリシンおよび内部のリジンの両方での)は、膜へのタンパク質の局在化およびタンパク質-タンパク相互作用の安定化を含む、より高度な真核生物における種々の役割を演ずることが示された[39, 43-45]。
【0106】
実施例7
バキュロ系において
昆虫細胞のバキュロウイルス感染は高次元の真核細胞系であるので、多くのタイプのより進化したタンパク質修飾を行うことができる。実際、この系は、いくつかのNミリストイル化されたタンパク質を産出するために特異的に使用されており、NAP-22が最も記述されている[46]。
【0107】
実施例8
抗体産生
ウサギポリクローナル抗血清は、組換えMSP4p40、MSP4p20およびMSP5(p45およびp35)にたいして、標準的な免疫化手順を使用して、EUROGENTECによって産出した。すべてのウサギは、免疫化(抗原あたり2羽のウサギ)前にMSP4またはMSP5について陰性であり、3または4回の免疫化の後のELISAによって決定されるように高力価の特異的な血清抗体を産生した。還元および非還元組換えタンパク質に対する最終時点の力価を図12に示してある。パネルBに示したように、ウサギNo.8からの抗体によるMSP4p20の認識は、抗原が還元される時に50%以上減少し、MSP4p20タンパク質のエピトープの多くが高次構造的であることを示している。還元は、主にジスルフィド結合構造に影響を及ぼすので、このデータは、MSP4p20のEGF様ドメインが無傷であり、少なくとも1つのB細胞エピトープを形成することに関与することを示す。
【0108】
実施例9
寄生虫由来MSP4
予備的な免疫ブロットデータは、前述したポリクローナル免疫血清を使用して作製されており、図13に示してあり。MSP4についてのデータは、40kDaのタンパク質が専ら後期寄生虫の調製物のTriton X100可溶性画分において検出されるというR. Coppelのグループにより公開された結果[9]と一致する(図13A)。MSP4、MSP4p20のC末端の領域の特性を調べるために、プールしたウサギ血清(R6-9)および2002年においてSenegal のDielmoで収集したヒト血清を、固定したMSP4p20に対してアフィニティー精製した(Amersham: NHS-activated Sepharose 4 fast flow)。非同調性寄生虫材料(SDS-PAGEによって分離し、ニトロセルロース膜へ転写した)をこれらの血清でプローブしたときに、以前に観察された40 kDaでのバンドに加えて、およそ18 kDaのバンドも見られ(図13B)、自然感染において、MSP4抗原がMSP1p42に類似のタンパク質分解のプロセシングを受け得るか、またはバキュロウイルス感染の後期において得られるような自己制御様式でのプロテアーゼによって分解され得ることを示唆している。熱帯熱マラリア原虫(P. falciparum)メロゾイトのMSP4p20エピトープの天然の存在をさらに確認するために、間接的な免疫蛍光アッセイ法(IFA)を、風乾した非同期成長の寄生虫培養で行った。メロゾイト特異的な染色は、ヒト抗MSP4p20で見られ、二次抗体単独またはマラリア未感染血清で何も染色されなかった(図13C)。
【0109】
実施例10
寄生虫由来MSP5
バキュロウイルスが発現したMSP5に対して生じたポリクローナル血清を使用して得られた免疫ブロットデータは、その他のグループによって公開されたもの[9]とは非常に異なるプロフィールを示し、彼らは、MSP5の検出に明らかな問題を抱えており、それが低レベルで発現されていることを示唆する。本発明の免疫ブロットデータは、抗MSP5特異的な血清で強力なシグナルを示す(図13D)。MSP4とは異なり、寄生虫由来MSP5は、また、粗雑に調製した寄生虫抽出物の膜画分にも存在しており、これはGPIアンカーを有するだけのタンパク質と変わらぬ様式で分画しないことを示唆する。実際に、還元した寄生虫材料に注目した場合、推定のNミリストイル化MSP5に対して調製したポリクローナル血清は、その他のNミリストイル化されたタンパク質とも交差反応し得ると結論づけることができ、さらに、これが熱帯熱マラリア原虫(P. falciparum)における重要かつ頻繁に使用される翻訳後修飾であるという考えを支持する。
【0110】
実施例11
ヒト免疫血清の解析
本発明のタンパク質産物がマラリアに感染した個体の抗体によって認識されることを確実にするために、Senegal(Dielmo)における風土病村で生活している9人のランダムに選択された準免疫成人の血清を、ELISAによって解析した(図14)。すべての個体は、すべてのタンパク質産物について陽性であった(標準偏差3倍に加えて対照血清より大きいOD値を有する)。相対力価は、個体間でおよび産物間で変化したが、全体の最終時点の力価は、MSP4について1x105およびMSP5について1x104であった。ウエスタンブロット解析(図15)は、MSP4について高度に陽性の2人の個体(個体3および9)並びにMSP5について高度に陽性の1人の個体(個体8)からの血清を使用して行った。以前に抗MSP1p19で見られたように、EGF含有メロゾイト表面タンパク質に対するヒト抗体は、凝集した材料に対して特に親和性を有していた(図15)。
【0111】
実施例12
Senegalにおける抗MSP4抗体;保護における役割
自然感染における抗MSP4抗体のための推定上の役割をさらに調べるために、疫学調査を行った。MSP4の3つの異なる精製された形態であるp40、p30およびp20は、他で考証された大規模な先見の明ある研究で評価されたており(参照[47]を参照されたい)、実施例16において下記に詳述してある。
【0112】
要約すると、組換えMSP4タンパク質のそれぞれの形態は、90%を超える血清有病率で強く認識されたが、有意に異なる平均のIgG OD比(p40 = 20±10、p30 = 12.3±7.3およびp20 17.3±10)を示した。すべてのマラリア抗原で見られるように、抗体力価は、年齢と共に増大し、保護との相関は、年齢で調整した進行モデルを使用して対処しなければならない。この形態の解析を使用して、MSP4p40抗体ではなく抗MSP4-p30および抗MSP4-p20は、臨床的なマラリア発症の減少した発病率に正に相関した(それぞれP = 0.036、P = 0.018+ 0.067)。これらの知見は、MSP4タンパク質のN末端領域内の潜在的にバランスをとっている多型の同定を強化し、MSP4に基づいたワクチン構築物においてタンパク質のこの領域を欠失させるという考えを特に支持する。
【0113】
実施例13
Senegalにおける抗MSP5抗体;保護における役割
自然感染における抗MSP5抗体の役割をさらに調べるために、疫学調査を行った。MSP5の2つの高分子量型を含む標品は、他に記述されたように解析した(参照[47]を参照されたい)。血清有病率は、MSP4に関するよりもMSP5に関してより低い(59%)ものの、臨床的なマラリア発症の減少数との高度に有意な統計的相関が、年齢を調整した進行モデルを使用して観察された(P = 0.0028)。以前に考察したように、この抗原は、GPI修飾について見られるように、脂質が誘導する免疫原性および抗感染免疫における免疫反応にとって潜在的な重要性を持つ脂肪酸修飾を有し得る[48];Bonnetら、2005(作成中の刊行物)。
【0114】
要約すると、PfMSP4は、C末端にEGF様ドメインを有し、GPIアンカーによってメロゾイト表面に結合する。その機能は、未知であるが、生存可能なPfMSP4「ノックアウト」変異体は観察されなかったため、寄生虫生存のために必須と思われる。3つの組換えPfMSP4構築物は、EGF様ドメインの適切な折りたたみを含む天然の抗原の複製を最適化するために、バキュロウイルス昆虫細胞発現系において、可溶性な分泌タンパク質として発現される。2つの構築物は、それぞれN末端の近くの多形領域からの30アミノ酸の欠失を伴い、または伴わないMSP4外抗原(分泌を可能にするC末端の疎水性残基なし)のMSP4p30およびMSP4p40に対応する。第3の20 kDaのタンパク質は、EGFドメインを含むMSP4のC末端半分にほぼ対応する(MSP4p20)。より直接的様式でのこの抗原の発現は、前述した構築物MSP4p21/Hisを使用して評価されている。3つの異なる抗原に反応する抗MSP4抗体は、マラリアに曝露されたヒトにおける自然獲得免疫に関して、伝播が季節的である中間的な風土病の村で生活している205個体の断面調査で比較した。
【0115】
伝播期以前には、3つの構築物を強く認識した:血清有病率は90%を超えないが、MSP4p40、MSP4p30およびMSP4p20についてそれぞれ20±10、12.3±7.3および17.3±10の有意に異なる平均IgG OD比であった(約5×10-3力価)。MSP4構築物に反応する抗体は、成人よりも有意に低いIgGレベルを有する15歳より下の個体と年齢相関した(P<0.001、Rho 0.25-0.33)。伝播期を含むその後の6月の期間の間の臨床的発病の解析は、年齢補正モデルにおいて、両者とも多形性の領域を欠いているMSP4p30およびMSP4p20の両方に反応するIgGの存在は、マラリアの臨床的発症の減少した発病率と有意に関連したが、多形領域を有するMSP4p40は、関連していなかった(P<0.05、比# 0.75)ことを示した。まとめると、これらの結果は:(i)バキュロウイルス組換え抗原を使用してモニターすると、PfMSP4に反応する強力な自然に獲得したIgGが存在する;(ii)抗MSP4 IgGは、臨床的なマラリアに対する保護と相関する、および;(iii)可変領域に反応するIgGは、別の保護に関連する抗MSP4 IgGを妨げるかもしれない。
【0116】
実施例14
PfMSP4モノクローナル抗体
PfMSP4に特異的な4種のモノクローナル抗体(mAb)は、3つの異なるエピトープを認識する。
【0117】
モノクローナル抗体L11-16は、p40(完全なPfMSP4の多形性外部ドメイン)によって誘導される2つのその他の代表である。還元非感受性のそのエピトープ(17B)は、PfMSP4のN末端の半分に位置する(non-p20;16A)。これは、特にPfMSP4の不均一な凝集体と反応し、その形成は、還元感受性であり(図16A)、非還元(nR)および部分的還元(R)SDS-PAGEにおける高分子量物質が、実際にPfMSP4であることを証明するために役立った(バキュロウイルスPfMSP1p19について以前に見られたとおり)。
【0118】
モノクローナル抗体F12-7は、PfMSP4 p30(多形性領域を欠く)によって誘導され、PfMSP4のp20のC末端半分における還元に反応しないエピトープを認識する(図16Bおよび17B)。L11-16と比べて、その明らかな減少した凝集体との反応性は、そのかなりより低い親和性のためであろう:
クローン アイソタイプ KD-MSP5 エピトープ
L11-19 IgG1 3.2 10-9 A
F12-7 IgG1 1.0 10-7 C
【0119】
実施例15
PfMSP5モノクローナル抗体
PfMSP5(p45+p35)に特異的な7種のモノクローナル抗体(mAb)は、5つの異なるエピトープを認識する。
【0120】
モノクローナル抗体G21-2は、p45およびp35のPfMSP5産物を認識し(図18A)、そのエピトープは、還元感受性でない(図18Aおよび19A)。別々のより高分子量の凝集体と反応し(図18A、nR)、その形成は、還元感受性である(図18A、R)。
【0121】
モノクローナル抗体J18-14は、p45のみを認識し、そのエピトープは、還元感受性である(図19B)。また、これは、凝集体と反応する。
【0122】
これらの抗原は、C末端のヘキサヒスチジンタグを介して精製したので、p45およびp35がN末端で異なることは考えられる。これらの特徴は、J18-14エピトープがC末端のEGFドメインおよびN末端の両方を含む高次構造に対応し得ることを示唆する。両方のモノクローナル抗体は、同様の親和性を有する:
クローン アイソタイプ KD-MSP5 エピトープ
G21-2 IgG1 2.8 10-8 B
J18-14 IgG1 4.2 10-8 D
これらの2つのモノクローナル抗体の反応性は、PfMSP5産物を定義するために使用することができる。
【0123】
実施例16
曝露された個体の免疫血清および集団
以下の結果は、(A)風土病の地域で生活している病気の症候が明らかでない個体から採った過免疫血清の対照プール、(B)2005年6月に収集したDielmo(完全な風土病の伝播)の村からの個体の血清および(C)伝播シーズンが始まる前の2000年7月および8月にNdiop(中間的な風土病の伝播)の村において収集した205血清の集団を使用して得られた。その集団は、3および75歳の年齢の間に、108人の男性および97人の女性を含んだ。臨床的発作を以降5月の間に活発に記録し、合計278の臨床的発症を治療した(Perraut et al, JID 2005 191 264-271)。
【0124】
解析プロトコル
(1)抗原コーティングの最適化、(2)試験される標準的な血清希釈の選択、(3)血清の集団の系統分析法、(4)中央OD比値の算出、(5)年齢による結果の層別化、(6)浸潤および増殖阻害についての選択した血清の機能解析、(7)選択した血清グループについてのアイソタイピング、(8)臨床発症データに対する中央OD比の関数として層別化される結果の統計分析。
【0125】
(1)較正:抗原をコーティングする2つの濃度の1および0.5μg/mlを使用して、「血清過剰免役」(SHI)およびDielmo(2005)の村からの選択された個体の希釈系列を試験する。
【0126】
結果:0.5μg/mlでのコーティングは、結果が村およびSHIからの両者の個体について、1μg/ml の抗原で見られたのと同じであったので、至適であることが分かった。選択した個体の反応は、NdiopよりもDielmoにおいて強力であり、MSP5は、MSP4より低い程度の抗体反応を示した。図20Aおよび20Bを参照されたい。
【0127】
(2)較正:SHIおよびDielmoからの陽性血清の滴定
結果:図21A、21B、21Cおよび21Dを参照すると、Dielmoからの血清(60605)を使用したときに、抗MSP4反応は、高度に陽性であった(>SHI)。しかし、ODは、2000年のNdiop集団で後に得られたODに比べて比較的低く、それにもかかわらず、SHIの力価は、同程度であった。一見すると、検出された抗体は、鋭角の曲線で示されていたので、以前の研究からのMSP1に対するものと比較して、相対的に低い親和性を有するように見えた。
【0128】
SHI力価に対して MSP5=1/1600
MSP4-40 MSP4-30 MSP4-20=1/3200。
【0129】
(3)Ndiop集団2000年の解析:一次結果
SHIは、それぞれのアッセイ法において系統的に滴定した。OD比を、それぞれのプレート上の一連のSHI対照に関して算出した。この集団については、1/200希釈でのOD比は、すべて高かったが、実際の力価は、平均的であった(4〜6の間のOD比値は、約1/2000の最終時点の力価を与える)。
【0130】
(下記の結果)
力価の結果/OD比
【表3】
【0131】
(4)反応者の発病率
以下の表には、2000年のNdiopからの集団の個体からの抗体反応の特徴を示してある。陽性反応者は、2を超えるOD比をもつとみなした。以下に示すように、MSP4は、ほとんどすべての個体(>90%)によって認められ、MSP5認識は、実質的により低かった(約60%)。
【表4】
【0132】
(5)年齢およびその他の抗MSP反応との相関
図22A、22B、22Cおよび22Dを参照すると、その他の抗MSP反応との有意な相関があった。
【0133】
- MSP1で、有意なRho # 0.27-0.36
-種々のMSP4抗原との間で、Rho >0.9
-MSP4およびMSP5との間で、Rho # 0.44-0.47
Ndiop集団において、個体の年齢との有意な相関があったが(P<0.001、Rho0.33〜0.22)、MSP1とのものよりもわずかに目立たなかった。抗MSP5抗体反応は、年齢と最も相関しなかったが、<15歳の個体は、有意に低いレベルの抗体を有した。
【0134】
MSP4に対する着目:MSP4抗原の相違
図23を参照すると、MSP4の異なる断片の認識のレベルには、高度に有意な相違がある(P<0.001)。逆説的に、最も小さなおよび最も大きな抗原(p20およびp40)は、抗原MSP4-30(またはMSP4md2)よりも優れて認識され、それ自体は、全長MSP4(MSP4-40)よりも小さい30残基だけである。この結果は、それが免疫回避ストラテジーの存在と一致するようであるので、興味深く、この場合、抗体反応は、可変ドメインに対して集中する。これにより、IgG反応と浸潤阻害との間の相関が無いことを説明することができるであろう(下記を参照されたい)。
【0135】
(6)機能試験との関係
図24Aおよび24Bを参照すると、MSP4p40、MSP4p20、MSP4_mod(p30)(<20>)またはMSP5(<2.4>)についての中央OD比値の間の二比較に基づいて、成長または浸潤阻害における有意差は無い。(この特性は、この集団における抗MSP1p19抗体について統計学的に有意であるだけのように見えた)。しかし、メロゾイト食作用に関しては、有意差がある。メロゾイト食作用とのこの機能的相関は、この現象が特異的なAb力価(したがって、OD比率)の全体の増大と関連し、これは、風土病の地域における年齢の関数として増大することが公知であることから、予想された。
【0136】
下記の図を参照されたい:
(7)アイソタイピング
解析は、MSP4-40およびMSP5についての限られた数の血清に限定された。結果は、図25Aおよび25Bに示してある。MSP4-p40については、アイソタイププロフィールは、IgG1+、IgG3+であり、ごくわずかな量のIgG4であった:
-力価と最近の循環寄生虫血症との間に相関はない。
【0137】
-IgG3レベルは、15歳>個体についてより高く(P=0.006)、最近マラリア発症にかかった人々と同じであり(P=0.03)、これは、発症が風土病の地域において年齢と関連した事象であるので、予想できる結果である。
【0138】
-IgG1およびIgG3は、IgG反応の有意な成分であり、それぞれIgGの40%および30%を表した。
【0139】
-強力な反応者については、IgG反応と浸潤阻害との間に有意な関係が存在する(IgGサブクラスに非依存的)。
【0140】
MSP-5については、アイソタイププロフィールは、主にIgG1である。
【0141】
-最近の循環寄生虫血症(有意性P=0.06近く)または年齢との関係において相違がない。
【0142】
-IgG1は、IgG反応の有意な主要構成要素であり、反応の75%の合計となった。
【0143】
-IgG反応と浸潤または増殖阻害との間に相関はなかった(特定のアイソタイプは関連しなかった)。
【0144】
(8)マラリア発症との相関
年齢補正したポアソン回帰モデルを使用して、追跡調査期間の間にAg構築物およびマラリア発症の発病率に対して抗体反応性との間の関係を解析した。これらの抗原は、2000年のNdiopの村からの205個体の先見の明ある集団研究において、臨床的発症に対する保護と有意な相関を示した。
【0145】
・MSP4に関して
MSP4-p40およびMSP4-p30に対するIgG力価の間に有意差が見られたという前述の観察と一致して、全長抗原(MSP4-p40)に対する抗体反応は、保護と相関しない。しかし、抗原の30残基の可変ドメインの無い構築物(MSP4-p30)および最も小さな抗原(MSP4p20)構築物に対して向けられた抗体の反応性は、保護と有意に関連する(P<0.05)。これらの知見は、タンパク質のN末端での残基多型が免疫回避に関与すること、およびタンパク質のこの領域に向けられた抗体は、(A)限られた親和性を有し(B)株の特異性を示し、したがって大多数の株に限定された効果を有し、または/および(C)APLの存在のために、親和性成熟を達成できないか、もしくは必要とされるT細胞補助を欠くという考えを補強する。関与するメカニズムに関係なく、このデータは、保存されたタンパク質サブユニットが、免疫回避において機能すると考えられる多型の領域を含むことが多い完全な実体よりも広範な特異性免疫を誘導するより良い機会を有するという考えを支持する。
【表5】
【表6】
【表7】
【0146】
高力価と疾患発症の減少との間に高度に有意な相関を見いだした。抗原によって保持されると予測される脂肪酸修飾からみて、これらの結果は、重症度および抗脂質反応に関して興味深い意味を有する。
【表8】
【0147】
上に示されたデータから描区ことができる主要な結論は、(1)バキュロウイルスで発現したMSP4 およびMSP5は、MSP1p19について見られたものと同様の活性を有するワクチンとして有用であり、(2)これらの抗原は、個体を免疫することによって強く認識され、(3)MSP4の保存されたドメイン(p30およびより相応しいp20)のIgG認識の中央値および高度に保存されたMSP5抗原のものよりも高いことは、臨床的なマラリアに対する天然の保護と相関する。
【0148】
実施例17
MSP4p20発現構築物
MSP4p20の発現のために、熱帯熱マラリア原虫(Plasmodium falciparum)MSP4合成の遺伝子配列から2つの構築物をデザインした。1つの構築物は、PfMSP4p21ss1と他に名付けられたMSP4p21ss1として同定される[配列番号:28]。構築物MPS4p21ss1によってコードされるオープンリーディングフレーム[配列番号:19]および予想アミノ酸配列[配列番号:20]を以下に記述してある:
構築butMPS4p21ss1のヌクレオチド配列
>MSP4p21 ss1 ATGTGGATCGTAAAGTTCTTGATTGTGGTCCACTTCTTCATCATATGCACCATCAACTTCGACAAGCTCTACATTAGTTACTCTTACAACATCGTCCCTGAAAACGGACGTATGCTTAACATGAGGATCTTGGGTGAAGAAAAGCCTAACGTTGACGGTGTGTCAACATCTctaGAAAAGAGTCCCAAGGAGAGTCAAATGGTCGACGACAAGAAGAAGACCGAGGCCATTCCAAAGAAAGTCGTGCAGCCAAGCTCGAGCAACTCTGGAGGTCACGTCGGTGAAGAAGAAGACCACAACGAAGGAGAGGGAGAGCACGAAGAGGAGGAAGAACACGAAGAAGACGATGACGACGAGGACGACGACACATACAACAAAGACGACTTGGAGGACGAAGATCTTTGCAAGCACAACAACGGAGGATGTGGAGATGACAAGCTCTGCGAGTACGTTGGAAACCGTCGCGTAAAATGTAAATGTAAGGAAGGATACAAGTTGGAAGGAATTGAGTGCGTTGAACACCACCACCACCATCACTAA[配列番号:28]
構築物MSP4p21 ss1によってコードされるオープンリーディングフレーム
>MSP4p21 ss1 MWIVKFLIVVHFFIICTINFDKLYISYSYNIVPENGRMLNMRILGEEKPNVDGVSTSLEKSPKESQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEEDHNEGEGEHEEEEEHEEDDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLCEYVGNRRVKCKCKEGYKLEGIECVEHHHHHH[配列番号:19]
明るい灰色(GEEKPNVDGVSTS): MSP4p40の配列における残基MRIL直後の付加されたアミノ酸
濃い灰色(L):前記明るい灰色で表示されたアミノ酸配列直後の、MSP4p40の配列におけるMSP4p20配列の開始に直接先行する付加されたアミノ酸L
予想されるタンパク質産物MSP4p21 ss1
MRILGEEKPNVDGVSTSLEKSPKESQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEEDHNEGEGEHEEEEEHEEDDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLCEYVGNRRVKCKCKEGYKLEGIECVEHHHHHH[配列番号:20]
その他の構築物は、PfMSP4p21ss2と他に名付けられていたMSP4p21ss2として同定される[配列番号:29]。構築物MPS4p21ss2によってコードされるオープンリーディングフレーム[配列番号:21]および予想アミノ酸配列[配列番号:22]を本明細書に記述してある。
【0149】
構築物MSP4p21 ss2のヌクレオチド配列
>MSP4p21 ss2 ATGTGGATCGTAAAGTTCTTGATTGTGGTCCACTTCTTCATCATATGCACCATCAACTTCGACAAGCTCTACATTAGTTACTCTTACAACATCGTCCCTGAAAACGGACGTATGCTTAACATGAGGATCTTGGGTGAAGAAAAGCCTCTAGAAAAGAGTCCCAAGGAGAGTCAAATGGTCGACGACAAGAAGAAGACCGAGGCCATTCCAAAGAAAGTCGTGCAGCCAAGCTCGAGCAACTCTGGAGGTCACGTCGGTGAAGAAGAAGACCACAACGAAGGAGAGGGAGAGCACGAAGAGGAGGAAGAACACGAAGAAGACGATGACGACGAGGACGACGACACATACAACAAAGACGACTTGGAGGACGAAGATCTTTGCAAGCACAACAACGGAGGATGTGGAGATGACAAGCTCTGCGAGTACGTTGGAAACCGTCGCGTAAAATGTAAATGTAAGGAAGGATACAAGTTGGAAGGAATTGAGTGCGTTGAACACCACCACCACCATCACTAA[配列番号:29]
構築物MSP4p21 ss2によってコードされるORF
>MSP4p21 ss2 MWIVKFLIVVHFFIICTINFDKLYISYSYNIVPENGRMLNMRILGEEKPLEKSPKESQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEEDHNEGEGEHEEEEEHEEDDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLCEYVGNRRVKCKCKEGYKLEGIECVEHHHHHH[配列番号:21]
明るい灰色(GEEKP): MSP4p40の配列の切断部位MRILの直後に付加されたアミノ酸
濃い灰色(L):前記明るい灰色で表示されたアミノ酸配列直後の、MSP4p40の配列におけるMSP4p20配列の開始に直接先行する付加されたアミノ酸L
予想されるタンパク質産物MSP4p21 ss2
MRILGEEKPLEKSPKESQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEEDHNEGEGEHEEEEEHEEDDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLCEYVGNRRVKCKCKEGYKLEGIECVEHHHHHH[配列番号:22]
【0150】
実施例18
MSP4特色の進化的保存
熱帯熱マラリア原虫(P. falciparum)および三日熱マラリア原虫(P. vivax)由来のMSP4のアミノ酸配列を、一般の整列プログラム(ClustalX)を使用して整列させられると、いくつかの著明な観察がなされ得る。図26を参照されたい。第1に、熱帯熱マラリア原虫(P. falciparum)のシグナル配列切断部位の下流の残基は(本明細書において定義したとおり)、その種の間でほとんど100%保存される(Pf MRILGEおよびPv GRILGE)。この知見は、成熟したPfMSP4タンパク質が41残基にて始まり、直前の残基は、保存された切断部位の一部分であるという考えをもう一度強く支持する。第2に、SALSAとして公知の配列エレメント(下線を引いてある)は、三日熱マラリア原虫(P. vivax)タンパク質に存在しない。これは、肝細胞結合(インビボではPfMSP4によって媒介される)は、このタンパク質の保存されておらず、したがって重要な機能ではないことを示唆するであろう。その代わりに、三日熱マラリア原虫(P. vivax)タンパク質のこの領域は、熱帯熱マラリア原虫(P. falciparum)のものとは完全に異なり、非常に低い複雑さの配列を含む。明らかに熱帯熱マラリア原虫(P. falciparum)の配列と同等ではないが、免疫系における効果が同様であると想像することができる。熱帯熱マラリア原虫(P. falciparum)には、タンパク質の最N末端に、N末端の高頻度可変領域がある。多型頻度の統計分析は、免疫回避に関与し得るこの領域内の2つの部位を示す。これは、変異体配列に曝露されたときに、株特異的反応および/または免疫寛容を引き起こすBおよびT細胞エピトープの修飾または切除を介して達成されるであろう。また、このタイプの免疫調整は、反復配列の特性である。(A)これらは、それ自体変化したペプチドリガンドを含み、(B)これらは、T細胞非依存的なB細胞活性化を誘導することができ、これが古典的に短命で低親和性の反応の産生を引き起こす。整列した配列からなされ得る第3の観察は、MSP4p20の一次配列エレメントが両方の種に存在していることである。驚くことではないが、コアEFG-ドメインおよびGPI-付着シグナルの両方が保存されており、より面白いことに、酸性リピートは、同等の様式で配置されるようにである。これにより、自然感染において、MSP4は、メロゾイトの表面上の膜結合の小構造実体を残す様に分解または切断されるという考えに重きが置かれる。この実体は、MSP1p19で見られるように、新たに侵入したRBCに入るかどうかはいまだ確認されないが、本モデルは適合するようである。この比較に基づいて、以下のタンパク質配列は、抗三日熱マラリア原虫(P. vivax)マラリアワクチンの候補として提唱することができよう。
【0151】
実施例19
多形核好中球によるメロゾイトの抗体依存的食作用におけるMSP4p20に対する抗体の役割
好中球は、病原体を内部に取り込み、活性酸素種(ROS)および顆粒加水分解性タンパク質を使用してこれらを破壊する。活性化された好中球は、NADPHオキシダーゼを介して作動する呼吸バーストとして公知のプロセスにより、O2・-、H2O2...のようなROSを産出する際に非常に効果的である。ROSは、化学発光色素によって検出され、抗体が誘導する食細胞活性の測定として使用される。化学発光色素、はルミノールであり、ROSの存在下において、これが光を放射し、コンピュータに連結したルミノメーター(Microlumat+)で定量化する。
【0152】
多形核好中球(PMN)によるメロゾイトの抗体依存的食作用(ADPm)に基づいた新たな機能的アッセイ法を使用して、自然に獲得され、およびワクチン接種誘導された抗体によって誘導される熱帯熱マラリア原虫(Plasmodium falciparum)メロゾイト食作用を測定した。マラリア風土病地域における個体由来の免疫血清および凍結したメロゾイト標品を使用して、この試験では、ごくわずかな非免疫性バックグラウンドシグナルおよび優れたアッセイ内再現性を有し、高度に特異的であることが示された。アッセイ間の比較は、標準的な正の対照を使用することによって行う。
【0153】
季節的な伝播の地域に生活している個体からのデータは、ADPm活性が、年齢、並びにELISAによって測定されるメロゾイトおよびMSP組換え抗原に対するIgG抗体の反応性と有意に相関したことを示した。SenegalにおけるDielmoおよびNdiop村の居住者の風土病の免疫血清から、バキュロウイルス組換えMSP1p19およびMSP4p20に特異的な抗体を取り除くことは、これらの抗原に対する抗体がADPm活性の実質的な成分であることを示した。
【0154】
より詳細には、熱帯熱マラリア原虫(P. falciparum)メロゾイトを培養液上清の遠心分離によって収集し、凍結した一定量として貯蔵した。新鮮な献血から得られた多形核好中球を単純なFicoll-Hystopaque 1077勾配で単離して、すぐにADPmアッセイ法に使用した。風土病免疫血清は、Ndiop(中間的風土病)およびDielmo(完全風土病)の住民から得た。C末端のヘキサヒスチジンタグを有する組換えMSP1p19およびMSP4p20抗原をバキュロウイルス発現系において産生して、固定化金属-アフィニティークロマトグラフィによって精製した。対応する組換え抗原を荷電したTALON金属親和性樹脂を使用して、血清の抗MSP1p19または/および抗MSP4p20抗体を除去した。反応を開始するためのルミノールおよびPMN(ウェルあたり5.106細胞)の添加の前に、メロゾイトを96ウェルプレートにおいて血清と共にインキュベートした。発光の出力は、Berthold MicroLumat Plus 96ウェルを使用して1時間測定した。標準的なヒト免疫血清の陽性対照(HIS)を、反応を定量化するために使用した:
メロゾイトの抗原特異的のIgGオプソニン作用(結合)(補体の有無にかかわらず)は、非働化血清(56℃にて30分間の処理)またはプロテインGで精製された総IgGが初期の血清に対する同様の反応を誘導し、総IgGを除去した血清がもはや反応を産生しない(図34)ため、このアッセイ法における陽性の読み出しのために必要であった。しかし、特異的IgGによって産出される化学発光効果の大きさは変動する。図35は、バキュロウイルスMSP4p20およびMSP1p19に特異的な抗体は、このアッセイ法において両方とも、同じ程度ではないが、機能的であることを示す。MSP1p19抗体の枯渇による34%の減少と比較して、MSP4p20抗体の枯渇の後では、化学発光シグナルは、67%減少する。MSP4p20およびMSP1p19の両方に特異的な抗体の枯渇は、化学発光シグナルを78%にまで減少させた。
【0155】
これらの結果は、バキュロウイルスMSP4p20およびMSP1p19の組換えタンパク質を認識する自然抗体は、PMN免疫エフェクターによるメロゾイト食作用および破壊を媒介する重要な成分であることを示し、ワクチンとしてのこれらの抗原の使用を強く支持する。
【0156】
実施例20
PfMSP5修飾
バキュロウイルスで発現したPfMSP5に存在するように見えるN末端遮断の修飾の性質に注目するために、放射性標識実験を行った。質量分析は、その実体が共有結合で相互作用したミリストイル基であることを示唆したので、ミリストイル基の組み込みをず最初に調べた。実施例1において前述した様に、昆虫細胞感染を、SF9細胞を使用してT25細胞培養フラスコにおいて行った。感染後24時間にて、BSAを含まない脂肪酸との複合体の200μCiの[9,10(n)-3H]ミリスチン酸(Amersham)を培養液上清に添加した。感染後70時間にて、培養上清を収集し、20 mM Tris pH8;500 mM NaClに対して透析し、Talon(登録商標)樹脂上でバッチ精製した。ブルー染色したSDS-PAGEゲルにおいて、タンパク質発現は、非ラベル対照および放射標識の存在下において行った培養と同一であることが分かった。オートラジオグラフィーによって解析したときに、両方のPfMSP5バキュロウイルス発現タンパク質(35および45kDa)は、トリチウム化されたミリストイルを組み込んでいた(図27)。バックグランド活性は、非標識タンパク質では見られず、2つのPfMSP5タンパク質バンドは、ウイルス感染無しでは見られなかった。
【0157】
これらの結果は、バキュロウイルスに発現される組換えPfMSP5タンパク質は、ミリストイル化されることを示す。これらのデータは、昆虫細胞におけるPfMSP5の発現に関連した、実施例4〜7における以前のデータを確認する。
【0158】
実施例21
PfMSP4の運命
PfMSP4の自然の運命を追跡するために、いくつかの異なる精製された血清を産生した。最初に、4羽のMSP4ワクチン接種したウサギから収集した抗血清のプールを、バキュロウイルスで発現したPfMSP4p20を固定したNHS活性化セファロースに対してアフィニティー精製した。この仮定をNdiopまたはDielmoからのPfMSP4について陽性であることが知られている5つのヒト免疫血清の2つの異なるプールを使用して繰り返した。これらの抗体試薬を使用して、PfMSP4の運命を、3D7株を適応した培養において、IFAおよびウエスタンブロットによって追跡し、その知見を、PfMSP1p19に特異的モノクローナル抗体(G17.12)で見られたものと比較した。図28Aに示したように、PfMSP4は、後期寄生虫および自由なメロゾイトの表面にて検出されたが、PfMSP1p19を同定することができたアセトン固定リング期の寄生虫では検出されなかった。加えて、ウエスタンブロットによる寄生虫抽出物の同等量の平行分析に注目すると(図28B)、PfMSP4レベルは、寄生虫培養上清をから収集した自由なメロゾイトにおいて、成熟シゾントにおけるよりもはるかにより少ないよう見え、該タンパク質は、リング期の寄生虫抽出物において検出されない。このデータは、MSP1p19についてみられたように、PfMSP4がタンパク質分解的にプロセスされたというよりむしろ、Apical Membrane Antigen-1と同様の様式でメロゾイト表面から分断され、新たに侵入されたRBCに運ばれるという考えに一致する。
【0159】
面白いことに、両方のアフィニティー精製した血清(ウサギおよびヒト)は、成熟したシゾント抽出物に限定的な複数の高分子量タンパク質バンドを認識する。これは、以前にMSP3およびMSP6で見いだされたような交差反応性エピトープを含む成熟したシゾントに存在する抗原であり得るか、または寄生虫抽出物が還元されていないために、これが天然のPfMSP4タンパク質三量体であり得る。
【0160】
肝臓段階前の寄生虫におけるPfMSP4発現を調べるために、抗PfMSP4p20のアフィニティー精製したウサギ血清を使用して、熱帯熱マラリア原虫(P. falciparum)スポロゾイトに対してIFAを行った。陽性対照の抗CSPモノクローナル抗体は、強力で明らかな表面染色を与えたが、抗PfMSP4p20抗体では、染色は達成されなかった。
【0161】
これらの結果は、天然MSP4タンパク質がメロゾイトの表面上に局在化しており、寄生虫による血液細胞の再侵略において、もはや存在していないことを示す(リング期に感染した血液細胞のIFAを参照されたい(図28A)。図28Bのウエスタンブロットは、天然のMSP4に対して向けられた抗体(ヒト血清)および組換えMSP4に対して向けられた抗体(ウサギ血清)で一致したパターンを示す。そして、バキュロウイルス組換え抗原に対して向けられた抗体は、天然の形態の抗原を認識することができることがこれから推論することができる。
【0162】
実施例22
p40と独立したp20サブドメインの発現
p20サブドメインの直接の発現を促進するために、一連の新たな発現構築物を構築し(図29および実施例17)、試験した。図30に示したように、最小のシグナル配列をコードする構築物(PfMSP4p21)は、タンパク質分泌に至らなかったが、高レベルの細胞内タンパク質が検出された(データ示さず)。より優れたレベルの発現は、図30に示すように構築物PfMSP4p21ss1およびss2で達成され、N末端のシーケンシング(図29参照されたい)によって明らかにされるように、両タンパク質は、迅速に残りのN末端の残基を失ってバキュロウイルスPfMSP4p40で見られるのと同じ安定なp20実体を産生する。しかし、感染後66時間でのタンパク質発現を、バキュロウイルスPfMSP4p40およびPfMSP4p21ss1を使用して行った並行感染と比較すると、オリジナルの構築物は、はるかに優れたタンパク質収率(2倍)を引き起こす。これらの結果は、上記の実施例17を補完する。
【0163】
実施例23
バキュロウイルス発現系におけるPvMSP4の発現
PvMSP4オープンリーディングフレーム(ORF)を得るために、ネスティッドPCRを、寄生虫株BelemからのゲノムDNAに対して行った。三日熱マラリア原虫(P. vivax)ゲノムDNAは、約50%がGCリッチであるので、合成遺伝子を作る必要はなかった。全てのORFは、pMosBLUEにクローニングして、シーケンスした。次に、両方のエキソンをPCRによって増幅し、エキソン2の3'末端のGPI付着シグナルをヘキサhisタグによって置換した。加えて、適切な制限部位をエキソン1および2の3'または5'末端にそれぞれ導入し、これらをpMosBLUEベクターに連続してクローニングし、his-タグにおける連続したORF終末を形成した(図31)。
【0164】
バキュロイルスをコードするPvMSP4は、標準的なプロトコルを使用して産生し、ある時間にわたったタンパク質発現を、前述したようにHigh Five昆虫細胞を使用して評価した。図32に示したように、タンパク質発現は、感染後約30時間で検出可能であった。
【0165】
このタンパク質バンドについて行ったN末端の配列解析により、GIAACのN末端が明らかになった。タンパク質発現は、感染後54時間ごろにピークに達し、この時点にて、いくつかのタンパク質分解が明白であった。しかし、PvMSP4は、PfMSP4と同様の様式では分解しない。最終産物のN末端のシーケンシングでは、該タンパク質が13または30〜32アミノ酸にまで分解されて、EGGEQのN末端の配列、並びにGDSSG、DSSGGおよびSSGGLを示すの混合シグナルを示した。検出される最も小さなタンパク質産物は、サイズで約8 kDaであり、N末端の配列LDNNGを保持している。
【0166】
実施例24
ヒト免疫血清のPvMSP4の認識
時間経過解析の最後に(図32で示す66時間)、130 mLの培養液上清をTalon樹脂上でのHPLCによって精製した。精製した産物は、図32において見られる30、25 kDaおよび8 kDaのバンドからなる。
【0167】
ヒト血清認識に注目するために、ELISAプレートをバキュロイルス発現したPvMSP4、0.5 g/mLでコートした。
【0168】
合計24の血清試料を、スリランカにおいて三日熱マラリア原虫(P. vivax)マラリアの少なくとも1回の発症を経験した個体から収集して、平行して試験した。これらの試料のうちの17は、PvMSP4に対して陽性の血清で、1/25,000〜1/200,000の間のエンドポイント力価であった(図33A)。これらの17の陽性血清を、DTTおよびヨードアセトアミドでの処理によって不可逆的に還元したバキュロウイルスPvMSP4をコートしたELISAプレート上で、1/2700希釈にて試験した。17の陽性血清のうち11は、1/2700の固定された希釈にて、またはそれぞれの曲線(図33B)の中点値に当てはまる希釈を使用して算出した場合に、12〜80%からの範囲で、ある程度の感受性の減少を示した。
【0169】
三日熱マラリア原虫(Plasmodium vivax)MSP4(PvMSP4)に関する実施例23および24は、実施例18を補完する。図33Bに示したように、血清番号2、3、4、6、7、8、9、10、11、14および17は、、DTTで還元した抗原に対して非還元抗原に対してよりも少ない親和性を有する一方、血清番号12および15は、還元した抗原に対して、非還元のものに対してよりも優れた親和性を有する。抗原の高次構造は、抗体-抗原結合に影響を及ぼすと結論することができるが、プラスモディウム属感染に対して試験した血清のそれぞれの保護能力が確立されていないので、還元または非還元の抗原に対する抗体親和性は、2つのタイプの抗体の保護値に相関させることができない。
【0170】
以下のPfMSP5/His、PfMSP4p30/HisおよびPfMSP4p40/Hisバキュロウイルスは、2005年11月10日に、Institut PasteurのCollection Nationale de Cultures de Microorganismes(C.N.C.M.)、28, rue du Docteur Roux, 75724 Paris, Cedex 15, FRANCEに寄託し、以下の受託番号(Accession number)が割り当てられた。
【0171】
材料 受託番号
PfMSP5/His I-3512
PfMSP4p30/His I-3513
PfMSP4p40/His I-3514。
【0172】
以下のF12-7、G21-2、J18-14、およびL11-6ハイブリドーマは、2005年11月16日に、Institut PasteurのCollection Nationale de Cultures de Microorganismes(C.N.C.M.)、28, rue du Docteur Roux, 75724 Paris, Cedex 15, FRANCEに寄託し、以下の受託番号が割り当てられた。
【0173】
材料 受託番号
F12-7 I-3517
G21-2 I-3518
J18-147 I-3519
L11-6 I-3520。
【0174】
以下のPfMSP4p21ss1/His、PfMSP4p21ss2/HisおよびPvMSP4/Hisは、2006年11月21日に、Institut PasteurのCollection Nationale de Cultures de Microorganismes(C.N.C.M.)、28, rue du Docteur Roux, 75724 Paris, Cedex 15, FRANCEに寄託し、以下の受託番号が割り当てられた。
【0175】
材料 受託番号
PfMSP4p21ss1/His I-3695
PfMSP4p21ss2/His I-3696
PvMSP4/His I-3694。
【0176】
以下は、上記に記述したペプチドおよびポリヌクレオチドのアミノ酸および核酸配列である:
MSP5p10(MSP5の分解生成物、186-253残基)YNKVEKNVTDEMLLYNMMSDQNRKSCAINNGGCSDDQICININNIGVKCICKDGYLLGTKCIHHHHHH[配列番号:16] 。
【0177】
MSP5p10をコードする核酸配列
>readseq-48009_tmp_1 207 bp TACAACAAGGTCGAGAAGAACGTAACCGACGAGATGCTCTTGTACAACATGATGTCCGACCAAAACCGCAAAAGCTGTGCTATCAACAACGGTGGCTGCAGTGACGACCAGATCTGCATCAACATCAACAACATCGGTGTGAAGTGCATTTGTAAGGATGGATACCTACTTGGTACCAAGTGCATTCACCACCACCACCACCACTGA[配列番号:5]。
【0178】
配列番号:1 =配列番号:9のMSP4p20の分解生成物をコードする核酸(132-251残基)
>readseq-41112_tmp_1 363 bp AAGAGTCCCAAGGAGAGTCAAATGGTCGACGACAAGAAGAAGACCGAGGCCATTCCAAAGAAAGTCGTGCAGCCAAGCTCGAGCAACTCTGGAGGTCACGTCGGTGAAGAAGAAGACCACAACGAAGGAGAGGGAGAGCACGAAGAGGAGGAAGAACACGAAGAAGACGATGACGACGAGGACGACGACACATACAACAAAGACGACTTGGAGGACGAAGATCTTTGCAAGCACAACAACGGAGGATGTGGAGATGACAAGCTCTGCGAGTACGTTGGAAACCGTCGCGTAAAATGTAAATGTAAGGAAGGATACAAGTTGGAAGGAATTGAGTGCGTTGAACACCACCACCACCATCACTAA[配列番号:1]。
【0179】
MSP5
>readseq.input(1)762 bases, 6761DE71 checksum.
ATGAACATTCTCTGTATTCTCAGCTACATTTACTTCTTCGTCATCTTCTA CAGTTTAAACCTCAACAACAAAAACGAGAACTTCTTGGTGGTCCGCAGAC TCATGAACGACGAAAAGGGAGAAGGTGGCTTCACTAGTAAGAACAAGGAA AACGGAAACAACAACAGGAACAACGAGAACGAACTCAAAGAAGAAGGATC TTTGCCCACTAAGATGAACGAGAAAAACAGTAACTCCGCGGATAAGCAAC CAAACGACATCTCCCACGACGAAAGCAAGAGCAACAGTAACAACGCCCAA AACATCCAAAAGGAACCTGAAGAGAAGGAAAACTCAAACCCCAACCTCGA CTCGAGTGAAAACTCCGCTGAAAGTGCTACTAGAAGCGTCGACATCAGTG AACACAACTCAAACAACCCCGAAACTAAAGAAGAAAACGGAGAAGAACCT CTAGACCTGGAAATTAACGAAAACGCAGAAATCGGCCAGGAACCTCCAAA CCGTCTTCACTTCGACAACGTTGACGACGAAGTACCACATTACTCAGCCC TGAGGTACAACAAGGTCGAGAAGAACGTAACCGACGAGATGCTCTTGTAC AACATGATGTCCGACCAAAACCGCAAAAGCTGTGCTATCAACAACGGTGG CTGCAGTGACGACCAGATCTGCATCAACATCAACAACATCGGTGTGAAGT GCATTTGTAAGGATGGATACCTACTTGGTACCAAGTGCATTCACCACCAC CACCACCACTGA[配列番号:4]。
【0180】
ORFによってコードされるタンパク質
>readseq.input(1)253 bases, 7870B947 checksum.
MNILCILSYIYFFVIFYSLNLNNKNENFLVVRRLMNDEKGEGGFTSKNKE NGNNNRNNENELKEEGSLPTKMNEKNSNSADKQPNDISHDESKSNSNNAQ NIQKEPEEKENSNPNLDSSENSAESATRSVDISEHNSNNPETKEENGEEP LDLEINENAEIGQEPPNRLHFDNVDDEVPHYSALRYNKVEKNVTDEMLLY NMMSDQNRKSCAINNGGCSDDQICININNIGVKCICKDGYLLGTKCIHHH HHH[配列番号:14]。
【0181】
MSP4p40
>readseq.input(1)756 bases, 733F5E48 checksum.
ATGTGGATCGTAAAGTTCTTGATTGTGGTCCACTTCTTCATCATATGCAC CATCAACTTCGACAAGCTCTACATTAGTTACTCTTACAACATCGTCCCTG AAAACGGACGTATGCTTAACATGAGGATCTTGGGTGAAGAAAAGCCTAAC GTTGACGGTGTGTCAACATCTAACACACCTGGCGGAAACGAGGCATCTAG TGCTTCTCCTAACCTTGCTGACGCTGCAGAAAAGAAGGACGAAAAGGAAG CAAGCGAGCAAGGCGAAGAATCCCACAAGAAGGAAAACTCTCAGGAATCT GCAAACGGAAAAGACGACGTTAAGGAGGAGAAGAAGACCAACGAGAAGAA GGACGACGGAAAGACTGACAAGGTACAAGAAAAGGTTCTAGAAAAGAGTC CCAAGGAGAGTCAAATGGTCGACGACAAGAAGAAGACCGAGGCCATTCCA AAGAAAGTCGTGCAGCCAAGCTCGAGCAACTCTGGAGGTCACGTCGGTGA AGAAGAAGACCACAACGAAGGAGAGGGAGAGCACGAAGAGGAGGAAGAAC ACGAAGAAGACGATGACGACGAGGACGACGACACATACAACAAAGACGAC TTGGAGGACGAAGATCTTTGCAAGCACAACAACGGAGGATGTGGAGATGA CAAGCTCTGCGAGTACGTTGGAAACCGTCGCGTAAAATGTAAATGTAAGG AAGGATACAAGTTGGAAGGAATTGAGTGCGTTGAACACCACCACCACCAT CACTAA[配列番号:3]。
【0182】
ORFによってコードされるタンパク質
>readseq.input(1)251 bases, 1EFAEF35 checksum.
MWIVKFLIVVHFFIICTINFDKLYISYSYNIVPENGRMLNMRILGEEKPN VDGVSTSNTPGGNEASSASPNLADAAEKKDEKEASEQGEESHKKENSQES ANGKDDVKEEKKTNEKKDDGKTDKVQEKVLEKSPKESQMVDDKKKTEAIP KKVVQPSSSNSGGHVGEEEDHNEGEGEHEEEEEHEEDDDDEDDDTYNKDD LEDEDLCKHNNGGCGDDKLCEYVGNRRVKCKCKEGYKLEGIECVEHHHHH H[配列番号:12]。
【0183】
最終的に産生されたタンパク質
>readseq.input(1)211 bases, 67EAFE8C checksum.
MRILGEEKPNVDGVSTSNTPGGNEASSASPNLADAAEKKDEKEASEQGEE SHKKENSQESANGKDDVKEEKKTNEKKDDGKTDKVQEKVLEKSPKESQMV DDKKKTEAIPKKVVQPSSSNSGGHVGEEEDHNEGEGEHEEEEEHEEDDDD EDDDTYNKDDLEDEDLCKHNNGGCGDDKLCEYVGNRRVKCKCKEGYKLEG IECVEHHHHHH[配列番号:13]。
【0184】
最終的に産生されたタンパク質で分泌されるP20タンパク質
>readseq.input(1)120 bases, 9358BD28 checksum.
KSPKESQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEEDHNEGEGEHEEE EEHEEDDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLCEYVGNRRVKCK CKEGYKLEGIECVEHHHHHH[配列番号:9]。
【0185】
MSP4p30
>readseq.input(1)666 bases, A9B2DCBC checksum.
ATGTGGATCGTAAAGTTCTTGATTGTGGTCCACTTCTTCATCATATGCAC CATCAACTTCGACAAGCTCTACATTAGTTACTCTTACAACATCGTCCCTG AAAACGGACGTATGCTTAACATGAGGATCTTGGCTGCAGAAAAGAAGGAC GAAAAGGAAGCAAGCGAGCAAGGCGAAGAATCCCACAAGAAGGAAAACTC TCAGGAATCTGCAAACGGAAAAGACGACGTTAAGGAGGAGAAGAAGACCA ACGAGAAGAAGGACGACGGAAAGACTGACAAGGTACAAGAAAAGGTTCTA GAAAAGAGTCCCAAGGAGAGTCAAATGGTCGACGACAAGAAGAAGACCGA GGCCATTCCAAAGAAAGTCGTGCAGCCAAGCTCGAGCAACTCTGGAGGTC ACGTCGGTGAAGAAGAAGACCACAACGAAGGAGAGGGAGAGCACGAAGAG GAGGAAGAACACGAAGAAGACGATGACGACGAGGACGACGACACATACAA CAAAGACGACTTGGAGGACGAAGATCTTTGCAAGCACAACAACGGAGGAT GTGGAGATGACAAGCTCTGCGAGTACGTTGGAAACCGTCGCGTAAAATGT AAATGTAAGGAAGGATACAAGTTGGAAGGAATTGAGTGCGTTGAACACCA CCACCACCATCACTAA[配列番号:2]。
【0186】
ORFによってコードされるタンパク質
>readseq.input(1)221 bases, F982D7F0 checksum.
MWIVKFLIVVHFFIICTINFDKLYISYSYNIVPENGRMLNMRILAAEKKD EKEASEQGEESHKKENSQESANGKDDVKEEKKTNEKKDDGKTDKVQEKVL EKSPKESQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEEDHNEGEGEHEE EEEHEEDDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLCEYVGNRRVKC KCKEGYKLEGIECVEHHHHHH[配列番号:10]。
【0187】
最終的に産生されたタンパク質
>readseq.input(1)181 bases, FB2C49DD checksum.
MRILAAEKKDEKEASEQGEESHKKENSQESANGKDDVKEEKKTNEKKDDG KTDKVQEKVLEKSPKESQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEED HNEGEGEHEEEEEHEEDDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLC EYVGNRRVKCKCKEGYKLEGIECVEHHHHHH[配列番号:11]。
【0188】
MSP4p21
>readseq.input(1)498 bases, C2453A47 checksum.
ATGTGGATCGTAAAGTTCTTGATTGTGGTCCACTTCTTCATCATATGCAC CATCAACTTCGACAAGCTCTACATTAGTTACTCTTACAACATCGTCCCTG AAAACGGACGTATGCTTAACATGAGGATTCTAGAAAAGAGTCCCAAGGAG AGTCAAATGGTCGACGACAAGAAGAAGACCGAGGCCATTCCAAAGAAAGT CGTGCAGCCAAGCTCGAGCAACTCTGGAGGTCACGTCGGTGAAGAAGAAG ACCACAACGAAGGAGAGGGAGAGCACGAAGAGGAGGAAGAACACGAAGAA GACGATGACGACGAGGACGACGACACATACAACAAAGACGACTTGGAGGA CGAAGATCTTTGCAAGCACAACAACGGAGGATGTGGAGATGACAAGCTCT GCGAGTACGTTGGAAACCGTCGCGTAAAATGTAAATGTAAGGAAGGATAC AAGTTGGAAGGAATTGAGTGCGTTGAACACCACCACCACCATCACTAA[配列番号:6]。
【0189】
ORFによってコードされるタンパク質
>readseq.input(1)165 bases, 27BB4F2E checksum.
MWIVKFLIVVHFFIICTINFDKLYISYSYNIVPENGRMLNMRILEKSPKE SQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEEDHNEGEGEHEEEEEHEE DDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLCEYVGNRRVKCKCKEGY KLEGIECVEHHHHHH[配列番号:17]。
【0190】
最終的に産生されたタンパク質
readseq.input(1)125 bases, 50D64DD2 checksum.
MRILEKSPKESQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEEDHNEGEG EHEEEEEHEEDDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLCEYVGNR RVKCKCKEGYKLEGIECVEHHHHHH[配列番号:18]。
【0191】
MSP4p21 ss1
>readseq.input(1)540 bases, 96DADE5 checksum.
ATGTGGATCGTAAAGTTCTTGATTGTGGTCCACTTCTTCATCATATGCAC CATCAACTTCGACAAGCTCTACATTAGTTACTCTTACAACATCGTCCCTG AAAACGGACGTATGCTTAACATGAGGATCTTGGGTGAAGAAAAGCCTAAC GTTGACGGTGTGTCAACATCTctaGAAAAGAGTCCCAAGGAGAGTCAAAT GGTCGACGACAAGAAGAAGACCGAGGCCATTCCAAAGAAAGTCGTGCAGC CAAGCTCGAGCAACTCTGGAGGTCACGTCGGTGAAGAAGAAGACCACAAC GAAGGAGAGGGAGAGCACGAAGAGGAGGAAGAACACGAAGAAGACGATGA CGACGAGGACGACGACACATACAACAAAGACGACTTGGAGGACGAAGATC TTTGCAAGCACAACAACGGAGGATGTGGAGATGACAAGCTCTGCGAGTAC GTTGGAAACCGTCGCGTAAAATGTAAATGTAAGGAAGGATACAAGTTGGA AGGAATTGAGTGCGTTGAACACCACCACCACCATCACTAA[配列番号:7]。
【0192】
ORFによってコードされるタンパク質
>readseq.input(1)179 bases, F0D6C58 checksum.
MWIVKFLIVVHFFIICTINFDKLYISYSYNIVPENGRMLNMRILGEEKPN VDGVSTSLEKSPKESQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEEDHN EGEGEHEEEEEHEEDDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLCEY VGNRRVKCKCKEGYKLEGIECVEHHHHHH[配列番号:19]。
【0193】
最終的に産生されたタンパク質
>readseq.input(1)139 bases, 4D5C3E0F checksum.
MRILGEEKPNVDGVSTSLEKSPKESQMVDDKKKTEAIPKKVVQPSSSNSG GHVGEEEDHNEGEGEHEEEEEHEEDDDDEDDDTYNKDDLEDEDLCKHNNG GCGDDKLCEYVGNRRVKCKCKEGYKLEGIECVEHHHHHH[配列番号:20]。
【0194】
MSP4p21 ss2
>readseq.input(1)516 bases, 1B3CABBA checksum.
ATGTGGATCGTAAAGTTCTTGATTGTGGTCCACTTCTTCATCATATGCAC CATCAACTTCGACAAGCTCTACATTAGTTACTCTTACAACATCGTCCCTG AAAACGGACGTATGCTTAACATGAGGATCTTGGGTGAAGAAAAGCCTCTA GAAAAGAGTCCCAAGGAGAGTCAAATGGTCGACGACAAGAAGAAGACCGA GGCCATTCCAAAGAAAGTCGTGCAGCCAAGCTCGAGCAACTCTGGAGGTC ACGTCGGTGAAGAAGAAGACCACAACGAAGGAGAGGGAGAGCACGAAGAG GAGGAAGAACACGAAGAAGACGATGACGACGAGGACGACGACACATACAA CAAAGACGACTTGGAGGACGAAGATCTTTGCAAGCACAACAACGGAGGAT GTGGAGATGACAAGCTCTGCGAGTACGTTGGAAACCGTCGCGTAAAATGT AAATGTAAGGAAGGATACAAGTTGGAAGGAATTGAGTGCGTTGAACACCA CCACCACCATCACTAA[配列番号:8]。
【0195】
ORFによってコードされる予想タンパク質
>readseq.input(1)171 bases, EB09D214 checksum.
MWIVKFLIVVHFFIICTINFDKLYISYSYNIVPENGRMLNMRILGEEKPL EKSPKESQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEEDHNEGEGEHEE EEEHEEDDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLCEYVGNRRVKC KCKEGYKLEGIECVEHHHHHH[配列番号:21]。
【0196】
最終的に産生されたタンパク質
>readseq.input(1)131 bases, 6B6B091D checksum.
MRILGEEKPLEKSPKESQMVDDKKKTEAIPKKVVQPSSSNSGGHVGEEED HNEGEGEHEEEEEHEEDDDDEDDDTYNKDDLEDEDLCKHNNGGCGDDKLC EYVGNRRVKCKCKEGYKLEGIECVEHHHHHH[配列番号:22]。
【0197】
PvMSP4/Hisタンパク質
MKVAYFLSVLDLLIIFSLYFDGRRSAFAGIAACIRHGRILGEGGEQSGGASGGSSGGSSGDSSGGLSGGSSGGPSPPAGSSGSGGSDPANSATGPQNSTPGSGGQTGDHSAEAENGDYNEQGDDHGDDHGDDHGDDHGDEQDGEDYDDAEDDDLYELSEVDENANLCLDNNGGCGDDKICENLGKGIVKCLCKPGYKLVGTECVESHHHHHH[配列番号:16]。
【0198】
PfMSP5/His、PfMSP4p30/His、PfMSP4p40/His、PfMSP4p21ss1/His、PfMSP4p21ss2/HisおよびPvMSP4/Hisとして開示した生物材料は、本出願において、特に実施例において、記述してある。それは、以下の要求に従う:
・ウイルスを繁殖させるための必要物:
-細胞型:Spodoptera frugiperda(Sf9)
-培地:
L-グルタミンを含むSF-900 II培地(GIBCO, Invitrogen Corporation)Cat番号10902-088
L-グルタミン200 mM(GIBCO, Invitrogen Corporation):2 mM終濃度Cat番号25030-024
ゲンタマイシン(50 mg/ml;GIBCO, Invitrogen Corporation)):50μg/ml終濃度Cat番号15750-037
-血清:
牛胎児血清5%
-pH 6.2
-温度 27-28℃
-気体相:大気(CO2濃縮のない)
・ウイルス懸濁液を得るために使用した方法:
-増殖についての詳細:新鮮な完全培地(5%FCSと共に)にSf9細胞の単層を有する容器(T-25、T-75またはT-150)における培養を、少量のウイルスの懸濁液(またはプラークもしくは限界希釈によるクローニング手順から得られるマイクロタイトレーションプレートのウェル)で播種する。培養は、すべての細胞の溶解まで、5〜6日ごろ、27〜28℃にてインキュベートする。
【0199】
-感染の証拠のための基準:伸長した核を有する細胞、続いて数日後に顆粒状の様相および細胞溶解となる。
【0200】
-ウイルス懸濁液を調製するための詳細
-細胞上清を、4000gで10分遠心分離し、細胞破片を除去する。
【0201】
・滴定のために使用した方法
限界希釈(1つのウェルには、96ウェルの陽性マイクロタイタープレートの約1/10を生じる希釈から開始して感染させる)。
結果を6日後に読み込み、予想される力価は、107〜108 pfu/mlであった。
【0202】
・貯蔵のための条件
長期の維持は、-80℃での凍結により可能となる。
懸濁液体は、標準的な培地である。
【0203】
上記のハイブリドーマF12-7、G21-2、J18-14およびL11-16は、マウス骨髄腫細胞(plasmocytome Ag8 X 63 6.5.3)と特定の抗原で免疫した動物の脾臓から得られるマウス免疫細胞との間の細胞融合の結果として得られた。
【0204】
・細胞の特徴付けおよび産生
-骨髄腫細胞の名称:X63-Ag8-6.5.3
-抗原特異性: F 12-7についてはMSP4 II、G21-2および518-14についてはMSP5、L11-16についてはMSP4。
【0205】
-抗体サブクラス:IgG1k
-分泌の安定性:良および限定的
・培養の条件:
-培地
Eagle MEM-Eurobio
ピルビン酸ナトリウム−Gibco BRL
グルタミン−Gibco BRL
抗生物質:ペニ-ストレプトマイシン−Gibco BRL
ウマ血清:10%
-血清炭酸水素塩2.2 g/l
血清:ウマ10%
pH7,4
至適温度:37℃
気体相:7% CO2
解凍のために講じた警戒:迅速に37℃にて非凍結し、培地+血清で即座に洗浄する。1 mlの24ウェルを有するプレート上で培養する。
【0206】
・培養のための条件:細胞懸濁液
集団倍加時間:15時間
最適な分割比:3日
予想される細胞密度:5.106
限られた寿命
ルーチンのサブカルチャー技術:分裂
・貯蔵のための条件
懸濁液:95% ウマ血清- 5% DMSO
細胞収集の技術:細胞懸濁液を取り、遠心分離し、ペレットを凍結培地に取り込む。
【0207】
凍結するための技術:1 mlの凍結培地における5.106の細胞を-20℃にて2時間、-80℃にて3-4日、そして液体窒素へ。懸濁液は、標準的な培地である。
【0208】
参照文献
以下の刊行物のそれぞれの全ての開示は、本明細書に参照に依拠し、および組み込まれる。
【0209】
1. Genton, B., Al-Yaman, F., Betuela, I. et al. Safety and immunogenicity of a three-component blood-stage malaria vaccine (MSP1, MSP2, RESA) against Plasmodium falciparum in Papua New Guinean children. Vaccine 2003, 22(1), 30-41.
2. Stowers, A.W., Kennedy, M.C., Keegan, B.P., Saul, A., Long, C.A. & Miller, L.H. Vaccination of monkeys with recombinant Plasmodium falciparum apical membrane antigen 1 confers protection against blood-stage malaria. Infect Immun 2002, 70(12), 6961-6967.
3. O'Donnell, R.A., de Koning-Ward, T.F., Burt, R.A. et al. Antibodies against merozoite surface protein (MSP)-1(19) are a major component of the invasion-inhibitory response in individuals immune to malaria. J Exp Med 2001, 193(12), 1403-1412.
4. Hughes, M.K. & Hughes, A.L. Natural selection on Plasmodium surface proteins. Mol Biochem Parasitol 1995, 71(1), 99-113.
5. Escalante, A.A., Lal, A.A. & Ayala, F.J. Genetic polymorphism and natural selection in the malaria parasite Plasmodium falciparum. Genetics 1998, 149(1), 189-202.
6. Volkman, S.K., Hartl, D.L., Wirth, D.F. et al. Excess polymorphisms in genes for membrane proteins in Plasmodium falciparum. Science 2002, 298(5591), 216-218.
7. Rayner, J.C., Corredor, V., Feldman, D. et al. Extensive polymorphism in the plasmodium vivax merozoite surface coat protein MSP-3alpha is limited to specific domains. Parasitology 2002, 125(Pt 5), 393-405.
8. Taylor, R.R., Smith, D.B., Robinson, V.J., McBride, J.S. & Riley, E.M. Human antibody response to Plasmodium falciparum merozoite surface protein 2 is serogroup specific and predominantly of the immunoglobulin G3 subclass. Infect Immun 1995, 63(11), 4382-4388.
9. Wang, L., Mohandas, N., Thomas, A. & Coppel, R.L. Detection of detergent-resistant membranes in asexual blood-stage parasites of Plasmodium falciparum. Mol Biochem Parasitol 2003, 130(2), 149-153.
10. Marshall, V.M., Silva, A., Foley, M. et al. A second merozoite surface protein (MSP-4) of Plasmodium falciparum that contains an epidermal growth factor-like domain. Infect Immun 1997, 65(11), 4460-4467.
11. Wu, T., Black, C.G., Wang, L., Hibbs, A.R. & Coppel, R.L. Lack of sequence diversity in the gene encoding merozoite surface protein 5 of Plasmodium falciparum. Mol Biochem Parasitol 1999, 103(2), 243-250.
12. Wang, L., Black, C.G., Marshall, V.M. & Coppel, R.L. Structural and antigenic properties of merozoite surface protein 4 of Plasmodium falciparum. Infect Immun 1999, 67(5), 2193-2200.
13. Wang, L., Richie, T.L., Stowers, A., Nhan, D.H. & Coppel, R.L. Naturally acquired antibody responses to Plasmodium falciparum merozoite surface protein 4 in a population living in an area of endemicity in Vietnam. Infect Immun 2001, 69(7), 4390-4397.
14. Black, C.G., Wang, L., Hibbs, A.R., Werner, E. & Coppel, R.L. Identification of the Plasmodium chabaudi homologue of merozoite surface proteins 4 and 5 of Plasmodium falciparum. Infect Immun 1999, 67(5), 2075-2081.
15. Black, C.G. & Coppel, R.L. Synonymous and non-synonymous mutations in a region of the Plasmodium chabaudi genome and evidence for selection acting on a malaria vaccine candidate. Mol Biochem Parasitol 2000, 111(2), 447-451.
16. Kedzierski, L., Black, C.G. & Coppel, R.L. Characterization of the merozoite surface protein 4/5 gene of Plasmodium berghei and Plasmodium yoelii. Mol Biochem Parasitol 2000, 105(1), 137-147.
17. Kedzierski, L., Black, C.G., Goschnick, M.W., Stowers, A.W. & Coppel, R.L. Immunization with a Combination of Merozoite Surface Proteins 4/5 and 1 Enhances Protection against Lethal Challenge with Plasmodium yoelii. Infect Immun 2002, 70(12), 6606-6613.
18. Kedzierski, L., Black, C.G. & Coppel, R.L. Immunization with recombinant Plasmodium yoelii merozoite surface protein 4/5 protects mice against lethal challenge. Infect Immun 2000, 68(10), 6034-6037.
19. Kedzierski, L., Black, C.G., Stowers, A.W., Goschnick, M.W., Kaslow, D.C. & Coppel, R.L. Comparison of the protective efficacy of yeast-derived and Escherichia coli-derived recombinant merozoite surface protein 4/5 against lethal challenge by Plasmodium yoelii. Vaccine 2001, 19(32), 4661-4668.
20. Wang, L., Goschnick, M.W. & Coppel, R.L. Oral Immunization with a Combination of Plasmodium yoelii Merozoite Surface Proteins 1 and 4/5 Enhances Protection against Lethal Malaria Challenge. Infect Immun 2004, 72(10), 6172-6175.
21. Goschnick, M.W., Black, C.G., Kedzierski, L., Holder, A.A. & Coppel, R.L. Merozoite Surface Protein 4/5 Provides Protection against Lethal Challenge with a Heterologous Malaria Parasite Strain. Infect Immun 2004, 72(10), 5840-5849.
22. Marshall, V.M., Tieqiao, W. & Coppel, R.L. Close linkage of three merozoite surface protein genes on chromosome 2 of Plasmodium falciparum. Mol Biochem Parasitol 1998, 94(1), 13-25.
23. Nishimura, Y., Chen, Y.Z., Uemura, Y. et al. Degenerate recognition and response of human CD4+ Th cell clones: implications for basic and applied immunology. Mol Immunol 2004, 40(14-15), 1089-1094.
24. Brady, C.P., Shimp, R.L., Miles, A.P., Whitmore, M. & Stowers, A.W. High-level production and purification of P30P2MSP1(19), an important vaccine antigen for malaria, expressed in the methylotropic yeast Pichia pastoris. Protein Expr Purif 2001, 23(3), 468-475.
25. Epp, C., Kauth, C.W., Bujard, H. & Lutz, R. Expression and purification of Plasmodium falciparum MSP-1(42): A malaria vaccine candidate. J Chromatogr B Analyt Technol Biomed Life Sci 2003, 786(1-2), 61-72.
26. Withers-Martinez, C., Saldanha, J.W., Ely, B. et al. Expression of recombinant Plasmodium falciparum subtilisin-like protease-1 in insect cells. Characterization, comparison with the parasite protease, and homology modeling PCR-based gene synthesis as an efficient approach for expression of the A+T-rich malaria genome. J Biol Chem 2002, 277(33), 29698-29709. Epub 22002 Jun 29696.
27. Zhou, Z., Schnake, P., Xiao, L. & Lal, A.A. Enhanced expression of a recombinant malaria candidate vaccine in Escherichia coli by codon optimization. Protein Expr Purif 2004, 34(1), 87-94.
28. Weber, J.L. Molecular Biology of Malaria Parasites. Exp. Parasitol. 1988, 66, 143-170.
29. Withers-Martinez, C., Carpenter, E.P., Hackett, F. et al. PCR-based gene synthesis as an efficient approach for expression of the A+T-rich malaria genome. Protein Eng 1999, 12(12), 1113-1120.
30. Ballou, W.R., Arevalo-Herrera, M., Carucci, D. et al. Update on the clinical development of candidate malaria vaccines. Am J Trop Med Hyg 2004, 71(2 Suppl), 239-247.
31. Wang, L., Marshall, V.M. & Coppel, R.L. Limited polymorphism of the vaccine candidate merozoite surface protein 4 of Plasmodium falciparum. Mol Biochem Parasitol 2002, 120(2), 301-303.
32. Jongwutiwes, S., Putaporntip, C., Friedman, R. & Hughes, A.L. The Extent of Nucleotide Polymorphism is Highly Variable Across a 3-kb Region on Plasmodium falciparum Chromosome 2. Mol Biol Evol 2002, 19(9), 1585-1590.
33. Polson, H.E., Conway, D.J., Fandeur, T., Mercereau-Puijalon, O. & Longacre, S. Gene polymorphism of Plasmodium falciparum merozoite surface proteins 4 and 5. Mol Biochem Parasitol 2005, 142(1), 110-115.
34. Puentes, A.G., J. Vera, R. Oopez, R. Suarez, J. Rodriguez, L. Curtidor, H. Sporozoite and Liver Stage Antigen Plasmodium falciparum peptides bind specifically to human hepatocytes. Vaccine 2004, 22, 1150-1156.
35. Hale, R.S. & Thompson, G. Codon optimization of the gene encoding a domain from human type 1 neurofibromin protein results in a threefold improvement in expression level in Escherichia coli. Protein Expr Purif 1998, 12(2), 185-188.
36. Longacre, S., Mendis, K.N. & David, P.H. Plasmodium vivax merozoite surface protein 1 C-terminal recombinant proteins in baculovirus. Mol Biochem Parasitol 1994, 64(2), 191-205.
37. Nacer, A., Berry, L., Slomianny, C. & Mattei, D. Plasmodium falciparum signal sequences: simply sequences or special signals? Int J Parasitol 2001, 31(12), 1371-1379.
38. Utsumi, T., Nakano, K., Funakoshi, T. et al. Vertical-scanning mutagenesis of amino acids in a model N-myristoylation motif reveals the major amino-terminal sequence requirements for protein N-myristoylation. Eur J Biochem 2004, 271(4), 863-874.
39. Resh, M.D. Fatty acylation of proteins: new insights into membrane targeting of myristoylated and palmitoylated proteins. Biochim Biophys Acta 1999, 1451(1), 1-16.
40. Gunaratne, R.S., Sajid, M., Ling, I.T., Tripathi, R., Pachebat, J.A. & Holder, A.A. Characterization of N-myristoyltransferase from Plasmodium falciparum. Biochem J 2000, 348 Pt 2, 459-463.
41. Price, H.P., Menon, M.R., Panethymitaki, C., Goulding, D., McKean, P.G. & Smith, D.F. Myristoyl-CoA:protein N-myristoyltransferase, an essential enzyme and potential drug target in kinetoplastid parasites. J Biol Chem 2003, 278(9), 7206-7214.
42. Stafford, W.H., Stockley, R.W., Ludbrook, S.B. & Holder, A.A. Isolation, expression and characterization of the gene for an ADP-ribosylation factor from the human malaria parasite, Plasmodium falciparum. Eur J Biochem 1996, 242(1), 104-113.
43. Stevenson, F.T., Bursten, S.L., Locksley, R.M. & Lovett, D.H. Myristyl acylation of the tumor necrosis factor alpha precursor on specific lysine residues. J Exp Med 1992, 176(4), 1053-1062.
44. Farazi, T.A., Waksman, G. & Gordon, J.I. The biology and enzymology of protein N-myristoylation. J Biol Chem 2001, 276(43), 39501-39504.
45. Hayashi, N., Nakagawa, C., Ito, Y. et al. Myristoylation-regulated direct interaction between calcium-bound calmodulin and N-terminal region of pp60v-src. J Mol Biol 2004, 338(1), 169-180.
46. Maekawa, S., Matsuura, Y. & Nakamura, S. Expression and myristoylation of NAP-22 using a baculovirus transfer vector system. Biochim Biophys Acta 1994, 1218(1), 119-122.
47. Perraut, R., Marrama, L., Diouf, B. et al. Distinct surrogate markers for protection against Plasmodium falciparum infection and clinical malaria identified in a Senegalese community after radical drug cure. J Infect Dis 2003, 188(12), 1940-1950.
48. Schofield, L., Hewitt, M.C., Evans, K., Siomos, M.A. & Seeberger, P.H. Synthetic GPI as a candidate anti-toxic vaccine in a model of malaria. Nature 2002, 418(6899), 785-789.
49. Bonnet S. et al, Vaccine 2006, vol. 24, p. 5997-6008, soluble and glyco-lipid modified Baculovirus Plasmodium falciparum C-terminal Merozoite Surface Protein 1, two forms of leading malaria vaccine candidate.
【特許請求の範囲】
【請求項1】
配列番号:1、2、3、4、5、6、7、28、29または8のDNA配列を含む精製された核酸分子。
【請求項2】
配列番号:9、10、11、12、13、14、15、16、17、18、19、20、21、30または22の配列を含むアミノ酸配列をコードする精製された核酸分子。
【請求項3】
42℃にて50%のホルムアミドおよび6×SSCにおける適度なストリンジェンシーの条件下において、60℃、0.5×SSC、0.1%のSDSの洗浄条件で、請求項1または2のいずれか1項の核酸配列を含む変性された二本鎖DNAいずれかの鎖にハイブリダイズする精製された核酸分子。
【請求項4】
前記単離された核酸分子が配列番号:1〜8からのインビトロ突然変異誘発によって誘導される、請求項3に記載の精製された核酸分子。
【請求項5】
遺伝暗号の結果として配列番号:1、2、3、4、5、6、7または8から縮重された、精製された核酸分子。
【請求項6】
組換えMSP4もしくはMSP5ポリペプチド、組換えMSP4もしくはMSP5ポリペプチドDNAの対立遺伝子変異体または組換えMSP4もしくはMSP5ポリペプチドDNAの相同体をコードする精製された核酸分子。
【請求項7】
請求項1、2、5および6の精製された核酸分子からなる群より選択される核酸分子の発現を指揮する組換えベクター。
【請求項8】
請求項3の核酸分子の発現を指揮する組換えベクター。
【請求項9】
請求項4の核酸分子の発現を指揮する組換えベクター。
【請求項10】
請求項1、2、5および6の精製された核酸分子からなる群より選択される核酸分子によってコードされる精製されたポリペプチド。
【請求項11】
非グリコシル化形態の請求項10に記載の精製されたポリペプチド。
【請求項12】
請求項3の核酸分子によってコードされる精製されたポリペプチド。
【請求項13】
非グリコシル化形態の請求項12に記載の精製されたポリペプチド。
【請求項14】
請求項4の核酸分子によってコードされる精製されたポリペプチド。
【請求項15】
非グリコシル化形態の請求項14に記載の精製されたポリペプチド。
【請求項16】
請求項10のポリペプチドに結合する精製された抗体。
【請求項17】
前記抗体がモノクローナル抗体である、請求項16に記載の精製された抗体。
【請求項18】
請求項12のポリペプチドに結合する精製された抗体。
【請求項19】
前記抗体がモノクローナル抗体である、請求項18に記載の精製された抗体。
【請求項20】
請求項14のポリペプチドに結合する精製された抗体。
【請求項21】
前記抗体がモノクローナル抗体である、請求項20に記載の精製された抗体。
【請求項22】
請求項7のベクターがトランスフェクトされ、形質導入され、または感染した宿主細胞。
【請求項23】
組換えMSP4またはMSP5ポリペプチドの産生のための方法であって、発現を促進する条件下で請求項22の宿主細胞を培養することと、および前記培地からポリペプチドを回収することとを含む方法。
【請求項24】
前記宿主細胞が昆虫細胞、細菌細胞、酵母細胞、植物細胞および動物細胞からなる群より選択される、請求項23に記載の方法。
【請求項25】
請求項8のベクターがトランスフェクトされ、感染され、または形質導入された宿主細胞。
【請求項26】
組換えMSP4およびMSP5ポリペプチドの産生のための方法であって、発現を促進する条件下で請求項25の宿主細胞を培養することと、および前記培地または細胞のペレットからポリペプチドを回収することとを含む方法。
【請求項27】
前記宿主細胞が昆虫細胞、細菌細胞、酵母細胞、植物細胞および動物細胞からなる群より選択される、請求項26に記載の方法。
【請求項28】
請求項9のベクターがトランスフェクトされ、感染され、または形質導入された宿主細胞。
【請求項29】
組換えMSP4およびMSP5ポリペプチドの産生のための方法であって、発現を促進する条件下で請求項28の宿主細胞を培養することと、および培地または細胞の凝集体からポリペプチドを回収することとを含む方法。
【請求項30】
前記宿主細胞が昆虫細胞、細菌細胞、酵母細胞、植物細胞および動物細胞からなる群より選択される、請求項29に記載の方法。
【請求項31】
I-3512、I-3513、I-3514、I-3694、I-3695およびI-3696からなる群より選択される組換えベクター。
【請求項32】
組換えMSP4およびMSP5ポリペプチドと前記ポリペプチドを特異的に認識する抗体とを含む免疫学的複合体。
【請求項33】
マラリア原虫による感染を検出するための方法であって、マラリア原虫に感染したことが疑われる生物材料を含む組成物を提供することと、および組換えMSP4またはMSP5ポリペプチドに対して特異的な抗体に結合するマラリア原虫の天然のMSP4および/またはMSP5ポリペプチドの存在についてアッセイすることとを含む方法。
【請求項34】
熱帯熱マラリア原虫(Plasmodium falciparum)を検出するために使用される、請求項33に記載の方法。
【請求項35】
三日熱マラリア原虫(Plasmodium vivax)を検出するために使用される、請求項33に記載の方法。
【請求項36】
前記天然のMSP4またはMSP5ポリペプチドが電気泳動法によって、または前記組換えMSP4およびMSP5ポリペプチドと免疫学的反応性である抗体との免疫アッセイ法によってアッセイされる、請求項33に記載の方法。
【請求項37】
組換えMSP4またはMSP5ポリペプチドを含む抗原に結合する抗体の有無の検出のためのインビトロでの診断法であって、前記抗原を、生物学的体液と、生物学的体液中の前記抗原および抗体が抗原抗体複合体を形成するために十分な時間の間および条件下で接触させることと、および複合体の形成を検出することとを含む方法。
【請求項38】
前記抗原抗体複合体の形成を測定することをさらに含む、請求項35に記載の方法。
【請求項39】
抗原抗体複合体の形成がウエスタンブロット法、ELISA、間接免疫蛍光法アッセイ法または免疫沈降アッセイ法に基づいた免疫アッセイによって検出される、請求項35に記載の方法。
【請求項40】
組換えMSP4もしくはMSP5ポリペプチドまたはこれらの混合物に結合する抗体の有無の検出のための診断キットであって、前期組換えMSP4もしくはMSP5ポリペプチドまたは組換えMSP4およびMSP5ポリペプチドの混合物を含む抗原と、抗原と抗体との間の免疫複合体の形成を検出するための手段とを含み、前記手段は、前記検出を行うために十分な量で存在するキット。
【請求項41】
インビボにおいて免疫原性反応または保護反応を誘導するために十分な量の少なくとも1つの組換えMSP4もしくはMSP5ポリペプチドまたはこれらの混合物と、薬学的に許容される担体とを含む免疫原性組成物。
【請求項42】
前記組成物がインビボにおいて中和抗体を誘導するために十分な量の少なくとも1つの組換えMSP4またはMSP5ポリペプチドを含む、請求項41に記載の免疫原性組成物。
【請求項43】
少なくとも組換えポリペプチドMSP4p40を含む、請求項41に記載の免疫原性組成物。
【請求項44】
少なくとも組換えポリペプチドMSP4p30を含む、請求項41に記載の免疫原性組成物。
【請求項45】
少なくとも組換えポリペプチドMSP4p20を含む、請求項41に記載の免疫原性組成物。
【請求項46】
少なくとも組換えポリペプチドMSP5p45およびMSP5p35を含む、請求項41に記載の免疫原性組成物。
【請求項47】
少なくとも組換えポリペプチドPvMSP4p20を含む、請求項39に記載の免疫原性組成物。
【請求項48】
配列番号:1、2、3、4、5、6、7または8から選択されるポリヌクレオチド。
【請求項49】
配列番号:9、10、11、12、13、14、15、16、17、18、19、20、21、22または30から選択されるポリヌクレオチド。
【請求項50】
少なくともPfMSP4p21ss1およびPfMSP4p21ss2から選択される組換えポリペプチドを含む、請求項41の免疫原性組成物。
【請求項51】
少なくとも組換えポリペプチドMSP4p40およびMSP4p20を含む、請求項39の免疫原性組成物。
【請求項52】
組換えMSP1p19ポリペプチドを含む抗原をさらに含む、請求項39に記載の免疫原性組成物。
【請求項53】
インビボにおいて保護反応を誘導するために十分な量の、少なくとも1つの組換えMSP4もしくはMSP5ポリペプチドまたはこれらの混合物と、薬学的に許容される担体とを含むワクチン組成物であって、前記少なくとも1つの組換えMSP4またはMSP5ポリペプチドは、MSP4p40、MSP4p30、MSP4p20、MSP4p21ss1、MSP4p21ss2、MSP5p45、MSP5p35およびPvMSP4p20から選択されるワクチン組成物。
【請求項54】
寄生虫による感染が疑われるヒト由来の生物材料の試料におけるプラスモディウム属感染をインビトロで診断するための方法であって、以下の工程を含む方法:
-組換えMSP4またはMSP5ポリペプチドを含む抗原を、前記試料と、前記試料中の前記抗原および抗体が抗原抗体複合体を形成するために十分な時間の間および条件下で接触させる工程、および、
-前記複合体の形成を検出する工程。
【請求項55】
前記抗原抗体複合体の形成を測定することをさらに含む、請求項54に記載の方法。
【請求項56】
前記抗原抗体複合体の形成がウエスタンブロット法、ELISA、間接免疫蛍光法アッセイ法または免疫沈降アッセイ法に基づいた免疫アッセイによって検出される、請求項54に記載の方法。
【請求項57】
前記天然のMSP4またはMSP5ポリペプチドが2005年11月16日に受託番号I-3517、I-3518、I-3519またはI-3520の下でC.N.C.M.に寄託されたハイブリドーマによって分泌された抗体での免疫アッセイ法によってアッセイされる、請求項36に記載の方法。
【請求項58】
2005年11月16日に受託番号I-3517、I-3518、I-3519またはI-3520の下でC.N.C.M.に寄託されたハイブリドーマから産生される、請求項17に記載の精製された抗体。
【請求項59】
2005年11月16日に受託番号I-3517、I-3518、I-3519またはI-3520下でC.N.C.M.に寄託されたハイブリドーマ。
【請求項60】
マラリア原虫を検出するための方法であって、マラリア原虫に感染したことが疑われる生物材料を含む組成物を提供することと、および組換えMSP4もしくはMSP5ポリペプチドに対して特異的な少なくとも1つの抗体または組換えMSP4ポリペプチドに対して特異的な少なくとも1つの抗体と組換えMSP5ポリペプチドに対して特異的な抗体との混合物に結合するマラリア原虫の天然のMSP4および/またはMSP5ポリペプチドの存在についてアッセイすることとを含む方法。
【請求項61】
前記組換えMSP4またはMSp5ポリペプチドが請求項10の精製されたポリペプチドの中から選択される、請求項60に記載の方法。
【請求項62】
前記組換えMSP4またはMSP5ポリペプチドに対して特異的な抗体が請求項58の精製された抗体の中から選択される、請求項60に記載の方法。
【請求項63】
熱帯熱マラリア原虫(Plasmodium falciparum)または三日熱マラリア原虫(Plasmodium vivax)を検出するために使用される、請求項60に記載の方法。
【請求項1】
配列番号:1、2、3、4、5、6、7、28、29または8のDNA配列を含む精製された核酸分子。
【請求項2】
配列番号:9、10、11、12、13、14、15、16、17、18、19、20、21、30または22の配列を含むアミノ酸配列をコードする精製された核酸分子。
【請求項3】
42℃にて50%のホルムアミドおよび6×SSCにおける適度なストリンジェンシーの条件下において、60℃、0.5×SSC、0.1%のSDSの洗浄条件で、請求項1または2のいずれか1項の核酸配列を含む変性された二本鎖DNAいずれかの鎖にハイブリダイズする精製された核酸分子。
【請求項4】
前記単離された核酸分子が配列番号:1〜8からのインビトロ突然変異誘発によって誘導される、請求項3に記載の精製された核酸分子。
【請求項5】
遺伝暗号の結果として配列番号:1、2、3、4、5、6、7または8から縮重された、精製された核酸分子。
【請求項6】
組換えMSP4もしくはMSP5ポリペプチド、組換えMSP4もしくはMSP5ポリペプチドDNAの対立遺伝子変異体または組換えMSP4もしくはMSP5ポリペプチドDNAの相同体をコードする精製された核酸分子。
【請求項7】
請求項1、2、5および6の精製された核酸分子からなる群より選択される核酸分子の発現を指揮する組換えベクター。
【請求項8】
請求項3の核酸分子の発現を指揮する組換えベクター。
【請求項9】
請求項4の核酸分子の発現を指揮する組換えベクター。
【請求項10】
請求項1、2、5および6の精製された核酸分子からなる群より選択される核酸分子によってコードされる精製されたポリペプチド。
【請求項11】
非グリコシル化形態の請求項10に記載の精製されたポリペプチド。
【請求項12】
請求項3の核酸分子によってコードされる精製されたポリペプチド。
【請求項13】
非グリコシル化形態の請求項12に記載の精製されたポリペプチド。
【請求項14】
請求項4の核酸分子によってコードされる精製されたポリペプチド。
【請求項15】
非グリコシル化形態の請求項14に記載の精製されたポリペプチド。
【請求項16】
請求項10のポリペプチドに結合する精製された抗体。
【請求項17】
前記抗体がモノクローナル抗体である、請求項16に記載の精製された抗体。
【請求項18】
請求項12のポリペプチドに結合する精製された抗体。
【請求項19】
前記抗体がモノクローナル抗体である、請求項18に記載の精製された抗体。
【請求項20】
請求項14のポリペプチドに結合する精製された抗体。
【請求項21】
前記抗体がモノクローナル抗体である、請求項20に記載の精製された抗体。
【請求項22】
請求項7のベクターがトランスフェクトされ、形質導入され、または感染した宿主細胞。
【請求項23】
組換えMSP4またはMSP5ポリペプチドの産生のための方法であって、発現を促進する条件下で請求項22の宿主細胞を培養することと、および前記培地からポリペプチドを回収することとを含む方法。
【請求項24】
前記宿主細胞が昆虫細胞、細菌細胞、酵母細胞、植物細胞および動物細胞からなる群より選択される、請求項23に記載の方法。
【請求項25】
請求項8のベクターがトランスフェクトされ、感染され、または形質導入された宿主細胞。
【請求項26】
組換えMSP4およびMSP5ポリペプチドの産生のための方法であって、発現を促進する条件下で請求項25の宿主細胞を培養することと、および前記培地または細胞のペレットからポリペプチドを回収することとを含む方法。
【請求項27】
前記宿主細胞が昆虫細胞、細菌細胞、酵母細胞、植物細胞および動物細胞からなる群より選択される、請求項26に記載の方法。
【請求項28】
請求項9のベクターがトランスフェクトされ、感染され、または形質導入された宿主細胞。
【請求項29】
組換えMSP4およびMSP5ポリペプチドの産生のための方法であって、発現を促進する条件下で請求項28の宿主細胞を培養することと、および培地または細胞の凝集体からポリペプチドを回収することとを含む方法。
【請求項30】
前記宿主細胞が昆虫細胞、細菌細胞、酵母細胞、植物細胞および動物細胞からなる群より選択される、請求項29に記載の方法。
【請求項31】
I-3512、I-3513、I-3514、I-3694、I-3695およびI-3696からなる群より選択される組換えベクター。
【請求項32】
組換えMSP4およびMSP5ポリペプチドと前記ポリペプチドを特異的に認識する抗体とを含む免疫学的複合体。
【請求項33】
マラリア原虫による感染を検出するための方法であって、マラリア原虫に感染したことが疑われる生物材料を含む組成物を提供することと、および組換えMSP4またはMSP5ポリペプチドに対して特異的な抗体に結合するマラリア原虫の天然のMSP4および/またはMSP5ポリペプチドの存在についてアッセイすることとを含む方法。
【請求項34】
熱帯熱マラリア原虫(Plasmodium falciparum)を検出するために使用される、請求項33に記載の方法。
【請求項35】
三日熱マラリア原虫(Plasmodium vivax)を検出するために使用される、請求項33に記載の方法。
【請求項36】
前記天然のMSP4またはMSP5ポリペプチドが電気泳動法によって、または前記組換えMSP4およびMSP5ポリペプチドと免疫学的反応性である抗体との免疫アッセイ法によってアッセイされる、請求項33に記載の方法。
【請求項37】
組換えMSP4またはMSP5ポリペプチドを含む抗原に結合する抗体の有無の検出のためのインビトロでの診断法であって、前記抗原を、生物学的体液と、生物学的体液中の前記抗原および抗体が抗原抗体複合体を形成するために十分な時間の間および条件下で接触させることと、および複合体の形成を検出することとを含む方法。
【請求項38】
前記抗原抗体複合体の形成を測定することをさらに含む、請求項35に記載の方法。
【請求項39】
抗原抗体複合体の形成がウエスタンブロット法、ELISA、間接免疫蛍光法アッセイ法または免疫沈降アッセイ法に基づいた免疫アッセイによって検出される、請求項35に記載の方法。
【請求項40】
組換えMSP4もしくはMSP5ポリペプチドまたはこれらの混合物に結合する抗体の有無の検出のための診断キットであって、前期組換えMSP4もしくはMSP5ポリペプチドまたは組換えMSP4およびMSP5ポリペプチドの混合物を含む抗原と、抗原と抗体との間の免疫複合体の形成を検出するための手段とを含み、前記手段は、前記検出を行うために十分な量で存在するキット。
【請求項41】
インビボにおいて免疫原性反応または保護反応を誘導するために十分な量の少なくとも1つの組換えMSP4もしくはMSP5ポリペプチドまたはこれらの混合物と、薬学的に許容される担体とを含む免疫原性組成物。
【請求項42】
前記組成物がインビボにおいて中和抗体を誘導するために十分な量の少なくとも1つの組換えMSP4またはMSP5ポリペプチドを含む、請求項41に記載の免疫原性組成物。
【請求項43】
少なくとも組換えポリペプチドMSP4p40を含む、請求項41に記載の免疫原性組成物。
【請求項44】
少なくとも組換えポリペプチドMSP4p30を含む、請求項41に記載の免疫原性組成物。
【請求項45】
少なくとも組換えポリペプチドMSP4p20を含む、請求項41に記載の免疫原性組成物。
【請求項46】
少なくとも組換えポリペプチドMSP5p45およびMSP5p35を含む、請求項41に記載の免疫原性組成物。
【請求項47】
少なくとも組換えポリペプチドPvMSP4p20を含む、請求項39に記載の免疫原性組成物。
【請求項48】
配列番号:1、2、3、4、5、6、7または8から選択されるポリヌクレオチド。
【請求項49】
配列番号:9、10、11、12、13、14、15、16、17、18、19、20、21、22または30から選択されるポリヌクレオチド。
【請求項50】
少なくともPfMSP4p21ss1およびPfMSP4p21ss2から選択される組換えポリペプチドを含む、請求項41の免疫原性組成物。
【請求項51】
少なくとも組換えポリペプチドMSP4p40およびMSP4p20を含む、請求項39の免疫原性組成物。
【請求項52】
組換えMSP1p19ポリペプチドを含む抗原をさらに含む、請求項39に記載の免疫原性組成物。
【請求項53】
インビボにおいて保護反応を誘導するために十分な量の、少なくとも1つの組換えMSP4もしくはMSP5ポリペプチドまたはこれらの混合物と、薬学的に許容される担体とを含むワクチン組成物であって、前記少なくとも1つの組換えMSP4またはMSP5ポリペプチドは、MSP4p40、MSP4p30、MSP4p20、MSP4p21ss1、MSP4p21ss2、MSP5p45、MSP5p35およびPvMSP4p20から選択されるワクチン組成物。
【請求項54】
寄生虫による感染が疑われるヒト由来の生物材料の試料におけるプラスモディウム属感染をインビトロで診断するための方法であって、以下の工程を含む方法:
-組換えMSP4またはMSP5ポリペプチドを含む抗原を、前記試料と、前記試料中の前記抗原および抗体が抗原抗体複合体を形成するために十分な時間の間および条件下で接触させる工程、および、
-前記複合体の形成を検出する工程。
【請求項55】
前記抗原抗体複合体の形成を測定することをさらに含む、請求項54に記載の方法。
【請求項56】
前記抗原抗体複合体の形成がウエスタンブロット法、ELISA、間接免疫蛍光法アッセイ法または免疫沈降アッセイ法に基づいた免疫アッセイによって検出される、請求項54に記載の方法。
【請求項57】
前記天然のMSP4またはMSP5ポリペプチドが2005年11月16日に受託番号I-3517、I-3518、I-3519またはI-3520の下でC.N.C.M.に寄託されたハイブリドーマによって分泌された抗体での免疫アッセイ法によってアッセイされる、請求項36に記載の方法。
【請求項58】
2005年11月16日に受託番号I-3517、I-3518、I-3519またはI-3520の下でC.N.C.M.に寄託されたハイブリドーマから産生される、請求項17に記載の精製された抗体。
【請求項59】
2005年11月16日に受託番号I-3517、I-3518、I-3519またはI-3520下でC.N.C.M.に寄託されたハイブリドーマ。
【請求項60】
マラリア原虫を検出するための方法であって、マラリア原虫に感染したことが疑われる生物材料を含む組成物を提供することと、および組換えMSP4もしくはMSP5ポリペプチドに対して特異的な少なくとも1つの抗体または組換えMSP4ポリペプチドに対して特異的な少なくとも1つの抗体と組換えMSP5ポリペプチドに対して特異的な抗体との混合物に結合するマラリア原虫の天然のMSP4および/またはMSP5ポリペプチドの存在についてアッセイすることとを含む方法。
【請求項61】
前記組換えMSP4またはMSp5ポリペプチドが請求項10の精製されたポリペプチドの中から選択される、請求項60に記載の方法。
【請求項62】
前記組換えMSP4またはMSP5ポリペプチドに対して特異的な抗体が請求項58の精製された抗体の中から選択される、請求項60に記載の方法。
【請求項63】
熱帯熱マラリア原虫(Plasmodium falciparum)または三日熱マラリア原虫(Plasmodium vivax)を検出するために使用される、請求項60に記載の方法。
【図1】

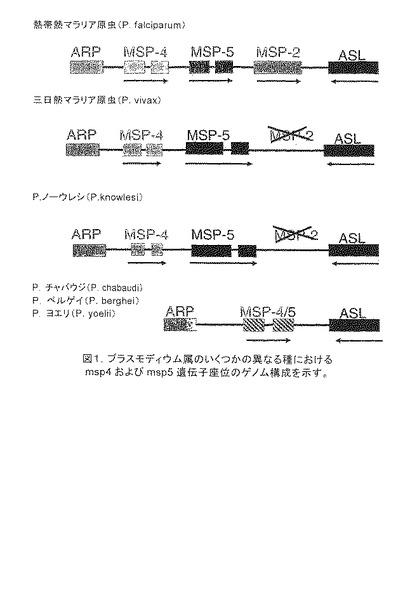
【図2】

【図3−1】
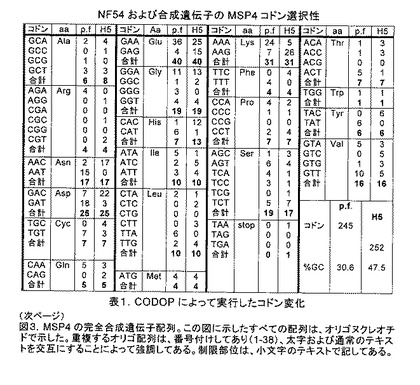
【図3−2】

【図4】
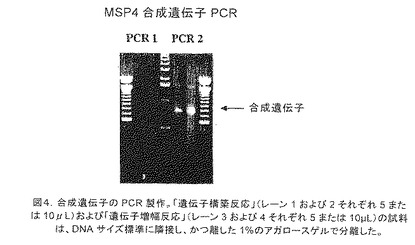
【図5】

【図6−1】
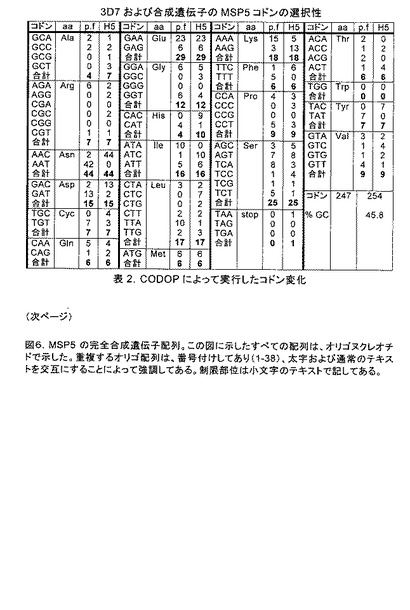
【図6−2】

【図7】
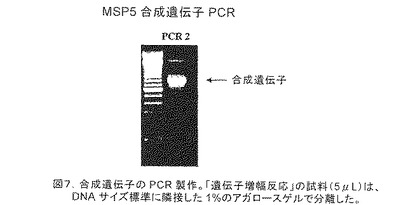
【図8】
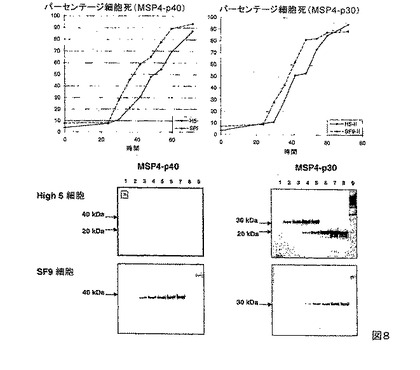
【図9】

【図10】
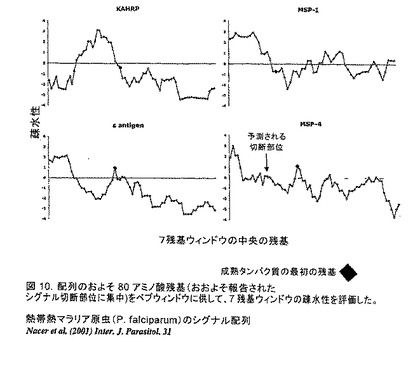
【図11】
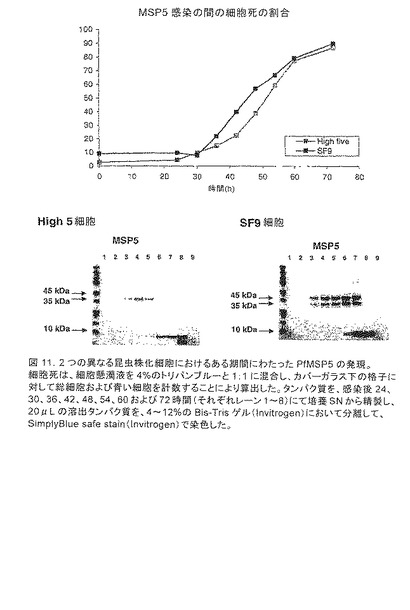
【図12】
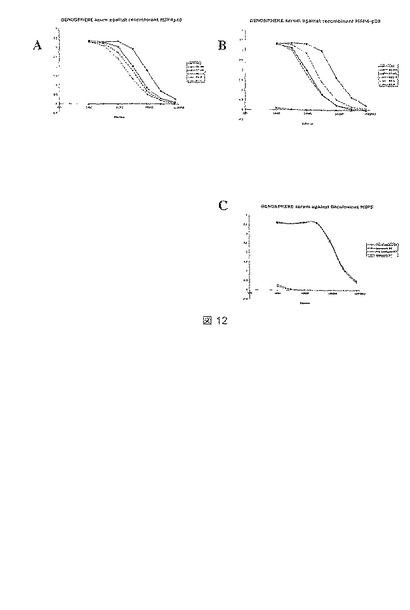
【図13】
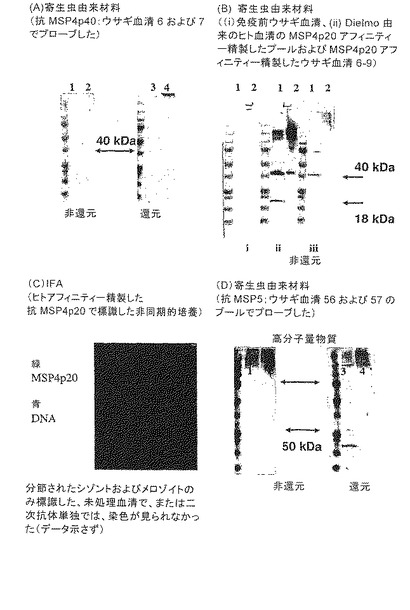
【図14】
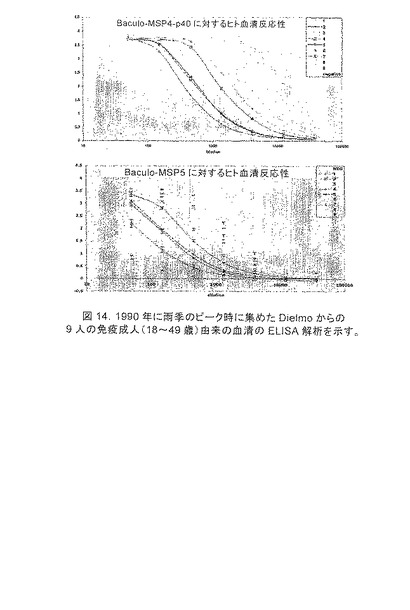
【図15】
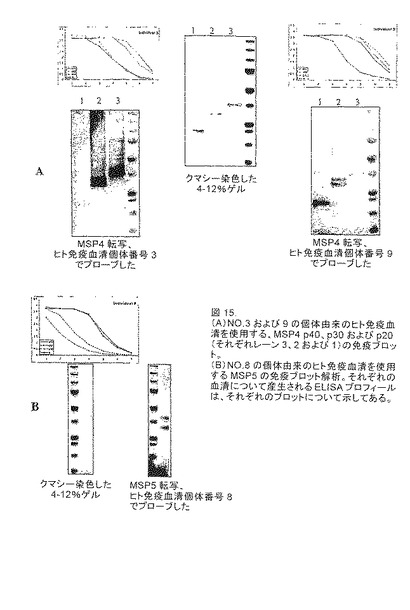
【図16】
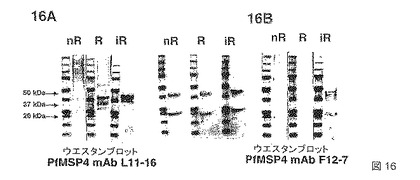
【図17】

【図18】
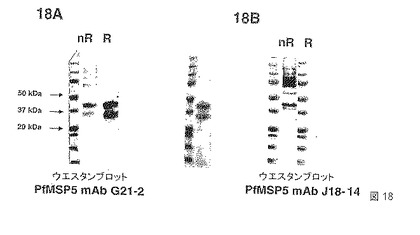
【図19】
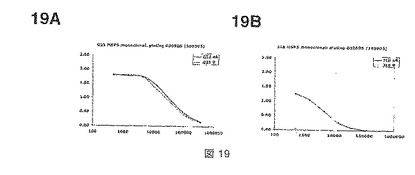
【図20】
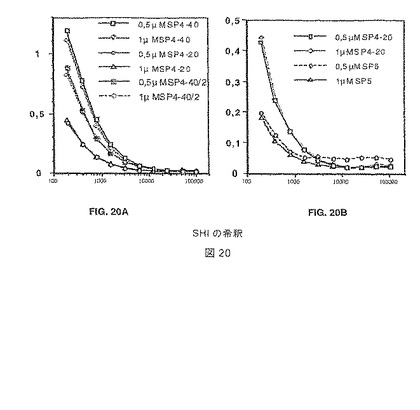
【図21】
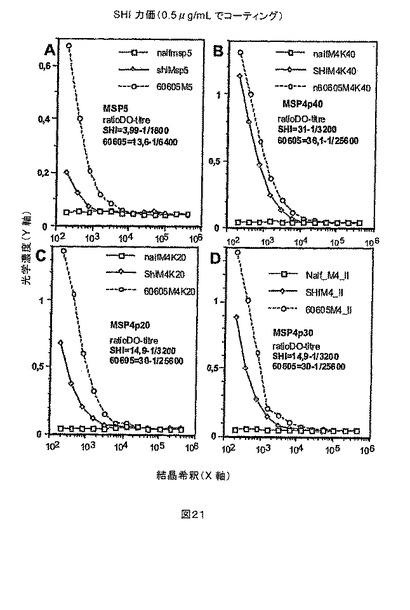
【図22】
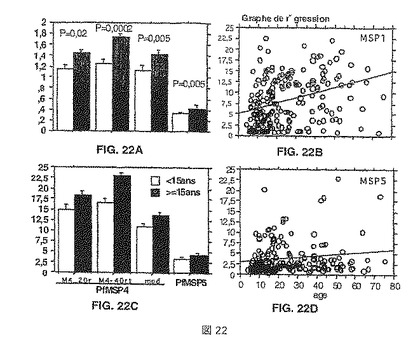
【図23】
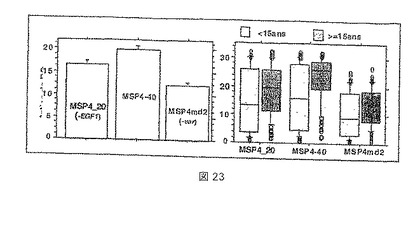
【図24】
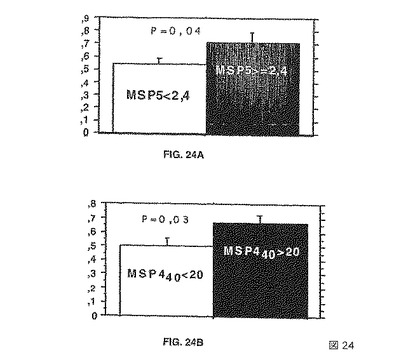
【図25】
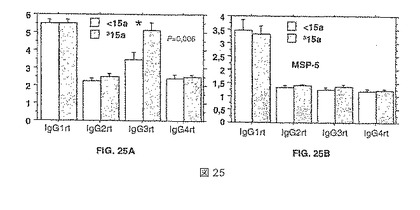
【図26】

【図27】
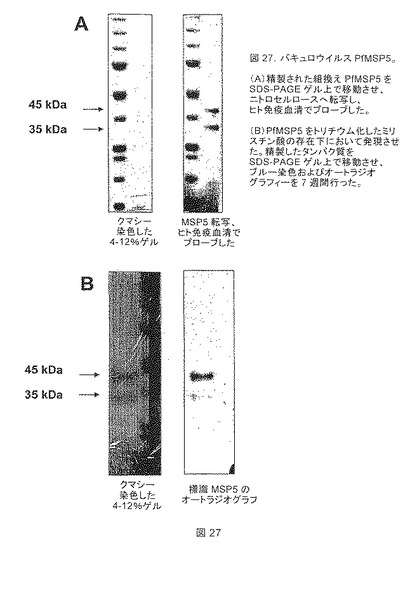
【図28】
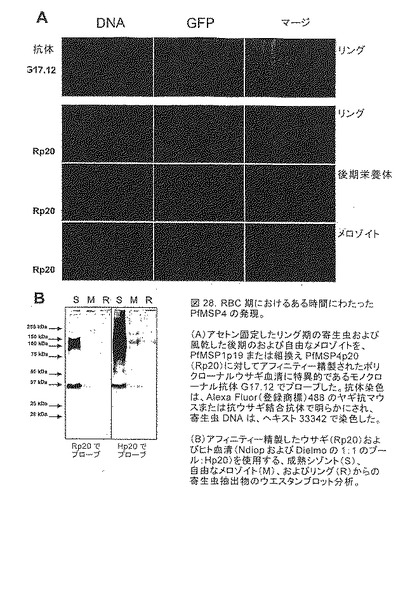
【図29】

【図30】
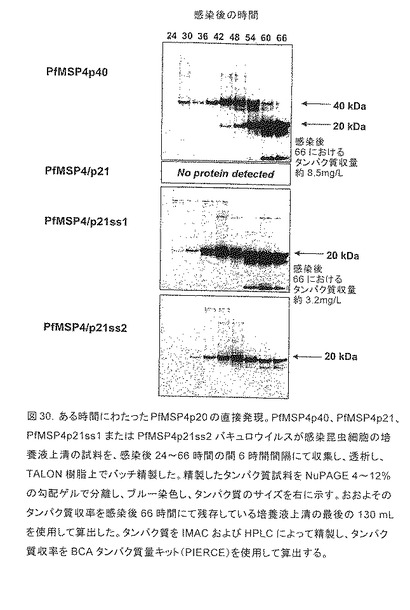
【図31】

【図32】

【図33】
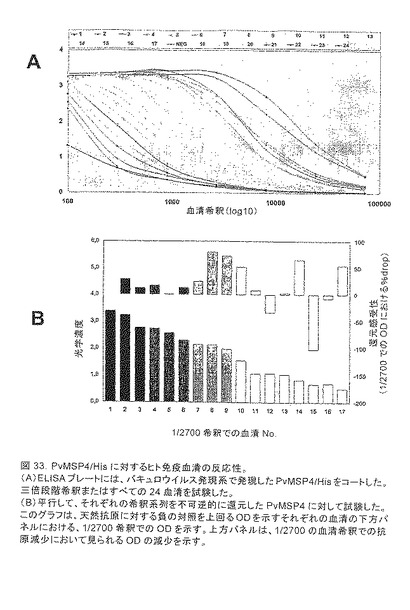
【図34】
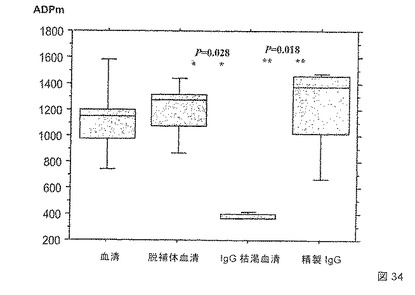
【図35】
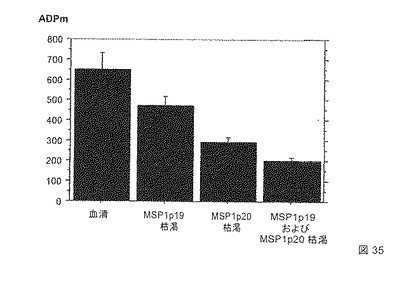
【図2】
【図3−1】
【図3−2】
【図4】
【図5】
【図6−1】
【図6−2】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31】
【図32】
【図33】
【図34】
【図35】
【公開番号】特開2012−254082(P2012−254082A)
【公開日】平成24年12月27日(2012.12.27)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−158992(P2012−158992)
【出願日】平成24年7月17日(2012.7.17)
【分割の表示】特願2008−541848(P2008−541848)の分割
【原出願日】平成18年11月23日(2006.11.23)
【出願人】(596009674)アンスティテュ・パストゥール (23)
【出願人】(500174661)サントル・ナショナル・ドゥ・ラ・レシェルシュ・サイエンティフィーク−セ・エン・エール・エス− (54)
【Fターム(参考)】
【公開日】平成24年12月27日(2012.12.27)
【国際特許分類】
【出願番号】特願2012−158992(P2012−158992)
【出願日】平成24年7月17日(2012.7.17)
【分割の表示】特願2008−541848(P2008−541848)の分割
【原出願日】平成18年11月23日(2006.11.23)
【出願人】(596009674)アンスティテュ・パストゥール (23)
【出願人】(500174661)サントル・ナショナル・ドゥ・ラ・レシェルシュ・サイエンティフィーク−セ・エン・エール・エス− (54)
【Fターム(参考)】
[ Back to top ]