
肝細胞成長因子(HGF)結合蛋白質
【課題】治療剤及び診断剤として使用できるさらなるHGF調節剤を提供すること。
【解決手段】本発明の一部は、HGF、特にヒトHGFを特異的に結合する一群の結合蛋白質の発見に基づいている。これらの結合蛋白質は、HGFを特異的に結合する一群の抗体のCDRに基づく抗原(即ち、HGF)結合部位を含有する限りにおいて、抗体系である。本願は肝細胞成長因子(HGF)、特にヒトHGFを結合し、その活性を中和する一群の結合蛋白質を提供する。これらの結合蛋白質は診断剤及び/又は治療剤として用いることができる。治療活性に関して言えば、これらの結合蛋白質は特定のHGF反応性疾患、例えば、特定のHGF反応性腫瘍を治療するのに用いることができる。
【解決手段】本発明の一部は、HGF、特にヒトHGFを特異的に結合する一群の結合蛋白質の発見に基づいている。これらの結合蛋白質は、HGFを特異的に結合する一群の抗体のCDRに基づく抗原(即ち、HGF)結合部位を含有する限りにおいて、抗体系である。本願は肝細胞成長因子(HGF)、特にヒトHGFを結合し、その活性を中和する一群の結合蛋白質を提供する。これらの結合蛋白質は診断剤及び/又は治療剤として用いることができる。治療活性に関して言えば、これらの結合蛋白質は特定のHGF反応性疾患、例えば、特定のHGF反応性腫瘍を治療するのに用いることができる。
【発明の詳細な説明】
【技術分野】
【0001】
関連する出願
本願は、2006年6月2日に出願された米国仮特許出願第60/810,714号、および2006年11月21日に出願された同第60/860,509号の利益およびそれらへの優先権を主張するものであり、それらの開示は本明細書中に参考として援用される。
【0002】
発明の分野
本発明の分野は、分子生物学、免疫学及び腫瘍学の分野である。より詳細には、この分野は、ヒト肝細胞成長因子(HGF)に結合する抗体系結合蛋白質の分野である。
【背景技術】
【0003】
背景
分散因子(SF:Scatter Factor)とも呼ばれる肝細胞成長因子(HGF:Hepatocyte Growth Factor)は、主として間葉細胞により産生される多機能性ヘテロ二量体蛋白質であり、Metチロシンキナーゼ受容体を発現する細胞のエフェクタである(非特許文献1、非特許文献2)。ヒトMet受容体は「c−Met」とも呼ばれる。成熟HGFは2本のポリペプチド鎖、α鎖及びβ鎖を含有している。公表された研究によれば、HGFのc−Met受容体結合ドメインを含有するのはα鎖であることが示唆されている。
【0004】
HGFは、その同族受容体に結合すると、多くの細胞活動を媒介する。HGF−Metシグナル伝達経路は、肝臓の再生、創傷治癒、神経再生、血管形成及び悪性腫瘍に関係している。例えば、非特許文献3、非特許文献4並びに特許文献1及び特許文献2を参照されたい。HGF活性が関係する各種の疾患、例えば、特定のHGF反応性癌、を治療するために、抗体を含む多くのHGF調節剤が研究者らにより開発されてきた。例えば、特許文献3を参照されたい。
【0005】
全ての抗体に共通する基本的構造を図1に摸式図的に示した。抗体は4本のポリペプチド鎖を含有する多量体蛋白質である。これらのポリペプチド鎖のうちの2本は重鎖又はH鎖と呼ばれ、他の2本は軽鎖又はL鎖と呼ばれている。これら免疫グロブリン重鎖及び軽鎖は、鎖間ジスルフィド結合によって連結されている。免疫グロブリン重鎖同士も多くの鎖間ジスルフィド結合により連結されている。軽鎖が1箇所の可変領域(図1中のVL)と1箇所の定常領域(図1中のCL)とで構成されるのに対して、重鎖は1箇所の可変領域(図1中のVH)と少なくとも3箇所の定常領域(図1中のCH1、CH2及びCH3)とで構成される。可変領域は抗体の特異性を決定し、定常領域は他の機能を有する。
【0006】
アミノ酸及び構造上の情報から、各可変領域は4箇所の比較的保存されたフレームワーク領域又はFRに隣接して存在する3箇所の超可変領域(相補性決定領域又はCDR(complementarity determining region)とも呼ばれる)を含むことが分かっている。CDR1、CDR2及びCDR3と称されるこれら3箇所のCDRは個々の抗体の結合特異性に関与している。抗体を診断剤及び治療剤として使用する場合には、通常、その標的分子に対して最も高い結合特異性及び親和性を有する抗体を作製することが望ましい。可変領域間の相違は、抗体の特異性及び親和性に対して大きな影響を与え得ると考えられている。
【0007】
特許文献4には、カポジ肉腫の治療における抗HGF抗体の使用について開示されている。同じように、特許文献5には、治療対象の患者に抗HGF抗体を投与して腫瘍における内因性HGFの血管形成促進能を妨げることによる腫瘍の治療について開示されている。最近になって、研究者らにより、HGFのβ鎖に結合する抗体がHGF依存性腫瘍の患者に対して治療剤となる可能性を有するとの提案がなされている(非特許文献4)。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】米国特許第5,997,868号明細書
【特許文献2】米国特許第5,707,624号明細書
【特許文献3】国際公開第2005/017107号パンフレット
【特許文献4】米国特許第5,707,624号明細書
【特許文献5】米国特許第5,997,868号明細書
【非特許文献】
【0009】
【非特許文献1】Bottaro et al. (1991年) SCIENCE 251: p.802−804
【非特許文献2】Rubin et al. (1993年) BIOCHIM. BIOPHYS. ACTA 1155: p.357−371
【非特許文献3】Cao et al.(2001年) PROC. NATL. ACAD. SCI. USA 98:p.7443−7448
【非特許文献4】Burgess et al.(2006年) CANCER RES. 66:p.1721−1729
【発明の概要】
【発明が解決しようとする課題】
【0010】
それでもなお、治療剤及び診断剤として使用できるHGF調節剤がさらに求められている。
【課題を解決するための手段】
【0011】
発明の要旨
本発明の一部は、HGF、特にヒトHGFを特異的に結合する一群の結合蛋白質の発見に基づいている。これらの結合蛋白質は、HGFを特異的に結合する一群の抗体のCDRに基づく抗原(即ち、HGF)結合部位を含有する限りにおいて、抗体系である。このCDRは結合蛋白質のHGFに対する結合特異性をもたらす。こうした結合蛋白質は診断剤及び治療剤として使用することができる。治療剤として用いる場合、結合蛋白質は、レシピエント(例えば、ヒト)に投与した時にこの結合蛋白質に対する免疫反応を誘発するリスクを低減又は排除できるように設計(例えば、ヒト化)する。
結合蛋白質は、HGF活性を中和するので、治療剤として用いることができる。一部の実施態様として、この結合蛋白質は、HGFがその同族受容体c−Metに結合するのを妨げることによってHGF活性を中和する。別の実施態様として、この結合蛋白質は、HGFに結合してその生物活性を中和するが、HGFがc−Met受容体に結合するのを妨げない。HGFが癌細胞の成長及び増殖に関与しているので、この結合蛋白質は癌細胞の増殖を抑制するのに用いることができる。さらに、この結合蛋白質は、哺乳動物に投与すると、その哺乳動物における腫瘍の成長を抑制又は低減することができる。
【0012】
本発明は、例えば、以下の項目を提供する。
(項目1)
ヒト肝細胞成長因子(HGF)に結合する単離結合蛋白質であって、
(a)構造CDRL1−CDRL2−CDRL3を含む免疫グロブリン軽鎖可変領域であって、
(i)CDRL1が配列番号18(2B8)のアミノ酸配列を含み、
(ii)CDRL2が配列番号206(LRMR2B8LC)のアミノ酸配列を含み、及び
(iii)CDRL3が配列番号20(2B8)のアミノ酸配列を含む
ものとする免疫グロブリン軽鎖可変領域、並びに
(b)免疫グロブリン重鎖可変領域
を含み、該免疫グロブリン軽鎖可変領域及び該免疫グロブリン重鎖可変領域が共同してヒトHGFに結合するための単一結合部位を規定するものとする結合蛋白質。
(項目2)
CDRL1、CDRL2及びCDRL3がヒト又はヒト化免疫グロブリンのフレームワーク領域間に挿入されている項目1の結合蛋白質。
(項目3)
前記結合蛋白質が抗体又はその抗原結合断片である項目1又は2の結合蛋白質。
(項目4)
配列番号193(LR2B8LC軽鎖可変領域)又は配列番号199(LRMR2B8LC軽鎖可変領域)の免疫グロブリン軽鎖可変領域を含む項目1乃至3のいずれか1項の単離結合蛋白質。
(項目5)
配列番号197(LR2B8LC+カッパ定常(km(3)アロタイプ)(対立遺伝子1))又は配列番号201(LRMR2B8LC+カッパ定常(km(3)アロタイプ)(対立遺伝子1))の免疫グロブリン軽鎖配列を含む項目4の単離結合蛋白質。
(項目6)
ヒト肝細胞成長因子(HGF)に結合する単離結合蛋白質であって、
(a)構造CDRH1−CDRH2−CDRH3を含む免疫グロブリン重鎖可変領域であって、
(i)CDRH1が配列番号15(2B8)のアミノ酸配列を含み、
(ii)CDRH2が配列番号204(LR2B8HC)及び配列番号205(LRMR2B8HC)からなる群から選ばれる配列を含み、及び
(iii)CDRH3が配列番号17(2B8)のアミノ酸配列を含む
ものとする免疫グロブリン重鎖可変領域、並びに
(b)免疫グロブリン軽鎖可変領域
を含み、該免疫グロブリン重鎖可変領域及び該免疫グロブリン軽鎖可変領域が共同してヒトHGFに結合するための単一結合部位を規定するものとする結合蛋白質。
(項目7)
CDRH1、CDRH2及びCDRH3がヒト又はヒト化免疫グロブリンの複数のフレームワーク領域間に挿入されている項目6の結合蛋白質。
(項目8)
前記結合蛋白質が抗体又はその抗原結合断片である項目7の結合蛋白質。
(項目9)
配列番号183(LR2B8HC重鎖可変領域)又は配列番号189(LRMR2B8HC重鎖可変領域)の免疫グロブリン重鎖可変領域を含む項目6乃至8のいずれか1項の単離結合蛋白質。
(項目10)
配列番号187(LR2B8HC+IgG1定常(G1m(3)アロタイプ)(対立遺伝子1))又は配列番号191(LRMR2B8HC+IgG1定常(Glm(3)アロタイプ)(対立遺伝子1))の免疫グロブリン重鎖配列を含む項目9の単離結合蛋白質。(項目11)
さらに、構造CDRL1−CDRL2−CDRL3を含む免疫グロブリン軽鎖可変領域を含み、
(i)CDRL1が配列番号18(2B8)のアミノ酸配列を含み、
(ii)CDRL2が配列番号19(2B8)又は配列番号206(LRMR2B8LC)のアミノ酸配列を含み、及び
(iii)CDRL3が配列番号20(2B8)のアミノ酸配列を含むものとし、該免疫グロブリン軽鎖可変領域及び前記免疫グロブリン重鎖可変領域が共同してヒトHGFに結合するための単一結合部位を規定するものとする項目6の単離結合蛋白質。
(項目12)
前記抗体がモノクローナル抗体である項目1、6又は11のいずれか1項の結合蛋白質。
(項目13)
項目1乃至5のいずれか1項の免疫グロブリン軽鎖可変領域をコードしているヌクレオチド配列を含む単離核酸。
(項目14)
項目6乃至10のいずれか1項の免疫グロブリン重鎖可変領域をコードしているヌクレオチド配列を含む単離核酸。
(項目15)
項目13又は14の核酸配列を含む発現ベクター。
(項目16)
項目15の発現ベクターを含む宿主細胞。
(項目17)
結合蛋白質を作製する方法であって、
(i)項目16の宿主細胞を該宿主細胞が前記免疫グロブリン軽鎖可変領域及び/又は前記免疫グロブリン重鎖可変領域を発現するような条件下で増殖させ、及び
(ii)該免疫グロブリン軽鎖可変領域及び/又は該免疫グロブリン重鎖可変領域を採取すること
を含む方法。
(項目18)
前記結合蛋白質が4.0×10−5s−1以下のkdでヒト肝細胞成長因子に結合する項目1乃至11のいずれか1項の単離結合蛋白質。
(項目19)
前記kdが3.0×10−5s−1以下である項目18の単離結合蛋白質。
(項目20)
前記kdが2.0×10−5s−1以下である項目19の単離結合蛋白質。
(項目21)
前記結合蛋白質が20pM以下のKDでヒト肝細胞成長因子に結合する項目1乃至11のいずれか1項の単離結合蛋白質。
(項目22)
前記KDが10pM以下である項目21の単離結合蛋白質。
(項目23)
前記KDが5pM以下である項目22の単離結合蛋白質。
(項目24)
腫瘍細胞に有効量の項目1乃至11のいずれか1項の結合蛋白質を作用させて該腫瘍細胞の増殖を抑制又は低減させることを含む腫瘍細胞の増殖を抑制又は低減させる方法。
(項目25)
前記腫瘍細胞がヒト腫瘍細胞である項目24の方法。
(項目26)
哺乳動物において腫瘍成長を抑制又は低減させる方法であって、該哺乳動物に有効量の項目1乃至11のいずれか1項の結合蛋白質を作用させて該腫瘍細胞の増殖を抑制又は低減させることを含む方法。
(項目27)
哺乳動物において腫瘍を治療する方法であって、有効量の項目1乃至11のいずれか1項の結合蛋白質を投与することを含む方法。
(項目28)
前記哺乳動物がヒトである項目26又は27の方法。
本発明の以上その他の態様及び効果は、以下の図、詳細な説明及び特許請求の範囲を考慮すれば明瞭に理解されよう。
【図面の簡単な説明】
【0013】
本発明は、以下の図面にてさらに完全な理解が可能となろう。
【図1】代表的な抗体を模式図化したものである。
【図2】1A3、1D3、1F3、2B8、2F8、3A12、3B6及び3D11で表した抗体の完全な免疫グロブリン重鎖可変領域を規定するアミノ酸配列を示す模式図である。各抗体のアミノ酸配列は全て相互に整列させてあり、シグナルペプチド、CDR1、CDR2及びCDR3を規定する領域は枠で囲んで識別されている。枠で囲んでない配列はFR配列を表す。
【図3】図2に示した免疫グロブリン重鎖可変領域配列のそれぞれのCDR1、CDR2及びCDR3を示す模式図である。
【図4】1A3、1D3、1F3、2B8、2F8、3A12、3B6及び3D11で表した抗体の完全な免疫グロブリン軽鎖可変領域を規定するアミノ酸配列を示す模式図である。各抗体のアミノ酸配列は全て相互に整列させてあり、シグナルペプチド、CDR1、CDR2及びCDR3を規定する領域は枠で囲んで識別されている。枠で囲んでない配列はFR配列を表す。
【図5】図4に示した免疫グロブリン軽鎖可変領域配列のそれぞれのCDR1、CDR2及びCDR3を示す模式図である。
【図6】U87MG異種移植モデルにおいて抗HGF抗体1D3、1F3、1A3及び2B8の腫瘍抑制活性を測定する実験からの結果をまとめたグラフである。菱形はPBS、三角形は抗HGF抗体1A3、Xは抗HGF抗体1D3、四角形は抗HGF抗体1F3、丸は抗HGF抗体2B8を示す。
【図7】U118異種移植モデルにおいて抗HGF抗体1D3、1F3、1A3及び2B8の腫瘍抑制活性を測定する実験からの結果をまとめたグラフである。菱形はIgG、四角形は抗HGF抗体1F3、三角形は抗HGF抗体1D3、Xは抗HGF抗体1A3、丸は抗HGF抗体2B8に相当する。
【図8】ヒトHGFとキメラ、キメラ/ヒト化又はヒト化2B8抗体との間の抗原結合親和性及び相互作用速度に関する表面プラスモン共鳴データをまとめた一覧表である。この表には試験したカッパ鎖とIgG1重鎖との対が示されている。標準偏差(STDEV)が示されている抗体は3回の独立した実験において解析したものである。
【図9】Hu2B8がマウスモノクローナル抗体2B8に対して互いに限定される抗原決定基に結合することを示す実験データをまとめた棒グラフである。ヒト化又はキメラ2B8を抗ヒトFcチップ上に捕捉した。次いで、HGFをこのヒト化又はキメラ2B8に結合させた。次に、マウス2B8又は対照抗体(ポリクロナールヤギ抗HGF抗体)の上記捕捉HGFへの結合能を測定した。その結果、ヒト化2B8抗体及びキメラ2B8はマウス2B8がHGFに結合するのを妨げた。白色棒はキメラ2B8抗体、灰色棒はヒト化Hu2B8抗体(カッパ可変領域Kv1−39.1及び重鎖可変領域Hv5−51.1)、黒色棒はヒト化Hu2B8抗体(カッパ可変領域Kv3−15.1及び重鎖可変領域Hv5−51.1)を示す。
【発明を実施するための形態】
【0014】
本発明の一部は、HGFを特異的に結合し、その活性を中和する一群の結合蛋白質、特にヒトHGFの発見に基づいている。こうした結合蛋白質は種々の診断的、治療的用途に使用することができる。この結合蛋白質は、HGFに結合し、その活性を中和する能力から選択された特定のモノクローナル抗体の抗原結合部位をベースとしている。具体的には、この結合蛋白質は、HGFに対する結合部位を共同して規定する免疫グロブリン可変領域CDR配列を含有している。
【0015】
こうした抗体は、その中和活性を考慮すると、HGF反応性細胞、例えば、癌細胞の成長及び/又は増殖を調節するのに特に有用である。この結合蛋白質は、治療剤として用いる場合には、レシピエントに投与した時にこの蛋白質に対する免疫反応を誘発するリスクをできる限り少なくしたり排除することができるように設計することができる。さらに、この結合蛋白質は、特定の用途に応じて、他の成分、例えば、検出可能な標識(例えば、放射能標識)並びにエフェクタ分子(例えば、他の蛋白質及び低分子系治療剤)に結合させることができることが企図されている。本発明のこれらの特徴及び態様については、それぞれ以下でさらに詳細に論じる。
【0016】
I. HGFに結合する結合蛋白質
一態様として、本発明はヒトHGFに結合する単離結合蛋白質を提供する。この結合蛋白質は、(i)CDRL1−CDRL2−CDRL3の構造を含む免疫グロブリン軽鎖可変領域及び(ii)3箇所の相補性決定領域(CDR)を含む免疫グロブリン重鎖可変領域を含む。この場合、免疫グロブリン軽鎖可変領域及び免疫グロブリン重鎖可変領域はヒトHGFに結合するための単一結合部位を共同して規定するものとする。CDRL1は、アミノ酸配列X1X2SerX4X5X6X7X8X9X10X11X12X13X14X15であって、アミノ酸X1がArg、Lys又はSerであり、X2がAla又はThrであり、X4がGlu、Gln又はSerであり、X5がAsn、Asp又はSerであり、X6がIle又はValであり、X7がAsp、Lys、Ser、Val又はTyrであり、X8がペプチド結合又はTyrであり、X9がペプチド結合又はAspであり、X10がペプチド結合又はGlyであり、X11がペプチド結合又はAsnであり、X12がペプチド結合、Ile又はSerであり、X13がAsn又はTyrであり、X14がIle、Leu、Met又はValであり、X15がAla、Asn、His又はSerであるものとするアミノ酸配列を含む。CDRL2は、アミノ酸配列X16X17X18X19X20X21X22であって、アミノ酸X16がAla、Asp、Arg、Gly又はValであり、X17がAla、Thr又はValであり、X18がAsn、Ser又はThrであり、X19がArg、Asn、Lys又はHisであり、X20がLeu又はArgであり、X21がAla、Asn、Glu、Val又はProであり、X22がAsp、Ser又はThrであるものとするアミノ酸配列を含む。CDRL3は、アミノ酸配列X23X24X25X26X27X28ProX30Thrであって、アミノ酸X23がLeu、Gly又はGlnであり、X24がHis又はGlnであり、X25がPhe、Ser、Trp又はTyrであり、X26がAsp、Ile、Ser、Trp又はTyrであり、X27がGly、Glu、Asn又はSerであり、X28がAsp、Asn、Phe、Thr又はTyrであり、X30がLeu、Phe、Pro又はTyrであるものとするアミノ酸配列を含む。
【0017】
別の態様として、本発明は、(i)CDRH1−CDRH2−CDRH3の構造を含む免疫グロブリン重鎖可変領域及び(ii)3箇所の相補性決定領域(CDR)を含む免疫グロブリン軽鎖可変領域を含む、ヒトHGFに結合する単離結合蛋白質であって、これら免疫グロブリン軽鎖可変領域及び免疫グロブリン重鎖可変領域がヒトHGFに結合するための単一結合部位を共同して規定するものとする単離結合蛋白質を提供する。CDRH1は、アミノ酸配列X1TyrX3X4X5であって、アミノ酸X1がAsp、Asn、Ser又はThrであり、X3がPhe、Ser、Trp又はTyrであり、X4がIle、Leu又はMetであり、X5がAsn、His又はSerであるものとするアミノ酸配列を含む。CDRH2は、アミノ酸配列X6IleX8X9X10X11GlyX13X14X15TyrX17X18X19X20X21X22であって、アミノ酸X6がLys、Gln、Glu、Val又はTyrであり、X8がAsn、Gly、Ser、Trp又はTyrであり、X9がAla、Pro又はSerであり、X10がGly又はThrであり、X11がペプチド結合、Asp、Asn、Gly又はSerであり、X13がAsp、Asn、His又はSerであり、X14がSer又はThrであり、X15がAsn又はTyrであり、X17がAsn又はProであり、X18がAla、Asp、Gly、Gln、Glu、Pro又Serであり、X19がAsn、Lys、Met又はSerであり、X20がLeu、Phe又はValであり、X21がLys、Met又はGlnであり、X22がAsp、Gly又はSerであるものとするアミノ酸配列を含む。CDRH3は、アミノ酸配列X23X24X25X26X27X28X29X30X31X32X33X34Tyrであって、アミノ酸X23がArg、Asn、Gln又はGluであり、X24がGly、Leu、Arg又はTyrであり、X25がペプチド結合、Asp又はGlyであり、X26がペプチド結合又はGlyであり、X27がペプチド結合又はTyrであり、X28がペプチド結合、Leu又はTyrであり、X29がペプチド結合、Gly、Leu、Arg又はValであり、X30がペプチド結合、Asp、Gly又はGluであり、X31がペプチド結合、Asn、Arg、Ser又はTyrであり、X32がペプチド結合、Ala、Gly、Ile又Tyrであり、X33がMet又はPheであり、X34がAla又はAspであるものとするアミノ酸配列を含む。
【0018】
上記結合蛋白質が上述の免疫グロブリン軽鎖及び免疫グロブリン重鎖の配列又はこれらの断片を共に含み得ることは理解されよう。さらに、上記結合蛋白質が完全な状態の抗体又はその抗原結合性断片或いは生合成抗体結合部位であってもよいことも理解されよう。
【0019】
一部の実施態様として、上記の免疫グロブリン軽鎖及び免疫グロブリン重鎖のCDR配列はフレームワーク領域(FR:framework region)と共に挿入されている。
【0020】
一部の実施態様として、上記の免疫グロブリン軽鎖及び免疫グロブリン重鎖のCDR配列は、複数のヒト又はヒト化フレームワーク領域間に挿入されている。
【0021】
別の態様として、本発明はヒトHGFに特異的に結合する単離結合蛋白質を提供する。この結合蛋白質は、(a)CDRL1−CDRL2−CDRL3の構造を有する免疫グロブリン軽鎖可変領域及び(b)免疫グロブリン重鎖可変領域を含む。この場合、免疫グロブリン軽鎖可変領域及び免疫グロブリン重鎖可変領域はヒトHGFに結合するための単一結合部位を共同して規定するものとする。上記CDRL1は、配列番号8(1A3)、配列番号18(2B8)、配列番号28(2F8)、配列番号38(3B6)、配列番号48(3D11)、配列番号58(1D3)、配列番号68(1F3)及び配列番号78(3A12)からなる群から選ばれる配列を含む。上記CDRL2は、配列番号9(1A3)、配列番号19(2B8)、配列番号29(2F8)、配列番号39(3B6)、配列番号49(3D11)、配列番号59(1D3)、配列番号69(1F3)、配列番号79(3A12)及び配列番号206(LRMR2B8LC)からなる群から選ばれる配列を含む。上記CDRL3は、配列番号10(1A3)、配列番号20(2B8)、配列番号30(2F8)、配列番号40(3B6)、配列番号50(3D11)、配列番号60(1D3)、配列番号70(1F3)及び配列番号80(3A12)からなる群から選ばれる配列を含む。本明細書及び特許請求の範囲の全体を通じて、特定の配列番号により示した配列の後に続けて、その特定の配列が由来する抗体を括弧内に示した。例えば、配列番号8(1A3)は、配列番号8の配列が抗体1A3に存在する配列に基づいていることを意味している。
【0022】
一実施態様として、上記結合蛋白質は、配列番号8(1A3)の配列を含むCDRL1、配列番号9(1A3)の配列を含むCDRL2及び配列番号10(1A3)を含むCDRL3を含む免疫グロブリン軽鎖可変領域を含む。
【0023】
別の実施態様として、上記結合蛋白質は、配列番号18(2B8)の配列を含むCDRL1、配列番号19(2B8)又は配列番号206(LRMR2B8LC)の配列を含むCDRL2及び配列番号20(2B8)を含むCDRL3を含む免疫グロブリン軽鎖可変領域を含む。
別の実施態様として、上記結合蛋白質は、配列番号28(2F8)の配列を含むCDRL1、配列番号29(2F8)の配列を含むCDRL2及び配列番号30(2F8)を含むCDRL3を含む免疫グロブリン軽鎖可変領域を含む。
【0024】
別の実施態様として、上記結合蛋白質は、配列番号38(3B6)の配列を含むCDRL1、配列番号39(3B6)配列を含むCDRL2及び配列番号40(3B6)を含むCDRL3を含む免疫グロブリン軽鎖可変領域を含む。
【0025】
別の実施態様として、上記結合蛋白質は、配列番号48(3D11)の配列を含むCDRL1、配列番号49(3D11)配列を含むCDRL2及び配列番号50(3D11)を含むCDRL3を含む免疫グロブリン軽鎖可変領域を含む。
【0026】
別の実施態様として、上記結合蛋白質は、配列番号58(1D3)の配列を含むCDRL1、配列番号59(1D3)配列を含むCDRL2及び配列番号60(1D3)を含むCDRL3を含む免疫グロブリン軽鎖可変領域を含む。
【0027】
別の実施態様として、上記結合蛋白質は、配列番号68(1F3)の配列を含むCDRL1、配列番号69(1F3)配列を含むCDRL2及び配列番号70(1F3)を含むCDRL3を含む免疫グロブリン軽鎖可変領域を含む。
【0028】
別の実施態様として、上記結合蛋白質は、配列番号78(3A12)の配列を含むCDRL1、配列番号79(3A12)配列を含むCDRL2及び配列番号80(3A12)を含むCDRL3を含む免疫グロブリン軽鎖可変領域を含む。
【0029】
前記の各実施態様において、CDRL1、CDRL2及びCDRL3の配列は、ヒト又はヒト化免疫グロブリンのFR間に挿入されていることが好ましい。上記結合蛋白質を完全な状態の抗体又はその抗原結合性断片或いは生合成抗体結合部位とすることができることは理解されよう。
【0030】
別の態様として、本発明はヒトHGFに結合する単離結合蛋白質を提供する。この結合蛋白質は、(a)CDRH1−CDRH2−CDRH3の構造を含む免疫グロブリン重鎖可変領域及び(b)免疫グロブリン軽鎖可変領域を含有する。この場合、これら免疫グロブリン重鎖可変領域及び免疫グロブリン軽鎖可変領域は、ヒトHGFに結合するための単一結合部位を共同して規定するものとする。上記CDRH1は、配列番号5(1A3)、配列番号15(2B8)、配列番号25(2F8)、配列番号35(3B6)、配列番号45(3D11)、配列番号55(1D3)、配列番号65(1F3)及び配列番号75(3A12)からなる群から選ばれる配列を含み、上記CDRH2は、配列番号6(1A3)、配列番号16(2B8)、配列番号26(2F8)、配列番号36(3B6)、配列番号46(3D11)、配列番号56(1D3)、配列番号66(1F3)、配列番号76(3A12)、配列番号202(Hu2B8 Hv1f.1)、配列番号203(Hu2B8 Hv5a.1又はHu2B8 Hv5−51.1)、配列番号204(LR2B8HC)及び配列番号205(LRMR2B8HC)からなる群から選ばれる配列を含み、上記CDRH3は、配列番号7(1A3)、配列番号17(2B8)、配列番号27(2F8)、配列番号37(3B6)、配列番号47(3D11)、配列番号57(1D3)、配列番号67(1F3)及び配列番号77(3A12)からなる群から選ばれる配列を含む。
【0031】
一実施態様として、上記結合蛋白質は、配列番号5(1A3)の配列を含むCDRH1、配列番号6(1A3)の配列を含むCDRH2及び配列番号7(1A3)を含むCDRH3を含む免疫グロブリン重鎖可変領域を含む。
【0032】
別の実施態様として、上記結合蛋白質は、配列番号15(2B8)の配列を含むCDRH1、配列番号16(2B8)、配列番号202(Hu2B8 Hv1f.1)、配列番号203(Hu2B8 Hv5a.1又はHu2B8 Hv5−51.1)、配列番号204(LR2B8HC)又は配列番号205(LRMR2B8HC)の配列を含むCDRH2及び配列番号17(2B8)を含むCDRH3を含む免疫グロブリン重鎖可変領域を含む。
【0033】
別の実施態様として、上記結合蛋白質は、配列番号25(2F8)の配列を含むCDRH1、配列番号26(2F8)の配列を含むCDRH2及び配列番号27(2F8)を含むCDRH3を含む免疫グロブリン重鎖可変領域を含む。
【0034】
別の実施態様として、上記結合蛋白質は、配列番号35(3B6)の配列を含むCDRH1、配列番号36(3B6)配列を含むCDRH2及び配列番号37(3B6)を含むCDRH3を含む免疫グロブリン重鎖可変領域を含む。
【0035】
別の実施態様として、上記結合蛋白質は、配列番号45(3D11)の配列を含むCDRH1、配列番号46(3D11)配列を含むCDRH2及び配列番号47(3D11)を含むCDRH3を含む免疫グロブリン重鎖可変領域を含む。
【0036】
別の実施態様として、上記結合蛋白質は、配列番号55(1D3)の配列を含むCDRH1、配列番号56(1D3)配列を含むCDRH2及び配列番号57(1D3)を含むCDRH3を含む免疫グロブリン重鎖可変領域を含む。
【0037】
別の実施態様として、上記結合蛋白質は、配列番号65(1F3)の配列を含むCDRH1、配列番号66(1F3)配列を含むCDRH2及び配列番号67(1F3)を含むCDRH3を含む免疫グロブリン重鎖可変領域を含む。
【0038】
別の実施態様として、上記結合蛋白質は、配列番号75(3A12)の配列を含むCDRH1、配列番号76(3A12)配列を含むCDRH2及び配列番号77(3A12)を含むCDRH3を含む免疫グロブリン重鎖可変領域を含む。
【0039】
前記の各実施態様において、CDRH1、CDRH2及びCDRH3の配列は、ヒト又はヒト化免疫グロブリンのFR間に挿入されていることが好ましい。上記結合蛋白質を完全な状態の抗体又はその抗原結合性断片或いは生合成抗体結合部位とすることができることは理解されよう。
【0040】
別の態様として、本発明はヒトHGFに結合する結合蛋白質を提供する。この結合蛋白質は、配列番号2(1A3)の20乃至141番目残基、配列番号12(2B8)の20乃至137番目残基、配列番号22(2F8)の20乃至137番目残基、配列番号32(3B6)の20乃至139番目残基、配列番号42(3D11)の20乃至132番目残基、配列番号52(1D3)の20乃至141番目残基、配列番号62(1F3)の20乃至141番目残基及び配列番号72(3A12)の20乃至141番目残基からなる群から選ばれる免疫グロブリン重鎖可変領域、並びに配列番号4(1A3)の21乃至127番目残基、配列番号14(2B8)の21乃至127番目残基、配列番号24(2F8)の20乃至131番目残基、配列番号34(3B6)の23乃至129番目残基、配列番号44(3D11)の23乃至128番目残基、配列番号54(1D3)の21乃至127番目残基、配列番号64(1F3)の21乃至127番目残基及び配列番号74(3A12)の21乃至127番目残基からなる群から選ばれる免疫グロブリン軽鎖可変領域を含む。
【0041】
別の実施態様として、上記結合蛋白質は、配列番号2(1A3)の20乃至141番目残基のアミノ酸配列を含む免疫グロブリン重鎖可変領域及び配列番号4(1A3)の21乃至127番目残基のアミノ酸配列を含む免疫グロブリン軽鎖可変領域を含む。
【0042】
一実施態様として、上記結合蛋白質は、配列番号12(2B8)の20乃至137番目残基のアミノ酸配列を含む免疫グロブリン重鎖可変領域及び配列番号14(2B8)の21乃至127番目残基のアミノ酸配列を含む免疫グロブリン軽鎖可変領域を含む。
【0043】
別の実施態様として、上記結合蛋白質は、配列番号22(2F8)の20乃至137番目残基のアミノ酸配列を含む免疫グロブリン重鎖可変領域及び配列番号24(2F8)の20乃至131番目残基のアミノ酸配列を含む免疫グロブリン軽鎖可変領域を含む。
【0044】
別の実施態様として、上記結合蛋白質は、配列番号32(3B6)の20乃至139番目残基のアミノ酸配列を含む免疫グロブリン重鎖可変領域及び配列番号34(3B6)の23乃至129番目残基のアミノ酸配列を含む免疫グロブリン軽鎖可変領域を含む。
【0045】
別の実施態様として、上記結合蛋白質は、配列番号42(3D11)の20乃至132番目残基のアミノ酸配列を含む免疫グロブリン重鎖可変領域及び配列番号44(3D11)の23乃至128番目残基のアミノ酸配列を含む免疫グロブリン軽鎖可変領域を含む。
【0046】
別の実施態様として、上記結合蛋白質は、配列番号52(1D3)の20乃至141番目残基のアミノ酸配列を含む免疫グロブリン重鎖可変領域及び配列番号54(1D3)の21乃至127番目残基のアミノ酸配列を含む免疫グロブリン軽鎖可変領域を含む。
【0047】
別の実施態様として、上記結合蛋白質は、配列番号62(1F3)の20乃至141番目残基のアミノ酸配列を含む免疫グロブリン重鎖可変領域及び配列番号64(1F3)の21乃至127番目残基のアミノ酸配列を含む免疫グロブリン軽鎖可変領域を含む。
【0048】
別の実施態様として、上記結合蛋白質は、配列番号72(3A12)の20乃至141番目残基のアミノ酸配列を含む免疫グロブリン重鎖可変領域及び配列番号74(3A12)の21乃至127番目の残基アミノ酸配列を含む免疫グロブリン軽鎖可変領域を含む。
【0049】
前記の各実施態様において、結合蛋白質は完全な状態の抗体又はその抗原結合性断片或いは生合成抗体結合部位とすることができる。
【0050】
別の態様として、本発明はヒトHGFに結合する単離結合蛋白質を提供する。この結合蛋白質は、(i)配列番号173(Hu2B8 Kv1−39.1軽鎖可変領域)、配列番号179(Hu2B8 Kv3−15.1軽鎖可変領域)、配列番号193(LR2B8LC軽鎖可変領域)及び配列番号199(LRMR2B8LC軽鎖可変領域)からなる群から選ばれる免疫グロブリン軽鎖可変領域、並びに(ii)配列番号159(Hu2B8 Hv1f.1重鎖可変領域)、配列番号165(Hu2B8 Hv5a.1重鎖可変領域)、配列番号169(Hu2B8 Hv5−51.1重鎖可変領域)、配列番号183(LR2B8HC重鎖可変領域)及び配列番号189(LRMR2B8LC軽鎖可変領域)からなる群から選ばれる免疫グロブリン重鎖可変領域を含有する。上記結合蛋白質は、完全な状態の抗体又はその抗原結合性断片或いは生合成抗体結合部位とすることができる。
【0051】
別の態様として、本発明はヒトHGFに結合する単離結合蛋白質を提供する。この結合蛋白質は、(i)配列番号177(Hu2B8 Kv1−39.1+カッパ定常(Km(3)アロタイプ(対立遺伝子2))、配列番号181(Hu2B8 Kv3−15.1+カッパ定常(Km(3)アロタイプ(対立遺伝子2))、配列番号197(LR2B8LC+カッパ定常(Km(3)アロタイプ(対立遺伝子1))及び配列番号201(LRMR2B8LC+カッパ定常(Km(3)アロタイプ(対立遺伝子1))からなる群から選ばれる免疫グロブリン軽鎖、並びに(ii)配列番号163(Hu2B8 Hv1f.1+IgG1定常(G1m(17,1)アロタイプ))、配列番号167(Hu2B8 Hv5a.1+IgG1定常領域(G1m(17,1)アロタイプ))、配列番号171(Hu2B8 Hv5−51.1+IgG1定常(G1m(17,1)アロタイプ))、配列番号187(LR2B8HC+IgG1定常(G1m(3)アロタイプ)(対立遺伝子1))及び配列番号191(LRMR2B8HC+IgG1定常(G1m(3)アロタイプ)(対立遺伝子1))からなる群から選ばれる免疫グロブリン重鎖を含む。上記結合蛋白質は、完全な状態の抗体又はその抗原結合性断片或いは生合成抗体結合部位とすることができる。
【0052】
別の態様として、本発明は還元型ヒトHGFに結合する単離結合蛋白質を提供する。この結合蛋白質は、(i)3箇所のCDRを含む免疫グロブリン軽鎖可変領域及び(ii)3箇所のCDRを含む免疫グロブリン重鎖可変領域を含む。これらのCDRは、通常FR間に挿入される。免疫グロブリン軽鎖及び免疫グロブリン重鎖のCDRは、還元型ヒトHGF、例えば、還元型HGFのα鎖に結合する結合部位を共同して規定する。還元型HGFとは、α鎖とβ鎖との間のジスルフィド結合を還元するのに十分な量の還元剤、例えば、ジチオスレイトール(DTT:dithiothreitol)、2−メルカプトエタノール又はグルタチオンで処理したHGFのことをいう。典型的な濃度としては、例えば、100mMのDTT及び5%の2−メルカプトエタノールが挙げられる。
【0053】
一部の実施態様として、上記結合蛋白質は、CDRL1、CDRL2及びCDRL3からなる群から選ばれる少なくとも1つのCDRを含む免疫グロブリン軽鎖可変領域を含む。上記結合蛋白質は、任意選択的に、2つのCDR、例えば、CDRL1とCDRL2又はCDRL1とCDRL3又はCDRL1とCDRL3を含む。上記結合蛋白質は、任意選択的に、3つのCDRの全て、即ち、CDRL1、CDRL2及びCDRL3を含む。CDRL1は、アミノ酸配列X1X2SerX4X5X6X7X8X9X10X11X12X13X14X15であって、アミノ酸X1がArg又はLysであり、X2がAla又はThrであり、X4がGlu又はGlnであり、X5がAsn、Ser又はAspであり、X6がIle又はValであり、X7がTyr、Asp又はLysであり、X8がペプチド結合又はTyrであり、X9がペプチド結合又はAspであり、X10がペプチド結合又はGlyであり、X11がペプチド結合又はAsnであり、X12がペプチド結合又はSerであり、X13がAsn又はTyrであり、X14がIle又はLeuであり、X15がAla、Asn又はSerであるものとする配列を含む。CDRL2は、アミノ酸配列X16X17X18X19LeuX21X22であって、アミノ酸X16がAla、Asp、Val又はArgであり、X17がAla又はValであり、X18がAsn、Ser又はThrであり、X19がArg、Asn又はHisであり、X21がAla、Glu、Val又はProであり、X22がAsp又はSerであるものとする配列を含む。CDRL3は、アミノ酸配列X23X24X25X26X27X28ProX30Thrであって、アミノ酸X23がLeu又はGlnであり、X24がHis又はGlnであり、X25がPhe、Ser又はTyrであり、X26がAsp、Ile又はTrpであり、X27がGly又はGluであり、X28がAsp、Phe又はThrであり、X30がPhe、Pro又はTyrであるものとする配列を含む。
【0054】
別の実施態様として、上記結合蛋白質は、CDRH1、CDRH2及びCDRH3からなる群から選ばれる少なくとも1つのCDRを含む免疫グロブリン重鎖可変領域を含む。上記結合蛋白質は、任意選択的に、2つのCDR、例えば、CDRH1とCDRH2又はCDRH1とCDRH3又はCDRH1とCDRH3を含む。上記結合蛋白質は、任意選択的に、3つのCDRの全て、即ち、CDRH1、CDRH2及びCDRH3を含む。CDRH1は、アミノ酸配列X1TyrX3X4X5であって、アミノ酸X1がAsp、Asn、Ser又はThrであり、X3がPhe、Trp又はTyrであり、X4がIle又はMetであり、X5がAsn、His又はSerであるものとする配列を含む。CDRH2は、アミノ酸配列X6IleX8X9GlyX11GlyX13X14X15TyrX17X18X19X20LysX22であって、アミノ酸X6がLys、Gln又はTyrであり、X8がGly、Ser又はTyrであり、X9がPro又はSerであり、X11がAsp、Gly又はSerであり、X13がAsp又はSerであり、X14がSer又はThrであり、X15がAsn又はTyrであり、X17がAsn又はProであり、X18がAla、Asp、Gly又はGluであり、X19がAsn、Met又はSerであり、X20がPhe又はValであり、X22がAsp又はGlyであるものとする配列を含む。CDRH3は、アミノ酸配列X23X24X25X26X27X28X29X30X31X32X33AspTyrであって、アミノ酸X23がArg又はGlnであり、X24がGly又はLeuであり、X25がAsp、Gly又はペプチド結合であり、X26がGly又はペプチド結合であり、X27がペプチド結合又はTyrであり、X28がLeu、ペプチド結合又はTyrであり、X29がGly、Arg又はLeuであり、X30がAsp、Gly又はGluであり、X31がTyr、Arg又はAsnであり、X32がAla、Gly又はTyrであり、X33がMet又はPheであるものとする配列を含む。
【0055】
上記結合蛋白質が上述の免疫グロブリン重鎖及び免疫グロブリン軽鎖の配列又はこれらの断片を含有し得ることは理解されよう。さらに、上記結合蛋白質は完全な状態の抗体又はその抗原結合性断片或いは生合成抗体結合部位とすることができることも理解されよう。
【0056】
一部の実施態様として、上記結合蛋白質は、(i)配列番号8(1A3)、配列番号28(2F8)、配列番号38(3B6)、配列番号58(1D3)及び配列番号68(1F3)からなる群から選ばれる配列を有するCDRL1、(ii)配列番号9(1A3)、配列番号29(2F8)、配列番号39(3B6)、配列番号59(1D3)及び配列番号69(1F3)からなる群から選ばれる配列を有するCDRL2並びに(iii)配列番号10(1A3)、配列番号30(2F8)、配列番号40(3B6)、配列番号60(1D3)及び配列番号70(1F3)からなる群から選ばれる配列を有するCDRL3を含む免疫グロブリン軽鎖可変領域を含む。これらのCDR配列はヒト又はヒト化FR間に挿入されていてもよい。他の実施態様として、上記結合蛋白質は、配列番号4(1A3)の21乃至127番目残基、配列番号24(2F8)の20乃至131番目残基、配列番号34(3B6)の23乃至129番目残基、配列番号54(1D3)の21乃至127番目残基及び配列番号64(1F3)の21乃至127番目残基からなる群から選ばれるアミノ酸配列を含む免疫グロブリン軽鎖可変領域を含む。
【0057】
一部の実施態様として、上記結合蛋白質は、(i)配列番号5(1A3)、配列番号25(2F8)、配列番号35(3B6)、配列番号55(1D3)及び配列番号65(1F3)からなる群から選ばれる配列を有するCDRH1、(ii)配列番号6(1A3)、配列番号26(2F8)、配列番号36(3B6)、配列番号56(1D3)及び配列番号66(1F3)からなる群から選ばれる配列を有するCDRH2並びに(iii)配列番号7(1A3)、配列番号27(2F8)、配列番号37(3B6)、配列番号57(1D3)及び配列番号67(1F3)からなる群から選ばれる配列を有するCDRH3を含む免疫グロブリン重鎖可変領域を含む。これらのCDR配列はヒト又はヒト化FR間に挿入されていてもよい。別の実施態様として、この免疫グロブリン重鎖可変領域は、配列番号2(1A3)の20乃至141番目残基、配列番号22(2F8)の20乃至137番目残基、配列番号32(3B6)の20乃至139番目残基、配列番号52(1D3)の20乃至141番目残基及び配列番号62(1F3)の20乃至141番目残基からなる群から選ばれるアミノ酸配列を含有する。
【0058】
別の態様として、本発明は、ヒトHGFに結合し、免疫グロブリン軽鎖可変領域及び免疫グロブリン重鎖可変領域を含む単離結合蛋白質を提供する。この単離結合蛋白質は、(i)配列番号24(2F8)の20乃至131番目残基の免疫グロブリン軽鎖可変領域及び配列番号22(2F8)の20乃至137番目残基の免疫グロブリン重鎖可変領域を有する抗体、(ii)配列番号34(3B6)の23乃至129番目残基の免疫グロブリン軽鎖可変領域及び配列番号32(3B6)の20乃至139番目残基の免疫グロブリン重鎖可変領域を有する抗体、並びに(iii)配列番号44(3D11)の23乃至128番目残基の免疫グロブリン軽鎖可変領域及び配列番号42(3D11)の20乃至132番目残基の免疫グロブリン重鎖可変領域を有する抗体からなる群から選ばれる少なくとも1種の対照抗体と、HGFへの結合に対して競合する。ある状況では、この結合蛋白質はこれらの対照抗体のうちの1種と同じHGFの抗原決定基に結合する。
【0059】
上述の各結合蛋白質が完全な状態の抗体、例えばモノクローナル抗体であってもよいことは理解されよう。或いは、上記結合蛋白質は、抗体の抗原結合性断片であってもよく、又は生合成抗体結合部位であってもよい。抗体断片としては、Fab、Fab’、(Fab’)2又はFv断片が挙げられる。このような抗体断片を作製するための技術は当業者に周知である。多くの生合成抗体結合部位が当該技術分野で周知であり、例えば、米国特許第5,476,786号明細書に記載されている単一Fv又はsFv分子が挙げられる。他の生合成抗体結合部位としては、二重特異性又は二機能性結合蛋白質、例えば、二重特異性又は二機能性抗体が挙げられ、これらは少なくとも2種の異なる抗原に結合する抗体又は抗体断片である。例えば、二重特異性結合蛋白質はHGF、例えばヒトHGFと別の対象とする抗原とに結合することができる。二重特異性抗体を作製するための方法は当該技術分野で周知であり、例えば、ハイブリドーマを融合させる方法又はFab’断片を連結させる方法が挙げられる。例えば、Songsivilai et al.(1990年) CLIN. Exp. IMMUNOL. 79:p.315−325及びKostelny et al.(1992年) J. IMMUNOL. 148:p.1547−1553を参照されたい。
【0060】
本発明の結合蛋白質は、561番目の位置にシステインからアルギニンへの置換及び555番目の位置にグリシンからグルタメートへの置換を含むhHGFに結合することができる。
【0061】
別の態様として、本発明は、4.0×10−5s−1以下、3.0×10−5s−1以下又は2.0×10−5s−1以下のKdでヒトHGFに結合する単離結合蛋白質を提供する。これらの単離結合蛋白質は、5.0×10−5s−1乃至0.5×10−5s−1又は4.0×10−5s−1乃至1.0×10−5s−1又は3.0×10−5s−1乃至1.5×10−5s−1のKdでヒトHGFに結合することができる。別の態様として、本発明は、100pM以下又は20pM以下又は10pM以下又は5pM以下のKDでヒトHGFに結合する単離結合蛋白質を提供する。これらの単離結合蛋白質は、100pM乃至5pM又は20pM乃至5pM又は15pM乃至10pM又は20pM乃至10pM乃至15pM乃至5pMのKDでヒトHGFに結合することができる。特に明記しない限り、KD値は実施例6に記載した方法及び条件下で測定する。
【0062】
別の態様として、本発明は、ヒトHGFに結合する単離結合蛋白質であって、この抗体が25℃よりも37℃でより低いKdでヒトHGFに結合するものとする結合蛋白質を提供する。この結合蛋白質結合は、任意選択的に、37℃において5pM未満のKdでヒトHGFに結合する。
【0063】
他の態様及び実施態様として、上記結合蛋白質は、hHGFがc−Metに結合するのを阻害することができる。例えば、上記結合蛋白質のIC50(最大阻害の50%を来す濃度)は、実施例7(a)に記載したプロトコルを用いて測定した場合、少なくとも約4.0、4.5、5.0、5.5、6.0、6.5及び7.0nMとすることができる。一部の他の実施態様として、上記結合蛋白質は、実施例7(b)に記載した方法を用いて4MBr−5細胞(ATCC、カタログ番号CCL208)におけるHGF BrdUの取り込みを中和することができる。
【0064】
上記結合蛋白質のIC50は、実施例7(b)に記載したプロトコルを用いて測定した場合、50nM以下、好ましくは、45、40、35、30、25、20、15、10、5、1又は0.5nM以下である。一部の他の実施態様として、上記結合蛋白質を用いて、実施例9に記載した測定法によるPC−3細胞(ATCC、マナサス、バージニア州、カタログ番号CCL1435)のHGF刺激c−Metリン酸化を抑制することができる。上記結合蛋白質は、実施例9に記載した測定法によるPC−3細胞におけるHGF刺激(1.25nM)c−Metリン酸化を2nM以下のIC50(表8)で抑制する。
【0065】
II.結合蛋白質の作製
本発明の結合蛋白質は、当該技術分野で周知の手法を用いる種々の方法で作製することができる。例えば、軽鎖可変領域及び重鎖可変領域をコードしているDNA分子は、市販の合成機及び本明細書に示した配列情報を用いて、化学的に合成することができる。このような合成DNA分子を、例えば、定常領域コード配列、発現制御配列などの他の適切なヌクレオチド配列に結合させて所望の結合蛋白質をコードしている通常の遺伝子発現構築物を作製することができる。特定の遺伝子構築物の作製は当該技術分野では日常的な技術の範囲内にある。或いは、本明細書に示した配列は、本明細書に示した配列情報、又はハイブリドーマ細胞中のマウス抗体の重及び軽鎖をコードしている遺伝子に関する従来技術の配列情報に基づいた配列を有する合成核酸プローブを用いて、通常のハイブリダイゼーション技術又はPCR技術によりハイブリドーマからクローニングすることができる。このようなプローブの作製及び使用法は、当該技術分野では日常的な技術の範囲内にある。
【0066】
この所望の結合蛋白質をコードしている核酸を発現ベクター中に導入(結合)して、これを、当該技術分野で周知の標準的な形質移入又は形質転換技術により宿主細胞中に導入することができる。典型的な宿主細胞としては、例えば、大腸菌細胞、チャイニーズハムスター卵巣(CHO:Chinese hamster ovary)細胞、ヒーラ(HeLa)細胞、仔ハムスター腎(BHK:baby hamster kidney)細胞、サル腎細胞(COS)、ヒト肝細胞癌細胞(例えば、Hep G2)及び普通なら免疫グロブリンを産生しない骨髄腫細胞が挙げられる。形質移入させた宿主細胞は、この宿主細胞が対象とする遺伝子、例えば免疫グロブリン軽又は重鎖可変領域をコードしている遺伝子を発現するのを可能にする条件下で増殖させることができる。得られる発現産物は当該技術分野で周知の技術を用いて採取することができる。
【0067】
特定の発現及び精製条件は、どんな発現系を用いるかによって異なるであろう。例えば、上記遺伝子を大腸菌で発現させる場合にはこれを先ず発現ベクター中にクローニングする。これは、好適な細菌プロモーター(例えばTrp又はTac)及びシグナル配列(例えばプロテインAのフラグメントB(FB)をコードしている配列)よりも下流に上記の設計遺伝子を配置することによって達成される。得られる発現融合蛋白質は、通常、細胞細胞質中の屈折体又は封入体に蓄積するので、フレンチプレス又は音波処理により細胞を破壊して採取することができる。次に、この屈折体を可溶化し、多くの他の組換え蛋白質について既に確立されている方法により発現蛋白質をリフォールディングし、切断する。
【0068】
上記設計遺伝子を真核宿主細胞、例えば骨髄腫細胞又はCHO細胞で発現させる場合には、先ず、これを、好適な真核生物プロモータ、分泌シグナル、免疫グロブリンエンハンサ及び各種イントロンを含有する発現ベクター中に挿入する。この発現ベクターには、任意選択的に定常領域の全て又は一部をコードしている配列を含有させることができ、これによって重又は軽鎖の全体又は一部の発現が可能となる。上記遺伝子構築物は、確立されている形質移入プロトコルを用いて骨髄腫細胞又はCHO細胞中に形質移入することができる。このような形質移入細胞は、VL又はVH断片、VL−VHヘテロ二量体、VH−VL又はVL−VH一本鎖ポリペプチド、完全な免疫グロブリン重又は軽鎖、或いはこれらの部分を発現することができ、これらは、それぞれ別の機能(例えば、細胞毒性)を有する蛋白質ドメインに結合させてもよい。
【0069】
III. 結合蛋白質に対する修飾
上記結合蛋白質を修飾してこの結合蛋白質の目的とする用途に応じて性能を最適化することができることは理解されよう。例えば、結合蛋白質を治療剤として用いようとする場合、この結合蛋白質を修飾して対象レシピエントにおける免疫原性を低減することができる。或いは、又はさらに、結合蛋白質を別の蛋白質又はペプチド、例えば、成長因子、サイトカインもしくは細胞毒と融合又は結合させることができる。このような修飾は、当該技術分野で周知の通常の遺伝子操作技術を用いることで達成することができる。
【0070】
抗体及び抗体断片の抗原性を低減させるための種々の技術が当該技術分野において周知である。これらの技術を用いて本発明の結合蛋白質の抗原性を低減又は除去することができる。例えば、上記結合蛋白質をヒトに投与しようとする場合、この結合蛋白質を改良してヒトにおけるその抗原性を低減させることが好ましい。このプロセスをヒト化と呼ぶことが多いが、このヒト化結合蛋白質は、それが由来する元の非ヒト化結合蛋白質と抗原に対する親和性が同じか実質的に同じであることが好ましい。
【0071】
ヒト化のための公知の一方法では、ある種、例えばマウスからの抗体の免疫グロブリン定常領域を別の異なる種、例えばヒトからの免疫グロブリン定常領域で置換したキメラ蛋白質を作製する。この例では、得られる抗体はマウス−ヒトキメラであり、このキメラのヒト定常領域の配列は、原理的に、対応するマウスの配列よりも免疫原性が少ない。この種の抗体改良については、例えば、Morrison,et al.(1984年) PROC.NAT.ACAD.SCI.81:p.6851−6855、Neuberger et al.(1984年)NATURE 312:p.604−608、並びに米国特許第6,893,625号明細書(Robinson)、同第5,500,362号明細書(Robinson)及び同第4,816,567号明細書(Cabilly)に記載されている。
【0072】
CDR移植と呼ばれる別の方法では、対象とする抗体の軽及び重鎖可変領域のCDRを別の種からのフレームワーク(FR)中に移植する。例えば、マウスCDRをヒトFR配列中に移植することができる。一部の実施態様として、抗HGF抗体の軽及び重鎖可変領域のCDRをヒトFR又はコンセンサスヒトFR中に移植する。コンセンサスヒトFRを作製するためには、いく種かのヒト重鎖又は軽鎖アミノ酸配列からのFRを整列させてコンセンサスアミノ酸配列を特定する。CDR移植については、例えば、米国特許第7,022,500号明細書(Queen)、同第6,982,321号明細書(Winter)、同第6,180,370号明細書(Queen)、同第6,054,297号明細書(Carter)、同第5,693,762号明細書(Queen)、同第5,859,205号明細書(Adair)、同第5,693,761号明細書(Queen)、同第5,565,332号明細書(Hoogenboom)、同第5,585,089号明細書(Queen)及び同第5,530,101号明細書(Queen)、Jones et al.(1986年)NATURE 321:p.522−525、Riechmann et al.(1988年)NATURE 332:p.323−327、Verhoeyen et al.(1988年)SCIENCE 239:p.1534−1536並びにWinter(1998年)FEBS LETT 430:p.92−94に記載されている。
【0073】
「超ヒト化」と呼ばれる方法では、ヒトにおける免疫原性が低減又は除去されている抗体を別のタイプの移植によって作製する。超ヒト化法では、ヒトCDRのヒト化対象マウス抗体のCDRに対する構造的類似性に基づいて一組のヒト生殖細胞遺伝子からヒトFR配列を選択する。この方法については、例えば、米国特許第6,881,557号明細書(Foote)及びTan et al.(2002年)J.IMMUNOL 169:p.1119−1125に記載されている。
【0074】
免疫原性を低減する他の方法としては、ヒト化抗体を作製するための「再成形(reshaping)」、「超キメラ化」又は「ベニア化(veneering)/再表面化(resurfacing)」と呼ばれる技術が挙げられる。例えば、Vaswami et al. (1998年) ANNALS OF ALLERGY, ASTHMA, & IMMUNOL. 81:p.105、Roguska et al.(1996年) PROT. ENGINEER 9:p.895−904及び米国特許第6,072,035号明細書(Hardman)を参照されたい。ベニア化/再表面化法では、マウス抗体の表面接近可能アミノ酸残基をヒト抗体の同一位置により多くみられるアミノ酸残基によって置換する。このタイプの抗体再表面化については、例えば、米国特許第5,639,641号明細書(Pedersen)に記載されている。
【0075】
マウス抗体をヒトにおける医学的用途に好適な構造に変換するための1つの典型的な方法は、ACTIVMAB(商標)技術(Vaccinex社、ロチェスター、ニューヨーク州)と呼ばれ、これは哺乳動物細胞で抗体を発現させるためのワクシニアウイルス系ベクターを必要とする。免疫グロブリン重及び軽鎖の高レベルの組み合わせ多様性がもたらされると言われている。例えば、米国特許第6,706,477号明細書(Zauderer)、同第6,800,442号明細書(Zauderer)及び同第6,872,518号明細書(Zauderer)を参照されたい。
【0076】
マウス抗体をヒトにおける用途に好適な構造に変換するための別の典型的な方法は、カロビオス・ファーマシューティカルズ社(KaloBios Pharmaceuticals, Inc.)(パロアルト、カリフォルニア州)により商業的に実施されている技術である。この技術は、特許権のあるヒト「アクセプター」ライブラリを用いて抗体選択用の「抗原決定基に焦点を合わせた」ライブラリを作製するものである。
【0077】
マウス抗体をヒトにおける医学的用途に好適な構造に修飾するための別の典型的な方法は、XOMA(米国)LLC社によって商業的に実施されているHUMAN ENGINEERING)(商標)(HE(商標))技術である。例えば、国際公開第93/11794号パンフレット並びに米国特許第5,766,886号明細書、同第5,770,196号明細書、同第5,821,123号明細書及び同第5,869,619号明細書を参照されたい。
【0078】
上記方法のいずれかを含む任意の適切な方法を用いて対象とする結合蛋白質のヒト免疫原性を低減又は除去することができる。
【0079】
さらに、マウスにおいて完全にヒト型の抗体を作製することが可能である。この方法では、マウスの抗体産生遺伝子がヒト抗体産生遺伝子の実質的な部分で置換されている遺伝子導入マウスを用いてヒト型抗体を調製する。このようなマウスは、マウスの免疫グロブリン分子ではなく、ヒト型の免疫グロブリンを産生する。例えば、国際公開第98/24893号パンフレット(Jacobovitz et al.)及びMendez et al. (1997年) NATURE GENETICS 15:p.146−156を参照されたい。完全にヒト型の抗HGFモノクローナル抗体は、以下の方法を用いて作製することができる。ヒト型免疫グロブリン遺伝子を有する遺伝子導入マウスを対象とする抗原、例えばHGFで免疫する。次いで、このマウスからマウスのリンパ細胞を得、これを脊髄系細胞株と融合させて不死化ハイブリドーマ細胞株を調製する。このハイブリドーマ細胞株をスクリーニングし、HGFに特異的な抗体を産生するハイブリドーマ細胞株を特定するための選択を行う。
【0080】
本発明の結合蛋白質は、その目的とする用途に応じて他の分子と結合させることができる。例えば、結合蛋白質を治療剤として用いようとする場合、この結合蛋白質を別の作用物質、例えば、その治療を調節するか、それとも促進するエフェクター分子と結合させることができる。このエフェクターが非蛋白質系の作用物質、例えば、低分子薬剤、放射標識又は毒素である限り、この物質を標準的なインビトロ結合化学反応を用いて結合蛋白質に化学的に結合させることができる。一方、エフェクター分子が蛋白質又はペプチド、例えば、酵素、受容体、毒素、成長因子、サイトカイン又は他の免疫調節因子である場合、結合蛋白質は、インビトロ結合化学反応を用いてこのエフェクターに化学的に結合させることができ、或いは、融合蛋白質としてこのエフェクターに結合させることができる。融合蛋白質は、セクションIIに記載したのと同様な技術を用いて構築し、発現させることができる。
【0081】
IV. 結合蛋白質の用途
本明細書に記載した結合蛋白質は、診断剤又は治療剤として用いることができる。
【0082】
(1)治療的用途
本発明の結合蛋白質は、HGFの活性を中和するので、種々の治療的用途に用いることができる。例えば、本発明の結合蛋白質の一部は、過剰増殖性疾病又は疾患、例えば、各種形態の癌の予防又は治療に有用である。
【0083】
上記結合蛋白質を用いることにより腫瘍細胞の増殖を抑制又は低減させることができる。このような方法では、腫瘍細胞に対して治療的有効量の上記結合蛋白質を作用させることによって腫瘍細胞の増殖を抑制又は低減させる。一部の実施態様として、上記結合蛋白質は、腫瘍細胞の増殖を少なくとも50%、60%、70%、80%、90%、95%又は100%抑制する。
【0084】
一部の実施態様として、上記結合蛋白質を用いることにより、結合蛋白質がhHGFのc−Metへの結合能を低減させることで腫瘍細胞の増殖を抑制又は低減させる。他の実施態様として、上記結合蛋白質を用いることにより、表5及び6において抗体3B6が示すように、この結合蛋白質がhHGFに結合するが、hHGFのc−Metへの結合を実質的に抑制しない場合にも腫瘍細胞の増殖は抑制され、又は低下する。
【0085】
さらに、上記結合蛋白質を用いることにより、哺乳動物における腫瘍の成長又は発症を抑制又は遅延させることができる。このような方法では、有効量の上記結合蛋白質を哺乳動物に投与することによってこの哺乳動物における腫瘍の成長を抑制又は遅延させる。従って、上記結合蛋白質を用いることによって、例えば、哺乳動物において腫瘍を治療することができる。この方法は哺乳動物に治療的有効量の上記結合蛋白質を投与するものである。上記結合蛋白質を単独又は別の医薬として有効な分子と併用して投与することにより腫瘍を治療することができる。
【0086】
本発明の結合蛋白質は、種々のHGF反応性疾患、例えば、肺癌、乳癌、大腸癌、前立腺癌、卵巣癌、頭頸部癌、卵巣癌、多発性骨髄腫、肝臓癌、胃癌、食道癌、腎癌、鼻咽腔癌、膵癌、中皮腫、黒色腫及びグリア芽細胞腫におけるHGF反応性腫瘍細胞などの治療に用いることができる。
【0087】
本明細書でいう「治療する」、「治療すること」及び「治療」とは、哺乳動物、特にヒトにおける病態の治療のことを指し、(a)その病態が哺乳動物において、特に、その哺乳動物がその病態に罹患する素因を有するが、まだこれを有していないと診断されている場合に発生するのを予防すること、(b)その病態を抑制する、即ち、その発症を阻止すること、及び/又は(c)その病態を緩和すること、即ち、その病態を軽減させること含む。
【0088】
一般に、活性成分の治療的有効量は、約0.1mg/kg乃至約100mg/kg、必要に応じて約1mg/kg乃至約100mg/kg、必要に応じて約1mg/kg乃至約10mg/kgの範囲となる。投与の量は、治療対象の疾患又は徴候の種類及び程度、特定の患者の全般的な健康状態、送達する結合蛋白質の相対的生物学的有効性、この結合蛋白質の剤型、製剤中の添加物の存在及び種類並びに投与経路などの変数によって決まることになる。所望の血液濃度又は組織濃度に急速に到達させるために、初期の投与用量を上記の上限を超えて増加させることができ、又は初期用量を最適用量より少なくすることができ、さらに、特定の状況に応じて治療の過程で初期用量を段階的に増加させることができる。ヒトにおける用量は、例えば、0.5mg/kg乃至20mg/kgで実施するようにデザインされた通常の第I相用量増加試験において最適化することができる。投与頻度については、投与経路、投与量、治療対象の疾患状態などの要因によって変更することができる。典型的な投与頻度は、1日当たり1回、1週当たり1回、2週ごとに1回である。好ましい投与経路は全身性投与、例えば、静脈内注入である。モノクローナル抗体系薬剤の製剤化については当該技術分野では日常的な技術の範囲内にある。本発明の一部の実施態様として、上記結合蛋白質、例えばモノクローナル抗体は、凍結乾燥され、投与時に緩衝生理食塩水で元に戻される。
【0089】
上記結合蛋白質は、単独又は他の医薬として有効な成分との併用で投与することができる。上記他の有効な成分、例えば、免疫調節剤はこの結合蛋白質と一緒に投与することができ、或いはこの結合蛋白質投与の前又は後で投与することができる。
【0090】
治療的用途の上記結合蛋白質含有製剤としては、典型的には、上記結合蛋白質を医薬用として許容可能な担体と組み合わせたものが挙げられる。本明細書でいう「医薬用として許容可能な担体」とは、正しい医学的判断の範囲内で、妥当な利益/危険度比に見合って、過度の毒性、刺激、アレルギー応答又は問題もしくは合併症を伴うことなくヒトその他の動物の組織と接触させて用いるのに適している緩衝剤、担体及び賦形剤を意味する。この担体は、製剤のその他の成分と適合し、レシピエントに対して有害でないという意味で「許容可能」でなければならない。なお、医薬用として許容可能な担体は、医薬用としての投与に適合するありとあらゆる緩衝剤、溶媒、分散媒、被覆剤、等張性吸収遅延剤などを含むものとする。医薬として有効な物質のためのこのような媒体及び剤については当該技術分野では周知である。
【0091】
こうした製剤は、投与単位形態で適宜提示することができ、薬学分野で公知の方法のいずれかを含め、任意の適切な方法によって調製することができる。本発明の医薬組成物は、対象とする投与経路に適合するように製剤化する必要がある。投与経路としては、例えば、全身性投与又は非全身性投与、例えば、静脈内、皮内、吸入、経皮(局所)、経粘膜及び直腸投与が挙げられる。経口又は全身性投与に有用な液剤は、例えば、「Remington’s Pharmaceutical Sciences」、第18版(マック出版(Mack Publishing Company)1990年)に記載されている、医薬分野で公知の方法のいずれかによって調製することができる。
【0092】
経口投与に適した製剤は、所定量の上記結合蛋白質を含有する、注射液、カプセル剤、ゼラチンカプセル剤、サッシェ剤、錠剤、トローチ剤又は舐剤などの個々の単位体;粉末状又は顆粒状組成物;水性液体又は非水性液体の液剤又は懸濁剤;或いは水中油型乳剤又は油中水型乳剤の形をとることができる。
【0093】
全身性投与に適した製剤は、例えば、以下の成分、即ち、注射用水などの滅菌希釈液、食塩水、固定油類、ポリエチレングリコール類、グリセリン、プロピレングリコール又は他の合成溶媒;ベンジルアルコール又はメチルパラベンなどの抗菌剤;アスコルビン酸又は亜硫酸水素ナトリウムなどの抗酸化剤;エチレンジアミン四酢酸などのキレート化剤;酢酸塩、クエン酸塩又はリン酸塩などの緩衝剤及び塩化ナトリウム又はデキストロースなどの等張性調整用剤を含む。pHは塩酸又は水酸化ナトリウムなどの酸又は塩基で調整することができる。全身性投与用製剤は、アンプル、使い捨て注射器、或いはガラス又はプラスチック製多回投与用バイアルに封入することができる。
【0094】
一般的に言って、注射剤用途に適した組成物としては、水溶液(水溶性の場合)又は分散液、及び滅菌注射液又は分散液の即時調整用粉末が挙げられる。静脈内投与の場合、好適な担体としては、生理食塩水、静菌水、クレモフォア(Cremophor)ELTM(BASF社、パーシパニ(Parsippany)、ニュージャージー州)又はリン酸緩衝生理食塩水(PBS:phosphate buffered saline)が挙げられる。これは作製及び貯蔵の条件下で安定である必要があり、細菌、真菌などの微生物の汚染作用に対して保護される必要がある。上記担体は、例えば水、エタノール、ポリオール(例えば、グリセロール、プロピレングリコール及び液状ポリエチレングリコール)並びにこれらの適切な混合物を含む溶媒又は分散媒とすることができる。
【0095】
医薬製剤は滅菌されていることが好ましい。滅菌は、例えば、滅菌濾過膜を通す濾過によって達成することができる。組成物が凍結乾燥される場合、この方法を用いた滅菌は凍結乾燥及び再形成の前又は後に行うことができる。医薬組成物は、ひとたび製剤化すれば、例えば、溶液、懸濁液、ゲル、乳液、固体として、又は脱水もしくは凍結乾燥粉末としてバイアルに貯蔵することができる。
【0096】
(2)診断用途
上記結合蛋白質を診断目的のためにインビトロ又はインビボで用いる場合はいつも、この結合蛋白質を検出可能な成分で直接又は間接に標識するのが通常である。この検出可能成分は、検出可能なシグナルを直接的又は間接的に発生することができる任意の成分とすることができる。例えば、この検出可能成分は、3水素(3H)、14炭素(14C)、32燐(32P)、35硫黄(35S)又は125ヨウ素(125I)などの放射性同位体;フルオレッセインイソチオシアネート、ローダミン又はルシフェリンなどの蛍光又は化学発光化合物;アルカリ性ホスファターゼ、ベータ−ガラクトシダーゼ又はホースラディッシュペルオキシダーゼなどの酵素;スピン標識などのスピンプローブ;或いは、カラー粒子、例えばラテックスもしくは金粒子であってもよい。上記結合蛋白質を、例えば、Hunter et al. (1962年) NATURE 144:p.945、David et a1. (1974年) BIOCHEMISTRY 13:p.1014、Pain et al. (I981年) J. IMMUNOL. METH. 40:p.219及びNygren (1982年) J. HISTOCHEM. AND CYTOCHEM. 30:p.407に記載されているような、当該技術分野において周知の多くの方法を用いて上記検出可能成分に結合させることができることは理解されよう。これらの標識は、例えば、目視により、又は分光光度計その他の検出器を用いて検出することができる。
【0097】
上記結合蛋白質は、当該技術分野で利用可能な広範囲の免疫測定技術において用いることができる。典型的な免疫測定法としては、例えば、サンドイッチ免疫測定法、競合免疫測定法、免疫組織化学的方法が挙げられる。
【0098】
サンドイッチ免疫測定法では、被分析物、即ち、対象とする抗原に結合する2種の抗体、例えば、固形担体上に固定化した1種と、溶液中に遊離し、検出可能成分で標識された1種とを用いる。抗原を含むサンプルをこの系に添加すると、抗原は、上記の固定化抗体及び標識抗体の両者に結合して担体の表面に「サンドイッチ」免疫複合体を形成する。結合しなかったサンプル成分及び過剰の標識抗体を洗い落とし、担体表面で蛋白質と複合体化した標識抗体の量を測定することにより上記複合体化蛋白質を検出する。或いは、上記の溶液中に遊離した抗体を、この遊離抗体に結合する検出可能成分で標識した第3の抗体によって検出することができる。免疫学的測定法のデザイン、理論及びプロトコルに関する詳細な総説は、Butt編、(1984年)PRACTICAL IMMUNOLOGY、マーセルデッカー社(Marcel Dekker)、ニューヨーク、Harlow et al.編(1988年)ANTIBODIES,A LABORATORY APPROACH、コールド・スプリング・ハーバー・ラボラトリ(Cold Spring Harbor Laboratory)及びDiamandis et al.編(1996年)IMMUNOASSAY、アカデミックプレス社(Academic Press)ボストンを含む多くの教科書に見出すことができる。
【0099】
上記標識結合蛋白質はインビボ造影剤として有用であり、これにより結合蛋白質はレシピエントの特定の対象組織を標的にしてこの造影剤を送達することができることが企図されている。インビボイメージング用の好ましい遠隔検出可能成分としては、約6時間の半減期を有するガンマ放出体である放射性原子テクネチウム-99m(99mTc)が挙げられる。また、インビボイメージングにおいて有用な非放射性成分としては、インサイツ(insitu)でプロトン緩和を誘導するニトロキシドスピンラベル並びにランタニド及び遷移金属イオンが挙げられる。免疫イメージングの他に、標的とする細胞を破壊するために、複合体化放射性成分を標準的な放射免疫治療プロトコルで用いることができる。高線量放射免疫治療用の好ましいヌクレオチド類は、放射性原子90イットリウム(90Yt)、131ヨウ素(131I)及び111インジウム(111In)を含む。上記結合蛋白質は、イメージング分野で周知の結合技術を用いて131I 、111In及び−99mTcで標識することができる。同様に、造影剤を調製し投与する方法及びイメージを捕捉し処理する方法もイメージング分野において公知であるので、本明細書では詳しく論じない。同様に、抗体を利用した免疫療法を実施する方法も当該分野では公知である。例えば、米国特許第5,534,254号を参照されたい。
【0100】
本説明の全体を通して、組成物が特定の成分を有する(having)、含む(including)又は含む(comprising)と記載されているところでは、組成物は、本質的にその列挙された成分からなる、又はこれらの成分からなることも企図されている。同様に、工程が特定の工程段階を有する(having)、含む(including)又は含む(comprising)と記載されているところでは、その工程は、また、本質的にその列挙された工程段階からなる、又はこれらの工程段階からなる。特に示さない限り、段階の順序又は特定の処置を行う順序は、本発明が実施可能である限り、重要ではない。さらに、特に断りのない限り、2つ以上の段階又は処置を同時に実施することができる。
【実施例】
【0101】
以下の実施例では、いくつかの抗hHGFモノクローナル抗体の作製及び特性化について検討する。
【0102】
(実施例1)
抗hHGFモノクローナル抗体の作製
この実施例ではいくつかの抗hHGFモノクローナル抗体の作製について説明する。
【0103】
免疫化、融合及び一次スクリーニングについては、反復性複数部位免疫化(RIMMS:Repetitive Immunization Multiple Sites)プロトコルに従ってMBS社(ポートランド、メイン州)で行われた。AJマウス5匹及びBalb/cマウス5匹を組換えヒトHGF(R&Dシステムズ社(R&D Systems)、ミネアポリス、ミネソタ州、カタログ番号294−HGN−025)で免疫化した。酵素結合免疫測定法(ELISA:Enzyme Linked Immunosorbent Assay)で最も高い抗HGF活性を示す血清を有するマウス2匹を後の融合のために選んだ。該当するマウスから脾臓及びリンパ節を採取した。次いで、B細胞を採取し、骨髄腫細胞株と融合させた。融合産物は1枚以上のプレート上でほぼクローン性になるまで連続希釈した。得られた融合産物からの上清をELISAによりそのhHGFへの結合についてスクリーニングした。HGFに対する抗体を含むとみなされた上清は、さらに、以下の実施例で検討したようにして、インビボ機能試験により特性を明らかにした。1パネルのハイブリドーマを選択し、このハイブリドーマをサブクローニングして増やした。次いで、得られたモノクローナル抗体を標準的な条件下でプロテインA/G樹脂を用いるアフィニティークロマトグラフィーにより精製した。
【0104】
(実施例2)
抗hHGFモノクローナル抗体の配列分析
この実施例では実施例1で作製した抗hHGFモノクローナル抗体のアイソタイプ及び配列の分析について説明する。
【0105】
a.HGFマウスモノクローナル抗体アイソタイプの決定
各モノクローナル抗体の軽鎖タイプ及び重鎖アイソタイプについて、イソストリップマウスモノクローナル抗体アイソタイピングキット(IsoStrip Mouse Monoclonal Antibody Isotyping Kit)を用い、そのメーカーの使用説明書(ロシュ・アプライド・サイエンス社(Roche Applied Science))に従って決定した。
【0106】
全ての抗体は、カッパ免疫グロブリン軽鎖及びIgG1免疫グロブリン重鎖を含むように決定した。
【0107】
b.免疫グロブリン重及び軽鎖可変領域をコードしているヌクレオチド配列の決定
各モノクローナルハイブリドーマ細胞株から、RNeasyミニプレップ(Miniprep)キットを用い、そのメーカーの使用説明書(キアゲン社(Qiagen)、ヴェンロ、オランダ)に従って全RNAを抽出した。BD SMART(商標)RACE cDNA増幅キットを用い、そのメーカーの使用説明書(クロンテック社(Clontech))に従い、5’RACE(cDNA末端の迅速増幅(Rapid Amplification of cDNA Ends))のためのオリゴヌクレオチドプライマBD SMART IIA(5’aagcagtggtatcaacgcagagtacgcggg 3’)(配列番号85)及び5’−RACE CDS Primer(5’ tttttttttttttttttttttttttvn 3’、但し、v=a、g又はc及びn=a、g、c又はt)(配列番号86)を用いて、完全長の第1鎖cDNAを作製した。
【0108】
エキスパンドハイフィデリティ(Expand High−Fidelity)PCRシステム(ロシュ・アプライド・サイエンス社)を用い、そのメーカーの使用説明書に従ってPCR(Polymerase Chain Reaction(ポリメラーゼ連鎖反応))により上記カッパ及び重(IgG1)免疫グロブリン鎖の可変領域を増幅させた。重鎖可変領域は、5’オリゴヌクレオチドプライマ混合物ユニバーサルプライマミックスA(Universal Primer Mix A)( 5’ctaatacgactcactatagggcaagcagtggtatcaacgcagagt 3’(配列番号87)と5’ctaatacgactcactatagggc 3’(配列番号88)との混合物)及び3’IgG1定常領域特異的プライマ5’tatgcaaggcttacaaccaca 3’(配列番号89)又は5’gccagtggatagacagatgggggtgtcg 3’(配列番号90)を用いて増幅した。カッパ鎖可変領域は、5’オリゴヌクレオチドプライマ混合物ユニバーサルプライマミックスA及び3’カッパ定常領域特異的プライマ5’ctcattcctgttgaagctcttgacaat 3’(配列番号91)又は5’cgactgaggcacctccagatgtt 3’(配列番号92)を用いて増幅した。
【0109】
個々のPCR産物をアガロースゲル電気泳動により分画し、キアクイック・ゲル精製(Qiaquick Gel Purification)キットを用い、そのメーカーの使用説明書(キアゲン社)に従って精製した。次いで、トポイソメラーゼ利用クローニングキットTOPO TAクローニング(登録商標)キット(pCR(登録商標)2.1−TOPO(登録商標)ベクターを含む)を用い、そのメーカーの使用説明書(インビトロジェン社(Invitrogen)、カールズバッド、カリフォルニア州)に従って、これらのPCR産物をpCR2.1 TOPOプラスミド中にクローニングした後、標準的な形質転換技術を用いて細菌DH5に形質転換した。形質転換した細菌のクローンから単離したプラスミドDNAについて、アジェンコートバイオサイエンス社(Agencourt Bioscience)のT7(5’TAATACGACTCACTATAGGG3’)(配列番号93)、M13順方向プライマ(5’GTAAAACGACGGCCAGT3’)(配列番号94)及びM13逆方向プライマ(5’CAGGAAACAGCTATGACC3’)(配列番号95)を用い、標準的なジデオキシDNA配列決定方法により配列を決定することにより可変領域配列の配列を特定した。ベクターNTI(Vector NTI)ソフトウェア(インビトロジェン社、カールズバッド、カリフォルニア州)及びIMGT/V−クエスト(Quest)ウェブサーバー(http://imgt.cines.fr/textes/vquest)を用いてこれらの配列を解析し、可変領域の配列を同定、確認した。
【0110】
c.1A3、1D3、1F3及び2B8カッパ及びIgG1鎖の免疫グロブリン重及び軽鎖定常領域の配列をコードしているヌクレオチド配列の決定
順方向プライマ5’ggggacaagtttgtacaaaaaagcaggctgccaccatgaactttgggctcagattgattttcc 3’(下線箇所:開始コドン)(配列番号96)及び逆方向プライマ5’ggggaccactttgtacaagaaagctgggttcatttaccaggagagtgggagagg3’(下線箇所:停止コドン)(配列番号97)を用いて上記で作製したcDNAから、1A3、1D3及び1F3 IgG1鎖の完全長cDNAをPCRにより増幅させた。順方向プライマ5’ggggacaagtttgtacaaaaaagcaggctgccaccatgggatggagctatatcatcctcttt3’(下線箇所:開始コドン)(配列番号98)及び逆方向プライマ5’ggggaccactttgtacaagaaagctgggttcatttaccaggagagtgggagag3’(下線箇所:停止コドン)(配列番号99)を用いて上記で作製したcDNAから、2B8IgG1鎖の完全長cDNAを増幅させた。
【0111】
順方向プライマ5’ggggacaagtttgtacaaaaaagcaggctgccaccatggaatcacagactctggtcttcata3’(下線箇所:開始コドン)(配列番号100)及び逆方向プライマ5’ggggaccactttgtacaagaaagctgggtctaacactcattcctgttgaagctc3’(下線箇所:停止コドン)(配列番号101)を用いて2B8カッパ鎖の完全長cDNAを増幅させた。PCR断片をゲートウェイ(Gateway)BP組換え反応によりpDONR221(インビトロジェン社、カールズバッド、カリフォルニア州)中にクローニングし、標準的なジデオキシDNA配列決定方法を用いてアジェンコートバイオサイエンス社で配列決定することによりその定常領域の配列を同定し、さらに可変領域の配列を確認した。
【0112】
d.配列の解析
可変領域(標準テキスト)は、IMGT/V−QUESTウェブサーバーソフトウェア(http://imgt.cines.fr/textes/vquest/)を用いて同定した。シグナルペプチド配列は、同定した可変領域の上流にあるインフレーム開始コドン(ATG)の同定に基づいて予測した。シグナルペプチド配列を同定し、下記に下線で示した。
【0113】
各可変領域の最後のヌクレオチドは、可変/定常領域結合により生じる次のコドンの最初の塩基である。このヌクレオチドは、そのエキソンの一部であるので、可変領域に含まれる。以下に列挙した定常領域のアミノ酸配列にはこの結合コドンの翻訳が含まれる。
【0114】
完全な重鎖又はカッパ鎖抗体配列を作製するためには、下記の可変領域の配列をそのそれぞれの定常領域の配列と結合させる(下線箇所:シグナル配列)。
【0115】
(1)1A3重鎖可変領域(配列番号1)
【0116】
【化1】

【0117】
(2)1A3カッパ軽鎖可変領域(配列番号3)
【0118】
【化2】
【0119】
(3)2B8重鎖可変領域(配列番号11)
【0120】
【化3】
【0121】
【化4】
【0122】
(4)2B8カッパ軽鎖可変領域(配列番号13)
【0123】
【化5】
【0124】
(5)2F8重鎖可変領域(配列番号21)
【0125】
【化6】
【0126】
(6)2F8カッパ軽鎖可変領域(配列番号23)
【0127】
【化7】
【0128】
(7)3B6重鎖可変領域(配列番号31)
【0129】
【化8】
【0130】
【化9】
【0131】
(8)3B6カッパ軽鎖可変領域(2個の可能なATG開始コドン(大文字))(配列番号33)
【0132】
【化10】
【0133】
(9)3D11重鎖可変領域(配列番号41)
【0134】
【化11】
【0135】
(10)3D11カッパ軽鎖可変領域(配列番号43)
【0136】
【化12】
【0137】
(11)1D3重鎖可変領域(配列番号51)
【0138】
【化13】
【0139】
(12)1D3カッパ軽鎖可変領域(配列番号53)
【0140】
【化14】
【0141】
(13)1F3重鎖可変領域(配列番号61)
【0142】
【化15】
【0143】
(14)1F3カッパ軽鎖可変領域(配列番号63)
【0144】
【化16】
【0145】
【化17】
【0146】
(15)3A12重鎖可変領域(配列番号71)
【0147】
【化18】
【0148】
(16)3A12カッパ軽鎖可変領域(配列番号73)
【0149】
【化19】
【0150】
(17)対照マウスIgG1重鎖定常領域(J00453)(配列番号81)
【0151】
【化20】
【0152】
【化21】
【0153】
(18)1A3、1D3、1F3及び2B8(AJ系マウス由来)について決定されたマウスIgG1重鎖定常領域(配列番号82)
【0154】
【化22】
【0155】
(19)対照マウスカッパ軽鎖定常領域(V00807)並びに1D3、1F3及び2B8(AJ系マウス由来)について決定されたマウスカッパ軽鎖定常領域(配列番号83)
【0156】
【化23】
【0157】
(20)1D3、1F3及び2B8に比し変更されたヌクレオチド(下線箇所)を1個含む1A3について決定されたマウスカッパ軽鎖定常領域(配列番号84)
【0158】
【化24】
【0159】
実施例1で作製した抗体の免疫グロブリン重鎖可変領域を規定するアミノ酸配列は、いずれも図2に記載されている。これらの配列は全て相互に整列させてあり、シグナルペプチド、CDR1、CDR2及びCDR3を規定する配列は枠で囲んで識別されている。図3は、全ての抗体のCDR1、CDR2及びCDR3配列を別々に整列させて示したものである。
【0160】
実施例1で作製した全ての抗体の免疫グロブリン軽鎖可変領域を規定するアミノ酸配列は、いずれも図4に記載されている。これらの配列は全て相互に整列させてあり、シグナルペプチド、CDR1、CDR2及びCDR3を規定する配列は枠で囲んで識別されている。図5は、全ての抗体のCDR1、CDR2及びCDR3配列を別々に整列させて示したものである。
【0161】
便宜のため、本実施例で検討した抗体配列と配列表に示したものとの対応関系を示すコンコーダンス表を表1に示した。
【0162】
【表1A】
【0163】
【表1B】
【0164】
また、便宜のため、下記の配列は、本実施例に記載した全ての抗体の実際の、又は企図された完全長の重及び軽鎖配列(即ち、可変領域及び定常領域の両者の配列を含む)を表したものである。マウス抗体2F8、3A12、3B6及び3D11の定常領域は配列決定を行わなかったが、これらが全てAJ系マウス由来の抗体であることから、配列決定した1D3、1F3及び2B8抗体と同一の定常領域配列を有すると推定されることは注目される。しかしながら、本明細書に記載した可変領域の配列を当業者に周知の他のいく種かの定常領域配列のそれぞれに結合させて活性のある完全長の免疫グロブリン重及び軽鎖を作製することができることは了解されよう。
【0165】
(1)完全長1A3重鎖配列(1A3重鎖可変領域及びIgG1定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号122)
【0166】
【化25】
【0167】
(2)完全長1A3重鎖配列(1A3重鎖可変領域及びIgG1定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号123)
【0168】
【化26】
【0169】
(3)完全長1A3軽鎖配列(1A3カッパ可変領域及び定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号124)
【0170】
【化27】
【0171】
【化28】
【0172】
(4)完全長1A3軽鎖配列(1A3カッパ可変領域及び定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号125)
【0173】
【化29】
【0174】
(5)完全長2B8重鎖配列(2B8重鎖可変領域及びIgG1定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号126)
【0175】
【化30】
【0176】
(6)完全長2B8重鎖配列(2B8重鎖可変領域及びIgG1定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号127)
【0177】
【化31】
【0178】
【化32】
【0179】
(7)完全長2B8軽鎖配列(2B8カッパ可変領域及び定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号128)
【0180】
【化33】
【0181】
(8)完全長2B8軽鎖配列(2B8カッパ可変領域及び定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号129)
【0182】
【化34】
【0183】
(9)完全長2F8重鎖配列(2F8重鎖可変領域及びIgG1定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号130)
【0184】
【化35】
【0185】
【化36】
【0186】
(10)完全長2F8重鎖配列(2F8重鎖可変領域及びIgG1定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号131)
【0187】
【化37】
【0188】
(11)完全長2F8軽鎖配列(2F8カッパ可変領域及び定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号132)
【0189】
【化38】
【0190】
(12)完全長2F8軽鎖配列(2F8カッパ可変領域及び定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号133)
【0191】
【化39】
【0192】
(13)完全長3B6重鎖配列(3B6重鎖可変領域及びIgG1定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号134)
【0193】
【化40】
【0194】
【化41】
【0195】
(14)完全長3B6重鎖配列(3B6重鎖可変領域及びIgG1定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号135)
【0196】
【化42】
【0197】
(15)完全長3B6軽鎖配列(3B6カッパ可変領域及び定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号136)
【0198】
【化43】
【0199】
(16)完全長3B6軽鎖配列(3B6カッパ可変領域及び定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号137)
【0200】
【化44】
【0201】
(17)完全長3D11重鎖配列(3D11重鎖可変領域及びIgG1定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号138)
【0202】
【化45】
【0203】
(18)完全長3D11重鎖配列(3D11重鎖可変領域及びIgG1定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号139)
【0204】
【化46】
【0205】
(19)完全長3D11軽鎖配列(3D11カッパ可変領域及び定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号140)
【0206】
【化47】
【0207】
(20)完全長3D11軽鎖配列(3D11カッパ可変領域及び定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号141)
【0208】
【化48】
【0209】
(21)完全長1D3重鎖配列(1D3重鎖可変領域及びIgG1定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号142)
【0210】
【化49】
【0211】
(22)完全長1D3重鎖配列(1D3重鎖可変領域及びIgG1定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号143)
【0212】
【化50】
【0213】
(23)完全長1D3軽鎖配列(1D3カッパ可変領域及び定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号144)
【0214】
【化51】
【0215】
(24)完全長1D3軽鎖配列(1D3カッパ可変領域及び定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号145)
【0216】
【化52】
【0217】
(25)完全長1F3重鎖配列(1F3重鎖可変領域及びIgG1定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号146)
【0218】
【化53】
【0219】
【化54】
【0220】
(26)完全長1F3重鎖配列(1F3重鎖可変領域及びIgG1定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号147)
【0221】
【化55】
【0222】
(27)完全長1F3軽鎖配列(1F3カッパ可変領域及び定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号148)
【0223】
【化56】
【0224】
(28)完全長1F3軽鎖配列(1F3カッパ可変領域及び定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号149)
【0225】
【化57】
【0226】
(29)完全長3A12重鎖配列(3A12重鎖可変領域及びIgG1定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号150)
【0227】
【化58】
【0228】
(30)完全長3A12重鎖配列(3A12重鎖可変領域及びIgG1定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号151)
【0229】
【化59】
【0230】
(31)完全長3A12軽鎖配列(3A12カッパ可変領域及び定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号152)
【0231】
【化60】
【0232】
(32)完全長3A12軽鎖配列(3A12カッパ可変領域及び定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号153)
【0233】
【化61】
【0234】
便宜のため、本実施例で検討した抗体の完全長配列と配列表に示したものとの対応関系を示すコンコーダンス表を表2に示した。
【0235】
【表2A】
【0236】
【表2B】
【0237】
(実施例3)
各種組換えhHGF蛋白質の作製
本実施例では、実施例1及び実施例14で作製された抗体の特性を明らかにするために用いるいくつかの組換え蛋白質のクローニング及び発現について説明する。具体的には、本実施例では、組換えhHGF蛋白質、555番目の位置にグリシンからグルタメートへの置換を含む組換えhHGF蛋白質(G555E)、561番目の位置にシステインからアルギニンへの置換を含む組換えhHGF蛋白質(C561R)、マウスHGF配列内に配置されたヒトV495乃至L585HGF配列を含む組換えマウス−ヒト−マウス(mhm)キメラHGF蛋白質、マウスHGF配列内に配置されたヒトI499乃至R566HGF配列を含む組換えmhmキメラHGF蛋白質及びマウスHGF配列内に配置されたヒトW507乃至L585HGF配列を含む組換えmhmキメラHGF蛋白質のクローニング及び発現について説明する。
【0238】
標準的な分子技術を用いて以下の発現構築物を作製し、得られたcDNA配列をDNA配列決定法によって確認した。
【0239】
a. hHGF−Fc
1回目のPCRでは、NotI部位を導入し、hHGFとhIgFcとの間に6×His標識をコードしている2本の重なり合うPCR断片を作製した。2回目に、これらの重なり合うPCR断片を鋳型としてhHGF−his−IgFcを増幅させた。得られた断片をNheI及びBamHIで消化し、pcDNA5/FRT(インビトロジェン社、#35−3014)中にクローニングした。次いで、インビトロジェン社のクローンID:IOH29794(ヒトHGF cDNA)からhHGFを増幅させた。その配列は、アクセッション番号NM_000601.4としてNCBIに寄託されている配列に相当することが分かった。
【0240】
(1) 5’hHGF NheIプライマー
【0241】
【化62】
【0242】
(2) 3’hHGF NotI His標識プライマー
【0243】
【化63】
【0244】
(3) 5’HisIgFcプライマー
【0245】
【化64】
【0246】
(4) 3’IgFc BamHIプライマー
【0247】
【化65】
【0248】
b. hHGF−Fc G555E及びhHGF−Fc C561R
hHGF−Fc突然変異体G555E及びC561Rは、クイックチェンジ(QuickChange)II XL部位特異的突然変異誘発キット(ストラタジーン社(Stratagene))をメーカーの使用説明書に従って用いた部位特異的突然変異誘発法により作製した。
【0249】
(1) hHGF−Fc(G555E)センスプライマ
【0250】
【化66】
【0251】
(2) hHGF−Fc(G555E)アンチセンスプライマ
【0252】
【化67】
【0253】
(3) hHGF−Fc(C561R)センスプライマ
【0254】
【化68】
【0255】
(4) hHGF−Fc(C561R)アンチセンスプライマ
【0256】
【化69】
【0257】
c. マウス−ヒト−マウスキメラFc
マウス−ヒト−マウスキメラIgFc構築物は、mHGFアルファ鎖hHGF、ヒトHGFのβ鎖アミノ酸Val495乃至Leu585、及びmHGF C末端β鎖とこれに続く6×His標識及びIgG−Fcを含む。
【0258】
アミノ酸V495乃至L585をコードしているヒトHGF cDNAは、インビトロジェン社のクローンID:IOH29794(ヒトHGF cDNA)から増幅させた。その配列は、アクセッション番号NM_000601.4としてNCBIに寄託されている配列に相当する。マウスHGF配列は、インビトロジェン社製のスーパースクリプトワンステップ(Super Script One Step)RT−PCRキット(#10928−034)をメーカーの使用説明書に従って用い、マウス肝の全RNA(クローンテック社、#636603)からRT−PCRによって増幅させた。そのmHGF cDNA配列は、アセッション番号D10213.1としてNCBIに寄託されている配列に相当する。
【0259】
重なり合うPCRプライマーを用いて断片1、2及び3と称する3本の断片を作製し、連続回のPCR増幅でアニーリングした。最終産物をNheI及びNotIで切断し、pcDNA5/FRT IgGFc中にクローニングした。
【0260】
(1) mHGFアルファ鎖5’NheIの断片1プライマー
【0261】
【化70】
【0262】
【化71】
【0263】
(2) hHGFベータ鎖aaV495乃至L585の断片2プライマー
【0264】
【化72】
【0265】
【化73】
【0266】
(3) mHGFベータ鎖C末端3’NotIの断片3プライマー
【0267】
【化74】
【0268】
【化75】
【0269】
d. hHGF及びmhmキメラの構築
hHGF及びmhmキメラ(V495乃至L585)をコードしているベクターであるpcDNA5/FRT hHGF及びpcDNA5/FRT−mhmキメラ(V495乃至L−L585)を、Fc標識を含ませずに、部位特異的突然変異誘発法により作製した。前記クイックチェンジII XL部位特異的突然変異誘発キット(ストラタジーン社)をメーカーの使用説明書に従って用い、上記6×His標識の3’末端に停止コドンを導入した。突然変異誘発プライマーには、プライマー1:
【0270】
【化76】
【0271】
及びプライマー2:
【0272】
【化77】
【0273】
を含めた。
【0274】
さらに、前記クイックチェンジII XL部位特異的突然変異誘発キット(ストラタジーン社)をメーカーの使用説明書に従って用い、部位特異的突然変異誘発法により上記pcDNA5/FRT−mhm(V495乃至L−L585)構築物から2種の別のmhmキメラを作製した。そのうちの1種のmhm構築物はマウス配列間に配置されたhHGFのI499乃至R556の領域を含んだ。もう一方のmhm構築物は、マウス配列間に配置されたhHGFのW507乃至L585の領域を含んだ。
【0275】
上記mhmキメラ(I499乃至R556)の場合、適切なオリゴヌクレオチド配列を用いて、鋳型pcDNA5/FRT−mhmキメラ(V495乃至L585)構築物中に、点突然変異D558E、C561R、V564I、V567I及びM583Lを順番に起こさせた。上記mhmキメラ(W507乃至L585)の場合、適切なオリゴヌクレオチド配列を用いて、鋳型pcDNA5/FRT−mhmキメラ(V495乃至L585)構築物中に、点突然変異Q502R、N504T及びI505Vを1ステップで導入した。
【0276】
得られたhHGF−Fc蛋白質のヌクレオチド配列は、シグナル配列(1乃至93番目ヌクレオチド)及びプロドメイン(94乃至162番目ヌクレオチド)を含めて、配列番号118として記載している。このhHGF−Fc蛋白質のアミノ酸配列は、配列番号119として記載している。
【0277】
得られたmhm(V495−L585)−Fcキメラ蛋白質をコードしているヌクレオチド配列は、シグナル配列(1乃至96番目ヌクレオチド)及びプロドメイン(97乃至165番目ヌクレオチド)を含めて、配列番号120として記載している。このmhm(V495−L585)−Fcキメラ蛋白質のアミノ酸配列は、配列番号121として記載している。
【0278】
得られたmhm(V495−L585)構築物をコードしているヌクレオチド配列及びこの構築物を規定する蛋白質配列は、それぞれ配列番号211及び212として記載している。配列番号211で表される核酸配列は、シグナル配列(1乃至96番目ヌクレオチド)及びプロドメイン(97乃至165番目ヌクレオチド)を含み、配列番号212で表される蛋白質配列は、(シグナル配列又はプロドメインを含まない)活性蛋白質配列を含む。得られたmhm(I499〜R556)構築物をコードしているヌクレオチド配列及びこの構築物を規定する蛋白質配列は、それぞれ配列番号213及び214として記載している。配列番号213で表される核酸配列は、シグナル配列(1乃至96番目ヌクレオチド)及びプロドメイン(97乃至165番目ヌクレオチド)を含み、配列番号214で表される蛋白質配列は、(シグナル配列又はプロドメインを含まない)活性蛋白質配列を含む。得られたmhm(W507〜L585)をコードしているヌクレオチド配列及びこの構築物を規定する蛋白質配列は、それぞれ配列番号215及び216として記載している。配列番号215で表される核酸配列は、シグナル配列(1乃至96番目ヌクレオチド)及びプロドメイン(97乃至165番目ヌクレオチド)を含み、配列番号216で表される蛋白質配列は、(シグナル配列又はプロドメインを含まない)活性蛋白質配列を含む。
【0279】
e. 蛋白質発現
(1) 細胞培養
CHO FIpIn細胞(インビトロジェン社、カタログ番号R758−07)をF12K培地(ATTC、カタログ番号30−2004)、10%FCS(インビトロジェン社、カタログ番号10438026)、1%ペニシリン(10,000単位/mL)/ストレプトマイシン(10,000μg/mL)(インビトロジェン社、カタログ番号15140−122)中、37℃、5% CO2、100μg/mLゼオシン(Zeocin)(インビトロジェン社、カタログ番号R250−01)の条件下で増殖させた。
【0280】
(2) 安定なCHO FIpIn細胞株の作製
リポフェクタミン2000(インビトロジェン社、カタログ番号11668−027)をそのメーカーの使用説明書に従って用い、発現プラスミドDNA pOG44及びpcDNA5/FRTを9:1の比率で用いてCHO FIpIn宿主細胞に形質移入した。対照として、空のpcDNA5/FRTベクター/pOG44及びpOG44プラスミド(インビトロジェン社、カタログ番号35−3018)単独を細胞に形質移入した。形質移入後24時間に、この細胞を分裂させ、48時間後に、0.5mg/mLのハイグロマイシンB(シグマ社(Sigma)、カタログ番号H0654−SPEC)を細胞に添加した。F12K、10%FCS、1%ペニシリン/ストレプトマイシン、0.5mg/mLハイグロマイシンB中で安定細胞の多クローン性選択を行った。
【0281】
(3) 安定なCHO FIpIn細胞株における蛋白質発現
約2×106個の細胞を15cmプレートに接種し、F12K(ATCC、カタログ番号30−2004)/DMEM高グルコース(インビトロジェン社、カタログ番号11995065)1:1、5%超低IgG FCS(インビトロジェン社、#16250−78)中、37℃、5%CO2で5乃至6日間増殖させた。上清を採取し、得られた蛋白質をELISA及び表面プラスモン共鳴により分析した。
【0282】
(実施例4)
抗hHGFモノクローナル抗体の結合特性
実施例1で作製したモノクローナル抗体の特徴は、これらがhHGF及び実施例3で作製した組換えHGF蛋白質の一部に結合することができることにあった。
【0283】
BlAcore T100装置を用いて表面プラスモン共鳴によりこれらの抗体を分析することによってHGF及び実施例3に記載した融合蛋白質の一部へのその結合能を評価した。各抗体は、標準的なカップリングプロトコルをメーカーの使用説明書に従って用いたアミンカップリング(BIAcore、カタログ番号BR−1000−50)によりカルボキシメチル化デキストランCM5センサーチップ(BIAcore、カタログ番号BR−1006−68)上に固定化した。
【0284】
分析は、0.05%界面活性剤P20(BIAcore、カタログ番号R−1000−54)、2mg/mLのBSA(EMD社カタログ番号2930)及び10mg/mLのCM−デキストランナトリウム塩(フルカ社(Fluka)、カタログ番号86524)を含有するPBS(GIBCO社、カタログ番号14040−133)を泳動用緩衝液として用い、25℃で行った。種々のHGF融合蛋白質を含む上清又は空のベクターを形質移入した細胞からの上清を各抗体上に30μL/分の流速で3分間注入した。その結果生じた結合は、注入終了後30秒のベースラインに対する共鳴単位(RU)として求めた。結合は、泳動用緩衝液で希釈したヒトHGF(R&Dシステムズ社(R&D Systems)、カタログ番号294−HGN−025)と比較した。非特異的な結合は、同じアミンカップリング法を用いてマウスIgG(ロックランド社(Rockland)、カタログ番号010−0102)を固定化した対照表面への結合を比較することにより測定した。
【0285】
以上の結果は表3にまとめた。
【0286】
【表3】
【0287】
表3の結果から明らかなように、上記の各抗体はrHGF及び精製ヒトHGFに結合する。さらに、これらの抗体は全て、点突然変異G555E及びC561Rを含むhHGFに結合する。総じて、1F3及び2F8を除く全ての抗体はマウスHGFに結合せず、このことから抗体1A3、1D3、2B8、3A12、3B6及び3D11がヒトHGFに特異的に結合することが明らかとなった。抗体1D3、1F3及び2B8がマウス−ヒト−マウスキメラに結合するのに対して、残りの抗体は結合しなかった。以上の結果から、抗体1D3及び2B8は、ヒトHGFの495乃至585番目残基に少なくとも部分的に結合することが分かる。抗体1A3、3A12、3B6及び3D11は、495乃至585番目残基以外のヒトhHGFの部分に結合するように思われる。2F8がhHGF及びmHGFのいずれにも結合するように思われるので、現在のところ、これがなぜmhmキメラに結合しないのか不明である。
【0288】
(実施例5)
抗hHGFモノクローナル抗体の還元及び非還元HGFへの結合能
本実施例では、実施例1で作製した抗hHGFモノクローナル抗体の還元及び非還元HGFへの結合能について分析した。
【0289】
抗HGF血清の上記組換えhHGFとの反応性を免疫ブロット法によって評価した。NuPAGEサンプル還元緩衝液(インビトロジェン社)を含む、又は含まないNuPAGE MOPS SDS泳動用緩衝液(インビトロジェン社)中8μgの組換えhHGF蛋白質を4乃至12%ビス−トリス1.0mm×2Dウェルゲル(インビトロジェン社、カールズバッド、カリフォルニア州)で分画した。次いで、分画した蛋白質を標準的な手順によりニトロセルロース膜上に移した。このニトロセルロース膜を、0.1% Tween20(登録商標)含有トリス緩衝食塩液(TBST)に溶かした脱脂乳粉の5%溶液でブロックした後、更なるブロックのために、ミニプロテアンIIマルチスクリーン(Mini Protean II Multi−Screen)装置(バイオラッド社(BioRad))に取り付けた。
【0290】
その結果得られた膜をマルチスクリーン装置上で精製抗体によりプローブした。精製抗体はブロッキング緩衝液で5μg/mLに希釈した。次いで、このニトロセルロース膜を装置から取り外し、ホースラディッシュペルオキシダーゼ標識抗マウスIgG抗体と共にインキュベートした。得られた結果は表4に示した。表中、数字は結合の程度を示し、−は最も弱い(殆どあるいは全く結合しない)、3+は最も強い結合を表している。
【0291】
【表4】
【0292】
表4のデータから明らかなように、抗体は全て、非還元rhHGFに結合する。これに対し、モノクローナル抗体1A3、1D3、1F3、2F8及び3B6は還元rhHGFに結合したが、抗体2B8、3A12及び3D11は還元rhHGFに結合しなかった。
【0293】
(実施例6)
結合親和性
実施例1で作製した各抗体のhHGFに対する結合親和性及び相互作用の速度を表面プラスモン共鳴によって測定した。
【0294】
ウサギ抗マウス免疫グロブリン(BIAcore、カタログ番号BR−1005−14)を、標準的なカップリングプロトコルをメーカーの使用説明書に従って用いたアミンカップリング(BIAcore、カタログ番号BR−1000−50)によりカルボキシメチル化デキストランCM5センサーチップ(BIAcore、カタログ番号BR−1006−68)上に固定化した。分析は、0.05%界面活性剤P20(BIAcore、カタログ番号R−1000−54)、2mg/mLのBSA(EMD社カタログ番号2930)及び10mg/mLのCM−デキストランナトリウム塩(フルカ社(Fluka)、カタログ番号86524)を含有するPBS(GIBCO社、カタログ番号14040−133)を泳動用緩衝液として用い、25℃で行った。
【0295】
前記抗体は、個々のフローセル内で10μL/分の流速で捕捉した。各抗体が各サイクルで捕捉される約20RUの抗体を生じるための注入時間にはばらつきがあった。緩衝液、又は泳動用緩衝液で希釈したHGF(R&Dシステムズ社、カタログ番号294−HGN−025)を対照表面(捕捉抗体なし)及び活性表面(試験対象抗体あり)上に60μL/分で2分間順次注入した。解離相を濃度に応じて15又は90分間モニターした。その後、上記表面は、別のサイクルが開始される前に、pH1.7の10mMグリシン−HCl(BIAcore社、カタログ番号BR−1003−54)を60μL/分の流速で3分間注入して再生させた。試験したHGF濃度は0.46nM乃至7.5nMであった。
【0296】
速度論的パラメータは、対照値減算を含むBIAエバリュエーション(BIAevalutation)ソフトウェアの速度関数を用いて求めた。各抗体の速度論的パラメータka(会合速度定数)、kd(解離速度定数)及びKD(平衡解離定数)を表5にまとめた。
【0297】
【表5】
【0298】
表5のデータから明らかなように、これらの抗体は、KD値約100pM以下、約50pM以下又は約20pM以下でhHGFに結合する。
【0299】
(実施例7)
抗hHGF抗体の中和活性
本実施例では、実施例1で作製した抗体について、(a)c−MetへのhHGFの結合を阻害し、(b)4MBr−5細胞におけるBrdU取り込みのHGFによる促進を阻害する能力を特性化した。
【0300】
a. HGF−Met結合阻害試験(中和試験)
上記抗体について、c−MetへのhHGF結合を阻害する能力をELISAにより試験した。
【0301】
すなわち、ワラック96穴DELFIAアッセイプレート(ワラック社、カタログ番号AAAND−0001)を6.25μg/mL HGF(R&Dシステムズ社、カタログ番号294−HGN−025)の炭酸コーティング緩衝液(15mM Na2CO3及び34mM NaHCO3、pH9.0)溶液100μLにより4℃で16時間コーティングした。次に、このプレートを室温で1時間5%脱脂粉乳のPBS溶液200μLでブロックした。抗体は、別のプレートにおいて5%脱脂粉乳のPBS溶液に溶解した2nMのc−Met(R&Dシステムズ社、カタログ番号358−MT/CF)に検討対象の抗体を漸増濃度(0.033乃至667nM、3倍段階希釈)で加えることにより調製した。ウェル当たり100μLのサンプルを上記アッセイプレートに移し、4℃で一夜インキュベートした。次いで、このアッセイプレートをPBS−0.1% Tween20で3回洗浄した後、5%脱脂粉乳のPBS溶液で調製した100μL/ウェルの2μg/mLビオチン化抗ヒトc−Met抗体(R&Dシステムズ社、カタログ番号BAF358)溶液と共に室温で2時間インキュベートした。
【0302】
その後、上記で得られたプレートをPBS−0.1% Tween20で3回洗浄した後、DELFIAアッセイ緩衝液(ワラック社、カタログ番号4002−0010)で1:1,000に希釈したEu標識ストレプトアビジン(ワラック社、カタログ番号1244−360)と共に室温で1時間インキュベートした。その結果得られたプレートをDELFIA洗浄液(ワラック社、カタログ番号4010−0010)で3回洗浄した後、100μL/ウェルのDELFIA強化溶液(ワラック社、#4001−0010)と共に攪拌しながら室温で15分間インキュベートした。
【0303】
上記プレートは、ユウロピウム法を用いるビクター3V装置(パーキンエルマー社(Perkin Elmer))で読み取らせた。IC50値を算出し、表6にまとめた。
【0304】
【表6】
【0305】
以上の結果から明らかなように、3B6以外の全ての抗体(即ち、1D3、1A3、2B8、3A12、1F3、3D11及び2F8)は、c−MetへのHGFの結合を効率的に中和する。
【0306】
b. 4MBr−5細胞におけるBrdU取り込みのHGFによる促進の中和
12.5nMのhHGF 10μLを96穴組織培養マイクロタイタープレート(コスター社(Costar)カタログ番号3903)の個々のウェルに分注した。次いで、6667、2222、740、247、82、27、9.1、3.0、1.0及び0.33nMの濃度に連続希釈した抗体10μLを各ウェルに添加した。次に、このHGF抗体混合液を室温で30分間インキュベートした。F−12K(ATCC、30−2004)、15%FBS(ギブコ(Gibco)10438−026)、30ng/mL EGF(シグマ(Sigma)E9644)、1%ペニシリン/ストレプトマイシン(PS、ギブコ社カタログ番号15140−122)中で培養したサル気管支上皮細胞4MBr−5(ATCC、CCL208)をトリプシン(Trypsin)(ギブコ社カタログ番号25200−056)で解離させ、75,000個/mLの濃度でアッセイ培地(F−12K、2.5% FBS、1% PS)に再懸濁し、この細胞懸濁液80μLを上記HGF抗体混合液に分注した。
【0307】
この細胞を37℃、5% CO2の条件下でインキュベートした。48時間後、100μM BrdU(ロシュ社(Roche)、カタログ番号1669915)を10μL添加した。72時間後、培地を除去し、プレートをヘアドライヤーで乾燥させた後、BrdU ELISAをメーカーの使用説明書(ロシュ社、カタログ番号1669915)に従って用いて処理した。
【0308】
発光シグナルは、シナジーHT(Synergy HT)プレートリーダー(バイオテック社(Bio−Tek))によって定量化した。このデータは、グラフパッドプリズム(GraphPad Prism)(グラフパッドソフトウェア社(GraphPad Software))において式y=ボトム+(トップ−ボトム)/(1+10^(log(EC50−x)*ヒル勾配))の可変勾配を有するS字形の用量反応に当てはめた。各実験は、2回ずつ、少なくとも3回繰り返した。表7にEC50の平均値を示した。
【0309】
【表7】
【0310】
表7の結果から明らかなように、抗体1A3、ID3、1F3、2B8、2F8、3A12、3B6及び3D11は全て、4MBr−5細胞のHGF誘発性増殖を抑制する。
【0311】
(実施例8)
抗hHGF抗体の抗散乱(Anti−Scatter)活性
本実施例では、実施例1で作製した抗体のHGF誘発性散乱活性を抑制する能力に関する特性化について説明する。HGFは、MDCK細胞(ATCC、マナッサス、バージニア州、カタログ番号CCL−34)においてクラスターの「散乱」(運動)を誘発する。
【0312】
MDCK細胞は、10%ウシ胎仔血清(インビトロジェン社、カタログ番号10438026)及び1%ペニシリン−ストレプトマイシン(インビトロジェン社カタログ番号15140122)を含有するMEM(ATCC、マナッサス、バージニア州、カタログ番号30−2003)80μL中ウェル当たり4×103個の密度で96穴Costar組織培養プレート(コーニング社(Corning Incorporated)、コーニング、ニューヨーク州、カタログ番号3595)に接種した。検討対象の各抗体は、10%ウシ胎仔血清及び1%ペニシリン−ストレプトマイシンを含有するMEMで6,667nMに希釈した。次いで、種々の抗体希釈液のそれぞれ、及び10%ウシ胎仔血清及び1%ペニシリン−ストレプトマイシンを含有し抗体を含まないMEMを別々に、10%ウシ胎仔血清及び1%ペニシリン−ストレプトマイシン並びに100ng/mlのHGF(R&Dシステムズ社、カタログ番号294−HGN−025)を含有する等容量のMEMと混合した。各抗体/HGF希釈液は、25℃で30分間インキュベートした。各抗体/HGF希釈液20μLを個別に個々のウェルに添加し、最終抗体濃度666.7nM及び最終HGF濃度10ng/mlを得た。次に、MDCK細胞を37℃、5%CO2で24時間インキュベートした。
【0313】
24時間のインキュベーション後、MDCK細胞をウェル当たり100μLの氷冷PBS(インビトロジェン社カタログ番号14190144)で一度注意深く洗い、25℃で10分間揺動させながらウェル当たり100μLの氷冷メタノールで固定した。次いで、プレートを一度蒸留水で注意深く洗浄した。蒸留水に溶解した0.5%クリスタルバイオレット(シグマ社、セントルイス、ミズーリ州、カタログ番号C3886)及び50%エタノールからなる100μL容のクリスタルバイオレット溶液を各ウェルに添加し、細胞を揺動させながら25℃で20分間インキュベートした。
【0314】
クリスタルバイオレット溶液による染色後、細胞を蒸留水で三度注意深く洗浄した。次に、各ウェルにPBSを添加してサンプルの乾燥を防いだ。ライカ(Leica)DMIRB顕微鏡(ライカマイクロシステムズ社、ウェッツラー、ドイツ)、DC500カメラ(ライカマイクロシステムズ社、ウェッツラー、ドイツ)及びマグナファイア(MagnaFire)2.1Cソフトウェア(Optronics、ゴレタ、カリフォルニア)を用いて細胞を撮像し、サンプルを散乱レベルについて評価した。表8にその結果をまとめた。
【0315】
【表8】
【0316】
表8の結果から明らかなように、抗体2B8は、その他の抗体よりも多くのHGF誘発性散乱を抑制した。抗体1D3及び3B6は中間的なレベルの抑制を示し、抗体1A3は低乃至中間レベルの抑制を示し、抗体1F3及び2F8は低レベルの抑制を示し、抗体3A12及び3D11はほとんど或いは全く検出可能な抑制を示さなかった。
【0317】
(実施例9)
HGF刺激c−Metリン酸化の阻害
本実施例では、実施例1で作製した抗体の、PC−3細胞におけるHGF刺激c−Metリン酸化を阻害する能力に関する特性化について説明する。HGFはPC−3細胞(ATCC番号CRL−1435)においてMetのリン酸化を誘発する。
【0318】
PC−3細胞は、10%ウシ胎仔血清(インビトロジェン社、カタログ番号10438026)及び1%ペニシリン−ストレプトマイシン(インビトロジェン社カタログ番号15140122)を含有するF−12K(ATCC、マナッサス、バージニア州、カタログ番号30−2004)100μL中ウェル当たり4.5×104個の密度で96穴Costar組織培養プレート(コーニング社カタログ番号3595)の個々のウェルに接種した。37℃、5% CO2で24時間培養後、培地を除去し、1% ペニシリン−ストレプトマイシンを含有する無血清F−12Kで細胞を一度洗った。次いで、細胞を1% ペニシリン−ストレプトマイシンを含有する無血清F−12K 100μL中で24時間インキュベートした。
【0319】
1% ペニシリン−ストレプトマイシンを含有する無血清F−12Kを用いて、検討対象の各抗体の10種の希釈液、即ち、6,667nM、2,222nM、741nM、247nM、82.3nM、27.4nM、9.1nM、3.0nM、1.0nM及び0.3nMの希釈液を調製した。各抗体希釈液、及び1% ペニシリン−ストレプトマイシンを含有するが抗体を含まない無血清F−12Kを別々に、1% ペニシリン−ストレプトマイシン及び500ng/mLのHGF(R&Dシステムズ社カタログ番号294−HGN−025)を含有する等容量の無血清F−12Kと混合した。次いで、これらの抗体/HGF希釈液を25℃で30分間インキュベートした。これによりHGFの最終濃度は1.25nMとなった。
【0320】
その後、1% ペニシリン−ストレプトマイシンを含有する無血清F−12Kで上記PC−3細胞を一度洗った。次に、この細胞に、1% ペニシリン−ストレプトマイシンを含有する無血清F−12Kを70μL添加した後、10mM Na3VO4(シグマ社カタログ番号S6508)の1% ペニシリン−ストレプトマイシン含有無血清F−12K溶液10μLを添加した。次いで、この細胞を37℃、5% CO2で60分間インキュベートした。このインキュベーションの後、各抗体/HGF希釈液20μLを別々に個別のウェルに添加し、最終HGF濃度50ng/mL並びに各抗体の最終濃度666.7nM、222.2nM、74.1nM、24.7nM、8.23nM、2.74nM、0.91nM、0.30nM、0.10nM及び0.03nMを得た。次に、この細胞を37℃、5%CO2で10分間インキュベートし、その時点後に培地/抗体/HGF混合物を除去してプレートを氷上に置いた。その後、細胞を、1mM Na3VO4を含有する100μL/ウェルの氷冷PBS(インビトロジェン社カタログ番号14190144)で一度洗った。次いで、この細胞を、1% OmniPur Triton X−100(ドイツメルク社(MERCK KGaA)、ダルムシュタット、ドイツ、カタログ番号9410)、50mMトリス−HCl pH8.0、100mM NaCl、0.3mM Na3VO4、1×プロテアーゼ阻害剤カクテル(シグマ社カタログ番号P8340)及び1×ホスファターゼ阻害剤カクテル2(シグマ社カタログ番号5726)からなる100μL/ウェルの氷冷溶解緩衝液中、4℃で、30分間インキュベートした。
【0321】
ビオチニル化抗ヒトHGF−R(c−met)抗体(R&Dシステムズ社カタログ番号BAF358)を、1%ウシ血清アルブミン(シグマ社カタログ番号A2153)を含有するDELFIAアッセイバッファ(Assay Buffer)(パーキンエルマー社(PerkinElmer)、トゥルク、フィンランド、カタログ番号4002−0010)で2μg/mLの濃度に希釈し、黄色ストレプトアビジンマイクロタイトレーションプレート(パーキンエルマー社カタログ番号AAAND−0005)のウェル当たり50μLのこの希釈液を添加した。次に、このプレートを揺動させながら25℃で30分間抗体と共にインキュベートした。インキュベーション後、このプレートをDELFIA洗浄液(パーキンエルマー社カタログ番号4010−0010)で洗浄し、各種PC−3細胞溶解物のそれぞれ80μLを別々に、洗浄したストレプトアビジンマイクロタイトレーションプレートの個々のウェルに添加した。
【0322】
上記のPC−3細胞溶解物を含むストレプトアビジンマイクロタイトレーションプレートを振盪しながら25℃で60分間インキュベートした後、DELFIA洗浄溶液で洗浄した。この前もってPC−3細胞溶解物とインキュベートした洗浄ストレプトアビジンマイクロタイトレーションプレートの各ウェルに、1%ウシ血清アルブミンを含有するDELFIAアッセイバッファーで希釈した600ng/mL DELFIA Eu−N1 P−tyr−100抗体(パーキンエルマー社、カタログ番号AD0159)溶液100μLを添加した。次いで、このプレートを揺動させながら25℃で60分間インキュベートした。最後に、このプレートをDELFIA洗浄溶液で洗浄した。次に、この洗浄ストレプトアビジンマイクロタイトレーションプレートの各ウェルにDELFIAエンハンスメントソリューション(Enhancement Solution)(パーキンエルマー社カタログ番号4001−0010)200μLを添加し、プレートを暗所で振盪しながら25℃で5分間インキュベートした。
【0323】
その後、ビクター3V(Victor3V)リーダ(パーキンエルマー社)でユウロピウム(Europium)プロトコルを用いてシグナルを測定した。ウィンドウズ(登録商標)(Windows(登録商標))用プリズム(Prism)4(グラフパッドソフトウェア社(GraphPad Software,Inc.)、サンディエゴ、カリフォルニア州)及びS字形用量反応式を用いてEC50値を算出した。
【0324】
nM単位のEC50としてまとめた結果を表9に示した。
【0325】
【表9】
【0326】
表9のデータから明らかなように、8種の抗体は全てPC−3細胞におけるHGF誘導性c−Metリン酸化の強力な阻害剤である。
【0327】
(実施例10)
U87MG異種移植モデルにおける腫瘍抑制
U87MG異種移植モデルを用い、本発明のマウスモノクローナル抗体の腫瘍増殖抑制能について試験した。U87MG細胞(ATCC)は、ダルベッコ改変イーグル培地(DMEM:Dulbecco’s Modified Eagle medium)を10%ウシ胎仔血清、100単位/mLペニシリン及び100μg/mLストレプトマイシンと共に含む培地を用いた、5%CO2及び95%空気を含有する雰囲気中37℃の培養で増殖させた。これらの細胞は、トリプシン−EDTAを用いて培養皿の壁から細胞を剥離することにより二次培養して維持した。
【0328】
ほぼコンフルエントの細胞をトリプシン処理により回収し、次いで、50%マトリゲル(Matrigel)(BDバイオサイエンシズ社(BD Biosciences)、カタログ番号356237)中の細胞5×106個を、7週齢雌性ICR SCIDマウス(タコニックラボラトリー社(Taconic Labs))の肩甲骨間の上背部に皮下注射した。キャリパで腫瘍の長径(L)及び短径(W)(mm)を測定した。腫瘍容積(vol.)は、容積(mm3)=L×W2/2として計算した。腫瘍が約200mm3にまで成長した時に、担腫瘍マウスを1群10匹の5群に無作為化した。1群にはPBSを投与した。その他の4群のそれぞれには、抗体1A3、1D3、1F3又は2B8のうちの1種を投与した。抗体は全て、5用量の腹腔内注射により週2回1mg/kg体重を投与した。腫瘍容積及びマウス体重を週2回記録した。スチューデントのt−検定を用いて腫瘍成長の抑制を解析した。結果は図6及び表10にまとめた。
【0329】
【表10】
【0330】
2B8投与群で部分的な退縮が達成された(図6)。1A3投与群及び1F3投与群では統計的に有意な成長抑制が認められた(表10)。1D3の場合、51%の腫瘍成長抑制(p値:0.075)が認められた。有意な体重減少は認められなかった。
【0331】
(実施例11)
U118異種移植モデルにおける腫瘍抑制
U118異種移植モデルを用い、抗体1A3、1D3、1F3及び2B8の腫瘍増殖抑制能について試験した。U118細胞(ATCC)は、U87MG細胞に関して(上記)実施例10に記載したのと同様にして増殖させた。
【0332】
使用マウスが7週齢雌性NCrヌードマウス(タコニック社)であること以外は上記実施例10に記載したのと同様にして皮下腫瘍を確立し、投与は、腫瘍が約80mm3にまで成長した時に開始した。U87MGモデルの場合と同様に、抗体は全て、4用量の腹腔内注射により週2回1mg/kg体重を投与した。腫瘍容積及びマウス体重を週2回記録した。スチューデントのt−検定を用いて腫瘍成長の抑制を解析した。結果は図7及び表11にまとめた。
【0333】
【表11】
【0334】
2B8投与群及び1A3投与群では統計的に有意な成長抑制が認められた(図7)。1F3群及び1D3群では中等度の腫瘍成長抑制が認められ、p値は、本試験で統計的に有意であると定義した0.05未満であった(表11)。有意な体重減少は認められなかった。
【0335】
(実施例12)
マウスモノクローナル抗体のヒト化
本実施例では、マウス2B8抗体のヒト化及び得られるヒト化抗体の特性化について説明する。マウス2B8重及び軽鎖可変領域は2通りの方法で「ヒト化」した。
【0336】
A. ヒト化の方法1
第一の方法では、Hwangほか (2005年) METHODS 36:p.35−42、Tanほか (2002年) J. IMMUNOL. 169:p.1119−1125及び米国特許第6,881,557号明細書に記載されている「超ヒト化」方法に基づいて、3つのヒト化重鎖可変領域及び2つのヒト化カッパ軽鎖可変領域を設計した。
【0337】
各マウス2B8CDRについてCDRの長さ及びアミノ酸組成に基づき、チョチア規定構造クラス(Chothia canonical structural class)を特定した。同じチョチア規定構造クラスの軽及び重鎖可変領域からなるヒト生殖細胞系可変領域は、国際免疫遺伝学情報システム(IMGT)ウェブサイトに記載されている既知ヒト生殖細胞系可変領域基準対立遺伝子(ワールドワイドウェブのimgt.cines.fr及びbiochem.unizh.ch/antibody/Sequences/index.htmlで入手可能)に基づいて確認した。同構造クラスのこれらヒト生殖細胞系可変領域は、CDRアミノ酸残基間の完全な同一性又は類似性を計算することによってマウス2B8可変領域と比較した。マウス2B8CDR残基と最も高い同一性及び/又は類似性を有するヒト生殖細胞系可変領域をCDR移植用に選んだ。これらのヒト生殖細胞系可変領域のフレームワーク残基を保存する一方、マウス2B8 CDR残基は、マウス2B8 CDRとヒト生殖細胞系CDRとの間で異なる対応するヒト生殖細胞系可変領域残基を置換するために用いた。次いで、2B8マウスJ領域に最も類似したヒトJ領域を上記「超ヒト化」可変領域のカルボキシル末端に付加した。次に、この「超ヒト化」可変領域のアミノ末端にシグナル配列を付加し、これらのアミノ酸配列を核酸配列に変換した。
【0338】
この完全な可変領域核酸配列は、遺伝子合成PCR法(Youngほか (2004年) NUCL. ACIDS RES. 32:e59)を用いて構築し、標準的な分子生物学的技術を使用して、(上記可変領域の下流にある)ヒト定常IgG1(G1m(17,1)アロタイプ)又はカッパ(Km(3)アロタイプ(対立遺伝子2))領域を含む(pcDNA3.2DEST(インビトロジェン社)を利用した)哺乳動物発現ベクター中にクローニングした。4種の重鎖IgG1抗体(キメラ2B8及び3種のヒト化重鎖(Hu2B8 Hv1−f.1、Hu2B8 Hv5−a.1及びHu2B8 Hv5−51.1))の全てを3種のカッパ鎖抗体(キメラ2B8及び2種のヒト化軽鎖(Hu2B8 Kv1−39.1及びHu2B8 Kv3−15.1))の全てとの考えられる全ての組合せで発現させ、12種の抗体蛋白質を作製した。次いで、これらキメラ、キメラ/ヒト化及びヒト化抗体のヒトHGFへの結合を下記のようにして測定し、その結果を図8にまとめた。免疫グロブリン重鎖可変領域と免疫グロブリン軽鎖可変領域との考えられる全ての組合せはいずれも下記の表12Aに記載した。
【0339】
【表12A】
【0340】
免疫グロブリン重鎖と免疫グロブリン軽鎖とのえられる全ての組合せはいずれも下記の表12Bに記載した。
【0341】
【表12B】
【0342】
ヒト化可変領域を含む完全長免疫グロブリン重及び軽鎖を含む考えられる抗体構築物のうちの2種について下記の通り命名した。
【0343】
sh2B8−9 (G1m(17,1))=hu2B8 Hv5−51.1(+IgG1定常領域(Glm(17,1)アロタイプ)(配列番号171)プラスhu2B8 Kv 1−39.1 (+カッパ定常領域(Km(3)アロタイプ(対立遺伝子2))) (配列番号177)
sh2B8−12 (Glm(17,1))=hu2B8 Hv5−51.1(+IgG1定常領域(Glm(l7,1)アロタイプ))(配列番号171)プラスhu2B8
Kv 3−15.1 (+カッパ定常領域(Km(3)アロタイプ(対立遺伝子2))) (配列番号181)
各ヒト化抗体をコードしている核酸配列及び各ヒト化抗体を規定する蛋白質配列を下記にまとめた。このセクションでは、各可変領域の最後のヌクレオチドは、可変/定常領域接合部によって生じる次のコドンの最初の塩基である。このヌクレオチドは、可変領域に含まれる。何故なら、これはそのエクソンの一部であるからである。下記に列挙した定常領域のアミノ酸配列はこの接合部コドンの翻訳物を含む。
【0344】
(1) 完全長キメラ2B8重鎖(マウス可変領域及びヒトIgG1定常領域)(アロタイプG1m(17,1))をコードしている核酸配列(下線:シグナル配列)(配列番号154)
【0345】
【化78】
【0346】
(2) 完全長キメラ2B8重鎖(キメラ2B8 IgG1(G1m(17,1)アロタイプ)を規定する蛋白質配列(シグナル配列なし)(配列番号155)
【0347】
【化79】
【0348】
(3) 完全長キメラ2B8軽鎖(マウス可変領域及びヒト定常領域)(キメラ2B8カッパ(Km(3)))をコードしている核酸配列(下線:シグナル配列)(配列番号156)
【0349】
【化80】
【0350】
【化81】
【0351】
(4) 完全長キメラ2B8軽鎖(キメラ2B8カッパ(Km(3)))を規定する蛋白質配列(シグナル配列なし)(配列番号157)
【0352】
【化82】
【0353】
(5) ヒト化Hu2B8 Hv1−f.1重鎖可変領域をコードしている核酸配列(下線:シグナル配列)(配列番号158)
【0354】
【化83】
【0355】
(6) ヒト化Hu2B8 Hv1−f.1重鎖可変領域を規定する蛋白質配列(シグナル配列なし)(配列番号159)
【0356】
【化84】
【0357】
(7) ヒトIgG1重鎖定常領域(G1m(17,1)アロタイプ)をコードしている核酸配列(配列番号160)
【0358】
【化85】
【0359】
(8) ヒトIgG1重鎖定常領域(G1m(17,1)アロタイプ)を規定する蛋白質配列(配列番号161) 最初のアミノ酸は、可変領域の最後のヌクレオチド及びIgG1重鎖配列の初めの2個のヌクレオチドの翻訳物に由来する。
【0360】
【化86】
【0361】
(9) 完全長重鎖ヒト化Hu2B8 Hv1f.1可変領域及びヒトIgG1(G1m(17,1)アロタイプ)重鎖定常領域をコードしている核酸配列(下線:シグナル配列)(配列番号162)
【0362】
【化87】
【0363】
(10) 完全長重鎖ヒト化Hu2B8 Hv1f.1可変領域及びヒトIgG1重鎖定常領域(G1m(17,1)アロタイプ)を規定する蛋白質配列(配列番号163)
【0364】
【化88】
【0365】
(11) ヒト化Hu2B8 Hv5a.1重鎖可変領域をコードしている核酸配列(下線:シグナル配列)(配列番号164)
【0366】
【化89】
【0367】
【化90】
【0368】
(12) ヒト化Hu2B8 Hv5a.1重鎖可変領域を規定する蛋白質配列(配列番号165)
【0369】
【化91】
【0370】
(13) 完全長ヒト化Hu2B8 Hv5a.1重鎖可変領域及びヒトIgG1(G1m(17,1)アロタイプ)重鎖定常領域をコードしている核酸配列(下線:シグナル配列)(配列番号166)
【0371】
【化92】
【0372】
(14) 完全長ヒト化Hu2B8 Hv5a.1重鎖可変領域及びヒトIgG1(G1m(17,1)アロタイプ)重鎖定常領域を規定する蛋白質配列(シグナル配列なし)(配列番号167)
【0373】
【化93】
【0374】
【化94】
【0375】
(15) ヒト化Hu2B8 Hv5−51.1重鎖可変領域をコードしている核酸配列(下線:シグナル配列)(配列番号168)
【0376】
【化95】
【0377】
(16) ヒト化Hu2B8 Hv5−51.1重鎖可変領域を規定する蛋白質配列(シグナル配列なし)(配列番号169)
【0378】
【化96】
【0379】
(17) 完全長重鎖ヒト化Hu2B8 Hv5−51.1重鎖可変領域及びヒトIgG1(G1m(17,1)アロタイプ)重鎖定常領域をコードしている核酸配列(下線:シグナル配列)(配列番号170)
【0380】
【化97】
【0381】
【化98】
【0382】
(18) 完全長重鎖ヒト化Hu2B8 Hv5−51.1重鎖可変領域及びヒトIgG1(G1m(17,1)アロタイプ)重鎖定常領域を規定する蛋白質配列(シグナル配列なし)(配列番号171)
【0383】
【化99】
【0384】
(19) ヒト化Hu2B8 Kv1−39.1カッパ鎖可変領域をコードしている核酸配列(下線:シグナル配列)(配列番号172)。考えられる2つの開始ATGを大文字で示した。
【0385】
【化100】
【0386】
(20) ヒト化Hu2B8 Kv1−39.1カッパ鎖可変領域を規定する蛋白質配列(シグナル配列なし)(配列番号173)
【0387】
【化101】
【0388】
(21) ヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子2)をコードしている核酸配列(配列番号174)
【0389】
【化102】
【0390】
【化103】
【0391】
(22) ヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子2)を規定する蛋白質配列 (配列番号175)
【0392】
【化104】
【0393】
(23) 完全長ヒト化Hu2B8 Kv1−39.1軽鎖可変領域及びヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子2)をコードしている核酸配列(下線:シグナル配列)(配列番号176)
【0394】
【化105】
【0395】
(24) 完全長ヒト化Hu2B8 Kv1−39.1軽鎖可変領域及びヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子1)を規定する蛋白質配列 (配列番号177)
【0396】
【化106】
【0397】
(25) ヒト化Hu2B8 Kv3−15.1軽鎖可変領域をコードしている核酸配列(下線:シグナル配列)(配列番号178)
【0398】
【化107】
【0399】
【化108】
【0400】
(26) ヒト化Hu2B8 Kv3−15.1軽鎖可変領域を規定する蛋白質配列(シグナル配列なし)(配列番号179)
【0401】
【化109】
【0402】
(27) 完全長ヒト化Hu2B8 Kv3−15.1軽鎖可変領域及びヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子2)をコードしている核酸配列(下線:シグナル配列)(配列番号180)
【0403】
【化110】
【0404】
(28) ヒト化Hu2B8 Kv3−15.1軽鎖可変領域及びヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子2)を規定する蛋白質配列(シグナル配列なし)(配列番号181)
【0405】
【化111】
【0406】
便宜のため、このセクションで説明した抗体の完全長配列と配列表に示したものとの対応関系を示すコンコーダンス表を表13に示した。
【0407】
【表13】
【0408】
B. ヒト化の方法2
マウス2B8抗体の免疫原性を低減させるのに用いる第二のヒト化方法は、Studnickaほか(1994年) PROTEIN ENG. 7:p.805−814に記載されている方法に基づくものである。重及びカッパヒト生殖細胞系可変領域のうち、マウス2B8のものに(アミノ酸レベルで)最も一致する領域を特定した。マウスとヒトとの間で異なる残基は、そのような変化が結合又は免疫原性に影響すると思われるリスクに応じてヒトの配列に変換した。低リスク残基(即ち、変化があっても、抗原結合に影響しないと考えられ、免疫原性を生じる可能性も少ない残基)は、ヒトアミノ酸への変更を重鎖可変領域(LR2B8HCを作製)及びカッパ可変領域(LR2B8LCを作製)において行った。さらに、低リスク及び中間リスク残基(即ち、変化があると、抗原結合に影響する可能性が若干高いが、免疫原性を生じる可能性も少ない残基)は、ヒトアミノ酸への変更を重鎖可変領域(LRMR2B8HCを作製)及びカッパ可変領域(LRMR2B8LCを作製)において行った。上記2種のヒト型に設計された重鎖可変領域のカルボキシル末端にヒトIgG1重鎖定常領域(G1m(3)アロタイプ(対立遺伝子1))を付加し、2種のヒト型に設計された軽鎖可変領域のカルボキシル末端にヒトカッパ定常領域(Km(3)アロタイプ(対立遺伝子1))を付加して、ヒト型に設計された抗体鎖を作製した。先ず、遺伝子合成法によって各可変領域の核酸配列を合成した後、これに各ヒト定常領域の配列に付加した。これらのヒト型に設計された抗体を哺乳動物の蛋白質発現ベクター中にクローニングし、重鎖プラス軽鎖の考えられる4種の組合せの蛋白質を発現させた。キメラ、キメラ/ヒト化又はヒト化抗体のヒトHGFへの結合については、下記のように、従来の技術を用いて測定した。
【0409】
各ヒト化抗体をコードしている核酸配列及び各ヒト化抗体を規定する蛋白質配列を下記にまとめた。このセクションでは、各可変領域の最後のヌクレオチドは、可変/定常領域接合部によって生じる次のコドンの最初の塩基である。このヌクレオチドは、可変領域に含まれる。何故なら、これはそのエクソンの一部であるからである。下記に列挙した定常領域のアミノ酸配列はこの接合部コドンの翻訳物を含む。
【0410】
(1) ヒト化LR2B8HC重鎖可変領域をコードしている核酸配列(下線:シグナル配列)(配列番号182)
【0411】
【化112】
【0412】
(2) ヒト化LR2B8HC重鎖可変領域を規定する蛋白質配列(シグナル配列なし)(配列番号183)
【0413】
【化113】
【0414】
(3) ヒトIgG1重鎖定常領域(G1m(3)アロタイプ)(対立遺伝子1)をコードしている核酸配列(配列番号184)
【0415】
【化114】
【0416】
(4) ヒトIgG1重鎖定常領域(G1m(3)アロタイプ)(対立遺伝子1又は2)を規定する蛋白質配列(配列番号185)。最初のアミノ酸は、可変領域の最後のヌクレオチド及びIgG1重鎖配列の初めの2個のヌクレオチドの翻訳物に由来する。
【0417】
【化115】
【0418】
【化116】
【0419】
(5) 完全長重鎖ヒト化LR2B8HC重鎖可変領域及びヒトIgG1重鎖定常領域(G1m(3)アロタイプ)(対立遺伝子1)(下線:シグナル配列)(配列番号186)
【0420】
【化117】
【0421】
(6) 完全長重鎖ヒト化LR2B8HC重鎖可変領域及びヒトIgG1重鎖定常領域(G1m(3)アロタイプ)(対立遺伝子1)を規定する蛋白質配列(シグナル配列なし)(配列番号187)
【0422】
【化118】
【0423】
(7) ヒト化LRMR2B8HC重鎖可変領域をコードしている核酸配列(下線:シグナル配列)(配列番号188)
【0424】
【化119】
【0425】
(8) ヒト化LRMR2B8HC重鎖可変領域を規定する蛋白質配列(シグナル配列なし)(配列番号189)
【0426】
【化120】
【0427】
(9) 完全長重鎖ヒト化LRMR2B8HC重鎖可変領域及びヒトIgG1重鎖定常領域(G1m(3)アロタイプ)(対立遺伝子1)をコードしている核酸配列(下線:シグナル配列)(配列番号190)
【0428】
【化121】
【0429】
(10) 完全長重鎖ヒト化LRMR2B8HC重鎖可変領域及びヒトIgG1重鎖定常領域(G1m(3)アロタイプ)(対立遺伝子1)を規定する蛋白質配列(シグナル配列なし)(配列番号191)
【0430】
【化122】
【0431】
(11) ヒト化LR2B8LC軽鎖可変領域をコードしている核酸配列(下線:シグナル配列)(配列番号192)
【0432】
【化123】
【0433】
(12) ヒト化LR2B8LC軽鎖可変領域を規定する蛋白質配列(シグナル配列なし)(配列番号193)
【0434】
【化124】
【0435】
(13) ヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子1)をコードしている核酸配列(配列番号194)。最初のアミノ酸は、可変領域の最後のヌクレオチドおよびκ軽鎖配列の開始2ヌクレオチドの翻訳物に由来する。
【0436】
【化125】
【0437】
(14) ヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子1)を規定する蛋白質配列(配列番号195)。最初のアミノ酸は、可変領域の最後のヌクレオチドおよびκ軽鎖配列の開始2ヌクレオチドの翻訳物に由来する。
【0438】
【化126】
【0439】
(15) 完全長LR2B8LC軽鎖可変領域及びヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子1)をコードしている核酸配列 (配列番号196)
【0440】
【化127】
【0441】
(16) 完全長LR2B8LC軽鎖可変領域及びヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子1)をコードしている蛋白質配列(配列番号197)
【0442】
【化128】
【0443】
(17) ヒト化LRMR2B8LC軽鎖可変領域をコードしている核酸配列(下線:シグナル配列)(配列番号198)
【0444】
【化129】
【0445】
(18) ヒト化LRMR2B8LC軽鎖可変領域を規定する蛋白質配列(シグナル配列なし)(配列番号199)
【0446】
【化130】
【0447】
(19) 完全長ヒト化LRMR2B8LC軽鎖可変領域及びヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子1)をコードしている核酸配列(下線:シグナル配列)(配列番号200)
【0448】
【化131】
【0449】
(20) 完全長ヒト化LRMR2B8LC軽鎖可変領域及びヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子1)を規定する蛋白質配列(配列番号201)
【0450】
【化132】
【0451】
便宜のため、このセクションで説明した抗体の完全長配列と配列表に示したものとの対応関系を示すコンコーダンス表を表14に示した。
【0452】
【表14】
【0453】
表15は、以上の本実施例で説明したヒト化の方法1及びヒト化の方法2により作製したヒト化2B8抗体の重鎖CDR配列(カバットの定義)をまとめたものである。
【0454】
【表15】
【0455】
表16は、以上の本実施例で説明したヒト化の方法1及びヒト化の方法2により作製したヒト化2B8抗体の軽鎖CDR配列(カバット(Kabat)の定義)をまとめたものである。
【0456】
【表16】
【0457】
C. ヒト化2B8抗体の結合親和性
BIAcoreT100装置を用いて表面プラスモン共鳴法により抗原結合親和性及び相互作用の速度を評価した。マウス抗ヒト免疫グロブリン(ジャクソン・イムノリサーチ・ラボラトリーズ社(Jackson ImmunoResearch Labs)、209−005−098)は、標準的なカップリングプロトコルをメーカーの推奨事項に従って用いたアミンカップリング(BIAcore、カタログ番号BR−1000−50)によりカルボキシメチル化デキストランCM4センサーチップ(BIAcore、カタログ番号BR−1005−34)上に固定化した。分析は、0.05%の界面活性剤P20(BIAcore、カタログ番号BR−1000−54)、2mg/mLのBSA(EMD社カタログ番号2930)及び10mg/mLのCM−デキストランナトリウム塩(フルカ社(Fluka)、カタログ番号86524)を含有するPBS(GIBCO社、カタログ番号14040−133)を泳動用緩衝液として用い、25℃で行った。
【0458】
前記抗体は、個々のフローセル上で10μL/分の流速で捕捉した。各抗体が各サイクルで捕捉される約20RUの抗体を生じるための注入時間にはばらつきがあった。緩衝液又は泳動用緩衝液で希釈したHGF(R&Dシステムズ社、カタログ番号294−HGN−025)を対照表面(捕捉抗体なし)及び活性表面(試験対象抗体あり)上に60μL/分で2分間順次注入した。解離相を濃度に応じて15又は90分間モニターした。その後、上記表面は、別のサイクルが開始される前に、pH2.0の10mMグリシン−塩酸(BIAcore社、カタログ番号BR−1003−55)を60μL/分の流速で3分間注入して再生させた。試験したHGF濃度は1.88、3.75及び7.5nMであった。速度論的パラメータは、対照値減算を含むBIAエバリュエーション(BIAevalutation)ソフトウェアの速度関数を用いて求めた。各抗体の速度論的パラメータka(会合速度定数)、kd(解離速度定数)及びKD(平衡解離定数)を図8にまとめた。
【0459】
図8にまとめた結果から、超ヒト化重鎖(Hu2B8 Hv5a.1、Hu2B8 Hv5−51.1又はHu2B8 Hvl−f.l)及び軽鎖(Hu2B8 Kvl−39.1又はHu2B8 Kv3−15.1)の一部の組合せは、キメラ2B8(マウス可変領域及びヒト定常領域)並びに2B8(表5)と同様なHGFとの結合親和性(KD)を保持することが分かる。
【0460】
D. 相互排他的結合
HGFへの相互排他的結合についてBIAcore T100装置を用いて表面プラスモン共鳴により評価した。マウス抗ヒト免疫グロブリン(ジャクソン・イムノリサーチ・ラボラトリーズ社(Jackson ImmunoResearch Labs)、209−005−098)は、標準的なカップリングプロトコルをメーカーの推奨事項に従って用いたアミンカップリング(BIAcore、カタログ番号BR−1000−50)によりカルボキシメチル化デキストランCM5センサーチップ(BIAcore、カタログ番号BR−1006−68)上に固定化した。分析は、0.05%の界面活性剤P20(BIAcore、BR−1000−54)、2mg/mLのBSA(EMD社カタログ番号2930)及び10mg/mLのCM−デキストランナトリウム塩(フルカ社(Fluka)、カタログ番号86524)を含有するPBS(GIBCO社、カタログ番号14040−133)を泳動用緩衝液として用い、25℃で行った。
【0461】
前記ヒト化抗体は、個々のフローセル上で30μL/分の流速で捕捉した。各抗体が各サイクルで捕捉される約150RUの抗体を生じるための注入時間にはばらつきがあった。7.5μg/mLの最終濃度に泳動用緩衝液で希釈したHGF(R&Dシステムズ社、カタログ番号294−HGN−025)を上記捕捉ヒト化抗体上に30μL/分で90秒間注入した。HGFの結合を測定した後、続いてマウス2B8抗体又はヤギポリクロナール抗HGF抗体(R&Dシステムズ社、AF294)を30μL/分で3分間注入した。その後、上記表面は、別の抗体が試験される前に、pH2.0の10mMグリシン−塩酸(BIAcore社、カタログ番号BR−1003−55)を60μL/分の流速で3分間注入して再生させた。得られた結果は図9にまとめた。
【0462】
図9にまとめた結果から、ヒト化2B8抗体及びキメラ2B8抗体はいずれもマウス2B8がHGFに結合するのを妨げることが分かる。以上の結果は、ヒト化抗体がそれでもなお元の2B8抗体と同じHGF抗原決定基に結合することを証明している。
【0463】
(実施例13)
ヒト化2B8変異体の作製
a. ヒューマン・エンジニアード(商標)抗体
ヒトカッパ及びガンマ−1定常領域モジュールを含むXOMA社の一過性抗体発現ベクター中に、コドン−及び発現適正化低リスク及び低プラス中等度リスクヒューマン・エンジニアード軽鎖(それぞれ、LR2B8LC及びLRMR2B8L)及び重鎖(それぞれ、LR2B8HC及びLRMR2BHC)をインフェーズでクローニングした。これら4種のヒューマン・エンジニアード2B8変異体は、HEK293E細胞への一過性形質移入によって作製した。以下の4種の抗体を作製した。
【0464】
HE2B8−1 = LR2B8HC(+IgG1定常領域(Glm(3)アロタイプ(対立遺伝子1))(配列番号187)プラスLR2B8LC(+カッパ鎖定常領域(Km(3)アロタイプ(対立遺伝子1)))(配列番号197)
HE2B8−2 = LR2B8HC(+IgGl定常領域(Glm(3)アロタイプ(対立遺伝子1))(配列番号187)プラスLRMR2B8LC(+カッパ鎖定常領域(Km(3)アロタイプ(対立遺伝子1)))(配列番号201)
HE2B8−3 = LRMR2B8HC(+IgG1定常領域(Glm(3)アロタイプ(対立遺伝子1))(配列番号191)プラスLR2B8LC(+カッパ鎖定常領域(Km(3)アロタイプ(対立遺伝子1)))(配列番号197)
HE2B8−4 = LRMR2B8HC(+IgG1定常領域(Glm(3)アロタイプ(対立遺伝子1))(配列番号191)プラスLRMR2B8LC(+カッパ鎖定常領域(Km(3)アロタイプ(対立遺伝子1)))(配列番号201)
これら軽及び重鎖は、2リットルの振盪フラスコを用いてIS293培地(アーバイン・サイエンティフィック社、アーバイン、カリフォルニア州)中で増殖させたXOMA社の懸濁適応HEK293E細胞中に同時形質移入した。振盪フラスコでの24時間の形質移入後、形質移入細胞200mLを遠心分離し、新鮮培地40mLに再懸濁し、産生用インテグラ(Integra)フラスコ(ウィルソン・ウォルフ・マニュファクチャリング社(Wilson Wolf Manufacturing Inc.)、ミネソタ州)に移した。数日間のインキュベーション後、細胞懸濁液をインテグラフラスコから取り出し、遠心分離して培養上清を残した。この培養上清中の抗体をプロテインAスピンカラム(プロ−ケム社)で精製し、PBSに対して透析し、濃縮して除菌した。
【0465】
b. スーパーヒューマナイズド(SUPERHUMANIZED)(商標)抗体
完全長Hu2B8_Hv5−51.1+ヒトIgG1定常ドメイン(G1m(3)アロタイプ)cDNAをHindIII及びEcoRI制限部位を利用してpEE6.4(ロンザバイオロジックス社(Lonza Biologics)、バークシャー、英国)中にクローニングした。完全長Hu2B8_Kv−39.1可変領域+ヒトカッパ鎖定常ドメインcDNA及び完全長Hu2B8_Kv3−15.1可変領域+ヒトカッパ鎖定常ドメインcDNAをそれぞれ、HindIII及びEcoRI制限部位を利用してpEE14.4(ロンザバイオロジックス社)中にクローニングした。(pEE6.4内の)hCMV−MIEプロモーター+完全長Hu2B8_Hv5−51.1+ヒトIgG1定常ドメイン(G1m(3)アロタイプ)cDNA+SV40ポリA断片をNotI/SalI消化により切り出し、どちらのカッパ鎖pEE14.4ベクター中にもNotI/SalI部位から挿入することにより、それぞれ重及び軽鎖を同時に発現する2種の発現ベクターを作り、以下の抗体を作製した。
【0466】
sh2B8−9(Glm(3)) = hu2B8 Hv5−51.1(+IgGl定常領域(Glm(3)アロタイプ)(対立遺伝子2))(配列番号210)プラスhu2B8 Kv 1−39.1(+カッパ鎖定常領域(Km(3)アロタイプ(対立遺伝子2)))(配列番号177)
sh2B8−12(Glm(3)) = hu2B8 Hv5−51.1(+IgG1定常領域(Glm(3)アロタイプ)(対立遺伝子 2))(配列番号210)プラスhu2B8 Kv 3−15.1(+カッパ鎖定常領域(Km(3)アロタイプ(対立遺伝子2)))(配列番号181)
上記ヒトIgG1重鎖定常領域G1m(3)アロタイプ(対立遺伝子2)及び上記各完全長重鎖配列をコードしている核酸配列並びにこれらを規定する蛋白質配列を以下に記載した。軽鎖の配列は実施例12に記載したのと同じであった。
【0467】
(1) ヒトIgG1重鎖定常領域(G1m(3)アロタイプ)(対立遺伝子2)をコードしている核酸配列(配列番号207)。
【0468】
【化133】
【0469】
【化134】
【0470】
(2) ヒトIgG1重鎖定常領域(G1m(3)アロタイプ)(対立遺伝子1又は2) を規定する蛋白質配列(配列番号208)。最初のアミノ酸は、可変領域の最後のヌクレオチドおよびIgG1重鎖配列の開始2ヌクレオチドの翻訳物に由来する。
【0471】
【化135】
【0472】
(3) ヒト化Hu2B8 Hv5−51.1重鎖可変領域及びヒトIgG1重鎖定常領域G1m(3)アロタイプ(対立遺伝子2)を含む完全長鎖をコードしている核酸配列(下線:シグナル配列)(配列番号209)
【0473】
【化136】
【0474】
【化137】
【0475】
(4) ヒト化Hu2B8 Hv5−51.1及びヒトIgG1重鎖定常領域G1m(3)アロタイプ(対立遺伝子2)を含む完全長鎖を規定する蛋白質配列(シグナル配列なし)(配列番号210)
【0476】
【化138】
【0477】
各二重発現ベクターを293T細胞中に形質移入し、DMEM 10%ウシ胎仔血清を用いて一過性に発現させた。形質移入後48時間に、無血清培地である、4mMのL−グルタミンを含有するIS GRO(商標)(アーバイン・サイエンティフィック社(Irvine Scientific)、サンタアナ、カリフォルニア州)で細胞を洗浄した後、これで置換した。上清を10日間毎日採取し、新鮮培地で置換した。この培養上清を遠心分離し、濾過(0.45μm)して10〜100倍に濃縮した。抗体は、プロステップ(ProSep)vA樹脂(ミリポア社(Millipore))で精製し、PBSに対して透析し、除菌した。
【0478】
(実施例14)
ヒト化2B8変異体の結合特性
実施例13で作製したヒト化抗体の特徴は、hHGF及び実施例3で作製した組換えHGF蛋白質に結合できることであった。
【0479】
BIAcore T100装置を用いて表面プラスモン共鳴によりこれらの抗体を分析することによってHGF及び実施例3に記載した融合蛋白質へのその結合能を評価した。各抗体は、標準的なカップリングプロトコルをメーカーの使用説明書に従って用いたアミンカップリング(BIAcore、カタログ番号BR−1000−50)によりカルボキシメチル化デキストランCM5センサーチップ(BIAcore、カタログ番号BR−1006−68)上に固定化した。
【0480】
分析は、0.05%の界面活性剤P20(BIAcore、カタログ番号R−1000−54)、2mg/mLのBSA(EMD社カタログ番号2930)及び10mg/mLのCM−デキストランナトリウム塩(フルカ社(Fluka)、カタログ番号86524)を含有するPBS(GIBCO社、カタログ番号14040−133)を泳動用緩衝液として用い、25℃で行った。種々のHGF融合蛋白質を含む上清又は空のベクターを形質移入した細胞からの上清を各抗体上に30μL/分の流速で3分間注入した。その結果生じた結合は、注入終了後30秒のベースラインに対する共鳴単位(RU)として求めた。結合は、泳動用緩衝液で希釈したヒトHGF(R&Dシステムズ社(R&D Systems)、カタログ番号294−HGN−025)と比較した。非特異的な結合は、対照表面への結合を比較することにより測定した。得られた結果は表17にまとめた。
【0481】
【表17】
【0482】
表17の結果から明らかなように、ヒト化2B8系抗体はrhHGF及び3種のマウス−ヒト−マウスキメラ抗体の全てに結合する。
【0483】
(実施例15)
ヒト化2B8変異体の結合親和性
表15に示した抗体の結合親和性及び相互作用の速度を表面プラスモン共鳴によって測定した。
【0484】
マウス抗ヒト免疫グロブリン(ジャクソン・ラボラトリーズ社(Jackson Labs)、209−005)は、標準的なカップリングプロトコルをメーカーの使用説明書に従って用いたアミンカップリング(BIAcore、カタログ番号BR−1000−50)によりカルボキシメチル化デキストランCM4センサーチップ(BIAcore、カタログ番号BR−1006−68)上に固定化した。分析は、0.05%の界面活性剤P20(BIAcore、カタログ番号BR−1000−54)及び2mg/mLのBSA(EMD社カタログ番号2930)を含有するPBS(GIBCO社、カタログ番号14040−133)を用い、25℃で行った。
【0485】
前記抗体は、個々のフローセル内で10μL/分の流速で捕捉した。各抗体が各サイクルで捕捉される約20RUの抗体を生じるための注入時間にはばらつきがあった。緩衝液又は泳動用緩衝液で希釈したHGF(R&Dシステムズ社、カタログ番号294−HGN−025)を対照表面(捕捉抗体なし)及び活性表面(試験対象抗体あり)上に60μL/分で2分間順次注入した。解離相を濃度に応じて15又は90分間モニターした。その後、上記表面は、別のサイクルが開始される前に、pH2.2の10mMグリシン−HCl(BIAcore社、カタログ番号BR−1003−54)を60μL/分の流速で3分間注入して再生させた。試験したHGF濃度は0.46nM及び7.5nMであった。
【0486】
速度論的パラメータは、対照値減算を含むBIAエバリュエーション(商標)ソフトウェアの速度関数を用いて求めた。各抗体の速度論的パラメータka(会合速度定数)、kd(解離速度定数)及びKD(平衡解離定数)を表18にまとめた。
【0487】
【表18】
【0488】
これらのデータから、これらのヒト化抗体は速い会合速度(ka)、極めて遅い解離速度(kd)及び極めて高い親和性(KD)を有することが分かる。具体的には、これらの抗体は2.0乃至12pMの範囲の親和性を有する。
【0489】
(実施例16)
25℃及び37℃における結合親和性の比較
抗体HE2B8−4、sh2B8−9、sh2B8−12及びマウス2B8の結合親和性及び相互作用の速度を種々の条件下で表面プラスモン共鳴により測定した。
【0490】
マウス抗ヒト免疫グロブリン(ジャクソン・ラボラトリーズ社(Jackson Labs)、209−005)又はウサギ抗マウス免疫グロブリン(BIAcore、カタログ番号BR−1005−14)は、標準的なカップリングプロトコルをメーカーの使用説明書に従って用いたアミンカップリング(BIAcore、カタログ番号BR−1000−50)によりカルボキシメチル化デキストランCM4センサーチップ(BIAcore、カタログ番号BR−1006−68)上に固定化した。sH2b8−9及びsh2B8−12についての25℃における測定の場合、CM5センサーチップ(BIAcore、カタログ番号BR−1006−68)を用いた。これらの分析は、0.05%の界面活性剤P20(BIAcore、カタログ番号BR−1000−54)及び2mg/mLのBSA(EMD社カタログ番号2930)を含有するPBS(GIBCO社、カタログ番号14040−133)を泳動用緩衝液として用い、25℃及び37℃で行った。
【0491】
前記抗体は、個々のフローセル内で10μL/分の流速で捕捉した。各抗体が各サイクルで捕捉される約20RUの抗体を生じるための注入時間にはばらつきがあった。緩衝液又は泳動用緩衝液で希釈したHGF(R&Dシステムズ社、カタログ番号294−HGN−025)を対照表面(捕捉抗体なし)及び活性表面(試験対象抗体あり)上に60μL/分で2分間順次注入した。解離相を濃度に応じて15又は90分間モニターした。その後、マウス抗ヒト免疫グロブリンセンサーチップの表面は、別のサイクルが開始される前に、pH2.2の10mMグリシン−HCl(BIAcore社、カタログ番号BR−1003−54)を60μL/分の流速で3分間注入して再生させた。ウサギ抗マウスグロブリンセンサーチップの表面は、別のサイクルが開始される前に、pH1.7の10mMグリシン−HCl(BIAcore社、カタログ番号BR−1003−54)を60μL/分の流速で3分間注入して再生させた。試験したHGF濃度は0.46nM及び7.5nMであった。
【0492】
速度論的パラメータは、対照値減算を含むBIAエバリュエーションソフトウェアの速度関数を用いて求めた。各抗体の速度論的パラメータka(会合速度定数)、kd(解離速度定数)及びKD(平衡解離定数)を下記の表19にまとめた。
【0493】
【表19】
【0494】
予想どおり、会合速度定数は温度の上昇と共に増大した。意外なことに、解離定数は温度の対応する上昇と共に増大することはなかった。その結果、全体としての平衡解離定数(KD)は、生理的温度(37℃)で約1.4乃至3倍小さい(高い親和性)ものであった。
【0495】
(実施例17)
ヒト化2B8変異体の中和活性
実施例14で作製した抗体について、(a)c−MetへのhHGFの結合を阻害し、(b)4MBr−5細胞におけるBrdU取り込みのHGFによる促進を阻害する能力を特性化した。
【0496】
HGF−Met結合阻害試験(中和試験)は以下のようにして実施した。c−MetへのhHGFの結合を阻害する抗体の能力についてはELISAにより試験した。すなわち、ワラック96穴DELFIAアッセイプレート(ワラック社、カタログ番号AAAND−0001)を6.25μg/mL HGF(R&Dシステムズ社、カタログ番号294−HGN−025)の炭酸コーティング緩衝液(15mM Na2CO3及び34mM NaHCO3、pH9.0)溶液100μLにより4℃で16時間コーティングした。次に、このプレートを室温で1時間5%脱脂粉乳のPBS溶液200μLでブロックした。抗体は、別のプレートにおいて5%脱脂粉乳のPBS溶液に溶解した2nMのビオチニル化c−Metに検討対象の抗体を漸増濃度(0.033乃至250nM、2倍段階希釈)で加えることにより調製した。c−Met(R&Dシステムズ社、カタログ番号358−MT/CF)のビオチニル化は、10:1のビオチン/c−Met比でメーカーの使用説明書に従って行う(ピアス社(Pierce)、カタログ番号21335)。ウェル当たり100μLのサンプルを上記アッセイプレートに移し、室温で2時間インキュベートした。次いで、上記で得られたプレートをPBS−0.1% Tween20で3回洗浄した後、DELFIAアッセイ緩衝液(ワラック社、カタログ番号4002−0010)で1:1,000に希釈したEu標識ストレプトアビジン(ワラック社、カタログ番号1244−360)と共に室温で1時間インキュベートした。その結果得られたプレートをDELFIA洗浄液(ワラック社、カタログ番号4010−0010)で3回洗浄した後、100μL/ウェルのDELFIA強化溶液(ワラック社、#4001−0010)と共に攪拌しながら室温で15分間インキュベートした。上記プレートは、ユウロピウム法を用いるビクター3V装置(パーキンエルマー社)で読み取らせた。IC50値はプリズム(Prism)を用いて算出した。
【0497】
得られたIC50値を表20に示した。
【0498】
【表20】
【0499】
この表20の結果から明らかなように、試験したヒト化抗体はc−MetへのHGFの結合を効率的に中和する。
【0500】
表17の抗体は、実施例7(b)で説明した細胞増殖アッセイにおいても試験した。その結果は下記の表21にまとめた。
【0501】
【表21】
【0502】
この表21の結果から明らかなように、試験したヒト化抗体は全て4MBr−5細胞のHGF誘発性増殖を抑制する。
【0503】
(実施例18)
ヒト化2B8変異体の抗散乱活性
表17の抗体を実施例8で説明した抗散乱アッセイにおいて試験した。その結果を下記の表22にまとめた。
【0504】
【表22】
【0505】
表22の結果から明らかなように、試験したヒト化抗体は全て、マウスモノクローナル抗体2B8と同程度にHGF誘発性散乱を抑制した。
【0506】
(実施例19)
HGF刺激c−Metリン酸化の阻害
表17の抗体を実施例9で説明したc−Metリン酸化アッセイにおいて試験した。その結果を下記の表23にまとめた。
【0507】
【表23】
【0508】
表23の結果から明らかなように、試験したヒト化抗体は、PC−3細胞においてHGF誘発性c−Metリン酸化の強力な阻害剤である。
【0509】
(実施例20)
U87MG異種移植モデルにおける腫瘍抑制
U87MG異種移植モデルを用い、本発明のヒト化モノクローナル抗体の腫瘍増殖抑制能について試験した。U87MG細胞(ATCC)は、ダルベッコ改変イーグル培地(DMEM:Dulbecco’s Modified Eagle medium)を10%ウシ胎仔血清、100単位/mLペニシリン及び100μg/mLストレプトマイシンと共に含む培地を用いた、5%CO2及び95%空気を含有する雰囲気中37℃の培養で増殖させた。これらの細胞は、トリプシン−EDTAを用いて培養皿の壁から細胞を剥離することにより二次培養して維持した。
【0510】
コンフルエントに近い細胞をトリプシン処理により回収し、次いで、50%マトリゲル(Matrigel)(BDバイオサイエンシズ社(BD Biosciences)、カタログ番号356237)中の細胞5×106個を、7週齢雌性ICR SCIDマウス(タコニックラボラトリー社(Taconic Labs))の肩甲骨間の上背部に皮下注射した。キャリパで腫瘍の長径(L)及び短径(W)(mm)を測定した。腫瘍容積(vol.)は、容積(mm3)=L×W2/2として計算した。腫瘍が約200mm3にまで成長した時に、担腫瘍マウスを1群10匹の5群に無作為化した。1群にはPBSを投与し、1群にはヒトIgG対照を投与した。その他の4群のそれぞれには、ヒト化抗体(HE2B8−1、HE2B8−2、HE2B8−3及びHE2B8−4)のうちの1種を投与した。抗体は全て、5用量の腹腔内注射により週2回0.25mg/kg体重を投与した。腫瘍容積及びマウス体重を週2回記録した。スチューデントのt−検定を用いて腫瘍成長の抑制を解析した。
【0511】
試験したヒト化抗体はインビボで活性があった。HE2B8−1では57%の腫瘍成長抑制(p値:0.02)、HE2B8−2では61%の腫瘍成長抑制(p値:0.02)、HE2B8−3では85%の腫瘍成長抑制(p値:0.0004)、HE2B8−4では74%の腫瘍成長抑制(p値:0.001)が認められた。有意な体重減少は認められなかった。
【0512】
その後の実験は、側腹部に接種された皮下U87MG腫瘍を有する雌性NCRヌードマウス(タコニックラボラトリー社)において前述のようにして実施した。各群(1群10匹)には0.5mg/kgの以下の処置剤、即ち、PBS溶媒対照、huIgG対照、HE2B8−4又はsh2B8−9のうちの1種を投与した。処置剤は、最低でも5週間、週2回腹腔内投与した。各投与群は、腫瘍成長の抑制がsh2B8−9で113%、HE2B8−4で115%と同様な腫瘍退縮を示し、最低腫瘍成長遅延日数が30日であった。両投与とも有意な体重減少がなく、忍容性良好であった。
引用による組込み
本明細書で触れた特許文献及び科学論文のそれぞれの開示内容はその全体があらゆる目的で引用により組み込まれている。
【0513】
均等物
本発明は, 当該発明の精神または主要な特徴から逸脱することなく, 他のさまざまな形式で実施することができる。そのため、前述の実施態様はあらゆる点で単なる例示にすぎず、本明細書に記載された発明に対して限定的に解釈してはならない。従って、本発明の範囲は、特許請求の範囲によって示すものであり、明細書の本文によって拘束されるものではない。また、特許請求の範囲の記載内容と同等の範囲に含まれる変形もしくは変更は、すべて本発明の範囲内のものである。
【技術分野】
【0001】
関連する出願
本願は、2006年6月2日に出願された米国仮特許出願第60/810,714号、および2006年11月21日に出願された同第60/860,509号の利益およびそれらへの優先権を主張するものであり、それらの開示は本明細書中に参考として援用される。
【0002】
発明の分野
本発明の分野は、分子生物学、免疫学及び腫瘍学の分野である。より詳細には、この分野は、ヒト肝細胞成長因子(HGF)に結合する抗体系結合蛋白質の分野である。
【背景技術】
【0003】
背景
分散因子(SF:Scatter Factor)とも呼ばれる肝細胞成長因子(HGF:Hepatocyte Growth Factor)は、主として間葉細胞により産生される多機能性ヘテロ二量体蛋白質であり、Metチロシンキナーゼ受容体を発現する細胞のエフェクタである(非特許文献1、非特許文献2)。ヒトMet受容体は「c−Met」とも呼ばれる。成熟HGFは2本のポリペプチド鎖、α鎖及びβ鎖を含有している。公表された研究によれば、HGFのc−Met受容体結合ドメインを含有するのはα鎖であることが示唆されている。
【0004】
HGFは、その同族受容体に結合すると、多くの細胞活動を媒介する。HGF−Metシグナル伝達経路は、肝臓の再生、創傷治癒、神経再生、血管形成及び悪性腫瘍に関係している。例えば、非特許文献3、非特許文献4並びに特許文献1及び特許文献2を参照されたい。HGF活性が関係する各種の疾患、例えば、特定のHGF反応性癌、を治療するために、抗体を含む多くのHGF調節剤が研究者らにより開発されてきた。例えば、特許文献3を参照されたい。
【0005】
全ての抗体に共通する基本的構造を図1に摸式図的に示した。抗体は4本のポリペプチド鎖を含有する多量体蛋白質である。これらのポリペプチド鎖のうちの2本は重鎖又はH鎖と呼ばれ、他の2本は軽鎖又はL鎖と呼ばれている。これら免疫グロブリン重鎖及び軽鎖は、鎖間ジスルフィド結合によって連結されている。免疫グロブリン重鎖同士も多くの鎖間ジスルフィド結合により連結されている。軽鎖が1箇所の可変領域(図1中のVL)と1箇所の定常領域(図1中のCL)とで構成されるのに対して、重鎖は1箇所の可変領域(図1中のVH)と少なくとも3箇所の定常領域(図1中のCH1、CH2及びCH3)とで構成される。可変領域は抗体の特異性を決定し、定常領域は他の機能を有する。
【0006】
アミノ酸及び構造上の情報から、各可変領域は4箇所の比較的保存されたフレームワーク領域又はFRに隣接して存在する3箇所の超可変領域(相補性決定領域又はCDR(complementarity determining region)とも呼ばれる)を含むことが分かっている。CDR1、CDR2及びCDR3と称されるこれら3箇所のCDRは個々の抗体の結合特異性に関与している。抗体を診断剤及び治療剤として使用する場合には、通常、その標的分子に対して最も高い結合特異性及び親和性を有する抗体を作製することが望ましい。可変領域間の相違は、抗体の特異性及び親和性に対して大きな影響を与え得ると考えられている。
【0007】
特許文献4には、カポジ肉腫の治療における抗HGF抗体の使用について開示されている。同じように、特許文献5には、治療対象の患者に抗HGF抗体を投与して腫瘍における内因性HGFの血管形成促進能を妨げることによる腫瘍の治療について開示されている。最近になって、研究者らにより、HGFのβ鎖に結合する抗体がHGF依存性腫瘍の患者に対して治療剤となる可能性を有するとの提案がなされている(非特許文献4)。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】米国特許第5,997,868号明細書
【特許文献2】米国特許第5,707,624号明細書
【特許文献3】国際公開第2005/017107号パンフレット
【特許文献4】米国特許第5,707,624号明細書
【特許文献5】米国特許第5,997,868号明細書
【非特許文献】
【0009】
【非特許文献1】Bottaro et al. (1991年) SCIENCE 251: p.802−804
【非特許文献2】Rubin et al. (1993年) BIOCHIM. BIOPHYS. ACTA 1155: p.357−371
【非特許文献3】Cao et al.(2001年) PROC. NATL. ACAD. SCI. USA 98:p.7443−7448
【非特許文献4】Burgess et al.(2006年) CANCER RES. 66:p.1721−1729
【発明の概要】
【発明が解決しようとする課題】
【0010】
それでもなお、治療剤及び診断剤として使用できるHGF調節剤がさらに求められている。
【課題を解決するための手段】
【0011】
発明の要旨
本発明の一部は、HGF、特にヒトHGFを特異的に結合する一群の結合蛋白質の発見に基づいている。これらの結合蛋白質は、HGFを特異的に結合する一群の抗体のCDRに基づく抗原(即ち、HGF)結合部位を含有する限りにおいて、抗体系である。このCDRは結合蛋白質のHGFに対する結合特異性をもたらす。こうした結合蛋白質は診断剤及び治療剤として使用することができる。治療剤として用いる場合、結合蛋白質は、レシピエント(例えば、ヒト)に投与した時にこの結合蛋白質に対する免疫反応を誘発するリスクを低減又は排除できるように設計(例えば、ヒト化)する。
結合蛋白質は、HGF活性を中和するので、治療剤として用いることができる。一部の実施態様として、この結合蛋白質は、HGFがその同族受容体c−Metに結合するのを妨げることによってHGF活性を中和する。別の実施態様として、この結合蛋白質は、HGFに結合してその生物活性を中和するが、HGFがc−Met受容体に結合するのを妨げない。HGFが癌細胞の成長及び増殖に関与しているので、この結合蛋白質は癌細胞の増殖を抑制するのに用いることができる。さらに、この結合蛋白質は、哺乳動物に投与すると、その哺乳動物における腫瘍の成長を抑制又は低減することができる。
【0012】
本発明は、例えば、以下の項目を提供する。
(項目1)
ヒト肝細胞成長因子(HGF)に結合する単離結合蛋白質であって、
(a)構造CDRL1−CDRL2−CDRL3を含む免疫グロブリン軽鎖可変領域であって、
(i)CDRL1が配列番号18(2B8)のアミノ酸配列を含み、
(ii)CDRL2が配列番号206(LRMR2B8LC)のアミノ酸配列を含み、及び
(iii)CDRL3が配列番号20(2B8)のアミノ酸配列を含む
ものとする免疫グロブリン軽鎖可変領域、並びに
(b)免疫グロブリン重鎖可変領域
を含み、該免疫グロブリン軽鎖可変領域及び該免疫グロブリン重鎖可変領域が共同してヒトHGFに結合するための単一結合部位を規定するものとする結合蛋白質。
(項目2)
CDRL1、CDRL2及びCDRL3がヒト又はヒト化免疫グロブリンのフレームワーク領域間に挿入されている項目1の結合蛋白質。
(項目3)
前記結合蛋白質が抗体又はその抗原結合断片である項目1又は2の結合蛋白質。
(項目4)
配列番号193(LR2B8LC軽鎖可変領域)又は配列番号199(LRMR2B8LC軽鎖可変領域)の免疫グロブリン軽鎖可変領域を含む項目1乃至3のいずれか1項の単離結合蛋白質。
(項目5)
配列番号197(LR2B8LC+カッパ定常(km(3)アロタイプ)(対立遺伝子1))又は配列番号201(LRMR2B8LC+カッパ定常(km(3)アロタイプ)(対立遺伝子1))の免疫グロブリン軽鎖配列を含む項目4の単離結合蛋白質。
(項目6)
ヒト肝細胞成長因子(HGF)に結合する単離結合蛋白質であって、
(a)構造CDRH1−CDRH2−CDRH3を含む免疫グロブリン重鎖可変領域であって、
(i)CDRH1が配列番号15(2B8)のアミノ酸配列を含み、
(ii)CDRH2が配列番号204(LR2B8HC)及び配列番号205(LRMR2B8HC)からなる群から選ばれる配列を含み、及び
(iii)CDRH3が配列番号17(2B8)のアミノ酸配列を含む
ものとする免疫グロブリン重鎖可変領域、並びに
(b)免疫グロブリン軽鎖可変領域
を含み、該免疫グロブリン重鎖可変領域及び該免疫グロブリン軽鎖可変領域が共同してヒトHGFに結合するための単一結合部位を規定するものとする結合蛋白質。
(項目7)
CDRH1、CDRH2及びCDRH3がヒト又はヒト化免疫グロブリンの複数のフレームワーク領域間に挿入されている項目6の結合蛋白質。
(項目8)
前記結合蛋白質が抗体又はその抗原結合断片である項目7の結合蛋白質。
(項目9)
配列番号183(LR2B8HC重鎖可変領域)又は配列番号189(LRMR2B8HC重鎖可変領域)の免疫グロブリン重鎖可変領域を含む項目6乃至8のいずれか1項の単離結合蛋白質。
(項目10)
配列番号187(LR2B8HC+IgG1定常(G1m(3)アロタイプ)(対立遺伝子1))又は配列番号191(LRMR2B8HC+IgG1定常(Glm(3)アロタイプ)(対立遺伝子1))の免疫グロブリン重鎖配列を含む項目9の単離結合蛋白質。(項目11)
さらに、構造CDRL1−CDRL2−CDRL3を含む免疫グロブリン軽鎖可変領域を含み、
(i)CDRL1が配列番号18(2B8)のアミノ酸配列を含み、
(ii)CDRL2が配列番号19(2B8)又は配列番号206(LRMR2B8LC)のアミノ酸配列を含み、及び
(iii)CDRL3が配列番号20(2B8)のアミノ酸配列を含むものとし、該免疫グロブリン軽鎖可変領域及び前記免疫グロブリン重鎖可変領域が共同してヒトHGFに結合するための単一結合部位を規定するものとする項目6の単離結合蛋白質。
(項目12)
前記抗体がモノクローナル抗体である項目1、6又は11のいずれか1項の結合蛋白質。
(項目13)
項目1乃至5のいずれか1項の免疫グロブリン軽鎖可変領域をコードしているヌクレオチド配列を含む単離核酸。
(項目14)
項目6乃至10のいずれか1項の免疫グロブリン重鎖可変領域をコードしているヌクレオチド配列を含む単離核酸。
(項目15)
項目13又は14の核酸配列を含む発現ベクター。
(項目16)
項目15の発現ベクターを含む宿主細胞。
(項目17)
結合蛋白質を作製する方法であって、
(i)項目16の宿主細胞を該宿主細胞が前記免疫グロブリン軽鎖可変領域及び/又は前記免疫グロブリン重鎖可変領域を発現するような条件下で増殖させ、及び
(ii)該免疫グロブリン軽鎖可変領域及び/又は該免疫グロブリン重鎖可変領域を採取すること
を含む方法。
(項目18)
前記結合蛋白質が4.0×10−5s−1以下のkdでヒト肝細胞成長因子に結合する項目1乃至11のいずれか1項の単離結合蛋白質。
(項目19)
前記kdが3.0×10−5s−1以下である項目18の単離結合蛋白質。
(項目20)
前記kdが2.0×10−5s−1以下である項目19の単離結合蛋白質。
(項目21)
前記結合蛋白質が20pM以下のKDでヒト肝細胞成長因子に結合する項目1乃至11のいずれか1項の単離結合蛋白質。
(項目22)
前記KDが10pM以下である項目21の単離結合蛋白質。
(項目23)
前記KDが5pM以下である項目22の単離結合蛋白質。
(項目24)
腫瘍細胞に有効量の項目1乃至11のいずれか1項の結合蛋白質を作用させて該腫瘍細胞の増殖を抑制又は低減させることを含む腫瘍細胞の増殖を抑制又は低減させる方法。
(項目25)
前記腫瘍細胞がヒト腫瘍細胞である項目24の方法。
(項目26)
哺乳動物において腫瘍成長を抑制又は低減させる方法であって、該哺乳動物に有効量の項目1乃至11のいずれか1項の結合蛋白質を作用させて該腫瘍細胞の増殖を抑制又は低減させることを含む方法。
(項目27)
哺乳動物において腫瘍を治療する方法であって、有効量の項目1乃至11のいずれか1項の結合蛋白質を投与することを含む方法。
(項目28)
前記哺乳動物がヒトである項目26又は27の方法。
本発明の以上その他の態様及び効果は、以下の図、詳細な説明及び特許請求の範囲を考慮すれば明瞭に理解されよう。
【図面の簡単な説明】
【0013】
本発明は、以下の図面にてさらに完全な理解が可能となろう。
【図1】代表的な抗体を模式図化したものである。
【図2】1A3、1D3、1F3、2B8、2F8、3A12、3B6及び3D11で表した抗体の完全な免疫グロブリン重鎖可変領域を規定するアミノ酸配列を示す模式図である。各抗体のアミノ酸配列は全て相互に整列させてあり、シグナルペプチド、CDR1、CDR2及びCDR3を規定する領域は枠で囲んで識別されている。枠で囲んでない配列はFR配列を表す。
【図3】図2に示した免疫グロブリン重鎖可変領域配列のそれぞれのCDR1、CDR2及びCDR3を示す模式図である。
【図4】1A3、1D3、1F3、2B8、2F8、3A12、3B6及び3D11で表した抗体の完全な免疫グロブリン軽鎖可変領域を規定するアミノ酸配列を示す模式図である。各抗体のアミノ酸配列は全て相互に整列させてあり、シグナルペプチド、CDR1、CDR2及びCDR3を規定する領域は枠で囲んで識別されている。枠で囲んでない配列はFR配列を表す。
【図5】図4に示した免疫グロブリン軽鎖可変領域配列のそれぞれのCDR1、CDR2及びCDR3を示す模式図である。
【図6】U87MG異種移植モデルにおいて抗HGF抗体1D3、1F3、1A3及び2B8の腫瘍抑制活性を測定する実験からの結果をまとめたグラフである。菱形はPBS、三角形は抗HGF抗体1A3、Xは抗HGF抗体1D3、四角形は抗HGF抗体1F3、丸は抗HGF抗体2B8を示す。
【図7】U118異種移植モデルにおいて抗HGF抗体1D3、1F3、1A3及び2B8の腫瘍抑制活性を測定する実験からの結果をまとめたグラフである。菱形はIgG、四角形は抗HGF抗体1F3、三角形は抗HGF抗体1D3、Xは抗HGF抗体1A3、丸は抗HGF抗体2B8に相当する。
【図8】ヒトHGFとキメラ、キメラ/ヒト化又はヒト化2B8抗体との間の抗原結合親和性及び相互作用速度に関する表面プラスモン共鳴データをまとめた一覧表である。この表には試験したカッパ鎖とIgG1重鎖との対が示されている。標準偏差(STDEV)が示されている抗体は3回の独立した実験において解析したものである。
【図9】Hu2B8がマウスモノクローナル抗体2B8に対して互いに限定される抗原決定基に結合することを示す実験データをまとめた棒グラフである。ヒト化又はキメラ2B8を抗ヒトFcチップ上に捕捉した。次いで、HGFをこのヒト化又はキメラ2B8に結合させた。次に、マウス2B8又は対照抗体(ポリクロナールヤギ抗HGF抗体)の上記捕捉HGFへの結合能を測定した。その結果、ヒト化2B8抗体及びキメラ2B8はマウス2B8がHGFに結合するのを妨げた。白色棒はキメラ2B8抗体、灰色棒はヒト化Hu2B8抗体(カッパ可変領域Kv1−39.1及び重鎖可変領域Hv5−51.1)、黒色棒はヒト化Hu2B8抗体(カッパ可変領域Kv3−15.1及び重鎖可変領域Hv5−51.1)を示す。
【発明を実施するための形態】
【0014】
本発明の一部は、HGFを特異的に結合し、その活性を中和する一群の結合蛋白質、特にヒトHGFの発見に基づいている。こうした結合蛋白質は種々の診断的、治療的用途に使用することができる。この結合蛋白質は、HGFに結合し、その活性を中和する能力から選択された特定のモノクローナル抗体の抗原結合部位をベースとしている。具体的には、この結合蛋白質は、HGFに対する結合部位を共同して規定する免疫グロブリン可変領域CDR配列を含有している。
【0015】
こうした抗体は、その中和活性を考慮すると、HGF反応性細胞、例えば、癌細胞の成長及び/又は増殖を調節するのに特に有用である。この結合蛋白質は、治療剤として用いる場合には、レシピエントに投与した時にこの蛋白質に対する免疫反応を誘発するリスクをできる限り少なくしたり排除することができるように設計することができる。さらに、この結合蛋白質は、特定の用途に応じて、他の成分、例えば、検出可能な標識(例えば、放射能標識)並びにエフェクタ分子(例えば、他の蛋白質及び低分子系治療剤)に結合させることができることが企図されている。本発明のこれらの特徴及び態様については、それぞれ以下でさらに詳細に論じる。
【0016】
I. HGFに結合する結合蛋白質
一態様として、本発明はヒトHGFに結合する単離結合蛋白質を提供する。この結合蛋白質は、(i)CDRL1−CDRL2−CDRL3の構造を含む免疫グロブリン軽鎖可変領域及び(ii)3箇所の相補性決定領域(CDR)を含む免疫グロブリン重鎖可変領域を含む。この場合、免疫グロブリン軽鎖可変領域及び免疫グロブリン重鎖可変領域はヒトHGFに結合するための単一結合部位を共同して規定するものとする。CDRL1は、アミノ酸配列X1X2SerX4X5X6X7X8X9X10X11X12X13X14X15であって、アミノ酸X1がArg、Lys又はSerであり、X2がAla又はThrであり、X4がGlu、Gln又はSerであり、X5がAsn、Asp又はSerであり、X6がIle又はValであり、X7がAsp、Lys、Ser、Val又はTyrであり、X8がペプチド結合又はTyrであり、X9がペプチド結合又はAspであり、X10がペプチド結合又はGlyであり、X11がペプチド結合又はAsnであり、X12がペプチド結合、Ile又はSerであり、X13がAsn又はTyrであり、X14がIle、Leu、Met又はValであり、X15がAla、Asn、His又はSerであるものとするアミノ酸配列を含む。CDRL2は、アミノ酸配列X16X17X18X19X20X21X22であって、アミノ酸X16がAla、Asp、Arg、Gly又はValであり、X17がAla、Thr又はValであり、X18がAsn、Ser又はThrであり、X19がArg、Asn、Lys又はHisであり、X20がLeu又はArgであり、X21がAla、Asn、Glu、Val又はProであり、X22がAsp、Ser又はThrであるものとするアミノ酸配列を含む。CDRL3は、アミノ酸配列X23X24X25X26X27X28ProX30Thrであって、アミノ酸X23がLeu、Gly又はGlnであり、X24がHis又はGlnであり、X25がPhe、Ser、Trp又はTyrであり、X26がAsp、Ile、Ser、Trp又はTyrであり、X27がGly、Glu、Asn又はSerであり、X28がAsp、Asn、Phe、Thr又はTyrであり、X30がLeu、Phe、Pro又はTyrであるものとするアミノ酸配列を含む。
【0017】
別の態様として、本発明は、(i)CDRH1−CDRH2−CDRH3の構造を含む免疫グロブリン重鎖可変領域及び(ii)3箇所の相補性決定領域(CDR)を含む免疫グロブリン軽鎖可変領域を含む、ヒトHGFに結合する単離結合蛋白質であって、これら免疫グロブリン軽鎖可変領域及び免疫グロブリン重鎖可変領域がヒトHGFに結合するための単一結合部位を共同して規定するものとする単離結合蛋白質を提供する。CDRH1は、アミノ酸配列X1TyrX3X4X5であって、アミノ酸X1がAsp、Asn、Ser又はThrであり、X3がPhe、Ser、Trp又はTyrであり、X4がIle、Leu又はMetであり、X5がAsn、His又はSerであるものとするアミノ酸配列を含む。CDRH2は、アミノ酸配列X6IleX8X9X10X11GlyX13X14X15TyrX17X18X19X20X21X22であって、アミノ酸X6がLys、Gln、Glu、Val又はTyrであり、X8がAsn、Gly、Ser、Trp又はTyrであり、X9がAla、Pro又はSerであり、X10がGly又はThrであり、X11がペプチド結合、Asp、Asn、Gly又はSerであり、X13がAsp、Asn、His又はSerであり、X14がSer又はThrであり、X15がAsn又はTyrであり、X17がAsn又はProであり、X18がAla、Asp、Gly、Gln、Glu、Pro又Serであり、X19がAsn、Lys、Met又はSerであり、X20がLeu、Phe又はValであり、X21がLys、Met又はGlnであり、X22がAsp、Gly又はSerであるものとするアミノ酸配列を含む。CDRH3は、アミノ酸配列X23X24X25X26X27X28X29X30X31X32X33X34Tyrであって、アミノ酸X23がArg、Asn、Gln又はGluであり、X24がGly、Leu、Arg又はTyrであり、X25がペプチド結合、Asp又はGlyであり、X26がペプチド結合又はGlyであり、X27がペプチド結合又はTyrであり、X28がペプチド結合、Leu又はTyrであり、X29がペプチド結合、Gly、Leu、Arg又はValであり、X30がペプチド結合、Asp、Gly又はGluであり、X31がペプチド結合、Asn、Arg、Ser又はTyrであり、X32がペプチド結合、Ala、Gly、Ile又Tyrであり、X33がMet又はPheであり、X34がAla又はAspであるものとするアミノ酸配列を含む。
【0018】
上記結合蛋白質が上述の免疫グロブリン軽鎖及び免疫グロブリン重鎖の配列又はこれらの断片を共に含み得ることは理解されよう。さらに、上記結合蛋白質が完全な状態の抗体又はその抗原結合性断片或いは生合成抗体結合部位であってもよいことも理解されよう。
【0019】
一部の実施態様として、上記の免疫グロブリン軽鎖及び免疫グロブリン重鎖のCDR配列はフレームワーク領域(FR:framework region)と共に挿入されている。
【0020】
一部の実施態様として、上記の免疫グロブリン軽鎖及び免疫グロブリン重鎖のCDR配列は、複数のヒト又はヒト化フレームワーク領域間に挿入されている。
【0021】
別の態様として、本発明はヒトHGFに特異的に結合する単離結合蛋白質を提供する。この結合蛋白質は、(a)CDRL1−CDRL2−CDRL3の構造を有する免疫グロブリン軽鎖可変領域及び(b)免疫グロブリン重鎖可変領域を含む。この場合、免疫グロブリン軽鎖可変領域及び免疫グロブリン重鎖可変領域はヒトHGFに結合するための単一結合部位を共同して規定するものとする。上記CDRL1は、配列番号8(1A3)、配列番号18(2B8)、配列番号28(2F8)、配列番号38(3B6)、配列番号48(3D11)、配列番号58(1D3)、配列番号68(1F3)及び配列番号78(3A12)からなる群から選ばれる配列を含む。上記CDRL2は、配列番号9(1A3)、配列番号19(2B8)、配列番号29(2F8)、配列番号39(3B6)、配列番号49(3D11)、配列番号59(1D3)、配列番号69(1F3)、配列番号79(3A12)及び配列番号206(LRMR2B8LC)からなる群から選ばれる配列を含む。上記CDRL3は、配列番号10(1A3)、配列番号20(2B8)、配列番号30(2F8)、配列番号40(3B6)、配列番号50(3D11)、配列番号60(1D3)、配列番号70(1F3)及び配列番号80(3A12)からなる群から選ばれる配列を含む。本明細書及び特許請求の範囲の全体を通じて、特定の配列番号により示した配列の後に続けて、その特定の配列が由来する抗体を括弧内に示した。例えば、配列番号8(1A3)は、配列番号8の配列が抗体1A3に存在する配列に基づいていることを意味している。
【0022】
一実施態様として、上記結合蛋白質は、配列番号8(1A3)の配列を含むCDRL1、配列番号9(1A3)の配列を含むCDRL2及び配列番号10(1A3)を含むCDRL3を含む免疫グロブリン軽鎖可変領域を含む。
【0023】
別の実施態様として、上記結合蛋白質は、配列番号18(2B8)の配列を含むCDRL1、配列番号19(2B8)又は配列番号206(LRMR2B8LC)の配列を含むCDRL2及び配列番号20(2B8)を含むCDRL3を含む免疫グロブリン軽鎖可変領域を含む。
別の実施態様として、上記結合蛋白質は、配列番号28(2F8)の配列を含むCDRL1、配列番号29(2F8)の配列を含むCDRL2及び配列番号30(2F8)を含むCDRL3を含む免疫グロブリン軽鎖可変領域を含む。
【0024】
別の実施態様として、上記結合蛋白質は、配列番号38(3B6)の配列を含むCDRL1、配列番号39(3B6)配列を含むCDRL2及び配列番号40(3B6)を含むCDRL3を含む免疫グロブリン軽鎖可変領域を含む。
【0025】
別の実施態様として、上記結合蛋白質は、配列番号48(3D11)の配列を含むCDRL1、配列番号49(3D11)配列を含むCDRL2及び配列番号50(3D11)を含むCDRL3を含む免疫グロブリン軽鎖可変領域を含む。
【0026】
別の実施態様として、上記結合蛋白質は、配列番号58(1D3)の配列を含むCDRL1、配列番号59(1D3)配列を含むCDRL2及び配列番号60(1D3)を含むCDRL3を含む免疫グロブリン軽鎖可変領域を含む。
【0027】
別の実施態様として、上記結合蛋白質は、配列番号68(1F3)の配列を含むCDRL1、配列番号69(1F3)配列を含むCDRL2及び配列番号70(1F3)を含むCDRL3を含む免疫グロブリン軽鎖可変領域を含む。
【0028】
別の実施態様として、上記結合蛋白質は、配列番号78(3A12)の配列を含むCDRL1、配列番号79(3A12)配列を含むCDRL2及び配列番号80(3A12)を含むCDRL3を含む免疫グロブリン軽鎖可変領域を含む。
【0029】
前記の各実施態様において、CDRL1、CDRL2及びCDRL3の配列は、ヒト又はヒト化免疫グロブリンのFR間に挿入されていることが好ましい。上記結合蛋白質を完全な状態の抗体又はその抗原結合性断片或いは生合成抗体結合部位とすることができることは理解されよう。
【0030】
別の態様として、本発明はヒトHGFに結合する単離結合蛋白質を提供する。この結合蛋白質は、(a)CDRH1−CDRH2−CDRH3の構造を含む免疫グロブリン重鎖可変領域及び(b)免疫グロブリン軽鎖可変領域を含有する。この場合、これら免疫グロブリン重鎖可変領域及び免疫グロブリン軽鎖可変領域は、ヒトHGFに結合するための単一結合部位を共同して規定するものとする。上記CDRH1は、配列番号5(1A3)、配列番号15(2B8)、配列番号25(2F8)、配列番号35(3B6)、配列番号45(3D11)、配列番号55(1D3)、配列番号65(1F3)及び配列番号75(3A12)からなる群から選ばれる配列を含み、上記CDRH2は、配列番号6(1A3)、配列番号16(2B8)、配列番号26(2F8)、配列番号36(3B6)、配列番号46(3D11)、配列番号56(1D3)、配列番号66(1F3)、配列番号76(3A12)、配列番号202(Hu2B8 Hv1f.1)、配列番号203(Hu2B8 Hv5a.1又はHu2B8 Hv5−51.1)、配列番号204(LR2B8HC)及び配列番号205(LRMR2B8HC)からなる群から選ばれる配列を含み、上記CDRH3は、配列番号7(1A3)、配列番号17(2B8)、配列番号27(2F8)、配列番号37(3B6)、配列番号47(3D11)、配列番号57(1D3)、配列番号67(1F3)及び配列番号77(3A12)からなる群から選ばれる配列を含む。
【0031】
一実施態様として、上記結合蛋白質は、配列番号5(1A3)の配列を含むCDRH1、配列番号6(1A3)の配列を含むCDRH2及び配列番号7(1A3)を含むCDRH3を含む免疫グロブリン重鎖可変領域を含む。
【0032】
別の実施態様として、上記結合蛋白質は、配列番号15(2B8)の配列を含むCDRH1、配列番号16(2B8)、配列番号202(Hu2B8 Hv1f.1)、配列番号203(Hu2B8 Hv5a.1又はHu2B8 Hv5−51.1)、配列番号204(LR2B8HC)又は配列番号205(LRMR2B8HC)の配列を含むCDRH2及び配列番号17(2B8)を含むCDRH3を含む免疫グロブリン重鎖可変領域を含む。
【0033】
別の実施態様として、上記結合蛋白質は、配列番号25(2F8)の配列を含むCDRH1、配列番号26(2F8)の配列を含むCDRH2及び配列番号27(2F8)を含むCDRH3を含む免疫グロブリン重鎖可変領域を含む。
【0034】
別の実施態様として、上記結合蛋白質は、配列番号35(3B6)の配列を含むCDRH1、配列番号36(3B6)配列を含むCDRH2及び配列番号37(3B6)を含むCDRH3を含む免疫グロブリン重鎖可変領域を含む。
【0035】
別の実施態様として、上記結合蛋白質は、配列番号45(3D11)の配列を含むCDRH1、配列番号46(3D11)配列を含むCDRH2及び配列番号47(3D11)を含むCDRH3を含む免疫グロブリン重鎖可変領域を含む。
【0036】
別の実施態様として、上記結合蛋白質は、配列番号55(1D3)の配列を含むCDRH1、配列番号56(1D3)配列を含むCDRH2及び配列番号57(1D3)を含むCDRH3を含む免疫グロブリン重鎖可変領域を含む。
【0037】
別の実施態様として、上記結合蛋白質は、配列番号65(1F3)の配列を含むCDRH1、配列番号66(1F3)配列を含むCDRH2及び配列番号67(1F3)を含むCDRH3を含む免疫グロブリン重鎖可変領域を含む。
【0038】
別の実施態様として、上記結合蛋白質は、配列番号75(3A12)の配列を含むCDRH1、配列番号76(3A12)配列を含むCDRH2及び配列番号77(3A12)を含むCDRH3を含む免疫グロブリン重鎖可変領域を含む。
【0039】
前記の各実施態様において、CDRH1、CDRH2及びCDRH3の配列は、ヒト又はヒト化免疫グロブリンのFR間に挿入されていることが好ましい。上記結合蛋白質を完全な状態の抗体又はその抗原結合性断片或いは生合成抗体結合部位とすることができることは理解されよう。
【0040】
別の態様として、本発明はヒトHGFに結合する結合蛋白質を提供する。この結合蛋白質は、配列番号2(1A3)の20乃至141番目残基、配列番号12(2B8)の20乃至137番目残基、配列番号22(2F8)の20乃至137番目残基、配列番号32(3B6)の20乃至139番目残基、配列番号42(3D11)の20乃至132番目残基、配列番号52(1D3)の20乃至141番目残基、配列番号62(1F3)の20乃至141番目残基及び配列番号72(3A12)の20乃至141番目残基からなる群から選ばれる免疫グロブリン重鎖可変領域、並びに配列番号4(1A3)の21乃至127番目残基、配列番号14(2B8)の21乃至127番目残基、配列番号24(2F8)の20乃至131番目残基、配列番号34(3B6)の23乃至129番目残基、配列番号44(3D11)の23乃至128番目残基、配列番号54(1D3)の21乃至127番目残基、配列番号64(1F3)の21乃至127番目残基及び配列番号74(3A12)の21乃至127番目残基からなる群から選ばれる免疫グロブリン軽鎖可変領域を含む。
【0041】
別の実施態様として、上記結合蛋白質は、配列番号2(1A3)の20乃至141番目残基のアミノ酸配列を含む免疫グロブリン重鎖可変領域及び配列番号4(1A3)の21乃至127番目残基のアミノ酸配列を含む免疫グロブリン軽鎖可変領域を含む。
【0042】
一実施態様として、上記結合蛋白質は、配列番号12(2B8)の20乃至137番目残基のアミノ酸配列を含む免疫グロブリン重鎖可変領域及び配列番号14(2B8)の21乃至127番目残基のアミノ酸配列を含む免疫グロブリン軽鎖可変領域を含む。
【0043】
別の実施態様として、上記結合蛋白質は、配列番号22(2F8)の20乃至137番目残基のアミノ酸配列を含む免疫グロブリン重鎖可変領域及び配列番号24(2F8)の20乃至131番目残基のアミノ酸配列を含む免疫グロブリン軽鎖可変領域を含む。
【0044】
別の実施態様として、上記結合蛋白質は、配列番号32(3B6)の20乃至139番目残基のアミノ酸配列を含む免疫グロブリン重鎖可変領域及び配列番号34(3B6)の23乃至129番目残基のアミノ酸配列を含む免疫グロブリン軽鎖可変領域を含む。
【0045】
別の実施態様として、上記結合蛋白質は、配列番号42(3D11)の20乃至132番目残基のアミノ酸配列を含む免疫グロブリン重鎖可変領域及び配列番号44(3D11)の23乃至128番目残基のアミノ酸配列を含む免疫グロブリン軽鎖可変領域を含む。
【0046】
別の実施態様として、上記結合蛋白質は、配列番号52(1D3)の20乃至141番目残基のアミノ酸配列を含む免疫グロブリン重鎖可変領域及び配列番号54(1D3)の21乃至127番目残基のアミノ酸配列を含む免疫グロブリン軽鎖可変領域を含む。
【0047】
別の実施態様として、上記結合蛋白質は、配列番号62(1F3)の20乃至141番目残基のアミノ酸配列を含む免疫グロブリン重鎖可変領域及び配列番号64(1F3)の21乃至127番目残基のアミノ酸配列を含む免疫グロブリン軽鎖可変領域を含む。
【0048】
別の実施態様として、上記結合蛋白質は、配列番号72(3A12)の20乃至141番目残基のアミノ酸配列を含む免疫グロブリン重鎖可変領域及び配列番号74(3A12)の21乃至127番目の残基アミノ酸配列を含む免疫グロブリン軽鎖可変領域を含む。
【0049】
前記の各実施態様において、結合蛋白質は完全な状態の抗体又はその抗原結合性断片或いは生合成抗体結合部位とすることができる。
【0050】
別の態様として、本発明はヒトHGFに結合する単離結合蛋白質を提供する。この結合蛋白質は、(i)配列番号173(Hu2B8 Kv1−39.1軽鎖可変領域)、配列番号179(Hu2B8 Kv3−15.1軽鎖可変領域)、配列番号193(LR2B8LC軽鎖可変領域)及び配列番号199(LRMR2B8LC軽鎖可変領域)からなる群から選ばれる免疫グロブリン軽鎖可変領域、並びに(ii)配列番号159(Hu2B8 Hv1f.1重鎖可変領域)、配列番号165(Hu2B8 Hv5a.1重鎖可変領域)、配列番号169(Hu2B8 Hv5−51.1重鎖可変領域)、配列番号183(LR2B8HC重鎖可変領域)及び配列番号189(LRMR2B8LC軽鎖可変領域)からなる群から選ばれる免疫グロブリン重鎖可変領域を含有する。上記結合蛋白質は、完全な状態の抗体又はその抗原結合性断片或いは生合成抗体結合部位とすることができる。
【0051】
別の態様として、本発明はヒトHGFに結合する単離結合蛋白質を提供する。この結合蛋白質は、(i)配列番号177(Hu2B8 Kv1−39.1+カッパ定常(Km(3)アロタイプ(対立遺伝子2))、配列番号181(Hu2B8 Kv3−15.1+カッパ定常(Km(3)アロタイプ(対立遺伝子2))、配列番号197(LR2B8LC+カッパ定常(Km(3)アロタイプ(対立遺伝子1))及び配列番号201(LRMR2B8LC+カッパ定常(Km(3)アロタイプ(対立遺伝子1))からなる群から選ばれる免疫グロブリン軽鎖、並びに(ii)配列番号163(Hu2B8 Hv1f.1+IgG1定常(G1m(17,1)アロタイプ))、配列番号167(Hu2B8 Hv5a.1+IgG1定常領域(G1m(17,1)アロタイプ))、配列番号171(Hu2B8 Hv5−51.1+IgG1定常(G1m(17,1)アロタイプ))、配列番号187(LR2B8HC+IgG1定常(G1m(3)アロタイプ)(対立遺伝子1))及び配列番号191(LRMR2B8HC+IgG1定常(G1m(3)アロタイプ)(対立遺伝子1))からなる群から選ばれる免疫グロブリン重鎖を含む。上記結合蛋白質は、完全な状態の抗体又はその抗原結合性断片或いは生合成抗体結合部位とすることができる。
【0052】
別の態様として、本発明は還元型ヒトHGFに結合する単離結合蛋白質を提供する。この結合蛋白質は、(i)3箇所のCDRを含む免疫グロブリン軽鎖可変領域及び(ii)3箇所のCDRを含む免疫グロブリン重鎖可変領域を含む。これらのCDRは、通常FR間に挿入される。免疫グロブリン軽鎖及び免疫グロブリン重鎖のCDRは、還元型ヒトHGF、例えば、還元型HGFのα鎖に結合する結合部位を共同して規定する。還元型HGFとは、α鎖とβ鎖との間のジスルフィド結合を還元するのに十分な量の還元剤、例えば、ジチオスレイトール(DTT:dithiothreitol)、2−メルカプトエタノール又はグルタチオンで処理したHGFのことをいう。典型的な濃度としては、例えば、100mMのDTT及び5%の2−メルカプトエタノールが挙げられる。
【0053】
一部の実施態様として、上記結合蛋白質は、CDRL1、CDRL2及びCDRL3からなる群から選ばれる少なくとも1つのCDRを含む免疫グロブリン軽鎖可変領域を含む。上記結合蛋白質は、任意選択的に、2つのCDR、例えば、CDRL1とCDRL2又はCDRL1とCDRL3又はCDRL1とCDRL3を含む。上記結合蛋白質は、任意選択的に、3つのCDRの全て、即ち、CDRL1、CDRL2及びCDRL3を含む。CDRL1は、アミノ酸配列X1X2SerX4X5X6X7X8X9X10X11X12X13X14X15であって、アミノ酸X1がArg又はLysであり、X2がAla又はThrであり、X4がGlu又はGlnであり、X5がAsn、Ser又はAspであり、X6がIle又はValであり、X7がTyr、Asp又はLysであり、X8がペプチド結合又はTyrであり、X9がペプチド結合又はAspであり、X10がペプチド結合又はGlyであり、X11がペプチド結合又はAsnであり、X12がペプチド結合又はSerであり、X13がAsn又はTyrであり、X14がIle又はLeuであり、X15がAla、Asn又はSerであるものとする配列を含む。CDRL2は、アミノ酸配列X16X17X18X19LeuX21X22であって、アミノ酸X16がAla、Asp、Val又はArgであり、X17がAla又はValであり、X18がAsn、Ser又はThrであり、X19がArg、Asn又はHisであり、X21がAla、Glu、Val又はProであり、X22がAsp又はSerであるものとする配列を含む。CDRL3は、アミノ酸配列X23X24X25X26X27X28ProX30Thrであって、アミノ酸X23がLeu又はGlnであり、X24がHis又はGlnであり、X25がPhe、Ser又はTyrであり、X26がAsp、Ile又はTrpであり、X27がGly又はGluであり、X28がAsp、Phe又はThrであり、X30がPhe、Pro又はTyrであるものとする配列を含む。
【0054】
別の実施態様として、上記結合蛋白質は、CDRH1、CDRH2及びCDRH3からなる群から選ばれる少なくとも1つのCDRを含む免疫グロブリン重鎖可変領域を含む。上記結合蛋白質は、任意選択的に、2つのCDR、例えば、CDRH1とCDRH2又はCDRH1とCDRH3又はCDRH1とCDRH3を含む。上記結合蛋白質は、任意選択的に、3つのCDRの全て、即ち、CDRH1、CDRH2及びCDRH3を含む。CDRH1は、アミノ酸配列X1TyrX3X4X5であって、アミノ酸X1がAsp、Asn、Ser又はThrであり、X3がPhe、Trp又はTyrであり、X4がIle又はMetであり、X5がAsn、His又はSerであるものとする配列を含む。CDRH2は、アミノ酸配列X6IleX8X9GlyX11GlyX13X14X15TyrX17X18X19X20LysX22であって、アミノ酸X6がLys、Gln又はTyrであり、X8がGly、Ser又はTyrであり、X9がPro又はSerであり、X11がAsp、Gly又はSerであり、X13がAsp又はSerであり、X14がSer又はThrであり、X15がAsn又はTyrであり、X17がAsn又はProであり、X18がAla、Asp、Gly又はGluであり、X19がAsn、Met又はSerであり、X20がPhe又はValであり、X22がAsp又はGlyであるものとする配列を含む。CDRH3は、アミノ酸配列X23X24X25X26X27X28X29X30X31X32X33AspTyrであって、アミノ酸X23がArg又はGlnであり、X24がGly又はLeuであり、X25がAsp、Gly又はペプチド結合であり、X26がGly又はペプチド結合であり、X27がペプチド結合又はTyrであり、X28がLeu、ペプチド結合又はTyrであり、X29がGly、Arg又はLeuであり、X30がAsp、Gly又はGluであり、X31がTyr、Arg又はAsnであり、X32がAla、Gly又はTyrであり、X33がMet又はPheであるものとする配列を含む。
【0055】
上記結合蛋白質が上述の免疫グロブリン重鎖及び免疫グロブリン軽鎖の配列又はこれらの断片を含有し得ることは理解されよう。さらに、上記結合蛋白質は完全な状態の抗体又はその抗原結合性断片或いは生合成抗体結合部位とすることができることも理解されよう。
【0056】
一部の実施態様として、上記結合蛋白質は、(i)配列番号8(1A3)、配列番号28(2F8)、配列番号38(3B6)、配列番号58(1D3)及び配列番号68(1F3)からなる群から選ばれる配列を有するCDRL1、(ii)配列番号9(1A3)、配列番号29(2F8)、配列番号39(3B6)、配列番号59(1D3)及び配列番号69(1F3)からなる群から選ばれる配列を有するCDRL2並びに(iii)配列番号10(1A3)、配列番号30(2F8)、配列番号40(3B6)、配列番号60(1D3)及び配列番号70(1F3)からなる群から選ばれる配列を有するCDRL3を含む免疫グロブリン軽鎖可変領域を含む。これらのCDR配列はヒト又はヒト化FR間に挿入されていてもよい。他の実施態様として、上記結合蛋白質は、配列番号4(1A3)の21乃至127番目残基、配列番号24(2F8)の20乃至131番目残基、配列番号34(3B6)の23乃至129番目残基、配列番号54(1D3)の21乃至127番目残基及び配列番号64(1F3)の21乃至127番目残基からなる群から選ばれるアミノ酸配列を含む免疫グロブリン軽鎖可変領域を含む。
【0057】
一部の実施態様として、上記結合蛋白質は、(i)配列番号5(1A3)、配列番号25(2F8)、配列番号35(3B6)、配列番号55(1D3)及び配列番号65(1F3)からなる群から選ばれる配列を有するCDRH1、(ii)配列番号6(1A3)、配列番号26(2F8)、配列番号36(3B6)、配列番号56(1D3)及び配列番号66(1F3)からなる群から選ばれる配列を有するCDRH2並びに(iii)配列番号7(1A3)、配列番号27(2F8)、配列番号37(3B6)、配列番号57(1D3)及び配列番号67(1F3)からなる群から選ばれる配列を有するCDRH3を含む免疫グロブリン重鎖可変領域を含む。これらのCDR配列はヒト又はヒト化FR間に挿入されていてもよい。別の実施態様として、この免疫グロブリン重鎖可変領域は、配列番号2(1A3)の20乃至141番目残基、配列番号22(2F8)の20乃至137番目残基、配列番号32(3B6)の20乃至139番目残基、配列番号52(1D3)の20乃至141番目残基及び配列番号62(1F3)の20乃至141番目残基からなる群から選ばれるアミノ酸配列を含有する。
【0058】
別の態様として、本発明は、ヒトHGFに結合し、免疫グロブリン軽鎖可変領域及び免疫グロブリン重鎖可変領域を含む単離結合蛋白質を提供する。この単離結合蛋白質は、(i)配列番号24(2F8)の20乃至131番目残基の免疫グロブリン軽鎖可変領域及び配列番号22(2F8)の20乃至137番目残基の免疫グロブリン重鎖可変領域を有する抗体、(ii)配列番号34(3B6)の23乃至129番目残基の免疫グロブリン軽鎖可変領域及び配列番号32(3B6)の20乃至139番目残基の免疫グロブリン重鎖可変領域を有する抗体、並びに(iii)配列番号44(3D11)の23乃至128番目残基の免疫グロブリン軽鎖可変領域及び配列番号42(3D11)の20乃至132番目残基の免疫グロブリン重鎖可変領域を有する抗体からなる群から選ばれる少なくとも1種の対照抗体と、HGFへの結合に対して競合する。ある状況では、この結合蛋白質はこれらの対照抗体のうちの1種と同じHGFの抗原決定基に結合する。
【0059】
上述の各結合蛋白質が完全な状態の抗体、例えばモノクローナル抗体であってもよいことは理解されよう。或いは、上記結合蛋白質は、抗体の抗原結合性断片であってもよく、又は生合成抗体結合部位であってもよい。抗体断片としては、Fab、Fab’、(Fab’)2又はFv断片が挙げられる。このような抗体断片を作製するための技術は当業者に周知である。多くの生合成抗体結合部位が当該技術分野で周知であり、例えば、米国特許第5,476,786号明細書に記載されている単一Fv又はsFv分子が挙げられる。他の生合成抗体結合部位としては、二重特異性又は二機能性結合蛋白質、例えば、二重特異性又は二機能性抗体が挙げられ、これらは少なくとも2種の異なる抗原に結合する抗体又は抗体断片である。例えば、二重特異性結合蛋白質はHGF、例えばヒトHGFと別の対象とする抗原とに結合することができる。二重特異性抗体を作製するための方法は当該技術分野で周知であり、例えば、ハイブリドーマを融合させる方法又はFab’断片を連結させる方法が挙げられる。例えば、Songsivilai et al.(1990年) CLIN. Exp. IMMUNOL. 79:p.315−325及びKostelny et al.(1992年) J. IMMUNOL. 148:p.1547−1553を参照されたい。
【0060】
本発明の結合蛋白質は、561番目の位置にシステインからアルギニンへの置換及び555番目の位置にグリシンからグルタメートへの置換を含むhHGFに結合することができる。
【0061】
別の態様として、本発明は、4.0×10−5s−1以下、3.0×10−5s−1以下又は2.0×10−5s−1以下のKdでヒトHGFに結合する単離結合蛋白質を提供する。これらの単離結合蛋白質は、5.0×10−5s−1乃至0.5×10−5s−1又は4.0×10−5s−1乃至1.0×10−5s−1又は3.0×10−5s−1乃至1.5×10−5s−1のKdでヒトHGFに結合することができる。別の態様として、本発明は、100pM以下又は20pM以下又は10pM以下又は5pM以下のKDでヒトHGFに結合する単離結合蛋白質を提供する。これらの単離結合蛋白質は、100pM乃至5pM又は20pM乃至5pM又は15pM乃至10pM又は20pM乃至10pM乃至15pM乃至5pMのKDでヒトHGFに結合することができる。特に明記しない限り、KD値は実施例6に記載した方法及び条件下で測定する。
【0062】
別の態様として、本発明は、ヒトHGFに結合する単離結合蛋白質であって、この抗体が25℃よりも37℃でより低いKdでヒトHGFに結合するものとする結合蛋白質を提供する。この結合蛋白質結合は、任意選択的に、37℃において5pM未満のKdでヒトHGFに結合する。
【0063】
他の態様及び実施態様として、上記結合蛋白質は、hHGFがc−Metに結合するのを阻害することができる。例えば、上記結合蛋白質のIC50(最大阻害の50%を来す濃度)は、実施例7(a)に記載したプロトコルを用いて測定した場合、少なくとも約4.0、4.5、5.0、5.5、6.0、6.5及び7.0nMとすることができる。一部の他の実施態様として、上記結合蛋白質は、実施例7(b)に記載した方法を用いて4MBr−5細胞(ATCC、カタログ番号CCL208)におけるHGF BrdUの取り込みを中和することができる。
【0064】
上記結合蛋白質のIC50は、実施例7(b)に記載したプロトコルを用いて測定した場合、50nM以下、好ましくは、45、40、35、30、25、20、15、10、5、1又は0.5nM以下である。一部の他の実施態様として、上記結合蛋白質を用いて、実施例9に記載した測定法によるPC−3細胞(ATCC、マナサス、バージニア州、カタログ番号CCL1435)のHGF刺激c−Metリン酸化を抑制することができる。上記結合蛋白質は、実施例9に記載した測定法によるPC−3細胞におけるHGF刺激(1.25nM)c−Metリン酸化を2nM以下のIC50(表8)で抑制する。
【0065】
II.結合蛋白質の作製
本発明の結合蛋白質は、当該技術分野で周知の手法を用いる種々の方法で作製することができる。例えば、軽鎖可変領域及び重鎖可変領域をコードしているDNA分子は、市販の合成機及び本明細書に示した配列情報を用いて、化学的に合成することができる。このような合成DNA分子を、例えば、定常領域コード配列、発現制御配列などの他の適切なヌクレオチド配列に結合させて所望の結合蛋白質をコードしている通常の遺伝子発現構築物を作製することができる。特定の遺伝子構築物の作製は当該技術分野では日常的な技術の範囲内にある。或いは、本明細書に示した配列は、本明細書に示した配列情報、又はハイブリドーマ細胞中のマウス抗体の重及び軽鎖をコードしている遺伝子に関する従来技術の配列情報に基づいた配列を有する合成核酸プローブを用いて、通常のハイブリダイゼーション技術又はPCR技術によりハイブリドーマからクローニングすることができる。このようなプローブの作製及び使用法は、当該技術分野では日常的な技術の範囲内にある。
【0066】
この所望の結合蛋白質をコードしている核酸を発現ベクター中に導入(結合)して、これを、当該技術分野で周知の標準的な形質移入又は形質転換技術により宿主細胞中に導入することができる。典型的な宿主細胞としては、例えば、大腸菌細胞、チャイニーズハムスター卵巣(CHO:Chinese hamster ovary)細胞、ヒーラ(HeLa)細胞、仔ハムスター腎(BHK:baby hamster kidney)細胞、サル腎細胞(COS)、ヒト肝細胞癌細胞(例えば、Hep G2)及び普通なら免疫グロブリンを産生しない骨髄腫細胞が挙げられる。形質移入させた宿主細胞は、この宿主細胞が対象とする遺伝子、例えば免疫グロブリン軽又は重鎖可変領域をコードしている遺伝子を発現するのを可能にする条件下で増殖させることができる。得られる発現産物は当該技術分野で周知の技術を用いて採取することができる。
【0067】
特定の発現及び精製条件は、どんな発現系を用いるかによって異なるであろう。例えば、上記遺伝子を大腸菌で発現させる場合にはこれを先ず発現ベクター中にクローニングする。これは、好適な細菌プロモーター(例えばTrp又はTac)及びシグナル配列(例えばプロテインAのフラグメントB(FB)をコードしている配列)よりも下流に上記の設計遺伝子を配置することによって達成される。得られる発現融合蛋白質は、通常、細胞細胞質中の屈折体又は封入体に蓄積するので、フレンチプレス又は音波処理により細胞を破壊して採取することができる。次に、この屈折体を可溶化し、多くの他の組換え蛋白質について既に確立されている方法により発現蛋白質をリフォールディングし、切断する。
【0068】
上記設計遺伝子を真核宿主細胞、例えば骨髄腫細胞又はCHO細胞で発現させる場合には、先ず、これを、好適な真核生物プロモータ、分泌シグナル、免疫グロブリンエンハンサ及び各種イントロンを含有する発現ベクター中に挿入する。この発現ベクターには、任意選択的に定常領域の全て又は一部をコードしている配列を含有させることができ、これによって重又は軽鎖の全体又は一部の発現が可能となる。上記遺伝子構築物は、確立されている形質移入プロトコルを用いて骨髄腫細胞又はCHO細胞中に形質移入することができる。このような形質移入細胞は、VL又はVH断片、VL−VHヘテロ二量体、VH−VL又はVL−VH一本鎖ポリペプチド、完全な免疫グロブリン重又は軽鎖、或いはこれらの部分を発現することができ、これらは、それぞれ別の機能(例えば、細胞毒性)を有する蛋白質ドメインに結合させてもよい。
【0069】
III. 結合蛋白質に対する修飾
上記結合蛋白質を修飾してこの結合蛋白質の目的とする用途に応じて性能を最適化することができることは理解されよう。例えば、結合蛋白質を治療剤として用いようとする場合、この結合蛋白質を修飾して対象レシピエントにおける免疫原性を低減することができる。或いは、又はさらに、結合蛋白質を別の蛋白質又はペプチド、例えば、成長因子、サイトカインもしくは細胞毒と融合又は結合させることができる。このような修飾は、当該技術分野で周知の通常の遺伝子操作技術を用いることで達成することができる。
【0070】
抗体及び抗体断片の抗原性を低減させるための種々の技術が当該技術分野において周知である。これらの技術を用いて本発明の結合蛋白質の抗原性を低減又は除去することができる。例えば、上記結合蛋白質をヒトに投与しようとする場合、この結合蛋白質を改良してヒトにおけるその抗原性を低減させることが好ましい。このプロセスをヒト化と呼ぶことが多いが、このヒト化結合蛋白質は、それが由来する元の非ヒト化結合蛋白質と抗原に対する親和性が同じか実質的に同じであることが好ましい。
【0071】
ヒト化のための公知の一方法では、ある種、例えばマウスからの抗体の免疫グロブリン定常領域を別の異なる種、例えばヒトからの免疫グロブリン定常領域で置換したキメラ蛋白質を作製する。この例では、得られる抗体はマウス−ヒトキメラであり、このキメラのヒト定常領域の配列は、原理的に、対応するマウスの配列よりも免疫原性が少ない。この種の抗体改良については、例えば、Morrison,et al.(1984年) PROC.NAT.ACAD.SCI.81:p.6851−6855、Neuberger et al.(1984年)NATURE 312:p.604−608、並びに米国特許第6,893,625号明細書(Robinson)、同第5,500,362号明細書(Robinson)及び同第4,816,567号明細書(Cabilly)に記載されている。
【0072】
CDR移植と呼ばれる別の方法では、対象とする抗体の軽及び重鎖可変領域のCDRを別の種からのフレームワーク(FR)中に移植する。例えば、マウスCDRをヒトFR配列中に移植することができる。一部の実施態様として、抗HGF抗体の軽及び重鎖可変領域のCDRをヒトFR又はコンセンサスヒトFR中に移植する。コンセンサスヒトFRを作製するためには、いく種かのヒト重鎖又は軽鎖アミノ酸配列からのFRを整列させてコンセンサスアミノ酸配列を特定する。CDR移植については、例えば、米国特許第7,022,500号明細書(Queen)、同第6,982,321号明細書(Winter)、同第6,180,370号明細書(Queen)、同第6,054,297号明細書(Carter)、同第5,693,762号明細書(Queen)、同第5,859,205号明細書(Adair)、同第5,693,761号明細書(Queen)、同第5,565,332号明細書(Hoogenboom)、同第5,585,089号明細書(Queen)及び同第5,530,101号明細書(Queen)、Jones et al.(1986年)NATURE 321:p.522−525、Riechmann et al.(1988年)NATURE 332:p.323−327、Verhoeyen et al.(1988年)SCIENCE 239:p.1534−1536並びにWinter(1998年)FEBS LETT 430:p.92−94に記載されている。
【0073】
「超ヒト化」と呼ばれる方法では、ヒトにおける免疫原性が低減又は除去されている抗体を別のタイプの移植によって作製する。超ヒト化法では、ヒトCDRのヒト化対象マウス抗体のCDRに対する構造的類似性に基づいて一組のヒト生殖細胞遺伝子からヒトFR配列を選択する。この方法については、例えば、米国特許第6,881,557号明細書(Foote)及びTan et al.(2002年)J.IMMUNOL 169:p.1119−1125に記載されている。
【0074】
免疫原性を低減する他の方法としては、ヒト化抗体を作製するための「再成形(reshaping)」、「超キメラ化」又は「ベニア化(veneering)/再表面化(resurfacing)」と呼ばれる技術が挙げられる。例えば、Vaswami et al. (1998年) ANNALS OF ALLERGY, ASTHMA, & IMMUNOL. 81:p.105、Roguska et al.(1996年) PROT. ENGINEER 9:p.895−904及び米国特許第6,072,035号明細書(Hardman)を参照されたい。ベニア化/再表面化法では、マウス抗体の表面接近可能アミノ酸残基をヒト抗体の同一位置により多くみられるアミノ酸残基によって置換する。このタイプの抗体再表面化については、例えば、米国特許第5,639,641号明細書(Pedersen)に記載されている。
【0075】
マウス抗体をヒトにおける医学的用途に好適な構造に変換するための1つの典型的な方法は、ACTIVMAB(商標)技術(Vaccinex社、ロチェスター、ニューヨーク州)と呼ばれ、これは哺乳動物細胞で抗体を発現させるためのワクシニアウイルス系ベクターを必要とする。免疫グロブリン重及び軽鎖の高レベルの組み合わせ多様性がもたらされると言われている。例えば、米国特許第6,706,477号明細書(Zauderer)、同第6,800,442号明細書(Zauderer)及び同第6,872,518号明細書(Zauderer)を参照されたい。
【0076】
マウス抗体をヒトにおける用途に好適な構造に変換するための別の典型的な方法は、カロビオス・ファーマシューティカルズ社(KaloBios Pharmaceuticals, Inc.)(パロアルト、カリフォルニア州)により商業的に実施されている技術である。この技術は、特許権のあるヒト「アクセプター」ライブラリを用いて抗体選択用の「抗原決定基に焦点を合わせた」ライブラリを作製するものである。
【0077】
マウス抗体をヒトにおける医学的用途に好適な構造に修飾するための別の典型的な方法は、XOMA(米国)LLC社によって商業的に実施されているHUMAN ENGINEERING)(商標)(HE(商標))技術である。例えば、国際公開第93/11794号パンフレット並びに米国特許第5,766,886号明細書、同第5,770,196号明細書、同第5,821,123号明細書及び同第5,869,619号明細書を参照されたい。
【0078】
上記方法のいずれかを含む任意の適切な方法を用いて対象とする結合蛋白質のヒト免疫原性を低減又は除去することができる。
【0079】
さらに、マウスにおいて完全にヒト型の抗体を作製することが可能である。この方法では、マウスの抗体産生遺伝子がヒト抗体産生遺伝子の実質的な部分で置換されている遺伝子導入マウスを用いてヒト型抗体を調製する。このようなマウスは、マウスの免疫グロブリン分子ではなく、ヒト型の免疫グロブリンを産生する。例えば、国際公開第98/24893号パンフレット(Jacobovitz et al.)及びMendez et al. (1997年) NATURE GENETICS 15:p.146−156を参照されたい。完全にヒト型の抗HGFモノクローナル抗体は、以下の方法を用いて作製することができる。ヒト型免疫グロブリン遺伝子を有する遺伝子導入マウスを対象とする抗原、例えばHGFで免疫する。次いで、このマウスからマウスのリンパ細胞を得、これを脊髄系細胞株と融合させて不死化ハイブリドーマ細胞株を調製する。このハイブリドーマ細胞株をスクリーニングし、HGFに特異的な抗体を産生するハイブリドーマ細胞株を特定するための選択を行う。
【0080】
本発明の結合蛋白質は、その目的とする用途に応じて他の分子と結合させることができる。例えば、結合蛋白質を治療剤として用いようとする場合、この結合蛋白質を別の作用物質、例えば、その治療を調節するか、それとも促進するエフェクター分子と結合させることができる。このエフェクターが非蛋白質系の作用物質、例えば、低分子薬剤、放射標識又は毒素である限り、この物質を標準的なインビトロ結合化学反応を用いて結合蛋白質に化学的に結合させることができる。一方、エフェクター分子が蛋白質又はペプチド、例えば、酵素、受容体、毒素、成長因子、サイトカイン又は他の免疫調節因子である場合、結合蛋白質は、インビトロ結合化学反応を用いてこのエフェクターに化学的に結合させることができ、或いは、融合蛋白質としてこのエフェクターに結合させることができる。融合蛋白質は、セクションIIに記載したのと同様な技術を用いて構築し、発現させることができる。
【0081】
IV. 結合蛋白質の用途
本明細書に記載した結合蛋白質は、診断剤又は治療剤として用いることができる。
【0082】
(1)治療的用途
本発明の結合蛋白質は、HGFの活性を中和するので、種々の治療的用途に用いることができる。例えば、本発明の結合蛋白質の一部は、過剰増殖性疾病又は疾患、例えば、各種形態の癌の予防又は治療に有用である。
【0083】
上記結合蛋白質を用いることにより腫瘍細胞の増殖を抑制又は低減させることができる。このような方法では、腫瘍細胞に対して治療的有効量の上記結合蛋白質を作用させることによって腫瘍細胞の増殖を抑制又は低減させる。一部の実施態様として、上記結合蛋白質は、腫瘍細胞の増殖を少なくとも50%、60%、70%、80%、90%、95%又は100%抑制する。
【0084】
一部の実施態様として、上記結合蛋白質を用いることにより、結合蛋白質がhHGFのc−Metへの結合能を低減させることで腫瘍細胞の増殖を抑制又は低減させる。他の実施態様として、上記結合蛋白質を用いることにより、表5及び6において抗体3B6が示すように、この結合蛋白質がhHGFに結合するが、hHGFのc−Metへの結合を実質的に抑制しない場合にも腫瘍細胞の増殖は抑制され、又は低下する。
【0085】
さらに、上記結合蛋白質を用いることにより、哺乳動物における腫瘍の成長又は発症を抑制又は遅延させることができる。このような方法では、有効量の上記結合蛋白質を哺乳動物に投与することによってこの哺乳動物における腫瘍の成長を抑制又は遅延させる。従って、上記結合蛋白質を用いることによって、例えば、哺乳動物において腫瘍を治療することができる。この方法は哺乳動物に治療的有効量の上記結合蛋白質を投与するものである。上記結合蛋白質を単独又は別の医薬として有効な分子と併用して投与することにより腫瘍を治療することができる。
【0086】
本発明の結合蛋白質は、種々のHGF反応性疾患、例えば、肺癌、乳癌、大腸癌、前立腺癌、卵巣癌、頭頸部癌、卵巣癌、多発性骨髄腫、肝臓癌、胃癌、食道癌、腎癌、鼻咽腔癌、膵癌、中皮腫、黒色腫及びグリア芽細胞腫におけるHGF反応性腫瘍細胞などの治療に用いることができる。
【0087】
本明細書でいう「治療する」、「治療すること」及び「治療」とは、哺乳動物、特にヒトにおける病態の治療のことを指し、(a)その病態が哺乳動物において、特に、その哺乳動物がその病態に罹患する素因を有するが、まだこれを有していないと診断されている場合に発生するのを予防すること、(b)その病態を抑制する、即ち、その発症を阻止すること、及び/又は(c)その病態を緩和すること、即ち、その病態を軽減させること含む。
【0088】
一般に、活性成分の治療的有効量は、約0.1mg/kg乃至約100mg/kg、必要に応じて約1mg/kg乃至約100mg/kg、必要に応じて約1mg/kg乃至約10mg/kgの範囲となる。投与の量は、治療対象の疾患又は徴候の種類及び程度、特定の患者の全般的な健康状態、送達する結合蛋白質の相対的生物学的有効性、この結合蛋白質の剤型、製剤中の添加物の存在及び種類並びに投与経路などの変数によって決まることになる。所望の血液濃度又は組織濃度に急速に到達させるために、初期の投与用量を上記の上限を超えて増加させることができ、又は初期用量を最適用量より少なくすることができ、さらに、特定の状況に応じて治療の過程で初期用量を段階的に増加させることができる。ヒトにおける用量は、例えば、0.5mg/kg乃至20mg/kgで実施するようにデザインされた通常の第I相用量増加試験において最適化することができる。投与頻度については、投与経路、投与量、治療対象の疾患状態などの要因によって変更することができる。典型的な投与頻度は、1日当たり1回、1週当たり1回、2週ごとに1回である。好ましい投与経路は全身性投与、例えば、静脈内注入である。モノクローナル抗体系薬剤の製剤化については当該技術分野では日常的な技術の範囲内にある。本発明の一部の実施態様として、上記結合蛋白質、例えばモノクローナル抗体は、凍結乾燥され、投与時に緩衝生理食塩水で元に戻される。
【0089】
上記結合蛋白質は、単独又は他の医薬として有効な成分との併用で投与することができる。上記他の有効な成分、例えば、免疫調節剤はこの結合蛋白質と一緒に投与することができ、或いはこの結合蛋白質投与の前又は後で投与することができる。
【0090】
治療的用途の上記結合蛋白質含有製剤としては、典型的には、上記結合蛋白質を医薬用として許容可能な担体と組み合わせたものが挙げられる。本明細書でいう「医薬用として許容可能な担体」とは、正しい医学的判断の範囲内で、妥当な利益/危険度比に見合って、過度の毒性、刺激、アレルギー応答又は問題もしくは合併症を伴うことなくヒトその他の動物の組織と接触させて用いるのに適している緩衝剤、担体及び賦形剤を意味する。この担体は、製剤のその他の成分と適合し、レシピエントに対して有害でないという意味で「許容可能」でなければならない。なお、医薬用として許容可能な担体は、医薬用としての投与に適合するありとあらゆる緩衝剤、溶媒、分散媒、被覆剤、等張性吸収遅延剤などを含むものとする。医薬として有効な物質のためのこのような媒体及び剤については当該技術分野では周知である。
【0091】
こうした製剤は、投与単位形態で適宜提示することができ、薬学分野で公知の方法のいずれかを含め、任意の適切な方法によって調製することができる。本発明の医薬組成物は、対象とする投与経路に適合するように製剤化する必要がある。投与経路としては、例えば、全身性投与又は非全身性投与、例えば、静脈内、皮内、吸入、経皮(局所)、経粘膜及び直腸投与が挙げられる。経口又は全身性投与に有用な液剤は、例えば、「Remington’s Pharmaceutical Sciences」、第18版(マック出版(Mack Publishing Company)1990年)に記載されている、医薬分野で公知の方法のいずれかによって調製することができる。
【0092】
経口投与に適した製剤は、所定量の上記結合蛋白質を含有する、注射液、カプセル剤、ゼラチンカプセル剤、サッシェ剤、錠剤、トローチ剤又は舐剤などの個々の単位体;粉末状又は顆粒状組成物;水性液体又は非水性液体の液剤又は懸濁剤;或いは水中油型乳剤又は油中水型乳剤の形をとることができる。
【0093】
全身性投与に適した製剤は、例えば、以下の成分、即ち、注射用水などの滅菌希釈液、食塩水、固定油類、ポリエチレングリコール類、グリセリン、プロピレングリコール又は他の合成溶媒;ベンジルアルコール又はメチルパラベンなどの抗菌剤;アスコルビン酸又は亜硫酸水素ナトリウムなどの抗酸化剤;エチレンジアミン四酢酸などのキレート化剤;酢酸塩、クエン酸塩又はリン酸塩などの緩衝剤及び塩化ナトリウム又はデキストロースなどの等張性調整用剤を含む。pHは塩酸又は水酸化ナトリウムなどの酸又は塩基で調整することができる。全身性投与用製剤は、アンプル、使い捨て注射器、或いはガラス又はプラスチック製多回投与用バイアルに封入することができる。
【0094】
一般的に言って、注射剤用途に適した組成物としては、水溶液(水溶性の場合)又は分散液、及び滅菌注射液又は分散液の即時調整用粉末が挙げられる。静脈内投与の場合、好適な担体としては、生理食塩水、静菌水、クレモフォア(Cremophor)ELTM(BASF社、パーシパニ(Parsippany)、ニュージャージー州)又はリン酸緩衝生理食塩水(PBS:phosphate buffered saline)が挙げられる。これは作製及び貯蔵の条件下で安定である必要があり、細菌、真菌などの微生物の汚染作用に対して保護される必要がある。上記担体は、例えば水、エタノール、ポリオール(例えば、グリセロール、プロピレングリコール及び液状ポリエチレングリコール)並びにこれらの適切な混合物を含む溶媒又は分散媒とすることができる。
【0095】
医薬製剤は滅菌されていることが好ましい。滅菌は、例えば、滅菌濾過膜を通す濾過によって達成することができる。組成物が凍結乾燥される場合、この方法を用いた滅菌は凍結乾燥及び再形成の前又は後に行うことができる。医薬組成物は、ひとたび製剤化すれば、例えば、溶液、懸濁液、ゲル、乳液、固体として、又は脱水もしくは凍結乾燥粉末としてバイアルに貯蔵することができる。
【0096】
(2)診断用途
上記結合蛋白質を診断目的のためにインビトロ又はインビボで用いる場合はいつも、この結合蛋白質を検出可能な成分で直接又は間接に標識するのが通常である。この検出可能成分は、検出可能なシグナルを直接的又は間接的に発生することができる任意の成分とすることができる。例えば、この検出可能成分は、3水素(3H)、14炭素(14C)、32燐(32P)、35硫黄(35S)又は125ヨウ素(125I)などの放射性同位体;フルオレッセインイソチオシアネート、ローダミン又はルシフェリンなどの蛍光又は化学発光化合物;アルカリ性ホスファターゼ、ベータ−ガラクトシダーゼ又はホースラディッシュペルオキシダーゼなどの酵素;スピン標識などのスピンプローブ;或いは、カラー粒子、例えばラテックスもしくは金粒子であってもよい。上記結合蛋白質を、例えば、Hunter et al. (1962年) NATURE 144:p.945、David et a1. (1974年) BIOCHEMISTRY 13:p.1014、Pain et al. (I981年) J. IMMUNOL. METH. 40:p.219及びNygren (1982年) J. HISTOCHEM. AND CYTOCHEM. 30:p.407に記載されているような、当該技術分野において周知の多くの方法を用いて上記検出可能成分に結合させることができることは理解されよう。これらの標識は、例えば、目視により、又は分光光度計その他の検出器を用いて検出することができる。
【0097】
上記結合蛋白質は、当該技術分野で利用可能な広範囲の免疫測定技術において用いることができる。典型的な免疫測定法としては、例えば、サンドイッチ免疫測定法、競合免疫測定法、免疫組織化学的方法が挙げられる。
【0098】
サンドイッチ免疫測定法では、被分析物、即ち、対象とする抗原に結合する2種の抗体、例えば、固形担体上に固定化した1種と、溶液中に遊離し、検出可能成分で標識された1種とを用いる。抗原を含むサンプルをこの系に添加すると、抗原は、上記の固定化抗体及び標識抗体の両者に結合して担体の表面に「サンドイッチ」免疫複合体を形成する。結合しなかったサンプル成分及び過剰の標識抗体を洗い落とし、担体表面で蛋白質と複合体化した標識抗体の量を測定することにより上記複合体化蛋白質を検出する。或いは、上記の溶液中に遊離した抗体を、この遊離抗体に結合する検出可能成分で標識した第3の抗体によって検出することができる。免疫学的測定法のデザイン、理論及びプロトコルに関する詳細な総説は、Butt編、(1984年)PRACTICAL IMMUNOLOGY、マーセルデッカー社(Marcel Dekker)、ニューヨーク、Harlow et al.編(1988年)ANTIBODIES,A LABORATORY APPROACH、コールド・スプリング・ハーバー・ラボラトリ(Cold Spring Harbor Laboratory)及びDiamandis et al.編(1996年)IMMUNOASSAY、アカデミックプレス社(Academic Press)ボストンを含む多くの教科書に見出すことができる。
【0099】
上記標識結合蛋白質はインビボ造影剤として有用であり、これにより結合蛋白質はレシピエントの特定の対象組織を標的にしてこの造影剤を送達することができることが企図されている。インビボイメージング用の好ましい遠隔検出可能成分としては、約6時間の半減期を有するガンマ放出体である放射性原子テクネチウム-99m(99mTc)が挙げられる。また、インビボイメージングにおいて有用な非放射性成分としては、インサイツ(insitu)でプロトン緩和を誘導するニトロキシドスピンラベル並びにランタニド及び遷移金属イオンが挙げられる。免疫イメージングの他に、標的とする細胞を破壊するために、複合体化放射性成分を標準的な放射免疫治療プロトコルで用いることができる。高線量放射免疫治療用の好ましいヌクレオチド類は、放射性原子90イットリウム(90Yt)、131ヨウ素(131I)及び111インジウム(111In)を含む。上記結合蛋白質は、イメージング分野で周知の結合技術を用いて131I 、111In及び−99mTcで標識することができる。同様に、造影剤を調製し投与する方法及びイメージを捕捉し処理する方法もイメージング分野において公知であるので、本明細書では詳しく論じない。同様に、抗体を利用した免疫療法を実施する方法も当該分野では公知である。例えば、米国特許第5,534,254号を参照されたい。
【0100】
本説明の全体を通して、組成物が特定の成分を有する(having)、含む(including)又は含む(comprising)と記載されているところでは、組成物は、本質的にその列挙された成分からなる、又はこれらの成分からなることも企図されている。同様に、工程が特定の工程段階を有する(having)、含む(including)又は含む(comprising)と記載されているところでは、その工程は、また、本質的にその列挙された工程段階からなる、又はこれらの工程段階からなる。特に示さない限り、段階の順序又は特定の処置を行う順序は、本発明が実施可能である限り、重要ではない。さらに、特に断りのない限り、2つ以上の段階又は処置を同時に実施することができる。
【実施例】
【0101】
以下の実施例では、いくつかの抗hHGFモノクローナル抗体の作製及び特性化について検討する。
【0102】
(実施例1)
抗hHGFモノクローナル抗体の作製
この実施例ではいくつかの抗hHGFモノクローナル抗体の作製について説明する。
【0103】
免疫化、融合及び一次スクリーニングについては、反復性複数部位免疫化(RIMMS:Repetitive Immunization Multiple Sites)プロトコルに従ってMBS社(ポートランド、メイン州)で行われた。AJマウス5匹及びBalb/cマウス5匹を組換えヒトHGF(R&Dシステムズ社(R&D Systems)、ミネアポリス、ミネソタ州、カタログ番号294−HGN−025)で免疫化した。酵素結合免疫測定法(ELISA:Enzyme Linked Immunosorbent Assay)で最も高い抗HGF活性を示す血清を有するマウス2匹を後の融合のために選んだ。該当するマウスから脾臓及びリンパ節を採取した。次いで、B細胞を採取し、骨髄腫細胞株と融合させた。融合産物は1枚以上のプレート上でほぼクローン性になるまで連続希釈した。得られた融合産物からの上清をELISAによりそのhHGFへの結合についてスクリーニングした。HGFに対する抗体を含むとみなされた上清は、さらに、以下の実施例で検討したようにして、インビボ機能試験により特性を明らかにした。1パネルのハイブリドーマを選択し、このハイブリドーマをサブクローニングして増やした。次いで、得られたモノクローナル抗体を標準的な条件下でプロテインA/G樹脂を用いるアフィニティークロマトグラフィーにより精製した。
【0104】
(実施例2)
抗hHGFモノクローナル抗体の配列分析
この実施例では実施例1で作製した抗hHGFモノクローナル抗体のアイソタイプ及び配列の分析について説明する。
【0105】
a.HGFマウスモノクローナル抗体アイソタイプの決定
各モノクローナル抗体の軽鎖タイプ及び重鎖アイソタイプについて、イソストリップマウスモノクローナル抗体アイソタイピングキット(IsoStrip Mouse Monoclonal Antibody Isotyping Kit)を用い、そのメーカーの使用説明書(ロシュ・アプライド・サイエンス社(Roche Applied Science))に従って決定した。
【0106】
全ての抗体は、カッパ免疫グロブリン軽鎖及びIgG1免疫グロブリン重鎖を含むように決定した。
【0107】
b.免疫グロブリン重及び軽鎖可変領域をコードしているヌクレオチド配列の決定
各モノクローナルハイブリドーマ細胞株から、RNeasyミニプレップ(Miniprep)キットを用い、そのメーカーの使用説明書(キアゲン社(Qiagen)、ヴェンロ、オランダ)に従って全RNAを抽出した。BD SMART(商標)RACE cDNA増幅キットを用い、そのメーカーの使用説明書(クロンテック社(Clontech))に従い、5’RACE(cDNA末端の迅速増幅(Rapid Amplification of cDNA Ends))のためのオリゴヌクレオチドプライマBD SMART IIA(5’aagcagtggtatcaacgcagagtacgcggg 3’)(配列番号85)及び5’−RACE CDS Primer(5’ tttttttttttttttttttttttttvn 3’、但し、v=a、g又はc及びn=a、g、c又はt)(配列番号86)を用いて、完全長の第1鎖cDNAを作製した。
【0108】
エキスパンドハイフィデリティ(Expand High−Fidelity)PCRシステム(ロシュ・アプライド・サイエンス社)を用い、そのメーカーの使用説明書に従ってPCR(Polymerase Chain Reaction(ポリメラーゼ連鎖反応))により上記カッパ及び重(IgG1)免疫グロブリン鎖の可変領域を増幅させた。重鎖可変領域は、5’オリゴヌクレオチドプライマ混合物ユニバーサルプライマミックスA(Universal Primer Mix A)( 5’ctaatacgactcactatagggcaagcagtggtatcaacgcagagt 3’(配列番号87)と5’ctaatacgactcactatagggc 3’(配列番号88)との混合物)及び3’IgG1定常領域特異的プライマ5’tatgcaaggcttacaaccaca 3’(配列番号89)又は5’gccagtggatagacagatgggggtgtcg 3’(配列番号90)を用いて増幅した。カッパ鎖可変領域は、5’オリゴヌクレオチドプライマ混合物ユニバーサルプライマミックスA及び3’カッパ定常領域特異的プライマ5’ctcattcctgttgaagctcttgacaat 3’(配列番号91)又は5’cgactgaggcacctccagatgtt 3’(配列番号92)を用いて増幅した。
【0109】
個々のPCR産物をアガロースゲル電気泳動により分画し、キアクイック・ゲル精製(Qiaquick Gel Purification)キットを用い、そのメーカーの使用説明書(キアゲン社)に従って精製した。次いで、トポイソメラーゼ利用クローニングキットTOPO TAクローニング(登録商標)キット(pCR(登録商標)2.1−TOPO(登録商標)ベクターを含む)を用い、そのメーカーの使用説明書(インビトロジェン社(Invitrogen)、カールズバッド、カリフォルニア州)に従って、これらのPCR産物をpCR2.1 TOPOプラスミド中にクローニングした後、標準的な形質転換技術を用いて細菌DH5に形質転換した。形質転換した細菌のクローンから単離したプラスミドDNAについて、アジェンコートバイオサイエンス社(Agencourt Bioscience)のT7(5’TAATACGACTCACTATAGGG3’)(配列番号93)、M13順方向プライマ(5’GTAAAACGACGGCCAGT3’)(配列番号94)及びM13逆方向プライマ(5’CAGGAAACAGCTATGACC3’)(配列番号95)を用い、標準的なジデオキシDNA配列決定方法により配列を決定することにより可変領域配列の配列を特定した。ベクターNTI(Vector NTI)ソフトウェア(インビトロジェン社、カールズバッド、カリフォルニア州)及びIMGT/V−クエスト(Quest)ウェブサーバー(http://imgt.cines.fr/textes/vquest)を用いてこれらの配列を解析し、可変領域の配列を同定、確認した。
【0110】
c.1A3、1D3、1F3及び2B8カッパ及びIgG1鎖の免疫グロブリン重及び軽鎖定常領域の配列をコードしているヌクレオチド配列の決定
順方向プライマ5’ggggacaagtttgtacaaaaaagcaggctgccaccatgaactttgggctcagattgattttcc 3’(下線箇所:開始コドン)(配列番号96)及び逆方向プライマ5’ggggaccactttgtacaagaaagctgggttcatttaccaggagagtgggagagg3’(下線箇所:停止コドン)(配列番号97)を用いて上記で作製したcDNAから、1A3、1D3及び1F3 IgG1鎖の完全長cDNAをPCRにより増幅させた。順方向プライマ5’ggggacaagtttgtacaaaaaagcaggctgccaccatgggatggagctatatcatcctcttt3’(下線箇所:開始コドン)(配列番号98)及び逆方向プライマ5’ggggaccactttgtacaagaaagctgggttcatttaccaggagagtgggagag3’(下線箇所:停止コドン)(配列番号99)を用いて上記で作製したcDNAから、2B8IgG1鎖の完全長cDNAを増幅させた。
【0111】
順方向プライマ5’ggggacaagtttgtacaaaaaagcaggctgccaccatggaatcacagactctggtcttcata3’(下線箇所:開始コドン)(配列番号100)及び逆方向プライマ5’ggggaccactttgtacaagaaagctgggtctaacactcattcctgttgaagctc3’(下線箇所:停止コドン)(配列番号101)を用いて2B8カッパ鎖の完全長cDNAを増幅させた。PCR断片をゲートウェイ(Gateway)BP組換え反応によりpDONR221(インビトロジェン社、カールズバッド、カリフォルニア州)中にクローニングし、標準的なジデオキシDNA配列決定方法を用いてアジェンコートバイオサイエンス社で配列決定することによりその定常領域の配列を同定し、さらに可変領域の配列を確認した。
【0112】
d.配列の解析
可変領域(標準テキスト)は、IMGT/V−QUESTウェブサーバーソフトウェア(http://imgt.cines.fr/textes/vquest/)を用いて同定した。シグナルペプチド配列は、同定した可変領域の上流にあるインフレーム開始コドン(ATG)の同定に基づいて予測した。シグナルペプチド配列を同定し、下記に下線で示した。
【0113】
各可変領域の最後のヌクレオチドは、可変/定常領域結合により生じる次のコドンの最初の塩基である。このヌクレオチドは、そのエキソンの一部であるので、可変領域に含まれる。以下に列挙した定常領域のアミノ酸配列にはこの結合コドンの翻訳が含まれる。
【0114】
完全な重鎖又はカッパ鎖抗体配列を作製するためには、下記の可変領域の配列をそのそれぞれの定常領域の配列と結合させる(下線箇所:シグナル配列)。
【0115】
(1)1A3重鎖可変領域(配列番号1)
【0116】
【化1】
【0117】
(2)1A3カッパ軽鎖可変領域(配列番号3)
【0118】
【化2】
【0119】
(3)2B8重鎖可変領域(配列番号11)
【0120】
【化3】
【0121】
【化4】
【0122】
(4)2B8カッパ軽鎖可変領域(配列番号13)
【0123】
【化5】
【0124】
(5)2F8重鎖可変領域(配列番号21)
【0125】
【化6】
【0126】
(6)2F8カッパ軽鎖可変領域(配列番号23)
【0127】
【化7】
【0128】
(7)3B6重鎖可変領域(配列番号31)
【0129】
【化8】
【0130】
【化9】
【0131】
(8)3B6カッパ軽鎖可変領域(2個の可能なATG開始コドン(大文字))(配列番号33)
【0132】
【化10】
【0133】
(9)3D11重鎖可変領域(配列番号41)
【0134】
【化11】
【0135】
(10)3D11カッパ軽鎖可変領域(配列番号43)
【0136】
【化12】
【0137】
(11)1D3重鎖可変領域(配列番号51)
【0138】
【化13】
【0139】
(12)1D3カッパ軽鎖可変領域(配列番号53)
【0140】
【化14】
【0141】
(13)1F3重鎖可変領域(配列番号61)
【0142】
【化15】
【0143】
(14)1F3カッパ軽鎖可変領域(配列番号63)
【0144】
【化16】
【0145】
【化17】
【0146】
(15)3A12重鎖可変領域(配列番号71)
【0147】
【化18】
【0148】
(16)3A12カッパ軽鎖可変領域(配列番号73)
【0149】
【化19】
【0150】
(17)対照マウスIgG1重鎖定常領域(J00453)(配列番号81)
【0151】
【化20】
【0152】
【化21】
【0153】
(18)1A3、1D3、1F3及び2B8(AJ系マウス由来)について決定されたマウスIgG1重鎖定常領域(配列番号82)
【0154】
【化22】
【0155】
(19)対照マウスカッパ軽鎖定常領域(V00807)並びに1D3、1F3及び2B8(AJ系マウス由来)について決定されたマウスカッパ軽鎖定常領域(配列番号83)
【0156】
【化23】
【0157】
(20)1D3、1F3及び2B8に比し変更されたヌクレオチド(下線箇所)を1個含む1A3について決定されたマウスカッパ軽鎖定常領域(配列番号84)
【0158】
【化24】
【0159】
実施例1で作製した抗体の免疫グロブリン重鎖可変領域を規定するアミノ酸配列は、いずれも図2に記載されている。これらの配列は全て相互に整列させてあり、シグナルペプチド、CDR1、CDR2及びCDR3を規定する配列は枠で囲んで識別されている。図3は、全ての抗体のCDR1、CDR2及びCDR3配列を別々に整列させて示したものである。
【0160】
実施例1で作製した全ての抗体の免疫グロブリン軽鎖可変領域を規定するアミノ酸配列は、いずれも図4に記載されている。これらの配列は全て相互に整列させてあり、シグナルペプチド、CDR1、CDR2及びCDR3を規定する配列は枠で囲んで識別されている。図5は、全ての抗体のCDR1、CDR2及びCDR3配列を別々に整列させて示したものである。
【0161】
便宜のため、本実施例で検討した抗体配列と配列表に示したものとの対応関系を示すコンコーダンス表を表1に示した。
【0162】
【表1A】
【0163】
【表1B】
【0164】
また、便宜のため、下記の配列は、本実施例に記載した全ての抗体の実際の、又は企図された完全長の重及び軽鎖配列(即ち、可変領域及び定常領域の両者の配列を含む)を表したものである。マウス抗体2F8、3A12、3B6及び3D11の定常領域は配列決定を行わなかったが、これらが全てAJ系マウス由来の抗体であることから、配列決定した1D3、1F3及び2B8抗体と同一の定常領域配列を有すると推定されることは注目される。しかしながら、本明細書に記載した可変領域の配列を当業者に周知の他のいく種かの定常領域配列のそれぞれに結合させて活性のある完全長の免疫グロブリン重及び軽鎖を作製することができることは了解されよう。
【0165】
(1)完全長1A3重鎖配列(1A3重鎖可変領域及びIgG1定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号122)
【0166】
【化25】
【0167】
(2)完全長1A3重鎖配列(1A3重鎖可変領域及びIgG1定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号123)
【0168】
【化26】
【0169】
(3)完全長1A3軽鎖配列(1A3カッパ可変領域及び定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号124)
【0170】
【化27】
【0171】
【化28】
【0172】
(4)完全長1A3軽鎖配列(1A3カッパ可変領域及び定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号125)
【0173】
【化29】
【0174】
(5)完全長2B8重鎖配列(2B8重鎖可変領域及びIgG1定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号126)
【0175】
【化30】
【0176】
(6)完全長2B8重鎖配列(2B8重鎖可変領域及びIgG1定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号127)
【0177】
【化31】
【0178】
【化32】
【0179】
(7)完全長2B8軽鎖配列(2B8カッパ可変領域及び定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号128)
【0180】
【化33】
【0181】
(8)完全長2B8軽鎖配列(2B8カッパ可変領域及び定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号129)
【0182】
【化34】
【0183】
(9)完全長2F8重鎖配列(2F8重鎖可変領域及びIgG1定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号130)
【0184】
【化35】
【0185】
【化36】
【0186】
(10)完全長2F8重鎖配列(2F8重鎖可変領域及びIgG1定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号131)
【0187】
【化37】
【0188】
(11)完全長2F8軽鎖配列(2F8カッパ可変領域及び定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号132)
【0189】
【化38】
【0190】
(12)完全長2F8軽鎖配列(2F8カッパ可変領域及び定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号133)
【0191】
【化39】
【0192】
(13)完全長3B6重鎖配列(3B6重鎖可変領域及びIgG1定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号134)
【0193】
【化40】
【0194】
【化41】
【0195】
(14)完全長3B6重鎖配列(3B6重鎖可変領域及びIgG1定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号135)
【0196】
【化42】
【0197】
(15)完全長3B6軽鎖配列(3B6カッパ可変領域及び定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号136)
【0198】
【化43】
【0199】
(16)完全長3B6軽鎖配列(3B6カッパ可変領域及び定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号137)
【0200】
【化44】
【0201】
(17)完全長3D11重鎖配列(3D11重鎖可変領域及びIgG1定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号138)
【0202】
【化45】
【0203】
(18)完全長3D11重鎖配列(3D11重鎖可変領域及びIgG1定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号139)
【0204】
【化46】
【0205】
(19)完全長3D11軽鎖配列(3D11カッパ可変領域及び定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号140)
【0206】
【化47】
【0207】
(20)完全長3D11軽鎖配列(3D11カッパ可変領域及び定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号141)
【0208】
【化48】
【0209】
(21)完全長1D3重鎖配列(1D3重鎖可変領域及びIgG1定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号142)
【0210】
【化49】
【0211】
(22)完全長1D3重鎖配列(1D3重鎖可変領域及びIgG1定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号143)
【0212】
【化50】
【0213】
(23)完全長1D3軽鎖配列(1D3カッパ可変領域及び定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号144)
【0214】
【化51】
【0215】
(24)完全長1D3軽鎖配列(1D3カッパ可変領域及び定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号145)
【0216】
【化52】
【0217】
(25)完全長1F3重鎖配列(1F3重鎖可変領域及びIgG1定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号146)
【0218】
【化53】
【0219】
【化54】
【0220】
(26)完全長1F3重鎖配列(1F3重鎖可変領域及びIgG1定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号147)
【0221】
【化55】
【0222】
(27)完全長1F3軽鎖配列(1F3カッパ可変領域及び定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号148)
【0223】
【化56】
【0224】
(28)完全長1F3軽鎖配列(1F3カッパ可変領域及び定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号149)
【0225】
【化57】
【0226】
(29)完全長3A12重鎖配列(3A12重鎖可変領域及びIgG1定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号150)
【0227】
【化58】
【0228】
(30)完全長3A12重鎖配列(3A12重鎖可変領域及びIgG1定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号151)
【0229】
【化59】
【0230】
(31)完全長3A12軽鎖配列(3A12カッパ可変領域及び定常領域)をコードしている核酸配列(下線箇所:シグナル配列)(配列番号152)
【0231】
【化60】
【0232】
(32)完全長3A12軽鎖配列(3A12カッパ可変領域及び定常領域)を規定している蛋白質配列(シグナル配列含まず)(配列番号153)
【0233】
【化61】
【0234】
便宜のため、本実施例で検討した抗体の完全長配列と配列表に示したものとの対応関系を示すコンコーダンス表を表2に示した。
【0235】
【表2A】
【0236】
【表2B】
【0237】
(実施例3)
各種組換えhHGF蛋白質の作製
本実施例では、実施例1及び実施例14で作製された抗体の特性を明らかにするために用いるいくつかの組換え蛋白質のクローニング及び発現について説明する。具体的には、本実施例では、組換えhHGF蛋白質、555番目の位置にグリシンからグルタメートへの置換を含む組換えhHGF蛋白質(G555E)、561番目の位置にシステインからアルギニンへの置換を含む組換えhHGF蛋白質(C561R)、マウスHGF配列内に配置されたヒトV495乃至L585HGF配列を含む組換えマウス−ヒト−マウス(mhm)キメラHGF蛋白質、マウスHGF配列内に配置されたヒトI499乃至R566HGF配列を含む組換えmhmキメラHGF蛋白質及びマウスHGF配列内に配置されたヒトW507乃至L585HGF配列を含む組換えmhmキメラHGF蛋白質のクローニング及び発現について説明する。
【0238】
標準的な分子技術を用いて以下の発現構築物を作製し、得られたcDNA配列をDNA配列決定法によって確認した。
【0239】
a. hHGF−Fc
1回目のPCRでは、NotI部位を導入し、hHGFとhIgFcとの間に6×His標識をコードしている2本の重なり合うPCR断片を作製した。2回目に、これらの重なり合うPCR断片を鋳型としてhHGF−his−IgFcを増幅させた。得られた断片をNheI及びBamHIで消化し、pcDNA5/FRT(インビトロジェン社、#35−3014)中にクローニングした。次いで、インビトロジェン社のクローンID:IOH29794(ヒトHGF cDNA)からhHGFを増幅させた。その配列は、アクセッション番号NM_000601.4としてNCBIに寄託されている配列に相当することが分かった。
【0240】
(1) 5’hHGF NheIプライマー
【0241】
【化62】
【0242】
(2) 3’hHGF NotI His標識プライマー
【0243】
【化63】
【0244】
(3) 5’HisIgFcプライマー
【0245】
【化64】
【0246】
(4) 3’IgFc BamHIプライマー
【0247】
【化65】
【0248】
b. hHGF−Fc G555E及びhHGF−Fc C561R
hHGF−Fc突然変異体G555E及びC561Rは、クイックチェンジ(QuickChange)II XL部位特異的突然変異誘発キット(ストラタジーン社(Stratagene))をメーカーの使用説明書に従って用いた部位特異的突然変異誘発法により作製した。
【0249】
(1) hHGF−Fc(G555E)センスプライマ
【0250】
【化66】
【0251】
(2) hHGF−Fc(G555E)アンチセンスプライマ
【0252】
【化67】
【0253】
(3) hHGF−Fc(C561R)センスプライマ
【0254】
【化68】
【0255】
(4) hHGF−Fc(C561R)アンチセンスプライマ
【0256】
【化69】
【0257】
c. マウス−ヒト−マウスキメラFc
マウス−ヒト−マウスキメラIgFc構築物は、mHGFアルファ鎖hHGF、ヒトHGFのβ鎖アミノ酸Val495乃至Leu585、及びmHGF C末端β鎖とこれに続く6×His標識及びIgG−Fcを含む。
【0258】
アミノ酸V495乃至L585をコードしているヒトHGF cDNAは、インビトロジェン社のクローンID:IOH29794(ヒトHGF cDNA)から増幅させた。その配列は、アクセッション番号NM_000601.4としてNCBIに寄託されている配列に相当する。マウスHGF配列は、インビトロジェン社製のスーパースクリプトワンステップ(Super Script One Step)RT−PCRキット(#10928−034)をメーカーの使用説明書に従って用い、マウス肝の全RNA(クローンテック社、#636603)からRT−PCRによって増幅させた。そのmHGF cDNA配列は、アセッション番号D10213.1としてNCBIに寄託されている配列に相当する。
【0259】
重なり合うPCRプライマーを用いて断片1、2及び3と称する3本の断片を作製し、連続回のPCR増幅でアニーリングした。最終産物をNheI及びNotIで切断し、pcDNA5/FRT IgGFc中にクローニングした。
【0260】
(1) mHGFアルファ鎖5’NheIの断片1プライマー
【0261】
【化70】
【0262】
【化71】
【0263】
(2) hHGFベータ鎖aaV495乃至L585の断片2プライマー
【0264】
【化72】
【0265】
【化73】
【0266】
(3) mHGFベータ鎖C末端3’NotIの断片3プライマー
【0267】
【化74】
【0268】
【化75】
【0269】
d. hHGF及びmhmキメラの構築
hHGF及びmhmキメラ(V495乃至L585)をコードしているベクターであるpcDNA5/FRT hHGF及びpcDNA5/FRT−mhmキメラ(V495乃至L−L585)を、Fc標識を含ませずに、部位特異的突然変異誘発法により作製した。前記クイックチェンジII XL部位特異的突然変異誘発キット(ストラタジーン社)をメーカーの使用説明書に従って用い、上記6×His標識の3’末端に停止コドンを導入した。突然変異誘発プライマーには、プライマー1:
【0270】
【化76】
【0271】
及びプライマー2:
【0272】
【化77】
【0273】
を含めた。
【0274】
さらに、前記クイックチェンジII XL部位特異的突然変異誘発キット(ストラタジーン社)をメーカーの使用説明書に従って用い、部位特異的突然変異誘発法により上記pcDNA5/FRT−mhm(V495乃至L−L585)構築物から2種の別のmhmキメラを作製した。そのうちの1種のmhm構築物はマウス配列間に配置されたhHGFのI499乃至R556の領域を含んだ。もう一方のmhm構築物は、マウス配列間に配置されたhHGFのW507乃至L585の領域を含んだ。
【0275】
上記mhmキメラ(I499乃至R556)の場合、適切なオリゴヌクレオチド配列を用いて、鋳型pcDNA5/FRT−mhmキメラ(V495乃至L585)構築物中に、点突然変異D558E、C561R、V564I、V567I及びM583Lを順番に起こさせた。上記mhmキメラ(W507乃至L585)の場合、適切なオリゴヌクレオチド配列を用いて、鋳型pcDNA5/FRT−mhmキメラ(V495乃至L585)構築物中に、点突然変異Q502R、N504T及びI505Vを1ステップで導入した。
【0276】
得られたhHGF−Fc蛋白質のヌクレオチド配列は、シグナル配列(1乃至93番目ヌクレオチド)及びプロドメイン(94乃至162番目ヌクレオチド)を含めて、配列番号118として記載している。このhHGF−Fc蛋白質のアミノ酸配列は、配列番号119として記載している。
【0277】
得られたmhm(V495−L585)−Fcキメラ蛋白質をコードしているヌクレオチド配列は、シグナル配列(1乃至96番目ヌクレオチド)及びプロドメイン(97乃至165番目ヌクレオチド)を含めて、配列番号120として記載している。このmhm(V495−L585)−Fcキメラ蛋白質のアミノ酸配列は、配列番号121として記載している。
【0278】
得られたmhm(V495−L585)構築物をコードしているヌクレオチド配列及びこの構築物を規定する蛋白質配列は、それぞれ配列番号211及び212として記載している。配列番号211で表される核酸配列は、シグナル配列(1乃至96番目ヌクレオチド)及びプロドメイン(97乃至165番目ヌクレオチド)を含み、配列番号212で表される蛋白質配列は、(シグナル配列又はプロドメインを含まない)活性蛋白質配列を含む。得られたmhm(I499〜R556)構築物をコードしているヌクレオチド配列及びこの構築物を規定する蛋白質配列は、それぞれ配列番号213及び214として記載している。配列番号213で表される核酸配列は、シグナル配列(1乃至96番目ヌクレオチド)及びプロドメイン(97乃至165番目ヌクレオチド)を含み、配列番号214で表される蛋白質配列は、(シグナル配列又はプロドメインを含まない)活性蛋白質配列を含む。得られたmhm(W507〜L585)をコードしているヌクレオチド配列及びこの構築物を規定する蛋白質配列は、それぞれ配列番号215及び216として記載している。配列番号215で表される核酸配列は、シグナル配列(1乃至96番目ヌクレオチド)及びプロドメイン(97乃至165番目ヌクレオチド)を含み、配列番号216で表される蛋白質配列は、(シグナル配列又はプロドメインを含まない)活性蛋白質配列を含む。
【0279】
e. 蛋白質発現
(1) 細胞培養
CHO FIpIn細胞(インビトロジェン社、カタログ番号R758−07)をF12K培地(ATTC、カタログ番号30−2004)、10%FCS(インビトロジェン社、カタログ番号10438026)、1%ペニシリン(10,000単位/mL)/ストレプトマイシン(10,000μg/mL)(インビトロジェン社、カタログ番号15140−122)中、37℃、5% CO2、100μg/mLゼオシン(Zeocin)(インビトロジェン社、カタログ番号R250−01)の条件下で増殖させた。
【0280】
(2) 安定なCHO FIpIn細胞株の作製
リポフェクタミン2000(インビトロジェン社、カタログ番号11668−027)をそのメーカーの使用説明書に従って用い、発現プラスミドDNA pOG44及びpcDNA5/FRTを9:1の比率で用いてCHO FIpIn宿主細胞に形質移入した。対照として、空のpcDNA5/FRTベクター/pOG44及びpOG44プラスミド(インビトロジェン社、カタログ番号35−3018)単独を細胞に形質移入した。形質移入後24時間に、この細胞を分裂させ、48時間後に、0.5mg/mLのハイグロマイシンB(シグマ社(Sigma)、カタログ番号H0654−SPEC)を細胞に添加した。F12K、10%FCS、1%ペニシリン/ストレプトマイシン、0.5mg/mLハイグロマイシンB中で安定細胞の多クローン性選択を行った。
【0281】
(3) 安定なCHO FIpIn細胞株における蛋白質発現
約2×106個の細胞を15cmプレートに接種し、F12K(ATCC、カタログ番号30−2004)/DMEM高グルコース(インビトロジェン社、カタログ番号11995065)1:1、5%超低IgG FCS(インビトロジェン社、#16250−78)中、37℃、5%CO2で5乃至6日間増殖させた。上清を採取し、得られた蛋白質をELISA及び表面プラスモン共鳴により分析した。
【0282】
(実施例4)
抗hHGFモノクローナル抗体の結合特性
実施例1で作製したモノクローナル抗体の特徴は、これらがhHGF及び実施例3で作製した組換えHGF蛋白質の一部に結合することができることにあった。
【0283】
BlAcore T100装置を用いて表面プラスモン共鳴によりこれらの抗体を分析することによってHGF及び実施例3に記載した融合蛋白質の一部へのその結合能を評価した。各抗体は、標準的なカップリングプロトコルをメーカーの使用説明書に従って用いたアミンカップリング(BIAcore、カタログ番号BR−1000−50)によりカルボキシメチル化デキストランCM5センサーチップ(BIAcore、カタログ番号BR−1006−68)上に固定化した。
【0284】
分析は、0.05%界面活性剤P20(BIAcore、カタログ番号R−1000−54)、2mg/mLのBSA(EMD社カタログ番号2930)及び10mg/mLのCM−デキストランナトリウム塩(フルカ社(Fluka)、カタログ番号86524)を含有するPBS(GIBCO社、カタログ番号14040−133)を泳動用緩衝液として用い、25℃で行った。種々のHGF融合蛋白質を含む上清又は空のベクターを形質移入した細胞からの上清を各抗体上に30μL/分の流速で3分間注入した。その結果生じた結合は、注入終了後30秒のベースラインに対する共鳴単位(RU)として求めた。結合は、泳動用緩衝液で希釈したヒトHGF(R&Dシステムズ社(R&D Systems)、カタログ番号294−HGN−025)と比較した。非特異的な結合は、同じアミンカップリング法を用いてマウスIgG(ロックランド社(Rockland)、カタログ番号010−0102)を固定化した対照表面への結合を比較することにより測定した。
【0285】
以上の結果は表3にまとめた。
【0286】
【表3】
【0287】
表3の結果から明らかなように、上記の各抗体はrHGF及び精製ヒトHGFに結合する。さらに、これらの抗体は全て、点突然変異G555E及びC561Rを含むhHGFに結合する。総じて、1F3及び2F8を除く全ての抗体はマウスHGFに結合せず、このことから抗体1A3、1D3、2B8、3A12、3B6及び3D11がヒトHGFに特異的に結合することが明らかとなった。抗体1D3、1F3及び2B8がマウス−ヒト−マウスキメラに結合するのに対して、残りの抗体は結合しなかった。以上の結果から、抗体1D3及び2B8は、ヒトHGFの495乃至585番目残基に少なくとも部分的に結合することが分かる。抗体1A3、3A12、3B6及び3D11は、495乃至585番目残基以外のヒトhHGFの部分に結合するように思われる。2F8がhHGF及びmHGFのいずれにも結合するように思われるので、現在のところ、これがなぜmhmキメラに結合しないのか不明である。
【0288】
(実施例5)
抗hHGFモノクローナル抗体の還元及び非還元HGFへの結合能
本実施例では、実施例1で作製した抗hHGFモノクローナル抗体の還元及び非還元HGFへの結合能について分析した。
【0289】
抗HGF血清の上記組換えhHGFとの反応性を免疫ブロット法によって評価した。NuPAGEサンプル還元緩衝液(インビトロジェン社)を含む、又は含まないNuPAGE MOPS SDS泳動用緩衝液(インビトロジェン社)中8μgの組換えhHGF蛋白質を4乃至12%ビス−トリス1.0mm×2Dウェルゲル(インビトロジェン社、カールズバッド、カリフォルニア州)で分画した。次いで、分画した蛋白質を標準的な手順によりニトロセルロース膜上に移した。このニトロセルロース膜を、0.1% Tween20(登録商標)含有トリス緩衝食塩液(TBST)に溶かした脱脂乳粉の5%溶液でブロックした後、更なるブロックのために、ミニプロテアンIIマルチスクリーン(Mini Protean II Multi−Screen)装置(バイオラッド社(BioRad))に取り付けた。
【0290】
その結果得られた膜をマルチスクリーン装置上で精製抗体によりプローブした。精製抗体はブロッキング緩衝液で5μg/mLに希釈した。次いで、このニトロセルロース膜を装置から取り外し、ホースラディッシュペルオキシダーゼ標識抗マウスIgG抗体と共にインキュベートした。得られた結果は表4に示した。表中、数字は結合の程度を示し、−は最も弱い(殆どあるいは全く結合しない)、3+は最も強い結合を表している。
【0291】
【表4】
【0292】
表4のデータから明らかなように、抗体は全て、非還元rhHGFに結合する。これに対し、モノクローナル抗体1A3、1D3、1F3、2F8及び3B6は還元rhHGFに結合したが、抗体2B8、3A12及び3D11は還元rhHGFに結合しなかった。
【0293】
(実施例6)
結合親和性
実施例1で作製した各抗体のhHGFに対する結合親和性及び相互作用の速度を表面プラスモン共鳴によって測定した。
【0294】
ウサギ抗マウス免疫グロブリン(BIAcore、カタログ番号BR−1005−14)を、標準的なカップリングプロトコルをメーカーの使用説明書に従って用いたアミンカップリング(BIAcore、カタログ番号BR−1000−50)によりカルボキシメチル化デキストランCM5センサーチップ(BIAcore、カタログ番号BR−1006−68)上に固定化した。分析は、0.05%界面活性剤P20(BIAcore、カタログ番号R−1000−54)、2mg/mLのBSA(EMD社カタログ番号2930)及び10mg/mLのCM−デキストランナトリウム塩(フルカ社(Fluka)、カタログ番号86524)を含有するPBS(GIBCO社、カタログ番号14040−133)を泳動用緩衝液として用い、25℃で行った。
【0295】
前記抗体は、個々のフローセル内で10μL/分の流速で捕捉した。各抗体が各サイクルで捕捉される約20RUの抗体を生じるための注入時間にはばらつきがあった。緩衝液、又は泳動用緩衝液で希釈したHGF(R&Dシステムズ社、カタログ番号294−HGN−025)を対照表面(捕捉抗体なし)及び活性表面(試験対象抗体あり)上に60μL/分で2分間順次注入した。解離相を濃度に応じて15又は90分間モニターした。その後、上記表面は、別のサイクルが開始される前に、pH1.7の10mMグリシン−HCl(BIAcore社、カタログ番号BR−1003−54)を60μL/分の流速で3分間注入して再生させた。試験したHGF濃度は0.46nM乃至7.5nMであった。
【0296】
速度論的パラメータは、対照値減算を含むBIAエバリュエーション(BIAevalutation)ソフトウェアの速度関数を用いて求めた。各抗体の速度論的パラメータka(会合速度定数)、kd(解離速度定数)及びKD(平衡解離定数)を表5にまとめた。
【0297】
【表5】
【0298】
表5のデータから明らかなように、これらの抗体は、KD値約100pM以下、約50pM以下又は約20pM以下でhHGFに結合する。
【0299】
(実施例7)
抗hHGF抗体の中和活性
本実施例では、実施例1で作製した抗体について、(a)c−MetへのhHGFの結合を阻害し、(b)4MBr−5細胞におけるBrdU取り込みのHGFによる促進を阻害する能力を特性化した。
【0300】
a. HGF−Met結合阻害試験(中和試験)
上記抗体について、c−MetへのhHGF結合を阻害する能力をELISAにより試験した。
【0301】
すなわち、ワラック96穴DELFIAアッセイプレート(ワラック社、カタログ番号AAAND−0001)を6.25μg/mL HGF(R&Dシステムズ社、カタログ番号294−HGN−025)の炭酸コーティング緩衝液(15mM Na2CO3及び34mM NaHCO3、pH9.0)溶液100μLにより4℃で16時間コーティングした。次に、このプレートを室温で1時間5%脱脂粉乳のPBS溶液200μLでブロックした。抗体は、別のプレートにおいて5%脱脂粉乳のPBS溶液に溶解した2nMのc−Met(R&Dシステムズ社、カタログ番号358−MT/CF)に検討対象の抗体を漸増濃度(0.033乃至667nM、3倍段階希釈)で加えることにより調製した。ウェル当たり100μLのサンプルを上記アッセイプレートに移し、4℃で一夜インキュベートした。次いで、このアッセイプレートをPBS−0.1% Tween20で3回洗浄した後、5%脱脂粉乳のPBS溶液で調製した100μL/ウェルの2μg/mLビオチン化抗ヒトc−Met抗体(R&Dシステムズ社、カタログ番号BAF358)溶液と共に室温で2時間インキュベートした。
【0302】
その後、上記で得られたプレートをPBS−0.1% Tween20で3回洗浄した後、DELFIAアッセイ緩衝液(ワラック社、カタログ番号4002−0010)で1:1,000に希釈したEu標識ストレプトアビジン(ワラック社、カタログ番号1244−360)と共に室温で1時間インキュベートした。その結果得られたプレートをDELFIA洗浄液(ワラック社、カタログ番号4010−0010)で3回洗浄した後、100μL/ウェルのDELFIA強化溶液(ワラック社、#4001−0010)と共に攪拌しながら室温で15分間インキュベートした。
【0303】
上記プレートは、ユウロピウム法を用いるビクター3V装置(パーキンエルマー社(Perkin Elmer))で読み取らせた。IC50値を算出し、表6にまとめた。
【0304】
【表6】
【0305】
以上の結果から明らかなように、3B6以外の全ての抗体(即ち、1D3、1A3、2B8、3A12、1F3、3D11及び2F8)は、c−MetへのHGFの結合を効率的に中和する。
【0306】
b. 4MBr−5細胞におけるBrdU取り込みのHGFによる促進の中和
12.5nMのhHGF 10μLを96穴組織培養マイクロタイタープレート(コスター社(Costar)カタログ番号3903)の個々のウェルに分注した。次いで、6667、2222、740、247、82、27、9.1、3.0、1.0及び0.33nMの濃度に連続希釈した抗体10μLを各ウェルに添加した。次に、このHGF抗体混合液を室温で30分間インキュベートした。F−12K(ATCC、30−2004)、15%FBS(ギブコ(Gibco)10438−026)、30ng/mL EGF(シグマ(Sigma)E9644)、1%ペニシリン/ストレプトマイシン(PS、ギブコ社カタログ番号15140−122)中で培養したサル気管支上皮細胞4MBr−5(ATCC、CCL208)をトリプシン(Trypsin)(ギブコ社カタログ番号25200−056)で解離させ、75,000個/mLの濃度でアッセイ培地(F−12K、2.5% FBS、1% PS)に再懸濁し、この細胞懸濁液80μLを上記HGF抗体混合液に分注した。
【0307】
この細胞を37℃、5% CO2の条件下でインキュベートした。48時間後、100μM BrdU(ロシュ社(Roche)、カタログ番号1669915)を10μL添加した。72時間後、培地を除去し、プレートをヘアドライヤーで乾燥させた後、BrdU ELISAをメーカーの使用説明書(ロシュ社、カタログ番号1669915)に従って用いて処理した。
【0308】
発光シグナルは、シナジーHT(Synergy HT)プレートリーダー(バイオテック社(Bio−Tek))によって定量化した。このデータは、グラフパッドプリズム(GraphPad Prism)(グラフパッドソフトウェア社(GraphPad Software))において式y=ボトム+(トップ−ボトム)/(1+10^(log(EC50−x)*ヒル勾配))の可変勾配を有するS字形の用量反応に当てはめた。各実験は、2回ずつ、少なくとも3回繰り返した。表7にEC50の平均値を示した。
【0309】
【表7】
【0310】
表7の結果から明らかなように、抗体1A3、ID3、1F3、2B8、2F8、3A12、3B6及び3D11は全て、4MBr−5細胞のHGF誘発性増殖を抑制する。
【0311】
(実施例8)
抗hHGF抗体の抗散乱(Anti−Scatter)活性
本実施例では、実施例1で作製した抗体のHGF誘発性散乱活性を抑制する能力に関する特性化について説明する。HGFは、MDCK細胞(ATCC、マナッサス、バージニア州、カタログ番号CCL−34)においてクラスターの「散乱」(運動)を誘発する。
【0312】
MDCK細胞は、10%ウシ胎仔血清(インビトロジェン社、カタログ番号10438026)及び1%ペニシリン−ストレプトマイシン(インビトロジェン社カタログ番号15140122)を含有するMEM(ATCC、マナッサス、バージニア州、カタログ番号30−2003)80μL中ウェル当たり4×103個の密度で96穴Costar組織培養プレート(コーニング社(Corning Incorporated)、コーニング、ニューヨーク州、カタログ番号3595)に接種した。検討対象の各抗体は、10%ウシ胎仔血清及び1%ペニシリン−ストレプトマイシンを含有するMEMで6,667nMに希釈した。次いで、種々の抗体希釈液のそれぞれ、及び10%ウシ胎仔血清及び1%ペニシリン−ストレプトマイシンを含有し抗体を含まないMEMを別々に、10%ウシ胎仔血清及び1%ペニシリン−ストレプトマイシン並びに100ng/mlのHGF(R&Dシステムズ社、カタログ番号294−HGN−025)を含有する等容量のMEMと混合した。各抗体/HGF希釈液は、25℃で30分間インキュベートした。各抗体/HGF希釈液20μLを個別に個々のウェルに添加し、最終抗体濃度666.7nM及び最終HGF濃度10ng/mlを得た。次に、MDCK細胞を37℃、5%CO2で24時間インキュベートした。
【0313】
24時間のインキュベーション後、MDCK細胞をウェル当たり100μLの氷冷PBS(インビトロジェン社カタログ番号14190144)で一度注意深く洗い、25℃で10分間揺動させながらウェル当たり100μLの氷冷メタノールで固定した。次いで、プレートを一度蒸留水で注意深く洗浄した。蒸留水に溶解した0.5%クリスタルバイオレット(シグマ社、セントルイス、ミズーリ州、カタログ番号C3886)及び50%エタノールからなる100μL容のクリスタルバイオレット溶液を各ウェルに添加し、細胞を揺動させながら25℃で20分間インキュベートした。
【0314】
クリスタルバイオレット溶液による染色後、細胞を蒸留水で三度注意深く洗浄した。次に、各ウェルにPBSを添加してサンプルの乾燥を防いだ。ライカ(Leica)DMIRB顕微鏡(ライカマイクロシステムズ社、ウェッツラー、ドイツ)、DC500カメラ(ライカマイクロシステムズ社、ウェッツラー、ドイツ)及びマグナファイア(MagnaFire)2.1Cソフトウェア(Optronics、ゴレタ、カリフォルニア)を用いて細胞を撮像し、サンプルを散乱レベルについて評価した。表8にその結果をまとめた。
【0315】
【表8】
【0316】
表8の結果から明らかなように、抗体2B8は、その他の抗体よりも多くのHGF誘発性散乱を抑制した。抗体1D3及び3B6は中間的なレベルの抑制を示し、抗体1A3は低乃至中間レベルの抑制を示し、抗体1F3及び2F8は低レベルの抑制を示し、抗体3A12及び3D11はほとんど或いは全く検出可能な抑制を示さなかった。
【0317】
(実施例9)
HGF刺激c−Metリン酸化の阻害
本実施例では、実施例1で作製した抗体の、PC−3細胞におけるHGF刺激c−Metリン酸化を阻害する能力に関する特性化について説明する。HGFはPC−3細胞(ATCC番号CRL−1435)においてMetのリン酸化を誘発する。
【0318】
PC−3細胞は、10%ウシ胎仔血清(インビトロジェン社、カタログ番号10438026)及び1%ペニシリン−ストレプトマイシン(インビトロジェン社カタログ番号15140122)を含有するF−12K(ATCC、マナッサス、バージニア州、カタログ番号30−2004)100μL中ウェル当たり4.5×104個の密度で96穴Costar組織培養プレート(コーニング社カタログ番号3595)の個々のウェルに接種した。37℃、5% CO2で24時間培養後、培地を除去し、1% ペニシリン−ストレプトマイシンを含有する無血清F−12Kで細胞を一度洗った。次いで、細胞を1% ペニシリン−ストレプトマイシンを含有する無血清F−12K 100μL中で24時間インキュベートした。
【0319】
1% ペニシリン−ストレプトマイシンを含有する無血清F−12Kを用いて、検討対象の各抗体の10種の希釈液、即ち、6,667nM、2,222nM、741nM、247nM、82.3nM、27.4nM、9.1nM、3.0nM、1.0nM及び0.3nMの希釈液を調製した。各抗体希釈液、及び1% ペニシリン−ストレプトマイシンを含有するが抗体を含まない無血清F−12Kを別々に、1% ペニシリン−ストレプトマイシン及び500ng/mLのHGF(R&Dシステムズ社カタログ番号294−HGN−025)を含有する等容量の無血清F−12Kと混合した。次いで、これらの抗体/HGF希釈液を25℃で30分間インキュベートした。これによりHGFの最終濃度は1.25nMとなった。
【0320】
その後、1% ペニシリン−ストレプトマイシンを含有する無血清F−12Kで上記PC−3細胞を一度洗った。次に、この細胞に、1% ペニシリン−ストレプトマイシンを含有する無血清F−12Kを70μL添加した後、10mM Na3VO4(シグマ社カタログ番号S6508)の1% ペニシリン−ストレプトマイシン含有無血清F−12K溶液10μLを添加した。次いで、この細胞を37℃、5% CO2で60分間インキュベートした。このインキュベーションの後、各抗体/HGF希釈液20μLを別々に個別のウェルに添加し、最終HGF濃度50ng/mL並びに各抗体の最終濃度666.7nM、222.2nM、74.1nM、24.7nM、8.23nM、2.74nM、0.91nM、0.30nM、0.10nM及び0.03nMを得た。次に、この細胞を37℃、5%CO2で10分間インキュベートし、その時点後に培地/抗体/HGF混合物を除去してプレートを氷上に置いた。その後、細胞を、1mM Na3VO4を含有する100μL/ウェルの氷冷PBS(インビトロジェン社カタログ番号14190144)で一度洗った。次いで、この細胞を、1% OmniPur Triton X−100(ドイツメルク社(MERCK KGaA)、ダルムシュタット、ドイツ、カタログ番号9410)、50mMトリス−HCl pH8.0、100mM NaCl、0.3mM Na3VO4、1×プロテアーゼ阻害剤カクテル(シグマ社カタログ番号P8340)及び1×ホスファターゼ阻害剤カクテル2(シグマ社カタログ番号5726)からなる100μL/ウェルの氷冷溶解緩衝液中、4℃で、30分間インキュベートした。
【0321】
ビオチニル化抗ヒトHGF−R(c−met)抗体(R&Dシステムズ社カタログ番号BAF358)を、1%ウシ血清アルブミン(シグマ社カタログ番号A2153)を含有するDELFIAアッセイバッファ(Assay Buffer)(パーキンエルマー社(PerkinElmer)、トゥルク、フィンランド、カタログ番号4002−0010)で2μg/mLの濃度に希釈し、黄色ストレプトアビジンマイクロタイトレーションプレート(パーキンエルマー社カタログ番号AAAND−0005)のウェル当たり50μLのこの希釈液を添加した。次に、このプレートを揺動させながら25℃で30分間抗体と共にインキュベートした。インキュベーション後、このプレートをDELFIA洗浄液(パーキンエルマー社カタログ番号4010−0010)で洗浄し、各種PC−3細胞溶解物のそれぞれ80μLを別々に、洗浄したストレプトアビジンマイクロタイトレーションプレートの個々のウェルに添加した。
【0322】
上記のPC−3細胞溶解物を含むストレプトアビジンマイクロタイトレーションプレートを振盪しながら25℃で60分間インキュベートした後、DELFIA洗浄溶液で洗浄した。この前もってPC−3細胞溶解物とインキュベートした洗浄ストレプトアビジンマイクロタイトレーションプレートの各ウェルに、1%ウシ血清アルブミンを含有するDELFIAアッセイバッファーで希釈した600ng/mL DELFIA Eu−N1 P−tyr−100抗体(パーキンエルマー社、カタログ番号AD0159)溶液100μLを添加した。次いで、このプレートを揺動させながら25℃で60分間インキュベートした。最後に、このプレートをDELFIA洗浄溶液で洗浄した。次に、この洗浄ストレプトアビジンマイクロタイトレーションプレートの各ウェルにDELFIAエンハンスメントソリューション(Enhancement Solution)(パーキンエルマー社カタログ番号4001−0010)200μLを添加し、プレートを暗所で振盪しながら25℃で5分間インキュベートした。
【0323】
その後、ビクター3V(Victor3V)リーダ(パーキンエルマー社)でユウロピウム(Europium)プロトコルを用いてシグナルを測定した。ウィンドウズ(登録商標)(Windows(登録商標))用プリズム(Prism)4(グラフパッドソフトウェア社(GraphPad Software,Inc.)、サンディエゴ、カリフォルニア州)及びS字形用量反応式を用いてEC50値を算出した。
【0324】
nM単位のEC50としてまとめた結果を表9に示した。
【0325】
【表9】
【0326】
表9のデータから明らかなように、8種の抗体は全てPC−3細胞におけるHGF誘導性c−Metリン酸化の強力な阻害剤である。
【0327】
(実施例10)
U87MG異種移植モデルにおける腫瘍抑制
U87MG異種移植モデルを用い、本発明のマウスモノクローナル抗体の腫瘍増殖抑制能について試験した。U87MG細胞(ATCC)は、ダルベッコ改変イーグル培地(DMEM:Dulbecco’s Modified Eagle medium)を10%ウシ胎仔血清、100単位/mLペニシリン及び100μg/mLストレプトマイシンと共に含む培地を用いた、5%CO2及び95%空気を含有する雰囲気中37℃の培養で増殖させた。これらの細胞は、トリプシン−EDTAを用いて培養皿の壁から細胞を剥離することにより二次培養して維持した。
【0328】
ほぼコンフルエントの細胞をトリプシン処理により回収し、次いで、50%マトリゲル(Matrigel)(BDバイオサイエンシズ社(BD Biosciences)、カタログ番号356237)中の細胞5×106個を、7週齢雌性ICR SCIDマウス(タコニックラボラトリー社(Taconic Labs))の肩甲骨間の上背部に皮下注射した。キャリパで腫瘍の長径(L)及び短径(W)(mm)を測定した。腫瘍容積(vol.)は、容積(mm3)=L×W2/2として計算した。腫瘍が約200mm3にまで成長した時に、担腫瘍マウスを1群10匹の5群に無作為化した。1群にはPBSを投与した。その他の4群のそれぞれには、抗体1A3、1D3、1F3又は2B8のうちの1種を投与した。抗体は全て、5用量の腹腔内注射により週2回1mg/kg体重を投与した。腫瘍容積及びマウス体重を週2回記録した。スチューデントのt−検定を用いて腫瘍成長の抑制を解析した。結果は図6及び表10にまとめた。
【0329】
【表10】
【0330】
2B8投与群で部分的な退縮が達成された(図6)。1A3投与群及び1F3投与群では統計的に有意な成長抑制が認められた(表10)。1D3の場合、51%の腫瘍成長抑制(p値:0.075)が認められた。有意な体重減少は認められなかった。
【0331】
(実施例11)
U118異種移植モデルにおける腫瘍抑制
U118異種移植モデルを用い、抗体1A3、1D3、1F3及び2B8の腫瘍増殖抑制能について試験した。U118細胞(ATCC)は、U87MG細胞に関して(上記)実施例10に記載したのと同様にして増殖させた。
【0332】
使用マウスが7週齢雌性NCrヌードマウス(タコニック社)であること以外は上記実施例10に記載したのと同様にして皮下腫瘍を確立し、投与は、腫瘍が約80mm3にまで成長した時に開始した。U87MGモデルの場合と同様に、抗体は全て、4用量の腹腔内注射により週2回1mg/kg体重を投与した。腫瘍容積及びマウス体重を週2回記録した。スチューデントのt−検定を用いて腫瘍成長の抑制を解析した。結果は図7及び表11にまとめた。
【0333】
【表11】
【0334】
2B8投与群及び1A3投与群では統計的に有意な成長抑制が認められた(図7)。1F3群及び1D3群では中等度の腫瘍成長抑制が認められ、p値は、本試験で統計的に有意であると定義した0.05未満であった(表11)。有意な体重減少は認められなかった。
【0335】
(実施例12)
マウスモノクローナル抗体のヒト化
本実施例では、マウス2B8抗体のヒト化及び得られるヒト化抗体の特性化について説明する。マウス2B8重及び軽鎖可変領域は2通りの方法で「ヒト化」した。
【0336】
A. ヒト化の方法1
第一の方法では、Hwangほか (2005年) METHODS 36:p.35−42、Tanほか (2002年) J. IMMUNOL. 169:p.1119−1125及び米国特許第6,881,557号明細書に記載されている「超ヒト化」方法に基づいて、3つのヒト化重鎖可変領域及び2つのヒト化カッパ軽鎖可変領域を設計した。
【0337】
各マウス2B8CDRについてCDRの長さ及びアミノ酸組成に基づき、チョチア規定構造クラス(Chothia canonical structural class)を特定した。同じチョチア規定構造クラスの軽及び重鎖可変領域からなるヒト生殖細胞系可変領域は、国際免疫遺伝学情報システム(IMGT)ウェブサイトに記載されている既知ヒト生殖細胞系可変領域基準対立遺伝子(ワールドワイドウェブのimgt.cines.fr及びbiochem.unizh.ch/antibody/Sequences/index.htmlで入手可能)に基づいて確認した。同構造クラスのこれらヒト生殖細胞系可変領域は、CDRアミノ酸残基間の完全な同一性又は類似性を計算することによってマウス2B8可変領域と比較した。マウス2B8CDR残基と最も高い同一性及び/又は類似性を有するヒト生殖細胞系可変領域をCDR移植用に選んだ。これらのヒト生殖細胞系可変領域のフレームワーク残基を保存する一方、マウス2B8 CDR残基は、マウス2B8 CDRとヒト生殖細胞系CDRとの間で異なる対応するヒト生殖細胞系可変領域残基を置換するために用いた。次いで、2B8マウスJ領域に最も類似したヒトJ領域を上記「超ヒト化」可変領域のカルボキシル末端に付加した。次に、この「超ヒト化」可変領域のアミノ末端にシグナル配列を付加し、これらのアミノ酸配列を核酸配列に変換した。
【0338】
この完全な可変領域核酸配列は、遺伝子合成PCR法(Youngほか (2004年) NUCL. ACIDS RES. 32:e59)を用いて構築し、標準的な分子生物学的技術を使用して、(上記可変領域の下流にある)ヒト定常IgG1(G1m(17,1)アロタイプ)又はカッパ(Km(3)アロタイプ(対立遺伝子2))領域を含む(pcDNA3.2DEST(インビトロジェン社)を利用した)哺乳動物発現ベクター中にクローニングした。4種の重鎖IgG1抗体(キメラ2B8及び3種のヒト化重鎖(Hu2B8 Hv1−f.1、Hu2B8 Hv5−a.1及びHu2B8 Hv5−51.1))の全てを3種のカッパ鎖抗体(キメラ2B8及び2種のヒト化軽鎖(Hu2B8 Kv1−39.1及びHu2B8 Kv3−15.1))の全てとの考えられる全ての組合せで発現させ、12種の抗体蛋白質を作製した。次いで、これらキメラ、キメラ/ヒト化及びヒト化抗体のヒトHGFへの結合を下記のようにして測定し、その結果を図8にまとめた。免疫グロブリン重鎖可変領域と免疫グロブリン軽鎖可変領域との考えられる全ての組合せはいずれも下記の表12Aに記載した。
【0339】
【表12A】
【0340】
免疫グロブリン重鎖と免疫グロブリン軽鎖とのえられる全ての組合せはいずれも下記の表12Bに記載した。
【0341】
【表12B】
【0342】
ヒト化可変領域を含む完全長免疫グロブリン重及び軽鎖を含む考えられる抗体構築物のうちの2種について下記の通り命名した。
【0343】
sh2B8−9 (G1m(17,1))=hu2B8 Hv5−51.1(+IgG1定常領域(Glm(17,1)アロタイプ)(配列番号171)プラスhu2B8 Kv 1−39.1 (+カッパ定常領域(Km(3)アロタイプ(対立遺伝子2))) (配列番号177)
sh2B8−12 (Glm(17,1))=hu2B8 Hv5−51.1(+IgG1定常領域(Glm(l7,1)アロタイプ))(配列番号171)プラスhu2B8
Kv 3−15.1 (+カッパ定常領域(Km(3)アロタイプ(対立遺伝子2))) (配列番号181)
各ヒト化抗体をコードしている核酸配列及び各ヒト化抗体を規定する蛋白質配列を下記にまとめた。このセクションでは、各可変領域の最後のヌクレオチドは、可変/定常領域接合部によって生じる次のコドンの最初の塩基である。このヌクレオチドは、可変領域に含まれる。何故なら、これはそのエクソンの一部であるからである。下記に列挙した定常領域のアミノ酸配列はこの接合部コドンの翻訳物を含む。
【0344】
(1) 完全長キメラ2B8重鎖(マウス可変領域及びヒトIgG1定常領域)(アロタイプG1m(17,1))をコードしている核酸配列(下線:シグナル配列)(配列番号154)
【0345】
【化78】
【0346】
(2) 完全長キメラ2B8重鎖(キメラ2B8 IgG1(G1m(17,1)アロタイプ)を規定する蛋白質配列(シグナル配列なし)(配列番号155)
【0347】
【化79】
【0348】
(3) 完全長キメラ2B8軽鎖(マウス可変領域及びヒト定常領域)(キメラ2B8カッパ(Km(3)))をコードしている核酸配列(下線:シグナル配列)(配列番号156)
【0349】
【化80】
【0350】
【化81】
【0351】
(4) 完全長キメラ2B8軽鎖(キメラ2B8カッパ(Km(3)))を規定する蛋白質配列(シグナル配列なし)(配列番号157)
【0352】
【化82】
【0353】
(5) ヒト化Hu2B8 Hv1−f.1重鎖可変領域をコードしている核酸配列(下線:シグナル配列)(配列番号158)
【0354】
【化83】
【0355】
(6) ヒト化Hu2B8 Hv1−f.1重鎖可変領域を規定する蛋白質配列(シグナル配列なし)(配列番号159)
【0356】
【化84】
【0357】
(7) ヒトIgG1重鎖定常領域(G1m(17,1)アロタイプ)をコードしている核酸配列(配列番号160)
【0358】
【化85】
【0359】
(8) ヒトIgG1重鎖定常領域(G1m(17,1)アロタイプ)を規定する蛋白質配列(配列番号161) 最初のアミノ酸は、可変領域の最後のヌクレオチド及びIgG1重鎖配列の初めの2個のヌクレオチドの翻訳物に由来する。
【0360】
【化86】
【0361】
(9) 完全長重鎖ヒト化Hu2B8 Hv1f.1可変領域及びヒトIgG1(G1m(17,1)アロタイプ)重鎖定常領域をコードしている核酸配列(下線:シグナル配列)(配列番号162)
【0362】
【化87】
【0363】
(10) 完全長重鎖ヒト化Hu2B8 Hv1f.1可変領域及びヒトIgG1重鎖定常領域(G1m(17,1)アロタイプ)を規定する蛋白質配列(配列番号163)
【0364】
【化88】
【0365】
(11) ヒト化Hu2B8 Hv5a.1重鎖可変領域をコードしている核酸配列(下線:シグナル配列)(配列番号164)
【0366】
【化89】
【0367】
【化90】
【0368】
(12) ヒト化Hu2B8 Hv5a.1重鎖可変領域を規定する蛋白質配列(配列番号165)
【0369】
【化91】
【0370】
(13) 完全長ヒト化Hu2B8 Hv5a.1重鎖可変領域及びヒトIgG1(G1m(17,1)アロタイプ)重鎖定常領域をコードしている核酸配列(下線:シグナル配列)(配列番号166)
【0371】
【化92】
【0372】
(14) 完全長ヒト化Hu2B8 Hv5a.1重鎖可変領域及びヒトIgG1(G1m(17,1)アロタイプ)重鎖定常領域を規定する蛋白質配列(シグナル配列なし)(配列番号167)
【0373】
【化93】
【0374】
【化94】
【0375】
(15) ヒト化Hu2B8 Hv5−51.1重鎖可変領域をコードしている核酸配列(下線:シグナル配列)(配列番号168)
【0376】
【化95】
【0377】
(16) ヒト化Hu2B8 Hv5−51.1重鎖可変領域を規定する蛋白質配列(シグナル配列なし)(配列番号169)
【0378】
【化96】
【0379】
(17) 完全長重鎖ヒト化Hu2B8 Hv5−51.1重鎖可変領域及びヒトIgG1(G1m(17,1)アロタイプ)重鎖定常領域をコードしている核酸配列(下線:シグナル配列)(配列番号170)
【0380】
【化97】
【0381】
【化98】
【0382】
(18) 完全長重鎖ヒト化Hu2B8 Hv5−51.1重鎖可変領域及びヒトIgG1(G1m(17,1)アロタイプ)重鎖定常領域を規定する蛋白質配列(シグナル配列なし)(配列番号171)
【0383】
【化99】
【0384】
(19) ヒト化Hu2B8 Kv1−39.1カッパ鎖可変領域をコードしている核酸配列(下線:シグナル配列)(配列番号172)。考えられる2つの開始ATGを大文字で示した。
【0385】
【化100】
【0386】
(20) ヒト化Hu2B8 Kv1−39.1カッパ鎖可変領域を規定する蛋白質配列(シグナル配列なし)(配列番号173)
【0387】
【化101】
【0388】
(21) ヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子2)をコードしている核酸配列(配列番号174)
【0389】
【化102】
【0390】
【化103】
【0391】
(22) ヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子2)を規定する蛋白質配列 (配列番号175)
【0392】
【化104】
【0393】
(23) 完全長ヒト化Hu2B8 Kv1−39.1軽鎖可変領域及びヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子2)をコードしている核酸配列(下線:シグナル配列)(配列番号176)
【0394】
【化105】
【0395】
(24) 完全長ヒト化Hu2B8 Kv1−39.1軽鎖可変領域及びヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子1)を規定する蛋白質配列 (配列番号177)
【0396】
【化106】
【0397】
(25) ヒト化Hu2B8 Kv3−15.1軽鎖可変領域をコードしている核酸配列(下線:シグナル配列)(配列番号178)
【0398】
【化107】
【0399】
【化108】
【0400】
(26) ヒト化Hu2B8 Kv3−15.1軽鎖可変領域を規定する蛋白質配列(シグナル配列なし)(配列番号179)
【0401】
【化109】
【0402】
(27) 完全長ヒト化Hu2B8 Kv3−15.1軽鎖可変領域及びヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子2)をコードしている核酸配列(下線:シグナル配列)(配列番号180)
【0403】
【化110】
【0404】
(28) ヒト化Hu2B8 Kv3−15.1軽鎖可変領域及びヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子2)を規定する蛋白質配列(シグナル配列なし)(配列番号181)
【0405】
【化111】
【0406】
便宜のため、このセクションで説明した抗体の完全長配列と配列表に示したものとの対応関系を示すコンコーダンス表を表13に示した。
【0407】
【表13】
【0408】
B. ヒト化の方法2
マウス2B8抗体の免疫原性を低減させるのに用いる第二のヒト化方法は、Studnickaほか(1994年) PROTEIN ENG. 7:p.805−814に記載されている方法に基づくものである。重及びカッパヒト生殖細胞系可変領域のうち、マウス2B8のものに(アミノ酸レベルで)最も一致する領域を特定した。マウスとヒトとの間で異なる残基は、そのような変化が結合又は免疫原性に影響すると思われるリスクに応じてヒトの配列に変換した。低リスク残基(即ち、変化があっても、抗原結合に影響しないと考えられ、免疫原性を生じる可能性も少ない残基)は、ヒトアミノ酸への変更を重鎖可変領域(LR2B8HCを作製)及びカッパ可変領域(LR2B8LCを作製)において行った。さらに、低リスク及び中間リスク残基(即ち、変化があると、抗原結合に影響する可能性が若干高いが、免疫原性を生じる可能性も少ない残基)は、ヒトアミノ酸への変更を重鎖可変領域(LRMR2B8HCを作製)及びカッパ可変領域(LRMR2B8LCを作製)において行った。上記2種のヒト型に設計された重鎖可変領域のカルボキシル末端にヒトIgG1重鎖定常領域(G1m(3)アロタイプ(対立遺伝子1))を付加し、2種のヒト型に設計された軽鎖可変領域のカルボキシル末端にヒトカッパ定常領域(Km(3)アロタイプ(対立遺伝子1))を付加して、ヒト型に設計された抗体鎖を作製した。先ず、遺伝子合成法によって各可変領域の核酸配列を合成した後、これに各ヒト定常領域の配列に付加した。これらのヒト型に設計された抗体を哺乳動物の蛋白質発現ベクター中にクローニングし、重鎖プラス軽鎖の考えられる4種の組合せの蛋白質を発現させた。キメラ、キメラ/ヒト化又はヒト化抗体のヒトHGFへの結合については、下記のように、従来の技術を用いて測定した。
【0409】
各ヒト化抗体をコードしている核酸配列及び各ヒト化抗体を規定する蛋白質配列を下記にまとめた。このセクションでは、各可変領域の最後のヌクレオチドは、可変/定常領域接合部によって生じる次のコドンの最初の塩基である。このヌクレオチドは、可変領域に含まれる。何故なら、これはそのエクソンの一部であるからである。下記に列挙した定常領域のアミノ酸配列はこの接合部コドンの翻訳物を含む。
【0410】
(1) ヒト化LR2B8HC重鎖可変領域をコードしている核酸配列(下線:シグナル配列)(配列番号182)
【0411】
【化112】
【0412】
(2) ヒト化LR2B8HC重鎖可変領域を規定する蛋白質配列(シグナル配列なし)(配列番号183)
【0413】
【化113】
【0414】
(3) ヒトIgG1重鎖定常領域(G1m(3)アロタイプ)(対立遺伝子1)をコードしている核酸配列(配列番号184)
【0415】
【化114】
【0416】
(4) ヒトIgG1重鎖定常領域(G1m(3)アロタイプ)(対立遺伝子1又は2)を規定する蛋白質配列(配列番号185)。最初のアミノ酸は、可変領域の最後のヌクレオチド及びIgG1重鎖配列の初めの2個のヌクレオチドの翻訳物に由来する。
【0417】
【化115】
【0418】
【化116】
【0419】
(5) 完全長重鎖ヒト化LR2B8HC重鎖可変領域及びヒトIgG1重鎖定常領域(G1m(3)アロタイプ)(対立遺伝子1)(下線:シグナル配列)(配列番号186)
【0420】
【化117】
【0421】
(6) 完全長重鎖ヒト化LR2B8HC重鎖可変領域及びヒトIgG1重鎖定常領域(G1m(3)アロタイプ)(対立遺伝子1)を規定する蛋白質配列(シグナル配列なし)(配列番号187)
【0422】
【化118】
【0423】
(7) ヒト化LRMR2B8HC重鎖可変領域をコードしている核酸配列(下線:シグナル配列)(配列番号188)
【0424】
【化119】
【0425】
(8) ヒト化LRMR2B8HC重鎖可変領域を規定する蛋白質配列(シグナル配列なし)(配列番号189)
【0426】
【化120】
【0427】
(9) 完全長重鎖ヒト化LRMR2B8HC重鎖可変領域及びヒトIgG1重鎖定常領域(G1m(3)アロタイプ)(対立遺伝子1)をコードしている核酸配列(下線:シグナル配列)(配列番号190)
【0428】
【化121】
【0429】
(10) 完全長重鎖ヒト化LRMR2B8HC重鎖可変領域及びヒトIgG1重鎖定常領域(G1m(3)アロタイプ)(対立遺伝子1)を規定する蛋白質配列(シグナル配列なし)(配列番号191)
【0430】
【化122】
【0431】
(11) ヒト化LR2B8LC軽鎖可変領域をコードしている核酸配列(下線:シグナル配列)(配列番号192)
【0432】
【化123】
【0433】
(12) ヒト化LR2B8LC軽鎖可変領域を規定する蛋白質配列(シグナル配列なし)(配列番号193)
【0434】
【化124】
【0435】
(13) ヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子1)をコードしている核酸配列(配列番号194)。最初のアミノ酸は、可変領域の最後のヌクレオチドおよびκ軽鎖配列の開始2ヌクレオチドの翻訳物に由来する。
【0436】
【化125】
【0437】
(14) ヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子1)を規定する蛋白質配列(配列番号195)。最初のアミノ酸は、可変領域の最後のヌクレオチドおよびκ軽鎖配列の開始2ヌクレオチドの翻訳物に由来する。
【0438】
【化126】
【0439】
(15) 完全長LR2B8LC軽鎖可変領域及びヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子1)をコードしている核酸配列 (配列番号196)
【0440】
【化127】
【0441】
(16) 完全長LR2B8LC軽鎖可変領域及びヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子1)をコードしている蛋白質配列(配列番号197)
【0442】
【化128】
【0443】
(17) ヒト化LRMR2B8LC軽鎖可変領域をコードしている核酸配列(下線:シグナル配列)(配列番号198)
【0444】
【化129】
【0445】
(18) ヒト化LRMR2B8LC軽鎖可変領域を規定する蛋白質配列(シグナル配列なし)(配列番号199)
【0446】
【化130】
【0447】
(19) 完全長ヒト化LRMR2B8LC軽鎖可変領域及びヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子1)をコードしている核酸配列(下線:シグナル配列)(配列番号200)
【0448】
【化131】
【0449】
(20) 完全長ヒト化LRMR2B8LC軽鎖可変領域及びヒトカッパ鎖定常領域(Km(3)アロタイプ)(対立遺伝子1)を規定する蛋白質配列(配列番号201)
【0450】
【化132】
【0451】
便宜のため、このセクションで説明した抗体の完全長配列と配列表に示したものとの対応関系を示すコンコーダンス表を表14に示した。
【0452】
【表14】
【0453】
表15は、以上の本実施例で説明したヒト化の方法1及びヒト化の方法2により作製したヒト化2B8抗体の重鎖CDR配列(カバットの定義)をまとめたものである。
【0454】
【表15】
【0455】
表16は、以上の本実施例で説明したヒト化の方法1及びヒト化の方法2により作製したヒト化2B8抗体の軽鎖CDR配列(カバット(Kabat)の定義)をまとめたものである。
【0456】
【表16】
【0457】
C. ヒト化2B8抗体の結合親和性
BIAcoreT100装置を用いて表面プラスモン共鳴法により抗原結合親和性及び相互作用の速度を評価した。マウス抗ヒト免疫グロブリン(ジャクソン・イムノリサーチ・ラボラトリーズ社(Jackson ImmunoResearch Labs)、209−005−098)は、標準的なカップリングプロトコルをメーカーの推奨事項に従って用いたアミンカップリング(BIAcore、カタログ番号BR−1000−50)によりカルボキシメチル化デキストランCM4センサーチップ(BIAcore、カタログ番号BR−1005−34)上に固定化した。分析は、0.05%の界面活性剤P20(BIAcore、カタログ番号BR−1000−54)、2mg/mLのBSA(EMD社カタログ番号2930)及び10mg/mLのCM−デキストランナトリウム塩(フルカ社(Fluka)、カタログ番号86524)を含有するPBS(GIBCO社、カタログ番号14040−133)を泳動用緩衝液として用い、25℃で行った。
【0458】
前記抗体は、個々のフローセル上で10μL/分の流速で捕捉した。各抗体が各サイクルで捕捉される約20RUの抗体を生じるための注入時間にはばらつきがあった。緩衝液又は泳動用緩衝液で希釈したHGF(R&Dシステムズ社、カタログ番号294−HGN−025)を対照表面(捕捉抗体なし)及び活性表面(試験対象抗体あり)上に60μL/分で2分間順次注入した。解離相を濃度に応じて15又は90分間モニターした。その後、上記表面は、別のサイクルが開始される前に、pH2.0の10mMグリシン−塩酸(BIAcore社、カタログ番号BR−1003−55)を60μL/分の流速で3分間注入して再生させた。試験したHGF濃度は1.88、3.75及び7.5nMであった。速度論的パラメータは、対照値減算を含むBIAエバリュエーション(BIAevalutation)ソフトウェアの速度関数を用いて求めた。各抗体の速度論的パラメータka(会合速度定数)、kd(解離速度定数)及びKD(平衡解離定数)を図8にまとめた。
【0459】
図8にまとめた結果から、超ヒト化重鎖(Hu2B8 Hv5a.1、Hu2B8 Hv5−51.1又はHu2B8 Hvl−f.l)及び軽鎖(Hu2B8 Kvl−39.1又はHu2B8 Kv3−15.1)の一部の組合せは、キメラ2B8(マウス可変領域及びヒト定常領域)並びに2B8(表5)と同様なHGFとの結合親和性(KD)を保持することが分かる。
【0460】
D. 相互排他的結合
HGFへの相互排他的結合についてBIAcore T100装置を用いて表面プラスモン共鳴により評価した。マウス抗ヒト免疫グロブリン(ジャクソン・イムノリサーチ・ラボラトリーズ社(Jackson ImmunoResearch Labs)、209−005−098)は、標準的なカップリングプロトコルをメーカーの推奨事項に従って用いたアミンカップリング(BIAcore、カタログ番号BR−1000−50)によりカルボキシメチル化デキストランCM5センサーチップ(BIAcore、カタログ番号BR−1006−68)上に固定化した。分析は、0.05%の界面活性剤P20(BIAcore、BR−1000−54)、2mg/mLのBSA(EMD社カタログ番号2930)及び10mg/mLのCM−デキストランナトリウム塩(フルカ社(Fluka)、カタログ番号86524)を含有するPBS(GIBCO社、カタログ番号14040−133)を泳動用緩衝液として用い、25℃で行った。
【0461】
前記ヒト化抗体は、個々のフローセル上で30μL/分の流速で捕捉した。各抗体が各サイクルで捕捉される約150RUの抗体を生じるための注入時間にはばらつきがあった。7.5μg/mLの最終濃度に泳動用緩衝液で希釈したHGF(R&Dシステムズ社、カタログ番号294−HGN−025)を上記捕捉ヒト化抗体上に30μL/分で90秒間注入した。HGFの結合を測定した後、続いてマウス2B8抗体又はヤギポリクロナール抗HGF抗体(R&Dシステムズ社、AF294)を30μL/分で3分間注入した。その後、上記表面は、別の抗体が試験される前に、pH2.0の10mMグリシン−塩酸(BIAcore社、カタログ番号BR−1003−55)を60μL/分の流速で3分間注入して再生させた。得られた結果は図9にまとめた。
【0462】
図9にまとめた結果から、ヒト化2B8抗体及びキメラ2B8抗体はいずれもマウス2B8がHGFに結合するのを妨げることが分かる。以上の結果は、ヒト化抗体がそれでもなお元の2B8抗体と同じHGF抗原決定基に結合することを証明している。
【0463】
(実施例13)
ヒト化2B8変異体の作製
a. ヒューマン・エンジニアード(商標)抗体
ヒトカッパ及びガンマ−1定常領域モジュールを含むXOMA社の一過性抗体発現ベクター中に、コドン−及び発現適正化低リスク及び低プラス中等度リスクヒューマン・エンジニアード軽鎖(それぞれ、LR2B8LC及びLRMR2B8L)及び重鎖(それぞれ、LR2B8HC及びLRMR2BHC)をインフェーズでクローニングした。これら4種のヒューマン・エンジニアード2B8変異体は、HEK293E細胞への一過性形質移入によって作製した。以下の4種の抗体を作製した。
【0464】
HE2B8−1 = LR2B8HC(+IgG1定常領域(Glm(3)アロタイプ(対立遺伝子1))(配列番号187)プラスLR2B8LC(+カッパ鎖定常領域(Km(3)アロタイプ(対立遺伝子1)))(配列番号197)
HE2B8−2 = LR2B8HC(+IgGl定常領域(Glm(3)アロタイプ(対立遺伝子1))(配列番号187)プラスLRMR2B8LC(+カッパ鎖定常領域(Km(3)アロタイプ(対立遺伝子1)))(配列番号201)
HE2B8−3 = LRMR2B8HC(+IgG1定常領域(Glm(3)アロタイプ(対立遺伝子1))(配列番号191)プラスLR2B8LC(+カッパ鎖定常領域(Km(3)アロタイプ(対立遺伝子1)))(配列番号197)
HE2B8−4 = LRMR2B8HC(+IgG1定常領域(Glm(3)アロタイプ(対立遺伝子1))(配列番号191)プラスLRMR2B8LC(+カッパ鎖定常領域(Km(3)アロタイプ(対立遺伝子1)))(配列番号201)
これら軽及び重鎖は、2リットルの振盪フラスコを用いてIS293培地(アーバイン・サイエンティフィック社、アーバイン、カリフォルニア州)中で増殖させたXOMA社の懸濁適応HEK293E細胞中に同時形質移入した。振盪フラスコでの24時間の形質移入後、形質移入細胞200mLを遠心分離し、新鮮培地40mLに再懸濁し、産生用インテグラ(Integra)フラスコ(ウィルソン・ウォルフ・マニュファクチャリング社(Wilson Wolf Manufacturing Inc.)、ミネソタ州)に移した。数日間のインキュベーション後、細胞懸濁液をインテグラフラスコから取り出し、遠心分離して培養上清を残した。この培養上清中の抗体をプロテインAスピンカラム(プロ−ケム社)で精製し、PBSに対して透析し、濃縮して除菌した。
【0465】
b. スーパーヒューマナイズド(SUPERHUMANIZED)(商標)抗体
完全長Hu2B8_Hv5−51.1+ヒトIgG1定常ドメイン(G1m(3)アロタイプ)cDNAをHindIII及びEcoRI制限部位を利用してpEE6.4(ロンザバイオロジックス社(Lonza Biologics)、バークシャー、英国)中にクローニングした。完全長Hu2B8_Kv−39.1可変領域+ヒトカッパ鎖定常ドメインcDNA及び完全長Hu2B8_Kv3−15.1可変領域+ヒトカッパ鎖定常ドメインcDNAをそれぞれ、HindIII及びEcoRI制限部位を利用してpEE14.4(ロンザバイオロジックス社)中にクローニングした。(pEE6.4内の)hCMV−MIEプロモーター+完全長Hu2B8_Hv5−51.1+ヒトIgG1定常ドメイン(G1m(3)アロタイプ)cDNA+SV40ポリA断片をNotI/SalI消化により切り出し、どちらのカッパ鎖pEE14.4ベクター中にもNotI/SalI部位から挿入することにより、それぞれ重及び軽鎖を同時に発現する2種の発現ベクターを作り、以下の抗体を作製した。
【0466】
sh2B8−9(Glm(3)) = hu2B8 Hv5−51.1(+IgGl定常領域(Glm(3)アロタイプ)(対立遺伝子2))(配列番号210)プラスhu2B8 Kv 1−39.1(+カッパ鎖定常領域(Km(3)アロタイプ(対立遺伝子2)))(配列番号177)
sh2B8−12(Glm(3)) = hu2B8 Hv5−51.1(+IgG1定常領域(Glm(3)アロタイプ)(対立遺伝子 2))(配列番号210)プラスhu2B8 Kv 3−15.1(+カッパ鎖定常領域(Km(3)アロタイプ(対立遺伝子2)))(配列番号181)
上記ヒトIgG1重鎖定常領域G1m(3)アロタイプ(対立遺伝子2)及び上記各完全長重鎖配列をコードしている核酸配列並びにこれらを規定する蛋白質配列を以下に記載した。軽鎖の配列は実施例12に記載したのと同じであった。
【0467】
(1) ヒトIgG1重鎖定常領域(G1m(3)アロタイプ)(対立遺伝子2)をコードしている核酸配列(配列番号207)。
【0468】
【化133】
【0469】
【化134】
【0470】
(2) ヒトIgG1重鎖定常領域(G1m(3)アロタイプ)(対立遺伝子1又は2) を規定する蛋白質配列(配列番号208)。最初のアミノ酸は、可変領域の最後のヌクレオチドおよびIgG1重鎖配列の開始2ヌクレオチドの翻訳物に由来する。
【0471】
【化135】
【0472】
(3) ヒト化Hu2B8 Hv5−51.1重鎖可変領域及びヒトIgG1重鎖定常領域G1m(3)アロタイプ(対立遺伝子2)を含む完全長鎖をコードしている核酸配列(下線:シグナル配列)(配列番号209)
【0473】
【化136】
【0474】
【化137】
【0475】
(4) ヒト化Hu2B8 Hv5−51.1及びヒトIgG1重鎖定常領域G1m(3)アロタイプ(対立遺伝子2)を含む完全長鎖を規定する蛋白質配列(シグナル配列なし)(配列番号210)
【0476】
【化138】
【0477】
各二重発現ベクターを293T細胞中に形質移入し、DMEM 10%ウシ胎仔血清を用いて一過性に発現させた。形質移入後48時間に、無血清培地である、4mMのL−グルタミンを含有するIS GRO(商標)(アーバイン・サイエンティフィック社(Irvine Scientific)、サンタアナ、カリフォルニア州)で細胞を洗浄した後、これで置換した。上清を10日間毎日採取し、新鮮培地で置換した。この培養上清を遠心分離し、濾過(0.45μm)して10〜100倍に濃縮した。抗体は、プロステップ(ProSep)vA樹脂(ミリポア社(Millipore))で精製し、PBSに対して透析し、除菌した。
【0478】
(実施例14)
ヒト化2B8変異体の結合特性
実施例13で作製したヒト化抗体の特徴は、hHGF及び実施例3で作製した組換えHGF蛋白質に結合できることであった。
【0479】
BIAcore T100装置を用いて表面プラスモン共鳴によりこれらの抗体を分析することによってHGF及び実施例3に記載した融合蛋白質へのその結合能を評価した。各抗体は、標準的なカップリングプロトコルをメーカーの使用説明書に従って用いたアミンカップリング(BIAcore、カタログ番号BR−1000−50)によりカルボキシメチル化デキストランCM5センサーチップ(BIAcore、カタログ番号BR−1006−68)上に固定化した。
【0480】
分析は、0.05%の界面活性剤P20(BIAcore、カタログ番号R−1000−54)、2mg/mLのBSA(EMD社カタログ番号2930)及び10mg/mLのCM−デキストランナトリウム塩(フルカ社(Fluka)、カタログ番号86524)を含有するPBS(GIBCO社、カタログ番号14040−133)を泳動用緩衝液として用い、25℃で行った。種々のHGF融合蛋白質を含む上清又は空のベクターを形質移入した細胞からの上清を各抗体上に30μL/分の流速で3分間注入した。その結果生じた結合は、注入終了後30秒のベースラインに対する共鳴単位(RU)として求めた。結合は、泳動用緩衝液で希釈したヒトHGF(R&Dシステムズ社(R&D Systems)、カタログ番号294−HGN−025)と比較した。非特異的な結合は、対照表面への結合を比較することにより測定した。得られた結果は表17にまとめた。
【0481】
【表17】
【0482】
表17の結果から明らかなように、ヒト化2B8系抗体はrhHGF及び3種のマウス−ヒト−マウスキメラ抗体の全てに結合する。
【0483】
(実施例15)
ヒト化2B8変異体の結合親和性
表15に示した抗体の結合親和性及び相互作用の速度を表面プラスモン共鳴によって測定した。
【0484】
マウス抗ヒト免疫グロブリン(ジャクソン・ラボラトリーズ社(Jackson Labs)、209−005)は、標準的なカップリングプロトコルをメーカーの使用説明書に従って用いたアミンカップリング(BIAcore、カタログ番号BR−1000−50)によりカルボキシメチル化デキストランCM4センサーチップ(BIAcore、カタログ番号BR−1006−68)上に固定化した。分析は、0.05%の界面活性剤P20(BIAcore、カタログ番号BR−1000−54)及び2mg/mLのBSA(EMD社カタログ番号2930)を含有するPBS(GIBCO社、カタログ番号14040−133)を用い、25℃で行った。
【0485】
前記抗体は、個々のフローセル内で10μL/分の流速で捕捉した。各抗体が各サイクルで捕捉される約20RUの抗体を生じるための注入時間にはばらつきがあった。緩衝液又は泳動用緩衝液で希釈したHGF(R&Dシステムズ社、カタログ番号294−HGN−025)を対照表面(捕捉抗体なし)及び活性表面(試験対象抗体あり)上に60μL/分で2分間順次注入した。解離相を濃度に応じて15又は90分間モニターした。その後、上記表面は、別のサイクルが開始される前に、pH2.2の10mMグリシン−HCl(BIAcore社、カタログ番号BR−1003−54)を60μL/分の流速で3分間注入して再生させた。試験したHGF濃度は0.46nM及び7.5nMであった。
【0486】
速度論的パラメータは、対照値減算を含むBIAエバリュエーション(商標)ソフトウェアの速度関数を用いて求めた。各抗体の速度論的パラメータka(会合速度定数)、kd(解離速度定数)及びKD(平衡解離定数)を表18にまとめた。
【0487】
【表18】
【0488】
これらのデータから、これらのヒト化抗体は速い会合速度(ka)、極めて遅い解離速度(kd)及び極めて高い親和性(KD)を有することが分かる。具体的には、これらの抗体は2.0乃至12pMの範囲の親和性を有する。
【0489】
(実施例16)
25℃及び37℃における結合親和性の比較
抗体HE2B8−4、sh2B8−9、sh2B8−12及びマウス2B8の結合親和性及び相互作用の速度を種々の条件下で表面プラスモン共鳴により測定した。
【0490】
マウス抗ヒト免疫グロブリン(ジャクソン・ラボラトリーズ社(Jackson Labs)、209−005)又はウサギ抗マウス免疫グロブリン(BIAcore、カタログ番号BR−1005−14)は、標準的なカップリングプロトコルをメーカーの使用説明書に従って用いたアミンカップリング(BIAcore、カタログ番号BR−1000−50)によりカルボキシメチル化デキストランCM4センサーチップ(BIAcore、カタログ番号BR−1006−68)上に固定化した。sH2b8−9及びsh2B8−12についての25℃における測定の場合、CM5センサーチップ(BIAcore、カタログ番号BR−1006−68)を用いた。これらの分析は、0.05%の界面活性剤P20(BIAcore、カタログ番号BR−1000−54)及び2mg/mLのBSA(EMD社カタログ番号2930)を含有するPBS(GIBCO社、カタログ番号14040−133)を泳動用緩衝液として用い、25℃及び37℃で行った。
【0491】
前記抗体は、個々のフローセル内で10μL/分の流速で捕捉した。各抗体が各サイクルで捕捉される約20RUの抗体を生じるための注入時間にはばらつきがあった。緩衝液又は泳動用緩衝液で希釈したHGF(R&Dシステムズ社、カタログ番号294−HGN−025)を対照表面(捕捉抗体なし)及び活性表面(試験対象抗体あり)上に60μL/分で2分間順次注入した。解離相を濃度に応じて15又は90分間モニターした。その後、マウス抗ヒト免疫グロブリンセンサーチップの表面は、別のサイクルが開始される前に、pH2.2の10mMグリシン−HCl(BIAcore社、カタログ番号BR−1003−54)を60μL/分の流速で3分間注入して再生させた。ウサギ抗マウスグロブリンセンサーチップの表面は、別のサイクルが開始される前に、pH1.7の10mMグリシン−HCl(BIAcore社、カタログ番号BR−1003−54)を60μL/分の流速で3分間注入して再生させた。試験したHGF濃度は0.46nM及び7.5nMであった。
【0492】
速度論的パラメータは、対照値減算を含むBIAエバリュエーションソフトウェアの速度関数を用いて求めた。各抗体の速度論的パラメータka(会合速度定数)、kd(解離速度定数)及びKD(平衡解離定数)を下記の表19にまとめた。
【0493】
【表19】
【0494】
予想どおり、会合速度定数は温度の上昇と共に増大した。意外なことに、解離定数は温度の対応する上昇と共に増大することはなかった。その結果、全体としての平衡解離定数(KD)は、生理的温度(37℃)で約1.4乃至3倍小さい(高い親和性)ものであった。
【0495】
(実施例17)
ヒト化2B8変異体の中和活性
実施例14で作製した抗体について、(a)c−MetへのhHGFの結合を阻害し、(b)4MBr−5細胞におけるBrdU取り込みのHGFによる促進を阻害する能力を特性化した。
【0496】
HGF−Met結合阻害試験(中和試験)は以下のようにして実施した。c−MetへのhHGFの結合を阻害する抗体の能力についてはELISAにより試験した。すなわち、ワラック96穴DELFIAアッセイプレート(ワラック社、カタログ番号AAAND−0001)を6.25μg/mL HGF(R&Dシステムズ社、カタログ番号294−HGN−025)の炭酸コーティング緩衝液(15mM Na2CO3及び34mM NaHCO3、pH9.0)溶液100μLにより4℃で16時間コーティングした。次に、このプレートを室温で1時間5%脱脂粉乳のPBS溶液200μLでブロックした。抗体は、別のプレートにおいて5%脱脂粉乳のPBS溶液に溶解した2nMのビオチニル化c−Metに検討対象の抗体を漸増濃度(0.033乃至250nM、2倍段階希釈)で加えることにより調製した。c−Met(R&Dシステムズ社、カタログ番号358−MT/CF)のビオチニル化は、10:1のビオチン/c−Met比でメーカーの使用説明書に従って行う(ピアス社(Pierce)、カタログ番号21335)。ウェル当たり100μLのサンプルを上記アッセイプレートに移し、室温で2時間インキュベートした。次いで、上記で得られたプレートをPBS−0.1% Tween20で3回洗浄した後、DELFIAアッセイ緩衝液(ワラック社、カタログ番号4002−0010)で1:1,000に希釈したEu標識ストレプトアビジン(ワラック社、カタログ番号1244−360)と共に室温で1時間インキュベートした。その結果得られたプレートをDELFIA洗浄液(ワラック社、カタログ番号4010−0010)で3回洗浄した後、100μL/ウェルのDELFIA強化溶液(ワラック社、#4001−0010)と共に攪拌しながら室温で15分間インキュベートした。上記プレートは、ユウロピウム法を用いるビクター3V装置(パーキンエルマー社)で読み取らせた。IC50値はプリズム(Prism)を用いて算出した。
【0497】
得られたIC50値を表20に示した。
【0498】
【表20】
【0499】
この表20の結果から明らかなように、試験したヒト化抗体はc−MetへのHGFの結合を効率的に中和する。
【0500】
表17の抗体は、実施例7(b)で説明した細胞増殖アッセイにおいても試験した。その結果は下記の表21にまとめた。
【0501】
【表21】
【0502】
この表21の結果から明らかなように、試験したヒト化抗体は全て4MBr−5細胞のHGF誘発性増殖を抑制する。
【0503】
(実施例18)
ヒト化2B8変異体の抗散乱活性
表17の抗体を実施例8で説明した抗散乱アッセイにおいて試験した。その結果を下記の表22にまとめた。
【0504】
【表22】
【0505】
表22の結果から明らかなように、試験したヒト化抗体は全て、マウスモノクローナル抗体2B8と同程度にHGF誘発性散乱を抑制した。
【0506】
(実施例19)
HGF刺激c−Metリン酸化の阻害
表17の抗体を実施例9で説明したc−Metリン酸化アッセイにおいて試験した。その結果を下記の表23にまとめた。
【0507】
【表23】
【0508】
表23の結果から明らかなように、試験したヒト化抗体は、PC−3細胞においてHGF誘発性c−Metリン酸化の強力な阻害剤である。
【0509】
(実施例20)
U87MG異種移植モデルにおける腫瘍抑制
U87MG異種移植モデルを用い、本発明のヒト化モノクローナル抗体の腫瘍増殖抑制能について試験した。U87MG細胞(ATCC)は、ダルベッコ改変イーグル培地(DMEM:Dulbecco’s Modified Eagle medium)を10%ウシ胎仔血清、100単位/mLペニシリン及び100μg/mLストレプトマイシンと共に含む培地を用いた、5%CO2及び95%空気を含有する雰囲気中37℃の培養で増殖させた。これらの細胞は、トリプシン−EDTAを用いて培養皿の壁から細胞を剥離することにより二次培養して維持した。
【0510】
コンフルエントに近い細胞をトリプシン処理により回収し、次いで、50%マトリゲル(Matrigel)(BDバイオサイエンシズ社(BD Biosciences)、カタログ番号356237)中の細胞5×106個を、7週齢雌性ICR SCIDマウス(タコニックラボラトリー社(Taconic Labs))の肩甲骨間の上背部に皮下注射した。キャリパで腫瘍の長径(L)及び短径(W)(mm)を測定した。腫瘍容積(vol.)は、容積(mm3)=L×W2/2として計算した。腫瘍が約200mm3にまで成長した時に、担腫瘍マウスを1群10匹の5群に無作為化した。1群にはPBSを投与し、1群にはヒトIgG対照を投与した。その他の4群のそれぞれには、ヒト化抗体(HE2B8−1、HE2B8−2、HE2B8−3及びHE2B8−4)のうちの1種を投与した。抗体は全て、5用量の腹腔内注射により週2回0.25mg/kg体重を投与した。腫瘍容積及びマウス体重を週2回記録した。スチューデントのt−検定を用いて腫瘍成長の抑制を解析した。
【0511】
試験したヒト化抗体はインビボで活性があった。HE2B8−1では57%の腫瘍成長抑制(p値:0.02)、HE2B8−2では61%の腫瘍成長抑制(p値:0.02)、HE2B8−3では85%の腫瘍成長抑制(p値:0.0004)、HE2B8−4では74%の腫瘍成長抑制(p値:0.001)が認められた。有意な体重減少は認められなかった。
【0512】
その後の実験は、側腹部に接種された皮下U87MG腫瘍を有する雌性NCRヌードマウス(タコニックラボラトリー社)において前述のようにして実施した。各群(1群10匹)には0.5mg/kgの以下の処置剤、即ち、PBS溶媒対照、huIgG対照、HE2B8−4又はsh2B8−9のうちの1種を投与した。処置剤は、最低でも5週間、週2回腹腔内投与した。各投与群は、腫瘍成長の抑制がsh2B8−9で113%、HE2B8−4で115%と同様な腫瘍退縮を示し、最低腫瘍成長遅延日数が30日であった。両投与とも有意な体重減少がなく、忍容性良好であった。
引用による組込み
本明細書で触れた特許文献及び科学論文のそれぞれの開示内容はその全体があらゆる目的で引用により組み込まれている。
【0513】
均等物
本発明は, 当該発明の精神または主要な特徴から逸脱することなく, 他のさまざまな形式で実施することができる。そのため、前述の実施態様はあらゆる点で単なる例示にすぎず、本明細書に記載された発明に対して限定的に解釈してはならない。従って、本発明の範囲は、特許請求の範囲によって示すものであり、明細書の本文によって拘束されるものではない。また、特許請求の範囲の記載内容と同等の範囲に含まれる変形もしくは変更は、すべて本発明の範囲内のものである。
【特許請求の範囲】
【請求項1】
ヒト肝細胞成長因子(HGF)に結合する単離された抗体、または、該抗体の抗原結合フラグメントであって、以下:
(a)構造CDRH1−CDRH2−CDRH3を含む免疫グロブリン重鎖可変領域であって、
(i)CDRH1が配列番号15のアミノ酸配列を含み、
(ii)CDRH2が配列番号205のアミノ酸配列を含み、かつ
(iii)CDRH3が配列番号17のアミノ酸配列を含む、
免疫グロブリン重鎖可変領域、および
(b)構造CDRL1−CDRL2−CDRL3を含む免疫グロブリン軽鎖可変領域であって、
(i)CDRL1が配列番号18のアミノ酸配列を含み、
(ii)CDRL2が配列番号206のアミノ酸配列を含み、かつ
(iii)CDRL3が配列番号20のアミノ酸配列を含む、
免疫グロブリン軽鎖可変領域
を含み、ただし、該抗体は、配列番号189のアミノ酸配列を含む免疫グロブリン重鎖可変領域と配列番号199のアミノ酸配列を含む免疫グロブリン軽鎖可変領域の両方を含む抗体を除く、抗体。
【請求項2】
前記相補性決定領域(CDR)がヒトまたはヒト化免疫グロブリンのフレームワーク領域間に挿入される、請求項1に記載の抗体。
【請求項3】
前記抗体がモノクローナル抗体である、請求項1に記載の抗体。
【請求項4】
前記抗体が4.0×10−5s−1以下のkdでヒト肝細胞成長因子に結合する、請求項1に記載の抗体。
【請求項5】
前記抗体が20pM以下のKDでヒト肝細胞成長因子に結合する、請求項1に記載の抗体。
【請求項6】
請求項1に記載の免疫グロブリン軽鎖可変領域をコードするヌクレオチド配列を含む、単離された核酸。
【請求項7】
請求項1に記載の免疫グロブリン重鎖可変領域をコードするヌクレオチド配列を含む、単離された核酸。
【請求項8】
請求項1に記載の免疫グロブリン軽鎖可変領域と、請求項1に記載の免疫グロブリン重鎖可変領域とをコードするヌクレオチド配列を含む、単離された核酸。
【請求項9】
前記免疫グロブリン軽鎖可変領域をコードするヌクレオチド配列が、配列番号199または配列番号201のアミノ酸配列を含む、請求項8に記載の単離された核酸。
【請求項10】
前記免疫グロブリン重鎖可変領域をコードするヌクレオチド配列が、配列番号189または配列番号191のアミノ酸配列を含む、請求項8に記載の単離された核酸。
【請求項11】
請求項6に記載の核酸を含む発現ベクター。
【請求項12】
請求項7に記載の核酸を含む発現ベクター。
【請求項13】
請求項8に記載の核酸を含む発現ベクター。
【請求項14】
請求項11に記載の発現ベクターを含む宿主細胞。
【請求項15】
請求項12に記載の発現ベクターを含む宿主細胞。
【請求項16】
請求項13に記載の発現ベクターを含む宿主細胞。
【請求項17】
少なくとも2つの発現ベクターを含む宿主細胞であって、第1の発現ベクターは請求項1に記載の免疫グロブリン軽鎖可変領域をコードする核酸配列を含み、そして、第2の発現ベクターは請求項1に記載の免疫グロブリン重鎖可変領域をコードする核酸配列を含む、宿主細胞。
【請求項18】
免疫グロブリン重鎖可変領域または免疫グロブリン軽鎖可変領域を含むポリペプチドを生成する方法であって、該方法は、以下:
(i)請求項14または15に記載の宿主細胞を、該宿主細胞が、該免疫グロブリン重鎖可変領域または免疫グロブリン軽鎖可変領域を含むポリペプチドを発現する条件下で増殖させる工程;および
(ii)該免疫グロブリン重鎖可変領域または免疫グロブリン軽鎖可変領域を含むポリペプチドを精製する工程
を包含する、方法。
【請求項19】
ヒト肝細胞成長因子(HGF)に結合するモノクローナル抗体または該抗体の抗原結合フラグメントを生成する方法であって、該方法は、以下:
(i)請求項16または17に記載の宿主細胞を、該免疫グロブリン重鎖可変領域を含むポリペプチドと、該免疫グロブリン軽鎖可変領域を含むポリペプチドの発現を可能にする条件下で増殖させ、それによって、該抗体または該抗体の抗原結合フラグメントを産生させる工程;および
(ii)該抗体または該抗体の抗原結合フラグメントを精製する工程
を包含する、方法。
【請求項20】
腫瘍細胞の増殖を抑制または低減するための組成物であって、該組成物は、ヒト肝細胞成長因子(HGF)に結合する抗体または該抗体の抗原結合フラグメントの、該腫瘍細胞の増殖を抑制または低減するために有効な量を含み、該抗体は、以下:
(i)配列番号18のアミノ酸配列を含むCDRL1、配列番号206のアミノ酸配列を含むCDRL2および配列番号20のアミノ酸配列を含むCDRL3を含む免疫グロブリン軽鎖可変領域;ならびに
(ii)配列番号15のアミノ酸配列を含むCDRH1、配列番号205のアミノ酸配列を含むCDRH2および配列番号17のアミノ酸配列を含むCDRH3を含む免疫グロブリン重鎖可変領域
を含む、組成物。
【請求項21】
哺乳動物における腫瘍の成長を抑制または低減するための組成物であって、該組成物は、ヒト肝細胞成長因子(HGF)に結合する抗体または該抗体の抗原結合フラグメントの、該腫瘍の成長を抑制または低減するために有効な量を含み、該抗体は、以下:
(i)配列番号18のアミノ酸配列を含むCDRL1、配列番号206のアミノ酸配列を含むCDRL2および配列番号20のアミノ酸配列を含むCDRL3を含む免疫グロブリン軽鎖可変領域;ならびに
(ii)配列番号15のアミノ酸配列を含むCDRH1、配列番号205のアミノ酸配列を含むCDRH2および配列番号17のアミノ酸配列を含むCDRH3を含む免疫グロブリン重鎖可変領域
を含む、組成物。
【請求項22】
哺乳動物における腫瘍を治療するための組成物であって、該組成物は、ヒト肝細胞成長因子(HGF)に結合する抗体または該抗体の抗原結合フラグメントの有効量を含み、該抗体は、以下:
(i)配列番号18のアミノ酸配列を含むCDRL1、配列番号206のアミノ酸配列を含むCDRL2および配列番号20のアミノ酸配列を含むCDRL3を含む免疫グロブリン軽鎖可変領域;ならびに
(ii)配列番号15のアミノ酸配列を含むCDRH1、配列番号205のアミノ酸配列を含むCDRH2および配列番号17のアミノ酸配列を含むCDRH3を含む免疫グロブリン重鎖可変領域
を含む、組成物。
【請求項23】
前記抗体が、配列番号199のアミノ酸配列を含む免疫グロブリン軽鎖可変領域および配列番号189のアミノ酸配列を含む免疫グロブリン重鎖可変領域、または該抗体の抗原結合フラグメントを含む、請求項20〜22のいずれか1項に記載の組成物。
【請求項24】
前記抗体が、配列番号201の免疫グロブリン軽鎖および配列番号191の免疫グロブリン重鎖、または該抗体の抗原結合フラグメントを含む、請求項20〜22のいずれか1項に記載の組成物。
【請求項25】
以下:
(a)(i)配列番号18のアミノ酸配列を含むCDRL1、配列番号19のアミノ酸配列を含むCDRL2および配列番号20のアミノ酸配列を含むCDRL3を含む免疫グロブリン軽鎖可変領域;ならびに
(ii)配列番号15のアミノ酸配列を含むCDRH1、配列番号204のアミノ酸配列を含むCDRH2および配列番号17のアミノ酸配列を含むCDRH3を含む免疫グロブリン重鎖可変領域;
(b)(i)配列番号18のアミノ酸配列を含むCDRL1、配列番号206のアミノ酸配列を含むCDRL2および配列番号20のアミノ酸配列を含むCDRL3を含む免疫グロブリン軽鎖可変領域;ならびに
(ii)配列番号15のアミノ酸配列を含むCDRH1、配列番号204のアミノ酸配列を含むCDRH2および配列番号17のアミノ酸配列を含むCDRH3を含む免疫グロブリン重鎖可変領域
(c)(i)配列番号18のアミノ酸配列を含むCDRL1、配列番号19のアミノ酸配列を含むCDRL2および配列番号20のアミノ酸配列を含むCDRL3を含む免疫グロブリン軽鎖可変領域;ならびに
(ii)配列番号15のアミノ酸配列を含むCDRH1、配列番号205のアミノ酸配列を含むCDRH2および配列番号17のアミノ酸配列を含むCDRH3を含む免疫グロブリン重鎖可変領域
からなる群より選択される免疫グロブリン軽鎖可変領域および免疫グロブリン重鎖可変領域を含む、ヒト肝細胞成長因子(HGF)に結合する単離された抗体またはその抗原結合フラグメント。
【請求項26】
前記相補性決定領域(CDR)の配列が、ヒトまたはヒト化フレームワーク配列の間に挿入される、請求項25に記載の抗体。
【請求項27】
以下:
(a)配列番号193のアミノ酸配列を含む免疫グロブリン軽鎖可変領域および配列番号183のアミノ酸配列を含む免疫グロブリン重鎖可変領域;
(b)配列番号199のアミノ酸配列を含む免疫グロブリン軽鎖可変領域および配列番号183のアミノ酸配列を含む免疫グロブリン重鎖可変領域;
(c)配列番号193のアミノ酸配列を含む免疫グロブリン軽鎖可変領域および配列番号189のアミノ酸配列を含む免疫グロブリン重鎖可変領域
からなる群より選択される免疫グロブリン軽鎖可変領域および免疫グロブリン重鎖可変領域を含む、ヒト肝細胞成長因子(HGF)に結合する単離された抗体またはその抗原結合フラグメント。
【請求項28】
以下:
(a)配列番号197のアミノ酸配列を含む免疫グロブリン軽鎖および配列番号187のアミノ酸配列を含む免疫グロブリン重鎖;
(b)配列番号201のアミノ酸配列を含む免疫グロブリン軽鎖および配列番号187のアミノ酸配列を含む免疫グロブリン重鎖;
(c)配列番号197のアミノ酸配列を含む免疫グロブリン軽鎖および配列番号191のアミノ酸配列を含む免疫グロブリン重鎖可変領域
からなる群より選択される免疫グロブリン軽鎖および免疫グロブリン重鎖を含む、ヒト肝細胞成長因子(HGF)に結合する単離された抗体またはその抗原結合フラグメント。
【請求項29】
前記抗体がモノクローナル抗体である、請求項25〜28のいずれか1項に記載の抗体。
【請求項30】
請求項25に記載の免疫グロブリン軽鎖可変領域をコードするヌクレオチド配列を含む、単離された核酸。
【請求項31】
請求項25に記載の免疫グロブリン重鎖可変領域をコードするヌクレオチド配列を含む、単離された核酸。
【請求項32】
請求項25に記載の免疫グロブリン軽鎖可変領域と、請求項25に記載の免疫グロブリン重鎖可変領域とをコードするヌクレオチド配列を含む、単離された核酸。
【請求項33】
請求項30に記載の核酸を含む発現ベクター。
【請求項34】
請求項31に記載の核酸を含む発現ベクター。
【請求項35】
請求項32に記載の核酸を含む発現ベクター。
【請求項36】
請求項33に記載の発現ベクターを含む宿主細胞。
【請求項37】
請求項34に記載の発現ベクターを含む宿主細胞。
【請求項38】
請求項35に記載の発現ベクターを含む宿主細胞。
【請求項39】
少なくとも2つの発現ベクターを含む宿主細胞であって、第1の発現ベクターは請求項25に記載の免疫グロブリン軽鎖可変領域をコードする核酸配列を含み、そして、第2の発現ベクターは請求項25に記載の免疫グロブリン重鎖可変領域をコードする核酸配列を含む、宿主細胞。
【請求項40】
免疫グロブリン重鎖可変領域または免疫グロブリン軽鎖可変領域を含むポリペプチドを生成する方法であって、該方法は、以下:
(i)請求項36または37に記載の宿主細胞を、該宿主細胞が、該免疫グロブリン重鎖可変領域または免疫グロブリン軽鎖可変領域を含むポリペプチドを発現する条件下で増殖させる工程;および
(ii)該免疫グロブリン重鎖可変領域または免疫グロブリン軽鎖可変領域を含むポリペプチドを精製する工程
を包含する、方法。
【請求項41】
ヒト肝細胞成長因子(HGF)に結合するモノクローナル抗体または該抗体の抗原結合フラグメントを生成する方法であって、該方法は、以下:
(i)請求項38または39に記載の宿主細胞を、該免疫グロブリン重鎖可変領域を含むポリペプチドと、該免疫グロブリン軽鎖可変領域を含むポリペプチドの発現を可能にする条件下で増殖させ、それによって、該抗体または該抗体の抗原結合フラグメントを産生させる工程;および
(ii)該抗体または該抗体の抗原結合フラグメントを精製する工程
を包含する、方法。
【請求項42】
腫瘍細胞の増殖を抑制または低減するための組成物であって、該組成物は、請求項25〜29のいずれか1項に記載の抗体の、該腫瘍細胞の増殖を抑制または低減するために有効な量を含む、組成物。
【請求項43】
哺乳動物における腫瘍の成長を抑制または低減するための組成物であって、該組成物は、請求項25〜29のいずれか1項に記載の抗体の、該腫瘍の成長を抑制または低減するために有効な量を含む、組成物。
【請求項44】
哺乳動物における腫瘍を治療するための組成物であって、該組成物は、請求項25〜29のいずれか1項に記載の抗体の有効量を含む、組成物。
【請求項45】
1.0×10−12M以下のKDでヒト肝細胞成長因子に結合する、請求項1に記載の抗体またはその抗原結合フラグメント。
【請求項46】
10pM以下のKDでヒト肝細胞成長因子に結合する、請求項1に記載の抗体またはその抗原結合フラグメント。
【請求項1】
ヒト肝細胞成長因子(HGF)に結合する単離された抗体、または、該抗体の抗原結合フラグメントであって、以下:
(a)構造CDRH1−CDRH2−CDRH3を含む免疫グロブリン重鎖可変領域であって、
(i)CDRH1が配列番号15のアミノ酸配列を含み、
(ii)CDRH2が配列番号205のアミノ酸配列を含み、かつ
(iii)CDRH3が配列番号17のアミノ酸配列を含む、
免疫グロブリン重鎖可変領域、および
(b)構造CDRL1−CDRL2−CDRL3を含む免疫グロブリン軽鎖可変領域であって、
(i)CDRL1が配列番号18のアミノ酸配列を含み、
(ii)CDRL2が配列番号206のアミノ酸配列を含み、かつ
(iii)CDRL3が配列番号20のアミノ酸配列を含む、
免疫グロブリン軽鎖可変領域
を含み、ただし、該抗体は、配列番号189のアミノ酸配列を含む免疫グロブリン重鎖可変領域と配列番号199のアミノ酸配列を含む免疫グロブリン軽鎖可変領域の両方を含む抗体を除く、抗体。
【請求項2】
前記相補性決定領域(CDR)がヒトまたはヒト化免疫グロブリンのフレームワーク領域間に挿入される、請求項1に記載の抗体。
【請求項3】
前記抗体がモノクローナル抗体である、請求項1に記載の抗体。
【請求項4】
前記抗体が4.0×10−5s−1以下のkdでヒト肝細胞成長因子に結合する、請求項1に記載の抗体。
【請求項5】
前記抗体が20pM以下のKDでヒト肝細胞成長因子に結合する、請求項1に記載の抗体。
【請求項6】
請求項1に記載の免疫グロブリン軽鎖可変領域をコードするヌクレオチド配列を含む、単離された核酸。
【請求項7】
請求項1に記載の免疫グロブリン重鎖可変領域をコードするヌクレオチド配列を含む、単離された核酸。
【請求項8】
請求項1に記載の免疫グロブリン軽鎖可変領域と、請求項1に記載の免疫グロブリン重鎖可変領域とをコードするヌクレオチド配列を含む、単離された核酸。
【請求項9】
前記免疫グロブリン軽鎖可変領域をコードするヌクレオチド配列が、配列番号199または配列番号201のアミノ酸配列を含む、請求項8に記載の単離された核酸。
【請求項10】
前記免疫グロブリン重鎖可変領域をコードするヌクレオチド配列が、配列番号189または配列番号191のアミノ酸配列を含む、請求項8に記載の単離された核酸。
【請求項11】
請求項6に記載の核酸を含む発現ベクター。
【請求項12】
請求項7に記載の核酸を含む発現ベクター。
【請求項13】
請求項8に記載の核酸を含む発現ベクター。
【請求項14】
請求項11に記載の発現ベクターを含む宿主細胞。
【請求項15】
請求項12に記載の発現ベクターを含む宿主細胞。
【請求項16】
請求項13に記載の発現ベクターを含む宿主細胞。
【請求項17】
少なくとも2つの発現ベクターを含む宿主細胞であって、第1の発現ベクターは請求項1に記載の免疫グロブリン軽鎖可変領域をコードする核酸配列を含み、そして、第2の発現ベクターは請求項1に記載の免疫グロブリン重鎖可変領域をコードする核酸配列を含む、宿主細胞。
【請求項18】
免疫グロブリン重鎖可変領域または免疫グロブリン軽鎖可変領域を含むポリペプチドを生成する方法であって、該方法は、以下:
(i)請求項14または15に記載の宿主細胞を、該宿主細胞が、該免疫グロブリン重鎖可変領域または免疫グロブリン軽鎖可変領域を含むポリペプチドを発現する条件下で増殖させる工程;および
(ii)該免疫グロブリン重鎖可変領域または免疫グロブリン軽鎖可変領域を含むポリペプチドを精製する工程
を包含する、方法。
【請求項19】
ヒト肝細胞成長因子(HGF)に結合するモノクローナル抗体または該抗体の抗原結合フラグメントを生成する方法であって、該方法は、以下:
(i)請求項16または17に記載の宿主細胞を、該免疫グロブリン重鎖可変領域を含むポリペプチドと、該免疫グロブリン軽鎖可変領域を含むポリペプチドの発現を可能にする条件下で増殖させ、それによって、該抗体または該抗体の抗原結合フラグメントを産生させる工程;および
(ii)該抗体または該抗体の抗原結合フラグメントを精製する工程
を包含する、方法。
【請求項20】
腫瘍細胞の増殖を抑制または低減するための組成物であって、該組成物は、ヒト肝細胞成長因子(HGF)に結合する抗体または該抗体の抗原結合フラグメントの、該腫瘍細胞の増殖を抑制または低減するために有効な量を含み、該抗体は、以下:
(i)配列番号18のアミノ酸配列を含むCDRL1、配列番号206のアミノ酸配列を含むCDRL2および配列番号20のアミノ酸配列を含むCDRL3を含む免疫グロブリン軽鎖可変領域;ならびに
(ii)配列番号15のアミノ酸配列を含むCDRH1、配列番号205のアミノ酸配列を含むCDRH2および配列番号17のアミノ酸配列を含むCDRH3を含む免疫グロブリン重鎖可変領域
を含む、組成物。
【請求項21】
哺乳動物における腫瘍の成長を抑制または低減するための組成物であって、該組成物は、ヒト肝細胞成長因子(HGF)に結合する抗体または該抗体の抗原結合フラグメントの、該腫瘍の成長を抑制または低減するために有効な量を含み、該抗体は、以下:
(i)配列番号18のアミノ酸配列を含むCDRL1、配列番号206のアミノ酸配列を含むCDRL2および配列番号20のアミノ酸配列を含むCDRL3を含む免疫グロブリン軽鎖可変領域;ならびに
(ii)配列番号15のアミノ酸配列を含むCDRH1、配列番号205のアミノ酸配列を含むCDRH2および配列番号17のアミノ酸配列を含むCDRH3を含む免疫グロブリン重鎖可変領域
を含む、組成物。
【請求項22】
哺乳動物における腫瘍を治療するための組成物であって、該組成物は、ヒト肝細胞成長因子(HGF)に結合する抗体または該抗体の抗原結合フラグメントの有効量を含み、該抗体は、以下:
(i)配列番号18のアミノ酸配列を含むCDRL1、配列番号206のアミノ酸配列を含むCDRL2および配列番号20のアミノ酸配列を含むCDRL3を含む免疫グロブリン軽鎖可変領域;ならびに
(ii)配列番号15のアミノ酸配列を含むCDRH1、配列番号205のアミノ酸配列を含むCDRH2および配列番号17のアミノ酸配列を含むCDRH3を含む免疫グロブリン重鎖可変領域
を含む、組成物。
【請求項23】
前記抗体が、配列番号199のアミノ酸配列を含む免疫グロブリン軽鎖可変領域および配列番号189のアミノ酸配列を含む免疫グロブリン重鎖可変領域、または該抗体の抗原結合フラグメントを含む、請求項20〜22のいずれか1項に記載の組成物。
【請求項24】
前記抗体が、配列番号201の免疫グロブリン軽鎖および配列番号191の免疫グロブリン重鎖、または該抗体の抗原結合フラグメントを含む、請求項20〜22のいずれか1項に記載の組成物。
【請求項25】
以下:
(a)(i)配列番号18のアミノ酸配列を含むCDRL1、配列番号19のアミノ酸配列を含むCDRL2および配列番号20のアミノ酸配列を含むCDRL3を含む免疫グロブリン軽鎖可変領域;ならびに
(ii)配列番号15のアミノ酸配列を含むCDRH1、配列番号204のアミノ酸配列を含むCDRH2および配列番号17のアミノ酸配列を含むCDRH3を含む免疫グロブリン重鎖可変領域;
(b)(i)配列番号18のアミノ酸配列を含むCDRL1、配列番号206のアミノ酸配列を含むCDRL2および配列番号20のアミノ酸配列を含むCDRL3を含む免疫グロブリン軽鎖可変領域;ならびに
(ii)配列番号15のアミノ酸配列を含むCDRH1、配列番号204のアミノ酸配列を含むCDRH2および配列番号17のアミノ酸配列を含むCDRH3を含む免疫グロブリン重鎖可変領域
(c)(i)配列番号18のアミノ酸配列を含むCDRL1、配列番号19のアミノ酸配列を含むCDRL2および配列番号20のアミノ酸配列を含むCDRL3を含む免疫グロブリン軽鎖可変領域;ならびに
(ii)配列番号15のアミノ酸配列を含むCDRH1、配列番号205のアミノ酸配列を含むCDRH2および配列番号17のアミノ酸配列を含むCDRH3を含む免疫グロブリン重鎖可変領域
からなる群より選択される免疫グロブリン軽鎖可変領域および免疫グロブリン重鎖可変領域を含む、ヒト肝細胞成長因子(HGF)に結合する単離された抗体またはその抗原結合フラグメント。
【請求項26】
前記相補性決定領域(CDR)の配列が、ヒトまたはヒト化フレームワーク配列の間に挿入される、請求項25に記載の抗体。
【請求項27】
以下:
(a)配列番号193のアミノ酸配列を含む免疫グロブリン軽鎖可変領域および配列番号183のアミノ酸配列を含む免疫グロブリン重鎖可変領域;
(b)配列番号199のアミノ酸配列を含む免疫グロブリン軽鎖可変領域および配列番号183のアミノ酸配列を含む免疫グロブリン重鎖可変領域;
(c)配列番号193のアミノ酸配列を含む免疫グロブリン軽鎖可変領域および配列番号189のアミノ酸配列を含む免疫グロブリン重鎖可変領域
からなる群より選択される免疫グロブリン軽鎖可変領域および免疫グロブリン重鎖可変領域を含む、ヒト肝細胞成長因子(HGF)に結合する単離された抗体またはその抗原結合フラグメント。
【請求項28】
以下:
(a)配列番号197のアミノ酸配列を含む免疫グロブリン軽鎖および配列番号187のアミノ酸配列を含む免疫グロブリン重鎖;
(b)配列番号201のアミノ酸配列を含む免疫グロブリン軽鎖および配列番号187のアミノ酸配列を含む免疫グロブリン重鎖;
(c)配列番号197のアミノ酸配列を含む免疫グロブリン軽鎖および配列番号191のアミノ酸配列を含む免疫グロブリン重鎖可変領域
からなる群より選択される免疫グロブリン軽鎖および免疫グロブリン重鎖を含む、ヒト肝細胞成長因子(HGF)に結合する単離された抗体またはその抗原結合フラグメント。
【請求項29】
前記抗体がモノクローナル抗体である、請求項25〜28のいずれか1項に記載の抗体。
【請求項30】
請求項25に記載の免疫グロブリン軽鎖可変領域をコードするヌクレオチド配列を含む、単離された核酸。
【請求項31】
請求項25に記載の免疫グロブリン重鎖可変領域をコードするヌクレオチド配列を含む、単離された核酸。
【請求項32】
請求項25に記載の免疫グロブリン軽鎖可変領域と、請求項25に記載の免疫グロブリン重鎖可変領域とをコードするヌクレオチド配列を含む、単離された核酸。
【請求項33】
請求項30に記載の核酸を含む発現ベクター。
【請求項34】
請求項31に記載の核酸を含む発現ベクター。
【請求項35】
請求項32に記載の核酸を含む発現ベクター。
【請求項36】
請求項33に記載の発現ベクターを含む宿主細胞。
【請求項37】
請求項34に記載の発現ベクターを含む宿主細胞。
【請求項38】
請求項35に記載の発現ベクターを含む宿主細胞。
【請求項39】
少なくとも2つの発現ベクターを含む宿主細胞であって、第1の発現ベクターは請求項25に記載の免疫グロブリン軽鎖可変領域をコードする核酸配列を含み、そして、第2の発現ベクターは請求項25に記載の免疫グロブリン重鎖可変領域をコードする核酸配列を含む、宿主細胞。
【請求項40】
免疫グロブリン重鎖可変領域または免疫グロブリン軽鎖可変領域を含むポリペプチドを生成する方法であって、該方法は、以下:
(i)請求項36または37に記載の宿主細胞を、該宿主細胞が、該免疫グロブリン重鎖可変領域または免疫グロブリン軽鎖可変領域を含むポリペプチドを発現する条件下で増殖させる工程;および
(ii)該免疫グロブリン重鎖可変領域または免疫グロブリン軽鎖可変領域を含むポリペプチドを精製する工程
を包含する、方法。
【請求項41】
ヒト肝細胞成長因子(HGF)に結合するモノクローナル抗体または該抗体の抗原結合フラグメントを生成する方法であって、該方法は、以下:
(i)請求項38または39に記載の宿主細胞を、該免疫グロブリン重鎖可変領域を含むポリペプチドと、該免疫グロブリン軽鎖可変領域を含むポリペプチドの発現を可能にする条件下で増殖させ、それによって、該抗体または該抗体の抗原結合フラグメントを産生させる工程;および
(ii)該抗体または該抗体の抗原結合フラグメントを精製する工程
を包含する、方法。
【請求項42】
腫瘍細胞の増殖を抑制または低減するための組成物であって、該組成物は、請求項25〜29のいずれか1項に記載の抗体の、該腫瘍細胞の増殖を抑制または低減するために有効な量を含む、組成物。
【請求項43】
哺乳動物における腫瘍の成長を抑制または低減するための組成物であって、該組成物は、請求項25〜29のいずれか1項に記載の抗体の、該腫瘍の成長を抑制または低減するために有効な量を含む、組成物。
【請求項44】
哺乳動物における腫瘍を治療するための組成物であって、該組成物は、請求項25〜29のいずれか1項に記載の抗体の有効量を含む、組成物。
【請求項45】
1.0×10−12M以下のKDでヒト肝細胞成長因子に結合する、請求項1に記載の抗体またはその抗原結合フラグメント。
【請求項46】
10pM以下のKDでヒト肝細胞成長因子に結合する、請求項1に記載の抗体またはその抗原結合フラグメント。
【図1】

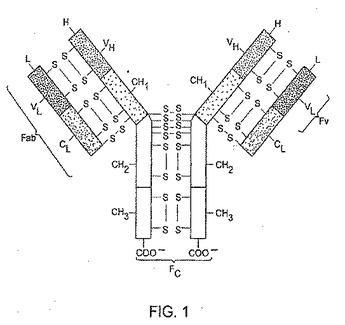
【図2】
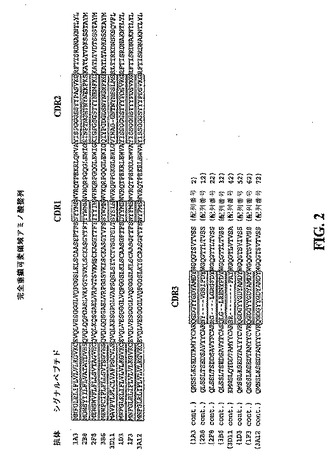
【図3】
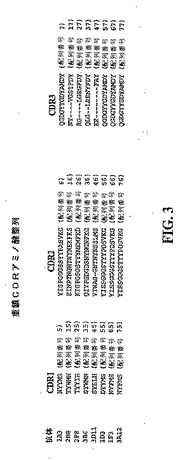
【図4】
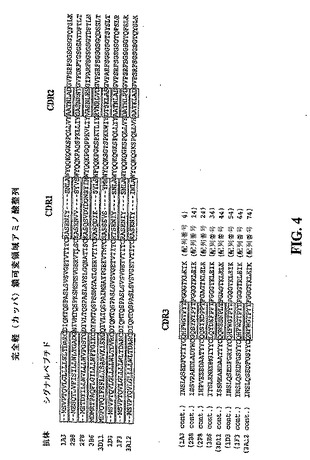
【図5】
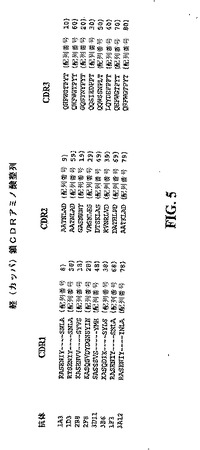
【図6】
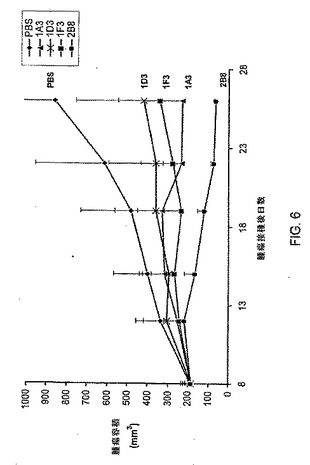
【図7】
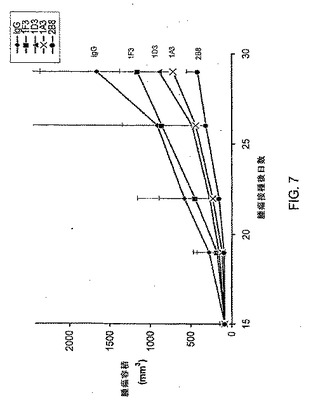
【図8】
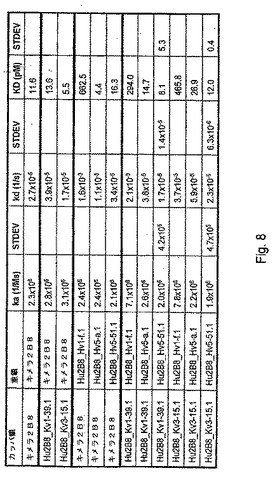
【図9】
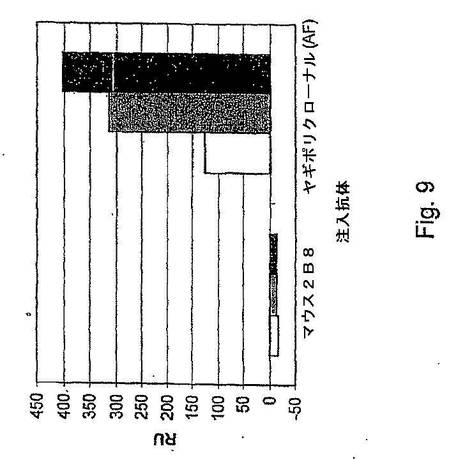
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公開番号】特開2011−72318(P2011−72318A)
【公開日】平成23年4月14日(2011.4.14)
【国際特許分類】
【出願番号】特願2010−294222(P2010−294222)
【出願日】平成22年12月28日(2010.12.28)
【分割の表示】特願2009−513295(P2009−513295)の分割
【原出願日】平成19年6月1日(2007.6.1)
【出願人】(506070224)アベオ ファーマシューティカルズ, インコーポレイテッド (14)
【出願人】(500570793)ゾーマ テクノロジー リミテッド (26)
【Fターム(参考)】
【公開日】平成23年4月14日(2011.4.14)
【国際特許分類】
【出願日】平成22年12月28日(2010.12.28)
【分割の表示】特願2009−513295(P2009−513295)の分割
【原出願日】平成19年6月1日(2007.6.1)
【出願人】(506070224)アベオ ファーマシューティカルズ, インコーポレイテッド (14)
【出願人】(500570793)ゾーマ テクノロジー リミテッド (26)
【Fターム(参考)】
[ Back to top ]