
耐病性キュウリ
【課題】キュウリに影響を及ぼすクロステロウイルスファミリーのウイルス(特にBPYV及びCYSDV)に対する耐性、S.fuliginea及び/又はE.cichoracearumにより生じるキュウリウドンコ病に対する抵抗性を有するキュウリの提供。
【解決手段】キュウリウドンコ病耐性と同時にキュウリクロステロウイルス耐性を有するキュウリ。かかる植物の好ましい実施形態では、クロステロウイルス耐性(QTL−1)に関与する1つ以上のゲノム領域、並びにウドンコ病耐性(pm−h及び/又はpm−l)に関与する1つ以上のゲノム領域が単一の染色体上に存在する。かかる植物のより好ましい実施形態では、pm−h及びpm−lの両方が存在し、かつQTL−1がpm−h及びpm−lの間に位置する。
【解決手段】キュウリウドンコ病耐性と同時にキュウリクロステロウイルス耐性を有するキュウリ。かかる植物の好ましい実施形態では、クロステロウイルス耐性(QTL−1)に関与する1つ以上のゲノム領域、並びにウドンコ病耐性(pm−h及び/又はpm−l)に関与する1つ以上のゲノム領域が単一の染色体上に存在する。かかる植物のより好ましい実施形態では、pm−h及びpm−lの両方が存在し、かつQTL−1がpm−h及びpm−lの間に位置する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、耐病性植物及び植物育種の分野、より詳細にはウドンコ病及びクロステロウイルス−耐性キュウリ植物の育種に関する。
【背景技術】
【0002】
キュウリ(Cucumis sativus)の農業生産においては、様々な植物病に見舞われることがしばしばある。黄化ウイルスによってそのような植物病が生じ、キュウリの生産において深刻な経済的ダメージが生じることもある。クロステロウイルス(Closteroviridae)(キュウリに悪影響を与える黄化ウイルスの中で最も上位の分類学上のクラスタを形成)のファミリーには、30種類以上の、曲状及び糸状の、昆虫により媒介される植物ウイルスが含まれる。上記ファミリーは3つの属からなり、特にモンシロチョウにより媒介されるCrinivirus属には、キュウリに深刻な影響を与える種が含まれる。この属には特に、ウリ科黄色萎縮ウイルス(CYSDV)、レタス伝染性黄斑ウイルス(LIYV)、及びビート偽黄化病ウイルス(BPYV)などの種が属する。CYSDV及びBPYVはキュウリ栽培者にとり最も脅威である。
【0003】
上記のウイルスは通常昆虫ベクター上に存在し、植物上の昆虫の摂食活動によって伝播する。したがって、キュウリクロステロウイルスは、殺虫剤の散布により抑制できる。しかしながら、好ましくは農業及び園芸におけるクロステロウイルスの制御は、寄主植物のウイルス耐性栽培品種を提供することにより行われる。現時点では、抵抗性のキュウリ種のゲノム上の少なくとも2つの量的形質遺伝子座又はQTLsが、キュウリのクロステロウイルス耐性と関連していることが知られている(特許文献1)。ゆえに子孫植物へのクロステロウイルス耐性関連の遺伝物質の適当な遺伝的浸透は、これらのQTLsに連結させた特異的なマーカーを検出することによってモニターできる。ゆえにQTLsについての見識を利用して、クロステロウイルス耐性栽培品種の育種を効率的に行うことが可能となる。
【0004】
キュウリの農業生産に影響を及ぼす他の重要な植物病として、キュウリウドンコ病(PM)が挙げられる。ウリ科のPMは、菌類のS.fuliginea及びE.cichoracearumによって生じうる。当該植物病は一年中発生し、その症状は、感染した葉の表面上における純粋な白い菌糸による小さなスポットの発生から始まり、そのスポットが次第に拡大し、最終的に茎及び葉が白い、粉末状の大量の胞子及び菌糸で覆われるに至る、というものである。重度の感染により葉の変色及び損失が生じ、その結果、果実の数及びサイズの減少に至る。ウドンコ病に対して効果的な多くの殺菌剤が存在するが、かかる化学物質に対する菌の耐性の問題が生じる。
【0005】
ある程度のウドンコ病耐性であれば、様々なキュウリ株において見られる。かかる株としては、例えばインドの野生のキュウリ:番号PI197088及び番号PI200815、PI200818、並びに栽培品種Natsufushinari及びAsomidori(モリシタら、2003)が挙げられる。また、PMに対する耐性と関連する様々な遺伝子が公知であり(Fanourakis、1984;Fujieda及びAkiya(1962);Kooistra、1968、1971;Shanmugasundarumら、1971、1972)、Natsufushinariのpm−1及びpm−2、PI200815及びPI200818のpm−3、及び栽培変種Wis.SMR18の「pm−h」(遺伝子「pm−h」は本願明細書においてpm−hとして示されるpm遺伝子座に相当する)などが挙げられる。栽培されている幾つかのキュウリの種類は、PMに対する部分的な耐性を示すものの、大部分の農業生産者は未だに葉への殺菌剤投与に依存している。この理由の1つとして、他のタイプの耐性を示す株に対してPM耐性を導入するのが困難であるという点が挙げられる。例えば、PM耐性対立遺伝子を得た株が、クロステロウイルスに対して感受性になる(それまではそうではなかった)、あるいはその逆という事態が育種の際に生じることが例として挙げられる。実際に、交配試験において、PM及びクロステロウイルス両方に対する耐性を示すキュウリの株はこれまで得られていない。すなわち有用な二重抵抗性の組換え体はこれまで得られていない。しかしながら驚くべきことに、比較的直接的な育種方法によっても、異なる特徴を単一のゲノムに取り込ませることが可能であることを見出した。
【特許文献1】国際公開第02/22836号パンフレット
【非特許文献1】Ausubel FM,Brent R,Kingston RE,Moore DD,Seidman JG,Smith JA,Struhl K(eds.)(1998)Current Protocols in Molecular Biology Wiley,New York.
【非特許文献2】Bai YL,Huang CC,van der Hulst R,Meijer Dekens F,Bonnema G,Lindhout P(2003)QTLs for tomato powdery mildew resistance(Oidium lycopersici)in Lycopersicon parviflorum Gl.1601 co−localize with two qualitative powdery mildew resistance genes.MoI.plant microbe interactions 16: 169−176.
【非特許文献3】Christou P,Murphy JE,and Swain WF(1987)Stable transformation of soybean by electroporation and root formation from transformed callus.Proc.Natl.Acad.Sd.USA 84:3962−3966.
【非特許文献4】Deshayes A,Herrera−Estrella L,Caboche M(1985)Liposome−mediated transformation of tobacco mesophyll protoplasts by an Escherichia coli plasmid.EMBO J.4:2731−2737.
【非特許文献5】D’Halluin K,Bonne E,Bossut M,De Beuckeleer M,Leemans J(1992)Plant.Cell 4: 1495−1505.
【非特許文献6】Draper J,Davey MR,Freeman JP,Cocking EC and Cox BJ(1982)Ti plasmid homologous sequences present in tissues from Agrobacterium plasmid− transformed Petunia protoplasts.Plant and Cell Physiol.23,451−458.
【非特許文献7】Fanourakis NE(1984)Inheritance and linkage studies of the fruit epidermis structure and investigation of linkage relations of several traits and of meiosis in cucumber.Ph.D.Diss.,Univ.of Wisconsin,Madison.
【非特許文献8】Fujieda K,and Akiya R(1962)Genetic study of powdery mildew resistance and spine color on fruit in cucumber.J.Jpn.Soc.HoH.Sd.31:30−32.
【非特許文献9】Ganal MW(1996)Isolation and analysis of high− molecular−weight DNA from plants,In: Nonmammalian Genomic Analysis: A Practical Guide.Academic Press Inc.,San Diego,61−73.
【非特許文献10】Gruber MY,Crosby WL(1993)Vectors for plant transformation.In BR Glick,JE Thompson,eds,Methods in Plant Molecular Biology and Biotechnology.CRC Press,Baton Rouge,LA,pp 89−119.
【非特許文献11】Hain R,Stabel P,Czernilofsky AP,Steinbliss HH,Herrera−Estrella L,Schell J(1985)Uptake,integration,expression and genetic transmission of a selectable chimaeric gene to plant protoplasts.MoI.Gen.Genet.199:161−168.
【非特許文献12】Hamilton CM.(1997)A binary−BAC system for plant transformation with high−molecular− weight DNA.Gene 200: 107−116.
【非特許文献13】Horejsi T,Staub JE and Thomas C.(2000)Linkage of random amplified polymorphic DNA markers to downy mildew resistance in cucumber(Cucumis sativus L.)Euphytica 115(2):105−113.
【非特許文献14】Horsch RB,Fraley RT,Rogers SG,Sanders PR,Lloyd A(1985)A simple and general method for transferring genes into plants.Science 227: 1229−1231.
【非特許文献15】Jiang J,Gill BS,Wang GL,Ronald PC and Ward DC(1995)Metaphase and interphase fluorescence in situ hybridization mapping of the rice genome with bacterial artificial chromosomes.Proc.Natl.Acad.Sci.USA 92:4487−4491.
【非特許文献16】Kado CI(1991): Molecular mechanisms of crown gall tumorigenesis.Crit.Rev.Plant Sci.10,1−32.
【非特許文献17】Klein TM,Gradziel T,Fromm ME,Sanford JC(1988). Factors influencing gene delivery into zea mays cells by high velocity microprojectiles.Biotechnology 6: 559−563.
【非特許文献18】Klein TM,Arentzen R,Lewis PA,and Fitzpatrick− McElligott S(1992)Transformation of microbes,plants and animals by particle bombardment.Bio /Technology 10:286−291.
【非特許文献19】Kleine M,Cai D,Hermann RG,and Jung C(1995)Physical mapping and cloning of a translocation in sugar beet(Beta vulgaris L.)carrying a gene for nematode(Heterodera schachtii)resistance from B.procumbens.Theor Appl Genet 90: 399−406.
【非特許文献20】Kooistra E(1968)Powdery mildew resistance in cucumber.Euphytica 17:236− 244.
【非特許文献21】Kooistra E(1971)Inheritance of flesh and skin colors in powdery mildew resistant cucumbers(Cucumis sativus L.).Euphytica 20:521−523.
【非特許文献22】Lui BH(1997)Statistical Genomics: Linkage Mapping and QTL Analysis.CRC Press,pg 18−19.
【非特許文献23】Miki BL,Fobert PF,Charest PJ,Iyer VN.1993.Procedures for Introducing Foreign DNA into Plants.In: Glick BR and Thompson JE(Eds.)Methods in Plant Molecular Biology & Biotechnology,CRC Press,pp.67−88.
【非特許文献24】Moloney MM,Walker JM,Sharma KK(1989)High efficiency transformation of Brassica napus using Agrobacteriuin vectors. Plant Cell Rep 8: 238− 242.
【非特許文献25】Morishita M,Sugiyama K,Saito T,and Sakata Y(2003)Review: Powdery Mildew Resistance in Cucumber.JARQ 37(1),7−14.
【非特許文献26】Nesbitt TC,Tanksley SD(2001)fw2.2 directly affects the size of developing tomato fruit,with secondary effects on fruit number and photosynthate distribution.Plant Physiol.127: 575−583.
【非特許文献27】Paterson AH(1996)Mapping genes responsible for differences in phenotype.p.41−54.In A.H.Paterson(ed.)Genome mapping in plants.R.G.Landes Company.
【非特許文献28】Phillips RL,Somers DA,Hibberd KA.1988.Cell/tissue culture and in vitro manipulation.In: G.F.Sprague & J.W.Dudley,eds.Corn and corn improvement,3rd ed.,p.345−387.Madison,WI,USA,American Society of Agronomy.
【非特許文献29】Pierik,R.L.M.(1999)In vitro Culture of Higher Plants,4th edition.Martinus Nijhoff Publishers,Dordrecht.
【非特許文献30】Sambrook J,and Russell DW(2001).Molecular Cloning: A Laboratory Manual.New York,NY,USA.,Cold Spring Harbor Laboratory Press.
【非特許文献31】Sanford JC,Klein TM,WoIf ED,Allen N(1987).Delivery of substances into cells and tissues using a particle bombardment process.J.Particulate Sd.Technol.5:27−37.
【非特許文献32】Sanford JC(1988)The biolistic process.Trends in Biotechnology 6:299−302.
【非特許文献33】Sanford JC(1990)Biolistic plant transformation.Physiologica Plantarum,79,206−209.
【非特許文献34】Sanford JC,Smith FD,and Russell JA(1993)Optimizing the biolistic process for different biological applications.Methods in Enzymology 217:483−509.
【非特許文献35】Shanmugasundarum S,Williams PH,and Peterson CE(1971)Inheritance of resistance to powdery mildew in cucumber.Phytopathology 61:1218−1221.
【非特許文献36】Shanmugasundarum S,Williams PH,and Peterson CE(1972)A recessive cotyledon marker gene in cucumber with pleiotropic effects.HortScience 7:555−556.
【非特許文献37】Tijssen P(1993)Hybridization with Nucleic Acid Probes.Part I.Theory and Nucleic Acid Preparation.In: Laboratory Techniques in Biochemistry and Molecular Biology.Elsevier.
【非特許文献38】Van Berloo R,Aalbers H,Werkman A,Niks RE(2001)Resistance QTL confirmed through development of QTL−NILs for barley leaf rust resistance.MoI.Breeding 8: 187−195.
【非特許文献39】Vos P,Hogers R,Bleeker M,Reijans M,van de Lee T,Homes M,Frijters A,Pot J,Peleman J,Kuiper M(1995)AFLP: a new technique for DNA fingerprinting.Nucl.Acids Res.23: 4407−4414.
【非特許文献40】Zhang L,Cheng L,Xu N,Zhao M,Li C,Yuan J,and Jia S(1991)Efficient transformation of tobacco by ultrasonication.Biotechnology.9:996−997.
【非特許文献41】Zhao S,and Stodolsky M(2004)Bacterial Artificial Chromosomes.Methods in Molecular Biology VoI 255.Humana Press Inc.Totowa,NJ,USA.
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明の目的は、キュウリに影響を及ぼすクロステロウイルスファミリーのウイルス(特にBPYV及びCYSDV)に対する耐性、S.fuliginea及び/又はE.cichoracearumにより生じるキュウリウドンコ病に対する抵抗性を有するキュウリの提供に関する。
【課題を解決するための手段】
【0007】
本発明の発明者らは鋭意研究の結果、CYSDV及びPMに対する耐性を示す多くのキュウリを見出した。更に、これらの二重抵抗性のキュウリが、ウドンコ病耐性と関連する2つの定量形質遺伝子座(QTLs)(本願明細書におけるpm−l及びpm−h)、並びにクロステロウイルス耐性に関連する主要なQTL(本願明細書におけるQTL−1)を含むことを発見した。更に重要なことに、全てのQTLsが単一の遺伝連鎖群に位置するということを発見した。すなわち本発明の発明者は基本的に、これらの植物ではクロステロウイルス及びウドンコ病に対する耐性遺伝子が単一の染色体に存在することを発見した。かかる植物は植物育種への使用に非常に適する。更に発明者らは、なぜこれまで二重抵抗性のキュウリが交配試験で得られなかったのかを示すメカニズムを発見した。特定の理論に束縛されるわけではないが、1つの植物病に対する耐性を与えるのに必要となる遺伝的要素の遺伝質浸透により、他の植物病に対する耐性を与えるのに必要となる遺伝的要素が損なわれるものと考えられる。この発見によって、発明者らはある特殊な育種方式を開発し、それにより上記の問題が解決され、所望の二重抵抗性を有する植物が得られるようになった。
【0008】
本発明の第1の態様は、キュウリウドンコ病耐性と同時にキュウリクロステロウイルス耐性を有するキュウリの提供に関する。
【0009】
本発明のかかる植物の好ましい実施形態では、クロステロウイルス耐性(QTL−1)に関与する1つ以上のゲノム領域、並びにウドンコ病耐性(pm−h及び/又はpm−l)に関与する1つ以上のゲノム領域は単一の染色体上に存在する。かかる植物のより好ましい実施形態では、pm−h及びpm−lの両方が存在し、かつQTL−1がpm−h及びpm−lの間に位置する。
【0010】
本発明の植物の更なる実施形態では、国際公開第02/22836号(本発明に援用する)において更に詳細に記載されているように、QTL−1の存在は、マーカーE16/M50−244、E16/M50−188及びE11/M48−251からなる群から選抜される少なくとも1つの隣接マーカーの存在の有無により示される。
【0011】
本発明の植物の他の実施形態では、pm−hと称されるQTLの存在は、前記植物のウドンコ病耐性を伴う単一のヌクレオチド多形(SNP)から生じる、少なくとも1つの突然変異を含んでなる核酸配列の存在に有無により示され、前記少なくとも1つの単一ヌクレオチド多形(SNP)は下記の表2に示すようにSNP1及びSNP2から選抜される。
【0012】
別の実施形態では、本発明の植物中の、pm−lと称されるQTLの存在は、前記植物のウドンコ病耐性と関連する、以下の表3中でSNP3として示される単一ヌクレオチド多形(SNP)から生じる少なくとも1つの突然変異を含んでなる核酸配列の存在の有無により示されるか、又は、前記植物のウドンコ病耐性と関連する、以下の表3における5bpの挿入部分(5’−AATTT−3’)として示す少なくとも1つの突然変異を含んでなる核酸配列の存在の有無により示される。
【0013】
本発明の好適な実施形態では、キュウリクロステロウイルス及びキュウリウドンコ病に対する抵抗性を示す植物の提供に関する。詳細には、前記植物はキュウリ(Cucumis sativus)であり、前記植物は単一の染色体上に、クロステロウイルス耐性を付与する少なくとも1つの染色体領域と、ウドンコ病耐性を付与する少なくとも1つの染色体領域を含んでなり、クロステロウイルス耐性を付与する前記少なくとも1つの領域が、E16/M50−244、E16/M50−188及びE11/M48−251からなる群から選択される少なくとも1つのマーカーと連結し、ウドンコ病耐性を付与する前記少なくとも1つの領域が、
− 配列番号1の単一ヌクレオチド多形マーカー 39T→G、
− 配列番号2の単一ヌクレオチド多形マーカー 29G→A、
− 配列番号3の単一ヌクレオチド多形マーカー 193C→T、
− 配列番号4の位置221における挿入突然変異5’−AATTT−3’、並びに−マーカーE16/M50−F−194、E11/M48−F−251、E23/M38−M001、E23/M40−M003、E24/M46−M002、E24/M46−M003、E12/M91−M003、E26/M43−M003、E14/M59−F−134及びE14/M59−F−200からなる群から選択される少なくとも1つのマーカーに連結している。
【0014】
本発明の植物は、クロステロウイルス耐性に関連する第2のQTLを任意に含んでもよい(国際公開第02/22836号に記載のQTL−2ように、このQTLの局在化及び特徴に関する詳細がこの特許文献の明細書に明確に示されている)。このQTLは、染色体上の別々の位置に配置していることが示されている。
【0015】
本発明の別の態様は、上記の本発明に係るキュウリの一部分の提供に関する。好ましくは、前記植物の一部分は、花粉、胚珠、葉、胚、根、根端、葯、花、果実、茎、芽、枝、根茎、種子、プロトプラスト及びカルスから選択され、最も好ましくは種子である。
【0016】
本発明の別の態様は、本発明に係る上記の二重抵抗性キュウリ(クロステロウイルス及びPMに耐性を示す[PM−葉(pm−l)及びPM−胚軸(pm−h)抵抗性の表現型を含む])と、農産物として望ましい特徴を有する第2のキュウリ又は他の種類の植物(好ましくはキュウリ株の植物)とを交配させることにより得られるF1種の提供に関する。好ましくは、前記第2のキュウリは最小限のpm−hを含む(劣性遺伝子であるため)。その場合、前記第2のキュウリは最小限のヘテロ型であり、好ましくは、劣性のpm−h特性を有するため、ホモ型である。他の好適な実施形態では、前記第2のキュウリはクロステロウイルスに感受性で、好ましくは近交系の植物である。
【0017】
本発明の別の態様は、本発明に係る上記F1種を栽培することによって得られる雑種植物の提供に関する。
【0018】
本発明の別の態様は、本発明にかかる上記ハイブリッド植物の一部分の提供に関する。好ましくは前記植物の一部分は、花粉、胚珠、葉、胚、根、根端、葯、花、果実、茎、芽、枝、根茎、種子、プロトプラスト及びカルスから選抜され、最も好ましくは果実である。
【0019】
上記したように、PM又はクロステロウイルスへの耐性に必要となる各々の遺伝要素を1つだけ有する抵抗性植物の交配による、当該遺伝要素の遺伝的浸透により、他の植物病に対する耐性に必要となる遺伝因子が損失すると考えられている。この発見により、発明者らはこの課題を解決できる育種方法の開発に至り、それにより所望の二重耐性植物を得るに至った。本発明の発明者は、耐性がQTL−1によって付与されるクロステロウイルス−耐性親キュウリ株と、耐性がpm−h及びpm−lによって付与されるPM耐性キュウリ株との交配により効果的に二重抵抗性のキュウリを作製する場合に、育種において、組換え体の作製(すなわち1つの染色体における相同組換えを発生させる)するステップと、それに続く、単一の染色体上にQTL−1及びpm−l、又はQTL−1及びpm−hを有する植物を選抜するステップを含める必要があることを見出した。好ましくは、前記選抜ステップは、単一の染色体上にQTL−1、pm−l及びpm−hを有する植物の選抜を含んでなる。より好ましくは、QTL−1はpm−h及びpm−lが配置されているゲノム領域の間に挿入され、それにより、クロステロウイルス耐性に必要となるQTL1がPM耐性に必要となる2つのQTLsの間に挟まれる位置関係をとる。
【0020】
PM又はクロステロウイルス耐性と関係する、本明細書で定義されるいずれかのQTLの存在の確認は、植物の染色体上の各々のQTLと連結した1つ以上の遺伝マーカー、植物の染色体上の各々のQTLに対して連鎖不平衡である遺伝マーカー、又はそれらの組み合わせを検出することによってなされる。
【0021】
前記植物の前記QTLsの存在及び/又は位置の適切な検出方法には、前記QTLsを特徴づけるAFLPマーカーの使用が含まれる。好ましくは、単一の染色体上におけるQTL−1、pm−l及び/又はpm−hを有する植物の上記選抜のためのステップは、p/n−4のための定量形質遺伝子座(QTL)を有する染色体上、又はPM−耐性と関連するpm−hのための定量形質遺伝子座(QTL)を有する染色体上、好ましくはpm−h及びpm−lのための定量形質遺伝子座を有する染色体上の、クロステロウイルス耐性関連の定量形質遺伝子座QTL−1を特徴づける少なくとも1つのAFLPマーカーの検出を含んでなり、その場合、前記QTL−1マーカーはQTL pm−hとQTL pm−lの間の染色体領域に位置する。すなわち、当該選抜プロセスは、表2、3及び4に記載のマーカーからなるマーカー群から選抜される1つ以上のマーカーの検出を含んでなってもよい。
【0022】
本発明の別の態様は、キュウリクロステロウイルス及びキュウリウドンコ病に対する耐性を示すキュウリの選抜方法であって、前記植物において、単一の染色体上のQTL−1、並びに少なくともQTLs pm−l及びpm−hのうちの1つを検出することを含んでなる方法の提供に関する。
【0023】
一実施形態では、当該方法は以下のステップを含んでなる:
a)キュウリからゲノムDNAのサンプルを得るステップと(当該サンプルは充分な長さのゲノムDNA断片を含んでなる)、
b)少なくとも1つの第1のQTL、又はそれに関連するQTL−1、pm−l及びpm−hからなる群から選抜される分子マーカーを含んでなる断片を選抜するための精製反応を実施するステップと、
c)前記選抜された核酸断片の増幅反応を実施するステップであって、少なくとも1つの第2のQTL、又はそれに関連するQTL−1、pm−l及びpm−hからなる群から選抜される分子マーカーを含んでなる断片を検出するステップと、
d)ステップc)の反応生成物中の、予測された長さ又は予測された核酸配列を有する増幅されたDNA断片を検出するステップ。
【0024】
好ましい実施形態では、前記ステップb)は、前記第1のQTLの分子マーカーを定義する少なくとも1つのプライマーセット、又は前記マーカーを定義する核酸配列と実質的に相補的である少なくとも1つの核酸プローブの使用を含んでなり、前記核酸プローブは前記1つ以上のマーカーを定義する核酸配列と、ストリンジェントなハイブリダイゼーション条件下で特異的にハイブリダイズする塩基配列を有する。
【0025】
他の好ましい例として、前記処理c)は、前記第2のQTLの分子マーカーを定義する少なくとも1つのプライマーセット、又は前記第2のQTLの分子マーカーの核酸配列とストリンジェントな条件下で特異的にハイブリダイズする少なくとも1つのプライマーセットの使用を含んでなる。
【0026】
本発明の方法の他の実施形態では、前記植物中の単一の染色体上の前記QTLsの存在を検出するステップは、in situハイブリダイゼーション技術、又はin situ増幅技術を用いて実施する。また、前記QTLsの分子マーカーを定義するか、又は前記QTLの分子マーカーの核酸配列とストリンジェントな条件下で特異的にハイブリダイズするプローブ及びプライマーを、前記方法で使用するのが好適である。
【0027】
本発明の幾つかの態様では、分子マーカーは好ましくは、SNP、挿入突然変異マーカー又はAFLPマーカーであり、好ましくは表2から4に列挙されるマーカーからなる群から選抜されるマーカー、並びに本願明細書に引用される文献において記載されたそれらである。
【0028】
本発明の別の態様は、キュウリクロステロウイルス、及びキュウリウドンコ病に対する耐性を示すキュウリ植物の作製方法の提供に関し、当該方法は以下のステップを含んでなる:
a)キュウリクロステロウイルス耐性を付与する染色体領域を含む第1のキュウリを選抜するステップであって、前記選抜が、前記植物のゲノムにおいて、マーカーE16/M50−244、E16/M50−188及びE11/M48−251により表される、キュウリクロステロウイルス耐性を付与するQTLに連結している少なくとも1つのマーカーの存在を検出することにより行われるステップと、
b)キュウリウドンコ病耐性を付与する少なくとも1つの染色体領域を含む第2のキュウリを選抜するステップであって、前記選抜が、前記植物のゲノムにおいて、
− 配列番号1の単一ヌクレオチド多形マーカー 39T→G、
− 配列番号2の単一ヌクレオチド多形マーカー 29G→A、並びに
− マーカーE16/M50−F−194、E11/M48−F−251及びE23/M38−M001により表される、第1のキュウリウドンコ病耐性を付与するQTLと連結している、少なくとも1つのマーカーの存在を検出することによって行われるステップか、又は、前記選抜が、前記植物のゲノムにおいて、
− 配列番号3の単一ヌクレオチド多形マーカー 193C→T、
− 配列番号4の位置221の挿入突然変異5’−AATTT−3’、並びに
− マーカーE23/M40−M003、E24/M46−M002、E24/M46−M003、E12/M91−M003、E26/M43−M003、E14/M59−F−134及びE14/M59−F−200により表される、第2のキュウリウドンコ病耐性を付与するQTLと連結している、少なくとも1つのマーカーの存在を検出することによって行われるステップによって行われるステップと、
c)ステップa)及びステップb)から得られた前記植物を交配してF1種子を得るステップと、
d)F1種子をF1植物に成長させ、交配又は自家交配することによって前記F1植物から更なる子孫を生じさせるステップと、
e)前記更なる子孫植物から、ステップa)において定義したキュウリクロステロウイルス耐性を付与するQTLに連結している少なくとも1つのマーカーと、ステップb)において定義したキュウリウドンコ病耐性を付与するQTLに連結している少なくとも1つのマーカーを含んでなる植物を選抜するステップ。
【0029】
当業者であれば、前記更なる子孫の集団は、その集団から単一の染色体上に少なくとも2つの相同組換えを生じさせている植物の存在を検出できるのに充分なサイズでなければならないことを理解するであろう。通常、1,000の植物からなる集団で充分である。本発明で提案するマーカー選抜技術による利点としては、かかる母集団のサイズであっても、目的の遺伝子型(表現型的として検知されなくてもよい)の存在を容易にスクリーニングできる点が挙げられる。
【0030】
本発明の方法の好ましい実施形態では、ステップb)の第2のキュウリを選抜するステップが、前記第1若しくは第2のキュウリウドンコ病耐性を付与するQTLsのうちの1つのみを有する第2のキュウリを選抜することを含んでなり、また前記方法は、以下のステップを更に含んでなる:
f)前記第1若しくは第2のキュウリウドンコ病耐性を付与するQTLs(すなわち前記第2のキュウリに存在しないQTL)のうちのもう1つを有する第3のキュウリを選抜するステップと、
g)ステップe)で得られたF1植物と、前記第3のキュウリとを交配させて、更なる子孫植物を得るステップと、
h)当該子孫植物から、ステップa)で定義されるキュウリクロステロウイルス耐性を付与するQTLと、ステップb)で定義されるキュウリウドンコ病耐性を付与する両方のQTLsを含む植物を選抜するステップ。
【0031】
好ましい実施形態では、ウドンコ病耐性を付与する遺伝子座prn−h及びpm−lの両方を含む植物が選抜され、詳細には、QTL−1によって特徴づけられるクロステロウイルス耐性を付与する遺伝子座が、ウドンコ病耐性を付与する遺伝子座pm−h及びpm−lの両方の間に配置されている。
【0032】
すなわち、キュウリクロステロウイルス及びキュウリウドンコ病耐性を示すキュウリの作製方法の他の好ましい実施態様では、ステップb)、e)、f)又はh)で定義されるキュウリウドンコ病耐性を付与する染色体領域を含むキュウリを選抜するステップは以下のステップを含んでなる:
−前記植物のゲノム中において、
配列番号1のSNPマーカー 39T→G、
配列番号2のSNPマーカー 29G→A、並びに
マーカーE16/M50−F−194、E11/M48−F−251及びE23/M38−M001で表される第1のキュウリウドンコ病耐性付与QTLに連結する少なくとも1つのマーカーの存在を検出するステップと、
−前記植物のゲノム中において、
配列番号3のSNPマーカー 193C→T、
配列番号4の位置221における挿入突然変異 5’−AATTT−3’、並びに
マーカーE23/M40−M003、E24/M46−M002、E24/M46−M003、E12/M91−M003、E26/M43−M003、E14/M59−F−134及びE14/M59−F−200で表される第2のキュウリウドンコ病耐性付与QTLに連結する少なくとも1つのマーカーの存在を検出するステップ。
【0033】
キュウリクロステロウイルス及びキュウリウドンコ病耐性を示すキュウリを作製する方法の他の好ましい実施形態では、ステップa)、b)、e)、f)又はh)のうちの少なくとも1つは、前記植物から得たゲノムDNAのサンプルを提供するステップと、前記ゲノムDNAのサンプル中から前記少なくとも1つのマーカーを検出するステップを含んでなる。
【0034】
他の好ましい実施形態では、ステップe)は、植物ゲノム中において、QTL−1と関連する少なくとも1つのマーカー、並びにpm−h若しくはpm−lに関連する少なくとも1つのマーカーを検出するステップを含んでなり、好ましくは、植物ゲノム中及び単一の染色体上において、QTL−1に関連する少なくとも1つのマーカー、pm−lに関連する少なくとも1つのマーカー、及びpm−hに関連する少なくとも1つのマーカーを検出するステップを含んでなり、好ましくは、表2、3及び4に列挙されるマーカー群から選択されるマーカーを含んでなる。
【0035】
他の好ましい実施形態では、ステップe)は、上記の二重抵抗性のキュウリを選抜するための本発明の方法により実施される。
【0036】
ステップc)からe)にて説明した方法であって、二重抵抗性のF1を生じさせる一度の交配において、単一の染色体上で少なくとも2つの相同組換えが生じる方法は、多くの世代を通じて実施可能であり、それにより、単一の染色体中における所望の(少なくとも2回の)相同組換え、及びそれによる二重抵抗性の植物の形成が、F2、F3、F4、F5又はいかなる世代においても実施可能となる。かかるバリエーションは本発明の範囲内に含まれ、当業者によって容易に実施される。
【0037】
キュウリクロステロウイルス及びキュウリウドンコ病に対する耐性を示すキュウリを作製する代替的な方法は、次のステップを含んでなる:
a)本発明の方法を実施して、キュウリクロステロウイルス及びキュウリウドンコ病に対する耐性を示す第1のキュウリを選抜するステップと、
b)前記植物を同系育種して、前記QTLsのホモ接合植物株を得るステップと、
c)ステップa)及びステップb)から得た前記植物を交配してF1種子を得るステップと、
d)前記F1種子をF1植物に成長させるステップ。
【0038】
本発明の別の態様は、本発明の方法で作製される植物又はその部分の提供に関する。
【0039】
本発明により子孫植物の非常に迅速なスクリーニングが可能となり、それにより二重耐性を示すと考えられる植物の選抜が可能となる。例えば、子孫植物の中から、少なくとも1つのpm−hマーカー、少なくとも1つのpm−lマーカー及び少なくとも1つのQTL−1マーカーを有する植物を検出することが可能となる。かかる植物は、望ましい遺伝的浸透が行われたと考えられる。更なるマーカーに関するスクリーニングを行うことにより、子孫植物が二重耐性を示すという予測上の信頼性を向上させることができる。3つのQTLsの各々の少なくとも1つのマーカーを示す植物の検出に基づき、植物を選抜することが可能となり、また育種プログラムあるいは更なる同定試験にも使用できる。
【0040】
本発明の植物の利点としては、上記の特性が同一の染色体に存在するため、従来の育種を使用したときでも、上記の両方の耐性を制御する遺伝子を(ハイブリッド)植物中へ容易に移動させることが可能となることであり、逆に従来の育種技術を用いた場合では、それらの特性が隔離した染色体上に存在するため、両方の特性を有するハイブリッドを作製するのがはるかに困難となる。
【発明を実施するための最良の形態】
【0041】
定義
本明細書で使用される「キュウリ」という用語には、一般にCucumberと呼ばれるプラント、アメリカガーキン、Cassabanana、Cuke、Gherkin、Hothouseキュウリ、Lemonキュウリ、Manderaキュウリ、Picklingキュウリ、Serpentキュウリ、Slicingキュウリ、Snakeキュウリ及び西インド諸島のガーキンなどが包含されるがこれらに限定されないキュウリ植物又はその一部を意味する。
【0042】
本発明の用語「植物部分」とは、単細胞及び細胞組織(例えば植物において、完全である植物細胞、細胞凝集塊及び植物が再生できる組織培養)などの植物の一部を意味する。植物部分の例としては、花粉、胚珠、葉、胚、根、根端、葯、花、果実、茎、芽及び種子に由来する単一の細胞及び組織、ならびに花粉、胚珠、葉、胚、根、根端、葯、花、果実、茎、芽、枝、根茎、種子、プロトプラスト、カルスなどが挙げられるが、これらに限定されない。
【0043】
本明細書で用いられる用語「クロステロウイルス」とはClosteriviridaeのファミリーのウイルスのことを指し、一般にCucurbit黄色萎縮障害ウイルス(CYSDV)、Lettuce伝染性黄化病ウイルス(LIYV)、及びBeet偽黄化病ウイルス(BPYV、またその同義語Cucumberクロロチックスポットウイルス(CCSV)、Cucumber黄化病ウイルス、Muskmelon黄化病ウイルス又はStrawberry pallidosisウイルスとして公知)と呼ばれるウイルス、好ましくはBPYV及びCYSDV、最も好ましくはCYSDVである。好ましくは、キュウリ植物に関する用語「closterovirus」とは、特にキュウリ(i.e.the Criniviideae)に影響を与える属を指す。
【0044】
本明細書で使用される「ウドンコ病」という用語は、真菌Sphaerotheca fuliginea(またその同義語Podosphaera黄色ii及びS.cucurbitaeとして公知)によって、及び/又は真菌Erysiphe cichoracearum(またその同義語Golovinomyces cichoracearumとして公知)によって、及び/又は真菌Leveillula taurica(またその同義語Oidiopsis taurica、Erysiphe taurica、Ovulariopsis cynarea、Leveillula solanacearumとして公知)によって、キュウリ(Cucumis sativus L.)において、生じる疾患を意味する。
【0045】
用語「QTL」は、本願明細書では、連続的に分散した(定量的な)表現型特徴の発現と関連する、対立遺伝子(例えば遺伝子又は制御配列の形)を含んでいる、当業者に公知の染色体領域を意味する用語として用いられる。「耐病性QTL」という用語は、特定の染色体に置かれる領域であって、すなわち、耐性遺伝子をコードする少なくとも1つの遺伝子に関連するか、又は、少なくとも1つの制御領域、(染色体領域)であって、耐性に関連する1つ以上の遺伝子の発現を制御するものを指す。耐性を伴うQTLsを指すものとして、より短い同義語:「耐性付与遺伝子座」を本願明細書では用いる。QTLsは、1つ以上の分子ゲノムマーカーを使用して、特異的なキュウリ継承のゲノムのそれらの遺伝子位置を示すことにより定義されてもよい。1つ以上のマーカーは、次に、特異的な遺伝子座を示す。遺伝子座の間の距離は、通常同じ染色体上の遺伝子座との間に、乗換えの頻度で測定される。2つの遺伝子座が遠くに離れるほど、より頻繁に交叉がそれらの間において、発生する。逆にいえば、2つの遺伝子座が間近である場合、それらの間での交叉は発生しにくい。概して、1センチモルガン、コーサンビー・マップ関数、cMは、遺伝子座(マーカー)との間に、1%の組換えにほぼ等しい(Lui、1997)。QTLが並列マーカーによって、示すことができる場合、エンドポイントマーカー間の遺伝的距離はQTLのサイズを表す。
【0046】
用語「染色体」は、本願明細書では、当業者に公知のように、細胞DNAを含有し、そのヌクレオチド配列に遺伝子の線形配列を有する、細胞核の自己再生する遺伝的構造のことを意味する用語として用いる。
【0047】
本発明で使用する「連鎖群」という用語は、同じ染色体に置かれる遺伝子又は遺伝形質の全てを指す。連鎖群の範囲内で、十分に間近であるそれらの遺伝子座は、遺伝的交雑の結合を示す。交差の確率が染色体上の遺伝子との間の物理的距離によって、増加するため、場所が連鎖群の中で各々からはるかに隔離した遺伝子は、遺伝子検査において、いかなる検出可能な直接的な結合も示すことはありえない。用語「連鎖群」は、大部分は染色体譲渡がまだなされなかった遺伝子のシステムの連結された挙動を示す遺伝子の遺伝子座を指すために用いる。したがって、本発明では、用語「連鎖群」とは、染色体(物理的存在)と同義である。
【0048】
本発明の用語「対立遺伝子」とは、遺伝子の1つ以上のいずれかの代替的な形を意味しその全ての対立遺伝子が少なくとも1つの特徴又は特徴に関する。二倍体細胞において、提供された遺伝子の2つの対立遺伝子は、一対の相同染色体上の対応する遺伝子座を占める。本発明はQTLs(すなわち1つ以上の遺伝子又は制御配列を含みうるゲノム領域)に関するため、若干の例においては「対立遺伝子」の代わりに「ハプロタイプ」(すなわち染色体部分の対立遺伝子)を指すのがより正確であるが、そのような場合、用語「対立遺伝子」は用語「ハプロタイプ」を含むものとして理解されるべきである。
【0049】
本願明細書における「遺伝子」とは、一連のDNAからなる遺伝単位として定義され、それは、染色体上の特定の位置を占め、また生物体の特定の形質又は特徴を制御する遺伝的な暗号を含んでなる。
【0050】
「遺伝子座」とは、提供された遺伝子又は制御配列が提供された種の染色体に占有する位置として、本願明細書に定義される。
【0051】
「相同組換え」とは、2つのDNA分子の間でのDNA断片の組換え(「クロスオーバー」)、又は同一のヌクレオチド配列の一定の領域における対染色体の染色分体間での組換えのことを指す。「組換え現象」は、本願明細書において、減数分裂のクロスオーバーを意味すると理解される。
【0052】
本発明で使用する「分子マーカー」という用語は、核酸配列の特徴の相違を視覚化する方法において、用いられる指標のことを指す。かかる指標の例としては、制限断片長多形性(RFLP)マーカー、増幅された断片長多型(AFLP)マーカー、単一ヌクレオチド多型(SNP)挿入変異、マイクロサテライトマーカー(SSRs)配列同定された増幅領域(SCARs)、切断された増幅された多形配列(CAPS)マーカー又はアイソザイムマーカー又は特異的な遺伝子の及び染色体位置を定義する本願明細書に記載されているマーカーの組合せが挙げられる。「QTLにリンクされる分子マーカー」は、本明細書で定義されるように、SNP、挿入変異、並びにAFLPマーカー又はフィールドにおいて、使用する通常の他のタイプのあらゆるマーカーのことを指してもよい。本願明細書におけるAFLPマーカーの前後関係において、マーカーとは、2つのAFLP−プライマーに隣接するキュウリ特異的なDNA塩基配列を指し、そのプライマーは、「コアプライマー」E及びM(制限酵素部位EcoRI及びMsel部位に相当)(Vosその他、1995、Baiその他。2003)、更に示すような2つ又は3つの選択的な塩基が続き、各々に選択的なヌクレオチドを確認する2桁のコードが続き、それによって、「コアプライマー」が延長される(コードに関しては表1を参照)。E16/M50−244は、増幅プライマーEcoRI + CC及びMsel + CATを用いて得られる、244bp.の全長を有する断片状のマーカーを意味する。断片の長さは断片の検出方法に依存すると考えられ、若干の塩基を増減させたものであっても、その正確な長さのものとさほど変わらない。本願明細書に記載のマーカーを定義する際、リケージマップの他のマーカーとの、そのマーカーの染色体上の相対的位置として参照する必要がある。すなわち、マーカーE16/M50−244は、そのプライマーの配列によって、増幅生成物の長さによって、並びに、E14/M59−F−200及び/又はE23/M38−M003との相対的位置によって、又は、本願明細書に記載のように、表4のマトリックスのcMの対応する距離を有する順序づけられたリストにて図示するように、他のマーカーとの相対的な位置により定義されてもよい。しかしながら、考慮が必要となることとして、植物間の交雑により、特定のマーカーが損なわれることもあることが挙げられるが、但し特定のマーカーの欠如が、耐病性を与え、マーカーがそこに連結する遺伝因子の存在を否定するわけではない。
【0053】
表1:通常、AFLP分析において適用され、本発明においても用いるプライマー伸長コード(Source:Keygene、Wageningen、オランダ)
【表1】

【0054】
用語「キュウリ特異的なDNA配列」とは、80%以上、好ましくは85%以上、好ましくは、90%以上、好ましくは、95%以上、好ましくは、97%以上、最も好ましくは99%以上、キュウリのゲノム配列とのヌクレオチド配列相同性を有するポリヌクレオチド配列を意味する。それは大きな類似性を示し、好ましくはQTL−1マーカーの場合、QTL−1マーカーに隣接するキュウリのアクセッションNo.PI 250147のDNA塩基配列の一部を構成する。
【0055】
本明細書で用いられる用語「ヌクレオチド配列相同性」とは、2つのポリヌクレオチド間の相同性を意味する。2つの配列のアラインメントを作成したときに最大の一致性でヌクレオチドの配列が同じである場合に、ポリヌクレオチドは「相同な」配列を有するという。2つ以上のポリヌクレオチドの間の配列比較は、通常、配列類似性を有する局部領域を同定し、比較ウインドウ上の2つの配列の部分を比較することにより実施される。比較ウインドウは通常、約20〜200の連続ヌクレオチドである。ポリヌクレオチドの「配列相同性のパーセンテージ」(例えば50、60、70、80、90、95、98、99又は100パーセント配列相同性)は、比較ウインドウの上の2つの最適に整列配置された配列を比較することによって、決定でき、比較ウインドウ中のポリヌクレオチド配列の部分には、最適な2つの配列の参照配列(付加又は欠失を含まない)と比較し添加又は欠失(すなわちギャップ)が存在する。パーセンテージの算出は、
(a)同一の核酸塩基が両方の配列において、存在する位置の数を勘定してマッチする位置の数を算出し、
(b)比較のウインドウの位置の総数で、上記のマッチする位置の数を除算し、
(c)結果に100を乗算して配列相同性パーセンテージとして示すことにより行われる。比較配列の最適配列は、周知のアルゴリズムを用いたコンピュータ化された実施態様で行ってもよく、又は視覚による検査によって、実施してもよい。直ちに利用できる配列比較及び並列配列アルゴリズムとしては、Basic Local Alignment Search Tool(BLAST)(Altschul et al.,1990、 Altschul et al,1997)、ClustalWプログラム(両方ともインターネットで利用可能)が挙げられる。他の適切なプログラムとしては、GAP、BestFit、PlotSimilarity及びFASTA(ウィスコンシンGenetics Software Package (Genetics Computer Group (GCG),Madison,WI,USA) (Devereux et al,1984))が挙げられるが、これらに限定されない。
【0056】
本明細書で使用される「pm−h」という用語は、下子葉部において、発現される推定のウドンコ病抵抗性遺伝子を意味し、好ましくは表2の特定のSNPの存在下で定義される。
【0057】
表2:ウドンコ病耐性表現型との完全な関連を示すSNPを、QTL pm−hとして本願明細書に示す。
【表2】
【0058】
本明細書で使用される「pm−l」という用語は、推定のウドンコ病抵抗性遺伝子pm−leafを意味し、好ましくは表3の特定の突然変異の存在下で定義される。
【0059】
表3:ウドンコ病耐性表現型との完全な関連を示している突然変異を、QTL pm−lとして本願明細書に示す。
【表3】
【0060】
「QTL−1」用語とは、クロステロウイルス耐性に連結するゲノム領域を指し、WO 02/22836において、詳細に記載されているマーカーE16/M50−244、E16/M50−188及び/又はE11/M48−251により定義される(上記開示内容を本発明に援用する)。
【0061】
「子孫」植物という用語は、1つ以上の親株からの無性生殖的若しくは有性生殖的な再生によって、得られる植物、又はその子孫としてのあらゆる植物のことを指す。例えば、子孫植物は、親植物のクローニング若しくは自家交配、又は2つの親植物を交配させることによって、得られ、F1若しくはF2、又はそれ以上の世代を含むものと解される。F1は、両親(その少なくとも1つが特徴ドナーとして初めて用いられ)から製作される第一世代の子であり、一方、第二世代(F2)又はそれ以降の世代(F3、F4など)の子孫がF1、F2などの自殖から作製される標本である。ゆえにF1は、2人の本物の繁殖両親(本当の繁殖は、特徴のためにホモ接合性である)のハイブリッドから生じているハイブリッドであると(通常)考えられ、F2が前記F1ハイブリッドの自家受粉から生じる子孫であると(通常)考えられる。
【0062】
本発明では、異なる対立遺伝子が相同染色体上の対応する場所に存在するとき、「ヘテロ接合体」という用語を、それを表す遺伝子的条件として用いる。
【0063】
本発明では、同一の対立遺伝子が相同染色体上の対応する場所に存在するとき、「ホモ接合性」という用語を、それを表す遺伝子的条件として用いる。
【0064】
植物育種の前後関係の「ハイブリッド」という用語は、異なる系列又は種類又は種の植物交配により作製される、遺伝的に異なる両親の子としての植物を意味し、2つの自殖系統の交配を含むがこれに限定されない。
【0065】
本発明で用いる「近交系」という用語は、実質的にホモ接合の個体又は株を意味する。
【0066】
本発明における用語「遺伝子移入」とは、天然の、及び人工的なプロセスであって、種を交配させることによって、1つの種、変異体多様性又は園芸品種のゲノム領域が他の種、変異体又は園芸品種のゲノムに移入することを指す。当該方法は、任意に再発する親に戻し交配して実施してもよい。
【0067】
本願明細書に用いられる「遺伝子工学」、「形質転換」及び「遺伝子組換え」の用語は、単離されクローニングされた遺伝子を、他の生物体のDNA(通常染色体DNA又はゲノム)へ移入することと、同義である。
【0068】
本発明の用語「集団」とは、共通の遺伝的な派生を有する、遺伝的に異質な植物のコレクションを意味する。
【0069】
本発明の用語「変異体」又は「栽培品種」とは、構造的若しくは遺伝子的特徴及び/又はパフォーマンスによって、同じ種の範囲内で他の種類と区別できる、類似する植物群を意味する。
【0070】
「抵抗性」及び「耐性」の用語は、感染に対する部分的及び完全な耐性のことを指す。感受性の植物は、感染に対して抵抗性を有さないか、又は耐性レベルが低いと考えられる。上記の用語には、「完全な耐性」、「免疫性」「中程度の耐性」、「部分的な耐性」、「過敏症」及び「許容性」などの、別に定義することが可能な形の抵抗性が包含される。
【0071】
「完全な耐性」とは、感染の後に病気が全く進行しない状態のことを指し、病原体が細胞内に入ること(第一の感染)がなされない結果によるものであってもよく、あるいは、細胞内で感染源が増殖できず、他の細胞にも感染しない(潜在的な感染ではなく、拡散することもない)結果によるものであってもよい。
【0072】
「感受性」という用語は、植物が疾患による影響を受け、病徴に至るような疾患に対する耐性を有しない植物を指すものとして、本願明細書で用いられる。ゆえに「感受性」という用語は、「非抵抗性」と同義語である。
【0073】
核酸に関する「ハイブリッド」という用語は、相補的なヌクレオチド塩基間の水素結合により形成される二本鎖核酸分子(又はデュプレックス)を指す。「ハイブリダイズする」又は「アニールする」という用語は、核酸配列の一本鎖が補完的な塩基との間で水素結合による二重螺旋部分を形成させる方法のことを指す。
【0074】
「プローブ」という用語は、目標核酸配列分析物又はその相補DNA誘導体の相補配列と共に水素結合デュプレックスを形成する、一本鎖オリゴヌクレオチド配列をいう。
【0075】
本明細書で使用される「プライマー」という用語は、増幅目標にアニーリングでき、DNAポリメラーゼの結合を可能にするオリゴヌクレオチドを意味し、それにより、プライマー伸長生成物の合成が誘導される条件(すなわちヌクレオチド及び例えばDNAポリメラーゼなどの酵素の存在)、並びに適切な温度及びpH条件下に設けられた場合に、DNA合成の開始位置として機能する。(増幅)プライマーは好ましくは増幅の最大効率のために一本鎖である。好ましくは、プライマーはオリゴデオキシリボヌクレオチドである。プライマーは、ポリメラーゼの存在下での伸長生成物の十分な合成を可能にするため、十分な長さを有する必要がある。プライマーの正確な長さは多くの因子に依存し、例えば温度及びプライマーの組成(A/T対G/C含量)などが挙げられる。例えばPCR増幅などのDNA増幅において、用いる、一対の双方向性プライマーは、1つの順方向及び1つの逆方向プライマーからなる。本明細書で用いられる「プライマー」は、複数のプライマーを指すこともあると理解され、特に増幅される目標領域の末端配列に関して情報の曖昧性が存在する場合には複数のプライマーが用いられる。それ故、「プライマー」には、配列又は中に若干の変異を有する配列を含んでなる、プライマーオリゴヌクレオチドのみならず、典型的な塩基対形成を可能にするプライマーも包含される。オリゴヌクレオチドプライマーは、いかなる適切な方法で調製してもよい。特異的な配列のオリゴヌクレオチドを調製する方法は公知であり、例えばクローニングや、配列の制限酵素処理、及び直接の化学合成などが挙げられる。化学合成の例としては、ホスホジ−又はトリエステル法、ジエチルホスホロアミデート法及び固相支持法(米国特許第4458066号で開示)などが挙げられる。プライマーは、必要に応じて標識してもよく、例えば分光、蛍光、光化学、生化学、免疫化学若しくは化学的な測定手段で検出される態様で標識してもよい。温度依存性のオリゴヌクレオチドプライマー伸長は、ポリメラーゼにより触媒され、4つのデオキシリボヌクレオチド三リン酸エステル(dATP、dGTP、dCTP及びdTTP(すなわちdNTPs)又はその類似体の十分な量の存在下、適当な塩、金属陽イオン及びpH緩衝システムを含有する反応溶媒中で当該触媒反応がなされる。適切なポリメラーゼは、プライマー−及び鋳型に依存してDNA合成反応を触媒することが知られている酵素である。公知のDNAポリメラーゼとしては、例えば大腸菌DNAポリメラーゼI又はそのクレノー断片、T4 DNAポリメラーゼ及びTaq DNAポリメラーゼなどが挙げられる。これらのDNAポリメラーゼを有するDNA合成に触媒作用を及ぼすための反応条件下は、従来技術において、周知である。合成の生成物は鋳型鎖及びプライマー拡張鎖からなる二重分子であり、標的配列を含む。これらの生成物は、次のラウンドで複製のための鋳型として役立つ。複製の2回において、第1のサイクルのプライマー伸長鎖は、その相補プライマーによって、アニールされる、合成により、「ショート」にプライマー配列又はそれらの補足によって、5’−及び3’末端に結合される生成物を与える。変性、プライマーアニーリング及び伸長の反復サイクルは、プライマーにより定義される目標領域の指数関数的な蓄積に結果としてなる。充分なサイクルは、核酸の目標領域を含んでいるポリヌクレオチドの所望の量を提供するために行われる。所望の量は異なる場合もあり、生成物ポリヌクレオチド実際に用いる目的により異なる。PCR方法は通常ハンドブックに記載され、当業者に知られている。PCRによる増幅の後、標的ポリヌクレオチドは、プローブポリヌクレオチドとのハイブリダイゼーションによって、検出できる。それはストリンジェント〜適度にストリンジェントなハイブリダイゼーション条件及び洗浄条件下で標的配列と共に安定ハイブリッドを形成する。プローブが標的配列と基本的に完全に相補的である(すなわち約99%以上)と考えられる場合、ストリンジェントな条件が用いられる。若干の不適当な組み合わせが予想される場合、例えばプローブが変異株と完全に相補的でないことが予想される場合、ハイブリダイゼーションのストリンジェンシーは減少すると考えられる。しかしながら、非特異/偶然の結合を排除する条件を選択する。ハイブリダイゼーションに影響を及ぼす、そして、非特異的結合に対して選択する条件は、従来技術において、周知で、例えば、Sambrook及びラッセル(2001)に記載されている。通常、低い塩濃度及びより高い温度は、ハイブリダイゼーション条件のストリンジェンシーを増加させる。
【0076】
用語「ストリンジェントなハイブリダイゼーション条件」とは、ポリヌクレオチドが、通常は核酸の複雑な混合物中でその標的とハイブリダイズするが、実質的に他の配列とはハイブリダイズしない条件のことを指す。ストリンジェントな条件は配列に依存し、また反応状況によっても異なる。長い配列は、高い温度で特異的にハイブリダイズする。核酸のハイブリダイゼーションの詳細にわたるガイドは、Tijssen(1993)を参照のこと。通常、ストリンジェントな条件は、一定のイオン強度pHにおいて、特定の配列の場合の熱融点(Tm)より5〜10℃低くなるように選択される。Tm値とは、所定のイオン強度、pH及び核酸濃度条件において、標的と相補的なプローブの50%が平衡状態において、標的配列とハイブリダイズする温度のことを指す(標的配列が過剰に存在するとき、Tm値では、プローブの50%が平衡状態にある)。ストリンジェントな条件は、塩濃度が約1.0M以下のナトリウムイオン濃度であり、通常はpH7.0〜8.3において、0.01〜1.0Mのナトリウムイオン(又は他の塩)濃度であり、温度が短いプローブ(例えば10〜50のヌクレオチド)では30℃以上、長いプローブ(例えば50超のヌクレオチド長)では約60℃以上である。ストリンジェントな条件は、不安定化剤(例えばホルムアミド)の添加によって、設定してもよい。選択的若しくは特異的なハイブリダイゼーションでは、陽性シグナルはバックグラウンドに対して少なくとも2倍(好ましくは10倍)のハイブリダイゼーションである。条件がしばしばそうである典型的な厳しいハイブリダイゼーション:50%ホルムアミド、5×SSC及び1%のSDS、42℃でインキュベート、又は、5×SSC、1%のSDS、65℃でインキュベートし、65℃で0.2×SSC及び0.1%のSDSで洗浄、である。PCRでは、アニーリング温度はプライマー長に応じて32℃〜48℃で変化しうるが、約36℃の温度が、低いストリンジェンシーによる増幅において、典型的である。ハイブリダイゼーションパラメータを決定するための更なるガイドラインは、多数の参考文献(例えばAusubelら、1999)において、開示されている。
【0077】
好ましい実施態様の説明二重耐性植物の生産農業及び園芸の繁殖プログラムの目的は、それらの遺伝組成を改良することによって、植物の性能を強化することである。本質的には、この改良は、目的の性能特性に影響している遺伝子に対して最も好ましい対立遺伝子の頻度を増加させることによって、なされる。野生の植物株を用い、遺伝子及び表現型バリエーションの豊富な資源とする。伝統的に、農業若しくは園芸分野においては、このような変種を用いて、所望の遺伝子型又は潜在的な表現型特性を有する野生植物系列又はその子孫を選択し、別の所望の遺伝子型又は潜在的な表現型特性を有する系列と交配し、所望の遺伝子型又は潜在的な表現型特性を示す(その頻度が増加した)植物を子孫植物から選択することを行う。メンデルの遺伝の法則の更なる理解及び利用(並びに分子遺伝学的ツールとの組み合わせ)によりこのような選抜プロセスが可能となった。例えば、所望の遺伝子型の又はポテンシャル表現型特性を有する植物を選択する方法は、前記植物を量的形質遺伝子座(QTL)の存在(すなわち連続的に分散(定量的に)した表現型特徴の発現と関連する対立遺伝子を含有する染色体領域の存在)の検出に基づいて行われる。通常、QTLは、表現型特徴における定量的変化に統計学的に結びつく1つ以上のマーカーによって、特徴づけられて、遺伝子と基本的に同義である。QTLマッピングは、目的の特徴の発現に影響を及ぼしている候補遺伝子座の同定を可能にする。植物繁殖においては、 マーカーによる選抜(MAS)が可能であり、すなわち植物中でQTL関連のマーカーを検出することによって、好ましい対立遺伝子を有する植物を選抜する。
【0078】
栽培植物の繁殖プログラムの大きな問題の1つは、別々の特徴間の負の遺伝相関の存在である。これは、例えばさまざまな疾患耐性植物系列の複製能及び産生間の負の遺伝相関を有するケースである。もう一方への1つの植物系列のゲノムからのDNAの遺伝子移入が基本的な繁殖形質の発現に妨げてもよいか又は影響を及ぼしうることが理解される。同様に、もう一方への1つの植物からの移入耐性を与えている遺伝子配列の試みにより、すでに宿主株に存在するに耐性特性が除去されうる。
【0079】
さまざまな特徴の遺伝についての知識により、例えば病害抵抗性と関連するQTLのためにホモ接合性株の選抜が可能となる。遺伝子の起源及び繁殖プログラムの所望の特徴の位置についての知識の中で使用法予測された繁殖している結果の精度を増加させ、従来の繁殖プログラムと比較し選抜速度が増加する。例えば、実際、所望の特徴が遺伝子的ベース(他の特徴に遺伝的にリンクされる)を有するとき、子孫中におけるそれらの2つの特徴の均一性の増加に役立つ。なぜなら、所望の対立遺伝子に関するホモ接合の親が、大部分の子孫に対して、分離の少ない状態でそれらを子孫に渡すからである。
【0080】
上記したように、本発明の発明者は、クロステロウイルス及びウドンコ病に耐性のある植物を見出した。かかる植物はこれまで公知ではなかった。すでにウドンコ病耐性植物系列へのクロステロウイルス耐性系列からのDNA移入(又はその逆)の試みはこれまで成功しなかった。
【0081】
本発明の発明者は、更に、二重耐性植物で、単一ユニットにつれて実質的に共遺伝するほど密接に、クロステロウイルス及びウドンコ病に対する耐性のための遺伝子が結合されるということを発見した。遺伝子(非常に密接に、連結される)間のかかる遺伝子の構成は、しばしば、カップリング期(シス)にある対立遺伝子(遺伝子)とも呼ばれる。そのような場合、遺伝子が単一染色体にあることが通常認められる。事実、単一染色体上に遺伝子又はQTLsが存在するとき、それらがカップリング期にあることが意味される。
【0082】
本発明の発明者は、二重耐性植物では、クロステロウイルス及びウドンコ病に対する耐性遺伝子が連鎖群4に存在することを発見した。Horejsiらによれば(2000)キュウリLの連鎖群4はまた、べと病(dm.)耐性遺伝子を有する。LG 4は更に、以下のRFLPマーカーのうちの1つ又は組合せ、(Software package INTMAP,Keygene,Wageningen,The Netherlandsによって、決定、マップ位置はcM、ブラケット):CsC032a/El(25.9)CsP357/H3(31.7)CsC588/H3(34.2)CsC477H3(35.3)CsC694/E5(38.5)CsP347/H3(38.5)CsC365/El(41)CsC386/El(41)CsC230/El(41.7)CsP064/El(45.5)により特徴付けられる。
【0083】
明確な染色体番号は、キュウリクロステロウイルス耐性QTL1、並びにウドンコ病耐性に関与するpm−l及びpm−hが位置する染色体には割り当てられていない。しかしながら、当該染色体はこれら及び他のゲノム領域が位置する連鎖群(LG4)を参照することにより示されてもよい。用語連鎖群とは、耐性を与えている対立遺伝子が位置し、染色体と同じ階層的なレベルを有する物理的ゲノム単位を指すものとして本願明細書で用いる。
【0084】
クロステロウイルス+pm耐性のための遺伝子が単一染色体に存在することを発見した利点として、2つ挙げられる。
【0085】
第1に、単一染色体上(好ましくは、小さい隣接する染色体部分)に遺伝子が存在するため、繁殖目的において、非常に有利であり、それらが共同で子孫植物に移入されるということである。ウドンコ病耐性及びクロステロウイルス耐性に関連する遺伝子との物理的な結合の結果として、交配による子孫植物(本発明の二重耐性植物を親株として用いた)では子孫植物における分離の頻度が低くなる。
【0086】
第2に、この知識は、2つの特徴を単一の植物系列内において、組み合わせるのが非常に困難である理由を説明するため、植物育種の助けとなる。いかなる理論にも束縛されないが、クロステロウイルス耐性に関与するQTLの特定の位置、及びそのQTLの選抜により、望ましい遺伝子移入の形成が妨げられたか、望ましい遺伝子移入の可能性が極めて低い確率に抑えられていた。ウドンコ病耐性及びクロステロウイルス耐性のためのQTLを伴うQTLsは近い位置に存在し、すでに第1の耐性(例えばウドンコ病)を有している植物系列への、第2の耐性特徴(例えばクロステロウイルス耐性)QTLの遺伝子移入により、第1の耐性特徴の(一部分の)損失が生じることが考えられる。植物系列がウドンコ病耐性(pm−l及びpm−h)のための両方のQTLsを有する場合、ウドンコ病耐性の基礎をなしている遺伝情報に「損傷を与え」ず、クロステロウイルス耐性のためのQTLの遺伝子移入が、ゲノムの非常に小さい部分において、のみ行われる。
【0087】
ゆえに、さまざまな抵抗性遺伝子が単一染色体に位置することを発見したこととは別に、両方のウドンコ病耐性対立遺伝子が二重耐性植物に存在するときに、クロステロウイルス耐性に関連するQTL(QTL−1)は、PM耐性(pm−h及びpm−l)を伴う2つのQTLsの間に存在することを見出した。この発見は、所望の遺伝子型を有する植物への組換え頻度が低いことを更に部分的に説明するものとも考えられる。また、かかる植物を作製するための、少なくとも3つの可能なシナリオが考えられ、それらを作製するいくつかの再現性のある方法が考案させる。
【0088】
第1の方法は、目的の第1の系列の植物に由来するクロステロウイルス耐性遺伝子座(本明細書でQTL−1として定義)を、2つのPM−耐性付与遺伝子座pm−h及びpm−lを有する、目的の第2の系列のウドンコ病耐性植物に移入することを含んでなる。これが目的の第2の系列の遺伝的バックグラウンドに結果としてなることである場合、当該方法は、QTL−1を、2つのPM−耐性付与遺伝子座pm−h及びpm−lの間のゲノム領域に遺伝子移入することを含んでなり、それにより、目的の第2の系列の遺伝的バックグラウンドを有する子孫植物中で、pm−l−QTL−1−pm−hの構成を有するゲノムDNA配列アセンブリとなる(図1を参照)。これに基づくと、単一染色体の特定の領域における二重の相同組換(すなわち各交差における1度の現象)のみにより、目的の第2の系列の遺伝的バックグラウンドにおいて、クロステロウイルス及びPM耐性の二重耐性の表現型が得られる子孫植物の適当な遺伝子移入を、QTL−1、pm−1及びpm−h特異的なマーカーを用いてモニターしてもよい。
【0089】
第2の方法では、異なる遺伝的バックグラウンド(すなわち第1の目的の株)において、耐性特徴を組み合わせることを望むかもしれないかかる方法は、目的の第2の系列の植物に由来するPM−耐性付与遺伝子座pm−h及びpm−lを、QTL−1場所を有するための第1の系列に係るクロステロウイルス耐性植物に移入することを含むと考えられる。この場合、当該方法は、pm−h及びpm−l遺伝子座を独立にQTL−1場所の両側に遺伝子移入することを含んでなり、それにより、目的の第1の系列の遺伝的バックグラウンドを有する子孫植物において、pm−l−QTL−1−pm−hの構成を有するゲノムDNA配列アセンブリとなる。(図2を参照)。この構成に基づくと、単一染色体の特定の領域において、少なくとも2つの二重相同組換(すなわち4回の交差)が、クロステロウイルス及びPM耐性の二重耐性表現型の獲得に必要であることが考えられる。また、子孫植物の適当な遺伝子移入を、QTL−1、pm−l及びpm−h特異的なマーカーを用いてモニターしてもよい。
【0090】
上記より、例えば野生型のCYSDV耐性の、例えば PM耐性の市販のキュウリへの遺伝子移入が非常に稀な現象であることが説明されると考えられる。同様に、例えばクロステロウイルス耐性の市販のキュウリへの、例えば野生型の両方のPM耐性遺伝子座の遺伝子移入が更に稀な現象である理由も説明ができる。
【0091】
組換えは、減数分裂の間の2つの相同染色体の間での情報の交換である。組換え植物において、最初は染色体の範囲内で特異的な位置に存在するDNAは、他の植物に由来するDNAと組み換えられる(父性から母性、あるいはその逆)。二重組換えでは、この組み換え反応は二度、例えば遺伝子/遺伝子座の両側において、行われる。必要な材料だけを組み換え、可能な限り染色体上の有益な第一の情報を維持するために、通常4度の交差現象(上記参照)が必要となる。かかる二重組換えを見つける通常の方法は、F2−植物の集団をスクリーニングすることである。この集団は、まれな(低頻度)二重組換えを検出するために充分なサイズでなければならない。二重組換えの頻度は、一重組換えの頻度に左右される。例えば、10cM領域の組換えは10%の頻度であった場合、二重組換えは、10%×10%=1%(1センチモルガンは、検定交雑の1%の組換え子孫として定義される)の頻度となる。
【0092】
低頻度のこの問題を迂回するための1つの方法は、「平行(パラレル)組換え」の実施である。これは基本的に、2つの組換えが別個に2つの異なる植物において、生じさせ、その後で、組換えを、得られるF2において、単純な交雑及び選抜を行うことによって、1つの植物で組み合わせることを意味する。これにより、スクリーニングする植物数が減少する。
【0093】
ゆえに、二重耐性植物を作製する第3の方法は、目的の第1の系列に由来する2つのPM−耐性付与遺伝子座pm−h又はpm−lのうちの1つを、QTL−1遺伝子座を有するための第2の系列のクロステロウイルス耐性植物に移入して、pm−l−QTL−1又はQTL−1−pm−hの構成を有するゲノムDNA配列アセンブリとすることと、目的の第3の系列に由来する2つのPM−耐性付与遺伝子座pm−h又はpm−lのうちの他方を、QTL−1遺伝子座を有するための第2の系列の他のクロステロウイルス耐性のある植物に移入して、QTL−1−pm−h又はpm−l−QTL−1の構成を有するゲノムDNA配列アセンブリとすることを含んでなる(図3を参照)。その後、個々の組換えから遺伝子移入を含んでいる子孫植物は、構成pm−lを有するゲノムDNA配列アセンブリに結果としてなるために交差する−QTL−1−pm−h(図3を参照)。また、さまざまな子孫植物への適当な遺伝子移入は、QTL−1、pm−l及びpm−hに特異的なマーカーを用いて、更に交雑された植物をモニターしてもよい。当業者であれば、図4−6を基にして他の方法を考案してもよい。
【0094】
スクリーニングに用いる植物数を制限する方法は、例えば以下の通りに実施してもよい:最初に、両方のPM−耐性対立遺伝子(pm−l及びpm−h)を有する個体と、QTL耐性対立遺伝子QTL−1を有する個体の2個体を交配する。次に、pm対立遺伝子の1つとQTL−1との間の1つの交差を有する、以下のような植物を選抜する:2pm対立遺伝子の2つのエンドポイント間の遺伝的距離が10cMであり、QTLlが中央(QTLlと、pm−l又はpm−hのいずれが1つの間が5cM)に位置する場合に、QTLlといずれかのpm−遺伝子座との間のかかる組換えが5%の頻度で生じる。それにより、最初に交配により得た200の子孫植物をスクリーニングする(10が組換えA型及び10の組換えB型である)。次に、このようにして同定された組換え体を自家交配し、10の植物をスクリーニングし、QTL−1及びpm−に隣接するマーカーを用いてホモ接合体の子孫を解析する。ホモ接合体A×B(10Aと10Bを交配)との間で10の交配を行い、得られるF2に関して10の植物をスクリーニングして1つの二重組換えを選抜する。最終的に、10の二重組換え体を得る。スクリーニングした植物数は全部で500であったが、従来の方法では合計4000の植物のスクリーニングが、10の独立した二重組換え体を得るのに必要であった(QTL−1及びいずれのpm−遺伝子座間も5cMの距離、0.05×0.05=0.25%の二重組換え頻度を有する結果となることが必要であった)。
【0095】
本発明は、マーカーにより補助される選抜方法(MAS)のより良好なモデルの提供に関する。ゆえに本発明は、植物の育種方法、植物(特にキュウリ)の選抜方法の提供に関する。特に繁殖プログラムに用いられる育種植物としての栽培用キュウリ、又は所望の遺伝子型又は潜在的表現型を有する栽培用キュウリの選抜方法の提供に関する。特に、商業的に有用な植物として本願明細書に記載した、有益なウリ科の果物の作製に際に応用できる。本願明細書における栽培用植物とは、故意に選抜されたか、又は所望の遺伝子型又は潜在的な表現型有するとして、農業若しくは園芸生産において故意に選抜された植物に由来する植物(特に同系交配によって得られた植物)として定義される。
【0096】
本発明の二重耐性植物は例えば、遺伝子型AABBcc、AABbcc、AaBBcc又はAaBbccを有してもよい。「A」とは優性のpm−l遺伝子座に基づく耐性遺伝子型であり、「a」はそれに対応する非抵抗性の対立遺伝子であり、「B」は優性のQTL−1遺伝子座に基づく耐性遺伝子型であり、「b」はその対応する感受性の対立遺伝子であり、「c」は劣性のpm−h遺伝子座に基づく耐性遺伝子型である。したがって、本発明の二重耐性植物には、ハイブリッド植物並びにホモ接合植物の両方が包含される。これらの遺伝子型は、以下の交配(親生殖体の遺伝子型)を行うことによって得られる:ABc×ABc、ABc×Abc、ABc×aBc、ABc×abc及びAbc×aBc。したがって、生殖体abcを作出する植物(pm−hを除き、耐性を有さない植物)であっても、本発明の二重耐性(ハイブリッド)植物の作製に使用できることが分かる。特に好ましい実施態様では、本発明の二重耐性植物は、近交系植物(耐性対立遺伝子がホモ接合)である。
【0097】
pm−h遺伝子は劣性遺伝子であるため、バイオアッセイを用いてF1又はBC1においてはモニターできない(すなわち感受性の親を用いてF1/BC1を作製した場合(繁殖の際に一般的))。したがって、本願明細書記載のように、子孫植物への適当な遺伝子移入が、QTL特異的なマーカーを用いてモニターできるというのが、本発明の顕著な効果である。MAS又はMAB方法を用いることにより、当業者は植物の選抜方法の提供をうけることになる。
【0098】
ゆえに本発明はまた、キュウリクロステロウイルス及びキュウリウドンコ病耐性を示すキュウリの選抜方法の提供に関し、当該方法は、前記植物において、本明細書で定義されるQTL−1の単一染色体上の存在、及び、本明細書で定義されるQTLsであるpm−l及びpm−hの少なくとも1つを検出することを含んでなる。かかる植物を選択するための本発明の好ましい方法は、a)キュウリからゲノムDNAのサンプルを調製するステップと、b)前記ゲノムDNAのサンプルにおいて、QTLに連結する、QTL−1、pm−l及びpm−hからなる群から選択される少なくとも1つの分子マーカーを検出する(好ましくは前記群から少なくとも2つの分子マーカーを検出するステップであって、1つのマーカーがクロステロウイルス耐性を検出し、他のマーカーがウドンコ病耐性を検出するステップを含んでなる。
【0099】
キュウリからゲノムDNAのサンプルを調製する処理は、公知の標準的なDNA単離方法により実施してもよい。
【0100】
分子マーカーを検出するステップ(ステップb)は、好ましい実施形態では、AFLP方法で用いられる双方向性プライマーセットを使用して、以下で適切なQTLのマーカーであると判明した増幅生成物を産生させることを含んでなる。かかるプライマーセットは、AFLPマーカーを定めるプライマー、又はマーカー特異的なプライマーとして本願明細書に記載する。双方向性とは、核酸の増幅反応において、1つが順方向プライマーとして機能し、もう1つが逆方向プライマーとして機能することを意味する。
【0101】
あるいは、分子マーカーを検出するステップ(ステップb)は、他の好ましい実施形態では、前記分子マーカーを定義する核酸配列と実質的に相補的な塩基配列を有する核酸プローブの使用を含んでなり、当該核酸プローブは、前記分子マーカーを定義する塩基配列と、ストリンジェントな条件下で特異的にハイブリダイズする。適切な核酸プローブは例えばマーカーに対応する増幅生成物の一本鎖でもよい。
【0102】
分子マーカーを検出するステップ(ステップb)は、前記ゲノムDNA上の核酸増幅反応を行わせて1つ以上のQTLsを検出するステップを含んでもよい。それは一組のマーカー特異的なプライマーを使用してPCR反応を実施することにより可能となる好ましい実施形態では、前記ステップb)は、前記QTLのAFLPマーカーを定義する少なくとも1つのプライマーセット、又は前記QTLのAFLPマーカーの核酸配列とストリンジェントな条件下で特異的にハイブリダイズするプライマーセットの使用を含んでなる。
【0103】
処理d)の、予測された長さ又は予測された核酸配列を有するDNAフラグメントを検出する処理は好ましくは、増幅されたDNAフラグメントが、最初にマーカーが検出された植物由来のDNAと同じプライマーを用いた同様の反応に基づいて増幅した場合の結果を基にした、予想される長さ(数塩基(例えば1、2又は3の塩基)長の増減があってもよい)に対応する長さを有する態様で実施するか、又は、前記増幅されたDNAフラグメントが、最初に前記マーカーが検出された植物中のQTLに関連したマーカーの配列に基づいて予想される配列に一致する核酸配列(80%以上、好ましくは90%以上、より好ましくは95%以上、更により好ましくは97%超、更に好ましくは99%を超の相同性を有しても良い)を有する態様で実施する。当業者に自明のように、感受性の親には存在するが耐性植物には存在しないマーカー(いわゆるトランスマーカー)はまた、子孫植物中の耐性を検出するアッセイにおいて有用であると考えられるが、但しマーカーの欠如を試験して特異的な特徴の存在を検出する場合には最適でない。
【0104】
予測された長さを又は予測された核酸配列を有する増幅されたDNAフラグメントを検出するステップは、標準的なゲル電気泳動、又は自動DNA解析装置を用いて実施してもよい。当該方法は無論、当業者にとり周知である。
【0105】
植物において単一染色体上に2つのQTLsの存在を検出する際、染色体ペインティング法を用いてもよい。かかる方法では、少なくとも第1のQTL及び少なくとも第2のQTLを、in situハイブリダイゼーション法又はin situPCR法で、同じ染色体上で検出してもよい。かかる技術を用いて、pm−l及びpm−hの位置とのQTL−1の前記染色体上の相対的位置を確認してもよく、またその方法は公知技術である(例えばvide Jiangら、1995を参照)。より簡便には、2つのQTLsが単一染色体に存在するという事実を、それらがカップリング期にあることに基づいて確認してもよい(すなわち、別々の染色体に個々の遺伝子が存在する場合と比較して分離が減少している)。
【0106】
分子マーカー及びQTLs:
分子マーカーを用いて、核酸配列の相違を視覚化してもよい。この視覚化は、制限酵素(RFLP)による切断後のDNA−DNAハイブリダイゼーション法、及び/又はポリメラーゼ連鎖反応を使用する方法(例えばSTS、マイクロサテライト、AFLP)において可能である。2つの親の遺伝子型の全ての違いは、これらの親の遺伝子型の交配に基づくマッピング集団(例えばBC1、F2、図2を参照)において分離している。異なるマーカーの分離を比較し、組換え頻度を算出してもよい。通常、異なる染色体上の分子マーカーの組換え頻度は50%である。同じ染色体に存在する分子マーカー間の組換え頻度は、マーカー間の距離に依存する。低い組換え頻度は、染色体上におけるマーカー間の低い遺伝的距離に対応する。全ての組換え頻度を比較することにより、染色体上の分子マーカーで最も論理的なオーダーが得られる。この最も論理的なオーダーは、連鎖地図(パターソン、1996)で表すことができる。耐病性の強化(例えば病原体による感染後の疾患を得ることの低下する発生率、及び/又は病原体との接触後の病変成長率の低下)と関連する連鎖地図上の隣接するマーカー群は、その疾患耐性に関連するQTLの位置をピンポイントで示すものである。
【0107】
本願明細書で同定されるマーカーを用いてもよく、様々な本発明の態様として本願明細書に示される。本発明の態様は、本願明細書で同定されるマーカーの使用に限定されるものではない。上記の態様は、本願明細書に開示されているマーカーの使用のみに限定されず、むしろ未同定のものも同様に含まれる。遺伝学的単位には、「遺伝子」(形質発現が予測できない多数の因子に依存する)以外にも遺伝学的単位「QTL」が存在し、それは定量可能な表現型に直接関与するゲノム領域のことを指す。したがって、遺伝子は植物育種にはほとんど関係しないが、QTLは植物育種に直接適用できるものである。本発明の発明者は、本明細書で定義されるキュウリのクロステロウイルス耐性及びウドンコ病耐性に関与するQTLsが、子孫植物のゲノムにおいて、各々特異的な構成を有しなければならないということを発見した。本発明者は、キュウリのゲノムの単一染色体上の、異なるQTLsに帰属する隣接するゲノムマーカーの一連の存在が、両方の耐性の存在と相関し、それにより、このゲノム構成が通常のメンデル式の遺伝形式で遺伝しうるという観察結果を基礎として、かかる新たな知見を得た。
【0108】
本発明で同定されたQTLsは、単一の染色体又は連鎖群上に位置し、それらの位置はそれ以外の多くの任意のマーカーによって十分に特徴づけられる。本明細書における試験では、増幅断片長多型(AFLP)マーカー、単一ヌクレオチド多型(SNP)及び挿入変異マーカーを用いたが、制限断片長多形性(RFLP)マーカー、マイクロサテライトマーカー(例えばSSRs)、配列依存的増幅領域(SCAR)マーカー、切断増幅多形配列(GAPS)マーカー若しくはアイソザイムマーカー、又はこれらのマーカーの組合せを用いてもよい。一般に、QTLは数百万塩基の領域にわたることもある。したがって、QTLの完全な配列情報を得ることは実際には不可能であり、また不必要でもある。むしろ、(一連の隣接するゲノムマーカーの存在と、特異的な表現型の存在との間の観察された相関によって)QTLを最初に検出する方法により、子孫植物の集団の中から、特異的な表現型特徴を示すポテンシャル遺伝子を有する植物を発見することが可能となる。したがって本発明において、非限定的なマーカーのリストを提供することにより、繁殖プログラムにおけるQTLsの有用性が提供される。
【0109】
マーカーは、ある特定の系列に対して特異的である。したがって、特異的な特徴は特異的なマーカーと関係している。本出願に示すマーカーはQTLの位置を示すだけでなく、それらはまた植物の特異的な表現型の存在との相関関係も示す。ゲノム上のQTLの位置を示す隣接ゲノムマーカーは専ら恣意的なものであり、非限定的であることに留意すべきである。一般に、QTLの位置は、一連の隣接するマーカーとして示され、それは表現型に対する統計学的な相関関係を示すものである。その一連の外でマーカーが見出されたとき(すなわち特異的な閾値以下のLOD−スコアを示し、当該マーカーが隔離され、その領域における当該マーカーとQTL間の組換えが頻繁に発生し、マーカーの存在が統計学的に有意に表現型の存在と相関しないとき)、QTLの境界がそこで設定される。したがって、その特定された領域に位置する他のマーカーを用いてQTLの位置を示すことも可能である。
【0110】
隣接するゲノムマーカーを用いて、個々の植物におけるQTL(すなわち表現型)の存在を示すことができる点は更に重要であり、すなわちマーカーアシスト選抜(MAS)方法に使用できる。原則として、多くの潜在的に有用なマーカーは限定されるが、多くのマーカーを用いてもよく、当業者であれば、本願明細書に記載のマーカー以外の他のマーカーを容易に同定することができる。QTLに連結したあらゆるマーカー(例えば、当該ゲノム領域の物理的に境界内に、ある一定の閾値を上回るLODスコアを示すマーカーが分布し、それにより交配の際、マーカーとQTLとの間で全く組換えが起こらないか、若しくはごくわずかな組換えしか起こらない)、並びにQTLに対して連鎖不平衡にあるいかなるマーカー、並びにQTL中で実際に突然変異を表すマーカーを、MAS方法に用いてもよい。このことは、本発明においてQTLsに関連するマーカーとして同定されたもの(例えばQTL−1に対するAFLPマーカーのE16/M50−244、E16/M50−188及びE11/M48−251)が、MAS方法の使用に適しているマーカーの単なる一例に過ぎないことを示すものである。更に、QTL(又は特異的な特徴を付与するその一部)が他の遺伝的バックグラウンド(すなわち他の植物のゲノム)に移入されると、若干のマーカーは、その特徴が表れるにもかかわらず、子孫内では存在しなくなることがある。すなわち、かかるマーカーは、ゲノム領域の外側に存在し、それはオリジナルの親系列においてのみQTLの特異的な特性を付与する部分であること、並びに、新規な遺伝的バックグラウンドは異なるゲノム構成を有することを示すものである。そのマーカーの不在が子孫への遺伝因子の良好な導入を示すかかるマーカーは「トランスマーカー」と称され、本発明におけるMAS法における使用も同様に好適である。
【0111】
QTLの同定後、QTLの効果(耐性)を、例えば試験対象のQTLsを分離させているBC2S1世代の耐性を評価することによって確認してもよい。耐性の評価は、周知のようにキュウリクロステロウイルス及びキュウリウドンコ病耐性をバイオアッセイするのが最適である。例えば、単離CYSDV(DSMZ−Deutsche Sammlung von Mikroorganismen及びZellkulturen社(ブラウンシュヴァイク、ドイツ)のコレクションno.PV−0592/EWSN_6として保管)を使用してもよく、実験的な感染は、例えばBemisia tabaci(Bバイオタイプ)による伝達により行ってもよい。また、自然感染条件下でのフィールド試験を行い、CYSDV又は他のクロステロウイルス耐性を評価してもよい。植物材料の人工接種によりウドンコ病耐性を評価してもよい。本発明に係るマーカーを最適に用いて、ウドンコ病及び/又はクロステロウイルス耐性を有すると思われるキュウリにおける、本発明の1つ以上のQTLsの存在を検出してもよく、またそれを用いて、マーカーアシスト選抜、並びにウドンコ病及びクロステロウイルス耐性キュウリの育種を実施してもよい。好ましくは、本発明のQTLの存在の検出は、本明細書で定義される少なくとも1つのQTLのマーカーを用いて実施される。ゆえに本発明の別の態様は、ウドンコ病及び/又はクロステロウイルス耐性を付与するQTLの存在の検出方法の提供に関する。当該方法は、ウドンコ病及び/又はクロステロウイルス耐性であると考えられるキュウリにおいて、前記QTLの核酸配列の存在を検出することを含んでなり、上述のマーカーを用いてその存在を検出することを特徴とする。
【0112】
本発明のQTLの核酸配列は、例えば前記QTLと関連する1つ以上のマーカーの核酸配列を決定し、前記マーカー配列の内部プライマーを設計し、更にそれを用いて前記マーカー配列の外のQTLの配列を決定して解析してもよい。例えば、表2、3及び4のAFLPマーカーの核酸配列は、対象となる植物のゲノム中の前記マーカーの存在の決定において使用する泳動ゲルから前記マーカーを単離し、例えば公知の方法であるジデオキシ末端停止法により、前記マーカーの核酸配列を決定してもよい。
【0113】
ウドンコ病及び/又はクロステロウイルス耐性を有すると思われるキュウリのQTLの存在を検出するかかる方法の実施形態では、当該方法は、前記QTL(好ましくは表2、3及び4のマーカーから選択)に連結したマーカーの核酸配列にストリンジェントなハイブリダイゼーション条件下でハイブリダイズできるオリゴヌクレオチド又はポリヌクレオチドを提供するステップと、ウドンコ病及び/又はクロステロウイルス耐性を有すると思われるキュウリのゲノム核酸の断片と、前記オリゴヌクレオチド又はポリヌクレオチドとを接触させるステップと、前記ゲノム核酸断片に対する、前記オリゴヌクレオチド又はポリヌクレオチドの特異的なハイブリダイゼーションの存在を測定するステップを含んでなる。
【0114】
好ましくは前記方法は前記ウドンコ病及び/又はクロステロウイルス耐性を有すると思われるキュウリから得られる核酸サンプルに対して実施するが、in situハイブリダイゼーション法を用いてもよい。あるいは、より好ましい実施形態ではあるが、当業者であれば、QTLの核酸配列が決定された後、前記QTLの核酸配列にストリンジェントなハイブリダイゼーション条件下でハイブリダイズできる特異的なハイブリッド形成用のプローブ若しくはオリゴヌクレオチドを設計し、かかるハイブリダイゼーションプローブを用いて、ウドンコ病及び/又はクロステロウイルス耐性を有すると思われるキュウリにおいて、本発明のQTLの存在を検出することができる。
【0115】
主要なクロステロウイルス抵抗性遺伝子の位置を示すマーカーは、親株から生じた耐性個体及び高感受性個体からのキュウリDNA断片の選択的増幅によって得てもよい。当該方法は、−前記断片を制限酵素(例えばEcoRI及びMsel)で処理し、更にライゲーションさせ、当該断片の端にプライマーに対応する補助アダプター(それらの端に1つ以上の特異的なヌクレオチドを有する)を連結させるステップであって、プライマー対のうちの1つのプライマーが検出目的で標識されているステップと、−変性条件下でゲル電気泳動を行い、増幅生成物を分離させるステップと、−抵抗性の子孫に由来する断片の混合物、及び感受性の子孫に由来する断片の混合物と、親株に由来する断片とを、それらの電気泳動プロファイルを比較することによって、抵抗性の遺伝子座と遺伝的に連結している多形バンドを同定するステップを含んでなる。前記同定の後、全ての個体に関して確証試験を行い、更にマーカーと耐性遺伝子座との間の遺伝子組換え率を算出する。
【0116】
原則として、本明細書で用いられる個々のAPLPマーカーは指定されたマーカーコードを有する。このコードは2つのプライマーを定義し、任意に、定義済みのアクセッションNo.(上記表1の説明を参照)のプライマーの増幅生成物の長さを示す図との組み合わせにより説明する。すなわちAFLPマーカーを用いることにより、キュウリゲノムDNAを鋳型とした増幅反応を実施し、一本鎖若しくは二本鎖DNAフラグメントを得、示されたアクセッションNo.の場合、示された長さの断片が得られる。更に当該マーカーは、5’−3’方向において、第1のプライマー配列、キュウリ特異的なDNA塩基配列、及び第2のプライマー配列からなる配列、更にはその補足配列を含んでなる。したがって、キュウリ特異的なDNA塩基配列には2つのプライマーが隣接している。用語「キュウリ特異的なDNA塩基配列」は、それぞれのプライマーが隣接する領域の核酸配列を意味し、国際公開第02/22836号にて説明したように、特定のプライマーを使用してキュウリ目録Khira PI250147から増幅される配列、又はそれに対して少なくとも90%、好ましくは少なくとも95%、最も好ましくは少なくとも98%の配列相同性を有する配列を表す。
【0117】
トランスジェニック技術によるウドンコ病及びクロステロウイルス耐性キュウリの育種:
本発明の別の態様では、少なくともQTL−1を含んでなる核酸(好ましくはDNA)配列、並びにpm−l及びpm−h又はその耐性付与部分のうちの1つ(好ましくは両方)を、本発明の二重耐性キュウリの育種に用いてもよい。本発明のこの態様では、本明細書で定義されるQTLs又はその耐性付与部分の使用により、二重耐性キュウリを作製し、当該使用においては、適切な宿主植物への前記QTLを含む核酸配列の導入を含んでなる。前述のように、前記核酸配列は、適切なクロステロウイルス及び/又はウドンコ病耐性ドナー植物に由来してもよい。QTL−1として本願明細書に同定されるクロステロウイルス耐性遺伝子座の適切な供給源として、キュウリランドレース Khira(PI250147)(パキスタン産)が挙げられる。それらの商業的な供給源、並びにその他の多くのPM耐性キュウリ園芸品種は、例えばhttp://cuke.hort.ncsu.edu/cucurbit/cuke/cukemain.html.に列挙されている。PM耐性植物は、例えばT.C.Wehner,cucumber gene curator for the Cucurbit Genetics Cooperative(CGC),Department of Horticultural Science,North Carolina State University,Ealeigh,NC27695−7609 U.S.A.から得られてもよい。本願明細書に記載する両方のpm遺伝子座の供給源としては、NPI(PI 200815を有する交雑Natsufushinari(PI 279465)から調製)が挙げられる。それらは例えば、Centre for Genetic Resources,the Netherlands(CGN),Wageningen,The Netherlandsから得てもよい。ECP/GR Cucurbits Database hosted by the Center for the Conservation and Breeding of Agrodiversity(COMAV) in Valencia,Spain、又はGermplasm Resources Information Network(GRIN) hosted by the USDA’s National Germplasm Resources Laboratory,Beltsville,Marylandなどの幾つかの植物データベースを用いて、登録株の検索を行ってもよい。本発明によるこれらの材料の同定方法において記載したように、クロステロウイルス又はウドンコ病耐性を示す他のキュウリを耐性ドナー植物として利用してもよい。
【0118】
適切なドナー植物を同定した後、クロステロウイルス又はウドンコ病耐性を付与するQTLを含む核酸配列、又はその耐性付与部分を、利用可能ないかなる方法によって適切な宿主植物へ導入してもよい。例えば、上述の核酸配列は、感受性の宿主植物と、クロステロウイルス又はウドンコ病耐性ドナー植物を交配すること(すなわち遺伝子移入)、形質転換、プロトプラスト融合、二重ハプロイド技術、又は胚レスキュー若しくは他のいかなる核酸伝達システムによって行ってもよく、任意に、QTLを含む耐性型の子孫植物の選抜をその後行ってもよい。クロステロウイルス又はウドンコ病耐性(又はその耐性付与部分)のためのQTLを含んでなる核酸配列を導入するトランスジェニック技術では、当該核酸を公知の方法を用いて前記ドナー植物から単離し、その単離された核酸をトランスジェニック技術を用いて宿主植物に導入する。当該導入方法としては、例えばベクター、生殖体、又は他の適切な媒体(例えば前記核酸配列でコーティングした粒子)の導入が挙げられる。
【0119】
植物の形質転換は一般に、植物細胞において機能する発現ベクターの構築を必要とする。本発明において、かかるベクターはクロステロウイルス又はウドンコ病耐性のためのQTL(又はその耐性付与部分)を含む核酸配列を含んでなり、当該ベクターにおいて、クロステロウイルス又はウドンコ病−耐性を与えている遺伝子は、調節エレメント(例えばプロモータ)と連結しているか、又は制御下にある。当該組合せに含まれる遺伝子の少なくとも1つがクロステロウイルス又はウドンコ病−耐性をコードする場合、当該発現ベクターは1つ以上のかかる制御可能に連結された遺伝子/制御エレメントの組合せを含んでもよい。当該ベクターはプラスミドの形でもよく、単独で使用してもよく、又は他のプラスミドとの組合せで使用してもよく、それらを用いて、公知技術の形質転換法(例えばアグロバクテリウム形質転換系)を用いて、クロステロウイルス耐性及びウドンコ病遺伝子を導入した植物を得る。
【0120】
発現ベクターは、使用可能な状態で調節エレメント(例えばプロモータ)と連結された少なくとも1つのマーカー遺伝子を含んでもよく、それにより、マーカーを含む形質転換細胞を、ネガティブ選抜(選択可能なマーカー遺伝子を含まない細胞生長を阻害)によって、又は、ポジティブ選抜(マーカー遺伝子によってコードされる生成物のスクリーニング)により回収することができる。植物の形質転換に一般的に用いられる選択可能な多くのマーカー遺伝子が従来技術において周知であり、例えば、選抜用の化学物質の毒性を代謝的に除去する酵素をコードする遺伝子(抗生物質又は除草剤)、又は阻害剤に対する感受性を喪失する形に変換された標的をコードする遺伝子などが挙げられる。マンノース選抜のようないくつかのポジティブ選抜方法が公知である。あるいは、マーカーを使用しない形質転換を行い、前記マーカー遺伝子のない植物を得てもよく、その技術は従来技術において周知である。
【0121】
発現ベクターを植物に導入する1つの方法は、アグロバクテリウム(例えばHorschら、1985)の自然形質転換システムに基づく。A.tumefaciens及びA.rhizogenesは遺伝的に植物細胞を形質転換する植物病原性土壌細菌である。A.tumefaciens及びA.rhizogenesのTi及びRiプラスミドはそれぞれ、植物の形質転換に関与する遺伝子を担持する(例えばKado(1991)を参照)。発現ベクターを植物組織に導入する方法としては、アグロバクテリウムチュメファシエンス、(Horschその他、1985)を用いた植物細胞の直接感染又は共存培養が挙げられる。グルーバー及びクロスビー(1993)及びマローニー(1989)は、アグロバクテリウムを媒介された遺伝子移入その他のアグロバクテリウムベクターシステムを開示している。また米国特許第5591616号を参照のこと。植物発現ベクター及びレポーター遺伝子及び変換プロトコルの一般的説明、並びにアグロバクテリウムの媒介による遺伝子移入ためのアグロバクテリウムベクターシステム及びそれを用いた方法の説明は、グルーバー及びクロスビー(1993)に記載されている。植物組織を培養する一般の方法は、三木ら(1993)による例、及びフィリップスら(1988)により開示されている。分子クローニング技術及び適切な発現ベクターのための適当な参照ハンドブックとしては、Sambrook及びRussell(2001)が挙げられる。
【0122】
発現ベクターを植物に導入する他の方法として、マイクロインジェクションにより媒介される形質転換に基づく方法が挙げられる。当該方法では、DNAをマイクロ発射体の表面上に担持させることを特徴とする。発現ベクターは、植物細胞壁及び膜を透過するのに十分な300〜600m/sの速度にマイクロ発射体を加速する装置を用いて植物組織に導入される(サンフォードその他、1987、1993;サンフォード、1988、1990;クラインら、1988、1992を参照)。植物にDNAを導入する他の方法としては、標的細胞(チャンその他は、知る、1991)の超音波処理を介する方法が挙げられる。あるいは、リポソーム又はスフェロプラスト融合を用いて、発現ベクターを植物に導入してもよい(例えばDeshayesら、1985、及びChristouら(1987を参照)。CaCl2沈殿、ポリビニルアルコール又はポリ−L−オルニチンを用いたプロトプラストへのDNAの直接の取り込みも報告されている(例えばHainら、1985、及びDraperら、1982を参照)。プロトプラスト及び全細胞及び組織へのエレクトロポーレーションに関しても開示されている(D’Halluinら、1992、及びラウルセン、1994を参照)。
【0123】
他の周知の技術(例えばBACの使用、キュウリゲノムの一部を細菌人工染色体(BAC)に導入(すなわちベクターを用いて大腸菌細胞のDNAフラグメント(100〜300kbのサイズを挿入、平均150kb)をクローニング)し、バクテリア大腸菌で見つかる天然のP因子プラスミドに基づく(Zhao及びStodolsky、2004)方法)、を例えばBIBACシステム(ハミルトン、1997)との組合せで使用し、遺伝子導入植物を作製してもよい。
【0124】
キュウリ標的組織の形質転換に続いて、上記した選択可能なマーカー遺伝子を発現させ、現在公知技術の再生及び選抜方法を用いて形質転換細胞、組織及び/又は植物の選抜を行う。
【0125】
非トランスジェニック方法によるクロステロウイルス及びウドンコ病耐性キュウリの育種:
クロステロウイルス及びウドンコ病耐性キュウリを作製するための別の実施形態では、プロトプラスト融合を用いて、ドナー植物から宿主植物への核酸導入を行ってもよい。プロトプラスト融合とは、2つ以上のプロトプラスト(細胞壁を酵素処理によって除去した細胞)間の誘発若しくは自然的な結合(例えば体細胞ハイブリダイゼーション)により、単一の二核若しくは多核細胞を作製する技術のことを指す。融合細胞(性質により交配できない植物種から得てもよい)を組織培養して、望ましい特徴の組合せを示すハイブリッド植物を形成できる。より詳しくは、第1のプロトプラストを、クロステロウイルス又はウドンコ病感染に対する耐性を示すにキュウリ又は他の植物系列から得てもよい。第2のプロトプラストは、例えば、病害抵抗性、虫害抵抗性、有益な果物特徴など(これらに限定されない)を有する第2のキュウリ又は他の植物変異体(好ましくは商業的に有益な特徴を有するキュウリ系列)から調製してもよい。次にプロトプラストを、従来公知のプロトプラスト融合法を用いて融合させる。
【0126】
あるいは、ドナー植物から宿主植物への胚レスキュー(embryo rescue)を採用し、本願明細書に記載の1つ以上のQTLsを含有する核酸を導入してもよい。胚レスキューは、植物が生存できる種子の作製に失敗した時、交配株から胚を単離するのに用いることができる。この方法では、植物の受精卵又は未熟種子を組織培養し、新規な植物を作製する(Pierik、1999)。
【0127】
本発明はまた、クロステロウイルス又はウドンコ病耐性キュウリの作製方法にも関する。当該方法は、上記したドナーキュウリ中において、クロステロウイルス又はウドンコ病耐性に関連する定量耐性遺伝子座(QTL)の存在を検出する方法を実施するステップと、少なくとも検出した当該1つ以上のQTL、又はクロステロウイルス若しくはウドンコ病耐性付与部分を含んでなる核酸配列を、前記ドナー植物からクロステロウイルス又はウドンコ病感受性の宿主キュウリに導入するステップを含んでなる。前記核酸配列の導入は本願明細書に上記したいかなる方法で実施してもよい。
【0128】
かかる方法の好ましい実施態様として、交雑によって、前記核酸配列を、クロステロウイルス又はウドンコ病耐性ドナーキュウリから、クロステロウイルス又はウドンコ病感受性の宿主キュウリへの遺伝子導入する態様が包含される。したがって、この遺伝子導入は従来の繁殖方法を用いて適宜実施してもよい。QTLsは好ましくはマーカーアシスト選抜(MAS)又はマーカーアシスト育種(MAB)を用いて商業的なキュウリ種へ移入される。MAS及びMABは、所望の特徴をコードする遺伝子を1つ以上含むそれらの子孫植物の同定及び選抜における、当該分子マーカーの1つ以上の使用を含んでなる。かかる例においては、その同定及び選抜は本発明に係るQTLs又はそれに連結するマーカーの選抜に基づく。MASを用いて、目的のQTLを有する近同質遺伝子系統(NIL)を育種してもよく、各QTLの効果の詳細な検討も可能となり、また戻し交配自殖系統(BIL)集団の育種にも応用することができる(例えばNesbittら、2001、van Berlooら、2001)。この実施形態により育種されるキュウリは、宿主植物から大多数のそれらの特徴を好適に引き継ぎ、またドナー植物からのクロステロウイルス及びウドンコ病耐性の特徴も有する。
【0129】
上記したように、従来の繁殖方法を用いて、クロステロウイルス又はウドンコ病耐性に関与する核酸配列を、クロステロウイルス及びウドンコ病感受性の宿主キュウリに導入することができる。系統育種と呼ばれる1つの方法では、クロステロウイルス又はウドンコ病に対する耐性を示し、クロステロウイルス又はウドンコ病耐性に関与する核酸コード配列を有するドナーキュウリを、クロステロウイルス若しくはウドンコ病感受性の宿主キュウリ(好ましくは商業的に望ましい特徴(限定されないが病害抵抗性、虫害抵抗性、有益な果物特徴など)を示す)と交配させる。得られる植物群(F1ハイブリッド)を更に自家授粉し、種子(F2種子)を得る。F2種子から成長させたF2植物を、更にクロステロウイルス及びウドンコ病耐性に関してスクリーニングする。当該集団を、様々な方法でスクリーニングしてもよい。
【0130】
第1に、従来の疾患スクリーンを使用して集団をスクリーニングしてもよい。かかる病態スクリーニングは公知である。好ましくは、定量的バイオアッセイを用いる。第2に、マーカーアシスト選抜を実施し、上記した分子マーカーの1つ以上を使用してクロステロウイルス又はウドンコ病耐性に関与する核酸コード配列を含むそれらの子孫を同定してもよい。上記した他の方法を用いて、QTLの存在を検出してもよい。また、マーカーアシスト選抜を用いて定量的バイオアッセイから得られる結果を確認してもよく、またその他の幾つかの方法を組み合わせて用いてもよい。
【0131】
近交系クロステロウイルス及びウドンコ病耐性キュウリ系列の親系列は、循環選択及び戻し交配の技術を使用して育種してもよく、自殖及び/又はジハプロイド又は他のあらゆる技術を用いて作製してもよい。循環選択及び戻し交配において、クロステロウイルス及びウドンコ病耐性は、循環親と、第1のドナー植物を交配させることにより標的宿主植物に移入することもできる。それは反復親とは異なり、本明細書において「一回親」と称される。循環親はクロステロウイルス及びウドンコ病に対する耐性を有さないか、又は低い耐性を有する植物であって、例えば商業的に望ましい特徴(病害抵抗性、虫害抵抗性、有益な果物特徴など)を有するものである。一回親は、クロステロウイルス及びウドンコ病耐性を示し、クロステロウイルス及びウドンコ病耐性に関与する核酸配列を含んでなる。一回親は、循環親との交配により得られる植物変異体又は近交系であってもよい。循環親及び一回親との間で交配して得た株を、循環親に戻し交配する。得られる植物群を更に所望の特徴に関してスクリーニングする。当該スクリーニング方法は特に限定されない。例えば、周知の表現型病理スクリーニング又は定量的バイオアッセイを用いて、集団をスクリーニングしてもよい。あるいは、バイオアッセイを使用する代わりに、マーカーアシスト選抜(MAS)を、上記の分子マーカー、ハイブリッド形成プローブ又はポリヌクレオチドの1つ以上を使用して、クロステロウイルス及びウドンコ病耐性に関与する核酸配列を含む子孫を同定してもよい。また、MASを用いて定量的バイオアッセイから得られる結果を確認してもよい。また、pm−hの劣性形質は、この遺伝子が表現型スクリーン(例えば耐性バイオアッセイ)を用いてF1又はBC1集団から選抜できないことを意味する。ゆえに本願明細書に係るマーカーは、遺伝子型のスクリーニングによって適当な子孫植物を選択するのに適切である。
【0132】
スクリーニングの後、クロステロウイルス及びウドンコ病耐性表現型を示すF1ハイブリッド植物、又は好ましくは遺伝子型は、クロステロウイルス及びウドンコ病耐性に関与する核酸配列を含んでなり、それを更に選抜して循環親に戻し交配し、キュウリを次第に近交系にする。このプロセスは、2〜5以上の世代にわたり実施することが好ましい。原則として、循環親と一回親との交配により得られる、クロステロウイルス及びウドンコ病耐性株の子孫は、クロステロウイルス及びウドンコ病耐性に関与する1つ以上の遺伝子に関してヘテロ接合体である。
【0133】
例えば、植物系列にpm−l及びpm−h遺伝子座を移入したとき、pm−h遺伝子座は劣性であり、その耐性表現型は、ホモ接合のpm−h植物を形成できる条件下においてのみ子孫植物において生じることに留意すべきである。
【0134】
一般に、所望の特徴をハイブリッドキュウリ変異体に導入する方法は、以下のステップを含んでなる:
(a)1つ以上の所望の特徴を含む他のキュウリと、近交系キュウリ親を交配して、F1子孫植物を作製するステップであって、当該所望の特徴が、クロステロウイルス耐性及びウドンコ病耐性からなる群から選択されるステップと、
(b)好ましくは本明細書で定義される分子マーカーを使用して、所望の特徴を有する前記F1子孫植物を選抜するステップと、
(c)前記近交系キュウリ親株と、前記選択された子孫との戻し交配して戻し交配植物を得るステップと、
(d)戻し交配子孫植物を、所望の特徴及び前記近交系キュウリ親株の形態的、生理的な特徴に関して選択するステップであって、前記選抜が、ゲノムDNAを単離し、上記DNAを、QTL−1、pm−l及び/又はpm−hの少なくとも1つの分子マーカー(好ましくは本願明細書に記載のもの)の存在に関して試験するステップと、
(e)ステップ(c)及び(d)を2回以上反復し、第3世代若しくはそれ以上の戻し交配子孫植物を作製するステップと、
(f)選抜した戻し交配子孫を任意に近交し、ホモ接合の植物を同定するステップと、
(g)最低1つの前記戻し交配子孫又は近交系植物と、他の近交系のキュウリ親株(好ましくはpm−hypo耐性、すなわちホモ接合のpm−h)とを交配し、ハイブリッドキュウリ変異体(同じ環境で成長させたとき、ハイブリッドキュウリ変異体の所望の特徴、並びに形態的及び生理的特徴の全てを有する)を得るステップを含んでなる。
【0135】
上記のように、最後の戻し交配生成は近交により行い、クロステロウイルス及びウドンコ病耐性に関してホモ接合の純粋株(近交系)子孫を得てもよい。したがって、循環選択、戻し交配及び近交により、商業的に有用な特徴と関連する他の遺伝子と共に、クロステロウイルス及びウドンコ病耐性に関連する遺伝子と相同な遺伝子を有する系列が得られる。
【0136】
クロステロウイルス及びウドンコ病耐性キュウリ及び種子:
植物育種の目的は、単一の変異体又はハイブリッド中で様々な望ましい特徴を組み合わせることである。商品作物の場合、これらの特徴としては、病気及び昆虫に対する耐性、熱及びかんばつに対する体制、成熟期間の短縮、収率の上昇及び農産物としての優れた品質などが挙げられる。植物特徴(例えば発芽及び成長、成長率、成熟及び草高の均一性など)が重要となることもある。
【0137】
商品作物は、花粉媒介による植物的な方法を利用して育種される。植物は、同一の花に由来する花粉が、その同一の花若しくは同じ植物の他の花へ移される場合に自家授粉される。同じファミリー又は系列の個体を花粉媒介に用いたとき、植物は兄弟授粉される。花粉が、異なるファミリー又は異なる系列の植物に存在する花に由来する場合、植物は他花受粉される。
【0138】
多世代にわたり自動授粉され、形態を基に選抜された植物は、全ての遺伝子座においてホモ接合性になっており、真の繁殖子孫としての同一集団を形成する。2つの異なるホモ接合性系列間での交配により、多くの遺伝子座に関してヘテロ接合体であるハイブリッド植物の同一集団を形成する。多くの遺伝子座でヘテロ接合体となっている2本の植物の交配により、遺伝的に多様な、同一でないヘテロな植物集団を形成する。
【0139】
キュウリ育種プログラムにおけるハイブリッドキュウリ多様性の成長は、以下の3つのステップを含んでなる:
(1)第一の育種交配のために、様々な生殖質プールから植物を選抜するステップと、
(2)繁殖交配から選抜された植物を数世代にわたり近交し、個々に繁殖した、非常に均一な一連の近交系を作製するステップと、
(3)ハイブリッド子孫(F1)の作製に無関係な近交系と、選抜された近交系とを交配させるステップ。同系交配を充分行った後、連続したハイブリッド世代は単に、育種された近交系の種子を増加させる目的でのみ用いる。好ましくは、近交系は、その遺伝子座の約95%以上でホモ接合性対立遺伝子を含まなければならない。
【0140】
近交系のホモ接合性及び均質性による顕著な効果として、確立された近交系のペアを交配して作製したハイブリッドは、常に同質であるということである。優れたハイブリッドを作製する近交系を同定した後、これらの近交系両親を使用してハイブリッド種を継続的に供給することができ、更にこのハイブリッド種子からハイブリッドキュウリを栽培することができる。
【0141】
本発明の方法で作製できるクロステロウイルス及びウドンコ病抵抗キュウリ、又はその部分は、本発明に包含される。
【0142】
本発明の別の態様は、上記にて説明したQTLsを何らかの構成で含んでなるクロステロウイルス及びウドンコ病耐性キュウリ又はその部分の提供に関し、詳細には、少なくとも1つの前記QTLsはその天然の遺伝的バックグラウンドに存在しない。本発明のクロステロウイルス及びウドンコ病耐性キュウリは、例えば近交系のいかなる遺伝子タイプ、ハイブリッド、ハプロイド、ジハプロイド、又はトランスジェニックであってもよい。更に、本発明の植物は、耐性特徴に関してヘテロ接合体であってもよく、またホモ接合性であってもよい(好ましくはホモ接合性である)。本発明のQTLs(並びにその耐性付与部分)をいかなる植物に導入してクロステロウイルス及びウドンコ病耐性植物を作製してもよいが、本発明に係る方法及び植物は、キュウリに関連する植物である。
【0143】
本願明細書に記載されているクロステロウイルス及びウドンコ病耐性近交系キュウリ系列を用いて、更に交配を行い、クロステロウイルス及びウドンコ病耐性ハイブリッド植物を作製してもよい。例えば、本発明の第1のクロステロウイルス及びウドンコ病耐性近交系キュウリは、商業的に望ましい特徴(例えば病害抵抗性、虫害抵抗性、望ましい果物特徴など)を有する第2の近交系キュウリと交配してもよい。この第2の近交系キュウリ系列は、クロステロウイルス又はウドンコ病耐性を有さなくてもよい。好ましくは、この系列は、この劣性形質がハイブリッドの子孫植物において発現されるために、pm−hに関してホモ接合性である。
【0144】
本発明の別の態様は、クロステロウイルス及びウドンコ病耐性キュウリを栽培する際の種子の作製方法に関する。一実施形態では、当該方法は、
クロステロウイルス及びウドンコ病耐性キュウリを準備するステップと、
前記クロステロウイルス及びウドンコ病耐性植物を他のキュウリと交配するステップと、
前記交配から得られる種子を収集するステップを含んでなる。更にそれを栽培し、クロステロウイルス及びウドンコ病耐性キュウリを作製する。
【0145】
別の実施形態では、当該方法は以下のステップを含んでなる:
本発明のクロステロウイルス及びウドンコ病耐性キュウリを準備するステップと、
前記クロステロウイルス及びウドンコ病耐性植物とキュウリとを交配するステップと、
前記交配から生じる種子を回収し、前記種を植物体に成長させるステップと、
本願明細書に記載のいずれかの方法で、クロステロウイルス及びウドンコ病耐性植物を選抜するステップと、
充分な世代数にわたり、選択された植物を自家授粉して繁殖させ、クロステロウイルス及びウドンコ病耐性を付与する対立遺伝子が安定した植物を得るステップと、
そのように得られた植物を、望ましい表現型を有するキュウリと十分な世代数にわたり戻し交配して、クロステロウイルス及びウドンコ病耐性、並びに望ましい表現型を有するキュウリを得るステップと、
最後の戻し交配から生じる植物から作製される種子を回収するステップ。上記の種子を栽培し、クロステロウイルス及びウドンコ病耐性を有するキュウリを作製する。
【0146】
表4のマトリックスにおいて、pm耐性若しくはクロステロウイルス耐性の親に由来する(耐性親の存在として示される)二重耐性植物における、マーカーの有無を示す。マトリックスは更に、二重(PM及びクロステロウイルス)耐性植物系列と関連するAFLPマーカーの起源及び相対的位置を示す。用語pm−l及びpm−hにより示される「マーカー」は、2つのそれぞれのウドンコ病耐性遺伝子座の中心位置を示す。QTL−1遺伝子座の位置は決定しなかったが、クロステロウイルス関連マーカーを使用することにより、その遺伝子座は2つのpm遺伝子座の間に存在することが示される。表より、プールnrs.1〜6では、プールnrs.10〜15より短い、クロステロウイルス耐性親からの遺伝子移入が可能であることが示唆される。マトリックスの解釈は、以下の通りである:例えば、マーカーE23/M40−M003(56.9cM)は、pm耐性親及び二重耐性子孫に存在したが、一方マーカーE11/M80−M003(79.1cM)は、クロステロウイルス耐性親には存在したものの、二重耐性子孫には存在しなかった。すなわち、マーカーE23/M40−M003はPM関連のマーカーであり(その存在は、テストした植物系列のpm−l関連遺伝子の領域の存在を示す)、一方、マーカーE11/M80−M003は、クロステロウイルス耐性に関連するQTL−1遺伝子座の近傍の遺伝的領域であるが、pm−hypo(pm−h)耐性の発現にとっては必須ではないことを示す。
【0147】
【表4】
+=ホモ接合性が存在(高密度のバンド)、
−=存在しない(バンドなし)、
+/−=ヘテロ接合体が(低密度バンド)、
D=中程度に存在(中程度のバンド)、
・=PM抵抗親におけるマーカースコア(両方存在:[+]、不在:[−])、
・=マーカースコア(クロステロウイルス耐性親)、
・=親系列における未同定マーカーの存在。
【図面の簡単な説明】
【0148】
【図1】考えられる組換えシナリオを示す図。2つの別々の植物から得たそうであるウドンコ病(pm)及びクロステロウイルス(QTL−1)耐性を、単一の子孫植物内で連結させた。多くの耐性遺伝子座を有するものとして示される様々な親植物は、それ自体交配若しくは自家交配の結果でありえることを理解すべきである。すなわち、QTLであるpm−l及びpm−hを有する植物は、単一のQTLを有する植物間での交配によって得られる。
【図2】同上。
【図3】同上。
【図4】同上。
【図5】同上。
【図6】同上。
【図7】単一の連鎖群における、キュウリNPI由来のウドンコ病耐性(pm)遺伝子、及びC.sativus Khira由来のクロステロウイルス耐性(QTL−1)遺伝子の様々な位置関係を示す。Pm−leaf1は、本明細書で用いられるpm−lに相当し、Pm hypoは本明細書で用いられるpm−hに相当する。
【図8】本願明細書の表2及び3中の各配列番号により示されるSNP及び挿入マーカー近傍のDNA配列を示す。
【技術分野】
【0001】
本発明は、耐病性植物及び植物育種の分野、より詳細にはウドンコ病及びクロステロウイルス−耐性キュウリ植物の育種に関する。
【背景技術】
【0002】
キュウリ(Cucumis sativus)の農業生産においては、様々な植物病に見舞われることがしばしばある。黄化ウイルスによってそのような植物病が生じ、キュウリの生産において深刻な経済的ダメージが生じることもある。クロステロウイルス(Closteroviridae)(キュウリに悪影響を与える黄化ウイルスの中で最も上位の分類学上のクラスタを形成)のファミリーには、30種類以上の、曲状及び糸状の、昆虫により媒介される植物ウイルスが含まれる。上記ファミリーは3つの属からなり、特にモンシロチョウにより媒介されるCrinivirus属には、キュウリに深刻な影響を与える種が含まれる。この属には特に、ウリ科黄色萎縮ウイルス(CYSDV)、レタス伝染性黄斑ウイルス(LIYV)、及びビート偽黄化病ウイルス(BPYV)などの種が属する。CYSDV及びBPYVはキュウリ栽培者にとり最も脅威である。
【0003】
上記のウイルスは通常昆虫ベクター上に存在し、植物上の昆虫の摂食活動によって伝播する。したがって、キュウリクロステロウイルスは、殺虫剤の散布により抑制できる。しかしながら、好ましくは農業及び園芸におけるクロステロウイルスの制御は、寄主植物のウイルス耐性栽培品種を提供することにより行われる。現時点では、抵抗性のキュウリ種のゲノム上の少なくとも2つの量的形質遺伝子座又はQTLsが、キュウリのクロステロウイルス耐性と関連していることが知られている(特許文献1)。ゆえに子孫植物へのクロステロウイルス耐性関連の遺伝物質の適当な遺伝的浸透は、これらのQTLsに連結させた特異的なマーカーを検出することによってモニターできる。ゆえにQTLsについての見識を利用して、クロステロウイルス耐性栽培品種の育種を効率的に行うことが可能となる。
【0004】
キュウリの農業生産に影響を及ぼす他の重要な植物病として、キュウリウドンコ病(PM)が挙げられる。ウリ科のPMは、菌類のS.fuliginea及びE.cichoracearumによって生じうる。当該植物病は一年中発生し、その症状は、感染した葉の表面上における純粋な白い菌糸による小さなスポットの発生から始まり、そのスポットが次第に拡大し、最終的に茎及び葉が白い、粉末状の大量の胞子及び菌糸で覆われるに至る、というものである。重度の感染により葉の変色及び損失が生じ、その結果、果実の数及びサイズの減少に至る。ウドンコ病に対して効果的な多くの殺菌剤が存在するが、かかる化学物質に対する菌の耐性の問題が生じる。
【0005】
ある程度のウドンコ病耐性であれば、様々なキュウリ株において見られる。かかる株としては、例えばインドの野生のキュウリ:番号PI197088及び番号PI200815、PI200818、並びに栽培品種Natsufushinari及びAsomidori(モリシタら、2003)が挙げられる。また、PMに対する耐性と関連する様々な遺伝子が公知であり(Fanourakis、1984;Fujieda及びAkiya(1962);Kooistra、1968、1971;Shanmugasundarumら、1971、1972)、Natsufushinariのpm−1及びpm−2、PI200815及びPI200818のpm−3、及び栽培変種Wis.SMR18の「pm−h」(遺伝子「pm−h」は本願明細書においてpm−hとして示されるpm遺伝子座に相当する)などが挙げられる。栽培されている幾つかのキュウリの種類は、PMに対する部分的な耐性を示すものの、大部分の農業生産者は未だに葉への殺菌剤投与に依存している。この理由の1つとして、他のタイプの耐性を示す株に対してPM耐性を導入するのが困難であるという点が挙げられる。例えば、PM耐性対立遺伝子を得た株が、クロステロウイルスに対して感受性になる(それまではそうではなかった)、あるいはその逆という事態が育種の際に生じることが例として挙げられる。実際に、交配試験において、PM及びクロステロウイルス両方に対する耐性を示すキュウリの株はこれまで得られていない。すなわち有用な二重抵抗性の組換え体はこれまで得られていない。しかしながら驚くべきことに、比較的直接的な育種方法によっても、異なる特徴を単一のゲノムに取り込ませることが可能であることを見出した。
【特許文献1】国際公開第02/22836号パンフレット
【非特許文献1】Ausubel FM,Brent R,Kingston RE,Moore DD,Seidman JG,Smith JA,Struhl K(eds.)(1998)Current Protocols in Molecular Biology Wiley,New York.
【非特許文献2】Bai YL,Huang CC,van der Hulst R,Meijer Dekens F,Bonnema G,Lindhout P(2003)QTLs for tomato powdery mildew resistance(Oidium lycopersici)in Lycopersicon parviflorum Gl.1601 co−localize with two qualitative powdery mildew resistance genes.MoI.plant microbe interactions 16: 169−176.
【非特許文献3】Christou P,Murphy JE,and Swain WF(1987)Stable transformation of soybean by electroporation and root formation from transformed callus.Proc.Natl.Acad.Sd.USA 84:3962−3966.
【非特許文献4】Deshayes A,Herrera−Estrella L,Caboche M(1985)Liposome−mediated transformation of tobacco mesophyll protoplasts by an Escherichia coli plasmid.EMBO J.4:2731−2737.
【非特許文献5】D’Halluin K,Bonne E,Bossut M,De Beuckeleer M,Leemans J(1992)Plant.Cell 4: 1495−1505.
【非特許文献6】Draper J,Davey MR,Freeman JP,Cocking EC and Cox BJ(1982)Ti plasmid homologous sequences present in tissues from Agrobacterium plasmid− transformed Petunia protoplasts.Plant and Cell Physiol.23,451−458.
【非特許文献7】Fanourakis NE(1984)Inheritance and linkage studies of the fruit epidermis structure and investigation of linkage relations of several traits and of meiosis in cucumber.Ph.D.Diss.,Univ.of Wisconsin,Madison.
【非特許文献8】Fujieda K,and Akiya R(1962)Genetic study of powdery mildew resistance and spine color on fruit in cucumber.J.Jpn.Soc.HoH.Sd.31:30−32.
【非特許文献9】Ganal MW(1996)Isolation and analysis of high− molecular−weight DNA from plants,In: Nonmammalian Genomic Analysis: A Practical Guide.Academic Press Inc.,San Diego,61−73.
【非特許文献10】Gruber MY,Crosby WL(1993)Vectors for plant transformation.In BR Glick,JE Thompson,eds,Methods in Plant Molecular Biology and Biotechnology.CRC Press,Baton Rouge,LA,pp 89−119.
【非特許文献11】Hain R,Stabel P,Czernilofsky AP,Steinbliss HH,Herrera−Estrella L,Schell J(1985)Uptake,integration,expression and genetic transmission of a selectable chimaeric gene to plant protoplasts.MoI.Gen.Genet.199:161−168.
【非特許文献12】Hamilton CM.(1997)A binary−BAC system for plant transformation with high−molecular− weight DNA.Gene 200: 107−116.
【非特許文献13】Horejsi T,Staub JE and Thomas C.(2000)Linkage of random amplified polymorphic DNA markers to downy mildew resistance in cucumber(Cucumis sativus L.)Euphytica 115(2):105−113.
【非特許文献14】Horsch RB,Fraley RT,Rogers SG,Sanders PR,Lloyd A(1985)A simple and general method for transferring genes into plants.Science 227: 1229−1231.
【非特許文献15】Jiang J,Gill BS,Wang GL,Ronald PC and Ward DC(1995)Metaphase and interphase fluorescence in situ hybridization mapping of the rice genome with bacterial artificial chromosomes.Proc.Natl.Acad.Sci.USA 92:4487−4491.
【非特許文献16】Kado CI(1991): Molecular mechanisms of crown gall tumorigenesis.Crit.Rev.Plant Sci.10,1−32.
【非特許文献17】Klein TM,Gradziel T,Fromm ME,Sanford JC(1988). Factors influencing gene delivery into zea mays cells by high velocity microprojectiles.Biotechnology 6: 559−563.
【非特許文献18】Klein TM,Arentzen R,Lewis PA,and Fitzpatrick− McElligott S(1992)Transformation of microbes,plants and animals by particle bombardment.Bio /Technology 10:286−291.
【非特許文献19】Kleine M,Cai D,Hermann RG,and Jung C(1995)Physical mapping and cloning of a translocation in sugar beet(Beta vulgaris L.)carrying a gene for nematode(Heterodera schachtii)resistance from B.procumbens.Theor Appl Genet 90: 399−406.
【非特許文献20】Kooistra E(1968)Powdery mildew resistance in cucumber.Euphytica 17:236− 244.
【非特許文献21】Kooistra E(1971)Inheritance of flesh and skin colors in powdery mildew resistant cucumbers(Cucumis sativus L.).Euphytica 20:521−523.
【非特許文献22】Lui BH(1997)Statistical Genomics: Linkage Mapping and QTL Analysis.CRC Press,pg 18−19.
【非特許文献23】Miki BL,Fobert PF,Charest PJ,Iyer VN.1993.Procedures for Introducing Foreign DNA into Plants.In: Glick BR and Thompson JE(Eds.)Methods in Plant Molecular Biology & Biotechnology,CRC Press,pp.67−88.
【非特許文献24】Moloney MM,Walker JM,Sharma KK(1989)High efficiency transformation of Brassica napus using Agrobacteriuin vectors. Plant Cell Rep 8: 238− 242.
【非特許文献25】Morishita M,Sugiyama K,Saito T,and Sakata Y(2003)Review: Powdery Mildew Resistance in Cucumber.JARQ 37(1),7−14.
【非特許文献26】Nesbitt TC,Tanksley SD(2001)fw2.2 directly affects the size of developing tomato fruit,with secondary effects on fruit number and photosynthate distribution.Plant Physiol.127: 575−583.
【非特許文献27】Paterson AH(1996)Mapping genes responsible for differences in phenotype.p.41−54.In A.H.Paterson(ed.)Genome mapping in plants.R.G.Landes Company.
【非特許文献28】Phillips RL,Somers DA,Hibberd KA.1988.Cell/tissue culture and in vitro manipulation.In: G.F.Sprague & J.W.Dudley,eds.Corn and corn improvement,3rd ed.,p.345−387.Madison,WI,USA,American Society of Agronomy.
【非特許文献29】Pierik,R.L.M.(1999)In vitro Culture of Higher Plants,4th edition.Martinus Nijhoff Publishers,Dordrecht.
【非特許文献30】Sambrook J,and Russell DW(2001).Molecular Cloning: A Laboratory Manual.New York,NY,USA.,Cold Spring Harbor Laboratory Press.
【非特許文献31】Sanford JC,Klein TM,WoIf ED,Allen N(1987).Delivery of substances into cells and tissues using a particle bombardment process.J.Particulate Sd.Technol.5:27−37.
【非特許文献32】Sanford JC(1988)The biolistic process.Trends in Biotechnology 6:299−302.
【非特許文献33】Sanford JC(1990)Biolistic plant transformation.Physiologica Plantarum,79,206−209.
【非特許文献34】Sanford JC,Smith FD,and Russell JA(1993)Optimizing the biolistic process for different biological applications.Methods in Enzymology 217:483−509.
【非特許文献35】Shanmugasundarum S,Williams PH,and Peterson CE(1971)Inheritance of resistance to powdery mildew in cucumber.Phytopathology 61:1218−1221.
【非特許文献36】Shanmugasundarum S,Williams PH,and Peterson CE(1972)A recessive cotyledon marker gene in cucumber with pleiotropic effects.HortScience 7:555−556.
【非特許文献37】Tijssen P(1993)Hybridization with Nucleic Acid Probes.Part I.Theory and Nucleic Acid Preparation.In: Laboratory Techniques in Biochemistry and Molecular Biology.Elsevier.
【非特許文献38】Van Berloo R,Aalbers H,Werkman A,Niks RE(2001)Resistance QTL confirmed through development of QTL−NILs for barley leaf rust resistance.MoI.Breeding 8: 187−195.
【非特許文献39】Vos P,Hogers R,Bleeker M,Reijans M,van de Lee T,Homes M,Frijters A,Pot J,Peleman J,Kuiper M(1995)AFLP: a new technique for DNA fingerprinting.Nucl.Acids Res.23: 4407−4414.
【非特許文献40】Zhang L,Cheng L,Xu N,Zhao M,Li C,Yuan J,and Jia S(1991)Efficient transformation of tobacco by ultrasonication.Biotechnology.9:996−997.
【非特許文献41】Zhao S,and Stodolsky M(2004)Bacterial Artificial Chromosomes.Methods in Molecular Biology VoI 255.Humana Press Inc.Totowa,NJ,USA.
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明の目的は、キュウリに影響を及ぼすクロステロウイルスファミリーのウイルス(特にBPYV及びCYSDV)に対する耐性、S.fuliginea及び/又はE.cichoracearumにより生じるキュウリウドンコ病に対する抵抗性を有するキュウリの提供に関する。
【課題を解決するための手段】
【0007】
本発明の発明者らは鋭意研究の結果、CYSDV及びPMに対する耐性を示す多くのキュウリを見出した。更に、これらの二重抵抗性のキュウリが、ウドンコ病耐性と関連する2つの定量形質遺伝子座(QTLs)(本願明細書におけるpm−l及びpm−h)、並びにクロステロウイルス耐性に関連する主要なQTL(本願明細書におけるQTL−1)を含むことを発見した。更に重要なことに、全てのQTLsが単一の遺伝連鎖群に位置するということを発見した。すなわち本発明の発明者は基本的に、これらの植物ではクロステロウイルス及びウドンコ病に対する耐性遺伝子が単一の染色体に存在することを発見した。かかる植物は植物育種への使用に非常に適する。更に発明者らは、なぜこれまで二重抵抗性のキュウリが交配試験で得られなかったのかを示すメカニズムを発見した。特定の理論に束縛されるわけではないが、1つの植物病に対する耐性を与えるのに必要となる遺伝的要素の遺伝質浸透により、他の植物病に対する耐性を与えるのに必要となる遺伝的要素が損なわれるものと考えられる。この発見によって、発明者らはある特殊な育種方式を開発し、それにより上記の問題が解決され、所望の二重抵抗性を有する植物が得られるようになった。
【0008】
本発明の第1の態様は、キュウリウドンコ病耐性と同時にキュウリクロステロウイルス耐性を有するキュウリの提供に関する。
【0009】
本発明のかかる植物の好ましい実施形態では、クロステロウイルス耐性(QTL−1)に関与する1つ以上のゲノム領域、並びにウドンコ病耐性(pm−h及び/又はpm−l)に関与する1つ以上のゲノム領域は単一の染色体上に存在する。かかる植物のより好ましい実施形態では、pm−h及びpm−lの両方が存在し、かつQTL−1がpm−h及びpm−lの間に位置する。
【0010】
本発明の植物の更なる実施形態では、国際公開第02/22836号(本発明に援用する)において更に詳細に記載されているように、QTL−1の存在は、マーカーE16/M50−244、E16/M50−188及びE11/M48−251からなる群から選抜される少なくとも1つの隣接マーカーの存在の有無により示される。
【0011】
本発明の植物の他の実施形態では、pm−hと称されるQTLの存在は、前記植物のウドンコ病耐性を伴う単一のヌクレオチド多形(SNP)から生じる、少なくとも1つの突然変異を含んでなる核酸配列の存在に有無により示され、前記少なくとも1つの単一ヌクレオチド多形(SNP)は下記の表2に示すようにSNP1及びSNP2から選抜される。
【0012】
別の実施形態では、本発明の植物中の、pm−lと称されるQTLの存在は、前記植物のウドンコ病耐性と関連する、以下の表3中でSNP3として示される単一ヌクレオチド多形(SNP)から生じる少なくとも1つの突然変異を含んでなる核酸配列の存在の有無により示されるか、又は、前記植物のウドンコ病耐性と関連する、以下の表3における5bpの挿入部分(5’−AATTT−3’)として示す少なくとも1つの突然変異を含んでなる核酸配列の存在の有無により示される。
【0013】
本発明の好適な実施形態では、キュウリクロステロウイルス及びキュウリウドンコ病に対する抵抗性を示す植物の提供に関する。詳細には、前記植物はキュウリ(Cucumis sativus)であり、前記植物は単一の染色体上に、クロステロウイルス耐性を付与する少なくとも1つの染色体領域と、ウドンコ病耐性を付与する少なくとも1つの染色体領域を含んでなり、クロステロウイルス耐性を付与する前記少なくとも1つの領域が、E16/M50−244、E16/M50−188及びE11/M48−251からなる群から選択される少なくとも1つのマーカーと連結し、ウドンコ病耐性を付与する前記少なくとも1つの領域が、
− 配列番号1の単一ヌクレオチド多形マーカー 39T→G、
− 配列番号2の単一ヌクレオチド多形マーカー 29G→A、
− 配列番号3の単一ヌクレオチド多形マーカー 193C→T、
− 配列番号4の位置221における挿入突然変異5’−AATTT−3’、並びに−マーカーE16/M50−F−194、E11/M48−F−251、E23/M38−M001、E23/M40−M003、E24/M46−M002、E24/M46−M003、E12/M91−M003、E26/M43−M003、E14/M59−F−134及びE14/M59−F−200からなる群から選択される少なくとも1つのマーカーに連結している。
【0014】
本発明の植物は、クロステロウイルス耐性に関連する第2のQTLを任意に含んでもよい(国際公開第02/22836号に記載のQTL−2ように、このQTLの局在化及び特徴に関する詳細がこの特許文献の明細書に明確に示されている)。このQTLは、染色体上の別々の位置に配置していることが示されている。
【0015】
本発明の別の態様は、上記の本発明に係るキュウリの一部分の提供に関する。好ましくは、前記植物の一部分は、花粉、胚珠、葉、胚、根、根端、葯、花、果実、茎、芽、枝、根茎、種子、プロトプラスト及びカルスから選択され、最も好ましくは種子である。
【0016】
本発明の別の態様は、本発明に係る上記の二重抵抗性キュウリ(クロステロウイルス及びPMに耐性を示す[PM−葉(pm−l)及びPM−胚軸(pm−h)抵抗性の表現型を含む])と、農産物として望ましい特徴を有する第2のキュウリ又は他の種類の植物(好ましくはキュウリ株の植物)とを交配させることにより得られるF1種の提供に関する。好ましくは、前記第2のキュウリは最小限のpm−hを含む(劣性遺伝子であるため)。その場合、前記第2のキュウリは最小限のヘテロ型であり、好ましくは、劣性のpm−h特性を有するため、ホモ型である。他の好適な実施形態では、前記第2のキュウリはクロステロウイルスに感受性で、好ましくは近交系の植物である。
【0017】
本発明の別の態様は、本発明に係る上記F1種を栽培することによって得られる雑種植物の提供に関する。
【0018】
本発明の別の態様は、本発明にかかる上記ハイブリッド植物の一部分の提供に関する。好ましくは前記植物の一部分は、花粉、胚珠、葉、胚、根、根端、葯、花、果実、茎、芽、枝、根茎、種子、プロトプラスト及びカルスから選抜され、最も好ましくは果実である。
【0019】
上記したように、PM又はクロステロウイルスへの耐性に必要となる各々の遺伝要素を1つだけ有する抵抗性植物の交配による、当該遺伝要素の遺伝的浸透により、他の植物病に対する耐性に必要となる遺伝因子が損失すると考えられている。この発見により、発明者らはこの課題を解決できる育種方法の開発に至り、それにより所望の二重耐性植物を得るに至った。本発明の発明者は、耐性がQTL−1によって付与されるクロステロウイルス−耐性親キュウリ株と、耐性がpm−h及びpm−lによって付与されるPM耐性キュウリ株との交配により効果的に二重抵抗性のキュウリを作製する場合に、育種において、組換え体の作製(すなわち1つの染色体における相同組換えを発生させる)するステップと、それに続く、単一の染色体上にQTL−1及びpm−l、又はQTL−1及びpm−hを有する植物を選抜するステップを含める必要があることを見出した。好ましくは、前記選抜ステップは、単一の染色体上にQTL−1、pm−l及びpm−hを有する植物の選抜を含んでなる。より好ましくは、QTL−1はpm−h及びpm−lが配置されているゲノム領域の間に挿入され、それにより、クロステロウイルス耐性に必要となるQTL1がPM耐性に必要となる2つのQTLsの間に挟まれる位置関係をとる。
【0020】
PM又はクロステロウイルス耐性と関係する、本明細書で定義されるいずれかのQTLの存在の確認は、植物の染色体上の各々のQTLと連結した1つ以上の遺伝マーカー、植物の染色体上の各々のQTLに対して連鎖不平衡である遺伝マーカー、又はそれらの組み合わせを検出することによってなされる。
【0021】
前記植物の前記QTLsの存在及び/又は位置の適切な検出方法には、前記QTLsを特徴づけるAFLPマーカーの使用が含まれる。好ましくは、単一の染色体上におけるQTL−1、pm−l及び/又はpm−hを有する植物の上記選抜のためのステップは、p/n−4のための定量形質遺伝子座(QTL)を有する染色体上、又はPM−耐性と関連するpm−hのための定量形質遺伝子座(QTL)を有する染色体上、好ましくはpm−h及びpm−lのための定量形質遺伝子座を有する染色体上の、クロステロウイルス耐性関連の定量形質遺伝子座QTL−1を特徴づける少なくとも1つのAFLPマーカーの検出を含んでなり、その場合、前記QTL−1マーカーはQTL pm−hとQTL pm−lの間の染色体領域に位置する。すなわち、当該選抜プロセスは、表2、3及び4に記載のマーカーからなるマーカー群から選抜される1つ以上のマーカーの検出を含んでなってもよい。
【0022】
本発明の別の態様は、キュウリクロステロウイルス及びキュウリウドンコ病に対する耐性を示すキュウリの選抜方法であって、前記植物において、単一の染色体上のQTL−1、並びに少なくともQTLs pm−l及びpm−hのうちの1つを検出することを含んでなる方法の提供に関する。
【0023】
一実施形態では、当該方法は以下のステップを含んでなる:
a)キュウリからゲノムDNAのサンプルを得るステップと(当該サンプルは充分な長さのゲノムDNA断片を含んでなる)、
b)少なくとも1つの第1のQTL、又はそれに関連するQTL−1、pm−l及びpm−hからなる群から選抜される分子マーカーを含んでなる断片を選抜するための精製反応を実施するステップと、
c)前記選抜された核酸断片の増幅反応を実施するステップであって、少なくとも1つの第2のQTL、又はそれに関連するQTL−1、pm−l及びpm−hからなる群から選抜される分子マーカーを含んでなる断片を検出するステップと、
d)ステップc)の反応生成物中の、予測された長さ又は予測された核酸配列を有する増幅されたDNA断片を検出するステップ。
【0024】
好ましい実施形態では、前記ステップb)は、前記第1のQTLの分子マーカーを定義する少なくとも1つのプライマーセット、又は前記マーカーを定義する核酸配列と実質的に相補的である少なくとも1つの核酸プローブの使用を含んでなり、前記核酸プローブは前記1つ以上のマーカーを定義する核酸配列と、ストリンジェントなハイブリダイゼーション条件下で特異的にハイブリダイズする塩基配列を有する。
【0025】
他の好ましい例として、前記処理c)は、前記第2のQTLの分子マーカーを定義する少なくとも1つのプライマーセット、又は前記第2のQTLの分子マーカーの核酸配列とストリンジェントな条件下で特異的にハイブリダイズする少なくとも1つのプライマーセットの使用を含んでなる。
【0026】
本発明の方法の他の実施形態では、前記植物中の単一の染色体上の前記QTLsの存在を検出するステップは、in situハイブリダイゼーション技術、又はin situ増幅技術を用いて実施する。また、前記QTLsの分子マーカーを定義するか、又は前記QTLの分子マーカーの核酸配列とストリンジェントな条件下で特異的にハイブリダイズするプローブ及びプライマーを、前記方法で使用するのが好適である。
【0027】
本発明の幾つかの態様では、分子マーカーは好ましくは、SNP、挿入突然変異マーカー又はAFLPマーカーであり、好ましくは表2から4に列挙されるマーカーからなる群から選抜されるマーカー、並びに本願明細書に引用される文献において記載されたそれらである。
【0028】
本発明の別の態様は、キュウリクロステロウイルス、及びキュウリウドンコ病に対する耐性を示すキュウリ植物の作製方法の提供に関し、当該方法は以下のステップを含んでなる:
a)キュウリクロステロウイルス耐性を付与する染色体領域を含む第1のキュウリを選抜するステップであって、前記選抜が、前記植物のゲノムにおいて、マーカーE16/M50−244、E16/M50−188及びE11/M48−251により表される、キュウリクロステロウイルス耐性を付与するQTLに連結している少なくとも1つのマーカーの存在を検出することにより行われるステップと、
b)キュウリウドンコ病耐性を付与する少なくとも1つの染色体領域を含む第2のキュウリを選抜するステップであって、前記選抜が、前記植物のゲノムにおいて、
− 配列番号1の単一ヌクレオチド多形マーカー 39T→G、
− 配列番号2の単一ヌクレオチド多形マーカー 29G→A、並びに
− マーカーE16/M50−F−194、E11/M48−F−251及びE23/M38−M001により表される、第1のキュウリウドンコ病耐性を付与するQTLと連結している、少なくとも1つのマーカーの存在を検出することによって行われるステップか、又は、前記選抜が、前記植物のゲノムにおいて、
− 配列番号3の単一ヌクレオチド多形マーカー 193C→T、
− 配列番号4の位置221の挿入突然変異5’−AATTT−3’、並びに
− マーカーE23/M40−M003、E24/M46−M002、E24/M46−M003、E12/M91−M003、E26/M43−M003、E14/M59−F−134及びE14/M59−F−200により表される、第2のキュウリウドンコ病耐性を付与するQTLと連結している、少なくとも1つのマーカーの存在を検出することによって行われるステップによって行われるステップと、
c)ステップa)及びステップb)から得られた前記植物を交配してF1種子を得るステップと、
d)F1種子をF1植物に成長させ、交配又は自家交配することによって前記F1植物から更なる子孫を生じさせるステップと、
e)前記更なる子孫植物から、ステップa)において定義したキュウリクロステロウイルス耐性を付与するQTLに連結している少なくとも1つのマーカーと、ステップb)において定義したキュウリウドンコ病耐性を付与するQTLに連結している少なくとも1つのマーカーを含んでなる植物を選抜するステップ。
【0029】
当業者であれば、前記更なる子孫の集団は、その集団から単一の染色体上に少なくとも2つの相同組換えを生じさせている植物の存在を検出できるのに充分なサイズでなければならないことを理解するであろう。通常、1,000の植物からなる集団で充分である。本発明で提案するマーカー選抜技術による利点としては、かかる母集団のサイズであっても、目的の遺伝子型(表現型的として検知されなくてもよい)の存在を容易にスクリーニングできる点が挙げられる。
【0030】
本発明の方法の好ましい実施形態では、ステップb)の第2のキュウリを選抜するステップが、前記第1若しくは第2のキュウリウドンコ病耐性を付与するQTLsのうちの1つのみを有する第2のキュウリを選抜することを含んでなり、また前記方法は、以下のステップを更に含んでなる:
f)前記第1若しくは第2のキュウリウドンコ病耐性を付与するQTLs(すなわち前記第2のキュウリに存在しないQTL)のうちのもう1つを有する第3のキュウリを選抜するステップと、
g)ステップe)で得られたF1植物と、前記第3のキュウリとを交配させて、更なる子孫植物を得るステップと、
h)当該子孫植物から、ステップa)で定義されるキュウリクロステロウイルス耐性を付与するQTLと、ステップb)で定義されるキュウリウドンコ病耐性を付与する両方のQTLsを含む植物を選抜するステップ。
【0031】
好ましい実施形態では、ウドンコ病耐性を付与する遺伝子座prn−h及びpm−lの両方を含む植物が選抜され、詳細には、QTL−1によって特徴づけられるクロステロウイルス耐性を付与する遺伝子座が、ウドンコ病耐性を付与する遺伝子座pm−h及びpm−lの両方の間に配置されている。
【0032】
すなわち、キュウリクロステロウイルス及びキュウリウドンコ病耐性を示すキュウリの作製方法の他の好ましい実施態様では、ステップb)、e)、f)又はh)で定義されるキュウリウドンコ病耐性を付与する染色体領域を含むキュウリを選抜するステップは以下のステップを含んでなる:
−前記植物のゲノム中において、
配列番号1のSNPマーカー 39T→G、
配列番号2のSNPマーカー 29G→A、並びに
マーカーE16/M50−F−194、E11/M48−F−251及びE23/M38−M001で表される第1のキュウリウドンコ病耐性付与QTLに連結する少なくとも1つのマーカーの存在を検出するステップと、
−前記植物のゲノム中において、
配列番号3のSNPマーカー 193C→T、
配列番号4の位置221における挿入突然変異 5’−AATTT−3’、並びに
マーカーE23/M40−M003、E24/M46−M002、E24/M46−M003、E12/M91−M003、E26/M43−M003、E14/M59−F−134及びE14/M59−F−200で表される第2のキュウリウドンコ病耐性付与QTLに連結する少なくとも1つのマーカーの存在を検出するステップ。
【0033】
キュウリクロステロウイルス及びキュウリウドンコ病耐性を示すキュウリを作製する方法の他の好ましい実施形態では、ステップa)、b)、e)、f)又はh)のうちの少なくとも1つは、前記植物から得たゲノムDNAのサンプルを提供するステップと、前記ゲノムDNAのサンプル中から前記少なくとも1つのマーカーを検出するステップを含んでなる。
【0034】
他の好ましい実施形態では、ステップe)は、植物ゲノム中において、QTL−1と関連する少なくとも1つのマーカー、並びにpm−h若しくはpm−lに関連する少なくとも1つのマーカーを検出するステップを含んでなり、好ましくは、植物ゲノム中及び単一の染色体上において、QTL−1に関連する少なくとも1つのマーカー、pm−lに関連する少なくとも1つのマーカー、及びpm−hに関連する少なくとも1つのマーカーを検出するステップを含んでなり、好ましくは、表2、3及び4に列挙されるマーカー群から選択されるマーカーを含んでなる。
【0035】
他の好ましい実施形態では、ステップe)は、上記の二重抵抗性のキュウリを選抜するための本発明の方法により実施される。
【0036】
ステップc)からe)にて説明した方法であって、二重抵抗性のF1を生じさせる一度の交配において、単一の染色体上で少なくとも2つの相同組換えが生じる方法は、多くの世代を通じて実施可能であり、それにより、単一の染色体中における所望の(少なくとも2回の)相同組換え、及びそれによる二重抵抗性の植物の形成が、F2、F3、F4、F5又はいかなる世代においても実施可能となる。かかるバリエーションは本発明の範囲内に含まれ、当業者によって容易に実施される。
【0037】
キュウリクロステロウイルス及びキュウリウドンコ病に対する耐性を示すキュウリを作製する代替的な方法は、次のステップを含んでなる:
a)本発明の方法を実施して、キュウリクロステロウイルス及びキュウリウドンコ病に対する耐性を示す第1のキュウリを選抜するステップと、
b)前記植物を同系育種して、前記QTLsのホモ接合植物株を得るステップと、
c)ステップa)及びステップb)から得た前記植物を交配してF1種子を得るステップと、
d)前記F1種子をF1植物に成長させるステップ。
【0038】
本発明の別の態様は、本発明の方法で作製される植物又はその部分の提供に関する。
【0039】
本発明により子孫植物の非常に迅速なスクリーニングが可能となり、それにより二重耐性を示すと考えられる植物の選抜が可能となる。例えば、子孫植物の中から、少なくとも1つのpm−hマーカー、少なくとも1つのpm−lマーカー及び少なくとも1つのQTL−1マーカーを有する植物を検出することが可能となる。かかる植物は、望ましい遺伝的浸透が行われたと考えられる。更なるマーカーに関するスクリーニングを行うことにより、子孫植物が二重耐性を示すという予測上の信頼性を向上させることができる。3つのQTLsの各々の少なくとも1つのマーカーを示す植物の検出に基づき、植物を選抜することが可能となり、また育種プログラムあるいは更なる同定試験にも使用できる。
【0040】
本発明の植物の利点としては、上記の特性が同一の染色体に存在するため、従来の育種を使用したときでも、上記の両方の耐性を制御する遺伝子を(ハイブリッド)植物中へ容易に移動させることが可能となることであり、逆に従来の育種技術を用いた場合では、それらの特性が隔離した染色体上に存在するため、両方の特性を有するハイブリッドを作製するのがはるかに困難となる。
【発明を実施するための最良の形態】
【0041】
定義
本明細書で使用される「キュウリ」という用語には、一般にCucumberと呼ばれるプラント、アメリカガーキン、Cassabanana、Cuke、Gherkin、Hothouseキュウリ、Lemonキュウリ、Manderaキュウリ、Picklingキュウリ、Serpentキュウリ、Slicingキュウリ、Snakeキュウリ及び西インド諸島のガーキンなどが包含されるがこれらに限定されないキュウリ植物又はその一部を意味する。
【0042】
本発明の用語「植物部分」とは、単細胞及び細胞組織(例えば植物において、完全である植物細胞、細胞凝集塊及び植物が再生できる組織培養)などの植物の一部を意味する。植物部分の例としては、花粉、胚珠、葉、胚、根、根端、葯、花、果実、茎、芽及び種子に由来する単一の細胞及び組織、ならびに花粉、胚珠、葉、胚、根、根端、葯、花、果実、茎、芽、枝、根茎、種子、プロトプラスト、カルスなどが挙げられるが、これらに限定されない。
【0043】
本明細書で用いられる用語「クロステロウイルス」とはClosteriviridaeのファミリーのウイルスのことを指し、一般にCucurbit黄色萎縮障害ウイルス(CYSDV)、Lettuce伝染性黄化病ウイルス(LIYV)、及びBeet偽黄化病ウイルス(BPYV、またその同義語Cucumberクロロチックスポットウイルス(CCSV)、Cucumber黄化病ウイルス、Muskmelon黄化病ウイルス又はStrawberry pallidosisウイルスとして公知)と呼ばれるウイルス、好ましくはBPYV及びCYSDV、最も好ましくはCYSDVである。好ましくは、キュウリ植物に関する用語「closterovirus」とは、特にキュウリ(i.e.the Criniviideae)に影響を与える属を指す。
【0044】
本明細書で使用される「ウドンコ病」という用語は、真菌Sphaerotheca fuliginea(またその同義語Podosphaera黄色ii及びS.cucurbitaeとして公知)によって、及び/又は真菌Erysiphe cichoracearum(またその同義語Golovinomyces cichoracearumとして公知)によって、及び/又は真菌Leveillula taurica(またその同義語Oidiopsis taurica、Erysiphe taurica、Ovulariopsis cynarea、Leveillula solanacearumとして公知)によって、キュウリ(Cucumis sativus L.)において、生じる疾患を意味する。
【0045】
用語「QTL」は、本願明細書では、連続的に分散した(定量的な)表現型特徴の発現と関連する、対立遺伝子(例えば遺伝子又は制御配列の形)を含んでいる、当業者に公知の染色体領域を意味する用語として用いられる。「耐病性QTL」という用語は、特定の染色体に置かれる領域であって、すなわち、耐性遺伝子をコードする少なくとも1つの遺伝子に関連するか、又は、少なくとも1つの制御領域、(染色体領域)であって、耐性に関連する1つ以上の遺伝子の発現を制御するものを指す。耐性を伴うQTLsを指すものとして、より短い同義語:「耐性付与遺伝子座」を本願明細書では用いる。QTLsは、1つ以上の分子ゲノムマーカーを使用して、特異的なキュウリ継承のゲノムのそれらの遺伝子位置を示すことにより定義されてもよい。1つ以上のマーカーは、次に、特異的な遺伝子座を示す。遺伝子座の間の距離は、通常同じ染色体上の遺伝子座との間に、乗換えの頻度で測定される。2つの遺伝子座が遠くに離れるほど、より頻繁に交叉がそれらの間において、発生する。逆にいえば、2つの遺伝子座が間近である場合、それらの間での交叉は発生しにくい。概して、1センチモルガン、コーサンビー・マップ関数、cMは、遺伝子座(マーカー)との間に、1%の組換えにほぼ等しい(Lui、1997)。QTLが並列マーカーによって、示すことができる場合、エンドポイントマーカー間の遺伝的距離はQTLのサイズを表す。
【0046】
用語「染色体」は、本願明細書では、当業者に公知のように、細胞DNAを含有し、そのヌクレオチド配列に遺伝子の線形配列を有する、細胞核の自己再生する遺伝的構造のことを意味する用語として用いる。
【0047】
本発明で使用する「連鎖群」という用語は、同じ染色体に置かれる遺伝子又は遺伝形質の全てを指す。連鎖群の範囲内で、十分に間近であるそれらの遺伝子座は、遺伝的交雑の結合を示す。交差の確率が染色体上の遺伝子との間の物理的距離によって、増加するため、場所が連鎖群の中で各々からはるかに隔離した遺伝子は、遺伝子検査において、いかなる検出可能な直接的な結合も示すことはありえない。用語「連鎖群」は、大部分は染色体譲渡がまだなされなかった遺伝子のシステムの連結された挙動を示す遺伝子の遺伝子座を指すために用いる。したがって、本発明では、用語「連鎖群」とは、染色体(物理的存在)と同義である。
【0048】
本発明の用語「対立遺伝子」とは、遺伝子の1つ以上のいずれかの代替的な形を意味しその全ての対立遺伝子が少なくとも1つの特徴又は特徴に関する。二倍体細胞において、提供された遺伝子の2つの対立遺伝子は、一対の相同染色体上の対応する遺伝子座を占める。本発明はQTLs(すなわち1つ以上の遺伝子又は制御配列を含みうるゲノム領域)に関するため、若干の例においては「対立遺伝子」の代わりに「ハプロタイプ」(すなわち染色体部分の対立遺伝子)を指すのがより正確であるが、そのような場合、用語「対立遺伝子」は用語「ハプロタイプ」を含むものとして理解されるべきである。
【0049】
本願明細書における「遺伝子」とは、一連のDNAからなる遺伝単位として定義され、それは、染色体上の特定の位置を占め、また生物体の特定の形質又は特徴を制御する遺伝的な暗号を含んでなる。
【0050】
「遺伝子座」とは、提供された遺伝子又は制御配列が提供された種の染色体に占有する位置として、本願明細書に定義される。
【0051】
「相同組換え」とは、2つのDNA分子の間でのDNA断片の組換え(「クロスオーバー」)、又は同一のヌクレオチド配列の一定の領域における対染色体の染色分体間での組換えのことを指す。「組換え現象」は、本願明細書において、減数分裂のクロスオーバーを意味すると理解される。
【0052】
本発明で使用する「分子マーカー」という用語は、核酸配列の特徴の相違を視覚化する方法において、用いられる指標のことを指す。かかる指標の例としては、制限断片長多形性(RFLP)マーカー、増幅された断片長多型(AFLP)マーカー、単一ヌクレオチド多型(SNP)挿入変異、マイクロサテライトマーカー(SSRs)配列同定された増幅領域(SCARs)、切断された増幅された多形配列(CAPS)マーカー又はアイソザイムマーカー又は特異的な遺伝子の及び染色体位置を定義する本願明細書に記載されているマーカーの組合せが挙げられる。「QTLにリンクされる分子マーカー」は、本明細書で定義されるように、SNP、挿入変異、並びにAFLPマーカー又はフィールドにおいて、使用する通常の他のタイプのあらゆるマーカーのことを指してもよい。本願明細書におけるAFLPマーカーの前後関係において、マーカーとは、2つのAFLP−プライマーに隣接するキュウリ特異的なDNA塩基配列を指し、そのプライマーは、「コアプライマー」E及びM(制限酵素部位EcoRI及びMsel部位に相当)(Vosその他、1995、Baiその他。2003)、更に示すような2つ又は3つの選択的な塩基が続き、各々に選択的なヌクレオチドを確認する2桁のコードが続き、それによって、「コアプライマー」が延長される(コードに関しては表1を参照)。E16/M50−244は、増幅プライマーEcoRI + CC及びMsel + CATを用いて得られる、244bp.の全長を有する断片状のマーカーを意味する。断片の長さは断片の検出方法に依存すると考えられ、若干の塩基を増減させたものであっても、その正確な長さのものとさほど変わらない。本願明細書に記載のマーカーを定義する際、リケージマップの他のマーカーとの、そのマーカーの染色体上の相対的位置として参照する必要がある。すなわち、マーカーE16/M50−244は、そのプライマーの配列によって、増幅生成物の長さによって、並びに、E14/M59−F−200及び/又はE23/M38−M003との相対的位置によって、又は、本願明細書に記載のように、表4のマトリックスのcMの対応する距離を有する順序づけられたリストにて図示するように、他のマーカーとの相対的な位置により定義されてもよい。しかしながら、考慮が必要となることとして、植物間の交雑により、特定のマーカーが損なわれることもあることが挙げられるが、但し特定のマーカーの欠如が、耐病性を与え、マーカーがそこに連結する遺伝因子の存在を否定するわけではない。
【0053】
表1:通常、AFLP分析において適用され、本発明においても用いるプライマー伸長コード(Source:Keygene、Wageningen、オランダ)
【表1】
【0054】
用語「キュウリ特異的なDNA配列」とは、80%以上、好ましくは85%以上、好ましくは、90%以上、好ましくは、95%以上、好ましくは、97%以上、最も好ましくは99%以上、キュウリのゲノム配列とのヌクレオチド配列相同性を有するポリヌクレオチド配列を意味する。それは大きな類似性を示し、好ましくはQTL−1マーカーの場合、QTL−1マーカーに隣接するキュウリのアクセッションNo.PI 250147のDNA塩基配列の一部を構成する。
【0055】
本明細書で用いられる用語「ヌクレオチド配列相同性」とは、2つのポリヌクレオチド間の相同性を意味する。2つの配列のアラインメントを作成したときに最大の一致性でヌクレオチドの配列が同じである場合に、ポリヌクレオチドは「相同な」配列を有するという。2つ以上のポリヌクレオチドの間の配列比較は、通常、配列類似性を有する局部領域を同定し、比較ウインドウ上の2つの配列の部分を比較することにより実施される。比較ウインドウは通常、約20〜200の連続ヌクレオチドである。ポリヌクレオチドの「配列相同性のパーセンテージ」(例えば50、60、70、80、90、95、98、99又は100パーセント配列相同性)は、比較ウインドウの上の2つの最適に整列配置された配列を比較することによって、決定でき、比較ウインドウ中のポリヌクレオチド配列の部分には、最適な2つの配列の参照配列(付加又は欠失を含まない)と比較し添加又は欠失(すなわちギャップ)が存在する。パーセンテージの算出は、
(a)同一の核酸塩基が両方の配列において、存在する位置の数を勘定してマッチする位置の数を算出し、
(b)比較のウインドウの位置の総数で、上記のマッチする位置の数を除算し、
(c)結果に100を乗算して配列相同性パーセンテージとして示すことにより行われる。比較配列の最適配列は、周知のアルゴリズムを用いたコンピュータ化された実施態様で行ってもよく、又は視覚による検査によって、実施してもよい。直ちに利用できる配列比較及び並列配列アルゴリズムとしては、Basic Local Alignment Search Tool(BLAST)(Altschul et al.,1990、 Altschul et al,1997)、ClustalWプログラム(両方ともインターネットで利用可能)が挙げられる。他の適切なプログラムとしては、GAP、BestFit、PlotSimilarity及びFASTA(ウィスコンシンGenetics Software Package (Genetics Computer Group (GCG),Madison,WI,USA) (Devereux et al,1984))が挙げられるが、これらに限定されない。
【0056】
本明細書で使用される「pm−h」という用語は、下子葉部において、発現される推定のウドンコ病抵抗性遺伝子を意味し、好ましくは表2の特定のSNPの存在下で定義される。
【0057】
表2:ウドンコ病耐性表現型との完全な関連を示すSNPを、QTL pm−hとして本願明細書に示す。
【表2】
【0058】
本明細書で使用される「pm−l」という用語は、推定のウドンコ病抵抗性遺伝子pm−leafを意味し、好ましくは表3の特定の突然変異の存在下で定義される。
【0059】
表3:ウドンコ病耐性表現型との完全な関連を示している突然変異を、QTL pm−lとして本願明細書に示す。
【表3】
【0060】
「QTL−1」用語とは、クロステロウイルス耐性に連結するゲノム領域を指し、WO 02/22836において、詳細に記載されているマーカーE16/M50−244、E16/M50−188及び/又はE11/M48−251により定義される(上記開示内容を本発明に援用する)。
【0061】
「子孫」植物という用語は、1つ以上の親株からの無性生殖的若しくは有性生殖的な再生によって、得られる植物、又はその子孫としてのあらゆる植物のことを指す。例えば、子孫植物は、親植物のクローニング若しくは自家交配、又は2つの親植物を交配させることによって、得られ、F1若しくはF2、又はそれ以上の世代を含むものと解される。F1は、両親(その少なくとも1つが特徴ドナーとして初めて用いられ)から製作される第一世代の子であり、一方、第二世代(F2)又はそれ以降の世代(F3、F4など)の子孫がF1、F2などの自殖から作製される標本である。ゆえにF1は、2人の本物の繁殖両親(本当の繁殖は、特徴のためにホモ接合性である)のハイブリッドから生じているハイブリッドであると(通常)考えられ、F2が前記F1ハイブリッドの自家受粉から生じる子孫であると(通常)考えられる。
【0062】
本発明では、異なる対立遺伝子が相同染色体上の対応する場所に存在するとき、「ヘテロ接合体」という用語を、それを表す遺伝子的条件として用いる。
【0063】
本発明では、同一の対立遺伝子が相同染色体上の対応する場所に存在するとき、「ホモ接合性」という用語を、それを表す遺伝子的条件として用いる。
【0064】
植物育種の前後関係の「ハイブリッド」という用語は、異なる系列又は種類又は種の植物交配により作製される、遺伝的に異なる両親の子としての植物を意味し、2つの自殖系統の交配を含むがこれに限定されない。
【0065】
本発明で用いる「近交系」という用語は、実質的にホモ接合の個体又は株を意味する。
【0066】
本発明における用語「遺伝子移入」とは、天然の、及び人工的なプロセスであって、種を交配させることによって、1つの種、変異体多様性又は園芸品種のゲノム領域が他の種、変異体又は園芸品種のゲノムに移入することを指す。当該方法は、任意に再発する親に戻し交配して実施してもよい。
【0067】
本願明細書に用いられる「遺伝子工学」、「形質転換」及び「遺伝子組換え」の用語は、単離されクローニングされた遺伝子を、他の生物体のDNA(通常染色体DNA又はゲノム)へ移入することと、同義である。
【0068】
本発明の用語「集団」とは、共通の遺伝的な派生を有する、遺伝的に異質な植物のコレクションを意味する。
【0069】
本発明の用語「変異体」又は「栽培品種」とは、構造的若しくは遺伝子的特徴及び/又はパフォーマンスによって、同じ種の範囲内で他の種類と区別できる、類似する植物群を意味する。
【0070】
「抵抗性」及び「耐性」の用語は、感染に対する部分的及び完全な耐性のことを指す。感受性の植物は、感染に対して抵抗性を有さないか、又は耐性レベルが低いと考えられる。上記の用語には、「完全な耐性」、「免疫性」「中程度の耐性」、「部分的な耐性」、「過敏症」及び「許容性」などの、別に定義することが可能な形の抵抗性が包含される。
【0071】
「完全な耐性」とは、感染の後に病気が全く進行しない状態のことを指し、病原体が細胞内に入ること(第一の感染)がなされない結果によるものであってもよく、あるいは、細胞内で感染源が増殖できず、他の細胞にも感染しない(潜在的な感染ではなく、拡散することもない)結果によるものであってもよい。
【0072】
「感受性」という用語は、植物が疾患による影響を受け、病徴に至るような疾患に対する耐性を有しない植物を指すものとして、本願明細書で用いられる。ゆえに「感受性」という用語は、「非抵抗性」と同義語である。
【0073】
核酸に関する「ハイブリッド」という用語は、相補的なヌクレオチド塩基間の水素結合により形成される二本鎖核酸分子(又はデュプレックス)を指す。「ハイブリダイズする」又は「アニールする」という用語は、核酸配列の一本鎖が補完的な塩基との間で水素結合による二重螺旋部分を形成させる方法のことを指す。
【0074】
「プローブ」という用語は、目標核酸配列分析物又はその相補DNA誘導体の相補配列と共に水素結合デュプレックスを形成する、一本鎖オリゴヌクレオチド配列をいう。
【0075】
本明細書で使用される「プライマー」という用語は、増幅目標にアニーリングでき、DNAポリメラーゼの結合を可能にするオリゴヌクレオチドを意味し、それにより、プライマー伸長生成物の合成が誘導される条件(すなわちヌクレオチド及び例えばDNAポリメラーゼなどの酵素の存在)、並びに適切な温度及びpH条件下に設けられた場合に、DNA合成の開始位置として機能する。(増幅)プライマーは好ましくは増幅の最大効率のために一本鎖である。好ましくは、プライマーはオリゴデオキシリボヌクレオチドである。プライマーは、ポリメラーゼの存在下での伸長生成物の十分な合成を可能にするため、十分な長さを有する必要がある。プライマーの正確な長さは多くの因子に依存し、例えば温度及びプライマーの組成(A/T対G/C含量)などが挙げられる。例えばPCR増幅などのDNA増幅において、用いる、一対の双方向性プライマーは、1つの順方向及び1つの逆方向プライマーからなる。本明細書で用いられる「プライマー」は、複数のプライマーを指すこともあると理解され、特に増幅される目標領域の末端配列に関して情報の曖昧性が存在する場合には複数のプライマーが用いられる。それ故、「プライマー」には、配列又は中に若干の変異を有する配列を含んでなる、プライマーオリゴヌクレオチドのみならず、典型的な塩基対形成を可能にするプライマーも包含される。オリゴヌクレオチドプライマーは、いかなる適切な方法で調製してもよい。特異的な配列のオリゴヌクレオチドを調製する方法は公知であり、例えばクローニングや、配列の制限酵素処理、及び直接の化学合成などが挙げられる。化学合成の例としては、ホスホジ−又はトリエステル法、ジエチルホスホロアミデート法及び固相支持法(米国特許第4458066号で開示)などが挙げられる。プライマーは、必要に応じて標識してもよく、例えば分光、蛍光、光化学、生化学、免疫化学若しくは化学的な測定手段で検出される態様で標識してもよい。温度依存性のオリゴヌクレオチドプライマー伸長は、ポリメラーゼにより触媒され、4つのデオキシリボヌクレオチド三リン酸エステル(dATP、dGTP、dCTP及びdTTP(すなわちdNTPs)又はその類似体の十分な量の存在下、適当な塩、金属陽イオン及びpH緩衝システムを含有する反応溶媒中で当該触媒反応がなされる。適切なポリメラーゼは、プライマー−及び鋳型に依存してDNA合成反応を触媒することが知られている酵素である。公知のDNAポリメラーゼとしては、例えば大腸菌DNAポリメラーゼI又はそのクレノー断片、T4 DNAポリメラーゼ及びTaq DNAポリメラーゼなどが挙げられる。これらのDNAポリメラーゼを有するDNA合成に触媒作用を及ぼすための反応条件下は、従来技術において、周知である。合成の生成物は鋳型鎖及びプライマー拡張鎖からなる二重分子であり、標的配列を含む。これらの生成物は、次のラウンドで複製のための鋳型として役立つ。複製の2回において、第1のサイクルのプライマー伸長鎖は、その相補プライマーによって、アニールされる、合成により、「ショート」にプライマー配列又はそれらの補足によって、5’−及び3’末端に結合される生成物を与える。変性、プライマーアニーリング及び伸長の反復サイクルは、プライマーにより定義される目標領域の指数関数的な蓄積に結果としてなる。充分なサイクルは、核酸の目標領域を含んでいるポリヌクレオチドの所望の量を提供するために行われる。所望の量は異なる場合もあり、生成物ポリヌクレオチド実際に用いる目的により異なる。PCR方法は通常ハンドブックに記載され、当業者に知られている。PCRによる増幅の後、標的ポリヌクレオチドは、プローブポリヌクレオチドとのハイブリダイゼーションによって、検出できる。それはストリンジェント〜適度にストリンジェントなハイブリダイゼーション条件及び洗浄条件下で標的配列と共に安定ハイブリッドを形成する。プローブが標的配列と基本的に完全に相補的である(すなわち約99%以上)と考えられる場合、ストリンジェントな条件が用いられる。若干の不適当な組み合わせが予想される場合、例えばプローブが変異株と完全に相補的でないことが予想される場合、ハイブリダイゼーションのストリンジェンシーは減少すると考えられる。しかしながら、非特異/偶然の結合を排除する条件を選択する。ハイブリダイゼーションに影響を及ぼす、そして、非特異的結合に対して選択する条件は、従来技術において、周知で、例えば、Sambrook及びラッセル(2001)に記載されている。通常、低い塩濃度及びより高い温度は、ハイブリダイゼーション条件のストリンジェンシーを増加させる。
【0076】
用語「ストリンジェントなハイブリダイゼーション条件」とは、ポリヌクレオチドが、通常は核酸の複雑な混合物中でその標的とハイブリダイズするが、実質的に他の配列とはハイブリダイズしない条件のことを指す。ストリンジェントな条件は配列に依存し、また反応状況によっても異なる。長い配列は、高い温度で特異的にハイブリダイズする。核酸のハイブリダイゼーションの詳細にわたるガイドは、Tijssen(1993)を参照のこと。通常、ストリンジェントな条件は、一定のイオン強度pHにおいて、特定の配列の場合の熱融点(Tm)より5〜10℃低くなるように選択される。Tm値とは、所定のイオン強度、pH及び核酸濃度条件において、標的と相補的なプローブの50%が平衡状態において、標的配列とハイブリダイズする温度のことを指す(標的配列が過剰に存在するとき、Tm値では、プローブの50%が平衡状態にある)。ストリンジェントな条件は、塩濃度が約1.0M以下のナトリウムイオン濃度であり、通常はpH7.0〜8.3において、0.01〜1.0Mのナトリウムイオン(又は他の塩)濃度であり、温度が短いプローブ(例えば10〜50のヌクレオチド)では30℃以上、長いプローブ(例えば50超のヌクレオチド長)では約60℃以上である。ストリンジェントな条件は、不安定化剤(例えばホルムアミド)の添加によって、設定してもよい。選択的若しくは特異的なハイブリダイゼーションでは、陽性シグナルはバックグラウンドに対して少なくとも2倍(好ましくは10倍)のハイブリダイゼーションである。条件がしばしばそうである典型的な厳しいハイブリダイゼーション:50%ホルムアミド、5×SSC及び1%のSDS、42℃でインキュベート、又は、5×SSC、1%のSDS、65℃でインキュベートし、65℃で0.2×SSC及び0.1%のSDSで洗浄、である。PCRでは、アニーリング温度はプライマー長に応じて32℃〜48℃で変化しうるが、約36℃の温度が、低いストリンジェンシーによる増幅において、典型的である。ハイブリダイゼーションパラメータを決定するための更なるガイドラインは、多数の参考文献(例えばAusubelら、1999)において、開示されている。
【0077】
好ましい実施態様の説明二重耐性植物の生産農業及び園芸の繁殖プログラムの目的は、それらの遺伝組成を改良することによって、植物の性能を強化することである。本質的には、この改良は、目的の性能特性に影響している遺伝子に対して最も好ましい対立遺伝子の頻度を増加させることによって、なされる。野生の植物株を用い、遺伝子及び表現型バリエーションの豊富な資源とする。伝統的に、農業若しくは園芸分野においては、このような変種を用いて、所望の遺伝子型又は潜在的な表現型特性を有する野生植物系列又はその子孫を選択し、別の所望の遺伝子型又は潜在的な表現型特性を有する系列と交配し、所望の遺伝子型又は潜在的な表現型特性を示す(その頻度が増加した)植物を子孫植物から選択することを行う。メンデルの遺伝の法則の更なる理解及び利用(並びに分子遺伝学的ツールとの組み合わせ)によりこのような選抜プロセスが可能となった。例えば、所望の遺伝子型の又はポテンシャル表現型特性を有する植物を選択する方法は、前記植物を量的形質遺伝子座(QTL)の存在(すなわち連続的に分散(定量的に)した表現型特徴の発現と関連する対立遺伝子を含有する染色体領域の存在)の検出に基づいて行われる。通常、QTLは、表現型特徴における定量的変化に統計学的に結びつく1つ以上のマーカーによって、特徴づけられて、遺伝子と基本的に同義である。QTLマッピングは、目的の特徴の発現に影響を及ぼしている候補遺伝子座の同定を可能にする。植物繁殖においては、 マーカーによる選抜(MAS)が可能であり、すなわち植物中でQTL関連のマーカーを検出することによって、好ましい対立遺伝子を有する植物を選抜する。
【0078】
栽培植物の繁殖プログラムの大きな問題の1つは、別々の特徴間の負の遺伝相関の存在である。これは、例えばさまざまな疾患耐性植物系列の複製能及び産生間の負の遺伝相関を有するケースである。もう一方への1つの植物系列のゲノムからのDNAの遺伝子移入が基本的な繁殖形質の発現に妨げてもよいか又は影響を及ぼしうることが理解される。同様に、もう一方への1つの植物からの移入耐性を与えている遺伝子配列の試みにより、すでに宿主株に存在するに耐性特性が除去されうる。
【0079】
さまざまな特徴の遺伝についての知識により、例えば病害抵抗性と関連するQTLのためにホモ接合性株の選抜が可能となる。遺伝子の起源及び繁殖プログラムの所望の特徴の位置についての知識の中で使用法予測された繁殖している結果の精度を増加させ、従来の繁殖プログラムと比較し選抜速度が増加する。例えば、実際、所望の特徴が遺伝子的ベース(他の特徴に遺伝的にリンクされる)を有するとき、子孫中におけるそれらの2つの特徴の均一性の増加に役立つ。なぜなら、所望の対立遺伝子に関するホモ接合の親が、大部分の子孫に対して、分離の少ない状態でそれらを子孫に渡すからである。
【0080】
上記したように、本発明の発明者は、クロステロウイルス及びウドンコ病に耐性のある植物を見出した。かかる植物はこれまで公知ではなかった。すでにウドンコ病耐性植物系列へのクロステロウイルス耐性系列からのDNA移入(又はその逆)の試みはこれまで成功しなかった。
【0081】
本発明の発明者は、更に、二重耐性植物で、単一ユニットにつれて実質的に共遺伝するほど密接に、クロステロウイルス及びウドンコ病に対する耐性のための遺伝子が結合されるということを発見した。遺伝子(非常に密接に、連結される)間のかかる遺伝子の構成は、しばしば、カップリング期(シス)にある対立遺伝子(遺伝子)とも呼ばれる。そのような場合、遺伝子が単一染色体にあることが通常認められる。事実、単一染色体上に遺伝子又はQTLsが存在するとき、それらがカップリング期にあることが意味される。
【0082】
本発明の発明者は、二重耐性植物では、クロステロウイルス及びウドンコ病に対する耐性遺伝子が連鎖群4に存在することを発見した。Horejsiらによれば(2000)キュウリLの連鎖群4はまた、べと病(dm.)耐性遺伝子を有する。LG 4は更に、以下のRFLPマーカーのうちの1つ又は組合せ、(Software package INTMAP,Keygene,Wageningen,The Netherlandsによって、決定、マップ位置はcM、ブラケット):CsC032a/El(25.9)CsP357/H3(31.7)CsC588/H3(34.2)CsC477H3(35.3)CsC694/E5(38.5)CsP347/H3(38.5)CsC365/El(41)CsC386/El(41)CsC230/El(41.7)CsP064/El(45.5)により特徴付けられる。
【0083】
明確な染色体番号は、キュウリクロステロウイルス耐性QTL1、並びにウドンコ病耐性に関与するpm−l及びpm−hが位置する染色体には割り当てられていない。しかしながら、当該染色体はこれら及び他のゲノム領域が位置する連鎖群(LG4)を参照することにより示されてもよい。用語連鎖群とは、耐性を与えている対立遺伝子が位置し、染色体と同じ階層的なレベルを有する物理的ゲノム単位を指すものとして本願明細書で用いる。
【0084】
クロステロウイルス+pm耐性のための遺伝子が単一染色体に存在することを発見した利点として、2つ挙げられる。
【0085】
第1に、単一染色体上(好ましくは、小さい隣接する染色体部分)に遺伝子が存在するため、繁殖目的において、非常に有利であり、それらが共同で子孫植物に移入されるということである。ウドンコ病耐性及びクロステロウイルス耐性に関連する遺伝子との物理的な結合の結果として、交配による子孫植物(本発明の二重耐性植物を親株として用いた)では子孫植物における分離の頻度が低くなる。
【0086】
第2に、この知識は、2つの特徴を単一の植物系列内において、組み合わせるのが非常に困難である理由を説明するため、植物育種の助けとなる。いかなる理論にも束縛されないが、クロステロウイルス耐性に関与するQTLの特定の位置、及びそのQTLの選抜により、望ましい遺伝子移入の形成が妨げられたか、望ましい遺伝子移入の可能性が極めて低い確率に抑えられていた。ウドンコ病耐性及びクロステロウイルス耐性のためのQTLを伴うQTLsは近い位置に存在し、すでに第1の耐性(例えばウドンコ病)を有している植物系列への、第2の耐性特徴(例えばクロステロウイルス耐性)QTLの遺伝子移入により、第1の耐性特徴の(一部分の)損失が生じることが考えられる。植物系列がウドンコ病耐性(pm−l及びpm−h)のための両方のQTLsを有する場合、ウドンコ病耐性の基礎をなしている遺伝情報に「損傷を与え」ず、クロステロウイルス耐性のためのQTLの遺伝子移入が、ゲノムの非常に小さい部分において、のみ行われる。
【0087】
ゆえに、さまざまな抵抗性遺伝子が単一染色体に位置することを発見したこととは別に、両方のウドンコ病耐性対立遺伝子が二重耐性植物に存在するときに、クロステロウイルス耐性に関連するQTL(QTL−1)は、PM耐性(pm−h及びpm−l)を伴う2つのQTLsの間に存在することを見出した。この発見は、所望の遺伝子型を有する植物への組換え頻度が低いことを更に部分的に説明するものとも考えられる。また、かかる植物を作製するための、少なくとも3つの可能なシナリオが考えられ、それらを作製するいくつかの再現性のある方法が考案させる。
【0088】
第1の方法は、目的の第1の系列の植物に由来するクロステロウイルス耐性遺伝子座(本明細書でQTL−1として定義)を、2つのPM−耐性付与遺伝子座pm−h及びpm−lを有する、目的の第2の系列のウドンコ病耐性植物に移入することを含んでなる。これが目的の第2の系列の遺伝的バックグラウンドに結果としてなることである場合、当該方法は、QTL−1を、2つのPM−耐性付与遺伝子座pm−h及びpm−lの間のゲノム領域に遺伝子移入することを含んでなり、それにより、目的の第2の系列の遺伝的バックグラウンドを有する子孫植物中で、pm−l−QTL−1−pm−hの構成を有するゲノムDNA配列アセンブリとなる(図1を参照)。これに基づくと、単一染色体の特定の領域における二重の相同組換(すなわち各交差における1度の現象)のみにより、目的の第2の系列の遺伝的バックグラウンドにおいて、クロステロウイルス及びPM耐性の二重耐性の表現型が得られる子孫植物の適当な遺伝子移入を、QTL−1、pm−1及びpm−h特異的なマーカーを用いてモニターしてもよい。
【0089】
第2の方法では、異なる遺伝的バックグラウンド(すなわち第1の目的の株)において、耐性特徴を組み合わせることを望むかもしれないかかる方法は、目的の第2の系列の植物に由来するPM−耐性付与遺伝子座pm−h及びpm−lを、QTL−1場所を有するための第1の系列に係るクロステロウイルス耐性植物に移入することを含むと考えられる。この場合、当該方法は、pm−h及びpm−l遺伝子座を独立にQTL−1場所の両側に遺伝子移入することを含んでなり、それにより、目的の第1の系列の遺伝的バックグラウンドを有する子孫植物において、pm−l−QTL−1−pm−hの構成を有するゲノムDNA配列アセンブリとなる。(図2を参照)。この構成に基づくと、単一染色体の特定の領域において、少なくとも2つの二重相同組換(すなわち4回の交差)が、クロステロウイルス及びPM耐性の二重耐性表現型の獲得に必要であることが考えられる。また、子孫植物の適当な遺伝子移入を、QTL−1、pm−l及びpm−h特異的なマーカーを用いてモニターしてもよい。
【0090】
上記より、例えば野生型のCYSDV耐性の、例えば PM耐性の市販のキュウリへの遺伝子移入が非常に稀な現象であることが説明されると考えられる。同様に、例えばクロステロウイルス耐性の市販のキュウリへの、例えば野生型の両方のPM耐性遺伝子座の遺伝子移入が更に稀な現象である理由も説明ができる。
【0091】
組換えは、減数分裂の間の2つの相同染色体の間での情報の交換である。組換え植物において、最初は染色体の範囲内で特異的な位置に存在するDNAは、他の植物に由来するDNAと組み換えられる(父性から母性、あるいはその逆)。二重組換えでは、この組み換え反応は二度、例えば遺伝子/遺伝子座の両側において、行われる。必要な材料だけを組み換え、可能な限り染色体上の有益な第一の情報を維持するために、通常4度の交差現象(上記参照)が必要となる。かかる二重組換えを見つける通常の方法は、F2−植物の集団をスクリーニングすることである。この集団は、まれな(低頻度)二重組換えを検出するために充分なサイズでなければならない。二重組換えの頻度は、一重組換えの頻度に左右される。例えば、10cM領域の組換えは10%の頻度であった場合、二重組換えは、10%×10%=1%(1センチモルガンは、検定交雑の1%の組換え子孫として定義される)の頻度となる。
【0092】
低頻度のこの問題を迂回するための1つの方法は、「平行(パラレル)組換え」の実施である。これは基本的に、2つの組換えが別個に2つの異なる植物において、生じさせ、その後で、組換えを、得られるF2において、単純な交雑及び選抜を行うことによって、1つの植物で組み合わせることを意味する。これにより、スクリーニングする植物数が減少する。
【0093】
ゆえに、二重耐性植物を作製する第3の方法は、目的の第1の系列に由来する2つのPM−耐性付与遺伝子座pm−h又はpm−lのうちの1つを、QTL−1遺伝子座を有するための第2の系列のクロステロウイルス耐性植物に移入して、pm−l−QTL−1又はQTL−1−pm−hの構成を有するゲノムDNA配列アセンブリとすることと、目的の第3の系列に由来する2つのPM−耐性付与遺伝子座pm−h又はpm−lのうちの他方を、QTL−1遺伝子座を有するための第2の系列の他のクロステロウイルス耐性のある植物に移入して、QTL−1−pm−h又はpm−l−QTL−1の構成を有するゲノムDNA配列アセンブリとすることを含んでなる(図3を参照)。その後、個々の組換えから遺伝子移入を含んでいる子孫植物は、構成pm−lを有するゲノムDNA配列アセンブリに結果としてなるために交差する−QTL−1−pm−h(図3を参照)。また、さまざまな子孫植物への適当な遺伝子移入は、QTL−1、pm−l及びpm−hに特異的なマーカーを用いて、更に交雑された植物をモニターしてもよい。当業者であれば、図4−6を基にして他の方法を考案してもよい。
【0094】
スクリーニングに用いる植物数を制限する方法は、例えば以下の通りに実施してもよい:最初に、両方のPM−耐性対立遺伝子(pm−l及びpm−h)を有する個体と、QTL耐性対立遺伝子QTL−1を有する個体の2個体を交配する。次に、pm対立遺伝子の1つとQTL−1との間の1つの交差を有する、以下のような植物を選抜する:2pm対立遺伝子の2つのエンドポイント間の遺伝的距離が10cMであり、QTLlが中央(QTLlと、pm−l又はpm−hのいずれが1つの間が5cM)に位置する場合に、QTLlといずれかのpm−遺伝子座との間のかかる組換えが5%の頻度で生じる。それにより、最初に交配により得た200の子孫植物をスクリーニングする(10が組換えA型及び10の組換えB型である)。次に、このようにして同定された組換え体を自家交配し、10の植物をスクリーニングし、QTL−1及びpm−に隣接するマーカーを用いてホモ接合体の子孫を解析する。ホモ接合体A×B(10Aと10Bを交配)との間で10の交配を行い、得られるF2に関して10の植物をスクリーニングして1つの二重組換えを選抜する。最終的に、10の二重組換え体を得る。スクリーニングした植物数は全部で500であったが、従来の方法では合計4000の植物のスクリーニングが、10の独立した二重組換え体を得るのに必要であった(QTL−1及びいずれのpm−遺伝子座間も5cMの距離、0.05×0.05=0.25%の二重組換え頻度を有する結果となることが必要であった)。
【0095】
本発明は、マーカーにより補助される選抜方法(MAS)のより良好なモデルの提供に関する。ゆえに本発明は、植物の育種方法、植物(特にキュウリ)の選抜方法の提供に関する。特に繁殖プログラムに用いられる育種植物としての栽培用キュウリ、又は所望の遺伝子型又は潜在的表現型を有する栽培用キュウリの選抜方法の提供に関する。特に、商業的に有用な植物として本願明細書に記載した、有益なウリ科の果物の作製に際に応用できる。本願明細書における栽培用植物とは、故意に選抜されたか、又は所望の遺伝子型又は潜在的な表現型有するとして、農業若しくは園芸生産において故意に選抜された植物に由来する植物(特に同系交配によって得られた植物)として定義される。
【0096】
本発明の二重耐性植物は例えば、遺伝子型AABBcc、AABbcc、AaBBcc又はAaBbccを有してもよい。「A」とは優性のpm−l遺伝子座に基づく耐性遺伝子型であり、「a」はそれに対応する非抵抗性の対立遺伝子であり、「B」は優性のQTL−1遺伝子座に基づく耐性遺伝子型であり、「b」はその対応する感受性の対立遺伝子であり、「c」は劣性のpm−h遺伝子座に基づく耐性遺伝子型である。したがって、本発明の二重耐性植物には、ハイブリッド植物並びにホモ接合植物の両方が包含される。これらの遺伝子型は、以下の交配(親生殖体の遺伝子型)を行うことによって得られる:ABc×ABc、ABc×Abc、ABc×aBc、ABc×abc及びAbc×aBc。したがって、生殖体abcを作出する植物(pm−hを除き、耐性を有さない植物)であっても、本発明の二重耐性(ハイブリッド)植物の作製に使用できることが分かる。特に好ましい実施態様では、本発明の二重耐性植物は、近交系植物(耐性対立遺伝子がホモ接合)である。
【0097】
pm−h遺伝子は劣性遺伝子であるため、バイオアッセイを用いてF1又はBC1においてはモニターできない(すなわち感受性の親を用いてF1/BC1を作製した場合(繁殖の際に一般的))。したがって、本願明細書記載のように、子孫植物への適当な遺伝子移入が、QTL特異的なマーカーを用いてモニターできるというのが、本発明の顕著な効果である。MAS又はMAB方法を用いることにより、当業者は植物の選抜方法の提供をうけることになる。
【0098】
ゆえに本発明はまた、キュウリクロステロウイルス及びキュウリウドンコ病耐性を示すキュウリの選抜方法の提供に関し、当該方法は、前記植物において、本明細書で定義されるQTL−1の単一染色体上の存在、及び、本明細書で定義されるQTLsであるpm−l及びpm−hの少なくとも1つを検出することを含んでなる。かかる植物を選択するための本発明の好ましい方法は、a)キュウリからゲノムDNAのサンプルを調製するステップと、b)前記ゲノムDNAのサンプルにおいて、QTLに連結する、QTL−1、pm−l及びpm−hからなる群から選択される少なくとも1つの分子マーカーを検出する(好ましくは前記群から少なくとも2つの分子マーカーを検出するステップであって、1つのマーカーがクロステロウイルス耐性を検出し、他のマーカーがウドンコ病耐性を検出するステップを含んでなる。
【0099】
キュウリからゲノムDNAのサンプルを調製する処理は、公知の標準的なDNA単離方法により実施してもよい。
【0100】
分子マーカーを検出するステップ(ステップb)は、好ましい実施形態では、AFLP方法で用いられる双方向性プライマーセットを使用して、以下で適切なQTLのマーカーであると判明した増幅生成物を産生させることを含んでなる。かかるプライマーセットは、AFLPマーカーを定めるプライマー、又はマーカー特異的なプライマーとして本願明細書に記載する。双方向性とは、核酸の増幅反応において、1つが順方向プライマーとして機能し、もう1つが逆方向プライマーとして機能することを意味する。
【0101】
あるいは、分子マーカーを検出するステップ(ステップb)は、他の好ましい実施形態では、前記分子マーカーを定義する核酸配列と実質的に相補的な塩基配列を有する核酸プローブの使用を含んでなり、当該核酸プローブは、前記分子マーカーを定義する塩基配列と、ストリンジェントな条件下で特異的にハイブリダイズする。適切な核酸プローブは例えばマーカーに対応する増幅生成物の一本鎖でもよい。
【0102】
分子マーカーを検出するステップ(ステップb)は、前記ゲノムDNA上の核酸増幅反応を行わせて1つ以上のQTLsを検出するステップを含んでもよい。それは一組のマーカー特異的なプライマーを使用してPCR反応を実施することにより可能となる好ましい実施形態では、前記ステップb)は、前記QTLのAFLPマーカーを定義する少なくとも1つのプライマーセット、又は前記QTLのAFLPマーカーの核酸配列とストリンジェントな条件下で特異的にハイブリダイズするプライマーセットの使用を含んでなる。
【0103】
処理d)の、予測された長さ又は予測された核酸配列を有するDNAフラグメントを検出する処理は好ましくは、増幅されたDNAフラグメントが、最初にマーカーが検出された植物由来のDNAと同じプライマーを用いた同様の反応に基づいて増幅した場合の結果を基にした、予想される長さ(数塩基(例えば1、2又は3の塩基)長の増減があってもよい)に対応する長さを有する態様で実施するか、又は、前記増幅されたDNAフラグメントが、最初に前記マーカーが検出された植物中のQTLに関連したマーカーの配列に基づいて予想される配列に一致する核酸配列(80%以上、好ましくは90%以上、より好ましくは95%以上、更により好ましくは97%超、更に好ましくは99%を超の相同性を有しても良い)を有する態様で実施する。当業者に自明のように、感受性の親には存在するが耐性植物には存在しないマーカー(いわゆるトランスマーカー)はまた、子孫植物中の耐性を検出するアッセイにおいて有用であると考えられるが、但しマーカーの欠如を試験して特異的な特徴の存在を検出する場合には最適でない。
【0104】
予測された長さを又は予測された核酸配列を有する増幅されたDNAフラグメントを検出するステップは、標準的なゲル電気泳動、又は自動DNA解析装置を用いて実施してもよい。当該方法は無論、当業者にとり周知である。
【0105】
植物において単一染色体上に2つのQTLsの存在を検出する際、染色体ペインティング法を用いてもよい。かかる方法では、少なくとも第1のQTL及び少なくとも第2のQTLを、in situハイブリダイゼーション法又はin situPCR法で、同じ染色体上で検出してもよい。かかる技術を用いて、pm−l及びpm−hの位置とのQTL−1の前記染色体上の相対的位置を確認してもよく、またその方法は公知技術である(例えばvide Jiangら、1995を参照)。より簡便には、2つのQTLsが単一染色体に存在するという事実を、それらがカップリング期にあることに基づいて確認してもよい(すなわち、別々の染色体に個々の遺伝子が存在する場合と比較して分離が減少している)。
【0106】
分子マーカー及びQTLs:
分子マーカーを用いて、核酸配列の相違を視覚化してもよい。この視覚化は、制限酵素(RFLP)による切断後のDNA−DNAハイブリダイゼーション法、及び/又はポリメラーゼ連鎖反応を使用する方法(例えばSTS、マイクロサテライト、AFLP)において可能である。2つの親の遺伝子型の全ての違いは、これらの親の遺伝子型の交配に基づくマッピング集団(例えばBC1、F2、図2を参照)において分離している。異なるマーカーの分離を比較し、組換え頻度を算出してもよい。通常、異なる染色体上の分子マーカーの組換え頻度は50%である。同じ染色体に存在する分子マーカー間の組換え頻度は、マーカー間の距離に依存する。低い組換え頻度は、染色体上におけるマーカー間の低い遺伝的距離に対応する。全ての組換え頻度を比較することにより、染色体上の分子マーカーで最も論理的なオーダーが得られる。この最も論理的なオーダーは、連鎖地図(パターソン、1996)で表すことができる。耐病性の強化(例えば病原体による感染後の疾患を得ることの低下する発生率、及び/又は病原体との接触後の病変成長率の低下)と関連する連鎖地図上の隣接するマーカー群は、その疾患耐性に関連するQTLの位置をピンポイントで示すものである。
【0107】
本願明細書で同定されるマーカーを用いてもよく、様々な本発明の態様として本願明細書に示される。本発明の態様は、本願明細書で同定されるマーカーの使用に限定されるものではない。上記の態様は、本願明細書に開示されているマーカーの使用のみに限定されず、むしろ未同定のものも同様に含まれる。遺伝学的単位には、「遺伝子」(形質発現が予測できない多数の因子に依存する)以外にも遺伝学的単位「QTL」が存在し、それは定量可能な表現型に直接関与するゲノム領域のことを指す。したがって、遺伝子は植物育種にはほとんど関係しないが、QTLは植物育種に直接適用できるものである。本発明の発明者は、本明細書で定義されるキュウリのクロステロウイルス耐性及びウドンコ病耐性に関与するQTLsが、子孫植物のゲノムにおいて、各々特異的な構成を有しなければならないということを発見した。本発明者は、キュウリのゲノムの単一染色体上の、異なるQTLsに帰属する隣接するゲノムマーカーの一連の存在が、両方の耐性の存在と相関し、それにより、このゲノム構成が通常のメンデル式の遺伝形式で遺伝しうるという観察結果を基礎として、かかる新たな知見を得た。
【0108】
本発明で同定されたQTLsは、単一の染色体又は連鎖群上に位置し、それらの位置はそれ以外の多くの任意のマーカーによって十分に特徴づけられる。本明細書における試験では、増幅断片長多型(AFLP)マーカー、単一ヌクレオチド多型(SNP)及び挿入変異マーカーを用いたが、制限断片長多形性(RFLP)マーカー、マイクロサテライトマーカー(例えばSSRs)、配列依存的増幅領域(SCAR)マーカー、切断増幅多形配列(GAPS)マーカー若しくはアイソザイムマーカー、又はこれらのマーカーの組合せを用いてもよい。一般に、QTLは数百万塩基の領域にわたることもある。したがって、QTLの完全な配列情報を得ることは実際には不可能であり、また不必要でもある。むしろ、(一連の隣接するゲノムマーカーの存在と、特異的な表現型の存在との間の観察された相関によって)QTLを最初に検出する方法により、子孫植物の集団の中から、特異的な表現型特徴を示すポテンシャル遺伝子を有する植物を発見することが可能となる。したがって本発明において、非限定的なマーカーのリストを提供することにより、繁殖プログラムにおけるQTLsの有用性が提供される。
【0109】
マーカーは、ある特定の系列に対して特異的である。したがって、特異的な特徴は特異的なマーカーと関係している。本出願に示すマーカーはQTLの位置を示すだけでなく、それらはまた植物の特異的な表現型の存在との相関関係も示す。ゲノム上のQTLの位置を示す隣接ゲノムマーカーは専ら恣意的なものであり、非限定的であることに留意すべきである。一般に、QTLの位置は、一連の隣接するマーカーとして示され、それは表現型に対する統計学的な相関関係を示すものである。その一連の外でマーカーが見出されたとき(すなわち特異的な閾値以下のLOD−スコアを示し、当該マーカーが隔離され、その領域における当該マーカーとQTL間の組換えが頻繁に発生し、マーカーの存在が統計学的に有意に表現型の存在と相関しないとき)、QTLの境界がそこで設定される。したがって、その特定された領域に位置する他のマーカーを用いてQTLの位置を示すことも可能である。
【0110】
隣接するゲノムマーカーを用いて、個々の植物におけるQTL(すなわち表現型)の存在を示すことができる点は更に重要であり、すなわちマーカーアシスト選抜(MAS)方法に使用できる。原則として、多くの潜在的に有用なマーカーは限定されるが、多くのマーカーを用いてもよく、当業者であれば、本願明細書に記載のマーカー以外の他のマーカーを容易に同定することができる。QTLに連結したあらゆるマーカー(例えば、当該ゲノム領域の物理的に境界内に、ある一定の閾値を上回るLODスコアを示すマーカーが分布し、それにより交配の際、マーカーとQTLとの間で全く組換えが起こらないか、若しくはごくわずかな組換えしか起こらない)、並びにQTLに対して連鎖不平衡にあるいかなるマーカー、並びにQTL中で実際に突然変異を表すマーカーを、MAS方法に用いてもよい。このことは、本発明においてQTLsに関連するマーカーとして同定されたもの(例えばQTL−1に対するAFLPマーカーのE16/M50−244、E16/M50−188及びE11/M48−251)が、MAS方法の使用に適しているマーカーの単なる一例に過ぎないことを示すものである。更に、QTL(又は特異的な特徴を付与するその一部)が他の遺伝的バックグラウンド(すなわち他の植物のゲノム)に移入されると、若干のマーカーは、その特徴が表れるにもかかわらず、子孫内では存在しなくなることがある。すなわち、かかるマーカーは、ゲノム領域の外側に存在し、それはオリジナルの親系列においてのみQTLの特異的な特性を付与する部分であること、並びに、新規な遺伝的バックグラウンドは異なるゲノム構成を有することを示すものである。そのマーカーの不在が子孫への遺伝因子の良好な導入を示すかかるマーカーは「トランスマーカー」と称され、本発明におけるMAS法における使用も同様に好適である。
【0111】
QTLの同定後、QTLの効果(耐性)を、例えば試験対象のQTLsを分離させているBC2S1世代の耐性を評価することによって確認してもよい。耐性の評価は、周知のようにキュウリクロステロウイルス及びキュウリウドンコ病耐性をバイオアッセイするのが最適である。例えば、単離CYSDV(DSMZ−Deutsche Sammlung von Mikroorganismen及びZellkulturen社(ブラウンシュヴァイク、ドイツ)のコレクションno.PV−0592/EWSN_6として保管)を使用してもよく、実験的な感染は、例えばBemisia tabaci(Bバイオタイプ)による伝達により行ってもよい。また、自然感染条件下でのフィールド試験を行い、CYSDV又は他のクロステロウイルス耐性を評価してもよい。植物材料の人工接種によりウドンコ病耐性を評価してもよい。本発明に係るマーカーを最適に用いて、ウドンコ病及び/又はクロステロウイルス耐性を有すると思われるキュウリにおける、本発明の1つ以上のQTLsの存在を検出してもよく、またそれを用いて、マーカーアシスト選抜、並びにウドンコ病及びクロステロウイルス耐性キュウリの育種を実施してもよい。好ましくは、本発明のQTLの存在の検出は、本明細書で定義される少なくとも1つのQTLのマーカーを用いて実施される。ゆえに本発明の別の態様は、ウドンコ病及び/又はクロステロウイルス耐性を付与するQTLの存在の検出方法の提供に関する。当該方法は、ウドンコ病及び/又はクロステロウイルス耐性であると考えられるキュウリにおいて、前記QTLの核酸配列の存在を検出することを含んでなり、上述のマーカーを用いてその存在を検出することを特徴とする。
【0112】
本発明のQTLの核酸配列は、例えば前記QTLと関連する1つ以上のマーカーの核酸配列を決定し、前記マーカー配列の内部プライマーを設計し、更にそれを用いて前記マーカー配列の外のQTLの配列を決定して解析してもよい。例えば、表2、3及び4のAFLPマーカーの核酸配列は、対象となる植物のゲノム中の前記マーカーの存在の決定において使用する泳動ゲルから前記マーカーを単離し、例えば公知の方法であるジデオキシ末端停止法により、前記マーカーの核酸配列を決定してもよい。
【0113】
ウドンコ病及び/又はクロステロウイルス耐性を有すると思われるキュウリのQTLの存在を検出するかかる方法の実施形態では、当該方法は、前記QTL(好ましくは表2、3及び4のマーカーから選択)に連結したマーカーの核酸配列にストリンジェントなハイブリダイゼーション条件下でハイブリダイズできるオリゴヌクレオチド又はポリヌクレオチドを提供するステップと、ウドンコ病及び/又はクロステロウイルス耐性を有すると思われるキュウリのゲノム核酸の断片と、前記オリゴヌクレオチド又はポリヌクレオチドとを接触させるステップと、前記ゲノム核酸断片に対する、前記オリゴヌクレオチド又はポリヌクレオチドの特異的なハイブリダイゼーションの存在を測定するステップを含んでなる。
【0114】
好ましくは前記方法は前記ウドンコ病及び/又はクロステロウイルス耐性を有すると思われるキュウリから得られる核酸サンプルに対して実施するが、in situハイブリダイゼーション法を用いてもよい。あるいは、より好ましい実施形態ではあるが、当業者であれば、QTLの核酸配列が決定された後、前記QTLの核酸配列にストリンジェントなハイブリダイゼーション条件下でハイブリダイズできる特異的なハイブリッド形成用のプローブ若しくはオリゴヌクレオチドを設計し、かかるハイブリダイゼーションプローブを用いて、ウドンコ病及び/又はクロステロウイルス耐性を有すると思われるキュウリにおいて、本発明のQTLの存在を検出することができる。
【0115】
主要なクロステロウイルス抵抗性遺伝子の位置を示すマーカーは、親株から生じた耐性個体及び高感受性個体からのキュウリDNA断片の選択的増幅によって得てもよい。当該方法は、−前記断片を制限酵素(例えばEcoRI及びMsel)で処理し、更にライゲーションさせ、当該断片の端にプライマーに対応する補助アダプター(それらの端に1つ以上の特異的なヌクレオチドを有する)を連結させるステップであって、プライマー対のうちの1つのプライマーが検出目的で標識されているステップと、−変性条件下でゲル電気泳動を行い、増幅生成物を分離させるステップと、−抵抗性の子孫に由来する断片の混合物、及び感受性の子孫に由来する断片の混合物と、親株に由来する断片とを、それらの電気泳動プロファイルを比較することによって、抵抗性の遺伝子座と遺伝的に連結している多形バンドを同定するステップを含んでなる。前記同定の後、全ての個体に関して確証試験を行い、更にマーカーと耐性遺伝子座との間の遺伝子組換え率を算出する。
【0116】
原則として、本明細書で用いられる個々のAPLPマーカーは指定されたマーカーコードを有する。このコードは2つのプライマーを定義し、任意に、定義済みのアクセッションNo.(上記表1の説明を参照)のプライマーの増幅生成物の長さを示す図との組み合わせにより説明する。すなわちAFLPマーカーを用いることにより、キュウリゲノムDNAを鋳型とした増幅反応を実施し、一本鎖若しくは二本鎖DNAフラグメントを得、示されたアクセッションNo.の場合、示された長さの断片が得られる。更に当該マーカーは、5’−3’方向において、第1のプライマー配列、キュウリ特異的なDNA塩基配列、及び第2のプライマー配列からなる配列、更にはその補足配列を含んでなる。したがって、キュウリ特異的なDNA塩基配列には2つのプライマーが隣接している。用語「キュウリ特異的なDNA塩基配列」は、それぞれのプライマーが隣接する領域の核酸配列を意味し、国際公開第02/22836号にて説明したように、特定のプライマーを使用してキュウリ目録Khira PI250147から増幅される配列、又はそれに対して少なくとも90%、好ましくは少なくとも95%、最も好ましくは少なくとも98%の配列相同性を有する配列を表す。
【0117】
トランスジェニック技術によるウドンコ病及びクロステロウイルス耐性キュウリの育種:
本発明の別の態様では、少なくともQTL−1を含んでなる核酸(好ましくはDNA)配列、並びにpm−l及びpm−h又はその耐性付与部分のうちの1つ(好ましくは両方)を、本発明の二重耐性キュウリの育種に用いてもよい。本発明のこの態様では、本明細書で定義されるQTLs又はその耐性付与部分の使用により、二重耐性キュウリを作製し、当該使用においては、適切な宿主植物への前記QTLを含む核酸配列の導入を含んでなる。前述のように、前記核酸配列は、適切なクロステロウイルス及び/又はウドンコ病耐性ドナー植物に由来してもよい。QTL−1として本願明細書に同定されるクロステロウイルス耐性遺伝子座の適切な供給源として、キュウリランドレース Khira(PI250147)(パキスタン産)が挙げられる。それらの商業的な供給源、並びにその他の多くのPM耐性キュウリ園芸品種は、例えばhttp://cuke.hort.ncsu.edu/cucurbit/cuke/cukemain.html.に列挙されている。PM耐性植物は、例えばT.C.Wehner,cucumber gene curator for the Cucurbit Genetics Cooperative(CGC),Department of Horticultural Science,North Carolina State University,Ealeigh,NC27695−7609 U.S.A.から得られてもよい。本願明細書に記載する両方のpm遺伝子座の供給源としては、NPI(PI 200815を有する交雑Natsufushinari(PI 279465)から調製)が挙げられる。それらは例えば、Centre for Genetic Resources,the Netherlands(CGN),Wageningen,The Netherlandsから得てもよい。ECP/GR Cucurbits Database hosted by the Center for the Conservation and Breeding of Agrodiversity(COMAV) in Valencia,Spain、又はGermplasm Resources Information Network(GRIN) hosted by the USDA’s National Germplasm Resources Laboratory,Beltsville,Marylandなどの幾つかの植物データベースを用いて、登録株の検索を行ってもよい。本発明によるこれらの材料の同定方法において記載したように、クロステロウイルス又はウドンコ病耐性を示す他のキュウリを耐性ドナー植物として利用してもよい。
【0118】
適切なドナー植物を同定した後、クロステロウイルス又はウドンコ病耐性を付与するQTLを含む核酸配列、又はその耐性付与部分を、利用可能ないかなる方法によって適切な宿主植物へ導入してもよい。例えば、上述の核酸配列は、感受性の宿主植物と、クロステロウイルス又はウドンコ病耐性ドナー植物を交配すること(すなわち遺伝子移入)、形質転換、プロトプラスト融合、二重ハプロイド技術、又は胚レスキュー若しくは他のいかなる核酸伝達システムによって行ってもよく、任意に、QTLを含む耐性型の子孫植物の選抜をその後行ってもよい。クロステロウイルス又はウドンコ病耐性(又はその耐性付与部分)のためのQTLを含んでなる核酸配列を導入するトランスジェニック技術では、当該核酸を公知の方法を用いて前記ドナー植物から単離し、その単離された核酸をトランスジェニック技術を用いて宿主植物に導入する。当該導入方法としては、例えばベクター、生殖体、又は他の適切な媒体(例えば前記核酸配列でコーティングした粒子)の導入が挙げられる。
【0119】
植物の形質転換は一般に、植物細胞において機能する発現ベクターの構築を必要とする。本発明において、かかるベクターはクロステロウイルス又はウドンコ病耐性のためのQTL(又はその耐性付与部分)を含む核酸配列を含んでなり、当該ベクターにおいて、クロステロウイルス又はウドンコ病−耐性を与えている遺伝子は、調節エレメント(例えばプロモータ)と連結しているか、又は制御下にある。当該組合せに含まれる遺伝子の少なくとも1つがクロステロウイルス又はウドンコ病−耐性をコードする場合、当該発現ベクターは1つ以上のかかる制御可能に連結された遺伝子/制御エレメントの組合せを含んでもよい。当該ベクターはプラスミドの形でもよく、単独で使用してもよく、又は他のプラスミドとの組合せで使用してもよく、それらを用いて、公知技術の形質転換法(例えばアグロバクテリウム形質転換系)を用いて、クロステロウイルス耐性及びウドンコ病遺伝子を導入した植物を得る。
【0120】
発現ベクターは、使用可能な状態で調節エレメント(例えばプロモータ)と連結された少なくとも1つのマーカー遺伝子を含んでもよく、それにより、マーカーを含む形質転換細胞を、ネガティブ選抜(選択可能なマーカー遺伝子を含まない細胞生長を阻害)によって、又は、ポジティブ選抜(マーカー遺伝子によってコードされる生成物のスクリーニング)により回収することができる。植物の形質転換に一般的に用いられる選択可能な多くのマーカー遺伝子が従来技術において周知であり、例えば、選抜用の化学物質の毒性を代謝的に除去する酵素をコードする遺伝子(抗生物質又は除草剤)、又は阻害剤に対する感受性を喪失する形に変換された標的をコードする遺伝子などが挙げられる。マンノース選抜のようないくつかのポジティブ選抜方法が公知である。あるいは、マーカーを使用しない形質転換を行い、前記マーカー遺伝子のない植物を得てもよく、その技術は従来技術において周知である。
【0121】
発現ベクターを植物に導入する1つの方法は、アグロバクテリウム(例えばHorschら、1985)の自然形質転換システムに基づく。A.tumefaciens及びA.rhizogenesは遺伝的に植物細胞を形質転換する植物病原性土壌細菌である。A.tumefaciens及びA.rhizogenesのTi及びRiプラスミドはそれぞれ、植物の形質転換に関与する遺伝子を担持する(例えばKado(1991)を参照)。発現ベクターを植物組織に導入する方法としては、アグロバクテリウムチュメファシエンス、(Horschその他、1985)を用いた植物細胞の直接感染又は共存培養が挙げられる。グルーバー及びクロスビー(1993)及びマローニー(1989)は、アグロバクテリウムを媒介された遺伝子移入その他のアグロバクテリウムベクターシステムを開示している。また米国特許第5591616号を参照のこと。植物発現ベクター及びレポーター遺伝子及び変換プロトコルの一般的説明、並びにアグロバクテリウムの媒介による遺伝子移入ためのアグロバクテリウムベクターシステム及びそれを用いた方法の説明は、グルーバー及びクロスビー(1993)に記載されている。植物組織を培養する一般の方法は、三木ら(1993)による例、及びフィリップスら(1988)により開示されている。分子クローニング技術及び適切な発現ベクターのための適当な参照ハンドブックとしては、Sambrook及びRussell(2001)が挙げられる。
【0122】
発現ベクターを植物に導入する他の方法として、マイクロインジェクションにより媒介される形質転換に基づく方法が挙げられる。当該方法では、DNAをマイクロ発射体の表面上に担持させることを特徴とする。発現ベクターは、植物細胞壁及び膜を透過するのに十分な300〜600m/sの速度にマイクロ発射体を加速する装置を用いて植物組織に導入される(サンフォードその他、1987、1993;サンフォード、1988、1990;クラインら、1988、1992を参照)。植物にDNAを導入する他の方法としては、標的細胞(チャンその他は、知る、1991)の超音波処理を介する方法が挙げられる。あるいは、リポソーム又はスフェロプラスト融合を用いて、発現ベクターを植物に導入してもよい(例えばDeshayesら、1985、及びChristouら(1987を参照)。CaCl2沈殿、ポリビニルアルコール又はポリ−L−オルニチンを用いたプロトプラストへのDNAの直接の取り込みも報告されている(例えばHainら、1985、及びDraperら、1982を参照)。プロトプラスト及び全細胞及び組織へのエレクトロポーレーションに関しても開示されている(D’Halluinら、1992、及びラウルセン、1994を参照)。
【0123】
他の周知の技術(例えばBACの使用、キュウリゲノムの一部を細菌人工染色体(BAC)に導入(すなわちベクターを用いて大腸菌細胞のDNAフラグメント(100〜300kbのサイズを挿入、平均150kb)をクローニング)し、バクテリア大腸菌で見つかる天然のP因子プラスミドに基づく(Zhao及びStodolsky、2004)方法)、を例えばBIBACシステム(ハミルトン、1997)との組合せで使用し、遺伝子導入植物を作製してもよい。
【0124】
キュウリ標的組織の形質転換に続いて、上記した選択可能なマーカー遺伝子を発現させ、現在公知技術の再生及び選抜方法を用いて形質転換細胞、組織及び/又は植物の選抜を行う。
【0125】
非トランスジェニック方法によるクロステロウイルス及びウドンコ病耐性キュウリの育種:
クロステロウイルス及びウドンコ病耐性キュウリを作製するための別の実施形態では、プロトプラスト融合を用いて、ドナー植物から宿主植物への核酸導入を行ってもよい。プロトプラスト融合とは、2つ以上のプロトプラスト(細胞壁を酵素処理によって除去した細胞)間の誘発若しくは自然的な結合(例えば体細胞ハイブリダイゼーション)により、単一の二核若しくは多核細胞を作製する技術のことを指す。融合細胞(性質により交配できない植物種から得てもよい)を組織培養して、望ましい特徴の組合せを示すハイブリッド植物を形成できる。より詳しくは、第1のプロトプラストを、クロステロウイルス又はウドンコ病感染に対する耐性を示すにキュウリ又は他の植物系列から得てもよい。第2のプロトプラストは、例えば、病害抵抗性、虫害抵抗性、有益な果物特徴など(これらに限定されない)を有する第2のキュウリ又は他の植物変異体(好ましくは商業的に有益な特徴を有するキュウリ系列)から調製してもよい。次にプロトプラストを、従来公知のプロトプラスト融合法を用いて融合させる。
【0126】
あるいは、ドナー植物から宿主植物への胚レスキュー(embryo rescue)を採用し、本願明細書に記載の1つ以上のQTLsを含有する核酸を導入してもよい。胚レスキューは、植物が生存できる種子の作製に失敗した時、交配株から胚を単離するのに用いることができる。この方法では、植物の受精卵又は未熟種子を組織培養し、新規な植物を作製する(Pierik、1999)。
【0127】
本発明はまた、クロステロウイルス又はウドンコ病耐性キュウリの作製方法にも関する。当該方法は、上記したドナーキュウリ中において、クロステロウイルス又はウドンコ病耐性に関連する定量耐性遺伝子座(QTL)の存在を検出する方法を実施するステップと、少なくとも検出した当該1つ以上のQTL、又はクロステロウイルス若しくはウドンコ病耐性付与部分を含んでなる核酸配列を、前記ドナー植物からクロステロウイルス又はウドンコ病感受性の宿主キュウリに導入するステップを含んでなる。前記核酸配列の導入は本願明細書に上記したいかなる方法で実施してもよい。
【0128】
かかる方法の好ましい実施態様として、交雑によって、前記核酸配列を、クロステロウイルス又はウドンコ病耐性ドナーキュウリから、クロステロウイルス又はウドンコ病感受性の宿主キュウリへの遺伝子導入する態様が包含される。したがって、この遺伝子導入は従来の繁殖方法を用いて適宜実施してもよい。QTLsは好ましくはマーカーアシスト選抜(MAS)又はマーカーアシスト育種(MAB)を用いて商業的なキュウリ種へ移入される。MAS及びMABは、所望の特徴をコードする遺伝子を1つ以上含むそれらの子孫植物の同定及び選抜における、当該分子マーカーの1つ以上の使用を含んでなる。かかる例においては、その同定及び選抜は本発明に係るQTLs又はそれに連結するマーカーの選抜に基づく。MASを用いて、目的のQTLを有する近同質遺伝子系統(NIL)を育種してもよく、各QTLの効果の詳細な検討も可能となり、また戻し交配自殖系統(BIL)集団の育種にも応用することができる(例えばNesbittら、2001、van Berlooら、2001)。この実施形態により育種されるキュウリは、宿主植物から大多数のそれらの特徴を好適に引き継ぎ、またドナー植物からのクロステロウイルス及びウドンコ病耐性の特徴も有する。
【0129】
上記したように、従来の繁殖方法を用いて、クロステロウイルス又はウドンコ病耐性に関与する核酸配列を、クロステロウイルス及びウドンコ病感受性の宿主キュウリに導入することができる。系統育種と呼ばれる1つの方法では、クロステロウイルス又はウドンコ病に対する耐性を示し、クロステロウイルス又はウドンコ病耐性に関与する核酸コード配列を有するドナーキュウリを、クロステロウイルス若しくはウドンコ病感受性の宿主キュウリ(好ましくは商業的に望ましい特徴(限定されないが病害抵抗性、虫害抵抗性、有益な果物特徴など)を示す)と交配させる。得られる植物群(F1ハイブリッド)を更に自家授粉し、種子(F2種子)を得る。F2種子から成長させたF2植物を、更にクロステロウイルス及びウドンコ病耐性に関してスクリーニングする。当該集団を、様々な方法でスクリーニングしてもよい。
【0130】
第1に、従来の疾患スクリーンを使用して集団をスクリーニングしてもよい。かかる病態スクリーニングは公知である。好ましくは、定量的バイオアッセイを用いる。第2に、マーカーアシスト選抜を実施し、上記した分子マーカーの1つ以上を使用してクロステロウイルス又はウドンコ病耐性に関与する核酸コード配列を含むそれらの子孫を同定してもよい。上記した他の方法を用いて、QTLの存在を検出してもよい。また、マーカーアシスト選抜を用いて定量的バイオアッセイから得られる結果を確認してもよく、またその他の幾つかの方法を組み合わせて用いてもよい。
【0131】
近交系クロステロウイルス及びウドンコ病耐性キュウリ系列の親系列は、循環選択及び戻し交配の技術を使用して育種してもよく、自殖及び/又はジハプロイド又は他のあらゆる技術を用いて作製してもよい。循環選択及び戻し交配において、クロステロウイルス及びウドンコ病耐性は、循環親と、第1のドナー植物を交配させることにより標的宿主植物に移入することもできる。それは反復親とは異なり、本明細書において「一回親」と称される。循環親はクロステロウイルス及びウドンコ病に対する耐性を有さないか、又は低い耐性を有する植物であって、例えば商業的に望ましい特徴(病害抵抗性、虫害抵抗性、有益な果物特徴など)を有するものである。一回親は、クロステロウイルス及びウドンコ病耐性を示し、クロステロウイルス及びウドンコ病耐性に関与する核酸配列を含んでなる。一回親は、循環親との交配により得られる植物変異体又は近交系であってもよい。循環親及び一回親との間で交配して得た株を、循環親に戻し交配する。得られる植物群を更に所望の特徴に関してスクリーニングする。当該スクリーニング方法は特に限定されない。例えば、周知の表現型病理スクリーニング又は定量的バイオアッセイを用いて、集団をスクリーニングしてもよい。あるいは、バイオアッセイを使用する代わりに、マーカーアシスト選抜(MAS)を、上記の分子マーカー、ハイブリッド形成プローブ又はポリヌクレオチドの1つ以上を使用して、クロステロウイルス及びウドンコ病耐性に関与する核酸配列を含む子孫を同定してもよい。また、MASを用いて定量的バイオアッセイから得られる結果を確認してもよい。また、pm−hの劣性形質は、この遺伝子が表現型スクリーン(例えば耐性バイオアッセイ)を用いてF1又はBC1集団から選抜できないことを意味する。ゆえに本願明細書に係るマーカーは、遺伝子型のスクリーニングによって適当な子孫植物を選択するのに適切である。
【0132】
スクリーニングの後、クロステロウイルス及びウドンコ病耐性表現型を示すF1ハイブリッド植物、又は好ましくは遺伝子型は、クロステロウイルス及びウドンコ病耐性に関与する核酸配列を含んでなり、それを更に選抜して循環親に戻し交配し、キュウリを次第に近交系にする。このプロセスは、2〜5以上の世代にわたり実施することが好ましい。原則として、循環親と一回親との交配により得られる、クロステロウイルス及びウドンコ病耐性株の子孫は、クロステロウイルス及びウドンコ病耐性に関与する1つ以上の遺伝子に関してヘテロ接合体である。
【0133】
例えば、植物系列にpm−l及びpm−h遺伝子座を移入したとき、pm−h遺伝子座は劣性であり、その耐性表現型は、ホモ接合のpm−h植物を形成できる条件下においてのみ子孫植物において生じることに留意すべきである。
【0134】
一般に、所望の特徴をハイブリッドキュウリ変異体に導入する方法は、以下のステップを含んでなる:
(a)1つ以上の所望の特徴を含む他のキュウリと、近交系キュウリ親を交配して、F1子孫植物を作製するステップであって、当該所望の特徴が、クロステロウイルス耐性及びウドンコ病耐性からなる群から選択されるステップと、
(b)好ましくは本明細書で定義される分子マーカーを使用して、所望の特徴を有する前記F1子孫植物を選抜するステップと、
(c)前記近交系キュウリ親株と、前記選択された子孫との戻し交配して戻し交配植物を得るステップと、
(d)戻し交配子孫植物を、所望の特徴及び前記近交系キュウリ親株の形態的、生理的な特徴に関して選択するステップであって、前記選抜が、ゲノムDNAを単離し、上記DNAを、QTL−1、pm−l及び/又はpm−hの少なくとも1つの分子マーカー(好ましくは本願明細書に記載のもの)の存在に関して試験するステップと、
(e)ステップ(c)及び(d)を2回以上反復し、第3世代若しくはそれ以上の戻し交配子孫植物を作製するステップと、
(f)選抜した戻し交配子孫を任意に近交し、ホモ接合の植物を同定するステップと、
(g)最低1つの前記戻し交配子孫又は近交系植物と、他の近交系のキュウリ親株(好ましくはpm−hypo耐性、すなわちホモ接合のpm−h)とを交配し、ハイブリッドキュウリ変異体(同じ環境で成長させたとき、ハイブリッドキュウリ変異体の所望の特徴、並びに形態的及び生理的特徴の全てを有する)を得るステップを含んでなる。
【0135】
上記のように、最後の戻し交配生成は近交により行い、クロステロウイルス及びウドンコ病耐性に関してホモ接合の純粋株(近交系)子孫を得てもよい。したがって、循環選択、戻し交配及び近交により、商業的に有用な特徴と関連する他の遺伝子と共に、クロステロウイルス及びウドンコ病耐性に関連する遺伝子と相同な遺伝子を有する系列が得られる。
【0136】
クロステロウイルス及びウドンコ病耐性キュウリ及び種子:
植物育種の目的は、単一の変異体又はハイブリッド中で様々な望ましい特徴を組み合わせることである。商品作物の場合、これらの特徴としては、病気及び昆虫に対する耐性、熱及びかんばつに対する体制、成熟期間の短縮、収率の上昇及び農産物としての優れた品質などが挙げられる。植物特徴(例えば発芽及び成長、成長率、成熟及び草高の均一性など)が重要となることもある。
【0137】
商品作物は、花粉媒介による植物的な方法を利用して育種される。植物は、同一の花に由来する花粉が、その同一の花若しくは同じ植物の他の花へ移される場合に自家授粉される。同じファミリー又は系列の個体を花粉媒介に用いたとき、植物は兄弟授粉される。花粉が、異なるファミリー又は異なる系列の植物に存在する花に由来する場合、植物は他花受粉される。
【0138】
多世代にわたり自動授粉され、形態を基に選抜された植物は、全ての遺伝子座においてホモ接合性になっており、真の繁殖子孫としての同一集団を形成する。2つの異なるホモ接合性系列間での交配により、多くの遺伝子座に関してヘテロ接合体であるハイブリッド植物の同一集団を形成する。多くの遺伝子座でヘテロ接合体となっている2本の植物の交配により、遺伝的に多様な、同一でないヘテロな植物集団を形成する。
【0139】
キュウリ育種プログラムにおけるハイブリッドキュウリ多様性の成長は、以下の3つのステップを含んでなる:
(1)第一の育種交配のために、様々な生殖質プールから植物を選抜するステップと、
(2)繁殖交配から選抜された植物を数世代にわたり近交し、個々に繁殖した、非常に均一な一連の近交系を作製するステップと、
(3)ハイブリッド子孫(F1)の作製に無関係な近交系と、選抜された近交系とを交配させるステップ。同系交配を充分行った後、連続したハイブリッド世代は単に、育種された近交系の種子を増加させる目的でのみ用いる。好ましくは、近交系は、その遺伝子座の約95%以上でホモ接合性対立遺伝子を含まなければならない。
【0140】
近交系のホモ接合性及び均質性による顕著な効果として、確立された近交系のペアを交配して作製したハイブリッドは、常に同質であるということである。優れたハイブリッドを作製する近交系を同定した後、これらの近交系両親を使用してハイブリッド種を継続的に供給することができ、更にこのハイブリッド種子からハイブリッドキュウリを栽培することができる。
【0141】
本発明の方法で作製できるクロステロウイルス及びウドンコ病抵抗キュウリ、又はその部分は、本発明に包含される。
【0142】
本発明の別の態様は、上記にて説明したQTLsを何らかの構成で含んでなるクロステロウイルス及びウドンコ病耐性キュウリ又はその部分の提供に関し、詳細には、少なくとも1つの前記QTLsはその天然の遺伝的バックグラウンドに存在しない。本発明のクロステロウイルス及びウドンコ病耐性キュウリは、例えば近交系のいかなる遺伝子タイプ、ハイブリッド、ハプロイド、ジハプロイド、又はトランスジェニックであってもよい。更に、本発明の植物は、耐性特徴に関してヘテロ接合体であってもよく、またホモ接合性であってもよい(好ましくはホモ接合性である)。本発明のQTLs(並びにその耐性付与部分)をいかなる植物に導入してクロステロウイルス及びウドンコ病耐性植物を作製してもよいが、本発明に係る方法及び植物は、キュウリに関連する植物である。
【0143】
本願明細書に記載されているクロステロウイルス及びウドンコ病耐性近交系キュウリ系列を用いて、更に交配を行い、クロステロウイルス及びウドンコ病耐性ハイブリッド植物を作製してもよい。例えば、本発明の第1のクロステロウイルス及びウドンコ病耐性近交系キュウリは、商業的に望ましい特徴(例えば病害抵抗性、虫害抵抗性、望ましい果物特徴など)を有する第2の近交系キュウリと交配してもよい。この第2の近交系キュウリ系列は、クロステロウイルス又はウドンコ病耐性を有さなくてもよい。好ましくは、この系列は、この劣性形質がハイブリッドの子孫植物において発現されるために、pm−hに関してホモ接合性である。
【0144】
本発明の別の態様は、クロステロウイルス及びウドンコ病耐性キュウリを栽培する際の種子の作製方法に関する。一実施形態では、当該方法は、
クロステロウイルス及びウドンコ病耐性キュウリを準備するステップと、
前記クロステロウイルス及びウドンコ病耐性植物を他のキュウリと交配するステップと、
前記交配から得られる種子を収集するステップを含んでなる。更にそれを栽培し、クロステロウイルス及びウドンコ病耐性キュウリを作製する。
【0145】
別の実施形態では、当該方法は以下のステップを含んでなる:
本発明のクロステロウイルス及びウドンコ病耐性キュウリを準備するステップと、
前記クロステロウイルス及びウドンコ病耐性植物とキュウリとを交配するステップと、
前記交配から生じる種子を回収し、前記種を植物体に成長させるステップと、
本願明細書に記載のいずれかの方法で、クロステロウイルス及びウドンコ病耐性植物を選抜するステップと、
充分な世代数にわたり、選択された植物を自家授粉して繁殖させ、クロステロウイルス及びウドンコ病耐性を付与する対立遺伝子が安定した植物を得るステップと、
そのように得られた植物を、望ましい表現型を有するキュウリと十分な世代数にわたり戻し交配して、クロステロウイルス及びウドンコ病耐性、並びに望ましい表現型を有するキュウリを得るステップと、
最後の戻し交配から生じる植物から作製される種子を回収するステップ。上記の種子を栽培し、クロステロウイルス及びウドンコ病耐性を有するキュウリを作製する。
【0146】
表4のマトリックスにおいて、pm耐性若しくはクロステロウイルス耐性の親に由来する(耐性親の存在として示される)二重耐性植物における、マーカーの有無を示す。マトリックスは更に、二重(PM及びクロステロウイルス)耐性植物系列と関連するAFLPマーカーの起源及び相対的位置を示す。用語pm−l及びpm−hにより示される「マーカー」は、2つのそれぞれのウドンコ病耐性遺伝子座の中心位置を示す。QTL−1遺伝子座の位置は決定しなかったが、クロステロウイルス関連マーカーを使用することにより、その遺伝子座は2つのpm遺伝子座の間に存在することが示される。表より、プールnrs.1〜6では、プールnrs.10〜15より短い、クロステロウイルス耐性親からの遺伝子移入が可能であることが示唆される。マトリックスの解釈は、以下の通りである:例えば、マーカーE23/M40−M003(56.9cM)は、pm耐性親及び二重耐性子孫に存在したが、一方マーカーE11/M80−M003(79.1cM)は、クロステロウイルス耐性親には存在したものの、二重耐性子孫には存在しなかった。すなわち、マーカーE23/M40−M003はPM関連のマーカーであり(その存在は、テストした植物系列のpm−l関連遺伝子の領域の存在を示す)、一方、マーカーE11/M80−M003は、クロステロウイルス耐性に関連するQTL−1遺伝子座の近傍の遺伝的領域であるが、pm−hypo(pm−h)耐性の発現にとっては必須ではないことを示す。
【0147】
【表4】
+=ホモ接合性が存在(高密度のバンド)、
−=存在しない(バンドなし)、
+/−=ヘテロ接合体が(低密度バンド)、
D=中程度に存在(中程度のバンド)、
・=PM抵抗親におけるマーカースコア(両方存在:[+]、不在:[−])、
・=マーカースコア(クロステロウイルス耐性親)、
・=親系列における未同定マーカーの存在。
【図面の簡単な説明】
【0148】
【図1】考えられる組換えシナリオを示す図。2つの別々の植物から得たそうであるウドンコ病(pm)及びクロステロウイルス(QTL−1)耐性を、単一の子孫植物内で連結させた。多くの耐性遺伝子座を有するものとして示される様々な親植物は、それ自体交配若しくは自家交配の結果でありえることを理解すべきである。すなわち、QTLであるpm−l及びpm−hを有する植物は、単一のQTLを有する植物間での交配によって得られる。
【図2】同上。
【図3】同上。
【図4】同上。
【図5】同上。
【図6】同上。
【図7】単一の連鎖群における、キュウリNPI由来のウドンコ病耐性(pm)遺伝子、及びC.sativus Khira由来のクロステロウイルス耐性(QTL−1)遺伝子の様々な位置関係を示す。Pm−leaf1は、本明細書で用いられるpm−lに相当し、Pm hypoは本明細書で用いられるpm−hに相当する。
【図8】本願明細書の表2及び3中の各配列番号により示されるSNP及び挿入マーカー近傍のDNA配列を示す。
【特許請求の範囲】
【請求項1】
キュウリクロステロウイルス及びキュウリウドンコ病に対する耐性を示す植物であって、前記植物がキュウリであり、前記植物が単一の染色体上に、クロステロウイルス耐性を付与する少なくとも1つの染色体領域と、ウドンコ病耐性を付与する少なくとも1つの染色体領域を含んでなり、クロステロウイルス耐性を付与する少なくとも1つの前記領域が、E16/M50−244、E16/M50−188及びE11/M48−251からなる群から選抜される少なくとも1つのマーカーと連結し、ウドンコ病耐性を付与する少なくとも1つの前記領域が、
− 配列番号1の単一ヌクレオチド多形マーカー 39T→G、
− 配列番号2の単一ヌクレオチド多形マーカー 29G→A、
− 配列番号3の単一ヌクレオチド多形マーカー 193C→T、
− 配列番号4の位置221における挿入突然変異5’−AATTT−3’、並びに−マーカーE16/M50−F−194、E11/M48−F−251、E23/M38−M001、E23/M40−M003、E24/M46−M002、E24/M46−M003、E12/M91−M003、E26/M43−M003、E14/M59−F−134及びE14/M59−F−200からなる群から選択される少なくとも1つのマーカーに連結している植物。
【請求項2】
請求項1記載の植物であって、前記植物が、ウドンコ病耐性を付与する少なくとも2つの染色体領域を含んでなり、
前記少なくとも2つの領域のうちの第1が、
− 配列番号1の単一ヌクレオチド多形マーカー 39T→G、
− 配列番号2の単一ヌクレオチド多形マーカー 29G→A、並びに
− マーカーE16/M50−F−194、E11/M48−F−251及びE23/M38−M001からなる群から選択される少なくとも1つのマーカーと連結し、
前記少なくとも2つの領域のうちの第2が、
− 配列番号3の単一ヌクレオチド多形マーカー 193C→T、
− 配列番号4の位置221の挿入突然変異5’−AATTT−3’、並びに
マーカーE23/M40−M003、E24/M46−M002、E24/M46−M003、E12/M91−M003、E26/M43−M003、E14/M59−F−134及びE14/M59−F−200からなる群から選択される少なくとも1つのマーカーに連結している植物。
【請求項3】
請求項2記載の植物であって、クロステロウイルス耐性を付与する少なくとも1つの前記領域が、請求項2記載のウドンコ病耐性を付与する少なくとも2つの前記領域の間に位置する植物。
【請求項4】
請求項1から3のいずれか1項記載のキュウリの部分。
【請求項5】
請求項4記載の部分であって、花粉、胚珠、葉、胚、根、根端、葯、花、果実、茎、芽、枝、根茎、種子、プロトプラスト及びカルスから選択される、前記部分。
【請求項6】
農産物として望ましい特徴を有する第2のキュウリ又は他の種類の植物(好ましくはキュウリ株)と、請求項1から3のいずれか1項記載のキュウリを交配させることにより得られるF1種子であって、前記種子が、マーカーE16/M50−244、E16/M50−188及びE11/M48−251からなる群から選抜されるクロステロウイルス耐性に関連する少なくとも1つのマーカーと、ウドンコ病耐性に関する、
− 配列番号1の単一ヌクレオチド多形マーカー 39T→G、
− 配列番号2の単一ヌクレオチド多形マーカー 29G→A、
− 配列番号3の単一ヌクレオチド多形マーカー 193C→T、
− 配列番号4の位置221における挿入突然変異5’−AATTT−3’、並びに
− マーカーE16/M50−F−194、E11/M48−F−251、E23/M38−M001、E23/M40−M003、E24/M46−M002、E24/M46−M003、E12/M91−M003、E26/M43−M003、E14/M59−F−134及びE14/M59−F−200からなる群から選択される少なくとも1つのマーカーとを含んでなる種子。
【請求項7】
請求項6記載の種子であって、前記第2のキュウリがクロステロウイルスに感受性であり、少なくとも1つの前記ウドンコ病耐性マーカーがホモ接合的に存在する種子。
【請求項8】
前記第2のキュウリが近交系の植物である、請求項6又は7記載の種子。
【請求項9】
請求項4から8のいずれか1項記載の種子を成長させることにより得られるハイブリッド植物。
【請求項10】
請求項9記載のハイブリッド植物の部分。
【請求項11】
請求項10記載の部分であって、前記部分が花粉、胚珠、葉、胚、根、根端、葯、花、果実、茎、芽、枝、根茎、種子、プロトプラスト及びカルスから選抜される部分。
【請求項12】
キュウリクロステロウイルス及びキュウリウドンコ病に対して抵抗性の植物の選抜方法であって、前記植物がキュウリであり、前記方法がマーカー検出アッセイを実施することを含んでなり、前記アッセイが、
a)前記植物のゲノムにおいて、マーカーE16/M50−244、E16/M50−188及びE11/M48−251からなる群から選抜されるクロステロウイルス耐性に関連する少なくとも1つのマーカーの存在を検出するステップと、
b)前記植物のゲノムにおいて、
− 配列番号1の単一ヌクレオチド多形マーカー 39T→G、
− 配列番号2の単一ヌクレオチド多形マーカー 29G→A、
− 配列番号3の単一ヌクレオチド多形マーカー 193C→T、
− 配列番号4の位置221における挿入突然変異5’−AATTT−3’、並びに
− マーカーE16/M50−F−194、E11/M48−F−251、E23/M38−M001、E23/M40−M003、E24/M46−M002、E24/M46−M003、E12/M91−M003、E26/M43−M003、E14/M59−F−134及びE14/M59−F−200からなる群から選択される、ウドンコ病耐性に関連する少なくとも1つのマーカーの存在を検出するステップを含んでなる方法。
【請求項13】
請求項12記載の方法であって、
−前記植物のゲノム中において、
配列番号1のSNPマーカー 39T→G、
配列番号2のSNPマーカー 29G→A、並びに
マーカーE16/M50−F−194、E11/M48−F−251及びE23/M38−M001で表される第1のキュウリウドンコ病耐性付与QTLに連結する少なくとも1つのマーカーの存在を検出するステップと、
−前記植物のゲノム中において、
配列番号3のSNPマーカー 193C→T、
配列番号4の位置221における挿入突然変異 5’−AATTT−3’、並びに
マーカーE23/M40−M003、E24/M46−M002、E24/M46−M003、E12/M91−M003、E26/M43−M003、E14/M59−F−134及びE14/M59−F−200で表される第2のキュウリウドンコ病耐性付与QTLに連結する少なくとも1つのマーカーの存在を検出するステップを含んでなる方法。
【請求項14】
請求項12又は13記載の方法であって、前記方法が、
a)前記植物からゲノムDNAのサンプルを得るステップと、
b)前記植物のゲノムDNAのサンプルにおいて、クロステロウイルス耐性に関連する少なくとも1つの前記マーカーと、ウドンコ病耐性に連結する少なくとも1つの前記マーカーを検出するステップを含んでなる方法。
【請求項15】
請求項14記載の方法であって、前記ステップb)が、前記マーカーを定義する少なくとも1つのプライマーセット、又は前記マーカーを定義する核酸配列と実質的に相補的である少なくとも1つの核酸プローブの使用を含んでなり、前記核酸プローブが1つ以上の前記マーカーを定義する核酸配列と、ストリンジェントなハイブリダイゼーション条件下で特異的にハイブリダイズする塩基配列を有する方法。
【請求項16】
請求項12から15のいずれか1項記載の方法であって、前記マーカーが、例えば交配試験において、カップリングしないマーカーの場合と比較しマーカー同士の分離が減少することにより、同一の染色体上において存在し、カップリング段階であることが示される方法。
【請求項17】
請求項12から16のいずれか1項記載の方法により選抜される植物。
【請求項18】
キュウリクロステロウイルス及びキュウリウドンコ病に対する耐性を示すキュウリの作製方法であって、
a)キュウリクロステロウイルス耐性を付与する染色体領域を含む第1のキュウリを選抜するステップであって、前記選抜が、前記植物のゲノムにおいて、マーカーE16/M50−244、E16/M50−188及びE11/M48−251により表される、キュウリクロステロウイルス耐性を付与するQTLに連結している少なくとも1つのマーカーの存在を検出することにより行われるステップと、
b)キュウリウドンコ病耐性を付与する少なくとも1つの染色体領域を含む第2のキュウリを選抜するステップであって、前記選抜が、前記植物のゲノムにおいて、
− 配列番号1の単一ヌクレオチド多形マーカー 39T→G、
− 配列番号2の単一ヌクレオチド多形マーカー 29G→A、並びに
− マーカーE16/M50−F−194、E11/M48−F−251及びE23/M38−M001により表される、第1のキュウリウドンコ病耐性を付与するQTLと連結している、少なくとも1つのマーカーの存在を検出することによって行われるステップか、
又は、前記選抜が、前記植物のゲノムにおいて、
− 配列番号3の単一ヌクレオチド多形マーカー 193C→T、
− 配列番号4の位置221の挿入突然変異5’−AATTT−3’、並びに
− マーカーE23/M40−M003、E24/M46−M002、E24/M46−M003、E12/M91−M003、E26/M43−M003、E14/M59−F−134及びE14/M59−F−200により表される、第2のキュウリウドンコ病耐性を付与するQTLと連結している、少なくとも1つのマーカーの存在を検出することによって行われるステップによって行われるステップと、
c)ステップa)及びステップb)から得られた前記植物を交配してF1種子を得るステップと、
d)F1種子をF1植物に成長させ、交配又は自家交配することによって前記F1植物から更なる子孫を生じさせるステップと、
e)前記更なる子孫植物から、ステップa)において定義したキュウリクロステロウイルス耐性を付与するQTLに連結している少なくとも1つのマーカーと、ステップb)において定義したキュウリウドンコ病耐性を付与するQTLに連結している少なくとも1つのマーカーを含んでなる植物を選抜するステップを含んでなる方法。
【請求項19】
請求項18記載の方法であって、ステップb)の第2のキュウリを選抜するステップが、前記第1若しくは第2のキュウリウドンコ病耐性を付与するQTLsのうちの1つのみを有する第2のキュウリを選抜することを含んでなり、
更に、
f)前記第1若しくは第2のキュウリウドンコ病耐性を付与するQTLs(すなわち前記第2のキュウリに存在しないQTL)のうちのもう1つを有する第3のキュウリを選抜するステップと、
g)ステップe)で得られたF1植物と、前記第3のキュウリとを交配させて、更なる子孫植物を得るステップと、
h)当該子孫植物から、ステップa)で定義されるキュウリクロステロウイルス耐性を付与するQTLと、ステップb)で定義されるキュウリウドンコ病耐性を付与する両方のQTLsを含む植物を選抜するステップを含んでなる方法。
【請求項20】
請求項17から19のいずれか1項記載の方法であって、ステップb)、e)、f)又はh)で定義されるキュウリウドンコ病耐性を付与する染色体領域を含むキュウリを選抜するステップが、
−前記植物のゲノム中において、
配列番号1のSNPマーカー 39T→G、
配列番号2のSNPマーカー 29G→A、並びに
マーカーE16/M50−F−194、E11/M48−F−251及びE23/M38−M001で表される第1のキュウリウドンコ病耐性付与QTLに連結する少なくとも1つのマーカーの存在を検出するステップと、
−前記植物のゲノム中において、
配列番号3のSNPマーカー 193C→T、
配列番号4の位置221における挿入突然変異 5’−AATTT−3’、並びに
マーカーE23/M40−M003、E24/M46−M002、E24/M46−M003、E12/M91−M003、E26/M43−M003、E14/M59−F−134及びE14/M59−F−200で表される第2のキュウリウドンコ病耐性付与QTLに連結する少なくとも1つのマーカーの存在を検出するステップを含んでなる方法。
【請求項21】
請求項17から20のいずれか1項記載の方法であって、ステップa)、b)、e)、f)又はh)のうちの少なくとも1つは、前記植物から得たゲノムDNAのサンプルを提供するステップと、前記ゲノムDNAのサンプル中から前記少なくとも1つのマーカーを検出するステップを含んでなる方法。
【請求項22】
キュウリクロステロウイルス及びキュウリウドンコ病に対する耐性を示すキュウリを作製する方法であって、
a)請求項12から16のいずれか1項記載の方法を実施して、キュウリクロステロウイルス及びキュウリウドンコ病に対する耐性を示す第1のキュウリを選抜するステップと、
b)前記植物を同系育種して、前記QTLsのホモ接合植物株を得るステップと、
c)ステップa)及びステップb)から得た前記植物を交配してF1種子を得るステップと、
d)前記F1種子をF1植物に成長させるステップを含んでなる方法。
【請求項23】
請求項18から22のいずれか1項記載の方法で得られる植物又はその部分。
【請求項1】
キュウリクロステロウイルス及びキュウリウドンコ病に対する耐性を示す植物であって、前記植物がキュウリであり、前記植物が単一の染色体上に、クロステロウイルス耐性を付与する少なくとも1つの染色体領域と、ウドンコ病耐性を付与する少なくとも1つの染色体領域を含んでなり、クロステロウイルス耐性を付与する少なくとも1つの前記領域が、E16/M50−244、E16/M50−188及びE11/M48−251からなる群から選抜される少なくとも1つのマーカーと連結し、ウドンコ病耐性を付与する少なくとも1つの前記領域が、
− 配列番号1の単一ヌクレオチド多形マーカー 39T→G、
− 配列番号2の単一ヌクレオチド多形マーカー 29G→A、
− 配列番号3の単一ヌクレオチド多形マーカー 193C→T、
− 配列番号4の位置221における挿入突然変異5’−AATTT−3’、並びに−マーカーE16/M50−F−194、E11/M48−F−251、E23/M38−M001、E23/M40−M003、E24/M46−M002、E24/M46−M003、E12/M91−M003、E26/M43−M003、E14/M59−F−134及びE14/M59−F−200からなる群から選択される少なくとも1つのマーカーに連結している植物。
【請求項2】
請求項1記載の植物であって、前記植物が、ウドンコ病耐性を付与する少なくとも2つの染色体領域を含んでなり、
前記少なくとも2つの領域のうちの第1が、
− 配列番号1の単一ヌクレオチド多形マーカー 39T→G、
− 配列番号2の単一ヌクレオチド多形マーカー 29G→A、並びに
− マーカーE16/M50−F−194、E11/M48−F−251及びE23/M38−M001からなる群から選択される少なくとも1つのマーカーと連結し、
前記少なくとも2つの領域のうちの第2が、
− 配列番号3の単一ヌクレオチド多形マーカー 193C→T、
− 配列番号4の位置221の挿入突然変異5’−AATTT−3’、並びに
マーカーE23/M40−M003、E24/M46−M002、E24/M46−M003、E12/M91−M003、E26/M43−M003、E14/M59−F−134及びE14/M59−F−200からなる群から選択される少なくとも1つのマーカーに連結している植物。
【請求項3】
請求項2記載の植物であって、クロステロウイルス耐性を付与する少なくとも1つの前記領域が、請求項2記載のウドンコ病耐性を付与する少なくとも2つの前記領域の間に位置する植物。
【請求項4】
請求項1から3のいずれか1項記載のキュウリの部分。
【請求項5】
請求項4記載の部分であって、花粉、胚珠、葉、胚、根、根端、葯、花、果実、茎、芽、枝、根茎、種子、プロトプラスト及びカルスから選択される、前記部分。
【請求項6】
農産物として望ましい特徴を有する第2のキュウリ又は他の種類の植物(好ましくはキュウリ株)と、請求項1から3のいずれか1項記載のキュウリを交配させることにより得られるF1種子であって、前記種子が、マーカーE16/M50−244、E16/M50−188及びE11/M48−251からなる群から選抜されるクロステロウイルス耐性に関連する少なくとも1つのマーカーと、ウドンコ病耐性に関する、
− 配列番号1の単一ヌクレオチド多形マーカー 39T→G、
− 配列番号2の単一ヌクレオチド多形マーカー 29G→A、
− 配列番号3の単一ヌクレオチド多形マーカー 193C→T、
− 配列番号4の位置221における挿入突然変異5’−AATTT−3’、並びに
− マーカーE16/M50−F−194、E11/M48−F−251、E23/M38−M001、E23/M40−M003、E24/M46−M002、E24/M46−M003、E12/M91−M003、E26/M43−M003、E14/M59−F−134及びE14/M59−F−200からなる群から選択される少なくとも1つのマーカーとを含んでなる種子。
【請求項7】
請求項6記載の種子であって、前記第2のキュウリがクロステロウイルスに感受性であり、少なくとも1つの前記ウドンコ病耐性マーカーがホモ接合的に存在する種子。
【請求項8】
前記第2のキュウリが近交系の植物である、請求項6又は7記載の種子。
【請求項9】
請求項4から8のいずれか1項記載の種子を成長させることにより得られるハイブリッド植物。
【請求項10】
請求項9記載のハイブリッド植物の部分。
【請求項11】
請求項10記載の部分であって、前記部分が花粉、胚珠、葉、胚、根、根端、葯、花、果実、茎、芽、枝、根茎、種子、プロトプラスト及びカルスから選抜される部分。
【請求項12】
キュウリクロステロウイルス及びキュウリウドンコ病に対して抵抗性の植物の選抜方法であって、前記植物がキュウリであり、前記方法がマーカー検出アッセイを実施することを含んでなり、前記アッセイが、
a)前記植物のゲノムにおいて、マーカーE16/M50−244、E16/M50−188及びE11/M48−251からなる群から選抜されるクロステロウイルス耐性に関連する少なくとも1つのマーカーの存在を検出するステップと、
b)前記植物のゲノムにおいて、
− 配列番号1の単一ヌクレオチド多形マーカー 39T→G、
− 配列番号2の単一ヌクレオチド多形マーカー 29G→A、
− 配列番号3の単一ヌクレオチド多形マーカー 193C→T、
− 配列番号4の位置221における挿入突然変異5’−AATTT−3’、並びに
− マーカーE16/M50−F−194、E11/M48−F−251、E23/M38−M001、E23/M40−M003、E24/M46−M002、E24/M46−M003、E12/M91−M003、E26/M43−M003、E14/M59−F−134及びE14/M59−F−200からなる群から選択される、ウドンコ病耐性に関連する少なくとも1つのマーカーの存在を検出するステップを含んでなる方法。
【請求項13】
請求項12記載の方法であって、
−前記植物のゲノム中において、
配列番号1のSNPマーカー 39T→G、
配列番号2のSNPマーカー 29G→A、並びに
マーカーE16/M50−F−194、E11/M48−F−251及びE23/M38−M001で表される第1のキュウリウドンコ病耐性付与QTLに連結する少なくとも1つのマーカーの存在を検出するステップと、
−前記植物のゲノム中において、
配列番号3のSNPマーカー 193C→T、
配列番号4の位置221における挿入突然変異 5’−AATTT−3’、並びに
マーカーE23/M40−M003、E24/M46−M002、E24/M46−M003、E12/M91−M003、E26/M43−M003、E14/M59−F−134及びE14/M59−F−200で表される第2のキュウリウドンコ病耐性付与QTLに連結する少なくとも1つのマーカーの存在を検出するステップを含んでなる方法。
【請求項14】
請求項12又は13記載の方法であって、前記方法が、
a)前記植物からゲノムDNAのサンプルを得るステップと、
b)前記植物のゲノムDNAのサンプルにおいて、クロステロウイルス耐性に関連する少なくとも1つの前記マーカーと、ウドンコ病耐性に連結する少なくとも1つの前記マーカーを検出するステップを含んでなる方法。
【請求項15】
請求項14記載の方法であって、前記ステップb)が、前記マーカーを定義する少なくとも1つのプライマーセット、又は前記マーカーを定義する核酸配列と実質的に相補的である少なくとも1つの核酸プローブの使用を含んでなり、前記核酸プローブが1つ以上の前記マーカーを定義する核酸配列と、ストリンジェントなハイブリダイゼーション条件下で特異的にハイブリダイズする塩基配列を有する方法。
【請求項16】
請求項12から15のいずれか1項記載の方法であって、前記マーカーが、例えば交配試験において、カップリングしないマーカーの場合と比較しマーカー同士の分離が減少することにより、同一の染色体上において存在し、カップリング段階であることが示される方法。
【請求項17】
請求項12から16のいずれか1項記載の方法により選抜される植物。
【請求項18】
キュウリクロステロウイルス及びキュウリウドンコ病に対する耐性を示すキュウリの作製方法であって、
a)キュウリクロステロウイルス耐性を付与する染色体領域を含む第1のキュウリを選抜するステップであって、前記選抜が、前記植物のゲノムにおいて、マーカーE16/M50−244、E16/M50−188及びE11/M48−251により表される、キュウリクロステロウイルス耐性を付与するQTLに連結している少なくとも1つのマーカーの存在を検出することにより行われるステップと、
b)キュウリウドンコ病耐性を付与する少なくとも1つの染色体領域を含む第2のキュウリを選抜するステップであって、前記選抜が、前記植物のゲノムにおいて、
− 配列番号1の単一ヌクレオチド多形マーカー 39T→G、
− 配列番号2の単一ヌクレオチド多形マーカー 29G→A、並びに
− マーカーE16/M50−F−194、E11/M48−F−251及びE23/M38−M001により表される、第1のキュウリウドンコ病耐性を付与するQTLと連結している、少なくとも1つのマーカーの存在を検出することによって行われるステップか、
又は、前記選抜が、前記植物のゲノムにおいて、
− 配列番号3の単一ヌクレオチド多形マーカー 193C→T、
− 配列番号4の位置221の挿入突然変異5’−AATTT−3’、並びに
− マーカーE23/M40−M003、E24/M46−M002、E24/M46−M003、E12/M91−M003、E26/M43−M003、E14/M59−F−134及びE14/M59−F−200により表される、第2のキュウリウドンコ病耐性を付与するQTLと連結している、少なくとも1つのマーカーの存在を検出することによって行われるステップによって行われるステップと、
c)ステップa)及びステップb)から得られた前記植物を交配してF1種子を得るステップと、
d)F1種子をF1植物に成長させ、交配又は自家交配することによって前記F1植物から更なる子孫を生じさせるステップと、
e)前記更なる子孫植物から、ステップa)において定義したキュウリクロステロウイルス耐性を付与するQTLに連結している少なくとも1つのマーカーと、ステップb)において定義したキュウリウドンコ病耐性を付与するQTLに連結している少なくとも1つのマーカーを含んでなる植物を選抜するステップを含んでなる方法。
【請求項19】
請求項18記載の方法であって、ステップb)の第2のキュウリを選抜するステップが、前記第1若しくは第2のキュウリウドンコ病耐性を付与するQTLsのうちの1つのみを有する第2のキュウリを選抜することを含んでなり、
更に、
f)前記第1若しくは第2のキュウリウドンコ病耐性を付与するQTLs(すなわち前記第2のキュウリに存在しないQTL)のうちのもう1つを有する第3のキュウリを選抜するステップと、
g)ステップe)で得られたF1植物と、前記第3のキュウリとを交配させて、更なる子孫植物を得るステップと、
h)当該子孫植物から、ステップa)で定義されるキュウリクロステロウイルス耐性を付与するQTLと、ステップb)で定義されるキュウリウドンコ病耐性を付与する両方のQTLsを含む植物を選抜するステップを含んでなる方法。
【請求項20】
請求項17から19のいずれか1項記載の方法であって、ステップb)、e)、f)又はh)で定義されるキュウリウドンコ病耐性を付与する染色体領域を含むキュウリを選抜するステップが、
−前記植物のゲノム中において、
配列番号1のSNPマーカー 39T→G、
配列番号2のSNPマーカー 29G→A、並びに
マーカーE16/M50−F−194、E11/M48−F−251及びE23/M38−M001で表される第1のキュウリウドンコ病耐性付与QTLに連結する少なくとも1つのマーカーの存在を検出するステップと、
−前記植物のゲノム中において、
配列番号3のSNPマーカー 193C→T、
配列番号4の位置221における挿入突然変異 5’−AATTT−3’、並びに
マーカーE23/M40−M003、E24/M46−M002、E24/M46−M003、E12/M91−M003、E26/M43−M003、E14/M59−F−134及びE14/M59−F−200で表される第2のキュウリウドンコ病耐性付与QTLに連結する少なくとも1つのマーカーの存在を検出するステップを含んでなる方法。
【請求項21】
請求項17から20のいずれか1項記載の方法であって、ステップa)、b)、e)、f)又はh)のうちの少なくとも1つは、前記植物から得たゲノムDNAのサンプルを提供するステップと、前記ゲノムDNAのサンプル中から前記少なくとも1つのマーカーを検出するステップを含んでなる方法。
【請求項22】
キュウリクロステロウイルス及びキュウリウドンコ病に対する耐性を示すキュウリを作製する方法であって、
a)請求項12から16のいずれか1項記載の方法を実施して、キュウリクロステロウイルス及びキュウリウドンコ病に対する耐性を示す第1のキュウリを選抜するステップと、
b)前記植物を同系育種して、前記QTLsのホモ接合植物株を得るステップと、
c)ステップa)及びステップb)から得た前記植物を交配してF1種子を得るステップと、
d)前記F1種子をF1植物に成長させるステップを含んでなる方法。
【請求項23】
請求項18から22のいずれか1項記載の方法で得られる植物又はその部分。
【図1】

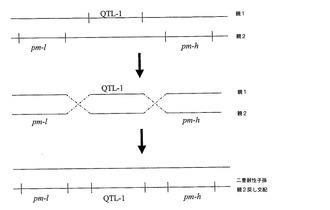
【図2】
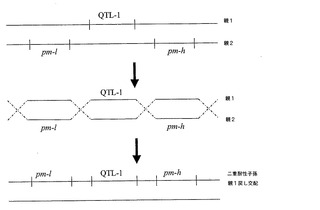
【図3】
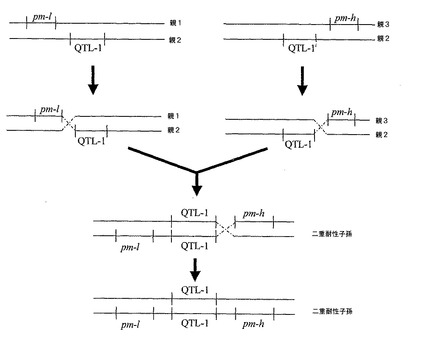
【図4】
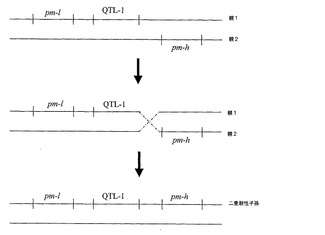
【図5】
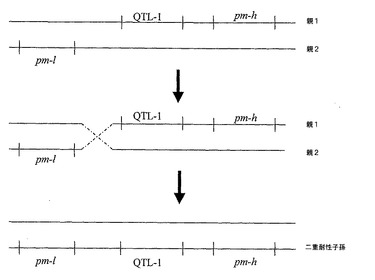
【図6】
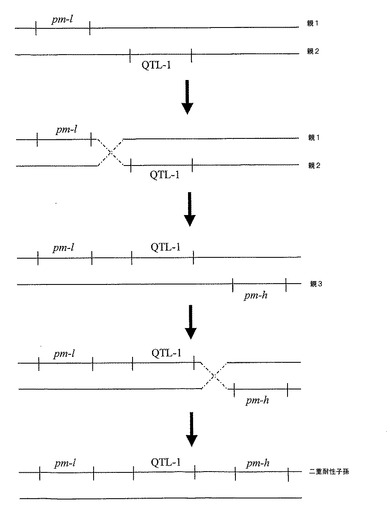
【図7】
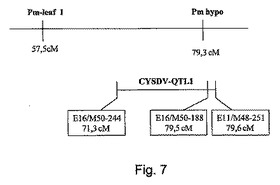
【図8】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公表番号】特表2009−514523(P2009−514523A)
【公表日】平成21年4月9日(2009.4.9)
【国際特許分類】
【出願番号】特願2008−538835(P2008−538835)
【出願日】平成18年11月6日(2006.11.6)
【国際出願番号】PCT/NL2006/000553
【国際公開番号】WO2007/053015
【国際公開日】平成19年5月10日(2007.5.10)
【出願人】(508135079)デ ルイター シーズ アール アンド ディー ビー.ヴイ. (1)
【Fターム(参考)】
【公表日】平成21年4月9日(2009.4.9)
【国際特許分類】
【出願日】平成18年11月6日(2006.11.6)
【国際出願番号】PCT/NL2006/000553
【国際公開番号】WO2007/053015
【国際公開日】平成19年5月10日(2007.5.10)
【出願人】(508135079)デ ルイター シーズ アール アンド ディー ビー.ヴイ. (1)
【Fターム(参考)】
[ Back to top ]