
耐病性植物
本発明は、向上した耐病性を有する植物を産生するための方法に関する。NRC1タンパク質およびこれらをコードする核酸配列、並びにNRC1タンパク質を産生するトランスジェニック植物が提供される。
【発明の詳細な説明】
【発明の分野】
【0001】
本発明は、ゲノムに統合されてNRC1タンパク質(NB-LRR Required for HR-associated Cell Death 1)をコードする遺伝子を含むトランスジェニック植物および植物細胞、並びにかかる植物および細胞を作成するための方法に関する。とりわけ、向上した耐病性を備えたナス科(Solanaceae)植物および植物の一部(種子、果実、葉など)が提供される。また、本発明のNRC1タンパク質をコードする単離された核酸分子、これらを含むベクター、並びに単離されたNRC1タンパク質自体が提供される。更に、植物および植物細胞に耐病性を付与する一または複数の突然変異を内在性NRC1対立遺伝子に含む植物細胞および植物が提供される。
【発明の背景】
【0002】
植物の積極的防御は、耐性遺伝子により仲介される病原体の無毒性因子の認識により誘発され、遺伝子対遺伝子(gene-for-gene)モデルに従う(Dangl and Jones, 2001, Nature 411, 826-833)。今日までに幾つかの植物耐性遺伝子(R遺伝子)がクローニングされており、これらは、コードするタンパク質の構造に基いて、幾つかのグループに分類される(Hammond-Kosack and Jones, 1997, Annu. Rev. Plant Physiol. Plant Molec. Biol. 48, 575-607)。ほとんどのR遺伝子は、ヌクレオチド結合部位(NB)およびロイシンリッチ繰返し配列(LRR)を含有する細胞質NB-LRRタンパク質をコードする。このグループは、二重コイル(coiled−coil)ドメインを含有するCC-NB-LRRタンパク質をコードする遺伝子、および哺乳類のTollおよびインターロイキン(IL)レセプターに類似のドメインを有するタンパク質、いわゆるTIR-NB-LRRタンパク質をコードする遺伝子からなる(Hammond-Kosack and Jones, 1997, supra)。
【0003】
このような特定の耐性遺伝子を永続的耐性のための育種プログラムで使用することは、病原体が、無毒性因子の突然変異により認識を容易に回避し、これにより積極的防御の誘導を妨害するため、問題がある(Westerink et al., 2004, Mol. Microbiol. 54, 533-545)。耐性タンパク質(Rタンパク質)の間の類似性は、共通の耐性経路の存在を示唆する(Shirasu and Schulze-Lefert, 2000, Plant Mol. Biol. 44, 371-385)。したがって、耐性に必要な追加の遺伝子を同定することにより、かかるシグナリング経路がどのように機能するかについて情報が提供されるだけでなく、我々が、耐性においてより一般的な役割を果たす遺伝子を同定することも可能になる。たとえば、Nicotiana benthamianaにおけるウイルス誘導性遺伝子サイレンシング(VIGS)により、SGT1が、複数の防御経路、たとえばN-、Rx-およびPto-介在性HRおよび耐性、並びにCf-4-およびCf-9-介在性HRに関与していることが示された(Peart et al., 2002, Proc. Natl. Acad. Sci. USA 99, 10865-10869; Zhang et al., 2004, Plant J. 40, 213-224)。SGT1は、SKP1の相互作用物質であり、タンパク質のユビキチン化、タンパク質を分解の標的にする修飾に関与するSCF E3-リガーゼ複合体の構成成分である(Schwechheimer and Schwager, 2004, Plant Cell Reports 23, 353-364)。このタンパク質分解システムの必須遺伝子をサイレンシングすることは、ユビキチン化プロセスを妨害し、これにより防御の活性化に必要な負の調節因子の分解を阻害すると仮定される(Azevedo et al., 2002, Science 295, 2073-2076)。
【0004】
幾つかの耐性経路において、MAPKs(マイトジェン活性化プロテインキナーゼ)が活性化される(Zhang and Klessig, 2001, Trends Plant Sci. 6, 520-527; Pedley and Martin, 2005, Curr. Opin. Plant Biol. 8, 541-547)。Cf-9-含有タバコ植物およびAvr-9で攻撃された(challenged)細胞培養物において、NtWIPK (傷害誘導性プロテインキナーゼ) およびNtSIPK (サリチル酸誘導性プロテインキナーゼ) が活性化される(Romeis et al., 1999, Plant Cell 11, 273-287)。N. benthamianaにおけるNtCDPK (カルシウム依存性プロテインキナーゼ) のVIGSは、Cf-9/Avr9-およびCf-4/Avr4-依存性HRを阻害し (Romeis et al., 2001, EMBO J. 20, 5556-5567)、トマトにおけるLeACIK1 (Avr/Cf-induced kinase 1) のVIGSは、C. fulvum耐性の減少という結果につながる (Rowland et al., 2005, Plant Cell 17, 295-310)。防御の間のキナーゼの活性化およびキナーゼをコードする遺伝子の「ノックダウン」による耐性の減少は、防御の活性化におけるキナーゼの機能をサポートする。
【0005】
偏りのあるアプローチに従って、防御関連のシグナリングに関与することが公知の21個の遺伝子が、トマトにおいてVIGSのために使用され、これらのうち9個の遺伝子が、Pto-介在性耐性に関与することが見出された。これらのなかに、MAPKKsをコードする2個の遺伝子(LeMEK1およびLeMEK2)、およびMAPKsをコードする2個の遺伝子(LeNTF6およびLeWIPK)が含まれる(Ekengren et al., 2003, Plant J. 36, 905-917)。別の研究において、N. benthamiana cDNAの標準化ライブラリー由来の2400を超えるcDNAをポテトウイルスX-ベースのベクターにクローニングし、N. benthamianaにおいてVIGSのために使用した。cDNAsの約3%が、サイレンシングによりPto-依存性HRに影響を及ぼした。これらのうち、MAPKKKαが、耐性と病気の両方の正の調節因子として同定された(Del Pozo et al., 2004, EMBO J. 23, 3072-3082)。
【0006】
Lu et al. (2003, EMBO J. 22, 5690-5699) は、PVXベクターにクローニングされた標準化N. benthamiana cDNAライブラリー由来の4992個のcDNAを用いてVIGSを行った。cDNAのうち、79個(1.6%)が、Pto-介在性HRに必要な遺伝子に相当し、それらの僅か6個のサイレンシングが、Pseudomonas syringaeに対するPto-介在性耐性を減少させた。HSP90に相当するcDNAを用いたVIGSは、Pto-介在性HRだけでなく、Pto-、Rx-およびN-介在性耐性を破壊し、このことは、HSP90が複数の耐病性経路に必要であることを示す。cDNAの同じセットを、N-トランスジェニックN. benthamianaにおいてVIGSのために使用し、その後、植物にTMVのGFP標識株を接種した。TMVに対する耐性は、NRG1 (N requirement gene 1) と称されるCC-NB-LRRコード遺伝子に由来するcDNAフラグメントを用いたサイレンシングにより最も顕著に抑制された(Peart et al., 2005, Curr. Biol. 15, 968-973)。NRG1は、N遺伝子の機能に特に必要であることが示され、このことは、CC-NB-LRRタンパク質が、無毒性因子の認識に関与する耐性タンパク質として機能するだけでなく、TIR-NB-LRRタンパク質Nにより開始され、最終的に耐性につながるシグナリング経路に関与することを示す(Peart et al., 2005, supra)。このように、タバコのNRG1タンパク質は、耐性タンパク質により開始される植物の防御シグナリングカスケードの下流で機能するが、タバコモザイクウイルス(TMV)に対するN-介在性耐性に特異的に関与し、耐病性の一般的な補助因子でない(PVXおよびPseudomonas syringaeに対するRx-およびPto-介在性耐性は、NRG1サイレンシングにより影響を受けない)という欠点を有し、これにより、トマトなどの作物において広範囲の病原体耐性をつくるのに適していないかもしれない。
【0007】
耐病性経路についての情報は増えているが、永続的で広範囲の耐病性を備えた植物をつくるために使用することができる遺伝子およびタンパク質を同定する需要がなお存在する。向上した耐病性を備えた植物、とりわけナス科に属する植物をつくるためのかかる核酸、タンパク質および方法を提供することが本発明の課題である。
【一般的な定義】
【0008】
“HR”は、高感受性応答、すなわち(Rivas and Thomas, 2005, Ann Rev Phytopath 43: 395-436により記載される)顕微鏡的な病変および/または肉眼的な病変として観察される局所的な細胞死を指す。高感受性細胞死は、通常、その他の植物応答、たとえば反応性酸素種の産生およびHR病変を囲む細胞における防御関連遺伝子の活性化などと関連がある。
【0009】
“植物病原体”は、植物に病気を引き起こすことができる生物薬剤、たとえば植物病原性の真菌、細菌、ウイルス、卵菌類、マイコプラズマ様生物、線虫、コナジラミおよびアブラムシなどを指す。一般に、宿主組織に病気を引き起こすことができる病原体種の株、品種またはパソバールのすべてが、本明細書において包含される。
【0010】
“生物栄養性(biotrophic)植物病原体”または“生物栄養体(biotroph)”は、植物細胞を生きたまま維持し、増殖および組織コロニー化のために生細胞に依存する病原体を指す。
【0011】
“半生物栄養性(hemibiotrophic)植物病原体”または“半生物栄養体(hemibiotroph)”は、少なくとも生活環の一部の期間、宿主細胞を生きたまま維持する植物病原体を指す。
【0012】
“壊死栄養性(necrotrophic)植物病原体”は、宿主細胞を殺傷する毒性酵素、タンパク質または代謝産物を産生することにより、組織コロニー化により植物細胞を積極的に殺傷する植物病原体を指す。
【0013】
“誘導因子非依存性HR”は、病原体または病原体の誘導因子(たとえば真菌のAvrタンパク質)の存在なしで発生する高感受性応答を指す。
【0014】
本発明のNRC1タンパク質(たとえば、構成的に活性なNRC1タンパク質)を発現する植物を参照する場合、病原体または病原体の誘導因子タンパク質の非存在下でのHR病変の発生を言及する“構成的HR”と、誘導性刺激の存在後(たとえば、NRC1タンパク質またはその変異体をコードする核酸配列の発現を駆動するプロモーターの誘導後)のHR病変の発生を言及する“誘導的HR”とを区別してもよい。
【0015】
“ナス科(Solanaceae)”は、本明細書において、ナス科に属する植物の属、種、およびその品種を指す。これらには、ナス属(Solanum)(以前はLycopersicon esculentumとして知られていたSolanum lycopersicumを含む)、タバコ属(Nicotiana)、トウガラシ属(Capsicum)、ペチュニア属(Petunia)、およびその他の属が含まれる。
【0016】
“耐病性”は、本明細書において種々のレベルの植物の病気抵抗性または耐性を指し、一または複数の病原体に対する中程度な抵抗性および高度な抵抗性または完全な抵抗性を含む。耐病性は、病原体により引き起こされる徴候(たとえばHR病変、真菌の菌糸体などの頻度および/またはサイズ)を、同じ病気プレッシャーの下で生育させたときの感受性コントロール植物で観察される徴候と比較することにより測定し、必要に応じて定量することができる。かかる病気バイオアッセイは、公知の方法を用いて行うことができる。また、耐病性は、病気プレッシャーの下で生育させたときの、感受性植物と比較して耐性植物の高い収量により間接的に測定することもできる。
【0017】
“向上した耐病性”は、適切なコントロールと比較して、植物または植物組織の耐病性が統計的に有意に増大することを指す。本明細書において、(たとえば感受性から抵抗性までの)定性的増大および定量的増大の両方が包含される。また、病気の発生率(感染している植物のパーセンテージ)の低下および/または病気の重症度の低下の両方が包含される。好ましくは、少なくとも一つの病原体に対して向上した耐病性を有する植物は、耐病性を評価するための適切なバイオアッセイおよび/またはフィールドアッセイを用いて、コントロール植物より病原体に対して少なくとも1%、2%、5%、10%、15%、20%、30%、50%、70%、80%、90%、または100%もの高いレベルの耐性を備えた植物である。
【0018】
“広範囲(broad spectrum)”の耐病性は、少なくとも2、3、4、またはそれ以上の異なる病原体種の病原体に対して向上した耐性を指す。たとえば、幾つかの生物栄養性および/または半生物栄養性および/または壊死栄養性の病原体種に対して向上した耐性を有する宿主植物は、広範囲の耐性を有すると考えられる。
【0019】
“病原体により引き起こされる徴候”には、病気の任意の徴候、たとえば宿主組織の上/中での菌糸体の増殖/バイオマス、細菌の増殖/バイオマス、植物組織上での壊死性または退緑性病変のサイズおよび/または頻度、潰瘍のサイズおよび/または頻度などが含まれる。
【0020】
“核酸配列”(または核酸分子)の用語は、一本鎖または二本鎖の標準的な形態のDNAまたはRNA分子、とりわけ本発明のタンパク質またはタンパク質フラグメントをコードするDNAを指す。“単離された核酸配列”は、自然環境から単離され、もはや自然環境に存在しない核酸配列、たとえば細菌の宿主細胞または植物の核もしくはプラスチドゲノムに存在する核酸配列を指す。
【0021】
“タンパク質”または“ポリペプチド”の用語は、交換可能に使用され、アミノ酸の鎖から成る分子を指し、具体的な作用形態、サイズ、3次元構造または由来を問わない。このため、タンパク質の“フラグメント”または“一部”は、なお“タンパク質”を指してもよい。“単離されたタンパク質”は、自然環境にもはや存在しない、たとえばインビトロまたは組換え細菌または植物の宿主細胞に存在する、タンパク質を指すために使用される。
【0022】
NRC1タンパク質(または変異体、たとえばオルソログまたは突然変異体、およびフラグメント)に関して“機能的”とは、植物においてNRC1コード遺伝子の発現レベルを改変することにより(たとえば過剰発現またはサイレンシングにより)、HR病変の(定量的および/または定性的)発生および/または耐病性のレベルを改変する能力を指す。たとえば、X植物種から得られる推定NRC1タンパク質の機能性は、種々の方法により試験することができる。当該タンパク質が機能的である場合、たとえばVIGSもしくは遺伝子サイレンシングベクターを用いて、X植物種で当該タンパク質をコードするNRC1遺伝子をサイレンシングさせると、トマトについての実施例で示されるとおり、病原体もしくは誘導因子誘発性HR病変の減少または抑制、および/または病原体耐性の低下という結果につながる。また、機能的なNRC1タンパク質を補足すると、HR病変および/または病原体耐性を回復させることができる。あるいは、(必要に応じて、転写後遺伝子サイレンシング阻害剤と一緒に)NRC1タンパク質をコードする遺伝子をX種で一過性もしくは安定(過剰)発現させると、誘導因子非依存性HR病変の発生、および/またはとりわけ生物栄養性および/または半生物栄養性病原体に対して向上した耐病性という結果につながる。実施例も参照されたい。
【0023】
“遺伝子”の用語は、適切な調節領域(たとえばプロモーター)に動作可能に連結された、細胞においてRNA分子(たとえばmRNA)に転写される領域(転写領域)を含むDNA配列を意味する。このため、遺伝子は、幾つかの動作可能に連結された配列、たとえば、プロモーター、転写開始に関与する配列などを含む5’リーダー配列、(タンパク質)コード領域(cDNAまたはゲノムDNA)、および転写終結部位などを含む3’非翻訳配列を含んでいてもよい。
【0024】
“キメラ遺伝子”(または組換え遺伝子)は、生物種において天然に通常見出されない任意の遺伝子、とりわけ、天然において互いに関連がない核酸配列の一または複数の部分が存在する遺伝子を指す。たとえば、プロモーターは、天然において、転写領域の一部もしくは全部、または別の調節領域と関連がない。“キメラ遺伝子”の用語は、プロモーターまたは転写調節配列が、一もしくは複数のコード配列、またはアンチセンス(センス鎖のリバース相補鎖)もしくは逆方向反復配列(RNA転写産物が転写により二本鎖RNAを形成するセンスとアンチセンス)に動作可能に連結された発現構築物を含むと理解される。
【0025】
“3’UTR”または“3’非翻訳配列”(しばしば3’非翻訳領域または3’端とも称される)は、遺伝子のコード鎖の下流に見出される核酸配列を指し、たとえば、転写終結部位および(ほとんどであるが全てではない真核生物mRNAにおいて)ポリアデニル化シグナル(たとえばAAUAAAまたはその変異体)を含む。mRNA転写産物は、転写終結後にポリアデニル化シグナルの下流で切断され、mRNAの(翻訳が起こる)細胞質への輸送に関与するポリ(A)テールが付加され得る。
【0026】
“遺伝子の発現”は、適切な調節領域、とりわけプロモーターに動作可能に連結されたDNA領域が、生物学的に活性なRNA、すなわち生物学的に活性なタンパク質またはペプチド(または活性なペプチドフラグメント)に翻訳され得るRNAまたは(たとえば転写後の遺伝子サイレンシングまたはRNAiにおいて)それ自体で活性なRNAに転写されるプロセスを指す。ある態様において活性なタンパク質は、構成的に活性なタンパク質を指す。コード配列は、好ましくはセンスの向きであり、所望の生物学的に活性なタンパク質またはペプチド、または活性なペプチドフラグメントをコードする。遺伝子サイレンシングアプローチにおいて、DNA配列は、好ましくは、ターゲット遺伝子の短い配列をアンチセンスの向きまたはセンスとアンチセンスの向きで含むアンチセンスDNAまたは逆方向反復DNAの形態で存在する。“異所性発現”は、遺伝子が通常発現しない組織での発現を指す。
【0027】
“転写調節配列”は、本明細書において、転写調節配列に動作可能に連結された(コード)配列の転写速度を調節することができる核酸配列と規定される。このため、本明細書で規定される転写調節配列は、転写の開始に必要な配列エレメント(プロモーターエレメント)、転写の維持および調節に必要な配列エレメント(たとえば、アテニュエーターまたはエンハンサー)の全てを含む。多くの場合、コード配列の上流(5’)転写調節配列が言及されるが、コード配列の下流(3’)に見出される調節配列も、この規定に包含される。
【0028】
本明細書で使用される“プロモーター”の用語は、遺伝子の転写開始部位の転写の方向に対して上流に位置して、一または複数の遺伝子の転写を制御するために機能する核酸フラグメントを指し、DNA依存性RNAポリメラーゼの結合部位、転写開始部位、並びに任意の他のDNA配列(転写因子結合部位、リプレッサーおよびアクチベータータンパク質結合部位、およびプロモーターからの転写量を調節するのに直接または間接的に作用することが当業者に公知の任意の他のヌクレオチド配列を含むがこれらに限定されない)の存在により構造的に同定される。“構成的”プロモーターは、ほとんどの組織でほとんどの生理および発生条件下において活性なプロモーターである。“誘導性”プロモーターは、生理的に(たとえば、ある化合物の外部からの適用により)または発生で調節されるプロモーターである。“組織特異的”プロモーターは、特定タイプの組織または細胞においてのみ活性である。“植物または植物細胞で活性なプロモーター”は、植物または植物細胞の中で転写を駆動するプロモーターの一般的能力を指す。これは、プロモーターの時空的活性について何の意味ももたない。
【0029】
本明細書で使用される“動作可能に連結された”の用語は、機能的関係におけるポリヌクレオチドエレメントの連結を指す。核酸が、別の核酸配列と機能的関係に配置されるとき、“動作可能に連結されている”。たとえば、プロモーター、正確にいえば転写調節配列が、コード配列の転写に影響を及ぼす場合、コード配列に動作可能に連結されている。動作可能に連結されているとは、連結されているDNA配列が、典型的には隣接していることを意味し、二つのタンパク質コード領域を結合する必要がある場合、“キメラタンパク質”を産生するようにリーディングフレームで隣接していることを意味する。“キメラタンパク質”または“ハイブリッドタンパク質”は、種々のタンパク質“ドメイン”(またはモチーフ)から構成されるタンパク質であり、それ自体天然には見出されないが、機能的タンパク質を形成するために結合され、結合されたドメインの機能性を示す(たとえば、二重コイル(coiled−coil)ドメイン、ヌクレオチド結合ドメイン(NB-ARC)およびロイシンリッチ繰返し配列(LRR)領域を組合せることができる)。また、キメラタンパク質は、天然に存在する二つ以上のタンパク質の融合タンパク質であってもよい。本明細書で使用される“ドメイン”の用語は、ドメインの機能的特徴を少なくとも備えた新規ハイブリッドタンパク質を提供するために別のタンパク質に転移させることができる特定の構造または機能を備えたタンパク質の任意の部分またはドメインを意味する。また、特定のドメインは、他のNRC1タンパク質、たとえば他の植物種に由来するNRC1オルソログを同定するために使用することもできる。
【0030】
“ターゲットペプチド”の用語は、タンパク質またはタンパク質フラグメントを、細胞内オルガネラ、たとえばプラスチド、好ましくは葉緑体、ミトコンドリアに向かわせるアミノ酸配列、または細胞外空間もしくはアドプラスド(adoplast)に向かわせるアミノ酸配列(分泌シグナルペプチド)を指す。ターゲットペプチドをコードする核酸配列は、タンパク質またはタンパク質フラグメントのアミノ末端(N-末端)をコードする核酸配列に(フレームで)融合してもよいし、天然のターゲッティングペプチドを置換するために使用してもよい。
【0031】
“核酸構築物”または“ベクター”は、組換えDNA技術の使用により得られ、外因性DNAを宿主細胞にデリバーするために使用される人工の核酸分子を意味すると本明細書において理解される。ベクターの骨格は、当該技術分野で公知であり、本明細書の他の箇所で記載される、たとえばバイナリーもしくはスーパーバイナリーベクター(たとえばUS 5591616、US 2002138879およびWO 95/06722を参照)、コインテグレート(co-integrate)ベクター、またはT-DNAベクターとすることができ、これにキメラ遺伝子が統合されるか、あるいは適切な転写調節配列が既に存在する場合には、所望の核酸配列(たとえばコード配列、アンチセンスまたは逆方向反復配列)のみが転写調節配列の下流に統合される。ベクターは、分子クローニングでの使用を容易にするための更なる遺伝的構成要素、たとえば選択マーカー、マルチクローニングサイトなどを通常含む(下記参照)。
【0032】
“宿主細胞”または“組換え宿主細胞”または“形質転換細胞”は、所望のタンパク質をコードするキメラ遺伝子またはターゲット遺伝子/遺伝子ファミリーのサイレンシングのためのアンチセンスRNAまたは逆方向反復RNA(またはヘアピンRNA)を転写により産生する核酸配列をとりわけ含む少なくとも一つの核酸分子を前記細胞に導入した結果生じる新規な個々の細胞(または生物)を指す用語である。宿主細胞は、好ましくは植物細胞または細菌細胞である。宿主細胞は、核酸構築物を染色体外(エピソーム)複製分子として含んでいてもよいし、より好ましくは、キメラ遺伝子を宿主細胞の核もしくはプラスチドゲノムに統合して含んでいてもよい。また本明細書を通して、“宿主”の用語は、病原体が侵入もしくは感染することが可能な宿主植物種を指してもよいが、このことは、本明細書の文脈から明らかである。植物種は、病原体との関係で、“宿主”または“非宿主”種に分類される。“非宿主”種は、病気発生に最適な条件下においても、病原体のあらゆる品種または株の病原体感染に対して完全に免疫性がある。“宿主”種は、病原体の“宿主範囲”とも呼ばれ、病原体の(全てではないが)ある品種に対して免疫性がある。
【0033】
“選択マーカー”の用語は、当業者によく知られた用語であり、発現されたときに、選択マーカーを含有する一つの細胞または複数の細胞を選択するために使用することができる任意の遺伝的構成要素(entity)を記載するために本明細書で使用される。選択マーカーの遺伝子産物は、たとえば抗生物質耐性、より好ましくは除草剤耐性または別の選択可能な特性、たとえば表現型の特性(たとえば色素形成の変化)または栄養要求性を付与する。“レポーター”の用語は、主に、可視マーカー、たとえば緑色蛍光タンパク質(GFP)、eGFP、ルシフェラーゼ、GUSなどを指すために使用される。
【0034】
遺伝子またはタンパク質の“オルソログ(ortholog)”の用語は、本明細書において、別の種で見出される相同な遺伝子またはタンパク質を指し、これは、当該遺伝子またはタンパク質と同じ機能を有するが、当該遺伝子を有する種が分岐した時点から配列が(通常)分岐している(すなわち、種分化により共通祖先から進化した遺伝子)。よって、トマトnrc1遺伝子のオルソログは、(たとえば全配列または特定ドメインにわたるパーセンテージ配列同一性に基く)配列の比較と機能分析の両方に基いて、他の植物種で同定され得る。
【0035】
“同種の(homologous)”および“異種の(heterologous)”は、とりわけトランスジェニック生物の文脈において、核酸またはアミノ酸配列とその宿主細胞または生物との関係を指す。よって、同種の配列は、宿主種(たとえばトマト遺伝子で形質転換されたトマト植物)で天然に見出されるが、異種の配列は、宿主細胞(たとえばポテト植物由来の配列で形質転換されたトマト植物)で天然に見出されない。あるいは、文脈に応じて“相同体(homolog)”または“相同な(homologous)”は、共通祖先の配列から伝わる配列を指してもよい(たとえば、これらはオルソログであり得る)。
【0036】
“ストリンジェントなハイブリダイゼーション条件”は、所定のヌクレオチド配列と実質的に同一のヌクレオチド配列を同定するために使用することができる。ストリンジェントな条件は、配列に依存し、異なる環境では異なる。一般に、ストリンジェントな条件は、規定のイオン強度とpHにおける特定の配列に対して、熱融点(Tm)より約5℃低くなるように選択される。Tmは、ターゲット配列の50%が完全に一致するプローブとハイブリダイズする(規定のイオン強度とpHの下での)温度である。典型的には、塩濃度がpH 7で約0.02モルであり、温度が少なくとも60℃であるストリンジェントな条件が選択される。塩濃度の低下および/または温度の増大は、ストリンジェンシーを増大させる。RNA-DNAハイブリダイゼーション(たとえば100ntのプローブを用いたノーザンブロット)のストリンジェントな条件は、たとえば、63℃で20分間の0.2×SSCでの少なくとも1回の洗浄を含む条件、または同等の条件である。DNA-DNAハイブリダイゼーション(たとえば100ntのプローブを用いたサザンブロット)のストリンジェントな条件は、たとえば、少なくとも50℃の温度(通常約55℃)で20分間の0.2×SSCでの少なくとも1回(通常2回)の洗浄を含む条件、または同等の条件である。Sambrook et al. (1989)およびSambrook and Russell (2001)も参照されたい。
【0037】
“配列同一性”および“配列類似性”は、グローバルまたはローカルアラインメントアルゴリズムを用いて、二つのペプチドまたは二つのヌクレオチドの配列を整列させること(alignment)により決定することができる。(デフォルトパラメーターを用いてたとえばプログラムGAPまたはBESTFITにより最適に整列させた際の)配列が、(以下で規定される)配列同一性の最小パーセンテージを少なくとも共有する場合、配列は、“実質的に同一である”または“本質的に類似している”ということができる。GAPは、Needleman and Wunschのグローバルアラインメントアルゴリズムを使用して、二つの配列を全長にわたって整列させ、一致の数を最大にし、ギャップの数を最小にする。一般に、GAPデフォルトパラメーターが使用され、ギャップ生成ペナルティー=50(ヌクレオチド)/8(タンパク質)であり、ギャップ伸長ペナルティー=3(ヌクレオチド)/2(タンパク質)である。ヌクレオチドについて、使用されるデフォルトスコアリングマトリクスは、nwsgapdnaであり、タンパク質について、デフォルトスコアリングマトリクスは、Blosum62である(Henikoff & Henikoff, 1992, PNAS 89, 915-919)。配列アラインメントおよびパーセンテージ配列同一性のスコアは、コンピュータープログラム、たとえばGCG Wisconsin Package, Version 10.3(Accelrys Inc., 9685 Scranton Road, San Diego, CA 92121-3752 USAから入手可能)を用いて、あるいはEmbossWIN version 2.10.0(プログラム“needle”を使用)を用いて決定され得る。あるいは、パーセンテージ類似性または同一性は、たとえばFASTA、BLASTなどのアルゴリズムを用いてデータベースに対してサーチすることにより決定してもよい。
【0038】
本明細書および特許請求の範囲において、動詞“含むこと(to comprise)”およびその語形変化は、非限定的な意味で使用され、そのワードの後につづく項目を含むが、具体的に挙げられていない項目を排除しないことを意味する。加えて、不定冠詞“a”または“an”によるエレメントの指示は、その文脈が、一つおよび唯一のエレメントが存在することを明らかに要求しない限り、二つ以上のエレメントが存在する可能性を排除しない。よって、不定冠詞“a”または“an”は、通常“少なくとも一つ”を意味する。更に、本明細書で“配列”を参照する場合、ある配列のサブユニット(たとえばアミノ酸)を備えた実際の物理的分子を指すと理解される。
【0039】
本明細書で使用される“植物”の用語は、植物細胞、植物組織または器官、植物プロトプラスト、植物を再生させることができる植物細胞組織培養物、植物カルス、植物細胞塊、および植物においてインタクトな植物細胞、または植物の部分、たとえば胚、花粉、胚珠、果実(たとえば収穫されたトマト)、花、葉、種子、根、根端などを含む。
【発明の詳細な説明】
【0040】
本発明者らは、VIGS(ウイルス誘導性遺伝子サイレンシング)と組合せてcDNA-AFLP分析を使用して、Cf-4/Avr4依存性HRおよび耐病性に関与する遺伝子を同定した。VIGSがAvr-4誘導性HRの抑制につながる遺伝子のうち、CC-NB-LRRタイプの耐性タンパク質類似体をコードする(本明細書においてNRC1と称される)一つのトマト遺伝子が同定された(本明細書では、NB-LRR protein Required for HR-associated Cell death 1のためNRC1と称される)。トマトにおけるNRC1のサイレンシングは、Avr4誘導性HRの発生だけでなく、トマト病原体Cladosporium fulvumに対する耐性を低下させた(compromise)。これにより、トマトのCf-4耐性タンパク質(細胞外レセプター様タンパク質)が、細胞質NB-LRRタンパク質を機能的にするのに必要であることが示された。
【0041】
更に、NRC1は、複数のHRおよび複数の耐病性/細胞死シグナリング経路、たとえばCf-9/Avr-9-、LeEix2/Eix-、Pto/AvrPto-およびRx/CP-開始HRに関与していることが驚くべきことに見出された(実施例参照)。更に、NRC1が他のHR、たとえば(線虫、コナジラミおよびアブラムシ-誘導性HRに対する耐性を付与する)Mi-介在性HRにも関与しているかどうかを決定するために試験が行われている(US 6613962およびEP0937155B1参照)。このため、NRC1は、様々なクラスに属する細胞外および細胞内の耐病性タンパク質:細胞外レセプター様タンパク質 (RLPs、たとえばCf-4、Cf-9およびLeEix2)、Ser/Thrプロテインキナーゼ、たとえばPtoおよびCC-NB-LRRタンパク質(Rx)(これらは、真菌(Cladosporium fulvumおよびTrichoderma viride)、細菌(Pseudomonas syringae pv tomato) またはウイルス(PVX)のそれぞれに耐性を付与する)により誘発されるHR経路に関与している。
【0042】
NRCタンパク質(およびそれをコードするNRC1遺伝子)は、種々の病原体、とりわけ生物栄養性および半生物栄養性植物病原体、更に壊死栄養性植物病原体、たとえばBotrytis種に対する植物耐性を付与するかまたは向上させるために使用することができる。特に、NRC1(または他の箇所で規定されるその変異体もしくはフラグメント)の発現は、とりわけ生物栄養性および/または半生物栄養性病原体、すなわち生細胞から栄養を得るあらゆる病原体に対する耐性の向上につながる。本発明の範囲を限定しないが、内在性NRC1遺伝子のノックダウン(遺伝子サイレンシング)または(たとえばTILLINGによる)ノックアウトは、壊死につながる経路が影響を受け、壊死栄養性病原体がこの経路を必要とするため、壊死栄養性病原体に対する耐性を付与するかまたは向上させるために使用することができると考えられる。このため、耐性を向上させたい病原体に応じて、NRC1発現レベルの増大または減少の何れかを、耐性を向上させるために使用することができる。必要に応じて、両方のアプローチを、たとえば異なるプロモーターの制御下において一つの植物で使用してもよい。たとえば、NRC1を、(半)生物栄養性病原体により誘導されるプロモーターの制御下において発現させて、生物栄養性および/または半生物栄養性の葉の病原体に対する耐性を付与することができ、同時に、内在性NRC1遺伝子(または遺伝子ファミリー)を、壊死栄養性病原体または傷害により誘導性であるプロモーターを用いて、ある組織で、または壊死栄養体による誘導によりサイレンシングさせることができる。
【0043】
構成的に活性なNRC1タンパク質(NRC1D481V)が、トマトにおいて一過性で産生されると、その植物組織は、誘導因子に依存しない細胞死(HR)を示すことが更に発見され、これは、機能的NRC1タンパク質の発現を、植物に耐病性を付与するかまたは向上させるために使用することができることを示す。
【0044】
本発明のタンパク質および核酸配列
トマトから得られたNRC1タンパク質は、タバコのNRG1に対して低い配列同一性(25%未満)を示す。またNRC1は、NRG1より多数のロイシンリッチ繰返し配列(LRR)を含有する。NRC1のタンパク質構造は、図1および配列番号2に示される。
【0045】
本発明の一つの態様において、NRC1タンパク質の核酸配列およびアミノ酸配列(オルソログを含む)が提供されるとともに、他の植物種、たとえば他のナス科、好ましくはポテトにおいてNRC1のオルソログを単離または同定するための方法が提供される。同様に、他のNRC1対立遺伝子、たとえば他のトマトの種、品種、系統またはアクセッション(accession)由来の対立遺伝子を単離または同定するための方法が本明細書において提供される。
【0046】
一つの態様において、NRC1タンパク質が提供される。“NRC1タンパク質”は、配列番号2(野生型)および配列番号4(構成的突然変異体)に示されるタンパク質、並びにそのフラグメントおよび変異体を含む。NRC1の変異体には、たとえば、配列番号2および/または4に対して(全長にわたって)少なくとも30、35、40、45、50、55、60、65、70、75、80、85、90、95、98、99%またはそれ以上のアミノ酸配列同一性を有するタンパク質が含まれる。アミノ酸配列同一性は、上述のとおり、Needleman and WunschアルゴリズムおよびGAPデフォルトパラメーターを用いたペアワイズアラインメントにより決定される。NRC1の変異体は、種々の供給源、たとえば既存の配列データベース、他の植物種(とりわけナス科の他の種、たとえばポテト)または他の品種から入手することができるし、あるいはデノボ合成、突然変異誘発などにより作成することもできる。たとえば、配列番号4、すなわち構成的に活性なNRC1突然変異体は、配列番号2の変異体であり、オーバーラップPCRを用いた標的突然変異誘発によって作成された(実施例参照)。このように、本発明のNRC1タンパク質は、天然源から単離されてもよいし、(たとえばApplied Biosystemsにより供給されるようなペプチド合成機を用いて)化学合成によりデノボ合成されてもよいし、組換え宿主細胞により、NRC1タンパク質、フラグメントまたは変異体をコードする核酸配列を発現させることにより産生してもよい。
【0047】
NRC1変異体は、塩基性(たとえばArg、His、Lys)、酸性(たとえばAsp、Glu)、非極性(たとえばAla、Val、Trp、Leu、Ile、Pro、Met、Phe、Trp)または極性(たとえばGly、Ser、Thr、Tyr、Cys、Asn、Gln)のカテゴリー内において保存的アミノ酸置換を含んでいてもよい。加えて、非保存的アミノ酸置換も本発明の範囲内である。
【0048】
NRCタンパク質、変異体またはフラグメントの機能性は、種々の方法を用いて決定することができる。たとえば、植物細胞における一過性または安定な過剰発現を用いて、当該タンパク質が植物体で(in planta)活性を有しているかどうか試験することができる。機能性は、好ましくは、タンパク質を入手したのと同一の植物種において試験する。このように、たとえば、一過性または安定な発現を用いて、HRが発生しているかどうか、および/または耐性が向上しているかどうかを決定し、機能性を示すことができる。あるいは、内在性遺伝子または遺伝子ファミリーのサイレンシングは、NRC1タンパク質が機能的であるかどうかを示す。たとえば、VIGSは、種々のナス科、たとえばポテト、トマトおよびタバコ(Brigneti et al., 2004, Plant Journal 39: 264; Faivre-Rampant et al. Plant Physiology 134: 1308-1316; Baulcombe 1999, Curr. Opinion. Plant Biol. 2: 109-113; Lu et al. 2003, EMBO J. 22: 5690-5699参照)、アラビドプシス(Arabidopsis)などのモデル生物(Turnage et al. 2002, Plant J. 30: 107-114)、オオムギなどの単子葉植物(Holzberg et al. 2002, Plant J. 30: 315-327)で使用することができる。あるいは、NRC1遺伝子のセンスおよび/またはアンチセンスフラグメントを含むサイレンシングベクターを使用して、植物細胞を形質転換し(下記参照)、その後アッセイにより、HR病変を発生する能力および/または耐病性が改変されているかどうかを決定することができる。
【0049】
好ましい態様において、NRC1の変異体には、植物細胞で構成的に活性なNRC1タンパク質、たとえばMHDドメインに単一のアミノ酸置換(D481V)を含む配列番号4で提供されるNRC1タンパク質が含まれる(図1参照)。構成的活性は、タンパク質が誘導因子の非存在下において植物組織でHRを誘導することができるかどうかを決定することにより試験することができる。たとえば、実施例に記載される35S:NRC1構築物のアグロインフィルトレーション(Agroinfiltration)は、葉組織に浸潤させる(infiltrate)ために使用することができる。構成的に活性な他のNRC1タンパク質は、ランダム突然変異誘発とその後の活性試験により(Bendahame et al., 2002, p196に記載される)、あるいはMHDドメイン(アミノ酸VHDまたはVHDMの何れか一つが別のアミノ酸に置換され得る)、NB-ARCドメイン、たとえばRNBS-Dドメイン(アミノ酸FLYFGTFPRGY)、または13のLRRドメインの一つにおける単一アミノ酸の位置指定突然変異誘発により(図1参照)、作成することができる。あるいは、構成的に活性なNRC1タンパク質をコードする核酸配列は、たとえば種子に突然変異を起こさせ、これら種子を自然発生的病変の表現型(たとえば顕微鏡的な病変)の存在についてスクリーニングすることにより、植物から入手することができる。たとえばSharino et al. (2002, The Plant Cell 14: 3149-3162)および後述の記載を参照されたい。
【0050】
一つの態様において、キメラNRC1タンパク質も提供される。かかるタンパク質は、少なくともCCドメイン、NB-ARCドメイン、および好ましくは少なくとも13のLRRを含む。CC-、NB-ARC-およびLRR-ドメインは、好ましくは、それぞれ配列番号2のアミノ酸1-150、アミノ酸151-508、またはアミノ酸509-846に対して少なくとも30、40、50、60、70、80、90、95、98、99%またはそれ以上のアミノ酸配列同一性を含むアミノ酸モチーフを指す。このため、ドメインは、得られるキメラタンパク質の機能性が、NRC1、または好ましくはNRC1D481Vと本質的に類似している限り、NRC1タンパク質の間、またはNRC1タンパク質と他のCC-NB-LRRもしくはTIR-NB-LRRタンパク質との間で交換(ドメインスワッピング)されてもよい。最も好ましくは、キメラタンパク質は、後述されるとおり、組換え植物細胞により産生されると、耐病性を付与するかまたは向上させる能力を保持する。
【0051】
上述のとおりNRC1タンパク質およびNRC1タンパク質の変異体の“フラグメント”は、100、150、200、300、400、500、600、700、800、850、855の連続アミノ酸またはそれ以上のフラグメントを含む。好ましくは、かかるフラグメントは、植物組織において機能的であり、すなわち植物細胞で産生されると病原体耐性を付与するかまたは向上させることができる。また、フラグメントは、上述のとおり、キメラタンパク質を作成するために使用してもよい。
【0052】
別の態様において、上記タンパク質、変異体またはフラグメントの何れかをコードする単離された核酸配列、たとえばcDNA、ゲノムDNAおよびRNAの配列が提供される。遺伝暗号の縮重のため、種々の核酸配列が、同一のアミノ酸配列をコードし得る。NRC1タンパク質または変異体をコードする任意の核酸配列が、本明細書において“NRC1”と称される。提供される核酸配列は、天然に存在する核酸配列、人工もしくは合成の核酸配列を含む。NRC1タンパク質をコードする核酸配列の例は、配列番号1および3において提供される。RNAが言及されているのにDNA配列として配列が示される場合、RNA分子の実際の塩基配列は、チミン(T)がウラシル(U)に置換されるという違いはあるが同一であることが理解される。
【0053】
また、NRC1核酸配列の変異体およびフラグメント、たとえば規定されるストリンジェントな条件下でNRC1核酸配列とハイブリダイズする核酸配列も包含される。NRC1核酸配列の変異体には、配列番号1または3に対して(全長にわたって)少なくとも50%以上、好ましくは少なくとも55%、60%、70%、80%、90%、95%、99%、99.5%、99.8%またはそれ以上の配列同一性を有する核酸配列も包含される。好ましい態様において、NRC1の変異体は、記載されるとおり、構成的に活性なNRC1タンパク質をコードする。NRC1核酸配列の変異体またはフラグメントを同定、合成または単離するために多くの方法が使用できること、たとえば核酸ハイブリダイゼーション、PCR技術、インシリコ(in silico)分析、および核酸合成などが使用できることは明らかである。
【0054】
本発明のNRC1タンパク質をコードする核酸配列、とりわけDNA配列は、後述のとおり、発現ベクターに挿入して、多量のNRC1タンパク質(またはたとえばキメラNRC1タンパク質)を産生することができる。宿主細胞での最適な発現のために、NRC1のDNA配列は、利用可能なコドン使用頻度表を用いて、コドン使用頻度を植物遺伝子で最も好ましいもの、とりわけ対象の植物属または種に生来備わっている遺伝子に適合させることにより、コドン最適化を行うことができる(Bennetzen & Hall, 1982, J. Biol. Chem. 257, 3026-3031; Itakura et al., 1977 Science 198, 1056-1063.)(たとえば、ワタ、ダイズ、トウモロコシまたはイネでの発現に対して適合させる)。種々の植物種についてのコドン使用頻度表は、たとえばIkemura (1993, In “Plant Molecular Biology Labfax”, Croy, ed., Bios Scientific Publishers Ltd.) およびNakamura et al. (2000, Nucl. Acids Res. 28, 292.)により、並びにメジャーなDNA配列データベース (たとえばEMBL at Heidelberg, Germany)で公開されている。したがって、合成DNA配列は、同一または実質的に同一のタンパク質が産生されるように構築することができる。コドン使用頻度を宿主細胞に好ましいものに改変するための幾つかの技術は、特許および科学文献に見出すことができる。コドン使用頻度の正確な改変方法は、本発明にとって重大ではない。
【0055】
上述されるようなDNA配列に対する小さな改変は、ルーチンで、すなわちPCRによる突然変異誘発により行うことができる(Ho et al., 1989, Gene 77, 51-59., White et al., 1989, Trends in Genet. 5, 185-189)。DNA配列に対する甚大な改変は、利用可能な技術を用いて所望のコード領域のデノボDNA合成によりルーチンで行うことができる。
【0056】
また、NRC1核酸配列は、NRC1タンパク質のN末端に一または複数のアミノ酸を付加するかまたは欠失させることにより、NRC1タンパク質のN末端が最適な翻訳開始の文脈情報(context)を有するように改変することができる。多くの場合、植物細胞で発現される本発明のタンパク質は、最適な翻訳開始のためにMet-AspまたはMet-Alaジペプチドで開始することが好ましい。このため、AspまたはAlaのコドンが、既存のMetの後に挿入されてもよいし、あるいは2番目のコドンValが、Asp(GATまたはGAC)またはAla(GCT、GCC、GCAまたはGCG)に置換されてもよい。またDNA配列は、異常なスプライス部位を除去するように改変されてもよい。
【0057】
NRC1核酸配列の“フラグメント”には、配列番号1もしくは3、または配列番号1もしくは3の変異体の少なくとも10、12、15、16、18、20、30、40、50、100、200、500、1000、1500、2000またはそれ以上の連続ヌクレオチドのフラグメントが含まれる。短いフラグメントは、たとえばPCRプライマーまたはハイブリダイゼーションプローブとして使用することができる。
【0058】
本発明の別の態様において、NRC1のDNAまたはRNA配列を検出するためのPCRプライマーおよび/またはプローブおよびキットが提供される。サンプルからNRC1のDNAを増幅するための縮重または特異的PCRプライマーのペアは、当該技術分野で公知のとおり、配列番号1または3(またはその変異体)に基いて合成することができる(Dieffenbach and Dveksler (1995) PCR Primer: A Laboratory Manual, Cold Spring Harbor Laboratory Press, and McPherson at al. (2000) PCR-Basics: From Background to Bench, First Edition, Springer Verlag, Germany参照)。たとえば、配列番号1または3(またはその相補鎖)の9、10、11、12、13、14、15、16、18またはそれ以上の連続ヌクレオチドの任意の長さが、プライマーまたはプローブとして使用され得る。同様に、配列番号1または3(またはその変異体)のDNAフラグメントを、ハイブリダイゼーションプローブとして使用することができる。NRC1検出キットは、NRC1特異的プライマーおよび/またはNRC1特異的プローブ、およびサンプル中のNRC1 DNAを検出するためにプライマーまたはプローブを使用するための関連プロトコールを含む。かかる検出キットは、たとえば、植物が本発明のNRC1遺伝子(またはその一部)で形質転換されたかどうかを決定するために使用され得る。遺伝暗号の縮重のため、幾つかのアミノ酸コドンは、タンパク質のアミノ酸配列を変更しないで他のものに置換することができる。
【0059】
更に別の態様において、トマトNRC1遺伝子(配列番号1および3)のオルソログまたは対立遺伝子を同定し、使用するための方法が提供される。この方法は、以下の工程を含む:
a)配列番号1および/または3に対して少なくとも70%の核酸同一性(または上述のより高いパーセンテージの配列同一性)を含む核酸配列を入手または同定する工程、
b)必要に応じて、構成的に活性なNRC1タンパク質をコードするように核酸配列を改変する工程、および
c)a)の核酸配列を使用して、発現および/またはサイレンシングベクターを作成する工程、またはb)の核酸配列を使用して発現ベクターを作成する工程、
d)一または複数のc)のベクターを使用して、好ましくは核酸を入手した植物種の、植物または植物細胞を形質転換する工程、
e)植物体において(in planta)遺伝子の機能を決定または検証するため、および/または向上した耐病性を有するトランスジェニック植物を作成するために、形質転換体のHR病変(すなわちHR病変の表現型、これは必要に応じて定量することができる)および/または耐病性を発生する形質転換植物/植物組織の能力を分析する工程、
f)必要に応じて、トランスジェニック植物に向上した耐病性を付与するが、発現により弱いHR表現型を付与する(すなわち、HR病変の表現型を引き起こさないかまたは減少したHR病変の表現型を引き起こす)対立遺伝子またはオルソログを、更なる使用のために選択する工程。
【0060】
このようにして、配列番号1または3の発現または形質転換される宿主種から得られた野生型NRC1対立遺伝子の発現により観察されるよりも少ないおよび/または小さいHR病変が植物での発現により得られるNRC1対立遺伝子またはオルソログを、この方法を用いて同定することができる。より好ましくは、発現によりHR病変を引き起こさないかまたは少なくとも肉眼で見えるHR病変を引き起こさないが、向上した耐病性を付与するNRC1対立遺伝子またはオルソログが同定される。
【0061】
同じプロモーターを使用して発現ベクターを作成し、これらを用いて宿主植物を形質転換し、これらトランスジェニック植物の間でHR病変の表現型を比較することにより、異なるNRC1対立遺伝子および/またはオルソログのHR表現型を比較することができる。異なる対立遺伝子をトランスジェニックナス科植物において(たとえば構成的または誘導性プロモーターの制御下で)比較する場合、好ましくは、配列番号1または3を発現する形質転換体のHR病変の表現型がレファレンスとして使用される。あるいは、形質転換される宿主種から得られた野生型対立遺伝子をレファレンスとして使用することができる。その後、配列番号1または3と比較して少ないおよび/または小さいHR病変、または形質転換される種から得られた野生型対立遺伝子の発現により引き起こされるよりも少ないおよび/または小さい病変を提供する対立遺伝子を、更なる使用のために選択することができる。たとえば、これらを発現するトランスジェニック植物は、後述されるとおり作成することができる。
【0062】
たとえばNRC1特異的PCRプライマーもしくはプローブ、またはインシリコでの(in silico)バイオインフォマティクス分析を用いて、とりわけ、トマト由来の対立遺伝子およびポテト由来のオルソログが入手または同定され得る。また、遺伝子マッピングを使用して、植物(たとえばトマトまたはポテト)ゲノムにおけるNRC1遺伝子座をマッピングすることができ、ゲノムマップを(たとえばトマトの配列決定プロジェクトで開発された)既存のゲノム配列決定データベースにリンクさせることにより配列を入手することができる。かかる対立遺伝子および/またはオルソログは、向上した耐病性を備えた植物を作成するために特に適している。
【0063】
ポテトのNRC1オルソログを上記方法で同定した場合、これらオルソログ(またはこれらオルソログの変異体)は、好ましくは、Phytophthora infestansに対して向上した耐性を有する植物を作成するために使用される。
【0064】
本発明のキメラ遺伝子、発現ベクターおよび組換え生物
本発明の一つの態様において、上述のNRC1タンパク質をコードする核酸配列(変異体またはフラグメントを含む)を使用して、キメラ遺伝子の宿主細胞への転移のため、およびNRC1タンパク質の宿主細胞(たとえば細胞、組織、器官または形質転換細胞に由来する生物)での産生のためにキメラ遺伝子およびこれらを含むベクターを作成する。NRC1タンパク質(またはタンパク質フラグメントまたは変異体)を植物細胞で産生するためのベクターは、本明細書において、すなわち“発現ベクター”と称される。宿主細胞は、好ましくは植物細胞であるが、微生物の宿主(細菌、たとえばアグロバクテリウム、酵母、真菌など)も想定される。
【0065】
任意の植物、たとえば単子葉植物または双子葉植物を適切な宿主としてもよいが、最も好ましくは、宿主植物はナス科に属する。たとえば、植物は、ナス属(Solanum)(Lycopersiconを含む)、タバコ属(Nicotiana)、トウガラシ属(Capsicum)、ペチュニア属(Petunia)および他の属に属する。以下の宿主種が、適切に使用され得る:タバコ(Nicotiana種、たとえばN. benthamiana、N. plumbaginifolia、N. tabacumなど)、野菜の種、たとえばトマト(L. esculentum, syn. Solanum lycopersicum)たとえばチェリートマト、var. cerasiformeまたはカレントトマト、var. pimpinellifolium)またはツリートマト(S. betaceum, syn. Cyphomandra betaceae)、ポテト(Solanum tuberosum)、ナス(Solanum melongena)、ペピーノ(Solanum muricatum)、ココナ(Solanum sessiliflorum)およびナランヒージャ(Solanum quitoense);コショウ(Capsicum annuum、Capsicum frutescens、Capsicum baccatum)、観賞植物種(たとえばPetunia hybrida、Petunia axillaries、P. integrifolia)。
【0066】
あるいは、植物は、その他の任意の科、たとえばウリ科(Cucurbitaceae)またはイネ科(Gramineae)に属していてもよい。適切な宿主植物には、たとえば以下のものが含まれる:トウモロコシ(maize/corn)(Zeaの種)、コムギ(Triticumの種)、オオムギ(たとえばHordeum vulgare)、カラスムギ(たとえばAvena sativa)、モロコシ(Sorghum bicolor)、ライムギ(Secale cereale)、ダイズ(Glycine spp、たとえばG. max)、ワタ(Gossypiumの種、たとえばG. hirsutum、G. barbadense)、アブラナ属の種(Brassica spp.)(たとえばB. napus、B. juncea、B. oleracea、B. rapaなど)、ヒマワリ(Helianthus annus)、ベニバナ、ヤム、キャッサバ、アルファルファ(Medicago sativa)、イネ(Oryzaの種、たとえばO. sativaインディカ栽培品種-グループまたはジャポニカ栽培品種-グループ)、まぐさ(forage grasses)、パールミレット(Pennisetumの種、たとえばP. glaucum)、樹木の種(マツ属(Pinus)、ポプラ、モミ、プランタン(plantain)など)、チャ、コーヒー、アブラヤシ、ココナッツ、野菜の種、たとえばエンドウ、ズッキーニ、マメ(たとえばPhaseolusの種)、キュウリ、アーティチョーク、アスパラガス、ブロッコリー、ニンニク、ニラ、レタス、タマネギ、ラディッシュ、カブ、芽キャベツ、ニンジン、カリフラワー、チコリ、セロリ、ホウレンソウ、エンダイブ、ウイキョウ、ビート、多肉質の果実をつける植物(グレープ、ピーチ、プラム、ストロベリー、マンゴー、リンゴ、プラム、チェリー、アプリコット、バナナ、ブラックベリー、ブルーベリー、シトラス、キウイ、イチジク、レモン、ライム、ネクタリン、ラズベリー、スイカ、オレンジ、グレープフルーツなど)、観賞植物種(たとえばバラ、ペチュニア、キク、ユリ、ガーベラの種)、ハーブ(ミント、パセリ、バジル、タイムなど)、木本(たとえばPopulus、Salix、Quercus、Eucalyptusの種)、繊維の種、たとえばアマ(Linum usitatissimum)およびアサ(Cannabis sativa)、またはモデル生物、たとえばシロイヌナズナ(Arabidopsis thaliana)。
【0067】
好ましい宿主は、“作物植物”、すなわちヒトにより栽培され、繁殖される植物種である。作物植物は、食物の目的のため(たとえば農作物など)、または装飾の目的のため(たとえば切花、芝生用の草の生産など)に栽培されてもよい。本明細書で規定される作物植物には、食物以外の生産物、たとえば燃料用オイル、プラスチックポリマー、医薬品、コルクなどが収穫される植物も含まれる。
【0068】
NRC1タンパク質コード核酸配列の宿主細胞ゲノムへの好ましくは安定した導入のためのキメラ遺伝子およびベクターの構築は、一般に当該技術分野で公知である。キメラ遺伝子を作成するために、NRC1タンパク質(または変異体もしくはフラグメント)をコードする核酸配列は、標準的な分子生物学技術を用いて、宿主細胞での発現に適したプロモーター配列に動作可能に連結される。プロモーター配列は、NRC1核酸配列が、ベクターにプロモーター配列の下流で簡単に挿入されるように、ベクターに既に存在していてもよい。その後、ベクターは、宿主細胞を形質転換するために使用され、キメラ遺伝子は、核ゲノム、またはプラスチド、ミトコンドリアまたは葉緑体のゲノムに挿入され、その場所で適切なプロモーターを用いて発現される(たとえば、Mc Bride et al., 1995 Bio/Technology 13, 362; US 5,693,507)。一つの態様において、キメラ遺伝子は、植物細胞または微生物細胞(たとえば細菌)での発現に適したプロモーターを含み、これに、本発明のNRC1タンパク質をコードする核酸配列が動作可能に連結され、必要に応じてその後に3’非翻訳核酸配列が続く。
【0069】
機能的なNRC1タンパク質(またはある態様において構成的に活性なNRC1タンパク質)をコードするNRC1核酸配列、好ましくはNRC1キメラ遺伝子は、慣用的なやり方で、単一の植物細胞の核ゲノムに安定に挿入することができ、このように形質転換された植物細胞を慣用的なやり方で使用して、ある細胞にある期間NRC1タンパク質が存在することにより変化した表現型を有する形質転換植物を産生することができる。これに関して、アグロバクテリウムツメファシエンス(Agrobacterium tumefaciens)内のNRC1タンパク質をコードする核酸配列を含むT-DNAベクターを、植物細胞を形質転換するために使用することができ、その後、たとえばEP 0 116 718、EP 0 270 822、PCT公報 WO 84/02913および公開された欧州特許出願EP 0 242 246、およびGould et al. (1991, Plant Physiol. 95,426-434) に記載される手法を用いて、形質転換された植物細胞から形質転換植物を再生させることができる。アグロバクテリウム媒介性の植物の形質転換のためのT-DNAベクターの構築は、当該技術分野で周知である。T-DNAベクターは、EP 0 120 561およびEP 0 120 515に記載されるバイナリーベクター、または相同組換えによりアグロバクテリウムTi-プラスミドに統合することができるEP 0 116 718に記載されるコインテグレート(co-integrate)ベクターの何れであってもよい。
【0070】
好ましいT-DNAベクターはそれぞれ、T-DNAのボーダー配列の間、または少なくとも右ボーダー配列の左側の位置に、NRC1コード核酸配列(たとえば配列番号2または配列番号4をコードする)に動作可能に連結されたプロモーターを含有する。ボーダー配列は、Gielen et al. (1984, EMBO J 3,835-845) に記載される。当然、他のタイプのベクターを使用して、直接的な遺伝子導入(たとえばEP 0 223 247に記載される)、花粉媒介性の形質転換(たとえばEP 0 270 356およびWO 85/01856に記載される)、プロトプラストの形質転換(たとえばUS 4,684,611に記載される)、植物RNAウイルス媒介性の形質転換(たとえばEP 0 067 553およびUS 4,407, 956に記載される)、リポソーム媒介性の形質転換(たとえばUS 4,536,475に記載される)などの手法を用いて、植物細胞を形質転換することができる。トマトまたはタバコの形質転換のためには、An G. et al., 1986, Plant Physiol. 81: 301-305; Horsch R.B. et al., 1988, In: Plant Molecular Biology Manual A5, Dordrecht, Netherlands, Kluwer Academic Publishers. pp 1-9; Koornneef M. et al., 1986, In: Nevins D.J. and R.A. Jones, eds. Tomato Biotechnology, New York, NY, USA, Alan R. Liss, Inc. pp 169-178も参照される。ポテトの形質転換のためには、たとえばSherman and Bevan (1988, Plant Cell Rep. 7: 13-16) が参照される。
【0071】
同様に、形質転換された細胞からの形質転換植物の選択および再生は、当該技術分野で周知である。明らかに、様々な種に対して、更に単一種の様々な品種または栽培品種に対しても、高い頻度で形質転換体を再生させるためにプロトコールは特異的に適合される。
【0072】
核ゲノムの形質転換に加えて、プラスチドのゲノム、好ましくは葉緑体のゲノムの形質転換も本発明に包含される。プラスチドのゲノムの形質転換は、導入遺伝子の蔓延のリスクを低減できるという利点がある。プラスチドのゲノムの形質転換は、当該技術分野で公知のとおり行うことができ、たとえばSidorov VA et al. 1999, Plant J.19: 209-216またはLutz KA et al. 2004, Plant J. 37(6): 906-13が参照される。
【0073】
得られた形質転換植物は、慣用的な植物育種スキームで使用して、導入遺伝子を含有する多くの形質転換植物を産生することができる。シングルコピーの形質転換体は、たとえばサザンブロット分析またはPCRベースの方法またはInvader(登録商標)テクノロジーアッセイ(Third Wave Technologies, Inc.)を用いて選択することができる。形質転換された細胞および植物は、キメラ遺伝子の存在により非形質転換体と容易に区別することができる。導入遺伝子の挿入部位に隣接する植物DNAの配列も配列決定することができ、これにより、ルーチン使用のために“イベント特異的な”検出方法を開発することができる。たとえば、WO 01/41558が参照され、これは、たとえば統合された配列および隣接する(ゲノム)配列に基く、選抜きのイベント検出キット(たとえばPCR検出キット)を記載する。
【0074】
NRC1核酸配列は、挿入されるコード配列が、植物細胞で発現を駆動することができるプロモーターの下流(すなわち3’)および制御下に位置するように植物細胞ゲノムに挿入される。これは、好ましくは、キメラ遺伝子を植物細胞ゲノム、とりわけ核またはプラスチド(たとえば葉緑体)ゲノムに挿入することにより達成される。
【0075】
NRC1タンパク質の構成的な産生は、細胞死の誘導(たとえば顕微鏡的な病変および/または肉眼的な病変)および/または低い収量につながり得るため、一つの態様において、活性が誘導性であるプロモーターを使用することが好ましい。誘導性プロモーターの例は、傷害誘導性プロモーター、たとえば、Cordera et al.(1994, The Plant Journal 6, 141) に記載される、傷害(たとえば昆虫または物理的傷害により引き起こされる)によって誘導されるMPIプロモーター、またはCOMPTIIプロモーター (WO 00/56897) またはUS 6031151に記載されるPR1プロモーターである。あるいは、プロモーターは、化学物質、たとえば、Aoyama and Chua (1997, Plant Journal 11: 605-612) およびUS 6063985に記載されるデキサメタゾン、またはテトラサイクリン (TOPFREEまたはTOP 10プロモーター、Gatz, 1997, Annu Rev Plant Physiol Plant Mol Biol. 48: 89-108 and Love et al. 2000, Plant J. 21: 579-88参照) により誘導されてもよい。他の誘導性プロモーターは、たとえば、温度の変化(たとえばUS 5,447,858に記載される熱ショックプロモーター)、嫌気的条件(たとえばトウモロコシADH1Sプロモーター)、光(US6455760)、病原体(たとえばEP759085のgst1プロモーターまたはEP309862のvst1プロモーター)、または老化(SAG12およびSAG13、US5689042参照)により誘導性である。明らかに、種々の他のプロモーターが利用可能である。
【0076】
一つの態様において、好ましくは、病原体誘導性プロモーターが使用され、これによりNRC1タンパク質(または変異体もしくはフラグメント)は、病原体が植物組織を攻撃した後にのみ産生される。とりわけ、病原体の攻撃の直後にアップレギュレートされる遺伝子のプロモーターが望ましい。病原体誘導性プロモーターには、たとえば、タバコ由来のhsr203J、str246Cおよびsgd24プロモーター、Yin et al. (1997, Plant Physiology 115(2): 437-51) に記載されるEAS4プロモーター、tap1またはtap2プロモーター (Mohan et al., 1993, Plant Mol Biol. 1993 22: 475-90)、gst1プロモーターまたはその変異体 (Martini et al. 1993, Mol. Gen. Gen. 236, 179-186; Hennin C., 1997, Afstudeerwerk, Faculteit Landbouwkundige en Toegepaste Biologische Wetenschappen, University of Gent, Belgium)、WRKYプロモーター (Eulgem et al., EMBO J., 1999, 18(17): 4689-99およびWO 00/29592に記載されるキメラプロモーター) が含まれる。また、特定の植物病原体により誘導性であるプロモーターは、公知の方法、たとえばcDNA-AFLP(登録商標)を用いて同定されてもよい。
【0077】
好ましくは、プロモーターは、複数の病原体により誘導され、すなわち宿主植物の広範囲の病原体により誘導性である。特定の宿主植物種のそれぞれに対して、異なるプロモーターが最適であってもよい。たとえば、トマトを宿主として使用する場合、プロモーターは、少なくとも一、好ましくは二以上のトマト病原体により誘導される。とりわけ、一または複数の真菌の植物病原体および/または細菌の植物病原体(とりわけ一または複数の生物栄養性および/または半生物栄養性植物病原体)により誘導性であるプロモーターが好ましい。
【0078】
植物病原体、それらにより引き起こされる病気の徴候、およびそれらの生活環の詳細な説明は、植物種のそれぞれに対して見出すことができる。たとえば、トマトの病原体は、“Compendium of Tomato Diseases”, Editors Jones, Jones, Stall and Zitter, ISBN 0-89054-120-5, APS Press (http:/www.shopapspress/org) に記載される。ポテトの病原体は、“Compendium of Potato Disease”, 2nd edition, Editors Stevenson, Franc and Weingartner, APS Press, ISBN 0-89054-275-9 に記載される。
【0079】
トマトの病原体には、たとえば、(非限定的な)以下の真菌および細菌種およびウイルスが含まれる:Botrytis cinerea (真菌/壊死栄養体); Colletotrichum coccodes (真菌/壊死栄養体); Alternaria alternata (真菌); Alternaria solani (真菌/壊死栄養体); Stemphylium solani; Phytophthora infestans (卵菌(oomycte)/半生物栄養体); Septoria lycopersici; Cladosporium fulvum (真菌/半生物栄養体); Phytophthora parasitica; Oidium lycopersicum (生物栄養体); Fusarium oxysporum; Sclerotium rolfsii; Pythium; Rhizoctonia (真菌/壊死栄養体); Corynebacterium michiganense (細菌); Pseudomonas syringae pv tomatoまたはpv syringae (細菌/生物栄養体); Pseudomonas solanacearum; Pseudomonas corrugate; Clavibacter Xanthomonas campestris (細菌/生物栄養体); Verticillium (真菌)、トマト・スポッテッド・ウイルト・ウイルスtomato spotted wilt virus (TSWV); タバコまたはトマト・モザイク・ウイルス (TobMV、TomMV)。
【0080】
ポテトの病原体には、たとえば、種々の真菌、細菌、線虫およびウイルス、たとえば以下のものが含まれる:Phytophthora infestans (oomycte /半生物栄養体), 線虫 (生物栄養体); Erwinia carotovora (細菌); Colletotrichum coccodes (真菌); Rhizoctonia solani (真菌/壊死栄養体); Verticillium dahliae (真菌); Streptomyces scabies; Alternaria solani (真菌/壊死栄養体); Pythium; Spongospora subterranean; PVXおよびPVY; ポテト・リーフロール・ウイルス (PLRV); など。
【0081】
種々の植物種の植物の病気については、http://www.apsnet.org/online/common/toc.aspも参照される。よって、一つの態様において、プロモーターは、好ましくは、上記病原体の一または複数により、最も好ましくは少なくとも上記生物栄養性および/または半生物栄養性病原体の一または複数により誘導性である。
【0082】
あるいは、宿主植物は、種々の病原体の攻撃後に確実にNRC1タンパク質が産生されるように、異なる病原体誘導性プロモーターの制御下に、種々のNRC1導入遺伝子をそれぞれ含んでいてもよい。たとえば、トマトの形質転換について、一つのプロモーターはPhytophthoraにより誘導性で、一つのプロモーターはCladosporiumにより誘導性であってもよい。
【0083】
“誘導性(inducible)”の用語は、プロモーターが、誘導物質刺激の非存在下において完全に不活性であることを必ずしも必要としない。低レベルの非特異的活性は、それが、植物の収量または品質の重いペナルティーという結果につながらない限り、存在していてもよい。よって、誘導性とは、好ましくは、誘導物質との接触後に、下流のNRC1コード領域の転写の増大につながるプロモーターの活性の増大を指す。
【0084】
本明細書において、最も好ましい組合せは、上述のとおり、構成的に活性なNRC1タンパク質をコードするNRC1核酸配列に動作可能に連結された病原体誘導性プロモーターの使用である。この場合、病原体の攻撃により、構成的に活性なNRC1が発現され、これが(病原体の攻撃部位に限定される)局所的なHRにつながり、(半)生物栄養性病原体の更なる増殖を妨害する。
【0085】
別の態様において、たとえば以下の構成的プロモーターが使用されてもよい:カリフラワーモザイクウイルス(CaMV)の単離体CM 1841 (Gardner et al., 1981, Nucleic Acids Research 9, 2871-2887)、CabbB-S (Franck et al., 1980, Cell 21, 285-294)、およびCabbB-JI (Hull and Howell, 1987, Virology 86, 482-493) の強力な構成的35Sプロモーターまたは増強(enhanced)35Sプロモーター(“35Sプロモーター”);Odell et al. (1985, Nature 313, 810-812) またはUS5164316に記載される35Sプロモーター、ユビキチンファミリー由来のプロモーター (たとえばChristensen et al., 1992, Plant Mol. Biol. 18, 675-689, EP 0 342 926のトウモロコシのユビキチンプロモーター、Cornejo et al. 1993, Plant Mol. Biol. 23, 567-581も参照)、gos2プロモーター (de Pater et al., 1992 Plant J. 2, 837-844)、emuプロモーター (Last et al., 1990, Theor. Appl. Genet. 81,581-588)、アラビドプシス(Arabidopsis)のアクチンプロモーター、たとえばAn et al. (1996, Plant J. 10, 107.)に記載されるプロモーター、イネのアクチンプロモーター、たとえばZhang et al.(1991, The Plant Cell 3, 1155-1165)に記載されるプロモーターおよびUS 5,641,876に記載されるプロモーターまたはWO 070067に記載されるイネのアクチン2プロモーター;キャッサバ葉脈モザイクウイルスのプロモーター (WO 97/48819, Verdaguer et al. 1998, Plant Mol. Biol. 37, 1055-1067)、サブタレニアン・クローバー萎縮ウイルス由来のpPLEXシリーズのプロモーター (WO 96/06932、とりわけS7プロモーター)、アルコールデヒドロゲナーゼプロモーター、たとえばpAdh1S (GenBankアクセッション番号X04049、X00581)、T-DNAのそれぞれ1’および2’遺伝子の発現を駆動するTR1’プロモーターおよびTR2’プロモーター(それぞれ“TR1’プロモーター”および“TR2’プロモーター”)(Velten et al., 1984, EMBO J 3, 2723-2730)、US6051753およびEP426641に記載されるゴマノハグサモザイクウイルスプロモーター、ヒストン遺伝子プロモーター、たとえばアラビドプシス(Arabidopsis)由来のPh4a748プロモーター (PMB 8: 179-191)、またはその他。好ましい態様において、PCT/NL2005/050083 (2005年12月16日出願) に記載されるAA6プロモーターが使用される。
【0086】
あるいは、構成的ではなく植物の一または複数の組織または器官に特異的なプロモーター(組織優先的/組織特異的、発生で調節されるプロモーターを含む、たとえば葉優先的、表皮優先的、根優先的、花組織、たとえば絨緞組織または葯優先的、種子優先的、莢優先的など)を使用することができ、これによりNRC1遺伝子は、特異的な組織または器官の細胞でのみ、および/またはある発生段階の間のみ発現される。たとえば、NRC1遺伝子は、光誘導性プロモーター(たとえば、植物それ自体または別の植物、たとえばUS 5,254,799に開示されるエンドウまたはUS5034322に開示されるアラビドプシスの、リブロース-1,5-ビスリン酸カルボキシラーゼの小サブユニット遺伝子のプロモーター)の制御下にコード配列を置くことにより、植物の葉で選択的に発現させることができる。
【0087】
一つの態様において、内在性NRC1遺伝子のプロモーターが使用される。たとえば、トマトNRC1遺伝子のプロモーターを単離し、配列番号2または4のNRC1タンパク質をコードするコード領域に動作可能に連結してもよい。NRC1プロモーター(配列番号1および3の上流の転写調節領域)は、公知の方法、たとえばTAIL-PCR(Liu et al. 1995, Genomics 25(3): 674-81; Liu et al. 2005, Methods Mol Biol. 286: 341-8)、Linker-PCR、またはInverse PCR (IPCR) を用いて、トマト植物から単離することができる。
【0088】
NRC1コード配列は、好ましくは、コード配列が、適切な3’端の非翻訳領域(“3’端”または3’UTR)の上流(すなわち5’)に位置するように植物ゲノムに挿入される。適切な3’端には、CaMV 35S遺伝子のもの(“3’35S”)、ノパリンシンターゼ遺伝子のもの(“3’nos”) (Depicker et al., 1982 J. Molec. Appl. Genetics 1, 561-573.)、オクトピンシンターゼ遺伝子のもの(“3’ocs”) (Gielen et al., 1984, EMBO J 3, 835-845)、およびT-DNA遺伝子7のもの (“3’遺伝子7”) (Velten and Schell, 1985, Nucleic Acids Research 13, 6981-6998) などが含まれ、これらは、形質転換された植物細胞において3’-非翻訳DNA配列として機能する。一つの態様において、配列番号3の2748ヌクレオチドから3168ヌクレオチドまで、および配列番号5に示される、トマトNRC1遺伝子の3’UTRが使用される。また、NRC1の3’UTRは、他のコード領域と組み合わせて3’UTRとして使用され得るため、本明細書においてそれ自体で一つの態様でもある。同様に、配列番号5の任意の変異体またはフラグメントが提供される。配列番号5の変異体には、配列番号5に対して少なくとも40、50、60、70、80、90、95、98、99%またはそれ以上の核酸配列同一性を含む核酸配列が含まれる(上記で規定されるNeedleman and WunschアルゴリズムおよびGAPペナルティーを用いて決定される)。フラグメントには、配列番号5または配列番号5の変異体の少なくとも30、50、100、150、200、300、400またはそれ以上の連続ヌクレオチドを含む任意のヌクレオチド配列が含まれる。
【0089】
T-DNAベクターのアグロバクテリウムへの導入は、公知の方法、たとえばエレクトロポレーションまたはトリペアレンタルメイティングを用いて行うことができる。
【0090】
NRC1コード核酸配列は、必要に応じて、ハイブリッド遺伝子配列として植物ゲノムに挿入することができ、これによりNRC1遺伝子は、容易に検出可能な融合タンパク質を植物が発現するように、選択またはスコアマーカーをコードする遺伝子、たとえばカナマイシン耐性をコードするneo(またはnptII)遺伝子(EP 0 242 236)にフレームで連結される(US 5,254, 799; Vaeck et al., 1987, Nature 328, 33-37)。
【0091】
NRC1タンパク質(または変異体もしくはフラグメント)をコードするNRC1核酸配列の全部または一部は、微生物、たとえば細菌(たとえば大腸菌(Escherichia coli)、シュードモナス属(Pseudomonas)、アグロバクテリウム属(Agrobacterium)、バチルス属(Bacillus)など)、真菌類、または藻類または昆虫を形質転換するため、あるいは組換えウイルスを作成するために使用することができる。適切なクローニング媒体に組み込まれた本発明のNRC1核酸配列の全部または一部による細菌の形質転換は、慣用的なやり方で、好ましくはMaillon et al. (1989, FEMS Microbiol. Letters 60, 205-210.) およびWO 90/06999に記載される慣用的なエレクトロポレーション技術を用いて行うことができる。原核宿主細胞での発現のために、核酸配列のコドン使用頻度は、(上記で植物について記載したとおり)適宜、最適化してもよい。イントロン配列は除去すべきであり、最適な発現のための他の適合を公知のとおり行ってもよい。
【0092】
NRC1核酸配列のDNA配列は、翻訳に中立的なやり方で、遺伝子部分に存在する抑制の可能性があるDNA配列を改変して、および/または上述のとおりコドン使用頻度に変化を導入することにより、たとえばコドン使用頻度を植物(好ましくは特定の関連の植物属)に最も好ましいものに適合させることにより、更に変化させることができる。
【0093】
本発明の一つの態様に従って、NRC1タンパク質(またはキメラタンパク質)は、細胞内オルガネラ、たとえばプラスチド、好ましくは葉緑体、ミトコンドリアをターゲットにするか、あるいは潜在的にタンパク質の安定性および/または発現を最適化して、細胞から分泌される。同様に、タンパク質は、液胞をターゲットにしてもよい。この目的のため、本発明の一つの態様において、本発明のキメラ遺伝子は、シグナルまたはターゲットペプチドをコードするコード領域を、本発明のNRC1タンパク質コード領域に連結して含む。本発明のタンパク質に含有される特に好ましいペプチドは、葉緑体または他のプラスチドターゲッティングのためのトランジットペプチド、とりわけ遺伝子産物がプラスチドをターゲットにする植物遺伝子由来の重複トランジットペプチド領域、Capellades et al. (US 5,635,618) の最適化されたトランジットペプチド、ホウレンソウ由来のフェレドキシン-NADP+オキシドレダクターゼのトランジットペプチド(Oelmuller et al., 1993, Mol. Gen. Genet. 237,261-272)、Wong et al. (1992, Plant Molec. Biol. 20, 81-93) に記載されるトランジットペプチド、およびPCT特許出願公開公報WO 00/26371のターゲッティングペプチドである。また、かかるペプチドに連結されたタンパク質の細胞外への分泌のシグナルを送るペプチド、たとえば、ポテトのプロテイナーゼインヒビターIIの分泌シグナル(Keil et al., 1986, Nucl. Acids Res. 14, 5641-5650)、イネのアルファーアミラーゼ3遺伝子の分泌シグナル(Sutliff et al., 1991, Plant Molec. Biol. 16, 579-591)、およびトマトのPR1タンパク質の分泌シグナル(Cornelissen et al., 1986, EMBO J. 5, 37-40)も好ましい。本発明に従って特に有用なシグナルペプチドには、葉緑体トランジットペプチド(たとえばVan Den Broeck et al., 1985, Nature 313, 358)、または葉緑体へのタンパク質の輸送を引き起こすUS 5,510,471およびUS 5,635,618の最適化された葉緑体トランジットペプチド、分泌シグナルペプチド、または他のプラスチド、ミトコンドリア、ER、または別のオルガネラをタンパク質のターゲットにするペプチドが含まれる。細胞内オルガネラへのターゲッティングまたは植物細胞の外もしくは細胞壁への分泌のためのシグナル配列は、天然においてターゲットにされるかまたは分泌されるタンパク質に見出され、好ましくは以下の文献に記載されるものである:Klosgen et al. (1989, Mol. Gen. Genet. 217, 155-161)、Klosgen and Weil (1991, Mol. Gen. Genet. 225, 297-304)、Neuhaus & Rogers (1998, Plant Mol. Biol. 38, 127-144)、Bih et al. (1999, J. Biol. Chem. 274, 22884-22894)、Morris et al. (1999, Biochem. Biophys. Res. Commun. 255, 328-333)、Hesse et al. (1989, EMBO J. 8, 2453-2461)、Tavladoraki et al. (1998, FEBS Lett. 426,62-66.)、Terashima et al. (1999, Appl. Microbiol. Biotechnol. 52,516-523)、Park et al. (1997, J.Biol. Chem. 272, 6876-6881)、Shcherban et al. (1995, Proc. Natl. Acad. Sci USA 92,9245-9249)。
【0094】
形質転換された宿主細胞の外へNRC1タンパク質を分泌するためには、適切な分泌シグナルが、NRC1タンパク質のアミノ末端(N-末端)に融合され得る。推定シグナルペプチドは、コンピューターベースの分析を用いて、プログラムSignal Peptide search (SignalP V1.1または2.0) などのプログラムを用いて検出することができる(Von Heijne, Gunnar, 1986 and Nielsen et al., 1996)。
【0095】
一つの態様において、幾つかのNRC1コード核酸配列を、単一の宿主において、必要に応じて異なるプロモーターの制御下で共発現させる。共発現する宿主植物は、本発明のNRC1タンパク質を既に発現している植物を形質転換することにより、あるいは本発明の異なるNRC1タンパク質で形質転換された植物を交配させることにより容易に得られる。あるいは、幾つかのNRC1タンパク質をコードする核酸配列を、単一の形質転換ベクター上に存在させることができるし、あるいは別々のベクターを用いて同時に共形質転換(co-transform)して、両方のキメラ遺伝子を含む形質転換体を選択することもできる。同様に、一または複数のNRC1コード遺伝子を、たとえば耐病性を向上させるかまたは耐病性シグナリング経路に関与する他のタンパク質などをコードする他のキメラ遺伝子と一緒に、単一の植物で発現させてもよい。
【0096】
異なるタンパク質を同一の植物で発現させることができること、あるいは各々のタンパク質を単一の植物で発現させた後に、当該植物を互いに交配することにより同一の植物で各タンパク質を組み合わせることができることが理解される。たとえば、ハイブリッド種子生産において、各々の親植物は、単一のタンパク質を発現することができる。親植物を交配してハイブリッドを産生すると、ハイブリッド植物において両方のタンパク質が組み合わされる。
【0097】
好ましくは、選択の目的のため、更に雑草防除オプションのために、本発明のトランスジェニック植物は、除草剤、たとえば広範囲の除草剤、たとえば有効成分としてグルホシネートアンモニウムを主成分とする除草剤(たとえばLiberty(登録商標)またはBASTA; 耐性は、PATまたはbar遺伝子により付与される; EP 0 242 236およびEP 0 242 246参照)またはグリホサートを主成分とする除草剤(たとえばRoundUp(登録商標); 耐性は、EPSPS遺伝子により付与される、たとえばEP 0 508 909およびEP 0 507 698参照)に対する耐性を付与するタンパク質をコードするDNAで形質転換される。更に、除草剤耐性遺伝子(または所望の表現型を付与する他の遺伝子)の選択マーカーとしての使用は、抗生物質耐性の導入を回避することができるという利点を有する。
【0098】
あるいは、他の選択マーカー遺伝子、たとえば抗生物質耐性遺伝子が使用されてもよい。一般に、形質転換宿主植物に抗生物質耐性遺伝子を保持することは受け入れられないため、これら遺伝子は、形質転換体の選択後に再度除去することができる。導入遺伝子を除去するための幾つかの技術が存在する。除去するための一つの方法は、キメラ遺伝子をlox部位と隣接させ、選択後、形質転換植物をCREリコンビナーゼ発現植物と交配することによるものである(たとえばEP506763B1参照)。部位特異的組換えの結果、マーカー遺伝子は切り出される。別の部位特異的組換えシステムは、EP686191およびUS5527695に記載されるFLP/FRTシステムである。遺伝子スタッキングの目的のために、CRE/LOXおよびFLP/FRTなどの部位特異的組換えシステムが使用されてもよい。更に、一成分(one-component)切出しシステムが記載されている(たとえばWO9737012またはWO9500555参照)。
【0099】
本発明の形質転換された植物細胞/植物/種子、並びに核酸配列およびタンパク質の使用
以下の記載において、向上した耐病性の表現型を備えたトランスジェニック植物細胞、植物、植物種子など、およびこれらの誘導体/子孫を作成するための本発明のNRC1配列の使用について説明される。
【0100】
向上した耐病性を備えたトランスジェニック植物は、上述のとおり、少なくとも一つのNRC1タンパク質を適切なプロモーターの制御下でコードする核酸配列を用いて植物宿主細胞を形質転換し、前記細胞からトランスジェニック植物を再生することにより作成することができる。
【0101】
好ましいプロモーターは、生物的および/または非生物的な外的刺激により誘導性であるプロモーターである。とりわけ病原体誘導性であるプロモーターが、上述のとおり好ましい。好ましいプロモーターとNRC1の組合せは、以下のとおりである:
a)病原体誘導性プロモーター − 構成的に活性なNRC1タンパク質をコードする核酸配列;
b)病原体誘導性プロモーター − 野生型NRC1タンパク質をコードする核酸配列;
c)植物NRC1遺伝子のプロモーター(好ましくは形質転換される同一種のもの) − 構成的に活性なNRC1タンパク質をコードする核酸配列;
d)植物NRC1遺伝子のプロモーター(好ましくは形質転換される同一種のもの) − 野生型NRC1タンパク質をコードする核酸配列;
e)生物的ストレス誘導性プロモーター(たとえば昆虫傷害誘導性、病原体誘導性など) − 構成的に活性なNRC1タンパク質をコードする核酸配列;
f)生物的ストレス誘導性プロモーター(たとえば昆虫傷害誘導性、病原体誘導性など) − 野生型NRC1タンパク質をコードする核酸配列;
g)構成的プロモーター(たとえば35Sプロモーター) − 野生型NRC1タンパク質をコードする核酸配列;
h)構成的プロモーター(たとえば35Sプロモーター) − 配列番号2に対して全長にわたって少なくとも70%のアミノ酸配列同一性を含むアミノ酸配列をコードする核酸配列;
i)病原体誘導性プロモーター − 配列番号2に対して全長にわたって少なくとも70%のアミノ酸配列同一性を含むアミノ酸配列をコードする核酸配列;
j)植物NRC1遺伝子のプロモーター − 配列番号2に対して全長にわたって少なくとも70%のアミノ酸配列同一性を含むアミノ酸配列をコードする核酸配列。
【0102】
一つの態様において、トランスジェニック植物は、構成的HR病変または誘導性HR病変の何れか、および一または複数の病原体に対して向上した耐病性を示すことができる。しかし、植物が向上した耐病性を示している間に、HR病変が発生しないか、または“弱い”HR病変(たとえば小さい病変、たとえばミクロ病変、および/または低い病変の頻度)が発生することも本明細書において想定される。宿主植物において同一プロモーターの制御下で発現されたときに、配列番号1または3より、あるいは形質転換されるのと同じ宿主種から得た野生型のNRC1対立遺伝子の発現より、少ないおよび/または小さいHR病変につながるNRC1対立遺伝子またはオルソログが、本明細書において、とりわけ上記アプローチg)およびh)において、特に好ましい。かかる対立遺伝子/オルソログは、所定の宿主に“弱いHR表現型”を付与するNRC1対立遺伝子ということもできる。かかるNRC1対立遺伝子またはオルソログは、上述のとおり同定および/または単離することができる。異なるNRC1対立遺伝子および/またはオルソログのHR表現型は、これらを用いて発現ベクターを作成し(好ましくは、比較される全ての核酸を同一のプロモーター、たとえば35Sに動作可能に連結し)、これらを用いて植物または植物組織を形質転換し、これら植物の間でHR表現型を比べることにより比較することができる。ナス科植物の形質転換体について、配列番号1または3を発現する形質転換体のHR病変の表現型が、好ましくはレファンレスとして使用され、同じプロモーターの制御下での発現により少ないおよび/または小さいHR病変が結果として得られる対立遺伝子が、弱いHR表現型を付与する対立遺伝子である。HR病変の表現型は、種々の方法を用いて、たとえば顕微鏡検査(必要に応じて死細胞の染色)、視覚的スコアリング、cm2あたりの数を計算するための病変の計数、HR病変の直径の測定などにより比較し、必要に応じて定量することができる。
【0103】
好ましくは、本発明のトランスジェニック植物は、一または複数の病原体、とりわけトランスジェニック植物種の生物栄養性および/または半生物栄養性病原体に対して向上した耐病性を含む。このため、たとえばトランスジェニックトマトまたはポテト植物は、少なくとも一つまたは複数の上述の真菌、細菌、線虫の種および/またはウイルス病原体に対して向上した耐性、最も好ましくは一つまたは幾つかの生物栄養性および/または半生物栄養性の種に対して少なくとも向上した耐性を含む。
【0104】
“耐病性”または“増大/向上した耐病性”は、一または複数の植物病原体の攻撃に耐える形質転換体の(野生型またはコントロール形質転換体と比較して)向上した能力を指すために本明細書において使用され、言い換えれば、非形質転換(または空ベクター形質転換)コントロールと比較して、形質転換体の病気の徴候が有意に減少したことを指す。耐病性または向上した耐病性は、種々の方法を用いて決定され得る。しばしば、病気の徴候は、病原体の接種または接触後の一または複数の時点において病気の徴候を評価することにより(バイオアッセイまたは野外研究で)視覚的にスコア化される。代わりの方法には、病原体を検出し、必要に応じて定量する方法が含まれる。このため、組織の中/上で検出される病原体の量がコントロールと比較して有意に少ないか、または病原体の広がりがコントロールより有意に遅い場合、トランスジェニック植物は、向上した耐病性を示すことができる。最終的には、同等の病気プレッシャーの下で生育させたときに、コントロールと比較して形質転換体の平均収量が有意に増大すること(たとえば少なくとも1%、2%、5%、10%またはそれ以上)が、向上した耐病性の間接的な測定(measurement)を提供する。
【0105】
このため、NRC1タンパク質(または構成的に活性なNRC1タンパク質)を発現する複数の形質転換植物が、非形質転換コントロールまたは空ベクター形質転換コントロールと比較して病気の徴候の有意な減少を示す場合、向上した耐病性を示す。明らかに、有意な差が存在するかどうかを決定するためには統計分析が必要である。好ましくは、一または複数の病気の徴候は、コントロール植物よりNRC1形質転換において、平均して少なくとも2%、5%、10%、15%、20%、30%、40%、50%、または100%も少ない。病気のアッセイは、宿主と病原体の組合せごとに異なるため、特定のプロトコールは提供されないが、当業者であれば、形質転換体が、一または複数の病原体に対して有意に向上した耐病性を示すかどうかを決定する方法が分かる。各々の植物と病原体の組合せについて当該技術分野で公知のバイオアッセイを使用して、トランスジェニック植物の耐性を適切なコントロールと比較することができる。
【0106】
NRC1タンパク質は、幾つかの態様において(たとえばNRC1遺伝子が構成的プロモーターの制御下にある場合)病原体の非存在下においてHR病変という結果につながるため、NRC1発現により引き起こされる徴候と病原体の感染および広がりにより引き起こされる徴候とを区別することが、ある態様においては重要であり得る。したがって、(植物組織上の壊死よりも)病原体それ自体を検出すること、または存在する病原体の量または病原体の広がりの速度を比較することが好ましい。たとえば、病原体を染色により検出することができるバイオアッセイが使用され得る。実施例では、GUSを発現するトランスジェニックC. fulvum品種が使用される。したがって、真菌の菌糸体は、接種された植物組織のX-gluc染色を用いて視覚化することができる。コントロールと比較してトランスジェニック植物において真菌の菌糸体が有意に減少することは、真菌に対して向上した耐性を示す。
【0107】
また、幾つかのNRC1タンパク質を、好ましくは異なるプロモーターの制御下、たとえば異なる病原体誘導性プロモーターの制御下で、発現するトランスジェニック植物を作成することも一つの態様である。
【0108】
耐病性の表現型は、適切な時および場所に適切な量のNRC1タンパク質を発現させることにより微調整することができる。かかる微調整は、特定の宿主−病原体の組合せに対して最も適切なプロモーターを決定し、所望の発現レベルを示すトランスジェニック“イベント”を選択することにより行うことができる。病原体の攻撃後の低すぎるレベルのNRC1タンパク質またはNRC1タンパク質生産の遅すぎる誘導は、耐病性を向上させるのに不十分であり得る。一方、病原体の攻撃のない時および場所における高すぎるタンパク質のレベルまたは発現は、農学的に望ましくない表現型、たとえば病原体の非存在下における葉または果実の病変および収量の損失という結果につながり得る。しかし、当業者であれば、向上した耐病性を有するが、同時に農学的に許容される植物を容易に作成することができる。最適なNRC1対立遺伝子、たとえば高い耐性レベルとわずかな弱いHR表現型を提供する対立遺伝子は、記載されるとおり単離または同定することができる。
【0109】
所望のレベルのNRC1タンパク質を発現する形質転換体は、たとえば、コピー数の分析(サザンブロット分析)、mRNA転写産物レベルの分析(たとえばNRC1プライマー対またはフランキングプライマーを用いたRT-PCR)、または種々の組織におけるNRC1タンパク質の存在およびレベルの分析(たとえばSDS-PAGE;ELISA分析など)により選択される。調節の理由のため、好ましくは単一コピーの形質転換体が、好ましくは選択され、キメラ遺伝子の挿入部位に隣接する配列が分析され、好ましくはその“イベント”を特徴づけるために配列決定される。安定なNRC1導入遺伝子を備えた高性能の選抜きのイベントが得られるまで、高いまたは適度なNRC1発現トランスジェニックイベントが、更なる交配/戻し交配/自家受粉のために選択される。
【0110】
また、本発明の一または複数のNRC1遺伝子を発現する形質転換体は、他の導入遺伝子、たとえば耐病性を付与する遺伝子または他の生物的および/または非生物的ストレスに対する耐性を付与する遺伝子を含んでいてもよい。“多重(stacked)”導入遺伝子を備えたかかる植物を得るために、他の導入遺伝子がNRC1形質転換体に遺伝子移入されてもよいし、あるいはNRC1形質転換体が、一または複数の他の遺伝子でその後に形質転換されてもよいし、あるいは植物系統または品種を形質転換するために幾つかのキメラ遺伝子が使用されてもよい。たとえば、幾つかのキメラ遺伝子が、単一のベクターに存在していてもよいし、あるいは共形質転換される(co-transform)異なるベクターに存在していてもよい。
【0111】
一つの態様において、本発明の一または複数のNRC1遺伝子と以下の遺伝子が組み合わせられる:公知の耐病性遺伝子、とりわけ壊死栄養性病原体に対して向上した耐性を付与する遺伝子、ウイルス耐性遺伝子、昆虫耐性遺伝子、非生物的ストレス耐性遺伝子(たとえば干ばつ耐性、塩耐性、熱もしくは低温耐性など)、殺虫剤耐性遺伝子など。このため、多重(stacked)形質転換体は、さらに広い生物的および/または非生物的ストレス耐性、病原体耐性、昆虫耐性、線虫耐性、塩度、低温ストレス、熱ストレス、水ストレス耐性などをもち得る。また、NRC1サイレンシングアプローチが、単一の植物においてNRC1発現アプローチと組み合わせられてもよい。たとえば、根または塊茎におけるNRC1過剰発現は、土壌病原体に対する根または塊茎の耐性を付与するかまたは向上させ得る。同時に、空気中の(aerial)部分におけるNRC1のダウンレギュレーションは、壊死栄養性病原体に対する耐性を付与するかまたは向上させ得る(または逆も同様である(vice versa))。
【0112】
また、NRC1遺伝子を、既にあるレベルの耐病性を有する植物育種系統に導入または遺伝子移入することも可能である。畑での耐病性の耐久性のため、好ましくは耐性の供給源が、異なる基本的分子メカニズムを有する、幾つかの耐病性メカニズムを植物で重ねることが望ましい。
【0113】
上述の任意の形質転換植物の植物全体、種子、細胞、組織および子孫(たとえばF1雑種、F2種子/植物など)は、本明細書に包含され、DNAにおける導入遺伝子の存在により、たとえば鋳型として全ゲノムDNAを用いてNRC1特異的PCRプライマー対を用いてPCR分析を行うことにより、同定することができる。また、PCRプライマーが、挿入されたキメラ遺伝子に隣接する植物DNAに基づいている“イベント特異的”PCR診断方法を開発することができる。US6563026参照。同様に、トランスジェニック植物、またはそれに由来する任意の植物、種子、組織または細胞を同定するイベント特異的AFLPフィンガープリントまたはRFLPフィンガープリントが開発され得る。
【0114】
本発明のトランスジェニック植物は、好ましくは、望ましくない表現型、たとえば収量の減少、病気(とりわけ壊死栄養体)に対して向上した感受性、または望ましくない構造の変化(矮性、変形)などを示さないこと、並びにかかる表現型が第一形質転換体で観察される場合、これらは、通常の育種と選抜法(交配/戻し交配/自家受粉など)により除去することができることが理解される。本明細書で記載される任意のトランスジェニック植物は、導入遺伝子についてホモ接合であっても半接合であってもよい。
【0115】
NRC1遺伝子サイレンシングアプローチおよび遺伝子サイレンシングベクター
NRC1遺伝子サイレンシングベクターで形質転換され、とりわけ壊死栄養性病原体に対して向上した耐病性を備えた植物を提供することが本発明の更なる態様である。本発明の範囲を限定しないが、内在性NRC1遺伝子または遺伝子ファミリーのサイレンシングは、トランスジェニック植物がHR応答を誘発および/または開始できないという結果につながると考えられる。壊死栄養性病原体は、増殖および発生のために細胞死を必要とするため、かかる植物は、一または複数の壊死栄養性病原体に対して向上した耐性を含み得る。
【0116】
“遺伝子サイレンシング”は、一または複数のターゲット遺伝子(たとえば内在性NRC1遺伝子)の遺伝子発現のダウンレギュレーションまたは完全な抑制を指す。遺伝子発現を低下または廃止する抑制性RNAの使用は、当該技術分野で十分に確立されており、幾つかのレビューの主題である(たとえばBaulcombe 1996, Stam et al. 1997, Depicker and Van Montagu, 1997)。植物で遺伝子サイレンシングを達成するために利用可能な幾つかの技術が存在し、たとえばターゲット遺伝子の全部または一部のアンチセンスRNAを産生するキメラ遺伝子(たとえばEP 0140308 B1、EP 0240208 B1およびEP 0223399 B1参照)、または(共抑制とも称される)センスRNAを産生するキメラ遺伝子(EP 0465572 B1参照)である。
【0117】
しかし、これまで最も成功したアプローチは、ターゲット遺伝子のセンスおよびアンチセンスRNAの両方の産生(“逆方向反復配列”)であり、これは、二本鎖RNA(dsRNA)を細胞で形成し、ターゲット遺伝子をサイレンシングする。dsRNAの産生および遺伝子サイレンシングのための方法およびベクターは、EP 1068311、EP 983370 A1、EP 1042462 A1、EP 1071762 A1およびEP 1080208 A1に記載されている。したがって、本発明のベクターは、本発明のNRC1遺伝子のセンスおよび/またはアンチセンスDNAフラグメントに動作可能に連結された植物細胞で活性な転写調節領域を含み得る。一般に、短い(センスおよびアンチセンス)長さのターゲット遺伝子配列、たとえば17、18、19、20、21、22または23ヌクレオチドのコードまたは非コード配列で十分である。より長い配列、たとえば50、100、200または250ヌクレオチドまたはそれ以上も使用することができる。好ましくは、短いセンスおよびアンチセンスのフラグメントは、スペーサー配列、たとえばイントロンにより分離され、これによりdsRNAの形成によりループ(またはヘアピン)が形成される。任意の短い長さの配列番号1または3、またはその変異体を使用して、NRC1遺伝子サイレンシングベクター、および一または複数のNRC1遺伝子が(使用されるプロモーターに依存して)全部または幾つかの組織または器官でサイレンシングされたトランスジェニック植物を作成してもよい。ヘアピン構築物を作成する簡便な方法は、ジェネリックベクター、たとえばpHANNIBALおよびpHELLSGATE、Gateway(登録商標)技術に基くベクターを使用することである(Wesley et al. 2004, Methods Mol Biol. 265:117-30; Wesley et al. 2003, Methods Mol Biol. 236:273-86 and Helliwell & Waterhouse 2003, Methods 30(4):289-95参照。全てが参照により本明細書に組み込まれる)。
【0118】
NRC1遺伝子の保存された核酸部分を選択することにより、宿主の植物または植物部分においてNRC1ファミリーメンバーをサイレンシングさせることができる。NRC1遺伝子のセンスおよび/またはアンチセンスDNAフラグメントに動作可能に連結された転写調節エレメントを含み、一または複数の病原体、とりわけ壊死栄養性病原体に対して向上した耐性を示すトランスジェニック植物も本明細書に包含される。
【0119】
また、一または複数の生物栄養性および/または半生物栄養性病原体および一または複数の壊死栄養性病原体に対して向上した耐性を有する植物も提供される。かかる植物は、適切なプロモーター−NRC1遺伝子の組合せを選択することにより作成することができる。たとえば、内在性NRC1遺伝子が、種々の組織および/または種々の時期に(たとえば芽生え、根または塊茎などにおいて)サイレンシングされ、一または複数の壊死栄養性病原体に対して耐性を提供している間に、機能的なNRC1タンパク質が、ある組織においてある時期に(たとえば誘導により、または空気中の(aerial)植物部分において)産生され、生物栄養性および/または半生物栄養性病原体に対して耐性を提供してもよい。したがって、単一の植物が、キメラNRC1発現導入遺伝子およびNRC1サイレンシング遺伝子の両方を含んでいてもよい。
【0120】
本発明の突然変異した対立遺伝子および植物
非トランスジェニック方法、たとえば、TILLINGなどの突然変異誘発システム(Targeting Induced Local Lesions IN Genomics; McCallum et al., 2000, Nat Biotech 18:455, and McCallum et al. 2000, Plant Physiol. 123, 439-442、何れも参照により本明細書に組み込まれる)を使用すること、および高レベルの本発明の一または複数のNRC1タンパク質を産生し、および/または構成的に活性なNRC1タンパク質を産生する植物系統を作成するための選択を使用することも本発明の一つの態様である。本発明の範囲を限定しないが、かかる植物は、宿主NRC1遺伝子の発現を構成的にするかまたは高くするリプレッサータンパク質の結合部位であるプロモーター内のポイント/欠失突然変異を含むことができると考えられる。構成的に活性なNRC1突然変異体は、コード領域、たとえばMHD領域に突然変異を含む。好ましくは、突然変異体または突然変異体の一部におけるNRC1タンパク質レベルは、突然変異体において、非突然変異体の植物と比較して少なくとも約2、5、10、15%、またはそれ以上増大している。TILLINGは、従来の化学的突然変異誘発(たとえばEMS突然変異誘発)と、その後の突然変異のハイスループットスクリーニング(たとえば突然変異体−野生型のDNAへテロ二本鎖の細胞分裂および配列決定ゲルシステムを用いた検出)を使用する(たとえばHenikoff et al. Plant Physiology Preview May 21, 2004参照)。このため、一または複数の組織で向上したNRC1遺伝子発現を含み、本発明の一または複数のNRC1表現型(向上した耐病性および/またはHR病変)を含む非トランスジェニック植物、種子および組織、並びにかかる植物を作成し、同定する方法が、本明細書に包含される。
【0121】
この方法は、一つの態様において、植物種子の突然変異誘発(たとえばEMS突然変異誘発)、植物個体またはDNAのプーリング、対象領域のPCR増幅、ヘテロ二本鎖の形成およびハイスループット検出、突然変異体植物の同定、突然変異体PCR産物の配列決定の工程を含む。他の突然変異誘発および選択方法が、かかる突然変異体植物を作成するために同様に使用され得ることが理解される。たとえば、種子を、放射線照射するかまたは化学的に処理し、植物を、改変された表現型、たとえば向上した耐病性および/またはHR病変についてスクリーニングしてもよい。
【0122】
本発明の別の態様において、植物材料は、NRC1オルソログのコード配列および/または調節配列のDNA配列に多型または変異を含む種または関連種の天然の集団である。NRC1遺伝子ターゲットにおける突然変異は、ECOTILLINGアプローチを使用してスクリーニングすることができる(Henikoff et al 2004, supra)。この方法において、育種系統または関連種における天然の多型は、上記TILLING方法によりスクリーニングされ、ここで植物の個体またはプールは、NRC1ターゲットのPCR増幅、ヘテロ二本鎖形成、およびハイスループット分析のために使用される。この後、必要な突然変異を有する個々の植物の選択を行うことができ、これは、所望のNRC1オルソログ対立遺伝子を組み込んで所望の特徴を備えた栽培品種を開発するための育種プログラムで引き続き使用することができる。
【0123】
突然変異体の植物は、分子的方法により、たとえばDNAに存在する突然変異、NRC1タンパク質レベル、NRC1 RNAレベルなどにより、および改変された表現型特性により、非突然変異体と区別することができる。
【0124】
非トランスジェニック突然変異体は、突然変異について、ホモ接合であってもヘテロ接合であってもよい。
【0125】
参照される配列
配列番号1:トマトNRC1遺伝子のコード領域
配列番号2:トマトNRC1タンパク質のアミノ酸配列
配列番号3:トマトNRC1遺伝子の全長cDNA(5’および3’UTRを含む)
配列番号4:トマトNRC1D481Vタンパク質のアミノ酸配列
配列番号5:トマトNRC1遺伝子の3’UTR
図説
図1 − NRC1タンパク質の推定配列
最初の150アミノ酸残基は、二重コイル(coiled-coil)(CC)ドメインを表し、CC構造を形成すると推定される残基に下線を付す。151〜508残基は、以下の(下線とラベルを付した)モチーフを備えたヌクレオチド結合(NB-ARC)ドメインを含む:Kinase1A (P loop)、RNBS-A、Kinase 2、RNBS-B、RNBS-C、GLPL、RNBS-DおよびMHD。509〜846残基は、13の不完全なロイシンリッチ繰返し配列(LRRs)を含み;保存された疎水性残基およびプロリン残基は太字で示す。タンパク質配列の下に、LRRコンセンサスモチーフを示す:‘l’は、保存された脂肪族残基を示し、‘c’は、保存された電荷を帯びた残基を示し、‘P’は、保存されたプロリン残基を示す。
【0126】
図2A − NRC1は、Cladosporium fulvumに対するトマトの完全なCf-4-介在性HRに必要である
Cf0トマトおよびCf-4-含有トマト植物に、表示のTRV構築物を接種し、植物をVIGSの開始から3週間分析した。TRV感染Cf-4-含有トマト植物の小葉にAvr4タンパク質を注入し、HRの発生について調査した。TRV:00-感染植物上でHRが載っている部位の数を100%とした。各々の誤差バーは、4つの独立実験に由来する標準偏差を示す。
【0127】
図2B − NRC1は、Cladosporium fulvumに対するトマトの完全なCf-4-介在性耐性に必要である
非TRV感染およびTRV感染Cf-4またはCf-0植物に、C. fulvum-pGPD::GUSを接種し、接種から2週間後に、小葉のコロニー形成をX-glucアッセイで調査した。
【0128】
図3 − N. benthamianaへのTRV:NRC1の接種は、Cf/Avr-、LeEix2/tvEix-、Pto/AvrPto-およびRx/CP-誘導性HRに影響を及ぼす
N. benthamianaに、TRV:00 (空ベクター)、TRV:NRC1およびTRV:SGT1を接種した。3週間後、HR-誘導性タンパク質を発現するアグロバクテリアを葉にインフィルトレーションし(infiltrate)、インフィルトレーションから4日後に写真を撮影した。第1、第2および第3カラム:Avr4、またはCf-9とAvr9の混合物、またはLeEix2とtvEixの混合物 (1:1の比で混合)をそれぞれアグロインフィルトレーションされた、Cf-4耐性遺伝子を発現するN. benthamianaの葉。第4カラム:AvrPtoをアグロインフィルトレーションされた、Pto耐性遺伝子を発現するトランスジェニックN. benthamianaの葉。第5カラム:PVX (CP)のコートタンパク質を発現する遺伝子でアグロインフィルトレーションされた、Rx耐性遺伝子を発現するトランスジェニックN. benthamianaの葉。濃い円はHRを示し、薄い円は低下した(compromised)HRを示す。
【0129】
図4 − 構成的に活性なNRC1は、誘導因子−非依存性HRを誘導し、細胞死シグナリング経路にNRC1を配置させる
Cf-4耐性遺伝子を発現するN. benthamianaに、表示の遺伝子をアグロインフィルトレーションした。パネルAとCについては、アグロインフィルトレーションの3週間前に、植物に表示のTRV構築物を接種した。濃い円はHRを示し、薄い円は低下した(compromised)HRを示す。
【0130】
(A) 構成的に活性なMAPKKおよびMAPKキナーゼをコードする遺伝子のアグロインフィルトレーション。第一カラム:LeMAPKKKαの構成的に活性なキナーゼドメイン (MAPKKK-KD) をコードする遺伝子を用いたアグロインフィルトレーション。第2カラム:LeMEK2の構成的に活性な型 (MEK2DD) をコードする遺伝子を用いたアグロインフィルトレーション。MAPKKK-KDまたはMEK2DDのインフィルトレーションから2日後、葉にエストラジオールをスプレーすることにより発現は誘導された。アグロインフィルトレーションから4日後に写真を撮影した。
【0131】
(B) 35S-プロモーターの制御下の野生型NRC1 (wt) および当該遺伝子の突然変異型のアグロインフィルトレーションであって、サイレンシングサプレッサーp19をコードする遺伝子の発現を駆動するアグロバクテリウムと1:1の比で混合されたアグロインフィルトレーション (左パネル)、または単独のアグロインフィルトレーション (右パネル)。NRC1K191R (K191R):NRC1の不活性なP-ループ突然変異体;NRC1D481V (D481V):構成的に活性なNRC1 (MHDモチーフでの突然変異);NRC1K191R/D481V (K191R/D481V):NRC1のダブル突然変異体。アグロインフィルトレーションから3日後に写真を撮影した。
【0132】
(C) Avr4および構成的に活性なNRC1D481V (D481V) をコードする遺伝子のアグロインフィルトレーション。アグロインフィルトレーションから3日後に写真を撮影した。
【0133】
図5 − NRC1介在性細胞死シグナリングのモデル
細胞死アッセイとN. benthamianaにおけるVIGSを組み合わせるエピスタシス実験に基くモデル。EDS1-、NRC1-、MEK2-、およびSGT1/RAR1依存的なやり方でのCf-4/Avr4介在性細胞死シグナル。
【0134】
以下の非限定的な実施例は、本発明の様々な態様を詳説する。実施例において特に記載しない限り、全ての組換えDNA技術は、Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press、およびSambrook and Russell (2001) Molecular Cloning: A Laboratory Manual, Third Edition, Cold Spring Harbor Laboratory Press, NY; およびAusubel et al. (1994) Current Protocols in Molecular Biology, Current Protocols, USAのVolumes 1 and 2に記載されるとおり、標準的なプロトコールに従って実施した。植物の分子的研究のための標準的な材料および方法は、Plant Molecular Biology Labfax (1993) by R.D.D. Croy, jointly published by BIOS Scientific Publications Ltd (UK) and Blackwell Scientific Publications, UKに記載される。
【0135】
実施例
1.材料および方法
1.1 N. benthamianaにおけるVIGS、アグロインフィルトレーション、HRおよび病気アッセイ
4週齢のN. benthamiana植物に、pTV00-由来の構築物 (バイナリーTRV RNA2ベクター) とpBintra6 (バイナリーTRV RNA1ベクター) の1:1混合物 (Ratcliff et al., 2001 Plant J. 25, 237-245)、またはpTRV-RNA2-由来の構築物とpTRV-RNA1の1:1 混合物 (Liu et al., 2002, Plant J. 31, 777-786; Liu et al., 2002, Plant J. 30, 415-429) をアグロインフィルトレーションした。以下のTRV構築物を使用した: TRV:NRC1、TRV:Cf-4 およびTRV:SGT1 (Peart et al., 2002, Proc. Natl. Acad. Sci. USA 99, 10865-10869)、Ratcliff et al. (2001, supra) に記載されるTRVベクターの全ておよびTRV:EDS1、TRV:MEK2、TRV:RAR1およびTRV:NDR1 (Ekengren et al., 2003, Plant J. 36, 905-917)、Liu et al. (2002, Plant J. 30, 415-429) に記載されるTRVベクターの全て。各TRV構築物について、各実験で4つの植物を使用した。AvrPtoおよびCPを、耐性遺伝子Ptoを発現するTRV-感染N. benthamiana (N. benthamiana:Pto (line 38-12 (Rommens et al., 1995, Plant Cell 7, 1537-1544)) (Pedley and Martin, 2003, Annu. Rev. Phytopathol. 41, 215-243) および耐性遺伝子Rxを発現するTRV-感染N. benthamiana (N. benthamiana:Rx (line Rx-18) (Bendahmane et al., 1999, Plant Cell 11, 781-791)に、それぞれアグロインフィルトレーションした。全ての他のケースにおいて、アグロインフィルトレーションは、耐性遺伝子Cf-4を発現するN. benthamiana (N. benthamiana:Cf-4) で行った。TRV接種から3週間後、接種された葉の上の第三葉、第四葉および第五葉を、以下の発現を駆動するAgrobacterium tumefaciensで攻撃した:AvrPto (OD600=0.06) (Tang et al., 1996, Science 274, 2060-2063)、CP (OD600=0.12) (Bendahmane et al., 1999, supra)、Avr4 (OD600=0.03)、Cf-9およびAvr9 (1:1の比で混合、OD600=0.2) (Van der Hoorn et al., 2000 Mol. Plant-Microbe Interact. 13, 439-446)、LeEix2およびtvEix (1:1の比で混合、OD600=1) (Ron and Avni, 2004, Plant Cell 16, 1604-1615)、β-グルクロニダーゼ (GUS) 遺伝子 (OD600=2) (Van der Hoorn et al., 2000, Mol. Plant-Microbe Interact. 13, 439-446)、NRC1およびp19 (1:1の比で混合、OD600=1) (Voinnet et al., 2003, Plant J. 33, 949-956)、構成的に活性なNRC1D481Vまたは不活性なNRC1K191R/D481V ダブル突然変異体 (OD600=2)、LeMAPKKKαKD、LeMAPKKKαKD- (Del Pozo et al., 2004, EMBO J. 23, 3072-3082) (何れもOD600=0.12) またはLeMEK2DDおよびLeMEK2 (Del Pozo et al., 2004, supra) (何れもOD600=0.25)。LeMAPKKKαKD、LeMAPKKKαKD-、LeMEK2DDまたはLeMEK2のインフィルトレーションから2日後に、silwet (4μl/100 ml) を含有する17-β-エストラジオールの7.5 μM水溶液を葉にスプレーした (Del Pozo et al., 2004, supra)。タンパク質の注入のために、Avr4-HIS-FLAG-標識タンパク質を、製造メーカー (Inv
itrogen, Breda, NL) の推奨に従ってエンテロキナーゼEK-maxで処理し、0.2% tween (v/v) を添加した水中の5μM Avr4タンパク質を、注入のために使用した。アグロインフィルトレーションまたはタンパク質の注入から3〜5日後、HRの発生について葉を調査するか、またはβ-グルクロニダーゼ (GUS) 活性について葉をアッセイした。
【0136】
1.2 トマトにおけるVIGS、HRおよび病気アッセイ
トマトにおけるVIGSのために、Liu et al. (2002, Plant J. 30, 415-429) に記載されるpTRV-RNA1およびpTRV-RNA2ベクターを使用した。Cf-4およびNRC1フラグメントは、BamH1/Asp718を用いた消化によりpTV00から切出し、BamH1/Asp718-消化pTRV-RNA2 (pYL156) に挿入した (Liu et al., 2002, Plant J. 31, 777-786)。TRV:222-UTRを構築するために、NRC1の3’-UTRの一部を、プライマー222-3’UTR-F (5’-GTGGATCCGCAGGTTCAACCAGCCTGGT-3’; BamH1部位に下線を付す) および222-3’UTR-R (5’-GTGGTACCCAAGTGACTTGTTCTGCTGT-3’; Asp718部位に下線を付す) を用いて増幅し、TRV:222-LRRを構築するために、LRRをコードするNRC1領域の一部を、プライマー222-LRR-F (5’-GTGGATCCGTTAAGAGGCTGCAATTTCT-3’; BamH1部位に下線を付す) および222-LRR-R (5’-GTGGTACCGATCTTCTCAAGTTTATCAC-3’; Asp718部位に下線を付す) を用いて増幅した。PCRフラグメントは、BamH1/Asp718で消化し、BamH1/Asp718-消化pTRV-RNA2に挿入した。TRV:Prfの構築は記載されている (Ekengren et al., 2003, Plant J. 36, 905-917)。全てのプラスミドは、エレクトロポレーションによりA. tumefaciens株 GV3101に形質転換した (Takken et al., 2000, Plant J. 24, 275-283)。トマトにおいてVIGSを確立するために、10〜12日齢のトマト芽生えの子葉に、pTRV-RNA1とpTRV-RNA2-由来構築物の混合物(1:1の比で混合)をアグロインフィルトレーションした (Liu et al., 2002, supra)。TRV構築物の各々について、Avr4を発現するC. fulvumに対して耐性のCf-4-含有トマト植物の4個体 (Hcr9-4D (Cf-4)で形質転換したCf0植物) (Thomas et al., 1997, Plant Cell 9, 2209-2224) またはAvr9を発現するC. fulvumに対して耐性のCf-9-含有トマト植物の4個体 (Hcr9-9C (Cf-9)で形質転換したCf0植物) (Jones et al., 1994, Science 266, 789-793) の何れかを使用した。コントロールとして、TRV:00- またはTRV:NRC1- の何れかを接種され、C. fulvumに完全に感受性を有するCf0トマト植物 (MM-Cf0) を使用した。病気アッセイのために、TRV接種から3週間後に、Cf0およびCf-4-含有植物にC. fulvumを接種した (De Wit, 1977, Neth. J. Plant Path. 83, 109-122)。C. fulvum品種5- pGPD::GUSを使用した (構成的GPDプロモーターの制御下でAvr4およびβ-グルクロニダーゼ遺伝子を発現する)。小葉のコロニー形成は、X-gluc 染色により2週間後に評価した。平行して、植物の第二、第三または第四複葉の小葉をRT-PCR分析のために使用して、対象遺伝子の‘ノックダウン’について試験した (下記参照)。HRアッセイのために、TRV-感染のCf-4-またはCf-9-含有植物の第三複葉の小葉に、それぞれ、Avr4またはAvr9を注入した。何れの誘導因子も、マイクロシリンジで小葉に注入した (Ito Corporation, Fuji, Japan)。Avr4は、10μMの濃度で、小葉あたり10の部位に、植物あたり4つの小葉に注入した。Avr9については、(C. fulvumの品種5とCf0植物との適合性相互作用から分離される)約10μMのAvr9を含有する8倍希釈のアポプラスト液を、小葉あたり8つの部位に、植物あたり4つの小葉に注入した。Pseudomonas syringae pv. tomatoに対する耐性は、TRV:00、TRV:PrfまたはTRV:NRC1を接種されたトマトRG-PtoR (Pto/Pto, Prf/Prf) においてアッセイした。接種の手順および葉の細菌コロニー形成の決定は、以前に記載されるとおり行った (Ekengren et al., 2003, supra)。
【0137】
1.3 バイナリー35S:NRC1ベクターの構築および突然変異誘発
全長NRC1 cDNAは、プライマー
222-Start-F (5’-GGGATCCATGGTTGATGTAGGGGTTGA-3’) および
222-Stop-R (5’-GTCACTGCAGACCTTTCTAAGAAGCTGTCTG-3’)
を使用してPCR増幅し、これによりそれぞれNcoIおよびPstI制限部位を導入した (制限部位に下線を付す)。PCRフラグメントは、NcoI/PstI-消化し、NcoI/PstI-消化pRH80に挿入した (Van der Hoorn et al., 2000, Mol. Plant-Microbe Interact. 13, 439-446)。その後、構築物をXbaI/KpnI-消化し、35Sプロモーター、NRC1オープンリーディングフレームおよびNOSターミネーター (tNOS) を含有する得られたフラグメントを、XbaI/KpnI-消化pMOG800バイナリーベクターにクローニングして (Honee et al., 1998, Plant Physiol. 117, 809-820)、プラスミドNRC1(wt)を作成した。構成的に活性なバイナリーNRC1D481Vを作成するため、D481V突然変異は、鋳型としてNRC1wtプラスミドを使用し、フランキングプライマー222-Start-Fおよび222-Stop-Rおよびミスマッチプライマー
222MHD-F (5’-CAAAACTTGTCGTGTTCATGTCATGTTGTATGAG-3’) および
222MHD-R (5’-CCAGCAAAACTCATACAACATGACATGAACACGAC-3’) (突然変異に下線を付す)
を使用して、オーバーラップ伸長PCRにより導入した (Higuchi et al., 1988, Nucleic Acids Res. 16, 7351-7367)。上述のとおり、フラグメントをNcoI/PstI-消化し、pRH80に挿入し、35S-NRC1D481V-tNOSフラグメントを切出し、その後pMOG800に挿入した。同様の方法で、P-ループ突然変異体NRC1K191Rおよび不活性なダブル突然変異体NRC1K191R/D481Vを作成した。ここで、K191R突然変異は、鋳型としてそれぞれプラスミドNRC1(wt)およびNRC1D481Vを使用し、ミスマッチプライマー
222Ploop-F (5’-GGAATGCCTGGTCTTGGCAGAACTACACTAGC-3’) および
222Ploop-R (5’- GCTAGTGTAGTTCTGCCAAGACCAGGCATTCC-3’) (突然変異に下線を付す)
を使用して導入した。全ての構築物は、配列を確認し、A. tumefaciens株GV3101に形質転換した。
【0138】
1.4 DNAゲル-ブロット分析
N. benthamiana由来のゲノムDNAは、QIA-Gen DNA抽出プロトコール (Qiagen, Venlo, NL) を使用して単離し、トマトについては、(Sambrook and Russell, 2001, Molecular cloning: A Laboratory Manual, 3rd ed. (Cold Spring Harbor, NY, U.S.A.: Cold Spring Harbor Laboratory Press) に記載される標準プロトコールを使用した。DNAは、BamHI、HindIII、EcoRI、EcoRVまたはXbaIで消化した。N. benthamianaのゲル-ブロットは、TRV:NRC1ベクターに存在するthe 32P-標識 (Prime-a-gene Labeling System, Promega, Madison, WI) 252 bpフラグメントとハイブリダイズさせ、トマトのDNAゲル-ブロットは、全長NRC1 cDNAの1876〜3168ヌクレオチドに対応する1293塩基の32P-標識プローブとハイブリダイズさせた。使用した制限酵素の部位は、プローブに存在しない。低ストリンジェンシーは、2x SSCおよび0.5% SDS中の55℃での洗浄を指す。高ストリンジェンシー条件は、0.5x SSCおよび0.5% SDS中の65℃での洗浄から構成される。
【0139】
1.5 トマトにおけるNRC1のサイレンシングを示すためのRT-PCR
4つの葉ディスク (トータルで約100 mgの組織) を、TRV-感染植物の第二、第三または第四複葉から集めた。トータルRNAを、QIA-Gen RNAeasy抽出プロトコール (Qiagen, Venlo, NL) を用いて抽出し、RNase-Free DNase (Bio-Rad, Veenendaal, NL) で処理した。第一鎖cDNAは、1μgのトータルRNAから、Bio-Rad cDNA合成キット (Bio-Rad, Veenendaal, NL) を用いて合成し、RT-PCRは、以下のサイクル:95℃で15秒、60℃で45秒および72℃で60秒を使用して行った。使用されたプライマー
(222F: 5’- TGAGGTATATTGCTTTCTCATCTGAC-3’ および
222R: 5’-AGCTATTTTCCCACGGATGCCCAG-3’)
は、TRV:NRC1に挿入されるフラグメントを対象としない。アクチンプライマー
(ActinFnr182: 5’-TATGGAAACATTGTGCTCAGTGG-3’ および
ActinRnr183: 5’-CCAGATTCGTCATACTCTGCC-3’)
は、PCR反応において等量のcDNAの存在をチェックするために使用した。
【0140】
例2 − 結果
2.1 トマトNRC1;CC-NB-LRRタンパク質
cDNA-AFLP分析を行い、その後、トマトの同定されたフラグメントのVIGSをN. benthamiana:Cf-4において行った。VIGSがCf-4/Avr4-誘導性HRに影響を及ぼす20のcDNAフラグメントが同定された。これらの一つであるNRC1について、配列番号3に示されるとおり、全長cDNAを単離した。オープンリーディングフレームは、配列番号1に示され、これは、配列番号2に示されるNRC1タンパク質をコードする。
【0141】
NRC1タンパク質(配列番号2)の推定される一次構造は、CC-NB-LRR耐性タンパク質のものと典型的に類似している(図1)。NRC1は、アミノ末端の二重コイル (CC) ドメイン、NB-ARC (Apaf-1、Rタンパク質およびCED4により共有されるヌクレオチド結合アダプター) ドメイン (Van der Biezen and Jones, 1998, Curr. Biol. 8, R226-R227; Aravind et al., 1999, Trends Biochem. Sci. 24, 47-53)、および13の不完全なロイシン-リッチ繰返し配列 (LRRs) を有する。図1に示されるとおり、相同なNB-ARCドメインとの比較により、Kinase1AまたはP-loopモチーフ、4つのRNBS (Resistance Nucleotide Binding Site) モチーフ、並びにGLPLおよびMHDモチーフの存在が明らかにされた (Meyers et al., 1999, Plant J. 20, 317-332; Meyers et al., 2003, Plant Cell 15, 809-834)。
【0142】
VIGS のために使用されるTRV:NRC1ベクターに存在する252 bpのcDNA-AFLPフラグメントは、599-681アミノ酸をコードし、これはLRRs 4〜7に位置する。
【0143】
BamHI-、HindIII-、EcoRI-、EcoRV-およびXbaI-で消化されたトマトのゲノムDNAの低ストリンジェンシーDNAゲル-ブロット分析は、プローブとして、TRV:NRC1-、TRV:NRC1-LRR-およびTRV:NRC1-UTR構築物 (下記参照) に存在するNRC1配列をカバーする1293 bpのNRC1 cDNAフラグメント (配列番号3の1876〜3168ヌクレオチド) とハイブリダイズさせた。このサザンブロットにより、高ストリンジェンシー洗浄後にたった一つの目立ったバンドが出現し、これは、NRC1がトマトにおいて単一コピー遺伝子であることを示す。
【0144】
BamHI-、HindIII-、EcoRI-、EcoRV-およびXbaI-で消化されたN. benthamianaのゲノムDNAのゲルブロットは、TRVベクターに存在するトマトNRC1 cDNA-AFLPフラグメントをプローブとし、2〜3のハイブリダイズするバンドが見出された (結果は示さず) (0.5 x SSC, 0.5% SDS, 65℃)。これにより、TRV:NRC1の接種によりサイレンシングされ得る少なくとも2〜3のNRC1オルソログが、N. benthamianaのゲノムに存在することが示唆される。
【0145】
2.2 NRC1-サイレンシングされたトマトは、Cf-4-介在性HRおよび耐病性に影響を受ける
HR-シグナリングおよびC.fulvumに対する耐性におけるNRC1の機能を調査するため、本発明者らは、この植物がこの真菌の唯一の宿主であるため、トマトにおいてVIGSを行った。10日齢のトマト芽生えに、TRV:NRC1をアグロインフィルトレーションし、インフィルトレーションから3週間後に、サイレンシングされた可能性のある小葉からRNAを単離し、RT-PCRにより分析した。NRC1転写産物レベルは、異なるTRV:NRC1-感染植物では変動するが、ほとんどの場合、TRV:00-感染植物より低く、これは、NRC1発現の‘ノックダウン’が起こったことを示す(データ示さず)。
【0146】
我々がトマトで観察する表現型が、別のNB-LRRタンパク質のサイレンシングにより引き起こされる可能性を排除するため、我々は、LRRs 8〜12をターゲットとするNRC1の360 bpフラグメント (TRV:NRC1-LRR)、およびNRC1の3’’-非翻訳領域 (UTR) の297 bpからなるフラグメント (TRV:NRC1-UTR) を用いて、トマトにおいてVIGSを更に行った。これらの構築物を用いて、我々は、TRV:222-LRRおよびTRV:222-UTR-感染Cf-4-含有トマト植物にAvr4タンパク質を注入することにより、NRC1が、トマトにおけるCf-4-介在性HRに必要であるかどうかを試験した。(3つの構築物の各々を用いた) NRC1のサイレンシングは、トマト植物が、TRV:00-またはTRV:Cf-4-感染植物よりいくらか小さかったため、軽い表現型という結果につながる (データ示さず)。コントロールとして、TRV:00-およびTRV:Cf-4-感染植物に、Avr4タンパク質を注入した。TRV:Cf-4-感染植物において、Avr4-注入部位の応答パーセンテージは52% (図2) であり、これは、Cf-4のサイレンシングによりHRが減少することを示した。TRV:222-LRRおよびTRV:222-UTR-感染植物において、このパーセンテージは類似しており (それぞれ56%および48%) (図2)、これにより、NRC1がCf-4/Avr4-誘導性HRに機能することがトマトにおいても確認された。Cf-9-含有トマトにおけるNrc1のVIGSおよびその後のAvr9含有アポプラスト液の注入により、同様の結果が得られた(図示せず)。404 bpのCf-4フラグメントは、高度に保存されたLRRs 15〜21をコードし、これが、Cf-4と相同Cf-9耐性遺伝子の両方のサイレンシングを可能にするため、Cf-9-含有トマトにおけるCf-9のVIGSのために、我々はTRV:Cf-4構築物を使用した (Van der Hoorn et al., 2001, supra)。
【0147】
更に、NRC1が、トマトのC. fulvumに対する完全な耐性にとっても必要であるかどうか調査した。Cf0およびCf-4-植物にTRV:00、TRV:Cf-4およびTRV:NRC1を接種し、3週間後、サイレンシングされた植物に、Avr4およびβ-グルクロニダーゼ (GUS) 遺伝子を発現するC. fulvumの株を接種し、これにより真菌の増殖を視覚化した。C. fulvumの接種から2週間後、葉をX-glucで染色した。TRV:00を感染させたCf-4植物の小葉では、C. fulvumの増殖は検出されなかったが、TRV:Cf-4-感染Cf-4植物において青色染色のパッチは、低下した(compromised)Cf-4-介在性耐性を示す (図示せず)。またTRV:NRC1-感染植物においても、青色染色の小さいパッチは、真菌に対する完全な耐性が失われたことを示す。顕微鏡分析により、TRV:Cf-4-およびTRV:NRC1-感染植物において真菌の菌糸の細胞間増殖が明らかにされたが、TRV:00-感染コントロール植物では明らかにされなかった。Cf0植物はすべて、C. fulvumによる広範なコロニー形成を示し、これは、TRV感染自体も、TRV:NRC1を用いたVIGSも、これら植物の真菌に対する感受性に影響を及ぼさないことを示す。
【0148】
2.3 NRC1のVIGSは、異なるマッチングR遺伝子/Avr遺伝子の組合せにより誘導されるHRに影響を及ぼす
NRC1を用いたVIGSによるCf-4/Avr4-誘導性HRの減少に加えて、卵菌の病原体Phytophthora infestansのInf1誘導因子により誘導されるHRも、N. benthamianaにおいて、NRC1を用いたVIGSにより減少することが見出された。防御シグナリングにおけるNRC1の特異性を更に調査するため、本発明者らは、別のR/Avrの組合せにより誘導されるHRにNRC1が必要であるかを試験した。SGT1は、幾つかのR/Avrの組合せにより誘導されるHRに必要であることが知られているため、コントロールとしてTRV:00 (空のベクター) およびTRV:SGT1を含めた (Peart et al., 2002, Proc. Natl. Acad. Sci. USA 99, 10865-10869)。
【0149】
TRV:NRC1-感染N. benthamianaに、Cf-9およびAvr9の混合物 (Van der Hoorn et al., 2000, supra)、またはLeEix2およびtvEixの混合物 (Ron and Avni, 2004, Plant Cell 16, 1604-1615) をアグロインフィルトレーションすると、HRは減少したが、TRV:00-感染植物では、HRは普通に発生した。TRV:SGT1-感染植物においてHRは完全になくなり、これは、Peart et al. (2002, supra) の観察を確認する (図3)。また、細菌の病原体Pseudomonas syringae pv tomatoに由来するAvrPtoおよびポテトウイルスX (PVX) のコートタンパク質 (CP) をコードする遺伝子を、耐性遺伝子Pto (Pedley and Martin, 2003, Annu. Rev. Phytopathol. 41, 215-243) およびRx (Bendahmane et al., 1999, Plant Cell 11, 781-791) をそれぞれ発現するTRV-感染N. benthamianaにアグロインフィルトレーションした。両ケースにおいて、TRV:00を感染させた植物はHRを示したが、TRV:SGT1-感染植物でHRはなくなった。TRV:NRC1-感染により、Pto/AvrPto-およびRx/CP-誘導性HRは大幅に抑制され、これは、N. benthamianaにおいてNRC1タンパク質が、幾つかのR/Avrの遺伝子対遺伝子(gene-for-gene)の組合せにより活性化されるHRシグナリングに必要であることを示す (図3)。
【0150】
TRV:NRC1-感染N. benthamianaにおいて低下した(compromised)HRは、アグロバクテリウムによる形質転換効率の減少に由来するという可能性を排除するため、本発明者らは、TRV:00-およびTRV:NRC1-感染N. benthamiana:Cf-4に、β-グルクロニダーゼ (GUS) 遺伝子を発現するアグロバクテリウムをインフィルトレーションした (Van der Hoorn et al., 2000, supra)。インフィルトレーションから3日後、TRV:00-およびTRV:NRC1-感染植物における同程度の強度の青色染色により、アグロバクテリウムによる植物の形質転換効率は影響を受けないことが明らかにされた (データ示さず)。更に、TRV:NRC1-感染植物も、Avr4タンパク質の注入によりHRの減少を示したが、TRV:00-感染植物では、明らかなHRが2日以内に発生した。
【0151】
2.4 NRC1は、細胞死シグナリング経路においてEDS1の下流およびMAPKカスケードの上流に作用する
NRC1は、Cf-4/Avr4-誘導性HRだけでなく、幾つかの別のR/Avrの組合せにより誘導されるHRにも必要であるため、NRC1は、共通のHRシグナリング経路に関与していると思われる。HRの開始前に起こる典型的な宿主応答には、MAPKカスケードの活性化が含まれる (Romeis et al., 2001, EMBO J. 20, 5556-5567; Del Pozo et al., 2004, EMBO J. 23, 3072-3082; Pedley and Martin, 2005, Plant Biol. 8, 541-547)。
【0152】
MAPKsにより開始されるHRにNRC1が必要であるか調査するため、N. benthamianaにおいてエピスタシス実験を行った。植物に、TRV:00, TRV:SGT1およびTRV:NRC1を接種し、その後、LeMAPKKKα (LeMAPKKKαKD) のキナーゼドメインまたは構成的に活性なLeMEK2 (LeMEK2DD) をコードする遺伝子をアグロインフィルトレーションした (Yang et al., 2001, Proc. Natl. Acad. Sci. USA 98, 741-746; Del Pozo et al., 2004, EMBO J. 23, 3072-3082.)。アグロインフィルトレーションから2日後、インフィルトレーションされた葉にエストラジオールをスプレーすることにより遺伝子の発現を誘導した。各遺伝子の一過性発現は、TRV:00-感染植物においてHRという結果につながるが、TRV:SGT1-感染植物においてHRは減少する (図4A)。TRV:NRC1-感染植物では、構成的に活性な何れのキナーゼにより引き起こされるHRも影響を受けない (図4A)。対応するネガティブコントロール、LeMAPKKKαKD- および野生型LeMEK2のアグロインフィルトレーションは、TRV-感染植物の何れにおいてもHRという結果につなからなかった (データ示さず)。これらの結果は、SGT1が、これらMAPKsの下流で機能するが、MAPKsは、NRC1の下流で作用するか、あるいはNRC1とは独立して作用することを示す。
【0153】
2.5 NRC1の一過性過剰発現および構成的に活性なNRC1タンパク質の構築
遺伝子が、CC-NB-LRRタンパク質によるHRシグナリングに必要であるか更に調査するため、NRC1の過剰発現の効果を調査した。したがって、cDNAのコード配列 (配列番号1) を、構成的35Sプロモーターに融合し、バイナリーベクターに挿入した。N. benthamianaにおいてこの構築物のアグロインフィルトレーションは、HRという結果につながらなかったが、NRC1およびp19サイレンシング阻害剤の混合物の発現 (Voinnet et al., 2003, Plant J. 33, 949-956) は、誘導因子-非依存性HRを引き起こした (図4B)。P-loopモチーフを破壊することによりATP加水分解に影響を及ぼすNRC1のP-loop突然変異体 (K191R) をコードする構築物のアグロインフィルトレーション (Tameling et al., 2002, Plant Cell 14, 2929-2939) は、p19が存在しても存在しなくても、HRという結果につながらなかった (図4B)。
【0154】
上記データは、NRC1遺伝子の翻訳後遺伝子サイレンシング (PTGS) が、NRC1過剰発現組織においてHRの発生を防ぎ得ることを示した。また、P-loopモチーフの破壊は、非機能的NRC1タンパク質という結果につながる。
【0155】
NB-LRR耐性タンパク質RxのMHDモチーフにおける突然変異 (D460V) (Bendahmane et al., 2002; Tameling et al., 2002) およびNB-LRR耐性タンパク質I-2のMHDモチーフにおける突然変異 (D495V) (Bendahmane et al., 2002, Plant J. 32, 195-204; Tameling et al., 2002, Plant Cell 14, 2929-2939; Van Bentem et al., 2005, Plant J. 43, 284-298) は、構成的活性という結果につながるため、本発明者らは、NRC1の同様の突然変異体 (NRC1D481V) を作成した。実際、NRC1D481Vのアグロインフィルトレーションは、インフィルトレーションから3日以内に、N. benthamianaの葉において誘導因子-非依存性HRという結果につながり、また、ダブル突然変異体NRC1K191R/D481V のアグロインフィルトレーションにより、HRは観察されなかった (図4B)。更に、SGT1-サイレンシング植物において、NRC1D481Vの発現によりHRは誘導されなかった (下記参照)。これらの結果は、NRC1D481Vのアグロインフィルトレーションにより誘導される応答は、特異的に、NRC1タンパク質の構成的活性によるものであること、並びにNRC1は、HRにつながるシグナル伝達カスケードにおいて機能することを示す。
【0156】
2.6 構成的に活性なNRC1タンパク質を用いたエピスタシス実験
NRC1D481Vを使用するエピスタシス実験を行い、このタンパク質によるHRシグナリングに遺伝子が必要であるか更に調査し、これにより、HR経路におけるその推定位置を決定する。HRシグナリングに一般に関与することが知られている遺伝子、たとえばSGT1およびRAR1 (Required for Mla12 resistance) (Shirasu and Schulze-Lefert, 2003, Trends Plant Sci. 8, 252-258) のVIGSに加えて、NDR1 (non race-specific disease resistance) (Century et al., 1995, Proc. Natl. Acad. Sci. USA 92, 6597-6601)、EDS1 (enhanced disease susceptibility) (Aarts et al., 1998, Proc. Natl. Acad. Sci. USA 95, 10306-10311)、およびMEK2 (a MAPKK) (Ekengren et al., 2003, Plant J. 36, 905-917) について、N. benthamiana:Cf-4をサイレンシングし、その後、NRC1D481VまたはAvr4を用いてアグロインフィルトレーションした。更に、TRV:00、TRV:Cf-4およびTRV:NRC1を用いたVIGSを、コントロールとして含めた。低下した(compromised)NRC1D481V-またはAvr4-誘導性HRは、NRC1-またはCf-4/Avr4-誘導性HRシグナリングのそれぞれに必要な遺伝子の‘ノックダウン’を示す。
【0157】
予測されるとおり、Avrのアグロインフィルトレーションにより誘導されるHRは、TRV:Cf-4-およびTRV:NRC1-感染植物において低下した(compromise)。また、この遺伝子についてサイレンシングされた植物が、それほどひどくないAvr4-誘導性HRを示すため、Cf-4-介在性シグナリングもEDS1を必要とする。更に、本発明者らは、MEK2、RAR1およびSGT1についてサイレンシングされた植物において、Avr4のアグロインフィルトレーションによりHRが減少することを見出した (図4C;薄い円)。Avr4-誘導性HRは、TRV:00-およびTRV:NDR1-感染植物において低下せず(not compromise)(図4C;濃い円)、これは、NDR1がCf-4-介在性シグナリングに必要でないことを示す。同様に、NRC1D481V-誘導性HRは、TRV:00-およびTRV:NDR1-感染植物において低下せず(not compromise)、TRV:Cf-4-感染植物においても低下しなかった。興味深いことに、Avr4とは対照的に、NRC1D481Vは、TRV:EDS1-感染植物においてもHRを誘導し、これは、NRC1がEDS1の下流で機能的であることを示す (図4C;濃い円)。NRC1D481V-誘導性HRは、MEK2についてサイレンシングされた植物において低下し(compromise)、これは、NRC1が、そのシグナリングのためにMAPキナーゼカスケードを必要とし、これらキナーゼの上流に位置することができることを示す。また、RAR1およびSGT1のVIGSも、Avr4により誘導されるHRと同様、D481V-誘導性HRを低下させる(compromise)(図4C;薄い円)。このため、NRC1は、Cf-4により開始されるHRシグナリングに必要であり、MAPKカスケードの上流およびEDS1の下流に位置することができる。
【0158】
NRC1介在性細胞シグナリングのモデルについては、図5を参照されたい。
【0159】
例3 − Mi-介在性耐性に対するNRC1の必要性
NRC1が、線虫、コナジラミおよびアブラムシに対するMi-介在性耐性に必要であるか否かを決定するため、Miの構成的に活性な型 (US 6613962およびEP0937155B1参照) を、NRC1サイレンシング植物にアグロインフィルトレーションする。NRC1サイレンシング植物におけるHRの減少は、NRC1が、Mi-介在性HRにも必要であること、並びにNRC1の(過剰)発現が、線虫、コナジラミおよびアブラムシに対して向上した耐性を有するトランスジェニック植物を作成するために使用することができることを示す。
【図面の簡単な説明】
【0160】
【図1】NRC1タンパク質の推定配列を示す図。
【図2a】NRC1は、Cladosporium fulvumに対するトマトの完全なCf-4-介在性HRに必要であることを示す図。
【図2b】NRC1は、Cladosporium fulvumに対するトマトの完全なCf-4-介在性耐性に必要であることを示す写真。
【図3】N. benthamianaへのTRV:NRC1の接種は、Cf/Avr-、LeEix2/tvEix-、Pto/AvrPto-およびRx/CP-誘導性HRに影響を及ぼすことを示す写真。
【図4a】構成的に活性なNRC1は、誘導因子−非依存性HRを誘導し、細胞死シグナリング経路にNRC1を配置させることを示す写真。構成的に活性なMAPKKおよびMAPKキナーゼをコードする遺伝子のアグロインフィルトレーション。
【図4b】構成的に活性なNRC1は、誘導因子−非依存性HRを誘導し、細胞死シグナリング経路にNRC1を配置させることを示す写真。35S-プロモーターの制御下の野生型NRC1 (wt) および当該遺伝子の突然変異型のアグロインフィルトレーションであって、サイレンシングサプレッサーp19をコードする遺伝子の発現を駆動するアグロバクテリウムと1:1の比で混合されたアグロインフィルトレーション (左パネル)、または単独のアグロインフィルトレーション (右パネル)。
【図4c】構成的に活性なNRC1は、誘導因子−非依存性HRを誘導し、細胞死シグナリング経路にNRC1を配置させることを示す写真。Avr4および構成的に活性なNRC1D481V (D481V) をコードする遺伝子のアグロインフィルトレーション。
【図5】NRC1介在性細胞死シグナリングのモデルを示す図。
【発明の分野】
【0001】
本発明は、ゲノムに統合されてNRC1タンパク質(NB-LRR Required for HR-associated Cell Death 1)をコードする遺伝子を含むトランスジェニック植物および植物細胞、並びにかかる植物および細胞を作成するための方法に関する。とりわけ、向上した耐病性を備えたナス科(Solanaceae)植物および植物の一部(種子、果実、葉など)が提供される。また、本発明のNRC1タンパク質をコードする単離された核酸分子、これらを含むベクター、並びに単離されたNRC1タンパク質自体が提供される。更に、植物および植物細胞に耐病性を付与する一または複数の突然変異を内在性NRC1対立遺伝子に含む植物細胞および植物が提供される。
【発明の背景】
【0002】
植物の積極的防御は、耐性遺伝子により仲介される病原体の無毒性因子の認識により誘発され、遺伝子対遺伝子(gene-for-gene)モデルに従う(Dangl and Jones, 2001, Nature 411, 826-833)。今日までに幾つかの植物耐性遺伝子(R遺伝子)がクローニングされており、これらは、コードするタンパク質の構造に基いて、幾つかのグループに分類される(Hammond-Kosack and Jones, 1997, Annu. Rev. Plant Physiol. Plant Molec. Biol. 48, 575-607)。ほとんどのR遺伝子は、ヌクレオチド結合部位(NB)およびロイシンリッチ繰返し配列(LRR)を含有する細胞質NB-LRRタンパク質をコードする。このグループは、二重コイル(coiled−coil)ドメインを含有するCC-NB-LRRタンパク質をコードする遺伝子、および哺乳類のTollおよびインターロイキン(IL)レセプターに類似のドメインを有するタンパク質、いわゆるTIR-NB-LRRタンパク質をコードする遺伝子からなる(Hammond-Kosack and Jones, 1997, supra)。
【0003】
このような特定の耐性遺伝子を永続的耐性のための育種プログラムで使用することは、病原体が、無毒性因子の突然変異により認識を容易に回避し、これにより積極的防御の誘導を妨害するため、問題がある(Westerink et al., 2004, Mol. Microbiol. 54, 533-545)。耐性タンパク質(Rタンパク質)の間の類似性は、共通の耐性経路の存在を示唆する(Shirasu and Schulze-Lefert, 2000, Plant Mol. Biol. 44, 371-385)。したがって、耐性に必要な追加の遺伝子を同定することにより、かかるシグナリング経路がどのように機能するかについて情報が提供されるだけでなく、我々が、耐性においてより一般的な役割を果たす遺伝子を同定することも可能になる。たとえば、Nicotiana benthamianaにおけるウイルス誘導性遺伝子サイレンシング(VIGS)により、SGT1が、複数の防御経路、たとえばN-、Rx-およびPto-介在性HRおよび耐性、並びにCf-4-およびCf-9-介在性HRに関与していることが示された(Peart et al., 2002, Proc. Natl. Acad. Sci. USA 99, 10865-10869; Zhang et al., 2004, Plant J. 40, 213-224)。SGT1は、SKP1の相互作用物質であり、タンパク質のユビキチン化、タンパク質を分解の標的にする修飾に関与するSCF E3-リガーゼ複合体の構成成分である(Schwechheimer and Schwager, 2004, Plant Cell Reports 23, 353-364)。このタンパク質分解システムの必須遺伝子をサイレンシングすることは、ユビキチン化プロセスを妨害し、これにより防御の活性化に必要な負の調節因子の分解を阻害すると仮定される(Azevedo et al., 2002, Science 295, 2073-2076)。
【0004】
幾つかの耐性経路において、MAPKs(マイトジェン活性化プロテインキナーゼ)が活性化される(Zhang and Klessig, 2001, Trends Plant Sci. 6, 520-527; Pedley and Martin, 2005, Curr. Opin. Plant Biol. 8, 541-547)。Cf-9-含有タバコ植物およびAvr-9で攻撃された(challenged)細胞培養物において、NtWIPK (傷害誘導性プロテインキナーゼ) およびNtSIPK (サリチル酸誘導性プロテインキナーゼ) が活性化される(Romeis et al., 1999, Plant Cell 11, 273-287)。N. benthamianaにおけるNtCDPK (カルシウム依存性プロテインキナーゼ) のVIGSは、Cf-9/Avr9-およびCf-4/Avr4-依存性HRを阻害し (Romeis et al., 2001, EMBO J. 20, 5556-5567)、トマトにおけるLeACIK1 (Avr/Cf-induced kinase 1) のVIGSは、C. fulvum耐性の減少という結果につながる (Rowland et al., 2005, Plant Cell 17, 295-310)。防御の間のキナーゼの活性化およびキナーゼをコードする遺伝子の「ノックダウン」による耐性の減少は、防御の活性化におけるキナーゼの機能をサポートする。
【0005】
偏りのあるアプローチに従って、防御関連のシグナリングに関与することが公知の21個の遺伝子が、トマトにおいてVIGSのために使用され、これらのうち9個の遺伝子が、Pto-介在性耐性に関与することが見出された。これらのなかに、MAPKKsをコードする2個の遺伝子(LeMEK1およびLeMEK2)、およびMAPKsをコードする2個の遺伝子(LeNTF6およびLeWIPK)が含まれる(Ekengren et al., 2003, Plant J. 36, 905-917)。別の研究において、N. benthamiana cDNAの標準化ライブラリー由来の2400を超えるcDNAをポテトウイルスX-ベースのベクターにクローニングし、N. benthamianaにおいてVIGSのために使用した。cDNAsの約3%が、サイレンシングによりPto-依存性HRに影響を及ぼした。これらのうち、MAPKKKαが、耐性と病気の両方の正の調節因子として同定された(Del Pozo et al., 2004, EMBO J. 23, 3072-3082)。
【0006】
Lu et al. (2003, EMBO J. 22, 5690-5699) は、PVXベクターにクローニングされた標準化N. benthamiana cDNAライブラリー由来の4992個のcDNAを用いてVIGSを行った。cDNAのうち、79個(1.6%)が、Pto-介在性HRに必要な遺伝子に相当し、それらの僅か6個のサイレンシングが、Pseudomonas syringaeに対するPto-介在性耐性を減少させた。HSP90に相当するcDNAを用いたVIGSは、Pto-介在性HRだけでなく、Pto-、Rx-およびN-介在性耐性を破壊し、このことは、HSP90が複数の耐病性経路に必要であることを示す。cDNAの同じセットを、N-トランスジェニックN. benthamianaにおいてVIGSのために使用し、その後、植物にTMVのGFP標識株を接種した。TMVに対する耐性は、NRG1 (N requirement gene 1) と称されるCC-NB-LRRコード遺伝子に由来するcDNAフラグメントを用いたサイレンシングにより最も顕著に抑制された(Peart et al., 2005, Curr. Biol. 15, 968-973)。NRG1は、N遺伝子の機能に特に必要であることが示され、このことは、CC-NB-LRRタンパク質が、無毒性因子の認識に関与する耐性タンパク質として機能するだけでなく、TIR-NB-LRRタンパク質Nにより開始され、最終的に耐性につながるシグナリング経路に関与することを示す(Peart et al., 2005, supra)。このように、タバコのNRG1タンパク質は、耐性タンパク質により開始される植物の防御シグナリングカスケードの下流で機能するが、タバコモザイクウイルス(TMV)に対するN-介在性耐性に特異的に関与し、耐病性の一般的な補助因子でない(PVXおよびPseudomonas syringaeに対するRx-およびPto-介在性耐性は、NRG1サイレンシングにより影響を受けない)という欠点を有し、これにより、トマトなどの作物において広範囲の病原体耐性をつくるのに適していないかもしれない。
【0007】
耐病性経路についての情報は増えているが、永続的で広範囲の耐病性を備えた植物をつくるために使用することができる遺伝子およびタンパク質を同定する需要がなお存在する。向上した耐病性を備えた植物、とりわけナス科に属する植物をつくるためのかかる核酸、タンパク質および方法を提供することが本発明の課題である。
【一般的な定義】
【0008】
“HR”は、高感受性応答、すなわち(Rivas and Thomas, 2005, Ann Rev Phytopath 43: 395-436により記載される)顕微鏡的な病変および/または肉眼的な病変として観察される局所的な細胞死を指す。高感受性細胞死は、通常、その他の植物応答、たとえば反応性酸素種の産生およびHR病変を囲む細胞における防御関連遺伝子の活性化などと関連がある。
【0009】
“植物病原体”は、植物に病気を引き起こすことができる生物薬剤、たとえば植物病原性の真菌、細菌、ウイルス、卵菌類、マイコプラズマ様生物、線虫、コナジラミおよびアブラムシなどを指す。一般に、宿主組織に病気を引き起こすことができる病原体種の株、品種またはパソバールのすべてが、本明細書において包含される。
【0010】
“生物栄養性(biotrophic)植物病原体”または“生物栄養体(biotroph)”は、植物細胞を生きたまま維持し、増殖および組織コロニー化のために生細胞に依存する病原体を指す。
【0011】
“半生物栄養性(hemibiotrophic)植物病原体”または“半生物栄養体(hemibiotroph)”は、少なくとも生活環の一部の期間、宿主細胞を生きたまま維持する植物病原体を指す。
【0012】
“壊死栄養性(necrotrophic)植物病原体”は、宿主細胞を殺傷する毒性酵素、タンパク質または代謝産物を産生することにより、組織コロニー化により植物細胞を積極的に殺傷する植物病原体を指す。
【0013】
“誘導因子非依存性HR”は、病原体または病原体の誘導因子(たとえば真菌のAvrタンパク質)の存在なしで発生する高感受性応答を指す。
【0014】
本発明のNRC1タンパク質(たとえば、構成的に活性なNRC1タンパク質)を発現する植物を参照する場合、病原体または病原体の誘導因子タンパク質の非存在下でのHR病変の発生を言及する“構成的HR”と、誘導性刺激の存在後(たとえば、NRC1タンパク質またはその変異体をコードする核酸配列の発現を駆動するプロモーターの誘導後)のHR病変の発生を言及する“誘導的HR”とを区別してもよい。
【0015】
“ナス科(Solanaceae)”は、本明細書において、ナス科に属する植物の属、種、およびその品種を指す。これらには、ナス属(Solanum)(以前はLycopersicon esculentumとして知られていたSolanum lycopersicumを含む)、タバコ属(Nicotiana)、トウガラシ属(Capsicum)、ペチュニア属(Petunia)、およびその他の属が含まれる。
【0016】
“耐病性”は、本明細書において種々のレベルの植物の病気抵抗性または耐性を指し、一または複数の病原体に対する中程度な抵抗性および高度な抵抗性または完全な抵抗性を含む。耐病性は、病原体により引き起こされる徴候(たとえばHR病変、真菌の菌糸体などの頻度および/またはサイズ)を、同じ病気プレッシャーの下で生育させたときの感受性コントロール植物で観察される徴候と比較することにより測定し、必要に応じて定量することができる。かかる病気バイオアッセイは、公知の方法を用いて行うことができる。また、耐病性は、病気プレッシャーの下で生育させたときの、感受性植物と比較して耐性植物の高い収量により間接的に測定することもできる。
【0017】
“向上した耐病性”は、適切なコントロールと比較して、植物または植物組織の耐病性が統計的に有意に増大することを指す。本明細書において、(たとえば感受性から抵抗性までの)定性的増大および定量的増大の両方が包含される。また、病気の発生率(感染している植物のパーセンテージ)の低下および/または病気の重症度の低下の両方が包含される。好ましくは、少なくとも一つの病原体に対して向上した耐病性を有する植物は、耐病性を評価するための適切なバイオアッセイおよび/またはフィールドアッセイを用いて、コントロール植物より病原体に対して少なくとも1%、2%、5%、10%、15%、20%、30%、50%、70%、80%、90%、または100%もの高いレベルの耐性を備えた植物である。
【0018】
“広範囲(broad spectrum)”の耐病性は、少なくとも2、3、4、またはそれ以上の異なる病原体種の病原体に対して向上した耐性を指す。たとえば、幾つかの生物栄養性および/または半生物栄養性および/または壊死栄養性の病原体種に対して向上した耐性を有する宿主植物は、広範囲の耐性を有すると考えられる。
【0019】
“病原体により引き起こされる徴候”には、病気の任意の徴候、たとえば宿主組織の上/中での菌糸体の増殖/バイオマス、細菌の増殖/バイオマス、植物組織上での壊死性または退緑性病変のサイズおよび/または頻度、潰瘍のサイズおよび/または頻度などが含まれる。
【0020】
“核酸配列”(または核酸分子)の用語は、一本鎖または二本鎖の標準的な形態のDNAまたはRNA分子、とりわけ本発明のタンパク質またはタンパク質フラグメントをコードするDNAを指す。“単離された核酸配列”は、自然環境から単離され、もはや自然環境に存在しない核酸配列、たとえば細菌の宿主細胞または植物の核もしくはプラスチドゲノムに存在する核酸配列を指す。
【0021】
“タンパク質”または“ポリペプチド”の用語は、交換可能に使用され、アミノ酸の鎖から成る分子を指し、具体的な作用形態、サイズ、3次元構造または由来を問わない。このため、タンパク質の“フラグメント”または“一部”は、なお“タンパク質”を指してもよい。“単離されたタンパク質”は、自然環境にもはや存在しない、たとえばインビトロまたは組換え細菌または植物の宿主細胞に存在する、タンパク質を指すために使用される。
【0022】
NRC1タンパク質(または変異体、たとえばオルソログまたは突然変異体、およびフラグメント)に関して“機能的”とは、植物においてNRC1コード遺伝子の発現レベルを改変することにより(たとえば過剰発現またはサイレンシングにより)、HR病変の(定量的および/または定性的)発生および/または耐病性のレベルを改変する能力を指す。たとえば、X植物種から得られる推定NRC1タンパク質の機能性は、種々の方法により試験することができる。当該タンパク質が機能的である場合、たとえばVIGSもしくは遺伝子サイレンシングベクターを用いて、X植物種で当該タンパク質をコードするNRC1遺伝子をサイレンシングさせると、トマトについての実施例で示されるとおり、病原体もしくは誘導因子誘発性HR病変の減少または抑制、および/または病原体耐性の低下という結果につながる。また、機能的なNRC1タンパク質を補足すると、HR病変および/または病原体耐性を回復させることができる。あるいは、(必要に応じて、転写後遺伝子サイレンシング阻害剤と一緒に)NRC1タンパク質をコードする遺伝子をX種で一過性もしくは安定(過剰)発現させると、誘導因子非依存性HR病変の発生、および/またはとりわけ生物栄養性および/または半生物栄養性病原体に対して向上した耐病性という結果につながる。実施例も参照されたい。
【0023】
“遺伝子”の用語は、適切な調節領域(たとえばプロモーター)に動作可能に連結された、細胞においてRNA分子(たとえばmRNA)に転写される領域(転写領域)を含むDNA配列を意味する。このため、遺伝子は、幾つかの動作可能に連結された配列、たとえば、プロモーター、転写開始に関与する配列などを含む5’リーダー配列、(タンパク質)コード領域(cDNAまたはゲノムDNA)、および転写終結部位などを含む3’非翻訳配列を含んでいてもよい。
【0024】
“キメラ遺伝子”(または組換え遺伝子)は、生物種において天然に通常見出されない任意の遺伝子、とりわけ、天然において互いに関連がない核酸配列の一または複数の部分が存在する遺伝子を指す。たとえば、プロモーターは、天然において、転写領域の一部もしくは全部、または別の調節領域と関連がない。“キメラ遺伝子”の用語は、プロモーターまたは転写調節配列が、一もしくは複数のコード配列、またはアンチセンス(センス鎖のリバース相補鎖)もしくは逆方向反復配列(RNA転写産物が転写により二本鎖RNAを形成するセンスとアンチセンス)に動作可能に連結された発現構築物を含むと理解される。
【0025】
“3’UTR”または“3’非翻訳配列”(しばしば3’非翻訳領域または3’端とも称される)は、遺伝子のコード鎖の下流に見出される核酸配列を指し、たとえば、転写終結部位および(ほとんどであるが全てではない真核生物mRNAにおいて)ポリアデニル化シグナル(たとえばAAUAAAまたはその変異体)を含む。mRNA転写産物は、転写終結後にポリアデニル化シグナルの下流で切断され、mRNAの(翻訳が起こる)細胞質への輸送に関与するポリ(A)テールが付加され得る。
【0026】
“遺伝子の発現”は、適切な調節領域、とりわけプロモーターに動作可能に連結されたDNA領域が、生物学的に活性なRNA、すなわち生物学的に活性なタンパク質またはペプチド(または活性なペプチドフラグメント)に翻訳され得るRNAまたは(たとえば転写後の遺伝子サイレンシングまたはRNAiにおいて)それ自体で活性なRNAに転写されるプロセスを指す。ある態様において活性なタンパク質は、構成的に活性なタンパク質を指す。コード配列は、好ましくはセンスの向きであり、所望の生物学的に活性なタンパク質またはペプチド、または活性なペプチドフラグメントをコードする。遺伝子サイレンシングアプローチにおいて、DNA配列は、好ましくは、ターゲット遺伝子の短い配列をアンチセンスの向きまたはセンスとアンチセンスの向きで含むアンチセンスDNAまたは逆方向反復DNAの形態で存在する。“異所性発現”は、遺伝子が通常発現しない組織での発現を指す。
【0027】
“転写調節配列”は、本明細書において、転写調節配列に動作可能に連結された(コード)配列の転写速度を調節することができる核酸配列と規定される。このため、本明細書で規定される転写調節配列は、転写の開始に必要な配列エレメント(プロモーターエレメント)、転写の維持および調節に必要な配列エレメント(たとえば、アテニュエーターまたはエンハンサー)の全てを含む。多くの場合、コード配列の上流(5’)転写調節配列が言及されるが、コード配列の下流(3’)に見出される調節配列も、この規定に包含される。
【0028】
本明細書で使用される“プロモーター”の用語は、遺伝子の転写開始部位の転写の方向に対して上流に位置して、一または複数の遺伝子の転写を制御するために機能する核酸フラグメントを指し、DNA依存性RNAポリメラーゼの結合部位、転写開始部位、並びに任意の他のDNA配列(転写因子結合部位、リプレッサーおよびアクチベータータンパク質結合部位、およびプロモーターからの転写量を調節するのに直接または間接的に作用することが当業者に公知の任意の他のヌクレオチド配列を含むがこれらに限定されない)の存在により構造的に同定される。“構成的”プロモーターは、ほとんどの組織でほとんどの生理および発生条件下において活性なプロモーターである。“誘導性”プロモーターは、生理的に(たとえば、ある化合物の外部からの適用により)または発生で調節されるプロモーターである。“組織特異的”プロモーターは、特定タイプの組織または細胞においてのみ活性である。“植物または植物細胞で活性なプロモーター”は、植物または植物細胞の中で転写を駆動するプロモーターの一般的能力を指す。これは、プロモーターの時空的活性について何の意味ももたない。
【0029】
本明細書で使用される“動作可能に連結された”の用語は、機能的関係におけるポリヌクレオチドエレメントの連結を指す。核酸が、別の核酸配列と機能的関係に配置されるとき、“動作可能に連結されている”。たとえば、プロモーター、正確にいえば転写調節配列が、コード配列の転写に影響を及ぼす場合、コード配列に動作可能に連結されている。動作可能に連結されているとは、連結されているDNA配列が、典型的には隣接していることを意味し、二つのタンパク質コード領域を結合する必要がある場合、“キメラタンパク質”を産生するようにリーディングフレームで隣接していることを意味する。“キメラタンパク質”または“ハイブリッドタンパク質”は、種々のタンパク質“ドメイン”(またはモチーフ)から構成されるタンパク質であり、それ自体天然には見出されないが、機能的タンパク質を形成するために結合され、結合されたドメインの機能性を示す(たとえば、二重コイル(coiled−coil)ドメイン、ヌクレオチド結合ドメイン(NB-ARC)およびロイシンリッチ繰返し配列(LRR)領域を組合せることができる)。また、キメラタンパク質は、天然に存在する二つ以上のタンパク質の融合タンパク質であってもよい。本明細書で使用される“ドメイン”の用語は、ドメインの機能的特徴を少なくとも備えた新規ハイブリッドタンパク質を提供するために別のタンパク質に転移させることができる特定の構造または機能を備えたタンパク質の任意の部分またはドメインを意味する。また、特定のドメインは、他のNRC1タンパク質、たとえば他の植物種に由来するNRC1オルソログを同定するために使用することもできる。
【0030】
“ターゲットペプチド”の用語は、タンパク質またはタンパク質フラグメントを、細胞内オルガネラ、たとえばプラスチド、好ましくは葉緑体、ミトコンドリアに向かわせるアミノ酸配列、または細胞外空間もしくはアドプラスド(adoplast)に向かわせるアミノ酸配列(分泌シグナルペプチド)を指す。ターゲットペプチドをコードする核酸配列は、タンパク質またはタンパク質フラグメントのアミノ末端(N-末端)をコードする核酸配列に(フレームで)融合してもよいし、天然のターゲッティングペプチドを置換するために使用してもよい。
【0031】
“核酸構築物”または“ベクター”は、組換えDNA技術の使用により得られ、外因性DNAを宿主細胞にデリバーするために使用される人工の核酸分子を意味すると本明細書において理解される。ベクターの骨格は、当該技術分野で公知であり、本明細書の他の箇所で記載される、たとえばバイナリーもしくはスーパーバイナリーベクター(たとえばUS 5591616、US 2002138879およびWO 95/06722を参照)、コインテグレート(co-integrate)ベクター、またはT-DNAベクターとすることができ、これにキメラ遺伝子が統合されるか、あるいは適切な転写調節配列が既に存在する場合には、所望の核酸配列(たとえばコード配列、アンチセンスまたは逆方向反復配列)のみが転写調節配列の下流に統合される。ベクターは、分子クローニングでの使用を容易にするための更なる遺伝的構成要素、たとえば選択マーカー、マルチクローニングサイトなどを通常含む(下記参照)。
【0032】
“宿主細胞”または“組換え宿主細胞”または“形質転換細胞”は、所望のタンパク質をコードするキメラ遺伝子またはターゲット遺伝子/遺伝子ファミリーのサイレンシングのためのアンチセンスRNAまたは逆方向反復RNA(またはヘアピンRNA)を転写により産生する核酸配列をとりわけ含む少なくとも一つの核酸分子を前記細胞に導入した結果生じる新規な個々の細胞(または生物)を指す用語である。宿主細胞は、好ましくは植物細胞または細菌細胞である。宿主細胞は、核酸構築物を染色体外(エピソーム)複製分子として含んでいてもよいし、より好ましくは、キメラ遺伝子を宿主細胞の核もしくはプラスチドゲノムに統合して含んでいてもよい。また本明細書を通して、“宿主”の用語は、病原体が侵入もしくは感染することが可能な宿主植物種を指してもよいが、このことは、本明細書の文脈から明らかである。植物種は、病原体との関係で、“宿主”または“非宿主”種に分類される。“非宿主”種は、病気発生に最適な条件下においても、病原体のあらゆる品種または株の病原体感染に対して完全に免疫性がある。“宿主”種は、病原体の“宿主範囲”とも呼ばれ、病原体の(全てではないが)ある品種に対して免疫性がある。
【0033】
“選択マーカー”の用語は、当業者によく知られた用語であり、発現されたときに、選択マーカーを含有する一つの細胞または複数の細胞を選択するために使用することができる任意の遺伝的構成要素(entity)を記載するために本明細書で使用される。選択マーカーの遺伝子産物は、たとえば抗生物質耐性、より好ましくは除草剤耐性または別の選択可能な特性、たとえば表現型の特性(たとえば色素形成の変化)または栄養要求性を付与する。“レポーター”の用語は、主に、可視マーカー、たとえば緑色蛍光タンパク質(GFP)、eGFP、ルシフェラーゼ、GUSなどを指すために使用される。
【0034】
遺伝子またはタンパク質の“オルソログ(ortholog)”の用語は、本明細書において、別の種で見出される相同な遺伝子またはタンパク質を指し、これは、当該遺伝子またはタンパク質と同じ機能を有するが、当該遺伝子を有する種が分岐した時点から配列が(通常)分岐している(すなわち、種分化により共通祖先から進化した遺伝子)。よって、トマトnrc1遺伝子のオルソログは、(たとえば全配列または特定ドメインにわたるパーセンテージ配列同一性に基く)配列の比較と機能分析の両方に基いて、他の植物種で同定され得る。
【0035】
“同種の(homologous)”および“異種の(heterologous)”は、とりわけトランスジェニック生物の文脈において、核酸またはアミノ酸配列とその宿主細胞または生物との関係を指す。よって、同種の配列は、宿主種(たとえばトマト遺伝子で形質転換されたトマト植物)で天然に見出されるが、異種の配列は、宿主細胞(たとえばポテト植物由来の配列で形質転換されたトマト植物)で天然に見出されない。あるいは、文脈に応じて“相同体(homolog)”または“相同な(homologous)”は、共通祖先の配列から伝わる配列を指してもよい(たとえば、これらはオルソログであり得る)。
【0036】
“ストリンジェントなハイブリダイゼーション条件”は、所定のヌクレオチド配列と実質的に同一のヌクレオチド配列を同定するために使用することができる。ストリンジェントな条件は、配列に依存し、異なる環境では異なる。一般に、ストリンジェントな条件は、規定のイオン強度とpHにおける特定の配列に対して、熱融点(Tm)より約5℃低くなるように選択される。Tmは、ターゲット配列の50%が完全に一致するプローブとハイブリダイズする(規定のイオン強度とpHの下での)温度である。典型的には、塩濃度がpH 7で約0.02モルであり、温度が少なくとも60℃であるストリンジェントな条件が選択される。塩濃度の低下および/または温度の増大は、ストリンジェンシーを増大させる。RNA-DNAハイブリダイゼーション(たとえば100ntのプローブを用いたノーザンブロット)のストリンジェントな条件は、たとえば、63℃で20分間の0.2×SSCでの少なくとも1回の洗浄を含む条件、または同等の条件である。DNA-DNAハイブリダイゼーション(たとえば100ntのプローブを用いたサザンブロット)のストリンジェントな条件は、たとえば、少なくとも50℃の温度(通常約55℃)で20分間の0.2×SSCでの少なくとも1回(通常2回)の洗浄を含む条件、または同等の条件である。Sambrook et al. (1989)およびSambrook and Russell (2001)も参照されたい。
【0037】
“配列同一性”および“配列類似性”は、グローバルまたはローカルアラインメントアルゴリズムを用いて、二つのペプチドまたは二つのヌクレオチドの配列を整列させること(alignment)により決定することができる。(デフォルトパラメーターを用いてたとえばプログラムGAPまたはBESTFITにより最適に整列させた際の)配列が、(以下で規定される)配列同一性の最小パーセンテージを少なくとも共有する場合、配列は、“実質的に同一である”または“本質的に類似している”ということができる。GAPは、Needleman and Wunschのグローバルアラインメントアルゴリズムを使用して、二つの配列を全長にわたって整列させ、一致の数を最大にし、ギャップの数を最小にする。一般に、GAPデフォルトパラメーターが使用され、ギャップ生成ペナルティー=50(ヌクレオチド)/8(タンパク質)であり、ギャップ伸長ペナルティー=3(ヌクレオチド)/2(タンパク質)である。ヌクレオチドについて、使用されるデフォルトスコアリングマトリクスは、nwsgapdnaであり、タンパク質について、デフォルトスコアリングマトリクスは、Blosum62である(Henikoff & Henikoff, 1992, PNAS 89, 915-919)。配列アラインメントおよびパーセンテージ配列同一性のスコアは、コンピュータープログラム、たとえばGCG Wisconsin Package, Version 10.3(Accelrys Inc., 9685 Scranton Road, San Diego, CA 92121-3752 USAから入手可能)を用いて、あるいはEmbossWIN version 2.10.0(プログラム“needle”を使用)を用いて決定され得る。あるいは、パーセンテージ類似性または同一性は、たとえばFASTA、BLASTなどのアルゴリズムを用いてデータベースに対してサーチすることにより決定してもよい。
【0038】
本明細書および特許請求の範囲において、動詞“含むこと(to comprise)”およびその語形変化は、非限定的な意味で使用され、そのワードの後につづく項目を含むが、具体的に挙げられていない項目を排除しないことを意味する。加えて、不定冠詞“a”または“an”によるエレメントの指示は、その文脈が、一つおよび唯一のエレメントが存在することを明らかに要求しない限り、二つ以上のエレメントが存在する可能性を排除しない。よって、不定冠詞“a”または“an”は、通常“少なくとも一つ”を意味する。更に、本明細書で“配列”を参照する場合、ある配列のサブユニット(たとえばアミノ酸)を備えた実際の物理的分子を指すと理解される。
【0039】
本明細書で使用される“植物”の用語は、植物細胞、植物組織または器官、植物プロトプラスト、植物を再生させることができる植物細胞組織培養物、植物カルス、植物細胞塊、および植物においてインタクトな植物細胞、または植物の部分、たとえば胚、花粉、胚珠、果実(たとえば収穫されたトマト)、花、葉、種子、根、根端などを含む。
【発明の詳細な説明】
【0040】
本発明者らは、VIGS(ウイルス誘導性遺伝子サイレンシング)と組合せてcDNA-AFLP分析を使用して、Cf-4/Avr4依存性HRおよび耐病性に関与する遺伝子を同定した。VIGSがAvr-4誘導性HRの抑制につながる遺伝子のうち、CC-NB-LRRタイプの耐性タンパク質類似体をコードする(本明細書においてNRC1と称される)一つのトマト遺伝子が同定された(本明細書では、NB-LRR protein Required for HR-associated Cell death 1のためNRC1と称される)。トマトにおけるNRC1のサイレンシングは、Avr4誘導性HRの発生だけでなく、トマト病原体Cladosporium fulvumに対する耐性を低下させた(compromise)。これにより、トマトのCf-4耐性タンパク質(細胞外レセプター様タンパク質)が、細胞質NB-LRRタンパク質を機能的にするのに必要であることが示された。
【0041】
更に、NRC1は、複数のHRおよび複数の耐病性/細胞死シグナリング経路、たとえばCf-9/Avr-9-、LeEix2/Eix-、Pto/AvrPto-およびRx/CP-開始HRに関与していることが驚くべきことに見出された(実施例参照)。更に、NRC1が他のHR、たとえば(線虫、コナジラミおよびアブラムシ-誘導性HRに対する耐性を付与する)Mi-介在性HRにも関与しているかどうかを決定するために試験が行われている(US 6613962およびEP0937155B1参照)。このため、NRC1は、様々なクラスに属する細胞外および細胞内の耐病性タンパク質:細胞外レセプター様タンパク質 (RLPs、たとえばCf-4、Cf-9およびLeEix2)、Ser/Thrプロテインキナーゼ、たとえばPtoおよびCC-NB-LRRタンパク質(Rx)(これらは、真菌(Cladosporium fulvumおよびTrichoderma viride)、細菌(Pseudomonas syringae pv tomato) またはウイルス(PVX)のそれぞれに耐性を付与する)により誘発されるHR経路に関与している。
【0042】
NRCタンパク質(およびそれをコードするNRC1遺伝子)は、種々の病原体、とりわけ生物栄養性および半生物栄養性植物病原体、更に壊死栄養性植物病原体、たとえばBotrytis種に対する植物耐性を付与するかまたは向上させるために使用することができる。特に、NRC1(または他の箇所で規定されるその変異体もしくはフラグメント)の発現は、とりわけ生物栄養性および/または半生物栄養性病原体、すなわち生細胞から栄養を得るあらゆる病原体に対する耐性の向上につながる。本発明の範囲を限定しないが、内在性NRC1遺伝子のノックダウン(遺伝子サイレンシング)または(たとえばTILLINGによる)ノックアウトは、壊死につながる経路が影響を受け、壊死栄養性病原体がこの経路を必要とするため、壊死栄養性病原体に対する耐性を付与するかまたは向上させるために使用することができると考えられる。このため、耐性を向上させたい病原体に応じて、NRC1発現レベルの増大または減少の何れかを、耐性を向上させるために使用することができる。必要に応じて、両方のアプローチを、たとえば異なるプロモーターの制御下において一つの植物で使用してもよい。たとえば、NRC1を、(半)生物栄養性病原体により誘導されるプロモーターの制御下において発現させて、生物栄養性および/または半生物栄養性の葉の病原体に対する耐性を付与することができ、同時に、内在性NRC1遺伝子(または遺伝子ファミリー)を、壊死栄養性病原体または傷害により誘導性であるプロモーターを用いて、ある組織で、または壊死栄養体による誘導によりサイレンシングさせることができる。
【0043】
構成的に活性なNRC1タンパク質(NRC1D481V)が、トマトにおいて一過性で産生されると、その植物組織は、誘導因子に依存しない細胞死(HR)を示すことが更に発見され、これは、機能的NRC1タンパク質の発現を、植物に耐病性を付与するかまたは向上させるために使用することができることを示す。
【0044】
本発明のタンパク質および核酸配列
トマトから得られたNRC1タンパク質は、タバコのNRG1に対して低い配列同一性(25%未満)を示す。またNRC1は、NRG1より多数のロイシンリッチ繰返し配列(LRR)を含有する。NRC1のタンパク質構造は、図1および配列番号2に示される。
【0045】
本発明の一つの態様において、NRC1タンパク質の核酸配列およびアミノ酸配列(オルソログを含む)が提供されるとともに、他の植物種、たとえば他のナス科、好ましくはポテトにおいてNRC1のオルソログを単離または同定するための方法が提供される。同様に、他のNRC1対立遺伝子、たとえば他のトマトの種、品種、系統またはアクセッション(accession)由来の対立遺伝子を単離または同定するための方法が本明細書において提供される。
【0046】
一つの態様において、NRC1タンパク質が提供される。“NRC1タンパク質”は、配列番号2(野生型)および配列番号4(構成的突然変異体)に示されるタンパク質、並びにそのフラグメントおよび変異体を含む。NRC1の変異体には、たとえば、配列番号2および/または4に対して(全長にわたって)少なくとも30、35、40、45、50、55、60、65、70、75、80、85、90、95、98、99%またはそれ以上のアミノ酸配列同一性を有するタンパク質が含まれる。アミノ酸配列同一性は、上述のとおり、Needleman and WunschアルゴリズムおよびGAPデフォルトパラメーターを用いたペアワイズアラインメントにより決定される。NRC1の変異体は、種々の供給源、たとえば既存の配列データベース、他の植物種(とりわけナス科の他の種、たとえばポテト)または他の品種から入手することができるし、あるいはデノボ合成、突然変異誘発などにより作成することもできる。たとえば、配列番号4、すなわち構成的に活性なNRC1突然変異体は、配列番号2の変異体であり、オーバーラップPCRを用いた標的突然変異誘発によって作成された(実施例参照)。このように、本発明のNRC1タンパク質は、天然源から単離されてもよいし、(たとえばApplied Biosystemsにより供給されるようなペプチド合成機を用いて)化学合成によりデノボ合成されてもよいし、組換え宿主細胞により、NRC1タンパク質、フラグメントまたは変異体をコードする核酸配列を発現させることにより産生してもよい。
【0047】
NRC1変異体は、塩基性(たとえばArg、His、Lys)、酸性(たとえばAsp、Glu)、非極性(たとえばAla、Val、Trp、Leu、Ile、Pro、Met、Phe、Trp)または極性(たとえばGly、Ser、Thr、Tyr、Cys、Asn、Gln)のカテゴリー内において保存的アミノ酸置換を含んでいてもよい。加えて、非保存的アミノ酸置換も本発明の範囲内である。
【0048】
NRCタンパク質、変異体またはフラグメントの機能性は、種々の方法を用いて決定することができる。たとえば、植物細胞における一過性または安定な過剰発現を用いて、当該タンパク質が植物体で(in planta)活性を有しているかどうか試験することができる。機能性は、好ましくは、タンパク質を入手したのと同一の植物種において試験する。このように、たとえば、一過性または安定な発現を用いて、HRが発生しているかどうか、および/または耐性が向上しているかどうかを決定し、機能性を示すことができる。あるいは、内在性遺伝子または遺伝子ファミリーのサイレンシングは、NRC1タンパク質が機能的であるかどうかを示す。たとえば、VIGSは、種々のナス科、たとえばポテト、トマトおよびタバコ(Brigneti et al., 2004, Plant Journal 39: 264; Faivre-Rampant et al. Plant Physiology 134: 1308-1316; Baulcombe 1999, Curr. Opinion. Plant Biol. 2: 109-113; Lu et al. 2003, EMBO J. 22: 5690-5699参照)、アラビドプシス(Arabidopsis)などのモデル生物(Turnage et al. 2002, Plant J. 30: 107-114)、オオムギなどの単子葉植物(Holzberg et al. 2002, Plant J. 30: 315-327)で使用することができる。あるいは、NRC1遺伝子のセンスおよび/またはアンチセンスフラグメントを含むサイレンシングベクターを使用して、植物細胞を形質転換し(下記参照)、その後アッセイにより、HR病変を発生する能力および/または耐病性が改変されているかどうかを決定することができる。
【0049】
好ましい態様において、NRC1の変異体には、植物細胞で構成的に活性なNRC1タンパク質、たとえばMHDドメインに単一のアミノ酸置換(D481V)を含む配列番号4で提供されるNRC1タンパク質が含まれる(図1参照)。構成的活性は、タンパク質が誘導因子の非存在下において植物組織でHRを誘導することができるかどうかを決定することにより試験することができる。たとえば、実施例に記載される35S:NRC1構築物のアグロインフィルトレーション(Agroinfiltration)は、葉組織に浸潤させる(infiltrate)ために使用することができる。構成的に活性な他のNRC1タンパク質は、ランダム突然変異誘発とその後の活性試験により(Bendahame et al., 2002, p196に記載される)、あるいはMHDドメイン(アミノ酸VHDまたはVHDMの何れか一つが別のアミノ酸に置換され得る)、NB-ARCドメイン、たとえばRNBS-Dドメイン(アミノ酸FLYFGTFPRGY)、または13のLRRドメインの一つにおける単一アミノ酸の位置指定突然変異誘発により(図1参照)、作成することができる。あるいは、構成的に活性なNRC1タンパク質をコードする核酸配列は、たとえば種子に突然変異を起こさせ、これら種子を自然発生的病変の表現型(たとえば顕微鏡的な病変)の存在についてスクリーニングすることにより、植物から入手することができる。たとえばSharino et al. (2002, The Plant Cell 14: 3149-3162)および後述の記載を参照されたい。
【0050】
一つの態様において、キメラNRC1タンパク質も提供される。かかるタンパク質は、少なくともCCドメイン、NB-ARCドメイン、および好ましくは少なくとも13のLRRを含む。CC-、NB-ARC-およびLRR-ドメインは、好ましくは、それぞれ配列番号2のアミノ酸1-150、アミノ酸151-508、またはアミノ酸509-846に対して少なくとも30、40、50、60、70、80、90、95、98、99%またはそれ以上のアミノ酸配列同一性を含むアミノ酸モチーフを指す。このため、ドメインは、得られるキメラタンパク質の機能性が、NRC1、または好ましくはNRC1D481Vと本質的に類似している限り、NRC1タンパク質の間、またはNRC1タンパク質と他のCC-NB-LRRもしくはTIR-NB-LRRタンパク質との間で交換(ドメインスワッピング)されてもよい。最も好ましくは、キメラタンパク質は、後述されるとおり、組換え植物細胞により産生されると、耐病性を付与するかまたは向上させる能力を保持する。
【0051】
上述のとおりNRC1タンパク質およびNRC1タンパク質の変異体の“フラグメント”は、100、150、200、300、400、500、600、700、800、850、855の連続アミノ酸またはそれ以上のフラグメントを含む。好ましくは、かかるフラグメントは、植物組織において機能的であり、すなわち植物細胞で産生されると病原体耐性を付与するかまたは向上させることができる。また、フラグメントは、上述のとおり、キメラタンパク質を作成するために使用してもよい。
【0052】
別の態様において、上記タンパク質、変異体またはフラグメントの何れかをコードする単離された核酸配列、たとえばcDNA、ゲノムDNAおよびRNAの配列が提供される。遺伝暗号の縮重のため、種々の核酸配列が、同一のアミノ酸配列をコードし得る。NRC1タンパク質または変異体をコードする任意の核酸配列が、本明細書において“NRC1”と称される。提供される核酸配列は、天然に存在する核酸配列、人工もしくは合成の核酸配列を含む。NRC1タンパク質をコードする核酸配列の例は、配列番号1および3において提供される。RNAが言及されているのにDNA配列として配列が示される場合、RNA分子の実際の塩基配列は、チミン(T)がウラシル(U)に置換されるという違いはあるが同一であることが理解される。
【0053】
また、NRC1核酸配列の変異体およびフラグメント、たとえば規定されるストリンジェントな条件下でNRC1核酸配列とハイブリダイズする核酸配列も包含される。NRC1核酸配列の変異体には、配列番号1または3に対して(全長にわたって)少なくとも50%以上、好ましくは少なくとも55%、60%、70%、80%、90%、95%、99%、99.5%、99.8%またはそれ以上の配列同一性を有する核酸配列も包含される。好ましい態様において、NRC1の変異体は、記載されるとおり、構成的に活性なNRC1タンパク質をコードする。NRC1核酸配列の変異体またはフラグメントを同定、合成または単離するために多くの方法が使用できること、たとえば核酸ハイブリダイゼーション、PCR技術、インシリコ(in silico)分析、および核酸合成などが使用できることは明らかである。
【0054】
本発明のNRC1タンパク質をコードする核酸配列、とりわけDNA配列は、後述のとおり、発現ベクターに挿入して、多量のNRC1タンパク質(またはたとえばキメラNRC1タンパク質)を産生することができる。宿主細胞での最適な発現のために、NRC1のDNA配列は、利用可能なコドン使用頻度表を用いて、コドン使用頻度を植物遺伝子で最も好ましいもの、とりわけ対象の植物属または種に生来備わっている遺伝子に適合させることにより、コドン最適化を行うことができる(Bennetzen & Hall, 1982, J. Biol. Chem. 257, 3026-3031; Itakura et al., 1977 Science 198, 1056-1063.)(たとえば、ワタ、ダイズ、トウモロコシまたはイネでの発現に対して適合させる)。種々の植物種についてのコドン使用頻度表は、たとえばIkemura (1993, In “Plant Molecular Biology Labfax”, Croy, ed., Bios Scientific Publishers Ltd.) およびNakamura et al. (2000, Nucl. Acids Res. 28, 292.)により、並びにメジャーなDNA配列データベース (たとえばEMBL at Heidelberg, Germany)で公開されている。したがって、合成DNA配列は、同一または実質的に同一のタンパク質が産生されるように構築することができる。コドン使用頻度を宿主細胞に好ましいものに改変するための幾つかの技術は、特許および科学文献に見出すことができる。コドン使用頻度の正確な改変方法は、本発明にとって重大ではない。
【0055】
上述されるようなDNA配列に対する小さな改変は、ルーチンで、すなわちPCRによる突然変異誘発により行うことができる(Ho et al., 1989, Gene 77, 51-59., White et al., 1989, Trends in Genet. 5, 185-189)。DNA配列に対する甚大な改変は、利用可能な技術を用いて所望のコード領域のデノボDNA合成によりルーチンで行うことができる。
【0056】
また、NRC1核酸配列は、NRC1タンパク質のN末端に一または複数のアミノ酸を付加するかまたは欠失させることにより、NRC1タンパク質のN末端が最適な翻訳開始の文脈情報(context)を有するように改変することができる。多くの場合、植物細胞で発現される本発明のタンパク質は、最適な翻訳開始のためにMet-AspまたはMet-Alaジペプチドで開始することが好ましい。このため、AspまたはAlaのコドンが、既存のMetの後に挿入されてもよいし、あるいは2番目のコドンValが、Asp(GATまたはGAC)またはAla(GCT、GCC、GCAまたはGCG)に置換されてもよい。またDNA配列は、異常なスプライス部位を除去するように改変されてもよい。
【0057】
NRC1核酸配列の“フラグメント”には、配列番号1もしくは3、または配列番号1もしくは3の変異体の少なくとも10、12、15、16、18、20、30、40、50、100、200、500、1000、1500、2000またはそれ以上の連続ヌクレオチドのフラグメントが含まれる。短いフラグメントは、たとえばPCRプライマーまたはハイブリダイゼーションプローブとして使用することができる。
【0058】
本発明の別の態様において、NRC1のDNAまたはRNA配列を検出するためのPCRプライマーおよび/またはプローブおよびキットが提供される。サンプルからNRC1のDNAを増幅するための縮重または特異的PCRプライマーのペアは、当該技術分野で公知のとおり、配列番号1または3(またはその変異体)に基いて合成することができる(Dieffenbach and Dveksler (1995) PCR Primer: A Laboratory Manual, Cold Spring Harbor Laboratory Press, and McPherson at al. (2000) PCR-Basics: From Background to Bench, First Edition, Springer Verlag, Germany参照)。たとえば、配列番号1または3(またはその相補鎖)の9、10、11、12、13、14、15、16、18またはそれ以上の連続ヌクレオチドの任意の長さが、プライマーまたはプローブとして使用され得る。同様に、配列番号1または3(またはその変異体)のDNAフラグメントを、ハイブリダイゼーションプローブとして使用することができる。NRC1検出キットは、NRC1特異的プライマーおよび/またはNRC1特異的プローブ、およびサンプル中のNRC1 DNAを検出するためにプライマーまたはプローブを使用するための関連プロトコールを含む。かかる検出キットは、たとえば、植物が本発明のNRC1遺伝子(またはその一部)で形質転換されたかどうかを決定するために使用され得る。遺伝暗号の縮重のため、幾つかのアミノ酸コドンは、タンパク質のアミノ酸配列を変更しないで他のものに置換することができる。
【0059】
更に別の態様において、トマトNRC1遺伝子(配列番号1および3)のオルソログまたは対立遺伝子を同定し、使用するための方法が提供される。この方法は、以下の工程を含む:
a)配列番号1および/または3に対して少なくとも70%の核酸同一性(または上述のより高いパーセンテージの配列同一性)を含む核酸配列を入手または同定する工程、
b)必要に応じて、構成的に活性なNRC1タンパク質をコードするように核酸配列を改変する工程、および
c)a)の核酸配列を使用して、発現および/またはサイレンシングベクターを作成する工程、またはb)の核酸配列を使用して発現ベクターを作成する工程、
d)一または複数のc)のベクターを使用して、好ましくは核酸を入手した植物種の、植物または植物細胞を形質転換する工程、
e)植物体において(in planta)遺伝子の機能を決定または検証するため、および/または向上した耐病性を有するトランスジェニック植物を作成するために、形質転換体のHR病変(すなわちHR病変の表現型、これは必要に応じて定量することができる)および/または耐病性を発生する形質転換植物/植物組織の能力を分析する工程、
f)必要に応じて、トランスジェニック植物に向上した耐病性を付与するが、発現により弱いHR表現型を付与する(すなわち、HR病変の表現型を引き起こさないかまたは減少したHR病変の表現型を引き起こす)対立遺伝子またはオルソログを、更なる使用のために選択する工程。
【0060】
このようにして、配列番号1または3の発現または形質転換される宿主種から得られた野生型NRC1対立遺伝子の発現により観察されるよりも少ないおよび/または小さいHR病変が植物での発現により得られるNRC1対立遺伝子またはオルソログを、この方法を用いて同定することができる。より好ましくは、発現によりHR病変を引き起こさないかまたは少なくとも肉眼で見えるHR病変を引き起こさないが、向上した耐病性を付与するNRC1対立遺伝子またはオルソログが同定される。
【0061】
同じプロモーターを使用して発現ベクターを作成し、これらを用いて宿主植物を形質転換し、これらトランスジェニック植物の間でHR病変の表現型を比較することにより、異なるNRC1対立遺伝子および/またはオルソログのHR表現型を比較することができる。異なる対立遺伝子をトランスジェニックナス科植物において(たとえば構成的または誘導性プロモーターの制御下で)比較する場合、好ましくは、配列番号1または3を発現する形質転換体のHR病変の表現型がレファレンスとして使用される。あるいは、形質転換される宿主種から得られた野生型対立遺伝子をレファレンスとして使用することができる。その後、配列番号1または3と比較して少ないおよび/または小さいHR病変、または形質転換される種から得られた野生型対立遺伝子の発現により引き起こされるよりも少ないおよび/または小さい病変を提供する対立遺伝子を、更なる使用のために選択することができる。たとえば、これらを発現するトランスジェニック植物は、後述されるとおり作成することができる。
【0062】
たとえばNRC1特異的PCRプライマーもしくはプローブ、またはインシリコでの(in silico)バイオインフォマティクス分析を用いて、とりわけ、トマト由来の対立遺伝子およびポテト由来のオルソログが入手または同定され得る。また、遺伝子マッピングを使用して、植物(たとえばトマトまたはポテト)ゲノムにおけるNRC1遺伝子座をマッピングすることができ、ゲノムマップを(たとえばトマトの配列決定プロジェクトで開発された)既存のゲノム配列決定データベースにリンクさせることにより配列を入手することができる。かかる対立遺伝子および/またはオルソログは、向上した耐病性を備えた植物を作成するために特に適している。
【0063】
ポテトのNRC1オルソログを上記方法で同定した場合、これらオルソログ(またはこれらオルソログの変異体)は、好ましくは、Phytophthora infestansに対して向上した耐性を有する植物を作成するために使用される。
【0064】
本発明のキメラ遺伝子、発現ベクターおよび組換え生物
本発明の一つの態様において、上述のNRC1タンパク質をコードする核酸配列(変異体またはフラグメントを含む)を使用して、キメラ遺伝子の宿主細胞への転移のため、およびNRC1タンパク質の宿主細胞(たとえば細胞、組織、器官または形質転換細胞に由来する生物)での産生のためにキメラ遺伝子およびこれらを含むベクターを作成する。NRC1タンパク質(またはタンパク質フラグメントまたは変異体)を植物細胞で産生するためのベクターは、本明細書において、すなわち“発現ベクター”と称される。宿主細胞は、好ましくは植物細胞であるが、微生物の宿主(細菌、たとえばアグロバクテリウム、酵母、真菌など)も想定される。
【0065】
任意の植物、たとえば単子葉植物または双子葉植物を適切な宿主としてもよいが、最も好ましくは、宿主植物はナス科に属する。たとえば、植物は、ナス属(Solanum)(Lycopersiconを含む)、タバコ属(Nicotiana)、トウガラシ属(Capsicum)、ペチュニア属(Petunia)および他の属に属する。以下の宿主種が、適切に使用され得る:タバコ(Nicotiana種、たとえばN. benthamiana、N. plumbaginifolia、N. tabacumなど)、野菜の種、たとえばトマト(L. esculentum, syn. Solanum lycopersicum)たとえばチェリートマト、var. cerasiformeまたはカレントトマト、var. pimpinellifolium)またはツリートマト(S. betaceum, syn. Cyphomandra betaceae)、ポテト(Solanum tuberosum)、ナス(Solanum melongena)、ペピーノ(Solanum muricatum)、ココナ(Solanum sessiliflorum)およびナランヒージャ(Solanum quitoense);コショウ(Capsicum annuum、Capsicum frutescens、Capsicum baccatum)、観賞植物種(たとえばPetunia hybrida、Petunia axillaries、P. integrifolia)。
【0066】
あるいは、植物は、その他の任意の科、たとえばウリ科(Cucurbitaceae)またはイネ科(Gramineae)に属していてもよい。適切な宿主植物には、たとえば以下のものが含まれる:トウモロコシ(maize/corn)(Zeaの種)、コムギ(Triticumの種)、オオムギ(たとえばHordeum vulgare)、カラスムギ(たとえばAvena sativa)、モロコシ(Sorghum bicolor)、ライムギ(Secale cereale)、ダイズ(Glycine spp、たとえばG. max)、ワタ(Gossypiumの種、たとえばG. hirsutum、G. barbadense)、アブラナ属の種(Brassica spp.)(たとえばB. napus、B. juncea、B. oleracea、B. rapaなど)、ヒマワリ(Helianthus annus)、ベニバナ、ヤム、キャッサバ、アルファルファ(Medicago sativa)、イネ(Oryzaの種、たとえばO. sativaインディカ栽培品種-グループまたはジャポニカ栽培品種-グループ)、まぐさ(forage grasses)、パールミレット(Pennisetumの種、たとえばP. glaucum)、樹木の種(マツ属(Pinus)、ポプラ、モミ、プランタン(plantain)など)、チャ、コーヒー、アブラヤシ、ココナッツ、野菜の種、たとえばエンドウ、ズッキーニ、マメ(たとえばPhaseolusの種)、キュウリ、アーティチョーク、アスパラガス、ブロッコリー、ニンニク、ニラ、レタス、タマネギ、ラディッシュ、カブ、芽キャベツ、ニンジン、カリフラワー、チコリ、セロリ、ホウレンソウ、エンダイブ、ウイキョウ、ビート、多肉質の果実をつける植物(グレープ、ピーチ、プラム、ストロベリー、マンゴー、リンゴ、プラム、チェリー、アプリコット、バナナ、ブラックベリー、ブルーベリー、シトラス、キウイ、イチジク、レモン、ライム、ネクタリン、ラズベリー、スイカ、オレンジ、グレープフルーツなど)、観賞植物種(たとえばバラ、ペチュニア、キク、ユリ、ガーベラの種)、ハーブ(ミント、パセリ、バジル、タイムなど)、木本(たとえばPopulus、Salix、Quercus、Eucalyptusの種)、繊維の種、たとえばアマ(Linum usitatissimum)およびアサ(Cannabis sativa)、またはモデル生物、たとえばシロイヌナズナ(Arabidopsis thaliana)。
【0067】
好ましい宿主は、“作物植物”、すなわちヒトにより栽培され、繁殖される植物種である。作物植物は、食物の目的のため(たとえば農作物など)、または装飾の目的のため(たとえば切花、芝生用の草の生産など)に栽培されてもよい。本明細書で規定される作物植物には、食物以外の生産物、たとえば燃料用オイル、プラスチックポリマー、医薬品、コルクなどが収穫される植物も含まれる。
【0068】
NRC1タンパク質コード核酸配列の宿主細胞ゲノムへの好ましくは安定した導入のためのキメラ遺伝子およびベクターの構築は、一般に当該技術分野で公知である。キメラ遺伝子を作成するために、NRC1タンパク質(または変異体もしくはフラグメント)をコードする核酸配列は、標準的な分子生物学技術を用いて、宿主細胞での発現に適したプロモーター配列に動作可能に連結される。プロモーター配列は、NRC1核酸配列が、ベクターにプロモーター配列の下流で簡単に挿入されるように、ベクターに既に存在していてもよい。その後、ベクターは、宿主細胞を形質転換するために使用され、キメラ遺伝子は、核ゲノム、またはプラスチド、ミトコンドリアまたは葉緑体のゲノムに挿入され、その場所で適切なプロモーターを用いて発現される(たとえば、Mc Bride et al., 1995 Bio/Technology 13, 362; US 5,693,507)。一つの態様において、キメラ遺伝子は、植物細胞または微生物細胞(たとえば細菌)での発現に適したプロモーターを含み、これに、本発明のNRC1タンパク質をコードする核酸配列が動作可能に連結され、必要に応じてその後に3’非翻訳核酸配列が続く。
【0069】
機能的なNRC1タンパク質(またはある態様において構成的に活性なNRC1タンパク質)をコードするNRC1核酸配列、好ましくはNRC1キメラ遺伝子は、慣用的なやり方で、単一の植物細胞の核ゲノムに安定に挿入することができ、このように形質転換された植物細胞を慣用的なやり方で使用して、ある細胞にある期間NRC1タンパク質が存在することにより変化した表現型を有する形質転換植物を産生することができる。これに関して、アグロバクテリウムツメファシエンス(Agrobacterium tumefaciens)内のNRC1タンパク質をコードする核酸配列を含むT-DNAベクターを、植物細胞を形質転換するために使用することができ、その後、たとえばEP 0 116 718、EP 0 270 822、PCT公報 WO 84/02913および公開された欧州特許出願EP 0 242 246、およびGould et al. (1991, Plant Physiol. 95,426-434) に記載される手法を用いて、形質転換された植物細胞から形質転換植物を再生させることができる。アグロバクテリウム媒介性の植物の形質転換のためのT-DNAベクターの構築は、当該技術分野で周知である。T-DNAベクターは、EP 0 120 561およびEP 0 120 515に記載されるバイナリーベクター、または相同組換えによりアグロバクテリウムTi-プラスミドに統合することができるEP 0 116 718に記載されるコインテグレート(co-integrate)ベクターの何れであってもよい。
【0070】
好ましいT-DNAベクターはそれぞれ、T-DNAのボーダー配列の間、または少なくとも右ボーダー配列の左側の位置に、NRC1コード核酸配列(たとえば配列番号2または配列番号4をコードする)に動作可能に連結されたプロモーターを含有する。ボーダー配列は、Gielen et al. (1984, EMBO J 3,835-845) に記載される。当然、他のタイプのベクターを使用して、直接的な遺伝子導入(たとえばEP 0 223 247に記載される)、花粉媒介性の形質転換(たとえばEP 0 270 356およびWO 85/01856に記載される)、プロトプラストの形質転換(たとえばUS 4,684,611に記載される)、植物RNAウイルス媒介性の形質転換(たとえばEP 0 067 553およびUS 4,407, 956に記載される)、リポソーム媒介性の形質転換(たとえばUS 4,536,475に記載される)などの手法を用いて、植物細胞を形質転換することができる。トマトまたはタバコの形質転換のためには、An G. et al., 1986, Plant Physiol. 81: 301-305; Horsch R.B. et al., 1988, In: Plant Molecular Biology Manual A5, Dordrecht, Netherlands, Kluwer Academic Publishers. pp 1-9; Koornneef M. et al., 1986, In: Nevins D.J. and R.A. Jones, eds. Tomato Biotechnology, New York, NY, USA, Alan R. Liss, Inc. pp 169-178も参照される。ポテトの形質転換のためには、たとえばSherman and Bevan (1988, Plant Cell Rep. 7: 13-16) が参照される。
【0071】
同様に、形質転換された細胞からの形質転換植物の選択および再生は、当該技術分野で周知である。明らかに、様々な種に対して、更に単一種の様々な品種または栽培品種に対しても、高い頻度で形質転換体を再生させるためにプロトコールは特異的に適合される。
【0072】
核ゲノムの形質転換に加えて、プラスチドのゲノム、好ましくは葉緑体のゲノムの形質転換も本発明に包含される。プラスチドのゲノムの形質転換は、導入遺伝子の蔓延のリスクを低減できるという利点がある。プラスチドのゲノムの形質転換は、当該技術分野で公知のとおり行うことができ、たとえばSidorov VA et al. 1999, Plant J.19: 209-216またはLutz KA et al. 2004, Plant J. 37(6): 906-13が参照される。
【0073】
得られた形質転換植物は、慣用的な植物育種スキームで使用して、導入遺伝子を含有する多くの形質転換植物を産生することができる。シングルコピーの形質転換体は、たとえばサザンブロット分析またはPCRベースの方法またはInvader(登録商標)テクノロジーアッセイ(Third Wave Technologies, Inc.)を用いて選択することができる。形質転換された細胞および植物は、キメラ遺伝子の存在により非形質転換体と容易に区別することができる。導入遺伝子の挿入部位に隣接する植物DNAの配列も配列決定することができ、これにより、ルーチン使用のために“イベント特異的な”検出方法を開発することができる。たとえば、WO 01/41558が参照され、これは、たとえば統合された配列および隣接する(ゲノム)配列に基く、選抜きのイベント検出キット(たとえばPCR検出キット)を記載する。
【0074】
NRC1核酸配列は、挿入されるコード配列が、植物細胞で発現を駆動することができるプロモーターの下流(すなわち3’)および制御下に位置するように植物細胞ゲノムに挿入される。これは、好ましくは、キメラ遺伝子を植物細胞ゲノム、とりわけ核またはプラスチド(たとえば葉緑体)ゲノムに挿入することにより達成される。
【0075】
NRC1タンパク質の構成的な産生は、細胞死の誘導(たとえば顕微鏡的な病変および/または肉眼的な病変)および/または低い収量につながり得るため、一つの態様において、活性が誘導性であるプロモーターを使用することが好ましい。誘導性プロモーターの例は、傷害誘導性プロモーター、たとえば、Cordera et al.(1994, The Plant Journal 6, 141) に記載される、傷害(たとえば昆虫または物理的傷害により引き起こされる)によって誘導されるMPIプロモーター、またはCOMPTIIプロモーター (WO 00/56897) またはUS 6031151に記載されるPR1プロモーターである。あるいは、プロモーターは、化学物質、たとえば、Aoyama and Chua (1997, Plant Journal 11: 605-612) およびUS 6063985に記載されるデキサメタゾン、またはテトラサイクリン (TOPFREEまたはTOP 10プロモーター、Gatz, 1997, Annu Rev Plant Physiol Plant Mol Biol. 48: 89-108 and Love et al. 2000, Plant J. 21: 579-88参照) により誘導されてもよい。他の誘導性プロモーターは、たとえば、温度の変化(たとえばUS 5,447,858に記載される熱ショックプロモーター)、嫌気的条件(たとえばトウモロコシADH1Sプロモーター)、光(US6455760)、病原体(たとえばEP759085のgst1プロモーターまたはEP309862のvst1プロモーター)、または老化(SAG12およびSAG13、US5689042参照)により誘導性である。明らかに、種々の他のプロモーターが利用可能である。
【0076】
一つの態様において、好ましくは、病原体誘導性プロモーターが使用され、これによりNRC1タンパク質(または変異体もしくはフラグメント)は、病原体が植物組織を攻撃した後にのみ産生される。とりわけ、病原体の攻撃の直後にアップレギュレートされる遺伝子のプロモーターが望ましい。病原体誘導性プロモーターには、たとえば、タバコ由来のhsr203J、str246Cおよびsgd24プロモーター、Yin et al. (1997, Plant Physiology 115(2): 437-51) に記載されるEAS4プロモーター、tap1またはtap2プロモーター (Mohan et al., 1993, Plant Mol Biol. 1993 22: 475-90)、gst1プロモーターまたはその変異体 (Martini et al. 1993, Mol. Gen. Gen. 236, 179-186; Hennin C., 1997, Afstudeerwerk, Faculteit Landbouwkundige en Toegepaste Biologische Wetenschappen, University of Gent, Belgium)、WRKYプロモーター (Eulgem et al., EMBO J., 1999, 18(17): 4689-99およびWO 00/29592に記載されるキメラプロモーター) が含まれる。また、特定の植物病原体により誘導性であるプロモーターは、公知の方法、たとえばcDNA-AFLP(登録商標)を用いて同定されてもよい。
【0077】
好ましくは、プロモーターは、複数の病原体により誘導され、すなわち宿主植物の広範囲の病原体により誘導性である。特定の宿主植物種のそれぞれに対して、異なるプロモーターが最適であってもよい。たとえば、トマトを宿主として使用する場合、プロモーターは、少なくとも一、好ましくは二以上のトマト病原体により誘導される。とりわけ、一または複数の真菌の植物病原体および/または細菌の植物病原体(とりわけ一または複数の生物栄養性および/または半生物栄養性植物病原体)により誘導性であるプロモーターが好ましい。
【0078】
植物病原体、それらにより引き起こされる病気の徴候、およびそれらの生活環の詳細な説明は、植物種のそれぞれに対して見出すことができる。たとえば、トマトの病原体は、“Compendium of Tomato Diseases”, Editors Jones, Jones, Stall and Zitter, ISBN 0-89054-120-5, APS Press (http:/www.shopapspress/org) に記載される。ポテトの病原体は、“Compendium of Potato Disease”, 2nd edition, Editors Stevenson, Franc and Weingartner, APS Press, ISBN 0-89054-275-9 に記載される。
【0079】
トマトの病原体には、たとえば、(非限定的な)以下の真菌および細菌種およびウイルスが含まれる:Botrytis cinerea (真菌/壊死栄養体); Colletotrichum coccodes (真菌/壊死栄養体); Alternaria alternata (真菌); Alternaria solani (真菌/壊死栄養体); Stemphylium solani; Phytophthora infestans (卵菌(oomycte)/半生物栄養体); Septoria lycopersici; Cladosporium fulvum (真菌/半生物栄養体); Phytophthora parasitica; Oidium lycopersicum (生物栄養体); Fusarium oxysporum; Sclerotium rolfsii; Pythium; Rhizoctonia (真菌/壊死栄養体); Corynebacterium michiganense (細菌); Pseudomonas syringae pv tomatoまたはpv syringae (細菌/生物栄養体); Pseudomonas solanacearum; Pseudomonas corrugate; Clavibacter Xanthomonas campestris (細菌/生物栄養体); Verticillium (真菌)、トマト・スポッテッド・ウイルト・ウイルスtomato spotted wilt virus (TSWV); タバコまたはトマト・モザイク・ウイルス (TobMV、TomMV)。
【0080】
ポテトの病原体には、たとえば、種々の真菌、細菌、線虫およびウイルス、たとえば以下のものが含まれる:Phytophthora infestans (oomycte /半生物栄養体), 線虫 (生物栄養体); Erwinia carotovora (細菌); Colletotrichum coccodes (真菌); Rhizoctonia solani (真菌/壊死栄養体); Verticillium dahliae (真菌); Streptomyces scabies; Alternaria solani (真菌/壊死栄養体); Pythium; Spongospora subterranean; PVXおよびPVY; ポテト・リーフロール・ウイルス (PLRV); など。
【0081】
種々の植物種の植物の病気については、http://www.apsnet.org/online/common/toc.aspも参照される。よって、一つの態様において、プロモーターは、好ましくは、上記病原体の一または複数により、最も好ましくは少なくとも上記生物栄養性および/または半生物栄養性病原体の一または複数により誘導性である。
【0082】
あるいは、宿主植物は、種々の病原体の攻撃後に確実にNRC1タンパク質が産生されるように、異なる病原体誘導性プロモーターの制御下に、種々のNRC1導入遺伝子をそれぞれ含んでいてもよい。たとえば、トマトの形質転換について、一つのプロモーターはPhytophthoraにより誘導性で、一つのプロモーターはCladosporiumにより誘導性であってもよい。
【0083】
“誘導性(inducible)”の用語は、プロモーターが、誘導物質刺激の非存在下において完全に不活性であることを必ずしも必要としない。低レベルの非特異的活性は、それが、植物の収量または品質の重いペナルティーという結果につながらない限り、存在していてもよい。よって、誘導性とは、好ましくは、誘導物質との接触後に、下流のNRC1コード領域の転写の増大につながるプロモーターの活性の増大を指す。
【0084】
本明細書において、最も好ましい組合せは、上述のとおり、構成的に活性なNRC1タンパク質をコードするNRC1核酸配列に動作可能に連結された病原体誘導性プロモーターの使用である。この場合、病原体の攻撃により、構成的に活性なNRC1が発現され、これが(病原体の攻撃部位に限定される)局所的なHRにつながり、(半)生物栄養性病原体の更なる増殖を妨害する。
【0085】
別の態様において、たとえば以下の構成的プロモーターが使用されてもよい:カリフラワーモザイクウイルス(CaMV)の単離体CM 1841 (Gardner et al., 1981, Nucleic Acids Research 9, 2871-2887)、CabbB-S (Franck et al., 1980, Cell 21, 285-294)、およびCabbB-JI (Hull and Howell, 1987, Virology 86, 482-493) の強力な構成的35Sプロモーターまたは増強(enhanced)35Sプロモーター(“35Sプロモーター”);Odell et al. (1985, Nature 313, 810-812) またはUS5164316に記載される35Sプロモーター、ユビキチンファミリー由来のプロモーター (たとえばChristensen et al., 1992, Plant Mol. Biol. 18, 675-689, EP 0 342 926のトウモロコシのユビキチンプロモーター、Cornejo et al. 1993, Plant Mol. Biol. 23, 567-581も参照)、gos2プロモーター (de Pater et al., 1992 Plant J. 2, 837-844)、emuプロモーター (Last et al., 1990, Theor. Appl. Genet. 81,581-588)、アラビドプシス(Arabidopsis)のアクチンプロモーター、たとえばAn et al. (1996, Plant J. 10, 107.)に記載されるプロモーター、イネのアクチンプロモーター、たとえばZhang et al.(1991, The Plant Cell 3, 1155-1165)に記載されるプロモーターおよびUS 5,641,876に記載されるプロモーターまたはWO 070067に記載されるイネのアクチン2プロモーター;キャッサバ葉脈モザイクウイルスのプロモーター (WO 97/48819, Verdaguer et al. 1998, Plant Mol. Biol. 37, 1055-1067)、サブタレニアン・クローバー萎縮ウイルス由来のpPLEXシリーズのプロモーター (WO 96/06932、とりわけS7プロモーター)、アルコールデヒドロゲナーゼプロモーター、たとえばpAdh1S (GenBankアクセッション番号X04049、X00581)、T-DNAのそれぞれ1’および2’遺伝子の発現を駆動するTR1’プロモーターおよびTR2’プロモーター(それぞれ“TR1’プロモーター”および“TR2’プロモーター”)(Velten et al., 1984, EMBO J 3, 2723-2730)、US6051753およびEP426641に記載されるゴマノハグサモザイクウイルスプロモーター、ヒストン遺伝子プロモーター、たとえばアラビドプシス(Arabidopsis)由来のPh4a748プロモーター (PMB 8: 179-191)、またはその他。好ましい態様において、PCT/NL2005/050083 (2005年12月16日出願) に記載されるAA6プロモーターが使用される。
【0086】
あるいは、構成的ではなく植物の一または複数の組織または器官に特異的なプロモーター(組織優先的/組織特異的、発生で調節されるプロモーターを含む、たとえば葉優先的、表皮優先的、根優先的、花組織、たとえば絨緞組織または葯優先的、種子優先的、莢優先的など)を使用することができ、これによりNRC1遺伝子は、特異的な組織または器官の細胞でのみ、および/またはある発生段階の間のみ発現される。たとえば、NRC1遺伝子は、光誘導性プロモーター(たとえば、植物それ自体または別の植物、たとえばUS 5,254,799に開示されるエンドウまたはUS5034322に開示されるアラビドプシスの、リブロース-1,5-ビスリン酸カルボキシラーゼの小サブユニット遺伝子のプロモーター)の制御下にコード配列を置くことにより、植物の葉で選択的に発現させることができる。
【0087】
一つの態様において、内在性NRC1遺伝子のプロモーターが使用される。たとえば、トマトNRC1遺伝子のプロモーターを単離し、配列番号2または4のNRC1タンパク質をコードするコード領域に動作可能に連結してもよい。NRC1プロモーター(配列番号1および3の上流の転写調節領域)は、公知の方法、たとえばTAIL-PCR(Liu et al. 1995, Genomics 25(3): 674-81; Liu et al. 2005, Methods Mol Biol. 286: 341-8)、Linker-PCR、またはInverse PCR (IPCR) を用いて、トマト植物から単離することができる。
【0088】
NRC1コード配列は、好ましくは、コード配列が、適切な3’端の非翻訳領域(“3’端”または3’UTR)の上流(すなわち5’)に位置するように植物ゲノムに挿入される。適切な3’端には、CaMV 35S遺伝子のもの(“3’35S”)、ノパリンシンターゼ遺伝子のもの(“3’nos”) (Depicker et al., 1982 J. Molec. Appl. Genetics 1, 561-573.)、オクトピンシンターゼ遺伝子のもの(“3’ocs”) (Gielen et al., 1984, EMBO J 3, 835-845)、およびT-DNA遺伝子7のもの (“3’遺伝子7”) (Velten and Schell, 1985, Nucleic Acids Research 13, 6981-6998) などが含まれ、これらは、形質転換された植物細胞において3’-非翻訳DNA配列として機能する。一つの態様において、配列番号3の2748ヌクレオチドから3168ヌクレオチドまで、および配列番号5に示される、トマトNRC1遺伝子の3’UTRが使用される。また、NRC1の3’UTRは、他のコード領域と組み合わせて3’UTRとして使用され得るため、本明細書においてそれ自体で一つの態様でもある。同様に、配列番号5の任意の変異体またはフラグメントが提供される。配列番号5の変異体には、配列番号5に対して少なくとも40、50、60、70、80、90、95、98、99%またはそれ以上の核酸配列同一性を含む核酸配列が含まれる(上記で規定されるNeedleman and WunschアルゴリズムおよびGAPペナルティーを用いて決定される)。フラグメントには、配列番号5または配列番号5の変異体の少なくとも30、50、100、150、200、300、400またはそれ以上の連続ヌクレオチドを含む任意のヌクレオチド配列が含まれる。
【0089】
T-DNAベクターのアグロバクテリウムへの導入は、公知の方法、たとえばエレクトロポレーションまたはトリペアレンタルメイティングを用いて行うことができる。
【0090】
NRC1コード核酸配列は、必要に応じて、ハイブリッド遺伝子配列として植物ゲノムに挿入することができ、これによりNRC1遺伝子は、容易に検出可能な融合タンパク質を植物が発現するように、選択またはスコアマーカーをコードする遺伝子、たとえばカナマイシン耐性をコードするneo(またはnptII)遺伝子(EP 0 242 236)にフレームで連結される(US 5,254, 799; Vaeck et al., 1987, Nature 328, 33-37)。
【0091】
NRC1タンパク質(または変異体もしくはフラグメント)をコードするNRC1核酸配列の全部または一部は、微生物、たとえば細菌(たとえば大腸菌(Escherichia coli)、シュードモナス属(Pseudomonas)、アグロバクテリウム属(Agrobacterium)、バチルス属(Bacillus)など)、真菌類、または藻類または昆虫を形質転換するため、あるいは組換えウイルスを作成するために使用することができる。適切なクローニング媒体に組み込まれた本発明のNRC1核酸配列の全部または一部による細菌の形質転換は、慣用的なやり方で、好ましくはMaillon et al. (1989, FEMS Microbiol. Letters 60, 205-210.) およびWO 90/06999に記載される慣用的なエレクトロポレーション技術を用いて行うことができる。原核宿主細胞での発現のために、核酸配列のコドン使用頻度は、(上記で植物について記載したとおり)適宜、最適化してもよい。イントロン配列は除去すべきであり、最適な発現のための他の適合を公知のとおり行ってもよい。
【0092】
NRC1核酸配列のDNA配列は、翻訳に中立的なやり方で、遺伝子部分に存在する抑制の可能性があるDNA配列を改変して、および/または上述のとおりコドン使用頻度に変化を導入することにより、たとえばコドン使用頻度を植物(好ましくは特定の関連の植物属)に最も好ましいものに適合させることにより、更に変化させることができる。
【0093】
本発明の一つの態様に従って、NRC1タンパク質(またはキメラタンパク質)は、細胞内オルガネラ、たとえばプラスチド、好ましくは葉緑体、ミトコンドリアをターゲットにするか、あるいは潜在的にタンパク質の安定性および/または発現を最適化して、細胞から分泌される。同様に、タンパク質は、液胞をターゲットにしてもよい。この目的のため、本発明の一つの態様において、本発明のキメラ遺伝子は、シグナルまたはターゲットペプチドをコードするコード領域を、本発明のNRC1タンパク質コード領域に連結して含む。本発明のタンパク質に含有される特に好ましいペプチドは、葉緑体または他のプラスチドターゲッティングのためのトランジットペプチド、とりわけ遺伝子産物がプラスチドをターゲットにする植物遺伝子由来の重複トランジットペプチド領域、Capellades et al. (US 5,635,618) の最適化されたトランジットペプチド、ホウレンソウ由来のフェレドキシン-NADP+オキシドレダクターゼのトランジットペプチド(Oelmuller et al., 1993, Mol. Gen. Genet. 237,261-272)、Wong et al. (1992, Plant Molec. Biol. 20, 81-93) に記載されるトランジットペプチド、およびPCT特許出願公開公報WO 00/26371のターゲッティングペプチドである。また、かかるペプチドに連結されたタンパク質の細胞外への分泌のシグナルを送るペプチド、たとえば、ポテトのプロテイナーゼインヒビターIIの分泌シグナル(Keil et al., 1986, Nucl. Acids Res. 14, 5641-5650)、イネのアルファーアミラーゼ3遺伝子の分泌シグナル(Sutliff et al., 1991, Plant Molec. Biol. 16, 579-591)、およびトマトのPR1タンパク質の分泌シグナル(Cornelissen et al., 1986, EMBO J. 5, 37-40)も好ましい。本発明に従って特に有用なシグナルペプチドには、葉緑体トランジットペプチド(たとえばVan Den Broeck et al., 1985, Nature 313, 358)、または葉緑体へのタンパク質の輸送を引き起こすUS 5,510,471およびUS 5,635,618の最適化された葉緑体トランジットペプチド、分泌シグナルペプチド、または他のプラスチド、ミトコンドリア、ER、または別のオルガネラをタンパク質のターゲットにするペプチドが含まれる。細胞内オルガネラへのターゲッティングまたは植物細胞の外もしくは細胞壁への分泌のためのシグナル配列は、天然においてターゲットにされるかまたは分泌されるタンパク質に見出され、好ましくは以下の文献に記載されるものである:Klosgen et al. (1989, Mol. Gen. Genet. 217, 155-161)、Klosgen and Weil (1991, Mol. Gen. Genet. 225, 297-304)、Neuhaus & Rogers (1998, Plant Mol. Biol. 38, 127-144)、Bih et al. (1999, J. Biol. Chem. 274, 22884-22894)、Morris et al. (1999, Biochem. Biophys. Res. Commun. 255, 328-333)、Hesse et al. (1989, EMBO J. 8, 2453-2461)、Tavladoraki et al. (1998, FEBS Lett. 426,62-66.)、Terashima et al. (1999, Appl. Microbiol. Biotechnol. 52,516-523)、Park et al. (1997, J.Biol. Chem. 272, 6876-6881)、Shcherban et al. (1995, Proc. Natl. Acad. Sci USA 92,9245-9249)。
【0094】
形質転換された宿主細胞の外へNRC1タンパク質を分泌するためには、適切な分泌シグナルが、NRC1タンパク質のアミノ末端(N-末端)に融合され得る。推定シグナルペプチドは、コンピューターベースの分析を用いて、プログラムSignal Peptide search (SignalP V1.1または2.0) などのプログラムを用いて検出することができる(Von Heijne, Gunnar, 1986 and Nielsen et al., 1996)。
【0095】
一つの態様において、幾つかのNRC1コード核酸配列を、単一の宿主において、必要に応じて異なるプロモーターの制御下で共発現させる。共発現する宿主植物は、本発明のNRC1タンパク質を既に発現している植物を形質転換することにより、あるいは本発明の異なるNRC1タンパク質で形質転換された植物を交配させることにより容易に得られる。あるいは、幾つかのNRC1タンパク質をコードする核酸配列を、単一の形質転換ベクター上に存在させることができるし、あるいは別々のベクターを用いて同時に共形質転換(co-transform)して、両方のキメラ遺伝子を含む形質転換体を選択することもできる。同様に、一または複数のNRC1コード遺伝子を、たとえば耐病性を向上させるかまたは耐病性シグナリング経路に関与する他のタンパク質などをコードする他のキメラ遺伝子と一緒に、単一の植物で発現させてもよい。
【0096】
異なるタンパク質を同一の植物で発現させることができること、あるいは各々のタンパク質を単一の植物で発現させた後に、当該植物を互いに交配することにより同一の植物で各タンパク質を組み合わせることができることが理解される。たとえば、ハイブリッド種子生産において、各々の親植物は、単一のタンパク質を発現することができる。親植物を交配してハイブリッドを産生すると、ハイブリッド植物において両方のタンパク質が組み合わされる。
【0097】
好ましくは、選択の目的のため、更に雑草防除オプションのために、本発明のトランスジェニック植物は、除草剤、たとえば広範囲の除草剤、たとえば有効成分としてグルホシネートアンモニウムを主成分とする除草剤(たとえばLiberty(登録商標)またはBASTA; 耐性は、PATまたはbar遺伝子により付与される; EP 0 242 236およびEP 0 242 246参照)またはグリホサートを主成分とする除草剤(たとえばRoundUp(登録商標); 耐性は、EPSPS遺伝子により付与される、たとえばEP 0 508 909およびEP 0 507 698参照)に対する耐性を付与するタンパク質をコードするDNAで形質転換される。更に、除草剤耐性遺伝子(または所望の表現型を付与する他の遺伝子)の選択マーカーとしての使用は、抗生物質耐性の導入を回避することができるという利点を有する。
【0098】
あるいは、他の選択マーカー遺伝子、たとえば抗生物質耐性遺伝子が使用されてもよい。一般に、形質転換宿主植物に抗生物質耐性遺伝子を保持することは受け入れられないため、これら遺伝子は、形質転換体の選択後に再度除去することができる。導入遺伝子を除去するための幾つかの技術が存在する。除去するための一つの方法は、キメラ遺伝子をlox部位と隣接させ、選択後、形質転換植物をCREリコンビナーゼ発現植物と交配することによるものである(たとえばEP506763B1参照)。部位特異的組換えの結果、マーカー遺伝子は切り出される。別の部位特異的組換えシステムは、EP686191およびUS5527695に記載されるFLP/FRTシステムである。遺伝子スタッキングの目的のために、CRE/LOXおよびFLP/FRTなどの部位特異的組換えシステムが使用されてもよい。更に、一成分(one-component)切出しシステムが記載されている(たとえばWO9737012またはWO9500555参照)。
【0099】
本発明の形質転換された植物細胞/植物/種子、並びに核酸配列およびタンパク質の使用
以下の記載において、向上した耐病性の表現型を備えたトランスジェニック植物細胞、植物、植物種子など、およびこれらの誘導体/子孫を作成するための本発明のNRC1配列の使用について説明される。
【0100】
向上した耐病性を備えたトランスジェニック植物は、上述のとおり、少なくとも一つのNRC1タンパク質を適切なプロモーターの制御下でコードする核酸配列を用いて植物宿主細胞を形質転換し、前記細胞からトランスジェニック植物を再生することにより作成することができる。
【0101】
好ましいプロモーターは、生物的および/または非生物的な外的刺激により誘導性であるプロモーターである。とりわけ病原体誘導性であるプロモーターが、上述のとおり好ましい。好ましいプロモーターとNRC1の組合せは、以下のとおりである:
a)病原体誘導性プロモーター − 構成的に活性なNRC1タンパク質をコードする核酸配列;
b)病原体誘導性プロモーター − 野生型NRC1タンパク質をコードする核酸配列;
c)植物NRC1遺伝子のプロモーター(好ましくは形質転換される同一種のもの) − 構成的に活性なNRC1タンパク質をコードする核酸配列;
d)植物NRC1遺伝子のプロモーター(好ましくは形質転換される同一種のもの) − 野生型NRC1タンパク質をコードする核酸配列;
e)生物的ストレス誘導性プロモーター(たとえば昆虫傷害誘導性、病原体誘導性など) − 構成的に活性なNRC1タンパク質をコードする核酸配列;
f)生物的ストレス誘導性プロモーター(たとえば昆虫傷害誘導性、病原体誘導性など) − 野生型NRC1タンパク質をコードする核酸配列;
g)構成的プロモーター(たとえば35Sプロモーター) − 野生型NRC1タンパク質をコードする核酸配列;
h)構成的プロモーター(たとえば35Sプロモーター) − 配列番号2に対して全長にわたって少なくとも70%のアミノ酸配列同一性を含むアミノ酸配列をコードする核酸配列;
i)病原体誘導性プロモーター − 配列番号2に対して全長にわたって少なくとも70%のアミノ酸配列同一性を含むアミノ酸配列をコードする核酸配列;
j)植物NRC1遺伝子のプロモーター − 配列番号2に対して全長にわたって少なくとも70%のアミノ酸配列同一性を含むアミノ酸配列をコードする核酸配列。
【0102】
一つの態様において、トランスジェニック植物は、構成的HR病変または誘導性HR病変の何れか、および一または複数の病原体に対して向上した耐病性を示すことができる。しかし、植物が向上した耐病性を示している間に、HR病変が発生しないか、または“弱い”HR病変(たとえば小さい病変、たとえばミクロ病変、および/または低い病変の頻度)が発生することも本明細書において想定される。宿主植物において同一プロモーターの制御下で発現されたときに、配列番号1または3より、あるいは形質転換されるのと同じ宿主種から得た野生型のNRC1対立遺伝子の発現より、少ないおよび/または小さいHR病変につながるNRC1対立遺伝子またはオルソログが、本明細書において、とりわけ上記アプローチg)およびh)において、特に好ましい。かかる対立遺伝子/オルソログは、所定の宿主に“弱いHR表現型”を付与するNRC1対立遺伝子ということもできる。かかるNRC1対立遺伝子またはオルソログは、上述のとおり同定および/または単離することができる。異なるNRC1対立遺伝子および/またはオルソログのHR表現型は、これらを用いて発現ベクターを作成し(好ましくは、比較される全ての核酸を同一のプロモーター、たとえば35Sに動作可能に連結し)、これらを用いて植物または植物組織を形質転換し、これら植物の間でHR表現型を比べることにより比較することができる。ナス科植物の形質転換体について、配列番号1または3を発現する形質転換体のHR病変の表現型が、好ましくはレファンレスとして使用され、同じプロモーターの制御下での発現により少ないおよび/または小さいHR病変が結果として得られる対立遺伝子が、弱いHR表現型を付与する対立遺伝子である。HR病変の表現型は、種々の方法を用いて、たとえば顕微鏡検査(必要に応じて死細胞の染色)、視覚的スコアリング、cm2あたりの数を計算するための病変の計数、HR病変の直径の測定などにより比較し、必要に応じて定量することができる。
【0103】
好ましくは、本発明のトランスジェニック植物は、一または複数の病原体、とりわけトランスジェニック植物種の生物栄養性および/または半生物栄養性病原体に対して向上した耐病性を含む。このため、たとえばトランスジェニックトマトまたはポテト植物は、少なくとも一つまたは複数の上述の真菌、細菌、線虫の種および/またはウイルス病原体に対して向上した耐性、最も好ましくは一つまたは幾つかの生物栄養性および/または半生物栄養性の種に対して少なくとも向上した耐性を含む。
【0104】
“耐病性”または“増大/向上した耐病性”は、一または複数の植物病原体の攻撃に耐える形質転換体の(野生型またはコントロール形質転換体と比較して)向上した能力を指すために本明細書において使用され、言い換えれば、非形質転換(または空ベクター形質転換)コントロールと比較して、形質転換体の病気の徴候が有意に減少したことを指す。耐病性または向上した耐病性は、種々の方法を用いて決定され得る。しばしば、病気の徴候は、病原体の接種または接触後の一または複数の時点において病気の徴候を評価することにより(バイオアッセイまたは野外研究で)視覚的にスコア化される。代わりの方法には、病原体を検出し、必要に応じて定量する方法が含まれる。このため、組織の中/上で検出される病原体の量がコントロールと比較して有意に少ないか、または病原体の広がりがコントロールより有意に遅い場合、トランスジェニック植物は、向上した耐病性を示すことができる。最終的には、同等の病気プレッシャーの下で生育させたときに、コントロールと比較して形質転換体の平均収量が有意に増大すること(たとえば少なくとも1%、2%、5%、10%またはそれ以上)が、向上した耐病性の間接的な測定(measurement)を提供する。
【0105】
このため、NRC1タンパク質(または構成的に活性なNRC1タンパク質)を発現する複数の形質転換植物が、非形質転換コントロールまたは空ベクター形質転換コントロールと比較して病気の徴候の有意な減少を示す場合、向上した耐病性を示す。明らかに、有意な差が存在するかどうかを決定するためには統計分析が必要である。好ましくは、一または複数の病気の徴候は、コントロール植物よりNRC1形質転換において、平均して少なくとも2%、5%、10%、15%、20%、30%、40%、50%、または100%も少ない。病気のアッセイは、宿主と病原体の組合せごとに異なるため、特定のプロトコールは提供されないが、当業者であれば、形質転換体が、一または複数の病原体に対して有意に向上した耐病性を示すかどうかを決定する方法が分かる。各々の植物と病原体の組合せについて当該技術分野で公知のバイオアッセイを使用して、トランスジェニック植物の耐性を適切なコントロールと比較することができる。
【0106】
NRC1タンパク質は、幾つかの態様において(たとえばNRC1遺伝子が構成的プロモーターの制御下にある場合)病原体の非存在下においてHR病変という結果につながるため、NRC1発現により引き起こされる徴候と病原体の感染および広がりにより引き起こされる徴候とを区別することが、ある態様においては重要であり得る。したがって、(植物組織上の壊死よりも)病原体それ自体を検出すること、または存在する病原体の量または病原体の広がりの速度を比較することが好ましい。たとえば、病原体を染色により検出することができるバイオアッセイが使用され得る。実施例では、GUSを発現するトランスジェニックC. fulvum品種が使用される。したがって、真菌の菌糸体は、接種された植物組織のX-gluc染色を用いて視覚化することができる。コントロールと比較してトランスジェニック植物において真菌の菌糸体が有意に減少することは、真菌に対して向上した耐性を示す。
【0107】
また、幾つかのNRC1タンパク質を、好ましくは異なるプロモーターの制御下、たとえば異なる病原体誘導性プロモーターの制御下で、発現するトランスジェニック植物を作成することも一つの態様である。
【0108】
耐病性の表現型は、適切な時および場所に適切な量のNRC1タンパク質を発現させることにより微調整することができる。かかる微調整は、特定の宿主−病原体の組合せに対して最も適切なプロモーターを決定し、所望の発現レベルを示すトランスジェニック“イベント”を選択することにより行うことができる。病原体の攻撃後の低すぎるレベルのNRC1タンパク質またはNRC1タンパク質生産の遅すぎる誘導は、耐病性を向上させるのに不十分であり得る。一方、病原体の攻撃のない時および場所における高すぎるタンパク質のレベルまたは発現は、農学的に望ましくない表現型、たとえば病原体の非存在下における葉または果実の病変および収量の損失という結果につながり得る。しかし、当業者であれば、向上した耐病性を有するが、同時に農学的に許容される植物を容易に作成することができる。最適なNRC1対立遺伝子、たとえば高い耐性レベルとわずかな弱いHR表現型を提供する対立遺伝子は、記載されるとおり単離または同定することができる。
【0109】
所望のレベルのNRC1タンパク質を発現する形質転換体は、たとえば、コピー数の分析(サザンブロット分析)、mRNA転写産物レベルの分析(たとえばNRC1プライマー対またはフランキングプライマーを用いたRT-PCR)、または種々の組織におけるNRC1タンパク質の存在およびレベルの分析(たとえばSDS-PAGE;ELISA分析など)により選択される。調節の理由のため、好ましくは単一コピーの形質転換体が、好ましくは選択され、キメラ遺伝子の挿入部位に隣接する配列が分析され、好ましくはその“イベント”を特徴づけるために配列決定される。安定なNRC1導入遺伝子を備えた高性能の選抜きのイベントが得られるまで、高いまたは適度なNRC1発現トランスジェニックイベントが、更なる交配/戻し交配/自家受粉のために選択される。
【0110】
また、本発明の一または複数のNRC1遺伝子を発現する形質転換体は、他の導入遺伝子、たとえば耐病性を付与する遺伝子または他の生物的および/または非生物的ストレスに対する耐性を付与する遺伝子を含んでいてもよい。“多重(stacked)”導入遺伝子を備えたかかる植物を得るために、他の導入遺伝子がNRC1形質転換体に遺伝子移入されてもよいし、あるいはNRC1形質転換体が、一または複数の他の遺伝子でその後に形質転換されてもよいし、あるいは植物系統または品種を形質転換するために幾つかのキメラ遺伝子が使用されてもよい。たとえば、幾つかのキメラ遺伝子が、単一のベクターに存在していてもよいし、あるいは共形質転換される(co-transform)異なるベクターに存在していてもよい。
【0111】
一つの態様において、本発明の一または複数のNRC1遺伝子と以下の遺伝子が組み合わせられる:公知の耐病性遺伝子、とりわけ壊死栄養性病原体に対して向上した耐性を付与する遺伝子、ウイルス耐性遺伝子、昆虫耐性遺伝子、非生物的ストレス耐性遺伝子(たとえば干ばつ耐性、塩耐性、熱もしくは低温耐性など)、殺虫剤耐性遺伝子など。このため、多重(stacked)形質転換体は、さらに広い生物的および/または非生物的ストレス耐性、病原体耐性、昆虫耐性、線虫耐性、塩度、低温ストレス、熱ストレス、水ストレス耐性などをもち得る。また、NRC1サイレンシングアプローチが、単一の植物においてNRC1発現アプローチと組み合わせられてもよい。たとえば、根または塊茎におけるNRC1過剰発現は、土壌病原体に対する根または塊茎の耐性を付与するかまたは向上させ得る。同時に、空気中の(aerial)部分におけるNRC1のダウンレギュレーションは、壊死栄養性病原体に対する耐性を付与するかまたは向上させ得る(または逆も同様である(vice versa))。
【0112】
また、NRC1遺伝子を、既にあるレベルの耐病性を有する植物育種系統に導入または遺伝子移入することも可能である。畑での耐病性の耐久性のため、好ましくは耐性の供給源が、異なる基本的分子メカニズムを有する、幾つかの耐病性メカニズムを植物で重ねることが望ましい。
【0113】
上述の任意の形質転換植物の植物全体、種子、細胞、組織および子孫(たとえばF1雑種、F2種子/植物など)は、本明細書に包含され、DNAにおける導入遺伝子の存在により、たとえば鋳型として全ゲノムDNAを用いてNRC1特異的PCRプライマー対を用いてPCR分析を行うことにより、同定することができる。また、PCRプライマーが、挿入されたキメラ遺伝子に隣接する植物DNAに基づいている“イベント特異的”PCR診断方法を開発することができる。US6563026参照。同様に、トランスジェニック植物、またはそれに由来する任意の植物、種子、組織または細胞を同定するイベント特異的AFLPフィンガープリントまたはRFLPフィンガープリントが開発され得る。
【0114】
本発明のトランスジェニック植物は、好ましくは、望ましくない表現型、たとえば収量の減少、病気(とりわけ壊死栄養体)に対して向上した感受性、または望ましくない構造の変化(矮性、変形)などを示さないこと、並びにかかる表現型が第一形質転換体で観察される場合、これらは、通常の育種と選抜法(交配/戻し交配/自家受粉など)により除去することができることが理解される。本明細書で記載される任意のトランスジェニック植物は、導入遺伝子についてホモ接合であっても半接合であってもよい。
【0115】
NRC1遺伝子サイレンシングアプローチおよび遺伝子サイレンシングベクター
NRC1遺伝子サイレンシングベクターで形質転換され、とりわけ壊死栄養性病原体に対して向上した耐病性を備えた植物を提供することが本発明の更なる態様である。本発明の範囲を限定しないが、内在性NRC1遺伝子または遺伝子ファミリーのサイレンシングは、トランスジェニック植物がHR応答を誘発および/または開始できないという結果につながると考えられる。壊死栄養性病原体は、増殖および発生のために細胞死を必要とするため、かかる植物は、一または複数の壊死栄養性病原体に対して向上した耐性を含み得る。
【0116】
“遺伝子サイレンシング”は、一または複数のターゲット遺伝子(たとえば内在性NRC1遺伝子)の遺伝子発現のダウンレギュレーションまたは完全な抑制を指す。遺伝子発現を低下または廃止する抑制性RNAの使用は、当該技術分野で十分に確立されており、幾つかのレビューの主題である(たとえばBaulcombe 1996, Stam et al. 1997, Depicker and Van Montagu, 1997)。植物で遺伝子サイレンシングを達成するために利用可能な幾つかの技術が存在し、たとえばターゲット遺伝子の全部または一部のアンチセンスRNAを産生するキメラ遺伝子(たとえばEP 0140308 B1、EP 0240208 B1およびEP 0223399 B1参照)、または(共抑制とも称される)センスRNAを産生するキメラ遺伝子(EP 0465572 B1参照)である。
【0117】
しかし、これまで最も成功したアプローチは、ターゲット遺伝子のセンスおよびアンチセンスRNAの両方の産生(“逆方向反復配列”)であり、これは、二本鎖RNA(dsRNA)を細胞で形成し、ターゲット遺伝子をサイレンシングする。dsRNAの産生および遺伝子サイレンシングのための方法およびベクターは、EP 1068311、EP 983370 A1、EP 1042462 A1、EP 1071762 A1およびEP 1080208 A1に記載されている。したがって、本発明のベクターは、本発明のNRC1遺伝子のセンスおよび/またはアンチセンスDNAフラグメントに動作可能に連結された植物細胞で活性な転写調節領域を含み得る。一般に、短い(センスおよびアンチセンス)長さのターゲット遺伝子配列、たとえば17、18、19、20、21、22または23ヌクレオチドのコードまたは非コード配列で十分である。より長い配列、たとえば50、100、200または250ヌクレオチドまたはそれ以上も使用することができる。好ましくは、短いセンスおよびアンチセンスのフラグメントは、スペーサー配列、たとえばイントロンにより分離され、これによりdsRNAの形成によりループ(またはヘアピン)が形成される。任意の短い長さの配列番号1または3、またはその変異体を使用して、NRC1遺伝子サイレンシングベクター、および一または複数のNRC1遺伝子が(使用されるプロモーターに依存して)全部または幾つかの組織または器官でサイレンシングされたトランスジェニック植物を作成してもよい。ヘアピン構築物を作成する簡便な方法は、ジェネリックベクター、たとえばpHANNIBALおよびpHELLSGATE、Gateway(登録商標)技術に基くベクターを使用することである(Wesley et al. 2004, Methods Mol Biol. 265:117-30; Wesley et al. 2003, Methods Mol Biol. 236:273-86 and Helliwell & Waterhouse 2003, Methods 30(4):289-95参照。全てが参照により本明細書に組み込まれる)。
【0118】
NRC1遺伝子の保存された核酸部分を選択することにより、宿主の植物または植物部分においてNRC1ファミリーメンバーをサイレンシングさせることができる。NRC1遺伝子のセンスおよび/またはアンチセンスDNAフラグメントに動作可能に連結された転写調節エレメントを含み、一または複数の病原体、とりわけ壊死栄養性病原体に対して向上した耐性を示すトランスジェニック植物も本明細書に包含される。
【0119】
また、一または複数の生物栄養性および/または半生物栄養性病原体および一または複数の壊死栄養性病原体に対して向上した耐性を有する植物も提供される。かかる植物は、適切なプロモーター−NRC1遺伝子の組合せを選択することにより作成することができる。たとえば、内在性NRC1遺伝子が、種々の組織および/または種々の時期に(たとえば芽生え、根または塊茎などにおいて)サイレンシングされ、一または複数の壊死栄養性病原体に対して耐性を提供している間に、機能的なNRC1タンパク質が、ある組織においてある時期に(たとえば誘導により、または空気中の(aerial)植物部分において)産生され、生物栄養性および/または半生物栄養性病原体に対して耐性を提供してもよい。したがって、単一の植物が、キメラNRC1発現導入遺伝子およびNRC1サイレンシング遺伝子の両方を含んでいてもよい。
【0120】
本発明の突然変異した対立遺伝子および植物
非トランスジェニック方法、たとえば、TILLINGなどの突然変異誘発システム(Targeting Induced Local Lesions IN Genomics; McCallum et al., 2000, Nat Biotech 18:455, and McCallum et al. 2000, Plant Physiol. 123, 439-442、何れも参照により本明細書に組み込まれる)を使用すること、および高レベルの本発明の一または複数のNRC1タンパク質を産生し、および/または構成的に活性なNRC1タンパク質を産生する植物系統を作成するための選択を使用することも本発明の一つの態様である。本発明の範囲を限定しないが、かかる植物は、宿主NRC1遺伝子の発現を構成的にするかまたは高くするリプレッサータンパク質の結合部位であるプロモーター内のポイント/欠失突然変異を含むことができると考えられる。構成的に活性なNRC1突然変異体は、コード領域、たとえばMHD領域に突然変異を含む。好ましくは、突然変異体または突然変異体の一部におけるNRC1タンパク質レベルは、突然変異体において、非突然変異体の植物と比較して少なくとも約2、5、10、15%、またはそれ以上増大している。TILLINGは、従来の化学的突然変異誘発(たとえばEMS突然変異誘発)と、その後の突然変異のハイスループットスクリーニング(たとえば突然変異体−野生型のDNAへテロ二本鎖の細胞分裂および配列決定ゲルシステムを用いた検出)を使用する(たとえばHenikoff et al. Plant Physiology Preview May 21, 2004参照)。このため、一または複数の組織で向上したNRC1遺伝子発現を含み、本発明の一または複数のNRC1表現型(向上した耐病性および/またはHR病変)を含む非トランスジェニック植物、種子および組織、並びにかかる植物を作成し、同定する方法が、本明細書に包含される。
【0121】
この方法は、一つの態様において、植物種子の突然変異誘発(たとえばEMS突然変異誘発)、植物個体またはDNAのプーリング、対象領域のPCR増幅、ヘテロ二本鎖の形成およびハイスループット検出、突然変異体植物の同定、突然変異体PCR産物の配列決定の工程を含む。他の突然変異誘発および選択方法が、かかる突然変異体植物を作成するために同様に使用され得ることが理解される。たとえば、種子を、放射線照射するかまたは化学的に処理し、植物を、改変された表現型、たとえば向上した耐病性および/またはHR病変についてスクリーニングしてもよい。
【0122】
本発明の別の態様において、植物材料は、NRC1オルソログのコード配列および/または調節配列のDNA配列に多型または変異を含む種または関連種の天然の集団である。NRC1遺伝子ターゲットにおける突然変異は、ECOTILLINGアプローチを使用してスクリーニングすることができる(Henikoff et al 2004, supra)。この方法において、育種系統または関連種における天然の多型は、上記TILLING方法によりスクリーニングされ、ここで植物の個体またはプールは、NRC1ターゲットのPCR増幅、ヘテロ二本鎖形成、およびハイスループット分析のために使用される。この後、必要な突然変異を有する個々の植物の選択を行うことができ、これは、所望のNRC1オルソログ対立遺伝子を組み込んで所望の特徴を備えた栽培品種を開発するための育種プログラムで引き続き使用することができる。
【0123】
突然変異体の植物は、分子的方法により、たとえばDNAに存在する突然変異、NRC1タンパク質レベル、NRC1 RNAレベルなどにより、および改変された表現型特性により、非突然変異体と区別することができる。
【0124】
非トランスジェニック突然変異体は、突然変異について、ホモ接合であってもヘテロ接合であってもよい。
【0125】
参照される配列
配列番号1:トマトNRC1遺伝子のコード領域
配列番号2:トマトNRC1タンパク質のアミノ酸配列
配列番号3:トマトNRC1遺伝子の全長cDNA(5’および3’UTRを含む)
配列番号4:トマトNRC1D481Vタンパク質のアミノ酸配列
配列番号5:トマトNRC1遺伝子の3’UTR
図説
図1 − NRC1タンパク質の推定配列
最初の150アミノ酸残基は、二重コイル(coiled-coil)(CC)ドメインを表し、CC構造を形成すると推定される残基に下線を付す。151〜508残基は、以下の(下線とラベルを付した)モチーフを備えたヌクレオチド結合(NB-ARC)ドメインを含む:Kinase1A (P loop)、RNBS-A、Kinase 2、RNBS-B、RNBS-C、GLPL、RNBS-DおよびMHD。509〜846残基は、13の不完全なロイシンリッチ繰返し配列(LRRs)を含み;保存された疎水性残基およびプロリン残基は太字で示す。タンパク質配列の下に、LRRコンセンサスモチーフを示す:‘l’は、保存された脂肪族残基を示し、‘c’は、保存された電荷を帯びた残基を示し、‘P’は、保存されたプロリン残基を示す。
【0126】
図2A − NRC1は、Cladosporium fulvumに対するトマトの完全なCf-4-介在性HRに必要である
Cf0トマトおよびCf-4-含有トマト植物に、表示のTRV構築物を接種し、植物をVIGSの開始から3週間分析した。TRV感染Cf-4-含有トマト植物の小葉にAvr4タンパク質を注入し、HRの発生について調査した。TRV:00-感染植物上でHRが載っている部位の数を100%とした。各々の誤差バーは、4つの独立実験に由来する標準偏差を示す。
【0127】
図2B − NRC1は、Cladosporium fulvumに対するトマトの完全なCf-4-介在性耐性に必要である
非TRV感染およびTRV感染Cf-4またはCf-0植物に、C. fulvum-pGPD::GUSを接種し、接種から2週間後に、小葉のコロニー形成をX-glucアッセイで調査した。
【0128】
図3 − N. benthamianaへのTRV:NRC1の接種は、Cf/Avr-、LeEix2/tvEix-、Pto/AvrPto-およびRx/CP-誘導性HRに影響を及ぼす
N. benthamianaに、TRV:00 (空ベクター)、TRV:NRC1およびTRV:SGT1を接種した。3週間後、HR-誘導性タンパク質を発現するアグロバクテリアを葉にインフィルトレーションし(infiltrate)、インフィルトレーションから4日後に写真を撮影した。第1、第2および第3カラム:Avr4、またはCf-9とAvr9の混合物、またはLeEix2とtvEixの混合物 (1:1の比で混合)をそれぞれアグロインフィルトレーションされた、Cf-4耐性遺伝子を発現するN. benthamianaの葉。第4カラム:AvrPtoをアグロインフィルトレーションされた、Pto耐性遺伝子を発現するトランスジェニックN. benthamianaの葉。第5カラム:PVX (CP)のコートタンパク質を発現する遺伝子でアグロインフィルトレーションされた、Rx耐性遺伝子を発現するトランスジェニックN. benthamianaの葉。濃い円はHRを示し、薄い円は低下した(compromised)HRを示す。
【0129】
図4 − 構成的に活性なNRC1は、誘導因子−非依存性HRを誘導し、細胞死シグナリング経路にNRC1を配置させる
Cf-4耐性遺伝子を発現するN. benthamianaに、表示の遺伝子をアグロインフィルトレーションした。パネルAとCについては、アグロインフィルトレーションの3週間前に、植物に表示のTRV構築物を接種した。濃い円はHRを示し、薄い円は低下した(compromised)HRを示す。
【0130】
(A) 構成的に活性なMAPKKおよびMAPKキナーゼをコードする遺伝子のアグロインフィルトレーション。第一カラム:LeMAPKKKαの構成的に活性なキナーゼドメイン (MAPKKK-KD) をコードする遺伝子を用いたアグロインフィルトレーション。第2カラム:LeMEK2の構成的に活性な型 (MEK2DD) をコードする遺伝子を用いたアグロインフィルトレーション。MAPKKK-KDまたはMEK2DDのインフィルトレーションから2日後、葉にエストラジオールをスプレーすることにより発現は誘導された。アグロインフィルトレーションから4日後に写真を撮影した。
【0131】
(B) 35S-プロモーターの制御下の野生型NRC1 (wt) および当該遺伝子の突然変異型のアグロインフィルトレーションであって、サイレンシングサプレッサーp19をコードする遺伝子の発現を駆動するアグロバクテリウムと1:1の比で混合されたアグロインフィルトレーション (左パネル)、または単独のアグロインフィルトレーション (右パネル)。NRC1K191R (K191R):NRC1の不活性なP-ループ突然変異体;NRC1D481V (D481V):構成的に活性なNRC1 (MHDモチーフでの突然変異);NRC1K191R/D481V (K191R/D481V):NRC1のダブル突然変異体。アグロインフィルトレーションから3日後に写真を撮影した。
【0132】
(C) Avr4および構成的に活性なNRC1D481V (D481V) をコードする遺伝子のアグロインフィルトレーション。アグロインフィルトレーションから3日後に写真を撮影した。
【0133】
図5 − NRC1介在性細胞死シグナリングのモデル
細胞死アッセイとN. benthamianaにおけるVIGSを組み合わせるエピスタシス実験に基くモデル。EDS1-、NRC1-、MEK2-、およびSGT1/RAR1依存的なやり方でのCf-4/Avr4介在性細胞死シグナル。
【0134】
以下の非限定的な実施例は、本発明の様々な態様を詳説する。実施例において特に記載しない限り、全ての組換えDNA技術は、Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press、およびSambrook and Russell (2001) Molecular Cloning: A Laboratory Manual, Third Edition, Cold Spring Harbor Laboratory Press, NY; およびAusubel et al. (1994) Current Protocols in Molecular Biology, Current Protocols, USAのVolumes 1 and 2に記載されるとおり、標準的なプロトコールに従って実施した。植物の分子的研究のための標準的な材料および方法は、Plant Molecular Biology Labfax (1993) by R.D.D. Croy, jointly published by BIOS Scientific Publications Ltd (UK) and Blackwell Scientific Publications, UKに記載される。
【0135】
実施例
1.材料および方法
1.1 N. benthamianaにおけるVIGS、アグロインフィルトレーション、HRおよび病気アッセイ
4週齢のN. benthamiana植物に、pTV00-由来の構築物 (バイナリーTRV RNA2ベクター) とpBintra6 (バイナリーTRV RNA1ベクター) の1:1混合物 (Ratcliff et al., 2001 Plant J. 25, 237-245)、またはpTRV-RNA2-由来の構築物とpTRV-RNA1の1:1 混合物 (Liu et al., 2002, Plant J. 31, 777-786; Liu et al., 2002, Plant J. 30, 415-429) をアグロインフィルトレーションした。以下のTRV構築物を使用した: TRV:NRC1、TRV:Cf-4 およびTRV:SGT1 (Peart et al., 2002, Proc. Natl. Acad. Sci. USA 99, 10865-10869)、Ratcliff et al. (2001, supra) に記載されるTRVベクターの全ておよびTRV:EDS1、TRV:MEK2、TRV:RAR1およびTRV:NDR1 (Ekengren et al., 2003, Plant J. 36, 905-917)、Liu et al. (2002, Plant J. 30, 415-429) に記載されるTRVベクターの全て。各TRV構築物について、各実験で4つの植物を使用した。AvrPtoおよびCPを、耐性遺伝子Ptoを発現するTRV-感染N. benthamiana (N. benthamiana:Pto (line 38-12 (Rommens et al., 1995, Plant Cell 7, 1537-1544)) (Pedley and Martin, 2003, Annu. Rev. Phytopathol. 41, 215-243) および耐性遺伝子Rxを発現するTRV-感染N. benthamiana (N. benthamiana:Rx (line Rx-18) (Bendahmane et al., 1999, Plant Cell 11, 781-791)に、それぞれアグロインフィルトレーションした。全ての他のケースにおいて、アグロインフィルトレーションは、耐性遺伝子Cf-4を発現するN. benthamiana (N. benthamiana:Cf-4) で行った。TRV接種から3週間後、接種された葉の上の第三葉、第四葉および第五葉を、以下の発現を駆動するAgrobacterium tumefaciensで攻撃した:AvrPto (OD600=0.06) (Tang et al., 1996, Science 274, 2060-2063)、CP (OD600=0.12) (Bendahmane et al., 1999, supra)、Avr4 (OD600=0.03)、Cf-9およびAvr9 (1:1の比で混合、OD600=0.2) (Van der Hoorn et al., 2000 Mol. Plant-Microbe Interact. 13, 439-446)、LeEix2およびtvEix (1:1の比で混合、OD600=1) (Ron and Avni, 2004, Plant Cell 16, 1604-1615)、β-グルクロニダーゼ (GUS) 遺伝子 (OD600=2) (Van der Hoorn et al., 2000, Mol. Plant-Microbe Interact. 13, 439-446)、NRC1およびp19 (1:1の比で混合、OD600=1) (Voinnet et al., 2003, Plant J. 33, 949-956)、構成的に活性なNRC1D481Vまたは不活性なNRC1K191R/D481V ダブル突然変異体 (OD600=2)、LeMAPKKKαKD、LeMAPKKKαKD- (Del Pozo et al., 2004, EMBO J. 23, 3072-3082) (何れもOD600=0.12) またはLeMEK2DDおよびLeMEK2 (Del Pozo et al., 2004, supra) (何れもOD600=0.25)。LeMAPKKKαKD、LeMAPKKKαKD-、LeMEK2DDまたはLeMEK2のインフィルトレーションから2日後に、silwet (4μl/100 ml) を含有する17-β-エストラジオールの7.5 μM水溶液を葉にスプレーした (Del Pozo et al., 2004, supra)。タンパク質の注入のために、Avr4-HIS-FLAG-標識タンパク質を、製造メーカー (Inv
itrogen, Breda, NL) の推奨に従ってエンテロキナーゼEK-maxで処理し、0.2% tween (v/v) を添加した水中の5μM Avr4タンパク質を、注入のために使用した。アグロインフィルトレーションまたはタンパク質の注入から3〜5日後、HRの発生について葉を調査するか、またはβ-グルクロニダーゼ (GUS) 活性について葉をアッセイした。
【0136】
1.2 トマトにおけるVIGS、HRおよび病気アッセイ
トマトにおけるVIGSのために、Liu et al. (2002, Plant J. 30, 415-429) に記載されるpTRV-RNA1およびpTRV-RNA2ベクターを使用した。Cf-4およびNRC1フラグメントは、BamH1/Asp718を用いた消化によりpTV00から切出し、BamH1/Asp718-消化pTRV-RNA2 (pYL156) に挿入した (Liu et al., 2002, Plant J. 31, 777-786)。TRV:222-UTRを構築するために、NRC1の3’-UTRの一部を、プライマー222-3’UTR-F (5’-GTGGATCCGCAGGTTCAACCAGCCTGGT-3’; BamH1部位に下線を付す) および222-3’UTR-R (5’-GTGGTACCCAAGTGACTTGTTCTGCTGT-3’; Asp718部位に下線を付す) を用いて増幅し、TRV:222-LRRを構築するために、LRRをコードするNRC1領域の一部を、プライマー222-LRR-F (5’-GTGGATCCGTTAAGAGGCTGCAATTTCT-3’; BamH1部位に下線を付す) および222-LRR-R (5’-GTGGTACCGATCTTCTCAAGTTTATCAC-3’; Asp718部位に下線を付す) を用いて増幅した。PCRフラグメントは、BamH1/Asp718で消化し、BamH1/Asp718-消化pTRV-RNA2に挿入した。TRV:Prfの構築は記載されている (Ekengren et al., 2003, Plant J. 36, 905-917)。全てのプラスミドは、エレクトロポレーションによりA. tumefaciens株 GV3101に形質転換した (Takken et al., 2000, Plant J. 24, 275-283)。トマトにおいてVIGSを確立するために、10〜12日齢のトマト芽生えの子葉に、pTRV-RNA1とpTRV-RNA2-由来構築物の混合物(1:1の比で混合)をアグロインフィルトレーションした (Liu et al., 2002, supra)。TRV構築物の各々について、Avr4を発現するC. fulvumに対して耐性のCf-4-含有トマト植物の4個体 (Hcr9-4D (Cf-4)で形質転換したCf0植物) (Thomas et al., 1997, Plant Cell 9, 2209-2224) またはAvr9を発現するC. fulvumに対して耐性のCf-9-含有トマト植物の4個体 (Hcr9-9C (Cf-9)で形質転換したCf0植物) (Jones et al., 1994, Science 266, 789-793) の何れかを使用した。コントロールとして、TRV:00- またはTRV:NRC1- の何れかを接種され、C. fulvumに完全に感受性を有するCf0トマト植物 (MM-Cf0) を使用した。病気アッセイのために、TRV接種から3週間後に、Cf0およびCf-4-含有植物にC. fulvumを接種した (De Wit, 1977, Neth. J. Plant Path. 83, 109-122)。C. fulvum品種5- pGPD::GUSを使用した (構成的GPDプロモーターの制御下でAvr4およびβ-グルクロニダーゼ遺伝子を発現する)。小葉のコロニー形成は、X-gluc 染色により2週間後に評価した。平行して、植物の第二、第三または第四複葉の小葉をRT-PCR分析のために使用して、対象遺伝子の‘ノックダウン’について試験した (下記参照)。HRアッセイのために、TRV-感染のCf-4-またはCf-9-含有植物の第三複葉の小葉に、それぞれ、Avr4またはAvr9を注入した。何れの誘導因子も、マイクロシリンジで小葉に注入した (Ito Corporation, Fuji, Japan)。Avr4は、10μMの濃度で、小葉あたり10の部位に、植物あたり4つの小葉に注入した。Avr9については、(C. fulvumの品種5とCf0植物との適合性相互作用から分離される)約10μMのAvr9を含有する8倍希釈のアポプラスト液を、小葉あたり8つの部位に、植物あたり4つの小葉に注入した。Pseudomonas syringae pv. tomatoに対する耐性は、TRV:00、TRV:PrfまたはTRV:NRC1を接種されたトマトRG-PtoR (Pto/Pto, Prf/Prf) においてアッセイした。接種の手順および葉の細菌コロニー形成の決定は、以前に記載されるとおり行った (Ekengren et al., 2003, supra)。
【0137】
1.3 バイナリー35S:NRC1ベクターの構築および突然変異誘発
全長NRC1 cDNAは、プライマー
222-Start-F (5’-GGGATCCATGGTTGATGTAGGGGTTGA-3’) および
222-Stop-R (5’-GTCACTGCAGACCTTTCTAAGAAGCTGTCTG-3’)
を使用してPCR増幅し、これによりそれぞれNcoIおよびPstI制限部位を導入した (制限部位に下線を付す)。PCRフラグメントは、NcoI/PstI-消化し、NcoI/PstI-消化pRH80に挿入した (Van der Hoorn et al., 2000, Mol. Plant-Microbe Interact. 13, 439-446)。その後、構築物をXbaI/KpnI-消化し、35Sプロモーター、NRC1オープンリーディングフレームおよびNOSターミネーター (tNOS) を含有する得られたフラグメントを、XbaI/KpnI-消化pMOG800バイナリーベクターにクローニングして (Honee et al., 1998, Plant Physiol. 117, 809-820)、プラスミドNRC1(wt)を作成した。構成的に活性なバイナリーNRC1D481Vを作成するため、D481V突然変異は、鋳型としてNRC1wtプラスミドを使用し、フランキングプライマー222-Start-Fおよび222-Stop-Rおよびミスマッチプライマー
222MHD-F (5’-CAAAACTTGTCGTGTTCATGTCATGTTGTATGAG-3’) および
222MHD-R (5’-CCAGCAAAACTCATACAACATGACATGAACACGAC-3’) (突然変異に下線を付す)
を使用して、オーバーラップ伸長PCRにより導入した (Higuchi et al., 1988, Nucleic Acids Res. 16, 7351-7367)。上述のとおり、フラグメントをNcoI/PstI-消化し、pRH80に挿入し、35S-NRC1D481V-tNOSフラグメントを切出し、その後pMOG800に挿入した。同様の方法で、P-ループ突然変異体NRC1K191Rおよび不活性なダブル突然変異体NRC1K191R/D481Vを作成した。ここで、K191R突然変異は、鋳型としてそれぞれプラスミドNRC1(wt)およびNRC1D481Vを使用し、ミスマッチプライマー
222Ploop-F (5’-GGAATGCCTGGTCTTGGCAGAACTACACTAGC-3’) および
222Ploop-R (5’- GCTAGTGTAGTTCTGCCAAGACCAGGCATTCC-3’) (突然変異に下線を付す)
を使用して導入した。全ての構築物は、配列を確認し、A. tumefaciens株GV3101に形質転換した。
【0138】
1.4 DNAゲル-ブロット分析
N. benthamiana由来のゲノムDNAは、QIA-Gen DNA抽出プロトコール (Qiagen, Venlo, NL) を使用して単離し、トマトについては、(Sambrook and Russell, 2001, Molecular cloning: A Laboratory Manual, 3rd ed. (Cold Spring Harbor, NY, U.S.A.: Cold Spring Harbor Laboratory Press) に記載される標準プロトコールを使用した。DNAは、BamHI、HindIII、EcoRI、EcoRVまたはXbaIで消化した。N. benthamianaのゲル-ブロットは、TRV:NRC1ベクターに存在するthe 32P-標識 (Prime-a-gene Labeling System, Promega, Madison, WI) 252 bpフラグメントとハイブリダイズさせ、トマトのDNAゲル-ブロットは、全長NRC1 cDNAの1876〜3168ヌクレオチドに対応する1293塩基の32P-標識プローブとハイブリダイズさせた。使用した制限酵素の部位は、プローブに存在しない。低ストリンジェンシーは、2x SSCおよび0.5% SDS中の55℃での洗浄を指す。高ストリンジェンシー条件は、0.5x SSCおよび0.5% SDS中の65℃での洗浄から構成される。
【0139】
1.5 トマトにおけるNRC1のサイレンシングを示すためのRT-PCR
4つの葉ディスク (トータルで約100 mgの組織) を、TRV-感染植物の第二、第三または第四複葉から集めた。トータルRNAを、QIA-Gen RNAeasy抽出プロトコール (Qiagen, Venlo, NL) を用いて抽出し、RNase-Free DNase (Bio-Rad, Veenendaal, NL) で処理した。第一鎖cDNAは、1μgのトータルRNAから、Bio-Rad cDNA合成キット (Bio-Rad, Veenendaal, NL) を用いて合成し、RT-PCRは、以下のサイクル:95℃で15秒、60℃で45秒および72℃で60秒を使用して行った。使用されたプライマー
(222F: 5’- TGAGGTATATTGCTTTCTCATCTGAC-3’ および
222R: 5’-AGCTATTTTCCCACGGATGCCCAG-3’)
は、TRV:NRC1に挿入されるフラグメントを対象としない。アクチンプライマー
(ActinFnr182: 5’-TATGGAAACATTGTGCTCAGTGG-3’ および
ActinRnr183: 5’-CCAGATTCGTCATACTCTGCC-3’)
は、PCR反応において等量のcDNAの存在をチェックするために使用した。
【0140】
例2 − 結果
2.1 トマトNRC1;CC-NB-LRRタンパク質
cDNA-AFLP分析を行い、その後、トマトの同定されたフラグメントのVIGSをN. benthamiana:Cf-4において行った。VIGSがCf-4/Avr4-誘導性HRに影響を及ぼす20のcDNAフラグメントが同定された。これらの一つであるNRC1について、配列番号3に示されるとおり、全長cDNAを単離した。オープンリーディングフレームは、配列番号1に示され、これは、配列番号2に示されるNRC1タンパク質をコードする。
【0141】
NRC1タンパク質(配列番号2)の推定される一次構造は、CC-NB-LRR耐性タンパク質のものと典型的に類似している(図1)。NRC1は、アミノ末端の二重コイル (CC) ドメイン、NB-ARC (Apaf-1、Rタンパク質およびCED4により共有されるヌクレオチド結合アダプター) ドメイン (Van der Biezen and Jones, 1998, Curr. Biol. 8, R226-R227; Aravind et al., 1999, Trends Biochem. Sci. 24, 47-53)、および13の不完全なロイシン-リッチ繰返し配列 (LRRs) を有する。図1に示されるとおり、相同なNB-ARCドメインとの比較により、Kinase1AまたはP-loopモチーフ、4つのRNBS (Resistance Nucleotide Binding Site) モチーフ、並びにGLPLおよびMHDモチーフの存在が明らかにされた (Meyers et al., 1999, Plant J. 20, 317-332; Meyers et al., 2003, Plant Cell 15, 809-834)。
【0142】
VIGS のために使用されるTRV:NRC1ベクターに存在する252 bpのcDNA-AFLPフラグメントは、599-681アミノ酸をコードし、これはLRRs 4〜7に位置する。
【0143】
BamHI-、HindIII-、EcoRI-、EcoRV-およびXbaI-で消化されたトマトのゲノムDNAの低ストリンジェンシーDNAゲル-ブロット分析は、プローブとして、TRV:NRC1-、TRV:NRC1-LRR-およびTRV:NRC1-UTR構築物 (下記参照) に存在するNRC1配列をカバーする1293 bpのNRC1 cDNAフラグメント (配列番号3の1876〜3168ヌクレオチド) とハイブリダイズさせた。このサザンブロットにより、高ストリンジェンシー洗浄後にたった一つの目立ったバンドが出現し、これは、NRC1がトマトにおいて単一コピー遺伝子であることを示す。
【0144】
BamHI-、HindIII-、EcoRI-、EcoRV-およびXbaI-で消化されたN. benthamianaのゲノムDNAのゲルブロットは、TRVベクターに存在するトマトNRC1 cDNA-AFLPフラグメントをプローブとし、2〜3のハイブリダイズするバンドが見出された (結果は示さず) (0.5 x SSC, 0.5% SDS, 65℃)。これにより、TRV:NRC1の接種によりサイレンシングされ得る少なくとも2〜3のNRC1オルソログが、N. benthamianaのゲノムに存在することが示唆される。
【0145】
2.2 NRC1-サイレンシングされたトマトは、Cf-4-介在性HRおよび耐病性に影響を受ける
HR-シグナリングおよびC.fulvumに対する耐性におけるNRC1の機能を調査するため、本発明者らは、この植物がこの真菌の唯一の宿主であるため、トマトにおいてVIGSを行った。10日齢のトマト芽生えに、TRV:NRC1をアグロインフィルトレーションし、インフィルトレーションから3週間後に、サイレンシングされた可能性のある小葉からRNAを単離し、RT-PCRにより分析した。NRC1転写産物レベルは、異なるTRV:NRC1-感染植物では変動するが、ほとんどの場合、TRV:00-感染植物より低く、これは、NRC1発現の‘ノックダウン’が起こったことを示す(データ示さず)。
【0146】
我々がトマトで観察する表現型が、別のNB-LRRタンパク質のサイレンシングにより引き起こされる可能性を排除するため、我々は、LRRs 8〜12をターゲットとするNRC1の360 bpフラグメント (TRV:NRC1-LRR)、およびNRC1の3’’-非翻訳領域 (UTR) の297 bpからなるフラグメント (TRV:NRC1-UTR) を用いて、トマトにおいてVIGSを更に行った。これらの構築物を用いて、我々は、TRV:222-LRRおよびTRV:222-UTR-感染Cf-4-含有トマト植物にAvr4タンパク質を注入することにより、NRC1が、トマトにおけるCf-4-介在性HRに必要であるかどうかを試験した。(3つの構築物の各々を用いた) NRC1のサイレンシングは、トマト植物が、TRV:00-またはTRV:Cf-4-感染植物よりいくらか小さかったため、軽い表現型という結果につながる (データ示さず)。コントロールとして、TRV:00-およびTRV:Cf-4-感染植物に、Avr4タンパク質を注入した。TRV:Cf-4-感染植物において、Avr4-注入部位の応答パーセンテージは52% (図2) であり、これは、Cf-4のサイレンシングによりHRが減少することを示した。TRV:222-LRRおよびTRV:222-UTR-感染植物において、このパーセンテージは類似しており (それぞれ56%および48%) (図2)、これにより、NRC1がCf-4/Avr4-誘導性HRに機能することがトマトにおいても確認された。Cf-9-含有トマトにおけるNrc1のVIGSおよびその後のAvr9含有アポプラスト液の注入により、同様の結果が得られた(図示せず)。404 bpのCf-4フラグメントは、高度に保存されたLRRs 15〜21をコードし、これが、Cf-4と相同Cf-9耐性遺伝子の両方のサイレンシングを可能にするため、Cf-9-含有トマトにおけるCf-9のVIGSのために、我々はTRV:Cf-4構築物を使用した (Van der Hoorn et al., 2001, supra)。
【0147】
更に、NRC1が、トマトのC. fulvumに対する完全な耐性にとっても必要であるかどうか調査した。Cf0およびCf-4-植物にTRV:00、TRV:Cf-4およびTRV:NRC1を接種し、3週間後、サイレンシングされた植物に、Avr4およびβ-グルクロニダーゼ (GUS) 遺伝子を発現するC. fulvumの株を接種し、これにより真菌の増殖を視覚化した。C. fulvumの接種から2週間後、葉をX-glucで染色した。TRV:00を感染させたCf-4植物の小葉では、C. fulvumの増殖は検出されなかったが、TRV:Cf-4-感染Cf-4植物において青色染色のパッチは、低下した(compromised)Cf-4-介在性耐性を示す (図示せず)。またTRV:NRC1-感染植物においても、青色染色の小さいパッチは、真菌に対する完全な耐性が失われたことを示す。顕微鏡分析により、TRV:Cf-4-およびTRV:NRC1-感染植物において真菌の菌糸の細胞間増殖が明らかにされたが、TRV:00-感染コントロール植物では明らかにされなかった。Cf0植物はすべて、C. fulvumによる広範なコロニー形成を示し、これは、TRV感染自体も、TRV:NRC1を用いたVIGSも、これら植物の真菌に対する感受性に影響を及ぼさないことを示す。
【0148】
2.3 NRC1のVIGSは、異なるマッチングR遺伝子/Avr遺伝子の組合せにより誘導されるHRに影響を及ぼす
NRC1を用いたVIGSによるCf-4/Avr4-誘導性HRの減少に加えて、卵菌の病原体Phytophthora infestansのInf1誘導因子により誘導されるHRも、N. benthamianaにおいて、NRC1を用いたVIGSにより減少することが見出された。防御シグナリングにおけるNRC1の特異性を更に調査するため、本発明者らは、別のR/Avrの組合せにより誘導されるHRにNRC1が必要であるかを試験した。SGT1は、幾つかのR/Avrの組合せにより誘導されるHRに必要であることが知られているため、コントロールとしてTRV:00 (空のベクター) およびTRV:SGT1を含めた (Peart et al., 2002, Proc. Natl. Acad. Sci. USA 99, 10865-10869)。
【0149】
TRV:NRC1-感染N. benthamianaに、Cf-9およびAvr9の混合物 (Van der Hoorn et al., 2000, supra)、またはLeEix2およびtvEixの混合物 (Ron and Avni, 2004, Plant Cell 16, 1604-1615) をアグロインフィルトレーションすると、HRは減少したが、TRV:00-感染植物では、HRは普通に発生した。TRV:SGT1-感染植物においてHRは完全になくなり、これは、Peart et al. (2002, supra) の観察を確認する (図3)。また、細菌の病原体Pseudomonas syringae pv tomatoに由来するAvrPtoおよびポテトウイルスX (PVX) のコートタンパク質 (CP) をコードする遺伝子を、耐性遺伝子Pto (Pedley and Martin, 2003, Annu. Rev. Phytopathol. 41, 215-243) およびRx (Bendahmane et al., 1999, Plant Cell 11, 781-791) をそれぞれ発現するTRV-感染N. benthamianaにアグロインフィルトレーションした。両ケースにおいて、TRV:00を感染させた植物はHRを示したが、TRV:SGT1-感染植物でHRはなくなった。TRV:NRC1-感染により、Pto/AvrPto-およびRx/CP-誘導性HRは大幅に抑制され、これは、N. benthamianaにおいてNRC1タンパク質が、幾つかのR/Avrの遺伝子対遺伝子(gene-for-gene)の組合せにより活性化されるHRシグナリングに必要であることを示す (図3)。
【0150】
TRV:NRC1-感染N. benthamianaにおいて低下した(compromised)HRは、アグロバクテリウムによる形質転換効率の減少に由来するという可能性を排除するため、本発明者らは、TRV:00-およびTRV:NRC1-感染N. benthamiana:Cf-4に、β-グルクロニダーゼ (GUS) 遺伝子を発現するアグロバクテリウムをインフィルトレーションした (Van der Hoorn et al., 2000, supra)。インフィルトレーションから3日後、TRV:00-およびTRV:NRC1-感染植物における同程度の強度の青色染色により、アグロバクテリウムによる植物の形質転換効率は影響を受けないことが明らかにされた (データ示さず)。更に、TRV:NRC1-感染植物も、Avr4タンパク質の注入によりHRの減少を示したが、TRV:00-感染植物では、明らかなHRが2日以内に発生した。
【0151】
2.4 NRC1は、細胞死シグナリング経路においてEDS1の下流およびMAPKカスケードの上流に作用する
NRC1は、Cf-4/Avr4-誘導性HRだけでなく、幾つかの別のR/Avrの組合せにより誘導されるHRにも必要であるため、NRC1は、共通のHRシグナリング経路に関与していると思われる。HRの開始前に起こる典型的な宿主応答には、MAPKカスケードの活性化が含まれる (Romeis et al., 2001, EMBO J. 20, 5556-5567; Del Pozo et al., 2004, EMBO J. 23, 3072-3082; Pedley and Martin, 2005, Plant Biol. 8, 541-547)。
【0152】
MAPKsにより開始されるHRにNRC1が必要であるか調査するため、N. benthamianaにおいてエピスタシス実験を行った。植物に、TRV:00, TRV:SGT1およびTRV:NRC1を接種し、その後、LeMAPKKKα (LeMAPKKKαKD) のキナーゼドメインまたは構成的に活性なLeMEK2 (LeMEK2DD) をコードする遺伝子をアグロインフィルトレーションした (Yang et al., 2001, Proc. Natl. Acad. Sci. USA 98, 741-746; Del Pozo et al., 2004, EMBO J. 23, 3072-3082.)。アグロインフィルトレーションから2日後、インフィルトレーションされた葉にエストラジオールをスプレーすることにより遺伝子の発現を誘導した。各遺伝子の一過性発現は、TRV:00-感染植物においてHRという結果につながるが、TRV:SGT1-感染植物においてHRは減少する (図4A)。TRV:NRC1-感染植物では、構成的に活性な何れのキナーゼにより引き起こされるHRも影響を受けない (図4A)。対応するネガティブコントロール、LeMAPKKKαKD- および野生型LeMEK2のアグロインフィルトレーションは、TRV-感染植物の何れにおいてもHRという結果につなからなかった (データ示さず)。これらの結果は、SGT1が、これらMAPKsの下流で機能するが、MAPKsは、NRC1の下流で作用するか、あるいはNRC1とは独立して作用することを示す。
【0153】
2.5 NRC1の一過性過剰発現および構成的に活性なNRC1タンパク質の構築
遺伝子が、CC-NB-LRRタンパク質によるHRシグナリングに必要であるか更に調査するため、NRC1の過剰発現の効果を調査した。したがって、cDNAのコード配列 (配列番号1) を、構成的35Sプロモーターに融合し、バイナリーベクターに挿入した。N. benthamianaにおいてこの構築物のアグロインフィルトレーションは、HRという結果につながらなかったが、NRC1およびp19サイレンシング阻害剤の混合物の発現 (Voinnet et al., 2003, Plant J. 33, 949-956) は、誘導因子-非依存性HRを引き起こした (図4B)。P-loopモチーフを破壊することによりATP加水分解に影響を及ぼすNRC1のP-loop突然変異体 (K191R) をコードする構築物のアグロインフィルトレーション (Tameling et al., 2002, Plant Cell 14, 2929-2939) は、p19が存在しても存在しなくても、HRという結果につながらなかった (図4B)。
【0154】
上記データは、NRC1遺伝子の翻訳後遺伝子サイレンシング (PTGS) が、NRC1過剰発現組織においてHRの発生を防ぎ得ることを示した。また、P-loopモチーフの破壊は、非機能的NRC1タンパク質という結果につながる。
【0155】
NB-LRR耐性タンパク質RxのMHDモチーフにおける突然変異 (D460V) (Bendahmane et al., 2002; Tameling et al., 2002) およびNB-LRR耐性タンパク質I-2のMHDモチーフにおける突然変異 (D495V) (Bendahmane et al., 2002, Plant J. 32, 195-204; Tameling et al., 2002, Plant Cell 14, 2929-2939; Van Bentem et al., 2005, Plant J. 43, 284-298) は、構成的活性という結果につながるため、本発明者らは、NRC1の同様の突然変異体 (NRC1D481V) を作成した。実際、NRC1D481Vのアグロインフィルトレーションは、インフィルトレーションから3日以内に、N. benthamianaの葉において誘導因子-非依存性HRという結果につながり、また、ダブル突然変異体NRC1K191R/D481V のアグロインフィルトレーションにより、HRは観察されなかった (図4B)。更に、SGT1-サイレンシング植物において、NRC1D481Vの発現によりHRは誘導されなかった (下記参照)。これらの結果は、NRC1D481Vのアグロインフィルトレーションにより誘導される応答は、特異的に、NRC1タンパク質の構成的活性によるものであること、並びにNRC1は、HRにつながるシグナル伝達カスケードにおいて機能することを示す。
【0156】
2.6 構成的に活性なNRC1タンパク質を用いたエピスタシス実験
NRC1D481Vを使用するエピスタシス実験を行い、このタンパク質によるHRシグナリングに遺伝子が必要であるか更に調査し、これにより、HR経路におけるその推定位置を決定する。HRシグナリングに一般に関与することが知られている遺伝子、たとえばSGT1およびRAR1 (Required for Mla12 resistance) (Shirasu and Schulze-Lefert, 2003, Trends Plant Sci. 8, 252-258) のVIGSに加えて、NDR1 (non race-specific disease resistance) (Century et al., 1995, Proc. Natl. Acad. Sci. USA 92, 6597-6601)、EDS1 (enhanced disease susceptibility) (Aarts et al., 1998, Proc. Natl. Acad. Sci. USA 95, 10306-10311)、およびMEK2 (a MAPKK) (Ekengren et al., 2003, Plant J. 36, 905-917) について、N. benthamiana:Cf-4をサイレンシングし、その後、NRC1D481VまたはAvr4を用いてアグロインフィルトレーションした。更に、TRV:00、TRV:Cf-4およびTRV:NRC1を用いたVIGSを、コントロールとして含めた。低下した(compromised)NRC1D481V-またはAvr4-誘導性HRは、NRC1-またはCf-4/Avr4-誘導性HRシグナリングのそれぞれに必要な遺伝子の‘ノックダウン’を示す。
【0157】
予測されるとおり、Avrのアグロインフィルトレーションにより誘導されるHRは、TRV:Cf-4-およびTRV:NRC1-感染植物において低下した(compromise)。また、この遺伝子についてサイレンシングされた植物が、それほどひどくないAvr4-誘導性HRを示すため、Cf-4-介在性シグナリングもEDS1を必要とする。更に、本発明者らは、MEK2、RAR1およびSGT1についてサイレンシングされた植物において、Avr4のアグロインフィルトレーションによりHRが減少することを見出した (図4C;薄い円)。Avr4-誘導性HRは、TRV:00-およびTRV:NDR1-感染植物において低下せず(not compromise)(図4C;濃い円)、これは、NDR1がCf-4-介在性シグナリングに必要でないことを示す。同様に、NRC1D481V-誘導性HRは、TRV:00-およびTRV:NDR1-感染植物において低下せず(not compromise)、TRV:Cf-4-感染植物においても低下しなかった。興味深いことに、Avr4とは対照的に、NRC1D481Vは、TRV:EDS1-感染植物においてもHRを誘導し、これは、NRC1がEDS1の下流で機能的であることを示す (図4C;濃い円)。NRC1D481V-誘導性HRは、MEK2についてサイレンシングされた植物において低下し(compromise)、これは、NRC1が、そのシグナリングのためにMAPキナーゼカスケードを必要とし、これらキナーゼの上流に位置することができることを示す。また、RAR1およびSGT1のVIGSも、Avr4により誘導されるHRと同様、D481V-誘導性HRを低下させる(compromise)(図4C;薄い円)。このため、NRC1は、Cf-4により開始されるHRシグナリングに必要であり、MAPKカスケードの上流およびEDS1の下流に位置することができる。
【0158】
NRC1介在性細胞シグナリングのモデルについては、図5を参照されたい。
【0159】
例3 − Mi-介在性耐性に対するNRC1の必要性
NRC1が、線虫、コナジラミおよびアブラムシに対するMi-介在性耐性に必要であるか否かを決定するため、Miの構成的に活性な型 (US 6613962およびEP0937155B1参照) を、NRC1サイレンシング植物にアグロインフィルトレーションする。NRC1サイレンシング植物におけるHRの減少は、NRC1が、Mi-介在性HRにも必要であること、並びにNRC1の(過剰)発現が、線虫、コナジラミおよびアブラムシに対して向上した耐性を有するトランスジェニック植物を作成するために使用することができることを示す。
【図面の簡単な説明】
【0160】
【図1】NRC1タンパク質の推定配列を示す図。
【図2a】NRC1は、Cladosporium fulvumに対するトマトの完全なCf-4-介在性HRに必要であることを示す図。
【図2b】NRC1は、Cladosporium fulvumに対するトマトの完全なCf-4-介在性耐性に必要であることを示す写真。
【図3】N. benthamianaへのTRV:NRC1の接種は、Cf/Avr-、LeEix2/tvEix-、Pto/AvrPto-およびRx/CP-誘導性HRに影響を及ぼすことを示す写真。
【図4a】構成的に活性なNRC1は、誘導因子−非依存性HRを誘導し、細胞死シグナリング経路にNRC1を配置させることを示す写真。構成的に活性なMAPKKおよびMAPKキナーゼをコードする遺伝子のアグロインフィルトレーション。
【図4b】構成的に活性なNRC1は、誘導因子−非依存性HRを誘導し、細胞死シグナリング経路にNRC1を配置させることを示す写真。35S-プロモーターの制御下の野生型NRC1 (wt) および当該遺伝子の突然変異型のアグロインフィルトレーションであって、サイレンシングサプレッサーp19をコードする遺伝子の発現を駆動するアグロバクテリウムと1:1の比で混合されたアグロインフィルトレーション (左パネル)、または単独のアグロインフィルトレーション (右パネル)。
【図4c】構成的に活性なNRC1は、誘導因子−非依存性HRを誘導し、細胞死シグナリング経路にNRC1を配置させることを示す写真。Avr4および構成的に活性なNRC1D481V (D481V) をコードする遺伝子のアグロインフィルトレーション。
【図5】NRC1介在性細胞死シグナリングのモデルを示す図。
【特許請求の範囲】
【請求項1】
非トランスジェニックコントロール植物と比べて向上した耐病性を有するトランスジェニック植物を産生するための方法であって、
(a)植物または植物細胞を、植物細胞で活性なプロモーターに動作可能に連結されたNRC1タンパク質をコードするヌクレオチド配列で形質転換する工程、
(b)植物を再生する工程
を含む方法。
【請求項2】
前記ヌクレオチド配列が前記植物のゲノムに統合されている、請求項1に記載の方法。
【請求項3】
(c)再生した植物、または再生した植物から自家受粉または異種交配により得られる植物を、一または複数の植物病原体に対する耐性についてスクリーニングし、前記植物病原体の一または複数に対して向上した耐性を含む植物を同定する工程
を更に含む、請求項1または2に記載の方法。
【請求項4】
前記プロモーターが病原体誘導性プロモーターである、前述の請求項の何れか一項に記載の方法。
【請求項5】
前記NRC1タンパク質が、配列番号2または配列番号4のアミノ酸配列、または配列番号2の全長に対して少なくとも70%のアミノ酸同一性を含むアミノ酸配列を含む、前述の請求項の何れか一項に記載の方法。
【請求項6】
前記植物がナス科(Solanaceae)に属する、前述の請求項の何れか一項に記載の方法。
【請求項7】
前記植物がナス属(Solanum)である、請求項6に記載の方法。
【請求項8】
前述の請求項の何れか一項に記載の方法により得られる、トランスジェニック植物、植物細胞、種子または果実。
【請求項9】
配列番号2または配列番号4のアミノ酸配列、または配列番号2の全長に対して少なくとも70%のアミノ酸配列同一性を含むアミノ酸配列を含む、単離されたタンパク質。
【請求項10】
請求項9に記載のタンパク質をコードする単離された核酸分子。
【請求項11】
請求項10に記載の核酸分子に動作可能に連結され、必要に応じて3’非翻訳核酸分子に更に動作可能に連結された、植物細胞で活性なプロモーターを含むキメラ遺伝子。
【請求項12】
請求項11に記載のキメラ遺伝子を含むベクター。
【請求項13】
配列番号2または配列番号4のアミノ酸配列、または配列番号2の全長に対して少なくとも70%のアミノ酸配列同一性を含むアミノ酸配列を含むNRC1タンパク質をコードする核酸分子の、耐病性植物の生成のための使用。
【請求項1】
非トランスジェニックコントロール植物と比べて向上した耐病性を有するトランスジェニック植物を産生するための方法であって、
(a)植物または植物細胞を、植物細胞で活性なプロモーターに動作可能に連結されたNRC1タンパク質をコードするヌクレオチド配列で形質転換する工程、
(b)植物を再生する工程
を含む方法。
【請求項2】
前記ヌクレオチド配列が前記植物のゲノムに統合されている、請求項1に記載の方法。
【請求項3】
(c)再生した植物、または再生した植物から自家受粉または異種交配により得られる植物を、一または複数の植物病原体に対する耐性についてスクリーニングし、前記植物病原体の一または複数に対して向上した耐性を含む植物を同定する工程
を更に含む、請求項1または2に記載の方法。
【請求項4】
前記プロモーターが病原体誘導性プロモーターである、前述の請求項の何れか一項に記載の方法。
【請求項5】
前記NRC1タンパク質が、配列番号2または配列番号4のアミノ酸配列、または配列番号2の全長に対して少なくとも70%のアミノ酸同一性を含むアミノ酸配列を含む、前述の請求項の何れか一項に記載の方法。
【請求項6】
前記植物がナス科(Solanaceae)に属する、前述の請求項の何れか一項に記載の方法。
【請求項7】
前記植物がナス属(Solanum)である、請求項6に記載の方法。
【請求項8】
前述の請求項の何れか一項に記載の方法により得られる、トランスジェニック植物、植物細胞、種子または果実。
【請求項9】
配列番号2または配列番号4のアミノ酸配列、または配列番号2の全長に対して少なくとも70%のアミノ酸配列同一性を含むアミノ酸配列を含む、単離されたタンパク質。
【請求項10】
請求項9に記載のタンパク質をコードする単離された核酸分子。
【請求項11】
請求項10に記載の核酸分子に動作可能に連結され、必要に応じて3’非翻訳核酸分子に更に動作可能に連結された、植物細胞で活性なプロモーターを含むキメラ遺伝子。
【請求項12】
請求項11に記載のキメラ遺伝子を含むベクター。
【請求項13】
配列番号2または配列番号4のアミノ酸配列、または配列番号2の全長に対して少なくとも70%のアミノ酸配列同一性を含むアミノ酸配列を含むNRC1タンパク質をコードする核酸分子の、耐病性植物の生成のための使用。
【図1】



【図2a】
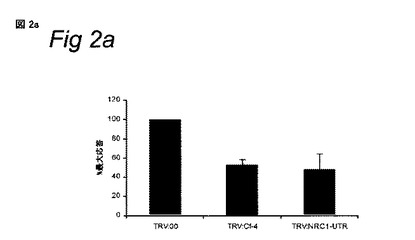
【図2b】

【図3】
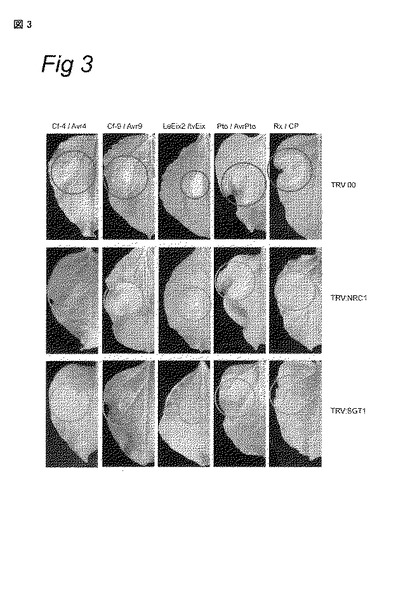
【図4a】
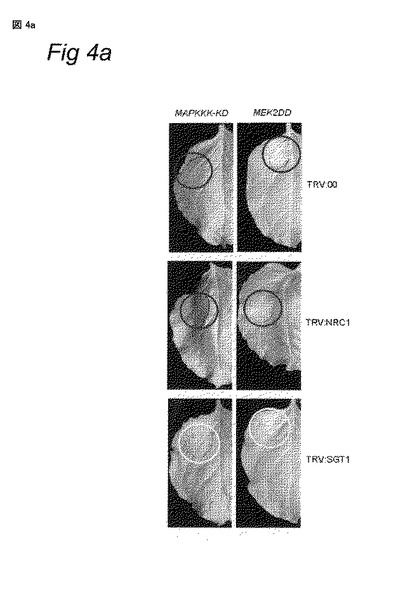
【図4b】
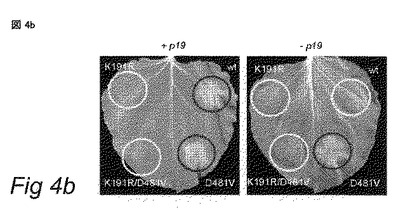
【図4c】
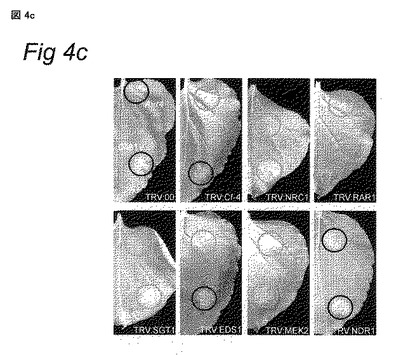
【図5】
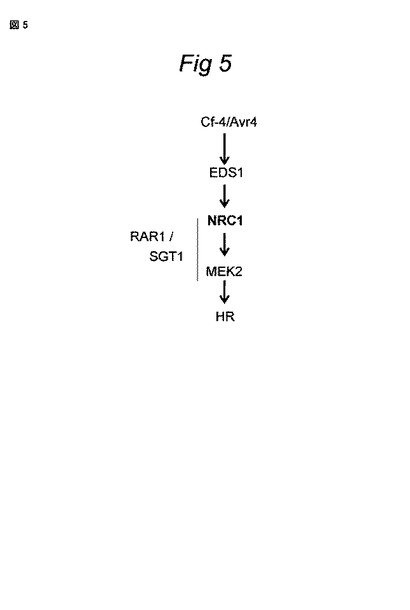
【図2a】
【図2b】
【図3】
【図4a】
【図4b】
【図4c】
【図5】
【公表番号】特表2009−528053(P2009−528053A)
【公表日】平成21年8月6日(2009.8.6)
【国際特許分類】
【出願番号】特願2008−557222(P2008−557222)
【出願日】平成18年2月28日(2006.2.28)
【国際出願番号】PCT/NL2006/050038
【国際公開番号】WO2007/100238
【国際公開日】平成19年9月7日(2007.9.7)
【出願人】(505477187)ケイヘーネ・エヌ・ブイ (10)
【出願人】(508261448)ワーヘニンヘン・ウニベルシティ (1)
【氏名又は名称原語表記】WAGENINGEN UNIVERSITY
【住所又は居所原語表記】Department of Plant Sciences, Laboratory of Phytopathology, Binnenhaven 5, NL−6709 PD WAGENINGEN, the Netherlands
【Fターム(参考)】
【公表日】平成21年8月6日(2009.8.6)
【国際特許分類】
【出願日】平成18年2月28日(2006.2.28)
【国際出願番号】PCT/NL2006/050038
【国際公開番号】WO2007/100238
【国際公開日】平成19年9月7日(2007.9.7)
【出願人】(505477187)ケイヘーネ・エヌ・ブイ (10)
【出願人】(508261448)ワーヘニンヘン・ウニベルシティ (1)
【氏名又は名称原語表記】WAGENINGEN UNIVERSITY
【住所又は居所原語表記】Department of Plant Sciences, Laboratory of Phytopathology, Binnenhaven 5, NL−6709 PD WAGENINGEN, the Netherlands
【Fターム(参考)】
[ Back to top ]