
耐熱性の植物および植物組織ならびにそれを作製および使用するための方法および材料
本発明は、熱条件に対する耐性が増大し、かつ/またはデンプン生合成が増大した植物または植物組織を提供するための材料および方法に関する。熱条件に対する植物または植物組織の耐性が増大すると、高温で一般に観察される収量減少と比べて少ない収量減少が実現される。本発明の1つの局面は、変異植物AGPase小サブユニットをコードするポリヌクレオチドに関する。本発明はまた、本発明のポリヌクレオチドにコードされる変異植物AGPase小サブユニットも含む。本発明はまた、本発明のポリヌクレオチドを含む植物およびこれらの植物を作製するための方法にも関する。
【発明の詳細な説明】
【背景技術】
【0001】
政府の支援
本出願の主題は、助成金番号IOS-0444031のもとで、米国国立科学財団(National Science Foundation)からの研究助成金によって支援されている。したがって、政府は、本発明において一定の権利を有する。
【0002】
発明の背景
熱ストレスは、トウモロコシ収量の減少をもたらす(Peters et al., 1971; Thompson, 1975; Chang, 1981; ChristyおよびWilliamson, 1985)。これは、光合成産物の利用可能度およびソース組織からシンク組織への輸送の減少、乏しい受粉、細胞および顆粒のサイズおよび数の減少、初期の種子発育不全(seed abortion)、ならびに/または登熟期間の短縮に帰され得る。内乳の成長は、細胞が活発に分裂する誘導期(lag phase)から始まり、細胞のサイズが増大し、デンプン合成が起こる直線期(linear phase)に続く。誘導期の間の温度上昇は、収量の減少をもたらした(Jones et al., 1984)。これらの研究者らは、収量の減少が細胞および顆粒の数およびサイズの減少ならびに種子発育不全に起因することを示唆した。さらに、直線期の間の温度上昇により、登熟期間が短くなり、続いて、穀粒が小さくなった(Jones et al., 1984)。同様の結果が、Hunter et al. (1977)ならびにTollenaarおよびBruulsema (1988)によって見出された。
【0003】
古くからUSトウモロコシの50%超を生産する5つの州での記録から、1日の平均温度は23.6℃であり、登熟の間の最適温度より約2℃高いことが示された(Singletary et al., 1994)。少なくともオオムギおよびコムギにおいて、登熟中の光合成産物の利用可能度は高温度で低下することはない。実際、オオムギ種子およびコムギ種子中のスクロース含有量は、高温度で不変であるか、または増加していた(BhullarおよびJenner, 1986; Wallwork et al., 1998)。また、トウモロコシの光合成は、32℃までは増加する(DuncanおよびHesketh, 1968; HofstraおよびHesketh, 1969; Christy et al., 1985)。さらに、CheikhnおよびJones (1995)は、トウモロコシ穀粒が14Cスクロースおよびヘキソースを固定する能力を様々な温度で調査した。彼らは、種子中のこれらの糖が高温で増加することを発見した。上記の証拠から、登熟の間に糖の利用可能度および穀粒中への輸送が限定されることが、温度によって誘発される収量減少の原因ではないことが示唆される。
【0004】
温度上昇の間に登熟に強い影響を与える生化学的経路を同定するために多大な努力が払われてきた。Singletary et al. (1993; 1994)は、インビトロで増殖させたトウモロコシ穀粒中のデンプン生合成酵素を高温(22℃〜36℃)において分析した。彼らは、ADPグルコースピロホスホリラーゼ(AGPase)および可溶性デンプン合成酵素(SSS)の方が、デンプン合成に関与する他の酵素と比べて熱不安定性であることを見出した。彼らは、AGPaseおよびSSSの熱不安定性が登熟停止に寄与していることを示唆した。DukeおよびDoehlert(1996)は、AGPaseをコードするものを含む、デンプン合成経路の酵素をコードするいくつかの遺伝子の転写物が、25℃と比較すると35℃では減少していることを発見した。しかしながら、酵素アッセイ法により、AGPase活性のみが目立って低いことが示された。これは他の酵素と比べて高いAGPaseの代謝回転率に起因し得ると彼らは示唆した。最後に、Wilhelm et al. (1999)は、Q10解析を通じて、他のいくつかの酵素と比べて、AGPaseの活性が最も顕著に低下することを示した。57℃で5分間加熱した際、トウモロコシAGPaseは実際にその活性の96%を失った(Hannah et al., 1980)。
【0005】
AGPaseは、デンプン(植物)合成およびグリコーゲン(細菌)合成において最初の方向付けされた(committed)段階を触媒する。これは、ADP-グルコースおよびピロリン酸(PPi)へのグルコース-1-P(G-1-P)およびATPの変換を伴う。AGPaseは、2つの同一な小サブユニットおよび2つの同一な大サブユニットからなる、植物中のヘテロ四量体である。これらの大サブユニットおよび小サブユニットは、それぞれ、トウモロコシ内乳中のしわ型(shrunken)-2(Sh2)およびカマイラズ(brittle)-2(Bt2)にコードされている。AGPaseは、細胞のエネルギー状態を示す小型エフェクター分子によってアロステリックに調節される。AGPaseは、藍藻類、緑藻類、および被子植物において、最初の炭素同化産物である3-PGAによって活性化され、無機リン酸(Pi)によって阻害/不活性化される。
【0006】
デンプン合成におけるトウモロコシ内乳AGPaseの重要性は、酵素のいずれかのサブユニットの変異体の穀粒表現型によって示されている。実際、このような変異体は、しわ型穀粒および内乳デンプン含有量の大幅な減少をもたらす(TsaiおよびNelson, 1966; HannahおよびNelson, 1976)。AGPaseがデンプン合成の律速段階を触媒するという証拠もある(Stark et al. 1992; Giroux et al. 1996; Greene et al. 1998; Sakulsingharoja et al. 2004; Obana et al. 2006; Wang et al. 2007)。
【0007】
GreeneおよびHannah (1998a)は、HS33と呼ばれる大サブユニット中に1つのアミノ酸変化を有する変異型トウモロコシAGPaseを単離した。彼らは、改変された酵素の方が熱安定性が高いこと、および安定性はサブユニット間のより強い相互作用の寄与によることを示した。AGPaseのアロステリック特性に影響を及ぼす変化(Rev6)(Giroux et al., 1996)と共にHS33変化を含むSh2バリアントでコムギおよびイネを形質転換した場合、収量はそれぞれ38%および23%増加した(Smidansky et al., 2002; 2003)。驚くべきことに、この増加は、個々の種子重量よりはむしろ、種子数の増加に起因した。
【0008】
また、Rev6変化およびHS33変化を含むSh2バリアントによるトウモロコシの形質転換も、種子数の増加をもたらす。トウモロコシの場合、種子収量/穂は最大で68%増加し得る。トウモロコシのトランスジェニック事象の詳細な特徴付けは進行中である(GreeneおよびHannah、出版準備中)。トウモロコシにおいてRev6を単独で発現させた場合、種子重量のみが増加するため、種子数の増加をRev6で説明することはできない(Hannah、未発表)。上記の研究から、穀物収量におけるAGPase熱安定性の重要性が示される。
【0009】
Cross et al. (2004)は、BT2の最初の200個のアミノ酸およびジャガイモ塊茎小サブユニットの最後の275個のアミノ酸からなるモザイク小サブユニット(MP)を作製した。SH2との複合体中のMPは、農学的利益をもたらし得るいくつかの特徴を有した(Cross et al., 2004; Boehlein et al., 2005)。これらの特徴のいくつかは、野生型トウモロコシ内乳AGPase(BT2/SH2)と比べて、アクチベーター3-PGAの不在下での増大した活性、3-PGAに対する増大した親和性、および上昇した熱安定性であった。予備データから、トウモロコシ内乳において発現されるAGPaseバリアントを含む、トランスジェニックMPを有するトウモロコシ植物が、デンプン収量の増加を実現することが示されている(Hannah、未発表データ)。
【発明の概要】
【0010】
本発明は、熱条件に対する耐性が増大し、かつ/またはデンプン生合成が増大した植物または植物組織を提供するための材料および方法に関する。熱条件に対する植物または植物組織の耐性が増大すると、高温で一般に観察される収量減少と比べて少ない収量減少が提供される。本発明の1つの局面は、変異植物AGPase小サブユニットをコードするポリヌクレオチドに関する。1つの態様において、本発明のポリヌクレオチドは、野生型トウモロコシAGPase小サブユニットのアミノ酸462位に対応するトレオニンアミノ酸が、増大した熱安定性を与えるアミノ酸によって置換されているアミノ酸変異を有する、植物AGPase小サブユニットをコードする。別の態様において、ポリヌクレオチドは、2種の異なる植物に由来する配列から構成され(米国特許第7,173,165号に記載されている)、かつ、野生型トウモロコシAGPase小サブユニットのアミノ酸462位に対応するトレオニンアミノ酸が、増大した熱安定性を与えるアミノ酸によって置換されている本発明のアミノ酸変異を含む、キメラ植物AGPase小サブユニットをコードする。キメラAGPaseの変異は、熱安定性を相乗的に高める。本発明はまた、本発明のポリヌクレオチドにコードされる、変異植物AGPase小サブユニットも含む。熱安定性ならびに動力学的特性およびアロステリック特性の特徴付けから、本発明のポリヌクレオチドが単子葉植物内乳など植物において発現される場合、デンプン収量の増加が実現されることが示される。
【図面の簡単な説明】
【0011】
【図1】SH2と共にBT2、BT2-TI、MP、MP-TIを発現する大腸菌(E. coli)細胞によって産生されたグリコーゲンを示す。SH2、BT2、BT2-TI、およびMPのみを単独で発現する細胞に由来するグリコーゲン。グリコーゲンはグルコース単位で測定される。エラーバーは標準偏差を示す(N=3)。
【図2】図2Aおよび図2Bは、相補的なサブユニットSH2と共にBT2、BT2-TI、MP、MP-TIを発現する大腸菌細胞に由来する粗製抽出物のドットブロットを示す。BT2に対するモノクローナル抗体を用いてAGPaseを可視化した。スポットの密度は、ImageJ(http://rsb.info.nih.gov/ij/)を用いて概算した。
【図3】誘導されていない大腸菌細胞に由来する粗製タンパク質抽出物および部分的に精製したタンパク質抽出物中のAGPaseバリアントの比活性を示す。活性は逆方向で測定した。n.d.=検出不可。エラーバーは標準偏差を示す(N=3)。
【図4】AGPaseの精製を示す。精製した組換えBT2/SH2、TI/SH2、MP/SH2、およびMP-TI/SH2のSDS-PAGE。Biorad社製のPrecision Plus Protein All Blue Standardをマーカーとして使用した。左側の上の矢印は、大サブユニットを示す。左側の下の矢印は、小サブユニットを示す。
【図5A】精製したBT2/SH2、BT2-TI/SH2、およびMP/SH2の熱安定性を示す。各AGPaseの半減期(T1/2)は、平均値±標準誤差として表される。p値は、Prizm (Graph pad, San Diego CA)を用いて実施されるF検定によって概算する。図5Aにおいて、このアッセイ法は順方向で実施した。
【図5B】精製したBT2/SH2、BT2-TI/SH2、およびMP/SH2の熱安定性を示す。各AGPaseの半減期(T1/2)は、平均値±標準誤差として表される。p値は、Prizm (Graph pad, San Diego CA)を用いて実施されるF検定によって概算する。図5Bにおいて、このアッセイ法は逆方向で実施した。
【図6A】精製したMP/SH2およびMP-TI/SH2の熱安定性を示す。各AGPaseの半減期(T1/2)は、平均値±標準誤差として表される。p値は、Prizm (Graph pad, San Diego CA)を用いて実施されるF検定によって概算する。図6Aにおいて、このアッセイ法は順方向で実施した。
【図6B】精製したMP/SH2およびMP-TI/SH2の熱安定性を示す。各AGPaseの半減期(T1/2)は、平均値±標準誤差として表される。p値は、Prizm (Graph pad, San Diego CA)を用いて実施されるF検定によって概算する。図6Bにおいて、このアッセイ法は逆方向で実施した。
【図7A】BT2およびTIの3Dモデリングを示す。図7Aは、BT2モノマーの予測される3D構造である。TI変化には赤い丸でマークを付けている。サブユニット間の相互作用に直接関与しているBT2の領域は、黄色い四角で強調している。
【図7B】BT2およびTIの3Dモデリングを示す。図7Bは、Pro60(4、5)およびLeu61(3)の炭素原子からのThr462(1、2)の炭素原子の距離を示す。Thr462の接触残基は、FirstGlance Jmolを用いて決定した。濃い灰色の球は、Thr462の炭素原子を示す。明るい灰色の球は、接触残基の炭素原子を示す。酸素原子および窒素原子は、それぞれ赤色および青色で示す。
【図7C】BT2およびTIの3Dモデリングを示す。図7Cは、Pro60(5、6)およびLeu61(7、8)の炭素原子からのIle462(1、2、3、4)の炭素原子の距離を示す。Ile462の接触残基は、FirstGlance Jmolを用いて決定した。濃い灰色の球は、Ile462の炭素原子を示す。明るい灰色の球は、接触残基の炭素原子を示す。酸素原子および窒素原子は、それぞれ赤色および青色で示す。
【図8】AGPaseのサブユニット間の相互作用の強さを示す。酵母ツーハイブリッドシステムにおいてSH2をベイトとして使用し、BT2、TI、およびMPをプレイとして使用した。定量的β-ガラクトシダーゼアッセイ法を用いて、ベイトとプレイの間の相互作用を定量した。エラーバーは、2×標準誤差を示す(N=4)。
【発明を実施するための形態】
【0012】
配列の簡単な説明
配列番号:1は、本発明の変異ポリペプチド(TI)をコードするヌクレオチド配列を含むポリヌクレオチド配列である。
【0013】
配列番号:2は、本発明の変異ポリペプチド(TI)のアミノ酸配列である。
【0014】
配列番号:3は、本発明の変異ポリペプチド(MP-TI)をコードするヌクレオチド配列を含むポリヌクレオチド配列である。
【0015】
配列番号:4は、本発明の変異ポリペプチド(MP-TI)のアミノ酸配列である。
【0016】
配列番号:5は、本発明の変異ポリペプチド(TI+YC)のアミノ酸配列である。
【0017】
配列番号:6は、本発明の変異ポリペプチド(TI+QTCL)のアミノ酸配列である。
【0018】
配列番号:7は、本発明の変異ポリペプチド(TI+ETCL)のアミノ酸配列である。
【0019】
配列番号:8は、本発明の変異ポリペプチド(MP-TI+YC)のアミノ酸配列である。
【0020】
配列番号:9は、本発明の変異ポリペプチド(MP-TI+QTCL)のアミノ酸配列である。
【0021】
配列番号:10は、本発明の変異ポリペプチド(MP-TI+ETCL)のアミノ酸配列である。
【0022】
配列番号:11は、本発明の変異ポリペプチド(配列番号:5)をコードするヌクレオチド配列を含むポリヌクレオチド配列である。
【0023】
配列番号:12は、本発明の変異ポリペプチド(配列番号:6)をコードするヌクレオチド配列を含むポリヌクレオチド配列である。
【0024】
配列番号:13は、本発明の変異ポリペプチド(配列番号:7)をコードするヌクレオチド配列を含むポリヌクレオチド配列である。
【0025】
配列番号:14は、本発明の変異ポリペプチド(配列番号:8)をコードするヌクレオチド配列を含むポリヌクレオチド配列である。
【0026】
配列番号:15は、本発明の変異ポリペプチド(配列番号:9)をコードするヌクレオチド配列を含むポリヌクレオチド配列である。
【0027】
配列番号:16は、本発明の変異ポリペプチド(配列番号:10)をコードするヌクレオチド配列を含むポリヌクレオチド配列である。
【0028】
配列番号:17は、本発明に従って使用することができるオリゴヌクレオチドである。
【0029】
配列番号:18は、本発明に従って使用することができるオリゴヌクレオチドである。
【0030】
配列番号:19は、本発明に従って使用することができるオリゴヌクレオチドである。
【0031】
配列番号:20は、本発明に従って使用することができるオリゴヌクレオチドである。
【0032】
配列番号:21は、本発明の変異ポリペプチド(TI)のアミノ酸配列である。
【0033】
配列番号:22は、本発明の変異ポリペプチド(TI)のアミノ酸配列である。
【0034】
配列番号:23は、本発明の変異ポリペプチド(TI)のアミノ酸配列である。
【0035】
配列番号:24は、本発明の変異ポリペプチド(TI)のアミノ酸配列である。
【0036】
配列番号:25は、本発明の変異ポリペプチド(TI)のアミノ酸配列である。
【0037】
配列番号:26は、ジャガイモ塊茎のAGPase小サブユニットタンパク質のアミノ酸配列である。
【0038】
発明の詳細な説明
本発明は、熱条件に対する耐性が増大し、かつ/またはデンプン生合成が増大した植物を提供するための材料および方法に関する。熱条件に対する植物の耐性が増大すると、高温で一般に観察される収量減少と比べて少ない収量減少が提供される。
【0039】
本発明の1つの局面は、変異植物AGPase小サブユニットをコードするポリヌクレオチドに関する。1つの態様において、本発明のポリヌクレオチドは、野生型トウモロコシ内乳AGPase小サブユニットのアミノ酸462位に対応するトレオニンアミノ酸が、増大した熱安定性を与えるアミノ酸によって置換されているアミノ酸変異を有する、植物AGPase小サブユニットをコードする。特定の態様において、置換アミノ酸(amino acid substituted)はイソロイシンである。1つの態様において、変異植物AGPase小サブユニットはトウモロコシ内乳AGPase小サブユニットである。例示的な態様において、変異植物AGPase小サブユニットは、配列番号:2に示すアミノ酸配列またはその断片もしくはバリアントを含む。特定の態様において、ポリヌクレオチドは、配列番号:1に示すヌクレオチド配列またはその断片もしくはバリアントを含む。別の態様において、変異植物AGPase小サブユニットは、オオムギ、コムギ、モロコシ、ジャガイモ、またはイネである。特定の態様において、オオムギ、コムギ、モロコシ、またはジャガイモの変異AGPase小サブユニットは、配列番号:21、配列番号:22、配列番号:23、配列番号:24、または配列番号:25にそれぞれ示すアミノ酸配列を含む。別の態様において、ポリヌクレオチドは、公開されている国際特許出願の国際公開公報第2005/019425号(HannahおよびLinebarger)に記載されているようなアミノ酸変異をさらに含んでよい変異植物AGPase小サブユニットをコードする。1つの態様において、このポリヌクレオチドにコードされる変異AGPase小サブユニットは、野生型トウモロコシ内乳AGPaseのアミノ酸36位に対応するチロシンがシステインで置換されているアミノ酸変異を含む。また、この変異AGPase小サブユニットは、野生型トウモロコシ内乳AGPaseのアミノ酸34位およびアミノ酸35位にそれぞれ対応するセリンアミノ酸とトレオニンアミノ酸の間に挿入されたアミノ酸を任意で含んでよい。特定の態様において、AGPase小サブユニットの34位および35位のアミノ酸の間に挿入されるアミノ酸は、グルタミン酸またはグルタミンである。例示的な態様において、変異植物AGPase小サブユニットは、配列番号:5、配列番号:6、もしくは配列番号:7に示すアミノ酸配列またはその断片もしくはバリアントを含む。特定の態様において、ポリヌクレオチドは、配列番号:11、配列番号:12、もしくは配列番号:13に示すヌクレオチド配列またはその断片もしくはバリアントを含む。
【0040】
別の態様において、ポリヌクレオチドは、2種の異なる植物に由来する配列から構成され(米国特許第7,173,165号に記載されている)、かつ、野生型トウモロコシ内乳AGPase小サブユニットのアミノ酸462位に対応するトレオニンアミノ酸が、増大した熱安定性を与えるアミノ酸によって置換されている本発明のアミノ酸変異もまた含む、キメラ植物AGPase小サブユニットをコードする。特定の態様において、置換アミノ酸はイソロイシンである。1つの態様において、キメラAGPase小サブユニットは、1つの植物に由来するC末端部分および別の植物に由来するN末端部分を含む。1つの態様において、本発明のキメラタンパク質は、第1の植物AGPase小サブユニットのN末端の最初の約150〜250個のアミノ酸を有するN末端配列および第2の植物AGPase小サブユニットのC末端の最後の約300個またはそれ以下の残基を含むC末端配列を含む。したがって、キメラサブユニットのC末端は、第2の植物のC末端の最後の300個、または299個、または298個、または297個、または296個、または295個などの残基を含み得る。サブユニット配列は、単子葉植物の、もしくは双子葉植物の、または単子葉植物と双子葉植物の両方のAGPaseに由来してよい。例えば、イネ、コムギ、オオムギ、エンバク、モロコシ、トウモロコシ、ユリ、およびキビ(millet)などの単子葉植物は、本発明の範囲内に含まれる。双子葉植物には、例えば、タバコ、ダイズ、ジャガイモ、サツマイモ、ラディッシュ、キャベツ、セイヨウアブラナ、リンゴの木、およびレタスが含まれ得る。1つの態様において、キメラタンパク質のN末端の最初の200個程度のアミノ酸はトウモロコシ内乳AGPase小サブユニットのN末端に由来し、C末端アミノ酸は、ジャガイモ塊茎AGPase小サブユニットのC末端に由来し、本発明のアミノ酸462位に対応する変異を含む。特定の態様において、本発明のキメラタンパク質のC末端領域は、ジャガイモ塊茎のAGPase小サブユニットの最後の276個のアミノ酸を含む。例示的な態様において、キメラタンパク質は、トウモロコシ内乳AGPaseの小サブユニットの一部分およびジャガイモ塊茎AGPaseの小サブユニットの一部分を含む。特定の態様において、キメラタンパク質は、a)トウモロコシ内乳AGPaseの小サブユニットに由来する最初の199個のアミノ酸(すなわち、アミノ酸1〜199)、およびアミノ酸246から始まる、ジャガイモ塊茎AGPaseの小サブユニットのカルボキシル末端(すなわち、アミノ酸246〜521)(Genbankアクセッション番号X61186として寄託されているタンパク質に関して示されているアミノ酸配列を用いた場合)(または、別法として、Hannah et al., 2001で記載されているジャガイモAGPaseサブユニットの番号付け方式を用いた場合、アミノ酸175から始まる)、ならびにb)本発明のアミノ酸462位に対応する変異を含む。例示的な態様において、植物キメラAGPase小サブユニットは、配列番号:4に示すアミノ酸配列またはその断片もしくはバリアントを含む。特定の態様において、ポリヌクレオチドは、配列番号:3に示すヌクレオチド配列またはその断片もしくはバリアントを含む。別の態様において、ポリヌクレオチドは、公開されている国際特許出願の国際公開公報第2005/019425号(HannahおよびLinebarger)に記載されているアミノ酸変異をさらに含んでよい変異植物AGPase小サブユニットをコードする。さらなる態様において、このポリヌクレオチドにコードされる変異AGPase小サブユニットはまた、36位のチロシンがシステインで置換されているアミノ酸変異を含む。また、この変異AGPase小サブユニットは、それぞれ34位および35位のセリンアミノ酸とトレオニンアミノ酸の間に挿入されたアミノ酸を任意で含んでよい。特定の態様において、変異AGPase小サブユニットの34位および35位のアミノ酸の間に挿入されるアミノ酸は、グルタミン酸またはグルタミンである。例示的な態様において、変異植物AGPase小サブユニットは、配列番号:8、配列番号:9、または配列番号:10に示すアミノ酸配列またはその断片もしくはバリアントを含む。特定の態様において、ポリヌクレオチドは、配列番号:14、配列番号:15、または配列番号:16に示すヌクレオチド配列またはその断片もしくはバリアントを含む。
【0041】
本発明はまた、植物または植物組織の熱安定性を増大させるため、および/またはデンプン生合成を増大させるため、ならびに収穫量を増加させるための方法にも関する。1つの態様において、本発明の方法は、本発明の1つまたは複数のポリヌクレオチドを植物中に導入する段階を含む。一定の態様において、植物中に導入されるポリヌクレオチドは、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、配列番号:10、配列番号:21、配列番号:22、配列番号:23、配列番号:24、もしくは配列番号:25のいずれかに示すアミノ酸配列またはその断片もしくはバリアントを含む1種または複数種のポリペプチドをコードする。特定の態様において、ポリヌクレオチドは、配列番号:1もしくは配列番号:3に示すヌクレオチド配列またはその断片もしくはバリアントを含む。さらなる特定の態様において、ポリヌクレオチドは、配列番号:11、配列番号:12、配列番号:13、配列番号:14、配列番号:15、もしくは配列番号:16に示すヌクレオチド配列またはその断片もしくはバリアントを含む。1つの態様において、ポリヌクレオチドは、植物または植物組織のゲノム中に安定に組み込まれる。ポリヌクレオチドは、ポリヌクレオチドおよび/またはそれにコードされるポリペプチドの発現を増大させる調節エレメント、例えばプロモーター配列および/またはエンハンサー配列を含んでよい。特定の態様において、プロモーター配列は、構成的発現または組織特異的(例えば内乳)発現をもたらすものである。このポリヌクレオチドを含む植物もしくは植物組織、またはこれらの植物の子孫は、本発明のポリヌクレオチドまたはポリペプチドの発現の増大について任意でスクリーニングすることができる。1つの態様において、本発明の1つまたは複数のポリヌクレオチドの複数のコピーは、植物または植物組織中に導入され、その植物のゲノム中に安定に組み込まれる。1つの態様において、本発明のポリヌクレオチドは、本明細書において説明する発現構築物において提供される。
【0042】
本発明はまた、本発明のポリヌクレオチドにコードされる、変異AGPase小サブユニットポリペプチドにも関する。1つの態様において、このポリペプチドは、配列番号:2に示すアミノ酸配列またはその断片もしくはバリアントを含む。他の態様において、このポリペプチドは、配列番号:21、配列番号:22、配列番号:23、配列番号:24、もしくは配列番号:25に示すアミノ酸配列またはその断片もしくはバリアントを含む。別の態様において、このポリペプチドは、配列番号:4に示すアミノ酸配列またはその断片もしくはバリアントを含む。さらに別の態様において、このポリヌクレオチドは、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、もしくは配列番号:10のいずれかに示すアミノ酸配列またはその断片もしくはバリアントを含む。
【0043】
本発明はまた、本発明の1つまたは複数の変異AGPase小サブユニットポリペプチドを含む変異植物AGPase酵素にも関する。変異植物AGPaseはまた、1つまたは複数の野生型AGPase大サブユニットポリペプチドも含んでよい。特定の態様において、変異植物AGPase酵素は、そのうちのいずれかが配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、配列番号:10、配列番号:21、配列番号:22、配列番号:23、配列番号:24、もしくは配列番号:25のアミノ酸配列または任意のこのような配列の断片もしくはバリアントを含んでよい1つまたは複数の変異AGPase小サブユニットポリペプチドを含み、この変異AGPase酵素は、野生型AGPase酵素と比べて増大した熱安定性を示す。1つの態様において、変異植物酵素は、本発明の2つの変異AGPase小サブユニットを含み、ここで、これらの変異ポリペプチドは同じ変異を有してもよく、または異なる変異を有してもよい。本発明はまた、本発明の1つまたは複数の変異AGPase小サブユニットポリペプチドおよび1つまたは複数の変異AGPase大サブユニットポリペプチドを含む、変異植物AGPase酵素にも関する。1つの態様において、変異AGPase大サブユニットポリペプチドは、米国特許第5,589,618号;同第5,650,557号;同第5,872,216号;同第6,069,300号;同第6,184,438号;同第6,403,863号;同第6,809,235号;同第7,173,165号;同第7,312,378号;および同第6,969,783号のいずれかに記載されているもののいずれかでよい。1つの態様において、変異AGPase大サブユニットポリペプチドは、Rev6変異を含む。別の態様において、変異AGPase大サブユニットポリペプチドは、米国特許第6,069,300号;同第6,403,863号;同第6,809,235号;同第7,312,378号;および同第6,969,783号、ならびに公開されている国際特許出願の国際公開公報第99/58698号;国際公開公報第2003/0070901号;国際公開公報第98/22601号;および国際公開公報第02/072784号に記載されているような1つまたは複数の熱安定性(HS)変異、例えば、HS33変異などを含む。1つの態様において、変異植物AGPase酵素は、本明細書において説明するように、同じ変異を有してもよく、または異なる変異を有してもよい本発明の2つの変異AGPase小サブユニットポリペプチドを含む。別の態様において、変異植物AGPase酵素は、同じ変異を有してもよく、または異なる変異を有してもよい2つの変異AGPase大サブユニットポリペプチドを含む。さらなる態様において、変異植物AGPase酵素は、本発明の2つの変異AGPase小サブユニットポリペプチドおよび2つの変異SH2ポリペプチドを含み、ここで、本明細書において説明するように、これらの変異AGPase小サブユニットポリペプチドおよび変異AGPase大サブユニットポリペプチドは、同じ変異を有してもよく、または異なる変異を有してもよい。
【0044】
本発明はまた、野生型植物AGPaseと比べて熱安定性が増大した変異植物AGPase酵素を提供するための方法にも関する。1つの態様において、この方法は、AGPase酵素において、本発明の1つまたは複数の変異AGPase小サブユニットポリペプチドを、野生型AGPase大サブユニットもしくは変異AGPase大サブユニットと合体させる段階、または野生型AGPase大サブユニットもしくは変異AGPase大サブユニットと共に提供する段階を含む。1つの態様において、AGPase酵素は、ポリペプチドサブユニットの四量体を含み、それらのサブユニットの内の1つ、2つ、またはそれ以上が本発明の変異ポリペプチドである。1つの態様において、AGPase酵素はまた、変異AGPase大サブユニットポリペプチドサブユニット、例えば、Rev6および/またはHS33のような熱安定性変異を含む変異大サブユニットを含む。
【0045】
本発明はまた、ポリヌクレオチドもしくは本発明のポリヌクレオチドにコードされるタンパク質を含むか、または本発明の変異ポリペプチドもしくはその断片もしくはバリアントを発現するか、または本発明の変異植物AGP酵素を含むかもしくは発現する、本発明の植物、植物組織、および植物細胞にも関する。植物組織には、種子、接ぎ穂、および台木が含まれるがこれらに限定されるわけではない。本発明の範囲内の植物には、例えば、イネ、コムギ、オオムギ、エンバク、ライムギ、モロコシ、トウモロコシ、サトウキビ、パイナップル、タマネギ、バナナ、ココナツ、ユリ、芝草(turfgrass)、およびキビなどの単子葉植物が含まれる。また、本発明の範囲内の植物には、例えば、トマト、キュウリ、カボチャ、エンドウマメ、アルファルファ、メロン、ヒヨコマメ、チコリ、クローバー、ケール、レンズマメ、ダイズ、マメ、タバコ、ジャガイモ、サツマイモ、ヤムイモ、カッサバ、ラディッシュ、ブロッコリー、ホウレンソウ、キャベツ、セイヨウアブラナ、リンゴの木、柑橘類(オレンジ、マンダリン、グレープフルーツ、レモン、およびライムなどを含む)、ブドウ、綿、ヒマワリ、イチゴ、レタス、およびホップなどの双子葉植物も含まれる。本発明のポリヌクレオチドを含むハーブ植物もまた、本発明の範囲内で企図される。ハーブ植物には、パセリ、セージ、ローズマリー、およびタイムなどが含まれる。1つの態様において、植物、植物組織、または植物細胞は、トウモロコシ(Zea mays)である。1つの態様において、植物、植物組織、または植物細胞は、トランスジェニックである植物、植物組織、または植物細胞である。別の態様において、植物、植物組織、または植物細胞は、育種プログラムを通じて得られたものである。
【0046】
本発明において有用なポリヌクレオチドを発現構築物において提供することができる。一般に、本発明の発現構築物は、その発現構築物が発現されることになっている意図された宿主細胞において機能的である調節エレメントを含む。したがって、当業者は、細菌宿主細胞、酵母宿主細胞、植物宿主細胞、昆虫宿主細胞、哺乳動物宿主細胞、およびヒト宿主細胞において使用するための調節エレメントを選択することができる。調節エレメントには、プロモーター、転写終結配列、翻訳終結配列、エンハンサー、およびポリアデニル化エレメントが含まれる。本明細書において使用される場合、「発現構築物」という用語は、機能的に連結された核酸配列の転写を実現する核酸配列の組み合わせを意味する。本明細書において使用される場合、「機能的に連結される」という用語は、記述された構成要素の並置を意味し、それらの構成要素は、意図された様式でそれらが機能することを可能にする関係にある。一般に、機能的に連結された構成要素は隣接関係にある。
【0047】
本発明の発現構築物は、本発明の変異ポリペプチドをコードするポリヌクレオチド配列に機能的に連結されたプロモーター配列を含んでよい。プロモーターは、当技術分野において公知の標準技術を用いてポリヌクレオチド中に組み入れることができる。プロモーターの複数のコピーまたは複数のプロモーターを本発明の発現構築物において使用することができる。好ましい態様において、プロモーターは、発現構築物中の転写開始部位からの距離が、天然の遺伝的環境における転写開始部位からの距離とほぼ同じになるように配置されてよい。プロモーター活性を実質的に低下させずに、この距離をいくらか変更することは許容される。転写開始部位は、典型的には発現構築物中に含まれる。
【0048】
発現構築物が植物細胞中で提供される予定であるか、または植物細胞中に導入される予定である場合には、植物ウイルスプロモーター、例えば、カリフラワーモザイクウイルス(CaMV) 35Sプロモーター(増強されたCaMV 35Sプロモーター(例えば米国特許第5,106,739号を参照されたい)を含む)もしくはCaMV 19Sプロモーターまたはキャッサバ葉脈モザイク(cassava vein mosaic)などが使用され得る。植物中の発現構築物のために使用され得る他のプロモーターには、例えば、プロリフェラ(prolifera)プロモーター、Ap3プロモーター、熱ショックプロモーター、A.ツメファシエンス(A.tumefaciens)のT-DNA 1'-プロモーターまたはT-DNA 2'-プロモーター、ポリガラクツロナーゼプロモーター、ペチュニア由来のカルコン合成酵素A(CHS-A)プロモーター、タバコPR-1aプロモーター、ユビキチンプロモーター、アクチンプロモーター、alcA遺伝子プロモーター、pin2プロモーター(Xu et al., 1993)、トウモロコシWipIプロモーター、トウモロコシtrpA遺伝子プロモーター(米国特許第5,625,136号)、トウモロコシCDPK遺伝子プロモーターが含まれ、また、RUBISCO SSU プロモーター(米国特許第5,034,322号)も使用され得る。組織特異的プロモーター、例えば、トマトのE8プロモーターのような果実特異的プロモーター(アクセッション番号: AF515784; Good et al. (1994))が使用され得る。花器官特異的プロモーターのような果実特異的プロモーターは、植物の花器官において本発明のポリヌクレオチドを発現させるために本発明の発現構築物と共に使用され得る。花器官特異的プロモーターの例には、米国特許第6,462,185号;同第5,639,948号;および同第5,589,610号で説明されているプロモーター配列のいずれかが含まれる。(例えばインゲンマメの)β-ファゼオリン遺伝子または(例えばダイズの)グリシニン遺伝子に由来するプロモーターのような種子特異的プロモーターおよび他のものもまた、使用され得る。内乳特異的プロモーターには、MEG1 (EPO出願番号EP1528104)ならびにWu et al. (1998)、Furtado et al. (2001)、およびHwang et al. (2002)によって説明されているものが含まれるが、それらに限定されるわけではない。根特異的プロモーター、例えば、米国特許第6,455,760号もしくは米国特許第6,696,623号;または米国特許出願公開第20040078841号;同第20040067506号;同第20040019934号;同第20030177536号;同第20030084486号;もしくは同第20040123349号で説明されているプロモーター配列のいずれかが、本発明の発現構築物と共に使用され得る。構成的プロモーター(CaMVプロモーター、ユビキチンプロモーター、アクチンプロモーター、またはNOSプロモーターなど)、発生に関連して調節されるプロモーター、および誘導性プロモーター(例えば、熱、光、ホルモン、または化学物質によって誘導され得るプロモーター類)もまた、本発明のポリヌクレオチド発現構築物と共に使用するために企図される。
【0049】
本発明の発現構築物は、転写終結配列、翻訳終結配列、シグナルペプチドをコードする配列、および/またはエンハンサーエレメントを任意で含んでよい。転写終結領域は、典型的には、真核生物またはウイルスの遺伝子配列の3'非翻訳領域から得ることができる。転写終結配列は、効率的な終結を実現するために、コード配列の下流に配置されてよい。シグナルペプチド配列は、特定の細胞小器官区画からタンパク質作用部位に及ぶ広範囲の翻訳後細胞内目的地および細胞外環境に、機能的に連結された成熟ポリペプチドを再配置するのを担当するタンパク質のアミノ末端に典型的に存在する短いアミノ酸配列である。機能的に連結されたシグナルペプチド配列を用いて、意図した細胞内目的地および/または細胞外目的地に遺伝子産物をターゲティングすることが、本発明のポリペプチドと共に使用するために企図される。古典的なエンハンサーは、遺伝子転写を増大させるシス作用エレメントであり、これもまた発現構築物中に含まれてよい。古典的なエンハンサーエレメントは当技術分野において公知であり、CaMV 35Sエンハンサーエレメント、サイトメガロウイルス(CMV)初期プロモーターエンハンサーエレメント、およびSV40エンハンサーエレメントが含まれるが、それらに限定されるわけではない。遺伝子発現を促進するイントロンを介したエンハンサーエレメントもまた、当技術分野において公知である。これらのエレメントは、転写領域内に存在しなければならず、配向依存性(orientation dependent)である。例には、トウモロコシしわ型-1エンハンサーエレメント(ClancyおよびHannah, 2002)が含まれる。
【0050】
発現構築物から転写されたmRNAのポリアデニル化を指示するDNA配列もまた、発現構築物中に含まれてよく、これらにはオクトピン合成酵素シグナルまたはノパリン合成酵素シグナルが含まれるが、それらに限定されるわけではない。本発明の発現構築物はまた、他の遺伝子の転位を指示するポリヌクレオチド配列、すなわちトランスポゾンも含んでよい。
【0051】
本発明のポリヌクレオチドは、RNAまたはDNAのいずれかから構成され得る。好ましくは、ポリヌクレオチドはDNAから構成される。本発明はまた、本明細書において開示するポリヌクレオチドに配列が相補的であるポリヌクレオチドを包含する。本発明のポリヌクレオチドおよびポリペプチドは、精製型または単離型で提供されてよい。
【0052】
遺伝コードの縮重があるため、様々な異なるポリヌクレオチド配列が、本発明の変異ポリペプチドをコードし得る。存在し得るすべての三つ組コドン(UはTも表す)および各コドンにコードされるアミノ酸を示す表がLewin (1985)に記載されている。さらに、本発明の同じまたは本質的に同じ変異ポリペプチドをコードする代替のポリヌクレオチド配列を作り出すことは、当業者の技能で十分に対応できる範囲である。これらの変異ポリヌクレオチド配列または代替ポリヌクレオチド配列は、本発明の範囲内である。本明細書において使用される場合、「本質的に同じ」配列への言及は、本発明のポリヌクレオチドにコードされるポリペプチドの機能活性を実質的に改変しないアミノ酸置換、欠失、付加、または挿入をコードする配列を意味する。本発明の野生型ポリペプチドまたは変異ポリペプチドをコードするヌクレオチド配列のアレルバリアントもまた、本発明の範囲内に含まれる。
【0053】
具体的に例示したもの、または本発明の野生型ポリペプチドもしくは変異ポリペプチドおよび/またはAGPase酵素中に天然に存在するもの以外のアミノ酸置換もまた、本発明の範囲内で企図される。例えば、置換されたアミノ酸を有する変異ポリペプチドが、アミノ酸が置換されていない変異ポリペプチドと実質的に同じ機能活性を保持する限りにおいて、変異AGPase小サブユニットポリペプチドのアミノ酸を非天然アミノ酸で置換してよい。非天然アミノ酸の例には、オルニチン、シトルリン、ヒドロキシプロリン、ホモセリン、フェニルグリシン、タウリン、ヨードチロシン、2,4-ジアミノ酪酸、a-アミノイソ酪酸、4-アミノ酪酸、2-アミノ酪酸、γ-アミノ酪酸、e-アミノヘキサン酸、6-アミノヘキサン酸、2-アミノイソ酪酸、3-アミノプロピオン酸、ノルロイシン、ノルバリン、サルコシン、ホモシトルリン、システイン酸、t-ブチルグリシン、t-ブチルアラニン、フェニルグリシン、シクロヘキシルアラニン、β-アラニン、フルオロアミノ酸、デザイナーアミノ酸、例えば、β-メチルアミノ酸、C-メチルアミノ酸、N-メチルアミノ酸、および一般的なアミノ酸類似体が含まれるが、それらに限定されるわけではない。また、非天然アミノ酸には、誘導体化された側鎖を有するアミノ酸も含まれる。さらに、タンパク質中のアミノ酸の任意のものは、D(右旋性)型またはL(左旋性)型でよい。本発明の野生型または変異型のAGPase小サブユニットポリペプチドまたはAGPase大サブユニットポリペプチドのタンパク質配列のアレルバリアントもまた、本発明の範囲内に含まれる。
【0054】
アミノ酸は一般に次のクラスに分類され得る:非極性、非荷電極性、塩基性、および酸性。あるクラスのアミノ酸を有する本発明の変異AGPase小サブユニットポリペプチドおよび/または野生型もしくは変異型のAGPase大サブユニットポリペプチドが同じクラスの別のアミノ酸で置換される保存的置換は、その置換を有するポリペプチドが、置換を有さないポリペプチドと実質的に同じ機能活性(例えば、AGPase酵素の酵素安定性および/または増大した熱安定性)を引き続き保持する限りにおいて、本発明の範囲内に含まれる。配列中に1つまたは複数のアミノ酸置換を有する変異AGPase小サブユニットポリペプチドおよび/または野生型もしくは変異型のAGPase大サブユニットポリペプチドをコードするポリヌクレオチドは、本発明の範囲内で企図される。下記の表1は、各クラスに属するアミノ酸の例のリストを提供する。
【0055】
【表1】

【0056】
本発明はまた、本発明の機能的な野生型または変異型のAGPase小サブユニットポリペプチドまたはAGPase大サブユニットポリペプチドをコードする本発明のポリヌクレオチドのバリアントにも関する。バリアント配列には、その配列の1つまたは複数のヌクレオチドが置換、欠失、および/または挿入された配列が含まれる。DNAの天然ヌクレオチドを置換できるヌクレオチドは塩基部分を有し、この塩基部分には、イノシン、5-フルオロウラシル、5-ブロモウラシル、ヒポキサンチン、1-メチルグアニン、5-メチルシトシン、およびトリチル化塩基が含まれ得るが、それらに限定されるわけではない。ある配列中のヌクレオチドの糖部分もまた修飾されてよく、アラビノース、キシルロース、およびヘキソースが含まれるが、それらに限定されるわけではない。さらに、ヌクレオチドのアデニン塩基、シトシン塩基、グアニン塩基、チミン塩基、およびウラシル塩基は、アセチル基、メチル基、および/またはチオ基で修飾されてもよい。ヌクレオチドの置換、欠失、および/または挿入を含む配列は、当技術分野において公知の標準技術を用いて調製し、試験することができる。
【0057】
本発明の変異ポリペプチドの断片およびバリアントは、本明細書において説明するようにして作製することができ、かつ、当技術分野において公知の標準技術を用いて、酵素機能および熱安定機能の存在に関して試験することができる。すなわち、当業者は、本発明の変異ポリペプチドの断片およびバリアントを容易に調製して試験し、その断片またはバリアントが完全長または非バリアント(non-variant)変異ポリペプチドと比べて機能活性を保持しているかどうかを判定することができる。
【0058】
本発明の範囲内で企図されるポリヌクレオチドおよびポリペプチドはまた、本明細書において具体的に例示する本発明の配列に対する同一性および/または類似性の範囲がより特定的であるという面から定義することもできる。配列同一性は、典型的には60%より高く、好ましくは75%より高く、より好ましくは80%より高く、さらにより好ましくは90%より高く、95%より高くでもよい。配列の同一性および/または類似性は、本明細書において例示する配列と比較して、49%、50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、または99%でよい。別段の指定が無い限り、本明細書において使用される場合、2つの配列の配列同一性パーセントおよび/または類似性パーセントは、KarlinおよびAltschul (1993)において改良された、KarlinおよびAltschul (1990)のアルゴリズムを用いて決定することができる。このようなアルゴリズムは、Altschul et al. (1990)のNBLASTプログラムおよびXBLASTプログラムに組み入れられている。NBLASTプログラム、スコア=100、ワード長=12を用いてBLAST探索を実施して、所望の配列同一性パーセントを有する配列を得ることができる。比較目的のギャップありアライメントを得るために、Altschul et al. (1997)で説明されているようにしてGapped BLASTを使用することができる。BLASTプログラムおよびGapped BLASTプログラムを使用する場合、各プログラム(NBLASTおよびXBLAST)の初期設定パラメーターを使用することができる。NCBI/NIHのウェブサイトを参照されたい。
【0059】
本発明はまた、本明細書において例示するポリヌクレオチド配列と十分に相同で、その結果、標準のストリンジェントな条件および標準的方法(Maniatis et al., 1982)のもとでその配列とハイブリダイゼーションすることを可能にする配列を有するポリヌクレオチド分子も企図する。本明細書において使用される場合、ハイブリダイゼーションのための「ストリンジェントな」条件とは、典型的には6×SSPE、5×デンハルト液、0.1% SDS、0.1mg/ml変性DNAにおいて、DNAハイブリッドの融解温度(Tm)より20〜25C低い温度で一晩、ハイブリダイゼーションを実施する条件を意味する。融解温度Tmは次の式によって説明される(Beltz et al., 1983):
Tm=81.5C+16.6Log[Na+]+0.41(%G+C)−0.61(%ホルムアミド)−600/二重鎖の長さ(単位:塩基対)。
【0060】
洗浄は典型的には以下のようにして実施する:
(1)室温で15分間、1×SSPE、0.1%SDS中で2回(低ストリンジェンシー洗浄)。
(2)Tm-20Cで15分間、0.2×SSPE、0.1%SDS中で1回(中程度のストリンジェンシー洗浄)。
【0061】
本明細書において使用される場合、「核酸」および「ポリヌクレオチド」という用語は、単鎖または二本鎖いずれかの形態のデオキシリボヌクレオチドポリマー、リボヌクレオチドポリマー、またはデオキシリボヌクレオチドとリボヌクレオチドの混合ポリマーを意味し、別段の制限がない限り、天然に存在するヌクレオチドと同様の様式で機能できる、天然ヌクレオチドの公知の類似体も包含する。これらのポリヌクレオチド配列には、RNAに転写されるDNA鎖配列および転写されるDNA鎖に相補的である鎖配列が含まれる。また、これらのポリヌクレオチド配列には、完全長配列ならびに完全長配列に由来する短い配列の両方が含まれる。例示する配列のアレル変種(variation)もまた、本発明の範囲内に入る。これらのポリヌクレオチド配列には、個々の鎖としてか、または二重鎖におけるセンス鎖およびアンチセンス鎖の両方が含まれる。
【0062】
植物細胞をある遺伝子で形質転換するための技術は当技術分野において公知であり、例えば、アグロバクテリウム(Agrobacterium)感染、バイオリスティック法、エレクトロポレーション、塩化カルシウム処理、PEGを介した形質転換などが含まれる。米国特許第5,661,017号では、異種ポリヌクレオチドで藻類細胞を形質転換するための方法および材料が教示されている。当技術分野において公知の標準的方法を用いて、形質転換細胞を選択し、再分化させ、本発明のポリヌクレオチドを含み発現する植物に成長させることができる。本発明の形質転換されたかまたはトランスジェニックである任意の植物細胞または植物の種子および他の植物組織ならびに子孫もまた、本発明の範囲内に含まれる。
【0063】
本発明はまた、野生型植物と比べて増大した熱安定性を示す植物を作製するための方法であって、本発明の変異AGPase小サブユニットポリペプチドをコードするポリヌクレオチドが植物細胞中に導入され、そのポリヌクレオチドにコードされるポリペプチドが発現される方法にも関する。1つの態様において、植物細胞は、野生型AGPase大サブユニットポリペプチドをコードする非変異遺伝子を含む。別の態様において、植物細胞は、変異AGPase大サブユニットポリペプチドをコードする少なくとも1つのポリヌクレオチドを含む。さらなる態様において、変異AGPase大サブユニットポリペプチドをコードするポリヌクレオチドはまた、変異AGPase小サブユニットポリペプチドをコードするポリヌクレオチドと共に植物細胞中に導入される。1つの態様において、1つまたは複数のポリヌクレオチドは、植物細胞のゲノム中に組み込まれ、その植物細胞から植物が成長させられる。好ましい態様において、植物細胞から成長させられた植物は、組み込まれた1つまたは複数のポリヌクレオチドを安定に発現する。
【0064】
本発明はまた、本発明のポリヌクレオチドのコード配列または非コード配列にハイブリダイズできるオリゴヌクレオチドプローブおよびオリゴヌクレオチドプライマー、例えばポリメラーゼ連鎖反応法(PCR)プライマーにも関する。本発明のオリゴヌクレオチドプローブは、本発明の変異AGPase小サブユニットポリペプチドをコードする核酸配列を検出および定量するための方法において使用され得る。本発明のオリゴヌクレオチドプライマーは、核酸増幅を伴うPCR法および他の方法において使用することができる。好ましい態様において、本発明のプローブまたはプライマーは、ストリンジェントな条件下で本発明のポリヌクレオチドにハイブリダイズすることができる。本発明のプローブおよびプライマーは、検出可能な標識またはレポーター分子、例えば、蛍光分子、酵素、および放射性部分(例えば、3H、35S、125Iなど)などを任意で含んでよい。本発明のプローブおよびプライマーは、それらが使用される方法またはアッセイ法に適した任意の長さのものでよい。典型的には、本発明のプローブおよびプライマーのヌクレオチド長は、10〜500またはそれ以上である。ヌクレオチド長が10〜20、21〜30、31〜40、41〜50、51〜60、61〜70、71〜80、81〜90、91〜100またはそれ以上であるプローブおよびプライマーは、本発明の範囲内で企図される。本発明のプローブおよびプライマーは、ポリヌクレオチド配列とのヌクレオチド配列同一性が完全(100%)でもよく、または配列同一性は100%未満でもよい。例えば、プローブまたはプライマーとある配列との配列同一性は、そのプローブまたはプライマーがストリンジェントな条件下で本発明のポリヌクレオチドのヌクレオチド配列にハイブリダイズできる限り、99%、98%、97%、96%、95%、90%、85%、80%、75%、70%または他の任意の配列同一率(%)でよい。1つの態様において、本発明のプローブおよびプライマーは、配列番号:1、配列番号:3、配列番号:11、配列番号:12、配列番号:13、配列番号:14、配列番号:15、もしくは配列番号:16、またはそれらの相補物のヌクレオチド配列に対して70%もしくはそれ以上、75%もしくはそれ以上、80%もしくはそれ以上、85%もしくはそれ以上、90%もしくはそれ以上、または95%〜100%の配列同一性を有する。
【0065】
本発明はまた、単離された変異AGPase小サブユニットポリペプチドにも関する。1つの態様において、変異AGPase小サブユニットポリペプチドはトウモロコシのAGPase小サブユニットポリペプチドである。特定の態様において、本発明のAGPase小サブユニットポリペプチドは、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、配列番号:10、配列番号:21、配列番号:22、配列番号:23、配列番号:24、もしくは配列番号:25に示すアミノ酸配列またはその機能的断片もしくはバリアントを有する。本発明のAGPase小サブユニットポリペプチドまたは酵素は、当技術分野において公知の標準技術を用いて精製することができる。1つの態様において、AGPase小サブユニットポリペプチドをコードする本発明のポリヌクレオチドは大腸菌のような微生物中に組み込まれ、AGPase小サブユニットポリペプチドはその微生物中で発現され、次いでそれから単離される。
【0066】
特定の態様において、本発明のポリペプチドおよびその機能的ペプチド断片は、本発明のポリペプチドに特異的に結合する抗体を作製するために使用され得、そのような抗体は本発明の範囲内で企図される。本発明の抗体は、ポリクローナルまたはモノクローナルでよく、当技術分野において公知の標準的方法を用いて作製および単離することができる。
【0067】
本発明によるポリペプチド断片は、典型的には、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、配列番号:10の少なくともまたは約
個のアミノ酸の連続スパン(contiguous span)を含む。
【0068】
一定の態様において、本発明のポリペプチド断片の長さは、少なくとも約25個の連続アミノ酸から、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、または配列番号:10に示す配列のような完全長配列よりアミノ酸1個分少ないものまでの任意の整数個でよい。したがって、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、または配列番号:10の場合、ポリペプチド断片はアミノ酸約25〜475個の、任意の整数個の連続アミノ酸でよい。「整数」という用語は、本明細書においてその数学的意味で使用され、したがって、代表的な整数には、
が含まれる。
【0069】
本発明の各ポリペプチド断片はまた、N末端位置およびC末端位置の観点から説明することもできる。例えば、約25個の連続アミノ酸から、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、または配列番号:10のポリペプチドのような完全長ポリペプチドよりアミノ酸1個分少ないものまでのN末端断片からC末端断片までの組み合わせが本発明に含まれる。したがって、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、または配列番号:10を例として用いると、25個の連続アミノ酸断片は、次からなる群より選択される、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、または配列番号:10のアミノ酸に対応し得る:
。同様に、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、または配列番号:10の26個の連続アミノ酸と474個(または475個)の連続アミノ酸の間のサイズである他のすべての断片に対応するアミノ酸も、本発明に含まれ、これらの例に基づいて直ちに予想することもできる。したがって、本明細書を不必要に長くするのを避けるために、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、または配列番号:10のポリペプチドの様々な断片を示すその他の例を本明細書で個別に列挙することはしない。
【0070】
配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、または配列番号:10の
個の連続アミノ酸を含むポリペプチド断片は、別法として、式「n〜c」(両端の数字を含む)(「n」はそのポリペプチドのN末端アミノ酸位置に等しく、「c」はC末端アミノ酸位置に等しい)を用いて説明することもできる。本発明のこの態様において、「n」は、下限値が1であり、上限値が完全長ポリペプチドのアミノ酸の総数-24(例えば、配列番号:2、配列番号:4、配列番号:5、または配列番号:8の場合、475−24=451;配列番号:6、配列番号:7、配列番号:9、または配列番号:10の場合、476−24=452)である整数である。「c」は、25と完全長ポリペプチド配列のアミノ酸の数(配列番号:2、配列番号:4、配列番号:5、または配列番号:8の場合、475;配列番号:6、配列番号:7、配列番号:9、または配列番号:10の場合、476)の間の整数であり、「n」は、「c」より少なくとも24少ない整数である。したがって、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、または配列番号:10の場合、「n」は、次からなるリストより選択される任意の整数であり:
;かつ「c」は、次からなる群より選択される任意の整数である:
。ただし、「n」は「c」はより少なくとも24少ない値であることを条件とする。「n」位置および「c」位置のあらゆる組み合わせが、本発明のポリペプチド断片の特定の態様として含まれる。本発明の任意のポリペプチド断片態様を説明するために使用される範囲はすべて、別段の具体的な説明が無い限り、両端の数字を含む(inclusive)。
【0071】
本発明の変異AGPase小サブユニットポリペプチドまたはAGPase大サブユニットポリペプチドの断片は、本明細書において説明するように、タンパク質分解酵素(トリプシン、キモトリプシン、もしくはコラゲナーゼなど)または臭化シアン(CNBr)のような化学試薬を用いて本発明のポリペプチドを切断することによって得ることができる。あるいは、強酸性環境、例えばpH2.5でポリペプチド断片を生成させることもできる。また、ポリペプチド断片は、化学合成によって、または本発明のAGPase大サブユニットポリペプチドの断片もしくは変異AGPase小サブユニットポリペプチドの断片、例えば、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、配列番号:10、配列番号:21、配列番号:22、配列番号:23、配列番号:24、もしくは配列番号:25に示すアミノ酸配列の断片である変異ポリペプチドをコードするポリヌクレオチドを含む発現ベクターで形質転換させた宿主細胞を用いて、調製することもできる。本明細書において同様に企図される本発明の変異AGPase大サブユニットポリペプチドまたは変異AGPase小サブユニットポリペプチドの断片には、そのポリペプチドのトランジット配列またはシグナル配列の全体または一部分が除去されているポリペプチド断片が含まれる。
【0072】
本発明はまた、本発明の変異AGPase小サブユニットポリペプチドをコードする本発明のポリヌクレオチドで形質転換させた細胞にも関する。1つの態様において、細胞は、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、配列番号:10、配列番号:21、配列番号:22、配列番号:23、配列番号:24、もしくは配列番号:25に示すアミノ酸配列またはその機能的な断片もしくはバリアントをコードする配列を含むポリヌクレオチド配列で形質転換される。特定の態様において、細胞は、配列番号:1、配列番号:3、配列番号:11、配列番号:12、配列番号:13、配列番号:14、配列番号:15、もしくは配列番号:16に示すポリヌクレオチド配列、または配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、配列番号:10、配列番号:21、配列番号:22、配列番号:23、配列番号:24、もしくは配列番号:25の機能的な断片もしくはバリアントをコードする配列で形質転換される。1つの態様において、細胞はまた、本明細書において説明する変異AGPase大サブユニットポリペプチドをコードするポリヌクレオチドで形質転換される。1つの態様において、このポリヌクレオチド配列は、本明細書の発現構築物において提供される。形質転換細胞は、原核細胞、例えば、大腸菌もしくは枯草菌(B.subtilis)などの細菌細胞でよく、または、形質転換細胞は、真核細胞、例えば、プロトプラストを含む植物細胞もしくは動物細胞でよい。植物細胞には双子葉植物細胞、単子葉植物細胞、および針葉樹細胞が含まれるが、それらに限定されるわけではない。1つの態様において、植物細胞は、トウモロコシ植物に由来する細胞である。動物細胞にはヒト細胞、哺乳動物細胞、鳥類細胞、および昆虫細胞が含まれる。哺乳動物細胞にはCOS細胞、3T3細胞、およびCHO細胞が含まれるが、それらに限定されるわけではない。
【0073】
本発明はまた、植物または植物組織(例えば植物種子もしくは内乳組織)におけるデンプン合成を増大させるための方法にも関する。1つの態様において、本発明の方法は、本発明の1つまたは複数のポリヌクレオチドを植物中に導入する段階を含む。一定の態様において、植物中に導入されるポリヌクレオチドは、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、配列番号:10、配列番号:21、配列番号:22、配列番号:23、配列番号:24、もしくは配列番号:25のいずれかに示すアミノ酸配列またはその断片もしくはバリアントを含む1種または複数種のポリペプチドをコードする。特定の態様において、ポリヌクレオチドは、配列番号:1もしくは配列番号:3に示すヌクレオチド配列またはその断片もしくはバリアントを含む。別の特定の態様において、ポリヌクレオチドは、配列番号:11、配列番号:12、配列番号:13、配列番号:14、配列番号:15、もしくは配列番号:16に示すヌクレオチド配列またはその断片もしくはバリアントを含む。1つの態様において、ポリヌクレオチドは、植物または植物組織のゲノム中に安定に組み込まれる。ポリヌクレオチドは、ポリヌクレオチドおよび/またはそれにコードされるポリペプチドの発現を増大させる調節エレメント、例えばプロモーター配列および/またはエンハンサー配列を含んでよい。特定の態様において、プロモーター配列は、構成的発現または組織特異的(例えば内乳)発現をもたらすものである。このポリヌクレオチドを含む植物もしくは植物組織、またはこれらの植物の子孫は、本発明のポリヌクレオチドまたはポリペプチドの発現の増大について任意でスクリーニングすることができる。1つの態様において、本発明の1つまたは複数のポリヌクレオチドの複数のコピーは、植物または植物組織中に導入され、その植物のゲノム中に安定に組み込まれる。1つの態様において、本発明のポリヌクレオチドは、本明細書において説明する発現構築物において提供される。
【0074】
一文字のアミノ酸略語を表2で定義する。
【0075】
【表2】
【0076】
材料および方法
ランダム変異誘発
PCRランダム変異誘発(GeneMorph II EZClone Domain Mutagenesis Kit, Stratagene)によって、Sh2およびBt2中に変異を導入した。バイアスがかかっていない誤りがちな(error-prone)DNAポリメラーゼの混合物を用いて点変異を導入した。それぞれpMONcSh2およびpMONcBt2(Giroux et al., 1996)中の野生型のSh2コード配列およびBt2コード配列をPCR用の鋳型として使用した。Sh2およびBt2に隣接する2対のプライマー
を誤りがちなPCRのために使用した。Stratagene社のプロトコールに従って、PCRによって作製した変異Sh2クローンをベクターpMONcSh2中にサブクローニングした。次いで、pMONcSh2を用いて、適合性のあるベクターpMONcBt2中に野生型Bt2を含む大腸菌株AC70R1-504を形質転換させた。PCRによって作製した変異Bt2クローンをベクターpMONcBt2中にサブクローニングした。次いで、pMONcBt2を用いて、適合性のあるベクターpMONcSh2中に野生型Sh2を含む大腸菌株AC70R1-504を形質転換させた。
【0077】
細菌発現系
細菌発現系(Iglesias et al., 1993)のおかげで、本発明者らは、トウモロコシ内乳AGPase遺伝子をランダムに変異誘発し、後述するようにヨウ素蒸気にプレートを曝露することによって迅速かつ効率的にAGPase活性を採点することが可能になった。大腸菌系は、Georgelis et al. (2007)において考察されているいくつかの理由から、植物AGPaseを研究するために理想的である。
【0078】
グリコーゲン検出
ヨウ素蒸気への曝露後に茶色に染色されたコロニーを得ることによって、グリコーゲン合成を検出した。75μg/mLのスペクチノマイシン、50μg/mLのカナマイシン、および1%(w/v)グルコースの存在下、37℃で16時間、コーンバーグ(Kornberg)培地上で大腸菌細胞を増殖させた(Govons et al., 1969)。これらのコロニーをヨウ素蒸気に1分間曝露した。不活性なAGPaseを有するコロニーはヨウ素蒸気への曝露後に発色しなかったのに対し、活性なAGPaseはグリコーゲンを産生しており、その結果、ヨウ素による茶色の染色を示した。野生型よりも濃く染色されたAGPaseバリアントを、さらに研究するために選択した。
【0079】
グリコーゲン定量
フェノール反応(HansonおよびPhillips, 1981)を用いてグリコーゲン定量を実施した。手短に言えば、50%(w/v)KOH中で3時間煮沸することによって、2%(w/v)グルコースを含むLB中で増殖させた大腸菌細胞(OD600=2.0)1.6mlからグリコーゲンを抽出した。次いで、70%(v/v)までエタノールを添加し、10000×gで10分間遠心分離することによってグリコーゲンを沈殿させた。沈殿物を乾燥した後に、200μlの脱イオン水、200μlの5%(w/v)フェノール、および1mLの濃硫酸を添加した。488nmでの吸光度に基づいてグリコーゲンを概算した。
【0080】
DNA配列決定
グリコーゲンの産生が増大したSh2変異体およびBt2変異体をフロリダ大学のInterdisciplinary Center for Biotechnology Research(バイオテクノロジー共同研究センター)のGenome Sequencing Services Laboratory (GSSL)によって2回(double-pass)配列決定した。Bioeditソフトウェア(Hall, 1999)によってデータ解析を実施した。
【0081】
AC70R1-504大腸菌細胞からのトウモロコシ内乳AGPaseの精製
トウモロコシ内乳AGPaseを発現するAC70R1-504大腸菌細胞を、75μg/mLのスペクチノマイシン、50μg/mLのカナマイシン、および2%(w/v)グルコースの存在下、37℃で16時間、振盪しながら、2Lのルリアブロス(LB)培地中で増殖させた。OD600=0.6で、0.2mMのイソプロピル-β-D-チオガラクトシド(IPTG)および0.02mg/mLのナリジクス酸を添加してタンパク質発現を誘導した。培養物を直ちに室温に移し、振盪しながら4時間増殖させた。下記の段階を4℃で実施した。3000×gで遠心分離することによって細胞を回収し、沈殿物を16mLの緩衝液A(50mM KH2PO4 pH 7.0、5mM MgCl2、0.5mM EDTA)およびプロテアーゼ阻害剤(1μg/mLのペプスタチン、0.1mMのPMSF、10μg/mLのキモスタチン、および1mMのベンズアミジン)中に再懸濁した。フレンチプレスを用いてこれらの細胞を溶解させ、26000×gで遠心分離した。緩衝液Aを添加することによって、上清のタンパク質濃度を30mg/mLに調整した。10分の3の体積の1%硫酸プロタミンを添加し、混合物を氷上で20分間撹拌し、次いで、26000×gで20分間遠心分離した。上清を硫酸アンモニウムで45%飽和にし、氷上で20分間撹拌し、26000×gで20分間遠心分離した。2〜2.5mLの緩衝液A中に沈殿物を再懸濁した。Boehlein et al. (2005)によって説明されているようにして、強陰イオン交換カラム(macro-prep High Q support, Biorad)およびEcono-pacヒドロキシアパタイトカートリッジ(Biorad)にこの混合物を通した。分析する前にZeba Micro Desalt Spin Columns (Pierce) を用いることによってAGPaseを脱塩した。AGPaseを50mM HEPES、5mM MgCl2、0.5mM EDTA、および0.5mg/mL BSA中に移した(exchanged)(安定性のため)。
【0082】
AGPaseの動力学的特徴付け
ATPおよびG-1-Pのkcat、Km、ならびに3-PGAおよびPiの親和性を推定するために、反応の順方向を使用した(G-1-P+ATP→ADP-グルコース+PPi)。より具体的には、総体積200μlの50mM HEPES pH 7.4、15mM MgCl2、1.0mM ATP、および2.0mM G-1-Pにおいて、37℃で10分間、0.04〜0.06μgの精製したAGPaseの比活性を分析した。ATPおよびG-1-PのKmならびに3-PGAのKaを決定するために、それぞれ様々な量のATP、G-1-P、および3-PGAを用いた。様々な量のPi、1mM ATP、2mM G-1-P、および2.5mM 3-PGAを添加することによって、PiのKiを概算した。2分間煮沸することによって反応を停止させた。カップリング試薬300μlを用いてNADH濃度の低下とこの反応を関連付けることによってPPiを概算した。カップリング試薬は、25mMイミダゾールpH 7.4、4mM MgCl2、1mM EDTA、0.2mM NADH、0.725Uアルドラーゼ、0.4Uトリオースリン酸イソメラーゼ、0.6Uグリセロリン酸デヒドロゲナーゼ、1mMフルクトース-6-リン酸、および0.8μgのピロリン酸依存性ホスホフルクトキナーゼ(PPi-PFK)を含んだ。これらの酵素はPPi-PFK以外はすべてSigma社から購入した。PPi-PFKは、Deng et al. 1999に従い、いくつかの改良を加えて作製し精製した(BoehleinおよびHannah、未発表データ)。340nmでの吸光度に基づいてNADH濃度を概算した。PPi濃度は、AGPaseの代わりに様々な量のPPiを用いることによって作成した検量線を用いて算出した。AGPaseによって産生されるPPiの量は、時間および酵素濃度に比例した。Prism 4.0 (Graph Pad, San Diego CA)によってAGPaseの動力学的定数を算出した。
【0083】
粗製タンパク質抽出物または部分的に精製したタンパク質抽出物に由来するAGPaseの比活性の測定
トウモロコシ内乳AGPaseを発現するAC70R1-504大腸菌細胞を、75μg/mLのスペクチノマイシン、50μg/mLのカナマイシン、および2%(w/v)グルコースの存在下、37℃で振盪しながら、OD600=2.0になるまで2Lのルリアブロス(LB)培地中で増殖させた。遺伝子発現は誘導しなかった。下記の段階を4℃で実施した。3000×gで遠心分離することによって細胞を回収し、沈殿物を16mLの緩衝液A(50mM KH2PO4 pH 7.0、5mM MgCl2、0.5mM EDTA)およびプロテアーゼ阻害剤(1μg/mLのペプスタチン、0.1mMのPMSF、10μg/mLのキモスタチン、および1mMのベンズアミジン)中に再懸濁した。フレンチプレスを用いてこれらの細胞を溶解させ、26000×gで遠心分離した。-80℃で保存した上清を用いて、粗製抽出物のAGPase活性を測定した。前述のようにして硫酸プロタミンおよび硫酸アンモニウムを用いて、上清の残りを部分的に精製した。アッセイ法の前にBoehlein et al. (2005)によって説明されているようにしてタンパク質抽出物を脱塩し、50mM HEPES、5mM MgCl2、および0.5mM EDTA中に移した。Boehlein et al. (2005)によって説明されているようにして、飽和量の基質およびアクチベーターを用いて、逆方向(ADP-グルコース+PPi→G-1-P+ATP)におけるAGPase比活性をモニターした。
【0084】
AGPaseの熱安定性の測定
AGPaseを前述のようにして精製した。50mM HEPES、5mM MgCl2、0.5mM EDTA、および0.5mg/mL BSA中でAGPaseをさらに1/100(v/v)希釈し、様々な時間に渡って42℃または53℃で熱処理し、次いで氷上で冷却した。飽和量のATP、G-1-P、および3-PGAを用いて、処理後に残っている活性を順方向および逆方向においてモニターした。熱処理の時間に対する残存活性率(%)の対数としてデータをプロットした。式t1/2=0.693/(-2.3×傾き)を用いて不活性化定数t1/2を算出した。
【0085】
AGPase純度の定性的測定
AGPaseの純度を以下のようにしてモニターした。6μgのAGPaseを変性溶液(100mM Tris-Cl pH 6.8、4% SDS、および8mM DTT)中で1:1希釈し、95℃で5分間加熱し、150Vで1時間、5% SDSポリアクリルアミドゲル上で電気泳動させ、クーマシーブリリアントブルー染色によって可視化した(Laemmli, 1970)。
【0086】
粗製抽出物のタンパク質ブロット解析
Hybri-Dotブロット装置(Life Technologies)を用いて、試料をPVDF膜(Biorad)上に真空ブロットした。PVDF膜はメタノール中で5分間、次いでトランスファー緩衝液[20%(v/v)メタノール、0.303%(w/v)Trisおよび1.44%(w/v)グリシン]中で10分間、予め浸しておいた。絶えず振盪しながら1時間、ブロッキング緩衝液[0.8%(w/v)NaCl、0.02%(w/v)KCl、0.144%(w/v)Na2HPO4、0.024%(w/v)KH2PO4、5%(w/v)ウシ血清アルブミン(BSA)、および0.05%(v/v)Tween-20]と共に膜をインキュベートした。振盪しながら1時間、BT2に対する1:10000(v/v)のモノクローナル抗体(Sue Boehleinの好意で提供された)を含むブロッキング緩衝液と共にブロットをインキュベートした。次いで、絶えず振盪しながら、そのブロットを洗浄緩衝液(ブロッキング緩衝液−BSA)で10分間×3回洗浄した。次いで、ホースラディッシュペルオキシダーゼと結合されたヤギ抗マウス二次抗体(Pierce)の1:60000希釈物と共に45分間、ブロットをインキュベートした。最後に、ブロットを10分間×3回洗浄した。増感された化学発光基質キット(Pierce)を用いてタンパク質を可視化した。
【0087】
3Dモデリング
最近公開されたジャガイモ塊茎ホモ四量体AGPase(RCSBタンパク質データバンク番号:1YP2)の三次構造におけるジャガイモ小ユニットにならって、BT2モノマーの構造のモデルを作った。SWISS MODELを用いてホモロジーモデリングを行った(Peitsch, 1995; GuexおよびPeitsch, 1997; Schwede et al. 2003; KoppおよびSchwede, 2004; Arnold et al, 2006)。Jmol、すなわちJavaによるオープンソースの三次元化学構造表示ソフト(open-source Java viewer for chemical structures in 3D)(http://www.jmol.org/)を用いて、アミノ酸462(ThrまたはIle)接触残基を決定した。
【0088】
酵母ツーハイブリッド
GreeneおよびHannah(1998b)によって説明されているようにして、酵母形質転換およびβ-ガラクトシダーゼアッセイ法を実施した。唯一変更した点は、ベイトおよびプレイ用のベクターとしてそれぞれpGBKT7およびpGADT7を使用したことであった。pGBKT7-53プラスミドおよびpGADT7-Tプラスミドを陽性対照として使用した。
【0089】
図面および表をすべて含む、本明細書において参照または引用される特許、特許出願、仮出願、および刊行物はすべて、本明細書の明確な教示と矛盾しない範囲で、そっくりそのまま参照により組み入れられる。
【0090】
以下は、本発明を実践するための手順を例示する実施例である。これらの実施例は、限定として解釈すべきではない。別段の注記の無い限り、パーセンテージはすべて重量に基づき、溶媒の混合比率はすべて体積に基づく。
【実施例】
【0091】
実施例1
誤りがちなPCRによって、変異Bt2ライブラリーを作り出した。突然変異荷重は、クローン1個当たり〜2個の非同義変異であった(Georgelis et al. 2007)。これらの変異体を野生型Sh2遺伝子と共に大腸菌において発現させた。約50,000個のコロニーをグリコーゲン産生についてスクリーニングした。濃く染色された10個のコロニーを選んだ。染色が最も濃いBt2変異体2つを配列決定した。両方とも、トレオニンからイソロイシンへのアミノ酸462の変化をもたらす同じ非同義変異(TI)を有していた。この位置のトレオニンは、高等植物小サブユニット間で絶対的に保存されている(データ不掲載)。BT2-TI/SH2(TI変異を含み、SH2と複合体を形成したBT2)は、BT2/SH2よりも多量のグリコーゲンを産生した(図1)。ホモ四量体としてBT2-TIおよびBT2を発現する細胞は、検出可能な量のグリコーゲンを産生しなかった(図1)。このことから、大腸菌によって合成されるグリコーゲンの量が、BT2-TIまたはBT2とSH2との複合体にもっぱら依存することが示される。
【0092】
BT2/SH2およびBT2-TI/SH2を発現する細胞に由来する粗製抽出物のドットブロットから、BT2-TIが大腸菌において多量に存在することが示された(図2A〜2B)。BT2/SH2およびBT2-TI/SH2を発現する誘導されていない細胞に由来する粗製抽出物のAGPase活性レベルは低すぎて検出できなかったのに対し、BT2-TI/SH2由来の部分的に精製した抽出物は、BT2/SH2由来の部分的に精製した抽出物よりも20倍高い活性を有していた(図3)。より効率的な転写/翻訳が理由となってBT2-TI/SHの方が多くのタンパク質および高い活性をもたらしたという可能性は存在しにくい。なぜならば、ACA(T)〜ATA(I)のコドンは大腸菌では同じ頻度で使用されているからである(それぞれ6.1%および5.0%)(Nakamura et al., 2000)。このことから、BT2-TI/SH2細胞の方がタンパク質が多量であり活性が高いのは、AGPaseの安定性の上昇に起因することが示唆される。
【0093】
BT2-TI/SH2の動力学的諸特性および熱安定性を決定し、大腸菌におけるグリコーゲン合成の増大の原因を解明するために、組換えBT2-TI/SH2 AGPaseおよびBT2/SH2 AGPaseを精製した(図4)。表3に要約するBT2-TI/SH2の動力学的諸特性から、G-1-PおよびATPのKm、3-PGAのKa、およびPiのKiがBT2/SH2の場合と区別不能であったことが示される。驚くべきことに、BT2-TIのkcatはBT2/SHのkcatより30%小さい。したがって、これらの動力学的諸特性では、大腸菌においてBT2-TI/SH2の方が染色が濃いことを説明することができない。しかしながら、BT2-TI/SH2がBT2/SH2よりも熱安定性であることは明らかである(図5A〜5B)。これらの結果から、BT2-TI/SH2の高い熱安定性が、大腸菌におけるグリコーゲンの量の増加の原因であることが強く示唆される。
【0094】
MPは、農学的利益をもたらし得る小サブユニットバリアントである。その特徴のいくつかには、BT2/SH2と比べて、アクチベーター3-PGAの不在下での増大した活性、3-PGAに対する増大した親和性、Piに対する減少した親和性(表3)、および上昇した熱安定性が含まれる(図3)(Cross et al., 2004; Boehlein et al., 2005)。BT2-TI/SH2はMP/SH2ほど熱安定性ではなかったため(図3)、MPの熱安定性をさらに高めるために、アミノ酸変化TIをMPに導入した(MP-TI)。
【0095】
MP-TI/SH2(TI変異を有し、SH2と複合体を形成したMP)を発現する細胞は、MP/SH2を発現する細胞と同じ量のグリコーゲンを産生した(図1)。しかしながら、BT2と比べてより多量のMP-TIタンパク質が、SH2を有する2種のタンパク質を発現する大腸菌の粗製抽出物で存在していた(図2A〜2B)。MP-TI/SH2に由来する粗製抽出物および部分的に精製した抽出物の活性は、MP/SH2の活性より2〜3倍高かった(図3)。純粋な形態のMP-TI/SH2(図4)は、MP/SH2と比べてkcatが〜30%減少した以外は、MP/SH2の好ましい動力学的諸特性を維持していた(表3)。さらに、MP-TI/SH2は、MP/SH2よりも高い熱安定性を示す(図6A〜6B)。
【0096】
トウモロコシ内乳AGPaseの結晶構造は解明されていない。唯一の関連する構造体は、ジャガイモ塊茎小サブユニットホモ四量体である(Jin et al., 2005)。ジャガイモ塊茎小サブユニットは、BT2に対して88%の同一性および96%の類似性を示す。解明されたジャガイモ塊茎小サブユニットの構造にならって、BT2モノマーの構造のモデルを作った(図7A)。TI中で変異した残基はβ-ヘリックスの一部分であり、これは、小サブユニットのN末端の2つの残基(Pro、Leu)との疎水的接触を作り出す(図7B)。TIにおいてThrからIleにアミノ酸が変化すると、前述のProおよびLeuからの距離が短くなる(図7C)。TI変異が小サブユニットのC末端とN末端の間の疎水性相互作用を強化し、その結果、より高い安定性をもたらすと推測したくなる。サブユニット同士の境界の/近くの残基に熱安定性が帰されるMPとは異なって、TIはサブユニット同士の境界から離れているため、サブユニット間の相互作用に直接影響を及ぼすことはできない(図7A)。
【0097】
TIがサブユニット間の相互作用の強度に影響を及ぼすかどうかを判定するために、酵母ツーハイブリッドシステム(Y2H)においてSH2をベイトとして使用し、BT2、TI、およびMPをプレイとして使用した。定量的β-ガラクトシダーゼアッセイ法により、MPとは対照的に、TIはサブユニット間の相互作用の強度を上昇させないことが示された(図8)。
【0098】
最後に、セリン、アラニン、およびグリシンなど側鎖の短いアミノ酸で元のトレオニンを置換しても、kcatは10分の1またはそれ以下に減少したにもかかわらず(データ不掲載)、AGPaseの熱安定性は影響を及ぼされなかった。このことから、462位の元のトレオニンはAGPase活性にとっては重要であるが熱安定性にとっては重要でないことが示唆される。
【0099】
実施例2
本発明は、ランダム変異誘発および異種細菌発現系を用いることによって、農学的に重要な植物AGPaseバリアントを提供する。SH2と共に発現された場合に大腸菌細胞によるグリコーゲン産生量を増加させる小サブユニットバリアントとしてBT2-TIを単離した。BT2-TI/SH2を発現する細胞は、BT2/SH2を発現する細胞よりも20倍高いAGPase活性を有していた。ドットブロットにより、BT2-TI/SH2を発現する細胞に由来する粗製タンパク質抽出物の方が、BT2/SH2を発現する細胞と比べて検出可能なBT2タンパク質を有することが示された。この結果は、より効率的な転写/翻訳またはより高いAGPase安定性および/もしくは溶解度に帰され得る。先に言及したように、より効率的な転写/翻訳は、コドン使用に基づくと、見込みは少ない。その一方で、精製型のBT2-TI/SH2は、精製型のBT2/SH2より有意に熱安定性が高いことが示された。このことにより、大腸菌においてBT2-TI/SH2複合体がBT2/SH2と比べてタンパク質分解および/または凝集を起こしにくくなる可能性がある。
【0100】
BT2-TI/SH2の動力学的諸特性およびアロステリック特性は、kcatが30%小さかった以外は、BT2/SH2と区別不能であった。BT2-TI/SH2発現細胞のAGPase活性が20倍上昇していることは、活性なAGPase分子の数がBT2/SH2発現細胞と比べて多いことと解釈される。これはまた、BT2/SH2において実際に機能しているのは潜在的AGPase分子の5%未満であることも意味する。
【0101】
ジャガイモ塊茎小サブユニットは、極めて多量のアクチベーター3-PGAを与えられた場合に、顕著な活性を有するホモ四量体を形成できることが報告されている(Ballicora et al., 1995)。本明細書において示すように、ホモ四量体としてBT2およびBT2-TIを発現する大腸菌細胞は、検出可能な量のグリコーゲンを産生しない。したがって、BT2-TI/SH2を発現する細胞において観察された、BT2/SH2を発現する細胞と比べて量の多いグリコーゲンは、BT2-TIホモ四量体ではなく、BT2-TIとSH2の複合体に起因する。
【0102】
SH2との複合体において熱安定性の上昇をもたらす別の小サブユニットバリアントはMPである。MPによって与えられる熱安定性は、サブユニット同士が相互作用する境界の近くまたは境界の残基にマッピングされている(Boehlein, Shaw, Stewartおよび Hannah, 未発表データ)。一方、BT2-TIのアミノ酸変化はこれらの境界から遠い。構造モデリングから、TI変化によってC末端とN末端の間のサブユニット内疎水性相互作用が強化されることが示唆される。Y2Hの結果は、MPと対照的にTIはサブユニット間の相互作用を強化しないという考えを裏付ける。しかしながら、立体構造的変化を通じてTIがサブユニット間の相互作用に間接的に影響を及ぼす可能性も切り捨てることはできない。Y2Hが、30℃という酵母増殖温度でのTI/SH2およびBT2/SH2におけるサブユニット間の相互作用の違いを明らかにするのに十分な感度を持ち得ないという可能性もある。
【0103】
純粋なBT2-TI/SH2およびMP/SH2の熱安定性の比較から、BT2-TI/SH2はMP/SH2ほど熱安定性ではないことが示された。さらに、MP/SH2はBT2-TI/SH2には無いいくつかの利点を有する。例えば、BT2/SH2と比べて、アクチベーター3-PGAの不在下での活性、3-PGAに対する高い親和性、阻害剤Piに対する低い親和性、および大きなkcatなどである。TI変化をMPに導入することによってMP/SH2の熱安定性がさらに改善され得るかどうかを調査した。結果として生じるバリアントMP-TIは、大腸菌においてSH2と共に発現させた場合、MP/SH2と等量のグリコーゲンを産生した。これは、MP-TI/SH2がMP/SH2以上に熱安定性ではないこと、またはAGPaseによって触媒されるADP-グルコース産生が大腸菌においてもはや律速的(limiting)ではないことのいずれかを意味し得る。MP-TI/SH2を発現する細胞の粗製抽出物および部分的に精製した抽出物におけるAGPase活性は、MP/SH2を発現する細胞より2〜3倍高かったため、後者の解釈が好まれ得る。MP-TI/SH2は、30%小さいkcat以外は、MP/SH2の動力学的諸特性およびアロステリック特性のすべてを維持していた。より重要なことには、MP-TI/SH2は、MP/SH2よりも熱安定性であった。MP-TI/SH2およびMP/SH2の両方において二相の熱安定性が観察された。これらの相は、おそらくは、AGPaseの異なる状態の結果であった。熱安定性のこの二相性の様式は、以前に観察されており、MP/SH2にもMP-TIにも特異的ではない(Boehlein et al. 2008)。第1の相は、第2の相よりも低い熱安定性を示す。MP-TI/SH2は両方の相においてMP/SH2と比べて熱安定性が高い。しかしながら、各相においてAGPaseの状態が正確にどのようであるかは、依然として謎である。熱安定性の二相性の様式は、BT2/SH2およびBT2-TI/SH2では観察されなかった。これは、これらの試料が第2の相に到達するのに十分な長さの期間加熱されず、MP/SH2およびMP-TI/SH2より低い温度(53℃の代わりに42℃)で加熱されたためである。
【0104】
表3は、精製された組換えAGPaseバリアントの動力学的諸特性を示す。AGPaseバリアントの動力学的特性およびアロステリック特性を順方向で決定した。G-1-Pの量を変更し、飽和量(1mM)のATPを維持することによって、kcat(s-1)(G-1-P)を概算した。ATPの量を変更し、飽和量(2mM)のG-1-Pを維持することによって、kcat(s-1)(ATP)を概算した。Kiの値(Ki's)は、平均値(95%信頼区間)として表している。他の値はすべて、平均値±標準偏差として表している。3-PGAの不在下でのAGPaseの比活性を、10mMの3-PGAの存在下での比活性に対する比率(%)として表している(平均値±標準誤差)。
【0105】
【表3】
【0106】
本明細書において説明される実施例および態様は、例示するためのものにすぎないこと、ならびにそれらを踏まえた様々な修正または変更が当業者に示唆され、かつそれらが本出願の精神および範囲内、ならびに添付の特許請求の範囲内に含まれ得ることが理解されるべきである。さらに、本明細書において開示される任意の発明またはその態様の任意の要素または制限は、本明細書において開示される他の任意の発明またはその態様の任意および/または他のすべての要素または制限と(個別または任意の組み合わせで)組み合わせられてよく、このような組み合わせはすべて、それらに限定されることなく、本発明の範囲内で企図される。
【0107】
参考文献
【背景技術】
【0001】
政府の支援
本出願の主題は、助成金番号IOS-0444031のもとで、米国国立科学財団(National Science Foundation)からの研究助成金によって支援されている。したがって、政府は、本発明において一定の権利を有する。
【0002】
発明の背景
熱ストレスは、トウモロコシ収量の減少をもたらす(Peters et al., 1971; Thompson, 1975; Chang, 1981; ChristyおよびWilliamson, 1985)。これは、光合成産物の利用可能度およびソース組織からシンク組織への輸送の減少、乏しい受粉、細胞および顆粒のサイズおよび数の減少、初期の種子発育不全(seed abortion)、ならびに/または登熟期間の短縮に帰され得る。内乳の成長は、細胞が活発に分裂する誘導期(lag phase)から始まり、細胞のサイズが増大し、デンプン合成が起こる直線期(linear phase)に続く。誘導期の間の温度上昇は、収量の減少をもたらした(Jones et al., 1984)。これらの研究者らは、収量の減少が細胞および顆粒の数およびサイズの減少ならびに種子発育不全に起因することを示唆した。さらに、直線期の間の温度上昇により、登熟期間が短くなり、続いて、穀粒が小さくなった(Jones et al., 1984)。同様の結果が、Hunter et al. (1977)ならびにTollenaarおよびBruulsema (1988)によって見出された。
【0003】
古くからUSトウモロコシの50%超を生産する5つの州での記録から、1日の平均温度は23.6℃であり、登熟の間の最適温度より約2℃高いことが示された(Singletary et al., 1994)。少なくともオオムギおよびコムギにおいて、登熟中の光合成産物の利用可能度は高温度で低下することはない。実際、オオムギ種子およびコムギ種子中のスクロース含有量は、高温度で不変であるか、または増加していた(BhullarおよびJenner, 1986; Wallwork et al., 1998)。また、トウモロコシの光合成は、32℃までは増加する(DuncanおよびHesketh, 1968; HofstraおよびHesketh, 1969; Christy et al., 1985)。さらに、CheikhnおよびJones (1995)は、トウモロコシ穀粒が14Cスクロースおよびヘキソースを固定する能力を様々な温度で調査した。彼らは、種子中のこれらの糖が高温で増加することを発見した。上記の証拠から、登熟の間に糖の利用可能度および穀粒中への輸送が限定されることが、温度によって誘発される収量減少の原因ではないことが示唆される。
【0004】
温度上昇の間に登熟に強い影響を与える生化学的経路を同定するために多大な努力が払われてきた。Singletary et al. (1993; 1994)は、インビトロで増殖させたトウモロコシ穀粒中のデンプン生合成酵素を高温(22℃〜36℃)において分析した。彼らは、ADPグルコースピロホスホリラーゼ(AGPase)および可溶性デンプン合成酵素(SSS)の方が、デンプン合成に関与する他の酵素と比べて熱不安定性であることを見出した。彼らは、AGPaseおよびSSSの熱不安定性が登熟停止に寄与していることを示唆した。DukeおよびDoehlert(1996)は、AGPaseをコードするものを含む、デンプン合成経路の酵素をコードするいくつかの遺伝子の転写物が、25℃と比較すると35℃では減少していることを発見した。しかしながら、酵素アッセイ法により、AGPase活性のみが目立って低いことが示された。これは他の酵素と比べて高いAGPaseの代謝回転率に起因し得ると彼らは示唆した。最後に、Wilhelm et al. (1999)は、Q10解析を通じて、他のいくつかの酵素と比べて、AGPaseの活性が最も顕著に低下することを示した。57℃で5分間加熱した際、トウモロコシAGPaseは実際にその活性の96%を失った(Hannah et al., 1980)。
【0005】
AGPaseは、デンプン(植物)合成およびグリコーゲン(細菌)合成において最初の方向付けされた(committed)段階を触媒する。これは、ADP-グルコースおよびピロリン酸(PPi)へのグルコース-1-P(G-1-P)およびATPの変換を伴う。AGPaseは、2つの同一な小サブユニットおよび2つの同一な大サブユニットからなる、植物中のヘテロ四量体である。これらの大サブユニットおよび小サブユニットは、それぞれ、トウモロコシ内乳中のしわ型(shrunken)-2(Sh2)およびカマイラズ(brittle)-2(Bt2)にコードされている。AGPaseは、細胞のエネルギー状態を示す小型エフェクター分子によってアロステリックに調節される。AGPaseは、藍藻類、緑藻類、および被子植物において、最初の炭素同化産物である3-PGAによって活性化され、無機リン酸(Pi)によって阻害/不活性化される。
【0006】
デンプン合成におけるトウモロコシ内乳AGPaseの重要性は、酵素のいずれかのサブユニットの変異体の穀粒表現型によって示されている。実際、このような変異体は、しわ型穀粒および内乳デンプン含有量の大幅な減少をもたらす(TsaiおよびNelson, 1966; HannahおよびNelson, 1976)。AGPaseがデンプン合成の律速段階を触媒するという証拠もある(Stark et al. 1992; Giroux et al. 1996; Greene et al. 1998; Sakulsingharoja et al. 2004; Obana et al. 2006; Wang et al. 2007)。
【0007】
GreeneおよびHannah (1998a)は、HS33と呼ばれる大サブユニット中に1つのアミノ酸変化を有する変異型トウモロコシAGPaseを単離した。彼らは、改変された酵素の方が熱安定性が高いこと、および安定性はサブユニット間のより強い相互作用の寄与によることを示した。AGPaseのアロステリック特性に影響を及ぼす変化(Rev6)(Giroux et al., 1996)と共にHS33変化を含むSh2バリアントでコムギおよびイネを形質転換した場合、収量はそれぞれ38%および23%増加した(Smidansky et al., 2002; 2003)。驚くべきことに、この増加は、個々の種子重量よりはむしろ、種子数の増加に起因した。
【0008】
また、Rev6変化およびHS33変化を含むSh2バリアントによるトウモロコシの形質転換も、種子数の増加をもたらす。トウモロコシの場合、種子収量/穂は最大で68%増加し得る。トウモロコシのトランスジェニック事象の詳細な特徴付けは進行中である(GreeneおよびHannah、出版準備中)。トウモロコシにおいてRev6を単独で発現させた場合、種子重量のみが増加するため、種子数の増加をRev6で説明することはできない(Hannah、未発表)。上記の研究から、穀物収量におけるAGPase熱安定性の重要性が示される。
【0009】
Cross et al. (2004)は、BT2の最初の200個のアミノ酸およびジャガイモ塊茎小サブユニットの最後の275個のアミノ酸からなるモザイク小サブユニット(MP)を作製した。SH2との複合体中のMPは、農学的利益をもたらし得るいくつかの特徴を有した(Cross et al., 2004; Boehlein et al., 2005)。これらの特徴のいくつかは、野生型トウモロコシ内乳AGPase(BT2/SH2)と比べて、アクチベーター3-PGAの不在下での増大した活性、3-PGAに対する増大した親和性、および上昇した熱安定性であった。予備データから、トウモロコシ内乳において発現されるAGPaseバリアントを含む、トランスジェニックMPを有するトウモロコシ植物が、デンプン収量の増加を実現することが示されている(Hannah、未発表データ)。
【発明の概要】
【0010】
本発明は、熱条件に対する耐性が増大し、かつ/またはデンプン生合成が増大した植物または植物組織を提供するための材料および方法に関する。熱条件に対する植物または植物組織の耐性が増大すると、高温で一般に観察される収量減少と比べて少ない収量減少が提供される。本発明の1つの局面は、変異植物AGPase小サブユニットをコードするポリヌクレオチドに関する。1つの態様において、本発明のポリヌクレオチドは、野生型トウモロコシAGPase小サブユニットのアミノ酸462位に対応するトレオニンアミノ酸が、増大した熱安定性を与えるアミノ酸によって置換されているアミノ酸変異を有する、植物AGPase小サブユニットをコードする。別の態様において、ポリヌクレオチドは、2種の異なる植物に由来する配列から構成され(米国特許第7,173,165号に記載されている)、かつ、野生型トウモロコシAGPase小サブユニットのアミノ酸462位に対応するトレオニンアミノ酸が、増大した熱安定性を与えるアミノ酸によって置換されている本発明のアミノ酸変異を含む、キメラ植物AGPase小サブユニットをコードする。キメラAGPaseの変異は、熱安定性を相乗的に高める。本発明はまた、本発明のポリヌクレオチドにコードされる、変異植物AGPase小サブユニットも含む。熱安定性ならびに動力学的特性およびアロステリック特性の特徴付けから、本発明のポリヌクレオチドが単子葉植物内乳など植物において発現される場合、デンプン収量の増加が実現されることが示される。
【図面の簡単な説明】
【0011】
【図1】SH2と共にBT2、BT2-TI、MP、MP-TIを発現する大腸菌(E. coli)細胞によって産生されたグリコーゲンを示す。SH2、BT2、BT2-TI、およびMPのみを単独で発現する細胞に由来するグリコーゲン。グリコーゲンはグルコース単位で測定される。エラーバーは標準偏差を示す(N=3)。
【図2】図2Aおよび図2Bは、相補的なサブユニットSH2と共にBT2、BT2-TI、MP、MP-TIを発現する大腸菌細胞に由来する粗製抽出物のドットブロットを示す。BT2に対するモノクローナル抗体を用いてAGPaseを可視化した。スポットの密度は、ImageJ(http://rsb.info.nih.gov/ij/)を用いて概算した。
【図3】誘導されていない大腸菌細胞に由来する粗製タンパク質抽出物および部分的に精製したタンパク質抽出物中のAGPaseバリアントの比活性を示す。活性は逆方向で測定した。n.d.=検出不可。エラーバーは標準偏差を示す(N=3)。
【図4】AGPaseの精製を示す。精製した組換えBT2/SH2、TI/SH2、MP/SH2、およびMP-TI/SH2のSDS-PAGE。Biorad社製のPrecision Plus Protein All Blue Standardをマーカーとして使用した。左側の上の矢印は、大サブユニットを示す。左側の下の矢印は、小サブユニットを示す。
【図5A】精製したBT2/SH2、BT2-TI/SH2、およびMP/SH2の熱安定性を示す。各AGPaseの半減期(T1/2)は、平均値±標準誤差として表される。p値は、Prizm (Graph pad, San Diego CA)を用いて実施されるF検定によって概算する。図5Aにおいて、このアッセイ法は順方向で実施した。
【図5B】精製したBT2/SH2、BT2-TI/SH2、およびMP/SH2の熱安定性を示す。各AGPaseの半減期(T1/2)は、平均値±標準誤差として表される。p値は、Prizm (Graph pad, San Diego CA)を用いて実施されるF検定によって概算する。図5Bにおいて、このアッセイ法は逆方向で実施した。
【図6A】精製したMP/SH2およびMP-TI/SH2の熱安定性を示す。各AGPaseの半減期(T1/2)は、平均値±標準誤差として表される。p値は、Prizm (Graph pad, San Diego CA)を用いて実施されるF検定によって概算する。図6Aにおいて、このアッセイ法は順方向で実施した。
【図6B】精製したMP/SH2およびMP-TI/SH2の熱安定性を示す。各AGPaseの半減期(T1/2)は、平均値±標準誤差として表される。p値は、Prizm (Graph pad, San Diego CA)を用いて実施されるF検定によって概算する。図6Bにおいて、このアッセイ法は逆方向で実施した。
【図7A】BT2およびTIの3Dモデリングを示す。図7Aは、BT2モノマーの予測される3D構造である。TI変化には赤い丸でマークを付けている。サブユニット間の相互作用に直接関与しているBT2の領域は、黄色い四角で強調している。
【図7B】BT2およびTIの3Dモデリングを示す。図7Bは、Pro60(4、5)およびLeu61(3)の炭素原子からのThr462(1、2)の炭素原子の距離を示す。Thr462の接触残基は、FirstGlance Jmolを用いて決定した。濃い灰色の球は、Thr462の炭素原子を示す。明るい灰色の球は、接触残基の炭素原子を示す。酸素原子および窒素原子は、それぞれ赤色および青色で示す。
【図7C】BT2およびTIの3Dモデリングを示す。図7Cは、Pro60(5、6)およびLeu61(7、8)の炭素原子からのIle462(1、2、3、4)の炭素原子の距離を示す。Ile462の接触残基は、FirstGlance Jmolを用いて決定した。濃い灰色の球は、Ile462の炭素原子を示す。明るい灰色の球は、接触残基の炭素原子を示す。酸素原子および窒素原子は、それぞれ赤色および青色で示す。
【図8】AGPaseのサブユニット間の相互作用の強さを示す。酵母ツーハイブリッドシステムにおいてSH2をベイトとして使用し、BT2、TI、およびMPをプレイとして使用した。定量的β-ガラクトシダーゼアッセイ法を用いて、ベイトとプレイの間の相互作用を定量した。エラーバーは、2×標準誤差を示す(N=4)。
【発明を実施するための形態】
【0012】
配列の簡単な説明
配列番号:1は、本発明の変異ポリペプチド(TI)をコードするヌクレオチド配列を含むポリヌクレオチド配列である。
【0013】
配列番号:2は、本発明の変異ポリペプチド(TI)のアミノ酸配列である。
【0014】
配列番号:3は、本発明の変異ポリペプチド(MP-TI)をコードするヌクレオチド配列を含むポリヌクレオチド配列である。
【0015】
配列番号:4は、本発明の変異ポリペプチド(MP-TI)のアミノ酸配列である。
【0016】
配列番号:5は、本発明の変異ポリペプチド(TI+YC)のアミノ酸配列である。
【0017】
配列番号:6は、本発明の変異ポリペプチド(TI+QTCL)のアミノ酸配列である。
【0018】
配列番号:7は、本発明の変異ポリペプチド(TI+ETCL)のアミノ酸配列である。
【0019】
配列番号:8は、本発明の変異ポリペプチド(MP-TI+YC)のアミノ酸配列である。
【0020】
配列番号:9は、本発明の変異ポリペプチド(MP-TI+QTCL)のアミノ酸配列である。
【0021】
配列番号:10は、本発明の変異ポリペプチド(MP-TI+ETCL)のアミノ酸配列である。
【0022】
配列番号:11は、本発明の変異ポリペプチド(配列番号:5)をコードするヌクレオチド配列を含むポリヌクレオチド配列である。
【0023】
配列番号:12は、本発明の変異ポリペプチド(配列番号:6)をコードするヌクレオチド配列を含むポリヌクレオチド配列である。
【0024】
配列番号:13は、本発明の変異ポリペプチド(配列番号:7)をコードするヌクレオチド配列を含むポリヌクレオチド配列である。
【0025】
配列番号:14は、本発明の変異ポリペプチド(配列番号:8)をコードするヌクレオチド配列を含むポリヌクレオチド配列である。
【0026】
配列番号:15は、本発明の変異ポリペプチド(配列番号:9)をコードするヌクレオチド配列を含むポリヌクレオチド配列である。
【0027】
配列番号:16は、本発明の変異ポリペプチド(配列番号:10)をコードするヌクレオチド配列を含むポリヌクレオチド配列である。
【0028】
配列番号:17は、本発明に従って使用することができるオリゴヌクレオチドである。
【0029】
配列番号:18は、本発明に従って使用することができるオリゴヌクレオチドである。
【0030】
配列番号:19は、本発明に従って使用することができるオリゴヌクレオチドである。
【0031】
配列番号:20は、本発明に従って使用することができるオリゴヌクレオチドである。
【0032】
配列番号:21は、本発明の変異ポリペプチド(TI)のアミノ酸配列である。
【0033】
配列番号:22は、本発明の変異ポリペプチド(TI)のアミノ酸配列である。
【0034】
配列番号:23は、本発明の変異ポリペプチド(TI)のアミノ酸配列である。
【0035】
配列番号:24は、本発明の変異ポリペプチド(TI)のアミノ酸配列である。
【0036】
配列番号:25は、本発明の変異ポリペプチド(TI)のアミノ酸配列である。
【0037】
配列番号:26は、ジャガイモ塊茎のAGPase小サブユニットタンパク質のアミノ酸配列である。
【0038】
発明の詳細な説明
本発明は、熱条件に対する耐性が増大し、かつ/またはデンプン生合成が増大した植物を提供するための材料および方法に関する。熱条件に対する植物の耐性が増大すると、高温で一般に観察される収量減少と比べて少ない収量減少が提供される。
【0039】
本発明の1つの局面は、変異植物AGPase小サブユニットをコードするポリヌクレオチドに関する。1つの態様において、本発明のポリヌクレオチドは、野生型トウモロコシ内乳AGPase小サブユニットのアミノ酸462位に対応するトレオニンアミノ酸が、増大した熱安定性を与えるアミノ酸によって置換されているアミノ酸変異を有する、植物AGPase小サブユニットをコードする。特定の態様において、置換アミノ酸(amino acid substituted)はイソロイシンである。1つの態様において、変異植物AGPase小サブユニットはトウモロコシ内乳AGPase小サブユニットである。例示的な態様において、変異植物AGPase小サブユニットは、配列番号:2に示すアミノ酸配列またはその断片もしくはバリアントを含む。特定の態様において、ポリヌクレオチドは、配列番号:1に示すヌクレオチド配列またはその断片もしくはバリアントを含む。別の態様において、変異植物AGPase小サブユニットは、オオムギ、コムギ、モロコシ、ジャガイモ、またはイネである。特定の態様において、オオムギ、コムギ、モロコシ、またはジャガイモの変異AGPase小サブユニットは、配列番号:21、配列番号:22、配列番号:23、配列番号:24、または配列番号:25にそれぞれ示すアミノ酸配列を含む。別の態様において、ポリヌクレオチドは、公開されている国際特許出願の国際公開公報第2005/019425号(HannahおよびLinebarger)に記載されているようなアミノ酸変異をさらに含んでよい変異植物AGPase小サブユニットをコードする。1つの態様において、このポリヌクレオチドにコードされる変異AGPase小サブユニットは、野生型トウモロコシ内乳AGPaseのアミノ酸36位に対応するチロシンがシステインで置換されているアミノ酸変異を含む。また、この変異AGPase小サブユニットは、野生型トウモロコシ内乳AGPaseのアミノ酸34位およびアミノ酸35位にそれぞれ対応するセリンアミノ酸とトレオニンアミノ酸の間に挿入されたアミノ酸を任意で含んでよい。特定の態様において、AGPase小サブユニットの34位および35位のアミノ酸の間に挿入されるアミノ酸は、グルタミン酸またはグルタミンである。例示的な態様において、変異植物AGPase小サブユニットは、配列番号:5、配列番号:6、もしくは配列番号:7に示すアミノ酸配列またはその断片もしくはバリアントを含む。特定の態様において、ポリヌクレオチドは、配列番号:11、配列番号:12、もしくは配列番号:13に示すヌクレオチド配列またはその断片もしくはバリアントを含む。
【0040】
別の態様において、ポリヌクレオチドは、2種の異なる植物に由来する配列から構成され(米国特許第7,173,165号に記載されている)、かつ、野生型トウモロコシ内乳AGPase小サブユニットのアミノ酸462位に対応するトレオニンアミノ酸が、増大した熱安定性を与えるアミノ酸によって置換されている本発明のアミノ酸変異もまた含む、キメラ植物AGPase小サブユニットをコードする。特定の態様において、置換アミノ酸はイソロイシンである。1つの態様において、キメラAGPase小サブユニットは、1つの植物に由来するC末端部分および別の植物に由来するN末端部分を含む。1つの態様において、本発明のキメラタンパク質は、第1の植物AGPase小サブユニットのN末端の最初の約150〜250個のアミノ酸を有するN末端配列および第2の植物AGPase小サブユニットのC末端の最後の約300個またはそれ以下の残基を含むC末端配列を含む。したがって、キメラサブユニットのC末端は、第2の植物のC末端の最後の300個、または299個、または298個、または297個、または296個、または295個などの残基を含み得る。サブユニット配列は、単子葉植物の、もしくは双子葉植物の、または単子葉植物と双子葉植物の両方のAGPaseに由来してよい。例えば、イネ、コムギ、オオムギ、エンバク、モロコシ、トウモロコシ、ユリ、およびキビ(millet)などの単子葉植物は、本発明の範囲内に含まれる。双子葉植物には、例えば、タバコ、ダイズ、ジャガイモ、サツマイモ、ラディッシュ、キャベツ、セイヨウアブラナ、リンゴの木、およびレタスが含まれ得る。1つの態様において、キメラタンパク質のN末端の最初の200個程度のアミノ酸はトウモロコシ内乳AGPase小サブユニットのN末端に由来し、C末端アミノ酸は、ジャガイモ塊茎AGPase小サブユニットのC末端に由来し、本発明のアミノ酸462位に対応する変異を含む。特定の態様において、本発明のキメラタンパク質のC末端領域は、ジャガイモ塊茎のAGPase小サブユニットの最後の276個のアミノ酸を含む。例示的な態様において、キメラタンパク質は、トウモロコシ内乳AGPaseの小サブユニットの一部分およびジャガイモ塊茎AGPaseの小サブユニットの一部分を含む。特定の態様において、キメラタンパク質は、a)トウモロコシ内乳AGPaseの小サブユニットに由来する最初の199個のアミノ酸(すなわち、アミノ酸1〜199)、およびアミノ酸246から始まる、ジャガイモ塊茎AGPaseの小サブユニットのカルボキシル末端(すなわち、アミノ酸246〜521)(Genbankアクセッション番号X61186として寄託されているタンパク質に関して示されているアミノ酸配列を用いた場合)(または、別法として、Hannah et al., 2001で記載されているジャガイモAGPaseサブユニットの番号付け方式を用いた場合、アミノ酸175から始まる)、ならびにb)本発明のアミノ酸462位に対応する変異を含む。例示的な態様において、植物キメラAGPase小サブユニットは、配列番号:4に示すアミノ酸配列またはその断片もしくはバリアントを含む。特定の態様において、ポリヌクレオチドは、配列番号:3に示すヌクレオチド配列またはその断片もしくはバリアントを含む。別の態様において、ポリヌクレオチドは、公開されている国際特許出願の国際公開公報第2005/019425号(HannahおよびLinebarger)に記載されているアミノ酸変異をさらに含んでよい変異植物AGPase小サブユニットをコードする。さらなる態様において、このポリヌクレオチドにコードされる変異AGPase小サブユニットはまた、36位のチロシンがシステインで置換されているアミノ酸変異を含む。また、この変異AGPase小サブユニットは、それぞれ34位および35位のセリンアミノ酸とトレオニンアミノ酸の間に挿入されたアミノ酸を任意で含んでよい。特定の態様において、変異AGPase小サブユニットの34位および35位のアミノ酸の間に挿入されるアミノ酸は、グルタミン酸またはグルタミンである。例示的な態様において、変異植物AGPase小サブユニットは、配列番号:8、配列番号:9、または配列番号:10に示すアミノ酸配列またはその断片もしくはバリアントを含む。特定の態様において、ポリヌクレオチドは、配列番号:14、配列番号:15、または配列番号:16に示すヌクレオチド配列またはその断片もしくはバリアントを含む。
【0041】
本発明はまた、植物または植物組織の熱安定性を増大させるため、および/またはデンプン生合成を増大させるため、ならびに収穫量を増加させるための方法にも関する。1つの態様において、本発明の方法は、本発明の1つまたは複数のポリヌクレオチドを植物中に導入する段階を含む。一定の態様において、植物中に導入されるポリヌクレオチドは、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、配列番号:10、配列番号:21、配列番号:22、配列番号:23、配列番号:24、もしくは配列番号:25のいずれかに示すアミノ酸配列またはその断片もしくはバリアントを含む1種または複数種のポリペプチドをコードする。特定の態様において、ポリヌクレオチドは、配列番号:1もしくは配列番号:3に示すヌクレオチド配列またはその断片もしくはバリアントを含む。さらなる特定の態様において、ポリヌクレオチドは、配列番号:11、配列番号:12、配列番号:13、配列番号:14、配列番号:15、もしくは配列番号:16に示すヌクレオチド配列またはその断片もしくはバリアントを含む。1つの態様において、ポリヌクレオチドは、植物または植物組織のゲノム中に安定に組み込まれる。ポリヌクレオチドは、ポリヌクレオチドおよび/またはそれにコードされるポリペプチドの発現を増大させる調節エレメント、例えばプロモーター配列および/またはエンハンサー配列を含んでよい。特定の態様において、プロモーター配列は、構成的発現または組織特異的(例えば内乳)発現をもたらすものである。このポリヌクレオチドを含む植物もしくは植物組織、またはこれらの植物の子孫は、本発明のポリヌクレオチドまたはポリペプチドの発現の増大について任意でスクリーニングすることができる。1つの態様において、本発明の1つまたは複数のポリヌクレオチドの複数のコピーは、植物または植物組織中に導入され、その植物のゲノム中に安定に組み込まれる。1つの態様において、本発明のポリヌクレオチドは、本明細書において説明する発現構築物において提供される。
【0042】
本発明はまた、本発明のポリヌクレオチドにコードされる、変異AGPase小サブユニットポリペプチドにも関する。1つの態様において、このポリペプチドは、配列番号:2に示すアミノ酸配列またはその断片もしくはバリアントを含む。他の態様において、このポリペプチドは、配列番号:21、配列番号:22、配列番号:23、配列番号:24、もしくは配列番号:25に示すアミノ酸配列またはその断片もしくはバリアントを含む。別の態様において、このポリペプチドは、配列番号:4に示すアミノ酸配列またはその断片もしくはバリアントを含む。さらに別の態様において、このポリヌクレオチドは、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、もしくは配列番号:10のいずれかに示すアミノ酸配列またはその断片もしくはバリアントを含む。
【0043】
本発明はまた、本発明の1つまたは複数の変異AGPase小サブユニットポリペプチドを含む変異植物AGPase酵素にも関する。変異植物AGPaseはまた、1つまたは複数の野生型AGPase大サブユニットポリペプチドも含んでよい。特定の態様において、変異植物AGPase酵素は、そのうちのいずれかが配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、配列番号:10、配列番号:21、配列番号:22、配列番号:23、配列番号:24、もしくは配列番号:25のアミノ酸配列または任意のこのような配列の断片もしくはバリアントを含んでよい1つまたは複数の変異AGPase小サブユニットポリペプチドを含み、この変異AGPase酵素は、野生型AGPase酵素と比べて増大した熱安定性を示す。1つの態様において、変異植物酵素は、本発明の2つの変異AGPase小サブユニットを含み、ここで、これらの変異ポリペプチドは同じ変異を有してもよく、または異なる変異を有してもよい。本発明はまた、本発明の1つまたは複数の変異AGPase小サブユニットポリペプチドおよび1つまたは複数の変異AGPase大サブユニットポリペプチドを含む、変異植物AGPase酵素にも関する。1つの態様において、変異AGPase大サブユニットポリペプチドは、米国特許第5,589,618号;同第5,650,557号;同第5,872,216号;同第6,069,300号;同第6,184,438号;同第6,403,863号;同第6,809,235号;同第7,173,165号;同第7,312,378号;および同第6,969,783号のいずれかに記載されているもののいずれかでよい。1つの態様において、変異AGPase大サブユニットポリペプチドは、Rev6変異を含む。別の態様において、変異AGPase大サブユニットポリペプチドは、米国特許第6,069,300号;同第6,403,863号;同第6,809,235号;同第7,312,378号;および同第6,969,783号、ならびに公開されている国際特許出願の国際公開公報第99/58698号;国際公開公報第2003/0070901号;国際公開公報第98/22601号;および国際公開公報第02/072784号に記載されているような1つまたは複数の熱安定性(HS)変異、例えば、HS33変異などを含む。1つの態様において、変異植物AGPase酵素は、本明細書において説明するように、同じ変異を有してもよく、または異なる変異を有してもよい本発明の2つの変異AGPase小サブユニットポリペプチドを含む。別の態様において、変異植物AGPase酵素は、同じ変異を有してもよく、または異なる変異を有してもよい2つの変異AGPase大サブユニットポリペプチドを含む。さらなる態様において、変異植物AGPase酵素は、本発明の2つの変異AGPase小サブユニットポリペプチドおよび2つの変異SH2ポリペプチドを含み、ここで、本明細書において説明するように、これらの変異AGPase小サブユニットポリペプチドおよび変異AGPase大サブユニットポリペプチドは、同じ変異を有してもよく、または異なる変異を有してもよい。
【0044】
本発明はまた、野生型植物AGPaseと比べて熱安定性が増大した変異植物AGPase酵素を提供するための方法にも関する。1つの態様において、この方法は、AGPase酵素において、本発明の1つまたは複数の変異AGPase小サブユニットポリペプチドを、野生型AGPase大サブユニットもしくは変異AGPase大サブユニットと合体させる段階、または野生型AGPase大サブユニットもしくは変異AGPase大サブユニットと共に提供する段階を含む。1つの態様において、AGPase酵素は、ポリペプチドサブユニットの四量体を含み、それらのサブユニットの内の1つ、2つ、またはそれ以上が本発明の変異ポリペプチドである。1つの態様において、AGPase酵素はまた、変異AGPase大サブユニットポリペプチドサブユニット、例えば、Rev6および/またはHS33のような熱安定性変異を含む変異大サブユニットを含む。
【0045】
本発明はまた、ポリヌクレオチドもしくは本発明のポリヌクレオチドにコードされるタンパク質を含むか、または本発明の変異ポリペプチドもしくはその断片もしくはバリアントを発現するか、または本発明の変異植物AGP酵素を含むかもしくは発現する、本発明の植物、植物組織、および植物細胞にも関する。植物組織には、種子、接ぎ穂、および台木が含まれるがこれらに限定されるわけではない。本発明の範囲内の植物には、例えば、イネ、コムギ、オオムギ、エンバク、ライムギ、モロコシ、トウモロコシ、サトウキビ、パイナップル、タマネギ、バナナ、ココナツ、ユリ、芝草(turfgrass)、およびキビなどの単子葉植物が含まれる。また、本発明の範囲内の植物には、例えば、トマト、キュウリ、カボチャ、エンドウマメ、アルファルファ、メロン、ヒヨコマメ、チコリ、クローバー、ケール、レンズマメ、ダイズ、マメ、タバコ、ジャガイモ、サツマイモ、ヤムイモ、カッサバ、ラディッシュ、ブロッコリー、ホウレンソウ、キャベツ、セイヨウアブラナ、リンゴの木、柑橘類(オレンジ、マンダリン、グレープフルーツ、レモン、およびライムなどを含む)、ブドウ、綿、ヒマワリ、イチゴ、レタス、およびホップなどの双子葉植物も含まれる。本発明のポリヌクレオチドを含むハーブ植物もまた、本発明の範囲内で企図される。ハーブ植物には、パセリ、セージ、ローズマリー、およびタイムなどが含まれる。1つの態様において、植物、植物組織、または植物細胞は、トウモロコシ(Zea mays)である。1つの態様において、植物、植物組織、または植物細胞は、トランスジェニックである植物、植物組織、または植物細胞である。別の態様において、植物、植物組織、または植物細胞は、育種プログラムを通じて得られたものである。
【0046】
本発明において有用なポリヌクレオチドを発現構築物において提供することができる。一般に、本発明の発現構築物は、その発現構築物が発現されることになっている意図された宿主細胞において機能的である調節エレメントを含む。したがって、当業者は、細菌宿主細胞、酵母宿主細胞、植物宿主細胞、昆虫宿主細胞、哺乳動物宿主細胞、およびヒト宿主細胞において使用するための調節エレメントを選択することができる。調節エレメントには、プロモーター、転写終結配列、翻訳終結配列、エンハンサー、およびポリアデニル化エレメントが含まれる。本明細書において使用される場合、「発現構築物」という用語は、機能的に連結された核酸配列の転写を実現する核酸配列の組み合わせを意味する。本明細書において使用される場合、「機能的に連結される」という用語は、記述された構成要素の並置を意味し、それらの構成要素は、意図された様式でそれらが機能することを可能にする関係にある。一般に、機能的に連結された構成要素は隣接関係にある。
【0047】
本発明の発現構築物は、本発明の変異ポリペプチドをコードするポリヌクレオチド配列に機能的に連結されたプロモーター配列を含んでよい。プロモーターは、当技術分野において公知の標準技術を用いてポリヌクレオチド中に組み入れることができる。プロモーターの複数のコピーまたは複数のプロモーターを本発明の発現構築物において使用することができる。好ましい態様において、プロモーターは、発現構築物中の転写開始部位からの距離が、天然の遺伝的環境における転写開始部位からの距離とほぼ同じになるように配置されてよい。プロモーター活性を実質的に低下させずに、この距離をいくらか変更することは許容される。転写開始部位は、典型的には発現構築物中に含まれる。
【0048】
発現構築物が植物細胞中で提供される予定であるか、または植物細胞中に導入される予定である場合には、植物ウイルスプロモーター、例えば、カリフラワーモザイクウイルス(CaMV) 35Sプロモーター(増強されたCaMV 35Sプロモーター(例えば米国特許第5,106,739号を参照されたい)を含む)もしくはCaMV 19Sプロモーターまたはキャッサバ葉脈モザイク(cassava vein mosaic)などが使用され得る。植物中の発現構築物のために使用され得る他のプロモーターには、例えば、プロリフェラ(prolifera)プロモーター、Ap3プロモーター、熱ショックプロモーター、A.ツメファシエンス(A.tumefaciens)のT-DNA 1'-プロモーターまたはT-DNA 2'-プロモーター、ポリガラクツロナーゼプロモーター、ペチュニア由来のカルコン合成酵素A(CHS-A)プロモーター、タバコPR-1aプロモーター、ユビキチンプロモーター、アクチンプロモーター、alcA遺伝子プロモーター、pin2プロモーター(Xu et al., 1993)、トウモロコシWipIプロモーター、トウモロコシtrpA遺伝子プロモーター(米国特許第5,625,136号)、トウモロコシCDPK遺伝子プロモーターが含まれ、また、RUBISCO SSU プロモーター(米国特許第5,034,322号)も使用され得る。組織特異的プロモーター、例えば、トマトのE8プロモーターのような果実特異的プロモーター(アクセッション番号: AF515784; Good et al. (1994))が使用され得る。花器官特異的プロモーターのような果実特異的プロモーターは、植物の花器官において本発明のポリヌクレオチドを発現させるために本発明の発現構築物と共に使用され得る。花器官特異的プロモーターの例には、米国特許第6,462,185号;同第5,639,948号;および同第5,589,610号で説明されているプロモーター配列のいずれかが含まれる。(例えばインゲンマメの)β-ファゼオリン遺伝子または(例えばダイズの)グリシニン遺伝子に由来するプロモーターのような種子特異的プロモーターおよび他のものもまた、使用され得る。内乳特異的プロモーターには、MEG1 (EPO出願番号EP1528104)ならびにWu et al. (1998)、Furtado et al. (2001)、およびHwang et al. (2002)によって説明されているものが含まれるが、それらに限定されるわけではない。根特異的プロモーター、例えば、米国特許第6,455,760号もしくは米国特許第6,696,623号;または米国特許出願公開第20040078841号;同第20040067506号;同第20040019934号;同第20030177536号;同第20030084486号;もしくは同第20040123349号で説明されているプロモーター配列のいずれかが、本発明の発現構築物と共に使用され得る。構成的プロモーター(CaMVプロモーター、ユビキチンプロモーター、アクチンプロモーター、またはNOSプロモーターなど)、発生に関連して調節されるプロモーター、および誘導性プロモーター(例えば、熱、光、ホルモン、または化学物質によって誘導され得るプロモーター類)もまた、本発明のポリヌクレオチド発現構築物と共に使用するために企図される。
【0049】
本発明の発現構築物は、転写終結配列、翻訳終結配列、シグナルペプチドをコードする配列、および/またはエンハンサーエレメントを任意で含んでよい。転写終結領域は、典型的には、真核生物またはウイルスの遺伝子配列の3'非翻訳領域から得ることができる。転写終結配列は、効率的な終結を実現するために、コード配列の下流に配置されてよい。シグナルペプチド配列は、特定の細胞小器官区画からタンパク質作用部位に及ぶ広範囲の翻訳後細胞内目的地および細胞外環境に、機能的に連結された成熟ポリペプチドを再配置するのを担当するタンパク質のアミノ末端に典型的に存在する短いアミノ酸配列である。機能的に連結されたシグナルペプチド配列を用いて、意図した細胞内目的地および/または細胞外目的地に遺伝子産物をターゲティングすることが、本発明のポリペプチドと共に使用するために企図される。古典的なエンハンサーは、遺伝子転写を増大させるシス作用エレメントであり、これもまた発現構築物中に含まれてよい。古典的なエンハンサーエレメントは当技術分野において公知であり、CaMV 35Sエンハンサーエレメント、サイトメガロウイルス(CMV)初期プロモーターエンハンサーエレメント、およびSV40エンハンサーエレメントが含まれるが、それらに限定されるわけではない。遺伝子発現を促進するイントロンを介したエンハンサーエレメントもまた、当技術分野において公知である。これらのエレメントは、転写領域内に存在しなければならず、配向依存性(orientation dependent)である。例には、トウモロコシしわ型-1エンハンサーエレメント(ClancyおよびHannah, 2002)が含まれる。
【0050】
発現構築物から転写されたmRNAのポリアデニル化を指示するDNA配列もまた、発現構築物中に含まれてよく、これらにはオクトピン合成酵素シグナルまたはノパリン合成酵素シグナルが含まれるが、それらに限定されるわけではない。本発明の発現構築物はまた、他の遺伝子の転位を指示するポリヌクレオチド配列、すなわちトランスポゾンも含んでよい。
【0051】
本発明のポリヌクレオチドは、RNAまたはDNAのいずれかから構成され得る。好ましくは、ポリヌクレオチドはDNAから構成される。本発明はまた、本明細書において開示するポリヌクレオチドに配列が相補的であるポリヌクレオチドを包含する。本発明のポリヌクレオチドおよびポリペプチドは、精製型または単離型で提供されてよい。
【0052】
遺伝コードの縮重があるため、様々な異なるポリヌクレオチド配列が、本発明の変異ポリペプチドをコードし得る。存在し得るすべての三つ組コドン(UはTも表す)および各コドンにコードされるアミノ酸を示す表がLewin (1985)に記載されている。さらに、本発明の同じまたは本質的に同じ変異ポリペプチドをコードする代替のポリヌクレオチド配列を作り出すことは、当業者の技能で十分に対応できる範囲である。これらの変異ポリヌクレオチド配列または代替ポリヌクレオチド配列は、本発明の範囲内である。本明細書において使用される場合、「本質的に同じ」配列への言及は、本発明のポリヌクレオチドにコードされるポリペプチドの機能活性を実質的に改変しないアミノ酸置換、欠失、付加、または挿入をコードする配列を意味する。本発明の野生型ポリペプチドまたは変異ポリペプチドをコードするヌクレオチド配列のアレルバリアントもまた、本発明の範囲内に含まれる。
【0053】
具体的に例示したもの、または本発明の野生型ポリペプチドもしくは変異ポリペプチドおよび/またはAGPase酵素中に天然に存在するもの以外のアミノ酸置換もまた、本発明の範囲内で企図される。例えば、置換されたアミノ酸を有する変異ポリペプチドが、アミノ酸が置換されていない変異ポリペプチドと実質的に同じ機能活性を保持する限りにおいて、変異AGPase小サブユニットポリペプチドのアミノ酸を非天然アミノ酸で置換してよい。非天然アミノ酸の例には、オルニチン、シトルリン、ヒドロキシプロリン、ホモセリン、フェニルグリシン、タウリン、ヨードチロシン、2,4-ジアミノ酪酸、a-アミノイソ酪酸、4-アミノ酪酸、2-アミノ酪酸、γ-アミノ酪酸、e-アミノヘキサン酸、6-アミノヘキサン酸、2-アミノイソ酪酸、3-アミノプロピオン酸、ノルロイシン、ノルバリン、サルコシン、ホモシトルリン、システイン酸、t-ブチルグリシン、t-ブチルアラニン、フェニルグリシン、シクロヘキシルアラニン、β-アラニン、フルオロアミノ酸、デザイナーアミノ酸、例えば、β-メチルアミノ酸、C-メチルアミノ酸、N-メチルアミノ酸、および一般的なアミノ酸類似体が含まれるが、それらに限定されるわけではない。また、非天然アミノ酸には、誘導体化された側鎖を有するアミノ酸も含まれる。さらに、タンパク質中のアミノ酸の任意のものは、D(右旋性)型またはL(左旋性)型でよい。本発明の野生型または変異型のAGPase小サブユニットポリペプチドまたはAGPase大サブユニットポリペプチドのタンパク質配列のアレルバリアントもまた、本発明の範囲内に含まれる。
【0054】
アミノ酸は一般に次のクラスに分類され得る:非極性、非荷電極性、塩基性、および酸性。あるクラスのアミノ酸を有する本発明の変異AGPase小サブユニットポリペプチドおよび/または野生型もしくは変異型のAGPase大サブユニットポリペプチドが同じクラスの別のアミノ酸で置換される保存的置換は、その置換を有するポリペプチドが、置換を有さないポリペプチドと実質的に同じ機能活性(例えば、AGPase酵素の酵素安定性および/または増大した熱安定性)を引き続き保持する限りにおいて、本発明の範囲内に含まれる。配列中に1つまたは複数のアミノ酸置換を有する変異AGPase小サブユニットポリペプチドおよび/または野生型もしくは変異型のAGPase大サブユニットポリペプチドをコードするポリヌクレオチドは、本発明の範囲内で企図される。下記の表1は、各クラスに属するアミノ酸の例のリストを提供する。
【0055】
【表1】
【0056】
本発明はまた、本発明の機能的な野生型または変異型のAGPase小サブユニットポリペプチドまたはAGPase大サブユニットポリペプチドをコードする本発明のポリヌクレオチドのバリアントにも関する。バリアント配列には、その配列の1つまたは複数のヌクレオチドが置換、欠失、および/または挿入された配列が含まれる。DNAの天然ヌクレオチドを置換できるヌクレオチドは塩基部分を有し、この塩基部分には、イノシン、5-フルオロウラシル、5-ブロモウラシル、ヒポキサンチン、1-メチルグアニン、5-メチルシトシン、およびトリチル化塩基が含まれ得るが、それらに限定されるわけではない。ある配列中のヌクレオチドの糖部分もまた修飾されてよく、アラビノース、キシルロース、およびヘキソースが含まれるが、それらに限定されるわけではない。さらに、ヌクレオチドのアデニン塩基、シトシン塩基、グアニン塩基、チミン塩基、およびウラシル塩基は、アセチル基、メチル基、および/またはチオ基で修飾されてもよい。ヌクレオチドの置換、欠失、および/または挿入を含む配列は、当技術分野において公知の標準技術を用いて調製し、試験することができる。
【0057】
本発明の変異ポリペプチドの断片およびバリアントは、本明細書において説明するようにして作製することができ、かつ、当技術分野において公知の標準技術を用いて、酵素機能および熱安定機能の存在に関して試験することができる。すなわち、当業者は、本発明の変異ポリペプチドの断片およびバリアントを容易に調製して試験し、その断片またはバリアントが完全長または非バリアント(non-variant)変異ポリペプチドと比べて機能活性を保持しているかどうかを判定することができる。
【0058】
本発明の範囲内で企図されるポリヌクレオチドおよびポリペプチドはまた、本明細書において具体的に例示する本発明の配列に対する同一性および/または類似性の範囲がより特定的であるという面から定義することもできる。配列同一性は、典型的には60%より高く、好ましくは75%より高く、より好ましくは80%より高く、さらにより好ましくは90%より高く、95%より高くでもよい。配列の同一性および/または類似性は、本明細書において例示する配列と比較して、49%、50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、または99%でよい。別段の指定が無い限り、本明細書において使用される場合、2つの配列の配列同一性パーセントおよび/または類似性パーセントは、KarlinおよびAltschul (1993)において改良された、KarlinおよびAltschul (1990)のアルゴリズムを用いて決定することができる。このようなアルゴリズムは、Altschul et al. (1990)のNBLASTプログラムおよびXBLASTプログラムに組み入れられている。NBLASTプログラム、スコア=100、ワード長=12を用いてBLAST探索を実施して、所望の配列同一性パーセントを有する配列を得ることができる。比較目的のギャップありアライメントを得るために、Altschul et al. (1997)で説明されているようにしてGapped BLASTを使用することができる。BLASTプログラムおよびGapped BLASTプログラムを使用する場合、各プログラム(NBLASTおよびXBLAST)の初期設定パラメーターを使用することができる。NCBI/NIHのウェブサイトを参照されたい。
【0059】
本発明はまた、本明細書において例示するポリヌクレオチド配列と十分に相同で、その結果、標準のストリンジェントな条件および標準的方法(Maniatis et al., 1982)のもとでその配列とハイブリダイゼーションすることを可能にする配列を有するポリヌクレオチド分子も企図する。本明細書において使用される場合、ハイブリダイゼーションのための「ストリンジェントな」条件とは、典型的には6×SSPE、5×デンハルト液、0.1% SDS、0.1mg/ml変性DNAにおいて、DNAハイブリッドの融解温度(Tm)より20〜25C低い温度で一晩、ハイブリダイゼーションを実施する条件を意味する。融解温度Tmは次の式によって説明される(Beltz et al., 1983):
Tm=81.5C+16.6Log[Na+]+0.41(%G+C)−0.61(%ホルムアミド)−600/二重鎖の長さ(単位:塩基対)。
【0060】
洗浄は典型的には以下のようにして実施する:
(1)室温で15分間、1×SSPE、0.1%SDS中で2回(低ストリンジェンシー洗浄)。
(2)Tm-20Cで15分間、0.2×SSPE、0.1%SDS中で1回(中程度のストリンジェンシー洗浄)。
【0061】
本明細書において使用される場合、「核酸」および「ポリヌクレオチド」という用語は、単鎖または二本鎖いずれかの形態のデオキシリボヌクレオチドポリマー、リボヌクレオチドポリマー、またはデオキシリボヌクレオチドとリボヌクレオチドの混合ポリマーを意味し、別段の制限がない限り、天然に存在するヌクレオチドと同様の様式で機能できる、天然ヌクレオチドの公知の類似体も包含する。これらのポリヌクレオチド配列には、RNAに転写されるDNA鎖配列および転写されるDNA鎖に相補的である鎖配列が含まれる。また、これらのポリヌクレオチド配列には、完全長配列ならびに完全長配列に由来する短い配列の両方が含まれる。例示する配列のアレル変種(variation)もまた、本発明の範囲内に入る。これらのポリヌクレオチド配列には、個々の鎖としてか、または二重鎖におけるセンス鎖およびアンチセンス鎖の両方が含まれる。
【0062】
植物細胞をある遺伝子で形質転換するための技術は当技術分野において公知であり、例えば、アグロバクテリウム(Agrobacterium)感染、バイオリスティック法、エレクトロポレーション、塩化カルシウム処理、PEGを介した形質転換などが含まれる。米国特許第5,661,017号では、異種ポリヌクレオチドで藻類細胞を形質転換するための方法および材料が教示されている。当技術分野において公知の標準的方法を用いて、形質転換細胞を選択し、再分化させ、本発明のポリヌクレオチドを含み発現する植物に成長させることができる。本発明の形質転換されたかまたはトランスジェニックである任意の植物細胞または植物の種子および他の植物組織ならびに子孫もまた、本発明の範囲内に含まれる。
【0063】
本発明はまた、野生型植物と比べて増大した熱安定性を示す植物を作製するための方法であって、本発明の変異AGPase小サブユニットポリペプチドをコードするポリヌクレオチドが植物細胞中に導入され、そのポリヌクレオチドにコードされるポリペプチドが発現される方法にも関する。1つの態様において、植物細胞は、野生型AGPase大サブユニットポリペプチドをコードする非変異遺伝子を含む。別の態様において、植物細胞は、変異AGPase大サブユニットポリペプチドをコードする少なくとも1つのポリヌクレオチドを含む。さらなる態様において、変異AGPase大サブユニットポリペプチドをコードするポリヌクレオチドはまた、変異AGPase小サブユニットポリペプチドをコードするポリヌクレオチドと共に植物細胞中に導入される。1つの態様において、1つまたは複数のポリヌクレオチドは、植物細胞のゲノム中に組み込まれ、その植物細胞から植物が成長させられる。好ましい態様において、植物細胞から成長させられた植物は、組み込まれた1つまたは複数のポリヌクレオチドを安定に発現する。
【0064】
本発明はまた、本発明のポリヌクレオチドのコード配列または非コード配列にハイブリダイズできるオリゴヌクレオチドプローブおよびオリゴヌクレオチドプライマー、例えばポリメラーゼ連鎖反応法(PCR)プライマーにも関する。本発明のオリゴヌクレオチドプローブは、本発明の変異AGPase小サブユニットポリペプチドをコードする核酸配列を検出および定量するための方法において使用され得る。本発明のオリゴヌクレオチドプライマーは、核酸増幅を伴うPCR法および他の方法において使用することができる。好ましい態様において、本発明のプローブまたはプライマーは、ストリンジェントな条件下で本発明のポリヌクレオチドにハイブリダイズすることができる。本発明のプローブおよびプライマーは、検出可能な標識またはレポーター分子、例えば、蛍光分子、酵素、および放射性部分(例えば、3H、35S、125Iなど)などを任意で含んでよい。本発明のプローブおよびプライマーは、それらが使用される方法またはアッセイ法に適した任意の長さのものでよい。典型的には、本発明のプローブおよびプライマーのヌクレオチド長は、10〜500またはそれ以上である。ヌクレオチド長が10〜20、21〜30、31〜40、41〜50、51〜60、61〜70、71〜80、81〜90、91〜100またはそれ以上であるプローブおよびプライマーは、本発明の範囲内で企図される。本発明のプローブおよびプライマーは、ポリヌクレオチド配列とのヌクレオチド配列同一性が完全(100%)でもよく、または配列同一性は100%未満でもよい。例えば、プローブまたはプライマーとある配列との配列同一性は、そのプローブまたはプライマーがストリンジェントな条件下で本発明のポリヌクレオチドのヌクレオチド配列にハイブリダイズできる限り、99%、98%、97%、96%、95%、90%、85%、80%、75%、70%または他の任意の配列同一率(%)でよい。1つの態様において、本発明のプローブおよびプライマーは、配列番号:1、配列番号:3、配列番号:11、配列番号:12、配列番号:13、配列番号:14、配列番号:15、もしくは配列番号:16、またはそれらの相補物のヌクレオチド配列に対して70%もしくはそれ以上、75%もしくはそれ以上、80%もしくはそれ以上、85%もしくはそれ以上、90%もしくはそれ以上、または95%〜100%の配列同一性を有する。
【0065】
本発明はまた、単離された変異AGPase小サブユニットポリペプチドにも関する。1つの態様において、変異AGPase小サブユニットポリペプチドはトウモロコシのAGPase小サブユニットポリペプチドである。特定の態様において、本発明のAGPase小サブユニットポリペプチドは、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、配列番号:10、配列番号:21、配列番号:22、配列番号:23、配列番号:24、もしくは配列番号:25に示すアミノ酸配列またはその機能的断片もしくはバリアントを有する。本発明のAGPase小サブユニットポリペプチドまたは酵素は、当技術分野において公知の標準技術を用いて精製することができる。1つの態様において、AGPase小サブユニットポリペプチドをコードする本発明のポリヌクレオチドは大腸菌のような微生物中に組み込まれ、AGPase小サブユニットポリペプチドはその微生物中で発現され、次いでそれから単離される。
【0066】
特定の態様において、本発明のポリペプチドおよびその機能的ペプチド断片は、本発明のポリペプチドに特異的に結合する抗体を作製するために使用され得、そのような抗体は本発明の範囲内で企図される。本発明の抗体は、ポリクローナルまたはモノクローナルでよく、当技術分野において公知の標準的方法を用いて作製および単離することができる。
【0067】
本発明によるポリペプチド断片は、典型的には、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、配列番号:10の少なくともまたは約
個のアミノ酸の連続スパン(contiguous span)を含む。
【0068】
一定の態様において、本発明のポリペプチド断片の長さは、少なくとも約25個の連続アミノ酸から、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、または配列番号:10に示す配列のような完全長配列よりアミノ酸1個分少ないものまでの任意の整数個でよい。したがって、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、または配列番号:10の場合、ポリペプチド断片はアミノ酸約25〜475個の、任意の整数個の連続アミノ酸でよい。「整数」という用語は、本明細書においてその数学的意味で使用され、したがって、代表的な整数には、
が含まれる。
【0069】
本発明の各ポリペプチド断片はまた、N末端位置およびC末端位置の観点から説明することもできる。例えば、約25個の連続アミノ酸から、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、または配列番号:10のポリペプチドのような完全長ポリペプチドよりアミノ酸1個分少ないものまでのN末端断片からC末端断片までの組み合わせが本発明に含まれる。したがって、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、または配列番号:10を例として用いると、25個の連続アミノ酸断片は、次からなる群より選択される、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、または配列番号:10のアミノ酸に対応し得る:
。同様に、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、または配列番号:10の26個の連続アミノ酸と474個(または475個)の連続アミノ酸の間のサイズである他のすべての断片に対応するアミノ酸も、本発明に含まれ、これらの例に基づいて直ちに予想することもできる。したがって、本明細書を不必要に長くするのを避けるために、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、または配列番号:10のポリペプチドの様々な断片を示すその他の例を本明細書で個別に列挙することはしない。
【0070】
配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、または配列番号:10の
個の連続アミノ酸を含むポリペプチド断片は、別法として、式「n〜c」(両端の数字を含む)(「n」はそのポリペプチドのN末端アミノ酸位置に等しく、「c」はC末端アミノ酸位置に等しい)を用いて説明することもできる。本発明のこの態様において、「n」は、下限値が1であり、上限値が完全長ポリペプチドのアミノ酸の総数-24(例えば、配列番号:2、配列番号:4、配列番号:5、または配列番号:8の場合、475−24=451;配列番号:6、配列番号:7、配列番号:9、または配列番号:10の場合、476−24=452)である整数である。「c」は、25と完全長ポリペプチド配列のアミノ酸の数(配列番号:2、配列番号:4、配列番号:5、または配列番号:8の場合、475;配列番号:6、配列番号:7、配列番号:9、または配列番号:10の場合、476)の間の整数であり、「n」は、「c」より少なくとも24少ない整数である。したがって、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、または配列番号:10の場合、「n」は、次からなるリストより選択される任意の整数であり:
;かつ「c」は、次からなる群より選択される任意の整数である:
。ただし、「n」は「c」はより少なくとも24少ない値であることを条件とする。「n」位置および「c」位置のあらゆる組み合わせが、本発明のポリペプチド断片の特定の態様として含まれる。本発明の任意のポリペプチド断片態様を説明するために使用される範囲はすべて、別段の具体的な説明が無い限り、両端の数字を含む(inclusive)。
【0071】
本発明の変異AGPase小サブユニットポリペプチドまたはAGPase大サブユニットポリペプチドの断片は、本明細書において説明するように、タンパク質分解酵素(トリプシン、キモトリプシン、もしくはコラゲナーゼなど)または臭化シアン(CNBr)のような化学試薬を用いて本発明のポリペプチドを切断することによって得ることができる。あるいは、強酸性環境、例えばpH2.5でポリペプチド断片を生成させることもできる。また、ポリペプチド断片は、化学合成によって、または本発明のAGPase大サブユニットポリペプチドの断片もしくは変異AGPase小サブユニットポリペプチドの断片、例えば、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、配列番号:10、配列番号:21、配列番号:22、配列番号:23、配列番号:24、もしくは配列番号:25に示すアミノ酸配列の断片である変異ポリペプチドをコードするポリヌクレオチドを含む発現ベクターで形質転換させた宿主細胞を用いて、調製することもできる。本明細書において同様に企図される本発明の変異AGPase大サブユニットポリペプチドまたは変異AGPase小サブユニットポリペプチドの断片には、そのポリペプチドのトランジット配列またはシグナル配列の全体または一部分が除去されているポリペプチド断片が含まれる。
【0072】
本発明はまた、本発明の変異AGPase小サブユニットポリペプチドをコードする本発明のポリヌクレオチドで形質転換させた細胞にも関する。1つの態様において、細胞は、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、配列番号:10、配列番号:21、配列番号:22、配列番号:23、配列番号:24、もしくは配列番号:25に示すアミノ酸配列またはその機能的な断片もしくはバリアントをコードする配列を含むポリヌクレオチド配列で形質転換される。特定の態様において、細胞は、配列番号:1、配列番号:3、配列番号:11、配列番号:12、配列番号:13、配列番号:14、配列番号:15、もしくは配列番号:16に示すポリヌクレオチド配列、または配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、配列番号:10、配列番号:21、配列番号:22、配列番号:23、配列番号:24、もしくは配列番号:25の機能的な断片もしくはバリアントをコードする配列で形質転換される。1つの態様において、細胞はまた、本明細書において説明する変異AGPase大サブユニットポリペプチドをコードするポリヌクレオチドで形質転換される。1つの態様において、このポリヌクレオチド配列は、本明細書の発現構築物において提供される。形質転換細胞は、原核細胞、例えば、大腸菌もしくは枯草菌(B.subtilis)などの細菌細胞でよく、または、形質転換細胞は、真核細胞、例えば、プロトプラストを含む植物細胞もしくは動物細胞でよい。植物細胞には双子葉植物細胞、単子葉植物細胞、および針葉樹細胞が含まれるが、それらに限定されるわけではない。1つの態様において、植物細胞は、トウモロコシ植物に由来する細胞である。動物細胞にはヒト細胞、哺乳動物細胞、鳥類細胞、および昆虫細胞が含まれる。哺乳動物細胞にはCOS細胞、3T3細胞、およびCHO細胞が含まれるが、それらに限定されるわけではない。
【0073】
本発明はまた、植物または植物組織(例えば植物種子もしくは内乳組織)におけるデンプン合成を増大させるための方法にも関する。1つの態様において、本発明の方法は、本発明の1つまたは複数のポリヌクレオチドを植物中に導入する段階を含む。一定の態様において、植物中に導入されるポリヌクレオチドは、配列番号:2、配列番号:4、配列番号:5、配列番号:6、配列番号:7、配列番号:8、配列番号:9、配列番号:10、配列番号:21、配列番号:22、配列番号:23、配列番号:24、もしくは配列番号:25のいずれかに示すアミノ酸配列またはその断片もしくはバリアントを含む1種または複数種のポリペプチドをコードする。特定の態様において、ポリヌクレオチドは、配列番号:1もしくは配列番号:3に示すヌクレオチド配列またはその断片もしくはバリアントを含む。別の特定の態様において、ポリヌクレオチドは、配列番号:11、配列番号:12、配列番号:13、配列番号:14、配列番号:15、もしくは配列番号:16に示すヌクレオチド配列またはその断片もしくはバリアントを含む。1つの態様において、ポリヌクレオチドは、植物または植物組織のゲノム中に安定に組み込まれる。ポリヌクレオチドは、ポリヌクレオチドおよび/またはそれにコードされるポリペプチドの発現を増大させる調節エレメント、例えばプロモーター配列および/またはエンハンサー配列を含んでよい。特定の態様において、プロモーター配列は、構成的発現または組織特異的(例えば内乳)発現をもたらすものである。このポリヌクレオチドを含む植物もしくは植物組織、またはこれらの植物の子孫は、本発明のポリヌクレオチドまたはポリペプチドの発現の増大について任意でスクリーニングすることができる。1つの態様において、本発明の1つまたは複数のポリヌクレオチドの複数のコピーは、植物または植物組織中に導入され、その植物のゲノム中に安定に組み込まれる。1つの態様において、本発明のポリヌクレオチドは、本明細書において説明する発現構築物において提供される。
【0074】
一文字のアミノ酸略語を表2で定義する。
【0075】
【表2】
【0076】
材料および方法
ランダム変異誘発
PCRランダム変異誘発(GeneMorph II EZClone Domain Mutagenesis Kit, Stratagene)によって、Sh2およびBt2中に変異を導入した。バイアスがかかっていない誤りがちな(error-prone)DNAポリメラーゼの混合物を用いて点変異を導入した。それぞれpMONcSh2およびpMONcBt2(Giroux et al., 1996)中の野生型のSh2コード配列およびBt2コード配列をPCR用の鋳型として使用した。Sh2およびBt2に隣接する2対のプライマー
を誤りがちなPCRのために使用した。Stratagene社のプロトコールに従って、PCRによって作製した変異Sh2クローンをベクターpMONcSh2中にサブクローニングした。次いで、pMONcSh2を用いて、適合性のあるベクターpMONcBt2中に野生型Bt2を含む大腸菌株AC70R1-504を形質転換させた。PCRによって作製した変異Bt2クローンをベクターpMONcBt2中にサブクローニングした。次いで、pMONcBt2を用いて、適合性のあるベクターpMONcSh2中に野生型Sh2を含む大腸菌株AC70R1-504を形質転換させた。
【0077】
細菌発現系
細菌発現系(Iglesias et al., 1993)のおかげで、本発明者らは、トウモロコシ内乳AGPase遺伝子をランダムに変異誘発し、後述するようにヨウ素蒸気にプレートを曝露することによって迅速かつ効率的にAGPase活性を採点することが可能になった。大腸菌系は、Georgelis et al. (2007)において考察されているいくつかの理由から、植物AGPaseを研究するために理想的である。
【0078】
グリコーゲン検出
ヨウ素蒸気への曝露後に茶色に染色されたコロニーを得ることによって、グリコーゲン合成を検出した。75μg/mLのスペクチノマイシン、50μg/mLのカナマイシン、および1%(w/v)グルコースの存在下、37℃で16時間、コーンバーグ(Kornberg)培地上で大腸菌細胞を増殖させた(Govons et al., 1969)。これらのコロニーをヨウ素蒸気に1分間曝露した。不活性なAGPaseを有するコロニーはヨウ素蒸気への曝露後に発色しなかったのに対し、活性なAGPaseはグリコーゲンを産生しており、その結果、ヨウ素による茶色の染色を示した。野生型よりも濃く染色されたAGPaseバリアントを、さらに研究するために選択した。
【0079】
グリコーゲン定量
フェノール反応(HansonおよびPhillips, 1981)を用いてグリコーゲン定量を実施した。手短に言えば、50%(w/v)KOH中で3時間煮沸することによって、2%(w/v)グルコースを含むLB中で増殖させた大腸菌細胞(OD600=2.0)1.6mlからグリコーゲンを抽出した。次いで、70%(v/v)までエタノールを添加し、10000×gで10分間遠心分離することによってグリコーゲンを沈殿させた。沈殿物を乾燥した後に、200μlの脱イオン水、200μlの5%(w/v)フェノール、および1mLの濃硫酸を添加した。488nmでの吸光度に基づいてグリコーゲンを概算した。
【0080】
DNA配列決定
グリコーゲンの産生が増大したSh2変異体およびBt2変異体をフロリダ大学のInterdisciplinary Center for Biotechnology Research(バイオテクノロジー共同研究センター)のGenome Sequencing Services Laboratory (GSSL)によって2回(double-pass)配列決定した。Bioeditソフトウェア(Hall, 1999)によってデータ解析を実施した。
【0081】
AC70R1-504大腸菌細胞からのトウモロコシ内乳AGPaseの精製
トウモロコシ内乳AGPaseを発現するAC70R1-504大腸菌細胞を、75μg/mLのスペクチノマイシン、50μg/mLのカナマイシン、および2%(w/v)グルコースの存在下、37℃で16時間、振盪しながら、2Lのルリアブロス(LB)培地中で増殖させた。OD600=0.6で、0.2mMのイソプロピル-β-D-チオガラクトシド(IPTG)および0.02mg/mLのナリジクス酸を添加してタンパク質発現を誘導した。培養物を直ちに室温に移し、振盪しながら4時間増殖させた。下記の段階を4℃で実施した。3000×gで遠心分離することによって細胞を回収し、沈殿物を16mLの緩衝液A(50mM KH2PO4 pH 7.0、5mM MgCl2、0.5mM EDTA)およびプロテアーゼ阻害剤(1μg/mLのペプスタチン、0.1mMのPMSF、10μg/mLのキモスタチン、および1mMのベンズアミジン)中に再懸濁した。フレンチプレスを用いてこれらの細胞を溶解させ、26000×gで遠心分離した。緩衝液Aを添加することによって、上清のタンパク質濃度を30mg/mLに調整した。10分の3の体積の1%硫酸プロタミンを添加し、混合物を氷上で20分間撹拌し、次いで、26000×gで20分間遠心分離した。上清を硫酸アンモニウムで45%飽和にし、氷上で20分間撹拌し、26000×gで20分間遠心分離した。2〜2.5mLの緩衝液A中に沈殿物を再懸濁した。Boehlein et al. (2005)によって説明されているようにして、強陰イオン交換カラム(macro-prep High Q support, Biorad)およびEcono-pacヒドロキシアパタイトカートリッジ(Biorad)にこの混合物を通した。分析する前にZeba Micro Desalt Spin Columns (Pierce) を用いることによってAGPaseを脱塩した。AGPaseを50mM HEPES、5mM MgCl2、0.5mM EDTA、および0.5mg/mL BSA中に移した(exchanged)(安定性のため)。
【0082】
AGPaseの動力学的特徴付け
ATPおよびG-1-Pのkcat、Km、ならびに3-PGAおよびPiの親和性を推定するために、反応の順方向を使用した(G-1-P+ATP→ADP-グルコース+PPi)。より具体的には、総体積200μlの50mM HEPES pH 7.4、15mM MgCl2、1.0mM ATP、および2.0mM G-1-Pにおいて、37℃で10分間、0.04〜0.06μgの精製したAGPaseの比活性を分析した。ATPおよびG-1-PのKmならびに3-PGAのKaを決定するために、それぞれ様々な量のATP、G-1-P、および3-PGAを用いた。様々な量のPi、1mM ATP、2mM G-1-P、および2.5mM 3-PGAを添加することによって、PiのKiを概算した。2分間煮沸することによって反応を停止させた。カップリング試薬300μlを用いてNADH濃度の低下とこの反応を関連付けることによってPPiを概算した。カップリング試薬は、25mMイミダゾールpH 7.4、4mM MgCl2、1mM EDTA、0.2mM NADH、0.725Uアルドラーゼ、0.4Uトリオースリン酸イソメラーゼ、0.6Uグリセロリン酸デヒドロゲナーゼ、1mMフルクトース-6-リン酸、および0.8μgのピロリン酸依存性ホスホフルクトキナーゼ(PPi-PFK)を含んだ。これらの酵素はPPi-PFK以外はすべてSigma社から購入した。PPi-PFKは、Deng et al. 1999に従い、いくつかの改良を加えて作製し精製した(BoehleinおよびHannah、未発表データ)。340nmでの吸光度に基づいてNADH濃度を概算した。PPi濃度は、AGPaseの代わりに様々な量のPPiを用いることによって作成した検量線を用いて算出した。AGPaseによって産生されるPPiの量は、時間および酵素濃度に比例した。Prism 4.0 (Graph Pad, San Diego CA)によってAGPaseの動力学的定数を算出した。
【0083】
粗製タンパク質抽出物または部分的に精製したタンパク質抽出物に由来するAGPaseの比活性の測定
トウモロコシ内乳AGPaseを発現するAC70R1-504大腸菌細胞を、75μg/mLのスペクチノマイシン、50μg/mLのカナマイシン、および2%(w/v)グルコースの存在下、37℃で振盪しながら、OD600=2.0になるまで2Lのルリアブロス(LB)培地中で増殖させた。遺伝子発現は誘導しなかった。下記の段階を4℃で実施した。3000×gで遠心分離することによって細胞を回収し、沈殿物を16mLの緩衝液A(50mM KH2PO4 pH 7.0、5mM MgCl2、0.5mM EDTA)およびプロテアーゼ阻害剤(1μg/mLのペプスタチン、0.1mMのPMSF、10μg/mLのキモスタチン、および1mMのベンズアミジン)中に再懸濁した。フレンチプレスを用いてこれらの細胞を溶解させ、26000×gで遠心分離した。-80℃で保存した上清を用いて、粗製抽出物のAGPase活性を測定した。前述のようにして硫酸プロタミンおよび硫酸アンモニウムを用いて、上清の残りを部分的に精製した。アッセイ法の前にBoehlein et al. (2005)によって説明されているようにしてタンパク質抽出物を脱塩し、50mM HEPES、5mM MgCl2、および0.5mM EDTA中に移した。Boehlein et al. (2005)によって説明されているようにして、飽和量の基質およびアクチベーターを用いて、逆方向(ADP-グルコース+PPi→G-1-P+ATP)におけるAGPase比活性をモニターした。
【0084】
AGPaseの熱安定性の測定
AGPaseを前述のようにして精製した。50mM HEPES、5mM MgCl2、0.5mM EDTA、および0.5mg/mL BSA中でAGPaseをさらに1/100(v/v)希釈し、様々な時間に渡って42℃または53℃で熱処理し、次いで氷上で冷却した。飽和量のATP、G-1-P、および3-PGAを用いて、処理後に残っている活性を順方向および逆方向においてモニターした。熱処理の時間に対する残存活性率(%)の対数としてデータをプロットした。式t1/2=0.693/(-2.3×傾き)を用いて不活性化定数t1/2を算出した。
【0085】
AGPase純度の定性的測定
AGPaseの純度を以下のようにしてモニターした。6μgのAGPaseを変性溶液(100mM Tris-Cl pH 6.8、4% SDS、および8mM DTT)中で1:1希釈し、95℃で5分間加熱し、150Vで1時間、5% SDSポリアクリルアミドゲル上で電気泳動させ、クーマシーブリリアントブルー染色によって可視化した(Laemmli, 1970)。
【0086】
粗製抽出物のタンパク質ブロット解析
Hybri-Dotブロット装置(Life Technologies)を用いて、試料をPVDF膜(Biorad)上に真空ブロットした。PVDF膜はメタノール中で5分間、次いでトランスファー緩衝液[20%(v/v)メタノール、0.303%(w/v)Trisおよび1.44%(w/v)グリシン]中で10分間、予め浸しておいた。絶えず振盪しながら1時間、ブロッキング緩衝液[0.8%(w/v)NaCl、0.02%(w/v)KCl、0.144%(w/v)Na2HPO4、0.024%(w/v)KH2PO4、5%(w/v)ウシ血清アルブミン(BSA)、および0.05%(v/v)Tween-20]と共に膜をインキュベートした。振盪しながら1時間、BT2に対する1:10000(v/v)のモノクローナル抗体(Sue Boehleinの好意で提供された)を含むブロッキング緩衝液と共にブロットをインキュベートした。次いで、絶えず振盪しながら、そのブロットを洗浄緩衝液(ブロッキング緩衝液−BSA)で10分間×3回洗浄した。次いで、ホースラディッシュペルオキシダーゼと結合されたヤギ抗マウス二次抗体(Pierce)の1:60000希釈物と共に45分間、ブロットをインキュベートした。最後に、ブロットを10分間×3回洗浄した。増感された化学発光基質キット(Pierce)を用いてタンパク質を可視化した。
【0087】
3Dモデリング
最近公開されたジャガイモ塊茎ホモ四量体AGPase(RCSBタンパク質データバンク番号:1YP2)の三次構造におけるジャガイモ小ユニットにならって、BT2モノマーの構造のモデルを作った。SWISS MODELを用いてホモロジーモデリングを行った(Peitsch, 1995; GuexおよびPeitsch, 1997; Schwede et al. 2003; KoppおよびSchwede, 2004; Arnold et al, 2006)。Jmol、すなわちJavaによるオープンソースの三次元化学構造表示ソフト(open-source Java viewer for chemical structures in 3D)(http://www.jmol.org/)を用いて、アミノ酸462(ThrまたはIle)接触残基を決定した。
【0088】
酵母ツーハイブリッド
GreeneおよびHannah(1998b)によって説明されているようにして、酵母形質転換およびβ-ガラクトシダーゼアッセイ法を実施した。唯一変更した点は、ベイトおよびプレイ用のベクターとしてそれぞれpGBKT7およびpGADT7を使用したことであった。pGBKT7-53プラスミドおよびpGADT7-Tプラスミドを陽性対照として使用した。
【0089】
図面および表をすべて含む、本明細書において参照または引用される特許、特許出願、仮出願、および刊行物はすべて、本明細書の明確な教示と矛盾しない範囲で、そっくりそのまま参照により組み入れられる。
【0090】
以下は、本発明を実践するための手順を例示する実施例である。これらの実施例は、限定として解釈すべきではない。別段の注記の無い限り、パーセンテージはすべて重量に基づき、溶媒の混合比率はすべて体積に基づく。
【実施例】
【0091】
実施例1
誤りがちなPCRによって、変異Bt2ライブラリーを作り出した。突然変異荷重は、クローン1個当たり〜2個の非同義変異であった(Georgelis et al. 2007)。これらの変異体を野生型Sh2遺伝子と共に大腸菌において発現させた。約50,000個のコロニーをグリコーゲン産生についてスクリーニングした。濃く染色された10個のコロニーを選んだ。染色が最も濃いBt2変異体2つを配列決定した。両方とも、トレオニンからイソロイシンへのアミノ酸462の変化をもたらす同じ非同義変異(TI)を有していた。この位置のトレオニンは、高等植物小サブユニット間で絶対的に保存されている(データ不掲載)。BT2-TI/SH2(TI変異を含み、SH2と複合体を形成したBT2)は、BT2/SH2よりも多量のグリコーゲンを産生した(図1)。ホモ四量体としてBT2-TIおよびBT2を発現する細胞は、検出可能な量のグリコーゲンを産生しなかった(図1)。このことから、大腸菌によって合成されるグリコーゲンの量が、BT2-TIまたはBT2とSH2との複合体にもっぱら依存することが示される。
【0092】
BT2/SH2およびBT2-TI/SH2を発現する細胞に由来する粗製抽出物のドットブロットから、BT2-TIが大腸菌において多量に存在することが示された(図2A〜2B)。BT2/SH2およびBT2-TI/SH2を発現する誘導されていない細胞に由来する粗製抽出物のAGPase活性レベルは低すぎて検出できなかったのに対し、BT2-TI/SH2由来の部分的に精製した抽出物は、BT2/SH2由来の部分的に精製した抽出物よりも20倍高い活性を有していた(図3)。より効率的な転写/翻訳が理由となってBT2-TI/SHの方が多くのタンパク質および高い活性をもたらしたという可能性は存在しにくい。なぜならば、ACA(T)〜ATA(I)のコドンは大腸菌では同じ頻度で使用されているからである(それぞれ6.1%および5.0%)(Nakamura et al., 2000)。このことから、BT2-TI/SH2細胞の方がタンパク質が多量であり活性が高いのは、AGPaseの安定性の上昇に起因することが示唆される。
【0093】
BT2-TI/SH2の動力学的諸特性および熱安定性を決定し、大腸菌におけるグリコーゲン合成の増大の原因を解明するために、組換えBT2-TI/SH2 AGPaseおよびBT2/SH2 AGPaseを精製した(図4)。表3に要約するBT2-TI/SH2の動力学的諸特性から、G-1-PおよびATPのKm、3-PGAのKa、およびPiのKiがBT2/SH2の場合と区別不能であったことが示される。驚くべきことに、BT2-TIのkcatはBT2/SHのkcatより30%小さい。したがって、これらの動力学的諸特性では、大腸菌においてBT2-TI/SH2の方が染色が濃いことを説明することができない。しかしながら、BT2-TI/SH2がBT2/SH2よりも熱安定性であることは明らかである(図5A〜5B)。これらの結果から、BT2-TI/SH2の高い熱安定性が、大腸菌におけるグリコーゲンの量の増加の原因であることが強く示唆される。
【0094】
MPは、農学的利益をもたらし得る小サブユニットバリアントである。その特徴のいくつかには、BT2/SH2と比べて、アクチベーター3-PGAの不在下での増大した活性、3-PGAに対する増大した親和性、Piに対する減少した親和性(表3)、および上昇した熱安定性が含まれる(図3)(Cross et al., 2004; Boehlein et al., 2005)。BT2-TI/SH2はMP/SH2ほど熱安定性ではなかったため(図3)、MPの熱安定性をさらに高めるために、アミノ酸変化TIをMPに導入した(MP-TI)。
【0095】
MP-TI/SH2(TI変異を有し、SH2と複合体を形成したMP)を発現する細胞は、MP/SH2を発現する細胞と同じ量のグリコーゲンを産生した(図1)。しかしながら、BT2と比べてより多量のMP-TIタンパク質が、SH2を有する2種のタンパク質を発現する大腸菌の粗製抽出物で存在していた(図2A〜2B)。MP-TI/SH2に由来する粗製抽出物および部分的に精製した抽出物の活性は、MP/SH2の活性より2〜3倍高かった(図3)。純粋な形態のMP-TI/SH2(図4)は、MP/SH2と比べてkcatが〜30%減少した以外は、MP/SH2の好ましい動力学的諸特性を維持していた(表3)。さらに、MP-TI/SH2は、MP/SH2よりも高い熱安定性を示す(図6A〜6B)。
【0096】
トウモロコシ内乳AGPaseの結晶構造は解明されていない。唯一の関連する構造体は、ジャガイモ塊茎小サブユニットホモ四量体である(Jin et al., 2005)。ジャガイモ塊茎小サブユニットは、BT2に対して88%の同一性および96%の類似性を示す。解明されたジャガイモ塊茎小サブユニットの構造にならって、BT2モノマーの構造のモデルを作った(図7A)。TI中で変異した残基はβ-ヘリックスの一部分であり、これは、小サブユニットのN末端の2つの残基(Pro、Leu)との疎水的接触を作り出す(図7B)。TIにおいてThrからIleにアミノ酸が変化すると、前述のProおよびLeuからの距離が短くなる(図7C)。TI変異が小サブユニットのC末端とN末端の間の疎水性相互作用を強化し、その結果、より高い安定性をもたらすと推測したくなる。サブユニット同士の境界の/近くの残基に熱安定性が帰されるMPとは異なって、TIはサブユニット同士の境界から離れているため、サブユニット間の相互作用に直接影響を及ぼすことはできない(図7A)。
【0097】
TIがサブユニット間の相互作用の強度に影響を及ぼすかどうかを判定するために、酵母ツーハイブリッドシステム(Y2H)においてSH2をベイトとして使用し、BT2、TI、およびMPをプレイとして使用した。定量的β-ガラクトシダーゼアッセイ法により、MPとは対照的に、TIはサブユニット間の相互作用の強度を上昇させないことが示された(図8)。
【0098】
最後に、セリン、アラニン、およびグリシンなど側鎖の短いアミノ酸で元のトレオニンを置換しても、kcatは10分の1またはそれ以下に減少したにもかかわらず(データ不掲載)、AGPaseの熱安定性は影響を及ぼされなかった。このことから、462位の元のトレオニンはAGPase活性にとっては重要であるが熱安定性にとっては重要でないことが示唆される。
【0099】
実施例2
本発明は、ランダム変異誘発および異種細菌発現系を用いることによって、農学的に重要な植物AGPaseバリアントを提供する。SH2と共に発現された場合に大腸菌細胞によるグリコーゲン産生量を増加させる小サブユニットバリアントとしてBT2-TIを単離した。BT2-TI/SH2を発現する細胞は、BT2/SH2を発現する細胞よりも20倍高いAGPase活性を有していた。ドットブロットにより、BT2-TI/SH2を発現する細胞に由来する粗製タンパク質抽出物の方が、BT2/SH2を発現する細胞と比べて検出可能なBT2タンパク質を有することが示された。この結果は、より効率的な転写/翻訳またはより高いAGPase安定性および/もしくは溶解度に帰され得る。先に言及したように、より効率的な転写/翻訳は、コドン使用に基づくと、見込みは少ない。その一方で、精製型のBT2-TI/SH2は、精製型のBT2/SH2より有意に熱安定性が高いことが示された。このことにより、大腸菌においてBT2-TI/SH2複合体がBT2/SH2と比べてタンパク質分解および/または凝集を起こしにくくなる可能性がある。
【0100】
BT2-TI/SH2の動力学的諸特性およびアロステリック特性は、kcatが30%小さかった以外は、BT2/SH2と区別不能であった。BT2-TI/SH2発現細胞のAGPase活性が20倍上昇していることは、活性なAGPase分子の数がBT2/SH2発現細胞と比べて多いことと解釈される。これはまた、BT2/SH2において実際に機能しているのは潜在的AGPase分子の5%未満であることも意味する。
【0101】
ジャガイモ塊茎小サブユニットは、極めて多量のアクチベーター3-PGAを与えられた場合に、顕著な活性を有するホモ四量体を形成できることが報告されている(Ballicora et al., 1995)。本明細書において示すように、ホモ四量体としてBT2およびBT2-TIを発現する大腸菌細胞は、検出可能な量のグリコーゲンを産生しない。したがって、BT2-TI/SH2を発現する細胞において観察された、BT2/SH2を発現する細胞と比べて量の多いグリコーゲンは、BT2-TIホモ四量体ではなく、BT2-TIとSH2の複合体に起因する。
【0102】
SH2との複合体において熱安定性の上昇をもたらす別の小サブユニットバリアントはMPである。MPによって与えられる熱安定性は、サブユニット同士が相互作用する境界の近くまたは境界の残基にマッピングされている(Boehlein, Shaw, Stewartおよび Hannah, 未発表データ)。一方、BT2-TIのアミノ酸変化はこれらの境界から遠い。構造モデリングから、TI変化によってC末端とN末端の間のサブユニット内疎水性相互作用が強化されることが示唆される。Y2Hの結果は、MPと対照的にTIはサブユニット間の相互作用を強化しないという考えを裏付ける。しかしながら、立体構造的変化を通じてTIがサブユニット間の相互作用に間接的に影響を及ぼす可能性も切り捨てることはできない。Y2Hが、30℃という酵母増殖温度でのTI/SH2およびBT2/SH2におけるサブユニット間の相互作用の違いを明らかにするのに十分な感度を持ち得ないという可能性もある。
【0103】
純粋なBT2-TI/SH2およびMP/SH2の熱安定性の比較から、BT2-TI/SH2はMP/SH2ほど熱安定性ではないことが示された。さらに、MP/SH2はBT2-TI/SH2には無いいくつかの利点を有する。例えば、BT2/SH2と比べて、アクチベーター3-PGAの不在下での活性、3-PGAに対する高い親和性、阻害剤Piに対する低い親和性、および大きなkcatなどである。TI変化をMPに導入することによってMP/SH2の熱安定性がさらに改善され得るかどうかを調査した。結果として生じるバリアントMP-TIは、大腸菌においてSH2と共に発現させた場合、MP/SH2と等量のグリコーゲンを産生した。これは、MP-TI/SH2がMP/SH2以上に熱安定性ではないこと、またはAGPaseによって触媒されるADP-グルコース産生が大腸菌においてもはや律速的(limiting)ではないことのいずれかを意味し得る。MP-TI/SH2を発現する細胞の粗製抽出物および部分的に精製した抽出物におけるAGPase活性は、MP/SH2を発現する細胞より2〜3倍高かったため、後者の解釈が好まれ得る。MP-TI/SH2は、30%小さいkcat以外は、MP/SH2の動力学的諸特性およびアロステリック特性のすべてを維持していた。より重要なことには、MP-TI/SH2は、MP/SH2よりも熱安定性であった。MP-TI/SH2およびMP/SH2の両方において二相の熱安定性が観察された。これらの相は、おそらくは、AGPaseの異なる状態の結果であった。熱安定性のこの二相性の様式は、以前に観察されており、MP/SH2にもMP-TIにも特異的ではない(Boehlein et al. 2008)。第1の相は、第2の相よりも低い熱安定性を示す。MP-TI/SH2は両方の相においてMP/SH2と比べて熱安定性が高い。しかしながら、各相においてAGPaseの状態が正確にどのようであるかは、依然として謎である。熱安定性の二相性の様式は、BT2/SH2およびBT2-TI/SH2では観察されなかった。これは、これらの試料が第2の相に到達するのに十分な長さの期間加熱されず、MP/SH2およびMP-TI/SH2より低い温度(53℃の代わりに42℃)で加熱されたためである。
【0104】
表3は、精製された組換えAGPaseバリアントの動力学的諸特性を示す。AGPaseバリアントの動力学的特性およびアロステリック特性を順方向で決定した。G-1-Pの量を変更し、飽和量(1mM)のATPを維持することによって、kcat(s-1)(G-1-P)を概算した。ATPの量を変更し、飽和量(2mM)のG-1-Pを維持することによって、kcat(s-1)(ATP)を概算した。Kiの値(Ki's)は、平均値(95%信頼区間)として表している。他の値はすべて、平均値±標準偏差として表している。3-PGAの不在下でのAGPaseの比活性を、10mMの3-PGAの存在下での比活性に対する比率(%)として表している(平均値±標準誤差)。
【0105】
【表3】
【0106】
本明細書において説明される実施例および態様は、例示するためのものにすぎないこと、ならびにそれらを踏まえた様々な修正または変更が当業者に示唆され、かつそれらが本出願の精神および範囲内、ならびに添付の特許請求の範囲内に含まれ得ることが理解されるべきである。さらに、本明細書において開示される任意の発明またはその態様の任意の要素または制限は、本明細書において開示される他の任意の発明またはその態様の任意および/または他のすべての要素または制限と(個別または任意の組み合わせで)組み合わせられてよく、このような組み合わせはすべて、それらに限定されることなく、本発明の範囲内で企図される。
【0107】
参考文献
【特許請求の範囲】
【請求項1】
変異植物AGPase小サブユニットタンパク質または該タンパク質の機能的断片をコードするポリヌクレオチドであって、
前記タンパク質が、
野生型トウモロコシ内乳AGPase小サブユニットタンパク質の462位のトレオニンアミノ酸に対応するアミノ酸が、変異AGPase小サブユニットが発現されてAGPase酵素を形成した場合に増大した熱安定性を与えるアミノ酸によって置換されている、アミノ酸変異
を含む、
前記ポリヌクレオチド。
【請求項2】
増大した熱安定性を与える置換アミノ酸が、イソロイシンである、請求項1記載のポリヌクレオチド。
【請求項3】
変異AGPase小サブユニットがトウモロコシ内乳AGPase小サブユニットである、請求項1または2記載のポリヌクレオチド。
【請求項4】
ポリヌクレオチドにコードされる変異植物AGPase小サブユニットタンパク質が、配列番号:2、配列番号:5、配列番号:6、配列番号:7、配列番号:21、配列番号:22、配列番号:23、配列番号:24、もしくは配列番号:25のいずれかに示すアミノ酸配列またはその機能的断片を含む、請求項1記載のポリヌクレオチド。
【請求項5】
配列番号:1、配列番号:11、配列番号:12、もしくは配列番号:13のいずれかに示すヌクレオチド配列またはその機能的断片を含む、請求項1記載のポリヌクレオチド。
【請求項6】
発現構築物において提供される、請求項1記載のポリヌクレオチド。
【請求項7】
キメラ植物AGPase小サブユニットタンパク質または該タンパク質の機能的断片をコードするポリヌクレオチドであって、
前記キメラAGPaseタンパク質が、
第1の植物に由来する植物AGPase小サブユニットのN末端領域に由来するN末端配列、および第2の植物に由来する植物AGPase小サブユニットのC末端領域に由来するC末端配列
を含み、かつ
前記キメラ植物AGPase小サブユニットタンパク質が、
野生型トウモロコシ内乳AGPase小サブユニットタンパク質の462位のトレオニンアミノ酸に対応するアミノ酸が、変異AGPase小サブユニットが発現されてAGPase酵素を形成した場合に増大した熱安定性を与えるアミノ酸によって置換されている、アミノ酸変異
を含む、
前記ポリヌクレオチド。
【請求項8】
N末端配列が、第1の植物に由来するAGPaseサブユニットのN末端領域の最初の150〜250個のアミノ酸を含み、かつC末端配列が、第2の植物に由来するAGPaseサブユニットのC末端領域の最後の300個またはそれ以下の残基を含む、請求項7記載のポリヌクレオチド。
【請求項9】
増大した熱安定性を与える置換アミノ酸が、イソロイシンである、請求項7記載のポリヌクレオチド。
【請求項10】
N末端領域がトウモロコシ内乳のAGPase小サブユニットに由来する、請求項7〜9のいずれか一項記載のポリヌクレオチド。
【請求項11】
C末端領域がジャガイモ塊茎のAGPase小サブユニットに由来する、請求項7〜9のいずれか一項記載のポリヌクレオチド。
【請求項12】
ポリヌクレオチドにコードされる植物AGPase小サブユニットタンパク質が、配列番号:4、配列番号:8、配列番号:9、もしくは配列番号:10のいずれかに示すアミノ酸配列またはその機能的断片を含む、請求項7記載のポリヌクレオチド。
【請求項13】
配列番号:3、配列番号:14、配列番号:15、もしくは配列番号:16のいずれかに示すヌクレオチド配列またはその機能的断片を含む、請求項7記載のポリヌクレオチド。
【請求項14】
発現構築物において提供される、請求項7〜13のいずれか一項記載のポリヌクレオチド。
【請求項15】
以下によってコードされるポリペプチド:
a)請求項1〜6のいずれか一項記載のポリヌクレオチド;または
b)請求項7〜14のいずれか一項記載のポリヌクレオチド。
【請求項16】
以下を含む、形質転換されたかまたはトランスジェニックである植物または植物組織もしくは植物細胞:
a)請求項1〜6のいずれか一項記載のポリヌクレオチド;もしくは
b)請求項7〜14のいずれか一項記載のポリヌクレオチド;または
c)a)およびb)の両方のポリヌクレオチド。
【請求項17】
トウモロコシAGPase大サブユニットもまた発現する、請求項16記載の植物または植物組織もしくは植物細胞。
【請求項18】
AGPaseの変異大サブユニットを発現する植物または植物組織もしくは植物細胞であって、前記変異大サブユニットが、増大した熱安定性を与える変異を含み、かつ/または前記変異大サブユニットが、増加した種子重量を与える変異を含む、請求項16記載の植物または植物組織もしくは植物細胞。
【請求項19】
変異大サブユニットがRev6変異を含む、請求項16記載の植物または植物組織もしくは植物細胞。
【請求項20】
単子葉植物のものである、請求項16記載の植物または植物組織もしくは植物細胞。
【請求項21】
単子葉の植物または植物組織もしくは植物細胞が、イネ、コムギ、オオムギ、エンバク、ライムギ、モロコシ、トウモロコシ、サトウキビ、パイナップル、タマネギ、バナナ、ココナツ、ユリ、芝草(turfgrass)、およびキビ(millet)からなる群より選択される、請求項20記載の植物または植物組織もしくは植物細胞。
【請求項22】
植物がトウモロコシ(Zea mays)であるか、または植物組織もしくは植物細胞がトウモロコシに由来する、請求項16記載の植物または植物組織もしくは植物細胞。
【請求項23】
双子葉植物のものである、請求項16記載の植物または植物組織もしくは植物細胞。
【請求項24】
双子葉の植物または植物組織もしくは植物細胞が、トマト、キュウリ、カボチャ、エンドウマメ、アルファルファ、メロン、ヒヨコマメ、チコリ、クローバー、ケール、レンズマメ、ダイズ、マメ、タバコ、ジャガイモ、サツマイモ、ヤムイモ、カッサバ、ラディッシュ、ブロッコリー、ホウレンソウ、キャベツ、セイヨウアブラナ、リンゴの木、柑橘類(オレンジ、マンダリン、グレープフルーツ、レモン、およびライムなどを含む)、ブドウ、綿、ヒマワリ、イチゴ、レタス、およびホップからなる群より選択される、請求項23記載の植物または植物組織もしくは植物細胞。
【請求項25】
植物組織が、種子、接ぎ穂、または台木である、請求項16記載の植物または植物組織もしくは植物細胞。
【請求項26】
請求項15記載の1つまたは複数の変異ポリペプチドを含む、変異植物AGPase酵素。
【請求項27】
植物のゲノム中に1つまたは複数のポリヌクレオチドを組み入れる段階、および前記ポリヌクレオチドにコードされるタンパク質を発現させる段階を含む、熱ストレス条件に対する植物の耐性を増大させ、かつ/または植物のデンプン生合成を増大させる方法であって、前記ポリヌクレオチドが、以下であるか、または以下を含む、前記方法:
a)請求項1〜6のいずれか一項記載の任意のポリヌクレオチド;および/または
b)請求項7〜14のいずれか一項記載の任意のポリヌクレオチド。
【請求項28】
植物がトウモロコシAGPase大サブユニットもまた発現する、請求項27記載の方法。
【請求項29】
植物が、AGPaseの変異大サブユニットを発現し、該変異大サブユニットが、増大した熱安定性を与える変異を含み、かつ/または該変異大サブユニットが、増加した種子重量を与える変異を含む、請求項27記載の方法。
【請求項30】
変異大サブユニットがRev6変異を含む、請求項27記載の方法。
【請求項31】
植物が単子葉植物である、請求項27記載の方法。
【請求項32】
単子葉植物が、イネ、コムギ、オオムギ、エンバク、ライムギ、モロコシ、トウモロコシ、サトウキビ、パイナップル、タマネギ、バナナ、ココナツ、ユリ、芝草、およびキビからなる群より選択される、請求項31記載の方法。
【請求項33】
植物がトウモロコシである、請求項27記載の方法。
【請求項34】
植物が双子葉植物である、請求項27記載の方法。
【請求項35】
双子葉植物が、トマト、キュウリ、カボチャ、エンドウマメ、アルファルファ、メロン、ヒヨコマメ、チコリ、クローバー、ケール、レンズマメ、ダイズ、マメ、タバコ、ジャガイモ、サツマイモ、ヤムイモ、カッサバ、ラディッシュ、ブロッコリー、ホウレンソウ、キャベツ、セイヨウアブラナ、リンゴの木、柑橘類(オレンジ、マンダリン、グレープフルーツ、レモン、およびライムなどを含む)、ブドウ、綿、ヒマワリ、イチゴ、レタス、およびホップからなる群より選択される、請求項34記載の方法。
【請求項36】
植物細胞中に1つまたは複数のポリヌクレオチドを導入する段階、および該植物細胞から植物を成長させる段階を含む、野生型AGPase酵素を有する植物と比べて増大した熱安定性および/または増大したデンプン生合成を植物において提供するAGPase酵素を有する植物を調製するための方法であって、前記ポリヌクレオチドが、以下であるか、または以下を含む、前記方法:
a)請求項1〜6のいずれか一項記載の任意のポリヌクレオチド;および/または
b)請求項7〜14のいずれか一項記載の任意のポリヌクレオチド。
【請求項37】
植物がトウモロコシAGPase大サブユニットもまた発現する、請求項36記載の方法。
【請求項38】
植物が、AGPaseの変異大サブユニットを発現し、該変異大サブユニットが、増大した熱安定性を与える変異を含み、かつ/または該変異大サブユニットが、増加した種子重量を与える変異を含む、請求項36記載の方法。
【請求項39】
変異大サブユニットがRev6変異を含む、請求項36記載の方法。
【請求項40】
植物が単子葉植物である、請求項36記載の方法。
【請求項41】
単子葉植物が、イネ、コムギ、オオムギ、エンバク、ライムギ、モロコシ、トウモロコシ、サトウキビ、パイナップル、タマネギ、バナナ、ココナツ、ユリ、芝草、およびキビからなる群より選択される、請求項40記載の方法。
【請求項42】
植物がトウモロコシである、請求項36記載の方法。
【請求項43】
植物が双子葉植物である、請求項36記載の方法。
【請求項44】
双子葉植物が、トマト、キュウリ、カボチャ、エンドウマメ、アルファルファ、メロン、ヒヨコマメ、チコリ、クローバー、ケール、レンズマメ、ダイズ、マメ、タバコ、ジャガイモ、サツマイモ、ヤムイモ、カッサバ、ラディッシュ、ブロッコリー、ホウレンソウ、キャベツ、セイヨウアブラナ、リンゴの木、柑橘類(オレンジ、マンダリン、グレープフルーツ、レモン、およびライムなどを含む)、ブドウ、綿、ヒマワリ、イチゴ、レタス、およびホップからなる群より選択される、請求項43記載の方法。
【請求項1】
変異植物AGPase小サブユニットタンパク質または該タンパク質の機能的断片をコードするポリヌクレオチドであって、
前記タンパク質が、
野生型トウモロコシ内乳AGPase小サブユニットタンパク質の462位のトレオニンアミノ酸に対応するアミノ酸が、変異AGPase小サブユニットが発現されてAGPase酵素を形成した場合に増大した熱安定性を与えるアミノ酸によって置換されている、アミノ酸変異
を含む、
前記ポリヌクレオチド。
【請求項2】
増大した熱安定性を与える置換アミノ酸が、イソロイシンである、請求項1記載のポリヌクレオチド。
【請求項3】
変異AGPase小サブユニットがトウモロコシ内乳AGPase小サブユニットである、請求項1または2記載のポリヌクレオチド。
【請求項4】
ポリヌクレオチドにコードされる変異植物AGPase小サブユニットタンパク質が、配列番号:2、配列番号:5、配列番号:6、配列番号:7、配列番号:21、配列番号:22、配列番号:23、配列番号:24、もしくは配列番号:25のいずれかに示すアミノ酸配列またはその機能的断片を含む、請求項1記載のポリヌクレオチド。
【請求項5】
配列番号:1、配列番号:11、配列番号:12、もしくは配列番号:13のいずれかに示すヌクレオチド配列またはその機能的断片を含む、請求項1記載のポリヌクレオチド。
【請求項6】
発現構築物において提供される、請求項1記載のポリヌクレオチド。
【請求項7】
キメラ植物AGPase小サブユニットタンパク質または該タンパク質の機能的断片をコードするポリヌクレオチドであって、
前記キメラAGPaseタンパク質が、
第1の植物に由来する植物AGPase小サブユニットのN末端領域に由来するN末端配列、および第2の植物に由来する植物AGPase小サブユニットのC末端領域に由来するC末端配列
を含み、かつ
前記キメラ植物AGPase小サブユニットタンパク質が、
野生型トウモロコシ内乳AGPase小サブユニットタンパク質の462位のトレオニンアミノ酸に対応するアミノ酸が、変異AGPase小サブユニットが発現されてAGPase酵素を形成した場合に増大した熱安定性を与えるアミノ酸によって置換されている、アミノ酸変異
を含む、
前記ポリヌクレオチド。
【請求項8】
N末端配列が、第1の植物に由来するAGPaseサブユニットのN末端領域の最初の150〜250個のアミノ酸を含み、かつC末端配列が、第2の植物に由来するAGPaseサブユニットのC末端領域の最後の300個またはそれ以下の残基を含む、請求項7記載のポリヌクレオチド。
【請求項9】
増大した熱安定性を与える置換アミノ酸が、イソロイシンである、請求項7記載のポリヌクレオチド。
【請求項10】
N末端領域がトウモロコシ内乳のAGPase小サブユニットに由来する、請求項7〜9のいずれか一項記載のポリヌクレオチド。
【請求項11】
C末端領域がジャガイモ塊茎のAGPase小サブユニットに由来する、請求項7〜9のいずれか一項記載のポリヌクレオチド。
【請求項12】
ポリヌクレオチドにコードされる植物AGPase小サブユニットタンパク質が、配列番号:4、配列番号:8、配列番号:9、もしくは配列番号:10のいずれかに示すアミノ酸配列またはその機能的断片を含む、請求項7記載のポリヌクレオチド。
【請求項13】
配列番号:3、配列番号:14、配列番号:15、もしくは配列番号:16のいずれかに示すヌクレオチド配列またはその機能的断片を含む、請求項7記載のポリヌクレオチド。
【請求項14】
発現構築物において提供される、請求項7〜13のいずれか一項記載のポリヌクレオチド。
【請求項15】
以下によってコードされるポリペプチド:
a)請求項1〜6のいずれか一項記載のポリヌクレオチド;または
b)請求項7〜14のいずれか一項記載のポリヌクレオチド。
【請求項16】
以下を含む、形質転換されたかまたはトランスジェニックである植物または植物組織もしくは植物細胞:
a)請求項1〜6のいずれか一項記載のポリヌクレオチド;もしくは
b)請求項7〜14のいずれか一項記載のポリヌクレオチド;または
c)a)およびb)の両方のポリヌクレオチド。
【請求項17】
トウモロコシAGPase大サブユニットもまた発現する、請求項16記載の植物または植物組織もしくは植物細胞。
【請求項18】
AGPaseの変異大サブユニットを発現する植物または植物組織もしくは植物細胞であって、前記変異大サブユニットが、増大した熱安定性を与える変異を含み、かつ/または前記変異大サブユニットが、増加した種子重量を与える変異を含む、請求項16記載の植物または植物組織もしくは植物細胞。
【請求項19】
変異大サブユニットがRev6変異を含む、請求項16記載の植物または植物組織もしくは植物細胞。
【請求項20】
単子葉植物のものである、請求項16記載の植物または植物組織もしくは植物細胞。
【請求項21】
単子葉の植物または植物組織もしくは植物細胞が、イネ、コムギ、オオムギ、エンバク、ライムギ、モロコシ、トウモロコシ、サトウキビ、パイナップル、タマネギ、バナナ、ココナツ、ユリ、芝草(turfgrass)、およびキビ(millet)からなる群より選択される、請求項20記載の植物または植物組織もしくは植物細胞。
【請求項22】
植物がトウモロコシ(Zea mays)であるか、または植物組織もしくは植物細胞がトウモロコシに由来する、請求項16記載の植物または植物組織もしくは植物細胞。
【請求項23】
双子葉植物のものである、請求項16記載の植物または植物組織もしくは植物細胞。
【請求項24】
双子葉の植物または植物組織もしくは植物細胞が、トマト、キュウリ、カボチャ、エンドウマメ、アルファルファ、メロン、ヒヨコマメ、チコリ、クローバー、ケール、レンズマメ、ダイズ、マメ、タバコ、ジャガイモ、サツマイモ、ヤムイモ、カッサバ、ラディッシュ、ブロッコリー、ホウレンソウ、キャベツ、セイヨウアブラナ、リンゴの木、柑橘類(オレンジ、マンダリン、グレープフルーツ、レモン、およびライムなどを含む)、ブドウ、綿、ヒマワリ、イチゴ、レタス、およびホップからなる群より選択される、請求項23記載の植物または植物組織もしくは植物細胞。
【請求項25】
植物組織が、種子、接ぎ穂、または台木である、請求項16記載の植物または植物組織もしくは植物細胞。
【請求項26】
請求項15記載の1つまたは複数の変異ポリペプチドを含む、変異植物AGPase酵素。
【請求項27】
植物のゲノム中に1つまたは複数のポリヌクレオチドを組み入れる段階、および前記ポリヌクレオチドにコードされるタンパク質を発現させる段階を含む、熱ストレス条件に対する植物の耐性を増大させ、かつ/または植物のデンプン生合成を増大させる方法であって、前記ポリヌクレオチドが、以下であるか、または以下を含む、前記方法:
a)請求項1〜6のいずれか一項記載の任意のポリヌクレオチド;および/または
b)請求項7〜14のいずれか一項記載の任意のポリヌクレオチド。
【請求項28】
植物がトウモロコシAGPase大サブユニットもまた発現する、請求項27記載の方法。
【請求項29】
植物が、AGPaseの変異大サブユニットを発現し、該変異大サブユニットが、増大した熱安定性を与える変異を含み、かつ/または該変異大サブユニットが、増加した種子重量を与える変異を含む、請求項27記載の方法。
【請求項30】
変異大サブユニットがRev6変異を含む、請求項27記載の方法。
【請求項31】
植物が単子葉植物である、請求項27記載の方法。
【請求項32】
単子葉植物が、イネ、コムギ、オオムギ、エンバク、ライムギ、モロコシ、トウモロコシ、サトウキビ、パイナップル、タマネギ、バナナ、ココナツ、ユリ、芝草、およびキビからなる群より選択される、請求項31記載の方法。
【請求項33】
植物がトウモロコシである、請求項27記載の方法。
【請求項34】
植物が双子葉植物である、請求項27記載の方法。
【請求項35】
双子葉植物が、トマト、キュウリ、カボチャ、エンドウマメ、アルファルファ、メロン、ヒヨコマメ、チコリ、クローバー、ケール、レンズマメ、ダイズ、マメ、タバコ、ジャガイモ、サツマイモ、ヤムイモ、カッサバ、ラディッシュ、ブロッコリー、ホウレンソウ、キャベツ、セイヨウアブラナ、リンゴの木、柑橘類(オレンジ、マンダリン、グレープフルーツ、レモン、およびライムなどを含む)、ブドウ、綿、ヒマワリ、イチゴ、レタス、およびホップからなる群より選択される、請求項34記載の方法。
【請求項36】
植物細胞中に1つまたは複数のポリヌクレオチドを導入する段階、および該植物細胞から植物を成長させる段階を含む、野生型AGPase酵素を有する植物と比べて増大した熱安定性および/または増大したデンプン生合成を植物において提供するAGPase酵素を有する植物を調製するための方法であって、前記ポリヌクレオチドが、以下であるか、または以下を含む、前記方法:
a)請求項1〜6のいずれか一項記載の任意のポリヌクレオチド;および/または
b)請求項7〜14のいずれか一項記載の任意のポリヌクレオチド。
【請求項37】
植物がトウモロコシAGPase大サブユニットもまた発現する、請求項36記載の方法。
【請求項38】
植物が、AGPaseの変異大サブユニットを発現し、該変異大サブユニットが、増大した熱安定性を与える変異を含み、かつ/または該変異大サブユニットが、増加した種子重量を与える変異を含む、請求項36記載の方法。
【請求項39】
変異大サブユニットがRev6変異を含む、請求項36記載の方法。
【請求項40】
植物が単子葉植物である、請求項36記載の方法。
【請求項41】
単子葉植物が、イネ、コムギ、オオムギ、エンバク、ライムギ、モロコシ、トウモロコシ、サトウキビ、パイナップル、タマネギ、バナナ、ココナツ、ユリ、芝草、およびキビからなる群より選択される、請求項40記載の方法。
【請求項42】
植物がトウモロコシである、請求項36記載の方法。
【請求項43】
植物が双子葉植物である、請求項36記載の方法。
【請求項44】
双子葉植物が、トマト、キュウリ、カボチャ、エンドウマメ、アルファルファ、メロン、ヒヨコマメ、チコリ、クローバー、ケール、レンズマメ、ダイズ、マメ、タバコ、ジャガイモ、サツマイモ、ヤムイモ、カッサバ、ラディッシュ、ブロッコリー、ホウレンソウ、キャベツ、セイヨウアブラナ、リンゴの木、柑橘類(オレンジ、マンダリン、グレープフルーツ、レモン、およびライムなどを含む)、ブドウ、綿、ヒマワリ、イチゴ、レタス、およびホップからなる群より選択される、請求項43記載の方法。
【図1】

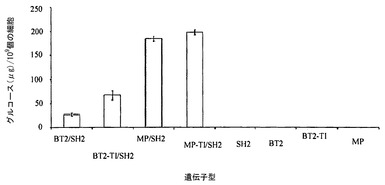
【図2】
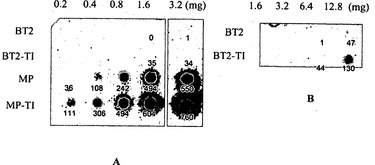
【図3】
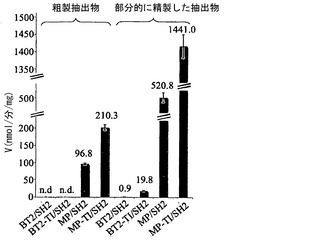
【図4】
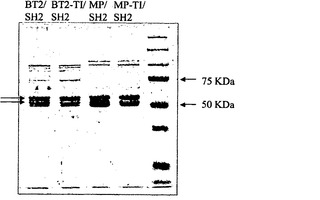
【図5A】
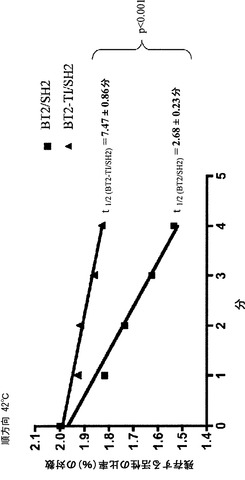
【図5B】
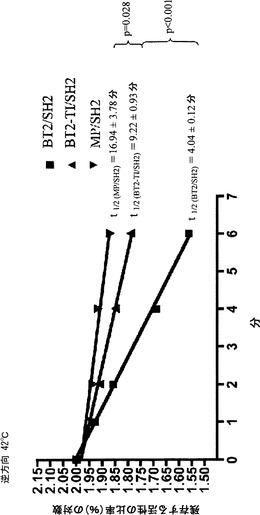
【図6A】
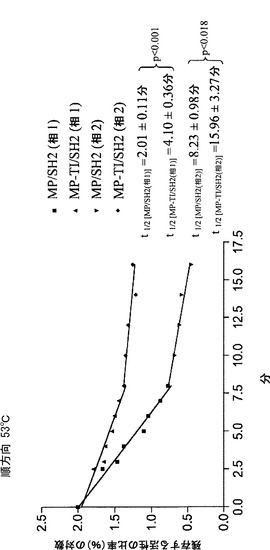
【図6B】
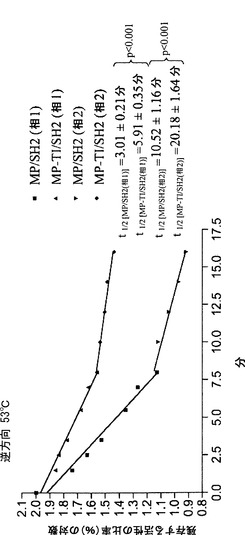
【図7A】
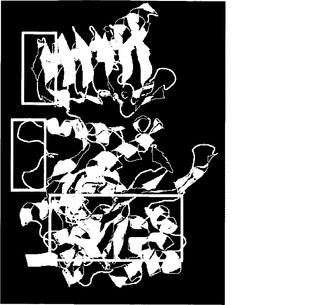
【図7B】
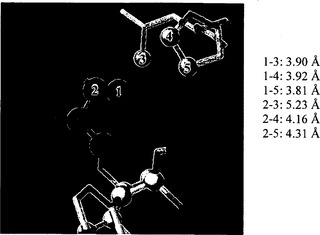
【図7C】
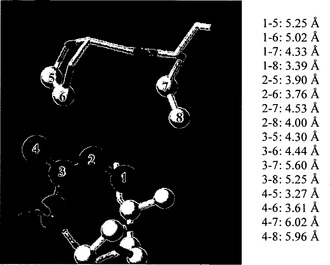
【図8】
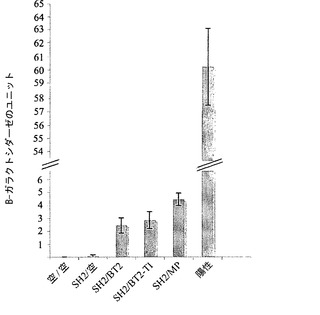
【図2】
【図3】
【図4】
【図5A】
【図5B】
【図6A】
【図6B】
【図7A】
【図7B】
【図7C】
【図8】
【公表番号】特表2011−519270(P2011−519270A)
【公表日】平成23年7月7日(2011.7.7)
【国際特許分類】
【出願番号】特願2011−503967(P2011−503967)
【出願日】平成21年3月26日(2009.3.26)
【国際出願番号】PCT/US2009/001903
【国際公開番号】WO2009/126208
【国際公開日】平成21年10月15日(2009.10.15)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.JAVA
【出願人】(507371168)ユニバーシティ オブ フロリダ リサーチ ファンデーション インコーポレーティッド (38)
【Fターム(参考)】
【公表日】平成23年7月7日(2011.7.7)
【国際特許分類】
【出願日】平成21年3月26日(2009.3.26)
【国際出願番号】PCT/US2009/001903
【国際公開番号】WO2009/126208
【国際公開日】平成21年10月15日(2009.10.15)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.JAVA
【出願人】(507371168)ユニバーシティ オブ フロリダ リサーチ ファンデーション インコーポレーティッド (38)
【Fターム(参考)】
[ Back to top ]