
組換えDsbCを発現し、低減したTsp活性を有する細菌宿主系統
本発明は、細胞が、a.DsbCをコードする組換えポリヌクレオチドを含んでおり、b.野生型細胞に比べて低減したTspタンパク質活性を有することを特徴とする、組換えグラム陰性細菌細胞を提供する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、組換え細菌宿主系統、特に大腸菌(E.coli)に関する。本発明は、このような細胞中で対象のタンパク質を生成するための方法にも関する。
【背景技術】
【0002】
大腸菌などの細菌細胞は、組換えタンパク質を生成するために一般的に用いられる。特に、プラスミドによって遺伝子の挿入を可能にする宿主細胞として細菌細胞が多用途であるという性質のため、組換えタンパク質を生成するのに大腸菌などの細菌細胞を用いると有利な点が多い。大腸菌は、ヒトインスリンを含めた多くの組換えタンパク質を生成するのに用いられている。
【0003】
組換えタンパク質を生成するのに細菌細胞を用いるのは利点が多いにもかかわらず、プロテアーゼ感受性のタンパク質を生成するのが困難であるなど、重大な制限が依然として存在する。プロテアーゼは、大腸菌のペリプラズム及び細胞質における古く、損傷を受け、又はミスフォールディングされたタンパク質を代謝回転する上で重要な役割を果たす。細菌のプロテアーゼは、対象の組換えタンパク質を分解するように作用し、それによって活性タンパク質の収率を大幅に低減することが多い。
【0004】
数々の細菌のプロテアーゼが同定されている。大腸菌において、ProteaseIII(ptr)、DegP、OmpT、Tsp、prlC、ptrA、ptrB、pepA−T、tsh、espc、eatA、clpP、及びlonを含めたプロテアーゼが同定されている。
【0005】
Tsp(Prcとしても知られている)は、60kDaのペリプラズムプロテアーゼである。Tspの最初に知られた基質はペニシリン結合性タンパク質−3(PBP3)であったが(「Determination of the cleavage site involved in C−terminal processing of penicillin−binding protein 3 of Escherichia coli」Nagasawa H、Sakagami Y、Suzuki A、Suzuki H、Hara H、Hirota Y.、J Bacteriol.、1989年11月、171巻(11)、5890〜3頁、及び「Cloning,mapping and characterization of the Escherichia coli Tsp gene which is involved in C−terminal processing of penicillin−binding protein 3」、Hara H、Yamamoto Y、Higashitani A、Suzuki H、Nishimura Y.、J Bacteriol.、1991年8月、173巻(15)、4799〜813頁)、Tspはファージテイルタンパク質を切断することもできることが後に発見され、したがってテイル特異的プロテアーゼ(Tail Specific Protease)(Tsp)と新たに命名された(Silberら、Proc.Natl.Acad.Sci.USA、89巻、295〜299頁(1992年))、Silberらは(「Deletion of the prc(tsp) gene provides evidence for additional tail−specific proteolytic activity in Escherichia coli K−12」、Silber、K.R.、Sauer、R.T.、Mol Gen Genet、1994年、242巻、237〜240頁)、prc遺伝子のセグメントをKanrマーカーを含むフラグメントで置換することによって変異が作り出されたprcの欠失系統(KS1000)について記載している。
【0006】
Tsp(prc)活性を低減することは、対象のタンパク質のタンパク質分解を低減するのに望ましい。しかし、プロテアーゼprcを欠く細胞は、低いモル浸透圧濃度で熱感受性の増殖を示すことが見出された。Haraらは、遺伝子外サプレッサー(spr)変異を含む熱耐性復帰変異体を単離した(Haraら、Microbial Drug Resistance、2巻、63〜72頁(1996年))。sprは、18kDaの膜結合ペリプラズムプロテアーゼであり、sprの基質は細胞分裂の間細胞壁の加水分解に関与する外膜におけるTsp及びペプチドグリカンである。spr遺伝子は、UniProtKB/Swiss−Prot P0AFV4(SPR_ECOLI)と呼ばれている。
【0007】
変異体spr遺伝子を含んでいる改善されたプロテアーゼ欠損系統は記載されている。Chenら(Chen C、Snedecor B、Nishihara JC、Joly JC、McFarland N、Andersen DC、Battersby JE、Champion KM.、Biotechnol Bioeng.、2004年3月5日、85巻(5)、463〜74頁)は、prc(Tsp)、並びに遺伝子の上流及び下流の領域を増幅し、選択マーカー及びsprW174R変異を含んでいるベクター上でこれらを一緒にライゲートすることによって作り出された別のプロテアーゼであるDegP(大腸菌のペリプラズムにおける組換え抗体フラグメントの高レベルの蓄積は三重変異体(ΔDegPΔprc sprW174R)宿主系統を必要とする)における変異の異なる組合せを有する大腸菌系統の構築について記載している。ΔDegP、Δprc、及びsprW174R変異の組合せは、最高レベルの抗体軽鎖、抗体重鎖、及びF(ab)’2−LZを提供することが見出された。EP1341899は、プロテアーゼDegP及びPrcをそれぞれコードする染色体DegP及びprcを欠いており、prc変異体を抱く系統が示す増殖表現型を抑制するタンパク質をコードする変異体spr遺伝子を抱く大腸菌系統を開示している。
【0008】
タンパク質ジスルフィドイソメラーゼは、タンパク質がフォールディングするときのタンパク質内のシステイン残基間のジスルフィド結合の形成及び切断を触媒する酵素である。宿主細胞中でタンパク質の発現を改善するジスルフィド結合の形成を触媒するタンパク質を同時発現することが知られている。WO98/56930は細菌細胞中で異種性のジスルフィド結合含有ポリペプチドを生成するための方法を開示しており、DsbC又はDsbGなどの原核生物のジスルフィドイソメラーゼが真核生物のポリペプチドと同時発現される。US6673569は、外来タンパク質を生成するのに用いるためのDsbA、DsbB、DsbC、及びDsbDの各々をコードするポリヌクレオチドを含む人工オペロンを開示している。EP0786009は、細菌において異種性のポリペプチドを生成するためのプロセスを開示しており、DsbA又はDsbCをコードする核酸の発現は、異種性のポリペプチドをコードする核酸の発現の誘導の前に誘発される。
【0009】
DsbCは、大腸菌におけるジスルフィド結合の形成を触媒する大腸菌のペリプラズムにおいて見出された原核生物のタンパク質である。DsbCは、236個(シグナルペプチドを含む)の長さのアミノ酸配列、及び分子量25.6KDaを有する(UniProt No.p0AEG6)。DsbCは、1994年に始めて同定された(Missiakas eら、「The Escherichia coli dsbC(xprA)gene encodes a periplasmic protein involved in disulfide bond formation」、The EMBO Journal、13巻8号、2013〜2020頁、1994年、及びShevchikら、「Characterization of DsbC、a periplasmic protein of Erwinia chrysanthemi and Escherichia coli with disulfide isomerase activity」、The EMBO Jounral、13巻8号、2007〜2012頁、1994年)。
【0010】
驚くべきことに、グラム陰性細菌細胞における組換えDsbCからのDsbCの過剰発現は、プロテアーゼTspを欠く細胞の細胞溶解表現型を改善することが見出されている。したがって、本発明者らは、対象のタンパク質を生成するための有利な性質を有する新規な系統を提供する。
【発明の概要】
【課題を解決するための手段】
【0011】
本発明は、
a.DsbCをコードする組換えポリヌクレオチドを含んでおり、
b.野生型細胞に比べて低減したTspタンパク質活性を有する
ことを特徴とする、組換えグラム陰性細菌細胞を提供する。
【0012】
一実施形態において、細胞は野生型spr遺伝子を含んでいる。この実施形態において、細胞のゲノムが、野生型細胞に比べてTspタンパク質活性を低減するのに必要とされる修飾以外、野生型細菌細胞に対して同質遺伝子であるのが好ましい。
【0013】
さらなる一実施形態において、本発明による細胞は、野生型細胞に比べて低減したTspタンパク質活性を有し、DsbC及び変異spr遺伝子をコードする組換えポリヌクレオチドを含んでいる。この実施形態において、細胞のゲノムが、変異spr遺伝子及び野生型細胞に比べてTspタンパク質活性を低減するのに必要とされる修飾以外、野生型細菌細胞に対して同質遺伝子であるのが好ましい。
【0014】
遺伝子修飾の上記の特異的な組合せを有するグラム陰性細菌細胞は、有利な増殖及びタンパク質生成の表現型を示す。
【0015】
本発明は、上記に規定した組換えグラム陰性細菌細胞において対象のタンパク質を発現することを含む、対象のタンパク質を生成するための方法も提供する。方法は、上記に規定した組換えグラム陰性細菌細胞を、培養培地中、対象のタンパク質及びDsbCをコードする組換えポリヌクレオチドを発現するのに効果的な条件下で培養し、対象のタンパク質を、組換えグラム陰性細菌細胞のペリプラズム及び/又は培養培地から回収することを含む。
【図面の簡単な説明】
【0016】
【図1】抗TNF Fab’発現系統MXE001 MXE008、並びに抗TNFαFab’及び組換えDsbC発現系統MXE001及びMXE008の増殖プロファイルを示す図である。
【図2】抗TNFα Fab’及び組換えDsbC発現性大腸菌系統であるMXE001、MXE008、及びMXE009からのペリプラズム(黒記号)及び上清(白記号)からの抗TNFFab’の収率を示す図である。
【図3】抗TNFαFab’発現性大腸菌系統であるMXE001及びMXE008からの、並びに、抗TNF Fab’及び組換えDsbC発現性大腸菌系統であるMXE001及びMXE008からの、ペリプラズム(黒記号)及び上清(白記号)からの、抗TNF Fab’の収率を示す図である。
【図4】FabA及びFabB発現性大腸菌系統であるW3110からの、並びにFabA及び組換えDsbC、又はFabB及び組換えDsbC発現性大腸菌系統であるMXE008からの、ペリプラズムからの抗TNF Fab’の収率を示す図である。
【図5a】野生型ptr(ProteaseIII)及びノックアウト変異ptr(ProteaseIII)の5’末端のタンパク質及び遺伝子配列を示す図である。
【図5b】野生型Tsp及びノックアウト変異Tspの5’末端のタンパク質及び遺伝子配列を示す図である。
【図5c】野生型DegP及び変異DegPタンパク質のある領域及び遺伝子配列を示す図である。
【図6】本発明の一実施形態による細胞を生成するのに用いるためのベクターの構築を示す図である。
【図7】抗TNF Fab’及び組換えDsbC発現性大腸菌系統MXE008の5L及び200Lの発酵の比較上の増殖プロファイルを示す図である。
【図8】抗TNF Fab’及び組換えDsbC発現性大腸菌系統MXE008の5L及び200Lの発酵の比較上のFab’力価を示す図である。
【図9】抗TNF Fab’及び組換えDsbC発現性大腸菌系統MXE008及びMXE009の発酵の比較上の増殖プロファイルを示す図である。
【図10】抗TNF Fab’及び組換えDsbC発現性大腸菌系統MXE008及びMXE009の発酵の比較上のFab’力価を示す図である。
【0017】
配列の簡単な説明
配列番号1は、開始コドンの上流の6個のヌクレオチドATGAACを含んでいる野生型Tsp遺伝子のDNA配列である。
【0018】
配列番号2は、野生型Tspタンパク質のアミノ酸配列である。
【0019】
配列番号3は、開始コドンの上流の6個のヌクレオチドATGAATを含んでいる変異ノックアウトTsp遺伝子のDNA配列である。
【0020】
配列番号4は、野生型ProteaseIII遺伝子のDNA配列である。
【0021】
配列番号5は、野生型ProteaseIIIタンパク質のアミノ酸配列である。
【0022】
配列番号6は、変異ノックアウトProteaseIII遺伝子のDNA配列である。
【0023】
配列番号7は、野生型DegP遺伝子のDNA配列である。
【0024】
配列番号8は、野生型DegPタンパク質のアミノ酸配列である。
【0025】
配列番号9は、変異DegP遺伝子のDNA配列である。
【0026】
配列番号10は、変異DegPタンパク質のアミノ酸配列である。
【0027】
配列番号11は、抗TNF抗体の軽鎖可変領域のアミノ酸配列である。
【0028】
配列番号12は、抗TNF抗体の重鎖可変領域のアミノ酸配列である。
【0029】
配列番号13は、抗TNF抗体の軽鎖のアミノ酸配列である。
【0030】
配列番号14は、抗TNF抗体の重鎖のアミノ酸配列である。
【0031】
配列番号15は、AseI制限部位を含んでいる変異Tsp遺伝子の領域に対する3’オリゴヌクレオチドプライマーの配列である。
【0032】
配列番号16は、AseI制限部位を含んでいる変異Tsp遺伝子の領域に対する5’オリゴヌクレオチドプライマーの配列である。
【0033】
配列番号17は、AseI制限部位を含んでいる変異ProteaseIII遺伝子の領域に対する3’オリゴヌクレオチドプライマーの配列である。
【0034】
配列番号18は、AseI制限部位を含んでいる変異ProteaseIII遺伝子の領域に対する5’オリゴヌクレオチドプライマーの配列である。
【0035】
配列番号19は、AseI制限部位を含んでいる変異DegP遺伝子の領域に対する5’オリゴヌクレオチドプライマーの配列である。
【0036】
配列番号20は、AseI制限部位を含んでいる変異DegP遺伝子の領域に対する3’オリゴヌクレオチドプライマーの配列である。
【0037】
配列番号21は、最初のアミノ酸残基26個であるシグナル配列を含んでいる野生型spr遺伝子の配列である。
【0038】
配列番号22は、シグナル配列のない野生型spr遺伝子の配列である。
【0039】
配列番号23は、D210A及びH212A変異を含んでいる変異OmpT配列のヌクレオチド配列である。
【0040】
配列番号24は、D210A及びH212A変異を含んでいる変異OmpT配列のアミノ酸配列である。
【0041】
配列番号25は、変異ノックアウトOmpT配列のヌクレオチド配列である。
【0042】
配列番号26は、hisタグ付けしたDsbCのヌクレオチド配列である。
【0043】
配列番号27は、hisタグ付けしたDsbCのアミノ酸配列である。
【0044】
配列番号28は、hTNF40のCDRH1のアミノ酸配列を示す。
【0045】
配列番号29は、ハイブリッドCDRであるhTNF40のCDRH2のアミノ酸配列を示し、C末端の6個のアミノ酸はヒトサブグループ3生殖系列抗体のH2 CDR配列からであり、このハイブリダイゼーションに起因する配列に対するアミノ酸の変化には以下の通り下線をつける:WINTYIGEPI YADSVKG。
【0046】
配列番号30は、hTNF40のCDRH3のアミノ酸配列を示す。
【0047】
配列番号31は、hTNF40のCDRL1のアミノ酸配列を示す。
【0048】
配列番号32は、hTNF40のCDRL2のアミノ酸配列を示す。
【0049】
配列番号33は、hTNF40のCDRL3のアミノ酸配列を示す。
【0050】
配列番号34は、hTNF40のCDRH2のアミノ酸配列を示す。
【0051】
配列番号35は、OmpAオリゴヌクレオチドアダプターの配列を示す。
【0052】
配列番号36は、大腸菌Fab発現に対する遺伝子間配列1(IGS1)をコードするオリゴヌクレオチドカセットを示す。
【0053】
配列番号37は、大腸菌Fab発現に対する遺伝子間配列2(IGS2)をコードするオリゴヌクレオチドカセットを示す。
【0054】
配列番号38は、大腸菌Fab発現に対する遺伝子間配列3(IGS3)をコードするオリゴヌクレオチドカセットを示す。
【0055】
配列番号39は、大腸菌Fab発現に対する遺伝子間配列4(IGS4)をコードするオリゴヌクレオチドカセットを示す。
【発明を実施するための形態】
【0056】
本発明を、次に、より詳しく記載する。
【0057】
「タンパク質」及び「ポリペプチド」の語は、文脈上別段の記載がなければ、本明細書において交換可能に用いられる。「ペプチド」は10個以下のアミノ酸を意味するものとされる。
【0058】
「ポリヌクレオチド」の語は、文脈上別段の記載がなければ、遺伝子、DNA、cDNA、RNA、mRNAなどを含む。
【0059】
本明細書で用いられる「含んでいる(comprising)」の語は、本明細書の文脈において「含んでいる(including)」と解釈されたい。
【0060】
非変異(non−mutated)細胞又はコントロール細胞(control cell)は、本発明の文脈において、本発明の組換えグラム陰性菌と同じ型の細胞を意味し、細胞はTspタンパク質活性を低減し、組換えDsbC配列及び任意選択によって変異体のspr遺伝子を有するように修飾されていない。例えば、非変異細胞は野生型細胞であってよく、あらゆる変異を導入する修飾の前の本発明の細胞と同じ宿主細胞の集団に由来してよい。
【0061】
「細胞」、「細胞系」、「細胞培養物」、及び「系統」の表現は交換可能に用いられる。
【0062】
「変異Tsp遺伝子を含んでいる細胞の表現型」の表現は、本発明の文脈において、変異体のTsp遺伝子を抱く細胞によって表される表現型を意味する。典型的に、変異体のTsp遺伝子を含んでいる細胞は、特に高い細胞密度で溶解することができる。これらの細胞の溶解は、あらゆる組換えタンパク質の上清中への漏出をもたらす。変異Tsp遺伝子を有する細胞は、低いモル浸透圧濃度では熱感受性の増殖も示すことがある。例えば、40℃以上などの高温では、細胞は増殖率を示さず、若しくは低減した増殖率を示し、又は細胞は低浸透圧性の培地中で死滅する。
【0063】
「同質遺伝子の」の語は、本発明の文脈において、細胞のゲノムが、細胞が由来する野生型の細胞に比べて、野生型細胞に比べてTspタンパク質活性を低減するのに必要とされる修飾、及び任意選択によって変異spr遺伝子以外、同じ、又は実質的に同じ核酸配列(単数若しくは複数)を含んでいることを意味する。この実施形態において、細胞のゲノムはさらなる天然に存在しない変異又は遺伝子操作された変異を含んでいない。一実施形態において、本発明の細胞のゲノムは、野生型細胞に比べてTspタンパク質活性を低減するのに必要とされる修飾、及び任意選択によって変異spr遺伝子以外、野生型細胞に比べて実質的に同じ又は同じゲノム配列を有する。一実施形態において、本発明による細胞は、起こり得るあらゆる天然に存在する変異を考慮に入れて、野生型細胞に比べてTspタンパク質活性を低減するのに必要とされる修飾、及び任意選択によって変異spr遺伝子以外、野生型細胞に比べて実質的に同じゲノム配列を有し得る。一実施形態において、本発明による細胞は、野生型細胞に比べてTspタンパク質活性を低減するのに必要とされる修飾、及び任意選択によって変異spr遺伝子以外、野生型細胞に比べて正確に同じゲノム配列を有し得る。
【0064】
DsbCをコードする組換えポリヌクレオチドは、細胞中に形質転換され、及び/又は宿主細胞のゲノム中に組み入れられた適切な発現ベクター上に存在していてよい。DsbCをコードするポリヌクレオチドが宿主のゲノム中に挿入される実施形態において、細胞のゲノムはDsbCをコードする挿入されたポリヌクレオチド配列のために、野生型細胞とやはり異なる。DsbCをコードするポリヌクレオチドが細胞中の発現ベクター中にあり、それによって宿主細胞のゲノムの最小のかく乱をもたらすのが好ましい。
【0065】
一実施形態において、本発明の細胞は、対象のタンパク質をコードするポリヌクレオチドを含んでいる。この実施形態において、対象のタンパク質をコードするポリヌクレオチドは、細胞に形質転換され、及び/又は宿主細胞のゲノム中に組み入れられる適切な発現ベクター内に含まれていてよい。対象のタンパク質をコードするポリヌクレオチドが宿主のゲノム中に挿入されている実施形態において、挿入されたポリヌクレオチド配列が対象のタンパク質をコードするため、ゲノムは野生型細胞とやはり異なる。対象のタンパク質をコードするポリヌクレオチドが細胞中の発現ベクター中にあり、それによって宿主細胞のゲノムの最小のかく乱をもたらすのが好ましい。
【0066】
「野生型」の語は、本発明の文脈において、天然に存在することがあり、又は環境から単離され得るグラム陰性細菌細胞の系統を意味し、いかなる遺伝子操作された変異も保有しない。大腸菌の野生型系統の一例にはW3110−12系統などのW3110がある。
【0067】
本発明は、野生型細胞に比べて低減したTspタンパク質活性を有し、DsbCをコードする組換えポリヌクレオチドを含んでいる、対象のタンパク質を発現するのに適する組換えグラム陰性細菌細胞を提供する。
【0068】
本発明による細胞は、DsbCをコードする組換えポリヌクレオチドを含んでいる。本明細書で用いられる「組換えポリペプチド」は、組換えDNA技術を用いて構築又は生成されるタンパク質を意味する。DsbCをコードするポリヌクレオチド配列は、細菌細胞中に見出されるDsbCをコードする内在性の配列に同一であってよい。或いは、DsbCをコードする組換えポリヌクレオチド配列は、EcoRI部位などの制限部位が除去された、及び/又はhisタグをコードする配列を有するなど、野生型DsbC配列の変異バージョンである。本発明に用いるための一例の修飾DsbCヌクレオチド配列を配列番号26に示すが、これは配列番号27に示すhisタグ付けしたDsbCアミノ酸配列をコードするものである。
【0069】
本発明者らは、野生型細胞に比べて低減したTspタンパク質活性を有する細菌細胞においてDsbCをコードする組換えポリヌクレオチド発現の特定の組合せ、及び好ましい実施形態において、さらに変異spr遺伝子は、対象のタンパク質を発現するために改善された宿主を提供することを見出した。上記の遺伝子変異の特定の組合せは、ノックアウト変異Tsp遺伝子を有する細胞に比べて、細胞の健康状態及び増殖の改善された表現型を有する新しい系統を提供する。変異Tsp遺伝子を有する細胞は細胞増殖速度が良好であることがあるが、これらの細胞の限界の一つは、特に高い細胞密度で溶解する傾向があることである。したがって、変異Tsp遺伝子を含んでいる細胞の表現型は、特に高い細胞密度で溶解する傾向がある。しかし、本発明の細胞中でのDsbCを発現させると、低減したTsp表現型を抑制し、したがって細胞は溶解の低減を示す。
【0070】
本発明による細胞は、ノックアウト変異Tsp遺伝子を有する細胞に比べて改善されたタンパク質生成収率を示す。改善されたタンパク質収率は、タンパク質生成速度であっても、及び/又は細胞からのタンパク質生成の期間であってもよい。改善されたタンパク質収率は、ペリプラズマのタンパク質収率でも、及び/又は上清のタンパク質収率でもよい。一実施形態において、本発明の細胞は、細胞からの漏出が低減するために、変異Tsp遺伝子を有する細胞に比べて改善されたペリプラズマタンパク質収率を示す。組換え細菌細胞は、対象のタンパク質の生成速度を速めることが可能であることがあり、したがって同じ量の対象のタンパク質が、変異Tsp遺伝子を含んでいる細胞に比べてより短時間で生成され得る。対象のタンパク質の生成速度がより速いのは、細胞の増殖の最初の期間にわたって、例えば、タンパク質発現の誘導後最初の5時間、10時間、20時間、又は30時間にわたって、特に著しいことがある。
【0071】
本発明による細胞が、ペリプラズム及び/又は培地において、対象のタンパク質のおよそ1.0g/L、1.5g/L、1.8g/L、2.0g/L、2.4g/L、2.5g/L、3.0g/L、3.5g/L、又は4.0g/Lの、最大の収率を発現するのが好ましい。
【0072】
本発明が提供する細胞は、野生型細胞に比べて低減したプロテアーゼ活性を有し、本発明が提供する細胞は対象の組換えタンパク質、特にTspプロテアーゼに対してタンパク質分解感受性である対象のタンパク質のタンパク質分解性を低減し得る。したがって、本発明が提供する細胞は、野生型の細菌細胞に比べて、インタクトなタンパク質、好ましくは対象のタンパク質をより高い収率で提供し、タンパク質、好ましくは対象のタンパク質のタンパク質分解性フラグメントをより低い収率で提供し、好ましくはタンパク質分解性フラグメントを全く提供しない。
【0073】
当業者であれば、発酵法、ELISA、及びタンパク質G HPLCを含めた当技術分野においてよく知られている方法を用いて、それが望ましい収率の対象のタンパク質を有するか否かを見るために、候補の細胞クローンを試験することが容易にできる。適切な発酵法は、Humphreys D Pら(1997年)、「Formation of dimeric Fabs in E. coli:effect of hinge size and isotype,presence of interchain disulphide bond,Fab’ expression levels,tail piece sequences and growth conditions.」、J.IMMUNOL.METH.、209巻、193〜202頁;Backlund E.、Reeks D.、Markland K.、Weir N.、Bowering L.、Larsson G.、「Fedbatch design for periplasmic product retention in Escherichia coli」、雑誌記事、Research Support Non−U.S. Gov’t Journal of Biotechnology.、135巻(4)、358〜65頁、2008年7月31日; Champion KM.、Nishihara JC.、Joly JC.、Arnott D.、「Similarity of the Escherichia coli proteome upon completion of different biopharmaceutical fermentation processes.」、[雑誌記事]Proteomics.、1巻(9)、1133〜48頁、2001年9月;及びHorn U.、Strittmatter W.、Krebber A.、Knupfer U.、Kujau M.、Wenderoth R.、Muller K.、Matzku S.、Pluckthun A.、Riesenberg D.、「High volumetric yields of functional dimeric miniantibodies in Escherichia coli,using an optimized expression vector and high−cell−density fermentation under non−limited growth conditions」、雑誌記事、Research Support、Non−U.S. Gov’t Applied Microbiology & Biotechnology.、46巻(5〜6)、524〜32頁、1996年12月に記載されている。当業者であれば、タンパク質G HPLC、円偏光二色性、NMR、X線結晶構造解析、及びエピトープ親和性測定方法など、当技術分野においてよく知られている方法を用いて、タンパク質が正確にフォールディングされているか否かを見るために分泌されたタンパク質を試験することも容易にできる。
【0074】
一実施形態において、本発明による細胞は、以下の1つ又は複数のさらなるタンパク質も発現する。
タンパク質のフォールディングを促進することができる1つ又は複数のタンパク質、例えば、FkpA、Skp、SirA、PPiA、及びPPid、並びに/或いは、
タンパク質の分泌若しくは翻訳を促進することができる1つ又は複数のタンパク質、例えば、SecY、SecE、SecG、SecYEG、SecA、SecB、FtsY、及びLep、並びに/或いは、
ジスルフィド結合の形成を促進することができる1つ又は複数のタンパク質、例えば、DsbA、DsbB、DsbD、DsbG。
【0075】
1つ又は複数の上記のタンパク質を、細胞のゲノム中に組み入れてもよく、且つ/又は発現ベクター中に挿入してもよい。
【0076】
一実施形態において、本発明による細胞は、1つ又は複数の以下のさらなるタンパク質をコードする組換えポリヌクレオチドを含んでいない。
タンパク質のフォールディングを促進することができる1つ又は複数のタンパク質、例えば、FkpA、Skp、SurA、PPiA、及びPPiD、
タンパク質の分泌又は翻訳を促進することができる1つ又は複数のタンパク質、例えば、SecY、SecE、SecG、SecYEG、SecA、SecB、FtsY、及びLep、並びに
ジスルフィド結合の形成を促進することができる1つ又は複数のタンパク質、例えば、DsbA、DsbB、DsbD、DsbG。
【0077】
本発明の実施形態において、細胞は変異spr遺伝子をさらに含んでいる。sprタンパク質は、大腸菌膜結合ペリプラズムプロテアーゼである。
【0078】
sprタンパク質の野生型アミノ酸配列を、N末端にシグナル配列を有する配列番号21、及びアミノ酸26個のシグナル配列のない配列番号22において示す(UniProt受諾番号P0AFV4による)。本発明におけるsprタンパク質配列のアミノ酸番号付けはシグナル配列を含んでいる。したがって、sprタンパク質のアミノ酸1は、配列番号21に示す最初のアミノ酸(Met)である。
【0079】
本発明による細胞が変異spr遺伝子を含んでいる実施形態において、変異spr遺伝子が細胞の染色体spr遺伝子であるのが好ましい。変異spr遺伝子は、変異Tsp遺伝子を含んでいる細胞の表現型を抑制することができるsprタンパク質をコードしている。変異Tsp遺伝子を有する細胞は、良好な細胞増殖速度を有し得るが、これらの細胞の一つの制限は、特に高い細胞濃度で溶解する傾向があることである。したがって、変異Tsp遺伝子を含んでいる細胞の表現型は、特に高い細胞濃度で溶解する傾向がある。変異Tsp遺伝子を有する細胞も、低いモル浸透圧濃度で熱感受性の増殖を示す。しかし、本発明の細胞が有するspr変異は、低減したTsp活性を有する細胞中に導入した場合、低減したTsp表現型を抑制し、したがって、細胞は、特に高い細胞密度で溶解の低減を表す。細胞の増殖の表現型は、当業者であれば、フラスコ振盪又は高い細胞密度の発酵技術の間に容易に測定することができる。細胞溶解の表現型の抑制は、特にペリプラズムにおいて増殖速度の改善及び/又は組換えタンパク質の生成から見ることができ、Tsp変異体及び野生型sprを有する細胞に比べて、spr変異体を有し、低減したTsp活性を有する細胞によって示される。
【0080】
本発明による細胞は、好ましくはN31、R62、I70、Q73、C94、S95、V98、Q99、R100、L108、Y115、D133、V135、L136、G140、R144、H145、G147、H157、及びW174から選択される1つ又は複数のアミノ酸、より好ましくはC94、S95、V98、Y115、D133、V135、H145、G147、H157、及びW174から選択される1つ又は複数のアミノ酸に変異を有するsprタンパク質をコードする変異体spr遺伝子を含んでいる。好ましくは、変異体spr遺伝子は、N31、R62、I70、Q73、C94、S95、V98、Q99、R100、L108、Y115、D133、V135、L136、G140、R144、H145、G147、及びH157から選択される1つ又は複数のアミノ酸、より好ましくはC94、S95、V98、Y115、D133、V135、H145、G147、及びH157から選択される1つ又は複数のアミノ酸に変異を有するsprタンパク質をコードしている。この実施形態において、sprタンパク質にいかなるさらなる変異もないのが好ましい。好ましくは、変異体のspr遺伝子は、N31、R62、I70、Q73、S95、V98、Q99、R100、L108、Y115、D133、V135、L136、G140、R144、及びG147から選択される1つ又は複数のアミノ酸に、より好ましくはS95、V98、Y115、D133、V135、及びG147から選択される1つ又は複数のアミノ酸に変異を有するsprタンパク質をコードする。この実施形態において、sprタンパク質にいかなるさらなる変異もないのが好ましい。
【0081】
本発明者らは、変異Tsp遺伝子を含んでいる細胞の増殖の表現型を抑制することができるspr変異を同定している。
【0082】
本発明者らはまた、驚くべきことに、組換えDsbC遺伝子、新たな変異spr遺伝子を有し、野生型細胞に比べて低減したTspタンパク質活性を有する細胞は、変異Tsp遺伝子を含んでいる細胞に比べて増大した細胞増殖速度、及び増大した細胞生存期間を示すことも見出した。特に、組換えDsbC遺伝子、spr変異を有し、低減したTspタンパク質活性を有する細胞は、変異Tsp遺伝子を有する細胞に比べて低減した細胞溶解の表現型を示す。
【0083】
上記のsprアミノ酸の1つ又は複数の変異は、アミノ酸をコードするヌクレオチドの1個、2個、又は3個に対するあらゆる適切なミスセンス変異であってよい。変異は、アミノ酸残基を、変異Tsp遺伝子を含んでいる細胞の表現型を抑制することができる変異sprタンパク質をもたらすあらゆる適切なアミノ酸へ変化させる。ミスセンス変異は、アミノ酸を、野生型アミノ酸に比べて異なるサイズであり、且つ/又は異なる化学性質を有するアミノ酸へ変化させることができる。
【0084】
一実施形態において、変異は、C94、H145、及びH157のアミノ酸残基の触媒三残基の1つ、2つ、又は3つに対するものである(「Solution NMR Structure of the NlpC/P60 Domain of Lipoprotein Spr from Escherichia coli Structural Evidence for a Novel Cysteine Peptidase Catalytic Triad」、Biochemistry、2008年、47巻、9715〜9717頁)。
【0085】
したがって、変異spr遺伝子は、
C94に対する変異、又は
H145に対する変異、又は
H157に対する変異、又は
C94及びH145に対する変異、又は
C94及びH157に対する変異、又は
H145及びH157に対する変異、又は
C94、H145、及びH157に対する変異
を含んでいてよい。
【0086】
この実施形態において、sprタンパク質がいかなるさらなる変異も有さないのが好ましい。
【0087】
C94、H145、及びH157の1つ、2つ、又は3つが、変異Tsp遺伝子を含んでいる細胞の表現型を抑制することができるsprタンパク質をもたらすあらゆる適切なアミノ酸に変異されてよい。例えば、C94、H145、及びH157の1つ、2つ、又は3つは、Gly又はAlaなどの小型のアミノ酸に変異されてよい。したがって、sprタンパク質は、C94、H145、及びH157の1つ、2つ、又は3つの変異を有することができる。spr遺伝子が、変異Tsp遺伝子を含んでいる細胞の表現型を抑制することができるsprタンパク質を生成することが見出されているミスセンス変異H145Aを含んでいるのが好ましい。
【0088】
本明細書の置換変異体に対する呼称は、1つの文字とそれに続く1つの数字とそれに続く1つの文字からなる。最初の文字は野生型タンパク質におけるアミノ酸を指定する。数字はアミノ酸置換が行われるアミノ酸位置を表し、2番目の文字は野生型アミノ酸を置換するのに用いられるアミノ酸を指定する。
【0089】
好ましい一実施形態において、変異体のsprタンパク質は、N31、R62、I70、Q73、S95、V98、Q99、R100、L108、Y115、D133、V135、L136、G140、R144、及びG147から選択される1つ又は複数のアミノ酸の変異、好ましくはS95、V98、Y115、D133、V135、及びG147から選択される1つ又は複数のアミノ酸の変異を含んでいる。この実施形態において、sprタンパク質がいかなるさらなる変異を有さないのが好ましい。したがって、変異spr遺伝子は、
N31に対する変異、又は
R62に対する変異、又は
I70に対する変異、又は
Q73に対する変異、又は
S95に対する変異、又は
V98に対する変異、又は
Q99に対する変異、又は
R100に対する変異、又は
L108に対する変異、又は
Y115に対する変異、又は
D133に対する変異、又は
V135に対する変異、又は
L136に対する変異、又は
G140に対する変異、又は
R144に対する変異、又は
G147に対する変異
を含んでいてよい。
【0090】
一実施形態において、変異体sprタンパク質は、アミノ酸:
S95及びY115、又は
N31、Q73、R100及びG140、又は
Q73、R100及びG140、又は
R100及びG140、又は
Q73及びG140、又は
Q73及びR100、又は
R62、Q99及びR144、又は
Q99及びR144
に対する複数の変異を含んでいる。
【0091】
N31、R62、I70、Q73、S95、V98、Q99、R100、L108、Y115、D133、V135、L136、G140、R144、及びG147の1つ又は複数のアミノ酸は、変異Tsp遺伝子を含んでいる細胞の表現型を抑制することができるsprタンパク質をもたらすあらゆる適切なアミノ酸に変異されてよい。例えば、N31、R62、I70、Q73、S95、V98、Q99、R100、L108、Y115、D133、V135、L136、G140、及びR144の1つ又は複数は、Gly又はAlaなどの小型のアミノ酸に変異されてよい。
【0092】
好ましい一実施形態において、sprタンパク質は、1つ又は複数の以下の変異を含んでいる:N31Y、R62C、I70T、Q73R、S95F、V98E、Q99P、R100G、L108S、Y115F、D133A、V135D又はV135G、L136P、G140C、R144C、及びG147C。sprタンパク質が1つ又は複数の以下の変異を含んでいるのが好ましい:S95F、V98E、Y115F、D133A、V135D又はV135G、及びG147C。この実施形態において、sprタンパク質にいかなるさらなる変異もないのが好ましい。
【0093】
一実施形態において、sprタンパク質は、N31Y、R62C、I70T、Q73R、S95F、V98E、Q99P、R100G、L108S、Y115F、D133A、V135D又はV135G、L136P、G140C、R144C、及びG147Cから選択される変異を1つ有する。この実施形態において、sprタンパク質にいかなるさらなる変異もないのが好ましい。
【0094】
さらなる一実施形態において、sprタンパク質は、以下から選択される複数の変異を有する:
S95F及びY115F
N31Y、Q73R、R100G、及びG140C、
Q73R、R100G、及びG140C、
R100G及びG140C、
Q73R及びG140C、
Q73R及びR100G、
R62C、Q99P及びR144C、又は
Q99P及びR144C。
【0095】
変異体のspr遺伝子が、C94A、D133A、H145A、及びH157Aから選択される変異を有するsprタンパク質をコードするのが好ましい。
【0096】
さらなる一実施形態において、変異spr遺伝子は変異W174Rを有するsprタンパク質をコードする。代替の一実施形態において、sprタンパク質は変異W174Rを有さない。
【0097】
本発明による細胞は、野生型細胞に比べて低減したTspタンパク質活性を有する。「野生型細胞に比べて低減したTspタンパク質活性」の表現は、細胞のTsp活性が野生型細胞のTsp活性に比べて低減していることを意味する。細胞を、Tspの活性を低減するのに適するあらゆる手段によって修飾することができる。
【0098】
一実施形態において、低減したTsp活性は、Tsp及び/又は関連の調節性の発現配列をコードする内因性ポリヌクレオチドの修飾による。修飾は、Tsp遺伝子の転写及び翻訳を低減又は停止することができ、或いは野生型Tspタンパク質に比べて低減したプロテアーゼ活性を有するTspタンパク質の発現を提供することができる。
【0099】
一実施形態において、関連の調節性の発現配列を、Tsp発現を低減するように修飾する。例えば、Tsp遺伝子に対するプロモーターを遺伝子の発現を防止するように変異させてもよい。
【0100】
好ましい一実施形態において、本発明による細胞は、低減したプロテアーゼ活性を有するTspタンパク質をコードする変異Tsp遺伝子、又はノックアウト変異Tsp遺伝子を有している。染色体のTsp遺伝子が変異しているのが好ましい。
【0101】
本明細書で用いられる「Tsp遺伝子」は、ペニシリン結合性タンパク質−3(PBP3)及びファージテイルタンパク質に対して作用することができるペリプラズムプロテアーゼであるプロテアーゼTsp(Prcとしても知られている)をコードする遺伝子を意味する。野生型Tsp遺伝子の配列を配列番号1に示し、野生型Tspタンパク質の配列を配列番号2に示す。
【0102】
変異Tsp遺伝子、又はTspをコードする変異Tsp遺伝子に対する言及は、別段の指摘がなければ、低減したプロテアーゼ活性を有するTspタンパク質をコードする変異Tsp遺伝子、又はノックアウト変異Tsp遺伝子のいずれかを意味する。
【0103】
「低減したプロテアーゼ活性を有するTspタンパク質をコードする変異Tsp遺伝子」の表現は、本発明の文脈において、変異Tsp遺伝子が野生型の非変異Tsp遺伝子に比べて完全なプロテアーゼ活性を有さないことを意味する。
【0104】
変異Tsp遺伝子が、野生型の非変異Tspタンパク質のプロテアーゼ活性の50%以下、40%以下、30%以下、20%以下、10%以下、又は5%以下を有するTspタンパク質をコードするのが好ましい。変異Tsp遺伝子が、プロテアーゼ活性を有さないTspタンパク質をコードするのがより好ましい。この実施形態において、細胞は染色体Tspを欠くのではなく、即ち、Tsp遺伝子配列はあらゆる形態のTspタンパク質の発現を防ぐように欠失又は変異されていない。
【0105】
低減したプロテアーゼ活性を有するタンパク質を生成するために、あらゆる適切な変異をTsp遺伝子中に導入することができる。グラム陰性細菌から発現されるTspタンパク質のプロテアーゼ活性は、当業者であれば、Tspのプロテアーゼ活性が試験されるKeilerら(「Identification of Active Site Residues of the Tsp Protease*」、THE JOURNAL OF BIOLOGICAL CHEMISTRY、270巻48号、12月1日発行、28864〜28868頁、1995年、Kenneth C.Keiler及びRobert T.Sauer)において記載されている方法など、当技術分野におけるあらゆる適切な方法によって容易に試験することができる。
【0106】
TspはKeilerら(上述)において、残基S430、D441、及びK455を含む活性部位を有すると報告されており、残基G375、G376、E433、及びT452はTspの構造を維持するのに重要である。Keilerら(上述)は、変異Tsp遺伝子S430A、D441A、K455A、K455H、K455R、G375A、G376A、E433A、及びT452Aには検出可能なプロテアーゼ活性がなかったという所見を報告している。変異Tsp遺伝子S430Cは野生型活性の約5〜10%を示したことがさらに報告されている。したがって、低減したプロテアーゼ活性を有するタンパク質を生成するためのTsp変異は、S430、D441、K455、G375、G376、E433、及びT452の1つ又は複数の残基に対するミスセンス変異などの変異を含むことがある。低減したプロテアーゼ活性を有するタンパク質を生成するためのTsp変異が、活性部位の残基S430、D441、及びK455の1つ、2つ、又は3つ全てに対するミスセンス変異などの変異を含むことができるのが好ましい。
【0107】
したがって、変異Tsp遺伝子は以下を含むことができる:
S430に対する変異、又は
D441に対する変異、又は
K455に対する変異、又は
S430及びD441に対する変異、又は
S430及びK455に対する変異、又は
D441及びK455に対する変異、又は
S430、D441、及びK455に対する変異。
【0108】
S430、D441、K455、G375、G376、E433、及びT452の1つ又は複数が、低減したプロテアーゼ活性を有するタンパク質をもたらすあらゆる適切なアミノ酸に変異していてよい。適切な変異の例には、S430A、S430C、D441A、K455A、K455H、K455R、G375A、G376A、E433A、及びT452Aがある。変異Tsp遺伝子は、活性部位残基に対する1つ、2つ、又は3つの変異を含むことができ、例えば、遺伝子は以下を含むことができる:
S430A、若しくはS430C、及び/又は
D441A及び/又は
K455A若しくはK455H、若しくはK455R。
【0109】
Tsp遺伝子が点変異S430A又はS430Cを有するのが好ましい。
【0110】
「ノックアウト変異Tsp遺伝子」の表現は、本発明の文脈において、Tsp遺伝子が、Tspタンパク質を欠く細胞を提供するために、野生型遺伝子によってコードされるTspタンパク質の発現を防ぐ1つ又は複数の変異を含んでいることを意味する。ノックアウト遺伝子は、部分的又は完全に転写され得るが、コードタンパク質に翻訳されない。ノックアウト変異Tsp遺伝子は、タンパク質の発現をもたらさないように、1つ又は複数の欠失、挿入、点、ミスセンス、ナンセンス、及びフレームシフトの変異によるなど、あらゆる適切な方法において変異され得る。例えば、遺伝子は、遺伝子コード配列中に、抗生物質耐性マーカーなどの外来のDNA配列を挿入することによってノックアウトされてよい。
【0111】
好ましい一実施形態において、Tsp遺伝子は、遺伝子コード配列中に、抗生物質耐性マーカーなどの外来のDNA配列を挿入することによって変異されない。一実施形態において、Tsp遺伝子は、遺伝子開始コドン、並びに/或いは遺伝子開始コドンの下流、及び遺伝子終止コドンの上流に位置する1つ又は複数の終止コドンに対する変異を含んでおり、それによってTspタンパク質の発現を防いでいる。開始コドンに対する変異は、開始コドンの1つ、2つ、又は3つ全てのヌクレオチドのミスセンス変異であってよい。或いは、又はさらに、開始コドンは、挿入又は欠失のフレームシフト変異によって変異されてよい。Tsp遺伝子は、コード配列の5’末端に2つのATGコドンを含んでおり、ATGコドンの1つ又は両方はミスセンス変異によって変異されてよい。Tsp遺伝子は、図5bに示す通り、第2のATGコドン(コドン3)からTCGコドンに変異されてよい。Tsp遺伝子は、或いは、又はさらに、遺伝子開始コドンの下流及び遺伝子終止コドンの上流に位置する1つ又は複数の終止コドンを含んでいてよい。ノックアウト変異Tsp遺伝子が、開始コドンに対するミスセンス変異、及び1つ又は複数の挿入された終止コドンの両方を含んでいるのが好ましい。好ましい一実施形態において、Tsp遺伝子は第5のコドンから「T」を欠失するように変異されており、それによって、図5bに示す通り、コドン11及び16に終止コドンをもたらすフレームシフトを生じている。好ましい一実施形態において、図5bに示す通り、コドン21に第3のインフレーム終止コドンを作り出すように、Tsp遺伝子はAseI制限部位を挿入するように変異される。
【0112】
好ましい一実施形態において、ノックアウト変異Tsp遺伝子は配列番号3のDNA配列を有し、これは開始コドンの上流にATGAATの6個のヌクレオチドを含んでいる。配列番号3のノックアウト変異Tsp配列においてなされた変異を図5bに示す。一実施形態において、変異Tsp遺伝子は、配列番号3のヌクレオチド7から2048のDNA配列を有する。
【0113】
本発明の好ましい一実施形態において、組換えグラム陰性細菌細胞は、シャペロン活性及び低減したプロテアーゼ活性を有するDegPタンパク質をコードする変異DegP遺伝子、並びに/或いは変異ptr遺伝子をさらに含んでおり、変異ptr遺伝子は低減したプロテアーゼ活性を有するProteaseIIIタンパク質をコードする、又はノックアウト変異ptr遺伝子及び/又は変異OmpT遺伝子であり、変異OmpT遺伝子は、低減したプロテアーゼ活性を有するOmpTタンパク質をコードする、又はノックアウト変異OmpT遺伝子である。
【0114】
一実施形態において、本発明は、
a.DsbCをコードする組換えポリヌクレオチド、
b.低減したプロテアーゼ活性を有するTspタンパク質をコードする変異Tsp遺伝子、又はノックアウト変異Tsp遺伝子、
c.シャペロン活性及び低減したプロテアーゼ活性、及び/又は変異OmpTを有するDegPタンパク質をコードする変異DegP遺伝子(変異OmpT遺伝子は、低減したプロテアーゼ活性を有するOmpTタンパク質をコードする、又はノックアウト変異OmpT遺伝子である)、並びに
d.任意選択によって変異ptr遺伝子
を含んでいる組換えグラム陰性細菌細胞を提供する。
【0115】
この実施形態において、細胞のゲノムが、上記の変異b、c、及びd以外は野生型細菌細胞に対して同質遺伝子であるのが好ましい。
【0116】
一実施形態において、本発明は、
a.DsbCをコードする組換えポリヌクレオチド、
b.低減したプロテアーゼ活性を有するTspタンパク質をコードする変異Tsp遺伝子、又はノックアウト変異Tsp遺伝子、
c.変異ptr遺伝子(変異ptr遺伝子は、低減したプロテアーゼ活性を有するProteaseIIIをコードする、又はノックアウト変異ptr遺伝子であり、且つ/又は変異OmpTであり、変異OmpT遺伝子は、低減したプロテアーゼ活性を有するOmpTタンパク質をコードする、又はノックアウト変異OmpT遺伝子である)、並びに
d.任意選択によって変異spr遺伝子
を含んでいる組換えグラム陰性細菌細胞を提供する。
【0117】
この実施形態において、細胞のゲノムが、上記の変異b、c、及びd以外は野生型細菌細胞に対して同質遺伝子であるのが好ましい。
【0118】
一実施形態において、本発明は、
a.DsbCをコードする組換えポリヌクレオチド、
b.低減したプロテアーゼ活性を有するTspタンパク質をコードする変異Tsp遺伝子、又はノックアウト変異Tsp遺伝子、
c.シャペロン活性及び低減したプロテアーゼ活性を有するDegPタンパク質をコードする変異DegP遺伝子、
d.変異ptr遺伝子(変異ptr遺伝子は、低減したプロテアーゼ活性を有するProteaseIIIタンパク質をコードする、又はノックアウト変異ptr遺伝子である)、
e.任意選択によって変異OmpT(変異Ompt遺伝子は、低減したプロテアーゼ活性を有するOmpTタンパク質をコードする、又はノックアウト変異OmpT遺伝子である)、並びに
f.任意選択によって変異spr遺伝子
を含んでいる細胞を提供する。
【0119】
この実施形態において、細胞のゲノムが、上記の変異b、c、d、e、及びf以外は野生型細菌細胞に対して同質遺伝子であるのが好ましい。
【0120】
本発明の実施形態において、細胞は変異DegP遺伝子を含んでいる。本明細書で用いられる「DegP」は、DegPタンパク質(HtrAとしても知られている)をコードする遺伝子を意味し、DegPタンパク質はシャペロン及びプロテアーゼとしての二重機能を有する(「Families of serine peptidases」、Rawlings ND、Barrett AJ.、Methods Enzymol.、1994年、244巻、19〜61頁)。非変異のDegP遺伝子の配列を配列番号7に示し、非変異のDegPタンパク質の配列を配列番号8に示す。
【0121】
低温でDegPはシャペロンとして機能し、高温でDegPはプロテアーゼとしての機能を優先する(「A Temperature−Dependent Switch from Chaperone to Protease in a Widely Conserved Heat Shock Protein.」Cell、97巻3号、339〜347頁、Spiess C、Beil A、Ehrmann M)、及び「The proteolytic activity of the HtrA (DegP) protein from Escherichia coli at low temperatures」、Skorko−Glonek Jら、Microbiology、2008年、154巻、3649〜3658頁)。
【0122】
細胞がDegP変異を含んでいる実施形態において、細胞におけるDegP変異は、シャペロン活性を有するが完全なプロテアーゼを有さないDegPタンパク質をコードする変異DegP遺伝子を提供する。
【0123】
「シャペロン活性を有する」の表現は、本発明の文脈において、変異DegPタンパク質が、野生型の非変異のDegPタンパク質に比べて同じ、又は実質的に同じシャペロン活性を有することを意味する。変異DegP遺伝子が、野生型の非変異のDegPタンパク質のシャペロン活性の50%以上、60%以上、70%以上、80%以上、90%以上、又は95%以上を有するDegPタンパク質をコードするのが好ましい。変異DegP遺伝子が、野生型DegPに比べて同じシャペロン活性を有するDegPタンパク質をコードするのがより好ましい。
【0124】
「低減したプロテアーゼ活性を有する」の表現は、本発明の文脈において、変異DegPタンパク質が、野生型の非変異のDegPタンパク質に比べて完全なプロテアーゼ活性を有さないことを意味する。変異DegP遺伝子が、野生型の非変異のDegPタンパク質のプロテアーゼ活性の50%以下、40%以下、30%以下、20%以下、10%以下、又は5%以下を有するDegPタンパク質をコードするのが好ましい。変異DegP遺伝子が、プロテアーゼ活性のないDegPタンパク質をコードするのがより好ましい。細胞は、染色体のDegPを欠くのではなく、即ち、DegP遺伝子配列は、あらゆる形態のDegPタンパク質の発現を防ぐように欠失又は変異されていない。
【0125】
シャペロン活性及び低減したプロテアーゼ活性を有するタンパク質を生成するために、あらゆる適切な変異をDegP遺伝子中に導入することができる。グラム陰性細菌から発現されるDegPタンパク質のプロテアーゼ及びシャペロンの活性は、当業者であれば、DegPのプロテアーゼ及びシャペロン活性がDegPの天然の基質であるMalS上で試験されるSpiessらにおいて記載されている方法(「A Temperature−Dependent Switch from Chaperone to Protease in a Widely Conserved Heat Shock Protein.」Cell、97巻3号、339〜347頁、Spiess C、Beil A、Ehrmann M)、及びまた「The proteolytic activity of the HtrA (DegP) protein from Escherichia coli at low temperatures」、Skorko−Glonek Jら、Microbiology、2008年、154巻、3649〜3658頁において記載されている方法などのあらゆる適切な方法によって容易に試験することができる。
【0126】
DegPはセリンプロテアーゼであり、アミノ酸残基His105、Asp135、及びSer210の触媒三塩基からなる活性中心を有する(「Families of serine peptidases」、Methods Enzymol.、1994年、244巻、19〜61頁、Rawlings N及びBarrett A)。シャペロン活性及び低減したプロテアーゼ活性を有するタンパク質を生成するためのDegP変異は、His105、Asp135、及びSer210の1つ、2つ、又は3つに対するミスセンス変異などの変異を含んでいてよい。
【0127】
したがって、変異DegP遺伝子は以下を含んでいてよい:
His105に対する変異、又は、
Asp135に対する変異、又は、
Ser210に対する変異、又は、
His105及びAsp135に対する変異、又は、
His105及びSer210に対する変異、又は、
Asp135及びSer210に対する変異、又は、
His105、Asp135、及びSer210に対する変異。
【0128】
His105、Asp135、及びSer210の1つ、2つ、又は3つは、シャペロン活性、及び低減したプロテアーゼ活性を有するタンパク質をもたらすあらゆる適切なアミノ酸に変異されていてよい。例えば、His105、Asp135、及びSer210の1つ、2つ、又は3つは、Gly又はAlaなどの小型のアミノ酸に変異されてよい。さらなる適する変異は、例えば、Asp135がLys又はArgに変異され、極性のHis105がGly、Ala、Val、又はLeuなどの非極性のアミノ酸に変異され、小型の親水性のSer210がVal、Leu、Phe、又はTyrなどの大型又は疎水性の残基に変異されるなど、His105、Asp135、及びSer210の1つ、2つ、又は3つの、反対の性質を有するアミノ酸への変化である。DegP遺伝子が、図5cに示す通り、シャペロン活性を有するがプロテアーゼを有さないタンパク質を生成することが見出されている点変異S210Aを含んでいるのが好ましい(「A Temperature−Dependent Switch from Chaperone to Protease in a Widely Conserved Heat Shock Protein.」Cell、97巻3号、339〜347頁、Spiess C、Beil A、Ehrmann M)。
【0129】
DegPは、タンパク質−タンパク質相互作用を媒介する、PDZ1(残基260〜358)及びPDZ2(残基359〜448)の2つのPDZドメインを有する(「A Temperature−Dependent Switch from Chaperone to Protease in a Widely Conserved Heat Shock Protein.」、Cell、97巻3号、339〜347頁、Spiess C、Beil A、Ehrmann M)。本発明の一実施形態において、degP遺伝子は、PDZ1ドメイン及び/又はPDZ2ドメインを欠失するように変異される。PDZ1及びPDZ2の欠失は、DegPタンパク質のプロテアーゼ活性の完全な喪失及び野生型DegPタンパク質に比べて低下したシャペロン活性をもたらし、PDZ1又はPDZ2いずれかの欠失は、野生型DegPタンパク質に比べて5%のプロテアーゼ活性及び同様のシャペロン活性をもたらす(「A Temperature−Dependent Switch from Chaperone to Protease in a Widely Conserved Heat Shock Protein.」、Cell、97巻3号、339〜347頁、Spiess C、Beil A、Ehrmann M)。
【0130】
変異DegP遺伝子は、例えば、図5cに示す通り、同定及びスクリーニング方法において助けとなるためにAseIなど、サイレントの天然に存在しない制限部位も含んでいてよい。
【0131】
点変異S210A及びAseI制限マーカー部位を含んでいる変異DegP遺伝子の好ましい配列を配列番号9に提供し、コードされるタンパク質配列を配列番号10に示す。配列番号9の変異DegP配列になされた変異を図5cに示す。
【0132】
細胞が、シャペロン活性及び低減したプロテアーゼ活性を有するDegPタンパク質をコードする変異DegP遺伝子を含んでいる本発明の実施形態において、本発明が提供する1つ又は複数の細胞は、変異細胞に比べて細胞からの正確にフォールディングされたタンパク質の収率の改善を提供することができ、DegP遺伝子は、染色体欠損DegPなど、DegP発現を防ぐノックアウトDegPに対して変異されている。DegPの発現を防ぐノックアウト変異DegP遺伝子を含んでいる細胞においてDegPのシャペロン活性は完全に失われるが、本発明による細胞においてDegPのシャペロン活性は保持され、完全なプロテアーゼ活性は失われる。これらの実施形態において、本発明による1つ又は複数の細胞は、タンパク質のタンパク質分解性を防ぐようにプロテアーゼ活性は低減されており、宿主細胞におけるタンパク質において正確なフォールディング及び輸送を可能にするようにシャペロン活性は維持されている。
【0133】
当業者であれば、タンパク質G HPLC、円偏光二色性、NMR、X線結晶構造解析、及びエピトープ親和性測定方法など、当技術分野においてよく知られている方法を用いて、タンパク質が正確にフォールディングされているか否かを見るために分泌されたタンパク質を試験することが容易にできる。
【0134】
これらの実施形態において、本発明による1つ又は複数の細胞は、DegP発現を防ぐ変異ノックアウトDegP遺伝子を有する細胞に比べて細胞の増殖が改善されていてよい。理論によって拘泥しようとするものではないが、シャペロン活性を必要とする全てのタンパク質をプロセシングする細胞の能力を増大し得るシャペロン活性をDegPプロテアーゼが保持するため、細胞増殖の改善が示され得る。したがって、細胞の増殖及び再生に必要な正確にフォールディングされたタンパク質の生成は、DegPノックアウト変異を有する細胞に比べて本発明の1つ又は複数の細胞において増大し、それによって増殖を制御する細胞の経路を改善し得る。さらに、知られているDegPプロテアーゼ欠損系統は一般に温度感受性であり、約28℃を超える温度では通常増殖しない。しかし、本発明による細胞は温度感受性ではなく、細菌からタンパク質を生成する工業規模に通常用いられる、およそ30℃からおよそ37℃の温度を含めた28℃以上の温度で増殖することができる。
【0135】
本発明の実施形態において、細胞は変異ptr遺伝子を含んでいる。本明細書で用いられる「ptr遺伝子」は、高分子量のタンパク質を分解するプロテアーゼであるProteaseIIIをコードする遺伝子を意味する。非変異のptr遺伝子の配列を配列番号4に示し、非変異のProteaseIIIタンパク質の配列を配列番号5に示す。
【0136】
変異ptr遺伝子、又はProteaseIIIをコードする変異ptr遺伝子に対する言及は、別段の指摘がなければ、低減したプロテアーゼ活性を有するProteaseIIIタンパク質をコードする変異ptr遺伝子、又はノックアウト変異ptr遺伝子のいずれかを意味する。
【0137】
「低減したプロテアーゼ活性を有するProteaseIIIタンパク質をコードする変異ptr遺伝子」の表現は、本発明の文脈において、変異ptr遺伝子が、野生型の非変異のptr遺伝子に比べて完全なプロテアーゼ活性を有さないことを意味する。
【0138】
変異ptr遺伝子が、野生型の非変異のProteaseIIIタンパク質のプロテアーゼ活性の50%以下、40%以下、30%以下、20%以下、10%以下、又は5%以下を有するProteaseIIIをコードするのが好ましい。変異ptr遺伝子がプロテアーゼ活性を有さないProteaseIIIタンパク質をコードするのがより好ましい。この実施形態において、細胞は染色体ptrを欠いておらず、即ち、ptr遺伝子配列は、あらゆる形態のProteaseIIIタンパク質の発現を防止するように欠失又は変異されていない。
【0139】
低減したプロテアーゼ活性を有するProteaseIIIタンパク質を生成するために、あらゆる適切な変異がptr遺伝子中に導入されてよい。グラム陰性細菌から発現されたProteaseIIIタンパク質のプロテアーゼ活性は、当業者であれば、当技術分野におけるあらゆる適切な方法によって容易に試験することができる。
【0140】
「ノックアウト変異ptr遺伝子」の表現は、本発明の文脈において、遺伝子が1つ又は複数の変異を含んでおり、それによって、ノックアウト変異遺伝子によってコードされるタンパク質を欠く細胞を提供するように、遺伝子によってコードされるタンパク質の発現を引き起こさないことを意味する。ノックアウト遺伝子は、部分的又は完全に転写され得るが、コードタンパク質に翻訳され得ない。ノックアウト変異ptr遺伝子は、タンパク質の発現を引き起こさないように、例えば、1つ又は複数の欠失、挿入、点、ミスセンス、ナンセンス、及びフレームシフトの変異によるなど、あらゆる適切な方法において変異されてよい。例えば、遺伝子は、遺伝子コード配列中に、抗生物質耐性マーカーなどの外来のDNA配列を挿入することによってノックアウトされてよい。
【0141】
好ましい一実施形態において、遺伝子は、遺伝子コード配列中に、抗生物質耐性マーカーなどの外来のDNA配列を挿入することによって変異されない。ProteaseIII遺伝子は、遺伝子開始コドン、並びに/或いは遺伝子開始コドンの下流及び遺伝子終止コドンの上流に位置する1つ又は複数の終止コドンに対する変異を含んでおり、それによってProteaseIIIタンパク質の発現を防いでいる。
【0142】
標的のノックアウト遺伝子開始コドンに対する変異は開始コドンの機能の喪失を引き起こし、それによって標的の遺伝子は確実に、コード配列の開始に適切な開始コドンを含んでいない。開始コドンに対する変異は開始コドンの1つ、2つ、又は3つ全てのヌクレオチドのミスセンス変異であってよい。或いは、又はさらに、開始コドンは挿入又は欠失のフレームシフト変異によって変異されてよい。
【0143】
好ましい一実施形態において、ptr遺伝子は、図5aに示す通り、ATG開始コドンがATTに変化するように変異されている。
【0144】
ノックアウト変異ptr遺伝子は、或いは、又はさらに、遺伝子開始コドンの下流及び遺伝子終止コドンの上流に位置する1つ又は複数の終止コドンを含んでいてよい。ノックアウト変異ptr遺伝子が、開始コドンに対するミスセンス変異、及び1つ又は複数の挿入された終止コドンの両方を含んでいるのが好ましい。
【0145】
1つ又は複数の挿入された終止コドンがインフレーム終止コドンであるのが好ましい。しかし、1つ又は複数の挿入された終止コドンは、或いは、又はさらに、フレーム外(out−of−frame)終止コドンであってよい。1つ又は複数のフレーム外終止コドンは、フレーム外開始コドンが挿入又は欠失のフレームシフト変異によってインフレーム開始コドンに変化される場合に翻訳を停止するのに必要とされ得る。1つ又は複数の終止コドンが、ナンセンス点変異及びフレームシフト変異を含めたあらゆる適切な変異によって導入されてよい。1つ又は複数の終止コドンがフレームシフト変異及び/又は挿入変異によって、好ましくは遺伝子配列のセグメントを、終止コドンを含んでいる配列で置換することによって導入されるのが好ましい。例えば、終止コドンTAAを含んでいるAseI制限部位を挿入してもよい。
【0146】
好ましい一実施形態において、ptr遺伝子は、図5aに示す通り、AseI制限部位の挿入によって、インフレーム終止コドンを挿入するように変異される。好ましい一実施形態において、ノックアウト変異ptr遺伝子は配列番号6のDNA配列を有する。配列番号6のノックアウト変異ptr遺伝子配列においてなされた変異を図5aに示す。
【0147】
上記に記載したノックアウト変異は、標的のノックアウト遺伝子部位の上流又は下流の染色体DNAに対して最小のかく乱を引き起こし、又はかく乱を引き起こさず、外来のDNA(例えば、対象のタンパク質、特に治療用タンパク質の発現に対する細胞の適合性に影響を及ぼし得る、抗生物質耐性マーカー)の挿入及び保持を必要としないので有利である。したがって、本発明による1つ又は複数の細胞は、増殖の特徴の改善及び/又は外来DNAの遺伝子コード配列中への挿入によってプロテアーゼ遺伝子がノックアウトされた細胞に比べてタンパク質発現の改善を示すことがある。
【0148】
本発明の実施形態において、細胞は変異OmpT遺伝子を有する。本明細書で用いられる「OmpT遺伝子」は、外膜プロテアーゼであるプロテアーゼOmpT(外膜プロテアーゼT)をコードする遺伝子を意味する。野生型の非変異OmpT遺伝子の配列は、SWISS−PROT P09169である。
【0149】
変異OmpT遺伝子、又はOmpTをコードする変異OmpT遺伝子に対する言及は、別段の指摘がなければ、低減したプロテアーゼ活性を有するOmpTタンパク質をコードする変異OmpT遺伝子、又はノックアウト変異OmpT遺伝子のいずれかを意味する。
【0150】
「低減したプロテアーゼ活性を有するOmpTタンパク質をコードする変異OmpT遺伝子」の表現は、本発明の文脈において、変異OmpT遺伝子が野生型の非変異OmpT遺伝子に比べて完全なプロテアーゼ活性を有さないことを意味する。変異OmpT遺伝子は、野生型の非変異OmpTタンパク質のプロテアーゼ活性の50%以下、40%以下、30%以下、20%以下、10%以下、又は5%以下を有するOmpTタンパク質をコードしていてよい。変異OmpT遺伝子は、プロテアーゼ活性を有さないOmpTタンパク質をコードしていてよい。この実施形態において、細胞は染色体OmpTを欠くのではなく、即ち、OmpT遺伝子配列はあらゆる形態のOmpTタンパク質の発現を防ぐように欠失又は変異されていない。
【0151】
低減したプロテアーゼ活性を有するタンパク質を生成するために、あらゆる適切な変異をOmpT遺伝子中に導入することができる。グラム陰性細菌から発現されるOmpTタンパク質のプロテアーゼ活性は、当業者であれば、Kramerら(「Identification of essential acidic residues of outer membrane protease OmpT supports a novel active site」、FEBS Letters、505巻(2001年) 426〜430頁)及びDekkerら(「Substrate Specitificity of the Integral Membrane Protease OmpT Determined by Spatially Addressed Peptide Libraries」、Biochemistry、2001年、40巻、1694〜1701頁)において記載されている方法など、当技術分野におけるあらゆる適切な方法によって容易に試験することができる。
【0152】
OmpTは、Kramerら(「Identification of active site serine and histidine residues in Escherichia coli outer membrane protease OmpT」、FEBS Letters、2000年、468巻、220〜224頁)において報告されており、これは、セリン、ヒスチジン、及び酸性残基をアラニンによって置換すると、Glu27、Asp97、Asp208、又はHis101ではおよそ10倍活性が低減し、Ser99ではおよそ500倍活性が低減し、Asp83、Asp85、Asp210、又はHis212ではおよそ10000倍活性が低減すると開示している。Vandeputte−Ruttenら(「Crystal Structure of the Outer Membrane Protease OmpT from Escherichia coli suggests a novel catalytic site」、The EMBO Journal、2001年、20巻18号、5033〜5039頁)は、Asp83−Asp85対及びHis212−Asp210対を含む活性部位を有すると開示している。さらに、Kramerら(「Lipopolysaccharide regions involved in the activation of Escherichia coli outer membrane protease OmpT」、Eur.J.Biochem.、FEBS、2002年、269巻、1746〜1752頁)は、ループL4における変異D208A、D210A、H212A、H212N、H212Q、G216K/K217G、K217T、及びR218Lは全て、酵素活性の部分的又はほぼ完全な喪失をもたらすことを開示している。
【0153】
したがって、低減したプロテアーゼ活性を有するタンパク質を生成するためのOmpT変異は、E27、D43、D83、D85、D97、S99、H101、E111、E136、E193、D206、D208、D210、H212、G216、K217、R218、及びE250の残基の1つ又は複数に対するミスセンス変異などの変異を含んでいてよい。
【0154】
E27、D43、D83、D85、D97、S99、H101、E111、E136、E193、D206、D208、D210、H212、G216、K217、R218、及びE250の1つ又は複数は、低減したプロテアーゼ活性を有するタンパク質をもたらすあらゆる適切なアミノ酸に変異していてよい。例えば、E27、D43、D83、D85、D97、S99、H101、E111、E136、E193、D206、D208、D210、H212、G216、K217、R218、及びE250の1つ又は複数は、アラニンに変異していてよい。適切な変異の例には、E27A、D43A、D83A、D85A、D97A、S99A、H101A、E111A、E136A、E193A、D206A、D208A、D210A、H212A、H212N、H212Q、G216K、K217G、K217T、R218L、及びE250Aがある。一実施形態において、変異OmpT遺伝子は、D210A及びH212A変異を含んでいる。D210A及びH212A変異を含んでいる適切な変異OmpT配列を配列番号23に示す。
【0155】
「ノックアウト変異OmpT遺伝子」の表現は、本発明の文脈において、遺伝子が1つ又は複数の変異を含んでおり、それによって、ノックアウト変異遺伝子によってコードされるタンパク質を欠く細胞を提供するように、遺伝子によってコードされるタンパク質の発現を引き起こさないことを意味する。ノックアウト遺伝子は、部分的又は完全に転写され得るが、コードタンパク質に翻訳され得ない。ノックアウト変異OmpT遺伝子は、タンパク質の発現を引き起こさないように、例えば、1つ又は複数の欠失、挿入、点、ミスセンス、ナンセンス、及びフレームシフトの変異によるなど、あらゆる適切な方法において変異されてよい。例えば、遺伝子は、遺伝子コード配列中に、抗生物質耐性マーカーなどの外来のDNA配列を挿入することによってノックアウトされてよい。
【0156】
一実施形態において、OmpT遺伝子は、遺伝子開始コドン、並びに/或いは遺伝子開始コドンの下流及び遺伝子終止コドンの上流に位置する1つ又は複数の終止コドンに対する変異を含んでおり、それによってOmpTタンパク質の発現を防いでいる。開始コドンに対する変異は開始コドンの1つ、2つ、又は3つ全てのヌクレオチドのミスセンス変異であってよい。或いは、又はさらに、開始コドンは挿入又は欠失のフレームシフト変異によって変異されてよい。
【0157】
適切な変異ノックアウトOmpT配列を、配列番号24に示す。
【0158】
一実施形態において、本発明によるグラム陰性細菌細胞は、染色体ompTを欠くなど、ノックアウト変異ompT遺伝子を有さない。
【0159】
一実施形態において、本発明によるグラム陰性細菌細胞は、染色体degPを欠くなど、ノックアウト変異degP遺伝子を有さない。一実施形態において、本発明によるグラム陰性細菌細胞は、変異degP遺伝子を有さない。
【0160】
一実施形態において、本発明によるグラム陰性細菌細胞は、染色体ptrを欠くなど、ノックアウト変異ptr遺伝子を有さない。
【0161】
ノックアウト変異を含めた多くの遺伝子操作された変異は、上首尾に変異させた細胞の選択及び同定を可能にする抗生物質耐性マーカーの使用を伴う。しかし、上記で論じたように、抗生物質耐性マーカーの使用には数々の不都合がある。
【0162】
本発明のさらなる一実施形態は、抗生物質耐性マーカーを使用する上記の不都合を克服するものであり、変異Tsp遺伝子、任意選択によって変異spr、任意選択によって変異DegP遺伝子、及び/又は変異ptr遺伝子、及び/又は変異OmpT遺伝子は1つ又は複数の制限マーカー部位を含むように変異される。制限部位は遺伝子中に遺伝子操作され、天然に存在しない。制限マーカー部位が有利であるのは、制限マーカー部位により、必要とされる染色体の変異を含んでいる正確に修飾されている細胞のスクリーニング及び同定が可能になるからである。1つ又は複数の変異プロテアーゼ遺伝子を有するように修飾された細胞は、天然に存在しない制限マーカー部位を含むゲノムDNAの領域を増幅するようにデザインされたオリゴヌクレオチド対を用いて、細胞溶解物からゲノムDNAのPCRによって分析することができる。増幅されたDNAを、次いで、天然に存在しない制限マーカー部位でDNAを消化することができる適切な制限酵素とインキュベートする前及び後にアガロースゲル電気泳動によって分析してもよい。制限酵素とインキュベートした後にDNAフラグメントが存在すれば、細胞が、1つ又は複数の変異遺伝子を有するように上首尾に修飾されたことを確認している。
【0163】
細胞が配列番号6のDNA配列を有するノックアウト変異ptr遺伝子を有する実施形態において、配列番号17及び配列番号18に示すオリゴヌクレオチドプライマー配列を用いて、形質転換した細胞のゲノムDNAから天然に存在しないAseI制限部位を含むDNAの領域を増幅してもよい。次いで、増幅したゲノムDNAを、AseI制限酵素とインキュベートし、ゲル電気泳動によって分析して、ゲノムDNA中の変異ptr遺伝子の存在を確認してもよい。
【0164】
細胞が、配列番号3のDNA配列又は配列番号3のヌクレオチド7から2048を有するノックアウト変異Tsp遺伝子を含んでいる実施形態において、配列番号15及び配列番号16に示すオリゴヌクレオチドプライマー配列を用いて、形質転換した細胞のゲノムDNAから天然に存在しないAseI制限部位を含んでいるDNAの領域を増幅してもよい。次いで、増幅したゲノムDNAを、AseI制限酵素とインキュベートし、ゲル電気泳動によって分析して、ゲノムDNA中の変異Tsp遺伝子の存在を確認してもよい。
【0165】
細胞が、配列番号9のDNA配列を有する変異DegP遺伝子を含んでいる実施形態において、配列番号19及び配列番号20に示すオリゴヌクレオチドプライマー配列を用いて、形質転換した細胞のゲノムDNAから天然に存在しないAseI制限部位を含んでいるDNAの領域を増幅してもよい。次いで、増幅したゲノムDNAを、AseI制限酵素とインキュベートし、ゲル電気泳動によって分析して、ゲノムDNA中の変異DegP遺伝子の存在を確認してもよい。
【0166】
1つ又は複数の制限部位を、1つ又は複数の欠失、挿入、点、ミスセンス、ナンセンス、及びフレームシフトの変異を含むあらゆる適切な変異によって導入することができる。制限部位を、上記に記載した通り、開始コドンの変異、及び/又は1つ若しくは複数の終止コドンを導入するような変異によって導入することができる。この実施形態が有利であるのは、制限マーカー部位が、導入されるノックアウト変異の直接的且つ独特なマーカーであるからである。
【0167】
AseI制限部位など、インフレームの終止コドンを含む制限マーカー部位を挿入してもよい。挿入される制限部位が制限マーカー部位及び終止コドンの両方として作用して遺伝子コード配列の完全な転写を防ぐので、これは特に有利である。例えば、終止コドンがAseI部位の導入によってptr遺伝子に導入される実施形態において、これは、図5aに示すように制限部位をまた作り出す。例えば、AseI部位の導入によって終止コドンがコドン21でTsp遺伝子に導入される実施形態において、これは、図5bに示すように制限部位をまた作り出す。
【0168】
制限マーカー部位を、開始コドンに対する変異、及び任意選択によって1つ又は複数のさらなる点変異によって挿入してもよい。この実施形態において、制限マーカー部位がEcoRI制限部位であるのが好ましい。開始コドンに対する変異は制限マーカー部位も作り出すので、これは特に有利である。例えば、ptr遺伝子の開始コドンがATTに変更される実施形態において、これは、図5aに示すようにEcoRIマーカー部位をまた作り出す。例えば、Tsp遺伝子の開始コドン(コドン3)がATGからTCGに変更される実施形態において、図1bに示すように、コドン2のAACからAATへのさらなる点変異、及びコドン3のATGからTCGへの変異は、図5bに示すように、EcoRI制限マーカー部位を作り出す。
【0169】
細胞が変異OmpT遺伝子を有する本発明の実施形態において、1つ又は複数の制限部位を、1つ又は複数の欠失、挿入、点、ミスセンス、ナンセンス、及びフレームシフトの変異を含むあらゆる適切な変異によって導入してもよい。例えば、OmpT遺伝子が変異D210A及びH212Aを含んでいる実施形態において、これらの変異は、選択マーカーとして用いることができるサイレントのHindIII制限部位を導入する。
【0170】
DegP遺伝子又はspr遺伝子において、マーカー制限部位を、サイレントコドン変化を用いて導入してもよい。例えば、AseI部位をサイレントの制限マーカー部位として用いてもよく、変異DegP遺伝子に対して図5cに示すように、TAA終止コドンはフレーム外である。
【0171】
ptr遺伝子及び/又はTsp遺伝子が低減したプロテアーゼ活性を有するProteaseIII又はTspをコードするように変異されている本発明の実施形態において、1つ又は複数のマーカー制限部位を、サイレントのコドン変化を用いて導入してもよい。
【0172】
あらゆる適切なグラム陰性細菌を、本発明の組換え細胞を生成するための親細胞として用いることができる。適切なグラム陰性細菌には、ネズミチフス菌(Salmonella typhimurium)、蛍光菌(Pseudomonas fluorescens)、軟腐病菌(Erwinia carotovora)、シゲラ(Shigella)、肺炎桿菌(Klebsiella pneumoniae)、レジオネラ・ニューモフィラ(Legionella pneumophila)、緑膿菌(Pseudomonas aeruginosa)、アシネトバクター・バウマンニイ(Acinetobacter baumannii)、及び大腸菌が含まれる。親細胞が大腸菌であるのが好ましい。本発明において大腸菌のあらゆる適切な系統を用いることができるが、K−12 W3110などの野生型W3110系統を用いるのが好ましい。
【0173】
予め作り出され、組換えタンパク質を発現するのに用いられるプロテアーゼ欠損細菌系統に付随する欠点は、これらが、大腸菌系統において、例えば、phoA、fhuA、lac、rec、gal、ara、arg、thi、及びproなど、細胞の代謝及びDNA複製に関与する遺伝子に対するさらなる変異を含むことである。これらの変異は、細胞の増殖、安定性、組換えタンパク質発現収率、及び毒性に対する効果を含めた、宿主細胞に対する多くの有害効果を有し得る。これらのゲノムの変異を1つ又は複数有する系統、特にこれらの変異を多数有する系統は、細菌の増殖速度を、産業的なタンパク質生成に適さないレベルまで低減する適応度の喪失を表し得る。さらに、あらゆる上記のゲノムの変異は、予測不可能な有害な方法で、シス及び/又はトランスにおいて他の遺伝子に影響を及ぼし、それによって系統の表現型、適応度、及びタンパク質のプロファイルを変更し得る。さらに、大幅に変異した細胞を使用することは、商用、特に治療剤に使用するための組換えタンパク質を生成するのに一般に適さない、というのは、このような系統は一般的に欠陥のある代謝経路を有しており、したがって最小の培地又は化学的に規定された培地において不十分にしか増殖せず、又は全く増殖しないことがあるからである。
【0174】
好ましい一実施形態において、細胞は、DsbCをコードする組換えポリヌクレオチドを導入するのに最小の変異だけを有するが、この修飾はTspタンパク質活性、及び任意選択によって変異spr遺伝子を低減するのに必要とされる。組換えポリヌクレオチド及び変異を導入するのに、最小の変異だけが細胞のゲノムに対して行われる。細胞は、細胞の増殖及び/又は対象のタンパク質を発現する能力に対して有害な効果を有し得るいかなる他の変異を有さない。したがって、本発明の1つ又は複数の組換え宿主細胞は、ゲノム配列に対してさらなる遺伝子操作した変異を含んでいる細胞に比べて、改善されたタンパク質発現及び/又は改善された増殖の特徴を表し得る。本発明が提供する細胞はまた、細胞のゲノムに対するさらなるかく乱を含んでいる細胞に比べて、治療用タンパク質の生成において用いるのにより適している。
【0175】
したがって、本発明は、Tspタンパク質活性、及び任意選択によって変異spr遺伝子を低減するのに必要とされる組換え、修飾以外、野生型細菌細胞に対して同質遺伝子であるゲノムを有するグラム陰性細菌細胞も提供する。細胞は、DsbCをコードする組換えポリヌクレオチド、及び任意選択によって対象のタンパク質をコードするポリヌクレオチド配列をさらに有する。
【0176】
好ましい一実施形態において、細胞は、野生型細胞及び任意選択によって変異spr遺伝子に比べてTspタンパク質活性を低減するのに必要とされる修飾以外、W3110系統などの野生型大腸菌細胞に対して同質遺伝子である。細胞は、DsbCをコードする組換えポリヌクレオチド、及び任意選択によって対象のタンパク質をコードするポリヌクレオチド配列をさらに有する。
【0177】
本発明による細胞は、対象のタンパク質をコードするポリヌクレオチド配列をさらに含んでいてよい。対象のタンパク質をコードするポリヌクレオチド配列は、外来性でも、又は内在性でもよい。対象のタンパク質をコードするポリヌクレオチド配列は、宿主の染色体中に組み込まれてもよく、又は通常はプラスミドであるベクター中に非組入れ(non−integrated)であってもよい。ポリヌクレオチドが細胞における発現ベクター中にあり、それによって宿主細胞のゲノムに対して最小のかく乱を引き起こすのが好ましい。
【0178】
「対象のタンパク質」は、本明細書の文脈において、通常は組換えポリペプチドである、発現用のポリペプチドを意味するものとされる。しかし、対象のタンパク質は、宿主細胞において内在性の遺伝子から発現される内在性タンパク質であってもよい。
【0179】
本明細書で用いられる、「組換えポリペプチド」は、組換えDNA技術を用いて構築又は生成されるタンパク質を意味する。対象のタンパク質は、外来性のベクターから発現された、内在性のタンパク質若しくはその変異バージョン(例えば、減弱された生物学的活性を有するもの)に同一の外来性の配列であってよく、又はそのフラグメントであってよい。或いは、対象のタンパク質は、宿主細胞によって通常発現されない、異種性のタンパク質であってよい。
【0180】
対象のタンパク質は、治療用、予防用、又は診断用のタンパク質を含めたあらゆる適切なタンパク質であってよい。
【0181】
一実施形態において、対象のタンパク質は、炎症性疾患及び障害、免疫疾患及び障害、線維性障害、並びに癌を含めた疾患又は障害の処置において有用である。
【0182】
「炎症性疾患」又は「障害」、及び「免疫疾患又は障害」の語は、関節リウマチ、乾癬性関節炎、スティル病、マックルウェルズ病、乾癬、クローン病、潰瘍性大腸炎、SLE(全身性エリテマトーデス)、喘息、アレルギー性鼻炎、アトピー性皮膚炎、多発性硬化症、血管炎、I型糖尿病、移植、及び移植片対宿主病を含む。
【0183】
「線維性障害」の語は、特発性肺線維症(IPF)、全身性硬化症(又は強皮症)、腎臓線維症、糖尿病性腎症、IgA腎症、高血圧、末期腎不全、腹膜線維症(連続の外来腹膜透析)、肝硬変、加齢黄斑変性(ARMD)、網膜症、反応性心臓線維症(cardiac reactive fibrosis)、瘢痕、ケロイド、火傷、皮膚潰瘍、血管形成、冠状動脈バイパス手術、関節形成術、及び白内障手術を含む。
【0184】
「癌」の語は、上皮から生じ、皮膚中又はより一般的には身体器官(例えば、乳房、卵巣、前立腺、肺、腎臓、膵臓、胃、膀胱、又は腸)の内膜に見出される悪性の新たな増殖を含む。癌は、隣接の組織中に浸潤し、例えば、骨、肝臓、肺、又は脳などの遠位の器官に広がる(転移)傾向がある。
【0185】
タンパク質は、タンパク質分解感受性のポリペプチド、即ち、自然状態又は分泌の間のいずれかにおいて、大腸菌などの1つ又は複数のグラム陰性細菌、プロテアーゼによって切断を受けやすく、切断に感受性であり、又は切断されるタンパク質であってよい。一実施形態において、対象のタンパク質は、DegP、ProteaseIII、及びTspから選択されるプロテアーゼに対してタンパク質分解感受性である。一実施形態において、対象のタンパク質は、プロテアーゼTspに対してタンパク質分解感受性である。一実施形態において、対象のタンパク質は、プロテアーゼであるDegP及びProteaseIIIに対してタンパク質分解感受性である。一実施形態において、対象のタンパク質は、プロテアーゼであるDegP及びTspに対してタンパク質分解感受性である。一実施形態において、対象のタンパク質は、プロテアーゼであるTsP及びProteaseIIIに対してタンパク質分解感受性である。一実施形態において、対象のタンパク質は、DegP、ProteaseIII、及びTsPに対してタンパク質分解感受性である。
【0186】
タンパク質が真核生物のポリペプチドであるのが好ましい。
【0187】
本発明による細胞によって発現される対象のタンパク質は、例えば免疫原、2つの異種性のタンパク質を含む融合タンパク質、又は抗体であってよい。対象のタンパク質として用いるための抗体には、モノクローナル抗体、多価抗体、多重特異性抗体、ヒト化抗体、完全ヒト抗体、又はキメラ抗体が含まれる。抗体はあらゆる種からのものであってよいが、モノクローナル抗体、ヒト抗体、又はヒト化フラグメントに由来しているのが好ましい。抗体は、免疫グロブリン分子のあらゆるクラス(例えば、IgG、IgE、IgM、IgD、若しくはIgA)又はサブクラスに由来していてよく、例えば、マウス、ラット、サメ、ウサギ、ブタ、ハムスター、ラクダ、ラマ、ヤギ、若しくはヒトを含めたあらゆる種から得ることができる。抗体フラグメントの部分は、1つを超える種から得ることができ、例えば、抗体フラグメントはキメラであってよい。一例において、定常領域はある種からであり、可変領域は別の種からである。
【0188】
抗体は、全長の重鎖及び軽鎖を有する完全な抗体分子であってよく、又はそのフラグメント、例えば、PCT/GB2008/003331に記載されている通り、VH、VL、VHH、Fab、修飾Fab’、Fab’、F(ab’)2、Fv、scFvフラグメント、Fab−Fv、又は二重特異性抗体、例えば、Fab−dAbであってよい。
【0189】
抗体はあらゆる標的の抗原に特異的であってよい。抗原は、細胞に付随するタンパク質、例えば、細菌細胞、酵母菌細胞、T細胞、内皮細胞、若しくは腫瘍細胞などの細胞状の細胞表面タンパク質であってよく、又は可溶性タンパク質であってよい。対象の抗原はまた、疾患若しくは感染の間に上方制御されるタンパク質などのあらゆる医学的に関連のあるタンパク質、例えば、受容体であってよく、且つ/又はこれらの対応するリガンドであってもよい。細胞表面タンパク質の特定の例には、接着分子、例えば、β1インテグリンなどのインテグリン、例えば、VLA−4、E−セレクチン、Pセレクチン、若しくはL−セレクチン、CD2、CD3、CD4、CD5、CD7、CD8、CD11a、CD11b、CD18、CD19、CD20、CD23、CD25、CD33、CD38、CD40、CD40L、CD45、CDW52、CD69、CD134(OX40)、ICOS、BCMP7、CD137、CD27L、CDCP1、CSF1、若しくはCSF1受容体、DPCR1、DPCR1、ヅヅリン(dudulin)2、FLJ20584、FLJ40787、HEK2、KIAA0634、KIAA0659、KIAA1246、KIAA1455、LTBP2、LTK、MAL2、MRP2、ネクチン様2、NKCC1、PTK7、RAIG1、TCAM1、SC6、BCMP101、BCMP84、BCMP11、DTD、癌胎児性抗原(CEA)、ヒト乳脂肪グロブリン(HMFG1及び2)、MHCクラスI及びMHCクラスII抗原、KDR及びVEGF、及び好適な場合にはこれらの受容体が含まれる。
【0190】
可溶性抗原には、インターロイキン、例えば、IL−1、IL−2、IL−3、IL−4、IL−5、IL−6、IL−8、IL−12、IL−13、IL−14、IL−16、又はIL−17、例えば、IL17A及び/又はIL17F、ウイルス抗原、例えば、呼吸器合胞体ウイルス抗原又はサイトメガロウイルス抗原、免疫グロブリン、例えば、IgE、インターフェロン、例えば、インターフェロンα、インターフェロンβ、又はインターフェロンγ、腫瘍壊死因子TNF(以前は腫瘍壊死因子−αとして知られていた)、腫瘍壊死因子−β、コロニー刺激因子、例えば、G−CSF又はGM−CSF、及び血小板由来増殖因子、例えば、PDGF−α及びPDGF−β、並びに好適な場合にはこれらの受容体が含まれる。他の抗原には、細菌の細胞表面抗原、細菌の毒素、ウイルス、例えば、インフルエンザ、EBV、HepA、B、及びC、生物テロ薬剤、放射性核種及び重金属、並びにヘビ毒及びクモ毒、及び毒素が含まれる。
【0191】
一実施形態において、対象の抗原の活性を機能的に変更するのに抗体を用いることができる。例えば、抗体は、前記抗原の活性を、直接的又は間接的に、中和し、拮抗する(antagonize)、又は作用し(アゴナイズ:agonise)得る。
【0192】
本発明は、DsbC、変異Tsp遺伝子をコードする組換えポリヌクレオチド、及びTNFに特異的な抗体又はその抗原結合性フラグメントをコードするポリヌクレオチド配列を含んでいる組換えグラム陰性細菌細胞も提供し、変異Tsp遺伝子は、低減したプロテアーゼ活性を有するTspタンパク質をコードする、又はノックアウト変異Tsp遺伝子である。好ましい一実施形態において、細胞は、変異体sprタンパク質をコードする変異体spr遺伝子をさらに含んでいる。
【0193】
好ましい一実施形態において、WO01/094585(この内容は参照によって本明細書に組み込まれる)に記載されている通り、本発明による細胞によって発現される対象のタンパク質は、抗TNF抗体、より好ましくは抗TNF Fab’である。
【0194】
一実施形態において、ヒトTNFαに対する特異性を有する抗体は重鎖を含んでおり、可変ドメインは、CDRH1に対して配列番号28に示す配列、CDRH2に対して配列番号29若しくは配列番号34に示す配列、又はCDRH3に対して配列番号30に示す配列を有するCDRを含んでいる。
【0195】
一実施形態において、抗体は軽鎖を含んでおり、可変ドメインは、CDRL1に対して配列番号31に示す配列、CDRL2に対して配列番号32に示す配列、又はCDRL3に対して配列番号33に示す配列を有するCDRを含んでいる。
【0196】
上記に言及した配列番号28、及び配列番号30から配列番号34に示したCDRは、マウスモノクローナル抗体hTNF40に由来する。しかし、配列番号29はハイブリッドCDRからなる。ハイブリッドCDRは、マウスモノクローナル抗体hTNF40(配列番号40)からの重鎖CDR2の部分、及びヒトグループ3生殖系列V領域配列からの重鎖CDR2の部分を含んでいる。
【0197】
一実施形態において、抗体は重鎖(可変ドメインは、CDRH1に対して配列番号28に示す配列、CDRH2に対して配列番号29若しくは配列番号34に示す配列、又はCDRH3に対して配列番号30に示す配列を有するCDRを含んでいる)、及び軽鎖(可変ドメインは、CDRL1に対して配列番号31に示す配列、CDRL2に対して配列番号32に示す配列、又はCDRL3に対して配列番号33に示す配列を有するCDRを含んでいる)を含んでいる。
【0198】
一実施形態において、抗体は、CDRH1に対して配列番号28、CDRH2に対して配列番号29又は配列番号34、CDRH3に対して配列番号30、CDRL1に対して配列番号31、CDRL2に対して配列番号32、及びCDRL3に対して配列番号33を含んでいる。抗体がCDRH2に対して配列番号29を含んでいるのが好ましい。
【0199】
抗TNF抗体がCDRグラフト化抗体分子であるのが好ましい。好ましい一実施形態において、可変ドメインは、ヒトアクセプターフレームワーク領域及び非ヒトドナーCDRを含んでいる。
【0200】
抗体分子がヒトTNF(以前はTNFαとして知られていた)に対して特異性を有するのが好ましく、軽鎖は配列番号11の軽鎖可変領域を含んでおり、重鎖は配列番号12の重鎖可変領域を含んでいる。
【0201】
抗TNF抗体がFab又はFab’のフラグメントであるのが好ましい。
【0202】
ヒトTNFに対して特異性を有する抗体分子がFab’であり、配列番号13を含んでいる、又はそれからなる軽鎖配列、及び配列番号14を含んでいる、又はそれからなる重鎖配列を有するのが好ましい。
【0203】
発現後、抗体フラグメントを、例えば、エフェクター分子などの別の実体に対してコンジュゲートすることによってさらにプロセシングしてもよい。
【0204】
本明細書で用いられるエフェクター分子の語には、例えば、抗悪性腫瘍薬、薬物、毒素(例えば、細菌若しくは植物起源の酵素的に活性な毒素及びこれらのフラグメント、例えば、リシン(ricin)及びそのフラグメント)生物学的に活性なタンパク質、例えば、酵素、他の抗体又は抗体フラグメント、合成の、又は天然に存在するポリマー、核酸及びこれらのフラグメント、例えば、DNA、RNA、及びこれらのフラグメント、放射性核種、特に放射性ヨウ化物、放射性同位元素、キレート化金属、ナノ粒子及びレポーターグループ、例えば、蛍光化合物又はNMR若しくはESR分光法によって検出され得る化合物が含まれる。エフェクター分子は、あらゆる適切な方法によって抗体又はそのフラグメントに付着していてよく、例えば、抗体フラグメントは、WO05/0031731又はWO05/003170(これらの内容は参照によって本明細書に組み入れられる)に記載されている通り、少なくとも1つのエフェクター分子で付着するように修飾されていてよい。WO05/003171又はWO05/003170も適切なエフェクター分子も記載している。
【0205】
一実施形態において、所望により、抗体又はそのフラグメント、例えばFabをPEG化して必要とされる性質、例えば、抗体全体に類似の性質を有する生成物を生成する。例えば、抗体は、WO01/094585に記載されている通り、PEG化した抗TNFα−Fab’であってよく、好ましくは、重鎖のC末端終端でリシル−マレイミド由来の基(lysyl−maleimide−derived group)でシステイン残基の1つに付着しており、リシル残基の2つのアミノ基の各々はこれを、分子量約20000Daを有するメトキシポリ(エチレングリコール)残基に共有結合させており、その結果メトキシポリ(エチレングリコール)残基の合計平均分子量は約40000Daであり、より好ましくはリシル−マレイミド由来の基は、[1−[[[2−[[3−(2,5−ジオキソ−1−ピロリジニル)−1−オキソプロピル]アミノ]エチル]アミノ]−カルボニル]−1,5−ペンタンジイル]ビス(イミノカルボニル)である。
【0206】
細胞はまた、1つ又は複数のさらなる対象のタンパク質をコードするさらなるポリヌクレオチド配列を含んでいてよい。
【0207】
一実施形態において、野生型が精製の間に対象の組換えタンパク質と同時精製することが知られている1つ又は複数の大腸菌宿主タンパク質は、Humphreysら、「Engineering of Escherichia coli to improve the purification of periplasmic Fab’ fragments:changing the pI of the chromosomally encoded PhoS/PstS protein」、Protein Expression and Purification、37巻(2004年)、109〜118頁及びWO04/035792(これらの内容は参照によって本明細書に組み入れられる)において記載されている通り、遺伝子修飾用に選択される。このような修飾された宿主タンパク質の使用により、これらが組換え抗体と最早同時精製されないように、選択された大腸菌タンパク質の物理的性質を変更することによって、大腸菌において生成された対象のタンパク質、特に抗体に対する精製プロセスが改善される。変更される大腸菌タンパク質が、リン酸結合性タンパク質(PhoS/PstS)、ジペプチド結合性タンパク質(DppA)、マルトース結合性タンパク質(MBP)、及びチオレドキシンの1つ又は複数から選択されるのが好ましい。
【0208】
一実施形態において、汚染性の宿主タンパク質の物理的性質は、アミノ酸タグをC末端又はN末端に添加することによって変更される。好ましい一実施形態において、変更される物理的性質は等電点であり、アミノ酸タグはC末端に付着するポリアスパラギン酸タグである。一実施形態において、前記タグの添加によって変更される大腸菌タンパク質は、ジペプチド結合性タンパク質(DppA)、マルトース結合性タンパク質(MBP)、チオレドキシン、及びリン酸結合性タンパク質(PhoS/PstS)である。詳しい一実施形態において、大腸菌のリン酸結合性タンパク質(PhoS/PstS)のpIは、アスパラギン酸残基を6個含んでいるポリアスパラギン酸タグ(PolyD)をC末端に加えることよって7.2から5.1に低減される。
【0209】
単独で、又はN末端若しくはC末端にタグを加えることとの組合せのいずれかで、その物理的性質を変更するための、汚染性の大腸菌タンパク質の特定の残基の修飾も好ましい。このような変更は、pI又は疎水性を変更するためのタンパク質又はアミノ酸置換基のサイズを変更するための挿入又は欠失を含むことができる。一実施形態において、これらの残基はタンパク質の表面上に位置する。好ましい一実施形態において、タンパク質のpIを低減するために、PhoSタンパク質の表面残基が変更される。機能的なPhoSタンパク質を維持するために、リン酸塩の結合において重要であると関係付けられている残基(Bass、US5,304,472)を回避するのが好ましい。タンパク質の表面から遠くはなれて突出している、又は塩基性残基の大型の基である、若しくは塩基性残基の大型の基の近くにあるリシン残基を標的にするのが好ましい。一実施形態において、PhoSタンパク質はC末端に付着しているヘキサポリアスパラギン酸タグを有し、分子の反対の終端にある表面残基が置換用に標的にされる。選択されたリシン残基が、中性残基から酸性残基に変わる場合よりも大きな潜在的なpI変化を付与するようにグルタミン酸又はアスパラギン酸を置換するのが好ましい。本明細書における置換変異体に対する呼称は、文字とその後に続く数字、その後に続く文字からなる。最初の文字は野生型タンパク質におけるアミノ酸を指定する。数字はアミノ酸置換が行われるアミノ酸位置を意味し、2番目の文字は野生型アミノ酸を置換するのに用いられるアミノ酸を指定する。本発明におけるPhoSの好ましい変異において、リシン残基(K)275、107、109、110、262、265、266、309、313は、単一変異又は複合変異としてグルタミン酸(E)又はグルタミン(Q)に対して置換され、さらにリシン(K)318は、単一変異又は複合変異としてアスパラギン酸(D)に対して置換されてよい。単独の変異がK262E、K265E、及びK266Eであるのが好ましい。複合変異がK265/266E及びK110/265/266Eであるのが好ましい。全ての変異が、C末端で付着しているポリアスパラギン酸(PolyD)タグと、任意選択によってK318D置換とも組み合わされるのがより好ましい。好ましい一実施形態において、変異は少なくとも2単位のpIにおける低減をもたらす。本発明の変異がPhoSのpIを7.2から約4と約5.5との間に低減するのが好ましい。本発明の一実施形態において、大腸菌のPhoSタンパク質のpIは、polyD K318D、polyD K265/266E、及びpolyD K110/265/266の変異を用いて、7.2からそれぞれ約4.9、約4.8、及び約4.5に低減される。
【0210】
本発明による組換えグラム陰性細菌細胞はあらゆる適切な手段によって生成され得る。
【0211】
染色体Tsp遺伝子が変異され、任意選択によって1つ又は複数の染色体spr遺伝子、Deg遺伝子、ptr遺伝子、及びOmpT遺伝子が変異される本発明の実施形態において、当業者であれば、染色体の遺伝子配列を、変異遺伝子配列で置換するのに用いることができる適切な技術を知っている。相同的組換えによって宿主の染色体中に組み込むのを可能にする適切なベクターを用いることができる。細胞が上記の変異遺伝子の2つ又は3つを含んでいる実施形態において、変異遺伝子を、同じ又は異なるベクター上のグラム陰性細菌中に導入してもよい。
【0212】
適切な遺伝子置換方法は、例えば、Hamiltonら(「New Method for Generating Deletions and Gene Replacements in Escherichia coli」、Hamilton C.M.ら、Journal of Bacteriology、1989年9月、171巻9号、4617〜4622頁)、Skorupskiら(「Positive selection vectors for allelic exchange」、Skorupski K及びTaylor R. K.、Gene、1996年、169巻、47〜52頁)、Kielら(「A general method for the construction of Escherichia coli mutants by homologous recombination and plasmid segregation」、Kiel J.A.K.W.ら、Mol Gen Genet、1987年、207巻、294〜301頁)、Blomfieldら(「Allelic exchange in Escherichia coli using the Bacillus subtilis sacB gene and a temperature sensitive pSC101 replicon」、Blomfield I.C.ら、Molecular Microbiology、1991年、5巻(6)、1447〜1457頁)、並びにRiedら(「An nptI−sacB−sacR cartridge for constructing directed, unmarked mutations in Gram−negative bacteria by marker exchange−eviction mutagenesis」、Ried J.L. 及びCollmer A.、Gene、57巻(1987年)239〜246頁)に記載されている。相同組換え/置換を可能にする適切なプラスミドはpKO3プラスミドである(Linkら、1997年、Journal of Bacteriology、179巻、6228〜6237頁)。
【0213】
当業者であれば、DsbCをコードする組換えポリヌクレオチドを挿入するのに用いることができる適切な技術を知っている。DsbCをコードする組換えポリヌクレオチドを、pKO3プラスミドなどの適切なベクターを用いて細胞のゲノム中に組み込むことができる。
【0214】
或いは、又はさらに、DsbCをコードする組換えポリヌクレオチドは、組換え発現カセット中に非組入れであってもよい。一実施形態において、DsbCをコードするタンパク質コード配列及び1つ又は複数の制御発現配列を典型的に含んでいる、DsbCをコードするポリヌクレオチドを有するために、本発明において発現カセットが用いられる。1つ又は複数の制御発現配列はプロモーターを含んでいてよい。1つ又は複数の制御発現配列は、終止配列などの3’非翻訳領域も含んでいてよい。適切なプロモーターを以下により詳しく論じる。
【0215】
一実施形態において、1つ若しくは複数の制御発現配列、1つ若しくは複数の対象のタンパク質をコードする1つ若しくは複数のコード配列、及び/又はDsbCをコードするコード配列を典型的に含んでいる、対象のタンパク質をコードするポリヌクレオチド及び/又はDsbCをコードする組換えポリヌクレオチドを有するために、本発明において発現カセットが用いられる。1つ又は複数の制御発現配列はプロモーターを含んでいてよい。1つ又は複数の制御発現配列は、終止配列などの3’非翻訳領域も含んでいてよい。適切なプロモーターを以下により詳しく論じる。
【0216】
一実施形態において、本発明による細胞は、プラスミドなどの1つ又は複数のベクターを含んでいる。ベクターが上記に規定した1つ又は複数の発現カセットを含んでいるのが好ましい。一実施形態において、対象のタンパク質をコードするポリヌクレオチド配列、及びDsbCをコードするポリヌクレオチドを1つのベクター中に挿入する。或いは、対象のタンパク質をコードするポリヌクレオチド配列、及びDsbCをコードするポリヌクレオチドを別々のベクター中に挿入する。
【0217】
対象のタンパク質が重鎖及び軽鎖の両方を含む抗体である実施形態において、第1のベクターが軽鎖ポリペプチドをコードし、第2のベクターが重鎖ポリペプチドをコードする、2つのベクターを細胞系にトランスフェクトしてもよい。或いは、ベクターが軽鎖及び重鎖のポリペプチドをコードする配列を含んでいる、単一のベクターを用いてもよい。或いは、抗体をコードするポリヌクレオチド配列及びDsbCをコードするポリヌクレオチドを1つのベクター中に挿入する。ベクターが、抗体の軽鎖及び重鎖のポリペプチドをコードする配列を含んでいるのが好ましい。
【0218】
細胞が以下の1つ又は複数のさらなるタンパク質:
タンパク質のフォールディングを促進することができる1つ又は複数のタンパク質、例えば、FkpA、Skp、SurA、PPiA、及びPPiD、並びに/又は
タンパク質の分泌又は転位置を促進することができる1つ又は複数のタンパク質、例えば、SecY、SecE、SecG、SecYEG、SecA、SecB、FtsY、及びLep、並びに/又は
ジスルフィド結合形成を促進することができる1つ又は複数のタンパク質、例えば、DsbA、DsbB、DsbD、DsbG
も発現する実施形態において、1つ又は複数のさらなるタンパク質を、DsbCをコードするポリヌクレオチド及び/又は対象のタンパク質をコードするポリヌクレオチド配列として同じベクター中に挿入された1つ又は複数のポリヌクレオチドから発現させてもよい。或いは、1つ又は複数のポリヌクレオチドを別々のベクター中に挿入してもよい。
【0219】
本発明において用いるためのベクターは、適切なベクター中に上記で定義した1つ又は複数の発現カセットを挿入することによって生成することができる。或いは、ポリヌクレオチド配列の発現を指示するための制御発現配列がベクターに含まれていてよく、したがってポリヌクレオチドのコード領域だけがベクターを完成するのに必要とされ得る。
【0220】
DsbCをコードするポリヌクレオチド及び/又は対象のタンパク質をコードするポリヌクレオチドを、細胞中で細胞に適するプロモーターの制御下で発現させるために、複製可能なベクター、典型的には自律的複製ベクター中に挿入するのが適切である。多くのベクターがこの目的で当技術分野において知られており、好適なベクターの選択は核酸のサイズ及び特定の細胞型に依存し得る。
【0221】
本発明によるポリヌクレオチドで宿主細胞を形質転換するのに用いることができるベクターの例には:
プラスミド、例えば、pBR322若しくはpACYC184、及び/又は
ウイルスのベクター、例えば、細菌のファージ
転位性遺伝要素、例えば、トランスポゾン
が含まれる。
【0222】
このようなベクターは、通常、プラスミドのDNA複製開始点、抗生物質の選択マーカー、プロモーター、及びマルチクローニング部位によって分離されている転写ターミネーター(発現カセット)、及びリボソーム結合部位をコードするDNA配列を含んでいる。
【0223】
本発明において用いられるプロモーターは、関連のポリヌクレオチドに直接連結していてよく、又は代替として好適な位置に、例えば、関連のポリペプチドが関連のプロモーターに挿入された場合に関連のプロモーターに対して作用することができるようなベクター中に位置していてよい。一実施形態において、プロモーターは、それに対してプロモーターが作用するポリヌクレオチドのコード部分の前に、例えば、ポリヌクレオチドの各コード部分の前の関連のプロモーターに位置している。本明細書で用いられる「前」はプロモーターが、コードするポリヌクレオチド部分に関して5プライムエンドに位置することを意味するものとされる。
【0224】
プロモーターは、宿主細胞に対して内在性でも、又は外因性でもよい。適切なプロモーターには、Lac、tac、trp、PhoA、Ipp、Arab、Tet、及びT7が含まれる。
【0225】
用いられる1つ又は複数のプロモーターは誘導プロモーターであってよい。
【0226】
DsbCをコードするポリヌクレオチド、及び対象のタンパク質をコードするポリヌクレオチドが1つのベクター中に挿入されている実施形態において、DsbC及び対象のタンパク質をコードするヌクレオチド配列は、単一のプロモーター又は別々のプロモーターの制御下にあってよい。DsbC及び対象のタンパク質をコードするヌクレオチド配列が別々のプロモーターの制御下にある実施形態において、プロモーターは独立に誘導プロモーターであってよい。
【0227】
細菌系において用いるためのプロモーターは、対象のポリペプチドをコードするDNAに作動可能にリンクしているシャインダルガノ(S.D.)配列も一般に含んでいる。プロモーターは、制限酵素の消化によって細菌の供給源のDNAから除去し、所望のDNAを含んでいるベクター中に挿入することができる。
【0228】
ポリヌクレオチド配列が、抗体軽鎖及び抗体重鎖など、2つ以上の対象のタンパク質に対するコード配列を2つ以上含んでいる本発明の実施形態において、ポリヌクレオチド配列は、mRNAの中央における翻訳開始を可能にする1つ又は複数の配列内リボソーム進入部位(IRES)配列を含んでいてよい。IRES配列は、コードされたポリペプチド配列を生成するためのmRNAの別々の翻訳を増強するためにコードするポリヌクレオチド配列間に位置していてよい。
【0229】
発現ベクターが、WO03/048208又はWO2007/039714(これらの内容は参照によって本明細書に組み入れられる)に記載されている抗体又はその抗原結合性フラグメントを生成するためのジシストロン性のメッセージも含んでいるのが好ましい。好ましくは、抗体の軽鎖をコードするDNAを含んでいる上流のシストロン、及び対応する重鎖をコードするDNAを含んでいる下流のシストロン、並びにジシストロン性の遺伝子間配列(IGS)が、IGS1(配列番号36)、IGS2(配列番号37)、IGS3(配列番号38)、及びIGS4(配列番号39)から選択される配列を含んでいるのが好ましい。
【0230】
ターミネーターは宿主細胞に対して内在性でも、又は外因性でもよい。適切なターミネーターはrrnBである。
【0231】
プロモーター及びターミネーターを含んでいるさらなる適切な転写レギュレーター、並びにタンパク質ターゲティング方法は、「Strategies for Achieving High−Level Expression of Genes in Escherichia coli」、Savvas C. Makrides、Microbiological Reviews、1996年9月、512〜538頁に見ることができる。
【0232】
発現ベクター中に挿入されたDsbCポリヌクレオチドが、DsbCシグナル配列及びDsbCコード配列をコードする核酸を含んでいるのが好ましい。ベクターは、好ましくは宿主の染色体を独立に複製するために、ベクターの1つ又は複数の選択された宿主細胞における複製を可能にする核酸配列を含んでいるのが好ましい。このような配列は様々な細菌ではよく知られている。
【0233】
一実施形態において、DsbC及び/又は対象のタンパク質は、N末端及び/又はC末端にヒスチジンタグを含んでいる。
【0234】
抗体分子は、細胞から分泌されることもあり、又は適切なシグナル配列によってペリプラズムに対して標的化することもある。或いは、抗体分子は細胞の原形質内に蓄積することもある。抗体分子がペリプラズムに対して標的化するのが好ましい。
【0235】
対象のタンパク質をコードするポリヌクレオチドは、別のポリペプチド、好ましくは、成熟ポリペプチドのN末端に特異的な切断部位を有するシグナル配列又は他のポリペプチドとの融合物として発現されてよい。選択される異種性のシグナル配列は、宿主細胞によって認識され、プロセシングされるものでなければならない。天然又は真核生物のポリペプチドシグナル配列を認識せず、プロセシングしない原核生物の宿主細胞では、シグナル配列は原核生物のシグナル配列によって置換される。適切なシグナル配列には、OmpA、PhoA、LamB、PelB、DsbA、及びDsbCが含まれる。
【0236】
上記に列挙した成分の1つ又は複数を含んでいる適切なベクターの構築は、標準のライゲーション技術を用いるものである。単離したプラスミド又はDNAフラグメントを、必要とするプラスミドを生成するのに望ましい形態において、切断し、誂え、再ライゲーション(re−ligated)する。
【0237】
本発明の好ましい一実施形態において、本発明は、DsbCをコードするポリヌクレオチド配列及び対象のタンパク質をコードするポリヌクレオチド配列を含んでいる多シストロン性の(multi−cistronic)ベクターを提供する。多シストロン性のベクターは、ポリヌクレオチド配列のベクター中への繰返し連続クローニングを可能にする有利なクローニング方法によって生成することができる。方法は、AseI及びNdeI制限部位の「AT」末端などの1対の制限部位の適合性の付着末端を用いるものである。コード配列を含んでおり、AseI−NdeIフラグメントなどの適合性の付着末端を有するポリヌクレオチド配列を、NdeIなどのベクターにおける制限部位中にクローニングしてもよい。ポリヌクレオチド配列の挿入は5’制限部位を破壊するが、NdeIなどの新たな3’制限部位を作り出し、次いでこの新たな制限部位を用いて、適合性の付着末端を含んでいるさらなるポリヌクレオチド配列を挿入してもよい。次いでこのプロセスを繰り返してさらなる配列を挿入する。ベクター中に挿入された各ポリヌクレオチド配列は、終止コドンに対して3’の非コード配列を含んでおり、終止コドンはスクリーニング用のSspI部位、シャインダルガノリボソーム結合性配列、Aリッチスペーサー、及び開始コドンをコードするNdeI部位を含んでいてよい。
【0238】
ある抗体の軽鎖(LC)、ある抗体の重鎖(HC)をコードするポリヌクレオチド配列、DsbCポリヌクレオチド配列、及びさらなるポリヌクレオチド配列を含んでいるベクターの創製を示す図を図6に示す。
【0239】
上首尾に変異させた系統を、コロニーPCR DNA配列決定及びコロニーPCR制限酵素マッピングを含めた当技術分野においてよく知られている方法を用いて同定してもよい。
【0240】
ポリヌクレオチドに関して本明細書に記載する本発明の実施形態は、ベクター、発現カセット、及び/又はこの中に用いられる成分を含んでいる宿主細胞など、関連する態様をこれらに適用することができる限り、本発明の代替の実施形態に等しく適用されるものである。
【0241】
本発明の第3の態様にしたがって、本発明の第1の態様又は第2の態様において上記に記載した組換えグラム陰性細菌細胞において、対象の組換えタンパク質を発現することを含む、対象の組換えタンパク質を生成するための方法を提供する。対象の組換えタンパク質を生成するための方法は、
a.対象の組換えタンパク質、及びDsbCをコードする組換えポリヌクレオチドを発現させるのに効果的な条件下で培養培地において上記に規定した組換えグラム陰性細菌細胞を培養するステップと、
b.組換えグラム陰性細菌細胞のペリプラズム及び/又は培養培地から対象の組換えタンパク質を回収するステップと
を含む。
【0242】
本発明の方法において用いるのが好ましいグラム陰性細菌細胞及び対象のタンパク質は、上記に詳しく記載したものである。
【0243】
対象のタンパク質をコードするポリヌクレオチドが外来性である場合、ポリヌクレオチドを当技術分野において知られているあらゆる適切な手段を用いて宿主細胞中に組み入れることができる。上記で論じた通り、ポリヌクレオチドが、細胞中に形質転換される発現ベクターの部分として組み入れられるのが典型的である。したがって、本発明による細胞の一態様は、対象のタンパク質をコードするポリヌクレオチドを含む発現カセット、及びDsbCをコードするポリヌクレオチドを含んでいる発現カセットを含む。
【0244】
対象のタンパク質をコードするポリヌクレオチド配列、及びDsbCをコードするポリヌクレオチド配列を、塩化ルビジウム、PEG、又は電気穿孔を用いるなど、標準の技術を用いて細胞中に形質転換してもよい。
【0245】
本発明による方法は、対象のタンパク質をコードするポリヌクレオチドで上首尾に形質転換されている安定な細胞の選択を促進する選択系も用いることができる。選択系は通常、選択マーカーをコードするポリヌクレオチド配列の同時形質転換を使用する。一実施形態において、細胞中に形質転換された各ポリヌクレオチドは、1つ又は複数の選択マーカーをコードするポリヌクレオチド配列をさらに含んでいる。したがって、DsbC及び対象のタンパク質をコードするポリヌクレオチドの形質転換、並びにマーカーをコードする1つ又は複数のポリヌクレオチドの形質転換は一緒に生じ、選択系を用いて所望のタンパク質を生成するこれらの細胞を選択することができる。
【0246】
1つ又は複数のマーカーを発現することができる細胞は、細胞中に組み入れられる選択系のポリペプチド/遺伝子又はポリペプチド成分によって付与される性質(例えば、抗生物質耐性)のために、例えば、毒素又は抗生物質の添加など、ある種の人工的に課せられた条件下で生存/増殖/繁殖することができる。1つ又は複数のマーカーを発現することができない細胞は、人工的に課せられた条件下で生存/増殖/繁殖することができない。人工的に課せられる条件は、所望により幾分強力であるように選択され得る。
【0247】
本発明においてあらゆる適切な選択系を用いることができる。典型的に、選択系は、例えば、テトラサイクリン、クロラムフェニコール、カナマイシン、又はアンピリシン耐性の遺伝子など、既知の抗生物質に対して耐性をもたらすベクターの1つ又は複数の遺伝子中に含まれていることに基づいていてよい。関連の抗生物質の存在下で増殖する細胞は、これらが抗生物質に対する耐性をもたらす遺伝子及び所望のタンパク質の両方を発現するように選択されてよい。
【0248】
一実施形態において、本発明による方法は、形質転換した細胞を培地中で培養し、それによって対象のタンパク質を発現させるステップをさらに含んでいる。
【0249】
対象のタンパク質及び/又はDsbCを発現させるために、誘導可能な発現系、又は構成的なプロモーターを本発明において用いることができる。適切な誘導可能な発現系及び構成的なプロモーターは、当技術分野においてよく知られている。
【0250】
DsbCをコードするポリヌクレオチド、及び対象のタンパク質をコードするポリヌクレオチドが、同じ又は別々の誘導プロモーターの制御下にある一実施形態において、対象のタンパク質をコードするポリヌクレオチド配列及びDsbCをコードする組換えポリヌクレオチドは、培養培地に誘導因子を加えることによって誘導される。
【0251】
形質転換細胞を培養するのにあらゆる適切な培地を用いることができる。例えば、培地中で増殖するように上首尾に形質転換されている細胞だけを可能にするように、培地は抗生物質を含み得るなど、培地を特定の選択系に適応させてもよい。
【0252】
培地から得た細胞を、所望により、さらにスクリーニング及び/又は精製にかけてもよい。方法は、所望により対象のタンパク質を抽出及び精製する1つ又は複数のステップをさらに含むことができる。
【0253】
ポリペプチドを、原形質、ペリプラズム、又は上清からを含む、系統から回収することができる。
【0254】
タンパク質を精製するために用いる特定の方法(単数又は複数)はタンパク質のタイプに依存する。適切な方法には、イムノ−アフィニティカラム、又はイオン交換カラム上での分画化、エタノール沈澱、逆相HPLC、疎水性−相互作用クロマトグラフィー、シリカ上クロマトグラフィー、SEPHAROSE及びDEAEなどのイオン交換樹脂上のクロマトグラフィー、クロマトフォーカシング、硫酸アンモニウム沈澱、及びゲルろ過が含まれる。
【0255】
一実施形態において、方法は、DsbCから対象の組換えタンパク質を分離することをさらに含む。
【0256】
抗体を、培養培地及び/又は原形質抽出物及び/又はペリプラズム抽出物から、従来の抗体精製手順、例えば、タンパク質A−セファロース、タンパク質Gクロマトグラフィー、タンパク質Lクロマトグラフィー、チオフィリック(thiophilic)、混合モード樹脂(mixed mode resins)、Hisタグ、FLAGタグ、ヒドロキシルアパタイトクロマトグラフィー、ゲル電気泳動、透析、アフィニティクロマトグラフィー、硫酸アンモニウム、エタノール、又はPEG分画/沈澱、イオン交換膜、膨張床吸着クロマトグラフィー(EBA)、又は擬似移動床クロマトグラフィーなどによって適切に分離することができる。
【0257】
方法は、対象のタンパク質の発現量を測定し、高発現レベルの対象のタンパク質を有する細胞を選択するさらなるステップも含むことができる。
【0258】
方法は、抗体又は抗体フラグメントなどの対象のタンパク質のペグ化などの1つ又は複数のさらなる下流のプロセシングのステップも含むことができる。
【0259】
本明細書に記載する1つ又は複数の方法のステップを、バイオリアクターなどの適切な容器において組み合わせて行ってもよい。
【実施例】
【0260】
細胞系
全ての実験に対して、親の野生型細胞系として大腸菌細胞系W3110を用いた。
【0261】
以下の変異を有する細胞系を作り出した:
a.変異Tsp遺伝子、
b.変異Tsp遺伝子及び組換えDsbCを有する、
c.変異Tsp遺伝子及び変異spr遺伝子、
d.変異Tsp遺伝子及び変異spr遺伝子及び組換えDsbCを有する。
【0262】
(例1)
変異Tsp遺伝子を有する細胞系MXE001(ΔTsp)の生成
MXR001系統を以下の通り生成した:
Tspカセットを、SalI、NotI制限フラグメントとして、同様に制限したpKO3プラスミド中に移動させた。pKO3プラスミドは、温度感受性変異体のpSC101複製開始点(RepA)を、染色体の組入れ事象に対して強化及び選択するためのクロラムフェニコールマーカーと一緒に用いる。レバンスクラーゼをコードするsacB遺伝子は、ショ糖上で増殖させた大腸菌に致死的であり、したがって(クロラムフェニコールマーカー及びpSC101開始点と一緒に)用いて脱組込み(de−integration)及びプラスミドキュアリング事象に対して強化及び選択するのに用いられる。この方法は以前に記載されている(Hamiltonら、1989年、Journal of Bacteriology、171巻、4617〜4622頁、及びBlomfieldら、1991年、Molecular Microbiology、5巻、1447〜1457頁)。pKO3系は全ての選択マーカーを、挿入された遺伝子以外の宿主ゲノムから除去する。
【0263】
以下のプラスミドを構築した。
EcoRI及びAseI制限マーカーを含んでいる、配列番号3に示すノックアウト変異Tsp遺伝子を含んでいるpMXE191。
【0264】
次いで、プラスミドを、Miller,E.M.及びNickoloff,J.A.における「Escherichia coli electrotransformation」、Methods in Molecular Biology、47巻、Nickoloff,J.A.(編集)、Humana Press、Totowa、NJ、105巻(1995年)に見られる方法を用いて調製した、電気的コンピテント大腸菌W3110細胞中に形質転換した。
【0265】
1日目 大腸菌細胞40μlを、冷却したBioRad0.2cm電気穿孔キュベット中、pKO3 DNA(10pg)1μgと混合し、その後2500V、25μF、及び200Ωで電気穿孔した。2×PY1000μlを直ちに加え、30℃のインキュベーター中で1時間、250rpmで振盪することによって細胞を回収した。細胞を2×PY中1/10に段階希釈し、その後100μlのアリコートを、クロラムフェニコール20μgを含み30℃及び43℃に予め温めた2×PY寒天平板上に塗抹した。平板を30℃及び43℃で一夜インキュベートした。
【0266】
2日目 30℃で増殖させたコロニーの数により電気穿孔の効果が推定されたが、43℃の増殖を生存したコロニーは潜在的な組入れ事象を表している。43℃の平板から単一のコロニーを拾い、2×PY10ml中に再懸濁した。この100μlを、ショ糖5%(w/v)を含み30℃に予め温めた2×PY寒天平板上に塗抹して単一のコロニーを生成させた。平板を30℃で一夜インキュベートした。
【0267】
3日目 ここでコロニーは、潜在的な同時の脱組込み及びプラスミドキュアリング事象を表す。脱組込み及びキュアリング事象が増殖における初期に生じた場合、大部分のコロニー塊はクローンとなる。単一のコロニーを拾い、クロラムフェニコール20μg/ml又は5%(w/v)ショ糖のいずれかを含む2×PY寒天上にレプリカプレーティングした。平板を30℃で一夜インキュベートした。
【0268】
4日目 ショ糖上で増殖し、且つクロラムフェニコール上で死滅した両方のコロニーは、潜在的な染色体の置換及びプラスミドキュアリング事象を表している。これらを拾い、変異特異的なオリゴヌクレオチドでPCRによってスクリーニングした。正確なサイズのPCR陽性のバンドを生じたコロニーを削除して、ショ糖5%(w/v)を含む2×PY寒天上で単一のコロニーを生成し、平板を30℃で一夜インキュベートした。
【0269】
5日目 PCR陽性、クロラムフェニコール感受性、及びショ糖耐性の大腸菌の単一のコロニーを用いてグリセロール株、化学的コンピテント細胞を作製し、5’及び3’に隣接するオリゴとのPCR反応用のPCR鋳型として作用させて、Taqポリメラーゼを用いた直接DNA配列決定用のPCR生成物を生成した。
【0270】
オリゴヌクレオチドプライマーを用いて、図1a、1b、及び1cに示す通り、天然に存在しないAseI制限部位を含んでいるTsp遺伝子の領域をPCR増幅することによって、変異Tsp遺伝子を有するゲノムDNAの上首尾な修飾を確認するために細胞系統MXE001を試験した。次いで、変異遺伝子における天然に存在しないAseI制限部位の存在を確認するために、AseI制限酵素でのインキュベートの前及び後に、DNAの増幅した領域をゲル電気泳動によって分析した。この方法を以下の通り行った。
【0271】
以下のオリゴを用いて、PCRを用い、MXE001及びW3110から調製した大腸菌細胞可溶化物からゲノムDNAを増幅した。
【化1】
【0272】
細胞のコロニー1個を、1×PCRバッファー20μl中95℃で10分間加熱することによって、可溶化物を調製した。混合物を室温に放冷し、次いで13200rpmで10分間遠心した。上清を除去し、「細胞可溶化物」のラベルを付けた。
【0273】
Tspオリゴ対を用いて各系統を増幅した。
【0274】
DNAを標準のPCR手順を用いて増幅した。
5μl バッファー×1 (Roche)
1μl dNTP混合物(Roche、10mM混合物)
1.5μl 5’オリゴ(5pmol)
1.5μl 3’オリゴ(5pmol)
2μl 細胞可溶化物
0.5μl Taq DNAポリメラーゼ(Roche 5U/μl)
38.5μl H2O
PCRサイクル
94℃ 1分
94℃ 1分)
55℃ 1分)30サイクル繰返し
72℃ 1分)
72℃ 10分
【0275】
反応が完了したら25μlをAseIでの消化用に新たな微量遠心管に移した。PCR反応25μlに、H2O19μl、バッファー3(NEB)5μl、AseI(NEB)1μlを加え、混合し、37℃で2時間インキュベートした。
【0276】
残りのPCR反応に、ローディングバッファー(×6)5μlを加え、20μlを0.8%TAE200mlアガロースゲル(Invitrogen)上にローディングし、エチジウムブロマイド(貯蔵液10ml/mlを5μl)を加え、100ボルトで1時間泳動させた。サイズマーカー(PerfectDNAマーカー0.1〜12kb、Novagen)10μlを最終レーンにローディングした。
【0277】
AseI消化が完了したら、ローディングバッファー(×6)10μlを加え、20μlを0.8%TAEアガロースゲル(Invitrogen)上にローディングし、エチジウムブロマイド(10ml/ml貯蔵液を5μl)を加え、100ボルトで1時間泳動した。サイズマーカー(PerfectDNAマーカー0.1〜12kb、Novagen)10μlを最終レーンにローディングした。ゲルを両方ともUVトランスイルミネーターを用いて可視化した。
【0278】
増幅したゲノムフラグメントは、Tspに対して2.8kbの正確なサイズバンドを示した。引き続きAseIで消化させると、Tsp欠損系統MXE001に導入されたAseI部位の存在が確認されたが、W3110コントロールでは確認されなかった。
【0279】
MXE001:Tspプライマーセットを用いてゲノムDNAを増幅し、得られたDNAをAseIで消化して2.2kbps及び0.6kbpsのバンドを生成した。
【0280】
W3110のPCR増幅したDNAは、AseI制限酵素によって消化されなかった。
【0281】
(例2)
変異spr遺伝子を有する細胞系統、並びに変異Tsp遺伝子及び変異spr遺伝子を有する細胞系統の生成
spr変異を、相補性アッセイを用いて生成し選択した。
【0282】
spr遺伝子を、1000bpあたり1つから2つの変異を導入するClontech(登録商標)ランダム変異ダイバーシティPCRキットを用いて変異させた。変異sprPCR DNAを、spr変異体と一緒にCDP870Fab’を発現する誘導発現ベクター[pTTO CDP870]中にクローニングした。次いで、このライゲーションを、Miller,E.M.及びNickoloff, J.A.、「Escherichia coli electrotransformation」、Methods in Molecular Biology、47巻、Nickoloff,J.A.(編集)、Humana Press、Totowa、NJ、105巻(1995年)において見られる方法を用いて、大腸菌系統MXE001(ΔTsp)中に電気的形質転換した。以下のプロトコールを用いた。電気的コンピテントMXE001 40μl、ライゲーション2.5μl(DNA100pg)を0.2cm電気穿孔キュベット中に加え、以下の条件、2500V、25μF、及び200ΩでBioRad Genepulser Xcellを用いて電気的形質転換を行った。電気的形質転換後、SOC(Invitrogen)1ml(37℃に予め加温)を加え、穏やかに振盪して細胞を37℃1時間回復させた。
【0283】
細胞をHypotonic寒天(酵母菌抽出物5g/L、トリプトン2.5g/L、寒天15g/L(全てDifco))上に塗抹し、40℃でインキュベートした。コロニーを形成した細胞をHLB上に43℃で再塗抹し、MXE001系統に対して低浸透圧条件下高温で増殖する能力の回復を確認した。選択したクローンからプラスミドDNAを調製し、配列決定してspr変異を同定した。
【0284】
この方法を用いて、sprタンパク質における、以下の通りΔTsp表現型を補足する8つの単一の変異、1つのダブル変異、及び2つの複数の変異を単離した:
1.V98E
2.D133A
3.V135D
4.V135G
5.G147C
6.S95F及びY115F
7.I70T
8.N31T、Q73R、R100G、G140C
9.R62C、Q99P、R144C
10.L108S
11.L136P
【0285】
上記で同定した1から5の個々の変異、並びにspr(C94A、H145A、H157A)及びW174Rの3つの触媒三残基の変異を用いて、複合のΔTsp/変異体spr系統を作製するための例1からのspr変異系統又はMXE001系統を作り出すために、いずれかの野生型W3110大腸菌系統(遺伝子型:F−LAM−IN(rrnD−RRnE)1rph1(ATCC no.27325))を用いて新たな系統を生成した。
【0286】
MXE001を作製するための例1に記載した通り、遺伝子置換ベクター系を用いて、pKO3相同組換え/置換プラスミドを用いて、以下の変異体の大腸菌細胞系統を作製した(Linkら、1997年、Journal of Bacteriology、179巻、6228〜6237頁)。
【表1】
【0287】
変異体spr組入れカセットを、SalI、NotI制限フラグメントとして、同様に制限したpKO3プラスミド中に移動した。
【0288】
プラスミドは、温度感受性変異体のpSC101複製開始点(RepA)を、染色体の組入れ事象に対して強化及び選択するためのクロラムフェニコールマーカーと一緒に用いている。レバンスクラーゼをコードするsacB遺伝子は、ショ糖上で増殖させた大腸菌に対して致死的であり、したがって(クロラムフェニコールマーカー及びpSC101開始点と一緒に)脱組込み及びプラスミドのキュアリングの事象に対して強化及び選択するのに用いられる。この方法は以前に記載されている(Hamiltonら、1989年、Journal of Bacteriology、171巻、4617〜4622頁、及びBlomfieldら、1991年、Molecular Microbiology、5巻、1447〜1457頁)。pKO3系は全ての選択マーカーを、挿入された遺伝子以外の宿主のゲノムから除去する。
【0289】
SalI制限部位をクローン同定用に除去するspr配列内にサイレント変異を含んでいる変異spr遺伝子を含んでいる、以下に列挙するpK03ベクターを構築した。
pMXE336、pK03 spr S95F (−SalI)
pMXE337、pK03 spr Y115F (−SalI)
pMXE338、pK03 spr G147C (−SalI)
pMXE339、pK03 spr D133A (−SalI)
pMXE340、pK03 spr V135D (−SalI)
pMXE341、pK03 spr V135G (−SalI)
pMXE342、pK03 spr V98E (−SalI)
pMXE343、pK03 spr C94A (−SalI)
pMXE344、pK03 spr H145A (−SalI)
pMXE345、pK03 spr H157A (−SalI)
pMXE346、pK03 spr W174R (−SalI)
【0290】
次いで、これらのプラスミドを、Miller,E.M.及びNickoloff,J.A.、「Escherichia coli electrotransformation」、Methods in Molecular Biology、47巻、Nickoloff,J.A.(編集)、Humana Press、Totowa、NJ、105巻(1995年)に見られる方法を用いて調製した、化学的コンピテント大腸菌W3110細胞中に、又は例1からのMXE001系統中に形質転換して、表1に示す通り、複合のΔTsp/変異体spr系統を作製した。
【0291】
1日目 電気的コンピテント大腸菌細胞又はMXE001細胞を、冷却したBioRad0.2cm電気穿孔キュベット中、pKO3 DNA(10pg)1μgと混合し、その後2500V、25μF、及び200Ωで電気穿孔した。2×PY1000μlを直ちに加え、30℃のインキュベーター中で1時間、250rpmで振盪することによって細胞を回収した。細胞を2×PY中1/10に段階希釈し、その後100μlのアリコートを、クロラムフェニコール20μgを含み30℃及び43℃に予め温めた2×PY寒天平板上に塗抹した。平板を30℃及び43℃で一夜インキュベートした。
【0292】
2日目 30℃で増殖させたコロニーの数により電気穿孔の効果が推定されたが、43℃の増殖を生存したコロニーは潜在的な組入れ事象を表している。43℃の平板から単一のコロニーを拾い、2×PY10ml中に再懸濁した。この100μlを、ショ糖5%(w/v)を含み30℃に予め温めた2×PY寒天平板上に塗抹して単一のコロニーを生成させた。平板を30℃で一夜インキュベートした。
【0293】
3日目 ここでコロニーは、潜在的な同時の脱組込み及びプラスミドキュアリング事象を表す。脱組込み及びキュアリング事象が増殖における初期に生じた場合、大部分のコロニー塊はクローンとなる。単一のコロニーを拾い、クロラムフェニコール20μg/ml又は5%(w/v)ショ糖のいずれかを含む2×PY寒天上にレプリカプレーティングした。平板を30℃で一夜インキュベートした。
【0294】
4日目 ショ糖上で増殖し、且つクロラムフェニコール上で死滅した両方のコロニーは潜在的な染色体の置換及びプラスミドキュアリング事象を表している。これらを拾い、SalI部位の喪失に対してPCRプラス制限消化によってスクリーニングした。正確なサイズのPCR陽性のバンド及びSalIによる消化に対する耐性を生じたコロニーを削除して、ショ糖5%(w/v)を含む2×PY寒天上で単一のコロニーを生成し、平板を30℃で一夜インキュベートした。
【0295】
5日目 PCR陽性、クロラムフェニコール感受性、及びショ糖耐性の大腸菌の単一のコロニーを用いてグリセロール株、化学的コンピテント細胞を作製し、5’及び3’に隣接するオリゴとのPCR反応用のPCRテンプレートとして作用させて、Taqポリメラーゼを用いた直接DNA配列決定用のPCR生成物を生成して正確な変異を確認した。
【0296】
(例3)
Fab’及びDsbC発現用のプラスミドの生成
抗TNF Fab’の重鎖及び軽鎖配列の両方、並びにDsbCをコードする配列を含んでいるプラスミドを構築した。
【0297】
WO01/94585に記載されている抗TNFαFab’フラグメント(CDP870と呼ぶ)のジシストロン性のメッセージを作成した。上流のシストロンは抗体の軽鎖(配列番号13)をコードし、下流のシストロンは抗体の重鎖(配列番号14)をコードしている。OmpAシグナルペプチドをコードするDNA配列は、軽鎖及び重鎖の各々をコードするDNAの5’末端に融合してペリプラズムへの効果的な分泌を可能にした。遺伝子間配列(IGS)2を、配列番号37に示す通りに用いた。
【0298】
CDP870Fab’(抗TNF Fab’)及びDsbC(ペリプラズムポリペプチド)に対する発現ベクターであるプラスミドpDPG358(pTTO40.4 CDP870 IGS2)を、Sambrookら、1989年、「Molecular cloning:a laboratory manual」、CSHL press、N.Yにおいて見ることができる従来の制限クローニング法を用いて構築した。プラスミドpDPH358は以下の特徴:強力なtacプロモーター及びlacオペレーター配列を含んでいた。図6に示す通り、プラスミドは独特のEcoRI制限部位をFab’重鎖のコード領域の後に含んでおり、その後に非コード配列、次いで独特のNdeI制限部位が続いた。DsbC遺伝子を、W3110粗製染色体DNAを鋳型として用いて、5’EcoRI部位をコードするPCR生成物の後に強力なリボソーム結合が続き、その後に天然の開始コドン、シグナル配列、及びDsbCの成熟配列、C末端Hisタグにおける終結、及び最後に非コードNdeI部位が続くようにPCRクローニングした。EcoRI−NdelI PCRフラグメントを制限し、3個全てのポリペプチド:Fab’軽鎖、Fab’重鎖、及びDsbCが単一のポリシストロン性のmRNA上にコードされるように発現ベクター中にライゲートした。
【0299】
Fab軽鎖、重鎖遺伝子、及びDcbC遺伝子を、単一のポリシストロン性メッセージとして転写した。大腸菌のペリプラズムに対するポリペプチドの転移を指示する大腸菌OmpAタンパク質からのシグナルペプチドをコードするDNAを軽鎖及び重鎖の遺伝子配列両方の5’末端に融合した。転写を二重転写ターミネーターrrnB t1t2を用いて終結させた。lacIq遺伝子は構成的に発現されるLacIリプレッサータンパク質をコードしていた。これは、アロラクトース又はIPTGの存在によって抑制解除が誘導されるまで、tacプロモーターからの転写を抑制した。用いた複製開始点はp15Aであり、これは低コピー数を維持した。プラスミドは、抗生物質選択用にテトラサイクリン耐性遺伝子を含んでいた。
【0300】
(例4)
変異細胞系における抗TNF Fab’及びDsbCの発現
抗TNF Fab’及びDsbCの発現
例1において提供したMXE001系統、及び例2において提供した変異系統であるMXE008及びMXE009を、例3において生成したプラスミドで形質転換した。
【0301】
系統の形質転換を、Chung C.Tら、「Transformation and storage of bacterial cells in the same solution.」PNAS、86巻、2172〜2175頁(1989年)に見られる方法を用いて行った。
【0302】
抗TNF Fab’の発現
MXE001、MXE008、及びMXE009の系統を、例3に記載した通り、CDP870Fab’に対する発現ベクター(配列番号13に示す軽鎖配列及び配列番号14に示す重鎖配列を有する抗TNF Fab’)であるプラスミドpMXE117(pTTO CDP870又は40.4 IGS2)で形質転換したが、この発現ベクターは、Sambrookら、1989年、「Molecular cloning:a laboratory manual.」、CSHL press、N.Y.に見ることができる従来の制限クローニング法を用いて構築されたものである。プラスミドpMXE117(pTTO CDP870又は40.4 IGS2)は以下の特徴:強力なtacプロモーター及びlacオペレーター配列を含んでいた。Fab軽鎖及び重鎖遺伝子を、単一のジシストロン性メッセージとして転写した。大腸菌OmpAタンパク質からのシグナルペプチドをコードするDNAを、ポリペプチドの大腸菌ペリプラズムへの移動を指示する軽鎖及び重鎖両方の遺伝子配列5’末端に融合させた。転写を二重転写ターミネーターrrnB t1t2を用いて終結させた。lacIq遺伝子は構成的に発現されるLacIリプレッサータンパク質をコードしていた。これは、アロラクトース又はIPTGの存在によって抑制解除が誘導されるまで、tacプロモーターからの転写を抑制した。用いた複製開始点はp15Aであり、低コピー数を維持した。プラスミドは、抗生物質選択用にテトラサイクリン耐性遺伝子を含んでいた。
【0303】
系統の形質転換を、Chung C.Tら、「Transformation and storage of bacterial cells in the same solution.」、PNAS、86巻、2172〜2175頁(1989年)に見られる方法を用いて行った。
【0304】
(例5)
高密度発酵を用いた変異大腸菌系統の増殖及び変異大腸菌系統における抗TNF Fab’の発現
例4において生成した以下の系統を、抗TNFαFab’の増殖及び発現を比べる発酵実験において試験した。
抗TNF Fab’を発現するMXE001
抗TNF Fab’を発現するMXE008
抗TNF Fab’を発現するMXE009
抗TNF Fab’及びDsbCを発現するMXE001
抗TNF Fab’及びDsbCを発現するMXE008
抗TNF Fab’及びDsbCを発現するMXE009
【0305】
増殖培地
発酵増殖培地は、NaH2PO4・H2O 3.86g/l及びグリセロール112g/lを有するSM6E培地(Humphreyら、2002年、Protein Expression and Purification、26巻、309〜320頁に記載)をベースとした。
【0306】
接種
接種培養物を、テトラサイクリン10μg/mlを補った同じ培地中で増殖させた。培養物をおよそ22時間、撹拌しながら30℃でインキュベートした。
【0307】
発酵
発酵槽(総容積2.5リットル)に、接種培養物を0.3〜0.5OD600に接種した。温度は増殖相の間は30℃に維持し、誘導前に25℃に下げた。溶存酸素濃度を、可変性の撹拌及びエアフローによって空気飽和を30%上回って維持した。15%(v/v)NH4OH及び10%(v/v)濃H2SO4で自動滴定することによって、培養物のpHを7.0で制御した。10%(v/v)Struktol J673溶液(Schill and Seilacher)を加えることによって泡立ちを制御した。発酵の様々な段階で数々の添加を行った。バイオマス濃度がおよそ40OD600に到達したらマグネシウム塩及びNaH2PO4・H2Oを加えた。誘導相の前及び間にNaH2PO4・H2Oのさらなる添加を行って、確実にリン酸塩が過剰に維持されるようにした。発酵の初期に存在したグリセロールが枯渇したら(およそ75OD600)、80%(w/w)の継続的なグリセロールの供給を適用した。発酵における同じ時点に170μMのIPTGの供給を適用した。IPTG供給の開始を誘導の開始と理解した。発酵は、典型的に、グリセロール供給速度(0.5から2.5ml/hの間の範囲)で64〜120時間行った。
【0308】
バイオマス濃度及び増殖速度の測定
600nmでの培養物の光学密度を測定することによってバイオマス濃度を決定した。
【0309】
ペリプラズム抽出
細胞を、遠心分離によって培養物試料から回収した。さらなる分析用に上清の分画を(−20℃で)保持した。細胞ペレット分画を、抽出バッファー(100mM Tris−HCl、10mM EDTA、pH7.4)中、元の培養体積に再懸濁した。およそ16時間、60℃でインキュベートした後、遠心分離によって抽出物を澄明にし、上清分画を分析用に(−20℃で)保持した。
【0310】
Fab’定量
ペリプラズム抽出物及び培養物上清中のFab’濃度を、Humphreysら、2002年、Protein Expression and Purification、26巻、309〜320に記載されている通り、Fab’アセンブリーELISAによって、タンパク質G HPLCを用いて決定した。HiTrap タンパク質G HP1mlカラム(GE−Healthcare又は同等物)に分析物(およそ中性のpH、30℃、0.2μmろ過)を2ml/分でローディングし、カラムを20mMリン酸塩、50mM NaCl、pH7.4で洗浄し、次いで50mMグリシン/HCl pH2.7の注入を用いてFab’を溶出した。溶出されたFab’をAgilient1100又は1200HPLCシステム上A280によって測定し、既知濃度の精製Fab’タンパク質の検量線を参照することによって定量した。
【0311】
図1は、抗TNF Fab’発現系統性MXE001及びMXE008の増殖プロファイル、並びに抗TNF Fab’及び組換えDsbC発現性系統MXE001及びMXE008の増殖プロファイルを示す。DsbCを発現する系統は、組換えDsbCを発現しない対応する細胞系統に比べて改善された増殖を表すことが分かる。系統におけるspr変異の存在が細胞増殖を改善することも分かる。
【0312】
図2は、抗TNF Fab’及び組換えDsbC発現性大腸菌系統MXE001、MXE008、及びMXE009からのペリプラズム(黒記号)及び上清(白記号)からのFabの収率を示す。このグラフから、3つの系統全てが高収率の抗TNFα Fab’を生成し、MXE008及びMXE009系統は2.2g/Lを超えるペリプラズム抗TNF Fab’を40時間で生成したことが分かる。変異体spr遺伝子を有するMXE008及びMXE009系統はMXE001(上清の抗TNFαFab’が少ないと分かる)に比べて低減した溶解を示したことも分かる(白記号)。
【0313】
図3は、抗TNF Fab’発現性大腸菌系統MXE001及びMXE008からの、並びに抗TNF Fab’及び組換えDsbC発現性大腸菌系統MXE001及びMXE008からの、ペリプラズム(黒記号)及び上清(白記号)からのFabの収率を示す。このグラフから、組換えDsbCを発現する系統は高収率の抗TNFαFab’を生成し、MXE008系統は2.4g/Lのペリプラズム抗TNF Fab’を40時間で生成したことが分かる。変異体spr遺伝子を有するMXE008系統はMXE001系統(上清の抗TNFαFab’が少ないと分かる)に比べて低減した溶解を示したことも分かる(白記号)。
【0314】
(例6)
高密度発酵を用いた変異大腸菌系統の増殖並びに変異大腸菌系統におけるFabA及びFabBの発現
本発明の細胞からのタンパク質生成の収率に対する効果も、抗原Aに対する特異性を有するFabA及び抗原Bに対する特異性を有するFabBである2つのさらなるFabタンパク質に対して、上記例5に記載したのと同じ方法を用いて行い、FabA及びFabBを発現するW3110系統に対して比較した。
【0315】
図4は、FabA及びFabB発現性大腸菌系統W3110からの、並びに、FabA及び組換えDsbC発現性又はFab及び組換えDsbC発現性大腸菌系統MXE008からの、ペリプラズムからのFab収率の合計を示す。このグラフから、組換えDsbCを発現する系統は、MXE008において、W3110に比べて有意に高い収率のFabA及びFabBを生成したことが分かる。
【0316】
(例7)
大規模発酵を用いた抗TNF Fab’及びDsbCを発現する大腸菌系統の増殖
例4によって生成した以下の系統を、抗TNFα Fab’の系統の増殖及び発現を比べる発酵実験において試験した:
抗TNF Fab’及びDsbCを発現するMXE008
【0317】
発酵を以下の通りに行った:
抗TNF Fab’及びDsbC細胞を発現するMXE008を、最初に、フラスコ振盪培養物中酵母菌抽出物及びペプトンの複合培地を用いて増殖させた。次いで、細胞を、化学的に規定された培地を用いて接種段階の発酵槽に移した。細胞を、規定された移行点まで栄養非制限の条件下で増殖させた。次いで、細胞を、同様の化学的に規定された培地を用いて、5L又は250Lの生成発酵槽にそれぞれおよそ3.3L又は230Lの最終体積で移した。培養物を最初に、炭素源が枯渇するまで、バッチモード中で増殖させた。炭素源が枯渇した後、炭素源を限定する供給を、指数関数的に増大する速度で供給した。規定された量の炭素源を加えた後、フィード溶液を添加する速度を下げ、IPTGを加えてFab’の発現を誘導した。次いで、発酵を続け、Fab’をペリプラズム中に蓄積させた。誘導後の規定された時点で、遠心分離によって培養物を回収し、回収した細胞をTris及びEDTAバッファー中に再懸濁し、59℃に加熱することによって、Fab’を細胞から抽出した。
【0318】
発酵の増殖プロファイルを、600nmの培養物の光学密度を測定することによって決定した。
【0319】
Fab’の力価を、上記例5に記載した通りタンパク質G HPLCによって決定したが、ペリプラズム抽出の間新鮮な細胞を用い、抽出バッファー1mLを細胞培養液に加える点が異なった。上清及びペリプラズムのFab’を、例5に記載した通りに測定した。図8はペリプラズムのFab’の力価を示す。
【0320】
図7は、抗TNF Fab’及び組換えDsbC発現性大腸菌系統MXE008の、5L及び200L発酵の、比較上の増殖プロファイルを示す。
【0321】
図8は、抗TNF Fab’及び組換えDsbC発現性大腸菌系統MXE008の、5L及び200L発酵の、比較上のFab’力価を示す。
【0322】
(例8)
大規模発酵を用いた抗TNF Fab’及びDsbCを発現する大腸菌系統の増殖
例4によって生成した以下の系統を、抗TNFα Fab’の系統の増殖及び発現を比べる発酵実験において試験した:
抗TNF Fab’及びDsbCを発現するMXE008
抗TNF Fab’及びDsbCを発現するMXE009。
【0323】
発酵を、20L生成発酵槽を用いて上記例7に記載した通りに行った。
【0324】
発酵の増殖プロファイルを、600nmの培養物の光学密度を測定することによって決定した。
【0325】
Fab’の力価を、上記例7に記載した通り、タンパク質G HPLCによって決定した。
【0326】
図9は、抗TNF Fab’及び組換えDsbC発現性大腸菌系統MXE008及びMXE009の、発酵の比較上の増殖プロファイルを示す。
【0327】
図10は、抗TNF Fab’及び組換えDsbC発現性大腸菌系統MXE008及びMXE009の、発酵の比較上のペリプラズマのFab’力価を示す。
【0328】
本発明を、好ましい実施形態を参照にして特に示し、記載してきたが、当業者であれば、添付の特許請求の範囲によって規定される通り、本発明の範囲から逸脱することなしに形態及び詳細において様々な変更を行うことができることが理解されよう。
【技術分野】
【0001】
本発明は、組換え細菌宿主系統、特に大腸菌(E.coli)に関する。本発明は、このような細胞中で対象のタンパク質を生成するための方法にも関する。
【背景技術】
【0002】
大腸菌などの細菌細胞は、組換えタンパク質を生成するために一般的に用いられる。特に、プラスミドによって遺伝子の挿入を可能にする宿主細胞として細菌細胞が多用途であるという性質のため、組換えタンパク質を生成するのに大腸菌などの細菌細胞を用いると有利な点が多い。大腸菌は、ヒトインスリンを含めた多くの組換えタンパク質を生成するのに用いられている。
【0003】
組換えタンパク質を生成するのに細菌細胞を用いるのは利点が多いにもかかわらず、プロテアーゼ感受性のタンパク質を生成するのが困難であるなど、重大な制限が依然として存在する。プロテアーゼは、大腸菌のペリプラズム及び細胞質における古く、損傷を受け、又はミスフォールディングされたタンパク質を代謝回転する上で重要な役割を果たす。細菌のプロテアーゼは、対象の組換えタンパク質を分解するように作用し、それによって活性タンパク質の収率を大幅に低減することが多い。
【0004】
数々の細菌のプロテアーゼが同定されている。大腸菌において、ProteaseIII(ptr)、DegP、OmpT、Tsp、prlC、ptrA、ptrB、pepA−T、tsh、espc、eatA、clpP、及びlonを含めたプロテアーゼが同定されている。
【0005】
Tsp(Prcとしても知られている)は、60kDaのペリプラズムプロテアーゼである。Tspの最初に知られた基質はペニシリン結合性タンパク質−3(PBP3)であったが(「Determination of the cleavage site involved in C−terminal processing of penicillin−binding protein 3 of Escherichia coli」Nagasawa H、Sakagami Y、Suzuki A、Suzuki H、Hara H、Hirota Y.、J Bacteriol.、1989年11月、171巻(11)、5890〜3頁、及び「Cloning,mapping and characterization of the Escherichia coli Tsp gene which is involved in C−terminal processing of penicillin−binding protein 3」、Hara H、Yamamoto Y、Higashitani A、Suzuki H、Nishimura Y.、J Bacteriol.、1991年8月、173巻(15)、4799〜813頁)、Tspはファージテイルタンパク質を切断することもできることが後に発見され、したがってテイル特異的プロテアーゼ(Tail Specific Protease)(Tsp)と新たに命名された(Silberら、Proc.Natl.Acad.Sci.USA、89巻、295〜299頁(1992年))、Silberらは(「Deletion of the prc(tsp) gene provides evidence for additional tail−specific proteolytic activity in Escherichia coli K−12」、Silber、K.R.、Sauer、R.T.、Mol Gen Genet、1994年、242巻、237〜240頁)、prc遺伝子のセグメントをKanrマーカーを含むフラグメントで置換することによって変異が作り出されたprcの欠失系統(KS1000)について記載している。
【0006】
Tsp(prc)活性を低減することは、対象のタンパク質のタンパク質分解を低減するのに望ましい。しかし、プロテアーゼprcを欠く細胞は、低いモル浸透圧濃度で熱感受性の増殖を示すことが見出された。Haraらは、遺伝子外サプレッサー(spr)変異を含む熱耐性復帰変異体を単離した(Haraら、Microbial Drug Resistance、2巻、63〜72頁(1996年))。sprは、18kDaの膜結合ペリプラズムプロテアーゼであり、sprの基質は細胞分裂の間細胞壁の加水分解に関与する外膜におけるTsp及びペプチドグリカンである。spr遺伝子は、UniProtKB/Swiss−Prot P0AFV4(SPR_ECOLI)と呼ばれている。
【0007】
変異体spr遺伝子を含んでいる改善されたプロテアーゼ欠損系統は記載されている。Chenら(Chen C、Snedecor B、Nishihara JC、Joly JC、McFarland N、Andersen DC、Battersby JE、Champion KM.、Biotechnol Bioeng.、2004年3月5日、85巻(5)、463〜74頁)は、prc(Tsp)、並びに遺伝子の上流及び下流の領域を増幅し、選択マーカー及びsprW174R変異を含んでいるベクター上でこれらを一緒にライゲートすることによって作り出された別のプロテアーゼであるDegP(大腸菌のペリプラズムにおける組換え抗体フラグメントの高レベルの蓄積は三重変異体(ΔDegPΔprc sprW174R)宿主系統を必要とする)における変異の異なる組合せを有する大腸菌系統の構築について記載している。ΔDegP、Δprc、及びsprW174R変異の組合せは、最高レベルの抗体軽鎖、抗体重鎖、及びF(ab)’2−LZを提供することが見出された。EP1341899は、プロテアーゼDegP及びPrcをそれぞれコードする染色体DegP及びprcを欠いており、prc変異体を抱く系統が示す増殖表現型を抑制するタンパク質をコードする変異体spr遺伝子を抱く大腸菌系統を開示している。
【0008】
タンパク質ジスルフィドイソメラーゼは、タンパク質がフォールディングするときのタンパク質内のシステイン残基間のジスルフィド結合の形成及び切断を触媒する酵素である。宿主細胞中でタンパク質の発現を改善するジスルフィド結合の形成を触媒するタンパク質を同時発現することが知られている。WO98/56930は細菌細胞中で異種性のジスルフィド結合含有ポリペプチドを生成するための方法を開示しており、DsbC又はDsbGなどの原核生物のジスルフィドイソメラーゼが真核生物のポリペプチドと同時発現される。US6673569は、外来タンパク質を生成するのに用いるためのDsbA、DsbB、DsbC、及びDsbDの各々をコードするポリヌクレオチドを含む人工オペロンを開示している。EP0786009は、細菌において異種性のポリペプチドを生成するためのプロセスを開示しており、DsbA又はDsbCをコードする核酸の発現は、異種性のポリペプチドをコードする核酸の発現の誘導の前に誘発される。
【0009】
DsbCは、大腸菌におけるジスルフィド結合の形成を触媒する大腸菌のペリプラズムにおいて見出された原核生物のタンパク質である。DsbCは、236個(シグナルペプチドを含む)の長さのアミノ酸配列、及び分子量25.6KDaを有する(UniProt No.p0AEG6)。DsbCは、1994年に始めて同定された(Missiakas eら、「The Escherichia coli dsbC(xprA)gene encodes a periplasmic protein involved in disulfide bond formation」、The EMBO Journal、13巻8号、2013〜2020頁、1994年、及びShevchikら、「Characterization of DsbC、a periplasmic protein of Erwinia chrysanthemi and Escherichia coli with disulfide isomerase activity」、The EMBO Jounral、13巻8号、2007〜2012頁、1994年)。
【0010】
驚くべきことに、グラム陰性細菌細胞における組換えDsbCからのDsbCの過剰発現は、プロテアーゼTspを欠く細胞の細胞溶解表現型を改善することが見出されている。したがって、本発明者らは、対象のタンパク質を生成するための有利な性質を有する新規な系統を提供する。
【発明の概要】
【課題を解決するための手段】
【0011】
本発明は、
a.DsbCをコードする組換えポリヌクレオチドを含んでおり、
b.野生型細胞に比べて低減したTspタンパク質活性を有する
ことを特徴とする、組換えグラム陰性細菌細胞を提供する。
【0012】
一実施形態において、細胞は野生型spr遺伝子を含んでいる。この実施形態において、細胞のゲノムが、野生型細胞に比べてTspタンパク質活性を低減するのに必要とされる修飾以外、野生型細菌細胞に対して同質遺伝子であるのが好ましい。
【0013】
さらなる一実施形態において、本発明による細胞は、野生型細胞に比べて低減したTspタンパク質活性を有し、DsbC及び変異spr遺伝子をコードする組換えポリヌクレオチドを含んでいる。この実施形態において、細胞のゲノムが、変異spr遺伝子及び野生型細胞に比べてTspタンパク質活性を低減するのに必要とされる修飾以外、野生型細菌細胞に対して同質遺伝子であるのが好ましい。
【0014】
遺伝子修飾の上記の特異的な組合せを有するグラム陰性細菌細胞は、有利な増殖及びタンパク質生成の表現型を示す。
【0015】
本発明は、上記に規定した組換えグラム陰性細菌細胞において対象のタンパク質を発現することを含む、対象のタンパク質を生成するための方法も提供する。方法は、上記に規定した組換えグラム陰性細菌細胞を、培養培地中、対象のタンパク質及びDsbCをコードする組換えポリヌクレオチドを発現するのに効果的な条件下で培養し、対象のタンパク質を、組換えグラム陰性細菌細胞のペリプラズム及び/又は培養培地から回収することを含む。
【図面の簡単な説明】
【0016】
【図1】抗TNF Fab’発現系統MXE001 MXE008、並びに抗TNFαFab’及び組換えDsbC発現系統MXE001及びMXE008の増殖プロファイルを示す図である。
【図2】抗TNFα Fab’及び組換えDsbC発現性大腸菌系統であるMXE001、MXE008、及びMXE009からのペリプラズム(黒記号)及び上清(白記号)からの抗TNFFab’の収率を示す図である。
【図3】抗TNFαFab’発現性大腸菌系統であるMXE001及びMXE008からの、並びに、抗TNF Fab’及び組換えDsbC発現性大腸菌系統であるMXE001及びMXE008からの、ペリプラズム(黒記号)及び上清(白記号)からの、抗TNF Fab’の収率を示す図である。
【図4】FabA及びFabB発現性大腸菌系統であるW3110からの、並びにFabA及び組換えDsbC、又はFabB及び組換えDsbC発現性大腸菌系統であるMXE008からの、ペリプラズムからの抗TNF Fab’の収率を示す図である。
【図5a】野生型ptr(ProteaseIII)及びノックアウト変異ptr(ProteaseIII)の5’末端のタンパク質及び遺伝子配列を示す図である。
【図5b】野生型Tsp及びノックアウト変異Tspの5’末端のタンパク質及び遺伝子配列を示す図である。
【図5c】野生型DegP及び変異DegPタンパク質のある領域及び遺伝子配列を示す図である。
【図6】本発明の一実施形態による細胞を生成するのに用いるためのベクターの構築を示す図である。
【図7】抗TNF Fab’及び組換えDsbC発現性大腸菌系統MXE008の5L及び200Lの発酵の比較上の増殖プロファイルを示す図である。
【図8】抗TNF Fab’及び組換えDsbC発現性大腸菌系統MXE008の5L及び200Lの発酵の比較上のFab’力価を示す図である。
【図9】抗TNF Fab’及び組換えDsbC発現性大腸菌系統MXE008及びMXE009の発酵の比較上の増殖プロファイルを示す図である。
【図10】抗TNF Fab’及び組換えDsbC発現性大腸菌系統MXE008及びMXE009の発酵の比較上のFab’力価を示す図である。
【0017】
配列の簡単な説明
配列番号1は、開始コドンの上流の6個のヌクレオチドATGAACを含んでいる野生型Tsp遺伝子のDNA配列である。
【0018】
配列番号2は、野生型Tspタンパク質のアミノ酸配列である。
【0019】
配列番号3は、開始コドンの上流の6個のヌクレオチドATGAATを含んでいる変異ノックアウトTsp遺伝子のDNA配列である。
【0020】
配列番号4は、野生型ProteaseIII遺伝子のDNA配列である。
【0021】
配列番号5は、野生型ProteaseIIIタンパク質のアミノ酸配列である。
【0022】
配列番号6は、変異ノックアウトProteaseIII遺伝子のDNA配列である。
【0023】
配列番号7は、野生型DegP遺伝子のDNA配列である。
【0024】
配列番号8は、野生型DegPタンパク質のアミノ酸配列である。
【0025】
配列番号9は、変異DegP遺伝子のDNA配列である。
【0026】
配列番号10は、変異DegPタンパク質のアミノ酸配列である。
【0027】
配列番号11は、抗TNF抗体の軽鎖可変領域のアミノ酸配列である。
【0028】
配列番号12は、抗TNF抗体の重鎖可変領域のアミノ酸配列である。
【0029】
配列番号13は、抗TNF抗体の軽鎖のアミノ酸配列である。
【0030】
配列番号14は、抗TNF抗体の重鎖のアミノ酸配列である。
【0031】
配列番号15は、AseI制限部位を含んでいる変異Tsp遺伝子の領域に対する3’オリゴヌクレオチドプライマーの配列である。
【0032】
配列番号16は、AseI制限部位を含んでいる変異Tsp遺伝子の領域に対する5’オリゴヌクレオチドプライマーの配列である。
【0033】
配列番号17は、AseI制限部位を含んでいる変異ProteaseIII遺伝子の領域に対する3’オリゴヌクレオチドプライマーの配列である。
【0034】
配列番号18は、AseI制限部位を含んでいる変異ProteaseIII遺伝子の領域に対する5’オリゴヌクレオチドプライマーの配列である。
【0035】
配列番号19は、AseI制限部位を含んでいる変異DegP遺伝子の領域に対する5’オリゴヌクレオチドプライマーの配列である。
【0036】
配列番号20は、AseI制限部位を含んでいる変異DegP遺伝子の領域に対する3’オリゴヌクレオチドプライマーの配列である。
【0037】
配列番号21は、最初のアミノ酸残基26個であるシグナル配列を含んでいる野生型spr遺伝子の配列である。
【0038】
配列番号22は、シグナル配列のない野生型spr遺伝子の配列である。
【0039】
配列番号23は、D210A及びH212A変異を含んでいる変異OmpT配列のヌクレオチド配列である。
【0040】
配列番号24は、D210A及びH212A変異を含んでいる変異OmpT配列のアミノ酸配列である。
【0041】
配列番号25は、変異ノックアウトOmpT配列のヌクレオチド配列である。
【0042】
配列番号26は、hisタグ付けしたDsbCのヌクレオチド配列である。
【0043】
配列番号27は、hisタグ付けしたDsbCのアミノ酸配列である。
【0044】
配列番号28は、hTNF40のCDRH1のアミノ酸配列を示す。
【0045】
配列番号29は、ハイブリッドCDRであるhTNF40のCDRH2のアミノ酸配列を示し、C末端の6個のアミノ酸はヒトサブグループ3生殖系列抗体のH2 CDR配列からであり、このハイブリダイゼーションに起因する配列に対するアミノ酸の変化には以下の通り下線をつける:WINTYIGEPI YADSVKG。
【0046】
配列番号30は、hTNF40のCDRH3のアミノ酸配列を示す。
【0047】
配列番号31は、hTNF40のCDRL1のアミノ酸配列を示す。
【0048】
配列番号32は、hTNF40のCDRL2のアミノ酸配列を示す。
【0049】
配列番号33は、hTNF40のCDRL3のアミノ酸配列を示す。
【0050】
配列番号34は、hTNF40のCDRH2のアミノ酸配列を示す。
【0051】
配列番号35は、OmpAオリゴヌクレオチドアダプターの配列を示す。
【0052】
配列番号36は、大腸菌Fab発現に対する遺伝子間配列1(IGS1)をコードするオリゴヌクレオチドカセットを示す。
【0053】
配列番号37は、大腸菌Fab発現に対する遺伝子間配列2(IGS2)をコードするオリゴヌクレオチドカセットを示す。
【0054】
配列番号38は、大腸菌Fab発現に対する遺伝子間配列3(IGS3)をコードするオリゴヌクレオチドカセットを示す。
【0055】
配列番号39は、大腸菌Fab発現に対する遺伝子間配列4(IGS4)をコードするオリゴヌクレオチドカセットを示す。
【発明を実施するための形態】
【0056】
本発明を、次に、より詳しく記載する。
【0057】
「タンパク質」及び「ポリペプチド」の語は、文脈上別段の記載がなければ、本明細書において交換可能に用いられる。「ペプチド」は10個以下のアミノ酸を意味するものとされる。
【0058】
「ポリヌクレオチド」の語は、文脈上別段の記載がなければ、遺伝子、DNA、cDNA、RNA、mRNAなどを含む。
【0059】
本明細書で用いられる「含んでいる(comprising)」の語は、本明細書の文脈において「含んでいる(including)」と解釈されたい。
【0060】
非変異(non−mutated)細胞又はコントロール細胞(control cell)は、本発明の文脈において、本発明の組換えグラム陰性菌と同じ型の細胞を意味し、細胞はTspタンパク質活性を低減し、組換えDsbC配列及び任意選択によって変異体のspr遺伝子を有するように修飾されていない。例えば、非変異細胞は野生型細胞であってよく、あらゆる変異を導入する修飾の前の本発明の細胞と同じ宿主細胞の集団に由来してよい。
【0061】
「細胞」、「細胞系」、「細胞培養物」、及び「系統」の表現は交換可能に用いられる。
【0062】
「変異Tsp遺伝子を含んでいる細胞の表現型」の表現は、本発明の文脈において、変異体のTsp遺伝子を抱く細胞によって表される表現型を意味する。典型的に、変異体のTsp遺伝子を含んでいる細胞は、特に高い細胞密度で溶解することができる。これらの細胞の溶解は、あらゆる組換えタンパク質の上清中への漏出をもたらす。変異Tsp遺伝子を有する細胞は、低いモル浸透圧濃度では熱感受性の増殖も示すことがある。例えば、40℃以上などの高温では、細胞は増殖率を示さず、若しくは低減した増殖率を示し、又は細胞は低浸透圧性の培地中で死滅する。
【0063】
「同質遺伝子の」の語は、本発明の文脈において、細胞のゲノムが、細胞が由来する野生型の細胞に比べて、野生型細胞に比べてTspタンパク質活性を低減するのに必要とされる修飾、及び任意選択によって変異spr遺伝子以外、同じ、又は実質的に同じ核酸配列(単数若しくは複数)を含んでいることを意味する。この実施形態において、細胞のゲノムはさらなる天然に存在しない変異又は遺伝子操作された変異を含んでいない。一実施形態において、本発明の細胞のゲノムは、野生型細胞に比べてTspタンパク質活性を低減するのに必要とされる修飾、及び任意選択によって変異spr遺伝子以外、野生型細胞に比べて実質的に同じ又は同じゲノム配列を有する。一実施形態において、本発明による細胞は、起こり得るあらゆる天然に存在する変異を考慮に入れて、野生型細胞に比べてTspタンパク質活性を低減するのに必要とされる修飾、及び任意選択によって変異spr遺伝子以外、野生型細胞に比べて実質的に同じゲノム配列を有し得る。一実施形態において、本発明による細胞は、野生型細胞に比べてTspタンパク質活性を低減するのに必要とされる修飾、及び任意選択によって変異spr遺伝子以外、野生型細胞に比べて正確に同じゲノム配列を有し得る。
【0064】
DsbCをコードする組換えポリヌクレオチドは、細胞中に形質転換され、及び/又は宿主細胞のゲノム中に組み入れられた適切な発現ベクター上に存在していてよい。DsbCをコードするポリヌクレオチドが宿主のゲノム中に挿入される実施形態において、細胞のゲノムはDsbCをコードする挿入されたポリヌクレオチド配列のために、野生型細胞とやはり異なる。DsbCをコードするポリヌクレオチドが細胞中の発現ベクター中にあり、それによって宿主細胞のゲノムの最小のかく乱をもたらすのが好ましい。
【0065】
一実施形態において、本発明の細胞は、対象のタンパク質をコードするポリヌクレオチドを含んでいる。この実施形態において、対象のタンパク質をコードするポリヌクレオチドは、細胞に形質転換され、及び/又は宿主細胞のゲノム中に組み入れられる適切な発現ベクター内に含まれていてよい。対象のタンパク質をコードするポリヌクレオチドが宿主のゲノム中に挿入されている実施形態において、挿入されたポリヌクレオチド配列が対象のタンパク質をコードするため、ゲノムは野生型細胞とやはり異なる。対象のタンパク質をコードするポリヌクレオチドが細胞中の発現ベクター中にあり、それによって宿主細胞のゲノムの最小のかく乱をもたらすのが好ましい。
【0066】
「野生型」の語は、本発明の文脈において、天然に存在することがあり、又は環境から単離され得るグラム陰性細菌細胞の系統を意味し、いかなる遺伝子操作された変異も保有しない。大腸菌の野生型系統の一例にはW3110−12系統などのW3110がある。
【0067】
本発明は、野生型細胞に比べて低減したTspタンパク質活性を有し、DsbCをコードする組換えポリヌクレオチドを含んでいる、対象のタンパク質を発現するのに適する組換えグラム陰性細菌細胞を提供する。
【0068】
本発明による細胞は、DsbCをコードする組換えポリヌクレオチドを含んでいる。本明細書で用いられる「組換えポリペプチド」は、組換えDNA技術を用いて構築又は生成されるタンパク質を意味する。DsbCをコードするポリヌクレオチド配列は、細菌細胞中に見出されるDsbCをコードする内在性の配列に同一であってよい。或いは、DsbCをコードする組換えポリヌクレオチド配列は、EcoRI部位などの制限部位が除去された、及び/又はhisタグをコードする配列を有するなど、野生型DsbC配列の変異バージョンである。本発明に用いるための一例の修飾DsbCヌクレオチド配列を配列番号26に示すが、これは配列番号27に示すhisタグ付けしたDsbCアミノ酸配列をコードするものである。
【0069】
本発明者らは、野生型細胞に比べて低減したTspタンパク質活性を有する細菌細胞においてDsbCをコードする組換えポリヌクレオチド発現の特定の組合せ、及び好ましい実施形態において、さらに変異spr遺伝子は、対象のタンパク質を発現するために改善された宿主を提供することを見出した。上記の遺伝子変異の特定の組合せは、ノックアウト変異Tsp遺伝子を有する細胞に比べて、細胞の健康状態及び増殖の改善された表現型を有する新しい系統を提供する。変異Tsp遺伝子を有する細胞は細胞増殖速度が良好であることがあるが、これらの細胞の限界の一つは、特に高い細胞密度で溶解する傾向があることである。したがって、変異Tsp遺伝子を含んでいる細胞の表現型は、特に高い細胞密度で溶解する傾向がある。しかし、本発明の細胞中でのDsbCを発現させると、低減したTsp表現型を抑制し、したがって細胞は溶解の低減を示す。
【0070】
本発明による細胞は、ノックアウト変異Tsp遺伝子を有する細胞に比べて改善されたタンパク質生成収率を示す。改善されたタンパク質収率は、タンパク質生成速度であっても、及び/又は細胞からのタンパク質生成の期間であってもよい。改善されたタンパク質収率は、ペリプラズマのタンパク質収率でも、及び/又は上清のタンパク質収率でもよい。一実施形態において、本発明の細胞は、細胞からの漏出が低減するために、変異Tsp遺伝子を有する細胞に比べて改善されたペリプラズマタンパク質収率を示す。組換え細菌細胞は、対象のタンパク質の生成速度を速めることが可能であることがあり、したがって同じ量の対象のタンパク質が、変異Tsp遺伝子を含んでいる細胞に比べてより短時間で生成され得る。対象のタンパク質の生成速度がより速いのは、細胞の増殖の最初の期間にわたって、例えば、タンパク質発現の誘導後最初の5時間、10時間、20時間、又は30時間にわたって、特に著しいことがある。
【0071】
本発明による細胞が、ペリプラズム及び/又は培地において、対象のタンパク質のおよそ1.0g/L、1.5g/L、1.8g/L、2.0g/L、2.4g/L、2.5g/L、3.0g/L、3.5g/L、又は4.0g/Lの、最大の収率を発現するのが好ましい。
【0072】
本発明が提供する細胞は、野生型細胞に比べて低減したプロテアーゼ活性を有し、本発明が提供する細胞は対象の組換えタンパク質、特にTspプロテアーゼに対してタンパク質分解感受性である対象のタンパク質のタンパク質分解性を低減し得る。したがって、本発明が提供する細胞は、野生型の細菌細胞に比べて、インタクトなタンパク質、好ましくは対象のタンパク質をより高い収率で提供し、タンパク質、好ましくは対象のタンパク質のタンパク質分解性フラグメントをより低い収率で提供し、好ましくはタンパク質分解性フラグメントを全く提供しない。
【0073】
当業者であれば、発酵法、ELISA、及びタンパク質G HPLCを含めた当技術分野においてよく知られている方法を用いて、それが望ましい収率の対象のタンパク質を有するか否かを見るために、候補の細胞クローンを試験することが容易にできる。適切な発酵法は、Humphreys D Pら(1997年)、「Formation of dimeric Fabs in E. coli:effect of hinge size and isotype,presence of interchain disulphide bond,Fab’ expression levels,tail piece sequences and growth conditions.」、J.IMMUNOL.METH.、209巻、193〜202頁;Backlund E.、Reeks D.、Markland K.、Weir N.、Bowering L.、Larsson G.、「Fedbatch design for periplasmic product retention in Escherichia coli」、雑誌記事、Research Support Non−U.S. Gov’t Journal of Biotechnology.、135巻(4)、358〜65頁、2008年7月31日; Champion KM.、Nishihara JC.、Joly JC.、Arnott D.、「Similarity of the Escherichia coli proteome upon completion of different biopharmaceutical fermentation processes.」、[雑誌記事]Proteomics.、1巻(9)、1133〜48頁、2001年9月;及びHorn U.、Strittmatter W.、Krebber A.、Knupfer U.、Kujau M.、Wenderoth R.、Muller K.、Matzku S.、Pluckthun A.、Riesenberg D.、「High volumetric yields of functional dimeric miniantibodies in Escherichia coli,using an optimized expression vector and high−cell−density fermentation under non−limited growth conditions」、雑誌記事、Research Support、Non−U.S. Gov’t Applied Microbiology & Biotechnology.、46巻(5〜6)、524〜32頁、1996年12月に記載されている。当業者であれば、タンパク質G HPLC、円偏光二色性、NMR、X線結晶構造解析、及びエピトープ親和性測定方法など、当技術分野においてよく知られている方法を用いて、タンパク質が正確にフォールディングされているか否かを見るために分泌されたタンパク質を試験することも容易にできる。
【0074】
一実施形態において、本発明による細胞は、以下の1つ又は複数のさらなるタンパク質も発現する。
タンパク質のフォールディングを促進することができる1つ又は複数のタンパク質、例えば、FkpA、Skp、SirA、PPiA、及びPPid、並びに/或いは、
タンパク質の分泌若しくは翻訳を促進することができる1つ又は複数のタンパク質、例えば、SecY、SecE、SecG、SecYEG、SecA、SecB、FtsY、及びLep、並びに/或いは、
ジスルフィド結合の形成を促進することができる1つ又は複数のタンパク質、例えば、DsbA、DsbB、DsbD、DsbG。
【0075】
1つ又は複数の上記のタンパク質を、細胞のゲノム中に組み入れてもよく、且つ/又は発現ベクター中に挿入してもよい。
【0076】
一実施形態において、本発明による細胞は、1つ又は複数の以下のさらなるタンパク質をコードする組換えポリヌクレオチドを含んでいない。
タンパク質のフォールディングを促進することができる1つ又は複数のタンパク質、例えば、FkpA、Skp、SurA、PPiA、及びPPiD、
タンパク質の分泌又は翻訳を促進することができる1つ又は複数のタンパク質、例えば、SecY、SecE、SecG、SecYEG、SecA、SecB、FtsY、及びLep、並びに
ジスルフィド結合の形成を促進することができる1つ又は複数のタンパク質、例えば、DsbA、DsbB、DsbD、DsbG。
【0077】
本発明の実施形態において、細胞は変異spr遺伝子をさらに含んでいる。sprタンパク質は、大腸菌膜結合ペリプラズムプロテアーゼである。
【0078】
sprタンパク質の野生型アミノ酸配列を、N末端にシグナル配列を有する配列番号21、及びアミノ酸26個のシグナル配列のない配列番号22において示す(UniProt受諾番号P0AFV4による)。本発明におけるsprタンパク質配列のアミノ酸番号付けはシグナル配列を含んでいる。したがって、sprタンパク質のアミノ酸1は、配列番号21に示す最初のアミノ酸(Met)である。
【0079】
本発明による細胞が変異spr遺伝子を含んでいる実施形態において、変異spr遺伝子が細胞の染色体spr遺伝子であるのが好ましい。変異spr遺伝子は、変異Tsp遺伝子を含んでいる細胞の表現型を抑制することができるsprタンパク質をコードしている。変異Tsp遺伝子を有する細胞は、良好な細胞増殖速度を有し得るが、これらの細胞の一つの制限は、特に高い細胞濃度で溶解する傾向があることである。したがって、変異Tsp遺伝子を含んでいる細胞の表現型は、特に高い細胞濃度で溶解する傾向がある。変異Tsp遺伝子を有する細胞も、低いモル浸透圧濃度で熱感受性の増殖を示す。しかし、本発明の細胞が有するspr変異は、低減したTsp活性を有する細胞中に導入した場合、低減したTsp表現型を抑制し、したがって、細胞は、特に高い細胞密度で溶解の低減を表す。細胞の増殖の表現型は、当業者であれば、フラスコ振盪又は高い細胞密度の発酵技術の間に容易に測定することができる。細胞溶解の表現型の抑制は、特にペリプラズムにおいて増殖速度の改善及び/又は組換えタンパク質の生成から見ることができ、Tsp変異体及び野生型sprを有する細胞に比べて、spr変異体を有し、低減したTsp活性を有する細胞によって示される。
【0080】
本発明による細胞は、好ましくはN31、R62、I70、Q73、C94、S95、V98、Q99、R100、L108、Y115、D133、V135、L136、G140、R144、H145、G147、H157、及びW174から選択される1つ又は複数のアミノ酸、より好ましくはC94、S95、V98、Y115、D133、V135、H145、G147、H157、及びW174から選択される1つ又は複数のアミノ酸に変異を有するsprタンパク質をコードする変異体spr遺伝子を含んでいる。好ましくは、変異体spr遺伝子は、N31、R62、I70、Q73、C94、S95、V98、Q99、R100、L108、Y115、D133、V135、L136、G140、R144、H145、G147、及びH157から選択される1つ又は複数のアミノ酸、より好ましくはC94、S95、V98、Y115、D133、V135、H145、G147、及びH157から選択される1つ又は複数のアミノ酸に変異を有するsprタンパク質をコードしている。この実施形態において、sprタンパク質にいかなるさらなる変異もないのが好ましい。好ましくは、変異体のspr遺伝子は、N31、R62、I70、Q73、S95、V98、Q99、R100、L108、Y115、D133、V135、L136、G140、R144、及びG147から選択される1つ又は複数のアミノ酸に、より好ましくはS95、V98、Y115、D133、V135、及びG147から選択される1つ又は複数のアミノ酸に変異を有するsprタンパク質をコードする。この実施形態において、sprタンパク質にいかなるさらなる変異もないのが好ましい。
【0081】
本発明者らは、変異Tsp遺伝子を含んでいる細胞の増殖の表現型を抑制することができるspr変異を同定している。
【0082】
本発明者らはまた、驚くべきことに、組換えDsbC遺伝子、新たな変異spr遺伝子を有し、野生型細胞に比べて低減したTspタンパク質活性を有する細胞は、変異Tsp遺伝子を含んでいる細胞に比べて増大した細胞増殖速度、及び増大した細胞生存期間を示すことも見出した。特に、組換えDsbC遺伝子、spr変異を有し、低減したTspタンパク質活性を有する細胞は、変異Tsp遺伝子を有する細胞に比べて低減した細胞溶解の表現型を示す。
【0083】
上記のsprアミノ酸の1つ又は複数の変異は、アミノ酸をコードするヌクレオチドの1個、2個、又は3個に対するあらゆる適切なミスセンス変異であってよい。変異は、アミノ酸残基を、変異Tsp遺伝子を含んでいる細胞の表現型を抑制することができる変異sprタンパク質をもたらすあらゆる適切なアミノ酸へ変化させる。ミスセンス変異は、アミノ酸を、野生型アミノ酸に比べて異なるサイズであり、且つ/又は異なる化学性質を有するアミノ酸へ変化させることができる。
【0084】
一実施形態において、変異は、C94、H145、及びH157のアミノ酸残基の触媒三残基の1つ、2つ、又は3つに対するものである(「Solution NMR Structure of the NlpC/P60 Domain of Lipoprotein Spr from Escherichia coli Structural Evidence for a Novel Cysteine Peptidase Catalytic Triad」、Biochemistry、2008年、47巻、9715〜9717頁)。
【0085】
したがって、変異spr遺伝子は、
C94に対する変異、又は
H145に対する変異、又は
H157に対する変異、又は
C94及びH145に対する変異、又は
C94及びH157に対する変異、又は
H145及びH157に対する変異、又は
C94、H145、及びH157に対する変異
を含んでいてよい。
【0086】
この実施形態において、sprタンパク質がいかなるさらなる変異も有さないのが好ましい。
【0087】
C94、H145、及びH157の1つ、2つ、又は3つが、変異Tsp遺伝子を含んでいる細胞の表現型を抑制することができるsprタンパク質をもたらすあらゆる適切なアミノ酸に変異されてよい。例えば、C94、H145、及びH157の1つ、2つ、又は3つは、Gly又はAlaなどの小型のアミノ酸に変異されてよい。したがって、sprタンパク質は、C94、H145、及びH157の1つ、2つ、又は3つの変異を有することができる。spr遺伝子が、変異Tsp遺伝子を含んでいる細胞の表現型を抑制することができるsprタンパク質を生成することが見出されているミスセンス変異H145Aを含んでいるのが好ましい。
【0088】
本明細書の置換変異体に対する呼称は、1つの文字とそれに続く1つの数字とそれに続く1つの文字からなる。最初の文字は野生型タンパク質におけるアミノ酸を指定する。数字はアミノ酸置換が行われるアミノ酸位置を表し、2番目の文字は野生型アミノ酸を置換するのに用いられるアミノ酸を指定する。
【0089】
好ましい一実施形態において、変異体のsprタンパク質は、N31、R62、I70、Q73、S95、V98、Q99、R100、L108、Y115、D133、V135、L136、G140、R144、及びG147から選択される1つ又は複数のアミノ酸の変異、好ましくはS95、V98、Y115、D133、V135、及びG147から選択される1つ又は複数のアミノ酸の変異を含んでいる。この実施形態において、sprタンパク質がいかなるさらなる変異を有さないのが好ましい。したがって、変異spr遺伝子は、
N31に対する変異、又は
R62に対する変異、又は
I70に対する変異、又は
Q73に対する変異、又は
S95に対する変異、又は
V98に対する変異、又は
Q99に対する変異、又は
R100に対する変異、又は
L108に対する変異、又は
Y115に対する変異、又は
D133に対する変異、又は
V135に対する変異、又は
L136に対する変異、又は
G140に対する変異、又は
R144に対する変異、又は
G147に対する変異
を含んでいてよい。
【0090】
一実施形態において、変異体sprタンパク質は、アミノ酸:
S95及びY115、又は
N31、Q73、R100及びG140、又は
Q73、R100及びG140、又は
R100及びG140、又は
Q73及びG140、又は
Q73及びR100、又は
R62、Q99及びR144、又は
Q99及びR144
に対する複数の変異を含んでいる。
【0091】
N31、R62、I70、Q73、S95、V98、Q99、R100、L108、Y115、D133、V135、L136、G140、R144、及びG147の1つ又は複数のアミノ酸は、変異Tsp遺伝子を含んでいる細胞の表現型を抑制することができるsprタンパク質をもたらすあらゆる適切なアミノ酸に変異されてよい。例えば、N31、R62、I70、Q73、S95、V98、Q99、R100、L108、Y115、D133、V135、L136、G140、及びR144の1つ又は複数は、Gly又はAlaなどの小型のアミノ酸に変異されてよい。
【0092】
好ましい一実施形態において、sprタンパク質は、1つ又は複数の以下の変異を含んでいる:N31Y、R62C、I70T、Q73R、S95F、V98E、Q99P、R100G、L108S、Y115F、D133A、V135D又はV135G、L136P、G140C、R144C、及びG147C。sprタンパク質が1つ又は複数の以下の変異を含んでいるのが好ましい:S95F、V98E、Y115F、D133A、V135D又はV135G、及びG147C。この実施形態において、sprタンパク質にいかなるさらなる変異もないのが好ましい。
【0093】
一実施形態において、sprタンパク質は、N31Y、R62C、I70T、Q73R、S95F、V98E、Q99P、R100G、L108S、Y115F、D133A、V135D又はV135G、L136P、G140C、R144C、及びG147Cから選択される変異を1つ有する。この実施形態において、sprタンパク質にいかなるさらなる変異もないのが好ましい。
【0094】
さらなる一実施形態において、sprタンパク質は、以下から選択される複数の変異を有する:
S95F及びY115F
N31Y、Q73R、R100G、及びG140C、
Q73R、R100G、及びG140C、
R100G及びG140C、
Q73R及びG140C、
Q73R及びR100G、
R62C、Q99P及びR144C、又は
Q99P及びR144C。
【0095】
変異体のspr遺伝子が、C94A、D133A、H145A、及びH157Aから選択される変異を有するsprタンパク質をコードするのが好ましい。
【0096】
さらなる一実施形態において、変異spr遺伝子は変異W174Rを有するsprタンパク質をコードする。代替の一実施形態において、sprタンパク質は変異W174Rを有さない。
【0097】
本発明による細胞は、野生型細胞に比べて低減したTspタンパク質活性を有する。「野生型細胞に比べて低減したTspタンパク質活性」の表現は、細胞のTsp活性が野生型細胞のTsp活性に比べて低減していることを意味する。細胞を、Tspの活性を低減するのに適するあらゆる手段によって修飾することができる。
【0098】
一実施形態において、低減したTsp活性は、Tsp及び/又は関連の調節性の発現配列をコードする内因性ポリヌクレオチドの修飾による。修飾は、Tsp遺伝子の転写及び翻訳を低減又は停止することができ、或いは野生型Tspタンパク質に比べて低減したプロテアーゼ活性を有するTspタンパク質の発現を提供することができる。
【0099】
一実施形態において、関連の調節性の発現配列を、Tsp発現を低減するように修飾する。例えば、Tsp遺伝子に対するプロモーターを遺伝子の発現を防止するように変異させてもよい。
【0100】
好ましい一実施形態において、本発明による細胞は、低減したプロテアーゼ活性を有するTspタンパク質をコードする変異Tsp遺伝子、又はノックアウト変異Tsp遺伝子を有している。染色体のTsp遺伝子が変異しているのが好ましい。
【0101】
本明細書で用いられる「Tsp遺伝子」は、ペニシリン結合性タンパク質−3(PBP3)及びファージテイルタンパク質に対して作用することができるペリプラズムプロテアーゼであるプロテアーゼTsp(Prcとしても知られている)をコードする遺伝子を意味する。野生型Tsp遺伝子の配列を配列番号1に示し、野生型Tspタンパク質の配列を配列番号2に示す。
【0102】
変異Tsp遺伝子、又はTspをコードする変異Tsp遺伝子に対する言及は、別段の指摘がなければ、低減したプロテアーゼ活性を有するTspタンパク質をコードする変異Tsp遺伝子、又はノックアウト変異Tsp遺伝子のいずれかを意味する。
【0103】
「低減したプロテアーゼ活性を有するTspタンパク質をコードする変異Tsp遺伝子」の表現は、本発明の文脈において、変異Tsp遺伝子が野生型の非変異Tsp遺伝子に比べて完全なプロテアーゼ活性を有さないことを意味する。
【0104】
変異Tsp遺伝子が、野生型の非変異Tspタンパク質のプロテアーゼ活性の50%以下、40%以下、30%以下、20%以下、10%以下、又は5%以下を有するTspタンパク質をコードするのが好ましい。変異Tsp遺伝子が、プロテアーゼ活性を有さないTspタンパク質をコードするのがより好ましい。この実施形態において、細胞は染色体Tspを欠くのではなく、即ち、Tsp遺伝子配列はあらゆる形態のTspタンパク質の発現を防ぐように欠失又は変異されていない。
【0105】
低減したプロテアーゼ活性を有するタンパク質を生成するために、あらゆる適切な変異をTsp遺伝子中に導入することができる。グラム陰性細菌から発現されるTspタンパク質のプロテアーゼ活性は、当業者であれば、Tspのプロテアーゼ活性が試験されるKeilerら(「Identification of Active Site Residues of the Tsp Protease*」、THE JOURNAL OF BIOLOGICAL CHEMISTRY、270巻48号、12月1日発行、28864〜28868頁、1995年、Kenneth C.Keiler及びRobert T.Sauer)において記載されている方法など、当技術分野におけるあらゆる適切な方法によって容易に試験することができる。
【0106】
TspはKeilerら(上述)において、残基S430、D441、及びK455を含む活性部位を有すると報告されており、残基G375、G376、E433、及びT452はTspの構造を維持するのに重要である。Keilerら(上述)は、変異Tsp遺伝子S430A、D441A、K455A、K455H、K455R、G375A、G376A、E433A、及びT452Aには検出可能なプロテアーゼ活性がなかったという所見を報告している。変異Tsp遺伝子S430Cは野生型活性の約5〜10%を示したことがさらに報告されている。したがって、低減したプロテアーゼ活性を有するタンパク質を生成するためのTsp変異は、S430、D441、K455、G375、G376、E433、及びT452の1つ又は複数の残基に対するミスセンス変異などの変異を含むことがある。低減したプロテアーゼ活性を有するタンパク質を生成するためのTsp変異が、活性部位の残基S430、D441、及びK455の1つ、2つ、又は3つ全てに対するミスセンス変異などの変異を含むことができるのが好ましい。
【0107】
したがって、変異Tsp遺伝子は以下を含むことができる:
S430に対する変異、又は
D441に対する変異、又は
K455に対する変異、又は
S430及びD441に対する変異、又は
S430及びK455に対する変異、又は
D441及びK455に対する変異、又は
S430、D441、及びK455に対する変異。
【0108】
S430、D441、K455、G375、G376、E433、及びT452の1つ又は複数が、低減したプロテアーゼ活性を有するタンパク質をもたらすあらゆる適切なアミノ酸に変異していてよい。適切な変異の例には、S430A、S430C、D441A、K455A、K455H、K455R、G375A、G376A、E433A、及びT452Aがある。変異Tsp遺伝子は、活性部位残基に対する1つ、2つ、又は3つの変異を含むことができ、例えば、遺伝子は以下を含むことができる:
S430A、若しくはS430C、及び/又は
D441A及び/又は
K455A若しくはK455H、若しくはK455R。
【0109】
Tsp遺伝子が点変異S430A又はS430Cを有するのが好ましい。
【0110】
「ノックアウト変異Tsp遺伝子」の表現は、本発明の文脈において、Tsp遺伝子が、Tspタンパク質を欠く細胞を提供するために、野生型遺伝子によってコードされるTspタンパク質の発現を防ぐ1つ又は複数の変異を含んでいることを意味する。ノックアウト遺伝子は、部分的又は完全に転写され得るが、コードタンパク質に翻訳されない。ノックアウト変異Tsp遺伝子は、タンパク質の発現をもたらさないように、1つ又は複数の欠失、挿入、点、ミスセンス、ナンセンス、及びフレームシフトの変異によるなど、あらゆる適切な方法において変異され得る。例えば、遺伝子は、遺伝子コード配列中に、抗生物質耐性マーカーなどの外来のDNA配列を挿入することによってノックアウトされてよい。
【0111】
好ましい一実施形態において、Tsp遺伝子は、遺伝子コード配列中に、抗生物質耐性マーカーなどの外来のDNA配列を挿入することによって変異されない。一実施形態において、Tsp遺伝子は、遺伝子開始コドン、並びに/或いは遺伝子開始コドンの下流、及び遺伝子終止コドンの上流に位置する1つ又は複数の終止コドンに対する変異を含んでおり、それによってTspタンパク質の発現を防いでいる。開始コドンに対する変異は、開始コドンの1つ、2つ、又は3つ全てのヌクレオチドのミスセンス変異であってよい。或いは、又はさらに、開始コドンは、挿入又は欠失のフレームシフト変異によって変異されてよい。Tsp遺伝子は、コード配列の5’末端に2つのATGコドンを含んでおり、ATGコドンの1つ又は両方はミスセンス変異によって変異されてよい。Tsp遺伝子は、図5bに示す通り、第2のATGコドン(コドン3)からTCGコドンに変異されてよい。Tsp遺伝子は、或いは、又はさらに、遺伝子開始コドンの下流及び遺伝子終止コドンの上流に位置する1つ又は複数の終止コドンを含んでいてよい。ノックアウト変異Tsp遺伝子が、開始コドンに対するミスセンス変異、及び1つ又は複数の挿入された終止コドンの両方を含んでいるのが好ましい。好ましい一実施形態において、Tsp遺伝子は第5のコドンから「T」を欠失するように変異されており、それによって、図5bに示す通り、コドン11及び16に終止コドンをもたらすフレームシフトを生じている。好ましい一実施形態において、図5bに示す通り、コドン21に第3のインフレーム終止コドンを作り出すように、Tsp遺伝子はAseI制限部位を挿入するように変異される。
【0112】
好ましい一実施形態において、ノックアウト変異Tsp遺伝子は配列番号3のDNA配列を有し、これは開始コドンの上流にATGAATの6個のヌクレオチドを含んでいる。配列番号3のノックアウト変異Tsp配列においてなされた変異を図5bに示す。一実施形態において、変異Tsp遺伝子は、配列番号3のヌクレオチド7から2048のDNA配列を有する。
【0113】
本発明の好ましい一実施形態において、組換えグラム陰性細菌細胞は、シャペロン活性及び低減したプロテアーゼ活性を有するDegPタンパク質をコードする変異DegP遺伝子、並びに/或いは変異ptr遺伝子をさらに含んでおり、変異ptr遺伝子は低減したプロテアーゼ活性を有するProteaseIIIタンパク質をコードする、又はノックアウト変異ptr遺伝子及び/又は変異OmpT遺伝子であり、変異OmpT遺伝子は、低減したプロテアーゼ活性を有するOmpTタンパク質をコードする、又はノックアウト変異OmpT遺伝子である。
【0114】
一実施形態において、本発明は、
a.DsbCをコードする組換えポリヌクレオチド、
b.低減したプロテアーゼ活性を有するTspタンパク質をコードする変異Tsp遺伝子、又はノックアウト変異Tsp遺伝子、
c.シャペロン活性及び低減したプロテアーゼ活性、及び/又は変異OmpTを有するDegPタンパク質をコードする変異DegP遺伝子(変異OmpT遺伝子は、低減したプロテアーゼ活性を有するOmpTタンパク質をコードする、又はノックアウト変異OmpT遺伝子である)、並びに
d.任意選択によって変異ptr遺伝子
を含んでいる組換えグラム陰性細菌細胞を提供する。
【0115】
この実施形態において、細胞のゲノムが、上記の変異b、c、及びd以外は野生型細菌細胞に対して同質遺伝子であるのが好ましい。
【0116】
一実施形態において、本発明は、
a.DsbCをコードする組換えポリヌクレオチド、
b.低減したプロテアーゼ活性を有するTspタンパク質をコードする変異Tsp遺伝子、又はノックアウト変異Tsp遺伝子、
c.変異ptr遺伝子(変異ptr遺伝子は、低減したプロテアーゼ活性を有するProteaseIIIをコードする、又はノックアウト変異ptr遺伝子であり、且つ/又は変異OmpTであり、変異OmpT遺伝子は、低減したプロテアーゼ活性を有するOmpTタンパク質をコードする、又はノックアウト変異OmpT遺伝子である)、並びに
d.任意選択によって変異spr遺伝子
を含んでいる組換えグラム陰性細菌細胞を提供する。
【0117】
この実施形態において、細胞のゲノムが、上記の変異b、c、及びd以外は野生型細菌細胞に対して同質遺伝子であるのが好ましい。
【0118】
一実施形態において、本発明は、
a.DsbCをコードする組換えポリヌクレオチド、
b.低減したプロテアーゼ活性を有するTspタンパク質をコードする変異Tsp遺伝子、又はノックアウト変異Tsp遺伝子、
c.シャペロン活性及び低減したプロテアーゼ活性を有するDegPタンパク質をコードする変異DegP遺伝子、
d.変異ptr遺伝子(変異ptr遺伝子は、低減したプロテアーゼ活性を有するProteaseIIIタンパク質をコードする、又はノックアウト変異ptr遺伝子である)、
e.任意選択によって変異OmpT(変異Ompt遺伝子は、低減したプロテアーゼ活性を有するOmpTタンパク質をコードする、又はノックアウト変異OmpT遺伝子である)、並びに
f.任意選択によって変異spr遺伝子
を含んでいる細胞を提供する。
【0119】
この実施形態において、細胞のゲノムが、上記の変異b、c、d、e、及びf以外は野生型細菌細胞に対して同質遺伝子であるのが好ましい。
【0120】
本発明の実施形態において、細胞は変異DegP遺伝子を含んでいる。本明細書で用いられる「DegP」は、DegPタンパク質(HtrAとしても知られている)をコードする遺伝子を意味し、DegPタンパク質はシャペロン及びプロテアーゼとしての二重機能を有する(「Families of serine peptidases」、Rawlings ND、Barrett AJ.、Methods Enzymol.、1994年、244巻、19〜61頁)。非変異のDegP遺伝子の配列を配列番号7に示し、非変異のDegPタンパク質の配列を配列番号8に示す。
【0121】
低温でDegPはシャペロンとして機能し、高温でDegPはプロテアーゼとしての機能を優先する(「A Temperature−Dependent Switch from Chaperone to Protease in a Widely Conserved Heat Shock Protein.」Cell、97巻3号、339〜347頁、Spiess C、Beil A、Ehrmann M)、及び「The proteolytic activity of the HtrA (DegP) protein from Escherichia coli at low temperatures」、Skorko−Glonek Jら、Microbiology、2008年、154巻、3649〜3658頁)。
【0122】
細胞がDegP変異を含んでいる実施形態において、細胞におけるDegP変異は、シャペロン活性を有するが完全なプロテアーゼを有さないDegPタンパク質をコードする変異DegP遺伝子を提供する。
【0123】
「シャペロン活性を有する」の表現は、本発明の文脈において、変異DegPタンパク質が、野生型の非変異のDegPタンパク質に比べて同じ、又は実質的に同じシャペロン活性を有することを意味する。変異DegP遺伝子が、野生型の非変異のDegPタンパク質のシャペロン活性の50%以上、60%以上、70%以上、80%以上、90%以上、又は95%以上を有するDegPタンパク質をコードするのが好ましい。変異DegP遺伝子が、野生型DegPに比べて同じシャペロン活性を有するDegPタンパク質をコードするのがより好ましい。
【0124】
「低減したプロテアーゼ活性を有する」の表現は、本発明の文脈において、変異DegPタンパク質が、野生型の非変異のDegPタンパク質に比べて完全なプロテアーゼ活性を有さないことを意味する。変異DegP遺伝子が、野生型の非変異のDegPタンパク質のプロテアーゼ活性の50%以下、40%以下、30%以下、20%以下、10%以下、又は5%以下を有するDegPタンパク質をコードするのが好ましい。変異DegP遺伝子が、プロテアーゼ活性のないDegPタンパク質をコードするのがより好ましい。細胞は、染色体のDegPを欠くのではなく、即ち、DegP遺伝子配列は、あらゆる形態のDegPタンパク質の発現を防ぐように欠失又は変異されていない。
【0125】
シャペロン活性及び低減したプロテアーゼ活性を有するタンパク質を生成するために、あらゆる適切な変異をDegP遺伝子中に導入することができる。グラム陰性細菌から発現されるDegPタンパク質のプロテアーゼ及びシャペロンの活性は、当業者であれば、DegPのプロテアーゼ及びシャペロン活性がDegPの天然の基質であるMalS上で試験されるSpiessらにおいて記載されている方法(「A Temperature−Dependent Switch from Chaperone to Protease in a Widely Conserved Heat Shock Protein.」Cell、97巻3号、339〜347頁、Spiess C、Beil A、Ehrmann M)、及びまた「The proteolytic activity of the HtrA (DegP) protein from Escherichia coli at low temperatures」、Skorko−Glonek Jら、Microbiology、2008年、154巻、3649〜3658頁において記載されている方法などのあらゆる適切な方法によって容易に試験することができる。
【0126】
DegPはセリンプロテアーゼであり、アミノ酸残基His105、Asp135、及びSer210の触媒三塩基からなる活性中心を有する(「Families of serine peptidases」、Methods Enzymol.、1994年、244巻、19〜61頁、Rawlings N及びBarrett A)。シャペロン活性及び低減したプロテアーゼ活性を有するタンパク質を生成するためのDegP変異は、His105、Asp135、及びSer210の1つ、2つ、又は3つに対するミスセンス変異などの変異を含んでいてよい。
【0127】
したがって、変異DegP遺伝子は以下を含んでいてよい:
His105に対する変異、又は、
Asp135に対する変異、又は、
Ser210に対する変異、又は、
His105及びAsp135に対する変異、又は、
His105及びSer210に対する変異、又は、
Asp135及びSer210に対する変異、又は、
His105、Asp135、及びSer210に対する変異。
【0128】
His105、Asp135、及びSer210の1つ、2つ、又は3つは、シャペロン活性、及び低減したプロテアーゼ活性を有するタンパク質をもたらすあらゆる適切なアミノ酸に変異されていてよい。例えば、His105、Asp135、及びSer210の1つ、2つ、又は3つは、Gly又はAlaなどの小型のアミノ酸に変異されてよい。さらなる適する変異は、例えば、Asp135がLys又はArgに変異され、極性のHis105がGly、Ala、Val、又はLeuなどの非極性のアミノ酸に変異され、小型の親水性のSer210がVal、Leu、Phe、又はTyrなどの大型又は疎水性の残基に変異されるなど、His105、Asp135、及びSer210の1つ、2つ、又は3つの、反対の性質を有するアミノ酸への変化である。DegP遺伝子が、図5cに示す通り、シャペロン活性を有するがプロテアーゼを有さないタンパク質を生成することが見出されている点変異S210Aを含んでいるのが好ましい(「A Temperature−Dependent Switch from Chaperone to Protease in a Widely Conserved Heat Shock Protein.」Cell、97巻3号、339〜347頁、Spiess C、Beil A、Ehrmann M)。
【0129】
DegPは、タンパク質−タンパク質相互作用を媒介する、PDZ1(残基260〜358)及びPDZ2(残基359〜448)の2つのPDZドメインを有する(「A Temperature−Dependent Switch from Chaperone to Protease in a Widely Conserved Heat Shock Protein.」、Cell、97巻3号、339〜347頁、Spiess C、Beil A、Ehrmann M)。本発明の一実施形態において、degP遺伝子は、PDZ1ドメイン及び/又はPDZ2ドメインを欠失するように変異される。PDZ1及びPDZ2の欠失は、DegPタンパク質のプロテアーゼ活性の完全な喪失及び野生型DegPタンパク質に比べて低下したシャペロン活性をもたらし、PDZ1又はPDZ2いずれかの欠失は、野生型DegPタンパク質に比べて5%のプロテアーゼ活性及び同様のシャペロン活性をもたらす(「A Temperature−Dependent Switch from Chaperone to Protease in a Widely Conserved Heat Shock Protein.」、Cell、97巻3号、339〜347頁、Spiess C、Beil A、Ehrmann M)。
【0130】
変異DegP遺伝子は、例えば、図5cに示す通り、同定及びスクリーニング方法において助けとなるためにAseIなど、サイレントの天然に存在しない制限部位も含んでいてよい。
【0131】
点変異S210A及びAseI制限マーカー部位を含んでいる変異DegP遺伝子の好ましい配列を配列番号9に提供し、コードされるタンパク質配列を配列番号10に示す。配列番号9の変異DegP配列になされた変異を図5cに示す。
【0132】
細胞が、シャペロン活性及び低減したプロテアーゼ活性を有するDegPタンパク質をコードする変異DegP遺伝子を含んでいる本発明の実施形態において、本発明が提供する1つ又は複数の細胞は、変異細胞に比べて細胞からの正確にフォールディングされたタンパク質の収率の改善を提供することができ、DegP遺伝子は、染色体欠損DegPなど、DegP発現を防ぐノックアウトDegPに対して変異されている。DegPの発現を防ぐノックアウト変異DegP遺伝子を含んでいる細胞においてDegPのシャペロン活性は完全に失われるが、本発明による細胞においてDegPのシャペロン活性は保持され、完全なプロテアーゼ活性は失われる。これらの実施形態において、本発明による1つ又は複数の細胞は、タンパク質のタンパク質分解性を防ぐようにプロテアーゼ活性は低減されており、宿主細胞におけるタンパク質において正確なフォールディング及び輸送を可能にするようにシャペロン活性は維持されている。
【0133】
当業者であれば、タンパク質G HPLC、円偏光二色性、NMR、X線結晶構造解析、及びエピトープ親和性測定方法など、当技術分野においてよく知られている方法を用いて、タンパク質が正確にフォールディングされているか否かを見るために分泌されたタンパク質を試験することが容易にできる。
【0134】
これらの実施形態において、本発明による1つ又は複数の細胞は、DegP発現を防ぐ変異ノックアウトDegP遺伝子を有する細胞に比べて細胞の増殖が改善されていてよい。理論によって拘泥しようとするものではないが、シャペロン活性を必要とする全てのタンパク質をプロセシングする細胞の能力を増大し得るシャペロン活性をDegPプロテアーゼが保持するため、細胞増殖の改善が示され得る。したがって、細胞の増殖及び再生に必要な正確にフォールディングされたタンパク質の生成は、DegPノックアウト変異を有する細胞に比べて本発明の1つ又は複数の細胞において増大し、それによって増殖を制御する細胞の経路を改善し得る。さらに、知られているDegPプロテアーゼ欠損系統は一般に温度感受性であり、約28℃を超える温度では通常増殖しない。しかし、本発明による細胞は温度感受性ではなく、細菌からタンパク質を生成する工業規模に通常用いられる、およそ30℃からおよそ37℃の温度を含めた28℃以上の温度で増殖することができる。
【0135】
本発明の実施形態において、細胞は変異ptr遺伝子を含んでいる。本明細書で用いられる「ptr遺伝子」は、高分子量のタンパク質を分解するプロテアーゼであるProteaseIIIをコードする遺伝子を意味する。非変異のptr遺伝子の配列を配列番号4に示し、非変異のProteaseIIIタンパク質の配列を配列番号5に示す。
【0136】
変異ptr遺伝子、又はProteaseIIIをコードする変異ptr遺伝子に対する言及は、別段の指摘がなければ、低減したプロテアーゼ活性を有するProteaseIIIタンパク質をコードする変異ptr遺伝子、又はノックアウト変異ptr遺伝子のいずれかを意味する。
【0137】
「低減したプロテアーゼ活性を有するProteaseIIIタンパク質をコードする変異ptr遺伝子」の表現は、本発明の文脈において、変異ptr遺伝子が、野生型の非変異のptr遺伝子に比べて完全なプロテアーゼ活性を有さないことを意味する。
【0138】
変異ptr遺伝子が、野生型の非変異のProteaseIIIタンパク質のプロテアーゼ活性の50%以下、40%以下、30%以下、20%以下、10%以下、又は5%以下を有するProteaseIIIをコードするのが好ましい。変異ptr遺伝子がプロテアーゼ活性を有さないProteaseIIIタンパク質をコードするのがより好ましい。この実施形態において、細胞は染色体ptrを欠いておらず、即ち、ptr遺伝子配列は、あらゆる形態のProteaseIIIタンパク質の発現を防止するように欠失又は変異されていない。
【0139】
低減したプロテアーゼ活性を有するProteaseIIIタンパク質を生成するために、あらゆる適切な変異がptr遺伝子中に導入されてよい。グラム陰性細菌から発現されたProteaseIIIタンパク質のプロテアーゼ活性は、当業者であれば、当技術分野におけるあらゆる適切な方法によって容易に試験することができる。
【0140】
「ノックアウト変異ptr遺伝子」の表現は、本発明の文脈において、遺伝子が1つ又は複数の変異を含んでおり、それによって、ノックアウト変異遺伝子によってコードされるタンパク質を欠く細胞を提供するように、遺伝子によってコードされるタンパク質の発現を引き起こさないことを意味する。ノックアウト遺伝子は、部分的又は完全に転写され得るが、コードタンパク質に翻訳され得ない。ノックアウト変異ptr遺伝子は、タンパク質の発現を引き起こさないように、例えば、1つ又は複数の欠失、挿入、点、ミスセンス、ナンセンス、及びフレームシフトの変異によるなど、あらゆる適切な方法において変異されてよい。例えば、遺伝子は、遺伝子コード配列中に、抗生物質耐性マーカーなどの外来のDNA配列を挿入することによってノックアウトされてよい。
【0141】
好ましい一実施形態において、遺伝子は、遺伝子コード配列中に、抗生物質耐性マーカーなどの外来のDNA配列を挿入することによって変異されない。ProteaseIII遺伝子は、遺伝子開始コドン、並びに/或いは遺伝子開始コドンの下流及び遺伝子終止コドンの上流に位置する1つ又は複数の終止コドンに対する変異を含んでおり、それによってProteaseIIIタンパク質の発現を防いでいる。
【0142】
標的のノックアウト遺伝子開始コドンに対する変異は開始コドンの機能の喪失を引き起こし、それによって標的の遺伝子は確実に、コード配列の開始に適切な開始コドンを含んでいない。開始コドンに対する変異は開始コドンの1つ、2つ、又は3つ全てのヌクレオチドのミスセンス変異であってよい。或いは、又はさらに、開始コドンは挿入又は欠失のフレームシフト変異によって変異されてよい。
【0143】
好ましい一実施形態において、ptr遺伝子は、図5aに示す通り、ATG開始コドンがATTに変化するように変異されている。
【0144】
ノックアウト変異ptr遺伝子は、或いは、又はさらに、遺伝子開始コドンの下流及び遺伝子終止コドンの上流に位置する1つ又は複数の終止コドンを含んでいてよい。ノックアウト変異ptr遺伝子が、開始コドンに対するミスセンス変異、及び1つ又は複数の挿入された終止コドンの両方を含んでいるのが好ましい。
【0145】
1つ又は複数の挿入された終止コドンがインフレーム終止コドンであるのが好ましい。しかし、1つ又は複数の挿入された終止コドンは、或いは、又はさらに、フレーム外(out−of−frame)終止コドンであってよい。1つ又は複数のフレーム外終止コドンは、フレーム外開始コドンが挿入又は欠失のフレームシフト変異によってインフレーム開始コドンに変化される場合に翻訳を停止するのに必要とされ得る。1つ又は複数の終止コドンが、ナンセンス点変異及びフレームシフト変異を含めたあらゆる適切な変異によって導入されてよい。1つ又は複数の終止コドンがフレームシフト変異及び/又は挿入変異によって、好ましくは遺伝子配列のセグメントを、終止コドンを含んでいる配列で置換することによって導入されるのが好ましい。例えば、終止コドンTAAを含んでいるAseI制限部位を挿入してもよい。
【0146】
好ましい一実施形態において、ptr遺伝子は、図5aに示す通り、AseI制限部位の挿入によって、インフレーム終止コドンを挿入するように変異される。好ましい一実施形態において、ノックアウト変異ptr遺伝子は配列番号6のDNA配列を有する。配列番号6のノックアウト変異ptr遺伝子配列においてなされた変異を図5aに示す。
【0147】
上記に記載したノックアウト変異は、標的のノックアウト遺伝子部位の上流又は下流の染色体DNAに対して最小のかく乱を引き起こし、又はかく乱を引き起こさず、外来のDNA(例えば、対象のタンパク質、特に治療用タンパク質の発現に対する細胞の適合性に影響を及ぼし得る、抗生物質耐性マーカー)の挿入及び保持を必要としないので有利である。したがって、本発明による1つ又は複数の細胞は、増殖の特徴の改善及び/又は外来DNAの遺伝子コード配列中への挿入によってプロテアーゼ遺伝子がノックアウトされた細胞に比べてタンパク質発現の改善を示すことがある。
【0148】
本発明の実施形態において、細胞は変異OmpT遺伝子を有する。本明細書で用いられる「OmpT遺伝子」は、外膜プロテアーゼであるプロテアーゼOmpT(外膜プロテアーゼT)をコードする遺伝子を意味する。野生型の非変異OmpT遺伝子の配列は、SWISS−PROT P09169である。
【0149】
変異OmpT遺伝子、又はOmpTをコードする変異OmpT遺伝子に対する言及は、別段の指摘がなければ、低減したプロテアーゼ活性を有するOmpTタンパク質をコードする変異OmpT遺伝子、又はノックアウト変異OmpT遺伝子のいずれかを意味する。
【0150】
「低減したプロテアーゼ活性を有するOmpTタンパク質をコードする変異OmpT遺伝子」の表現は、本発明の文脈において、変異OmpT遺伝子が野生型の非変異OmpT遺伝子に比べて完全なプロテアーゼ活性を有さないことを意味する。変異OmpT遺伝子は、野生型の非変異OmpTタンパク質のプロテアーゼ活性の50%以下、40%以下、30%以下、20%以下、10%以下、又は5%以下を有するOmpTタンパク質をコードしていてよい。変異OmpT遺伝子は、プロテアーゼ活性を有さないOmpTタンパク質をコードしていてよい。この実施形態において、細胞は染色体OmpTを欠くのではなく、即ち、OmpT遺伝子配列はあらゆる形態のOmpTタンパク質の発現を防ぐように欠失又は変異されていない。
【0151】
低減したプロテアーゼ活性を有するタンパク質を生成するために、あらゆる適切な変異をOmpT遺伝子中に導入することができる。グラム陰性細菌から発現されるOmpTタンパク質のプロテアーゼ活性は、当業者であれば、Kramerら(「Identification of essential acidic residues of outer membrane protease OmpT supports a novel active site」、FEBS Letters、505巻(2001年) 426〜430頁)及びDekkerら(「Substrate Specitificity of the Integral Membrane Protease OmpT Determined by Spatially Addressed Peptide Libraries」、Biochemistry、2001年、40巻、1694〜1701頁)において記載されている方法など、当技術分野におけるあらゆる適切な方法によって容易に試験することができる。
【0152】
OmpTは、Kramerら(「Identification of active site serine and histidine residues in Escherichia coli outer membrane protease OmpT」、FEBS Letters、2000年、468巻、220〜224頁)において報告されており、これは、セリン、ヒスチジン、及び酸性残基をアラニンによって置換すると、Glu27、Asp97、Asp208、又はHis101ではおよそ10倍活性が低減し、Ser99ではおよそ500倍活性が低減し、Asp83、Asp85、Asp210、又はHis212ではおよそ10000倍活性が低減すると開示している。Vandeputte−Ruttenら(「Crystal Structure of the Outer Membrane Protease OmpT from Escherichia coli suggests a novel catalytic site」、The EMBO Journal、2001年、20巻18号、5033〜5039頁)は、Asp83−Asp85対及びHis212−Asp210対を含む活性部位を有すると開示している。さらに、Kramerら(「Lipopolysaccharide regions involved in the activation of Escherichia coli outer membrane protease OmpT」、Eur.J.Biochem.、FEBS、2002年、269巻、1746〜1752頁)は、ループL4における変異D208A、D210A、H212A、H212N、H212Q、G216K/K217G、K217T、及びR218Lは全て、酵素活性の部分的又はほぼ完全な喪失をもたらすことを開示している。
【0153】
したがって、低減したプロテアーゼ活性を有するタンパク質を生成するためのOmpT変異は、E27、D43、D83、D85、D97、S99、H101、E111、E136、E193、D206、D208、D210、H212、G216、K217、R218、及びE250の残基の1つ又は複数に対するミスセンス変異などの変異を含んでいてよい。
【0154】
E27、D43、D83、D85、D97、S99、H101、E111、E136、E193、D206、D208、D210、H212、G216、K217、R218、及びE250の1つ又は複数は、低減したプロテアーゼ活性を有するタンパク質をもたらすあらゆる適切なアミノ酸に変異していてよい。例えば、E27、D43、D83、D85、D97、S99、H101、E111、E136、E193、D206、D208、D210、H212、G216、K217、R218、及びE250の1つ又は複数は、アラニンに変異していてよい。適切な変異の例には、E27A、D43A、D83A、D85A、D97A、S99A、H101A、E111A、E136A、E193A、D206A、D208A、D210A、H212A、H212N、H212Q、G216K、K217G、K217T、R218L、及びE250Aがある。一実施形態において、変異OmpT遺伝子は、D210A及びH212A変異を含んでいる。D210A及びH212A変異を含んでいる適切な変異OmpT配列を配列番号23に示す。
【0155】
「ノックアウト変異OmpT遺伝子」の表現は、本発明の文脈において、遺伝子が1つ又は複数の変異を含んでおり、それによって、ノックアウト変異遺伝子によってコードされるタンパク質を欠く細胞を提供するように、遺伝子によってコードされるタンパク質の発現を引き起こさないことを意味する。ノックアウト遺伝子は、部分的又は完全に転写され得るが、コードタンパク質に翻訳され得ない。ノックアウト変異OmpT遺伝子は、タンパク質の発現を引き起こさないように、例えば、1つ又は複数の欠失、挿入、点、ミスセンス、ナンセンス、及びフレームシフトの変異によるなど、あらゆる適切な方法において変異されてよい。例えば、遺伝子は、遺伝子コード配列中に、抗生物質耐性マーカーなどの外来のDNA配列を挿入することによってノックアウトされてよい。
【0156】
一実施形態において、OmpT遺伝子は、遺伝子開始コドン、並びに/或いは遺伝子開始コドンの下流及び遺伝子終止コドンの上流に位置する1つ又は複数の終止コドンに対する変異を含んでおり、それによってOmpTタンパク質の発現を防いでいる。開始コドンに対する変異は開始コドンの1つ、2つ、又は3つ全てのヌクレオチドのミスセンス変異であってよい。或いは、又はさらに、開始コドンは挿入又は欠失のフレームシフト変異によって変異されてよい。
【0157】
適切な変異ノックアウトOmpT配列を、配列番号24に示す。
【0158】
一実施形態において、本発明によるグラム陰性細菌細胞は、染色体ompTを欠くなど、ノックアウト変異ompT遺伝子を有さない。
【0159】
一実施形態において、本発明によるグラム陰性細菌細胞は、染色体degPを欠くなど、ノックアウト変異degP遺伝子を有さない。一実施形態において、本発明によるグラム陰性細菌細胞は、変異degP遺伝子を有さない。
【0160】
一実施形態において、本発明によるグラム陰性細菌細胞は、染色体ptrを欠くなど、ノックアウト変異ptr遺伝子を有さない。
【0161】
ノックアウト変異を含めた多くの遺伝子操作された変異は、上首尾に変異させた細胞の選択及び同定を可能にする抗生物質耐性マーカーの使用を伴う。しかし、上記で論じたように、抗生物質耐性マーカーの使用には数々の不都合がある。
【0162】
本発明のさらなる一実施形態は、抗生物質耐性マーカーを使用する上記の不都合を克服するものであり、変異Tsp遺伝子、任意選択によって変異spr、任意選択によって変異DegP遺伝子、及び/又は変異ptr遺伝子、及び/又は変異OmpT遺伝子は1つ又は複数の制限マーカー部位を含むように変異される。制限部位は遺伝子中に遺伝子操作され、天然に存在しない。制限マーカー部位が有利であるのは、制限マーカー部位により、必要とされる染色体の変異を含んでいる正確に修飾されている細胞のスクリーニング及び同定が可能になるからである。1つ又は複数の変異プロテアーゼ遺伝子を有するように修飾された細胞は、天然に存在しない制限マーカー部位を含むゲノムDNAの領域を増幅するようにデザインされたオリゴヌクレオチド対を用いて、細胞溶解物からゲノムDNAのPCRによって分析することができる。増幅されたDNAを、次いで、天然に存在しない制限マーカー部位でDNAを消化することができる適切な制限酵素とインキュベートする前及び後にアガロースゲル電気泳動によって分析してもよい。制限酵素とインキュベートした後にDNAフラグメントが存在すれば、細胞が、1つ又は複数の変異遺伝子を有するように上首尾に修飾されたことを確認している。
【0163】
細胞が配列番号6のDNA配列を有するノックアウト変異ptr遺伝子を有する実施形態において、配列番号17及び配列番号18に示すオリゴヌクレオチドプライマー配列を用いて、形質転換した細胞のゲノムDNAから天然に存在しないAseI制限部位を含むDNAの領域を増幅してもよい。次いで、増幅したゲノムDNAを、AseI制限酵素とインキュベートし、ゲル電気泳動によって分析して、ゲノムDNA中の変異ptr遺伝子の存在を確認してもよい。
【0164】
細胞が、配列番号3のDNA配列又は配列番号3のヌクレオチド7から2048を有するノックアウト変異Tsp遺伝子を含んでいる実施形態において、配列番号15及び配列番号16に示すオリゴヌクレオチドプライマー配列を用いて、形質転換した細胞のゲノムDNAから天然に存在しないAseI制限部位を含んでいるDNAの領域を増幅してもよい。次いで、増幅したゲノムDNAを、AseI制限酵素とインキュベートし、ゲル電気泳動によって分析して、ゲノムDNA中の変異Tsp遺伝子の存在を確認してもよい。
【0165】
細胞が、配列番号9のDNA配列を有する変異DegP遺伝子を含んでいる実施形態において、配列番号19及び配列番号20に示すオリゴヌクレオチドプライマー配列を用いて、形質転換した細胞のゲノムDNAから天然に存在しないAseI制限部位を含んでいるDNAの領域を増幅してもよい。次いで、増幅したゲノムDNAを、AseI制限酵素とインキュベートし、ゲル電気泳動によって分析して、ゲノムDNA中の変異DegP遺伝子の存在を確認してもよい。
【0166】
1つ又は複数の制限部位を、1つ又は複数の欠失、挿入、点、ミスセンス、ナンセンス、及びフレームシフトの変異を含むあらゆる適切な変異によって導入することができる。制限部位を、上記に記載した通り、開始コドンの変異、及び/又は1つ若しくは複数の終止コドンを導入するような変異によって導入することができる。この実施形態が有利であるのは、制限マーカー部位が、導入されるノックアウト変異の直接的且つ独特なマーカーであるからである。
【0167】
AseI制限部位など、インフレームの終止コドンを含む制限マーカー部位を挿入してもよい。挿入される制限部位が制限マーカー部位及び終止コドンの両方として作用して遺伝子コード配列の完全な転写を防ぐので、これは特に有利である。例えば、終止コドンがAseI部位の導入によってptr遺伝子に導入される実施形態において、これは、図5aに示すように制限部位をまた作り出す。例えば、AseI部位の導入によって終止コドンがコドン21でTsp遺伝子に導入される実施形態において、これは、図5bに示すように制限部位をまた作り出す。
【0168】
制限マーカー部位を、開始コドンに対する変異、及び任意選択によって1つ又は複数のさらなる点変異によって挿入してもよい。この実施形態において、制限マーカー部位がEcoRI制限部位であるのが好ましい。開始コドンに対する変異は制限マーカー部位も作り出すので、これは特に有利である。例えば、ptr遺伝子の開始コドンがATTに変更される実施形態において、これは、図5aに示すようにEcoRIマーカー部位をまた作り出す。例えば、Tsp遺伝子の開始コドン(コドン3)がATGからTCGに変更される実施形態において、図1bに示すように、コドン2のAACからAATへのさらなる点変異、及びコドン3のATGからTCGへの変異は、図5bに示すように、EcoRI制限マーカー部位を作り出す。
【0169】
細胞が変異OmpT遺伝子を有する本発明の実施形態において、1つ又は複数の制限部位を、1つ又は複数の欠失、挿入、点、ミスセンス、ナンセンス、及びフレームシフトの変異を含むあらゆる適切な変異によって導入してもよい。例えば、OmpT遺伝子が変異D210A及びH212Aを含んでいる実施形態において、これらの変異は、選択マーカーとして用いることができるサイレントのHindIII制限部位を導入する。
【0170】
DegP遺伝子又はspr遺伝子において、マーカー制限部位を、サイレントコドン変化を用いて導入してもよい。例えば、AseI部位をサイレントの制限マーカー部位として用いてもよく、変異DegP遺伝子に対して図5cに示すように、TAA終止コドンはフレーム外である。
【0171】
ptr遺伝子及び/又はTsp遺伝子が低減したプロテアーゼ活性を有するProteaseIII又はTspをコードするように変異されている本発明の実施形態において、1つ又は複数のマーカー制限部位を、サイレントのコドン変化を用いて導入してもよい。
【0172】
あらゆる適切なグラム陰性細菌を、本発明の組換え細胞を生成するための親細胞として用いることができる。適切なグラム陰性細菌には、ネズミチフス菌(Salmonella typhimurium)、蛍光菌(Pseudomonas fluorescens)、軟腐病菌(Erwinia carotovora)、シゲラ(Shigella)、肺炎桿菌(Klebsiella pneumoniae)、レジオネラ・ニューモフィラ(Legionella pneumophila)、緑膿菌(Pseudomonas aeruginosa)、アシネトバクター・バウマンニイ(Acinetobacter baumannii)、及び大腸菌が含まれる。親細胞が大腸菌であるのが好ましい。本発明において大腸菌のあらゆる適切な系統を用いることができるが、K−12 W3110などの野生型W3110系統を用いるのが好ましい。
【0173】
予め作り出され、組換えタンパク質を発現するのに用いられるプロテアーゼ欠損細菌系統に付随する欠点は、これらが、大腸菌系統において、例えば、phoA、fhuA、lac、rec、gal、ara、arg、thi、及びproなど、細胞の代謝及びDNA複製に関与する遺伝子に対するさらなる変異を含むことである。これらの変異は、細胞の増殖、安定性、組換えタンパク質発現収率、及び毒性に対する効果を含めた、宿主細胞に対する多くの有害効果を有し得る。これらのゲノムの変異を1つ又は複数有する系統、特にこれらの変異を多数有する系統は、細菌の増殖速度を、産業的なタンパク質生成に適さないレベルまで低減する適応度の喪失を表し得る。さらに、あらゆる上記のゲノムの変異は、予測不可能な有害な方法で、シス及び/又はトランスにおいて他の遺伝子に影響を及ぼし、それによって系統の表現型、適応度、及びタンパク質のプロファイルを変更し得る。さらに、大幅に変異した細胞を使用することは、商用、特に治療剤に使用するための組換えタンパク質を生成するのに一般に適さない、というのは、このような系統は一般的に欠陥のある代謝経路を有しており、したがって最小の培地又は化学的に規定された培地において不十分にしか増殖せず、又は全く増殖しないことがあるからである。
【0174】
好ましい一実施形態において、細胞は、DsbCをコードする組換えポリヌクレオチドを導入するのに最小の変異だけを有するが、この修飾はTspタンパク質活性、及び任意選択によって変異spr遺伝子を低減するのに必要とされる。組換えポリヌクレオチド及び変異を導入するのに、最小の変異だけが細胞のゲノムに対して行われる。細胞は、細胞の増殖及び/又は対象のタンパク質を発現する能力に対して有害な効果を有し得るいかなる他の変異を有さない。したがって、本発明の1つ又は複数の組換え宿主細胞は、ゲノム配列に対してさらなる遺伝子操作した変異を含んでいる細胞に比べて、改善されたタンパク質発現及び/又は改善された増殖の特徴を表し得る。本発明が提供する細胞はまた、細胞のゲノムに対するさらなるかく乱を含んでいる細胞に比べて、治療用タンパク質の生成において用いるのにより適している。
【0175】
したがって、本発明は、Tspタンパク質活性、及び任意選択によって変異spr遺伝子を低減するのに必要とされる組換え、修飾以外、野生型細菌細胞に対して同質遺伝子であるゲノムを有するグラム陰性細菌細胞も提供する。細胞は、DsbCをコードする組換えポリヌクレオチド、及び任意選択によって対象のタンパク質をコードするポリヌクレオチド配列をさらに有する。
【0176】
好ましい一実施形態において、細胞は、野生型細胞及び任意選択によって変異spr遺伝子に比べてTspタンパク質活性を低減するのに必要とされる修飾以外、W3110系統などの野生型大腸菌細胞に対して同質遺伝子である。細胞は、DsbCをコードする組換えポリヌクレオチド、及び任意選択によって対象のタンパク質をコードするポリヌクレオチド配列をさらに有する。
【0177】
本発明による細胞は、対象のタンパク質をコードするポリヌクレオチド配列をさらに含んでいてよい。対象のタンパク質をコードするポリヌクレオチド配列は、外来性でも、又は内在性でもよい。対象のタンパク質をコードするポリヌクレオチド配列は、宿主の染色体中に組み込まれてもよく、又は通常はプラスミドであるベクター中に非組入れ(non−integrated)であってもよい。ポリヌクレオチドが細胞における発現ベクター中にあり、それによって宿主細胞のゲノムに対して最小のかく乱を引き起こすのが好ましい。
【0178】
「対象のタンパク質」は、本明細書の文脈において、通常は組換えポリペプチドである、発現用のポリペプチドを意味するものとされる。しかし、対象のタンパク質は、宿主細胞において内在性の遺伝子から発現される内在性タンパク質であってもよい。
【0179】
本明細書で用いられる、「組換えポリペプチド」は、組換えDNA技術を用いて構築又は生成されるタンパク質を意味する。対象のタンパク質は、外来性のベクターから発現された、内在性のタンパク質若しくはその変異バージョン(例えば、減弱された生物学的活性を有するもの)に同一の外来性の配列であってよく、又はそのフラグメントであってよい。或いは、対象のタンパク質は、宿主細胞によって通常発現されない、異種性のタンパク質であってよい。
【0180】
対象のタンパク質は、治療用、予防用、又は診断用のタンパク質を含めたあらゆる適切なタンパク質であってよい。
【0181】
一実施形態において、対象のタンパク質は、炎症性疾患及び障害、免疫疾患及び障害、線維性障害、並びに癌を含めた疾患又は障害の処置において有用である。
【0182】
「炎症性疾患」又は「障害」、及び「免疫疾患又は障害」の語は、関節リウマチ、乾癬性関節炎、スティル病、マックルウェルズ病、乾癬、クローン病、潰瘍性大腸炎、SLE(全身性エリテマトーデス)、喘息、アレルギー性鼻炎、アトピー性皮膚炎、多発性硬化症、血管炎、I型糖尿病、移植、及び移植片対宿主病を含む。
【0183】
「線維性障害」の語は、特発性肺線維症(IPF)、全身性硬化症(又は強皮症)、腎臓線維症、糖尿病性腎症、IgA腎症、高血圧、末期腎不全、腹膜線維症(連続の外来腹膜透析)、肝硬変、加齢黄斑変性(ARMD)、網膜症、反応性心臓線維症(cardiac reactive fibrosis)、瘢痕、ケロイド、火傷、皮膚潰瘍、血管形成、冠状動脈バイパス手術、関節形成術、及び白内障手術を含む。
【0184】
「癌」の語は、上皮から生じ、皮膚中又はより一般的には身体器官(例えば、乳房、卵巣、前立腺、肺、腎臓、膵臓、胃、膀胱、又は腸)の内膜に見出される悪性の新たな増殖を含む。癌は、隣接の組織中に浸潤し、例えば、骨、肝臓、肺、又は脳などの遠位の器官に広がる(転移)傾向がある。
【0185】
タンパク質は、タンパク質分解感受性のポリペプチド、即ち、自然状態又は分泌の間のいずれかにおいて、大腸菌などの1つ又は複数のグラム陰性細菌、プロテアーゼによって切断を受けやすく、切断に感受性であり、又は切断されるタンパク質であってよい。一実施形態において、対象のタンパク質は、DegP、ProteaseIII、及びTspから選択されるプロテアーゼに対してタンパク質分解感受性である。一実施形態において、対象のタンパク質は、プロテアーゼTspに対してタンパク質分解感受性である。一実施形態において、対象のタンパク質は、プロテアーゼであるDegP及びProteaseIIIに対してタンパク質分解感受性である。一実施形態において、対象のタンパク質は、プロテアーゼであるDegP及びTspに対してタンパク質分解感受性である。一実施形態において、対象のタンパク質は、プロテアーゼであるTsP及びProteaseIIIに対してタンパク質分解感受性である。一実施形態において、対象のタンパク質は、DegP、ProteaseIII、及びTsPに対してタンパク質分解感受性である。
【0186】
タンパク質が真核生物のポリペプチドであるのが好ましい。
【0187】
本発明による細胞によって発現される対象のタンパク質は、例えば免疫原、2つの異種性のタンパク質を含む融合タンパク質、又は抗体であってよい。対象のタンパク質として用いるための抗体には、モノクローナル抗体、多価抗体、多重特異性抗体、ヒト化抗体、完全ヒト抗体、又はキメラ抗体が含まれる。抗体はあらゆる種からのものであってよいが、モノクローナル抗体、ヒト抗体、又はヒト化フラグメントに由来しているのが好ましい。抗体は、免疫グロブリン分子のあらゆるクラス(例えば、IgG、IgE、IgM、IgD、若しくはIgA)又はサブクラスに由来していてよく、例えば、マウス、ラット、サメ、ウサギ、ブタ、ハムスター、ラクダ、ラマ、ヤギ、若しくはヒトを含めたあらゆる種から得ることができる。抗体フラグメントの部分は、1つを超える種から得ることができ、例えば、抗体フラグメントはキメラであってよい。一例において、定常領域はある種からであり、可変領域は別の種からである。
【0188】
抗体は、全長の重鎖及び軽鎖を有する完全な抗体分子であってよく、又はそのフラグメント、例えば、PCT/GB2008/003331に記載されている通り、VH、VL、VHH、Fab、修飾Fab’、Fab’、F(ab’)2、Fv、scFvフラグメント、Fab−Fv、又は二重特異性抗体、例えば、Fab−dAbであってよい。
【0189】
抗体はあらゆる標的の抗原に特異的であってよい。抗原は、細胞に付随するタンパク質、例えば、細菌細胞、酵母菌細胞、T細胞、内皮細胞、若しくは腫瘍細胞などの細胞状の細胞表面タンパク質であってよく、又は可溶性タンパク質であってよい。対象の抗原はまた、疾患若しくは感染の間に上方制御されるタンパク質などのあらゆる医学的に関連のあるタンパク質、例えば、受容体であってよく、且つ/又はこれらの対応するリガンドであってもよい。細胞表面タンパク質の特定の例には、接着分子、例えば、β1インテグリンなどのインテグリン、例えば、VLA−4、E−セレクチン、Pセレクチン、若しくはL−セレクチン、CD2、CD3、CD4、CD5、CD7、CD8、CD11a、CD11b、CD18、CD19、CD20、CD23、CD25、CD33、CD38、CD40、CD40L、CD45、CDW52、CD69、CD134(OX40)、ICOS、BCMP7、CD137、CD27L、CDCP1、CSF1、若しくはCSF1受容体、DPCR1、DPCR1、ヅヅリン(dudulin)2、FLJ20584、FLJ40787、HEK2、KIAA0634、KIAA0659、KIAA1246、KIAA1455、LTBP2、LTK、MAL2、MRP2、ネクチン様2、NKCC1、PTK7、RAIG1、TCAM1、SC6、BCMP101、BCMP84、BCMP11、DTD、癌胎児性抗原(CEA)、ヒト乳脂肪グロブリン(HMFG1及び2)、MHCクラスI及びMHCクラスII抗原、KDR及びVEGF、及び好適な場合にはこれらの受容体が含まれる。
【0190】
可溶性抗原には、インターロイキン、例えば、IL−1、IL−2、IL−3、IL−4、IL−5、IL−6、IL−8、IL−12、IL−13、IL−14、IL−16、又はIL−17、例えば、IL17A及び/又はIL17F、ウイルス抗原、例えば、呼吸器合胞体ウイルス抗原又はサイトメガロウイルス抗原、免疫グロブリン、例えば、IgE、インターフェロン、例えば、インターフェロンα、インターフェロンβ、又はインターフェロンγ、腫瘍壊死因子TNF(以前は腫瘍壊死因子−αとして知られていた)、腫瘍壊死因子−β、コロニー刺激因子、例えば、G−CSF又はGM−CSF、及び血小板由来増殖因子、例えば、PDGF−α及びPDGF−β、並びに好適な場合にはこれらの受容体が含まれる。他の抗原には、細菌の細胞表面抗原、細菌の毒素、ウイルス、例えば、インフルエンザ、EBV、HepA、B、及びC、生物テロ薬剤、放射性核種及び重金属、並びにヘビ毒及びクモ毒、及び毒素が含まれる。
【0191】
一実施形態において、対象の抗原の活性を機能的に変更するのに抗体を用いることができる。例えば、抗体は、前記抗原の活性を、直接的又は間接的に、中和し、拮抗する(antagonize)、又は作用し(アゴナイズ:agonise)得る。
【0192】
本発明は、DsbC、変異Tsp遺伝子をコードする組換えポリヌクレオチド、及びTNFに特異的な抗体又はその抗原結合性フラグメントをコードするポリヌクレオチド配列を含んでいる組換えグラム陰性細菌細胞も提供し、変異Tsp遺伝子は、低減したプロテアーゼ活性を有するTspタンパク質をコードする、又はノックアウト変異Tsp遺伝子である。好ましい一実施形態において、細胞は、変異体sprタンパク質をコードする変異体spr遺伝子をさらに含んでいる。
【0193】
好ましい一実施形態において、WO01/094585(この内容は参照によって本明細書に組み込まれる)に記載されている通り、本発明による細胞によって発現される対象のタンパク質は、抗TNF抗体、より好ましくは抗TNF Fab’である。
【0194】
一実施形態において、ヒトTNFαに対する特異性を有する抗体は重鎖を含んでおり、可変ドメインは、CDRH1に対して配列番号28に示す配列、CDRH2に対して配列番号29若しくは配列番号34に示す配列、又はCDRH3に対して配列番号30に示す配列を有するCDRを含んでいる。
【0195】
一実施形態において、抗体は軽鎖を含んでおり、可変ドメインは、CDRL1に対して配列番号31に示す配列、CDRL2に対して配列番号32に示す配列、又はCDRL3に対して配列番号33に示す配列を有するCDRを含んでいる。
【0196】
上記に言及した配列番号28、及び配列番号30から配列番号34に示したCDRは、マウスモノクローナル抗体hTNF40に由来する。しかし、配列番号29はハイブリッドCDRからなる。ハイブリッドCDRは、マウスモノクローナル抗体hTNF40(配列番号40)からの重鎖CDR2の部分、及びヒトグループ3生殖系列V領域配列からの重鎖CDR2の部分を含んでいる。
【0197】
一実施形態において、抗体は重鎖(可変ドメインは、CDRH1に対して配列番号28に示す配列、CDRH2に対して配列番号29若しくは配列番号34に示す配列、又はCDRH3に対して配列番号30に示す配列を有するCDRを含んでいる)、及び軽鎖(可変ドメインは、CDRL1に対して配列番号31に示す配列、CDRL2に対して配列番号32に示す配列、又はCDRL3に対して配列番号33に示す配列を有するCDRを含んでいる)を含んでいる。
【0198】
一実施形態において、抗体は、CDRH1に対して配列番号28、CDRH2に対して配列番号29又は配列番号34、CDRH3に対して配列番号30、CDRL1に対して配列番号31、CDRL2に対して配列番号32、及びCDRL3に対して配列番号33を含んでいる。抗体がCDRH2に対して配列番号29を含んでいるのが好ましい。
【0199】
抗TNF抗体がCDRグラフト化抗体分子であるのが好ましい。好ましい一実施形態において、可変ドメインは、ヒトアクセプターフレームワーク領域及び非ヒトドナーCDRを含んでいる。
【0200】
抗体分子がヒトTNF(以前はTNFαとして知られていた)に対して特異性を有するのが好ましく、軽鎖は配列番号11の軽鎖可変領域を含んでおり、重鎖は配列番号12の重鎖可変領域を含んでいる。
【0201】
抗TNF抗体がFab又はFab’のフラグメントであるのが好ましい。
【0202】
ヒトTNFに対して特異性を有する抗体分子がFab’であり、配列番号13を含んでいる、又はそれからなる軽鎖配列、及び配列番号14を含んでいる、又はそれからなる重鎖配列を有するのが好ましい。
【0203】
発現後、抗体フラグメントを、例えば、エフェクター分子などの別の実体に対してコンジュゲートすることによってさらにプロセシングしてもよい。
【0204】
本明細書で用いられるエフェクター分子の語には、例えば、抗悪性腫瘍薬、薬物、毒素(例えば、細菌若しくは植物起源の酵素的に活性な毒素及びこれらのフラグメント、例えば、リシン(ricin)及びそのフラグメント)生物学的に活性なタンパク質、例えば、酵素、他の抗体又は抗体フラグメント、合成の、又は天然に存在するポリマー、核酸及びこれらのフラグメント、例えば、DNA、RNA、及びこれらのフラグメント、放射性核種、特に放射性ヨウ化物、放射性同位元素、キレート化金属、ナノ粒子及びレポーターグループ、例えば、蛍光化合物又はNMR若しくはESR分光法によって検出され得る化合物が含まれる。エフェクター分子は、あらゆる適切な方法によって抗体又はそのフラグメントに付着していてよく、例えば、抗体フラグメントは、WO05/0031731又はWO05/003170(これらの内容は参照によって本明細書に組み入れられる)に記載されている通り、少なくとも1つのエフェクター分子で付着するように修飾されていてよい。WO05/003171又はWO05/003170も適切なエフェクター分子も記載している。
【0205】
一実施形態において、所望により、抗体又はそのフラグメント、例えばFabをPEG化して必要とされる性質、例えば、抗体全体に類似の性質を有する生成物を生成する。例えば、抗体は、WO01/094585に記載されている通り、PEG化した抗TNFα−Fab’であってよく、好ましくは、重鎖のC末端終端でリシル−マレイミド由来の基(lysyl−maleimide−derived group)でシステイン残基の1つに付着しており、リシル残基の2つのアミノ基の各々はこれを、分子量約20000Daを有するメトキシポリ(エチレングリコール)残基に共有結合させており、その結果メトキシポリ(エチレングリコール)残基の合計平均分子量は約40000Daであり、より好ましくはリシル−マレイミド由来の基は、[1−[[[2−[[3−(2,5−ジオキソ−1−ピロリジニル)−1−オキソプロピル]アミノ]エチル]アミノ]−カルボニル]−1,5−ペンタンジイル]ビス(イミノカルボニル)である。
【0206】
細胞はまた、1つ又は複数のさらなる対象のタンパク質をコードするさらなるポリヌクレオチド配列を含んでいてよい。
【0207】
一実施形態において、野生型が精製の間に対象の組換えタンパク質と同時精製することが知られている1つ又は複数の大腸菌宿主タンパク質は、Humphreysら、「Engineering of Escherichia coli to improve the purification of periplasmic Fab’ fragments:changing the pI of the chromosomally encoded PhoS/PstS protein」、Protein Expression and Purification、37巻(2004年)、109〜118頁及びWO04/035792(これらの内容は参照によって本明細書に組み入れられる)において記載されている通り、遺伝子修飾用に選択される。このような修飾された宿主タンパク質の使用により、これらが組換え抗体と最早同時精製されないように、選択された大腸菌タンパク質の物理的性質を変更することによって、大腸菌において生成された対象のタンパク質、特に抗体に対する精製プロセスが改善される。変更される大腸菌タンパク質が、リン酸結合性タンパク質(PhoS/PstS)、ジペプチド結合性タンパク質(DppA)、マルトース結合性タンパク質(MBP)、及びチオレドキシンの1つ又は複数から選択されるのが好ましい。
【0208】
一実施形態において、汚染性の宿主タンパク質の物理的性質は、アミノ酸タグをC末端又はN末端に添加することによって変更される。好ましい一実施形態において、変更される物理的性質は等電点であり、アミノ酸タグはC末端に付着するポリアスパラギン酸タグである。一実施形態において、前記タグの添加によって変更される大腸菌タンパク質は、ジペプチド結合性タンパク質(DppA)、マルトース結合性タンパク質(MBP)、チオレドキシン、及びリン酸結合性タンパク質(PhoS/PstS)である。詳しい一実施形態において、大腸菌のリン酸結合性タンパク質(PhoS/PstS)のpIは、アスパラギン酸残基を6個含んでいるポリアスパラギン酸タグ(PolyD)をC末端に加えることよって7.2から5.1に低減される。
【0209】
単独で、又はN末端若しくはC末端にタグを加えることとの組合せのいずれかで、その物理的性質を変更するための、汚染性の大腸菌タンパク質の特定の残基の修飾も好ましい。このような変更は、pI又は疎水性を変更するためのタンパク質又はアミノ酸置換基のサイズを変更するための挿入又は欠失を含むことができる。一実施形態において、これらの残基はタンパク質の表面上に位置する。好ましい一実施形態において、タンパク質のpIを低減するために、PhoSタンパク質の表面残基が変更される。機能的なPhoSタンパク質を維持するために、リン酸塩の結合において重要であると関係付けられている残基(Bass、US5,304,472)を回避するのが好ましい。タンパク質の表面から遠くはなれて突出している、又は塩基性残基の大型の基である、若しくは塩基性残基の大型の基の近くにあるリシン残基を標的にするのが好ましい。一実施形態において、PhoSタンパク質はC末端に付着しているヘキサポリアスパラギン酸タグを有し、分子の反対の終端にある表面残基が置換用に標的にされる。選択されたリシン残基が、中性残基から酸性残基に変わる場合よりも大きな潜在的なpI変化を付与するようにグルタミン酸又はアスパラギン酸を置換するのが好ましい。本明細書における置換変異体に対する呼称は、文字とその後に続く数字、その後に続く文字からなる。最初の文字は野生型タンパク質におけるアミノ酸を指定する。数字はアミノ酸置換が行われるアミノ酸位置を意味し、2番目の文字は野生型アミノ酸を置換するのに用いられるアミノ酸を指定する。本発明におけるPhoSの好ましい変異において、リシン残基(K)275、107、109、110、262、265、266、309、313は、単一変異又は複合変異としてグルタミン酸(E)又はグルタミン(Q)に対して置換され、さらにリシン(K)318は、単一変異又は複合変異としてアスパラギン酸(D)に対して置換されてよい。単独の変異がK262E、K265E、及びK266Eであるのが好ましい。複合変異がK265/266E及びK110/265/266Eであるのが好ましい。全ての変異が、C末端で付着しているポリアスパラギン酸(PolyD)タグと、任意選択によってK318D置換とも組み合わされるのがより好ましい。好ましい一実施形態において、変異は少なくとも2単位のpIにおける低減をもたらす。本発明の変異がPhoSのpIを7.2から約4と約5.5との間に低減するのが好ましい。本発明の一実施形態において、大腸菌のPhoSタンパク質のpIは、polyD K318D、polyD K265/266E、及びpolyD K110/265/266の変異を用いて、7.2からそれぞれ約4.9、約4.8、及び約4.5に低減される。
【0210】
本発明による組換えグラム陰性細菌細胞はあらゆる適切な手段によって生成され得る。
【0211】
染色体Tsp遺伝子が変異され、任意選択によって1つ又は複数の染色体spr遺伝子、Deg遺伝子、ptr遺伝子、及びOmpT遺伝子が変異される本発明の実施形態において、当業者であれば、染色体の遺伝子配列を、変異遺伝子配列で置換するのに用いることができる適切な技術を知っている。相同的組換えによって宿主の染色体中に組み込むのを可能にする適切なベクターを用いることができる。細胞が上記の変異遺伝子の2つ又は3つを含んでいる実施形態において、変異遺伝子を、同じ又は異なるベクター上のグラム陰性細菌中に導入してもよい。
【0212】
適切な遺伝子置換方法は、例えば、Hamiltonら(「New Method for Generating Deletions and Gene Replacements in Escherichia coli」、Hamilton C.M.ら、Journal of Bacteriology、1989年9月、171巻9号、4617〜4622頁)、Skorupskiら(「Positive selection vectors for allelic exchange」、Skorupski K及びTaylor R. K.、Gene、1996年、169巻、47〜52頁)、Kielら(「A general method for the construction of Escherichia coli mutants by homologous recombination and plasmid segregation」、Kiel J.A.K.W.ら、Mol Gen Genet、1987年、207巻、294〜301頁)、Blomfieldら(「Allelic exchange in Escherichia coli using the Bacillus subtilis sacB gene and a temperature sensitive pSC101 replicon」、Blomfield I.C.ら、Molecular Microbiology、1991年、5巻(6)、1447〜1457頁)、並びにRiedら(「An nptI−sacB−sacR cartridge for constructing directed, unmarked mutations in Gram−negative bacteria by marker exchange−eviction mutagenesis」、Ried J.L. 及びCollmer A.、Gene、57巻(1987年)239〜246頁)に記載されている。相同組換え/置換を可能にする適切なプラスミドはpKO3プラスミドである(Linkら、1997年、Journal of Bacteriology、179巻、6228〜6237頁)。
【0213】
当業者であれば、DsbCをコードする組換えポリヌクレオチドを挿入するのに用いることができる適切な技術を知っている。DsbCをコードする組換えポリヌクレオチドを、pKO3プラスミドなどの適切なベクターを用いて細胞のゲノム中に組み込むことができる。
【0214】
或いは、又はさらに、DsbCをコードする組換えポリヌクレオチドは、組換え発現カセット中に非組入れであってもよい。一実施形態において、DsbCをコードするタンパク質コード配列及び1つ又は複数の制御発現配列を典型的に含んでいる、DsbCをコードするポリヌクレオチドを有するために、本発明において発現カセットが用いられる。1つ又は複数の制御発現配列はプロモーターを含んでいてよい。1つ又は複数の制御発現配列は、終止配列などの3’非翻訳領域も含んでいてよい。適切なプロモーターを以下により詳しく論じる。
【0215】
一実施形態において、1つ若しくは複数の制御発現配列、1つ若しくは複数の対象のタンパク質をコードする1つ若しくは複数のコード配列、及び/又はDsbCをコードするコード配列を典型的に含んでいる、対象のタンパク質をコードするポリヌクレオチド及び/又はDsbCをコードする組換えポリヌクレオチドを有するために、本発明において発現カセットが用いられる。1つ又は複数の制御発現配列はプロモーターを含んでいてよい。1つ又は複数の制御発現配列は、終止配列などの3’非翻訳領域も含んでいてよい。適切なプロモーターを以下により詳しく論じる。
【0216】
一実施形態において、本発明による細胞は、プラスミドなどの1つ又は複数のベクターを含んでいる。ベクターが上記に規定した1つ又は複数の発現カセットを含んでいるのが好ましい。一実施形態において、対象のタンパク質をコードするポリヌクレオチド配列、及びDsbCをコードするポリヌクレオチドを1つのベクター中に挿入する。或いは、対象のタンパク質をコードするポリヌクレオチド配列、及びDsbCをコードするポリヌクレオチドを別々のベクター中に挿入する。
【0217】
対象のタンパク質が重鎖及び軽鎖の両方を含む抗体である実施形態において、第1のベクターが軽鎖ポリペプチドをコードし、第2のベクターが重鎖ポリペプチドをコードする、2つのベクターを細胞系にトランスフェクトしてもよい。或いは、ベクターが軽鎖及び重鎖のポリペプチドをコードする配列を含んでいる、単一のベクターを用いてもよい。或いは、抗体をコードするポリヌクレオチド配列及びDsbCをコードするポリヌクレオチドを1つのベクター中に挿入する。ベクターが、抗体の軽鎖及び重鎖のポリペプチドをコードする配列を含んでいるのが好ましい。
【0218】
細胞が以下の1つ又は複数のさらなるタンパク質:
タンパク質のフォールディングを促進することができる1つ又は複数のタンパク質、例えば、FkpA、Skp、SurA、PPiA、及びPPiD、並びに/又は
タンパク質の分泌又は転位置を促進することができる1つ又は複数のタンパク質、例えば、SecY、SecE、SecG、SecYEG、SecA、SecB、FtsY、及びLep、並びに/又は
ジスルフィド結合形成を促進することができる1つ又は複数のタンパク質、例えば、DsbA、DsbB、DsbD、DsbG
も発現する実施形態において、1つ又は複数のさらなるタンパク質を、DsbCをコードするポリヌクレオチド及び/又は対象のタンパク質をコードするポリヌクレオチド配列として同じベクター中に挿入された1つ又は複数のポリヌクレオチドから発現させてもよい。或いは、1つ又は複数のポリヌクレオチドを別々のベクター中に挿入してもよい。
【0219】
本発明において用いるためのベクターは、適切なベクター中に上記で定義した1つ又は複数の発現カセットを挿入することによって生成することができる。或いは、ポリヌクレオチド配列の発現を指示するための制御発現配列がベクターに含まれていてよく、したがってポリヌクレオチドのコード領域だけがベクターを完成するのに必要とされ得る。
【0220】
DsbCをコードするポリヌクレオチド及び/又は対象のタンパク質をコードするポリヌクレオチドを、細胞中で細胞に適するプロモーターの制御下で発現させるために、複製可能なベクター、典型的には自律的複製ベクター中に挿入するのが適切である。多くのベクターがこの目的で当技術分野において知られており、好適なベクターの選択は核酸のサイズ及び特定の細胞型に依存し得る。
【0221】
本発明によるポリヌクレオチドで宿主細胞を形質転換するのに用いることができるベクターの例には:
プラスミド、例えば、pBR322若しくはpACYC184、及び/又は
ウイルスのベクター、例えば、細菌のファージ
転位性遺伝要素、例えば、トランスポゾン
が含まれる。
【0222】
このようなベクターは、通常、プラスミドのDNA複製開始点、抗生物質の選択マーカー、プロモーター、及びマルチクローニング部位によって分離されている転写ターミネーター(発現カセット)、及びリボソーム結合部位をコードするDNA配列を含んでいる。
【0223】
本発明において用いられるプロモーターは、関連のポリヌクレオチドに直接連結していてよく、又は代替として好適な位置に、例えば、関連のポリペプチドが関連のプロモーターに挿入された場合に関連のプロモーターに対して作用することができるようなベクター中に位置していてよい。一実施形態において、プロモーターは、それに対してプロモーターが作用するポリヌクレオチドのコード部分の前に、例えば、ポリヌクレオチドの各コード部分の前の関連のプロモーターに位置している。本明細書で用いられる「前」はプロモーターが、コードするポリヌクレオチド部分に関して5プライムエンドに位置することを意味するものとされる。
【0224】
プロモーターは、宿主細胞に対して内在性でも、又は外因性でもよい。適切なプロモーターには、Lac、tac、trp、PhoA、Ipp、Arab、Tet、及びT7が含まれる。
【0225】
用いられる1つ又は複数のプロモーターは誘導プロモーターであってよい。
【0226】
DsbCをコードするポリヌクレオチド、及び対象のタンパク質をコードするポリヌクレオチドが1つのベクター中に挿入されている実施形態において、DsbC及び対象のタンパク質をコードするヌクレオチド配列は、単一のプロモーター又は別々のプロモーターの制御下にあってよい。DsbC及び対象のタンパク質をコードするヌクレオチド配列が別々のプロモーターの制御下にある実施形態において、プロモーターは独立に誘導プロモーターであってよい。
【0227】
細菌系において用いるためのプロモーターは、対象のポリペプチドをコードするDNAに作動可能にリンクしているシャインダルガノ(S.D.)配列も一般に含んでいる。プロモーターは、制限酵素の消化によって細菌の供給源のDNAから除去し、所望のDNAを含んでいるベクター中に挿入することができる。
【0228】
ポリヌクレオチド配列が、抗体軽鎖及び抗体重鎖など、2つ以上の対象のタンパク質に対するコード配列を2つ以上含んでいる本発明の実施形態において、ポリヌクレオチド配列は、mRNAの中央における翻訳開始を可能にする1つ又は複数の配列内リボソーム進入部位(IRES)配列を含んでいてよい。IRES配列は、コードされたポリペプチド配列を生成するためのmRNAの別々の翻訳を増強するためにコードするポリヌクレオチド配列間に位置していてよい。
【0229】
発現ベクターが、WO03/048208又はWO2007/039714(これらの内容は参照によって本明細書に組み入れられる)に記載されている抗体又はその抗原結合性フラグメントを生成するためのジシストロン性のメッセージも含んでいるのが好ましい。好ましくは、抗体の軽鎖をコードするDNAを含んでいる上流のシストロン、及び対応する重鎖をコードするDNAを含んでいる下流のシストロン、並びにジシストロン性の遺伝子間配列(IGS)が、IGS1(配列番号36)、IGS2(配列番号37)、IGS3(配列番号38)、及びIGS4(配列番号39)から選択される配列を含んでいるのが好ましい。
【0230】
ターミネーターは宿主細胞に対して内在性でも、又は外因性でもよい。適切なターミネーターはrrnBである。
【0231】
プロモーター及びターミネーターを含んでいるさらなる適切な転写レギュレーター、並びにタンパク質ターゲティング方法は、「Strategies for Achieving High−Level Expression of Genes in Escherichia coli」、Savvas C. Makrides、Microbiological Reviews、1996年9月、512〜538頁に見ることができる。
【0232】
発現ベクター中に挿入されたDsbCポリヌクレオチドが、DsbCシグナル配列及びDsbCコード配列をコードする核酸を含んでいるのが好ましい。ベクターは、好ましくは宿主の染色体を独立に複製するために、ベクターの1つ又は複数の選択された宿主細胞における複製を可能にする核酸配列を含んでいるのが好ましい。このような配列は様々な細菌ではよく知られている。
【0233】
一実施形態において、DsbC及び/又は対象のタンパク質は、N末端及び/又はC末端にヒスチジンタグを含んでいる。
【0234】
抗体分子は、細胞から分泌されることもあり、又は適切なシグナル配列によってペリプラズムに対して標的化することもある。或いは、抗体分子は細胞の原形質内に蓄積することもある。抗体分子がペリプラズムに対して標的化するのが好ましい。
【0235】
対象のタンパク質をコードするポリヌクレオチドは、別のポリペプチド、好ましくは、成熟ポリペプチドのN末端に特異的な切断部位を有するシグナル配列又は他のポリペプチドとの融合物として発現されてよい。選択される異種性のシグナル配列は、宿主細胞によって認識され、プロセシングされるものでなければならない。天然又は真核生物のポリペプチドシグナル配列を認識せず、プロセシングしない原核生物の宿主細胞では、シグナル配列は原核生物のシグナル配列によって置換される。適切なシグナル配列には、OmpA、PhoA、LamB、PelB、DsbA、及びDsbCが含まれる。
【0236】
上記に列挙した成分の1つ又は複数を含んでいる適切なベクターの構築は、標準のライゲーション技術を用いるものである。単離したプラスミド又はDNAフラグメントを、必要とするプラスミドを生成するのに望ましい形態において、切断し、誂え、再ライゲーション(re−ligated)する。
【0237】
本発明の好ましい一実施形態において、本発明は、DsbCをコードするポリヌクレオチド配列及び対象のタンパク質をコードするポリヌクレオチド配列を含んでいる多シストロン性の(multi−cistronic)ベクターを提供する。多シストロン性のベクターは、ポリヌクレオチド配列のベクター中への繰返し連続クローニングを可能にする有利なクローニング方法によって生成することができる。方法は、AseI及びNdeI制限部位の「AT」末端などの1対の制限部位の適合性の付着末端を用いるものである。コード配列を含んでおり、AseI−NdeIフラグメントなどの適合性の付着末端を有するポリヌクレオチド配列を、NdeIなどのベクターにおける制限部位中にクローニングしてもよい。ポリヌクレオチド配列の挿入は5’制限部位を破壊するが、NdeIなどの新たな3’制限部位を作り出し、次いでこの新たな制限部位を用いて、適合性の付着末端を含んでいるさらなるポリヌクレオチド配列を挿入してもよい。次いでこのプロセスを繰り返してさらなる配列を挿入する。ベクター中に挿入された各ポリヌクレオチド配列は、終止コドンに対して3’の非コード配列を含んでおり、終止コドンはスクリーニング用のSspI部位、シャインダルガノリボソーム結合性配列、Aリッチスペーサー、及び開始コドンをコードするNdeI部位を含んでいてよい。
【0238】
ある抗体の軽鎖(LC)、ある抗体の重鎖(HC)をコードするポリヌクレオチド配列、DsbCポリヌクレオチド配列、及びさらなるポリヌクレオチド配列を含んでいるベクターの創製を示す図を図6に示す。
【0239】
上首尾に変異させた系統を、コロニーPCR DNA配列決定及びコロニーPCR制限酵素マッピングを含めた当技術分野においてよく知られている方法を用いて同定してもよい。
【0240】
ポリヌクレオチドに関して本明細書に記載する本発明の実施形態は、ベクター、発現カセット、及び/又はこの中に用いられる成分を含んでいる宿主細胞など、関連する態様をこれらに適用することができる限り、本発明の代替の実施形態に等しく適用されるものである。
【0241】
本発明の第3の態様にしたがって、本発明の第1の態様又は第2の態様において上記に記載した組換えグラム陰性細菌細胞において、対象の組換えタンパク質を発現することを含む、対象の組換えタンパク質を生成するための方法を提供する。対象の組換えタンパク質を生成するための方法は、
a.対象の組換えタンパク質、及びDsbCをコードする組換えポリヌクレオチドを発現させるのに効果的な条件下で培養培地において上記に規定した組換えグラム陰性細菌細胞を培養するステップと、
b.組換えグラム陰性細菌細胞のペリプラズム及び/又は培養培地から対象の組換えタンパク質を回収するステップと
を含む。
【0242】
本発明の方法において用いるのが好ましいグラム陰性細菌細胞及び対象のタンパク質は、上記に詳しく記載したものである。
【0243】
対象のタンパク質をコードするポリヌクレオチドが外来性である場合、ポリヌクレオチドを当技術分野において知られているあらゆる適切な手段を用いて宿主細胞中に組み入れることができる。上記で論じた通り、ポリヌクレオチドが、細胞中に形質転換される発現ベクターの部分として組み入れられるのが典型的である。したがって、本発明による細胞の一態様は、対象のタンパク質をコードするポリヌクレオチドを含む発現カセット、及びDsbCをコードするポリヌクレオチドを含んでいる発現カセットを含む。
【0244】
対象のタンパク質をコードするポリヌクレオチド配列、及びDsbCをコードするポリヌクレオチド配列を、塩化ルビジウム、PEG、又は電気穿孔を用いるなど、標準の技術を用いて細胞中に形質転換してもよい。
【0245】
本発明による方法は、対象のタンパク質をコードするポリヌクレオチドで上首尾に形質転換されている安定な細胞の選択を促進する選択系も用いることができる。選択系は通常、選択マーカーをコードするポリヌクレオチド配列の同時形質転換を使用する。一実施形態において、細胞中に形質転換された各ポリヌクレオチドは、1つ又は複数の選択マーカーをコードするポリヌクレオチド配列をさらに含んでいる。したがって、DsbC及び対象のタンパク質をコードするポリヌクレオチドの形質転換、並びにマーカーをコードする1つ又は複数のポリヌクレオチドの形質転換は一緒に生じ、選択系を用いて所望のタンパク質を生成するこれらの細胞を選択することができる。
【0246】
1つ又は複数のマーカーを発現することができる細胞は、細胞中に組み入れられる選択系のポリペプチド/遺伝子又はポリペプチド成分によって付与される性質(例えば、抗生物質耐性)のために、例えば、毒素又は抗生物質の添加など、ある種の人工的に課せられた条件下で生存/増殖/繁殖することができる。1つ又は複数のマーカーを発現することができない細胞は、人工的に課せられた条件下で生存/増殖/繁殖することができない。人工的に課せられる条件は、所望により幾分強力であるように選択され得る。
【0247】
本発明においてあらゆる適切な選択系を用いることができる。典型的に、選択系は、例えば、テトラサイクリン、クロラムフェニコール、カナマイシン、又はアンピリシン耐性の遺伝子など、既知の抗生物質に対して耐性をもたらすベクターの1つ又は複数の遺伝子中に含まれていることに基づいていてよい。関連の抗生物質の存在下で増殖する細胞は、これらが抗生物質に対する耐性をもたらす遺伝子及び所望のタンパク質の両方を発現するように選択されてよい。
【0248】
一実施形態において、本発明による方法は、形質転換した細胞を培地中で培養し、それによって対象のタンパク質を発現させるステップをさらに含んでいる。
【0249】
対象のタンパク質及び/又はDsbCを発現させるために、誘導可能な発現系、又は構成的なプロモーターを本発明において用いることができる。適切な誘導可能な発現系及び構成的なプロモーターは、当技術分野においてよく知られている。
【0250】
DsbCをコードするポリヌクレオチド、及び対象のタンパク質をコードするポリヌクレオチドが、同じ又は別々の誘導プロモーターの制御下にある一実施形態において、対象のタンパク質をコードするポリヌクレオチド配列及びDsbCをコードする組換えポリヌクレオチドは、培養培地に誘導因子を加えることによって誘導される。
【0251】
形質転換細胞を培養するのにあらゆる適切な培地を用いることができる。例えば、培地中で増殖するように上首尾に形質転換されている細胞だけを可能にするように、培地は抗生物質を含み得るなど、培地を特定の選択系に適応させてもよい。
【0252】
培地から得た細胞を、所望により、さらにスクリーニング及び/又は精製にかけてもよい。方法は、所望により対象のタンパク質を抽出及び精製する1つ又は複数のステップをさらに含むことができる。
【0253】
ポリペプチドを、原形質、ペリプラズム、又は上清からを含む、系統から回収することができる。
【0254】
タンパク質を精製するために用いる特定の方法(単数又は複数)はタンパク質のタイプに依存する。適切な方法には、イムノ−アフィニティカラム、又はイオン交換カラム上での分画化、エタノール沈澱、逆相HPLC、疎水性−相互作用クロマトグラフィー、シリカ上クロマトグラフィー、SEPHAROSE及びDEAEなどのイオン交換樹脂上のクロマトグラフィー、クロマトフォーカシング、硫酸アンモニウム沈澱、及びゲルろ過が含まれる。
【0255】
一実施形態において、方法は、DsbCから対象の組換えタンパク質を分離することをさらに含む。
【0256】
抗体を、培養培地及び/又は原形質抽出物及び/又はペリプラズム抽出物から、従来の抗体精製手順、例えば、タンパク質A−セファロース、タンパク質Gクロマトグラフィー、タンパク質Lクロマトグラフィー、チオフィリック(thiophilic)、混合モード樹脂(mixed mode resins)、Hisタグ、FLAGタグ、ヒドロキシルアパタイトクロマトグラフィー、ゲル電気泳動、透析、アフィニティクロマトグラフィー、硫酸アンモニウム、エタノール、又はPEG分画/沈澱、イオン交換膜、膨張床吸着クロマトグラフィー(EBA)、又は擬似移動床クロマトグラフィーなどによって適切に分離することができる。
【0257】
方法は、対象のタンパク質の発現量を測定し、高発現レベルの対象のタンパク質を有する細胞を選択するさらなるステップも含むことができる。
【0258】
方法は、抗体又は抗体フラグメントなどの対象のタンパク質のペグ化などの1つ又は複数のさらなる下流のプロセシングのステップも含むことができる。
【0259】
本明細書に記載する1つ又は複数の方法のステップを、バイオリアクターなどの適切な容器において組み合わせて行ってもよい。
【実施例】
【0260】
細胞系
全ての実験に対して、親の野生型細胞系として大腸菌細胞系W3110を用いた。
【0261】
以下の変異を有する細胞系を作り出した:
a.変異Tsp遺伝子、
b.変異Tsp遺伝子及び組換えDsbCを有する、
c.変異Tsp遺伝子及び変異spr遺伝子、
d.変異Tsp遺伝子及び変異spr遺伝子及び組換えDsbCを有する。
【0262】
(例1)
変異Tsp遺伝子を有する細胞系MXE001(ΔTsp)の生成
MXR001系統を以下の通り生成した:
Tspカセットを、SalI、NotI制限フラグメントとして、同様に制限したpKO3プラスミド中に移動させた。pKO3プラスミドは、温度感受性変異体のpSC101複製開始点(RepA)を、染色体の組入れ事象に対して強化及び選択するためのクロラムフェニコールマーカーと一緒に用いる。レバンスクラーゼをコードするsacB遺伝子は、ショ糖上で増殖させた大腸菌に致死的であり、したがって(クロラムフェニコールマーカー及びpSC101開始点と一緒に)用いて脱組込み(de−integration)及びプラスミドキュアリング事象に対して強化及び選択するのに用いられる。この方法は以前に記載されている(Hamiltonら、1989年、Journal of Bacteriology、171巻、4617〜4622頁、及びBlomfieldら、1991年、Molecular Microbiology、5巻、1447〜1457頁)。pKO3系は全ての選択マーカーを、挿入された遺伝子以外の宿主ゲノムから除去する。
【0263】
以下のプラスミドを構築した。
EcoRI及びAseI制限マーカーを含んでいる、配列番号3に示すノックアウト変異Tsp遺伝子を含んでいるpMXE191。
【0264】
次いで、プラスミドを、Miller,E.M.及びNickoloff,J.A.における「Escherichia coli electrotransformation」、Methods in Molecular Biology、47巻、Nickoloff,J.A.(編集)、Humana Press、Totowa、NJ、105巻(1995年)に見られる方法を用いて調製した、電気的コンピテント大腸菌W3110細胞中に形質転換した。
【0265】
1日目 大腸菌細胞40μlを、冷却したBioRad0.2cm電気穿孔キュベット中、pKO3 DNA(10pg)1μgと混合し、その後2500V、25μF、及び200Ωで電気穿孔した。2×PY1000μlを直ちに加え、30℃のインキュベーター中で1時間、250rpmで振盪することによって細胞を回収した。細胞を2×PY中1/10に段階希釈し、その後100μlのアリコートを、クロラムフェニコール20μgを含み30℃及び43℃に予め温めた2×PY寒天平板上に塗抹した。平板を30℃及び43℃で一夜インキュベートした。
【0266】
2日目 30℃で増殖させたコロニーの数により電気穿孔の効果が推定されたが、43℃の増殖を生存したコロニーは潜在的な組入れ事象を表している。43℃の平板から単一のコロニーを拾い、2×PY10ml中に再懸濁した。この100μlを、ショ糖5%(w/v)を含み30℃に予め温めた2×PY寒天平板上に塗抹して単一のコロニーを生成させた。平板を30℃で一夜インキュベートした。
【0267】
3日目 ここでコロニーは、潜在的な同時の脱組込み及びプラスミドキュアリング事象を表す。脱組込み及びキュアリング事象が増殖における初期に生じた場合、大部分のコロニー塊はクローンとなる。単一のコロニーを拾い、クロラムフェニコール20μg/ml又は5%(w/v)ショ糖のいずれかを含む2×PY寒天上にレプリカプレーティングした。平板を30℃で一夜インキュベートした。
【0268】
4日目 ショ糖上で増殖し、且つクロラムフェニコール上で死滅した両方のコロニーは、潜在的な染色体の置換及びプラスミドキュアリング事象を表している。これらを拾い、変異特異的なオリゴヌクレオチドでPCRによってスクリーニングした。正確なサイズのPCR陽性のバンドを生じたコロニーを削除して、ショ糖5%(w/v)を含む2×PY寒天上で単一のコロニーを生成し、平板を30℃で一夜インキュベートした。
【0269】
5日目 PCR陽性、クロラムフェニコール感受性、及びショ糖耐性の大腸菌の単一のコロニーを用いてグリセロール株、化学的コンピテント細胞を作製し、5’及び3’に隣接するオリゴとのPCR反応用のPCR鋳型として作用させて、Taqポリメラーゼを用いた直接DNA配列決定用のPCR生成物を生成した。
【0270】
オリゴヌクレオチドプライマーを用いて、図1a、1b、及び1cに示す通り、天然に存在しないAseI制限部位を含んでいるTsp遺伝子の領域をPCR増幅することによって、変異Tsp遺伝子を有するゲノムDNAの上首尾な修飾を確認するために細胞系統MXE001を試験した。次いで、変異遺伝子における天然に存在しないAseI制限部位の存在を確認するために、AseI制限酵素でのインキュベートの前及び後に、DNAの増幅した領域をゲル電気泳動によって分析した。この方法を以下の通り行った。
【0271】
以下のオリゴを用いて、PCRを用い、MXE001及びW3110から調製した大腸菌細胞可溶化物からゲノムDNAを増幅した。
【化1】
【0272】
細胞のコロニー1個を、1×PCRバッファー20μl中95℃で10分間加熱することによって、可溶化物を調製した。混合物を室温に放冷し、次いで13200rpmで10分間遠心した。上清を除去し、「細胞可溶化物」のラベルを付けた。
【0273】
Tspオリゴ対を用いて各系統を増幅した。
【0274】
DNAを標準のPCR手順を用いて増幅した。
5μl バッファー×1 (Roche)
1μl dNTP混合物(Roche、10mM混合物)
1.5μl 5’オリゴ(5pmol)
1.5μl 3’オリゴ(5pmol)
2μl 細胞可溶化物
0.5μl Taq DNAポリメラーゼ(Roche 5U/μl)
38.5μl H2O
PCRサイクル
94℃ 1分
94℃ 1分)
55℃ 1分)30サイクル繰返し
72℃ 1分)
72℃ 10分
【0275】
反応が完了したら25μlをAseIでの消化用に新たな微量遠心管に移した。PCR反応25μlに、H2O19μl、バッファー3(NEB)5μl、AseI(NEB)1μlを加え、混合し、37℃で2時間インキュベートした。
【0276】
残りのPCR反応に、ローディングバッファー(×6)5μlを加え、20μlを0.8%TAE200mlアガロースゲル(Invitrogen)上にローディングし、エチジウムブロマイド(貯蔵液10ml/mlを5μl)を加え、100ボルトで1時間泳動させた。サイズマーカー(PerfectDNAマーカー0.1〜12kb、Novagen)10μlを最終レーンにローディングした。
【0277】
AseI消化が完了したら、ローディングバッファー(×6)10μlを加え、20μlを0.8%TAEアガロースゲル(Invitrogen)上にローディングし、エチジウムブロマイド(10ml/ml貯蔵液を5μl)を加え、100ボルトで1時間泳動した。サイズマーカー(PerfectDNAマーカー0.1〜12kb、Novagen)10μlを最終レーンにローディングした。ゲルを両方ともUVトランスイルミネーターを用いて可視化した。
【0278】
増幅したゲノムフラグメントは、Tspに対して2.8kbの正確なサイズバンドを示した。引き続きAseIで消化させると、Tsp欠損系統MXE001に導入されたAseI部位の存在が確認されたが、W3110コントロールでは確認されなかった。
【0279】
MXE001:Tspプライマーセットを用いてゲノムDNAを増幅し、得られたDNAをAseIで消化して2.2kbps及び0.6kbpsのバンドを生成した。
【0280】
W3110のPCR増幅したDNAは、AseI制限酵素によって消化されなかった。
【0281】
(例2)
変異spr遺伝子を有する細胞系統、並びに変異Tsp遺伝子及び変異spr遺伝子を有する細胞系統の生成
spr変異を、相補性アッセイを用いて生成し選択した。
【0282】
spr遺伝子を、1000bpあたり1つから2つの変異を導入するClontech(登録商標)ランダム変異ダイバーシティPCRキットを用いて変異させた。変異sprPCR DNAを、spr変異体と一緒にCDP870Fab’を発現する誘導発現ベクター[pTTO CDP870]中にクローニングした。次いで、このライゲーションを、Miller,E.M.及びNickoloff, J.A.、「Escherichia coli electrotransformation」、Methods in Molecular Biology、47巻、Nickoloff,J.A.(編集)、Humana Press、Totowa、NJ、105巻(1995年)において見られる方法を用いて、大腸菌系統MXE001(ΔTsp)中に電気的形質転換した。以下のプロトコールを用いた。電気的コンピテントMXE001 40μl、ライゲーション2.5μl(DNA100pg)を0.2cm電気穿孔キュベット中に加え、以下の条件、2500V、25μF、及び200ΩでBioRad Genepulser Xcellを用いて電気的形質転換を行った。電気的形質転換後、SOC(Invitrogen)1ml(37℃に予め加温)を加え、穏やかに振盪して細胞を37℃1時間回復させた。
【0283】
細胞をHypotonic寒天(酵母菌抽出物5g/L、トリプトン2.5g/L、寒天15g/L(全てDifco))上に塗抹し、40℃でインキュベートした。コロニーを形成した細胞をHLB上に43℃で再塗抹し、MXE001系統に対して低浸透圧条件下高温で増殖する能力の回復を確認した。選択したクローンからプラスミドDNAを調製し、配列決定してspr変異を同定した。
【0284】
この方法を用いて、sprタンパク質における、以下の通りΔTsp表現型を補足する8つの単一の変異、1つのダブル変異、及び2つの複数の変異を単離した:
1.V98E
2.D133A
3.V135D
4.V135G
5.G147C
6.S95F及びY115F
7.I70T
8.N31T、Q73R、R100G、G140C
9.R62C、Q99P、R144C
10.L108S
11.L136P
【0285】
上記で同定した1から5の個々の変異、並びにspr(C94A、H145A、H157A)及びW174Rの3つの触媒三残基の変異を用いて、複合のΔTsp/変異体spr系統を作製するための例1からのspr変異系統又はMXE001系統を作り出すために、いずれかの野生型W3110大腸菌系統(遺伝子型:F−LAM−IN(rrnD−RRnE)1rph1(ATCC no.27325))を用いて新たな系統を生成した。
【0286】
MXE001を作製するための例1に記載した通り、遺伝子置換ベクター系を用いて、pKO3相同組換え/置換プラスミドを用いて、以下の変異体の大腸菌細胞系統を作製した(Linkら、1997年、Journal of Bacteriology、179巻、6228〜6237頁)。
【表1】
【0287】
変異体spr組入れカセットを、SalI、NotI制限フラグメントとして、同様に制限したpKO3プラスミド中に移動した。
【0288】
プラスミドは、温度感受性変異体のpSC101複製開始点(RepA)を、染色体の組入れ事象に対して強化及び選択するためのクロラムフェニコールマーカーと一緒に用いている。レバンスクラーゼをコードするsacB遺伝子は、ショ糖上で増殖させた大腸菌に対して致死的であり、したがって(クロラムフェニコールマーカー及びpSC101開始点と一緒に)脱組込み及びプラスミドのキュアリングの事象に対して強化及び選択するのに用いられる。この方法は以前に記載されている(Hamiltonら、1989年、Journal of Bacteriology、171巻、4617〜4622頁、及びBlomfieldら、1991年、Molecular Microbiology、5巻、1447〜1457頁)。pKO3系は全ての選択マーカーを、挿入された遺伝子以外の宿主のゲノムから除去する。
【0289】
SalI制限部位をクローン同定用に除去するspr配列内にサイレント変異を含んでいる変異spr遺伝子を含んでいる、以下に列挙するpK03ベクターを構築した。
pMXE336、pK03 spr S95F (−SalI)
pMXE337、pK03 spr Y115F (−SalI)
pMXE338、pK03 spr G147C (−SalI)
pMXE339、pK03 spr D133A (−SalI)
pMXE340、pK03 spr V135D (−SalI)
pMXE341、pK03 spr V135G (−SalI)
pMXE342、pK03 spr V98E (−SalI)
pMXE343、pK03 spr C94A (−SalI)
pMXE344、pK03 spr H145A (−SalI)
pMXE345、pK03 spr H157A (−SalI)
pMXE346、pK03 spr W174R (−SalI)
【0290】
次いで、これらのプラスミドを、Miller,E.M.及びNickoloff,J.A.、「Escherichia coli electrotransformation」、Methods in Molecular Biology、47巻、Nickoloff,J.A.(編集)、Humana Press、Totowa、NJ、105巻(1995年)に見られる方法を用いて調製した、化学的コンピテント大腸菌W3110細胞中に、又は例1からのMXE001系統中に形質転換して、表1に示す通り、複合のΔTsp/変異体spr系統を作製した。
【0291】
1日目 電気的コンピテント大腸菌細胞又はMXE001細胞を、冷却したBioRad0.2cm電気穿孔キュベット中、pKO3 DNA(10pg)1μgと混合し、その後2500V、25μF、及び200Ωで電気穿孔した。2×PY1000μlを直ちに加え、30℃のインキュベーター中で1時間、250rpmで振盪することによって細胞を回収した。細胞を2×PY中1/10に段階希釈し、その後100μlのアリコートを、クロラムフェニコール20μgを含み30℃及び43℃に予め温めた2×PY寒天平板上に塗抹した。平板を30℃及び43℃で一夜インキュベートした。
【0292】
2日目 30℃で増殖させたコロニーの数により電気穿孔の効果が推定されたが、43℃の増殖を生存したコロニーは潜在的な組入れ事象を表している。43℃の平板から単一のコロニーを拾い、2×PY10ml中に再懸濁した。この100μlを、ショ糖5%(w/v)を含み30℃に予め温めた2×PY寒天平板上に塗抹して単一のコロニーを生成させた。平板を30℃で一夜インキュベートした。
【0293】
3日目 ここでコロニーは、潜在的な同時の脱組込み及びプラスミドキュアリング事象を表す。脱組込み及びキュアリング事象が増殖における初期に生じた場合、大部分のコロニー塊はクローンとなる。単一のコロニーを拾い、クロラムフェニコール20μg/ml又は5%(w/v)ショ糖のいずれかを含む2×PY寒天上にレプリカプレーティングした。平板を30℃で一夜インキュベートした。
【0294】
4日目 ショ糖上で増殖し、且つクロラムフェニコール上で死滅した両方のコロニーは潜在的な染色体の置換及びプラスミドキュアリング事象を表している。これらを拾い、SalI部位の喪失に対してPCRプラス制限消化によってスクリーニングした。正確なサイズのPCR陽性のバンド及びSalIによる消化に対する耐性を生じたコロニーを削除して、ショ糖5%(w/v)を含む2×PY寒天上で単一のコロニーを生成し、平板を30℃で一夜インキュベートした。
【0295】
5日目 PCR陽性、クロラムフェニコール感受性、及びショ糖耐性の大腸菌の単一のコロニーを用いてグリセロール株、化学的コンピテント細胞を作製し、5’及び3’に隣接するオリゴとのPCR反応用のPCRテンプレートとして作用させて、Taqポリメラーゼを用いた直接DNA配列決定用のPCR生成物を生成して正確な変異を確認した。
【0296】
(例3)
Fab’及びDsbC発現用のプラスミドの生成
抗TNF Fab’の重鎖及び軽鎖配列の両方、並びにDsbCをコードする配列を含んでいるプラスミドを構築した。
【0297】
WO01/94585に記載されている抗TNFαFab’フラグメント(CDP870と呼ぶ)のジシストロン性のメッセージを作成した。上流のシストロンは抗体の軽鎖(配列番号13)をコードし、下流のシストロンは抗体の重鎖(配列番号14)をコードしている。OmpAシグナルペプチドをコードするDNA配列は、軽鎖及び重鎖の各々をコードするDNAの5’末端に融合してペリプラズムへの効果的な分泌を可能にした。遺伝子間配列(IGS)2を、配列番号37に示す通りに用いた。
【0298】
CDP870Fab’(抗TNF Fab’)及びDsbC(ペリプラズムポリペプチド)に対する発現ベクターであるプラスミドpDPG358(pTTO40.4 CDP870 IGS2)を、Sambrookら、1989年、「Molecular cloning:a laboratory manual」、CSHL press、N.Yにおいて見ることができる従来の制限クローニング法を用いて構築した。プラスミドpDPH358は以下の特徴:強力なtacプロモーター及びlacオペレーター配列を含んでいた。図6に示す通り、プラスミドは独特のEcoRI制限部位をFab’重鎖のコード領域の後に含んでおり、その後に非コード配列、次いで独特のNdeI制限部位が続いた。DsbC遺伝子を、W3110粗製染色体DNAを鋳型として用いて、5’EcoRI部位をコードするPCR生成物の後に強力なリボソーム結合が続き、その後に天然の開始コドン、シグナル配列、及びDsbCの成熟配列、C末端Hisタグにおける終結、及び最後に非コードNdeI部位が続くようにPCRクローニングした。EcoRI−NdelI PCRフラグメントを制限し、3個全てのポリペプチド:Fab’軽鎖、Fab’重鎖、及びDsbCが単一のポリシストロン性のmRNA上にコードされるように発現ベクター中にライゲートした。
【0299】
Fab軽鎖、重鎖遺伝子、及びDcbC遺伝子を、単一のポリシストロン性メッセージとして転写した。大腸菌のペリプラズムに対するポリペプチドの転移を指示する大腸菌OmpAタンパク質からのシグナルペプチドをコードするDNAを軽鎖及び重鎖の遺伝子配列両方の5’末端に融合した。転写を二重転写ターミネーターrrnB t1t2を用いて終結させた。lacIq遺伝子は構成的に発現されるLacIリプレッサータンパク質をコードしていた。これは、アロラクトース又はIPTGの存在によって抑制解除が誘導されるまで、tacプロモーターからの転写を抑制した。用いた複製開始点はp15Aであり、これは低コピー数を維持した。プラスミドは、抗生物質選択用にテトラサイクリン耐性遺伝子を含んでいた。
【0300】
(例4)
変異細胞系における抗TNF Fab’及びDsbCの発現
抗TNF Fab’及びDsbCの発現
例1において提供したMXE001系統、及び例2において提供した変異系統であるMXE008及びMXE009を、例3において生成したプラスミドで形質転換した。
【0301】
系統の形質転換を、Chung C.Tら、「Transformation and storage of bacterial cells in the same solution.」PNAS、86巻、2172〜2175頁(1989年)に見られる方法を用いて行った。
【0302】
抗TNF Fab’の発現
MXE001、MXE008、及びMXE009の系統を、例3に記載した通り、CDP870Fab’に対する発現ベクター(配列番号13に示す軽鎖配列及び配列番号14に示す重鎖配列を有する抗TNF Fab’)であるプラスミドpMXE117(pTTO CDP870又は40.4 IGS2)で形質転換したが、この発現ベクターは、Sambrookら、1989年、「Molecular cloning:a laboratory manual.」、CSHL press、N.Y.に見ることができる従来の制限クローニング法を用いて構築されたものである。プラスミドpMXE117(pTTO CDP870又は40.4 IGS2)は以下の特徴:強力なtacプロモーター及びlacオペレーター配列を含んでいた。Fab軽鎖及び重鎖遺伝子を、単一のジシストロン性メッセージとして転写した。大腸菌OmpAタンパク質からのシグナルペプチドをコードするDNAを、ポリペプチドの大腸菌ペリプラズムへの移動を指示する軽鎖及び重鎖両方の遺伝子配列5’末端に融合させた。転写を二重転写ターミネーターrrnB t1t2を用いて終結させた。lacIq遺伝子は構成的に発現されるLacIリプレッサータンパク質をコードしていた。これは、アロラクトース又はIPTGの存在によって抑制解除が誘導されるまで、tacプロモーターからの転写を抑制した。用いた複製開始点はp15Aであり、低コピー数を維持した。プラスミドは、抗生物質選択用にテトラサイクリン耐性遺伝子を含んでいた。
【0303】
系統の形質転換を、Chung C.Tら、「Transformation and storage of bacterial cells in the same solution.」、PNAS、86巻、2172〜2175頁(1989年)に見られる方法を用いて行った。
【0304】
(例5)
高密度発酵を用いた変異大腸菌系統の増殖及び変異大腸菌系統における抗TNF Fab’の発現
例4において生成した以下の系統を、抗TNFαFab’の増殖及び発現を比べる発酵実験において試験した。
抗TNF Fab’を発現するMXE001
抗TNF Fab’を発現するMXE008
抗TNF Fab’を発現するMXE009
抗TNF Fab’及びDsbCを発現するMXE001
抗TNF Fab’及びDsbCを発現するMXE008
抗TNF Fab’及びDsbCを発現するMXE009
【0305】
増殖培地
発酵増殖培地は、NaH2PO4・H2O 3.86g/l及びグリセロール112g/lを有するSM6E培地(Humphreyら、2002年、Protein Expression and Purification、26巻、309〜320頁に記載)をベースとした。
【0306】
接種
接種培養物を、テトラサイクリン10μg/mlを補った同じ培地中で増殖させた。培養物をおよそ22時間、撹拌しながら30℃でインキュベートした。
【0307】
発酵
発酵槽(総容積2.5リットル)に、接種培養物を0.3〜0.5OD600に接種した。温度は増殖相の間は30℃に維持し、誘導前に25℃に下げた。溶存酸素濃度を、可変性の撹拌及びエアフローによって空気飽和を30%上回って維持した。15%(v/v)NH4OH及び10%(v/v)濃H2SO4で自動滴定することによって、培養物のpHを7.0で制御した。10%(v/v)Struktol J673溶液(Schill and Seilacher)を加えることによって泡立ちを制御した。発酵の様々な段階で数々の添加を行った。バイオマス濃度がおよそ40OD600に到達したらマグネシウム塩及びNaH2PO4・H2Oを加えた。誘導相の前及び間にNaH2PO4・H2Oのさらなる添加を行って、確実にリン酸塩が過剰に維持されるようにした。発酵の初期に存在したグリセロールが枯渇したら(およそ75OD600)、80%(w/w)の継続的なグリセロールの供給を適用した。発酵における同じ時点に170μMのIPTGの供給を適用した。IPTG供給の開始を誘導の開始と理解した。発酵は、典型的に、グリセロール供給速度(0.5から2.5ml/hの間の範囲)で64〜120時間行った。
【0308】
バイオマス濃度及び増殖速度の測定
600nmでの培養物の光学密度を測定することによってバイオマス濃度を決定した。
【0309】
ペリプラズム抽出
細胞を、遠心分離によって培養物試料から回収した。さらなる分析用に上清の分画を(−20℃で)保持した。細胞ペレット分画を、抽出バッファー(100mM Tris−HCl、10mM EDTA、pH7.4)中、元の培養体積に再懸濁した。およそ16時間、60℃でインキュベートした後、遠心分離によって抽出物を澄明にし、上清分画を分析用に(−20℃で)保持した。
【0310】
Fab’定量
ペリプラズム抽出物及び培養物上清中のFab’濃度を、Humphreysら、2002年、Protein Expression and Purification、26巻、309〜320に記載されている通り、Fab’アセンブリーELISAによって、タンパク質G HPLCを用いて決定した。HiTrap タンパク質G HP1mlカラム(GE−Healthcare又は同等物)に分析物(およそ中性のpH、30℃、0.2μmろ過)を2ml/分でローディングし、カラムを20mMリン酸塩、50mM NaCl、pH7.4で洗浄し、次いで50mMグリシン/HCl pH2.7の注入を用いてFab’を溶出した。溶出されたFab’をAgilient1100又は1200HPLCシステム上A280によって測定し、既知濃度の精製Fab’タンパク質の検量線を参照することによって定量した。
【0311】
図1は、抗TNF Fab’発現系統性MXE001及びMXE008の増殖プロファイル、並びに抗TNF Fab’及び組換えDsbC発現性系統MXE001及びMXE008の増殖プロファイルを示す。DsbCを発現する系統は、組換えDsbCを発現しない対応する細胞系統に比べて改善された増殖を表すことが分かる。系統におけるspr変異の存在が細胞増殖を改善することも分かる。
【0312】
図2は、抗TNF Fab’及び組換えDsbC発現性大腸菌系統MXE001、MXE008、及びMXE009からのペリプラズム(黒記号)及び上清(白記号)からのFabの収率を示す。このグラフから、3つの系統全てが高収率の抗TNFα Fab’を生成し、MXE008及びMXE009系統は2.2g/Lを超えるペリプラズム抗TNF Fab’を40時間で生成したことが分かる。変異体spr遺伝子を有するMXE008及びMXE009系統はMXE001(上清の抗TNFαFab’が少ないと分かる)に比べて低減した溶解を示したことも分かる(白記号)。
【0313】
図3は、抗TNF Fab’発現性大腸菌系統MXE001及びMXE008からの、並びに抗TNF Fab’及び組換えDsbC発現性大腸菌系統MXE001及びMXE008からの、ペリプラズム(黒記号)及び上清(白記号)からのFabの収率を示す。このグラフから、組換えDsbCを発現する系統は高収率の抗TNFαFab’を生成し、MXE008系統は2.4g/Lのペリプラズム抗TNF Fab’を40時間で生成したことが分かる。変異体spr遺伝子を有するMXE008系統はMXE001系統(上清の抗TNFαFab’が少ないと分かる)に比べて低減した溶解を示したことも分かる(白記号)。
【0314】
(例6)
高密度発酵を用いた変異大腸菌系統の増殖並びに変異大腸菌系統におけるFabA及びFabBの発現
本発明の細胞からのタンパク質生成の収率に対する効果も、抗原Aに対する特異性を有するFabA及び抗原Bに対する特異性を有するFabBである2つのさらなるFabタンパク質に対して、上記例5に記載したのと同じ方法を用いて行い、FabA及びFabBを発現するW3110系統に対して比較した。
【0315】
図4は、FabA及びFabB発現性大腸菌系統W3110からの、並びに、FabA及び組換えDsbC発現性又はFab及び組換えDsbC発現性大腸菌系統MXE008からの、ペリプラズムからのFab収率の合計を示す。このグラフから、組換えDsbCを発現する系統は、MXE008において、W3110に比べて有意に高い収率のFabA及びFabBを生成したことが分かる。
【0316】
(例7)
大規模発酵を用いた抗TNF Fab’及びDsbCを発現する大腸菌系統の増殖
例4によって生成した以下の系統を、抗TNFα Fab’の系統の増殖及び発現を比べる発酵実験において試験した:
抗TNF Fab’及びDsbCを発現するMXE008
【0317】
発酵を以下の通りに行った:
抗TNF Fab’及びDsbC細胞を発現するMXE008を、最初に、フラスコ振盪培養物中酵母菌抽出物及びペプトンの複合培地を用いて増殖させた。次いで、細胞を、化学的に規定された培地を用いて接種段階の発酵槽に移した。細胞を、規定された移行点まで栄養非制限の条件下で増殖させた。次いで、細胞を、同様の化学的に規定された培地を用いて、5L又は250Lの生成発酵槽にそれぞれおよそ3.3L又は230Lの最終体積で移した。培養物を最初に、炭素源が枯渇するまで、バッチモード中で増殖させた。炭素源が枯渇した後、炭素源を限定する供給を、指数関数的に増大する速度で供給した。規定された量の炭素源を加えた後、フィード溶液を添加する速度を下げ、IPTGを加えてFab’の発現を誘導した。次いで、発酵を続け、Fab’をペリプラズム中に蓄積させた。誘導後の規定された時点で、遠心分離によって培養物を回収し、回収した細胞をTris及びEDTAバッファー中に再懸濁し、59℃に加熱することによって、Fab’を細胞から抽出した。
【0318】
発酵の増殖プロファイルを、600nmの培養物の光学密度を測定することによって決定した。
【0319】
Fab’の力価を、上記例5に記載した通りタンパク質G HPLCによって決定したが、ペリプラズム抽出の間新鮮な細胞を用い、抽出バッファー1mLを細胞培養液に加える点が異なった。上清及びペリプラズムのFab’を、例5に記載した通りに測定した。図8はペリプラズムのFab’の力価を示す。
【0320】
図7は、抗TNF Fab’及び組換えDsbC発現性大腸菌系統MXE008の、5L及び200L発酵の、比較上の増殖プロファイルを示す。
【0321】
図8は、抗TNF Fab’及び組換えDsbC発現性大腸菌系統MXE008の、5L及び200L発酵の、比較上のFab’力価を示す。
【0322】
(例8)
大規模発酵を用いた抗TNF Fab’及びDsbCを発現する大腸菌系統の増殖
例4によって生成した以下の系統を、抗TNFα Fab’の系統の増殖及び発現を比べる発酵実験において試験した:
抗TNF Fab’及びDsbCを発現するMXE008
抗TNF Fab’及びDsbCを発現するMXE009。
【0323】
発酵を、20L生成発酵槽を用いて上記例7に記載した通りに行った。
【0324】
発酵の増殖プロファイルを、600nmの培養物の光学密度を測定することによって決定した。
【0325】
Fab’の力価を、上記例7に記載した通り、タンパク質G HPLCによって決定した。
【0326】
図9は、抗TNF Fab’及び組換えDsbC発現性大腸菌系統MXE008及びMXE009の、発酵の比較上の増殖プロファイルを示す。
【0327】
図10は、抗TNF Fab’及び組換えDsbC発現性大腸菌系統MXE008及びMXE009の、発酵の比較上のペリプラズマのFab’力価を示す。
【0328】
本発明を、好ましい実施形態を参照にして特に示し、記載してきたが、当業者であれば、添付の特許請求の範囲によって規定される通り、本発明の範囲から逸脱することなしに形態及び詳細において様々な変更を行うことができることが理解されよう。
【特許請求の範囲】
【請求項1】
a.DsbCをコードする組換えポリヌクレオチドを含んでおり、
b.野生型細胞に比べて低減したTspタンパク質活性を有する
ことを特徴とする、組換えグラム陰性細菌細胞。
【請求項2】
変異体sprタンパク質をコードする変異spr遺伝子をさらに含んでいる、請求項1に記載の細胞。
【請求項3】
変異spr遺伝子が、H157、N31、R62、I70、Q73、C94、S95、V98、Q99、R100、L108、Y115、D133、V135、L136、G140、R144、H145、及びG147から選択される1つ又は複数のアミノ酸に変異を有するsprタンパク質をコードする、請求項2に記載の細胞。
【請求項4】
変異体spr遺伝子が、H157A、N31Y、R62C、I70T、Q73R、C94A、S95F、V98E、Q99P、R100G、L108S、Y115F、D133A、V135D、V135G、L136P、G140C、R144C、H145A、及びG147Cから選択される1つ又は複数の変異を有するsprタンパク質をコードする、請求項3に記載の細胞。
【請求項5】
1つ又は複数のsprタンパク質の変異が、S95F、V98E、Y115F、D133A、V135D、V135G、及びG147Cから選択される、請求項4に記載の細胞。
【請求項6】
変異体spr遺伝子が、S95F及びY115Fの変異を有するsprタンパク質をコードする、請求項5に記載の細胞。
【請求項7】
変異体spr遺伝子が、C94A、D133A、H145A、及びH157Aから選択される変異を有するsprタンパク質をコードする、請求項4に記載の細胞。
【請求項8】
以下の変異遺伝子:
a.シャペロン活性及び低減したプロテアーゼ活性を有するDegPタンパク質をコードする変異DegP遺伝子、
b.低減したプロテアーゼ活性を有するProtease IIIタンパク質をコードする、又はノックアウト変異ptr遺伝子である、変異ptr遺伝子、
c.低減したプロテアーゼ活性を有するOmpTタンパク質をコードする、又はノックアウト変異OmpT遺伝子である、変異OmpT遺伝子
の1つ又は複数をさらに含んでいる、請求項1から7までのいずれか一項に記載の細胞。
【請求項9】
低減したプロテアーゼ活性を有するTspタンパク質をコードする変異Tsp遺伝子を含んでいる、又はノックアウト変異Tsp遺伝子である、請求項1から8までのいずれか一項に記載の細胞。
【請求項10】
細胞のゲノムが、変異Tsp遺伝子及び任意選択によって変異spr遺伝子以外は野生型細菌細胞に対して同質遺伝子である、請求項9に記載の細胞。
【請求項11】
遺伝子開始コドン、並びに/或いは遺伝子開始コドンの下流及び遺伝子終止コドンの上流に位置する1つ又は複数の終止コドンに対する変異を含んでいるノックアウト変異遺伝子を有する、請求項9又は請求項10に記載の細胞。
【請求項12】
ノックアウト変異Tsp遺伝子が、遺伝子開始コドンに対するミスセンス変異によって作り出された制限マーカー部位、及び任意選択によって1つ又は複数のさらなる点変異を含んでいる、請求項11に記載の細胞。
【請求項13】
ノックアウト変異Tsp遺伝子が配列番号3を含んでいる、請求項12に記載の細胞。
【請求項14】
大腸菌(E.coli)である、請求項1から13までのいずれか一項に記載の細胞。
【請求項15】
対象のタンパク質をコードするポリヌクレオチド配列を含んでいる、請求項1から14までのいずれか一項に記載の細胞。
【請求項16】
DsbCをコードする組換えポリヌクレオチド及び対象のタンパク質をコードするポリヌクレオチド配列を含むベクターを含んでいる、請求項15に記載の細胞。
【請求項17】
ベクターが、DsbCをコードする組換えポリヌクレオチドの発現を制御するプロモーター、及び対象のタンパク質をコードするポリヌクレオチド配列を含んでいる、請求項16に記載の細胞。
【請求項18】
対象のタンパク質が、抗体又は抗体の抗原結合性フラグメントである、請求項15から17までのいずれか一項に記載の細胞。
【請求項19】
抗体又は抗体の抗原結合性フラグメントがTNFに特異的である、請求項18に記載の細胞。
【請求項20】
a.請求項1から19までのいずれか一項に記載の組換えグラム陰性細菌細胞を、培養培地中、対象の組換えタンパク質及びDsbCをコードする組換えポリヌクレオチドを発現するのに効果的な条件下で培養するステップと、
b.対象のタンパク質を、組換えグラム陰性細菌細胞のペリプラズム及び/又は培養培地から回収するステップと
を含む、対象のタンパク質を生成するための方法。
【請求項21】
対象のタンパク質をコードするポリヌクレオチド配列、及びDsbCをコードする組換えポリヌクレオチドの発現が、培養培地に誘導因子を加えることによって誘発される、請求項20に記載の方法。
【請求項22】
対象のタンパク質をDsbCから分離するステップをさらに含む、請求項20又は請求項21に記載の方法。
【請求項1】
a.DsbCをコードする組換えポリヌクレオチドを含んでおり、
b.野生型細胞に比べて低減したTspタンパク質活性を有する
ことを特徴とする、組換えグラム陰性細菌細胞。
【請求項2】
変異体sprタンパク質をコードする変異spr遺伝子をさらに含んでいる、請求項1に記載の細胞。
【請求項3】
変異spr遺伝子が、H157、N31、R62、I70、Q73、C94、S95、V98、Q99、R100、L108、Y115、D133、V135、L136、G140、R144、H145、及びG147から選択される1つ又は複数のアミノ酸に変異を有するsprタンパク質をコードする、請求項2に記載の細胞。
【請求項4】
変異体spr遺伝子が、H157A、N31Y、R62C、I70T、Q73R、C94A、S95F、V98E、Q99P、R100G、L108S、Y115F、D133A、V135D、V135G、L136P、G140C、R144C、H145A、及びG147Cから選択される1つ又は複数の変異を有するsprタンパク質をコードする、請求項3に記載の細胞。
【請求項5】
1つ又は複数のsprタンパク質の変異が、S95F、V98E、Y115F、D133A、V135D、V135G、及びG147Cから選択される、請求項4に記載の細胞。
【請求項6】
変異体spr遺伝子が、S95F及びY115Fの変異を有するsprタンパク質をコードする、請求項5に記載の細胞。
【請求項7】
変異体spr遺伝子が、C94A、D133A、H145A、及びH157Aから選択される変異を有するsprタンパク質をコードする、請求項4に記載の細胞。
【請求項8】
以下の変異遺伝子:
a.シャペロン活性及び低減したプロテアーゼ活性を有するDegPタンパク質をコードする変異DegP遺伝子、
b.低減したプロテアーゼ活性を有するProtease IIIタンパク質をコードする、又はノックアウト変異ptr遺伝子である、変異ptr遺伝子、
c.低減したプロテアーゼ活性を有するOmpTタンパク質をコードする、又はノックアウト変異OmpT遺伝子である、変異OmpT遺伝子
の1つ又は複数をさらに含んでいる、請求項1から7までのいずれか一項に記載の細胞。
【請求項9】
低減したプロテアーゼ活性を有するTspタンパク質をコードする変異Tsp遺伝子を含んでいる、又はノックアウト変異Tsp遺伝子である、請求項1から8までのいずれか一項に記載の細胞。
【請求項10】
細胞のゲノムが、変異Tsp遺伝子及び任意選択によって変異spr遺伝子以外は野生型細菌細胞に対して同質遺伝子である、請求項9に記載の細胞。
【請求項11】
遺伝子開始コドン、並びに/或いは遺伝子開始コドンの下流及び遺伝子終止コドンの上流に位置する1つ又は複数の終止コドンに対する変異を含んでいるノックアウト変異遺伝子を有する、請求項9又は請求項10に記載の細胞。
【請求項12】
ノックアウト変異Tsp遺伝子が、遺伝子開始コドンに対するミスセンス変異によって作り出された制限マーカー部位、及び任意選択によって1つ又は複数のさらなる点変異を含んでいる、請求項11に記載の細胞。
【請求項13】
ノックアウト変異Tsp遺伝子が配列番号3を含んでいる、請求項12に記載の細胞。
【請求項14】
大腸菌(E.coli)である、請求項1から13までのいずれか一項に記載の細胞。
【請求項15】
対象のタンパク質をコードするポリヌクレオチド配列を含んでいる、請求項1から14までのいずれか一項に記載の細胞。
【請求項16】
DsbCをコードする組換えポリヌクレオチド及び対象のタンパク質をコードするポリヌクレオチド配列を含むベクターを含んでいる、請求項15に記載の細胞。
【請求項17】
ベクターが、DsbCをコードする組換えポリヌクレオチドの発現を制御するプロモーター、及び対象のタンパク質をコードするポリヌクレオチド配列を含んでいる、請求項16に記載の細胞。
【請求項18】
対象のタンパク質が、抗体又は抗体の抗原結合性フラグメントである、請求項15から17までのいずれか一項に記載の細胞。
【請求項19】
抗体又は抗体の抗原結合性フラグメントがTNFに特異的である、請求項18に記載の細胞。
【請求項20】
a.請求項1から19までのいずれか一項に記載の組換えグラム陰性細菌細胞を、培養培地中、対象の組換えタンパク質及びDsbCをコードする組換えポリヌクレオチドを発現するのに効果的な条件下で培養するステップと、
b.対象のタンパク質を、組換えグラム陰性細菌細胞のペリプラズム及び/又は培養培地から回収するステップと
を含む、対象のタンパク質を生成するための方法。
【請求項21】
対象のタンパク質をコードするポリヌクレオチド配列、及びDsbCをコードする組換えポリヌクレオチドの発現が、培養培地に誘導因子を加えることによって誘発される、請求項20に記載の方法。
【請求項22】
対象のタンパク質をDsbCから分離するステップをさらに含む、請求項20又は請求項21に記載の方法。
【図1】

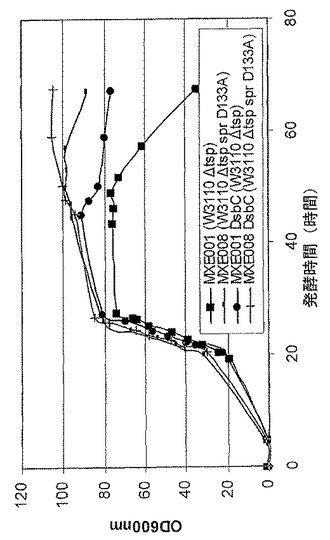
【図2】
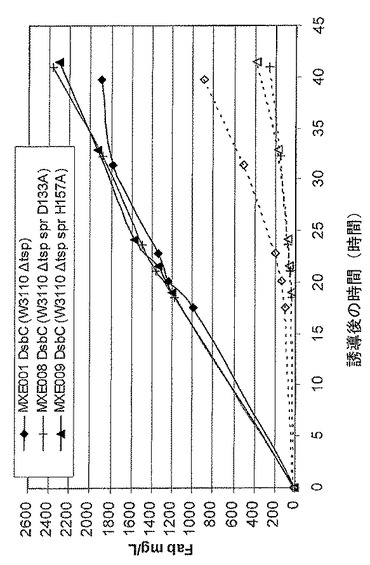
【図3】
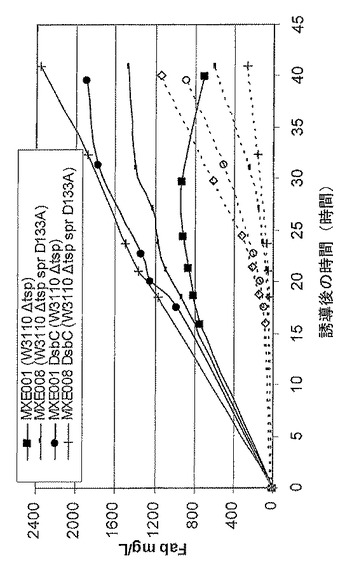
【図4】
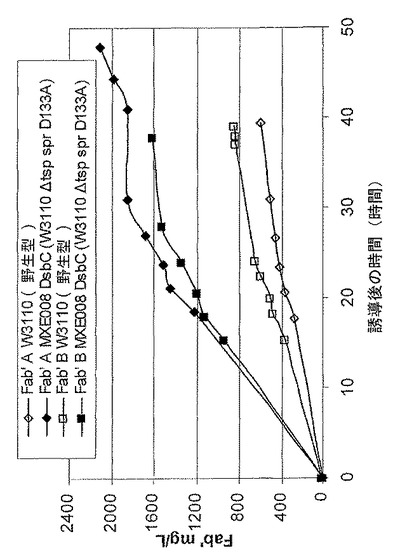
【図5a】

【図5b】
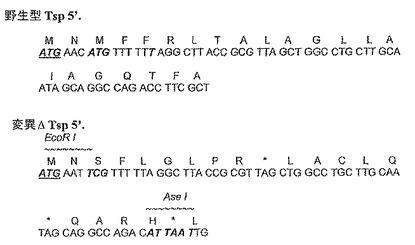
【図5c】
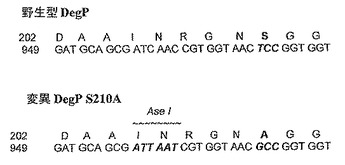
【図6】
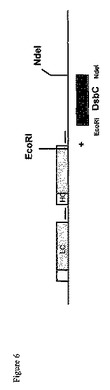
【図7】
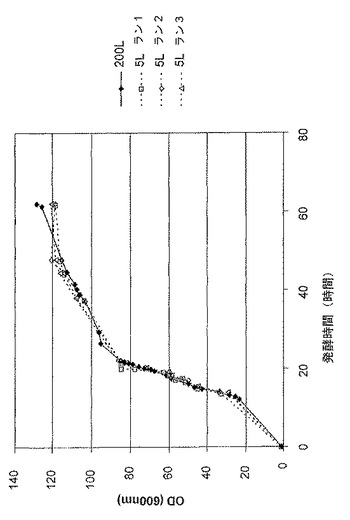
【図8】
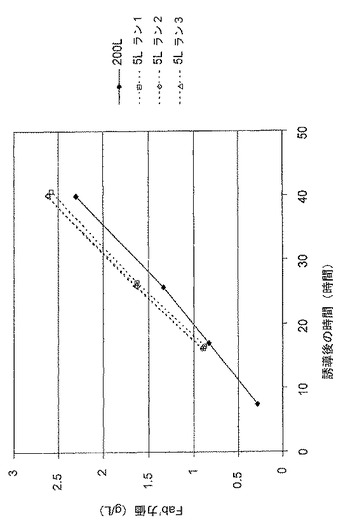
【図9】
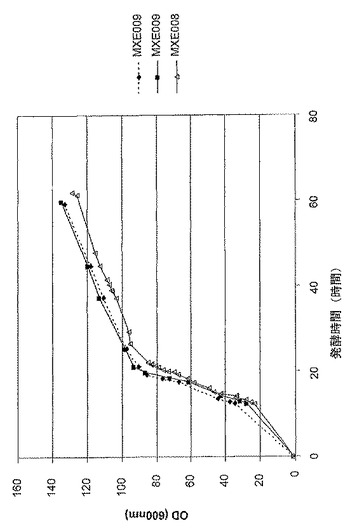
【図10】
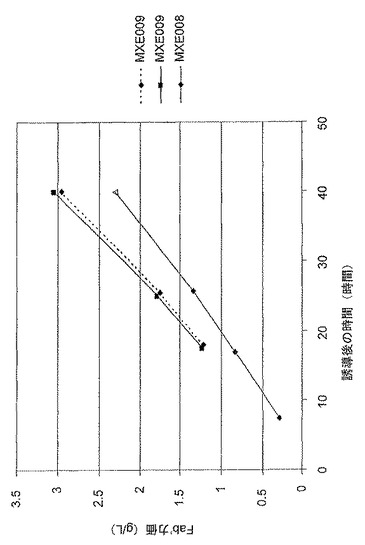
【図2】
【図3】
【図4】
【図5a】
【図5b】
【図5c】
【図6】
【図7】
【図8】
【図9】
【図10】
【公表番号】特表2013−516966(P2013−516966A)
【公表日】平成25年5月16日(2013.5.16)
【国際特許分類】
【出願番号】特願2012−548441(P2012−548441)
【出願日】平成23年1月13日(2011.1.13)
【国際出願番号】PCT/EP2011/050416
【国際公開番号】WO2011/086139
【国際公開日】平成23年7月21日(2011.7.21)
【出願人】(507073918)ユセベ ファルマ ソシエテ アノニム (70)
【Fターム(参考)】
【公表日】平成25年5月16日(2013.5.16)
【国際特許分類】
【出願日】平成23年1月13日(2011.1.13)
【国際出願番号】PCT/EP2011/050416
【国際公開番号】WO2011/086139
【国際公開日】平成23年7月21日(2011.7.21)
【出願人】(507073918)ユセベ ファルマ ソシエテ アノニム (70)
【Fターム(参考)】
[ Back to top ]