
肝特異的有機アニオントランスポーターとその遺伝子
【課題】肝臓における有機アニオン性物質の取り込み排出の制御をする蛋白質として有用な肝特異的有機アニオントランスポーターOAT2、それをコードする塩基配列を有する核酸、及びそれに対する抗体を提供する。
【解決手段】特定のアミノ酸配列を有する、肝特異的有機アニオントランスポーターOAT2、前記肝特異的有機アニオントランスポーターOAT2のアミノ酸配列をコードする核酸、及び前記肝特異的有機アニオントランスポーターOAT2を認識しうる抗体。
【解決手段】特定のアミノ酸配列を有する、肝特異的有機アニオントランスポーターOAT2、前記肝特異的有機アニオントランスポーターOAT2のアミノ酸配列をコードする核酸、及び前記肝特異的有機アニオントランスポーターOAT2を認識しうる抗体。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は有機陰イオン(有機アニオン)輸送に関与する遺伝子と、その遺伝子がコードするポリペプチドに関する。
【背景技術】
【0002】
肝臓は、生体異物や薬物の代謝および体外排出に関して重要な役割を果たしている。肝細胞は極性を有する上皮細胞であり、血液とは側底膜、また胆汁とは胆管側膜を介して接している。陰イオン(アニオン)性の薬物は、輸送担体(トランスポーター)により側底膜を介して肝臓中に取り込まれ、また肝細胞内で代謝により産生された有機アニオンもトランスポーターにより肝細胞より排出される。
【0003】
肝細胞の側底膜を介した有機アニオンの取り込み(即ち血液と肝細胞間での有機アニオンの輸送)については、これまで摘出臓器灌流法や単離細胞膜小胞系などを用いた実験系により研究されてきた。これらの研究により主に、生理的有機アニオンの一つである胆汁酸の輸送について多くの知見が得られてきた。
【0004】
しかし従来の手法では、肝細胞の特に側底膜を介した物質輸送について詳細に解析することは困難であり、トランスポーターそのものを単離して詳細に解析することが望まれてきた。
【0005】
これまでの分子生物学的な研究から、肝臓の側底膜に発現している有機アニオントランスポーターとしては2種類のものがクローニングされている(Jacquemin,E.ら、 Proc.Natl.Acad.Sci.USA、91巻、133−7頁、1994年、Hagenbuch,B.ら、Proc.Natl.Acad.Sci.USA、88巻、10629−33頁、1991年)。
【0006】
しかしながら、これらのトランスポーターのみでは肝臓の側底膜における多様な有機アニオン輸送を説明することが出来ず、新規有機アニオントランスポーターの存在が予想されていた。
【0007】
腎臓は肝臓とならび、有機アニオン輸送を最も活発におこなっている臓器である。腎臓における(特に側底膜を介した)有機アニオン輸送の研究は100年近い歴史を持ち、その輸送特性については膨大な検討がなされてきた。我々は最近、この腎臓での有機アニオン輸送において最も重要な役割を果たしている有機アニオントランスポーターOAT1の単離に成功し(Sekine,T.ら、J.Biol.Chem.、272巻、18526−9頁、1997)、既に特許出願済みである。OAT1は基質選択性の極めて広い、即ち化学構造の異なる多くの有機アニオンを輸送することの出来るトランスポーターである。また多くの薬物が有機アニオンに属することからOAT1は薬物トランスポーターとしても認識されている。OAT1の発現は腎臓に限られており、肝臓には全くその発現が認められない。
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明の目的は、肝臓における有機アニオン輸送に関与する新規な有機アニオンントランスポーター遺伝子およびその遺伝子がコードするポリペプチドである有機アニオントランスポーターを同定し、提供することにある。その他の目的については以下の記載より明白である。
【課題を解決するための手段】
【0009】
本発明者らは、腎臓の有機アニオントランスポーターOAT1との構造上の類似性から、肝細胞に発現している肝特異的有機アニオントランスポーター(OAT2)を同定した。
【0010】
本発明の有機アニオントランスポーターOAT2は、ジカルボン酸、プロスタグランジン、非ステロイド系抗炎症薬や抗腫瘍薬等の異なる化学構造を持った薬物や内因性物質に対してこれらを輸送する(取り込む能力を有する)、非常に広い範囲の基質選択性を有するものである。
【0011】
本発明のタンパク質としては、配列番号1で示されたアミノ酸配列を有するもののほか、例えば、配列番号1で示されたアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列を有するものが挙げられる。アミノ酸の欠失、置換もしくは付加は、有機アニオン輸送活性が失われない程度であればよく、通常1〜約110個、好ましくは1〜約55個である。このようなタンパク質は、配列番号1で示されたアミノ酸配列と通常、1〜80%、好ましくは1〜90%のアミノ酸配列のホモロジーを有する。
【0012】
本発明において、ストリンジェントな条件下でのハイブリダイゼーションは、通常、ハイブリダイゼーションを、5×SSC又はこれと同等の塩濃度のハイブリダイゼーション溶液中、37〜42℃の温度条件下、約12時間行い、5×SSCまたはこれと同等の塩濃度の溶液等で予備洗浄を行った後に、1×SSC又はこれと同等の塩濃度で洗浄を行うことにより実施できる。また、より高いストリンジェンシーを得るためには、洗浄を0.1×SSC又はこれと同等の塩濃度の溶液中で洗浄を行うことにより実施できる。
【0013】
本発明の有機アニオントランスポーター遺伝子は、適当な哺乳動物の肝臓の組織や細胞を遺伝子源として用いてスクリーニングを行うことにより単離取得できる。哺乳動物としては、イヌ、ウシ、ウマ、ヤギ、ヒツジ、サル、ブタ、ウサギ、ラット、マウスなどの非ヒト動物のほか、ヒトが挙げられる。
【0014】
遺伝子のスクリーニングおよび単離は、ホモロジースクリーニングおよびPCR(Polymerase Chain Reaction)スクリーニングなどにより好適に実施できる。
【0015】
得られたcDNAについては、常法により塩基配列を決定し、翻訳領域を解析
して、これにコードされるタンパク質、即ちOAT2のアミノ酸配列を決定することができる。
【0016】
得られたcDNAが、有機アニオントランスポーター遺伝子のcDNAであること、即ちcDNAにコードされた遺伝子産物が有機アニオントランスポーターであることは、例えば次のようにして検証することができる。即ち、得られたOAT2遺伝子から調製したcRNAを卵母細胞に導入して発現させ、有機アニオンを細胞内に輸送する(取り込む)能力を、適当な有機アニオンを基質とする通常の取り込み実験(Sekine,T.ら、J.Biol.Chem.、272巻、18526−9頁、1997)により、細胞内への基質取り込みを測定することにより確認できる。
【0017】
また、OAT2発現細胞について、同様の取り込み実験を応用して、OAT2の輸送特性や基質特異性などを調べることができる。
【0018】
得られたOAT1遺伝子のcDNAを用いて、異なる遺伝子源で作製された適当なcDNAライブラリー又はゲノミックDNAライブラリーをスクリーニングすることにより、異なる組織、異なる生物由来の相同遺伝子や染色体遺伝子等を単離することができる。
【0019】
また、開示された本発明の遺伝子の塩基配列(配列番号2)に示された塩基配列、もしくはその一部の情報に基づいて設計された合成プライマーを用い、通常のPCR法によりcDNAライブラリーから遺伝子を単離することが出来る。
【0020】
cDNAライブラリー及びゲノミックDNAライブラリー等のDNAライブラリーは例えば、「Molecular Cloning;Sambrook, J.,Fritsh,E.F.およびManiatis,T.著、Cold Spring Harbor Laboratory Pressより、1989年に発刊」に記載の方法により調製することができる。あるいは、市販のライブラリーがある場合にはこれを用いてもよい。
【0021】
本発明の有機アニオントランスポーター(OAT2)は、例えば、有機アニオントランスポーターをコードするcDNAを用い、遺伝子組み換え技術により生産することができる。例えば、有機アニオントランスポーターをコードするDNA(cDNA等)を適当な発現ベクターに組み込み、得られた組み換えDNAを適当な宿主細胞に導入することができる。ポリペプチドを生産するための発現系としては、例えば、細菌、酵母、昆虫細胞および哺乳動物細胞の発現系等が挙げられる。このうち、機能タンパクを得るためには、昆虫細胞および哺乳動物細胞を用いることが望ましい。
【0022】
例えば、ポリペプチドを哺乳動物で発現させる場合には、有機アニオントランスポーターをコードするDNAを、適当な発現ベクター(例えば、レトロウイルス系ベクター、パピローマウイルスベクター、ワクシニアウイルスベクター、SV40系ベクター等)中の適当なプロモーター(例えばSV40プロモーター、LTRプロモーター、エロンゲーション1αプロモーター等)の下流に挿入して発現ベクターを構築する。次に、得られた発現ベクターで適当な動物細胞を形質転換して、形質転換体を適当な培地で培養することによって、目的とするポリペプチドが生産される。宿主とする哺乳動物細胞としては、サルCOS−7細胞、チャイニーズハムスターCHO細胞、ヒトHela細胞または、腎臓組織由来の初代培養細胞やブタ腎由来LLC−PK1細胞、フクロネズミ腎由来OK細胞等の細胞株が挙げられる。
【0023】
有機アニオントランスポーターOAT2をコードするcDNAとしては、例えば、配列番号2に示される塩基配列を有するcDNAを用いることが出来るほか、前記のcDNAに限定されることなく、アミノ酸配列に対応するDNAを設計し、ポリペプチドをコードするDNAをもちいることもできる。この場合、一つのアミノ酸をコードするコドンは各々1〜6種類知られており、用いるコドンの選択は任意でよいが、例えば発現に利用する宿主のコドン使用頻度を考慮して、より発現の高い配列を設計することができる。設計した塩基配列をもつDNAはDNAの化学合成、前記cDNAの断片化と結合、塩基配列の一部改変等によって取得できる。人為的な塩基配列の一部改変、変異導入は、所望の改変をコードする合成オリゴヌクレオチドからなるプライマーを利用して部位変異導入法(site specific mutagenesis)「Mark,D.F.ら、Proc Natl Acad Sci USA 第18巻、5662−5666頁、1984年」等により実施できる。
【0024】
本発明の有機アニオントランスポーター遺伝子にストリンジェントな条件下でハイブリダイズするヌクレオチド(オリゴヌクレオチドもしくはポリヌクレオチド)は、有機アニオントランスポーター遺伝子を検出するためのプローブとして使用できるほか、有機アニオントランスポーターの発現を変調させるために、例えばアンチセンスオリゴヌクレオチドやリボザイム、デコイとして使用することもできる。このようなヌクレオチドとしては、例えば、配列番号2で示される塩基配列の中の通常、連続する14塩基以上の部分配列もしくはその相補的な配列を含むヌクレオチドを用いることができ、ハイブリダイズをより特異的とするためには、部分配列としてより長い配列、例えば20塩基以上あるいは30塩基以上の配列を用いても良い。
【0025】
また、本発明の有機アニオントランスポーターまたは、これと免疫学的同等性を有するポリペプチドを用いて、その抗体を取得することが出来、抗体は、有機アニオントランスポーター検出や精製などに利用できる。抗体は、本発明の有機アニオントランスポーター、その断片、またはその部分配列を有する合成ペプチド等を抗原として用いて製造できる。ポリクロナール抗体は、宿主動物(たとえば、ラットやウサギ)に抗原を接種し、免疫血清を回収する通常の方法により製造することができ、モノクロナール抗体は、通常ハイブリドーマ法などの技術により製造できる。
【図面の簡単な説明】
【0026】
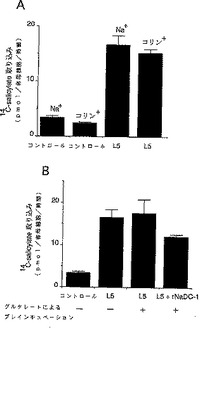
【図1】図1は、卵母細胞におけるサリチル酸の取り込みを示したものである。図1のBはグルタル酸とのプレインキュベーションの影響(図1Bの「+」)をみたものである。図1中の「L5」はL5を発現させせた卵母細胞を用いた系である。
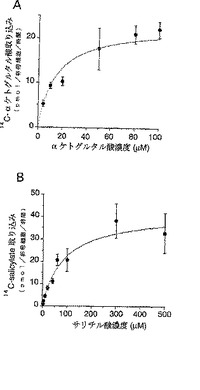
【図2】図2は、本発明の有機アニオントランスポーターにおける基質α−ケトグルタル酸(図2A)とサリチル酸(図2B)の動力学試験の結果を示すものである。
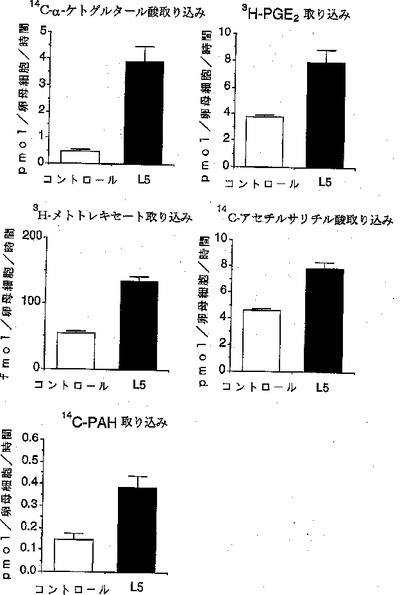
【図3】図3は、本発明の有機アニオントランスポーターの各種薬剤および内因性有機アニオンの取り込みを示すものである。図3中の「3H−PGE2」は3H標識化プロスタグランジンE2を示し、「14C−PAH」は14C標識化パラアミノ馬尿酸を示す。
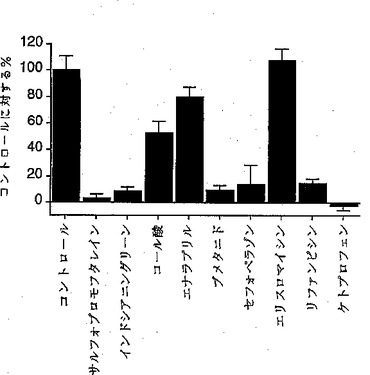
【図4】図4は、各種のアニオン物質の存在下における、本発明の有機アニオントランスポーターのサリチル酸の取り込みの影響を調べたものである。
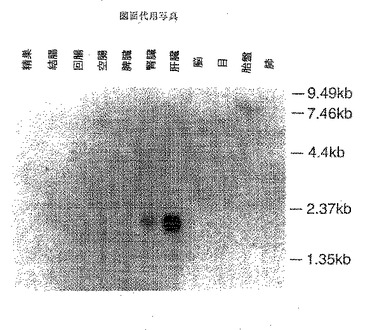
【図5】図5は、本発明の遺伝子のノーザンブロッティング解析の結果を示すものである。
【0027】
以下、実施例をもって本説明をさらに詳しく説明するが、これらの実施例は本発明を制限するものではない。
【0028】
なお下記実施例において、各操作は特に断りがない限り、「Molecular Cloning:Sambrook,J.,Fritsh,E.F.およびManiatis,T.著、Cold Spring HarborPressより、1989年に発刊」に記載の方法により行うか、または、市販のキットを用いる場合には市販品の指示書に従って使用した。
【実施例】
【0029】
実施例1 ラット肝特異的有機アニオントランスポーター(NLT:novel liber transporter)cDNAの単離とその解析
(1)腎特異的有機アニオントランスポーターOAT1の塩基配列情報を基にした、関連遺伝子の検索
【0030】
既に我々が単離したOAT1(Sekine,T.ら、J Biol Chem 272巻、18526−9頁、1997年)の塩基配列情報をもとにDNAデータベースを検索したところ、機能未知として報告されていた膜タンパクNLT(Simonson,G.D.ら、J.Cell.Sci.、107巻、1065−1072頁、1994)がOAT1と弱い相同性(アミノ酸レベルで42%)を有することが判明した。またNLTは肝細胞の側底膜に限局して発現していると考えられた。我々は、NLTが肝細胞側底膜に存在する多選択性有機アニオントランスポーターであると仮定してその単離と機能解析を行った。
【0031】
cDNAライブラリーは、ラット肝ポリ(A)+RNAから、cDNA合成キット(商品名:Superscript Choice System,ギブコ社製)を使用して作製し、ファージベクターλZipLox(ギブコ社製)の制限酵素EcoRI切断部位に組み込んだ。PCR法にて、既に報告されているNLTの 131−673塩基に相当する部位を単離し、32P−dCTPでラベルし、これをプローブとして用いてラットのcDNAライブラリーをスクリーニングした。ハイブリダイゼーションは、37℃のハイブリダイゼーション用溶液中で一昼夜行い、その後フィルター膜は、37℃で0.1×SSC/0.1%SDSで洗浄した。ハイブリダイゼーション溶液としては、50% ホルムアミド、5×standard saline citrate(SSC)、3×デンハード液、0.2% SDS、10%硫酸デキストラン、0.2mg/ml変性サーモン精子DNA、2.5mMピロリン酸ナトリウム、25mM MES、0.01% Antifoam B(シグマ社製)を含むpH6.5の緩衝液を用いた。λZipLox中に単離されたクローンは、in vitro excision法によりプラスミドベクターpZLにサブクローン化した。
【0032】
上記により得られたクローン(L5)の塩基配列の決定は、NLTに対する特異的オリゴヌクレオチドプライマーを合成し、Sequenase ver.2.0(アマシャム社製)を用いておこなった。
【0033】
これにより、NLTと相同なクローン(L5)遺伝子のcDNA塩基配列が得られた。
(2)NLTの機能の特定
【0034】
上記により得られたクローン(L5)を含むプラスミドから、T7 RNAポリメラーゼを用いて、cRNA(cDNAに相補的なRNA)を調製した。
【0035】
得られたcRNAを、既に報告されている方法に従い(Sekine,T.,et al.J Biol Chem 272巻、18526−9頁、1997年)、アフリカツメガエルの卵母細胞に注入し、この卵母細胞についてグルタル酸およびサリチル酸を基質として用いた取り込み実験をおこなった。実験には、放射能ラベルした基質(14C−グルタル酸および14C−サリチル酸)を用いた。その結果、L5を発現させた卵母細胞は14C−グルタル酸および14C−サリチル酸の取り込みを示すことが判明した。
【0036】
L5の輸送活性における細胞外Naイオンの影響を検討する実験を行った。サリチル酸の取り込み実験は前記に記載の方法に準じて実施した。但し、incubation溶液はND96溶液および、塩化コリン溶液(ND96溶液の96mMの塩化ナトリウムを96mMの塩化コリンで置換し、pHを7.4に調製した)を用いておこなった。その結果、細胞外のナトリウムをコリンと置換しても、サリチル酸取り込みに何ら影響を与えなかった(図1A)。このことから、NLTはNaイオン非依存的に働くトランスポーターであることが示された。
【0037】
NLT cRNAを注入した卵母細胞によるサリチル酸の取り込み実験においてグルタル酸とのプレインキュベーションの影響を調べた。サリチル酸の取り込み実験は、NLT cRNAもしくはNLT cRNAとラットNaDC−1(Na−ジカルボン酸トランスポーター)cRNAを注入した卵母細胞を1mMグルタル酸添加もしくは無添加のND96溶液で2時間培養した後、14C−サリチル酸を添加して室温で1時間培養し放射能ラベルされた基質の取り込みを調べた。その結果1mMグルタル酸添加した溶液で前処置しても14C−サリチル酸の取り込みは変わらなかった(図1B)。この結果はNLTが交換輸送体ではなく、促進拡散型輸送体であることを示唆するものである。
【0038】
NLTの有機アニオン輸送のミカエリスーメンテンの動力学試験をおこなった。
基質サリチル酸およびα−ケトグルタル酸の濃度の違いによるそれぞれの取り込み率の変化を調べることにより、NLTのミカエリスーメンテンの動力学試験をおこなった。サリチル酸およびα−ケトグルタル酸の取り込み実験は、NLT cRNAを注入した卵母細胞を用い、前記記載方法に準じて実施した。この結果(図2)、サリチル酸およびα−ケトグルタル酸のKm値はそれぞれ 88.8 ± 23.4 μMおよび17.8 ± 2.9μMであった。
【0039】
NLTの基質選択性を検討するために、各種アニオン性物質を基質とする取り込み試験を行った。取り込み実験は、NLT cRNAを注入した卵母細胞を用い、ND96溶液中で前記記載方法に準じて実施した。但し、基質としては、放射能でラベルした各種化合物を用いた。その結果、α−ケトグルタレート(14C標識)、メトトレキセート(3H標識)、プロスタグランジンE2(3H標識)、アセチルサリチル酸(14C標識)、パラアミノ馬尿酸(14C標識)を基質とした場合に、卵母細胞への取り込みが認められた(図3)。一方、テトラエチルアンモニウム(14C標識)、タウロコール酸(14C標識)およびコール酸(14C標識)では取り込みを示さなかった。
【0040】
NLTの基質選択性をさらに検討するために、NLT cRNAを注入した卵母細胞によるサリチル酸の取り込み実験において、系へ各種アニオン性物質を添加し、その影響を調べた(阻害実験)。サリチル酸の取り込み実験は、NLT cRNAを注入した卵母細胞を用い、前記に記載の方法に準じて実施した。但し、ND96溶液を用い、1mMの各種化合物(非標識)の存在下および非存在下で、サリチル酸の取り込みを測定した。その結果、構造的に無関係のアニオン性物質の添加で、cis−阻害効害効果が観察された(図4)。
【0041】
種々の組織におけるNLT遺伝子の発現(ノーザンブロッティング)の解析を行った。NLT cDNAの全長を32P−dCTPでラベルし、これをプローブとして用いてラットの種々の組織から抽出したRNAに対してノーザンブロッティングを以下のように行った。3μgのポリ(A)+ RNAを1%アガロース/ホルムアルデヒドゲルで電気泳動した後にニトロセルロースフィルターにトランスファーした。このフィルターを42℃で、32P−dCTPでラベルしたNLT cDNA全長を含んだハイブリダイゼーション液で一晩ハイブリダイセーションを行った。フィルターを65℃にて、0.1%SDSを含む0.1xSSCで洗浄した。
ノーザンブロットの結果(図5)、肝臓において、2.0Kb付近に強いバンドが検出され、また腎臓に弱い発現が観察された。
【技術分野】
【0001】
本発明は有機陰イオン(有機アニオン)輸送に関与する遺伝子と、その遺伝子がコードするポリペプチドに関する。
【背景技術】
【0002】
肝臓は、生体異物や薬物の代謝および体外排出に関して重要な役割を果たしている。肝細胞は極性を有する上皮細胞であり、血液とは側底膜、また胆汁とは胆管側膜を介して接している。陰イオン(アニオン)性の薬物は、輸送担体(トランスポーター)により側底膜を介して肝臓中に取り込まれ、また肝細胞内で代謝により産生された有機アニオンもトランスポーターにより肝細胞より排出される。
【0003】
肝細胞の側底膜を介した有機アニオンの取り込み(即ち血液と肝細胞間での有機アニオンの輸送)については、これまで摘出臓器灌流法や単離細胞膜小胞系などを用いた実験系により研究されてきた。これらの研究により主に、生理的有機アニオンの一つである胆汁酸の輸送について多くの知見が得られてきた。
【0004】
しかし従来の手法では、肝細胞の特に側底膜を介した物質輸送について詳細に解析することは困難であり、トランスポーターそのものを単離して詳細に解析することが望まれてきた。
【0005】
これまでの分子生物学的な研究から、肝臓の側底膜に発現している有機アニオントランスポーターとしては2種類のものがクローニングされている(Jacquemin,E.ら、 Proc.Natl.Acad.Sci.USA、91巻、133−7頁、1994年、Hagenbuch,B.ら、Proc.Natl.Acad.Sci.USA、88巻、10629−33頁、1991年)。
【0006】
しかしながら、これらのトランスポーターのみでは肝臓の側底膜における多様な有機アニオン輸送を説明することが出来ず、新規有機アニオントランスポーターの存在が予想されていた。
【0007】
腎臓は肝臓とならび、有機アニオン輸送を最も活発におこなっている臓器である。腎臓における(特に側底膜を介した)有機アニオン輸送の研究は100年近い歴史を持ち、その輸送特性については膨大な検討がなされてきた。我々は最近、この腎臓での有機アニオン輸送において最も重要な役割を果たしている有機アニオントランスポーターOAT1の単離に成功し(Sekine,T.ら、J.Biol.Chem.、272巻、18526−9頁、1997)、既に特許出願済みである。OAT1は基質選択性の極めて広い、即ち化学構造の異なる多くの有機アニオンを輸送することの出来るトランスポーターである。また多くの薬物が有機アニオンに属することからOAT1は薬物トランスポーターとしても認識されている。OAT1の発現は腎臓に限られており、肝臓には全くその発現が認められない。
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明の目的は、肝臓における有機アニオン輸送に関与する新規な有機アニオンントランスポーター遺伝子およびその遺伝子がコードするポリペプチドである有機アニオントランスポーターを同定し、提供することにある。その他の目的については以下の記載より明白である。
【課題を解決するための手段】
【0009】
本発明者らは、腎臓の有機アニオントランスポーターOAT1との構造上の類似性から、肝細胞に発現している肝特異的有機アニオントランスポーター(OAT2)を同定した。
【0010】
本発明の有機アニオントランスポーターOAT2は、ジカルボン酸、プロスタグランジン、非ステロイド系抗炎症薬や抗腫瘍薬等の異なる化学構造を持った薬物や内因性物質に対してこれらを輸送する(取り込む能力を有する)、非常に広い範囲の基質選択性を有するものである。
【0011】
本発明のタンパク質としては、配列番号1で示されたアミノ酸配列を有するもののほか、例えば、配列番号1で示されたアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列を有するものが挙げられる。アミノ酸の欠失、置換もしくは付加は、有機アニオン輸送活性が失われない程度であればよく、通常1〜約110個、好ましくは1〜約55個である。このようなタンパク質は、配列番号1で示されたアミノ酸配列と通常、1〜80%、好ましくは1〜90%のアミノ酸配列のホモロジーを有する。
【0012】
本発明において、ストリンジェントな条件下でのハイブリダイゼーションは、通常、ハイブリダイゼーションを、5×SSC又はこれと同等の塩濃度のハイブリダイゼーション溶液中、37〜42℃の温度条件下、約12時間行い、5×SSCまたはこれと同等の塩濃度の溶液等で予備洗浄を行った後に、1×SSC又はこれと同等の塩濃度で洗浄を行うことにより実施できる。また、より高いストリンジェンシーを得るためには、洗浄を0.1×SSC又はこれと同等の塩濃度の溶液中で洗浄を行うことにより実施できる。
【0013】
本発明の有機アニオントランスポーター遺伝子は、適当な哺乳動物の肝臓の組織や細胞を遺伝子源として用いてスクリーニングを行うことにより単離取得できる。哺乳動物としては、イヌ、ウシ、ウマ、ヤギ、ヒツジ、サル、ブタ、ウサギ、ラット、マウスなどの非ヒト動物のほか、ヒトが挙げられる。
【0014】
遺伝子のスクリーニングおよび単離は、ホモロジースクリーニングおよびPCR(Polymerase Chain Reaction)スクリーニングなどにより好適に実施できる。
【0015】
得られたcDNAについては、常法により塩基配列を決定し、翻訳領域を解析
して、これにコードされるタンパク質、即ちOAT2のアミノ酸配列を決定することができる。
【0016】
得られたcDNAが、有機アニオントランスポーター遺伝子のcDNAであること、即ちcDNAにコードされた遺伝子産物が有機アニオントランスポーターであることは、例えば次のようにして検証することができる。即ち、得られたOAT2遺伝子から調製したcRNAを卵母細胞に導入して発現させ、有機アニオンを細胞内に輸送する(取り込む)能力を、適当な有機アニオンを基質とする通常の取り込み実験(Sekine,T.ら、J.Biol.Chem.、272巻、18526−9頁、1997)により、細胞内への基質取り込みを測定することにより確認できる。
【0017】
また、OAT2発現細胞について、同様の取り込み実験を応用して、OAT2の輸送特性や基質特異性などを調べることができる。
【0018】
得られたOAT1遺伝子のcDNAを用いて、異なる遺伝子源で作製された適当なcDNAライブラリー又はゲノミックDNAライブラリーをスクリーニングすることにより、異なる組織、異なる生物由来の相同遺伝子や染色体遺伝子等を単離することができる。
【0019】
また、開示された本発明の遺伝子の塩基配列(配列番号2)に示された塩基配列、もしくはその一部の情報に基づいて設計された合成プライマーを用い、通常のPCR法によりcDNAライブラリーから遺伝子を単離することが出来る。
【0020】
cDNAライブラリー及びゲノミックDNAライブラリー等のDNAライブラリーは例えば、「Molecular Cloning;Sambrook, J.,Fritsh,E.F.およびManiatis,T.著、Cold Spring Harbor Laboratory Pressより、1989年に発刊」に記載の方法により調製することができる。あるいは、市販のライブラリーがある場合にはこれを用いてもよい。
【0021】
本発明の有機アニオントランスポーター(OAT2)は、例えば、有機アニオントランスポーターをコードするcDNAを用い、遺伝子組み換え技術により生産することができる。例えば、有機アニオントランスポーターをコードするDNA(cDNA等)を適当な発現ベクターに組み込み、得られた組み換えDNAを適当な宿主細胞に導入することができる。ポリペプチドを生産するための発現系としては、例えば、細菌、酵母、昆虫細胞および哺乳動物細胞の発現系等が挙げられる。このうち、機能タンパクを得るためには、昆虫細胞および哺乳動物細胞を用いることが望ましい。
【0022】
例えば、ポリペプチドを哺乳動物で発現させる場合には、有機アニオントランスポーターをコードするDNAを、適当な発現ベクター(例えば、レトロウイルス系ベクター、パピローマウイルスベクター、ワクシニアウイルスベクター、SV40系ベクター等)中の適当なプロモーター(例えばSV40プロモーター、LTRプロモーター、エロンゲーション1αプロモーター等)の下流に挿入して発現ベクターを構築する。次に、得られた発現ベクターで適当な動物細胞を形質転換して、形質転換体を適当な培地で培養することによって、目的とするポリペプチドが生産される。宿主とする哺乳動物細胞としては、サルCOS−7細胞、チャイニーズハムスターCHO細胞、ヒトHela細胞または、腎臓組織由来の初代培養細胞やブタ腎由来LLC−PK1細胞、フクロネズミ腎由来OK細胞等の細胞株が挙げられる。
【0023】
有機アニオントランスポーターOAT2をコードするcDNAとしては、例えば、配列番号2に示される塩基配列を有するcDNAを用いることが出来るほか、前記のcDNAに限定されることなく、アミノ酸配列に対応するDNAを設計し、ポリペプチドをコードするDNAをもちいることもできる。この場合、一つのアミノ酸をコードするコドンは各々1〜6種類知られており、用いるコドンの選択は任意でよいが、例えば発現に利用する宿主のコドン使用頻度を考慮して、より発現の高い配列を設計することができる。設計した塩基配列をもつDNAはDNAの化学合成、前記cDNAの断片化と結合、塩基配列の一部改変等によって取得できる。人為的な塩基配列の一部改変、変異導入は、所望の改変をコードする合成オリゴヌクレオチドからなるプライマーを利用して部位変異導入法(site specific mutagenesis)「Mark,D.F.ら、Proc Natl Acad Sci USA 第18巻、5662−5666頁、1984年」等により実施できる。
【0024】
本発明の有機アニオントランスポーター遺伝子にストリンジェントな条件下でハイブリダイズするヌクレオチド(オリゴヌクレオチドもしくはポリヌクレオチド)は、有機アニオントランスポーター遺伝子を検出するためのプローブとして使用できるほか、有機アニオントランスポーターの発現を変調させるために、例えばアンチセンスオリゴヌクレオチドやリボザイム、デコイとして使用することもできる。このようなヌクレオチドとしては、例えば、配列番号2で示される塩基配列の中の通常、連続する14塩基以上の部分配列もしくはその相補的な配列を含むヌクレオチドを用いることができ、ハイブリダイズをより特異的とするためには、部分配列としてより長い配列、例えば20塩基以上あるいは30塩基以上の配列を用いても良い。
【0025】
また、本発明の有機アニオントランスポーターまたは、これと免疫学的同等性を有するポリペプチドを用いて、その抗体を取得することが出来、抗体は、有機アニオントランスポーター検出や精製などに利用できる。抗体は、本発明の有機アニオントランスポーター、その断片、またはその部分配列を有する合成ペプチド等を抗原として用いて製造できる。ポリクロナール抗体は、宿主動物(たとえば、ラットやウサギ)に抗原を接種し、免疫血清を回収する通常の方法により製造することができ、モノクロナール抗体は、通常ハイブリドーマ法などの技術により製造できる。
【図面の簡単な説明】
【0026】
【図1】図1は、卵母細胞におけるサリチル酸の取り込みを示したものである。図1のBはグルタル酸とのプレインキュベーションの影響(図1Bの「+」)をみたものである。図1中の「L5」はL5を発現させせた卵母細胞を用いた系である。
【図2】図2は、本発明の有機アニオントランスポーターにおける基質α−ケトグルタル酸(図2A)とサリチル酸(図2B)の動力学試験の結果を示すものである。
【図3】図3は、本発明の有機アニオントランスポーターの各種薬剤および内因性有機アニオンの取り込みを示すものである。図3中の「3H−PGE2」は3H標識化プロスタグランジンE2を示し、「14C−PAH」は14C標識化パラアミノ馬尿酸を示す。
【図4】図4は、各種のアニオン物質の存在下における、本発明の有機アニオントランスポーターのサリチル酸の取り込みの影響を調べたものである。
【図5】図5は、本発明の遺伝子のノーザンブロッティング解析の結果を示すものである。
【0027】
以下、実施例をもって本説明をさらに詳しく説明するが、これらの実施例は本発明を制限するものではない。
【0028】
なお下記実施例において、各操作は特に断りがない限り、「Molecular Cloning:Sambrook,J.,Fritsh,E.F.およびManiatis,T.著、Cold Spring HarborPressより、1989年に発刊」に記載の方法により行うか、または、市販のキットを用いる場合には市販品の指示書に従って使用した。
【実施例】
【0029】
実施例1 ラット肝特異的有機アニオントランスポーター(NLT:novel liber transporter)cDNAの単離とその解析
(1)腎特異的有機アニオントランスポーターOAT1の塩基配列情報を基にした、関連遺伝子の検索
【0030】
既に我々が単離したOAT1(Sekine,T.ら、J Biol Chem 272巻、18526−9頁、1997年)の塩基配列情報をもとにDNAデータベースを検索したところ、機能未知として報告されていた膜タンパクNLT(Simonson,G.D.ら、J.Cell.Sci.、107巻、1065−1072頁、1994)がOAT1と弱い相同性(アミノ酸レベルで42%)を有することが判明した。またNLTは肝細胞の側底膜に限局して発現していると考えられた。我々は、NLTが肝細胞側底膜に存在する多選択性有機アニオントランスポーターであると仮定してその単離と機能解析を行った。
【0031】
cDNAライブラリーは、ラット肝ポリ(A)+RNAから、cDNA合成キット(商品名:Superscript Choice System,ギブコ社製)を使用して作製し、ファージベクターλZipLox(ギブコ社製)の制限酵素EcoRI切断部位に組み込んだ。PCR法にて、既に報告されているNLTの 131−673塩基に相当する部位を単離し、32P−dCTPでラベルし、これをプローブとして用いてラットのcDNAライブラリーをスクリーニングした。ハイブリダイゼーションは、37℃のハイブリダイゼーション用溶液中で一昼夜行い、その後フィルター膜は、37℃で0.1×SSC/0.1%SDSで洗浄した。ハイブリダイゼーション溶液としては、50% ホルムアミド、5×standard saline citrate(SSC)、3×デンハード液、0.2% SDS、10%硫酸デキストラン、0.2mg/ml変性サーモン精子DNA、2.5mMピロリン酸ナトリウム、25mM MES、0.01% Antifoam B(シグマ社製)を含むpH6.5の緩衝液を用いた。λZipLox中に単離されたクローンは、in vitro excision法によりプラスミドベクターpZLにサブクローン化した。
【0032】
上記により得られたクローン(L5)の塩基配列の決定は、NLTに対する特異的オリゴヌクレオチドプライマーを合成し、Sequenase ver.2.0(アマシャム社製)を用いておこなった。
【0033】
これにより、NLTと相同なクローン(L5)遺伝子のcDNA塩基配列が得られた。
(2)NLTの機能の特定
【0034】
上記により得られたクローン(L5)を含むプラスミドから、T7 RNAポリメラーゼを用いて、cRNA(cDNAに相補的なRNA)を調製した。
【0035】
得られたcRNAを、既に報告されている方法に従い(Sekine,T.,et al.J Biol Chem 272巻、18526−9頁、1997年)、アフリカツメガエルの卵母細胞に注入し、この卵母細胞についてグルタル酸およびサリチル酸を基質として用いた取り込み実験をおこなった。実験には、放射能ラベルした基質(14C−グルタル酸および14C−サリチル酸)を用いた。その結果、L5を発現させた卵母細胞は14C−グルタル酸および14C−サリチル酸の取り込みを示すことが判明した。
【0036】
L5の輸送活性における細胞外Naイオンの影響を検討する実験を行った。サリチル酸の取り込み実験は前記に記載の方法に準じて実施した。但し、incubation溶液はND96溶液および、塩化コリン溶液(ND96溶液の96mMの塩化ナトリウムを96mMの塩化コリンで置換し、pHを7.4に調製した)を用いておこなった。その結果、細胞外のナトリウムをコリンと置換しても、サリチル酸取り込みに何ら影響を与えなかった(図1A)。このことから、NLTはNaイオン非依存的に働くトランスポーターであることが示された。
【0037】
NLT cRNAを注入した卵母細胞によるサリチル酸の取り込み実験においてグルタル酸とのプレインキュベーションの影響を調べた。サリチル酸の取り込み実験は、NLT cRNAもしくはNLT cRNAとラットNaDC−1(Na−ジカルボン酸トランスポーター)cRNAを注入した卵母細胞を1mMグルタル酸添加もしくは無添加のND96溶液で2時間培養した後、14C−サリチル酸を添加して室温で1時間培養し放射能ラベルされた基質の取り込みを調べた。その結果1mMグルタル酸添加した溶液で前処置しても14C−サリチル酸の取り込みは変わらなかった(図1B)。この結果はNLTが交換輸送体ではなく、促進拡散型輸送体であることを示唆するものである。
【0038】
NLTの有機アニオン輸送のミカエリスーメンテンの動力学試験をおこなった。
基質サリチル酸およびα−ケトグルタル酸の濃度の違いによるそれぞれの取り込み率の変化を調べることにより、NLTのミカエリスーメンテンの動力学試験をおこなった。サリチル酸およびα−ケトグルタル酸の取り込み実験は、NLT cRNAを注入した卵母細胞を用い、前記記載方法に準じて実施した。この結果(図2)、サリチル酸およびα−ケトグルタル酸のKm値はそれぞれ 88.8 ± 23.4 μMおよび17.8 ± 2.9μMであった。
【0039】
NLTの基質選択性を検討するために、各種アニオン性物質を基質とする取り込み試験を行った。取り込み実験は、NLT cRNAを注入した卵母細胞を用い、ND96溶液中で前記記載方法に準じて実施した。但し、基質としては、放射能でラベルした各種化合物を用いた。その結果、α−ケトグルタレート(14C標識)、メトトレキセート(3H標識)、プロスタグランジンE2(3H標識)、アセチルサリチル酸(14C標識)、パラアミノ馬尿酸(14C標識)を基質とした場合に、卵母細胞への取り込みが認められた(図3)。一方、テトラエチルアンモニウム(14C標識)、タウロコール酸(14C標識)およびコール酸(14C標識)では取り込みを示さなかった。
【0040】
NLTの基質選択性をさらに検討するために、NLT cRNAを注入した卵母細胞によるサリチル酸の取り込み実験において、系へ各種アニオン性物質を添加し、その影響を調べた(阻害実験)。サリチル酸の取り込み実験は、NLT cRNAを注入した卵母細胞を用い、前記に記載の方法に準じて実施した。但し、ND96溶液を用い、1mMの各種化合物(非標識)の存在下および非存在下で、サリチル酸の取り込みを測定した。その結果、構造的に無関係のアニオン性物質の添加で、cis−阻害効害効果が観察された(図4)。
【0041】
種々の組織におけるNLT遺伝子の発現(ノーザンブロッティング)の解析を行った。NLT cDNAの全長を32P−dCTPでラベルし、これをプローブとして用いてラットの種々の組織から抽出したRNAに対してノーザンブロッティングを以下のように行った。3μgのポリ(A)+ RNAを1%アガロース/ホルムアルデヒドゲルで電気泳動した後にニトロセルロースフィルターにトランスファーした。このフィルターを42℃で、32P−dCTPでラベルしたNLT cDNA全長を含んだハイブリダイゼーション液で一晩ハイブリダイセーションを行った。フィルターを65℃にて、0.1%SDSを含む0.1xSSCで洗浄した。
ノーザンブロットの結果(図5)、肝臓において、2.0Kb付近に強いバンドが検出され、また腎臓に弱い発現が観察された。
【特許請求の範囲】
【請求項1】
肝特異的有機アニオントランスポーターOAT2。
【請求項2】
配列表の配列番号1に示されるアミノ酸配列、又は、その一部のアミノ酸配列が欠失し、他のアミノ酸で置換若しくは付加されていてもよいアミノ酸配列を有する請求項1に記載の肝特異的有機アニオントランスポーターOAT2。
【請求項3】
配列表の配列番号1に示されるアミノ酸配列、又は、その一部のアミノ酸配列が欠失し、他のアミノ酸で置換若しくは付加されていてもよいアミノ酸配列をコードする核酸。
【請求項4】
配列表の配列番号2で示される塩基配列を有するDNAである請求項3に記載の核酸。
【請求項5】
配列表の配列番号2で示される塩基配列を有するDNAの連続する少なくとも14塩基又はその相補鎖からなる核酸。
【請求項6】
塩基の数が20以上である請求項5に記載の核酸。
【請求項7】
請求項1又は2に記載の肝特異的有機アニオントランスポーターを認識し得る抗体。
【請求項1】
肝特異的有機アニオントランスポーターOAT2。
【請求項2】
配列表の配列番号1に示されるアミノ酸配列、又は、その一部のアミノ酸配列が欠失し、他のアミノ酸で置換若しくは付加されていてもよいアミノ酸配列を有する請求項1に記載の肝特異的有機アニオントランスポーターOAT2。
【請求項3】
配列表の配列番号1に示されるアミノ酸配列、又は、その一部のアミノ酸配列が欠失し、他のアミノ酸で置換若しくは付加されていてもよいアミノ酸配列をコードする核酸。
【請求項4】
配列表の配列番号2で示される塩基配列を有するDNAである請求項3に記載の核酸。
【請求項5】
配列表の配列番号2で示される塩基配列を有するDNAの連続する少なくとも14塩基又はその相補鎖からなる核酸。
【請求項6】
塩基の数が20以上である請求項5に記載の核酸。
【請求項7】
請求項1又は2に記載の肝特異的有機アニオントランスポーターを認識し得る抗体。
【図1】


【図2】
【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2012−139231(P2012−139231A)
【公開日】平成24年7月26日(2012.7.26)
【国際特許分類】
【出願番号】特願2012−55037(P2012−55037)
【出願日】平成24年3月12日(2012.3.12)
【分割の表示】特願平10−169174の分割
【原出願日】平成10年6月3日(1998.6.3)
【出願人】(503360115)独立行政法人科学技術振興機構 (1,734)
【Fターム(参考)】
【公開日】平成24年7月26日(2012.7.26)
【国際特許分類】
【出願日】平成24年3月12日(2012.3.12)
【分割の表示】特願平10−169174の分割
【原出願日】平成10年6月3日(1998.6.3)
【出願人】(503360115)独立行政法人科学技術振興機構 (1,734)
【Fターム(参考)】
[ Back to top ]