
肥満細胞の脱顆粒抑制剤
【課題】新たな作用機序に基づく肥満細胞の活性化の抑制手段、具体的には肥満細胞の脱顆粒抑制剤等を提供する。
【解決手段】STIM1阻害物質を有効成分として含有する肥満細胞の脱顆粒抑制剤、STIM1阻害物質を有効成分として含有するアレルギーの予防または治療剤、STIM1阻害物質を有効成分として含有するアナフィラキシーの予防または治療剤、STIM1阻害物質を有効成分として含有する肥満細胞の炎症性サイトカインの産生抑制剤、および被験物質がSTIM1遺伝子の発現または機能を抑制し得るか否かを評価することを含む、肥満細胞の脱顆粒を抑制し得る物質のスクリーニング方法。
【解決手段】STIM1阻害物質を有効成分として含有する肥満細胞の脱顆粒抑制剤、STIM1阻害物質を有効成分として含有するアレルギーの予防または治療剤、STIM1阻害物質を有効成分として含有するアナフィラキシーの予防または治療剤、STIM1阻害物質を有効成分として含有する肥満細胞の炎症性サイトカインの産生抑制剤、および被験物質がSTIM1遺伝子の発現または機能を抑制し得るか否かを評価することを含む、肥満細胞の脱顆粒を抑制し得る物質のスクリーニング方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、肥満細胞の脱顆粒抑制剤に関し、アレルギーの治療分野に属する。
【背景技術】
【0002】
肥満細胞はイムノグロブリンE(IgE)関連Th2ヘルパー細胞依存性の即時型過剰反応およびアレルギー疾患ならびにある種の自然免疫応答において、主要なエフェクター細胞である(非特許文献1〜3)。高親和性IgEレセプター(FcεRI)に結合したIgE分子への多価抗原の結合によって、肥満細胞上に発現するFcεRIが凝集し、かかる凝集が肥満細胞の活性化を開始する(非特許文献4、5)。活性化した肥満細胞は、プロテアーゼや、ヒスタミン等の血管作動性アミンを含む前もって作られた(細胞質内の顆粒に貯蔵されている)メディエーターを分泌する。また、肥満細胞の活性化は、前炎症性脂質メディエーターおよびサイトカインのde novo合成につながる。FcεRI凝集に応答したこのようなメディエーターの合成および放出は、多数のプロテインチロシンキナーゼカスケードを含むシグナル伝達経路を介して調節される。
【0003】
これらのカスケードの下流の重要な標的の1つは、ホスホリパーゼC−γ(PLCγ)である。PLCγは、ホスファチジルイノシトール−4,5−ビスホスフェートのジアシルグリセロールおよびイノシトール−1,4,5−トリホスフェート(IP3)への加水分解を触媒する。IP3は、小胞体(ER)膜のその受容体に結合し、それにより、Ca2+が迅速ではあるが一過性にERストアから放出される。カルシウム動員の第2ステージでは、形質膜(PM)を横切って細胞外Ca2+が持続的に流入する。この第2段階で、ERプール内のCa2+ストアが空になることそれ自体がPMのCa2+チャネルを活性化すると考えられている(いわゆるストア作動性Ca2+(SOC)流入メカニズム)(非特許文献6、7)。しかしながら、ストア枯渇に依存しない、PM中のCa2+チャネルを活性化する他のメカニズムも報告されている。
【0004】
最近の研究により、SOC流入メカニズムにおける主要なプレーヤーが明確にされてきた(非特許文献8〜11)。Ca2+流入のゲノムワイドRNAiスクリーニングにより、ER膜タンパク質STIM1が同定された。STIM1は、そのEFハンドCa2+結合モチーフにより、内腔のCa2+量の枯渇を検出するCa2+センサーである(非特許文献8、9)。ストア枯渇の際に、STIM1はPM下部の点状に移動し、その後PMのSOCチャネルが活性化される(非特許文献9、12、13)。
【0005】
肥満細胞活性化および脱顆粒におけるCa2+の重要性は、薬理学的試験に基づいて広く信じられている。EGTA処理は、FcεRIが介在するカルシウム動員および脱顆粒をほぼ完全に抑制した(非特許文献14、15)。しかし、最近の知見によると(非特許文献16〜19)、Lyn欠損肥満細胞はカルシウムの動員が減弱しているにも関らず、脱顆粒を誘導可能であり、カルシウム動員の第2ステージ(主としてカルシウム流入)が実際に肥満細胞の活性化および脱顆粒に必須であるのか否か不明である。さらに、もしそうだとしても、SOC依存性および非依存性経路の両方が共存していると思われるので、SOC経路が肥満細胞応答に主要な役割を果たしているのか未だに不明である。
【非特許文献1】Wedemeyer, J. et al. Curr Opin Immunol 12, 624-631 (2000)
【非特許文献2】Bischoff, S. C. Nat Rev Immunol 7, 93-104 (2007)
【非特許文献3】Galli, S. J. et al. Nat Immunol 6, 135-142 (2005)
【非特許文献4】Kraft, S. & Kinet, J. P. Nat Rev Immunol 7, 365-378 (2007)
【非特許文献5】Gilfillan, A. M. & Tkaczyk, C. Nat Rev Immunol 6, 218-230 (2006)
【非特許文献6】Venkatachalam, K. et al. Nat Cell Biol 4, E263-272 (2002)
【非特許文献7】Parekh, A. B. & Putney, J. W. Physiol Rev 85, 757-810 (2005)
【非特許文献8】Roos, J. et al. J Cell Biol 169, 435-445 (2005)
【非特許文献9】Liou, J. et al. Curr Biol 15, 1235-1241 (2005)
【非特許文献10】Putney, J. W., Jr. J Cell Sci 120, 1959-1965 (2007)
【非特許文献11】Hogan, P. G. & Rao, A. Trends Biochem Sci 32, 235-245 (2007)
【非特許文献12】Baba, Y. et al. Proc Natl Acad Sci USA 103, 16704-16709 (2006)
【非特許文献13】Luik, R. M. & Lewis, R. S. Trends Mol Med 13, 103-107 (2007)
【非特許文献14】Ozawa, K. et al. J Biol Chem 268, 1749-1756 (1993)
【非特許文献15】Nishida, K. et al. J Cell Biol 170, 115-126 (2005)
【非特許文献16】Odom, S. et al. J Exp Med 199, 1491-1502 (2004)
【非特許文献17】Parravicini, V. et al. Nat Immunol 3, 741-748 (2002)
【非特許文献18】Nishizumi, H. & Yamamoto, T. J Immunol 158, 2350-2355 (1997)
【非特許文献19】Hernandez-Hansen, V. et al. J Immunol 173, 100-112 (2004)
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明の目的は、新たな作用機序に基づく肥満細胞の活性化の抑制手段、具体的には肥満細胞の脱顆粒抑制剤等を提供することにある。
【課題を解決するための手段】
【0007】
肥満細胞活性化におけるSOC流入の関与に直接取り組むために、本発明者らは、STIM1欠損マウスを作出した。その結果、STIM1がFcεRI誘導性Ca2+流入を厳密に調節し、NF−κBおよびNFAT活性化を介し、脱顆粒ならびにin vivoアナフィラキシー応答を調節していることを解明し、本発明を完成するに至った。これは、肥満細胞活性化におけるSTIM1の重要性を直接遺伝学的に証明したものである。即ち、本願発明は、以下に示す通りである。
【0008】
〔1〕 STIM1阻害物質を有効成分として含有する肥満細胞の脱顆粒抑制剤。
〔2〕 STIM1阻害物質を有効成分として含有するアレルギーの予防または治療剤。
〔3〕 STIM1阻害物質を有効成分として含有するアナフィラキシーの予防または治療剤。
〔4〕 STIM1阻害物質を有効成分として含有する肥満細胞の炎症性サイトカインの産生抑制剤。
〔5〕 STIM1阻害物質が、RNAi誘導性核酸、アンチセンス核酸もしくはリボザイムまたはそれらの発現ベクターである、前記〔1〕〜〔4〕いずれかに記載の剤。
〔6〕 RNAi誘導性核酸がsiRNAである、前記〔5〕に記載の剤。
〔7〕 siRNAが、配列番号1の塩基配列に対応するmRNAにおける19〜25個の連続する塩基配列を含むセンス鎖と、その相補配列を含むアンチセンス鎖からなるものである、前記〔6〕に記載の剤。
〔8〕 被験物質がSTIM1遺伝子の発現または機能を抑制し得るか否かを評価することを含む、肥満細胞の脱顆粒を抑制し得る物質のスクリーニング方法。
〔9〕 下記の工程(a)〜(c)を含む、前記〔8〕記載の方法:
(a)被験物質とSTIM1遺伝子の発現を測定可能な細胞とを接触させる工程;
(b)被験物質を接触させた細胞におけるSTIM1遺伝子の発現量を測定し、該発現量を被験物質を接触させない対照細胞におけるSTIM1遺伝子の発現量と比較する工程;および
(c)上記(b)の比較結果に基づいて、STIM1遺伝子の発現量を抑制する被験物質を選択する工程。
〔10〕 下記の工程(a)〜(c)を含む、前記〔8〕記載の方法:
(a)被験物質をSTIM1蛋白質に接触させる工程;
(b)被験物質のSTIM1蛋白質に対する結合能を測定する工程;および
(c)上記(b)の結果に基づいて、STIM1蛋白質に結合能を有する被験物質を選択する工程。
【発明の効果】
【0009】
本発明の肥満細胞の脱顆粒抑制剤によれば、STIM1阻害物質を有効成分として含むことから、これまでにない作用機序に基づいた分子標的医薬品の開発に貢献することができる。STIM1阻害物質を有効成分として含む医薬は、アレルギーまたはアナフィラキシーの予防または治療薬として有用である。また、本発明のスクリーニング方法によれば、STIM1を分子標的とする候補物質を迅速にスクリーニングすることができる。
【発明を実施するための最良の形態】
【0010】
本発明は、STIM1阻害物質を有効成分として含有する肥満細胞の脱顆粒抑制剤を提供する。
【0011】
本発明において、STIM1とは、1つの膜貫通ドメインと1つのEF−ハンドカルシウム結合モチーフを有する間質細胞相互作用分子1をいい、小胞体膜に存在するカルシウムセンサータンパク質である。
【0012】
本発明において、STIM1は、任意の哺乳動物由来の膜タンパク質である。哺乳動物としては、ヒトおよびヒトを除く哺乳動物が挙げられ、ヒトを除く哺乳動物としては、例えば、マウス、ラット、ハムスター、モルモット等のげっ歯類やウサギ等の実験動物、ブタ、ウシ、ヤギ、ウマ、ヒツジ等の家畜、イヌ、ネコ等のペット、サル、オランウータン、チンパンジー等の霊長類が挙げられる。ヒトのアレルギー疾患の治療に用いるためには、ヒト由来のSTIM1が好ましい。ヒトSTIM1の塩基配列およびアミノ酸配列は公知であり、例えば、STIM1の塩基配列(配列番号1)およびアミノ酸配列(配列番号2)(GenBank Accession No. NM_003156)などがGenBankに登録され、公表されている。また、マウスSTIM1の塩基配列およびアミノ酸配列も公知であり、例えば、マウスSTIM1の塩基配列(配列番号3)およびアミノ酸配列(配列番号4)(GenBank Accession No. NM_009287)などがGenBankに登録され、公表されている。
【0013】
本発明において、STIM1阻害物質とは、STIM1の発現を阻害する物質およびSTIM1の機能(作用)を阻害する物質の両方を含む。
【0014】
本発明の薬剤に有効成分として含まれるSTIM1の発現を阻害する物質は、STIM1の転写過程に作用してその発現を阻害する物質であれば特に限定されるものではない。かかる阻害物質としては、RNAi誘導性核酸、アンチセンス核酸もしくはリボザイムまたはそれらの発現ベクターが挙げられる。
【0015】
前記RNAi誘導性核酸とは、細胞内に導入されることにより、RNA干渉を誘導し得るポリヌクレオチドをいい、好ましくはRNAまたはRNAとDNAのキメラ分子である。RNA干渉とは、mRNAと同一の塩基配列(またはその部分配列)を含む2本鎖構造のRNAが、当該mRNAの発現を抑制する効果をいう。このRNAi効果を得るには、例えば、少なくとも19の連続する標的mRNAと同一の塩基配列(またはその部分配列)を有する2本鎖構造のRNAを用いることが好ましい。ただし、STIM1の発現阻害作用を有していれば数塩基置換されているものであってもよく、19塩基長よりも短いRNAであってもよい。2本鎖構造は、センス鎖とアンチセンス鎖の異なるストランドで構成されていてもよいし、一つのRNAのステムループ構造によって与えられる2本鎖(shRNA)であってもよい。RNAi誘導性核酸としては、例えばsiRNA、miRNAなどが挙げられる。
【0016】
RNAi誘導性核酸は、転写抑制活性が強いという観点から、siRNAが好ましい。STIM1に対するsiRNAは、STIM1のmRNAの任意の部分を標的とすることができる。STIM1に対するsiRNA分子は、RNAi効果を誘導できる限り特に制限されないが、例えば19〜27塩基長、好ましくは21〜25塩基長である。STIM1に対するsiRNAは、センス鎖およびアンチセンス鎖を含む二重鎖である。具体的には、STIM1に対するsiRNAは、配列番号1または3の塩基配列に対応するmRNAにおける19〜25個の連続する塩基配列を含むセンス鎖と、その相補配列を含むアンチセンス鎖からなるものである。STIM1に対するsiRNAは、センス鎖、アンチセンス鎖の一方または双方の5’末端または3’末端においてオーバーハング(overhang)を有していてもよい。オーバーハングは、センス鎖および/またはアンチセンス鎖の末端における1〜数個(例、1、2または3個)の塩基の付加により形成されるものである。siRNAの設計方法は、当業者に公知であり、siRNAの様々な設計ソフトウエアまたはアルゴリズムを用いて、上記塩基配列から適切なsiRNAの塩基配列を選択することができる。
【0017】
STIM1に対するsiRNAのセンス鎖の一具体例としては、下記配列:
5’-AAGGCTCTGGATACAGTGCTC-3’(配列番号5)
5’-AAGAAGCTGCGCGATGAGATC-3’(配列番号6)
が挙げられる。
【0018】
STIM1に対するアンチセンス核酸は、STIM1の転写産物(mRNAまたは初期転写産物)を発現する細胞の生理的条件下で該転写産物とハイブリダイズし得る塩基配列からなり、且つハイブリダイズした状態で該転写産物にコードされるポリペプチドの翻訳を阻害し得るポリヌクレオチドをいう。アンチセンス核酸の種類はDNAであってもRNAであってもよいし、あるいはDNA/RNAキメラであってもよい。アンチセンス核酸は、天然型のリン酸ジエステル結合を有するものであっても、分解酵素に安定なチオリン酸型(リン酸結合のP=OをP=Sに置換)や2’-O-メチル型等の修飾ヌクレオチドであってもよい。アンチセンス核酸の設計に重要な他の要素として、水溶性および細胞膜透過性を高めること等が挙げられるが、これらはリポソームやマイクロスフェアを使用するなどの剤形の工夫によっても克服できる。アンチセンス核酸の長さは、STIM1の転写産物(例、配列番号1または3の塩基配列に対応するmRNA)と特異的にハイブリダイズし得る限り特に制限はなく、短いもので約15塩基程度、長いもので転写産物の全配列に相補的な配列を含むような配列であってもよい。合成の容易さや抗原性の問題等から、例えば約15塩基以上、好ましくは約15〜約30塩基、より好ましくは約18塩基〜約30塩基からなるオリゴヌクレオチドが例示される。さらに、アンチセンス核酸は、STIM1の転写産物とハイブリダイズして翻訳を阻害するだけでなく、二本鎖DNAと結合して三重鎖(トリプレックス)を形成し、mRNAへの転写を阻害し得るものであってもよい。
【0019】
本明細書において、「相補的である」とは、塩基配列間で約70%以上、好ましくは約80%以上、より好ましくは約90%以上、更に好ましくは約95%以上、最も好ましくは100%の相補性を有することをいう。本明細書における塩基配列の相同性は、相同性計算アルゴリズムNCBI BLAST(NationalCenter for Biotechnology Information Basic Local Alignment Search Tool)を用い、以下の条件(期待値=10;ギャップを許す;フィルタリング=ON;マッチスコア=1;ミスマッチスコア=-3)にて計算することができる。
【0020】
前記「リボザイム」とは核酸を切断する酵素活性を有するRNAをいうが、最近では当該酵素活性部位の塩基配列を有するオリゴDNAも同様に核酸切断活性を有することが明らかになっているので、本明細書では配列特異的な核酸切断活性を有する限りDNAをも包含する概念として用いる。具体的には、リボザイムは、STIM1をコードするmRNAまたは初期転写産物を、コード領域の内部(初期転写産物の場合はイントロン部分を含む)で特異的に切断し得る。リボザイムとして最も汎用性の高いものとしては、ウイロイドやウイルソイド等の感染性RNAに見られるセルフスプライシングRNAがあり、ハンマーヘッド型やヘアピン型等が知られている。ハンマーヘッド型は約40塩基程度で酵素活性を発揮し、ハンマーヘッド構造をとる部分に隣接する両端の数塩基ずつ(合わせて約10塩基程度)をmRNAの所望の切断部位と相補的な配列にすることにより、標的mRNAのみを特異的に切断することが可能である。さらに、リボザイムを、それをコードするDNAを含む発現ベクターの形態で使用する場合には、転写産物の細胞質への移行を促進するために、tRNAを改変した配列をさらに連結したハイブリッドリボザイムとすることもできる(Nucleic Acids Res., 29(13): 2780-2788 (2001))。
【0021】
STIM1の機能(作用)を阻害する物質としては、STIM1に対する抗体があげられる。該抗体は、ポリクローナル抗体、モノクローナル抗体のいずれであってもよく、周知の免疫学的手法により作製できる。また、該抗体は、抗体のフラグメント(例えば、Fab、F(ab’)2)、組換え抗体(例えば、単鎖抗体)であってもよく、さらに、該抗体をコードする核酸(プロモーター活性を有する核酸に機能可能に連結されたもの)であってもよい。
【0022】
前記抗体は、ヒトにおける治療効果と安全性を考慮すると、キメラ抗体、ヒト化またはヒト型抗体であってもよい。キメラ抗体は、例えば「実験医学(臨時増刊号), Vol.6, No.10, 1988」、特公平3-73280号公報等を、ヒト化抗体は、例えば特表平4-506458号公報、特開昭62-296890号公報等を、ヒト抗体は、例えば「Nature Genetics, Vol.15, p.146-156, 1997」、「Nature Genetics, Vol.7, p.13-21, 1994」、特表平4-504365号公報、国際出願公開WO94/25585号公報、「日経サイエンス、6月号、第40〜第50頁、1995年」、「Nature, Vol.368, p.856-859, 1994」、特表平6-500233号公報等を参考にそれぞれ作製することができる。
【0023】
さらに別の実施形態では、STIM1の機能(作用)を阻害する物質は、STIM1のドミナントネガティブ変異体、該変異体をコードする核酸を含む発現ベクターが例示される。
【0024】
STIM1のドミナントネガティブ変異体とは、STIM1に対する変異の導入によりその活性が低減したものをいう。該ドミナントネガティブ変異体は、天然のSTIM1と競合することで間接的にその機能を阻害することができる。該ドミナントネガティブ変異体は、STIM1遺伝子をコードする核酸に変異を導入することによって作製することができる。変異としては、例えば、機能性部位における、当該部位が担う機能の低下をもたらすようなアミノ酸の変異(例えば、1以上のアミノ酸の欠失、置換、付加)が挙げられる。ドミナントネガティブ変異体は、PCRや公知のキットを用いる自体公知の方法により作製できる。
【0025】
STIM1阻害物質は、発現ベクターとしても提供され得る。かかる発現ベクターは、STIM1阻害物質をコードするポリヌクレオチド、および当該ポリヌクレオチドに機能可能に連結されたプロモーターを含む。
【0026】
前記プロモーターは、その制御下にある発現対象の核酸の種類により適宜選択され得るが、例えば、polIIIプロモーター(例、tRNAプロモーター、U6プロモーター、H1プロモーター)、哺乳動物用プロモーター(例、CMVプロモーター、CAGプロモーター、SV40プロモーター)が挙げられる。
【0027】
本発明の発現ベクターはさらに、選択マーカー遺伝子(テトラサイクリン、アンピシリン、カナマイシン、ハイグロマイシン、ホスフィノスリシン等の薬剤に対する抵抗性を付与する遺伝子、栄養要求性変異を相補する遺伝子等)をさらに含んでいてもよい。
【0028】
本発明の発現ベクターのバックボーン(backbone)としては、ヒト等の哺乳動物細胞中でSTIM1阻害物質を産生できるものであれば特に制限されないが、例えば、プラスミドベクター、ウイルスベクターが挙げられる。哺乳動物への投与に好適なベクターとしては、レトロウイルス、アデノウイルス、アデノ随伴ウイルス、ヘルペスウイルス、ワクシニアウイルス、ポックスウイルス、ポリオウイルス、シンドビスウイルス、センダイウイルス等のウイルスベクターが挙げられる。なかでも、レトロウイルス、アデノウイルス、アデノ随伴ウイルス、ワクシニアウイルス由来のウイルスベクターが好ましい。
【0029】
本発明の肥満細胞の脱顆粒抑制剤は、肥満細胞からのヒスタミン、サイトカイン、ケモカインおよびプロテアーゼ等の遊離物質の分泌を抑制することが可能であり、かかる遊離物質に起因する様々なアレルギー疾患の予防または治療剤としても有用である。アレルギー疾患としては、喘息、アレルギー性鼻炎、アレルギー性結膜炎、アトピー性皮膚炎、アレルギー性胃腸炎などが挙げられる。
【0030】
本発明において、STIM1が受動皮膚アナフィラキシー反応(PCA)に必要であることを生体内で確認したことにより、本発明は、STIM1阻害物質を有効成分として含有するアナフィラキシーの予防または治療剤を提供することができる。
【0031】
また、本発明において、肥満細胞からの炎症性サイトカインの産生および分泌にSTIM1が必須の役割を果たしていることを確認したことにより、本発明は、STIM1阻害物質を有効成分として含有する肥満細胞の炎症性サイトカインの産生抑制剤を提供することができる。
【0032】
前記炎症性サイトカインとしては、IL−3、IL−6、TNF−αなどが挙げられる。
【0033】
前記脱顆粒抑制剤、アレルギー疾患の予防または治療剤、アナフィラキシーの予防または治療剤および炎症性サイトカインの産生抑制剤を総称して、本発明の薬剤と略す場合がある。
【0034】
本発明の薬剤の投与量は、有効成分の種類もしくは活性、投与対象となる動物種、投与対象の病気の重篤度、薬物受容性、体重、年齢等によって異なるが、通常、成人1日あたり有効成分量として約0.0001〜約1000mg/kgが例示される。
【0035】
本発明の薬剤は、患者に対して経口的または非経口的に投与することができ、投与形態としては、経口投与、局所投与(点鼻投与、点眼投与、吸入投与等)、静脈内投与、経皮投与などが挙げられ、必要に応じて、製薬学的に許容され得る添加剤と共に、投与に適した剤型に製剤化される。経口投与に適した剤型としては、例えば、錠剤、カプセル剤、顆粒剤、散剤などが挙げられ、非経口投与に適した剤型としては、例えば、点鼻剤、点眼剤、注射剤、貼付剤、ローション剤、クリーム剤などが挙げられる。これらは当該分野で汎用されている通常の技術を用い、調製することができる。本発明の薬剤は、上述の治療効果を奏する限りその投与経路および剤形は特に限定されない。
【0036】
例えば、本発明の薬剤を注射剤として用いる場合、安定剤(例えば、亜硫酸水素ナトリウム、チオ硫酸ナトリウム、エデト酸ナトリウム、クエン酸ナトリウム、アスコルビン酸、ジブチルヒドロキシトルエンなど)、溶解補助剤(例えば、グリセリン、プロピレングリコール、マクロゴール、ポリオキシエチレン硬化ヒマシ油など)、懸濁化剤(例えば、ポリビニルピロリドン、ヒドロキシプロピルメチルセルロース、ヒドロキシメチルセルロース、カルボキシメチルセルロースナトリウムなど)、乳化剤(例えば、ポリビニルピロリドン、大豆レシチン、卵黄レシチン、ポリオキシエチレン硬化ヒマシ油、ポリソルベート80など)、緩衝剤(例えば、リン酸緩衝液、酢酸緩衝液、ホウ酸緩衝液、炭酸緩衝液、クエン酸緩衝液、トリス緩衝液、グルタミン酸、イプシロンアミノカプロン酸など)、粘稠剤(例えば、メチルセルロース、ヒドロキシエチルセルロース、ヒドロキシプロピルメチルセルロース、カルボキシメチルセルロースなどの水溶性セルロース誘導体、コンドロイチン硫酸ナトリウム、ヒアルロン酸ナトリウム、カルボキシビニルポリマー、ポリビニルアルコール、ポリビニルピロリドン、マクロゴールなど)、保存剤(例えば、塩化ベンザルコニウム、塩化ベンゼトニウム、グルコン酸クロルヘキシジン、クロロブタノール、ベンジルアルコール、デヒドロ酢酸ナトリウム、パラオキシ安息香酸エステル類、エデト酸ナトリウム、ホウ酸など)、等張化剤(例えば、塩化ナトリウム、塩化カリウム、グリセリン、マンニトール、ソルビトール、ホウ酸、ブドウ糖、プロピレングリコールなど)、pH調整剤(例えば、塩酸、水酸化ナトリウム、リン酸、酢酸など)、清涼化剤(例えば、l−メントール、d−カンフル、d−ボルネオール、ハッカ油など)、軟膏基剤(白色ワセリン、精製ラノリン、流動パラフィン、植物油(オリーブ油、椿油、落花生油など)など)などを添加剤として加えることができる。これら添加剤の添加量は、添加する添加剤の種類、用途などによって異なるが、添加剤の目的を達成し得る濃度を添加すればよい。
【0037】
本発明の薬剤は、siRNA等の核酸をリポフェクション法を用いて製剤化することもできる。
【0038】
本発明はまた、被験物質がSTIM1遺伝子の発現または機能を抑制し得るか否かを評価することを含む、肥満細胞の脱顆粒抑制物質のスクリーニング方法、ならびに当該スクリーニング方法により得られる物質、および当該物質を含有してなる脱顆粒抑制剤を提供する。
【0039】
スクリーニング方法に供される被験物質は、いかなる公知化合物および新規化合物であってもよく、例えば、核酸、糖質、脂質、蛋白質、ペプチド、有機低分子化合物、コンビナトリアルケミストリー技術を用いて作製された化合物ライブラリー、固相合成やファージディスプレイ法により作製されたランダムペプチドライブラリー、あるいは微生物、動植物、海洋生物等由来の天然成分等が挙げられる。
【0040】
一実施形態では、本発明のスクリーニング方法は、下記の工程(a)〜(c)を含む:
(a)被験物質とSTIM1遺伝子の発現を測定可能な細胞とを接触させる工程;
(b)被験物質を接触させた細胞におけるSTIM1遺伝子の発現量を測定し、該発現量を被験物質を接触させない対照細胞におけるSTIM1遺伝子の発現量と比較する工程;
(c)上記(b)の比較結果に基づいて、STIM1遺伝子の発現量を抑制する被験物質を選択する工程。
【0041】
上記方法の工程(a)では、被験物質がSTIM1遺伝子の発現を測定可能な細胞と接触条件下におかれる。STIM1遺伝子の発現を測定可能な細胞に対する被験物質の接触は、培養培地中で行われ得る。
【0042】
STIM1遺伝子の発現を測定可能な細胞とは、STIM1遺伝子の産物、例えば、転写産物、翻訳産物の発現レベルを直接的または間接的に評価可能な細胞をいう。STIM1遺伝子の産物の発現レベルを直接的に評価可能な細胞は、STIM1遺伝子を天然で発現可能な細胞であり得、一方、STIM1遺伝子の産物の発現レベルを間接的に評価可能な細胞は、STIM1遺伝子転写調節領域についてレポーターアッセイを可能とする細胞であり得る。STIM1遺伝子の発現を測定可能な細胞は、動物細胞、例えばマウス、ラット、ハムスター、モルモット、ウサギ、イヌ、サル、ヒト等の哺乳動物細胞であり得る。
【0043】
STIM1遺伝子を天然で発現可能な細胞は、STIM1遺伝子を潜在的に発現するものである限り特に限定されない。かかる細胞は、当業者であれば容易に同定でき、初代培養細胞、当該初代培養細胞から誘導された細胞株、市販の細胞株、セルバンクより入手可能な細胞株などを使用できる。
【0044】
STIM1遺伝子転写調節領域についてレポーターアッセイを可能とする細胞は、STIM1遺伝子転写調節領域、当該領域に機能可能に連結されたレポーター遺伝子を含む細胞である。STIM1遺伝子転写調節領域およびレポーター遺伝子は、発現ベクター中に挿入され得る。STIM1遺伝子転写調節領域は、STIM1遺伝子の発現を制御し得る領域である限り特に限定されないが、例えば、転写開始点から上流約2kbpまでの領域、あるいは該領域の塩基配列において1以上の塩基が欠失、置換若しくは付加された塩基配列からなり、且つSTIM1遺伝子の転写を制御する能力を有する領域などが挙げられる。レポーター遺伝子は、検出可能な蛋白質または検出可能な物質を生成する酵素をコードする遺伝子であればよく、例えばGFP(緑色蛍光蛋白質)遺伝子、GUS(β−グルクロニダーゼ)遺伝子、LUC(ルシフェラーゼ)遺伝子、CAT(クロラムフェニコルアセチルトランスフェラーゼ)遺伝子等が挙げられる。
【0045】
STIM1遺伝子転写調節領域、当該領域に機能可能に連結されたレポーター遺伝子が導入される細胞は、STIM1遺伝子転写調節機能を評価できる限り、即ち、該レポーター遺伝子の発現量が定量的に解析可能である限り特に限定されない。しかしながら、STIM1遺伝子に対する生理的な転写調節因子を発現し、STIM1遺伝子の発現調節の評価により適切であると考えられることから、該導入される細胞としては、STIM1遺伝子を天然で発現可能な細胞が好ましい。
【0046】
被験物質とSTIM1遺伝子の発現を測定可能な細胞とが接触される培養培地は、用いられる細胞の種類などに応じて適宜選択されるが、例えば、約5〜20%のウシ胎仔血清を含む最少必須培地(MEM)、ダルベッコ改変最少必須培地(DMEM)、RPMI1640培地、199培地などである。培養条件もまた、用いられる細胞の種類などに応じて適宜決定されるが、例えば、培地のpHは約6〜約8であり、培養温度は通常約30〜約40℃であり、培養時間は約12〜約72時間である。
【0047】
上記方法の工程(b)では、先ず、被験物質を接触させた細胞におけるSTIM1遺伝子の発現量が測定される。発現量の測定は、用いた細胞の種類などを考慮し、自体公知の方法により行われ得る。例えば、STIM1遺伝子の発現を測定可能な細胞として、STIM1遺伝子を天然で発現可能な細胞を用いた場合、発現量は、STIM1遺伝子の産物、例えば、転写産物または翻訳産物を対象として自体公知の方法により測定できる。例えば、転写産物の発現量は、細胞からtotal RNAを調製し、RT−PCR、ノーザンブロッティング等により測定され得る。また、翻訳産物の発現量は、細胞から抽出液を調製し、免疫学的手法により測定され得る。免疫学的手法としては、放射性同位元素免疫測定法(RIA法)、ELISA法(Methods in Enzymol. 70: 419-439 (1980))、蛍光抗体法などが使用できる。一方、STIM1遺伝子の発現を測定可能な細胞として、STIM1遺伝子転写調節領域についてレポーターアッセイを可能とする細胞を用いた場合、発現量は、レポーターのシグナル強度に基づき測定され得る。
【0048】
次いで、被験物質を接触させた細胞におけるSTIM1遺伝子の発現量が、被験物質を接触させない対照細胞におけるSTIM1遺伝子の発現量と比較される。発現量の比較は、好ましくは、有意差の有無に基づいて行なわれる。被験物質を接触させない対照細胞におけるSTIM1遺伝子の発現量は、被験物質を接触させた細胞におけるSTIM1遺伝子の発現量の測定に対し、事前に測定した発現量であっても、同時に測定した発現量であってもよいが、実験の精度、再現性の観点から同時に測定した発現量であることが好ましい。
【0049】
上記方法の工程(c)では、STIM1遺伝子の発現量を抑制する被験物質が選択される。STIM1遺伝子の発現量の抑制は、発現量の減少であり得る。STIM1遺伝子の発現量を減少させる(発現を抑制する)被験物質は、脱顆粒機能を抑制する作用を有し得、アレルギー疾患またはアナフィラキシーの予防・治療薬となり得る。従って、STIM1遺伝子の発現量を指標として、アレルギー疾患の予防・治療剤等の医薬、または研究用試薬のための候補物質を選択することが可能となる。
【0050】
別の実施形態では、本発明のスクリーニング方法は、下記の工程(a)〜(c)を含む:
(a)被験物質をSTIM1蛋白質に接触させる工程;
(b)被験物質のSTIM1蛋白質に対する結合能を測定する工程;
(c)上記(b)の結果に基づいて、STIM1蛋白質に結合能を有する被験物質を選択する工程。
【0051】
上記方法の工程(a)では、被験物質がSTIM1蛋白質と接触条件下におかれる。被験物質の該蛋白質に対する接触は、溶液中での被験物質とSTIM1蛋白質との混合により行われ得る。
【0052】
STIM1蛋白質は自体公知の方法により調製できる。例えば、遺伝子組換え技術により組換えSTIM1蛋白質を調製するのが好ましい。組換え蛋白質は、細胞系、無細胞系のいずれで調製したものでもよい。
【0053】
上記方法の工程(b)では、STIM1蛋白質に対する被験物質の結合能が測定される。結合能の測定は、自体公知の方法、例えば、バインディングアッセイ、表面プラズモン共鳴を利用する方法(例えば、Biacore(登録商標)の使用)により行われ得る。STIM1蛋白質に対する被験物質の結合能はまた、STIM1蛋白質に対する拮抗体(例えば、STIM1蛋白質の共役因子)の存在下で測定してもよい。この場合、例えば、STIM1蛋白質に対する拮抗体の結合量の変化を指標に、STIM1蛋白質に対する被験物質の結合能が測定され得る。
【0054】
上記方法の工程(c)では、STIM1蛋白質に結合能を有する被験物質が選択される。STIM1蛋白質に結合能を有する被験物質は、STIM1遺伝子の機能を抑制し得る物質となる可能性がある。従って、本方法論は、例えば、STIM1遺伝子の機能を抑制し得る物質の一次スクリーニングとして有用である。
【0055】
本発明のスクリーニング方法はまた、被験物質の動物への投与により行われ得る。該動物としては、例えば、マウス、ラット、ハムスター、モルモット、ウサギ、イヌ、サル等の哺乳動物が挙げられる。動物を用いて本発明のスクリーニング方法が行われる場合、例えば、STIM1遺伝子の発現量を抑制する被験物質が選択され得る。
【0056】
本発明のスクリーニング方法は、脱顆粒を抑制し得る物質のスクリーニングを可能とする。従って、本発明のスクリーニング方法は、上述の薬剤または研究用試薬の開発などに有用である。
【実施例】
【0057】
以下、実施例により本発明を詳細に説明するが、本発明はこれらの実施例により何ら限定されるものではない。
【0058】
実施例1
STIM1標的化マウスの作出
STIM1標的化マウスを作出するためのターゲティングベクターは、STIM1の膜貫通領域をコードする配列を含むエキソン6の両端にそれぞれloxP部位を有するように2つのloxP部位を隣接させて設計した。かかるコンストラクトのために、ゲノムDNA断片をC57BL/6起源のBruce-4 ES細胞(ハーバードメディカルスクール、CBRバイオメディカルリサーチ研究所のK. Rajewsky博士より供与)から増幅させた。KSTKNEOLOXPベクター(K. Rajewsky博士より供与)中の第1のloxP部位を有するネオマイシン耐性カセットを、Stim1のイントロン5を含む1.7-kbの5'ホモロジー領域の下流にクローニングし、エキソン6を含む断片を2つのloxP部位の間に挿入した。エキソン7を含む4.5-kbの3'ホモロジー領域の後に、ジフテリアトキシンA(DTA)カセット(Yagi, T. et al. Proc Natl Acad Sci U S A 87, 9918-9922 (1990))を連結した。DTA 遺伝子は、ランダムインテグレーションを有するクローンのネガティブセレクションに使用した。ターゲティングコンストラクトを直鎖状にし、Bruce-4 ES細胞にエレクトロポレーションにより導入した。形質転換細胞は、G418で選択した。ターゲットクローンをサザンブロット解析により同定し、C57BL/6の胚盤胞にマイクロインジェクションし、キメラを生成させた。キメラ雄性マウスとC57BL/6雌性マウスとの交配により、flox-neoアレルが生殖系列へ伝達された。得られたStim1flox/+マウスを、サイトメガロウイルス即時型初期エンハンサー−ニワトリアクチンハイブリッド(CAG)プロモーターの制御下にCreトランスジーンを保持するトランスジェニックマウス系列と交配して、CAGCre/+Stim1flox/+(遺伝子型;Stim1+/-)を作出した。次いで、このマウスを交雑させて、Stim1-/-マウスを作出した。すべての実験は、理化学研究所の動物実験委員会のガイドラインに遵守してなされた。
【0059】
抗体
Syk、LAT、JNK、p38、IκBα、p65、ActinおよびNFAT1 (NFATc2)に対する抗体は、Santa Cruz社から購入した。抗リン酸化JNK抗体、抗リン酸化p38抗体および抗Erk1/2抗体は、Promega社から購入した。PLCγ1、リン酸化PLCγ1、リン酸化Syk、リン酸化IκBα、リン酸化LATおよびリン酸化Erk(p42/44)に対する抗体は、Cell Signaling Technology社から購入し、リン酸化NFAT1に対する抗体は、Abcam社から購入した。抗STIM1抗体は、既報(Baba, Y. et al. Proc Natl Acad Sci USA103, 16704-16709 (2006))に従って製造した。FITC-結合抗CD45.2 mAbおよびPE-結合抗FcεRI mAb、APC-結合抗c-Kit mAbは、それぞれ、BD社およびeBioscience社から購入した。
【0060】
実施例2
肥満細胞の培養
胎仔肝由来肥満細胞(FLMCs)の調製のため、STIM1-/-マウスおよび同腹子野生型マウス由来のE15.5胎仔肝から懸濁細胞を取り出し、10% FCS、β-メルカプトエタノール、抗生物質および20ng/ml組換えマウス(rm)IL-3(R&D systems)を補足したRPMI1640で培養した。培養中、培地を3〜4日毎に交換した。培養後4〜5週に、当該細胞がその細胞表面にFcεRIおよびc-Kitを発現していることを確認し、実験に使用した(< 98% 肥満細胞)。
【0061】
実施例3
組織染色
E15.5の胎仔の5μmパラフィン切片を固定し、ニュークレアファストレッドおよびアルシャンブルーで染色した。肥満細胞を観察するために、各試料中のアルシャンブルー染色細胞を計数した。
【0062】
実施例4
脱顆粒およびサイトカイン産生の測定
細胞(1 x 106/ml)を、1μg/ml 抗DNP IgE (SPE-7; Sigma)を用いて37℃で6時間感作し、次いで、感作した細胞を、Tyrode’s緩衝液(10 mM HEPES pH 7.4、130 mM NaCl、5 mM KCl、1.4 mM CaCl2、1 mM MgCl2、5.6 mM グルコース、0.1% BSA)中、多価ジニトロフェニル-ヒト血清アルブミン(DNP-HSA, Sigma)で30分間刺激した。β-ヘキソサミニダーゼ反応に関しては、50μlの細胞上清または細胞溶解液を、100μlの1.3mg/ml p-ニトロフェニル-N-アセチル-D-グルコサミド(0.1M クエン酸塩(pH 4.5)中)とともにインキュベートし、37℃で60分発色させた。次いで、0.2M グリシン-NaOH(pH 10.2)を150μl添加することにより酵素反応を停止させ、405nmでの吸光度をマイクロプレートリーダー(Bio-Rad)で測定した。細胞を、1%Triton X-100を含むTyrode’s緩衝液で溶解し、β-ヘキソサミニダーゼ活性を測定した。放出されたβ-ヘキソサミニダーゼの割合を、下記式:
放出(%) = 上清 / (上清 + 細胞溶解液) x 100
を用いて計算した。サイトカイン放出の測定のため、細胞培養上清中のIL-6、TNF-αおよびIL-13を、既報(Kabu, K. et al. J Immunol 177, 1296-1305 (2006))に記載のELISAアッセイにより検出した。
【0063】
実施例5
カルシウム濃度の測定
細胞質ゾルのカルシウム濃度は、既報(Baba, Y. et al. Proc Natl Acad Sci USA103, 16704-16709 (2006))に従って測定した。簡潔に記載すると、胎仔肝肥満細胞を、RPMI1640/ 1% FCS中、37℃で45分、Indo-1-AMおよびPluronic F-127 (Invitrogen)を負荷した。FcεRI刺激に関しては、胎仔肝肥満細胞を抗DNP IgE(2μg/ml)で同時に感作した。蛍光強度の変化を、LSRII(BD Biosciences)でモニターした。FL4(Ca2+-フリーindo-1)の蛍光に対するFL5(Ca2+-結合indo-1)の蛍光の比として、値をプロットした。データは、Flowjo software(Treestar)で解析した。
【0064】
実施例6
細胞の刺激およびイムノブロッティング
細胞(1 x 106/ml)を、1μg/ml IgEで37℃、6時間感作し、次いで、Tyrode’s緩衝液中50 ng/ml DNP-HASで刺激した。あるいは、未感作細胞を1μM イオノマイシンで刺激した。刺激後、前記細胞を、10 mM Tris-HCl pH 7.4、150 mM NaCl、1 % NP-40、0.1 % SDS、0.5 mM EDTA、プロテアーゼ阻害剤(Roche)およびホスファターゼ(Nakarai)阻害剤カクテルを含む溶解緩衝液中で溶解し、清澄化させた細胞溶解液を、5 x SDSサンプル緩衝液とともに煮沸した。5 x 105〜1 x 106 細胞分の試料を、PVDF膜に電気泳動で転写し、適切な抗体でブロットした。核抽出物は、Extraction Kit (Pierce)を用いて製造業者のプロトコルに従って調製し、イムノブロッティングに供した。
【0065】
実施例7
イムノフルオレッセンスおよび共焦点顕微鏡観察
細胞を、余熱した4 %パラホルムアルデヒドで15分間固定し、0.2 % Triton X-100で5分間透過処理した。前記細胞を2 %ヤギ血清でブロッキングした後、2 %ヤギ血清を含むPBSで1:50希釈した抗p65 Abとともに1時間インキュベートした。前記一次抗体を、Alexa Fluor 488で標識した抗ウサギ二次抗体(Invitrogen)を用いて検出した。核は、DAPIで共染色した。染色した細胞は、共焦点蛍光顕微鏡(DMIRE2; Leica Microsystems) 下で観察した。
【0066】
実施例8
RT-PCR解析
MMCP-5、MMCP-6、アクチン、IL-6、TNF-α、IL-13およびGAPDHの発現を評価するためのRT-PCRは、既報(Kabu, K. et al. J Immunol 177, 1296-1305 (2006))に記載された遺伝子特異的プライマーを用いて行った。
【0067】
実施例9
胎仔肝細胞の移植
STIM1-/-または野生型同腹子のE15.5胎仔由来の2 x 106個の胎仔肝細胞を、致死量(950rad)を照射したコンジェニックマウス(CD45.1)に静脈内に移植した。10週間後、前記マウスから腹膜腔を回収し、フローサイトメトリーにより解析した。CD45.2を用いて、宿主細胞とドナー細胞とを区別した。
【0068】
実施例10
レトロウイルスの産生および感染
GFPタグ化したSTIM1 cDNA(Baba, Y. et al. Proc Natl Acad Sci USA103, 16704-16709 (2006))を、pMX-puroレトロウイルスベクター(東京大学T. Kitamura博士より供与)にクローニングした。得られたベクターおよび対照のGFP単独mockベクターを、FuGENE6(Roche)を用いてEcoPack2(BD Biosciences)にトランスフェクションし、形質転換した細胞を、1.5 μg/mlピューロマイシンで選択した。培地交換後24時間で培養上清を回収し、直ちに感染に用いた。胎仔肝由来肥満細胞の感染のため、当該細胞を20ng/ml rmIL-3およびrmSCF(R&D systems)の存在下で16〜18時間、前刺激した。次いで、等量のウイルス上清および10ng/ml DOTAP(Roche)を添加した後、32℃で2時間、2000rpmでのスピン感染を行い、プレートを37℃で一晩インキュベートした。翌日、rmIL-3を含む新鮮な培地と交換した。
【0069】
実施例11
受動皮膚アナフィラキシー(PCA)
PCAは、既報(Kabu, K. et al. J Immunol 177, 1296-1305 (2006))に従って行った。動物の右耳に、0.5μgの抗DNP IgEを含む20μl生理食塩水を皮下に注射した。左耳には、20μlの生理食塩水を注射した。16〜18時間後、前記動物を、250μgの多価ジニトロフェニル-ウシ血清アルブミン(DNP-BSA)および5mg/mlエバンスブルーダイ(Sigma)を含む250μlの生理食塩水でチャレンジした。前記耳におけるエバンスブルーの血管外溢出を30分間モニターし、耳バイオプシーを、700μlホルムアミド中、63℃で一晩インキュベートした。ホルムアミド抽出物の定量解析は、620nmでのエバンスブルーの吸光度を測定することにより決定した。
【0070】
結果
STIM1欠損マウスでの肥満細胞の発生
肥満細胞およびアレルギー応答におけるSTIM1の生理学的機能を研究するため、マウスのStim1遺伝子を遺伝的に欠失させた。Stim1遺伝子を、Cre-loxP-介在遺伝子ターゲティングストラテジーを用いて、C57B6/Cマウスで不活性化した(図1a)。サザンブロッティングにより、組換えおよびSTIM1の膜貫通領域をコードするエキソン6のCreによる除去を確認した(図1b)。胎仔肝細胞および胎仔肝由来肥満細胞(FLMCs)におけるSTIM1タンパク質の破壊は、イムノブロット解析により確認した。STIM1+/+、STIM1+/-およびSTIM1-/-細胞の細胞溶解液を、抗STIM1抗体でブロットした。STIM1+/+細胞は、STIM1タンパク質を発現していたが、STIM1-/-細胞ではSTIM1タンパク質が検出されず、STIM1+/-細胞ではその発現レベルが著しく低下していた(図1c、d)。ホモ接合性STIM1-/-マウスは、原因は不明であるが、周産期死亡を示した。E18.5まで、STIM1-/-胎仔は、明らかな異常は示さなかった。STIM1-/-マウスは稀に出生する場合があり、新生STIM1-/-はSTIM1+/+マウスよりも若干小さく、呼吸障害により一日以内に死亡した。
【0071】
胎生致死のため、E15.5胎仔由来の肝臓から肥満細胞前駆細胞を収集し、IL-3を含む培地中で高度に純粋な肥満細胞を生成させた。細胞の純度は、FcεRIおよびc-Kitの細胞表面の発現で評価した(図2a)。サイトスピンした試料のアルシャンブルー染色により、STIM1-/-肥満細胞の形態は野生型肥満細胞の形態とは異ならないことが示された(図2b)。MMCP-5およびMMCP-6等の肥満細胞に特異的な遺伝子は、野生型およびSTIM1欠損FLMCsで同程度のレベルで発現していた(図2c)。
肥満細胞のin vivo の分化状態を調べるため、野生型およびSTIM1欠損胎仔での組織肥満細胞の数を決定した。STIM1欠損E15.5胎仔の背の皮膚における肥満細胞の密度は、野生型の密度と同程度であった(図2d)。さらに、野生型およびSTIM1欠損マウスの胎仔肝細胞(CD45.2)を、致死量放射線を照射したコンジェニックマウス(CD45.1)に適応移植し、フローサイトメトリーにより再構成の有無を解析した。ドナー型細胞を、CD45マーカーにより区別した。再構成したマウス末梢血中のSTIM1+/+およびSTIM1-/- 細胞によるキメリズムは、ほとんど同等であった(>90%;データは示さず)。このことから、STIM1欠損マウス由来の造血幹細胞の正着能力は正常であることが示唆された。腹腔内のフローサイトメトリー解析により、肥満細胞の発生も無傷であることがわかった(図2e)。前記結果をまとめると、肥満細胞の分化はSTIM1非存在下でも障害されないことが示された。
【0072】
STIM1欠損FLMCsにおける障害されたストア感受性Ca2+流入
STIM1は、RNAiに基づいたノックダウンHeLa細胞およびJurkat細胞、ならびにノックアウトDT40 B細胞におけるSOC流入の重要な分子であることが示されている。肥満細胞におけるSOC流入に対するSTIM1の効果を決定するために、0.5 mM EGTAの存在下で小胞体Ca2+ポンプ阻害剤であるタプシガルギン(TG)処理でCa2+ストアを枯渇させ、FLMCsの細胞質内Ca2+をモニターし、次いで、2 mM Ca2+ を戻し入れてCa2+ 流入をモニターした。図3aに示すように、STIM1欠損FLMCsにおいて、Ca2+流入の有意な抑制が観察された。このSTIM1依存性SOC流入メカニズムは、FcεRIシグナル伝達コンテクストによって優位に利用されており、STIM1の非存在下ではFcεRIにより誘導されたストア枯渇からCa2+流入の障害も生じた(図3a)。Ca2+動員におけるSTIM1の効果をさらに評価するために、細胞外Ca2+が存在する条件下で、IgEおよび抗原によるFcεRI刺激を行った。野生型FLMCsは、シャープなスパイクを示した後、持続的なCa2+流入を示した(図3b)。対照的に、STIM欠損FLMCsでは細胞質内Ca2+レベルが有意に低下していた。イオノマイシン処理野生型FLMCsは、連続的に高いレベルのCa2+増加を示した。この持続したCa2+は、STIM1欠損FLMCsでは劇的に低下した(図3b)。これらの結果より、イオノマイシンはCa2+流入の引き金となり得ることを示唆し、これは全体的にSTIM1に依存する。イオノマイシンが内部のCa2+ストアを枯渇させることによりPMにおけるSOCチャネルを活性化できるという以前の証拠があることから、STIM1は、このイオノマイシン依存性Ca2+枯渇のセンサーとしても機能し、Ca2+流入を活性化すると思われる。
【0073】
Ca2+流入の上記した欠損がSTIM1が存在しないことによるものであるか否かを検証するために、GFP標識したSTIM1をSTIM1欠損FLMCsにレトロウイルスにより導入し、細胞質内Ca2+プロフィールを分析した。モックGFPを導入したSTIM1欠損FLMCsは、Ca2+流入を誘導する能力がなかったが、GFP-STIM1のバックトランスフェクションにより、FcεRIおよびTG刺激後、Ca2+流入を明確にレスキューした(図3c)。一貫して、2 mM Ca2+ の存在下でIgE凝集またはイオノマイシンによって媒介されるCa2+の動員は、GFP-STIM1導入細胞において十分回復した。まとめると、これらの結果から、STIM1が肥満細胞においてSOC流入活性化に関与すること、およびこのメカニズムはFcεRIおよびイオノマイシン刺激により誘導されるCa2+流入において重要な役割を果たすことを明確に示している。
【0074】
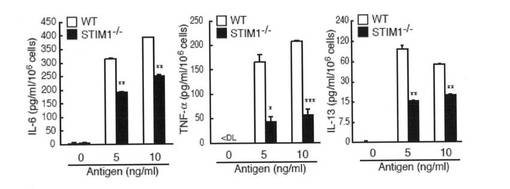
STIM1は肥満細胞脱顆粒およびサイトカイン産生に必須である
FcεRIが介在する肥満細胞活性化は、迅速な脱顆粒ならびに代謝産物およびサイトカインのde novoの合成を誘導する。このことは、アレルギー反応に重要な役割を果たす。これらの肥満細胞の機能にとってSTIM1が必要であるか否かを探究するために、FcεRI刺激後の野生型およびSTIM1欠損FLMCs由来のβ-ヘキソサミニダーゼの放出を測定した。STIM1の喪失により、抗原により誘発される脱顆粒の顕著な抑制が生じた(図4a)。また、STIM1欠損FLMCsがイオノマシンにより誘導される脱顆粒の極度の低下を示したことを見出した。イオノマイシンは、FcεRI近位のシグナル伝達をバイパスし、遊離Ca2+イオンの動員を通じて直接細胞を刺激することが知られている。β-ヘキソサミニダーゼの総含量は野生型FLMCsとSTIM1欠損FLMCsとで相違しなかった点に注目すべきである (データ示さず)。したがって、STIM1は肥満細胞の脱顆粒にとって必須であることが結論される。これらの知見は、一過性のCa2+ の上昇では脱顆粒を有効に行うには不十分であることも示唆する。
【0075】
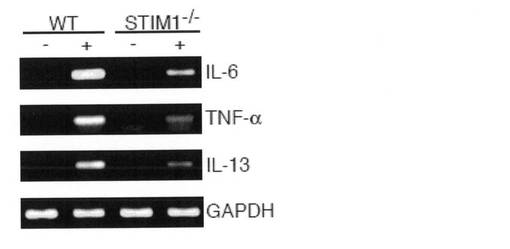
サイトカイン産生を評価するために、FcεRIライゲーション後の野生型およびSTIM1欠損FLMCsにおけるELISAアッセイを行った。IL-6、TNF-αおよびIL-13の分泌は、野生型FLMCsに比べてSTIM1欠損FLMCsで顕著に低下していた(図4b)。半定量的RT-PCRにより、これらのサイトカインのmRNA合成もSTIM1の非存在下で顕著に抑制されていたことが示された(図4c)。これらの結果は、STIM1がIL-6、TNF-αおよびIL-13の遺伝子発現ならびに当該サイトカインの分泌に必須であることを示している。
【0076】
STIM1欠損FLMCsにおける正常なFcεRI介在近位シグナル伝達およびMAPKs活性化
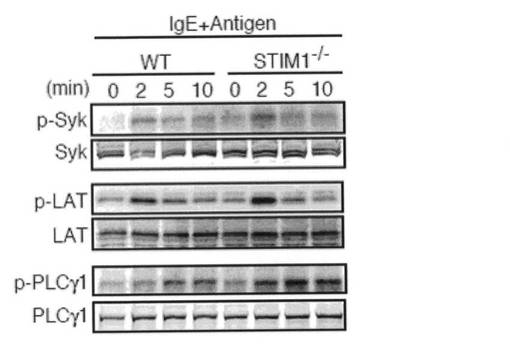
上記したデータは、STIM1依存性の持続的Ca2+ 障害が肥満細胞の機能に応答可能な細胞内シグナル伝達事象の欠損に導く可能性を示唆した。どの細胞内シグナル伝達経路が攪乱されるのかを決定するために、野生型およびSTIM1欠損FLMCsにおける様々なFcεRIシグナル経路を調べた。感作したFLMCsを所定の時間抗原刺激し、イムノブロッティングによりSyk、LATおよびPLCγ1等の近位分子の活性化状態を調べた。STIM1欠損FLMCsは、Syk、LATおよびPLCγ1の正常なチロシンリン酸化を示した(図5a)。このことは、FcεRI近位のシグナル伝達は一般にSTIM1により影響されないことを示す。MAPKs カスケードは、サイトカイン産生を調節していることが知られている。しかしながら、FcεRIが介在するJNK、p38およびErkの活性化は、STIM1の不在によって変化しなかった(図5b)。イオノマイシンによる細胞質内Ca2+濃度の強制的上昇は、野生型FLMCs においてJNK、p38およびErkを活性化した(図5b)。対照的に、イオノマイシンで誘導されたこれらのMAPKsの活性化は、STIM1欠損FLMCsで消滅した。このことは、STIM1はイオノマイシンで誘導されたCa2+流入により誘発されるJNK、p38およびErkの活性化に不可欠であることを示唆する。
【0077】
NF-κBおよびNFATシグナル伝達経路に対するSTIM1の必須の役割
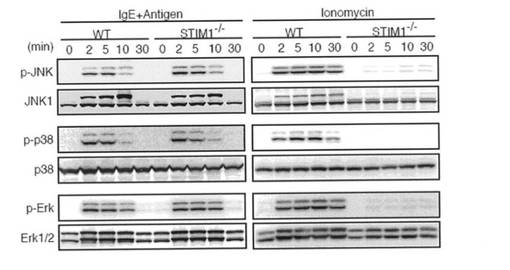
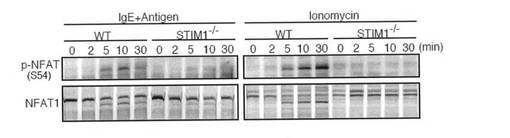
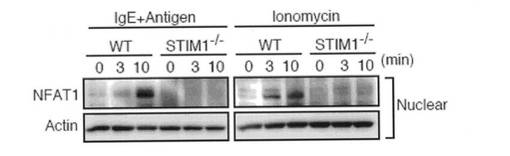
FcεRIの刺激は、2つの転写因子NF-κBおよびNFATを活性化させる。当該転写因子は、サイトカイン産生等の肥満細胞の機能を調節していることが知られている。以前の研究により、NF-κBは大きな一過性のCa2+上昇により選択的に活性化され、他方、NFATは低い持続したCa2+プラトーにより活性化されることが示されている。そこで、本発明者らは、NF-κBおよびNFAT活性化に対するSTIM1の寄与を評価した。STIM1欠損FLMCsにおいて、FcεRIおよびイオノマイシン刺激後のIκBαのリン酸化および分解は、野生型FLMCsと比較して有意に低下していた(図6a)。さらに、FcεRIが介在するp65の核への移行は、STIM1欠損FLMCsにおいて顕著に低下していた(図6b)。NFAT活性化を探究するために、NFAT1のSer54におけるリン酸化の状況を調べた。リン酸化がNFAT活性を抑制する他の部位とは異なり、Ser54をリン酸化されたNFATは、自身の活性を高める。図6cに示すように、STIM1欠損FLMCsでは、FcεRIおよびイオノマイシン刺激後にNFATのSer54はほとんどリン酸化されていなかった。さらに、NFAT1の脱リン酸化および核への移行がSTIM1欠損FLMCsで非常に障害されていたことを見出した(図6c、下部パネル、および図6d)。以上の結果をまとめると、NF-κBおよびNFAT活性化は両方とも、STIM1依存性Ca2+流入により調節されていると結論付けられる。肥満細胞におけるIL-6/TNF-αおよびTNF-α/IL-13産生はそれぞれ、NF-κBおよびNFATにより調節されていることを考慮すると、STIM1不在によるNF-κBおよびNFAT活性化の低下は、かかるサイトカインの産生を不十分にさせるものと思われる。
【0078】
STIM1はin vivoでのアナフィラキシー応答に必要である
STIM1がin vivoの肥満細胞において機能的な役割を果たしているかどうかを評価するために、FcεRIが介在する受動皮膚アナフィラキシー(PCA)反応を行った。STIM1欠損マウスは胎生致死であるので、成体のSTIM1+/-マウスを解析した。図1に示すように、STIM1+/-マウス由来のFLMCsでは、STIM1+/+マウス由来のものに比べて、STIM1タンパク質が非常に低量であった。PCAは、抗DNP IgEを右耳の耳介に皮内注射することにより誘導した。内部標準として、左耳に同量の生理食塩水を注射した。16〜18時間後、マウスを抗原DNP-HSAで静脈内注射によりチャレンジし、エバンスブルー染色を行った。PCA誘導後最初の1時間の間のエバンスブルー染色の血管外溢出は、主として、活性化した肥満細胞の迅速なヒスタミンおよびセロトニン放出を伴う脱顆粒と、それによる局所的に血管の透過性が上昇することに依存している。血管外溢出は、生理食塩水処理耳と比較して、IgE処理耳由来の組織バイオプシーにおける染色のレベルを決定することにより定量した。対照条件として、STIM1+/+マウスとSTIM1+/- マウスとの間で、エバンスブルーのレベルは相違しなかった。しかしながら、STIM1+/- マウスのIgE処理耳介における血管外溢出は、STIM1+/+マウスよりも有意に低く、このことから、STIM1は抗原で誘導された肥満細胞が介在するPCA反応に必要であることが示唆された。
【0079】
配列表のフリーテキスト
〔配列番号:5〕siRNAセンス鎖
〔配列番号:6〕siRNAセンス鎖
【産業上の利用可能性】
【0080】
本発明によれば、新たな作用機序に基づく肥満細胞の脱顆粒抑制剤が提供される。当該作用機序を指標とした本発明のスクリーニング方法は、STIM1に特異的に作用する薬剤の開発に貢献することができる。
【図面の簡単な説明】
【0081】
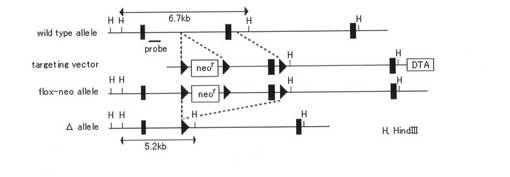
【図1a】図1aは、Stim1遺伝子の標的化戦略を示す概略図である。標的ベクターは、Stim1遺伝子のエキソン6およびneor遺伝子を2つのloxP部位(黒三角)に隣接するように設計した。Cre介在の遺伝子除去を行い、欠損したアレル(D)を生成させた。関連するHind III認識部位(H)を示す。
【図1b】図1bは、E15.5胎仔由来のHind III消化ゲノムDNAのサザンブロット解析を示す。使用したプローブは、図1aに示す。
【図1c】図1cは、E15.5胎仔由来の肝臓の全細胞抽出物の、STIM1およびActinに特異的な抗体を用いたイムノブロット解析を示す。標準の分子量をkDaで示す。
【図1d】図1dは、E15.5胎仔肝臓由来の肥満細胞(FLMCs)の全細胞抽出物の、STIM1およびActinに特異的な抗体を用いたイムノブロット解析を示す。標準の分子量をkDaで示す。
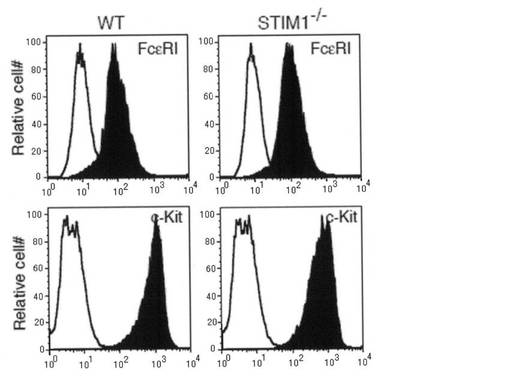
【図2a】図2aは、野生型(WT)およびSTIM1-/-マウス由来のFLMCsにおけるFcεRIおよびc-Kitの細胞表面発現を調べたフローサイトメトリーを示す。
【図2b】図2bは、サイトスピン標品のニュークレアファストレッド染色およびアルシャンブルー染色の代表例を示す。
【図2c】図2cは、WTおよびSTIM1-/-FLMCsにおけるRT-PCRによるMMCP-5およびMMCP-6 mRNA発現を調べた結果を示す。
【図2d】図2dは、WTおよびSTIM1-/-E15.5 胎仔の皮膚組織の肥満細胞の組織学的解析を示す。切片を、ニュークレアファストレッドおよびアルシャンブルーで染色した。矢頭は、アルシャンブルーで染色された肥満細胞を示す。肥満細胞の絶対数(mm2当たり)を、右側のグラフに示す。データは平均+ s.d.で示す。
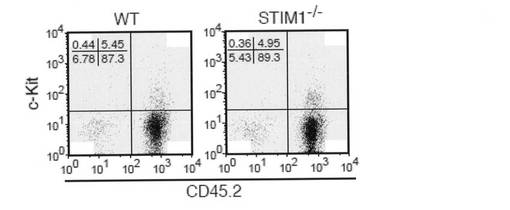
【図2e】図2eは、WTおよびSTIM1-/-マウス(CD45.2)のE15.5 胎仔肝臓由来の2 x 106個の細胞を、致死量を照射したコンジェニックマウス(CD45.1)に注入した結果を示す。10週後、細胞を腹腔から単離し、フローサイトメトリーにより解析した。ドナー型細胞は、CD45.2マーカーに従って同定した。
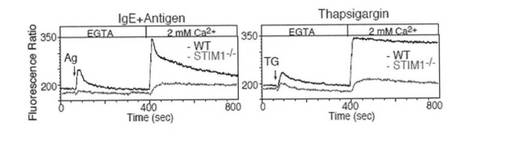
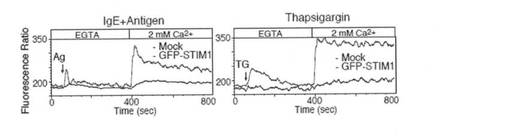
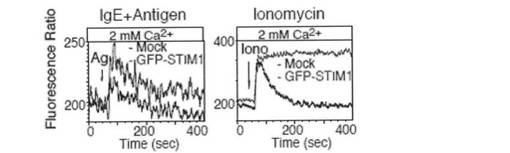
【図3a】図3aは、Indo-1イメージングを用いて、STIM1欠損FLMCs におけるCa2+ 動員プロファイルをモニターしたグラフである。まず、抗DNP IgEで感作したFLMCsを、DNP-HSA抗原(Ag; 50ng/ml)またはタプシガルギン(TG; 2μM)の刺激によりCa2+フリー条件下(0.5 mM EGTA)でCa2+放出を惹起し、野生型(WT; 黒)およびSTIM1欠損(STIM1-/-; 赤)FLMCsにおいて細胞外Ca2+ 濃度を2mMに回復させることによってCa2+流入を誘導した。解析結果は、少なくとも3つの独立した実験の代表例である。
【図3b】図3bは、2mM Ca2+ 存在下、WTおよびSTIM1-/- マウス由来の抗DNP IgEで感作したFLMCsをDNP-HAS抗原(50 ng/ml)での刺激後、またはイオノマイシン(Iono; 1μM)刺激後の未感作FLMCsにおいて、Ca2+動員プロファイルをモニターしたグラフである。解析結果は、少なくとも3つの独立した実験の代表例である。
【図3c】図3cは、GFP-STIM1を発現するSTIM1欠損FLMCsがCa2+流入を回復したことを示すグラフである。GFP単独(Mock; 黒)またはGFP-STIM1(青)をレトロウイルスにより導入したSTIM1-/- FLMCs におけるCa2+ 動員プロファイルを、図3aと同様に示す。データは、GFP陽性ゲートで解析した。値は、FL5/FL4 (FL4= 500-520 nm、FL5= 400-420 nm)での蛍光比としてプロットした。解析結果は、少なくとも3つの独立した実験の代表例である。
【図3d】図3dは、GFP-STIM1を発現するSTIM1欠損FLMCsがCa2+流入を回復したことを示すグラフである。GFP単独(Mock; 黒)またはGFP-STIM1(青)をレトロウイルスにより導入したSTIM1-/-FLMCsにおけるCa2+ 動員プロファイルを、図3bと同様に示す。データは、GFP陽性ゲートで解析した。値は、FL5/FL4 (FL4= 500-520 nm、FL5= 400-420 nm)での蛍光比としてプロットした。すべての解析結果は、少なくとも3つの独立した実験の代表例である。
【図4a】図4aは、野生型(WT)およびSTIM1-/-マウス由来のFLMCsの脱顆粒について、抗DNP IgEで感作した細胞を所定の用量のDNP-HAS抗原で30分間刺激した後、β-ヘキサミニダーゼ放出を測定することによって評価した結果を示す。未感作細胞は、1μM イオノマイシンで刺激した。結果は、三連の試料からの平均 + s.d.であり、3つの異なるペアを用いた3つの独立した実験の代表例である。*, P < 0.05、**, P < 0.01、***, P < 0.001、野生型コントロールに対する。
【図4b】図4bは、野生型(WT)およびSTIM1-/- マウス由来のFLMCsを3時間抗原刺激後、IL-6、TNF-αおよびIL-13の放出を測定したELISA法によるサイトカイン産生の評価を示す。結果は、3つの独立した実験の代表例である(平均 + s.d.)。*, P < 0.05、**, P < 0.01、***, P < 0.001、野生型コントロールに対する。< DL, 検出限界未満。
【図4c】図4cは、感作したFLMCsを抗原で1時間刺激し、サイトカインおよびGAPDHのmRNA発現を半定量的RT-PCRにより検出した結果を示す。結果は、3つの独立した実験の代表例である。
【図5a】図5aは、野生型(WT)およびSTIM1欠損FLMCsにおけるFcεRI近位シグナル伝達の活性化を示す。抗DNP IgEで感作したWTおよびSTIM1-/-FLMCsを、50 ng/ml DNP-HAS抗原で所定の間隔を置いて刺激した。全細胞溶解液をSDS-PAGEで分画し、所定の抗体でイムノブロットした。結果は、3つの独立した実験の代表例である。
【図5b】図5bは、野生型(WT)およびSTIM1欠損FLMCsにおけるMAPKsの活性化を示す。抗DNP IgEで感作したWTおよびSTIM1-/-FLMCsを、50ng/ml DNP-HAS抗原で所定の間隔を置いて刺激した(図5bの左パネル)。あるいは、未感作のWTおよびSTIM1-/-FLMCsを、1μM イオノマイシンで所定の間隔を置いて刺激した(図5bの右パネル)。全細胞溶解液をSDS-PAGEで分画し、所定の抗体でイムノブロットした。結果は、3つの独立した実験の代表例である。
【図6a】図6aは、野生型(WT)およびSTIM1-/- マウス由来のFLMCsを抗DNP IgEで感作し、次いで、所定の間隔を置いて50 ng/ml DNP-HAS抗原で刺激した場合のNF-κBおよびNFATの活性化を示す。あるいは、未感作WTおよびSTIM1-/-FLMCsを1μM イオノマイシンで刺激した場合のNF-κBおよびNFATの活性化を示す。全細胞溶解液を、SDS-PAGEで分画した。IκBαのリン酸化および分解を、イムノブロッティングにより決定した。イムノブロット解析は、少なくとも3つの独立した実験の代表例である。
【図6b】図6bは、共焦点顕微鏡によるNF-κBの核移行を評価した結果を示す。抗DNP IgEで感作したWTおよびSTIM1-/- FLMCsを、100 ng/ml DNP-HAS抗原で10分間刺激し、次いで固定し、抗p65 Ab (緑)およびDAPI (青)での免疫蛍光染色に供した。スケールバー、5μm。各試料中の全部で100個の細胞をランダムにカウントし、p65が核に移行した細胞の頻度を計算した。データは平均 + s.d.である。***, P < 0.001、野生型コントロールに対する。
【図6c】図6cは、FLMCsを所定の間隔を置いて図6aと同様に刺激した結果を示す。全細胞溶解液を、SDS-PAGEにより分画し、リン酸化NFAT (Ser54)およびNFAT1に対する特異的抗体でイムノブロットした。イムノブロット解析は、少なくとも3つの独立した実験の代表例である。
【図6d】図6dは、NFAT1の核移行をイムノブロットにより決定した結果を示す。刺激した細胞由来の核抽出物を、抗NFAT1抗体および抗actin抗体でブロットした。イムノブロット解析は、少なくとも3つの独立した実験の代表例である。
【図7】図7は、STIM1+/-マウスにおいてin vivoアナフィラキシーが減少することを示すグラフである。PCA反応のため、マウスの右耳に抗DNP IgEを皮下注射し、対照として、左耳に生理食塩水を皮下注射した(STIM1+/+; n=10, STIM1+/-; n=8)。16〜18時間後、DNP-HASをエバンスブルー色素とともに静脈内投与し、エバンスブルーの両耳への血管外溢出を測定した。データは、エバンスブルー色素の量を示す(平均 + s.d.)。*, P < 0.05、**, P < 0.01、対応する対照に対する。n.s., 有意差なし。
【技術分野】
【0001】
本発明は、肥満細胞の脱顆粒抑制剤に関し、アレルギーの治療分野に属する。
【背景技術】
【0002】
肥満細胞はイムノグロブリンE(IgE)関連Th2ヘルパー細胞依存性の即時型過剰反応およびアレルギー疾患ならびにある種の自然免疫応答において、主要なエフェクター細胞である(非特許文献1〜3)。高親和性IgEレセプター(FcεRI)に結合したIgE分子への多価抗原の結合によって、肥満細胞上に発現するFcεRIが凝集し、かかる凝集が肥満細胞の活性化を開始する(非特許文献4、5)。活性化した肥満細胞は、プロテアーゼや、ヒスタミン等の血管作動性アミンを含む前もって作られた(細胞質内の顆粒に貯蔵されている)メディエーターを分泌する。また、肥満細胞の活性化は、前炎症性脂質メディエーターおよびサイトカインのde novo合成につながる。FcεRI凝集に応答したこのようなメディエーターの合成および放出は、多数のプロテインチロシンキナーゼカスケードを含むシグナル伝達経路を介して調節される。
【0003】
これらのカスケードの下流の重要な標的の1つは、ホスホリパーゼC−γ(PLCγ)である。PLCγは、ホスファチジルイノシトール−4,5−ビスホスフェートのジアシルグリセロールおよびイノシトール−1,4,5−トリホスフェート(IP3)への加水分解を触媒する。IP3は、小胞体(ER)膜のその受容体に結合し、それにより、Ca2+が迅速ではあるが一過性にERストアから放出される。カルシウム動員の第2ステージでは、形質膜(PM)を横切って細胞外Ca2+が持続的に流入する。この第2段階で、ERプール内のCa2+ストアが空になることそれ自体がPMのCa2+チャネルを活性化すると考えられている(いわゆるストア作動性Ca2+(SOC)流入メカニズム)(非特許文献6、7)。しかしながら、ストア枯渇に依存しない、PM中のCa2+チャネルを活性化する他のメカニズムも報告されている。
【0004】
最近の研究により、SOC流入メカニズムにおける主要なプレーヤーが明確にされてきた(非特許文献8〜11)。Ca2+流入のゲノムワイドRNAiスクリーニングにより、ER膜タンパク質STIM1が同定された。STIM1は、そのEFハンドCa2+結合モチーフにより、内腔のCa2+量の枯渇を検出するCa2+センサーである(非特許文献8、9)。ストア枯渇の際に、STIM1はPM下部の点状に移動し、その後PMのSOCチャネルが活性化される(非特許文献9、12、13)。
【0005】
肥満細胞活性化および脱顆粒におけるCa2+の重要性は、薬理学的試験に基づいて広く信じられている。EGTA処理は、FcεRIが介在するカルシウム動員および脱顆粒をほぼ完全に抑制した(非特許文献14、15)。しかし、最近の知見によると(非特許文献16〜19)、Lyn欠損肥満細胞はカルシウムの動員が減弱しているにも関らず、脱顆粒を誘導可能であり、カルシウム動員の第2ステージ(主としてカルシウム流入)が実際に肥満細胞の活性化および脱顆粒に必須であるのか否か不明である。さらに、もしそうだとしても、SOC依存性および非依存性経路の両方が共存していると思われるので、SOC経路が肥満細胞応答に主要な役割を果たしているのか未だに不明である。
【非特許文献1】Wedemeyer, J. et al. Curr Opin Immunol 12, 624-631 (2000)
【非特許文献2】Bischoff, S. C. Nat Rev Immunol 7, 93-104 (2007)
【非特許文献3】Galli, S. J. et al. Nat Immunol 6, 135-142 (2005)
【非特許文献4】Kraft, S. & Kinet, J. P. Nat Rev Immunol 7, 365-378 (2007)
【非特許文献5】Gilfillan, A. M. & Tkaczyk, C. Nat Rev Immunol 6, 218-230 (2006)
【非特許文献6】Venkatachalam, K. et al. Nat Cell Biol 4, E263-272 (2002)
【非特許文献7】Parekh, A. B. & Putney, J. W. Physiol Rev 85, 757-810 (2005)
【非特許文献8】Roos, J. et al. J Cell Biol 169, 435-445 (2005)
【非特許文献9】Liou, J. et al. Curr Biol 15, 1235-1241 (2005)
【非特許文献10】Putney, J. W., Jr. J Cell Sci 120, 1959-1965 (2007)
【非特許文献11】Hogan, P. G. & Rao, A. Trends Biochem Sci 32, 235-245 (2007)
【非特許文献12】Baba, Y. et al. Proc Natl Acad Sci USA 103, 16704-16709 (2006)
【非特許文献13】Luik, R. M. & Lewis, R. S. Trends Mol Med 13, 103-107 (2007)
【非特許文献14】Ozawa, K. et al. J Biol Chem 268, 1749-1756 (1993)
【非特許文献15】Nishida, K. et al. J Cell Biol 170, 115-126 (2005)
【非特許文献16】Odom, S. et al. J Exp Med 199, 1491-1502 (2004)
【非特許文献17】Parravicini, V. et al. Nat Immunol 3, 741-748 (2002)
【非特許文献18】Nishizumi, H. & Yamamoto, T. J Immunol 158, 2350-2355 (1997)
【非特許文献19】Hernandez-Hansen, V. et al. J Immunol 173, 100-112 (2004)
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明の目的は、新たな作用機序に基づく肥満細胞の活性化の抑制手段、具体的には肥満細胞の脱顆粒抑制剤等を提供することにある。
【課題を解決するための手段】
【0007】
肥満細胞活性化におけるSOC流入の関与に直接取り組むために、本発明者らは、STIM1欠損マウスを作出した。その結果、STIM1がFcεRI誘導性Ca2+流入を厳密に調節し、NF−κBおよびNFAT活性化を介し、脱顆粒ならびにin vivoアナフィラキシー応答を調節していることを解明し、本発明を完成するに至った。これは、肥満細胞活性化におけるSTIM1の重要性を直接遺伝学的に証明したものである。即ち、本願発明は、以下に示す通りである。
【0008】
〔1〕 STIM1阻害物質を有効成分として含有する肥満細胞の脱顆粒抑制剤。
〔2〕 STIM1阻害物質を有効成分として含有するアレルギーの予防または治療剤。
〔3〕 STIM1阻害物質を有効成分として含有するアナフィラキシーの予防または治療剤。
〔4〕 STIM1阻害物質を有効成分として含有する肥満細胞の炎症性サイトカインの産生抑制剤。
〔5〕 STIM1阻害物質が、RNAi誘導性核酸、アンチセンス核酸もしくはリボザイムまたはそれらの発現ベクターである、前記〔1〕〜〔4〕いずれかに記載の剤。
〔6〕 RNAi誘導性核酸がsiRNAである、前記〔5〕に記載の剤。
〔7〕 siRNAが、配列番号1の塩基配列に対応するmRNAにおける19〜25個の連続する塩基配列を含むセンス鎖と、その相補配列を含むアンチセンス鎖からなるものである、前記〔6〕に記載の剤。
〔8〕 被験物質がSTIM1遺伝子の発現または機能を抑制し得るか否かを評価することを含む、肥満細胞の脱顆粒を抑制し得る物質のスクリーニング方法。
〔9〕 下記の工程(a)〜(c)を含む、前記〔8〕記載の方法:
(a)被験物質とSTIM1遺伝子の発現を測定可能な細胞とを接触させる工程;
(b)被験物質を接触させた細胞におけるSTIM1遺伝子の発現量を測定し、該発現量を被験物質を接触させない対照細胞におけるSTIM1遺伝子の発現量と比較する工程;および
(c)上記(b)の比較結果に基づいて、STIM1遺伝子の発現量を抑制する被験物質を選択する工程。
〔10〕 下記の工程(a)〜(c)を含む、前記〔8〕記載の方法:
(a)被験物質をSTIM1蛋白質に接触させる工程;
(b)被験物質のSTIM1蛋白質に対する結合能を測定する工程;および
(c)上記(b)の結果に基づいて、STIM1蛋白質に結合能を有する被験物質を選択する工程。
【発明の効果】
【0009】
本発明の肥満細胞の脱顆粒抑制剤によれば、STIM1阻害物質を有効成分として含むことから、これまでにない作用機序に基づいた分子標的医薬品の開発に貢献することができる。STIM1阻害物質を有効成分として含む医薬は、アレルギーまたはアナフィラキシーの予防または治療薬として有用である。また、本発明のスクリーニング方法によれば、STIM1を分子標的とする候補物質を迅速にスクリーニングすることができる。
【発明を実施するための最良の形態】
【0010】
本発明は、STIM1阻害物質を有効成分として含有する肥満細胞の脱顆粒抑制剤を提供する。
【0011】
本発明において、STIM1とは、1つの膜貫通ドメインと1つのEF−ハンドカルシウム結合モチーフを有する間質細胞相互作用分子1をいい、小胞体膜に存在するカルシウムセンサータンパク質である。
【0012】
本発明において、STIM1は、任意の哺乳動物由来の膜タンパク質である。哺乳動物としては、ヒトおよびヒトを除く哺乳動物が挙げられ、ヒトを除く哺乳動物としては、例えば、マウス、ラット、ハムスター、モルモット等のげっ歯類やウサギ等の実験動物、ブタ、ウシ、ヤギ、ウマ、ヒツジ等の家畜、イヌ、ネコ等のペット、サル、オランウータン、チンパンジー等の霊長類が挙げられる。ヒトのアレルギー疾患の治療に用いるためには、ヒト由来のSTIM1が好ましい。ヒトSTIM1の塩基配列およびアミノ酸配列は公知であり、例えば、STIM1の塩基配列(配列番号1)およびアミノ酸配列(配列番号2)(GenBank Accession No. NM_003156)などがGenBankに登録され、公表されている。また、マウスSTIM1の塩基配列およびアミノ酸配列も公知であり、例えば、マウスSTIM1の塩基配列(配列番号3)およびアミノ酸配列(配列番号4)(GenBank Accession No. NM_009287)などがGenBankに登録され、公表されている。
【0013】
本発明において、STIM1阻害物質とは、STIM1の発現を阻害する物質およびSTIM1の機能(作用)を阻害する物質の両方を含む。
【0014】
本発明の薬剤に有効成分として含まれるSTIM1の発現を阻害する物質は、STIM1の転写過程に作用してその発現を阻害する物質であれば特に限定されるものではない。かかる阻害物質としては、RNAi誘導性核酸、アンチセンス核酸もしくはリボザイムまたはそれらの発現ベクターが挙げられる。
【0015】
前記RNAi誘導性核酸とは、細胞内に導入されることにより、RNA干渉を誘導し得るポリヌクレオチドをいい、好ましくはRNAまたはRNAとDNAのキメラ分子である。RNA干渉とは、mRNAと同一の塩基配列(またはその部分配列)を含む2本鎖構造のRNAが、当該mRNAの発現を抑制する効果をいう。このRNAi効果を得るには、例えば、少なくとも19の連続する標的mRNAと同一の塩基配列(またはその部分配列)を有する2本鎖構造のRNAを用いることが好ましい。ただし、STIM1の発現阻害作用を有していれば数塩基置換されているものであってもよく、19塩基長よりも短いRNAであってもよい。2本鎖構造は、センス鎖とアンチセンス鎖の異なるストランドで構成されていてもよいし、一つのRNAのステムループ構造によって与えられる2本鎖(shRNA)であってもよい。RNAi誘導性核酸としては、例えばsiRNA、miRNAなどが挙げられる。
【0016】
RNAi誘導性核酸は、転写抑制活性が強いという観点から、siRNAが好ましい。STIM1に対するsiRNAは、STIM1のmRNAの任意の部分を標的とすることができる。STIM1に対するsiRNA分子は、RNAi効果を誘導できる限り特に制限されないが、例えば19〜27塩基長、好ましくは21〜25塩基長である。STIM1に対するsiRNAは、センス鎖およびアンチセンス鎖を含む二重鎖である。具体的には、STIM1に対するsiRNAは、配列番号1または3の塩基配列に対応するmRNAにおける19〜25個の連続する塩基配列を含むセンス鎖と、その相補配列を含むアンチセンス鎖からなるものである。STIM1に対するsiRNAは、センス鎖、アンチセンス鎖の一方または双方の5’末端または3’末端においてオーバーハング(overhang)を有していてもよい。オーバーハングは、センス鎖および/またはアンチセンス鎖の末端における1〜数個(例、1、2または3個)の塩基の付加により形成されるものである。siRNAの設計方法は、当業者に公知であり、siRNAの様々な設計ソフトウエアまたはアルゴリズムを用いて、上記塩基配列から適切なsiRNAの塩基配列を選択することができる。
【0017】
STIM1に対するsiRNAのセンス鎖の一具体例としては、下記配列:
5’-AAGGCTCTGGATACAGTGCTC-3’(配列番号5)
5’-AAGAAGCTGCGCGATGAGATC-3’(配列番号6)
が挙げられる。
【0018】
STIM1に対するアンチセンス核酸は、STIM1の転写産物(mRNAまたは初期転写産物)を発現する細胞の生理的条件下で該転写産物とハイブリダイズし得る塩基配列からなり、且つハイブリダイズした状態で該転写産物にコードされるポリペプチドの翻訳を阻害し得るポリヌクレオチドをいう。アンチセンス核酸の種類はDNAであってもRNAであってもよいし、あるいはDNA/RNAキメラであってもよい。アンチセンス核酸は、天然型のリン酸ジエステル結合を有するものであっても、分解酵素に安定なチオリン酸型(リン酸結合のP=OをP=Sに置換)や2’-O-メチル型等の修飾ヌクレオチドであってもよい。アンチセンス核酸の設計に重要な他の要素として、水溶性および細胞膜透過性を高めること等が挙げられるが、これらはリポソームやマイクロスフェアを使用するなどの剤形の工夫によっても克服できる。アンチセンス核酸の長さは、STIM1の転写産物(例、配列番号1または3の塩基配列に対応するmRNA)と特異的にハイブリダイズし得る限り特に制限はなく、短いもので約15塩基程度、長いもので転写産物の全配列に相補的な配列を含むような配列であってもよい。合成の容易さや抗原性の問題等から、例えば約15塩基以上、好ましくは約15〜約30塩基、より好ましくは約18塩基〜約30塩基からなるオリゴヌクレオチドが例示される。さらに、アンチセンス核酸は、STIM1の転写産物とハイブリダイズして翻訳を阻害するだけでなく、二本鎖DNAと結合して三重鎖(トリプレックス)を形成し、mRNAへの転写を阻害し得るものであってもよい。
【0019】
本明細書において、「相補的である」とは、塩基配列間で約70%以上、好ましくは約80%以上、より好ましくは約90%以上、更に好ましくは約95%以上、最も好ましくは100%の相補性を有することをいう。本明細書における塩基配列の相同性は、相同性計算アルゴリズムNCBI BLAST(NationalCenter for Biotechnology Information Basic Local Alignment Search Tool)を用い、以下の条件(期待値=10;ギャップを許す;フィルタリング=ON;マッチスコア=1;ミスマッチスコア=-3)にて計算することができる。
【0020】
前記「リボザイム」とは核酸を切断する酵素活性を有するRNAをいうが、最近では当該酵素活性部位の塩基配列を有するオリゴDNAも同様に核酸切断活性を有することが明らかになっているので、本明細書では配列特異的な核酸切断活性を有する限りDNAをも包含する概念として用いる。具体的には、リボザイムは、STIM1をコードするmRNAまたは初期転写産物を、コード領域の内部(初期転写産物の場合はイントロン部分を含む)で特異的に切断し得る。リボザイムとして最も汎用性の高いものとしては、ウイロイドやウイルソイド等の感染性RNAに見られるセルフスプライシングRNAがあり、ハンマーヘッド型やヘアピン型等が知られている。ハンマーヘッド型は約40塩基程度で酵素活性を発揮し、ハンマーヘッド構造をとる部分に隣接する両端の数塩基ずつ(合わせて約10塩基程度)をmRNAの所望の切断部位と相補的な配列にすることにより、標的mRNAのみを特異的に切断することが可能である。さらに、リボザイムを、それをコードするDNAを含む発現ベクターの形態で使用する場合には、転写産物の細胞質への移行を促進するために、tRNAを改変した配列をさらに連結したハイブリッドリボザイムとすることもできる(Nucleic Acids Res., 29(13): 2780-2788 (2001))。
【0021】
STIM1の機能(作用)を阻害する物質としては、STIM1に対する抗体があげられる。該抗体は、ポリクローナル抗体、モノクローナル抗体のいずれであってもよく、周知の免疫学的手法により作製できる。また、該抗体は、抗体のフラグメント(例えば、Fab、F(ab’)2)、組換え抗体(例えば、単鎖抗体)であってもよく、さらに、該抗体をコードする核酸(プロモーター活性を有する核酸に機能可能に連結されたもの)であってもよい。
【0022】
前記抗体は、ヒトにおける治療効果と安全性を考慮すると、キメラ抗体、ヒト化またはヒト型抗体であってもよい。キメラ抗体は、例えば「実験医学(臨時増刊号), Vol.6, No.10, 1988」、特公平3-73280号公報等を、ヒト化抗体は、例えば特表平4-506458号公報、特開昭62-296890号公報等を、ヒト抗体は、例えば「Nature Genetics, Vol.15, p.146-156, 1997」、「Nature Genetics, Vol.7, p.13-21, 1994」、特表平4-504365号公報、国際出願公開WO94/25585号公報、「日経サイエンス、6月号、第40〜第50頁、1995年」、「Nature, Vol.368, p.856-859, 1994」、特表平6-500233号公報等を参考にそれぞれ作製することができる。
【0023】
さらに別の実施形態では、STIM1の機能(作用)を阻害する物質は、STIM1のドミナントネガティブ変異体、該変異体をコードする核酸を含む発現ベクターが例示される。
【0024】
STIM1のドミナントネガティブ変異体とは、STIM1に対する変異の導入によりその活性が低減したものをいう。該ドミナントネガティブ変異体は、天然のSTIM1と競合することで間接的にその機能を阻害することができる。該ドミナントネガティブ変異体は、STIM1遺伝子をコードする核酸に変異を導入することによって作製することができる。変異としては、例えば、機能性部位における、当該部位が担う機能の低下をもたらすようなアミノ酸の変異(例えば、1以上のアミノ酸の欠失、置換、付加)が挙げられる。ドミナントネガティブ変異体は、PCRや公知のキットを用いる自体公知の方法により作製できる。
【0025】
STIM1阻害物質は、発現ベクターとしても提供され得る。かかる発現ベクターは、STIM1阻害物質をコードするポリヌクレオチド、および当該ポリヌクレオチドに機能可能に連結されたプロモーターを含む。
【0026】
前記プロモーターは、その制御下にある発現対象の核酸の種類により適宜選択され得るが、例えば、polIIIプロモーター(例、tRNAプロモーター、U6プロモーター、H1プロモーター)、哺乳動物用プロモーター(例、CMVプロモーター、CAGプロモーター、SV40プロモーター)が挙げられる。
【0027】
本発明の発現ベクターはさらに、選択マーカー遺伝子(テトラサイクリン、アンピシリン、カナマイシン、ハイグロマイシン、ホスフィノスリシン等の薬剤に対する抵抗性を付与する遺伝子、栄養要求性変異を相補する遺伝子等)をさらに含んでいてもよい。
【0028】
本発明の発現ベクターのバックボーン(backbone)としては、ヒト等の哺乳動物細胞中でSTIM1阻害物質を産生できるものであれば特に制限されないが、例えば、プラスミドベクター、ウイルスベクターが挙げられる。哺乳動物への投与に好適なベクターとしては、レトロウイルス、アデノウイルス、アデノ随伴ウイルス、ヘルペスウイルス、ワクシニアウイルス、ポックスウイルス、ポリオウイルス、シンドビスウイルス、センダイウイルス等のウイルスベクターが挙げられる。なかでも、レトロウイルス、アデノウイルス、アデノ随伴ウイルス、ワクシニアウイルス由来のウイルスベクターが好ましい。
【0029】
本発明の肥満細胞の脱顆粒抑制剤は、肥満細胞からのヒスタミン、サイトカイン、ケモカインおよびプロテアーゼ等の遊離物質の分泌を抑制することが可能であり、かかる遊離物質に起因する様々なアレルギー疾患の予防または治療剤としても有用である。アレルギー疾患としては、喘息、アレルギー性鼻炎、アレルギー性結膜炎、アトピー性皮膚炎、アレルギー性胃腸炎などが挙げられる。
【0030】
本発明において、STIM1が受動皮膚アナフィラキシー反応(PCA)に必要であることを生体内で確認したことにより、本発明は、STIM1阻害物質を有効成分として含有するアナフィラキシーの予防または治療剤を提供することができる。
【0031】
また、本発明において、肥満細胞からの炎症性サイトカインの産生および分泌にSTIM1が必須の役割を果たしていることを確認したことにより、本発明は、STIM1阻害物質を有効成分として含有する肥満細胞の炎症性サイトカインの産生抑制剤を提供することができる。
【0032】
前記炎症性サイトカインとしては、IL−3、IL−6、TNF−αなどが挙げられる。
【0033】
前記脱顆粒抑制剤、アレルギー疾患の予防または治療剤、アナフィラキシーの予防または治療剤および炎症性サイトカインの産生抑制剤を総称して、本発明の薬剤と略す場合がある。
【0034】
本発明の薬剤の投与量は、有効成分の種類もしくは活性、投与対象となる動物種、投与対象の病気の重篤度、薬物受容性、体重、年齢等によって異なるが、通常、成人1日あたり有効成分量として約0.0001〜約1000mg/kgが例示される。
【0035】
本発明の薬剤は、患者に対して経口的または非経口的に投与することができ、投与形態としては、経口投与、局所投与(点鼻投与、点眼投与、吸入投与等)、静脈内投与、経皮投与などが挙げられ、必要に応じて、製薬学的に許容され得る添加剤と共に、投与に適した剤型に製剤化される。経口投与に適した剤型としては、例えば、錠剤、カプセル剤、顆粒剤、散剤などが挙げられ、非経口投与に適した剤型としては、例えば、点鼻剤、点眼剤、注射剤、貼付剤、ローション剤、クリーム剤などが挙げられる。これらは当該分野で汎用されている通常の技術を用い、調製することができる。本発明の薬剤は、上述の治療効果を奏する限りその投与経路および剤形は特に限定されない。
【0036】
例えば、本発明の薬剤を注射剤として用いる場合、安定剤(例えば、亜硫酸水素ナトリウム、チオ硫酸ナトリウム、エデト酸ナトリウム、クエン酸ナトリウム、アスコルビン酸、ジブチルヒドロキシトルエンなど)、溶解補助剤(例えば、グリセリン、プロピレングリコール、マクロゴール、ポリオキシエチレン硬化ヒマシ油など)、懸濁化剤(例えば、ポリビニルピロリドン、ヒドロキシプロピルメチルセルロース、ヒドロキシメチルセルロース、カルボキシメチルセルロースナトリウムなど)、乳化剤(例えば、ポリビニルピロリドン、大豆レシチン、卵黄レシチン、ポリオキシエチレン硬化ヒマシ油、ポリソルベート80など)、緩衝剤(例えば、リン酸緩衝液、酢酸緩衝液、ホウ酸緩衝液、炭酸緩衝液、クエン酸緩衝液、トリス緩衝液、グルタミン酸、イプシロンアミノカプロン酸など)、粘稠剤(例えば、メチルセルロース、ヒドロキシエチルセルロース、ヒドロキシプロピルメチルセルロース、カルボキシメチルセルロースなどの水溶性セルロース誘導体、コンドロイチン硫酸ナトリウム、ヒアルロン酸ナトリウム、カルボキシビニルポリマー、ポリビニルアルコール、ポリビニルピロリドン、マクロゴールなど)、保存剤(例えば、塩化ベンザルコニウム、塩化ベンゼトニウム、グルコン酸クロルヘキシジン、クロロブタノール、ベンジルアルコール、デヒドロ酢酸ナトリウム、パラオキシ安息香酸エステル類、エデト酸ナトリウム、ホウ酸など)、等張化剤(例えば、塩化ナトリウム、塩化カリウム、グリセリン、マンニトール、ソルビトール、ホウ酸、ブドウ糖、プロピレングリコールなど)、pH調整剤(例えば、塩酸、水酸化ナトリウム、リン酸、酢酸など)、清涼化剤(例えば、l−メントール、d−カンフル、d−ボルネオール、ハッカ油など)、軟膏基剤(白色ワセリン、精製ラノリン、流動パラフィン、植物油(オリーブ油、椿油、落花生油など)など)などを添加剤として加えることができる。これら添加剤の添加量は、添加する添加剤の種類、用途などによって異なるが、添加剤の目的を達成し得る濃度を添加すればよい。
【0037】
本発明の薬剤は、siRNA等の核酸をリポフェクション法を用いて製剤化することもできる。
【0038】
本発明はまた、被験物質がSTIM1遺伝子の発現または機能を抑制し得るか否かを評価することを含む、肥満細胞の脱顆粒抑制物質のスクリーニング方法、ならびに当該スクリーニング方法により得られる物質、および当該物質を含有してなる脱顆粒抑制剤を提供する。
【0039】
スクリーニング方法に供される被験物質は、いかなる公知化合物および新規化合物であってもよく、例えば、核酸、糖質、脂質、蛋白質、ペプチド、有機低分子化合物、コンビナトリアルケミストリー技術を用いて作製された化合物ライブラリー、固相合成やファージディスプレイ法により作製されたランダムペプチドライブラリー、あるいは微生物、動植物、海洋生物等由来の天然成分等が挙げられる。
【0040】
一実施形態では、本発明のスクリーニング方法は、下記の工程(a)〜(c)を含む:
(a)被験物質とSTIM1遺伝子の発現を測定可能な細胞とを接触させる工程;
(b)被験物質を接触させた細胞におけるSTIM1遺伝子の発現量を測定し、該発現量を被験物質を接触させない対照細胞におけるSTIM1遺伝子の発現量と比較する工程;
(c)上記(b)の比較結果に基づいて、STIM1遺伝子の発現量を抑制する被験物質を選択する工程。
【0041】
上記方法の工程(a)では、被験物質がSTIM1遺伝子の発現を測定可能な細胞と接触条件下におかれる。STIM1遺伝子の発現を測定可能な細胞に対する被験物質の接触は、培養培地中で行われ得る。
【0042】
STIM1遺伝子の発現を測定可能な細胞とは、STIM1遺伝子の産物、例えば、転写産物、翻訳産物の発現レベルを直接的または間接的に評価可能な細胞をいう。STIM1遺伝子の産物の発現レベルを直接的に評価可能な細胞は、STIM1遺伝子を天然で発現可能な細胞であり得、一方、STIM1遺伝子の産物の発現レベルを間接的に評価可能な細胞は、STIM1遺伝子転写調節領域についてレポーターアッセイを可能とする細胞であり得る。STIM1遺伝子の発現を測定可能な細胞は、動物細胞、例えばマウス、ラット、ハムスター、モルモット、ウサギ、イヌ、サル、ヒト等の哺乳動物細胞であり得る。
【0043】
STIM1遺伝子を天然で発現可能な細胞は、STIM1遺伝子を潜在的に発現するものである限り特に限定されない。かかる細胞は、当業者であれば容易に同定でき、初代培養細胞、当該初代培養細胞から誘導された細胞株、市販の細胞株、セルバンクより入手可能な細胞株などを使用できる。
【0044】
STIM1遺伝子転写調節領域についてレポーターアッセイを可能とする細胞は、STIM1遺伝子転写調節領域、当該領域に機能可能に連結されたレポーター遺伝子を含む細胞である。STIM1遺伝子転写調節領域およびレポーター遺伝子は、発現ベクター中に挿入され得る。STIM1遺伝子転写調節領域は、STIM1遺伝子の発現を制御し得る領域である限り特に限定されないが、例えば、転写開始点から上流約2kbpまでの領域、あるいは該領域の塩基配列において1以上の塩基が欠失、置換若しくは付加された塩基配列からなり、且つSTIM1遺伝子の転写を制御する能力を有する領域などが挙げられる。レポーター遺伝子は、検出可能な蛋白質または検出可能な物質を生成する酵素をコードする遺伝子であればよく、例えばGFP(緑色蛍光蛋白質)遺伝子、GUS(β−グルクロニダーゼ)遺伝子、LUC(ルシフェラーゼ)遺伝子、CAT(クロラムフェニコルアセチルトランスフェラーゼ)遺伝子等が挙げられる。
【0045】
STIM1遺伝子転写調節領域、当該領域に機能可能に連結されたレポーター遺伝子が導入される細胞は、STIM1遺伝子転写調節機能を評価できる限り、即ち、該レポーター遺伝子の発現量が定量的に解析可能である限り特に限定されない。しかしながら、STIM1遺伝子に対する生理的な転写調節因子を発現し、STIM1遺伝子の発現調節の評価により適切であると考えられることから、該導入される細胞としては、STIM1遺伝子を天然で発現可能な細胞が好ましい。
【0046】
被験物質とSTIM1遺伝子の発現を測定可能な細胞とが接触される培養培地は、用いられる細胞の種類などに応じて適宜選択されるが、例えば、約5〜20%のウシ胎仔血清を含む最少必須培地(MEM)、ダルベッコ改変最少必須培地(DMEM)、RPMI1640培地、199培地などである。培養条件もまた、用いられる細胞の種類などに応じて適宜決定されるが、例えば、培地のpHは約6〜約8であり、培養温度は通常約30〜約40℃であり、培養時間は約12〜約72時間である。
【0047】
上記方法の工程(b)では、先ず、被験物質を接触させた細胞におけるSTIM1遺伝子の発現量が測定される。発現量の測定は、用いた細胞の種類などを考慮し、自体公知の方法により行われ得る。例えば、STIM1遺伝子の発現を測定可能な細胞として、STIM1遺伝子を天然で発現可能な細胞を用いた場合、発現量は、STIM1遺伝子の産物、例えば、転写産物または翻訳産物を対象として自体公知の方法により測定できる。例えば、転写産物の発現量は、細胞からtotal RNAを調製し、RT−PCR、ノーザンブロッティング等により測定され得る。また、翻訳産物の発現量は、細胞から抽出液を調製し、免疫学的手法により測定され得る。免疫学的手法としては、放射性同位元素免疫測定法(RIA法)、ELISA法(Methods in Enzymol. 70: 419-439 (1980))、蛍光抗体法などが使用できる。一方、STIM1遺伝子の発現を測定可能な細胞として、STIM1遺伝子転写調節領域についてレポーターアッセイを可能とする細胞を用いた場合、発現量は、レポーターのシグナル強度に基づき測定され得る。
【0048】
次いで、被験物質を接触させた細胞におけるSTIM1遺伝子の発現量が、被験物質を接触させない対照細胞におけるSTIM1遺伝子の発現量と比較される。発現量の比較は、好ましくは、有意差の有無に基づいて行なわれる。被験物質を接触させない対照細胞におけるSTIM1遺伝子の発現量は、被験物質を接触させた細胞におけるSTIM1遺伝子の発現量の測定に対し、事前に測定した発現量であっても、同時に測定した発現量であってもよいが、実験の精度、再現性の観点から同時に測定した発現量であることが好ましい。
【0049】
上記方法の工程(c)では、STIM1遺伝子の発現量を抑制する被験物質が選択される。STIM1遺伝子の発現量の抑制は、発現量の減少であり得る。STIM1遺伝子の発現量を減少させる(発現を抑制する)被験物質は、脱顆粒機能を抑制する作用を有し得、アレルギー疾患またはアナフィラキシーの予防・治療薬となり得る。従って、STIM1遺伝子の発現量を指標として、アレルギー疾患の予防・治療剤等の医薬、または研究用試薬のための候補物質を選択することが可能となる。
【0050】
別の実施形態では、本発明のスクリーニング方法は、下記の工程(a)〜(c)を含む:
(a)被験物質をSTIM1蛋白質に接触させる工程;
(b)被験物質のSTIM1蛋白質に対する結合能を測定する工程;
(c)上記(b)の結果に基づいて、STIM1蛋白質に結合能を有する被験物質を選択する工程。
【0051】
上記方法の工程(a)では、被験物質がSTIM1蛋白質と接触条件下におかれる。被験物質の該蛋白質に対する接触は、溶液中での被験物質とSTIM1蛋白質との混合により行われ得る。
【0052】
STIM1蛋白質は自体公知の方法により調製できる。例えば、遺伝子組換え技術により組換えSTIM1蛋白質を調製するのが好ましい。組換え蛋白質は、細胞系、無細胞系のいずれで調製したものでもよい。
【0053】
上記方法の工程(b)では、STIM1蛋白質に対する被験物質の結合能が測定される。結合能の測定は、自体公知の方法、例えば、バインディングアッセイ、表面プラズモン共鳴を利用する方法(例えば、Biacore(登録商標)の使用)により行われ得る。STIM1蛋白質に対する被験物質の結合能はまた、STIM1蛋白質に対する拮抗体(例えば、STIM1蛋白質の共役因子)の存在下で測定してもよい。この場合、例えば、STIM1蛋白質に対する拮抗体の結合量の変化を指標に、STIM1蛋白質に対する被験物質の結合能が測定され得る。
【0054】
上記方法の工程(c)では、STIM1蛋白質に結合能を有する被験物質が選択される。STIM1蛋白質に結合能を有する被験物質は、STIM1遺伝子の機能を抑制し得る物質となる可能性がある。従って、本方法論は、例えば、STIM1遺伝子の機能を抑制し得る物質の一次スクリーニングとして有用である。
【0055】
本発明のスクリーニング方法はまた、被験物質の動物への投与により行われ得る。該動物としては、例えば、マウス、ラット、ハムスター、モルモット、ウサギ、イヌ、サル等の哺乳動物が挙げられる。動物を用いて本発明のスクリーニング方法が行われる場合、例えば、STIM1遺伝子の発現量を抑制する被験物質が選択され得る。
【0056】
本発明のスクリーニング方法は、脱顆粒を抑制し得る物質のスクリーニングを可能とする。従って、本発明のスクリーニング方法は、上述の薬剤または研究用試薬の開発などに有用である。
【実施例】
【0057】
以下、実施例により本発明を詳細に説明するが、本発明はこれらの実施例により何ら限定されるものではない。
【0058】
実施例1
STIM1標的化マウスの作出
STIM1標的化マウスを作出するためのターゲティングベクターは、STIM1の膜貫通領域をコードする配列を含むエキソン6の両端にそれぞれloxP部位を有するように2つのloxP部位を隣接させて設計した。かかるコンストラクトのために、ゲノムDNA断片をC57BL/6起源のBruce-4 ES細胞(ハーバードメディカルスクール、CBRバイオメディカルリサーチ研究所のK. Rajewsky博士より供与)から増幅させた。KSTKNEOLOXPベクター(K. Rajewsky博士より供与)中の第1のloxP部位を有するネオマイシン耐性カセットを、Stim1のイントロン5を含む1.7-kbの5'ホモロジー領域の下流にクローニングし、エキソン6を含む断片を2つのloxP部位の間に挿入した。エキソン7を含む4.5-kbの3'ホモロジー領域の後に、ジフテリアトキシンA(DTA)カセット(Yagi, T. et al. Proc Natl Acad Sci U S A 87, 9918-9922 (1990))を連結した。DTA 遺伝子は、ランダムインテグレーションを有するクローンのネガティブセレクションに使用した。ターゲティングコンストラクトを直鎖状にし、Bruce-4 ES細胞にエレクトロポレーションにより導入した。形質転換細胞は、G418で選択した。ターゲットクローンをサザンブロット解析により同定し、C57BL/6の胚盤胞にマイクロインジェクションし、キメラを生成させた。キメラ雄性マウスとC57BL/6雌性マウスとの交配により、flox-neoアレルが生殖系列へ伝達された。得られたStim1flox/+マウスを、サイトメガロウイルス即時型初期エンハンサー−ニワトリアクチンハイブリッド(CAG)プロモーターの制御下にCreトランスジーンを保持するトランスジェニックマウス系列と交配して、CAGCre/+Stim1flox/+(遺伝子型;Stim1+/-)を作出した。次いで、このマウスを交雑させて、Stim1-/-マウスを作出した。すべての実験は、理化学研究所の動物実験委員会のガイドラインに遵守してなされた。
【0059】
抗体
Syk、LAT、JNK、p38、IκBα、p65、ActinおよびNFAT1 (NFATc2)に対する抗体は、Santa Cruz社から購入した。抗リン酸化JNK抗体、抗リン酸化p38抗体および抗Erk1/2抗体は、Promega社から購入した。PLCγ1、リン酸化PLCγ1、リン酸化Syk、リン酸化IκBα、リン酸化LATおよびリン酸化Erk(p42/44)に対する抗体は、Cell Signaling Technology社から購入し、リン酸化NFAT1に対する抗体は、Abcam社から購入した。抗STIM1抗体は、既報(Baba, Y. et al. Proc Natl Acad Sci USA103, 16704-16709 (2006))に従って製造した。FITC-結合抗CD45.2 mAbおよびPE-結合抗FcεRI mAb、APC-結合抗c-Kit mAbは、それぞれ、BD社およびeBioscience社から購入した。
【0060】
実施例2
肥満細胞の培養
胎仔肝由来肥満細胞(FLMCs)の調製のため、STIM1-/-マウスおよび同腹子野生型マウス由来のE15.5胎仔肝から懸濁細胞を取り出し、10% FCS、β-メルカプトエタノール、抗生物質および20ng/ml組換えマウス(rm)IL-3(R&D systems)を補足したRPMI1640で培養した。培養中、培地を3〜4日毎に交換した。培養後4〜5週に、当該細胞がその細胞表面にFcεRIおよびc-Kitを発現していることを確認し、実験に使用した(< 98% 肥満細胞)。
【0061】
実施例3
組織染色
E15.5の胎仔の5μmパラフィン切片を固定し、ニュークレアファストレッドおよびアルシャンブルーで染色した。肥満細胞を観察するために、各試料中のアルシャンブルー染色細胞を計数した。
【0062】
実施例4
脱顆粒およびサイトカイン産生の測定
細胞(1 x 106/ml)を、1μg/ml 抗DNP IgE (SPE-7; Sigma)を用いて37℃で6時間感作し、次いで、感作した細胞を、Tyrode’s緩衝液(10 mM HEPES pH 7.4、130 mM NaCl、5 mM KCl、1.4 mM CaCl2、1 mM MgCl2、5.6 mM グルコース、0.1% BSA)中、多価ジニトロフェニル-ヒト血清アルブミン(DNP-HSA, Sigma)で30分間刺激した。β-ヘキソサミニダーゼ反応に関しては、50μlの細胞上清または細胞溶解液を、100μlの1.3mg/ml p-ニトロフェニル-N-アセチル-D-グルコサミド(0.1M クエン酸塩(pH 4.5)中)とともにインキュベートし、37℃で60分発色させた。次いで、0.2M グリシン-NaOH(pH 10.2)を150μl添加することにより酵素反応を停止させ、405nmでの吸光度をマイクロプレートリーダー(Bio-Rad)で測定した。細胞を、1%Triton X-100を含むTyrode’s緩衝液で溶解し、β-ヘキソサミニダーゼ活性を測定した。放出されたβ-ヘキソサミニダーゼの割合を、下記式:
放出(%) = 上清 / (上清 + 細胞溶解液) x 100
を用いて計算した。サイトカイン放出の測定のため、細胞培養上清中のIL-6、TNF-αおよびIL-13を、既報(Kabu, K. et al. J Immunol 177, 1296-1305 (2006))に記載のELISAアッセイにより検出した。
【0063】
実施例5
カルシウム濃度の測定
細胞質ゾルのカルシウム濃度は、既報(Baba, Y. et al. Proc Natl Acad Sci USA103, 16704-16709 (2006))に従って測定した。簡潔に記載すると、胎仔肝肥満細胞を、RPMI1640/ 1% FCS中、37℃で45分、Indo-1-AMおよびPluronic F-127 (Invitrogen)を負荷した。FcεRI刺激に関しては、胎仔肝肥満細胞を抗DNP IgE(2μg/ml)で同時に感作した。蛍光強度の変化を、LSRII(BD Biosciences)でモニターした。FL4(Ca2+-フリーindo-1)の蛍光に対するFL5(Ca2+-結合indo-1)の蛍光の比として、値をプロットした。データは、Flowjo software(Treestar)で解析した。
【0064】
実施例6
細胞の刺激およびイムノブロッティング
細胞(1 x 106/ml)を、1μg/ml IgEで37℃、6時間感作し、次いで、Tyrode’s緩衝液中50 ng/ml DNP-HASで刺激した。あるいは、未感作細胞を1μM イオノマイシンで刺激した。刺激後、前記細胞を、10 mM Tris-HCl pH 7.4、150 mM NaCl、1 % NP-40、0.1 % SDS、0.5 mM EDTA、プロテアーゼ阻害剤(Roche)およびホスファターゼ(Nakarai)阻害剤カクテルを含む溶解緩衝液中で溶解し、清澄化させた細胞溶解液を、5 x SDSサンプル緩衝液とともに煮沸した。5 x 105〜1 x 106 細胞分の試料を、PVDF膜に電気泳動で転写し、適切な抗体でブロットした。核抽出物は、Extraction Kit (Pierce)を用いて製造業者のプロトコルに従って調製し、イムノブロッティングに供した。
【0065】
実施例7
イムノフルオレッセンスおよび共焦点顕微鏡観察
細胞を、余熱した4 %パラホルムアルデヒドで15分間固定し、0.2 % Triton X-100で5分間透過処理した。前記細胞を2 %ヤギ血清でブロッキングした後、2 %ヤギ血清を含むPBSで1:50希釈した抗p65 Abとともに1時間インキュベートした。前記一次抗体を、Alexa Fluor 488で標識した抗ウサギ二次抗体(Invitrogen)を用いて検出した。核は、DAPIで共染色した。染色した細胞は、共焦点蛍光顕微鏡(DMIRE2; Leica Microsystems) 下で観察した。
【0066】
実施例8
RT-PCR解析
MMCP-5、MMCP-6、アクチン、IL-6、TNF-α、IL-13およびGAPDHの発現を評価するためのRT-PCRは、既報(Kabu, K. et al. J Immunol 177, 1296-1305 (2006))に記載された遺伝子特異的プライマーを用いて行った。
【0067】
実施例9
胎仔肝細胞の移植
STIM1-/-または野生型同腹子のE15.5胎仔由来の2 x 106個の胎仔肝細胞を、致死量(950rad)を照射したコンジェニックマウス(CD45.1)に静脈内に移植した。10週間後、前記マウスから腹膜腔を回収し、フローサイトメトリーにより解析した。CD45.2を用いて、宿主細胞とドナー細胞とを区別した。
【0068】
実施例10
レトロウイルスの産生および感染
GFPタグ化したSTIM1 cDNA(Baba, Y. et al. Proc Natl Acad Sci USA103, 16704-16709 (2006))を、pMX-puroレトロウイルスベクター(東京大学T. Kitamura博士より供与)にクローニングした。得られたベクターおよび対照のGFP単独mockベクターを、FuGENE6(Roche)を用いてEcoPack2(BD Biosciences)にトランスフェクションし、形質転換した細胞を、1.5 μg/mlピューロマイシンで選択した。培地交換後24時間で培養上清を回収し、直ちに感染に用いた。胎仔肝由来肥満細胞の感染のため、当該細胞を20ng/ml rmIL-3およびrmSCF(R&D systems)の存在下で16〜18時間、前刺激した。次いで、等量のウイルス上清および10ng/ml DOTAP(Roche)を添加した後、32℃で2時間、2000rpmでのスピン感染を行い、プレートを37℃で一晩インキュベートした。翌日、rmIL-3を含む新鮮な培地と交換した。
【0069】
実施例11
受動皮膚アナフィラキシー(PCA)
PCAは、既報(Kabu, K. et al. J Immunol 177, 1296-1305 (2006))に従って行った。動物の右耳に、0.5μgの抗DNP IgEを含む20μl生理食塩水を皮下に注射した。左耳には、20μlの生理食塩水を注射した。16〜18時間後、前記動物を、250μgの多価ジニトロフェニル-ウシ血清アルブミン(DNP-BSA)および5mg/mlエバンスブルーダイ(Sigma)を含む250μlの生理食塩水でチャレンジした。前記耳におけるエバンスブルーの血管外溢出を30分間モニターし、耳バイオプシーを、700μlホルムアミド中、63℃で一晩インキュベートした。ホルムアミド抽出物の定量解析は、620nmでのエバンスブルーの吸光度を測定することにより決定した。
【0070】
結果
STIM1欠損マウスでの肥満細胞の発生
肥満細胞およびアレルギー応答におけるSTIM1の生理学的機能を研究するため、マウスのStim1遺伝子を遺伝的に欠失させた。Stim1遺伝子を、Cre-loxP-介在遺伝子ターゲティングストラテジーを用いて、C57B6/Cマウスで不活性化した(図1a)。サザンブロッティングにより、組換えおよびSTIM1の膜貫通領域をコードするエキソン6のCreによる除去を確認した(図1b)。胎仔肝細胞および胎仔肝由来肥満細胞(FLMCs)におけるSTIM1タンパク質の破壊は、イムノブロット解析により確認した。STIM1+/+、STIM1+/-およびSTIM1-/-細胞の細胞溶解液を、抗STIM1抗体でブロットした。STIM1+/+細胞は、STIM1タンパク質を発現していたが、STIM1-/-細胞ではSTIM1タンパク質が検出されず、STIM1+/-細胞ではその発現レベルが著しく低下していた(図1c、d)。ホモ接合性STIM1-/-マウスは、原因は不明であるが、周産期死亡を示した。E18.5まで、STIM1-/-胎仔は、明らかな異常は示さなかった。STIM1-/-マウスは稀に出生する場合があり、新生STIM1-/-はSTIM1+/+マウスよりも若干小さく、呼吸障害により一日以内に死亡した。
【0071】
胎生致死のため、E15.5胎仔由来の肝臓から肥満細胞前駆細胞を収集し、IL-3を含む培地中で高度に純粋な肥満細胞を生成させた。細胞の純度は、FcεRIおよびc-Kitの細胞表面の発現で評価した(図2a)。サイトスピンした試料のアルシャンブルー染色により、STIM1-/-肥満細胞の形態は野生型肥満細胞の形態とは異ならないことが示された(図2b)。MMCP-5およびMMCP-6等の肥満細胞に特異的な遺伝子は、野生型およびSTIM1欠損FLMCsで同程度のレベルで発現していた(図2c)。
肥満細胞のin vivo の分化状態を調べるため、野生型およびSTIM1欠損胎仔での組織肥満細胞の数を決定した。STIM1欠損E15.5胎仔の背の皮膚における肥満細胞の密度は、野生型の密度と同程度であった(図2d)。さらに、野生型およびSTIM1欠損マウスの胎仔肝細胞(CD45.2)を、致死量放射線を照射したコンジェニックマウス(CD45.1)に適応移植し、フローサイトメトリーにより再構成の有無を解析した。ドナー型細胞を、CD45マーカーにより区別した。再構成したマウス末梢血中のSTIM1+/+およびSTIM1-/- 細胞によるキメリズムは、ほとんど同等であった(>90%;データは示さず)。このことから、STIM1欠損マウス由来の造血幹細胞の正着能力は正常であることが示唆された。腹腔内のフローサイトメトリー解析により、肥満細胞の発生も無傷であることがわかった(図2e)。前記結果をまとめると、肥満細胞の分化はSTIM1非存在下でも障害されないことが示された。
【0072】
STIM1欠損FLMCsにおける障害されたストア感受性Ca2+流入
STIM1は、RNAiに基づいたノックダウンHeLa細胞およびJurkat細胞、ならびにノックアウトDT40 B細胞におけるSOC流入の重要な分子であることが示されている。肥満細胞におけるSOC流入に対するSTIM1の効果を決定するために、0.5 mM EGTAの存在下で小胞体Ca2+ポンプ阻害剤であるタプシガルギン(TG)処理でCa2+ストアを枯渇させ、FLMCsの細胞質内Ca2+をモニターし、次いで、2 mM Ca2+ を戻し入れてCa2+ 流入をモニターした。図3aに示すように、STIM1欠損FLMCsにおいて、Ca2+流入の有意な抑制が観察された。このSTIM1依存性SOC流入メカニズムは、FcεRIシグナル伝達コンテクストによって優位に利用されており、STIM1の非存在下ではFcεRIにより誘導されたストア枯渇からCa2+流入の障害も生じた(図3a)。Ca2+動員におけるSTIM1の効果をさらに評価するために、細胞外Ca2+が存在する条件下で、IgEおよび抗原によるFcεRI刺激を行った。野生型FLMCsは、シャープなスパイクを示した後、持続的なCa2+流入を示した(図3b)。対照的に、STIM欠損FLMCsでは細胞質内Ca2+レベルが有意に低下していた。イオノマイシン処理野生型FLMCsは、連続的に高いレベルのCa2+増加を示した。この持続したCa2+は、STIM1欠損FLMCsでは劇的に低下した(図3b)。これらの結果より、イオノマイシンはCa2+流入の引き金となり得ることを示唆し、これは全体的にSTIM1に依存する。イオノマイシンが内部のCa2+ストアを枯渇させることによりPMにおけるSOCチャネルを活性化できるという以前の証拠があることから、STIM1は、このイオノマイシン依存性Ca2+枯渇のセンサーとしても機能し、Ca2+流入を活性化すると思われる。
【0073】
Ca2+流入の上記した欠損がSTIM1が存在しないことによるものであるか否かを検証するために、GFP標識したSTIM1をSTIM1欠損FLMCsにレトロウイルスにより導入し、細胞質内Ca2+プロフィールを分析した。モックGFPを導入したSTIM1欠損FLMCsは、Ca2+流入を誘導する能力がなかったが、GFP-STIM1のバックトランスフェクションにより、FcεRIおよびTG刺激後、Ca2+流入を明確にレスキューした(図3c)。一貫して、2 mM Ca2+ の存在下でIgE凝集またはイオノマイシンによって媒介されるCa2+の動員は、GFP-STIM1導入細胞において十分回復した。まとめると、これらの結果から、STIM1が肥満細胞においてSOC流入活性化に関与すること、およびこのメカニズムはFcεRIおよびイオノマイシン刺激により誘導されるCa2+流入において重要な役割を果たすことを明確に示している。
【0074】
STIM1は肥満細胞脱顆粒およびサイトカイン産生に必須である
FcεRIが介在する肥満細胞活性化は、迅速な脱顆粒ならびに代謝産物およびサイトカインのde novoの合成を誘導する。このことは、アレルギー反応に重要な役割を果たす。これらの肥満細胞の機能にとってSTIM1が必要であるか否かを探究するために、FcεRI刺激後の野生型およびSTIM1欠損FLMCs由来のβ-ヘキソサミニダーゼの放出を測定した。STIM1の喪失により、抗原により誘発される脱顆粒の顕著な抑制が生じた(図4a)。また、STIM1欠損FLMCsがイオノマシンにより誘導される脱顆粒の極度の低下を示したことを見出した。イオノマイシンは、FcεRI近位のシグナル伝達をバイパスし、遊離Ca2+イオンの動員を通じて直接細胞を刺激することが知られている。β-ヘキソサミニダーゼの総含量は野生型FLMCsとSTIM1欠損FLMCsとで相違しなかった点に注目すべきである (データ示さず)。したがって、STIM1は肥満細胞の脱顆粒にとって必須であることが結論される。これらの知見は、一過性のCa2+ の上昇では脱顆粒を有効に行うには不十分であることも示唆する。
【0075】
サイトカイン産生を評価するために、FcεRIライゲーション後の野生型およびSTIM1欠損FLMCsにおけるELISAアッセイを行った。IL-6、TNF-αおよびIL-13の分泌は、野生型FLMCsに比べてSTIM1欠損FLMCsで顕著に低下していた(図4b)。半定量的RT-PCRにより、これらのサイトカインのmRNA合成もSTIM1の非存在下で顕著に抑制されていたことが示された(図4c)。これらの結果は、STIM1がIL-6、TNF-αおよびIL-13の遺伝子発現ならびに当該サイトカインの分泌に必須であることを示している。
【0076】
STIM1欠損FLMCsにおける正常なFcεRI介在近位シグナル伝達およびMAPKs活性化
上記したデータは、STIM1依存性の持続的Ca2+ 障害が肥満細胞の機能に応答可能な細胞内シグナル伝達事象の欠損に導く可能性を示唆した。どの細胞内シグナル伝達経路が攪乱されるのかを決定するために、野生型およびSTIM1欠損FLMCsにおける様々なFcεRIシグナル経路を調べた。感作したFLMCsを所定の時間抗原刺激し、イムノブロッティングによりSyk、LATおよびPLCγ1等の近位分子の活性化状態を調べた。STIM1欠損FLMCsは、Syk、LATおよびPLCγ1の正常なチロシンリン酸化を示した(図5a)。このことは、FcεRI近位のシグナル伝達は一般にSTIM1により影響されないことを示す。MAPKs カスケードは、サイトカイン産生を調節していることが知られている。しかしながら、FcεRIが介在するJNK、p38およびErkの活性化は、STIM1の不在によって変化しなかった(図5b)。イオノマイシンによる細胞質内Ca2+濃度の強制的上昇は、野生型FLMCs においてJNK、p38およびErkを活性化した(図5b)。対照的に、イオノマイシンで誘導されたこれらのMAPKsの活性化は、STIM1欠損FLMCsで消滅した。このことは、STIM1はイオノマイシンで誘導されたCa2+流入により誘発されるJNK、p38およびErkの活性化に不可欠であることを示唆する。
【0077】
NF-κBおよびNFATシグナル伝達経路に対するSTIM1の必須の役割
FcεRIの刺激は、2つの転写因子NF-κBおよびNFATを活性化させる。当該転写因子は、サイトカイン産生等の肥満細胞の機能を調節していることが知られている。以前の研究により、NF-κBは大きな一過性のCa2+上昇により選択的に活性化され、他方、NFATは低い持続したCa2+プラトーにより活性化されることが示されている。そこで、本発明者らは、NF-κBおよびNFAT活性化に対するSTIM1の寄与を評価した。STIM1欠損FLMCsにおいて、FcεRIおよびイオノマイシン刺激後のIκBαのリン酸化および分解は、野生型FLMCsと比較して有意に低下していた(図6a)。さらに、FcεRIが介在するp65の核への移行は、STIM1欠損FLMCsにおいて顕著に低下していた(図6b)。NFAT活性化を探究するために、NFAT1のSer54におけるリン酸化の状況を調べた。リン酸化がNFAT活性を抑制する他の部位とは異なり、Ser54をリン酸化されたNFATは、自身の活性を高める。図6cに示すように、STIM1欠損FLMCsでは、FcεRIおよびイオノマイシン刺激後にNFATのSer54はほとんどリン酸化されていなかった。さらに、NFAT1の脱リン酸化および核への移行がSTIM1欠損FLMCsで非常に障害されていたことを見出した(図6c、下部パネル、および図6d)。以上の結果をまとめると、NF-κBおよびNFAT活性化は両方とも、STIM1依存性Ca2+流入により調節されていると結論付けられる。肥満細胞におけるIL-6/TNF-αおよびTNF-α/IL-13産生はそれぞれ、NF-κBおよびNFATにより調節されていることを考慮すると、STIM1不在によるNF-κBおよびNFAT活性化の低下は、かかるサイトカインの産生を不十分にさせるものと思われる。
【0078】
STIM1はin vivoでのアナフィラキシー応答に必要である
STIM1がin vivoの肥満細胞において機能的な役割を果たしているかどうかを評価するために、FcεRIが介在する受動皮膚アナフィラキシー(PCA)反応を行った。STIM1欠損マウスは胎生致死であるので、成体のSTIM1+/-マウスを解析した。図1に示すように、STIM1+/-マウス由来のFLMCsでは、STIM1+/+マウス由来のものに比べて、STIM1タンパク質が非常に低量であった。PCAは、抗DNP IgEを右耳の耳介に皮内注射することにより誘導した。内部標準として、左耳に同量の生理食塩水を注射した。16〜18時間後、マウスを抗原DNP-HSAで静脈内注射によりチャレンジし、エバンスブルー染色を行った。PCA誘導後最初の1時間の間のエバンスブルー染色の血管外溢出は、主として、活性化した肥満細胞の迅速なヒスタミンおよびセロトニン放出を伴う脱顆粒と、それによる局所的に血管の透過性が上昇することに依存している。血管外溢出は、生理食塩水処理耳と比較して、IgE処理耳由来の組織バイオプシーにおける染色のレベルを決定することにより定量した。対照条件として、STIM1+/+マウスとSTIM1+/- マウスとの間で、エバンスブルーのレベルは相違しなかった。しかしながら、STIM1+/- マウスのIgE処理耳介における血管外溢出は、STIM1+/+マウスよりも有意に低く、このことから、STIM1は抗原で誘導された肥満細胞が介在するPCA反応に必要であることが示唆された。
【0079】
配列表のフリーテキスト
〔配列番号:5〕siRNAセンス鎖
〔配列番号:6〕siRNAセンス鎖
【産業上の利用可能性】
【0080】
本発明によれば、新たな作用機序に基づく肥満細胞の脱顆粒抑制剤が提供される。当該作用機序を指標とした本発明のスクリーニング方法は、STIM1に特異的に作用する薬剤の開発に貢献することができる。
【図面の簡単な説明】
【0081】
【図1a】図1aは、Stim1遺伝子の標的化戦略を示す概略図である。標的ベクターは、Stim1遺伝子のエキソン6およびneor遺伝子を2つのloxP部位(黒三角)に隣接するように設計した。Cre介在の遺伝子除去を行い、欠損したアレル(D)を生成させた。関連するHind III認識部位(H)を示す。
【図1b】図1bは、E15.5胎仔由来のHind III消化ゲノムDNAのサザンブロット解析を示す。使用したプローブは、図1aに示す。
【図1c】図1cは、E15.5胎仔由来の肝臓の全細胞抽出物の、STIM1およびActinに特異的な抗体を用いたイムノブロット解析を示す。標準の分子量をkDaで示す。
【図1d】図1dは、E15.5胎仔肝臓由来の肥満細胞(FLMCs)の全細胞抽出物の、STIM1およびActinに特異的な抗体を用いたイムノブロット解析を示す。標準の分子量をkDaで示す。
【図2a】図2aは、野生型(WT)およびSTIM1-/-マウス由来のFLMCsにおけるFcεRIおよびc-Kitの細胞表面発現を調べたフローサイトメトリーを示す。
【図2b】図2bは、サイトスピン標品のニュークレアファストレッド染色およびアルシャンブルー染色の代表例を示す。
【図2c】図2cは、WTおよびSTIM1-/-FLMCsにおけるRT-PCRによるMMCP-5およびMMCP-6 mRNA発現を調べた結果を示す。
【図2d】図2dは、WTおよびSTIM1-/-E15.5 胎仔の皮膚組織の肥満細胞の組織学的解析を示す。切片を、ニュークレアファストレッドおよびアルシャンブルーで染色した。矢頭は、アルシャンブルーで染色された肥満細胞を示す。肥満細胞の絶対数(mm2当たり)を、右側のグラフに示す。データは平均+ s.d.で示す。
【図2e】図2eは、WTおよびSTIM1-/-マウス(CD45.2)のE15.5 胎仔肝臓由来の2 x 106個の細胞を、致死量を照射したコンジェニックマウス(CD45.1)に注入した結果を示す。10週後、細胞を腹腔から単離し、フローサイトメトリーにより解析した。ドナー型細胞は、CD45.2マーカーに従って同定した。
【図3a】図3aは、Indo-1イメージングを用いて、STIM1欠損FLMCs におけるCa2+ 動員プロファイルをモニターしたグラフである。まず、抗DNP IgEで感作したFLMCsを、DNP-HSA抗原(Ag; 50ng/ml)またはタプシガルギン(TG; 2μM)の刺激によりCa2+フリー条件下(0.5 mM EGTA)でCa2+放出を惹起し、野生型(WT; 黒)およびSTIM1欠損(STIM1-/-; 赤)FLMCsにおいて細胞外Ca2+ 濃度を2mMに回復させることによってCa2+流入を誘導した。解析結果は、少なくとも3つの独立した実験の代表例である。
【図3b】図3bは、2mM Ca2+ 存在下、WTおよびSTIM1-/- マウス由来の抗DNP IgEで感作したFLMCsをDNP-HAS抗原(50 ng/ml)での刺激後、またはイオノマイシン(Iono; 1μM)刺激後の未感作FLMCsにおいて、Ca2+動員プロファイルをモニターしたグラフである。解析結果は、少なくとも3つの独立した実験の代表例である。
【図3c】図3cは、GFP-STIM1を発現するSTIM1欠損FLMCsがCa2+流入を回復したことを示すグラフである。GFP単独(Mock; 黒)またはGFP-STIM1(青)をレトロウイルスにより導入したSTIM1-/- FLMCs におけるCa2+ 動員プロファイルを、図3aと同様に示す。データは、GFP陽性ゲートで解析した。値は、FL5/FL4 (FL4= 500-520 nm、FL5= 400-420 nm)での蛍光比としてプロットした。解析結果は、少なくとも3つの独立した実験の代表例である。
【図3d】図3dは、GFP-STIM1を発現するSTIM1欠損FLMCsがCa2+流入を回復したことを示すグラフである。GFP単独(Mock; 黒)またはGFP-STIM1(青)をレトロウイルスにより導入したSTIM1-/-FLMCsにおけるCa2+ 動員プロファイルを、図3bと同様に示す。データは、GFP陽性ゲートで解析した。値は、FL5/FL4 (FL4= 500-520 nm、FL5= 400-420 nm)での蛍光比としてプロットした。すべての解析結果は、少なくとも3つの独立した実験の代表例である。
【図4a】図4aは、野生型(WT)およびSTIM1-/-マウス由来のFLMCsの脱顆粒について、抗DNP IgEで感作した細胞を所定の用量のDNP-HAS抗原で30分間刺激した後、β-ヘキサミニダーゼ放出を測定することによって評価した結果を示す。未感作細胞は、1μM イオノマイシンで刺激した。結果は、三連の試料からの平均 + s.d.であり、3つの異なるペアを用いた3つの独立した実験の代表例である。*, P < 0.05、**, P < 0.01、***, P < 0.001、野生型コントロールに対する。
【図4b】図4bは、野生型(WT)およびSTIM1-/- マウス由来のFLMCsを3時間抗原刺激後、IL-6、TNF-αおよびIL-13の放出を測定したELISA法によるサイトカイン産生の評価を示す。結果は、3つの独立した実験の代表例である(平均 + s.d.)。*, P < 0.05、**, P < 0.01、***, P < 0.001、野生型コントロールに対する。< DL, 検出限界未満。
【図4c】図4cは、感作したFLMCsを抗原で1時間刺激し、サイトカインおよびGAPDHのmRNA発現を半定量的RT-PCRにより検出した結果を示す。結果は、3つの独立した実験の代表例である。
【図5a】図5aは、野生型(WT)およびSTIM1欠損FLMCsにおけるFcεRI近位シグナル伝達の活性化を示す。抗DNP IgEで感作したWTおよびSTIM1-/-FLMCsを、50 ng/ml DNP-HAS抗原で所定の間隔を置いて刺激した。全細胞溶解液をSDS-PAGEで分画し、所定の抗体でイムノブロットした。結果は、3つの独立した実験の代表例である。
【図5b】図5bは、野生型(WT)およびSTIM1欠損FLMCsにおけるMAPKsの活性化を示す。抗DNP IgEで感作したWTおよびSTIM1-/-FLMCsを、50ng/ml DNP-HAS抗原で所定の間隔を置いて刺激した(図5bの左パネル)。あるいは、未感作のWTおよびSTIM1-/-FLMCsを、1μM イオノマイシンで所定の間隔を置いて刺激した(図5bの右パネル)。全細胞溶解液をSDS-PAGEで分画し、所定の抗体でイムノブロットした。結果は、3つの独立した実験の代表例である。
【図6a】図6aは、野生型(WT)およびSTIM1-/- マウス由来のFLMCsを抗DNP IgEで感作し、次いで、所定の間隔を置いて50 ng/ml DNP-HAS抗原で刺激した場合のNF-κBおよびNFATの活性化を示す。あるいは、未感作WTおよびSTIM1-/-FLMCsを1μM イオノマイシンで刺激した場合のNF-κBおよびNFATの活性化を示す。全細胞溶解液を、SDS-PAGEで分画した。IκBαのリン酸化および分解を、イムノブロッティングにより決定した。イムノブロット解析は、少なくとも3つの独立した実験の代表例である。
【図6b】図6bは、共焦点顕微鏡によるNF-κBの核移行を評価した結果を示す。抗DNP IgEで感作したWTおよびSTIM1-/- FLMCsを、100 ng/ml DNP-HAS抗原で10分間刺激し、次いで固定し、抗p65 Ab (緑)およびDAPI (青)での免疫蛍光染色に供した。スケールバー、5μm。各試料中の全部で100個の細胞をランダムにカウントし、p65が核に移行した細胞の頻度を計算した。データは平均 + s.d.である。***, P < 0.001、野生型コントロールに対する。
【図6c】図6cは、FLMCsを所定の間隔を置いて図6aと同様に刺激した結果を示す。全細胞溶解液を、SDS-PAGEにより分画し、リン酸化NFAT (Ser54)およびNFAT1に対する特異的抗体でイムノブロットした。イムノブロット解析は、少なくとも3つの独立した実験の代表例である。
【図6d】図6dは、NFAT1の核移行をイムノブロットにより決定した結果を示す。刺激した細胞由来の核抽出物を、抗NFAT1抗体および抗actin抗体でブロットした。イムノブロット解析は、少なくとも3つの独立した実験の代表例である。
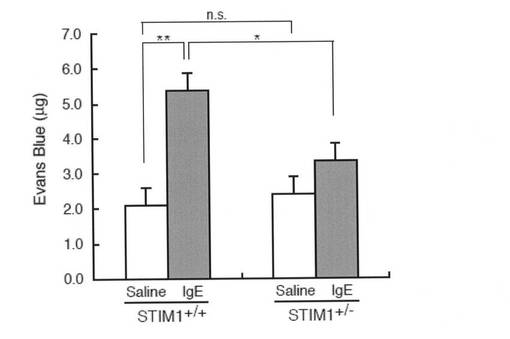
【図7】図7は、STIM1+/-マウスにおいてin vivoアナフィラキシーが減少することを示すグラフである。PCA反応のため、マウスの右耳に抗DNP IgEを皮下注射し、対照として、左耳に生理食塩水を皮下注射した(STIM1+/+; n=10, STIM1+/-; n=8)。16〜18時間後、DNP-HASをエバンスブルー色素とともに静脈内投与し、エバンスブルーの両耳への血管外溢出を測定した。データは、エバンスブルー色素の量を示す(平均 + s.d.)。*, P < 0.05、**, P < 0.01、対応する対照に対する。n.s., 有意差なし。
【特許請求の範囲】
【請求項1】
STIM1阻害物質を有効成分として含有する肥満細胞の脱顆粒抑制剤。
【請求項2】
STIM1阻害物質を有効成分として含有するアレルギーの予防または治療剤。
【請求項3】
STIM1阻害物質を有効成分として含有するアナフィラキシーの予防または治療剤。
【請求項4】
STIM1阻害物質を有効成分として含有する肥満細胞の炎症性サイトカインの産生抑制剤。
【請求項5】
STIM1阻害物質が、RNAi誘導性核酸、アンチセンス核酸もしくはリボザイムまたはそれらの発現ベクターである、請求項1〜4いずれかに記載の剤。
【請求項6】
RNAi誘導性核酸がsiRNAである、請求項5に記載の剤。
【請求項7】
siRNAが、配列番号1の塩基配列に対応するmRNAにおける19〜25個の連続する塩基配列を含むセンス鎖と、その相補配列を含むアンチセンス鎖からなるものである、請求項6に記載の剤。
【請求項8】
被験物質がSTIM1遺伝子の発現または機能を抑制し得るか否かを評価することを含む、肥満細胞の脱顆粒を抑制し得る物質のスクリーニング方法。
【請求項9】
下記の工程(a)〜(c)を含む、請求項8記載の方法:
(a)被験物質とSTIM1遺伝子の発現を測定可能な細胞とを接触させる工程;
(b)被験物質を接触させた細胞におけるSTIM1遺伝子の発現量を測定し、該発現量を被験物質を接触させない対照細胞におけるSTIM1遺伝子の発現量と比較する工程;および
(c)上記(b)の比較結果に基づいて、STIM1遺伝子の発現量を抑制する被験物質を選択する工程。
【請求項10】
下記の工程(a)〜(c)を含む、請求項8記載の方法:
(a)被験物質をSTIM1蛋白質に接触させる工程;
(b)被験物質のSTIM1蛋白質に対する結合能を測定する工程;および
(c)上記(b)の結果に基づいて、STIM1蛋白質に結合能を有する被験物質を選択する工程。
【請求項1】
STIM1阻害物質を有効成分として含有する肥満細胞の脱顆粒抑制剤。
【請求項2】
STIM1阻害物質を有効成分として含有するアレルギーの予防または治療剤。
【請求項3】
STIM1阻害物質を有効成分として含有するアナフィラキシーの予防または治療剤。
【請求項4】
STIM1阻害物質を有効成分として含有する肥満細胞の炎症性サイトカインの産生抑制剤。
【請求項5】
STIM1阻害物質が、RNAi誘導性核酸、アンチセンス核酸もしくはリボザイムまたはそれらの発現ベクターである、請求項1〜4いずれかに記載の剤。
【請求項6】
RNAi誘導性核酸がsiRNAである、請求項5に記載の剤。
【請求項7】
siRNAが、配列番号1の塩基配列に対応するmRNAにおける19〜25個の連続する塩基配列を含むセンス鎖と、その相補配列を含むアンチセンス鎖からなるものである、請求項6に記載の剤。
【請求項8】
被験物質がSTIM1遺伝子の発現または機能を抑制し得るか否かを評価することを含む、肥満細胞の脱顆粒を抑制し得る物質のスクリーニング方法。
【請求項9】
下記の工程(a)〜(c)を含む、請求項8記載の方法:
(a)被験物質とSTIM1遺伝子の発現を測定可能な細胞とを接触させる工程;
(b)被験物質を接触させた細胞におけるSTIM1遺伝子の発現量を測定し、該発現量を被験物質を接触させない対照細胞におけるSTIM1遺伝子の発現量と比較する工程;および
(c)上記(b)の比較結果に基づいて、STIM1遺伝子の発現量を抑制する被験物質を選択する工程。
【請求項10】
下記の工程(a)〜(c)を含む、請求項8記載の方法:
(a)被験物質をSTIM1蛋白質に接触させる工程;
(b)被験物質のSTIM1蛋白質に対する結合能を測定する工程;および
(c)上記(b)の結果に基づいて、STIM1蛋白質に結合能を有する被験物質を選択する工程。
【図1a】


【図1b】

【図1c】

【図1d】

【図2a】
【図2b】

【図2c】

【図2d】

【図2e】
【図3a】
【図3b】

【図3c】
【図3d】
【図4a】

【図4b】
【図4c】
【図5a】
【図5b】
【図6a】

【図6b】

【図6c】
【図6d】
【図7】
【図1b】
【図1c】
【図1d】
【図2a】
【図2b】
【図2c】
【図2d】
【図2e】
【図3a】
【図3b】
【図3c】
【図3d】
【図4a】
【図4b】
【図4c】
【図5a】
【図5b】
【図6a】
【図6b】
【図6c】
【図6d】
【図7】
【公開番号】特開2009−102231(P2009−102231A)
【公開日】平成21年5月14日(2009.5.14)
【国際特許分類】
【出願番号】特願2007−273055(P2007−273055)
【出願日】平成19年10月19日(2007.10.19)
【出願人】(503359821)独立行政法人理化学研究所 (1,056)
【Fターム(参考)】
【公開日】平成21年5月14日(2009.5.14)
【国際特許分類】
【出願日】平成19年10月19日(2007.10.19)
【出願人】(503359821)独立行政法人理化学研究所 (1,056)
【Fターム(参考)】
[ Back to top ]