
緑膿菌に対して感染防御能を誘導できる医薬組成物
【課題】 緑膿菌に対して効果的に感染防御能を誘導できる医薬組成物の提供。
【解決手段】 下記(1)〜(3)等のいずれかの成分を含む医薬組成物。(1) 緑膿菌の細胞外膜蛋白OprFの全長(一部)をコ-ドするDNA、緑膿菌の細胞外膜蛋白OprIの全長(一部)をコ-ドするDNA、緑膿菌のIII型分泌機構制御蛋白PcrVの全長(一部)をコ-ドするDNA、及び緑膿菌の毛状付属器蛋白PilAの全長(一部)をコ-ドするDNAをそれぞれ含むベクタ-の混合物(2) 緑膿菌の細胞外膜蛋白OprFの全長(一部)、緑膿菌の細胞外膜蛋白OprIの全長(一部)、III型分泌機構制御蛋白PcrVの全長(一部)、及び毛状付属器蛋白PilAの全長(一部)の融合物をコ-ドするDNAを含むベクタ-(3) 緑膿菌の細胞外膜蛋白OprFの全長(一部)、緑膿菌の細胞外膜蛋白OprIの全長(一部)、緑膿菌のIII型分泌機構制御蛋白PcrVの全長(一部)、及び緑膿菌の毛状付属器蛋白PilAの全長(一部)の混合物
【解決手段】 下記(1)〜(3)等のいずれかの成分を含む医薬組成物。(1) 緑膿菌の細胞外膜蛋白OprFの全長(一部)をコ-ドするDNA、緑膿菌の細胞外膜蛋白OprIの全長(一部)をコ-ドするDNA、緑膿菌のIII型分泌機構制御蛋白PcrVの全長(一部)をコ-ドするDNA、及び緑膿菌の毛状付属器蛋白PilAの全長(一部)をコ-ドするDNAをそれぞれ含むベクタ-の混合物(2) 緑膿菌の細胞外膜蛋白OprFの全長(一部)、緑膿菌の細胞外膜蛋白OprIの全長(一部)、III型分泌機構制御蛋白PcrVの全長(一部)、及び毛状付属器蛋白PilAの全長(一部)の融合物をコ-ドするDNAを含むベクタ-(3) 緑膿菌の細胞外膜蛋白OprFの全長(一部)、緑膿菌の細胞外膜蛋白OprIの全長(一部)、緑膿菌のIII型分泌機構制御蛋白PcrVの全長(一部)、及び緑膿菌の毛状付属器蛋白PilAの全長(一部)の混合物
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、緑膿菌に対して感染防御能を誘導できる医薬組成物に関する。
【背景技術】
【0002】
緑膿菌は、慢性疾患に伴う日和見感染症の原因菌として、薬剤耐性株の出現などから臨床上非常に問題となっている。様々なワクチンが検討されてきたが、臨床応用に至っているものは未だ開発されていない(非特許文献1)。例えば、細胞外膜タンパクOprF, OprIはワクチン標的分子として検討されている。マウスにおいてOprF/Iワクチンは、肺感染症・敗血症モデルでの有効性が確認されている。OprF/I融合分子(B細胞エピト-プ)は第III相臨床治験が行われ、安全性と有効性が確認されている(非特許文献2〜6)。III型分泌機構制御タンパクPcrVもワクチン標的分子として検討されている。特異抗体は菌毒素を宿主細胞内に注入するのを抑制するため、感染に伴う組織障害を抑制すると考えられている。特異抗体のみで感染を抑制することが報告されている(非特許文献7〜9)。毛状付属器タンパクPilAは細胞外膜に存在する繊毛で、菌の運動や粘膜への接着に係わる他、IV型分泌機構に重要である。ワクチン標的分子として有効性が確認されている(非特許文献10〜13)。
【0003】
【非特許文献1】Holder IA. Pseudomonas immunotherapy: a histrical overview. Vaccine 2004;22(7):831-9
【非特許文献2】Infect Immun, 63:1855, 1995
【非特許文献3】Vaccine, 17:1663, 1999
【非特許文献4】Infect Immun, 67:1461, 1999
【非特許文献5】FEMS Immunol Med Microbiol, 33:89, 2002
【非特許文献6】Vaccine, 24:6240, 2006
【非特許文献7】Nat Med, 5:392, 1999
【非特許文献8】J Immunol, 167:5880, 2001
【非特許文献9】Vaccine, 24:6240, 2006
【非特許文献10】Infect Immun, 63:1278, 1995
【非特許文献11】Vaccine, 22:831, 2004
【非特許文献12】FEMS Immunol Med Microbiol, 47:107, 2006
【非特許文献13】Vaccine, 24:6240, 2006
【発明の開示】
【発明が解決しようとする課題】
【0004】
本発明は、緑膿菌に対して効果的に感染防御能を誘導する医薬組成物を提供することを目的とする。
【課題を解決するための手段】
【0005】
本発明者らは、OprF/I、PcrV及びPilAを混合して免疫することで、多価かつ相乗的なワクチン効果を賦与できることを見出した。しかし、個々の標的抗原は、分子量が大きいため、遺伝子ワクチンとして応用した際に発現効率が悪く、病原因子としての危険性を孕んでいる。そこで、上記混合ワクチンの効果を効率化させ、病原因子の危険性を軽減させるために、各標的タンパクの抗原性を有する領域(エピト-プ)のみを連結し、抗原の分子量を縮小させた抗原エピト-プ融合ワクチンを作製した。この抗原を標的とするDNAワクチンを免疫すると、前記混合ワクチンと同等の防御的抗緑膿菌免疫応答を誘導することが確認された。このことから、抗原エピト-プ融合ワクチンを用いることで、幅広いスペクトラムの緑膿菌に対して効率的かつ安全に感染防御能を賦与できるワクチンの開発が可能となった。
【0006】
本発明の要旨は以下の通りである。
(1)下記(i)〜(v)のいずれかの成分を含む、緑膿菌に対して感染防御能を誘導できる医薬組成物。
(i) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部をコ-ドするDNA、緑膿菌の細胞外膜タンパクOprIの全長又はその一部をコ-ドするDNA、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部をコ-ドするDNA、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部をコ-ドするDNAをそれぞれ含むベクタ-の混合物
(ii) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、III型分泌機構制御タンパクPcrVの全長又はその一部、及び毛状付属器タンパクPilAの全長又はその一部の融合物をコ-ドするDNAを含むベクタ-
(iii) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部の混合物
(iv) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部の融合物
(v) (iii)の混合物又は(iv)の融合物に対して誘導された抗体
【0007】
(2)緑膿菌の細胞外膜タンパクOprFの全長又はその一部が下記(A)又は(B)のタンパク質であり、緑膿菌の細胞外膜タンパクOprIの全長又はその一部が下記(C)又は(D)のペプチドであり、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部が下記(E)又は(F)のタンパク質であり、緑膿菌の毛状付属器タンパクPilAの全長又はその一部が下記(G)又は(H)のペプチドである(1)記載の医薬組成物。
(A)配列番号2のアミノ酸配列からなるタンパク質
(B)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の細胞外膜タンパクOprFの抗原活性を有するタンパク質
(C)配列番号4のアミノ酸配列からなるペプチド
(D)配列番号4のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の細胞外膜タンパクOprIの抗原活性を有するペプチド
(E)配列番号6のアミノ酸配列からなるタンパク質
(F)配列番号6のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌のIII型分泌機構制御タンパクPcrVの抗原活性を有するタンパク質
(G)配列番号8のアミノ酸配列からなるペプチド
(H)配列番号8のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の毛状付属器タンパクPilAの抗原活性を有するペプチド
【0008】
(3)緑膿菌の細胞外膜タンパクOprFの全長又はその一部をコ-ドするDNAが下記(a)又は(b)のDNAであり、緑膿菌の細胞外膜タンパクOprIの全長又はその一部をコ-ドするDNAが下記(c)又は(d)のDNAであり、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部をコ-ドするDNAが下記(e)又は(f)のDNAであり、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部をコ-ドするDNAが下記(g)又は(h)のDNAである(1)又は(2)記載の医薬組成物。
(a)配列番号1のヌクレオチド配列で表されるDNA
(b)配列番号1のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の細胞外膜タンパクOprFの抗原活性を有するタンパク質をコ-ドするDNA
(c) 配列番号3のヌクレオチド配列で表されるDNA
(d) 配列番号3のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の細胞外膜タンパクOprIの抗原活性を有するペプチドをコ-ドするDNA
(e)配列番号5のヌクレオチド配列で表されるDNA
(f)配列番号5のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌のIII型分泌機構制御タンパクPcrVの抗原活性を有するタンパク質をコ-ドするDNA
(g)配列番号7のヌクレオチド配列で表されるDNA
(h)配列番号7のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の毛状付属器タンパクPilAの抗原活性を有するタンパク質をコ-ドするDNA
【0009】
(4)(i)〜(iv)のいずれかの成分を含むワクチンである(1)〜(3)のいずれかに記載の医薬組成物。
【0010】
(5)(i)のベクタ-混合物又は(ii)のベクタ-を含む遺伝子ワクチンである(4)記載の医薬組成物。
【0011】
(6)ベクタ-がプラスミド又はウイルスベクタ-である(5)記載の医薬組成物。
【0012】
(7)下記(I)の混合物又は(II)の融合物に対して誘導された抗体。
(I) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部の混合物
(II) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部の融合物
【発明の効果】
【0013】
本発明の医薬組成物は、異種の抗原若しくは抗原エピト-プを混合又は融合させたものを標的分子とするので、幅広いスペクトラムの緑膿菌に対して効率的かつ安全に感染防御能を賦与できる。
【発明を実施するための最良の形態】
【0014】
以下、本発明の実施の形態についてより詳細に説明する。
本発明は、緑膿菌に対して感染防御能を誘導できる医薬組成物を提供する。
本発明の医薬組成物は、ワクチン(遺伝子ワクチン、タンパクワクチン)又は抗体医薬として利用できる。
【0015】
本発明のワクチンが標的とする分子は、緑膿菌の細胞外膜タンパクOprFの全長若しくはその一部、緑膿菌の細胞外膜タンパクOprIの全長若しくはその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長若しくはその一部、及び緑膿菌の毛状付属器タンパクPilAの全長若しくはその一部の混合物又は融合物である。すなわち、本発明のワクチンは、緑膿菌の細胞外膜タンパクOprFの全長若しくはその一部、緑膿菌の細胞外膜タンパクOprIの全長若しくはその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長若しくはその一部、及び緑膿菌の毛状付属器タンパクPilAの全長若しくはその一部を抗原又は抗原エピト-プとして利用するものである。ワクチンは標的病原体に対する特異的抗原を標的に能動的に免疫応答を誘導することを目的とするが、病原因子のような抗原の場合(エンドトキシン、ペプチドグリカン、リポタンパク、ゲノムDNAなど)、投与することで生体に有害な作用(例えば、ショック症状、免疫応答の過剰活性化)を引き起こすことがあるため、免疫応答を誘導するのに必要な最小領域をもちいるのが望ましい。
【0016】
本発明のワクチンは、下記(i)〜(iv)のいずれかの成分を含む。
(i) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部をコ-ドするDNA、緑膿菌の細胞外膜タンパクOprIの全長又はその一部をコ-ドするDNA、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部をコ-ドするDNA、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部をコ-ドするDNAをそれぞれ含むベクタ-の混合物
(ii) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、III型分泌機構制御タンパクPcrVの全長又はその一部、及び毛状付属器タンパクPilAの全長又はその一部の融合物をコ-ドするDNAを含むベクタ-
(iii) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部の混合物
(iv) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部の融合物
緑膿菌(Pseudomonas aeruginosa)PA01株の全ゲノム配列は解読されており、NCBI gene bank にaccession #AE004091の登録番号で登録されている。従って、上記の(i)〜(iv)の成分は、この配列情報を利用して調製することができる。また、PA01株以外の他の株(PAK、D4など)の配列情報を利用してもよい。
【0017】
緑膿菌の細胞外膜タンパクOprFの全長又はその一部は、抗原又は抗原エピト-プとして機能するものであれば、特に限定されないが、例えば、PA01株のOprF190-342、そのホモロ-グなどを挙げることができ、具体的には、下記(A)又は(B)のタンパク質を例示することができる。
【0018】
(A)配列番号2のアミノ酸配列からなるタンパク質
(B)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の細胞外膜タンパクOprFの抗原活性を有するタンパク質
配列番号2は、PA01株のOprF190-342のアミノ酸配列を示す。また、PA01株のOprFの全長(OprF1-350)のアミノ酸配列は配列番号10に示す。緑膿菌の細胞外膜タンパクOprFの全長としては、下記(A1)又は(B1)のタンパク質を例示することができる。
【0019】
(A1)配列番号10のアミノ酸配列からなるタンパク質
(B1)配列番号10のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の細胞外膜タンパクOprFの抗原活性を有するタンパク質
緑膿菌の細胞外膜タンパクOprIの全長又はその一部は、抗原又は抗原エピト-プとして機能するものであれば、特に限定されないが、例えば、PA01株のOprI21-83、そのホモロ-グなどを挙げることができ、具体的には、下記(C)又は(D)のペプチドを例示することができる。
【0020】
(C)配列番号4のアミノ酸配列からなるペプチド
(D)配列番号4のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の細胞外膜タンパクOprIの抗原活性を有するペプチド
配列番号4は、PA01株のOprI21-83のアミノ酸配列を示す。また、PA01株のOprIの全長(OprI1-83)のアミノ酸配列は配列番号12に示す。緑膿菌の細胞外膜タンパクOprIの全長としては、下記(C1)又は(D1)のタンパク質を例示することができる。
【0021】
(C1)配列番号12のアミノ酸配列からなるペプチド
(D1)配列番号12のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の細胞外膜タンパクOprIの抗原活性を有するペプチド
緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部は、抗原又は抗原エピト-プとして機能するものであれば、特に限定されないが、例えば、PA01株のPcrV1-294、そのホモロ-グなどを挙げることができ、具体的には、下記(E)又は(F)のタンパク質を例示することができる。
【0022】
(E)配列番号6のアミノ酸配列からなるタンパク質
(F)配列番号6のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌のIII型分泌機構制御タンパクPcrVの抗原活性を有するタンパク質
配列番号6は、PA01株のPcrV1-294(全長)のアミノ酸配列を示す。
緑膿菌の毛状付属器タンパクPilAの全長又はその一部は、抗原又は抗原エピト-プとして機能するものであれば、特に限定されないが、例えば、PA01株のPilA136-149、そのホモロ-グなどを挙げることができ、具体的には、下記(G)又は(H)のペプチドを例示することができる。
【0023】
(G)配列番号8のアミノ酸配列からなるペプチド
(H)配列番号8のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の毛状付属器タンパクPilAの抗原活性を有するペプチド
配列番号8は、PA01株のPilA136-149のアミノ酸配列を示す。また、PA01株のPilA1-149の(全長)のアミノ酸配列は配列番号14に示す。緑膿菌の細胞外膜タンパクPilAの全長としては、下記(G1)又は(H1)のタンパク質を例示することができる。
【0024】
(G1)配列番号14のアミノ酸配列からなるタンパク質
(H1)配列番号14のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の毛状付属器タンパクPilAの抗原活性を有するタンパク質
緑膿菌の細胞外膜タンパクOprFの全長又はその一部をコ-ドするDNAとしては、例えば、PA01株のOprF190-342、そのホモロ-グなどをコ-ドするDNAを挙げることができ、具体的には、下記(a)又は(b)のDNAを例示することができる。
【0025】
(a)配列番号1のヌクレオチド配列で表されるDNA
(b)配列番号1のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の細胞外膜タンパクOprFの抗原活性を有するタンパク質をコ-ドするDNA
配列番号1は、PA01株のOprF190-342をコ-ドするDNAのヌクレオチド配列を示す。また、PA01株のOprFの全長をコ-ドするDNAのヌクレオチド配列は配列番号9に示す。緑膿菌の細胞外膜タンパクOprFの全長をコ-ドするDNAとしては、下記(a1)又は(b1)のDNAを例示することができる。
【0026】
(a1)配列番号9のヌクレオチド配列で表されるDNA
(b1)配列番号9のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の細胞外膜タンパクOprFの抗原活性を有するタンパク質をコ-ドするDNA
緑膿菌の細胞外膜タンパクOprIの全長又はその一部をコ-ドするDNAとしては、例えば、PA01株のOprI21-83、そのホモロ-グなどをコ-ドするDNAを挙げることができ、具体的には、下記(c)又は(d)のDNAを例示することができる。
【0027】
(c) 配列番号3のヌクレオチド配列で表されるDNA
(d) 配列番号3のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の細胞外膜タンパクOprIの抗原活性を有するペプチドをコ-ドするDNA
配列番号3は、PA01株のOprI21-83をコ-ドするDNAのヌクレオチド配列を示す。また、PA01株のOprIの全長をコ-ドするDNAのヌクレオチド配列は配列番号11に示す。緑膿菌の細胞外膜タンパクOprIの全長をコ-ドするDNAとしては、下記(c1)又は(d1)のDNAを例示することができる。
【0028】
(c1)配列番号11のヌクレオチド配列で表されるDNA
(d1)配列番号11のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の細胞外膜タンパクOprFの抗原活性を有するタンパク質をコ-ドするDNA
緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部をコ-ドするDNAとしては、例えば、PA01株のPcrV1-294、そのホモロ-グなどをコ-ドするDNAを挙げることができ、具体的には、下記(e)又は(f)のDNAを例示することができる。
【0029】
(e)配列番号5のヌクレオチド配列で表されるDNA
(f)配列番号5のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌のIII型分泌機構制御タンパクPcrVの抗原活性を有するタンパク質をコ-ドするDNA
配列番号5は、PA01株のPcrV1-294(全長)をコ-ドするDNAのヌクレオチド配列を示す。
緑膿菌の毛状付属器タンパクPilAの全長又はその一部をコ-ドするDNAとしては、例えば、PA01株のPilA136-149、そのホモロ-グなどをコ-ドするDNAを挙げることができ、具体的には、下記(g)又は(h)のDNAを例示することができる。
【0030】
(g)配列番号7のヌクレオチド配列で表されるDNA
(h)配列番号7のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の毛状付属器タンパクPilAの抗原活性を有するタンパク質をコ-ドするDNA
配列番号7は、PA01株のPilA136-149をコ-ドするDNAのヌクレオチド配列を示す。また、PA01株のPilAの全長(PilA1-149)をコ-ドするDNAのヌクレオチド配列は配列番号13に示す。緑膿菌の毛状付属器タンパクPilAの全長をコ-ドするDNAとしては、下記(g1)又は(h1)のDNAを例示することができる。
【0031】
(g1)配列番号13のヌクレオチド配列で表されるDNA
(h1)配列番号13のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の細胞外膜タンパクOprFの抗原活性を有するタンパク質をコ-ドするDNA
配列番号1、3、5、7、9、11又は13のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズするDNAは、配列番号1、3、5、7、9、11又は13のヌクレオチド配列で表されるDNAに相補的なDNAの全部又は一部と少なくとも80%(好ましくは、少なくとも95%、より好ましくは、少なくとも98%)の同一性があるとよい。「同一性」という用語は当業者に周知の用語であり、配列の同一性は入手可能な配列解析ソフトウェア(例えば、GCG Wisconsin Package (Accelrys INC.提供、Nucleic Acid Research 12 (1):387, 1984)、BLAST Nプログラム(NCBI提供、J Mol Biol 215: 403-410, 1990)などを用いて決定することができる。ハイブリダイゼ-ションはストリンジェントな条件下で行われる。核酸二本鎖又はハイブリッドの安定性は、融解温度Tm(プロ-ブが標的DNAから解離する温度)で表される。この融解温度はストリンジェントな条件を定義するために用いられる。1%のミスマッチによりTmが1℃低下すると仮定すると、ハイブリダイゼ-ション反応の最終洗浄の温度を低くしなければならない。例えば、プロ-ブと95%以上の同一性を有する配列を求める場合には、最終洗浄温度を5℃低くしなければならない。実際、1%のミスマッチにつき、0.5〜1.5℃の間でTmが変わることになる。ストリンジェントな条件の例としては、5x SSC/5x デンハルト溶液/1.0% SDS中68℃でハイブリダイズさせ、0.2x SSC/0.1%SDS中室温で洗浄することである。中程度にストリンジェントな条件の例としては、3x SSC中42℃で洗浄することである。塩濃度や温度は、プロ-ブと標的核酸との同一性の最適なレベルを達成するために変更されうる。このような条件に関するさらなる指針として、Sambrook et al., 1989, Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Press, N.Y.; and Ausubel et al. (eds.), 1995, Current Protocols in Molecular Biology, (John Wiley & Sons. N.Y.) at Unit 2.10を参照されたい。
【0032】
緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部、緑膿菌の毛状付属器タンパクPilAの全長又はその一部(以下、「抗原又は抗原エピト-プ」と呼ぶことがある)、及びそれらの融合物をコ-ドするDNAは、例えば、以下のようにして製造することができる。
【0033】
緑膿菌からゲノムDNAを抽出し、目的とする抗原又は抗原エピト-プのコ-ド領域をPCRによって増幅する。得られたPCR産物が目的の抗原又は抗原エピト-プをコ-ドするDNAである。抗原又は抗原エピト-プの融合物は、抗原又は抗原エピト-プをコ-ドするDNAをライゲートすることにより得られる。抗原若しくは抗原エピト-プ又はそれらの融合物をコ-ドするDNAを含有する組換えベクタ-は、公知の方法(例えば、Molecular Cloning2nd Edition, J. Sambrook et al., Cold Spring Harbor Lab. Press, 1989に記載の方法)により、抗原若しくは抗原エピト-プ又はそれらの融合物をコ-ドするDNAを適当な発現ベクタ-に挿入することにより得られる。
【0034】
発現ベクタ-としては、大腸菌由来のプラスミド(例、pBR322,pBR325,pUC12,pUC13, pGACAG)、枯草菌由来のプラスミド(例、pUB110,pTP5,pC194)、酵母由来プラスミド(例、pSH19,pSH15)、λファ-ジなどのバクテリオファ-ジ、レトロウイルス,アデノウイルス、ワクシニアウイルスなどの動物ウイルス、バキュロウイルスなどの昆虫病原ウイルスなどを用いることができる。
【0035】
発現ベクタ-には、プロモ-タ-(例えば、CAGプロモ-タ-、CMVプロモーター、SV40プロモーター、EF-1プロモーター))、イントロン(例えば、IntronA、b-actinイントロン))、エンハンサ-、スプライシングシグナル、ポリA付加シグナル、選択マ-カ-、タグ(例えば、FLAG配列、HA配列、c-myc配列、IgFc配列、6xHis配列、GST配列))、SV40複製オリジンなどを付加してもよい。
【0036】
また、発現ベクタ-は、融合タンパク質発現ベクタ-であってもよい。種々の融合タンパク質発現ベクタ-が市販されており、pGEXシリ-ズ(アマシャムファルマシアバイオテク社)、pET CBD Fusion System 34b-38b(Novagen社)、pET Dsb Fusion Systems 39b and 40b(Novagen社)、pET GST Fusion System 41 and 42(Novagen社)、pFLAGシリ-ズ(シグマアルドリッチ社)などを例示することができる。
【0037】
組換えベクタ-を遺伝子ワクチン(プラスミドワクチン・ウイルスベクタ-ワクチン)として使用する場合には、すでに遺伝子ワクチン用に開発したpGACAGプラスミドベクタ-に標的遺伝子を導入しプラスミドワクチンを作製するとよい。
抗原若しくは抗原エピト-プ又はそれらの融合物をコ-ドするDNAを含有する組換えベクタ-を宿主に導入することにより、形質転換体を得ることができる。
【0038】
宿主としては、細菌細胞(例えば、エシェリヒア属菌、バチルス属菌、枯草菌など)、真菌細胞(例えば、酵母、アスペルギルスなど)、昆虫細胞(例えば、S2細胞、Sf細胞など)、動物細胞(例えば、CHO細胞、COS細胞、HeLa細胞、C127細胞、3T3細胞、BHK細胞、HEK293細胞など)、植物細胞などを例示することができる。
組換えベクタ-を宿主に導入するには、Molecular Cloning2nd Edition, J. Sambrook et al., Cold Spring Harbor Lab. Press, 1989に記載の方法(例えば、リン酸カルシウム法、DEAE-デキストラン法、トランスフェクション法、マイクロインジェクション法、リポフェクション法、エレクロトポレ-ション法、形質導入法、スクレ-プロ-ディング法、ショットガン法など)または感染により行うことができる。
【0039】
形質転換体を培地で培養し、培養物から抗原若しくは抗原エピト-プ又はそれらの融合物を採取することができる。抗原若しくは抗原エピト-プ又はそれらの融合物が培地に分泌される場合には、培地を回収し、その培地から抗原若しくは抗原エピト-プ又はそれらの融合物を分離し、精製すればよい。抗原若しくは抗原エピト-プ又はそれらの融合物が形質転換された細胞内に産生される場合には、その細胞を溶解し、その溶解物から抗原若しくは抗原エピト-プ又はそれらの融合物を分離し、精製すればよい。
【0040】
抗原若しくは抗原エピト-プ又はそれらの融合物が別のタンパク質(タグとして機能する)との融合タンパク質の形態で発現される場合には、融合タンパク質を分離及び精製した後に、FactorXaや酵素(エンテロキナ-ゼ)処理をすることにより、別のタンパク質を切断し、目的とする抗原若しくは抗原エピト-プ又はそれらの融合物を得ることができる。
【0041】
抗原若しくは抗原エピト-プ又はそれらの融合物の分離及び精製は、公知の方法により行うことができる。公知の分離、精製法としては、塩析や溶媒沈澱法などの溶解度の差を利用する方法、透析法、限外ろ過法、ゲルろ過法、およびSDS-ポリアクリルアミドゲル電気泳動法などの分子量の差を利用する方法、イオン交換クロマトグラフィ-などの荷電の差を利用する方法、アフィニティ-クロマトグラフィ-などの特異的親和性を利用する方法、逆相高速液体クロマトグラフィ-などの疎水性の差を利用する方法、等電点電気泳動法などの等電点の差を利用する方法などが用いられる。
【0042】
抗原及び抗原エピト-プ並びにそれらの融合物は公知のペプチド合成法に従って製造してもよい。
【0043】
また、抗原若しくは抗原エピト-プに変異が導入されているもの(例えば、天然型の抗原又は抗原エピト-プのアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ前記抗原又は抗原エピト-プの抗原活性を有するタンパク質又はペプチド)又はそれらの融合物をコ-ドするDNAは、天然型の抗原又は抗原エピト-プのコ-ド領域を点突然変異誘発法により変異させることにより作製することができる。変異させたコ-ド領域をPCRによって増幅する。得られたPCR産物が抗原若しくは抗原エピト-プの変異体をコ-ドするDNAである。また、この変異体をコ-ドするDNAをライゲ-トすることにより、変異体の融合物をコ-ドするDNAを得ることができる。得られたDNAを適当な発現ベクタ-に組み込んだ後、適当な宿主に導入し、組換え蛋白質として生産させる(例えば、西郷薫、佐野弓子共訳、CURRENT PROTOCOLSコンパクト版、分子生物学実験プロトコ-ル、I、II、III、丸善株式会社:原著、Ausubel,F.M.等, Short Protocols in Molecular Biology, Third Edition, John Wiley & Sons, Inc., New Yorkを参照のこと)ことにより、抗原若しくは抗原エピト-プの変異体又はそれらの融合物を得ることができる。
【0044】
抗原若しくは抗原エピト-プの混合物又は融合物、あるいは抗原若しくは抗原エピト-プをコ-ドするDNAの混合物又は融合物を含有するベクタ-は、緑膿菌に対して感染防御能を誘導するためのワクチンの有効成分として利用することができる。例えば、抗原若しくは抗原エピト-プの混合物又は融合物、あるいは抗原若しくは抗原エピト-プをコ-ドするDNAの混合物又は融合物を含有するベクタ-をPBSなどの緩衝液、生理食塩水、滅菌水などに溶解し、必要に応じてフィルタ-などで濾過滅菌した後、注射により被験者に投与されるとよい。また、この溶液には、添加剤(例えば、不活化剤、保存剤、アジュバント、乳化剤など)などを添加してもよい。投与経路としては、静脈、筋肉、腹腔、皮下、皮内投与などが可能であり、また、経鼻、経口投与してもよい。
【0045】
抗原若しくは抗原エピト-プの混合物又は融合物、あるいは抗原若しくは抗原エピト-プをコ-ドするDNAの混合物又は融合物を含有するベクタ-の投与量、投与の回数及び頻度は、被験者の症状、年齢、体重、投与方法、投与形態などにより異なるが、例えば、通常、成人一人当たり0.01〜1mg/kg体重、好ましくは、0.01〜0.1mg/kg体重の抗原若しくは抗原エピト-プの混合物又は融合物を、少なくとも1回、所望の効果が持続する頻度で投与するとよい。抗原若しくは抗原エピト-プをコ-ドするDNAの混合物又は融合物を含有するプラスミドベクタ-を投与する場合には、例えば、通常、成人一人当たり1〜100mg、好ましくは、1〜10mgの投与量で少なくとも1回、所望の効果が持続する頻度で投与するとよい。ベクタ-がウイルスの場合には、成人一人当たり109〜1012ウイルス粒子、好ましくは1011〜1012ウイルス粒子の投与量で少なくとも1回、所望の効果が持続する頻度で投与するとよい。
【0046】
プラスミドワクチンはエンドトキシン除去した後投与するとよい。
筋肉内投与後エレクトロポレ-ション法で細胞内に導入する方法やリポソ-ムなどトランスフェクション増強剤との複合体を作製後投与する方法によりワクチン効果を高める方法も併用するとよい。
【0047】
本発明は、抗原若しくは抗原エピト-プの混合物又はそれらの融合物に対して誘導された抗体も提供する。本発明の抗体は緑膿菌に対して特異的に反応することができる。
本発明の抗体は、慣用のプロトコ-ルを用いて、抗原若しくは抗原エピト-プの混合物又はそれらの融合物を動物に投与することにより得られる。
本発明の抗体は、ポリクロ-ナル抗体、モノクロ-ナル抗体、キメラ抗体、一本鎖抗体、ヒト化抗体のいずれであってもよい。
【0048】
ポリクロ-ナル抗体を作製するには、公知あるいはそれに準じる方法にしたがって製造することができる。例えば、免疫抗原(タンパク質抗原)をコ-ドするDNAフラグメントを発現ベクタ-に導入し、これを動物に投与(免疫)し、該免疫動物からタンパク質に対する抗体含有物を採取して、抗体の分離精製を行なうことにより製造できる。投与に際して抗体産生能を高めるため、完全フロイントアジュバントや不完全フロイントアジュバントを投与してもよい。投与は、通常約2〜6週毎に1回ずつ、計約2〜10回程度行なわれる。ポリクロ-ナル抗体は、免疫動物の血液、腹水など、好ましくは血液から採取する(抗血清)ことができる。ポリクロ-ナル抗体の分離精製は、免疫グロブリンの分離精製法(例えば、塩析法、アルコ-ル沈殿法、等電点沈殿法、電気泳動法、イオン交換体による吸脱着法、超遠心法、ゲルろ過法、抗原結合固相あるいはプロテインAあるいはプロテインGなどの活性吸着剤により抗体のみを採取し、結合を解離させて抗体を得る特異的精製法)に従って行なうことができる。
【0049】
モノクロ-ナル抗体は、Nature (1975) 256: 495、Science (1980) 208: 692-に記載されている、G. Koehler及びC. Milsteinのハイブリド-マ法により作製することができる。すなわち、動物を免疫した後、免疫動物の脾臓から抗体産生細胞を単離し、これを骨髄腫細胞と融合させることによりモノクロ-ナル抗体産生細胞を調製する。さらに、抗原若しくは抗原エピト-プの混合物又はそれらの融合物と特異的に反応するが、他の抗原タンパク質とは実質的に交差反応しない抗体を産生する細胞系を単離するとよい。この細胞系を培養し、培養物から所望のモノクロ-ナル抗体を取得することができる。モノクロ-ナル抗体の精製は、上記の免疫グロブリンの分離精製法に従って行なうことができる。
【0050】
一本鎖抗体を作製する技法は、米国特許第4,946,778号に記載されている。
ヒト化抗体を作製する技法は、Biotechnology 10, 1121-, 1992; Biotechnology 10, 169-, 1992に記載されている。
【0051】
本発明の抗体は、緑膿菌に対する感染防御能を誘導するための医薬として利用することができる。この医薬の利点は、宿主の免疫系が活性化され、十分に高い防御レベルに達するまで待つ必要がなく、即座に効果が期待できることである。特に、感染が成立している場合や疾患が成立している場合に、抗体を投与すると、病原体や病原体が生産する毒素から患者を即座に回復させることができる。例えば、本発明の抗体をPBSなどの緩衝液、生理食塩水、滅菌水などに溶解し、必要に応じてフィルタ-などで濾過滅菌した後、注射により被験者に投与するとよい。また、この溶液には、添加剤(例えば、着色剤、乳化剤、懸濁剤、界面活性剤、溶解補助剤、安定化剤、保存剤、酸化防止剤、緩衝剤、等張化剤など)などを添加してもよい。投与経路としては、静脈、筋肉、腹腔、皮下、皮内投与などが可能であり、また、経鼻、経口投与してもよい。
【0052】
本発明の抗体の投与量、投与の回数及び頻度は、被験者の症状、年齢、体重、投与方法、投与形態などにより異なるが、例えば、通常、成人一人当たり1,000〜10,000 mg/kg体重、好ましくは、 2,000〜5,000 mg/kg体重の抗体を、少なくとも1回、所望の効果が持続する頻度で投与するとよい。
【実施例】
【0053】
以下、実施例に基づいて本発明を詳細に説明するが、本発明はこれらの実施例に限定されるものではない。
【0054】
〔実施例1〕
1.はじめに
緑膿菌(P. Aeruginosa)は、嚢胞性線維症に関連する罹患と死亡率に寄与する。なぜならば、その感染が直接肺実質にダメ-ジを与え、進行性の閉塞症、小気道の線維症及び肺機能の悪化に寄与するからである[1]。この感染の処置については抗生物質による治療が利用できるが、薬剤耐性分離株が現れ、これが慢性的に緑膿菌のコロニ-を形成する患者に重大な治療を提起する。緑膿菌に対する効果的なワクチンが開発されれば、このような感染の予防及び治療が促進されることになる[2]。
遺伝子免疫は、広範なウイルス、細菌及び寄生虫の病原体に対する保護的な免疫応答を誘導するための有望なストラテジ-である[3]。動物モデルにおいて、DNAワクチンは、AIDS様レトロウイルス、インフルエンザ、マラリア、結核及び緑膿菌感染に対する保護を与える[4-8]。免疫原性の他に、DNAワクチンは、通常の生又はサブユニットワクチンと比較したとき、いくつかの他の潜在的な利点を有する。DNAワクチンは製造が容易であり、安定性が高く、潜在的に安全である[9, 10]。DNAワクチンの主な潜在的利点は、単独の病原体の異なる株由来の抗原又は複数の病原体由来の抗原をコ-ドするいくつかのプラスミドを混合すること、多価抗原に対する免疫応答を誘導することができ、その結果、多種の感染に対して効果的に保護できることである。マラリア又は結核のマウスモデルにおいて、いくつかの異なる標的を包含するDNAプラスミドの組合せにより、いずれかのプラスミドを単独で用いたときに観察された保護の遺伝子的制限が抑制されるか、あるいは抗原の競合を見せることなく保護が増強された[11, 12]。異なるグレ-ド由来の複数のHIV-1遺伝子を発現するプラスミドの組合せで免疫すると、個々のプラスミドによって誘発されたものに匹敵する体液性及び細胞性免疫応答が誘発され、その結果、免疫干渉なしに抗ウイルス免疫応答が拡大した[13]。
本実施例では、異なる抗原をコ-ドするプラスミドからなる多価DNAワクチンの免疫原性を評価した。これまでに報告された緑膿菌抗原の可能性のある候補の中で、細胞外膜タンパクの融合物(OprF/I)、III型分泌機構制御タンパク(PcrV)、及び毛状付属器タンパク(PilA)を選択した。筋肉内エレクトロポ-レイション(imEPT)又は遺伝子銃(GG)によりプラスミドワクチン接種の有効性を比較し、これらの2つの方法を用いた場合に免疫原性が変化するかどうかを調べた。OprF/I、PcrV又はPilAを単独(一価)又は組合せ(多価)で発現するプラスミドでマウスを免疫した後、個々の抗原に対する血清抗体価と全細菌細胞に対する血清抗体価をモニタ-した。鼻腔内に細菌を投与した後、サイトカイン/ケモカイン mRNA産生レベル、細菌の生存数、好中球数及びマクロファ-ジ数をワクチン接種マウスの気管支肺胞洗浄(BAL)において評価した。最後に、致死量の緑膿菌を投与することにより、多価DNAワクチンの保護の可能性を評価した。
【0055】
2.材料及び方法
2.1.細菌
緑膿菌株PAO1、PAK及びD4を本実施例に用いた。PAO1及びPAKは、線毛、鞭毛及びIII型分泌細胞外酵素のような異なる病原性因子を発現する[14, 15]。D4は、好中球減少性の菌血症患者の血液から単離した[16]。すべての菌株はTerrific broth(Sigma, St. Louis, MO)中37℃で16時間インキュベ-トした。鼻腔内投与のために、細菌培養物を3000Xgで10分間遠心し、ペレットをPBS中で2回洗浄し、550 nmでの光学濃度1.0でPBSに再懸濁した。この懸濁液30又は40μlは、それぞれ、PAK又はD4のほぼ2LD50に相当した。
【0056】
2.2.ワクチン接種のための哺乳類発現プラスミドの構築(図8)
緑膿菌株PAO1のゲノムDNAを既報のように調製した[17]。以下のプライマ-セットを用いて、oprF、oprI、pcrV及びpilA遺伝子をPCR増幅した。
oprFセンス: 5’-GGA ATT CGC CAT GGC TCC GGC TCC GGA ACC GG-3’(配列番号15)
oprFアンチセンス: 5’-ACT GAG CTC TCA ACG CGA CGG TTG ATA GC-3’(配列番号16)
oprIセンス: 5’-TTG GAG CTT CCA CTC CAA AGA AAC CGA A-3’(配列番号17)
oprIアンチセンス: 5’-CCG GAT CCC TTG CGG CTG GCT TTT TCC AGC-3’(配列番号18)
pcrVセンス: 5’-GGA ATT CGC CAT GGA AGT CAG AAA CCT TAA-3’(配列番号19)
pcrVアンチセンス: 5’-CCG GAT TCG ATC GCG CTG AGA ATG TCG CGC-3’(配列番号20)
pilAセンス: 5’-GGA ATT CGG CAT GAA AGC TCA AAA AGG CTT-3’(配列番号21)
pilAアンチセンス: 5’-ACG CGT CGA CGT TAT CAC AAC CTT TCG GAG-3’(配列番号22)
融合遺伝子oprF/Iを構築するために、OprFのアミノ酸190-342及びOprIのアミノ酸21-83をコ-ドするDNAフラグメントを既報のようにタンデムにライゲ-トした[18]。プラスミドpGACAG-OprF/I、pGACAG-PcrV又はpGACAG-PilAを得るために、各遺伝子フラグメントにFLAGシグナル配列をタグ付けしてから、pGACAGベクタ-(DNAワクチンの臨床応用のために最適化された哺乳類発現ベクタ-[19])のEcoRI部位に導入した。すべてのプラスミドをE.coli DH5αに形質転換し、Endo-free Maxiprep kit(Quigen, Hilden, Germany)を用いて精製した。
【0057】
2.3.組み換えタンパク質の精製
組み換えタンパク質は、既報のように、E.coli DH5α中に発現させ、精製した[20]。
【0058】
2.4.免疫スケジュ-ル
8週令の雌BALB/cマウスをSLC(静岡県)から購入し、特定病原体除去条件下にある動物施設に入れた。これらのマウスをケタミン/キシラジン混合物で麻酔した後、100 μgのpGACAG(対照プラスミド)、pGACAG-OprF/I、pGACAG-PcrV、pGACAG-PilA又は33μgの各プラスミド(pGACAG-OprF/I+pGACAG-PcrV+pGACAG-PilA)の混合物のいずれかを含むプラスミド溶液(1 μg/μl食塩水、50 μl/筋肉)をマウス(各グル-プ8匹)の四頭筋に注射した。30 V/cmの一定の電場強度で、50 ms X 3パルスにて、CUY21EDIT(NepaGene、東京都、日本)を用いて、注入部位に電気穿孔した。最初の免疫化の後4週間目に追加免疫を行った。あるいはまた、微粒子銃粒子注入法(Helios Gene Gun kit, Bio-Rad, Richmond, CA)を用いて、0.5 mg金粒子にコ-ティングしたpGACAG-OprF/I+pGACAG-PcrV+pGACAG-PilAのプラスミド混合物1 μgでマウスを免疫した。マウスの腹部皮膚に2回の照射を14日の間隔で全3回行った。すべての動物実験は、動物実験委員会による承認を得、マウスはNIH animal care guidelineに従って取り扱われた。
【0059】
2.5.ワクチン接種したマウス由来の血清の特異性
PAO1、PAK又はD4から調製した細胞溶解物の免疫ブロットにより、抗体特異性を分析した。膜をブロックし、ワクチン接種した種々のグル-プのマウス由来の血清と反応させた。HRP標識抗マウスIgG抗体と共にインキュベ-トした後、ブロットをECL puls基板(Amersham Bioscience)と反応させてから、オ-トラジオグラフィ-で分析した。
【0060】
2.6.ELISA
血清抗体価を既報のようにELISAで測定した[19]。
【0061】
2.7.肺及び血液中の緑膿菌の定量
最終の免疫化後2週間目に、マウスを麻酔し、20μlのPBS中の5 X 105 CFUのD4を鼻腔内接種した。感染後(p.i.)24時間で再度マウスを麻酔した。マウスの眼を2回70%エタノ-ルで洗浄した後、眼窩後穿刺により無菌的に採血した。CO2に富む雰囲気下でマウスを屠殺した後、肺を除去し、4 ml PBS中、無菌条件下でホモジナイズした。血液と肺ホモジネ-トの10倍段階希釈液をLB寒天プレ-ト(Sigma)上にまき、このプレ-トを37℃で24時間インキュベ-トした。コロニ-数を二重盲検でマニュアルで数えた。
【0062】
2.8.BAL液中の好中球及びマクロファ-ジの定量
5 X 105 CFUのD4を鼻腔内接種したマウスを感染後3及び6時間目にCO2に富む雰囲気下で屠殺した。1.7 mm-ODポリエチレンカテ-テルを用い、0.5 mlのPBSで連続して2回洗浄することにより、BALFを回収した。試料を遠心し、20 μlのPBS中に再懸濁させた。5 μlのスメアがスライドグラス上に調製され、これをギムザ染色した。0.37 μm X 0.37 μmに広がる無作為抽出の10個の領域に存在する好中球及びマクロファ-ジの数を顕微鏡下でカウントしてから、全細胞数を算出した。
【0063】
2.9.マクロファ-ジ炎症性タンパク質2(MIP-2)及びIFN-γ mRNAのリアルタイムPCR分析
上述のように5 X 105 CFUの緑膿菌D4を用いて、感染後2、12及び24時間目にBAL細胞を調製した。細胞をTRIzo試薬(Invitrogen, Carlsbad, CA)に再懸濁し、全RNAを単離し、既報のように逆転写した[21]。ABI PRISM 7700 sequence detection system(Applied Biosystems)を用い、製造者の指示に従い、MIP-2、IFN-γ又は18S rRNAに特異的なTaqMan probe(Applied Biosystems, Foster City, CA)を使用して、リアルタイムPCRを行った。同じ試料の18S rRNAのレベルをノ-マライズすることにより、試料中のMIP-2及びIFN-γ mRNAの反応発現レベルを算出した。
【0064】
2.10.保護テスト
最終の免疫化の後2週間目に、マウスを麻酔して、2LD50のPAK又はD4を鼻腔内に投与した。このマウスの致死率をその後10日間モニタ-した。
【0065】
2.11.受動保護テスト
6週令の雌BALB/cマウスを用いた。PAK及びD4を16時間TB broth中で培養し、PBS中で3回洗浄し、ペレットをPBSに再懸濁し、550 nmの光学密度を1.0に調整し、所望の接種菌液を調製した。種々のグル-プの免疫マウス由来の血清のIgG画分をProtein G sepharose column(Amersham Biosciences)で精製した。各IgG画分10 μgを2LD50の細菌と混合し、37℃で30分間インキュベ-トした。これらの懸濁液を各グル-プ10匹のマウスの鼻腔内に投与し(全50 μl/マウス)、致死率をその後10日間モニタ-した。
【0066】
2.12.統計分析
すべての実験を少なくとも2回繰り返した。統計的な有意差は、ANOVAボンフェロ-ニ多重比較検定を用いて評価した。マウスのグル-プ間の生存率の差は、Mantel-Cox log 順位検定を用いて評価した。p値<0.05は、統計的な有意差があるとみなした。
【0067】
3.結果
3.1.DNAワクチンにより誘発された抗体産生のキャラクタリゼ-ション
緑膿菌を標的とする多価DNAワクチンの免疫原性が、一価DNAワクチンにより誘発された免疫原性より大きいかどうかを調べるために、3つの異なるプラスミド(各々は、oprF/I、pcrV又はpilA遺伝子を包含する)を調製した。その後、対照プラスミド、各単独プラスミド(一価)又は3つの異なるプラスミドの混合物(多価)で、imEPTにてマウスを免疫した。PAO1、PAK及びD4細胞溶解物の免疫ブロットを用いて、各標的抗原に特異的な抗体応答を調べた。pGACAG-OprF/Iで免疫したマウスから得られた血清は、実際に、すべての細胞溶解物中に存在する〜36及び〜12 kDaタンパク質と反応した(図1A)。〜36及び〜12 kDaタンパク質は、それぞれ、OprF及びOprIであると推定された。pGACAG-PcrVで免疫したマウス由来の血清は、D4及びPAK細胞溶解物中に存在する〜32 kDaのタンパク質(PcrVと同じ大きさ)と反応したが、pGACAG-PilAで免疫したマウス由来の血清は、PAO1及びD4中に存在する〜15 kDaのタンパク質(PilAと同じ大きさ)と反応した(図1B及びC)。これらの結果は、各DNAワクチンにより生じた抗体は、緑膿菌の異なる株により発現される内在性の抗原標的に特異的に結合することを保証するものである。
imEPT又はGGのいずれかにより多価DNAワクチンで免疫されたマウスから得られた血清は、いくつかのタンパク質、おそらくはOprF、OprI、PcrV及びPilAと反応したが、OprFとPcrVを標的とする抗体レベルは、分子サイズが近いために、免疫ブロットにより区別することはできなかった(〜36及び〜32 kDa、図1D及びE)。全IgG、IgG1、及びIgG2aサブタイプを含む血清抗体価をELISAにより規則的な時間間隔でモニタ-した。対照プラスミド又は単独プラスミドのいずれかでワクチン接種したマウスから得られた血清は、他の抗原標的と交叉反応をしなかった(デ-タは示さず)。図2Aに示されるように、各一価又は多価DNAワクチンにより誘導されたIgGは緑膿菌D4株の全細胞と特異的に反応した。各組み換え標的タンパク質に対する全IgG産生のレベルは、一価及び多価ワクチン接種グル-プ間で匹敵するものであり、このことは、OprF/I、PcrV及びPilAを標的とする多価DNAワクチンが、抗原間の免疫干渉なしに、容易に多価抗体応答を誘導することを示唆している(図2B-D)。IgG1及びIgG2サブタイプを調べたときに、同様の結果が得られた(図3)。多価DNAワクチンを用いて、imEPT及びGGにより誘発される免疫応答を比較したところ、各標的タンパク質に対する全IgG及びIgG1サブタイプの匹敵するレベルがどちらの方法でも誘導された(図2及び3)。しかしながら、GGに仲介された免疫化は、すべての標的抗原に対する体液性応答のIgG2aサブタイプは誘導しなかった(図3D-F, p<0.01)。
【0068】
3.2.肺及び血液からの細菌のクリアランス
DNAワクチンにより生じた免疫が緑膿菌感染を排除する能力を調べるために、D4を鼻腔内投与した後に肺ホモジネ-ト又は血液中の生存細菌の数を調べた。一価DNAワクチンの各々でワクチン接種すると、対照プラスミド処理と比較して、肺及び血液の両方で細菌の数はより少なくなった(図4)。重大なことは、imEPTによる多価DNAワクチン接種により肺中の細菌が有意に清掃されたことである(図4A、p<0.05)。非常に重要な点は、このグル-プのみに菌血症の完全な保護が観察されたことである(図4、p<0.01)。これらの結果は、明らかに、OprF/I、PcrV及びPilAからなる多価DNAワクチン接種によって、免疫応答の付与に成功し、緑膿菌の肺感染から保護されたことを示している。しかしながら、GGによる多価DNAワクチン接種が誘発した免疫応答は、緑膿菌肺炎に対する保護効果が不十分であった。
【0069】
3.3.BAL細胞のキャラクタリゼ-ション
多価DNAワクチンによる保護免疫応答の根底にある機構を明らかにするために、免疫マウスのBAL細胞をキャラクタライズした。他のグル-プと比較したときに、imEPTにより多価DNAワクチンが投与されたマウスから得られたBAL試料中の好中球及びマクロファ-ジの両方の数が有意に増加したことがギムザ染色により示された(図5、p<0.05)。この迅速な炎症反応の基礎を明らかにするために、BAL細胞中のいくつかのサイトカイン及びケモカインのmRNAレベルをリアルタイムPCRで定量した。ケモカインの中でも、MIP-2は気管支肺胞での好中球動員の中心的役割を果たす[22]。最も高いレベルのMIP-2 mRNAが、imEPTによる多価ワクチン接種を受けたマウスに観察された(図6A、p<0.05)が、このことは、MIP-2の発現が増加することにより、気管支肺胞空間への強い好中球動員が誘発されたことを示唆している。IFN-γ mRNAレベルは、すべてのグル-プにおいて、中程度だが、持続的に、感染後24時間まで増加した(図6B)。最も高いレベルのIFN-γ mRNAが、imEPTにより多価ワクチン接種を受けたマウスにおいて、感染後24時間で検出された(図6B、p<0.01)が、このことは、IFN-γの産生が増加したことにより、好中球及びマクロファ-ジにより貪食された細菌の自然な死滅がさらに活性化されたことを示唆している。これらのデ-タにより、多価DNAワクチンはMIP-2産生を通して好中球動員を優先的に誘導し、IFN-γ産生により好中球及びマクロファ-ジの殺菌活性を増強したことを暗に意味している。一価及び多価ワクチン接種グル-プ間で、IL-4、IL-6、TNF-α及びJE/MCP-1のような他のサイトカイン及びケモカインのレベルの有意差はなかった(デ-タは示さず)。
【0070】
3.4.imEPTによる多価DNAワクチン接種はD4又はPAKの致死的投与からマウスを保護した
ワクチン接種したマウスの鼻腔内に2LD50投与量のPAK又はD4を投与した。接種マウスの致死率をその後10日間モニタ-した。imEPTにより多価DNAをワクチン接種したすべてのマウスは、PAKの感染後10日間以上生存した(図7A)。一方、GGにより、対照プラスミド、PilAワクチン又は多価ワクチンで免疫したマウスの70%は感染後5日以内に死亡した(p<0.01)。OprF/I又はPcrVを標的とする一価DNAワクチン接種でさえ、80%のマウスを感染後4日目で保護することができた(p<0.01)。D4株を用いたときも、同様の結果が得られた(図7B)が、このことは、OprF/I、PcrV及びPilAを標的とするimEPTによる多価DNAワクチン接種により、緑膿菌感染に対して顕著な保護免疫応答を誘導することを示唆している。
【0071】
3.5.多価DNAワクチンで免疫したマウス由来の抗血清による受動保護
受動保護の検討を行い、抗体が保護能力に重大であるかどうかを評価した。ワクチン接種したマウスから得られた血清の精製IgG画分とともにD4又はPAKを前培養し、その後、この混合物を未処理マウスの鼻腔内に接種した。ワクチン接種グル-プの中で、imEPTにより多価DNAワクチンを接種したマウス由来の血清が、PAK株の致死的な感染から未処理マウスを保護する効果が高かった(図7C、p<0.01)。もう一つのD4株を用いたときも同様の結果が得られた(図7D)。
【0072】
4.考察
緑膿菌は院内肺炎の原因となる主要な病原体である。この病原体は、LPS、外毒素、線毛、鞭毛、エラスタ-ゼ、及びプロテア-ゼを含む数々の病原性因子を保有する[14,15,20,23-26]。しかしながら、これらの病原性因子はすべてのタイプの緑膿菌感染に関連しているわけではない。緑膿菌の複雑な病原性とその病原性因子の多様な機能が、効果的で汎用性があるワクチンの開発に大きな障害となっている[27]。従って、細菌の異なる成分を標的とするDNAワクチンが協同的に作用して、緑膿菌に対する宿主の保護免疫の範囲を拡大することができるかどうかを調べた。本実施例において、比較的広い範囲の臨床株に保存されている3つの主要な抗原を用いた。また、免疫応答を拡大するために、細菌器官の異なる成分由来のこれらの抗原を選択した。OprF及びOprIのような外膜タンパクでワクチン接種すると、いくつかのげっ歯類モデルにおいて、緑膿菌感染に対して効果的であることが示された[28,29]。PcrVは、III型分泌-中毒機構のエフェクタ-の転座に重要な役割を果たす。PcrVに対する能動及び受動免疫により、緑膿菌により引き起こされる急性及び慢性感染の両方での生存が有意に増加した。PcrVに対する抗体はエフェクタ-の転座を防止し、次に、肺胞マクロファ-ジ生存率と食細胞機能を増強する[30,31]。線毛は、細菌の付属器として機能するが、上皮細胞への接着に利用される。事実、線毛を有しない株は病原性が減少し、抗線毛血清は宿主細胞への細菌の結合を抑制することが実証されている[32,33]。また、異なる株を起源とする線毛に対する抗体は、他の株由来の線毛にも広く交叉反応する[15]。従って、我々のワクチン製剤は、宿主の生体防御から細菌が逃れることを防ぐための3つの独立な作用機序を有するのかもしれない。
【0073】
いくつかの研究では、DNAワクチンの組合せでは、単一のDNAワクチンと比較して、個々の抗原に対する免疫原性が減少することが示され、このことは、「抗原競合」と呼ばれている[34]。しかしながら、本実施例により、多価緑膿菌DNAワクチンが、imEPTにより免疫したマウスにおいて細菌に対して強力な保護免疫応答を誘発することが明らかに示された。imEPT技術は、トランスフェクション効率と炎症を増強することにより、DNAワクチンの免疫原性を強化することが実証されている[35]。事実、免疫化がimEPTにより行われると、同じレベルの最適免疫応答を誘導するには、20分の1程度の低量のプラスミドで十分であった(デ-タは示さず)。従って、この現象が不十分なトランスフェクション効率では起こらない場合、imEPTによる効率的なトランスフェクション及びその後の免疫原性の増強により、「抗原競合」が抑制されたのかもしれない。ワクチンの多様性については、使用されたDNAの量がimEPT法とGG法とで異なったのであるが、GGと比較して、imEPTは、緑膿菌に対する保護免疫の誘導においてより顕著な効果を示した。
【0074】
好中球及びマクロファ-ジは、急性肺細菌感染に対する保護において鍵となる役割を果たす[36]。本実施例において、多価DNAワクチンで免疫したマウスにおける緑膿菌の肺クリアランスの増強は、BALにおける好中球及びマクロファ-ジの数の増加と関連があった。MIP-2はヒトIL-8の機能的ホモログであるが、これは強力な化学誘引物質であり、好中球の活性化因子である。予想されるように、BAL細胞におけるMIP-2 mRNAレベルのアップレギュレ-ションは、好中球の流入量と関連があった。IFN-γは、肺胞マクロファ-ジと好中球の両方において細胞内殺菌活性を制御する鍵となる因子である[37]。IFN-γ mRNAは、多価DNAワクチン接種グル-プにおいて遅延相(>12h)で有意に誘導されたが、このことは、図5Bに観察されるように、強力なIFN-γが細菌転移の完全な保護の要因となっている可能性があることを示唆している、この観察結果は、IFN-γ仲介Th1免疫応答が緑膿菌に対する肺宿主生体防御と慢性緑膿菌感染の予後を改善することを示した以前の報告[38-40]と一致している。
【0075】
受動保護に関する本実施例の結果は、多価抗体応答が緑膿菌に対する保護免疫応答の主要な因子であることを示している。非常に重要なことは、IgG2aが保護に重要であるように思われることである(図4)。事実、IgG2aは、Fcγ受容体に最も強い親和性を持ち、IFN-γが仲介するB細胞のアイソタイプスイッチングの結果として産生される[41]。さらに、これらの結果は、抗体が仲介する免疫は保護の主要な因子であるが、より高いTh2免疫応答(IgG1)が、Th1免疫応答(IgG2a)よりも緑膿菌感染におけるより重篤な肺病理に関連性があることを示す以前の研究をさらに進めるものである[42,43]。
【0076】
以上を要約すると、本実施例の結果は、OprF/I、PcrV、及びPilAを標的とする多価DNAワクチンが協同的に緑膿菌肺炎のマウスモデルにおける保護効率を強めることを直接的に実証した。imEPT技術は、保護機構を誘発するという点で、GGよりかなり優れている。我々の進行中の研究は、抗原インサ-トのサイズを最小化し、OprF、OprI、PcrV、及びPilA由来の入れ子状態のエピト-プからなる単一の融合抗原を理想的に設計することに焦点を当てている。そのような研究は、緑膿菌多価DNAワクチンの臨床応用を支持するであろう。
【0077】
〔実施例2〕
図9に示すように、OprF190-342アミノ酸領域、OprI20-83アミノ酸領域、PcrV1-294アミノ酸領域、PilA136-149アミノ酸領域を直列に連結した抗原エピトープ融合DNAワクチンpGACAG-OprF/I-PcrV-PilAを実施例1と同様の方法で作製した。このワクチンを実施例1と同様に、筋注エレクトロポレーション法で導入し(0週と4週に1回ずつ両側大腿四頭筋に25μg)、6週目の血清を用いてELISA法により抗OprF/I、PcrV、PilA抗体価を測定した。その結果、それぞれの抗原に対するIgG抗体が誘導されていることが確認された(図10)。同様に、異なる菌株由来の抗原に対して多価の抗体が誘導されているか確認するため、各菌株(PAO1, PAK, PA103, D4)の菌体溶解物をSDS-PAGE法で分画し、PVDF膜に転写後、抗原エピトープ融合ワクチンで免疫したマウス血清と反応させ、抗マウスIgG抗体を用いてimmunoblottingを行った。その結果、各菌株由来のOprF,PvrV, PilA, OprIに対して反応する特異抗体が誘導されたことが確認された(図11)。実際に、抗原エピトープ融合ワクチンpGACAG-OprF/I-PcrV-PilA(Multiepitope)、実施例1で至適化した混合ワクチンpGACAG-OprF/I+pGACAG-PcrV+pGACAG-PilA(Mix)、もしくは空ベクターpGACAG(Empty)を筋注エレクトロポレーション法で導入し(0週と4週に1回ずつ両側大腿四頭筋に25μg)、6週目に2LD50量の緑膿菌株(PAO1, PAK, D4)を経鼻感染させてその後の生存率を観察した。その結果、Multiepitope投与群においてはMix群と同等の生存率を示し、抗原エピトープ融合ワクチンの有効性が確認された(図12)。実際に、PAO1感染後12および24時間後の肺内残存生菌数を数えると、Multiepitope投与群においてはMix群と同等の菌数であり、抗原エピトープ融合ワクチンにより緑膿菌感染に対する防御的免疫応答を誘導できたことが確認された(図13)。抗原エピトープ融合ワクチンによる緑膿菌感染に対する防御的免疫応答について、腸管感染由来敗血症モデル(易感染モデル)でも評価した(図14)。飲料水中に緑膿菌PAO1株を入れ(1 x 107/ml)、アンピシリン(1回/日、200 mg/kg)を3日間腹腔内に投与した。4日目に無菌の飲料水に交換して飼育し、4日目、6日目に1回ずつシクロフォスファミド(200 mg/kg)を腹腔内に投与して易感染状態とし、その後の生存率を観察した(図14)。その結果、抗原エピトープ融合ワクチンによる、易感染状態における抗緑膿菌感染防御能の誘導が確認され、その効果はわれわれが検討してきた至適化混合ワクチンとほぼ同等であることがあきらかとなった。抗原エピトープ融合ワクチンにより誘導される抗体の抗緑膿菌感染防御効果について検討した(図15)。実施例1と同様に抗原エピトープ融合ワクチン(Multiepitope)、至適化混合ワクチン(Mix)、もしくは空ベクター(Empty)を筋注エレクトロポレーション法で導入し(0週と4週に1回ずつ両側大腿四頭筋に25μg)、6週目に血清を採取し、IgG分画を精製した。15μgの精製IgGと2LD50量のPAO1株またはPAK株と37℃で30分間反応された後、ナイーブマウスに経鼻感染させ、その後の生存率を観察した。その結果、これまでの結果と同様に、ワクチン免疫により誘導されたIgG分画に緑膿菌感染を抑制する作用があることが確認され、抗原エピトープ融合ワクチンによる効果は、至適化混合ワクチンと同等であった。
【0078】
引用文献
[1] Pier GB. CFTR mutations and host susceptibility to Pseudomonas aeruginosa lung infection. Curr Opin Microbiol 2002;5(1): 81-6.
[2] Lang AB, Horn MP, Imboden MA, Zuercher AW. Prophylaxis and therapy of Pseudomonas aeruginosa infection in cystic fibrosis and immunocompromised patients. Vaccine 2004;22(Suppl 1):S44-8.
[3] Srivastava IK, Liu MA. Gene vaccines. Ann Intern Med 2003; 138(7):550-9.
[4] Ha SJ, Jeon BY, Youn JI, Kim SC, Cho SN, Sung YC. Protective effect of DNA vaccine during chemotherapy on reactivation and reinfection of Mycobacterium tuberculosis. Gene Ther 2005;12(7): 634-8.
[5] Okuda K, Xin KQ, Haruki A, et al. Transplacental genetic immunization after intravenous delivery of plasmid DNA to pregnant mice. J Immunol 2001;167(9):5478-84.
[6] Sasaki S, Amara RR, Oran AE, Smith JM, Robinson HL. Apoptosismediated enhancement of DNA-raised immune responses by mutant caspases. Nat Biotechnol 2001;19(6):543-7.
[7] Takakura M, Okuda K, Matsuda T, et al. Combination of DNA vaccine and adenovirus vector by cutaneous administration induced strong HIV-specific cellular immune responses in mice. Vaccine 2005;23(7):847-8.
[8] Vuola JM, Keating S, Webster DP, et al. Differential immunogenicity of various heterologous prime-boost vaccine regimens using DNA and viral vectors in healthy volunteers. J Immunol 2005;174(1): 449-55.
[9] Smith HA, Klinman DM. The regulation of DNA vaccines. Curr Opin Biotechnol 2001;12(3):299-303.
[10] Staczek J, Gilleland LB, van der Heyde HC, Gilleland HE. DNA vaccines against chronic lung infections by Pseudomonas aeruginosa. FEMS Immunol Med Microbiol 2003;37(2/3):147-53.
[11] Doolan DL, Aguiar JC, Weiss WR, et al. Utilization of genomic sequence information to develop malaria vaccines. J Exp Biol 2003;206(Pt 21):3789-802.
[12] Morris S, Kelley C, Howard A, Li Z, Collins F. The immunogenicity of single and combination DNA vaccines against tuberculosis. Vaccine 2000;18(20):2155-63.
[13] Kong WP, Huang Y, Yang ZY, Chakrabarti BK, Moodie Z, Nabel GJ. Immunogenicity of multiple gene and clade human immunode-ficiency virus type 1 DNA vaccines. J Virol 2003;77(23):12764-72.
[14] Hertle R, Mrsny R, Fitzgerald DJ. Dual-function vaccine for Pseudomonas aeruginosa: characterization of chimeric exotoxin A-pilin protein. Infect Immun 2001;69(11):6962-9.
[15] Sheth HB, Glasier LM, Ellert NW, et al. Development of an anti-adhesive vaccine for Pseudomonas aeruginosa targeting the Cterminal region of the pilin structural protein. Biomed Pept Proteins Nucl Acids 1995;1(3):141-8.
[16] Matsumoto T, Furuya N, Tateda K, et al. Effect of passive immunotherapy on murine gut-derived sepsis caused by Pseudomonas aeruginosa. J Med Microbiol 1999;48(8):765-70.
[17] Fukushima J, Yamamoto S, Morihara K, et al. Structural gene and complete amino acid sequence of Pseudomonas aeruginosa IFO 3455 elastase. J Bacteriol 1989;171(3):1698-704.
[18] Larbig M, Mansouri E, Freihorst J, et al. Safety and immunogenicity of an intranasal Pseudomonas aeruginosa hybrid outer membrane protein F-I vaccine in human volunteers. Vaccine 2001;19(17-19): 2291-7.
[19] Sasaki S, Xin KQ, Okudela K, Okuda K, Ishii N. Immunomodulation by apoptosis-inducing caspases for an influenza DNA vaccine delivered by gene gun. Gene Ther 2002;9(12):828-31.
[20] Kawamoto S, Shibano Y, Fukushima J, Ishii N, Morihara K, Okuda K. Site-directed mutagenesis of Glu-141 and His-223 in Pseudomonas aeruginosa elastase: catalytic activity, processing, and protective activity of the elastase against Pseudomonas infection. Infect Immun 1993;61(4):1400-5.
[21] Takeshita F, Suzuki K, Sasaki S, Ishii N, Klinman DM, Ishii KJ. Transcriptional regulation of the human TLR9 gene. J Immunol 2004;173(4):2552-61.
[22] Tsai WC, Strieter RM, Mehrad B, Newstead MW, Zeng X, Standiford TJ. CXC chemokine receptor CXCR2 is essential for protective innate host response in murine Pseudomonas aeruginosa pneumonia. Infect Immun 2000;68(7):4289-96.
[23] Atsumi Y, Yamamoto S, Morihara K, et al. Cloning and expression of the alkaline proteinase gene from Pseudomonas aeruginosa IFO 3455. J Bacteriol 1989;171(9):5173-5.
[24] Matsumoto T, Tateda K, Furuya N, et al. Efficacies of alkaline protease, elastase and exotoxin A toxoid vaccines against gutderived Pseudomonas aeruginosa sepsis in mice. J Med Microbiol 1998;47(4):303-8.
[25] Pier GB. Promises and pitfalls of Pseudomonas aeruginosa lipopolysaccharide as a vaccine antigen. Carbohydr Res 2003;338(23):2549-56.
[26] Tanaka E, Kawamoto S, Fukushima J, et al. Detection of elastase production in Escherichia coli with the elastase structural gene from several non-elastase-producing strains of Pseudomonas aeruginosa. J Bacteriol 1991;173(19):6153-8.
[27] Holder IA. Pseudomonas immunotherapy: a historical overview. Vaccine 2004;22(7):831-9.
[28] von Specht BU, Knapp B, Muth G, et al. Protection of immunocompromised mice against lethal infection with Pseudomonas aeruginosa by active or passive immunization with recombinant P. aeruginosa outer membrane protein F and outer membrane protein I fusion proteins. Infect Immun 1995;63(5):1855-62.
[29] Worgall S, Krause A, Rivara M, et al. Protection against P. aeruginosa with an adenovirus vector containing an OprF epitope in the capsid. J Clin Invest 2005;115(5):1281-9.
[30] Sawa T, Yahr TL, Ohara M, et al. Active and passive immunization with the Pseudomonas V antigen protects against type III intoxication and lung injury. Nat Med 1999;5(4):392-8.
[31] Shime N, Sawa T, Fujimoto J, et al. Therapeutic administration of anti-PcrV F(ab$)(2) in sepsis associated with Pseudomonas aeruginosa. J Immunol 2001;167(10):5880-6.
[32] Farinha MA, Conway BD, Glasier LM, et al. Alteration of the pilin adhesin of Pseudomonas aeruginosa PAO results in normal pilus biogenesis but a loss of adherence to human pneumocyte cells and decreased virulence in mice. Infect Immun 1994;62(10): 4118-23.
[33] Tang H, Kays M, Prince A. Role of Pseudomonas aeruginosa pili in acute pulmonary infection. Infect Immun 1995;63(4):1278-85.
[34] Sedegah M, Charoenvit Y, Minh L, et al. Reduced immunogenicity of DNA vaccine plasmids in mixtures. Gene Ther 2004;11(5): 448-56.
[35] Widera G, Austin M, Rabussay D, et al. Increased DNA vaccine delivery and immunogenicity by electroporation in vivo. J Immunol 2000;164(9):4635-40.
[36] McClellan SA, Huang X, Barrett RP, van Rooijen N, Hazlett LD. Macrophages restrict Pseudomonas aeruginosa growth, regulate polymorphonuclear neutrophil influx, and balance pro- and anti-inflammatory cytokines in BALB/c mice. J Immunol 2003;170(10): 5219-27.
[37] Ojielo CI, Cooke K, Mancuso P, et al. Defective phagocytosis and clearance of Pseudomonas aeruginosa in the lung following bone marrow transplantation. J Immunol 2003;171(8):4416-24.
[38] Cripps AW, Dunkley ML, Clancy RL, Kyd J. Vaccine strategies against Pseudomonas aeruginosa infection in the lung. Behring Inst Mitt 1997;98:262-8.
[39] Johansen HK, Cryz Jr SJ, Hougen HP, Moser C, Hoiby N. Vaccination promotes TH1-like inflammation and survival in chronic Pseudomonas aeruginosa pneumonia. A new prophylactic principle. Behring Inst Mitt 1997;98:269-73.
[40] Lei D, Lancaster Jr JR, Joshi MS, et al. Activation of alveolar macrophages and lung host defenses using transfer of the interferon-! gene. Am J Physiol 1997;272(5 Pt 1):L852-9.
[41] Hamaguchi Y, Xiu Y, Komura K, Nimmerjahn F, Tedder TF. Antibody isotype-specific engagement of Fc{gamma} receptors regulates B lymphocyte depletion during CD20 immunotherapy. J Exp Med 2006;203(3):743-53.
[42] Price BM, Barten Legutki J, Galloway DR, et al. Enhancement of the protective efficacy of an oprF DNA vaccine against Pseudomonas aeruginosa. FEMS Immunol Med Microbiol 2002;33(2):89-99.
[43] Song Z, Kharazmi A, Wu H, et al. Effects of ginseng treatment on neutrophil chemiluminescence and immunoglobulin G subclasses in a rat model of chronic Pseudomonas aeruginosa pneumonia. Clin Diagn Lab Immunol 1998;5(6):882-7.
【産業上の利用可能性】
【0079】
本発明の医薬組成物は、院内感染の原因菌である緑膿菌に対して効果的に感染防御能を誘導することができる。
【図面の簡単な説明】
【0080】
【図1】DNAワクチンは緑膿菌標的抗原に結合する特異的IgGを産生させた。PAO1、D4、及びPAKから調製された細胞溶解物、及び一価(pGACAG-OprF/I、pGACAG-PcrV又はpGACAG-PilA)又は多価(pGACAG-OprF/I+pGACAG-PcrV+pGACAG-PilA)DNAワクチンにより生じた血清を用いて、免疫ブロッティング分析を行った。OprF(36 kDa)、OprI(12 kDa)、PcrV(32 kDa)及びPilA(15 kDa)に相当するバンドを示す。3つの独立する実験から同じ結果が得られた。n.s.:非特異的バンド。
【図2】DNAワクチン接種により生じた緑膿菌抗原に対する血清IgG力値。100 μgのpGACAG(対照プラスミド)、pGACAG-OrrF/OprI、pGACAG-PcrV、pGACAG-PilA、又は33 μgの個々のプラスミド(pGACAG-OprF/I+pGACAG-PcrV+pGACAG-PilA)の混合物のいずれかでimEPTにより、マウスを2回免疫した(0及び4週)。あるいは、0.5 mgの金粒子にコ-ティングしたpGACAG-OprF/I+pGACAG-PcrV+pGACAG-PilA の混合物1 μgでGGにより、マウスを免疫した。血液は、最初の免疫化の後、0、2、4又は6週目に採取した。緑膿菌D4、rOprF/I、rPcrV、及びrPilAの全細胞に対する血清全IgG力値をELISAで測定した。グラフは、平均値±S.D.値(n=8匹のマウス/グル-プ)を示す。
【図3】DNAワクチンにより生じた緑膿菌抗原に対する血清IgG1及びIgG2a力値。マウスは図2で記述したように免疫した。rOprF/I、rPcrV、及びrPilAに対するIgG1又はIgG2a力値をELISAで測定した。グラフは、平均値±S.D.値(n=8匹のマウス/グル-プ)を示す。同様の結果が2つの独立な実験で得られた。** ANOVAボンフェロ-ニ多重比較検定によりp<0.01。
【図4】緑膿菌D4を鼻腔内投与した後にワクチン接種したマウスの肺ホモゲネ-ト及び血液中の生存細菌の数。マウスは図2で記述したように免疫した。感染後24時間で、マウスを屠殺し、希釈試料をLBプレ-トに広げて、(A)肺ホモゲネ-ト及び(B)血液中の全細菌数をカウントした。各LBプレ-ト上のコロニ-数を数えた後、CFUを算出した。グラフは、平均値±S.D.値(n=5匹のマウス/グル-プ)を示す。同様の結果が2つの独立な実験で得られた。* ANOVAボンフェロ-ニ多重比較検定によりp<0.05、** p<0.01。
【図5】緑膿菌D4を鼻腔内投与した後にワクチン接種したマウスの気管支肺胞洗浄(BAL)における好中球及びマクロファ-ジの数。マウスは図2で記述したように免疫した。5X105 CFUの細菌で感染させた後3及び6時間で、マウスを屠殺した後、BALを回収した。BAL中の細胞集団をギムザ染色によりキャラクタライズした。0.37 μm X 0.37 μmに広がる無作為抽出した10個の領域に存在する好中球及びマクロファ-ジの数を顕微鏡下でカウントしてから、全細胞数を算出した。グラフは、平均値±S.D.値(n=3匹のマウス/条件)を示す。同様の結果が2つの独立な実験で得られた。* ANOVAボンフェロ-ニ多重比較検定によりp<0.05、** p<0.01。
【図6】緑膿菌D4を鼻腔内投与した後にワクチン接種したマウスの気管支肺胞洗浄(BAL)細胞におけるマクロファ-ジ炎症性タンパク2(MIP-2)及びIFN-γ mRNAの生成。マウスは図2で記述したように免疫した。5 X 105 CFUの細菌で感染させた後2、12及び24時間で、マウスを屠殺した後、BALを回収した。全RNAを単離し、逆転写し、MIP-2、IFN-γ又は18S rRNAを標的とするリアルタイムPCRに各試料のアリコ-トをかけ、標的cDNAのレベルを定量した。18S rRNAのレベルでノ-マライズすることにより、相対的発現レベルを算出した。グラフは、平均値±S.D.値(n=3匹のマウス/条件)を示す。同様の結果が2つの独立な実験で得られた。* ANOVAボンフェロ-ニ多重比較検定によりp<0.05。
【図7】緑膿菌PAK又はD4を鼻腔内投与したマウスの生存率。マウスは図2で記述したように免疫した。2LD50の投与量の(A)PAK又は(B)D4(n=16-20匹のマウス/グル-プ)を鼻腔内に投与した後、生存率をモニタ-した。免疫したマウスの血清から精製したIgG画分(10 μg)を2LD50の投与量の(C)PAK又は(D)D4とともに37℃で30分間インキュベ-トした。その後、懸濁液を未処理マウスの鼻腔内に接種した(10匹のマウス/グル-プ)。生存率はその後10日間モニタ-した。同様の結果が2つの独立な実験で得られた。** Mantel-Cox log順位検定によりp<0.01。
【図8】プラスミドDNAワクチンの構成を示す。
【図9】抗原エピト-プを融合させたプラスミドDNAワクチンの構成を示す。
【図10】抗原エピトープ融合ワクチンにより誘導される多価抗体。抗原エピトープ融合ワクチンを筋注エレクトロポレーション法で免疫し(0、4週目、50μg/mouse)、血清(6週目)を採取した。リコンビナントOprF/I, PcrV, PilAをコーティングし、ELISA法で血清中のIgGを定量した。
【図11】抗原エピトープ融合ワクチンにより誘導される多価抗体。各菌株(PAO1, PAK, PA103, D4)をSDSサンプルバッファーで溶解し、SDS-PAGE法で分画し、PVDF膜に転写した。抗原エピトープ融合ワクチンで免疫したマウス血清と反応させ、抗マウスIgG抗体を用いてimmunoblottingを行った。
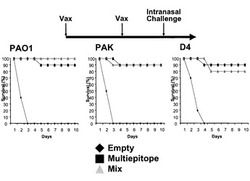
【図12】抗原エピト-プ融合ワクチンにより誘導される抗緑膿菌防御的免疫応答の評価。抗原エピト-プ融合ワクチン(Multiepitope)、至適化混合ワクチン(Mix)、空ベクター(Empty)を筋注エレクトロポレーション法で2回接種した後に2LD50量の緑膿菌(PAO1)を経鼻投与した。投与後のマウス生存率を示す。N = 10/グループ。
【図13】抗原エピト-プ融合ワクチンにより誘導される抗緑膿菌防御的免疫応答の評価。抗原エピト-プ融合ワクチン(Multiepitope)、至適化混合ワクチン(Mix)、空ベクター(Empty)を筋注エレクトロポレーション法で2回接種した後に2LD50量の緑膿菌(PAO1)を経鼻投与した。投与後12時間後と24時間後のマウス肺中の生存緑膿菌数を示す。N = 10/グループ。
【図14】抗原エピトープ融合ワクチンによる緑膿菌感染に対する防御的免疫応答の評価。抗原エピト-プ融合ワクチン(Multiepitope)、至適化混合ワクチン(Mix)、空ベクター(Empty)を筋注エレクトロポレーション法で2回接種した。6週目に、飲料水中に緑膿菌PAO1株を入れ(1 x 107/ml)、アンピシリン(1回/日、200 mg/kg)を3日間腹腔内に投与した。4日目に無菌の飲料水に交換して飼育し、4日目、6日目に1回ずつシクロフォスファミド(200 mg/kg)を腹腔内に投与して腸管感染由来敗血症モデル(易感染モデル)を作製した。図は7日以降の生存率を示す。N = 10/グループ。
【図15】抗原エピトープ融合ワクチンにより誘導された抗体の緑膿菌感染に対する防御能の評価。抗原エピト-プ融合ワクチン(Multiepitope)、至適化混合ワクチン(Mix)、空ベクター(Empty)を筋注エレクトロポレーション法で2回接種した。6週目に、採血し、IgG分画を精製した。15μgのIgGと2LD50のPAO1またはPAKを37℃で30分間反応させた後、ナイーブマウスに経鼻投与した。投与後の生存率を示す。N = 10/グループ。
【配列表フリーテキスト】
【0081】
<配列番号1>
配列番号1は、緑膿菌(PA01)由来の細胞外膜タンパクOprF190-342の塩基配列を示す。
<配列番号2>
配列番号2は、緑膿菌(PA01)由来の細胞外膜タンパクOprF190-342のアミノ酸配列を示す。
<配列番号3>
配列番号3は、緑膿菌(PA01)由来の細胞外膜タンパクOprI21-83の塩基配列を示す。
<配列番号4>
配列番号4は、緑膿菌(PA01)由来の細胞外膜タンパクOprI21-83のアミノ酸配列を示す。
<配列番号5>
配列番号5は、緑膿菌(PA01)由来のIII型分泌機構制御タンパクPcrV1-293(全長)(全長)の塩基配列を示す。
<配列番号6>
配列番号6は、緑膿菌(PA01)由来のIII型分泌機構制御タンパクPcrV1-293のアミノ酸配列を示す。
<配列番号7>
配列番号7は、緑膿菌(PA01)由来の毛状付属器タンパクPilA 136-149の塩基配列を示す。
<配列番号8>
配列番号9は、緑膿菌(PA01)由来の毛状付属器タンパクPilA 136-149のアミノ酸配列を示す。
<配列番号9>
配列番号9は、緑膿菌(PA01)由来の細胞外膜タンパクOprF全長の塩基配列を示す。
<配列番号10>
配列番号10は、緑膿菌(PA01)由来の細胞外膜タンパクOprF全長のアミノ酸配列を示す。
<配列番号11>
配列番号11は、緑膿菌(PA01)由来の細胞外膜タンパクOprI全長の塩基配列を示す。
<配列番号12>
配列番号12は、緑膿菌(PA01)由来の細胞外膜タンパクOprI全長のアミノ酸配列を示す。
<配列番号13>
配列番号13は、緑膿菌(PA01)由来の毛状付属器タンパクPilA全長(PilA 1-149)の塩基配列を示す。
<配列番号14>
配列番号14は、緑膿菌(PA01)由来の毛状付属器タンパクPilA全長(PilA 1-149)のアミノ酸配列を示す。
<配列番号15>
配列番号15は、oprFセンスプライマ-のDNA配列を示す。
<配列番号16>
配列番号16は、oprFアンチセンスプライマ-のDNA配列を示す。
<配列番号17>
配列番号17は、oprIセンスプライマ-のDNA配列を示す。
<配列番号18>
配列番号18は、oprIアンチセンスプライマ-のDNA配列を示す。
<配列番号19>
配列番号19は、pcrVセンスプライマ-のDNA配列を示す。
<配列番号20>
配列番号20は、pcrVアンチセンスプライマ-のDNA配列を示す。
<配列番号21>
配列番号21は、pilAセンスプライマ-のDNA配列を示す。
<配列番号22>
配列番号22は、pilAアンチセンスプライマ-のDNA配列を示す。
【技術分野】
【0001】
本発明は、緑膿菌に対して感染防御能を誘導できる医薬組成物に関する。
【背景技術】
【0002】
緑膿菌は、慢性疾患に伴う日和見感染症の原因菌として、薬剤耐性株の出現などから臨床上非常に問題となっている。様々なワクチンが検討されてきたが、臨床応用に至っているものは未だ開発されていない(非特許文献1)。例えば、細胞外膜タンパクOprF, OprIはワクチン標的分子として検討されている。マウスにおいてOprF/Iワクチンは、肺感染症・敗血症モデルでの有効性が確認されている。OprF/I融合分子(B細胞エピト-プ)は第III相臨床治験が行われ、安全性と有効性が確認されている(非特許文献2〜6)。III型分泌機構制御タンパクPcrVもワクチン標的分子として検討されている。特異抗体は菌毒素を宿主細胞内に注入するのを抑制するため、感染に伴う組織障害を抑制すると考えられている。特異抗体のみで感染を抑制することが報告されている(非特許文献7〜9)。毛状付属器タンパクPilAは細胞外膜に存在する繊毛で、菌の運動や粘膜への接着に係わる他、IV型分泌機構に重要である。ワクチン標的分子として有効性が確認されている(非特許文献10〜13)。
【0003】
【非特許文献1】Holder IA. Pseudomonas immunotherapy: a histrical overview. Vaccine 2004;22(7):831-9
【非特許文献2】Infect Immun, 63:1855, 1995
【非特許文献3】Vaccine, 17:1663, 1999
【非特許文献4】Infect Immun, 67:1461, 1999
【非特許文献5】FEMS Immunol Med Microbiol, 33:89, 2002
【非特許文献6】Vaccine, 24:6240, 2006
【非特許文献7】Nat Med, 5:392, 1999
【非特許文献8】J Immunol, 167:5880, 2001
【非特許文献9】Vaccine, 24:6240, 2006
【非特許文献10】Infect Immun, 63:1278, 1995
【非特許文献11】Vaccine, 22:831, 2004
【非特許文献12】FEMS Immunol Med Microbiol, 47:107, 2006
【非特許文献13】Vaccine, 24:6240, 2006
【発明の開示】
【発明が解決しようとする課題】
【0004】
本発明は、緑膿菌に対して効果的に感染防御能を誘導する医薬組成物を提供することを目的とする。
【課題を解決するための手段】
【0005】
本発明者らは、OprF/I、PcrV及びPilAを混合して免疫することで、多価かつ相乗的なワクチン効果を賦与できることを見出した。しかし、個々の標的抗原は、分子量が大きいため、遺伝子ワクチンとして応用した際に発現効率が悪く、病原因子としての危険性を孕んでいる。そこで、上記混合ワクチンの効果を効率化させ、病原因子の危険性を軽減させるために、各標的タンパクの抗原性を有する領域(エピト-プ)のみを連結し、抗原の分子量を縮小させた抗原エピト-プ融合ワクチンを作製した。この抗原を標的とするDNAワクチンを免疫すると、前記混合ワクチンと同等の防御的抗緑膿菌免疫応答を誘導することが確認された。このことから、抗原エピト-プ融合ワクチンを用いることで、幅広いスペクトラムの緑膿菌に対して効率的かつ安全に感染防御能を賦与できるワクチンの開発が可能となった。
【0006】
本発明の要旨は以下の通りである。
(1)下記(i)〜(v)のいずれかの成分を含む、緑膿菌に対して感染防御能を誘導できる医薬組成物。
(i) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部をコ-ドするDNA、緑膿菌の細胞外膜タンパクOprIの全長又はその一部をコ-ドするDNA、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部をコ-ドするDNA、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部をコ-ドするDNAをそれぞれ含むベクタ-の混合物
(ii) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、III型分泌機構制御タンパクPcrVの全長又はその一部、及び毛状付属器タンパクPilAの全長又はその一部の融合物をコ-ドするDNAを含むベクタ-
(iii) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部の混合物
(iv) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部の融合物
(v) (iii)の混合物又は(iv)の融合物に対して誘導された抗体
【0007】
(2)緑膿菌の細胞外膜タンパクOprFの全長又はその一部が下記(A)又は(B)のタンパク質であり、緑膿菌の細胞外膜タンパクOprIの全長又はその一部が下記(C)又は(D)のペプチドであり、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部が下記(E)又は(F)のタンパク質であり、緑膿菌の毛状付属器タンパクPilAの全長又はその一部が下記(G)又は(H)のペプチドである(1)記載の医薬組成物。
(A)配列番号2のアミノ酸配列からなるタンパク質
(B)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の細胞外膜タンパクOprFの抗原活性を有するタンパク質
(C)配列番号4のアミノ酸配列からなるペプチド
(D)配列番号4のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の細胞外膜タンパクOprIの抗原活性を有するペプチド
(E)配列番号6のアミノ酸配列からなるタンパク質
(F)配列番号6のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌のIII型分泌機構制御タンパクPcrVの抗原活性を有するタンパク質
(G)配列番号8のアミノ酸配列からなるペプチド
(H)配列番号8のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の毛状付属器タンパクPilAの抗原活性を有するペプチド
【0008】
(3)緑膿菌の細胞外膜タンパクOprFの全長又はその一部をコ-ドするDNAが下記(a)又は(b)のDNAであり、緑膿菌の細胞外膜タンパクOprIの全長又はその一部をコ-ドするDNAが下記(c)又は(d)のDNAであり、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部をコ-ドするDNAが下記(e)又は(f)のDNAであり、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部をコ-ドするDNAが下記(g)又は(h)のDNAである(1)又は(2)記載の医薬組成物。
(a)配列番号1のヌクレオチド配列で表されるDNA
(b)配列番号1のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の細胞外膜タンパクOprFの抗原活性を有するタンパク質をコ-ドするDNA
(c) 配列番号3のヌクレオチド配列で表されるDNA
(d) 配列番号3のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の細胞外膜タンパクOprIの抗原活性を有するペプチドをコ-ドするDNA
(e)配列番号5のヌクレオチド配列で表されるDNA
(f)配列番号5のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌のIII型分泌機構制御タンパクPcrVの抗原活性を有するタンパク質をコ-ドするDNA
(g)配列番号7のヌクレオチド配列で表されるDNA
(h)配列番号7のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の毛状付属器タンパクPilAの抗原活性を有するタンパク質をコ-ドするDNA
【0009】
(4)(i)〜(iv)のいずれかの成分を含むワクチンである(1)〜(3)のいずれかに記載の医薬組成物。
【0010】
(5)(i)のベクタ-混合物又は(ii)のベクタ-を含む遺伝子ワクチンである(4)記載の医薬組成物。
【0011】
(6)ベクタ-がプラスミド又はウイルスベクタ-である(5)記載の医薬組成物。
【0012】
(7)下記(I)の混合物又は(II)の融合物に対して誘導された抗体。
(I) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部の混合物
(II) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部の融合物
【発明の効果】
【0013】
本発明の医薬組成物は、異種の抗原若しくは抗原エピト-プを混合又は融合させたものを標的分子とするので、幅広いスペクトラムの緑膿菌に対して効率的かつ安全に感染防御能を賦与できる。
【発明を実施するための最良の形態】
【0014】
以下、本発明の実施の形態についてより詳細に説明する。
本発明は、緑膿菌に対して感染防御能を誘導できる医薬組成物を提供する。
本発明の医薬組成物は、ワクチン(遺伝子ワクチン、タンパクワクチン)又は抗体医薬として利用できる。
【0015】
本発明のワクチンが標的とする分子は、緑膿菌の細胞外膜タンパクOprFの全長若しくはその一部、緑膿菌の細胞外膜タンパクOprIの全長若しくはその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長若しくはその一部、及び緑膿菌の毛状付属器タンパクPilAの全長若しくはその一部の混合物又は融合物である。すなわち、本発明のワクチンは、緑膿菌の細胞外膜タンパクOprFの全長若しくはその一部、緑膿菌の細胞外膜タンパクOprIの全長若しくはその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長若しくはその一部、及び緑膿菌の毛状付属器タンパクPilAの全長若しくはその一部を抗原又は抗原エピト-プとして利用するものである。ワクチンは標的病原体に対する特異的抗原を標的に能動的に免疫応答を誘導することを目的とするが、病原因子のような抗原の場合(エンドトキシン、ペプチドグリカン、リポタンパク、ゲノムDNAなど)、投与することで生体に有害な作用(例えば、ショック症状、免疫応答の過剰活性化)を引き起こすことがあるため、免疫応答を誘導するのに必要な最小領域をもちいるのが望ましい。
【0016】
本発明のワクチンは、下記(i)〜(iv)のいずれかの成分を含む。
(i) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部をコ-ドするDNA、緑膿菌の細胞外膜タンパクOprIの全長又はその一部をコ-ドするDNA、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部をコ-ドするDNA、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部をコ-ドするDNAをそれぞれ含むベクタ-の混合物
(ii) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、III型分泌機構制御タンパクPcrVの全長又はその一部、及び毛状付属器タンパクPilAの全長又はその一部の融合物をコ-ドするDNAを含むベクタ-
(iii) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部の混合物
(iv) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部の融合物
緑膿菌(Pseudomonas aeruginosa)PA01株の全ゲノム配列は解読されており、NCBI gene bank にaccession #AE004091の登録番号で登録されている。従って、上記の(i)〜(iv)の成分は、この配列情報を利用して調製することができる。また、PA01株以外の他の株(PAK、D4など)の配列情報を利用してもよい。
【0017】
緑膿菌の細胞外膜タンパクOprFの全長又はその一部は、抗原又は抗原エピト-プとして機能するものであれば、特に限定されないが、例えば、PA01株のOprF190-342、そのホモロ-グなどを挙げることができ、具体的には、下記(A)又は(B)のタンパク質を例示することができる。
【0018】
(A)配列番号2のアミノ酸配列からなるタンパク質
(B)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の細胞外膜タンパクOprFの抗原活性を有するタンパク質
配列番号2は、PA01株のOprF190-342のアミノ酸配列を示す。また、PA01株のOprFの全長(OprF1-350)のアミノ酸配列は配列番号10に示す。緑膿菌の細胞外膜タンパクOprFの全長としては、下記(A1)又は(B1)のタンパク質を例示することができる。
【0019】
(A1)配列番号10のアミノ酸配列からなるタンパク質
(B1)配列番号10のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の細胞外膜タンパクOprFの抗原活性を有するタンパク質
緑膿菌の細胞外膜タンパクOprIの全長又はその一部は、抗原又は抗原エピト-プとして機能するものであれば、特に限定されないが、例えば、PA01株のOprI21-83、そのホモロ-グなどを挙げることができ、具体的には、下記(C)又は(D)のペプチドを例示することができる。
【0020】
(C)配列番号4のアミノ酸配列からなるペプチド
(D)配列番号4のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の細胞外膜タンパクOprIの抗原活性を有するペプチド
配列番号4は、PA01株のOprI21-83のアミノ酸配列を示す。また、PA01株のOprIの全長(OprI1-83)のアミノ酸配列は配列番号12に示す。緑膿菌の細胞外膜タンパクOprIの全長としては、下記(C1)又は(D1)のタンパク質を例示することができる。
【0021】
(C1)配列番号12のアミノ酸配列からなるペプチド
(D1)配列番号12のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の細胞外膜タンパクOprIの抗原活性を有するペプチド
緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部は、抗原又は抗原エピト-プとして機能するものであれば、特に限定されないが、例えば、PA01株のPcrV1-294、そのホモロ-グなどを挙げることができ、具体的には、下記(E)又は(F)のタンパク質を例示することができる。
【0022】
(E)配列番号6のアミノ酸配列からなるタンパク質
(F)配列番号6のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌のIII型分泌機構制御タンパクPcrVの抗原活性を有するタンパク質
配列番号6は、PA01株のPcrV1-294(全長)のアミノ酸配列を示す。
緑膿菌の毛状付属器タンパクPilAの全長又はその一部は、抗原又は抗原エピト-プとして機能するものであれば、特に限定されないが、例えば、PA01株のPilA136-149、そのホモロ-グなどを挙げることができ、具体的には、下記(G)又は(H)のペプチドを例示することができる。
【0023】
(G)配列番号8のアミノ酸配列からなるペプチド
(H)配列番号8のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の毛状付属器タンパクPilAの抗原活性を有するペプチド
配列番号8は、PA01株のPilA136-149のアミノ酸配列を示す。また、PA01株のPilA1-149の(全長)のアミノ酸配列は配列番号14に示す。緑膿菌の細胞外膜タンパクPilAの全長としては、下記(G1)又は(H1)のタンパク質を例示することができる。
【0024】
(G1)配列番号14のアミノ酸配列からなるタンパク質
(H1)配列番号14のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の毛状付属器タンパクPilAの抗原活性を有するタンパク質
緑膿菌の細胞外膜タンパクOprFの全長又はその一部をコ-ドするDNAとしては、例えば、PA01株のOprF190-342、そのホモロ-グなどをコ-ドするDNAを挙げることができ、具体的には、下記(a)又は(b)のDNAを例示することができる。
【0025】
(a)配列番号1のヌクレオチド配列で表されるDNA
(b)配列番号1のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の細胞外膜タンパクOprFの抗原活性を有するタンパク質をコ-ドするDNA
配列番号1は、PA01株のOprF190-342をコ-ドするDNAのヌクレオチド配列を示す。また、PA01株のOprFの全長をコ-ドするDNAのヌクレオチド配列は配列番号9に示す。緑膿菌の細胞外膜タンパクOprFの全長をコ-ドするDNAとしては、下記(a1)又は(b1)のDNAを例示することができる。
【0026】
(a1)配列番号9のヌクレオチド配列で表されるDNA
(b1)配列番号9のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の細胞外膜タンパクOprFの抗原活性を有するタンパク質をコ-ドするDNA
緑膿菌の細胞外膜タンパクOprIの全長又はその一部をコ-ドするDNAとしては、例えば、PA01株のOprI21-83、そのホモロ-グなどをコ-ドするDNAを挙げることができ、具体的には、下記(c)又は(d)のDNAを例示することができる。
【0027】
(c) 配列番号3のヌクレオチド配列で表されるDNA
(d) 配列番号3のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の細胞外膜タンパクOprIの抗原活性を有するペプチドをコ-ドするDNA
配列番号3は、PA01株のOprI21-83をコ-ドするDNAのヌクレオチド配列を示す。また、PA01株のOprIの全長をコ-ドするDNAのヌクレオチド配列は配列番号11に示す。緑膿菌の細胞外膜タンパクOprIの全長をコ-ドするDNAとしては、下記(c1)又は(d1)のDNAを例示することができる。
【0028】
(c1)配列番号11のヌクレオチド配列で表されるDNA
(d1)配列番号11のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の細胞外膜タンパクOprFの抗原活性を有するタンパク質をコ-ドするDNA
緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部をコ-ドするDNAとしては、例えば、PA01株のPcrV1-294、そのホモロ-グなどをコ-ドするDNAを挙げることができ、具体的には、下記(e)又は(f)のDNAを例示することができる。
【0029】
(e)配列番号5のヌクレオチド配列で表されるDNA
(f)配列番号5のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌のIII型分泌機構制御タンパクPcrVの抗原活性を有するタンパク質をコ-ドするDNA
配列番号5は、PA01株のPcrV1-294(全長)をコ-ドするDNAのヌクレオチド配列を示す。
緑膿菌の毛状付属器タンパクPilAの全長又はその一部をコ-ドするDNAとしては、例えば、PA01株のPilA136-149、そのホモロ-グなどをコ-ドするDNAを挙げることができ、具体的には、下記(g)又は(h)のDNAを例示することができる。
【0030】
(g)配列番号7のヌクレオチド配列で表されるDNA
(h)配列番号7のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の毛状付属器タンパクPilAの抗原活性を有するタンパク質をコ-ドするDNA
配列番号7は、PA01株のPilA136-149をコ-ドするDNAのヌクレオチド配列を示す。また、PA01株のPilAの全長(PilA1-149)をコ-ドするDNAのヌクレオチド配列は配列番号13に示す。緑膿菌の毛状付属器タンパクPilAの全長をコ-ドするDNAとしては、下記(g1)又は(h1)のDNAを例示することができる。
【0031】
(g1)配列番号13のヌクレオチド配列で表されるDNA
(h1)配列番号13のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の細胞外膜タンパクOprFの抗原活性を有するタンパク質をコ-ドするDNA
配列番号1、3、5、7、9、11又は13のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズするDNAは、配列番号1、3、5、7、9、11又は13のヌクレオチド配列で表されるDNAに相補的なDNAの全部又は一部と少なくとも80%(好ましくは、少なくとも95%、より好ましくは、少なくとも98%)の同一性があるとよい。「同一性」という用語は当業者に周知の用語であり、配列の同一性は入手可能な配列解析ソフトウェア(例えば、GCG Wisconsin Package (Accelrys INC.提供、Nucleic Acid Research 12 (1):387, 1984)、BLAST Nプログラム(NCBI提供、J Mol Biol 215: 403-410, 1990)などを用いて決定することができる。ハイブリダイゼ-ションはストリンジェントな条件下で行われる。核酸二本鎖又はハイブリッドの安定性は、融解温度Tm(プロ-ブが標的DNAから解離する温度)で表される。この融解温度はストリンジェントな条件を定義するために用いられる。1%のミスマッチによりTmが1℃低下すると仮定すると、ハイブリダイゼ-ション反応の最終洗浄の温度を低くしなければならない。例えば、プロ-ブと95%以上の同一性を有する配列を求める場合には、最終洗浄温度を5℃低くしなければならない。実際、1%のミスマッチにつき、0.5〜1.5℃の間でTmが変わることになる。ストリンジェントな条件の例としては、5x SSC/5x デンハルト溶液/1.0% SDS中68℃でハイブリダイズさせ、0.2x SSC/0.1%SDS中室温で洗浄することである。中程度にストリンジェントな条件の例としては、3x SSC中42℃で洗浄することである。塩濃度や温度は、プロ-ブと標的核酸との同一性の最適なレベルを達成するために変更されうる。このような条件に関するさらなる指針として、Sambrook et al., 1989, Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Press, N.Y.; and Ausubel et al. (eds.), 1995, Current Protocols in Molecular Biology, (John Wiley & Sons. N.Y.) at Unit 2.10を参照されたい。
【0032】
緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部、緑膿菌の毛状付属器タンパクPilAの全長又はその一部(以下、「抗原又は抗原エピト-プ」と呼ぶことがある)、及びそれらの融合物をコ-ドするDNAは、例えば、以下のようにして製造することができる。
【0033】
緑膿菌からゲノムDNAを抽出し、目的とする抗原又は抗原エピト-プのコ-ド領域をPCRによって増幅する。得られたPCR産物が目的の抗原又は抗原エピト-プをコ-ドするDNAである。抗原又は抗原エピト-プの融合物は、抗原又は抗原エピト-プをコ-ドするDNAをライゲートすることにより得られる。抗原若しくは抗原エピト-プ又はそれらの融合物をコ-ドするDNAを含有する組換えベクタ-は、公知の方法(例えば、Molecular Cloning2nd Edition, J. Sambrook et al., Cold Spring Harbor Lab. Press, 1989に記載の方法)により、抗原若しくは抗原エピト-プ又はそれらの融合物をコ-ドするDNAを適当な発現ベクタ-に挿入することにより得られる。
【0034】
発現ベクタ-としては、大腸菌由来のプラスミド(例、pBR322,pBR325,pUC12,pUC13, pGACAG)、枯草菌由来のプラスミド(例、pUB110,pTP5,pC194)、酵母由来プラスミド(例、pSH19,pSH15)、λファ-ジなどのバクテリオファ-ジ、レトロウイルス,アデノウイルス、ワクシニアウイルスなどの動物ウイルス、バキュロウイルスなどの昆虫病原ウイルスなどを用いることができる。
【0035】
発現ベクタ-には、プロモ-タ-(例えば、CAGプロモ-タ-、CMVプロモーター、SV40プロモーター、EF-1プロモーター))、イントロン(例えば、IntronA、b-actinイントロン))、エンハンサ-、スプライシングシグナル、ポリA付加シグナル、選択マ-カ-、タグ(例えば、FLAG配列、HA配列、c-myc配列、IgFc配列、6xHis配列、GST配列))、SV40複製オリジンなどを付加してもよい。
【0036】
また、発現ベクタ-は、融合タンパク質発現ベクタ-であってもよい。種々の融合タンパク質発現ベクタ-が市販されており、pGEXシリ-ズ(アマシャムファルマシアバイオテク社)、pET CBD Fusion System 34b-38b(Novagen社)、pET Dsb Fusion Systems 39b and 40b(Novagen社)、pET GST Fusion System 41 and 42(Novagen社)、pFLAGシリ-ズ(シグマアルドリッチ社)などを例示することができる。
【0037】
組換えベクタ-を遺伝子ワクチン(プラスミドワクチン・ウイルスベクタ-ワクチン)として使用する場合には、すでに遺伝子ワクチン用に開発したpGACAGプラスミドベクタ-に標的遺伝子を導入しプラスミドワクチンを作製するとよい。
抗原若しくは抗原エピト-プ又はそれらの融合物をコ-ドするDNAを含有する組換えベクタ-を宿主に導入することにより、形質転換体を得ることができる。
【0038】
宿主としては、細菌細胞(例えば、エシェリヒア属菌、バチルス属菌、枯草菌など)、真菌細胞(例えば、酵母、アスペルギルスなど)、昆虫細胞(例えば、S2細胞、Sf細胞など)、動物細胞(例えば、CHO細胞、COS細胞、HeLa細胞、C127細胞、3T3細胞、BHK細胞、HEK293細胞など)、植物細胞などを例示することができる。
組換えベクタ-を宿主に導入するには、Molecular Cloning2nd Edition, J. Sambrook et al., Cold Spring Harbor Lab. Press, 1989に記載の方法(例えば、リン酸カルシウム法、DEAE-デキストラン法、トランスフェクション法、マイクロインジェクション法、リポフェクション法、エレクロトポレ-ション法、形質導入法、スクレ-プロ-ディング法、ショットガン法など)または感染により行うことができる。
【0039】
形質転換体を培地で培養し、培養物から抗原若しくは抗原エピト-プ又はそれらの融合物を採取することができる。抗原若しくは抗原エピト-プ又はそれらの融合物が培地に分泌される場合には、培地を回収し、その培地から抗原若しくは抗原エピト-プ又はそれらの融合物を分離し、精製すればよい。抗原若しくは抗原エピト-プ又はそれらの融合物が形質転換された細胞内に産生される場合には、その細胞を溶解し、その溶解物から抗原若しくは抗原エピト-プ又はそれらの融合物を分離し、精製すればよい。
【0040】
抗原若しくは抗原エピト-プ又はそれらの融合物が別のタンパク質(タグとして機能する)との融合タンパク質の形態で発現される場合には、融合タンパク質を分離及び精製した後に、FactorXaや酵素(エンテロキナ-ゼ)処理をすることにより、別のタンパク質を切断し、目的とする抗原若しくは抗原エピト-プ又はそれらの融合物を得ることができる。
【0041】
抗原若しくは抗原エピト-プ又はそれらの融合物の分離及び精製は、公知の方法により行うことができる。公知の分離、精製法としては、塩析や溶媒沈澱法などの溶解度の差を利用する方法、透析法、限外ろ過法、ゲルろ過法、およびSDS-ポリアクリルアミドゲル電気泳動法などの分子量の差を利用する方法、イオン交換クロマトグラフィ-などの荷電の差を利用する方法、アフィニティ-クロマトグラフィ-などの特異的親和性を利用する方法、逆相高速液体クロマトグラフィ-などの疎水性の差を利用する方法、等電点電気泳動法などの等電点の差を利用する方法などが用いられる。
【0042】
抗原及び抗原エピト-プ並びにそれらの融合物は公知のペプチド合成法に従って製造してもよい。
【0043】
また、抗原若しくは抗原エピト-プに変異が導入されているもの(例えば、天然型の抗原又は抗原エピト-プのアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ前記抗原又は抗原エピト-プの抗原活性を有するタンパク質又はペプチド)又はそれらの融合物をコ-ドするDNAは、天然型の抗原又は抗原エピト-プのコ-ド領域を点突然変異誘発法により変異させることにより作製することができる。変異させたコ-ド領域をPCRによって増幅する。得られたPCR産物が抗原若しくは抗原エピト-プの変異体をコ-ドするDNAである。また、この変異体をコ-ドするDNAをライゲ-トすることにより、変異体の融合物をコ-ドするDNAを得ることができる。得られたDNAを適当な発現ベクタ-に組み込んだ後、適当な宿主に導入し、組換え蛋白質として生産させる(例えば、西郷薫、佐野弓子共訳、CURRENT PROTOCOLSコンパクト版、分子生物学実験プロトコ-ル、I、II、III、丸善株式会社:原著、Ausubel,F.M.等, Short Protocols in Molecular Biology, Third Edition, John Wiley & Sons, Inc., New Yorkを参照のこと)ことにより、抗原若しくは抗原エピト-プの変異体又はそれらの融合物を得ることができる。
【0044】
抗原若しくは抗原エピト-プの混合物又は融合物、あるいは抗原若しくは抗原エピト-プをコ-ドするDNAの混合物又は融合物を含有するベクタ-は、緑膿菌に対して感染防御能を誘導するためのワクチンの有効成分として利用することができる。例えば、抗原若しくは抗原エピト-プの混合物又は融合物、あるいは抗原若しくは抗原エピト-プをコ-ドするDNAの混合物又は融合物を含有するベクタ-をPBSなどの緩衝液、生理食塩水、滅菌水などに溶解し、必要に応じてフィルタ-などで濾過滅菌した後、注射により被験者に投与されるとよい。また、この溶液には、添加剤(例えば、不活化剤、保存剤、アジュバント、乳化剤など)などを添加してもよい。投与経路としては、静脈、筋肉、腹腔、皮下、皮内投与などが可能であり、また、経鼻、経口投与してもよい。
【0045】
抗原若しくは抗原エピト-プの混合物又は融合物、あるいは抗原若しくは抗原エピト-プをコ-ドするDNAの混合物又は融合物を含有するベクタ-の投与量、投与の回数及び頻度は、被験者の症状、年齢、体重、投与方法、投与形態などにより異なるが、例えば、通常、成人一人当たり0.01〜1mg/kg体重、好ましくは、0.01〜0.1mg/kg体重の抗原若しくは抗原エピト-プの混合物又は融合物を、少なくとも1回、所望の効果が持続する頻度で投与するとよい。抗原若しくは抗原エピト-プをコ-ドするDNAの混合物又は融合物を含有するプラスミドベクタ-を投与する場合には、例えば、通常、成人一人当たり1〜100mg、好ましくは、1〜10mgの投与量で少なくとも1回、所望の効果が持続する頻度で投与するとよい。ベクタ-がウイルスの場合には、成人一人当たり109〜1012ウイルス粒子、好ましくは1011〜1012ウイルス粒子の投与量で少なくとも1回、所望の効果が持続する頻度で投与するとよい。
【0046】
プラスミドワクチンはエンドトキシン除去した後投与するとよい。
筋肉内投与後エレクトロポレ-ション法で細胞内に導入する方法やリポソ-ムなどトランスフェクション増強剤との複合体を作製後投与する方法によりワクチン効果を高める方法も併用するとよい。
【0047】
本発明は、抗原若しくは抗原エピト-プの混合物又はそれらの融合物に対して誘導された抗体も提供する。本発明の抗体は緑膿菌に対して特異的に反応することができる。
本発明の抗体は、慣用のプロトコ-ルを用いて、抗原若しくは抗原エピト-プの混合物又はそれらの融合物を動物に投与することにより得られる。
本発明の抗体は、ポリクロ-ナル抗体、モノクロ-ナル抗体、キメラ抗体、一本鎖抗体、ヒト化抗体のいずれであってもよい。
【0048】
ポリクロ-ナル抗体を作製するには、公知あるいはそれに準じる方法にしたがって製造することができる。例えば、免疫抗原(タンパク質抗原)をコ-ドするDNAフラグメントを発現ベクタ-に導入し、これを動物に投与(免疫)し、該免疫動物からタンパク質に対する抗体含有物を採取して、抗体の分離精製を行なうことにより製造できる。投与に際して抗体産生能を高めるため、完全フロイントアジュバントや不完全フロイントアジュバントを投与してもよい。投与は、通常約2〜6週毎に1回ずつ、計約2〜10回程度行なわれる。ポリクロ-ナル抗体は、免疫動物の血液、腹水など、好ましくは血液から採取する(抗血清)ことができる。ポリクロ-ナル抗体の分離精製は、免疫グロブリンの分離精製法(例えば、塩析法、アルコ-ル沈殿法、等電点沈殿法、電気泳動法、イオン交換体による吸脱着法、超遠心法、ゲルろ過法、抗原結合固相あるいはプロテインAあるいはプロテインGなどの活性吸着剤により抗体のみを採取し、結合を解離させて抗体を得る特異的精製法)に従って行なうことができる。
【0049】
モノクロ-ナル抗体は、Nature (1975) 256: 495、Science (1980) 208: 692-に記載されている、G. Koehler及びC. Milsteinのハイブリド-マ法により作製することができる。すなわち、動物を免疫した後、免疫動物の脾臓から抗体産生細胞を単離し、これを骨髄腫細胞と融合させることによりモノクロ-ナル抗体産生細胞を調製する。さらに、抗原若しくは抗原エピト-プの混合物又はそれらの融合物と特異的に反応するが、他の抗原タンパク質とは実質的に交差反応しない抗体を産生する細胞系を単離するとよい。この細胞系を培養し、培養物から所望のモノクロ-ナル抗体を取得することができる。モノクロ-ナル抗体の精製は、上記の免疫グロブリンの分離精製法に従って行なうことができる。
【0050】
一本鎖抗体を作製する技法は、米国特許第4,946,778号に記載されている。
ヒト化抗体を作製する技法は、Biotechnology 10, 1121-, 1992; Biotechnology 10, 169-, 1992に記載されている。
【0051】
本発明の抗体は、緑膿菌に対する感染防御能を誘導するための医薬として利用することができる。この医薬の利点は、宿主の免疫系が活性化され、十分に高い防御レベルに達するまで待つ必要がなく、即座に効果が期待できることである。特に、感染が成立している場合や疾患が成立している場合に、抗体を投与すると、病原体や病原体が生産する毒素から患者を即座に回復させることができる。例えば、本発明の抗体をPBSなどの緩衝液、生理食塩水、滅菌水などに溶解し、必要に応じてフィルタ-などで濾過滅菌した後、注射により被験者に投与するとよい。また、この溶液には、添加剤(例えば、着色剤、乳化剤、懸濁剤、界面活性剤、溶解補助剤、安定化剤、保存剤、酸化防止剤、緩衝剤、等張化剤など)などを添加してもよい。投与経路としては、静脈、筋肉、腹腔、皮下、皮内投与などが可能であり、また、経鼻、経口投与してもよい。
【0052】
本発明の抗体の投与量、投与の回数及び頻度は、被験者の症状、年齢、体重、投与方法、投与形態などにより異なるが、例えば、通常、成人一人当たり1,000〜10,000 mg/kg体重、好ましくは、 2,000〜5,000 mg/kg体重の抗体を、少なくとも1回、所望の効果が持続する頻度で投与するとよい。
【実施例】
【0053】
以下、実施例に基づいて本発明を詳細に説明するが、本発明はこれらの実施例に限定されるものではない。
【0054】
〔実施例1〕
1.はじめに
緑膿菌(P. Aeruginosa)は、嚢胞性線維症に関連する罹患と死亡率に寄与する。なぜならば、その感染が直接肺実質にダメ-ジを与え、進行性の閉塞症、小気道の線維症及び肺機能の悪化に寄与するからである[1]。この感染の処置については抗生物質による治療が利用できるが、薬剤耐性分離株が現れ、これが慢性的に緑膿菌のコロニ-を形成する患者に重大な治療を提起する。緑膿菌に対する効果的なワクチンが開発されれば、このような感染の予防及び治療が促進されることになる[2]。
遺伝子免疫は、広範なウイルス、細菌及び寄生虫の病原体に対する保護的な免疫応答を誘導するための有望なストラテジ-である[3]。動物モデルにおいて、DNAワクチンは、AIDS様レトロウイルス、インフルエンザ、マラリア、結核及び緑膿菌感染に対する保護を与える[4-8]。免疫原性の他に、DNAワクチンは、通常の生又はサブユニットワクチンと比較したとき、いくつかの他の潜在的な利点を有する。DNAワクチンは製造が容易であり、安定性が高く、潜在的に安全である[9, 10]。DNAワクチンの主な潜在的利点は、単独の病原体の異なる株由来の抗原又は複数の病原体由来の抗原をコ-ドするいくつかのプラスミドを混合すること、多価抗原に対する免疫応答を誘導することができ、その結果、多種の感染に対して効果的に保護できることである。マラリア又は結核のマウスモデルにおいて、いくつかの異なる標的を包含するDNAプラスミドの組合せにより、いずれかのプラスミドを単独で用いたときに観察された保護の遺伝子的制限が抑制されるか、あるいは抗原の競合を見せることなく保護が増強された[11, 12]。異なるグレ-ド由来の複数のHIV-1遺伝子を発現するプラスミドの組合せで免疫すると、個々のプラスミドによって誘発されたものに匹敵する体液性及び細胞性免疫応答が誘発され、その結果、免疫干渉なしに抗ウイルス免疫応答が拡大した[13]。
本実施例では、異なる抗原をコ-ドするプラスミドからなる多価DNAワクチンの免疫原性を評価した。これまでに報告された緑膿菌抗原の可能性のある候補の中で、細胞外膜タンパクの融合物(OprF/I)、III型分泌機構制御タンパク(PcrV)、及び毛状付属器タンパク(PilA)を選択した。筋肉内エレクトロポ-レイション(imEPT)又は遺伝子銃(GG)によりプラスミドワクチン接種の有効性を比較し、これらの2つの方法を用いた場合に免疫原性が変化するかどうかを調べた。OprF/I、PcrV又はPilAを単独(一価)又は組合せ(多価)で発現するプラスミドでマウスを免疫した後、個々の抗原に対する血清抗体価と全細菌細胞に対する血清抗体価をモニタ-した。鼻腔内に細菌を投与した後、サイトカイン/ケモカイン mRNA産生レベル、細菌の生存数、好中球数及びマクロファ-ジ数をワクチン接種マウスの気管支肺胞洗浄(BAL)において評価した。最後に、致死量の緑膿菌を投与することにより、多価DNAワクチンの保護の可能性を評価した。
【0055】
2.材料及び方法
2.1.細菌
緑膿菌株PAO1、PAK及びD4を本実施例に用いた。PAO1及びPAKは、線毛、鞭毛及びIII型分泌細胞外酵素のような異なる病原性因子を発現する[14, 15]。D4は、好中球減少性の菌血症患者の血液から単離した[16]。すべての菌株はTerrific broth(Sigma, St. Louis, MO)中37℃で16時間インキュベ-トした。鼻腔内投与のために、細菌培養物を3000Xgで10分間遠心し、ペレットをPBS中で2回洗浄し、550 nmでの光学濃度1.0でPBSに再懸濁した。この懸濁液30又は40μlは、それぞれ、PAK又はD4のほぼ2LD50に相当した。
【0056】
2.2.ワクチン接種のための哺乳類発現プラスミドの構築(図8)
緑膿菌株PAO1のゲノムDNAを既報のように調製した[17]。以下のプライマ-セットを用いて、oprF、oprI、pcrV及びpilA遺伝子をPCR増幅した。
oprFセンス: 5’-GGA ATT CGC CAT GGC TCC GGC TCC GGA ACC GG-3’(配列番号15)
oprFアンチセンス: 5’-ACT GAG CTC TCA ACG CGA CGG TTG ATA GC-3’(配列番号16)
oprIセンス: 5’-TTG GAG CTT CCA CTC CAA AGA AAC CGA A-3’(配列番号17)
oprIアンチセンス: 5’-CCG GAT CCC TTG CGG CTG GCT TTT TCC AGC-3’(配列番号18)
pcrVセンス: 5’-GGA ATT CGC CAT GGA AGT CAG AAA CCT TAA-3’(配列番号19)
pcrVアンチセンス: 5’-CCG GAT TCG ATC GCG CTG AGA ATG TCG CGC-3’(配列番号20)
pilAセンス: 5’-GGA ATT CGG CAT GAA AGC TCA AAA AGG CTT-3’(配列番号21)
pilAアンチセンス: 5’-ACG CGT CGA CGT TAT CAC AAC CTT TCG GAG-3’(配列番号22)
融合遺伝子oprF/Iを構築するために、OprFのアミノ酸190-342及びOprIのアミノ酸21-83をコ-ドするDNAフラグメントを既報のようにタンデムにライゲ-トした[18]。プラスミドpGACAG-OprF/I、pGACAG-PcrV又はpGACAG-PilAを得るために、各遺伝子フラグメントにFLAGシグナル配列をタグ付けしてから、pGACAGベクタ-(DNAワクチンの臨床応用のために最適化された哺乳類発現ベクタ-[19])のEcoRI部位に導入した。すべてのプラスミドをE.coli DH5αに形質転換し、Endo-free Maxiprep kit(Quigen, Hilden, Germany)を用いて精製した。
【0057】
2.3.組み換えタンパク質の精製
組み換えタンパク質は、既報のように、E.coli DH5α中に発現させ、精製した[20]。
【0058】
2.4.免疫スケジュ-ル
8週令の雌BALB/cマウスをSLC(静岡県)から購入し、特定病原体除去条件下にある動物施設に入れた。これらのマウスをケタミン/キシラジン混合物で麻酔した後、100 μgのpGACAG(対照プラスミド)、pGACAG-OprF/I、pGACAG-PcrV、pGACAG-PilA又は33μgの各プラスミド(pGACAG-OprF/I+pGACAG-PcrV+pGACAG-PilA)の混合物のいずれかを含むプラスミド溶液(1 μg/μl食塩水、50 μl/筋肉)をマウス(各グル-プ8匹)の四頭筋に注射した。30 V/cmの一定の電場強度で、50 ms X 3パルスにて、CUY21EDIT(NepaGene、東京都、日本)を用いて、注入部位に電気穿孔した。最初の免疫化の後4週間目に追加免疫を行った。あるいはまた、微粒子銃粒子注入法(Helios Gene Gun kit, Bio-Rad, Richmond, CA)を用いて、0.5 mg金粒子にコ-ティングしたpGACAG-OprF/I+pGACAG-PcrV+pGACAG-PilAのプラスミド混合物1 μgでマウスを免疫した。マウスの腹部皮膚に2回の照射を14日の間隔で全3回行った。すべての動物実験は、動物実験委員会による承認を得、マウスはNIH animal care guidelineに従って取り扱われた。
【0059】
2.5.ワクチン接種したマウス由来の血清の特異性
PAO1、PAK又はD4から調製した細胞溶解物の免疫ブロットにより、抗体特異性を分析した。膜をブロックし、ワクチン接種した種々のグル-プのマウス由来の血清と反応させた。HRP標識抗マウスIgG抗体と共にインキュベ-トした後、ブロットをECL puls基板(Amersham Bioscience)と反応させてから、オ-トラジオグラフィ-で分析した。
【0060】
2.6.ELISA
血清抗体価を既報のようにELISAで測定した[19]。
【0061】
2.7.肺及び血液中の緑膿菌の定量
最終の免疫化後2週間目に、マウスを麻酔し、20μlのPBS中の5 X 105 CFUのD4を鼻腔内接種した。感染後(p.i.)24時間で再度マウスを麻酔した。マウスの眼を2回70%エタノ-ルで洗浄した後、眼窩後穿刺により無菌的に採血した。CO2に富む雰囲気下でマウスを屠殺した後、肺を除去し、4 ml PBS中、無菌条件下でホモジナイズした。血液と肺ホモジネ-トの10倍段階希釈液をLB寒天プレ-ト(Sigma)上にまき、このプレ-トを37℃で24時間インキュベ-トした。コロニ-数を二重盲検でマニュアルで数えた。
【0062】
2.8.BAL液中の好中球及びマクロファ-ジの定量
5 X 105 CFUのD4を鼻腔内接種したマウスを感染後3及び6時間目にCO2に富む雰囲気下で屠殺した。1.7 mm-ODポリエチレンカテ-テルを用い、0.5 mlのPBSで連続して2回洗浄することにより、BALFを回収した。試料を遠心し、20 μlのPBS中に再懸濁させた。5 μlのスメアがスライドグラス上に調製され、これをギムザ染色した。0.37 μm X 0.37 μmに広がる無作為抽出の10個の領域に存在する好中球及びマクロファ-ジの数を顕微鏡下でカウントしてから、全細胞数を算出した。
【0063】
2.9.マクロファ-ジ炎症性タンパク質2(MIP-2)及びIFN-γ mRNAのリアルタイムPCR分析
上述のように5 X 105 CFUの緑膿菌D4を用いて、感染後2、12及び24時間目にBAL細胞を調製した。細胞をTRIzo試薬(Invitrogen, Carlsbad, CA)に再懸濁し、全RNAを単離し、既報のように逆転写した[21]。ABI PRISM 7700 sequence detection system(Applied Biosystems)を用い、製造者の指示に従い、MIP-2、IFN-γ又は18S rRNAに特異的なTaqMan probe(Applied Biosystems, Foster City, CA)を使用して、リアルタイムPCRを行った。同じ試料の18S rRNAのレベルをノ-マライズすることにより、試料中のMIP-2及びIFN-γ mRNAの反応発現レベルを算出した。
【0064】
2.10.保護テスト
最終の免疫化の後2週間目に、マウスを麻酔して、2LD50のPAK又はD4を鼻腔内に投与した。このマウスの致死率をその後10日間モニタ-した。
【0065】
2.11.受動保護テスト
6週令の雌BALB/cマウスを用いた。PAK及びD4を16時間TB broth中で培養し、PBS中で3回洗浄し、ペレットをPBSに再懸濁し、550 nmの光学密度を1.0に調整し、所望の接種菌液を調製した。種々のグル-プの免疫マウス由来の血清のIgG画分をProtein G sepharose column(Amersham Biosciences)で精製した。各IgG画分10 μgを2LD50の細菌と混合し、37℃で30分間インキュベ-トした。これらの懸濁液を各グル-プ10匹のマウスの鼻腔内に投与し(全50 μl/マウス)、致死率をその後10日間モニタ-した。
【0066】
2.12.統計分析
すべての実験を少なくとも2回繰り返した。統計的な有意差は、ANOVAボンフェロ-ニ多重比較検定を用いて評価した。マウスのグル-プ間の生存率の差は、Mantel-Cox log 順位検定を用いて評価した。p値<0.05は、統計的な有意差があるとみなした。
【0067】
3.結果
3.1.DNAワクチンにより誘発された抗体産生のキャラクタリゼ-ション
緑膿菌を標的とする多価DNAワクチンの免疫原性が、一価DNAワクチンにより誘発された免疫原性より大きいかどうかを調べるために、3つの異なるプラスミド(各々は、oprF/I、pcrV又はpilA遺伝子を包含する)を調製した。その後、対照プラスミド、各単独プラスミド(一価)又は3つの異なるプラスミドの混合物(多価)で、imEPTにてマウスを免疫した。PAO1、PAK及びD4細胞溶解物の免疫ブロットを用いて、各標的抗原に特異的な抗体応答を調べた。pGACAG-OprF/Iで免疫したマウスから得られた血清は、実際に、すべての細胞溶解物中に存在する〜36及び〜12 kDaタンパク質と反応した(図1A)。〜36及び〜12 kDaタンパク質は、それぞれ、OprF及びOprIであると推定された。pGACAG-PcrVで免疫したマウス由来の血清は、D4及びPAK細胞溶解物中に存在する〜32 kDaのタンパク質(PcrVと同じ大きさ)と反応したが、pGACAG-PilAで免疫したマウス由来の血清は、PAO1及びD4中に存在する〜15 kDaのタンパク質(PilAと同じ大きさ)と反応した(図1B及びC)。これらの結果は、各DNAワクチンにより生じた抗体は、緑膿菌の異なる株により発現される内在性の抗原標的に特異的に結合することを保証するものである。
imEPT又はGGのいずれかにより多価DNAワクチンで免疫されたマウスから得られた血清は、いくつかのタンパク質、おそらくはOprF、OprI、PcrV及びPilAと反応したが、OprFとPcrVを標的とする抗体レベルは、分子サイズが近いために、免疫ブロットにより区別することはできなかった(〜36及び〜32 kDa、図1D及びE)。全IgG、IgG1、及びIgG2aサブタイプを含む血清抗体価をELISAにより規則的な時間間隔でモニタ-した。対照プラスミド又は単独プラスミドのいずれかでワクチン接種したマウスから得られた血清は、他の抗原標的と交叉反応をしなかった(デ-タは示さず)。図2Aに示されるように、各一価又は多価DNAワクチンにより誘導されたIgGは緑膿菌D4株の全細胞と特異的に反応した。各組み換え標的タンパク質に対する全IgG産生のレベルは、一価及び多価ワクチン接種グル-プ間で匹敵するものであり、このことは、OprF/I、PcrV及びPilAを標的とする多価DNAワクチンが、抗原間の免疫干渉なしに、容易に多価抗体応答を誘導することを示唆している(図2B-D)。IgG1及びIgG2サブタイプを調べたときに、同様の結果が得られた(図3)。多価DNAワクチンを用いて、imEPT及びGGにより誘発される免疫応答を比較したところ、各標的タンパク質に対する全IgG及びIgG1サブタイプの匹敵するレベルがどちらの方法でも誘導された(図2及び3)。しかしながら、GGに仲介された免疫化は、すべての標的抗原に対する体液性応答のIgG2aサブタイプは誘導しなかった(図3D-F, p<0.01)。
【0068】
3.2.肺及び血液からの細菌のクリアランス
DNAワクチンにより生じた免疫が緑膿菌感染を排除する能力を調べるために、D4を鼻腔内投与した後に肺ホモジネ-ト又は血液中の生存細菌の数を調べた。一価DNAワクチンの各々でワクチン接種すると、対照プラスミド処理と比較して、肺及び血液の両方で細菌の数はより少なくなった(図4)。重大なことは、imEPTによる多価DNAワクチン接種により肺中の細菌が有意に清掃されたことである(図4A、p<0.05)。非常に重要な点は、このグル-プのみに菌血症の完全な保護が観察されたことである(図4、p<0.01)。これらの結果は、明らかに、OprF/I、PcrV及びPilAからなる多価DNAワクチン接種によって、免疫応答の付与に成功し、緑膿菌の肺感染から保護されたことを示している。しかしながら、GGによる多価DNAワクチン接種が誘発した免疫応答は、緑膿菌肺炎に対する保護効果が不十分であった。
【0069】
3.3.BAL細胞のキャラクタリゼ-ション
多価DNAワクチンによる保護免疫応答の根底にある機構を明らかにするために、免疫マウスのBAL細胞をキャラクタライズした。他のグル-プと比較したときに、imEPTにより多価DNAワクチンが投与されたマウスから得られたBAL試料中の好中球及びマクロファ-ジの両方の数が有意に増加したことがギムザ染色により示された(図5、p<0.05)。この迅速な炎症反応の基礎を明らかにするために、BAL細胞中のいくつかのサイトカイン及びケモカインのmRNAレベルをリアルタイムPCRで定量した。ケモカインの中でも、MIP-2は気管支肺胞での好中球動員の中心的役割を果たす[22]。最も高いレベルのMIP-2 mRNAが、imEPTによる多価ワクチン接種を受けたマウスに観察された(図6A、p<0.05)が、このことは、MIP-2の発現が増加することにより、気管支肺胞空間への強い好中球動員が誘発されたことを示唆している。IFN-γ mRNAレベルは、すべてのグル-プにおいて、中程度だが、持続的に、感染後24時間まで増加した(図6B)。最も高いレベルのIFN-γ mRNAが、imEPTにより多価ワクチン接種を受けたマウスにおいて、感染後24時間で検出された(図6B、p<0.01)が、このことは、IFN-γの産生が増加したことにより、好中球及びマクロファ-ジにより貪食された細菌の自然な死滅がさらに活性化されたことを示唆している。これらのデ-タにより、多価DNAワクチンはMIP-2産生を通して好中球動員を優先的に誘導し、IFN-γ産生により好中球及びマクロファ-ジの殺菌活性を増強したことを暗に意味している。一価及び多価ワクチン接種グル-プ間で、IL-4、IL-6、TNF-α及びJE/MCP-1のような他のサイトカイン及びケモカインのレベルの有意差はなかった(デ-タは示さず)。
【0070】
3.4.imEPTによる多価DNAワクチン接種はD4又はPAKの致死的投与からマウスを保護した
ワクチン接種したマウスの鼻腔内に2LD50投与量のPAK又はD4を投与した。接種マウスの致死率をその後10日間モニタ-した。imEPTにより多価DNAをワクチン接種したすべてのマウスは、PAKの感染後10日間以上生存した(図7A)。一方、GGにより、対照プラスミド、PilAワクチン又は多価ワクチンで免疫したマウスの70%は感染後5日以内に死亡した(p<0.01)。OprF/I又はPcrVを標的とする一価DNAワクチン接種でさえ、80%のマウスを感染後4日目で保護することができた(p<0.01)。D4株を用いたときも、同様の結果が得られた(図7B)が、このことは、OprF/I、PcrV及びPilAを標的とするimEPTによる多価DNAワクチン接種により、緑膿菌感染に対して顕著な保護免疫応答を誘導することを示唆している。
【0071】
3.5.多価DNAワクチンで免疫したマウス由来の抗血清による受動保護
受動保護の検討を行い、抗体が保護能力に重大であるかどうかを評価した。ワクチン接種したマウスから得られた血清の精製IgG画分とともにD4又はPAKを前培養し、その後、この混合物を未処理マウスの鼻腔内に接種した。ワクチン接種グル-プの中で、imEPTにより多価DNAワクチンを接種したマウス由来の血清が、PAK株の致死的な感染から未処理マウスを保護する効果が高かった(図7C、p<0.01)。もう一つのD4株を用いたときも同様の結果が得られた(図7D)。
【0072】
4.考察
緑膿菌は院内肺炎の原因となる主要な病原体である。この病原体は、LPS、外毒素、線毛、鞭毛、エラスタ-ゼ、及びプロテア-ゼを含む数々の病原性因子を保有する[14,15,20,23-26]。しかしながら、これらの病原性因子はすべてのタイプの緑膿菌感染に関連しているわけではない。緑膿菌の複雑な病原性とその病原性因子の多様な機能が、効果的で汎用性があるワクチンの開発に大きな障害となっている[27]。従って、細菌の異なる成分を標的とするDNAワクチンが協同的に作用して、緑膿菌に対する宿主の保護免疫の範囲を拡大することができるかどうかを調べた。本実施例において、比較的広い範囲の臨床株に保存されている3つの主要な抗原を用いた。また、免疫応答を拡大するために、細菌器官の異なる成分由来のこれらの抗原を選択した。OprF及びOprIのような外膜タンパクでワクチン接種すると、いくつかのげっ歯類モデルにおいて、緑膿菌感染に対して効果的であることが示された[28,29]。PcrVは、III型分泌-中毒機構のエフェクタ-の転座に重要な役割を果たす。PcrVに対する能動及び受動免疫により、緑膿菌により引き起こされる急性及び慢性感染の両方での生存が有意に増加した。PcrVに対する抗体はエフェクタ-の転座を防止し、次に、肺胞マクロファ-ジ生存率と食細胞機能を増強する[30,31]。線毛は、細菌の付属器として機能するが、上皮細胞への接着に利用される。事実、線毛を有しない株は病原性が減少し、抗線毛血清は宿主細胞への細菌の結合を抑制することが実証されている[32,33]。また、異なる株を起源とする線毛に対する抗体は、他の株由来の線毛にも広く交叉反応する[15]。従って、我々のワクチン製剤は、宿主の生体防御から細菌が逃れることを防ぐための3つの独立な作用機序を有するのかもしれない。
【0073】
いくつかの研究では、DNAワクチンの組合せでは、単一のDNAワクチンと比較して、個々の抗原に対する免疫原性が減少することが示され、このことは、「抗原競合」と呼ばれている[34]。しかしながら、本実施例により、多価緑膿菌DNAワクチンが、imEPTにより免疫したマウスにおいて細菌に対して強力な保護免疫応答を誘発することが明らかに示された。imEPT技術は、トランスフェクション効率と炎症を増強することにより、DNAワクチンの免疫原性を強化することが実証されている[35]。事実、免疫化がimEPTにより行われると、同じレベルの最適免疫応答を誘導するには、20分の1程度の低量のプラスミドで十分であった(デ-タは示さず)。従って、この現象が不十分なトランスフェクション効率では起こらない場合、imEPTによる効率的なトランスフェクション及びその後の免疫原性の増強により、「抗原競合」が抑制されたのかもしれない。ワクチンの多様性については、使用されたDNAの量がimEPT法とGG法とで異なったのであるが、GGと比較して、imEPTは、緑膿菌に対する保護免疫の誘導においてより顕著な効果を示した。
【0074】
好中球及びマクロファ-ジは、急性肺細菌感染に対する保護において鍵となる役割を果たす[36]。本実施例において、多価DNAワクチンで免疫したマウスにおける緑膿菌の肺クリアランスの増強は、BALにおける好中球及びマクロファ-ジの数の増加と関連があった。MIP-2はヒトIL-8の機能的ホモログであるが、これは強力な化学誘引物質であり、好中球の活性化因子である。予想されるように、BAL細胞におけるMIP-2 mRNAレベルのアップレギュレ-ションは、好中球の流入量と関連があった。IFN-γは、肺胞マクロファ-ジと好中球の両方において細胞内殺菌活性を制御する鍵となる因子である[37]。IFN-γ mRNAは、多価DNAワクチン接種グル-プにおいて遅延相(>12h)で有意に誘導されたが、このことは、図5Bに観察されるように、強力なIFN-γが細菌転移の完全な保護の要因となっている可能性があることを示唆している、この観察結果は、IFN-γ仲介Th1免疫応答が緑膿菌に対する肺宿主生体防御と慢性緑膿菌感染の予後を改善することを示した以前の報告[38-40]と一致している。
【0075】
受動保護に関する本実施例の結果は、多価抗体応答が緑膿菌に対する保護免疫応答の主要な因子であることを示している。非常に重要なことは、IgG2aが保護に重要であるように思われることである(図4)。事実、IgG2aは、Fcγ受容体に最も強い親和性を持ち、IFN-γが仲介するB細胞のアイソタイプスイッチングの結果として産生される[41]。さらに、これらの結果は、抗体が仲介する免疫は保護の主要な因子であるが、より高いTh2免疫応答(IgG1)が、Th1免疫応答(IgG2a)よりも緑膿菌感染におけるより重篤な肺病理に関連性があることを示す以前の研究をさらに進めるものである[42,43]。
【0076】
以上を要約すると、本実施例の結果は、OprF/I、PcrV、及びPilAを標的とする多価DNAワクチンが協同的に緑膿菌肺炎のマウスモデルにおける保護効率を強めることを直接的に実証した。imEPT技術は、保護機構を誘発するという点で、GGよりかなり優れている。我々の進行中の研究は、抗原インサ-トのサイズを最小化し、OprF、OprI、PcrV、及びPilA由来の入れ子状態のエピト-プからなる単一の融合抗原を理想的に設計することに焦点を当てている。そのような研究は、緑膿菌多価DNAワクチンの臨床応用を支持するであろう。
【0077】
〔実施例2〕
図9に示すように、OprF190-342アミノ酸領域、OprI20-83アミノ酸領域、PcrV1-294アミノ酸領域、PilA136-149アミノ酸領域を直列に連結した抗原エピトープ融合DNAワクチンpGACAG-OprF/I-PcrV-PilAを実施例1と同様の方法で作製した。このワクチンを実施例1と同様に、筋注エレクトロポレーション法で導入し(0週と4週に1回ずつ両側大腿四頭筋に25μg)、6週目の血清を用いてELISA法により抗OprF/I、PcrV、PilA抗体価を測定した。その結果、それぞれの抗原に対するIgG抗体が誘導されていることが確認された(図10)。同様に、異なる菌株由来の抗原に対して多価の抗体が誘導されているか確認するため、各菌株(PAO1, PAK, PA103, D4)の菌体溶解物をSDS-PAGE法で分画し、PVDF膜に転写後、抗原エピトープ融合ワクチンで免疫したマウス血清と反応させ、抗マウスIgG抗体を用いてimmunoblottingを行った。その結果、各菌株由来のOprF,PvrV, PilA, OprIに対して反応する特異抗体が誘導されたことが確認された(図11)。実際に、抗原エピトープ融合ワクチンpGACAG-OprF/I-PcrV-PilA(Multiepitope)、実施例1で至適化した混合ワクチンpGACAG-OprF/I+pGACAG-PcrV+pGACAG-PilA(Mix)、もしくは空ベクターpGACAG(Empty)を筋注エレクトロポレーション法で導入し(0週と4週に1回ずつ両側大腿四頭筋に25μg)、6週目に2LD50量の緑膿菌株(PAO1, PAK, D4)を経鼻感染させてその後の生存率を観察した。その結果、Multiepitope投与群においてはMix群と同等の生存率を示し、抗原エピトープ融合ワクチンの有効性が確認された(図12)。実際に、PAO1感染後12および24時間後の肺内残存生菌数を数えると、Multiepitope投与群においてはMix群と同等の菌数であり、抗原エピトープ融合ワクチンにより緑膿菌感染に対する防御的免疫応答を誘導できたことが確認された(図13)。抗原エピトープ融合ワクチンによる緑膿菌感染に対する防御的免疫応答について、腸管感染由来敗血症モデル(易感染モデル)でも評価した(図14)。飲料水中に緑膿菌PAO1株を入れ(1 x 107/ml)、アンピシリン(1回/日、200 mg/kg)を3日間腹腔内に投与した。4日目に無菌の飲料水に交換して飼育し、4日目、6日目に1回ずつシクロフォスファミド(200 mg/kg)を腹腔内に投与して易感染状態とし、その後の生存率を観察した(図14)。その結果、抗原エピトープ融合ワクチンによる、易感染状態における抗緑膿菌感染防御能の誘導が確認され、その効果はわれわれが検討してきた至適化混合ワクチンとほぼ同等であることがあきらかとなった。抗原エピトープ融合ワクチンにより誘導される抗体の抗緑膿菌感染防御効果について検討した(図15)。実施例1と同様に抗原エピトープ融合ワクチン(Multiepitope)、至適化混合ワクチン(Mix)、もしくは空ベクター(Empty)を筋注エレクトロポレーション法で導入し(0週と4週に1回ずつ両側大腿四頭筋に25μg)、6週目に血清を採取し、IgG分画を精製した。15μgの精製IgGと2LD50量のPAO1株またはPAK株と37℃で30分間反応された後、ナイーブマウスに経鼻感染させ、その後の生存率を観察した。その結果、これまでの結果と同様に、ワクチン免疫により誘導されたIgG分画に緑膿菌感染を抑制する作用があることが確認され、抗原エピトープ融合ワクチンによる効果は、至適化混合ワクチンと同等であった。
【0078】
引用文献
[1] Pier GB. CFTR mutations and host susceptibility to Pseudomonas aeruginosa lung infection. Curr Opin Microbiol 2002;5(1): 81-6.
[2] Lang AB, Horn MP, Imboden MA, Zuercher AW. Prophylaxis and therapy of Pseudomonas aeruginosa infection in cystic fibrosis and immunocompromised patients. Vaccine 2004;22(Suppl 1):S44-8.
[3] Srivastava IK, Liu MA. Gene vaccines. Ann Intern Med 2003; 138(7):550-9.
[4] Ha SJ, Jeon BY, Youn JI, Kim SC, Cho SN, Sung YC. Protective effect of DNA vaccine during chemotherapy on reactivation and reinfection of Mycobacterium tuberculosis. Gene Ther 2005;12(7): 634-8.
[5] Okuda K, Xin KQ, Haruki A, et al. Transplacental genetic immunization after intravenous delivery of plasmid DNA to pregnant mice. J Immunol 2001;167(9):5478-84.
[6] Sasaki S, Amara RR, Oran AE, Smith JM, Robinson HL. Apoptosismediated enhancement of DNA-raised immune responses by mutant caspases. Nat Biotechnol 2001;19(6):543-7.
[7] Takakura M, Okuda K, Matsuda T, et al. Combination of DNA vaccine and adenovirus vector by cutaneous administration induced strong HIV-specific cellular immune responses in mice. Vaccine 2005;23(7):847-8.
[8] Vuola JM, Keating S, Webster DP, et al. Differential immunogenicity of various heterologous prime-boost vaccine regimens using DNA and viral vectors in healthy volunteers. J Immunol 2005;174(1): 449-55.
[9] Smith HA, Klinman DM. The regulation of DNA vaccines. Curr Opin Biotechnol 2001;12(3):299-303.
[10] Staczek J, Gilleland LB, van der Heyde HC, Gilleland HE. DNA vaccines against chronic lung infections by Pseudomonas aeruginosa. FEMS Immunol Med Microbiol 2003;37(2/3):147-53.
[11] Doolan DL, Aguiar JC, Weiss WR, et al. Utilization of genomic sequence information to develop malaria vaccines. J Exp Biol 2003;206(Pt 21):3789-802.
[12] Morris S, Kelley C, Howard A, Li Z, Collins F. The immunogenicity of single and combination DNA vaccines against tuberculosis. Vaccine 2000;18(20):2155-63.
[13] Kong WP, Huang Y, Yang ZY, Chakrabarti BK, Moodie Z, Nabel GJ. Immunogenicity of multiple gene and clade human immunode-ficiency virus type 1 DNA vaccines. J Virol 2003;77(23):12764-72.
[14] Hertle R, Mrsny R, Fitzgerald DJ. Dual-function vaccine for Pseudomonas aeruginosa: characterization of chimeric exotoxin A-pilin protein. Infect Immun 2001;69(11):6962-9.
[15] Sheth HB, Glasier LM, Ellert NW, et al. Development of an anti-adhesive vaccine for Pseudomonas aeruginosa targeting the Cterminal region of the pilin structural protein. Biomed Pept Proteins Nucl Acids 1995;1(3):141-8.
[16] Matsumoto T, Furuya N, Tateda K, et al. Effect of passive immunotherapy on murine gut-derived sepsis caused by Pseudomonas aeruginosa. J Med Microbiol 1999;48(8):765-70.
[17] Fukushima J, Yamamoto S, Morihara K, et al. Structural gene and complete amino acid sequence of Pseudomonas aeruginosa IFO 3455 elastase. J Bacteriol 1989;171(3):1698-704.
[18] Larbig M, Mansouri E, Freihorst J, et al. Safety and immunogenicity of an intranasal Pseudomonas aeruginosa hybrid outer membrane protein F-I vaccine in human volunteers. Vaccine 2001;19(17-19): 2291-7.
[19] Sasaki S, Xin KQ, Okudela K, Okuda K, Ishii N. Immunomodulation by apoptosis-inducing caspases for an influenza DNA vaccine delivered by gene gun. Gene Ther 2002;9(12):828-31.
[20] Kawamoto S, Shibano Y, Fukushima J, Ishii N, Morihara K, Okuda K. Site-directed mutagenesis of Glu-141 and His-223 in Pseudomonas aeruginosa elastase: catalytic activity, processing, and protective activity of the elastase against Pseudomonas infection. Infect Immun 1993;61(4):1400-5.
[21] Takeshita F, Suzuki K, Sasaki S, Ishii N, Klinman DM, Ishii KJ. Transcriptional regulation of the human TLR9 gene. J Immunol 2004;173(4):2552-61.
[22] Tsai WC, Strieter RM, Mehrad B, Newstead MW, Zeng X, Standiford TJ. CXC chemokine receptor CXCR2 is essential for protective innate host response in murine Pseudomonas aeruginosa pneumonia. Infect Immun 2000;68(7):4289-96.
[23] Atsumi Y, Yamamoto S, Morihara K, et al. Cloning and expression of the alkaline proteinase gene from Pseudomonas aeruginosa IFO 3455. J Bacteriol 1989;171(9):5173-5.
[24] Matsumoto T, Tateda K, Furuya N, et al. Efficacies of alkaline protease, elastase and exotoxin A toxoid vaccines against gutderived Pseudomonas aeruginosa sepsis in mice. J Med Microbiol 1998;47(4):303-8.
[25] Pier GB. Promises and pitfalls of Pseudomonas aeruginosa lipopolysaccharide as a vaccine antigen. Carbohydr Res 2003;338(23):2549-56.
[26] Tanaka E, Kawamoto S, Fukushima J, et al. Detection of elastase production in Escherichia coli with the elastase structural gene from several non-elastase-producing strains of Pseudomonas aeruginosa. J Bacteriol 1991;173(19):6153-8.
[27] Holder IA. Pseudomonas immunotherapy: a historical overview. Vaccine 2004;22(7):831-9.
[28] von Specht BU, Knapp B, Muth G, et al. Protection of immunocompromised mice against lethal infection with Pseudomonas aeruginosa by active or passive immunization with recombinant P. aeruginosa outer membrane protein F and outer membrane protein I fusion proteins. Infect Immun 1995;63(5):1855-62.
[29] Worgall S, Krause A, Rivara M, et al. Protection against P. aeruginosa with an adenovirus vector containing an OprF epitope in the capsid. J Clin Invest 2005;115(5):1281-9.
[30] Sawa T, Yahr TL, Ohara M, et al. Active and passive immunization with the Pseudomonas V antigen protects against type III intoxication and lung injury. Nat Med 1999;5(4):392-8.
[31] Shime N, Sawa T, Fujimoto J, et al. Therapeutic administration of anti-PcrV F(ab$)(2) in sepsis associated with Pseudomonas aeruginosa. J Immunol 2001;167(10):5880-6.
[32] Farinha MA, Conway BD, Glasier LM, et al. Alteration of the pilin adhesin of Pseudomonas aeruginosa PAO results in normal pilus biogenesis but a loss of adherence to human pneumocyte cells and decreased virulence in mice. Infect Immun 1994;62(10): 4118-23.
[33] Tang H, Kays M, Prince A. Role of Pseudomonas aeruginosa pili in acute pulmonary infection. Infect Immun 1995;63(4):1278-85.
[34] Sedegah M, Charoenvit Y, Minh L, et al. Reduced immunogenicity of DNA vaccine plasmids in mixtures. Gene Ther 2004;11(5): 448-56.
[35] Widera G, Austin M, Rabussay D, et al. Increased DNA vaccine delivery and immunogenicity by electroporation in vivo. J Immunol 2000;164(9):4635-40.
[36] McClellan SA, Huang X, Barrett RP, van Rooijen N, Hazlett LD. Macrophages restrict Pseudomonas aeruginosa growth, regulate polymorphonuclear neutrophil influx, and balance pro- and anti-inflammatory cytokines in BALB/c mice. J Immunol 2003;170(10): 5219-27.
[37] Ojielo CI, Cooke K, Mancuso P, et al. Defective phagocytosis and clearance of Pseudomonas aeruginosa in the lung following bone marrow transplantation. J Immunol 2003;171(8):4416-24.
[38] Cripps AW, Dunkley ML, Clancy RL, Kyd J. Vaccine strategies against Pseudomonas aeruginosa infection in the lung. Behring Inst Mitt 1997;98:262-8.
[39] Johansen HK, Cryz Jr SJ, Hougen HP, Moser C, Hoiby N. Vaccination promotes TH1-like inflammation and survival in chronic Pseudomonas aeruginosa pneumonia. A new prophylactic principle. Behring Inst Mitt 1997;98:269-73.
[40] Lei D, Lancaster Jr JR, Joshi MS, et al. Activation of alveolar macrophages and lung host defenses using transfer of the interferon-! gene. Am J Physiol 1997;272(5 Pt 1):L852-9.
[41] Hamaguchi Y, Xiu Y, Komura K, Nimmerjahn F, Tedder TF. Antibody isotype-specific engagement of Fc{gamma} receptors regulates B lymphocyte depletion during CD20 immunotherapy. J Exp Med 2006;203(3):743-53.
[42] Price BM, Barten Legutki J, Galloway DR, et al. Enhancement of the protective efficacy of an oprF DNA vaccine against Pseudomonas aeruginosa. FEMS Immunol Med Microbiol 2002;33(2):89-99.
[43] Song Z, Kharazmi A, Wu H, et al. Effects of ginseng treatment on neutrophil chemiluminescence and immunoglobulin G subclasses in a rat model of chronic Pseudomonas aeruginosa pneumonia. Clin Diagn Lab Immunol 1998;5(6):882-7.
【産業上の利用可能性】
【0079】
本発明の医薬組成物は、院内感染の原因菌である緑膿菌に対して効果的に感染防御能を誘導することができる。
【図面の簡単な説明】
【0080】
【図1】DNAワクチンは緑膿菌標的抗原に結合する特異的IgGを産生させた。PAO1、D4、及びPAKから調製された細胞溶解物、及び一価(pGACAG-OprF/I、pGACAG-PcrV又はpGACAG-PilA)又は多価(pGACAG-OprF/I+pGACAG-PcrV+pGACAG-PilA)DNAワクチンにより生じた血清を用いて、免疫ブロッティング分析を行った。OprF(36 kDa)、OprI(12 kDa)、PcrV(32 kDa)及びPilA(15 kDa)に相当するバンドを示す。3つの独立する実験から同じ結果が得られた。n.s.:非特異的バンド。
【図2】DNAワクチン接種により生じた緑膿菌抗原に対する血清IgG力値。100 μgのpGACAG(対照プラスミド)、pGACAG-OrrF/OprI、pGACAG-PcrV、pGACAG-PilA、又は33 μgの個々のプラスミド(pGACAG-OprF/I+pGACAG-PcrV+pGACAG-PilA)の混合物のいずれかでimEPTにより、マウスを2回免疫した(0及び4週)。あるいは、0.5 mgの金粒子にコ-ティングしたpGACAG-OprF/I+pGACAG-PcrV+pGACAG-PilA の混合物1 μgでGGにより、マウスを免疫した。血液は、最初の免疫化の後、0、2、4又は6週目に採取した。緑膿菌D4、rOprF/I、rPcrV、及びrPilAの全細胞に対する血清全IgG力値をELISAで測定した。グラフは、平均値±S.D.値(n=8匹のマウス/グル-プ)を示す。
【図3】DNAワクチンにより生じた緑膿菌抗原に対する血清IgG1及びIgG2a力値。マウスは図2で記述したように免疫した。rOprF/I、rPcrV、及びrPilAに対するIgG1又はIgG2a力値をELISAで測定した。グラフは、平均値±S.D.値(n=8匹のマウス/グル-プ)を示す。同様の結果が2つの独立な実験で得られた。** ANOVAボンフェロ-ニ多重比較検定によりp<0.01。
【図4】緑膿菌D4を鼻腔内投与した後にワクチン接種したマウスの肺ホモゲネ-ト及び血液中の生存細菌の数。マウスは図2で記述したように免疫した。感染後24時間で、マウスを屠殺し、希釈試料をLBプレ-トに広げて、(A)肺ホモゲネ-ト及び(B)血液中の全細菌数をカウントした。各LBプレ-ト上のコロニ-数を数えた後、CFUを算出した。グラフは、平均値±S.D.値(n=5匹のマウス/グル-プ)を示す。同様の結果が2つの独立な実験で得られた。* ANOVAボンフェロ-ニ多重比較検定によりp<0.05、** p<0.01。
【図5】緑膿菌D4を鼻腔内投与した後にワクチン接種したマウスの気管支肺胞洗浄(BAL)における好中球及びマクロファ-ジの数。マウスは図2で記述したように免疫した。5X105 CFUの細菌で感染させた後3及び6時間で、マウスを屠殺した後、BALを回収した。BAL中の細胞集団をギムザ染色によりキャラクタライズした。0.37 μm X 0.37 μmに広がる無作為抽出した10個の領域に存在する好中球及びマクロファ-ジの数を顕微鏡下でカウントしてから、全細胞数を算出した。グラフは、平均値±S.D.値(n=3匹のマウス/条件)を示す。同様の結果が2つの独立な実験で得られた。* ANOVAボンフェロ-ニ多重比較検定によりp<0.05、** p<0.01。
【図6】緑膿菌D4を鼻腔内投与した後にワクチン接種したマウスの気管支肺胞洗浄(BAL)細胞におけるマクロファ-ジ炎症性タンパク2(MIP-2)及びIFN-γ mRNAの生成。マウスは図2で記述したように免疫した。5 X 105 CFUの細菌で感染させた後2、12及び24時間で、マウスを屠殺した後、BALを回収した。全RNAを単離し、逆転写し、MIP-2、IFN-γ又は18S rRNAを標的とするリアルタイムPCRに各試料のアリコ-トをかけ、標的cDNAのレベルを定量した。18S rRNAのレベルでノ-マライズすることにより、相対的発現レベルを算出した。グラフは、平均値±S.D.値(n=3匹のマウス/条件)を示す。同様の結果が2つの独立な実験で得られた。* ANOVAボンフェロ-ニ多重比較検定によりp<0.05。
【図7】緑膿菌PAK又はD4を鼻腔内投与したマウスの生存率。マウスは図2で記述したように免疫した。2LD50の投与量の(A)PAK又は(B)D4(n=16-20匹のマウス/グル-プ)を鼻腔内に投与した後、生存率をモニタ-した。免疫したマウスの血清から精製したIgG画分(10 μg)を2LD50の投与量の(C)PAK又は(D)D4とともに37℃で30分間インキュベ-トした。その後、懸濁液を未処理マウスの鼻腔内に接種した(10匹のマウス/グル-プ)。生存率はその後10日間モニタ-した。同様の結果が2つの独立な実験で得られた。** Mantel-Cox log順位検定によりp<0.01。
【図8】プラスミドDNAワクチンの構成を示す。
【図9】抗原エピト-プを融合させたプラスミドDNAワクチンの構成を示す。
【図10】抗原エピトープ融合ワクチンにより誘導される多価抗体。抗原エピトープ融合ワクチンを筋注エレクトロポレーション法で免疫し(0、4週目、50μg/mouse)、血清(6週目)を採取した。リコンビナントOprF/I, PcrV, PilAをコーティングし、ELISA法で血清中のIgGを定量した。
【図11】抗原エピトープ融合ワクチンにより誘導される多価抗体。各菌株(PAO1, PAK, PA103, D4)をSDSサンプルバッファーで溶解し、SDS-PAGE法で分画し、PVDF膜に転写した。抗原エピトープ融合ワクチンで免疫したマウス血清と反応させ、抗マウスIgG抗体を用いてimmunoblottingを行った。
【図12】抗原エピト-プ融合ワクチンにより誘導される抗緑膿菌防御的免疫応答の評価。抗原エピト-プ融合ワクチン(Multiepitope)、至適化混合ワクチン(Mix)、空ベクター(Empty)を筋注エレクトロポレーション法で2回接種した後に2LD50量の緑膿菌(PAO1)を経鼻投与した。投与後のマウス生存率を示す。N = 10/グループ。
【図13】抗原エピト-プ融合ワクチンにより誘導される抗緑膿菌防御的免疫応答の評価。抗原エピト-プ融合ワクチン(Multiepitope)、至適化混合ワクチン(Mix)、空ベクター(Empty)を筋注エレクトロポレーション法で2回接種した後に2LD50量の緑膿菌(PAO1)を経鼻投与した。投与後12時間後と24時間後のマウス肺中の生存緑膿菌数を示す。N = 10/グループ。
【図14】抗原エピトープ融合ワクチンによる緑膿菌感染に対する防御的免疫応答の評価。抗原エピト-プ融合ワクチン(Multiepitope)、至適化混合ワクチン(Mix)、空ベクター(Empty)を筋注エレクトロポレーション法で2回接種した。6週目に、飲料水中に緑膿菌PAO1株を入れ(1 x 107/ml)、アンピシリン(1回/日、200 mg/kg)を3日間腹腔内に投与した。4日目に無菌の飲料水に交換して飼育し、4日目、6日目に1回ずつシクロフォスファミド(200 mg/kg)を腹腔内に投与して腸管感染由来敗血症モデル(易感染モデル)を作製した。図は7日以降の生存率を示す。N = 10/グループ。
【図15】抗原エピトープ融合ワクチンにより誘導された抗体の緑膿菌感染に対する防御能の評価。抗原エピト-プ融合ワクチン(Multiepitope)、至適化混合ワクチン(Mix)、空ベクター(Empty)を筋注エレクトロポレーション法で2回接種した。6週目に、採血し、IgG分画を精製した。15μgのIgGと2LD50のPAO1またはPAKを37℃で30分間反応させた後、ナイーブマウスに経鼻投与した。投与後の生存率を示す。N = 10/グループ。
【配列表フリーテキスト】
【0081】
<配列番号1>
配列番号1は、緑膿菌(PA01)由来の細胞外膜タンパクOprF190-342の塩基配列を示す。
<配列番号2>
配列番号2は、緑膿菌(PA01)由来の細胞外膜タンパクOprF190-342のアミノ酸配列を示す。
<配列番号3>
配列番号3は、緑膿菌(PA01)由来の細胞外膜タンパクOprI21-83の塩基配列を示す。
<配列番号4>
配列番号4は、緑膿菌(PA01)由来の細胞外膜タンパクOprI21-83のアミノ酸配列を示す。
<配列番号5>
配列番号5は、緑膿菌(PA01)由来のIII型分泌機構制御タンパクPcrV1-293(全長)(全長)の塩基配列を示す。
<配列番号6>
配列番号6は、緑膿菌(PA01)由来のIII型分泌機構制御タンパクPcrV1-293のアミノ酸配列を示す。
<配列番号7>
配列番号7は、緑膿菌(PA01)由来の毛状付属器タンパクPilA 136-149の塩基配列を示す。
<配列番号8>
配列番号9は、緑膿菌(PA01)由来の毛状付属器タンパクPilA 136-149のアミノ酸配列を示す。
<配列番号9>
配列番号9は、緑膿菌(PA01)由来の細胞外膜タンパクOprF全長の塩基配列を示す。
<配列番号10>
配列番号10は、緑膿菌(PA01)由来の細胞外膜タンパクOprF全長のアミノ酸配列を示す。
<配列番号11>
配列番号11は、緑膿菌(PA01)由来の細胞外膜タンパクOprI全長の塩基配列を示す。
<配列番号12>
配列番号12は、緑膿菌(PA01)由来の細胞外膜タンパクOprI全長のアミノ酸配列を示す。
<配列番号13>
配列番号13は、緑膿菌(PA01)由来の毛状付属器タンパクPilA全長(PilA 1-149)の塩基配列を示す。
<配列番号14>
配列番号14は、緑膿菌(PA01)由来の毛状付属器タンパクPilA全長(PilA 1-149)のアミノ酸配列を示す。
<配列番号15>
配列番号15は、oprFセンスプライマ-のDNA配列を示す。
<配列番号16>
配列番号16は、oprFアンチセンスプライマ-のDNA配列を示す。
<配列番号17>
配列番号17は、oprIセンスプライマ-のDNA配列を示す。
<配列番号18>
配列番号18は、oprIアンチセンスプライマ-のDNA配列を示す。
<配列番号19>
配列番号19は、pcrVセンスプライマ-のDNA配列を示す。
<配列番号20>
配列番号20は、pcrVアンチセンスプライマ-のDNA配列を示す。
<配列番号21>
配列番号21は、pilAセンスプライマ-のDNA配列を示す。
<配列番号22>
配列番号22は、pilAアンチセンスプライマ-のDNA配列を示す。
【特許請求の範囲】
【請求項1】
下記(i)〜(v)のいずれかの成分を含む、緑膿菌に対して感染防御能を誘導できる医薬組成物。
(i) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部をコ-ドするDNA、緑膿菌の細胞外膜タンパクOprIの全長又はその一部をコ-ドするDNA、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部をコ-ドするDNA、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部をコ-ドするDNAをそれぞれ含むベクタ-の混合物
(ii) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、III型分泌機構制御タンパクPcrVの全長又はその一部、及び毛状付属器タンパクPilAの全長又はその一部の融合物をコ-ドするDNAを含むベクタ-
(iii) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部の混合物
(iv) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部の融合物
(v) (iii)の混合物又は(iv)の融合物に対して誘導された抗体
【請求項2】
緑膿菌の細胞外膜タンパクOprFの全長又はその一部が下記(A)又は(B)のタンパク質であり、緑膿菌の細胞外膜タンパクOprIの全長又はその一部が下記(C)又は(D)のペプチドであり、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部が下記(E)又は(F)のタンパク質であり、緑膿菌の毛状付属器タンパクPilAの全長又はその一部が下記(G)又は(H)のペプチドである請求項1記載の医薬組成物。
(A)配列番号2のアミノ酸配列からなるタンパク質
(B)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の細胞外膜タンパクOprFの抗原活性を有するタンパク質
(C)配列番号4のアミノ酸配列からなるペプチド
(D)配列番号4のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の細胞外膜タンパクOprIの抗原活性を有するペプチド
(E)配列番号6のアミノ酸配列からなるタンパク質
(F)配列番号6のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌のIII型分泌機構制御タンパクPcrVの抗原活性を有するタンパク質
(G)配列番号8のアミノ酸配列からなるペプチド
(H)配列番号8のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の毛状付属器タンパクPilAの抗原活性を有するペプチド
【請求項3】
緑膿菌の細胞外膜タンパクOprFの全長又はその一部をコ-ドするDNAが下記(a)又は(b)のDNAであり、緑膿菌の細胞外膜タンパクOprIの全長又はその一部をコ-ドするDNAが下記(c)又は(d)のDNAであり、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部をコ-ドするDNAが下記(e)又は(f)のDNAであり、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部をコ-ドするDNAが下記(g)又は(h)のDNAである請求項1又は2記載の医薬組成物。
(a)配列番号1のヌクレオチド配列で表されるDNA
(b)配列番号1のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の細胞外膜タンパクOprFの抗原活性を有するタンパク質をコ-ドするDNA
(c) 配列番号3のヌクレオチド配列で表されるDNA
(d) 配列番号3のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の細胞外膜タンパクOprIの抗原活性を有するペプチドをコ-ドするDNA
(e)配列番号5のヌクレオチド配列で表されるDNA
(f)配列番号5のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌のIII型分泌機構制御タンパクPcrVの抗原活性を有するタンパク質をコ-ドするDNA
(g)配列番号7のヌクレオチド配列で表されるDNA
(h)配列番号7のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の毛状付属器タンパクPilAの抗原活性を有するタンパク質をコ-ドするDNA
【請求項4】
(i)〜(iv)のいずれかの成分を含むワクチンである請求項1〜3のいずれかに記載の医薬組成物。
【請求項5】
(i)のベクタ-混合物又は(ii)のベクタ-を含む遺伝子ワクチンである請求項4記載の医薬組成物。
【請求項6】
ベクタ-がプラスミド又はウイルスベクタ-である請求項5記載の医薬組成物。
【請求項7】
下記(I)の混合物又は(II)の融合物に対して誘導された抗体。
(I) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部の混合物
(II) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部の融合物
【請求項1】
下記(i)〜(v)のいずれかの成分を含む、緑膿菌に対して感染防御能を誘導できる医薬組成物。
(i) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部をコ-ドするDNA、緑膿菌の細胞外膜タンパクOprIの全長又はその一部をコ-ドするDNA、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部をコ-ドするDNA、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部をコ-ドするDNAをそれぞれ含むベクタ-の混合物
(ii) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、III型分泌機構制御タンパクPcrVの全長又はその一部、及び毛状付属器タンパクPilAの全長又はその一部の融合物をコ-ドするDNAを含むベクタ-
(iii) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部の混合物
(iv) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部の融合物
(v) (iii)の混合物又は(iv)の融合物に対して誘導された抗体
【請求項2】
緑膿菌の細胞外膜タンパクOprFの全長又はその一部が下記(A)又は(B)のタンパク質であり、緑膿菌の細胞外膜タンパクOprIの全長又はその一部が下記(C)又は(D)のペプチドであり、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部が下記(E)又は(F)のタンパク質であり、緑膿菌の毛状付属器タンパクPilAの全長又はその一部が下記(G)又は(H)のペプチドである請求項1記載の医薬組成物。
(A)配列番号2のアミノ酸配列からなるタンパク質
(B)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の細胞外膜タンパクOprFの抗原活性を有するタンパク質
(C)配列番号4のアミノ酸配列からなるペプチド
(D)配列番号4のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の細胞外膜タンパクOprIの抗原活性を有するペプチド
(E)配列番号6のアミノ酸配列からなるタンパク質
(F)配列番号6のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌のIII型分泌機構制御タンパクPcrVの抗原活性を有するタンパク質
(G)配列番号8のアミノ酸配列からなるペプチド
(H)配列番号8のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ緑膿菌の毛状付属器タンパクPilAの抗原活性を有するペプチド
【請求項3】
緑膿菌の細胞外膜タンパクOprFの全長又はその一部をコ-ドするDNAが下記(a)又は(b)のDNAであり、緑膿菌の細胞外膜タンパクOprIの全長又はその一部をコ-ドするDNAが下記(c)又は(d)のDNAであり、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部をコ-ドするDNAが下記(e)又は(f)のDNAであり、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部をコ-ドするDNAが下記(g)又は(h)のDNAである請求項1又は2記載の医薬組成物。
(a)配列番号1のヌクレオチド配列で表されるDNA
(b)配列番号1のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の細胞外膜タンパクOprFの抗原活性を有するタンパク質をコ-ドするDNA
(c) 配列番号3のヌクレオチド配列で表されるDNA
(d) 配列番号3のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の細胞外膜タンパクOprIの抗原活性を有するペプチドをコ-ドするDNA
(e)配列番号5のヌクレオチド配列で表されるDNA
(f)配列番号5のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌のIII型分泌機構制御タンパクPcrVの抗原活性を有するタンパク質をコ-ドするDNA
(g)配列番号7のヌクレオチド配列で表されるDNA
(h)配列番号7のヌクレオチド配列で表されるDNAに相補的なDNAとストリンジェントな条件下でハイブリダイズし、かつ緑膿菌の毛状付属器タンパクPilAの抗原活性を有するタンパク質をコ-ドするDNA
【請求項4】
(i)〜(iv)のいずれかの成分を含むワクチンである請求項1〜3のいずれかに記載の医薬組成物。
【請求項5】
(i)のベクタ-混合物又は(ii)のベクタ-を含む遺伝子ワクチンである請求項4記載の医薬組成物。
【請求項6】
ベクタ-がプラスミド又はウイルスベクタ-である請求項5記載の医薬組成物。
【請求項7】
下記(I)の混合物又は(II)の融合物に対して誘導された抗体。
(I) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部の混合物
(II) 緑膿菌の細胞外膜タンパクOprFの全長又はその一部、緑膿菌の細胞外膜タンパクOprIの全長又はその一部、緑膿菌のIII型分泌機構制御タンパクPcrVの全長又はその一部、及び緑膿菌の毛状付属器タンパクPilAの全長又はその一部の融合物
【図2】


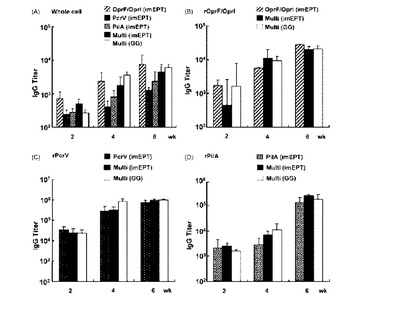
【図3】
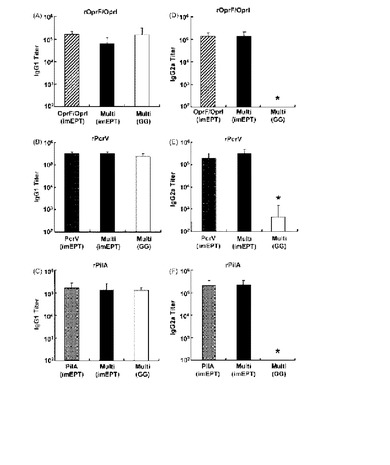
【図4】
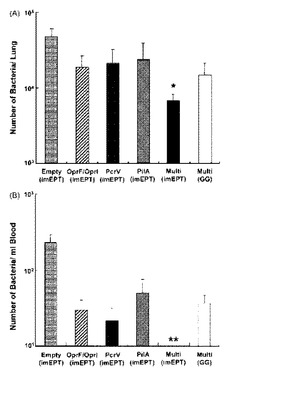
【図5】
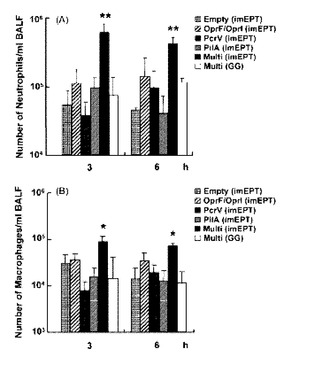
【図6】
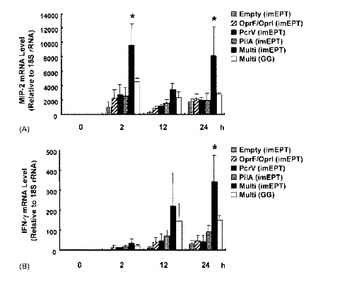
【図8】

【図9】
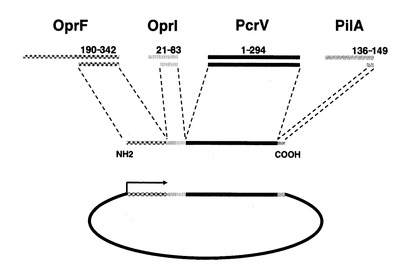
【図10】
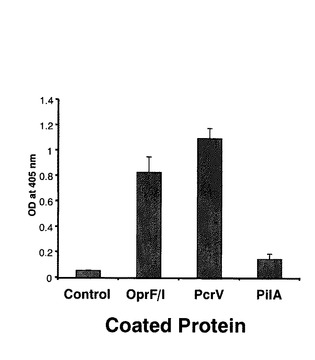
【図11】
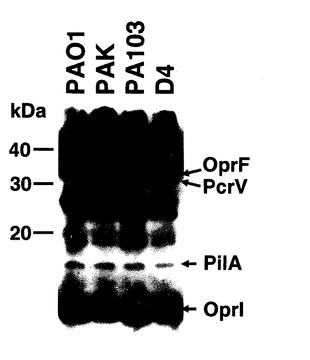
【図12】
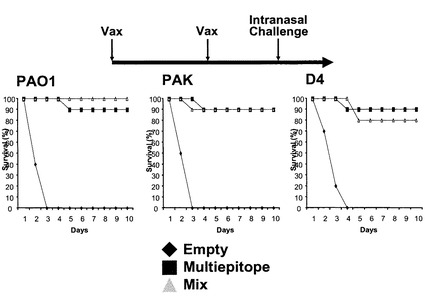
【図13】
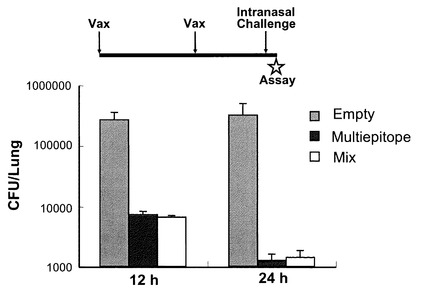
【図14】
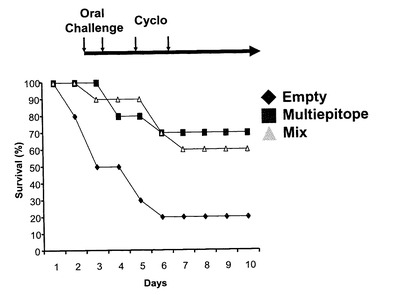
【図15】
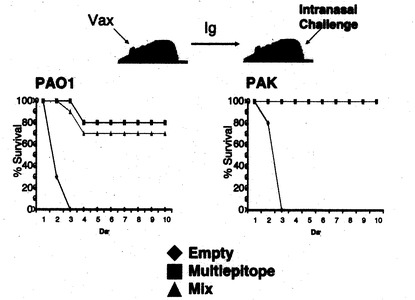
【図1】
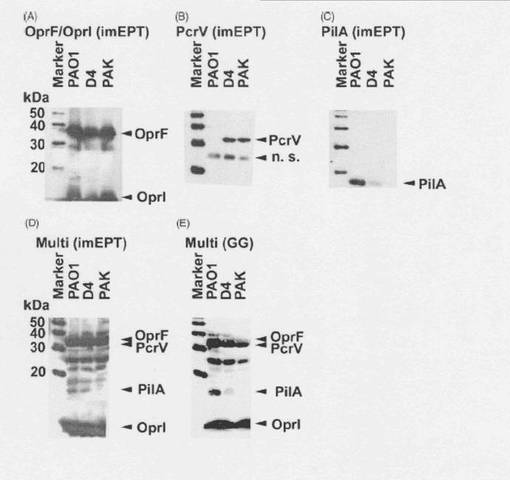
【図7】
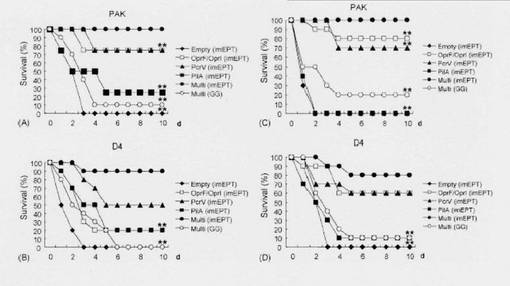
【図3】
【図4】
【図5】
【図6】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図1】
【図7】
【公開番号】特開2008−133206(P2008−133206A)
【公開日】平成20年6月12日(2008.6.12)
【国際特許分類】
【出願番号】特願2006−319535(P2006−319535)
【出願日】平成18年11月28日(2006.11.28)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 2006年6月9日 Elsevier Ltd.(Web公開)の「Vaccine 第24巻 2006年」に発表
【出願人】(505155528)公立大学法人横浜市立大学 (101)
【Fターム(参考)】
【公開日】平成20年6月12日(2008.6.12)
【国際特許分類】
【出願日】平成18年11月28日(2006.11.28)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 2006年6月9日 Elsevier Ltd.(Web公開)の「Vaccine 第24巻 2006年」に発表
【出願人】(505155528)公立大学法人横浜市立大学 (101)
【Fターム(参考)】
[ Back to top ]