
緑のコーヒー粒中のコーヒーフレーバー前駆体レベルの調節
本発明は、システインプロテイナーゼ;システインプロテイナーゼインヒビター;及びアスパラギン酸エンドプロテイナーゼをコードする単離されたポリヌクレオチドに関する。本発明はまた、形質転換された宿主細胞(特に植物細胞)に関し、この細胞中、これらのポリヌクレオチドの過剰発現又は過少発現は、緑のコーヒー粒中のコーヒーフレーバー前駆体レベル、特に、アミノ基含有分子(例えば、アミノ酸、ペプチド及びタンパク質)のレベルの変更を生じる。
【発明の詳細な説明】
【技術分野】
【0001】
コーヒーは、フレーバー分子の高度に複雑な混合物を含む。インスタント及び新鮮な粉砕されたコーヒー飲料の組成についての広範な研究は、今日までに850を超える化合物を同定しており、これらのうち多くがフレーバー活性分子である(Flament,I(2002)、コーヒーフレーバー化学(Coffee Flavor Chemistry)、John Wiley and Sons、UK)。しかし、1杯のコーヒー中に見出される最終コーヒーフレーバー分子は、原材料(植物種Coffea arabica又はCoffea canephora(robusta)の緑の粒(緑の豆))中にはほとんど存在しない。実際、コーヒーフレーバー化合物の大多数は、熟した赤いコーヒーチェリーの収穫から最終的な焙煎され粉砕されたコーヒー製品又はその抽出物(例えば、可溶性コーヒー製品)までに生じる複数の加工ステップの1つ又は複数の間に生成される。
【背景技術】
【0002】
コーヒーの生産における種々のステップが、Smith,A.W.、コーヒー;第1巻:化学(Coffee;Volume 1:Chemistry)1−41頁、Clark,R.J.及びMacrea,R.編、Elsevier Applied Science London and New York、1985;Clarke,R.J.、コーヒー:植物学、生化学並びに豆及び飲料の生産(Coffee:Botany,Biochemistry,and Production of Beans and Beverage)、230−250頁及び375−393頁;並びにClifford,M.N.及びWillson,K.C.編、Croom Helm Ltd、London中に記載されている。簡潔に述べると、このプロセスは、成熟して熟した赤いチェリーの収集で開始される。次いで、最も外側の層(すなわち果皮)が、乾式プロセス又は湿式プロセスのいずれかを使用して除去できる。乾式プロセスは最も単純であり、以下を含む:1)チェリーの等級分け(classification)及び洗浄;2)等級付け(grading)後のチェリーの乾燥(空気乾燥又は機械的乾燥のいずれか)、及び3)乾燥した果皮を除去するための乾燥したチェリーの脱穀(dehusking)。湿式プロセスは僅かにより複雑であり、より高い品質の緑の豆の生産を一般に導く。湿式プロセスは、C.arabicaチェリーとより頻繁に関連する。湿式プロセスは以下からなる:1)チェリーの等級分け、2)チェリーの脱肉(パルピング)(このステップは、収穫直後に実施され、一般に、成熟チェリーの「パルプ」すなわち果皮の機械的除去を含む)、3)脱肉後にチェリーの粒に付着したままの粘液が、バッチプロセスを使用して粒+付着した粘液をタンク中で水と共にインキュベートさせることによって除去される、「発酵」。この「発酵」プロセスは80時間まで継続されるが、発酵、4)乾燥(このステップは、発酵したコーヒー粒の空気乾燥又は機械的な熱気乾燥のいずれかを含む)及び5)「ハリング(hulling)」(このステップは、乾燥したコーヒー粒(乾燥したパーチメントコーヒー)の「パーチ(parch)」の機械的除去を含み、シルバースキン(銀皮)もまたしばしばこの段階で除去される)の間に増殖する微生物の種々の酵素活性及び代謝作用に起因して、しばしば24時間が、受容可能な発酵を可能にしかつ約6.8〜6.9から4.2〜4.6までpHを低下させるのに一般に充分である。湿式加工又は乾式加工の後に、得られた緑のコーヒー粒はしばしば分類され、ほとんどの分類手順は、粒のサイズ及び/又は形状に基づく。
【0003】
コーヒー加工における次のステップは、乾式加工したコーヒー又は湿式加工したコーヒーそれぞれの、脱穀(dehusking)又は脱穀(dehulling)後の緑の粒の焙煎である。これは、豆における有意な化学変化を誘導する時間依存的なプロセスである。焙煎の第1段階は、供給された熱が粒中に残留する水を追い出す場合に生じる。水の大部分がなくなると、温度が190℃〜200℃まで上昇して、適切な焙煎が開始される。豆の発色によって通常モニタリングされる焙煎の程度は、最終飲料製品のフレーバー特徴を決定するのに主要な役割を果たす。従って、焙煎の時間及び温度は、所望のコーヒーフレーバープロフィールを達成するために厳密に制御される。焙煎後、コーヒーは、コーヒー飲料又はコーヒー抽出物(後者は、インスタントコーヒー製品を製造するために使用される)の生産の間の抽出を容易にするために粉砕される。再度、粉砕のタイプは、飲料の最終的なフレーバーに影響を与え得る。
【0004】
かなりの量の研究がコーヒー中のフレーバー分子の同定について実施されているが、加工ステップの各々の間にコーヒー粒内で生じる物理反応及び化学反応に関して実施された研究はかなり少ない。この後者の点は焙煎反応について特に明白であり、多数の粒の成分が、熱により誘導される反応の非常に複雑な連続を経る(Homma,S.2001、「コーヒー:新事情(Coffee:Recent Developments)」.R.J.Clarke及びO.G.Vitzthum編、Blackwell Science、London;Yeretzian,C.ら((2002)Eur.Food Res.Technol.214、92−104;Flament,I(2002)、コーヒーフレーバー化学(Coffee Flavor Chemistry)、John Wiley and Sons、UK;Reineccius,G.A.、「メイラード反応及びコーヒーフレーバー(The Maillard Reaction and Coffee Flavor)」Conference Proceedings of ASIC、16th Colloque、Kyoto、Japan 1995)。
【0005】
コーヒー加工の異なるステップの間に生じる反応のほとんどの詳細は比較的不明のままであるが、コーヒーの芳香に関連するフレーバーの多数を担う重要なフレーバー生成反応は、コーヒー焙煎の間の「メイラード」反応であると考えられている。活発なメイラード反応は、焙煎ステップの間に、粒の還元糖/多糖分解産物とアミノ基含有分子(特に、タンパク質、ペプチド及びアミノ酸)との間で生じる。
【0006】
メイラード反応は、コーヒー焙煎の間のコーヒーのフレーバー分子及び芳香分子の生成に重要な貢献を明らかに果しているので、緑の豆中の1次メイラード反応のレベルと焙煎後に発生したフレーバー/芳香の品質との間には関連性がある可能性がある。
【0007】
上記のように、メイラード反応における基質の重要な群は、アミノ酸、ペプチド及びタンパク質である。2次元電気泳動を使用して、arabica及びrobustaの緑のコーヒー豆中の主要な貯蔵タンパク質のレベル及び量に差異が存在することが示されている。しかし、これらの保存タンパク質の差異とフレーバーの品質との間の関連性は注目されていない(Rogersら、1999、Plant Physiol.Biochem.第37巻、261−272)。未熟なコーヒー豆と成熟したコーヒー豆(これらは異なるフレーバー品質を有する)との間には貯蔵タンパク質に小さい差異が存在することもまた、最近見出された(Montavon,P.ら、2003、J.Agric and Food Chemistry 第51巻、2328−2334)。種子の成熟の間には多数の変化が生じるので、この後者の研究は、成熟によって引き起こされる品質の改善と主要なコーヒーの貯蔵タンパク質の2次元ゲルパターンにおいて見られる差異との間に関連性が存在し得ることを示唆している。
【0008】
arabica及びrobustaの緑の豆から単離されたペプチドのプロフィールにおいて差異が存在することが最近示された(Ludwigら、2000、Eur.Food Res Technol.、第211巻、111−116)。これらの結果は、arabica及びrobustaのペプチド抽出物がその芳香前駆体プロフィールにおいて異なることを示しているが、この報告中に示されるデータは、抽出物中のどの成分がこれらの芳香プロフィールの差異を担うかを同定していない。これらの研究者はまた、緑のコーヒーの粗製抽出物中に少なくとも2つの異なるプロテイナーゼ活性を検出したが、彼らは、いずれの比活性をも芳香/フレーバーの品質と相関させなかった(Ludwigら、2000、Eur.Food Res Technol.、第211巻、111−116)。最後に、緑のコーヒー粒の焙煎の後期段階の間に使用される非常に高い温度は、このコーヒー粒中に存在するタンパク質の引き続く切断を引き起こすとも考えられている(Homma,S.2001、「コーヒー:新事情(Coffee:Recent Developments)」.R.J.Clarke及びO.G.Vitzthum編、Blackwell Science、London;Montavon,P.ら、2003、「焙煎の間の緑のコーヒーのタンパク質プロフィールの変化(Changes in green coffee protein profiles during roasting)」、J.Agric.Food Chem.51、2335−2343)。しかし、このタンパク質分解についての全体的スキームは非常に不十分にしか理解されていないが、これはとりわけ、焙煎の開始前の原材料中の主要なコーヒータンパク質の正確な状態におそらく依存する。本発明者らが知る限り、コーヒー中のペプチドプロフィールがコーヒーの芳香/フレーバーの生成に関与する可能性を扱っている他の有意な報告は存在しない。
【0009】
Theobroma cacao(カカオ豆)の発酵した種子の焙煎において、メイラード反応の芳香/フレーバーの発生には、種子のアミノ酸及びペプチドが関与するようである。他の種子と比較すると、T.cacaoの種子は、異常に高いレベルのアスパラギン酸プロテイナーゼ活性を有することが示されている(Biehl,B.、Voigt,J.、Voigt,G.、Heinrichs,H.、Senyuk,V.及びBytof,G.(1994)「生のカカオ豆における芳香前駆体であるオリゴペプチド及びアミノ酸のpH依存的な酵素的形成(pH dependent enzymatic formation of oligopeptides and amino acids,the aroma precursors in raw cocoa beans)」.The Proceedings of the 11th International Cocoa Research Conference、1993年7月18日〜24日、Yamoussoukro、Ivory Coast)。高レベルのココアフレーバー前駆体を有するカカオ豆を生産するために、天然の発酵ステップを実施することが必要である(未発酵の豆は、焙煎したときにほとんどフレーバーを発生させない)。この発酵ステップの間に、パルプ中の糖が発酵し、高レベルの酸(特に酢酸)を生成する(Carr,J.G.(1982)ココア−発酵食品−経済学的微生物学(Cocoa.In Fermented Foods.Economic Microbiology.)第7巻.275−292頁.(A.H.Rose編).Academic Press)。発酵が継続するにつれて種子中のpHは低下し、細胞構造が破壊される。低いpHは、豊富なカカオ種子のアスパラギン酸プロテイナーゼを起動及び/又は活性化させ、細胞性タンパク質の大量分解を生じる(Biehl,B.、Passern,D.及びSagemann,W.(1982)「カカオ豆子葉の細胞内構造に対する酢酸の影響(Effect of Acetic Acid on Subcellular Structures of Cocoa Bean Cotylydons)」.J.Sci.Food Agric.33、1101−1109;Biehl,B.、Brunner,E.、Passern,D.、Quesnel,V.C.及びAdomako,D.(1985)「カカオ豆の発酵における酸性化、タンパク質分解及びフレーバー潜在能力(Acidification,proteolysis and flavour potential in fermenting cocoa beans)」.J.Sci.Food Agric.36、583−598)。ペプチド及びアミノ酸は、ココアフレーバー前駆体であることが示されている(Rohan,T.(1964)「チョコレート芳香の前駆体:発酵したカカオ豆及び未発酵のカカオ豆の比較研究(The precursors of chocolate aroma:a comparative study of fermented and unfermented cocoa beans)」.J.Food Sci.、29、456−459;Voigt,J.及びBiehl,B.(1995)「ココア特異的芳香成分の前駆体は、タンパク質分解加工によってココア種子のビシリンクラス(7S)グロブリンから誘導される(Precursors of the cocoa specific aroma components are derived from the vicilin−class(7S)globulin of the cocoa seeds by proteolytic processing)」.Bot.Acta 108、283−289)。従って、T.cacao種子のアスパラギン酸プロテイナーゼは、種子のセリンカルボキシペプチダーゼと共に、発酵の間のココアフレーバー前駆体の生成に重要であることが提唱されている(Voigt,J.及びBiehl,B.(1995)「ココア特異的芳香成分の前駆体は、タンパク質分解加工によってココア種子のビシリンクラス(7S)グロブリンから誘導される(Precursors of the cocoa specific aroma components are derived from the vicilin−class(7S)globulin of the cocoa seeds by proteolytic processing)」.Bot.Acta 108、283−289;Voigt,J.、Heinrichs,H.、Voigt,G.及びBiehl,B.(1994)「ココア特異的芳香前駆体は、ココア種子のビシリン様グロブリンのタンパク質分解消化によって生成される(Cocoa−specific aroma precursors are generated by proteolytic digestion of the vicilin−like globulin of cocoa seeds)」.Food Chemistry、50、177−184)。豊富なココア種子のアスパラギン酸プロテイナーゼをコードする遺伝子が同定されており、発酵したカカオ豆中のココアフレーバー前駆体のアミノ酸及びペプチドのレベルを増大させることができる、カカオ種子中でこのタンパク質を過剰発現させる方法は、国際特許公開第02/04617号(この全内容は、本明細書中に参照として組み込まれる)中に最近記載された。しかし、国際特許公開第02/04617号の教示は、酸発酵ステップを受けないコーヒー粒とは異なり、特定の長さの酸発酵ステップを受けるカカオ種子に関するものである。
【0010】
重要な液胞システインプロテイナーゼ(CP)は、KDEL含有システインプロテイナーゼである。このタイプのプロテイナーゼは、いくつかの植物において特徴付けられている。今日まで、C末端KDEL配列を有するシステインプロテイナーゼをコードする3つの遺伝子がarabidopsisにおいて見出されている(Gietl,C.及びSchmid,M.2001、Naturwissenschaften 88、49−58)。1つは成熟中の胚珠中で発現され、1つは導管中で発現され、3番目は成熟中の長角果において発現される。しかし、このタンパク質についてのより詳細な研究は、他の植物において実施されてきた。例えば、スルフヒドリル−エンドプロテイナーゼ(SH−EP)と称されるCPが、Vigna mungo種子の子葉において特徴付けられている(Toyooka,K.、Okamoto,T.及びMinamikawa,T.(200)J.Cell Biol.148、453−463)。SH−EPは、V.mungoの発芽中の子葉において新たに発現され、タンパク質貯蔵液胞中に蓄積した貯蔵タンパク質の分解に関与すると提唱されている(Okamato,T及びMinamikawa,T.J.Plant Physiol.152、675−682)。SH−EPポリペプチドの重要な特徴は、小胞体(ER)からタンパク質貯蔵液胞までのこのタンパク質の輸送を指図する特異的COOH末端配列KDELをこれが保有することである(Toyookaら、2000)。このSH−EPタンパク質は、そのKDEL配列の存在を介して、以前に記載されていない小胞輸送システムにおけるKV(KDEL Vesicles)と称される特定の小胞の形成に実際に関与することもまた、最近提唱されている(Okamato,T.、Shimada,T.、Hara−Nishimura,I.、Nishimura,M.及びMinamikawa,T.(2003)Plant Physiology、132、1892−1900)。
【0011】
関連の提案が、発芽中のトウゴマ(Ricinus communis)の実の子葉において見出されたKDEL含有CPタンパク質についてなされている。この植物において、著者らは、内胚乳のプログラム細胞死において、発芽中のトウゴマの実の胚に栄養を供給し続けるKDELプロテイナーゼを示している(Gietl,C.及びSchmid,M.2001、Naturwissenschaften 88、49−58)。これらの著者らは、トウゴマの実において、このKDELプロテイナーゼが3日目の前に発芽中の種子のERにおいて作製されることを提唱している。種子の皮が脱げると、3日目付近で、KDEL含有CPは、リシノソーム(ricinosome)と称される特定の小胞中に詰め込まれる。後に、内胚乳が4日目〜5日目の間に軟化するにつれて、KDEL−CPは、そのアンカー配列(KDEL)が切断して除かれ、このプロテイナーゼは、細胞性タンパク質の全般的な分解においてそれが補助する場所である細胞質に移動する。
【発明の開示】
【発明が解決しようとする課題】
【0012】
本発明の目的は、コーヒー中のタンパク質/ペプチド/アミノ酸のフレーバー前駆体プールを改変することである。
【0013】
より具体的には、本発明の目的は、収穫後の処理及び焙煎加工の後に変更されたフレーバーが達成できるように、原材料(緑の粒)中のフレーバー前駆体のレベルを改変することである。理論によって束縛されないが、有意に異なるフレーバーを有するコーヒー間のペプチド及びタンパク質の分解のレベルにバリエーションが存在する場合、これらのバリエーションは、これらの異なる粒中の内因性プロテイナーゼ活性の差異に起因することがあると考えられる。この差異は、特定の種子プロテイナーゼ遺伝子についての発現レベルのバリエーションによって、mRNA発現のレベルで検出可能である場合がある。
【課題を解決するための手段】
【0014】
従って本発明は、コーヒー粒(種子)特異的プロテイナーゼをコードする遺伝子配列を同定すること、並びにarabica及びrobustaにおけるこれらの遺伝子の発現にバリエーションが実際に存在することを示すことを含む。
【0015】
より具体的には、本発明は、2つの主要なコーヒーシステインプロテイナーゼ(CcCP−1及びCcCP−4)、4つの主要なコーヒーシステインプロテイナーゼインヒビター(CcCPI−1、CcCPI−2、CcCPI−3及びCcCPI−4)並びに2つのコーヒーアスパラギン酸プロテイナーゼ(CcAP−1及びCcAP−2)を開示し、これらは全てコーヒーの種子中で発現される。本発明者らは、種子発達における特に後期のこれらのタンパク質の過剰発現又は後期種子発達の間のこれらのタンパク質の低下した発現のいずれかが、成熟豆のアミノ酸/ペプチド/タンパク質のプロフィールを如何に変更できるかをさらに示す。成熟豆のアミノ酸/ペプチド/タンパク質のプロフィールを変更するために開示された遺伝子配列及び遺伝子構築物の1つ又は複数を使用することによって、本発明者らは、成熟コーヒー豆のフレーバー前駆体プロフィールを変更するための新たな方法を開示する。
【0016】
第1の態様において、本発明は、システインプロテイナーゼ活性を有するポリペプチドをコードするヌクレオチド配列又はこのヌクレオチド配列の相補体を含む単離されたポリヌクレオチドを提供し、ここで、このポリペプチドのアミノ酸配列と配列番号2又は16から選択されるアミノ酸配列とは、ClustalWアラインメント法に基づいて、少なくとも70%、好ましくは少なくとも80%の配列同一性を有し、この相補体はこのヌクレオチド配列と同数のヌクレオチドを含み、この相補体とこのヌクレオチド配列とは100%相補的である。好ましくは、このポリペプチドのアミノ酸配列と配列番号2又は16のアミノ酸配列とは、ClustalWアラインメント法に基づいて、少なくとも85%、好ましくは少なくとも90%、任意選択で少なくとも95%の配列同一性を有する。好ましくは、このヌクレオチド配列は、配列番号1又は15のヌクレオチド配列を含む。好ましくは、このポリペプチドは、配列番号2又は16のアミノ酸配列を含む。
【0017】
第2の態様において、システインプロテイナーゼインヒビター活性を有するポリペプチドをコードするヌクレオチド配列又はこのヌクレオチド配列の相補体を含む単離されたポリヌクレオチドが提供され、このポリペプチドのアミノ酸配列と配列番号4、10、12及び14から選択されるアミノ酸配列とは、ClustalWアラインメント法に基づいて、少なくとも70%、好ましくは少なくとも80%の配列同一性を有し、この相補体はこのヌクレオチド配列と同数のヌクレオチドを含み、この相補体とこのヌクレオチド配列とは100%相補的である。好ましくは、このポリペプチドのアミノ酸配列と配列番号4、10、12及び14から選択されるアミノ酸配列とは、ClustalWアラインメント法に基づいて、少なくとも85%、好ましくは少なくとも90%、任意選択で少なくとも95%の配列同一性を有する。好ましくは、このヌクレオチド配列は、配列番号3、9、11又は13から、任意選択で配列番号9、11又は13から、さらに任意選択で配列番号9又は13から選択される;なおさらに任意選択で配列番号9であるヌクレオチド配列を含む。好ましくは、このポリペプチドは、配列番号4、10、12及び14から、任意選択で配列番号10、12及び14から選択される、さらに任意選択で配列番号10又は14から;なおさらに任意選択で配列番号10であるアミノ酸配列を含む。
【0018】
第3の態様において、アスパラギン酸エンドプロテイナーゼ活性を有するポリペプチドをコードするヌクレオチド配列又はこのヌクレオチド配列の相補体を含む単離されたポリヌクレオチドが提供され、このポリペプチドのアミノ酸配列と配列番号6又は8、好ましくは配列番号8から選択されるアミノ酸配列とは、ClustalWアラインメント法に基づいて、少なくとも75%、好ましくは少なくとも80%の配列同一性を有し、この相補体はこのヌクレオチド配列と同数のヌクレオチドを含み、この相補体とこのヌクレオチド配列とは100%相補的である。好ましくは、このポリペプチドのアミノ酸配列と配列番号6又は8、好ましくは配列番号8から選択されるアミノ酸配列とは、ClustalWアラインメント法に基づいて、少なくとも85%、好ましくは少なくとも90%、任意選択で少なくとも95%の配列同一性を有する。好ましくは、このヌクレオチド配列は、配列番号5又は7、好ましくは配列番号7のヌクレオチド配列を含む。好ましくは、このポリペプチドは、配列番号6又は8、好ましくは配列番号8のアミノ酸配列を含む。
【0019】
さらなる態様において、本発明の第1の態様〜第3の態様のいずれか1つに記載のポリヌクレオチドを含むベクターが提供される。
【0020】
さらなる態様において、調節配列に作動可能に連結された本発明の第1の態様〜第3の態様のいずれか1つに記載のポリヌクレオチドを含む、非ネイティブの組換えDNA構築物が提供される。この非ネイティブの構築物において、このポリヌクレオチドが非ネイティブであるか、調節配列が非ネイティブであるか、又は両方が非ネイティブであるかのいずれかであることが理解される。
【0021】
さらなる態様において、本発明の第1の態様〜第3の態様のいずれか1つに記載のポリヌクレオチドで細胞を形質転換するステップを含む、細胞を形質転換するための方法が提供される。
【0022】
さらなる態様において、上記の非ネイティブの組換えDNA構築物を含む細胞が提供され、この細胞は好ましくは、原核細胞、真核細胞又は植物細胞、好ましくはコーヒー細胞である。
【0023】
さらなる態様において、このような形質転換された細胞を含むトランスジェニック植物が提供される。
【0024】
本願において、コーヒーチェリーとの用語は以下のように規定される:コーヒーチェリー;果実全体;外果皮、皮;果皮、チェリーの多肉質の主要な外側層;及び粒、コーヒーの種子。これらの用語のより完全な説明については、Clarke,R.J.、コーヒー:植物学、生化学並びに豆及び飲料の生産(Coffee:Botany,Biochemistry,and Production of Beans and Beverage)、230頁、Clifford,M.N.及びWillson,K.C.編、Croom Helm Ltd、London(その内容は、その全体が参照として組み込まれる)を参照のこと。
【発明を実施するための最良の形態】
【0025】
本発明は、以下の詳細な説明及び本願の一部をなす添付の配列表から理解できる。
【0026】
表1は、添付の表において使用される通りの対応する配列同定子(配列番号)と共に、本明細書中に記載されるポリペプチドを本明細書中以下に列挙する。
【0027】
表1:
配列番号1(CcCP1:システインプロテイナーゼ、核酸及びその対応するアミノ酸)
配列番号2(CcCP1:システインプロテイナーゼ、アミノ酸)
配列番号3(CcCPI−1:システインプロテイナーゼインヒビター、核酸及びその対応するアミノ酸)
配列番号4(CcCPI−1:システインプロテイナーゼインヒビター、アミノ酸)
配列番号5(CcAP1:アスパラギン酸エンドプロテイナーゼ1、核酸及びその対応するアミノ酸)
配列番号6(CcAP1:アスパラギン酸エンドプロテイナーゼ1、アミノ酸)
配列番号7(CcAP2:アスパラギン酸プロテイナーゼ2、核酸及びその対応するアミノ酸)
配列番号8(CcAP2:アスパラギン酸プロテイナーゼ2、アミノ酸)
配列番号9(CcCPI−2:システインプロテイナーゼインヒビター、核酸及びその対応するアミノ酸)
配列番号10(CcCPI−2:システインプロテイナーゼインヒビター、アミノ酸)
配列番号11(CcCPI−3:システインプロテイナーゼインヒビター、核酸及びその対応するアミノ酸)
配列番号12(CcCPI−3:システインプロテイナーゼインヒビター、アミノ酸)
配列番号13(CcCPI−4:システインプロテイナーゼインヒビター、核酸及びその対応するアミノ酸)
配列番号14(CcCPI−4:システインプロテイナーゼインヒビター、アミノ酸)
配列番号15(CcCP−4:システインプロテイナーゼ、核酸及びその対応するアミノ酸)
配列番号16(CcCP−4:システインプロテイナーゼ、アミノ酸)
【0028】
この配列表は、ヌクレオチド配列特徴について1文字コードを使用し、アミノ酸について、IUPAC−IUBMB Standardsのために規定され、Nucleic Acids Research 13:3021−3030(1985)(これは、本明細書中に参照として組み込まれる)中に記載されるような3文字コードを使用する。
【0029】
本明細書中で使用する場合、「ポリヌクレオチド」は、核酸フラグメントのようなヌクレオチド配列である。ポリヌクレオチドは、一本鎖又は二本鎖の任意選択で合成、非天然又は変更されたヌクレオチド塩基を含むRNA又はDNAのポリマーであってもよい。DNAのポリマーの形態のポリヌクレオチドは、cDNA、ゲノムDNA、合成DNA又はそれらの混合物の1つ又は複数のセグメントを含んでもよい。
【0030】
本発明において、当業者に一般に使用されるアルゴリズムによって決定されるような、本明細書中に開示されるアミノ酸配列に対する、類似の核酸フラグメントがコードするアミノ酸配列の%同一性によって、この類似の核酸フラグメントが特徴付けられる。適切な核酸フラグメント(すなわち、本発明の第1の態様〜第3の態様に記載の単離されたポリヌクレオチド)は、本明細書中に開示されるアミノ酸配列に対して少なくとも70%同一、好ましくは少なくとも80%同一であるポリペプチドをコードする。好ましい核酸フラグメントは、本明細書中に開示されるアミノ酸配列に対して少なくとも85%同一であるアミノ酸配列をコードする。より好ましい核酸フラグメントは、本明細書中に開示されるアミノ酸配列に対して少なくとも90%同一であるアミノ酸配列をコードする。本明細書中に開示されるアミノ酸配列に対して少なくとも95%同一であるアミノ酸配列をコードする核酸フラグメントが、なおより好ましい。複数の配列アラインメントは、ClustalW法のアラインメントを使用して実施されるべきである(Thompsonら、1994、Nucleic Acids Research、第22巻、4673−4680頁;Higgins及びSharp 1989 Cabios.5:151−153)。
【0031】
本明細書中で使用される場合、用語「類似の核酸フラグメント」とは、1つ又は複数のヌクレオチド塩基の変化が1つ又は複数のアミノ酸の置換を生じるが、その変化がこのヌクレオチド配列によってコードされるポリペプチドの機能に影響を与えないか、又は例えばアンチセンス技術若しくは同時発現技術を介した遺伝子サイレンシングによって核酸フラグメントが遺伝子発現を媒介する能力に影響を与えないかのいずれかである、ポリヌクレオチド配列をいう。用語「類似の核酸フラグメント」とはまた、1つ又は複数のヌクレオチド塩基が欠失又は挿入された改変ポリヌクレオチド配列をいい、但しこの改変は、このヌクレオチド配列によってコードされるポリペプチドの機能に影響を与えないか、又は遺伝子サイレンシングによって核酸フラグメントが遺伝子発現を媒介する能力に影響を与えないかのいずれかである。従って、本発明の範囲が、本明細書中に具体的に開示されたポリヌクレオチド配列及びポリペプチド配列を超えて広がることが理解される。類似の核酸フラグメントは、植物又は植物細胞中の改変核酸フラグメントによってコードされるポリペプチドのレベルに影響を与えるそれらの能力について、部分フラグメント又は改変核酸フラグメントの形態の核酸フラグメントをスクリーニングすることによって選択できる。
【0032】
用語「作動可能に連結された」とは、一方の核酸フラグメントの機能が他方の核酸フラグメントによって影響されるような、単一の核酸フラグメントに対する2つ以上の核酸フラグメントの結合をいう。「調節配列」とは、コード配列の上流、その内部又はその下流に位置し、それに結合したコード配列の転写、RNAのプロセシング若しくは安定性又は翻訳に影響を与えるヌクレオチド配列をいう。調節配列には、プロモーター、翻訳リーダー配列、イントロン、転写終結配列及びポリアデニル化認識配列が含まれてもよい。プロモーターの形態の調節配列がコード配列に作動可能に連結される場合、この調節配列は、コード配列の発現に影響を与えることがある。コード配列は、センス又はアンチセンスの配向で、調節配列に作動可能に連結できる。
【0033】
用語「発現」とは、本発明の核酸フラグメント由来のセンスRNA(mRNA)又はアンチセンスRNAの転写及び安定な蓄積をいう。発現はまた、mRNAのポリペプチドへの翻訳を指すこともある。過剰発現とは、正常細胞すなわち形質転換されていない細胞における産生レベルを超える、トランスジェニック細胞における遺伝子産物の産生をいう。「変更されたレベル」とは、正常細胞すなわち形質転換されていない細胞のものとは異なる量又は割合の、トランスジェニック細胞における遺伝子産物の産生をいう。
【0034】
「形質転換」とは、遺伝的に安定な継承を生じる、宿主細胞のゲノムへの核酸フラグメントの移入をいう。形質転換された核酸フラグメントを含む宿主細胞は、本明細書中で「トランスジェニック細胞」と称される。
【0035】
本明細書中で使用されるような標準的な組換えDNA技術及び分子クローニング技術は当該分野で周知であり、Sambrookら「分子クローニング:ラボラトリーマニュアル(Molecular Cloning:A Laboratory Manual)」;Cold Spring Harbor Laboratory Press:Cold Spring Harbor、1989(これは、本明細書中に参照として組み込まれる)中により完全に記載されている。
【実施例】
【0036】
以下の実施例は、本発明をこれらの実施例に限定することなく、本発明を例示する。これらの実施例において、他のように特定されない限り、全ての部及び百分率は重量により、度数はセルシウスである。
【0037】
以下の実施例において、これらの略号が使用されている:
PCR:ポリメラーゼ連鎖反応
RACE:cDNA末端の迅速増幅
【0038】
上記の考察及び下記の実施例から、当業者は、本発明の本質的な特徴を確認することができ、本発明の範囲から逸脱することなく本発明の本質的な特徴を様々に変化及び改変して、本発明を所望に応じて種々の用途及び条件に適合させることができる。
【0039】
cDNAライブラリーの生成及びスクリーニング
種子特異的RNAの生成
Robusta変種Q121のコーヒーチェリーを、ICCRI(インドネシア)において30WAF(開花後の週)で収穫した。次いで、これらのチェリーの果皮を除去し、残留する外胚乳/内胚乳材料を凍結し、液体窒素中で粉末になるまで粉砕した。RNAを、カカオ種子のRNA抽出のための以前に記載された方法を使用して、この凍結粉末材料から抽出した(Guilloteau,M.ら、2003、Theobroma cacao種子中の油体:15.8kDa及び16.9kDaのオレオシンをコードするcDNAのクローニング及び特徴付け(Oil bodies in Theobroma cacao seeds:cloning and characterisation of cDNA encoding the 15.8 and 16.9 kDa oleosins).Plant Science 第164巻、597−606)。ポリA+RNAを、AMBION(Ambion,Inc.製)の「PolyA Purist(商標)」キットをキットの指示書に従って使用して、約250μgの総RNAから調製した。
【0040】
種子cDNAクローンの第1セットの生成
次いで、約50ng〜100ngのこのポリA+RNAを、「SuperScript(商標)II RNaseH−逆転写酵素(GIBCOBRL(商標))及び以下のようなSMART(商標)PCR cDNA合成キット(Clontech)を使用して、第1鎖cDNAの合成において使用した。2μlの30WAF ポリA+RNA、1μLのCDSオリゴ(SMART(商標)PCR cDNAキット、Clontech)、1μLのSmart IIオリゴ(SMART(商標)PCR cDNAキット、Clontech)及び8μLの脱イオンH2Oを含む反応。この混合物を72℃で5分間加熱し、次いで氷上に置いた。次いで以下を添加した;1μLの10mM dNTP、4μLのSuperScriptII(商標)第1鎖緩衝液及び2μLのDTT。この混合物を42℃で2分間置き、次いで1μLのSuperScriptII(商標)RNaseH−逆転写酵素(200単位/μL、GIBCO BRL(商標))を添加し、この混合物を空気循環インキュベータ中で42℃でさらに50分間インキュベートした。
【0041】
逆転写反応後、以下のPCR反応を実施した。Advantage(商標)2ポリメラーゼ(Advantage(商標)2 PCRキット、ClonTech)を含むSMART(商標)PCR cDNAキット(Clontech)中に記載される98μLのMaster Mixを氷上に設置し、次いで3μLの上記第1鎖cDNA合成反応を添加した。次いで、この100μLのPCR反応を、MJ Research PTC−150 HB装置中に配置し、以下のPCR条件を実施した:95℃で1分間、次いで(95℃で15秒間、65℃で30秒間、68℃で6分間)×16サイクル。増幅されたDNAを、Strataprep(商標)PCR Purification Kit(Stratagene)を供給者の指示に従って使用して精製した。次いで、50μLの脱イオン水中に溶出されたDNAを、以下のようなPCR−Script(商標)Ampクローニングキット(Stratagene)中に含まれるPfu−1ポリメラーゼ試薬を使用して「研磨」した;50μLのDNA、5μLの10mM dNTP、6.5μLの10×Pfu−1研磨緩衝液、5μLのクローニングされたPfu−1 DNAポリメラーゼ(0.5U/μl)。次いでこの反応を、加熱されたカバーを有するPCR装置(Perkin Elmer)中で72℃で30分間インキュベートした。pPCR−Script(商標)Ampキット(Stratagene)中に記載されるプロトコルを使用して、研磨した(平滑化した)PCR産物を、Srf−1酵素の存在下でSrf−1消化したpPCR−Script(商標)Amp SK(+)ベクター中に連結し、この連結反応産物を、XL−10 Gold(商標)Kan ultracompetent E.coli細胞中に形質転換した。挿入配列を含むプラスミドを有する形質転換についての選択を、LB−Ampプレート並びにその表面上に広げられたIPTG及びXgalを使用して、pPCR−Script(商標)Ampキット中に記載されるように実施した。白色コロニーを選択し、これらのクローンをDavl−1などと命名した。
【0042】
サイズ選択されたcDNAを有する種子cDNAクローンの第2セットの生成
種子は、少数のタンパク質(例えば、種子の貯蔵タンパク質)を高度に発現する(Whiteら、2000、Plant Physiology、第124巻、1582−1594)。cDNAがこのような組織から調製される場合、非常に高いレベルの貯蔵タンパク質及び他の種子特異的タンパク質が、高レベルのcDNAの「重複」を導く。すなわち、生成されたcDNA集団は、高い割合の同じcDNAを含む。コーヒー種子のmRNAから作製されたcDNAの重複を低下させるため及び弱く発現された長いcDNAを選択的に特徴付けるために、第2のcDNAクローニングストラテジーもまた使用した。上記の逆転写酵素反応の産物を使用して、以下のPCR反応を、Advantage(商標)2 PCRキット(ClonTech):3μLの逆転写酵素反応、5μLの10×Advantage(商標)2 PCR緩衝液、1μLのdNTP(各々10mM)、2μLのPCRプライマー(SMART(商標)PCR cDNAキット、Clontech)、39μLの脱イオン水及び1μLの50×Advantage(商標)2ポリメラーゼミックスを使用して設定した。次いで、このPCR反応を、MJ Research PTC−150 HB装置中に配置し、以下のPCR条件を実施した:95℃で1分間、次いで(95℃で15秒間、65℃で30秒間、68℃で6分間)×16サイクル。PCRの終了時に、1μLの10% SDSをゲルローディング緩衝液と共に添加し、このサンプルを37℃で10分間加熱した。次いで、このサンプルを、エチジウムブロマイドを含まない0.7%アガロースゲル上にロードするために分割し、10%をDNAマーカーレーンの脇の小さいウェル中にロードし、他の90%を隣接する大きい調製スケールのウェル中にロードした。このゲルを泳動した後に、サイズマーカーを有するゲル区画+10%反応サンプルを、エチジウムブロマイドで染色した。次いで、この染色したゲル区画をテンプレートとして使用して、このゲルの残りの未染色の(調製)部分中に存在するcDNAとは異なるサイズのPCR増幅されたcDNAを含むゲルスライスを生成した。示されたサイズ範囲のPCRフラグメントを有する6つのゲルスライスを生成した;A1A(0.8kb〜1kb)、A1B(1kb〜1.5kb)、A2(1.5kb〜2.25kb)、A3(2.25kb〜3.25kb)、A4(3.25kb〜4kb)及びA5(4kb〜6.5kb)。
【0043】
各ゲルスライス中のDNAを、供給業者の指示に従ってQiagenのQIAEX IIキットを使用してアガロースから溶出した(サンプル3A、4A及び5Aについては50℃で10分間加熱し、サンプル1A、1B及び2Aについては室温で10分間加熱した)。次いで、精製した二本鎖cDNAを、3’Tオーバーハングを有するフラグメントを作製する以下のようなTAQ酵素ミックスを用いたPCRによってさらに再増幅した:30μLのゲル単離された二本鎖cDNA、5μLの10×TAQ緩衝液(TAQ PLUS精密ポリメラーゼミックスを補充したもの、Stratagene)、1μLの40mM dNTP(各々10mM)、2μLのPCRプライマー(SMART(商標)PCR cDNAキット、Clontech)、0.5μLのTAQ PLUS精密ポリメラーゼミックス(Stratagene)及び11.5μLの脱イオン水。PCR反応条件は以下の通りであった:95℃で1分間、次いで(95℃で15秒間、65℃で1分間、72℃で8分間)×7サイクル、次いで(95℃で15秒間、65℃で1分間、72℃で10分間)×1サイクル。
【0044】
次いで、生成されたPCR増幅されたDNAを、ベクターpCR(商標)−TOPO(商標)中に連結し、TOPO(商標)TAキット(Invitrogen)を供給業者によって記載される通りに使用して、TOP10 E.coli細胞中にクローニングした。これらのクローンを、その単離の順番及びサイジングゲルにおけるそれらの位置によって命名した(例えば、A2−1、A2−2など)。
【0045】
種子cDNAのスクリーニング及び予備的同定
Dav−1ライブラリーにおいて得られた白色コロニーの第1セットを、使用したクローニング部位に隣接したT3プライマー及びT7プライマーを使用して挿入配列をPCR増幅し、ゲル上のPCR増幅されたフラグメントを試験することによって、各挿入配列のサイズを最初に決定することによってスクリーニングした。
【0046】
各白色コロニーを200μlの滅菌水中に再懸濁し、このうち10μl〜30μlを、5μlの10×Taqポリメラーゼ緩衝液(Stratagene)、1μlの10mM dNTPミックス、2.5μlの20μM T3プライマー、2.5μlの20μM T7プライマー、1μlのDMSO、0.5μlのTaqポリメラーゼ(Stratagene)及び50μlの最終容量までのH2Oに添加した。使用したPCR反応プログラムは、94℃で1分間、次いで(94℃で1分間、55℃で1.5分間及び72℃で3.5分間)×30サイクル、及び72℃で7分間の最終サイクルであった。重複を低下させるために、類似のサイズのPCR挿入配列を、制限酵素Hae IIIによる消化に供した。同じHae III制限パターンを有するPCRフラグメントをさらに研究することはしなかった。次いで、>500bpでありかつ独自のHae III制限パターンを有したPCRフラグメントを有するクローンのプラスミドを、隣接するベクター配列中にコードされた適切なT7配列決定プライマー又はT3配列決定プライマーを使用する5’末端ジデオキシ配列決定のために、Qiawall 8ウルトラプラスミドキット(Qiagen)を使用することによって精製した。これらの挿入配列は方向性をもった様式でクローニングされるわけではないので、精製されたプラスミドDNAのSca 1消化によって各クローンの5’末端を決定することが最初に必要であった(CDS SMARTプライマーは、挿入配列の配向の決定を可能にするSca 1部位を含む)。得られたDNA配列データを、プログラムBLASTX(商標)を使用してGENEBANK中の非重複データベースタンパク質に対して引き続いて書き込み、各cDNAクローンの予備的注釈を得た。
【0047】
種子cDNAバンクは、高レベルの重複を有する。すなわち、少数の種子mRNA(例えば、種子の貯蔵タンパク質をコードするmRNA)が異常に高いレベルの発現を有し、従って、それらのcDNAは、種子cDNAバンク中に非常に豊富である(Whiteら、2000、Plant Physiology、第124巻、1582−1594)。従って、コーヒー種子のcDNAを配列決定する第1ラウンドにおいて主要な重複cDNAを同定した直後に、プレスクリーニングステップを、挿入配列のサイズ決定の前に、白色の挿入配列含有コロニーのために付加した。4つの配列が非常に高度に発現され、以下の特異的プライマーセットを、これらの重複配列の各々について作製した。

【0048】
このプレスクリーニングステップのためのPCR反応は、以下のように実施した:滅菌H2O中の白色コロニー10μl〜30μl、5μlの10×Taq緩衝液(Stratagene)、1μlの10mM dNTP、2.5μlの各プライマー(20μM)、1μlのDMSO、0.5μlのTaqポリメラーゼ(Stratagene 10U/μl)及び滅菌H2Oを添加して、50μlの最終反応総容量にした。PCRプログラムは、94℃で1分間、次いで(94℃で1分間、各プライマー対に特異的な温度で1.5分間、72℃で2.5分間)×30サイクル、その後の72℃で7分間であった。
【0049】
全長cDNA挿入配列の配列決定及び配列分析
その部分配列がプロテイナーゼ及びプロテイナーゼインヒビターに対して最初の相同性を示したcDNAクローンを、標準的なジデオキシプライマーウォーキングストラテジーを使用して、両方の鎖について完全に配列決定した。これらの配列は、配列番号1、3、5、7、9、11、13及び15の下に示される。得られた全長配列を、BLASTXを使用してGanBank非重複タンパク質データベースに対して再度書き込み、予備的注釈を強化した。
【0050】
配列対の配列の同一性を、Lasergene(商標)ソフトウェアパッケージ(DNASTAR Inc)中のMegAlign(商標)モジュール中に含まれるClustalW(商標)プログラムを使用して計算した。デフォルトパラメータは以下のように選択した:(1−MULTIPLE ALIGNMENT PARAMETERS−Gap penalty 15.00、Gap length penalty 6.66、Delay divergent Seqs(%)30、DNA transition weight 0.5、Protein Weight Matrix−Gonnet Series、DNA Weight Matrix IUB。2−PAIRWISE ALIGNMENT PARAMETERS−Slow/Accurate(Gap Penalty 15.00、Gap Length Penalty 6.66)、Protein Weight Matrix−Gonnet 250、DNA Weight Matrix−IUB)。使用した配列は、各cDNA又は各cDNAの完全ORF(オープンリーディングフレーム)の全長ヌクレオチド配列のいずれかであった。
【0051】
表2:CcCP−1、CcCPI−1、CcAP−1及びCcAP−2とGenBankの非重複タンパク質データベース中に見出される関連遺伝子及びWO 02/04617号の関連遺伝子との間の、核酸配列及びアミノ酸配列の同一性の値。
【表1】
【0052】
5’RACE PCR
クローンA5−812のcDNA挿入配列はイントロンを含むことが見出された。従って、このタンパク質のコード配列を確認するために、完全コード配列を含む新たなcDNAを単離することが必要であった。これを、SMART(商標)RACE cDNA増幅キット(Clontech)を使用することによって達成した。5’RACEに使用した第1鎖cDNAを、上記のcDNAライブラリーについてすでに記載した通りに作製した。遺伝子特異的プライマーrAP2(5’ CATATAATATTAAAAGCACCACCCATAA 3’)を設計した。この配列は、A5−812クローンのポリ(A)テイルから92pbに位置する。次いで、この特異的プライマーを、以下の条件下でPCR反応においてCLONTECHキット中のUniversal Primer Mix(UPM)と共に使用した;2.5μlの第1鎖cDNA産物、5μlの10×Advantage 2 PCR Buffer(CLONTECH)、1μlのdNTP Mix(10mM)、1μlの50×Advantage 2 Polymerase Mix(CLONTECH)、5μlの「Universal Primer A Mix」(10×)(CLONTECH)、1μlのrAP2(10μM)及び滅菌水を添加して、50μlの最終容量にした。PCRサイクル条件は、(94℃で30秒間、68℃で30秒間及び72℃で3分間)×20サイクルとその後の72℃で5分間の最終伸張反応であった。約1700pbのフラグメントが得られ、「CONCERT(商標)Rapid Gel Extractionキット」(GibcoBRL)を使用してこれをゲルから切り出した。単離したフラグメントをpCR4−TOPOベクター中にクローニングし、Topo−TAクローニングキット(Invitorogen)を使用してEscherichia coli中に形質転換した。次いで、得られたプラスミドを、プラスミド抽出キット(QIAfilter Plasmid Midi Kit、Qiagen、France)を使用して精製し、このプラスミドの挿入配列を二本鎖配列決定した。
【0053】
クローンA5−442(AP1)のDNAは、cDNAの5’領域を欠くことが見出された。この領域を単離するために、5’RACEを、SMART(商標)RACE cDNA増幅Kit(Clontech)を使用して実施した。ポリ(A)テイルから396pbに位置する配列特異的プライマーrAP1(5’−TGGAGTCACAAGATGTCTCGACGAACTG−3’)を設計した。次いで、この特異的プライマーを、以下の条件下でPCR反応においてCLONTECHキット中のUniversal Primer Mix(UPM)と共に使用した;2.5μlの第1鎖cDNA、5μlの10×Advantage 2 PCR Buffer(CLONTECH)、1μlのdNTP Mix(10mM)、1μlの50×Advantage 2 Polymerase Mix(CLONTECH)、5μlの「Universal Primer A Mix」(10×)(CLONTECH)、1μlのrAP1及び滅菌水を添加して、50μlの最終容量にした。PCRサイクル条件は、(94℃で30秒間、68℃で30秒間及び72℃で3分間)×20サイクルとその後の72℃で5分間の最終伸張反応であった。約2,000bpのフラグメントが得られ、「CONCERT(商標)Rapid Gel Extractionキット」(GibcoBRL)を使用してこれをゲルから切り出した。単離したフラグメントをpCR4−TOPOベクター中にクローニングし、Topo−TAクローニングキット(Invitorogen)を使用してEscherichia coli中に形質転換した。次いで、得られたプラスミドを、プラスミド抽出キット(QIAfilter Plasmid Midi Kit、Qiagen、France)を使用して精製し、このプラスミドの挿入配列を二本鎖配列決定した。
【0054】
大きいEstライブラリーのためのRNA調製:
RNAを、以前に記載された方法を使用して、種々の発達段階の解剖した粒組織及び果皮組織並びに若葉から単離した。異なるEstライブラリーを生成するためのRNAを調製するために使用した変種及び組織は以下の通りであった:(1)若葉、1つの変種(FRT−32);(2)5つの変種(FRT32、FRT−31、FRT−400、FRT−4001及びQ121)由来の果皮(8つの異なる発達段階);(3)1つの変種(FRT−31)由来のチェリー全体、受精後22週(WAF);(4)5つの変種(FRT32、FRT−31、FRT−400、FRT−4001及びQ121)由来の粒、18+22WAF;(5)5つの変種(FRT32、FRT−31、FRT−400、FRT−4001及びQ121)由来の粒、30WAF;(6)5つの変種(FRT32、FRT−31、FRT−400、FRT−4001及びQ121)由来の粒、42WAF;並びに(7)2つの変種(FRT−32及びQ121)由来の粒、46WAF。
【0055】
cDNAクローンの生成及びDNA配列分析。
種々のEstライブラリーのためのcDNAクローンを、以下のように調製した:ポリA+mRNAを、小規模の単離のための製造業者の指示に従ってPolyATrack(商標)mRNA Isolation System(System IV、Promega)を使用して単離した。次いで、精製したポリA+mRNAを使用して、ZAP−cDNA(商標)ライブラリー構築キット(カタログ番号200450 Stratagene)中に記載されるように、λファージ中への一方向クローニングのためのcDNAを調製した。マス切り出しプロトコルは、Uni−ZAP XRベクターからpBlueScriptファージミドを切り出すためのものであり、80μlのx−gal(20mg/ml)及び16μlのIPTG(0.5M)を含む150mmのLB−アンピシリン寒天プレート上にプレートした後に白色コロニーが得られた。単一コロニーをランダムに選択して、cDNA挿入配列の5’末端を配列決定するために次いで使用されるプラスミドDNAを生成した。
【0056】
得られたDNA配列は、各クローンについてのEST配列(Expressed Sequence Tag)を生成した。次いで、7つのライブラリー由来の全てのEst配列データを「in−silico」でクラスター化し、「ユニジーン(unigene)」配列セットと称される独自の群の配列を生成した。従って、各「ユニジーン」配列は、理論的に別個の遺伝子産物に対応する。しかし、多数のユニジーンは部分的cDNA配列を示しているだけなので、いくつかの遺伝子が2つ以上のユニジーンによって示される可能性があることに留意すべきである。次いで、ユニジーンセットの予備的注釈を、各ユニジーン配列が非重複GenBankタンパク質データベースに対して検索される自動BLAST検索を用いて実施した。このBLAST検索アプローチは、「ユニジーン注釈」と称される5つの最良のBLAST「ヒット」(最低のe値を有する「ヒット」)を生じた。
【0057】
ノザンブロット分析
新たに採取した根、若葉、茎、花並びに異なる発達段階の果実(小さい緑の果実(SG)、大きい緑の果実(LG)、黄色い果実(Y)及び赤い果実(R))を、トゥール(フランス)において温室条件下(25℃、70RH)で成長したCoffea arabica CCCA2及びエクアドル又はICCRI(インドネシア)のいずれかで成長したCoffea canephora FRT32から採取した。新たな組織を液体窒素中で即座に凍結し、上記抽出手段を使用して各組織から総RNAを単離した。合計5μgのRNAを、ホルムアルデヒドを含む1.2%(w/v)の変性RNAゲル上で泳動した。各植物組織由来の総RNAサンプルを、7μLの「RNA Sample Loading Buffer」(エチジウムブロマイドを含まない、Sigma)の存在下で65℃で15分間加熱し、次いで直後に氷上に2分間置き、その後1.2%のRNAゲル上にロードした。このゲルを、60ボルトで5時間泳動した。次いで、このゲルを10×SSC中に20分間にわたって2回浸した。ゲル中のRNAを、10×SSC中で「Positive TM Membrane」(Qbiogene)にキャピラリートランスファーによって一晩移し、このRNAを、このブロットを80℃で30分間加熱することによって固定した。プローブを、(P32)dCTPの存在下で「Rediprime(商標)IIランダムプライム標識システム」キット(Amersham)を使用して生成した。ハイブリダイゼーションを、ハイブリダイゼーション溶液(5×SSC、40μg/mlの変性サケ精子DNA、5%[w/v]のSDS及び5×Denhardt’s溶液)中で65℃で24時間実施した。次いでこのメンブレンを、各々30分間の間に、2×SSC、0.1% SDS[w/v]及び1×SSC、0.1% SDS[w/v]を使用して65℃で2回洗浄した。
【0058】
図1中に示されるノザンブロット分析は、コーヒーのシステインプロテイナーゼ遺伝子CcCp−1遺伝子が、試験した全ての段階でC.arabicaのコーヒーチェリーにおいて発現され、黄色いチェリーが他の段階よりも僅かに高い発現レベルを示すことを実証する。C.arabicaの根でも茎でも葉でも、この遺伝子の発現は検出されなかった。図2は、RNAの新たな調製物を使用した、C.arabicaにおけるCcCP−1の発現を試験する別のノザンブロット実験を示す。この実験のために、4つの段階についてチェリーを解剖して、各段階のチェリー発達について果皮組織及び粒組織を生成した。次いで、総RNAをこれらの組織から抽出した。得られた結果は、チェリー発達の間のCcCP−1の発現の同じ一時的パターンを示すが、この新たな実験は、CcCP−1が、チェリーの粒組織中のみにおいて高レベルで主に発現されることをさらに示す。CcCP−1遺伝子の有意な発現は、コーヒーチェリーの果皮においては見られない。この後者の結果は、通常の成長条件下でのコーヒー粒のタンパク質、ペプチド及びアミノ酸のプロフィールの全面的な変更におけるこの遺伝子産物の役割を支持する。
【0059】
本発明者らは、コーヒーの葉並びに異なる段階の発達中のコーヒーチェリーから解剖した種子組織及び果皮組織から、ESTライブラリーを生成した。異なるライブラリー中のCcCP−1 ESTの検出(以下に示される−表3を参照のこと)もまた、この遺伝子が粒において強く発現されるが、果皮においても葉においても有意には発現されないことを実証する。種子の発達の間のCcCP−1の発現パターンは、Vicia sativaのその提唱された相同配列(CPR4遺伝子:Fischer,J.ら、2000.Plant Molecular Biology、43、83−101)について見られる発現パターンと類似である。これらの著者らは、CPR4が葉においても根においても茎においてもノザンブロッティングによって検出されないことを示しており、CcCP−1が粒特異的であるという主張をさらに強化している。例えば、CcCP−1のアンチセンス構築物又はCcCP−1の過剰発現構築物のために粒特異的プロモーターを使用することによって、本明細書中で示唆されるような粒におけるCcCP−1の発現を特異的に変更することは、他の組織において代謝を妨害しないと予測される。
【0060】
【表2】
【0061】
CcCP−1についての任意選択のアラインメント(図2A)は、このcDNAがシステインプロテイナーゼをコードすることを示す。
【0062】
図3中に示されるノザンブロット分析は、コーヒーのシステインプロテイナーゼインヒビター遺伝子CcCPI−1遺伝子が、試験した全ての段階でC.arabicaのコーヒーチェリーにおいて発現されることを実証する。しかし、システインプロテイナーゼCcCP−1について見られた発現とは対照的に、CcCPI−1は、2つの初期段階のコーヒーチェリー発達(小さい緑及び大きい緑)においてより高い発現を示し、この遺伝子は、2つの後期段階のチェリー発達においてより低いレベルで発現される。この発現パターンは、システインプロテイナーゼインヒビタータンパク質(CcCPI−1)が、種子において特異的に発現されるシステインプロテイナーゼ(例えば、コーヒーチェリー中のCcCP−1)の活性レベルを制御するという本発明の仮説と一致する。制御タンパク質(例えば、システインプロテイナーゼインヒビタータンパク質)は、標的タンパク質が発現される時点から連続的にその標的タンパク質の活性レベルを制御する必要がある場合、その標的タンパク質よりも早く発現されることが予測できる。C.arabicaの根でも茎でも葉でも、この遺伝子の発現は検出されなかった。CcCP−1及びCcCPI−1の発現パターンの類似性は、これらのタンパク質が機能的に相互作用し得るという本発明の仮説と一致することが留意される。
【0063】
ノザンブロッティングの結果(図3)は、CcCPI−1がコーヒーチェリーにおいて全ての段階で発現されることを示した。しかし、この実験は、この発現がチェリー全体中に存在するのか、又は果皮若しくは粒の中にのみ存在するのかを決定しなかった。葉における発現もまた、試験しなかった。しかし、異なるEstライブラリーにおけるCcCPI−1の発現(以下の表4中に示される)は、この遺伝子が粒においてのみ特異的に発現され、果皮でも葉でも発現が検出されなかったことを実証する。この結果は、CcCPI−1が種子において特異的に発現されるシステインプロテイナーゼ(例えば、CcCP−1)の活性レベルを制御することをさらに示唆する。
【0064】
【表3】
【0065】
図4中に示されるノザンブロット分析は、コーヒーのシステインプロテイナーゼインヒビター遺伝子CcCPI−1遺伝子が、C.canephora(robusta)のチェリー 対C.arabicaのチェリーにおいて差示的に発現されることを実証する。第1に、図4のデータは、このCcCPI−1遺伝子が、C.arabicaにおいて僅かにより早く発現されることを示す。第2に、そしてより重要なことに、このCcCPI−1遺伝子は、C.canephoraのチェリーにおいて有意により高いレベルで発現される。発現におけるこの差異は、C.arabicaのチェリー 対C.canephoraのチェリーにおいて見出されるシステインプロテイナーゼ活性のレベルにおそらく影響を与える。このクラスのタンパク質は植物における耐虫性と広く関連しているので、C.canephoraにおけるCcCPI−1遺伝子の高い発現は、robusta変種 対arabica変種についてしばしば見られるより高い耐病性に寄与する可能性もある。
【0066】
粒の発芽の間のCcCP−1発現のRT−PCR分析
コーヒー粒の発芽の間のCcCP−1の発現を決定するために、コーヒーの果実を成熟段階で収穫し、水でリンスし、その果皮を除去した(各果実は通常2つの粒を含む)。得られた粒を、室温で戸外にて1週間乾燥させた。発芽前に、各粒のパーチメント及び銀皮(種皮)を手動で除去し、次いで粒を1%(w/v)の次亜塩素酸ナトリウム中に1時間配置することによって滅菌し、次いで滅菌脱イオン水によって2回洗浄した。発芽のために、150個の滅菌した粒を、10mlの固体Heller成長培地H15(Hellerの塩(Heller、1953)を含む)及び7g/lの寒天を含む試験管中に個々に配置し、次いで1日8時間の明期で25℃でインキュベートした。
【0067】
10個の粒の3つのセットを、発芽の2日後、3日後、5日後、1ヵ月後及び2ヵ月後に採取し、液体窒素中で即座に凍結し、RNA抽出まで−80℃で保存した。1ヶ月及び2ヶ月の発芽サンプルについて、これらのサンプルに付随した幼根をサンプリング時に摘出し、粒とは別に凍結した。30個の滅菌した粒を、T(0)コントロールとして使用するためにT=0で採取して凍結した。
【0068】
各サンプルから抽出した4μgのDNase処理した総RNAを使用して、Superscript II Reverse Transcriptase(Invitrogen、Carlsbad、CA)のプロトコルに従ってヘキサマーオリゴヌクレオチドを使用してcDNAを合成した。コーヒーのリボソームタンパク質L39遺伝子のフラグメントを、cDNA合成ステップのためのコントロールとして各cDNAサンプルについて増幅した。これらのPCR反応を、cDNAの1/100希釈物10μl、1μMの各プライマー、5μlの10×ThermoPol PCR緩衝液(10mMの(NH4)2SO4、2mMのMgSO4、20mMのTris−HCl(25℃でpH8.8)、10mMのKCl及び0.1%のTriton X−100)及び2.5単位のTaqポリメラーゼ(New England Biolabs、Beverly、MA)を含む50μlの反応を使用して実施した。サイクル条件は、94℃で2分間、その後の(94℃で1分間、60℃で1.5分間及び72℃で2.5分間)×35サイクルであった。最終伸長ステップは72℃で7分間であった。以下のプライマーをPCRによる増幅のために使用し、726bpのcDNA産物を得た:CcCP−1 up 5’ ACCGAGGAGGAGTTTGAGGCTACG 3’及びCcCP−1 low:5’ ACGCTTCCCCCATGAGTTCTTGA 3’。RPL39タンパク質のためのプライマーは、
であった。
【0069】
RT−PCRを使用して、発芽の異なる段階の間のCcCP−1の発現を決定した。得られた結果は、CcCP−1転写物が試験した全ての発芽時点において粒全体中で検出されることを実証する(図5)。Fischer,J.ら、2000(Plant Molecular Biology、43、83−101)によって、V.sativa由来の提唱されたCcCP−1相同体CPR4のRNAもまた、発芽の間にV.sativa種子の胚軸及び子葉の両方において発現されることが以前に示されている。
【0070】
ウエスタンブロット分析
分析した葉組織及びチェリー組織は、Coffea arabica CCCA2由来であり、使用前にこれらの組織を−80℃で凍結保存した。異なる発達段階のチェリーの粒組織及び果皮組織を、可能な限り小さい果皮の解凍で、別々に解剖した。次いで、これらの異なる組織を迅速に粉砕して微細粉末にした。これは、例えば、予め凍結した乳鉢及び乳棒を用いて液体窒素を使用して実施できる。タンパク質抽出物を、Tanakaら、1986(Plant Physiology、81 802−806)によって記載された抽出手順の改変バージョンを使用してこの組織から調製した。使用した緩衝液は以下であった:
Tanaka緩衝液:
・スクロース 0.7M
・Tris−HCl(pH8) 0.5M
・β−メルカプト−エタノール 2%(v/v)
・NaCl 0.1M
そしてこの緩衝液を使用する直前に以下を添加した:
・EDTA 5mM
・PMSF 2mM
ゲルローディング緩衝液
・グリセロール 15%(v/v)
・β−メルカプト−エタノール 2%(v/v)
・SDS 3%(v/v)
・Tris−HCl(pH6.8) 62.5mM
【0071】
数百ミリグラムの凍結粉砕粉末を、650μlのTanaka緩衝液に添加した。タンパク質を、1体積のTris飽和フェノール(pH8)(すなわち、10mMのTris−HCl(pH8)で飽和した)の添加によって抽出した。各サンプルを20分間激しく混合し、次いで室温で13000gで20分間遠心分離した。遠心分離後、これらのタンパク質はフェノール相中に存在する。20μlのサンプルを分析(以下を参照のこと)のために維持し、フェノール相中の残留タンパク質を、0.1Mの酢酸アンモニウムを含む5体積のメタノールの添加後に、−20℃で一晩沈殿させた。引き続いて、これらのサンプルを室温で13000gで20分間遠心分離し、得られたペレットを、0.1Mの酢酸アンモニウムを含む500μlのメタノール中で2回洗浄した。得られたペレットを、タンパク質定量まで、30μlのゲルローディング緩衝液中に再懸濁した。
【0072】
フェノール相の20μlのサンプル中のタンパク質もまた上記のように沈殿させ、最終ペレットをBioRad DC ProteinアッセイKitのサンプル緩衝液中に再懸濁した。このサンプル中の総タンパク質の定量を、供給業者によって記載されるようにBioRad DC Proteinアッセイキットを使用して実施した。引き続いて、全ての主要なサンプルを、ゲルローディング緩衝液の添加によって調整して、5μg/μlにした。
【0073】
各サンプルの約50μgのタンパク質を含むサンプルを、SDS−ポリアクリルアミドゲル(12%のトリス−グリシン(Novex(登録商標)Invitrogen(商標))における電気泳動によって分離した。次いで、これらのタンパク質を、標準的なプロトコルを使用したエレクトロブロッティングによってPVDFメンブレンに移した。このメンブレン上の非特異的結合部位を、TBS緩衝液(BioRad(商標))中の10%脱脂乾燥ミルク中でこのメンブレンを室温で1時間又は4℃で一晩インキュベートすることによってブロッキングした。ブロットされたタンパク質を、Vicia sativa由来のCPR4の予測された相同体に対して惹起されたポリクローナル抗体(TBS 10%脱脂乾燥ミルク中、1/5000e希釈)(A.Schlereth及びK.Muntz、Institut fur Pflanzengenetik und kulturpflanzenforchung(IPK)、Germany(A.Schlereth、C.Becker、C.Horstmann、J.Tiedmann及びK.Muntz、2000、Journal of Experimental Botany、51:1423−1433)の厚意により寄贈されたもの)を用いて室温で2時間又は4℃で一晩プローブした。次いで、このメンブレンを、TBS+0.1%のTween 20緩衝液中で20分間にわたって3回洗浄した。このメンブレンを引き続き、西洋ワサビペルオキシダーゼで標識された2次抗体(ヤギ抗ウサギIg、Immunopure(登録商標)、Pierce(商標))と共に1時間インキュベートした。次いで、このメンブレンをTBS+0.1%のTween 20緩衝液中で20分間にわたって2回洗浄し、次いでTBS中で20分間にわたって1回洗浄した。この2次抗体に連結された酵素の存在を、増強されたECL+(登録商標)システム(Amersham Life Science)を供給業者によって記載される通りに使用して、化学ルミネセンス検出によって可視化した。
【0074】
得られた結果は、約41kDaのポリペプチド(これは、CcCP−1前駆体ポリペプチドの予測された分子量(43735Da)と密接に対応する)が、試験した粒成熟段階の全てにおいて検出されるが、果皮組織においては検出されないことを示す(図6)。このタンパク質発現パターンは、CcCP−1 mRNAについて見られた発現パターン(図2)と類似である。約22kDaの別のポリペプチドもまた、黄色い段階及び赤い段階の粒において検出されるが、その量は、41kDaのポリペプチドよりも少ない。この第2のポリペプチドのサイズは、CcCP−1の成熟形態の予測されたサイズ(25,239Da)と一致する。プロセシング後の成熟CcCP−1の予測されたサイズを、CcCP1の完全ORF配列とCPR4の予測された成熟形態の配列(Vicia sativa−登録番号Z99172、CcCP1と60.9%の同一性)との間のタンパク質アラインメントによって決定した。成熟形態を生成するためのCPR4ポリペプチドのプロセシングのN末端部位を、他のパパイン様CPRポリペプチドとの配列比較によって予測した(J.Fisher、C.Becker、S.Hillmer、C.Horstmann、B.Neubohn、A.Schlereth、V.Senyuk、A.Shutov及びK.Muntz.2000 Plant molecular biology 43:83−101)。興味深いことに、CcCP−1の前駆体及び成熟形態の両方が粒の発達の間に検出される本明細書中に示される結果とは対照的に、CPR4ポリペプチドの成熟形態のみが発達中の種子において検出され、V.sativa種子の発芽の間にも検出された(Fisherら、2000)。
【0075】
robusta変種FRT−32についての遺伝子発現のRT−PCR分析。
FRT−32の異なる組織を調製し、総RNAを、以前に記載された方法によってこれらの組織から抽出した。cDNAを、DNase処理した総RNAから、arabica cDNAを用いたRT−PCR実験について上記したように調製した。次いで、特異的PCR反応を、arabica cDNAを用いたRT−PCR実験のための上記の反応条件を使用して実施した。使用した特異的増幅条件及びオリゴヌクレオチドプライマーは、各実験について図の説明文中に与えられる。
【0076】
CcCPI−1
a)このcDNAがシステインプロテイナーゼインヒビターをコードすることを示す、CcCPI−1についての最適なアラインメント(図6A)。
【0077】
b)arabica及びrobustaにおけるCcCPI−1についてのRT−PCR発現データ(図7)。PCR反応を以前に記載されたように実施した。使用したサイクル条件及びPCRプライマーは、図の説明文中に与えられる。これらのデータは、CcCPI−1が粒においてのみ発現され、果皮においては発現されないことを示すので、arabica発現について以前に示されたデータを補足及び拡張する。この遺伝子の弱い発現が花においても検出され、これは、ノザンブロット分析によって以前に見られなかった結果である。robustaにおけるRT−PCR発現もまた決定した(図7)。これは、花においても小さい緑の粒においても発現が検出されなかったこと以外、arabicaについて見られたのと同じ発現パターンである。robustaの小さい緑の段階について見られた発現の非存在は他の遺伝子についても見られ、従って、これはCcCPI−1遺伝子に独自のことではない。
【0078】
表5:異なるEstライブラリーにおけるシステインプロテイナーゼインヒビター遺伝子CPI−2、CPI−3及びCPI−4についてのEstの発生。
【表4】
【0079】
CcCPI−2
a)このcDNAがシステインプロテイナーゼインヒビターをコードすることを示す、CcCPI−2についての最適なアラインメント(図8)。
【0080】
b)arabica及びrobustaにおけるCcCPI−2についてのRT−PCR発現データ(図9)。PCR反応を以前に記載されたように実施した。使用したPCRプライマーは図の説明文中に与えられる。これらのデータは、CcCP−2が全ての組織において発現されることを示し、従って、この遺伝子のタンパク質産物が、これらの組織中に存在する1つ又は複数のシステインプロテイナーゼを制御することにおいて重要な役割をおそらく果たすことを示す。上記表5中に見られる各ライブラリー中のEstの数は、CPI−2が、葉、果皮又は受粉後46週の種子よりも、受粉後30週の粒(種子)においてより多く発現できることを示唆する。
【0081】
CcCPI−3
a)このcDNAがシステインプロテイナーゼインヒビターをコードすることを示す、CcCPI−3についての最適なアラインメント(図10)。
【0082】
b)このシステインプロテイナーゼインヒビターについて現在入手可能なRT−PCR発現データは存在しない。しかし、各ライブラリー中に出現するEstの数(上記表5)によって決定されるようなこの遺伝子の「in silico」発現は、CcCPI−3が、コーヒー粒において発現される(「種子30w」及び「種子46w」、すなわち30週及び46週の種子ライブラリー中に存在する)ことを示す。果皮、葉又はチェリー全体におけるこの遺伝子についてのEstの非存在は、この遺伝子が粒特異的遺伝子であり得ることを示唆する。
【0083】
CcCPI−4
a)このcDNAがシステインプロテイナーゼインヒビターをコードすることを示す、CcCPI−4についての最適なアラインメント(図11)。
【0084】
b)arabica及びrobustaにおけるCcCPI−4についてのRT−PCR発現データ(図12)。PCR反応を以前に記載されたように実施した。使用したPCRプライマーは図の説明文中に与えられる。得られたデータは、この遺伝子がarabicaの葉、花及び赤い段階の粒において有意に発現されることを示す。最初のゲルの綿密な試験(パネルA:arabica)は、小さい緑の粒及び大きい緑の果皮のレーンにおいても弱いバンドが検出されることを示すので、この遺伝子はまた、arabicaの研究した全てのチェリー発達段階の粒及び果皮において弱く発現できる。robustaについて得られたデータは、この遺伝子が、葉、花、小さい緑の粒及び大きい緑の粒において有意に発現されることを示す。CcCPI−4についてたった1つのEstが種子又は果皮のライブラリーにおいて見出され(上記の表5)、このことは、粒及び/又は果皮におけるこの遺伝子の発現が比較的低いか、或いはこれら2つの組織の小さい規定された領域に限定されることを示す。
【0085】
システインプロテイナーゼインヒビター(CPI)遺伝子についての各々の場合において、粒発達の間のこれらの遺伝子の過剰発現又は発現の阻害(すなわち、非常に強い粒特異的プロモーター(例えば、コーヒー11Sプロモーター)の制御下での)は、成熟粒におけるタンパク質、ペプチド及びアミノ酸のプロフィール(並びに従って、フレーバー前駆体のレベル)を変更することが予測される。
【0086】
発芽及びRT−PCR分析
滅菌した乾燥C.arabica CCCA2の粒(パーチメント及び銀皮を除去したもの)を、10mlの固体Heller成長培地H15及び7g/lの寒天を含む試験管中に個々に配置し、1日8時間の明期で25℃でインキュベートした。発芽の2日後、3日後、5日後、1ヵ月後及び2ヵ月後に3個の粒を採取し、存在する場合には幼根を除去し、粒及び幼根の両方を液体窒素中で即座に凍結し、RNA抽出まで−80℃で保存した。同様に乾燥及び滅菌した非発芽粒(T0)をコントロールとして使用した。RNAを、以前に記載されたように粒サンプルから抽出した。各サンプルから抽出したDNase処理した総RNAを使用して、Superscript II Reverse Transcriptaseキット(Invitrogen、Carlsbad、CA)のプロトコルに従って、プライマーとしてオリゴ(dT)20を使用してcDNAを合成した。次いで、PCR反応を、各cDNA反応のアリコート(10μlの1/10希釈したcDNA、1μMの各プライマー、5μlの10×ThermoPol PCR緩衝液、200μmのdNTP及び2単位のTaqポリメラーゼ(New England Biolabs、Beverly、MA)を含む50μlの反応)を使用して実施した。サイクル条件は、94℃で2分間、(94℃で1分間、54℃で1.5分間及び72℃で2.5分間)×40サイクルであった。最終伸長ステップは72℃で7分間であった。PCRプライマーは以下であった:
【0087】
ゲノムPCR及び精製されたPCRフラグメントのDNA配列決定
5つの異なるコーヒー変種(FRT−07、FRT−19、FRT−32、CCCA2及びGPFA57)のゲノムDNAを、発芽RT−PCR発現研究のために上記のPCR反応において使用した。予測されたサイズのPCR産物が得られ、これらのフラグメントをゲルから精製した。次いで、このPCR増幅されたDNAを第2ラウンドのPCR増幅に供し、次いで、この配列決定反応から得られたDNAを、増幅のために使用したのと同じプライマーを使用して配列決定した。
【0088】
システインプロテイナーゼCcCP−4の単離及び特徴付け
1)異なる発達段階のコーヒー粒、異なる発達段階のコーヒーの果皮組織、及び葉から単離されたRNAを用いて作製されたEst(Expressed Sequence Tags)の収集を使用して、本発明者らは、C末端KDDL配列を有するコーヒーシステインプロテイナーゼをコードする全長cDNAを単離した。本発明者らは、このcDNAをCcCP−4(KDDL)と命名した(図14)。このcDNAによってコードされるタンパク質と他の高度に相同な植物システインプロテイナーゼとのアラインメントが図15中に示される。このアラインメントデータ及び関連のBlast検索は、コーヒーCcCP−4(KDDL)配列によってコードされるタンパク質が、植物のKDEL含有システインプロテイナーゼファミリーのメンバーであることを明らかに示す(図15)。CcCP−4(KDDL)と最も相同なデータベース配列との間の正確な同一性は、表6A及び6B中に与えられる。
【0089】
表6A:Coffea canephoraシステインプロテイナーゼCcCP−4(KDDL)のアミノ酸配列と、最も相同なGenBank配列のアミノ酸配列との同一性
【表5】
【0090】
表6B:Coffea canephoraシステインプロテイナーゼCcCP−4(KDDL)の核酸(cDNA)配列と、最も相同なGenBank配列の核酸配列との同一性
【表6】
【0091】
明白に、得られたコーヒーCcCP−4 KDDL配列は、図15中に示される他のほぼ全ての配列とは異なる1つの重要な差異を有する。すなわち、この配列は、予測された小胞体(ER)残留配列(C末端KDEL配列)を有さないが、この配列の変異体(すなわちKDDL)を有する。C末端KDEL配列におけるバリエーションが植物細胞中のERにおける残留を指向する能力を試験することによって、Deneckeら(Denecke,J.、De Rycke,R.及びBotterman,J.1992 EMBO J.11、2345−2355)は、C末端変異体(例えば、SDEL、KDDL、KDEI及びKDEV)が、小胞体残留機能を完全に喪失することもあることを以前に示している。従って、植物KDELシステインプロテイナーゼのコーヒー相同体中のKDDL配列の存在は予測されなかった。表7は、cDNA CcCP−4(KDDL)を含むユニジーンが21個のEstを有することを示す。従って、次いで本発明者らは、このユニジーン中の他のEstの配列を試験し、これらのEstのうち7つが、KDDL領域についての良好な配列データを含むことを見出した。これら7つのcDNA配列のうち、6つはKDDL配列を有し、1つはKDEL配列を有した。本発明者らは、KDEL C末端配列を有するcDNAクローンを引き続いて単離し、この部分的cDNAクローンについての完全配列を得た。得られたDNA配列及びタンパク質配列はそれぞれ、図16及び図17中に示される。
【0092】
表7.全長cDNA CcCP−4(KDDL)を含むユニジーン中のEstの数。
【表7】
【0093】
図16中に示されるCcCP−4(KDEL)の配列をコードするcDNAは、部分的cDNAに過ぎない。すなわち、これは、全長cDNAクローンCcCP−4(KDDL)についての1336bp長に対してたった817bp長である。部分的cDNA CcCP−4(KDEL)は、cDNAクローンCcCP−4(KDDL)中に見出される等価な配列からの8個の単一ヌクレオチド残基変化を有するが、これらのヌクレオチド変化のうち2つだけがオープンリーディングフレームのアミノ酸配列の変化を引き起こす(図17)。3’非翻訳領域中に、3つの明白なヌクレオチド変化が存在する。さらに、マイクロサテライト領域内であるように思われるCcCP−4(KDEL)cDNA配列の3’非翻訳領域中に、12ヌクレオチドの挿入もまた存在する。このデータは、コーヒーCcCP−4遺伝子のこれら2つの対立遺伝子についての未発見の2つの異なる重要な分子マーカーを示したに過ぎず、そのうち一方は、機能的に重要なKDEL部位と関連するSNPであり、他方は、この遺伝子の3’非翻訳領域と関連するマイクロサテライトマーカーである。後者の点は、マイクロサテライト配列が高い変動性を有する遺伝子マーカーであると通常みなされるので重要であり、従って、この遺伝子の他の対立遺伝子は、このマイクロサテライト含有領域を使用して見出される可能性がある。
【0094】
arabica及びrobustaの異なる変種において上で同定されたCcCP−4遺伝子の2つの対立遺伝子の分布を試験するために、CcCP−4遺伝子を保有するゲノム配列の小さい領域を、5つの異なる遺伝子型からPCRによって増幅した。予測されたサイズ(207塩基対)のPCRフラグメントが各ゲノムDNAサンプルから得られ、これらのPCR産物をゲル精製し、次いで再増幅して、このPCR産物の直接的DNA配列決定に充分なDNAを生成した。配列決定反応から得られた結果が図18中に示される。5つの配列についての配列決定クロマトグラムは、試験した2つのarabica変種がKDEL配列を明らかに有し、試験した3つのrobusta変種がKDDL配列を有することを示す。この結果は、KDDL対立遺伝子がrobusta変種に限定できること及びこれがarabica変種においては見出されないことを示す。KDEL配列は本明細書中で研究した3つのrobusta変種においては見出されなかったが、Cornell Estライブラリーにおける1つのKDEL配列の発見は、この対立遺伝子が少なくともいくつかのrobustaクローン中に存在できることを示す。
【0095】
CcCP−4遺伝子の発現を、ノザンブロット及びRT−PCR分析を使用して研究した。図19は、arabica変種の異なる発達段階のコーヒーの粒及び果皮から抽出したRNA並びに根、若葉、茎及び花から単離したRNAを使用したノザンブロット実験から得られた結果を示す。CcCP−4(KDEL)対立遺伝子の既知のDNA配列との約98%の相同性を有するCcCP−4(KDDL)プローブを使用して得られたデータは、CcCP−4が粒においてのみ発現されることを示した。発現は、果皮においても、根、茎、花及び葉においても検出されなかった。2つの対立遺伝子間の非常に高いレベルの同一性に起因して、このCcCP−4(KDDL)プローブは、両方の対立遺伝子由来の転写物にハイブリダイズすると予測される。RT−PCR分析を使用した類似の実験もまた、CcCP−4遺伝子について同じ発現プロフィールを示した。
【0096】
CcCP−4の発現を、RT−PCR分析を使用して、発芽の間の種子全体においても研究した。この実験は、CcCP−4 KDEL対立遺伝子及びCcCP−4 KDDL対立遺伝子の両方に共通するプライマーを使用した。この実験の結果は、図20中に示される。CcCP−4転写物は試験した全ての発芽段階で検出されたが、これらの転写物のレベルは、3日目に僅かに低下するようであり、次いで発芽が進行するにつれて再度上昇し始めた(1ヶ月目及び2ヶ月目のサンプルにおいて最高レベル)。
【0097】
Lingら(Ling,J.−Q.、Kojima,T.、Shiraiwa,M.及びTakahara,H.、2003 Biochim.Biophys.Acta 1627、129−139)は、KDEL含有システインプロテイナーゼをコードする2つのcDNAを、ダイズ子葉から単離した。これら2つのcDNAは、DNAレベルで93.5%の類似性を有し、根、花及び種子発達の間に発現された。発達中の種子においても成熟種子においてもノザンブロッティングによって発現は検出されなかったが、成熟した莢において発現が検出された。KDEL含有システインプロテイナーゼをコードするcDNAは、ニンジンからも単離された(Sakuta,C.、Oda,A.、Konishi,M.、Yamakawa,S.、Kamada,H.及びSatoh,S.2001 Biosci.Biotechnol.Biochem.65、2243−2248)。この遺伝子の転写物は、成熟した乾燥種子並びに吸水の2日後及び3日後の発芽中の種子全体において検出された。他のニンジン組織におけるこの遺伝子の発現又は種子発達の間のこの遺伝子の発現は示されなかった。別のKDEL含有タンパク質及びその対応するcDNAがV.sativaから単離されている(Fischer,J、Becker,C.、Hillmer,S.、Horstmann,C.、Neubohn,B、Schlereth,A.、Senyuk,V.、Shutov,A.及びMuntz,K.(2000)Plant Molecular Biol.43、83−101)。ノザンブロッティングを使用して、この遺伝子の転写物は、発芽の間に子葉において検出されたが、発芽中の種子の胚軸においては検出されなかった。成熟中の種子においても、成熟種子においても、葉及び根においても、転写物は検出されなかった。
【0098】
本明細書中に示される結果は、コーヒーのKDEL型システインプロテイナーゼが、いくつかの新規かつ予測されなかった特徴を示すことを示す。第1に、本発明者らは、robustaコーヒー粒が、KDEL領域をコードする配列中にKDELからKDDLへの変化を生じる単一変異を有するKDEL型CP遺伝子を発現することを発見した。Deneckeら(1992)のデータに基づいて、この残留配列におけるこの特定の変更は、robusta CcCP−4(KDDL)タンパク質の細胞局在及び/又は制御を変更することが予測される。本発明者らは、CcCP−4 KDDL遺伝子の転写されたコピーが存在することにより、CcCP−4 KDEL配列を有する変種と比較して、コーヒー粒におけるペプチド/アミノ酸のプロフィールの有意な変化を生成できることを提唱する。本発明者らはまた、コーヒーのKDEL型システインプロテイナーゼが、粒の発芽の間に予測された発現を示しつつ、研究した全ての粒の発達段階の間にも予測に反して発現されることを本明細書中で示した。上記のように、これまで、他の植物における種子発達の間のKDEL型システインプロテイナーゼの有意な発現を実証する明確なデータは刊行された文献中には存在しないが、その転写物は、成熟ニンジン種子において検出されている(Sakutaら、2001)。
【0099】
上で示されたコーヒーのKDEL型システインプロテイナーゼの新規特性は、arabica及びrobustaの成熟粒におけるペプチドプロフィール及びアミノ酸プロフィールに対して重要な影響をおそらく有し、従って、重要なコーヒーフレーバー前駆体のプールを変更する。KDEL型システインプロテイナーゼの転写物が成熟粒中に存在することを考慮すると、このKDEL型タンパク質はコーヒーの湿式加工の間に活性化でき、それによって湿式プロセスのコーヒー粒のペプチド/アミノ酸のプロフィールをさらに変更することもまた可能である。記載された研究は、KDEL型システインプロテイナーゼ遺伝子の特定の対立遺伝子を有するコーヒー変種(これは、タンパク質/ペプチド/アミノ酸のプロフィールにおける付随した変更を有する)を得るために古典的選択及び交配作業において使用できる分子マーカー(SNP及びマイクロサテライトマーカー)を生成している。例えば、CcCP−4 KDEL対立遺伝子のみを有するか又は低い発現レベルのCcCP−4 KDDL対立遺伝子のみを有するrobustaの変種が、選択/交配できる。さらに、遺伝子改変技術を使用して、KDEL型又はKDDL型のシステインプロテイナーゼの種子特異的な過剰発現によってコーヒー又は他の植物におけるKDEL型システインプロテイナーゼ活性を変更することが想定できる。或いは、KDEL型システインプロテイナーゼのレベルは、アンチセンス、センス又はRNAiの技術を使用して低下できる。両方の場合において、得られた形質転換された植物中のタンパク質/ペプチド/アミノ酸のプールが変更されて、タンパク質/ペプチド/アミノ酸のフレーバー前駆体プールの新たなプロフィールが導かれる。
【0100】
図13中に示されるノザンブロット分析は、コーヒーのアスパラギン酸プロテイナーゼCcAP−2遺伝子が、試験した全てのチェリー発達段階で、C.arabicaコーヒーチェリーの粒及び果皮の両方において発現されることを実証する。このCcAP−2遺伝子はまた、根において比較的高い発現を有する。フィルムをより長く露光させると、CcAP−2発現は、C.arabicaの茎、葉及び花の組織においても検出された。
【0101】
CcAP−1及びCcAP−2
図21及び22は、CcAP−1及びCcAP−2の各々がアスパラギン酸プロテイナーゼをコードすることを示す。
【0102】
コーヒー種子におけるCcCP−1、CcCP−4、CcAP−1及びCcAP−2のプロテイナーゼ遺伝子配列並びにCcCPI−1、CcCPI−2、CcCPI−3及びCcCPI−4のプロテイナーゼインヒビターの過剰発現及び過少発現。
主要な貯蔵タンパク質のプロフィール及びアミノ酸/ペプチドのプロフィールは、本明細書中に開示される1つ又は複数の遺伝子の発現を上方又は下方のいずれかで変更することによって、成熟コーヒー粒において変化できることが予測される。
【0103】
目的の遺伝子の過剰発現のための方法は当該分野で周知である。このような方法は、以下の3つの主要な成分のキメラ遺伝子を生成するステップからなる:1)遺伝子の5’末端のプロモーター配列、好ましくは本願においてはMarracciniら、1999(Marracciniら、1999、Coffea arabicaの完全11S種子貯蔵タンパク質遺伝子の分子クローニング及びトランスジェニックタバコ植物におけるプロモーター分析(Molecular cloning of the complete 11S seed storage protein gene of Coffea arabica and promoter analysis in transgenic tobacco plants)、Plant Physiol.Biochem.第37巻、273−282及びWO 99/02688号)中に記載されるコーヒー種子特異的プロモーターのような種子特異的プロモーター、2)発現されるべき遺伝子の全コード配列、並びに3)Agrobacterium tumefaciensのTiプラスミドのT−DNA由来のノパリンシンターゼ遺伝子由来の3’領域のような3’制御領域。次いで、このキメラ遺伝子は、Agrobacterium tumefaciens形質転換ベクター中にクローニングでき、このベクターは、Leroyら、2000、(Leroyら、2000、リーフマイナーに対する抵抗性についてのBacillus thuringiensis cry1Ac遺伝子を発現する、遺伝的に変更されたコーヒー植物(Genetically modified coffee plants expressing the Bacillus thuringiensis cry1Ac gene for resistance to leaf minor).Plant Cell Reports 2000、19、382−389)によって詳細に記載されているコーヒー形質転換において使用するためのAgrobacterium tumefaciens株中に形質転換できる。安定な形質転換挿入配列を有する植物を次いでスクリーニングして、遺伝子の過剰発現又はタンパク質活性の過剰発現の検出のような方法を使用して、モック形質転換した植物由来の種子に対して、形質転換実験において使用する特定の遺伝子を成熟種子において特異的に過剰発現する植物をスクリーニングすることができる。
【0104】
例えば、当業者は、以下からなる組換え構築物を生成できる:1)Marracciniら(1999)中に記載される最長のコーヒー11S遺伝子プロモーター配列、2)ポリAテイルを有さない、CcCP−1又はCcCP−4(KDDL)の全長cDNA配列、及び3)既知の転写ターミネーター配列(例えば、充分研究されたノパリンターミネーター)。CcCP4若しくは他のcDNA配列の5’非コード領域の、コーヒー11S遺伝子の5’非コード領域又はコーヒー若しくは他の関連の植物種のいずれかの他の強力な種子特異的プロモーターの5’非コード領域による置換に起因して、より高いレベルの組換え構築物の過剰発現が生じることもまた可能である。次いで、この組換え遺伝子配列を、Leroyらにおいて記載されるAgrobacterium T−DNAベクターの適切な部位中に挿入できる。このように構築されたT−DNAベクターは、適切なAgrobacterium株(例えば、Leroyらにおいて記載される株)中に配置することができ、このT−DNA含有Agrobacteriumは、Leroyらにおいて詳述される方法に従ってコーヒーを形質転換するために使用できる。
【0105】
既知の遺伝子配列の発現が、アンチセンス抑制によって、遺伝子の全コード領域未満を示す核酸フラグメントを使用した遺伝子発現によって、抑制されるべき遺伝子と100%の配列同一性を共有しない核酸によって、低下又は完全に遮断できることは、当該分野で周知である。この場合、特定のアンチセンス抑制又は共抑制実験のために選択された配列は、上に示したキメラ遺伝子構築スキームにおいて全長遺伝子を置換する。得られたアンチセンス抑制又は共抑制キメラ構築物は、Agrobacterium tumefaciens形質転換ベクター中に再度クローニングされ、上記のようにコーヒー形質転換において使用するためにAgrobacterium tumefaciens株中に形質転換される。次いで、安定な形質転換挿入配列を有する植物は、形質転換された植物の種子において使用される特定の遺伝子配列の発現を低下させたものについてスクリーニングできる。低下された発現は、ノザンブロッティング;半定量的RT−PCR及び/又は定量的RT−PCRのような技術によって検出できる。
【0106】
植物中の遺伝子発現を低下又は排除するための別の方法は、本明細書中に開示される遺伝子配列の小部分を使用して、RNAiの使用を介したRNAサイレンシングを生じることである(Hannon,G.J.、2002、Nature、第418巻、244−251;Tangら、2003、Genes Dev、第17巻、49−63)。このアプローチにおいて、本明細書中に開示される1つ又は複数の配列の小領域が、種子特異的プロモーター及び適切な3’調節領域を有する上記のようなAgrobacterium tumefaciens形質転換ベクター中にクローニングされる。RNAiのためのこの挿入された新たな配列は、生成されたRNAがin vivoでRNA構造を形成して、形質転換された細胞において小さい二本鎖RNAの生成を生じ、それによってこれらの小さい二本鎖RNA配列がこれらの形質転換された細胞において相同なmRNAの分解を誘発するように構築されるべきである。
【0107】
CcCP−1遺伝子、CcCP−4遺伝子、CcAP−1遺伝子、CcAP−2遺伝子、CcCPI−1遺伝子、CcCPI−2遺伝子、CcCPI−3遺伝子、CcCPI−4遺伝子における天然に存在するバリエーションについてのスクリーニング及びこれらの遺伝子における新たな変異の生成。
本明細書中に開示される配列は、これらの遺伝子における対立遺伝子変異体について天然の集団をスクリーニングするために使用できる。これは、CcCP−1配列、CcCP−4配列、CcAP−1配列、CcAP−2配列、CcCPI−1配列、CcCPI−2配列、CcCPI−3配列及びCcCPI−4配列を、異なるコーヒー植物変種由来のゲノムDNA中の天然に存在するRFLP(制限酵素断片長多型)についての研究においてプローブとして使用することによって達成できる。対立遺伝子変異体を見出すためのより強力な方法は、TILLING法(Till,B.J.ら、2003、高スループットTILLINGを用いた、誘導された点変異の大規模発見(Large scale discovery of induced point mutations with high−thruput TILLING).Genome Research 第13巻、524−530)に関連する変異スクリーニング技術を使用することである。この場合、特定の遺伝子配列(例えば、本明細書中のCcCP−1配列、CcCP−4配列、CcAP−1配列、CcAP−2配列、CcCPI−1配列、CcCPI−2配列、CcCPI−3配列及びCcCPI−4配列)が一旦単離及びクローニングされると、TILLING法に関連する変異スクリーニング技術は、クローニングされた配列と異なる変種中の対応するcDNA又はゲノム配列との間で配列変異体を同定するために使用できる。700塩基対〜1000塩基対のDNAセグメントをコードするPCRプライマー対を使用して、既知のクローニングされた遺伝子が、異なる変種中の天然に存在する配列バリエーションについてスキャンできる。理想的な状況において、1つ又は複数の配列変異体はまた、特定の表現型バリエーションと相関させることができ、それによってこの表現型変異体に対する遺伝子マーカーを同定する。
【0108】
さらに、CcCP−1、CcCP−4、CcAP−1、CcAP−2、CcCPI−1、CcCPI−2、CcCPI−3及びCcCPI−4についての本明細書中に開示される配列を使用して、完全なTILLING法の適用は、これらの遺伝子中の新たな変異を生成及び検出するため、従って、これらの特定の変異を含む植物を生成するために使用できる。例えば、完全なTILLING法を使用して、特定の変異(例えば、目的の遺伝子標的を不活化する、コード配列中のミスセンス変異)を有するコーヒー植物が生成できる。
【図面の簡単な説明】
【0109】
【図1】Coffea arabicaの異なる組織におけるシステインプロテイナーゼ遺伝子のノザンブロット分析を示す図であり、図中、レーンは以下で標識される:R:根、S:茎、L:若葉;並びにSG、LG、Y及びRedはそれぞれ、小さい緑の果実、大きい緑の果実、黄色い果実及び赤い果実由来の粒である。各レーンにおいて5μgの総RNAをロードした。MWはRNAサイズラダーである。パネルBは、試験した組織中のCcCP−1 mRNAの出現を示す、24時間の露光後のオートラジオグラムを示し、パネルAは、ブロッティング前のゲルのエチジウムブロマイド染色を実証する。
【図2】Coffea arabicaの異なる組織におけるシステインプロテイナーゼCcCP−1遺伝子の発現のノザンブロット分析を示す図であり、図中、レーンは以下で標識される:R、根;S、茎;L、若葉;F、花。SG(G)、LG(G)、Y(G)及びRed(G)はそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリーの粒から単離されたRNAに対応し、SG(P)、LG(P)、Y(P)及びRed(P)で標識されたレーンはそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリーの果皮組織から単離されたRNAに対応する。各レーンにおいて5μgの総RNAをロードした。パネルAは、ローディングコントロールとしてのブロッティング前の大きいリボソームRNAのエチジウムブロマイド染色を実証し、パネルBは、試験した特定の組織中のCcCP−1 mRNAの出現を示すオートラジオグラムである。
【図2A】CcCP−1 cDNAによってコードされるタンパク質の完全配列と、NCBIデータベースにおいて入手可能な他の全長システインプロテイナーゼとのアラインメントの図である。これは、MegAlign(DNASTAR)中のCLUSTAL法によって、Megalignにおいて実施した。影付のブロックは同一のアミノ酸を示す。EMBLデータベースの登録番号は括弧内に与えられる。Arabidopsis thaliana(AY070063);Vicia sativa(Z99172);Glycine max GMCP3(Z32795);Glycine max GmPM33(AF167986);Phaseolus vulgaris Moldavain(Z99955);Solanum melongena(AF082181);Nicotiana tabacum(AJ242994);Lycopersicon esculentum(Z14028);Vicia faba(AY161277)。
【図3】Coffea arabicaの異なる組織におけるシステインプロテイナーゼインヒビター(CcCPI−1)遺伝子のノザンブロット分析を示す図であり、図中、レーンは以下で標識される:R:根、S:茎、L:若葉;並びにSG、LG、Y及びRedはそれぞれ、小さい緑の果実、大きい緑の果実、黄色い果実及び赤い果実由来の粒について。各レーンにおいて5μgの総RNAをロードした。MWはRNAサイズラダーである。パネルBは、24時間の露光後のオートラジオグラムを示し、パネルAは、ブロッティング前のゲルのエチジウムブロマイド染色を実証する。
【図4】Coffea arabica(ARA)及びCoffea robusta(ROB)の果実の異なる発達段階におけるシステインプロテイナーゼインヒビター(CcCPI−1)遺伝子のノザンブロット分析を示す図である。レーンは、小さい緑の果実(SG)、大きい緑の果実(LG)、黄色い果実(Y)及び赤い果実(Red)でそれぞれ標識される。各レーンにおいて5μgの総RNAをロードした。MWはRNAサイズラダーである。パネルBは、試験した特定の組織中のCcCPI−1 mRNAの出現を示す、24時間の露光後のオートラジオグラムを示す。パネルAは、ブロッティング前のゲルのエチジウムブロマイド染色を実証する。
【図5】Coffea arabica粒の出芽の間のCcCP−1の発現のRT−PCR分析を示す図である。PCR反応を、1/100希釈した各cDNA10μlを使用して実施した。サイクル条件は、94℃で2分間、(94℃で1分間、61℃で1.5分間及び72℃で2.5分間)×35サイクルであった。最終伸長ステップは72℃で7分間であった。PCRプライマーは、A4-43-upper:5'-ACCGAGGAGGAGTTTGAGGCTACG-3'、A4-43-lower:5'-ACGCTTCCCCCATGAGTTCTTGA-3'であった。mRNAを、異なるテンプレート(それぞれ、滅菌した種子(T0)並びに出芽の2日後(2d)、3日後(3d)、5日後(5d)、1ヵ月後(1m)及び2ヵ月後(2m)に採取した種子由来のcDNA)に対して特異的プライマー(CcCP−1 up/CcCP−1 low)を使用するRT−PCRによって増幅した。これらのPCR産物を、1%(w/v)アガロースゲル中で分離し、エチジウムブロマイドで染色した。RPL39;60Sリボソーム大サブユニットのL39タンパク質をコードするcDNAの増幅されたフラグメント。
【図6】CcCP1タンパク質の発現のウエスタンブロット分析を示す図である(A)。総タンパク質を、小さい緑(SG)、大きい緑(LG)、黄色(Y)及び赤(Red)の段階の、発達中のコーヒーチェリーから収集した粒(g)及び果皮(p)から抽出した。パネルB−クマシーブルーで染色した12% SDS−PAGEゲル上での50μgの総タンパク質の分離。パネルA−タンパク質検出を、これらの方法において記載されるように、抗CRP4ポリクローナル抗体(ウサギ)を使用して実施した。パネルB中のバンドのおおよそのサイズを左側に矢印で示す。各パネル内の大きい矢印は、これらの抗体のうち1つと交差反応する主要な貯蔵タンパク質の存在を示す。
【図6A】CcCPI−1 cDNAによってコードされる完全タンパク質と、NCBIにおいて入手可能な他の相同な全長システインプロテイナーゼとの最適なアラインメントを示す図である。影付のブロックは同一のアミノ酸を示す。EMBLデータベースの登録番号及び%同一性は、括弧内に与えられる。Malus x domestica(AAO18638;42.3%の同一性)、Common sunflower(JE0308;41.5%の同一性)、Arabidopsis thaliana(AAM64985;30%の同一性)及びRumex obtusifolius(CAD21441;29.3%の同一性)。
【図7】Coffea arabica CCCA2(A)及びCoffea robusta FRT−32(B)の異なる組織におけるCcCPI−1遺伝子の発現のRT−PCR分析を示す図である。PCR反応を、1/1000希釈した各cDNA 10μlを使用して実施した。サイクル条件は、94℃で2分間、(94℃で1分間、60℃で1.5分間及び72℃で1分間)×40サイクルであった。最終伸長ステップは72℃で7分間であった。PCRプライマーは、CcCPI-1(up)5'AGGAAAGTGGGAGCAAGGGAGAAGA3'、CcCPI-1(low)5'TAGTATGAACCCAAGGCCGAACCAC3'.であった。レーンは以下のように標識される:M、マーカー;+P、CcCPI−1遺伝子を含む希釈したプラスミド;R、根;S、茎;L、若葉;F、花。SG(G)、LG(G)、Y(G)及びRed(G)はそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリーから単離された粒である。SG(P)、LG(P)、Y(P)及びRed(P)はそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリーから単離された果皮組織である。
【図8】CcCPI−2 cDNAによってコードされる完全タンパク質と、NCBIにおいて入手可能な他の相同な全長システインプロテイナーゼとの最適なアラインメントを示す図である。影付のブロックは同一のアミノ酸を示す。EMBLデータベースの登録番号及び%同一性は、括弧内に与えられる。Rumex obtusifolius(CAD21441;66.7%の同一性)、Dianthus caryophyllus(AAK30004;71.7%の同一性)、Manihot esculenta(AAF72202;65.2%の同一性)。
【図9】Coffea arabica CCCA2(A)及びCoffea robusta FRT−32(B)の異なる組織におけるCcCPI−2遺伝子の発現のRT−PCR分析を示す図である。PCR反応を、1/1000希釈した各cDNA10μlを使用して実施した。サイクル条件は、94℃で2分間、(94℃で1分間、57℃で1.5分間及び72℃で1分間)×40サイクルであった。最終伸長ステップは72℃で7分間であった。PCRプライマーは、CcCPI-2(up)5'GTGAAGCCATGGTTGAACTT3'、CcCPI-2(low)5'GTAATGATACCTCAAGCCAGA3'.であった。レーンは以下のように標識される:M、マーカー;+P、CcCPI−2遺伝子を含む希釈したプラスミド;R、根;S、茎;L、若葉;F、花。SG(G)、LG(G)、Y(G)及びRed(G)はそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリーから単離された粒である。SG(P)、LG(P)、Y(P)及びRed(P)はそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリーから単離された果皮組織である。
【図10】CcCPI−3 cDNAによってコードされる完全タンパク質と、NCBIにおいて入手可能な他の相同な全長システインプロテイナーゼとの最適なアラインメントを示す図である。影付のブロックは同一のアミノ酸を示す。EMBLデータベースの登録番号及び%同一性は、括弧内に与えられる。Citrus x paradisi(AAG38521;42.4%の同一性)、Actinidia deliciosa(AAR92223;44.4%の同一性)及びArabidopsis thaliana(AAM64661;44%の同一性)。
【図11】CcCPI−4 cDNAによってコードされる完全タンパク質と、NCBIにおいて入手可能な他の相同な全長システインプロテイナーゼとの最適なアラインメントを示す図である。影付のブロックは同一のアミノ酸を示す。EMBLデータベースの登録番号及び%同一性は、括弧内に与えられる。Citrus x paradisi(AAG38521;23.6%の同一性)及びArabidopsis thaliana(AAM64661;20%の同一性)。
【図12】Coffea arabica CCCA2(A)及びCoffea robusta FRT−32(B)の異なる組織におけるCcCPI−4遺伝子の発現のRT−PCR分析を示す図である。PCR反応を、1/100希釈した各cDNA10μlを使用して実施した。サイクル条件は、94℃で2分間、(94℃で1分間、60℃で1.5分間及び72℃で1分間)×40サイクルであった。最終伸長ステップは72℃で7分間であった。PCRプライマーは、CcCPI-4(up)5'CTACGGTCGCAGCCAAATC3'、CcCPI-4(low)5'ACAACTGCACCTTCAATGTAC3'.であった。レーンは以下のように標識される:M、マーカー;+P、CcCPI−4遺伝子を含む希釈したプラスミド;R、根;S、茎;L、若葉;F、花。SG(G)、LG(G)、Y(G)及びRed(G)はそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリーから単離された粒である。SG(P)、LG(P)、Y(P)及びRed(P)はそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリーから単離された果皮組織である。
【図13】Coffea arabicaの異なる組織におけるアスパラギン酸プロテイナーゼ2(CcAP2)遺伝子のノザンブロット分析を示す図であり、図中、レーンは以下で標識される:R:根、S:茎、L:若葉;F:花;SG(G)及びSG(P)、LG(G)及びLG(P)、Y(G)及びY(P)並びにRed(G)及びRed(P)はそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリー由来の粒及び果皮についてであり、SG(G)、LG(G)、Y(G)及びR(G)はそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリー由来の果皮についてである。各レーンにおいて5μgの総RNAをロードした。パネルAは、ローディングコントロールとしてのブロッティング前の大きいリボソームRNAのエチジウムブロマイド染色を実証し、パネルBは、試験した特定の組織中のCcAP2 mRNAの出現を示すオートラジオグラムである。
【図14】CcCP−4のcDNA配列及び推定アミノ酸配列を示す図である。小文字:5’非翻訳領域及び3’非翻訳領域;大文字:オープンリーディングフレーム;太字:アミノ酸配列;*:終止コドン。
【図15】CcCP−4 cDNAによってコードされるタンパク質の完全配列と、NCBIデータベースにおいて入手可能な他の全長システインプロテイナーゼとのアラインメントを示す図である。これは、MegAlignソフトウェア(Lasergeneパッケージ、DNASTAR)中のCLUSTAL Wプログラムを使用して実施した。影付のブロックは同一のアミノ酸を示す。登録番号は括弧内に与えられる。Dacus carrota(JC7787);Ricinus communis(AF050756);Vicia sativa(Z34895);Phaseolus vulgaris(X56753);Helianthus annuus(AB109188);Glycine max Cys1(AB092555);Glycine max Cys2(AB092557);Canavalia ensiformis(P49046);Oryza sativa(AB004648);Vigna mungo(P12412);Pisum sativum(AJ004985)。
【図16】Megalign中のプログラムClustalWを使用して整列された全長cDNA配列CcCP−4 KDDL及び部分的cDNA配列CcCP−4(KDEL)を示す図である。
【図17】Megalign中のプログラムClustalWを使用して整列されたCcCP−4(KDDL)の完全オープンリーディングフレーム及びCcCP−4(KDEL)の部分的オープンリーディングフレームを示す図である。
【図18】CcCP−4遺伝子のKDEL/KDDL領域をコードするPCR増幅されたゲノムDNAについてのDNA配列クロマトグラムを示す図である。Robはrobusta変種を示し、Arabはarabica変種を示す。
【図19】Coffea arabicaの異なる組織におけるシステインプロテイナーゼCcCP−4遺伝子の発現のノザンブロット分析を示す図である。レーンは以下で標識される:R、根;S、茎;L、若葉;F、花。SG(G)、LG(G)、Y(G)及びRed(G)はそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリーから単離された粒である。SG(P)、LG(P)、Y(P)及びRed(P)はそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリーから単離された果皮組織である。各レーンにおいて5μgの総RNAをロードした。パネルAは、ローディングコントロールとしてのブロッティング前の大きいリボソームRNAのエチジウムブロマイド染色を実証し、パネルBは、試験した特定の組織中のCcCP−3 mRNAの出現を示すオートラジオグラムである。
【図20】発芽の間の粒全体におけるCcCP−4の発現のRT−PCR分析を示す図である。サンプリング時間は以下であった:0、滅菌処理の直後;2D、処理の2日後;3D、処理の3日後;5D、処理の5日後;1M、処理の1ヵ月後;2M、処理の2ヵ月後;−、DNAなしのコントロール;+P、希釈したCcCP−4プラスミドDNA;M、分子量マーカー。
【図21】CcAP−1 cDNAによってコードされる完全タンパク質と、NCBIにおいて入手可能な他の相同な全長アスパラギン酸プロテイナーゼ配列との最適なアラインメントを示す図である。影付のブロックは同一のアミノ酸を示す。データベースの登録番号は括弧内に与えられる。Arabidopsis thaliana(AY099617)及びArabidopsis thaliana(BAB09366)。
【図22】CcAP−2 cDNAによってコードされる完全タンパク質と、NCBIにおいて入手可能な他の相同な全長アスパラギン酸プロテイナーゼ配列との最適なアラインメントを示す図である。影付のブロックは同一のアミノ酸を示す。データベースの登録番号は括弧内に与えられる。Glycine max(BAB64296)、Ipomoea batatas(AAK48494)、Lycopersicon esculentum(S71591)及びNepenthes alata(BAB20972)。
【技術分野】
【0001】
コーヒーは、フレーバー分子の高度に複雑な混合物を含む。インスタント及び新鮮な粉砕されたコーヒー飲料の組成についての広範な研究は、今日までに850を超える化合物を同定しており、これらのうち多くがフレーバー活性分子である(Flament,I(2002)、コーヒーフレーバー化学(Coffee Flavor Chemistry)、John Wiley and Sons、UK)。しかし、1杯のコーヒー中に見出される最終コーヒーフレーバー分子は、原材料(植物種Coffea arabica又はCoffea canephora(robusta)の緑の粒(緑の豆))中にはほとんど存在しない。実際、コーヒーフレーバー化合物の大多数は、熟した赤いコーヒーチェリーの収穫から最終的な焙煎され粉砕されたコーヒー製品又はその抽出物(例えば、可溶性コーヒー製品)までに生じる複数の加工ステップの1つ又は複数の間に生成される。
【背景技術】
【0002】
コーヒーの生産における種々のステップが、Smith,A.W.、コーヒー;第1巻:化学(Coffee;Volume 1:Chemistry)1−41頁、Clark,R.J.及びMacrea,R.編、Elsevier Applied Science London and New York、1985;Clarke,R.J.、コーヒー:植物学、生化学並びに豆及び飲料の生産(Coffee:Botany,Biochemistry,and Production of Beans and Beverage)、230−250頁及び375−393頁;並びにClifford,M.N.及びWillson,K.C.編、Croom Helm Ltd、London中に記載されている。簡潔に述べると、このプロセスは、成熟して熟した赤いチェリーの収集で開始される。次いで、最も外側の層(すなわち果皮)が、乾式プロセス又は湿式プロセスのいずれかを使用して除去できる。乾式プロセスは最も単純であり、以下を含む:1)チェリーの等級分け(classification)及び洗浄;2)等級付け(grading)後のチェリーの乾燥(空気乾燥又は機械的乾燥のいずれか)、及び3)乾燥した果皮を除去するための乾燥したチェリーの脱穀(dehusking)。湿式プロセスは僅かにより複雑であり、より高い品質の緑の豆の生産を一般に導く。湿式プロセスは、C.arabicaチェリーとより頻繁に関連する。湿式プロセスは以下からなる:1)チェリーの等級分け、2)チェリーの脱肉(パルピング)(このステップは、収穫直後に実施され、一般に、成熟チェリーの「パルプ」すなわち果皮の機械的除去を含む)、3)脱肉後にチェリーの粒に付着したままの粘液が、バッチプロセスを使用して粒+付着した粘液をタンク中で水と共にインキュベートさせることによって除去される、「発酵」。この「発酵」プロセスは80時間まで継続されるが、発酵、4)乾燥(このステップは、発酵したコーヒー粒の空気乾燥又は機械的な熱気乾燥のいずれかを含む)及び5)「ハリング(hulling)」(このステップは、乾燥したコーヒー粒(乾燥したパーチメントコーヒー)の「パーチ(parch)」の機械的除去を含み、シルバースキン(銀皮)もまたしばしばこの段階で除去される)の間に増殖する微生物の種々の酵素活性及び代謝作用に起因して、しばしば24時間が、受容可能な発酵を可能にしかつ約6.8〜6.9から4.2〜4.6までpHを低下させるのに一般に充分である。湿式加工又は乾式加工の後に、得られた緑のコーヒー粒はしばしば分類され、ほとんどの分類手順は、粒のサイズ及び/又は形状に基づく。
【0003】
コーヒー加工における次のステップは、乾式加工したコーヒー又は湿式加工したコーヒーそれぞれの、脱穀(dehusking)又は脱穀(dehulling)後の緑の粒の焙煎である。これは、豆における有意な化学変化を誘導する時間依存的なプロセスである。焙煎の第1段階は、供給された熱が粒中に残留する水を追い出す場合に生じる。水の大部分がなくなると、温度が190℃〜200℃まで上昇して、適切な焙煎が開始される。豆の発色によって通常モニタリングされる焙煎の程度は、最終飲料製品のフレーバー特徴を決定するのに主要な役割を果たす。従って、焙煎の時間及び温度は、所望のコーヒーフレーバープロフィールを達成するために厳密に制御される。焙煎後、コーヒーは、コーヒー飲料又はコーヒー抽出物(後者は、インスタントコーヒー製品を製造するために使用される)の生産の間の抽出を容易にするために粉砕される。再度、粉砕のタイプは、飲料の最終的なフレーバーに影響を与え得る。
【0004】
かなりの量の研究がコーヒー中のフレーバー分子の同定について実施されているが、加工ステップの各々の間にコーヒー粒内で生じる物理反応及び化学反応に関して実施された研究はかなり少ない。この後者の点は焙煎反応について特に明白であり、多数の粒の成分が、熱により誘導される反応の非常に複雑な連続を経る(Homma,S.2001、「コーヒー:新事情(Coffee:Recent Developments)」.R.J.Clarke及びO.G.Vitzthum編、Blackwell Science、London;Yeretzian,C.ら((2002)Eur.Food Res.Technol.214、92−104;Flament,I(2002)、コーヒーフレーバー化学(Coffee Flavor Chemistry)、John Wiley and Sons、UK;Reineccius,G.A.、「メイラード反応及びコーヒーフレーバー(The Maillard Reaction and Coffee Flavor)」Conference Proceedings of ASIC、16th Colloque、Kyoto、Japan 1995)。
【0005】
コーヒー加工の異なるステップの間に生じる反応のほとんどの詳細は比較的不明のままであるが、コーヒーの芳香に関連するフレーバーの多数を担う重要なフレーバー生成反応は、コーヒー焙煎の間の「メイラード」反応であると考えられている。活発なメイラード反応は、焙煎ステップの間に、粒の還元糖/多糖分解産物とアミノ基含有分子(特に、タンパク質、ペプチド及びアミノ酸)との間で生じる。
【0006】
メイラード反応は、コーヒー焙煎の間のコーヒーのフレーバー分子及び芳香分子の生成に重要な貢献を明らかに果しているので、緑の豆中の1次メイラード反応のレベルと焙煎後に発生したフレーバー/芳香の品質との間には関連性がある可能性がある。
【0007】
上記のように、メイラード反応における基質の重要な群は、アミノ酸、ペプチド及びタンパク質である。2次元電気泳動を使用して、arabica及びrobustaの緑のコーヒー豆中の主要な貯蔵タンパク質のレベル及び量に差異が存在することが示されている。しかし、これらの保存タンパク質の差異とフレーバーの品質との間の関連性は注目されていない(Rogersら、1999、Plant Physiol.Biochem.第37巻、261−272)。未熟なコーヒー豆と成熟したコーヒー豆(これらは異なるフレーバー品質を有する)との間には貯蔵タンパク質に小さい差異が存在することもまた、最近見出された(Montavon,P.ら、2003、J.Agric and Food Chemistry 第51巻、2328−2334)。種子の成熟の間には多数の変化が生じるので、この後者の研究は、成熟によって引き起こされる品質の改善と主要なコーヒーの貯蔵タンパク質の2次元ゲルパターンにおいて見られる差異との間に関連性が存在し得ることを示唆している。
【0008】
arabica及びrobustaの緑の豆から単離されたペプチドのプロフィールにおいて差異が存在することが最近示された(Ludwigら、2000、Eur.Food Res Technol.、第211巻、111−116)。これらの結果は、arabica及びrobustaのペプチド抽出物がその芳香前駆体プロフィールにおいて異なることを示しているが、この報告中に示されるデータは、抽出物中のどの成分がこれらの芳香プロフィールの差異を担うかを同定していない。これらの研究者はまた、緑のコーヒーの粗製抽出物中に少なくとも2つの異なるプロテイナーゼ活性を検出したが、彼らは、いずれの比活性をも芳香/フレーバーの品質と相関させなかった(Ludwigら、2000、Eur.Food Res Technol.、第211巻、111−116)。最後に、緑のコーヒー粒の焙煎の後期段階の間に使用される非常に高い温度は、このコーヒー粒中に存在するタンパク質の引き続く切断を引き起こすとも考えられている(Homma,S.2001、「コーヒー:新事情(Coffee:Recent Developments)」.R.J.Clarke及びO.G.Vitzthum編、Blackwell Science、London;Montavon,P.ら、2003、「焙煎の間の緑のコーヒーのタンパク質プロフィールの変化(Changes in green coffee protein profiles during roasting)」、J.Agric.Food Chem.51、2335−2343)。しかし、このタンパク質分解についての全体的スキームは非常に不十分にしか理解されていないが、これはとりわけ、焙煎の開始前の原材料中の主要なコーヒータンパク質の正確な状態におそらく依存する。本発明者らが知る限り、コーヒー中のペプチドプロフィールがコーヒーの芳香/フレーバーの生成に関与する可能性を扱っている他の有意な報告は存在しない。
【0009】
Theobroma cacao(カカオ豆)の発酵した種子の焙煎において、メイラード反応の芳香/フレーバーの発生には、種子のアミノ酸及びペプチドが関与するようである。他の種子と比較すると、T.cacaoの種子は、異常に高いレベルのアスパラギン酸プロテイナーゼ活性を有することが示されている(Biehl,B.、Voigt,J.、Voigt,G.、Heinrichs,H.、Senyuk,V.及びBytof,G.(1994)「生のカカオ豆における芳香前駆体であるオリゴペプチド及びアミノ酸のpH依存的な酵素的形成(pH dependent enzymatic formation of oligopeptides and amino acids,the aroma precursors in raw cocoa beans)」.The Proceedings of the 11th International Cocoa Research Conference、1993年7月18日〜24日、Yamoussoukro、Ivory Coast)。高レベルのココアフレーバー前駆体を有するカカオ豆を生産するために、天然の発酵ステップを実施することが必要である(未発酵の豆は、焙煎したときにほとんどフレーバーを発生させない)。この発酵ステップの間に、パルプ中の糖が発酵し、高レベルの酸(特に酢酸)を生成する(Carr,J.G.(1982)ココア−発酵食品−経済学的微生物学(Cocoa.In Fermented Foods.Economic Microbiology.)第7巻.275−292頁.(A.H.Rose編).Academic Press)。発酵が継続するにつれて種子中のpHは低下し、細胞構造が破壊される。低いpHは、豊富なカカオ種子のアスパラギン酸プロテイナーゼを起動及び/又は活性化させ、細胞性タンパク質の大量分解を生じる(Biehl,B.、Passern,D.及びSagemann,W.(1982)「カカオ豆子葉の細胞内構造に対する酢酸の影響(Effect of Acetic Acid on Subcellular Structures of Cocoa Bean Cotylydons)」.J.Sci.Food Agric.33、1101−1109;Biehl,B.、Brunner,E.、Passern,D.、Quesnel,V.C.及びAdomako,D.(1985)「カカオ豆の発酵における酸性化、タンパク質分解及びフレーバー潜在能力(Acidification,proteolysis and flavour potential in fermenting cocoa beans)」.J.Sci.Food Agric.36、583−598)。ペプチド及びアミノ酸は、ココアフレーバー前駆体であることが示されている(Rohan,T.(1964)「チョコレート芳香の前駆体:発酵したカカオ豆及び未発酵のカカオ豆の比較研究(The precursors of chocolate aroma:a comparative study of fermented and unfermented cocoa beans)」.J.Food Sci.、29、456−459;Voigt,J.及びBiehl,B.(1995)「ココア特異的芳香成分の前駆体は、タンパク質分解加工によってココア種子のビシリンクラス(7S)グロブリンから誘導される(Precursors of the cocoa specific aroma components are derived from the vicilin−class(7S)globulin of the cocoa seeds by proteolytic processing)」.Bot.Acta 108、283−289)。従って、T.cacao種子のアスパラギン酸プロテイナーゼは、種子のセリンカルボキシペプチダーゼと共に、発酵の間のココアフレーバー前駆体の生成に重要であることが提唱されている(Voigt,J.及びBiehl,B.(1995)「ココア特異的芳香成分の前駆体は、タンパク質分解加工によってココア種子のビシリンクラス(7S)グロブリンから誘導される(Precursors of the cocoa specific aroma components are derived from the vicilin−class(7S)globulin of the cocoa seeds by proteolytic processing)」.Bot.Acta 108、283−289;Voigt,J.、Heinrichs,H.、Voigt,G.及びBiehl,B.(1994)「ココア特異的芳香前駆体は、ココア種子のビシリン様グロブリンのタンパク質分解消化によって生成される(Cocoa−specific aroma precursors are generated by proteolytic digestion of the vicilin−like globulin of cocoa seeds)」.Food Chemistry、50、177−184)。豊富なココア種子のアスパラギン酸プロテイナーゼをコードする遺伝子が同定されており、発酵したカカオ豆中のココアフレーバー前駆体のアミノ酸及びペプチドのレベルを増大させることができる、カカオ種子中でこのタンパク質を過剰発現させる方法は、国際特許公開第02/04617号(この全内容は、本明細書中に参照として組み込まれる)中に最近記載された。しかし、国際特許公開第02/04617号の教示は、酸発酵ステップを受けないコーヒー粒とは異なり、特定の長さの酸発酵ステップを受けるカカオ種子に関するものである。
【0010】
重要な液胞システインプロテイナーゼ(CP)は、KDEL含有システインプロテイナーゼである。このタイプのプロテイナーゼは、いくつかの植物において特徴付けられている。今日まで、C末端KDEL配列を有するシステインプロテイナーゼをコードする3つの遺伝子がarabidopsisにおいて見出されている(Gietl,C.及びSchmid,M.2001、Naturwissenschaften 88、49−58)。1つは成熟中の胚珠中で発現され、1つは導管中で発現され、3番目は成熟中の長角果において発現される。しかし、このタンパク質についてのより詳細な研究は、他の植物において実施されてきた。例えば、スルフヒドリル−エンドプロテイナーゼ(SH−EP)と称されるCPが、Vigna mungo種子の子葉において特徴付けられている(Toyooka,K.、Okamoto,T.及びMinamikawa,T.(200)J.Cell Biol.148、453−463)。SH−EPは、V.mungoの発芽中の子葉において新たに発現され、タンパク質貯蔵液胞中に蓄積した貯蔵タンパク質の分解に関与すると提唱されている(Okamato,T及びMinamikawa,T.J.Plant Physiol.152、675−682)。SH−EPポリペプチドの重要な特徴は、小胞体(ER)からタンパク質貯蔵液胞までのこのタンパク質の輸送を指図する特異的COOH末端配列KDELをこれが保有することである(Toyookaら、2000)。このSH−EPタンパク質は、そのKDEL配列の存在を介して、以前に記載されていない小胞輸送システムにおけるKV(KDEL Vesicles)と称される特定の小胞の形成に実際に関与することもまた、最近提唱されている(Okamato,T.、Shimada,T.、Hara−Nishimura,I.、Nishimura,M.及びMinamikawa,T.(2003)Plant Physiology、132、1892−1900)。
【0011】
関連の提案が、発芽中のトウゴマ(Ricinus communis)の実の子葉において見出されたKDEL含有CPタンパク質についてなされている。この植物において、著者らは、内胚乳のプログラム細胞死において、発芽中のトウゴマの実の胚に栄養を供給し続けるKDELプロテイナーゼを示している(Gietl,C.及びSchmid,M.2001、Naturwissenschaften 88、49−58)。これらの著者らは、トウゴマの実において、このKDELプロテイナーゼが3日目の前に発芽中の種子のERにおいて作製されることを提唱している。種子の皮が脱げると、3日目付近で、KDEL含有CPは、リシノソーム(ricinosome)と称される特定の小胞中に詰め込まれる。後に、内胚乳が4日目〜5日目の間に軟化するにつれて、KDEL−CPは、そのアンカー配列(KDEL)が切断して除かれ、このプロテイナーゼは、細胞性タンパク質の全般的な分解においてそれが補助する場所である細胞質に移動する。
【発明の開示】
【発明が解決しようとする課題】
【0012】
本発明の目的は、コーヒー中のタンパク質/ペプチド/アミノ酸のフレーバー前駆体プールを改変することである。
【0013】
より具体的には、本発明の目的は、収穫後の処理及び焙煎加工の後に変更されたフレーバーが達成できるように、原材料(緑の粒)中のフレーバー前駆体のレベルを改変することである。理論によって束縛されないが、有意に異なるフレーバーを有するコーヒー間のペプチド及びタンパク質の分解のレベルにバリエーションが存在する場合、これらのバリエーションは、これらの異なる粒中の内因性プロテイナーゼ活性の差異に起因することがあると考えられる。この差異は、特定の種子プロテイナーゼ遺伝子についての発現レベルのバリエーションによって、mRNA発現のレベルで検出可能である場合がある。
【課題を解決するための手段】
【0014】
従って本発明は、コーヒー粒(種子)特異的プロテイナーゼをコードする遺伝子配列を同定すること、並びにarabica及びrobustaにおけるこれらの遺伝子の発現にバリエーションが実際に存在することを示すことを含む。
【0015】
より具体的には、本発明は、2つの主要なコーヒーシステインプロテイナーゼ(CcCP−1及びCcCP−4)、4つの主要なコーヒーシステインプロテイナーゼインヒビター(CcCPI−1、CcCPI−2、CcCPI−3及びCcCPI−4)並びに2つのコーヒーアスパラギン酸プロテイナーゼ(CcAP−1及びCcAP−2)を開示し、これらは全てコーヒーの種子中で発現される。本発明者らは、種子発達における特に後期のこれらのタンパク質の過剰発現又は後期種子発達の間のこれらのタンパク質の低下した発現のいずれかが、成熟豆のアミノ酸/ペプチド/タンパク質のプロフィールを如何に変更できるかをさらに示す。成熟豆のアミノ酸/ペプチド/タンパク質のプロフィールを変更するために開示された遺伝子配列及び遺伝子構築物の1つ又は複数を使用することによって、本発明者らは、成熟コーヒー豆のフレーバー前駆体プロフィールを変更するための新たな方法を開示する。
【0016】
第1の態様において、本発明は、システインプロテイナーゼ活性を有するポリペプチドをコードするヌクレオチド配列又はこのヌクレオチド配列の相補体を含む単離されたポリヌクレオチドを提供し、ここで、このポリペプチドのアミノ酸配列と配列番号2又は16から選択されるアミノ酸配列とは、ClustalWアラインメント法に基づいて、少なくとも70%、好ましくは少なくとも80%の配列同一性を有し、この相補体はこのヌクレオチド配列と同数のヌクレオチドを含み、この相補体とこのヌクレオチド配列とは100%相補的である。好ましくは、このポリペプチドのアミノ酸配列と配列番号2又は16のアミノ酸配列とは、ClustalWアラインメント法に基づいて、少なくとも85%、好ましくは少なくとも90%、任意選択で少なくとも95%の配列同一性を有する。好ましくは、このヌクレオチド配列は、配列番号1又は15のヌクレオチド配列を含む。好ましくは、このポリペプチドは、配列番号2又は16のアミノ酸配列を含む。
【0017】
第2の態様において、システインプロテイナーゼインヒビター活性を有するポリペプチドをコードするヌクレオチド配列又はこのヌクレオチド配列の相補体を含む単離されたポリヌクレオチドが提供され、このポリペプチドのアミノ酸配列と配列番号4、10、12及び14から選択されるアミノ酸配列とは、ClustalWアラインメント法に基づいて、少なくとも70%、好ましくは少なくとも80%の配列同一性を有し、この相補体はこのヌクレオチド配列と同数のヌクレオチドを含み、この相補体とこのヌクレオチド配列とは100%相補的である。好ましくは、このポリペプチドのアミノ酸配列と配列番号4、10、12及び14から選択されるアミノ酸配列とは、ClustalWアラインメント法に基づいて、少なくとも85%、好ましくは少なくとも90%、任意選択で少なくとも95%の配列同一性を有する。好ましくは、このヌクレオチド配列は、配列番号3、9、11又は13から、任意選択で配列番号9、11又は13から、さらに任意選択で配列番号9又は13から選択される;なおさらに任意選択で配列番号9であるヌクレオチド配列を含む。好ましくは、このポリペプチドは、配列番号4、10、12及び14から、任意選択で配列番号10、12及び14から選択される、さらに任意選択で配列番号10又は14から;なおさらに任意選択で配列番号10であるアミノ酸配列を含む。
【0018】
第3の態様において、アスパラギン酸エンドプロテイナーゼ活性を有するポリペプチドをコードするヌクレオチド配列又はこのヌクレオチド配列の相補体を含む単離されたポリヌクレオチドが提供され、このポリペプチドのアミノ酸配列と配列番号6又は8、好ましくは配列番号8から選択されるアミノ酸配列とは、ClustalWアラインメント法に基づいて、少なくとも75%、好ましくは少なくとも80%の配列同一性を有し、この相補体はこのヌクレオチド配列と同数のヌクレオチドを含み、この相補体とこのヌクレオチド配列とは100%相補的である。好ましくは、このポリペプチドのアミノ酸配列と配列番号6又は8、好ましくは配列番号8から選択されるアミノ酸配列とは、ClustalWアラインメント法に基づいて、少なくとも85%、好ましくは少なくとも90%、任意選択で少なくとも95%の配列同一性を有する。好ましくは、このヌクレオチド配列は、配列番号5又は7、好ましくは配列番号7のヌクレオチド配列を含む。好ましくは、このポリペプチドは、配列番号6又は8、好ましくは配列番号8のアミノ酸配列を含む。
【0019】
さらなる態様において、本発明の第1の態様〜第3の態様のいずれか1つに記載のポリヌクレオチドを含むベクターが提供される。
【0020】
さらなる態様において、調節配列に作動可能に連結された本発明の第1の態様〜第3の態様のいずれか1つに記載のポリヌクレオチドを含む、非ネイティブの組換えDNA構築物が提供される。この非ネイティブの構築物において、このポリヌクレオチドが非ネイティブであるか、調節配列が非ネイティブであるか、又は両方が非ネイティブであるかのいずれかであることが理解される。
【0021】
さらなる態様において、本発明の第1の態様〜第3の態様のいずれか1つに記載のポリヌクレオチドで細胞を形質転換するステップを含む、細胞を形質転換するための方法が提供される。
【0022】
さらなる態様において、上記の非ネイティブの組換えDNA構築物を含む細胞が提供され、この細胞は好ましくは、原核細胞、真核細胞又は植物細胞、好ましくはコーヒー細胞である。
【0023】
さらなる態様において、このような形質転換された細胞を含むトランスジェニック植物が提供される。
【0024】
本願において、コーヒーチェリーとの用語は以下のように規定される:コーヒーチェリー;果実全体;外果皮、皮;果皮、チェリーの多肉質の主要な外側層;及び粒、コーヒーの種子。これらの用語のより完全な説明については、Clarke,R.J.、コーヒー:植物学、生化学並びに豆及び飲料の生産(Coffee:Botany,Biochemistry,and Production of Beans and Beverage)、230頁、Clifford,M.N.及びWillson,K.C.編、Croom Helm Ltd、London(その内容は、その全体が参照として組み込まれる)を参照のこと。
【発明を実施するための最良の形態】
【0025】
本発明は、以下の詳細な説明及び本願の一部をなす添付の配列表から理解できる。
【0026】
表1は、添付の表において使用される通りの対応する配列同定子(配列番号)と共に、本明細書中に記載されるポリペプチドを本明細書中以下に列挙する。
【0027】
表1:
配列番号1(CcCP1:システインプロテイナーゼ、核酸及びその対応するアミノ酸)
配列番号2(CcCP1:システインプロテイナーゼ、アミノ酸)
配列番号3(CcCPI−1:システインプロテイナーゼインヒビター、核酸及びその対応するアミノ酸)
配列番号4(CcCPI−1:システインプロテイナーゼインヒビター、アミノ酸)
配列番号5(CcAP1:アスパラギン酸エンドプロテイナーゼ1、核酸及びその対応するアミノ酸)
配列番号6(CcAP1:アスパラギン酸エンドプロテイナーゼ1、アミノ酸)
配列番号7(CcAP2:アスパラギン酸プロテイナーゼ2、核酸及びその対応するアミノ酸)
配列番号8(CcAP2:アスパラギン酸プロテイナーゼ2、アミノ酸)
配列番号9(CcCPI−2:システインプロテイナーゼインヒビター、核酸及びその対応するアミノ酸)
配列番号10(CcCPI−2:システインプロテイナーゼインヒビター、アミノ酸)
配列番号11(CcCPI−3:システインプロテイナーゼインヒビター、核酸及びその対応するアミノ酸)
配列番号12(CcCPI−3:システインプロテイナーゼインヒビター、アミノ酸)
配列番号13(CcCPI−4:システインプロテイナーゼインヒビター、核酸及びその対応するアミノ酸)
配列番号14(CcCPI−4:システインプロテイナーゼインヒビター、アミノ酸)
配列番号15(CcCP−4:システインプロテイナーゼ、核酸及びその対応するアミノ酸)
配列番号16(CcCP−4:システインプロテイナーゼ、アミノ酸)
【0028】
この配列表は、ヌクレオチド配列特徴について1文字コードを使用し、アミノ酸について、IUPAC−IUBMB Standardsのために規定され、Nucleic Acids Research 13:3021−3030(1985)(これは、本明細書中に参照として組み込まれる)中に記載されるような3文字コードを使用する。
【0029】
本明細書中で使用する場合、「ポリヌクレオチド」は、核酸フラグメントのようなヌクレオチド配列である。ポリヌクレオチドは、一本鎖又は二本鎖の任意選択で合成、非天然又は変更されたヌクレオチド塩基を含むRNA又はDNAのポリマーであってもよい。DNAのポリマーの形態のポリヌクレオチドは、cDNA、ゲノムDNA、合成DNA又はそれらの混合物の1つ又は複数のセグメントを含んでもよい。
【0030】
本発明において、当業者に一般に使用されるアルゴリズムによって決定されるような、本明細書中に開示されるアミノ酸配列に対する、類似の核酸フラグメントがコードするアミノ酸配列の%同一性によって、この類似の核酸フラグメントが特徴付けられる。適切な核酸フラグメント(すなわち、本発明の第1の態様〜第3の態様に記載の単離されたポリヌクレオチド)は、本明細書中に開示されるアミノ酸配列に対して少なくとも70%同一、好ましくは少なくとも80%同一であるポリペプチドをコードする。好ましい核酸フラグメントは、本明細書中に開示されるアミノ酸配列に対して少なくとも85%同一であるアミノ酸配列をコードする。より好ましい核酸フラグメントは、本明細書中に開示されるアミノ酸配列に対して少なくとも90%同一であるアミノ酸配列をコードする。本明細書中に開示されるアミノ酸配列に対して少なくとも95%同一であるアミノ酸配列をコードする核酸フラグメントが、なおより好ましい。複数の配列アラインメントは、ClustalW法のアラインメントを使用して実施されるべきである(Thompsonら、1994、Nucleic Acids Research、第22巻、4673−4680頁;Higgins及びSharp 1989 Cabios.5:151−153)。
【0031】
本明細書中で使用される場合、用語「類似の核酸フラグメント」とは、1つ又は複数のヌクレオチド塩基の変化が1つ又は複数のアミノ酸の置換を生じるが、その変化がこのヌクレオチド配列によってコードされるポリペプチドの機能に影響を与えないか、又は例えばアンチセンス技術若しくは同時発現技術を介した遺伝子サイレンシングによって核酸フラグメントが遺伝子発現を媒介する能力に影響を与えないかのいずれかである、ポリヌクレオチド配列をいう。用語「類似の核酸フラグメント」とはまた、1つ又は複数のヌクレオチド塩基が欠失又は挿入された改変ポリヌクレオチド配列をいい、但しこの改変は、このヌクレオチド配列によってコードされるポリペプチドの機能に影響を与えないか、又は遺伝子サイレンシングによって核酸フラグメントが遺伝子発現を媒介する能力に影響を与えないかのいずれかである。従って、本発明の範囲が、本明細書中に具体的に開示されたポリヌクレオチド配列及びポリペプチド配列を超えて広がることが理解される。類似の核酸フラグメントは、植物又は植物細胞中の改変核酸フラグメントによってコードされるポリペプチドのレベルに影響を与えるそれらの能力について、部分フラグメント又は改変核酸フラグメントの形態の核酸フラグメントをスクリーニングすることによって選択できる。
【0032】
用語「作動可能に連結された」とは、一方の核酸フラグメントの機能が他方の核酸フラグメントによって影響されるような、単一の核酸フラグメントに対する2つ以上の核酸フラグメントの結合をいう。「調節配列」とは、コード配列の上流、その内部又はその下流に位置し、それに結合したコード配列の転写、RNAのプロセシング若しくは安定性又は翻訳に影響を与えるヌクレオチド配列をいう。調節配列には、プロモーター、翻訳リーダー配列、イントロン、転写終結配列及びポリアデニル化認識配列が含まれてもよい。プロモーターの形態の調節配列がコード配列に作動可能に連結される場合、この調節配列は、コード配列の発現に影響を与えることがある。コード配列は、センス又はアンチセンスの配向で、調節配列に作動可能に連結できる。
【0033】
用語「発現」とは、本発明の核酸フラグメント由来のセンスRNA(mRNA)又はアンチセンスRNAの転写及び安定な蓄積をいう。発現はまた、mRNAのポリペプチドへの翻訳を指すこともある。過剰発現とは、正常細胞すなわち形質転換されていない細胞における産生レベルを超える、トランスジェニック細胞における遺伝子産物の産生をいう。「変更されたレベル」とは、正常細胞すなわち形質転換されていない細胞のものとは異なる量又は割合の、トランスジェニック細胞における遺伝子産物の産生をいう。
【0034】
「形質転換」とは、遺伝的に安定な継承を生じる、宿主細胞のゲノムへの核酸フラグメントの移入をいう。形質転換された核酸フラグメントを含む宿主細胞は、本明細書中で「トランスジェニック細胞」と称される。
【0035】
本明細書中で使用されるような標準的な組換えDNA技術及び分子クローニング技術は当該分野で周知であり、Sambrookら「分子クローニング:ラボラトリーマニュアル(Molecular Cloning:A Laboratory Manual)」;Cold Spring Harbor Laboratory Press:Cold Spring Harbor、1989(これは、本明細書中に参照として組み込まれる)中により完全に記載されている。
【実施例】
【0036】
以下の実施例は、本発明をこれらの実施例に限定することなく、本発明を例示する。これらの実施例において、他のように特定されない限り、全ての部及び百分率は重量により、度数はセルシウスである。
【0037】
以下の実施例において、これらの略号が使用されている:
PCR:ポリメラーゼ連鎖反応
RACE:cDNA末端の迅速増幅
【0038】
上記の考察及び下記の実施例から、当業者は、本発明の本質的な特徴を確認することができ、本発明の範囲から逸脱することなく本発明の本質的な特徴を様々に変化及び改変して、本発明を所望に応じて種々の用途及び条件に適合させることができる。
【0039】
cDNAライブラリーの生成及びスクリーニング
種子特異的RNAの生成
Robusta変種Q121のコーヒーチェリーを、ICCRI(インドネシア)において30WAF(開花後の週)で収穫した。次いで、これらのチェリーの果皮を除去し、残留する外胚乳/内胚乳材料を凍結し、液体窒素中で粉末になるまで粉砕した。RNAを、カカオ種子のRNA抽出のための以前に記載された方法を使用して、この凍結粉末材料から抽出した(Guilloteau,M.ら、2003、Theobroma cacao種子中の油体:15.8kDa及び16.9kDaのオレオシンをコードするcDNAのクローニング及び特徴付け(Oil bodies in Theobroma cacao seeds:cloning and characterisation of cDNA encoding the 15.8 and 16.9 kDa oleosins).Plant Science 第164巻、597−606)。ポリA+RNAを、AMBION(Ambion,Inc.製)の「PolyA Purist(商標)」キットをキットの指示書に従って使用して、約250μgの総RNAから調製した。
【0040】
種子cDNAクローンの第1セットの生成
次いで、約50ng〜100ngのこのポリA+RNAを、「SuperScript(商標)II RNaseH−逆転写酵素(GIBCOBRL(商標))及び以下のようなSMART(商標)PCR cDNA合成キット(Clontech)を使用して、第1鎖cDNAの合成において使用した。2μlの30WAF ポリA+RNA、1μLのCDSオリゴ(SMART(商標)PCR cDNAキット、Clontech)、1μLのSmart IIオリゴ(SMART(商標)PCR cDNAキット、Clontech)及び8μLの脱イオンH2Oを含む反応。この混合物を72℃で5分間加熱し、次いで氷上に置いた。次いで以下を添加した;1μLの10mM dNTP、4μLのSuperScriptII(商標)第1鎖緩衝液及び2μLのDTT。この混合物を42℃で2分間置き、次いで1μLのSuperScriptII(商標)RNaseH−逆転写酵素(200単位/μL、GIBCO BRL(商標))を添加し、この混合物を空気循環インキュベータ中で42℃でさらに50分間インキュベートした。
【0041】
逆転写反応後、以下のPCR反応を実施した。Advantage(商標)2ポリメラーゼ(Advantage(商標)2 PCRキット、ClonTech)を含むSMART(商標)PCR cDNAキット(Clontech)中に記載される98μLのMaster Mixを氷上に設置し、次いで3μLの上記第1鎖cDNA合成反応を添加した。次いで、この100μLのPCR反応を、MJ Research PTC−150 HB装置中に配置し、以下のPCR条件を実施した:95℃で1分間、次いで(95℃で15秒間、65℃で30秒間、68℃で6分間)×16サイクル。増幅されたDNAを、Strataprep(商標)PCR Purification Kit(Stratagene)を供給者の指示に従って使用して精製した。次いで、50μLの脱イオン水中に溶出されたDNAを、以下のようなPCR−Script(商標)Ampクローニングキット(Stratagene)中に含まれるPfu−1ポリメラーゼ試薬を使用して「研磨」した;50μLのDNA、5μLの10mM dNTP、6.5μLの10×Pfu−1研磨緩衝液、5μLのクローニングされたPfu−1 DNAポリメラーゼ(0.5U/μl)。次いでこの反応を、加熱されたカバーを有するPCR装置(Perkin Elmer)中で72℃で30分間インキュベートした。pPCR−Script(商標)Ampキット(Stratagene)中に記載されるプロトコルを使用して、研磨した(平滑化した)PCR産物を、Srf−1酵素の存在下でSrf−1消化したpPCR−Script(商標)Amp SK(+)ベクター中に連結し、この連結反応産物を、XL−10 Gold(商標)Kan ultracompetent E.coli細胞中に形質転換した。挿入配列を含むプラスミドを有する形質転換についての選択を、LB−Ampプレート並びにその表面上に広げられたIPTG及びXgalを使用して、pPCR−Script(商標)Ampキット中に記載されるように実施した。白色コロニーを選択し、これらのクローンをDavl−1などと命名した。
【0042】
サイズ選択されたcDNAを有する種子cDNAクローンの第2セットの生成
種子は、少数のタンパク質(例えば、種子の貯蔵タンパク質)を高度に発現する(Whiteら、2000、Plant Physiology、第124巻、1582−1594)。cDNAがこのような組織から調製される場合、非常に高いレベルの貯蔵タンパク質及び他の種子特異的タンパク質が、高レベルのcDNAの「重複」を導く。すなわち、生成されたcDNA集団は、高い割合の同じcDNAを含む。コーヒー種子のmRNAから作製されたcDNAの重複を低下させるため及び弱く発現された長いcDNAを選択的に特徴付けるために、第2のcDNAクローニングストラテジーもまた使用した。上記の逆転写酵素反応の産物を使用して、以下のPCR反応を、Advantage(商標)2 PCRキット(ClonTech):3μLの逆転写酵素反応、5μLの10×Advantage(商標)2 PCR緩衝液、1μLのdNTP(各々10mM)、2μLのPCRプライマー(SMART(商標)PCR cDNAキット、Clontech)、39μLの脱イオン水及び1μLの50×Advantage(商標)2ポリメラーゼミックスを使用して設定した。次いで、このPCR反応を、MJ Research PTC−150 HB装置中に配置し、以下のPCR条件を実施した:95℃で1分間、次いで(95℃で15秒間、65℃で30秒間、68℃で6分間)×16サイクル。PCRの終了時に、1μLの10% SDSをゲルローディング緩衝液と共に添加し、このサンプルを37℃で10分間加熱した。次いで、このサンプルを、エチジウムブロマイドを含まない0.7%アガロースゲル上にロードするために分割し、10%をDNAマーカーレーンの脇の小さいウェル中にロードし、他の90%を隣接する大きい調製スケールのウェル中にロードした。このゲルを泳動した後に、サイズマーカーを有するゲル区画+10%反応サンプルを、エチジウムブロマイドで染色した。次いで、この染色したゲル区画をテンプレートとして使用して、このゲルの残りの未染色の(調製)部分中に存在するcDNAとは異なるサイズのPCR増幅されたcDNAを含むゲルスライスを生成した。示されたサイズ範囲のPCRフラグメントを有する6つのゲルスライスを生成した;A1A(0.8kb〜1kb)、A1B(1kb〜1.5kb)、A2(1.5kb〜2.25kb)、A3(2.25kb〜3.25kb)、A4(3.25kb〜4kb)及びA5(4kb〜6.5kb)。
【0043】
各ゲルスライス中のDNAを、供給業者の指示に従ってQiagenのQIAEX IIキットを使用してアガロースから溶出した(サンプル3A、4A及び5Aについては50℃で10分間加熱し、サンプル1A、1B及び2Aについては室温で10分間加熱した)。次いで、精製した二本鎖cDNAを、3’Tオーバーハングを有するフラグメントを作製する以下のようなTAQ酵素ミックスを用いたPCRによってさらに再増幅した:30μLのゲル単離された二本鎖cDNA、5μLの10×TAQ緩衝液(TAQ PLUS精密ポリメラーゼミックスを補充したもの、Stratagene)、1μLの40mM dNTP(各々10mM)、2μLのPCRプライマー(SMART(商標)PCR cDNAキット、Clontech)、0.5μLのTAQ PLUS精密ポリメラーゼミックス(Stratagene)及び11.5μLの脱イオン水。PCR反応条件は以下の通りであった:95℃で1分間、次いで(95℃で15秒間、65℃で1分間、72℃で8分間)×7サイクル、次いで(95℃で15秒間、65℃で1分間、72℃で10分間)×1サイクル。
【0044】
次いで、生成されたPCR増幅されたDNAを、ベクターpCR(商標)−TOPO(商標)中に連結し、TOPO(商標)TAキット(Invitrogen)を供給業者によって記載される通りに使用して、TOP10 E.coli細胞中にクローニングした。これらのクローンを、その単離の順番及びサイジングゲルにおけるそれらの位置によって命名した(例えば、A2−1、A2−2など)。
【0045】
種子cDNAのスクリーニング及び予備的同定
Dav−1ライブラリーにおいて得られた白色コロニーの第1セットを、使用したクローニング部位に隣接したT3プライマー及びT7プライマーを使用して挿入配列をPCR増幅し、ゲル上のPCR増幅されたフラグメントを試験することによって、各挿入配列のサイズを最初に決定することによってスクリーニングした。
【0046】
各白色コロニーを200μlの滅菌水中に再懸濁し、このうち10μl〜30μlを、5μlの10×Taqポリメラーゼ緩衝液(Stratagene)、1μlの10mM dNTPミックス、2.5μlの20μM T3プライマー、2.5μlの20μM T7プライマー、1μlのDMSO、0.5μlのTaqポリメラーゼ(Stratagene)及び50μlの最終容量までのH2Oに添加した。使用したPCR反応プログラムは、94℃で1分間、次いで(94℃で1分間、55℃で1.5分間及び72℃で3.5分間)×30サイクル、及び72℃で7分間の最終サイクルであった。重複を低下させるために、類似のサイズのPCR挿入配列を、制限酵素Hae IIIによる消化に供した。同じHae III制限パターンを有するPCRフラグメントをさらに研究することはしなかった。次いで、>500bpでありかつ独自のHae III制限パターンを有したPCRフラグメントを有するクローンのプラスミドを、隣接するベクター配列中にコードされた適切なT7配列決定プライマー又はT3配列決定プライマーを使用する5’末端ジデオキシ配列決定のために、Qiawall 8ウルトラプラスミドキット(Qiagen)を使用することによって精製した。これらの挿入配列は方向性をもった様式でクローニングされるわけではないので、精製されたプラスミドDNAのSca 1消化によって各クローンの5’末端を決定することが最初に必要であった(CDS SMARTプライマーは、挿入配列の配向の決定を可能にするSca 1部位を含む)。得られたDNA配列データを、プログラムBLASTX(商標)を使用してGENEBANK中の非重複データベースタンパク質に対して引き続いて書き込み、各cDNAクローンの予備的注釈を得た。
【0047】
種子cDNAバンクは、高レベルの重複を有する。すなわち、少数の種子mRNA(例えば、種子の貯蔵タンパク質をコードするmRNA)が異常に高いレベルの発現を有し、従って、それらのcDNAは、種子cDNAバンク中に非常に豊富である(Whiteら、2000、Plant Physiology、第124巻、1582−1594)。従って、コーヒー種子のcDNAを配列決定する第1ラウンドにおいて主要な重複cDNAを同定した直後に、プレスクリーニングステップを、挿入配列のサイズ決定の前に、白色の挿入配列含有コロニーのために付加した。4つの配列が非常に高度に発現され、以下の特異的プライマーセットを、これらの重複配列の各々について作製した。
【0048】
このプレスクリーニングステップのためのPCR反応は、以下のように実施した:滅菌H2O中の白色コロニー10μl〜30μl、5μlの10×Taq緩衝液(Stratagene)、1μlの10mM dNTP、2.5μlの各プライマー(20μM)、1μlのDMSO、0.5μlのTaqポリメラーゼ(Stratagene 10U/μl)及び滅菌H2Oを添加して、50μlの最終反応総容量にした。PCRプログラムは、94℃で1分間、次いで(94℃で1分間、各プライマー対に特異的な温度で1.5分間、72℃で2.5分間)×30サイクル、その後の72℃で7分間であった。
【0049】
全長cDNA挿入配列の配列決定及び配列分析
その部分配列がプロテイナーゼ及びプロテイナーゼインヒビターに対して最初の相同性を示したcDNAクローンを、標準的なジデオキシプライマーウォーキングストラテジーを使用して、両方の鎖について完全に配列決定した。これらの配列は、配列番号1、3、5、7、9、11、13及び15の下に示される。得られた全長配列を、BLASTXを使用してGanBank非重複タンパク質データベースに対して再度書き込み、予備的注釈を強化した。
【0050】
配列対の配列の同一性を、Lasergene(商標)ソフトウェアパッケージ(DNASTAR Inc)中のMegAlign(商標)モジュール中に含まれるClustalW(商標)プログラムを使用して計算した。デフォルトパラメータは以下のように選択した:(1−MULTIPLE ALIGNMENT PARAMETERS−Gap penalty 15.00、Gap length penalty 6.66、Delay divergent Seqs(%)30、DNA transition weight 0.5、Protein Weight Matrix−Gonnet Series、DNA Weight Matrix IUB。2−PAIRWISE ALIGNMENT PARAMETERS−Slow/Accurate(Gap Penalty 15.00、Gap Length Penalty 6.66)、Protein Weight Matrix−Gonnet 250、DNA Weight Matrix−IUB)。使用した配列は、各cDNA又は各cDNAの完全ORF(オープンリーディングフレーム)の全長ヌクレオチド配列のいずれかであった。
【0051】
表2:CcCP−1、CcCPI−1、CcAP−1及びCcAP−2とGenBankの非重複タンパク質データベース中に見出される関連遺伝子及びWO 02/04617号の関連遺伝子との間の、核酸配列及びアミノ酸配列の同一性の値。
【表1】
【0052】
5’RACE PCR
クローンA5−812のcDNA挿入配列はイントロンを含むことが見出された。従って、このタンパク質のコード配列を確認するために、完全コード配列を含む新たなcDNAを単離することが必要であった。これを、SMART(商標)RACE cDNA増幅キット(Clontech)を使用することによって達成した。5’RACEに使用した第1鎖cDNAを、上記のcDNAライブラリーについてすでに記載した通りに作製した。遺伝子特異的プライマーrAP2(5’ CATATAATATTAAAAGCACCACCCATAA 3’)を設計した。この配列は、A5−812クローンのポリ(A)テイルから92pbに位置する。次いで、この特異的プライマーを、以下の条件下でPCR反応においてCLONTECHキット中のUniversal Primer Mix(UPM)と共に使用した;2.5μlの第1鎖cDNA産物、5μlの10×Advantage 2 PCR Buffer(CLONTECH)、1μlのdNTP Mix(10mM)、1μlの50×Advantage 2 Polymerase Mix(CLONTECH)、5μlの「Universal Primer A Mix」(10×)(CLONTECH)、1μlのrAP2(10μM)及び滅菌水を添加して、50μlの最終容量にした。PCRサイクル条件は、(94℃で30秒間、68℃で30秒間及び72℃で3分間)×20サイクルとその後の72℃で5分間の最終伸張反応であった。約1700pbのフラグメントが得られ、「CONCERT(商標)Rapid Gel Extractionキット」(GibcoBRL)を使用してこれをゲルから切り出した。単離したフラグメントをpCR4−TOPOベクター中にクローニングし、Topo−TAクローニングキット(Invitorogen)を使用してEscherichia coli中に形質転換した。次いで、得られたプラスミドを、プラスミド抽出キット(QIAfilter Plasmid Midi Kit、Qiagen、France)を使用して精製し、このプラスミドの挿入配列を二本鎖配列決定した。
【0053】
クローンA5−442(AP1)のDNAは、cDNAの5’領域を欠くことが見出された。この領域を単離するために、5’RACEを、SMART(商標)RACE cDNA増幅Kit(Clontech)を使用して実施した。ポリ(A)テイルから396pbに位置する配列特異的プライマーrAP1(5’−TGGAGTCACAAGATGTCTCGACGAACTG−3’)を設計した。次いで、この特異的プライマーを、以下の条件下でPCR反応においてCLONTECHキット中のUniversal Primer Mix(UPM)と共に使用した;2.5μlの第1鎖cDNA、5μlの10×Advantage 2 PCR Buffer(CLONTECH)、1μlのdNTP Mix(10mM)、1μlの50×Advantage 2 Polymerase Mix(CLONTECH)、5μlの「Universal Primer A Mix」(10×)(CLONTECH)、1μlのrAP1及び滅菌水を添加して、50μlの最終容量にした。PCRサイクル条件は、(94℃で30秒間、68℃で30秒間及び72℃で3分間)×20サイクルとその後の72℃で5分間の最終伸張反応であった。約2,000bpのフラグメントが得られ、「CONCERT(商標)Rapid Gel Extractionキット」(GibcoBRL)を使用してこれをゲルから切り出した。単離したフラグメントをpCR4−TOPOベクター中にクローニングし、Topo−TAクローニングキット(Invitorogen)を使用してEscherichia coli中に形質転換した。次いで、得られたプラスミドを、プラスミド抽出キット(QIAfilter Plasmid Midi Kit、Qiagen、France)を使用して精製し、このプラスミドの挿入配列を二本鎖配列決定した。
【0054】
大きいEstライブラリーのためのRNA調製:
RNAを、以前に記載された方法を使用して、種々の発達段階の解剖した粒組織及び果皮組織並びに若葉から単離した。異なるEstライブラリーを生成するためのRNAを調製するために使用した変種及び組織は以下の通りであった:(1)若葉、1つの変種(FRT−32);(2)5つの変種(FRT32、FRT−31、FRT−400、FRT−4001及びQ121)由来の果皮(8つの異なる発達段階);(3)1つの変種(FRT−31)由来のチェリー全体、受精後22週(WAF);(4)5つの変種(FRT32、FRT−31、FRT−400、FRT−4001及びQ121)由来の粒、18+22WAF;(5)5つの変種(FRT32、FRT−31、FRT−400、FRT−4001及びQ121)由来の粒、30WAF;(6)5つの変種(FRT32、FRT−31、FRT−400、FRT−4001及びQ121)由来の粒、42WAF;並びに(7)2つの変種(FRT−32及びQ121)由来の粒、46WAF。
【0055】
cDNAクローンの生成及びDNA配列分析。
種々のEstライブラリーのためのcDNAクローンを、以下のように調製した:ポリA+mRNAを、小規模の単離のための製造業者の指示に従ってPolyATrack(商標)mRNA Isolation System(System IV、Promega)を使用して単離した。次いで、精製したポリA+mRNAを使用して、ZAP−cDNA(商標)ライブラリー構築キット(カタログ番号200450 Stratagene)中に記載されるように、λファージ中への一方向クローニングのためのcDNAを調製した。マス切り出しプロトコルは、Uni−ZAP XRベクターからpBlueScriptファージミドを切り出すためのものであり、80μlのx−gal(20mg/ml)及び16μlのIPTG(0.5M)を含む150mmのLB−アンピシリン寒天プレート上にプレートした後に白色コロニーが得られた。単一コロニーをランダムに選択して、cDNA挿入配列の5’末端を配列決定するために次いで使用されるプラスミドDNAを生成した。
【0056】
得られたDNA配列は、各クローンについてのEST配列(Expressed Sequence Tag)を生成した。次いで、7つのライブラリー由来の全てのEst配列データを「in−silico」でクラスター化し、「ユニジーン(unigene)」配列セットと称される独自の群の配列を生成した。従って、各「ユニジーン」配列は、理論的に別個の遺伝子産物に対応する。しかし、多数のユニジーンは部分的cDNA配列を示しているだけなので、いくつかの遺伝子が2つ以上のユニジーンによって示される可能性があることに留意すべきである。次いで、ユニジーンセットの予備的注釈を、各ユニジーン配列が非重複GenBankタンパク質データベースに対して検索される自動BLAST検索を用いて実施した。このBLAST検索アプローチは、「ユニジーン注釈」と称される5つの最良のBLAST「ヒット」(最低のe値を有する「ヒット」)を生じた。
【0057】
ノザンブロット分析
新たに採取した根、若葉、茎、花並びに異なる発達段階の果実(小さい緑の果実(SG)、大きい緑の果実(LG)、黄色い果実(Y)及び赤い果実(R))を、トゥール(フランス)において温室条件下(25℃、70RH)で成長したCoffea arabica CCCA2及びエクアドル又はICCRI(インドネシア)のいずれかで成長したCoffea canephora FRT32から採取した。新たな組織を液体窒素中で即座に凍結し、上記抽出手段を使用して各組織から総RNAを単離した。合計5μgのRNAを、ホルムアルデヒドを含む1.2%(w/v)の変性RNAゲル上で泳動した。各植物組織由来の総RNAサンプルを、7μLの「RNA Sample Loading Buffer」(エチジウムブロマイドを含まない、Sigma)の存在下で65℃で15分間加熱し、次いで直後に氷上に2分間置き、その後1.2%のRNAゲル上にロードした。このゲルを、60ボルトで5時間泳動した。次いで、このゲルを10×SSC中に20分間にわたって2回浸した。ゲル中のRNAを、10×SSC中で「Positive TM Membrane」(Qbiogene)にキャピラリートランスファーによって一晩移し、このRNAを、このブロットを80℃で30分間加熱することによって固定した。プローブを、(P32)dCTPの存在下で「Rediprime(商標)IIランダムプライム標識システム」キット(Amersham)を使用して生成した。ハイブリダイゼーションを、ハイブリダイゼーション溶液(5×SSC、40μg/mlの変性サケ精子DNA、5%[w/v]のSDS及び5×Denhardt’s溶液)中で65℃で24時間実施した。次いでこのメンブレンを、各々30分間の間に、2×SSC、0.1% SDS[w/v]及び1×SSC、0.1% SDS[w/v]を使用して65℃で2回洗浄した。
【0058】
図1中に示されるノザンブロット分析は、コーヒーのシステインプロテイナーゼ遺伝子CcCp−1遺伝子が、試験した全ての段階でC.arabicaのコーヒーチェリーにおいて発現され、黄色いチェリーが他の段階よりも僅かに高い発現レベルを示すことを実証する。C.arabicaの根でも茎でも葉でも、この遺伝子の発現は検出されなかった。図2は、RNAの新たな調製物を使用した、C.arabicaにおけるCcCP−1の発現を試験する別のノザンブロット実験を示す。この実験のために、4つの段階についてチェリーを解剖して、各段階のチェリー発達について果皮組織及び粒組織を生成した。次いで、総RNAをこれらの組織から抽出した。得られた結果は、チェリー発達の間のCcCP−1の発現の同じ一時的パターンを示すが、この新たな実験は、CcCP−1が、チェリーの粒組織中のみにおいて高レベルで主に発現されることをさらに示す。CcCP−1遺伝子の有意な発現は、コーヒーチェリーの果皮においては見られない。この後者の結果は、通常の成長条件下でのコーヒー粒のタンパク質、ペプチド及びアミノ酸のプロフィールの全面的な変更におけるこの遺伝子産物の役割を支持する。
【0059】
本発明者らは、コーヒーの葉並びに異なる段階の発達中のコーヒーチェリーから解剖した種子組織及び果皮組織から、ESTライブラリーを生成した。異なるライブラリー中のCcCP−1 ESTの検出(以下に示される−表3を参照のこと)もまた、この遺伝子が粒において強く発現されるが、果皮においても葉においても有意には発現されないことを実証する。種子の発達の間のCcCP−1の発現パターンは、Vicia sativaのその提唱された相同配列(CPR4遺伝子:Fischer,J.ら、2000.Plant Molecular Biology、43、83−101)について見られる発現パターンと類似である。これらの著者らは、CPR4が葉においても根においても茎においてもノザンブロッティングによって検出されないことを示しており、CcCP−1が粒特異的であるという主張をさらに強化している。例えば、CcCP−1のアンチセンス構築物又はCcCP−1の過剰発現構築物のために粒特異的プロモーターを使用することによって、本明細書中で示唆されるような粒におけるCcCP−1の発現を特異的に変更することは、他の組織において代謝を妨害しないと予測される。
【0060】
【表2】
【0061】
CcCP−1についての任意選択のアラインメント(図2A)は、このcDNAがシステインプロテイナーゼをコードすることを示す。
【0062】
図3中に示されるノザンブロット分析は、コーヒーのシステインプロテイナーゼインヒビター遺伝子CcCPI−1遺伝子が、試験した全ての段階でC.arabicaのコーヒーチェリーにおいて発現されることを実証する。しかし、システインプロテイナーゼCcCP−1について見られた発現とは対照的に、CcCPI−1は、2つの初期段階のコーヒーチェリー発達(小さい緑及び大きい緑)においてより高い発現を示し、この遺伝子は、2つの後期段階のチェリー発達においてより低いレベルで発現される。この発現パターンは、システインプロテイナーゼインヒビタータンパク質(CcCPI−1)が、種子において特異的に発現されるシステインプロテイナーゼ(例えば、コーヒーチェリー中のCcCP−1)の活性レベルを制御するという本発明の仮説と一致する。制御タンパク質(例えば、システインプロテイナーゼインヒビタータンパク質)は、標的タンパク質が発現される時点から連続的にその標的タンパク質の活性レベルを制御する必要がある場合、その標的タンパク質よりも早く発現されることが予測できる。C.arabicaの根でも茎でも葉でも、この遺伝子の発現は検出されなかった。CcCP−1及びCcCPI−1の発現パターンの類似性は、これらのタンパク質が機能的に相互作用し得るという本発明の仮説と一致することが留意される。
【0063】
ノザンブロッティングの結果(図3)は、CcCPI−1がコーヒーチェリーにおいて全ての段階で発現されることを示した。しかし、この実験は、この発現がチェリー全体中に存在するのか、又は果皮若しくは粒の中にのみ存在するのかを決定しなかった。葉における発現もまた、試験しなかった。しかし、異なるEstライブラリーにおけるCcCPI−1の発現(以下の表4中に示される)は、この遺伝子が粒においてのみ特異的に発現され、果皮でも葉でも発現が検出されなかったことを実証する。この結果は、CcCPI−1が種子において特異的に発現されるシステインプロテイナーゼ(例えば、CcCP−1)の活性レベルを制御することをさらに示唆する。
【0064】
【表3】
【0065】
図4中に示されるノザンブロット分析は、コーヒーのシステインプロテイナーゼインヒビター遺伝子CcCPI−1遺伝子が、C.canephora(robusta)のチェリー 対C.arabicaのチェリーにおいて差示的に発現されることを実証する。第1に、図4のデータは、このCcCPI−1遺伝子が、C.arabicaにおいて僅かにより早く発現されることを示す。第2に、そしてより重要なことに、このCcCPI−1遺伝子は、C.canephoraのチェリーにおいて有意により高いレベルで発現される。発現におけるこの差異は、C.arabicaのチェリー 対C.canephoraのチェリーにおいて見出されるシステインプロテイナーゼ活性のレベルにおそらく影響を与える。このクラスのタンパク質は植物における耐虫性と広く関連しているので、C.canephoraにおけるCcCPI−1遺伝子の高い発現は、robusta変種 対arabica変種についてしばしば見られるより高い耐病性に寄与する可能性もある。
【0066】
粒の発芽の間のCcCP−1発現のRT−PCR分析
コーヒー粒の発芽の間のCcCP−1の発現を決定するために、コーヒーの果実を成熟段階で収穫し、水でリンスし、その果皮を除去した(各果実は通常2つの粒を含む)。得られた粒を、室温で戸外にて1週間乾燥させた。発芽前に、各粒のパーチメント及び銀皮(種皮)を手動で除去し、次いで粒を1%(w/v)の次亜塩素酸ナトリウム中に1時間配置することによって滅菌し、次いで滅菌脱イオン水によって2回洗浄した。発芽のために、150個の滅菌した粒を、10mlの固体Heller成長培地H15(Hellerの塩(Heller、1953)を含む)及び7g/lの寒天を含む試験管中に個々に配置し、次いで1日8時間の明期で25℃でインキュベートした。
【0067】
10個の粒の3つのセットを、発芽の2日後、3日後、5日後、1ヵ月後及び2ヵ月後に採取し、液体窒素中で即座に凍結し、RNA抽出まで−80℃で保存した。1ヶ月及び2ヶ月の発芽サンプルについて、これらのサンプルに付随した幼根をサンプリング時に摘出し、粒とは別に凍結した。30個の滅菌した粒を、T(0)コントロールとして使用するためにT=0で採取して凍結した。
【0068】
各サンプルから抽出した4μgのDNase処理した総RNAを使用して、Superscript II Reverse Transcriptase(Invitrogen、Carlsbad、CA)のプロトコルに従ってヘキサマーオリゴヌクレオチドを使用してcDNAを合成した。コーヒーのリボソームタンパク質L39遺伝子のフラグメントを、cDNA合成ステップのためのコントロールとして各cDNAサンプルについて増幅した。これらのPCR反応を、cDNAの1/100希釈物10μl、1μMの各プライマー、5μlの10×ThermoPol PCR緩衝液(10mMの(NH4)2SO4、2mMのMgSO4、20mMのTris−HCl(25℃でpH8.8)、10mMのKCl及び0.1%のTriton X−100)及び2.5単位のTaqポリメラーゼ(New England Biolabs、Beverly、MA)を含む50μlの反応を使用して実施した。サイクル条件は、94℃で2分間、その後の(94℃で1分間、60℃で1.5分間及び72℃で2.5分間)×35サイクルであった。最終伸長ステップは72℃で7分間であった。以下のプライマーをPCRによる増幅のために使用し、726bpのcDNA産物を得た:CcCP−1 up 5’ ACCGAGGAGGAGTTTGAGGCTACG 3’及びCcCP−1 low:5’ ACGCTTCCCCCATGAGTTCTTGA 3’。RPL39タンパク質のためのプライマーは、
であった。
【0069】
RT−PCRを使用して、発芽の異なる段階の間のCcCP−1の発現を決定した。得られた結果は、CcCP−1転写物が試験した全ての発芽時点において粒全体中で検出されることを実証する(図5)。Fischer,J.ら、2000(Plant Molecular Biology、43、83−101)によって、V.sativa由来の提唱されたCcCP−1相同体CPR4のRNAもまた、発芽の間にV.sativa種子の胚軸及び子葉の両方において発現されることが以前に示されている。
【0070】
ウエスタンブロット分析
分析した葉組織及びチェリー組織は、Coffea arabica CCCA2由来であり、使用前にこれらの組織を−80℃で凍結保存した。異なる発達段階のチェリーの粒組織及び果皮組織を、可能な限り小さい果皮の解凍で、別々に解剖した。次いで、これらの異なる組織を迅速に粉砕して微細粉末にした。これは、例えば、予め凍結した乳鉢及び乳棒を用いて液体窒素を使用して実施できる。タンパク質抽出物を、Tanakaら、1986(Plant Physiology、81 802−806)によって記載された抽出手順の改変バージョンを使用してこの組織から調製した。使用した緩衝液は以下であった:
Tanaka緩衝液:
・スクロース 0.7M
・Tris−HCl(pH8) 0.5M
・β−メルカプト−エタノール 2%(v/v)
・NaCl 0.1M
そしてこの緩衝液を使用する直前に以下を添加した:
・EDTA 5mM
・PMSF 2mM
ゲルローディング緩衝液
・グリセロール 15%(v/v)
・β−メルカプト−エタノール 2%(v/v)
・SDS 3%(v/v)
・Tris−HCl(pH6.8) 62.5mM
【0071】
数百ミリグラムの凍結粉砕粉末を、650μlのTanaka緩衝液に添加した。タンパク質を、1体積のTris飽和フェノール(pH8)(すなわち、10mMのTris−HCl(pH8)で飽和した)の添加によって抽出した。各サンプルを20分間激しく混合し、次いで室温で13000gで20分間遠心分離した。遠心分離後、これらのタンパク質はフェノール相中に存在する。20μlのサンプルを分析(以下を参照のこと)のために維持し、フェノール相中の残留タンパク質を、0.1Mの酢酸アンモニウムを含む5体積のメタノールの添加後に、−20℃で一晩沈殿させた。引き続いて、これらのサンプルを室温で13000gで20分間遠心分離し、得られたペレットを、0.1Mの酢酸アンモニウムを含む500μlのメタノール中で2回洗浄した。得られたペレットを、タンパク質定量まで、30μlのゲルローディング緩衝液中に再懸濁した。
【0072】
フェノール相の20μlのサンプル中のタンパク質もまた上記のように沈殿させ、最終ペレットをBioRad DC ProteinアッセイKitのサンプル緩衝液中に再懸濁した。このサンプル中の総タンパク質の定量を、供給業者によって記載されるようにBioRad DC Proteinアッセイキットを使用して実施した。引き続いて、全ての主要なサンプルを、ゲルローディング緩衝液の添加によって調整して、5μg/μlにした。
【0073】
各サンプルの約50μgのタンパク質を含むサンプルを、SDS−ポリアクリルアミドゲル(12%のトリス−グリシン(Novex(登録商標)Invitrogen(商標))における電気泳動によって分離した。次いで、これらのタンパク質を、標準的なプロトコルを使用したエレクトロブロッティングによってPVDFメンブレンに移した。このメンブレン上の非特異的結合部位を、TBS緩衝液(BioRad(商標))中の10%脱脂乾燥ミルク中でこのメンブレンを室温で1時間又は4℃で一晩インキュベートすることによってブロッキングした。ブロットされたタンパク質を、Vicia sativa由来のCPR4の予測された相同体に対して惹起されたポリクローナル抗体(TBS 10%脱脂乾燥ミルク中、1/5000e希釈)(A.Schlereth及びK.Muntz、Institut fur Pflanzengenetik und kulturpflanzenforchung(IPK)、Germany(A.Schlereth、C.Becker、C.Horstmann、J.Tiedmann及びK.Muntz、2000、Journal of Experimental Botany、51:1423−1433)の厚意により寄贈されたもの)を用いて室温で2時間又は4℃で一晩プローブした。次いで、このメンブレンを、TBS+0.1%のTween 20緩衝液中で20分間にわたって3回洗浄した。このメンブレンを引き続き、西洋ワサビペルオキシダーゼで標識された2次抗体(ヤギ抗ウサギIg、Immunopure(登録商標)、Pierce(商標))と共に1時間インキュベートした。次いで、このメンブレンをTBS+0.1%のTween 20緩衝液中で20分間にわたって2回洗浄し、次いでTBS中で20分間にわたって1回洗浄した。この2次抗体に連結された酵素の存在を、増強されたECL+(登録商標)システム(Amersham Life Science)を供給業者によって記載される通りに使用して、化学ルミネセンス検出によって可視化した。
【0074】
得られた結果は、約41kDaのポリペプチド(これは、CcCP−1前駆体ポリペプチドの予測された分子量(43735Da)と密接に対応する)が、試験した粒成熟段階の全てにおいて検出されるが、果皮組織においては検出されないことを示す(図6)。このタンパク質発現パターンは、CcCP−1 mRNAについて見られた発現パターン(図2)と類似である。約22kDaの別のポリペプチドもまた、黄色い段階及び赤い段階の粒において検出されるが、その量は、41kDaのポリペプチドよりも少ない。この第2のポリペプチドのサイズは、CcCP−1の成熟形態の予測されたサイズ(25,239Da)と一致する。プロセシング後の成熟CcCP−1の予測されたサイズを、CcCP1の完全ORF配列とCPR4の予測された成熟形態の配列(Vicia sativa−登録番号Z99172、CcCP1と60.9%の同一性)との間のタンパク質アラインメントによって決定した。成熟形態を生成するためのCPR4ポリペプチドのプロセシングのN末端部位を、他のパパイン様CPRポリペプチドとの配列比較によって予測した(J.Fisher、C.Becker、S.Hillmer、C.Horstmann、B.Neubohn、A.Schlereth、V.Senyuk、A.Shutov及びK.Muntz.2000 Plant molecular biology 43:83−101)。興味深いことに、CcCP−1の前駆体及び成熟形態の両方が粒の発達の間に検出される本明細書中に示される結果とは対照的に、CPR4ポリペプチドの成熟形態のみが発達中の種子において検出され、V.sativa種子の発芽の間にも検出された(Fisherら、2000)。
【0075】
robusta変種FRT−32についての遺伝子発現のRT−PCR分析。
FRT−32の異なる組織を調製し、総RNAを、以前に記載された方法によってこれらの組織から抽出した。cDNAを、DNase処理した総RNAから、arabica cDNAを用いたRT−PCR実験について上記したように調製した。次いで、特異的PCR反応を、arabica cDNAを用いたRT−PCR実験のための上記の反応条件を使用して実施した。使用した特異的増幅条件及びオリゴヌクレオチドプライマーは、各実験について図の説明文中に与えられる。
【0076】
CcCPI−1
a)このcDNAがシステインプロテイナーゼインヒビターをコードすることを示す、CcCPI−1についての最適なアラインメント(図6A)。
【0077】
b)arabica及びrobustaにおけるCcCPI−1についてのRT−PCR発現データ(図7)。PCR反応を以前に記載されたように実施した。使用したサイクル条件及びPCRプライマーは、図の説明文中に与えられる。これらのデータは、CcCPI−1が粒においてのみ発現され、果皮においては発現されないことを示すので、arabica発現について以前に示されたデータを補足及び拡張する。この遺伝子の弱い発現が花においても検出され、これは、ノザンブロット分析によって以前に見られなかった結果である。robustaにおけるRT−PCR発現もまた決定した(図7)。これは、花においても小さい緑の粒においても発現が検出されなかったこと以外、arabicaについて見られたのと同じ発現パターンである。robustaの小さい緑の段階について見られた発現の非存在は他の遺伝子についても見られ、従って、これはCcCPI−1遺伝子に独自のことではない。
【0078】
表5:異なるEstライブラリーにおけるシステインプロテイナーゼインヒビター遺伝子CPI−2、CPI−3及びCPI−4についてのEstの発生。
【表4】
【0079】
CcCPI−2
a)このcDNAがシステインプロテイナーゼインヒビターをコードすることを示す、CcCPI−2についての最適なアラインメント(図8)。
【0080】
b)arabica及びrobustaにおけるCcCPI−2についてのRT−PCR発現データ(図9)。PCR反応を以前に記載されたように実施した。使用したPCRプライマーは図の説明文中に与えられる。これらのデータは、CcCP−2が全ての組織において発現されることを示し、従って、この遺伝子のタンパク質産物が、これらの組織中に存在する1つ又は複数のシステインプロテイナーゼを制御することにおいて重要な役割をおそらく果たすことを示す。上記表5中に見られる各ライブラリー中のEstの数は、CPI−2が、葉、果皮又は受粉後46週の種子よりも、受粉後30週の粒(種子)においてより多く発現できることを示唆する。
【0081】
CcCPI−3
a)このcDNAがシステインプロテイナーゼインヒビターをコードすることを示す、CcCPI−3についての最適なアラインメント(図10)。
【0082】
b)このシステインプロテイナーゼインヒビターについて現在入手可能なRT−PCR発現データは存在しない。しかし、各ライブラリー中に出現するEstの数(上記表5)によって決定されるようなこの遺伝子の「in silico」発現は、CcCPI−3が、コーヒー粒において発現される(「種子30w」及び「種子46w」、すなわち30週及び46週の種子ライブラリー中に存在する)ことを示す。果皮、葉又はチェリー全体におけるこの遺伝子についてのEstの非存在は、この遺伝子が粒特異的遺伝子であり得ることを示唆する。
【0083】
CcCPI−4
a)このcDNAがシステインプロテイナーゼインヒビターをコードすることを示す、CcCPI−4についての最適なアラインメント(図11)。
【0084】
b)arabica及びrobustaにおけるCcCPI−4についてのRT−PCR発現データ(図12)。PCR反応を以前に記載されたように実施した。使用したPCRプライマーは図の説明文中に与えられる。得られたデータは、この遺伝子がarabicaの葉、花及び赤い段階の粒において有意に発現されることを示す。最初のゲルの綿密な試験(パネルA:arabica)は、小さい緑の粒及び大きい緑の果皮のレーンにおいても弱いバンドが検出されることを示すので、この遺伝子はまた、arabicaの研究した全てのチェリー発達段階の粒及び果皮において弱く発現できる。robustaについて得られたデータは、この遺伝子が、葉、花、小さい緑の粒及び大きい緑の粒において有意に発現されることを示す。CcCPI−4についてたった1つのEstが種子又は果皮のライブラリーにおいて見出され(上記の表5)、このことは、粒及び/又は果皮におけるこの遺伝子の発現が比較的低いか、或いはこれら2つの組織の小さい規定された領域に限定されることを示す。
【0085】
システインプロテイナーゼインヒビター(CPI)遺伝子についての各々の場合において、粒発達の間のこれらの遺伝子の過剰発現又は発現の阻害(すなわち、非常に強い粒特異的プロモーター(例えば、コーヒー11Sプロモーター)の制御下での)は、成熟粒におけるタンパク質、ペプチド及びアミノ酸のプロフィール(並びに従って、フレーバー前駆体のレベル)を変更することが予測される。
【0086】
発芽及びRT−PCR分析
滅菌した乾燥C.arabica CCCA2の粒(パーチメント及び銀皮を除去したもの)を、10mlの固体Heller成長培地H15及び7g/lの寒天を含む試験管中に個々に配置し、1日8時間の明期で25℃でインキュベートした。発芽の2日後、3日後、5日後、1ヵ月後及び2ヵ月後に3個の粒を採取し、存在する場合には幼根を除去し、粒及び幼根の両方を液体窒素中で即座に凍結し、RNA抽出まで−80℃で保存した。同様に乾燥及び滅菌した非発芽粒(T0)をコントロールとして使用した。RNAを、以前に記載されたように粒サンプルから抽出した。各サンプルから抽出したDNase処理した総RNAを使用して、Superscript II Reverse Transcriptaseキット(Invitrogen、Carlsbad、CA)のプロトコルに従って、プライマーとしてオリゴ(dT)20を使用してcDNAを合成した。次いで、PCR反応を、各cDNA反応のアリコート(10μlの1/10希釈したcDNA、1μMの各プライマー、5μlの10×ThermoPol PCR緩衝液、200μmのdNTP及び2単位のTaqポリメラーゼ(New England Biolabs、Beverly、MA)を含む50μlの反応)を使用して実施した。サイクル条件は、94℃で2分間、(94℃で1分間、54℃で1.5分間及び72℃で2.5分間)×40サイクルであった。最終伸長ステップは72℃で7分間であった。PCRプライマーは以下であった:
【0087】
ゲノムPCR及び精製されたPCRフラグメントのDNA配列決定
5つの異なるコーヒー変種(FRT−07、FRT−19、FRT−32、CCCA2及びGPFA57)のゲノムDNAを、発芽RT−PCR発現研究のために上記のPCR反応において使用した。予測されたサイズのPCR産物が得られ、これらのフラグメントをゲルから精製した。次いで、このPCR増幅されたDNAを第2ラウンドのPCR増幅に供し、次いで、この配列決定反応から得られたDNAを、増幅のために使用したのと同じプライマーを使用して配列決定した。
【0088】
システインプロテイナーゼCcCP−4の単離及び特徴付け
1)異なる発達段階のコーヒー粒、異なる発達段階のコーヒーの果皮組織、及び葉から単離されたRNAを用いて作製されたEst(Expressed Sequence Tags)の収集を使用して、本発明者らは、C末端KDDL配列を有するコーヒーシステインプロテイナーゼをコードする全長cDNAを単離した。本発明者らは、このcDNAをCcCP−4(KDDL)と命名した(図14)。このcDNAによってコードされるタンパク質と他の高度に相同な植物システインプロテイナーゼとのアラインメントが図15中に示される。このアラインメントデータ及び関連のBlast検索は、コーヒーCcCP−4(KDDL)配列によってコードされるタンパク質が、植物のKDEL含有システインプロテイナーゼファミリーのメンバーであることを明らかに示す(図15)。CcCP−4(KDDL)と最も相同なデータベース配列との間の正確な同一性は、表6A及び6B中に与えられる。
【0089】
表6A:Coffea canephoraシステインプロテイナーゼCcCP−4(KDDL)のアミノ酸配列と、最も相同なGenBank配列のアミノ酸配列との同一性
【表5】
【0090】
表6B:Coffea canephoraシステインプロテイナーゼCcCP−4(KDDL)の核酸(cDNA)配列と、最も相同なGenBank配列の核酸配列との同一性
【表6】
【0091】
明白に、得られたコーヒーCcCP−4 KDDL配列は、図15中に示される他のほぼ全ての配列とは異なる1つの重要な差異を有する。すなわち、この配列は、予測された小胞体(ER)残留配列(C末端KDEL配列)を有さないが、この配列の変異体(すなわちKDDL)を有する。C末端KDEL配列におけるバリエーションが植物細胞中のERにおける残留を指向する能力を試験することによって、Deneckeら(Denecke,J.、De Rycke,R.及びBotterman,J.1992 EMBO J.11、2345−2355)は、C末端変異体(例えば、SDEL、KDDL、KDEI及びKDEV)が、小胞体残留機能を完全に喪失することもあることを以前に示している。従って、植物KDELシステインプロテイナーゼのコーヒー相同体中のKDDL配列の存在は予測されなかった。表7は、cDNA CcCP−4(KDDL)を含むユニジーンが21個のEstを有することを示す。従って、次いで本発明者らは、このユニジーン中の他のEstの配列を試験し、これらのEstのうち7つが、KDDL領域についての良好な配列データを含むことを見出した。これら7つのcDNA配列のうち、6つはKDDL配列を有し、1つはKDEL配列を有した。本発明者らは、KDEL C末端配列を有するcDNAクローンを引き続いて単離し、この部分的cDNAクローンについての完全配列を得た。得られたDNA配列及びタンパク質配列はそれぞれ、図16及び図17中に示される。
【0092】
表7.全長cDNA CcCP−4(KDDL)を含むユニジーン中のEstの数。
【表7】
【0093】
図16中に示されるCcCP−4(KDEL)の配列をコードするcDNAは、部分的cDNAに過ぎない。すなわち、これは、全長cDNAクローンCcCP−4(KDDL)についての1336bp長に対してたった817bp長である。部分的cDNA CcCP−4(KDEL)は、cDNAクローンCcCP−4(KDDL)中に見出される等価な配列からの8個の単一ヌクレオチド残基変化を有するが、これらのヌクレオチド変化のうち2つだけがオープンリーディングフレームのアミノ酸配列の変化を引き起こす(図17)。3’非翻訳領域中に、3つの明白なヌクレオチド変化が存在する。さらに、マイクロサテライト領域内であるように思われるCcCP−4(KDEL)cDNA配列の3’非翻訳領域中に、12ヌクレオチドの挿入もまた存在する。このデータは、コーヒーCcCP−4遺伝子のこれら2つの対立遺伝子についての未発見の2つの異なる重要な分子マーカーを示したに過ぎず、そのうち一方は、機能的に重要なKDEL部位と関連するSNPであり、他方は、この遺伝子の3’非翻訳領域と関連するマイクロサテライトマーカーである。後者の点は、マイクロサテライト配列が高い変動性を有する遺伝子マーカーであると通常みなされるので重要であり、従って、この遺伝子の他の対立遺伝子は、このマイクロサテライト含有領域を使用して見出される可能性がある。
【0094】
arabica及びrobustaの異なる変種において上で同定されたCcCP−4遺伝子の2つの対立遺伝子の分布を試験するために、CcCP−4遺伝子を保有するゲノム配列の小さい領域を、5つの異なる遺伝子型からPCRによって増幅した。予測されたサイズ(207塩基対)のPCRフラグメントが各ゲノムDNAサンプルから得られ、これらのPCR産物をゲル精製し、次いで再増幅して、このPCR産物の直接的DNA配列決定に充分なDNAを生成した。配列決定反応から得られた結果が図18中に示される。5つの配列についての配列決定クロマトグラムは、試験した2つのarabica変種がKDEL配列を明らかに有し、試験した3つのrobusta変種がKDDL配列を有することを示す。この結果は、KDDL対立遺伝子がrobusta変種に限定できること及びこれがarabica変種においては見出されないことを示す。KDEL配列は本明細書中で研究した3つのrobusta変種においては見出されなかったが、Cornell Estライブラリーにおける1つのKDEL配列の発見は、この対立遺伝子が少なくともいくつかのrobustaクローン中に存在できることを示す。
【0095】
CcCP−4遺伝子の発現を、ノザンブロット及びRT−PCR分析を使用して研究した。図19は、arabica変種の異なる発達段階のコーヒーの粒及び果皮から抽出したRNA並びに根、若葉、茎及び花から単離したRNAを使用したノザンブロット実験から得られた結果を示す。CcCP−4(KDEL)対立遺伝子の既知のDNA配列との約98%の相同性を有するCcCP−4(KDDL)プローブを使用して得られたデータは、CcCP−4が粒においてのみ発現されることを示した。発現は、果皮においても、根、茎、花及び葉においても検出されなかった。2つの対立遺伝子間の非常に高いレベルの同一性に起因して、このCcCP−4(KDDL)プローブは、両方の対立遺伝子由来の転写物にハイブリダイズすると予測される。RT−PCR分析を使用した類似の実験もまた、CcCP−4遺伝子について同じ発現プロフィールを示した。
【0096】
CcCP−4の発現を、RT−PCR分析を使用して、発芽の間の種子全体においても研究した。この実験は、CcCP−4 KDEL対立遺伝子及びCcCP−4 KDDL対立遺伝子の両方に共通するプライマーを使用した。この実験の結果は、図20中に示される。CcCP−4転写物は試験した全ての発芽段階で検出されたが、これらの転写物のレベルは、3日目に僅かに低下するようであり、次いで発芽が進行するにつれて再度上昇し始めた(1ヶ月目及び2ヶ月目のサンプルにおいて最高レベル)。
【0097】
Lingら(Ling,J.−Q.、Kojima,T.、Shiraiwa,M.及びTakahara,H.、2003 Biochim.Biophys.Acta 1627、129−139)は、KDEL含有システインプロテイナーゼをコードする2つのcDNAを、ダイズ子葉から単離した。これら2つのcDNAは、DNAレベルで93.5%の類似性を有し、根、花及び種子発達の間に発現された。発達中の種子においても成熟種子においてもノザンブロッティングによって発現は検出されなかったが、成熟した莢において発現が検出された。KDEL含有システインプロテイナーゼをコードするcDNAは、ニンジンからも単離された(Sakuta,C.、Oda,A.、Konishi,M.、Yamakawa,S.、Kamada,H.及びSatoh,S.2001 Biosci.Biotechnol.Biochem.65、2243−2248)。この遺伝子の転写物は、成熟した乾燥種子並びに吸水の2日後及び3日後の発芽中の種子全体において検出された。他のニンジン組織におけるこの遺伝子の発現又は種子発達の間のこの遺伝子の発現は示されなかった。別のKDEL含有タンパク質及びその対応するcDNAがV.sativaから単離されている(Fischer,J、Becker,C.、Hillmer,S.、Horstmann,C.、Neubohn,B、Schlereth,A.、Senyuk,V.、Shutov,A.及びMuntz,K.(2000)Plant Molecular Biol.43、83−101)。ノザンブロッティングを使用して、この遺伝子の転写物は、発芽の間に子葉において検出されたが、発芽中の種子の胚軸においては検出されなかった。成熟中の種子においても、成熟種子においても、葉及び根においても、転写物は検出されなかった。
【0098】
本明細書中に示される結果は、コーヒーのKDEL型システインプロテイナーゼが、いくつかの新規かつ予測されなかった特徴を示すことを示す。第1に、本発明者らは、robustaコーヒー粒が、KDEL領域をコードする配列中にKDELからKDDLへの変化を生じる単一変異を有するKDEL型CP遺伝子を発現することを発見した。Deneckeら(1992)のデータに基づいて、この残留配列におけるこの特定の変更は、robusta CcCP−4(KDDL)タンパク質の細胞局在及び/又は制御を変更することが予測される。本発明者らは、CcCP−4 KDDL遺伝子の転写されたコピーが存在することにより、CcCP−4 KDEL配列を有する変種と比較して、コーヒー粒におけるペプチド/アミノ酸のプロフィールの有意な変化を生成できることを提唱する。本発明者らはまた、コーヒーのKDEL型システインプロテイナーゼが、粒の発芽の間に予測された発現を示しつつ、研究した全ての粒の発達段階の間にも予測に反して発現されることを本明細書中で示した。上記のように、これまで、他の植物における種子発達の間のKDEL型システインプロテイナーゼの有意な発現を実証する明確なデータは刊行された文献中には存在しないが、その転写物は、成熟ニンジン種子において検出されている(Sakutaら、2001)。
【0099】
上で示されたコーヒーのKDEL型システインプロテイナーゼの新規特性は、arabica及びrobustaの成熟粒におけるペプチドプロフィール及びアミノ酸プロフィールに対して重要な影響をおそらく有し、従って、重要なコーヒーフレーバー前駆体のプールを変更する。KDEL型システインプロテイナーゼの転写物が成熟粒中に存在することを考慮すると、このKDEL型タンパク質はコーヒーの湿式加工の間に活性化でき、それによって湿式プロセスのコーヒー粒のペプチド/アミノ酸のプロフィールをさらに変更することもまた可能である。記載された研究は、KDEL型システインプロテイナーゼ遺伝子の特定の対立遺伝子を有するコーヒー変種(これは、タンパク質/ペプチド/アミノ酸のプロフィールにおける付随した変更を有する)を得るために古典的選択及び交配作業において使用できる分子マーカー(SNP及びマイクロサテライトマーカー)を生成している。例えば、CcCP−4 KDEL対立遺伝子のみを有するか又は低い発現レベルのCcCP−4 KDDL対立遺伝子のみを有するrobustaの変種が、選択/交配できる。さらに、遺伝子改変技術を使用して、KDEL型又はKDDL型のシステインプロテイナーゼの種子特異的な過剰発現によってコーヒー又は他の植物におけるKDEL型システインプロテイナーゼ活性を変更することが想定できる。或いは、KDEL型システインプロテイナーゼのレベルは、アンチセンス、センス又はRNAiの技術を使用して低下できる。両方の場合において、得られた形質転換された植物中のタンパク質/ペプチド/アミノ酸のプールが変更されて、タンパク質/ペプチド/アミノ酸のフレーバー前駆体プールの新たなプロフィールが導かれる。
【0100】
図13中に示されるノザンブロット分析は、コーヒーのアスパラギン酸プロテイナーゼCcAP−2遺伝子が、試験した全てのチェリー発達段階で、C.arabicaコーヒーチェリーの粒及び果皮の両方において発現されることを実証する。このCcAP−2遺伝子はまた、根において比較的高い発現を有する。フィルムをより長く露光させると、CcAP−2発現は、C.arabicaの茎、葉及び花の組織においても検出された。
【0101】
CcAP−1及びCcAP−2
図21及び22は、CcAP−1及びCcAP−2の各々がアスパラギン酸プロテイナーゼをコードすることを示す。
【0102】
コーヒー種子におけるCcCP−1、CcCP−4、CcAP−1及びCcAP−2のプロテイナーゼ遺伝子配列並びにCcCPI−1、CcCPI−2、CcCPI−3及びCcCPI−4のプロテイナーゼインヒビターの過剰発現及び過少発現。
主要な貯蔵タンパク質のプロフィール及びアミノ酸/ペプチドのプロフィールは、本明細書中に開示される1つ又は複数の遺伝子の発現を上方又は下方のいずれかで変更することによって、成熟コーヒー粒において変化できることが予測される。
【0103】
目的の遺伝子の過剰発現のための方法は当該分野で周知である。このような方法は、以下の3つの主要な成分のキメラ遺伝子を生成するステップからなる:1)遺伝子の5’末端のプロモーター配列、好ましくは本願においてはMarracciniら、1999(Marracciniら、1999、Coffea arabicaの完全11S種子貯蔵タンパク質遺伝子の分子クローニング及びトランスジェニックタバコ植物におけるプロモーター分析(Molecular cloning of the complete 11S seed storage protein gene of Coffea arabica and promoter analysis in transgenic tobacco plants)、Plant Physiol.Biochem.第37巻、273−282及びWO 99/02688号)中に記載されるコーヒー種子特異的プロモーターのような種子特異的プロモーター、2)発現されるべき遺伝子の全コード配列、並びに3)Agrobacterium tumefaciensのTiプラスミドのT−DNA由来のノパリンシンターゼ遺伝子由来の3’領域のような3’制御領域。次いで、このキメラ遺伝子は、Agrobacterium tumefaciens形質転換ベクター中にクローニングでき、このベクターは、Leroyら、2000、(Leroyら、2000、リーフマイナーに対する抵抗性についてのBacillus thuringiensis cry1Ac遺伝子を発現する、遺伝的に変更されたコーヒー植物(Genetically modified coffee plants expressing the Bacillus thuringiensis cry1Ac gene for resistance to leaf minor).Plant Cell Reports 2000、19、382−389)によって詳細に記載されているコーヒー形質転換において使用するためのAgrobacterium tumefaciens株中に形質転換できる。安定な形質転換挿入配列を有する植物を次いでスクリーニングして、遺伝子の過剰発現又はタンパク質活性の過剰発現の検出のような方法を使用して、モック形質転換した植物由来の種子に対して、形質転換実験において使用する特定の遺伝子を成熟種子において特異的に過剰発現する植物をスクリーニングすることができる。
【0104】
例えば、当業者は、以下からなる組換え構築物を生成できる:1)Marracciniら(1999)中に記載される最長のコーヒー11S遺伝子プロモーター配列、2)ポリAテイルを有さない、CcCP−1又はCcCP−4(KDDL)の全長cDNA配列、及び3)既知の転写ターミネーター配列(例えば、充分研究されたノパリンターミネーター)。CcCP4若しくは他のcDNA配列の5’非コード領域の、コーヒー11S遺伝子の5’非コード領域又はコーヒー若しくは他の関連の植物種のいずれかの他の強力な種子特異的プロモーターの5’非コード領域による置換に起因して、より高いレベルの組換え構築物の過剰発現が生じることもまた可能である。次いで、この組換え遺伝子配列を、Leroyらにおいて記載されるAgrobacterium T−DNAベクターの適切な部位中に挿入できる。このように構築されたT−DNAベクターは、適切なAgrobacterium株(例えば、Leroyらにおいて記載される株)中に配置することができ、このT−DNA含有Agrobacteriumは、Leroyらにおいて詳述される方法に従ってコーヒーを形質転換するために使用できる。
【0105】
既知の遺伝子配列の発現が、アンチセンス抑制によって、遺伝子の全コード領域未満を示す核酸フラグメントを使用した遺伝子発現によって、抑制されるべき遺伝子と100%の配列同一性を共有しない核酸によって、低下又は完全に遮断できることは、当該分野で周知である。この場合、特定のアンチセンス抑制又は共抑制実験のために選択された配列は、上に示したキメラ遺伝子構築スキームにおいて全長遺伝子を置換する。得られたアンチセンス抑制又は共抑制キメラ構築物は、Agrobacterium tumefaciens形質転換ベクター中に再度クローニングされ、上記のようにコーヒー形質転換において使用するためにAgrobacterium tumefaciens株中に形質転換される。次いで、安定な形質転換挿入配列を有する植物は、形質転換された植物の種子において使用される特定の遺伝子配列の発現を低下させたものについてスクリーニングできる。低下された発現は、ノザンブロッティング;半定量的RT−PCR及び/又は定量的RT−PCRのような技術によって検出できる。
【0106】
植物中の遺伝子発現を低下又は排除するための別の方法は、本明細書中に開示される遺伝子配列の小部分を使用して、RNAiの使用を介したRNAサイレンシングを生じることである(Hannon,G.J.、2002、Nature、第418巻、244−251;Tangら、2003、Genes Dev、第17巻、49−63)。このアプローチにおいて、本明細書中に開示される1つ又は複数の配列の小領域が、種子特異的プロモーター及び適切な3’調節領域を有する上記のようなAgrobacterium tumefaciens形質転換ベクター中にクローニングされる。RNAiのためのこの挿入された新たな配列は、生成されたRNAがin vivoでRNA構造を形成して、形質転換された細胞において小さい二本鎖RNAの生成を生じ、それによってこれらの小さい二本鎖RNA配列がこれらの形質転換された細胞において相同なmRNAの分解を誘発するように構築されるべきである。
【0107】
CcCP−1遺伝子、CcCP−4遺伝子、CcAP−1遺伝子、CcAP−2遺伝子、CcCPI−1遺伝子、CcCPI−2遺伝子、CcCPI−3遺伝子、CcCPI−4遺伝子における天然に存在するバリエーションについてのスクリーニング及びこれらの遺伝子における新たな変異の生成。
本明細書中に開示される配列は、これらの遺伝子における対立遺伝子変異体について天然の集団をスクリーニングするために使用できる。これは、CcCP−1配列、CcCP−4配列、CcAP−1配列、CcAP−2配列、CcCPI−1配列、CcCPI−2配列、CcCPI−3配列及びCcCPI−4配列を、異なるコーヒー植物変種由来のゲノムDNA中の天然に存在するRFLP(制限酵素断片長多型)についての研究においてプローブとして使用することによって達成できる。対立遺伝子変異体を見出すためのより強力な方法は、TILLING法(Till,B.J.ら、2003、高スループットTILLINGを用いた、誘導された点変異の大規模発見(Large scale discovery of induced point mutations with high−thruput TILLING).Genome Research 第13巻、524−530)に関連する変異スクリーニング技術を使用することである。この場合、特定の遺伝子配列(例えば、本明細書中のCcCP−1配列、CcCP−4配列、CcAP−1配列、CcAP−2配列、CcCPI−1配列、CcCPI−2配列、CcCPI−3配列及びCcCPI−4配列)が一旦単離及びクローニングされると、TILLING法に関連する変異スクリーニング技術は、クローニングされた配列と異なる変種中の対応するcDNA又はゲノム配列との間で配列変異体を同定するために使用できる。700塩基対〜1000塩基対のDNAセグメントをコードするPCRプライマー対を使用して、既知のクローニングされた遺伝子が、異なる変種中の天然に存在する配列バリエーションについてスキャンできる。理想的な状況において、1つ又は複数の配列変異体はまた、特定の表現型バリエーションと相関させることができ、それによってこの表現型変異体に対する遺伝子マーカーを同定する。
【0108】
さらに、CcCP−1、CcCP−4、CcAP−1、CcAP−2、CcCPI−1、CcCPI−2、CcCPI−3及びCcCPI−4についての本明細書中に開示される配列を使用して、完全なTILLING法の適用は、これらの遺伝子中の新たな変異を生成及び検出するため、従って、これらの特定の変異を含む植物を生成するために使用できる。例えば、完全なTILLING法を使用して、特定の変異(例えば、目的の遺伝子標的を不活化する、コード配列中のミスセンス変異)を有するコーヒー植物が生成できる。
【図面の簡単な説明】
【0109】
【図1】Coffea arabicaの異なる組織におけるシステインプロテイナーゼ遺伝子のノザンブロット分析を示す図であり、図中、レーンは以下で標識される:R:根、S:茎、L:若葉;並びにSG、LG、Y及びRedはそれぞれ、小さい緑の果実、大きい緑の果実、黄色い果実及び赤い果実由来の粒である。各レーンにおいて5μgの総RNAをロードした。MWはRNAサイズラダーである。パネルBは、試験した組織中のCcCP−1 mRNAの出現を示す、24時間の露光後のオートラジオグラムを示し、パネルAは、ブロッティング前のゲルのエチジウムブロマイド染色を実証する。
【図2】Coffea arabicaの異なる組織におけるシステインプロテイナーゼCcCP−1遺伝子の発現のノザンブロット分析を示す図であり、図中、レーンは以下で標識される:R、根;S、茎;L、若葉;F、花。SG(G)、LG(G)、Y(G)及びRed(G)はそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリーの粒から単離されたRNAに対応し、SG(P)、LG(P)、Y(P)及びRed(P)で標識されたレーンはそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリーの果皮組織から単離されたRNAに対応する。各レーンにおいて5μgの総RNAをロードした。パネルAは、ローディングコントロールとしてのブロッティング前の大きいリボソームRNAのエチジウムブロマイド染色を実証し、パネルBは、試験した特定の組織中のCcCP−1 mRNAの出現を示すオートラジオグラムである。
【図2A】CcCP−1 cDNAによってコードされるタンパク質の完全配列と、NCBIデータベースにおいて入手可能な他の全長システインプロテイナーゼとのアラインメントの図である。これは、MegAlign(DNASTAR)中のCLUSTAL法によって、Megalignにおいて実施した。影付のブロックは同一のアミノ酸を示す。EMBLデータベースの登録番号は括弧内に与えられる。Arabidopsis thaliana(AY070063);Vicia sativa(Z99172);Glycine max GMCP3(Z32795);Glycine max GmPM33(AF167986);Phaseolus vulgaris Moldavain(Z99955);Solanum melongena(AF082181);Nicotiana tabacum(AJ242994);Lycopersicon esculentum(Z14028);Vicia faba(AY161277)。
【図3】Coffea arabicaの異なる組織におけるシステインプロテイナーゼインヒビター(CcCPI−1)遺伝子のノザンブロット分析を示す図であり、図中、レーンは以下で標識される:R:根、S:茎、L:若葉;並びにSG、LG、Y及びRedはそれぞれ、小さい緑の果実、大きい緑の果実、黄色い果実及び赤い果実由来の粒について。各レーンにおいて5μgの総RNAをロードした。MWはRNAサイズラダーである。パネルBは、24時間の露光後のオートラジオグラムを示し、パネルAは、ブロッティング前のゲルのエチジウムブロマイド染色を実証する。
【図4】Coffea arabica(ARA)及びCoffea robusta(ROB)の果実の異なる発達段階におけるシステインプロテイナーゼインヒビター(CcCPI−1)遺伝子のノザンブロット分析を示す図である。レーンは、小さい緑の果実(SG)、大きい緑の果実(LG)、黄色い果実(Y)及び赤い果実(Red)でそれぞれ標識される。各レーンにおいて5μgの総RNAをロードした。MWはRNAサイズラダーである。パネルBは、試験した特定の組織中のCcCPI−1 mRNAの出現を示す、24時間の露光後のオートラジオグラムを示す。パネルAは、ブロッティング前のゲルのエチジウムブロマイド染色を実証する。
【図5】Coffea arabica粒の出芽の間のCcCP−1の発現のRT−PCR分析を示す図である。PCR反応を、1/100希釈した各cDNA10μlを使用して実施した。サイクル条件は、94℃で2分間、(94℃で1分間、61℃で1.5分間及び72℃で2.5分間)×35サイクルであった。最終伸長ステップは72℃で7分間であった。PCRプライマーは、A4-43-upper:5'-ACCGAGGAGGAGTTTGAGGCTACG-3'、A4-43-lower:5'-ACGCTTCCCCCATGAGTTCTTGA-3'であった。mRNAを、異なるテンプレート(それぞれ、滅菌した種子(T0)並びに出芽の2日後(2d)、3日後(3d)、5日後(5d)、1ヵ月後(1m)及び2ヵ月後(2m)に採取した種子由来のcDNA)に対して特異的プライマー(CcCP−1 up/CcCP−1 low)を使用するRT−PCRによって増幅した。これらのPCR産物を、1%(w/v)アガロースゲル中で分離し、エチジウムブロマイドで染色した。RPL39;60Sリボソーム大サブユニットのL39タンパク質をコードするcDNAの増幅されたフラグメント。
【図6】CcCP1タンパク質の発現のウエスタンブロット分析を示す図である(A)。総タンパク質を、小さい緑(SG)、大きい緑(LG)、黄色(Y)及び赤(Red)の段階の、発達中のコーヒーチェリーから収集した粒(g)及び果皮(p)から抽出した。パネルB−クマシーブルーで染色した12% SDS−PAGEゲル上での50μgの総タンパク質の分離。パネルA−タンパク質検出を、これらの方法において記載されるように、抗CRP4ポリクローナル抗体(ウサギ)を使用して実施した。パネルB中のバンドのおおよそのサイズを左側に矢印で示す。各パネル内の大きい矢印は、これらの抗体のうち1つと交差反応する主要な貯蔵タンパク質の存在を示す。
【図6A】CcCPI−1 cDNAによってコードされる完全タンパク質と、NCBIにおいて入手可能な他の相同な全長システインプロテイナーゼとの最適なアラインメントを示す図である。影付のブロックは同一のアミノ酸を示す。EMBLデータベースの登録番号及び%同一性は、括弧内に与えられる。Malus x domestica(AAO18638;42.3%の同一性)、Common sunflower(JE0308;41.5%の同一性)、Arabidopsis thaliana(AAM64985;30%の同一性)及びRumex obtusifolius(CAD21441;29.3%の同一性)。
【図7】Coffea arabica CCCA2(A)及びCoffea robusta FRT−32(B)の異なる組織におけるCcCPI−1遺伝子の発現のRT−PCR分析を示す図である。PCR反応を、1/1000希釈した各cDNA 10μlを使用して実施した。サイクル条件は、94℃で2分間、(94℃で1分間、60℃で1.5分間及び72℃で1分間)×40サイクルであった。最終伸長ステップは72℃で7分間であった。PCRプライマーは、CcCPI-1(up)5'AGGAAAGTGGGAGCAAGGGAGAAGA3'、CcCPI-1(low)5'TAGTATGAACCCAAGGCCGAACCAC3'.であった。レーンは以下のように標識される:M、マーカー;+P、CcCPI−1遺伝子を含む希釈したプラスミド;R、根;S、茎;L、若葉;F、花。SG(G)、LG(G)、Y(G)及びRed(G)はそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリーから単離された粒である。SG(P)、LG(P)、Y(P)及びRed(P)はそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリーから単離された果皮組織である。
【図8】CcCPI−2 cDNAによってコードされる完全タンパク質と、NCBIにおいて入手可能な他の相同な全長システインプロテイナーゼとの最適なアラインメントを示す図である。影付のブロックは同一のアミノ酸を示す。EMBLデータベースの登録番号及び%同一性は、括弧内に与えられる。Rumex obtusifolius(CAD21441;66.7%の同一性)、Dianthus caryophyllus(AAK30004;71.7%の同一性)、Manihot esculenta(AAF72202;65.2%の同一性)。
【図9】Coffea arabica CCCA2(A)及びCoffea robusta FRT−32(B)の異なる組織におけるCcCPI−2遺伝子の発現のRT−PCR分析を示す図である。PCR反応を、1/1000希釈した各cDNA10μlを使用して実施した。サイクル条件は、94℃で2分間、(94℃で1分間、57℃で1.5分間及び72℃で1分間)×40サイクルであった。最終伸長ステップは72℃で7分間であった。PCRプライマーは、CcCPI-2(up)5'GTGAAGCCATGGTTGAACTT3'、CcCPI-2(low)5'GTAATGATACCTCAAGCCAGA3'.であった。レーンは以下のように標識される:M、マーカー;+P、CcCPI−2遺伝子を含む希釈したプラスミド;R、根;S、茎;L、若葉;F、花。SG(G)、LG(G)、Y(G)及びRed(G)はそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリーから単離された粒である。SG(P)、LG(P)、Y(P)及びRed(P)はそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリーから単離された果皮組織である。
【図10】CcCPI−3 cDNAによってコードされる完全タンパク質と、NCBIにおいて入手可能な他の相同な全長システインプロテイナーゼとの最適なアラインメントを示す図である。影付のブロックは同一のアミノ酸を示す。EMBLデータベースの登録番号及び%同一性は、括弧内に与えられる。Citrus x paradisi(AAG38521;42.4%の同一性)、Actinidia deliciosa(AAR92223;44.4%の同一性)及びArabidopsis thaliana(AAM64661;44%の同一性)。
【図11】CcCPI−4 cDNAによってコードされる完全タンパク質と、NCBIにおいて入手可能な他の相同な全長システインプロテイナーゼとの最適なアラインメントを示す図である。影付のブロックは同一のアミノ酸を示す。EMBLデータベースの登録番号及び%同一性は、括弧内に与えられる。Citrus x paradisi(AAG38521;23.6%の同一性)及びArabidopsis thaliana(AAM64661;20%の同一性)。
【図12】Coffea arabica CCCA2(A)及びCoffea robusta FRT−32(B)の異なる組織におけるCcCPI−4遺伝子の発現のRT−PCR分析を示す図である。PCR反応を、1/100希釈した各cDNA10μlを使用して実施した。サイクル条件は、94℃で2分間、(94℃で1分間、60℃で1.5分間及び72℃で1分間)×40サイクルであった。最終伸長ステップは72℃で7分間であった。PCRプライマーは、CcCPI-4(up)5'CTACGGTCGCAGCCAAATC3'、CcCPI-4(low)5'ACAACTGCACCTTCAATGTAC3'.であった。レーンは以下のように標識される:M、マーカー;+P、CcCPI−4遺伝子を含む希釈したプラスミド;R、根;S、茎;L、若葉;F、花。SG(G)、LG(G)、Y(G)及びRed(G)はそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリーから単離された粒である。SG(P)、LG(P)、Y(P)及びRed(P)はそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリーから単離された果皮組織である。
【図13】Coffea arabicaの異なる組織におけるアスパラギン酸プロテイナーゼ2(CcAP2)遺伝子のノザンブロット分析を示す図であり、図中、レーンは以下で標識される:R:根、S:茎、L:若葉;F:花;SG(G)及びSG(P)、LG(G)及びLG(P)、Y(G)及びY(P)並びにRed(G)及びRed(P)はそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリー由来の粒及び果皮についてであり、SG(G)、LG(G)、Y(G)及びR(G)はそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリー由来の果皮についてである。各レーンにおいて5μgの総RNAをロードした。パネルAは、ローディングコントロールとしてのブロッティング前の大きいリボソームRNAのエチジウムブロマイド染色を実証し、パネルBは、試験した特定の組織中のCcAP2 mRNAの出現を示すオートラジオグラムである。
【図14】CcCP−4のcDNA配列及び推定アミノ酸配列を示す図である。小文字:5’非翻訳領域及び3’非翻訳領域;大文字:オープンリーディングフレーム;太字:アミノ酸配列;*:終止コドン。
【図15】CcCP−4 cDNAによってコードされるタンパク質の完全配列と、NCBIデータベースにおいて入手可能な他の全長システインプロテイナーゼとのアラインメントを示す図である。これは、MegAlignソフトウェア(Lasergeneパッケージ、DNASTAR)中のCLUSTAL Wプログラムを使用して実施した。影付のブロックは同一のアミノ酸を示す。登録番号は括弧内に与えられる。Dacus carrota(JC7787);Ricinus communis(AF050756);Vicia sativa(Z34895);Phaseolus vulgaris(X56753);Helianthus annuus(AB109188);Glycine max Cys1(AB092555);Glycine max Cys2(AB092557);Canavalia ensiformis(P49046);Oryza sativa(AB004648);Vigna mungo(P12412);Pisum sativum(AJ004985)。
【図16】Megalign中のプログラムClustalWを使用して整列された全長cDNA配列CcCP−4 KDDL及び部分的cDNA配列CcCP−4(KDEL)を示す図である。
【図17】Megalign中のプログラムClustalWを使用して整列されたCcCP−4(KDDL)の完全オープンリーディングフレーム及びCcCP−4(KDEL)の部分的オープンリーディングフレームを示す図である。
【図18】CcCP−4遺伝子のKDEL/KDDL領域をコードするPCR増幅されたゲノムDNAについてのDNA配列クロマトグラムを示す図である。Robはrobusta変種を示し、Arabはarabica変種を示す。
【図19】Coffea arabicaの異なる組織におけるシステインプロテイナーゼCcCP−4遺伝子の発現のノザンブロット分析を示す図である。レーンは以下で標識される:R、根;S、茎;L、若葉;F、花。SG(G)、LG(G)、Y(G)及びRed(G)はそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリーから単離された粒である。SG(P)、LG(P)、Y(P)及びRed(P)はそれぞれ、小さい緑のチェリー、大きい緑のチェリー、黄色いチェリー及び赤いチェリーから単離された果皮組織である。各レーンにおいて5μgの総RNAをロードした。パネルAは、ローディングコントロールとしてのブロッティング前の大きいリボソームRNAのエチジウムブロマイド染色を実証し、パネルBは、試験した特定の組織中のCcCP−3 mRNAの出現を示すオートラジオグラムである。
【図20】発芽の間の粒全体におけるCcCP−4の発現のRT−PCR分析を示す図である。サンプリング時間は以下であった:0、滅菌処理の直後;2D、処理の2日後;3D、処理の3日後;5D、処理の5日後;1M、処理の1ヵ月後;2M、処理の2ヵ月後;−、DNAなしのコントロール;+P、希釈したCcCP−4プラスミドDNA;M、分子量マーカー。
【図21】CcAP−1 cDNAによってコードされる完全タンパク質と、NCBIにおいて入手可能な他の相同な全長アスパラギン酸プロテイナーゼ配列との最適なアラインメントを示す図である。影付のブロックは同一のアミノ酸を示す。データベースの登録番号は括弧内に与えられる。Arabidopsis thaliana(AY099617)及びArabidopsis thaliana(BAB09366)。
【図22】CcAP−2 cDNAによってコードされる完全タンパク質と、NCBIにおいて入手可能な他の相同な全長アスパラギン酸プロテイナーゼ配列との最適なアラインメントを示す図である。影付のブロックは同一のアミノ酸を示す。データベースの登録番号は括弧内に与えられる。Glycine max(BAB64296)、Ipomoea batatas(AAK48494)、Lycopersicon esculentum(S71591)及びNepenthes alata(BAB20972)。
【特許請求の範囲】
【請求項1】
システインプロテイナーゼ活性を有するポリペプチドをコードするヌクレオチド配列又は前記ヌクレオチド配列の相補体を含む単離されたポリヌクレオチドであって、前記ポリペプチドのアミノ酸配列と配列番号2のアミノ酸配列とは、ClustalWアラインメント法に基づいて、少なくとも70%、好ましくは少なくとも80%の配列同一性を有し、前記相補体は前記ヌクレオチド配列と同数のヌクレオチドを含み、前記相補体と前記ヌクレオチド配列とは100%相補的であるポリヌクレオチド。
【請求項2】
前記ポリペプチドのアミノ酸配列と配列番号2のアミノ酸配列とが、ClustalWアラインメント法に基づいて、少なくとも85%、好ましくは少なくとも90%、任意選択で少なくとも95%の配列同一性を有する、請求項1に記載のポリヌクレオチド。
【請求項3】
前記ヌクレオチド配列が配列番号1のヌクレオチド配列を含む、請求項1に記載のポリヌクレオチド。
【請求項4】
前記ポリペプチドが配列番号2のアミノ酸配列を含む、請求項1に記載のポリヌクレオチド。
【請求項5】
システインプロテイナーゼインヒビター活性を有するポリペプチドをコードするヌクレオチド配列又は前記ヌクレオチド配列の相補体を含む単離されたポリヌクレオチドであって、前記ポリペプチドのアミノ酸配列と配列番号4、10、12及び14からなる群から選択されるアミノ酸配列とは、ClustalWアラインメント法に基づいて、少なくとも70%、好ましくは少なくとも80%の配列同一性を有し、前記相補体は前記ヌクレオチド配列と同数のヌクレオチドを含み、前記相補体と前記ヌクレオチド配列とは100%相補的であるポリヌクレオチド。
【請求項6】
前記ポリペプチドのアミノ酸配列と配列番号4、10、12及び14からなる群から選択されるアミノ酸配列とが、ClustalWアラインメント法に基づいて、少なくとも85%、好ましくは少なくとも90%、任意選択で少なくとも95%の配列同一性を有する、請求項5に記載のポリヌクレオチド。
【請求項7】
前記ヌクレオチド配列が、配列番号3、9、11及び13からなる群から選択されるヌクレオチド配列を含む、請求項5に記載のポリヌクレオチド。
【請求項8】
前記ポリペプチドが、配列番号4、10、12及び14からなる群から選択されるアミノ酸配列を含む、請求項5に記載のポリヌクレオチド。
【請求項9】
アスパラギン酸エンドプロテイナーゼ活性を有するポリペプチドをコードするヌクレオチド配列又は前記ヌクレオチド配列の相補体を含む単離されたポリヌクレオチドであって、前記ポリペプチドのアミノ酸配列と配列番号6又は8から選択されるアミノ酸配列、好ましくは配列番号8のアミノ酸配列とは、ClustalWアラインメント法に基づいて、少なくとも75%、好ましくは少なくとも80%の配列同一性を有し、前記相補体は前記ヌクレオチド配列と同数のヌクレオチドを含み、前記相補体と前記ヌクレオチド配列とは100%相補的であるポリヌクレオチド。
【請求項10】
前記ポリペプチドのアミノ酸配列と配列番号6又は8、好ましくは配列番号8から選択されるアミノ酸配列とが、ClustalWアラインメント法に基づいて、少なくとも85%、好ましくは少なくとも90%、任意選択で少なくとも95%の配列同一性を有する、請求項9に記載のポリヌクレオチド。
【請求項11】
前記ヌクレオチド配列が、配列番号5又は7、好ましくは配列番号7のヌクレオチド配列を含む、請求項9に記載のポリヌクレオチド。
【請求項12】
前記ポリペプチドが、配列番号6又は8、好ましくは配列番号8のアミノ酸配列を含む、請求項9に記載のポリヌクレオチド。
【請求項13】
システインプロテイナーゼ活性を有するポリペプチドをコードするヌクレオチド配列又は前記ヌクレオチド配列の相補体を含む単離されたポリヌクレオチドであって、前記ポリペプチドのアミノ酸配列と配列番号16のアミノ酸配列とは、ClustalWアラインメント法に基づいて、少なくとも70%、好ましくは少なくとも80%の配列同一性を有し、前記相補体は前記ヌクレオチド配列と同数のヌクレオチドを含み、前記相補体と前記ヌクレオチド配列とは100%相補的であるポリヌクレオチド。
【請求項14】
前記ポリペプチドのアミノ酸配列と配列番号16のアミノ酸配列とが、ClustalWアラインメント法に基づいて、少なくとも85%、好ましくは少なくとも90%、任意選択で少なくとも95%の配列同一性を有する、請求項13に記載のポリヌクレオチド。
【請求項15】
前記ヌクレオチド配列が配列番号15のヌクレオチド配列を含む、請求項13に記載のポリヌクレオチド。
【請求項16】
前記ポリペプチドが配列番号16のアミノ酸配列を含む、請求項13に記載のポリヌクレオチド。
【請求項17】
請求項1から16までのいずれか一項に記載のポリヌクレオチドを含むベクター。
【請求項18】
調節配列に作動可能に連結された請求項1から16までのいずれか一項に記載のポリヌクレオチドを含む、非ネイティブの組換えDNA構築物。
【請求項19】
請求項1から16までのいずれか一項に記載のポリヌクレオチドで細胞を形質転換するステップを含む、細胞を形質転換するための方法。
【請求項20】
請求項18に記載の非ネイティブの組換えDNA構築物を含む細胞。
【請求項21】
原核細胞、真核細胞又は植物細胞、好ましくはコーヒー細胞である、請求項20に記載の細胞。
【請求項22】
請求項20又は21に記載の細胞を含むトランスジェニック植物。
【請求項23】
緑のコーヒー粒中のコーヒーフレーバー前駆体レベルを調節するための方法であって、請求項18に記載の非ネイティブの組換えDNA構築物をコーヒー植物中に導入するステップを含む方法。
【請求項1】
システインプロテイナーゼ活性を有するポリペプチドをコードするヌクレオチド配列又は前記ヌクレオチド配列の相補体を含む単離されたポリヌクレオチドであって、前記ポリペプチドのアミノ酸配列と配列番号2のアミノ酸配列とは、ClustalWアラインメント法に基づいて、少なくとも70%、好ましくは少なくとも80%の配列同一性を有し、前記相補体は前記ヌクレオチド配列と同数のヌクレオチドを含み、前記相補体と前記ヌクレオチド配列とは100%相補的であるポリヌクレオチド。
【請求項2】
前記ポリペプチドのアミノ酸配列と配列番号2のアミノ酸配列とが、ClustalWアラインメント法に基づいて、少なくとも85%、好ましくは少なくとも90%、任意選択で少なくとも95%の配列同一性を有する、請求項1に記載のポリヌクレオチド。
【請求項3】
前記ヌクレオチド配列が配列番号1のヌクレオチド配列を含む、請求項1に記載のポリヌクレオチド。
【請求項4】
前記ポリペプチドが配列番号2のアミノ酸配列を含む、請求項1に記載のポリヌクレオチド。
【請求項5】
システインプロテイナーゼインヒビター活性を有するポリペプチドをコードするヌクレオチド配列又は前記ヌクレオチド配列の相補体を含む単離されたポリヌクレオチドであって、前記ポリペプチドのアミノ酸配列と配列番号4、10、12及び14からなる群から選択されるアミノ酸配列とは、ClustalWアラインメント法に基づいて、少なくとも70%、好ましくは少なくとも80%の配列同一性を有し、前記相補体は前記ヌクレオチド配列と同数のヌクレオチドを含み、前記相補体と前記ヌクレオチド配列とは100%相補的であるポリヌクレオチド。
【請求項6】
前記ポリペプチドのアミノ酸配列と配列番号4、10、12及び14からなる群から選択されるアミノ酸配列とが、ClustalWアラインメント法に基づいて、少なくとも85%、好ましくは少なくとも90%、任意選択で少なくとも95%の配列同一性を有する、請求項5に記載のポリヌクレオチド。
【請求項7】
前記ヌクレオチド配列が、配列番号3、9、11及び13からなる群から選択されるヌクレオチド配列を含む、請求項5に記載のポリヌクレオチド。
【請求項8】
前記ポリペプチドが、配列番号4、10、12及び14からなる群から選択されるアミノ酸配列を含む、請求項5に記載のポリヌクレオチド。
【請求項9】
アスパラギン酸エンドプロテイナーゼ活性を有するポリペプチドをコードするヌクレオチド配列又は前記ヌクレオチド配列の相補体を含む単離されたポリヌクレオチドであって、前記ポリペプチドのアミノ酸配列と配列番号6又は8から選択されるアミノ酸配列、好ましくは配列番号8のアミノ酸配列とは、ClustalWアラインメント法に基づいて、少なくとも75%、好ましくは少なくとも80%の配列同一性を有し、前記相補体は前記ヌクレオチド配列と同数のヌクレオチドを含み、前記相補体と前記ヌクレオチド配列とは100%相補的であるポリヌクレオチド。
【請求項10】
前記ポリペプチドのアミノ酸配列と配列番号6又は8、好ましくは配列番号8から選択されるアミノ酸配列とが、ClustalWアラインメント法に基づいて、少なくとも85%、好ましくは少なくとも90%、任意選択で少なくとも95%の配列同一性を有する、請求項9に記載のポリヌクレオチド。
【請求項11】
前記ヌクレオチド配列が、配列番号5又は7、好ましくは配列番号7のヌクレオチド配列を含む、請求項9に記載のポリヌクレオチド。
【請求項12】
前記ポリペプチドが、配列番号6又は8、好ましくは配列番号8のアミノ酸配列を含む、請求項9に記載のポリヌクレオチド。
【請求項13】
システインプロテイナーゼ活性を有するポリペプチドをコードするヌクレオチド配列又は前記ヌクレオチド配列の相補体を含む単離されたポリヌクレオチドであって、前記ポリペプチドのアミノ酸配列と配列番号16のアミノ酸配列とは、ClustalWアラインメント法に基づいて、少なくとも70%、好ましくは少なくとも80%の配列同一性を有し、前記相補体は前記ヌクレオチド配列と同数のヌクレオチドを含み、前記相補体と前記ヌクレオチド配列とは100%相補的であるポリヌクレオチド。
【請求項14】
前記ポリペプチドのアミノ酸配列と配列番号16のアミノ酸配列とが、ClustalWアラインメント法に基づいて、少なくとも85%、好ましくは少なくとも90%、任意選択で少なくとも95%の配列同一性を有する、請求項13に記載のポリヌクレオチド。
【請求項15】
前記ヌクレオチド配列が配列番号15のヌクレオチド配列を含む、請求項13に記載のポリヌクレオチド。
【請求項16】
前記ポリペプチドが配列番号16のアミノ酸配列を含む、請求項13に記載のポリヌクレオチド。
【請求項17】
請求項1から16までのいずれか一項に記載のポリヌクレオチドを含むベクター。
【請求項18】
調節配列に作動可能に連結された請求項1から16までのいずれか一項に記載のポリヌクレオチドを含む、非ネイティブの組換えDNA構築物。
【請求項19】
請求項1から16までのいずれか一項に記載のポリヌクレオチドで細胞を形質転換するステップを含む、細胞を形質転換するための方法。
【請求項20】
請求項18に記載の非ネイティブの組換えDNA構築物を含む細胞。
【請求項21】
原核細胞、真核細胞又は植物細胞、好ましくはコーヒー細胞である、請求項20に記載の細胞。
【請求項22】
請求項20又は21に記載の細胞を含むトランスジェニック植物。
【請求項23】
緑のコーヒー粒中のコーヒーフレーバー前駆体レベルを調節するための方法であって、請求項18に記載の非ネイティブの組換えDNA構築物をコーヒー植物中に導入するステップを含む方法。
【図1】

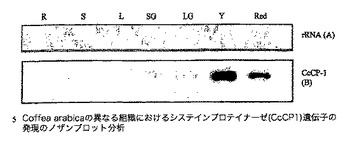
【図2】
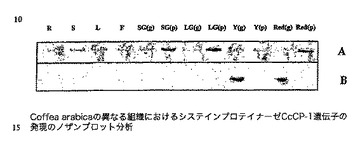
【図2A】

【図3】
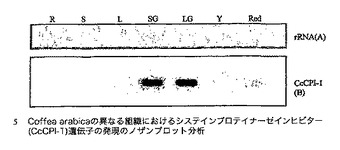
【図4】

【図5】

【図6】
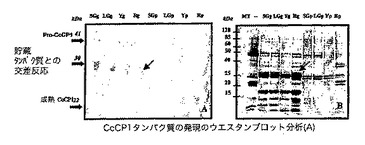
【図6A】

【図7】

【図8】
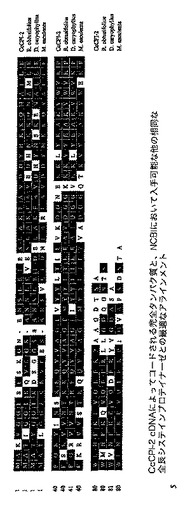
【図9】
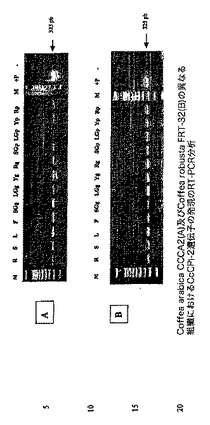
【図10】

【図11】

【図12】
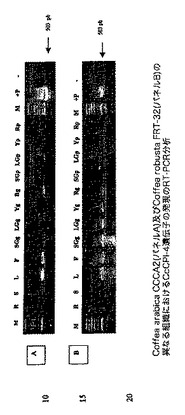
【図13】
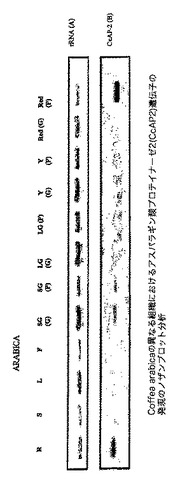
【図14】

【図15】

【図16】
【図17】
【図18】
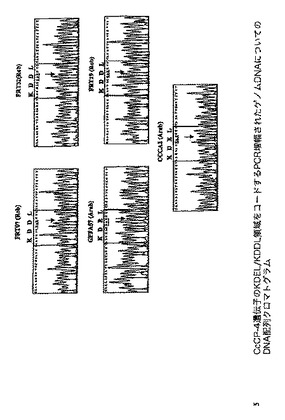
【図19】

【図20】
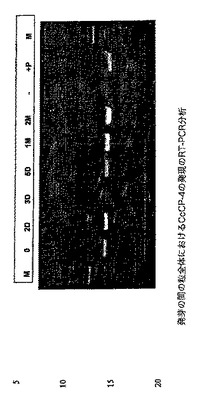
【図21】

【図22】

【図2】
【図2A】
【図3】
【図4】
【図5】
【図6】
【図6A】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【公表番号】特表2009−513102(P2009−513102A)
【公表日】平成21年4月2日(2009.4.2)
【国際特許分類】
【出願番号】特願2006−516040(P2006−516040)
【出願日】平成16年6月18日(2004.6.18)
【国際出願番号】PCT/EP2004/006805
【国際公開番号】WO2004/113520
【国際公開日】平成16年12月29日(2004.12.29)
【出願人】(599132904)ネステク ソシエテ アノニム (637)
【出願人】(505294425)コーネル リサーチ ファウンデイション インコーポレイテッド (11)
【Fターム(参考)】
【公表日】平成21年4月2日(2009.4.2)
【国際特許分類】
【出願日】平成16年6月18日(2004.6.18)
【国際出願番号】PCT/EP2004/006805
【国際公開番号】WO2004/113520
【国際公開日】平成16年12月29日(2004.12.29)
【出願人】(599132904)ネステク ソシエテ アノニム (637)
【出願人】(505294425)コーネル リサーチ ファウンデイション インコーポレイテッド (11)
【Fターム(参考)】
[ Back to top ]