
維管束優先的発現プロモーター
本発明はポリヌクレオチドの転写および/または発現の調節に関する。特に、本発明は、植物細胞での維管束優先的なポリヌクレオチド転写を可能にするEucalyptus grandisおよびPinus radiataから単離されたポリヌクレオチド調節配列に関する。内在的および/または異種由来のポリヌクレオチドの転写を改変するために本発明の調節配列を利用するコンストラクトおよび方法も本発明に含まれる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明はポリヌクレオチド転写および/または発現の調節に関する。特に本発明は、ユーカリ・グランディスまたはユーカリ(Eucalyptus grandis)およびラジアータマツまたはマツ(Pinus radiata)から単離され、植物細胞内におけるポリヌクレオチドの維管束優先発現的転写を行うポリヌクレオチド調節配列に関する。内在性および/または異種由来のポリヌクレオチドの転写を改変するために本発明の調節配列を用いるコンストラクトおよび方法も、本発明に含まれる。
【背景技術】
【0002】
本出願は、米国仮出願第60/428,287号(2002年9月22日出願)に基づく優先権主張を伴う。
【0003】
林業および製紙産業の大きな目標は、植物のリグニン含量の制御である。リグニンは、木材の機械強度、着色、防腐性に関与するシンナミルアルコールの複合ポリマーである。木本種は、木材の乾燥重量の20%ないし30%を占める大量のリグニンを合成する。剛性を与えることに加えて、リグニンは、細胞壁に疎水性および非透水性を与えて、水の輸送を助ける。したがって、樹木のリグニン濃度を増大させることは、耐病性および建築用の強度の改善を木に与えることのような、一部の用途で有利になることがわかることになる。また、リグニンは燃料としても有用であり、木炭製造用材と、トウモロコシ稈と、バイオ燃料として用いられる柳および成長の早いポプラ雑種とのような燃料用の木材その他のバイオマスにおけるセルロース含量の増大したリグニンは好ましい。
【0004】
逆に樹木におけるリグニン含量の増大は、製紙産業にとって重大な問題をもたらし、製紙産業はセルロース繊維からリグニンを除去するために多大な資源を費やさなければならない。米国内だけでも、年間約2千万トンのリグニンが除去される。さらにリグニン含量は、飼草作物の消化性の良否の大きな原因であり、植物のリグニン含量のわずかな上昇が消化性の比較的大きな低下につながる。例えば、リグニン含量が低い作物は家畜にとってより効率的な飼草を提供し、飼草作物の消費量に対する乳および肉の生産収量が向上する。リグニン含量は植物の成育中に増大するので、農家は早めの収穫を行って少収量で消化の良い作物を得るか、あるいは遅めに収穫を行って多収量で消化の悪い作物を得るか、のいずれかを選択しなければならない。
【0005】
かかる理由から、植物の遺伝子改変によるリグニン含量または組成の制御は望ましい。この目標に向けて、リグニン生合成を担う遺伝子の同定および特徴付けと、その発現を調節する配列の決定とに多大な努力が注がれてきた。マツ由来のシンナミルアルコールデヒドロゲナーゼ(CAD)、シンナメート4−ヒドロキシラーゼ(C4H)、クマレート3−ヒドロキシラーゼ(C3H)、フェノラーゼ(PNL)、O−メチルトランスフェラーゼ(OMT)、シンナモイル−CoAレダクターゼ(CCR)、フェニルアラニンアンモニアリアーゼ(PAL)、4−クマレート:CoAリガーゼ(4CL)およびペルオキシダーゼ(POX)を含む、リグニン生合成に関与する酵素の多くをエンコードするポリヌクレオチドがクローニングされてきた(特許文献1)。
【特許文献1】米国特許第6,204,434号明細書
【0006】
これらの遺伝子の発現操作がリグニン含量の改変に利用されてきた。かかる実験は、CAD、コニフェリンβ−グルコシダーゼ(CBG)、およびカフェイン酸3−O−メチルトランスフェラーゼ(COMT)をエンコードする遺伝子のコピー数の変更を含む(特許文献2、特許文献3および非特許文献1)。さらに、ポプラ、タバコ(N. tabacum)およびマツにおいてCADをエンコードする配列のアンチセンス発現は、組成の異なるリグニン産生を起こす(それぞれ、非特許文献2、3および4を参照)。
【特許文献2】米国特許第5,451,514号明細書
【特許文献3】国際公開第WO94/23044号公報
【非特許文献1】Dharmawardhanaら、Plant Mol.Biol.、40巻、365−72頁(1999年)
【非特許文献2】Grandら、Planta(Berl.)、163巻、232−37頁(1985年)
【非特許文献3】Yahiaouiら、Phytochemistry、49巻、295−306頁(1998年)
【非特許文献4】Baucherら、Plant Physiol.、112巻、1479頁(1996年)
【0007】
林産品、紙、植物バイオマスおよび飼草産業のもう1つの大目標は、茎からのセルロース回収を容易化するために、茎のサイズを増大させ、茎のセルロース含量を操作し、あるいは細胞壁の特性を操作することである。例えばセルロースは、パルプ生産では木部繊維から回収され、木部繊維数および管状要素数、細胞壁の厚さ、管腔径、繊維長、セルロース微繊維の角度その他のこれら木部の特性が、回収されるセルロースの質および量を決定する。セルロース生合成に関与する遺伝子の操作は、植物のバイオマス総量および植物からのセルロース収率の増大に有用であり、かかる遺伝子のアンチセンス発現は細胞壁の発達に影響を及ぼすことが示されている(非特許文献5)。また、エンド−1,4−β−グルカナーゼとセルロース結合ドメインとによる多糖類と植物細胞壁の改変が報告されている(非特許文献6)。したがって本発明のポリヌクレオチドは、セルロース生合成を改変し、これによって植物の生育とバイオマスとに影響を及ぼすために、維管束組織内でヌクレオチド配列を発現するのに用いられる場合がある。
【非特許文献5】Shani Z.、Shpigel,E.、Roiz L.、Goren R.、Vinocur B.、Tzfira T.、 Altman A.およびShoseyov O.、Cellulose−binding domain increases cellulose synthase activity in Acetobacter xylinum,and biomass of transgenic plants、A.Altman、M.Ziv、S.Izhar共編、Plant Biotechnology and In Vitro Biology in the 21st Century、213−218頁、Kluwer Academic Publishers(1999年)
【非特許文献6】Levy,I.、Shani,Z.およびShoseyov O.、Biomol.Eng.、19巻、17−30頁(2002年)
【発明の開示】
【発明が解決しようとする課題】
【0008】
前記改変植物への全身的な有害な影響を回避するために、生合成経路の遺伝子的調節は狭く限定された組織タイプにおいて実行することが好ましい。例えば、特定の遺伝子産物の発現によりリグニンの含量または組成が特定の遺伝子産物の発現によって影響を受ける場合、前記植物の病気に対する耐性の低下を避けるために、前記遺伝子産物の発現を前記植物の一部のセグメントまたは一部の生育段階に限定することが好ましい場合がある。異種遺伝子は、組織優先発現的プロモーターと操作可能に連結することにより、選択された組織において発現される場合がある。好適な組織優先発現的プロモーターは、維管束組織の原生木部管状要素内で特異的な活性があるマメgrb1.8プロモーターがある(非特許文献7)。これらのプロモーターは木質化部にて優先的に発現するユーカリCADプロモーターも含む(非特許文献8)。かかる組織優先発現的プロモーターは、特異的な組織におけるアンチセンス分子の遺伝子発現の調節に用いられてきた(非特許文献9−11)。
【非特許文献7】Kellerら、EMBO J.、8巻、1309頁(1989年)
【非特許文献8】Feuilletら、Plant Mol.Biol.、27巻、651頁(1995年)
【非特許文献9】Van der Meerら、Plant Cell、4巻、253頁(1992年)
【非特許文献10】Salehuzzamanら、Plant Mol.Biol.、 23巻、947頁(1993年)
【非特許文献11】Matsudaら、Plant Cell Physiol.、37巻、215頁(1996年)
【0009】
組織優先発現的プロモーターは、異なる環境では活性が弱い場合があり、構成的プロモーターで達成されるレベルと常に同じレベルで遺伝子を発現させるとは限らない(非特許文献12)。さらに、これらのプロモーターが活性を示す発生学的なウィンドウ、またはそれらの活性の空間分布が、それらの有用性を制限する場合がある。したがって、本技術分野では、所望の空間的・時間的な発現パターンを有するさらなる組織優先発現的プロモーター、特に、維管束優先発現的プロモーターの開発する継続的な必要性が存在する。非特許文献13に総説される。
【非特許文献12】Yahiaouiら、Phytochemistry、49巻、295−306頁(1998年)
【非特許文献13】Grima−Pettenatiら、Plant Science、145巻、51−65頁(1999年)
【課題を解決するための手段】
【0010】
発明の概要
本発明は、維管束優先発現的遺伝子発現に関与する単離ポリヌクレオチド調節配列に関する。本発明のポリヌクレオチドは、植物のリグニン含量、セルロース含量、個体のサイズまたは細胞壁の成長の調節に用いることができる。本発明のポリヌクレオチドはまた、リグニン、セルロース、および植物細胞壁の生合成にも用いられる場合がある。
【0011】
1つの局面において、本発明は、維管束優先発現的ポリヌクレオチド転写をさせることが可能なポリヌクレオチドを含む単離核酸分子を提供する。1つの実施態様において、前記単離核酸分子は配列番号1−85のいずれか1つとその機能変異体とから選択されるポリヌクレオチドを含む。別の実施態様において前記機能変異体は、配列番号1−85のいずれかの配列との同一性が99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、89%、88%、87%、86%、85%、84%、83%、82%、81%、80%、79%、78%、77%、76%、75%、74%、73%、72%、71%、70%、69%、68%、67%、66%、65%、64%、63%、62%、61%または60%以上である。
【0012】
他の局面において、本発明は、(a)配列番号1−85およびその機能変異体のいずれか1つに対する相補配列と、(b)配列番号1−85の配列のいずれかに対する逆相補体の配列と、(c)前記(a)または(b)のポリヌクレオチドのいずれかと雑種形成する、少なくとも20個の連続する塩基を含む配列とから選択される配列を有する単離ポリヌクレオチドを提供する。1つの実施態様において、本発明は、前記ポリヌクレオチドが植物細胞における木部優先発現的の遺伝子発現を実現できるようにすることを意図する。別の実施態様において、本発明は、植物細胞における操作可能に連結した遺伝子の発現を上向き調節または下向き調節することが可能なポリヌクレオチドを提供する。
【0013】
別の局面において、本発明は、(a)配列番号1−85のいずれかの配列を有する少なくとも1個のポリヌクレオチド配列と、(b)所望の遺伝子とを含み、前記ポリヌクレオチドと前記所望の遺伝子とが操作可能に連結している植物細胞を提供する。1つの実施態様において、前記所望の遺伝子はポリペプチドまたはタンパクをエンコードする。別の実施態様において、前記タンパクは細胞壁の生合成に関与する酵素である。さらなる実施態様において、前記タンパクはリグニンの生合成に関与する酵素である。さらに別の実施態様において、前記所望の遺伝子はRNA転写産物を生成する。さらに別の実施態様において、前記RNA転写産物は植物細胞に内在する遺伝子のアンチセンス配列を有する。さらに別の実施態様において、前記RNA転写産物は植物細胞中で正常に発現される遺伝子のRNA干渉を誘導する。
【0014】
別の局面において、本発明は、(a)配列番号1−85のいずれかの配列を有する少なくとも1個のポリヌクレオチド配列と、(b)所望の遺伝子とを含み、前記ポリヌクレオチドと前記所望の遺伝子とが操作可能に連結している、植物細胞を含む植物を提供する。1つの実施態様において、前記植物は被子植物および裸子植物から選択される。
【0015】
別の実施態様において、本発明は、(a)配列番号1−85のいずれかの配列を有する少なくとも1個のポリヌクレオチド配列と、(b)所望の遺伝子とを含み、前記ポリヌクレオチドと前記所望の遺伝子とが操作可能に連結しているDNAコンストラクトを含む、植物細胞を含む植物を栽培することを含む、植物のリグニン含量の調節方法を意図する。
【0016】
別の局面において、本発明は、(a)配列番号1−85のいずれかの配列を有する少なくとも1個のポリヌクレオチド配列と、(b)所望の遺伝子とを含み、前記ポリヌクレオチドと前記所望の遺伝子とが操作可能に連結しているDNAコンストラクトを含む、植物細胞を含む植物を栽培することを含む、植物の細胞壁の発生の調節方法を意図する。
【0017】
他の局面において、本発明は、配列番号1−85およびその機能変異体のいずれかから選択されるポリヌクレオチド配列を含む、トランスジェニック植物を提供する。
【0018】
1つの局面において、本発明は、(a)(i)配列番号1ないし85およびその機能変異体のいずれかの配列を有するプロモーターと、(ii)所望の核酸とを含み、前記プロモーターは前記所望の核酸の発現を調節するDNAコンストラクトを木本植物の植物細胞に導入すること、(b)前記形質転換された植物細胞を植物の生育を促進する条件下で培養すること、および、(c)前記植物から木材を得ることとを含む、木材を得る方法を提供する。1つの実施態様において、前記木本植物は、ユーカリ属またはマツ属の植物種から選択される。
【0019】
本発明はまた、配列番号1−85からなる群から選択される単離ポリヌクレオチド配列と、これらの配列のいずれかに対して少なくとも60%の同一性を有するポリヌクレオチドとを意図する。本発明はまた、配列番号1−85のいずれかに対して少なくとも65%の配列同一性を有するポリヌクレオチドを意図する。本発明はまた、配列番号1−85のいずれかに対して少なくとも70%の配列同一性を有するポリヌクレオチドを意図する。本発明はまた、配列番号1−85のいずれかに対して少なくとも75%の配列同一性を有するポリヌクレオチドを提供する。本発明はまた、配列番号1−85のいずれかに対して少なくとも80%の配列同一性を有するポリヌクレオチドを意図する。また、配列番号1−85のいずれかに対して少なくとも85%の配列同一性を有するポリヌクレオチドが提供される。本発明はまた、配列番号1−85のいずれかに対して少なくとも90%の配列同一性を有するポリヌクレオチドを意図する。配列番号1−85のいずれかに対して少なくとも95%の配列同一性を有するポリヌクレオチドが提供される。
【0020】
本発明は、表8に挙げたポリヌクレオチド配列のいずれかからなる群から選択される配列に対して少なくとも95%の同一性がある配列を有するポリヌクレオチドを含む単離核酸分子に関する。本発明はまた、表8に開示されるポリヌクレオチド配列の機能断片も提供する。本発明はさらに、表8に列挙されるポリヌクレオチド配列のいずれかに対する相補的な核酸またはその断片と、表8に開示されるポリヌクレオチド配列のいずれかと雑種形成し、少なくとも15個の連続する塩基を含む核酸とを提供する。
【0021】
本発明の核酸のヌクレオチド配列は配列表に示される。ここで、「配列番号(2N+1)」とは、例えば1、3、5、7…等の、すべての奇数番号の配列をいう。「配列番号2N」とは、すべての偶数番号の配列をいう。ある奇数番号の配列とすぐ次の偶数番号の配列(例えば、配列番号1と配列番号2)との間の違いは、前記偶数番号の配列は追加の塩基を有する点である。偶数番号の配列の末端の3塩基は通常ATGで、(通常、メチオニンをコードする)翻訳の開始コドンを表す。
【0022】
本発明は当業者に周知の用語および表現を用いる。別の定義が与えられないかぎり、本明細書中で用いられるすべての科学技術用語は、本発明が属する技術分野において通常の知識を有する者によって普通に理解されるのと同じ意味を有する。一般に、ここで用いる用語法と、ここで説明される細胞培養、分子遺伝学、核酸化学および雑種形成法における各実験手順とは、当該技術分野において周知慣用のものである。核酸組換法、ポリヌクレオチド合成、微生物培養、細胞培養、組織培養、形質転換、トランスフェクション、トランスダクション、分析化学、有機合成化学、化学合成、化学分析および医薬品処方および薬物送達には、標準的な手法が用いられる。一般に、酵素反応、精製および/または単離のステップは製造者の仕様にしたがって行われる。手法および手順は従来の方法論にしたがって実行されるのが一般的である。例えば、Sambrook & Russel、MOLECULAR CLONING:A LABORATORY MANUAL第3版、Cold Spring Harbor Laboratory Press刊、Cold Spring Harbor、NY、2001年を参照せよ。
【0023】
アグロバクテリア:当技術分野で周知のとおり、植物細胞の形質転換に用いられるアグロ細菌は、ベクターを含む、アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)またはアグロバクテリウム・リゾゲネス(Agrobacterium rhizogenes)の無毒化された(disarmed)ビルレントな誘導体であることが一般である。前記ベクターは、T−DNAの境界の間に配置された所望のポリヌクレオチドを含むことが典型的である。
【0024】
被子植物:種子が子房に被覆されている維管束植物。被子植物は、花を付け、実を結ぶ種子植物である。被子植物は、双子葉植物と単子葉植物とに分類される。
【0025】
所望のポリヌクレオチド:本発明の所望のポリヌクレオチドとは、プロモーター、エンハンサーまたはターミネーターのような遺伝子エレメントか、ゲノム中に前記所望のポリヌクレオチドを含む形質転換細胞で転写および/または翻訳されるべき遺伝子またはポリヌクレオチドかをいう。前記所望のポリヌクレオチドがタンパク産物をエンコードする配列を含む場合、そのコーディング領域は、当該所望のポリヌクレオチドによってエンコードされるメッセンジャーRNA転写産物および/またはタンパク産物の発現をもたらす、プロモーターおよびターミネーターのような調節エレメントと操作可能に連結している場合がある。したがって、「所望のポリヌクレオチド」は、5’から3’への配向で、プロモーターと、タンパクをエンコードする遺伝子と、ターミネーターとに操作可能に連結する遺伝子を含む場合がある。代替的には、前記所望のポリヌクレオチドは、翻訳されると植物細胞内での内在性遺伝子の発現に影響を与える2次構造を形成するかもしれない核酸を産生するような、「アンチセンス」配向の遺伝子またはその断片を含む場合がある。所望のポリヌクレオチドはまた、転写されると前記所望のポリヌクレオチドが会合する遺伝子のRNA干渉を引き起こすような、2本鎖RNAを産生する場合がある。本発明の所望のポリヌクレオチドは、T−DNAの左右の境界配列が前記所望のポリヌクレオチドを挟むか、あるいは所望のポリヌクレオチドのいずれか片側に配置されるように、T−DNA内に配置される場合がある。本発明は、少なくとも1個の植物細胞のゲノムに1個または2個以上の所望のポリヌクレオチドを安定的に組み込むことを可能にする。所望のポリヌクレオチドは、突然変異処理されていても、その野生型配列の変異体であってもよい。前記所望のポリヌクレオチドの全部または一部が植物ゲノムに組み込まれ場合があるものと理解されている。また、「所望のポリヌクレオチド」という用語は、かかるポリヌクレオチドの1個または2個以上を包含するものと理解されている。したがって、本発明のT−DNAは、1個、2個、3個、4個、5個、6個、7個、8個、9個、10個または11個以上の所望のポリヌクレオチドを含む場合がある。
【0026】
双子葉植物:胚が2分割状の種子、すなわち子葉と、分岐した葉脈と、4または5の倍数に分割される花とを持つ顕花植物。双子葉植物の例は、ユーカリ属、ハコヤナギ属、フウ(Liquidamber),アカシア属、チーク、マホガニー、ワタ、タバコ、シロイヌナズナ(Arabidopsis)、トマト、ジャガイモ、テンサイ、ブロッコリ、キャッサバ、サツマイモ、コショウ、ポインセチア、マメ、アルファルファ、ダイズ、ニンジン、イチゴ、レタス、カシ、カエデ、クルミ、バラ、ミント、カボチャ、ヒナギク、ゼラニウム、アボカド、サボテンがを含むが、これらに限定されない。
【0027】
内在性とは、ある植物ゲノムに元々存在する遺伝子を指す。
【0028】
繊維組成:本明細書で用いられる繊維組成とは、繊維の構造、外観または用途を変更するために改変可能な形質をいう。繊維組成を決定する形質は、繊維長、粗さ、強度、色、断面幅、繊維密度を含むが、これらに限定されない。例えば、繊維長は強度を与え、繊維の粗さは風合いおよび可撓性を決定する要因であることが知られている。
【0029】
外来性:ある核酸に関して「外来性」とは、その核酸が植物以外の生物に由来するか、形質転換されるべき植物とは異なる植物に由来するか、形質転換されるべき植物と交配できない植物に由来するか、あるいは標的植物の種に属さないことを意味する。本発明によれば、外来性のDNAまたはRNAは、真菌、細菌、ウィルス、哺乳類、魚類または鳥類の遺伝物質中に天然に存在するが、形質転換されるべき植物中には天然には存在しない核酸を含む場合がある。したがって、ある外来性の核酸は、例えば、形質転換された植物によって天然には生産されないポリぺプチドをエンコードするものである。外来性の核酸は、タンパク産物をエンコードする必要はない。
【0030】
遺伝子:産物、すなわちポリペプチド鎖またはRNA分子の合成に必要な情報のすべてを含むDNA分子のセグメントであり、コーディング配列およびノンコーディング配列の双方を含む。
【0031】
遺伝子エレメント:「遺伝子エレメント」とは、プロモーター、遺伝子、ターミネーター、イントロン、エンハンサー、スペーサー、5’−非翻訳領域、3’−非翻訳領域、またはリコンビナーゼ認識部位を含むがこれらに限定されない、離散的なヌクレオチド配列のいずれかをいう。
【0032】
遺伝子改変:分子細胞生物学的手法を適用することによって、ある生物のゲノムにDNAを安定的に導入すること。
【0033】
裸子植物:本明細書で用いられる場合には、子房を持たず、種子が露出している種子植物をいう。裸子植物の例は、針葉樹、ソテツ、イチョウおよびマオウを含む。
【0034】
導入:本明細書で用いられる場合には、感染、トランスフェクション、形質転換またはトランスダクション等の方法により、細胞中に核酸配列を挿入することをいう。
【0035】
幼若性:若木と成木との間の生理学的差異を説明する。本発明では、幼若性とは、若木と成木との間の微繊維角度、木材密度、セルロース収量、再生能力および生殖能力の差異をいう。例えば、木本植物組織は成熟するにしたがって再生能力を失うことが知られている。
【0036】
リグニン:本明細書で用いられる場合には、モノリグノールである、コニフェリルアルコール、クマリルアルコールおよびシナピルアルコールの重合誘導体を含む、フェニルプロパノイド単位で構成されるポリマー組成物をいう。リグニンの品質とは、細胞壁マトリクスに強度を与え、水分輸送を助け、および/または細胞壁多糖類の分解に抵抗する、リグニン組成物の能力をいう。リグニン組成物またはリグニン構造物は、各モノリグノールの相対量を変更するか、リグニンのタイプを変更することによって変化される場合がある。例えば、(フェルラ酸から誘導される)グアイアシルリグニンは軟木種では顕著だが、(フェルラ酸およびシナピン酸から誘導される)グアイアシル−シリンギルリグニンは硬木種に特徴的である。マツのような軟木由来のリグニンの分解には、硬木由来のリグニンの除去に比べてかなり大量のアルカリと長いインキュべーション時間とを要する。リグニン組成は、リグニン生合成に関与する酵素の上向き調節または下向き調節のいずれかにより調節される場合がある。例えば、重要なリグニン生合成酵素は、4−クマリン酸:補酵素Aリガーゼ(4CL)、シンナミルアルコールデヒドロゲナーゼ(CAD)およびシナピルアルコールデヒドロゲナーゼ(SAD)を含むが、これらに限定されない。
【0037】
単子葉植物:1枚の子葉または種葉(seedleaf)を有する胚と、平行な葉脈と、3の倍数に分割される花を有する顕花植物である。単子葉植物の例は、シバ、トウモロコシ、コメ、エンバク、コムギ、オオムギ、モロコシ、ラン、アヤメ、ユリ、タマネギ、ヤシを含むが、これらに限定されない。シバの例としては、Agrostis spp.(コロニアルベントグラスおよびクリーピングベントグラスを含むベントグラス種)、Poa pratensis(ケンタッキーブルーグラス)、Lolium spp.(1年生ライグラスおよび多年生ライグラス等のライグラス種)、Festuca arundiytacea(トールフェスク)、Festuca rubra commutata(ファインフェスク)、Cynodon dactylos(ティフグリーン、ティフウェイIIおよびサンタアナ等のコモンバミューダグラスの変種、並びにその雑種形成種)、Pennisetum clandestinum(アフリカチカラシバ)、Stenotaphrum secundatum(イヌシバ)、Zoysia japonica(シバ)、およびDichondra micranthaを含むが、これらに限定されない。
【0038】
操作可能に連結した:2個または3個以上の分子を、それらがある植物細胞内で組み合わせて適切に機能するように組み合わせること。例えば、プロモーターが構造遺伝子の転写を制御するとき、プロモーターは構造遺伝子に操作可能に連結している。
【0039】
表現型:表現型は、区別できる植物の特徴または特性であって、本発明では1個または2個以上の「所望のポリヌクレオチド」および/またはスクリーニング/選択可能なマーカーを、形質転換された植物の少なくとも1個の細胞のゲノムに組み込むことによって変化させられる場合がある。「(単数または複数の)所望のポリヌクレオチド」および/またはマーカーは、形質転換された植物細胞または植物個体全体の多数の遺伝的、分子的、生化学的、生理学的、形態学的、農業経済学的特徴または特性を改変することにより、形質転換された植物の表現型の変化を可能にする場合がある。したがって、植物ゲノム中に安定に組み込まれた1個または2個以上の所望のポリヌクレオチドの発現は、例えば、干魃耐性の向上、耐寒性および耐霜性強化、活力の向上、色彩の強化、保健栄養的な特性の強化、保存性の向上、収量の増大、耐塩性向上、重金属耐性向上、耐病性の向上、耐虫性の向上、耐乾性の強化、甘味の増強、活力の向上、味の改善、食感の改善、リン酸含量の低減、発芽率の増大、微量栄養素の取込みの増大、デンプン組成の改善、開花寿命の改善、密度の変更、茎強度または茎剛性の変化、寸法安定性の向上およびセルロースまたはリグニン含量の変化からなる群から選択される表現型を与える場合がある。
【0040】
植物組織:「植物」とは、胚を生産し、葉緑体を含み、セルロース細胞壁を有することを特徴とする、光合成能を有する植物界の真核多細胞生物のいずれかをいう。植物の一部、すなわち「植物組織」は、トランスジェニック植物を作出するために、本発明の方法により形質転換される場合がある。多くの適当な植物組織が、本発明の方法によって形質転換可能で、体細胞胚、花粉、葉、茎、カルス、走根、マイクロチューバーおよび苗条を含むが、これらに限定されない。このように、本発明はシバ、コムギ、トウモロコシ、コメ、オオムギ、エンバク、テンサイ、ジャガイモ、トマト、タバコ、アルファルファ、レタス、ニンジン、イチゴ、キャッサバ、サツマイモ、ゼラニウム、ダイズ、カシ、リンゴ、ブドウ、マツ、モミ、アカシア、ユーカリ、クルミおよびヤシのような被子植物および裸子植物の形質転換を意図する。本発明によると、「植物組織」は植物細胞も包む。植物細胞は、懸濁培養、カルス、胚、成長点領域、カルス組織、葉、根、苗条、配偶子、胞子体、花粉、種子および小胞子を含む。植物組織は成熟のいかなる段階にあるものでもよく、液体または固体培地か、土壌か、植木鉢、温室または圃場の適当な培地かで生育されてもよい。植物組織とは、有性的、無性的のいずれによって生じたかを問わず、かかる植物個体、種子、子孫、ムカゴのいずれかのクローンと、切断片や種子のような、これらのいずれかの派生物をもいう。特に興味深いのは、マツ、モミおよびトウヒのような針葉樹と、ケンタッキーブルーグラス、クリーピングベントグラス、トウモロコシおよびコムギのような単子葉植物と、ワタ、トマト、レタス、シロイヌナズナ、タバコ、リンゴおよびゼラニウムのような双子葉植物とである。
【0041】
植物の形質転換および細胞培養:広義には、植物細胞が遺伝子的に改変され、維持、さらなる成長および/またはさらなる発生のために適当な植物培地に移されるプロセスをいう。これらの方法は、当業者に周知である。
【0042】
子孫:トランスジェニック植物の子孫のような、本発明の「子孫」とは、植物またはトランスジェニック植物から生まれたか、これらの間に授かったか、これらに由来するものをいう。したがって、「子孫」植物、すなわち「F1」世代の植物は、本発明の方法により作出されたトランスジェニック植物の子、または子孫である。トランスジェニック植物の子孫は、少なくとも1個、一部、あるいは全ての細胞ゲノムに、本明細書に説明される方法によって親のトランスジェニック植物に組み込まれた所望のポリヌクレオチドを含む場合がある。したがって、所望のポリヌクレオチドは、子孫の植物によって「伝達される(transmitted)」か、「受け継がれる(inherited)」。そのように子孫植物に受け継がれる所望のポリヌクレオチドは、T−DNAコンストラクト中に存在する場合があるが、T−DNAコンストラクトもまたその親植物から子孫植物へと受け継がれる。本明細書で用いられる「子孫(progeny)」という用語は、ある群の植物の子(offspring)、または子孫(descendants)と考えられる場合もある。本発明のトランスジェニック植物は、子孫植物を作出するために無性生殖により繁殖される場合がある。
【0043】
プロモーター:RNAポリメラーゼおよび/または他の転写調節エレメントに結合する核酸、好ましくはDNAを指す意味で用いられる。あらゆるプロモーターと同様、本発明のプロモーター配列は、該プロモーターと操作可能に連結した核酸分子からmRNAを生成するために、DNAまたはRNAの転写を促進または制御する。上述したように、生成したRNAはタンパクまたはポリペプチドをコードする場合があり、RNA干渉分子またはアンチセンス分子をコードする場合がある。
【0044】
本明細書で用いられるプロモーターとは、調節エレメントを含む場合もある。逆に、ある調節因子がプロモーターから分離される場合もある。調節因子は、1つのプロモーター領域に対して多数の重要な性質を与える。ある種の要素は、操作可能に連結した核酸の転写速度を上げる転写因子に結合する。また、別の要素は、転写活性を阻害するレプレッサーに結合する。プロモーター活性に及ぼす転写因子の影響は、そのプロモーター活性が高いか低いか、すなわち、プロモーターが「強い」か「弱い」かを決定する。
【0045】
植物プロモーターとは、植物細胞由来であるか否かによらず、植物細胞中における転写を開始することができるプロモーターをいう。代表的な植物プロモーターは、植物と、植物ウィルスと、植物細胞で発現される遺伝子を含む、アグロバクテリウム属またはリゾビウム属のような細菌とから得られるものを含むが、これらに限定されない。発生学的な制御下にあるプロモーターの例は、木部、葉、根または種子のような特定の組織で転写を優先的に開始するプロモーターを含む。かかるプロモーターが、組織優先発現的(tissue−preferred)プロモーターと呼ばれる。特定の組織中でのみ転写を開始するプロモーターは、組織特異的プロモーターと呼ばれる。細胞タイプ特異的プロモーターは、例えば根または葉の維管束系細胞のような1種類または2種類以上の器官における特定の細胞タイプでの発現を主として推進する。誘導可能または抑制可能なプロモーターは、環境による制御の下にあるプロモーターである。誘導可能なプロモータによって転写に影響を及ぼし得る環境条件の例は、嫌気的条件および光の存在を含む。組織特異的、組織優先発現的、細胞タイプ特異的、および誘導可能プロモーターは、非構成的プロモーターのクラスを構成する。構成的プロモーターは、ほとんどの環境条件の下で、かつ、ほとんどの植物の部分で活性があるプロモーターである。
【0046】
ポリヌクレオチドとは、遺伝子をコードする配列または(少なくとも15個の連続したヌクレオチド、少なくとも30個の連続したヌクレオチド、または少なくとも50個の連続したヌクレオチドを含む)その断片と、プロモーターと、イントロンと、エンハンサー領域と、ポリアデニル化部位と、翻訳開始部位と、5’または3’非翻訳部位と、レポーター遺伝子と、選択可能マーカー等とを含むヌクレオチド配列をいう。ポリヌクレオチドは、1本鎖または2本鎖のDNAまたはRNAを含む場合がある。ポリヌクレオチドは、修飾塩基または修飾されたバックボーンを含む場合がある。ポリヌクレオチドは、ゲノム由来か、(mRNAのような)RNA転写産物か、(cDNAのような)プロセシングを経たヌクレオチド配列かの場合がある。ポリヌクレオチドは、センス配向またはアンチセンス配向を有する配列を含む場合がある。
【0047】
単離ポリヌクレオチドは、天然状態ではないポリヌクレオチド配列である。例えば、前記ポリヌクレオチドは、自然界には存在しないヌクレオチド配列で構成されるか、その近傍に存在するのが典型的なヌクレオチド配列から切り離されているか、あるいは、その近傍には存在しないのが典型的なヌクレオチド配列の近傍に存在する。
【0048】
再生能力:本明細書で用いられるところでは、脱分化組織から植物が再分化する能力をいう。
【0049】
種子:「種子」とは、胚を含む熟した植物の胚珠と、塊茎または胞子のような植物の増殖部位として考えられる場合がある。種子は、発芽促進のために、アグロバクテリアを介する形質転換の前に、例えば暗所でインキュベーションされる場合がある。種子はまた、インキュベーションの前に、短時間の漂白剤処理のような処理によって滅菌される場合がある。得られた実生はアグロバクテリアの所望の菌種に曝露できる。
【0050】
選択可能なマーカー/スクリーニング可能なマーカー:ある遺伝子が植物個体または植物組織において発現している場合に、該植物個体または植物組織を前記遺伝子が発現していない他の植物個体または植物組織と区別することを可能にする遺伝子をいう。スクリーニング手順は、前記スクーニング可能なマーカー遺伝子にエンコードされるタンパクの発現についてのアッセイが必要な場合がある。かかるマーカーの例は、ベータ・グルクロニダーゼ(GUS)遺伝子と、ルシフェラーゼ(LUX)遺伝子とを含む。選択可能なマーカーの例は、カナマイシンおよびジェネチシン耐性をエンコードするネオマイシンホスホトランスフェラーゼ(NPTII)遺伝子と、ハイグロマイシン耐性をエンコードするハイグロマイシンホスホトランスフェラーゼ(HPTまたはAPHIV)遺伝子と、スルホニル尿素系除草剤耐性をエンコードするアセトラクテートシンターゼ(als)遺伝子と、グルタミンシンターゼの活性を阻害するように作用する、ホスフィノトリシン(LibertyまたはBasta)のような除草剤への耐性をコードする遺伝子(BARおよび/またはPAT)と、その他の本技術分野で知られた類似の遺伝子を含む。
【0051】
配列の同一性:本明細書で用いられる、2個の核酸配列またはポリペプチド配列について「配列の同一性」または「同一性」は、ある特定の領域にわたって前記2個の配列を最大限対応するようにアライメントをとるときに、前記2個の配列中で同一となる残基をさすことを含む。
【0052】
本明細書で用いられる同一性の百分率は、ある比較ウィンドウにわたって最適なアライメントをとった2個の配列を比較することによって決定される値を意味するが、ここで、前記2個の配列の最適なアライメントのための((例えばギャップのような)付加または欠失を含まない)基準配列と比較するとき、前記比較ウィンドウ内の前記ポリヌクレオチド配列の一部は付加または欠失を含む場合がある。前記百分率は、両方の配列中に同一の核酸塩基が出現する塩基数を数えることにより一致塩基数を算出し、この一致塩基数を比較ウィンドウ中の総塩基数で除し、この商に100を乗ずることにより算出される。
【0053】
転写因子:転写因子とは、遺伝子コード配列を伴う1個または2個以上のヌクレオチド配列に直接結合するか、あるいは、遺伝子をコードする配列を伴う1個または2個以上のヌクレオデチド配列に直接結合する他の(単数または複数の)ポリペプチドの活性に間接的に影響を及ぼすかのいずれかによって、単数または複数の遺伝子の発現を調節するポリペプチド配列をいう。転写因子は、単数または複数の遺伝子の発現を活性化(上向き調節)または抑制(下向き調節)する場合がある。転写因子は、DNA結合ドメインと、活性化ドメインまたはタンパク−タンパク相互作用ドメインとを含む場合がある。本発明では、転写因子は、(1)核酸配列に結合すること、あるいは(2)植物における遺伝子の発現を調節すること、の少なくともいずれかを行うことができる。さらに、本発明のポリヌクレオチド配列およびこれに対応するポリペプチド配列は、被子植物および裸子植物を含む、いずれかの植物種において転写因子として機能する。
【0054】
転写および翻訳のターミネーター:本発明の発現DNAコンストラクトには、典型的には、転写開始調節エレメントの反対側に転写終結領域がある。この転写終結領域は、発現を増強するmRNAの安定、および/または、遺伝子転写産物にポリアデニル鎖の尾部の付加のために選択される場合がある。
【0055】
転移DNA(T−DNA):アグロバクテリアのT−DNAは、その境界内に含まれるヌクレオチド配列を別のゲノムに組み入れることが可能な因子として周知の遺伝子エレメントである。この点で、T−DNAは典型的には2つの「境界」配列に挟まれている。本発明の所望のポリヌクレオチドおよび選択可能なマーカーは、あるT−DNAの左側の境界様配列と右側の境界様配列との間に位置する場合がある。T−DNAに含まれる前記所望のポリヌクレオチドおよび選択可能なマーカーは、その発現、すなわち前記所望のポリヌクレオチドまたは選択可能なマーカーによってエンコードされるDNA配列の転写および/または翻訳を促進するプロモーターおよびターミネーターの調節エレメントのような、さまざまな異なる植物特異的な(すなわち、天然の)、または外来の核酸と操作可能に連結している場合がある。
【0056】
植物細胞の形質転換:植物細胞のゲノムに核酸を安定に挿入するプロセスをいう。形質転換は当業者に周知のさまざまな方法を用い、自然または人工的な条件下で起こる場合がある。形質転換は、原核または真核宿主細胞へ核酸配列を挿入するいずれかの知られた方法によって行われる場合があり、これら方法としては、アグロバクテリアを介する形質転換法、ウィルス感染法、ウィスカー法、エレクトロポレーション法、顕微注入法、ポリエチレングリコール処理法、熱ショック法、リポフェクション法および微粒子銃法を含む。
【0057】
トランスジェニック植物:本発明のトランスジェニック植物とは、外来性の核酸が安定に組み込まれた少なくとも1個の細胞ゲノムを含むものをいう。本発明によると、トランスジェニック植物は、遺伝的に改変された細胞および細胞ゲノムを1つだけ含む場合があるか、複数ないし多数の遺伝的に改変された細胞を含む場合があるか、全ての細胞が遺伝的に改変されている場合がある。本発明のトランスジェニック植物は、所望のポリヌクレオチド、すなわち外来性の核酸の発現が、植物個体の特定の部分のみで起こる場合がある。したがって、トランスジェニック植物は、その構造の特定の部分には遺伝的に改変された細胞のみを含む場合がある。
【0058】
変異体:ここで言う「変異体(variant)」とは、ある特定の遺伝子の基準(すなわち天然、標準、所与の)ヌクレオチド配列から逸脱(deviate)したヌクレオチド配列をいう。「アイソフォーム」、「アイソタイプ」および「類似体」という用語もヌクレオチド配列の「変異」した類型を指す。
【0059】
「変異体」には、Maxygenに付与された特許に記載されているような「シャッフルされた(shuffled)遺伝子」を指す場合もある。例えば、本発明で言う変異体は、引用によりここに取り込まれる米国特許第6,132,970号明細書に開示された方法および原理にしたがって改変された配列および所望のポリヌクレオチドの変異体も含む場合がある。
【0060】
本明細書で用いられるところの木材組成は、木材の構造、外観または用途を変更するために改変可能な形質を指す。木材組成を決定する特性は、細胞壁の厚さと、細胞の長さと、細胞のサイズと、内腔の大きさと、細胞密度と、微繊維の角度と、引張強度と、引裂強度と、木材の色と、細胞分裂の所用時間および頻度とを含むが、これらに限定されない。
【0061】
木材パルプとは、さまざまな純度の木材から得られる繊維をいう。木材パルプは、紙、板紙、化学製品の製造に用いることができる。
【0062】
本発明は、本明細書で説明する特定の方法論、プロトコール、ベクター、試薬等に限定されるものではないことが理解されるが、それは変更が可能だからである。また、本明細書で用いる用語法は特定の実施態様のみを説明する目的で用いられており、本発明の範囲を限定することを意図するものではないことが理解される。また、本明細書および添付する請求の範囲で用いられる単数形の「a」、「an」、「the」は、その文脈の中で特に明言しない限り、複数形も包含することに留意しなければならない。したがって例えば、「遺伝子」というときは、1個または2個以上の遺伝子を指し、当業者等に知られた均等物等を含む。実際、当業者は、植物宿主系でいずれかの天然の(現在または将来知られる)遺伝子を発現させるためにここに説明する方法を利用することができる。
【0063】
(単数または複数の)「単離された」核酸分子とは、天然の環境から移された核酸分子である、DNAまたはRNAを指す。例えば、あるベクターに含まれる組換えDNA分子は、本発明の目的のために単離されたものと考えられる。単離DNA分子の他の例は、異種宿主細胞内で維持される組み替えDNA分子か、(部分的または実質的に)精製された溶液中のDNA分子かを含む。単離RNA分子は、本発明のDNA分子の試験管内RNA転写産物を含む。本発明の単離核酸分子は、人工合成で製造された分子も含む。
【0064】
本発明の核酸分子は、mRNAのようなRNAの形態か、クローニングや人工合成によって製造されたcDNAおよびゲノムDNAのようなDNAの形態かの場合がある。前記DNAまたはRNAは2本鎖または1本鎖の場合がある。1本鎖DNAは、コーディング鎖、別名センス鎖か、ノンコーディング鎖、別名アンチセンス鎖かの場合がある。
【0065】
特に断らない限り、本明細書でDNA分子の配列決定により決定されたすべてのヌクレオチド配列は、(アプライド・バイオシステムズ社製、モデル3700のような)自動シーケンサを用いて決定され、また本明細書で決定されたDNA分子にエンコードされたポリペプチドのすべてのアミノ酸配列は、上記のとおり決定されたDNA配列を翻訳することによって予測された。したがって、かかる自動化手法により決定されたいずれかのDNA配列について当業者に知られているとおり、本明細書で決定されたヌクレオチド配列にはなんらかのエラーが含まれている場合がある。自動的に決定されたヌクレオチド配列は、配列決定されたDNA分子の実際のヌクレオチド配列に対して典型的には少なくとも約95%、より典型的には少なくとも約96%ないし約99.9%の同一性を有する。前記実際の配列は、当業者に周知の手作業によるDNA配列決定を含むの他のアプローチにより、より正確に決定することができる。当業者に知られているとおり、決定されたヌクレオチド配列中に実際の配列と比較してたった1個の挿入または欠失は、ヌクレオチド配列の翻訳でフレームシフトを起こし、決定されたヌクレオチド配列によってエンコードされる予測アミノ酸配列が、このような挿入または欠失が生じた塩基から後は、配列決定されたDNA分子によって実際にエンコードされるアミノ酸配列とは全く異なったものになる。
【0066】
特に断らない限り、ここに示す「ヌクレオチド配列」のそれぞれは、デオキシリボヌクレオチド(A、G、C、Tと略記)の配列として提供される。しかし、核酸分子またはポリヌクレオチドの「ヌクレオチド配列」の語は、DNA分子またはポリヌクレオチドの場合はデオキシリボヌクレオチド配列を指し、RNA分子またはポリヌクレオチドの場合は、対応するデオキシヌクレオチド配列中における各チミジンデオキシヌクレオチド(T)がウリジンリボヌクレオチド(U)に置換された、対応するリボヌクレオチド(A、G、C、U)配列を指す。例えば、デオキシリボヌクレオチドの省略記号を用いて示される配列番号1の配列を有するRNA分子への言及は、配列番号1の各デオキシヌクレオチドA、GまたはCが対応するリボヌクレオチドA、GまたはCに置換され、各デオキシヌクレオチドTがリボヌクレオチドUに置換された配列を有するRNA分子を示すことを意図する。
【0067】
本発明は、本明細書で説明される単離核酸分子の断片にも向けられる。表8に示されるヌクレオチド配列を有する単離DNA分子の断片は、以下に詳しく説明される診断プローブおよびプライマーとして有用な、少なくとも15ヌクレオチド、少なくとも20ヌクレオチド、少なくとも30ヌクレオチドの長さを有するDNA断片を指す。もちろん、従来の雑種形成手法によれば、本発明の核酸分子の全長までの長さを有するより大きな核酸断片もプローブとして診断上有用であり、また、例えば、引用によりここにその開示内容全体が取り込まれるMolecular Cloning、A Laboratory Manual第3版、Sambrook、JおよびRussel、D.W.共編(2001年)、Cold Spring Harbor Laboratory Press、Cold Spring Harbor、New Yorkに説明されるポリメラーゼ連鎖反応(PCR)による標的配列の増幅用のプライマーとしても有用である。
【0068】
例えば、少なくとも20ヌクレオチドの長さを有する断片とは、表8に示したヌクレオチド配列由来の20個以上の連続した塩基を有する断片をいう。表8に列挙されたヌクレオチド配列を含む核酸は、当業者にはルーティン作業であろう従来のDNA合成法を用いて作成することができる。例えば、制限エンドヌクレアーゼによる切断または超音波処理による剪断は、さまざまなサイズの断片を作成するために簡単に利用することができる場合がある。代替的に、本発明のDNA断片は、既知の方法にしたがって、人工合成によって作成できる場合がある。
【0069】
別の局面において、本発明は、ストリンジェントな雑種形成条件下で上記の本発明の核酸分子におけるポリヌクレオチドの一部と雑種形成するポリヌクレオチドを含む単離核酸分子を提供する。あるポリヌクレオチドの「部分」と雑種形成するポリヌクレオチドとは、前記基準ポリヌクレオチドの少なくとも約15ヌクレオチド、少なくとも約20ヌクレオチド、少なくとも約30ヌクレオチド、および、31ヌクレオチド以上と雑種形成する(DNAまたはRNAのいずれかの)ポリヌクレオチドをいう。前記基準断片と雑種形成するこれらの断片は、診断プローブおよびプライマーとして有用である。本明細書で用いられるプローブは、表8の1個の核酸の少なくとも約100個の連続する塩基として定義される。本発明の目的には、6X SSC、0.5% SDS、5X Denhardt溶液および非特異的キャリアDNA100μgを含む雑種形成溶液中で2個の配列が2本鎖複合体を形成するとき、前記2個の配列は雑種形成するといえる。Ausubelら、第2.9節、補遺27(1994年)を参照せよ。配列が「中程度のストリンジェンシー」で雑種形成するとは、6X SSC、0.5% SDS、5X Denhardt溶液および非特異的キャリアDNA100μgを含む雑種形成溶液中で、60°Cの条件をいう。「高度なストリンジェンシー」で雑種形成を行うには、温度を68°Cに上昇させる。かかる中程度のストリンジェンシーの雑種形成反応を行った後、2X SSCおよび0.05% SDSの溶液中で室温にて5回、前記ヌクレオチドは洗浄され、さらに0.1X SSCおよび0.1% SDS溶液を用いて60°Cで1時間洗浄される。「高度なストリンジェンシー」用には、洗浄温度を68°Cに上昇させる。本発明の目的には、雑種形成したヌクレオチドとは、比活性10,000cpm/ngの放射能標識プローブ1ngを用いて検出されるものであり、前記雑種形成ヌクレオチドは−70°C、72時間以内のX線フィルムへの露出の後明瞭に視認できる。
【0070】
上記のとおり、本出願は、上記の核酸配列との同一性が、少なくとも65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%または100%である核酸分子に向けられる。1つの実施態様は、表8に示される核酸配列との同一性が少なくとも95%、96%、97%、98%、99%または100%である核酸分子を包含する。ある基準ヌクレオチド配列に対して例えば少なくとも95%「同一性がある」ヌクレオチド配列を有するポリヌクレオチドとは、前記ポリヌクレオチドのヌクレオチド配列が、前記基準ヌクレオチド配列の100個のヌクレオチドあたり最大5個の点突然変異を含むかもしれないことを除いて、前記基準配列と同一であることを意味する。換言すれば、基準ヌクレオチド配列に対して少なくとも95%の同一性があるヌクレオチド配列を有するポリヌクレオチドを得るためには、前記基準配列のヌクレオチドの最大5%が欠失するかまたは別のヌクレオチドに置換されている場合があるか、前記基準配列のヌクレオチド総数の最大5%の数のヌクレオチドが前記基準配列に挿入されている場合がある。前記基準配列のこれらの突然変異は、前記基準ヌクレオチド配列の5’末端または3’末端に起こるか、あるいは、前記基準配列中のヌクレオチドの間に1個ずつ分散するか、または前記基準配列中の1個または2個以上のグループに分かれて分散してこれら両末端の間の任意の場所で起こる場合がある。
【0071】
実際問題として、ある特定の核酸分子が基準ヌクレオチド配列に対して少なくとも95%、96%、97%、98%または99%の同一性を有するか否かは、当該技術分野で周知の標準的なアルゴリズムを用いて2個の分子の間の比較を行うことで判る。配列の同一性の決定にはいかなる配列決定アルゴリズムを用いてもよいが、明確にするために、本発明はBasis Local Alignment Search Tool(BLAST)アルゴリズム(Altschulら、J.Mol.Biol.、215巻、403−410頁(1990年))を用いて同一性を決定し、本開示内容に示されるプロモーター配列が、その全長にわたってポリヌクレオチド相同体の同一性の百分率を決定するための基準配列として用いられる。より生物学的に現実性のある結果をもたらすパラメータはいくつか知られているが、一致、不一致、および挿入または欠失のパラメータ値の選択は任意である。
【0072】
本発明にしたがって、ある特定の配列が基準配列に対して、例えば、95%の同一性を有しているか否かを決定するためにBLASTその他の配列アライメントプログラムを用いるときには、同一性の百分率が前記基準ヌクレオチド配列の全長にわたって計算され、前記基準配列中のヌクレオチド総数の5%までの相同性のギャップが許容されるように、パラメータが設定される。
【0073】
2個のポリヌクレオチドの間の関連度(relatedness)は、前記ポリヌクレオチドが雑種形成して相補的塩基対の形成によって2本鎖複合体を形成する能力に関連づけて説明することができる。雑種形成条件については既に説明した。温度の上昇はこれらの複合体を解離するのに利用することができる。2つの配列の構造的な同一性が大きいほど、両者を解離または「融解」させるのに必要な温度は高くなる。2本鎖複合体を融解させるのに必要な温度は「Tm」と呼ばれる。Tmと他の雑種形成パラメータとの関係は、次式で表される。
Tm(°C)=81.5+16.6(log10[Na+])+0.41(G+C分画)−0.63(%ホルムアミド)−(600/l)
前記式で、Tmはプローブとその標的とからなるDNA2本鎖複合体の融解温度であり、lは雑種形成体の長さ(単位:塩基対)、但しl>100塩基対である。Boltonら、Proc.Natl.Acad.Sci.、48巻、1390頁(1962年)を参照せよ。一般に、融解温度が1°C変化することは、DNA配列の類似度に0.7%から3.2%までの相違があることを表す。Bonnerら、Journal of Molecular Biology、81巻、123−35頁(1973年);McCarthyら、EVOLUTION OF GENETIC SYSTEMS、H.H.Smith編、Brookhaven Symposium in Biology、No.23、Gordon and Breach、New York、1−43頁(1972年)。安定なDNA2本鎖複合体を60°Cで形成するには、配列の間に少なくとも80%の配列同一性が必要である。Sibleyら、ACTA、1巻、83−121頁(Proceedings of the 18th International Ornithological Congress、Moscow、8月16〜24日、1982年、ソ連科学アカデミー)。
【0074】
本発明の核酸は、植物の維管束組織においてポリペプチドまたはタンパクを優先的に発現させる。また本発明の核酸は、標的遺伝子の発現を阻害または完全にブロックする目的で使用可能な、アンチセンスRNAの発現、または、低分子干渉RNA(siRNA)のような、RNA干渉(RNAi)に関与するRNAの発現を植物の維管束組織において優先的に行わせることができる。
【0075】
本明細書で用いられる維管束植物組織とは、木部、篩部または維管束の形成層組織をいう。1つの実施態様では、本発明のプロモーターは「木部特異的」、「形成層特異的」および/または「篩部特異的」のいずれかであり、それぞれ木部、形成層および/または篩部におけるある操作可能に連結した核酸セグメントの発現を指令する。本明細書で用いられる「コード産物(coding product)」とは、前記プロモーターに操作可能に連結した核酸の最終産物を意味する。例えば、タンパクまたはポリペプチドと、アンチセンスRNAまたはsiRNAとはコード産物であり、siRNAは前記アンチセンスRNAの核酸コーディングの最終産物である。またコード産物は非翻訳mRNAの場合がある。本明細書では、ポリペプチドおよびタンパクという用語を互換的に用いる。例えば木部特異的とは、本発明の核酸分子が他のいかなる植物組織におけるよりも木部において活性が高いことを意味する。1つの実施態様において本発明の核酸は、それぞれ植物の木部、形成層、および/または篩部組織でのみ活性があるという意味で、木部、形成層および/または篩部で特異的に活性を示すプロモーターである。換言すれば、例えば「木部特異的」プロモーターは、検出可能なレベルのコード産物は植物の木部組織においてのみ発現するように、前記コード産物を発現させる。しかし、植物個体内での溶質輸送のために、木部、篩部または形成層で特異的に発現するコード産物が植物個体内の他の場所で見出される場合があるため、その存在は必ずしも木部組織には限定されない。一方、維管束優先発現的プロモーターは、木部、篩部または形成層組織のいずれかにおいて、あるいはこれら3つの組織タイプのうち少なくとも2つにおいて優先的に活性を示すことができる。維管束特異的プロモーターは、木部、篩部または形成層のいずれかにおいて、あるいはこれら3つの組織タイプのうち少なくとも2つにおいて特異的に活性を示すことができる。
【0076】
本明細書で用いられる維管束系は、高等植物において水および光合成産物の輸送に用いられる木部および篩部の各組織と、該木部と篩部との間に位置する維管束形成層とから構成される。維管束系の構成要素としては、木部と篩部とをまとめて維管束組織と呼ぶ。維管束組織は、2つの発生学的に区別される段階で形成される。胚形成段階および/または胚形成後の段階では、頂端分裂組織および前形成層の分化から一次維管束組織が発生する。二次維管束組織は、維管束形成層の分化から発生する。
【0077】
維管束形成層は、木部と篩部との間に位置する側生分裂組織であり、分化して二次維管束組織を生成する。維管束形成層は、紡錘形原始細胞および木質線原始細胞という2つの細胞タイプで構成され、これらは形成層原始細胞と総称される。紡錘形原始細胞の分裂は、木部および篩部の長手方向、すなわち軸系を形成し、線原始細胞の分裂は、横方向、すなわち線系の発達をもたらす。形成層原始細胞は、木部細胞が長軸の内側に向かって凝集し、篩部細胞が周辺へ向かって凝集するように、軸の接線方向に分裂する。
【0078】
本明細書で用いられる木部とは、水やミネラルイオンを根系から植物の空中部分へと輸送する伝導組織をいう。一次および二次木部組織の両方とも、異なる3つの細胞タイプ、すなわち、導管要素、繊維および柔細胞を含む。導管要素は、無孔仮導管および有孔導管要素を含み、水の導通機能を果たす。木部繊維は、繊維仮導管および靱皮状繊維を含み、木部の構造に強度および剛性を与える。柔細胞は、軸細胞および放射細胞を含み、デンプン、油その他のエネルギーの豊富な分子の貯蔵に重要である。したがって、本発明の木部特異的プロモーターは、一次および/または二次木部の導管要素、繊維および/または柔細胞で活性がある。
【0079】
篩部は、光合成産物を成熟した葉から成長および貯蔵を担う領域へと移動させる植物組織である。一次および二次篩部組織の両方を含む主な細胞は、篩要素、柔細胞および繊維を含む。篩要素は、篩部の伝導要素であって、裸子植物に存在する篩細胞か、被子植物に存在する篩管かを含む。篩管要素は篩管と呼ばれる固有の構造を形成し、篩管は、縦方向に配列され、篩板と呼ばれる細胞壁の一部を通じて相互に連絡する一連の篩管要素でできている。これに対し、裸子植物や下等維管束植物に存在する特殊化の程度の低い篩細胞は、相互に連絡して篩板を形成することはない。篩細胞は、篩域と呼ばれる拡大した孔部を通じて伝導が容易になるように、重複したアレイ状に配置される。篩要素は、被子植物では伴細胞、裸子植物では胚乳細胞と呼ばれる特殊な柔細胞と連携し、かつ、依存している。伴細胞および胚乳細胞の両方とも、篩要素の機能に必要なタンパクおよび代謝産物の貯蔵を担う。前記特殊な柔細胞に加え、篩部は、デンプン、脂肪その他の化合物を蓄積し貯蔵する機能がある柔細胞を含む。篩部繊維は篩部構造に強度を与え、場合によりデンプンの貯蔵に寄与する。したがって、本発明の篩部特異的プロモーターは、一次および/または二次篩部の篩要素、柔細胞、および/または繊維で活性がある。
【0080】
ほとんどの高等植物において維管束組織は、基本組織間に位置する円筒状をしている。根と茎の両方で、髄と皮層とが基本組織を構成する。髄は維管柱の内部に位置し、皮層は表皮と維管束組織との間に位置する。維管束系とこれに付随する基本組織を、維管柱と総称する。
【0081】
本明細書で用いられるプロモーターは、RNAポリメラーゼおよび/または他の転写調節エレメントに結合する核酸、好ましくはDNAを意味する。他のいかなるプロモーターと同様、本発明のプロモーターは、該プロモーターと操作可能に連結している核酸分子からmRNA分子を生成するために、DNAまたはRNAを転写することを促進または制御する。前記のとおり、生成されたRNAはタンパクまたはポリペプチドをコードするか、あるいはRNA干渉、またはアンチセンス分子をコードする場合がある。本明細書で用いられる「操作可能に連結した」とは、RNAセグメントに転写されるべき核酸配列に対してプロモーター−核酸配列の組合せが適切な配向で形成されるような、化学的融合、連結、またはDNA合成をいう。本発明のプロモーターは、生成するmRNA転写産物の5’非翻訳領域(5’UTR)の一部または全部を含む場合がある。一方、本発明のプロモーターは、必ずしも5’UTRを有している必要はない。
【0082】
本明細書で用いられるプロモーターは調節エレメントを含む場合もある。逆に、調節エレメントがプロモーターから分離される場合がある。調節エレメントはプロモーター領域に多数の重要な特徴を与える。一部のエレメントは、前記操作可能に連結した核酸の転写速度を増大させる転写因子と結合する。他のエレメントは、転写活性を阻害するリプレッサーと結合する。転写因子がプロモーター活性に総合的に及ぼす影響は、該プロモーター活性が高いか低いか、すなわち、プロモーターが「強い」か「弱い」かを決定する場合がある。調節エレメントと結合する転写因子自身が、他の結合タンパクとの相互作用または細胞外刺激に応答する共有結合による修飾、例えばリン酸化によって調節される場合がある。一部の転写因子は、細胞内代謝産物か、その生物個体にとって外来性の化学物質かのような細胞核と情報交換する情報伝達分子によってモジュレーションされる。細胞環境の変化によって影響を受けないプロモーターは、構成的プロモーターと呼ばれる。
【0083】
本発明は、本発明の単離核酸分子を含むベクターも提供する。1つの実施態様において、本発明のベクターはA.tumefaciens由来のTi−プラスミドである。
【0084】
本発明の核酸コンストラクトを開発するとき、前記コンストラクトまたはその断片のさまざまな構成要素を便利なクローニングベクター、例えば大腸菌のような細菌宿主内で自己複製能を有するプラスミドに通常の手法で挿入されるのが普通であろう。多数のベクターが文献に記載され、その多くは市販されている。それぞれのクローニングが終了した後、所望のインサートを有するクローニングベクターが単離され、所望の配列の構成要素を仕立てるために、制限消化、新しい断片またはヌクレオチドの挿入、連結、欠失、突然変異、再切断等の他の操作に供される。前記コンストラクトは、完成すると、宿主細胞の形質転換のやり方にしたがってさらなる操作を行うために適当なベクターに移植される場合がある。
【0085】
本発明の組換えDNA分子は、形質転換された細胞を容易に同定し、形質転換されていない細胞から選別できるように、選択可能なマーカーを含むのが典型的である。かかるマーカーの例は、カナマイシン耐性を可能にするネオマイシン・ホスホトランスフェラーゼ(nptII)遺伝子(Potrykusら、(1985年)、Mol.Gen.Genet.、199巻、183−188頁)を含むが、これに限定されない。nptII遺伝子を発現する細胞は、カナマイシンまたはG418のような適当な抗生物質を用いて選択できる。他の慣用される選択可能なマーカーは、ビアラホス耐性を可能にするbar遺伝子と、グリホセート耐性を可能にするEPSPシンターゼ遺伝子突然変異体(Hincheeら、Bio/Technology、6巻、915−922頁(1988年))と、ブロモキシニル耐性を可能にするニトリラーゼ遺伝子(Stalkerら、(1988年)、J.Biol.Chem.、263巻、6310−6314頁)と、イミダゾリノンまたはスルホニル尿素耐性を可能にするアセトラクテートシンターゼ遺伝子(ALS)突然変異体(欧州特許第154,204号、1985年)と、メトトレキセート耐性DHFR遺伝子(Thilletら、(1988年)、J.Biol.Chem.、263巻、12500−12508頁)とを含む。
【0086】
さらにベクターは、特定の宿主細胞の複製開始点(レプリコン)を含む場合がある。さまざまな原核生物のレプリコンが当業者に知られており、原核宿主細胞中における組換え分子の自己複製と維持を指令する役割を果たす。
【0087】
ベクターは、選択可能なマーカーを含むことが好ましい。トランスフェクションされた植物の選択に用いるための多数の選択可能なマーカーはカナマイシン、グリホセート耐性遺伝子を、大腸菌、A.tumefaciensその他の細菌中で培養するための選択可能なマーカーはテトラサイクリンまたはアンピシリン耐性遺伝子をそれぞれ含むが、これらに限定されない。
【0088】
本発明の核酸を微粒子銃法によって単子葉植物に導入するのに適したプラスミドベクターは、選択したプロモーターと、Hsp70イントロン(国際公開第WO93/19189号公報)のような遺伝子発現を促進するスプライシング部位を提供するイントロンと、ノパリンシンターゼ3’配列(NOS3’、Fraleyら(1983年)、Proc.Natl.Acad.Sci.、USA、80巻、4803−4807頁)のような3’ポリアデニル化配列とで構成される。この発現カセットは、DNAの大量生産に適したハイコピーレプリコン上に構築される場合がある。
【0089】
特に有用な双子葉植物の形質転換用アグロバクテリア利用植物形質転換ベクターは、プラスミドベクターpMON530(Rogersら、(1987年)「Improved vectors for plant transformation:expresssion cassette vectors and new selectable markers」、Methods in Enzymology.、R.Wuおよび L.Grossman共編、253−277頁、San Diego、Academic Press)である。プラスミドpMON530は、pMON316(Rogersら、(1987年)「Improved vectors for plant transformation:expression cassette vectors and new selectable markers」、Methods in Enzymology.、R.Wuおよび L.Grossman共編、253−277頁、San Diego、Academic Press)のStuI−HindIII断片(2.3kb)をpMON526に移植して調製されたpMON505の誘導体である。プラスミドpMON526は、前記SmaI部位がXmaI消化、Klenowポリメラーゼ処理および連結によって除去されたpMON505の単純な誘導体である。プラスミドpMON530は、pMON505およびCaMV35S−NOS発現カセットの特性のすべてを保持し、プロモーターとポリアデニル化シグナルとの間に唯一のSmaI切断部位を持つことになる。
【0090】
バイナリーベクターpMON505は、Tiプラスミド相同領域LIHが、ミニRK2プラスミドpTJS75(Schmidhauser and Helinski.、(1985年)、J.Bacteriol.、164−155頁)の3.8kbのHindIII−SmaIセグメントに置換されたpMON200(Rogersら、1987年)の誘導体である。このセグメントは、トリペアレンタル(tri−parental)交配法によるアグロバクテリアへの接合に用いられるRK2複製開始点oriVと転移起点oriTとを含む(HorschおよびKlee、(1986年)、Proc.Natl.Acad.Sci.、USA.、83巻、4428−4432頁)。プラスミドpMON505は、所望のDNA断片を挿入するための合成マルチリンカーと、植物細胞におけるカナマイシン耐性NOS/NPTII’/NOSキメラ遺伝子と、大腸菌およびA.tumefaciensにおける選択用のスペクチノマイシン/ストレプトマイシン耐性決定因子と、形質転換体のスコアリングおよび後代での遺伝子保持の確認を容易化するインタクトなノパリンシンターゼ遺伝子と、大腸菌体内でベクターの大量生産を可能とするpBR322複製開始点とを含む、pMON200の重要な特徴をすべて保持する。プラスミドpMON505は、pTiT37ノパリン型T−DNAの右端に由来する単一のT−DNA境界を含む。サザンブロット分析は、プラスミドpMON505およびこれに含まれるすべてのDNAが植物ゲノムに組み込まれていること、すなわち、プラスミド全体が植物ゲノムに組み込まれたT−DNAであることを示す。組込みDNAの一端は、右境界配列とノパリンシンターゼ遺伝子との間に位置し、他端は前記境界配列とpBR322配列との間に位置する。
【0091】
別の特に有用なTiプラスミドカセットベクターは、pMON17227である。このベクターは、国際公開第WO92/04449号公報に記載されており、ジャガイモ、トマトを含む多くの植物の優れた選択マーカーである、グリホセート耐性を可能にする酵素(CP4と命名)をエンコードする遺伝子を含む。この遺伝子はシロイヌナズナ(Arabidopsis)のEPSPS葉緑体輸送ペプチド(CTP2)と融合し、選択したプロモーターによって発現する。
【0092】
小胞体ルーメン、ペリプラズム間隙または細胞外環境へ翻訳されたタンパクを分泌するために、適当な分泌シグナルが発現したポリペプチドに組み込まれる場合がある。前記シグナルは、前記ポリペプチドに内在するものであっても、あるいは、異種シグナルであってもよい。
【0093】
1つの実施形態では本発明のベクターは、本明細書で説明される核酸が、関心のあるポリペプチドをエンコードするDNAに操作可能に連結した組織特異的プロモーターとなるように設計される。別の実施態様では、関心のあるポリペプチドは植物個体におけるリグニン生合成に関与する酵素である。リグニン生合成に関与する多くの酵素をエンコードするポリヌクレオチドは、シンナミルアルコールデヒドロゲナーゼ(CAD)、シンナメート4−ヒドロキシラーゼ(C4H)、クマレート3−ヒドロキシラーゼ(C3H)、フェノラーゼ(PNL)、O−メチルランスフェラーゼ(OMT)、シンナモイル−CoAレダクターゼ(CCR)、フェニルアラニンアンモニア−リアーゼ(PAL)、4−クマレート:CoAリガーゼ(4CL)およびマツ由来のペルオキシダーゼ(POX)(米国特許第6,204,434号)を含むが、これらに限定されない。他の酵素は、コニフェリンβ−グルコシダーゼ(CBG)およびカフェイン酸3−O−メチルトランスフェラーゼ(COMT)を含む(米国特許第5,451,514号明細書、国際公開第WO94/23044号明細書、およびDharmawardhanaら、Plant Mol.Biol.、40巻、365−72頁(1999年))。
【0094】
別の実施態様では、前記プロモーターに操作可能に連結されたコーディング配列は、リグニン生合成に関与する酵素の発現または活性を阻害する遺伝子産物をコードする場合がある。例えば、リグニン生合成の制御にとって特に関心があるのは、シンナミルアルコールデヒドロゲナーゼ(CAD)、シンナメート4−ヒドロキシラーゼ(C4H)、クマレート3−ヒドロキシラーゼ(C3H)、フェノラーゼ(PNL)、O−メチルトランスフェラーゼ(OMT)、シンナモイル−CoAレダクターゼ(CCR)、フェニルアラニンアンモニアリアーゼ(PAL)、4−クマレート:CoAリガーゼ(4CL)およびマツ由来のペルオキシダーゼ(POX)に対するアンチセンス遺伝子である。
【0095】
さらに別の実施態様では、本発明のベクターは、本発明の核酸が、関心のあるポリペプチドをコードする遺伝子に対応するアンチセンスRNAまたは干渉RNAをエンコードするDNAまたはRNAに操作可能に連結し、これによって標的遺伝子産物の発現を弱めるように設計される。抑制の標的となる遺伝子産物は、上記のとおり、リグニン生合成に関与する酵素であることが好ましい。遺伝子発現のRNAi阻害の利用は、Paddisonら、Genes & Dev.、16巻、948−958頁(2002年)に概説されており、また遺伝子発現を阻害するためのRNAiの利用は、国際公開第WO99/61631号公報に具体的に記載されており、両文献は、引用によりここに取り込まれる。
【0096】
特定の植物遺伝子の発現を低減または阻害するためのアンチセンス技術の利用は、例えば欧州出願公開第271988号公報に記載されている。遺伝子発現の低減は、植物の表現型に変化をもたらし、それは例えば、トマト果実中のリコピン合成欠損のため赤くなるはずの実が黄色くなるといった目に見えるレベルの表現型の差異か、あるいは例えばトマト果実の成熟過程におけるポリガラクトウロナーゼ量の変化およびペクチンの脱重合の低減のような、より微小な生化学的レベルかのいずれかで現れる(Smithら、Nature、334巻、724−726頁(1988年);Smithら、Plant Mol.Biol.、14巻、369−379頁(1990年))。このように、アンチセンスRNAは植物における遺伝子発現を低下させる上で有用であることが実証されている。
【0097】
本発明の植物の作成方法の1つの実施態様では、植物個体内で転写されてアンチセンスRNA転写産物を生成する外来性のDNAは、植物個体内、例えば植物細胞内に導入される。前記外来性DNAは、例えばそのプロモーターに対する遺伝子配列の配向を反転することにより調製することができる。植物細胞内における前記外来性DNAの転写は、その遺伝子に対して「アンチセンス」な細胞内RNA転写産物を生成する。
【0098】
例えばアンチセンス技術は、4CLのようなリグニン生合成に関与する酵素をエンコードする遺伝子の発現を抑制するために用いられる場合がある。4CLのアンチセンスDNA配列を含むコンストラクトは、プロモーターに操作可能に連結された4CLをエンコードする外来性DNAの配向を反転することにより調製することができる。前記外来性の4CLDNAの植物細胞における転写は、アンチセンス配向の4CLRNA転写産物を生成する。
【0099】
また本発明は、本発明のベクターを含む宿主細胞を提供する。本明細書で用いられる宿主細胞とは、コード産物が最終的に発現される細胞をいう。したがって宿主細胞は、個々の細胞、細胞培養、または生物個体の一部である複数の細胞であり得る。また宿主細胞は、胚、胚乳、精子または卵子、または受精卵の場合がある。
【0100】
本発明のポリヌクレオチドを含むポリヌクレオチドまたはベクターは、標的宿主生物への組換えベクターDNAを導入するための当該技術分野で知られた標準的な手順によって宿主細胞に導入される。かかる手順は、トランスフェクション、感染、形質転換、自然取込み、エレクトロポレーション法、微粒子銃法およびアグロバクテリアの使用を含むが、これらに限定されない。生物学的および物理的な植物形質転換プロトコールを含む、外来遺伝子を植物に導入する方法は当該技術分野で知られており、本発明の遺伝子コンストラクトを植物宿主に挿入するために用いられる場合がある。例えば、Mikiら、1993年、「Procedure for Introducing Foreign DNA Into Plants」、Methods in Plant Molecular Biology and Biotechnology、GlickおよびThompson共編、CRC Press Inc.、Boca Raton、67−88頁を参照せよ。選択される方法は宿主植物によって異なり、リン酸カルシウムのような化学的トランスフェクション法、アグロバクテリアのような微生物を介した遺伝子導入(Horschら、Science、227巻、1229−31頁、1985年)、エレクトロポレーション法、顕微注入法、および微粒子銃法を含む。
【0101】
したがって本発明は、本発明のポリヌクレオチドを含むポリヌクレオチドまたはベクターを含む、植物個体または植物細胞も提供する。1つの実施態様では前記植物は、被子植物または裸子植物である。別の実施態様ではこれら植物は、ユーカリ属およびマツ属の種から選択される。特にトランスジェニック植物は、ユーカリ・グランディス(Eucalyptus grandis)およびその交雑種、ラジアータマツ(Pinus radiata)、テーダマツ(Pinus taeda L、loblolly pine)、セイヨウハコヤナギ(Populus nigra)、カロリナポプラ(Populus deltoides)またはモミジバフウ(Liquidamber styraciflua)に属する植物種の場合がある。通常の植物の意味を超え、本明細書では「植物」の語を植物の果実、種子、花、胞子嚢等も指すものとする。本発明の植物は、直接トランスフェクション体、すなわちベクターがアグロバクテリアを介するやり方のように直接的に導入されるか、あるいはトランスフェクションされた植物の子孫植物の場合がある。前記子孫は、トランスフェクションされた植物の無性生殖によって得られる場合がある。第2世代またはその以降の世代の植物は、有性生殖すなわち受精によって作成される場合も、そうでない場合もある。さらに前記植物個体は、配偶体(単相期)または胞子体(複相期)の場合がある。
【0102】
本発明はまた、本発明のベクターを含む植物または種子を栽培することを含む、植物のリグニン含量の制御方法を提供する。本発明の植物または種子の成長または発芽を誘導または維持する適切な栽培法は種特異的であり、本発明の属する技術分野の通常の知識レベルの範囲内である。栽培場所は、植物個体または種子の成長または発芽を促進できるいかなる場所でもよい。さらに、栽培には、葯および/または小胞子内に胚形成を誘導できるストレス処理(例えば、窒素欠乏、熱ショック、ショ糖欠乏)が含まれていてもよいが、ストレス処理はこれらに限定されない。
【0103】
本発明はさらに、転写因子と結合し、組織優先発現的または組織特異的な発現を調節する能力を有する単離調節エレメントも提供する。前記調節エレメントによって実現される調節の程度は、完全、すなわち、前記転写因子無しでは転写が検出できない場合もあるが、部分的、すなわち、前記転写因子の存在下で転写が増強される場合もある。1つの実施態様では、複合プロモーターを用意するために、少なくとも1個の調節エレメントが異種プロモーターに連結される。前記複合プロモーターは、維管束組織内で優先的または特異的に発現させる。本明細書で用いられる異種プロモーターは、前記調節エレメントとの関係で定義される。自然環境で調節エレメントとプロモーターとが互いに協働していない場合は、そのプロモーターはその調節エレメントに対して異種とされる。典型的には、ある調節エレメントのプロモーター領域内での正確な配向はその活性に影響しない。さらに調節エレメントは、異種プロモーター領域に挿入されたときでも正常に機能することができる。したがって、例えば、木部特異的発現または木部優先的発現を実現するために、木部優先発現的調節エレメントが本来のプロモーターから取り出されて異種プロモーター領域に挿入される場合がある。同様に、篩部特異的発現または篩部優先的発現を実現するために、篩部優先発現的調節エレメントがその本来のプロモーターから取り出されて異種プロモーター領域に挿入される場合がある。同様に、形成層特異的発現または形成層優先的発現を実現するために、形成層優先発現的の調節エレメントがその本来のプロモーターから取り出されて異種プロモーター領域に挿入される場合がある。前記異種プロモーターは、例えば最小CaMV35Sプロモーターの場合がある。最小プロモーターに改変するのに適した植物細胞内での発現を指令するプロモーターは、カリフラワーウィルス(CaMV)35Sプロモーター(Jeffersonら、EMBO J.、6巻、3901−07頁(1987年))、コメのアクチンプロモーター(McElroyら、Plant Cell、2巻、163−71頁(1990年))、トウモロコシのユビキチン−1プロモーター(Christensenら、Transgenic Research、5巻、213−18頁(1996)年)、およびノパリンシンターゼプロモーター(Kononowicsら、Plant Cell、4巻、17−27頁(1992))を含む。
【0104】
本発明の核酸を調製するにあたり、さまざまな制限エンドヌクレアーゼを用いてゲノムをばらばらの断片に消化し、ラジアータマツ(Pinus radiata)およびユーカリ・グランディス(Eucalyptus grandis)からゲノムライブラリが作成された。ゲノムライブラリは、組織選択的プロモーターを得るのに適したいかなる植物種からも同様に構築することができる。Clontech社(カリフォルニア州、パロアルト)のGenomeWalkerシステム(登録商標)用に提供される手順にしたがって、アダプターがこれらゲノム配列の各々に連結された。次にプロモーター配列は、アダプター特異的プライマーと「遺伝子特異的プライマー」とを用いてPCR法で増幅された。代替的には、このPCR増幅のステップは、任意的に長鎖でバックグランドが最小の反応生成物を産生する方法であって、引用によりここに取り込まれる米国特許第5,565,340号および第5,759,822号明細書に記載される方法により実行される場合がある。この一般的なPCR増幅法を用いることにより、本発明のプロモーターの同定およびその組織選択的プロモーターとしての同定は、前記「遺伝子特異的プライマー」の選択よって左右される。
【0105】
遺伝子特異的プライマーは、関心のある組織内で高度に発現されるいずれかの転写配列である。本発明では、遺伝子特異的プライマーは、維管束組織中で高度に発現されるmRNAの断片か、あるいは該mRNAに対して相補的である。1つの実施態様では、前記遺伝子特異的プライマーは、特定の維管束組織のタイプにおいて特異的に発現することが知られている遺伝子に対する相同性によって選択される。特に関心のある遺伝子は、ある特定の維管束組織中で高度に発現される遺伝子であり、これは対応するプローモーターの維管束選択的活性の指標となるのが典型的である。
【0106】
発現配列タグ(EST)は遺伝子特異的プライマーの別の供給源である。ESTは、あるライブラリ中に存在する対応mRNAのcDNA断片である。いずれかの植物ESTデータベースが、所望の組織タイプで特異的に発現することが知られている遺伝子のセグメントとの同一性があるESTを捜すために、電子的に検索される場合がある(「インシリコ(in silico)スクリーニング」)。したがってこれらのESTは、あるゲノムライブラリ内の対応する遺伝子のプロモーターの増幅に用いられる遺伝子特異的プライマーを提供する。増幅された遺伝子プロモーターは、ESTデータベースの構築の元となる植物種と同じ植物種に由来するものである必要はない。前記ESTが、関心のある遺伝子プロモーターに対して、前記遺伝子の標的セグメントのPCR増幅用プライマーとして作用するのに十分な配列類似性を備えていることだけが必要である。
【0107】
組織特異的プロモーターを同定する代替的な方法は、ある組織タイプ内では発現するが他の組織内では発現しないmRNAは組織特異的プロモーターから転写されたことを意味するので、かかるmRNAの検出に基づく。mRNAの集団は、例えばサブトラクティブハイブリダイゼーション法により、この原理で区別することができる。かかるサブトラクティブハイブリダイゼーション法の1つの例は、ClontechによるPCR−Select(登録商標)法である。
【0108】
代替的には、組織特異的mRNAの分布は、植物組織の薄切片を放射標識プローブとin situで雑種形成させることにより、決定することができる。特定の組織タイプを放射能で反応するプローブは、以下に説明する方法を用いるゲノムライブラリのサザン分析により、mRNAと関連するプロモーターを検出するのに用いられる。前記の技術は全て関心のある組織、ここでは維管束組織からのmRNAライブラリの作成を要する。例えば、木部組織からのcDNAライブラリの調製は、Dharmawardhanaら、Plant Mol.Biol.、40巻、365−72頁(1999年)およびLoopstraら、Plant Mol.Biol.、27巻、277−91頁(1995年)に記載されている。簡潔には、まず伐採した木から活発に分化している木部を剥ぎ取る。常法にしたがって全RNAが単離され、ポリ(A)RNAが単離され、木部組織cDNAライブラリを構築するために逆転写される。例えばDharmawardhana(1999年)に説明されているように、前記cDNAライブラリは、Strategene cDNA合成およびGigapakII Gold(登録商標)パッケージングキットを用いて、λZAP−XRベクターで構築された。木部特異的プロモーターは、遺伝子特異的プローブと前記プロモーターの5’末端の配列を認識するプライマーとを用いて、PCR法により、かかるcDNAライブラリからを単離することができる。遺伝子特異的プローブは、前記インシリコのアプローチによって、あるいは、もしmRNA配列が既知の場合には、該mRNA配列にもとづく特異的プローブを設計することによって得られる場合がある。さらに、所望の標的遺伝子の5’UTRに相補的なプライマーが合成できる。代替的には、前記プライマーは、エンコードされたタンパクのアミノ酸配列の一部から、いわゆる縮重プライマーとして設計できる(Dharmawardhana(1999年)を参照せよ。)。
【0109】
関心のあるプロモーターを単離した後は、さまざまな手法がその組織特異的発現パターンやプロモーターの強さを調べるために利用できる。慣用される方法の1つは、前記プロモーターをアッセイの容易なレポーター遺伝子と操作可能に連結させることである。例えば、木部優先発現的プロモーターと、グニーユーカリ(Eucalyptus gunnii)シンナモイルCoAレダクターゼ(EgCCR)遺伝子(位置:−1448〜+200、ただし+1が転写開始部位)の5’UTRとが、β−グルクロニダーゼ(GUS)をエンコードする遺伝子と操作可能に連結される(Lacombeら、Plant J.、23巻、663−76頁(2000年))。適切な発現コンストラクトは、周知の方法にしたがって作出することができる。
【0110】
植物の形質転換は、米国特許第6,051,757号明細書に説明されるアグロバクテリアを介した形質転換法を含む多数の適切な方法のいずれかを用いて行うことができる。樹木を形質転換する当該技術分野における他の方法が、加速粒子による裸子植物の体細胞胚の形質転換を開示する米国特許第6,518,485号明細書に代表されるように、本発明の技術分野で知られている。他の形質転換法は、微粒子銃法(Kleinら、Biotechnology、6巻、559−63頁(1988年))、エレクトロポレーション法(Dhalluinら、Plant Cell、4巻、1495−1505頁(1992年))、およびポリエチレングリコール処理法(Golovkinら、Plant Sci.、90巻、41−52頁(1993年))を含む。さらに、米国特許第6,187,994号明細書は、リコンビナーゼに支援される形で発現コストラクトを植物ゲノム中の特異的選択部位に挿入する方法を提供する。上記の特許および文献はすべて引用によりここに含まれる。
【0111】
本発明のDNA分子は、いずれかの適当な方法を用いて植物のゲノムに挿入できる。好適な植物形質転換ベクターは、アグロバクテリウム・ツメファシエンスのTiプラスミドに由来するものと、例えばHerrera−Estrellaら、Nature、303巻、209頁(1983年)と、Bevan、Nucleic Acid Res.、12巻(22号)(1984年)、8711−8721頁と、Kleeら、Bio/Technology、3巻(7号)、637−642頁(1985年)と、欧州特許出願公開第120,516号公報に開示されるものとを含む。アグロバクテリアのTiまたは根誘導(Ri)プラスミドから誘導される植物形質転換ベクターに加えて、代替的な方法が、本発明のDNAコンストラクトを植物細胞に挿入するために利用可能である。かかる方法は、例えば、リポソームの使用と、エレクトロポレーション法と、遊離DNAの取込みを増大させる化学物質と、微粒子銃による遊離DNAの送達と、ウィルスまたは花粉を用いる形質転換とを含む場合がある。DNAは、葉緑体のゲノムに挿入される場合もある(Daniellら(1998年)、Nature Biotechnology、16巻、345−348頁)。
【0112】
関心のある核酸を含む細胞(またはプロトプラスト)が適当な数だけ得られると、該細胞(またはプロトプラスト)は完全な植物個体に再生される。再生ステップのための方法論の選択は重要ではなく、マメ科(アルファルファ、ダイズ、クローバー等)と、セリ科(ニンジン、セロリ、パースニップ)と、アブラナ科(キャベツ、ダイコン、ナタネ等)と、ウリ科(メロンおよびキュウリ)と、イネ科(コムギ、オオムギ、コメ、トウモロコシ等)と、ナス科(ジャガイモ、タバコ、トマト、コショウ)と、ヒマワリのようなさまざまな花卉作物と、アーモンド、カシュー、クルミ、ペカンのようなナッツを産する樹木とに由来する宿主について適切なプロトコルは入手可能である。例えば、Ammiratoら、(1984年)Handbook of Plant Cell Culture−Crop Species、Macmillan Publ.Co.と、Fromm,M.(1990年)UCLA Symposium on Molecular Strategies for Crop Improvement、4月16〜22日、1990年、コロラド州、キーストーンと、Vasilら、(1990年)Bio/Technology、8巻、429−434頁と、Vasilら、(1992年)Bio/Technology、10巻、667−674頁と、Hayashimotoら(1990年)Plant Physiol.、93巻、857−863頁と、Dattaら(1990年)とを参照せよ。
【0113】
プロモーターとレポーター遺伝子とを含むベクターは、該ベクターで形質転換に成功した植物細胞を選択するメカニズムを備え、例えばカナマイシン耐性を有する場合がある。形質転換体中のGUS遺伝子の有無は、GUS特異的PCRプライマー(Clontech、パロアルト)を用いてPCR法で確認される場合がある。形質転換された植物細胞の子孫におけるカナマイシン耐性の分離は、安定に挿入された前記ベクターを宿す遺伝子座の数を決定するためにサザン分析と組み合わせて利用される場合がある。プロモーター発現の時間的および空間的パターンは、Jeffersonら、EMBO J.、6巻、3901−07頁(1987年)の記載にしたがって、レポーター遺伝子発現の定量化から推測される。一般にGUSの発現は、先ず90%アセトンで固定した後、GUSの基質である5−ブロモ−4−クロロ−3−インドイル−β−D−グルクロン酸(X−Gluc)を含む緩衝液中で固定された植物組織の薄切片中で組織化学的に決定される。GUS発現産物の有無は、X−Glucを用いる比色定量反応により示される。
【0114】
例えば木部特異的な発現は、木部組織中で転写因子と特異的に結合する調節エレメントの存在によって実現される場合がある。木部特異的な発現の程度に影響する転写因子の間の相互作用は、調節エレメントの塩基対のサブセットと前記転写因子のアミノ酸残基とのアライメントに依存する。同様に、例えば篩部特異的または形成層特異的な発現は、それぞれ篩部および形成層組織中で転写因子を特異的に結合する調節エレメントの存在によって実現される場合がある。結合した転写因子と相互作用を行わない塩基対は、組織特異的転写因子を特異的に結合する調節エレメントの全体的な能力は維持したまま、他の塩基対に置換される場合がある。
【0115】
いったんプロモータが組織優先発現的または組織特異的と同定された後は、組織優先発現的または組織特異的プロモーターの活性に影響を与える調節エレメントの同定や特徴付けをするためにさまざまな方法論が利用できる。1つの方法では、前記プロモーター領域は5’末端が徐々に短くなるように切断され、切断された一連のプロモーターはそれぞれレポーター遺伝子に操作可能に連結される。調節エレメントが欠失すると、レポーター遺伝子の組織特異的発現が失われることからプロモーター活性への影響が推測される。代替的には、CaMV35S最小プロモーターのような最小プロモーターを含む発現コンストラクトに調節エレメントと推定される配列を挿入し(Kellerら、Plant Mol.Biol.、26巻、747−56頁)、前記調節エレメントと推定される配列が組織特異的発現を与えるか否かを確認することができる。最小プロモーターは、RNAポリメラーゼ結合部位のような、プロモーター活性に不可欠な要素のみを含む。推定的な調節エレメントの解明についての追加例は、L−フェニルアラニンアンモニアリアーゼ(PAL)および4−クマレートCoAリガーゼ(4CL)をエンコードする遺伝子の転写を協調的に調節する組織特異的調節エレメントの研究によりもたらされている(Hattonら、Plant J.、7巻、859−76頁(1995年); Leyvaら、Plant Cell、4巻、263−71頁(1992年); Hauffeら、Plant J.、4巻、235−53頁(1993年); Neustaedterら、Plant J.、18巻、77−88頁(1999年))。これらの文献はすべて、引用により本発明に含まれる。
【0116】
推定的な調節エレメントの位置決定を行う別の方法は、既知の維管束優先発現的または維管束特異的な調節エレメントの間で配列を比較することである。ヌクレオチド比較により、例えば維管束優先発現的調節エレメントのような既知の組織優先発現的調節エレメントと類似する領域が同定される。例えば、シンナミルCoAレダクターゼと、フェニルプロパノイド産生に関与する他のプロモーターとの間でプロモーター配列を比較することにより、配列CCCACCTACCを有する保存されたACリッチな領域の存在が明らかになる。Lacombe(2000年)を参照せよ。フェニルプロパノイド合成に関与するプロモーター中に前記配列が保存されていることは、保存されている前記配列が、複数の植物種でこれまでに同定されている、MYBのような協調的活性化転写因子に対する結合部位であることを意味する。Martin、Trends Genet.、13巻、67−73頁(1997年)を参照せよ。例えば、MYB結合部位は、トウモロコシでは([C/A]TCC[T/A]ACC)、ペチュニアでは(TAAC[C/G]GTTまたはTAACTAAC)と同定されている。同上。
【0117】
本発明の核酸配列にもとづいてこのような配列比較を行うことにより、以下の表1に示される一連のモチーフが明らかになる。表1のモチーフは、組織優先発現的または組織特異的転写因子に結合し、異種最小プロモーターの組織優先発現的または組織特異的発現のモジュレーションに利用できることが期待される。
【0118】
【表1】

【0119】
例えば、bZIPタンパクと維管束特異的または維管束優先発現的なプロモーターとの相互作用は、当業者がどのように調節エレメントの特徴付けを行えるかについて、別の方法を提供する。例えば、グリシンリッチなタンパクであるgrpl.8をエンコードする遺伝子は、GRP1.8−β−グルクロニダーゼ融合タンパクの組織化学的染色から決定されるとおり、木部特異的に発現される。Kellerら、EMBO J.、8巻、1309−14頁(1989年)。grpl.8プロモーターの5’欠損試験は、維管束の特異的発現に必須な20塩基対からなる負の調節エレメントを明らかにした。Kellerら、Plant Cell、3巻、1051−61頁。プロモーターの負の組織特異的調節によって作用する類似の木部特異的調節エレメントが、マメのpal2と、パセリの4CL−1とで同定されている。それぞれ、Leyvaら、Plant Cell、4巻、263−71頁(1992年)およびHauffeら、Plant J.、4巻、235−53頁(1993年)を参照せよ。木部の転写因子VSF−1は、grpl.8遺伝子の20塩基対の前記調節エレメント配列と特異的な相互作用を行う。VSF−1と調節エレメントとの相互作用は、結合特異性がVSF−1結合に関して競合する無標識DNA標的によって示される電気泳動移動度シフトアッセイを用いて、VSF−1と調節エレメントとによる高分子複合体の形成によって実証された。Torres−Schumannら、Plant J.、 9巻、283−96頁(1996年)。さらにVSF−1が相互作用を行う正確な残基は、DNaseIに対する調節エレメントの感受性を測定することによって解明された。簡潔には、VSF−1と調節エレメントとの相互作用は、結合した前記転写因子の立体障害のために、DNaseIがタンパクに接触した一部の残基を分解するのを防止する。典型的には、前記DNAaseI分析は、前記転写因子が結合するときに過敏になる残基も明らかにする。前記推定的な調節エレメントが木部特異的であることを確認するために、前記推定調節領域の単量体または多量体が最小CaMV35Sまたは最小grp1.8プロモーターの上流に融合される。改変プロモーターに操作可能に連結され、タバコに安定的にトランスフェクションされた融合β−グルクロニダーゼ遺伝子を用いた組織化学的分析により、前記推定調節領域はこれらのプロモーターに木部特異的発現を可能にすることがわかった。Torres−Schumann(1996年)を参照せよ。
【0120】
本発明のプロモーターの機能変異体または断片
本発明のプロモーターのさらに別の変異体または断片は、前記配列全体にわたって散在する改変を有するものである。本明細書で用いられる機能変異体または断片とは、基準の核酸に対して少なくとも約70%の同一性を有し、しかもコード産物の組織特異的発現を依然として可能にする核酸をいう。前記機能変異体の組織特異性または優先発現性は、基準の核酸と同じ組織に対するものでなければならない。しかし、前記機能変異体が前記基準の核酸と同様の優先発現性または特異性を有していない場合でも、前記変異体は依然として本明細書で用いられる機能変異体と考えられる。1つの実施態様では、前記機能変異体または断片の配列は、前記基準の核酸に対して少なくとも約75%の同一性を有する。別の実施態様では、前記機能変異体または断片の配列は、基準の核酸に対して少なくとも約80%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%または99%の同一性を有する。
【0121】
機能変異体を作成できる改変は、残基の5’末端からの逐次欠失、または5’UTRの3’末端からの欠失により作出される場合がある。代替的に内部残基が改変される場合もある。前記プロモーター領域の機能に影響を与えない改変は、転写因子の結合に影響を与えない改変である可能性が最も高い。本発明に含まれる改変は、本発明のプロモーターの対立遺伝子の変異体の形で天然に存在する改変を含む。本発明の変異体は、本明細書に説明される方法を用いてその器官または組織特異的活性、あるいは器官または組織優先発現的活性についてテストされる。
【0122】
本発明の核酸の作成方法
本発明の核酸は、周知の合成技術、標準的な組換え方法、精製技術、またはこれらの組合せを用いて得ることができる。例えば本発明の単離ポリヌクレオチドは、固相ホスホアミダイトトリエステル法(Beaucageら、Tetra.Letts.、22巻、1859−1862頁(1981年))、自動化合成装置(Van Devanterら、Nucleic Acids Res.、12巻、6159−6168頁(1984年))、または米国特許第4,458,066号明細書に記載の固体支持法を用いて、直接的な化学合成により調製することができる。化学合成は1本鎖オリゴヌクレオチドを製造するのが一般的で、該1本鎖オリゴヌクレオチドは、相補的な配列との雑種形成か、あるいは、前記1本鎖オリゴヌクレオチドをテンプレートとして用いる重合かによって、2本鎖オリゴヌクレオチドに変換される場合がある。また、もっと長い配列は、短い配列を連結することによって得られる場合もある。
【0123】
代替的には、本発明の核酸は、互いにプライマーとして使用可能なオリゴヌクレオチドを用いる組換え法により得られる場合がある。例えば、Ausubelら編、CURRENT PROTOCOLS IN MOLECULAR BIOLOGY(John Wiley & Sons,Inc.1990)を参照せよ。また、Wosnickら、Gene、60巻、115頁(1987年)およびAusubelら編、SHORT PROTOCOLS IN MOLECULAR BIOLOGY、第3版(John Wiley & Sons,Inc.1995)も参照せよ。ポリメラーゼ連鎖反応を利用する確立された技術は、少なくとも2キロベースの長さを有するポリヌクレオチドを合成する能力を提供する。Adangら、Plant Mol.Biol.、21巻、1131頁(1993年)、Bambotら、PCR Metods and Applications、2巻、266頁(1993年)、Dillonら、「Use of the Polymerase Chain Reaction for the Rapid Construction of Synthetic Genes」、METHODS IN MOLECULAR BIOLOGY、15巻:PCR PROTOCOLS:CURRENT METHODS AND APPLICATIONS、White編、263−268頁(Humana Press,Inc.、1993年)、Holowachukら、PCR Methods Appl.、4巻、299頁(1995年)。
【0124】
本発明の核酸の使用方法
本発明の核酸は、植物の性質を変化させるのに有用である。前記核酸は、維管束組織に見出される分子のレベルを増大させるために、関心のある遺伝子と操作可能に連結される場合がある。代替的には、関心のある遺伝子は、最終生成物、例えばセルロースの形成を阻害する場合があるので、本発明の核酸は、維管束組織中の所定の分子のレベルを減小させるために用いられる場合がある。
【0125】
かかる発現操作の主な標的の1つは、リグニンの生合成経路である。上記の理由のため、植物個体のリグニン含量を正負いずれかに調節することに大きな関心が寄せられており、前記経路に影響を与える遺伝子産物の発現を通じて達成できる。例えば、米国特許第5,451,514号明細書および国際公開第WO94/23044号公報に記載されているように、CADおよびCOMTのコピー数を操作すると、リグニン含量が改変される。さらに、ポプラまたはマツのCADをエンコードする配列のアンチセンス発現は、リグニン組成の改変をもたらす。それぞれ、Grandら、Planta(Berl.)、163巻、232−37頁(1985年);Baucherら、Plant Physiol.、112巻、1479頁(1996年)。
【0126】
変更が重要であるかもしれない他の性質には糖が含まれる。糖産生を担う酵素のレベルを例えば篩部組織で上昇させると、植物個体内の糖含量が増大する可能性がある。糖産生を増大または減少させる酵素は、β−グルコシダーゼ、β−1,3−グルカナーゼ、6−ホスホフルクト−1−キナーゼ、スクロースシンターゼ、UDP−グルコースピロホスホリラーゼ、ヘキソキナーゼ、ホスホグルコムターゼ、スクロース輸送体およびインベルターゼを含むが、これらに限定されない。
【0127】
さらなる性質は、植物の葉に存在する殺昆虫性、抗ウィルス性または抗菌性タンパクの増加を含むが、これらに限定されない。かかるタンパクは、セクロピン、チオニン、ディフェンシン、脂質輸送タンパク、ヘベイン様ペプチドおよびGASAペプチドを含むが、これらに限定されない。さらに他の性質は、干魃耐性、耐寒性、ペルオキシダーゼ活性、耐塩性および窒素輸送を含む。
【発明を実施するための最良の形態】
【0128】
以上概説された本発明は、以下の実施例を参照することにより、一層容易に理解されるであろう。ただし、これらの実施例は例示を目的として提供されるものであって、本発明を限定する意図はない。
【実施例1】
【0129】
維管束特異的プロモーターの単離
ラジアータマツおよびユーカリ・グランディスのcDNAライブラリが以下のとおり構築され、スクリーニングされた。若干改変されたChangら、Plant Molecular Biology Reporter、11巻、113−116頁、(1993年)のプロトコルを用いて、植物組織からmRNAが抽出された。具体的には、試料がCPC−RNAXB(100mM Tris−Cl、pH8.0、25mM EDTA、2.0M NaCl、2%CTAB、2%PVPおよび0.05%スペルミジン*3HCl)に溶解され、クロロホルム:イソアミルアルコール(24:1)で抽出された。
【0130】
RNAはエタノールで沈殿され、MessageMakerキット(Life Techologies社)を用いて精製された。精製されたmRNAから、逆転写酵素(Loopstraら、Plant Mol.Biol.、27巻、277−291頁(1995年))合成の後、得られたcDNAクローンをメーカー指定のプロトコルにしたがってZAP Express cDNA Synthesisキット(Stratagene)を用いてLambda ZAPに挿入して、cDNA発現ライブラリが構築された。得られたcDNAは、5μlのライゲーションミックスからの試料DNAを1μl用いて、Gigapack II Packaging Extra(Stratagene)でパッケージングされた。ライブラリの大量切り出しは、XL1−Blue MRF'細胞およびXLOLR細胞(Stratagene)を、ExAssistヘルパーファージ(Stratagene社)と併用して行われた。切り出されたファージミドは、NZYブロス(Gibco BRL社、メリーランド州、Gaithersburg)で希釈され、5−ブロモ−4−クロロ−3−インドイル−β−ガラクトシド(X−gal)とイソプロピルチオ−β−ガラクトシド(IPTG)とを含むLB−カナマイシン寒天プレートにプレーティングされた。
【0131】
コロニーを採取し、標準的なミニプレップ手法によりDNAを調製した。正しい導入断片を含むコロニーをカナマイシン含有NZYブロス中で培養し、cDNAをアルカリ法とポリエチレングリコール(PEG)沈殿法とにより精製した。1%アガロースゲルを用い、シーケンシングテンプレートへの染色体の混入の有無をスクリーニングした。Big Dye Chemistry(Applied Biosystems社製)を用い、メーカー指定のプロトコルにしたがってダイターミネーター化学反応を準備した。
【0132】
陽性クローンのDNA配列は、3700 Capillary Machine(Applied Biosystems社)またはPrism377シーケンサー(Perkin Elmer/Applied Biosystems部門)を用いて得た。cDNAクローンは、まず5’末端側から、そして場合によっては3'末端側からも配列決定した。一部のクローンについては、サブクローニングされた断片を用いて内部配列を得た。サブクローニングは、制限マッピングおよびpBluescript II SK+ベクターへのサブクローニングという標準的な手順にしたがって行った。
【0133】
選択された遺伝子と相同な植物ESTを、ラジアータマツ(Pinus radiata)またはユーカリ・グランディス(Eucalyptus grandis)のcDNA発現ライブラリのいずれかから同定し、表2に示した。
【0134】
【表2】
【0135】
ポリヌクレオチドまたはポリペプチド配列は、他のポリヌクレオチドおよび/またはポリペプチド配列とアライメントをとることができ、同一の配列を共有する割合は公的に利用可能なコンピューターアルゴリズムを用いて決定することができる。BLASTN、BLASTPおよびBLASTXを含むアルゴリズムのファミリー、BLASTが、Altschulらの文献「Gapped BLAST and PSI−BLAST:a new generation of protein database search programs」、Nucleic Acids Res.、25巻、3389−3402頁、1997年に記載されている。本発明のポリヌクレオチド変異体の決定には、上記アルゴリズムの添付書類に記載されたデフォルトパラメータに設定されているBLASTNアルゴリズムバージョン2.0.11(2000年1月20日)を用いることが好ましい。
【0136】
SWISSPROT−TREMBLE配列(2002年7月9日版)を使用し、2002年11月15日にBLASTXサーチを実施した。 BLASTXを用い、E値とポリペプチド配列の一致度とに寄与するアライメントと同一性とを決定するには、下記のランニングパラメータが好ましい。blastall −p blastx −d swissprotdb −e10 −G0 −E0 −FF −v30 −b 30 −iqueryseq −o results。また、パラメーターは以下のとおりである。−p プログラム名[文字列];−d データベース[文字列];−e 期待値(E)[実数];−G ギャップを空けるためのコスト(ゼロはデフォルト挙動)[整数];−E ギャップを拡張するためのコスト(ゼロはデフォルト挙動)[整数];−F フィルタークェリー配列[文字列];−v 1行説明の数(v)[整数];−b 表示するアライメントの数(b)[整数];−I クェリーファイル[File In];−o BLASTレポートアウトプットファイル[File Out]オプション。
【0137】
プロモーターは、Genome Walkerキット(Clontech社製、Palo Alto、CA)を用いてクローニングした。これはPCR法にもとづく方法であり、4本のプライマーを作成することが必要で、その中の2本は遺伝子特異的でなければならない。この遺伝子特異的プライマーは通常、その遺伝子の5’UTR内に設計される。断片が増幅され、GUSレポーター遺伝子の手前のT終端ベクター内にクローニングされる。
【実施例2】
【0138】
プロモーターの組織特異性の決定方法
実施例1で概説した手順によりプロモーターの同定およびクローニングが終了した後、プロモーターがレポーター遺伝子と操作可能に連結され、プロモーターが活性を示す組織の種類が決定される。このために、エレクトロポレーション法により、本発明のプロモーターを含むコンストラクトがアグロバクテリウム・ツメファシエンスに形質転換される。簡単に述べると、40μlの希釈AgL−1コンピテント細胞を氷上に置き、当該プロモーター配列を含むpART27ベクター約10ngと接触させた。エレクトロポレーション法は、下記のパラメーターの設定下で行った。
抵抗=129Ω
印加電圧=1.44kV
電界強度=14.4kV/cm
パルス持続時間=5.0ms
【0139】
エレクトロポレーション終了後、400μlのYEP液体培地を添加し、細胞を1時間、室温にて回復させる。次に、細胞を6000rpm、3分間遠心し、約50μlのYEPに再懸濁する。細胞試料をYEP Kan50/Rif50プレート上にスプレッドし、パラフィルムで密閉し、29°Cで2日間インキュベーションしコロニーを成長させる。
【0140】
次に、関心のあるコンストラクトを含むアグロバクテリウムを用い、フローラルディップ浸潤法(floral dip infiltration)により野生型シロイヌナズナ「コロンビア−0」株の植物個体を形質転換する。簡潔には、アグロバクテリウム培養を20°Cにて約8600rcfで10分間遠心し、光学濃度が約0.7〜0.8となるように再懸濁する。アグロバクテリウムを含む浸潤液に植物個体を5秒間浸漬する。植物個体から余分な液を水切りし、大気条件で生育光下に置く。24時間後、植物個体を噴霧し、種子を産生できる状態に維持する。T1種子の表面を市販の5%漂白液で滅菌し、カナマイシン(50mg/l)とチメンチン(250mg/l)とを含むMS培地上にスプレッドし、推定される形質転換体を選別する。
【0141】
アグロバクテリウムを介した葉組織形質転換により、タバコ(N.benthamiana)植物個体を、関心のあるコンストラクトを用いて形質転換する(Burowら、Plant Mol.Biol.Rep.、8巻、124−139頁、1990年)。
【0142】
形質転換に成功した植物個体は、操作可能に連結されたレポーター遺伝子が発現しているか否か分析される。葉、茎、根および花の各部分を染色液(50mM NaPO4、pH7.2、0.5% Triton X−100、1mM X−グルクロニド、シクロヘキシミド塩(Ducheffa))に浸漬される。5分間の真空吸引を2回行うことによって組織に染色液を浸透させる。前記組織は37°Cで終夜振盪され、呈色させる。前記組織は、組織の呈色を確認するために途中で3回または4回確認され、試料が早期に呈色する場合には、組織の一片が70%エタノール中で脱染される。次に、光学顕微鏡を用いてGUSの発現について調べるためにこの組織の写真が撮影される。
【実施例3】
【0143】
導管要素(TE)誘導性培地(FKH)および非誘導性培地(FK)中における百日草(Zinnia elegans)葉肉細胞の単離および培養
百日草の実生の初生葉および第二葉のペアが8パネットから採集された。葉は0.175%次亜塩素酸ナトリウム溶液500ml中で10秒間滅菌された。次に、葉は500mlの滅菌水中で2回リンスされた。1回当たり20〜30枚の葉を用いて、乳鉢および乳棒で25〜30mlのFK培地中で粉砕された。細胞が40μmのナイロンメッシュで濾過され、このやり方で合計90mlの葉肉細胞が得られた。前記細胞は20°Cで200×g、2分間遠心され、生成したペレットは、再度等量のFK培地を用いて洗浄された。2回目の洗浄が終了した後、ペレットは2等分され、一方の半分は45mlのFK培地で洗浄され、他方の半分は45mlのFKH培地で洗浄された。各ペレットは60mlのFK培地およびFKH培地にそれぞれ再懸濁された。これらは暗所にて、120rpmに設定されたロータリーシェーカー上の2枚の6ウェルプレートで培養された。
【実施例4】
【0144】
FK培地およびFKH培地中で終夜ないし3日間培養した葉または葉肉細胞からの百日草(Zinnia elegans)プロトプラストの単離
滅菌した百日草の初生葉(6〜8枚)は1mmの細片状に切断され、15mlの細胞壁消化酵素混合液(1%セルラーゼ オノヅカR−10および0.2%ペクトリアーゼ Y23を含むプロトプラスト単離緩衝液)中に添加された。FK培地(40ml)またはFKH培地(40ml)中で培養された葉肉細胞は、20°Cにて200×g、2分間遠心してペレット化された。各ペレットは、200mgのセルラーゼ オノヅカR−10と40mgのペクトリアーゼY23とを含む滅菌プロトプラスト単離緩衝液20ml中に再懸濁された。前記プロトプラストは、前記細胞懸濁液を約70rpm23°Cに設定されたロータリーシェーカー上のCellStar培養プレートで2〜4時間インキュベーションすることによって、単離された。前記プロトプラストは、各内容物を200×g、2分間遠心してペレット化された。各ぺレットは20mlの24%ショ糖溶液中に再懸濁された。
【0145】
Fukuda−Komamine培地保存液
【0146】
【表3】
【0147】
【表4】
【実施例5】
【0148】
百日草(Zinnia elegans)プロトプラストのトランスフェクション
24%ショ糖溶液中に懸濁させた百日草プロトプラスト上に1mlのW5溶液(150mM NaCl、125mM CaCl2・2H2O、5mM KC1、5mM ショ糖、pH5.6−6)を重層し、20°Cにて70×g、10分間の遠心を行い、ブレーキをかけずに停止させた。浮遊するプロトプラストを回収し、W5溶液10ml中に再懸濁させ、20°Cにて70×g、10分間の遠心を行い、ペレット化した。プロトプラストのペレットはプロトプラスト濃度が約5×106個/mlとなるようにMaMg培地(450mMマンニトール、15mM MgCl2、0.1% MES、pH5.6)中に再懸濁され、一定量ずつ別々の15ml試験管に分注された(300μl:プロトプラスト1.5×106個)。各コンストラクトにつき、上記プロトプラスト懸濁液に5μgのDNAおよび50μgのサケ精巣DNAが添加、混合され、20°Cで5分間インキューベートされた。300μlの40%PEG溶液(40%PEG3340、100mM Ca(NO3)2・4H2O、0.45Mマンニトール、pH9.0)が各分注プロトプラストに添加され、20°Cで20分間インキュベーションされた。5mlのK3/0.4Mショ糖(溶液1L当たり:4.3gムラシゲ・スクーグ植物培地用混合塩、100mgミオイノシトール、250mgキシロース、10mgチアミン塩酸塩、1mgニコチン酸、1mgピリドキシン塩酸塩、1mg NAA、0.2mgカイネチン、137gショ糖、pH5.6(KOHで調整))が、葉由来のトランスフェクションされたプロトプラスト、またはFK培地中で培養した葉肉細胞由来のトランスフェクションされたプロトプラストの各分注に添加、混合された。同様に、5mlのK3/0.4Mショ糖+0.lppm NAA+0.2ppm BAが、FKH培地で培養された葉肉細胞由来のトランスフェクションされた細胞の各分注に添加、混合された。これらトランスフェクションされたプロトプラスト懸濁液は、暗所で23°Cで終夜インキュベーションされた。
【実施例6】
【0149】
トランスフェクション百日草(Zinnia elegans)プロトプラストの回収およびレポーター遺伝子解析
トランスフェクションされた百日草プロトプラストを個々に回収するため、実施例5で説明したとおり調製された懸濁液にそれぞれ9.5mlのW5溶液が添加され、各試験管の内容物が混合され、20°Cにて70×g、10分間遠心された。上清の大部分がデカンテーションにより捨てられ、プロトプラストの体積が900μlとされた。900μlの各試料のうち、300μlのプロトプラストが5mlポリスチレン丸底試験管に分注され、500μlのW5培地中に再懸濁され、蛍光レポータ遺伝子発現および細胞生存度の分析用に取り置かれた。残り600μlのプロトプラストおよび溶液は個々のマイクロチューブに移されて、20°Cにて420×g、2分間の遠心によってペレット化された。このプロトプラストペレットは、Jefferson,R.A.、1987年、Plant Mol.Biol.Rep.5,387頁に記載される方法にしたがって、GUSレポーター遺伝子発現について検定された。GUS(MUG)の検定は、Wallac社(テュルク、フィンランド)のVictor21420マルチラベルカウンターを用いて行われた。ウンベリフェロンは355nm励起光フィルターおよび460nm発光フィルターを用いて1秒間検出された。
【実施例7】
【0150】
植物個体内における発現データ
土壌移植3週間後に、形質転換シロイヌナズナおよびタバコ組織はGUSレポーター遺伝子発現について解析される。GUS発現を検定するために、実施例2で説明したとおり、葉、根および花の材料がGUS溶液に浸漬される。組織に染色液を浸透させるために5分間の真空吸引を2回行い、前記組織は発色のため37°Cで終夜インキュベーションされる。終夜のインキュベーションの後、前記組織試料は70%エタノール中で脱染され、光学顕微鏡下でGUS発現について検査される。表5は、形質転換されたシロイヌナズナおよびタバコのGUS発現率(%)を含む。
【0151】
土壌移植3ヶ月後のタバコのT1植物に由来する組織がパラプラストに包埋され、ミクロトームで切片が作成され、光学顕微鏡下でGUS発現が検査される。表5に示したGUSの局在と組織鏡検結果(microtome results)は、本願明細書に開示された単離ヌクレオチド配列が、維管束の形成層、木部および/または篩部組織に優先的なレポーター遺伝子発現を可能にすることを証明する。
【0152】
【表5】
【0153】
タバコ組織の試料は、固定液(100%エタノール、氷酢酸、37%ホルムアルデヒド、ミリQ水)を含むバイアルに入れられ、15分間の真空吸引が2回行われる。前記試料は、確実に組織に浸透させるために固定液中で2時間インキュベーションされる。組織浸透の後、再び15分間の真空吸引を行い、前記組織は4°Cで終夜放置した。
【0154】
組織試料は、終夜のインキュベーションの後、エタノール系列のインキュベーションにより脱水されるが、ここでのすべてのインキュベーションは室温で行われる。前記終夜固定された固定液は組織試料を入れたバイアルから除去され、50%エタノールに置換される。30分後、前記50%エタノールはデカンテーションにより除去され、前記組織は50%エタノールの新しい分注液でインキュベーションされる。50%エタノールで30分間経過後、前記溶液は除去されて60%エタノールに置換される。30分間のインキュベーション後、60%エタノールは70%エタノールに置換される。30分後にこの70%エタノールはデカンテーションにより除去され、85%エタノールで置換される。30分後、前記85%エタノールは除去され、前記試料は95%エタノール中で終夜インキュベーションされる。
【0155】
組織の脱水の後、該組織は室温でキシレン溶液系列中でインキュベーションされる。前記終夜処理された95%エタノールが除去され、上記組織は100%エタノール中で30分間インキュベーションされる。30分後、100%エタノールが除去され、前記組織は25%キシレン:75%エタノール中に懸濁される。30分間のインキュベーション後、前記溶液は50%キシレン:50%エタノールに置換される。次に、前記溶液はデカンテーションにより除去され、75%キシレン:25%エタノールに置換される。30分後、前記組織は100%キシレンで60分のインキュベーションを3回される。次に前記組織は、キシレンと20個のパラプラストチップとを入れたバイアル中で、終夜インキュベーションされる。
【0156】
パラフィンを浸潤させるため、前記バイアルは、前記パラプラストチップが溶解するまで42°Cのハイブリダイゼーションオーブンで保温される。8時間の間に、合計60〜80個のパラフィンチップがバイアルに添加され、溶解される。前記試料は62°Cのハイブリダイゼーションオーブンで終夜保温される。次の2日間にわたり、パラプラストは12時間間隔で4回交換される。前記組織を包埋するために、流動パラフィンがカセット内に注入され、組織を適当な向きに配置される。前記カセットは4°Cで終夜静置され、パラフィンが硬化される。
【0157】
表5に示すとおり、本願明細書に開示される単離ヌクレオチド配列は、維管束の形成層、木部、および/または篩部組織で優先的なGUSレポーター遺伝子発現を可能にする。特に注目すべきは、配列番号21と配列番号13とが、配列番号13の5’末端に40bpの断片(AAATATAACATAATCTAACTATTGATGTACATTATTCGCC)の挿入を含むことを除いて、実質的な配列相同性を有する点である。この40bp断片(配列番号85)は、配列番号13では不完全な反復配列として存在するが、配列番号21には存在しない。興味深いことに、前記40bp断片が存在すると(例えば、配列番号13)GUS発現が可能になり、前記断片が存在しないと(例えば、配列番号21)GUS発現は起こらない。このデータは、本願明細書に開示される40bp断片(配列番号85)が維管束優先発現的な遺伝子発現に必要十分な要素を構成する可能性があることを示唆するため、意義深い。
【実施例8】
【0158】
維管束特異的プロモーターの利用方法
適当な組織特異的かつ発生段階特異的な発現パターンを有するプロモーターがいったん見つかると、このプロモーターは、トランスジェニック植物で所望の形質を調節するために利用可能である。1つの実施態様では、木部特異的プロモーターが植物のリグニン組成と含量とを調節するために利用される。本実施例では、本発明の木部特異的プロモーターがシンナミルアルコールデヒドロゲナーゼ(CAD)のコーディング領域の一部に対応するRNA干渉(RNAi)分子をエンコードする遺伝子と操作可能に連結される。CADは、リグニンモノマー合成の最終段階を触媒し、他のプロモーターを用いてアンチセンス配列を介した植物個体内のリグニンの下向き調節に成功した標的である。Yahiaouiら、Phytochemistry、49巻、295−306頁(1998年)と、これに引用されている各文献を参照せよ。CADの一部に対応するRNAi分子が発現すると、酵素活性が低下し、これに対応してトランスジェニック植物個体内のリグニンのシンナミルアルデヒドの割合が増大する。トランスジェニック植物個体におけるCAD活性は、Baucherら(Plant Physiol.、112巻、1479−90頁(1996年))によって改良された、Wyrambikら(Eur.J.Biochem.、59巻、9−15頁(1975年))の方法によって分析された。リグニン含量および組成は、Baucher(1996年)の方法によって測定された。CADの下向き調節は、木材の色の変化と、抽出がより容易なシンナミルアルデヒドの割合の増大につながり、このことはパルプ産業用の植物にとって商業上重要である。例えば、CAD活性を低減させたポプラは赤みを帯び、リグニンがより抽出しやすく、製紙産業の用途でのポプラ材の特性が改善される(Baucherら(1996年))を参照せよ。
【0159】
別の実施例では、ユーカリおよびマツが形質転換される。ユーカリおよびマツは、リグニン含量が多いため、リグニン含量および組成に対する遺伝子調節の効果を研究するための有用なモデル系を提供する。さらに、ユーカリおよびマツは、リグニン含量または組成のモジュレーションが製紙または木材産業にとって有用な特性を与えるような植物の代表例である。
【0160】
これらのトランスジェニック植物を作出するために、CADのcDNAの断片が、本発明の木部特異的プロモーターと、ノパリンシンターゼ3’ターミネーターとに適当な配向で操作可能に連結される。前記CADのcDNAは、例えば、遺伝子配列をそのプロモーターに対して反転させることにより調製できる。植物細胞内における前記CADのcDNAの転写は、CAD遺伝子に対して「アンチセンス」な細胞内RNA転写産物を生成する。コンストラクト全体が、制限断片としてバイナリーベクターpBI101.1(Clontech、カリフォルニア州、パロアルト)に挿入される。ベクターは、ユーカリまたはマツの形質転換を行うために、それぞれアグロバクテリウム・ツメファシエンス(A.tumefaciens)の菌株LBA4404またはC58pMP90にエレクトロポレーション法により導入される。一般的にはNoら、Plant Science、 160巻、77−86頁(2000年)を参照せよ。カナマイシン耐性形質転換体は、CAD活性について測定され、導入遺伝子のコピー数がサザン解析により決定され、CADのmRNA発現がYahiaoui(1998年)に説明されるとおりに決定される。好適な形質転換体は発根させて、温室に移植される。
【実施例9】
【0161】
インシリコ発現
コンピュータによる遺伝子発現は、コンセンシ(consensi)ESTライブラリのメンバー資格を決定するために利用できる。各ライブラリについて、いずれかの組織クラスのESTの数をそのクラスのESTの総数で割った商に1000を乗じ、コンセンサスが決定される。これらの値は、あるライブラリからの配列決定の程度に影響されない規格化値である。形成層、木部、篩部、栄養芽、根、生殖組織、葉、茎および果実のライブラリを含む複数のライブラリが、コンセンサス値についてサンプル調査された。下記の表6および表7に示すとおり、多数の本発明のプロモーター配列が維管束優先的発現を示し、樹木関連の発生過程に関与する可能性がある。
【0162】
【表6】
【0163】
【表7】
【0164】
【表8】
【図面の簡単な説明】
【0165】
【図1】3種類のプロモーターが培養3日目に同様の発現プロファイルを示すことを証明する百日草TE検定結果を示すグラフ。
【図2A】2種類のプロモーターが培養3日目に同様の発現プロファイルを示すことを証明する百日草TE検定結果を示すグラフ。
【図2B】2種類のプロモーターが培養3日目に同様の発現プロファイルを示すことを証明する百日草TE検定結果を示すグラフ。
【図3】2種類のプロモーターが培養3日目に同様の発現プロファイルを示すことを証明する百日草TE検定結果を示すグラフ。
【技術分野】
【0001】
本発明はポリヌクレオチド転写および/または発現の調節に関する。特に本発明は、ユーカリ・グランディスまたはユーカリ(Eucalyptus grandis)およびラジアータマツまたはマツ(Pinus radiata)から単離され、植物細胞内におけるポリヌクレオチドの維管束優先発現的転写を行うポリヌクレオチド調節配列に関する。内在性および/または異種由来のポリヌクレオチドの転写を改変するために本発明の調節配列を用いるコンストラクトおよび方法も、本発明に含まれる。
【背景技術】
【0002】
本出願は、米国仮出願第60/428,287号(2002年9月22日出願)に基づく優先権主張を伴う。
【0003】
林業および製紙産業の大きな目標は、植物のリグニン含量の制御である。リグニンは、木材の機械強度、着色、防腐性に関与するシンナミルアルコールの複合ポリマーである。木本種は、木材の乾燥重量の20%ないし30%を占める大量のリグニンを合成する。剛性を与えることに加えて、リグニンは、細胞壁に疎水性および非透水性を与えて、水の輸送を助ける。したがって、樹木のリグニン濃度を増大させることは、耐病性および建築用の強度の改善を木に与えることのような、一部の用途で有利になることがわかることになる。また、リグニンは燃料としても有用であり、木炭製造用材と、トウモロコシ稈と、バイオ燃料として用いられる柳および成長の早いポプラ雑種とのような燃料用の木材その他のバイオマスにおけるセルロース含量の増大したリグニンは好ましい。
【0004】
逆に樹木におけるリグニン含量の増大は、製紙産業にとって重大な問題をもたらし、製紙産業はセルロース繊維からリグニンを除去するために多大な資源を費やさなければならない。米国内だけでも、年間約2千万トンのリグニンが除去される。さらにリグニン含量は、飼草作物の消化性の良否の大きな原因であり、植物のリグニン含量のわずかな上昇が消化性の比較的大きな低下につながる。例えば、リグニン含量が低い作物は家畜にとってより効率的な飼草を提供し、飼草作物の消費量に対する乳および肉の生産収量が向上する。リグニン含量は植物の成育中に増大するので、農家は早めの収穫を行って少収量で消化の良い作物を得るか、あるいは遅めに収穫を行って多収量で消化の悪い作物を得るか、のいずれかを選択しなければならない。
【0005】
かかる理由から、植物の遺伝子改変によるリグニン含量または組成の制御は望ましい。この目標に向けて、リグニン生合成を担う遺伝子の同定および特徴付けと、その発現を調節する配列の決定とに多大な努力が注がれてきた。マツ由来のシンナミルアルコールデヒドロゲナーゼ(CAD)、シンナメート4−ヒドロキシラーゼ(C4H)、クマレート3−ヒドロキシラーゼ(C3H)、フェノラーゼ(PNL)、O−メチルトランスフェラーゼ(OMT)、シンナモイル−CoAレダクターゼ(CCR)、フェニルアラニンアンモニアリアーゼ(PAL)、4−クマレート:CoAリガーゼ(4CL)およびペルオキシダーゼ(POX)を含む、リグニン生合成に関与する酵素の多くをエンコードするポリヌクレオチドがクローニングされてきた(特許文献1)。
【特許文献1】米国特許第6,204,434号明細書
【0006】
これらの遺伝子の発現操作がリグニン含量の改変に利用されてきた。かかる実験は、CAD、コニフェリンβ−グルコシダーゼ(CBG)、およびカフェイン酸3−O−メチルトランスフェラーゼ(COMT)をエンコードする遺伝子のコピー数の変更を含む(特許文献2、特許文献3および非特許文献1)。さらに、ポプラ、タバコ(N. tabacum)およびマツにおいてCADをエンコードする配列のアンチセンス発現は、組成の異なるリグニン産生を起こす(それぞれ、非特許文献2、3および4を参照)。
【特許文献2】米国特許第5,451,514号明細書
【特許文献3】国際公開第WO94/23044号公報
【非特許文献1】Dharmawardhanaら、Plant Mol.Biol.、40巻、365−72頁(1999年)
【非特許文献2】Grandら、Planta(Berl.)、163巻、232−37頁(1985年)
【非特許文献3】Yahiaouiら、Phytochemistry、49巻、295−306頁(1998年)
【非特許文献4】Baucherら、Plant Physiol.、112巻、1479頁(1996年)
【0007】
林産品、紙、植物バイオマスおよび飼草産業のもう1つの大目標は、茎からのセルロース回収を容易化するために、茎のサイズを増大させ、茎のセルロース含量を操作し、あるいは細胞壁の特性を操作することである。例えばセルロースは、パルプ生産では木部繊維から回収され、木部繊維数および管状要素数、細胞壁の厚さ、管腔径、繊維長、セルロース微繊維の角度その他のこれら木部の特性が、回収されるセルロースの質および量を決定する。セルロース生合成に関与する遺伝子の操作は、植物のバイオマス総量および植物からのセルロース収率の増大に有用であり、かかる遺伝子のアンチセンス発現は細胞壁の発達に影響を及ぼすことが示されている(非特許文献5)。また、エンド−1,4−β−グルカナーゼとセルロース結合ドメインとによる多糖類と植物細胞壁の改変が報告されている(非特許文献6)。したがって本発明のポリヌクレオチドは、セルロース生合成を改変し、これによって植物の生育とバイオマスとに影響を及ぼすために、維管束組織内でヌクレオチド配列を発現するのに用いられる場合がある。
【非特許文献5】Shani Z.、Shpigel,E.、Roiz L.、Goren R.、Vinocur B.、Tzfira T.、 Altman A.およびShoseyov O.、Cellulose−binding domain increases cellulose synthase activity in Acetobacter xylinum,and biomass of transgenic plants、A.Altman、M.Ziv、S.Izhar共編、Plant Biotechnology and In Vitro Biology in the 21st Century、213−218頁、Kluwer Academic Publishers(1999年)
【非特許文献6】Levy,I.、Shani,Z.およびShoseyov O.、Biomol.Eng.、19巻、17−30頁(2002年)
【発明の開示】
【発明が解決しようとする課題】
【0008】
前記改変植物への全身的な有害な影響を回避するために、生合成経路の遺伝子的調節は狭く限定された組織タイプにおいて実行することが好ましい。例えば、特定の遺伝子産物の発現によりリグニンの含量または組成が特定の遺伝子産物の発現によって影響を受ける場合、前記植物の病気に対する耐性の低下を避けるために、前記遺伝子産物の発現を前記植物の一部のセグメントまたは一部の生育段階に限定することが好ましい場合がある。異種遺伝子は、組織優先発現的プロモーターと操作可能に連結することにより、選択された組織において発現される場合がある。好適な組織優先発現的プロモーターは、維管束組織の原生木部管状要素内で特異的な活性があるマメgrb1.8プロモーターがある(非特許文献7)。これらのプロモーターは木質化部にて優先的に発現するユーカリCADプロモーターも含む(非特許文献8)。かかる組織優先発現的プロモーターは、特異的な組織におけるアンチセンス分子の遺伝子発現の調節に用いられてきた(非特許文献9−11)。
【非特許文献7】Kellerら、EMBO J.、8巻、1309頁(1989年)
【非特許文献8】Feuilletら、Plant Mol.Biol.、27巻、651頁(1995年)
【非特許文献9】Van der Meerら、Plant Cell、4巻、253頁(1992年)
【非特許文献10】Salehuzzamanら、Plant Mol.Biol.、 23巻、947頁(1993年)
【非特許文献11】Matsudaら、Plant Cell Physiol.、37巻、215頁(1996年)
【0009】
組織優先発現的プロモーターは、異なる環境では活性が弱い場合があり、構成的プロモーターで達成されるレベルと常に同じレベルで遺伝子を発現させるとは限らない(非特許文献12)。さらに、これらのプロモーターが活性を示す発生学的なウィンドウ、またはそれらの活性の空間分布が、それらの有用性を制限する場合がある。したがって、本技術分野では、所望の空間的・時間的な発現パターンを有するさらなる組織優先発現的プロモーター、特に、維管束優先発現的プロモーターの開発する継続的な必要性が存在する。非特許文献13に総説される。
【非特許文献12】Yahiaouiら、Phytochemistry、49巻、295−306頁(1998年)
【非特許文献13】Grima−Pettenatiら、Plant Science、145巻、51−65頁(1999年)
【課題を解決するための手段】
【0010】
発明の概要
本発明は、維管束優先発現的遺伝子発現に関与する単離ポリヌクレオチド調節配列に関する。本発明のポリヌクレオチドは、植物のリグニン含量、セルロース含量、個体のサイズまたは細胞壁の成長の調節に用いることができる。本発明のポリヌクレオチドはまた、リグニン、セルロース、および植物細胞壁の生合成にも用いられる場合がある。
【0011】
1つの局面において、本発明は、維管束優先発現的ポリヌクレオチド転写をさせることが可能なポリヌクレオチドを含む単離核酸分子を提供する。1つの実施態様において、前記単離核酸分子は配列番号1−85のいずれか1つとその機能変異体とから選択されるポリヌクレオチドを含む。別の実施態様において前記機能変異体は、配列番号1−85のいずれかの配列との同一性が99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、89%、88%、87%、86%、85%、84%、83%、82%、81%、80%、79%、78%、77%、76%、75%、74%、73%、72%、71%、70%、69%、68%、67%、66%、65%、64%、63%、62%、61%または60%以上である。
【0012】
他の局面において、本発明は、(a)配列番号1−85およびその機能変異体のいずれか1つに対する相補配列と、(b)配列番号1−85の配列のいずれかに対する逆相補体の配列と、(c)前記(a)または(b)のポリヌクレオチドのいずれかと雑種形成する、少なくとも20個の連続する塩基を含む配列とから選択される配列を有する単離ポリヌクレオチドを提供する。1つの実施態様において、本発明は、前記ポリヌクレオチドが植物細胞における木部優先発現的の遺伝子発現を実現できるようにすることを意図する。別の実施態様において、本発明は、植物細胞における操作可能に連結した遺伝子の発現を上向き調節または下向き調節することが可能なポリヌクレオチドを提供する。
【0013】
別の局面において、本発明は、(a)配列番号1−85のいずれかの配列を有する少なくとも1個のポリヌクレオチド配列と、(b)所望の遺伝子とを含み、前記ポリヌクレオチドと前記所望の遺伝子とが操作可能に連結している植物細胞を提供する。1つの実施態様において、前記所望の遺伝子はポリペプチドまたはタンパクをエンコードする。別の実施態様において、前記タンパクは細胞壁の生合成に関与する酵素である。さらなる実施態様において、前記タンパクはリグニンの生合成に関与する酵素である。さらに別の実施態様において、前記所望の遺伝子はRNA転写産物を生成する。さらに別の実施態様において、前記RNA転写産物は植物細胞に内在する遺伝子のアンチセンス配列を有する。さらに別の実施態様において、前記RNA転写産物は植物細胞中で正常に発現される遺伝子のRNA干渉を誘導する。
【0014】
別の局面において、本発明は、(a)配列番号1−85のいずれかの配列を有する少なくとも1個のポリヌクレオチド配列と、(b)所望の遺伝子とを含み、前記ポリヌクレオチドと前記所望の遺伝子とが操作可能に連結している、植物細胞を含む植物を提供する。1つの実施態様において、前記植物は被子植物および裸子植物から選択される。
【0015】
別の実施態様において、本発明は、(a)配列番号1−85のいずれかの配列を有する少なくとも1個のポリヌクレオチド配列と、(b)所望の遺伝子とを含み、前記ポリヌクレオチドと前記所望の遺伝子とが操作可能に連結しているDNAコンストラクトを含む、植物細胞を含む植物を栽培することを含む、植物のリグニン含量の調節方法を意図する。
【0016】
別の局面において、本発明は、(a)配列番号1−85のいずれかの配列を有する少なくとも1個のポリヌクレオチド配列と、(b)所望の遺伝子とを含み、前記ポリヌクレオチドと前記所望の遺伝子とが操作可能に連結しているDNAコンストラクトを含む、植物細胞を含む植物を栽培することを含む、植物の細胞壁の発生の調節方法を意図する。
【0017】
他の局面において、本発明は、配列番号1−85およびその機能変異体のいずれかから選択されるポリヌクレオチド配列を含む、トランスジェニック植物を提供する。
【0018】
1つの局面において、本発明は、(a)(i)配列番号1ないし85およびその機能変異体のいずれかの配列を有するプロモーターと、(ii)所望の核酸とを含み、前記プロモーターは前記所望の核酸の発現を調節するDNAコンストラクトを木本植物の植物細胞に導入すること、(b)前記形質転換された植物細胞を植物の生育を促進する条件下で培養すること、および、(c)前記植物から木材を得ることとを含む、木材を得る方法を提供する。1つの実施態様において、前記木本植物は、ユーカリ属またはマツ属の植物種から選択される。
【0019】
本発明はまた、配列番号1−85からなる群から選択される単離ポリヌクレオチド配列と、これらの配列のいずれかに対して少なくとも60%の同一性を有するポリヌクレオチドとを意図する。本発明はまた、配列番号1−85のいずれかに対して少なくとも65%の配列同一性を有するポリヌクレオチドを意図する。本発明はまた、配列番号1−85のいずれかに対して少なくとも70%の配列同一性を有するポリヌクレオチドを意図する。本発明はまた、配列番号1−85のいずれかに対して少なくとも75%の配列同一性を有するポリヌクレオチドを提供する。本発明はまた、配列番号1−85のいずれかに対して少なくとも80%の配列同一性を有するポリヌクレオチドを意図する。また、配列番号1−85のいずれかに対して少なくとも85%の配列同一性を有するポリヌクレオチドが提供される。本発明はまた、配列番号1−85のいずれかに対して少なくとも90%の配列同一性を有するポリヌクレオチドを意図する。配列番号1−85のいずれかに対して少なくとも95%の配列同一性を有するポリヌクレオチドが提供される。
【0020】
本発明は、表8に挙げたポリヌクレオチド配列のいずれかからなる群から選択される配列に対して少なくとも95%の同一性がある配列を有するポリヌクレオチドを含む単離核酸分子に関する。本発明はまた、表8に開示されるポリヌクレオチド配列の機能断片も提供する。本発明はさらに、表8に列挙されるポリヌクレオチド配列のいずれかに対する相補的な核酸またはその断片と、表8に開示されるポリヌクレオチド配列のいずれかと雑種形成し、少なくとも15個の連続する塩基を含む核酸とを提供する。
【0021】
本発明の核酸のヌクレオチド配列は配列表に示される。ここで、「配列番号(2N+1)」とは、例えば1、3、5、7…等の、すべての奇数番号の配列をいう。「配列番号2N」とは、すべての偶数番号の配列をいう。ある奇数番号の配列とすぐ次の偶数番号の配列(例えば、配列番号1と配列番号2)との間の違いは、前記偶数番号の配列は追加の塩基を有する点である。偶数番号の配列の末端の3塩基は通常ATGで、(通常、メチオニンをコードする)翻訳の開始コドンを表す。
【0022】
本発明は当業者に周知の用語および表現を用いる。別の定義が与えられないかぎり、本明細書中で用いられるすべての科学技術用語は、本発明が属する技術分野において通常の知識を有する者によって普通に理解されるのと同じ意味を有する。一般に、ここで用いる用語法と、ここで説明される細胞培養、分子遺伝学、核酸化学および雑種形成法における各実験手順とは、当該技術分野において周知慣用のものである。核酸組換法、ポリヌクレオチド合成、微生物培養、細胞培養、組織培養、形質転換、トランスフェクション、トランスダクション、分析化学、有機合成化学、化学合成、化学分析および医薬品処方および薬物送達には、標準的な手法が用いられる。一般に、酵素反応、精製および/または単離のステップは製造者の仕様にしたがって行われる。手法および手順は従来の方法論にしたがって実行されるのが一般的である。例えば、Sambrook & Russel、MOLECULAR CLONING:A LABORATORY MANUAL第3版、Cold Spring Harbor Laboratory Press刊、Cold Spring Harbor、NY、2001年を参照せよ。
【0023】
アグロバクテリア:当技術分野で周知のとおり、植物細胞の形質転換に用いられるアグロ細菌は、ベクターを含む、アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)またはアグロバクテリウム・リゾゲネス(Agrobacterium rhizogenes)の無毒化された(disarmed)ビルレントな誘導体であることが一般である。前記ベクターは、T−DNAの境界の間に配置された所望のポリヌクレオチドを含むことが典型的である。
【0024】
被子植物:種子が子房に被覆されている維管束植物。被子植物は、花を付け、実を結ぶ種子植物である。被子植物は、双子葉植物と単子葉植物とに分類される。
【0025】
所望のポリヌクレオチド:本発明の所望のポリヌクレオチドとは、プロモーター、エンハンサーまたはターミネーターのような遺伝子エレメントか、ゲノム中に前記所望のポリヌクレオチドを含む形質転換細胞で転写および/または翻訳されるべき遺伝子またはポリヌクレオチドかをいう。前記所望のポリヌクレオチドがタンパク産物をエンコードする配列を含む場合、そのコーディング領域は、当該所望のポリヌクレオチドによってエンコードされるメッセンジャーRNA転写産物および/またはタンパク産物の発現をもたらす、プロモーターおよびターミネーターのような調節エレメントと操作可能に連結している場合がある。したがって、「所望のポリヌクレオチド」は、5’から3’への配向で、プロモーターと、タンパクをエンコードする遺伝子と、ターミネーターとに操作可能に連結する遺伝子を含む場合がある。代替的には、前記所望のポリヌクレオチドは、翻訳されると植物細胞内での内在性遺伝子の発現に影響を与える2次構造を形成するかもしれない核酸を産生するような、「アンチセンス」配向の遺伝子またはその断片を含む場合がある。所望のポリヌクレオチドはまた、転写されると前記所望のポリヌクレオチドが会合する遺伝子のRNA干渉を引き起こすような、2本鎖RNAを産生する場合がある。本発明の所望のポリヌクレオチドは、T−DNAの左右の境界配列が前記所望のポリヌクレオチドを挟むか、あるいは所望のポリヌクレオチドのいずれか片側に配置されるように、T−DNA内に配置される場合がある。本発明は、少なくとも1個の植物細胞のゲノムに1個または2個以上の所望のポリヌクレオチドを安定的に組み込むことを可能にする。所望のポリヌクレオチドは、突然変異処理されていても、その野生型配列の変異体であってもよい。前記所望のポリヌクレオチドの全部または一部が植物ゲノムに組み込まれ場合があるものと理解されている。また、「所望のポリヌクレオチド」という用語は、かかるポリヌクレオチドの1個または2個以上を包含するものと理解されている。したがって、本発明のT−DNAは、1個、2個、3個、4個、5個、6個、7個、8個、9個、10個または11個以上の所望のポリヌクレオチドを含む場合がある。
【0026】
双子葉植物:胚が2分割状の種子、すなわち子葉と、分岐した葉脈と、4または5の倍数に分割される花とを持つ顕花植物。双子葉植物の例は、ユーカリ属、ハコヤナギ属、フウ(Liquidamber),アカシア属、チーク、マホガニー、ワタ、タバコ、シロイヌナズナ(Arabidopsis)、トマト、ジャガイモ、テンサイ、ブロッコリ、キャッサバ、サツマイモ、コショウ、ポインセチア、マメ、アルファルファ、ダイズ、ニンジン、イチゴ、レタス、カシ、カエデ、クルミ、バラ、ミント、カボチャ、ヒナギク、ゼラニウム、アボカド、サボテンがを含むが、これらに限定されない。
【0027】
内在性とは、ある植物ゲノムに元々存在する遺伝子を指す。
【0028】
繊維組成:本明細書で用いられる繊維組成とは、繊維の構造、外観または用途を変更するために改変可能な形質をいう。繊維組成を決定する形質は、繊維長、粗さ、強度、色、断面幅、繊維密度を含むが、これらに限定されない。例えば、繊維長は強度を与え、繊維の粗さは風合いおよび可撓性を決定する要因であることが知られている。
【0029】
外来性:ある核酸に関して「外来性」とは、その核酸が植物以外の生物に由来するか、形質転換されるべき植物とは異なる植物に由来するか、形質転換されるべき植物と交配できない植物に由来するか、あるいは標的植物の種に属さないことを意味する。本発明によれば、外来性のDNAまたはRNAは、真菌、細菌、ウィルス、哺乳類、魚類または鳥類の遺伝物質中に天然に存在するが、形質転換されるべき植物中には天然には存在しない核酸を含む場合がある。したがって、ある外来性の核酸は、例えば、形質転換された植物によって天然には生産されないポリぺプチドをエンコードするものである。外来性の核酸は、タンパク産物をエンコードする必要はない。
【0030】
遺伝子:産物、すなわちポリペプチド鎖またはRNA分子の合成に必要な情報のすべてを含むDNA分子のセグメントであり、コーディング配列およびノンコーディング配列の双方を含む。
【0031】
遺伝子エレメント:「遺伝子エレメント」とは、プロモーター、遺伝子、ターミネーター、イントロン、エンハンサー、スペーサー、5’−非翻訳領域、3’−非翻訳領域、またはリコンビナーゼ認識部位を含むがこれらに限定されない、離散的なヌクレオチド配列のいずれかをいう。
【0032】
遺伝子改変:分子細胞生物学的手法を適用することによって、ある生物のゲノムにDNAを安定的に導入すること。
【0033】
裸子植物:本明細書で用いられる場合には、子房を持たず、種子が露出している種子植物をいう。裸子植物の例は、針葉樹、ソテツ、イチョウおよびマオウを含む。
【0034】
導入:本明細書で用いられる場合には、感染、トランスフェクション、形質転換またはトランスダクション等の方法により、細胞中に核酸配列を挿入することをいう。
【0035】
幼若性:若木と成木との間の生理学的差異を説明する。本発明では、幼若性とは、若木と成木との間の微繊維角度、木材密度、セルロース収量、再生能力および生殖能力の差異をいう。例えば、木本植物組織は成熟するにしたがって再生能力を失うことが知られている。
【0036】
リグニン:本明細書で用いられる場合には、モノリグノールである、コニフェリルアルコール、クマリルアルコールおよびシナピルアルコールの重合誘導体を含む、フェニルプロパノイド単位で構成されるポリマー組成物をいう。リグニンの品質とは、細胞壁マトリクスに強度を与え、水分輸送を助け、および/または細胞壁多糖類の分解に抵抗する、リグニン組成物の能力をいう。リグニン組成物またはリグニン構造物は、各モノリグノールの相対量を変更するか、リグニンのタイプを変更することによって変化される場合がある。例えば、(フェルラ酸から誘導される)グアイアシルリグニンは軟木種では顕著だが、(フェルラ酸およびシナピン酸から誘導される)グアイアシル−シリンギルリグニンは硬木種に特徴的である。マツのような軟木由来のリグニンの分解には、硬木由来のリグニンの除去に比べてかなり大量のアルカリと長いインキュべーション時間とを要する。リグニン組成は、リグニン生合成に関与する酵素の上向き調節または下向き調節のいずれかにより調節される場合がある。例えば、重要なリグニン生合成酵素は、4−クマリン酸:補酵素Aリガーゼ(4CL)、シンナミルアルコールデヒドロゲナーゼ(CAD)およびシナピルアルコールデヒドロゲナーゼ(SAD)を含むが、これらに限定されない。
【0037】
単子葉植物:1枚の子葉または種葉(seedleaf)を有する胚と、平行な葉脈と、3の倍数に分割される花を有する顕花植物である。単子葉植物の例は、シバ、トウモロコシ、コメ、エンバク、コムギ、オオムギ、モロコシ、ラン、アヤメ、ユリ、タマネギ、ヤシを含むが、これらに限定されない。シバの例としては、Agrostis spp.(コロニアルベントグラスおよびクリーピングベントグラスを含むベントグラス種)、Poa pratensis(ケンタッキーブルーグラス)、Lolium spp.(1年生ライグラスおよび多年生ライグラス等のライグラス種)、Festuca arundiytacea(トールフェスク)、Festuca rubra commutata(ファインフェスク)、Cynodon dactylos(ティフグリーン、ティフウェイIIおよびサンタアナ等のコモンバミューダグラスの変種、並びにその雑種形成種)、Pennisetum clandestinum(アフリカチカラシバ)、Stenotaphrum secundatum(イヌシバ)、Zoysia japonica(シバ)、およびDichondra micranthaを含むが、これらに限定されない。
【0038】
操作可能に連結した:2個または3個以上の分子を、それらがある植物細胞内で組み合わせて適切に機能するように組み合わせること。例えば、プロモーターが構造遺伝子の転写を制御するとき、プロモーターは構造遺伝子に操作可能に連結している。
【0039】
表現型:表現型は、区別できる植物の特徴または特性であって、本発明では1個または2個以上の「所望のポリヌクレオチド」および/またはスクリーニング/選択可能なマーカーを、形質転換された植物の少なくとも1個の細胞のゲノムに組み込むことによって変化させられる場合がある。「(単数または複数の)所望のポリヌクレオチド」および/またはマーカーは、形質転換された植物細胞または植物個体全体の多数の遺伝的、分子的、生化学的、生理学的、形態学的、農業経済学的特徴または特性を改変することにより、形質転換された植物の表現型の変化を可能にする場合がある。したがって、植物ゲノム中に安定に組み込まれた1個または2個以上の所望のポリヌクレオチドの発現は、例えば、干魃耐性の向上、耐寒性および耐霜性強化、活力の向上、色彩の強化、保健栄養的な特性の強化、保存性の向上、収量の増大、耐塩性向上、重金属耐性向上、耐病性の向上、耐虫性の向上、耐乾性の強化、甘味の増強、活力の向上、味の改善、食感の改善、リン酸含量の低減、発芽率の増大、微量栄養素の取込みの増大、デンプン組成の改善、開花寿命の改善、密度の変更、茎強度または茎剛性の変化、寸法安定性の向上およびセルロースまたはリグニン含量の変化からなる群から選択される表現型を与える場合がある。
【0040】
植物組織:「植物」とは、胚を生産し、葉緑体を含み、セルロース細胞壁を有することを特徴とする、光合成能を有する植物界の真核多細胞生物のいずれかをいう。植物の一部、すなわち「植物組織」は、トランスジェニック植物を作出するために、本発明の方法により形質転換される場合がある。多くの適当な植物組織が、本発明の方法によって形質転換可能で、体細胞胚、花粉、葉、茎、カルス、走根、マイクロチューバーおよび苗条を含むが、これらに限定されない。このように、本発明はシバ、コムギ、トウモロコシ、コメ、オオムギ、エンバク、テンサイ、ジャガイモ、トマト、タバコ、アルファルファ、レタス、ニンジン、イチゴ、キャッサバ、サツマイモ、ゼラニウム、ダイズ、カシ、リンゴ、ブドウ、マツ、モミ、アカシア、ユーカリ、クルミおよびヤシのような被子植物および裸子植物の形質転換を意図する。本発明によると、「植物組織」は植物細胞も包む。植物細胞は、懸濁培養、カルス、胚、成長点領域、カルス組織、葉、根、苗条、配偶子、胞子体、花粉、種子および小胞子を含む。植物組織は成熟のいかなる段階にあるものでもよく、液体または固体培地か、土壌か、植木鉢、温室または圃場の適当な培地かで生育されてもよい。植物組織とは、有性的、無性的のいずれによって生じたかを問わず、かかる植物個体、種子、子孫、ムカゴのいずれかのクローンと、切断片や種子のような、これらのいずれかの派生物をもいう。特に興味深いのは、マツ、モミおよびトウヒのような針葉樹と、ケンタッキーブルーグラス、クリーピングベントグラス、トウモロコシおよびコムギのような単子葉植物と、ワタ、トマト、レタス、シロイヌナズナ、タバコ、リンゴおよびゼラニウムのような双子葉植物とである。
【0041】
植物の形質転換および細胞培養:広義には、植物細胞が遺伝子的に改変され、維持、さらなる成長および/またはさらなる発生のために適当な植物培地に移されるプロセスをいう。これらの方法は、当業者に周知である。
【0042】
子孫:トランスジェニック植物の子孫のような、本発明の「子孫」とは、植物またはトランスジェニック植物から生まれたか、これらの間に授かったか、これらに由来するものをいう。したがって、「子孫」植物、すなわち「F1」世代の植物は、本発明の方法により作出されたトランスジェニック植物の子、または子孫である。トランスジェニック植物の子孫は、少なくとも1個、一部、あるいは全ての細胞ゲノムに、本明細書に説明される方法によって親のトランスジェニック植物に組み込まれた所望のポリヌクレオチドを含む場合がある。したがって、所望のポリヌクレオチドは、子孫の植物によって「伝達される(transmitted)」か、「受け継がれる(inherited)」。そのように子孫植物に受け継がれる所望のポリヌクレオチドは、T−DNAコンストラクト中に存在する場合があるが、T−DNAコンストラクトもまたその親植物から子孫植物へと受け継がれる。本明細書で用いられる「子孫(progeny)」という用語は、ある群の植物の子(offspring)、または子孫(descendants)と考えられる場合もある。本発明のトランスジェニック植物は、子孫植物を作出するために無性生殖により繁殖される場合がある。
【0043】
プロモーター:RNAポリメラーゼおよび/または他の転写調節エレメントに結合する核酸、好ましくはDNAを指す意味で用いられる。あらゆるプロモーターと同様、本発明のプロモーター配列は、該プロモーターと操作可能に連結した核酸分子からmRNAを生成するために、DNAまたはRNAの転写を促進または制御する。上述したように、生成したRNAはタンパクまたはポリペプチドをコードする場合があり、RNA干渉分子またはアンチセンス分子をコードする場合がある。
【0044】
本明細書で用いられるプロモーターとは、調節エレメントを含む場合もある。逆に、ある調節因子がプロモーターから分離される場合もある。調節因子は、1つのプロモーター領域に対して多数の重要な性質を与える。ある種の要素は、操作可能に連結した核酸の転写速度を上げる転写因子に結合する。また、別の要素は、転写活性を阻害するレプレッサーに結合する。プロモーター活性に及ぼす転写因子の影響は、そのプロモーター活性が高いか低いか、すなわち、プロモーターが「強い」か「弱い」かを決定する。
【0045】
植物プロモーターとは、植物細胞由来であるか否かによらず、植物細胞中における転写を開始することができるプロモーターをいう。代表的な植物プロモーターは、植物と、植物ウィルスと、植物細胞で発現される遺伝子を含む、アグロバクテリウム属またはリゾビウム属のような細菌とから得られるものを含むが、これらに限定されない。発生学的な制御下にあるプロモーターの例は、木部、葉、根または種子のような特定の組織で転写を優先的に開始するプロモーターを含む。かかるプロモーターが、組織優先発現的(tissue−preferred)プロモーターと呼ばれる。特定の組織中でのみ転写を開始するプロモーターは、組織特異的プロモーターと呼ばれる。細胞タイプ特異的プロモーターは、例えば根または葉の維管束系細胞のような1種類または2種類以上の器官における特定の細胞タイプでの発現を主として推進する。誘導可能または抑制可能なプロモーターは、環境による制御の下にあるプロモーターである。誘導可能なプロモータによって転写に影響を及ぼし得る環境条件の例は、嫌気的条件および光の存在を含む。組織特異的、組織優先発現的、細胞タイプ特異的、および誘導可能プロモーターは、非構成的プロモーターのクラスを構成する。構成的プロモーターは、ほとんどの環境条件の下で、かつ、ほとんどの植物の部分で活性があるプロモーターである。
【0046】
ポリヌクレオチドとは、遺伝子をコードする配列または(少なくとも15個の連続したヌクレオチド、少なくとも30個の連続したヌクレオチド、または少なくとも50個の連続したヌクレオチドを含む)その断片と、プロモーターと、イントロンと、エンハンサー領域と、ポリアデニル化部位と、翻訳開始部位と、5’または3’非翻訳部位と、レポーター遺伝子と、選択可能マーカー等とを含むヌクレオチド配列をいう。ポリヌクレオチドは、1本鎖または2本鎖のDNAまたはRNAを含む場合がある。ポリヌクレオチドは、修飾塩基または修飾されたバックボーンを含む場合がある。ポリヌクレオチドは、ゲノム由来か、(mRNAのような)RNA転写産物か、(cDNAのような)プロセシングを経たヌクレオチド配列かの場合がある。ポリヌクレオチドは、センス配向またはアンチセンス配向を有する配列を含む場合がある。
【0047】
単離ポリヌクレオチドは、天然状態ではないポリヌクレオチド配列である。例えば、前記ポリヌクレオチドは、自然界には存在しないヌクレオチド配列で構成されるか、その近傍に存在するのが典型的なヌクレオチド配列から切り離されているか、あるいは、その近傍には存在しないのが典型的なヌクレオチド配列の近傍に存在する。
【0048】
再生能力:本明細書で用いられるところでは、脱分化組織から植物が再分化する能力をいう。
【0049】
種子:「種子」とは、胚を含む熟した植物の胚珠と、塊茎または胞子のような植物の増殖部位として考えられる場合がある。種子は、発芽促進のために、アグロバクテリアを介する形質転換の前に、例えば暗所でインキュベーションされる場合がある。種子はまた、インキュベーションの前に、短時間の漂白剤処理のような処理によって滅菌される場合がある。得られた実生はアグロバクテリアの所望の菌種に曝露できる。
【0050】
選択可能なマーカー/スクリーニング可能なマーカー:ある遺伝子が植物個体または植物組織において発現している場合に、該植物個体または植物組織を前記遺伝子が発現していない他の植物個体または植物組織と区別することを可能にする遺伝子をいう。スクリーニング手順は、前記スクーニング可能なマーカー遺伝子にエンコードされるタンパクの発現についてのアッセイが必要な場合がある。かかるマーカーの例は、ベータ・グルクロニダーゼ(GUS)遺伝子と、ルシフェラーゼ(LUX)遺伝子とを含む。選択可能なマーカーの例は、カナマイシンおよびジェネチシン耐性をエンコードするネオマイシンホスホトランスフェラーゼ(NPTII)遺伝子と、ハイグロマイシン耐性をエンコードするハイグロマイシンホスホトランスフェラーゼ(HPTまたはAPHIV)遺伝子と、スルホニル尿素系除草剤耐性をエンコードするアセトラクテートシンターゼ(als)遺伝子と、グルタミンシンターゼの活性を阻害するように作用する、ホスフィノトリシン(LibertyまたはBasta)のような除草剤への耐性をコードする遺伝子(BARおよび/またはPAT)と、その他の本技術分野で知られた類似の遺伝子を含む。
【0051】
配列の同一性:本明細書で用いられる、2個の核酸配列またはポリペプチド配列について「配列の同一性」または「同一性」は、ある特定の領域にわたって前記2個の配列を最大限対応するようにアライメントをとるときに、前記2個の配列中で同一となる残基をさすことを含む。
【0052】
本明細書で用いられる同一性の百分率は、ある比較ウィンドウにわたって最適なアライメントをとった2個の配列を比較することによって決定される値を意味するが、ここで、前記2個の配列の最適なアライメントのための((例えばギャップのような)付加または欠失を含まない)基準配列と比較するとき、前記比較ウィンドウ内の前記ポリヌクレオチド配列の一部は付加または欠失を含む場合がある。前記百分率は、両方の配列中に同一の核酸塩基が出現する塩基数を数えることにより一致塩基数を算出し、この一致塩基数を比較ウィンドウ中の総塩基数で除し、この商に100を乗ずることにより算出される。
【0053】
転写因子:転写因子とは、遺伝子コード配列を伴う1個または2個以上のヌクレオチド配列に直接結合するか、あるいは、遺伝子をコードする配列を伴う1個または2個以上のヌクレオデチド配列に直接結合する他の(単数または複数の)ポリペプチドの活性に間接的に影響を及ぼすかのいずれかによって、単数または複数の遺伝子の発現を調節するポリペプチド配列をいう。転写因子は、単数または複数の遺伝子の発現を活性化(上向き調節)または抑制(下向き調節)する場合がある。転写因子は、DNA結合ドメインと、活性化ドメインまたはタンパク−タンパク相互作用ドメインとを含む場合がある。本発明では、転写因子は、(1)核酸配列に結合すること、あるいは(2)植物における遺伝子の発現を調節すること、の少なくともいずれかを行うことができる。さらに、本発明のポリヌクレオチド配列およびこれに対応するポリペプチド配列は、被子植物および裸子植物を含む、いずれかの植物種において転写因子として機能する。
【0054】
転写および翻訳のターミネーター:本発明の発現DNAコンストラクトには、典型的には、転写開始調節エレメントの反対側に転写終結領域がある。この転写終結領域は、発現を増強するmRNAの安定、および/または、遺伝子転写産物にポリアデニル鎖の尾部の付加のために選択される場合がある。
【0055】
転移DNA(T−DNA):アグロバクテリアのT−DNAは、その境界内に含まれるヌクレオチド配列を別のゲノムに組み入れることが可能な因子として周知の遺伝子エレメントである。この点で、T−DNAは典型的には2つの「境界」配列に挟まれている。本発明の所望のポリヌクレオチドおよび選択可能なマーカーは、あるT−DNAの左側の境界様配列と右側の境界様配列との間に位置する場合がある。T−DNAに含まれる前記所望のポリヌクレオチドおよび選択可能なマーカーは、その発現、すなわち前記所望のポリヌクレオチドまたは選択可能なマーカーによってエンコードされるDNA配列の転写および/または翻訳を促進するプロモーターおよびターミネーターの調節エレメントのような、さまざまな異なる植物特異的な(すなわち、天然の)、または外来の核酸と操作可能に連結している場合がある。
【0056】
植物細胞の形質転換:植物細胞のゲノムに核酸を安定に挿入するプロセスをいう。形質転換は当業者に周知のさまざまな方法を用い、自然または人工的な条件下で起こる場合がある。形質転換は、原核または真核宿主細胞へ核酸配列を挿入するいずれかの知られた方法によって行われる場合があり、これら方法としては、アグロバクテリアを介する形質転換法、ウィルス感染法、ウィスカー法、エレクトロポレーション法、顕微注入法、ポリエチレングリコール処理法、熱ショック法、リポフェクション法および微粒子銃法を含む。
【0057】
トランスジェニック植物:本発明のトランスジェニック植物とは、外来性の核酸が安定に組み込まれた少なくとも1個の細胞ゲノムを含むものをいう。本発明によると、トランスジェニック植物は、遺伝的に改変された細胞および細胞ゲノムを1つだけ含む場合があるか、複数ないし多数の遺伝的に改変された細胞を含む場合があるか、全ての細胞が遺伝的に改変されている場合がある。本発明のトランスジェニック植物は、所望のポリヌクレオチド、すなわち外来性の核酸の発現が、植物個体の特定の部分のみで起こる場合がある。したがって、トランスジェニック植物は、その構造の特定の部分には遺伝的に改変された細胞のみを含む場合がある。
【0058】
変異体:ここで言う「変異体(variant)」とは、ある特定の遺伝子の基準(すなわち天然、標準、所与の)ヌクレオチド配列から逸脱(deviate)したヌクレオチド配列をいう。「アイソフォーム」、「アイソタイプ」および「類似体」という用語もヌクレオチド配列の「変異」した類型を指す。
【0059】
「変異体」には、Maxygenに付与された特許に記載されているような「シャッフルされた(shuffled)遺伝子」を指す場合もある。例えば、本発明で言う変異体は、引用によりここに取り込まれる米国特許第6,132,970号明細書に開示された方法および原理にしたがって改変された配列および所望のポリヌクレオチドの変異体も含む場合がある。
【0060】
本明細書で用いられるところの木材組成は、木材の構造、外観または用途を変更するために改変可能な形質を指す。木材組成を決定する特性は、細胞壁の厚さと、細胞の長さと、細胞のサイズと、内腔の大きさと、細胞密度と、微繊維の角度と、引張強度と、引裂強度と、木材の色と、細胞分裂の所用時間および頻度とを含むが、これらに限定されない。
【0061】
木材パルプとは、さまざまな純度の木材から得られる繊維をいう。木材パルプは、紙、板紙、化学製品の製造に用いることができる。
【0062】
本発明は、本明細書で説明する特定の方法論、プロトコール、ベクター、試薬等に限定されるものではないことが理解されるが、それは変更が可能だからである。また、本明細書で用いる用語法は特定の実施態様のみを説明する目的で用いられており、本発明の範囲を限定することを意図するものではないことが理解される。また、本明細書および添付する請求の範囲で用いられる単数形の「a」、「an」、「the」は、その文脈の中で特に明言しない限り、複数形も包含することに留意しなければならない。したがって例えば、「遺伝子」というときは、1個または2個以上の遺伝子を指し、当業者等に知られた均等物等を含む。実際、当業者は、植物宿主系でいずれかの天然の(現在または将来知られる)遺伝子を発現させるためにここに説明する方法を利用することができる。
【0063】
(単数または複数の)「単離された」核酸分子とは、天然の環境から移された核酸分子である、DNAまたはRNAを指す。例えば、あるベクターに含まれる組換えDNA分子は、本発明の目的のために単離されたものと考えられる。単離DNA分子の他の例は、異種宿主細胞内で維持される組み替えDNA分子か、(部分的または実質的に)精製された溶液中のDNA分子かを含む。単離RNA分子は、本発明のDNA分子の試験管内RNA転写産物を含む。本発明の単離核酸分子は、人工合成で製造された分子も含む。
【0064】
本発明の核酸分子は、mRNAのようなRNAの形態か、クローニングや人工合成によって製造されたcDNAおよびゲノムDNAのようなDNAの形態かの場合がある。前記DNAまたはRNAは2本鎖または1本鎖の場合がある。1本鎖DNAは、コーディング鎖、別名センス鎖か、ノンコーディング鎖、別名アンチセンス鎖かの場合がある。
【0065】
特に断らない限り、本明細書でDNA分子の配列決定により決定されたすべてのヌクレオチド配列は、(アプライド・バイオシステムズ社製、モデル3700のような)自動シーケンサを用いて決定され、また本明細書で決定されたDNA分子にエンコードされたポリペプチドのすべてのアミノ酸配列は、上記のとおり決定されたDNA配列を翻訳することによって予測された。したがって、かかる自動化手法により決定されたいずれかのDNA配列について当業者に知られているとおり、本明細書で決定されたヌクレオチド配列にはなんらかのエラーが含まれている場合がある。自動的に決定されたヌクレオチド配列は、配列決定されたDNA分子の実際のヌクレオチド配列に対して典型的には少なくとも約95%、より典型的には少なくとも約96%ないし約99.9%の同一性を有する。前記実際の配列は、当業者に周知の手作業によるDNA配列決定を含むの他のアプローチにより、より正確に決定することができる。当業者に知られているとおり、決定されたヌクレオチド配列中に実際の配列と比較してたった1個の挿入または欠失は、ヌクレオチド配列の翻訳でフレームシフトを起こし、決定されたヌクレオチド配列によってエンコードされる予測アミノ酸配列が、このような挿入または欠失が生じた塩基から後は、配列決定されたDNA分子によって実際にエンコードされるアミノ酸配列とは全く異なったものになる。
【0066】
特に断らない限り、ここに示す「ヌクレオチド配列」のそれぞれは、デオキシリボヌクレオチド(A、G、C、Tと略記)の配列として提供される。しかし、核酸分子またはポリヌクレオチドの「ヌクレオチド配列」の語は、DNA分子またはポリヌクレオチドの場合はデオキシリボヌクレオチド配列を指し、RNA分子またはポリヌクレオチドの場合は、対応するデオキシヌクレオチド配列中における各チミジンデオキシヌクレオチド(T)がウリジンリボヌクレオチド(U)に置換された、対応するリボヌクレオチド(A、G、C、U)配列を指す。例えば、デオキシリボヌクレオチドの省略記号を用いて示される配列番号1の配列を有するRNA分子への言及は、配列番号1の各デオキシヌクレオチドA、GまたはCが対応するリボヌクレオチドA、GまたはCに置換され、各デオキシヌクレオチドTがリボヌクレオチドUに置換された配列を有するRNA分子を示すことを意図する。
【0067】
本発明は、本明細書で説明される単離核酸分子の断片にも向けられる。表8に示されるヌクレオチド配列を有する単離DNA分子の断片は、以下に詳しく説明される診断プローブおよびプライマーとして有用な、少なくとも15ヌクレオチド、少なくとも20ヌクレオチド、少なくとも30ヌクレオチドの長さを有するDNA断片を指す。もちろん、従来の雑種形成手法によれば、本発明の核酸分子の全長までの長さを有するより大きな核酸断片もプローブとして診断上有用であり、また、例えば、引用によりここにその開示内容全体が取り込まれるMolecular Cloning、A Laboratory Manual第3版、Sambrook、JおよびRussel、D.W.共編(2001年)、Cold Spring Harbor Laboratory Press、Cold Spring Harbor、New Yorkに説明されるポリメラーゼ連鎖反応(PCR)による標的配列の増幅用のプライマーとしても有用である。
【0068】
例えば、少なくとも20ヌクレオチドの長さを有する断片とは、表8に示したヌクレオチド配列由来の20個以上の連続した塩基を有する断片をいう。表8に列挙されたヌクレオチド配列を含む核酸は、当業者にはルーティン作業であろう従来のDNA合成法を用いて作成することができる。例えば、制限エンドヌクレアーゼによる切断または超音波処理による剪断は、さまざまなサイズの断片を作成するために簡単に利用することができる場合がある。代替的に、本発明のDNA断片は、既知の方法にしたがって、人工合成によって作成できる場合がある。
【0069】
別の局面において、本発明は、ストリンジェントな雑種形成条件下で上記の本発明の核酸分子におけるポリヌクレオチドの一部と雑種形成するポリヌクレオチドを含む単離核酸分子を提供する。あるポリヌクレオチドの「部分」と雑種形成するポリヌクレオチドとは、前記基準ポリヌクレオチドの少なくとも約15ヌクレオチド、少なくとも約20ヌクレオチド、少なくとも約30ヌクレオチド、および、31ヌクレオチド以上と雑種形成する(DNAまたはRNAのいずれかの)ポリヌクレオチドをいう。前記基準断片と雑種形成するこれらの断片は、診断プローブおよびプライマーとして有用である。本明細書で用いられるプローブは、表8の1個の核酸の少なくとも約100個の連続する塩基として定義される。本発明の目的には、6X SSC、0.5% SDS、5X Denhardt溶液および非特異的キャリアDNA100μgを含む雑種形成溶液中で2個の配列が2本鎖複合体を形成するとき、前記2個の配列は雑種形成するといえる。Ausubelら、第2.9節、補遺27(1994年)を参照せよ。配列が「中程度のストリンジェンシー」で雑種形成するとは、6X SSC、0.5% SDS、5X Denhardt溶液および非特異的キャリアDNA100μgを含む雑種形成溶液中で、60°Cの条件をいう。「高度なストリンジェンシー」で雑種形成を行うには、温度を68°Cに上昇させる。かかる中程度のストリンジェンシーの雑種形成反応を行った後、2X SSCおよび0.05% SDSの溶液中で室温にて5回、前記ヌクレオチドは洗浄され、さらに0.1X SSCおよび0.1% SDS溶液を用いて60°Cで1時間洗浄される。「高度なストリンジェンシー」用には、洗浄温度を68°Cに上昇させる。本発明の目的には、雑種形成したヌクレオチドとは、比活性10,000cpm/ngの放射能標識プローブ1ngを用いて検出されるものであり、前記雑種形成ヌクレオチドは−70°C、72時間以内のX線フィルムへの露出の後明瞭に視認できる。
【0070】
上記のとおり、本出願は、上記の核酸配列との同一性が、少なくとも65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%または100%である核酸分子に向けられる。1つの実施態様は、表8に示される核酸配列との同一性が少なくとも95%、96%、97%、98%、99%または100%である核酸分子を包含する。ある基準ヌクレオチド配列に対して例えば少なくとも95%「同一性がある」ヌクレオチド配列を有するポリヌクレオチドとは、前記ポリヌクレオチドのヌクレオチド配列が、前記基準ヌクレオチド配列の100個のヌクレオチドあたり最大5個の点突然変異を含むかもしれないことを除いて、前記基準配列と同一であることを意味する。換言すれば、基準ヌクレオチド配列に対して少なくとも95%の同一性があるヌクレオチド配列を有するポリヌクレオチドを得るためには、前記基準配列のヌクレオチドの最大5%が欠失するかまたは別のヌクレオチドに置換されている場合があるか、前記基準配列のヌクレオチド総数の最大5%の数のヌクレオチドが前記基準配列に挿入されている場合がある。前記基準配列のこれらの突然変異は、前記基準ヌクレオチド配列の5’末端または3’末端に起こるか、あるいは、前記基準配列中のヌクレオチドの間に1個ずつ分散するか、または前記基準配列中の1個または2個以上のグループに分かれて分散してこれら両末端の間の任意の場所で起こる場合がある。
【0071】
実際問題として、ある特定の核酸分子が基準ヌクレオチド配列に対して少なくとも95%、96%、97%、98%または99%の同一性を有するか否かは、当該技術分野で周知の標準的なアルゴリズムを用いて2個の分子の間の比較を行うことで判る。配列の同一性の決定にはいかなる配列決定アルゴリズムを用いてもよいが、明確にするために、本発明はBasis Local Alignment Search Tool(BLAST)アルゴリズム(Altschulら、J.Mol.Biol.、215巻、403−410頁(1990年))を用いて同一性を決定し、本開示内容に示されるプロモーター配列が、その全長にわたってポリヌクレオチド相同体の同一性の百分率を決定するための基準配列として用いられる。より生物学的に現実性のある結果をもたらすパラメータはいくつか知られているが、一致、不一致、および挿入または欠失のパラメータ値の選択は任意である。
【0072】
本発明にしたがって、ある特定の配列が基準配列に対して、例えば、95%の同一性を有しているか否かを決定するためにBLASTその他の配列アライメントプログラムを用いるときには、同一性の百分率が前記基準ヌクレオチド配列の全長にわたって計算され、前記基準配列中のヌクレオチド総数の5%までの相同性のギャップが許容されるように、パラメータが設定される。
【0073】
2個のポリヌクレオチドの間の関連度(relatedness)は、前記ポリヌクレオチドが雑種形成して相補的塩基対の形成によって2本鎖複合体を形成する能力に関連づけて説明することができる。雑種形成条件については既に説明した。温度の上昇はこれらの複合体を解離するのに利用することができる。2つの配列の構造的な同一性が大きいほど、両者を解離または「融解」させるのに必要な温度は高くなる。2本鎖複合体を融解させるのに必要な温度は「Tm」と呼ばれる。Tmと他の雑種形成パラメータとの関係は、次式で表される。
Tm(°C)=81.5+16.6(log10[Na+])+0.41(G+C分画)−0.63(%ホルムアミド)−(600/l)
前記式で、Tmはプローブとその標的とからなるDNA2本鎖複合体の融解温度であり、lは雑種形成体の長さ(単位:塩基対)、但しl>100塩基対である。Boltonら、Proc.Natl.Acad.Sci.、48巻、1390頁(1962年)を参照せよ。一般に、融解温度が1°C変化することは、DNA配列の類似度に0.7%から3.2%までの相違があることを表す。Bonnerら、Journal of Molecular Biology、81巻、123−35頁(1973年);McCarthyら、EVOLUTION OF GENETIC SYSTEMS、H.H.Smith編、Brookhaven Symposium in Biology、No.23、Gordon and Breach、New York、1−43頁(1972年)。安定なDNA2本鎖複合体を60°Cで形成するには、配列の間に少なくとも80%の配列同一性が必要である。Sibleyら、ACTA、1巻、83−121頁(Proceedings of the 18th International Ornithological Congress、Moscow、8月16〜24日、1982年、ソ連科学アカデミー)。
【0074】
本発明の核酸は、植物の維管束組織においてポリペプチドまたはタンパクを優先的に発現させる。また本発明の核酸は、標的遺伝子の発現を阻害または完全にブロックする目的で使用可能な、アンチセンスRNAの発現、または、低分子干渉RNA(siRNA)のような、RNA干渉(RNAi)に関与するRNAの発現を植物の維管束組織において優先的に行わせることができる。
【0075】
本明細書で用いられる維管束植物組織とは、木部、篩部または維管束の形成層組織をいう。1つの実施態様では、本発明のプロモーターは「木部特異的」、「形成層特異的」および/または「篩部特異的」のいずれかであり、それぞれ木部、形成層および/または篩部におけるある操作可能に連結した核酸セグメントの発現を指令する。本明細書で用いられる「コード産物(coding product)」とは、前記プロモーターに操作可能に連結した核酸の最終産物を意味する。例えば、タンパクまたはポリペプチドと、アンチセンスRNAまたはsiRNAとはコード産物であり、siRNAは前記アンチセンスRNAの核酸コーディングの最終産物である。またコード産物は非翻訳mRNAの場合がある。本明細書では、ポリペプチドおよびタンパクという用語を互換的に用いる。例えば木部特異的とは、本発明の核酸分子が他のいかなる植物組織におけるよりも木部において活性が高いことを意味する。1つの実施態様において本発明の核酸は、それぞれ植物の木部、形成層、および/または篩部組織でのみ活性があるという意味で、木部、形成層および/または篩部で特異的に活性を示すプロモーターである。換言すれば、例えば「木部特異的」プロモーターは、検出可能なレベルのコード産物は植物の木部組織においてのみ発現するように、前記コード産物を発現させる。しかし、植物個体内での溶質輸送のために、木部、篩部または形成層で特異的に発現するコード産物が植物個体内の他の場所で見出される場合があるため、その存在は必ずしも木部組織には限定されない。一方、維管束優先発現的プロモーターは、木部、篩部または形成層組織のいずれかにおいて、あるいはこれら3つの組織タイプのうち少なくとも2つにおいて優先的に活性を示すことができる。維管束特異的プロモーターは、木部、篩部または形成層のいずれかにおいて、あるいはこれら3つの組織タイプのうち少なくとも2つにおいて特異的に活性を示すことができる。
【0076】
本明細書で用いられる維管束系は、高等植物において水および光合成産物の輸送に用いられる木部および篩部の各組織と、該木部と篩部との間に位置する維管束形成層とから構成される。維管束系の構成要素としては、木部と篩部とをまとめて維管束組織と呼ぶ。維管束組織は、2つの発生学的に区別される段階で形成される。胚形成段階および/または胚形成後の段階では、頂端分裂組織および前形成層の分化から一次維管束組織が発生する。二次維管束組織は、維管束形成層の分化から発生する。
【0077】
維管束形成層は、木部と篩部との間に位置する側生分裂組織であり、分化して二次維管束組織を生成する。維管束形成層は、紡錘形原始細胞および木質線原始細胞という2つの細胞タイプで構成され、これらは形成層原始細胞と総称される。紡錘形原始細胞の分裂は、木部および篩部の長手方向、すなわち軸系を形成し、線原始細胞の分裂は、横方向、すなわち線系の発達をもたらす。形成層原始細胞は、木部細胞が長軸の内側に向かって凝集し、篩部細胞が周辺へ向かって凝集するように、軸の接線方向に分裂する。
【0078】
本明細書で用いられる木部とは、水やミネラルイオンを根系から植物の空中部分へと輸送する伝導組織をいう。一次および二次木部組織の両方とも、異なる3つの細胞タイプ、すなわち、導管要素、繊維および柔細胞を含む。導管要素は、無孔仮導管および有孔導管要素を含み、水の導通機能を果たす。木部繊維は、繊維仮導管および靱皮状繊維を含み、木部の構造に強度および剛性を与える。柔細胞は、軸細胞および放射細胞を含み、デンプン、油その他のエネルギーの豊富な分子の貯蔵に重要である。したがって、本発明の木部特異的プロモーターは、一次および/または二次木部の導管要素、繊維および/または柔細胞で活性がある。
【0079】
篩部は、光合成産物を成熟した葉から成長および貯蔵を担う領域へと移動させる植物組織である。一次および二次篩部組織の両方を含む主な細胞は、篩要素、柔細胞および繊維を含む。篩要素は、篩部の伝導要素であって、裸子植物に存在する篩細胞か、被子植物に存在する篩管かを含む。篩管要素は篩管と呼ばれる固有の構造を形成し、篩管は、縦方向に配列され、篩板と呼ばれる細胞壁の一部を通じて相互に連絡する一連の篩管要素でできている。これに対し、裸子植物や下等維管束植物に存在する特殊化の程度の低い篩細胞は、相互に連絡して篩板を形成することはない。篩細胞は、篩域と呼ばれる拡大した孔部を通じて伝導が容易になるように、重複したアレイ状に配置される。篩要素は、被子植物では伴細胞、裸子植物では胚乳細胞と呼ばれる特殊な柔細胞と連携し、かつ、依存している。伴細胞および胚乳細胞の両方とも、篩要素の機能に必要なタンパクおよび代謝産物の貯蔵を担う。前記特殊な柔細胞に加え、篩部は、デンプン、脂肪その他の化合物を蓄積し貯蔵する機能がある柔細胞を含む。篩部繊維は篩部構造に強度を与え、場合によりデンプンの貯蔵に寄与する。したがって、本発明の篩部特異的プロモーターは、一次および/または二次篩部の篩要素、柔細胞、および/または繊維で活性がある。
【0080】
ほとんどの高等植物において維管束組織は、基本組織間に位置する円筒状をしている。根と茎の両方で、髄と皮層とが基本組織を構成する。髄は維管柱の内部に位置し、皮層は表皮と維管束組織との間に位置する。維管束系とこれに付随する基本組織を、維管柱と総称する。
【0081】
本明細書で用いられるプロモーターは、RNAポリメラーゼおよび/または他の転写調節エレメントに結合する核酸、好ましくはDNAを意味する。他のいかなるプロモーターと同様、本発明のプロモーターは、該プロモーターと操作可能に連結している核酸分子からmRNA分子を生成するために、DNAまたはRNAを転写することを促進または制御する。前記のとおり、生成されたRNAはタンパクまたはポリペプチドをコードするか、あるいはRNA干渉、またはアンチセンス分子をコードする場合がある。本明細書で用いられる「操作可能に連結した」とは、RNAセグメントに転写されるべき核酸配列に対してプロモーター−核酸配列の組合せが適切な配向で形成されるような、化学的融合、連結、またはDNA合成をいう。本発明のプロモーターは、生成するmRNA転写産物の5’非翻訳領域(5’UTR)の一部または全部を含む場合がある。一方、本発明のプロモーターは、必ずしも5’UTRを有している必要はない。
【0082】
本明細書で用いられるプロモーターは調節エレメントを含む場合もある。逆に、調節エレメントがプロモーターから分離される場合がある。調節エレメントはプロモーター領域に多数の重要な特徴を与える。一部のエレメントは、前記操作可能に連結した核酸の転写速度を増大させる転写因子と結合する。他のエレメントは、転写活性を阻害するリプレッサーと結合する。転写因子がプロモーター活性に総合的に及ぼす影響は、該プロモーター活性が高いか低いか、すなわち、プロモーターが「強い」か「弱い」かを決定する場合がある。調節エレメントと結合する転写因子自身が、他の結合タンパクとの相互作用または細胞外刺激に応答する共有結合による修飾、例えばリン酸化によって調節される場合がある。一部の転写因子は、細胞内代謝産物か、その生物個体にとって外来性の化学物質かのような細胞核と情報交換する情報伝達分子によってモジュレーションされる。細胞環境の変化によって影響を受けないプロモーターは、構成的プロモーターと呼ばれる。
【0083】
本発明は、本発明の単離核酸分子を含むベクターも提供する。1つの実施態様において、本発明のベクターはA.tumefaciens由来のTi−プラスミドである。
【0084】
本発明の核酸コンストラクトを開発するとき、前記コンストラクトまたはその断片のさまざまな構成要素を便利なクローニングベクター、例えば大腸菌のような細菌宿主内で自己複製能を有するプラスミドに通常の手法で挿入されるのが普通であろう。多数のベクターが文献に記載され、その多くは市販されている。それぞれのクローニングが終了した後、所望のインサートを有するクローニングベクターが単離され、所望の配列の構成要素を仕立てるために、制限消化、新しい断片またはヌクレオチドの挿入、連結、欠失、突然変異、再切断等の他の操作に供される。前記コンストラクトは、完成すると、宿主細胞の形質転換のやり方にしたがってさらなる操作を行うために適当なベクターに移植される場合がある。
【0085】
本発明の組換えDNA分子は、形質転換された細胞を容易に同定し、形質転換されていない細胞から選別できるように、選択可能なマーカーを含むのが典型的である。かかるマーカーの例は、カナマイシン耐性を可能にするネオマイシン・ホスホトランスフェラーゼ(nptII)遺伝子(Potrykusら、(1985年)、Mol.Gen.Genet.、199巻、183−188頁)を含むが、これに限定されない。nptII遺伝子を発現する細胞は、カナマイシンまたはG418のような適当な抗生物質を用いて選択できる。他の慣用される選択可能なマーカーは、ビアラホス耐性を可能にするbar遺伝子と、グリホセート耐性を可能にするEPSPシンターゼ遺伝子突然変異体(Hincheeら、Bio/Technology、6巻、915−922頁(1988年))と、ブロモキシニル耐性を可能にするニトリラーゼ遺伝子(Stalkerら、(1988年)、J.Biol.Chem.、263巻、6310−6314頁)と、イミダゾリノンまたはスルホニル尿素耐性を可能にするアセトラクテートシンターゼ遺伝子(ALS)突然変異体(欧州特許第154,204号、1985年)と、メトトレキセート耐性DHFR遺伝子(Thilletら、(1988年)、J.Biol.Chem.、263巻、12500−12508頁)とを含む。
【0086】
さらにベクターは、特定の宿主細胞の複製開始点(レプリコン)を含む場合がある。さまざまな原核生物のレプリコンが当業者に知られており、原核宿主細胞中における組換え分子の自己複製と維持を指令する役割を果たす。
【0087】
ベクターは、選択可能なマーカーを含むことが好ましい。トランスフェクションされた植物の選択に用いるための多数の選択可能なマーカーはカナマイシン、グリホセート耐性遺伝子を、大腸菌、A.tumefaciensその他の細菌中で培養するための選択可能なマーカーはテトラサイクリンまたはアンピシリン耐性遺伝子をそれぞれ含むが、これらに限定されない。
【0088】
本発明の核酸を微粒子銃法によって単子葉植物に導入するのに適したプラスミドベクターは、選択したプロモーターと、Hsp70イントロン(国際公開第WO93/19189号公報)のような遺伝子発現を促進するスプライシング部位を提供するイントロンと、ノパリンシンターゼ3’配列(NOS3’、Fraleyら(1983年)、Proc.Natl.Acad.Sci.、USA、80巻、4803−4807頁)のような3’ポリアデニル化配列とで構成される。この発現カセットは、DNAの大量生産に適したハイコピーレプリコン上に構築される場合がある。
【0089】
特に有用な双子葉植物の形質転換用アグロバクテリア利用植物形質転換ベクターは、プラスミドベクターpMON530(Rogersら、(1987年)「Improved vectors for plant transformation:expresssion cassette vectors and new selectable markers」、Methods in Enzymology.、R.Wuおよび L.Grossman共編、253−277頁、San Diego、Academic Press)である。プラスミドpMON530は、pMON316(Rogersら、(1987年)「Improved vectors for plant transformation:expression cassette vectors and new selectable markers」、Methods in Enzymology.、R.Wuおよび L.Grossman共編、253−277頁、San Diego、Academic Press)のStuI−HindIII断片(2.3kb)をpMON526に移植して調製されたpMON505の誘導体である。プラスミドpMON526は、前記SmaI部位がXmaI消化、Klenowポリメラーゼ処理および連結によって除去されたpMON505の単純な誘導体である。プラスミドpMON530は、pMON505およびCaMV35S−NOS発現カセットの特性のすべてを保持し、プロモーターとポリアデニル化シグナルとの間に唯一のSmaI切断部位を持つことになる。
【0090】
バイナリーベクターpMON505は、Tiプラスミド相同領域LIHが、ミニRK2プラスミドpTJS75(Schmidhauser and Helinski.、(1985年)、J.Bacteriol.、164−155頁)の3.8kbのHindIII−SmaIセグメントに置換されたpMON200(Rogersら、1987年)の誘導体である。このセグメントは、トリペアレンタル(tri−parental)交配法によるアグロバクテリアへの接合に用いられるRK2複製開始点oriVと転移起点oriTとを含む(HorschおよびKlee、(1986年)、Proc.Natl.Acad.Sci.、USA.、83巻、4428−4432頁)。プラスミドpMON505は、所望のDNA断片を挿入するための合成マルチリンカーと、植物細胞におけるカナマイシン耐性NOS/NPTII’/NOSキメラ遺伝子と、大腸菌およびA.tumefaciensにおける選択用のスペクチノマイシン/ストレプトマイシン耐性決定因子と、形質転換体のスコアリングおよび後代での遺伝子保持の確認を容易化するインタクトなノパリンシンターゼ遺伝子と、大腸菌体内でベクターの大量生産を可能とするpBR322複製開始点とを含む、pMON200の重要な特徴をすべて保持する。プラスミドpMON505は、pTiT37ノパリン型T−DNAの右端に由来する単一のT−DNA境界を含む。サザンブロット分析は、プラスミドpMON505およびこれに含まれるすべてのDNAが植物ゲノムに組み込まれていること、すなわち、プラスミド全体が植物ゲノムに組み込まれたT−DNAであることを示す。組込みDNAの一端は、右境界配列とノパリンシンターゼ遺伝子との間に位置し、他端は前記境界配列とpBR322配列との間に位置する。
【0091】
別の特に有用なTiプラスミドカセットベクターは、pMON17227である。このベクターは、国際公開第WO92/04449号公報に記載されており、ジャガイモ、トマトを含む多くの植物の優れた選択マーカーである、グリホセート耐性を可能にする酵素(CP4と命名)をエンコードする遺伝子を含む。この遺伝子はシロイヌナズナ(Arabidopsis)のEPSPS葉緑体輸送ペプチド(CTP2)と融合し、選択したプロモーターによって発現する。
【0092】
小胞体ルーメン、ペリプラズム間隙または細胞外環境へ翻訳されたタンパクを分泌するために、適当な分泌シグナルが発現したポリペプチドに組み込まれる場合がある。前記シグナルは、前記ポリペプチドに内在するものであっても、あるいは、異種シグナルであってもよい。
【0093】
1つの実施形態では本発明のベクターは、本明細書で説明される核酸が、関心のあるポリペプチドをエンコードするDNAに操作可能に連結した組織特異的プロモーターとなるように設計される。別の実施態様では、関心のあるポリペプチドは植物個体におけるリグニン生合成に関与する酵素である。リグニン生合成に関与する多くの酵素をエンコードするポリヌクレオチドは、シンナミルアルコールデヒドロゲナーゼ(CAD)、シンナメート4−ヒドロキシラーゼ(C4H)、クマレート3−ヒドロキシラーゼ(C3H)、フェノラーゼ(PNL)、O−メチルランスフェラーゼ(OMT)、シンナモイル−CoAレダクターゼ(CCR)、フェニルアラニンアンモニア−リアーゼ(PAL)、4−クマレート:CoAリガーゼ(4CL)およびマツ由来のペルオキシダーゼ(POX)(米国特許第6,204,434号)を含むが、これらに限定されない。他の酵素は、コニフェリンβ−グルコシダーゼ(CBG)およびカフェイン酸3−O−メチルトランスフェラーゼ(COMT)を含む(米国特許第5,451,514号明細書、国際公開第WO94/23044号明細書、およびDharmawardhanaら、Plant Mol.Biol.、40巻、365−72頁(1999年))。
【0094】
別の実施態様では、前記プロモーターに操作可能に連結されたコーディング配列は、リグニン生合成に関与する酵素の発現または活性を阻害する遺伝子産物をコードする場合がある。例えば、リグニン生合成の制御にとって特に関心があるのは、シンナミルアルコールデヒドロゲナーゼ(CAD)、シンナメート4−ヒドロキシラーゼ(C4H)、クマレート3−ヒドロキシラーゼ(C3H)、フェノラーゼ(PNL)、O−メチルトランスフェラーゼ(OMT)、シンナモイル−CoAレダクターゼ(CCR)、フェニルアラニンアンモニアリアーゼ(PAL)、4−クマレート:CoAリガーゼ(4CL)およびマツ由来のペルオキシダーゼ(POX)に対するアンチセンス遺伝子である。
【0095】
さらに別の実施態様では、本発明のベクターは、本発明の核酸が、関心のあるポリペプチドをコードする遺伝子に対応するアンチセンスRNAまたは干渉RNAをエンコードするDNAまたはRNAに操作可能に連結し、これによって標的遺伝子産物の発現を弱めるように設計される。抑制の標的となる遺伝子産物は、上記のとおり、リグニン生合成に関与する酵素であることが好ましい。遺伝子発現のRNAi阻害の利用は、Paddisonら、Genes & Dev.、16巻、948−958頁(2002年)に概説されており、また遺伝子発現を阻害するためのRNAiの利用は、国際公開第WO99/61631号公報に具体的に記載されており、両文献は、引用によりここに取り込まれる。
【0096】
特定の植物遺伝子の発現を低減または阻害するためのアンチセンス技術の利用は、例えば欧州出願公開第271988号公報に記載されている。遺伝子発現の低減は、植物の表現型に変化をもたらし、それは例えば、トマト果実中のリコピン合成欠損のため赤くなるはずの実が黄色くなるといった目に見えるレベルの表現型の差異か、あるいは例えばトマト果実の成熟過程におけるポリガラクトウロナーゼ量の変化およびペクチンの脱重合の低減のような、より微小な生化学的レベルかのいずれかで現れる(Smithら、Nature、334巻、724−726頁(1988年);Smithら、Plant Mol.Biol.、14巻、369−379頁(1990年))。このように、アンチセンスRNAは植物における遺伝子発現を低下させる上で有用であることが実証されている。
【0097】
本発明の植物の作成方法の1つの実施態様では、植物個体内で転写されてアンチセンスRNA転写産物を生成する外来性のDNAは、植物個体内、例えば植物細胞内に導入される。前記外来性DNAは、例えばそのプロモーターに対する遺伝子配列の配向を反転することにより調製することができる。植物細胞内における前記外来性DNAの転写は、その遺伝子に対して「アンチセンス」な細胞内RNA転写産物を生成する。
【0098】
例えばアンチセンス技術は、4CLのようなリグニン生合成に関与する酵素をエンコードする遺伝子の発現を抑制するために用いられる場合がある。4CLのアンチセンスDNA配列を含むコンストラクトは、プロモーターに操作可能に連結された4CLをエンコードする外来性DNAの配向を反転することにより調製することができる。前記外来性の4CLDNAの植物細胞における転写は、アンチセンス配向の4CLRNA転写産物を生成する。
【0099】
また本発明は、本発明のベクターを含む宿主細胞を提供する。本明細書で用いられる宿主細胞とは、コード産物が最終的に発現される細胞をいう。したがって宿主細胞は、個々の細胞、細胞培養、または生物個体の一部である複数の細胞であり得る。また宿主細胞は、胚、胚乳、精子または卵子、または受精卵の場合がある。
【0100】
本発明のポリヌクレオチドを含むポリヌクレオチドまたはベクターは、標的宿主生物への組換えベクターDNAを導入するための当該技術分野で知られた標準的な手順によって宿主細胞に導入される。かかる手順は、トランスフェクション、感染、形質転換、自然取込み、エレクトロポレーション法、微粒子銃法およびアグロバクテリアの使用を含むが、これらに限定されない。生物学的および物理的な植物形質転換プロトコールを含む、外来遺伝子を植物に導入する方法は当該技術分野で知られており、本発明の遺伝子コンストラクトを植物宿主に挿入するために用いられる場合がある。例えば、Mikiら、1993年、「Procedure for Introducing Foreign DNA Into Plants」、Methods in Plant Molecular Biology and Biotechnology、GlickおよびThompson共編、CRC Press Inc.、Boca Raton、67−88頁を参照せよ。選択される方法は宿主植物によって異なり、リン酸カルシウムのような化学的トランスフェクション法、アグロバクテリアのような微生物を介した遺伝子導入(Horschら、Science、227巻、1229−31頁、1985年)、エレクトロポレーション法、顕微注入法、および微粒子銃法を含む。
【0101】
したがって本発明は、本発明のポリヌクレオチドを含むポリヌクレオチドまたはベクターを含む、植物個体または植物細胞も提供する。1つの実施態様では前記植物は、被子植物または裸子植物である。別の実施態様ではこれら植物は、ユーカリ属およびマツ属の種から選択される。特にトランスジェニック植物は、ユーカリ・グランディス(Eucalyptus grandis)およびその交雑種、ラジアータマツ(Pinus radiata)、テーダマツ(Pinus taeda L、loblolly pine)、セイヨウハコヤナギ(Populus nigra)、カロリナポプラ(Populus deltoides)またはモミジバフウ(Liquidamber styraciflua)に属する植物種の場合がある。通常の植物の意味を超え、本明細書では「植物」の語を植物の果実、種子、花、胞子嚢等も指すものとする。本発明の植物は、直接トランスフェクション体、すなわちベクターがアグロバクテリアを介するやり方のように直接的に導入されるか、あるいはトランスフェクションされた植物の子孫植物の場合がある。前記子孫は、トランスフェクションされた植物の無性生殖によって得られる場合がある。第2世代またはその以降の世代の植物は、有性生殖すなわち受精によって作成される場合も、そうでない場合もある。さらに前記植物個体は、配偶体(単相期)または胞子体(複相期)の場合がある。
【0102】
本発明はまた、本発明のベクターを含む植物または種子を栽培することを含む、植物のリグニン含量の制御方法を提供する。本発明の植物または種子の成長または発芽を誘導または維持する適切な栽培法は種特異的であり、本発明の属する技術分野の通常の知識レベルの範囲内である。栽培場所は、植物個体または種子の成長または発芽を促進できるいかなる場所でもよい。さらに、栽培には、葯および/または小胞子内に胚形成を誘導できるストレス処理(例えば、窒素欠乏、熱ショック、ショ糖欠乏)が含まれていてもよいが、ストレス処理はこれらに限定されない。
【0103】
本発明はさらに、転写因子と結合し、組織優先発現的または組織特異的な発現を調節する能力を有する単離調節エレメントも提供する。前記調節エレメントによって実現される調節の程度は、完全、すなわち、前記転写因子無しでは転写が検出できない場合もあるが、部分的、すなわち、前記転写因子の存在下で転写が増強される場合もある。1つの実施態様では、複合プロモーターを用意するために、少なくとも1個の調節エレメントが異種プロモーターに連結される。前記複合プロモーターは、維管束組織内で優先的または特異的に発現させる。本明細書で用いられる異種プロモーターは、前記調節エレメントとの関係で定義される。自然環境で調節エレメントとプロモーターとが互いに協働していない場合は、そのプロモーターはその調節エレメントに対して異種とされる。典型的には、ある調節エレメントのプロモーター領域内での正確な配向はその活性に影響しない。さらに調節エレメントは、異種プロモーター領域に挿入されたときでも正常に機能することができる。したがって、例えば、木部特異的発現または木部優先的発現を実現するために、木部優先発現的調節エレメントが本来のプロモーターから取り出されて異種プロモーター領域に挿入される場合がある。同様に、篩部特異的発現または篩部優先的発現を実現するために、篩部優先発現的調節エレメントがその本来のプロモーターから取り出されて異種プロモーター領域に挿入される場合がある。同様に、形成層特異的発現または形成層優先的発現を実現するために、形成層優先発現的の調節エレメントがその本来のプロモーターから取り出されて異種プロモーター領域に挿入される場合がある。前記異種プロモーターは、例えば最小CaMV35Sプロモーターの場合がある。最小プロモーターに改変するのに適した植物細胞内での発現を指令するプロモーターは、カリフラワーウィルス(CaMV)35Sプロモーター(Jeffersonら、EMBO J.、6巻、3901−07頁(1987年))、コメのアクチンプロモーター(McElroyら、Plant Cell、2巻、163−71頁(1990年))、トウモロコシのユビキチン−1プロモーター(Christensenら、Transgenic Research、5巻、213−18頁(1996)年)、およびノパリンシンターゼプロモーター(Kononowicsら、Plant Cell、4巻、17−27頁(1992))を含む。
【0104】
本発明の核酸を調製するにあたり、さまざまな制限エンドヌクレアーゼを用いてゲノムをばらばらの断片に消化し、ラジアータマツ(Pinus radiata)およびユーカリ・グランディス(Eucalyptus grandis)からゲノムライブラリが作成された。ゲノムライブラリは、組織選択的プロモーターを得るのに適したいかなる植物種からも同様に構築することができる。Clontech社(カリフォルニア州、パロアルト)のGenomeWalkerシステム(登録商標)用に提供される手順にしたがって、アダプターがこれらゲノム配列の各々に連結された。次にプロモーター配列は、アダプター特異的プライマーと「遺伝子特異的プライマー」とを用いてPCR法で増幅された。代替的には、このPCR増幅のステップは、任意的に長鎖でバックグランドが最小の反応生成物を産生する方法であって、引用によりここに取り込まれる米国特許第5,565,340号および第5,759,822号明細書に記載される方法により実行される場合がある。この一般的なPCR増幅法を用いることにより、本発明のプロモーターの同定およびその組織選択的プロモーターとしての同定は、前記「遺伝子特異的プライマー」の選択よって左右される。
【0105】
遺伝子特異的プライマーは、関心のある組織内で高度に発現されるいずれかの転写配列である。本発明では、遺伝子特異的プライマーは、維管束組織中で高度に発現されるmRNAの断片か、あるいは該mRNAに対して相補的である。1つの実施態様では、前記遺伝子特異的プライマーは、特定の維管束組織のタイプにおいて特異的に発現することが知られている遺伝子に対する相同性によって選択される。特に関心のある遺伝子は、ある特定の維管束組織中で高度に発現される遺伝子であり、これは対応するプローモーターの維管束選択的活性の指標となるのが典型的である。
【0106】
発現配列タグ(EST)は遺伝子特異的プライマーの別の供給源である。ESTは、あるライブラリ中に存在する対応mRNAのcDNA断片である。いずれかの植物ESTデータベースが、所望の組織タイプで特異的に発現することが知られている遺伝子のセグメントとの同一性があるESTを捜すために、電子的に検索される場合がある(「インシリコ(in silico)スクリーニング」)。したがってこれらのESTは、あるゲノムライブラリ内の対応する遺伝子のプロモーターの増幅に用いられる遺伝子特異的プライマーを提供する。増幅された遺伝子プロモーターは、ESTデータベースの構築の元となる植物種と同じ植物種に由来するものである必要はない。前記ESTが、関心のある遺伝子プロモーターに対して、前記遺伝子の標的セグメントのPCR増幅用プライマーとして作用するのに十分な配列類似性を備えていることだけが必要である。
【0107】
組織特異的プロモーターを同定する代替的な方法は、ある組織タイプ内では発現するが他の組織内では発現しないmRNAは組織特異的プロモーターから転写されたことを意味するので、かかるmRNAの検出に基づく。mRNAの集団は、例えばサブトラクティブハイブリダイゼーション法により、この原理で区別することができる。かかるサブトラクティブハイブリダイゼーション法の1つの例は、ClontechによるPCR−Select(登録商標)法である。
【0108】
代替的には、組織特異的mRNAの分布は、植物組織の薄切片を放射標識プローブとin situで雑種形成させることにより、決定することができる。特定の組織タイプを放射能で反応するプローブは、以下に説明する方法を用いるゲノムライブラリのサザン分析により、mRNAと関連するプロモーターを検出するのに用いられる。前記の技術は全て関心のある組織、ここでは維管束組織からのmRNAライブラリの作成を要する。例えば、木部組織からのcDNAライブラリの調製は、Dharmawardhanaら、Plant Mol.Biol.、40巻、365−72頁(1999年)およびLoopstraら、Plant Mol.Biol.、27巻、277−91頁(1995年)に記載されている。簡潔には、まず伐採した木から活発に分化している木部を剥ぎ取る。常法にしたがって全RNAが単離され、ポリ(A)RNAが単離され、木部組織cDNAライブラリを構築するために逆転写される。例えばDharmawardhana(1999年)に説明されているように、前記cDNAライブラリは、Strategene cDNA合成およびGigapakII Gold(登録商標)パッケージングキットを用いて、λZAP−XRベクターで構築された。木部特異的プロモーターは、遺伝子特異的プローブと前記プロモーターの5’末端の配列を認識するプライマーとを用いて、PCR法により、かかるcDNAライブラリからを単離することができる。遺伝子特異的プローブは、前記インシリコのアプローチによって、あるいは、もしmRNA配列が既知の場合には、該mRNA配列にもとづく特異的プローブを設計することによって得られる場合がある。さらに、所望の標的遺伝子の5’UTRに相補的なプライマーが合成できる。代替的には、前記プライマーは、エンコードされたタンパクのアミノ酸配列の一部から、いわゆる縮重プライマーとして設計できる(Dharmawardhana(1999年)を参照せよ。)。
【0109】
関心のあるプロモーターを単離した後は、さまざまな手法がその組織特異的発現パターンやプロモーターの強さを調べるために利用できる。慣用される方法の1つは、前記プロモーターをアッセイの容易なレポーター遺伝子と操作可能に連結させることである。例えば、木部優先発現的プロモーターと、グニーユーカリ(Eucalyptus gunnii)シンナモイルCoAレダクターゼ(EgCCR)遺伝子(位置:−1448〜+200、ただし+1が転写開始部位)の5’UTRとが、β−グルクロニダーゼ(GUS)をエンコードする遺伝子と操作可能に連結される(Lacombeら、Plant J.、23巻、663−76頁(2000年))。適切な発現コンストラクトは、周知の方法にしたがって作出することができる。
【0110】
植物の形質転換は、米国特許第6,051,757号明細書に説明されるアグロバクテリアを介した形質転換法を含む多数の適切な方法のいずれかを用いて行うことができる。樹木を形質転換する当該技術分野における他の方法が、加速粒子による裸子植物の体細胞胚の形質転換を開示する米国特許第6,518,485号明細書に代表されるように、本発明の技術分野で知られている。他の形質転換法は、微粒子銃法(Kleinら、Biotechnology、6巻、559−63頁(1988年))、エレクトロポレーション法(Dhalluinら、Plant Cell、4巻、1495−1505頁(1992年))、およびポリエチレングリコール処理法(Golovkinら、Plant Sci.、90巻、41−52頁(1993年))を含む。さらに、米国特許第6,187,994号明細書は、リコンビナーゼに支援される形で発現コストラクトを植物ゲノム中の特異的選択部位に挿入する方法を提供する。上記の特許および文献はすべて引用によりここに含まれる。
【0111】
本発明のDNA分子は、いずれかの適当な方法を用いて植物のゲノムに挿入できる。好適な植物形質転換ベクターは、アグロバクテリウム・ツメファシエンスのTiプラスミドに由来するものと、例えばHerrera−Estrellaら、Nature、303巻、209頁(1983年)と、Bevan、Nucleic Acid Res.、12巻(22号)(1984年)、8711−8721頁と、Kleeら、Bio/Technology、3巻(7号)、637−642頁(1985年)と、欧州特許出願公開第120,516号公報に開示されるものとを含む。アグロバクテリアのTiまたは根誘導(Ri)プラスミドから誘導される植物形質転換ベクターに加えて、代替的な方法が、本発明のDNAコンストラクトを植物細胞に挿入するために利用可能である。かかる方法は、例えば、リポソームの使用と、エレクトロポレーション法と、遊離DNAの取込みを増大させる化学物質と、微粒子銃による遊離DNAの送達と、ウィルスまたは花粉を用いる形質転換とを含む場合がある。DNAは、葉緑体のゲノムに挿入される場合もある(Daniellら(1998年)、Nature Biotechnology、16巻、345−348頁)。
【0112】
関心のある核酸を含む細胞(またはプロトプラスト)が適当な数だけ得られると、該細胞(またはプロトプラスト)は完全な植物個体に再生される。再生ステップのための方法論の選択は重要ではなく、マメ科(アルファルファ、ダイズ、クローバー等)と、セリ科(ニンジン、セロリ、パースニップ)と、アブラナ科(キャベツ、ダイコン、ナタネ等)と、ウリ科(メロンおよびキュウリ)と、イネ科(コムギ、オオムギ、コメ、トウモロコシ等)と、ナス科(ジャガイモ、タバコ、トマト、コショウ)と、ヒマワリのようなさまざまな花卉作物と、アーモンド、カシュー、クルミ、ペカンのようなナッツを産する樹木とに由来する宿主について適切なプロトコルは入手可能である。例えば、Ammiratoら、(1984年)Handbook of Plant Cell Culture−Crop Species、Macmillan Publ.Co.と、Fromm,M.(1990年)UCLA Symposium on Molecular Strategies for Crop Improvement、4月16〜22日、1990年、コロラド州、キーストーンと、Vasilら、(1990年)Bio/Technology、8巻、429−434頁と、Vasilら、(1992年)Bio/Technology、10巻、667−674頁と、Hayashimotoら(1990年)Plant Physiol.、93巻、857−863頁と、Dattaら(1990年)とを参照せよ。
【0113】
プロモーターとレポーター遺伝子とを含むベクターは、該ベクターで形質転換に成功した植物細胞を選択するメカニズムを備え、例えばカナマイシン耐性を有する場合がある。形質転換体中のGUS遺伝子の有無は、GUS特異的PCRプライマー(Clontech、パロアルト)を用いてPCR法で確認される場合がある。形質転換された植物細胞の子孫におけるカナマイシン耐性の分離は、安定に挿入された前記ベクターを宿す遺伝子座の数を決定するためにサザン分析と組み合わせて利用される場合がある。プロモーター発現の時間的および空間的パターンは、Jeffersonら、EMBO J.、6巻、3901−07頁(1987年)の記載にしたがって、レポーター遺伝子発現の定量化から推測される。一般にGUSの発現は、先ず90%アセトンで固定した後、GUSの基質である5−ブロモ−4−クロロ−3−インドイル−β−D−グルクロン酸(X−Gluc)を含む緩衝液中で固定された植物組織の薄切片中で組織化学的に決定される。GUS発現産物の有無は、X−Glucを用いる比色定量反応により示される。
【0114】
例えば木部特異的な発現は、木部組織中で転写因子と特異的に結合する調節エレメントの存在によって実現される場合がある。木部特異的な発現の程度に影響する転写因子の間の相互作用は、調節エレメントの塩基対のサブセットと前記転写因子のアミノ酸残基とのアライメントに依存する。同様に、例えば篩部特異的または形成層特異的な発現は、それぞれ篩部および形成層組織中で転写因子を特異的に結合する調節エレメントの存在によって実現される場合がある。結合した転写因子と相互作用を行わない塩基対は、組織特異的転写因子を特異的に結合する調節エレメントの全体的な能力は維持したまま、他の塩基対に置換される場合がある。
【0115】
いったんプロモータが組織優先発現的または組織特異的と同定された後は、組織優先発現的または組織特異的プロモーターの活性に影響を与える調節エレメントの同定や特徴付けをするためにさまざまな方法論が利用できる。1つの方法では、前記プロモーター領域は5’末端が徐々に短くなるように切断され、切断された一連のプロモーターはそれぞれレポーター遺伝子に操作可能に連結される。調節エレメントが欠失すると、レポーター遺伝子の組織特異的発現が失われることからプロモーター活性への影響が推測される。代替的には、CaMV35S最小プロモーターのような最小プロモーターを含む発現コンストラクトに調節エレメントと推定される配列を挿入し(Kellerら、Plant Mol.Biol.、26巻、747−56頁)、前記調節エレメントと推定される配列が組織特異的発現を与えるか否かを確認することができる。最小プロモーターは、RNAポリメラーゼ結合部位のような、プロモーター活性に不可欠な要素のみを含む。推定的な調節エレメントの解明についての追加例は、L−フェニルアラニンアンモニアリアーゼ(PAL)および4−クマレートCoAリガーゼ(4CL)をエンコードする遺伝子の転写を協調的に調節する組織特異的調節エレメントの研究によりもたらされている(Hattonら、Plant J.、7巻、859−76頁(1995年); Leyvaら、Plant Cell、4巻、263−71頁(1992年); Hauffeら、Plant J.、4巻、235−53頁(1993年); Neustaedterら、Plant J.、18巻、77−88頁(1999年))。これらの文献はすべて、引用により本発明に含まれる。
【0116】
推定的な調節エレメントの位置決定を行う別の方法は、既知の維管束優先発現的または維管束特異的な調節エレメントの間で配列を比較することである。ヌクレオチド比較により、例えば維管束優先発現的調節エレメントのような既知の組織優先発現的調節エレメントと類似する領域が同定される。例えば、シンナミルCoAレダクターゼと、フェニルプロパノイド産生に関与する他のプロモーターとの間でプロモーター配列を比較することにより、配列CCCACCTACCを有する保存されたACリッチな領域の存在が明らかになる。Lacombe(2000年)を参照せよ。フェニルプロパノイド合成に関与するプロモーター中に前記配列が保存されていることは、保存されている前記配列が、複数の植物種でこれまでに同定されている、MYBのような協調的活性化転写因子に対する結合部位であることを意味する。Martin、Trends Genet.、13巻、67−73頁(1997年)を参照せよ。例えば、MYB結合部位は、トウモロコシでは([C/A]TCC[T/A]ACC)、ペチュニアでは(TAAC[C/G]GTTまたはTAACTAAC)と同定されている。同上。
【0117】
本発明の核酸配列にもとづいてこのような配列比較を行うことにより、以下の表1に示される一連のモチーフが明らかになる。表1のモチーフは、組織優先発現的または組織特異的転写因子に結合し、異種最小プロモーターの組織優先発現的または組織特異的発現のモジュレーションに利用できることが期待される。
【0118】
【表1】
【0119】
例えば、bZIPタンパクと維管束特異的または維管束優先発現的なプロモーターとの相互作用は、当業者がどのように調節エレメントの特徴付けを行えるかについて、別の方法を提供する。例えば、グリシンリッチなタンパクであるgrpl.8をエンコードする遺伝子は、GRP1.8−β−グルクロニダーゼ融合タンパクの組織化学的染色から決定されるとおり、木部特異的に発現される。Kellerら、EMBO J.、8巻、1309−14頁(1989年)。grpl.8プロモーターの5’欠損試験は、維管束の特異的発現に必須な20塩基対からなる負の調節エレメントを明らかにした。Kellerら、Plant Cell、3巻、1051−61頁。プロモーターの負の組織特異的調節によって作用する類似の木部特異的調節エレメントが、マメのpal2と、パセリの4CL−1とで同定されている。それぞれ、Leyvaら、Plant Cell、4巻、263−71頁(1992年)およびHauffeら、Plant J.、4巻、235−53頁(1993年)を参照せよ。木部の転写因子VSF−1は、grpl.8遺伝子の20塩基対の前記調節エレメント配列と特異的な相互作用を行う。VSF−1と調節エレメントとの相互作用は、結合特異性がVSF−1結合に関して競合する無標識DNA標的によって示される電気泳動移動度シフトアッセイを用いて、VSF−1と調節エレメントとによる高分子複合体の形成によって実証された。Torres−Schumannら、Plant J.、 9巻、283−96頁(1996年)。さらにVSF−1が相互作用を行う正確な残基は、DNaseIに対する調節エレメントの感受性を測定することによって解明された。簡潔には、VSF−1と調節エレメントとの相互作用は、結合した前記転写因子の立体障害のために、DNaseIがタンパクに接触した一部の残基を分解するのを防止する。典型的には、前記DNAaseI分析は、前記転写因子が結合するときに過敏になる残基も明らかにする。前記推定的な調節エレメントが木部特異的であることを確認するために、前記推定調節領域の単量体または多量体が最小CaMV35Sまたは最小grp1.8プロモーターの上流に融合される。改変プロモーターに操作可能に連結され、タバコに安定的にトランスフェクションされた融合β−グルクロニダーゼ遺伝子を用いた組織化学的分析により、前記推定調節領域はこれらのプロモーターに木部特異的発現を可能にすることがわかった。Torres−Schumann(1996年)を参照せよ。
【0120】
本発明のプロモーターの機能変異体または断片
本発明のプロモーターのさらに別の変異体または断片は、前記配列全体にわたって散在する改変を有するものである。本明細書で用いられる機能変異体または断片とは、基準の核酸に対して少なくとも約70%の同一性を有し、しかもコード産物の組織特異的発現を依然として可能にする核酸をいう。前記機能変異体の組織特異性または優先発現性は、基準の核酸と同じ組織に対するものでなければならない。しかし、前記機能変異体が前記基準の核酸と同様の優先発現性または特異性を有していない場合でも、前記変異体は依然として本明細書で用いられる機能変異体と考えられる。1つの実施態様では、前記機能変異体または断片の配列は、前記基準の核酸に対して少なくとも約75%の同一性を有する。別の実施態様では、前記機能変異体または断片の配列は、基準の核酸に対して少なくとも約80%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%または99%の同一性を有する。
【0121】
機能変異体を作成できる改変は、残基の5’末端からの逐次欠失、または5’UTRの3’末端からの欠失により作出される場合がある。代替的に内部残基が改変される場合もある。前記プロモーター領域の機能に影響を与えない改変は、転写因子の結合に影響を与えない改変である可能性が最も高い。本発明に含まれる改変は、本発明のプロモーターの対立遺伝子の変異体の形で天然に存在する改変を含む。本発明の変異体は、本明細書に説明される方法を用いてその器官または組織特異的活性、あるいは器官または組織優先発現的活性についてテストされる。
【0122】
本発明の核酸の作成方法
本発明の核酸は、周知の合成技術、標準的な組換え方法、精製技術、またはこれらの組合せを用いて得ることができる。例えば本発明の単離ポリヌクレオチドは、固相ホスホアミダイトトリエステル法(Beaucageら、Tetra.Letts.、22巻、1859−1862頁(1981年))、自動化合成装置(Van Devanterら、Nucleic Acids Res.、12巻、6159−6168頁(1984年))、または米国特許第4,458,066号明細書に記載の固体支持法を用いて、直接的な化学合成により調製することができる。化学合成は1本鎖オリゴヌクレオチドを製造するのが一般的で、該1本鎖オリゴヌクレオチドは、相補的な配列との雑種形成か、あるいは、前記1本鎖オリゴヌクレオチドをテンプレートとして用いる重合かによって、2本鎖オリゴヌクレオチドに変換される場合がある。また、もっと長い配列は、短い配列を連結することによって得られる場合もある。
【0123】
代替的には、本発明の核酸は、互いにプライマーとして使用可能なオリゴヌクレオチドを用いる組換え法により得られる場合がある。例えば、Ausubelら編、CURRENT PROTOCOLS IN MOLECULAR BIOLOGY(John Wiley & Sons,Inc.1990)を参照せよ。また、Wosnickら、Gene、60巻、115頁(1987年)およびAusubelら編、SHORT PROTOCOLS IN MOLECULAR BIOLOGY、第3版(John Wiley & Sons,Inc.1995)も参照せよ。ポリメラーゼ連鎖反応を利用する確立された技術は、少なくとも2キロベースの長さを有するポリヌクレオチドを合成する能力を提供する。Adangら、Plant Mol.Biol.、21巻、1131頁(1993年)、Bambotら、PCR Metods and Applications、2巻、266頁(1993年)、Dillonら、「Use of the Polymerase Chain Reaction for the Rapid Construction of Synthetic Genes」、METHODS IN MOLECULAR BIOLOGY、15巻:PCR PROTOCOLS:CURRENT METHODS AND APPLICATIONS、White編、263−268頁(Humana Press,Inc.、1993年)、Holowachukら、PCR Methods Appl.、4巻、299頁(1995年)。
【0124】
本発明の核酸の使用方法
本発明の核酸は、植物の性質を変化させるのに有用である。前記核酸は、維管束組織に見出される分子のレベルを増大させるために、関心のある遺伝子と操作可能に連結される場合がある。代替的には、関心のある遺伝子は、最終生成物、例えばセルロースの形成を阻害する場合があるので、本発明の核酸は、維管束組織中の所定の分子のレベルを減小させるために用いられる場合がある。
【0125】
かかる発現操作の主な標的の1つは、リグニンの生合成経路である。上記の理由のため、植物個体のリグニン含量を正負いずれかに調節することに大きな関心が寄せられており、前記経路に影響を与える遺伝子産物の発現を通じて達成できる。例えば、米国特許第5,451,514号明細書および国際公開第WO94/23044号公報に記載されているように、CADおよびCOMTのコピー数を操作すると、リグニン含量が改変される。さらに、ポプラまたはマツのCADをエンコードする配列のアンチセンス発現は、リグニン組成の改変をもたらす。それぞれ、Grandら、Planta(Berl.)、163巻、232−37頁(1985年);Baucherら、Plant Physiol.、112巻、1479頁(1996年)。
【0126】
変更が重要であるかもしれない他の性質には糖が含まれる。糖産生を担う酵素のレベルを例えば篩部組織で上昇させると、植物個体内の糖含量が増大する可能性がある。糖産生を増大または減少させる酵素は、β−グルコシダーゼ、β−1,3−グルカナーゼ、6−ホスホフルクト−1−キナーゼ、スクロースシンターゼ、UDP−グルコースピロホスホリラーゼ、ヘキソキナーゼ、ホスホグルコムターゼ、スクロース輸送体およびインベルターゼを含むが、これらに限定されない。
【0127】
さらなる性質は、植物の葉に存在する殺昆虫性、抗ウィルス性または抗菌性タンパクの増加を含むが、これらに限定されない。かかるタンパクは、セクロピン、チオニン、ディフェンシン、脂質輸送タンパク、ヘベイン様ペプチドおよびGASAペプチドを含むが、これらに限定されない。さらに他の性質は、干魃耐性、耐寒性、ペルオキシダーゼ活性、耐塩性および窒素輸送を含む。
【発明を実施するための最良の形態】
【0128】
以上概説された本発明は、以下の実施例を参照することにより、一層容易に理解されるであろう。ただし、これらの実施例は例示を目的として提供されるものであって、本発明を限定する意図はない。
【実施例1】
【0129】
維管束特異的プロモーターの単離
ラジアータマツおよびユーカリ・グランディスのcDNAライブラリが以下のとおり構築され、スクリーニングされた。若干改変されたChangら、Plant Molecular Biology Reporter、11巻、113−116頁、(1993年)のプロトコルを用いて、植物組織からmRNAが抽出された。具体的には、試料がCPC−RNAXB(100mM Tris−Cl、pH8.0、25mM EDTA、2.0M NaCl、2%CTAB、2%PVPおよび0.05%スペルミジン*3HCl)に溶解され、クロロホルム:イソアミルアルコール(24:1)で抽出された。
【0130】
RNAはエタノールで沈殿され、MessageMakerキット(Life Techologies社)を用いて精製された。精製されたmRNAから、逆転写酵素(Loopstraら、Plant Mol.Biol.、27巻、277−291頁(1995年))合成の後、得られたcDNAクローンをメーカー指定のプロトコルにしたがってZAP Express cDNA Synthesisキット(Stratagene)を用いてLambda ZAPに挿入して、cDNA発現ライブラリが構築された。得られたcDNAは、5μlのライゲーションミックスからの試料DNAを1μl用いて、Gigapack II Packaging Extra(Stratagene)でパッケージングされた。ライブラリの大量切り出しは、XL1−Blue MRF'細胞およびXLOLR細胞(Stratagene)を、ExAssistヘルパーファージ(Stratagene社)と併用して行われた。切り出されたファージミドは、NZYブロス(Gibco BRL社、メリーランド州、Gaithersburg)で希釈され、5−ブロモ−4−クロロ−3−インドイル−β−ガラクトシド(X−gal)とイソプロピルチオ−β−ガラクトシド(IPTG)とを含むLB−カナマイシン寒天プレートにプレーティングされた。
【0131】
コロニーを採取し、標準的なミニプレップ手法によりDNAを調製した。正しい導入断片を含むコロニーをカナマイシン含有NZYブロス中で培養し、cDNAをアルカリ法とポリエチレングリコール(PEG)沈殿法とにより精製した。1%アガロースゲルを用い、シーケンシングテンプレートへの染色体の混入の有無をスクリーニングした。Big Dye Chemistry(Applied Biosystems社製)を用い、メーカー指定のプロトコルにしたがってダイターミネーター化学反応を準備した。
【0132】
陽性クローンのDNA配列は、3700 Capillary Machine(Applied Biosystems社)またはPrism377シーケンサー(Perkin Elmer/Applied Biosystems部門)を用いて得た。cDNAクローンは、まず5’末端側から、そして場合によっては3'末端側からも配列決定した。一部のクローンについては、サブクローニングされた断片を用いて内部配列を得た。サブクローニングは、制限マッピングおよびpBluescript II SK+ベクターへのサブクローニングという標準的な手順にしたがって行った。
【0133】
選択された遺伝子と相同な植物ESTを、ラジアータマツ(Pinus radiata)またはユーカリ・グランディス(Eucalyptus grandis)のcDNA発現ライブラリのいずれかから同定し、表2に示した。
【0134】
【表2】
【0135】
ポリヌクレオチドまたはポリペプチド配列は、他のポリヌクレオチドおよび/またはポリペプチド配列とアライメントをとることができ、同一の配列を共有する割合は公的に利用可能なコンピューターアルゴリズムを用いて決定することができる。BLASTN、BLASTPおよびBLASTXを含むアルゴリズムのファミリー、BLASTが、Altschulらの文献「Gapped BLAST and PSI−BLAST:a new generation of protein database search programs」、Nucleic Acids Res.、25巻、3389−3402頁、1997年に記載されている。本発明のポリヌクレオチド変異体の決定には、上記アルゴリズムの添付書類に記載されたデフォルトパラメータに設定されているBLASTNアルゴリズムバージョン2.0.11(2000年1月20日)を用いることが好ましい。
【0136】
SWISSPROT−TREMBLE配列(2002年7月9日版)を使用し、2002年11月15日にBLASTXサーチを実施した。 BLASTXを用い、E値とポリペプチド配列の一致度とに寄与するアライメントと同一性とを決定するには、下記のランニングパラメータが好ましい。blastall −p blastx −d swissprotdb −e10 −G0 −E0 −FF −v30 −b 30 −iqueryseq −o results。また、パラメーターは以下のとおりである。−p プログラム名[文字列];−d データベース[文字列];−e 期待値(E)[実数];−G ギャップを空けるためのコスト(ゼロはデフォルト挙動)[整数];−E ギャップを拡張するためのコスト(ゼロはデフォルト挙動)[整数];−F フィルタークェリー配列[文字列];−v 1行説明の数(v)[整数];−b 表示するアライメントの数(b)[整数];−I クェリーファイル[File In];−o BLASTレポートアウトプットファイル[File Out]オプション。
【0137】
プロモーターは、Genome Walkerキット(Clontech社製、Palo Alto、CA)を用いてクローニングした。これはPCR法にもとづく方法であり、4本のプライマーを作成することが必要で、その中の2本は遺伝子特異的でなければならない。この遺伝子特異的プライマーは通常、その遺伝子の5’UTR内に設計される。断片が増幅され、GUSレポーター遺伝子の手前のT終端ベクター内にクローニングされる。
【実施例2】
【0138】
プロモーターの組織特異性の決定方法
実施例1で概説した手順によりプロモーターの同定およびクローニングが終了した後、プロモーターがレポーター遺伝子と操作可能に連結され、プロモーターが活性を示す組織の種類が決定される。このために、エレクトロポレーション法により、本発明のプロモーターを含むコンストラクトがアグロバクテリウム・ツメファシエンスに形質転換される。簡単に述べると、40μlの希釈AgL−1コンピテント細胞を氷上に置き、当該プロモーター配列を含むpART27ベクター約10ngと接触させた。エレクトロポレーション法は、下記のパラメーターの設定下で行った。
抵抗=129Ω
印加電圧=1.44kV
電界強度=14.4kV/cm
パルス持続時間=5.0ms
【0139】
エレクトロポレーション終了後、400μlのYEP液体培地を添加し、細胞を1時間、室温にて回復させる。次に、細胞を6000rpm、3分間遠心し、約50μlのYEPに再懸濁する。細胞試料をYEP Kan50/Rif50プレート上にスプレッドし、パラフィルムで密閉し、29°Cで2日間インキュベーションしコロニーを成長させる。
【0140】
次に、関心のあるコンストラクトを含むアグロバクテリウムを用い、フローラルディップ浸潤法(floral dip infiltration)により野生型シロイヌナズナ「コロンビア−0」株の植物個体を形質転換する。簡潔には、アグロバクテリウム培養を20°Cにて約8600rcfで10分間遠心し、光学濃度が約0.7〜0.8となるように再懸濁する。アグロバクテリウムを含む浸潤液に植物個体を5秒間浸漬する。植物個体から余分な液を水切りし、大気条件で生育光下に置く。24時間後、植物個体を噴霧し、種子を産生できる状態に維持する。T1種子の表面を市販の5%漂白液で滅菌し、カナマイシン(50mg/l)とチメンチン(250mg/l)とを含むMS培地上にスプレッドし、推定される形質転換体を選別する。
【0141】
アグロバクテリウムを介した葉組織形質転換により、タバコ(N.benthamiana)植物個体を、関心のあるコンストラクトを用いて形質転換する(Burowら、Plant Mol.Biol.Rep.、8巻、124−139頁、1990年)。
【0142】
形質転換に成功した植物個体は、操作可能に連結されたレポーター遺伝子が発現しているか否か分析される。葉、茎、根および花の各部分を染色液(50mM NaPO4、pH7.2、0.5% Triton X−100、1mM X−グルクロニド、シクロヘキシミド塩(Ducheffa))に浸漬される。5分間の真空吸引を2回行うことによって組織に染色液を浸透させる。前記組織は37°Cで終夜振盪され、呈色させる。前記組織は、組織の呈色を確認するために途中で3回または4回確認され、試料が早期に呈色する場合には、組織の一片が70%エタノール中で脱染される。次に、光学顕微鏡を用いてGUSの発現について調べるためにこの組織の写真が撮影される。
【実施例3】
【0143】
導管要素(TE)誘導性培地(FKH)および非誘導性培地(FK)中における百日草(Zinnia elegans)葉肉細胞の単離および培養
百日草の実生の初生葉および第二葉のペアが8パネットから採集された。葉は0.175%次亜塩素酸ナトリウム溶液500ml中で10秒間滅菌された。次に、葉は500mlの滅菌水中で2回リンスされた。1回当たり20〜30枚の葉を用いて、乳鉢および乳棒で25〜30mlのFK培地中で粉砕された。細胞が40μmのナイロンメッシュで濾過され、このやり方で合計90mlの葉肉細胞が得られた。前記細胞は20°Cで200×g、2分間遠心され、生成したペレットは、再度等量のFK培地を用いて洗浄された。2回目の洗浄が終了した後、ペレットは2等分され、一方の半分は45mlのFK培地で洗浄され、他方の半分は45mlのFKH培地で洗浄された。各ペレットは60mlのFK培地およびFKH培地にそれぞれ再懸濁された。これらは暗所にて、120rpmに設定されたロータリーシェーカー上の2枚の6ウェルプレートで培養された。
【実施例4】
【0144】
FK培地およびFKH培地中で終夜ないし3日間培養した葉または葉肉細胞からの百日草(Zinnia elegans)プロトプラストの単離
滅菌した百日草の初生葉(6〜8枚)は1mmの細片状に切断され、15mlの細胞壁消化酵素混合液(1%セルラーゼ オノヅカR−10および0.2%ペクトリアーゼ Y23を含むプロトプラスト単離緩衝液)中に添加された。FK培地(40ml)またはFKH培地(40ml)中で培養された葉肉細胞は、20°Cにて200×g、2分間遠心してペレット化された。各ペレットは、200mgのセルラーゼ オノヅカR−10と40mgのペクトリアーゼY23とを含む滅菌プロトプラスト単離緩衝液20ml中に再懸濁された。前記プロトプラストは、前記細胞懸濁液を約70rpm23°Cに設定されたロータリーシェーカー上のCellStar培養プレートで2〜4時間インキュベーションすることによって、単離された。前記プロトプラストは、各内容物を200×g、2分間遠心してペレット化された。各ぺレットは20mlの24%ショ糖溶液中に再懸濁された。
【0145】
Fukuda−Komamine培地保存液
【0146】
【表3】
【0147】
【表4】
【実施例5】
【0148】
百日草(Zinnia elegans)プロトプラストのトランスフェクション
24%ショ糖溶液中に懸濁させた百日草プロトプラスト上に1mlのW5溶液(150mM NaCl、125mM CaCl2・2H2O、5mM KC1、5mM ショ糖、pH5.6−6)を重層し、20°Cにて70×g、10分間の遠心を行い、ブレーキをかけずに停止させた。浮遊するプロトプラストを回収し、W5溶液10ml中に再懸濁させ、20°Cにて70×g、10分間の遠心を行い、ペレット化した。プロトプラストのペレットはプロトプラスト濃度が約5×106個/mlとなるようにMaMg培地(450mMマンニトール、15mM MgCl2、0.1% MES、pH5.6)中に再懸濁され、一定量ずつ別々の15ml試験管に分注された(300μl:プロトプラスト1.5×106個)。各コンストラクトにつき、上記プロトプラスト懸濁液に5μgのDNAおよび50μgのサケ精巣DNAが添加、混合され、20°Cで5分間インキューベートされた。300μlの40%PEG溶液(40%PEG3340、100mM Ca(NO3)2・4H2O、0.45Mマンニトール、pH9.0)が各分注プロトプラストに添加され、20°Cで20分間インキュベーションされた。5mlのK3/0.4Mショ糖(溶液1L当たり:4.3gムラシゲ・スクーグ植物培地用混合塩、100mgミオイノシトール、250mgキシロース、10mgチアミン塩酸塩、1mgニコチン酸、1mgピリドキシン塩酸塩、1mg NAA、0.2mgカイネチン、137gショ糖、pH5.6(KOHで調整))が、葉由来のトランスフェクションされたプロトプラスト、またはFK培地中で培養した葉肉細胞由来のトランスフェクションされたプロトプラストの各分注に添加、混合された。同様に、5mlのK3/0.4Mショ糖+0.lppm NAA+0.2ppm BAが、FKH培地で培養された葉肉細胞由来のトランスフェクションされた細胞の各分注に添加、混合された。これらトランスフェクションされたプロトプラスト懸濁液は、暗所で23°Cで終夜インキュベーションされた。
【実施例6】
【0149】
トランスフェクション百日草(Zinnia elegans)プロトプラストの回収およびレポーター遺伝子解析
トランスフェクションされた百日草プロトプラストを個々に回収するため、実施例5で説明したとおり調製された懸濁液にそれぞれ9.5mlのW5溶液が添加され、各試験管の内容物が混合され、20°Cにて70×g、10分間遠心された。上清の大部分がデカンテーションにより捨てられ、プロトプラストの体積が900μlとされた。900μlの各試料のうち、300μlのプロトプラストが5mlポリスチレン丸底試験管に分注され、500μlのW5培地中に再懸濁され、蛍光レポータ遺伝子発現および細胞生存度の分析用に取り置かれた。残り600μlのプロトプラストおよび溶液は個々のマイクロチューブに移されて、20°Cにて420×g、2分間の遠心によってペレット化された。このプロトプラストペレットは、Jefferson,R.A.、1987年、Plant Mol.Biol.Rep.5,387頁に記載される方法にしたがって、GUSレポーター遺伝子発現について検定された。GUS(MUG)の検定は、Wallac社(テュルク、フィンランド)のVictor21420マルチラベルカウンターを用いて行われた。ウンベリフェロンは355nm励起光フィルターおよび460nm発光フィルターを用いて1秒間検出された。
【実施例7】
【0150】
植物個体内における発現データ
土壌移植3週間後に、形質転換シロイヌナズナおよびタバコ組織はGUSレポーター遺伝子発現について解析される。GUS発現を検定するために、実施例2で説明したとおり、葉、根および花の材料がGUS溶液に浸漬される。組織に染色液を浸透させるために5分間の真空吸引を2回行い、前記組織は発色のため37°Cで終夜インキュベーションされる。終夜のインキュベーションの後、前記組織試料は70%エタノール中で脱染され、光学顕微鏡下でGUS発現について検査される。表5は、形質転換されたシロイヌナズナおよびタバコのGUS発現率(%)を含む。
【0151】
土壌移植3ヶ月後のタバコのT1植物に由来する組織がパラプラストに包埋され、ミクロトームで切片が作成され、光学顕微鏡下でGUS発現が検査される。表5に示したGUSの局在と組織鏡検結果(microtome results)は、本願明細書に開示された単離ヌクレオチド配列が、維管束の形成層、木部および/または篩部組織に優先的なレポーター遺伝子発現を可能にすることを証明する。
【0152】
【表5】
【0153】
タバコ組織の試料は、固定液(100%エタノール、氷酢酸、37%ホルムアルデヒド、ミリQ水)を含むバイアルに入れられ、15分間の真空吸引が2回行われる。前記試料は、確実に組織に浸透させるために固定液中で2時間インキュベーションされる。組織浸透の後、再び15分間の真空吸引を行い、前記組織は4°Cで終夜放置した。
【0154】
組織試料は、終夜のインキュベーションの後、エタノール系列のインキュベーションにより脱水されるが、ここでのすべてのインキュベーションは室温で行われる。前記終夜固定された固定液は組織試料を入れたバイアルから除去され、50%エタノールに置換される。30分後、前記50%エタノールはデカンテーションにより除去され、前記組織は50%エタノールの新しい分注液でインキュベーションされる。50%エタノールで30分間経過後、前記溶液は除去されて60%エタノールに置換される。30分間のインキュベーション後、60%エタノールは70%エタノールに置換される。30分後にこの70%エタノールはデカンテーションにより除去され、85%エタノールで置換される。30分後、前記85%エタノールは除去され、前記試料は95%エタノール中で終夜インキュベーションされる。
【0155】
組織の脱水の後、該組織は室温でキシレン溶液系列中でインキュベーションされる。前記終夜処理された95%エタノールが除去され、上記組織は100%エタノール中で30分間インキュベーションされる。30分後、100%エタノールが除去され、前記組織は25%キシレン:75%エタノール中に懸濁される。30分間のインキュベーション後、前記溶液は50%キシレン:50%エタノールに置換される。次に、前記溶液はデカンテーションにより除去され、75%キシレン:25%エタノールに置換される。30分後、前記組織は100%キシレンで60分のインキュベーションを3回される。次に前記組織は、キシレンと20個のパラプラストチップとを入れたバイアル中で、終夜インキュベーションされる。
【0156】
パラフィンを浸潤させるため、前記バイアルは、前記パラプラストチップが溶解するまで42°Cのハイブリダイゼーションオーブンで保温される。8時間の間に、合計60〜80個のパラフィンチップがバイアルに添加され、溶解される。前記試料は62°Cのハイブリダイゼーションオーブンで終夜保温される。次の2日間にわたり、パラプラストは12時間間隔で4回交換される。前記組織を包埋するために、流動パラフィンがカセット内に注入され、組織を適当な向きに配置される。前記カセットは4°Cで終夜静置され、パラフィンが硬化される。
【0157】
表5に示すとおり、本願明細書に開示される単離ヌクレオチド配列は、維管束の形成層、木部、および/または篩部組織で優先的なGUSレポーター遺伝子発現を可能にする。特に注目すべきは、配列番号21と配列番号13とが、配列番号13の5’末端に40bpの断片(AAATATAACATAATCTAACTATTGATGTACATTATTCGCC)の挿入を含むことを除いて、実質的な配列相同性を有する点である。この40bp断片(配列番号85)は、配列番号13では不完全な反復配列として存在するが、配列番号21には存在しない。興味深いことに、前記40bp断片が存在すると(例えば、配列番号13)GUS発現が可能になり、前記断片が存在しないと(例えば、配列番号21)GUS発現は起こらない。このデータは、本願明細書に開示される40bp断片(配列番号85)が維管束優先発現的な遺伝子発現に必要十分な要素を構成する可能性があることを示唆するため、意義深い。
【実施例8】
【0158】
維管束特異的プロモーターの利用方法
適当な組織特異的かつ発生段階特異的な発現パターンを有するプロモーターがいったん見つかると、このプロモーターは、トランスジェニック植物で所望の形質を調節するために利用可能である。1つの実施態様では、木部特異的プロモーターが植物のリグニン組成と含量とを調節するために利用される。本実施例では、本発明の木部特異的プロモーターがシンナミルアルコールデヒドロゲナーゼ(CAD)のコーディング領域の一部に対応するRNA干渉(RNAi)分子をエンコードする遺伝子と操作可能に連結される。CADは、リグニンモノマー合成の最終段階を触媒し、他のプロモーターを用いてアンチセンス配列を介した植物個体内のリグニンの下向き調節に成功した標的である。Yahiaouiら、Phytochemistry、49巻、295−306頁(1998年)と、これに引用されている各文献を参照せよ。CADの一部に対応するRNAi分子が発現すると、酵素活性が低下し、これに対応してトランスジェニック植物個体内のリグニンのシンナミルアルデヒドの割合が増大する。トランスジェニック植物個体におけるCAD活性は、Baucherら(Plant Physiol.、112巻、1479−90頁(1996年))によって改良された、Wyrambikら(Eur.J.Biochem.、59巻、9−15頁(1975年))の方法によって分析された。リグニン含量および組成は、Baucher(1996年)の方法によって測定された。CADの下向き調節は、木材の色の変化と、抽出がより容易なシンナミルアルデヒドの割合の増大につながり、このことはパルプ産業用の植物にとって商業上重要である。例えば、CAD活性を低減させたポプラは赤みを帯び、リグニンがより抽出しやすく、製紙産業の用途でのポプラ材の特性が改善される(Baucherら(1996年))を参照せよ。
【0159】
別の実施例では、ユーカリおよびマツが形質転換される。ユーカリおよびマツは、リグニン含量が多いため、リグニン含量および組成に対する遺伝子調節の効果を研究するための有用なモデル系を提供する。さらに、ユーカリおよびマツは、リグニン含量または組成のモジュレーションが製紙または木材産業にとって有用な特性を与えるような植物の代表例である。
【0160】
これらのトランスジェニック植物を作出するために、CADのcDNAの断片が、本発明の木部特異的プロモーターと、ノパリンシンターゼ3’ターミネーターとに適当な配向で操作可能に連結される。前記CADのcDNAは、例えば、遺伝子配列をそのプロモーターに対して反転させることにより調製できる。植物細胞内における前記CADのcDNAの転写は、CAD遺伝子に対して「アンチセンス」な細胞内RNA転写産物を生成する。コンストラクト全体が、制限断片としてバイナリーベクターpBI101.1(Clontech、カリフォルニア州、パロアルト)に挿入される。ベクターは、ユーカリまたはマツの形質転換を行うために、それぞれアグロバクテリウム・ツメファシエンス(A.tumefaciens)の菌株LBA4404またはC58pMP90にエレクトロポレーション法により導入される。一般的にはNoら、Plant Science、 160巻、77−86頁(2000年)を参照せよ。カナマイシン耐性形質転換体は、CAD活性について測定され、導入遺伝子のコピー数がサザン解析により決定され、CADのmRNA発現がYahiaoui(1998年)に説明されるとおりに決定される。好適な形質転換体は発根させて、温室に移植される。
【実施例9】
【0161】
インシリコ発現
コンピュータによる遺伝子発現は、コンセンシ(consensi)ESTライブラリのメンバー資格を決定するために利用できる。各ライブラリについて、いずれかの組織クラスのESTの数をそのクラスのESTの総数で割った商に1000を乗じ、コンセンサスが決定される。これらの値は、あるライブラリからの配列決定の程度に影響されない規格化値である。形成層、木部、篩部、栄養芽、根、生殖組織、葉、茎および果実のライブラリを含む複数のライブラリが、コンセンサス値についてサンプル調査された。下記の表6および表7に示すとおり、多数の本発明のプロモーター配列が維管束優先的発現を示し、樹木関連の発生過程に関与する可能性がある。
【0162】
【表6】
【0163】
【表7】
【0164】
【表8】
【図面の簡単な説明】
【0165】
【図1】3種類のプロモーターが培養3日目に同様の発現プロファイルを示すことを証明する百日草TE検定結果を示すグラフ。
【図2A】2種類のプロモーターが培養3日目に同様の発現プロファイルを示すことを証明する百日草TE検定結果を示すグラフ。
【図2B】2種類のプロモーターが培養3日目に同様の発現プロファイルを示すことを証明する百日草TE検定結果を示すグラフ。
【図3】2種類のプロモーターが培養3日目に同様の発現プロファイルを示すことを証明する百日草TE検定結果を示すグラフ。
【特許請求の範囲】
【請求項1】
維管束優先的なポリヌクレオチド転写を可能にするポリヌクレオチドを含む、単離核酸分子。
【請求項2】
前記ポリヌクレオチドは配列番号1−85のいずれかと、その機能変異体とから選択される、請求項1に記載の単離核酸分子。
【請求項3】
前記機能変異体は、配列番号1−85の配列のいずれかとの同一性が、99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、89%、88%、87%、86%、85%、84%、83%、82%、81%、80%、79%、78%、77%、76%、75%、74%、73%、72%、71%、70%、69%、68%、67%、66%、65%、64%、63%、62%、61%または60%以上である、請求項2に記載の単離核酸分子。
【請求項4】
(a)請求項2に記載の配列のいずれかに対して相補的な配列と、
(b)請求項2に記載の配列のいずれかに対する逆相補体の配列と、
(c)前記(a)または(b)のポリヌクレオチドのいずれかと雑種形成する、少なくとも20個の連続する塩基を含む配列とから選択される配列を有する単離ポリヌクレオチド。
【請求項5】
前記ポリヌクレオチドは植物細胞内で木部優先的な遺伝子発現を可能にする、請求項1に記載の単離核酸分子。
【請求項6】
前記ポリヌクレオチドは操作可能に連結した遺伝子の植物細胞内での発現を上向き調節または下向き調節することができる、請求項1に記載の単離核酸分子。
【請求項7】
(a)配列番号1−85のいずれかの配列を有する少なくとも1個のポリヌクレオチド配列と、(b)所望の遺伝子とを含み、前記ポリヌクレオチドと前記所望の遺伝子とは操作可能に連結されている、植物細胞。
【請求項8】
前記所望の遺伝子はポリペプチドまたはタンパクをエンコードする、請求項7に記載の植物細胞。
【請求項9】
前記タンパクは細胞壁の生合成に関与する酵素である、請求項7に記載の植物細胞。
【請求項10】
前記タンパクはリグニンの生合成に関与する酵素である、請求項8に記載の植物細胞。
【請求項11】
前記所望の遺伝子はRNA転写産物を生成する、請求項7に記載の植物細胞。
【請求項12】
前記RNA転写産物は植物細胞に内在する遺伝子のアンチセンス配列を有する、請求項11に記載の植物細胞。
【請求項13】
前記RNA転写産物は植物細胞で正常に発現される遺伝子のRNA干渉を誘導する、請求項12に記載の植物細胞。
【請求項14】
請求項7に記載の植物細胞を含む植物。
【請求項15】
前記植物は被子植物および裸子植物から選択される、請求項14に記載の植物。
【請求項16】
請求項14に記載の植物を栽培することを含む、植物のリグニン含量を調節する方法。
【請求項17】
請求項14に記載の植物を栽培することを含む、植物の細胞壁の形成を調節する方法。
【請求項18】
配列番号1−85のいずれかと、その機能変異体とから選択されるポリヌクレオチド配列を含む、トランスジェニック植物。
【請求項19】
(a)木本植物の植物細胞に、(i)配列番号1−85のいずれかまたはその機能変異体の配列を有するプロモーターと、(ii)所望の核酸とを含み、前記プロモーターは前記所望の核酸の発現を調節するDNAコンストラクトを導入すること、(b)形質転換された前記植物細胞を植物の生育を促進する条件下で培養すること、および、(c)前記植物から木材を得ることを含む、木材を得る方法。
【請求項20】
前記木本植物は、ユーカリ属またはマツ属の植物種から選択される、請求項19に記載の方法。
【請求項1】
維管束優先的なポリヌクレオチド転写を可能にするポリヌクレオチドを含む、単離核酸分子。
【請求項2】
前記ポリヌクレオチドは配列番号1−85のいずれかと、その機能変異体とから選択される、請求項1に記載の単離核酸分子。
【請求項3】
前記機能変異体は、配列番号1−85の配列のいずれかとの同一性が、99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、89%、88%、87%、86%、85%、84%、83%、82%、81%、80%、79%、78%、77%、76%、75%、74%、73%、72%、71%、70%、69%、68%、67%、66%、65%、64%、63%、62%、61%または60%以上である、請求項2に記載の単離核酸分子。
【請求項4】
(a)請求項2に記載の配列のいずれかに対して相補的な配列と、
(b)請求項2に記載の配列のいずれかに対する逆相補体の配列と、
(c)前記(a)または(b)のポリヌクレオチドのいずれかと雑種形成する、少なくとも20個の連続する塩基を含む配列とから選択される配列を有する単離ポリヌクレオチド。
【請求項5】
前記ポリヌクレオチドは植物細胞内で木部優先的な遺伝子発現を可能にする、請求項1に記載の単離核酸分子。
【請求項6】
前記ポリヌクレオチドは操作可能に連結した遺伝子の植物細胞内での発現を上向き調節または下向き調節することができる、請求項1に記載の単離核酸分子。
【請求項7】
(a)配列番号1−85のいずれかの配列を有する少なくとも1個のポリヌクレオチド配列と、(b)所望の遺伝子とを含み、前記ポリヌクレオチドと前記所望の遺伝子とは操作可能に連結されている、植物細胞。
【請求項8】
前記所望の遺伝子はポリペプチドまたはタンパクをエンコードする、請求項7に記載の植物細胞。
【請求項9】
前記タンパクは細胞壁の生合成に関与する酵素である、請求項7に記載の植物細胞。
【請求項10】
前記タンパクはリグニンの生合成に関与する酵素である、請求項8に記載の植物細胞。
【請求項11】
前記所望の遺伝子はRNA転写産物を生成する、請求項7に記載の植物細胞。
【請求項12】
前記RNA転写産物は植物細胞に内在する遺伝子のアンチセンス配列を有する、請求項11に記載の植物細胞。
【請求項13】
前記RNA転写産物は植物細胞で正常に発現される遺伝子のRNA干渉を誘導する、請求項12に記載の植物細胞。
【請求項14】
請求項7に記載の植物細胞を含む植物。
【請求項15】
前記植物は被子植物および裸子植物から選択される、請求項14に記載の植物。
【請求項16】
請求項14に記載の植物を栽培することを含む、植物のリグニン含量を調節する方法。
【請求項17】
請求項14に記載の植物を栽培することを含む、植物の細胞壁の形成を調節する方法。
【請求項18】
配列番号1−85のいずれかと、その機能変異体とから選択されるポリヌクレオチド配列を含む、トランスジェニック植物。
【請求項19】
(a)木本植物の植物細胞に、(i)配列番号1−85のいずれかまたはその機能変異体の配列を有するプロモーターと、(ii)所望の核酸とを含み、前記プロモーターは前記所望の核酸の発現を調節するDNAコンストラクトを導入すること、(b)形質転換された前記植物細胞を植物の生育を促進する条件下で培養すること、および、(c)前記植物から木材を得ることを含む、木材を得る方法。
【請求項20】
前記木本植物は、ユーカリ属またはマツ属の植物種から選択される、請求項19に記載の方法。
【図1】

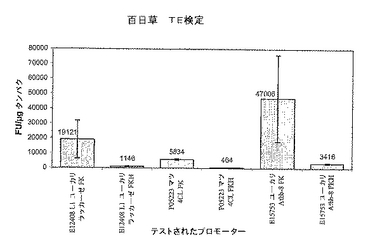
【図2A】
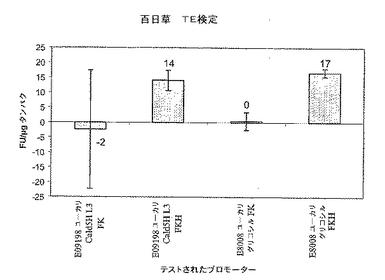
【図2B】
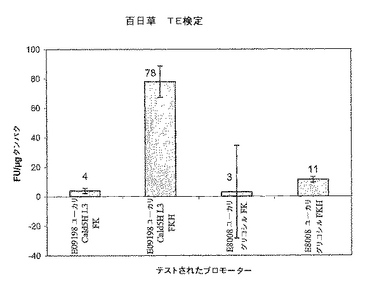
【図3】
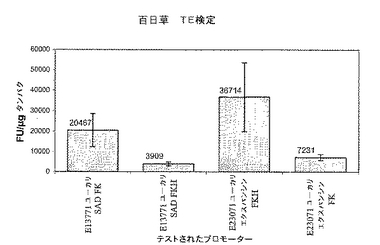
【図2A】
【図2B】
【図3】
【公表番号】特表2006−515509(P2006−515509A)
【公表日】平成18年6月1日(2006.6.1)
【国際特許分類】
【出願番号】特願2004−555614(P2004−555614)
【出願日】平成15年11月21日(2003.11.21)
【国際出願番号】PCT/US2003/037412
【国際公開番号】WO2004/048595
【国際公開日】平成16年6月10日(2004.6.10)
【出願人】(505186603)アーバーゲン エルエルシー (1)
【Fターム(参考)】
【公表日】平成18年6月1日(2006.6.1)
【国際特許分類】
【出願日】平成15年11月21日(2003.11.21)
【国際出願番号】PCT/US2003/037412
【国際公開番号】WO2004/048595
【国際公開日】平成16年6月10日(2004.6.10)
【出願人】(505186603)アーバーゲン エルエルシー (1)
【Fターム(参考)】
[ Back to top ]