
肝細胞増殖因子(HGF)の生物活性に関する評価
本発明では、治療の要素として使用するための有効な生物活性のある肝細胞増殖因子の識別法について示す。生物活性は、i)損傷細胞の回復のin vitroモデル、ならびに/あるいはii)C−met受容体、抗HGF抗体またはデキストランなどの多糖類に対して、SPRを使用する親和性測定またはエピトープマッピング、ならびに/あるいはiii)発毛速度に関する動物モデルを使用して、測定することができる。同様に、これらの方法によって、身体の分泌物におけるHGFの活性を評価することができる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、肝細胞増殖因子(HGF)の生物活性についての評価法に関する。本法は、例えば、組換え技術による、バッチ生産(batch production)後のHGFの品質管理のため、およびHGF製剤の評価のために、治療の必要な患者への投与前に使用することができる。また、本評価法は、HGF関連疾患の診断およびこれに続く治療選択を向上させるために使用することができる。したがって、本発明は、HGFの役割を評価するために、潰瘍分泌物、尿、糞便、唾液、胸膜における損傷部位の内因性HGFの活性を評価し、外因性HGFの損傷部位への適用が必要かどうかを判定する方法にも関する。
【背景技術】
【0002】
肝細胞増殖因子(HGF)は、細胞損傷に反応して分泌され、特定の器官の再生および創傷治癒のために重要と思われる増殖因子である(Arakaki Nら、 Hepatology 221, 1995)。HGFは、H鎖が約60kDaおよびL鎖が約30kDaのヘテロ二量体で、最初に、不活性前駆体として合成される(Miyazawa Kら、J Biol Chem 268, 1994)。この前駆体は、損傷器官で特定の活性化因子によって活性タンパク質へ切断される(Naka Dら、 J Biol Chem 276, 1992、Nishizaki Tら、J Am Coll Surg 181, 1995)。HGFは、内分泌作用だけでなく、パラクリン作用を有する(Yanagita K.ら、Biochem Biophys Res Commun 182, 1992、Kono Sら、Biochem Biophys Res Commun 186, 1992)。HGFの標的細胞は、完全に発育した上皮細胞である(Matsumoto K およびNakamura T. J Gastroenterology Hepatology 6, 1991)。HGFは、器官損傷部位において産生され、高濃度で存在する(Nayeri Fら、Scand J Infect Dis, 34, 2002)。我々は、以前に、様々な感染症でHGFの全身的産生および局所産生について検討し、胃腸炎などの急性感染症の間に、高濃度の血清HGFを観察している(Nayeri Fら、Scand J Infect Dis, 34, 2002)。我々は、髄膜炎中に、HGFの全身的産生の亢進と同時に、脳脊椎液で高濃度のHGFを発見した(Nayeri Fら、 J Infect Dis, 181, 2000)。肺炎患者における呼気凝縮物(exhaled breath condensate)中のHGF濃度の上昇(Nayeri Fら、Respir Med, 96, 2002)は、HGFの血清濃度とは相関がなく、肺炎中のHGFの局所産生を示した。さらに、我々は、血清におけるHGFの安定性について研究しており(Nayeri Fら、CYTOLIKE, 2002)、HGFが試料中(血清/血漿、糞便、呼気凝縮物)で非常に安定しており、数回にわたる凍結−解凍の繰り返し、異なる緩衝液または−20℃での数年間の保管は、HFG濃度に著しい影響を及ぼさないことを発見した。
【0003】
HGFは多機能の増殖因子であり、HGFの再生特性は、肝臓(Ishikiら、1992)、腎臓(Igawaら、1993)、肺(Yanagitaら、1993)および敗血症中(Kondoら、1999)の動物モデルで研究されている。HGFの標的細胞に対する分裂促進作用が、組織再生および増殖を生じさせるには十分でないことが示唆されている。細胞遊走促進作用および形態形成作用などのHGFの他の機能が再生過程を完了するために必要である(Montesanoら、1991)。HGFの切断種(truncated species)は、組織再生および修復の最終段階で、HGFの増殖促進作用に対するアンタゴニストとして機能する可能性があると仮定されている(Chanら、1991)。
【発明の開示】
【発明が解決しようとする課題】
【0004】
以前に我々は、外因性HGF投与により、治療が困難な慢性の下肢潰瘍の患者を治癒することができることを示している(Nayeriら、Dermatological Treatment, 2002)。最近の二重盲検試験の間、我々は、後に欠陥があることが明らかになった市販の組換えHGFを使用している。試験製剤における、この欠陥成分の使用により、潰瘍の十分な治癒が得られなかった。1年後、これらの患者の一部(n=4)に活性のある組換えHGFを投与したところ、最初の1週間以内に、かれらの潰瘍に著明な治癒が見られた(表1)。感染症治療におけるHGFの、将来の指標に加えて、抗生物質(EP01963646.3として)の治療的可能性により、治療の要素として使用されるHGFの生物活性について評価することは非常に重要である。
【0005】
本発明は、生物学的生産物と共に、組換え製剤におけるHGF活性の評価法について部分的に述べる。
【課題を解決するための手段】
【0006】
(発明の概要)
本発明の目的は、体液で検出されるHGFに加えて、組換えHGFの生物活性の評価法を提供することである。
【0007】
本発明のさまざまな他の目的および利点は、本発明の図および以下の記述から明らかになるであろう。
【0008】
一実施形態では、本発明は、活性HGFと相互に作用するHGF受容体を発現する上皮または内皮の細胞株を使用するHGF活性の評価のためのin vitroモデルについて述べる。特に、マウス皮膚上皮細胞株(CCL−53.1)が使用される。他の実施形態では、本発明は、HGFの生物活性についてのin vitroの評価法に関する。HGFに対するモノクローナル抗体またはポリクローナル抗体による1つまたは複数の相互作用の生物活性に関する評価において、C−met受容体および多糖類の特にデキストランを使用する。これらの相互作用の研究のために、多くの方法が利用可能で、当業者によく知られている。本発明の好ましい実施形態では、BIACORE(スウェーデン、ウプサラ)によって提供された器具を使用する。この技術は、表面プラズモン共鳴法に基づいており、多くの相互作用を並行して調べることができるという点で、非常に効果的である。しかし、もちろん、本発明による方法は、明確にするための記載が、この実施形態に焦点を合わせているとしても、特定の器具に限定されるものではない。
【0009】
本発明の他の実施形態では、HGFの生物活性は、マウス(例えば、C57BL/6)のin−vivoモデル利用する方法を使用して評価する。
【0010】
さらなる実施形態では、本発明は、HGFの生物活性の評価用キットに関する。この種のキットでは、細胞株CCL−53.1は、培養細胞株が損傷し、損傷部位に向けた隣接細胞の運動性がHGFの量および活性に用量依存的に関連する場合に使用する。各々の細胞に対する細胞の粘着性減少による形態形成変化、および細胞の細胞骨格特性の変化による損傷した無毛部位の増大に加えて、損傷部位への細胞の運動性の亢進が見られる。この反応は、HGFに特異的であって、抗HGF抗体によって止めることができる。線維芽細胞増殖因子、上皮増殖因子、血小板由来増殖因子およびケラチノサイト増殖因子の使用では、そのような影響を細胞に及ぼすことはない。不活性HGFまたは1ng/mLより低い用量で活性HGFを使用することで、細胞の形態学的活性または細胞遊走促進活性を誘発することはない。
【0011】
他の実施形態では、本発明は、HGFに対する少なくとも1つのリガンド(ポリクローナルまたはモノクローナルの抗HGF抗体またはc−metプロトオンコジーン受容体)で固定化されるいくつかのチャネルを有するセンサチップ、および不活性デキストランを有する少なくとも1つのチャンネルからなるプラズモン共鳴バイオセンサー(BIACORE)法を使用するHGFの生物活性評価のためのキットに関する。生物活性のあるHGFは、デキストランおよびアルブミンにだけでなくそのリガンドに高い親和性を有し、一方、生物活性のないHGFは、そのリガンドと相互に作用する場合と作用しない場合があり、デキストランまたはアルブミンとは相互に作用しない。
【発明の効果】
【0012】
(発明の詳細な説明)
HGFの変異体:
肝細胞増殖因子(HGF)は、分子量が約90kDaの大きなタンパク質である。活性化されると、このタンパク質は、α鎖およびβ鎖に切断される。以前の試験は、HGF分子内のN末端のヘアピンドメイン、最初のクリングルドメイン、2番目のクリングルドメインまたはセリンプロテイナーゼドメインの欠失が生物活性の全面的な喪失を招くことを示している(Matsumotoら、1991b)。HGF分子のα鎖またはβ鎖のいずれかの欠失は、HGFの活性の完全な喪失を招く(Matsumotoら、1991)。c−met受容体結合ドメインは、HGFのα鎖のNH2末端で同定されている(Mizunoら、1994)。また、HGFの受容体結合および活性で重要な役割を果たす27残基のヘアピンループの三次元(3D)モデルは、タンパク質工学および核磁気共鳴(NMR)技術(Zhouら、1998)によって得られている。ドメイン欠失実験の結果および個々のドメインの3Dモデルの有用性と共に、HGF配列の特徴から、いくつかの試験につながり、ここでは、特定の残基または残基のクラスターが、HGF活性におけるこれらの役割を明らかにするために置換されている(Chirgadzeら、1998)。Lokkerら(1994)は、Nドメイン内で、受容体結合のために重要な残基について研究している。今まで、多くの個々および群の突然変異について、ヘアピンループ自体の範囲内で、受容体結合にとって重要な残基を特定できなかった。しかし、彼らは、NドメインのC末端(His−114、Glu−115、Asp−171)に位置する残基のクラスターが、HGFの受容体結合および生物活性の両方に関わることを発見した(Lokkerら、1994)。また、彼らは、最初のクリングル内で、少なくとも7種のアミノ酸(Arg−197、Glu−189、Tyr−198、Glu−195、Asp−171、Gln−173およびSer−161)の置換が、HGFの受容体結合および生物活性に対して明らかな影響を有することを示している。最も大きな影響は、Arg−197およびGlu−159の置換で見られ、受容体結合および生物活性が50倍を超えて減少する(Lokkerら、1994)。HGFのセリンプロテイナーゼドメインにおけるいくつかの点変異(残基のGln−534およびTyr−673が、活性セリンプロテイナーゼのHisおよびSer残基に復帰した)は、受容体結合には影響を及ぼさないが、生物活性を著明に減少させた。同様の結果が、Val692Serの突然変異で得られた(Lokkerら、1992)。C末端のクリングル(クリングル4)とセリンプロテイナーゼドメインの間に位置するトリプシン様切断部位での重要な残基の突然変異は、HGFの単鎖前駆体型(pro−HGF)の転換を消失させた。結果として生じる単鎖タンパク質は、HGFの受容体結合活性を維持するものの、標的細胞に対する生物活性を欠いていた(Lokkerら、1992、Nakaら、1992、Gakら、1992)。
【0013】
HGFの生物活性についての評価の重要性
以前のパイロット研究では、高齢患者11例を対象に、19の慢性下肢潰瘍うちの15に対して、局所にゲル状HGFを1日1回、7日間塗布した。全例が以前に従来の方法によって治療されており、彼らの下肢潰瘍は、1年から14年の期間でやや安定した状態であった。アレルギー、不快感または疼痛に関するあらゆる徴候が毎日報告された。隣接する無傷の皮膚と比較して、潰瘍の微小循環灌流(microcirculation perfusion)について、(患者7例で)本研究の開始時、1週間後、および再度3カ月後にレーザードップラーによって測定した。また、潰瘍の大きさおよび特徴についても詳細に記録した。我々は、HGFの血管形成作用に反映する可能性がある微少循環灌流が、治療した潰瘍における潰瘍部位の縮小と統計的に有意な相関を示すことを観察した(r=0.94、p<0.002)。十分良好な治癒(84〜100%の領域縮小)または部分的治癒(58〜59%)が患者11例中8例で認められた。
【0014】
このパイロット研究中に観察された慢性下肢潰瘍に対する肝細胞増殖因子の正の効果を評価するために、患者20例を対象にして、市販の組換えHGFを使用した1週間の二重盲検プラセボ対照試験を開始した。本試験は、患者12例の組み入れ後、SDS−PAGEおよびウエスタンブロットにより、使用されたタンパク質に欠陥があった(わずか1本の約60〜67kDaの鎖が発見された)ことが明らかにされた時点で中断された。潰瘍部位由来の皮膚生検材料の免疫組織化学染色は、正常健常対照(n=10)と比較して、患者(n=12)では、c−met受容体の発現が有意に高いことを示した(p<0.001)。市販の組換えHGFを含む製剤を投与した患者(第1群、n=6)で、c−met発現が減少する傾向が観察されたが(タンパク質のc−met受容体との相互作用を示している可能性がある)、本群と塩化ナトリウム製剤を投与した群(第2群、n=6)との間の差は有意ではなかった(表1)。第1群では、毛細管増殖の亢進が観察されたが(有意ではない)、劇的な臨床的改善が認められたのは、わずか1例に過ぎなかった。にもかかわらず、第1群の3例および第2群の2例が9カ月以内に治癒した。他の患者7例には、生物活性のある組換えHGFによる治療を申し出て、4例(各群2例)が受け入れた(第3群)。これら当初不成功の全例において、潰瘍の状態の改善が、治療開始から1週間以内に観察された。潰瘍部位のc−met発現が、患者2例で確認され、2例とも1週間の治療後に減少した。
【0015】
我々は、生物活性のあるHGFが慢性下肢潰瘍の治癒に対して、プラセボ効果によるものでない正の効果を有すると結論を下した。欠陥のある組換えHGFと活性のある組換えHGFの特性について、患者に対する治療効果を有するHGFの生物活性型を特徴付けるために調査した。上記のように、HGFの分裂促進効果および細胞遊走促進効果は、効果的な治癒プロセスにとって必要とされる。
【0016】
本発明は、in−vitroにおいてHGFの免疫学的および生物学的特性を調査することによるHGFの生物活性の測定について述べる。
【実施例】
【0017】
本発明では、治療的処置用として適した生物活性のあるHGFを定義するために、以下の基準を採用している。本発明の方法を使用することで、生物活性のあるHGFは以下の通り。
【0018】
・CCL−53.1モデルで、18時間以内に少なくとも50%の損傷部位の減少を引き起こすはずである。
【0019】
・SPRベースの方法において、(20μg/mL)c−metプロトオンコジーンHGF受容体だけでなく、(50μg/mL)固定モノクローナルおよびポリクローナル抗HGF抗体に対して同等の親和性を有するはずである。
【0020】
・SPRベースの方法において、モノクローナル抗HGF抗体に対する親和性より低く、デキストランに対して陰性対照より高い親和性を有するはずである。この親和性は、試料にデキストランを添加すると、消失するはずである。
【0021】
・免疫組織化学染色によって、またはマウスが無毛部位で完全な発毛を示す時間(15日未満)を判定することによって判定して、HGF治療(少なくとも100ng/マウス/日を5日間)の1週間後、無毛部位から採取した皮膚生検材料で、PCNA発現によって判定したマウスの刈毛した皮膚部位より、2倍速い発毛を起こすはずである。
【0022】
以下の実施例は、本発明の実施形態をさらに説明することを意図するものであって、決して本発明の範囲を制限することを意図していない。
【0023】
(実施例1:in vitroでのマウスの損傷メラニン形成細胞の回復を加速する肝細胞増殖因子の効果)
以下の材料および条件を使用した。
【0024】
細胞:マウスの皮膚由来のCCL−53.1付着上皮細胞、メラニン形成細胞、黒色腫は、アメリカン・タイプ・カルチャー・コレクション(American Tissue Cell Collection:ATCC、米国バージニア州Manassas)から入手し、継代数3〜32で使用した。
【0025】
増殖因子:組換え肝細胞増殖因子(HGF)は、好意によりT. Nakamura教授(日本、大阪)から寄贈された。また、HGF(RD Systems、ロット:QF031062)、線維芽細胞増殖因子(FGF、RD Systems、ロット:EX164011)、血小板由来増殖因子(PDGF、RD Systems、ロット:D332071)、ケラチノサイト増殖因子(KGF、RD Systems、ロット番号:J0164021)、上皮増殖因子(EGF、RD Systems、ロット:HLM114031)は市販のものを購入した。
【0026】
Nakamura教授から入手した組換えHGFは、患者の慢性潰瘍治療に非常に有効であることが示されたので(実施例4)、本研究では、生物活性のあるHGFの陽性対照として使用している。
【0027】
細胞培養:CCL−53.1細胞は、37℃の5%CO2および95%空気の雰囲気中で、15%ウマ血清および2.5%ウシ胎児血清(Sigma-Aldrich、スウェーデン、ストックホルム)を補充したKaighn改変Ham’s F−12K培地(ATCC)で増殖した。細胞が集密に達した後、非酵素細胞解離液(non-enzymatic cell dissociation solution)(1×)(Sigma-Aldrich)により分離し、15%ウマ血清および2.5%ウシ胎児血清を補充したF−12K培地で懸濁し、24ウェル培養プレート(Nunc Brand Products、デンマーク、ロスキレ(Roskilde))に播種し、集密に達するまで、同一条件下で24〜48時間培養した。
【0028】
無血清培地:75−cm2組織培養フラスコ(Nuncolone)で増殖させたCCL−53.1細胞の集密状態の培養物をリン酸塩緩衝液(Sigma-Aldrich)で洗い、非酵素細胞解離液により分離し、無血清培地のNutridoma−NS培地(Roche Diagnostics Scandinavia AB、スウェーデン、ブロンマ(Bromma))で再懸濁し、24ウェル培養プレートに播種し、集密に達するまで、上記と同一条件下で24〜48時間培養した。
【0029】
損傷アッセイ:集密状態の単層を無菌のスチール器具によってこする。分離細胞をPBSで洗い、ウェルに新鮮培地を加える。細胞に覆われていない正方形部分(mm2)を顕微鏡(オリンパス)で測定し、各ウェルに記録した。次に、HGF(1、5、10ng/ml)を細胞に加え、5%CO2を含む加湿した雰囲気において、37℃でインキュベートした。24および48時間後、単層に覆われていない部分を再度測定し、記録した。実験は、3通り実施した。3通りの中央値を、13の個別の実験の平均値の計算に使用した。ヘパリン(Leo Pharma AB、スウェーデン、マルメー(Malmo)、SE−20124、Box404)を、異なる濃度(範囲1〜100IE/ml)で、HGF添加または不添加の傷つけた単層に加えた。
【0030】
細胞増殖:CCL−53.1細胞は、24ウェルプレートで、前述のようにウェルあたり1×106細胞の密度で培養した(細胞計数マルチサイザー[Cell Counter Multisizer]、KEBO Lab、スウェーデン、スパンガ(Spanga))。集密後、細胞を非酵素細胞解離培地(Sigma Aldrich)で分離した。DNA合成は、3H−チミジンの取り込みを測定することによって判定した。分離CCL−53.1細胞は、24ウェル培養プレートに播種し、15%ウマ血清および2.5%ウシ胎児血清を補充したF−12K培地で、24時間培養した。その翌日、この培地を新鮮培地に変えて、HGF(5ng/ml)を3つ一組のウェルに加え、同時に50μlの[メチル3H]チミジン(Amersham Pharmacia Biotech Europe GmbH、テュスクラン)を各ウェルに加えた。前述のように(Betsholtz CおよびWestermark 1984)DNAアッセイ法を実施する24時間前に、細胞をインキュベートした。
【0031】
遊走アッセイ:細胞は、集密になるまで、24ウェルディッシュの底面に置かれた無菌の円形カバーガラス上で培養し、24時間後に取り除き、新鮮培地5mlを含むペトリ皿(Nuncolone)にさかさまにして置かれた。HGF(5ng/ml)をペトリ皿に加え、5%CO2および95%の空気の雰囲気において、夜間に37℃でインキュベートした。次にペトリ皿を光学顕微鏡(オリンパス、東京)で検査し、ガラス周囲の細胞の遊走の長さを7つの部位で測定した。統計解析のために、7回の測定の中央値を、10回の実験の中央値の平均算出に使用している。
【0032】
細胞骨格および共焦点顕微鏡:細胞をカバーガラス上で培養した。前述のように、集密な単層を傷つけ、HGF10ng/mlで24時間インキュベートした。固定後(4%パラホルムアルデヒド/PBSで、室温で30分間)、細胞をKRGで3回洗い、1単位のAlexa488−ファロイジン(Molecular Probes、オレゴン州)/200μlKRGを、細胞入りの各ガラスに加えた。標識は、Bellydancer(Stovall Life Science INC、米国ノースカロライナ州Greensbro)上で、遮光下において、室温で20分間継続した。標識後、Alexa488−ファロイジンは除去し、ガラスをKRGで3×5分間洗い、ProLong(Molecular Probes、オレゴン州)にマウントした。細胞は、励起用の波長488nmのアルゴンイオンレーザーおよび×60の対物レンズ(N.A.1.4)を使用したSarastro 2000共焦点レーザー走査顕微鏡(Molecular Dynamics、カリフォルニア州Sunnyvale)で調査した。一連の水平x−y断面(断面間1.4μmの距離で)および垂直x−z断面を収集した。3Dルックスルー投射を異なる角度で実施し、90度投射を使用して、対照とHGF処理細胞の高さを比較した。
【0033】
免疫蛍光によるC−met/HGF/SF受容体の検出:前述のように、集密な単層を傷つけ、HGF10ng/mlで24時間インキュベートした。実験の日、細胞を4%パラホルムアルデヒド/PBSにより室温で30分間固定した後、PBSで2回完全に洗浄し、抗体の非特異的結合を遮断するために、室温で1時間、5%ウシ血清アルブミン(BSA)/PBSからなるブロッキングバッファーでプレインキュベートした。次に、マウスHGF受容体(Sigma-Aldrich)の細胞外ドメインに対するウサギのポリクローナル抗体(ブロッキングバッファーで1:50に希釈)により、室温で60分間単層をインキュベートした。細胞を上記のように洗い、フルオレセインイソチオシアネート(FITC)結合ヤギ抗ウサギIgG(1:50)(Sigma-Aldrich)により標識した。室温で60分間のインキュベーション後、細胞を洗浄し、17% Gelvatol/PBSでマウントし、落射蛍光顕微鏡使用(epifluorescence microscopy)(Zeiss Axioskop)で検査した。
【0034】
(実施例1a):細胞増殖:対照およびHGF処理ウェルにおけるCPM(カウント/毎分)比は以下のようであった。2ngHGF/対照=1.1(0.95〜1.20)、5ngHGF/対照=1.1(1.04〜1.18)および15ngHGF/対照=1.15(0.99〜1.23)。したがって、上皮細胞株(CCL−53.1)における細胞増殖は、[3H]チミジン取り込み測定による評価で、組換えHGFにより著しく増加しなかった。
【0035】
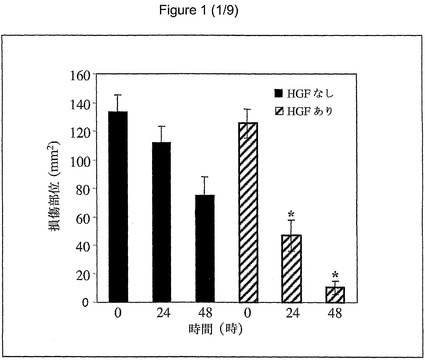
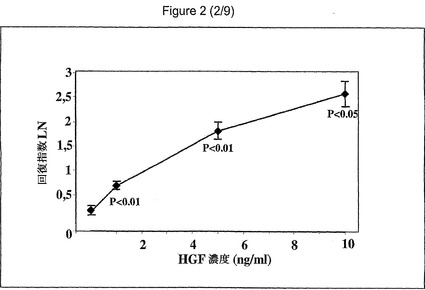
(実施例1b):損傷部位の回復:損傷した単層の回復に対するHGFの効果を、in−vitroの損傷モデルを使用して評価した。むきだし部分は、24時間および48時間後に減少した。損傷時と24時間後との測定されたむきだし部分の比は、HGF処理の方が(損傷時=125.25±10.08mm2、24時間後=46.73±11.03mm2、比=2.65、p<0.01、48時間後=10.26±4.76mm2、比=11.91)対照(損傷時=133.77±11.7mm2、24時間後=112.23±11.11mm2、比=1.15、48時間後=75±13.17mm2、比=2.43)と比較して、有意に高かった(図1)。従来の培地または血清除去した培地との間に、HGF添加後の損傷細胞の回復において有意差は認められなかった。上皮回復の有意な亢進が、低濃度(1ng/ml)のHGFで観察された。10ng/mlまでのHGFの用量増加は、さらなる回復の増進を引き起こした(対照と1ng/mlの間でp<0.01、1ng/mlと5ng/mlの間でp<0.01、5ng/mlと10ng/mlの間でp=0.05)(図2)。FGF(1〜15ng/ml)、EGF(2〜60ng/ml)、PDGF(1〜35ng/ml)、またはFGF/KGF(1〜15ng/ml)の添加は、損傷細胞の回復に有意な影響を及ぼさなかった。
【0036】
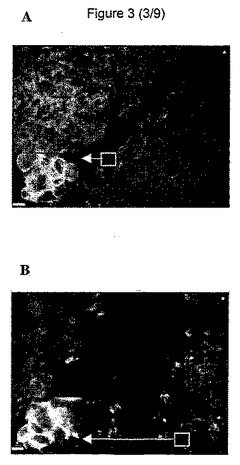
c−Met受容体を、ウェル内で免疫蛍光によって視覚化した。HGFの添加後または未処理の対照において、損傷部から遠位の他の細胞と比較して、むきだし部位に隣接する細胞でのc−Met発現における本法による有意差は認められなかった(図3)。
【0037】
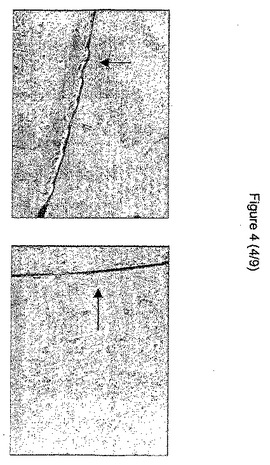
(実施例1c):細胞運動性:HGF添加後のガラス層からの遊走距離(mm)について、未処理の対照と比較した。HGF添加後の上皮細胞の遊走亢進が示された(HGF処理ペトリ皿で平均10.6±1.66mm、対照で1.2±0.51mm)(p<0.01)(図4)。
【0038】
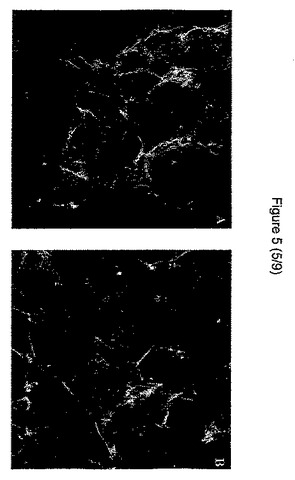
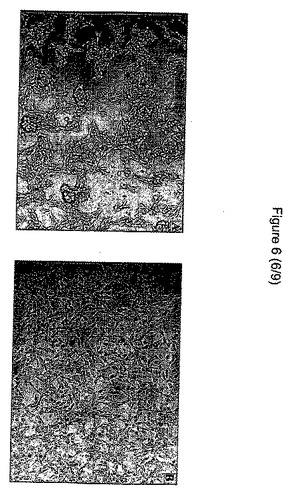
(実施例1d):形態および細胞骨格:上皮細胞CCL−53.1の形態は、HGF処理細胞において異なった。未処理のウェルでは、細胞は、より小さい細胞内空間で、円形の高密度の団集花序状に集まったが、HGF処理ウェルでは、平らになり、広がった形態であった。共焦点顕微鏡技術によって作られる3D再構築によって、我々は、HGF添加後のアクチン構造に著しい変化が認められ、未処理細胞と比較して、高密度の末梢構造でF−アクチンがより少ないことを発見した(図5および6)。
【0039】
上記の例は、どの程度損傷したCCL−53.1上皮細胞を、HGFの生物活性を検査するために用いることができるかを示している。生物活性のあるHGFは、損傷後の損傷CCL−53.1上皮細胞の回復を用量依存的に亢進する。これらの結果は、HGF処理後に細胞遊走促進効果が亢進し、最終的に、その結果として、細胞骨格構造が弱まることによって部分的に説明することができる。上記の知見は、活性HGFに特有であって、非活性HGFまたはEGF、PDGF、FGF、KGFは、細胞に対して同一な効果を誘発することができなかった。
【0040】
(実施例2:表面プラズモン共鳴法(Biacore)によって示される生物活性のある肝細胞増殖因子(HGF)の特性)
以下の材料および条件を使用した。
【0041】
SPR測定およびリガンド固定化手順。SPR測定は、それぞれ4つおよび2つのフローセルを備えた全自動Biacore 2000測定器および半自動Bicaore X測定器(Biacore AB、スウェーデン、ウプサラ)によって、760nmで実施した。フローセルの温度は、すべての実験で25℃であった。使用した試料表面は、Biacore AB(スウェーデン、ウプサラ)から購入したカルボキシメチル化デキストランCM5チップであった。デキストランヒドロゲルのカルボン酸基に対するリガンドの結合は、200mM EDC(N−エチル−N’−(3−ジエチルアミノプロピル)カルボジイミド)および50mM NHS(N−ヒドロキシスクシンイミド)を使用した従来のカルボジイミドの化学的作用によって実施した。活性化にかかる時間は7分で、2〜7分後にリガンドの注入を行った。残留する活性エステルの不活性化は、pH8.5のエタノールアミン/塩酸塩を7分注入することによって実施した。5μl/分の流速をすべての固定化の間に使用した。すべてのリガンドは、pH4.5の10mMアセテート緩衝液で希釈した(すなわち、タンパク質の等電点を下回るので、デキストランマトリックスとリガンドの間の静電的相互作用を亢進する)。モノクローナル抗HGF(500μg/ml)を1:10に希釈し、およびポリクローナル抗HGFも。組換えMetプロトオンコジーン受容体(100μg/ml)は1:5に希釈し、HGF組換え体(5μg/ml)は1:3に希釈した。接触時間は、2分から7分の間で変えて、8,000〜30,000RU(反応単位)の固定化レベルを得た。不活性化後、表面は、その後に、pH2.0の5mMグリシン緩衝液と1M塩化ナトリウムを1分間、5回注入することにより洗浄した。フローセルの1つは、デキストランに対するHGFの特異的な相互作用だけでなく、緩衝液および非特異的な相互作用による反応を監視するための基準として使用した。このフローセルは、固定化処置の間は、他のフローセルと同様に処理を行ったが、リガンドの固定化工程は除外した。
【0042】
(実施例2a:デキストランに対するHGFの結合)
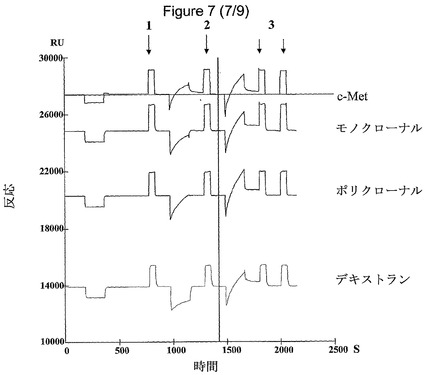
生物活性のあるHGFは、デキストランに対して高い親和性を持ち、試料へのデキストラン(0.05%)添加により、固定化されたモノクローナルおよびポリクローナル抗HGF抗体との、フローセルならびにデキストラン経路におけるシグナル応答が有意に減少した(p<0.05)(図7および8)。
【0043】
(実施例2b:リガンドへの結合)
生物活性のあるHGFは、生物活性のないHGFと比較して、固定化されたモノクローナル抗HGF抗体(p=0.005)またはc−Met受容体(p=0.02)とのフローセルにおけるシグナル応答が有意に高い(図7および8)。
【0044】
(実施例2c:ELISAとの相関)
免疫反応性HGFは、市販のキット(Quantikine HGF Immunoassy、R&D Systems Inc.、米国Minneapolis)を使用してELISAによって判定した。生物活性のあるHGFをSPRによる判定を用いて検査すると、結果は、ELISAによって測定した水準と有意に相関した。不活性HGFを使用すると、ELISAおよびSPRの結果は相関しなかった。
【0045】
(実施例2d:固定化レベルを変える影響)
固定化されたモノクローナル抗HGF抗体の量によるシグナル応答の影響について、異なる試料で測定した。固定化中に接触時間を増加することにより(1、5および10分)固定化レベルを増やすことで、生物活性のあるHGFを含むすべての試料の反応に有意な増加を生じた。流速を変更することでは(5、10および15μl/分)、試料の反応に有意な影響を及ぼさなかった。
【0046】
上記の実施例は、生物活性のあるHGFがデキストランに対して高い親和性を有すると思われ、これは細胞表面ヘパラン硫酸プロテオグリカン結合部位に対するそのような親和性に類似している可能性のあることを示している。これらの結合部位は、抗体結合エピトープと重複するか、デキストランと結合するとすぐに、HGFのコンホメーション変化を誘発し、この結果、立体的障害になり、したがって、抗体への結合を妨げる。
【0047】
(実施例3:マウスの正常な無毛の皮膚に対する生物活性のある組換えヒト肝細胞増殖因子(rHGF)の局所投与の影響)
マウス18匹(C57/B6ブラック、14〜16週齢、体重21〜29g、全マウスが雌)の背部を剪毛し、これにより、右側および左側に、2つの1×1cmの部分ができた。次に、マウスを3群に割り付けた。第1群(n=6)は、0.70μg/kg(12〜18ng/cm2無毛の皮膚)の組換えヒト肝細胞増殖因子(rHGF)を、右側に毎日5日間皮下に処置した。利用されたHGFは、試験前にCCL−53.1細胞の損傷分析(wound assay)に加えて、FLISA、表面プラズモン共鳴(SPR)法によって検査した。このHGF型は、SPRよる判定で、デキストランおよびHGFに対する抗体に高い親和性を持ち、ELISAによって得た濃度と相関した。組換えHGFは、CCL−53.1細胞の損傷部位への用量依存的な遊走を引き起こした。この効果は、抗HGF抗体によって停止させることができた。第2群(n=6)には、SPRによってデキストランに対する親和性が認められなかった組換えHGF 0.70μg/kgを5日間皮下投与したが、損傷分析において、CCL−53.1の遊走は生じなかった。第3群(n=6)には、無菌の生理的塩化ナトリウムを5日間皮下投与した。本群は対照群とした。治療コース中または後に、非忍容性、または疾患の何らかの徴候はいずれの治療群においても示されなかった。治療下で、発毛におけるいくつかの有意差は、明らかに見られなかったが、我々は、12日後、第1群のマウス5匹が、両側で完全に発毛したことを見て驚いた(図9)。第2群および第3群では、同じプロセスに非常に長い時間がかかった(20〜35日)。したがって我々は、マウスの正常な、無毛の皮膚に対する適量の生物活性のあるHGFによる局所治療は、発毛を促進し、この局所治療が、問題のない動物では忍容性を有すると結論する。
【0048】
(実施例4:静脈血流不全症患者の慢性下肢潰瘍に対する組換え肝細胞増殖因子の治療効果−症例報告)
最近のパイロット研究期間中に観察された慢性下肢潰瘍に対する肝細胞増殖因子の正の効果を評価するために、患者20例を対象にして、市販の組換えHGFを1週間使用する二重盲検プラセボ対照試験を開始した。本試験は、患者12例の組み入れ後、SDS−PAGEおよびウエスタンブロットにより、使用されたタンパク質に欠陥があったことが明らかにされた時点で中断された。潰瘍部位由来の生検材料の免疫組織化学染色は、HGF受容体(c−met)の発現が、塩化ナトリウム製剤を投与した患者(第2群、n=6)と比較して、市販の組換えHGFを含む製剤を投与した患者(第1群、n=6)で有意ではないが減少することを示した。このことはタンパク質のc−met受容体との相互作用を示す可能性がある。第1群で、毛細管増殖の増進が観察されたが、劇的な臨床的改善が認められたのは、わずか1例に過ぎなかった。にもかかわらず、第1群の3例および第2群の2例が9カ月以内に治癒した。他の患者7例には、生物活性のある組換えHGFによる治療を申し出て、4例(各群2例)が受け入れた(第3群)。これら当初不成功の全例で、潰瘍の状態の改善が、治療開始から1週間以内に観察された。我々は、生物活性のあるHGFが慢性下肢潰瘍の治癒に対して、プラセボ効果によるものでない正の効果を有すると結論を下した。
【0049】
(実施例5:SPRによるHGFエピトープマッピング)
(実施例5a:抗体親和性)
HGF(Santa Cruz)に対するエピトープ特異的ないくつかの抗体をSPRのフローセルに固定化することによって、我々は、生物活性のあるHGFまたは生物的に不活性な組換えHGFの多くの特性を判定することに成功している。
【0050】
1−生物活性のあるHGFは、HGFのα鎖およびβ鎖で異なるエピトープに対して一定の親和性を有する。
【0051】
2−生物活性のないHGFでは、N−17(ヒト由来のHGFαのN末端におけるペプチドマッピングに対する抗体)により固定化されたフローセルに対する親和性は、減少または無効になった。
【0052】
H−170(ヒト由来のHGFβのアミノ酸1〜170に対して作られる抗体)に対する親和性は減少した。
【0053】
c−met受容体およびH−145(ヒト由来のHGFαのアミノ酸32〜176に対する抗体)の親和性は維持した。
【0054】
(実施例5b:HGF受容体結合エピトープ)
SPRを使用して、我々は、HGFのc−met受容体への結合において重要なエピトープを確認することに成功している。我々は、HGFがc−met受容体と結合する時、H−145(ヒト由来のHGFαのアミノ酸32〜176に対する抗体)およびC−20(ヒト由来のHGFαのC末端におけるペプチドマッピングに対して作られた抗体)に結合するHGFのエピトープが関与していることを明らかにしている。依然として、HGFがc−met受容体と相互作用を持つ間に、N−17(ヒト由来のHGFαのN末端におけるペプチドマッピングに対する抗体)、N−19(ヒト由来のHGFβのN末端におけるペプチドマッピングに対する抗体)およびH−170(ヒト由来のHGFβのアミノ酸1〜170に対して作られる抗体)と相互に作用する他のエピトープが利用可能である。
【0055】
我々が試験した不活性HGF組換え型において、エピトープに結合する受容体は、損傷がなかったが、α鎖のN−17およびβ鎖のH−170と相互に作用するドメインは欠陥があった。
【図面の簡単な説明】
【0056】
【図1】傷害後の24時間および48時間以内の対照(単色)および活性HGFで治療した細胞における回復を示す図である。
【図2】異なるHGF濃度(1、5および10ng/ml)および24時間以内の回復の相違を示す図である。
【図3】免疫蛍光法によるCCL−53.1細胞におけるc−Met受容体の局在化を示す図である。本図で見られるように、免疫蛍光による受容体の提示で、生物活性のあるHGFでの治療(A)と対照(B)の間に相違が認められない。しかし、損傷したむきだし部分の減少が、(B)と比較して、(A)で明白に見られる。
【図4】遊走アッセイを示す図である。右側の写真は、生物活性のあるHGFの添加後の細胞の遊走を示し、左側の写真は、対照細胞を示している。矢印は、円形のカバーガラスの縁を示している。
【図5】アクチン構造のファロイジン染色後の対照(左側)と活性HGF処理細胞(右側)を比較した共焦点顕微鏡写真および3Dルックスルー投射図(3D look-through projection)である。両群間の細胞壁径の差異に留意のこと。
【図6】生物活性のない細胞(左側)と対照細胞(右側)の間の形態学的相違を示す図である。
【図7】デキストラン(0.05%)の添加前(左側の最初のシグナル)および添加後(2番目のシグナル)の生物活性のあるHGFのSPR反応を示す図である。再生パルスは、各シグナルの後に観察される。
【図8】生物活性のあるHGFおよび生物活性のないHGFからのSPRシグナル間の相違を示すヒストグラムである。
【図9】生物活性のあるHGFを5日間皮下投与したマウス(左側および右側)および無菌塩化ナトリウムを5日間皮下注射した対照マウス(中央)の発毛の相違を示す図である。写真は、最初の注射の12日後に撮影された。
【技術分野】
【0001】
本発明は、肝細胞増殖因子(HGF)の生物活性についての評価法に関する。本法は、例えば、組換え技術による、バッチ生産(batch production)後のHGFの品質管理のため、およびHGF製剤の評価のために、治療の必要な患者への投与前に使用することができる。また、本評価法は、HGF関連疾患の診断およびこれに続く治療選択を向上させるために使用することができる。したがって、本発明は、HGFの役割を評価するために、潰瘍分泌物、尿、糞便、唾液、胸膜における損傷部位の内因性HGFの活性を評価し、外因性HGFの損傷部位への適用が必要かどうかを判定する方法にも関する。
【背景技術】
【0002】
肝細胞増殖因子(HGF)は、細胞損傷に反応して分泌され、特定の器官の再生および創傷治癒のために重要と思われる増殖因子である(Arakaki Nら、 Hepatology 221, 1995)。HGFは、H鎖が約60kDaおよびL鎖が約30kDaのヘテロ二量体で、最初に、不活性前駆体として合成される(Miyazawa Kら、J Biol Chem 268, 1994)。この前駆体は、損傷器官で特定の活性化因子によって活性タンパク質へ切断される(Naka Dら、 J Biol Chem 276, 1992、Nishizaki Tら、J Am Coll Surg 181, 1995)。HGFは、内分泌作用だけでなく、パラクリン作用を有する(Yanagita K.ら、Biochem Biophys Res Commun 182, 1992、Kono Sら、Biochem Biophys Res Commun 186, 1992)。HGFの標的細胞は、完全に発育した上皮細胞である(Matsumoto K およびNakamura T. J Gastroenterology Hepatology 6, 1991)。HGFは、器官損傷部位において産生され、高濃度で存在する(Nayeri Fら、Scand J Infect Dis, 34, 2002)。我々は、以前に、様々な感染症でHGFの全身的産生および局所産生について検討し、胃腸炎などの急性感染症の間に、高濃度の血清HGFを観察している(Nayeri Fら、Scand J Infect Dis, 34, 2002)。我々は、髄膜炎中に、HGFの全身的産生の亢進と同時に、脳脊椎液で高濃度のHGFを発見した(Nayeri Fら、 J Infect Dis, 181, 2000)。肺炎患者における呼気凝縮物(exhaled breath condensate)中のHGF濃度の上昇(Nayeri Fら、Respir Med, 96, 2002)は、HGFの血清濃度とは相関がなく、肺炎中のHGFの局所産生を示した。さらに、我々は、血清におけるHGFの安定性について研究しており(Nayeri Fら、CYTOLIKE, 2002)、HGFが試料中(血清/血漿、糞便、呼気凝縮物)で非常に安定しており、数回にわたる凍結−解凍の繰り返し、異なる緩衝液または−20℃での数年間の保管は、HFG濃度に著しい影響を及ぼさないことを発見した。
【0003】
HGFは多機能の増殖因子であり、HGFの再生特性は、肝臓(Ishikiら、1992)、腎臓(Igawaら、1993)、肺(Yanagitaら、1993)および敗血症中(Kondoら、1999)の動物モデルで研究されている。HGFの標的細胞に対する分裂促進作用が、組織再生および増殖を生じさせるには十分でないことが示唆されている。細胞遊走促進作用および形態形成作用などのHGFの他の機能が再生過程を完了するために必要である(Montesanoら、1991)。HGFの切断種(truncated species)は、組織再生および修復の最終段階で、HGFの増殖促進作用に対するアンタゴニストとして機能する可能性があると仮定されている(Chanら、1991)。
【発明の開示】
【発明が解決しようとする課題】
【0004】
以前に我々は、外因性HGF投与により、治療が困難な慢性の下肢潰瘍の患者を治癒することができることを示している(Nayeriら、Dermatological Treatment, 2002)。最近の二重盲検試験の間、我々は、後に欠陥があることが明らかになった市販の組換えHGFを使用している。試験製剤における、この欠陥成分の使用により、潰瘍の十分な治癒が得られなかった。1年後、これらの患者の一部(n=4)に活性のある組換えHGFを投与したところ、最初の1週間以内に、かれらの潰瘍に著明な治癒が見られた(表1)。感染症治療におけるHGFの、将来の指標に加えて、抗生物質(EP01963646.3として)の治療的可能性により、治療の要素として使用されるHGFの生物活性について評価することは非常に重要である。
【0005】
本発明は、生物学的生産物と共に、組換え製剤におけるHGF活性の評価法について部分的に述べる。
【課題を解決するための手段】
【0006】
(発明の概要)
本発明の目的は、体液で検出されるHGFに加えて、組換えHGFの生物活性の評価法を提供することである。
【0007】
本発明のさまざまな他の目的および利点は、本発明の図および以下の記述から明らかになるであろう。
【0008】
一実施形態では、本発明は、活性HGFと相互に作用するHGF受容体を発現する上皮または内皮の細胞株を使用するHGF活性の評価のためのin vitroモデルについて述べる。特に、マウス皮膚上皮細胞株(CCL−53.1)が使用される。他の実施形態では、本発明は、HGFの生物活性についてのin vitroの評価法に関する。HGFに対するモノクローナル抗体またはポリクローナル抗体による1つまたは複数の相互作用の生物活性に関する評価において、C−met受容体および多糖類の特にデキストランを使用する。これらの相互作用の研究のために、多くの方法が利用可能で、当業者によく知られている。本発明の好ましい実施形態では、BIACORE(スウェーデン、ウプサラ)によって提供された器具を使用する。この技術は、表面プラズモン共鳴法に基づいており、多くの相互作用を並行して調べることができるという点で、非常に効果的である。しかし、もちろん、本発明による方法は、明確にするための記載が、この実施形態に焦点を合わせているとしても、特定の器具に限定されるものではない。
【0009】
本発明の他の実施形態では、HGFの生物活性は、マウス(例えば、C57BL/6)のin−vivoモデル利用する方法を使用して評価する。
【0010】
さらなる実施形態では、本発明は、HGFの生物活性の評価用キットに関する。この種のキットでは、細胞株CCL−53.1は、培養細胞株が損傷し、損傷部位に向けた隣接細胞の運動性がHGFの量および活性に用量依存的に関連する場合に使用する。各々の細胞に対する細胞の粘着性減少による形態形成変化、および細胞の細胞骨格特性の変化による損傷した無毛部位の増大に加えて、損傷部位への細胞の運動性の亢進が見られる。この反応は、HGFに特異的であって、抗HGF抗体によって止めることができる。線維芽細胞増殖因子、上皮増殖因子、血小板由来増殖因子およびケラチノサイト増殖因子の使用では、そのような影響を細胞に及ぼすことはない。不活性HGFまたは1ng/mLより低い用量で活性HGFを使用することで、細胞の形態学的活性または細胞遊走促進活性を誘発することはない。
【0011】
他の実施形態では、本発明は、HGFに対する少なくとも1つのリガンド(ポリクローナルまたはモノクローナルの抗HGF抗体またはc−metプロトオンコジーン受容体)で固定化されるいくつかのチャネルを有するセンサチップ、および不活性デキストランを有する少なくとも1つのチャンネルからなるプラズモン共鳴バイオセンサー(BIACORE)法を使用するHGFの生物活性評価のためのキットに関する。生物活性のあるHGFは、デキストランおよびアルブミンにだけでなくそのリガンドに高い親和性を有し、一方、生物活性のないHGFは、そのリガンドと相互に作用する場合と作用しない場合があり、デキストランまたはアルブミンとは相互に作用しない。
【発明の効果】
【0012】
(発明の詳細な説明)
HGFの変異体:
肝細胞増殖因子(HGF)は、分子量が約90kDaの大きなタンパク質である。活性化されると、このタンパク質は、α鎖およびβ鎖に切断される。以前の試験は、HGF分子内のN末端のヘアピンドメイン、最初のクリングルドメイン、2番目のクリングルドメインまたはセリンプロテイナーゼドメインの欠失が生物活性の全面的な喪失を招くことを示している(Matsumotoら、1991b)。HGF分子のα鎖またはβ鎖のいずれかの欠失は、HGFの活性の完全な喪失を招く(Matsumotoら、1991)。c−met受容体結合ドメインは、HGFのα鎖のNH2末端で同定されている(Mizunoら、1994)。また、HGFの受容体結合および活性で重要な役割を果たす27残基のヘアピンループの三次元(3D)モデルは、タンパク質工学および核磁気共鳴(NMR)技術(Zhouら、1998)によって得られている。ドメイン欠失実験の結果および個々のドメインの3Dモデルの有用性と共に、HGF配列の特徴から、いくつかの試験につながり、ここでは、特定の残基または残基のクラスターが、HGF活性におけるこれらの役割を明らかにするために置換されている(Chirgadzeら、1998)。Lokkerら(1994)は、Nドメイン内で、受容体結合のために重要な残基について研究している。今まで、多くの個々および群の突然変異について、ヘアピンループ自体の範囲内で、受容体結合にとって重要な残基を特定できなかった。しかし、彼らは、NドメインのC末端(His−114、Glu−115、Asp−171)に位置する残基のクラスターが、HGFの受容体結合および生物活性の両方に関わることを発見した(Lokkerら、1994)。また、彼らは、最初のクリングル内で、少なくとも7種のアミノ酸(Arg−197、Glu−189、Tyr−198、Glu−195、Asp−171、Gln−173およびSer−161)の置換が、HGFの受容体結合および生物活性に対して明らかな影響を有することを示している。最も大きな影響は、Arg−197およびGlu−159の置換で見られ、受容体結合および生物活性が50倍を超えて減少する(Lokkerら、1994)。HGFのセリンプロテイナーゼドメインにおけるいくつかの点変異(残基のGln−534およびTyr−673が、活性セリンプロテイナーゼのHisおよびSer残基に復帰した)は、受容体結合には影響を及ぼさないが、生物活性を著明に減少させた。同様の結果が、Val692Serの突然変異で得られた(Lokkerら、1992)。C末端のクリングル(クリングル4)とセリンプロテイナーゼドメインの間に位置するトリプシン様切断部位での重要な残基の突然変異は、HGFの単鎖前駆体型(pro−HGF)の転換を消失させた。結果として生じる単鎖タンパク質は、HGFの受容体結合活性を維持するものの、標的細胞に対する生物活性を欠いていた(Lokkerら、1992、Nakaら、1992、Gakら、1992)。
【0013】
HGFの生物活性についての評価の重要性
以前のパイロット研究では、高齢患者11例を対象に、19の慢性下肢潰瘍うちの15に対して、局所にゲル状HGFを1日1回、7日間塗布した。全例が以前に従来の方法によって治療されており、彼らの下肢潰瘍は、1年から14年の期間でやや安定した状態であった。アレルギー、不快感または疼痛に関するあらゆる徴候が毎日報告された。隣接する無傷の皮膚と比較して、潰瘍の微小循環灌流(microcirculation perfusion)について、(患者7例で)本研究の開始時、1週間後、および再度3カ月後にレーザードップラーによって測定した。また、潰瘍の大きさおよび特徴についても詳細に記録した。我々は、HGFの血管形成作用に反映する可能性がある微少循環灌流が、治療した潰瘍における潰瘍部位の縮小と統計的に有意な相関を示すことを観察した(r=0.94、p<0.002)。十分良好な治癒(84〜100%の領域縮小)または部分的治癒(58〜59%)が患者11例中8例で認められた。
【0014】
このパイロット研究中に観察された慢性下肢潰瘍に対する肝細胞増殖因子の正の効果を評価するために、患者20例を対象にして、市販の組換えHGFを使用した1週間の二重盲検プラセボ対照試験を開始した。本試験は、患者12例の組み入れ後、SDS−PAGEおよびウエスタンブロットにより、使用されたタンパク質に欠陥があった(わずか1本の約60〜67kDaの鎖が発見された)ことが明らかにされた時点で中断された。潰瘍部位由来の皮膚生検材料の免疫組織化学染色は、正常健常対照(n=10)と比較して、患者(n=12)では、c−met受容体の発現が有意に高いことを示した(p<0.001)。市販の組換えHGFを含む製剤を投与した患者(第1群、n=6)で、c−met発現が減少する傾向が観察されたが(タンパク質のc−met受容体との相互作用を示している可能性がある)、本群と塩化ナトリウム製剤を投与した群(第2群、n=6)との間の差は有意ではなかった(表1)。第1群では、毛細管増殖の亢進が観察されたが(有意ではない)、劇的な臨床的改善が認められたのは、わずか1例に過ぎなかった。にもかかわらず、第1群の3例および第2群の2例が9カ月以内に治癒した。他の患者7例には、生物活性のある組換えHGFによる治療を申し出て、4例(各群2例)が受け入れた(第3群)。これら当初不成功の全例において、潰瘍の状態の改善が、治療開始から1週間以内に観察された。潰瘍部位のc−met発現が、患者2例で確認され、2例とも1週間の治療後に減少した。
【0015】
我々は、生物活性のあるHGFが慢性下肢潰瘍の治癒に対して、プラセボ効果によるものでない正の効果を有すると結論を下した。欠陥のある組換えHGFと活性のある組換えHGFの特性について、患者に対する治療効果を有するHGFの生物活性型を特徴付けるために調査した。上記のように、HGFの分裂促進効果および細胞遊走促進効果は、効果的な治癒プロセスにとって必要とされる。
【0016】
本発明は、in−vitroにおいてHGFの免疫学的および生物学的特性を調査することによるHGFの生物活性の測定について述べる。
【実施例】
【0017】
本発明では、治療的処置用として適した生物活性のあるHGFを定義するために、以下の基準を採用している。本発明の方法を使用することで、生物活性のあるHGFは以下の通り。
【0018】
・CCL−53.1モデルで、18時間以内に少なくとも50%の損傷部位の減少を引き起こすはずである。
【0019】
・SPRベースの方法において、(20μg/mL)c−metプロトオンコジーンHGF受容体だけでなく、(50μg/mL)固定モノクローナルおよびポリクローナル抗HGF抗体に対して同等の親和性を有するはずである。
【0020】
・SPRベースの方法において、モノクローナル抗HGF抗体に対する親和性より低く、デキストランに対して陰性対照より高い親和性を有するはずである。この親和性は、試料にデキストランを添加すると、消失するはずである。
【0021】
・免疫組織化学染色によって、またはマウスが無毛部位で完全な発毛を示す時間(15日未満)を判定することによって判定して、HGF治療(少なくとも100ng/マウス/日を5日間)の1週間後、無毛部位から採取した皮膚生検材料で、PCNA発現によって判定したマウスの刈毛した皮膚部位より、2倍速い発毛を起こすはずである。
【0022】
以下の実施例は、本発明の実施形態をさらに説明することを意図するものであって、決して本発明の範囲を制限することを意図していない。
【0023】
(実施例1:in vitroでのマウスの損傷メラニン形成細胞の回復を加速する肝細胞増殖因子の効果)
以下の材料および条件を使用した。
【0024】
細胞:マウスの皮膚由来のCCL−53.1付着上皮細胞、メラニン形成細胞、黒色腫は、アメリカン・タイプ・カルチャー・コレクション(American Tissue Cell Collection:ATCC、米国バージニア州Manassas)から入手し、継代数3〜32で使用した。
【0025】
増殖因子:組換え肝細胞増殖因子(HGF)は、好意によりT. Nakamura教授(日本、大阪)から寄贈された。また、HGF(RD Systems、ロット:QF031062)、線維芽細胞増殖因子(FGF、RD Systems、ロット:EX164011)、血小板由来増殖因子(PDGF、RD Systems、ロット:D332071)、ケラチノサイト増殖因子(KGF、RD Systems、ロット番号:J0164021)、上皮増殖因子(EGF、RD Systems、ロット:HLM114031)は市販のものを購入した。
【0026】
Nakamura教授から入手した組換えHGFは、患者の慢性潰瘍治療に非常に有効であることが示されたので(実施例4)、本研究では、生物活性のあるHGFの陽性対照として使用している。
【0027】
細胞培養:CCL−53.1細胞は、37℃の5%CO2および95%空気の雰囲気中で、15%ウマ血清および2.5%ウシ胎児血清(Sigma-Aldrich、スウェーデン、ストックホルム)を補充したKaighn改変Ham’s F−12K培地(ATCC)で増殖した。細胞が集密に達した後、非酵素細胞解離液(non-enzymatic cell dissociation solution)(1×)(Sigma-Aldrich)により分離し、15%ウマ血清および2.5%ウシ胎児血清を補充したF−12K培地で懸濁し、24ウェル培養プレート(Nunc Brand Products、デンマーク、ロスキレ(Roskilde))に播種し、集密に達するまで、同一条件下で24〜48時間培養した。
【0028】
無血清培地:75−cm2組織培養フラスコ(Nuncolone)で増殖させたCCL−53.1細胞の集密状態の培養物をリン酸塩緩衝液(Sigma-Aldrich)で洗い、非酵素細胞解離液により分離し、無血清培地のNutridoma−NS培地(Roche Diagnostics Scandinavia AB、スウェーデン、ブロンマ(Bromma))で再懸濁し、24ウェル培養プレートに播種し、集密に達するまで、上記と同一条件下で24〜48時間培養した。
【0029】
損傷アッセイ:集密状態の単層を無菌のスチール器具によってこする。分離細胞をPBSで洗い、ウェルに新鮮培地を加える。細胞に覆われていない正方形部分(mm2)を顕微鏡(オリンパス)で測定し、各ウェルに記録した。次に、HGF(1、5、10ng/ml)を細胞に加え、5%CO2を含む加湿した雰囲気において、37℃でインキュベートした。24および48時間後、単層に覆われていない部分を再度測定し、記録した。実験は、3通り実施した。3通りの中央値を、13の個別の実験の平均値の計算に使用した。ヘパリン(Leo Pharma AB、スウェーデン、マルメー(Malmo)、SE−20124、Box404)を、異なる濃度(範囲1〜100IE/ml)で、HGF添加または不添加の傷つけた単層に加えた。
【0030】
細胞増殖:CCL−53.1細胞は、24ウェルプレートで、前述のようにウェルあたり1×106細胞の密度で培養した(細胞計数マルチサイザー[Cell Counter Multisizer]、KEBO Lab、スウェーデン、スパンガ(Spanga))。集密後、細胞を非酵素細胞解離培地(Sigma Aldrich)で分離した。DNA合成は、3H−チミジンの取り込みを測定することによって判定した。分離CCL−53.1細胞は、24ウェル培養プレートに播種し、15%ウマ血清および2.5%ウシ胎児血清を補充したF−12K培地で、24時間培養した。その翌日、この培地を新鮮培地に変えて、HGF(5ng/ml)を3つ一組のウェルに加え、同時に50μlの[メチル3H]チミジン(Amersham Pharmacia Biotech Europe GmbH、テュスクラン)を各ウェルに加えた。前述のように(Betsholtz CおよびWestermark 1984)DNAアッセイ法を実施する24時間前に、細胞をインキュベートした。
【0031】
遊走アッセイ:細胞は、集密になるまで、24ウェルディッシュの底面に置かれた無菌の円形カバーガラス上で培養し、24時間後に取り除き、新鮮培地5mlを含むペトリ皿(Nuncolone)にさかさまにして置かれた。HGF(5ng/ml)をペトリ皿に加え、5%CO2および95%の空気の雰囲気において、夜間に37℃でインキュベートした。次にペトリ皿を光学顕微鏡(オリンパス、東京)で検査し、ガラス周囲の細胞の遊走の長さを7つの部位で測定した。統計解析のために、7回の測定の中央値を、10回の実験の中央値の平均算出に使用している。
【0032】
細胞骨格および共焦点顕微鏡:細胞をカバーガラス上で培養した。前述のように、集密な単層を傷つけ、HGF10ng/mlで24時間インキュベートした。固定後(4%パラホルムアルデヒド/PBSで、室温で30分間)、細胞をKRGで3回洗い、1単位のAlexa488−ファロイジン(Molecular Probes、オレゴン州)/200μlKRGを、細胞入りの各ガラスに加えた。標識は、Bellydancer(Stovall Life Science INC、米国ノースカロライナ州Greensbro)上で、遮光下において、室温で20分間継続した。標識後、Alexa488−ファロイジンは除去し、ガラスをKRGで3×5分間洗い、ProLong(Molecular Probes、オレゴン州)にマウントした。細胞は、励起用の波長488nmのアルゴンイオンレーザーおよび×60の対物レンズ(N.A.1.4)を使用したSarastro 2000共焦点レーザー走査顕微鏡(Molecular Dynamics、カリフォルニア州Sunnyvale)で調査した。一連の水平x−y断面(断面間1.4μmの距離で)および垂直x−z断面を収集した。3Dルックスルー投射を異なる角度で実施し、90度投射を使用して、対照とHGF処理細胞の高さを比較した。
【0033】
免疫蛍光によるC−met/HGF/SF受容体の検出:前述のように、集密な単層を傷つけ、HGF10ng/mlで24時間インキュベートした。実験の日、細胞を4%パラホルムアルデヒド/PBSにより室温で30分間固定した後、PBSで2回完全に洗浄し、抗体の非特異的結合を遮断するために、室温で1時間、5%ウシ血清アルブミン(BSA)/PBSからなるブロッキングバッファーでプレインキュベートした。次に、マウスHGF受容体(Sigma-Aldrich)の細胞外ドメインに対するウサギのポリクローナル抗体(ブロッキングバッファーで1:50に希釈)により、室温で60分間単層をインキュベートした。細胞を上記のように洗い、フルオレセインイソチオシアネート(FITC)結合ヤギ抗ウサギIgG(1:50)(Sigma-Aldrich)により標識した。室温で60分間のインキュベーション後、細胞を洗浄し、17% Gelvatol/PBSでマウントし、落射蛍光顕微鏡使用(epifluorescence microscopy)(Zeiss Axioskop)で検査した。
【0034】
(実施例1a):細胞増殖:対照およびHGF処理ウェルにおけるCPM(カウント/毎分)比は以下のようであった。2ngHGF/対照=1.1(0.95〜1.20)、5ngHGF/対照=1.1(1.04〜1.18)および15ngHGF/対照=1.15(0.99〜1.23)。したがって、上皮細胞株(CCL−53.1)における細胞増殖は、[3H]チミジン取り込み測定による評価で、組換えHGFにより著しく増加しなかった。
【0035】
(実施例1b):損傷部位の回復:損傷した単層の回復に対するHGFの効果を、in−vitroの損傷モデルを使用して評価した。むきだし部分は、24時間および48時間後に減少した。損傷時と24時間後との測定されたむきだし部分の比は、HGF処理の方が(損傷時=125.25±10.08mm2、24時間後=46.73±11.03mm2、比=2.65、p<0.01、48時間後=10.26±4.76mm2、比=11.91)対照(損傷時=133.77±11.7mm2、24時間後=112.23±11.11mm2、比=1.15、48時間後=75±13.17mm2、比=2.43)と比較して、有意に高かった(図1)。従来の培地または血清除去した培地との間に、HGF添加後の損傷細胞の回復において有意差は認められなかった。上皮回復の有意な亢進が、低濃度(1ng/ml)のHGFで観察された。10ng/mlまでのHGFの用量増加は、さらなる回復の増進を引き起こした(対照と1ng/mlの間でp<0.01、1ng/mlと5ng/mlの間でp<0.01、5ng/mlと10ng/mlの間でp=0.05)(図2)。FGF(1〜15ng/ml)、EGF(2〜60ng/ml)、PDGF(1〜35ng/ml)、またはFGF/KGF(1〜15ng/ml)の添加は、損傷細胞の回復に有意な影響を及ぼさなかった。
【0036】
c−Met受容体を、ウェル内で免疫蛍光によって視覚化した。HGFの添加後または未処理の対照において、損傷部から遠位の他の細胞と比較して、むきだし部位に隣接する細胞でのc−Met発現における本法による有意差は認められなかった(図3)。
【0037】
(実施例1c):細胞運動性:HGF添加後のガラス層からの遊走距離(mm)について、未処理の対照と比較した。HGF添加後の上皮細胞の遊走亢進が示された(HGF処理ペトリ皿で平均10.6±1.66mm、対照で1.2±0.51mm)(p<0.01)(図4)。
【0038】
(実施例1d):形態および細胞骨格:上皮細胞CCL−53.1の形態は、HGF処理細胞において異なった。未処理のウェルでは、細胞は、より小さい細胞内空間で、円形の高密度の団集花序状に集まったが、HGF処理ウェルでは、平らになり、広がった形態であった。共焦点顕微鏡技術によって作られる3D再構築によって、我々は、HGF添加後のアクチン構造に著しい変化が認められ、未処理細胞と比較して、高密度の末梢構造でF−アクチンがより少ないことを発見した(図5および6)。
【0039】
上記の例は、どの程度損傷したCCL−53.1上皮細胞を、HGFの生物活性を検査するために用いることができるかを示している。生物活性のあるHGFは、損傷後の損傷CCL−53.1上皮細胞の回復を用量依存的に亢進する。これらの結果は、HGF処理後に細胞遊走促進効果が亢進し、最終的に、その結果として、細胞骨格構造が弱まることによって部分的に説明することができる。上記の知見は、活性HGFに特有であって、非活性HGFまたはEGF、PDGF、FGF、KGFは、細胞に対して同一な効果を誘発することができなかった。
【0040】
(実施例2:表面プラズモン共鳴法(Biacore)によって示される生物活性のある肝細胞増殖因子(HGF)の特性)
以下の材料および条件を使用した。
【0041】
SPR測定およびリガンド固定化手順。SPR測定は、それぞれ4つおよび2つのフローセルを備えた全自動Biacore 2000測定器および半自動Bicaore X測定器(Biacore AB、スウェーデン、ウプサラ)によって、760nmで実施した。フローセルの温度は、すべての実験で25℃であった。使用した試料表面は、Biacore AB(スウェーデン、ウプサラ)から購入したカルボキシメチル化デキストランCM5チップであった。デキストランヒドロゲルのカルボン酸基に対するリガンドの結合は、200mM EDC(N−エチル−N’−(3−ジエチルアミノプロピル)カルボジイミド)および50mM NHS(N−ヒドロキシスクシンイミド)を使用した従来のカルボジイミドの化学的作用によって実施した。活性化にかかる時間は7分で、2〜7分後にリガンドの注入を行った。残留する活性エステルの不活性化は、pH8.5のエタノールアミン/塩酸塩を7分注入することによって実施した。5μl/分の流速をすべての固定化の間に使用した。すべてのリガンドは、pH4.5の10mMアセテート緩衝液で希釈した(すなわち、タンパク質の等電点を下回るので、デキストランマトリックスとリガンドの間の静電的相互作用を亢進する)。モノクローナル抗HGF(500μg/ml)を1:10に希釈し、およびポリクローナル抗HGFも。組換えMetプロトオンコジーン受容体(100μg/ml)は1:5に希釈し、HGF組換え体(5μg/ml)は1:3に希釈した。接触時間は、2分から7分の間で変えて、8,000〜30,000RU(反応単位)の固定化レベルを得た。不活性化後、表面は、その後に、pH2.0の5mMグリシン緩衝液と1M塩化ナトリウムを1分間、5回注入することにより洗浄した。フローセルの1つは、デキストランに対するHGFの特異的な相互作用だけでなく、緩衝液および非特異的な相互作用による反応を監視するための基準として使用した。このフローセルは、固定化処置の間は、他のフローセルと同様に処理を行ったが、リガンドの固定化工程は除外した。
【0042】
(実施例2a:デキストランに対するHGFの結合)
生物活性のあるHGFは、デキストランに対して高い親和性を持ち、試料へのデキストラン(0.05%)添加により、固定化されたモノクローナルおよびポリクローナル抗HGF抗体との、フローセルならびにデキストラン経路におけるシグナル応答が有意に減少した(p<0.05)(図7および8)。
【0043】
(実施例2b:リガンドへの結合)
生物活性のあるHGFは、生物活性のないHGFと比較して、固定化されたモノクローナル抗HGF抗体(p=0.005)またはc−Met受容体(p=0.02)とのフローセルにおけるシグナル応答が有意に高い(図7および8)。
【0044】
(実施例2c:ELISAとの相関)
免疫反応性HGFは、市販のキット(Quantikine HGF Immunoassy、R&D Systems Inc.、米国Minneapolis)を使用してELISAによって判定した。生物活性のあるHGFをSPRによる判定を用いて検査すると、結果は、ELISAによって測定した水準と有意に相関した。不活性HGFを使用すると、ELISAおよびSPRの結果は相関しなかった。
【0045】
(実施例2d:固定化レベルを変える影響)
固定化されたモノクローナル抗HGF抗体の量によるシグナル応答の影響について、異なる試料で測定した。固定化中に接触時間を増加することにより(1、5および10分)固定化レベルを増やすことで、生物活性のあるHGFを含むすべての試料の反応に有意な増加を生じた。流速を変更することでは(5、10および15μl/分)、試料の反応に有意な影響を及ぼさなかった。
【0046】
上記の実施例は、生物活性のあるHGFがデキストランに対して高い親和性を有すると思われ、これは細胞表面ヘパラン硫酸プロテオグリカン結合部位に対するそのような親和性に類似している可能性のあることを示している。これらの結合部位は、抗体結合エピトープと重複するか、デキストランと結合するとすぐに、HGFのコンホメーション変化を誘発し、この結果、立体的障害になり、したがって、抗体への結合を妨げる。
【0047】
(実施例3:マウスの正常な無毛の皮膚に対する生物活性のある組換えヒト肝細胞増殖因子(rHGF)の局所投与の影響)
マウス18匹(C57/B6ブラック、14〜16週齢、体重21〜29g、全マウスが雌)の背部を剪毛し、これにより、右側および左側に、2つの1×1cmの部分ができた。次に、マウスを3群に割り付けた。第1群(n=6)は、0.70μg/kg(12〜18ng/cm2無毛の皮膚)の組換えヒト肝細胞増殖因子(rHGF)を、右側に毎日5日間皮下に処置した。利用されたHGFは、試験前にCCL−53.1細胞の損傷分析(wound assay)に加えて、FLISA、表面プラズモン共鳴(SPR)法によって検査した。このHGF型は、SPRよる判定で、デキストランおよびHGFに対する抗体に高い親和性を持ち、ELISAによって得た濃度と相関した。組換えHGFは、CCL−53.1細胞の損傷部位への用量依存的な遊走を引き起こした。この効果は、抗HGF抗体によって停止させることができた。第2群(n=6)には、SPRによってデキストランに対する親和性が認められなかった組換えHGF 0.70μg/kgを5日間皮下投与したが、損傷分析において、CCL−53.1の遊走は生じなかった。第3群(n=6)には、無菌の生理的塩化ナトリウムを5日間皮下投与した。本群は対照群とした。治療コース中または後に、非忍容性、または疾患の何らかの徴候はいずれの治療群においても示されなかった。治療下で、発毛におけるいくつかの有意差は、明らかに見られなかったが、我々は、12日後、第1群のマウス5匹が、両側で完全に発毛したことを見て驚いた(図9)。第2群および第3群では、同じプロセスに非常に長い時間がかかった(20〜35日)。したがって我々は、マウスの正常な、無毛の皮膚に対する適量の生物活性のあるHGFによる局所治療は、発毛を促進し、この局所治療が、問題のない動物では忍容性を有すると結論する。
【0048】
(実施例4:静脈血流不全症患者の慢性下肢潰瘍に対する組換え肝細胞増殖因子の治療効果−症例報告)
最近のパイロット研究期間中に観察された慢性下肢潰瘍に対する肝細胞増殖因子の正の効果を評価するために、患者20例を対象にして、市販の組換えHGFを1週間使用する二重盲検プラセボ対照試験を開始した。本試験は、患者12例の組み入れ後、SDS−PAGEおよびウエスタンブロットにより、使用されたタンパク質に欠陥があったことが明らかにされた時点で中断された。潰瘍部位由来の生検材料の免疫組織化学染色は、HGF受容体(c−met)の発現が、塩化ナトリウム製剤を投与した患者(第2群、n=6)と比較して、市販の組換えHGFを含む製剤を投与した患者(第1群、n=6)で有意ではないが減少することを示した。このことはタンパク質のc−met受容体との相互作用を示す可能性がある。第1群で、毛細管増殖の増進が観察されたが、劇的な臨床的改善が認められたのは、わずか1例に過ぎなかった。にもかかわらず、第1群の3例および第2群の2例が9カ月以内に治癒した。他の患者7例には、生物活性のある組換えHGFによる治療を申し出て、4例(各群2例)が受け入れた(第3群)。これら当初不成功の全例で、潰瘍の状態の改善が、治療開始から1週間以内に観察された。我々は、生物活性のあるHGFが慢性下肢潰瘍の治癒に対して、プラセボ効果によるものでない正の効果を有すると結論を下した。
【0049】
(実施例5:SPRによるHGFエピトープマッピング)
(実施例5a:抗体親和性)
HGF(Santa Cruz)に対するエピトープ特異的ないくつかの抗体をSPRのフローセルに固定化することによって、我々は、生物活性のあるHGFまたは生物的に不活性な組換えHGFの多くの特性を判定することに成功している。
【0050】
1−生物活性のあるHGFは、HGFのα鎖およびβ鎖で異なるエピトープに対して一定の親和性を有する。
【0051】
2−生物活性のないHGFでは、N−17(ヒト由来のHGFαのN末端におけるペプチドマッピングに対する抗体)により固定化されたフローセルに対する親和性は、減少または無効になった。
【0052】
H−170(ヒト由来のHGFβのアミノ酸1〜170に対して作られる抗体)に対する親和性は減少した。
【0053】
c−met受容体およびH−145(ヒト由来のHGFαのアミノ酸32〜176に対する抗体)の親和性は維持した。
【0054】
(実施例5b:HGF受容体結合エピトープ)
SPRを使用して、我々は、HGFのc−met受容体への結合において重要なエピトープを確認することに成功している。我々は、HGFがc−met受容体と結合する時、H−145(ヒト由来のHGFαのアミノ酸32〜176に対する抗体)およびC−20(ヒト由来のHGFαのC末端におけるペプチドマッピングに対して作られた抗体)に結合するHGFのエピトープが関与していることを明らかにしている。依然として、HGFがc−met受容体と相互作用を持つ間に、N−17(ヒト由来のHGFαのN末端におけるペプチドマッピングに対する抗体)、N−19(ヒト由来のHGFβのN末端におけるペプチドマッピングに対する抗体)およびH−170(ヒト由来のHGFβのアミノ酸1〜170に対して作られる抗体)と相互に作用する他のエピトープが利用可能である。
【0055】
我々が試験した不活性HGF組換え型において、エピトープに結合する受容体は、損傷がなかったが、α鎖のN−17およびβ鎖のH−170と相互に作用するドメインは欠陥があった。
【図面の簡単な説明】
【0056】
【図1】傷害後の24時間および48時間以内の対照(単色)および活性HGFで治療した細胞における回復を示す図である。
【図2】異なるHGF濃度(1、5および10ng/ml)および24時間以内の回復の相違を示す図である。
【図3】免疫蛍光法によるCCL−53.1細胞におけるc−Met受容体の局在化を示す図である。本図で見られるように、免疫蛍光による受容体の提示で、生物活性のあるHGFでの治療(A)と対照(B)の間に相違が認められない。しかし、損傷したむきだし部分の減少が、(B)と比較して、(A)で明白に見られる。
【図4】遊走アッセイを示す図である。右側の写真は、生物活性のあるHGFの添加後の細胞の遊走を示し、左側の写真は、対照細胞を示している。矢印は、円形のカバーガラスの縁を示している。
【図5】アクチン構造のファロイジン染色後の対照(左側)と活性HGF処理細胞(右側)を比較した共焦点顕微鏡写真および3Dルックスルー投射図(3D look-through projection)である。両群間の細胞壁径の差異に留意のこと。
【図6】生物活性のない細胞(左側)と対照細胞(右側)の間の形態学的相違を示す図である。
【図7】デキストラン(0.05%)の添加前(左側の最初のシグナル)および添加後(2番目のシグナル)の生物活性のあるHGFのSPR反応を示す図である。再生パルスは、各シグナルの後に観察される。
【図8】生物活性のあるHGFおよび生物活性のないHGFからのSPRシグナル間の相違を示すヒストグラムである。
【図9】生物活性のあるHGFを5日間皮下投与したマウス(左側および右側)および無菌塩化ナトリウムを5日間皮下注射した対照マウス(中央)の発毛の相違を示す図である。写真は、最初の注射の12日後に撮影された。
【特許請求の範囲】
【請求項1】
肝細胞増殖因子(HGF)の生物活性の測定のための方法であって、HGFの生物活性が、
i)損傷細胞の回復のin vitroモデル、および/または
ii)c−met受容体、モノクローナル抗HGF、ポリクローナル抗HGFおよび/または多糖類に対して、SPRを使用する親和性測定および/またはエピトープマッピング、ならびに/あるいは
iii)発毛速度に関する動物モデル
を使用して測定される方法。
【請求項2】
前記in vitroモデルi)がCCL53.1細胞を含む請求項1に記載の方法。
【請求項3】
ii)の前記多糖類がデキストランである請求項1に記載の方法。
【請求項4】
前記動物モデルiii)がC57/B6ブラックマウス系統である請求項1に記載の方法。
【請求項5】
HGFの生物活性の測定が、例えば、生物活性のあるHGFを含む医薬組成物の調製のための組換え技術による、バッチ生産後のHGFの品質管理に使用される前記請求項のいずれかに記載の方法。
【請求項6】
HGFの生物活性の測定が、治療上活性のあるHGFによる治療が必要な患者への投与前に、HGF製剤の評価に使用される請求項1から4のいずれかに記載の方法。
【請求項7】
外因性の治療上活性のあるHGFの損傷部位への適用が必要かどうか判定するために、HGFの生物活性の測定が、病変部位の、例えば、潰瘍分泌物、尿、糞便、唾液、胸膜などの体組織または体液で内因性HGFの活性の評価に使用される請求項1から4のいずれかに記載の方法。
【請求項8】
HGFの生物活性の測定が、さまざまなHGF関連疾患の診断をさらに正確にするために使用される請求項1から4のいずれかに記載の方法。
【請求項9】
前記請求項のいずれかに記載のHGFの生物活性に関する評価のためのキット。
【請求項1】
肝細胞増殖因子(HGF)の生物活性の測定のための方法であって、HGFの生物活性が、
i)損傷細胞の回復のin vitroモデル、および/または
ii)c−met受容体、モノクローナル抗HGF、ポリクローナル抗HGFおよび/または多糖類に対して、SPRを使用する親和性測定および/またはエピトープマッピング、ならびに/あるいは
iii)発毛速度に関する動物モデル
を使用して測定される方法。
【請求項2】
前記in vitroモデルi)がCCL53.1細胞を含む請求項1に記載の方法。
【請求項3】
ii)の前記多糖類がデキストランである請求項1に記載の方法。
【請求項4】
前記動物モデルiii)がC57/B6ブラックマウス系統である請求項1に記載の方法。
【請求項5】
HGFの生物活性の測定が、例えば、生物活性のあるHGFを含む医薬組成物の調製のための組換え技術による、バッチ生産後のHGFの品質管理に使用される前記請求項のいずれかに記載の方法。
【請求項6】
HGFの生物活性の測定が、治療上活性のあるHGFによる治療が必要な患者への投与前に、HGF製剤の評価に使用される請求項1から4のいずれかに記載の方法。
【請求項7】
外因性の治療上活性のあるHGFの損傷部位への適用が必要かどうか判定するために、HGFの生物活性の測定が、病変部位の、例えば、潰瘍分泌物、尿、糞便、唾液、胸膜などの体組織または体液で内因性HGFの活性の評価に使用される請求項1から4のいずれかに記載の方法。
【請求項8】
HGFの生物活性の測定が、さまざまなHGF関連疾患の診断をさらに正確にするために使用される請求項1から4のいずれかに記載の方法。
【請求項9】
前記請求項のいずれかに記載のHGFの生物活性に関する評価のためのキット。
【図1】


【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】

【図9】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公表番号】特表2008−516629(P2008−516629A)
【公表日】平成20年5月22日(2008.5.22)
【国際特許分類】
【出願番号】特願2007−537847(P2007−537847)
【出願日】平成17年10月20日(2005.10.20)
【国際出願番号】PCT/SE2005/001583
【国際公開番号】WO2006/043892
【国際公開日】平成18年4月27日(2006.4.27)
【出願人】(307035066)ペアス インスティテュート アーベー (1)
【Fターム(参考)】
【公表日】平成20年5月22日(2008.5.22)
【国際特許分類】
【出願日】平成17年10月20日(2005.10.20)
【国際出願番号】PCT/SE2005/001583
【国際公開番号】WO2006/043892
【国際公開日】平成18年4月27日(2006.4.27)
【出願人】(307035066)ペアス インスティテュート アーベー (1)
【Fターム(参考)】
[ Back to top ]