
線維性状態およびその他の疾患の治療のための、オリゴリボヌクレオチドおよびその使用方法
本発明は、転写後レベルにてヒトTGaseII遺伝子の発現をダウンレギュレートする二本鎖化合物、好ましくはオリゴリボヌクレオチド(siRNA)に関する。また、本発明は、化合物またはオリゴリボヌクレオチド化合物を発現することができるベクターと薬学的に許容されるキャリアとを含む薬学的組成物に関する。また、本発明は、肺、腎臓および肝臓線維症などの線維性疾患または眼瘢痕に罹患した患者を治療する方法であって、これにより患者を治療するために、患者に対して治療的に有効な用量の薬学的組成物を投与することを含む方法を提供する。また、本発明は、TGaseIIポリペプチドに対する抗体を使用することによる線維性疾患およびその他の疾患の治療に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本出願は、2004年1月30日に出願された米国仮特許出願第60/540,687号および2005年1月4日に出願された第60/641,522号の優先権を主張し、両方とも、参照によりこれらの全体が本明細書に援用される。
【0002】
本出願の全体を通じて、種々の特許および科学的刊行物を引用してある。これらの刊行物の開示は、本発明が属する技術の水準をより完全に記述するために、その全体が参照により本明細書によって本出願に援用される。
【背景技術】
【0003】
siRNAおよびRNA干渉
RNA干渉(RNAi)は、二本鎖(ds)RNA依存的な遺伝子特異的な転写後サイレンシングを含む現象である。元来、この現象を研究して、実験的に哺乳動物細胞を操る試みは、長鎖dsRNA分子に応答して活性化された活性な非特異的抗ウイルス防衛機構によって頓挫していた;Gil et al. 2000、Apoptosis、5:107-114を参照されたい。後に、21ヌクレオチドRNAの合成二重鎖が、一般的な抗ウイルス防衛機構を刺激することなく哺乳動物細胞の遺伝子特異的RNAiを媒介することができることが発見された。Elbashir et al. Nature 2001、411:494-498およびCaplen et al. Proc Natl Acad Sci 2001、98:9742-9747を参照されたい。その結果、短い二重鎖RNAである短鎖干渉RNA(siRNA)は、遺伝子機能を理解することを試みる際の強力なツールとなった。
【0004】
したがって、RNA干渉(RNAi)は、短鎖干渉RNA(siRNA)(Fire et al, 1998, Nature 391, 806)またはミクロRNA(miRNA)(Ambros V. Nature 431:7006,350-355(2004);およびBartel DP. Cell. 2004 an 23; 116(2): 281-97 MicroRNAs: genomics, biogenesis, mechanism, and function)によって媒介される、哺乳類の配列特異的な転写後遺伝子サイレンシングの過程をいう。植物の対応する過程は、一般に特異的な転写後遺伝子サイレンシングまたはRNAサイレンシングといわれ、真菌では、クエリングともいわれる。siRNAは、その内因性の(細胞の)対応物の遺伝子/mRNAの発現をダウンレギュレートし、またはサイレンスする(防げる)二重鎖RNA分子である。RNA干渉は、dsRNA種が特定のタンパク質複合体に入り、次いでこれが相補的な細胞RNAにターゲットされて、これを特異的に分解する能力に基づいている。したがって、RNA干渉反応は、一般にRNA誘導サイレンシング複合体(RISC)と称される、siRNA二重鎖のアンチセンス鎖に対して相補的な配列を有する一本鎖RNAの切断を媒介するsiRNAを含むエンドヌクレアーゼ複合体を特徴とする。標的RNAの切断は、siRNA二重鎖のアンチセンス鎖に対して相補的な領域の中央で生じ得る(Elbashir et al 2001、Genes Dev., 15、188)。さらに詳細には、より長いdsRNAが、III型RNAses(DICER、DROSHA、その他、Bernstein et al., Nature、2001、v.409、p.363-6;Lee et al., Nature、2003、425、p.415-9)によって短い(17〜29bp)dsRNA断片(短鎖阻害RNA−「siRNA」とも称される)に消化される。RISCタンパク質複合体は、これらの断片および相補的mRNAを認識する。標的mRNAのエンドヌクレアーゼ切断により、全ての過程が完了する(McManus&Sharp、Nature Rev Genet、2002、v.3、p.737-47;Paddison &Hannon、Curr Opinmol Ther. 2003の6月;5(3):217-24)。これらの用語および提唱されたメカニズムについての情報に関しては、Bernstein E., Denli AM. Hannon GJ: 2001 The rest is silence. RNA. I; 7(11): 1509-21; Nishikura K.: 2001 A short primer on RNAi: RNA-directed RNA polymerase acts as a key catalyst. Cell. I 16; 107(4): 415-8 およびPCT公開WO01/36646号(Glover et al)を参照されたい。
【0005】
既知の遺伝子に対応するsiRNAの選択および合成は、広く報告されてきた;たとえば、Chalk AM, Wahlestedt C, Sonnhammer EL. 2004 Improved and automated prediction of effective siRNA Biochem. Biophys. Res. Commun. Jun 18; 319(1): 264-74; Sioud M, Leirdal M., 2004, Potential design rules and enzymatic synthesis of siRNAs, Methods Mol Biol.; 252:457-69; Levenkova N, Gu Q, Rux JJ. 2004 ,Gene specific siRNA selector Bioinformatics. I 12; 20(3): 430-2. and Ui-Tei K, Naito Y, Takahashi F, Haraguchi T, Ohki-Hamazaki H, Juni A, Ueda R, Saigo K., Guidelines for the selection of highly effective siRNA sequences for mammalian and chick RNA interference Nucleic Acids Res. 2004 I 9;32(3):936-48.Se also Liu Y, Braasch DA, Nulf CJ, Corey DR. Efficient and isoform-selective inhibition of cellular gene expression by peptide nucleic acids, Biochemistry, 2004 I 24;43(7):1921-7を参照されたい。また、修飾された/より安定なsiRNAの産生については、PCT公開WO2004/015107号(Atugen)およびWO02/44321号(Tuschl et al)、並びにChiu YL, Rana TM. siRNA function in RNAi: a chemical modification analysis, RNA 2003 Sep;9(9):1034-48およびI 特許第5898031号および6107094号(Crooke)も参照されたい。
【0006】
いくつかのグループは、細胞内でsiRNAを産生することができるDNAに基づいたベクターの開発を記述した。該方法は、一般に、細胞内で効率的にプロセスされてsiRNAを形成する短いヘアピンRNAの転写を含む。Paddison et al. PNAS 2002, 99:1443-1448; Paddison et al. Genes & Dev 2002, 16:948-958; Sui et al. PNAS 2002, 8:5515-5520; and Brummelkamp et al. Science 2002, 296:550-553。これらの報告は、多数の内因的および外因的に発現された遺伝子を特異的にターゲットすることができるsiRNAを作製するための方法を記述する。
【0007】
siRNAは、最近、霊長類における阻害のために首尾よく使用されている;詳しくは、Tolentino et al., Retina 24(1) February 2004 I 132-138を参照されたい。
【0008】
トランスグルタミナーゼ(TGase)ファミリー
トランスグルタミナーゼ(EC 2.3.2.13)は、イプシロン-γグルタミルリジンイソペプチド結合によるタンパク質の架橋を触媒する酵素のファミリーである。本ファミリーは、9つの異なる酵素を含み、その中には、第XIIIa因子(血漿トランスグルタミナーゼ)、ケラチノサイトトランスグルタミナーゼ(TGaseI)、表皮トランスグルタミナーゼ(TGaseIII)、前立腺トランスグルタミナーゼ(TGaseIV)および組織型トランスグルタミナーゼ(TGaseII)がある。これらの酵素の全体的な一次構造は異なるが、これらは全て、活性部位にて共通のアミノ酸配列(Y-G-Q-CW)と、これらの活性のための厳密なカルシウム依存性とを共有する(Lesort M, Tucholski J, Miller ML, Johnson GV, Tissue transglutaminase: a possible role in neurodegenerative diseases. Prog Neurobiol. 2000 Aug; 61(5):439-63)。
【0009】
トランスグルタミナーゼII
トランスグルタミナーゼII(TGaseII)(タンパク質-グルタミンγ-グルタミルトランスフェラーゼ、TGase C、TGC、TG(C)および組織型トランスグルタミナーゼとしても知られる)は、二重酵素活性をもつ独特の多機能酵素である:
1)本酵素は、架橋活性をもつTGaseタンパク質(Ca2+で活性化され、GTPで不活性化される)として作用する(すなわち、これは、タンパク質架橋および/または生体アミンの共有結合性の組み込みを生じる反応を触媒する)。さらに、TGaseは、共有結合性のグルタミルリジル結合(タンパク質分解および変性剤に高度に耐性であり、任意の既知の脊椎動物のエンドペプチダーゼによって崩壊することができない独特のイソペプチド結合)の形成を触媒する。
【0010】
2)また本酵素は、α1Bおよびα1dアドレナリン作動性受容体からの、TPαトロンボキサンA2受容体からの、およびオキシトシン受容体からホスホリパーゼCδ1への、活性化シグナルを伝達するGTP結合タンパク質として作用する(すなわち、これは、イノシトールリン酸産生、カルシウム動員などを活性化する)。α1Dアドレナリン作動性受容体およびPLCは、両方ともトランスグルタミナーゼIIのためにグアニンヌクレオチド交換因子として作用することが示された(Baek KJ, Kang S, Damron D, Im M, Phospholipase C-delta1 is a guanine nucleotide exchanging factor for transglutaminase II (G alpha h) and promotes alpha 1B-adrenoreceptor-mediated GTP binding and intracellular calcium release. J Biol Chem. 2001 Feb 23;276(8):5591-7)。
【0011】
この酵素の二つの機能は、分離しており、また活性部位は、異なる位置に位置しており(Im MJ, Russell MA, Feng JF, Transglutaminase II: a new class of GTP-binding protein with new biological functions. Cell Signal. 1997 Nov;9(7):477-82)、TGaseIIの架橋活性の阻害は、そのGタンパク質機能を妨げることなく達成することができる。
【0012】
TGaseII:細胞下局在および線維症関連細胞内シグナリング活性
TGaseIIは、一様に発現されており、ECMと会合して細胞内にて膜結合型として、およびサイトゾルタンパク質として見いだすことができる。GTP-結合活性は、TGaseの膜画分でより高く、架橋活性は、サイトゾルおよび細胞外画分でより高い。細胞表面上において、TGaseは、その42kDaのゼラチン結合ドメインを経てフィブロネクチンと結合する。コラーゲンとフィブロネクチンの架橋により、ECMの構造安定化に寄与し、マトリックスメタロプロテイナーゼ(mettaloproteinases)によるタンパク質分解に耐性のマトリックスになる。したがって、TGaseIIは、細胞外タンパク質を沈殿の方に有利にして、前記タンパク質の破壊を阻害することによって、組織線維症の形成の中心的現象であるECMの蓄積に寄与する。
【0013】
加えて、TGaseは、この前線維性因子のその後の適当なプロセシングのために必要とされる工程であるECMに対する潜在性TGF-βの結合を媒介し、最終的に、活性なTGF-βを放出することとなる(Le et al., Connect Tissue Res. 2001;42(4):245-53. Rosenthal et al., Arthritis Rheum. 2000 Aug;43(8):1729-33)。
【0014】
細胞質において、TGaseIIは、細胞骨格再配列、細胞形態制御および分化において重要な役割を有することが既知であるrasファミリーの小Gタンパク質であるRhoAを活性化する。TGaseIIの活性化により、RhoAのアミド基転移の増大を生じ、次いで構成的に活性なGタンパク質として機能して、その下流の標的ROCK-2(Rho関連キナーゼ)の結合の増大、およびその活性化を示す(Singh et al. EMBO J. 2001 May 15;20(10):2413-23)。特異的ROCKキナーゼ阻害剤のY-27632は、腎線維症のUUOモデルにおいて、並びに肝臓または肺線維症のモデルにおいて、抗線維性効果を示した(Nagatoya K, Moriyama T, Kawada N, Takeji M, Oseto S, Murozono T, Ando A, Imai E, Hori M. Y-27632 was found to prevent tubulointerstitial fibrosis in mouse kidneys with unilateral urethral obstruction. Kidney Int. 2002 May;61(5):1684-95; Murata T, Arii S, Nakamura T, Mori A, Kaido T, Furuyama H, Furumoto K, Nakao T, Isobe N, Imamura M, Inhibitory effect of Y-27632, a ROCK inhibitor, on progression of rat liver fibrosis in association with inactivation of hepatic stellate cells. J Hepatol. 2001 Oct;35(4):474-81; Shimizu Y, Dobashi K, Iizuka K, Horie T, Suzuki K, Tukagoshi H, Nakazawa T, Nakazato Y, Mori M, Contribution of small GTPase Rho and its target protein ROCK in a murine model of lung fibrosis. Am J Respir Crit Care Med. 2001 Jan;163(1):210-7)。最近、十分に実証された前線維性活性をもつ転写因子であるNFκBの活性化に寄与するTGaseIIの独特の機能が記述された(Baud et al., J Soc Biol. 2002;196(4):269-73, Lee at al. J Biol Chem. 2004 Dec 17;279(51):53725-35)、
線維性疾患
線維性疾患は全て、細胞外基質の繊維状物質の過剰な堆積によって特徴づけられ、これが組織構築における異常な変化に寄与し、正常な器官機能を妨げる。残念なことに、線維症は、広く一般的で、衰弱して生命が脅かされることが多いが、現在、有効な治療を提供されていない。
【0015】
外傷による損傷を受けた全ての組織は、創傷治癒プログラムの開始によって応答する。創傷治癒応答の正常な自己制御過程が妨げられると、過剰の瘢痕によって特徴づけられる障害の一種である線維症が生じ、コラーゲンの過剰産生および堆積を生じさせる。その結果、正常な器官組織は、瘢痕組織に置き換わり、これが最終的に器官の機能不全に至る。
【0016】
線維症は、多様な原因によって、および種々の器官において開始する可能性がある。肝硬変、肺線維症、サルコイドーシス、ケロイドおよび腎臓線維症は、全て進行性線維症と関連する慢性状態であり、これにより正常組織機能の持続的な喪失が生じる。
【0017】
急性線維症(通常、突然および重篤で、かつ短期間に発症する)は、偶発性傷害(特に、脊椎および中枢神経系に対する傷害)、感染、外科手術、虚血性の疾病(たとえば、心臓麻痺後の心臓瘢痕)、やけど、環境汚染物、アルコールおよびその他のタイプの毒素、急性呼吸不全症候群、放射線、並びに化学療法治療を含む外傷の種々の形態に共通の応答として生じる。
【0018】
線維症の種々のタイプについてのさらなる情報は、Molina V, Blank M, Shoenfeld Y. (2002), “Fibrotic diseases”, Harefuah, 141(11): 973-8, 1009; Yu L, Noble NA, Border WA (2002), “Therapeutic strategies to halt renal fibrosis", Curr Opin Pharmacol. 2(2):177-81; Keane WF, Lyle PA. (2003), "Recent advances in management of type 2 diabetes and nephropathy: lessons from the RENAAL study", Am J Kidney Dis. 41(3 Suppl 2): S22-5; Bohle A, Kressel G, Muller CA, Muller GA. (1989), "The pathogenesis of chronic renal failure", Pathol Res Pract. 185(4):421-40; Kikkawa R, Togawa M, Isono M, Isshiki K, Haneda M. (1997), "Mechanism of the progression of diabetic nephropathy to renal failure", Kidney Int Suppl. 62:S39-40; Bataller R, Brenner DA. (2001), "Hepatic stellate cells as a target for the treatment of liver fibrosis", Semin Liver Dis. 21(3):437-51; Gross TJ, Hunninghake GW, (2001) "Idiopathic pulmonary fibrosis", N Engl J Med. 345(7):517-25; Frohlich ED. (2001) "Fibrosis and ischemia: the real risks in hypertensive heart disease", Am J Hypertens;14(6 Pt 2):194S-199Sを参照されたい。
【0019】
肝臓線維症
肝臓線維症(LF)は、一般に回復不能ないくつかの病因の肝臓損傷の結果である。西洋世界では、主な病因カテゴリーは:アルコール性肝疾患(30〜50%)、ウイルス性肝炎(30%)、胆汁疾患(5〜10%)、原発性血色素症(5%)および病因が未知の薬物関連および原因不明の肝硬変(10〜15%)。また、ウイルソン病、α1-アンチトリプシン欠損およびその他のまれな疾患では、症候Scheinberg IH、Sternlieb I、ウイルソン病および特発性銅中毒症のうちの1つとして肝臓線維症を有する。Am J Clin Nutr 1996 May;63(5):842S-5S; Parfrey H, Mahadeva R, Lomas DA., Alpha(1)-antitrypsin deficiency, liver disease and emphysema. Int J Biochem Cell Biol. 2003 Jul;35(7):1009-14。
【0020】
肝臓線維症の最終段階である肝硬変では、頻繁な肝移植が必要であり、西洋世界における上位10位の死因の一つである。
【0021】
肝臓星状細胞の活性化の阻害、肝細胞の増殖刺激およびコラーゲンの翻訳後修飾の阻害を生じさせる抗炎症薬は、全て肝臓線維症を治療するために使用されてきた。しかし、選択的ターゲティングが不十分であるために、とりわけこれらの治療では、重篤な副作用という欠点を被ってしまう。
【0022】
詳細は、Friedman SL. (2003), “Liver fibrosis - from bench to bedside”, J Hepatol. 38 Suppl 1:S38-53; Albanis E, Safadi R, Friedman SL. (2003), "Treatment of hepatic fibrosis: almost there", Curr Gastroenterol Rep. 5(1):48-56. See also Grenard P, Bresson-Hadni S, El Alaoui S, Chevallier M, Vuitton DA, Ricard-Blum S., Transglutaminase-mediated cross-linking is involved in the stabilization of extracellular matrix in human liver fibrosis. J Hepatol. 2001 Sep;35(3):367-75 ;Mirza A, Liu SL, Frizell E, Zhu J, Maddukuri S, Martinez J, Davies P, Schwarting R, Norton P, Zern MA., A role for tissue transglutaminase in hepatic injury and fibrogenesis, and its regulation by NF-kappaB. Am J Physiol. 1997 Feb;272(2 Pt 1):G281-8)を参照されたい。
【0023】
腎臓線維症および関連状態
慢性腎不全(CRF)
慢性腎不全は、腎臓が排泄物を排出し、尿を濃縮し、および電解質を保存する能力が徐々に、かつ進行性に失われる。CRFは、ゆっくりと進行する。これは、多くの場合、腎機能の段階的な喪失を生じさせる何らかの疾患によって生じ、線維症は、CRFを生じる主な病態である。
【0024】
糖尿病性腎障害
糖尿病性腎障害(その特徴は、糸球体硬化症および尿細管間質性線維症である)は、現代世界において、唯一末期腎疾患に多い原因であり、糖尿病患者は、透析を行う最大集団を構成する。このような療法は、費用がかかり、決して最適とはいえない。移植は、より優れた結果をもたらすが、ドナーの極度な不足に悩まされている。糖尿病性腎障害に対する(並びに、腎病態のその他のタイプに対する)よりターゲットされた療法は、これらの病態の基礎をなす分子機構の大部分が未知であるために、開発されていない。疾患において調整され、かつ糖尿病腎症の結果の重症度に影響を及ぼす必須な機能的標的遺伝子の同定は、診断的、並びに治療的に高い価値を有する。
【0025】
腎臓病態の起源
腎臓における多くの病理学的過程(たとえば糸球体腎炎、物理的な閉塞、中毒性傷害、代謝性および免疫学的疾患)は、最終的に同様の、または同一の形態学的変化、すなわち糸球体硬化症および尿細管間質性線維症となる。したがって、異なるタイプの傷害により、同じ単一の遺伝プログラムに収束して2つの線維症の特徴(線維芽細胞の増殖およびこれらによる結合組織の種々のタンパク質成分の過剰産生)を生じる。加えて、糸球体における基底膜の肥厚は、間質性線維症を伴い、結果的に糸球体硬化症になる。また、Johnson TS, Skill NJ, El Nahas AM, Oldroyd SD, Thomas GL, Douthwaite JA, Haylor JL, Griffin M, Transglutaminase transcription and antigen translocation in experimental renal scarring. J Am Soc Nephrol. 1999 Oct;10(10):2146-57; Johnson TS, Griffin M, Thomas GL, Skill J, Cox A, Yang B, Nicholas B, Birckbichler PJ, Muchaneta-Kubara C, Meguid El Nahas A, The role of transglutaminase in the rat subtotal nephrectomy model of renal fibrosis. J Clin Invest. 1997 Jun 15;99(12):2950-60)を参照されたい。
【0026】
肺線維症
間質性肺線維症(IPF)は、ミネラル粒子、有機性粉末およびオキシダントガスを含む種々の吸入によって、または未知の原因によって(特発性肺線維症)生じる肺の瘢痕である。本疾患は、世界的に何百万もの個体を苦しめ、有効な治療手段はない。有用な治療法がない主な理由は、疾患の分子機構が、療法のための適切な標的をデザインするほど十分に定義されていないことである(Lasky JA., Brody AR. (2000), “Interstitial fibrosis and growth factors”, Environ Health Perspect.;108 Suppl 4:751-62)。
【0027】
肺線維症の病原の細胞基質は、内皮および上皮細胞障害、炎症細胞およびこれらのメディエータの産生、並びに線維芽細胞活性化を含む。線維症は、種々のサイトカイン、ケモカインおよび成長因子によって媒介される炎症性細胞と構造細胞との間のクロストーク(これは、組織恒常性維持の役割を担い、かつ傷害に対する応答を調整する)の調節不全に関連があると考えられている(Kelly M, Kolb M, Bonniaud P, Gauldie J. (2003), “Re-evaluation of fibrogenic cytokines in lung fibrosis”, Curr Pharm Des. 9(1):39-49 )。
【0028】
糖質コルチコイドまたは細胞毒からなる従来の療法は、通常、疾患の進行の予防には効果がない。肺線維症に対する有効な治療の開発のためには、内皮細胞および上皮細胞傷害、炎症反応、線維芽細胞増殖、コラーゲン沈着および肺修復の分子機構をさらに理解することが必要であると考えられている(Kuwano K, Hagimoto N, Hara N. (2001), “Molecular mechanisms of pulmonary fibrosis and current treatment”, Curr Mol Med. 1(5):551-73)。また、Griffin M, Smith LL, Wynne J., Changes in transglutaminase activity in an experimental model of pulmonary fibrosis induced by Paraquat. Br. J. Exp. Pathol. 1979 Dec;60(6):653-61 )も参照されたい。
【0029】
心臓線維症
心不全は、それ単独での有病率が増大しているものの、一方でその他の状態は、著明に減少してきたという点で、主要な心血管障害の中では独特である。この一部には、米国および欧州の人口の老化に起因すると考えることができる。また、これらの患者は、心臓の有害な再造形によって左心室機能不全の進行を発症する可能性があるので、心筋障害をもつ患者を救う能力が主要な要因となる。
【0030】
正常な心筋は、種々の細胞、心臓筋細胞および非心筋細胞で構成され、これらには内皮および血管平滑筋細胞、並びに線維芽細胞を含む。(Weber KT. (2000), “Fibrosis and hypertensive heart disease”, Curr Opin Cardiol. 15(4):264-72)。
【0031】
心室壁の構造の再造形は、心疾患における臨床結果の重要な決定因子である。このような再造形は、細胞外基質タンパク質、細胞増殖および遊走、並びにアポトーシス性および壊死性細胞死の発生や破壊を含む。心臓線維芽細胞は、これらの過程に決定的に関与し、自己分泌因子およびパラ分泌因子として作用する成長因子およびサイトカイン、並びに細胞外基質タンパク質およびプロテイナーゼを産生する。最近の研究では、心臓線維芽細胞と心筋細胞との間の相互作用が、最終的効果が心機能の悪化および心不全の発症である心臓再造形の進行に必須であることが示された(Manabe I, Shindo T, Nagai R. (2002), “Gene expression in fibroblasts and fibrosis: involvement in cardiac hypertrophy”, Circ Res. 13;91(12):1103-13)。
【0032】
レニン-アンジオテンシン-アルドステロンおよび交感神経系を遮断する薬剤の使用により、心臓再造形を阻害し(および、時には逆転さえも)、心臓機能障害患者の臨床経過を改善することが示された。しかし、線維症の直接の抑制または減少に狙った薬物は、いまだ利用できない。また、Greenberg B. (2001), “Treatment of heart failure: state of the art and prospectives", J Cardiovasc Pharmacol. 38 Suppl 2:S59-63, Zhang Z, Vezza R, Plappert T, McNamara P, Lawson JA, Austin S, Pratico D, Sutton MS, FitzGerald GA, COX-2-dependent cardiac failure in Gh/tTG transgenic mice. Circ Res. 2003 May 30;92(10):1153-61. Epub 2003 Apr 17; Hwang KC, Gray CD, Sweet WE, Moravec CS, Im MJ., Alpha 1-adrenergic receptor coupling with Gh in the failing human heart. Circulation. 1996 Aug 15;94(4):718-26を参照されたい。
【0033】
神経系疾患
ポリグルタミン疾患は、ポリグルタミン挿入をコードするCAGトリヌクレオチド・リピートの増大することによって生じる一群の神経系疾患である。ポリグルタミン疾患は、ハンチントン病(HD)、球脊髄性筋肉萎縮症、歯状核赤核-淡蒼球ルイ体萎縮症および脊髄小脳失調(SCA)1、2、3、6、7および17を含む。これらの疾患は全て、ポリグルタミンの一続き(35〜40グルタミンを上回る)の増大が存在し、したがって、核内凝集体を形成して、ニューロンの死に至ることによって特徴づけられる。アルツハイマー病(AD)は、老齢患者の認知障害において最も一般的な原因であり、有病率が非常に増大することが予想される。対になったらせん状フィラメント(PHF)の形成と関連する神経原線維変性は、アルツハイマー病(AD)の重要な神経病理学的特徴のうちの1つである。パーキンソン病は、黒質内のドーパミン作動性ニューロンの選択的かつ進行性の喪失によって特徴づけられる老化における神経変性障害である。また、Mastroberardino PG, Iannicola C, Nardacci R, Bernassola F, De Laurenzi V, Melino G, Moreno S, Pavone F, Oliverio S, Fesus L, Piacentini M. Tissue transglutaminase ablation reduces neuronal death and prolongs survival in a mouse model of Huntington's disease. Cell Death Differ. 2002 Sep;9(9):873-80; Karpuj MV, Becher MW, Springer JE, Chabas D, Youssef S, Pedotti R, Mitchell D, Steinman L., Prolonged survival and decreased abnormal movements in transgenic model of Huntington disease, with administration of the transglutaminase inhibitor cystamine. Nat Med. 2002 Feb;8(2):143-9; Citron BA, Suo Z, SantaCruz K, Davies PJ, Qin F, Festoff BW., Protein crosslinking, tissue transglutaminase, alternative splicing and neurodegeneration. Neurochem Int. 2002 Jan;40(1):69-78; Chen JS, Mehta K., Tissue transglutaminase: an enzyme with a split personality. Int J Biochem Cell Biol. 1999 Aug;31(8):817-36を参照されたい。
【0034】
骨関節炎
骨関節炎の主な特徴の中には、関節軟骨の分解および関節の縁、いわゆる骨増殖体における新たな骨形成がある。Van den Berg WB., Growth factors in experimental osteoarthritis: transforming growth factor beta pathogenic? J Rheumatol Suppl. 1995 Feb;43:143-5; Scharstuhl A, Glansbeek HL, Van Beuningen HM, Vitters EL, Van der Kraan PM, Van den Berg WB., Inhibition of endogenous TGF-beta during experimental osteoarthritis prevents osteophyte formation and impairs cartilage repair. J Immunol. 2002 Jul 1;169(1):507-14; Karpouzas GA, Terkeltaub RA., New developments in the pathogenesis of articular cartilage calcification. Curr Rheumatol Rep. 1999 Dec;1(2):121-7 を参照されたい。
【0035】
眼疾患
年齢関連性白内障:白内障は、眼のレンズの主要な構造タンパク質であるクリスタリンの凝集および共有結合性架橋によって特徴づけられ、光散乱の増大、不透明化および白内障を生じる。組織におけるカルシウムホメオスタシスの障害は、白内障発生に関係する要因のうちの1つである。Shridas et al., FEBS Lett. 2001 Jun 22;499(3):245-50; Shin et al, J Biol Chem. 2004 Apr 9;279(15):15032-9; Wan et al., Br J Ophthalmol. 2002 Nov;86(11):1293-8. and Takeuchi N, Kamei A. Biol Pharm Bull. 2000 Mar;23(3):283-90を参照されたい。
【0036】
増殖性硝子体網膜症
増殖性硝子体網膜症(PVR)は、網膜剥離後の最も一般的な合併症であり、網膜の穴または破壊が付随する。PVRは、硝子体腔内の、並びに網膜色素上皮(RPE)細胞を含む網膜の正面上の、および裏手表面の細胞膜の増殖をいう。本質的に瘢痕組織であるこれらの膜は、網膜に対して牽引する作用があり、最初の良好な網膜剥離手順の後でさえも、網膜剥離の再発を生じ得る。
【0037】
網膜色素上皮(RPE)細胞の位置を変えるフィブロネクチンリッチな細胞外基質への遊走および接着は、増殖性硝子体網膜症(PVR)の初期工程である。また、Casaroli-Marano RP et al Invest Ophthalmol Vis Sci. 1999 Aug;40(9):2062-72). and Priglinger S et al, Invest Ophthalmol Vis Sci. 2004 Mar;45(3):955-63; Priglinger et al., Invest Ophthalmol Vis Sci. 2003 Jan;44(1):355-64を参照されたい。
【0038】
線維症に続く緑内障濾過手術
緑内障濾過術の目標は、眼内部の水性流体を逃がすことができる新たな通路を作製することより、圧力を下げることである。したがって、濾過により、眼の前房内部から、眼の最も外側の被覆である結膜と眼の根底にある白い解剖学的構造である強膜との間の「ポケット」へ液体を排水することができる。液体は、最終的に血管によって吸収される。都合の悪いことに、術後の瘢痕のために、作製された排水系が遮断され、さらなる外科的介入必要でとなる可能性もある。
【0039】
現在の抗瘢痕療法(マイトマイシンCまたは5FU)は、関連した合併症(たとえば、失明)のために、制限される。たとえば、Cordeiro MF, Gay JA, Khaw PT., Human anti-transforming growth factor-beta2 antibody: a new glaucoma anti-scarring agent Invest Ophthalmol Vis Sci. 1999 Sep;40(10):2225-34を参照されたい。
【0040】
結論として、上記した疾患のための療法の有効な方法がなく、したがって、これらの目的のための治療の新規の有効な化合物および方法を開発する需要がある。
【発明の開示】
【0041】
本発明は、新規二本鎖オリゴリボヌクレオチドを提供する。これらのオリゴリボヌクレオチドは、RNA干渉のメカニズムを経てヒトTGaseIIを阻害する。また、本発明は、このようなオリゴリボヌクレオチドを含む薬学的組成物およびオリゴリボヌクレオチドを発現することができるベクターを提供する。また、本発明は、線維症関連病態に罹患した患者を治療する方法であって、これにより患者を治療するために、患者に対して、典型的には薬学的組成物として治療的に有効な用量のオリゴリボヌクレオチドを投与することを含む方法を提供する。また、本発明は、その他の疾患および状態を治療することも想定する。本発明は、TGaseIIポリペプチドに対する抗体を使用することによる、線維性疾患およびその他の疾患の治療にも関する。
【発明を実施するための最良の形態】
【0042】
発明の詳細な説明
本発明は、下記に詳述したような種々の病態の、このような治療を必要とする被検者における治療のための方法であって、これにより被検者を治療するために、被検者に対してTGaseII活性の実質的阻害を生じさせるために十分なTGaseIIポリペプチドの阻害剤の量を投与することを含む方法を提供する。TGaseII阻害剤は、L683685、TGaseIIポリペプチドに対する抗体もしくはTGaseII RNAに対するsiRNA、または当技術分野において既知のTGaseII阻害剤のいずれかであってもよい(このような阻害剤についての詳細は、たとえば、米国特許番号5021440号;第4968713号および第5098707号を参照されたい)。特に、本発明は、TGaseII RNAをターゲットする特異的siRNAおよびこれらの使用に関する。また、TGaseIIに対する中和抗体の使用も開示してある。
【0043】
本発明は、一般に、ヒトTGaseII遺伝子の発現をダウンレギュレートする化合物に、特に新規短鎖干渉RNA(siRNA)に、種々の疾患および医学的状態、特に線維性疾患、線維性瘢痕に関連した疾患、およびその他のトランスグルタミナーゼIIの異常な発現に関連した疾患の治療におけるこれらの新規siRNA特定の使用に関する。
【0044】
本発明は、インビボで標的TGaseII遺伝子の発現を阻害するための方法および組成物を提供する。一般に、本方法は、生物学的条件下で(細胞内で)特定のmRNAにターゲットされて、これとハイブリダイズするか、もしくは相互作用する短鎖干渉RNA(すなわち、siRNA)か、または細胞においてsiRNAを産生することができる核酸材料などのオリゴリボヌクレオチドを、RNA干渉メカニズムによって標的遺伝子の発現をダウンレギュレートするために十分な量で投与することを含む。特に、本方法は、疾患治療のために、TGaseII遺伝子の発現を阻害するために使用することができる。
【0045】
したがって、TGaseII発現(転写または翻訳)またはポリペプチド活性の阻害剤は、とりわけ、単鎖抗体、アンチセンスオリゴヌクレオチド、アンチセンスDNAまたはRNA分子、タンパク質、ペプトイド擬態を含むポリペプチドおよびペプチド、並びにドミナントネガティブを含むsiRNA、抗体、好ましくは中和抗体またはこれらの断片、さらには上記の全てを発現する発現ベクターであってもよい。さらなる阻害剤は、一般に2000ダルトン未満、より好ましくは1000ダルトン未満、さらに好ましくは500ダルトン未満の分子量を有する小さな化学分子であってもよい。これらの阻害剤は、以下のとおりに作用し得る:小分子は、発現および/または活性に影響を及ぼし得るし;抗体は、活性に影響を及ぼし得るし;各種アンチセンスは、TGaseII発現に影響を及ぼし得るし;ドミナントネガティブ・ポリペプチドおよびペプトイド擬態は、活性に影響を及ぼし得るし;発現ベクターは、とりわけアンチセンスまたはドミナントネガティブ・ポリペプチドまたは抗体の送達のために使用し得る。
【0046】
本発明によれば、siRNA分子または抗体などのトランスグルタミナーゼIIの阻害剤は、線維症関連病態(下記に定義したとおり)を含む種々の病態を治療するための、さらに白内障を含む眼疾患、心臓血管疾患、神経系疾患、ポリグルタミン疾患(ハンチントン病(HD)、球脊髄性筋肉萎縮症、歯状核赤核-淡蒼球ルイ体萎縮症、並びに脊髄小脳失調(SCA)1、2、3、6、7、および17を含む)、アルツハイマー病およびパーキンソン病、並びに骨関節炎を治療する薬物として使用してもよい。
【0047】
本明細書に使用される「トランスグルタミナーゼII遺伝子」または「TGaseII遺伝子」、または「TGase遺伝子」という用語は、配列番号:1のアミノ酸コードする領域に対して好ましくは90%の相同性、より好ましくは95%の相同性、およびさらにより好ましくは98%の相同性か、または高度にストリンジェントなハイブリダイゼーション条件下でTGaseII遺伝子と結合する核酸配列を有するTGaseII遺伝子の相同体のいずれかとして定義され、これらは、当技術分野において周知である(たとえば、Ausubel et al., Current Protocols in Molecular Biology, John Wiley and Sons, Baltimore, Maryland (1988), updated in 1995 and 1998を参照されたい)。
【0048】
本明細書に使用される「トランスグルタミナーゼIIポリペプチド」または「TGaseIIポリペプチド」、または「TGase」という用語は、配列番号:2に対して好ましくは90%の相同性、より好ましくは、95%の相同性およびさらにより好ましくは98%の相同性を有するTGaseIIポリペプチドの相同体のいずれかとして、全長またはこれらの断片もしくはドメインのいずれかとして、スプライス変異体核酸配列によってコードされるポリペプチドの突然変異体として、その他のポリペプチドとのキメラとして定義されるが、ただし、上記のいずれも、TGaseIIポリペプチドと同じか、または実質的に同じ生物学的機能を有することを条件とする。TGaseIIポリペプチドまたはTGaseIIポリペプチド相同体は、可溶性タンパク質、膜結合(精製された膜標品で、または細胞表面上のいずれか)、ビーズ結合、またはTGaseIIタンパク質またはこれらの断片および誘導体ポリペプチドを提示するその他の何らかの形態含む(しかし、これらに限定されない)、種々の形態で存在してもよい。
【0049】
本明細書に使用される、「インタラクター」は、性質上、TGaseIIが結合し、または相互作用し、または活性化する分子;たとえば、TGaseIIポリペプチドを発現する細胞の表面上の分子、第二の細胞表面上の分子、または細胞質分子である。インタラクターは、TGaseII単独によって、またはその他の成分との複合体の一部としてのTGaseIIによって活性化されるリガンドであってもよい。インタラクターは、TGaseIIから細胞膜を通って細胞内への細胞外シグナルの伝達を容易にするシグナル伝達経路の成分であってもよい。インタラクターは、たとえばTGaseIIからの下流のシグナリングを媒介する第二の細胞間のタンパク質であることができる。インタラクターは、TGaseが既知のTGase基質(たとえば、フィブロネクチン)と競合して結合する分子である。
【0050】
本明細書に使用される「リジル・ドナー」または「Kドナー」という用語は、リジル側鎖を供与してグルタミン転移過程の間にγ-グルタミル-リジン結合を形成させる能力を有する何らかのポリペプチドとして定義される。
【0051】
本明細書に使用される「グルタミル・ドナー」または「Qドナー」という用語は、グルタミン側鎖を供与してグルタミン転移過程の間にγ-グルタミル-リジン結合を形成させる能力を有する何らかのポリペプチドとして定義される。
【0052】
本発明は、TGaseIIの発現をダウンレギュレートする二本鎖オリゴリボヌクレオチド(siRNA)を提供する。トランスグルタミナーゼIIの発現のダウンレギュレーションは、たとえばsiRNAの存在下において生じるリジル-グルタミル架橋された材料の量を測定することによって、またはTGaseII mRNAまたはポリペプチドの量を直接評価することによって測定することができる。TGaseII mRNAの量は、たとえばノーザンブロッティング、RNase保護、RT-PCRまたはリアルタイムPCRによって測定してもよい。TGaseIIポリペプチドの量は、免疫ブロットによって、または免疫沈降によって、またはTGaseII特異的抗体でのELISAによって測定してもよい。
【0053】
本発明のsiRNAは、センス鎖が遺伝子TGaseIIのmRNA配列に由来する二重鎖オリゴリボヌクレオチドであり、かつアンチセンス鎖がセンス鎖に対して相補的である。一般に、標的mRNA配列からのいくらかずれがあっても、siRNA活性を損なわずに、許容される(たとえばCzauderna et al 2003 Nucleic Acids Research 31(11), 2705-2716)。本発明のsiRNAは、mRNAを破壊すると共に、または破壊することなく、転写後レベルの遺伝子発現を阻害する。理論に拘束されないが、siRNAは、特異的切断および分解のためにmRNAをターゲットし得るし、および/またはターゲットされたメッセージからの翻訳を阻害し得る。
【0054】
少なくとも2つの変異体TGaseIIポリペプチドがあり、これらに関して、GeneBank参照は、変異体2 NM_198951.1 GI:39777598および変異体1NM_004613.2 GI:39777596である図1に示した配列は、変異体2(GI:39777598)のヌクレオチド配列である。両変異体およびその他の何らかの同様の軽微な変異体も、TGaseIIポリペプチドの定義に、およびこれらをコードするTGaseII遺伝子の定義に含まれる。
【0055】
本明細書に使用される「TGaseII遺伝子」という用語は、天然に存在するヒト遺伝子(これらの対立遺伝子変異体のいずれか、または配列番号:1のアミノ酸コードする領域に対して好ましくは90%の相同性、より好ましくは95%の相同性、およびさらに好ましくは98%の相同性か、もしくは高度にストリンジェントなハイブリダイゼーション条件下でTGaseII遺伝子と結合する核酸配列を有するTGaseII遺伝子の相同体のいずれかを含む)として定義され、これらは、当技術分野において周知である(たとえば、Ausubel et al., Current Protocols in Molecular Biology, John Wiley and Sons, Baltimore, Maryland (1988), updated in 1995 and 1998を参照されたい。
【0056】
本明細書に使用される「TGaseII」または「TGaseIIポリペプチド」という用語は、配列番号:2に対して好ましくは90%の相同性、より好ましくは、95%の相同性およびさらにより好ましくは98%の相同性を有するTGaseIIポリペプチドの相同体のいずれかとして、全長またはこれらの断片もしくはドメインのいずれかとして、スプライス変異体核酸配列によってコードされるポリペプチドの突然変異体として、その他のポリペプチドとのキメラとして定義されるが、ただし、上記のいずれも、TGaseIIポリペプチドと同じか、または実質的に同じ生物学的機能を有することを条件とする。
【0057】
より詳しくは、本発明は、以下の構造を有する化合物を提供する:
5'(N)x-Z3'(アンチセンス鎖)
3'Z'-(N’)y5'(センス鎖)
式中、それぞれのNおよびN’は、その糖残基が修飾されていても、または修飾されていなくてもよいリボヌクレオチドであり、かつ(N)xおよび(N’)yは、それぞれの連続したNまたはN’が、共有結合によって次のNまたはN’に連結されているオリゴマーであり;
式中、それぞれのxおよびyは、19〜40の間の整数であり;
式中、それぞれのZおよびZ’は、存在しても、または存在しなくてもよいが、存在する場合、dTdTであり、かつそれが存在する鎖の3'末端にて共有結合で付着されており;および、
式中、(N)xの配列は、表A、BおよびCに存在するアンチセンス配列のいずれか一つを含む。
【0058】
本発明の化合物が、共有結合を介して連結された複数のヌクレオチドからなることは、当業者には容易に理解されるであろう。このようなそれぞれの共有結合は、個々の鎖のヌクレオチド配列に沿って、ホスホジエステル結合、ホスホチオアート結合、または両方の組み合わせであってもよい。その他の可能性バックボーン修飾は、とりわけ米国特許第5,587,361号;第6,242,589号;第6,277,967号;第6,326,358号;第5,399,676号;第5,489,677号;および第5,596,086号に記述されている。
【0059】
特定の態様において、xおよびyは、好ましくは約19〜約27の間、最も好ましくは約19〜約23の整数である。本発明の化合物の特定の態様において、xは、yに等しく(すなわちx=y)、好ましい態様において、x=y=19またはx=y=21である。特に好ましい態様において、x=y=19である。
【0060】
一つの態様において、本発明の化合物の中で、ZおよびZ’は、両方とも存在せず;もう一つの態様において、ZまたはZ’のうちの1つが存在する。
【0061】
一つの態様において本発明の化合物の中で、化合物のリボヌクレオチドの全てにおいて、これらの糖残基が修飾されていない。
【0062】
本発明の化合物のいくつかの態様において、少なくとも1つのリボヌクレオチドは、その糖残基が修飾されている(好ましくは2’位の修飾)。2’位の修飾により、アミノ基、フルオロ基、メトキシ基、アルコキシ基およびアルキル基を含む群から選択される部分が存在することとなる。現在最も好ましい態様において、2’位の部分は、メトキシ(2’-0-メチル)である。
【0063】
本発明のいくつかの態様において、交互に現れるリボヌクレオチドが、アンチセンス鎖およびセンス鎖において修飾されている。
【0064】
本発明の特に好ましい態様において、アンチセンス鎖は、5'末端にてリン酸化されており(phophorylated)、かつ3'末端にてリン酸化されていても、またはリン酸化されていなくてもよく、かつセンス鎖は、5'末端にて、および3'末端にてリン酸化されていても、またはリン酸化されていなくてもよい。
【0065】
もう一つの態様において、本発明の化合物の中で、アンチセンス鎖の5'および3'末端のリボヌクレオチドは、これらの糖残基が修飾されており、かつセンス鎖の5'および3'末端のリボヌクレオチドは、これらの糖残基が修飾されていない。
【0066】
本発明は、細胞内にて無修飾形態で上述したオリゴリボヌクレオチドのいずれかを発現し、その後に適切な修飾を生じることができるベクターをさらに提供する。
【0067】
また、本発明は、本発明の化合物の1つまたは複数を、キャリア(好ましくは薬学的に許容されるキャリア)中に含む組成物を提供する。
【0068】
また、本発明は、キャリアと、ヒトTGaseII遺伝子の細胞における発現をダウンレギュレートするために有効な量で、実質的に(N)xの配列に相補的な配列を含む本発明の化合物の1つまたは複数を含む組成物を提供する。
【0069】
また、本発明は、対照と比較して少なくとも50%ヒトTGaseII遺伝子の発現をダウンレギュレートする方法であって、遺伝子のmRNA転写物を本発明の化合物の1つまたは複数と接触させることを含む方法を提供する。
【0070】
一つの態様において、化合物は、TGaseIIをダウンレギュレートし、TGaseIIのダウンレギュレーションは、TGaseII機能のダウンレギュレーション(これは、とりわけ、天然の遺伝子/ポリペプチドの既知のインタラクターでの酵素アッセイ法または結合アッセイによって調べてもよい)、TGaseIIタンパク質のダウンレギュレーション(これは、とりわけウエスタンブロット法、ELISAまたは免疫沈降法によって調べてもよい)およびTGaseII mRNA発現のダウンレギュレーション(これは、とりわけノーザンブロッティング、定量的RT-PCR、インサイチュー・ハイブリダイゼーションまたはマイクロアレイ・ハイブリダイゼーションによって調べてもよい)を含む群から選択されるものによる。
【0071】
また、本発明は、線維症または線維症関連病態に罹患した患者を治療する方法であって、これにより患者を治療するために、患者に治療的に有効な用量の本発明の組成物を投与することを含む方法を提供する。
【0072】
また、本発明は、トランスグルタミナーゼIIを経た細胞タンパク質の異常な架橋に関連した病態に罹患した患者を治療する方法であって、これにより患者を治療するために、患者に治療的に有効な用量の本発明の組成物を投与することを含む方法を提供する。
【0073】
また、本発明は、線維症または線維症関連病態またはトランスグルタミナーゼIIを経た細胞タンパク質の異常な架橋に関連した病態に罹患した患者における回復を促進するための組成物の調製のための、本発明の1つまたは複数の化合物の治療的に有効な用量の使用を提供する。
【0074】
線維症が明らかである線維性疾患(線維症関連病態)は、以下の全ての病原学的変種を含む器官における線維症の急性および慢性形態を含む:間質性肺疾患および線維性肺疾患を含む肺線維症、肝臓線維症、心臓線維症を含む心筋線維症、慢性腎不全を含む腎臓線維症、強皮症を含む皮膚線維症、ケロイドおよび肥厚性瘢痕;骨髄線維化(骨髄線維症);増殖性硝子体網膜症(PVR)および白内障または緑内障を治療する外科手術によって生じる全種類の眼の瘢痕を含む瘢痕;病因不定の炎症性腸疾患、黄斑変性症、グレーブス眼障害、薬物で誘導される麦角中毒、乾癬、リ-フラウメニ症候群におけるグリア芽細胞腫、散発性グリア芽細胞腫、骨髄球性白血病、急性骨髄性白血病、骨髄異形成症候群、骨髄増殖症候群、婦人科学的癌、カポジ肉腫、ハンセン病およびコラーゲン性大腸炎。
【0075】
本発明の化合物は、線維性疾患とは異なる多くのその他の疾患および状態を治療するために使用してもよい。その他の適応症は、白内障を含む眼疾患、心臓血管疾患、特に心肥大、アテローム性動脈硬化症/再狭窄、神経系疾患、封入ポリグルタミン疾患(ハンチントン病など)、球脊髄性筋肉萎縮症、歯状核赤核-淡蒼球ルイ体萎縮症、および脊髄小脳失調(SCA)1、2、3、6、7および17、アルツハイマー病、並びにパーキンソン病であってもよい。
【0076】
化合物は、それぞれの末端領域のリボヌクレオチドのうちの2つまで塩基が変更された相同体を有していてもよく;末端領域とは、4つの末端リボヌクレオチドをいいたとえば19mer配列の塩基1〜4および/または16〜19および21mer配列の塩基1〜4および/または18〜21をいう。
【0077】
本発明の好ましいオリゴヌクレオチドは、表A、BおよびCに一覧が記載されたオリゴヌクレオチド、好ましくはヒトcDNAをターゲットするオリゴヌクレオチドである。本発明の最も好ましいオリゴヌクレオチドは、表Dに示したように、阻害活性を有するオリゴヌクレオチド、好ましくはヒトTGaseII cDNAをターゲットするオリゴヌクレオチドである。
【0078】
現在、本発明の最も好ましい化合物は、平滑末端19merオリゴヌクレオチド、すなわちx=y=19かつZおよびZ’の両方が存在せず;オリゴヌクレオチドは、アンチセンス鎖の5'位にて、およびセンス鎖の3'位にてリン酸化されており、交互に現れるリボヌクレオチドは、アンチセンスおよびセンス鎖の2’位にて修飾されており、2’位の部分は、メトキシ(2’-0-メチル)であり、かつアンチセンス鎖の5'および3'末端のリボヌクレオチドは、これらの糖残基が修飾されており、かつセンス鎖の5'および3'末端のリボヌクレオチドは、これらの糖残基が修飾されていないオリゴヌクレオチドである。現在、最も好ましいこのような化合物は、TG_HMRG1.である。TG_HMRG1のアンチセンス鎖は、配列番号:22を有し、センス鎖は、配列番号:4を有する。その他の好ましい化合物は、TG_HMG1およびTG_HM1であり、これらは、それぞれ配列番号:23および25によって表されるアンチセンス鎖を有し、センス鎖は、それぞれ配列番号:5および7によって表される
本発明の一つの局面において、オリゴヌクレオチドは、二本鎖構造を含み、このような二本鎖構造は、
第一の鎖および第二の鎖を含み、
第一の鎖は、隣接するヌクレオチドの第一のひと配列を含み、第二の鎖は、隣接するヌクレオチドの第二のひと配列を含み、
第一のひと配列は、TGaseIIをコードする核酸配列に相補的か、または同一であり、第二のひと配列は、TGaseIIをコードする核酸配列と同一か、または相補的である。
【0079】
ある態様において、第一のひと配列および/または第二のひと配列は、約14〜40ヌクレオチド、好ましくは約18〜30ヌクレオチド、より好ましくは約19〜27ヌクレオチド、最も好ましくは約19〜21ヌクレオチド、特に約19〜23ヌクレオチドを含む。このような局面において、オリゴヌクレオチドは、17〜40ヌクレオチドの長さであってもよい。加えて、本発明によるさらなる核酸は、配列番号:3〜配列番号:416の一つの少なくとも14の隣接するヌクレオチド、より好ましくは上記のとおりの第一のひと配列および第二のひと配列で構成される二本鎖構造のいずれかの末端の14の隣接するヌクレオチド塩基対を含む。
【0080】
本明細書に使用される「治療」という用語は、疾患もしくは状態と関連する症候を寛解させるため、疾患もしくは状態の重症度を軽くするため、もしくは治癒するため、または疾患または状態が生じるのを予防するために有効な治療物質を投与することをいう。
【0081】
特定の態様において、投与は、静脈内投与を含む。もう一つの特定の態様において、投与は、局所的または局部的な投与を含む。
【0082】
加えて、本発明は、患者に対して核酸配列に対する少なくとも1つのアンチセンス(AS)オリゴヌクレオチドもしくは少なくとも1つのsiRNA、またはTGaseII配列もしくはTGaseIIタンパク質に対して向けられたドミナントネガティブ・ペプチド、TGaseIIポリペプチドに対して向けられた抗体の治療的に有効な用量を投与することによって、病態または疾患(上記で詳述したとおり)を、このような治療を必要とする患者において調節する方法を提供する。
【0083】
送達:哺乳類細胞へのsiRNAの送達が増強され、および改善されることを特異的に狙った送達系が開発されており、たとえば、Shen et al (FEBS letters 539: 111-114 (2003)), Xia et al., Nature Biotechnology 20: 1006-1010 (2002), Reich et al., Molecular Vision 9: 210-216 (2003), Sorensen et al. (J.Mol.Biol. 327: 761-766 (2003), Lewis et al., Nature Genetics 32: 107-108 (2002) and Simeoni et al., Nucleic Acids Research 31, 11: 2717-2724 (2003)を参照されたい。siRNAは、霊長類における阻害に関して、最近首尾よく使用されてきた;詳しくは、Tolentino et al., Retina 24(1) February 2004 I 132-138を参照されたい。siRNAのための呼吸器製剤は、Davis et al.の米国特許出願第2004/0063654号に記述されている。コレステロール抱合されたsiRNA(および、その他のステロイドおよび脂質抱合されたsiRNA)は、送達のために使用されてきた(Soutschek et al Nature 432: 173-177(2004) Therapeutic silencing of an endogenous gene by systemic administration of modified siRNAs; and Lorenz et al. Bioorg. Med. Chemistry. Lett. 14:4975-4977 (2004) Steroid and lipid conjugates of siRNAs to enhance cellular uptake and gene silencing in liver cellsを参照されたい。
【0084】
本発明のsiRNAまたは薬学的組成物は、個々の患者の臨床状態、治療される疾患、投与の部位および方法、投与のスケジューリング、患者の年齢、性、体重、並びに開業医にとって既知であるその他の因子を考慮して、優れた医事に従って施され、および投与される。
【0085】
したがって、本明細書における目的のための「治療的に有効な用量」は、当技術分野において既知である考察などによって決定される。用量は、生存率またはより迅速な回復を含む(しかし、これらに限定されない)改善、または当業者による適切な測定として選択される症候およびその他の指標の改善もしくは除去を達成するために有効でなければならない。本発明の化合物は、従来の投与経路のいずれによっても投与することができる。化合物は、化合物として、または薬学的に許容される塩として投与することができ、また単独で、または活性成分として薬学的に許容されるキャリア、溶媒、希釈剤、賦形剤、アジュバントおよび媒体と組み合わせて投与することができることに留意すべきである。化合物は、腹腔内、静脈内、動脈内、筋肉内および鼻腔内投与、並びに髄腔内および注入技術を含む経口的に、皮下に、または非経口的に投与することができる。また、化合物の移植も有用である。液体形態を注射のために調製してもよく、本用語には、皮下、経皮、経静脈、筋肉内、髄腔内およびその他の非経口(parental)投与経路を含む。液体組成物は、有機共溶媒を伴い、または伴わない水溶液、水性または油懸濁液、食用油との乳剤、並びに同様の薬学的媒体を含む。加えて、一定の環境下において、本発明の新規治療に使用するための組成物は、鼻腔等の投与のためおエアロゾルとして形成してもよい。治療される患者は、温血動物および、特にヒトを含む哺乳動物である。薬学的に許容されるキャリア、溶媒、希釈剤、賦形剤、アジュバントおよび媒体、並びに移植キャリアは、一般に本発明の活性成分と反応することのない不活性な無毒の固体または液体の充填剤、希釈剤または封入材料をいい、これらには、リポソームおよび微粒子を含む。本発明に有用な送達系の例は、米国特許第5,225,182号;第5,169,383号;第5,167,616号;第4,959,217号;第4,925,678号;第4,487,603;号;第,486,194号;第4,447,233号;第4,447,224号;第4,439,196号;および第4,475,196号を含む。多くのその他のこのようなインプラント、送達系およびモジュールが当業者に周知である。本発明の1つの特定の態様において、局所的および経皮の製剤が特に好ましい。
【0086】
一般に、ヒトのための化合物の活性な用量は、1日あたり1ng/kg〜約20〜100mg/kg体重に、好ましくは1日あたり約0.01mg〜約2〜10mg/kg体重の範囲で、少なくとも1〜4週またはそれ以上の期間の間に1日あたり1用量、または1日あたり2回もしくは3回以上の処方計画である。また、より長い年月の治療または生涯の治療さえも、本明細書に開示したいくつかの適応症にについて想定される。
【0087】
また、本発明は、薬学的組成物製剤を調製する方法であって:
提供する本発明の少なくとも1つの二本鎖siRNA化合物を得ることと;および、
前記化合物を薬学的に許容されるキャリアと混合することと、
を含む方法を提供する。
【0088】
また、本発明は、薬学的組成物を調製する方法であって、本発明の化合物を薬学的に許容されるキャリアと混合することを含む方法を提供する。
【0089】
好ましい態様において、薬学的組成物の調製に使用される化合物は、薬学的に有効な用量のキャリアと混合される。特定の態様において、本発明の化合物は、ステロイドに対して、または脂質に対して、または別の適切な分子に対して、たとえばコレステロールに対して抱合される。
【0090】
ヌクレオチドの治療特性を改善するために、ヌクレオチドの修飾または類似体を導入することができる。改善される特性は、増大されたヌクレアーゼ耐性および/または増大された細胞膜に浸透する能力を含む。
【0091】
したがって、本発明は、また、実質的にポリヌクレオチドまたはオリゴヌクレオチドの機能に影響を及ぼさない全ての本発明のオリゴヌクレオチドの類似体、またはオリゴヌクレオチドに対する修飾を含む。好ましい態様において、このような修飾は、ヌクレオチドの塩基部分に、ヌクレオチドの糖残基に、および/またはヌクレオチドのリン酸部分に関連する。
【0092】
本発明の態様において、ヌクレオチドは、天然に存在するか、または合成的に修飾された塩基から選択することができる。天然に存在する塩基は、アデニン、グアニン、シトシン、チミンおよびウラシルを含む。オリゴヌクレオチドの修飾塩基は、イノシン、キサンチン、ヒポキサンチン、2-アミノアデニン、6-メチル-、2-プロピル-およびその他のアルキル-アデニン、5-ハロウラシル、5-ハロシトシン、6-アザシトシンおよび6-アザチミン、偽ウラシル、4-チウラシル、8-ハロアデニン、8-アミノアデニン、8-チオールアデニン、8-チオアルキルアデニン、8-ヒドロキシルアデニンおよびその他の8置換されたアデニン、8-ハログアニン、8-アミノグアニン、8-チオールグアニン、8-チオアルキルグアニン、8-ヒドロキシルグアニンおよびその他の置換されたグアニン、その他のアザおよびデアザアデニン、その他のアザおよびデアザグアニン、5-トリフルオロメチルウラシル、並びに5-トリフルオロシトシンを含む。
【0093】
加えて、ヌクレオチドの構造が基本的に変更されており、かつ治療的または実験的試薬としてより適しているヌクレオチドの類似体を調製することができる。ヌクレオチド類似体の例は、DNA(またはRNA)のデオキシリボース(またはリボース)リン酸バックボーンがペプチドに見いだされるものと同様のポリアミドバックボーンで置換されているペプチド核酸(PNA)である。PNA類似体は、酵素による分解に耐性を示し、インビボおよびインビトロでの生存が延長された。さらに、PNAは、DNA分子よりも相補的DNA配列に対して強く結合することを示した。この観察は、PNA鎖とDNA鎖との間の電荷相反がないことに起因する。オリゴヌクレオチドに対して作製することができるその他の修飾は、ポリマー主鎖、環状バックボーンまたは非環状バックボーンを含む。
【0094】
一つの態様において、修飾は、リン酸部分の修飾であり、修飾されたリン酸部分は、ホスホチオアート基から選択される。
【0095】
本発明の化合物は、リボ核(または、デオキシリボ核)オリゴヌクレオチドの合成のための技術分野において周知の方法のいずれによっても合成することができる。このような合成は、とりわけ、Beaucage S.L. and Iyer R.P., Tetrahedron 1992; 48: 2223-2311, Beaucage S.L. and Iyer R.P., Tetrahedron 1993; 49: 6123-6194 and Caruthers M.H. et. al., Methods Enzymol. 1987; 154: 287-313に記述されており、チオアートの合成は、とりわけ、Eckstein F., Annu. Rev. Biochem. 1985; 54: 367-402, the synthesis of RNA molecules is described in Sproat B., in Humana Press 2005 Edited by Herdewijn P.; Kap. 2: 17-31に記述されており、それぞれの 顆粒のプロセスは、とりわけPingoud A. et. al., in IRL Press 1989 Edited by Oliver R.W.A.; Kap. 7: 183-208 and Sproat B., in Humana Press 2005 Edited by Herdewijn P.; Kap. 2: 17-31(前記)に記述されている。
【0096】
その他の合成手順は、当技術分野において既知であり、たとえばUsman et al., 1987, J. Am. Chem. Soc., 109, 7845; Scaringe et al., 1990, Nucleic Acids Res., 18, 5433; Wincott et al., 1995, Nucleic Acids Res. 23, 2677-2684; and Wincott et al., 1997, Methods Mol. Bio., 74, 59に記載されているような手順であり、これらの手順には、5'末端のジメトキシトリチルおよび3'末端のホスホラミダイトなどの共通の核酸保護基およびカップリング基を利用してもよい。修飾された(たとえば2’-O-メチル化された)ヌクレオチドおよび無修飾ヌクレオチドが、所望のとおりに取り込まれる。
【0097】
本発明のオリゴヌクレオチドは、別々に合成することができ、たとえばライゲーションによって、(Moore et al., 1992, Science 256, 9923; Draper et al., International PCT publication No. WO93/23569; Shabarova et al., 1991, Nucleic Acids Research 19, 4247; Bellon et al., 1997, Nucleosides & Nucleotides, 16, 951; Bellon et al., 1997, Bioconjugate Chem. 8, 204)または合成および/または脱保護後のハイブリダイゼーションによって合成後に共に連結することができる。
【0098】
市販の機械(とりわけ、Applied Biosystemsから利用できる)を使用することができる点;オリゴヌクレオチドは、本明細書に開示された配列に従って調製される点に留意されたい。化学的に合成された断片の重なり合う対は、当技術分野において周知の方法を使用して結合することができる(たとえば、米国特許第6,121,426号を参照されたい)。鎖を別々に合成して、次いで、チューブ内で互いにアニールさせる。次いで、二本鎖siRNAを、HPLCによってアニールされなかった一本鎖オリゴヌクレオチド(たとえば、これらのうちの一方が過剰なことを原因として生じるので)から分離する。本発明のsiRNAまたはsiRNA断片に関して、2つ以上のこのような配列を合成して、本発明に使用するために共に連結させることができる。また、本発明の化合物は、米国特許出願公開番号US2004/0019001号(McSwiggen)に記載されているように、両方のsiRNA鎖を、切断可能なリンカーによって分離された単一の隣接するオリゴヌクレオチド断片または鎖として合成し、これをその後に切断して、ハイブリダイズしてsiRNA二重鎖の精製を可能にする別々のsiRNA断片または鎖を提供するという、直列型合成法を経て合成することができる。リンカーは、ポリヌクレオチドリンカーまたは非ヌクレオチドリンカーであることができる。
【0099】
本発明の化合物は、直接、またはウイルスまたは非ウイルスベクターと共に送達することができる。直接送達されるときは、一般に配列をヌクレアーゼ耐性にする。あるいは、本明細書において下記に論議したように、配列は、配列が細胞に発現されるように、発現カセットまたは構築物に取り込むことができる。一般に、構築物は、配列をターゲットされる細胞にて発現することができるように、適当な制御配列またはプロモーターを含む。本発明の化合物の送達のために任意に使用されるベクターは、市販されており、当業者に既知の方法によって本発明の化合物を送達するために修飾してもよい。
【0100】
また、本発明のオリゴヌクレオチドの配列の1つまたは複数を含む長い二本鎖オリゴヌクレオチド(典型的には、25〜500ヌクレオチドの長さ)を送達してもよく、内因性細胞複合体によって(たとえば、上記のとおりのDICERによって)細胞内にてプロセスされて、本発明のオリゴヌクレオチドであるより小さな二本鎖オリゴヌクレオチドを産生し得ることが想定される。
【0101】
本明細書に使用される「ポリペプチド」という用語は、ポリペプチドに加えて、オリゴペプチド、ペプチドおよび完全なタンパク質をいう。
【0102】
動物モデル系:本発明の活性なsiRNAの試験は、予測的動物モデルで行ってもよい。腎臓線維症のためのいくつかのモデルを実施例3に記述してある。
【0103】
ラットの肝臓線維症の2つのモデルは、以下の参照に記載したように、対照として偽手術したBile Duct Ligation(BDL、)対照としてのオリーブ油を与えた動物であるCCl4中毒である:Lotersztajn S, Julien B, Teixeira-Clerc F, Grenard P, Mallat A, Hepatic Fibrosis: Molecular Mechanisms and Drug Targets. Annu Rev Pharmacol Toxicol. 2004 Oct 07; Uchio K, Graham M, Dean NM, Rosenbaum J, Desmouliere A., Down-regulation of connective tissue growth factor and type I collagen mRNA expression by connective tissue growth factor antisense oligonucleotide during experimental liver fibrosis. Wound Repair Regen. 2004 Jan-Feb;12(1):60-6. ;and. Xu XQ, Leow CK, Lu X, Zhang X, Liu JS, Wong WH, Asperger A, Deininger S, Eastwood Leung HC., Molecular classification of liver cirrhosis in a rat model by proteomics and bioinformatics Proteomics. 2004 Oct;4(10):3235-45。
【0104】
瘢痕組織阻害剤を評価するためのモデルは、当技術分野において周知であり、たとえば、Sherwood MB et al., J Glaucoma. 2004 Oct;13(5):407-12. A new model of glaucoma filtering surgery in the rat; Miller MH et al., Ophthalmic Surg. 1989 May;20(5):350-7. Wound healing in an animal model of glaucoma fistulizing surgery in the rabbit; vanBockxmeer FM et al., Retina. 1985 Fall-Winter; 5(4): 239-52. Models for assessing scar tissue inhibitors; Wiedemann P et al., J Pharmacol Methods. 1984 Aug; 12(1): 69-78. Proliferative vitreoretinopathy: the rabbit cell injection model for screening of antiproliferative drugs。
【0105】
白内障のモデルは、以下の刊行物に記述されている:The role of Src family kinases in cortical cataract formation. Zhou J, Menko AS.Invest Ophthalmol Vis Sci. 2002 Jul;43(7):2293-300; Bioavailability and anticataract effects of a topical ocular drug delivery system containing disulfiram and hydroxypropyl-beta-cyclodextrin on selenite-treated rats.Wang S, Li D, Ito Y, Nabekura T, Wang S, Zhang J, Wu C. Curr Eye Res. 2004 Jul;29(1):51-8; and Long-term organ culture system to study the effects of UV-Airradiation on lens transglutaminase. Weinreb O, Dovrat A.; Curr Eye Res. 2004 Jul;29(1):51-8。
【0106】
抗体産生
本発明に使用される「抗体」という用語は、ポリクローナルおよびモノクローナルの両方の完全抗体、並びにFab、F(ab')2およびFvなどのエピトープの決定因子に結合することができるこれらの断片を意味する。これらの抗体断片は、その抗原または受容体と選択的に結合する能力を保持し、とりわけ、以下のとおりに例示される:
(1)Fab、酵素パパインで抗体全体を消化して、軽鎖および一部の重鎖を得ることによって産生することができる抗体分子の一価の抗体結合性断片を含む断片;
(2)(Fab')2、酵素ペプシンで抗体全体を処置することによってその後に還元することなく得ることができる抗体の断片;F(ab'2)は、2つのジスルフィド結合によって共に保持された2つのFab断片の二量体である;
(3)Fv、2つの鎖として発現された軽鎖の可変領域と重鎖の可変領域とを含む遺伝的に操作された断片として定義される;および
(4)単鎖抗体(SCA)、遺伝的に融合された単一の鎖状分子として、適切なポリペプチド・リンカーによって連結された軽鎖の可変領域と重鎖の可変領域とを含む遺伝的に操作された分子として定義される。
【0107】
抗体機能的活性を有するこのような断片は、当業者に既知の方法によって調製することができる(たとえば、Bird et al.(1988)Science 242:423-426)。 都合よくは、抗体は、免疫原またはこれらの部分、たとえば配列に基づいた合成ペプチドに対して調製しても、もしくはクローン技術によって組換えで調製してもよく、または天然の遺伝子産物および/またはこれらの部分を単離して免疫原として使用してもよい。免疫原は、Harlow and Lane (1988), Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, and Borrebaeck (1992), Antibody Engineering - A Practical Guide, W.H. Freeman and Co., NYに一般に記述されているように、当業者に周知の標準的な抗体産生技術によって抗体を産生するために使用することができる。
【0108】
ポリクローナル抗体を産生するためには、ウサギまたはヤギなどの宿主を、一般にアジュバントと共に、必要に応じて、キャリアに結合させて免疫原または免疫原断片で免疫し;免疫原に対する抗体を血清から収集する。さらに、ポリクローナル抗体は、それが単一特異性であるように吸収することができ;すなわち、交差反応を起こし得る抗体が血清に残らないように、血清を関連免疫原に対して吸収して、それを単一特異性にすることができる。
【0109】
モノクローナル抗体を産生するためには、本技術には、適切なドナー、一般にマウスの免疫原での高度免疫処置、および脾臓抗体産生細胞の単離を含む。これらの細胞を骨髄腫細胞などの不死細胞に融合させて、不死で、かつ必要とされる抗体を分泌する融合細胞ハイブリッドを提供する。次いで、細胞を大量に培養し、モノクローナル抗体を使用のために培地から収集する。
【0110】
組換えを抗体産生については、一般にHuston et al. (1991) "Protein engineering of single-chain Fv analogs and fusion proteins" in Methods in Enzymology (JJ Langone, ed., Academic Press, New York, NY) 203:46-88; Johnson and Bird (1991) "Construction of single-chain Fvb derivatives of monoclonal antibodies and their production in Escherichia coli in Methods in Enzymology (JJ Langone, ed.; Academic Press, New York, NY) 203:88-99; Mernaugh and Mernaugh (1995) "An overview of phage-displayed recombinant antibodies" in Molecular Methods In Plant Pathology (RP Singh and US Singh, eds.; CRC Press Inc., Boca Raton, FL:359-365)を参照されたい。特に、scFv抗体は、国際公開公報第2004/007553号(TedescoおよびMarzari)に記述されている。加えて、動物の抗体産生Bリンパ球またはハイブリドーマからのメッセンジャーRNAを逆転写して相補的DNA(cDNA)を得ることができる。抗体cDNA(これは、完、または部分的な長さであることができる)を増幅し、ファージまたはプラスミドにクローン化する。cDNAは、分離されたか、またはリンカーによって接続された重鎖および軽鎖cDNAの部分的な長さであることができる。抗体または抗体断片を適切な発現系を使用して発現させて、組換え抗体を得る。また、抗体cDNAは、関連する発現ライブラリをスクリーニングすることによって得ることができる。
【0111】
抗体は、周知のように、固体支持体基体に結合し、または検出可能な部分と抱合し、または結合および抱合を両方することができる。(蛍光または酵素部分の抱合に関する一般的な議論については、Johnstone & Thorpe (1982.), Immunochemistry in Practice, Blackwell Scientific Publications, Oxfordを参照されたい)。また、固体支持体基体に対する抗体の結合は、当技術分野において周知である(一般的な考察については、Harlow & Lane (1988) Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Publications, New York; and Borrebaeck (1992), Antibody Engineering - A Practical Guide, W.H. Freeman and Co.を参照されたい)。本発明で想定される検出可能部分は、ビオチン、金、フェリチン、アルカリホスファターゼ、β-ガラクトシダーゼ、ペルオキシダーゼ、ウレアーゼ、フルオレッセイン、ローダミン、トリチウム、14C、およびヨウ素化などの蛍光、金属、酵素、および放射性のマーカーを含むことができるが、これらに限定されるわけではない。
【0112】
本明細書で論議される疾患および障害の治療に有用であると考えられるさらなる化合物は、アンチセンスDNA分子(これは、当技術分野において既知の方法によって、図1の配列を使用して作製することができる)、リボザイムなどの触媒RNA、ドミナントネガティブ・ペプチド(これは、当技術分野において既知の方法によって、図2の配列を使用して作製することができる)などのポリペプチドまたはその他のポリペプチド阻害剤であってもよい。本明細書に開示したsiRNA配列を含むアンチセンスDNA分子(DNAとRNAとの間の相違から生じる適切な核酸修飾をもつ)が特に望ましく、本明細書に開示した全ての使用および方法に対して、これらの対応するsiRNAと同じ能力で使用し得る。
【0113】
TGase不活性化化合物のスクリーニング
本発明のいくつかの化合物および組成物は、TGaseIIの活性を調整する化合物、特に線維性疾患または線維症関連病態を調整する化合物を同定し、および単離するためのスクリーニングアッセイ法に使用してもよい。スクリーニングされる化合物は、とりわけ小さな化学分子、抗体、特に中和抗体、阻害ポリペプチドおよびドミナントネガティブなどの物質を含む。
【0114】
TGaseIIポリペプチド酵素活性に対する本発明の化合物の阻害活性またはTGaseIIに対する本発明の化合物の結合は、TGaseIIポリペプチドとさらなる化合物の相互作用、たとえばさらなる化合物が、本発明の抗体またはドミナントネガティブ・ペプチドとTGaseII阻害に関して競合するかどうか、またはさらなる化合物が該阻害をレスキューするかどうかを決定するために使用してもよい。阻害または活性化は、とりわけTGaseIIポリペプチドの活性の産生または放射性または蛍光性競合アッセイ法においてTGaseIIポリペプチドからの結合化合物の置換についてアッセイすることなど、種々の手段によって試験することができる。
【0115】
本発明は加えて、TGaseIIポリペプチド、好ましくはヒトTGaseIIポリペプチドの酵素活性を阻害することができる化合物を得る方法であって:
(i)TGaseIIポリペプチドまたはTGaseIIポリペプチドを発現する細胞をリジル・ドナーである化合物と、および生体分子であるグルタミル・ドナーと接触させる工程と;
(ii)工程(i)の混合物から過剰なリジル・ドナーを除去する工程と;
(iii)化合物の存在下において産生されるリジル-グルタミル架橋物質の量を測定する工程と;
(iv)工程(iii)で測定したリジル-グルタミル架橋された物質の量を、一定条件下で化合物の非存在下において測定したものと比較する工程であって、リジル-グルタミル架橋された物質の量の減少は、化合物によるTGaseIIポリペプチド活性の阻害を指し示す工程と;および、
(v)TGaseIIポリペプチド活性の阻害剤として化合物を同定する工程と、
の工程を含む方法を提供する。
【0116】
一つの態様において、本明細書に記述した全ての方法の中で、このような方法に使用されるTGaseIIポリペプチドは、配列番号:2に記載された配列の連続したアミノ酸を含む。
【0117】
もう一つの態様において、工程(i)のリジル・ドナーにタグを付けてあり、工程(iii)の測定は、タグ関連読み出しであり、読み出しレベルの減少は、化合物によるTGaseIIポリペプチド活性の阻害を指し示す。
【0118】
生体分子であるグルタミル・ドナーは、たとえばフィブロネクチンまたはコラーゲンであってもよい。
【0119】
もう一つの態様において、このような方法に使用されるリジル・ドナーは、ダンシル蛍光タグでタグを付けたカダベリンである。
【0120】
もう一つの態様において、このような方法に使用されるリジル・ドナーは、ビオチンでタグを付けてあり、特定の態様において、リジル・ドナーは、ビオチン・カダベリンである。ビオチンは、当技術分野において既知の読み出し(またはヌトラビジン(nutravidin)などの修飾されたストレプトアビジン)に抱合されたストレプトアビジンによって検出される(たとえば、西洋ワサビペルオキシダーゼに抱合されており、西洋ワサビペルオキシダーゼの量は、基質としてテトラメチルベンジジンを使用して定量化する)。
【0121】
本発明のさらにもう一つの態様において、提供した方法におけるグルタミル・ドナーは、細胞外基質タンパク質または細胞タンパク質であってもよい。細胞外基質タンパク質は、以下からなる群より選択され:フィブロネクチン、コラーゲン、オステオネクチン、ECM-ゲル(たとえばSigma E1270)、また細胞内タンパク質は、とりわけ以下からなる群より選択される:RhoA、フィアルミン(fialmin)、スペクトリン、ビメンチン、HSP90、HSP60(Orru S, Caputo I, D'Amato A, Ruoppolo M, Esposito C Proteomics identification of acyl-acceptor and acyl-donor substrates for transglutaminase in a human intestinal epithelial cell line. Implications for celiac disease. J Biol Chem. 2003 Aug 22;278(34):31766-73を参照されたい)。
【0122】
異なる態様において、化合物を、TGaseIIポリペプチドを発現する細胞と接触させる。このような方法に使用してもよい細胞は、分離されていても、または組織に提示されていてもよい。細胞は、TGaseIIポリペプチドを天然に、またはTGaseII遺伝子で、一過性もしくは安定にトランスフェクトした結果として発現していてもよく、またはTGaseII遺伝子を過剰発現してもよく、過剰発現と正常TGaseII発現細胞との間でのTGaseIIの活性を比較する。さらなる態様において、グルタミル・ドナーは、天然物であるか、またはこれは、細胞外から添加される。
【0123】
異なる態様において、TGaseIIポリペプチドを化合物と接触させる。TGaseIIポリペプチドは、固定され、または溶液中で遊離していることができる。提供したスクリーニング法の態様では、グルタミル・ドナーを固定してある。
【0124】
さらなる態様において、ヒトTGaseIIの活性を阻害することができる化合物を得るために提供した方法は、工程(i)の前にTGaseIIを、TGaseIIに結合することが既知の第二の化合物と接触させるというさらなる工程を含んでいてもよい。
【0125】
本発明の一つの態様において、本明細書に記述した提供した方法におけるタグ読み出しでは、西洋ワサビペルオキシダーゼに抱合したヌトラビジンとビオチンの相互作用がなされ、細胞に結合した西洋ワサビペルオキシダーゼの量を、基質としてTMB(テトラメチルベンジジン)を使用して定量化する。
【0126】
なおさらなる態様において、提供した方法によって得られた化合物は、これがTGaseファミリーの少なくとも1つの他のメンバーの活性を阻害するよりも少なくとも2倍有効にTGaseIIポリペプチドの活性を阻害し、より好ましくは、阻害は、少なくともよりも10倍、50倍、さらに100倍有効である。このような方法では、TGaseIIポリペプチドまたは第二の化合物が固定されていてもよい。
【0127】
もう一つの態様において、提供した方法のいずれかによって得られた化合物は、抗体である。
【0128】
本発明は、さらに、TGaseIIポリペプチド、好ましくはヒトTGaseIIポリペプチドの活性を調整する化合物を得る方法であって:
(i)TGaseIIポリペプチドを、TGaseIIポリペプチドがインビボにおいて特異的に相互作用するインタラクターと接触させる工程と;
(ii)TGaseIIポリペプチドまたはインタラクターを該化合物と接触させる工程と;および、
(iii)線維症に関連したパラメーターを測定することにより、TGaseIIポリペプチドとインタラクターとの間の相互作用に対する化合物の効果を測定する工程と;および、
(iv)工程(iii)で測定された効果を、化合物の非存在下において測定された効果と比較する工程であって。測定された効果の変化は、化合物がヒトTGaseIIポリペプチドの活性を調整することを示す工程と、
の工程を含む方法を提供する。
【0129】
本発明の一つの態様において、提供された方法によって得られた化合物は、ヒトTGaseIIポリペプチドの活性を阻害する。
【0130】
本発明のもう一つの態様において、このような方法にTGaseIIポリペプチドは、配列番号:2に記載された配列の連続したアミノ酸を含む。さらなる態様において、TGaseIIポリペプチドまたはインタラクターは、固定されている。
【0131】
本発明のさらなる態様において、提供した方法のいずれかにおいて測定されるパラメーターは、一般のTGaseIIアップレギュレーションによって特徴づけられる病態に、上で定義したような線維症関連病態に、さらに白内障、心臓血管疾患、神経疾患、ハンチントン病(HD)を含むポリグルタミン疾患、球脊髄性筋肉萎縮症、歯状核赤核-淡蒼球ルイ体萎縮症、並びに脊髄小脳失調(SCA)1、2、3、6、7および17、アルツハイマー病およびパーキンソン病、小児脂肪便症および骨関節炎に関連がある。本発明のもう一つの目的は、ヒトTGaseIIポリペプチドの活性を阻害することができる化合物を得る方法であって:
(i)TGaseIIポリペプチドを発現するTGaseIIポリペプチドまたは細胞を、複数のタグを付けたリジル・ドナーである化合物と、生体分子であるグルタミル・ドナーと接触させる工程と、
(ii)工程(i)の混合物から過剰なタグを付けたリジル・ドナー洗浄する工程と;および、
(iii)複数の化合物の存在下においてタグに関連した読み出しを測定する工程と;
(iv)工程(iii)で測定されたタグ読み出しを、一定条件下で、複数の化合物の非存在下において測定されたものと比較する工程であって、読み出しレベルの減少は、複数の化合物によるTGaseIIポリペプチド活性の阻害を指し示す工程と;
(v)多数のうちのどの化合物がヒトTGaseIIの活性を阻害するかを別々に決定する工程と、
の工程を含む方法を提供することである。
【0132】
もう一つの本発明の局面は、TGaseIIポリペプチド、好ましくはヒトTGaseIIポリペプチドの活性を阻害することができる化合物を得る細胞に基づいた方法であって:
(i)活性なTGaseIIポリペプチドを発現する細胞を、化合物と、およびリジル・ドナーと接触させる工程と;
(ii)工程(i)の混合物から過剰なリジル・ドナーを除去する工程と;
(iii)化合物の存在下において産生されたリジル-グルタミル架橋された物質の量を測定する工程と;
(iv)工程(iii)で測定されたリジル-グルタミル架橋された物質の量を、一定条件下で、化合物の非存在下において測定されたものと比較する工程であって、リジル-グルタミル架橋された物質の量の減少は、化合物によってTGaseIIポリペプチド活性の阻害を指し示す工程と;および
(v)化合物をTGaseIIポリペプチドの阻害剤として同定する工程と、
の工程を含む方法を提供することである。
【0133】
もう一つの本発明の局面は、ヒトTGaseIIポリペプチドの活性を阻害することができる化合物を得る細胞に基づいた方法であって:
a. TGaseIIポリペプチドを発現する細胞を、複数の化合物と、およびリジル・ドナーと接触させる工程と;
b. 工程(i)の混合物から過剰なリジル・ドナーを除去する工程と;および、
c. 複数の化合物の存在下において産生されるリジル-グルタミル架橋された物質の量を測定する工程と;
d. 工程(iii)で測定されたリジル-グルタミル架橋された物質の量を、一定条件下で、複数の化合物の非存在下で測定されたものと比較する工程であって、リジル-グルタミル架橋された物質の量の減少は、複数の化合物によるTGaseIIポリペプチド活性の阻害を指し示す工程と;および、
(e)多数のうちのどの化合物がヒトTGaseIIの活性を阻害するかを別々に決定する工程と、
の工程を含む方法を提供することである。
【0134】
方法
分子生物学における一般的な方法
当技術分野において既知であり、かつ特に記述されていない標準的分子生物学技術は、一般に、Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, New York (1989)に、およびAusubel et al., Current Protocols in Molecular Biology, John Wiley and Sons, Baltimore, Maryland (1989)に、およびPerbal, A Practical Guide to Molecular Cloning, John Wiley & Sons, New York (1988)に、およびWatson et al., Recombinant DNA, Scientific American Books, New Yorkに、およびBirren et al (eds) Genome Analysis: A Laboratory Manual Series, Vols. 1-4 Cold Spring Harbor Laboratory Press, New York (1998)に記載されたとおりであり、並びに方法論は、米国特許第4,666,828号;第4,683,202号;第4,801,531号;第5,192,659号;および第5,272,057号に記載されており、これらは、参照により本明細書に援用される。ポリメラーゼ連鎖反応法(PCR)は、一般にPCR Protocols: A Guide To Methods And Applications, Academic Press, San Diego, CA (1990)と同様に行った。フローサイトメトリーと組み合わせたインサイチュウ(細胞における)PCRは、特異的DNAおよびmRNA配列を含む細胞の検出のために使用することができる(たとえばTestoni et al., 1996, Blood 87:3822)。また、RT-PCRを行う方法も、当技術分野において周知である。
【0135】
本発明は、実施例に関して、以下に詳細を例証してあるが、これらに限定されるものとして解釈されることはない。
【実施例】
【0136】
実施例1
活性なsiRNA化合物のデザイン
独占権下にあるアルゴリズムおよびTGaseIIのcDNAの既知の配列(配列番号:1)を使用して、多くの潜在性siRNAの配列を作製した。これらを下記の表に示してある。種のための略語は:H:ヒト;M:マウス;R:ラット;G:モルモット;C:ウシであることに留意されたい。
【0137】
表Aは、今までに化学的合成されており、かつ活性に関して試験された18個のsiRNAを示す(実施例2を参照されたい)。これらのsiRNAの全てが19merである。表Aにおいて、siRNA 1〜18のセンス鎖は、それぞれ配列番号:3〜20を有し、siRNA 1〜18のアンチセンス鎖は、それぞれ配列番号:21〜38を有する。
【0138】
下記の表Bは、独占権下にあるアルゴリズムによって作製されたが、いまだ活性について試験されていない153個のさらなる19mer siRNAを示す。表Bにおいて、siRNA 1〜153のセンス鎖は、それぞれ配列番号:39〜191を有し、siRNA 1〜153のアンチセンス鎖は、それぞれ配列番号:192〜344を有する。
【0139】
下記の表Cは、独占権下にあるアルゴリズムによって作製された36個のさらなる21mer siRNAを示す。表Cにおいて、siRNA 1〜36のセンス鎖は、それぞれ配列番号:345〜380を有し、siRNA 1-36のアンチセンス鎖は、それぞれ配列番号:381〜416を有する。
【表1】

【表2−1】
【表2−2】
【表2−3】
【表2−4】
【表2−5】
【表2−6】
【表3−1】
【表3−2】
実施例2
siRNA化合物の抗TGaseII活性についての試験
I. siRNA(二本鎖オリゴヌクレオチド)の使用液の調製
凍結乾燥したオリゴヌクレオチドを、RNAseを含まない二回蒸留水に溶解して100μMの終濃度に作製する。希釈したオリゴヌクレオチドを室温で15分間保持して、液体窒素中で直ちに凍結させた。オリゴヌクレオチドは、-80℃に貯蔵し、使用前にPBSで希釈した。
【0140】
II. ラットTGaseIIを安定に過剰発現するNRK49株化細胞の確立
PLXSNベクター(CLONTECH)にクローン化したラットTGaseIIの全長cDNAを、Rat1線維芽細胞株化細胞へのトランスフェクションに使用した。リポフェクトアミン試薬(Invitrogen)を、下記に記述されるプロトコルを使用するトランスフェクション試薬として使用した。トランスフェクションの48時間後、G418抗生物質(ネオマイシン)を使用して安定なクローンの選択を行った。このポリクローナル集団におけるTGaseIIの安定な過剰発現を、ウエスタンブロット法およびTGaseII活性アッセイ法によって検証した。細胞は、NRK49-TGaseII(TGASE-OE(登録商標))と命名した。
【0141】
III.NRK49におけるTGaseIIに関する活性のアッセイ法
4×103 NRK49-ラットTGASE-OE細胞を96ウェル・プレートにウェル毎に播種した。TGaseII活性のための細胞に基づいたアッセイ法を使用してTGaseII活性を測定した。簡潔には、増殖培地を細胞標識化培地(0.1%のBSA、25μM BC、10μM Calcimycin、16.2mM CaCl2のDMEM溶液)で37℃にて45分間置き換えた。45分後に、細胞を4%のホルムアルデヒドで固定して、ビオチン標識されたカダベリンの架橋を試験するために、0.6%のH2O2で処置して、ネトラビジン-HRP Abで染色した。吸光度をλ=450nmにて読み込んだ。規準化のために、細胞をヘキスト色素溶液で染色した。TGaseIIの活性は、NRK49対照細胞と比較して、NRK49-TGaseII細胞におけるよりも75倍高い。
【0142】
IV. リポフェクトアミン2000試薬を使用するsiRNAオリゴヌクレオチドによるトランスフェクション
2×105細胞を6ウェル・プレートのウェル毎に播種した。24時間後、細胞を以下の手順に従って、リポフェクトアミン2000試薬(Invitrogen)を使用してTGaseII特異的なsiRNAオリゴヌクレオチドをトランスフェクトした:
1. トランスフェクションの前に、細胞培地を抗生物質を含まない1500μlの新鮮な培地に置き換えた。
【0143】
2. 無菌プラスチックチューブに、リポフェクトアミン2000試薬(量は、1ウェルあたり5μlに従って算出する)を250μlの無血清培地に添加して、室温で5分間インキュベートした。
【0144】
3. もう一つのチューブに、siRNAオリゴヌクレオチド(ウェルあたりに所望の終濃度に適合するように様々の量)を250μlの無血清培地に添加した。
【0145】
4. リポフェクトアミン2000複合体をsiRNA溶液と合わせて、室温で20分間インキュベートした。
【0146】
5. 生じる混合物を細胞に滴下して、細胞を37℃にてsiRNA活性の解析までインキュベートした。
【0147】
V.トランスフェクトしたヒト、マウスおよびラット細胞における抗TGaseII siRNAの活性の試験
表Aの一覧表に記載したTGaseII siRNAの活性を異なる種に由来する3つの異なる株化細胞で試験した:
1. マウス特異的TGaseII siRNAは、マウスTGaseII NMUMG細胞を発現するマウス細胞において試験した。
【0148】
2. ヒト特異的TGaseII siRNAは、ヒトTgaseIIを発現するヒトHeLa細胞において試験した;図3を参照されたい。
【0149】
3. ラット特異的TGaseII siRNAは、ラットTGaseIIを過剰発現するNRK49(TGase-OE(登録商標)細胞)において試験した。
【0150】
siRNAのトランスフェクションの48時間〜6日後に、細胞を収集して、TGaseIIタンパク質の発現を特異的抗体でウエスタンブロット解析によって調べた。対照と比較して、TGaseIIに特異的なsiRNAオリゴヌクレオチドをトランスフェクトした細胞におけるTGaseIIポリペプチド発現の減少は、siRNAで誘発されるTGaseIIタンパク質発現に対する阻害作用を示し、これを定量化することができる。
【0151】
下記の表Dには、それぞれのsiRNAオリゴヌクレオチドのTGaseII阻害活性に関する情報を要約してある。表Dのオリゴヌクレオチドの名称は、表A.に使用した名称に対応する。比較は、異なるsiRNAによってトランスフェクトされたそれぞれの株化細胞における活性であり、株化細胞間の活性ではない。現在最も好ましいヒトsiRNAは、高いTGaseII阻害作用を誘発して、さらにマウス、ラットおよびモルモットTGaseIIを阻害したTG_HMRG1である。その他の好ましいヒトsiRNAは、TG_HMG1およびTG_HM1である。
【表4】
【0152】
実施例3
腎臓線維症の動物モデル
抗TGaseII siRNAの、もしくはTGaseIIに対する中和抗体の、またはその他のTGaseII阻害剤のインビボでの治療活性の試験は、疾患に関連したTGaseIIの過剰発現が見いだされた(インシチューハイブリダイゼイションによって検出された;下記参照)腎臓線維症の以下の動物モデルで行ってもよい。
【0153】
A. 腎臓線維症のRat糖尿病患者および老化モデル
A1. ZDFラット
9月齢のZDFラットの腎臓試料(Zucker糖尿病脂肪ラット)は、拡張型腎杯をもつ水腎症様変化を提示した。顕微鏡によって、これらの試料は、糸球体硬化症および尿細管間質性線維症の特色を示した。これらの形態学的変化に従って、インシチューハイブリダイゼイションによって測定される線維症マーカー遺伝子(オステオポンチン(OPN)、トランスフォーミング成長因子β1(TGF-β1)およびプロコラーゲン1(I)(Col1))の発現は、正常な腎臓と比較して有意に増大された。強力なOPN発現が、腎臓皮質および髄質の全ての管状構造で検出可能であった。TGF-β1発現は、間質細胞の全体にわたって広範囲におよんだ。また、一部の上皮細胞もTGF-β1発現を示した。Col1発現は、髄質内の大部分の間質細胞でインシチューハイブリダイゼイションによって検出可能であったが、皮質発現は「限局的」であった。
【0154】
A2. 高齢fa/fa(肥満Zucker)ラット
Fa/faラットは、遺伝的にレプチン受容体を欠損しており;その結果、進行性の糖尿病性腎障害を伴ったインスリン耐性糖尿病(II型糖尿病)を発病する。12月齢のfa/faラットの腎臓試料は、皮質および髄質の全体にわたって高度の糸球体硬化症および散在性の尿細管間質性線維症を呈する。線維症マーカー遺伝子発現のパターンは、形態学的変化に対応した。OPNは、皮質の管状構造に、および髄質に発現された。複数の間質細胞が、TGF-β1を発現した。複数の病巣および単一の間質細胞が、有意に、皮質および髄質における強力なCol1発現を示し、その結果、Col1発現細胞の数は、ZDF試料におけるよりもfa/fa腎臓試料において高いようであった。
【0155】
A3. 高齢SD系の(SD)(正常)ラット
また、高齢SD系ラットの腎臓試料は、糸球体および間隙におけるコラーゲンの蓄積が増大されたこと、および線維症マーカー遺伝子の発現が増加したことを示した。線維性変化の強度は、有意に、試料間で変化した。したがって、研究した4つの試料のうちの1つでは、幼若動物と比較して極めて少ない変化を示し;第二の試料では、線維性変化は、「極所」領域のみに限定されており、残りの2つの試料では、コラーゲンの一様な蓄積を示し、切片全体にわたってマーカー遺伝子の発現が上昇した。
【0156】
A4. 48週齢のGoto Kakizaki(GK)ラット
GKラットは、ウィスターラットに由来した、NIDDM(糖尿病II型)の自発的発症に関して選択された近交系である。GKおよび対照ウィスター48週齢ラットからの腎臓試料では、糸球体および間隙におけるコラーゲンの蓄積を示した。この蓄積は、GK試料においてよりはっきりしていた。
【0157】
TGaseIIハイブリダイゼーション・パターン
TGaseII特異的ハイブリダイゼーション・シグナルは、健康なラットおよび糖尿病ラットの腎臓における線維性表現型(散在性または局所性)と関連していた。非線維性腎臓試料は、近位尿細管全体に散在性で、低いが、検出可能なTGaseII特異的ハイブリダイゼーション・シグナルを示す。12月齢までに、より強力なシグナルが、明白な萎縮症の徴候を示す膨張する尿細管の間質細胞および上皮裏打ち全体に濃縮される。慢性腎不全試料(2歳のラットからの腎臓)では、強力なTGaseII発現が、間質を囲む範囲内の萎縮性または増殖性の上皮細胞に沿って並んだ管状プロフィールに見られる。同様に、ZDF試料では、線維症(または老化)の進行と共にハイブリダイゼーション・シグナルの増大を示した。
【0158】
B. 腎臓線維症誘導モデル。
【0159】
B1. 持続性一側性尿管閉塞(UUO)モデル
持続性一側性尿管閉塞(UUO)は、健康な非糖尿病動物に生じる急性間質性線維症のモデルである。腎臓線維症は、閉塞後数日以内に発病する。
【0160】
モデルを作製するために、ラットをケタミン/キシラジンで麻酔して、腹腔を開いた。曝露後、右の輸尿管を低い腎臓極レベルにて縫合により結紮した。見せかけの手術をしたラットでは、輸尿管を曝露したが、結紮しなかった。研究は、尿管閉塞後の種々の時点にて(たとえば、24時間、5日、10日、15日、20日および25日にて)終了し、腎臓を取り出して調べた。
【0161】
組織学的検査では、持続性UUOが、髄質および皮質の間質細胞によるコラーゲン合成の迅速な活性化を生じることが明らかになった。UUOの20〜25日までに、間質コラーゲンの有意な量が間隙に堆積したが、コラーゲンの糸球体蓄積は、外側のカプセルに限定された。
【0162】
閉塞腎臓試料のインシチューハイブリダイゼイション解析では、UUOの期間に依存したTGaseII発現のパターンの迅速かつ有意な変化を示した。UUOの24時間後、非常に強力なハイブリダイゼーション・シグナルが、皮質管状上皮全体に検出されたが、延髄上皮細胞は、非常に弱いシグナルを示した。UUOの5および10日目には、強力なハイブリダイゼーション・シグナルの「病巣」は、尿細管間質性の線維症を発症している領域と関連しており、ここでは、シグナルが上皮および間隙細胞に位置していた。同様の結果が、同じUUOモデルに曝露したマウスにおいても観察された。
【0163】
B2. 5/6腎摘除
5/6腎摘除は、明らかな線維症である慢性腎不全(CRI)のためのもう一つの有用な動物モデルである。
【0164】
要約すると、TGaseII発現および活性レベルの増加は、腎臓における線維性変化を伴うことを結果が示唆する。
【0165】
実施例4
TGaseIIノックアウト・マウスを使用するUUOモデルを使用する腎臓線維症におけるTGaseII役割の確立
ノックアウト・マウスは、Dr. Melino (De Laurenzi V, Melino G., Gene disruption of tissue transglutaminase, Mol Cell Biol. 2001 Jan;21(1):148-55)から得た。マウスは、表現型的に正常のように見えた。また、これらのマウスから得られる腎臓において、形態学的変化は観察されなかった。
【0166】
次いで、正常マウスと比較して腎臓コラーゲン蓄積を評価することによって、UUOの誘導後の腎臓線維症の発症の解析するために、TGaseII KOマウスを使用した。結果は、TGaseIIの破壊は、野生型マウスの閉塞腎臓におけるコラーゲン蓄積と比較すると、閉塞の三(3)および七(7)日に後の両方において、統計学的に有意なコラーゲン蓄積の減少を生じることを明らかに示し、したがって、TGaseIIは、腎臓線維症における機能的に重要な因子として関係づけられた。したがって、TGaseII活性の阻害は、抗線維治療効果を有し得ることが示唆される。
【0167】
実施例5
モノクローナル中和抗体を使用するTGaseII活性の阻害
TGaseIIに対する組換えモノクローナル・ヒト-抗ヒト抗体は、ファージディスプレイ・ライブラリーのスクリーニングによって得た。720クローンをヒトTGaseII結合についてスクリーニングした。酵素に対する強い結合を示した七(7)つのクローンを選択した。活性アッセイでは、これらの抗体が濃度範囲(1〜5nM)にてヒトTGaseII酵素の架橋活性を阻害することを示した。
【図面の簡単な説明】
【0168】
【図1】この図は、ヒトTGaseII cDNA(gi|13653650|ref|XM_009482.3| ORF )−配列番号:1のヌクレオチド配列を記載する。
【図2】この図は、ヒトTGaseIIに対応するポリペプチド−配列番号:2のアミノ酸配列を記載する。
【図3】ヒトTGaseIIポリペプチド発現に対する種々のTGaseII siRNAの効果を証明するウエスタンブロットの結果。図は、siRNAトランスフェクションの72時間後のHeLa細胞におけるヒトTGaseポリペプチドの発現を減少させる際の種々のsiRNAの活性を証明する。GAPDH発現は、タンパク質使用量のための対照として役立つ。数15および30は、オリゴヌクレオチドのnM濃度を表す。「細胞」(それぞれのパネルにおける第一のレーン)は、非トランスフェクト対照細胞を表す。それぞれのパネルの最後のレーンは、TGaseII発現における減少が抗TGase siRNA活性に特異的なことを保証するための、非関連(NR)siRNAをトランスフェクトしたHeLa細胞からのタンパク質抽出物を含む。
【技術分野】
【0001】
本出願は、2004年1月30日に出願された米国仮特許出願第60/540,687号および2005年1月4日に出願された第60/641,522号の優先権を主張し、両方とも、参照によりこれらの全体が本明細書に援用される。
【0002】
本出願の全体を通じて、種々の特許および科学的刊行物を引用してある。これらの刊行物の開示は、本発明が属する技術の水準をより完全に記述するために、その全体が参照により本明細書によって本出願に援用される。
【背景技術】
【0003】
siRNAおよびRNA干渉
RNA干渉(RNAi)は、二本鎖(ds)RNA依存的な遺伝子特異的な転写後サイレンシングを含む現象である。元来、この現象を研究して、実験的に哺乳動物細胞を操る試みは、長鎖dsRNA分子に応答して活性化された活性な非特異的抗ウイルス防衛機構によって頓挫していた;Gil et al. 2000、Apoptosis、5:107-114を参照されたい。後に、21ヌクレオチドRNAの合成二重鎖が、一般的な抗ウイルス防衛機構を刺激することなく哺乳動物細胞の遺伝子特異的RNAiを媒介することができることが発見された。Elbashir et al. Nature 2001、411:494-498およびCaplen et al. Proc Natl Acad Sci 2001、98:9742-9747を参照されたい。その結果、短い二重鎖RNAである短鎖干渉RNA(siRNA)は、遺伝子機能を理解することを試みる際の強力なツールとなった。
【0004】
したがって、RNA干渉(RNAi)は、短鎖干渉RNA(siRNA)(Fire et al, 1998, Nature 391, 806)またはミクロRNA(miRNA)(Ambros V. Nature 431:7006,350-355(2004);およびBartel DP. Cell. 2004 an 23; 116(2): 281-97 MicroRNAs: genomics, biogenesis, mechanism, and function)によって媒介される、哺乳類の配列特異的な転写後遺伝子サイレンシングの過程をいう。植物の対応する過程は、一般に特異的な転写後遺伝子サイレンシングまたはRNAサイレンシングといわれ、真菌では、クエリングともいわれる。siRNAは、その内因性の(細胞の)対応物の遺伝子/mRNAの発現をダウンレギュレートし、またはサイレンスする(防げる)二重鎖RNA分子である。RNA干渉は、dsRNA種が特定のタンパク質複合体に入り、次いでこれが相補的な細胞RNAにターゲットされて、これを特異的に分解する能力に基づいている。したがって、RNA干渉反応は、一般にRNA誘導サイレンシング複合体(RISC)と称される、siRNA二重鎖のアンチセンス鎖に対して相補的な配列を有する一本鎖RNAの切断を媒介するsiRNAを含むエンドヌクレアーゼ複合体を特徴とする。標的RNAの切断は、siRNA二重鎖のアンチセンス鎖に対して相補的な領域の中央で生じ得る(Elbashir et al 2001、Genes Dev., 15、188)。さらに詳細には、より長いdsRNAが、III型RNAses(DICER、DROSHA、その他、Bernstein et al., Nature、2001、v.409、p.363-6;Lee et al., Nature、2003、425、p.415-9)によって短い(17〜29bp)dsRNA断片(短鎖阻害RNA−「siRNA」とも称される)に消化される。RISCタンパク質複合体は、これらの断片および相補的mRNAを認識する。標的mRNAのエンドヌクレアーゼ切断により、全ての過程が完了する(McManus&Sharp、Nature Rev Genet、2002、v.3、p.737-47;Paddison &Hannon、Curr Opinmol Ther. 2003の6月;5(3):217-24)。これらの用語および提唱されたメカニズムについての情報に関しては、Bernstein E., Denli AM. Hannon GJ: 2001 The rest is silence. RNA. I; 7(11): 1509-21; Nishikura K.: 2001 A short primer on RNAi: RNA-directed RNA polymerase acts as a key catalyst. Cell. I 16; 107(4): 415-8 およびPCT公開WO01/36646号(Glover et al)を参照されたい。
【0005】
既知の遺伝子に対応するsiRNAの選択および合成は、広く報告されてきた;たとえば、Chalk AM, Wahlestedt C, Sonnhammer EL. 2004 Improved and automated prediction of effective siRNA Biochem. Biophys. Res. Commun. Jun 18; 319(1): 264-74; Sioud M, Leirdal M., 2004, Potential design rules and enzymatic synthesis of siRNAs, Methods Mol Biol.; 252:457-69; Levenkova N, Gu Q, Rux JJ. 2004 ,Gene specific siRNA selector Bioinformatics. I 12; 20(3): 430-2. and Ui-Tei K, Naito Y, Takahashi F, Haraguchi T, Ohki-Hamazaki H, Juni A, Ueda R, Saigo K., Guidelines for the selection of highly effective siRNA sequences for mammalian and chick RNA interference Nucleic Acids Res. 2004 I 9;32(3):936-48.Se also Liu Y, Braasch DA, Nulf CJ, Corey DR. Efficient and isoform-selective inhibition of cellular gene expression by peptide nucleic acids, Biochemistry, 2004 I 24;43(7):1921-7を参照されたい。また、修飾された/より安定なsiRNAの産生については、PCT公開WO2004/015107号(Atugen)およびWO02/44321号(Tuschl et al)、並びにChiu YL, Rana TM. siRNA function in RNAi: a chemical modification analysis, RNA 2003 Sep;9(9):1034-48およびI 特許第5898031号および6107094号(Crooke)も参照されたい。
【0006】
いくつかのグループは、細胞内でsiRNAを産生することができるDNAに基づいたベクターの開発を記述した。該方法は、一般に、細胞内で効率的にプロセスされてsiRNAを形成する短いヘアピンRNAの転写を含む。Paddison et al. PNAS 2002, 99:1443-1448; Paddison et al. Genes & Dev 2002, 16:948-958; Sui et al. PNAS 2002, 8:5515-5520; and Brummelkamp et al. Science 2002, 296:550-553。これらの報告は、多数の内因的および外因的に発現された遺伝子を特異的にターゲットすることができるsiRNAを作製するための方法を記述する。
【0007】
siRNAは、最近、霊長類における阻害のために首尾よく使用されている;詳しくは、Tolentino et al., Retina 24(1) February 2004 I 132-138を参照されたい。
【0008】
トランスグルタミナーゼ(TGase)ファミリー
トランスグルタミナーゼ(EC 2.3.2.13)は、イプシロン-γグルタミルリジンイソペプチド結合によるタンパク質の架橋を触媒する酵素のファミリーである。本ファミリーは、9つの異なる酵素を含み、その中には、第XIIIa因子(血漿トランスグルタミナーゼ)、ケラチノサイトトランスグルタミナーゼ(TGaseI)、表皮トランスグルタミナーゼ(TGaseIII)、前立腺トランスグルタミナーゼ(TGaseIV)および組織型トランスグルタミナーゼ(TGaseII)がある。これらの酵素の全体的な一次構造は異なるが、これらは全て、活性部位にて共通のアミノ酸配列(Y-G-Q-CW)と、これらの活性のための厳密なカルシウム依存性とを共有する(Lesort M, Tucholski J, Miller ML, Johnson GV, Tissue transglutaminase: a possible role in neurodegenerative diseases. Prog Neurobiol. 2000 Aug; 61(5):439-63)。
【0009】
トランスグルタミナーゼII
トランスグルタミナーゼII(TGaseII)(タンパク質-グルタミンγ-グルタミルトランスフェラーゼ、TGase C、TGC、TG(C)および組織型トランスグルタミナーゼとしても知られる)は、二重酵素活性をもつ独特の多機能酵素である:
1)本酵素は、架橋活性をもつTGaseタンパク質(Ca2+で活性化され、GTPで不活性化される)として作用する(すなわち、これは、タンパク質架橋および/または生体アミンの共有結合性の組み込みを生じる反応を触媒する)。さらに、TGaseは、共有結合性のグルタミルリジル結合(タンパク質分解および変性剤に高度に耐性であり、任意の既知の脊椎動物のエンドペプチダーゼによって崩壊することができない独特のイソペプチド結合)の形成を触媒する。
【0010】
2)また本酵素は、α1Bおよびα1dアドレナリン作動性受容体からの、TPαトロンボキサンA2受容体からの、およびオキシトシン受容体からホスホリパーゼCδ1への、活性化シグナルを伝達するGTP結合タンパク質として作用する(すなわち、これは、イノシトールリン酸産生、カルシウム動員などを活性化する)。α1Dアドレナリン作動性受容体およびPLCは、両方ともトランスグルタミナーゼIIのためにグアニンヌクレオチド交換因子として作用することが示された(Baek KJ, Kang S, Damron D, Im M, Phospholipase C-delta1 is a guanine nucleotide exchanging factor for transglutaminase II (G alpha h) and promotes alpha 1B-adrenoreceptor-mediated GTP binding and intracellular calcium release. J Biol Chem. 2001 Feb 23;276(8):5591-7)。
【0011】
この酵素の二つの機能は、分離しており、また活性部位は、異なる位置に位置しており(Im MJ, Russell MA, Feng JF, Transglutaminase II: a new class of GTP-binding protein with new biological functions. Cell Signal. 1997 Nov;9(7):477-82)、TGaseIIの架橋活性の阻害は、そのGタンパク質機能を妨げることなく達成することができる。
【0012】
TGaseII:細胞下局在および線維症関連細胞内シグナリング活性
TGaseIIは、一様に発現されており、ECMと会合して細胞内にて膜結合型として、およびサイトゾルタンパク質として見いだすことができる。GTP-結合活性は、TGaseの膜画分でより高く、架橋活性は、サイトゾルおよび細胞外画分でより高い。細胞表面上において、TGaseは、その42kDaのゼラチン結合ドメインを経てフィブロネクチンと結合する。コラーゲンとフィブロネクチンの架橋により、ECMの構造安定化に寄与し、マトリックスメタロプロテイナーゼ(mettaloproteinases)によるタンパク質分解に耐性のマトリックスになる。したがって、TGaseIIは、細胞外タンパク質を沈殿の方に有利にして、前記タンパク質の破壊を阻害することによって、組織線維症の形成の中心的現象であるECMの蓄積に寄与する。
【0013】
加えて、TGaseは、この前線維性因子のその後の適当なプロセシングのために必要とされる工程であるECMに対する潜在性TGF-βの結合を媒介し、最終的に、活性なTGF-βを放出することとなる(Le et al., Connect Tissue Res. 2001;42(4):245-53. Rosenthal et al., Arthritis Rheum. 2000 Aug;43(8):1729-33)。
【0014】
細胞質において、TGaseIIは、細胞骨格再配列、細胞形態制御および分化において重要な役割を有することが既知であるrasファミリーの小Gタンパク質であるRhoAを活性化する。TGaseIIの活性化により、RhoAのアミド基転移の増大を生じ、次いで構成的に活性なGタンパク質として機能して、その下流の標的ROCK-2(Rho関連キナーゼ)の結合の増大、およびその活性化を示す(Singh et al. EMBO J. 2001 May 15;20(10):2413-23)。特異的ROCKキナーゼ阻害剤のY-27632は、腎線維症のUUOモデルにおいて、並びに肝臓または肺線維症のモデルにおいて、抗線維性効果を示した(Nagatoya K, Moriyama T, Kawada N, Takeji M, Oseto S, Murozono T, Ando A, Imai E, Hori M. Y-27632 was found to prevent tubulointerstitial fibrosis in mouse kidneys with unilateral urethral obstruction. Kidney Int. 2002 May;61(5):1684-95; Murata T, Arii S, Nakamura T, Mori A, Kaido T, Furuyama H, Furumoto K, Nakao T, Isobe N, Imamura M, Inhibitory effect of Y-27632, a ROCK inhibitor, on progression of rat liver fibrosis in association with inactivation of hepatic stellate cells. J Hepatol. 2001 Oct;35(4):474-81; Shimizu Y, Dobashi K, Iizuka K, Horie T, Suzuki K, Tukagoshi H, Nakazawa T, Nakazato Y, Mori M, Contribution of small GTPase Rho and its target protein ROCK in a murine model of lung fibrosis. Am J Respir Crit Care Med. 2001 Jan;163(1):210-7)。最近、十分に実証された前線維性活性をもつ転写因子であるNFκBの活性化に寄与するTGaseIIの独特の機能が記述された(Baud et al., J Soc Biol. 2002;196(4):269-73, Lee at al. J Biol Chem. 2004 Dec 17;279(51):53725-35)、
線維性疾患
線維性疾患は全て、細胞外基質の繊維状物質の過剰な堆積によって特徴づけられ、これが組織構築における異常な変化に寄与し、正常な器官機能を妨げる。残念なことに、線維症は、広く一般的で、衰弱して生命が脅かされることが多いが、現在、有効な治療を提供されていない。
【0015】
外傷による損傷を受けた全ての組織は、創傷治癒プログラムの開始によって応答する。創傷治癒応答の正常な自己制御過程が妨げられると、過剰の瘢痕によって特徴づけられる障害の一種である線維症が生じ、コラーゲンの過剰産生および堆積を生じさせる。その結果、正常な器官組織は、瘢痕組織に置き換わり、これが最終的に器官の機能不全に至る。
【0016】
線維症は、多様な原因によって、および種々の器官において開始する可能性がある。肝硬変、肺線維症、サルコイドーシス、ケロイドおよび腎臓線維症は、全て進行性線維症と関連する慢性状態であり、これにより正常組織機能の持続的な喪失が生じる。
【0017】
急性線維症(通常、突然および重篤で、かつ短期間に発症する)は、偶発性傷害(特に、脊椎および中枢神経系に対する傷害)、感染、外科手術、虚血性の疾病(たとえば、心臓麻痺後の心臓瘢痕)、やけど、環境汚染物、アルコールおよびその他のタイプの毒素、急性呼吸不全症候群、放射線、並びに化学療法治療を含む外傷の種々の形態に共通の応答として生じる。
【0018】
線維症の種々のタイプについてのさらなる情報は、Molina V, Blank M, Shoenfeld Y. (2002), “Fibrotic diseases”, Harefuah, 141(11): 973-8, 1009; Yu L, Noble NA, Border WA (2002), “Therapeutic strategies to halt renal fibrosis", Curr Opin Pharmacol. 2(2):177-81; Keane WF, Lyle PA. (2003), "Recent advances in management of type 2 diabetes and nephropathy: lessons from the RENAAL study", Am J Kidney Dis. 41(3 Suppl 2): S22-5; Bohle A, Kressel G, Muller CA, Muller GA. (1989), "The pathogenesis of chronic renal failure", Pathol Res Pract. 185(4):421-40; Kikkawa R, Togawa M, Isono M, Isshiki K, Haneda M. (1997), "Mechanism of the progression of diabetic nephropathy to renal failure", Kidney Int Suppl. 62:S39-40; Bataller R, Brenner DA. (2001), "Hepatic stellate cells as a target for the treatment of liver fibrosis", Semin Liver Dis. 21(3):437-51; Gross TJ, Hunninghake GW, (2001) "Idiopathic pulmonary fibrosis", N Engl J Med. 345(7):517-25; Frohlich ED. (2001) "Fibrosis and ischemia: the real risks in hypertensive heart disease", Am J Hypertens;14(6 Pt 2):194S-199Sを参照されたい。
【0019】
肝臓線維症
肝臓線維症(LF)は、一般に回復不能ないくつかの病因の肝臓損傷の結果である。西洋世界では、主な病因カテゴリーは:アルコール性肝疾患(30〜50%)、ウイルス性肝炎(30%)、胆汁疾患(5〜10%)、原発性血色素症(5%)および病因が未知の薬物関連および原因不明の肝硬変(10〜15%)。また、ウイルソン病、α1-アンチトリプシン欠損およびその他のまれな疾患では、症候Scheinberg IH、Sternlieb I、ウイルソン病および特発性銅中毒症のうちの1つとして肝臓線維症を有する。Am J Clin Nutr 1996 May;63(5):842S-5S; Parfrey H, Mahadeva R, Lomas DA., Alpha(1)-antitrypsin deficiency, liver disease and emphysema. Int J Biochem Cell Biol. 2003 Jul;35(7):1009-14。
【0020】
肝臓線維症の最終段階である肝硬変では、頻繁な肝移植が必要であり、西洋世界における上位10位の死因の一つである。
【0021】
肝臓星状細胞の活性化の阻害、肝細胞の増殖刺激およびコラーゲンの翻訳後修飾の阻害を生じさせる抗炎症薬は、全て肝臓線維症を治療するために使用されてきた。しかし、選択的ターゲティングが不十分であるために、とりわけこれらの治療では、重篤な副作用という欠点を被ってしまう。
【0022】
詳細は、Friedman SL. (2003), “Liver fibrosis - from bench to bedside”, J Hepatol. 38 Suppl 1:S38-53; Albanis E, Safadi R, Friedman SL. (2003), "Treatment of hepatic fibrosis: almost there", Curr Gastroenterol Rep. 5(1):48-56. See also Grenard P, Bresson-Hadni S, El Alaoui S, Chevallier M, Vuitton DA, Ricard-Blum S., Transglutaminase-mediated cross-linking is involved in the stabilization of extracellular matrix in human liver fibrosis. J Hepatol. 2001 Sep;35(3):367-75 ;Mirza A, Liu SL, Frizell E, Zhu J, Maddukuri S, Martinez J, Davies P, Schwarting R, Norton P, Zern MA., A role for tissue transglutaminase in hepatic injury and fibrogenesis, and its regulation by NF-kappaB. Am J Physiol. 1997 Feb;272(2 Pt 1):G281-8)を参照されたい。
【0023】
腎臓線維症および関連状態
慢性腎不全(CRF)
慢性腎不全は、腎臓が排泄物を排出し、尿を濃縮し、および電解質を保存する能力が徐々に、かつ進行性に失われる。CRFは、ゆっくりと進行する。これは、多くの場合、腎機能の段階的な喪失を生じさせる何らかの疾患によって生じ、線維症は、CRFを生じる主な病態である。
【0024】
糖尿病性腎障害
糖尿病性腎障害(その特徴は、糸球体硬化症および尿細管間質性線維症である)は、現代世界において、唯一末期腎疾患に多い原因であり、糖尿病患者は、透析を行う最大集団を構成する。このような療法は、費用がかかり、決して最適とはいえない。移植は、より優れた結果をもたらすが、ドナーの極度な不足に悩まされている。糖尿病性腎障害に対する(並びに、腎病態のその他のタイプに対する)よりターゲットされた療法は、これらの病態の基礎をなす分子機構の大部分が未知であるために、開発されていない。疾患において調整され、かつ糖尿病腎症の結果の重症度に影響を及ぼす必須な機能的標的遺伝子の同定は、診断的、並びに治療的に高い価値を有する。
【0025】
腎臓病態の起源
腎臓における多くの病理学的過程(たとえば糸球体腎炎、物理的な閉塞、中毒性傷害、代謝性および免疫学的疾患)は、最終的に同様の、または同一の形態学的変化、すなわち糸球体硬化症および尿細管間質性線維症となる。したがって、異なるタイプの傷害により、同じ単一の遺伝プログラムに収束して2つの線維症の特徴(線維芽細胞の増殖およびこれらによる結合組織の種々のタンパク質成分の過剰産生)を生じる。加えて、糸球体における基底膜の肥厚は、間質性線維症を伴い、結果的に糸球体硬化症になる。また、Johnson TS, Skill NJ, El Nahas AM, Oldroyd SD, Thomas GL, Douthwaite JA, Haylor JL, Griffin M, Transglutaminase transcription and antigen translocation in experimental renal scarring. J Am Soc Nephrol. 1999 Oct;10(10):2146-57; Johnson TS, Griffin M, Thomas GL, Skill J, Cox A, Yang B, Nicholas B, Birckbichler PJ, Muchaneta-Kubara C, Meguid El Nahas A, The role of transglutaminase in the rat subtotal nephrectomy model of renal fibrosis. J Clin Invest. 1997 Jun 15;99(12):2950-60)を参照されたい。
【0026】
肺線維症
間質性肺線維症(IPF)は、ミネラル粒子、有機性粉末およびオキシダントガスを含む種々の吸入によって、または未知の原因によって(特発性肺線維症)生じる肺の瘢痕である。本疾患は、世界的に何百万もの個体を苦しめ、有効な治療手段はない。有用な治療法がない主な理由は、疾患の分子機構が、療法のための適切な標的をデザインするほど十分に定義されていないことである(Lasky JA., Brody AR. (2000), “Interstitial fibrosis and growth factors”, Environ Health Perspect.;108 Suppl 4:751-62)。
【0027】
肺線維症の病原の細胞基質は、内皮および上皮細胞障害、炎症細胞およびこれらのメディエータの産生、並びに線維芽細胞活性化を含む。線維症は、種々のサイトカイン、ケモカインおよび成長因子によって媒介される炎症性細胞と構造細胞との間のクロストーク(これは、組織恒常性維持の役割を担い、かつ傷害に対する応答を調整する)の調節不全に関連があると考えられている(Kelly M, Kolb M, Bonniaud P, Gauldie J. (2003), “Re-evaluation of fibrogenic cytokines in lung fibrosis”, Curr Pharm Des. 9(1):39-49 )。
【0028】
糖質コルチコイドまたは細胞毒からなる従来の療法は、通常、疾患の進行の予防には効果がない。肺線維症に対する有効な治療の開発のためには、内皮細胞および上皮細胞傷害、炎症反応、線維芽細胞増殖、コラーゲン沈着および肺修復の分子機構をさらに理解することが必要であると考えられている(Kuwano K, Hagimoto N, Hara N. (2001), “Molecular mechanisms of pulmonary fibrosis and current treatment”, Curr Mol Med. 1(5):551-73)。また、Griffin M, Smith LL, Wynne J., Changes in transglutaminase activity in an experimental model of pulmonary fibrosis induced by Paraquat. Br. J. Exp. Pathol. 1979 Dec;60(6):653-61 )も参照されたい。
【0029】
心臓線維症
心不全は、それ単独での有病率が増大しているものの、一方でその他の状態は、著明に減少してきたという点で、主要な心血管障害の中では独特である。この一部には、米国および欧州の人口の老化に起因すると考えることができる。また、これらの患者は、心臓の有害な再造形によって左心室機能不全の進行を発症する可能性があるので、心筋障害をもつ患者を救う能力が主要な要因となる。
【0030】
正常な心筋は、種々の細胞、心臓筋細胞および非心筋細胞で構成され、これらには内皮および血管平滑筋細胞、並びに線維芽細胞を含む。(Weber KT. (2000), “Fibrosis and hypertensive heart disease”, Curr Opin Cardiol. 15(4):264-72)。
【0031】
心室壁の構造の再造形は、心疾患における臨床結果の重要な決定因子である。このような再造形は、細胞外基質タンパク質、細胞増殖および遊走、並びにアポトーシス性および壊死性細胞死の発生や破壊を含む。心臓線維芽細胞は、これらの過程に決定的に関与し、自己分泌因子およびパラ分泌因子として作用する成長因子およびサイトカイン、並びに細胞外基質タンパク質およびプロテイナーゼを産生する。最近の研究では、心臓線維芽細胞と心筋細胞との間の相互作用が、最終的効果が心機能の悪化および心不全の発症である心臓再造形の進行に必須であることが示された(Manabe I, Shindo T, Nagai R. (2002), “Gene expression in fibroblasts and fibrosis: involvement in cardiac hypertrophy”, Circ Res. 13;91(12):1103-13)。
【0032】
レニン-アンジオテンシン-アルドステロンおよび交感神経系を遮断する薬剤の使用により、心臓再造形を阻害し(および、時には逆転さえも)、心臓機能障害患者の臨床経過を改善することが示された。しかし、線維症の直接の抑制または減少に狙った薬物は、いまだ利用できない。また、Greenberg B. (2001), “Treatment of heart failure: state of the art and prospectives", J Cardiovasc Pharmacol. 38 Suppl 2:S59-63, Zhang Z, Vezza R, Plappert T, McNamara P, Lawson JA, Austin S, Pratico D, Sutton MS, FitzGerald GA, COX-2-dependent cardiac failure in Gh/tTG transgenic mice. Circ Res. 2003 May 30;92(10):1153-61. Epub 2003 Apr 17; Hwang KC, Gray CD, Sweet WE, Moravec CS, Im MJ., Alpha 1-adrenergic receptor coupling with Gh in the failing human heart. Circulation. 1996 Aug 15;94(4):718-26を参照されたい。
【0033】
神経系疾患
ポリグルタミン疾患は、ポリグルタミン挿入をコードするCAGトリヌクレオチド・リピートの増大することによって生じる一群の神経系疾患である。ポリグルタミン疾患は、ハンチントン病(HD)、球脊髄性筋肉萎縮症、歯状核赤核-淡蒼球ルイ体萎縮症および脊髄小脳失調(SCA)1、2、3、6、7および17を含む。これらの疾患は全て、ポリグルタミンの一続き(35〜40グルタミンを上回る)の増大が存在し、したがって、核内凝集体を形成して、ニューロンの死に至ることによって特徴づけられる。アルツハイマー病(AD)は、老齢患者の認知障害において最も一般的な原因であり、有病率が非常に増大することが予想される。対になったらせん状フィラメント(PHF)の形成と関連する神経原線維変性は、アルツハイマー病(AD)の重要な神経病理学的特徴のうちの1つである。パーキンソン病は、黒質内のドーパミン作動性ニューロンの選択的かつ進行性の喪失によって特徴づけられる老化における神経変性障害である。また、Mastroberardino PG, Iannicola C, Nardacci R, Bernassola F, De Laurenzi V, Melino G, Moreno S, Pavone F, Oliverio S, Fesus L, Piacentini M. Tissue transglutaminase ablation reduces neuronal death and prolongs survival in a mouse model of Huntington's disease. Cell Death Differ. 2002 Sep;9(9):873-80; Karpuj MV, Becher MW, Springer JE, Chabas D, Youssef S, Pedotti R, Mitchell D, Steinman L., Prolonged survival and decreased abnormal movements in transgenic model of Huntington disease, with administration of the transglutaminase inhibitor cystamine. Nat Med. 2002 Feb;8(2):143-9; Citron BA, Suo Z, SantaCruz K, Davies PJ, Qin F, Festoff BW., Protein crosslinking, tissue transglutaminase, alternative splicing and neurodegeneration. Neurochem Int. 2002 Jan;40(1):69-78; Chen JS, Mehta K., Tissue transglutaminase: an enzyme with a split personality. Int J Biochem Cell Biol. 1999 Aug;31(8):817-36を参照されたい。
【0034】
骨関節炎
骨関節炎の主な特徴の中には、関節軟骨の分解および関節の縁、いわゆる骨増殖体における新たな骨形成がある。Van den Berg WB., Growth factors in experimental osteoarthritis: transforming growth factor beta pathogenic? J Rheumatol Suppl. 1995 Feb;43:143-5; Scharstuhl A, Glansbeek HL, Van Beuningen HM, Vitters EL, Van der Kraan PM, Van den Berg WB., Inhibition of endogenous TGF-beta during experimental osteoarthritis prevents osteophyte formation and impairs cartilage repair. J Immunol. 2002 Jul 1;169(1):507-14; Karpouzas GA, Terkeltaub RA., New developments in the pathogenesis of articular cartilage calcification. Curr Rheumatol Rep. 1999 Dec;1(2):121-7 を参照されたい。
【0035】
眼疾患
年齢関連性白内障:白内障は、眼のレンズの主要な構造タンパク質であるクリスタリンの凝集および共有結合性架橋によって特徴づけられ、光散乱の増大、不透明化および白内障を生じる。組織におけるカルシウムホメオスタシスの障害は、白内障発生に関係する要因のうちの1つである。Shridas et al., FEBS Lett. 2001 Jun 22;499(3):245-50; Shin et al, J Biol Chem. 2004 Apr 9;279(15):15032-9; Wan et al., Br J Ophthalmol. 2002 Nov;86(11):1293-8. and Takeuchi N, Kamei A. Biol Pharm Bull. 2000 Mar;23(3):283-90を参照されたい。
【0036】
増殖性硝子体網膜症
増殖性硝子体網膜症(PVR)は、網膜剥離後の最も一般的な合併症であり、網膜の穴または破壊が付随する。PVRは、硝子体腔内の、並びに網膜色素上皮(RPE)細胞を含む網膜の正面上の、および裏手表面の細胞膜の増殖をいう。本質的に瘢痕組織であるこれらの膜は、網膜に対して牽引する作用があり、最初の良好な網膜剥離手順の後でさえも、網膜剥離の再発を生じ得る。
【0037】
網膜色素上皮(RPE)細胞の位置を変えるフィブロネクチンリッチな細胞外基質への遊走および接着は、増殖性硝子体網膜症(PVR)の初期工程である。また、Casaroli-Marano RP et al Invest Ophthalmol Vis Sci. 1999 Aug;40(9):2062-72). and Priglinger S et al, Invest Ophthalmol Vis Sci. 2004 Mar;45(3):955-63; Priglinger et al., Invest Ophthalmol Vis Sci. 2003 Jan;44(1):355-64を参照されたい。
【0038】
線維症に続く緑内障濾過手術
緑内障濾過術の目標は、眼内部の水性流体を逃がすことができる新たな通路を作製することより、圧力を下げることである。したがって、濾過により、眼の前房内部から、眼の最も外側の被覆である結膜と眼の根底にある白い解剖学的構造である強膜との間の「ポケット」へ液体を排水することができる。液体は、最終的に血管によって吸収される。都合の悪いことに、術後の瘢痕のために、作製された排水系が遮断され、さらなる外科的介入必要でとなる可能性もある。
【0039】
現在の抗瘢痕療法(マイトマイシンCまたは5FU)は、関連した合併症(たとえば、失明)のために、制限される。たとえば、Cordeiro MF, Gay JA, Khaw PT., Human anti-transforming growth factor-beta2 antibody: a new glaucoma anti-scarring agent Invest Ophthalmol Vis Sci. 1999 Sep;40(10):2225-34を参照されたい。
【0040】
結論として、上記した疾患のための療法の有効な方法がなく、したがって、これらの目的のための治療の新規の有効な化合物および方法を開発する需要がある。
【発明の開示】
【0041】
本発明は、新規二本鎖オリゴリボヌクレオチドを提供する。これらのオリゴリボヌクレオチドは、RNA干渉のメカニズムを経てヒトTGaseIIを阻害する。また、本発明は、このようなオリゴリボヌクレオチドを含む薬学的組成物およびオリゴリボヌクレオチドを発現することができるベクターを提供する。また、本発明は、線維症関連病態に罹患した患者を治療する方法であって、これにより患者を治療するために、患者に対して、典型的には薬学的組成物として治療的に有効な用量のオリゴリボヌクレオチドを投与することを含む方法を提供する。また、本発明は、その他の疾患および状態を治療することも想定する。本発明は、TGaseIIポリペプチドに対する抗体を使用することによる、線維性疾患およびその他の疾患の治療にも関する。
【発明を実施するための最良の形態】
【0042】
発明の詳細な説明
本発明は、下記に詳述したような種々の病態の、このような治療を必要とする被検者における治療のための方法であって、これにより被検者を治療するために、被検者に対してTGaseII活性の実質的阻害を生じさせるために十分なTGaseIIポリペプチドの阻害剤の量を投与することを含む方法を提供する。TGaseII阻害剤は、L683685、TGaseIIポリペプチドに対する抗体もしくはTGaseII RNAに対するsiRNA、または当技術分野において既知のTGaseII阻害剤のいずれかであってもよい(このような阻害剤についての詳細は、たとえば、米国特許番号5021440号;第4968713号および第5098707号を参照されたい)。特に、本発明は、TGaseII RNAをターゲットする特異的siRNAおよびこれらの使用に関する。また、TGaseIIに対する中和抗体の使用も開示してある。
【0043】
本発明は、一般に、ヒトTGaseII遺伝子の発現をダウンレギュレートする化合物に、特に新規短鎖干渉RNA(siRNA)に、種々の疾患および医学的状態、特に線維性疾患、線維性瘢痕に関連した疾患、およびその他のトランスグルタミナーゼIIの異常な発現に関連した疾患の治療におけるこれらの新規siRNA特定の使用に関する。
【0044】
本発明は、インビボで標的TGaseII遺伝子の発現を阻害するための方法および組成物を提供する。一般に、本方法は、生物学的条件下で(細胞内で)特定のmRNAにターゲットされて、これとハイブリダイズするか、もしくは相互作用する短鎖干渉RNA(すなわち、siRNA)か、または細胞においてsiRNAを産生することができる核酸材料などのオリゴリボヌクレオチドを、RNA干渉メカニズムによって標的遺伝子の発現をダウンレギュレートするために十分な量で投与することを含む。特に、本方法は、疾患治療のために、TGaseII遺伝子の発現を阻害するために使用することができる。
【0045】
したがって、TGaseII発現(転写または翻訳)またはポリペプチド活性の阻害剤は、とりわけ、単鎖抗体、アンチセンスオリゴヌクレオチド、アンチセンスDNAまたはRNA分子、タンパク質、ペプトイド擬態を含むポリペプチドおよびペプチド、並びにドミナントネガティブを含むsiRNA、抗体、好ましくは中和抗体またはこれらの断片、さらには上記の全てを発現する発現ベクターであってもよい。さらなる阻害剤は、一般に2000ダルトン未満、より好ましくは1000ダルトン未満、さらに好ましくは500ダルトン未満の分子量を有する小さな化学分子であってもよい。これらの阻害剤は、以下のとおりに作用し得る:小分子は、発現および/または活性に影響を及ぼし得るし;抗体は、活性に影響を及ぼし得るし;各種アンチセンスは、TGaseII発現に影響を及ぼし得るし;ドミナントネガティブ・ポリペプチドおよびペプトイド擬態は、活性に影響を及ぼし得るし;発現ベクターは、とりわけアンチセンスまたはドミナントネガティブ・ポリペプチドまたは抗体の送達のために使用し得る。
【0046】
本発明によれば、siRNA分子または抗体などのトランスグルタミナーゼIIの阻害剤は、線維症関連病態(下記に定義したとおり)を含む種々の病態を治療するための、さらに白内障を含む眼疾患、心臓血管疾患、神経系疾患、ポリグルタミン疾患(ハンチントン病(HD)、球脊髄性筋肉萎縮症、歯状核赤核-淡蒼球ルイ体萎縮症、並びに脊髄小脳失調(SCA)1、2、3、6、7、および17を含む)、アルツハイマー病およびパーキンソン病、並びに骨関節炎を治療する薬物として使用してもよい。
【0047】
本明細書に使用される「トランスグルタミナーゼII遺伝子」または「TGaseII遺伝子」、または「TGase遺伝子」という用語は、配列番号:1のアミノ酸コードする領域に対して好ましくは90%の相同性、より好ましくは95%の相同性、およびさらにより好ましくは98%の相同性か、または高度にストリンジェントなハイブリダイゼーション条件下でTGaseII遺伝子と結合する核酸配列を有するTGaseII遺伝子の相同体のいずれかとして定義され、これらは、当技術分野において周知である(たとえば、Ausubel et al., Current Protocols in Molecular Biology, John Wiley and Sons, Baltimore, Maryland (1988), updated in 1995 and 1998を参照されたい)。
【0048】
本明細書に使用される「トランスグルタミナーゼIIポリペプチド」または「TGaseIIポリペプチド」、または「TGase」という用語は、配列番号:2に対して好ましくは90%の相同性、より好ましくは、95%の相同性およびさらにより好ましくは98%の相同性を有するTGaseIIポリペプチドの相同体のいずれかとして、全長またはこれらの断片もしくはドメインのいずれかとして、スプライス変異体核酸配列によってコードされるポリペプチドの突然変異体として、その他のポリペプチドとのキメラとして定義されるが、ただし、上記のいずれも、TGaseIIポリペプチドと同じか、または実質的に同じ生物学的機能を有することを条件とする。TGaseIIポリペプチドまたはTGaseIIポリペプチド相同体は、可溶性タンパク質、膜結合(精製された膜標品で、または細胞表面上のいずれか)、ビーズ結合、またはTGaseIIタンパク質またはこれらの断片および誘導体ポリペプチドを提示するその他の何らかの形態含む(しかし、これらに限定されない)、種々の形態で存在してもよい。
【0049】
本明細書に使用される、「インタラクター」は、性質上、TGaseIIが結合し、または相互作用し、または活性化する分子;たとえば、TGaseIIポリペプチドを発現する細胞の表面上の分子、第二の細胞表面上の分子、または細胞質分子である。インタラクターは、TGaseII単独によって、またはその他の成分との複合体の一部としてのTGaseIIによって活性化されるリガンドであってもよい。インタラクターは、TGaseIIから細胞膜を通って細胞内への細胞外シグナルの伝達を容易にするシグナル伝達経路の成分であってもよい。インタラクターは、たとえばTGaseIIからの下流のシグナリングを媒介する第二の細胞間のタンパク質であることができる。インタラクターは、TGaseが既知のTGase基質(たとえば、フィブロネクチン)と競合して結合する分子である。
【0050】
本明細書に使用される「リジル・ドナー」または「Kドナー」という用語は、リジル側鎖を供与してグルタミン転移過程の間にγ-グルタミル-リジン結合を形成させる能力を有する何らかのポリペプチドとして定義される。
【0051】
本明細書に使用される「グルタミル・ドナー」または「Qドナー」という用語は、グルタミン側鎖を供与してグルタミン転移過程の間にγ-グルタミル-リジン結合を形成させる能力を有する何らかのポリペプチドとして定義される。
【0052】
本発明は、TGaseIIの発現をダウンレギュレートする二本鎖オリゴリボヌクレオチド(siRNA)を提供する。トランスグルタミナーゼIIの発現のダウンレギュレーションは、たとえばsiRNAの存在下において生じるリジル-グルタミル架橋された材料の量を測定することによって、またはTGaseII mRNAまたはポリペプチドの量を直接評価することによって測定することができる。TGaseII mRNAの量は、たとえばノーザンブロッティング、RNase保護、RT-PCRまたはリアルタイムPCRによって測定してもよい。TGaseIIポリペプチドの量は、免疫ブロットによって、または免疫沈降によって、またはTGaseII特異的抗体でのELISAによって測定してもよい。
【0053】
本発明のsiRNAは、センス鎖が遺伝子TGaseIIのmRNA配列に由来する二重鎖オリゴリボヌクレオチドであり、かつアンチセンス鎖がセンス鎖に対して相補的である。一般に、標的mRNA配列からのいくらかずれがあっても、siRNA活性を損なわずに、許容される(たとえばCzauderna et al 2003 Nucleic Acids Research 31(11), 2705-2716)。本発明のsiRNAは、mRNAを破壊すると共に、または破壊することなく、転写後レベルの遺伝子発現を阻害する。理論に拘束されないが、siRNAは、特異的切断および分解のためにmRNAをターゲットし得るし、および/またはターゲットされたメッセージからの翻訳を阻害し得る。
【0054】
少なくとも2つの変異体TGaseIIポリペプチドがあり、これらに関して、GeneBank参照は、変異体2 NM_198951.1 GI:39777598および変異体1NM_004613.2 GI:39777596である図1に示した配列は、変異体2(GI:39777598)のヌクレオチド配列である。両変異体およびその他の何らかの同様の軽微な変異体も、TGaseIIポリペプチドの定義に、およびこれらをコードするTGaseII遺伝子の定義に含まれる。
【0055】
本明細書に使用される「TGaseII遺伝子」という用語は、天然に存在するヒト遺伝子(これらの対立遺伝子変異体のいずれか、または配列番号:1のアミノ酸コードする領域に対して好ましくは90%の相同性、より好ましくは95%の相同性、およびさらに好ましくは98%の相同性か、もしくは高度にストリンジェントなハイブリダイゼーション条件下でTGaseII遺伝子と結合する核酸配列を有するTGaseII遺伝子の相同体のいずれかを含む)として定義され、これらは、当技術分野において周知である(たとえば、Ausubel et al., Current Protocols in Molecular Biology, John Wiley and Sons, Baltimore, Maryland (1988), updated in 1995 and 1998を参照されたい。
【0056】
本明細書に使用される「TGaseII」または「TGaseIIポリペプチド」という用語は、配列番号:2に対して好ましくは90%の相同性、より好ましくは、95%の相同性およびさらにより好ましくは98%の相同性を有するTGaseIIポリペプチドの相同体のいずれかとして、全長またはこれらの断片もしくはドメインのいずれかとして、スプライス変異体核酸配列によってコードされるポリペプチドの突然変異体として、その他のポリペプチドとのキメラとして定義されるが、ただし、上記のいずれも、TGaseIIポリペプチドと同じか、または実質的に同じ生物学的機能を有することを条件とする。
【0057】
より詳しくは、本発明は、以下の構造を有する化合物を提供する:
5'(N)x-Z3'(アンチセンス鎖)
3'Z'-(N’)y5'(センス鎖)
式中、それぞれのNおよびN’は、その糖残基が修飾されていても、または修飾されていなくてもよいリボヌクレオチドであり、かつ(N)xおよび(N’)yは、それぞれの連続したNまたはN’が、共有結合によって次のNまたはN’に連結されているオリゴマーであり;
式中、それぞれのxおよびyは、19〜40の間の整数であり;
式中、それぞれのZおよびZ’は、存在しても、または存在しなくてもよいが、存在する場合、dTdTであり、かつそれが存在する鎖の3'末端にて共有結合で付着されており;および、
式中、(N)xの配列は、表A、BおよびCに存在するアンチセンス配列のいずれか一つを含む。
【0058】
本発明の化合物が、共有結合を介して連結された複数のヌクレオチドからなることは、当業者には容易に理解されるであろう。このようなそれぞれの共有結合は、個々の鎖のヌクレオチド配列に沿って、ホスホジエステル結合、ホスホチオアート結合、または両方の組み合わせであってもよい。その他の可能性バックボーン修飾は、とりわけ米国特許第5,587,361号;第6,242,589号;第6,277,967号;第6,326,358号;第5,399,676号;第5,489,677号;および第5,596,086号に記述されている。
【0059】
特定の態様において、xおよびyは、好ましくは約19〜約27の間、最も好ましくは約19〜約23の整数である。本発明の化合物の特定の態様において、xは、yに等しく(すなわちx=y)、好ましい態様において、x=y=19またはx=y=21である。特に好ましい態様において、x=y=19である。
【0060】
一つの態様において、本発明の化合物の中で、ZおよびZ’は、両方とも存在せず;もう一つの態様において、ZまたはZ’のうちの1つが存在する。
【0061】
一つの態様において本発明の化合物の中で、化合物のリボヌクレオチドの全てにおいて、これらの糖残基が修飾されていない。
【0062】
本発明の化合物のいくつかの態様において、少なくとも1つのリボヌクレオチドは、その糖残基が修飾されている(好ましくは2’位の修飾)。2’位の修飾により、アミノ基、フルオロ基、メトキシ基、アルコキシ基およびアルキル基を含む群から選択される部分が存在することとなる。現在最も好ましい態様において、2’位の部分は、メトキシ(2’-0-メチル)である。
【0063】
本発明のいくつかの態様において、交互に現れるリボヌクレオチドが、アンチセンス鎖およびセンス鎖において修飾されている。
【0064】
本発明の特に好ましい態様において、アンチセンス鎖は、5'末端にてリン酸化されており(phophorylated)、かつ3'末端にてリン酸化されていても、またはリン酸化されていなくてもよく、かつセンス鎖は、5'末端にて、および3'末端にてリン酸化されていても、またはリン酸化されていなくてもよい。
【0065】
もう一つの態様において、本発明の化合物の中で、アンチセンス鎖の5'および3'末端のリボヌクレオチドは、これらの糖残基が修飾されており、かつセンス鎖の5'および3'末端のリボヌクレオチドは、これらの糖残基が修飾されていない。
【0066】
本発明は、細胞内にて無修飾形態で上述したオリゴリボヌクレオチドのいずれかを発現し、その後に適切な修飾を生じることができるベクターをさらに提供する。
【0067】
また、本発明は、本発明の化合物の1つまたは複数を、キャリア(好ましくは薬学的に許容されるキャリア)中に含む組成物を提供する。
【0068】
また、本発明は、キャリアと、ヒトTGaseII遺伝子の細胞における発現をダウンレギュレートするために有効な量で、実質的に(N)xの配列に相補的な配列を含む本発明の化合物の1つまたは複数を含む組成物を提供する。
【0069】
また、本発明は、対照と比較して少なくとも50%ヒトTGaseII遺伝子の発現をダウンレギュレートする方法であって、遺伝子のmRNA転写物を本発明の化合物の1つまたは複数と接触させることを含む方法を提供する。
【0070】
一つの態様において、化合物は、TGaseIIをダウンレギュレートし、TGaseIIのダウンレギュレーションは、TGaseII機能のダウンレギュレーション(これは、とりわけ、天然の遺伝子/ポリペプチドの既知のインタラクターでの酵素アッセイ法または結合アッセイによって調べてもよい)、TGaseIIタンパク質のダウンレギュレーション(これは、とりわけウエスタンブロット法、ELISAまたは免疫沈降法によって調べてもよい)およびTGaseII mRNA発現のダウンレギュレーション(これは、とりわけノーザンブロッティング、定量的RT-PCR、インサイチュー・ハイブリダイゼーションまたはマイクロアレイ・ハイブリダイゼーションによって調べてもよい)を含む群から選択されるものによる。
【0071】
また、本発明は、線維症または線維症関連病態に罹患した患者を治療する方法であって、これにより患者を治療するために、患者に治療的に有効な用量の本発明の組成物を投与することを含む方法を提供する。
【0072】
また、本発明は、トランスグルタミナーゼIIを経た細胞タンパク質の異常な架橋に関連した病態に罹患した患者を治療する方法であって、これにより患者を治療するために、患者に治療的に有効な用量の本発明の組成物を投与することを含む方法を提供する。
【0073】
また、本発明は、線維症または線維症関連病態またはトランスグルタミナーゼIIを経た細胞タンパク質の異常な架橋に関連した病態に罹患した患者における回復を促進するための組成物の調製のための、本発明の1つまたは複数の化合物の治療的に有効な用量の使用を提供する。
【0074】
線維症が明らかである線維性疾患(線維症関連病態)は、以下の全ての病原学的変種を含む器官における線維症の急性および慢性形態を含む:間質性肺疾患および線維性肺疾患を含む肺線維症、肝臓線維症、心臓線維症を含む心筋線維症、慢性腎不全を含む腎臓線維症、強皮症を含む皮膚線維症、ケロイドおよび肥厚性瘢痕;骨髄線維化(骨髄線維症);増殖性硝子体網膜症(PVR)および白内障または緑内障を治療する外科手術によって生じる全種類の眼の瘢痕を含む瘢痕;病因不定の炎症性腸疾患、黄斑変性症、グレーブス眼障害、薬物で誘導される麦角中毒、乾癬、リ-フラウメニ症候群におけるグリア芽細胞腫、散発性グリア芽細胞腫、骨髄球性白血病、急性骨髄性白血病、骨髄異形成症候群、骨髄増殖症候群、婦人科学的癌、カポジ肉腫、ハンセン病およびコラーゲン性大腸炎。
【0075】
本発明の化合物は、線維性疾患とは異なる多くのその他の疾患および状態を治療するために使用してもよい。その他の適応症は、白内障を含む眼疾患、心臓血管疾患、特に心肥大、アテローム性動脈硬化症/再狭窄、神経系疾患、封入ポリグルタミン疾患(ハンチントン病など)、球脊髄性筋肉萎縮症、歯状核赤核-淡蒼球ルイ体萎縮症、および脊髄小脳失調(SCA)1、2、3、6、7および17、アルツハイマー病、並びにパーキンソン病であってもよい。
【0076】
化合物は、それぞれの末端領域のリボヌクレオチドのうちの2つまで塩基が変更された相同体を有していてもよく;末端領域とは、4つの末端リボヌクレオチドをいいたとえば19mer配列の塩基1〜4および/または16〜19および21mer配列の塩基1〜4および/または18〜21をいう。
【0077】
本発明の好ましいオリゴヌクレオチドは、表A、BおよびCに一覧が記載されたオリゴヌクレオチド、好ましくはヒトcDNAをターゲットするオリゴヌクレオチドである。本発明の最も好ましいオリゴヌクレオチドは、表Dに示したように、阻害活性を有するオリゴヌクレオチド、好ましくはヒトTGaseII cDNAをターゲットするオリゴヌクレオチドである。
【0078】
現在、本発明の最も好ましい化合物は、平滑末端19merオリゴヌクレオチド、すなわちx=y=19かつZおよびZ’の両方が存在せず;オリゴヌクレオチドは、アンチセンス鎖の5'位にて、およびセンス鎖の3'位にてリン酸化されており、交互に現れるリボヌクレオチドは、アンチセンスおよびセンス鎖の2’位にて修飾されており、2’位の部分は、メトキシ(2’-0-メチル)であり、かつアンチセンス鎖の5'および3'末端のリボヌクレオチドは、これらの糖残基が修飾されており、かつセンス鎖の5'および3'末端のリボヌクレオチドは、これらの糖残基が修飾されていないオリゴヌクレオチドである。現在、最も好ましいこのような化合物は、TG_HMRG1.である。TG_HMRG1のアンチセンス鎖は、配列番号:22を有し、センス鎖は、配列番号:4を有する。その他の好ましい化合物は、TG_HMG1およびTG_HM1であり、これらは、それぞれ配列番号:23および25によって表されるアンチセンス鎖を有し、センス鎖は、それぞれ配列番号:5および7によって表される
本発明の一つの局面において、オリゴヌクレオチドは、二本鎖構造を含み、このような二本鎖構造は、
第一の鎖および第二の鎖を含み、
第一の鎖は、隣接するヌクレオチドの第一のひと配列を含み、第二の鎖は、隣接するヌクレオチドの第二のひと配列を含み、
第一のひと配列は、TGaseIIをコードする核酸配列に相補的か、または同一であり、第二のひと配列は、TGaseIIをコードする核酸配列と同一か、または相補的である。
【0079】
ある態様において、第一のひと配列および/または第二のひと配列は、約14〜40ヌクレオチド、好ましくは約18〜30ヌクレオチド、より好ましくは約19〜27ヌクレオチド、最も好ましくは約19〜21ヌクレオチド、特に約19〜23ヌクレオチドを含む。このような局面において、オリゴヌクレオチドは、17〜40ヌクレオチドの長さであってもよい。加えて、本発明によるさらなる核酸は、配列番号:3〜配列番号:416の一つの少なくとも14の隣接するヌクレオチド、より好ましくは上記のとおりの第一のひと配列および第二のひと配列で構成される二本鎖構造のいずれかの末端の14の隣接するヌクレオチド塩基対を含む。
【0080】
本明細書に使用される「治療」という用語は、疾患もしくは状態と関連する症候を寛解させるため、疾患もしくは状態の重症度を軽くするため、もしくは治癒するため、または疾患または状態が生じるのを予防するために有効な治療物質を投与することをいう。
【0081】
特定の態様において、投与は、静脈内投与を含む。もう一つの特定の態様において、投与は、局所的または局部的な投与を含む。
【0082】
加えて、本発明は、患者に対して核酸配列に対する少なくとも1つのアンチセンス(AS)オリゴヌクレオチドもしくは少なくとも1つのsiRNA、またはTGaseII配列もしくはTGaseIIタンパク質に対して向けられたドミナントネガティブ・ペプチド、TGaseIIポリペプチドに対して向けられた抗体の治療的に有効な用量を投与することによって、病態または疾患(上記で詳述したとおり)を、このような治療を必要とする患者において調節する方法を提供する。
【0083】
送達:哺乳類細胞へのsiRNAの送達が増強され、および改善されることを特異的に狙った送達系が開発されており、たとえば、Shen et al (FEBS letters 539: 111-114 (2003)), Xia et al., Nature Biotechnology 20: 1006-1010 (2002), Reich et al., Molecular Vision 9: 210-216 (2003), Sorensen et al. (J.Mol.Biol. 327: 761-766 (2003), Lewis et al., Nature Genetics 32: 107-108 (2002) and Simeoni et al., Nucleic Acids Research 31, 11: 2717-2724 (2003)を参照されたい。siRNAは、霊長類における阻害に関して、最近首尾よく使用されてきた;詳しくは、Tolentino et al., Retina 24(1) February 2004 I 132-138を参照されたい。siRNAのための呼吸器製剤は、Davis et al.の米国特許出願第2004/0063654号に記述されている。コレステロール抱合されたsiRNA(および、その他のステロイドおよび脂質抱合されたsiRNA)は、送達のために使用されてきた(Soutschek et al Nature 432: 173-177(2004) Therapeutic silencing of an endogenous gene by systemic administration of modified siRNAs; and Lorenz et al. Bioorg. Med. Chemistry. Lett. 14:4975-4977 (2004) Steroid and lipid conjugates of siRNAs to enhance cellular uptake and gene silencing in liver cellsを参照されたい。
【0084】
本発明のsiRNAまたは薬学的組成物は、個々の患者の臨床状態、治療される疾患、投与の部位および方法、投与のスケジューリング、患者の年齢、性、体重、並びに開業医にとって既知であるその他の因子を考慮して、優れた医事に従って施され、および投与される。
【0085】
したがって、本明細書における目的のための「治療的に有効な用量」は、当技術分野において既知である考察などによって決定される。用量は、生存率またはより迅速な回復を含む(しかし、これらに限定されない)改善、または当業者による適切な測定として選択される症候およびその他の指標の改善もしくは除去を達成するために有効でなければならない。本発明の化合物は、従来の投与経路のいずれによっても投与することができる。化合物は、化合物として、または薬学的に許容される塩として投与することができ、また単独で、または活性成分として薬学的に許容されるキャリア、溶媒、希釈剤、賦形剤、アジュバントおよび媒体と組み合わせて投与することができることに留意すべきである。化合物は、腹腔内、静脈内、動脈内、筋肉内および鼻腔内投与、並びに髄腔内および注入技術を含む経口的に、皮下に、または非経口的に投与することができる。また、化合物の移植も有用である。液体形態を注射のために調製してもよく、本用語には、皮下、経皮、経静脈、筋肉内、髄腔内およびその他の非経口(parental)投与経路を含む。液体組成物は、有機共溶媒を伴い、または伴わない水溶液、水性または油懸濁液、食用油との乳剤、並びに同様の薬学的媒体を含む。加えて、一定の環境下において、本発明の新規治療に使用するための組成物は、鼻腔等の投与のためおエアロゾルとして形成してもよい。治療される患者は、温血動物および、特にヒトを含む哺乳動物である。薬学的に許容されるキャリア、溶媒、希釈剤、賦形剤、アジュバントおよび媒体、並びに移植キャリアは、一般に本発明の活性成分と反応することのない不活性な無毒の固体または液体の充填剤、希釈剤または封入材料をいい、これらには、リポソームおよび微粒子を含む。本発明に有用な送達系の例は、米国特許第5,225,182号;第5,169,383号;第5,167,616号;第4,959,217号;第4,925,678号;第4,487,603;号;第,486,194号;第4,447,233号;第4,447,224号;第4,439,196号;および第4,475,196号を含む。多くのその他のこのようなインプラント、送達系およびモジュールが当業者に周知である。本発明の1つの特定の態様において、局所的および経皮の製剤が特に好ましい。
【0086】
一般に、ヒトのための化合物の活性な用量は、1日あたり1ng/kg〜約20〜100mg/kg体重に、好ましくは1日あたり約0.01mg〜約2〜10mg/kg体重の範囲で、少なくとも1〜4週またはそれ以上の期間の間に1日あたり1用量、または1日あたり2回もしくは3回以上の処方計画である。また、より長い年月の治療または生涯の治療さえも、本明細書に開示したいくつかの適応症にについて想定される。
【0087】
また、本発明は、薬学的組成物製剤を調製する方法であって:
提供する本発明の少なくとも1つの二本鎖siRNA化合物を得ることと;および、
前記化合物を薬学的に許容されるキャリアと混合することと、
を含む方法を提供する。
【0088】
また、本発明は、薬学的組成物を調製する方法であって、本発明の化合物を薬学的に許容されるキャリアと混合することを含む方法を提供する。
【0089】
好ましい態様において、薬学的組成物の調製に使用される化合物は、薬学的に有効な用量のキャリアと混合される。特定の態様において、本発明の化合物は、ステロイドに対して、または脂質に対して、または別の適切な分子に対して、たとえばコレステロールに対して抱合される。
【0090】
ヌクレオチドの治療特性を改善するために、ヌクレオチドの修飾または類似体を導入することができる。改善される特性は、増大されたヌクレアーゼ耐性および/または増大された細胞膜に浸透する能力を含む。
【0091】
したがって、本発明は、また、実質的にポリヌクレオチドまたはオリゴヌクレオチドの機能に影響を及ぼさない全ての本発明のオリゴヌクレオチドの類似体、またはオリゴヌクレオチドに対する修飾を含む。好ましい態様において、このような修飾は、ヌクレオチドの塩基部分に、ヌクレオチドの糖残基に、および/またはヌクレオチドのリン酸部分に関連する。
【0092】
本発明の態様において、ヌクレオチドは、天然に存在するか、または合成的に修飾された塩基から選択することができる。天然に存在する塩基は、アデニン、グアニン、シトシン、チミンおよびウラシルを含む。オリゴヌクレオチドの修飾塩基は、イノシン、キサンチン、ヒポキサンチン、2-アミノアデニン、6-メチル-、2-プロピル-およびその他のアルキル-アデニン、5-ハロウラシル、5-ハロシトシン、6-アザシトシンおよび6-アザチミン、偽ウラシル、4-チウラシル、8-ハロアデニン、8-アミノアデニン、8-チオールアデニン、8-チオアルキルアデニン、8-ヒドロキシルアデニンおよびその他の8置換されたアデニン、8-ハログアニン、8-アミノグアニン、8-チオールグアニン、8-チオアルキルグアニン、8-ヒドロキシルグアニンおよびその他の置換されたグアニン、その他のアザおよびデアザアデニン、その他のアザおよびデアザグアニン、5-トリフルオロメチルウラシル、並びに5-トリフルオロシトシンを含む。
【0093】
加えて、ヌクレオチドの構造が基本的に変更されており、かつ治療的または実験的試薬としてより適しているヌクレオチドの類似体を調製することができる。ヌクレオチド類似体の例は、DNA(またはRNA)のデオキシリボース(またはリボース)リン酸バックボーンがペプチドに見いだされるものと同様のポリアミドバックボーンで置換されているペプチド核酸(PNA)である。PNA類似体は、酵素による分解に耐性を示し、インビボおよびインビトロでの生存が延長された。さらに、PNAは、DNA分子よりも相補的DNA配列に対して強く結合することを示した。この観察は、PNA鎖とDNA鎖との間の電荷相反がないことに起因する。オリゴヌクレオチドに対して作製することができるその他の修飾は、ポリマー主鎖、環状バックボーンまたは非環状バックボーンを含む。
【0094】
一つの態様において、修飾は、リン酸部分の修飾であり、修飾されたリン酸部分は、ホスホチオアート基から選択される。
【0095】
本発明の化合物は、リボ核(または、デオキシリボ核)オリゴヌクレオチドの合成のための技術分野において周知の方法のいずれによっても合成することができる。このような合成は、とりわけ、Beaucage S.L. and Iyer R.P., Tetrahedron 1992; 48: 2223-2311, Beaucage S.L. and Iyer R.P., Tetrahedron 1993; 49: 6123-6194 and Caruthers M.H. et. al., Methods Enzymol. 1987; 154: 287-313に記述されており、チオアートの合成は、とりわけ、Eckstein F., Annu. Rev. Biochem. 1985; 54: 367-402, the synthesis of RNA molecules is described in Sproat B., in Humana Press 2005 Edited by Herdewijn P.; Kap. 2: 17-31に記述されており、それぞれの 顆粒のプロセスは、とりわけPingoud A. et. al., in IRL Press 1989 Edited by Oliver R.W.A.; Kap. 7: 183-208 and Sproat B., in Humana Press 2005 Edited by Herdewijn P.; Kap. 2: 17-31(前記)に記述されている。
【0096】
その他の合成手順は、当技術分野において既知であり、たとえばUsman et al., 1987, J. Am. Chem. Soc., 109, 7845; Scaringe et al., 1990, Nucleic Acids Res., 18, 5433; Wincott et al., 1995, Nucleic Acids Res. 23, 2677-2684; and Wincott et al., 1997, Methods Mol. Bio., 74, 59に記載されているような手順であり、これらの手順には、5'末端のジメトキシトリチルおよび3'末端のホスホラミダイトなどの共通の核酸保護基およびカップリング基を利用してもよい。修飾された(たとえば2’-O-メチル化された)ヌクレオチドおよび無修飾ヌクレオチドが、所望のとおりに取り込まれる。
【0097】
本発明のオリゴヌクレオチドは、別々に合成することができ、たとえばライゲーションによって、(Moore et al., 1992, Science 256, 9923; Draper et al., International PCT publication No. WO93/23569; Shabarova et al., 1991, Nucleic Acids Research 19, 4247; Bellon et al., 1997, Nucleosides & Nucleotides, 16, 951; Bellon et al., 1997, Bioconjugate Chem. 8, 204)または合成および/または脱保護後のハイブリダイゼーションによって合成後に共に連結することができる。
【0098】
市販の機械(とりわけ、Applied Biosystemsから利用できる)を使用することができる点;オリゴヌクレオチドは、本明細書に開示された配列に従って調製される点に留意されたい。化学的に合成された断片の重なり合う対は、当技術分野において周知の方法を使用して結合することができる(たとえば、米国特許第6,121,426号を参照されたい)。鎖を別々に合成して、次いで、チューブ内で互いにアニールさせる。次いで、二本鎖siRNAを、HPLCによってアニールされなかった一本鎖オリゴヌクレオチド(たとえば、これらのうちの一方が過剰なことを原因として生じるので)から分離する。本発明のsiRNAまたはsiRNA断片に関して、2つ以上のこのような配列を合成して、本発明に使用するために共に連結させることができる。また、本発明の化合物は、米国特許出願公開番号US2004/0019001号(McSwiggen)に記載されているように、両方のsiRNA鎖を、切断可能なリンカーによって分離された単一の隣接するオリゴヌクレオチド断片または鎖として合成し、これをその後に切断して、ハイブリダイズしてsiRNA二重鎖の精製を可能にする別々のsiRNA断片または鎖を提供するという、直列型合成法を経て合成することができる。リンカーは、ポリヌクレオチドリンカーまたは非ヌクレオチドリンカーであることができる。
【0099】
本発明の化合物は、直接、またはウイルスまたは非ウイルスベクターと共に送達することができる。直接送達されるときは、一般に配列をヌクレアーゼ耐性にする。あるいは、本明細書において下記に論議したように、配列は、配列が細胞に発現されるように、発現カセットまたは構築物に取り込むことができる。一般に、構築物は、配列をターゲットされる細胞にて発現することができるように、適当な制御配列またはプロモーターを含む。本発明の化合物の送達のために任意に使用されるベクターは、市販されており、当業者に既知の方法によって本発明の化合物を送達するために修飾してもよい。
【0100】
また、本発明のオリゴヌクレオチドの配列の1つまたは複数を含む長い二本鎖オリゴヌクレオチド(典型的には、25〜500ヌクレオチドの長さ)を送達してもよく、内因性細胞複合体によって(たとえば、上記のとおりのDICERによって)細胞内にてプロセスされて、本発明のオリゴヌクレオチドであるより小さな二本鎖オリゴヌクレオチドを産生し得ることが想定される。
【0101】
本明細書に使用される「ポリペプチド」という用語は、ポリペプチドに加えて、オリゴペプチド、ペプチドおよび完全なタンパク質をいう。
【0102】
動物モデル系:本発明の活性なsiRNAの試験は、予測的動物モデルで行ってもよい。腎臓線維症のためのいくつかのモデルを実施例3に記述してある。
【0103】
ラットの肝臓線維症の2つのモデルは、以下の参照に記載したように、対照として偽手術したBile Duct Ligation(BDL、)対照としてのオリーブ油を与えた動物であるCCl4中毒である:Lotersztajn S, Julien B, Teixeira-Clerc F, Grenard P, Mallat A, Hepatic Fibrosis: Molecular Mechanisms and Drug Targets. Annu Rev Pharmacol Toxicol. 2004 Oct 07; Uchio K, Graham M, Dean NM, Rosenbaum J, Desmouliere A., Down-regulation of connective tissue growth factor and type I collagen mRNA expression by connective tissue growth factor antisense oligonucleotide during experimental liver fibrosis. Wound Repair Regen. 2004 Jan-Feb;12(1):60-6. ;and. Xu XQ, Leow CK, Lu X, Zhang X, Liu JS, Wong WH, Asperger A, Deininger S, Eastwood Leung HC., Molecular classification of liver cirrhosis in a rat model by proteomics and bioinformatics Proteomics. 2004 Oct;4(10):3235-45。
【0104】
瘢痕組織阻害剤を評価するためのモデルは、当技術分野において周知であり、たとえば、Sherwood MB et al., J Glaucoma. 2004 Oct;13(5):407-12. A new model of glaucoma filtering surgery in the rat; Miller MH et al., Ophthalmic Surg. 1989 May;20(5):350-7. Wound healing in an animal model of glaucoma fistulizing surgery in the rabbit; vanBockxmeer FM et al., Retina. 1985 Fall-Winter; 5(4): 239-52. Models for assessing scar tissue inhibitors; Wiedemann P et al., J Pharmacol Methods. 1984 Aug; 12(1): 69-78. Proliferative vitreoretinopathy: the rabbit cell injection model for screening of antiproliferative drugs。
【0105】
白内障のモデルは、以下の刊行物に記述されている:The role of Src family kinases in cortical cataract formation. Zhou J, Menko AS.Invest Ophthalmol Vis Sci. 2002 Jul;43(7):2293-300; Bioavailability and anticataract effects of a topical ocular drug delivery system containing disulfiram and hydroxypropyl-beta-cyclodextrin on selenite-treated rats.Wang S, Li D, Ito Y, Nabekura T, Wang S, Zhang J, Wu C. Curr Eye Res. 2004 Jul;29(1):51-8; and Long-term organ culture system to study the effects of UV-Airradiation on lens transglutaminase. Weinreb O, Dovrat A.; Curr Eye Res. 2004 Jul;29(1):51-8。
【0106】
抗体産生
本発明に使用される「抗体」という用語は、ポリクローナルおよびモノクローナルの両方の完全抗体、並びにFab、F(ab')2およびFvなどのエピトープの決定因子に結合することができるこれらの断片を意味する。これらの抗体断片は、その抗原または受容体と選択的に結合する能力を保持し、とりわけ、以下のとおりに例示される:
(1)Fab、酵素パパインで抗体全体を消化して、軽鎖および一部の重鎖を得ることによって産生することができる抗体分子の一価の抗体結合性断片を含む断片;
(2)(Fab')2、酵素ペプシンで抗体全体を処置することによってその後に還元することなく得ることができる抗体の断片;F(ab'2)は、2つのジスルフィド結合によって共に保持された2つのFab断片の二量体である;
(3)Fv、2つの鎖として発現された軽鎖の可変領域と重鎖の可変領域とを含む遺伝的に操作された断片として定義される;および
(4)単鎖抗体(SCA)、遺伝的に融合された単一の鎖状分子として、適切なポリペプチド・リンカーによって連結された軽鎖の可変領域と重鎖の可変領域とを含む遺伝的に操作された分子として定義される。
【0107】
抗体機能的活性を有するこのような断片は、当業者に既知の方法によって調製することができる(たとえば、Bird et al.(1988)Science 242:423-426)。 都合よくは、抗体は、免疫原またはこれらの部分、たとえば配列に基づいた合成ペプチドに対して調製しても、もしくはクローン技術によって組換えで調製してもよく、または天然の遺伝子産物および/またはこれらの部分を単離して免疫原として使用してもよい。免疫原は、Harlow and Lane (1988), Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, and Borrebaeck (1992), Antibody Engineering - A Practical Guide, W.H. Freeman and Co., NYに一般に記述されているように、当業者に周知の標準的な抗体産生技術によって抗体を産生するために使用することができる。
【0108】
ポリクローナル抗体を産生するためには、ウサギまたはヤギなどの宿主を、一般にアジュバントと共に、必要に応じて、キャリアに結合させて免疫原または免疫原断片で免疫し;免疫原に対する抗体を血清から収集する。さらに、ポリクローナル抗体は、それが単一特異性であるように吸収することができ;すなわち、交差反応を起こし得る抗体が血清に残らないように、血清を関連免疫原に対して吸収して、それを単一特異性にすることができる。
【0109】
モノクローナル抗体を産生するためには、本技術には、適切なドナー、一般にマウスの免疫原での高度免疫処置、および脾臓抗体産生細胞の単離を含む。これらの細胞を骨髄腫細胞などの不死細胞に融合させて、不死で、かつ必要とされる抗体を分泌する融合細胞ハイブリッドを提供する。次いで、細胞を大量に培養し、モノクローナル抗体を使用のために培地から収集する。
【0110】
組換えを抗体産生については、一般にHuston et al. (1991) "Protein engineering of single-chain Fv analogs and fusion proteins" in Methods in Enzymology (JJ Langone, ed., Academic Press, New York, NY) 203:46-88; Johnson and Bird (1991) "Construction of single-chain Fvb derivatives of monoclonal antibodies and their production in Escherichia coli in Methods in Enzymology (JJ Langone, ed.; Academic Press, New York, NY) 203:88-99; Mernaugh and Mernaugh (1995) "An overview of phage-displayed recombinant antibodies" in Molecular Methods In Plant Pathology (RP Singh and US Singh, eds.; CRC Press Inc., Boca Raton, FL:359-365)を参照されたい。特に、scFv抗体は、国際公開公報第2004/007553号(TedescoおよびMarzari)に記述されている。加えて、動物の抗体産生Bリンパ球またはハイブリドーマからのメッセンジャーRNAを逆転写して相補的DNA(cDNA)を得ることができる。抗体cDNA(これは、完、または部分的な長さであることができる)を増幅し、ファージまたはプラスミドにクローン化する。cDNAは、分離されたか、またはリンカーによって接続された重鎖および軽鎖cDNAの部分的な長さであることができる。抗体または抗体断片を適切な発現系を使用して発現させて、組換え抗体を得る。また、抗体cDNAは、関連する発現ライブラリをスクリーニングすることによって得ることができる。
【0111】
抗体は、周知のように、固体支持体基体に結合し、または検出可能な部分と抱合し、または結合および抱合を両方することができる。(蛍光または酵素部分の抱合に関する一般的な議論については、Johnstone & Thorpe (1982.), Immunochemistry in Practice, Blackwell Scientific Publications, Oxfordを参照されたい)。また、固体支持体基体に対する抗体の結合は、当技術分野において周知である(一般的な考察については、Harlow & Lane (1988) Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Publications, New York; and Borrebaeck (1992), Antibody Engineering - A Practical Guide, W.H. Freeman and Co.を参照されたい)。本発明で想定される検出可能部分は、ビオチン、金、フェリチン、アルカリホスファターゼ、β-ガラクトシダーゼ、ペルオキシダーゼ、ウレアーゼ、フルオレッセイン、ローダミン、トリチウム、14C、およびヨウ素化などの蛍光、金属、酵素、および放射性のマーカーを含むことができるが、これらに限定されるわけではない。
【0112】
本明細書で論議される疾患および障害の治療に有用であると考えられるさらなる化合物は、アンチセンスDNA分子(これは、当技術分野において既知の方法によって、図1の配列を使用して作製することができる)、リボザイムなどの触媒RNA、ドミナントネガティブ・ペプチド(これは、当技術分野において既知の方法によって、図2の配列を使用して作製することができる)などのポリペプチドまたはその他のポリペプチド阻害剤であってもよい。本明細書に開示したsiRNA配列を含むアンチセンスDNA分子(DNAとRNAとの間の相違から生じる適切な核酸修飾をもつ)が特に望ましく、本明細書に開示した全ての使用および方法に対して、これらの対応するsiRNAと同じ能力で使用し得る。
【0113】
TGase不活性化化合物のスクリーニング
本発明のいくつかの化合物および組成物は、TGaseIIの活性を調整する化合物、特に線維性疾患または線維症関連病態を調整する化合物を同定し、および単離するためのスクリーニングアッセイ法に使用してもよい。スクリーニングされる化合物は、とりわけ小さな化学分子、抗体、特に中和抗体、阻害ポリペプチドおよびドミナントネガティブなどの物質を含む。
【0114】
TGaseIIポリペプチド酵素活性に対する本発明の化合物の阻害活性またはTGaseIIに対する本発明の化合物の結合は、TGaseIIポリペプチドとさらなる化合物の相互作用、たとえばさらなる化合物が、本発明の抗体またはドミナントネガティブ・ペプチドとTGaseII阻害に関して競合するかどうか、またはさらなる化合物が該阻害をレスキューするかどうかを決定するために使用してもよい。阻害または活性化は、とりわけTGaseIIポリペプチドの活性の産生または放射性または蛍光性競合アッセイ法においてTGaseIIポリペプチドからの結合化合物の置換についてアッセイすることなど、種々の手段によって試験することができる。
【0115】
本発明は加えて、TGaseIIポリペプチド、好ましくはヒトTGaseIIポリペプチドの酵素活性を阻害することができる化合物を得る方法であって:
(i)TGaseIIポリペプチドまたはTGaseIIポリペプチドを発現する細胞をリジル・ドナーである化合物と、および生体分子であるグルタミル・ドナーと接触させる工程と;
(ii)工程(i)の混合物から過剰なリジル・ドナーを除去する工程と;
(iii)化合物の存在下において産生されるリジル-グルタミル架橋物質の量を測定する工程と;
(iv)工程(iii)で測定したリジル-グルタミル架橋された物質の量を、一定条件下で化合物の非存在下において測定したものと比較する工程であって、リジル-グルタミル架橋された物質の量の減少は、化合物によるTGaseIIポリペプチド活性の阻害を指し示す工程と;および、
(v)TGaseIIポリペプチド活性の阻害剤として化合物を同定する工程と、
の工程を含む方法を提供する。
【0116】
一つの態様において、本明細書に記述した全ての方法の中で、このような方法に使用されるTGaseIIポリペプチドは、配列番号:2に記載された配列の連続したアミノ酸を含む。
【0117】
もう一つの態様において、工程(i)のリジル・ドナーにタグを付けてあり、工程(iii)の測定は、タグ関連読み出しであり、読み出しレベルの減少は、化合物によるTGaseIIポリペプチド活性の阻害を指し示す。
【0118】
生体分子であるグルタミル・ドナーは、たとえばフィブロネクチンまたはコラーゲンであってもよい。
【0119】
もう一つの態様において、このような方法に使用されるリジル・ドナーは、ダンシル蛍光タグでタグを付けたカダベリンである。
【0120】
もう一つの態様において、このような方法に使用されるリジル・ドナーは、ビオチンでタグを付けてあり、特定の態様において、リジル・ドナーは、ビオチン・カダベリンである。ビオチンは、当技術分野において既知の読み出し(またはヌトラビジン(nutravidin)などの修飾されたストレプトアビジン)に抱合されたストレプトアビジンによって検出される(たとえば、西洋ワサビペルオキシダーゼに抱合されており、西洋ワサビペルオキシダーゼの量は、基質としてテトラメチルベンジジンを使用して定量化する)。
【0121】
本発明のさらにもう一つの態様において、提供した方法におけるグルタミル・ドナーは、細胞外基質タンパク質または細胞タンパク質であってもよい。細胞外基質タンパク質は、以下からなる群より選択され:フィブロネクチン、コラーゲン、オステオネクチン、ECM-ゲル(たとえばSigma E1270)、また細胞内タンパク質は、とりわけ以下からなる群より選択される:RhoA、フィアルミン(fialmin)、スペクトリン、ビメンチン、HSP90、HSP60(Orru S, Caputo I, D'Amato A, Ruoppolo M, Esposito C Proteomics identification of acyl-acceptor and acyl-donor substrates for transglutaminase in a human intestinal epithelial cell line. Implications for celiac disease. J Biol Chem. 2003 Aug 22;278(34):31766-73を参照されたい)。
【0122】
異なる態様において、化合物を、TGaseIIポリペプチドを発現する細胞と接触させる。このような方法に使用してもよい細胞は、分離されていても、または組織に提示されていてもよい。細胞は、TGaseIIポリペプチドを天然に、またはTGaseII遺伝子で、一過性もしくは安定にトランスフェクトした結果として発現していてもよく、またはTGaseII遺伝子を過剰発現してもよく、過剰発現と正常TGaseII発現細胞との間でのTGaseIIの活性を比較する。さらなる態様において、グルタミル・ドナーは、天然物であるか、またはこれは、細胞外から添加される。
【0123】
異なる態様において、TGaseIIポリペプチドを化合物と接触させる。TGaseIIポリペプチドは、固定され、または溶液中で遊離していることができる。提供したスクリーニング法の態様では、グルタミル・ドナーを固定してある。
【0124】
さらなる態様において、ヒトTGaseIIの活性を阻害することができる化合物を得るために提供した方法は、工程(i)の前にTGaseIIを、TGaseIIに結合することが既知の第二の化合物と接触させるというさらなる工程を含んでいてもよい。
【0125】
本発明の一つの態様において、本明細書に記述した提供した方法におけるタグ読み出しでは、西洋ワサビペルオキシダーゼに抱合したヌトラビジンとビオチンの相互作用がなされ、細胞に結合した西洋ワサビペルオキシダーゼの量を、基質としてTMB(テトラメチルベンジジン)を使用して定量化する。
【0126】
なおさらなる態様において、提供した方法によって得られた化合物は、これがTGaseファミリーの少なくとも1つの他のメンバーの活性を阻害するよりも少なくとも2倍有効にTGaseIIポリペプチドの活性を阻害し、より好ましくは、阻害は、少なくともよりも10倍、50倍、さらに100倍有効である。このような方法では、TGaseIIポリペプチドまたは第二の化合物が固定されていてもよい。
【0127】
もう一つの態様において、提供した方法のいずれかによって得られた化合物は、抗体である。
【0128】
本発明は、さらに、TGaseIIポリペプチド、好ましくはヒトTGaseIIポリペプチドの活性を調整する化合物を得る方法であって:
(i)TGaseIIポリペプチドを、TGaseIIポリペプチドがインビボにおいて特異的に相互作用するインタラクターと接触させる工程と;
(ii)TGaseIIポリペプチドまたはインタラクターを該化合物と接触させる工程と;および、
(iii)線維症に関連したパラメーターを測定することにより、TGaseIIポリペプチドとインタラクターとの間の相互作用に対する化合物の効果を測定する工程と;および、
(iv)工程(iii)で測定された効果を、化合物の非存在下において測定された効果と比較する工程であって。測定された効果の変化は、化合物がヒトTGaseIIポリペプチドの活性を調整することを示す工程と、
の工程を含む方法を提供する。
【0129】
本発明の一つの態様において、提供された方法によって得られた化合物は、ヒトTGaseIIポリペプチドの活性を阻害する。
【0130】
本発明のもう一つの態様において、このような方法にTGaseIIポリペプチドは、配列番号:2に記載された配列の連続したアミノ酸を含む。さらなる態様において、TGaseIIポリペプチドまたはインタラクターは、固定されている。
【0131】
本発明のさらなる態様において、提供した方法のいずれかにおいて測定されるパラメーターは、一般のTGaseIIアップレギュレーションによって特徴づけられる病態に、上で定義したような線維症関連病態に、さらに白内障、心臓血管疾患、神経疾患、ハンチントン病(HD)を含むポリグルタミン疾患、球脊髄性筋肉萎縮症、歯状核赤核-淡蒼球ルイ体萎縮症、並びに脊髄小脳失調(SCA)1、2、3、6、7および17、アルツハイマー病およびパーキンソン病、小児脂肪便症および骨関節炎に関連がある。本発明のもう一つの目的は、ヒトTGaseIIポリペプチドの活性を阻害することができる化合物を得る方法であって:
(i)TGaseIIポリペプチドを発現するTGaseIIポリペプチドまたは細胞を、複数のタグを付けたリジル・ドナーである化合物と、生体分子であるグルタミル・ドナーと接触させる工程と、
(ii)工程(i)の混合物から過剰なタグを付けたリジル・ドナー洗浄する工程と;および、
(iii)複数の化合物の存在下においてタグに関連した読み出しを測定する工程と;
(iv)工程(iii)で測定されたタグ読み出しを、一定条件下で、複数の化合物の非存在下において測定されたものと比較する工程であって、読み出しレベルの減少は、複数の化合物によるTGaseIIポリペプチド活性の阻害を指し示す工程と;
(v)多数のうちのどの化合物がヒトTGaseIIの活性を阻害するかを別々に決定する工程と、
の工程を含む方法を提供することである。
【0132】
もう一つの本発明の局面は、TGaseIIポリペプチド、好ましくはヒトTGaseIIポリペプチドの活性を阻害することができる化合物を得る細胞に基づいた方法であって:
(i)活性なTGaseIIポリペプチドを発現する細胞を、化合物と、およびリジル・ドナーと接触させる工程と;
(ii)工程(i)の混合物から過剰なリジル・ドナーを除去する工程と;
(iii)化合物の存在下において産生されたリジル-グルタミル架橋された物質の量を測定する工程と;
(iv)工程(iii)で測定されたリジル-グルタミル架橋された物質の量を、一定条件下で、化合物の非存在下において測定されたものと比較する工程であって、リジル-グルタミル架橋された物質の量の減少は、化合物によってTGaseIIポリペプチド活性の阻害を指し示す工程と;および
(v)化合物をTGaseIIポリペプチドの阻害剤として同定する工程と、
の工程を含む方法を提供することである。
【0133】
もう一つの本発明の局面は、ヒトTGaseIIポリペプチドの活性を阻害することができる化合物を得る細胞に基づいた方法であって:
a. TGaseIIポリペプチドを発現する細胞を、複数の化合物と、およびリジル・ドナーと接触させる工程と;
b. 工程(i)の混合物から過剰なリジル・ドナーを除去する工程と;および、
c. 複数の化合物の存在下において産生されるリジル-グルタミル架橋された物質の量を測定する工程と;
d. 工程(iii)で測定されたリジル-グルタミル架橋された物質の量を、一定条件下で、複数の化合物の非存在下で測定されたものと比較する工程であって、リジル-グルタミル架橋された物質の量の減少は、複数の化合物によるTGaseIIポリペプチド活性の阻害を指し示す工程と;および、
(e)多数のうちのどの化合物がヒトTGaseIIの活性を阻害するかを別々に決定する工程と、
の工程を含む方法を提供することである。
【0134】
方法
分子生物学における一般的な方法
当技術分野において既知であり、かつ特に記述されていない標準的分子生物学技術は、一般に、Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, New York (1989)に、およびAusubel et al., Current Protocols in Molecular Biology, John Wiley and Sons, Baltimore, Maryland (1989)に、およびPerbal, A Practical Guide to Molecular Cloning, John Wiley & Sons, New York (1988)に、およびWatson et al., Recombinant DNA, Scientific American Books, New Yorkに、およびBirren et al (eds) Genome Analysis: A Laboratory Manual Series, Vols. 1-4 Cold Spring Harbor Laboratory Press, New York (1998)に記載されたとおりであり、並びに方法論は、米国特許第4,666,828号;第4,683,202号;第4,801,531号;第5,192,659号;および第5,272,057号に記載されており、これらは、参照により本明細書に援用される。ポリメラーゼ連鎖反応法(PCR)は、一般にPCR Protocols: A Guide To Methods And Applications, Academic Press, San Diego, CA (1990)と同様に行った。フローサイトメトリーと組み合わせたインサイチュウ(細胞における)PCRは、特異的DNAおよびmRNA配列を含む細胞の検出のために使用することができる(たとえばTestoni et al., 1996, Blood 87:3822)。また、RT-PCRを行う方法も、当技術分野において周知である。
【0135】
本発明は、実施例に関して、以下に詳細を例証してあるが、これらに限定されるものとして解釈されることはない。
【実施例】
【0136】
実施例1
活性なsiRNA化合物のデザイン
独占権下にあるアルゴリズムおよびTGaseIIのcDNAの既知の配列(配列番号:1)を使用して、多くの潜在性siRNAの配列を作製した。これらを下記の表に示してある。種のための略語は:H:ヒト;M:マウス;R:ラット;G:モルモット;C:ウシであることに留意されたい。
【0137】
表Aは、今までに化学的合成されており、かつ活性に関して試験された18個のsiRNAを示す(実施例2を参照されたい)。これらのsiRNAの全てが19merである。表Aにおいて、siRNA 1〜18のセンス鎖は、それぞれ配列番号:3〜20を有し、siRNA 1〜18のアンチセンス鎖は、それぞれ配列番号:21〜38を有する。
【0138】
下記の表Bは、独占権下にあるアルゴリズムによって作製されたが、いまだ活性について試験されていない153個のさらなる19mer siRNAを示す。表Bにおいて、siRNA 1〜153のセンス鎖は、それぞれ配列番号:39〜191を有し、siRNA 1〜153のアンチセンス鎖は、それぞれ配列番号:192〜344を有する。
【0139】
下記の表Cは、独占権下にあるアルゴリズムによって作製された36個のさらなる21mer siRNAを示す。表Cにおいて、siRNA 1〜36のセンス鎖は、それぞれ配列番号:345〜380を有し、siRNA 1-36のアンチセンス鎖は、それぞれ配列番号:381〜416を有する。
【表1】
【表2−1】
【表2−2】
【表2−3】
【表2−4】
【表2−5】
【表2−6】
【表3−1】
【表3−2】
実施例2
siRNA化合物の抗TGaseII活性についての試験
I. siRNA(二本鎖オリゴヌクレオチド)の使用液の調製
凍結乾燥したオリゴヌクレオチドを、RNAseを含まない二回蒸留水に溶解して100μMの終濃度に作製する。希釈したオリゴヌクレオチドを室温で15分間保持して、液体窒素中で直ちに凍結させた。オリゴヌクレオチドは、-80℃に貯蔵し、使用前にPBSで希釈した。
【0140】
II. ラットTGaseIIを安定に過剰発現するNRK49株化細胞の確立
PLXSNベクター(CLONTECH)にクローン化したラットTGaseIIの全長cDNAを、Rat1線維芽細胞株化細胞へのトランスフェクションに使用した。リポフェクトアミン試薬(Invitrogen)を、下記に記述されるプロトコルを使用するトランスフェクション試薬として使用した。トランスフェクションの48時間後、G418抗生物質(ネオマイシン)を使用して安定なクローンの選択を行った。このポリクローナル集団におけるTGaseIIの安定な過剰発現を、ウエスタンブロット法およびTGaseII活性アッセイ法によって検証した。細胞は、NRK49-TGaseII(TGASE-OE(登録商標))と命名した。
【0141】
III.NRK49におけるTGaseIIに関する活性のアッセイ法
4×103 NRK49-ラットTGASE-OE細胞を96ウェル・プレートにウェル毎に播種した。TGaseII活性のための細胞に基づいたアッセイ法を使用してTGaseII活性を測定した。簡潔には、増殖培地を細胞標識化培地(0.1%のBSA、25μM BC、10μM Calcimycin、16.2mM CaCl2のDMEM溶液)で37℃にて45分間置き換えた。45分後に、細胞を4%のホルムアルデヒドで固定して、ビオチン標識されたカダベリンの架橋を試験するために、0.6%のH2O2で処置して、ネトラビジン-HRP Abで染色した。吸光度をλ=450nmにて読み込んだ。規準化のために、細胞をヘキスト色素溶液で染色した。TGaseIIの活性は、NRK49対照細胞と比較して、NRK49-TGaseII細胞におけるよりも75倍高い。
【0142】
IV. リポフェクトアミン2000試薬を使用するsiRNAオリゴヌクレオチドによるトランスフェクション
2×105細胞を6ウェル・プレートのウェル毎に播種した。24時間後、細胞を以下の手順に従って、リポフェクトアミン2000試薬(Invitrogen)を使用してTGaseII特異的なsiRNAオリゴヌクレオチドをトランスフェクトした:
1. トランスフェクションの前に、細胞培地を抗生物質を含まない1500μlの新鮮な培地に置き換えた。
【0143】
2. 無菌プラスチックチューブに、リポフェクトアミン2000試薬(量は、1ウェルあたり5μlに従って算出する)を250μlの無血清培地に添加して、室温で5分間インキュベートした。
【0144】
3. もう一つのチューブに、siRNAオリゴヌクレオチド(ウェルあたりに所望の終濃度に適合するように様々の量)を250μlの無血清培地に添加した。
【0145】
4. リポフェクトアミン2000複合体をsiRNA溶液と合わせて、室温で20分間インキュベートした。
【0146】
5. 生じる混合物を細胞に滴下して、細胞を37℃にてsiRNA活性の解析までインキュベートした。
【0147】
V.トランスフェクトしたヒト、マウスおよびラット細胞における抗TGaseII siRNAの活性の試験
表Aの一覧表に記載したTGaseII siRNAの活性を異なる種に由来する3つの異なる株化細胞で試験した:
1. マウス特異的TGaseII siRNAは、マウスTGaseII NMUMG細胞を発現するマウス細胞において試験した。
【0148】
2. ヒト特異的TGaseII siRNAは、ヒトTgaseIIを発現するヒトHeLa細胞において試験した;図3を参照されたい。
【0149】
3. ラット特異的TGaseII siRNAは、ラットTGaseIIを過剰発現するNRK49(TGase-OE(登録商標)細胞)において試験した。
【0150】
siRNAのトランスフェクションの48時間〜6日後に、細胞を収集して、TGaseIIタンパク質の発現を特異的抗体でウエスタンブロット解析によって調べた。対照と比較して、TGaseIIに特異的なsiRNAオリゴヌクレオチドをトランスフェクトした細胞におけるTGaseIIポリペプチド発現の減少は、siRNAで誘発されるTGaseIIタンパク質発現に対する阻害作用を示し、これを定量化することができる。
【0151】
下記の表Dには、それぞれのsiRNAオリゴヌクレオチドのTGaseII阻害活性に関する情報を要約してある。表Dのオリゴヌクレオチドの名称は、表A.に使用した名称に対応する。比較は、異なるsiRNAによってトランスフェクトされたそれぞれの株化細胞における活性であり、株化細胞間の活性ではない。現在最も好ましいヒトsiRNAは、高いTGaseII阻害作用を誘発して、さらにマウス、ラットおよびモルモットTGaseIIを阻害したTG_HMRG1である。その他の好ましいヒトsiRNAは、TG_HMG1およびTG_HM1である。
【表4】
【0152】
実施例3
腎臓線維症の動物モデル
抗TGaseII siRNAの、もしくはTGaseIIに対する中和抗体の、またはその他のTGaseII阻害剤のインビボでの治療活性の試験は、疾患に関連したTGaseIIの過剰発現が見いだされた(インシチューハイブリダイゼイションによって検出された;下記参照)腎臓線維症の以下の動物モデルで行ってもよい。
【0153】
A. 腎臓線維症のRat糖尿病患者および老化モデル
A1. ZDFラット
9月齢のZDFラットの腎臓試料(Zucker糖尿病脂肪ラット)は、拡張型腎杯をもつ水腎症様変化を提示した。顕微鏡によって、これらの試料は、糸球体硬化症および尿細管間質性線維症の特色を示した。これらの形態学的変化に従って、インシチューハイブリダイゼイションによって測定される線維症マーカー遺伝子(オステオポンチン(OPN)、トランスフォーミング成長因子β1(TGF-β1)およびプロコラーゲン1(I)(Col1))の発現は、正常な腎臓と比較して有意に増大された。強力なOPN発現が、腎臓皮質および髄質の全ての管状構造で検出可能であった。TGF-β1発現は、間質細胞の全体にわたって広範囲におよんだ。また、一部の上皮細胞もTGF-β1発現を示した。Col1発現は、髄質内の大部分の間質細胞でインシチューハイブリダイゼイションによって検出可能であったが、皮質発現は「限局的」であった。
【0154】
A2. 高齢fa/fa(肥満Zucker)ラット
Fa/faラットは、遺伝的にレプチン受容体を欠損しており;その結果、進行性の糖尿病性腎障害を伴ったインスリン耐性糖尿病(II型糖尿病)を発病する。12月齢のfa/faラットの腎臓試料は、皮質および髄質の全体にわたって高度の糸球体硬化症および散在性の尿細管間質性線維症を呈する。線維症マーカー遺伝子発現のパターンは、形態学的変化に対応した。OPNは、皮質の管状構造に、および髄質に発現された。複数の間質細胞が、TGF-β1を発現した。複数の病巣および単一の間質細胞が、有意に、皮質および髄質における強力なCol1発現を示し、その結果、Col1発現細胞の数は、ZDF試料におけるよりもfa/fa腎臓試料において高いようであった。
【0155】
A3. 高齢SD系の(SD)(正常)ラット
また、高齢SD系ラットの腎臓試料は、糸球体および間隙におけるコラーゲンの蓄積が増大されたこと、および線維症マーカー遺伝子の発現が増加したことを示した。線維性変化の強度は、有意に、試料間で変化した。したがって、研究した4つの試料のうちの1つでは、幼若動物と比較して極めて少ない変化を示し;第二の試料では、線維性変化は、「極所」領域のみに限定されており、残りの2つの試料では、コラーゲンの一様な蓄積を示し、切片全体にわたってマーカー遺伝子の発現が上昇した。
【0156】
A4. 48週齢のGoto Kakizaki(GK)ラット
GKラットは、ウィスターラットに由来した、NIDDM(糖尿病II型)の自発的発症に関して選択された近交系である。GKおよび対照ウィスター48週齢ラットからの腎臓試料では、糸球体および間隙におけるコラーゲンの蓄積を示した。この蓄積は、GK試料においてよりはっきりしていた。
【0157】
TGaseIIハイブリダイゼーション・パターン
TGaseII特異的ハイブリダイゼーション・シグナルは、健康なラットおよび糖尿病ラットの腎臓における線維性表現型(散在性または局所性)と関連していた。非線維性腎臓試料は、近位尿細管全体に散在性で、低いが、検出可能なTGaseII特異的ハイブリダイゼーション・シグナルを示す。12月齢までに、より強力なシグナルが、明白な萎縮症の徴候を示す膨張する尿細管の間質細胞および上皮裏打ち全体に濃縮される。慢性腎不全試料(2歳のラットからの腎臓)では、強力なTGaseII発現が、間質を囲む範囲内の萎縮性または増殖性の上皮細胞に沿って並んだ管状プロフィールに見られる。同様に、ZDF試料では、線維症(または老化)の進行と共にハイブリダイゼーション・シグナルの増大を示した。
【0158】
B. 腎臓線維症誘導モデル。
【0159】
B1. 持続性一側性尿管閉塞(UUO)モデル
持続性一側性尿管閉塞(UUO)は、健康な非糖尿病動物に生じる急性間質性線維症のモデルである。腎臓線維症は、閉塞後数日以内に発病する。
【0160】
モデルを作製するために、ラットをケタミン/キシラジンで麻酔して、腹腔を開いた。曝露後、右の輸尿管を低い腎臓極レベルにて縫合により結紮した。見せかけの手術をしたラットでは、輸尿管を曝露したが、結紮しなかった。研究は、尿管閉塞後の種々の時点にて(たとえば、24時間、5日、10日、15日、20日および25日にて)終了し、腎臓を取り出して調べた。
【0161】
組織学的検査では、持続性UUOが、髄質および皮質の間質細胞によるコラーゲン合成の迅速な活性化を生じることが明らかになった。UUOの20〜25日までに、間質コラーゲンの有意な量が間隙に堆積したが、コラーゲンの糸球体蓄積は、外側のカプセルに限定された。
【0162】
閉塞腎臓試料のインシチューハイブリダイゼイション解析では、UUOの期間に依存したTGaseII発現のパターンの迅速かつ有意な変化を示した。UUOの24時間後、非常に強力なハイブリダイゼーション・シグナルが、皮質管状上皮全体に検出されたが、延髄上皮細胞は、非常に弱いシグナルを示した。UUOの5および10日目には、強力なハイブリダイゼーション・シグナルの「病巣」は、尿細管間質性の線維症を発症している領域と関連しており、ここでは、シグナルが上皮および間隙細胞に位置していた。同様の結果が、同じUUOモデルに曝露したマウスにおいても観察された。
【0163】
B2. 5/6腎摘除
5/6腎摘除は、明らかな線維症である慢性腎不全(CRI)のためのもう一つの有用な動物モデルである。
【0164】
要約すると、TGaseII発現および活性レベルの増加は、腎臓における線維性変化を伴うことを結果が示唆する。
【0165】
実施例4
TGaseIIノックアウト・マウスを使用するUUOモデルを使用する腎臓線維症におけるTGaseII役割の確立
ノックアウト・マウスは、Dr. Melino (De Laurenzi V, Melino G., Gene disruption of tissue transglutaminase, Mol Cell Biol. 2001 Jan;21(1):148-55)から得た。マウスは、表現型的に正常のように見えた。また、これらのマウスから得られる腎臓において、形態学的変化は観察されなかった。
【0166】
次いで、正常マウスと比較して腎臓コラーゲン蓄積を評価することによって、UUOの誘導後の腎臓線維症の発症の解析するために、TGaseII KOマウスを使用した。結果は、TGaseIIの破壊は、野生型マウスの閉塞腎臓におけるコラーゲン蓄積と比較すると、閉塞の三(3)および七(7)日に後の両方において、統計学的に有意なコラーゲン蓄積の減少を生じることを明らかに示し、したがって、TGaseIIは、腎臓線維症における機能的に重要な因子として関係づけられた。したがって、TGaseII活性の阻害は、抗線維治療効果を有し得ることが示唆される。
【0167】
実施例5
モノクローナル中和抗体を使用するTGaseII活性の阻害
TGaseIIに対する組換えモノクローナル・ヒト-抗ヒト抗体は、ファージディスプレイ・ライブラリーのスクリーニングによって得た。720クローンをヒトTGaseII結合についてスクリーニングした。酵素に対する強い結合を示した七(7)つのクローンを選択した。活性アッセイでは、これらの抗体が濃度範囲(1〜5nM)にてヒトTGaseII酵素の架橋活性を阻害することを示した。
【図面の簡単な説明】
【0168】
【図1】この図は、ヒトTGaseII cDNA(gi|13653650|ref|XM_009482.3| ORF )−配列番号:1のヌクレオチド配列を記載する。
【図2】この図は、ヒトTGaseIIに対応するポリペプチド−配列番号:2のアミノ酸配列を記載する。
【図3】ヒトTGaseIIポリペプチド発現に対する種々のTGaseII siRNAの効果を証明するウエスタンブロットの結果。図は、siRNAトランスフェクションの72時間後のHeLa細胞におけるヒトTGaseポリペプチドの発現を減少させる際の種々のsiRNAの活性を証明する。GAPDH発現は、タンパク質使用量のための対照として役立つ。数15および30は、オリゴヌクレオチドのnM濃度を表す。「細胞」(それぞれのパネルにおける第一のレーン)は、非トランスフェクト対照細胞を表す。それぞれのパネルの最後のレーンは、TGaseII発現における減少が抗TGase siRNA活性に特異的なことを保証するための、非関連(NR)siRNAをトランスフェクトしたHeLa細胞からのタンパク質抽出物を含む。
【特許請求の範囲】
【請求項1】
以下の構造を有する化合物:
5'(N)x-Z3'(アンチセンス鎖)
3'Z'-(N’)y5'(センス鎖)
式中、それぞれのNおよびN’は、その糖残基が修飾されていても、または修飾されていなくてもよいリボヌクレオチドであり、かつ(N)xおよび(N’)yは、それぞれの連続したNまたはN’が、共有結合によって次のNまたはN’に連結されているオリゴマーであり;
式中、それぞれのxおよびyは、19〜40の間の整数であり;
式中、それぞれのZおよびZ’は、存在しても、または存在しなくてもよいが、存在する場合、dTdTであり、かつそれが存在する鎖の3'末端にて共有結合で付着されており;および、
式中、(N)xの配列は、配列番号:21〜38、192〜344および381〜416に記載した配列のいずれか1つを含む。
【請求項2】
前記共有結合がホスホジエステル結合である、請求項1に記載の化合物。
【請求項3】
x=yである、請求項2に記載の化合物。
【請求項4】
x=y=19である、請求項3に記載の化合物。
【請求項5】
ZおよびZ’が両方とも存在しない、請求項1、2、3または4に記載の化合物。
【請求項6】
ZまたはZ’のうちの1つが存在する、請求項1、2、3または4に記載の化合物。
【請求項7】
前記リボヌクレオチドの全てにおいて、これらの糖残基が修飾されていない、請求項1〜6のいずれか1項に記載に記載の化合物。
【請求項8】
少なくとも1つのリボヌクレオチドにおいて、その糖残基が修飾されている、請求項1〜6のいずれか1項に記載に記載の化合物。
【請求項9】
前記糖残基の修飾が、2’位における修飾を含む、請求項8に記載の化合物。
【請求項10】
前記2’位の修飾により、アミノ基、フルオロ基、メトキシ基、アルコキシ基およびアルキル基を含む群から選択される部分が存在することとなる、請求項9に記載の化合物。
【請求項11】
前記2’位の部分が、メトキシ(2’-0-メチル)である、請求項10に記載の化合物。
【請求項12】
交互に現れるリボヌクレオチドが、アンチセンス鎖およびセンス鎖において修飾されている、請求項1〜6または8〜11のいずれか1項に記載に記載の化合物。
【請求項13】
請求項1〜6または8〜12のいずれか1項に記載の化合物であって、前記アンチセンス鎖の5'および3'末端の前記リボヌクレオチドは、これらの糖残基が修飾されており、かつ前記センス鎖の5'および3'末端の前記リボヌクレオチドは、これらの糖残基が修飾されていない化合物。
【請求項14】
請求項1〜13のいずれか1項に記載の化合物であって、前記アンチセンス鎖は、5'末端にてリン酸化されており(phophorylated)、かつ3'末端にてリン酸化されていても、またはリン酸化されていなくてもよく;および、
前記センス鎖が5'末端にて、および3'末端にてリン酸化されていても、またはリン酸化されていなくてもよい化合物。
【請求項15】
請求項1〜7のいずれか1項に記載の化合物を発現することができるベクター。
【請求項16】
ヒトTGaseIIを阻害するために有効な量の請求項1〜14のいずれか1項に記載の化合物または請求項15記載のベクターとキャリアとを含む組成物。
【請求項17】
障害に罹患している患者を治療する方法であって、これにより患者を治療するために、患者に対して治療的に有効な用量のヒトTGaseIIの阻害剤を投与することを含む方法。
【請求項18】
前記阻害剤がsiRNAである、請求項17記載の方法。
【請求項19】
前記阻害剤が抗体である、請求項17記載の方法。
【請求項20】
前記siRNAが、請求項1〜14のいずれか1項に記載の化合物または請求項15に記載に記載のベクターを含む、請求項18記載の方法。
【請求項21】
前記障害が線維症に関連した病態である、請求項17記載の方法。
【請求項22】
前記線維症に関連した病が、腎臓線維症、肝臓線維症、肺線維症または眼の瘢痕である、請求項21記載の方法。
【請求項23】
前記障害が、眼疾患、特に白内障、心臓血管疾患、特に心肥大、アテローム性動脈硬化症/再狭窄、神経系疾患、封入ポリグルタミン疾患、球脊髄性筋肉萎縮症、歯状核赤核-淡蒼球ルイ体萎縮症、脊髄小脳失調(SCA)1、2、3、6、7および17、アルツハイマー病またはパーキンソン病のいずれか一つである、請求項17記載の方法。
【請求項1】
以下の構造を有する化合物:
5'(N)x-Z3'(アンチセンス鎖)
3'Z'-(N’)y5'(センス鎖)
式中、それぞれのNおよびN’は、その糖残基が修飾されていても、または修飾されていなくてもよいリボヌクレオチドであり、かつ(N)xおよび(N’)yは、それぞれの連続したNまたはN’が、共有結合によって次のNまたはN’に連結されているオリゴマーであり;
式中、それぞれのxおよびyは、19〜40の間の整数であり;
式中、それぞれのZおよびZ’は、存在しても、または存在しなくてもよいが、存在する場合、dTdTであり、かつそれが存在する鎖の3'末端にて共有結合で付着されており;および、
式中、(N)xの配列は、配列番号:21〜38、192〜344および381〜416に記載した配列のいずれか1つを含む。
【請求項2】
前記共有結合がホスホジエステル結合である、請求項1に記載の化合物。
【請求項3】
x=yである、請求項2に記載の化合物。
【請求項4】
x=y=19である、請求項3に記載の化合物。
【請求項5】
ZおよびZ’が両方とも存在しない、請求項1、2、3または4に記載の化合物。
【請求項6】
ZまたはZ’のうちの1つが存在する、請求項1、2、3または4に記載の化合物。
【請求項7】
前記リボヌクレオチドの全てにおいて、これらの糖残基が修飾されていない、請求項1〜6のいずれか1項に記載に記載の化合物。
【請求項8】
少なくとも1つのリボヌクレオチドにおいて、その糖残基が修飾されている、請求項1〜6のいずれか1項に記載に記載の化合物。
【請求項9】
前記糖残基の修飾が、2’位における修飾を含む、請求項8に記載の化合物。
【請求項10】
前記2’位の修飾により、アミノ基、フルオロ基、メトキシ基、アルコキシ基およびアルキル基を含む群から選択される部分が存在することとなる、請求項9に記載の化合物。
【請求項11】
前記2’位の部分が、メトキシ(2’-0-メチル)である、請求項10に記載の化合物。
【請求項12】
交互に現れるリボヌクレオチドが、アンチセンス鎖およびセンス鎖において修飾されている、請求項1〜6または8〜11のいずれか1項に記載に記載の化合物。
【請求項13】
請求項1〜6または8〜12のいずれか1項に記載の化合物であって、前記アンチセンス鎖の5'および3'末端の前記リボヌクレオチドは、これらの糖残基が修飾されており、かつ前記センス鎖の5'および3'末端の前記リボヌクレオチドは、これらの糖残基が修飾されていない化合物。
【請求項14】
請求項1〜13のいずれか1項に記載の化合物であって、前記アンチセンス鎖は、5'末端にてリン酸化されており(phophorylated)、かつ3'末端にてリン酸化されていても、またはリン酸化されていなくてもよく;および、
前記センス鎖が5'末端にて、および3'末端にてリン酸化されていても、またはリン酸化されていなくてもよい化合物。
【請求項15】
請求項1〜7のいずれか1項に記載の化合物を発現することができるベクター。
【請求項16】
ヒトTGaseIIを阻害するために有効な量の請求項1〜14のいずれか1項に記載の化合物または請求項15記載のベクターとキャリアとを含む組成物。
【請求項17】
障害に罹患している患者を治療する方法であって、これにより患者を治療するために、患者に対して治療的に有効な用量のヒトTGaseIIの阻害剤を投与することを含む方法。
【請求項18】
前記阻害剤がsiRNAである、請求項17記載の方法。
【請求項19】
前記阻害剤が抗体である、請求項17記載の方法。
【請求項20】
前記siRNAが、請求項1〜14のいずれか1項に記載の化合物または請求項15に記載に記載のベクターを含む、請求項18記載の方法。
【請求項21】
前記障害が線維症に関連した病態である、請求項17記載の方法。
【請求項22】
前記線維症に関連した病が、腎臓線維症、肝臓線維症、肺線維症または眼の瘢痕である、請求項21記載の方法。
【請求項23】
前記障害が、眼疾患、特に白内障、心臓血管疾患、特に心肥大、アテローム性動脈硬化症/再狭窄、神経系疾患、封入ポリグルタミン疾患、球脊髄性筋肉萎縮症、歯状核赤核-淡蒼球ルイ体萎縮症、脊髄小脳失調(SCA)1、2、3、6、7および17、アルツハイマー病またはパーキンソン病のいずれか一つである、請求項17記載の方法。
【図1】


【図2】

【図3】
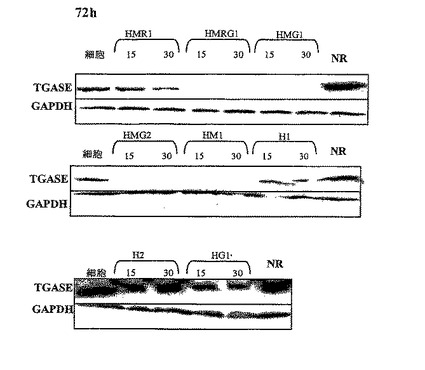
【図2】
【図3】
【公表番号】特表2007−528216(P2007−528216A)
【公表日】平成19年10月11日(2007.10.11)
【国際特許分類】
【出願番号】特願2006−550491(P2006−550491)
【出願日】平成17年1月27日(2005.1.27)
【国際出願番号】PCT/IL2005/000102
【国際公開番号】WO2005/072057
【国際公開日】平成17年8月11日(2005.8.11)
【出願人】(502337583)クアーク・ファーマスーティカルス、インコーポレイテッド (12)
【氏名又は名称原語表記】Quark Pharmaceuticals,Inc.
【Fターム(参考)】
【公表日】平成19年10月11日(2007.10.11)
【国際特許分類】
【出願日】平成17年1月27日(2005.1.27)
【国際出願番号】PCT/IL2005/000102
【国際公開番号】WO2005/072057
【国際公開日】平成17年8月11日(2005.8.11)
【出願人】(502337583)クアーク・ファーマスーティカルス、インコーポレイテッド (12)
【氏名又は名称原語表記】Quark Pharmaceuticals,Inc.
【Fターム(参考)】
[ Back to top ]