
肝細胞癌に関連する遺伝子およびポリペプチド、ならびに肝細胞癌を検出する方法
【課題】肝細胞癌で発現が上昇する遺伝子、この遺伝子によりコードされるポリペプチド、および肝細胞癌を検出する方法を提供する。
【解決手段】DDEFL1、VANGL1、LGN遺伝子、およびこれらの遺伝子によってコードされるポリペプチド、発現ベクター、形質転換体。また、これらの遺伝子配列を含むプローブ、プライマーならびにこれらのポリペプチドに対する抗体を用いた肝細胞癌検出法。肝細胞癌の成長を抑制するために使用することができるこれらの遺伝子のアンチセンスヌクレオチド配列。
【解決手段】DDEFL1、VANGL1、LGN遺伝子、およびこれらの遺伝子によってコードされるポリペプチド、発現ベクター、形質転換体。また、これらの遺伝子配列を含むプローブ、プライマーならびにこれらのポリペプチドに対する抗体を用いた肝細胞癌検出法。肝細胞癌の成長を抑制するために使用することができるこれらの遺伝子のアンチセンスヌクレオチド配列。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、肝細胞癌で発現が上昇する遺伝子、これらの遺伝子によりコードされるポリペプチド、および肝細胞癌を検出する方法に関する。
【背景技術】
【0002】
cDNAマイクロアレイ技術により、正常細胞と悪性細胞における遺伝子発現の包括的なプロファイルが得られるようになった(Perou, C. M.ら、Nature. 406:747-752、2000(非特許文献1);Clark, E. A.ら、Nature. 406:532-535、2000(非特許文献2);Okabe, H.ら、Cancer Res. 61:2129-2137、2001(非特許文献3))。この方法は、癌細胞の複雑な性質を明らかにしており、発癌の理解を進める一助となっている。腫瘍において調節が解除された遺伝子の同定が、個々の癌のさらに正確かつ精密な診断、および新たな治療標的の開発につながる可能性がある(Golub, T. R.ら、Science 286:531-537、1999(非特許文献4))。
【0003】
肝細胞癌(HCC)は、全世界での癌による死亡原因の第1位である。治療法が最近進歩しているにもかかわらず、進行期のHCC患者の予後は依然として悪い。分子的研究でTP53遺伝子、CTNNB1遺伝子、および/またはAXIN1遺伝子の変化が肝癌発生に関与する可能性があることが報告されているが(Perou, C. M.ら、Nature. 406:747-752、2000(非特許文献5);Satoh, S.ら、Nat Genet. 24:245-250、2000(非特許文献6))、このような変化は一部のHCCにしか関連していないようである。したがって、癌の治療の新たな診断マーカーおよび/または薬剤標的となりうる究極の遺伝子が求められている。
【0004】
本発明者らは過去に、新たな遺伝子VANGL1をHCCのゲノム全体にわたる解析により同定したことを報告した(Yagyu, R.ら、International Journal of Oncology 20:1173-1178、2002(非特許文献7))。
【非特許文献1】Perou, C. M.ら、Nature. 406:747-752、2000
【非特許文献2】Clark, E. A.ら、Nature. 406:532-535、2000
【非特許文献3】Okabe, H.ら、Cancer Res. 61:2129-2137、2001
【非特許文献4】Golub, T. R.ら、Science 286:531-537、1999
【非特許文献5】Perou, C. M.ら、Nature. 406:747-752、2000
【非特許文献6】Satoh, S.ら、Nat Genet. 24:245-250、2000
【非特許文献7】Yagyu, R.ら、International Journal of Oncology 20:1173-1178、2002
【発明の開示】
【0005】
本発明の目的は、肝細胞癌で発現が上昇する遺伝子、この遺伝子によりコードされるポリペプチド、および肝細胞癌を検出する方法を提供することである。
【0006】
本発明者らは、23,040遺伝子に相当するcDNAマイクロアレイを用いてHCCの発現プロファイルの解析を行った。この解析により、対応する非癌性肝組織と比較して、癌組織において高頻度で発現が上昇していると考えられる、69のESTを含む165の遺伝子が特定された。本発明者らは、発現がHCCで高頻度で上昇している転写物から3つの遺伝子を単離した。これらの遺伝子は、センタウリン(centaurin)ファミリーのタンパク質と構造的特徴を共有する産物をコードしている。
【0007】
3つの遺伝子の1つは、UniGeneクラスターのEST(Hs.44579)に対応し、染色体バンド1p36.13で過剰に発現されている新たな遺伝子であることが判明した。この遺伝子のオープンリーディングフレームは、発生・分化促進因子2(DDEF2)と約60%同一なタンパク質をコードしていたため、本発明者らはこの遺伝子を発生・分化促進様因子1(DDEFL1)と命名した。
【0008】
HCCで発現が上昇している別の遺伝子は、UniGeneクラスターのEST(Hs.122730)に対応する。この推定アミノ酸配列は、ショウジョウバエにおける細胞極性および細胞運命の決定に関与するstrabismus(Van Gogh)、およびVan Gogh Like 2(VANGL2)と40%および63%の同一性を有していた。したがって、この遺伝子をVan Gogh Like 1(VANGL1)と命名した。
【0009】
HCCで発現が上昇している別の遺伝子は、LGN(GenBankアクセッション番号U54999)であることが判明した。LGNタンパク質は、抑制性ヘテロ三量体Gタンパク質のαサブユニット(Gαi2)と相互作用する。
【0010】
DDEFL1またはLGNの遺伝子移入により、これら3つのいずれかの遺伝子の内因性発現を欠く細胞の増殖が促進された。また、これらの遺伝子に特異的なアンチセンスS-オリゴヌクレオチドをトランスフェクトしてDDEFL1、VANGL1、またはLGNの発現を低下させたところ、肝細胞癌細胞の成長が阻害された。
【0011】
この結果は、HCCの機構を明らかにするために、かつHCCの診断および治療の新たな戦略を開発するために寄与すると考えられる。
【0012】
本発明は特に以下を提供する:
(1)以下からなる群より選択される、単離された核酸:
(a)配列番号:1または配列番号:3のヌクレオチド配列を含む核酸;
(b)配列番号:2または配列番号:4のアミノ酸配列を含むポリペプチドをコードする核酸;
(c)配列番号:1もしくは配列番号:3、またはその相補物からなるヌクレオチド配列と高ストリンジェントな条件下でハイブリダイズする鎖を含む核酸、
(2)以下からなる群より選択される、単離されたポリペプチド:
(a)配列番号:1または配列番号:3のヌクレオチド配列によりコードされるポリペプチド;
(b)配列番号:2または配列番号:4のアミノ酸配列を含むポリペプチド;
(c)配列番号:2または配列番号:4と少なくとも65%の同一性を有するポリペプチド、
(3)(1)の核酸を有するベクター、
(4)(1)の核酸または(3)のベクターを有する形質転換体、
(5)(4)の形質転換体を培養物中で培養する段階、形質転換体中でポリペプチドを発現させる段階、およびポリペプチドを培養物から回収する段階を含む、ポリペプチドを作製する方法、
(6)(2)のポリペプチドに特異的に結合する抗体、
(7)以下の段階を含む、肝細胞癌を検出する方法:
(a)被験者から生物試料を調製する段階;
(b)配列番号:1のポリペプチド、配列番号:3のポリペプチド、および配列番号:5のポリペプチドからなる群より選択される、少なくとも1つのポリペプチドの発現レベルを測定する段階;
(c)この発現レベルを、非癌性試料で測定された発現レベルと比較する段階;ならびに
(d)被験者における癌の有無を判定する段階、
(8)配列番号:1、配列番号:3、もしくは配列番号:5、またはその相補物からなるヌクレオチド配列と、高ストリンジェントな条件下でハイブリダイズする鎖を含む核酸を含む、肝細胞癌を検出するための試薬、
(9)(6)の抗体を含む、肝細胞癌を検出するための試薬、ならびに
(10)配列番号:1、配列番号:3、または配列番号:5のヌクレオチド配列とハイブリダイズする、少なくとも1つのアンチセンスオリゴヌクレオチドを肝細胞癌に導入する段階を含む、肝細胞癌の成長を阻害する方法。
【0013】
本発明を以下でさらに詳細に説明する。
【0014】
核酸
本発明は、肝細胞癌で発現が上昇する遺伝子を提供する。
【0015】
DDEFL1のヌクレオチド配列およびアミノ酸配列をそれぞれ配列番号:1および配列番号:2に示す。DDEFL1の完全なcDNAは、903アミノ酸のタンパク質をコードする2712ヌクレオチドのオープンリーディングフレームを含む、4050ヌクレオチドからなる(GenBankアクセッション番号AB051853)。DDFEL1のアミノ酸配列はDDEFL2と60%の同一性を、DDEF/ASAP1と46%の同一性を示し、かつArf GTPアーゼ活性化タンパク質(ArfGAP)ドメイン、および2つのアンキリン反復配列を含んでいる。
【0016】
DDEFL1は、アクチン細胞骨格の再編成を調節するタンパク質であるセンタウリンファミリーのメンバーであるDDEF2と60%の同一性を示した。これは、DDEFL1も細胞構造の編成に重要な役割を果たす可能性を示唆している(Randazzo, P. A.ら、The Arf GTPase-activating protein ASAP1 regulates the actin cytoskeleton、Proc. Natl. Acad. Sci. U S A 97:4011-4016、2000)。DDEFL1は、PHドメインおよびArfGAPモチーフも保存しているため、GAP活性によってArf低分子GTPアーゼを調節する、センタウリンファミリーの新たなメンバーであると考えられる。GEFのDblファミリーに属する分子の多くに認められるPHドメインは、特定の標的分子と相互作用することにより、および/または触媒ドメインを直接調節することにより、タンパク質の再配置に重要な役割を果たすと考えられている(Jackson, T. R.ら、Trends Biochem Sci. 25:489-495、2000;Cerione, R. A.、およびZheng, Y.、Curr. Opin. Cell. Biol. 8:216-222、1996;Chardin, P.ら、Nature 384:481-484、1996)。DDEF2は、周辺の接着斑(focal adhesion)に局在するが、本発明者らは、myc標識DDEFL1タンパク質が細胞内に拡散していることを発見した。
【0017】
Arfタンパク質は、小胞の膜輸送、小胞体区画およびゴルジ区画の完全性の維持、ならびに周辺の細胞骨格の調節などの重要な細胞過程に関係している(Cukierman, E.ら、Science 270:1999-2002、1995)。Arfファミリーの6つのメンバー(Arf1〜Arf6)、およびこれらの機能は既に同定されている(Moss, J.およびVaughan, M.、J. Biol. Chem. 270:12327-12330、1995)。例えばArf6タンパク質は、接着斑の形態を変化させ、細胞の拡散を阻害する細胞骨格の調節因子とみなされており、DDEF2はArf1に対してGAP活性を示す。
【0018】
DDEFL1の過剰発現により、細胞の成長および生存が低血清条件下で促進された。これは、DDEFL1が貧栄養および低酸素の条件で癌細胞に成長上の利点をもたらす可能性があることを示唆する。HCCでDDEFL1の発現が高頻度で上昇することは、肝癌発生におけるこの遺伝子の重要性を強調している。
【0019】
VANGL1のヌクレオチド配列およびアミノ酸配列を、それぞれ配列番号:3および配列番号:4に示す。決定されたcDNA配列は、524アミノ酸のタンパク質をコードする1572ヌクレオチドのオープンリーディングフレームを含む、1879ヌクレオチドからなっていた(GenBankアクセッション番号AB057596)。
【0020】
strabismus(stbm)は、rough eyeの表現型をもつ変異ショウジョウバエの原因遺伝子として同定された(Wolff T.およびRubin G.M.、Development 125:1149-1159、1998)。この遺伝子は、ショウジョウバエの眼、脚、および剛毛の極性を維持するために、かつR3およびR4光受容器の細胞運命を決定するために必要である。stbmのマウス相同遺伝子Ltapは、神経管変異マウスLoop-tailにおいて変化しており、神経管欠損(NTD)のヒトモデルとなっている(Kibar Zら、Nat Genet. 28:251-255、2001)。したがってVANGL1は、細胞極性、細胞運命の決定、および/または組織の編成に重要な役割を果たす可能性がある。VANGL1の発現はHCCにおいて高頻度で上昇しており、この発現を抑制したところ、癌細胞の成長または生存率が有意に低下したことから、VANGL1は癌細胞に生存率の上昇および/または脱極性化した成長をもたらす可能性がある。
【0021】
LGNのヌクレオチド配列およびアミノ酸配列をそれぞれ配列番号:5および配列番号:6に示す。LGNのcDNAは2336ヌクレオチドを含み、677アミノ酸のペプチドをコードする。
【0022】
LGNタンパク質は、抑制性ヘテロ三量体Gタンパク質のαサブユニット(Gαi2)と相互作用するタンパク質として以前に報告された(Mochizuki, N.ら、Gene 181:39-43、1996)。Gαi2の活性化変異は、下垂体腫瘍および他の内分泌腫瘍において報告されている(Hermouet, S.ら、Proc. Natl. Acad. Sci. USA 88:10455-10459、1991;Pace, A. M.ら、Proc. Natl. Acad. Sci. USA. 88:7031-7035、1991;Lyons, J.ら、Science 249:655-659、1990)。しかし、腫瘍形成または発癌へのLGNの関与については未だ報告はない。コロニー形成アッセイ法により、LGNが発癌活性をもつ可能性が示唆された。LGNの発現の促進はGαi2を活性化し、肝癌発生における発癌性シグナルを介在しうる。
【0023】
本発明の核酸は、cDNA、ゲノムDNA、化学的に合成されたDNA、およびRNAを含む。本発明の核酸は1本鎖または2本鎖でありうる。
【0024】
本明細書で用いる「単離された核酸」という表現は、構造が任意の天然の核酸の構造と同一ではないか、または3つより多い別個の遺伝子にわたる天然のゲノム核酸の任意の断片と同一ではない構造をとる核酸を意味する。したがってこの表現は例えば、(a)生物のゲノム中に天然に存在する、天然のゲノムDNA分子の一部の配列を有するDNA;(b)得られる分子が任意の天然のベクターもしくはゲノムDNAと同一ではないように、ベクター中にまたは原核生物もしくは真核生物のゲノムDNA中に組み込まれる核酸;(c)cDNA、ゲノム断片、ポリメラーゼ連鎖反応(PCR)により作製された断片、または制限酵素切断断片などの分離した分子;ならびに(d)ハイブリッド遺伝子(融合タンパク質をコードする遺伝子)の一部である組換えヌクレオチド配列を含む。具体的には、様々な(i)DNA分子、(ii)トランスフェクトされた細胞、または(iii)細胞クローンの混合物中に存在するDNA分子(例えば、cDNAライブラリーまたはゲノムDNAライブラリーなどのDNAライブラリー中に存在するDNA分子)の核酸は、この定義から除外される。
【0025】
1つの態様では、本発明の核酸はDDEFL1またはVANGL1のヌクレオチド配列(特に配列番号:1または配列番号:3)を含む核酸を含む。
【0026】
別の態様では、本発明の核酸は、DDEFL1またはVANGL1のアミノ酸配列(特に配列番号:2または配列番号:4)を含むポリペプチドをコードする核酸を含む。したがって、遺伝コードの縮重に基づく任意の配列を含む核酸が含まれる。
【0027】
さらに別の態様では、本発明の核酸は、配列番号:1または配列番号:3のバリアント核酸を含む。このバリアントは、配列番号:1もしくは配列番号:3、またはその相補物からなるヌクレオチド配列と、高ストリンジェントな条件下でハイブリダイズする鎖を含む核酸を含む。
【0028】
本明細書で用いる「相補物」という表現は、全ての塩基がもう一方の鎖の塩基の配列と塩基対を形成することができる、2本鎖核酸の一方の鎖を意味する。また「相補的である」という表現は、少なくとも15残基の連続ヌクレオチドの連続領域内で完全にマッチするものだけでなく、この領域内で少なくとも65%、好ましくは70%、より好ましくは80%、さらにより好ましくは90%、また最も好ましくは95%またはそれ以上の同一性を有するものも意味する。
【0029】
本明細書で用いる、2つの核酸の「同一性%」は、KarlinおよびAltschul(Proc. Natl. Acad. Sci. USA 90:5873-5877、1993)のように改変された、KarlinおよびAltschulのアルゴリズム(Proc. Natl. Acad. Sci. USA 87:2264-2268、1990)を用いて決定される。このようなアルゴリズムは、Altschulら(J. Mol. Biol. 215:403-410、1990)のNBLASTプログラムおよびXBLASTプログラムに組み込まれている。BLASTによるヌクレオチド検索はNBLASTプログラムを用いて実施される(スコア=100、ワード長=12)。タンパク質の相同性検索は、例えば日本のDNAデータバンク(DDBJ)において、FASTAプログラムやBLASTプログラムなどを用いて容易に実施することができる。BLASTによるタンパク質検索はXBLASTプログラムを用いて実施される(スコア=50、ワード長=3)。2つの配列間にギャップが存在する場合は、Gapped BLASTをAltsuchlら(Nucleic Acids Res. 25:3389-3402、1997)に記載されたように用いる。BLASTプログラムおよびGapped BLASTプログラムを使用する場合は、個々のプログラム(例えばXBLASTおよびNBLAST)のデフォルトのパラメータを用いる。
【0030】
好ましくは、バリアントは配列番号:1または配列番号:3のヌクレオチド配列と少なくとも65%同一なヌクレオチド配列を含む。より好ましくは、バリアントは配列番号:1または配列番号:3のヌクレオチド配列と少なくとも65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%、またはそれ以上同一である。標準配列(例えば配列番号:1または配列番号:3)より長いかまたは同じ長さのバリアントの場合、比較は全長の標準配列に対して行われる。バリアントが標準配列より短い場合(例えば配列番号:1または配列番号:3より短い場合)、比較は同じ長さの標準配列のセグメント(相同性の計算に必要な任意のループを除く)に対して行われる。
【0031】
ハイブリダイゼーションのストリンジェンシーは、以下の条件下での平衡ハイブリダイゼーションとして定義される:42℃、2×SSC、0.1%SDS(低ストリンジェンシー);50℃、2×SSC、0.1%SDS(中ストリンジェンシー);および65℃、2×SSC、0.1%SDS(高ストリンジェンシー)。平衡に達するために洗浄が必要な場合は、所望の特定のストリンジェンシーのハイブリダイゼーション溶液で洗浄を行う。一般に、温度が高くなるほど、平衡時にハイブリダイズする2本鎖の間の相同性は高くなる。
【0032】
本発明の核酸の長さに制限はないが、好ましくは少なくとも15ヌクレオチド、20ヌクレオチド、30ヌクレオチド、40ヌクレオチド、50ヌクレオチド、100ヌクレオチド、150ヌクレオチド、200ヌクレオチド、300ヌクレオチド、400ヌクレオチド、500ヌクレオチド、1000ヌクレオチド、1500ヌクレオチド、2000ヌクレオチド、2500ヌクレオチド、または3000ヌクレオチドを含む。
【0033】
本発明の核酸は、配列番号:1もしくは配列番号:3またはその相補物のヌクレオチド配列と特異的にハイブリダイズするプローブまたはプライマーとして使用される、ポリヌクレオチドを含む。「特異的にハイブリダイズする」という表現は、通常のハイブリダイゼーション条件(好ましくはストリンジェントな条件)で、配列番号:1または配列番号:3のヌクレオチド配列とハイブリダイズするが、他のポリペプチドをコードするDNAとクロスハイブリダイズしないことを意味する。
【0034】
プライマーおよびプローブは、配列番号:1もしくは配列番号:3のヌクレオチド配列内に少なくとも15の連続ヌクレオチドを含むか、またはその配列に相補的である。一般にプライマーは、15〜100ヌクレオチド、好ましくは15〜35ヌクレオチドを含み、プローブは、配列番号:1もしくは配列番号:3の全配列の少なくとも一部または全体を含む、少なくとも15ヌクレオチド、好ましくは少なくとも30ヌクレオチドを含む。プライマーは、本発明のポリペプチドをコードする核酸の増幅に使用することができ、プローブは、本発明のポリペプチドをコードする核酸の単離または検出に使用することができる。本発明のプライマーおよびプローブは、例えば市販のオリゴヌクレオチド合成装置で調製することができる。プローブは、制限酵素処理などにより得られる2本鎖DNA断片として調製することもできる。
【0035】
本発明の核酸は、配列番号:1または配列番号:3のヌクレオチド配列内の任意の部位とハイブリダイズするアンチセンスオリゴヌクレオチドを含む。本明細書で用いる「アンチセンスオリゴヌクレオチド」という表現は、DNAまたはmRNAの特定の領域を構成する部分に対応するヌクレオチド全体が相補的なオリゴヌクレオチドだけでなく、DNAまたはmRNAおよびオリゴヌクレオチドが配列番号:1または配列番号:3のヌクレオチド配列と特異的にハイブリダイズすることが可能な限りは、1個または複数のヌクレオチドのミスマッチを有するオリゴヌクレオチドも意味する。
【0036】
アンチセンスオリゴヌクレオチドは好ましくは、配列番号:1または配列番号:3のヌクレオチド配列中の、少なくとも15の連続ヌクレオチドに対するものである。上述の少なくとも15の連続ヌクレオチド中に開始コドンを含む上述のアンチセンスオリゴヌクレオチドがさらにより好ましい。
【0037】
本発明のアンチセンスオリゴヌクレオチドは、低級アルキルホスホネート(例えばメチルホスホネートまたはエチルホスホネート)、ホスホチオエート、およびホスホアミデートを含む類似体を含む。
【0038】
本発明のアンチセンスオリゴヌクレオチドは、本発明のポリペプチドをコードするDNAまたはmRNAに結合することによって、本発明のポリペプチドを産生する細胞に作用し、その転写または翻訳を阻害し、mRNAの分解を促進し、本発明のポリペプチドの発現を阻害する。
【0039】
本発明の核酸は以下のように調製することができる。本発明のポリペプチドをコードするcDNAは、例えば本発明のポリペプチドをコードするDNAのヌクレオチド情報(例えば配列番号:1または配列番号:3)を基にプライマーを作製し、プラークPCRを行うことにより調製することができる(Affara NAら (1994)Genomics 22、205-210)。ゲノムDNAは、例えば市販の「QiagenゲノムDNAキット」(Qiagen、ヒルデン、ドイツ)を用いる方法で調製することができる。得られたDNAのヌクレオチド配列は、例えば市販の「ダイターミネーターシーケンシングキット」(Applied Biosystems)を用いて、当技術分野での通常の方法で決定することができる。本発明の核酸は、後述するように、組換えタンパク質の作製および肝細胞癌の検出に使用することができる。
【0040】
ベクター、形質転換体、および組換えポリペプチドの作製
本発明は、本発明の核酸が挿入されたベクターも特徴とする。
【0041】
本発明のベクターは、本発明の組換えポリペプチドを作製するためのベクターを含む。本発明のポリペプチドの発現を可能とする限りは、任意のベクターを使用することができる。
【0042】
発現ベクターの例には、細菌(例えば大腸菌(Escherichia coli))発現ベクター、酵母発現ベクター、昆虫発現ベクター、および哺乳類発現ベクターなどがある。本発明では、pcDNA 3.1-myc/HisまたはpcDNA 3.1ベクター(Invitrogen)などの哺乳類発現ベクターを使用することができる。本発明の核酸のベクターへの挿入は、当技術分野での通常の方法で実施することができる。
【0043】
本発明のベクターは、本発明のポリペプチドを(特に遺伝子治療を目的として)インビボで発現させるためのベクターも含む。本発明のポリペプチドをインビボで発現可能な限りは、様々なウイルスベクターおよび非ウイルスベクターを使用することができる。ウイルスベクターの例には、アデノウイルスベクターやレトロウイルスベクターなどがある。陽イオンリポソームは非ウイルスベクターの一例でありうる。
【0044】
本発明は、本発明の核酸を発現可能な状態で有する形質転換体も提供する。本発明の形質転換体は、本発明の核酸が挿入された上述の発現ベクターを有する形質転換体、および本発明の核酸が組み込まれた宿主ゲノムを有する形質転換体を含む。本発明の核酸は、形質転換体が核酸を発現できる限りは、形質転換体内に任意の状態で保持される。
【0045】
ベクターが細胞内で機能して本発明の核酸を発現する限りは、ベクターが挿入される細胞に関して特別の制限はない。例えば大腸菌(E. coli)、酵母、哺乳類細胞、および昆虫細胞を宿主として使用することができる。好ましくはCOS7細胞やNIH3T3細胞などの哺乳類細胞を使用する。ベクターの細胞への導入は、エレクトロポレーションおよびリン酸カルシウム法などの既知の方法を用いて行うことができる。
【0046】
当技術分野で用いられる一般的な方法を、形質転換体から対象組換えポリペプチドを単離および精製する際に使用することができる。例えば、形質転換体を回収して抽出物を得た後に、イオン交換クロマトグラフィー、逆相クロマトグラフィー、ゲル濾過、もしくは本発明のポリペプチドに対する抗体がカラムに固定されたアフィニティクロマトグラフィーにより、またはこのようなカラムをいくつか組み合わせることにより、目的のポリペプチドを精製および調製することができる。
【0047】
また本発明のポリペプチドをグルタチオン-S-トランスフェラーゼタンパク質との融合タンパク質として、または複数のヒスチジンを追加した組換えポリペプチドとして、宿主細胞(例えば動物細胞や大腸菌(E. coli))内で発現させる場合には、発現された組換えポリペプチドを、グルタチオンカラムまたはニッケルカラムを用いて精製することができる。融合タンパク質を精製後、トロンビンまたは第Xa因子で切断することにより、目的のポリペプチド以外の領域を除くことも可能である。
【0048】
ポリペプチド
本発明は、DDEFL1またはVANGL1(例えば配列番号:1または配列番号:3)によりコードされる単離されたポリペプチドを提供する。特定の態様では、本発明のポリペプチドは、配列番号:1または配列番号:3のヌクレオチド配列によりコードされるポリペプチド、ならびに配列番号:2または配列番号:4のアミノ酸配列を含むポリペプチドを含む。
【0049】
本明細書で用いる「単離されたポリペプチド」は、実質的に純粋で他の生体高分子を含まないポリペプチドを意味する。実質的に純粋なポリペプチドは、乾燥重量で少なくとも75%(例えば少なくとも80%、85%、95%、または99%)純粋である。純度は、任意の適切な標準的な方法(例えばカラムクロマトグラフィー、ポリアクリルアミドゲル電気泳動、またはHPLC解析)で測定することができる。
【0050】
本発明のポリペプチドは、バリアントが配列番号:2または配列番号:4と少なくとも65%同一である限りは、配列番号:2または配列番号:4のバリアントを含む。バリアントは、1つまたは複数のアミノ酸が置換、欠失、付加、および/または挿入されている、配列番号:2または配列番号:4のアミノ酸配列を含むポリペプチドでありうる。バリアントはまた、配列番号:1または配列番号:3からなるヌクレオチド配列と高ストリンジェントな条件下でハイブリダイズする鎖を含む核酸によりコードされるポリペプチドでありうる。
【0051】
あるアミノ酸配列の1つまたは複数のアミノ酸残基が欠失、付加、および/または置換により改変されたアミノ酸配列を有するポリペプチドは、当初の生物学的活性を保持することが知られている(Mark, D. F.ら、Proc. Natl. Acad. Sci. USA(1984)81、5662-5666、Zoller, M. J.およびSmith, M.、Nucleic Acids Research(1982)10、6487-6500、Wang, A.ら、Science 224、1431-1433、Dalbadie-McFarland, G.ら、Proc. Natl. Acad. Sci. USA(1982)79、6409-6413)。
【0052】
置換、欠失、付加、および/または挿入によって変異が導入されるアミノ酸の数には特に制限はない。通常、全アミノ酸残基の10%またはそれ未満、好ましくは5%またはそれ未満、より好ましくは1%またはそれ未満である。
【0053】
変異が導入されるアミノ酸残基に関しては、アミノ酸の側鎖の性質が保存されている、異なるアミノ酸に変化させることが好ましい。アミノ酸側鎖の性質の例は、疎水性アミノ酸(A、I、L、M、F、P、W、Y、V)、親水性アミノ酸(R、D、N、C、E、Q、G、H、K、S、T)、ならびに以下の側鎖を含むアミノ酸である:脂肪族側鎖(G、A、V、L、I、P);ヒドロキシル基を含む側鎖(S、T、Y);硫黄原子を含む側鎖(C、M);カルボン酸およびアミドを含む側鎖(D、N、E、Q);塩基を含む側鎖(R、K、H);ならびに芳香族官能基を含む側鎖(H、F、Y、W)(括弧内の文字は、アミノ酸の1文字コードを意味する)。「保存的アミノ酸置換」は、上記グループの1つに属する1つのアミノ酸と、同じグループの別のアミノ酸との置換である。
【0054】
欠失バリアントは、配列番号:1または配列番号:3のアミノ酸配列の断片を含む。このような断片は、本発明の上記のポリペプチドと(全体的ではなく)部分的に同一なアミノ酸配列を有するポリペプチドである。本発明のポリペプチド断片は通常、8アミノ酸残基またはそれ以上、好ましくは12アミノ酸残基またはそれ以上(例えば15アミノ酸残基またはそれ以上)を含む。好ましい断片の例は、アミノ末端またはカルボキシル末端のいずれかを含むひと続きのアミノ酸残基、または2つの連続するアミノ酸残基(一方はアミノ末端を含み、もう一方はカルボキシル末端を含む)を欠くアミノ酸配列を有する、切断ポリペプチドを含む。さらに、αヘリックスおよびαヘリックス形成領域、βシートおよびβシート形成領域、ターンおよびターン形成領域、コイルおよびコイル形成領域、親水性領域、疎水性領域、α両親媒性領域、β両親媒性領域、可変領域、表面形成領域、基質結合領域、および高抗原性指標領域を有する断片を含む、構造的または機能的な特徴をもつ断片も好ましい。生物学的な活性を有する断片もまた好ましい。類似の活性もしくは改良された活性を有する、または望ましくない活性を低下させた、生物学的な活性を有する断片は、本発明のポリペプチドの活性を介在する。例えば、リガンドの結合を介してシグナルを細胞に伝達する活性を有する断片、および動物(特にヒト)における抗原性または免疫原性を有する断片が含まれる。これらのポリペプチド断片は好ましくは、本発明のポリペプチドの抗原性を有する。
【0055】
また別のバリアントは、本発明のポリペプチドと別のペプチドまたはポリペプチドとの融合タンパク質を含む。融合タンパク質は、本発明のポリペプチドをコードするDNAを、他のペプチドまたはポリペプチドをコードするDNAに、フレームが一致するように連結させ、これを発現ベクターに挿入し、宿主内で発現させるような、当業者に周知の技術で作製することができる。本発明のポリペプチドに融合させるペプチドまたはポリペプチドに関して制限はない。
【0056】
既知のペプチド、例えばFLAG(Hopp, T.P.ら、Biotechnology(1988)6、1204-1210)、6個のHis(ヒスチジン)残基を含む6×His、10×His、インフルエンザ凝集素(HA)、ヒトc-myc断片、VSP-GP断片、p18 HIV断片、T7タグ、HSVタグ、Eタグ、SV40のT抗原断片、lckタグ、α-チューブリン断片、Bタグ、タンパク質C断片などを、本発明のポリペプチドに融合させるペプチドとして使用することができる。本発明のポリペプチドに融合させるポリペプチドの例には、GST(グルタチオン-S-トランスフェラーゼ)、インフルエンザ凝集素(HA)、免疫グロブリンの定常領域、β-ガラクトシダーゼ、MBP(マルトース結合タンパク質)などがある。
【0057】
融合タンパク質は、これらのペプチドまたはポリペプチドをコードする市販のDNAを、本発明のポリペプチドをコードするDNAと融合し、調製した融合DNAを発現させることにより、作製することができる。
【0058】
バリアントポリペプチドは、配列番号:2または配列番号:4のアミノ酸配列と好ましくは少なくとも65%同一である。具体的には、改変ポリペプチドは、配列番号:2または配列番号:4のアミノ酸配列と少なくとも65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%、またはそれ以上同一である。改変ポリペプチドが、標準配列(例えば配列番号:2または配列番号:4)より長いかまたは同じ長さの場合は、比較は全長の標準配列に対して行われる。改変ポリペプチドが、標準配列より短い(例えば配列番号:2または配列番号:4より短い)場合は、比較は同じ長さの標準配列のセグメントに対して行われる。
【0059】
本明細書で用いる、2つのアミノ酸配列の「同一性%」は、核酸に関する上記の手順と同様に決定される。
【0060】
本発明のポリペプチドは、天然のポリペプチド、または上述の遺伝子工学技術を用いて作製された組換えポリペプチドとして、当業者に公知の方法で調製することができる。例えば、天然のポリペプチドは、小動物を組換えポリペプチドで免疫化することにより得られる抗体を結合させたカラムを調製し、本発明のポリペプチドを高レベルで発現する、肝臓組織または細胞の抽出物に対してアフィニティクロマトグラフィーを行うことによって得られる。組換えポリペプチドは、本発明のポリペプチドをコードするDNA(例えば配列番号:1または配列番号:3のヌクレオチド配列を含むDNA)を適切な発現ベクターに挿入し、このベクターを宿主細胞に導入し、結果として得られる形質転換体にポリペプチドを発現させ、発現されたポリペプチドを回収することにより調製することができる。
【0061】
バリアントポリペプチドは例えば、PCRによる部位特異的変異導入システム(GIBCO-BRL、ゲイサーズバーグ、メリーランド)や、オリゴヌクレオチドによる部位特異的変異誘発法(Kramer, W.およびFritz, HJ(1987)Methods in Enzymol. 154:350-367)などの公知の方法により、配列番号:1または配列番号:3のアミノ酸配列に変異を挿入することで調製することができる。
【0062】
抗体
本発明は、本発明のポリペプチドに特異的に結合する抗体も特徴とする。本発明の抗体の形状に関して特に制限はなく、ポリクローナル抗体およびモノクローナル抗体が含まれる。ウサギなどの動物を本発明のポリペプチドで免疫化することによって得られる抗血清、あらゆるクラスのポリクローナル抗体およびモノクローナル抗体、遺伝子工学により作製されたヒト化抗体、ヒト抗体も含まれる。
【0063】
ポリクローナル抗体は、本発明のポリペプチドで免疫化したウサギなどの小動物の血清を得て、本発明のポリペプチドのみを認識する画分を、本発明のポリペプチドを結合させたアフィニティカラムを用いて入手し、免疫グロブリンGまたはMを、この画分からプロテインGまたはプロテインAカラムを用いて精製することにより、作製することができる。
【0064】
モノクローナル抗体は、マウスなどの小動物を本発明のポリペプチドで免疫化し、この動物から脾臓を切除し、細胞へと組織をホモジナイズし、ポリエチレングリコールなどの試薬を用いて細胞をマウスのミエローマ細胞と融合し、本発明のポリペプチドに対する抗体を産生するクローンを融合細胞(ハイブリドーマ)から選択し、得られたハイブリドーマをマウスの腹腔に移植し、腹水を抽出することで作製することができる。得られたモノクローナル抗体は例えば、硫酸アンモニウム沈殿法、プロテインAもしくはプロテインGカラム、DEAEイオン交換クロマトグラフィー、または本発明のポリペプチドを結合させたアフィニティカラムで精製することができる。本発明の抗体は、本発明のポリペプチドの精製および検出に使用することができる。特に本発明の抗体は、肝細胞癌を検出するために使用することができる。
【0065】
ヒト抗体またはヒト化抗体は、当業者に公知の方法で調製することができる。例えば、ヒト抗体は、免疫系をヒトの免疫系に変化させたマウスを本発明のポリペプチドで免疫化することにより作製することができる。またヒト化抗体は、例えば、モノクローナル抗体産生細胞から抗体遺伝子をクローニングし、遺伝子の抗原認識部位を公知のヒト抗体に移すCDRグラフト法を用いて調製することができる。
【0066】
検出法
本発明はさらに、DDEFL1、VANGL1、またはLGNのポリペプチドをマーカーとして用いて肝細胞癌を検出する方法を提供する。
【0067】
検出は、被験者由来の生物試料中のDDEFL1、VANGL1、およびLGNのポリペプチドの少なくとも1つの発現レベルを測定し、非癌性試料における発現レベルと比較し、被験者における癌の有無を判定することにより行うことができる。
【0068】
本発明で用いる生物試料は、肝細胞癌の検出を必要とする被験者から得られる任意の肝臓組織または細胞を含む。特に、肝生検標本を使用することができる。このような生物試料は、肝臓組織または細胞から調製されたmRNA試料、cRNA試料、またはcDNA試料も含む。mRNA試料およびcDNA試料は従来の方法で調製することができる。cRNAは、テンプレートのcDNAからRNAポリメラーゼを用いて転写されたRNAを意味する。cRNAは、T7 RNAポリメラーゼを用いて、テンプレートとしてT7プロモーターを結合したcDNAから合成することができる。DNAチップベースの発現プロファイリング用の市販のcRNA転写キットを使用することができる。
【0069】
特定の態様では、DDEFL1、VANGL1、またはLGNのポリペプチドの発現レベルは、RNA、cDNA、またはポリペプチドのレベルで測定することができる。
【0070】
mRNAの発現レベルは、例えばDDEFL1、VANGL1、またはLGNのヌクレオチド配列とハイブリダイズするプローブを用いるノーザンブロッティング法、DDEFL1、VANGL1、またはLGNのヌクレオチド配列とハイブリダイズするプライマーを用いるRT-PCR法などで測定することができる。
【0071】
本発明の検出法で使用されるプローブまたはプライマーは、配列番号:1、配列番号:3、もしくは配列番号:5のヌクレオチド配列、またはその相補物と特異的にハイブリダイズする核酸を含む。「特異的にハイブリダイズする」という表現は、通常のハイブリダイゼーション条件(好ましくはストリンジェントな条件)下で、配列番号:1、配列番号:3、または配列番号:5のヌクレオチド配列とハイブリダイズするが、他のポリペプチドをコードするDNAとはクロスハイブリダイズしないことを意味する。
【0072】
プライマーおよびプローブは、配列番号:1、配列番号:3、または配列番号:5のヌクレオチド配列内に、またはこの配列に相補的な配列内にある少なくとも15の連続ヌクレオチドを含む。一般にプライマーは15〜100ヌクレオチド、また好ましくは15〜35ヌクレオチドを含み、プローブは、配列番号:1、配列番号:3、または配列番号:5の配列の少なくとも一部または全体を含む、少なくとも15ヌクレオチド、好ましくは少なくとも30ヌクレオチドを含む。プライマーおよびプローブは、例えば市販のオリゴヌクレオチド合成装置で作製することができる。プローブは、制限酵素処理などで得られる2本鎖DNA断片として調製することもできる。
【0073】
cDNAの発現レベルは、例えばDNAアレイを用いる方法で測定することができる(村松正実および山本雅、新遺伝子工学ハンドブック、pp.280〜284、羊土社)。具体的にはまず、被験者から調製されたcDNA試料、およびDDEFL1、VANGL1、またはLGNのヌクレオチド配列とハイブリダイズするポリヌクレオチドプローブが固定された固相支持体が提供される。プローブとして、上述のプローブを使用することができる。複数の種類の標的ポリヌクレオチドを検出するために、複数の種類のプローブを固相支持体に固定することができる。cDNA試料を必要に応じて検出用に標識する。標識は、検出可能である限りは特に制限されず、例えば蛍光標識や放射性標識などを含む。標識は従来の方法で実施することができる(L. Luoら、「Gene expression profiles of laser-captured adjacent neuronal subtypes」、 Nat. Med.(1999)pp.117-122)。
【0074】
次にcDNA試料を固相支持体上のプローブと接触させて、cDNA試料をプローブとハイブリダイズさせる。ハイブリダイゼーションの反応溶液および反応条件は、プローブ長などの様々な要素に依存して変動するが、当業者に周知の通常の方法に従って決定することができる。
【0075】
固相支持体上におけるcDNA試料とプローブ間のハイブリダイゼーションの強度は、cDNA試料の標識の種類に応じて測定される。例えば蛍光標識は、蛍光シグナルをスキャナーで読み取ることで検出可能である。
【0076】
被験cDNA試料および対照cDNA試料(例えば非癌性の組織または細胞に由来するcDNA)のハイブリダイゼーションの強度を、これらを異なる蛍光標識で標識することにより1回の測定で同時に測定することができる。例えば上述のcDNA試料の1つをCy5で、もう1つをCy3で標識することができる。Cy5およびCy3の蛍光シグナルの強度は、各cDNA試料の発現レベルを示す(Dugganら、Nat. Genet. 21:10〜14、1999)。
【0077】
この方法で、cDNAに代えてcRNAを測定することができる。
【0078】
さらに、ポリペプチドの発現レベルは、DDEFL1、VANGL1、またはLGNのポリペプチドに対する抗体を用いて、例えばSDSポリアクリルアミド電気泳動、ウエスタンブロッティング、ドットブロッティング、免疫沈降法、蛍光免疫アッセイ法、放射免疫アッセイ法、酵素免疫アッセイ法(例えば酵素結合免疫吸着アッセイ法(ELISA))、ならびに免疫組織化学染色などの免疫アッセイ法により測定することができる。
【0079】
特定の態様では、固相支持体上に固定されたDDEFL1、VANGL1、またはLGNのポリペプチドに対する抗体に生物試料を接触させ、固相支持体上の抗体-抗原複合体を、検出可能な標識で標識した二次抗体に接触させ、この標識を適切な方法で検出する。
【0080】
本発明の検出法で使用される抗体には、ウサギなどの動物をDDEFL1、VANGL1、またはLGNのポリペプチドで免疫化することにより得られる抗血清、あらゆるクラスのポリクローナル抗体およびモノクローナル抗体、遺伝子工学により作製されたヒト化抗体、ならびにヒト抗体を含む、DDEFL1、VANGL1、またはLGNのポリペプチド(特に配列番号:2、配列番号:4、または配列番号:6のアミノ酸配列を有するポリペプチド)に結合する任意の抗体が含まれる。これらの抗体は、上述のように調製することができる。
【0081】
上述のように測定された発現レベルを、非癌性試料で測定された発現レベルと比較して、被験者における肝細胞癌の有無を判定する。被験者由来の試料で測定された発現レベルが、非癌性試料で測定された発現レベルより高い場合は、被験者は癌であるか、または癌の危険性があると判断される。一方、被験者の試料における発現レベルが、非癌性試料における発現レベルより高くない場合は、被験者は癌ではないと判断される。具体的には、被験者の試料中の発現レベルが、非癌性試料における発現レベルと比べて高いか否かは、相対発現比(被験者試料/非癌性試料)を基に判定することができる。発現レベルは、相対発現比が2.0を上回る場合に高いと判定される。
【0082】
検出試薬
本発明は、肝細胞癌の検出試薬を提供する。
【0083】
ある態様では、本発明の検出試薬は、DDEFL1、VANGL1、またはLGN(具体的には配列番号:1、配列番号:3、または配列番号:5)とハイブリダイズする、少なくとも15ヌクレオチドを有するポリヌクレオチドを含む。このポリヌクレオチドを、本発明の上述の検出法においてプローブまたはプライマーとして使用することができる。プローブとして使用する場合は、本発明の検出試薬に含まれるポリヌクレオチドを標識することができる。標識法には例えば、T4ポリヌクレオチドキナーゼを用いてポリヌクレオチドの5'末端を32Pでリン酸化する標識法や、ランダムヘキサマーオリゴヌクレオチドなどをプライマーとして、およびクレノー酵素などのDNAポリメラーゼを用いて、32Pなどの同位元素、蛍光色素、ビオチンなどで標識された基質塩基を導入する方法(ランダムプライム法など)がある。
【0084】
別の態様では、本発明の検出試薬は、DDEFL1、VANGL1、またはLGNのポリペプチド(具体的には特に配列番号:2、配列番号:4、または配列番号:6のアミノ酸配列を有するポリペプチド)に結合する抗体を含む。これらの抗体を用いて、本発明のポリペプチドを上述の本発明の検出法で検出する。抗体は検出法に従って標識することができる。また、抗体を固相支持体上に固定することができる。
【0085】
本発明の検出試薬はさらに、滅菌水、生理食塩水、植物油、界面活性剤、脂質、可溶化剤、緩衝液、タンパク質安定剤(ウシ血清アルブミンやゼラチンなど)、保存剤などを含む溶媒または添加物を、これらが本発明の検出法で用いられる反応に影響を及ぼさない限り、含むことができる。
【0086】
肝細胞癌の成長を阻害する方法
本発明はさらに、肝細胞癌の成長を阻害する方法を提供する。特定の態様では、この方法は、DDEFL1、VANGL1、またはLGNのアンチセンスオリゴヌクレオチドを標的細胞に導入することにより実施することができる。
【0087】
この方法で使用されるアンチセンスオリゴヌクレオチドは、配列番号:1、配列番号:3、または配列番号:5のヌクレオチド配列内の任意の部位とハイブリダイズする。このアンチセンスオリゴヌクレオチドは、DNAまたはmRNA、およびオリゴヌクレオチドが、配列番号:1、配列番号:3、または配列番号:5のヌクレオチド配列と特異的にハイブリダイズすることができる限りは、DNAまたはmRNAの特定の領域を構成する部分に対応するヌクレオチド全体が相補的なオリゴヌクレオチドだけでなく、1つまたは複数ヌクレオチドのミスマッチを有するオリゴヌクレオチドも含む。
【0088】
このようなアンチセンスオリゴヌクレオチドは好ましくは、配列番号:1、配列番号:3、または配列番号:5のヌクレオチド配列中の少なくとも15の連続ヌクレオチドに対するオリゴヌクレオチドである。上述の少なくとも15の連続ヌクレオチド中に開始コドンを含む、上述のアンチセンスオリゴヌクレオチドが、さらにより好ましい。
【0089】
アンチセンスオリゴヌクレオチドは、低級アルキルホスホネート(例えばメチルホスホネートまたはエチルホスホネート)、ホスホチオエート、およびホスホアミデートを含む類似体を含む。
【0090】
本明細書では、標的細胞は哺乳類細胞、好ましくはヒト細胞でありうる。
【0091】
導入法は、インビトロ、インビボ、またはエキソビボの移入法でありうる。ある態様では、アンチセンスオリゴヌクレオチドを、従来のトランスフェクション法で標的細胞に導入することができる。または、導入はアデノウイルスベクター、レトロウイルスベクター、または陽イオンリポソームなどのアンチセンスオリゴヌクレオチドを有するベクターを用いる、従来の遺伝子移入技術により行うことができる。
【0092】
本明細書で引用されるあらゆる特許、特許出願、および刊行物は参照として組み入れられる。
【0093】
発明を実施するための最良の形態
本発明を、以下の実施例を参照して説明するが、本発明がそれらに制限されるとは解釈されない。
【0094】
実施例1
1-1. ヒトの肝細胞癌において発現が一般的に上昇するDDEFL1の同定
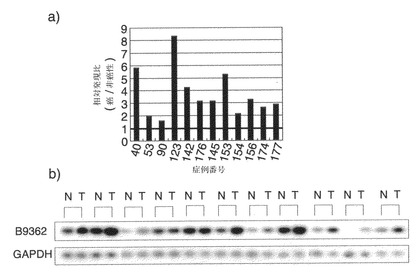
23040個の遺伝子を含むゲノム全体にわたるcDNAマイクロアレイによって、20例の肝細胞癌(HCC)の発現プロファイルを、対応する非癌性肝組織と比較した。全てのHCCの組織および対応する非癌性組織は、肝切除を受け、同意を得た患者の外科標本から入手した。UniGeneクラスターのEST(Hs.44579)に対応する施設内アクセッション番号B9362の遺伝子は、1.57〜5.83の範囲で過剰に発現されることが見出された(図1a)。この発現上昇(Cy3:Cy5強度比が2.0を上回る)は、カットオフフィルターを通過した(Cy3およびCy5の両シグナルが25,000を上回る)12例のHCCのうち11例で認められた。この遺伝子のオープンリーディングフレームは、発生・分化促進因子2(DDEF2)と約60%同一なタンパク質をコードしていたため、この遺伝子を「発生・分化促進因子様1」(DDEFL1)と命名した。cDNAマイクロアレイの結果を明らかにするために、別の11例のHCCにおいて、この転写物の発現を半定量的RT-PCRで調べた。GAPDHの発現を内部対照として用いた。RT-PCRは以下のように実施した。全RNAをQiagen RNeasyキット(Qiagen)、またはTrizol試薬(Life Technologies社)を用いて製造業者のプロトコールにしたがって抽出した。10 μgの全RNAを、ポリdT12-18プライマー(Amersham Pharmacia Biotech)を用いてSuperscript II逆転写酵素(Life Technologies)で逆転写して1本鎖のcDNAを得た。1本鎖cDNA調製物を、20 μlのPCR緩衝液(TAKARA)中で行う標準的なRT-PCR実験による、次のPCR増幅用に希釈した。増幅は、94℃で4分間処理して変性した後に、94℃で30秒間、56℃で30秒間、および72℃で45秒間を20サイクル(GAPDHの場合)または33サイクル(DDEFL1の場合)、GeneAmp PCR system 9700(Perkin-Elmer、フォスターシティ、カリフォルニア)で行った。プライマーの配列は以下であった;GAPDH用:フォワード、5'-ACAACAGCCTCAAGATCATCAG (配列番号: 7) およびリバース、5'-GGTCCACCACTGACACGTTG (配列番号: 8); DDEFL1用:フォワード、5'-AGCTGAGACATTTGTTCTCTTG (配列番号: 9)およびリバース、5'-TATAAACCAGCTGAGTCCAGAG (配列番号: 10)。結果から、DDEFL1の発現がこれらの腫瘍のうち9例で上昇することが確認された(図1b)。
【0095】
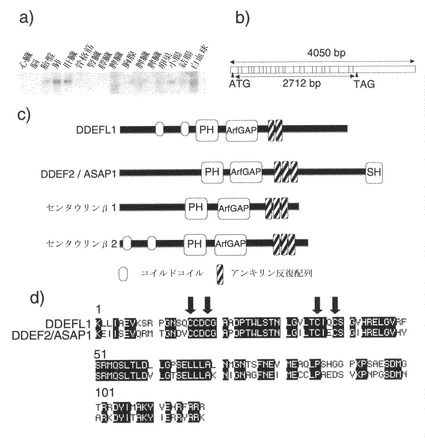
1-2. 新たな遺伝子DDEFL1の単離および構造
DDEFL1の発現を、DDEFL1のPCR産物をプローブとして用いた多組織ノーザンブロット解析で解析した。ヒト多組織ブロット(Clontech、パロアルト、カリフォルニア)を、32P標識DDEFL1 cDNAとハイブリダイズさせた。プレハイブリダイゼーション、ハイブリダイゼーション、および洗浄は、製造業者の推奨にしたがって行った。このブロットのオートラジオグラフを増感スクリーンを用いて、-80℃で72時間かけて得た。この結果、4 kbの転写物が、肺、肝臓、小腸、胎盤、および末梢血白血球で発現していることが判明した(図2a)。
【0096】
B9362は、ノーザンブロットで検出されたものより小さかったため、5'RACE実験を行って、この遺伝子のコード配列全体を決定した。5'RACE実験は、Marathon cDNA増幅キット(Clontech、パロアルト、カリフォルニア)を製造業者の指示書通りに用いて実施した。DDEFL1 cDNAの5'側部分の増幅には、遺伝子に特異的なリバースプライマー(5'-CTCACTTGGCACGTCAGCAGGG (配列番号: 11))およびキットに同梱されているAP-1プライマーを使用した。cDNAテンプレートは、ヒト肝臓のmRNAから合成した。PCR産物をTAクローニングキット(Invitrogen)を用いてクローニングし、配列をABI PRISM 3700 DNAシーケンサー(Applied Biosystems)で決定した。
【0097】
完全なcDNAは、903アミノ酸のタンパク質をコードする2712ヌクレオチドのオープンリーディングフレームを有する、4050ヌクレオチドからなる(GenBankアクセッション番号AB051853)。最初のATGは、真核生物における翻訳開始のコンセンサス配列と一致する配列(CCCGCCATGC(配列番号:12))に隣接しており、上流にインフレームの停止コドンをもっていた。NCBI(米国バイオテクノロジー情報センター)データベース上の、相同性探索用のBLASTプログラムによって、染色体バンド1p36.12に位置するGenBankアクセッション番号AL357134のゲノム配列が同定された。cDNA配列とゲノム配列の比較の結果、DDEFL1が25個のエキソンからなることが判明した(図2b)。
【0098】
Simple Modular Architecture Research Tool(SMART)によるタンパク質モチーフの探索により、推定タンパク質が2つのコイルドコイル領域(コドン141〜172および241〜278)、PH(プレックストリン(Pleckstrin)相同性)モチーフ(コドン303〜396)、ArfGAPのモチーフ(ArfのGTPアーゼ活性化タンパク質)(コドン426〜551)、ならびに2つのアンキリン反復配列(コドン585〜617および621〜653)を含むことが判明した。この構造は、センタウリンβ1およびβ2(図2c)と似ていた。特にDDEFL1は、ホスファチジルイノシトール3,4,5-三リン酸の標的であるPHドメイン、およびArfGAPのモチーフなどの、センタウリンファミリーのタンパク質の特徴を有していた。DDEFL1のArfGAPモチーフのアミノ酸配列は、DDEF2のアミノ酸配列と67.8%同一であった(図2d)。特に、GAP活性に不可欠なジンクフィンガー構造を示すCXXCX16CXXCモチーフは完全に保存されていた。
【0099】
1-3. DDEFL1の細胞内局在
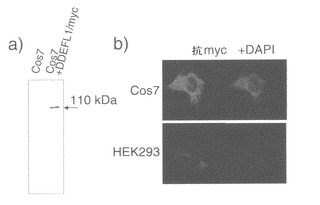
DDEFL1のコード配列をpcDNA3.1-myc/Hisベクター(Invitrogen)にクローニングした。myc標識DDEFL1タンパク質を発現する、得られたプラスミド(pDNA-myc/His-DDEFL1)を、COS7細胞(米国菌株保存機関(ATCC))に一過性にトランスフェクトした。推定myc標識タンパク質を、以下に示す免疫ブロッティング(ウエスタンブロッティング)で検出した。pcDNA3.1-myc/His-DDEFL1でトランスフェクトした細胞をPBSで2回洗浄し、溶解緩衝液(150 mM NaCl、1%トリトン X-100、50 mM Tris-HCl pH 7.4、1 mM DTT、および1×コンプリートプロテアーゼインヒビターカクテル(Boehringer))中に回収した。細胞をホモジナイズして10,000×gで30分間遠心した後に、上清をタンパク質濃度に関してBradfordアッセイ法(Bio-Rad)で標準化した。タンパク質を10%SDS-PAGEで分離し、マウス抗myc抗体で免疫ブロットを行った。HRP結合ヤギ抗マウスIgG(Amersham)をECL検出システム(Amersham)用の二次抗体として用いた。結果的に、DDEFL1タンパク質が抗myc抗体を用いたウエスタンブロットで検出された(図3a)。
【0100】
また、免疫細胞化学染色を以下のように行った。最初に細胞を4%パラホルムアルデヒドを含むPBSで15分間固定し、次に0.1%トリトン X-100を含むPBSで室温で2.5分間処理して透過性にした。次に、細胞を2%BSA(PBS中)で覆い、24時間4℃で処理して、非特異的なハイブリダイゼーションをブロックした。マウス抗mycモノクローナル抗体(Sigma)を1:1000の希釈率で、またはマウス抗FLAG抗体(Sigma)を1:2000の希釈率で一次抗体用に使用し、ローダミンを結合した抗マウス二次抗体(Leinco and ICN)とインキュベートした後、反応を可視化した。核を4',6'-ジアミン-2'-フェニルインドールジヒドロクロライド(DAPI)で対比染色した。蛍光像をECLIPSE E800顕微鏡下で得た。顕微鏡解析により、タンパク質が主に細胞質に存在することが示された(図3b)。DDEFL1もヒト胚腎(HEK293)細胞(ATCC)の細胞質に局在していた。
【0101】
1-4. DDEFL1が細胞成長に及ぼす作用
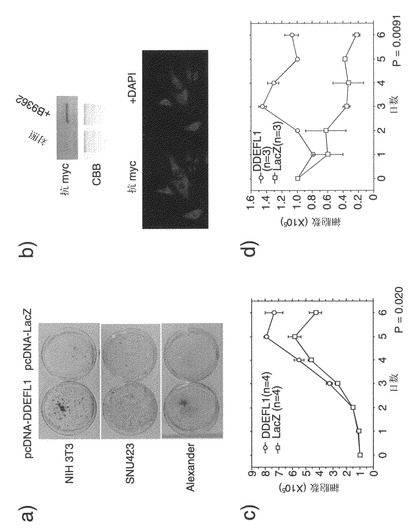
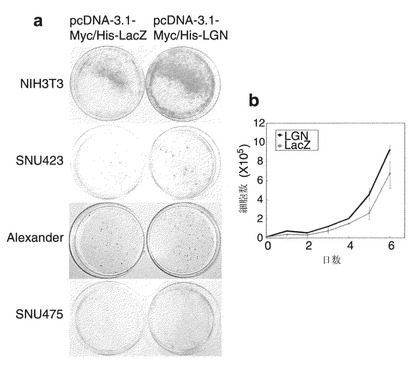
DDEFL1のコード配列をpcDNA 3.1ベクター(Invitrogen)にクローニングした。10 cmのディッシュに播いたNIH3T3細胞(ATCC)(2×105細胞/ディッシュ)に、DDEFL1を発現する得られたプラスミド(pcDNA-DDEFL1)、および対照プラスミド(pcDNA-LacZ)でトランスフェクトし、10%ウシ胎児血清および1%抗生物質/抗真菌剤溶液(Sigma)を添加したダルベッコ改変イーグル培地に、さらに適切な濃度のジェネティシンを添加した培地中で2週間培養した。次にこの細胞を100%メタノールで固定し、ギムザ溶液で染色した。pcDNA-DDEFL1でトランスフェクトした細胞は、対照細胞より著しく多数のコロニーを生じた。コロニー形成の増加は、DDEFL1の内因性発現が極めて低い、ヒト肝癌SNU423(韓国細胞株バンク)細胞、およびAlexander(ATCC)細胞をトランスフェクトした場合に同様に認められた(図4a)。
【0102】
この成長促進作用を詳細に調べるために、外因性DDEFL1を安定に発現するNIH3T3細胞を確立した。pDNA-myc/His-DDEFL1を、FuGENE6試薬(Boehringer)を製造業者の推奨通りに用いてNIH3T3細胞にトランスフェクトした。トランスフェクションから24時間後に、培養物にジェネティシンを添加し、トランスフェクションから2週間後に単一コロニーを選択した。DDEFL1の発現を、半定量的RT-PCRで決定した(図4b)。NIH3T3-DDEFL1細胞の成長速度は、擬似(NIH3T3-LacZ)細胞の成長速度より、10%FBSを含む培地中において統計的に速かった(P<0.05)(図4c)。0.1%FBSのみを含む培地中では、NIH3T3-DDEFL1細胞は6日間生存したが、対照NIH3T3細胞は同条件下で6日以内に死滅した。この場合、NIH3T3-DDEFL1細胞の成長は、0.1%FBSを含む培地における擬似処理細胞の成長より統計的に速かった(P<0.01)(図4d)。
【0103】
1-5. ヒト肝癌SNU475細胞におけるアンチセンスS-オリゴヌクレオチドによるDDEFL1発現の抑制
DDEFL1遺伝子に対応する、以下の6組の対照(センス)およびアンチセンスのS-オリゴヌクレオチドを合成した。
アンチセンス:
DDEFL1-AS1 5'-TGCTCCGGCATGGCGG-3' (配列番号: 13);
DDEFL1-AS2 5'-GCTGAACTGCTCCGGC-3' (配列番号: 14);
DDEFL1-AS3 5'-TCCAAGATCTCCTCCC-3' (配列番号: 15);
DDEFL1-AS4 5'-TCTCCTTCCAAGATCT-3' (配列番号: 16);
DDEFL1-AS5 5'-GCGCTGAGCCGGCCTC-3' (配列番号: 17);および
DDEFL1-AS6 5'-CCTCACCTCCTCCCGC-3' (配列番号: 18)
対照:
DDEFL1-S1 5'-CCGCCATGCCGGAGCA-3' (配列番号: 19);
DDEFL1-S2 5'-GCCGGAGCAGTTCAGC-3' (配列番号: 20);
DDEFL1-S3 5'-GGGAGGAGATCTTGGA-3' (配列番号: 21);
DDEFL1-S4 5'-AGATCTTGGAAGGAGA-3' (配列番号: 22);
DDEFL1-S5 5'-GAGGCCGGCTCAGCGC-3' (配列番号: 23); および
DDEFL1-S6 5'-GCGGGAGGAGGTGAGG-3' (配列番号: 24)
【0104】
LIPOFECTIN試薬(GIBCO BRL)を用いて、DDEFL1の発現レベルが、本発明者らが調べた6つの肝癌細胞株中で最高であった(データは示していない)、SNU475細胞(韓国細胞株バンク)に、合成S-オリゴヌクレオチドをトランスフェクトした。トランスフェクションから12時間および24時間後に、アンチセンスS-オリゴヌクレオチドAS1およびAS5は、DDEFL1の発現を各対照S-オリゴヌクレオチドS1およびS5と比較して有意に抑制した(図5a)。アンチセンスS-オリゴヌクレオチドAS1またはAS5によりトランスフェクトされた生存細胞の数は、トランスフェクションから6日後の時点で、各対照S-オリゴヌクレオチドS1またはS5によりトランスフェクトされた細胞と比較して著しく少なかった(図5b)。3回の独立した実験で一貫した結果が得られた。
【0105】
実施例2
2-1. ヒト肝細胞癌で一般的に発現が上昇しているVANGL1の同定
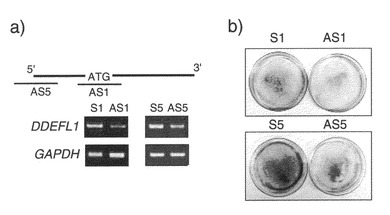
実施例1で実施したゲノム全体にわたるcDNAマイクロアレイ解析の結果では、UniGeneクラスターのEST(Hs.122730)に対応する施設内アクセッション番号D3244の遺伝子が、12例の臨床HCCのうち10例で、対応する非癌性肝組織と比較して有意に発現が上昇していることも判明した。これら12例の腫瘍の、対応する非癌性組織と比較した相対発現比の範囲は1.5〜16.0であった(図6a)。発現の上昇(Cy3:Cy5強度比が2.0を上回る)は、カットオフフィルターを通過した(Cy3およびCy5の両シグナルが25,000を上回る)12例のHCCのうち10例で認められた。D3244の発現上昇は、別の10例のHCC症例において、プライマー対、フォワード: 5'- GAGTTGTATTATGAAGAGGCCGA (配列番号: 25); リバース: 5'- ATGTCTCAGACTGTAAGCGAAGG (配列番号: 26)を用いて、実施例1-1と同様に実施した半定量的RT-PCRにより確認された。(図6b)
【0106】
2-2. ヒト成体組織におけるVANGL1の発現
D3244 cDNAをプローブとして用いた多組織ノーザンブロット解析を、実施例1-2と同様の手順で行い、その結果、1.9 kbの転写物が、精巣および卵巣で組織特異的に多量に発現されることが判明した(図7a)。D3244に対応するゲノム配列のNCBIデータベースの探索により、2つの配列(GenBankアクセッション番号:AL450389およびAL592436)が染色体バンド1p22に位置することが判明した。GENSCAN、および「Gene Recognition and Assembly Internet Link」プログラムを用いて候補エキソン配列を推定し、エキソンの連結を行った。またリバースプライマー(5'-TGTCAGCTCTCCGCTTGCGGAAAAAAAG (配列番号: 27))を用いて5'RACEを行い、転写物の5'領域の配列を実施例1-2と同様の手順で決定した。その結果、1572ヌクレオチドのオープンリーディングフレームを含む、1879ヌクレオチドのヒトcDNA配列がアセンブルして得られた(GenBankアクセッション番号:AB057596)。この推定アミノ酸配列は、strabismus(Van Gogh)と40%、VANGL2と63%同一であった。したがってD3244に対応する遺伝子をVan Gogh Like 1(VANGL1)と命名した。Simple Modular Architecture Research Toolにより、推定タンパク質が4つの推定膜貫通ドメイン(コドン111〜133、148〜170、182〜204、219〜241)を含むことが示唆された(図7b)。
【0107】
2-3. VANGL1の細胞内局在
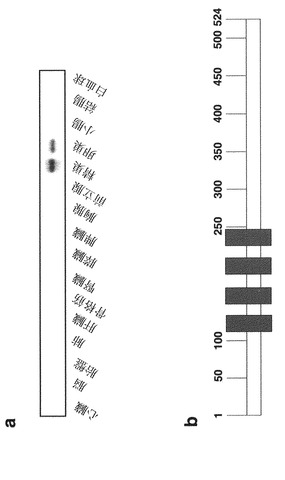
c-myc標識VANGL1タンパク質を発現するpcDNA3.1-myc/His-VANGL1プラスミドを、SNU475細胞(韓国細胞株バンク)に一過性にトランスフェクトした。免疫組織化学染色を実施例1-3と同様の手順で行った。この結果から、標識VANGL1タンパク質が細胞質に存在することが判明した(図8aおよび8b)。
【0108】
2-4. VANGL1の発現を低下させるように指定したアンチセンスS-オリゴヌクレオチドによる肝癌細胞の成長抑制
VANGL1の抑制がHCC細胞の細胞周期の停止および/または細胞死をもたらすかどうかを試験するために、以下の4組のアンチセンスおよび対照(センス)のS-オリゴヌクレオチドを合成し、SNU475細胞にトランスフェクトした。
アンチセンス:
アンチセンス1 5'-GTATCCATAGCAATGG-3' (配列番号: 28);
アンチセンス2 5'-TGGATTGGGTATCCAT-3' (配列番号: 29);
アンチセンス3 5'-TAAGTGGATTGGGTAT-3' (配列番号: 30); および
アンチセンス4 5'-ACTCCTACCTGCCTGT-3' (配列番号: 31)
対照:
センス1 5'-CCATTGCTATGGATAC-3' (配列番号: 32);
センス2 5'-ATGGATACCCAATCCA-3' (配列番号: 33);
センス3 5'-ATACCCAATCCACTTA-3' (配列番号: 34); および
センス4 5'-ACAGGCAGGTAGGAGT-3' (配列番号: 35)
【0109】
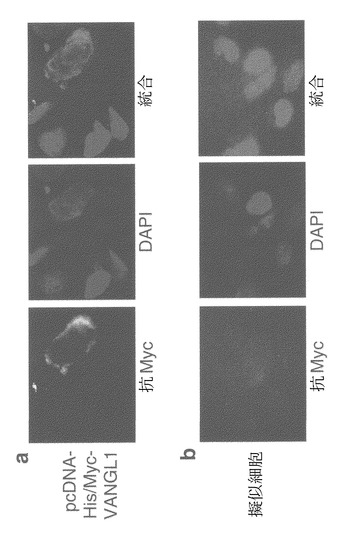
開始コドンを含むアンチセンスS-オリゴヌクレオチド(アンチセンス3)は、SNU475細胞におけるVANGL1の内因性発現を有意に低下させた(図9a)。
【0110】
細胞の生存は、3-(4,5-ジメチル-チアゾール-2-イル)-2,5-ジフェニルテトラゾリウムブロマイド(MTT)アッセイ法により、以下のように評価した。細胞を5×105細胞/100 mm ディッシュの密度で播いた。播種してから24時間後に、細胞を、センスまたはVANGL1を抑制するように指定したアンチセンスのS-オリゴヌクレオチドにより3回トランスフェクトした。トランスフェクションから72時間後に、培地を500 μg/mlの3-(4,5-ジメチルチアゾール-2-イル)-2,5-ジフェニルテトラゾリウムブロマイド(MTT)(Sigma)を含む新鮮な培地と交換し、プレートを37℃で4時間インキュベートした。続いて、1 mlの0.01 N HCl/10%SDSを添加して細胞を溶解し、溶解液の吸光度をELISAプレートリーダーで測定した(試験波長=570 nm、標準波長=630 nm)。細胞生存率を、対照細胞と比較した吸光度で表した。
【0111】
アンチセンスS-オリゴヌクレオチド、アンチセンス3のトランスフェクションにより、対照のセンスS-オリゴヌクレオチド、センス3の使用時と比較して、生存細胞数が有意に減少した(図9bおよび9c)。この結果は、3回の独立した実験で確認された。
【0112】
さらにフローサイトメトリー解析を以下のように行った。細胞を、1×105細胞/100 mm ディッシュの密度で播き、任意の経過時間後にトリプシン処理し、続いて70%冷エタノール中で固定した。RNA分解酵素処理後に、細胞をヨウ化プロピジウム(50 μg/ml、PBS中)で染色した。フローサイトメトリーをBecton Dickinson FACScanで行い、CellQuestおよびModFitソフトウェア(Verity Software House)で解析した。細胞周期のG0/G1期、S期、およびG2/M期にある核の割合、および任意のサブG1集団を、少なくとも20,000個の非同期細胞を対象に決定した。
【0113】
FACS解析の結果、VANGL1を阻害すると、サブG1期の細胞数が有意に増加することが判明した(図9d)。これらの結果は、VANGL1が、肝細胞癌細胞の成長および/または生存に重要な役割を果たす可能性があることを示唆する。
【0114】
実施例3
3-1. LGNは一般的にヒトの肝細胞癌で増加している
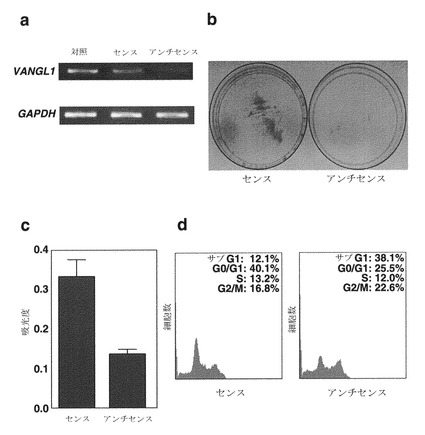
実施例1-1で実施したマイクロアレイ解析によって明らかになった、一般的に発現が上昇している遺伝子の中で、LGN(GenBankアクセッション番号:U54999)に対応する遺伝子D3636を選択した。というのはD3636の発現が、対応する非癌性肝組織と比較して、12例の臨床HCCのうち10例で有意に上昇していたためである。これら12例の腫瘍の、対応する非癌性組織と比較時の相対発現比は0.7〜16.0であった。LGNの発現の上昇(Cy3:Cy5強度比が2.0を上回る)は、カットオフフィルターを通過した(Cy3およびCy5の両シグナルが25,000を上回る)12例のHCCのうち10例で認められた(図10a)。LGNの発現の上昇は、別の10例のHCCでも、プライマー対、フォワード: 5'-ATCTGAAGCACTTAGCAATTGC (配列番号: 36)、リバース: 5'-CTGTAGCTCAGACCAAGAACC (配列番号: 37)を用いて、実施例1-1と同様に実施した半定量的RT-PCRにより確認された(図10b)。
【0115】
3-2. LGNのゲノム構造
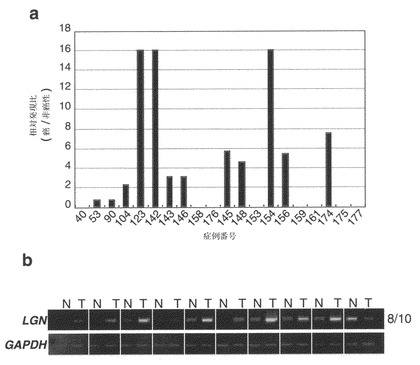
LGNのcDNAは2,336ヌクレオチドからなり、677アミノ酸のペプチドをコードする。cDNA配列とゲノム配列との比較により、LGN遺伝子が14個のエキソンからなることが判明した(図11)。
【0116】
3-3. LGNの細胞内局在
c-myc標識LGNタンパク質を発現するpcDNA3.1-myc/His-LGNプラスミドをCOS7細胞に一過性にトランスフェクトした。myc標識LGNタンパク質に対応する72 kDaのバンドが、実施例1-3と同様の手順で行った免疫ブロット解析により検出された(図12)。同様に、実施例1-3のように免疫細胞化学染色を行い、その結果、標識LGNタンパク質が、細胞の細胞質および核に存在することが判明した。
【0117】
3-4. LGN遺伝子の移入は細胞成長を促進することができる
LGNが細胞成長に及ぼす作用を解析するために、実施例1-4のように、NIH3T3細胞、SNU423細胞、Alexander細胞、およびSNU475細胞を、LGNを発現するプラスミド(pcDNA3.1-myc/His-LGN)によりトランスフェクトして、コロニー形成アッセイ法を実施した。対照プラスミド(pcDNA3.1-myc/His-LacZ)と比較して、pcDNA3.1-myc/His-LGNは、これらの細胞で著しく多数のコロニーを生じた(図13a)。この結果は3回の独立した実験で確認された。
【0118】
LGNが細胞成長に及ぼす作用をさらに調べるために、外因性LGNを安定に発現するNIH3T3細胞(NIH3T3-LGN細胞)を確立した。NIH3T3-LGN細胞は、対照のNIH3T3-LacZ細胞より速い成長速度を示した(図13b)。
【0119】
3-5. LGNのアンチセンスS-オリゴヌクレオチドはヒト肝癌SNU475細胞の成長を抑制した
LGNに対応する、以下の5組の対照(センス)およびアンチセンスのS-オリゴヌクレオチドを合成してSNU423細胞にトランスフェクトした。
アンチセンス:
アンチセンス1 5'-CCATCGAGTCATATTA-3'(配列番号: 38);
アンチセンス2 5'-TTCCTCCATCGAGTCA-3'(配列番号: 39);
アンチセンス3 5'-AAATTTTCCTCCATCG-3'(配列番号: 40);
アンチセンス4 5'-AGTCTTACCTGTAACG-3'(配列番号: 41); および
アンチセンス5 5'-GCTTCCATTCTACAAA-3'(配列番号: 42)
センス:
センス1 5'-TAATATGACTCGATGG-3'(配列番号: 43);
センス2 5'-TGACTCGATGGAGGAA-3'(配列番号: 44);
センス3 5'-CGATGGAGGAAAATTT-3'(配列番号: 45);
センス4 5'-CGTTACAGGTAAGACT-3'(配列番号: 46); および
センス5 5'-TTTGTAGAATGGAAGC-3'(配列番号: 47)
【0120】
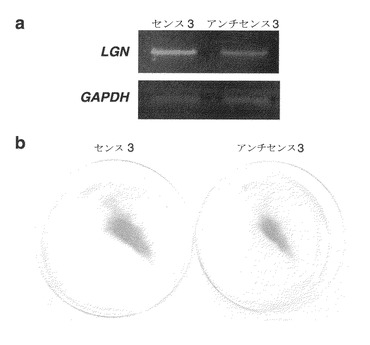
開始コドンを含むアンチセンスS-オリゴヌクレオチド(アンチセンス3)は、トランスフェクションから12時間後の対照S-オリゴヌクレオチド(センス3)と比較して、LGNの発現を有意に抑制した(図14a)。トランスフェクションから6日後に、アンチセンス3でトランスフェクトした生存細胞数は、対照のセンス3でトランスフェクトしたものより著しく少なかった(図14b)。3回の独立した実験で一貫した結果が得られた。
【0121】
産業上の利用可能性
本発明は、肝細胞癌で一般に発現が上昇することが見出されたDDEFL1、VANGL1、またはLGNの、cDNAヌクレオチド配列およびポリペプチドのアミノ酸配列を提供する。したがって、これらのポリペプチドは、肝臓癌の有無を判定するためのマーカーとして使用することができる。これらのヌクレオチド配列の情報により、DDEFL1、VANGL1、またはLGNの遺伝子を検出または増幅するためのプローブおよびプライマーを設計することができる。またDDEFL1、VANGL1、またはLGNのポリペプチドの発現を阻害するアンチセンスヌクレオチド配列を合成することも可能となる。アミノ酸配列情報を用いることで、DDEFL1、VANGL1、またはLGNのポリペプチドに結合する抗体を調製することができる。このようなプローブおよびプライマーならびに抗体は、肝細胞癌を検出するための試薬として有用である。さらに本発明者らは、アンチセンスオリゴヌクレオチドでDDEFL1、VANGL1、またはLGNの発現を抑制すると、HCC細胞の成長が著しく低下することを立証した。したがって、アンチセンスオリゴヌクレオチドを用いてHCC細胞の成長を阻害することができる。本発明は、肝細胞の発癌機構をさらに明らかにし、肝臓癌を治療するための有効な薬剤の開発に向けた分子標的を発見することにも寄与する。
【図面の簡単な説明】
【0122】
【図1】B9362と命名された遺伝子の、HCCにおける発現を示す。図1aは、20例の原発性HCCにおいてcDNAマイクロアレイを用いて調べたB9362の相対発現比(癌/非癌性)を示す。図1bは、別の11例のHCCを半定量的RT-PCRで解析したB9362の発現を示す写真である。GAPDHの発現を内部対照として用いた。
【図2】DDEFL1の同定の結果を示す。図2aは、ヒトの様々な組織におけるDDEFL1のノーザンブロット解析の結果を示す写真である。図2bはDDEFL1の構造を示す。図2cは、推定DDEFL1タンパク質とArfGAPファミリーのメンバーとの類似性を示す。図2dは、DDEFL1とDDEF2におけるArfGAPモチーフのアミノ酸配列間の同一性を示す。矢印は、GAP活性に不可欠なジンクフィンガー構造を示す、CXXCX16CXXCモチーフを示す。
【図3】DDEFL1の細胞内局在を示す。図3aは、cMyc標識DDEFL1タンパク質が、pcDNA-DDEFL1-mycプラスミドをトランスフェクトしたCOS7細胞で発現していることを示す、ウエスタンブロット解析の結果を示す写真である。図3bは、cMyc標識DDEFL1タンパク質が細胞質に局在することを示唆する、細胞の免疫組織化学染色を示す写真である。
【図4】DDEFL1の成長促進作用を示す。図4aは、DDEFL1がNIH3T3細胞、SNU423細胞、およびAlexander細胞の成長を促進することを示す、コロニー形成アッセイの結果を示す写真である。図4bは、NIH3T3-DDEFL1細胞による外因性DDEFL1の安定な発現を示す写真である。図4cは、10%FBSを含む培地中で外因性DDEFL1を安定に発現している、NIH3T3-DDEFL1細胞の成長を示すグラフである。図4dは、0.1%FBSを含む培地中におけるNIH3T3-DDEFL1細胞の成長を示すグラフである。(P<0.01)
【図5】SNU475細胞でDDEFL1を抑制するように指定したアンチセンスS-オリゴヌクレオチドによる、成長抑制を示す。図5aは、アンチセンスS-オリゴヌクレオチドを明示し、AS1またはAS5アンチセンスS-オリゴヌクレオチドのトランスフェクションによってDDEFL1の発現が低下したことを示す写真である。図5bは、AS1およびAS5がSNU475細胞の成長を抑制したことを示す写真である。
【図6】HCCにおけるVANGL1の発現を示す。図6aは、cDNAマイクロアレイで調べた、20例の原発性HCCにおけるVANGL1の相対発現比(癌/非癌性)を示す。図6bは、別の10例のHCCを用いて半定量的RT-PCRにより解析した、D3244の発現を示す写真である。T=腫瘍組織;N=正常組織。GAPDHの発現を内部対照として用いた。
【図7】VANGL1の同定の結果を示す。図7aは、ヒトの様々な組織におけるVANGL1の多組織ノーザンブロット解析の結果を示す写真である。図7bはVANGL1の推定タンパク質の構造を示す。
【図8】VANGL1の細胞内局在を示す。図8aは、pcDNA3.1-myc/His-VANGL1でトランスフェクトしたSNU475細胞をマウス抗mycモノクローナル抗体で染色し、ローダミンを結合した抗マウスIgG二次抗体で可視化した写真である。核はDAPIで対比染色した。図8bは、同様に染色して可視化した擬似細胞の写真である。
【図9】VANGL1を抑制するように指定したアンチセンスS-オリゴヌクレオチドの成長抑制作用を示す。図9aは、対照またはアンチセンスオリゴヌクレオチドのいずれかで12時間処理したSNU475細胞におけるVANGL1の発現を示す写真である。図9bは、S-オリゴヌクレオチドがSNU423細胞の成長を抑制したことを示す写真である。図9cは、MTTアッセイ法による細胞生存率解析の結果を示すグラフである。図9dは、センスまたはアンチセンスのオリゴヌクレオチドで処理した細胞の、蛍光活性化細胞選別(FACS)解析の結果を示す。
【図10】対応する非癌性肝組織と比較した、HCCにおけるLGN遺伝子の発現を示す。図10aは、20例の原発性HCCにおいてcDNAマイクロアレイにより調べたLGNの相対発現比(癌/非癌性)を示す。図10bは、別の10例のHCCを用いて半定量的RT-PCRで解析した、LGNの発現を示す写真である。GAPDHの発現を内部対照として用いた。T=腫瘍組織;N=正常組織。
【図11】LGNのゲノム構造を示す。
【図12】LGNの細胞内局在を示す。図12aは、pcDNA3.1-myc/His-LGNでトランスフェクトし、核をDAPIで対比染色したCOS7細胞の写真である。図12bは、pcDNA3.1-myc/His-LGNでトランスフェクトし、マウス抗c-myc抗体で染色し、ローダミンを結合した抗マウスIgG二次抗体で可視化したCOS7細胞の写真である。図12cはaとbの合成写真である。
【図13】LGNの成長促進作用を示す。図13aは、LGNがNIH3T3細胞、SNU423細胞、Alexander細胞、およびSNU475細胞の成長を促進することを示す、コロニー形成アッセイ法の結果を示す写真である。図13bは、外因性LGNを安定に発現しているNIH3T3-LGN細胞の成長が、10%FBSを含む培地中における擬似(NIH3T3-LacZ)細胞の成長を上回ることを示すグラフである。
【図14】ヒト肝癌SNU423細胞において、LGN発現を抑制するように指定したアンチセンスS-オリゴヌクレオチドによる成長抑制を示す。図14aは、アンチセンスS-オリゴヌクレオチド、アンチセンス3のトランスフェクションによる、LGNの発現低下を示す写真である。図14bは、アンチセンス3がSNU423細胞の成長を抑制したことを示す写真である。
【技術分野】
【0001】
本発明は、肝細胞癌で発現が上昇する遺伝子、これらの遺伝子によりコードされるポリペプチド、および肝細胞癌を検出する方法に関する。
【背景技術】
【0002】
cDNAマイクロアレイ技術により、正常細胞と悪性細胞における遺伝子発現の包括的なプロファイルが得られるようになった(Perou, C. M.ら、Nature. 406:747-752、2000(非特許文献1);Clark, E. A.ら、Nature. 406:532-535、2000(非特許文献2);Okabe, H.ら、Cancer Res. 61:2129-2137、2001(非特許文献3))。この方法は、癌細胞の複雑な性質を明らかにしており、発癌の理解を進める一助となっている。腫瘍において調節が解除された遺伝子の同定が、個々の癌のさらに正確かつ精密な診断、および新たな治療標的の開発につながる可能性がある(Golub, T. R.ら、Science 286:531-537、1999(非特許文献4))。
【0003】
肝細胞癌(HCC)は、全世界での癌による死亡原因の第1位である。治療法が最近進歩しているにもかかわらず、進行期のHCC患者の予後は依然として悪い。分子的研究でTP53遺伝子、CTNNB1遺伝子、および/またはAXIN1遺伝子の変化が肝癌発生に関与する可能性があることが報告されているが(Perou, C. M.ら、Nature. 406:747-752、2000(非特許文献5);Satoh, S.ら、Nat Genet. 24:245-250、2000(非特許文献6))、このような変化は一部のHCCにしか関連していないようである。したがって、癌の治療の新たな診断マーカーおよび/または薬剤標的となりうる究極の遺伝子が求められている。
【0004】
本発明者らは過去に、新たな遺伝子VANGL1をHCCのゲノム全体にわたる解析により同定したことを報告した(Yagyu, R.ら、International Journal of Oncology 20:1173-1178、2002(非特許文献7))。
【非特許文献1】Perou, C. M.ら、Nature. 406:747-752、2000
【非特許文献2】Clark, E. A.ら、Nature. 406:532-535、2000
【非特許文献3】Okabe, H.ら、Cancer Res. 61:2129-2137、2001
【非特許文献4】Golub, T. R.ら、Science 286:531-537、1999
【非特許文献5】Perou, C. M.ら、Nature. 406:747-752、2000
【非特許文献6】Satoh, S.ら、Nat Genet. 24:245-250、2000
【非特許文献7】Yagyu, R.ら、International Journal of Oncology 20:1173-1178、2002
【発明の開示】
【0005】
本発明の目的は、肝細胞癌で発現が上昇する遺伝子、この遺伝子によりコードされるポリペプチド、および肝細胞癌を検出する方法を提供することである。
【0006】
本発明者らは、23,040遺伝子に相当するcDNAマイクロアレイを用いてHCCの発現プロファイルの解析を行った。この解析により、対応する非癌性肝組織と比較して、癌組織において高頻度で発現が上昇していると考えられる、69のESTを含む165の遺伝子が特定された。本発明者らは、発現がHCCで高頻度で上昇している転写物から3つの遺伝子を単離した。これらの遺伝子は、センタウリン(centaurin)ファミリーのタンパク質と構造的特徴を共有する産物をコードしている。
【0007】
3つの遺伝子の1つは、UniGeneクラスターのEST(Hs.44579)に対応し、染色体バンド1p36.13で過剰に発現されている新たな遺伝子であることが判明した。この遺伝子のオープンリーディングフレームは、発生・分化促進因子2(DDEF2)と約60%同一なタンパク質をコードしていたため、本発明者らはこの遺伝子を発生・分化促進様因子1(DDEFL1)と命名した。
【0008】
HCCで発現が上昇している別の遺伝子は、UniGeneクラスターのEST(Hs.122730)に対応する。この推定アミノ酸配列は、ショウジョウバエにおける細胞極性および細胞運命の決定に関与するstrabismus(Van Gogh)、およびVan Gogh Like 2(VANGL2)と40%および63%の同一性を有していた。したがって、この遺伝子をVan Gogh Like 1(VANGL1)と命名した。
【0009】
HCCで発現が上昇している別の遺伝子は、LGN(GenBankアクセッション番号U54999)であることが判明した。LGNタンパク質は、抑制性ヘテロ三量体Gタンパク質のαサブユニット(Gαi2)と相互作用する。
【0010】
DDEFL1またはLGNの遺伝子移入により、これら3つのいずれかの遺伝子の内因性発現を欠く細胞の増殖が促進された。また、これらの遺伝子に特異的なアンチセンスS-オリゴヌクレオチドをトランスフェクトしてDDEFL1、VANGL1、またはLGNの発現を低下させたところ、肝細胞癌細胞の成長が阻害された。
【0011】
この結果は、HCCの機構を明らかにするために、かつHCCの診断および治療の新たな戦略を開発するために寄与すると考えられる。
【0012】
本発明は特に以下を提供する:
(1)以下からなる群より選択される、単離された核酸:
(a)配列番号:1または配列番号:3のヌクレオチド配列を含む核酸;
(b)配列番号:2または配列番号:4のアミノ酸配列を含むポリペプチドをコードする核酸;
(c)配列番号:1もしくは配列番号:3、またはその相補物からなるヌクレオチド配列と高ストリンジェントな条件下でハイブリダイズする鎖を含む核酸、
(2)以下からなる群より選択される、単離されたポリペプチド:
(a)配列番号:1または配列番号:3のヌクレオチド配列によりコードされるポリペプチド;
(b)配列番号:2または配列番号:4のアミノ酸配列を含むポリペプチド;
(c)配列番号:2または配列番号:4と少なくとも65%の同一性を有するポリペプチド、
(3)(1)の核酸を有するベクター、
(4)(1)の核酸または(3)のベクターを有する形質転換体、
(5)(4)の形質転換体を培養物中で培養する段階、形質転換体中でポリペプチドを発現させる段階、およびポリペプチドを培養物から回収する段階を含む、ポリペプチドを作製する方法、
(6)(2)のポリペプチドに特異的に結合する抗体、
(7)以下の段階を含む、肝細胞癌を検出する方法:
(a)被験者から生物試料を調製する段階;
(b)配列番号:1のポリペプチド、配列番号:3のポリペプチド、および配列番号:5のポリペプチドからなる群より選択される、少なくとも1つのポリペプチドの発現レベルを測定する段階;
(c)この発現レベルを、非癌性試料で測定された発現レベルと比較する段階;ならびに
(d)被験者における癌の有無を判定する段階、
(8)配列番号:1、配列番号:3、もしくは配列番号:5、またはその相補物からなるヌクレオチド配列と、高ストリンジェントな条件下でハイブリダイズする鎖を含む核酸を含む、肝細胞癌を検出するための試薬、
(9)(6)の抗体を含む、肝細胞癌を検出するための試薬、ならびに
(10)配列番号:1、配列番号:3、または配列番号:5のヌクレオチド配列とハイブリダイズする、少なくとも1つのアンチセンスオリゴヌクレオチドを肝細胞癌に導入する段階を含む、肝細胞癌の成長を阻害する方法。
【0013】
本発明を以下でさらに詳細に説明する。
【0014】
核酸
本発明は、肝細胞癌で発現が上昇する遺伝子を提供する。
【0015】
DDEFL1のヌクレオチド配列およびアミノ酸配列をそれぞれ配列番号:1および配列番号:2に示す。DDEFL1の完全なcDNAは、903アミノ酸のタンパク質をコードする2712ヌクレオチドのオープンリーディングフレームを含む、4050ヌクレオチドからなる(GenBankアクセッション番号AB051853)。DDFEL1のアミノ酸配列はDDEFL2と60%の同一性を、DDEF/ASAP1と46%の同一性を示し、かつArf GTPアーゼ活性化タンパク質(ArfGAP)ドメイン、および2つのアンキリン反復配列を含んでいる。
【0016】
DDEFL1は、アクチン細胞骨格の再編成を調節するタンパク質であるセンタウリンファミリーのメンバーであるDDEF2と60%の同一性を示した。これは、DDEFL1も細胞構造の編成に重要な役割を果たす可能性を示唆している(Randazzo, P. A.ら、The Arf GTPase-activating protein ASAP1 regulates the actin cytoskeleton、Proc. Natl. Acad. Sci. U S A 97:4011-4016、2000)。DDEFL1は、PHドメインおよびArfGAPモチーフも保存しているため、GAP活性によってArf低分子GTPアーゼを調節する、センタウリンファミリーの新たなメンバーであると考えられる。GEFのDblファミリーに属する分子の多くに認められるPHドメインは、特定の標的分子と相互作用することにより、および/または触媒ドメインを直接調節することにより、タンパク質の再配置に重要な役割を果たすと考えられている(Jackson, T. R.ら、Trends Biochem Sci. 25:489-495、2000;Cerione, R. A.、およびZheng, Y.、Curr. Opin. Cell. Biol. 8:216-222、1996;Chardin, P.ら、Nature 384:481-484、1996)。DDEF2は、周辺の接着斑(focal adhesion)に局在するが、本発明者らは、myc標識DDEFL1タンパク質が細胞内に拡散していることを発見した。
【0017】
Arfタンパク質は、小胞の膜輸送、小胞体区画およびゴルジ区画の完全性の維持、ならびに周辺の細胞骨格の調節などの重要な細胞過程に関係している(Cukierman, E.ら、Science 270:1999-2002、1995)。Arfファミリーの6つのメンバー(Arf1〜Arf6)、およびこれらの機能は既に同定されている(Moss, J.およびVaughan, M.、J. Biol. Chem. 270:12327-12330、1995)。例えばArf6タンパク質は、接着斑の形態を変化させ、細胞の拡散を阻害する細胞骨格の調節因子とみなされており、DDEF2はArf1に対してGAP活性を示す。
【0018】
DDEFL1の過剰発現により、細胞の成長および生存が低血清条件下で促進された。これは、DDEFL1が貧栄養および低酸素の条件で癌細胞に成長上の利点をもたらす可能性があることを示唆する。HCCでDDEFL1の発現が高頻度で上昇することは、肝癌発生におけるこの遺伝子の重要性を強調している。
【0019】
VANGL1のヌクレオチド配列およびアミノ酸配列を、それぞれ配列番号:3および配列番号:4に示す。決定されたcDNA配列は、524アミノ酸のタンパク質をコードする1572ヌクレオチドのオープンリーディングフレームを含む、1879ヌクレオチドからなっていた(GenBankアクセッション番号AB057596)。
【0020】
strabismus(stbm)は、rough eyeの表現型をもつ変異ショウジョウバエの原因遺伝子として同定された(Wolff T.およびRubin G.M.、Development 125:1149-1159、1998)。この遺伝子は、ショウジョウバエの眼、脚、および剛毛の極性を維持するために、かつR3およびR4光受容器の細胞運命を決定するために必要である。stbmのマウス相同遺伝子Ltapは、神経管変異マウスLoop-tailにおいて変化しており、神経管欠損(NTD)のヒトモデルとなっている(Kibar Zら、Nat Genet. 28:251-255、2001)。したがってVANGL1は、細胞極性、細胞運命の決定、および/または組織の編成に重要な役割を果たす可能性がある。VANGL1の発現はHCCにおいて高頻度で上昇しており、この発現を抑制したところ、癌細胞の成長または生存率が有意に低下したことから、VANGL1は癌細胞に生存率の上昇および/または脱極性化した成長をもたらす可能性がある。
【0021】
LGNのヌクレオチド配列およびアミノ酸配列をそれぞれ配列番号:5および配列番号:6に示す。LGNのcDNAは2336ヌクレオチドを含み、677アミノ酸のペプチドをコードする。
【0022】
LGNタンパク質は、抑制性ヘテロ三量体Gタンパク質のαサブユニット(Gαi2)と相互作用するタンパク質として以前に報告された(Mochizuki, N.ら、Gene 181:39-43、1996)。Gαi2の活性化変異は、下垂体腫瘍および他の内分泌腫瘍において報告されている(Hermouet, S.ら、Proc. Natl. Acad. Sci. USA 88:10455-10459、1991;Pace, A. M.ら、Proc. Natl. Acad. Sci. USA. 88:7031-7035、1991;Lyons, J.ら、Science 249:655-659、1990)。しかし、腫瘍形成または発癌へのLGNの関与については未だ報告はない。コロニー形成アッセイ法により、LGNが発癌活性をもつ可能性が示唆された。LGNの発現の促進はGαi2を活性化し、肝癌発生における発癌性シグナルを介在しうる。
【0023】
本発明の核酸は、cDNA、ゲノムDNA、化学的に合成されたDNA、およびRNAを含む。本発明の核酸は1本鎖または2本鎖でありうる。
【0024】
本明細書で用いる「単離された核酸」という表現は、構造が任意の天然の核酸の構造と同一ではないか、または3つより多い別個の遺伝子にわたる天然のゲノム核酸の任意の断片と同一ではない構造をとる核酸を意味する。したがってこの表現は例えば、(a)生物のゲノム中に天然に存在する、天然のゲノムDNA分子の一部の配列を有するDNA;(b)得られる分子が任意の天然のベクターもしくはゲノムDNAと同一ではないように、ベクター中にまたは原核生物もしくは真核生物のゲノムDNA中に組み込まれる核酸;(c)cDNA、ゲノム断片、ポリメラーゼ連鎖反応(PCR)により作製された断片、または制限酵素切断断片などの分離した分子;ならびに(d)ハイブリッド遺伝子(融合タンパク質をコードする遺伝子)の一部である組換えヌクレオチド配列を含む。具体的には、様々な(i)DNA分子、(ii)トランスフェクトされた細胞、または(iii)細胞クローンの混合物中に存在するDNA分子(例えば、cDNAライブラリーまたはゲノムDNAライブラリーなどのDNAライブラリー中に存在するDNA分子)の核酸は、この定義から除外される。
【0025】
1つの態様では、本発明の核酸はDDEFL1またはVANGL1のヌクレオチド配列(特に配列番号:1または配列番号:3)を含む核酸を含む。
【0026】
別の態様では、本発明の核酸は、DDEFL1またはVANGL1のアミノ酸配列(特に配列番号:2または配列番号:4)を含むポリペプチドをコードする核酸を含む。したがって、遺伝コードの縮重に基づく任意の配列を含む核酸が含まれる。
【0027】
さらに別の態様では、本発明の核酸は、配列番号:1または配列番号:3のバリアント核酸を含む。このバリアントは、配列番号:1もしくは配列番号:3、またはその相補物からなるヌクレオチド配列と、高ストリンジェントな条件下でハイブリダイズする鎖を含む核酸を含む。
【0028】
本明細書で用いる「相補物」という表現は、全ての塩基がもう一方の鎖の塩基の配列と塩基対を形成することができる、2本鎖核酸の一方の鎖を意味する。また「相補的である」という表現は、少なくとも15残基の連続ヌクレオチドの連続領域内で完全にマッチするものだけでなく、この領域内で少なくとも65%、好ましくは70%、より好ましくは80%、さらにより好ましくは90%、また最も好ましくは95%またはそれ以上の同一性を有するものも意味する。
【0029】
本明細書で用いる、2つの核酸の「同一性%」は、KarlinおよびAltschul(Proc. Natl. Acad. Sci. USA 90:5873-5877、1993)のように改変された、KarlinおよびAltschulのアルゴリズム(Proc. Natl. Acad. Sci. USA 87:2264-2268、1990)を用いて決定される。このようなアルゴリズムは、Altschulら(J. Mol. Biol. 215:403-410、1990)のNBLASTプログラムおよびXBLASTプログラムに組み込まれている。BLASTによるヌクレオチド検索はNBLASTプログラムを用いて実施される(スコア=100、ワード長=12)。タンパク質の相同性検索は、例えば日本のDNAデータバンク(DDBJ)において、FASTAプログラムやBLASTプログラムなどを用いて容易に実施することができる。BLASTによるタンパク質検索はXBLASTプログラムを用いて実施される(スコア=50、ワード長=3)。2つの配列間にギャップが存在する場合は、Gapped BLASTをAltsuchlら(Nucleic Acids Res. 25:3389-3402、1997)に記載されたように用いる。BLASTプログラムおよびGapped BLASTプログラムを使用する場合は、個々のプログラム(例えばXBLASTおよびNBLAST)のデフォルトのパラメータを用いる。
【0030】
好ましくは、バリアントは配列番号:1または配列番号:3のヌクレオチド配列と少なくとも65%同一なヌクレオチド配列を含む。より好ましくは、バリアントは配列番号:1または配列番号:3のヌクレオチド配列と少なくとも65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%、またはそれ以上同一である。標準配列(例えば配列番号:1または配列番号:3)より長いかまたは同じ長さのバリアントの場合、比較は全長の標準配列に対して行われる。バリアントが標準配列より短い場合(例えば配列番号:1または配列番号:3より短い場合)、比較は同じ長さの標準配列のセグメント(相同性の計算に必要な任意のループを除く)に対して行われる。
【0031】
ハイブリダイゼーションのストリンジェンシーは、以下の条件下での平衡ハイブリダイゼーションとして定義される:42℃、2×SSC、0.1%SDS(低ストリンジェンシー);50℃、2×SSC、0.1%SDS(中ストリンジェンシー);および65℃、2×SSC、0.1%SDS(高ストリンジェンシー)。平衡に達するために洗浄が必要な場合は、所望の特定のストリンジェンシーのハイブリダイゼーション溶液で洗浄を行う。一般に、温度が高くなるほど、平衡時にハイブリダイズする2本鎖の間の相同性は高くなる。
【0032】
本発明の核酸の長さに制限はないが、好ましくは少なくとも15ヌクレオチド、20ヌクレオチド、30ヌクレオチド、40ヌクレオチド、50ヌクレオチド、100ヌクレオチド、150ヌクレオチド、200ヌクレオチド、300ヌクレオチド、400ヌクレオチド、500ヌクレオチド、1000ヌクレオチド、1500ヌクレオチド、2000ヌクレオチド、2500ヌクレオチド、または3000ヌクレオチドを含む。
【0033】
本発明の核酸は、配列番号:1もしくは配列番号:3またはその相補物のヌクレオチド配列と特異的にハイブリダイズするプローブまたはプライマーとして使用される、ポリヌクレオチドを含む。「特異的にハイブリダイズする」という表現は、通常のハイブリダイゼーション条件(好ましくはストリンジェントな条件)で、配列番号:1または配列番号:3のヌクレオチド配列とハイブリダイズするが、他のポリペプチドをコードするDNAとクロスハイブリダイズしないことを意味する。
【0034】
プライマーおよびプローブは、配列番号:1もしくは配列番号:3のヌクレオチド配列内に少なくとも15の連続ヌクレオチドを含むか、またはその配列に相補的である。一般にプライマーは、15〜100ヌクレオチド、好ましくは15〜35ヌクレオチドを含み、プローブは、配列番号:1もしくは配列番号:3の全配列の少なくとも一部または全体を含む、少なくとも15ヌクレオチド、好ましくは少なくとも30ヌクレオチドを含む。プライマーは、本発明のポリペプチドをコードする核酸の増幅に使用することができ、プローブは、本発明のポリペプチドをコードする核酸の単離または検出に使用することができる。本発明のプライマーおよびプローブは、例えば市販のオリゴヌクレオチド合成装置で調製することができる。プローブは、制限酵素処理などにより得られる2本鎖DNA断片として調製することもできる。
【0035】
本発明の核酸は、配列番号:1または配列番号:3のヌクレオチド配列内の任意の部位とハイブリダイズするアンチセンスオリゴヌクレオチドを含む。本明細書で用いる「アンチセンスオリゴヌクレオチド」という表現は、DNAまたはmRNAの特定の領域を構成する部分に対応するヌクレオチド全体が相補的なオリゴヌクレオチドだけでなく、DNAまたはmRNAおよびオリゴヌクレオチドが配列番号:1または配列番号:3のヌクレオチド配列と特異的にハイブリダイズすることが可能な限りは、1個または複数のヌクレオチドのミスマッチを有するオリゴヌクレオチドも意味する。
【0036】
アンチセンスオリゴヌクレオチドは好ましくは、配列番号:1または配列番号:3のヌクレオチド配列中の、少なくとも15の連続ヌクレオチドに対するものである。上述の少なくとも15の連続ヌクレオチド中に開始コドンを含む上述のアンチセンスオリゴヌクレオチドがさらにより好ましい。
【0037】
本発明のアンチセンスオリゴヌクレオチドは、低級アルキルホスホネート(例えばメチルホスホネートまたはエチルホスホネート)、ホスホチオエート、およびホスホアミデートを含む類似体を含む。
【0038】
本発明のアンチセンスオリゴヌクレオチドは、本発明のポリペプチドをコードするDNAまたはmRNAに結合することによって、本発明のポリペプチドを産生する細胞に作用し、その転写または翻訳を阻害し、mRNAの分解を促進し、本発明のポリペプチドの発現を阻害する。
【0039】
本発明の核酸は以下のように調製することができる。本発明のポリペプチドをコードするcDNAは、例えば本発明のポリペプチドをコードするDNAのヌクレオチド情報(例えば配列番号:1または配列番号:3)を基にプライマーを作製し、プラークPCRを行うことにより調製することができる(Affara NAら (1994)Genomics 22、205-210)。ゲノムDNAは、例えば市販の「QiagenゲノムDNAキット」(Qiagen、ヒルデン、ドイツ)を用いる方法で調製することができる。得られたDNAのヌクレオチド配列は、例えば市販の「ダイターミネーターシーケンシングキット」(Applied Biosystems)を用いて、当技術分野での通常の方法で決定することができる。本発明の核酸は、後述するように、組換えタンパク質の作製および肝細胞癌の検出に使用することができる。
【0040】
ベクター、形質転換体、および組換えポリペプチドの作製
本発明は、本発明の核酸が挿入されたベクターも特徴とする。
【0041】
本発明のベクターは、本発明の組換えポリペプチドを作製するためのベクターを含む。本発明のポリペプチドの発現を可能とする限りは、任意のベクターを使用することができる。
【0042】
発現ベクターの例には、細菌(例えば大腸菌(Escherichia coli))発現ベクター、酵母発現ベクター、昆虫発現ベクター、および哺乳類発現ベクターなどがある。本発明では、pcDNA 3.1-myc/HisまたはpcDNA 3.1ベクター(Invitrogen)などの哺乳類発現ベクターを使用することができる。本発明の核酸のベクターへの挿入は、当技術分野での通常の方法で実施することができる。
【0043】
本発明のベクターは、本発明のポリペプチドを(特に遺伝子治療を目的として)インビボで発現させるためのベクターも含む。本発明のポリペプチドをインビボで発現可能な限りは、様々なウイルスベクターおよび非ウイルスベクターを使用することができる。ウイルスベクターの例には、アデノウイルスベクターやレトロウイルスベクターなどがある。陽イオンリポソームは非ウイルスベクターの一例でありうる。
【0044】
本発明は、本発明の核酸を発現可能な状態で有する形質転換体も提供する。本発明の形質転換体は、本発明の核酸が挿入された上述の発現ベクターを有する形質転換体、および本発明の核酸が組み込まれた宿主ゲノムを有する形質転換体を含む。本発明の核酸は、形質転換体が核酸を発現できる限りは、形質転換体内に任意の状態で保持される。
【0045】
ベクターが細胞内で機能して本発明の核酸を発現する限りは、ベクターが挿入される細胞に関して特別の制限はない。例えば大腸菌(E. coli)、酵母、哺乳類細胞、および昆虫細胞を宿主として使用することができる。好ましくはCOS7細胞やNIH3T3細胞などの哺乳類細胞を使用する。ベクターの細胞への導入は、エレクトロポレーションおよびリン酸カルシウム法などの既知の方法を用いて行うことができる。
【0046】
当技術分野で用いられる一般的な方法を、形質転換体から対象組換えポリペプチドを単離および精製する際に使用することができる。例えば、形質転換体を回収して抽出物を得た後に、イオン交換クロマトグラフィー、逆相クロマトグラフィー、ゲル濾過、もしくは本発明のポリペプチドに対する抗体がカラムに固定されたアフィニティクロマトグラフィーにより、またはこのようなカラムをいくつか組み合わせることにより、目的のポリペプチドを精製および調製することができる。
【0047】
また本発明のポリペプチドをグルタチオン-S-トランスフェラーゼタンパク質との融合タンパク質として、または複数のヒスチジンを追加した組換えポリペプチドとして、宿主細胞(例えば動物細胞や大腸菌(E. coli))内で発現させる場合には、発現された組換えポリペプチドを、グルタチオンカラムまたはニッケルカラムを用いて精製することができる。融合タンパク質を精製後、トロンビンまたは第Xa因子で切断することにより、目的のポリペプチド以外の領域を除くことも可能である。
【0048】
ポリペプチド
本発明は、DDEFL1またはVANGL1(例えば配列番号:1または配列番号:3)によりコードされる単離されたポリペプチドを提供する。特定の態様では、本発明のポリペプチドは、配列番号:1または配列番号:3のヌクレオチド配列によりコードされるポリペプチド、ならびに配列番号:2または配列番号:4のアミノ酸配列を含むポリペプチドを含む。
【0049】
本明細書で用いる「単離されたポリペプチド」は、実質的に純粋で他の生体高分子を含まないポリペプチドを意味する。実質的に純粋なポリペプチドは、乾燥重量で少なくとも75%(例えば少なくとも80%、85%、95%、または99%)純粋である。純度は、任意の適切な標準的な方法(例えばカラムクロマトグラフィー、ポリアクリルアミドゲル電気泳動、またはHPLC解析)で測定することができる。
【0050】
本発明のポリペプチドは、バリアントが配列番号:2または配列番号:4と少なくとも65%同一である限りは、配列番号:2または配列番号:4のバリアントを含む。バリアントは、1つまたは複数のアミノ酸が置換、欠失、付加、および/または挿入されている、配列番号:2または配列番号:4のアミノ酸配列を含むポリペプチドでありうる。バリアントはまた、配列番号:1または配列番号:3からなるヌクレオチド配列と高ストリンジェントな条件下でハイブリダイズする鎖を含む核酸によりコードされるポリペプチドでありうる。
【0051】
あるアミノ酸配列の1つまたは複数のアミノ酸残基が欠失、付加、および/または置換により改変されたアミノ酸配列を有するポリペプチドは、当初の生物学的活性を保持することが知られている(Mark, D. F.ら、Proc. Natl. Acad. Sci. USA(1984)81、5662-5666、Zoller, M. J.およびSmith, M.、Nucleic Acids Research(1982)10、6487-6500、Wang, A.ら、Science 224、1431-1433、Dalbadie-McFarland, G.ら、Proc. Natl. Acad. Sci. USA(1982)79、6409-6413)。
【0052】
置換、欠失、付加、および/または挿入によって変異が導入されるアミノ酸の数には特に制限はない。通常、全アミノ酸残基の10%またはそれ未満、好ましくは5%またはそれ未満、より好ましくは1%またはそれ未満である。
【0053】
変異が導入されるアミノ酸残基に関しては、アミノ酸の側鎖の性質が保存されている、異なるアミノ酸に変化させることが好ましい。アミノ酸側鎖の性質の例は、疎水性アミノ酸(A、I、L、M、F、P、W、Y、V)、親水性アミノ酸(R、D、N、C、E、Q、G、H、K、S、T)、ならびに以下の側鎖を含むアミノ酸である:脂肪族側鎖(G、A、V、L、I、P);ヒドロキシル基を含む側鎖(S、T、Y);硫黄原子を含む側鎖(C、M);カルボン酸およびアミドを含む側鎖(D、N、E、Q);塩基を含む側鎖(R、K、H);ならびに芳香族官能基を含む側鎖(H、F、Y、W)(括弧内の文字は、アミノ酸の1文字コードを意味する)。「保存的アミノ酸置換」は、上記グループの1つに属する1つのアミノ酸と、同じグループの別のアミノ酸との置換である。
【0054】
欠失バリアントは、配列番号:1または配列番号:3のアミノ酸配列の断片を含む。このような断片は、本発明の上記のポリペプチドと(全体的ではなく)部分的に同一なアミノ酸配列を有するポリペプチドである。本発明のポリペプチド断片は通常、8アミノ酸残基またはそれ以上、好ましくは12アミノ酸残基またはそれ以上(例えば15アミノ酸残基またはそれ以上)を含む。好ましい断片の例は、アミノ末端またはカルボキシル末端のいずれかを含むひと続きのアミノ酸残基、または2つの連続するアミノ酸残基(一方はアミノ末端を含み、もう一方はカルボキシル末端を含む)を欠くアミノ酸配列を有する、切断ポリペプチドを含む。さらに、αヘリックスおよびαヘリックス形成領域、βシートおよびβシート形成領域、ターンおよびターン形成領域、コイルおよびコイル形成領域、親水性領域、疎水性領域、α両親媒性領域、β両親媒性領域、可変領域、表面形成領域、基質結合領域、および高抗原性指標領域を有する断片を含む、構造的または機能的な特徴をもつ断片も好ましい。生物学的な活性を有する断片もまた好ましい。類似の活性もしくは改良された活性を有する、または望ましくない活性を低下させた、生物学的な活性を有する断片は、本発明のポリペプチドの活性を介在する。例えば、リガンドの結合を介してシグナルを細胞に伝達する活性を有する断片、および動物(特にヒト)における抗原性または免疫原性を有する断片が含まれる。これらのポリペプチド断片は好ましくは、本発明のポリペプチドの抗原性を有する。
【0055】
また別のバリアントは、本発明のポリペプチドと別のペプチドまたはポリペプチドとの融合タンパク質を含む。融合タンパク質は、本発明のポリペプチドをコードするDNAを、他のペプチドまたはポリペプチドをコードするDNAに、フレームが一致するように連結させ、これを発現ベクターに挿入し、宿主内で発現させるような、当業者に周知の技術で作製することができる。本発明のポリペプチドに融合させるペプチドまたはポリペプチドに関して制限はない。
【0056】
既知のペプチド、例えばFLAG(Hopp, T.P.ら、Biotechnology(1988)6、1204-1210)、6個のHis(ヒスチジン)残基を含む6×His、10×His、インフルエンザ凝集素(HA)、ヒトc-myc断片、VSP-GP断片、p18 HIV断片、T7タグ、HSVタグ、Eタグ、SV40のT抗原断片、lckタグ、α-チューブリン断片、Bタグ、タンパク質C断片などを、本発明のポリペプチドに融合させるペプチドとして使用することができる。本発明のポリペプチドに融合させるポリペプチドの例には、GST(グルタチオン-S-トランスフェラーゼ)、インフルエンザ凝集素(HA)、免疫グロブリンの定常領域、β-ガラクトシダーゼ、MBP(マルトース結合タンパク質)などがある。
【0057】
融合タンパク質は、これらのペプチドまたはポリペプチドをコードする市販のDNAを、本発明のポリペプチドをコードするDNAと融合し、調製した融合DNAを発現させることにより、作製することができる。
【0058】
バリアントポリペプチドは、配列番号:2または配列番号:4のアミノ酸配列と好ましくは少なくとも65%同一である。具体的には、改変ポリペプチドは、配列番号:2または配列番号:4のアミノ酸配列と少なくとも65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%、またはそれ以上同一である。改変ポリペプチドが、標準配列(例えば配列番号:2または配列番号:4)より長いかまたは同じ長さの場合は、比較は全長の標準配列に対して行われる。改変ポリペプチドが、標準配列より短い(例えば配列番号:2または配列番号:4より短い)場合は、比較は同じ長さの標準配列のセグメントに対して行われる。
【0059】
本明細書で用いる、2つのアミノ酸配列の「同一性%」は、核酸に関する上記の手順と同様に決定される。
【0060】
本発明のポリペプチドは、天然のポリペプチド、または上述の遺伝子工学技術を用いて作製された組換えポリペプチドとして、当業者に公知の方法で調製することができる。例えば、天然のポリペプチドは、小動物を組換えポリペプチドで免疫化することにより得られる抗体を結合させたカラムを調製し、本発明のポリペプチドを高レベルで発現する、肝臓組織または細胞の抽出物に対してアフィニティクロマトグラフィーを行うことによって得られる。組換えポリペプチドは、本発明のポリペプチドをコードするDNA(例えば配列番号:1または配列番号:3のヌクレオチド配列を含むDNA)を適切な発現ベクターに挿入し、このベクターを宿主細胞に導入し、結果として得られる形質転換体にポリペプチドを発現させ、発現されたポリペプチドを回収することにより調製することができる。
【0061】
バリアントポリペプチドは例えば、PCRによる部位特異的変異導入システム(GIBCO-BRL、ゲイサーズバーグ、メリーランド)や、オリゴヌクレオチドによる部位特異的変異誘発法(Kramer, W.およびFritz, HJ(1987)Methods in Enzymol. 154:350-367)などの公知の方法により、配列番号:1または配列番号:3のアミノ酸配列に変異を挿入することで調製することができる。
【0062】
抗体
本発明は、本発明のポリペプチドに特異的に結合する抗体も特徴とする。本発明の抗体の形状に関して特に制限はなく、ポリクローナル抗体およびモノクローナル抗体が含まれる。ウサギなどの動物を本発明のポリペプチドで免疫化することによって得られる抗血清、あらゆるクラスのポリクローナル抗体およびモノクローナル抗体、遺伝子工学により作製されたヒト化抗体、ヒト抗体も含まれる。
【0063】
ポリクローナル抗体は、本発明のポリペプチドで免疫化したウサギなどの小動物の血清を得て、本発明のポリペプチドのみを認識する画分を、本発明のポリペプチドを結合させたアフィニティカラムを用いて入手し、免疫グロブリンGまたはMを、この画分からプロテインGまたはプロテインAカラムを用いて精製することにより、作製することができる。
【0064】
モノクローナル抗体は、マウスなどの小動物を本発明のポリペプチドで免疫化し、この動物から脾臓を切除し、細胞へと組織をホモジナイズし、ポリエチレングリコールなどの試薬を用いて細胞をマウスのミエローマ細胞と融合し、本発明のポリペプチドに対する抗体を産生するクローンを融合細胞(ハイブリドーマ)から選択し、得られたハイブリドーマをマウスの腹腔に移植し、腹水を抽出することで作製することができる。得られたモノクローナル抗体は例えば、硫酸アンモニウム沈殿法、プロテインAもしくはプロテインGカラム、DEAEイオン交換クロマトグラフィー、または本発明のポリペプチドを結合させたアフィニティカラムで精製することができる。本発明の抗体は、本発明のポリペプチドの精製および検出に使用することができる。特に本発明の抗体は、肝細胞癌を検出するために使用することができる。
【0065】
ヒト抗体またはヒト化抗体は、当業者に公知の方法で調製することができる。例えば、ヒト抗体は、免疫系をヒトの免疫系に変化させたマウスを本発明のポリペプチドで免疫化することにより作製することができる。またヒト化抗体は、例えば、モノクローナル抗体産生細胞から抗体遺伝子をクローニングし、遺伝子の抗原認識部位を公知のヒト抗体に移すCDRグラフト法を用いて調製することができる。
【0066】
検出法
本発明はさらに、DDEFL1、VANGL1、またはLGNのポリペプチドをマーカーとして用いて肝細胞癌を検出する方法を提供する。
【0067】
検出は、被験者由来の生物試料中のDDEFL1、VANGL1、およびLGNのポリペプチドの少なくとも1つの発現レベルを測定し、非癌性試料における発現レベルと比較し、被験者における癌の有無を判定することにより行うことができる。
【0068】
本発明で用いる生物試料は、肝細胞癌の検出を必要とする被験者から得られる任意の肝臓組織または細胞を含む。特に、肝生検標本を使用することができる。このような生物試料は、肝臓組織または細胞から調製されたmRNA試料、cRNA試料、またはcDNA試料も含む。mRNA試料およびcDNA試料は従来の方法で調製することができる。cRNAは、テンプレートのcDNAからRNAポリメラーゼを用いて転写されたRNAを意味する。cRNAは、T7 RNAポリメラーゼを用いて、テンプレートとしてT7プロモーターを結合したcDNAから合成することができる。DNAチップベースの発現プロファイリング用の市販のcRNA転写キットを使用することができる。
【0069】
特定の態様では、DDEFL1、VANGL1、またはLGNのポリペプチドの発現レベルは、RNA、cDNA、またはポリペプチドのレベルで測定することができる。
【0070】
mRNAの発現レベルは、例えばDDEFL1、VANGL1、またはLGNのヌクレオチド配列とハイブリダイズするプローブを用いるノーザンブロッティング法、DDEFL1、VANGL1、またはLGNのヌクレオチド配列とハイブリダイズするプライマーを用いるRT-PCR法などで測定することができる。
【0071】
本発明の検出法で使用されるプローブまたはプライマーは、配列番号:1、配列番号:3、もしくは配列番号:5のヌクレオチド配列、またはその相補物と特異的にハイブリダイズする核酸を含む。「特異的にハイブリダイズする」という表現は、通常のハイブリダイゼーション条件(好ましくはストリンジェントな条件)下で、配列番号:1、配列番号:3、または配列番号:5のヌクレオチド配列とハイブリダイズするが、他のポリペプチドをコードするDNAとはクロスハイブリダイズしないことを意味する。
【0072】
プライマーおよびプローブは、配列番号:1、配列番号:3、または配列番号:5のヌクレオチド配列内に、またはこの配列に相補的な配列内にある少なくとも15の連続ヌクレオチドを含む。一般にプライマーは15〜100ヌクレオチド、また好ましくは15〜35ヌクレオチドを含み、プローブは、配列番号:1、配列番号:3、または配列番号:5の配列の少なくとも一部または全体を含む、少なくとも15ヌクレオチド、好ましくは少なくとも30ヌクレオチドを含む。プライマーおよびプローブは、例えば市販のオリゴヌクレオチド合成装置で作製することができる。プローブは、制限酵素処理などで得られる2本鎖DNA断片として調製することもできる。
【0073】
cDNAの発現レベルは、例えばDNAアレイを用いる方法で測定することができる(村松正実および山本雅、新遺伝子工学ハンドブック、pp.280〜284、羊土社)。具体的にはまず、被験者から調製されたcDNA試料、およびDDEFL1、VANGL1、またはLGNのヌクレオチド配列とハイブリダイズするポリヌクレオチドプローブが固定された固相支持体が提供される。プローブとして、上述のプローブを使用することができる。複数の種類の標的ポリヌクレオチドを検出するために、複数の種類のプローブを固相支持体に固定することができる。cDNA試料を必要に応じて検出用に標識する。標識は、検出可能である限りは特に制限されず、例えば蛍光標識や放射性標識などを含む。標識は従来の方法で実施することができる(L. Luoら、「Gene expression profiles of laser-captured adjacent neuronal subtypes」、 Nat. Med.(1999)pp.117-122)。
【0074】
次にcDNA試料を固相支持体上のプローブと接触させて、cDNA試料をプローブとハイブリダイズさせる。ハイブリダイゼーションの反応溶液および反応条件は、プローブ長などの様々な要素に依存して変動するが、当業者に周知の通常の方法に従って決定することができる。
【0075】
固相支持体上におけるcDNA試料とプローブ間のハイブリダイゼーションの強度は、cDNA試料の標識の種類に応じて測定される。例えば蛍光標識は、蛍光シグナルをスキャナーで読み取ることで検出可能である。
【0076】
被験cDNA試料および対照cDNA試料(例えば非癌性の組織または細胞に由来するcDNA)のハイブリダイゼーションの強度を、これらを異なる蛍光標識で標識することにより1回の測定で同時に測定することができる。例えば上述のcDNA試料の1つをCy5で、もう1つをCy3で標識することができる。Cy5およびCy3の蛍光シグナルの強度は、各cDNA試料の発現レベルを示す(Dugganら、Nat. Genet. 21:10〜14、1999)。
【0077】
この方法で、cDNAに代えてcRNAを測定することができる。
【0078】
さらに、ポリペプチドの発現レベルは、DDEFL1、VANGL1、またはLGNのポリペプチドに対する抗体を用いて、例えばSDSポリアクリルアミド電気泳動、ウエスタンブロッティング、ドットブロッティング、免疫沈降法、蛍光免疫アッセイ法、放射免疫アッセイ法、酵素免疫アッセイ法(例えば酵素結合免疫吸着アッセイ法(ELISA))、ならびに免疫組織化学染色などの免疫アッセイ法により測定することができる。
【0079】
特定の態様では、固相支持体上に固定されたDDEFL1、VANGL1、またはLGNのポリペプチドに対する抗体に生物試料を接触させ、固相支持体上の抗体-抗原複合体を、検出可能な標識で標識した二次抗体に接触させ、この標識を適切な方法で検出する。
【0080】
本発明の検出法で使用される抗体には、ウサギなどの動物をDDEFL1、VANGL1、またはLGNのポリペプチドで免疫化することにより得られる抗血清、あらゆるクラスのポリクローナル抗体およびモノクローナル抗体、遺伝子工学により作製されたヒト化抗体、ならびにヒト抗体を含む、DDEFL1、VANGL1、またはLGNのポリペプチド(特に配列番号:2、配列番号:4、または配列番号:6のアミノ酸配列を有するポリペプチド)に結合する任意の抗体が含まれる。これらの抗体は、上述のように調製することができる。
【0081】
上述のように測定された発現レベルを、非癌性試料で測定された発現レベルと比較して、被験者における肝細胞癌の有無を判定する。被験者由来の試料で測定された発現レベルが、非癌性試料で測定された発現レベルより高い場合は、被験者は癌であるか、または癌の危険性があると判断される。一方、被験者の試料における発現レベルが、非癌性試料における発現レベルより高くない場合は、被験者は癌ではないと判断される。具体的には、被験者の試料中の発現レベルが、非癌性試料における発現レベルと比べて高いか否かは、相対発現比(被験者試料/非癌性試料)を基に判定することができる。発現レベルは、相対発現比が2.0を上回る場合に高いと判定される。
【0082】
検出試薬
本発明は、肝細胞癌の検出試薬を提供する。
【0083】
ある態様では、本発明の検出試薬は、DDEFL1、VANGL1、またはLGN(具体的には配列番号:1、配列番号:3、または配列番号:5)とハイブリダイズする、少なくとも15ヌクレオチドを有するポリヌクレオチドを含む。このポリヌクレオチドを、本発明の上述の検出法においてプローブまたはプライマーとして使用することができる。プローブとして使用する場合は、本発明の検出試薬に含まれるポリヌクレオチドを標識することができる。標識法には例えば、T4ポリヌクレオチドキナーゼを用いてポリヌクレオチドの5'末端を32Pでリン酸化する標識法や、ランダムヘキサマーオリゴヌクレオチドなどをプライマーとして、およびクレノー酵素などのDNAポリメラーゼを用いて、32Pなどの同位元素、蛍光色素、ビオチンなどで標識された基質塩基を導入する方法(ランダムプライム法など)がある。
【0084】
別の態様では、本発明の検出試薬は、DDEFL1、VANGL1、またはLGNのポリペプチド(具体的には特に配列番号:2、配列番号:4、または配列番号:6のアミノ酸配列を有するポリペプチド)に結合する抗体を含む。これらの抗体を用いて、本発明のポリペプチドを上述の本発明の検出法で検出する。抗体は検出法に従って標識することができる。また、抗体を固相支持体上に固定することができる。
【0085】
本発明の検出試薬はさらに、滅菌水、生理食塩水、植物油、界面活性剤、脂質、可溶化剤、緩衝液、タンパク質安定剤(ウシ血清アルブミンやゼラチンなど)、保存剤などを含む溶媒または添加物を、これらが本発明の検出法で用いられる反応に影響を及ぼさない限り、含むことができる。
【0086】
肝細胞癌の成長を阻害する方法
本発明はさらに、肝細胞癌の成長を阻害する方法を提供する。特定の態様では、この方法は、DDEFL1、VANGL1、またはLGNのアンチセンスオリゴヌクレオチドを標的細胞に導入することにより実施することができる。
【0087】
この方法で使用されるアンチセンスオリゴヌクレオチドは、配列番号:1、配列番号:3、または配列番号:5のヌクレオチド配列内の任意の部位とハイブリダイズする。このアンチセンスオリゴヌクレオチドは、DNAまたはmRNA、およびオリゴヌクレオチドが、配列番号:1、配列番号:3、または配列番号:5のヌクレオチド配列と特異的にハイブリダイズすることができる限りは、DNAまたはmRNAの特定の領域を構成する部分に対応するヌクレオチド全体が相補的なオリゴヌクレオチドだけでなく、1つまたは複数ヌクレオチドのミスマッチを有するオリゴヌクレオチドも含む。
【0088】
このようなアンチセンスオリゴヌクレオチドは好ましくは、配列番号:1、配列番号:3、または配列番号:5のヌクレオチド配列中の少なくとも15の連続ヌクレオチドに対するオリゴヌクレオチドである。上述の少なくとも15の連続ヌクレオチド中に開始コドンを含む、上述のアンチセンスオリゴヌクレオチドが、さらにより好ましい。
【0089】
アンチセンスオリゴヌクレオチドは、低級アルキルホスホネート(例えばメチルホスホネートまたはエチルホスホネート)、ホスホチオエート、およびホスホアミデートを含む類似体を含む。
【0090】
本明細書では、標的細胞は哺乳類細胞、好ましくはヒト細胞でありうる。
【0091】
導入法は、インビトロ、インビボ、またはエキソビボの移入法でありうる。ある態様では、アンチセンスオリゴヌクレオチドを、従来のトランスフェクション法で標的細胞に導入することができる。または、導入はアデノウイルスベクター、レトロウイルスベクター、または陽イオンリポソームなどのアンチセンスオリゴヌクレオチドを有するベクターを用いる、従来の遺伝子移入技術により行うことができる。
【0092】
本明細書で引用されるあらゆる特許、特許出願、および刊行物は参照として組み入れられる。
【0093】
発明を実施するための最良の形態
本発明を、以下の実施例を参照して説明するが、本発明がそれらに制限されるとは解釈されない。
【0094】
実施例1
1-1. ヒトの肝細胞癌において発現が一般的に上昇するDDEFL1の同定
23040個の遺伝子を含むゲノム全体にわたるcDNAマイクロアレイによって、20例の肝細胞癌(HCC)の発現プロファイルを、対応する非癌性肝組織と比較した。全てのHCCの組織および対応する非癌性組織は、肝切除を受け、同意を得た患者の外科標本から入手した。UniGeneクラスターのEST(Hs.44579)に対応する施設内アクセッション番号B9362の遺伝子は、1.57〜5.83の範囲で過剰に発現されることが見出された(図1a)。この発現上昇(Cy3:Cy5強度比が2.0を上回る)は、カットオフフィルターを通過した(Cy3およびCy5の両シグナルが25,000を上回る)12例のHCCのうち11例で認められた。この遺伝子のオープンリーディングフレームは、発生・分化促進因子2(DDEF2)と約60%同一なタンパク質をコードしていたため、この遺伝子を「発生・分化促進因子様1」(DDEFL1)と命名した。cDNAマイクロアレイの結果を明らかにするために、別の11例のHCCにおいて、この転写物の発現を半定量的RT-PCRで調べた。GAPDHの発現を内部対照として用いた。RT-PCRは以下のように実施した。全RNAをQiagen RNeasyキット(Qiagen)、またはTrizol試薬(Life Technologies社)を用いて製造業者のプロトコールにしたがって抽出した。10 μgの全RNAを、ポリdT12-18プライマー(Amersham Pharmacia Biotech)を用いてSuperscript II逆転写酵素(Life Technologies)で逆転写して1本鎖のcDNAを得た。1本鎖cDNA調製物を、20 μlのPCR緩衝液(TAKARA)中で行う標準的なRT-PCR実験による、次のPCR増幅用に希釈した。増幅は、94℃で4分間処理して変性した後に、94℃で30秒間、56℃で30秒間、および72℃で45秒間を20サイクル(GAPDHの場合)または33サイクル(DDEFL1の場合)、GeneAmp PCR system 9700(Perkin-Elmer、フォスターシティ、カリフォルニア)で行った。プライマーの配列は以下であった;GAPDH用:フォワード、5'-ACAACAGCCTCAAGATCATCAG (配列番号: 7) およびリバース、5'-GGTCCACCACTGACACGTTG (配列番号: 8); DDEFL1用:フォワード、5'-AGCTGAGACATTTGTTCTCTTG (配列番号: 9)およびリバース、5'-TATAAACCAGCTGAGTCCAGAG (配列番号: 10)。結果から、DDEFL1の発現がこれらの腫瘍のうち9例で上昇することが確認された(図1b)。
【0095】
1-2. 新たな遺伝子DDEFL1の単離および構造
DDEFL1の発現を、DDEFL1のPCR産物をプローブとして用いた多組織ノーザンブロット解析で解析した。ヒト多組織ブロット(Clontech、パロアルト、カリフォルニア)を、32P標識DDEFL1 cDNAとハイブリダイズさせた。プレハイブリダイゼーション、ハイブリダイゼーション、および洗浄は、製造業者の推奨にしたがって行った。このブロットのオートラジオグラフを増感スクリーンを用いて、-80℃で72時間かけて得た。この結果、4 kbの転写物が、肺、肝臓、小腸、胎盤、および末梢血白血球で発現していることが判明した(図2a)。
【0096】
B9362は、ノーザンブロットで検出されたものより小さかったため、5'RACE実験を行って、この遺伝子のコード配列全体を決定した。5'RACE実験は、Marathon cDNA増幅キット(Clontech、パロアルト、カリフォルニア)を製造業者の指示書通りに用いて実施した。DDEFL1 cDNAの5'側部分の増幅には、遺伝子に特異的なリバースプライマー(5'-CTCACTTGGCACGTCAGCAGGG (配列番号: 11))およびキットに同梱されているAP-1プライマーを使用した。cDNAテンプレートは、ヒト肝臓のmRNAから合成した。PCR産物をTAクローニングキット(Invitrogen)を用いてクローニングし、配列をABI PRISM 3700 DNAシーケンサー(Applied Biosystems)で決定した。
【0097】
完全なcDNAは、903アミノ酸のタンパク質をコードする2712ヌクレオチドのオープンリーディングフレームを有する、4050ヌクレオチドからなる(GenBankアクセッション番号AB051853)。最初のATGは、真核生物における翻訳開始のコンセンサス配列と一致する配列(CCCGCCATGC(配列番号:12))に隣接しており、上流にインフレームの停止コドンをもっていた。NCBI(米国バイオテクノロジー情報センター)データベース上の、相同性探索用のBLASTプログラムによって、染色体バンド1p36.12に位置するGenBankアクセッション番号AL357134のゲノム配列が同定された。cDNA配列とゲノム配列の比較の結果、DDEFL1が25個のエキソンからなることが判明した(図2b)。
【0098】
Simple Modular Architecture Research Tool(SMART)によるタンパク質モチーフの探索により、推定タンパク質が2つのコイルドコイル領域(コドン141〜172および241〜278)、PH(プレックストリン(Pleckstrin)相同性)モチーフ(コドン303〜396)、ArfGAPのモチーフ(ArfのGTPアーゼ活性化タンパク質)(コドン426〜551)、ならびに2つのアンキリン反復配列(コドン585〜617および621〜653)を含むことが判明した。この構造は、センタウリンβ1およびβ2(図2c)と似ていた。特にDDEFL1は、ホスファチジルイノシトール3,4,5-三リン酸の標的であるPHドメイン、およびArfGAPのモチーフなどの、センタウリンファミリーのタンパク質の特徴を有していた。DDEFL1のArfGAPモチーフのアミノ酸配列は、DDEF2のアミノ酸配列と67.8%同一であった(図2d)。特に、GAP活性に不可欠なジンクフィンガー構造を示すCXXCX16CXXCモチーフは完全に保存されていた。
【0099】
1-3. DDEFL1の細胞内局在
DDEFL1のコード配列をpcDNA3.1-myc/Hisベクター(Invitrogen)にクローニングした。myc標識DDEFL1タンパク質を発現する、得られたプラスミド(pDNA-myc/His-DDEFL1)を、COS7細胞(米国菌株保存機関(ATCC))に一過性にトランスフェクトした。推定myc標識タンパク質を、以下に示す免疫ブロッティング(ウエスタンブロッティング)で検出した。pcDNA3.1-myc/His-DDEFL1でトランスフェクトした細胞をPBSで2回洗浄し、溶解緩衝液(150 mM NaCl、1%トリトン X-100、50 mM Tris-HCl pH 7.4、1 mM DTT、および1×コンプリートプロテアーゼインヒビターカクテル(Boehringer))中に回収した。細胞をホモジナイズして10,000×gで30分間遠心した後に、上清をタンパク質濃度に関してBradfordアッセイ法(Bio-Rad)で標準化した。タンパク質を10%SDS-PAGEで分離し、マウス抗myc抗体で免疫ブロットを行った。HRP結合ヤギ抗マウスIgG(Amersham)をECL検出システム(Amersham)用の二次抗体として用いた。結果的に、DDEFL1タンパク質が抗myc抗体を用いたウエスタンブロットで検出された(図3a)。
【0100】
また、免疫細胞化学染色を以下のように行った。最初に細胞を4%パラホルムアルデヒドを含むPBSで15分間固定し、次に0.1%トリトン X-100を含むPBSで室温で2.5分間処理して透過性にした。次に、細胞を2%BSA(PBS中)で覆い、24時間4℃で処理して、非特異的なハイブリダイゼーションをブロックした。マウス抗mycモノクローナル抗体(Sigma)を1:1000の希釈率で、またはマウス抗FLAG抗体(Sigma)を1:2000の希釈率で一次抗体用に使用し、ローダミンを結合した抗マウス二次抗体(Leinco and ICN)とインキュベートした後、反応を可視化した。核を4',6'-ジアミン-2'-フェニルインドールジヒドロクロライド(DAPI)で対比染色した。蛍光像をECLIPSE E800顕微鏡下で得た。顕微鏡解析により、タンパク質が主に細胞質に存在することが示された(図3b)。DDEFL1もヒト胚腎(HEK293)細胞(ATCC)の細胞質に局在していた。
【0101】
1-4. DDEFL1が細胞成長に及ぼす作用
DDEFL1のコード配列をpcDNA 3.1ベクター(Invitrogen)にクローニングした。10 cmのディッシュに播いたNIH3T3細胞(ATCC)(2×105細胞/ディッシュ)に、DDEFL1を発現する得られたプラスミド(pcDNA-DDEFL1)、および対照プラスミド(pcDNA-LacZ)でトランスフェクトし、10%ウシ胎児血清および1%抗生物質/抗真菌剤溶液(Sigma)を添加したダルベッコ改変イーグル培地に、さらに適切な濃度のジェネティシンを添加した培地中で2週間培養した。次にこの細胞を100%メタノールで固定し、ギムザ溶液で染色した。pcDNA-DDEFL1でトランスフェクトした細胞は、対照細胞より著しく多数のコロニーを生じた。コロニー形成の増加は、DDEFL1の内因性発現が極めて低い、ヒト肝癌SNU423(韓国細胞株バンク)細胞、およびAlexander(ATCC)細胞をトランスフェクトした場合に同様に認められた(図4a)。
【0102】
この成長促進作用を詳細に調べるために、外因性DDEFL1を安定に発現するNIH3T3細胞を確立した。pDNA-myc/His-DDEFL1を、FuGENE6試薬(Boehringer)を製造業者の推奨通りに用いてNIH3T3細胞にトランスフェクトした。トランスフェクションから24時間後に、培養物にジェネティシンを添加し、トランスフェクションから2週間後に単一コロニーを選択した。DDEFL1の発現を、半定量的RT-PCRで決定した(図4b)。NIH3T3-DDEFL1細胞の成長速度は、擬似(NIH3T3-LacZ)細胞の成長速度より、10%FBSを含む培地中において統計的に速かった(P<0.05)(図4c)。0.1%FBSのみを含む培地中では、NIH3T3-DDEFL1細胞は6日間生存したが、対照NIH3T3細胞は同条件下で6日以内に死滅した。この場合、NIH3T3-DDEFL1細胞の成長は、0.1%FBSを含む培地における擬似処理細胞の成長より統計的に速かった(P<0.01)(図4d)。
【0103】
1-5. ヒト肝癌SNU475細胞におけるアンチセンスS-オリゴヌクレオチドによるDDEFL1発現の抑制
DDEFL1遺伝子に対応する、以下の6組の対照(センス)およびアンチセンスのS-オリゴヌクレオチドを合成した。
アンチセンス:
DDEFL1-AS1 5'-TGCTCCGGCATGGCGG-3' (配列番号: 13);
DDEFL1-AS2 5'-GCTGAACTGCTCCGGC-3' (配列番号: 14);
DDEFL1-AS3 5'-TCCAAGATCTCCTCCC-3' (配列番号: 15);
DDEFL1-AS4 5'-TCTCCTTCCAAGATCT-3' (配列番号: 16);
DDEFL1-AS5 5'-GCGCTGAGCCGGCCTC-3' (配列番号: 17);および
DDEFL1-AS6 5'-CCTCACCTCCTCCCGC-3' (配列番号: 18)
対照:
DDEFL1-S1 5'-CCGCCATGCCGGAGCA-3' (配列番号: 19);
DDEFL1-S2 5'-GCCGGAGCAGTTCAGC-3' (配列番号: 20);
DDEFL1-S3 5'-GGGAGGAGATCTTGGA-3' (配列番号: 21);
DDEFL1-S4 5'-AGATCTTGGAAGGAGA-3' (配列番号: 22);
DDEFL1-S5 5'-GAGGCCGGCTCAGCGC-3' (配列番号: 23); および
DDEFL1-S6 5'-GCGGGAGGAGGTGAGG-3' (配列番号: 24)
【0104】
LIPOFECTIN試薬(GIBCO BRL)を用いて、DDEFL1の発現レベルが、本発明者らが調べた6つの肝癌細胞株中で最高であった(データは示していない)、SNU475細胞(韓国細胞株バンク)に、合成S-オリゴヌクレオチドをトランスフェクトした。トランスフェクションから12時間および24時間後に、アンチセンスS-オリゴヌクレオチドAS1およびAS5は、DDEFL1の発現を各対照S-オリゴヌクレオチドS1およびS5と比較して有意に抑制した(図5a)。アンチセンスS-オリゴヌクレオチドAS1またはAS5によりトランスフェクトされた生存細胞の数は、トランスフェクションから6日後の時点で、各対照S-オリゴヌクレオチドS1またはS5によりトランスフェクトされた細胞と比較して著しく少なかった(図5b)。3回の独立した実験で一貫した結果が得られた。
【0105】
実施例2
2-1. ヒト肝細胞癌で一般的に発現が上昇しているVANGL1の同定
実施例1で実施したゲノム全体にわたるcDNAマイクロアレイ解析の結果では、UniGeneクラスターのEST(Hs.122730)に対応する施設内アクセッション番号D3244の遺伝子が、12例の臨床HCCのうち10例で、対応する非癌性肝組織と比較して有意に発現が上昇していることも判明した。これら12例の腫瘍の、対応する非癌性組織と比較した相対発現比の範囲は1.5〜16.0であった(図6a)。発現の上昇(Cy3:Cy5強度比が2.0を上回る)は、カットオフフィルターを通過した(Cy3およびCy5の両シグナルが25,000を上回る)12例のHCCのうち10例で認められた。D3244の発現上昇は、別の10例のHCC症例において、プライマー対、フォワード: 5'- GAGTTGTATTATGAAGAGGCCGA (配列番号: 25); リバース: 5'- ATGTCTCAGACTGTAAGCGAAGG (配列番号: 26)を用いて、実施例1-1と同様に実施した半定量的RT-PCRにより確認された。(図6b)
【0106】
2-2. ヒト成体組織におけるVANGL1の発現
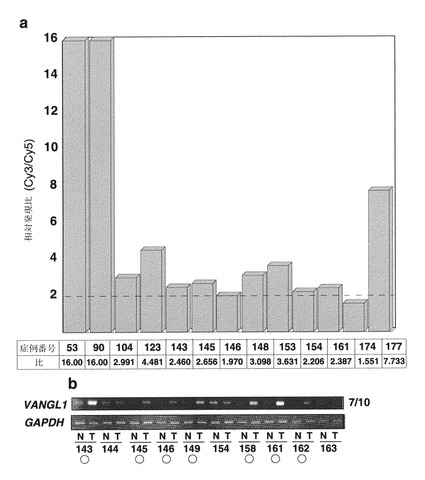
D3244 cDNAをプローブとして用いた多組織ノーザンブロット解析を、実施例1-2と同様の手順で行い、その結果、1.9 kbの転写物が、精巣および卵巣で組織特異的に多量に発現されることが判明した(図7a)。D3244に対応するゲノム配列のNCBIデータベースの探索により、2つの配列(GenBankアクセッション番号:AL450389およびAL592436)が染色体バンド1p22に位置することが判明した。GENSCAN、および「Gene Recognition and Assembly Internet Link」プログラムを用いて候補エキソン配列を推定し、エキソンの連結を行った。またリバースプライマー(5'-TGTCAGCTCTCCGCTTGCGGAAAAAAAG (配列番号: 27))を用いて5'RACEを行い、転写物の5'領域の配列を実施例1-2と同様の手順で決定した。その結果、1572ヌクレオチドのオープンリーディングフレームを含む、1879ヌクレオチドのヒトcDNA配列がアセンブルして得られた(GenBankアクセッション番号:AB057596)。この推定アミノ酸配列は、strabismus(Van Gogh)と40%、VANGL2と63%同一であった。したがってD3244に対応する遺伝子をVan Gogh Like 1(VANGL1)と命名した。Simple Modular Architecture Research Toolにより、推定タンパク質が4つの推定膜貫通ドメイン(コドン111〜133、148〜170、182〜204、219〜241)を含むことが示唆された(図7b)。
【0107】
2-3. VANGL1の細胞内局在
c-myc標識VANGL1タンパク質を発現するpcDNA3.1-myc/His-VANGL1プラスミドを、SNU475細胞(韓国細胞株バンク)に一過性にトランスフェクトした。免疫組織化学染色を実施例1-3と同様の手順で行った。この結果から、標識VANGL1タンパク質が細胞質に存在することが判明した(図8aおよび8b)。
【0108】
2-4. VANGL1の発現を低下させるように指定したアンチセンスS-オリゴヌクレオチドによる肝癌細胞の成長抑制
VANGL1の抑制がHCC細胞の細胞周期の停止および/または細胞死をもたらすかどうかを試験するために、以下の4組のアンチセンスおよび対照(センス)のS-オリゴヌクレオチドを合成し、SNU475細胞にトランスフェクトした。
アンチセンス:
アンチセンス1 5'-GTATCCATAGCAATGG-3' (配列番号: 28);
アンチセンス2 5'-TGGATTGGGTATCCAT-3' (配列番号: 29);
アンチセンス3 5'-TAAGTGGATTGGGTAT-3' (配列番号: 30); および
アンチセンス4 5'-ACTCCTACCTGCCTGT-3' (配列番号: 31)
対照:
センス1 5'-CCATTGCTATGGATAC-3' (配列番号: 32);
センス2 5'-ATGGATACCCAATCCA-3' (配列番号: 33);
センス3 5'-ATACCCAATCCACTTA-3' (配列番号: 34); および
センス4 5'-ACAGGCAGGTAGGAGT-3' (配列番号: 35)
【0109】
開始コドンを含むアンチセンスS-オリゴヌクレオチド(アンチセンス3)は、SNU475細胞におけるVANGL1の内因性発現を有意に低下させた(図9a)。
【0110】
細胞の生存は、3-(4,5-ジメチル-チアゾール-2-イル)-2,5-ジフェニルテトラゾリウムブロマイド(MTT)アッセイ法により、以下のように評価した。細胞を5×105細胞/100 mm ディッシュの密度で播いた。播種してから24時間後に、細胞を、センスまたはVANGL1を抑制するように指定したアンチセンスのS-オリゴヌクレオチドにより3回トランスフェクトした。トランスフェクションから72時間後に、培地を500 μg/mlの3-(4,5-ジメチルチアゾール-2-イル)-2,5-ジフェニルテトラゾリウムブロマイド(MTT)(Sigma)を含む新鮮な培地と交換し、プレートを37℃で4時間インキュベートした。続いて、1 mlの0.01 N HCl/10%SDSを添加して細胞を溶解し、溶解液の吸光度をELISAプレートリーダーで測定した(試験波長=570 nm、標準波長=630 nm)。細胞生存率を、対照細胞と比較した吸光度で表した。
【0111】
アンチセンスS-オリゴヌクレオチド、アンチセンス3のトランスフェクションにより、対照のセンスS-オリゴヌクレオチド、センス3の使用時と比較して、生存細胞数が有意に減少した(図9bおよび9c)。この結果は、3回の独立した実験で確認された。
【0112】
さらにフローサイトメトリー解析を以下のように行った。細胞を、1×105細胞/100 mm ディッシュの密度で播き、任意の経過時間後にトリプシン処理し、続いて70%冷エタノール中で固定した。RNA分解酵素処理後に、細胞をヨウ化プロピジウム(50 μg/ml、PBS中)で染色した。フローサイトメトリーをBecton Dickinson FACScanで行い、CellQuestおよびModFitソフトウェア(Verity Software House)で解析した。細胞周期のG0/G1期、S期、およびG2/M期にある核の割合、および任意のサブG1集団を、少なくとも20,000個の非同期細胞を対象に決定した。
【0113】
FACS解析の結果、VANGL1を阻害すると、サブG1期の細胞数が有意に増加することが判明した(図9d)。これらの結果は、VANGL1が、肝細胞癌細胞の成長および/または生存に重要な役割を果たす可能性があることを示唆する。
【0114】
実施例3
3-1. LGNは一般的にヒトの肝細胞癌で増加している
実施例1-1で実施したマイクロアレイ解析によって明らかになった、一般的に発現が上昇している遺伝子の中で、LGN(GenBankアクセッション番号:U54999)に対応する遺伝子D3636を選択した。というのはD3636の発現が、対応する非癌性肝組織と比較して、12例の臨床HCCのうち10例で有意に上昇していたためである。これら12例の腫瘍の、対応する非癌性組織と比較時の相対発現比は0.7〜16.0であった。LGNの発現の上昇(Cy3:Cy5強度比が2.0を上回る)は、カットオフフィルターを通過した(Cy3およびCy5の両シグナルが25,000を上回る)12例のHCCのうち10例で認められた(図10a)。LGNの発現の上昇は、別の10例のHCCでも、プライマー対、フォワード: 5'-ATCTGAAGCACTTAGCAATTGC (配列番号: 36)、リバース: 5'-CTGTAGCTCAGACCAAGAACC (配列番号: 37)を用いて、実施例1-1と同様に実施した半定量的RT-PCRにより確認された(図10b)。
【0115】
3-2. LGNのゲノム構造
LGNのcDNAは2,336ヌクレオチドからなり、677アミノ酸のペプチドをコードする。cDNA配列とゲノム配列との比較により、LGN遺伝子が14個のエキソンからなることが判明した(図11)。
【0116】
3-3. LGNの細胞内局在
c-myc標識LGNタンパク質を発現するpcDNA3.1-myc/His-LGNプラスミドをCOS7細胞に一過性にトランスフェクトした。myc標識LGNタンパク質に対応する72 kDaのバンドが、実施例1-3と同様の手順で行った免疫ブロット解析により検出された(図12)。同様に、実施例1-3のように免疫細胞化学染色を行い、その結果、標識LGNタンパク質が、細胞の細胞質および核に存在することが判明した。
【0117】
3-4. LGN遺伝子の移入は細胞成長を促進することができる
LGNが細胞成長に及ぼす作用を解析するために、実施例1-4のように、NIH3T3細胞、SNU423細胞、Alexander細胞、およびSNU475細胞を、LGNを発現するプラスミド(pcDNA3.1-myc/His-LGN)によりトランスフェクトして、コロニー形成アッセイ法を実施した。対照プラスミド(pcDNA3.1-myc/His-LacZ)と比較して、pcDNA3.1-myc/His-LGNは、これらの細胞で著しく多数のコロニーを生じた(図13a)。この結果は3回の独立した実験で確認された。
【0118】
LGNが細胞成長に及ぼす作用をさらに調べるために、外因性LGNを安定に発現するNIH3T3細胞(NIH3T3-LGN細胞)を確立した。NIH3T3-LGN細胞は、対照のNIH3T3-LacZ細胞より速い成長速度を示した(図13b)。
【0119】
3-5. LGNのアンチセンスS-オリゴヌクレオチドはヒト肝癌SNU475細胞の成長を抑制した
LGNに対応する、以下の5組の対照(センス)およびアンチセンスのS-オリゴヌクレオチドを合成してSNU423細胞にトランスフェクトした。
アンチセンス:
アンチセンス1 5'-CCATCGAGTCATATTA-3'(配列番号: 38);
アンチセンス2 5'-TTCCTCCATCGAGTCA-3'(配列番号: 39);
アンチセンス3 5'-AAATTTTCCTCCATCG-3'(配列番号: 40);
アンチセンス4 5'-AGTCTTACCTGTAACG-3'(配列番号: 41); および
アンチセンス5 5'-GCTTCCATTCTACAAA-3'(配列番号: 42)
センス:
センス1 5'-TAATATGACTCGATGG-3'(配列番号: 43);
センス2 5'-TGACTCGATGGAGGAA-3'(配列番号: 44);
センス3 5'-CGATGGAGGAAAATTT-3'(配列番号: 45);
センス4 5'-CGTTACAGGTAAGACT-3'(配列番号: 46); および
センス5 5'-TTTGTAGAATGGAAGC-3'(配列番号: 47)
【0120】
開始コドンを含むアンチセンスS-オリゴヌクレオチド(アンチセンス3)は、トランスフェクションから12時間後の対照S-オリゴヌクレオチド(センス3)と比較して、LGNの発現を有意に抑制した(図14a)。トランスフェクションから6日後に、アンチセンス3でトランスフェクトした生存細胞数は、対照のセンス3でトランスフェクトしたものより著しく少なかった(図14b)。3回の独立した実験で一貫した結果が得られた。
【0121】
産業上の利用可能性
本発明は、肝細胞癌で一般に発現が上昇することが見出されたDDEFL1、VANGL1、またはLGNの、cDNAヌクレオチド配列およびポリペプチドのアミノ酸配列を提供する。したがって、これらのポリペプチドは、肝臓癌の有無を判定するためのマーカーとして使用することができる。これらのヌクレオチド配列の情報により、DDEFL1、VANGL1、またはLGNの遺伝子を検出または増幅するためのプローブおよびプライマーを設計することができる。またDDEFL1、VANGL1、またはLGNのポリペプチドの発現を阻害するアンチセンスヌクレオチド配列を合成することも可能となる。アミノ酸配列情報を用いることで、DDEFL1、VANGL1、またはLGNのポリペプチドに結合する抗体を調製することができる。このようなプローブおよびプライマーならびに抗体は、肝細胞癌を検出するための試薬として有用である。さらに本発明者らは、アンチセンスオリゴヌクレオチドでDDEFL1、VANGL1、またはLGNの発現を抑制すると、HCC細胞の成長が著しく低下することを立証した。したがって、アンチセンスオリゴヌクレオチドを用いてHCC細胞の成長を阻害することができる。本発明は、肝細胞の発癌機構をさらに明らかにし、肝臓癌を治療するための有効な薬剤の開発に向けた分子標的を発見することにも寄与する。
【図面の簡単な説明】
【0122】
【図1】B9362と命名された遺伝子の、HCCにおける発現を示す。図1aは、20例の原発性HCCにおいてcDNAマイクロアレイを用いて調べたB9362の相対発現比(癌/非癌性)を示す。図1bは、別の11例のHCCを半定量的RT-PCRで解析したB9362の発現を示す写真である。GAPDHの発現を内部対照として用いた。
【図2】DDEFL1の同定の結果を示す。図2aは、ヒトの様々な組織におけるDDEFL1のノーザンブロット解析の結果を示す写真である。図2bはDDEFL1の構造を示す。図2cは、推定DDEFL1タンパク質とArfGAPファミリーのメンバーとの類似性を示す。図2dは、DDEFL1とDDEF2におけるArfGAPモチーフのアミノ酸配列間の同一性を示す。矢印は、GAP活性に不可欠なジンクフィンガー構造を示す、CXXCX16CXXCモチーフを示す。
【図3】DDEFL1の細胞内局在を示す。図3aは、cMyc標識DDEFL1タンパク質が、pcDNA-DDEFL1-mycプラスミドをトランスフェクトしたCOS7細胞で発現していることを示す、ウエスタンブロット解析の結果を示す写真である。図3bは、cMyc標識DDEFL1タンパク質が細胞質に局在することを示唆する、細胞の免疫組織化学染色を示す写真である。
【図4】DDEFL1の成長促進作用を示す。図4aは、DDEFL1がNIH3T3細胞、SNU423細胞、およびAlexander細胞の成長を促進することを示す、コロニー形成アッセイの結果を示す写真である。図4bは、NIH3T3-DDEFL1細胞による外因性DDEFL1の安定な発現を示す写真である。図4cは、10%FBSを含む培地中で外因性DDEFL1を安定に発現している、NIH3T3-DDEFL1細胞の成長を示すグラフである。図4dは、0.1%FBSを含む培地中におけるNIH3T3-DDEFL1細胞の成長を示すグラフである。(P<0.01)
【図5】SNU475細胞でDDEFL1を抑制するように指定したアンチセンスS-オリゴヌクレオチドによる、成長抑制を示す。図5aは、アンチセンスS-オリゴヌクレオチドを明示し、AS1またはAS5アンチセンスS-オリゴヌクレオチドのトランスフェクションによってDDEFL1の発現が低下したことを示す写真である。図5bは、AS1およびAS5がSNU475細胞の成長を抑制したことを示す写真である。
【図6】HCCにおけるVANGL1の発現を示す。図6aは、cDNAマイクロアレイで調べた、20例の原発性HCCにおけるVANGL1の相対発現比(癌/非癌性)を示す。図6bは、別の10例のHCCを用いて半定量的RT-PCRにより解析した、D3244の発現を示す写真である。T=腫瘍組織;N=正常組織。GAPDHの発現を内部対照として用いた。
【図7】VANGL1の同定の結果を示す。図7aは、ヒトの様々な組織におけるVANGL1の多組織ノーザンブロット解析の結果を示す写真である。図7bはVANGL1の推定タンパク質の構造を示す。
【図8】VANGL1の細胞内局在を示す。図8aは、pcDNA3.1-myc/His-VANGL1でトランスフェクトしたSNU475細胞をマウス抗mycモノクローナル抗体で染色し、ローダミンを結合した抗マウスIgG二次抗体で可視化した写真である。核はDAPIで対比染色した。図8bは、同様に染色して可視化した擬似細胞の写真である。
【図9】VANGL1を抑制するように指定したアンチセンスS-オリゴヌクレオチドの成長抑制作用を示す。図9aは、対照またはアンチセンスオリゴヌクレオチドのいずれかで12時間処理したSNU475細胞におけるVANGL1の発現を示す写真である。図9bは、S-オリゴヌクレオチドがSNU423細胞の成長を抑制したことを示す写真である。図9cは、MTTアッセイ法による細胞生存率解析の結果を示すグラフである。図9dは、センスまたはアンチセンスのオリゴヌクレオチドで処理した細胞の、蛍光活性化細胞選別(FACS)解析の結果を示す。
【図10】対応する非癌性肝組織と比較した、HCCにおけるLGN遺伝子の発現を示す。図10aは、20例の原発性HCCにおいてcDNAマイクロアレイにより調べたLGNの相対発現比(癌/非癌性)を示す。図10bは、別の10例のHCCを用いて半定量的RT-PCRで解析した、LGNの発現を示す写真である。GAPDHの発現を内部対照として用いた。T=腫瘍組織;N=正常組織。
【図11】LGNのゲノム構造を示す。
【図12】LGNの細胞内局在を示す。図12aは、pcDNA3.1-myc/His-LGNでトランスフェクトし、核をDAPIで対比染色したCOS7細胞の写真である。図12bは、pcDNA3.1-myc/His-LGNでトランスフェクトし、マウス抗c-myc抗体で染色し、ローダミンを結合した抗マウスIgG二次抗体で可視化したCOS7細胞の写真である。図12cはaとbの合成写真である。
【図13】LGNの成長促進作用を示す。図13aは、LGNがNIH3T3細胞、SNU423細胞、Alexander細胞、およびSNU475細胞の成長を促進することを示す、コロニー形成アッセイ法の結果を示す写真である。図13bは、外因性LGNを安定に発現しているNIH3T3-LGN細胞の成長が、10%FBSを含む培地中における擬似(NIH3T3-LacZ)細胞の成長を上回ることを示すグラフである。
【図14】ヒト肝癌SNU423細胞において、LGN発現を抑制するように指定したアンチセンスS-オリゴヌクレオチドによる成長抑制を示す。図14aは、アンチセンスS-オリゴヌクレオチド、アンチセンス3のトランスフェクションによる、LGNの発現低下を示す写真である。図14bは、アンチセンス3がSNU423細胞の成長を抑制したことを示す写真である。
【特許請求の範囲】
【請求項1】
以下からなる群より選択される、単離された核酸:
(a)配列番号:1または配列番号:3のヌクレオチド配列を含む核酸;
(b)配列番号:2または配列番号:4のアミノ酸配列を含むポリペプチドをコードする核酸;
(c)配列番号:1もしくは配列番号:3またはその相補物からなるヌクレオチド配列と、高ストリンジェントな条件下でハイブリダイズする鎖を含む核酸。
【請求項2】
以下からなる群より選択される、単離されたポリペプチド:
(d)配列番号:1または配列番号:3のヌクレオチド配列によりコードされるポリペプチド;
(e)配列番号:2または配列番号:4のアミノ酸配列を含むポリペプチド;
(f)配列番号:2または配列番号:4と少なくとも65%の同一性を有するポリペプチド。
【請求項3】
請求項1記載の核酸を有するベクター。
【請求項4】
請求項1記載の核酸または請求項3記載のベクターを有する形質転換体。
【請求項5】
請求項4記載の形質転換体を培地中で培養する段階、該形質転換体においてポリペプチドを発現させる段階、および該ポリペプチドを培地から回収する段階を含む、ポリペプチドを作製する方法。
【請求項6】
請求項2のポリペプチドに特異的に結合する抗体。
【請求項7】
以下の段階を含む、肝細胞癌を検出する方法:
(a)被験者から生物試料を調製する段階;
(b)配列番号:1記載のポリペプチド、配列番号:3記載のポリペプチド、および配列番号:5記載のポリペプチドからなる群より選択される、少なくとも1つのポリペプチドの発現レベルを測定する段階;
(c)発現レベルを、非癌性試料における発現レベルと比較する段階;ならびに
(d)被験者における癌の有無を判定する段階。
【請求項8】
配列番号:1、配列番号:3、もしくは配列番号:5、またはその相補物からなるヌクレオチド配列と、高ストリンジェントな条件下でハイブリダイズする鎖を含む核酸を含む、肝細胞癌を検出するための試薬。
【請求項9】
請求項6記載の抗体を含む、肝細胞癌を検出するための試薬。
【請求項10】
配列番号:1、配列番号:3、または配列番号:5のヌクレオチド配列とハイブリダイズする、少なくとも1つのアンチセンスオリゴヌクレオチドを肝細胞癌に導入する段階を含む、肝細胞癌の成長を阻害する方法。
【請求項1】
以下からなる群より選択される、単離された核酸:
(a)配列番号:1または配列番号:3のヌクレオチド配列を含む核酸;
(b)配列番号:2または配列番号:4のアミノ酸配列を含むポリペプチドをコードする核酸;
(c)配列番号:1もしくは配列番号:3またはその相補物からなるヌクレオチド配列と、高ストリンジェントな条件下でハイブリダイズする鎖を含む核酸。
【請求項2】
以下からなる群より選択される、単離されたポリペプチド:
(d)配列番号:1または配列番号:3のヌクレオチド配列によりコードされるポリペプチド;
(e)配列番号:2または配列番号:4のアミノ酸配列を含むポリペプチド;
(f)配列番号:2または配列番号:4と少なくとも65%の同一性を有するポリペプチド。
【請求項3】
請求項1記載の核酸を有するベクター。
【請求項4】
請求項1記載の核酸または請求項3記載のベクターを有する形質転換体。
【請求項5】
請求項4記載の形質転換体を培地中で培養する段階、該形質転換体においてポリペプチドを発現させる段階、および該ポリペプチドを培地から回収する段階を含む、ポリペプチドを作製する方法。
【請求項6】
請求項2のポリペプチドに特異的に結合する抗体。
【請求項7】
以下の段階を含む、肝細胞癌を検出する方法:
(a)被験者から生物試料を調製する段階;
(b)配列番号:1記載のポリペプチド、配列番号:3記載のポリペプチド、および配列番号:5記載のポリペプチドからなる群より選択される、少なくとも1つのポリペプチドの発現レベルを測定する段階;
(c)発現レベルを、非癌性試料における発現レベルと比較する段階;ならびに
(d)被験者における癌の有無を判定する段階。
【請求項8】
配列番号:1、配列番号:3、もしくは配列番号:5、またはその相補物からなるヌクレオチド配列と、高ストリンジェントな条件下でハイブリダイズする鎖を含む核酸を含む、肝細胞癌を検出するための試薬。
【請求項9】
請求項6記載の抗体を含む、肝細胞癌を検出するための試薬。
【請求項10】
配列番号:1、配列番号:3、または配列番号:5のヌクレオチド配列とハイブリダイズする、少なくとも1つのアンチセンスオリゴヌクレオチドを肝細胞癌に導入する段階を含む、肝細胞癌の成長を阻害する方法。
【図1】


【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【公開番号】特開2008−301816(P2008−301816A)
【公開日】平成20年12月18日(2008.12.18)
【国際特許分類】
【出願番号】特願2008−131279(P2008−131279)
【出願日】平成20年5月19日(2008.5.19)
【分割の表示】特願2003−530887(P2003−530887)の分割
【原出願日】平成14年9月25日(2002.9.25)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 2002年8月25日 日本癌学会発行の「第61回 日本癌学会総会記事」に発表
【出願人】(502240113)オンコセラピー・サイエンス株式会社 (142)
【Fターム(参考)】
【公開日】平成20年12月18日(2008.12.18)
【国際特許分類】
【出願日】平成20年5月19日(2008.5.19)
【分割の表示】特願2003−530887(P2003−530887)の分割
【原出願日】平成14年9月25日(2002.9.25)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 2002年8月25日 日本癌学会発行の「第61回 日本癌学会総会記事」に発表
【出願人】(502240113)オンコセラピー・サイエンス株式会社 (142)
【Fターム(参考)】
[ Back to top ]