
結晶性タンパク質VAP−1その用途
本発明は結晶性血管接着タンパク質-1(VAP-1)、および特に、結晶性ヒトVAP-1の構造情報の、リガンドおよび/または阻害剤の同定、設計および製造のため、ならびに該リガンドおよび/または阻害剤の検出のためのスクリーニングアッセイのための用途に関する。本発明はさらに、本発明のアッセイにより同定される阻害剤に関する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、結晶性ヒト血管接着タンパク質-1(VAP-1)、および特に、結晶性ヒトVAP-1の構造情報の、リガンドおよび/または阻害剤の同定、設計および製造のため、ならびに、かかるリガンドおよび/または阻害剤の検出のための、インシリコおよびインビトロのスクリーニングアッセイのための用途に関する。本発明はさらに、本発明のアッセイにより同定される阻害剤であって、急性および慢性の炎症性症状、糖質代謝関連疾患、脂肪細胞または平滑筋細胞機能における異常、および種々の血管疾患の治療において有用な阻害剤に関する。
【背景技術】
【0002】
発明の背景
生理的免疫監視機構は、血液と、種々のリンパ器官との間のリンパ球の継続的な監視に依存している。正常な非リンパ組織には、リンパ球は存在しないか、または非常に低レベルに存在するが、多くの炎症性性の疾病状態においては、莫大な数のリンパ球が種々の冒された組織および器官中に集積することがある。血管からのリンパ球の退出を制御している重要な分子の一つは、米国特許第5,580,780号に開示された血管接着タンパク質-1(VAP-1)である。VAP-1は、ホモダイマーの170〜180kDaの内皮糖タンパク質である。VAP-1は、ヒトの組織切片において、細静脈へのリンパ球の結合を仲介する。VAP-1は、重度にグリコシル化されており、糖部分は接着機能にとり重要である(サルミ(Salmi)ら、1996年)。VAP-1の接着機能を阻止すると、炎症性組織に浸潤する細胞の数が減少して炎症を散らすことができる。VAP-1はしたがって、抗炎症薬開発のターゲットである。
【0003】
ヒトの血管接着タンパク質-1(VAP-1)は、接着性および酵素的性質の双方をもつ、膜結合多機能糖タンパク質である。VAP-1のクローニングは、驚いたことに、それがセミカルバジド感受性モノアミンオキシダーゼ(SSAO;EC 1.4.3.6)に属することを明らかにした(国際特許公開 WO 98/53049)。VAP-1は、触媒活性のある大きい細胞外ドメインをもつII型の主要膜タンパク質である。したがって、VAP-1は細胞外酵素である。白血球-内皮相互作用におけるSSAO活性の役割も、その生理的基質も、どちらも充分には定義されていない。VAP-1は、哺乳類におけるこの酵素群のうちで初めて分子的に定義された膜貫通メンバーであり、細胞のSSAO活性の90%を占めている。とりわけ、SSAOは、細胞内局在、基質、補因子、阻害剤、およびタンパク質配列の点において、充分に特徴づけされたモノアミンオキシダーゼAおよびBとは異なっている。
【0004】
SSAO反応は生化学用語においては、1950年代から知られているが、これらの酵素の生理的機能は謎のままであった。SSAOの生理的基質は分かっていない。しかしながら二つの可能性のある候補、メチルアミンおおよびアミノアセトンは、ヒトでは中間代謝を通じて形成され、かつin vitroおよびin vivoにおいてSSAOにより脱アミノ化されることが可能である。
【0005】
VAP-1 SSAO活性は、白血球の表面に発現されたVAP-1リガンド上に呈示されたアミン基質との直接的な相互作用を含む新規な機構により、内皮細胞への白血球接着の経路に直接的に関与していると提案されていた(サルミ(salmi)ら、2001年)。この発表は、内皮への白血球の接着プロセスにおける、VAP-1 SSAO活性の直接的な関与を述べている。したがって、VAP-1 SSAO活性の阻害剤は、炎症領域における白血球接着を減少させ、それにより炎症領域内への白血球の通行を、それゆえ炎症プロセスそのものを低減することが期待できた。
【0006】
ヒトの臨床組織サンプルにおいては、VAP-1の発現は炎症の部位において誘導される。この増大されたVAP-1レベルは、血中に存在するモノアミンに対する、VAP-1SSAO細胞外ドメインの作用から発生される、H2O2の増大された産生をもたらすことが可能である。内皮細胞という局部的な環境におけるこのH2O2の発生は、別の細胞事象を開始することも可能であろう。H2O2は、他の接着性分子をアップレギュレートすることができるシグナリング分子として知られており、この増大された接着分子の発現が、VAP-1の発現されている領域内への白血球の交通の増強をもたらすのかもしれない。他のVAP-1 SSAO反応産物もまた、炎症プロセスにも寄与する生物学的作用を有しているのかもしれない。したがって、VAP-1 SSAO活性の産物は、炎症プロセスのエスカレーションに関与してよく、それは特異的なSSAO阻害剤によって阻止されることも可能である。
【0007】
VAP-1 SSAOは、VAP-1 SSAOの循環性アミン基質の増大されたレベルに関連する多くの他の病的症状に関係し得る。これらの基質の酸化的脱アミノ化は、内皮細胞という局所環境における毒性アルデヒドおよび酸素ラジカルのレベルの上昇をもたらすこととなり、そのことが細胞を損傷して血管損傷をもたらすことも可能であろう。メチルアミンおよびアミノアセトンの増大されたレベルは、I型およびII型の糖尿病をもつ患者において報告されており、後期糖尿病に見られる網膜症、神経障害、腎症のような血管症は、VAP-1SSAOの特異害剤を用いて治療されることも可能である。
【0008】
VAP-1活性を調整する特異的なVAP-1SSAO阻害剤の開発は、慢性関節炎、炎症性腸疾患、および皮膚疾患のような、急性および慢性の炎症症状または疾患、ならびに糖質代謝に関連した疾患(糖尿病および、糖尿病から結果として生じる血管症のような合併症を含めて)の治療に有用であろう。加えて、脂肪細胞の分化または機能、および平滑筋細胞の機能(特にアテローム硬化症における)、および種々の血管性疾患における異常は、VAP-1SSAO阻害剤による治療に適している可能性がある。
【0009】
国際特許公開WO 03/006003は、炭素環式ヒドラジノ化合物、ならびにそれらの、ヒト血管接着タンパク質-1(VAP-1)を含めた、セミカルバジド感受性アミンオキシダーゼ(SSAO)阻害剤としての用途を開示している。
【0010】
銅含有アミンオキシダーゼ(CAO;EC1.4.3.6)は、機能的に多様なアミンオキシダーゼのスーパーファミリーに属する(ドークス(Dawkes)ら、2001年)。それらはまた、その酵素活性がカルボニル反応性化合物、セミカルバジドにより阻止され得ることから、セミカルバジド感受性アミンオキシダーゼとしても知られている。それらは、第一級アミンの、対応するアルデヒドへの酸化的脱アミノ化を銅依存性の反応において触媒し、分子状酸素が消費され、かつアンモニアが放出される。すべてのCAOに特徴的な性質は、2,4,5-トリヒドロキシフェニルアラニルキノン、トパキノン(TPQ)の酸化還元補因子としての使用である。CAOは、細菌、真菌、植物、および哺乳類を含めたいくつかの異な生物から単離されている。植物においては、CAOは、たとえば損傷治癒に関与しており、一方原核生物においては、CAOはその生物が窒素および炭素の供給源として種々のアミンを代謝的に利用できるようにする。より高等な真核生物では、生体アミンおよび他のアミンの代謝におけるそれらの役割の他には、CAOの生物学的機能についてはほとんどわかっていない。
【0011】
シェパード(Shapard)ら、2002年、は、6つの既知の銅含有アミンオキシダーゼに対して阻害剤を検査した際の、選択性および不活性化の速度における著しい差異を報告している。
【0012】
CAOの結晶構造は、4つの異なる種:大腸菌(ECAO:たとえば、タンパク質データバンク(Protein Data Bank)、PDBコード1oac)(パーソンズ(Parsons)ら、1995年)、エンドウ(Pisum sativum)(PSAO;PDBコード1ksi)(クマー(Kumar)ら、1996年)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)(HPAO;たとえば、PDBコード1a2v)(リ(Li)ら、1998年)、およびアルスロバクター・グロビフォルミス(Arthobacter globiformis)(AGAO;PDBコード1av4)(ウィルス(Wilce)ら、1997年)から解明されてきた。これらのホモダイマー構造はすべて、一つの類似した全体的な基本構造を有しており、それはドメインD1-D4に分けられることが可能であり、そのうちのD1ドメインは大腸菌においてのみ見出されている。ドメインD2およびD3は、各々〜100アミノ酸であって、α/β型の基本構造を有しており、一方最大のC末端ドメインD4は、〜400アミノ酸長であって、ダイマー化に必要な独特のβ-サンドイッチ基本構造を有する。D4ドメイン中に局在する活性部位は、CAOファミリー中に高度に保存されている。それはタンパク質内に深く埋められており、主としてD3およびD4ドメインからのアミノ酸によって囲まれた長いチャンネルを通してのみ接近することが可能である。D3ドメインのアミノ酸残基は、実際の活性部位よりも軽度に保存されており、活性部位へ導くキャビティがCAOの基質特異性の決定において非常に重要であることを示唆している。
【0013】
既知のCAOタンパク質の構造が全く類似しているとはいえ、アミノ酸レベルでの配列同一性はわずか25〜35%である。構造が分かっているCAOファミリーのメンバーに対するVAP-1の進化的関係は特徴づけられていないが、VAP-1のN-末端における膜貫通ドメインの存在は、可溶性CAOからの実質的な相違を示唆している。
【発明の開示】
【発明が解決しようとする課題】
【0014】
本発明はヒトVAP-1の結晶化およびX線分析を提供する。これは結晶化された最初の哺乳類のCAOである。
【課題を解決するための手段】
【0015】
発明の詳細な説明
本発明は、結晶性血管接着タンパク質-1(VAP-1)、特にヒトVAP-1に関する。
【0016】
本発明は、精製されたVAP-1を結晶化し、得られたVAP-1結晶を分析し、結晶パラメータおよびX線回折データを得るための、手段および方法を提供する。
【0017】
本発明はさらに、結晶性VAP-1を含んでいる組成物、およびかかる組成物の用途に関する。
【0018】
本発明はさらに、結晶性のヒトVAP-1の構造情報、より具体的には、ヒトVAP-1の活性部位キャビティについての情報を提供しており、前記キャビティは、表面では幅約20Å×約10Åであり、かつ深さ約15Åであり、そしてアミノ酸86-87、97、168-173、176-177、180、184、205-212、216、227、232-234、236-239、344、388-390、393-397、415-419、421、421-426、467-470、647-651、および758-761はヒトVAP-1の一つのモノマーに由来し、アミノ酸433-449、および451はヒトVAP-1の別のモノマーに由来する。さらに具体的には、前記活性部位は、アミノ酸残基Ala370、Tyr384、Asp386、Asn470、Tpq471、およびTyr473によって裏打ちされた、該部位の底の狭い約4.5×約4.5のキャビティの上端にアミノ酸Leu469をさらに含む。
【0019】
本発明はさらに、ヒトVAP-1タンパク質の三次元構造を規定する原子座標/X線回折データをその上に記憶した、コンピュータ可読媒体であって、適当な機械によって読取られ、かつ分子構造を決定するためのコンピュータプログラムによって処理されたとき、ヒトVAP-1タンパク質のフラグメントを含んでいる分子の結晶の三次元表現をディスプレイすることができる媒体を提供する。
【0020】
本発明はさらに、リガンドおよび/または阻害剤のde novo設計のためのin sillicoアッセイであって、(i)VAP-1の活性部位内の箇所と相互作用することができる官能基または低分子フラグメントの同定、および(ii)これらを一つの化合物中に結合すること、を含むアッセイを提供する。
【0021】
本発明はさらに、既知の化合物および化合物ライブラリを、VAP-1活性を阻害する能力についてスクリーニングするための、in sillicoのアッセイを提供する。
【0022】
本発明はさらに、慢性関節炎、炎症性腸疾患、および皮膚疾患、多発性硬化症といった、急性および慢性の炎症性症状および疾患、ならびに糖質代謝に関連した疾患(糖尿病および、糖尿病から結果として生じる血管症のような合併症を含めて)の治療に有用な、新規VAP-1阻害剤を提供する。加えて、かかる阻害剤は、脂肪細胞の分化または機能、および平滑筋細胞の機能(特にアテローム硬化症における)、および種々の血管性疾患における異常を治療するために有用であってよい。
【発明を実施するための最良の形態】
【0023】
以下において、本発明は、好ましい態様により、添付の図面を参照して、さらに詳細に記述される。
【0024】
発明の詳細な説明
本発明は、哺乳類の銅含有アミンオキシダーゼ、CAOの結晶、特にヒトVAP-1の結晶を提供する。
【0025】
本発明の結晶を成長させるためには、N-末端膜貫通領域を含めた完全長タンパク質は、全タンパク質の80%を超え、さらに好ましくは全タンパク質の90%を越え、最も好ましくは全タンパク質の95%を超えるように精製されねばならない。発現および精製目的のためには、完全長VAP-1をコードしている配列(配列番号1)が使用される。選んだ精製法は、精製されたタンパク質がそのCAO活性を保持しているようにすることが重要であり、それはベンジルアミンを基質として用いることにより測定されてよい。
【0026】
当業界で公知の多くのベクター-宿主系は、結晶化プロセスのためのタンパク質の組換え体産生に使用されてよい。可能なベクターは、これに制限されないが、プラスミドまたは修飾されたウイルスを含むが、ベクター系は用いた宿主細胞と適合性でなければならない。ヒトVAP-1は重度にグリコシル化されているため、酵母または動物細胞の宿主のような、真核生物の宿主が好ましい。チャイニーズ・ハムスター・オバリー(CHO)細胞は、最も好ましい宿主細胞を形成しており、これらの細胞は完全なグリコシル化が可能である。
【0027】
本発明の結晶を得るためには、これに制限されないが、バッチ晶析法、蒸気拡散法、およびマイクロ透析法を含めて、この技術において周知の任意の結晶化技術が使用されてよい。X線用の品質の結晶を得ることが必要な場合には、標準的なミクロおよび/またはマクロの種晶の添加が使用されて良い。
【0028】
本発明の結晶は、非対称ユニット中に二つの分子をもち、a=b=225.9Å、c=218.7Å、α=β=90°、γ=120°のユニットディメンションをもつ空間群P6522を作る(以下の実施例2を参照)。しかしながら、本発明はP6522を含むがこれに制限されない任意の空間群を形成する結晶を検討している。結晶は、4Åを超える、好ましくは3Åを超える、最も好ましくは2.8Åを超える分解能までを回折する。
【0029】
本発明の結晶から回折データを収集するべく、結晶は、グリセロールのような凍結保護物質、および窒素流中での瞬間凍結を用いて保護されてよい。X線回折データは、XDSプログラム(カブシュ(Kabsch)ら、1993年)により処理されてよいが、当業者に周知の任意の方法がX線回折データの処理に使用されてもよい。
【0030】
本発明によるヒトVAP-1の原子構造を決定するためには、分子置換(MR)、モデル構築、およびリファインメントが行なわれてもよい。
【0031】
VAP-1構造の決定には、当業界で公知のCAOの既知の構造または、上記および以下の実施例3のように決定され得る他のCAO構造を用いた、分子置換が使用されてよい。
【0032】
当業者には周知の任意の方法が、本発明のタンパク質の三次元構造を分子置換により決定するべく使用されてよい。たとえば、CCO4iプログラムスイートのプログラムAMOREは、ヒトVAP-1の構造を、本文に記述されたように導出された原子座標表を用いて決定するべく使用されてよい。
【0033】
原子座標は、コンピュータ可読媒体上に与えられてよい。かかる記憶媒体は、好ましくはランダムアクセスメモリ(RAM)、読み出し専用メモリ(たとえば、CD-ROM)、またはディスケットである。記憶媒体は、ローカルアクセス可能なコンピュータ上か、またはインターネットを通じたリモートアクセス可能なものでよい。
【0034】
三次元構造の最初のモデルは、プログラムO(ジョーンズ(Jones)ら、1991年)を用いて構築されてよく、たとえばCCP4iプログラムスイートのREFMACプログラムを用いてリファインされる。
【0035】
リファインされた三次元の本発明のVAP-1構造は、表1に示された原子座標および構造決定統計学によって表される。リファインされたVAP-1ダイマーのX線構造は、モノマーA中にはモデリングされない残基A1-A54、A202-A204、およびA762-A763をもつ残基A55-A761と、モノマーB中にはモデリングされない残基B1-B56、B203、B742-B746、およびB762-B763をもつ残基B57-B761からなる。残基A471およびB471は、トパキノン(Tpq471)であり、それらは内在性チロシン残基の翻訳後修飾によって形成され、触媒反応に関係する。双方のモノマーは、活性部位に一つの銅イオンを含有する。
【0036】
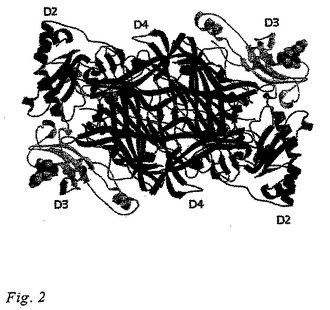
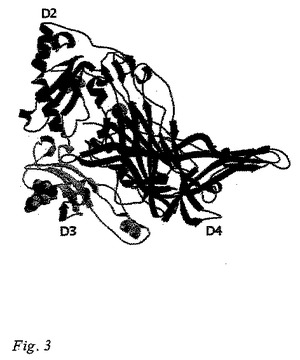
本発明の三次元構造から得られる情報は、ヒトVAP-1の活性部位(基質結合部位)が(1)銅イオンを配位している3つのヒスチジン、His520、His522、およびHis684、(2)触媒ベース、Asp386、ならびに(3)Tpq471のポジショニングに関係する、Tyr372およびAsn470、および(4)活性部位ゲート残基、Tyr384を含んでいることを示している。Tpq471は、VAP-1のX線構造では「オン・カッパー(on-copper)」(不活性な)立体配座にある。ヒトVAP-1構造の三次元表示は図2および3に示されている。
【0037】
VAP-1活性部位は深く埋められており、Tpq471は分子の表面から約22Åに位置している。この活性部位キャビティは、表面において幅約20Å×約10Åであり、深さ約15Åである。VAP-1のX線構造においては、キャビティの底では、Tpq471の上部に位置するLeu469が活性部位への入口をブロックしている。キャビティの一つの壁は、たとえば86-87、97、168-173、176-177、180,184、205-212、216、227、232-234、および236-239を含んでいるより小さなD2およびおD3ドメインに由来する残基と、443-449、および451を含んでいる他のモノマーから突出た長いβヘアピンアームに由来する残基とからなる。チャンネルの別の壁は、たとえば344、388-390、393-397、415-419、421,421-426、467-470、647-651、758-761を含んでいる触媒D4ドメインに由来する残基からなる。活性部位チャンネルの入口において、Asn232に付着したN-グリカンの一つの糖単位がX線構造中で見られる。Leu469の下では、約7Å長の活性部位キャビティは表面におけるよりもずっと狭く、約4.5×4.5Åのディメンションをもつほぼ円形である。チャンネルのこの部分は、残基Ala370、Tyr384、Asp386、Asn470、Tpq471、およびTyr473によって裏打ちされている。
【0038】
VAP-1X線構造は、驚いたことに、活性部位キャビティの独特の構造を明らかにしている。キャビティは、ECAO、HPAO、PSAO、およびAGAO構造内の狭い活性部位チャンネルに比較して、極めて幅広にマウントされかつオープンである。それゆえVAP-1の活性部位キャビティは、ECAO、HPAO、PSAO、およびAGAOの活性VAP-1キャビティよりもはるかに大きいリガンドおよび阻害剤構造物を収容することが可能である。VAP-1においてLeu469に相当する残基は、グリシン(ECAO、HPAO、およびAGAO)またはアラニン(PSAO)であり、したがって、それはECAO、HPAO、PSAO、およびAGAO構造物においては活性部位をブロックすることができない。この独特の構造についての知見は、ヒトVAP-1のファーマコフォアおよび、このキャビティにフィットする特異的リガンドおよび阻害剤構造物の設計に必要である。
【0039】
この情報は、ヒトVAP-1のファーマコフォア、すなわち生物活性に必要な特別の機能を表している化学的特徴および三次元的制約のコレクション、を定義するべく使用されることが可能である。ファーマコフォアは、好ましくは、水素結合供与体および受容体、荷電基、または疎水性部位といった、表面でアクセス可能な特徴を含む。かかる特徴は、生物活性に対するそれらの相対的重要性に基づき、ファーマコフォアモデルに包含されてよい。
【0040】
ファーマコフォアは、CATALYST、CERIUSといった可用性のコンピュータソフトウエアを用いるか、またはリード化合物についての既知の立体配座に基づく手動モデリングを使用することにより決定されてよい。ファーマコフォアは、以下にさらに詳細に記述されるような、可用性のコンピュータソフトウエアを用いて、in sillicoの化合物ライブラリをスクリーンするべく使用されてよい。
【0041】
したがって、本発明の一つの態様においては、分子モデリング技術がde novoの化合設計に使用される。デノボ化合物設計は、VAP-1のようなターゲット高分子の結合表面または活性部位が決定され、かつ、前記結合表面または活性部位と相互作用する化合物の合理的な設計のためのベースとして使用されるプロセスを指す。本発明による分子モデリングの段階は、ヒトVAP-1の原子座標および活性部位のモデルを利用する。好ましい態様においては、de novoの薬物設計は、ヒトVAP-1の結合表面における活性部位と相互作用することができる官能基または小さい分子フラグメントの同定、およびこれらの基またはフラグメントを結合して単一の化合物にすることを含む。ヒトVAP-1の特異的活性部位と相互作用する官能基または小さい分子フラグメントがひとたび同定されれば、好適なサイズを有するフラグメントをブリッジングすること、および活性部位にフィットするためのジオメトリの双方を用いて、単一の化合物へ結合されることが可能である。
【0042】
適切な官能基およびフラグメントの結合は手動で行なわれてもよいが、QUANTAまたはSYBYLといった適当なソフトウエアを使用することが好ましい。この技術において周知のさらなるソフトウエアは、たとえば、多数の官能基をデータベースからの分子鋳型と結びつけるHOOK、および制約された非環式分子に対して連結ユニットを設計するためのCAVEATである。de novoの化合物設計のための、他のコンピュータ・ベースのアプローチは、LUDI、SPROUT、およびLEAP-FROGを含む。
【0043】
本発明は、阻害性化合物を含め、ヒトVAP-1の活性部位に対して結合することが可能な、化学的実体および化合物を設計、同定、および合成するための、分子設計技術の使用を可能にする。ヒトVAP-1の原子座標は、ヒトVAP-1の潜在的な阻害剤を同定するべく、たとえば、GOLD、GRAM、DOCK、またはAUTODOCKといったドッキングプログラムを用いたコンピュータモデリングと一緒に使用されてよい。この手法は、潜在的阻害剤の形状および化学構造が、いかに充分に活性部位に相補的であるかを確認するための、VAP-1の活性部位に対する潜在的阻害剤のコンピュータフィッティングを含むことが可能である。ドッキングプログラムを用いたモデリングにより設計された潜在的阻害剤の実例は、以下に記述される一般式(I)に適合してもよい。
【0044】
本発明はさらに、ヒトVAP-1の活性部位と相互作用する化合物を同定するためのin sillicoの方法であって、(a)ヒトVAP-1のリガンド結合ドメインの原子座標をコンピュータの記憶媒体に供給すること、および(b)該コンピュータを使用して分子モデリング技術を該座標に適用すること、の段階を含む方法を提供する。
【0045】
上記の構造モデルはさらに、構築された分子のライブラリおよびSybyl v.6.9による最小化されたエネルギーの検査に使用され、VAP-1阻害剤として作用するそれらの能力について評価された。in sillicoスクリーニングに先立ち、VAP-1のX線構造は実施例4に記述されたように、CAOの「オフ・カッパー(off-copper)」の活性のある立体配座を模すべく修飾された。
【0046】
本発明のさらなる態様においては、in sillico法はヒトVAP-1の酵素活性を特異的に阻害する化合物の同定に使用されており、以下の段階:前記分子モデリング技術により同定される化合物を提供すること、前記化合物をヒトVAP-1と接触させること、および、相互作用を検出すること、を特徴とする。
【0047】
三次元構造情報および、VAP-1の前記構造情報に関連した原子座標は、合理的な薬物設計において有用であり、VAP-1に結合しかつVAP-1の酵素活性を阻害する阻害性化合物の同定法を提供する。SSAO活性を有する酵素のための、前記潜在的阻害剤を同定するための方法は、(a)表1にリストされたその原子座標をベースとしたVAP-1の三次元構造を用いること;(b)前記三次元構造を用いて前記潜在的阻害剤を設計または選択すること;(c)前記潜在的阻害剤を合成すること;(d)前記潜在的阻害剤を前記酵素と接触させること;および(e)前記SSAO活性を阻害する前記阻害剤の能力を測定すること、の段階を含む。
【0048】
本発明はさらに、ヒトVAP-1の酵素活性を阻害する化合物か、またはVAP-1の活性部位と相互作用する化合物を同定するための、本発明のin sillico法を用いて同定された化合物を包含する。
【0049】
本発明の方法により同定可能な潜在的VAP-1阻害剤の実例は、式(I):
【化1】
[式中、R1は水素、低級アルキル、または任意に置換されたフェニルまたはヘテロアリール基であり;
R2は水素または低級アルキルであるか、または
R1およびR2は、それらが結合している窒素原子と一緒になって飽和複素環を形成してもよく;
R3-R5は各々独立して水素、低級アルキル、アラルキル、任意に置換されたフェニルまたはヘテロアリール基を表しているか、または
R2およびR3は、それらが結合している原子と一緒になって飽和複素環を形成してもよく、あるいは
R3およびR5は、それらが結合している炭素原子と一緒になって飽和炭素環を形成してもよく;
R6はナフチル、フェニル、置換されたフェニルまたはヘテロアリール基であり;
R7は水素、低級アルキルまたはアラルキルであり;
nは1、2または3であり;かつ
X=O、S、SO、SO2またはNR2である]
により表されてよい。
【0050】
他のタイプの化合物も同様に、本発明による方法およびモデルを用いて充分に同定されてよい。当業者は、本文に記述された原子座標およびファーマコフォアに基づき、ヒトVAP-1の活性部位と相互作用する化合物を同定してよい。一般に、かかる潜在的阻害剤化合物は、前文に例示されたような、アミンまたはヒドラジンのようなカルボニル基反応因子によって特徴づけられてよいが、活性部位キャビティの4.5×4.5Åの底部分にはまりかつ活性部位の底部においてTPQに結合することが可能な、細い突出した部分をもつ他の構造物も、同様に潜在的阻害剤である。本発明の化合物のこの突出した部分は、この化合物の約7Å長の比較的疎水性の中央部分へ連結され、ともに「リンカー」として作用して、突出部分が活性部位へアクセスすることを可能にし、キャビティを裏打ちするアミノ酸残基Ala370、Tyr384、Asp386、Asn470、Tpq471、およびTyr473と相互作用する。最後に、本発明の化合物は、広いキャビティをふさぎ、かつ活性部位キャビティーの壁の代表的なアミノ酸残86-87、97、168-173、176-177、180、184、205-212、216、227、232-234、236-239、344、388-390、393-397、415-419、421、421-426、467-470、647-651、758-761、443-449、および451と相互作用する、よりかさばった部分を含む。
【0051】
さらに、本発明は同定された化合物の予想された活性を確かめるためのアッセイおよび手段を提供する。
【0052】
本発明はさらに、本発明のスクリーニングアッセイにより同定された化合物の、慢性関節炎、炎症性腸疾患、皮膚疾患、および多発性硬化症、のような慢性または急性の炎症性症状または疾患、ならびに糖質代謝に関連した疾患(糖尿病および、糖尿病から結果として生じる合併症、たとえば網膜症、腎症、および神経障害のような血管症を含めて)の治療のための薬の調製のための用途を包含する。加えて、かかる阻害剤は、脂肪細胞の分化または機能、および平滑筋細胞の機能(特にアテローム硬化症における)、および種々の血管性疾患における異常の治療に有用であってよい。
【0053】
本発明はさらに、本スクリーニングアッセイによって同定された化合物を含有する薬剤組成物を包含する。
【0054】
以下の実施例は、本発明を例証する。
【実施例1】
【0055】
ヒトVAP-1の産生および精製
N-末端膜貫通ドメインをもつ完全長のタンパク質は、スミス(Smith)ら、1998年、において記述されたように、グリコシル化コンピテントCHO細胞において発現された。収穫された細胞は、溶解緩衝液(150mM NaCl、10mM トリス塩基pH7.2、1.5mMのMgCl2、1%NP40)を用いて溶解された。浄化された細胞溶解物は、モノクローナル抗体アフィニティカラムに基づき、かつAKTATM精製システム(アマシャム・バイオテック(Amersham Biotech))を用いた、HVAP-1の精製に使用された。タンパク質は、アフィニティクロマトグラフィーを用いて均一(>95%)まで精製され、精製の後、VAP-1タンパク質の存在、90-および170〜180kDのバンドは、スミスら、1998年、により記述されたように、銀染色SDS-pageによって確認された。
【0056】
精製されたタンパク質は、基質としてベンジルアミンを用いて測定されるような、そのCAO活性を保持していた。アミンオキシダーゼ活性は、記載のスペクトロメトリー法(Haltら、1997)、基質として200μl体積及び1mMのベンジルアミンを使用することで測定された。吸光度の変化は、490nmにおいて、VictorTMマルチラベルプレートカウンタ(パーキンエルマー・ライフ・サイエンシズ(PerkinElmer Life Sciences))においてモニターされた。
【0057】
ヒトVAP-1のコード領域のヌクレオチド配列は、配列番号1として列挙されている配列に示されており、そして配列番号2のようなアミノ酸配列(763アミノ酸)に相当する。
【実施例2】
【0058】
結晶化および予備的分析
hVAP-1の最初の結晶化条件は、ウィザード(Wizard)Iランダムスパース行列結晶化スクリーン(エメラルド・バイオストラクチャーズ・インク(Emerald BioStructures, Inc., USA))および蒸気拡散法を用いて、室温でスクリーンされた。小さい六方晶の結晶が、1.0M酒石酸K/Na、100mMイミダゾール(pH8.0)、および200mM NaClを含有する条件において、数ヶ月間のインキュベーションの後に得られた。懸滴は、10mMリン酸カリウム緩衝液(pH7.2)中の2μlのタンパク質試料(1.0mg/ml)、および2μlの貯蔵溶液を含有していた。最適化の後、1.0M酒石酸K/Na、100mMイミダゾール(pH7.8)、および沈殿剤としての150〜250mM NaClからなる貯蔵溶液を用いて、最良の結晶が得られた。結晶は数日間で形成され、約0.15×0.15×0.1mmの最終サイズに成長した(図1)。
【0059】
一つの結晶がキャピラリーにマウントされ、予備的X線分析が、回転陽極放射線源(Cu Kα放射線、50kV、150mA)、およびMAR345イメージングプレート検出器を用いて、インハウスで行なわれた。結晶は、しかしながら、8Åまでの分解能しか回折せず、空間群は正確には測定できなかった。さらなるX線分析ならびにデータ収集はすべて、MARリサーチCCD検出器を具備したビームライン(beamline)X11(EMBL/DESY、ドイツ、ハンブルグ)において、シンクロトロン照射を用いて行なわれた。データ収集には、結晶は20%(v/v)グリセロールを用いて凍結保護され、100Kの窒素流中で瞬間凍結された。3つの異なる結晶から収集された回折データは、プログラムXDSにより処理された。溶媒含有率およびマシューズ(Matthews)係数は、分子量をモノマーあたり90kDと仮定し、CCP4スイートを用いて計算された。
【0060】
最良に見える六方晶結晶は、1.0M酒石酸K/Na、100mMイミダゾール(pH7.8)、および沈殿剤として150〜250mMのNaClを用いて得られた。結晶は、a,b=225.9Å、C=218.7Å、α,β=90°、およびγ=120°のユニットセルディメンションセルをもつ、典型的なサイズ0.15×0.15×0.1mmに成長した(図1a)。回折データ統計(表1)によれば、いくつかのフレームでは3.0Åをこえる分解能に相当する反射が観察されたにもかかわらず、VAP-1触媒の回折限界は、3.2Åであった(図1b)。結晶は、空間群P6522に属する。非対称ユニットあたり一つのダイマー(180kD)の存在を仮定すると、マシューズ係数は4.5Å3/Da、および溶媒含有率は72%である。結晶パラメータおよび回折データ統計は、表1に要約されている。
【0061】
【表1】
【実施例3】
【0062】
構造決定
VAP-1の構造は、CCP4iプログラムスイート(コラボラティブ・コンピュテイショナル・プロジェクト(Collaborative Computational Project(共同計算プロジェクト))、1994年)のプログラムAMORE(ナバザ(Navaza)、1994年)を用いた分子置換により解明された。この方法は、VAP-1の空間群が、非対称ユニットあたり一つの生物学的単位である一つのダイマーをもつ、P6522であったことを立証した。分子置換において検査された大腸菌(Escherichia coli)CAO(残基93-720;PDBコード10AC)、エンドウ(Pisum sativum)CAO(残基7-634;PDBコード1KSI)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)CAO(残基22-655;PDBコード1A2V)、およびアルスロバクター・グロビフォルミス(Arthobacter globiformis)CAO(残基9-623;PDBコード1AV4)の二量体ポリアラニン主鎖のうち、エンドウの構造は最良の相関係数(44.1%)およびR因子(53.4%)を与え、サーチモデルが使用された。
【0063】
電子密度マップはCCP4iスイートのFFTを用いて計算され、構築の初期においてであっても、いくつかのフラグメントにおいては、VAP-1ポリペプチドをトレースするのに充分に予測可能であった。モデルはプログラムOを用いて手動により構築され、CCP4iスイートのREFMAC5.1.24(ムルシュドフ(Murshudov)ら、1997年)により、分解能3.2Åまでリファインされた。
【0064】
リファインメントの多数のサイクルの間に、構造に側鎖が逐次的に加えられた。最終モデルにおいては、以下のアミノ酸だけはトレースできず、したがって、この構造には含まれていない:A1-A54、A202-A204、A762-A763、B1-B56、B203、B742-B746、およびB762-B763。トパキノン残基のための座標は、ヘテロコンパウンド・インフォメーションセンター(Hetero-compoun Information Centre)、ウプサラ(クレイウェグト&ジョーンズ(Kleywegt and Jones)、1998年)から取得され、そのための辞書はプログラムPRODRG(ヴァンアールテン(van Aalten)ら、1996年)を用いて生成された。最終的なVAP-1モデルの立体化学的な質は、PROCHECK(ラスコウスキー(Laskowski)ら、1993年)を用いて査定され、モデル中の1401個のアミノ酸のうち、84.0%がラマチャンドラン・プロット(ラマチャンドラン&サジーセクハラン(Ramachandran and Sasisekharan)、1968年)中の最も好ましい領域に生じ、わずか0.5%が寛大に許されるか、または許されない領域に生じた。構造決定統計の要約は、表1に呈示されている。ヒトVAP-1結晶構造の原子座標(および構造因子)は、PDBに登録コード1PU4および1US1で寄託されている。
【0065】
ヒトVAP-1の結晶形状はホモダイマーであり、各サブユニットはドメインD2、D3、およびD4を含有している。VAP-1構造の概略説明図は、図2に示されている。結晶構造に見られるD2、D3、およびD4ドメインは、各々残基約55-169、170-300、および301-761(各々、配列番号3、配列番号4、および配列番号5)からなる。他の既知のアミンオキシダーゼの構造に比較しての、ヒトVAP-1の構造における主要な差異は、ドメインD4にも差異は存在するが、ドメインD2およびD3において見られる。
【実施例4】
【0066】
潜在的阻害剤のインシリコスクリーニング法
本実施例は、アリールオキシメチル置換ヒドラジンアルコール化合物のライブラリが、実施例3に記述されたVAP-1の結合キャビティ内へドッキングすることにより、in sillicoにおいていかにしてスクリーンされるかを示している。スクリーンされた化合物は、式
【化2】
[式中、R1は水素、低級アルキルまたは任意に置換されたフェニルまたはヘテロアリール基であり;
R2は水素または低級アルキルであるか、または
R1およびR2は、それらが結合している窒素原子と一緒になって飽和複素環を形成してもよく;
R3-R5は各々独立して水素、低級アルキル、アラルキル、任意に置換されたフェニルまたはヘテロアリール基を表しているか、または
R2およびR3は、それらが結合している原子と一緒になって飽和複素環を形成してもよく、あるいは
R3およびR5は、それらが結合している炭素原子と一緒になって飽和炭素環を形成してもよく;
R6はナフチル、フェニル、置換フェニルまたはヘテロアリール基であり;
R7は水素、低級アルキルまたはアラルキルであり;
nは1、2または3であり;かつ
X=O、S、SO、SO2またはNR2である]
を有する。
【0067】
合成および/または構築され、かつin sillicoでスクリーンされた化合物の実例は、表2に示されている。
【0068】
【表2】
【0069】
X線構造の修飾およびリガンドドッキング
VAP-1X線構造は、以下に記述されたように、CAOの「オフ・カッパー(off-copper)」の活性のある立体配座を模すべく修飾された。最初に、トパキノンは、ECAO構造(PDBコード1D6Z(ウィルモット(wilmot)ら、1999年)にしたがって、活性のあるイミノキノン形へ修飾された。第二に、VAP-1構造の活性部位キャビティは、ドッキング研究に先立ち、ボディル(Bodil)モデリングパッケージ(http://www/abo.f//fak/mnf/reseach/jonnson/bodil.html)において側鎖ロータマー(Phe389、Tyr394、Asp386、およびLeu469)を選ぶことにより修飾され、それにより活性部位のイミノキノンをリガンドに関してさらに近づきやすくした。水素原子は、スクリーニングおよびドッキングに使用されたVAP-1構造に対し、何ら側鎖調整なしに、プログラムReduce v.2.15を用いて付加された(ワード(Word)ら、1999年)。
【0070】
潜在的リガンドのR-およびS-エナンチオマーが構築され、Sybyl v.6.9(トリポス・アソシエイツ(Tripos Associates)、米国、セントルイス)によりエネルギー最小化された。リガンドはイミノキノン残基へ共有結合により結合され、手動により結合キャビティ内へドッキングされた。
【0071】
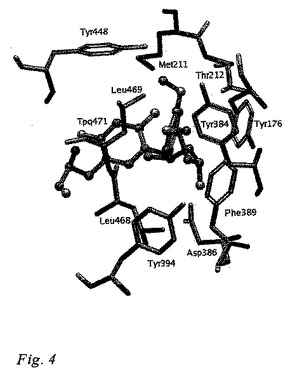
リガンド結合モード
図4は、スクリーンされたリガンドの実例を示している。ボディル・モデリングパッケージを用いて作成された図3には、BTT-2071(化合物9)のS-エナンチオマーの結合モードが存在する。BTT-2071(化合物9)のS-エナンチオマーは、ドッキングシミュレーションに基づけば、VAP-1構造と最も広範囲な相互作用を形成することができることから、図中の実例リガンドとなるべく選ばれた。
【0072】
VAP-1構造に基づけば、Xが酸素原子のかわりに硫黄原子であるリガンドは、VAP-1の相互作用表面が疎水性であることから、より有利にVAP-1と相互作用する。
【0073】
硫黄原子を含んでいるリガンドの疎水性部分は、親水性残基(Leu468、Leu469、Phe389、およびMet211)に対して固く締まる(pack)。リガンドのパラメトキシ基中の酸素原子は、側鎖のThr212の酸素に対して水素結合距離にあり、一方、リガンドのメタメトキシル基中の酸素原子は、側鎖のTyr394の水酸基に対して水素結合距離にある。Phe389は、リガンドの芳香環と容易に相互作用することができる位置に局在する。パラ-およびメタ-メトキシル基中のメチル基は、各々Tyr448およびTyr176と接触している。S-エナンチオマー中の水酸基は、Asp386と最適な水素結合を形成することが可能であるのに対し、R-エナンチオマーにおいては、水素結合距離およびジオメトリは最適ではない。
【実施例5】
【0074】
同定された潜在的阻害剤のVAP-1阻害効果を立証するためのインビトロアッセイ
VAP-1SSAO活性は、本質的にはモノアミンオキシダーゼおよび類縁酵素について記述されたような組合わされた比色法を用いて測定された。チャイニーズハムスターオバリー(CHO)において発現された細胞組換えヒトVAP-1 SSAOが、活性測定のためのVAP-1 SSAOの供給源として使用された。天然のCHO細胞は、無視できる程度のSSAO活性を有する。これらの細胞およびその培養物は、従来に記述されている(スミス(Smith D J)ら、1998年)。
【0075】
細胞溶解物は、約3.6×108個の細胞を25mlの溶解緩衝液(150mM NaCl、10mMトリス-塩基 pH7.2、1.5mMのMgCl2、1%NP40)中に懸諾すること、および回転台の上で4℃において一晩インキュベートすることにより調製された。溶解物は、室温において18000gで5分間遠心分離することにより透明にされ、上清はアッセイに直接使用された。
【0076】
VAP-1 SSAOアッセイは、96穴マイクロタイタープレートにおいて、以下のように行なわれた。各ウェルに対し、必要であれば、あらかじめ決められた量の阻害剤が添加された。阻害剤の量は各アッセイで異なったが、全般的には1nM〜50μMの濃度であった。対照は阻害剤を欠いていた。阻害剤は、水中で、全体積中に20:1であった。次に、以下の試薬が添加された。全反応体積200μlに対し、0.2Mリン酸カリウム緩衝液pH7.6、1mMの2,4-ジクロロフェノール、500μMの4-アミノアンチピリン、および4U/mlの西洋ワサビペルオキシダーゼを含有する、45μlの新たに作成された色素溶液、および、1時間当たり0.6のA490の変化を引き起こした量の、VAP-1 SSAOを含有しているCHO細胞溶解物。
【0077】
プレートは37℃で30分間インキュベートされ、バックグラウンド吸光度は、ウォラク・ビクター(Walac Victor)IIマルチラベルカウンタを用いて、490nmにおいて測定された。酵素反応を開始するためには、20μlの10mMベンジルアミン(最終濃度=1mM)が添加され、そしてプレートは37℃において1時間インキュベートされた。
【0078】
VAP-1 SSAO活性を反映する吸光度の増大は、490nmにおいて測定された。阻害は、バックグラウンド吸収について補正した後に、対照と比較されたパーセント阻害として表され、IC50値はグラフパッド・プリズム(GraphPad Prism)を用いて計算された。
【実施例6】
【0079】
VAP-1 SSAO活性対全ラットMAO活性の比較
ラットMAOは、ラット肝より、1gの肝試料を14mlのKCl-EDTA溶液中で数回洗浄して血液をすべて除去することにより調製された。次いで、1gの肝試料は、4mlの氷冷リン酸カリウム緩衝液(0.1M、pH7.4)中で、ウルトラ・トラックス(Ultra-Turrax)ホモジナイザ(設定11000rpm、4×10秒)によりホモジナイズされた。4℃において500gで10分間の遠心分離の後、上清は注意深く取出され、4℃において12300gで15分間遠心分離された。上清は排出され、沈降されたミトコンドリアは4mlの新鮮なリン酸緩衝液中に再懸濁され、前と同様に遠心分離された。ミトコンドリアは4mlの辛酸緩衝液中に懸濁され、ウルトラ・トラックス(Ultra-Turrax)ホモジナイザ(設定11000rpm、2×10秒)によりホモジナイズされた。ミトコンドリアプレパラートはアリコートに分けられ、-70℃に貯蔵された。
【0080】
全MAO活性は、2,4-ジクロロフェノールが1mMバニリン酸で置換えられたこと以外は、VAP-1 SSAOと類似の方法で測定された。各ウエルに対し、必要な場合には、あらかじめ決められた量の阻害剤が添加された。阻害剤の量は各アッセイで異なったが、全般的には10nM〜800mMの濃度であった。対照は阻害剤を欠いていた。阻害剤は、水中で、全体積中に20:1であった。次に、以下の試薬が添加された。全反応体積300μlに対し、0.2Mリン酸カリウム緩衝液pH7.6、50μlの新たに作成された色素溶液(上記と同じ)、および50μlのMAO標品。
【0081】
プレートは37℃で30分間インキュベートされ、バックグラウンド吸光度は、ウォラク・ビクター(Walac Victor)IIマルチラベルカウンタを用いて、490nmにおいて測定された。酵素反応を開始するためには、20μlの5mMトリアミン(最終濃度=0.5mM)が添加され、プレートは37℃において1時間インキュベートされた。MAO活性を反映する吸光度の増大は、490nmにおいて測定された。阻害は、バックグラウンド吸収について修正した後に、対照と比較されたパーセント阻害として表され、IC50値はグラフパッド・プリズム(GraphPad Prism)を用いて計算された。MAO阻害のための陽性対照として、クロルギリンおよぎパルギリン(各々MAO-AおよびMAO-Bの阻害剤)が0.5μMにて、いくつかのウエルに添加された。
【0082】
表3の化合物の、VAP-1 SSAOに対してMAOを超える特異性をもってVAP-1 SSAOを阻害する能力は、表2に示されている。結果は、本発明の化合物がヒトVAP-1 SSAO活性の特異阻害剤であることを示している。本発明の化合物は、そえ故、ヒトの接着性分子VAP-1のSSAO活性が役割を果たしている疾病および症状の治療において、治療上の有用性をもつことが期待される。
【0083】
【表3】
【0084】
当業者には、技術が進歩するにつれて、発明の概念が様々な方法で遂行され得ることが明らかであろう。本発明およびその態様は、上記の実施例に制限されず、特許請求の範囲内で変更されてよい。
【0085】
参考文献
コラボラティブ・コンピュテイショナル・プロジェクト(Collaborative Computational Project(共同計算プロジェクト))、Acta Cryst、1994年、第D51巻、p.760-763。
ドークス(Dawkes, H. C.)&フィリップス(Phillips, S. E.)著、Curr Opin Struct Biol、2001年、第11巻、p.666-673。
カブシュ(Kabsch, W.)著、Journal of Applied Crystallography、1993年、第26巻、p.795-800。
クマー(Kumar, V.)、ドゥーリー(Dooley, D. M.)、フリーマン(Freeman, H. G.)、ガス(Guss, J. M.)、ハーベイ(Harvey, I.)、マクグリル(McGuril, M. A.)、ウィルス(Wilce, M. C.)、およびズバク(Zubak, V. M.)著、Structure、1996年、第4巻、p.943-955。
リ(Li, R.)、クリンマン(Klinman, J. P.)&マテウス(Mathews, F. S.)著、Structure、1998年、第6巻、p.293-307。
パーソンズ(Parsons, M. R.)、コンベリー(Convery, M. A.)、ウイルモット(Wilmot, C. M.)、ヤダブ(Yadav, K. D.)、ブレーカリー(Blakeley, V.)、コーナー(Corner, A. S.)、フィリップス(Phillips, S. E.)、マクファーソン(McPharson, M. J.)、およびノウルズ(Knowles)著、Structure、1995年、第3巻、p.1171-1184。
サルミ(Salmi, M.)&ヤルカネン(Jalkanen, S.)著、J Exp Med、1996年、第183巻、p.569-579。
サルミ(Salmi, M.)&ヤルカネン(Jalkanen, S.)著、Trends Immunol、2001年、第22巻、p.211-216。
スミス(Smith, D. J.)、サルミ(Salmi, M.)、ボノ(Bono, P.)、ヘルマン(Hellman, J.)、ロイ(Leu, T.)、およびヤルカネン(Jalkanen, S.)著、J Exp Med、1998年、第188巻、p.17-27。
ウィルス(Wilce, M. C.)、ドゥーリー(Dooley, D. M.)、フリーマン(Freeman, H. C.)、ガス(Guss, J. M.)、マツナミ(Matsunami, H.)、マクインタイヤー(McIntire, W.S.)、ルギエロ(Ruggiero, C. E.)、タニザキ(Tanizaki, K.)、およびヤマグチ(Yamaguchi, H.)著、Biochemistry、1997年、第36巻、p.16116-16133。
ジョーンズ(Jones, T. A.)、ズー(Zou, J. Y.)、カウマン(Cowman, S. W.)、およびケルドガード(Kjeldgaard)著、Acta Crystallogr、1991年、第A47巻、パート2、p.110-119。
クレイウェグト(Kleywegt、G. J.)&ジョーンズ(Jones, T. A.)著、Acta Cryst、1998年、第D54巻、p.1119-1131。
ラスコウスキー(Laskowski, R. A.)、マッカーサー(Macarthur, M. W.)、モス(Moss, D. S.)、およびソーントン(Thornton, J. M.)著、Journal of Applied Crystallography、1993年、第26巻、p.283-291。
ムルシュドフ(Murshudov, G. N.)、ヴァギン(Vagin, A. A.)、ドドゥソン(Dodson, E. J.)著、Acta Crystallography、1997年、第D53巻、p.240-255。
ナバザ(Navaza, J.)著、Amore - an Automated Package for Molecular Replacement(AMoRe:分子置換用の自動化されたパッケージ). Acta Cryst、1994年、第A50巻、p.157-163。
ラマチャンドラン(Ramachandran, G. H.)&サジーセクハラン(Sasisekharan, V.)著、Adv Protein Chem、1968年、第23巻、p.283-438。
シェパード(Shepard, E. M.)、スミス(Smith, J.)、ブラッドリー(Bradley, O.E. )、クーチャー(Kuchar, J. A.)、ローレンス(Lawrence, M. S.)、およびドゥーリー(Dooley, D. M.)著、Eurl J. Biochem.、2002年、第269巻、p.3645-3658。
ヴァンアールテン(van Aalten, D. M.)、バイウォーター(Bywater, R.)、ファインドレイ(Findlay, J. B.)、へンドリッヒ(Hendlich, M.)、ホフト(Hooft, R. M.)、およびブリエンド(Vriend, G.)著、J Comput Aided Mol Des、1996年、第10巻、p.255-262。
ウィルモット(Wilmot, C. M.)、ホーイド(Hajdu, J.)、マクファーソン(McPherson, M. J.)、ノウルズ(Knowles, P. F.)、およびフィリップス(Phillips, S. E.)著、Science、1999年、第286巻、第5445号、p.1724-8。
ワード(Word, J. M.)、ローベル(Lovell, S. C.)、リチャードソン(Richardson, J. S.)、およびリチャードソン(Richardson, D. C.)著、J Mol Biol、1999年、第285巻、p.1735-47。
【図面の簡単な説明】
【0086】
【図1】パネルAにおいて六方晶のVAP-1結晶を、そしてパネルBにおいて典型的な回折パターンを示している。
【図2】ヒトVAP-1ダイマーの結晶構造を示しており、糖単位は区間充填モデルとして描かれており、双方のモノマー中のTpg471はD4ドメイン中の球として示されている。
【図3】VAP-1ダイマーの結晶構造の一つのモノマーを示している。
【図4】VAP-1構造におけるVAP-1阻害剤の結合モードを示している。視点は、表面から活性部位へ下っている。
【図1A】
【図1B】
【技術分野】
【0001】
本発明は、結晶性ヒト血管接着タンパク質-1(VAP-1)、および特に、結晶性ヒトVAP-1の構造情報の、リガンドおよび/または阻害剤の同定、設計および製造のため、ならびに、かかるリガンドおよび/または阻害剤の検出のための、インシリコおよびインビトロのスクリーニングアッセイのための用途に関する。本発明はさらに、本発明のアッセイにより同定される阻害剤であって、急性および慢性の炎症性症状、糖質代謝関連疾患、脂肪細胞または平滑筋細胞機能における異常、および種々の血管疾患の治療において有用な阻害剤に関する。
【背景技術】
【0002】
発明の背景
生理的免疫監視機構は、血液と、種々のリンパ器官との間のリンパ球の継続的な監視に依存している。正常な非リンパ組織には、リンパ球は存在しないか、または非常に低レベルに存在するが、多くの炎症性性の疾病状態においては、莫大な数のリンパ球が種々の冒された組織および器官中に集積することがある。血管からのリンパ球の退出を制御している重要な分子の一つは、米国特許第5,580,780号に開示された血管接着タンパク質-1(VAP-1)である。VAP-1は、ホモダイマーの170〜180kDaの内皮糖タンパク質である。VAP-1は、ヒトの組織切片において、細静脈へのリンパ球の結合を仲介する。VAP-1は、重度にグリコシル化されており、糖部分は接着機能にとり重要である(サルミ(Salmi)ら、1996年)。VAP-1の接着機能を阻止すると、炎症性組織に浸潤する細胞の数が減少して炎症を散らすことができる。VAP-1はしたがって、抗炎症薬開発のターゲットである。
【0003】
ヒトの血管接着タンパク質-1(VAP-1)は、接着性および酵素的性質の双方をもつ、膜結合多機能糖タンパク質である。VAP-1のクローニングは、驚いたことに、それがセミカルバジド感受性モノアミンオキシダーゼ(SSAO;EC 1.4.3.6)に属することを明らかにした(国際特許公開 WO 98/53049)。VAP-1は、触媒活性のある大きい細胞外ドメインをもつII型の主要膜タンパク質である。したがって、VAP-1は細胞外酵素である。白血球-内皮相互作用におけるSSAO活性の役割も、その生理的基質も、どちらも充分には定義されていない。VAP-1は、哺乳類におけるこの酵素群のうちで初めて分子的に定義された膜貫通メンバーであり、細胞のSSAO活性の90%を占めている。とりわけ、SSAOは、細胞内局在、基質、補因子、阻害剤、およびタンパク質配列の点において、充分に特徴づけされたモノアミンオキシダーゼAおよびBとは異なっている。
【0004】
SSAO反応は生化学用語においては、1950年代から知られているが、これらの酵素の生理的機能は謎のままであった。SSAOの生理的基質は分かっていない。しかしながら二つの可能性のある候補、メチルアミンおおよびアミノアセトンは、ヒトでは中間代謝を通じて形成され、かつin vitroおよびin vivoにおいてSSAOにより脱アミノ化されることが可能である。
【0005】
VAP-1 SSAO活性は、白血球の表面に発現されたVAP-1リガンド上に呈示されたアミン基質との直接的な相互作用を含む新規な機構により、内皮細胞への白血球接着の経路に直接的に関与していると提案されていた(サルミ(salmi)ら、2001年)。この発表は、内皮への白血球の接着プロセスにおける、VAP-1 SSAO活性の直接的な関与を述べている。したがって、VAP-1 SSAO活性の阻害剤は、炎症領域における白血球接着を減少させ、それにより炎症領域内への白血球の通行を、それゆえ炎症プロセスそのものを低減することが期待できた。
【0006】
ヒトの臨床組織サンプルにおいては、VAP-1の発現は炎症の部位において誘導される。この増大されたVAP-1レベルは、血中に存在するモノアミンに対する、VAP-1SSAO細胞外ドメインの作用から発生される、H2O2の増大された産生をもたらすことが可能である。内皮細胞という局部的な環境におけるこのH2O2の発生は、別の細胞事象を開始することも可能であろう。H2O2は、他の接着性分子をアップレギュレートすることができるシグナリング分子として知られており、この増大された接着分子の発現が、VAP-1の発現されている領域内への白血球の交通の増強をもたらすのかもしれない。他のVAP-1 SSAO反応産物もまた、炎症プロセスにも寄与する生物学的作用を有しているのかもしれない。したがって、VAP-1 SSAO活性の産物は、炎症プロセスのエスカレーションに関与してよく、それは特異的なSSAO阻害剤によって阻止されることも可能である。
【0007】
VAP-1 SSAOは、VAP-1 SSAOの循環性アミン基質の増大されたレベルに関連する多くの他の病的症状に関係し得る。これらの基質の酸化的脱アミノ化は、内皮細胞という局所環境における毒性アルデヒドおよび酸素ラジカルのレベルの上昇をもたらすこととなり、そのことが細胞を損傷して血管損傷をもたらすことも可能であろう。メチルアミンおよびアミノアセトンの増大されたレベルは、I型およびII型の糖尿病をもつ患者において報告されており、後期糖尿病に見られる網膜症、神経障害、腎症のような血管症は、VAP-1SSAOの特異害剤を用いて治療されることも可能である。
【0008】
VAP-1活性を調整する特異的なVAP-1SSAO阻害剤の開発は、慢性関節炎、炎症性腸疾患、および皮膚疾患のような、急性および慢性の炎症症状または疾患、ならびに糖質代謝に関連した疾患(糖尿病および、糖尿病から結果として生じる血管症のような合併症を含めて)の治療に有用であろう。加えて、脂肪細胞の分化または機能、および平滑筋細胞の機能(特にアテローム硬化症における)、および種々の血管性疾患における異常は、VAP-1SSAO阻害剤による治療に適している可能性がある。
【0009】
国際特許公開WO 03/006003は、炭素環式ヒドラジノ化合物、ならびにそれらの、ヒト血管接着タンパク質-1(VAP-1)を含めた、セミカルバジド感受性アミンオキシダーゼ(SSAO)阻害剤としての用途を開示している。
【0010】
銅含有アミンオキシダーゼ(CAO;EC1.4.3.6)は、機能的に多様なアミンオキシダーゼのスーパーファミリーに属する(ドークス(Dawkes)ら、2001年)。それらはまた、その酵素活性がカルボニル反応性化合物、セミカルバジドにより阻止され得ることから、セミカルバジド感受性アミンオキシダーゼとしても知られている。それらは、第一級アミンの、対応するアルデヒドへの酸化的脱アミノ化を銅依存性の反応において触媒し、分子状酸素が消費され、かつアンモニアが放出される。すべてのCAOに特徴的な性質は、2,4,5-トリヒドロキシフェニルアラニルキノン、トパキノン(TPQ)の酸化還元補因子としての使用である。CAOは、細菌、真菌、植物、および哺乳類を含めたいくつかの異な生物から単離されている。植物においては、CAOは、たとえば損傷治癒に関与しており、一方原核生物においては、CAOはその生物が窒素および炭素の供給源として種々のアミンを代謝的に利用できるようにする。より高等な真核生物では、生体アミンおよび他のアミンの代謝におけるそれらの役割の他には、CAOの生物学的機能についてはほとんどわかっていない。
【0011】
シェパード(Shapard)ら、2002年、は、6つの既知の銅含有アミンオキシダーゼに対して阻害剤を検査した際の、選択性および不活性化の速度における著しい差異を報告している。
【0012】
CAOの結晶構造は、4つの異なる種:大腸菌(ECAO:たとえば、タンパク質データバンク(Protein Data Bank)、PDBコード1oac)(パーソンズ(Parsons)ら、1995年)、エンドウ(Pisum sativum)(PSAO;PDBコード1ksi)(クマー(Kumar)ら、1996年)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)(HPAO;たとえば、PDBコード1a2v)(リ(Li)ら、1998年)、およびアルスロバクター・グロビフォルミス(Arthobacter globiformis)(AGAO;PDBコード1av4)(ウィルス(Wilce)ら、1997年)から解明されてきた。これらのホモダイマー構造はすべて、一つの類似した全体的な基本構造を有しており、それはドメインD1-D4に分けられることが可能であり、そのうちのD1ドメインは大腸菌においてのみ見出されている。ドメインD2およびD3は、各々〜100アミノ酸であって、α/β型の基本構造を有しており、一方最大のC末端ドメインD4は、〜400アミノ酸長であって、ダイマー化に必要な独特のβ-サンドイッチ基本構造を有する。D4ドメイン中に局在する活性部位は、CAOファミリー中に高度に保存されている。それはタンパク質内に深く埋められており、主としてD3およびD4ドメインからのアミノ酸によって囲まれた長いチャンネルを通してのみ接近することが可能である。D3ドメインのアミノ酸残基は、実際の活性部位よりも軽度に保存されており、活性部位へ導くキャビティがCAOの基質特異性の決定において非常に重要であることを示唆している。
【0013】
既知のCAOタンパク質の構造が全く類似しているとはいえ、アミノ酸レベルでの配列同一性はわずか25〜35%である。構造が分かっているCAOファミリーのメンバーに対するVAP-1の進化的関係は特徴づけられていないが、VAP-1のN-末端における膜貫通ドメインの存在は、可溶性CAOからの実質的な相違を示唆している。
【発明の開示】
【発明が解決しようとする課題】
【0014】
本発明はヒトVAP-1の結晶化およびX線分析を提供する。これは結晶化された最初の哺乳類のCAOである。
【課題を解決するための手段】
【0015】
発明の詳細な説明
本発明は、結晶性血管接着タンパク質-1(VAP-1)、特にヒトVAP-1に関する。
【0016】
本発明は、精製されたVAP-1を結晶化し、得られたVAP-1結晶を分析し、結晶パラメータおよびX線回折データを得るための、手段および方法を提供する。
【0017】
本発明はさらに、結晶性VAP-1を含んでいる組成物、およびかかる組成物の用途に関する。
【0018】
本発明はさらに、結晶性のヒトVAP-1の構造情報、より具体的には、ヒトVAP-1の活性部位キャビティについての情報を提供しており、前記キャビティは、表面では幅約20Å×約10Åであり、かつ深さ約15Åであり、そしてアミノ酸86-87、97、168-173、176-177、180、184、205-212、216、227、232-234、236-239、344、388-390、393-397、415-419、421、421-426、467-470、647-651、および758-761はヒトVAP-1の一つのモノマーに由来し、アミノ酸433-449、および451はヒトVAP-1の別のモノマーに由来する。さらに具体的には、前記活性部位は、アミノ酸残基Ala370、Tyr384、Asp386、Asn470、Tpq471、およびTyr473によって裏打ちされた、該部位の底の狭い約4.5×約4.5のキャビティの上端にアミノ酸Leu469をさらに含む。
【0019】
本発明はさらに、ヒトVAP-1タンパク質の三次元構造を規定する原子座標/X線回折データをその上に記憶した、コンピュータ可読媒体であって、適当な機械によって読取られ、かつ分子構造を決定するためのコンピュータプログラムによって処理されたとき、ヒトVAP-1タンパク質のフラグメントを含んでいる分子の結晶の三次元表現をディスプレイすることができる媒体を提供する。
【0020】
本発明はさらに、リガンドおよび/または阻害剤のde novo設計のためのin sillicoアッセイであって、(i)VAP-1の活性部位内の箇所と相互作用することができる官能基または低分子フラグメントの同定、および(ii)これらを一つの化合物中に結合すること、を含むアッセイを提供する。
【0021】
本発明はさらに、既知の化合物および化合物ライブラリを、VAP-1活性を阻害する能力についてスクリーニングするための、in sillicoのアッセイを提供する。
【0022】
本発明はさらに、慢性関節炎、炎症性腸疾患、および皮膚疾患、多発性硬化症といった、急性および慢性の炎症性症状および疾患、ならびに糖質代謝に関連した疾患(糖尿病および、糖尿病から結果として生じる血管症のような合併症を含めて)の治療に有用な、新規VAP-1阻害剤を提供する。加えて、かかる阻害剤は、脂肪細胞の分化または機能、および平滑筋細胞の機能(特にアテローム硬化症における)、および種々の血管性疾患における異常を治療するために有用であってよい。
【発明を実施するための最良の形態】
【0023】
以下において、本発明は、好ましい態様により、添付の図面を参照して、さらに詳細に記述される。
【0024】
発明の詳細な説明
本発明は、哺乳類の銅含有アミンオキシダーゼ、CAOの結晶、特にヒトVAP-1の結晶を提供する。
【0025】
本発明の結晶を成長させるためには、N-末端膜貫通領域を含めた完全長タンパク質は、全タンパク質の80%を超え、さらに好ましくは全タンパク質の90%を越え、最も好ましくは全タンパク質の95%を超えるように精製されねばならない。発現および精製目的のためには、完全長VAP-1をコードしている配列(配列番号1)が使用される。選んだ精製法は、精製されたタンパク質がそのCAO活性を保持しているようにすることが重要であり、それはベンジルアミンを基質として用いることにより測定されてよい。
【0026】
当業界で公知の多くのベクター-宿主系は、結晶化プロセスのためのタンパク質の組換え体産生に使用されてよい。可能なベクターは、これに制限されないが、プラスミドまたは修飾されたウイルスを含むが、ベクター系は用いた宿主細胞と適合性でなければならない。ヒトVAP-1は重度にグリコシル化されているため、酵母または動物細胞の宿主のような、真核生物の宿主が好ましい。チャイニーズ・ハムスター・オバリー(CHO)細胞は、最も好ましい宿主細胞を形成しており、これらの細胞は完全なグリコシル化が可能である。
【0027】
本発明の結晶を得るためには、これに制限されないが、バッチ晶析法、蒸気拡散法、およびマイクロ透析法を含めて、この技術において周知の任意の結晶化技術が使用されてよい。X線用の品質の結晶を得ることが必要な場合には、標準的なミクロおよび/またはマクロの種晶の添加が使用されて良い。
【0028】
本発明の結晶は、非対称ユニット中に二つの分子をもち、a=b=225.9Å、c=218.7Å、α=β=90°、γ=120°のユニットディメンションをもつ空間群P6522を作る(以下の実施例2を参照)。しかしながら、本発明はP6522を含むがこれに制限されない任意の空間群を形成する結晶を検討している。結晶は、4Åを超える、好ましくは3Åを超える、最も好ましくは2.8Åを超える分解能までを回折する。
【0029】
本発明の結晶から回折データを収集するべく、結晶は、グリセロールのような凍結保護物質、および窒素流中での瞬間凍結を用いて保護されてよい。X線回折データは、XDSプログラム(カブシュ(Kabsch)ら、1993年)により処理されてよいが、当業者に周知の任意の方法がX線回折データの処理に使用されてもよい。
【0030】
本発明によるヒトVAP-1の原子構造を決定するためには、分子置換(MR)、モデル構築、およびリファインメントが行なわれてもよい。
【0031】
VAP-1構造の決定には、当業界で公知のCAOの既知の構造または、上記および以下の実施例3のように決定され得る他のCAO構造を用いた、分子置換が使用されてよい。
【0032】
当業者には周知の任意の方法が、本発明のタンパク質の三次元構造を分子置換により決定するべく使用されてよい。たとえば、CCO4iプログラムスイートのプログラムAMOREは、ヒトVAP-1の構造を、本文に記述されたように導出された原子座標表を用いて決定するべく使用されてよい。
【0033】
原子座標は、コンピュータ可読媒体上に与えられてよい。かかる記憶媒体は、好ましくはランダムアクセスメモリ(RAM)、読み出し専用メモリ(たとえば、CD-ROM)、またはディスケットである。記憶媒体は、ローカルアクセス可能なコンピュータ上か、またはインターネットを通じたリモートアクセス可能なものでよい。
【0034】
三次元構造の最初のモデルは、プログラムO(ジョーンズ(Jones)ら、1991年)を用いて構築されてよく、たとえばCCP4iプログラムスイートのREFMACプログラムを用いてリファインされる。
【0035】
リファインされた三次元の本発明のVAP-1構造は、表1に示された原子座標および構造決定統計学によって表される。リファインされたVAP-1ダイマーのX線構造は、モノマーA中にはモデリングされない残基A1-A54、A202-A204、およびA762-A763をもつ残基A55-A761と、モノマーB中にはモデリングされない残基B1-B56、B203、B742-B746、およびB762-B763をもつ残基B57-B761からなる。残基A471およびB471は、トパキノン(Tpq471)であり、それらは内在性チロシン残基の翻訳後修飾によって形成され、触媒反応に関係する。双方のモノマーは、活性部位に一つの銅イオンを含有する。
【0036】
本発明の三次元構造から得られる情報は、ヒトVAP-1の活性部位(基質結合部位)が(1)銅イオンを配位している3つのヒスチジン、His520、His522、およびHis684、(2)触媒ベース、Asp386、ならびに(3)Tpq471のポジショニングに関係する、Tyr372およびAsn470、および(4)活性部位ゲート残基、Tyr384を含んでいることを示している。Tpq471は、VAP-1のX線構造では「オン・カッパー(on-copper)」(不活性な)立体配座にある。ヒトVAP-1構造の三次元表示は図2および3に示されている。
【0037】
VAP-1活性部位は深く埋められており、Tpq471は分子の表面から約22Åに位置している。この活性部位キャビティは、表面において幅約20Å×約10Åであり、深さ約15Åである。VAP-1のX線構造においては、キャビティの底では、Tpq471の上部に位置するLeu469が活性部位への入口をブロックしている。キャビティの一つの壁は、たとえば86-87、97、168-173、176-177、180,184、205-212、216、227、232-234、および236-239を含んでいるより小さなD2およびおD3ドメインに由来する残基と、443-449、および451を含んでいる他のモノマーから突出た長いβヘアピンアームに由来する残基とからなる。チャンネルの別の壁は、たとえば344、388-390、393-397、415-419、421,421-426、467-470、647-651、758-761を含んでいる触媒D4ドメインに由来する残基からなる。活性部位チャンネルの入口において、Asn232に付着したN-グリカンの一つの糖単位がX線構造中で見られる。Leu469の下では、約7Å長の活性部位キャビティは表面におけるよりもずっと狭く、約4.5×4.5Åのディメンションをもつほぼ円形である。チャンネルのこの部分は、残基Ala370、Tyr384、Asp386、Asn470、Tpq471、およびTyr473によって裏打ちされている。
【0038】
VAP-1X線構造は、驚いたことに、活性部位キャビティの独特の構造を明らかにしている。キャビティは、ECAO、HPAO、PSAO、およびAGAO構造内の狭い活性部位チャンネルに比較して、極めて幅広にマウントされかつオープンである。それゆえVAP-1の活性部位キャビティは、ECAO、HPAO、PSAO、およびAGAOの活性VAP-1キャビティよりもはるかに大きいリガンドおよび阻害剤構造物を収容することが可能である。VAP-1においてLeu469に相当する残基は、グリシン(ECAO、HPAO、およびAGAO)またはアラニン(PSAO)であり、したがって、それはECAO、HPAO、PSAO、およびAGAO構造物においては活性部位をブロックすることができない。この独特の構造についての知見は、ヒトVAP-1のファーマコフォアおよび、このキャビティにフィットする特異的リガンドおよび阻害剤構造物の設計に必要である。
【0039】
この情報は、ヒトVAP-1のファーマコフォア、すなわち生物活性に必要な特別の機能を表している化学的特徴および三次元的制約のコレクション、を定義するべく使用されることが可能である。ファーマコフォアは、好ましくは、水素結合供与体および受容体、荷電基、または疎水性部位といった、表面でアクセス可能な特徴を含む。かかる特徴は、生物活性に対するそれらの相対的重要性に基づき、ファーマコフォアモデルに包含されてよい。
【0040】
ファーマコフォアは、CATALYST、CERIUSといった可用性のコンピュータソフトウエアを用いるか、またはリード化合物についての既知の立体配座に基づく手動モデリングを使用することにより決定されてよい。ファーマコフォアは、以下にさらに詳細に記述されるような、可用性のコンピュータソフトウエアを用いて、in sillicoの化合物ライブラリをスクリーンするべく使用されてよい。
【0041】
したがって、本発明の一つの態様においては、分子モデリング技術がde novoの化合設計に使用される。デノボ化合物設計は、VAP-1のようなターゲット高分子の結合表面または活性部位が決定され、かつ、前記結合表面または活性部位と相互作用する化合物の合理的な設計のためのベースとして使用されるプロセスを指す。本発明による分子モデリングの段階は、ヒトVAP-1の原子座標および活性部位のモデルを利用する。好ましい態様においては、de novoの薬物設計は、ヒトVAP-1の結合表面における活性部位と相互作用することができる官能基または小さい分子フラグメントの同定、およびこれらの基またはフラグメントを結合して単一の化合物にすることを含む。ヒトVAP-1の特異的活性部位と相互作用する官能基または小さい分子フラグメントがひとたび同定されれば、好適なサイズを有するフラグメントをブリッジングすること、および活性部位にフィットするためのジオメトリの双方を用いて、単一の化合物へ結合されることが可能である。
【0042】
適切な官能基およびフラグメントの結合は手動で行なわれてもよいが、QUANTAまたはSYBYLといった適当なソフトウエアを使用することが好ましい。この技術において周知のさらなるソフトウエアは、たとえば、多数の官能基をデータベースからの分子鋳型と結びつけるHOOK、および制約された非環式分子に対して連結ユニットを設計するためのCAVEATである。de novoの化合物設計のための、他のコンピュータ・ベースのアプローチは、LUDI、SPROUT、およびLEAP-FROGを含む。
【0043】
本発明は、阻害性化合物を含め、ヒトVAP-1の活性部位に対して結合することが可能な、化学的実体および化合物を設計、同定、および合成するための、分子設計技術の使用を可能にする。ヒトVAP-1の原子座標は、ヒトVAP-1の潜在的な阻害剤を同定するべく、たとえば、GOLD、GRAM、DOCK、またはAUTODOCKといったドッキングプログラムを用いたコンピュータモデリングと一緒に使用されてよい。この手法は、潜在的阻害剤の形状および化学構造が、いかに充分に活性部位に相補的であるかを確認するための、VAP-1の活性部位に対する潜在的阻害剤のコンピュータフィッティングを含むことが可能である。ドッキングプログラムを用いたモデリングにより設計された潜在的阻害剤の実例は、以下に記述される一般式(I)に適合してもよい。
【0044】
本発明はさらに、ヒトVAP-1の活性部位と相互作用する化合物を同定するためのin sillicoの方法であって、(a)ヒトVAP-1のリガンド結合ドメインの原子座標をコンピュータの記憶媒体に供給すること、および(b)該コンピュータを使用して分子モデリング技術を該座標に適用すること、の段階を含む方法を提供する。
【0045】
上記の構造モデルはさらに、構築された分子のライブラリおよびSybyl v.6.9による最小化されたエネルギーの検査に使用され、VAP-1阻害剤として作用するそれらの能力について評価された。in sillicoスクリーニングに先立ち、VAP-1のX線構造は実施例4に記述されたように、CAOの「オフ・カッパー(off-copper)」の活性のある立体配座を模すべく修飾された。
【0046】
本発明のさらなる態様においては、in sillico法はヒトVAP-1の酵素活性を特異的に阻害する化合物の同定に使用されており、以下の段階:前記分子モデリング技術により同定される化合物を提供すること、前記化合物をヒトVAP-1と接触させること、および、相互作用を検出すること、を特徴とする。
【0047】
三次元構造情報および、VAP-1の前記構造情報に関連した原子座標は、合理的な薬物設計において有用であり、VAP-1に結合しかつVAP-1の酵素活性を阻害する阻害性化合物の同定法を提供する。SSAO活性を有する酵素のための、前記潜在的阻害剤を同定するための方法は、(a)表1にリストされたその原子座標をベースとしたVAP-1の三次元構造を用いること;(b)前記三次元構造を用いて前記潜在的阻害剤を設計または選択すること;(c)前記潜在的阻害剤を合成すること;(d)前記潜在的阻害剤を前記酵素と接触させること;および(e)前記SSAO活性を阻害する前記阻害剤の能力を測定すること、の段階を含む。
【0048】
本発明はさらに、ヒトVAP-1の酵素活性を阻害する化合物か、またはVAP-1の活性部位と相互作用する化合物を同定するための、本発明のin sillico法を用いて同定された化合物を包含する。
【0049】
本発明の方法により同定可能な潜在的VAP-1阻害剤の実例は、式(I):
【化1】
[式中、R1は水素、低級アルキル、または任意に置換されたフェニルまたはヘテロアリール基であり;
R2は水素または低級アルキルであるか、または
R1およびR2は、それらが結合している窒素原子と一緒になって飽和複素環を形成してもよく;
R3-R5は各々独立して水素、低級アルキル、アラルキル、任意に置換されたフェニルまたはヘテロアリール基を表しているか、または
R2およびR3は、それらが結合している原子と一緒になって飽和複素環を形成してもよく、あるいは
R3およびR5は、それらが結合している炭素原子と一緒になって飽和炭素環を形成してもよく;
R6はナフチル、フェニル、置換されたフェニルまたはヘテロアリール基であり;
R7は水素、低級アルキルまたはアラルキルであり;
nは1、2または3であり;かつ
X=O、S、SO、SO2またはNR2である]
により表されてよい。
【0050】
他のタイプの化合物も同様に、本発明による方法およびモデルを用いて充分に同定されてよい。当業者は、本文に記述された原子座標およびファーマコフォアに基づき、ヒトVAP-1の活性部位と相互作用する化合物を同定してよい。一般に、かかる潜在的阻害剤化合物は、前文に例示されたような、アミンまたはヒドラジンのようなカルボニル基反応因子によって特徴づけられてよいが、活性部位キャビティの4.5×4.5Åの底部分にはまりかつ活性部位の底部においてTPQに結合することが可能な、細い突出した部分をもつ他の構造物も、同様に潜在的阻害剤である。本発明の化合物のこの突出した部分は、この化合物の約7Å長の比較的疎水性の中央部分へ連結され、ともに「リンカー」として作用して、突出部分が活性部位へアクセスすることを可能にし、キャビティを裏打ちするアミノ酸残基Ala370、Tyr384、Asp386、Asn470、Tpq471、およびTyr473と相互作用する。最後に、本発明の化合物は、広いキャビティをふさぎ、かつ活性部位キャビティーの壁の代表的なアミノ酸残86-87、97、168-173、176-177、180、184、205-212、216、227、232-234、236-239、344、388-390、393-397、415-419、421、421-426、467-470、647-651、758-761、443-449、および451と相互作用する、よりかさばった部分を含む。
【0051】
さらに、本発明は同定された化合物の予想された活性を確かめるためのアッセイおよび手段を提供する。
【0052】
本発明はさらに、本発明のスクリーニングアッセイにより同定された化合物の、慢性関節炎、炎症性腸疾患、皮膚疾患、および多発性硬化症、のような慢性または急性の炎症性症状または疾患、ならびに糖質代謝に関連した疾患(糖尿病および、糖尿病から結果として生じる合併症、たとえば網膜症、腎症、および神経障害のような血管症を含めて)の治療のための薬の調製のための用途を包含する。加えて、かかる阻害剤は、脂肪細胞の分化または機能、および平滑筋細胞の機能(特にアテローム硬化症における)、および種々の血管性疾患における異常の治療に有用であってよい。
【0053】
本発明はさらに、本スクリーニングアッセイによって同定された化合物を含有する薬剤組成物を包含する。
【0054】
以下の実施例は、本発明を例証する。
【実施例1】
【0055】
ヒトVAP-1の産生および精製
N-末端膜貫通ドメインをもつ完全長のタンパク質は、スミス(Smith)ら、1998年、において記述されたように、グリコシル化コンピテントCHO細胞において発現された。収穫された細胞は、溶解緩衝液(150mM NaCl、10mM トリス塩基pH7.2、1.5mMのMgCl2、1%NP40)を用いて溶解された。浄化された細胞溶解物は、モノクローナル抗体アフィニティカラムに基づき、かつAKTATM精製システム(アマシャム・バイオテック(Amersham Biotech))を用いた、HVAP-1の精製に使用された。タンパク質は、アフィニティクロマトグラフィーを用いて均一(>95%)まで精製され、精製の後、VAP-1タンパク質の存在、90-および170〜180kDのバンドは、スミスら、1998年、により記述されたように、銀染色SDS-pageによって確認された。
【0056】
精製されたタンパク質は、基質としてベンジルアミンを用いて測定されるような、そのCAO活性を保持していた。アミンオキシダーゼ活性は、記載のスペクトロメトリー法(Haltら、1997)、基質として200μl体積及び1mMのベンジルアミンを使用することで測定された。吸光度の変化は、490nmにおいて、VictorTMマルチラベルプレートカウンタ(パーキンエルマー・ライフ・サイエンシズ(PerkinElmer Life Sciences))においてモニターされた。
【0057】
ヒトVAP-1のコード領域のヌクレオチド配列は、配列番号1として列挙されている配列に示されており、そして配列番号2のようなアミノ酸配列(763アミノ酸)に相当する。
【実施例2】
【0058】
結晶化および予備的分析
hVAP-1の最初の結晶化条件は、ウィザード(Wizard)Iランダムスパース行列結晶化スクリーン(エメラルド・バイオストラクチャーズ・インク(Emerald BioStructures, Inc., USA))および蒸気拡散法を用いて、室温でスクリーンされた。小さい六方晶の結晶が、1.0M酒石酸K/Na、100mMイミダゾール(pH8.0)、および200mM NaClを含有する条件において、数ヶ月間のインキュベーションの後に得られた。懸滴は、10mMリン酸カリウム緩衝液(pH7.2)中の2μlのタンパク質試料(1.0mg/ml)、および2μlの貯蔵溶液を含有していた。最適化の後、1.0M酒石酸K/Na、100mMイミダゾール(pH7.8)、および沈殿剤としての150〜250mM NaClからなる貯蔵溶液を用いて、最良の結晶が得られた。結晶は数日間で形成され、約0.15×0.15×0.1mmの最終サイズに成長した(図1)。
【0059】
一つの結晶がキャピラリーにマウントされ、予備的X線分析が、回転陽極放射線源(Cu Kα放射線、50kV、150mA)、およびMAR345イメージングプレート検出器を用いて、インハウスで行なわれた。結晶は、しかしながら、8Åまでの分解能しか回折せず、空間群は正確には測定できなかった。さらなるX線分析ならびにデータ収集はすべて、MARリサーチCCD検出器を具備したビームライン(beamline)X11(EMBL/DESY、ドイツ、ハンブルグ)において、シンクロトロン照射を用いて行なわれた。データ収集には、結晶は20%(v/v)グリセロールを用いて凍結保護され、100Kの窒素流中で瞬間凍結された。3つの異なる結晶から収集された回折データは、プログラムXDSにより処理された。溶媒含有率およびマシューズ(Matthews)係数は、分子量をモノマーあたり90kDと仮定し、CCP4スイートを用いて計算された。
【0060】
最良に見える六方晶結晶は、1.0M酒石酸K/Na、100mMイミダゾール(pH7.8)、および沈殿剤として150〜250mMのNaClを用いて得られた。結晶は、a,b=225.9Å、C=218.7Å、α,β=90°、およびγ=120°のユニットセルディメンションセルをもつ、典型的なサイズ0.15×0.15×0.1mmに成長した(図1a)。回折データ統計(表1)によれば、いくつかのフレームでは3.0Åをこえる分解能に相当する反射が観察されたにもかかわらず、VAP-1触媒の回折限界は、3.2Åであった(図1b)。結晶は、空間群P6522に属する。非対称ユニットあたり一つのダイマー(180kD)の存在を仮定すると、マシューズ係数は4.5Å3/Da、および溶媒含有率は72%である。結晶パラメータおよび回折データ統計は、表1に要約されている。
【0061】
【表1】
【実施例3】
【0062】
構造決定
VAP-1の構造は、CCP4iプログラムスイート(コラボラティブ・コンピュテイショナル・プロジェクト(Collaborative Computational Project(共同計算プロジェクト))、1994年)のプログラムAMORE(ナバザ(Navaza)、1994年)を用いた分子置換により解明された。この方法は、VAP-1の空間群が、非対称ユニットあたり一つの生物学的単位である一つのダイマーをもつ、P6522であったことを立証した。分子置換において検査された大腸菌(Escherichia coli)CAO(残基93-720;PDBコード10AC)、エンドウ(Pisum sativum)CAO(残基7-634;PDBコード1KSI)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)CAO(残基22-655;PDBコード1A2V)、およびアルスロバクター・グロビフォルミス(Arthobacter globiformis)CAO(残基9-623;PDBコード1AV4)の二量体ポリアラニン主鎖のうち、エンドウの構造は最良の相関係数(44.1%)およびR因子(53.4%)を与え、サーチモデルが使用された。
【0063】
電子密度マップはCCP4iスイートのFFTを用いて計算され、構築の初期においてであっても、いくつかのフラグメントにおいては、VAP-1ポリペプチドをトレースするのに充分に予測可能であった。モデルはプログラムOを用いて手動により構築され、CCP4iスイートのREFMAC5.1.24(ムルシュドフ(Murshudov)ら、1997年)により、分解能3.2Åまでリファインされた。
【0064】
リファインメントの多数のサイクルの間に、構造に側鎖が逐次的に加えられた。最終モデルにおいては、以下のアミノ酸だけはトレースできず、したがって、この構造には含まれていない:A1-A54、A202-A204、A762-A763、B1-B56、B203、B742-B746、およびB762-B763。トパキノン残基のための座標は、ヘテロコンパウンド・インフォメーションセンター(Hetero-compoun Information Centre)、ウプサラ(クレイウェグト&ジョーンズ(Kleywegt and Jones)、1998年)から取得され、そのための辞書はプログラムPRODRG(ヴァンアールテン(van Aalten)ら、1996年)を用いて生成された。最終的なVAP-1モデルの立体化学的な質は、PROCHECK(ラスコウスキー(Laskowski)ら、1993年)を用いて査定され、モデル中の1401個のアミノ酸のうち、84.0%がラマチャンドラン・プロット(ラマチャンドラン&サジーセクハラン(Ramachandran and Sasisekharan)、1968年)中の最も好ましい領域に生じ、わずか0.5%が寛大に許されるか、または許されない領域に生じた。構造決定統計の要約は、表1に呈示されている。ヒトVAP-1結晶構造の原子座標(および構造因子)は、PDBに登録コード1PU4および1US1で寄託されている。
【0065】
ヒトVAP-1の結晶形状はホモダイマーであり、各サブユニットはドメインD2、D3、およびD4を含有している。VAP-1構造の概略説明図は、図2に示されている。結晶構造に見られるD2、D3、およびD4ドメインは、各々残基約55-169、170-300、および301-761(各々、配列番号3、配列番号4、および配列番号5)からなる。他の既知のアミンオキシダーゼの構造に比較しての、ヒトVAP-1の構造における主要な差異は、ドメインD4にも差異は存在するが、ドメインD2およびD3において見られる。
【実施例4】
【0066】
潜在的阻害剤のインシリコスクリーニング法
本実施例は、アリールオキシメチル置換ヒドラジンアルコール化合物のライブラリが、実施例3に記述されたVAP-1の結合キャビティ内へドッキングすることにより、in sillicoにおいていかにしてスクリーンされるかを示している。スクリーンされた化合物は、式
【化2】
[式中、R1は水素、低級アルキルまたは任意に置換されたフェニルまたはヘテロアリール基であり;
R2は水素または低級アルキルであるか、または
R1およびR2は、それらが結合している窒素原子と一緒になって飽和複素環を形成してもよく;
R3-R5は各々独立して水素、低級アルキル、アラルキル、任意に置換されたフェニルまたはヘテロアリール基を表しているか、または
R2およびR3は、それらが結合している原子と一緒になって飽和複素環を形成してもよく、あるいは
R3およびR5は、それらが結合している炭素原子と一緒になって飽和炭素環を形成してもよく;
R6はナフチル、フェニル、置換フェニルまたはヘテロアリール基であり;
R7は水素、低級アルキルまたはアラルキルであり;
nは1、2または3であり;かつ
X=O、S、SO、SO2またはNR2である]
を有する。
【0067】
合成および/または構築され、かつin sillicoでスクリーンされた化合物の実例は、表2に示されている。
【0068】
【表2】
【0069】
X線構造の修飾およびリガンドドッキング
VAP-1X線構造は、以下に記述されたように、CAOの「オフ・カッパー(off-copper)」の活性のある立体配座を模すべく修飾された。最初に、トパキノンは、ECAO構造(PDBコード1D6Z(ウィルモット(wilmot)ら、1999年)にしたがって、活性のあるイミノキノン形へ修飾された。第二に、VAP-1構造の活性部位キャビティは、ドッキング研究に先立ち、ボディル(Bodil)モデリングパッケージ(http://www/abo.f//fak/mnf/reseach/jonnson/bodil.html)において側鎖ロータマー(Phe389、Tyr394、Asp386、およびLeu469)を選ぶことにより修飾され、それにより活性部位のイミノキノンをリガンドに関してさらに近づきやすくした。水素原子は、スクリーニングおよびドッキングに使用されたVAP-1構造に対し、何ら側鎖調整なしに、プログラムReduce v.2.15を用いて付加された(ワード(Word)ら、1999年)。
【0070】
潜在的リガンドのR-およびS-エナンチオマーが構築され、Sybyl v.6.9(トリポス・アソシエイツ(Tripos Associates)、米国、セントルイス)によりエネルギー最小化された。リガンドはイミノキノン残基へ共有結合により結合され、手動により結合キャビティ内へドッキングされた。
【0071】
リガンド結合モード
図4は、スクリーンされたリガンドの実例を示している。ボディル・モデリングパッケージを用いて作成された図3には、BTT-2071(化合物9)のS-エナンチオマーの結合モードが存在する。BTT-2071(化合物9)のS-エナンチオマーは、ドッキングシミュレーションに基づけば、VAP-1構造と最も広範囲な相互作用を形成することができることから、図中の実例リガンドとなるべく選ばれた。
【0072】
VAP-1構造に基づけば、Xが酸素原子のかわりに硫黄原子であるリガンドは、VAP-1の相互作用表面が疎水性であることから、より有利にVAP-1と相互作用する。
【0073】
硫黄原子を含んでいるリガンドの疎水性部分は、親水性残基(Leu468、Leu469、Phe389、およびMet211)に対して固く締まる(pack)。リガンドのパラメトキシ基中の酸素原子は、側鎖のThr212の酸素に対して水素結合距離にあり、一方、リガンドのメタメトキシル基中の酸素原子は、側鎖のTyr394の水酸基に対して水素結合距離にある。Phe389は、リガンドの芳香環と容易に相互作用することができる位置に局在する。パラ-およびメタ-メトキシル基中のメチル基は、各々Tyr448およびTyr176と接触している。S-エナンチオマー中の水酸基は、Asp386と最適な水素結合を形成することが可能であるのに対し、R-エナンチオマーにおいては、水素結合距離およびジオメトリは最適ではない。
【実施例5】
【0074】
同定された潜在的阻害剤のVAP-1阻害効果を立証するためのインビトロアッセイ
VAP-1SSAO活性は、本質的にはモノアミンオキシダーゼおよび類縁酵素について記述されたような組合わされた比色法を用いて測定された。チャイニーズハムスターオバリー(CHO)において発現された細胞組換えヒトVAP-1 SSAOが、活性測定のためのVAP-1 SSAOの供給源として使用された。天然のCHO細胞は、無視できる程度のSSAO活性を有する。これらの細胞およびその培養物は、従来に記述されている(スミス(Smith D J)ら、1998年)。
【0075】
細胞溶解物は、約3.6×108個の細胞を25mlの溶解緩衝液(150mM NaCl、10mMトリス-塩基 pH7.2、1.5mMのMgCl2、1%NP40)中に懸諾すること、および回転台の上で4℃において一晩インキュベートすることにより調製された。溶解物は、室温において18000gで5分間遠心分離することにより透明にされ、上清はアッセイに直接使用された。
【0076】
VAP-1 SSAOアッセイは、96穴マイクロタイタープレートにおいて、以下のように行なわれた。各ウェルに対し、必要であれば、あらかじめ決められた量の阻害剤が添加された。阻害剤の量は各アッセイで異なったが、全般的には1nM〜50μMの濃度であった。対照は阻害剤を欠いていた。阻害剤は、水中で、全体積中に20:1であった。次に、以下の試薬が添加された。全反応体積200μlに対し、0.2Mリン酸カリウム緩衝液pH7.6、1mMの2,4-ジクロロフェノール、500μMの4-アミノアンチピリン、および4U/mlの西洋ワサビペルオキシダーゼを含有する、45μlの新たに作成された色素溶液、および、1時間当たり0.6のA490の変化を引き起こした量の、VAP-1 SSAOを含有しているCHO細胞溶解物。
【0077】
プレートは37℃で30分間インキュベートされ、バックグラウンド吸光度は、ウォラク・ビクター(Walac Victor)IIマルチラベルカウンタを用いて、490nmにおいて測定された。酵素反応を開始するためには、20μlの10mMベンジルアミン(最終濃度=1mM)が添加され、そしてプレートは37℃において1時間インキュベートされた。
【0078】
VAP-1 SSAO活性を反映する吸光度の増大は、490nmにおいて測定された。阻害は、バックグラウンド吸収について補正した後に、対照と比較されたパーセント阻害として表され、IC50値はグラフパッド・プリズム(GraphPad Prism)を用いて計算された。
【実施例6】
【0079】
VAP-1 SSAO活性対全ラットMAO活性の比較
ラットMAOは、ラット肝より、1gの肝試料を14mlのKCl-EDTA溶液中で数回洗浄して血液をすべて除去することにより調製された。次いで、1gの肝試料は、4mlの氷冷リン酸カリウム緩衝液(0.1M、pH7.4)中で、ウルトラ・トラックス(Ultra-Turrax)ホモジナイザ(設定11000rpm、4×10秒)によりホモジナイズされた。4℃において500gで10分間の遠心分離の後、上清は注意深く取出され、4℃において12300gで15分間遠心分離された。上清は排出され、沈降されたミトコンドリアは4mlの新鮮なリン酸緩衝液中に再懸濁され、前と同様に遠心分離された。ミトコンドリアは4mlの辛酸緩衝液中に懸濁され、ウルトラ・トラックス(Ultra-Turrax)ホモジナイザ(設定11000rpm、2×10秒)によりホモジナイズされた。ミトコンドリアプレパラートはアリコートに分けられ、-70℃に貯蔵された。
【0080】
全MAO活性は、2,4-ジクロロフェノールが1mMバニリン酸で置換えられたこと以外は、VAP-1 SSAOと類似の方法で測定された。各ウエルに対し、必要な場合には、あらかじめ決められた量の阻害剤が添加された。阻害剤の量は各アッセイで異なったが、全般的には10nM〜800mMの濃度であった。対照は阻害剤を欠いていた。阻害剤は、水中で、全体積中に20:1であった。次に、以下の試薬が添加された。全反応体積300μlに対し、0.2Mリン酸カリウム緩衝液pH7.6、50μlの新たに作成された色素溶液(上記と同じ)、および50μlのMAO標品。
【0081】
プレートは37℃で30分間インキュベートされ、バックグラウンド吸光度は、ウォラク・ビクター(Walac Victor)IIマルチラベルカウンタを用いて、490nmにおいて測定された。酵素反応を開始するためには、20μlの5mMトリアミン(最終濃度=0.5mM)が添加され、プレートは37℃において1時間インキュベートされた。MAO活性を反映する吸光度の増大は、490nmにおいて測定された。阻害は、バックグラウンド吸収について修正した後に、対照と比較されたパーセント阻害として表され、IC50値はグラフパッド・プリズム(GraphPad Prism)を用いて計算された。MAO阻害のための陽性対照として、クロルギリンおよぎパルギリン(各々MAO-AおよびMAO-Bの阻害剤)が0.5μMにて、いくつかのウエルに添加された。
【0082】
表3の化合物の、VAP-1 SSAOに対してMAOを超える特異性をもってVAP-1 SSAOを阻害する能力は、表2に示されている。結果は、本発明の化合物がヒトVAP-1 SSAO活性の特異阻害剤であることを示している。本発明の化合物は、そえ故、ヒトの接着性分子VAP-1のSSAO活性が役割を果たしている疾病および症状の治療において、治療上の有用性をもつことが期待される。
【0083】
【表3】
【0084】
当業者には、技術が進歩するにつれて、発明の概念が様々な方法で遂行され得ることが明らかであろう。本発明およびその態様は、上記の実施例に制限されず、特許請求の範囲内で変更されてよい。
【0085】
参考文献
コラボラティブ・コンピュテイショナル・プロジェクト(Collaborative Computational Project(共同計算プロジェクト))、Acta Cryst、1994年、第D51巻、p.760-763。
ドークス(Dawkes, H. C.)&フィリップス(Phillips, S. E.)著、Curr Opin Struct Biol、2001年、第11巻、p.666-673。
カブシュ(Kabsch, W.)著、Journal of Applied Crystallography、1993年、第26巻、p.795-800。
クマー(Kumar, V.)、ドゥーリー(Dooley, D. M.)、フリーマン(Freeman, H. G.)、ガス(Guss, J. M.)、ハーベイ(Harvey, I.)、マクグリル(McGuril, M. A.)、ウィルス(Wilce, M. C.)、およびズバク(Zubak, V. M.)著、Structure、1996年、第4巻、p.943-955。
リ(Li, R.)、クリンマン(Klinman, J. P.)&マテウス(Mathews, F. S.)著、Structure、1998年、第6巻、p.293-307。
パーソンズ(Parsons, M. R.)、コンベリー(Convery, M. A.)、ウイルモット(Wilmot, C. M.)、ヤダブ(Yadav, K. D.)、ブレーカリー(Blakeley, V.)、コーナー(Corner, A. S.)、フィリップス(Phillips, S. E.)、マクファーソン(McPharson, M. J.)、およびノウルズ(Knowles)著、Structure、1995年、第3巻、p.1171-1184。
サルミ(Salmi, M.)&ヤルカネン(Jalkanen, S.)著、J Exp Med、1996年、第183巻、p.569-579。
サルミ(Salmi, M.)&ヤルカネン(Jalkanen, S.)著、Trends Immunol、2001年、第22巻、p.211-216。
スミス(Smith, D. J.)、サルミ(Salmi, M.)、ボノ(Bono, P.)、ヘルマン(Hellman, J.)、ロイ(Leu, T.)、およびヤルカネン(Jalkanen, S.)著、J Exp Med、1998年、第188巻、p.17-27。
ウィルス(Wilce, M. C.)、ドゥーリー(Dooley, D. M.)、フリーマン(Freeman, H. C.)、ガス(Guss, J. M.)、マツナミ(Matsunami, H.)、マクインタイヤー(McIntire, W.S.)、ルギエロ(Ruggiero, C. E.)、タニザキ(Tanizaki, K.)、およびヤマグチ(Yamaguchi, H.)著、Biochemistry、1997年、第36巻、p.16116-16133。
ジョーンズ(Jones, T. A.)、ズー(Zou, J. Y.)、カウマン(Cowman, S. W.)、およびケルドガード(Kjeldgaard)著、Acta Crystallogr、1991年、第A47巻、パート2、p.110-119。
クレイウェグト(Kleywegt、G. J.)&ジョーンズ(Jones, T. A.)著、Acta Cryst、1998年、第D54巻、p.1119-1131。
ラスコウスキー(Laskowski, R. A.)、マッカーサー(Macarthur, M. W.)、モス(Moss, D. S.)、およびソーントン(Thornton, J. M.)著、Journal of Applied Crystallography、1993年、第26巻、p.283-291。
ムルシュドフ(Murshudov, G. N.)、ヴァギン(Vagin, A. A.)、ドドゥソン(Dodson, E. J.)著、Acta Crystallography、1997年、第D53巻、p.240-255。
ナバザ(Navaza, J.)著、Amore - an Automated Package for Molecular Replacement(AMoRe:分子置換用の自動化されたパッケージ). Acta Cryst、1994年、第A50巻、p.157-163。
ラマチャンドラン(Ramachandran, G. H.)&サジーセクハラン(Sasisekharan, V.)著、Adv Protein Chem、1968年、第23巻、p.283-438。
シェパード(Shepard, E. M.)、スミス(Smith, J.)、ブラッドリー(Bradley, O.E. )、クーチャー(Kuchar, J. A.)、ローレンス(Lawrence, M. S.)、およびドゥーリー(Dooley, D. M.)著、Eurl J. Biochem.、2002年、第269巻、p.3645-3658。
ヴァンアールテン(van Aalten, D. M.)、バイウォーター(Bywater, R.)、ファインドレイ(Findlay, J. B.)、へンドリッヒ(Hendlich, M.)、ホフト(Hooft, R. M.)、およびブリエンド(Vriend, G.)著、J Comput Aided Mol Des、1996年、第10巻、p.255-262。
ウィルモット(Wilmot, C. M.)、ホーイド(Hajdu, J.)、マクファーソン(McPherson, M. J.)、ノウルズ(Knowles, P. F.)、およびフィリップス(Phillips, S. E.)著、Science、1999年、第286巻、第5445号、p.1724-8。
ワード(Word, J. M.)、ローベル(Lovell, S. C.)、リチャードソン(Richardson, J. S.)、およびリチャードソン(Richardson, D. C.)著、J Mol Biol、1999年、第285巻、p.1735-47。
【図面の簡単な説明】
【0086】
【図1】パネルAにおいて六方晶のVAP-1結晶を、そしてパネルBにおいて典型的な回折パターンを示している。
【図2】ヒトVAP-1ダイマーの結晶構造を示しており、糖単位は区間充填モデルとして描かれており、双方のモノマー中のTpg471はD4ドメイン中の球として示されている。
【図3】VAP-1ダイマーの結晶構造の一つのモノマーを示している。
【図4】VAP-1構造におけるVAP-1阻害剤の結合モードを示している。視点は、表面から活性部位へ下っている。
【図1A】
【図1B】
【特許請求の範囲】
【請求項1】
結晶性ヒト血管接着タンパク質1(VAP-1)。
【請求項2】
前記結晶が、二つの分子を非対称ユニットにもち、a=b=225.9Å、c=218.7Å、α=β=90°、γ=120°のユニットディメンションをもつ、空間群P6522の結晶として定義される、請求項1に記載の結晶性ヒトVAP-1。
【請求項3】
アミノ酸配列、配列番号3、配列番号4および配列番号5によって各々特徴づけられる、ドメインD2、D3、およびD4を含んでいる、請求項2に記載の結晶性VAP-1、またはその同族体。
【請求項4】
表面において幅約20Å×約10Å、かつ深さ約15Åである活性部位キャビティを含んでいる、請求項3に記載の結晶性VAP-1タンパク質。
【請求項5】
前記活性部位キャビティが、ヒトVAP-1の一つのモノマーに由来する配列番号2のアミノ酸86-87、97、168-173、176-177、180、184、205-212、216、227、232-234、236-239、344、388-390、393-397、415-419、421、421-426、467-470、647-651および758-761、およびヒトVAP-1の別のモノマーに由来する配列番号2のアミノ酸433-449および451を含んでいる、請求項4に記載の結晶性VAP-1タンパク質。
【請求項6】
前記活性部位が、前記キャビティの底の狭い約4.5Å×約4.5Åのキャビティの上端に、アミノ酸Leu469をさらに含む、請求項5に記載の結晶性VAP-1タンパク質。
【請求項7】
前記活性部位キャビティの底部が、アミノ酸残基Ala370、Tyr384、Asp386、Asn470、Tpq471およびTyr473によって裏打ちされた、請求項6に記載の結晶性VAP-1タンパク質。
【請求項8】
請求項1〜7のいずれか1項に記載の結晶性VAP-1を含んでいる組成物。
【請求項9】
ヒトVAP-1を結晶化するための方法であって、
a)ヒトVAP-1タンパク質を含んでいる水溶液を供給すること;
b)沈殿剤を含んでいる貯蔵溶液を供給すること;
c)ある体積の前記水溶液とある体積の前記保存溶液と混合して、混合液を形成すること;および
d)前記混合液の少なくとも一部を結晶化すること、
の段階を含む方法。
【請求項10】
段階a)で供給された前記水溶液が、1mg/mlのVAP-1濃度を有する、請求項4に記載の方法。
【請求項11】
前記沈殿剤が酒石酸K/Naである、請求項5に記載の方法。
【請求項12】
段階d)を蒸気拡散によって行う、請求項6に記載の方法。
【請求項13】
ヒトVAP-1タンパク質の三次元構造を規定する原子座標/X線回折データをその上に記憶した、機械可読データをコードしたデータ記憶装置を含んでいるコンピュータ可読媒体であって、分子構造を決定するための適当な機械によって読取られかつコンピュータプログラムによって処理されたとき、ヒトVAP-1タンパク質のフラグメントを含んでいる分子の結晶の三次元表現をディスプレイすることができる媒体。
【請求項14】
前記データが、ダイマーのヒトVAP-1タンパク質の活性部位キャビティを規定する、請求項13に記載のコンピュータ可読媒体。
【請求項15】
前記活性部位キャビティが、表面では幅約20Å×約10Åかつ深さ約15Åである、請求項14に記載のコンピュータ可読媒体。
【請求項16】
前記活性部位キャビティが、ヒトVAP-1の一つのモノマーに由来するアミノ酸86-87、97、168-173、176-177、180、184、205-212、216、227、232-234、236-239、344、388-390、393-397、415-419、421、421-426、467-470、647-651、および758-761、およびヒトVAP-1の別のモノマーに由来するアミノ酸433-449、および451を含んでいる、請求項15に記載のコンピュータ可読媒体。
【請求項17】
前記活性部位が、前記部位の底の狭い約4.5Å×約4.5Åのキャビティの上端に、アミノ酸Leu469をさらに含む、請求項16のコンピュータ可読媒体。
【請求項18】
前記活性部位キャビティの底部が、アミノ酸残基Ala370、Tyr384、Asp386、Asn470、Tpq471およびTyr473によって裏打ちされた、請求項17に記載のコンピュータ可読媒体。
【請求項19】
ヒトVAP-1タンパク質と相互作用する化合物を同定するための方法であって:
a)コンピュータ可読媒体上に前記タンパク質の原子座標を供給すること、およびb)コンピュータを用いて、前記座標に分子モデリング技術を適用すること、
の段階を含む方法。
【請求項20】
前記原子座標が、ヒトVAP-1の一つのモノマーに由来するアミノ酸86-87、97、168-173、176-177、180、184、205-212、216、227、232-234、236-239、334、388-390、393-397、415-419、421、421-426、467-470、647-651、および758-761、およびヒトVAP-1の別のモノマーに由来するアミノ酸433-449、および451のものである、請求項19に記載の方法。
【請求項21】
前記原子座標が、アミノ酸残基Ala370、Tyr384、Asp386、Asn470、Tpq471およびTyr473のものである、請求項19に記載の方法。
【請求項22】
前記分子モデリング技術が、de novoの化合物設計を含んでいる、請求項20〜21のいずれか1項に記載の方法。
【請求項23】
前記de novoの化合物設計が、(i)VAP-1の活性部位内の箇所と相互作用することができる官能基または低分子フラグメントの同定、および(ii)これらを一つの化合物中に結合すること、を含む請求項22に記載の方法。
【請求項24】
前記分子モデリング技術が、VAP-1のファーマコフォアを使用する、請求項19〜23のいずれか1項に記載の方法。
【請求項25】
前記分子モデリング技術が、自動化されたドッキングアルゴリズムを使用する、請求項19〜24のいずれか1項に記載の方法。
【請求項26】
更なる段階(c)前記分子モデリング技術により同定された化合物を供給すること;および(d)前記化合物をヒトVAP-1と接触させること、およびVAP-1のSSAO活性に対する前記化合物の阻害効果を検出すること、を含む請求項19〜25のいずれか1項に記載の方法。
【請求項27】
請求項19〜26のいずれか1項に記載の方法によって同定可能なヒトVAP-1阻害剤。
【請求項28】
式:
【化1】
[式中、
R1は水素、低級アルキルまたは任意に置換されたフェニルまたはヘテロアリール基であり;
R2は水素または低級アルキルであるか、または
R1およびR2は、それらが結合している窒素原子と一緒になって飽和複素環を形成してもよく;
R3-R5は各々独立して水素、低級アルキル、アラルキル、任意に置換されたフェニルまたはヘテロアリール基を表しているか、または
R2およびR3は、それらが結合している原子と一緒になって飽和複素環を形成してもよく、あるいは
R3およびR5は、それらが結合している炭素原子と一緒になって飽和炭素環を形成してもよく;
R6はナフチル、フェニル、置換フェニルまたはヘテロアリール基であり;
R7は水素、低級アルキルまたはアラルキルであり;
nは1、2または3であり;かつ
X=O、S、SO、SO2またはNR2である]
を特徴とする、請求項27に記載の阻害剤。
【請求項29】
慢性関節炎、炎症性腸疾患、および皮膚疾患及び多発性硬化症のような、急性および慢性の炎症症状または疾患、糖尿病および糖尿病から結果として生じる、たとえば網膜症、神経障害、腎症といった血管症のような合併症を含めた、糖質代謝に関連した疾患、脂肪細胞の分化または機能、および平滑筋細胞の機能、特にアテローム硬化症における異常、および種々の血管性疾患からなる群より選ばれる疾病の治療における使用のための薬剤製造のための、請求項27〜28のいずれか1項に記載の化合物の使用。
【請求項30】
請求項27〜28のいずれか1項に記載の化合物を含んでいる薬剤組成物。
【請求項1】
結晶性ヒト血管接着タンパク質1(VAP-1)。
【請求項2】
前記結晶が、二つの分子を非対称ユニットにもち、a=b=225.9Å、c=218.7Å、α=β=90°、γ=120°のユニットディメンションをもつ、空間群P6522の結晶として定義される、請求項1に記載の結晶性ヒトVAP-1。
【請求項3】
アミノ酸配列、配列番号3、配列番号4および配列番号5によって各々特徴づけられる、ドメインD2、D3、およびD4を含んでいる、請求項2に記載の結晶性VAP-1、またはその同族体。
【請求項4】
表面において幅約20Å×約10Å、かつ深さ約15Åである活性部位キャビティを含んでいる、請求項3に記載の結晶性VAP-1タンパク質。
【請求項5】
前記活性部位キャビティが、ヒトVAP-1の一つのモノマーに由来する配列番号2のアミノ酸86-87、97、168-173、176-177、180、184、205-212、216、227、232-234、236-239、344、388-390、393-397、415-419、421、421-426、467-470、647-651および758-761、およびヒトVAP-1の別のモノマーに由来する配列番号2のアミノ酸433-449および451を含んでいる、請求項4に記載の結晶性VAP-1タンパク質。
【請求項6】
前記活性部位が、前記キャビティの底の狭い約4.5Å×約4.5Åのキャビティの上端に、アミノ酸Leu469をさらに含む、請求項5に記載の結晶性VAP-1タンパク質。
【請求項7】
前記活性部位キャビティの底部が、アミノ酸残基Ala370、Tyr384、Asp386、Asn470、Tpq471およびTyr473によって裏打ちされた、請求項6に記載の結晶性VAP-1タンパク質。
【請求項8】
請求項1〜7のいずれか1項に記載の結晶性VAP-1を含んでいる組成物。
【請求項9】
ヒトVAP-1を結晶化するための方法であって、
a)ヒトVAP-1タンパク質を含んでいる水溶液を供給すること;
b)沈殿剤を含んでいる貯蔵溶液を供給すること;
c)ある体積の前記水溶液とある体積の前記保存溶液と混合して、混合液を形成すること;および
d)前記混合液の少なくとも一部を結晶化すること、
の段階を含む方法。
【請求項10】
段階a)で供給された前記水溶液が、1mg/mlのVAP-1濃度を有する、請求項4に記載の方法。
【請求項11】
前記沈殿剤が酒石酸K/Naである、請求項5に記載の方法。
【請求項12】
段階d)を蒸気拡散によって行う、請求項6に記載の方法。
【請求項13】
ヒトVAP-1タンパク質の三次元構造を規定する原子座標/X線回折データをその上に記憶した、機械可読データをコードしたデータ記憶装置を含んでいるコンピュータ可読媒体であって、分子構造を決定するための適当な機械によって読取られかつコンピュータプログラムによって処理されたとき、ヒトVAP-1タンパク質のフラグメントを含んでいる分子の結晶の三次元表現をディスプレイすることができる媒体。
【請求項14】
前記データが、ダイマーのヒトVAP-1タンパク質の活性部位キャビティを規定する、請求項13に記載のコンピュータ可読媒体。
【請求項15】
前記活性部位キャビティが、表面では幅約20Å×約10Åかつ深さ約15Åである、請求項14に記載のコンピュータ可読媒体。
【請求項16】
前記活性部位キャビティが、ヒトVAP-1の一つのモノマーに由来するアミノ酸86-87、97、168-173、176-177、180、184、205-212、216、227、232-234、236-239、344、388-390、393-397、415-419、421、421-426、467-470、647-651、および758-761、およびヒトVAP-1の別のモノマーに由来するアミノ酸433-449、および451を含んでいる、請求項15に記載のコンピュータ可読媒体。
【請求項17】
前記活性部位が、前記部位の底の狭い約4.5Å×約4.5Åのキャビティの上端に、アミノ酸Leu469をさらに含む、請求項16のコンピュータ可読媒体。
【請求項18】
前記活性部位キャビティの底部が、アミノ酸残基Ala370、Tyr384、Asp386、Asn470、Tpq471およびTyr473によって裏打ちされた、請求項17に記載のコンピュータ可読媒体。
【請求項19】
ヒトVAP-1タンパク質と相互作用する化合物を同定するための方法であって:
a)コンピュータ可読媒体上に前記タンパク質の原子座標を供給すること、およびb)コンピュータを用いて、前記座標に分子モデリング技術を適用すること、
の段階を含む方法。
【請求項20】
前記原子座標が、ヒトVAP-1の一つのモノマーに由来するアミノ酸86-87、97、168-173、176-177、180、184、205-212、216、227、232-234、236-239、334、388-390、393-397、415-419、421、421-426、467-470、647-651、および758-761、およびヒトVAP-1の別のモノマーに由来するアミノ酸433-449、および451のものである、請求項19に記載の方法。
【請求項21】
前記原子座標が、アミノ酸残基Ala370、Tyr384、Asp386、Asn470、Tpq471およびTyr473のものである、請求項19に記載の方法。
【請求項22】
前記分子モデリング技術が、de novoの化合物設計を含んでいる、請求項20〜21のいずれか1項に記載の方法。
【請求項23】
前記de novoの化合物設計が、(i)VAP-1の活性部位内の箇所と相互作用することができる官能基または低分子フラグメントの同定、および(ii)これらを一つの化合物中に結合すること、を含む請求項22に記載の方法。
【請求項24】
前記分子モデリング技術が、VAP-1のファーマコフォアを使用する、請求項19〜23のいずれか1項に記載の方法。
【請求項25】
前記分子モデリング技術が、自動化されたドッキングアルゴリズムを使用する、請求項19〜24のいずれか1項に記載の方法。
【請求項26】
更なる段階(c)前記分子モデリング技術により同定された化合物を供給すること;および(d)前記化合物をヒトVAP-1と接触させること、およびVAP-1のSSAO活性に対する前記化合物の阻害効果を検出すること、を含む請求項19〜25のいずれか1項に記載の方法。
【請求項27】
請求項19〜26のいずれか1項に記載の方法によって同定可能なヒトVAP-1阻害剤。
【請求項28】
式:
【化1】
[式中、
R1は水素、低級アルキルまたは任意に置換されたフェニルまたはヘテロアリール基であり;
R2は水素または低級アルキルであるか、または
R1およびR2は、それらが結合している窒素原子と一緒になって飽和複素環を形成してもよく;
R3-R5は各々独立して水素、低級アルキル、アラルキル、任意に置換されたフェニルまたはヘテロアリール基を表しているか、または
R2およびR3は、それらが結合している原子と一緒になって飽和複素環を形成してもよく、あるいは
R3およびR5は、それらが結合している炭素原子と一緒になって飽和炭素環を形成してもよく;
R6はナフチル、フェニル、置換フェニルまたはヘテロアリール基であり;
R7は水素、低級アルキルまたはアラルキルであり;
nは1、2または3であり;かつ
X=O、S、SO、SO2またはNR2である]
を特徴とする、請求項27に記載の阻害剤。
【請求項29】
慢性関節炎、炎症性腸疾患、および皮膚疾患及び多発性硬化症のような、急性および慢性の炎症症状または疾患、糖尿病および糖尿病から結果として生じる、たとえば網膜症、神経障害、腎症といった血管症のような合併症を含めた、糖質代謝に関連した疾患、脂肪細胞の分化または機能、および平滑筋細胞の機能、特にアテローム硬化症における異常、および種々の血管性疾患からなる群より選ばれる疾病の治療における使用のための薬剤製造のための、請求項27〜28のいずれか1項に記載の化合物の使用。
【請求項30】
請求項27〜28のいずれか1項に記載の化合物を含んでいる薬剤組成物。
【図2】

【図3】
【図4】
【図3】
【図4】
【公表番号】特表2007−509605(P2007−509605A)
【公表日】平成19年4月19日(2007.4.19)
【国際特許分類】
【出願番号】特願2006−530308(P2006−530308)
【出願日】平成16年5月25日(2004.5.25)
【国際出願番号】PCT/FI2004/000318
【国際公開番号】WO2004/104191
【国際公開日】平成16年12月2日(2004.12.2)
【出願人】(500021583)バイオティ セラピーズ コーポレイション (7)
【Fターム(参考)】
【公表日】平成19年4月19日(2007.4.19)
【国際特許分類】
【出願日】平成16年5月25日(2004.5.25)
【国際出願番号】PCT/FI2004/000318
【国際公開番号】WO2004/104191
【国際公開日】平成16年12月2日(2004.12.2)
【出願人】(500021583)バイオティ セラピーズ コーポレイション (7)
【Fターム(参考)】
[ Back to top ]