
組織中の無酸素的運動能力を高めるための方法及び組成物
【課題】筋肉及びその他の組織の無酸素的運動能力を高めるための方法及び組成物を提供する。
【解決手段】組織中におけるβ-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のβ-アラニンを血液または血漿に供給し、組織を前記血液または血漿にさらし、それによりβ-アラニルヒスチジンの濃度を前記組織中で増加させる、ことを含んでなる、組織中のヒドロニウムイオン濃度を制御する方法、ならびに組織の無酸素的運動能力を高めるための組成物。
【解決手段】組織中におけるβ-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のβ-アラニンを血液または血漿に供給し、組織を前記血液または血漿にさらし、それによりβ-アラニルヒスチジンの濃度を前記組織中で増加させる、ことを含んでなる、組織中のヒドロニウムイオン濃度を制御する方法、ならびに組織の無酸素的運動能力を高めるための組成物。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、筋肉及びその他の組織の無酸素的運動能力を高めるための方法及び組成物に関する。
【背景技術】
【0002】
発明の背景
天然食品の補助物質(supplement)は、典型的には現代のヒト及び動物における食餌の栄養素のレベル低下を補償するように設計される。特に、有用な補助物質は摂取したとき組織の機能を高めるものである。とりわけ、特定の種類の動物の食餌(その通常の食餌は肉や動物性食品のみから利用できる栄養素が不足する可能性がある)を補給することは重要である(例えば、ヒト菜食主義者やその他の動物は草食性食品を摂取する)。
【0003】
例えば、スポーツ及び競技の社会においては、レジャーや労働の目的で身体的頑強さを促進し強化する補助物質のような、特定的に運動能力を改善する自然食品補助物質の重要性が高まっている。別の例としては、無酸素的(例えば乳酸生成)ストレスが加齢とともに経験し得る疲労および不快感の発生を引き起し得る。また無酸素的ストレスは、筋肉内圧力の増加により(例えばロッククライミング、フリーダイビング、シンクロナイズドスイミング等の間に)局所的な循環が部分的あるいは完全に閉塞された場合の長時間の最大下の等尺運動からも起こり得る。過剰な乳酸の生成は細胞内環境の酸性化を起こし得る。
【0004】
クレアチン(すなわちN-(アミノイミノメチル)-N-グリシン、N-アミジノサルコシン、N-メチル-N-グアニルグリシン、またはメチルグリコシアミン)は、骨格筋及び高い可変的なエネルギー要求の能力を特徴とするその他の「励起可能な」組織(例えば平滑筋、心筋、あるいは精子)において多量に見られる。クレアチンは細胞内のエネルギー生成生化学的経路においてホスホリルクレアチンに変換される。哺乳動物の骨格筋においては、典型的なクレアチンの合計含量(すなわちクレアチンとホスホリルクレアチン)は、新鮮な筋肉1キログラムあたり25mmol未満から50mmol(すなわち筋肉1キログラムあたり3.2〜6.5グラム)まで変化し得る。
【0005】
クレアチンは肝臓で生成され、活性輸送系により例えば筋肉のような組織に取り込まれる。体内でのクレアチン合成は食肉中に存在するクレアチンの摂取(例えば平均的な肉食のヒトにおいては1日に体重1キログラムあたり5〜10ミリグラム、菜食主義者の食事においてはほぼゼロである)によっても増加される。
【0006】
集中的な運動の継続、あるいは局所的な低酸素状態での運動の継続の間に、解糖中に形成されたヒドロニウムイオンの蓄積および乳酸の蓄積(嫌気代謝)が細胞内のpHを著しく低下させることがある。低下したpHはクレアチン-ホスホリルクレアチン系の機能を低下させ得る。細胞内pHの低下は、例えば筋繊維中の収縮タンパク質の機能のような、細胞内のその他の機能に影響し得る。
【0007】
β-アラニンとヒスチジンのジペプチド及びそのメチル化類似体には、カルノシン(β-アラニル-L-ヒスチジン)、アンセリン(β-アラニル-L-1-メチルヒスチジン)、あるいはバレニン(β-アラニル-L-3-メチルヒスチジン)が含まれる。これらのジペプチドはヒトやその他の脊椎動物の筋肉中に存在する。カルノシンは例えばヒト及びウマ科の動物の筋肉中にかなりの量が見られる。アンセリン及びカルノシンは例えばイヌ、ラクダ、及び多くの鳥類の種の筋肉中に見られる。アンセリンは多くの魚類において見られる主要なβ-アラニルヒスチジンジペプチドである。バレニンは、水生哺乳動物及び爬虫類のいくつかの種に見られる主要なβ-アラニルヒスチジンジペプチドである。ヒト、ウマ及びラクダでは、β-アラニルヒスチジンジペプチドの最大濃度は、激しい運動の間に集中的に使用される急速収縮性解糖筋繊維(IIA及びIIB型)中に見られる。より低い濃度は、酸化的緩徐収縮性筋繊維(I型)中に見られる。例えば、Dunnett,M.& Harris,R.C.Equine Vet.J.Suppl.18,214-217(1995)を参照のこと。カルノシンは種々の筋繊維型においてヒドロニウムイオン緩衝能に貢献していると推定され、ウマII型繊維においては全体の最高50%である。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】Dunnett,M.& Harris,R.C.Equine Vet.J.Suppl.18,214-217(1995)
【発明の概要】
【課題を解決するための手段】
【0009】
発明の概要
一般に、本発明は、筋肉及びその他の組織の無酸素的運動能力を高めるための方法及び組成物に関する。前記方法は、体内の組織中でのクレアチン及びβ-アラニルヒスチジンジペプチド、あるいはβ-アラニン及びL-ヒスチジン類似体の同時蓄積を含む。前記方法は組成物を体内に摂取させること、あるいは注入することを含む。前記組成物は、ヒトあるいは動物の筋肉において、β-アラニルヒスチジンジペプチドの合成及び蓄積のためにクレアチン及び前駆体の利用可能性及び吸収を増加させることができる化合物の混合物である。前記組成物は、それが体内に導入されたときに、ヒトあるいは動物体内におけるβ-アラニルヒスチジンジペプチドの合成および蓄積を引き出すものである。
【0010】
前記組成物は、クレアチン及びβ-アラニン、クレアチン、β-アラニン及びL-ヒスチジン、またはクレアチン及びβ-アラニンもしくはL-ヒスチジンの活性誘導体の混合物を含む。β-アラニンまたはL-ヒスチジンのそれぞれは個々のアミノ酸であってもよく、あるいはジペプチド、オリゴペプチドまたはポリペプチドの成分であってもよい。β-アラニン及びL-ヒスチジンは活性誘導体であってもよい。活性誘導体は、体内でその物質と同じあるいは類似の方法で機能する物質から誘導される化合物もしくはその前駆体、あるいはそのような物質にプロセシングされて体内に配置される化合物である。例としては、エステル及びアミドが挙げられる。
【0011】
一つの面において、本発明は、組織中のヒドロニウムイオン濃度を制御する方法に関する。前記方法は、組織中のβ-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のβ-アラニンを血液あるいは血漿に供給し、組織をその血液あるいは血漿にさらし、それによりβ-アラニルヒスチジンの濃度を前記組織中で増加させるステップを含む。前記方法は、β-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のL-ヒスチジンを血液あるいは血漿に供給するステップを含むことができる。
【0012】
別の面においては、本発明は、組織の無酸素的運動能力を高める方法に関する。前記方法は、組織中のβ-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のβ-アラニンを血液あるいは血漿に供給し、組織中のβ-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のL-ヒスチジンを血液あるいは血漿に供給し、そして組織をその血液あるいは血漿にさらすステップを含む。β-アラニルヒスチジンの濃度が前記組織中で増加する。
【0013】
ある実施態様においては、前記方法は組織中のクレアチンの濃度を増加させるステップを含む。この増加ステップは、組織中のクレアチン濃度を増加させるのに有効な量のクレアチンを血液あるいは血漿に供給する(例えば前記量のクレアチンを血液あるいは血漿に供給することによる)ことを含み得る。
【0014】
前記方法の供給ステップは、前記量のβ-アラニンまたは前記量のβ-アラニン及びL-ヒスチジンを含む組成物の摂取あるいは注入(例えば注射)、または摂取と注入の組合せを含み得る。
【0015】
前記方法は、血液あるいは血漿中のインスリン濃度を増加させることを含み得る。インスリンの濃度は、例えばインスリンの注射により増加させることができる。
【0016】
前記組織は骨格筋であり得る。
【0017】
別の面においては、本発明は、β-アラニンを含むペプチド源、約39〜約99重量%の炭水化物、及び約60重量%までの水から実質的になる組成物に関する。前記組成物は約1〜約20重量%のβ-アラニンを含む。前記ペプチド源はL-ヒスチジンを含むことができる。前記組成物は約1〜約20重量%のL-ヒスチジンを含むことができる。
【0018】
前記炭水化物は単純な炭水化物(例えばグルコース)であってよい。別の面においては、本発明は、β-アラニンを含むペプチド源、約1〜約98重量%のクレアチン源、及び約97重量%までの水から実質的になる組成物に関する。前記組成物は約1〜約98重量%のβ-アラニンを含む。前記ペプチド源はL-ヒスチジンを含むことができ、前記組成物は約1〜約98重量%のL-ヒスチジンを含む。
【0019】
前記ペプチド源はアミノ酸、ジペプチド、オリゴペプチド、ポリペプチド、あるいはそれらの活性誘導体の混合物であり得る。
【0020】
前記組成物は食品補助物質とすることができる。前記クレアチン源はクレアチン一水和物であってよい。
【0021】
血液または血漿中の成分の濃度は、血漿濃度の増加を起こすように機能できる物質の注入(例えば注射)あるいは摂取により増加させることができる。前記組成物は1日あたり約10グラム〜約800グラムの間の量で摂取させることができる。この量は1日1回で、あるいは複数回に分けて投与することができる。
【0022】
クレアチン及びβ-アラニルヒスチジンジペプチドの筋内含量が増加することにより、無酸素的運動に伴うヒドロニウムイオン生成の増加に対する細胞の許容度が増加し、疲労が発生するまでの運動の持続期間を増加させることができる。
【0023】
前記組成物及び方法は、調理あるいは加工中のβ-アラニン、L-ヒスチジンあるいはクレアチンの分解または浸出(leaching)によるこれらの成分の損失を補充するのに有用であり得る。また前記組成物及び方法は、菜食主義者の食事におけるこれらの成分の欠損を補給するのにも有用であり得る。
【0024】
前記方法及び組成物は、例えば、スポーツマン、競技者、ボディビルダー、シンクロナイズドスイミングのスイマー、兵士、老年者、競争馬、作業犬及びレース犬、競技用の鳥等においてβ-アラニルヒスチジンジペプチドを増加させるのに使用し、筋肉疲労の発生を防止し、遅延させることができる。
【0025】
本発明のその他の利点及び特徴は詳細な説明及び請求の範囲から明らかになるであろう。
【図面の簡単な説明】
【0026】
図面の簡単な説明
【図1】β-アラニン及びL-ヒスチジンを30日間にわたって(それぞれ体重1キログラムあたり100ミリグラム及び体重1キログラムあたり12.5ミリグラム、1日3回)給餌した後の、5頭のウマの給餌前及び給餌後2時間おきの血漿中のβ-アラニンの濃度の変化を示すグラフである。
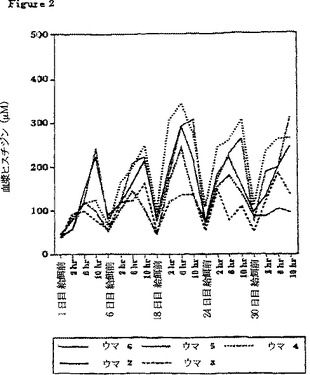
【図2】β-アラニン及びL-ヒスチジンを30日間にわたって(それぞれ体重1キログラムあたり100ミリグラム及び体重1キログラムあたり12.5ミリグラム、1日3回)給餌した後の、5頭のウマの給餌前及び給餌後2時間おきの血漿中のL-ヒスチジンの濃度の変化を示すグラフである。
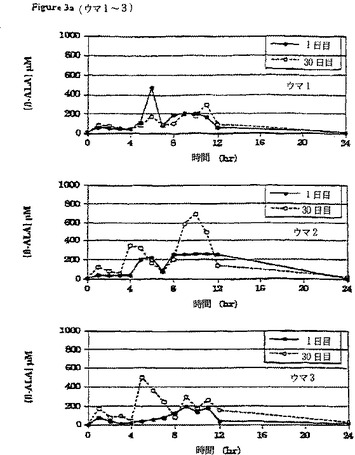
【図3a】β-アラニン及びL-ヒスチジンの30日間の食餌補給(それぞれ体重1キログラムあたり100ミリグラム及び体重1キログラムあたり12.5ミリグラム、1日3回)の初日及び最終日の、6頭のウマのうちのウマ1〜3の給餌前及び給餌後時間毎の血漿中のβ-アラニンの濃度の変化を比較して示すグラフである。
【図3b】β-アラニン及びL-ヒスチジンの30日間の食餌補給(それぞれ体重1キログラムあたり100ミリグラム及び体重1キログラムあたり12.5ミリグラム、1日3回)の初日及び最終日の、6頭のウマのうちのウマ4〜6の給餌前及び給餌後時間毎の血漿中のβ-アラニンの濃度の変化を比較して示すグラフである。
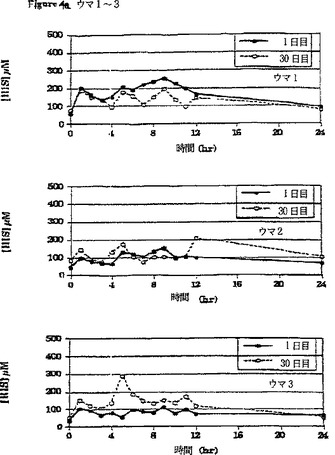
【図4a】β-アラニン及びL-ヒスチジンの30日間の食餌補給(それぞれ体重1キログラムあたり100ミリグラム及び体重1キログラムあたり12.5ミリグラム、1日3回)の初日及び最終日の、6頭のウマのうちのウマ1〜3の給餌前及び給餌後時間毎の血漿中のL-ヒスチジンの濃度の変化を比較して表すグラフである。
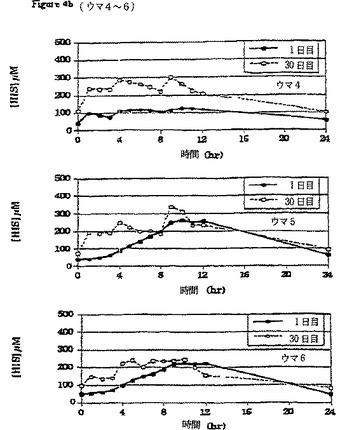
【図4b】β-アラニン及びL-ヒスチジンの30日間の食餌補給(それぞれ体重1キログラムあたり100ミリグラム及び体重1キログラムあたり12.5ミリグラム、1日3回)の初日及び最終日の、6頭のウマのうちのウマ4〜6の給餌前及び給餌後時間毎の血漿中のL-ヒスチジンの濃度の変化を比較して表すグラフである。
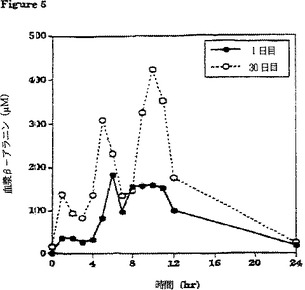
【図5】β-アラニン及びL-ヒスチジンの30日間の食餌補給(それぞれ体重1キログラムあたり100ミリグラム及び体重1キログラムあたり12.5ミリグラム、1日3回)の初日及び最終日の、給餌前及び給餌後時間毎のウマ血漿中のβ-アラニンの平均濃度(n=6)の変化を比較して示すグラフである。
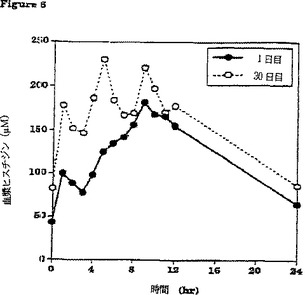
【図6】β-アラニン及びL-ヒスチジンの30日間の食餌補給(それぞれ体重1キログラムあたり100ミリグラム及び体重1キログラムあたり12.5ミリグラム、1日3回)の初日及び最終日の、給餌前及び給餌後時間毎のウマ血漿中のL-ヒスチジンの平均濃度(n=6)の変化を比較して示すグラフである。
【図7】6頭の純血種ウマのII型骨格筋繊維におけるカルノシン濃度の増加(IIA及びIIB型繊維の合計の平均)と、補給の1日目と30日目の間の血漿β-アラニン濃度-時間曲線下の当日の最初の12時間にわたる面積(AUC(0-12hr))の増加と、の相関関係を示すグラフである。
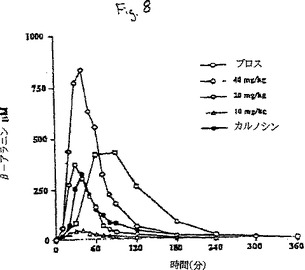
【図8】β-アラニン、ブロス、あるいはカルノシンの被験者への投与の平均の結果を示すグラフである。
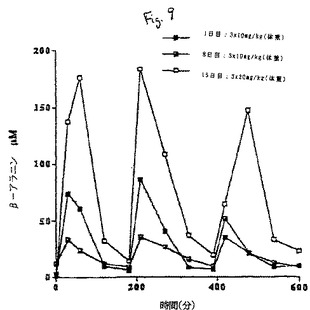
【図9】9時間の処置にわたる血漿β-アラニンの平均変化を示すグラフである。
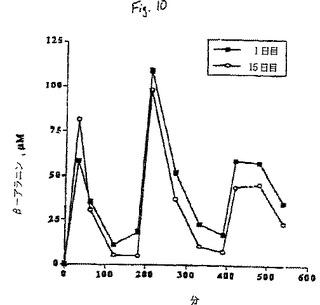
【図10】体重1キログラムあたり10ミリグラムのβ-アラニンの経口摂取後の9時間にわたる血漿β-アラニンの平均変化を示すグラフである。
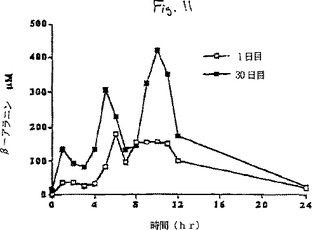
【図11】処置期間の1日目及び30日目の24時間にわたる平均(n=6)血漿β-アラニン濃度を示すグラフである。
【発明を実施するための形態】
【0027】
好ましい実施態様の説明
カルノシン、アンセリン及びバレニン等のβ-アラニルヒスチジンジペプチドは約6.8〜7.1の間のpKa値を有し、筋収縮及び疲労発生中の細胞内pHの恒常性の制御に関与している。ヒドロニウムイオンの緩衝に関与しているその他の物質、例えばタンパク質中のアミノ酸残基、無機及び有機のリン酸及び炭酸水素塩等の含量は、その他の細胞機能へのそれらの関与により拘束される。β-アラニルヒスチジンジペプチドは、細胞中にpH感受性ヒスチジン残基を蓄積する有効な方法を提供する。筋肉β-アラニルヒスチジンジペプチド濃度の変化は個々の競技者の無酸素的運動能力に影響する。
【0028】
β-アラニルヒスチジンジペプチドはβ-アラニンとL-ヒスチジンから体内で合成される。これらの前駆体は体内で生成されたり、摂取されたβ-アラニルヒスチジンジペプチドの分解からのものを含め、食餌から利用できるようにされる。
【0029】
体内のβ-アラニンは筋肉等の組織に輸送される。典型的な給餌状態では、ヒト及びウマ血漿中のL-ヒスチジンの濃度と比較してβ-アラニンの濃度は低い。これらの濃度は、カルノシン合成酵素であるカルノシンシンテターゼの、ミカエリス-メンテン定数(Km)により決定される、その基質に対する親和性と関連付けて考えなければならない。ヒスチジンについてのKmは約16.8μMである。β-アラニンについてのKmは約1000〜2300μMの間である。β-アラニンに対するカルノシンシンテターゼの低い親和性、及び筋肉中のβ-アラニンの低い濃度は、筋肉中のβ-アラニンの濃度によりβ-アラニルヒスチジンジペプチドの合成が制限されていることを示す。
【0030】
筋肉内のβ-アラニルヒスチジンジペプチド量を増加させると、筋肉の性能及びその筋肉により達成される運動量に好都合に影響する。従って、β-アラニルヒスチジンジペプチドの合成及び蓄積がヒトあるいは動物体内の組織において増加される。
【0031】
ヒトあるいは動物体内におけるβ-アラニルヒスチジンジペプチドの合成及び蓄積は、β-アラニンの血液もしくは血漿濃度の増加、β-アラニン及びクレアチンの血液もしくは血漿濃度の増加、あるいはβ-アラニン、L-ヒスチジン及びクレアチンの血液もしくは血漿濃度の増加により、体内のクレアチン含量の増加とともに増加させることができる。ジペプチドの増加は、β-アラニン濃度の増加と同時に起こり得る。
【0032】
β-アラニン、L-ヒスチジン及びクレアチンの血漿濃度は、β-アラニン、L-ヒスチジン及びクレアチンあるいはそれらの活性誘導体の摂取または注入により増加させることができる。前記組成物は、経口、経腸、あるいは非経口的に投与することができる。β-アラニン及びクレアチン、あるいはβ-アラニン、L-ヒスチジン及びクレアチンは好ましくは経口的に摂取される。
【0033】
前記組成物は炭水化物(例えば単純な炭水化物)、インスリン、またはインスリンの生成を刺激する物質を含んでもよい。
【0034】
前記組成物は食品の補助物質として摂取させることができる。好ましくは前記組成物は1日に1回以上投与される。β-アラニンの投与量は体重1キログラムあたり約5ミリグラムから約200ミリグラムの間とすることができる。クレアチン(例えばクレアチン一水和物)の投与量は体重1キログラムあたり約5ミリグラムから200ミリグラムの間とすることができる。L-ヒスチジンの投与量は体重1キログラムあたり約1ミリグラムから100ミリグラムの間とすることができる。
【0035】
単純な炭水化物(例えばグルコース)の投与量は体重1キログラムあたり約0.5グラムから2.0グラムの間とすることができる。
【0036】
80キログラムのヒトにおいては、1日あたりの適当な投与量は、0.4グラム〜16.0グラムのβ-アラニン、0.4グラム〜16.0グラムのクレアチン一水和物、0.08グラム〜8.0グラムのL-ヒスチジン、あるいは40グラム〜160グラムのグルコースもしくはその他の単純な炭水化物とすることができる。前記組成物は摂取するための固体もしくは液体の剤形、あるいは懸濁体の剤形、あるいは体内に注入するための液体もしくは懸濁体の剤形とすることができる。前記組成物は、1日あたり2グラム〜1000グラムの間の量(例えば10グラム〜800グラムの間の量)でヒトに摂取させるが、これは1日のうちに小分けして摂取させることができる。動物における1日の摂取量は体重に関して調整されるだろう。
【0037】
ヒト及び動物について、前記組成物は、(a) 1重量%〜99重量%のβ-アラニン、 1重量%〜99重量%のクレアチン一水和物、及び 0重量%〜98重量%の水;(b) 1重量%〜98重量%のβ-アラニン、 1重量%〜98重量%のL-ヒスチジン、 1重量%〜98重量%のクレアチン一水和物、及び0重量%〜97重量%の水;(c) 1重量%〜20重量%のβ-アラニン、 39重量%〜99重量%のグルコースまたはその他の単純な炭水化物、及び 0重量%〜60重量%の水、あるいは(d) 1重量%〜20重量%のβ-アラニン、 1重量%〜20重量%のL-ヒスチジン、 39重量%〜99重量%のグルコースまたはその他の単純な炭水化物、及び 0重量%〜60重量%の水、とすることができる。
【実施例】
【0038】
以下は、筋肉及びその他の組織の無酸素的運動能力を高めるための方法及び組成物の方法の具体例である。
【0039】
実施例1
標準飼料への1日複数回のβ-アラニンとL-ヒスチジンの補給が純血種のウマのI、IIAおよびIIB型の骨格筋繊維中のカルノシン濃度に及ぼす効果を評価した。普通の健康状態の4〜9歳の実験用純血種ウマ(若い雌ウマ3頭と去勢したウマ3頭)に、補給期間の開始に先立って1ヵ月(30日)(補給前期間)の給餌コンディショニングをおこなった。給餌コンディショニング期間中、各ウマに、複合および単純炭水化物の供給源として1キログラムの成形飼料(Spillers競争馬キューブ)と1キログラムの浸潰したサトウダイコンパルプを含む飼料を1日3回(それぞれ08:30、12:30および16:30)与えた。浸潰した干し草(乾燥重量で3キログラム)も1日2回(09:00および17:00に)与えた。水は自由に飲めるようにした。
【0040】
補給期間中は同一の給餌レジメを実施した。しかし、硬質飼料を与える際にはβ-アラニンとL-ヒスチジン(遊離塩基)を補給した。β-アラニンとL-ヒスチジンを標準飼料に直接混合した。β-アラニンとL-ヒスチジンの個々の投与量は体重に応じて算出した。β-アラニンは体重1キログラムにつき100ミリグラムを、L-ヒスチジンは体重1キログラムにつき12.5ミリグラムを投与した。餌の補給はこのプロトコールの1日目に開始し、30日目に終わらせた。1、6、18、24および30日目に血液サンプル(5ミリリットル)を採取し、ヘパリンを添加した。1日目と30日目には、1回目の給餌前と合計12時間にわたり1時間おきに血液サンプルを採取した。残りの3日の採血日には、1回目の給餌前とその後の各給餌の2時間後に血液を集めた。補給開始の前日(0日目)には、皮膚の局所麻酔後にBergstrom-Stille経皮生検針を使って各ウマの右側の中間臀部筋(m.gluteusmedius)から筋肉の生検材料を採取した。その後の筋生検材料はもとの採取部位に可能なかぎり接近させて補給期間の終了直後(31日目)に採取した。ウマの臨床的監視を毎日おこなった。この監視は肉眼による検査と体重測定、直腸温度の1日2回の測定、それに臨床生化学および血液学のための週1度の採血を含んでいた。実験期間中、ウマには正式な訓練や運動を施さなかったが、毎日1時間自由に運動させた。
【0041】
凍結乾燥した筋生検材料から切り裂いた個々の筋繊維の断片は、KaiserおよびBrcok,Arch.Neurol.,23:369-379(1970)に記載される方法の変法により、pH4.50でプレインキュベートした後にpH9.6でミオシンATPアーゼ活性を組織化学的に染色することで、I、IIAまたはIIB型のいずれかであると特性づけられた。
【0042】
ヘパリン添加血漿サンプルを抽出し、高性能液体クロマトグラフィー(HPLC)でβ-アラニンとL-ヒスチジンの濃度を分析した。計量した個々の筋繊維を抽出し、DunnettおよびHarris,「筋肉および個々の筋繊維中のイミダゾールジペプチド、ヒスチジン、1-メチルヒスチジン、3-メチルヒスチジンの高性能液体クロマトグラフィー測定」,J.Chromatogr.B.Biomed.Appl.,688:47-55(1997)に記載される方法に従ってHPLCでカルノシンを分析した。
【0043】
補給前後の繊維型に含まれるカルノシン濃度の差は、一方向分散分析(ANOVA)を用いてウマにおいて決定した。差が認められた場合には、多重比較検定(Fisher's PLSD)により有意差を求めた。
【0044】
飼料にβ-アラニンとL-ヒスチジンを添加しても味覚上の問題はまったく生起しなかった。30日の補給期間中、補給餌の有害な生理または行動作用はいずれのウマにも観察されなかった。体重の顕著な変化は記録されず、直腸温度も正常範囲内にとどまった。臨床生化学または血液学においても急性または慢性の変化が認められなかった。補給開始前のウマの血漿中にはβ-アラニンは検出されなかった。用いたアッセイによる血漿中のβ-アラニンの定量の下限は3マイクロモル(μM)であった。補給開始前の6頭のウマの血漿L-ヒスチジン濃度は36.6〜54.4μMであった。
【0045】
すべての採血日にわたる血漿β-アラニンおよびL-ヒスチジン濃度の個々の変化を、6頭のウマのうち5頭についてそれぞれ図1および2に示す。補給期間が増すにつれて、血漿β-アラニンおよびL-ヒスチジンの給餌前濃度は増加する傾向にあった。さらに、30日の補給期間にわたって、補給に対する血漿濃度応答も増加した。この応答はβ-アラニンの場合に、より大きかった。
【0046】
最初の補給の前日と、その後補給期間の初日と最終日の時間ごとの血漿β-アラニンおよびL-ヒスチジン濃度の変化の比較を、6頭の各ウマについてそれぞれ図3aと3b、および図4aと4bに示す。補給期間の初日(1日目)と最終日(30日目)の24時間にわたる経時的血漿β-アラニン濃度の平均(±SD)変化(n=6)を図5に対比して示す。24時間にわたる時間曲線に対する平均血漿β-アラニン濃度下の面積(AUC(0-24hr))は補給30日目に一層大きかった。
【0047】
補給期間の初日(1日目)と最終日(30日目)の24時間にわたる経時的血漿L-ヒスチジン濃度の平均(±SD)変化(n=6)を図6に対比して示す。24時間にわたる時間曲線に対する平均血漿β-アラニン濃度下の面積(AUC(0-24hr))は補給30日目により大きかった。補給初日(1日目)に対比させたときの補給最終日(30日目)の血漿β-アラニンのより大きいAUCは、補給が進行するにつれてウマの胃腸管からのβ-アラニンの吸収が増加することを示唆している。同様の効果が補給期間中の血漿L-ヒスチジン濃度の変化に関しても認められた。β-アラニンおよびL-ヒスチジンの最大血漿濃度はそれぞれの場合に給餌の約1〜2時間後に出現した。
【0048】
6頭のウマから合計397本の骨格筋繊維(補給前 192本;補給後 205本)を切除し、カルノシンについて分析した。6頭の各ウマから得られた補給前および補給後のI、IIAおよびIIB型骨格筋繊維中の平均(±SD)カルノシン濃度(乾燥重量1キログラムあたりのミリモル数(mmol kg-1dw)で表す)を表1に示してあるが、ここでnは分析した個々の筋繊維の数である。β-アラニンおよびL-ヒスチジンの補給30日後に、6頭すべてのウマにおいてIIAおよびIIB型の繊維中の平均カルノシン濃度が増加していた。これらの増加は7つの事例において統計的に有意であった。IIB型骨格筋繊維中の平均カルノシン濃度の増加は6頭のウマのうち5頭において統計的に有意であった。IIA型骨格筋繊維中の平均カルノシン濃度の増加は6頭のウマのうち2頭において統計的に有意であった。
【表1】

* 補給前と有意に異なる、p<0.05
** 補給前と有意に異なる、p<0.01
*** 補給前と有意に異なる、p<0.005
【0049】
6頭のウマから得られたIIAおよびIIB型骨格筋繊維中の平均カルノシン濃度の絶対増加(例えば、mmol kg-1dw)および増加パーセントを表2に示す。
【表2】
【0050】
30日の補給後に筋カルノシン濃度のより大きい増加を示したウマはまた、補給期間の1日目と30日目の間で血漿β-アラニンAUCのより大きい増加を示したことが観察された。図7を参照すると、IIAおよびIIB型骨格筋繊維間で平均した平均カルノシン濃度の増加と、補給1日目と30日目との最初の12時間にわたる血漿β-アラニンAUC(AUC(0-12hr))の増加と、の間には有意な相関関係(r=0.986,p=0.005)が6頭のウマのうち5頭について認められた。回帰線を求めるために5頭のウマだけを使用した。ウマ6(黒丸)は、1日目に観察された血漿β-アラニン濃度の増加より高い、評価できる増加を補給最終日まで示さなかった。このウマはそれぞれの採血日について漸増的増加を示した他の5頭のウマと相違していた。こうした理由のため、回帰方程式を求めることからウマ6を除外した。
【0051】
β-アラニンおよびL-ヒスチジンを30日間補給した後の筋カルノシン濃度の増加は全体的な筋緩衝能の直接的増加を引き起こすだろう。この増加はHenderson-Hasselbach式を使って計算することができる。6頭の純血種ウマにおけるIIAおよびIIB型骨格筋繊維の筋緩衝能の増加に関する計算値を表3に示す。
【表3】
【0052】
実施例2
常食への1日複数回のβ-アラニンとL-ヒスチジンの補給がヒトのI、IIAおよびIIB型の骨格筋繊維のカルノシン含量に及ぼす効果を評価した。約40mg/kg(体重)のβ-アラニンを供給するブロスを摂取した後の6人の正常被験者におけるβ-アラニンの血漿濃度をモニターした。10および20mg/kg(体重)の用量のβ-アラニンも投与した。
【0053】
ブロスは次のように調製した。新鮮な鶏むね肉(皮と骨を取ったもの)を細かく切り刻み、水(鶏肉1.5kgにつき1リットル)と共に15分間ボイルした。残存する鶏肉を目の粗い濾過にかけて取り除いた。濾液にニンジン、タマネギ、セロリ、塩、コショウ、バジル、パセリ、トマトピューレを加えて味付けし、さらに15分間再ボイルし、冷ました後に4℃で微細なモスリンに通して最終濾過にかけた。1.5kgの鶏肉と1Lの水から870mLのブロスが得られた。このストックの一部について、その全β-アラニル-ジペプチド含量(例えば、カルノシンおよびアンセリン)およびβ-アラニンをアッセイした。典型的な分析は以下のとおりであった。
【0054】
総β-アラニル-ジペプチド 74.5mM
遊離β-アラニン 5.7mM
6人の男性被験者は普通の健康状態にあり、表4に示すとおり年齢は25歳から53歳であった。実験を一夜の絶食(例えば、肉を含む最後の食事をとった後最低12時間)後に開始した。被験者には実験開始前に少量の温水を飲む自由が与えられた。カテーテル法を08:30に始め、実験を09:00に開始した。
【0055】
対照として、8mL/kg(体重)の水(例えば、体重が75kgの被験者では600mL)を摂取させた。
【0056】
一つのセッションでは、約40mg/kg(体重)のβ-アラニン(例えば、アンセリンおよびカルノシンの形のもの)を含む8mL/kg(体重)のブロスを摂取させた。体重が75kgの被験者において、これは3gのβ-アラニンを含むブロスを600mL摂取することに相当する。別のセッションでは、試験量のβ-アラニンを含む3mL/kg(体重)の液体を追加の5mL/kg(体重)の水と共に摂取させた。すべてのセッションにおいて、被験者は摂取後1〜2時間のうちにさらに8mL/kg(体重)の水(50mLずつ)を飲んだ。6時間後に菜食主義者用のピザを与えた。8時間後に常食に戻った。
【0057】
2.5mLの静脈血サンプルを、最初の90分間は10分おきに、その後は120、180、240および360分後に留置カテーテルから採取した。抗凝固剤としてリチウム-ヘパリンを含むチューブ中に血液サンプルを分配した。カテーテルは食塩水で洗い流すことで維持した。Jones & Gilligan(1983)J.Chromatogr.266:471-482(1983)に記載される方法に従ってHPLCで血漿サンプルを分析した。
【0058】
表4にはβ-アラニン吸収実験中の処置の割当てがまとめてある。概算された等量のβ-アラニンを表3に示す。
【表4】
【0059】
それぞれの処置後の血漿濃度曲線を図8にグラフで示す。表4の処置計画に従うβ-アラニン、ブロスまたはカルノシンの投与の平均結果。鶏肉ブロスの摂取後にまたは他の処置後にカルノシンもアンセリンも血漿中に検出されなかった。
【0060】
ブロスの摂取は最大血漿濃度427.9(SD 161.8)μMをもたらした。一人の被験者に20mg/kg(体重)のβ-アラニンに等しいカルノシンを投与すると、血漿β-アラニン濃度が同様に増加した。
【0061】
対照を除く全ての処置の投与は血漿タウリン濃度を増加させた。タウリン濃度の変化はβ-アラニンの濃度変化を密に反映していた。自然食品であるブロスの投与は血漿タウリンの同様の増加を引き起こし、このことは、こうした応答がほとんどの食事の摂取後に普通に起こっていることを示している。
【0062】
実施例3
10mg/kg(体重)のβ−アラニンを1日3回(すなわち、朝、昼、及び夜)、7日間投与した場合に、β−アラニン及びタウリンの血漿濃度プロフィールにどのような影響があるかを調べた。3人の被験者に、10mg/kg(体重)のβ−アラニンを1日3回、7日間投与し、この7日間の最初と最後における血漿濃度プロフィールを調べた。
【0063】
被験者は、年齢33〜53歳の正常健康体の3人の男性とした。被験者に、10mg/kg(体重)のβ−アラニンを1日3回、8日間投与した。被験者のうちの2人には、これに続けて、20mg/kg(体重)のβ−アラニンを1日3回、さらに7日間(9日目から15日目まで)投与した。被験者は、一晩絶食させた後、1日目(いかなる処置も施す前)、8日目、及び15日目の午前8時に血液採取室に出向かせた。被験者には、この試験の前12時間にはいかなる肉を含んだ食事もとらないように頼んでおいた。これらの3つの各試験日において、被験者にカテーテルをいれ、午前9時、正午及び午後3時丁度又はその近くにβ−アラニンを投与したとき、最初の血液サンプルを採取した。血液サンプルを、30分、60分、120分及び180分後に採取し、β−アラニン及びタウリンの血漿濃度の変化を分析した。各試験日に24時間尿サンプルを採取し、HPLCで分析してβ−アラニン及びタウリンの排泄量を測定した。処置の概要を表5に示す。
【表5】
【0064】
血漿β−アラニン濃度は、図9に要約する。各回の投与により、投与後1/2時間又は1時間で最大β−アラニン濃度に達し、その後、3時間(すなわち、次回の投与直前)で0−10μMの基底レベルまで減少した。血漿濃度曲線下面積(AUC)によって示されるように、処置8日目の応答は、1日目よりも低下する傾向にあった。
【0065】
実施例4
40mg/kg(体重)のβ−アラニンを1日3回(すなわち、朝、昼、及び夜)、2週間投与した場合に、筋肉のカルノシン含量及び最大随意収縮力の66%における等尺性持久力にどのような影響があるかを調べた。
【0066】
被験者として、代謝又は筋肉疾患の形跡のない、年齢25〜32歳の正常な6人の男性を募集した。被験者には、最近の食事及び補給物質の摂取習慣に関して質問した。被験者の中には、最近、クレアチンを含有する補給物質(supplement)を摂取した者はおらず、また最近の試験的補給法でそのようにした者もいなかった。被験者の身体的特徴を表6に要約する。
【表6】
【0067】
処置の2日前に、座らせた状態における被験者の膝伸筋の最大随意(等尺性)収縮力(MVC)を予備測定した。MVC出力を即時にディスプレイ表示することにより被験者を誘導しながら、Macflexシステムを用いてMVCを測定した。各被験者において、2回の試行を実施し、声援に関わらずもはや標的の力を維持することができなくなるまで66%MVCを持続させて、66%MVCにおける持久力を測定した。この最初の収縮の後、被験者を等尺性椅子に座らせたまま、60秒間の休憩時間を与えた。休憩時間の後、第2の収縮を疲労するまで継続させた。第2の休憩を60秒間与えた後、第3の収縮を疲労するまで継続させた。
【0068】
処置の1日前に、被験者を午前8〜10時の間に等尺性試験室に出向かせた。MVCを測定するとともに、上記のように60秒間の休憩間隔をとりながら3回の収縮を実施し、66%MVCにおける持久力を測定した。測定は被験者の利き足を用いて実施した。外側広筋の外側部分の生検材料を再度利き足から採取した。
【0069】
一晩絶食させるとともに最後にとった肉を含んだ食事から少なくとも12時間経過させた後、処置の1日目に、被験者を午前8時に血液採取室に出向かせた。各被験者にカテーテルを入れて最初の血液サンプルを採取した後、実施例3に記載する補給及び血液採取プロトコールを実施した。10mg/kg(体重)のβ−アラニンを0時間目(午前9時)、3時間目及び6時間目に投与した。
【0070】
2−15日目では、被験者に10mg/kg(体重)のβ−アラニンを3回投与し続けた。
【0071】
14日目の朝に、処置後における等尺性運動試験を利き足において実施し、MVCを測定するとともに、処置前に測定した66%MVCに対する相対66%MVCにおける持久力を測定した。その日の午後に、処置前に生検材料を採取した部位の近辺から外側広筋の生検材料を採取した。
【0072】
15日目に、1日目に実施した手順を繰り返し、補給15日間にわたるβ−アラニン及びタウリンの血漿濃度プロフィールの全体的な変化を求めた。10mg/kg(体重)量を1日3回投与した1日目及び15日目の0、3及び6時間目に10mg/kg(体重)のβ−アラニンを経口投与した後、9時間にわたる血漿β−アラニンの平均変化を図10に示す。
【0073】
1人の追加被験者(No.7)には、10mg/kg(体重)の用量を3回、7日間投与した後、20mg/kg(体重)の用量を3回、7日間投与する試験を実施した。この被験者からは、筋肉生検材料を採取しなかった。
【0074】
6人の被験者から採取した筋肉生検材料中の筋肉カルノシン含量には、明らかな変化はなかった。実施例2で述べたように、6人の被験者における血漿タウリン濃度の変化は、血漿β−アラニン濃度を反映していた。
【0075】
10mg/kg(体重)のβ−アラニンを3回投与する1日前及び投与した14日後におけるMVC及び66%MVCにおける持久力の測定値を表7に示す。66%MVCにおける平均持久時間は、6人の被験者のうち5人で増加した。また、より多くの用量を投与した被験者7においても増加が見られた。
【表7】
【0076】
その他の実施態様は請求の範囲内のものである。
【技術分野】
【0001】
本発明は、筋肉及びその他の組織の無酸素的運動能力を高めるための方法及び組成物に関する。
【背景技術】
【0002】
発明の背景
天然食品の補助物質(supplement)は、典型的には現代のヒト及び動物における食餌の栄養素のレベル低下を補償するように設計される。特に、有用な補助物質は摂取したとき組織の機能を高めるものである。とりわけ、特定の種類の動物の食餌(その通常の食餌は肉や動物性食品のみから利用できる栄養素が不足する可能性がある)を補給することは重要である(例えば、ヒト菜食主義者やその他の動物は草食性食品を摂取する)。
【0003】
例えば、スポーツ及び競技の社会においては、レジャーや労働の目的で身体的頑強さを促進し強化する補助物質のような、特定的に運動能力を改善する自然食品補助物質の重要性が高まっている。別の例としては、無酸素的(例えば乳酸生成)ストレスが加齢とともに経験し得る疲労および不快感の発生を引き起し得る。また無酸素的ストレスは、筋肉内圧力の増加により(例えばロッククライミング、フリーダイビング、シンクロナイズドスイミング等の間に)局所的な循環が部分的あるいは完全に閉塞された場合の長時間の最大下の等尺運動からも起こり得る。過剰な乳酸の生成は細胞内環境の酸性化を起こし得る。
【0004】
クレアチン(すなわちN-(アミノイミノメチル)-N-グリシン、N-アミジノサルコシン、N-メチル-N-グアニルグリシン、またはメチルグリコシアミン)は、骨格筋及び高い可変的なエネルギー要求の能力を特徴とするその他の「励起可能な」組織(例えば平滑筋、心筋、あるいは精子)において多量に見られる。クレアチンは細胞内のエネルギー生成生化学的経路においてホスホリルクレアチンに変換される。哺乳動物の骨格筋においては、典型的なクレアチンの合計含量(すなわちクレアチンとホスホリルクレアチン)は、新鮮な筋肉1キログラムあたり25mmol未満から50mmol(すなわち筋肉1キログラムあたり3.2〜6.5グラム)まで変化し得る。
【0005】
クレアチンは肝臓で生成され、活性輸送系により例えば筋肉のような組織に取り込まれる。体内でのクレアチン合成は食肉中に存在するクレアチンの摂取(例えば平均的な肉食のヒトにおいては1日に体重1キログラムあたり5〜10ミリグラム、菜食主義者の食事においてはほぼゼロである)によっても増加される。
【0006】
集中的な運動の継続、あるいは局所的な低酸素状態での運動の継続の間に、解糖中に形成されたヒドロニウムイオンの蓄積および乳酸の蓄積(嫌気代謝)が細胞内のpHを著しく低下させることがある。低下したpHはクレアチン-ホスホリルクレアチン系の機能を低下させ得る。細胞内pHの低下は、例えば筋繊維中の収縮タンパク質の機能のような、細胞内のその他の機能に影響し得る。
【0007】
β-アラニンとヒスチジンのジペプチド及びそのメチル化類似体には、カルノシン(β-アラニル-L-ヒスチジン)、アンセリン(β-アラニル-L-1-メチルヒスチジン)、あるいはバレニン(β-アラニル-L-3-メチルヒスチジン)が含まれる。これらのジペプチドはヒトやその他の脊椎動物の筋肉中に存在する。カルノシンは例えばヒト及びウマ科の動物の筋肉中にかなりの量が見られる。アンセリン及びカルノシンは例えばイヌ、ラクダ、及び多くの鳥類の種の筋肉中に見られる。アンセリンは多くの魚類において見られる主要なβ-アラニルヒスチジンジペプチドである。バレニンは、水生哺乳動物及び爬虫類のいくつかの種に見られる主要なβ-アラニルヒスチジンジペプチドである。ヒト、ウマ及びラクダでは、β-アラニルヒスチジンジペプチドの最大濃度は、激しい運動の間に集中的に使用される急速収縮性解糖筋繊維(IIA及びIIB型)中に見られる。より低い濃度は、酸化的緩徐収縮性筋繊維(I型)中に見られる。例えば、Dunnett,M.& Harris,R.C.Equine Vet.J.Suppl.18,214-217(1995)を参照のこと。カルノシンは種々の筋繊維型においてヒドロニウムイオン緩衝能に貢献していると推定され、ウマII型繊維においては全体の最高50%である。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】Dunnett,M.& Harris,R.C.Equine Vet.J.Suppl.18,214-217(1995)
【発明の概要】
【課題を解決するための手段】
【0009】
発明の概要
一般に、本発明は、筋肉及びその他の組織の無酸素的運動能力を高めるための方法及び組成物に関する。前記方法は、体内の組織中でのクレアチン及びβ-アラニルヒスチジンジペプチド、あるいはβ-アラニン及びL-ヒスチジン類似体の同時蓄積を含む。前記方法は組成物を体内に摂取させること、あるいは注入することを含む。前記組成物は、ヒトあるいは動物の筋肉において、β-アラニルヒスチジンジペプチドの合成及び蓄積のためにクレアチン及び前駆体の利用可能性及び吸収を増加させることができる化合物の混合物である。前記組成物は、それが体内に導入されたときに、ヒトあるいは動物体内におけるβ-アラニルヒスチジンジペプチドの合成および蓄積を引き出すものである。
【0010】
前記組成物は、クレアチン及びβ-アラニン、クレアチン、β-アラニン及びL-ヒスチジン、またはクレアチン及びβ-アラニンもしくはL-ヒスチジンの活性誘導体の混合物を含む。β-アラニンまたはL-ヒスチジンのそれぞれは個々のアミノ酸であってもよく、あるいはジペプチド、オリゴペプチドまたはポリペプチドの成分であってもよい。β-アラニン及びL-ヒスチジンは活性誘導体であってもよい。活性誘導体は、体内でその物質と同じあるいは類似の方法で機能する物質から誘導される化合物もしくはその前駆体、あるいはそのような物質にプロセシングされて体内に配置される化合物である。例としては、エステル及びアミドが挙げられる。
【0011】
一つの面において、本発明は、組織中のヒドロニウムイオン濃度を制御する方法に関する。前記方法は、組織中のβ-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のβ-アラニンを血液あるいは血漿に供給し、組織をその血液あるいは血漿にさらし、それによりβ-アラニルヒスチジンの濃度を前記組織中で増加させるステップを含む。前記方法は、β-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のL-ヒスチジンを血液あるいは血漿に供給するステップを含むことができる。
【0012】
別の面においては、本発明は、組織の無酸素的運動能力を高める方法に関する。前記方法は、組織中のβ-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のβ-アラニンを血液あるいは血漿に供給し、組織中のβ-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のL-ヒスチジンを血液あるいは血漿に供給し、そして組織をその血液あるいは血漿にさらすステップを含む。β-アラニルヒスチジンの濃度が前記組織中で増加する。
【0013】
ある実施態様においては、前記方法は組織中のクレアチンの濃度を増加させるステップを含む。この増加ステップは、組織中のクレアチン濃度を増加させるのに有効な量のクレアチンを血液あるいは血漿に供給する(例えば前記量のクレアチンを血液あるいは血漿に供給することによる)ことを含み得る。
【0014】
前記方法の供給ステップは、前記量のβ-アラニンまたは前記量のβ-アラニン及びL-ヒスチジンを含む組成物の摂取あるいは注入(例えば注射)、または摂取と注入の組合せを含み得る。
【0015】
前記方法は、血液あるいは血漿中のインスリン濃度を増加させることを含み得る。インスリンの濃度は、例えばインスリンの注射により増加させることができる。
【0016】
前記組織は骨格筋であり得る。
【0017】
別の面においては、本発明は、β-アラニンを含むペプチド源、約39〜約99重量%の炭水化物、及び約60重量%までの水から実質的になる組成物に関する。前記組成物は約1〜約20重量%のβ-アラニンを含む。前記ペプチド源はL-ヒスチジンを含むことができる。前記組成物は約1〜約20重量%のL-ヒスチジンを含むことができる。
【0018】
前記炭水化物は単純な炭水化物(例えばグルコース)であってよい。別の面においては、本発明は、β-アラニンを含むペプチド源、約1〜約98重量%のクレアチン源、及び約97重量%までの水から実質的になる組成物に関する。前記組成物は約1〜約98重量%のβ-アラニンを含む。前記ペプチド源はL-ヒスチジンを含むことができ、前記組成物は約1〜約98重量%のL-ヒスチジンを含む。
【0019】
前記ペプチド源はアミノ酸、ジペプチド、オリゴペプチド、ポリペプチド、あるいはそれらの活性誘導体の混合物であり得る。
【0020】
前記組成物は食品補助物質とすることができる。前記クレアチン源はクレアチン一水和物であってよい。
【0021】
血液または血漿中の成分の濃度は、血漿濃度の増加を起こすように機能できる物質の注入(例えば注射)あるいは摂取により増加させることができる。前記組成物は1日あたり約10グラム〜約800グラムの間の量で摂取させることができる。この量は1日1回で、あるいは複数回に分けて投与することができる。
【0022】
クレアチン及びβ-アラニルヒスチジンジペプチドの筋内含量が増加することにより、無酸素的運動に伴うヒドロニウムイオン生成の増加に対する細胞の許容度が増加し、疲労が発生するまでの運動の持続期間を増加させることができる。
【0023】
前記組成物及び方法は、調理あるいは加工中のβ-アラニン、L-ヒスチジンあるいはクレアチンの分解または浸出(leaching)によるこれらの成分の損失を補充するのに有用であり得る。また前記組成物及び方法は、菜食主義者の食事におけるこれらの成分の欠損を補給するのにも有用であり得る。
【0024】
前記方法及び組成物は、例えば、スポーツマン、競技者、ボディビルダー、シンクロナイズドスイミングのスイマー、兵士、老年者、競争馬、作業犬及びレース犬、競技用の鳥等においてβ-アラニルヒスチジンジペプチドを増加させるのに使用し、筋肉疲労の発生を防止し、遅延させることができる。
【0025】
本発明のその他の利点及び特徴は詳細な説明及び請求の範囲から明らかになるであろう。
【図面の簡単な説明】
【0026】
図面の簡単な説明
【図1】β-アラニン及びL-ヒスチジンを30日間にわたって(それぞれ体重1キログラムあたり100ミリグラム及び体重1キログラムあたり12.5ミリグラム、1日3回)給餌した後の、5頭のウマの給餌前及び給餌後2時間おきの血漿中のβ-アラニンの濃度の変化を示すグラフである。
【図2】β-アラニン及びL-ヒスチジンを30日間にわたって(それぞれ体重1キログラムあたり100ミリグラム及び体重1キログラムあたり12.5ミリグラム、1日3回)給餌した後の、5頭のウマの給餌前及び給餌後2時間おきの血漿中のL-ヒスチジンの濃度の変化を示すグラフである。
【図3a】β-アラニン及びL-ヒスチジンの30日間の食餌補給(それぞれ体重1キログラムあたり100ミリグラム及び体重1キログラムあたり12.5ミリグラム、1日3回)の初日及び最終日の、6頭のウマのうちのウマ1〜3の給餌前及び給餌後時間毎の血漿中のβ-アラニンの濃度の変化を比較して示すグラフである。
【図3b】β-アラニン及びL-ヒスチジンの30日間の食餌補給(それぞれ体重1キログラムあたり100ミリグラム及び体重1キログラムあたり12.5ミリグラム、1日3回)の初日及び最終日の、6頭のウマのうちのウマ4〜6の給餌前及び給餌後時間毎の血漿中のβ-アラニンの濃度の変化を比較して示すグラフである。
【図4a】β-アラニン及びL-ヒスチジンの30日間の食餌補給(それぞれ体重1キログラムあたり100ミリグラム及び体重1キログラムあたり12.5ミリグラム、1日3回)の初日及び最終日の、6頭のウマのうちのウマ1〜3の給餌前及び給餌後時間毎の血漿中のL-ヒスチジンの濃度の変化を比較して表すグラフである。
【図4b】β-アラニン及びL-ヒスチジンの30日間の食餌補給(それぞれ体重1キログラムあたり100ミリグラム及び体重1キログラムあたり12.5ミリグラム、1日3回)の初日及び最終日の、6頭のウマのうちのウマ4〜6の給餌前及び給餌後時間毎の血漿中のL-ヒスチジンの濃度の変化を比較して表すグラフである。
【図5】β-アラニン及びL-ヒスチジンの30日間の食餌補給(それぞれ体重1キログラムあたり100ミリグラム及び体重1キログラムあたり12.5ミリグラム、1日3回)の初日及び最終日の、給餌前及び給餌後時間毎のウマ血漿中のβ-アラニンの平均濃度(n=6)の変化を比較して示すグラフである。
【図6】β-アラニン及びL-ヒスチジンの30日間の食餌補給(それぞれ体重1キログラムあたり100ミリグラム及び体重1キログラムあたり12.5ミリグラム、1日3回)の初日及び最終日の、給餌前及び給餌後時間毎のウマ血漿中のL-ヒスチジンの平均濃度(n=6)の変化を比較して示すグラフである。
【図7】6頭の純血種ウマのII型骨格筋繊維におけるカルノシン濃度の増加(IIA及びIIB型繊維の合計の平均)と、補給の1日目と30日目の間の血漿β-アラニン濃度-時間曲線下の当日の最初の12時間にわたる面積(AUC(0-12hr))の増加と、の相関関係を示すグラフである。
【図8】β-アラニン、ブロス、あるいはカルノシンの被験者への投与の平均の結果を示すグラフである。
【図9】9時間の処置にわたる血漿β-アラニンの平均変化を示すグラフである。
【図10】体重1キログラムあたり10ミリグラムのβ-アラニンの経口摂取後の9時間にわたる血漿β-アラニンの平均変化を示すグラフである。
【図11】処置期間の1日目及び30日目の24時間にわたる平均(n=6)血漿β-アラニン濃度を示すグラフである。
【発明を実施するための形態】
【0027】
好ましい実施態様の説明
カルノシン、アンセリン及びバレニン等のβ-アラニルヒスチジンジペプチドは約6.8〜7.1の間のpKa値を有し、筋収縮及び疲労発生中の細胞内pHの恒常性の制御に関与している。ヒドロニウムイオンの緩衝に関与しているその他の物質、例えばタンパク質中のアミノ酸残基、無機及び有機のリン酸及び炭酸水素塩等の含量は、その他の細胞機能へのそれらの関与により拘束される。β-アラニルヒスチジンジペプチドは、細胞中にpH感受性ヒスチジン残基を蓄積する有効な方法を提供する。筋肉β-アラニルヒスチジンジペプチド濃度の変化は個々の競技者の無酸素的運動能力に影響する。
【0028】
β-アラニルヒスチジンジペプチドはβ-アラニンとL-ヒスチジンから体内で合成される。これらの前駆体は体内で生成されたり、摂取されたβ-アラニルヒスチジンジペプチドの分解からのものを含め、食餌から利用できるようにされる。
【0029】
体内のβ-アラニンは筋肉等の組織に輸送される。典型的な給餌状態では、ヒト及びウマ血漿中のL-ヒスチジンの濃度と比較してβ-アラニンの濃度は低い。これらの濃度は、カルノシン合成酵素であるカルノシンシンテターゼの、ミカエリス-メンテン定数(Km)により決定される、その基質に対する親和性と関連付けて考えなければならない。ヒスチジンについてのKmは約16.8μMである。β-アラニンについてのKmは約1000〜2300μMの間である。β-アラニンに対するカルノシンシンテターゼの低い親和性、及び筋肉中のβ-アラニンの低い濃度は、筋肉中のβ-アラニンの濃度によりβ-アラニルヒスチジンジペプチドの合成が制限されていることを示す。
【0030】
筋肉内のβ-アラニルヒスチジンジペプチド量を増加させると、筋肉の性能及びその筋肉により達成される運動量に好都合に影響する。従って、β-アラニルヒスチジンジペプチドの合成及び蓄積がヒトあるいは動物体内の組織において増加される。
【0031】
ヒトあるいは動物体内におけるβ-アラニルヒスチジンジペプチドの合成及び蓄積は、β-アラニンの血液もしくは血漿濃度の増加、β-アラニン及びクレアチンの血液もしくは血漿濃度の増加、あるいはβ-アラニン、L-ヒスチジン及びクレアチンの血液もしくは血漿濃度の増加により、体内のクレアチン含量の増加とともに増加させることができる。ジペプチドの増加は、β-アラニン濃度の増加と同時に起こり得る。
【0032】
β-アラニン、L-ヒスチジン及びクレアチンの血漿濃度は、β-アラニン、L-ヒスチジン及びクレアチンあるいはそれらの活性誘導体の摂取または注入により増加させることができる。前記組成物は、経口、経腸、あるいは非経口的に投与することができる。β-アラニン及びクレアチン、あるいはβ-アラニン、L-ヒスチジン及びクレアチンは好ましくは経口的に摂取される。
【0033】
前記組成物は炭水化物(例えば単純な炭水化物)、インスリン、またはインスリンの生成を刺激する物質を含んでもよい。
【0034】
前記組成物は食品の補助物質として摂取させることができる。好ましくは前記組成物は1日に1回以上投与される。β-アラニンの投与量は体重1キログラムあたり約5ミリグラムから約200ミリグラムの間とすることができる。クレアチン(例えばクレアチン一水和物)の投与量は体重1キログラムあたり約5ミリグラムから200ミリグラムの間とすることができる。L-ヒスチジンの投与量は体重1キログラムあたり約1ミリグラムから100ミリグラムの間とすることができる。
【0035】
単純な炭水化物(例えばグルコース)の投与量は体重1キログラムあたり約0.5グラムから2.0グラムの間とすることができる。
【0036】
80キログラムのヒトにおいては、1日あたりの適当な投与量は、0.4グラム〜16.0グラムのβ-アラニン、0.4グラム〜16.0グラムのクレアチン一水和物、0.08グラム〜8.0グラムのL-ヒスチジン、あるいは40グラム〜160グラムのグルコースもしくはその他の単純な炭水化物とすることができる。前記組成物は摂取するための固体もしくは液体の剤形、あるいは懸濁体の剤形、あるいは体内に注入するための液体もしくは懸濁体の剤形とすることができる。前記組成物は、1日あたり2グラム〜1000グラムの間の量(例えば10グラム〜800グラムの間の量)でヒトに摂取させるが、これは1日のうちに小分けして摂取させることができる。動物における1日の摂取量は体重に関して調整されるだろう。
【0037】
ヒト及び動物について、前記組成物は、(a) 1重量%〜99重量%のβ-アラニン、 1重量%〜99重量%のクレアチン一水和物、及び 0重量%〜98重量%の水;(b) 1重量%〜98重量%のβ-アラニン、 1重量%〜98重量%のL-ヒスチジン、 1重量%〜98重量%のクレアチン一水和物、及び0重量%〜97重量%の水;(c) 1重量%〜20重量%のβ-アラニン、 39重量%〜99重量%のグルコースまたはその他の単純な炭水化物、及び 0重量%〜60重量%の水、あるいは(d) 1重量%〜20重量%のβ-アラニン、 1重量%〜20重量%のL-ヒスチジン、 39重量%〜99重量%のグルコースまたはその他の単純な炭水化物、及び 0重量%〜60重量%の水、とすることができる。
【実施例】
【0038】
以下は、筋肉及びその他の組織の無酸素的運動能力を高めるための方法及び組成物の方法の具体例である。
【0039】
実施例1
標準飼料への1日複数回のβ-アラニンとL-ヒスチジンの補給が純血種のウマのI、IIAおよびIIB型の骨格筋繊維中のカルノシン濃度に及ぼす効果を評価した。普通の健康状態の4〜9歳の実験用純血種ウマ(若い雌ウマ3頭と去勢したウマ3頭)に、補給期間の開始に先立って1ヵ月(30日)(補給前期間)の給餌コンディショニングをおこなった。給餌コンディショニング期間中、各ウマに、複合および単純炭水化物の供給源として1キログラムの成形飼料(Spillers競争馬キューブ)と1キログラムの浸潰したサトウダイコンパルプを含む飼料を1日3回(それぞれ08:30、12:30および16:30)与えた。浸潰した干し草(乾燥重量で3キログラム)も1日2回(09:00および17:00に)与えた。水は自由に飲めるようにした。
【0040】
補給期間中は同一の給餌レジメを実施した。しかし、硬質飼料を与える際にはβ-アラニンとL-ヒスチジン(遊離塩基)を補給した。β-アラニンとL-ヒスチジンを標準飼料に直接混合した。β-アラニンとL-ヒスチジンの個々の投与量は体重に応じて算出した。β-アラニンは体重1キログラムにつき100ミリグラムを、L-ヒスチジンは体重1キログラムにつき12.5ミリグラムを投与した。餌の補給はこのプロトコールの1日目に開始し、30日目に終わらせた。1、6、18、24および30日目に血液サンプル(5ミリリットル)を採取し、ヘパリンを添加した。1日目と30日目には、1回目の給餌前と合計12時間にわたり1時間おきに血液サンプルを採取した。残りの3日の採血日には、1回目の給餌前とその後の各給餌の2時間後に血液を集めた。補給開始の前日(0日目)には、皮膚の局所麻酔後にBergstrom-Stille経皮生検針を使って各ウマの右側の中間臀部筋(m.gluteusmedius)から筋肉の生検材料を採取した。その後の筋生検材料はもとの採取部位に可能なかぎり接近させて補給期間の終了直後(31日目)に採取した。ウマの臨床的監視を毎日おこなった。この監視は肉眼による検査と体重測定、直腸温度の1日2回の測定、それに臨床生化学および血液学のための週1度の採血を含んでいた。実験期間中、ウマには正式な訓練や運動を施さなかったが、毎日1時間自由に運動させた。
【0041】
凍結乾燥した筋生検材料から切り裂いた個々の筋繊維の断片は、KaiserおよびBrcok,Arch.Neurol.,23:369-379(1970)に記載される方法の変法により、pH4.50でプレインキュベートした後にpH9.6でミオシンATPアーゼ活性を組織化学的に染色することで、I、IIAまたはIIB型のいずれかであると特性づけられた。
【0042】
ヘパリン添加血漿サンプルを抽出し、高性能液体クロマトグラフィー(HPLC)でβ-アラニンとL-ヒスチジンの濃度を分析した。計量した個々の筋繊維を抽出し、DunnettおよびHarris,「筋肉および個々の筋繊維中のイミダゾールジペプチド、ヒスチジン、1-メチルヒスチジン、3-メチルヒスチジンの高性能液体クロマトグラフィー測定」,J.Chromatogr.B.Biomed.Appl.,688:47-55(1997)に記載される方法に従ってHPLCでカルノシンを分析した。
【0043】
補給前後の繊維型に含まれるカルノシン濃度の差は、一方向分散分析(ANOVA)を用いてウマにおいて決定した。差が認められた場合には、多重比較検定(Fisher's PLSD)により有意差を求めた。
【0044】
飼料にβ-アラニンとL-ヒスチジンを添加しても味覚上の問題はまったく生起しなかった。30日の補給期間中、補給餌の有害な生理または行動作用はいずれのウマにも観察されなかった。体重の顕著な変化は記録されず、直腸温度も正常範囲内にとどまった。臨床生化学または血液学においても急性または慢性の変化が認められなかった。補給開始前のウマの血漿中にはβ-アラニンは検出されなかった。用いたアッセイによる血漿中のβ-アラニンの定量の下限は3マイクロモル(μM)であった。補給開始前の6頭のウマの血漿L-ヒスチジン濃度は36.6〜54.4μMであった。
【0045】
すべての採血日にわたる血漿β-アラニンおよびL-ヒスチジン濃度の個々の変化を、6頭のウマのうち5頭についてそれぞれ図1および2に示す。補給期間が増すにつれて、血漿β-アラニンおよびL-ヒスチジンの給餌前濃度は増加する傾向にあった。さらに、30日の補給期間にわたって、補給に対する血漿濃度応答も増加した。この応答はβ-アラニンの場合に、より大きかった。
【0046】
最初の補給の前日と、その後補給期間の初日と最終日の時間ごとの血漿β-アラニンおよびL-ヒスチジン濃度の変化の比較を、6頭の各ウマについてそれぞれ図3aと3b、および図4aと4bに示す。補給期間の初日(1日目)と最終日(30日目)の24時間にわたる経時的血漿β-アラニン濃度の平均(±SD)変化(n=6)を図5に対比して示す。24時間にわたる時間曲線に対する平均血漿β-アラニン濃度下の面積(AUC(0-24hr))は補給30日目に一層大きかった。
【0047】
補給期間の初日(1日目)と最終日(30日目)の24時間にわたる経時的血漿L-ヒスチジン濃度の平均(±SD)変化(n=6)を図6に対比して示す。24時間にわたる時間曲線に対する平均血漿β-アラニン濃度下の面積(AUC(0-24hr))は補給30日目により大きかった。補給初日(1日目)に対比させたときの補給最終日(30日目)の血漿β-アラニンのより大きいAUCは、補給が進行するにつれてウマの胃腸管からのβ-アラニンの吸収が増加することを示唆している。同様の効果が補給期間中の血漿L-ヒスチジン濃度の変化に関しても認められた。β-アラニンおよびL-ヒスチジンの最大血漿濃度はそれぞれの場合に給餌の約1〜2時間後に出現した。
【0048】
6頭のウマから合計397本の骨格筋繊維(補給前 192本;補給後 205本)を切除し、カルノシンについて分析した。6頭の各ウマから得られた補給前および補給後のI、IIAおよびIIB型骨格筋繊維中の平均(±SD)カルノシン濃度(乾燥重量1キログラムあたりのミリモル数(mmol kg-1dw)で表す)を表1に示してあるが、ここでnは分析した個々の筋繊維の数である。β-アラニンおよびL-ヒスチジンの補給30日後に、6頭すべてのウマにおいてIIAおよびIIB型の繊維中の平均カルノシン濃度が増加していた。これらの増加は7つの事例において統計的に有意であった。IIB型骨格筋繊維中の平均カルノシン濃度の増加は6頭のウマのうち5頭において統計的に有意であった。IIA型骨格筋繊維中の平均カルノシン濃度の増加は6頭のウマのうち2頭において統計的に有意であった。
【表1】
* 補給前と有意に異なる、p<0.05
** 補給前と有意に異なる、p<0.01
*** 補給前と有意に異なる、p<0.005
【0049】
6頭のウマから得られたIIAおよびIIB型骨格筋繊維中の平均カルノシン濃度の絶対増加(例えば、mmol kg-1dw)および増加パーセントを表2に示す。
【表2】
【0050】
30日の補給後に筋カルノシン濃度のより大きい増加を示したウマはまた、補給期間の1日目と30日目の間で血漿β-アラニンAUCのより大きい増加を示したことが観察された。図7を参照すると、IIAおよびIIB型骨格筋繊維間で平均した平均カルノシン濃度の増加と、補給1日目と30日目との最初の12時間にわたる血漿β-アラニンAUC(AUC(0-12hr))の増加と、の間には有意な相関関係(r=0.986,p=0.005)が6頭のウマのうち5頭について認められた。回帰線を求めるために5頭のウマだけを使用した。ウマ6(黒丸)は、1日目に観察された血漿β-アラニン濃度の増加より高い、評価できる増加を補給最終日まで示さなかった。このウマはそれぞれの採血日について漸増的増加を示した他の5頭のウマと相違していた。こうした理由のため、回帰方程式を求めることからウマ6を除外した。
【0051】
β-アラニンおよびL-ヒスチジンを30日間補給した後の筋カルノシン濃度の増加は全体的な筋緩衝能の直接的増加を引き起こすだろう。この増加はHenderson-Hasselbach式を使って計算することができる。6頭の純血種ウマにおけるIIAおよびIIB型骨格筋繊維の筋緩衝能の増加に関する計算値を表3に示す。
【表3】
【0052】
実施例2
常食への1日複数回のβ-アラニンとL-ヒスチジンの補給がヒトのI、IIAおよびIIB型の骨格筋繊維のカルノシン含量に及ぼす効果を評価した。約40mg/kg(体重)のβ-アラニンを供給するブロスを摂取した後の6人の正常被験者におけるβ-アラニンの血漿濃度をモニターした。10および20mg/kg(体重)の用量のβ-アラニンも投与した。
【0053】
ブロスは次のように調製した。新鮮な鶏むね肉(皮と骨を取ったもの)を細かく切り刻み、水(鶏肉1.5kgにつき1リットル)と共に15分間ボイルした。残存する鶏肉を目の粗い濾過にかけて取り除いた。濾液にニンジン、タマネギ、セロリ、塩、コショウ、バジル、パセリ、トマトピューレを加えて味付けし、さらに15分間再ボイルし、冷ました後に4℃で微細なモスリンに通して最終濾過にかけた。1.5kgの鶏肉と1Lの水から870mLのブロスが得られた。このストックの一部について、その全β-アラニル-ジペプチド含量(例えば、カルノシンおよびアンセリン)およびβ-アラニンをアッセイした。典型的な分析は以下のとおりであった。
【0054】
総β-アラニル-ジペプチド 74.5mM
遊離β-アラニン 5.7mM
6人の男性被験者は普通の健康状態にあり、表4に示すとおり年齢は25歳から53歳であった。実験を一夜の絶食(例えば、肉を含む最後の食事をとった後最低12時間)後に開始した。被験者には実験開始前に少量の温水を飲む自由が与えられた。カテーテル法を08:30に始め、実験を09:00に開始した。
【0055】
対照として、8mL/kg(体重)の水(例えば、体重が75kgの被験者では600mL)を摂取させた。
【0056】
一つのセッションでは、約40mg/kg(体重)のβ-アラニン(例えば、アンセリンおよびカルノシンの形のもの)を含む8mL/kg(体重)のブロスを摂取させた。体重が75kgの被験者において、これは3gのβ-アラニンを含むブロスを600mL摂取することに相当する。別のセッションでは、試験量のβ-アラニンを含む3mL/kg(体重)の液体を追加の5mL/kg(体重)の水と共に摂取させた。すべてのセッションにおいて、被験者は摂取後1〜2時間のうちにさらに8mL/kg(体重)の水(50mLずつ)を飲んだ。6時間後に菜食主義者用のピザを与えた。8時間後に常食に戻った。
【0057】
2.5mLの静脈血サンプルを、最初の90分間は10分おきに、その後は120、180、240および360分後に留置カテーテルから採取した。抗凝固剤としてリチウム-ヘパリンを含むチューブ中に血液サンプルを分配した。カテーテルは食塩水で洗い流すことで維持した。Jones & Gilligan(1983)J.Chromatogr.266:471-482(1983)に記載される方法に従ってHPLCで血漿サンプルを分析した。
【0058】
表4にはβ-アラニン吸収実験中の処置の割当てがまとめてある。概算された等量のβ-アラニンを表3に示す。
【表4】
【0059】
それぞれの処置後の血漿濃度曲線を図8にグラフで示す。表4の処置計画に従うβ-アラニン、ブロスまたはカルノシンの投与の平均結果。鶏肉ブロスの摂取後にまたは他の処置後にカルノシンもアンセリンも血漿中に検出されなかった。
【0060】
ブロスの摂取は最大血漿濃度427.9(SD 161.8)μMをもたらした。一人の被験者に20mg/kg(体重)のβ-アラニンに等しいカルノシンを投与すると、血漿β-アラニン濃度が同様に増加した。
【0061】
対照を除く全ての処置の投与は血漿タウリン濃度を増加させた。タウリン濃度の変化はβ-アラニンの濃度変化を密に反映していた。自然食品であるブロスの投与は血漿タウリンの同様の増加を引き起こし、このことは、こうした応答がほとんどの食事の摂取後に普通に起こっていることを示している。
【0062】
実施例3
10mg/kg(体重)のβ−アラニンを1日3回(すなわち、朝、昼、及び夜)、7日間投与した場合に、β−アラニン及びタウリンの血漿濃度プロフィールにどのような影響があるかを調べた。3人の被験者に、10mg/kg(体重)のβ−アラニンを1日3回、7日間投与し、この7日間の最初と最後における血漿濃度プロフィールを調べた。
【0063】
被験者は、年齢33〜53歳の正常健康体の3人の男性とした。被験者に、10mg/kg(体重)のβ−アラニンを1日3回、8日間投与した。被験者のうちの2人には、これに続けて、20mg/kg(体重)のβ−アラニンを1日3回、さらに7日間(9日目から15日目まで)投与した。被験者は、一晩絶食させた後、1日目(いかなる処置も施す前)、8日目、及び15日目の午前8時に血液採取室に出向かせた。被験者には、この試験の前12時間にはいかなる肉を含んだ食事もとらないように頼んでおいた。これらの3つの各試験日において、被験者にカテーテルをいれ、午前9時、正午及び午後3時丁度又はその近くにβ−アラニンを投与したとき、最初の血液サンプルを採取した。血液サンプルを、30分、60分、120分及び180分後に採取し、β−アラニン及びタウリンの血漿濃度の変化を分析した。各試験日に24時間尿サンプルを採取し、HPLCで分析してβ−アラニン及びタウリンの排泄量を測定した。処置の概要を表5に示す。
【表5】
【0064】
血漿β−アラニン濃度は、図9に要約する。各回の投与により、投与後1/2時間又は1時間で最大β−アラニン濃度に達し、その後、3時間(すなわち、次回の投与直前)で0−10μMの基底レベルまで減少した。血漿濃度曲線下面積(AUC)によって示されるように、処置8日目の応答は、1日目よりも低下する傾向にあった。
【0065】
実施例4
40mg/kg(体重)のβ−アラニンを1日3回(すなわち、朝、昼、及び夜)、2週間投与した場合に、筋肉のカルノシン含量及び最大随意収縮力の66%における等尺性持久力にどのような影響があるかを調べた。
【0066】
被験者として、代謝又は筋肉疾患の形跡のない、年齢25〜32歳の正常な6人の男性を募集した。被験者には、最近の食事及び補給物質の摂取習慣に関して質問した。被験者の中には、最近、クレアチンを含有する補給物質(supplement)を摂取した者はおらず、また最近の試験的補給法でそのようにした者もいなかった。被験者の身体的特徴を表6に要約する。
【表6】
【0067】
処置の2日前に、座らせた状態における被験者の膝伸筋の最大随意(等尺性)収縮力(MVC)を予備測定した。MVC出力を即時にディスプレイ表示することにより被験者を誘導しながら、Macflexシステムを用いてMVCを測定した。各被験者において、2回の試行を実施し、声援に関わらずもはや標的の力を維持することができなくなるまで66%MVCを持続させて、66%MVCにおける持久力を測定した。この最初の収縮の後、被験者を等尺性椅子に座らせたまま、60秒間の休憩時間を与えた。休憩時間の後、第2の収縮を疲労するまで継続させた。第2の休憩を60秒間与えた後、第3の収縮を疲労するまで継続させた。
【0068】
処置の1日前に、被験者を午前8〜10時の間に等尺性試験室に出向かせた。MVCを測定するとともに、上記のように60秒間の休憩間隔をとりながら3回の収縮を実施し、66%MVCにおける持久力を測定した。測定は被験者の利き足を用いて実施した。外側広筋の外側部分の生検材料を再度利き足から採取した。
【0069】
一晩絶食させるとともに最後にとった肉を含んだ食事から少なくとも12時間経過させた後、処置の1日目に、被験者を午前8時に血液採取室に出向かせた。各被験者にカテーテルを入れて最初の血液サンプルを採取した後、実施例3に記載する補給及び血液採取プロトコールを実施した。10mg/kg(体重)のβ−アラニンを0時間目(午前9時)、3時間目及び6時間目に投与した。
【0070】
2−15日目では、被験者に10mg/kg(体重)のβ−アラニンを3回投与し続けた。
【0071】
14日目の朝に、処置後における等尺性運動試験を利き足において実施し、MVCを測定するとともに、処置前に測定した66%MVCに対する相対66%MVCにおける持久力を測定した。その日の午後に、処置前に生検材料を採取した部位の近辺から外側広筋の生検材料を採取した。
【0072】
15日目に、1日目に実施した手順を繰り返し、補給15日間にわたるβ−アラニン及びタウリンの血漿濃度プロフィールの全体的な変化を求めた。10mg/kg(体重)量を1日3回投与した1日目及び15日目の0、3及び6時間目に10mg/kg(体重)のβ−アラニンを経口投与した後、9時間にわたる血漿β−アラニンの平均変化を図10に示す。
【0073】
1人の追加被験者(No.7)には、10mg/kg(体重)の用量を3回、7日間投与した後、20mg/kg(体重)の用量を3回、7日間投与する試験を実施した。この被験者からは、筋肉生検材料を採取しなかった。
【0074】
6人の被験者から採取した筋肉生検材料中の筋肉カルノシン含量には、明らかな変化はなかった。実施例2で述べたように、6人の被験者における血漿タウリン濃度の変化は、血漿β−アラニン濃度を反映していた。
【0075】
10mg/kg(体重)のβ−アラニンを3回投与する1日前及び投与した14日後におけるMVC及び66%MVCにおける持久力の測定値を表7に示す。66%MVCにおける平均持久時間は、6人の被験者のうち5人で増加した。また、より多くの用量を投与した被験者7においても増加が見られた。
【表7】
【0076】
その他の実施態様は請求の範囲内のものである。
【特許請求の範囲】
【請求項1】
組織中におけるβ-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のβ-アラニンを血液または血漿に供給し、組織を前記血液または血漿にさらし、それによりβ-アラニルヒスチジンの濃度を前記組織中で増加させる、ことを含んでなる、組織中のヒドロニウムイオン濃度を制御する方法。
【請求項2】
組織中のβ-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のL-ヒスチジンを血液または血漿に供給することをさらに含む、請求項1に記載の方法。
【請求項3】
組織中のクレアチンの濃度を増加させることをさらに含む、請求項1に記載の方法。
【請求項4】
組織中のβ-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のL-ヒスチジンを血液または血漿に供給することをさらに含む、請求項3に記載の方法。
【請求項5】
組織中のクレアチンの濃度を増加させることが、組織中のクレアチン濃度を増加させるのに有効な量のクレアチンを血液または血漿に供給することを含む、請求項3に記載の方法。
【請求項6】
組織中のβ-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のL-ヒスチジンを血液または血漿に供給することをさらに含む、請求項5に記載の方法。
【請求項7】
供給段階が前記量のβ-アラニンを含有する組成物の摂取を含んでなる、請求項1に記載の方法。
【請求項8】
供給段階が前記量のβ-アラニンを含有する組成物の注入を含んでなる、請求項1に記載の方法。
【請求項9】
血液または血漿中のインスリンの濃度を増加させることをさらに含む、請求項1に記載の方法。
【請求項10】
組織中のβ-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のL-ヒスチジンを血液または血漿に供給することをさらに含む、請求項9に記載の方法。
【請求項11】
組織が骨格筋である、請求項1に記載の方法。
【請求項12】
組織がヒト組織である、請求項1に記載の方法。
【請求項13】
組織が動物組織である、請求項1に記載の方法。
【請求項14】
組織中におけるβ-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のβ-アラニンを血液または血漿に供給し、β-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のL-ヒスチジンを血液または血漿に供給し、組織を前記血液または血漿にさらし、それによりβ-アラニルヒスチジンの濃度を前記組織中で増加させる、ことを含んでなる、組織の無酸素的運動能力を高める方法。
【請求項15】
組織中のクレアチンの濃度を増加させることをさらに含む、請求項14に記載の方法。
【請求項16】
供給段階が前記量のβ-アラニン及び前記量のL-ヒスチジンを含有する組成物の摂取を含む、請求項14に記載の方法。
【請求項17】
供給段階が前記量のβ-アラニン及び前記量のL-ヒスチジンを含有する組成物の注入を含む、請求項14に記載の方法。
【請求項18】
血液または血漿中のインスリンの濃度を増加させることをさらに含む、請求項14に記載の方法。
【請求項19】
組織が骨格筋である、請求項14に記載の方法。
【請求項20】
組織がヒト組織である、請求項14に記載の方法。
【請求項21】
組織が動物組織である、請求項14に記載の方法。
【請求項22】
β-アラニンを含むペプチド源、約39〜約99重量%の炭水化物、及び約60重量%までの水、から実質的になる組成物であって、約1〜約20重量%のβ-アラニンを含むことを特徴とする前記組成物。
【請求項23】
ペプチド源がL-ヒスチジンを含み、前記組成物が約1〜約20重量%のL-ヒスチジンを含む、請求項22に記載の組成物。
【請求項24】
β-アラニンを含むペプチド源、約1〜約98重量%のクレアチン源、及び約97重量%までの水、から実質的になる組成物であって、約1〜約98重量%のβ-アラニンを含むことを特徴とする前記組成物。
【請求項25】
ペプチド源がL-ヒスチジンを含み、前記組成物が約1〜約98重量%のL-ヒスチジンを含む、請求項24に記載の組成物。
【請求項1】
組織中におけるβ-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のβ-アラニンを血液または血漿に供給し、組織を前記血液または血漿にさらし、それによりβ-アラニルヒスチジンの濃度を前記組織中で増加させる、ことを含んでなる、組織中のヒドロニウムイオン濃度を制御する方法。
【請求項2】
組織中のβ-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のL-ヒスチジンを血液または血漿に供給することをさらに含む、請求項1に記載の方法。
【請求項3】
組織中のクレアチンの濃度を増加させることをさらに含む、請求項1に記載の方法。
【請求項4】
組織中のβ-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のL-ヒスチジンを血液または血漿に供給することをさらに含む、請求項3に記載の方法。
【請求項5】
組織中のクレアチンの濃度を増加させることが、組織中のクレアチン濃度を増加させるのに有効な量のクレアチンを血液または血漿に供給することを含む、請求項3に記載の方法。
【請求項6】
組織中のβ-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のL-ヒスチジンを血液または血漿に供給することをさらに含む、請求項5に記載の方法。
【請求項7】
供給段階が前記量のβ-アラニンを含有する組成物の摂取を含んでなる、請求項1に記載の方法。
【請求項8】
供給段階が前記量のβ-アラニンを含有する組成物の注入を含んでなる、請求項1に記載の方法。
【請求項9】
血液または血漿中のインスリンの濃度を増加させることをさらに含む、請求項1に記載の方法。
【請求項10】
組織中のβ-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のL-ヒスチジンを血液または血漿に供給することをさらに含む、請求項9に記載の方法。
【請求項11】
組織が骨格筋である、請求項1に記載の方法。
【請求項12】
組織がヒト組織である、請求項1に記載の方法。
【請求項13】
組織が動物組織である、請求項1に記載の方法。
【請求項14】
組織中におけるβ-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のβ-アラニンを血液または血漿に供給し、β-アラニルヒスチジンジペプチド合成を増加させるのに有効な量のL-ヒスチジンを血液または血漿に供給し、組織を前記血液または血漿にさらし、それによりβ-アラニルヒスチジンの濃度を前記組織中で増加させる、ことを含んでなる、組織の無酸素的運動能力を高める方法。
【請求項15】
組織中のクレアチンの濃度を増加させることをさらに含む、請求項14に記載の方法。
【請求項16】
供給段階が前記量のβ-アラニン及び前記量のL-ヒスチジンを含有する組成物の摂取を含む、請求項14に記載の方法。
【請求項17】
供給段階が前記量のβ-アラニン及び前記量のL-ヒスチジンを含有する組成物の注入を含む、請求項14に記載の方法。
【請求項18】
血液または血漿中のインスリンの濃度を増加させることをさらに含む、請求項14に記載の方法。
【請求項19】
組織が骨格筋である、請求項14に記載の方法。
【請求項20】
組織がヒト組織である、請求項14に記載の方法。
【請求項21】
組織が動物組織である、請求項14に記載の方法。
【請求項22】
β-アラニンを含むペプチド源、約39〜約99重量%の炭水化物、及び約60重量%までの水、から実質的になる組成物であって、約1〜約20重量%のβ-アラニンを含むことを特徴とする前記組成物。
【請求項23】
ペプチド源がL-ヒスチジンを含み、前記組成物が約1〜約20重量%のL-ヒスチジンを含む、請求項22に記載の組成物。
【請求項24】
β-アラニンを含むペプチド源、約1〜約98重量%のクレアチン源、及び約97重量%までの水、から実質的になる組成物であって、約1〜約98重量%のβ-アラニンを含むことを特徴とする前記組成物。
【請求項25】
ペプチド源がL-ヒスチジンを含み、前記組成物が約1〜約98重量%のL-ヒスチジンを含む、請求項24に記載の組成物。
【図1】


【図2】
【図3a】
【図3b】

【図4a】
【図4b】
【図5】
【図6】
【図7】

【図8】
【図9】
【図10】
【図11】
【図2】
【図3a】
【図3b】
【図4a】
【図4b】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公開番号】特開2010−31035(P2010−31035A)
【公開日】平成22年2月12日(2010.2.12)
【国際特許分類】
【出願番号】特願2009−247709(P2009−247709)
【出願日】平成21年10月28日(2009.10.28)
【分割の表示】特願平10−509546の分割
【原出願日】平成9年8月12日(1997.8.12)
【出願人】(507387491)
【出願人】(509298964)
【出願人】(509299031)
【Fターム(参考)】
【公開日】平成22年2月12日(2010.2.12)
【国際特許分類】
【出願日】平成21年10月28日(2009.10.28)
【分割の表示】特願平10−509546の分割
【原出願日】平成9年8月12日(1997.8.12)
【出願人】(507387491)
【出願人】(509298964)
【出願人】(509299031)
【Fターム(参考)】
[ Back to top ]