
肺臓の組織工学
本発明は脱細胞化組織を含む組成物に関する。本発明は、天然肺臓組織の特徴を示す人工三次元肺臓組織も提供する。この人工組織は肺臓の発生生物学および病理学の研究ならびに創薬に有用である。
【発明の詳細な説明】
【背景技術】
【0001】
発明の背景
毎年400,000人のアメリカ人が肺臓疾患で死亡する。さらに憂慮されることに、他の主要疾患カテゴリについては死亡率が減少しつつあるのに(心臓疾患、がんおよび脳卒中)、肺臓疾患による死亡率は増加しつつある。嚢胞性線維症、気腫/COPD、および特発性肺線維症を含むいくつかの肺臓疾患については、肺臓移植(lung transplantation)が今なお唯一の根治的処置である。しかし、肺臓移植後の患者生存率は、5年時点で50%、10年時点で24%しかない[Mondrinos et al., 2008, Tissue Eng 14:361-8(非特許文献1)]。したがって、移植に使用することができる人工(engineered)肺臓組織の開発が、強く求められている。人工肺臓組織の利点の一つは、患者自身の細胞を使って組織を成長させることができ、そうすることによって、現在の肺臓移植で要求されるような強い免疫抑制の必要を回避することができるという点である。免疫抑制は移植された器官の拒絶を防ぐために必要であるが、感染、悪性疾患、腎機能障害、心血管障害、および神経障害を含む広範な問題につながり得る[Pietra et al., 2000, J Clin Invest 106:1003-10(非特許文献2);Christie et al., 2009, J Heart Lung Transplant 28:1031-49(非特許文献3)]。
【0002】
組織工学は、細胞学的、分子学的、技術的および医学的進歩を組み合わせて埋植(implantation)に適した代用組織を作出しようとする成長分野である。前途有望な研究が、血管、膀胱、心臓弁、および心臓組織を含むさまざまな組織について行われている[Nichols et al., 2008, Proc Am Thor Soc 5:723-30(非特許文献4);Satchell et al., 2004, J Am Soc Nephrol 15:566-74(非特許文献5);Atala et al., 2006, Lancet 367:1241-6(非特許文献6);Orfanos et al., 2004, Intensive Care Med 30:1702-14(非特許文献7)]。しかし、肺臓は、実験室で工学的に操作(engineer)することが困難な組織である。呼吸の機械的圧力に耐えることができ、内皮、上皮および間葉細胞の成長を支持することができ、しかも、極めて異なっているが密接に近接している2つのコンパートメント間のガス交換のための手段を提供する複雑なマトリックスを、肺臓は必要とする。
【0003】
臨床現場における潜在的な患者使用の他に、人工肺臓組織は、肺の生物学および生理学の広範囲にわたる種々の重要な局面を研究するために、実験室で使用することもできる。インビトロ三次元肺臓培養モデルはごくわずかしかない[Vandenbroucke et al., 2008, Ann N Y Acad Sci 1123:134-4(非特許文献8)]。そのうえ、肺の内皮および上皮細胞は他の多くの細胞タイプよりも実験室で培養することが困難であり[Malda et al., 2004, Biomaterials 25:5773-80(非特許文献9);Reichenspurner, 2005, J Heart Lung Transplant 24:119-30(非特許文献10)]、肺の前駆細胞および幹細胞生物学の分野は進展が比較的遅かったBlaisdell et al., 2009, Stem Cells 27:2263-70(非特許文献11);Muratore et al., 2008, J Surg Res 155(2):225-30(非特許文献12)]。したがって、当技術分野では、未変性(native)肺環境の重要な特質を再現するインビトロ肺臓組織の開発が必要とされている。本発明は当技術分野におけるこの必要を満たすものである。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】Mondrinos et al., 2008, Tissue Eng 14:361-8
【非特許文献2】Pietra et al., 2000, J Clin Invest 106:1003-10
【非特許文献3】Christie et al., 2009, J Heart Lung Transplant 28:1031-49
【非特許文献4】Nichols et al., 2008, Proc Am Thor Soc 5:723-30
【非特許文献5】Satchell et al., 2004, J Am Soc Nephrol 15:566-74
【非特許文献6】Atala et al., 2006, Lancet 367:1241-6
【非特許文献7】Orfanos et al., 2004, Intensive Care Med 30:1702-14
【非特許文献8】Vandenbroucke et al., 2008, Ann N Y Acad Sci 1123:134-4
【非特許文献9】Malda et al., 2004, Biomaterials 25:5773-80
【非特許文献10】Reichenspurner, 2005, J Heart Lung Transplant 24:119-30
【非特許文献11】Blaisdell et al., 2009, Stem Cells 27:2263-70
【非特許文献12】Muratore et al., 2008, J Surg Res 155(2):225-30
【発明の概要】
【0005】
発明の概略
本発明は、細胞成長を支持する能力を有する脱細胞化組織を提供する。好ましくは、脱細胞化組織は、脱細胞化前の対応する天然組織の特徴を示す。より好ましくは、組織は肺臓である。
【0006】
ある態様では、脱細胞化組織が、他の点では同一な脱細胞化前の組織の形態と実質的に類似する形態を示す。
【0007】
別の態様において、請求項1の脱細胞化組織は、対応する天然組織の細胞外マトリックスを保っており、当該細胞外マトリックスは、実質的にインタクト(intact)な外面を含む。
【0008】
別の態様では、免疫原性マーカーが脱細胞化組織から実質的に除去されている。
【0009】
ある態様では、脱細胞化組織が、対応する天然組織の機械的性質と実質的に類似する機械的性質を示す。
【0010】
本発明は、三次元スキャフォールド(scaffold)と細胞の集団とを含む組成物を提供する。好ましくは、組成物は、肺臓細胞の分化状態を支持し維持する能力を有する。
【0011】
ある態様では、三次元スキャフォールドが脱細胞化組織である。
【0012】
別の態様では、組成物が、インタクトな気道樹および血管網を示す。
【0013】
別の態様では、細胞の集団が幹細胞を含む。
【0014】
別の態様では、細胞の集団が上皮細胞および内皮細胞を含む。
【0015】
別の態様では、細胞が遺伝子改変されている。ある態様では、細胞がCFTR遺伝子を発現するように遺伝子改変されている。
【0016】
ある態様では、組成物が、肺胞上皮細胞の分化状態を支持し維持する能力を有する。
【0017】
別の態様では、スキャフォールドが、フィブロネクチン、ラミニン、コラーゲン、糖タンパク質、トロンボスポンジン、エラスチン、フィブリリン、ムコ多糖、糖脂質、ヘパリン硫酸、コンドロイチン硫酸、ケラチン硫酸、グリコサミノグリカン、ヒアルロン酸、プロテオグリカン、ビトロネクチン、ポリ-D-リジン、多糖、およびそれらの組合せからなる群より選択される生体適合性材料を含む。
【0018】
ある態様では、細胞が、分枝形態形成の誘導に関連する遺伝子発現を示す。
【0019】
別の態様では、組成物が肺臓組織の特徴を含む。いくつかの例では、特徴が、分枝形態形成、遠位肺臓上皮細胞分化、上皮成長、血管発生、およびそれらの組合せからなる群より選択される。
【0020】
本発明は、肺臓細胞の分化状態を支持し維持する能力を有する人工三次元組織を製作する方法を提供する。本方法は、播種済スキャフォールドを作製するために脱細胞化スキャフォールドに細胞の集団を播種する工程を含む。
【0021】
本発明は、肺臓組織の健康状態を調整する試験作用因子の能力に関して試験作用因子をスクリーニングするためのインビトロ法を提供する。本方法は、試験作用因子を人工三次元肺臓組織モデルに接触させる工程と、試験作用因子がモデルに及ぼす効果を測定する工程とを含む。モデルに加えられる変更はいずれも、その試験作用因子が肺臓組織の健康状態を調整できることのしるしである。
【0022】
ある態様では、試験作用因子が、化学剤、医薬、ペプチド、核酸、および放射線からなる群より選択される。
【0023】
別の態様では、試験作用因子が治療剤のための送達媒体である。
【0024】
ある態様において、本方法は、細胞数、面積、体積、形状、形態、マーカー発現または染色体断片化に対する試験作用因子の効果を決定する工程を含む。
【0025】
別の態様において、本方法は、肺臓組織モデルに対して所望の効果を持つ作用因子を選択する工程を含む。
【0026】
本発明は、哺乳動物における肺臓欠陥を軽減または処置する方法を提供する。本方法は、肺臓細胞の分化状態を支持し維持する能力を有する三次元構築物を含む組成物の治療有効量を哺乳動物に投与し、それによってその哺乳動物における肺臓欠陥を軽減または処置する工程を含む。
【0027】
本発明は、細胞成長を支持する能力を有する脱細胞化組織を含む埋植可能な組成物を提供する。好ましくは、脱細胞化組織は、脱細胞化前の対応する天然組織の特徴を示す。
【0028】
ある態様において、埋植可能な組成物は細胞の集団を含む。好ましくは、埋植可能な組成物は、肺臓組織の分化状態を支持し維持する能力を有する。
【図面の簡単な説明】
【0029】
本発明を例証するために、本発明の一定の態様を図面に図示する。ただし、本発明は、図面に図示する態様の厳密な配置および道具に限定されるわけではない。
【0030】
【図1A】図1A〜1Dを含む図1は、未変性肺臓および脱細胞化肺臓のH&E染色および定量的DNAアッセイを図示する一連の画像である。細胞材料は除去されたが、それでもスキャフォールドのアーキテクチャは保たれたことを、図1は証明している。DNAは未変性レベルの約1.2%まで除去された。*はp<0.01を示す。
【図1B】図1A〜1Dを含む図1は、未変性肺臓および脱細胞化肺臓のH&E染色および定量的DNAアッセイを図示する一連の画像である。細胞材料は除去されたが、それでもスキャフォールドのアーキテクチャは保たれたことを、図1は証明している。DNAは未変性レベルの約1.2%まで除去された。*はp<0.01を示す。
【図1C】図1A〜1Dを含む図1は、未変性肺臓および脱細胞化肺臓のH&E染色および定量的DNAアッセイを図示する一連の画像である。細胞材料は除去されたが、それでもスキャフォールドのアーキテクチャは保たれたことを、図1は証明している。DNAは未変性レベルの約1.2%まで除去された。*はp<0.01を示す。
【図1D】図1A〜1Dを含む図1は、未変性肺臓および脱細胞化肺臓のH&E染色および定量的DNAアッセイを図示する一連の画像である。細胞材料は除去されたが、それでもスキャフォールドのアーキテクチャは保たれたことを、図1は証明している。DNAは未変性レベルの約1.2%まで除去された。*はp<0.01を示す。図1Dは脱細胞化肺臓の画像である。
【図2】図2Aおよび図2Bを含む図2は、脱細胞化スキャフォールド中の残存DNAに関する染色を図示する一連の画像である。DNAがDAPIで染色されている。比較が可能なように、画像は同じ時間、露出した。
【図3】図3は、MHCクラスIおよびII抗原に関するウェスタンブロットであり、脱細胞化スキャフォールドにおけるMHCクラスIまたはII抗原の欠如を証明している。
【図4】図4Aおよび4Bを含む図4は、未変性肺臓および脱細胞化肺臓におけるコラーゲン染色を図示する一連の画像である。コラーゲンIは緑色に染色され、コラーゲンIVは赤色に染色され、核はDAPIで青色に対比染色される。コラーゲンIは大血管の近くに見いだされ、コラーゲンIVは実質全体に分布している。未変性肺臓では、実質中の赤血球が自己蛍光ゆえに緑色に見えることに注意されたい。
【図5】図5Aおよび5Bを含む図5は、未変性肺臓および脱細胞化肺臓の走査型EMを図示する一連の画像である。肺胞中隔はインタクトである。スケールバーは、左側のパネルでは100μm、右側のパネルでは20μmである。
【図6】図6A〜6Cを含む図6は、未変性肺臓および脱細胞化肺臓の透過型EMを図示する一連の画像である。脱細胞化灌流圧を30mmHg未満に維持すると、肺胞基底膜が保たれる。Cは毛細管を示し、Aは肺胞を示し、Sは肺胞中隔を示す。スケールバーは全てのパネルにおいて2μmである。
【図7】図7は、脱細胞化肺臓の透過型EMを図示する画像であり、保存された毛細管を証明している。脱細胞化のための灌流圧は20mmHg未満だった。Cは毛細管を示し、Aは肺胞を示す。肺胞および毛細管の寸法は、脱細胞化マトリックスの圧縮ゆえに、インビボにある時よりも小さく見える。スケールバーは、上側のパネルでは2μm、左下のパネルでは1μm、右下のパネルでは500nmである。
【図8】脱細胞化スキャフォールドによる5μmマイクロスフェアの保持を図示するグラフである。マイクロスフェアアッセイは、脱細胞化時の低い灌流圧(<30mmHg)によって、気道コンパートメントにおける5μm粒子の95%の保持が可能になることを証明している。*は、未変性物との比較で、p<0.05を示す。
【図9】図9Aおよび9Bを含む図9は、未変性肺臓および脱細胞化肺臓の脈管構造のマイクロCTの一連の画像である。全体として、58μmの解像度でイメージングした場合、脱細胞化スキャフォールドは未変性物と類似しているようである。
【図10】図10Aおよび10Bを含む図10は、未変性肺臓および脱細胞化肺臓の脈管構造の高解像度マイクロCTを図示する一連の画像である。これらのスキャンの解像度は6.5μmである。
【図11】図11は、機械的試験プロトコールを図示する図解である。簡単に述べると、肺臓組織の条片を上側のプレートに取り付け、次にそれを下げて、組織を下側のプレートに取り付ける。組織を20%歪みまで周期的に伸張した後、破損するまで伸張する。
【図12】図12A〜12Cを含む図12は、未変性肺臓および脱細胞化肺臓のコラーゲン染色およびコラーゲン含量を図示する一連の画像である。Massonのトリクローム染色により、未変性肺臓および脱細胞化肺臓の両方に、波状の暗青色線維が明らかになる。定量的アッセイは、未変性肺臓および脱細胞化肺臓ではコラーゲンが保存されるが、ドデシル硫酸ナトリウム(SDS)を使った脱細胞化ではコラーゲンが失われることを証明している。*はp<0.01を示す。
【図13】図13A〜13Cを含む図13は、未変性肺臓および脱細胞化肺臓に関するエラスチン組織化学(Verhoff-van Geison)を図示する一連の画像である。図13は、未変性肺臓および脱細胞化肺臓の両方において、波状の暗いエラスチン線維を図示している。脱細胞化肺臓では、未変性物と比較して、エラスチンがいくらか保存されることが、定量的アッセイによって証明される。*はp<0.01を示す。
【図14】図14A〜14Cを含む図14は、未変性肺臓および脱細胞化肺臓に関するGAG組織化学(アルシアンブルー)を図示する一連の画像である。未変性肺臓では青色のGAG染色が図示されるが、脱細胞化肺臓にはそれらが存在しない。脱細胞化肺臓では、未変性肺臓と比較して、硫酸化GAGが失われることが、定量的アッセイによって証明される。*はp<0.01を示す。
【図15】図15は、未変性肺臓および脱細胞化肺臓の応力-歪み曲線を図示する画像である。SDSは、ドデシル硫酸ナトリウムで処理した肺臓を示す。
【図16】図16は、未変性、脱細胞化およびSDS脱細胞化肺臓の極限引張強さを図示する図表である。SDSはドデシル硫酸ナトリウムを使って脱細胞化した肺臓を示す。*は、未変性物との比較で、p<0.01を示す。
【図17】図17Aおよび17Bを含む図17は、インビトロ肺臓培養に使用されるバイオリアクタの概略図を図示する一連の画像である。
【図18】図18Aおよび18Bを含む図18は、インビトロ肺臓培養時の肺動脈圧および気管圧を図示する一連の画像である。灌流量は約5ml/分である。
【図19】図19A〜19Cを含む図19は、肺臓アーキテクチャ(architecture)および気道上皮に対する空気による換気と液体による換気の効果を対比して図示する一連の画像である。空気換気は3日間の培養後に気道の拡張と気道上皮の破壊を引き起こす。
【図20】図20Aおよび20Bを含む図20は、未変性肺臓培養時の細胞アポトーシスおよび細胞数に対する血管灌流および圧力の効果を図示する一連の画像である。未変性物との比較で、*はp<0.01を示し、#はp<0.05を示す。
【図21】図21A〜21Dを含む図21は、未変性肺臓および灌流培養肺臓におけるCCSPおよびSPC発現の比較を図示する一連の画像である。CCSPおよびSPCは赤く染色され、核はDAPIで青く対比染色される。
【図22】図22Aおよび22Bを含む図22は、未変性肺臓および灌流培養肺臓におけるPECAM発現の比較を図示する一連の画像である。灌流肺臓培養(30mmHg)ではPECAM発現が依然として認められる。PECAMは赤く染色され、核はDAPIで青く対比染色される。
【図23】図23Aおよび23Bを含む図23は、未変性肺臓培養時の細胞アポトーシスおよび細胞数に対する換気の効果を図示する一連の画像である。未変性物との比較で、*はp<0.01を示し、#はp<0.05を示す。
【図24】図24A〜24Cを含む図24は、未変性肺臓および換気培養肺臓におけるアポトーシス核を図示する一連の画像である。単一接続による換気は、未変性肺臓または気道「ループ」による換気と比較して、はるかに高い比率のアポトーシス核をもたらした。アポトーシス核はTUNELによって褐色に染色され、正常な核は緑色に対比染色される。
【図25−1】図25A〜25Jを含む図25は、未変性肺臓および7日間培養した肺臓における肺胞構造を図示する一連の画像である。細胞形態、肺胞構造、および中隔アーキテクチャは、未変性肺臓と培養換気肺臓の間で、類似しているようである。
【図25−2】図25A〜25Jを含む図25は、未変性肺臓および7日間培養した肺臓における肺胞構造を図示する一連の画像である。細胞形態、肺胞構造、および中隔アーキテクチャは、未変性肺臓と培養換気肺臓の間で、類似しているようである。図25C〜25Jは、7日間のインビトロ換気肺臓培養後の肺細胞分化の維持を図示している。
【図26】図26は、換気が肺脈管構造の受動的灌流を可能にすることを証明する画像である。マイクロスフェアは、もっぱらインビトロ培養時の肺臓の換気運動のみによって、血管および毛細管中に見いだされる。
【図27】図27Aおよび27Bを含む図27は、脱細胞化スキャフォールド上で培養した不死化上皮細胞株MLE-12のH&E染色を図示する一連の画像である。
【図28A】図28A〜図28Fを含む図28は、単離された新生児肺細胞の肺マーカーパネルのフローサイトメトリー染色を図示する一連の画像である。青色または緑色の曲線はアイソタイプ対照染色であり、赤色は表示した抗原である。
【図28B】図28A〜図28Fを含む図28は、単離された新生児肺細胞の肺マーカーパネルのフローサイトメトリー染色を図示する一連の画像である。青色または緑色の曲線はアイソタイプ対照染色であり、赤色は表示した抗原である。
【図28C】図28A〜図28Fを含む図28は、単離された新生児肺細胞の肺マーカーパネルのフローサイトメトリー染色を図示する一連の画像である。青色または緑色の曲線はアイソタイプ対照染色であり、赤色は表示した抗原である。
【図28D】図28A〜図28Fを含む図28は、単離された新生児肺細胞の肺マーカーパネルのフローサイトメトリー染色を図示する一連の画像である。青色または緑色の曲線はアイソタイプ対照染色であり、赤色は表示した抗原である。
【図28E】図28A〜図28Fを含む図28は、単離された新生児肺細胞の肺マーカーパネルのフローサイトメトリー染色を図示する一連の画像である。青色または緑色の曲線はアイソタイプ対照染色であり、赤色は表示した抗原である。
【図28F】図28A〜図28Fを含む図28は、単離された新生児肺細胞の肺マーカーパネルのフローサイトメトリー染色を図示する一連の画像である。青色または緑色の曲線はアイソタイプ対照染色であり、赤色は表示した抗原である。
【図29】図29は、培養8日時点の人工肺臓のH&E染色を図示する画像である。ここでは条件が上皮細胞成長に最適化されている。
【図30】図30Aおよび30Bを含む図30は、培養4日および8日時点の人工肺臓のPCNA染色を図示する一連の画像である。増殖性核はPCNAに関して褐色に染まり、陰性核はヘマトキシリンで対比染色される。
【図31】図31Aおよび31Bを含む図31は、培養4日および8日時点の人工肺臓のTUNEL染色を図示する一連の画像である。陽性核は褐色であり、陰性核はメチルグリーンで対比染色される。
【図32】図32A〜32Bを含む図32は、4日時点の未変性肺臓および人工肺臓のクララ細胞分泌タンパク質(CCSP)染色を図示する一連の画像である。CCSPは赤色に染色され、核はDAPIで青色に対比染色される。
【図33】図33A〜33Cを含む図33は、4日および8日時点の未変性肺臓および人工肺臓のサーファクタントタンパク質C染色を図示する一連の画像である。SPCは赤色に染色され、核はDAPIで青色に対比染色される。
【図34】図34A〜34Cを含む図34は、4日時点の未変性肺臓および人工肺臓のアクアポリン5染色を図示する一連の画像である。AQPは赤色に染色され、核はDAPIで青色に対比染色される。
【図35】図35Aおよび35Bを含む図35は、人工肺臓組織におけるSPCおよびCCSPに関する二重染色を図示する一連の画像である。SPCは緑色に染色され、CCSPは赤色に染色され、核はDAPIで青色に対比染色される。SPC-CCSP二重陽性細胞は黄色に見える。
【図36】図36A〜36Cを含む図36は、未変性肺臓および人工肺臓における基底細胞に関するサイトケラチン14染色を図示する一連の画像である。サイトケラチンは赤色に染まり、核はDAPIで青色に対比染色される。
【図37】図37は、人工肺臓におけるサイトケラチン14およびCCSPに関する二重染色を図示する画像である。サイトケラチン14は赤色に染色され、CCSPは緑色に染色され、核はDAPIで青色に対比染色される。
【図38】図38Aおよび38Bを含む図38は、未変性肺臓および人工肺臓のα-アクチン染色を図示する一連の画像である。α-アクチンは緑色に染色され、核はDAPIで青色に対比染色される。
【図39】図39A〜39Fを含む図39は、上皮発生に対する培地組成の効果を図示する一連の画像である。BGJb培地中で培養すると、上皮構造は、SPC顆粒の頂端部発現を起こし、CCSP発現の喪失を伴う。DMEM培地では、細胞がSPCとCCSPの発現をどちらも保ち、SPC発現は散在性に細胞質発現である。
【図40】図40は、人工上皮組織におけるサーファクタント発現を図示する画像である。「Lad」はタンパク質ラダーであり;表示のバンドは20および25kDaであり;「Nat」は未変性肺臓組織であり;「Vent」はDMEM培地で換気した人工肺臓組織であり;「Perf」はDMEM培地で灌流した人工肺臓組織であり;「DMEM」はDMEM培地中の静置培養人工肺臓であり;「BGJb」はBGJb培地中の静置培養人工肺臓であり;「ALI」は空気で換気した人工肺臓であり;「Decell」は脱細胞化スキャフォールドである。
【図41】図41A〜41Cを含む図41は、人工肺臓組織における上皮発生に対する空気による換気の効果を図示する一連の画像である。AQP発現は、実質細胞(これはSPCについても陽性である(図41B))中に認められると共に(左上)、立方上皮細胞にも時折強い発現が認められる(右上)。立方上皮のCCSP発現も認められる(図41C)。
【図42】図42Aおよび42Bを含む図42は、未変性肺臓および人工肺臓における繊毛上皮を図示する一連の画像である。人工肺臓については繊毛細胞が赤色で強調されている。
【図43】図43Aおよび43Bを含む図43は、人工肺臓培養に対する灌流および換気の効果を図示する一連の画像である。
【図44】図44A〜44Dを含む図44は、人工肺臓培養における細胞増殖およびアポトーシスに対する灌流および換気の効果を図示する一連の画像である。
【図45】図45Aおよび45Bを含む図45は、人工肺臓組織におけるCCSP発現に対する灌流および換気の効果を図示する一連の画像である。CCSPは赤色に染色され、核はDAPIで青色に対比染色される。
【図46】図46A〜46Bを含む図46は、人工肺臓組織におけるSPC発現に対する灌流および換気の効果を図示する一連の画像である。SPCは赤色に染色され、核はDAPIで青色に対比染色される。
【図47】図47は、ラット肺臓微小血管内皮細胞を播種したフィブロネクチン被覆脱細胞化スキャフォールドのH&E染色を図示する画像である。
【図48】図48Aおよび48Bを含む図48は、灌流人工肺臓内皮と換気人工肺臓内皮のH&E染色を対比して図示する一連の画像である。
【図49】図49Aおよび49Bを含む図49は、灌流人工肺臓内皮と換気人工肺臓内皮のTUNEL染色を対比して図示する一連の画像である。換気だけで培養されたECは、灌流肺臓よりも、実質的に多くアポトーシスを起こす。アポトーシス核はTUNELによって褐色に染まり、陰性核はメチルグリーンで対比染色される。
【図50】図50は、人工肺臓組織における内皮細胞間の密着結合形成の画像による証明である。内皮細胞には星印を付けてあり、それらは広範な細胞間結合によって分離されている。スケールバーは500nmである。
【図51】図51Aおよび51Bを含む図51は、未変性肺臓および人工肺臓におけるVE-カドヘリンの発現を図示する一連の画像である。VE-カドヘリンは赤色に染色され、核はDAPIで青色に対比染色される。
【図52】図52は、2メガダルトンFITC標識デキストランに対する、内皮細胞のみを播種した人工肺臓の透過性を図示する図表である。*は、脱細胞化スキャフォールドとの比較で、p<0.05を示す。
【図53】図53は、人工組織の極限引張強さを図示する図表である。未変性肺臓および脱細胞化肺臓の強さも示している。
【図54】図54A〜54Cを含む図54は、人工内皮組織の成長に対する培地の影響を図示する一連の画像である。人工灌流内皮を表示の培地タイプ中で培養した。H&E組織像を左側のパネルに示す。一方、右側のパネルは褐色(TUNELによる)のアポトーシス核を示し、正常な核はメチルグリーンで対比染色される。
【図55】図55は、CHAPS緩衝液中で4〜8時間インキュベートすることによって調製された脱細胞化気管が、コラーゲンマトリックスを維持し、大半の細胞の、組織からの除去を示したことを証明する一連の画像である。
【図56】図56は、脱細胞化気管が未変性気管に見られる3タイプのCOLの全てを含有していたことを証明する一連の画像である。
【図57】図57は、脱細胞化気管がNHBEの接着および成長を支持したことを証明する一連の画像である。
【図58】図58は、脱細胞化気管がSAECの接着および成長を支持したことを証明する一連の画像である。
【図59】図59は、GFPレンチウイルスに感染したNHBEが6時間後に明白な形態変化を示さなかったことを証明する一連の画像である。
【図60】図60は、気道への点滴注入による送達後に、かなりの数のマイクロスフェアが、マウス肺臓の全ての葉に存在していたことを証明する一連の画像である。
【図61】図61は、肺臓への細胞の注入の成功と、導入遺伝子(GFP)を形質導入されたヒト上皮細胞が肺臓上皮に接着したこととを証明する一連の画像である。
【図62】図62は、トリプシン処理前に見られる注入に使用したGFP細胞を図示する一連の画像である。
【図63A】図63A〜63Cを含む図63は、GFP陽性ヒト気道上皮細胞(NHBEとSAECの両方)が、気道への点滴注入後、何日間も、マウス肺臓に見いだされたことを証明する一連の画像である。
【図63B】図63A〜63Cを含む図63は、GFP陽性ヒト気道上皮細胞(NHBEとSAECの両方)が、気道への点滴注入後、何日間も、マウス肺臓に見いだされたことを証明する一連の画像である。
【図63C】図63A〜63Cを含む図63は、GFP陽性ヒト気道上皮細胞(NHBEとSAECの両方)が、気道への点滴注入後、何日間も、マウス肺臓に見いだされたことを証明する一連の画像である。
【図64】図64Aおよび64Bを含む図64は、換気サイクル中の拡張時および収縮時における、埋植された人工肺臓を示す一連の画像である。
【発明を実施するための形態】
【0031】
発明の詳細な説明
本発明は人工肺臓組織を提供する。本発明は、一つには、天然肺臓組織の特徴を示すように三次元肺臓組織を生成させることができるという発見に基づいている。
【0032】
ある態様では、人工肺臓組織が脱細胞化未変性肺臓組織から誘導される。脱細胞化組織は細胞およびDNAを実質的に欠いている。好ましくは、脱細胞化組織が免疫原性分子も欠いている。より好ましくは、脱細胞化組織は、細胞の付着と増殖にとって大切であるいくつかの重要な細胞外マトリックス分子を保っている。
【0033】
本発明は組織を脱細胞化する方法を包含する。この脱細胞化方法は、肺臓の細胞外マトリックスの重要な局面を保つと共に、肺臓の細胞外マトリックスに対するいかなる損傷も最小限に抑えつつ、組織から細胞材料および核材料を除去する工程を含む。ある態様では、脱細胞化プロセスが、組織から抗原性分子を除去し、それによって組織を非免疫原性にする工程も含む。ある態様では、本発明の脱細胞化プロセスが、細胞培養と完全に適合すると同時に障壁機能を提供する脱細胞化スキャフォールドを生成させる工程を含む。好ましくは、脱細胞化スキャフォールドは、インタクトな気道樹および血管網を持つ肺臓スキャフォールドである。
【0034】
本発明はバイオリアクタも包含する。好ましくは、バイオリアクタは、任意の三次元組織のインビトロ培養を支持する能力を有する。ある態様では、バイオリアクタが、陰圧によって肺臓を換気する能力を有するだけでなく、生理的速度および圧力で血管灌流および換気を提供する能力も持つ。バイオリアクタは、例えば脈管構造を通した培地の灌流、気道を出入りする培地または空気の動き、陰(ならびに陽)圧による肺臓の換気などを可能にする。
【0035】
本発明の肺臓組織のインビトロ三次元モデルは、肺臓発生生物学を調べるのに役立つ。また、本モデルは、その他、例えば創薬、毒性試験、疾患病理学などにも役立つ。
【0036】
本発明は、肺臓組織をインビトロで生成させることができるという発見にも関係する。インビトロモデルは、宿主循環と密接に連係された導管上皮から構成される肺胞形成単位を連想させる構造の形成を再現する。したがって本発明は、再生医学の一形態として、血管化された肺組織を生成させるための方法および組成物を提供する。
【0037】
本発明は、哺乳動物、好ましくはヒトにおける肺臓欠陥を軽減または処置する方法も提供する。本方法は、それを必要とする哺乳動物に、本発明の三次元構築物を含む組成物の治療有効量を投与し、それによってその哺乳動物における肺臓欠陥を軽減または処置する工程を含む。
【0038】
定義
別段の定義がない限り、本明細書において使用する技術用語および科学用語は全て、一般に、本発明が属する技術分野の当業者が通常理解しているものと同じ意味を持つ。一般に、本明細書において使用する用語体系、ならびに細胞培養、分子遺伝学、有機化学、ならびに核酸化学およびハイブリダイゼーションにおける実験手法は、当技術分野において周知であり、通常使用されているものである。
【0039】
核酸とペプチドの合成には標準的な技法を使用する。これらの技法および手法は、一般に、本明細書の至るところに記載する当技術分野における常法や種々の一般的参考文献(例えばSambrookおよびRussell(2001)「Molecular Cloning, A Laboratory Approach」(Cold Spring Harbor Laboratory Press、ニューヨーク州コールドスプリングハーバー)ならびにAusubel et al.(2002)「Current Protocols in Molecular Biology」(John Wiley & Sons、ニューヨーク州ニューヨーク))に従って行われる。
【0040】
冠詞「a」および「an」(「ある」「1つの」)は、本明細書では、1つの、または1つより多くの(すなわち少なくとも1つの)、その冠詞の文法上の対象を指すために使用される。例えば、「ある要素」とは、1つの要素または1つより多くの要素を意味する。
【0041】
「約」という用語は当業者には理解され、それが使用される文脈に基づいて、ある程度は変動するであろう。
【0042】
「前駆体細胞」、「前駆細胞」、および「幹細胞」という用語は、当技術分野では互換的に使用され、本明細書において使用される場合は、無限回にわたって有糸分裂して自分自身を再生するか、所望の細胞タイプへと分化するであろう子孫細胞を生み出す能力を、潜在的に有する、多能性の、または分化拘束されていない(lineage-uncommitted)前駆細胞を指す。多能性幹細胞とは対照的に、分化拘束された(lineage-committed)前駆細胞は、一般に、表現型が互いに異なる多数の細胞タイプを生じる能力は持たないと考えられる。その代わりに、前駆細胞は、1つか、場合によっては2つの、分化拘束された細胞タイプを生じる。
【0043】
本明細書において使用する「脱分化」という用語は、細胞が、専門性の低い状態に戻ることを指す。脱分化後、そのような細胞は、再プログラミング前に可能であったものと比較して、より多くの、または異なる細胞タイプへと、分化する能力を有するであろう。逆分化(すなわち脱分化)のプロセスは分化よりも複雑である可能性が高く、より原始的になるように細胞を「再プログラムする」ことが必要である。
【0044】
本明細書にいう「スキャフォールド」は、細胞の接着と増殖に適した表面を提供する生体適合性材料を含む構造を指す。スキャフォールドは、さらに、機械的安定性および機械的支持も提供し得る。スキャフォールドは、三次元形状に影響を及ぼすようにまたは三次元形状の境界を定めるように特定の形状または形態をとるか、増殖細胞の集団が呈する形態をとり得る。そのような形状または形態には、薄膜(例えば第3の次元よりも実質的に大きな2つの次元を持つ形態)、リボン、ひも、シート、平らな円板、円柱、球、三次元無定形の形状などがあるが、これらに限定されるわけではない。
【0045】
本明細書にいう「生体適合性」とは、哺乳動物に埋植された時にその哺乳動物において有害な応答を惹起しない、任意の材料を指す。生体適合性材料は、個体中に導入された場合に、その個体にとって毒性または有害でなく、その哺乳動物におけるその材料の免疫学的拒絶を誘導することもない。
【0046】
本明細書にいう「自家」とは、後にその材料が再導入されることになる個体と同じ個体に由来する、生物学的材料を指す。
【0047】
本明細書にいう「同種異系」とは、その材料が導入されることになる個体と同じ種の遺伝学的に異なる個体に由来する、生物学的材料を指す。
【0048】
本明細書において、「グラフト(graft)」とは、典型的には欠陥を置き換え、矯正し、または他の方法で克服するために個体に埋植される、細胞、組織または器官を指す。グラフトはさらにスキャフォールドを含んでもよい。組織または器官は、同じ個体から派生する細胞からなることができ、このグラフトを、本明細書では、以下の交換可能な用語で表す:「オートグラフト(autograft)」、「自家トランスプラント(autologous transplant)」、「自家インプラント(autologous implant)」および「自家グラフト」。同じ種の遺伝学的に異なる個体に由来する細胞を含むグラフトを、本明細書では、以下の交換可能な用語で表す:「アログラフト(allograft)」、「同種異系トランスプラント」、「同種異系インプラント」および「同種異系グラフト」。ある個体の一卵性双生児である兄妹から得られるグラフトを、本明細書では、「アイソグラフト(isograft)」、「同種同系トランスプラント」、「同種同系インプラント」または「同種同系グラフト」という。「キセノグラフト(xenograft)」、「異種トランスプラント」または「異種インプラント」とは、ある個体から、種が異なる別の個体へのグラフトを指す。
【0049】
本明細書において使用する「組織グラフティング(tissue grafting)」および「組織再建(tissue reconstructing)」という用語は、どちらも、肺臓欠陥または軟部組織欠陥などの組織欠陥を処置または軽減するために、グラフトを個体に埋植することを指す。
【0050】
本明細書において、疾患、欠陥、障害または状態を「軽減する」とは、その疾患、欠陥、障害または状態の1つまたは複数の症状の重症度を低減することを意味する。
【0051】
本明細書にいう「処置する」とは、ある疾患、欠陥、障害、または有害な状態などの症状を患者が経験する頻度を低減することを意味する。
【0052】
本明細書にいう「治療有効量」は、その組成物の投与を受ける個体に有益な効果を与えるのに十分であるような、本発明の組成物の量である。
【0053】
本明細書において使用する「成長培地」という用語は、細胞の成長を促進する培養培地を指すものとする。成長培地は、一般に、動物血清を含有するであろう。場合によっては、成長培地が動物血清を含有しないこともあるだろう。
【0054】
「分化培地」は、本明細書では、完全には分化していない幹細胞、胎児肺細胞または他のそのような前駆細胞が、その培地中でインキュベートされた時に、分化細胞の特徴の一部または全部を持つ細胞へと発生するように添加物を含むまたは添加物を欠く、細胞成長培地を指すために使用される。
【0055】
本明細書において使用する「成長因子産物」という用語は、細胞に対して成長効果、増殖効果、分化効果、または栄養効果を有する、タンパク質、ペプチド、マイトジェン、または他の分子を指す。成長因子には、線維芽細胞成長因子(FGF)、塩基性線維芽細胞成長因子(bFGF)、酸性線維芽細胞成長因子(aFGF)、上皮成長因子(EGF)、インスリン様成長因子I(IGF-T)、インスリン様成長因子II(IGF-II)、血小板由来成長因子(PDGF)、血管内皮細胞成長因子(VEGF)、アクチビンA、骨形態形成タンパク質(BMP)、インスリン、成長ホルモン、エリスロポエチン、トロンボポエチン、インターロイキン3(IL-3)、インターロイキン6(IL-6)、インターロイキン7(IL-7)、マクロファージコロニー刺激因子、c-kitリガンド/幹細胞因子、オステオプロテゲリンリガンド、インスリン、神経成長因子、毛様体神経栄養因子、サイトカイン、ケモカイン、モルフォゲン、中和抗体、他のタンパク質、および小分子などが含まれるが、これらに限定されるわけではない。好ましくは、FGFは、FGF2、FGF7、FGF10、およびその任意の組合せから選択される群から選択される。
【0056】
「単離された細胞」とは、組織または哺乳動物においてその単離された細胞に天然に付随している他の構成要素および/または細胞から分離された細胞を指す。
【0057】
本明細書にいう「胎児肺細胞」(FPC)は、胚の肺臓組織から単離された細胞を指す。FPCの混合集団は、上皮、間葉および内皮細胞を含み得るが、これらに限るわけではない。
【0058】
本明細書にいう「上皮細胞」とは、身体の外面を形成し、器官、腔および粘膜表面を裏打ちする細胞を意味する。
【0059】
本明細書にいう「内皮細胞」とは、血管およびリンパ管ならびに他のさまざまな体腔を裏打ちする細胞を意味する。
【0060】
本明細書にいう「実質的に精製された」細胞は、本質的に他の細胞タイプを含まない細胞である。したがって、実質的に精製された細胞とは、その天然状態ではそれに通常付随している他の細胞タイプから精製された細胞を指す。
【0061】
「拡大性(expandability)」は、本明細書では、増殖して、例えば数を拡大するか、細胞の集団の場合であれば、集団倍加を起こすという、細胞の能力を指すために使用される。
【0062】
「肺臓特異的」という用語は、身体の他の組織と比較して肺臓において優勢に発現する核酸分子またはポリペプチドを指す。ある好ましい態様では、「肺臓特異的」核酸分子またはポリペプチドが、身体の他のどの組織よりも5倍高いレベルで発現する。より好ましい態様では、「肺臓特異的」核酸分子またはポリペプチドが、身体の他のどの組織よりも10倍高いレベルで発現し、より好ましくは、身体の他のどの組織よりも少なくとも15倍、20倍、25倍、50倍または100倍高いレベルで発現する。核酸分子レベルは、ノーザンブロットハイブリダイゼーションなどの核酸ハイブリダイゼーションまたは定量PCRによって測定することができる。ポリペプチドレベルは、タンパク質レベルを正確に測定することが知られている任意の方法、例えばウェスタンブロット分析などによって測定することができる。
【0063】
「増殖」は、本明細書では、類似する形態の、特に細胞の、複製または増倍を指すために使用される。すなわち増殖は、より多数の細胞の産生を包含し、例えば、細胞の数を単純に数え上げること、3H-チミジンの細胞への取り込みを測定することなどによって測定することができる。
【0064】
本明細書にいう「組織工学」とは、組織代用(tissue replacement)または組織再建に使用するためにエクスビボで組織を生成させるプロセスを指す。組織工学は「再生医学」の一例であり、これは、細胞、遺伝子または他の生物学的構成単位の組込みと、生物工学的材料および技術とによる、組織および器官の修復または代用への取り組みを包含する。
【0065】
本明細書にいう「内在性」とは、ある生物、細胞または系に由来する、またはそれらの内部で産生された、任意の材料を指す。
【0066】
「外因性」とは、ある生物、細胞、または系に導入された、またはそれらの外で産生された、任意の材料を指す。
【0067】
「コードする」とは、所定の配列のヌクレオチド(すなわちrRNA、tRNAおよびmRNA)または所定の配列のアミノ酸とその結果生じる生物学的性質とを有する他のポリマーおよび高分子を生物学的プロセスにおいて合成するための鋳型として役立つという、遺伝子、cDNA、またはmRNAなどのポリヌクレオチドにおけるヌクレオチドの特異的配列が持つ固有の性質を指す。したがって、ある遺伝子に対応するmRNAの転写および翻訳が、細胞内または他の生物学的系内で、あるタンパク質を産生するのであれば、その遺伝子はそのタンパク質をコードしている。コード鎖(mRNA配列と同一のヌクレオチド配列を持ち、配列表に通常記載されるもの)と、遺伝子またはcDNAの転写のための鋳型として使用される非コード鎖は、どちらも、タンパク質、またはその遺伝子もしくはcDNAの他の産物をコードしているということができる。
【0068】
別段の指定がない限り、「アミノ酸配列をコードするヌクレオチド配列」には、互いの縮重型であって同じアミノ酸配列をコードする、全てのヌクレオチド配列が含まれる。タンパク質およびRNAをコードするヌクレオチド配列はイントロンを含み得る。
【0069】
「単離された核酸」とは、天然の状態でそれに隣接している配列から分離されている核酸セグメントまたは核酸フラグメント、すなわち通常はそのフラグメントに近接している配列(すなわち、それが天然に存在するゲノム中でそのフラグメントに近接している配列)から取り出されたDNAフラグメントを指す。この用語は、その核酸に天然に付随する他の構成要素、すなわち、細胞内でそれに天然に付随しているRNAまたはDNAまたはタンパク質から実質的に精製された核酸にも適用される。したがってこの用語には、例えば、ベクターに組み込まれているか、自律的に複製するプラスミドもしくはウイルスに組み込まれているか、または原生生物もしくは真核生物のゲノムDNAに組み込まれている組換えDNA、または他の配列から独立して別個の分子(すなわちcDNAまたはPCRもしくは制限酵素消化によって作り出されるゲノムフラグメントもしくはcDNAフラグメント)として存在する組換えDNAが含まれる。また、追加のポリペプチド配列をコードするハイブリッド遺伝子の一部である組換えDNAも含まれる。
【0070】
本発明については、よく見られる核酸塩基に対して、以下の略号を使用する。「A」はアデノシンを指し、「C」はシトシンを指し、「G」はグアノシンを指し、「T」はチミジンを指し、「U」はウリジンを指す。
【0071】
本明細書において使用する「転写制御下にある」または「作動的に連結された」という表現は、そのプロモーターがそのポリヌクレオチドに対して正しい位置と向きにあって、RNAポリメラーゼの開始およびポリヌクレオチドの発現を制御することを意味する。
【0072】
本明細書において使用する「プロモーター/調節配列」という用語は、そのプロモーター/制御配列に機能的に連結される遺伝子産物の発現にとって必要な核酸配列を意味する。いくつかの例では、この配列はコアプロモーター配列であることができ、他の例では、この配列はエンハンサー配列および遺伝子産物の発現に必要な他の調節要素も含むことができる。プロモーター/調節配列は、例えば、遺伝子産物を組織特異的に発現させるものであることができる。
【0073】
「構成的」プロモーターは、ある遺伝子産物をコードまたは指定するポリヌクレオチドと機能的に連結された場合に、ある細胞において、その細胞の大半のまたは全ての生理的条件下で、その遺伝子産物を産生させる、ヌクレオチド配列である。
【0074】
「誘導性」プロモーターは、ある遺伝子産物をコードまたは指定するポリヌクレオチドと機能的に連結された場合に、ある細胞において、実質上、そのプロモーターに対応する誘導因子がその細胞中に存在する場合にのみ、その遺伝子産物を産生させる、ヌクレオチド配列である。
【0075】
本明細書において使用する「組織」という用語には、例えば骨、神経組織、腱および靱帯を含む線維性結合組織、軟骨、硬膜、心膜、筋、肺臓、心臓弁、静脈および動脈ならびに他の脈管構造、真皮、脂肪組織、または腺組織が含まれるが、これらに限定されるわけではない。
【0076】
「組織特異的」プロモーターは、ある遺伝子産物をコードまたは指定するポリヌクレオチドと機能的に連結された場合に、ある細胞において、実質上、その細胞がそのプロモーターに対応する組織タイプの細胞である場合にのみ、その遺伝子産物を産生させる、ヌクレオチド配列である。
【0077】
「ベクター」は、単離された核酸を含み、その単離された核酸を細胞の内部に送達するために使用することができる、組成物である。限定するわけではないが、線状ポリヌクレオチド、イオン性または両親媒性化合物と会合したポリヌクレオチド、プラスミド、およびウイルスなど、数多くのベクターが当技術分野において公知である。したがって「ベクター」という用語には、自律的に複製するプラスミドまたはウイルスが含まれる。また、この用語は、細胞への核酸の導入を容易にする非プラスミドおよび非ウイルス化合物、例えばポリリジン化合物、リポソームなどを包含すると解釈すべきでもある。ウイルスベクターの例には、アデノウイルスベクター、アデノ随伴ウイルスベクター、レトロウイルスベクターなどがあるが、これらに限定されるわけではない。
【0078】
「発現ベクター」とは、発現させようとするヌクレオチド配列に作動的に連結された発現制御配列を含む組換えポリヌクレオチドを含むベクターを指す。発現ベクターは、発現にとって十分なシス作用要素を含み、発現のための他の要素は、宿主細胞によって供給されるか、インビトロ発現系で供給され得る。発現ベクターには、当技術分野において公知であるもの、例えばコスミド、プラスミド(すなわち裸のプラスミドまたはリポソームに含有されたプラスミド)およびウイルスなどであって、組換えポリヌクレオチドが組み込まれているものが、全て含まれる。
【0079】
本明細書において使用する「対象」および「患者」という用語は、互換的に使用される。本明細書にいう対象は、好ましくは、哺乳動物、例えば非霊長類(例えばウシ、ブタ、ウマ、ネコ、イヌ、ラットなど)および霊長類(例えばサルおよびヒト)、最も好ましくはヒトである。
【0080】
説明
本発明は、人工三次元肺組織、およびその三次元肺組織を製作する方法を提供する。好ましくは、肺組織は肺臓組織である。ある態様において、人工肺組織は、天然肺組織によって例示される分枝形態形成を示す。したがって本発明は、天然肺組織を模倣するインビトロモデルを提供する。インビトロ三次元肺組織モデルは、例えば創薬、毒性試験、疾患病理学などに役立つ。
【0081】
本発明は、細胞材料は除去するが細胞外マトリックスの重要な構成要素は保つ技法を使って肺臓組織を脱細胞化するのに有用な手法の発見に基づく。脱細胞化肺臓マトリックスの開発は、組織工学的応用のためのスキャフォールドとして重要である。したがって本発明は、組織または器官を実質的に脱細胞化するための方法を包含する。好ましくは、本方法は、組織または器官が、移植時に、レシピエントの免疫系によって拒絶されないように、組織または器官の免疫原性を著しく低減または排除する。本方法は、ドナーから組織を取り出し、組織または器官の細胞の実質上全てを除去するために組織を加工する工程を含む。本方法はさらに、レシピエントに埋植するために、幹細胞や胎児細胞など(ただしこれらに限るわけではない)の細胞を播種することによって、脱細胞化スキャフォールドに再集合させる工程を含む。好ましくは、脱細胞化スキャフォールドに、非免疫原性細胞を播種する。ある態様では、脱細胞化スキャフォールドに、意図するレシピエントにとって自家である細胞を播種する。処理し、置換しようとする組織のタイプに依存して、播種済インプラントをレシピエントに埋植した時に適当な組織が形成されるように、当技術分野において公知であるか今後公知になる異なる幹細胞が選択される。
【0082】
いくつかの例では、人工三次元肺組織が、組織上で培養された細胞を含む。本発明の脱細胞化組織上での培養には、任意の適切な細胞を使用することができる。いくつかの例では、肺臓組織を再生させるために、幹細胞を脱細胞化組織上で培養する。いくつかの例では、胎児または新生児肺細胞(NPC)を脱細胞化組織上で培養する。いくつかの例では、上皮細胞、間葉細胞、および内皮細胞(ただしこれらに限るわけではない)を含むNPCの混合集団を使用する。
【0083】
播種後に、スキャフォールド上の細胞は、任意で、拡大培地または分化培地に供されるか、組織特異的成長因子の存在下で培養される。次に、その組成物を、それを必要とする対象に埋植する。対象は哺乳動物であることができるが、好ましくはヒトであり、成長および埋植のための細胞の供給源は任意の哺乳動物、好ましくはヒトである。埋植された組成物はインビボでさらなる細胞成長を支持するので、組織再建をもたらす。したがって、本発明は、組織グラフティング療法のための人工三次元肺組織の使用を提供する。
【0084】
本発明は、インビボでの肺組織の生成も包含する。好ましくは、血管化した肺組織を、インビボで生成させる。ある局面では、インビボ肺組織形成を容易にするために、哺乳動物に、脱細胞化組織との関連において、胎児肺細胞が投与される。
【0085】
本発明では、脱細胞化組織に、新生児肺細胞または成人肺細胞などの適切な細胞を播種することができ、その結果生じる組成物は、血管化した三次元肺組織モデルとして、前臨床インビトロ薬理試験、生理学的試験および科学的試験に使用できることが証明される。また、脱細胞化組織には、新生児肺細胞または自家肺細胞などの適切な細胞を播種することができ、その結果生じる組成物は、インビボでの組織再建に使用することができる。
【0086】
本発明の組成物および方法には無数の有用な応用がある。本組成物は、個体における組織欠陥を軽減または処置するための治療方法に使用することができる。本組成物は治療化合物を同定するためにインビトロまたはインビボで使用することもでき、したがって治療能力を有し得る。
【0087】
脱細胞化
本発明は、当技術分野において公知の組織工学的技法を上回る進歩を提供する。具体的には、本発明は、脱細胞化組織(好ましくは哺乳動物から得られる脱細胞化天然組織)を出発源として使用して、人工組織スキャフォールドを製作する方法を提供する。
【0088】
脱細胞化プロセスは化学的方法に依拠する。ある局面では、脱細胞化に使用される化学溶液(脱細胞化溶液ともいう)は一般に、少なくとも高張溶液、界面活性剤、およびキレート剤を含む。好ましくは、高張溶液は高張塩化ナトリウム溶液である。好ましくは、界面活性剤はCHAPSなどの両性イオン性界面活性剤である。好ましくは、キレート剤はEDTAである。
【0089】
ある態様において、脱細胞化溶液は、細胞との浸透圧適合性のための緩衝液(例えばPBS)を含むことができる。いくつかの例では、脱細胞化溶液が、1つまたは複数のコラゲナーゼ、1つまたは複数のディスパーゼ、1つまたは複数のDNアーゼ、またはトリプシンなどのプロテアーゼなどの酵素(ただしこれらに限定されるわけではない)も含むことができる。いくつかの例では、脱細胞化溶液が、上記に加えて、または上記の代わりに、1つまたは複数の酵素の阻害剤(例えばプロテアーゼ阻害剤、ヌクレアーゼ阻害剤、および/またはコラゲナーゼ阻害剤)を含むことができる。
【0090】
ある態様において、組織を脱細胞化するための本発明の方法は、組織を脱細胞化溶液で灌流する工程を含む。脱細胞化溶液を組織に灌流させる圧力は、望ましい圧力に調節することができる。好ましくは、脱細胞化溶液を組織に約30mmHg未満の灌流圧で灌流させる。より好ましくは、脱細胞化溶液を組織に約20mmHg未満の圧力で灌流させる。
【0091】
ある態様では、細胞除去を達成するために脱細胞化溶液を肺臓組織の気道中に導入することができる。
【0092】
ある態様では、本発明の脱細胞化組織が、本質的に、血管樹のECM構成要素を含む、その組織の全てまたは大半の領域の細胞外マトリックス(ECM)構成要素からなる。ECM構成要素は、以下の物質をどれでも含むことができ、または以下の物質の全てを含むことができる:基底膜などの所定の構造として組織化された状態を保つことができる、フィブロネクチン、フィブリリン、ラミニン、エラスチン、コラーゲンファミリーのメンバー(例えばコラーゲンI、III、およびIV)、グリコサミノグリカン、礎質(ground substance)、細網線維およびトロンボスポンジン。脱細胞化の成功は、標準的な組織染色手法を使って組織切片中に検出可能な筋フィラメント、内皮細胞、平滑筋細胞、上皮細胞および核が存在しないことと定義される。そうでない場合もあるが、好ましくは、残留細胞片も脱細胞化組織から除去された。
【0093】
ある態様では、天然組織の脱細胞化プロセスが、組織の未変性三次元構造を保存する。すなわち、ECM構成要素を含む組織の形態およびアーキテクチャは、脱細胞化のプロセス中およびプロセス後に維持される。ECMの形態およびアーキテクチャは、視覚的および/または組織学的に調べることができる。例えば、固形臓器(solid organ)の外面上または器官もしくは組織の脈管構造内の基底膜は、脱細胞化によって除去されたり、著しく損傷されたりすべきでない。また、ECMの原線維は、脱細胞化されていない器官または臓器のそれと類似しているか、著しくは変化していないべきである。
【0094】
ある態様では、例えば脱細胞化組織を保存するために、または再細胞化に備えて脱細胞化組織を調製するために、かつ/または再細胞化プロセス中の細胞を補助もしくは刺激するために、1つまたは複数の化合物を脱細胞化組織中または脱細胞化組織上に適用することができる。そのような化合物には、1つまたは複数の成長因子(例えばVEGF、DKK-1、FGF、BMP-1、BMP-4、SDF-1、IGF、およびHGF)、免疫調整剤(例えばサイトカイン、グルココルチコイド、IL2Rアンタゴニスト、ロイコトリエンアンタゴニスト)、および/または凝固カスケードを修飾する因子(例えばアスピリン、ヘパリン結合タンパク質、およびヘパリン)などがあるが、これらに限定されるわけではない。また、脱細胞化された器官または組織をさらに、例えば照射(例えばUV、γ線)処理して、脱細胞化組織上または脱細胞化組織中に残っているあらゆるタイプの微生物を減少させ、またはその存在を排除することもできる。
【0095】
脱細胞化組織を生成させるための本発明の脱細胞化溶液の使用は、脈管構造を含む基礎をなすECMと元の組織の他の総体的な形態上の特質とをインタクトな状態で残しつつ、組織の細胞を破壊するための制御された正確な方法を提供する。したがって、脱細胞化されたスキャフォールドは、適当な細胞による播種に適している。このプロセスがインビトロで行われる場合、播種された組織は、代用組織として、レシピエントへの埋植に適している。脱細胞化組織そのものに加えて、本発明は、そのようなスキャフォールドから構築される人工組織の製造方法も包含する。
【0096】
本発明は、組織工学において使用するための組織スキャフォールドを作製するのに適した方法を提供する。組織の供給源は限定されないが、例示的な態様では、組織が、比較的大きな動物、または(関心対象の組織に関して)ヒトと類似する解剖学的形態を持つと認識されている動物、例えばブタ、ウシ、ウマ、サル、または類人猿に由来する。いくつかの態様では、組織の供給源がヒトであり、その使用により、当該スキャフォールドに基づく人工組織の拒絶の可能性を低減することができる。好ましい態様において、本方法は、肺胞中隔が保存された肺胞アーキテクチャなど、組織のインタクトな血管構造を残す。本明細書において使用する「インタクト」という用語は、ある要素がその要素の元の機能を相当程度果たす能力を有するような状態を指す。
【0097】
ある態様では、脱細胞化肺臓が、正常肺臓マトリックスのいくつかの重要な特徴を保っている。例えば、脱細胞化肺臓は、コラーゲン、エラスチン、フィブロネクチン、およびプロテオグリカンの少なくとも1つまたは複数を含む。
【0098】
脱細胞化組織は、主要組織適合遺伝子複合体(MHC)クラスIまたはII抗原を保っていないので、組織は、レシピエントに投与された時に、有害な免疫応答を引き起こさない。
【0099】
脱細胞化組織は正常な未変性肺臓の機械的性質を保っている。脱細胞化組織は正常な未変性肺臓の障壁機能の一部も保っている。
【0100】
バイオリアクタ
本発明は、組織を脱細胞化しかつ/または再細胞化するためのシステム(例えばバイオリアクタ)を提供する。バイオリアクタは、細胞生存性、細胞の分化状態、および肺臓形態の維持を可能にする。脱細胞化スキャフォールドは、適切な細胞源と共にバイオリアクタ中で培養すると、肺の内皮、上皮、および間葉細胞を含む広範な細胞タイプの接着および増殖を支持することができる。本発明のバイオリアクタには、生体(vivo)環境の重要な特質が組み込まれている。本バイオリアクタは、脱細胞化および/または再細胞化プロセスを最適化するための変更が可能なように設計された。ある態様では、バイオリアクタが、使用者の指定した流量かつ哺乳動物の生理的流量レベルおよび生理的圧力レベル内で、脈管構造を通して培地を灌流させる能力を有する。別の態様では、バイオリアクタが、気管を通して空気または培地で組織(例えば肺臓)を換気する能力を有する。好ましくは、正常な生理学的条件と合致するように陰圧換気法が使用されるが、陽圧を使った換気を行うこともできる。さらに別の態様において、バイオリアクタは、組織の血管コンパートメントと気道コンパートメントを異なる培地タイプで浸すことを可能にする能力を有する。別の態様において、バイオリアクタは、培養培地へのガス交換を可能にすると同時に、換気に関する所望の要件を満たす。別の態様では、バイオリアクタが、圧力測定(例えば肺動脈圧および気管圧の測定)を可能にするためのポート(port)を持つ。好ましくは、圧力は正常な生理学的値の範囲内にある。別の態様では、バイオリアクタが、定期的な培地交換を可能にする手段を持つ。
【0101】
本発明のバイオリアクタは、一般に、組織にカニューレを挿入するための少なくとも1つのカニューレ挿入装置、カニューレを通して培地を灌流させるための灌流器具、および器官または組織の滅菌環境を維持するための手段(例えば封じ込めシステム)を含む。カニューレ挿入装置は、一般に、組織の血管、導管、および/または腔中に導入するための適当なサイズの中空チューブを含む。典型的には、組織中の1つまたは複数の血管、導管、および/または腔にカニューレが挿入される。灌流器具は、液体(例えば細胞破壊培地)のための保持容器および1つまたは複数のカニューレを通して器官越しに液体を移動させるための機構(例えばポンプ、空気圧、重力)を含むことができる。脱細胞化および/または再細胞化中の組織の滅菌性は、本明細書の他の項で論じる方法を使って維持することができる。
【0102】
バイオリアクタは、本明細書に記載するとおり、組織を脱細胞化および再細胞化するために使用することができる。本プロセスは、一定の灌流特徴(例えば圧力、体積、フローパターン、温度、ガス、pH)、機械的力(例えば心室壁運動および心室壁応力)、および電気刺激(例えばペーシング)について監視することができる。灌流の有効性は流出液および組織切片において評価することができる。灌流体積、フローパターン、温度、O2およびCO2分圧、ならびにpHは、標準的な方法を使って監視することができる。
【0103】
バイオリアクタおよび/または組織を監視するために、センサを使用することができる。ソノミクロメトリー、微小検圧法、および/またはコンダクタンス測定を使って、圧力-体積を取得することができる。例えば、センサを使って、カニューレが挿入された器官または組織を通って移動する液体の圧力;系内の周囲温度および/または器官もしくは組織の温度;カニューレが挿入された器官または組織を通って移動する液体のpHおよび/または流量;および/または再細胞化組織の生物学的活性を監視することができる。そのような特質を監視するためのセンサを持つことに加えて、組織を脱細胞化および/または再細胞化するためのシステムは、そのような特質を維持または調節するための手段も含むことができる。そのような特質を維持または調節するための手段は、温度計、サーモスタット、電極、圧力センサ、オーバーフロー弁、液体の流量を変化させるための弁、溶液のpHを変化させるために使用される溶液への流体接続を開け閉めするための弁、風船、外部ペースメーカー、および/またはコンプライアンスチャンバなどの構成要素を含むことができる。安定した状態(例えば温度)の確保に役立つように、チャンバ、リザーバ、およびチューブには水ジャケットを装着することができる。
【0104】
バイオリアクタは、細胞の生存および分化を支持するために、肺臓組織に十分な栄養素供給および機械的刺激を与える能力を有する。バイオリアクタは、インビトロ肺臓組織培養および人工肺臓組織培養に使用することができる。好ましくは、本発明の脱細胞化肺臓スキャフォールドを使って人工肺臓組織を培養するために、バイオリアクタを使用することができる。
【0105】
肺臓組織の真の3次元セグメントをインビトロ培養する能力を有するバイオリアクタの開発は、臨床的に有用な人工肺臓組織の開発における重要な一工程である。例えば、人工肺臓をレシピエントに埋植する前に、人工肺臓組織の成長および成熟をバイオリアクタ中で行うことにより、最終的な埋植肺臓組織のインビボでの機能性を強化することができる。また、肺の生物学、生理学および発生の研究を支援するために、インビトロ肺臓培養のためのバイオリアクタを使用することもできる。すなわち、肺臓内皮および上皮細胞の相互作用による肺胞-毛細管障壁の形成を、本発明の人工肺臓組織およびバイオリアクタを使って研究することができる。当業者は、現在使用されているさまざまな動物モデルよりもよく制御された環境で、肺臓の挙動を研究することが可能になるだろう。人工肺臓組織およびバイオリアクタは、時間も費用もかかるヒト治験または動物治験に進む前に、ヒト組織および動物組織における薬理学的な試験および調査を行うために使用することもできる。
【0106】
組成物
本発明の組成物は、人工肺臓組織を含む。好ましくは、人工肺臓組織は、以下に挙げる性質の任意の1つまたは複数を示す:1)開存性の灌流された脈管構造と、換気することができる開存性の気道樹とが存在する、脈管構造および気道;2)人工肺臓が、レシピエントの生理学的必要を支えるために、気道と血管コンパートメントとの間で十分な気体を交換する能力を有し、最も好ましくは、肺静脈中の酸素分圧が少なくとも50mmHgであるような、ガス交換;3)人工組織が、必要とされる全ての動き(特に呼吸運動および血管灌流)にも外科的埋植中の操作にも耐え得るほど十分に強いような、力学的性質;4)レシピエントに埋植された時に人工肺臓組織が免疫応答を惹起しないような、免疫原性。
【0107】
本発明の組成物および方法は、任意の適切な細胞を使って実施することができる。好ましくは、適切な細胞または細胞群は再生性であり、本発明の脱細胞化組織を再細胞化するために使用することができる。再生性細胞の一例には、幹細胞、胚性幹細胞、成人幹細胞、臍帯血細胞、組織由来の幹細胞または前駆細胞、骨髄由来の幹細胞または前駆細胞、血液由来の幹細胞または前駆細胞、間葉系幹細胞(MSC)、骨格筋由来の細胞、複能性成人前駆細胞(MAPC)、胎児肺細胞、分化肺上皮細胞、肺前駆細胞、血管前駆細胞、分化血管細胞などがあるが、これらに限定されるわけではない。使用することができる他の再生性細胞には、骨髄由来の幹細胞、例えば骨髄単核球(BM-MNC)、内皮または血管幹細胞または前駆細胞、および末梢血由来の幹細胞、例えば内皮前駆細胞(EPC)などがある。
【0108】
好ましくは、適切な細胞は、哺乳動物、より好ましくは霊長類、さらに好ましくはヒトから単離される。本発明の方法において有用な細胞は、本明細書において(例えば実施例の項で)論じる方法を使って、または当技術分野で公知の任意の方法によって、単離される。単離に続いて、その適切な細胞を、培養培地中で培養する。
【0109】
限定ではない一例として、新生児肺細胞(NPC)を細胞の培養に関して詳述する。しかし、適切な細胞に合わせて培養条件を変更できることは、当業者にはわかるだろう。肺細胞の成長を支持する培地製剤には、イーグル最小必須培地、ADC-1、LPM(ウシ血清アルブミン非含有)、F10(HAM)、F12(HAM)、DCCM1、DCCM2、RPMI1640、BGJ培地(フィトン-ジャクソンの改良を含むものおよび含まないもの)、イーグル基礎培地(アールの塩類ベース(salt base)が添加されたBME)、ダルベッコ変法イーグル培地(血清を含まないDMEM)、ヤマネ(Yamane)、IMEM-20、グラスゴー変法イーグル培地(GMEM)、リーボビッツL-15培地、マッコイの5A培地、M199培地(アールの塩類ベースを含むM199E)、M199培地(ハンクスの塩類ベースを含むM199H)、イーグル最小必須培地(アールの塩類ベースを含むMEM-E)、イーグル最小必須培地(ハンクスの塩類ベースを含むMEM-H)およびイーグル最小必須培地(非必須アミノ酸を含むMEM-NAA)などがあるが、これらに限定されるわけではない。
【0110】
本発明の方法に役立つ培地のさらなる限定でない例は、ウシまたは他の種の胎児血清を、少なくとも1%〜約30%、好ましくは少なくとも約5%〜15%、最も好ましくは約10%の濃度で含有し得る。ウシまたは他の種の胎児抽出物は、約1%〜30%、好ましくは少なくとも約5%〜15%、最も好ましくは約10%の濃度で存在することができる。
【0111】
典型的には、NPC培養培地は、基礎培地、血清および抗生物質/抗真菌物質を含む。好ましい基礎培地の一つは、DMEM/F12(1:1)である。好ましい血清はウシ胎児血清(FBS)であるが、ウマ血清またはヒト血清を含む他の血清も使用することができる。好ましくは、NPCの成長を支持するために、20%までのFBSが上記の培地に加えられるであろう。しかし、NPC成長にとって必要なFBS中の成長因子、サイトカインおよびホルモンが同定され、それらが成長培地に適当な濃度で与えられるのであれば、合成培地を使用することもできる。さらに、追加の構成要素を培養培地に加え得ることも、理解される。そのような構成要素には、抗生物質、抗真菌物質、アルブミン、成長因子、アミノ酸、および細胞の培養に関して当技術分野で公知である他の構成要素などが含まれるが、これらに限定されるわけではない。培地に加えることができる抗生物質には、ペニシリンおよびストレプトマイシンがあるが、これらに限定されるわけではない。培養培地中のペニシリンの濃度は1mlあたり約10〜約200単位である。培養培地中のストレプトマイシンの濃度は約10〜約200μg/mlである。しかし本発明は、決して、NPCの培養に関して、どの一培地にも限定されると解釈されるべきではない。むしろ、組織培養において肺細胞を支持する能力を有する任意の培地を使用することができる。
【0112】
また、NPC培養培地には、少なくとも1つの成長因子を補足することもできる。好ましくは、成長因子は線維芽細胞成長因子(FGF)である。例えば、FGF10、FGF7、FGF2の任意の組合せを、NPC培養培地に補足することができる。FGF7の好ましい濃度は、約0.1〜100ng/ml(およびその中間にある任意の整数)であり、より好ましくは、濃度は約10ng/mlである。FGF10の好ましい濃度は約1〜200ng/ml(およびその中間にある任意の整数)であり、より好ましくは、濃度は約25ng/mlである。FGF2の好ましい濃度は約1〜200ng/ml(およびその中間にある任意の整数)であり、より好ましくは、濃度は約25ng/mlである。
【0113】
単離に続いて、NPCは、培養器具中の培養培地において、一定の期間、または細胞がコンフルエントに達するまで、培養した後、細胞を別の培養器具に移すことができる。最初のプレーティングに続いて、細胞を約6日間培養維持することにより、継代第0代(P0)の集団を得ることができる。細胞は、無限回にわたって継代することができ、各継代は、細胞を約6〜7日間培養することを含み、その間、細胞倍加時間は約3〜約5日の範囲に及び得る。培養器具は、細胞をインビトロで培養するために一般に使用される任意の培養器具であることができる。
【0114】
NPCは、ある期間にわたって、または細胞が一定レベルのコンフルエンスに達するまで、FGFを補足した培養培地中で培養することができる。好ましくは、コンフルエンスのレベルは70%より高い。より好ましくはコンフルエンスのレベルは、90%より高い。期間は、インビトロでの細胞の培養に適した任意の期間であることができる。NPC培養培地は、NPCの培養中はいつでも取り替えることができる。好ましくは、3〜4日ごとに培養培地を取り替える。次に、NPCを培養器具から収集し、その後、直ちにそれらを使用するか、低温保存して、後に使用するまで貯蔵しておくことができる。NPCはトリプシン処理、EDTA処理、または培養器具から細胞を収集するために使用される他の任意の手法によって、収集することができる。
【0115】
本明細書において記載するNPCは、常法に従って低温保存することができる。好ましくは、約100万〜1000万個の細胞を、10%DMSOを含有する培養培地に入れて、液体N2の気相中で低温保存する。凍結細胞は、37℃の浴中で旋回させることによって融解し、新鮮な成長培地中で再懸濁し、上述のように拡大することができる。
【0116】
本発明は、スキャフォールドに「播種(seed)」する細胞も提供する。NPCはスキャフォールド上で培養することができる。細胞は、細胞を分化培地中で培養することにより、インビトロで分化することもできる。あるいは、細胞が哺乳動物内で組織と接触するか、細胞が、組織から放出される物質(例えば成長因子、酵素、またはホルモン)による影響を受けるように、組織の十分近くにある場合には、細胞はインビボで分化することもできる。言い換えると、マトリックスのNPCは、組織からのシグナルを受け取ることにより、肺臓などの組織と接触することができる。そのようなシグナリングは、例えば、NPCの表面上またはNPCに由来する細胞の表面上にある受容体が、哺乳動物内で組織によって放出された成長因子、酵素、またはホルモンなどの分子を結合し、その分子からのシグナルを伝達する場合に、起こるだろう。これらの作用物質は分化を誘導するので、NPCは、それらが置かれた組織中の分化細胞によって通常発現されるものと同じタンパク質の一部を、そして場合によっては(全てではないとしても)大半を、発現するようになる。
【0117】
上記の代わりに、または上記に加えて、マトリックスのNPCは、その細胞の環境に、ある物質(例えば成長因子、酵素、ホルモン、または他のシグナリング分子)を加えることによって、分化するように誘導することができる。例えば、ある物質を、本発明の生物学的足場に加えることができる。
【0118】
NPCとそれに付随する細胞マトリックスは最終的に完全に分化した状態になることができ、状況によっては(例えば組織学的に成熟した完全な組織を再現するために細胞を使用する場合には)それが望ましい場合もあるが、処置を成功させるには、投与された細胞の全てが完全に分化する必要はなく、細胞マトリックスのNPCは、哺乳動物を処置するのに十分な点まで分化するだけでよい。その点には、マトリックスを患者に投与する前に到達するか、マトリックスを患者に投与した後に到達することができる。
【0119】
分化は、マトリックスの細胞が、埋植部位にある成熟細胞と基本的に同じ表現型を発現する場合に起こる。例えば、本発明の定義を目的とする場合、肺臓中に埋植された細胞マトリックスのNPCは、それが肺臓(例えば肺胞上皮細胞)によって発現されるタンパク質と本質的に同じタンパク質を発現する場合に、分化しているという。肺臓マーカーに対する抗体は市販されているか、そうでなくても、容易に取得することができる。
【0120】
分化細胞は、その外部形態によって、およびそれらが他の細胞と形成する接続によって、同定することもできる。例えば、肺臓細胞に分化する細胞は、細気管支に似た複雑な形態を発生することができる。例えば本発明は、三次元スキャフォールド上でNPCを培養することにより、成熟した肺臓細胞の特徴を示すことができるという、新規な発見に基づいている。
【0121】
器官または組織を生成させるために脱細胞化器官中および脱細胞化器官上に導入される細胞の数は、器官(例えば器官の種類、器官のサイズおよび重量)または組織にも、再生性細胞のタイプおよび発生段階にも依存する。異なるタイプの細胞は、その細胞が到達するであろう集団密度に関して、異なる傾向を持ち得る。同様に、異なる器官または組織は異なる密度で細胞化され得る。例えば、脱細胞化器官または組織には、少なくとも約1,000(例えば少なくとも10,000、100,000、1,000,000、10,000,000、または100,000,000)個の再生性細胞を播種することができ、あるいは、脱細胞化器官または組織は、そこに付着した約1,000細胞/mg組織(湿重量、すなわち脱細胞化前)〜約10,000,000細胞/mg組織(湿重量)を持つことができる。
【0122】
細胞は、1つまたは複数の場所への注入によって、脱細胞化器官または組織に導入することができる。また、2タイプ以上の細胞(すなわち細胞のカクテル)を脱細胞化器官または組織に導入することができる。例えば、細胞のカクテルを脱細胞化器官または組織中の複数の場所に注入するか、異なる細胞タイプを脱細胞化器官または組織の異なる部分に注入することができる。注入の代わりに、または注入に加えて、再生性細胞または細胞のカクテルは、カニューレを挿入した脱細胞化器官または組織への灌流によって導入することができる。例えば、細胞は灌流培地を使って脱細胞化器官中に灌流することができ、次にその灌流培地を拡大培地および/または分化培地に変えて、再生性細胞の成長および/または分化を誘導することができる。肺臓組織の場合、細胞は、気管を通して気道コンパートメントに導入するか、肺動脈もしくは肺静脈を通して血管コンパートメントに導入するか、またはその両方に導入することができる。
【0123】
再細胞化中、器官または組織は、再生性細胞の少なくとも一部が脱細胞化器官または組織内および脱細胞化器官または組織上で増倍しかつ/または分化することができるような条件下に維持される。それらの条件には、適当な温度および/または圧力、電気的および/または機械的活動、力、適当な量のO2および/またはCO2、適当な量の湿気、および滅菌またはほぼ滅菌状態が含まれるが、これらに限定されるわけではない。再細胞化中、脱細胞化器官または組織およびそこに付着した細胞は、適切な環境に維持される。例えば細胞は、栄養補給(例えば栄養素および/またはグルコースなどの炭素源)、外因性のホルモンもしくは成長因子、および/または特定のpHを必要とし得る。
【0124】
細胞は、脱細胞化器官または組織にとって同種異系(例えばヒト細胞を播種されたヒト脱細胞化器官または組織)であるか、再生性細胞は脱細胞化器官または組織にとって異種(例えばヒト細胞を播種されたブタ脱細胞化器官または組織)であることができる。
【0125】
いくつかの例では、本明細書に記載する方法によって生成させる器官または組織が、患者に移植されるものである。これらの場合、脱細胞化器官または組織を再細胞化するために使用される細胞は、再生性細胞が患者にとって自家細胞になるように、患者から得ることができる。患者の細胞は、例えば、当技術分野において公知の方法を使って、血液、骨髄、組織、または器官から、人生の異なる段階で(例えば、出生前または周産期に、思春期中に、または成人として)得ることができる。あるいは、脱細胞化器官または組織を再細胞化するために使用される細胞は、患者にとって同種同系の細胞(すなわち、一卵性双生児の一方から得られるもの)であるか、細胞は、例えば患者の親類から得られる、または患者とは親類関係にないHLA適合個体から得られる、ヒトリンパ球抗原(HLA)適合細胞であるか、または細胞は、例えば非HLA適合ドナーから得られる、患者にとって同種異系の細胞であることができる。
【0126】
細胞の供給源(例えば自家であるかないか)とは関係なく、脱細胞化固形臓器は、患者にとって、自家、同種異系または異種であることができる。
【0127】
一定の例では、脱細胞化組織が、インビボで(例えば組織が個体中に移植された後で)、細胞によって再細胞化され得る。インビボ再細胞化は、例えば本明細書に記載する細胞のいずれかを使って、上述のように行うことができる(例えば注入および/または灌流)。上記に代えて、または上記に加えて、内在性細胞による脱細胞化器官または組織のインビボ播種も、自然に起こるか、または再細胞化組織に送達される因子によって媒介され得る。
【0128】
遺伝子改変
本発明は、哺乳動物における肺臓細胞治療を容易にするために本発明の脱細胞化組織を使用することができるという発見に関する。
【0129】
別の態様において、脱細胞化肺臓組織は、肺上皮細胞などの所望の肺臓細胞を培養するために使用することができる。細胞は、遺伝子改変されていてもされていなくても、気腫、閉塞性細気管支炎、および嚢胞性線維症を含む(ただしこれらに限定されるわけではない)肺臓疾患を処置するために使用することができる。例えば、本発明の脱細胞化組織は、ヒト肺気道上皮細胞を培養するための基質として使用することができる。次に、培養ヒト気道上皮細胞を、例えば気管点滴注入、吸入または注入などによって、レシピエントに送達することができる。培養下に拡大されるそのような細胞は、レシピエントにおける治療を達成するために使用することができる。脱細胞化肺臓組織(例えば気管)は、組織培養プラスチック製品などの典型的な細胞培養環境では成長させることが通常は極めて困難な肺上皮細胞を培養し拡大するための傑出したプラットフォームになる。
【0130】
遺伝子治療との関連では、レシピエントの肺臓への細胞の送達に先だって、脱細胞化組織上で培養される細胞を、関心対象の遺伝子で処理することができる。いくつかの例では、そのような細胞ベースの遺伝子送達が、アデノウイルス遺伝子送達ベクターの吸入などといった肺臓への他の遺伝子送達手段に対して、著しい利点を示すことができる。宿主への細胞ベースの遺伝子送達が持つこの優位性は、吸入された遺伝子ベクターが典型的には、粘液層および宿主免疫系によって課される障壁により、低い細胞形質導入効率をもたらすという知見に由来する。細胞中に前もって挿入された治療遺伝子の送達では、レシピエント肺臓細胞への遺伝子治療ベクターの貫入に付随する問題が回避される。
【0131】
本発明の脱細胞化肺臓組織は、標準的な細胞培養プラスチック製品での培養と比較して、上皮細胞などの肺臓細胞を高度に生存可能かつ分化した状態で成長させるための便利で効率のよい手段を提供する。そして、脱細胞化マトリックス上での肺上皮細胞などの肺細胞の拡大は、細胞治療にとって有効であるような十分に多数の細胞を与える。また、脱細胞化マトリックス上での肺臓上皮細胞の拡大は、インビトロで培養細胞を遺伝子治療ベクターで処理するためのプラットフォームにもなる。選ばれた遺伝子をインビトロでトランスフェクトされた細胞は、次に、任意で、関心対象の導入遺伝子を発現する細胞だけを選択するために精製し、次いで、そのような細胞治療を必要とするレシピエントに導入することができる。そのようなアプローチは、嚢胞性線維症などの遺伝的肺臓疾患の処置において、とりわけ価値があるだろう。
【0132】
ある態様において、本発明は、嚢胞性線維症を処置する方法を提供する。本方法は、上皮細胞などの関心対象の細胞に、正常型のCFTR遺伝子(この遺伝子の突然変異型が嚢胞性線維症の原因遺伝子である)をトランスフェクトする工程を含む。そのようなトランスフェクト細胞を、気管への点滴注入、吸入、または他の導入手段によって患者に送達すれば、これらの患者への遺伝子ベクターの送達に付随してきた著しい困難が軽減される。こうして、嚢胞性線維症における有効な細胞治療および遺伝子送達を実現することができる。しかし本発明はCFTR遺伝子をトランスフェクトした細胞による嚢胞性線維症の処置だけに限定されるべきではない。むしろ本発明は、肺臓細胞に関連する任意の疾患または障害の処置を包含する。
【0133】
したがって本発明は、本発明の脱細胞化組織上で培養された遺伝子改変細胞(例えば肺細胞)の使用を提供する。遺伝子改変は、例えば外因性遺伝子(「導入遺伝子」)の発現をもたらすか、内在性遺伝子の発現の変化をもたらし得る。そのような遺伝子改変は治療上の利益を持ち得る。あるいは、遺伝子改変は、例えば本発明の組成物を個体に埋植した後に、そのように修飾された細胞を追跡しまたは同定するための手段にもなり得る。細胞の追跡には、移植された遺伝子改変細胞の移動、同化および生存を追跡することが含まれ得る。遺伝子改変は少なくとも第2の遺伝子を含むこともできる。第2の遺伝子は、例えば、選択可能な抗生物質耐性遺伝子または他の選択可能マーカーをコードし得る。
【0134】
細胞を追跡するのに有用なタンパク質には、緑色蛍光タンパク質(GFP)、他の蛍光タンパク質のいずれか(例えば強化緑色、シアン、黄色、青色および赤色蛍光タンパク質;Clontech、カリフォルニア州パロアルト)または他のタグタンパク質(例えばLacZ、FLAG-tag、Myc、His6など)などがあるが、これらに限定されるわけではない。
【0135】
細胞の遺伝子改変の目的が生物学的活性物質の産生にある場合、その物質は一般に、所与の障害の処置にとって有用なものであるだろう。例えば、細胞が骨または軟部組織形成に関連する一定の成長因子産物を分泌するように、細胞を遺伝子改変することが望ましい場合があるだろう。組織修復に関係する他の内在性細胞タイプの成長を誘導するための成長因子産物も有用である。例えば、内在性毛細管および/または微小血管内皮細胞を刺激するための成長因子は、軟部組織欠陥の修復において、特に大体積の欠陥に、役立ち得る。
【0136】
本発明の細胞は、細胞の培養にとって有益な分子、例えば栄養因子、成長因子、サイトカインなどを産生するように外因性遺伝物質を細胞中に導入することにより、遺伝子改変することができる。また、そのような分子を産生するように細胞を遺伝子改変することにより、細胞は、それを必要とする哺乳動物中に移植された時に、哺乳動物に対して追加の治療効果を与えることができる。例えば遺伝子改変細胞は、哺乳動物中の移植部位に近接する細胞にとって有益な分子を分泌することができる。
【0137】
肺細胞は、当業者に知られる任意の方法を使って遺伝子改変することができる。例えばSambrook et al.(2001)「Molecular Cloning: A Laboratory Manual」(Cold Spring Harbor Laboratory Press、ニューヨーク州コールドスプリングハーバー)およびAusubel et al.編(1997)「Current Protocols in Molecular Biology」(John Wiley & Sons、ニューヨーク州ニューヨーク)を参照されたい。例えば肺細胞を、導入遺伝子を含む核酸が含まれている発現ベクターに暴露して、導入遺伝子が細胞内で発現されるのに適当な条件下で核酸が細胞内に導入されるようにする。導入遺伝子は一般に、適切なプロモーターに機能的に連結されたポリヌクレオチドを含む発現カセットである。ポリヌクレオチドはタンパク質をコードするか、生物学的に活性なRNA(例えばアンチセンスRNAまたはリボザイム)をコードすることができる。したがって、例えばポリヌクレオチドは、毒素に対する耐性を付与する遺伝子、ホルモン(例えばペプチド成長ホルモン、ホルモン放出因子、性ホルモン、副腎皮質刺激ホルモン、サイトカイン(例えばインターフェリン(interferin)、インターロイキン、リンフォカイン)など)、細胞表面結合型細胞内シグナリング部分(例えば細胞接着分子、ホルモン受容体など)、所与の系譜の分化を促進する因子(例えば骨形態形成タンパク質(BMP))などをコードすることができる。
【0138】
発現カセット内では、コードポリヌクレオチドが適切なプロモーターに機能的に連結される。適切なプロモーターの例には、原核生物プロモーターおよびウイルスプロモーター(例えばレトロウイルスITR、LTR、前初期ウイルスプロモーター(IEp)、例えばヘルペスウイルスIEp(例えばICP4-IEpおよびICP0-IEEp)、サイトメガロウイルス(CMV)IEp、および他のウイルスプロモーター、例えばラウス肉腫ウイルス(RSV)プロモーター、およびマウス白血病ウイルス(MLV)プロモーター)が含まれる。他の適切なプロモーターは、真核生物プロモーター、例えばエンハンサー(例えばウサギβ-グロビン調節要素)、構成的に活性なプロモーター(例えばβ-アクチンプロモーターなど)、シグナル特異的プロモーター(例えばRU486に応答するプロモーターなどの誘導性プロモーター)、組織特異的プロモーターである。事前に定められた細胞との関連で遺伝子発現を駆動するのに適したプロモーターを選択することは、当技術分野の技能で十分に可能である。発現カセットは、2つ以上のコードポリヌクレオチドを含むことができ、所望どおりに他の要素(例えばポリアデニル化配列、膜挿入シグナルまたは分泌リーダーをコードする配列、リボソームエントリー配列、転写調節要素(例えばエンハンサー、サイレンサーなど)など)を含むことができる。
【0139】
導入遺伝子を含有する発現カセットは、細胞に導入遺伝子を送達するのに適した遺伝子ベクター中に組み込まれるべきである。所望する最終用途に応じて、任意のそれらベクターを、細胞を遺伝子改変するために、そのように使用することができる(例えばプラスミド、裸のDNA、アデノウイルス、アデノ随伴ウイルス、ヘルペスウイルス、レンチウイルス、パピローマウイルス、レトロウイルスなどのウイルスなど)。そのようなベクター内に所望の発現カセットを構築する任意の方法を使用することができ、それらの多くは当技術分野において周知である(例えば直接クローニング、相同組換えなど)。ベクターの選択が、細胞中にベクターを導入するために使用される一般に当技術分野において公知である方法(例えばプロトプラスト融合による方法、リン酸カルシウム沈殿法、遺伝子銃、エレクトロポレーション、DEAEデキストランまたは脂質担体によるトランスフェクション、ウイルスベクターによる感染など)を、おおむね決めることになる。
【0140】
ポリメラーゼ連鎖反応(PCR)、リガーゼ連鎖反応(LCR)、および他のDNAまたはRNAポリメラーゼ媒介技法を含むインビトロ増幅法を当業者に実行させるのに足りる技法の例が、Sambrook et al.「MOLECULAR CLONING: A LABORATORY MANUAL」第1〜3巻(第3版、Cold Spring Harbor Press、NY 2001)に見いだされる。
【0141】
いったんあるタンパク質に関して核酸をクローニングしたら、当業者は、さまざまな肺臓細胞中で組換え遺伝子を発現させることができる。当業者は、所望の導入遺伝子を発現させるために利用することができる数多くの発現系を知っていると予期される。
【0142】
本発明は、天然肺臓組織を模倣する人工三次元組織を提供する。天然肺臓組織を模倣する複合材料およびスキャフォールドを作り出す能力は、先行技術の方法より高度な組織および組織集合体の修復および再生を可能にし、先行技術の方法を使って達成できるものよりも、より正確な組織学的構造および機能を示す。例えば、人工肺臓組織は、出芽構造および伸長する管状構造を示す細胞を含む。そのうえ、細胞は、形態形成および肺臓上皮分化に関与する遺伝子を発現する。形態形成および肺臓上皮分化に関与する遺伝子には、遠位上皮マーカー遺伝子SpCおよびSpB、間葉由来モルフォゲンFGF10、FGFr2、および血管内皮成長因子A(VEGF)などがあるが、これらに限定されるわけではない。
【0143】
投与
本発明では、インビトロ環境とインビボ環境の両方での人工組織の使用が考えられる。したがって本発明は、研究目的および治療目的もしくは医学的/獣医学的目的での人工組織の使用を提供する。研究環境では、この技術に関して、数多くの実用的応用が存在する。そのような応用の一例は、エクスビボがんモデル(例えばさまざまなアブレーション技法(例えば放射線処置、化学療法処置、またはそれらの組合せを含む)の有効性を実験室で調べ、そうすることによって処置方法を最適化するために病気の患者を使用することを避けるためのものなど)における人工組織の使用である。例えば、新しく摘出した肺臓をバイオリアクタに取り付け、その肺臓を処理して組織をアブレートすることができる。インビボ使用の別の例は、組織工学のための使用である。
【0144】
本発明の人工組織はインビボでも役立つ。数ある用途のなかでも、対象(本明細書においては「患者」と互換的に使用され、ヒトと動物の両方を包含するものとする)のインビボ処置法を挙げることができる。一般に、一定の態様に関して、対象の処置方法は、対象中または対象の表面に本発明の人工組織を埋植する工程を含み、組織の埋植は対象における検出可能な変化をもたらす。検出可能な変化は、自然の知覚を使ってまたは人造の装置を使って検出することができる任意の変化であることができる。本発明では任意のタイプの処置(例えば疾患または障害の治療的処置、皮膚の斑点の美容的処置など)が想定されるが、多くの態様では、処置が、対象の疾患、障害、または他の苦痛の原因の治療的処置である。したがって検出可能な変化は、対象を冒す疾患または障害の少なくとも1つの臨床症状における変化(好ましくは改善)の検出であることができる。例示的なインビボ治療法には、腫瘍処置後の器官の再生、医学的装置を埋植するための手術部位の調製、皮膚グラフティング、および組織または器官(疾患または障害によって損傷を受けたか破壊されたものなど)の一部または全部の代用が含まれる。例示的な器官または組織には、以下に挙げるものが含まれる:心臓、肺臓、肝臓、腎臓、膀胱、脳、耳、目、または皮膚。対象はヒトまたは動物であることができるという事実を考慮すると、本発明は医学的応用と獣医学的応用の両方を持つ。
【0145】
ある態様において、本方法は、処理される組織の細胞を殺し、組織スキャフォールドを作り出すために、組織を本発明の脱細胞化方法に暴露する工程を含む。本方法はさらに、組織スキャフォールドに細胞を播種する工程、および播種された細胞を組織スキャフォールドの中または上で増殖させる工程を含むことができる。増殖により、健常で機能的な細胞を含有する再生組織が作製される。
【0146】
本発明は、人工肺臓組織を、それを必要とする哺乳動物中に埋植することによって、患者を処置する方法も提供する。いくつかの例では、人工肺臓組織が、適切な細胞、例えばNPCを含む。しかし本発明は、いかなる特定タイプの細胞にも限定されるべきでない。埋植後、グラフトされた細胞は、それに内在性組織の特徴を発生させる環境キューに応答することができる。好ましくは、細胞は、導管構造を形成する分化遠位上皮細胞(proSpC発現性)から構成される組織型(histiotypic)肺胞様構造を形成する。このように、埋植された細胞は、それを周囲の組織に似せる特徴を発生させるであろう。これらの方法を使って、生物学的足場は組織を増強することができ、本発明の生物学的足場は、組織工学に使用することができ、従来のいかなる組織工学的背景でも使用することができる。
【0147】
したがって本発明は組織再生への応用を包含する。組織再生治療アプローチの目的は、高密度の修復コンピテント細胞(または局所環境による影響を受けたときにコンピテントになることができる細胞)を、初期の創傷力学と最終的な新組織生成の両方を最適化するような形で、欠陥部位に送達することである。本発明の組成物は、個体における肺臓組織欠陥を軽減または処置するのにとりわけ有用である。有利なことに、本発明の組成物は、改善された肺臓組織再生をもたらす。具体的には、本発明の組成物の結果として、組織再生が、より迅速に達成される。
【0148】
有利なことに、本発明の組成物および方法は、先行技術の方法に対する改善を表す。好ましくは、肺臓欠陥を処置するために使用される組成物は、本明細書の他の項で説明するように、NPCを含み、より好ましくは、3次元培養物を生成するようにスキャフォールド上に播種されてインビトロで培養されたNPCを含む。
【0149】
創薬のためのモデル
本発明は、肺臓の疾患または障害に関して、試験化合物の治療活性の評価を可能にするのに適したインビトロ法を提供する。好ましくは、本方法は、人工三次元肺臓組織の使用を含む。
【0150】
本方法は、脱細胞化組織を使って開発されたモデルに基づく。いくつかの例では、脱細胞化組織に適切な細胞を播種することができる。いくつかの例では、上皮、間葉、および内皮細胞を含有するNPCの混合集団を使って、三次元人工肺臓組織を生成させる。例えば、NPCを三次元脱細胞化肺臓組織内に置く。こうして、本モデルには、成長および近接細胞との細胞間コミュニケーションに対するNPCの影響が組み入れられる。三次元肺臓組織は天然肺臓組織を模倣し、例えば人工肺臓組織は天然肺臓組織によって例示される分枝形態形成を示す。
【0151】
本モデルは肺臓組織の病変に対して薬物を試験するのに役立つ。また、本モデルは、肺臓組織の病変に対する、特定の治療剤送達媒体の効果を調べて、例えば異なる送達系によって投与された同じ薬剤の効果を比較するために使用するか、または単に送達媒体そのもの(例えばウイルスベクター)が肺臓の病変に影響を及ぼす能力を有するかどうかを評価するために使用することができる。
【0152】
ある態様において、本発明は、肺臓組織の健康状態を調整する試験作用因子の能力について、試験作用因子をスクリーニングするためのインビトロ法を提供する。本方法は、試験作用因子を人工三次元肺臓組織モデルと接触させる工程、および試験作用因子が肺臓組織モデルに対して持つ効果を測定する工程を含む。試験作用因子の存在下でモデルに加わる変化はいずれも、その試験作用因子が肺臓組織の健康状態を調整できることのしるしである。
【0153】
別の態様において、本発明は、ある試験作用因子が肺臓組織に対して持つ効果を観察するためのインビトロ法であって、以下の工程を含む方法を提供する:
a)正常肺臓組織をモデル化することを意図した少なくとも1つの三次元肺臓組織モデルを用意する工程;
b)試験作用因子を肺臓組織モデルと接触させる工程;および
c)試験作用因子が肺臓組織モデルに対して持つ効果を観察する工程。
【0154】
本組織モデルは、スキャフォールド(例えばコラーゲンマトリックス)上の三次元的細胞アレイと、少なくとも1つの試験細胞とを含む構築物である。本方法は、肺臓組織の病変に対する試験作用因子の効果を観察する工程を含む。しかし、本方法は、肺臓組織上の個々の細胞タイプに対する試験作用因子の効果を観察する工程をさらに含んでもよい。
【0155】
試験作用因子は、化学剤(例えば毒素)、医薬、ペプチド、タンパク質(例えば抗体、サイトカイン、酵素など)および核酸(タンパク質、アンチセンス剤(すなわち、ターゲット細胞タイプ中で発現するターゲットRNAに相補的な配列を含む核酸、例えばRNAiまたはsiRNA)、リボザイムなどの治療剤をコードし得る、遺伝子医薬品および導入された遺伝子を含む)を含む、任意の作用因子であることができる。上記に加えて、または上記に代えて、試験作用因子は、放射線(例えば電離放射線、UV光または熱)などの物理的作用因子であってもよく、これらは単独で試験するか、化学剤および他の作用因子と組合せて試験することができる。
【0156】
本モデルは送達媒体を試験するためにも使用することができる。これらは、従来の医薬製剤から遺伝子送達媒体まで、任意の形態をとり得る。例えば本モデルは、2つまたはそれ以上の異なる送達系(例えばデポー製剤と放出制御製剤)によって投与された同じ薬剤の治療効果に対する効果を比較するために使用することができる。ある特定媒体がそれだけで肺臓組織に対して効果を持つかどうかを調べるためにも使用することができる。遺伝子ベースの治療薬の使用が増加するにつれて、考え得る種々の送達系に付随する安全性の問題が、ますます重要になる。したがって本発明のモデルは、例えば裸のDNAまたはRNAなどの核酸治療薬のための送達系、ウイルスベクター(例えばレトロウイルスベクターまたはアデノウイルスベクター)、リポソームなどの性質を調べるために使用することができる。したがって試験作用因子は、任意の治療剤が付随しているまたは付随していない、任意の適当なタイプの送達媒体であることができる。
【0157】
試験作用因子は、試験されるモデルに、任意の適切な手段を使って加えることができる。例えば、試験作用因子をモデルの表面上に滴下して、拡散または他の方法でモデルに進入させるか、試験作用因子を栄養培地に加え、コラーゲンゲルを通して拡散させることができる。本モデルは、電離放射線、UV光または熱などの物理的作用因子の効果を、単独で、または化学剤と組み合わせて(例えば光線力学的治療において)、試験するのにも適している。
【0158】
試験作用因子がモデルに対して持つ効果の観察は、さまざまな方法を使って達成することができる。例えば、ある特定の作用因子は、アポトーシスに入るように細胞を誘導するかもしれない。細胞における検出可能な変化には、細胞の面積、体積、形状、形態、マーカー発現(例えば細胞表面マーカー発現)または他の適切な特徴、例えば染色体断片化が含まれ得る。細胞増殖に対する試験作用因子の効果を観察するために細胞数を監視することもでき、これは、存在する特定細胞タイプの数を計数することなどによって直接的に分析するか、特定細胞塊のサイズを測定することなどによって間接的に分析することができる。これらは、例えば適切な蛍光細胞染色などを使って、インタクトなモデル上で直接的または間接的に観察することができる。これは、生きているモデルを連続分析するための、生体染色色素または遺伝子的に導入された蛍光マーカー(例えば緑色蛍光タンパク質)による細胞の事前標識(pre-labeling)によって行うか、固定化と、ヨウ化プロピジウムまたは蛍光標識抗体などの蛍光物質による事後標識(post-labeling)とによって行うことができる。あるいは、特定のmRNA種の発現について試験するために、通常の組織学的方法、例えば適切な細胞ターゲットに対する抗体を使用する免疫組織化学、またはインサイチューハイブリダイゼーションによって、モデルを処理することもできる。さらにまた、コンピュータシステムおよびソフトウェアを使って細胞をさまざまな時点でイメージングし、例えば細胞の密度、場所および/または形態などの任意の変化を検出することによって、これを自動/ロボット的または半自動的に行うこともできる。特に、共焦点レーザー走査型顕微鏡法は、インタクトなモデルの三次元解析を可能にする。したがって、通常は従来の二次元培養中の細胞に対してのみ可能である細胞挙動の定量的解析を、インタクトな三次元肺臓組織モデルに直接応用することが可能である。このようにして、細胞の増殖、アポトーシス、壊死、移動およびマトリックス浸潤、その他の定量的連続分析が、従来の二次元細胞培養と生きている動物モデルの間の間隙を埋める三次元肺臓組織モデルで得られる。
【0159】
実験例
以下に実験例を挙げて本発明をさらに詳しく説明する。これらの実施例は、例示のために記載するに過ぎず、別段の指定がない限り、限定を意図するものではない。したがって本発明は、決して、以下の実施例に限定されると解釈されるべきではなく、本発明は、本明細書が提供する教示の結果として明白になる、ありとあらゆる変形を包含すると解釈すべきである。
【実施例】
【0160】
実施例1:ラット肺臓の脱細胞化および脱細胞化スキャフォールドの形態学的特徴付け
脱細胞化器官は、組織工学スキャフォールドとしての使用に、いくつかの利点をもたらす。ある局面において、脱細胞化スキャフォールドは、肺臓の場合であれば血管系および気道網を含む組織機能のための適当な三次元構成を持つ。また、細胞外マトリックス(ECM)構成要素は、種を超えて広く保存されているので、異種埋植時に脱細胞化スキャフォールドが免疫応答を誘導する可能性は低い[Bernard et al., 1983, Biochemistry 1983;22:5213-23]。別の局面において、未変性ECMは、細胞の付着、伝播、成長および分化にとって、最適な基質を提供する。
【0161】
本発明の脱細胞化プロセスの目標は、肺臓のECMの重要な局面を保ちかつ肺臓のECMに対するいかなる損傷も最小限に抑えつつ、細胞材料および核材料を除去することである。本明細書に提示する結果は、未変性肺臓組織を脱細胞化して、細胞構成要素および抗原性分子を除去し、しかも重要な細胞外マトリックス分子は保つことが可能であることを証明している。ある局面において、本発明の脱細胞化プロセスの目標は、細胞培養と完全に適合すると同時に障壁機能を提供する脱細胞化肺臓スキャフォールドを生成させることである。また、脱細胞化肺臓スキャフォールドは、インタクトな気道樹および血管網を持つことが望ましい。
【0162】
この研究では化学的な脱細胞化法を使用した。この研究で使用した化学薬品には、塩化ナトリウム、CHAPS、およびEDTAが含まれる。高張塩化ナトリウム溶液は、細胞を効率よく溶解することができるが、組織からの細胞構成要素の除去を補助することはない。CHAPSは両性イオン性界面活性剤であり、これは細胞材料の効率の良い可溶化を可能にし、それゆえに細胞材料の効率の良い除去を可能にする。EDTAは、ECMへの細胞付着の破壊を助ける重要な二価イオン(すなわちCa2+)を結合するキレート剤である。また、溶液は高いアルカリ性を持ち、それが、細胞質細胞構成要素と、可溶化しなければマトリックスを詰まらせるGAGとを、可溶化するのに役立つ[Gilbert et al., 2008 J Surg Res 152(l):135-9]]。
【0163】
これらの実験で使用した材料および方法を以下に説明する。
【0164】
材料および方法
器官収集
肺臓を若年成体(3ヶ月齢)雄フィッシャー344ラットから収集した。動物実験の作業は全て、イェール大学施設内動物実験委員会(Yale University Institutional Animal Care and Use Committee)からの承認を受けて行われた。動物をペントバルビタールナトリウム(Sigma、40mg/kg)の腹腔内注射によって麻酔した。麻酔の誘導後に、肋骨縁の直下で横切開によって腹部に入った。横隔膜を穿刺し、胸郭を切って、肺臓を露出させた。50U/mlヘパリン(Sigma)を含有するPBSで右心室を通して肺臓を灌流した。灌流が完了した後、心臓、肺臓および気管を切り離し、ひとまとめにして取り出した。
【0165】
バイオリアクタ構成要素
バイオリアクタ構成要素はCole-Parmer(イリノイ州バーノンヒルズ)から入手した。シリコーン栓と500mlのガラス製広口瓶とがバイオリアクタの基礎をなした。灌流ループおよび空気換気を含む、肺臓への必要な接続が可能になるように、シリコーン栓を通して、サイズL/S14およびL/S16のPharMedチューブ(オハイオ州ウェストレーク)を挿入した。圧力は、灌流ポンプと肺動脈への接続との間のTruWave圧変換器(Edwards Lifesciences、カリフォルニア州アービン)を使って監視した。灌流はMasterflex L/S可変速ローラーポンプ(Masterflex、イリノイ州バーノンヒルズ)を使って達成した。
【0166】
脱細胞化プロセス
脱細胞化に使用した流体は、PBS中の8mM CHAPS、1M NaCl、25mM EDTAだった。化学薬品は全てSigmaから入手し、PBSはGibcoから入手した。バイオリアクタを脱細胞化流体で満たし、バイオリアクタを37℃に保たれたインキュベータに移した。灌流圧は肺動脈幹への流入時に監視し、30または20mmHg未満に維持した。脱細胞化流体は以下の時点で新鮮な流体と交換した:30分、1時間、2時間、4時間、6時間。大半の条件について、4時間後または6時間後に脱細胞化を停止した。
【0167】
DNAアッセイ
Quant-iT PicoGreen dsDNAアッセイキット(Invitrogen、オレゴン州ユージーン)を、製造者の説明書に従って使用することにより、組織のDNA含量を定量した。簡単に述べると、組織試料を重量測定し、凍結乾燥し、TE緩衝液で希釈し、Quant-iT PicoGreen試薬と混合した。蛍光を485nmで励起して535nmで測定し、標準曲線を使ってDNA含量を定量した。未変性試料と脱細胞化試料の両方について、少なくとも4つの試料を測定した。
【0168】
ウェスタンブロット
ウェスタンブロッティング用の組織を、プロテアーゼ阻害剤(Sigma)を添加した冷RIPA緩衝液(Boston Bioproducts)中で消化し、15,000rpmで30秒間ホモジナイズした。4℃で1時間インキュベートした後、14,000gで25分間の遠心分離によって、不溶性粒子を除去した。タンパク質濃度をBradfordアッセイ[Bradford, 1976, Anal Biochem 72:248-54]によって定量した後、Laemmli還元緩衝液(Boston Bioproducts)中、65℃で25分間、煮沸した。試料を分析まで-80℃で保存した。25〜30μgのタンパク質を使って、さまざまなパーセンテージのポリアクリルアミドゲルで、試料を泳動した。電気泳動後に、タンパク質をニトロセルロース膜に転写した。膜をTBS中ですすいだ後、0.05%Tween-20を含むTBS(TBS-T)中の5%脱脂粉乳(NFDM)または3%ウシ血清アルブミン中で、1時間ブロッキングした。一次抗体を、TBS-T中の2%NFDMまたは3%BSA中で、一晩適用した。二次抗体は、ロバまたはヤギにおいて産生させたものを、Santa Cruzから入手し、1:2000の希釈度で室温において1時間適用した。Supersignal West Picoから得た基質(これを5分間適用してから、フィルム現像した)を使ってタンパク質を検出した。
【0169】
免疫蛍光
組織ブロックを3.7%ホルムアルデヒド(Sigma)中で4時間固定した後、70%エタノールに移し、パラフィンに包埋した。イェール大学組織学中核施設(Yale University Histology core facility)によって薄い(5μm)切片が調製された。組織切片をキシレン中で脱パラフィンし、エタノール勾配によって再水和し、緩衝液(PBS+0.2%Triton-X)中で15分間すすいだ。抗原回復(antigen retrieval)を0.01Mクエン酸、pH6.0中、70℃で20分間行った。室温まで冷却した後、切片を緩衝液中ですすぎ、次にPBS中、5%ウシ血清アルブミン(BSA)および0.75%グリシンで、室温において1時間ブロッキングした。一次抗体を、ブロッキング緩衝液中、適当な濃度で、4℃において一晩適用した。スライドを緩衝液中で3回すすいだ後、二次抗体をブロッキング緩衝液中、1:500希釈で、室温において1時間適用した。二次抗体は、Invitrogenから入手したAlexFluor555ロバ抗ヤギまたは抗ウサギ抗体およびAlexaFluor488ニワトリ抗ウサギ抗体とした。DAPI含有封入剤(Vector Labs)を使ってスライドを封入し、Zeiss Axiovert 200M倒立蛍光顕微鏡を使って画像を取得した。
【0170】
走査型電子顕微鏡法
0.1Mカコジル酸緩衝液(EMD Biosciences、ニュージャージー州ギブスタウン)中の2%グルタルアルデヒドおよび2.5%パラホルムアルデヒドを使って試料を室温で2時間固定した後、カコジル酸緩衝液中ですすぎ、薄切し、エタノール勾配によって脱水した。試料をヘキサメチルジシラザン中で10分間さらに脱水し、一晩乾燥した後、金でスパッタコーティングし、イェール大学地質学および地球物理学施設(Yale University Geology and Geophysics facility)においてJOEL JXA-8600を使って分析した。
【0171】
透過型電子顕微鏡法
PBS中の4%パラホルムアルデヒドを使って試料を固定した後、0.1Mカコジル酸ナトリウム緩衝固定剤(pH7.4)中の2%グルタルアルデヒドおよび2.5%パラホルムアルデヒドに、室温で2時間入れておいた。試料を0.1Mカコジル酸ナトリウム緩衝液中で3回すすぎ、1%四酸化オスミウム中で1時間、後固定し、次に、マレイン酸緩衝液(pH5.2)中の2%酢酸ウランにおいて、さらに1時間、一括して染色した。次に、試料をすすぎ、段階的エタノール系列によって脱水し、エポン樹脂を浸透させ、60℃で一晩ベーキングした。硬化したブロックを、Leica UltraCut UCTを使って切断し、ニッケルグリッド上に60nm切片を集め、2%酢酸ウランおよびクエン酸鉛を使って染色した。80kVのFEI Tencai Biotwin TEMで試料を観察した。Morada CCDデジタルカメラを使用し、iTEM(オリンパス)ソフトウェアを使って、画像を撮影した。
【0172】
マイクロスフェア保持
脱細胞化肺臓または未変性肺臓を、本明細書の他の項で説明するように、カニューレに取り付け、その肺臓を、気管を通して、5μmのマイクロスフェアを含有するPBSで膨らませた。次に、脈管構造を、PBS各10mlのすすぎ液で3回フラッシングした。マイクロスフェアをdH2O中で2回洗浄して、細片を除去し、溶解しなければ未変性肺臓の読みに影響を及ぼすであろう細胞を全て溶解した。4.9μm〜5.1μmの粒子を測定するように設定したCoulter計数器を使って、各試料中のマイクロスフェア濃度を定量し、マイクロスフェア注入前に測定したベースラインの読みと比較した。
【0173】
マイクロCTイメージング
未変性肺臓または脱細胞化肺臓を10%中性緩衝ホルマリン(Sigma)中で固定し、気道または脈管構造を通して造影剤を注入した。造影剤はPBS中の20%ビスマスおよび5%ゼラチン(Sigma)とした。造影剤の注入後、肺臓を氷浴で冷却してゼラチンを重合させた。
【0174】
肺臓全体に関して、有効検出器ピクセルサイズ0.029mmに設定したマイクロCTイメージングシステム(GE eXplore Locus SP、GE Healthcare)で肺脈管構造をイメージングした。マイクロCTは、ピークX線管電圧60kV、管電流80mA、1600ミリ秒/フレーム、22検出器ビニングモデル、720ビュー、および0.5°刻み/ビューで作動させた。1つの葉(右上葉)の高解像度イメージングのために、試料をコンピュータ制御された回転ステージ上に配置し、垂直軸の周りを、0.4°の回転間隔で、360°スキャンした。管はピーク値80kVおよび80mAで作動させる。1×1に設定した検出器ビニングモデルおよび0.0065mmの解像度で、各ビューごとの露出時間は、典型的には3000ミリ秒とした。どちらの取得も、肺臓または1つの葉を貫く連続的な軸方向VFFフォーマット画像のセットをもたらした。
【0175】
Microview Software(GE Healthcare)を使用することにより、生データを補正し、ボクセル寸法58μm×58μm×58μmで再構築して、肺臓中の血管樹全体を可視化した。高品質の血管樹(1つの葉)については、ボクセル寸法を6.5μm×6.5μm×6.5μmに設定した。このソフトウェアは、生データから最大値投影像を再構築するためにも使用した。
【0176】
多平面再構成、空間フィルタリング、およびボリュームレンダリング技法により、データセットを、横断面、矢状断面、前頭断面、ハイブリッド断面、および3Dフォーマットで見ることを可能にした。対象抽出および関心領域測定には二値画像を使用した。三次元ボリューム画像は、改変Feldkampフィルタ補正逆投影アルゴリズムを使用することにより、斜視図から再構築される。しかしこの系では、ラット肺臓全体(視野、約3.0cm)を、58μmという小さな典型的立方ボクセル寸法を持つ画像で、調べることができる。各ボクセルの不透明度は16ビットグレースケール値によって表される。
【0177】
実験の結果を以下に説明する。
【0178】
脱細胞化方法
本発明に提示する結果は、インタクトな齧歯類肺臓の完全な葉から細胞材料を除去する脱細胞化方法を証明している。1M NaCl、8mM CHAPSおよび25mM EDTAを使った脱細胞化は、細胞材料を除去するのに最適であり、しかもコラーゲンまたはエラスチン線維を除去せず(組織像に基づく)、マトリックスの構造的完全性も損傷しないらしい(機械的試験に基づく)ことが観察された。これに比して、SDSを含有する溶液による脱細胞化は、マトリックスの機械的強度を損なうことが見いだされた。他の条件は、細胞材料を効率よく除去しないか、マトリックスの完全性の著しい低下を引き起こすことが見いだされた。
【0179】
組織学的分析
組織学的検査を使って数多くの脱細胞化肺臓スキャフォールドを特徴付けた。核およびDNAに関するH&E染色およびDAPI染色によれば、脱細胞化肺臓は、インタクトな細胞をただ一つも示さなかった。時折、巻き戻されたDNAまたは細胞抗原が観察されたが、インタクトな細胞は観察されなかった。図1は、未変性肺臓および脱細胞化肺臓のH&E染色を示し、一方、図2は、残存DNAに関するDAPI染色を示している。標準的組織切片では肺胞中隔がインタクトに見え、大きい気道および血管もそうであったという事実に基づいて、肺構造の保存も観察された。
【0180】
DNA含量
細胞材料の完全な除去は、いくつかの理由で重要である。第1に、スキャフォールドを組織工学的応用に使用するつもりである場合は、スキャフォールドに新しい細胞源を播種する前に、全ての細胞がスキャフォールドから除去されることを確実にしなければならない。人工組織がインビボ応用に使用される場合は、いかなる残存細胞材料も、再播種済スキャフォールドの評価を複雑にするだけでなく、免疫合併症を引き起こすだろう[Conconi et al., 2005, Transpl Int 18:727-34;Macchiarini et al., 2008, Lancet 372(9655):2023-30;Alexander et al., 2009, Cell Transplant 18:255-9]。そのため、本発明のスキャフォールドは、MHCクラスIおよびII抗原がどちらも脱細胞化スキャフォールド中に存在しないことが確認された。第2に、細胞外マトリックスの肺臓力学への寄与を個別に評価するために、全ての細胞構成要素を除去すべきである。末梢肺臓力学に寄与し得る2クラスの構成要素は、細胞材料と細胞外マトリックスである。細胞外マトリックスはさらに、主としてコラーゲン、エラスチン、およびプロテオグリカンに分割することができる[Cavalcante et al., 2005, J Appl Physiol 98:672-9;Dunsmore et al., 1996, Am J Physiol 270:L3-27;Ito et al., 2005, J Appl Physiol 98:503-11;Suki et al., 2005, J Appl Physiol 98:1892-9]。脱細胞化スキャフォールドからの細胞構成要素の除去を確実にすることにより、スキャフォールドの機械的性質を評価することができる。
【0181】
細胞材料の除去を証拠付けるために、定量的DNAアッセイを行った。未変性肺臓と比較して脱細胞化スキャフォールドではDNA含量の激しい減少が観察された(図1C)。脱細胞化スキャフォールドは未変性肺臓中に見いだされるDNAの約1.2%を含有し、それは乾燥重量1mgあたり1.83±0.29ngのDNAに相当した。これに比して、未変性肺臓の場合は、38.7±5.8ng/mgである。残存DNAを最小限に抑えるためにスキャフォールドの十分な洗浄を一般に使用することができるが、図2においてDAPI染色が巻き戻されたDNAの小さなクラスターを示していることから証明されるように、全てのDNAの完全な除去は難しく、少量のDNAが残った。DNA含量の激しい低下は細胞除去を示し、組織学的所見と共に、生存可能な細胞材料の全てがスキャフォールドから欠けていることを確認した。
【0182】
脱細胞化スキャフォールドでは、DNAのほぼ99%が除去されていることが証明された。少量のDNAがマトリックス中に残るが、それは、図2に示すようにDNAの細長い鎖として存在する。DAPI染色によれば、核構造におけるこの残存DNAの組織化は観察されなかった。
【0183】
未変性肺臓と比較して98.8%のDNAが除去され、残留DNA濃度は組織1mg(乾燥重量)あたり1.83ngのDNAであることが観察された。これは、他の脱細胞化心臓組織について他の研究者らが観察した16.6ng/mgという残存DNAレベルと比較して優れており[Ott et al., 2008, Nat Med 14:213-21]、このレベルが、本研究および他の研究[Gilbert et al., 2008 J Surg Res 152(l):135-9]の場合のように乾燥重量ではなく、湿重量に規格化されていることを考えると、とりわけそうである。しかし、観察される残存DNAのレベルは、皮膚グラフトに使用される市販のおよび実験室で作製されたECMスキャフォールド(一部のスキャフォールドは1.13ng/mgもの残存DNAを持っていたものの[Gilbert et al., 2008 J Surg Res 152(l):135-9]、大半のスキャフォールドは乾燥重量1mgあたり0.2ng未満のDNAを示す)に見られるものよりも高い。
【0184】
免疫原性
脱細胞化スキャフォールドの免疫原性を、主要組織適合遺伝子複合体(MHC)クラスIおよびII抗原に関する染色によって特徴付けた。MHCクラスIおよびIIタンパク質は、抗原-特異的免疫応答において重要な膜糖タンパク質である。MHCクラスI抗原は全ての有核細胞において発現し、MHCクラスII抗原は免疫系の特殊化した細胞上に見いだされる。MHCクラスI抗原は生物が「自己」を「非自己」と識別することを可能にするので、将来的に動物モデル中に人工肺臓組織を埋植した際の免疫問題を回避するには、脱細胞化スキャフォールドから除去することが重要である。図3に、MHCクラスIおよびII抗原ならびにβ-アクチンに関するウェスタンブロッティングの結果を図示する。免疫ブロッティングによってMHCクラスIおよびII抗原の両方の完全な喪失が観察されたことから、脱細胞化スキャフォールドは、組織工学的応用に使用しても、著しい免疫応答を惹起しないと予想されることが確認された。β-アクチンも失われたことは、細胞材料の非存在と合致する。宿主に埋植された場合にスキャフォールドが免疫応答を惹起する可能性は低いと考えられる。
【0185】
細胞外マトリックスの特徴付け
コラーゲン:コラーゲンは、肺臓の最も重要な構造的構成要素であり、組織の総合的な機械的強度を、主に担っている。免疫蛍光を使って、図4に示すように、未変性肺臓および脱細胞化肺臓におけるコラーゲンIおよびIVの分布を特徴付けた。コラーゲンIおよびIVはどちらも脱細胞化マトリックスによって保持されており、コラーゲンIは主により大きな気道および脈管構造付近に認められ、コラーゲンIVは実質全体に認められる。類似する染色パターンが、未変性肺臓にも脱細胞化肺臓にも認められた。これらのコラーゲンサブタイプがそれぞれ解剖学上適切な場所に保存されることにより、人工肺臓組織の開発時に、細胞タイプの選択的沈着が可能になるだろう。
【0186】
脱細胞化スキャフォールドの走査型EM評価
走査型電子顕微鏡法(SEM)を使って、脱細胞化肺臓スキャフォールドの微細構造を評価した。図5に、細胞が除去され、しかも肺胞アーキテクチャが全体的に維持されていることを証明するサンプル画像を示す。脱細胞化肺臓中の肺胞はわずかにしぼんでいるように見えたが、これは固定の人為的結果である。未変性肺臓は、固定剤で肺臓を膨らませることによって固定したが、脱細胞化肺臓は、加圧時に肺胞コンパートメント内に固定剤流体を含有することができないので、しぼんだ外観の肺臓を与える。しかし、肺胞アーキテクチャは概して類似しており、肺胞中隔が保存されている。これらの結果は、組織学的研究による知見と共に、全体的な肺の気道アーキテクチャおよび肺胞中隔を含む肺胞構造が、脱細胞化スキャフォールドにおいてインタクトであったことを示している。
【0187】
スキャフォールド超微細構造に対する灌流圧の影響
走査型EM研究に加えて、透過型EM(TEM)も使って、毛細管-肺胞基底膜を調べた。インタクトな毛細管網の存在は、脱細胞化スキャフォールドが、肺胞腔への高分子の移行に対抗することを可能にし、人工肺臓組織における毛細管内皮の成長にとって適切な基質にもなるので、これは、脱細胞化スキャフォールドの決定的に重要な特質である。
【0188】
図7Aおよび7Bに、未変性肺臓と、血管灌流圧の制御なしで脱細胞化した肺臓のTEM像を図示する。このような条件下では、肺胞基底膜は時折、同定することができず、毛細管を見いだすことができなかった。いかなる特定の理論にも束縛されることは望まないが、基底膜および超微細構造の損傷は、脱細胞化プロセス中の灌流圧を最小限に抑え、脱細胞化を開始する前に脈管構造を最大限に血管拡張しておくことによって減少させることができると考えられる。脱細胞化流体は生理的流量未満の流量で脈管構造に灌流させたが、大量の細胞溶解ならびに脈管構造における細胞タンパク質およびDNAの蓄積ゆえに、血管灌流圧は脱細胞化中に超生理的になり得る。そこで、肺動脈圧を注意深く監視し、この圧力が約20〜30mmHg未満に厳密に保たれるように、脱細胞化バイオリアクタおよび灌流量を調整した。血管拡張剤ニトロプルシドナトリウムを利用して、初期灌流圧を最低限に抑えた。
【0189】
図6Cに、圧力を約30mmHg未満に保って脱細胞化したスキャフォールドのTEM像を示す。これらの条件下では、インタクトな連続的肺胞基底膜が観察された。コラーゲン線維および他のマトリックス構成要素は、肺胞中隔内に保たれている。しかし、肺胞を取り囲んで豊富に存在するはずの透明な毛細管構造の存在は一切認められない。
【0190】
脱細胞化スキャフォールドにおける毛細管構造の保持
齧歯類の肺血管系における典型的な圧力は15mmHg未満であり[Lee et al., 1999, Cell 99:301-12]、これは上記の研究で利用した30mmHgよりもかなり低い。灌流圧を下げるために灌流の流量を低下させ、血管拡張剤を使用したにもかかわらず、脱細胞化灌流圧を30mmHg未満に維持することは困難だった。しかし、脱細胞化プロトコールをわずかに変更すれば脱細胞化時の灌流を約20mmHg未満の圧力で行うことが可能になることが、発見された。重要なことに、これは毛細管構造の保持を可能にした。この変更には、脈管構造を通して脱細胞化流体の灌流を始める前に、気道コンパートメントを脱細胞化流体で洗浄することが含まれた。その結果、血管灌流圧が、特に脱細胞化プロセスの開始時において、有意に低下した。図8に示すように、この技法は、脱細胞化スキャフォールドにおける毛細管構造の保持を可能にした。毛細管の保持は、脱細胞化肺臓スキャフォールドの作出における重要な進展であると考えられる。
【0191】
スキャフォールドはインタクトな気道樹および血管網を保持するべきである。マイクロCTイメージングに加えて走査型および透過型電子顕微鏡法を使用することにより、全体として、スキャフォールドは脱細胞化プロセス後に極めてよく保存されていることが証明された。走査型EMは、所定の組織学的検査と共に、スキャフォールドが総体的にインタクトであり、大きな欠陥を持たない(すなわち肺胞および肺胞中隔はインタクトに見える)ことを証明した。透過型EMは、肺胞基底膜がよく保存されること、および少なくとも一部の毛細管が保たれることを証明した。マイクロCTイメージングは、脈管構造が、下は直径100μmの血管まで、インタクトであることを証明した。
【0192】
透過性評価
肺臓がインビボで機能するには、肺胞および間質腔への大量の血液損失を回避するために、肺臓が、開存性で漏出しにくい連続した脈管構造を持たなければならない。5μmマイクロスフェアを気道コンパートメント中に保持して脈管構造へのこれらの高分子の輸送を許さない脱細胞化肺臓スキャフォールドの能力を評価した。脈管構造中に保たれる必要があるであろう血液の主要構成要素である赤血球のサイズを模倣するために、5μm粒子を使用した。したがって、気道から脈管構造への5μm粒子の漏出を、脱細胞化膜を横切るそのような粒子の移動に著しい指向性はないという仮定の下で評価した。
【0193】
未変性肺臓、灌流圧を制御せずに(一定の灌流流量で)脱細胞化した肺臓、および血管拡張後に制御された灌流圧(30mmHg未満)で脱細胞化した肺臓の透過性を決定した。結果を図8に示す。この結果は、TEM所見を、より大規模に確認するものだった。高い(制御されない)灌流圧による脱細胞化が39%の漏出につながるのに対し、低圧脱細胞化の場合は5.7%、未変性肺臓の場合は2.1%だった。
【0194】
マイクロCTイメージング
マイクロCTイメージングを使って、脱細胞化肺臓スキャフォールドの気道および血管コンパートメントの開存性を評価した。この技法により、肺臓スキャフォールドの三次元像を得ることが可能になり、気道および血管コンパートメントの開存度の同定が容易になる。
【0195】
図9に58μmの解像度で脈管構造の画像を示す。この解像度で、大血管はインタクトであることが示され(図9の最上段のパネル)、未変性試料および脱細胞化試料が概して類似していることが、下段および中段のパネルに示されている。脈管構造の高解像度画像(6.8μm)を図10に示す。ここでは血管が三次元投影像(最大値投影像)として示されている。これらの画像では、わずかな血管漏出が、脱細胞化スキャフォールドの一部の領域に示される曇りとして、同定された。
【0196】
脱細胞化マトリックスの決定的に重要な特質は未変性三次元構造の保存である。脱細胞化スキャフォールドの構造がどの程度保存されているかを評価するために、走査型および透過型EM、マイクロCT、およびマイクロスフェア透過性アッセイの組合せを使用した。脱細胞化肺臓の超微細構造特徴をSEMで調べたところ、肺胞アーキテクチャおよび肺胞中隔の維持が証明された。透過型EMにより、完全にインタクトな肺胞基底膜ならびにコラーゲンおよびエラスチン線維が証明された。これらのEM所見は、肺臓マトリックスを脱細胞化する他の研究であって、そのような構造が保たれるもの[Lwebuga-Mukasa et al., 1986, Exp Cell Res 162:423-35]と合致している。脱細胞化時の血管灌流圧を厳密に制御することにより、本明細書に提示する結果は、毛細管の保持を証明している。マイクロCTイメージングにより、控えめに見積もっても下は直径100μmの血管までの血管網の保持が証明され、それより小さな血管もかなりの数がインタクトである。
【0197】
実施例2:脱細胞化肺臓組織の機械的完全性に対する細胞外マトリックス構成要素の寄与
スキャフォールドの機械的性質に焦点を絞って脱細胞化スキャフォールドの組成をさらに詳しく評価するために、以下の実験を計画した。いかなる特定の理論にも束縛されることは望まないが、脱細胞化肺臓スキャフォールドは、主にコラーゲンおよびエラスチンからの寄与によって、未変性肺臓の顕著な機械的特質を保っていると考えられる。本明細書に提示する結果は、細胞の寄与には依存しない肺臓力学を研究するためのプラットフォームとしての脱細胞化肺臓組織の有用性を証明している。
【0198】
本明細書に提示する結果は、コラーゲン含量が保たれ、エラスチン含量が未変性レベルの約40%のレベルで保たれ、一方、グリコサミノグリカンは、大部分が、脱細胞化スキャフォールドから失われることを証明している。
【0199】
以下にこれらの実験で使用した材料および方法を説明する。
【0200】
材料および方法
器官収集および脱細胞化
本明細書の他の項で説明したように肺臓組織を収集し、脱細胞化した。
【0201】
組織学的分析
組織学的検査を使って数多くの脱細胞化肺臓スキャフォールドを特徴付け、細胞材料の除去を確認した。組織を固定し、パラフィン包埋し、薄切りした。分析は、標準的なヘマトキシリンおよびエオシン染色(H&E)、コラーゲンに関するMassonのトリクローム染色、エラスチンに関するVerhoff van Gieson染色、およびプロテオグリカンに関するアルシアンブルー染色、ならびに4'6-ジアミジノ-2-フェニルインドール(DAPI)を用いるDNAの染色によって行った。
【0202】
コラーゲンアッセイ
改変されたGrantの方法[Grant 1964, 1964, J Clin Pathol 17:685-6]を使ってOH-プロリンを検出する比色アッセイで、コラーゲンを定量した。肺臓試料を凍結乾燥し、重量測定した後、パパイン(140μg/ml)中、60℃で一晩インキュベートした(Sigma)。パパイン消化した試料を6N HCl中、115℃で18時間インキュベートし、中和し、クロラミンTで酸化し、p-ジメチルアミノベンズアルデヒドと反応させた。吸光度を波長550nmで測定し、1:10(w/w)というヒドロキシプロリン対コラーゲンの比を使って、組織のコラーゲン含量を算出した。未変性試料および脱細胞化試料について少なくとも4つの試料を測定した。
【0203】
エラスチンアッセイ
Fastinエラスチン・アッセイ・キット(Biocolor、北アイルランド・ベルファスト)を使ってエラスチンを定量した。肺臓試料をまず凍結乾燥し、重量を測定してから、Foronjy et al.[Foronjy et al., 2008, Am J Physiol Lung Cell Mol Physiol 294:L1149-57]に記載の方法に従って、エラスチンを抽出した。試料を0.25Mシュウ酸と共に100℃でインキュベートした後、10,000gで遠心分離し、上清を取り置いた。5回の抽出から得られた上清をプールし、6回目の抽出から得られた上清も、組織中にもうエラスチンが残っていないことを確かめるために測定した。10,000分子量カットオフフィルタ(Millipore)を使ってシュウ酸を除去した後、dH2Oに再懸濁し、Fastinエラスチンキットを使って、製造者の説明書に従って分析した。未変性試料および脱細胞化試料について、少なくとも4つの試料を測定した。
【0204】
硫酸化グリコサミノグリカンアッセイ
Blyscan GAGアッセイキットを使って、コンドロイチン、デルマタン、ヘパランおよびケラタン硫酸を含む硫酸化グリコサミノグリカン(sGAG)を定量した。パパイン消化した試料(上記コラーゲンアッセイについて説明したように調製したもの)を、製造者の説明書に従ってアッセイした。簡単に述べると、硫酸化GAGを1,9-ジメチル-メチレンブルー色素で標識し、吸光度を650nmで測定した。
【0205】
機械的試験
10Nロードセルを装着したInstron 5848を使って未変性肺臓および脱細胞化肺臓試料を分析した。既知寸法の組織切片を20%歪みまで10サイクルにわたって周期的にプレ伸張して弾性的性質を調べた後、破損するまで伸張して、極限引張強さ(UTS)を評価した。試験プロトコールの図解については図11を参照されたい。組織寸法を使って、工学応力および工学歪みを力と距離から算出した。
【0206】
実験の結果を以下に説明する。
【0207】
コラーゲンおよびエラスチン含量
図12Cに示すように、脱細胞化スキャフォールド中のコラーゲン含量は、未変性肺臓と識別できなかった。コラーゲンは肺臓の機械的強度に重要な役割を果たすので、コラーゲンのこの保存は重要である。コラーゲン含量は、図12に示すMassonのトリクロームによる組織化学的染色でも維持されていた。同様に図12Cに示すように、適切でないことがわかった脱細胞化法の一つであるSDSで脱細胞化したスキャフォールドでは、コラーゲン含量が減少した。コラーゲンのこの喪失は、SDS脱細胞化スキャフォールドにおける低下した機械的完全性と相関すると考えられる。
【0208】
定量的アッセイでも組織染色でも証明されるとおり(図13)、脱細胞化スキャフォールドでは、エラスチン含量も(減少はするが)保存される。エラスチン線維は肺臓に弾性を与え、吸入後の肺臓の弛緩(したがって呼気)において重要な役割を果たす組織の自然反跳にとって、決定的に重要である。脱細胞化プロセス中にこれらの線維が保持されることは、決定的に重要である。というのも、それは、肺細胞集団をスキャフォールドに再播種しようとする努力において、肺臓スキャフォールドが適切に換気されることを可能にするからである。スキャフォールドは未変性エラスチン含量の60%を失ったが、残ったエラスチンは、本明細書の他の項で論じる機械的試験結果からわかるように、肺臓に弾性機能を与えるのに十分だった。
【0209】
全体として、これらの重要なECM構成要素の保持は、スキャフォールドが生理的レベルの機械適応力を経験することを可能にした。これは、重要なことである。というのも、さまざまな発生および細胞分化プロセスが機械的刺激に依拠するからである。また、ECMは、マトリックスへの細胞付着を補助するのに決定的に重要であり、これらの未変性ECM構成要素の保持は細胞の付着と伝播を容易にし、したがって生体工学的(bioengineered)肺臓組織の開発を容易にする。
【0210】
プロテオグリカン含量
プロテオグリカンは、1つまたは複数のグリコサミノグリカン(GAG)鎖に連結したコアタンパク質からなる。大半のGAGは硫酸化されており、それが定量的アッセイによるそれらの検出を可能にする。その結果を図14に示す。脱細胞化スキャフォールドのGAG含量は、未変性肺臓より有意に低かった(未変性肺臓レベルの約6%)。プロテオグリカンは、細胞表面上または細胞外マトリックス内に見いだされ[Ferdous et al., 2007, Tissue Engineering 13:1893-904]、それらの除去は、一つには、細胞結合型GAGの除去によるものである。しかし、ECM内に見いだされるGAGも、脱細胞化溶液によって可溶化され得る。図14は、プロテオグリカンに関するアルシアンブルー組織染色であり、これは、脱細胞化肺臓スキャフォールド中に残っているGAGの量が未変性肺臓と比較して減少したことを示しており、定量的アッセイの結果が確認された。
【0211】
機械的特徴付け
末梢肺臓条片の機械的試験を使って、未変性肺臓試料および脱細胞化肺臓試料の両方の準静的力学を評価した。応力-歪み曲線の弾性域は、未変性試料と脱細胞化試料がどちらも履歴挙動を表したことを示している。履歴現象は肺臓が粘弾性材料であることを証明しており、拡大曲線と弛緩曲線の間の相違は、弛緩中に回復されないエネルギーを表す。また、図15に示すように、試料はクリープを起こさなかった。仮に肺臓組織がクリープを起こすとすると、拡張後に元の位置に収縮することはなく、それゆえに肺臓は決して完全には収縮せず、ガス交換が損なわれるであろう。この適当な弾性肺臓挙動の保存は、肺臓スキャフォールドにとって重要である。なぜなら、肺弾性の喪失は、いくつかの疾患状態、とりわけ肺気腫に見られるからである[Gelb et al., 2002, Chest 121:715-21]。
【0212】
極限引張強さ(UTS)は破損時の試料上の応力であり、材料の強さの尺度である。図16に示すように、脱細胞化試料のUTSは、未変性試料のそれと識別できなかった。しかし、ドデシル硫酸ナトリウム(SDS)を含有する緩衝液中で試料を脱細胞化した場合は、UTSの減少によって証明されるように、機械的完全性が損なわれた。SDSはコラーゲンを分解して、組織の断片化および腫脹を引き起こすことができ[Bodnar et al., 1986, Thorac Cardiovasc Surg. 34(2):82-5;Gilbert et al., 2006, Biomaterials 27:3675-83]、組織の伸展性(extensibility)を増加させることも示されている[Mirsadraee et al., 2006, Tissue Eng 12:763-73]。SDSはイオン性が高い両親媒性界面活性剤であり、その疎水領域はタンパク質と相互作用することができ、一方、親水性部分は、特に負に帯電している場合には、水と結合して組織腫脹を引き起こす[Bodnar et al., 1986, Thorac Cardiovasc Surg. 34(2):82-5]。他の研究ではSDS処理によるUTSの減少が常に見られるわけではなかったが[Mirsadraee et al., 2006, Tissue Eng 12:763-73]、これは、組織の相違によるものだろう。Mirsadraee et al.は心臓周囲組織を研究したが、これは肺臓よりはるかに密に充填されたコラーゲン線維を含有している。肺臓では、組織の幾何学ゆえに、コラーゲン線維が高度に分散されており、SDSが誘発する腫脹は、定量的コラーゲンアッセイで見られるように、はるかに容易に、コラーゲンの除去につながり得る。
【0213】
本明細書に提示する結果は、脱細胞化スキャフォールドが、関連するインビボでの生理的力に耐え得ることを証明している。
【0214】
本明細書に提示する結果により、コラーゲンとエラスチンはどちらも機能的レベルで保存されることが確認された。これらの知見により、肺臓力学への主な寄与は、コラーゲンとエラスチンによってなされ、細胞構成成分またはプロテオグリカンによるものではないことが確認された。本明細書に提示する結果により、未変性肺臓の特徴を示す脱細胞化肺臓スキャフォールド(これらの特徴は、肺臓スキャフォールドを、組織工学的応用のための前途有望な基質にすると共に、詳細なマトリックス力学ならびに肺臓の生物学、発生および生理学を研究するためのプラットフォームにする)の作製が証明された。
【0215】
実施例3:3次元肺臓組織のインビトロ培養のためのバイオリアクタの設計と検証
バイオリアクタを使って3次元肺臓組織をインビトロで培養することができる。そのようなバイオリアクタの開発は、人工肺臓組織の成長に関する研究を行うのに有益であるばかりでなく、肺生物学の研究にも有益であるだろう。成人肺臓細胞の長期インビトロ培養を可能にするシステムは、今のところない。
【0216】
肺臓組織全体をインビトロ培養するためのバイオリアクタを設計するために、以下の実験を行った。細胞の生存および分化を支持するために肺臓組織に十分な栄養素供給および機械的刺激を与え得ることを目指して、一連の設計制約を満たすように、バイオリアクタを設計した。細胞の生存性および分化状態の維持によって証明される肺臓組織の全葉のインビトロ培養をバイオリアクタが支持できるかどうかを評価するために、実験を設計した。バイオリアクタを評価するための過程では、バイオリアクタにおける肺臓の生存に対する灌流および換気の効果を評価した。本明細書に提示する結果は、バイオリアクタがインビトロ肺臓組織培養に使用することができ、それゆえに肺臓組織のエンジニアリングに応用可能であることを証明している。
【0217】
これらの実験で使用した材料および方法を以下に説明する。
【0218】
材料および方法
全肺臓培養
肺臓を若年成体(3ヶ月齢)雄フィッシャー344ラットから収集した。動物実験の作業は全て、イェール大学施設内動物実験委員会からの承認を受けて行われた。動物をペントバルビタールナトリウム(Sigma、40mg/kg)の腹腔内注射によって麻酔した。麻酔の誘導後に、胸部および腹部にエタノールを噴霧し、肋骨縁の直下で横切開を施し、腹腔に入った。横隔膜を穿刺し、肺臓に触れないように注意しながら肋骨を後退させた。下大静脈を切断し、50U/mlヘパリン(Sigma)および1μg/mlニトロプルシドナトリウム(Sigma)を含有する20〜30mlのPBSで右心室を通して肺臓を灌流した。次に気管を切り離し、できるだけ高く切った。心臓および肺臓への残りの接続を全て切り離して、心臓、肺臓および気管をひとまとめにして動物から取り出せるようにした。
【0219】
カニューレ装着
器官の除去後に、気管と、心臓の右側を通して肺動脈幹とに、カニューレを接続した。心尖を円刃刀で切り落とし、直角カニューレを右心室を通して肺動脈幹へと挿入した。このカニューレにシリンジを取り付け、漏出を伴わない適正なカニューレ設置と肺臓の十分な灌流とを確かめるために、5〜10mlのヘパリン化食塩水を注入した。次に、このカニューレを心臓に縫合糸で固定した。別の真っ直ぐな返し付(barb-end)カニューレを気管に挿入し、縫合糸で固定した。次に、肺臓をバイオリアクタに接続し、本明細書の他の項で説明するプロトコールに従って、脱細胞化した。
【0220】
カニューレを、右心室を通して肺動脈と、気管とに取り付け、肺臓をバイオリアクタに接続した。PBS中の2%アンホテリシン、ペニシリンおよびストレプトマイシンで気道を洗浄した後、PBSで2回洗浄し、次にバイオリアクタを培地で満たして、培養を開始した。血管灌流および換気を実験条件の指図どおりに行った。
【0221】
バイオリアクタの構成要素と組み立て
バイオリアクタの構成要素は、別段の注記がない限り、Cole-Parmer(イリノイ州バーノンヒルズ)から入手した。シリコーン栓と500mlのガラス製広口瓶とがバイオリアクタの基礎をなした。灌流ループ、気管接続、空気換気、および培地交換ポートを含む肺臓への必要な接続が可能になるように、シリコーン栓を通して、サイズL/S14およびL/S16のPharMedチューブ(オハイオ州ウェストレーク)を挿入した。圧力は、灌流ポンプと肺動脈への接続との間のTruWave圧変換器(Edwards Lifesciences、カリフォルニア州アービン)を使って監視した。灌流はMasterflex L/S可変速ローラーポンプ(Masterflex、イリノイ州バーノンヒルズ)を使って達成した。換気は、多チャンネルプログラマブルシリンジポンプ(Cole Parmer)を使って行い、吸気と呼気をそれぞれ10mlの体積で30秒間かけて行った。バイオリアクタの略図を図17に示す。
【0222】
組織学的検査および免疫蛍光法
所望の培養期間後に、肺臓を固定し、パラフィン包埋し、薄切りした。所定の組織学的検査(H&E)を、アクアポリン5(I型上皮)、サーファクタントタンパク質C(II型上皮)、CCSP(クララ細胞)、およびPECAM-1(内皮)に関する免疫蛍光法と共に行った。切片をキシレン中で脱パラフィンし、再水和し、0.2%Triton-Xを含むPBS(緩衝液)と共に15分間インキュベートした。PBS中の0.02Mクエン酸を使って、75〜85℃で20分間、抗原回復を行った後、切片を緩衝液中ですすいだ。ブロッキングは、PBS+1%ウシ血清アルブミンおよび0.75%グリシンを使って室温で1時間行った。一次抗体を緩衝液ですすぎ落とし、二次抗体を1:500の希釈度で室温において1時間適用した。二次抗体はInvitrogenから入手した(AlexaFluor555またはAlexaFluor488x)。画像はZeiss Axiovert 200M倒立蛍光顕微鏡を使って取得した。
【0223】
細胞増殖は、増殖性細胞核抗原(PCNA)に関する染色によって評価し(Zymed、カリフォルニア州サンフランシスコ)、アポトーシス核を、末端デオキシヌクレオチジルトランスフェラーゼdUTPニック末端ラベリング(TUNEL)染色(Calbiochem、カリフォルニア州サンディエゴ)で検出した。どちらのアッセイも製造者の説明書に従って行った。
【0224】
マイクロスフェア換気アッセイ
バイオリアクタにおける肺臓の換気が、培地の移動を誘発して脈管構造を灌流するのに十分であるかどうかを決定するために、5μmポリスチレンマイクロスフェア(SPI Supplies、ペンシルバニア州ウェストチェスター)を使って簡単なアッセイを開発した。肺臓を、本明細書の他の項で説明するように、バイオリアクタに接続し、換気したが、灌流はしなかった。バイオリアクタチャンバを、1000万個のマイクロスフェアを含有する100mlの培地(培地1mlあたり10万個のマイクロスフェア)で満たした。培養を換気だけで3時間続けた。次に肺臓を固定し、パラフィン包埋し、薄切し、所定の組織学的検査(H&E)を使って分析した。
【0225】
実験の結果を以下に説明する。
【0226】
バイオリアクタ設計要件
バイオリアクタには、齧歯類インビボ環境の重要な特質を組み入れると共に、所望する条件に応じて使用者がいくつかの重要なパラメータを調整できるようにも設計した。設計目標は次のとおりである。
・システムは、使用者が指定した、生理的レベルの範囲内にある速度で、脈管構造を通して培地を灌流させる能力をもたなければならない。
・システムは、空気または培地を使って気管を通して肺臓を換気する能力をもたなければならない。正常な生理的条件と合致するように、陰圧換気が好ましく、肺臓を絶えず換気できることが好ましい。
・バイオリアクタは、好ましくは、肺臓の血管コンパートメントと気道コンパートメントとを異なる培地タイプで浸すことを可能にするべきである。
・バイオリアクタは、換気に関する上記の要件を満たすと同時に、培養培地へのガス交換も可能としなければならない。
・バイオリアクタは、肺動脈および気管圧の圧力測定を可能にするポートを持たなければならない。圧力は、理想的には、正常な生理学的値の範囲内にあるべきであり、肺動脈圧は15〜30mmHg未満である[Li et al., 2004, Proc Natl Acad Sci USA 101:11488-93]。
・バイオリアクタは、定期的な培地交換を可能にする手段を持たなければならない。
・バイオリアクタは、標準的な組織培養インキュベータの物理的境界内に収まり得るように、小さくかつ自給式でなければならない。
・全てのバイオリアクタ構成要素は、安価かつ入手が容易でなければならない。
・バイオリアクタおよび全ての構成要素は(好ましくはオートクレーブによる)滅菌が、可能でなければならない。
【0227】
上記の基準を満たすバイオリアクタを設計し、構築した。そのバイオリアクタの図解を図17に示す。
【0228】
バイオリアクタ灌流システム
肺臓への灌流は、バイオリアクタ本体から肺動脈へと培地を循環させるローラーポンプによって行った。使用者は灌流量を指定することができる。心臓の右心室を通した肺動脈幹へのカニューレの接続が容易になるように、ラットの心臓を肺臓に取り付けたままにしておく。ただし肺静脈は灌流ループに直接的には接続しなかった。むしろ肺静脈は心臓の左側からメインバイオリアクタリザーバに直接的に排液した。肺臓の静脈排出路はメインバイオリアクタ中に直接出る。
【0229】
肺臓を通る灌流量は使用者の指定に合わせて設定することができる。成体ラットにおける生理的流量は40〜80ml/分であるが、人工組織培養の場合、典型的には、流量はこの値よりはるかに低い。成体ラットの場合、酸素化されるには、全血液量が肺臓を通過しなければならないのに対し、人工組織培養中は、肺細胞の成長を支持するのに十分な培地を灌流するだけでよい。したがって、人工的培養(engineered culture)時の灌流量は、正常な生理学的シャントゆえに肺臓の血流量が心拍出量の8〜10%しかないラット胎児におけるそれに近い[Hislop et al., 2000, Ped Resp Rev 1:321-7]。圧力プロファイルは、肺血管圧を低下させるために使用することができるニトロプルシドナトリウムなどの血管拡張剤を使って、ある程度制御することができる。典型的には、灌流圧は、肺動脈系に見られる最大値である約30mmHg未満に保たれる[Li et al., 2004, Proc Natl Acad Sci USA 101:11488-93]。
【0230】
バイオリアクタ換気システム
バイオリアクタは陽圧換気と陰圧換気がどちらも可能だった。インビボでは、呼吸は通常、陰圧換気によって達成される。横隔膜が収縮し、胸郭が拡大して、胸腔内に陰圧を生じ、それが、この圧力不均衡を緩和するべく肺臓内に空気を流入させる。吸気後に、呼吸筋が弛緩し、肺臓は受動的にしぼむ。
【0231】
陰圧換気はバイオリアクタにおける換気の主要モードである。肺臓を取り巻く陰圧を達成するために、バイオリアクタのメインチャンバは完全に気密でなければならない。これは、全ての空気口および圧力監視口を閉じることによって達成される。次に、シリンジポンプを使って、バイオリアクタ本体から所定の体積の空気を抜いて、陰圧を生じさせる。この圧力を緩和させるための唯一の経路は、別個のリザーバに接続されている気管を通して培地(または空気)が肺臓に流入する経路である。次に、バイオリアクタ本体中に空気を押し戻す方向に、シリンジポンプを逆転させる。これはチャンバー内での陰圧の強化を逆戻りさせ、培地(または空気)は気管リザーバに向かって逆流する。この時、肺臓は受動的にしぼむ。
【0232】
気管カニューレは一方向弁を利用する:
図17に図示するように、気管への接続には、Y字連結器と、バイオリアクタ本体に向かって開く一方向弁が必要である。このタイプの接続は、流体の気道コンパートメントからの漏出ゆえに必要になる。吸気時は、ある体積の培地が肺臓に入る。しかし、この培地の一部は肺胞膜を横切って間質腔または脈管構造へと漏出する。それゆえ、吸気時に肺臓に進入した培地の全てが、呼気時に気管リザーバに戻ることができるわけではない。図17に示す設計には、吸気時には全ての培地が肺臓に進入することを許すという特徴が組み込まれている。しかし呼気時には、培地は肺臓から、またはバイオリアクタ本体から一方向弁を通して、気管リザーバに戻ることができる。
【0233】
バイオリアクタは、シリンジポンプを気管カニューレまたは気管リザーバに直接接続することによって、陽圧換気を利用することもできる。
【0234】
気管入口の改造
バイオリアクタに関して、換気時に、肺臓気道コンパートメントに十分な新鮮培地が供給されないことが観察された。いかなる特定の理論にも束縛されることは望まないが、これは、主として、気管と別個の気管リザーバとの間のチューブ内に含まれる培地の体積ゆえに、同じ培地が気管を出入りして、気管に進入する新鮮培地が不十分になっているからであったと考えられる。気道培地流ループ中の「死腔」が、呼吸時に新鮮培地が肺臓組織に到達することを妨げた。そのため、図17に概説するように、換気時に培地が異なる経路を辿って肺臓に出入りするように、バイオリアクタを改造した。この改造により、各吸息と共に気管に入る培地の大半は気管リザーバから直接供給される(したがって気管から出てくる培地と比較して「新鮮」である)ことになった。
【0235】
バイオリアクタ培養中の酸素供給:
十分な酸素含量があることを保証するために、肺臓培養中のバイオリアクタにおける組織培養培地の酸素含量を測定した。特に、陰圧換気(この間、バイオリアクタ本体は気密であり、酸素進入のための唯一の入口は気管リザーバを介したものである)中に、十分な酸素送達があることを保証する必要がある。酸素分圧は、培養の間、著しくは低下せず、正常組織培養培地におけるレベルと同じ6.0〜7.0mg/Lを保つことがわかった。これらのレベルは、80〜100mmHgという正常な生理的レベルを上回っている(6〜7mg/Lは分圧137〜159mmHgに相当する)。
【0236】
バイオリアクタ圧力プロファイル
圧力が予想限度内または生理学的限度内にあることを保証するために、バイオリアクタ中で培養されている人工肺臓組織の気管および肺動脈における圧力プロファイルを測定した。図18に代表的プロファイルを示す。灌流圧は、典型的には、約2〜30mmHgに保たれた。記載の例では、ベースライン灌流圧が10〜17mmHgで変動した。しかし、陰圧換気の効果がこのプロファイルに重ね合わされて、陰圧「吸息」時には灌流圧が0〜7mmHgまで低下した。この効果は生理学的に見られ、肺脈管構造の圧力は息の吸い込みに伴って低下する。バイオリアクタでは、肺静脈がメインチャンバに直接排液し、このメインチャンバは、「胸腔」としても機能し、肺臓を換気するために陰圧を生じさせた場所である。これが、バイオリアクタから灌流脈管構造への陰圧の伝達を増加させる役割を果たした。
【0237】
灌流圧プロファイルから、最大負「胸腔」圧は約-12mmHgであり、生理学的値とほぼ一致した。それゆえに、吸気中は、この陰圧が気道に加えられ、流体(または空気)を気管リザーバから肺臓へと動かす。この圧力は、気道樹を上がるにつれて徐々に減少し、気管の入口では-3mmHgだった。注目すべきことに、気管への入口における圧力は、大気圧に拘束されるので、生理的には本質的にゼロだった。しかし、バイオリアクタでは、気管と気管リザーバの間の長さ(ここで圧力が0に達する)により、この圧力が吸気中はわずかに負のままだった。
【0238】
培地および酸素要求量
以下の結果は、バイオリアクタ中で培養される齧歯類肺臓に要求される培地および空気の体積の決定に役立たせることを目的とした一連の計算を示す。
【0239】
組織培養比較:
インビトロ組織培養時は、500万個の細胞に、3日ごとに12mlの培地を供給することが一般的である。成体齧歯類肺臓が1億個の細胞を含有すると仮定すると、これは、3日ごとに多くて240mlという培地要求量に相当する。しかしこれは過剰評価であるだろう。というのも、組織培養中の細胞が一般に活発に複製するのに対して、インタクトな齧歯類肺臓中の多く細胞は休止状態にあり、それゆえに培地要求量が低いからである。
【0240】
グルコース消費要求量:
灌流ラット肺臓のグルコース消費量は、1時間あたり乾燥重量1グラムあたり43μmolであることが証明されている[Kerr et al., 1979, Am J Physiol 236:E229-33]。成体ラットの肺臓は約150〜250mgの乾燥重量を持ち[Inokawa et al., 2006, Ann Thorac Surg 82:1219-25]、組織培養培地は、典型的には、5.5mmol/Lのグルコースを含有する。それゆえに、成体ラットの肺臓は、そのグルコース消費要求量を供給するために、1日あたり28〜47mlの組織培養培地を要求するだろう。
【0241】
酸素要求量:
肺動脈内皮細胞は、1分あたり細胞100万個あたり6nmolの酸素を消費し[Xu et al., 2007, Proc Natl Acad Sci USA 104:1342-7]、一方、ラットII型上皮細胞は1分あたり1.25nmolを消費する[Dobbs et al., 1980, Biochim Biophys Acta 618:510-23]。成体ラット肺臓には1億個の細胞があり、肺臓中の全ての細胞が高い方の速度で酸素を消費すると仮定すると、ラット肺臓は1日あたり多くて26mgの酸素を要求するだろう。組織培養培地は、1リットルあたり約6mgの酸素を含有し、バイオリアクタは約300mlの培地を含有する。したがって培地は、新鮮培地の交換1回(3日ごと)につき1.8mgの酸素を供給することができる。さらに、酸素はバイオリアクタ中の空気にも含まれる。バイオリアクタ本体には約200mlの空気がある。培養器中の空気は約20%のO2を含有し、これは海面位、37℃で、空気1リットルあたり約260mgの酸素に相当する。したがってバイオリアクタ中の空気は約52mgの酸素を含有する。
【0242】
本発明のバイオリアクタは、上記の計算に基づいて、培養齧歯類肺臓の培地および酸素要求量を供給する。通例、バイオリアクタには合計240mlの培地を供給することができ(バイオリアクタ本体に180mlおよび気管リザーバに60ml)、バイオリアクタ中の空気は毎日交換することができる。これらの条件は、培養肺臓に必要以上の栄養素および酸素を提供するのに足りると考えられる。
【0243】
全肺臓培養
肺臓バイオリアクタの設計を検証し最適化するために、全未変性齧歯類肺臓のインビトロ培養を使用した。肺臓をバイオリアクタ中で最長7日間培養した。バイオリアクタは細胞の生存および分化ならびに肺臓形態を維持するのに十分な栄養素供給および機械的刺激を提供することが証明された。
【0244】
細胞生存、肺臓形態および細胞分化状態の維持に対するバイオリアクタ条件の効果を調べるためにも、バイオリアクタにおける未変性肺臓の培養を使用した。まず最初に、肺臓形態に対する空気換気と液体(培地)換気の効果を比較した。次に、細胞生存に対する換気技法および栄養素送達の効果を評価した。細胞の生存および分化に対する血管灌流圧の効果も評価した。7日間の培養中に細胞分化を維持するバイオリアクタの能力も評価した。
【0245】
全体的肺臓形態に対する空気換気と培地換気の効果:
バイオリアクタ中で培養される肺臓を培地または大気(約20% O2)で換気する効果を評価した。培地による換気は、改善された栄養素送達(これは、バイオリアクタでは、大きな気道に供給する灌流された気管支循環がないので、より一層重要になり得る)を提供するだろうから、改善された細胞生存をもたらすだろうと考えられる。しかし、空気による換気は、成体肺臓が条件づけられている条件であり、肺上皮は気液界面の存在下で培養されることが多く、それはラット胎児肺臓における適当な肺発生を可能にすることが示されている[Funkhouser et al., 1976, Biochem Biophys Res Comm 70:630-7]。そこで、培地による換気が、気液界面の欠如による上皮分化状態の喪失をもたらすかどうかを調べるための実験も計画した。
【0246】
3日間の培養後に、培地で換気した肺臓と空気で換気した肺臓との間に、有意な相違が認められた。図19に示すように、培地換気は未変性肺臓と同じように見えたが、空気換気肺臓は著しく拡張した気道を示し、気道中の細胞片が明らかだった(図19C)。そのうえ、図19Cの中央のパネルは、空気換気肺臓の気管支上皮および細気管支上皮が完全になくなっていることを示し、この所見は、肺臓全体にわたって一貫していた。さらにまた、図19Cの右側のパネルに示すように、拡張した末梢気腔も明白だった。
【0247】
(空気による換気に加えて)脈管構造に培地を灌流させても気道上皮は剥皮され、換気を行わずに脈管構造に培地を灌流させた場合は気道上皮がインタクトであることも観察された。これは、気道上皮の喪失が十分な培地の欠如によるものではなく、空気換気の効果に関係することを示唆している。気管支循環はどの培養でも灌流されないことが観察された。
【0248】
上皮細胞は、その生理的位置と同じく気液界面で培養されることが多い。気液界面(ALI)は上皮分化を誘導するために利用されることが多く[Gruenert et al., 1995, Am J Physiol 268:L347-60;Wong et al., 2009, J Clin Invest 1 19:336-48;Hosokawa et al., 2007, Connect Tissue Res 48:9-18]、気液界面の欠如は絨毛形成の減少につながる可能性がある[Ostrowski et al., 1995, Exp Lung Res 21:957-70;Yeh et al., 2007, Laryngoscope 117:1439-44]。さらにまた、気液界面は、インビトロ培養時に、II型上皮によるサーファクタントの分泌が維持されることを可能にする[Mason et al., 2002, Am J Physiol Lung Cell Mol Physiol 282:L249-58]。したがって、ALIの非存在下では、細胞分化状態の相違が生じるだろうと予想された。しかし、7日まで行った培養物について図25に示すように、培地で換気した肺臓におけるクララ細胞分泌タンパク質(CCSP)、サーファクタントタンパク質C(SPC)、またはアクアポリン(AQP)の発現パターンには、有意な変化が観察されなかった。
【0249】
細胞生存に対する灌流の効果:
インビトロ肺臓培養を灌流だけで支持することができるか、そしてもしそうなら、どの灌流圧が最適であるかを決定するために、バイオリアクタ中の培養未変性肺臓における細胞生存および細胞分化に対する血管灌流の効果を調べた。これらの実験を複雑にするのは、肺臓の外植後に血管透過性が急速に増加するという事実だった。10mmHgの圧力を使った単離肺臓の灌流は、10分以内に肺臓水腫を引き起こし得る[Wierup et al., 2005, J Heart Lung Transplant 24:379-85]。外植から5〜10分以内に血管漏出が観察され、小粒子(半径28nm)の3〜4%が肺胞-毛細管膜を横切って漏出した。広範な肺微小血管漏出は、遠位毛細管および静脈構造への培地送達を減少させるか、無くしてしまうことさえあり得る。したがって、流れを遠方まで送達し、遠位毛細管を開存させておくには、約1〜10mmHgという生理的レベルより高い血管灌流圧が要求されるだろう[Li et al., 2004, Proc Natl Acad Sci USA 101:11488-93]。
【0250】
バイオリアクタにおける3日間の未変性肺臓培養時の細胞生存に対する血管灌流圧の効果を調べた。図20に示すように、最高30mmHgの高い灌流圧は、10または20mmHgの圧力と比較して、少ないアポトーシス細胞と、高い細胞密度とをもたらした。しかし、灌流圧とは無関係に、血管灌流による細胞分化の維持は不十分だった。実質的に低いCCSPレベルおよびSPCレベルが観察され(図21)、アクアポリン発現はほとんど全く存在しなかった。図22に示すように、PECAM発現は、脈管構造の大きな血管では観察されたが、毛細管では、減少した発現が観察された。これらの実験により、灌流だけでは、十分な細胞生存または細胞分化を維持するのに十分でないことが証明された。
【0251】
気道コンパートメントにおける培地流路が細胞生存に及ぼす効果:
培地による換気は肺臓形態および細胞分化の維持を可能にしたが、換気培養肺臓では、未変性肺臓と比較して、有意に高いアポトーシス細胞率が観察された(図23および28参照)。これは、新鮮培地の送達が不十分であったためと考えられるので、換気中の気道コンパートメントへの新鮮培地の送達を増加させるためにバイオリアクタを改造する実験を計画した。図17に示し、本明細書の他の項で説明するように、バイオリアクタ本体を気管リザーバに接続する1本のラインがある。このチューブの長さは約40〜45cmであり、3〜3.5mlの培地を含有する。換気中は、陰圧吸気時に約2.5〜3.0mlの培地が肺臓に引き込まれ、同じ体積の培地がチューブを通って気管広口瓶に戻る。それゆえに、各「吸息」時に肺臓に進入する培地のうち、この約2.5〜3.0mlは新鮮ではなく、ただ単にチューブから肺臓へと戻るにすぎない。したがって、肺臓への実際の培地送達は、新鮮な培地を使った換気によって送達されるであろうものよりはるかに少ない。
【0252】
バイオリアクタ本体と気管リザーバとの間に第2の接続が付加されるように、バイオリアクタの設計を改造した。一方向逆止弁を使って、一方の接続を吸気中の培地送達に使用し、他方の接続を呼気中の培地還流に使用した。この改造により、「再利用される」培地が各換気サイクルにつき約2.5〜3.0mlからわずか約0.75mlに減少し、それゆえに新鮮培地の送達量が著しく増加した。
【0253】
このバイオリアクタ改造の効果を図24に示す。この図では、追加の呼吸ラインが細胞の生存を改善することが示された。アポトーシス細胞のパーセンテージは、1本のラインで換気した場合の21.5%(図27の「換気のみ」)から、「ループ」換気では3.9%まで低下した。これに対し、未変性肺臓では0.5%である。
【0254】
「ループ」換気は「再利用される」培地の量を減らすことによって肺臓への培地の送達を増加させるが、血管還流の追加によって培地送達を増加させることもできる。灌流は、換気と共同して、細胞アポトーシスを、単一ラインのみの場合の21.5%から7.9%まで低下させた(図23)。「ループ」換気への改造は単一ライン換気と比較して総細胞数をわずかに増加させたが(図23B)、これは有意な差ではなかった。単一ライン換気でも「ループ」換気でも、未変性肺臓と比較すると細胞数は減少したが、統計的に有意ではなかった。
【0255】
本明細書に提示する結果は、「ループ」換気への改造または補足的血液灌流を使って十分な新鮮培地が肺臓に送達されるのであれば、換気だけでも、3日間の培養中はバイオリアクタにおける未変性肺臓組織の生存が可能になることを証明している。ループ換気は最善の総合的結果を示し、細胞のアポトーシスを最小限に抑えると同時に、培養肺臓組織における細胞数を最大にする。
【0256】
細胞形態、細胞分化および肺胞構造:
バイオリアクタの設計をさらに詳しく検証するために、未変性肺臓の7日間培養を行った。これらの培養では、本明細書の他の項で説明する「ループ」改造を施した、培地による換気を利用し、いかなる血管灌流も行わなかった。血管灌流は利用しなかったが、今後の研究では、「ループ」換気を使った長期換気培養への灌流の付加を探究するかもしれない。
【0257】
肺臓を細胞増殖、アポトーシスおよび細胞分化について、アクアポリン5(I型上皮)、サーファクタントタンパク質C(II型上皮)、クララ細胞分泌タンパク質(クララ細胞)、およびPECAM-1(内皮)に関する染色により、組織学的検査で評価した。図29Aおよび29Bに示すように、全体的な肺アーキテクチャは肺胞構造を含めて保存された。低倍率画像は培地呼吸に関して図19に示したものと識別できなかった。さらにまた、図29C〜29Jに示すように、細胞マーカーの発現パターンは、未変性肺臓と実質的に異ならなかった。
【0258】
換気は単独で肺臓の脈管構造の受動的灌流を可能にする:
換気は単独で、内皮を含むいくつかの重要な肺臓細胞タイプの細胞生存と細胞表現型の維持とを、7日間まで可能にできることが証明された。しかしこれは、脈管構造を通した能動的灌流が存在しない状態でのことであり、それが最初は意外であった。灌流の欠如が内皮の生存または分化に影響を及ぼさない理由を調べるために、5μmマイクロスフェアを使って、脈管構造への流体の移動に換気が及ぼす影響を調べる実験を行った。換気によって誘発される物理的動きは、バイオリアクタ中の培地に向かって開いている脈管構造に入ったり出たりする培地の受動的移動を引き起こすのに十分であったと考えられる。この実験では、5μmマイクロスフェアを含有する培地で満たしたバイオリアクタ中で肺臓を3時間換気した。もし肺臓の脈管構造内にマイクロスフェアが観察されたら、それは、換気によって受動的灌流が誘導されたことを示す。図26に示すように、マイクロスフェアは大血管にも一部の毛細管にも見いだされた。これらの結果は、灌流はそれだけで、脈管構造における培地の移動を誘発するのに十分であり、したがって灌流の欠如にもかかわらず内皮の維持を可能にすることを示している。
【0259】
脈管構造における培地のこの移動は、換気による肺臓の物理的運動の結果である。このような大きな粒子を脈管構造中に移動させるには、拡散だけでは不十分であるだろう。拡散による脈管構造中へのマイクロスフェアの予想される移動は、フィックの第二法則を使って概算することができる。
【0260】
細胞の生存および分化の支持
本明細書に提示する結果は、血管灌流だけでは、細胞生存や、II型上皮によるサーファクタント産生を含む細胞分化を支持するには、十分でないことを証明した。しかし、培地による陰圧換気は、広範な細胞生存(未変性レベルの95.1%まで)を支持し、上皮および内皮の分化を維持するのに十分であった。
【0261】
本実施例の実験の全体的目的は、人工肺臓組織の今後の培養にこのバイオリアクタを使用するために、全齧歯類肺臓をインビトロで長期間にわたって培養する能力を有するバイオリアクタの根拠ある設計を証明することだった。上記の結果は、人工肺臓培養における使用に適した設計の成功を証明している。
【0262】
実施例4:人工肺臓組織における上皮発生
本明細書に提示する結果は、脱細胞化スキャフォールドが細胞毒性を持たず、上皮、内皮、および間葉細胞を含む広範な肺細胞タイプの接着と増殖を支持することを証明している。いくつかの例では、肺臓組織のエンジニアリングに、幹細胞を使用することができる。人工肺臓組織が有用であるためには、それを脈管構造および気道に接続することができなければならない。気道は肺胞とつながっていなくてはならず、血管接続は肺胞を取り巻く緻密な毛細管網に通じなくてはならない。
【0263】
本明細書に提示する結果は、人工肺臓組織の発生が、培地タイプ、灌流、換気、および気液界面の存在を含む重要なバイオリアクタ条件によって左右されることを証明している。いくつかの例では、培養培地による換気(すなわち「液体換気」)が、気道構造および上皮細胞の分化を助ける。さらにまた、気液界面における人工組織の小片の静置培養は、組織成長に対して有意な効果を示し、バイオリアクタでは、空気による換気が細胞の分化および上皮構造の発生に影響を及ぼすことを示した。
【0264】
これらの実験で使用した材料および方法を以下に説明する。
【0265】
材料および方法
スキャフォールド調製
成体Fischer 344ラットの肺臓を収集し、本明細書の他の項で説明するように脱細胞化した。脱細胞化操作に続いて、スキャフォールドを滅菌水(10回交換)中ですすぎ、次に、PBS中の10%ペニシリン/ストレプトマイシンにおいて少なくとも12時間すすいだ。後の培養では、DNA残存物の除去を助けるために、10%FBS中でもすすいだ。次に、完全な灌流および呼吸システムを取り付けた新しい滅菌バイオリアクタに、肺臓を移した。次に肺臓をPBSで2回すすぎ、培養に使用する培地で1回すすいだ。
【0266】
新生児細胞単離
肺臓を本明細書の他の項で論じるように新生児(約7日齢)ラットから単離した。次に肺臓を70%エタノール中で10秒間すすぎ、ダルベッコ変法イーグル培地(DMEM、Gibco)中で2回すすいだ後、滅菌乾燥ペトリ皿に移した。肺臓を円刃刀で5分間切り刻んでから、エラスターゼ消化用の円錐管に移した。DNアーゼ、コラゲナーゼおよびエラスターゼはWorthington Biochemical(ニュージャージー州レークウッド)から入手した。エラスターゼ消化は、DMEM中の4U/mlエラスターゼを100U/ml DNアーゼと共に使用し、撹拌しながら室温で20分間行った。次に、組織塊を70μmナイロンフィルタで濾過し、DMEMですすいだ。消化されなかった塊を新しいチューブに移し、Ca2+およびMg2+を含む1:1 DMEM:PBS中の1mg/mlコラゲナーゼ溶液において、撹拌しながら室温で1時間、コラゲナーゼ消化した。コラゲナーゼ消化組織を再び70μmフィルタで濾過し、消化されなかった断片をシリンジプランジャーを使って物理的に破砕した。残った組織をDMEMですすぎ、70μmフィルタで濾過した。コラゲナーゼ消化およびエラスターゼ消化から得た細胞を合わせてから、DMEM中で3回洗浄し、培養に使用する培地中で1回洗浄した。トリパンブルー色素排除法を使って細胞の生存性を評価した後、本明細書の他の項で説明するように、細胞を脱細胞化スキャフォールドに播種した。
【0267】
新生児細胞播種
肺細胞単離と脱細胞化スキャフォールドの調製後に、単離された細胞を、培養に使用する培地に懸濁した。気道コンパートメントの播種には、バイオリアクタあたり15mlの細胞懸濁液を気管リザーバに注入し、陰圧換気で肺臓の気道コンパートメント中に細胞を移動させることによって、細胞を播種した。脈管構造の播種には、バイオリアクタあたり3mlの細胞懸濁液を肺動脈中に注入した。灌流も換気も行わずに細胞を終夜付着させた後、実験条件に応じて、灌流および/または換気を開始した。
【0268】
人工組織培養
播種後、肺臓を一晩静置培養してから、灌流または換気を開始した。灌流および換気は、実験条件に応じて変更した。培養培地は週に2回入れ換えた。換気条件については、バイオリアクタ中の空気を手作業で交換できるように毎日1回、短時間停止した時間を除けば、肺臓を絶えず換気した。人工組織の断片の培養については、一晩播種した後、スキャフォールドをバイオリアクタから取り出し、滅菌剪刀を使って小さい(1〜3mm)断片に切断した。その断片を培養用ペトリ皿に移し、表示がある場合には、後に、気液界面培養のための0.4μmフィルタインサートを含むペトリ皿に移した。
【0269】
フローサイトメトリー
細胞単離後に、細胞を緩衝液(2mM EDTAおよび0.5%ウシ血清アルブミンを含むPBS)中ですすいだ。細胞内抗原を染色するために、細胞を室温において1%ホルムアルデヒドで15分間固定し、次にPBS中の0.2%Triton-Xで透過処理した。一次抗体を、1:100の希釈度で、室温にて30分間、緩衝液中で適用した。緩衝液中で3回すすいだ後、二次抗体を1:100の希釈度で室温にて20分間適用した。細胞は、イェール大学医学部細胞選別施設(Yale School of Medicine Cell Sorting Facility)のBecton-Dickinson FACSCalibur装置で分析した。
【0270】
実験の結果を以下に説明する。
【0271】
スキャフォールドには細胞毒性はない
まず最初に、腫瘍由来肺臓上皮細胞株MLE-12を、脱細胞化肺臓スキャフォールドにおける予備培養実験に使用した。これらの実験は、スキャフォールドに細胞毒性がないことを証明するため、および脱細胞化スキャフォールドを利用する培養のためのバイオリアクタシステムの一階妥当性(first-order validity)を証明するために行った。MLE-12細胞株は、脈管構造を通した培地の灌流を伴うバイオリアクタにおける脱細胞化スキャフォールド上での10日までの培養期間中、ロバストな細胞成長を示すことが観察された。組織像を図27に示す。細胞は、3日の時点では極めて原始的な肺胞構造を形成するようだったが、その後は著しく増殖し、7日までに無制御な細胞成長を示す。腫瘍由来の細胞株であるから、これは予想される帰結である。これらの実験は、バイオリアクタおよび脱細胞化肺臓スキャフォールドの検証における予備段階であり、これにより、新鮮単離新生児肺細胞を使った以後の実験が正当化された。
【0272】
新生児肺細胞の収集
齧歯類肺臓上皮はインビトロで培養することが難しいので、また新生児ラットの肺細胞は若く比較的可塑性が高いので[Massaro et al., 1985, J Clin Invest 76:1297-305;Meyrick et al., 1982, Am Rev Respir Dis 125:468-73]、肺細胞タイプの不均質な混合物を表す多数の細胞を単離できることを含むいくつかの理由から、新生児肺細胞を選択した。
【0273】
細胞数、生存性、およびフローサイトメトリーに基づく細胞タイプの分布に基づいて、細胞単離の条件を最適化した。使用した主なマーカーは、サーファクタントタンパク質C(SPC;II型肺細胞)、アクアポリン5(AQP;I型肺細胞)、クララ細胞分泌タンパク質(CCSP;クララ細胞)、および血小板内皮細胞接着分子1(PECAM-1;内皮細胞)である。細胞単離の条件は、総合的な細胞数および生存性を最適化する反復実験の結果として選択した。酵素およびインキュベーション条件の選択は、総細胞数、トリパンブルー色素排除法およびフローサイトメトリー分析によって評価される細胞の収量および生存性に基づいて最適化した。
【0274】
「最適化された」単離レジメンの下で得られた試料肺臓単離のフローサイトメトリーデータを図28に示す。典型的な単離では、細胞の5〜10%がCCSP陽性であり、細胞の40〜60%がSPC陽性であり、細胞の2〜8%がAQP陽性であり、細胞の1〜2%がサイトケラチン14陽性であり、細胞の10〜30%がPECAM-1陽性であり、細胞の5〜10%がα-アクチン陽性である。サイトスピン調製物を使って、CCSPおよびSPCに関する染色を確認した。これらのパーセンテージの大半は予想される範囲内にあるが、I型肺細胞(AQP陽性)については、未変性肺臓におけるその割合だけに基づけば、もっと高い収量が予想されるだろう。しかし、I型肺細胞は非常にもろく、それらの多くは細胞収集プロセスに耐え抜く可能性が低い。ひと腹の子(7〜12匹)からの総細胞収量は細胞約1億個であり、生存率は75〜85%だった。
【0275】
人工肺臓培養のためのバイオリアクタ条件の予備的確認
主に、組織学的検査による細胞密度、生存性および形態、ならびにいくらかのタンパク質発現評価に基づいて、さまざまな変量および適切な条件を探究するために、実験を計画した。評価した条件を以下に簡単に述べる。
【0276】
培地選択:基礎培地と血清濃度をどちらも変化させて、いくつかの培地組成を評価した。BGJb(無血清)、10%FBSを含むDMEM、15%FBSを含むEGM-2、およびEGM-2+15%FBSとBGJbの3対1混合物を含むいくつかの培地条件を使って、上皮再集合(repopulation)、主にII型上皮を観察した。組織学的検査および免疫蛍光染色によれば、培地タイプBGJbおよびDMEM+10%FBSは、総合的上皮成長にとって優れた条件をもたらした。その結果、以後の実験にはこれらの培地を使用した。ただし本発明はいかなる具体的培地にも限定されるべきでない。これは、所望する増殖および分化を促進する任意の培地を使用することができるからである。
【0277】
灌流および換気:予備実験では灌流と換気の両方を使用した。例えば、栄養素を供給するために培地を2〜5ml/分の流速で灌流した。1回の吸息による1日1回の換気の効果も評価した。しかし、その結果得られる人工肺臓培養に有意差は検出されなかった。1日1回の換気では明確な利益が観察されなかったにもかかわらず、この最低レベルの換気は、より生理的な培養環境を与えるであろうと推定された。それゆえに、以降の実験の大部分では、培養物を灌流し、1回の吸息により、1日1回、換気した。
【0278】
脱細胞化スキャフォールドは上皮、内皮および間葉細胞の成長を支持する
以下の実験は、発生工学的(development engineered)肺臓組織にとっての、脱細胞化スキャフォールド、肺臓バイオリアクタ、および単離新生児肺細胞集団の妥当性を証明するために計画した。これらの人工組織の培養に使用した詳細な条件を表1に示す。表には、これらの条件が人工組織の成長に及ぼす効果を評価するために具体的に調べた条件も記載する。ただし本発明はこれらの条件に限定されるわけではない。むしろ、本発明には、適用可能な条件であれば、その条件がバイオリアクタとの関連で人工肺臓の生成を促進する限り、いずれも包含される。
【0279】
(表1)人工肺臓培養のためのバイオリアクタ条件

【0280】
上皮細胞再集合の証明
本明細書に提示する結果は、脱細胞化肺臓スキャフォールドへの上皮細胞の接着および増殖を証明する。これらの実験には、DMEM+10%FBS培地、2ml/分での脈管構造の灌流、1日1回の換気、非被覆(uncoated)脱細胞化スキャフォールド、および無選別新生児肺細胞集団(本明細書の他の項で説明する「最適化された」条件)を使用した。
【0281】
図29に、人工肺臓組織のH&E染色を示す。4日時点で、著しい数の、組織化され立方上皮で裏打ちされた発生中の上皮構造が観察され、8日時点では、観察されるそのような構造が少なくなって、多くの細胞がより成熟した表現型をとった。4日時点では、多くの細胞が増殖性であったが、8日時点では、観察される増殖性細胞は少なくなった(図30)。4日時点でも、8日時点でも、著しい数のアポトーシス細胞は観察されなかった(図31)。
【0282】
免疫蛍光法を使って、I型肺細胞にはアクアポリン5を、II型肺細胞にはサーファクタントタンパク質Cを、そしてクララ細胞にはクララ細胞分泌タンパク質を使用して、重要な上皮細胞マーカーの発現を証拠立てた。図33に示すように、培養物ではII型上皮細胞が一般に優勢であり、後期の時点では特にそうだった。クララ細胞は4日時点で最も高密度に観察され、8日時点では少なくなった(図32)。4日時点ではアクアポリンに関する染色も観察されたが、それより遅い時点では、観察されるアクアポリン染色が著しく減少した(図34)。
【0283】
I型上皮に関するアクアポリン染色は、培養4日の時点では、形状が立方形である細胞を表すことが観察された。これは、機能している肺臓において肺胞を裏打ちする細胞としてのそれらの通常の扁平な形態とは相容れない。さらにまた、これらの立方形細胞は、図36および図37に見られるように、しばしばSPCまたはCCSPのいずれかについても陽性に染色された。したがって、4日時点でアクアポリンを発現する細胞が、平坦な形態と他のマーカーを伴わないアクアポリンの発現とによって示唆されるであろう成熟したI型上皮である可能性は低い。
【0284】
I型肺細胞はインビボでは、肺胞上皮の局所常在前駆体細胞であるII型上皮細胞に由来する。発生中の未変性肺臓では、胎児呼吸運動がなければI型肺細胞が最終分化状態を獲得せず[Inanlou et al., 2005, Dev Dyn 233:772-82]、組織像およびTEMでは形状が立方形のままである[Inanlou et al., 2005, Histol Histopathol 20:1261-6]。
【0285】
I型肺細胞分化の欠如は、定期的に換気されないこれらの培養物では、意外なことではなかった。アクアポリンを発現する細胞は、おそらく、I型上皮に完全には分化していなかった局所前駆体細胞から生じるのだろう。したがって脱細胞化スキャフォールドは肺上皮の付着および増殖を支持することができる。II型上皮、ならびにクララ細胞、および完全に分化したI型上皮細胞の前駆体である可能性が高い細胞のロバストな成長が観察された。本発明者らは、これらの所見を、脈管構造を通した培地灌流下に、呼吸運動を時折行うだけで観察している。
【0286】
上皮前駆細胞再集合
脱細胞化スキャフォールドにおける2タイプの肺上皮前駆細胞の成長が観察された。CCSPとSPCについて二重に陽性である細胞は、気管支肺胞幹細胞と呼ばれる局所前駆細胞であると報告されており、これはクララ細胞にもII型肺細胞にも分化することができ、気管支肺胞管接合部に見いだされる[Lane et al., 2007, Regenerative Medicine 2:407-15;Kim et al., 2005, Cell 121:823-35]。図35に、これらの細胞の予想される生理学的位置である末端細気管支の外観と合致する構造中に見いだされる、そのような二重陽性細胞を示す。
【0287】
基底細胞は肺気道の局所幹細胞であり、それらは円柱上皮の下に存在して、近位気道の上皮にとって再生性細胞源として役立つ。図36は、基底細胞マーカーであるサイトケラチン14について陽性である細胞を示している。比較のために、未変性肺臓を図36Aに示す。さらにまた、サイトケラチン14とCCSPに関する二重染色を図37に示す。図37は、クララ細胞が気道を裏打ちし、基底細胞がそれらの下に横たわっていることも示しており、これらは、その正常な解剖学的位置に合致している。これらの細胞は、時には、未変性肺臓におけるそれらの位置(図36B)と合致して、より大きい気道構造の下に見いだされるが、大きな気道には関係しないと思われるクラスター中にも見いだされる(図36C)。
【0288】
上皮細胞および内皮細胞の成長に加えて、間葉細胞も、脱細胞化肺臓スキャフォールドに再集合することができる。図38は、平滑筋および筋線維芽細胞を染色するα-アクチンの免疫蛍光染色を示す。これらの人工組織は、上皮成長にとって有利であることが示された同じ条件下で培養された。間葉細胞が、未変性肺臓におけるそれらの位置と合致して、発生中の上皮構造の下および間に位置することも見いだされた。したがって、本明細書に提示する結果は、脱細胞化スキャフォールドが間葉細胞の成長にとっても適切な基質であることと、新生児肺細胞の集団には生きた間葉細胞が含まれていたことを証明している。
【0289】
培地組成が上皮分化に及ぼす効果
培地タイプは、細胞の成長と分化に、したがって人工肺臓組織の発生に、著しい影響を持つことができる。これらの相違の一部をより詳しく調べるために、上皮分化に関して無血清培地(BGJb)と血清含有培地(DMEM+10%FBS)を使って人工肺臓培養の成長を比較した。これらの実験では、まず最初に、細胞をDMEM+10%FBS中のスキャフォールド上に播種し、この培地中で2日間培養した後、BGJb(無血清)に4日間かけて徐々に移行し、最後に純粋なBGJb培地中で2日間培養した。無血清培地への移行は、サーファクタントの発現に対する実質的な影響を引き起こした。無血清培地は、DMEM+10%FBSと比較して、より多くのサーファクタント(SPC)の頂端部発現につながることが観察された(図39Cおよび39D)。これは、無血清培地(BGJb)では、ウェスタンブロット上で、サーファクタントの発現が有意に増加することと一致する(図40;「DMEM」および「BGJb」と記したレーンを比較のこと)。さらにまた、サーファクタントの形態が、未変性肺臓と、はるかによく合致していた(BGJb培地では大半のサーファクタントが21kDaプロSPC形として認められた)。
【0290】
さらにまた、無血清培地への移行は、図43Fおよび43Eに記載の免疫蛍光によって認められるCCSP発現の現象につながる。どちらの培地タイプでも、散在性のCCSP発現が発生中の上皮構造の管腔に観察された。これらの培養は小さい組織切片で行われたので、この表現型は、灌流または換気の欠如によるものであったと考えられる。
【0291】
バイオリアクタにおける肺臓発生に気液界面が及ぼす効果
気液界面を作り出すために、人工肺臓をまず、培地による換気下で4日間培養して、細胞を付着させ増殖させた。培養の最後の4日間は換気を培地から濾過大気に切り換えた。
【0292】
空気による換気は、気道上皮に重度の損傷を引き起こすと共に、一部の肺胞壁に破壊的な変化を引き起こした(図19)。そこで、空気換気によって未変性肺臓に引き起こされる損傷を減少させる努力として、空気換気に使用する一回換気量を、液体または空気呼吸に関する約2.0〜2.5mlという先の値から、約50%減少させた。
【0293】
バイオリアクタにおける空気換気の存在に起因する細胞付着または細胞形態の変化は観察されなかった。しかし、バイオリアクタで培養された人工肺臓における空気による換気は、I型上皮の分化マーカーであるアクアポリンの発現の誘導につながることが観察された。これは、典型的にはサーファクタントタンパク質C(II型上皮を示す)についてのみ染色される実質の細胞でも、発生中の上皮構造における立方形細胞の偶発的染色でも認められた。観察された染色パターンは、アクアポリン発現細胞の大半がSPCも発現する可能性が極めて高いことを示している。図42Bからわかるように、実質中の細胞の事実上全てがSPCを発現し、一方、実質細胞のサブセットはAQPを発現する(図41A)。
【0294】
これらの知見は、気液界面がII型上皮からI型上皮への分化を誘導していることを、強く示唆している。II型上皮は、I型上皮にとって公知の局所前駆細胞であるので、この分化は意外なことではない[Adamson et al., 1974, Lab Invest 30:35-42;Fehrenbach, 2001, Respir Res 2:33-46]。そのうえ、図40に示すように、空気換気人工肺臓では、サーファクタントタンパク質Cの発現量の減少が観察された(レーン「ALI」を、培地で換気(「Vent」)および灌流(「Perf」)した肺と比較されたい)。これは、II型上皮からI型上皮への分化とも合致する。さらにまた、図42に示すように、絨毛上皮細胞の数の増加も観察された。これは空気界面を導入した結果である可能性が高い。気道上皮をインビトロで培養すると、液体から空気界面への細胞の移行が絨毛発現を誘導する[You et al., 2002, Am J Physiol Lung Cell Mol Physiol 283:L1315-21;Davidson et al., 2004, J Cyst Fibros 3 Suppl 2:59-62]。そのうえ、気液界面の欠如は絨毛形成の減少につながり得る[Ostrowski et al., 1995, Exp Lung Res 21 :957-70;Yeh et al., 2007, Laryngoscope 117:1439-44]。したがっていくつかの例では、空気界面が、インビトロでの肺臓組織の再生に、重要な影響を持つ。
【0295】
灌流および換気が人工肺臓組織における上皮構造の発生に及ぼす効果
灌流および換気は、バイオリアクタにおける肺臓組織の培養に著しい影響を及ぼすことができる。換気は、I型およびII型肺細胞の分化を含む肺臓上皮発生にも著しい影響を持つ[Inanlou et al., 2005, Histol Histopathol 20:1261-6;Inanlou et al., 2005, Dev Dyn 233:772-82;Inanlou et al., 2005, Dev Dyn 232:43-54]。そこで、バイオリアクタにおける8日間の培養中に灌流および換気が人工肺臓発生に及ぼす効果を比較した。これらの実験では、条件を、他の項で論じる換気実験時に利用したものと同じ条件とし、DMEM+10%FBSの培養培地、非被覆スキャフォールド、および無選別新生児肺細胞集団を使用した。ただし、培養物は、脈管構造を通して2ml/分で灌流するか、培地を使って1吸息/分で連続的に換気した。
【0296】
図50に示すように、上皮構造を形成する細胞の多くが、CCSPに関して陽性に染色される。換気した培養物ではCCSPに関する染色が観察されたが、細胞はより平坦な形態を獲得した。これは、換気が、肺胞上皮(I型またはII型肺細胞)に向かって分化するようにCCSP発現細胞を誘導していることを示すのだろう。
【0297】
換気が存在しない場合は、図43のH&E染色で目に見えるエオシン好性の物質、したがってタンパク質性である可能性が高い物質で、発生中の上皮構造の管腔が満たされた。この物質は、図45に見られるように、CCSPについても陽性に染色された。このCCSPの蓄積は、これらの上皮構造を裏打ちするクララ細胞がCCSPを産生していることを示す。さらにまた、換気によるこの物質の除去は、気道樹が依然としてインタクトであって流体を通すことを示し、これら発生中の上皮構造が気道樹の一部であって、マトリックス内でランダムには増殖しないことも示唆する。
【0298】
図46に示すように、灌流と換気は、SPCの発現に対して、有意な効果を持たなかった。細胞の大半は、換気条件下でも、灌流条件下でも、SPCについて陽性だった。
【0299】
実施例5:人工肺臓組織における内皮発生
脱細胞化スキャフォールドが人工肺臓内皮の成長を支持することができるかどうかを決定するために、また、いくつかの具体的因子が人工内皮の発生に及ぼす効果を、血管コンパートメントと気道コンパートメントの間の機能的な内皮障壁の形成に影響を及ぼすそれらの因子の能力に焦点を絞って評価するために、以下の実験を計画した。
【0300】
これらの実験で使用した材料および方法を以下に説明する。
【0301】
材料および方法
スキャフォールドの調製
脱細胞化スキャフォールドは、本明細書の他の項で説明するように調製した。
【0302】
新生児細胞の単離と播種
新生児ラット肺細胞は、本明細書の他の項で説明するように単離した。細胞を、本明細書の他の項で説明するように、スキャフォールドに播種した。
【0303】
内皮細胞培養
ラット肺臓微小血管内皮細胞をVEC Technologies(ニューヨーク州レンセリア)から入手し、フィブロネクチン被覆(約1μg/cm2、Gibco)組織培養器上、10%FBSと追加成長因子(supplemental growth factors)(VEC Technologies)を含むMCDB-131完全培地中で成長させた。
【0304】
人工内皮培養のための最適化された条件
スキャフォールドを1mgのフィブロネクチン(Gibco)で被覆し、37℃のPBS 60ml中、脈管構造を通して灌流した後、PBSおよび培地ですすいだ。培養0日目と培養2日目または3日目の2回、各回800万〜1000万個のラット肺臓微小血管ECを播種した(2回の播種のそれぞれについて、肺臓1つあたり2つのT150培養フラスコを使用した)。0.25%トリプシン(Gibco)を使って細胞を組織培養プレートからトリプシン処理し、細胞塊を除去するために40μmフィルタで濾過し、約3mlの培地に入れた単回ボーラス注射として、肺動脈に注入した。細胞を1時間接着させた後、脈管構造を通して約1.5ml/分で灌流を開始した。1〜2時間後に、7〜10日の培養期間の残りの部分は、灌流量を3ml/分に増加させた。培地は3〜4日ごとに換えた。
【0305】
免疫蛍光法
本明細書の他の項で説明するように、組織試料を調製し、染色した。
【0306】
透過型電子顕微鏡法(TEM)
本明細書の他の項で説明するように、試料を調製し、分析した。
【0307】
微粒子保持
巨大高分子程度のサイズを持つ小さな粒子に対する全ラット肺臓の透過性を評価するために、アッセイを開発した。このアッセイでは、気道-血管障壁を横切って起こるFITC標識デキストラン溶液の漏出を定量した。2,000,000Daの分子量を持つFITC標識デキストランをSigma(ミズーリ州セントルイス)から入手した。未変性肺臓および0.025%トリプシンで2分間処理した未変性肺臓の透過性を測定することによってアッセイの検証を行った。肺臓をヘパリン化PBSで灌流し、通常のバイオリアクタカニューレに接続した。ベースライン洗浄液試料を得た後、トリプシン処理肺臓は、PBS中の0.025%トリプシン10mlで灌流し、室温で2分間留まらせてから、10mlのPBSですすいだ。FITC標識デキストラン溶液(1mg/ml)を肺動脈に注入し、次に20mlのPBSでフラッシングした。次に、2つの洗浄液試料を直ちに、気管から連続して採取した。蛍光プレートリーダーを使って蛍光を測定し、データを標準曲線に当てはめた。脱細胞化肺臓または人工肺臓で行う場合は、マイクロスフェアアッセイ(3.2.9章参照)の場合と同様に、このアッセイを気道によって行った。したがって、FITC-デキストランを気道に注入し、脈管構造をPBSでフラッシングした。
【0308】
実験の結果を以下に説明する。
【0309】
細胞供給源:市販されているラット肺臓微小血管EC源を使用した。これらの細胞を、フィブロネクチン被覆したスキャフォールド中に播種して、EC特異的培地の存在下で培養すると、図47に示すように、内皮細胞の実質的成長が観察された。
【0310】
内皮スクリーニング実験の要約:本明細書の他の項で論じるスクリーニング実験により、人工内皮培養に適合する一組の条件を同定することができた。帰結は、主に、細胞生存性に関する組織学的検査と、免疫蛍光法でのPECAMの発現とによって評価した。これらのパイロット試験の結果、個別の条件が人工肺臓内皮に及ぼす影響を体系的に評価することができるように、スキャフォールド内部での内皮細胞成長を可能にする一組の条件が得られた。
【0311】
人工内皮の培養に適していると同定された条件は、次のとおりだった:精製されインビトロで拡大されたラット肺臓微小血管EC集団の使用;フィブロネクチン被覆スキャフォールド;およびEC特異的培地(10%FBSおよび追加成長因子を含むMCDB-131)の使用。
【0312】
FITC-デキストラン透過性アッセイの検証
巨大高分子程度のサイズを持つ小さな粒子に対する全ラット肺臓の透過性を評価するためのアッセイを開発した。このアッセイでは、気道-血管障壁を横切って起こるFITC標識デキストラン溶液の漏出を定量した。このアッセイは、培養の過程で繰り返し使用することができ、細胞培養に害を及ぼさない材料を使用し、全肺臓の透過性の測定をもたらす。さらにまた、アッセイを固定直前に行えば、FITC-デキストランを、抗FITC抗体を使って組織切片上で同定することができるだろう。
【0313】
FITC標識デキストランは2,000,000Daの分子量を持つ。単分散デキストランの場合、ストークス-アインシュタイン半径(nm)は、rs=0.0488(MW)0.437によって分子量と関連づけられる[Venturoli et al., 2005, Am J Physiol Renal Physiol 288:F605-13;Oliver et al., 1992, J Am Soc Nephrol 3:214-28]。2MDaデキストランの場合は、これにより、27.7nmという半径が得られる。未変性肺臓と、脈管構造を希釈トリプシンで短時間灌流することによって「漏出しやすく」した未変性肺臓の透過性を評価することによって、このアッセイを検証した。FITC標識デキストラン溶液を肺動脈に注入した後、食塩水でフラッシングした。次に、2つの洗浄液試料を直ちに、気管から連続して採取した。図48に示すように、肺臓の透過性は、基底膜への内皮付着が破壊されることにより、予想どおり、トリプシン処理によって増加する。しかし、未変性肺臓でも、このアッセイでは測定可能な漏出を与えた。この血管透過性の劣化は、動物の屠殺とデキストラン注入との間の遅延、ならびに肺臓組織の取扱の結果である。
【0314】
脱細胞化肺臓または人工肺臓で行う場合は、マイクロスフェアアッセイの場合と同様に、このアッセイを気道によって行った。したがって、FITC-デキストランを気道に注入し、脈管構造を食塩水でフラッシングした。血管コンパートメントに移動したデキストランを漏出として測定した。脱細胞化肺臓または人工肺臓では、組織が流体に対して高い透過性を持ち、気道洗浄後に還流試料を得ることができないので、アッセイをこの方法で行った。したがって、デキストランを単回ボーラス洗浄液として気道に注入し、脈管構造をフラッシングして、気道-血管障壁を横切って起こるFITC-デキストランの流出を測定した。
【0315】
人工肺臓内皮に対する還流と換気の効果
具体的条件が人工内皮組織の発生に及ぼす効果を、機能的内皮障壁の形成に焦点を絞って評価するために、以下の実験を計画した。灌流しながら人工肺臓内皮を培養することの効果と換気しながら人工肺臓内皮を培養することの効果を比較した。換気と灌流の両方を、内皮細胞の生存および増殖に関して評価し、透過型EMを使って細胞間結合の形成についても評価した。
【0316】
図48の組織像に示されるように、灌流は人工肺臓内皮の成長を実質的に改善することが観察された。また、図49に示すように、またH&E組織像におけるそれらの外観不良と合致して、換気では、より多くのアポトーシス細胞が観察された。
【0317】
また、灌流培養物と換気培養物を、細胞間結合の存在について、透過型EMおよびVE-カドヘリン染色を使って分析した。内皮細胞観の密着結合は、隣接する細胞を互いに堅く連結し、よってこれらの傍細胞間隙間での流体の動きを阻害するので、障壁機能の重要な手段である[Majno et al., 1961, J Biophys Biochem Cytol. 11:571-605]。これらの細胞結合が弱いか存在しない場合は、脈管構造外への流体の漏出が起こって、それが肺水腫を引き起こし得る[Orfanos et al., 2004, Intensive Care Med 30:1702-14;Maniatis et al., 2008, Vascular Pharmacology 49:119-33]。
【0318】
図50に示すように、TEMを使って、灌流人工組織における細胞間結合を観察した。全ての細胞が密着結合形成を示したわけではなかった。
【0319】
VE-カドヘリンに関する免疫蛍光法を使って細胞間結合形成を評価した。図51に示すように、灌流人工肺臓内皮には、VE-カドヘリンに関してロバストな染色が見いだされた。換気組織は、組織像およびTEMでのそれらの外観不良ゆえに、VE-カドヘリンについて染色しなかった。
【0320】
人工肺臓内皮の障壁機能の評価
血管床と気腔の間に機能的障壁を形成する人工肺臓内皮の能力を評価するために以下の実験を計画した。これは、肺胞への流体漏出を減少させ、よってガス交換を可能にするために重要であり、人工肺臓組織にとっての目標の重要な構成要素である。肺臓内皮によって提供される障壁機能を評価するために、透過性アッセイを使って、気腔から脈管構造への小さい(55nm)FITC-デキストラン粒子の移動を測定した。このアッセイは、本明細書の他の項で説明するように開発され、検証された。
【0321】
このアッセイでは、FITC-デキストランを気道コンパートメントに注入し、脈管構造を食塩水でフラッシングすることにより、肺胞-血管障壁を横切って漏出した量を測定した。脱細胞化スキャフォールドの場合、そのような小粒子に対する障壁機能は本質的に存在せず、事実上全て(98.4%)のデキストランが肺胞から脈管構造中に移動して、血管すすぎ液と共に回収された。これに対し、未変性肺臓は、同様に処理した場合に、12.9%の漏出を示す(図52)。
【0322】
灌流された人工肺臓内皮組織では、気道コンパートメントにおける30%までのデキストランの保持が、7〜10日の培養期間後に証明された。換気内皮組織は87%の透過性を示した。
【0323】
灌流人工肺臓内皮に関するこれらの知見は、特に蛍光免疫法によるロバストなVE-カドヘリン発現(図51)およびTEMによる細胞間結合形成という知見と合わせて考えると、人工内皮組織における機能的内皮障壁の形成が起こったことを示している。
【0324】
図63は人工組織の極限引張強さを図示する図表である。未変性肺臓と脱細胞化肺臓の強さも示されている。この図は、人工組織の極限引張応力が未変性肺臓とも脱細胞化マトリックスとも同等であることを示している。したがって、組織工学的肺臓の機械的性質は再細胞化プロセス後も維持される。
【0325】
実施例6:肺臓細胞治療
本明細書に提示する結果は、哺乳動物における肺臓細胞治療を達成するための脱細胞化肺臓組織の使用を証明する。一般に、工程には、気管の脱細胞化、脱細胞化気管組織内の細胞外マトリックス構成要素の検出、脱細胞化気管マトリックスにおけるヒト気管支(大気道)および小気道肺上皮細胞の培養、所望の遺伝子によるヒト肺上皮細胞の遺伝子治療、および点滴注入による肺臓へのヒト肺上皮細胞の点滴注入が含まれ、レシピエント肺臓における細胞の付着および生存が確認される。
【0326】
気管の脱細胞化
ブタ気管を収集し、血液を除去するためにPBS中ですすいだ。断片を切断し、10%中性緩衝ホルマリンで固定し、パラフィンに包埋し、5mm切片に切断した。残りを5つの環に切断し、CHAPS緩衝液(pH13.5)中で撹拌しながら、2時間、4時間および8時間の時点でCHAPS緩衝液を交換して、2〜24時間インキュベートした。表示した時点において、組織をCHAPS緩衝液から取り出し、PBSですすいだ。断片を切断し、10%ホルマリンで固定し、脱細胞化を確認するための組織学的分析用に加工した。
【0327】
CHAPS緩衝液中で4〜8時間インキュベートして調製した脱細胞化気管は、コラーゲンマトリックスを維持しており、細胞の大半が組織から除去されている(軟骨層を除く)ことが観察された。図55参照。以下の実験は、撹拌しながら6時間のCHAPSインキュベーションを行うことによる脱細胞化気管の調製に基づいた。
【0328】
脱細胞化の前後に気管中の細胞外マトリックスを検出するために、これらの実験を計画した。簡単に述べると、組織の断片を未変性気管および脱細胞化気管から切り取り、10%ホルマリンで固定した。5μmパラフィン包埋切片をH&EおよびMassonのトリクロームで染色した。パラフィン包埋切片を、コラーゲン(COL)、フィブロネクチン(FN)およびラミニン(LN)を含む細胞外マトリックスタンパク質についても免疫染色した。切片を脱パラフィンし、再水和し、抗原回復(プロテイナーゼKを使用)を行い、ブロッキングした。自家製のウサギ抗ヒトFN、COL I、COL III、COL IV、COL VまたはLN抗体を切片に適用し、次にAlexa Fluor 546コンジュゲートヤギ抗ウサギIgGを適用した。スライドをDAPIで対比染色した。未変性気管は、COL I、COL III、COL Vについて陽性に染色されるが、他のECMタンパク質については陽性に染色されないことが観察された。脱細胞化気管は、未変性気管に見られる3タイプのCOLを全て含有していた。図56参照。これらの結果は、脱細胞化気管組織が、インビトロで気道上皮細胞の接着、成長および分化を支持することを示唆している。
【0329】
脱細胞化気管組織上での気道上皮細胞の成長
脱細胞化組織上で細胞を成長させるために、次の一組の実験を計画した。簡単に述べると、ヒト気管支/気管上皮細胞(NHBE)を気管支上皮成長培地(BEGM)中で培養した。脱細胞化気管(6時間CHAPS緩衝液インキュベーション)を、滅菌PBSで十分に(少なくとも24時間は)すすいだ。軟骨層(ならびに外膜層)をはがして、後続の細胞播種のために気管粘膜および粘膜下層を残した。組織を約5×5cm2のサイズに切断し、6ウェルプレート中のトランスウェルインサート(孔径0.4μm)上に、上皮面を上向きにして置いた。
【0330】
細胞を脱細胞化気管組織に次のように播種した。50μlの細胞(密度3.5×106細胞/ml)を脱細胞化気管組織の上皮面に加え、37℃で30分間インキュベートした。次に、新鮮な培養培地をトランスウェルインサートの上側と下側の両方に加え、細胞をさらにインキュベートした。表示した時間で、組織をトランスウェルから取り出し、10%ホルマリンで固定した。パラフィン包埋した5μm切片をH&Eで染色した。脱細胞化気管はNHBEの接着と成長を支持することが観察された。図57参照。
【0331】
導入遺伝子でトランスフェクトされたヒト小気道上皮細胞(SAEC)を脱細胞化組織上で成長させるために、次の一組の実験を計画した。簡単に述べると、ヒトSAECに、GFPレトロウイルスを6時間トランスフェクトした。細胞を6ウェルプレート中の脱細胞化気管組織に次のように播種した。50μlの細胞(密度5×106細胞/ml)を脱細胞化気管組織の上皮面(約0.5×1.0cm2)に加え、37℃で30分間インキュベートした。次に、新鮮な培地を加え、細胞をさらにインキュベートした。脱細胞化気管はSAECの接着と成長も支持することが観察された。図58参照。さらにまた、脱細胞化気管は、関心対象の遺伝子がトランスフェクトされたヒト肺上皮細胞を培養することの原理証明として、GFPがトランスフェクトされたSAECの接着および成長も支持した。
【0332】
気道上皮細胞の遺伝子治療
肺臓細胞の遺伝子治療に脱細胞化組織を使用することの実現可能性を決定するために、次の一組の実験を計画した。遺伝子改変は次のように行った。Phoenixパッケージング細胞株を使ってEGFP(強化GFP)レトロウイルス上清を調製した。EGFP DNAをLZRSpBMNベクターに挿入した。NHBEおよびSAECを80%コンフルエントを超えるまで成長させた。感染当日に細胞を数回すすぎ、ウイルス上清(8μg/mlポリブレンを含有するもの)を37℃で6時間接種した。細胞を数回すすぎ、新鮮培地中で一晩インキュベートした。次に、フローサイトメトリーを使ってGFPについて分析した。感染細胞はGFPについて陽性に染色されることが観察された。例えばNHBE細胞は非感染細胞と比較してGFPについて18.5%の陽性染色を示した。SAEC細胞は、非感染細胞と比較してGFPについて16%の陽性染色を示した。本明細書に提示する結果は、レトロウイルスを使ってこの培養系でヒト肺上皮細胞に関心対象の遺伝子を感染させ得ることを証明している。しかし、導入遺伝子を送達するための他の手段も本発明には包含される。例えば、レンチウイルス系を使用することの実現可能性を調べるために、次の一組の実験を計画した。
【0333】
GFPレンチウイルスによる感染は次のように行った。EGFPレンチウイルス上清を無血清培地中の293T細胞で調製した。CMVプロモーターを持つpSicoRベクターにGFP DNAを挿入した。NHBEを1×105細胞/ウェルの密度で6ウェルプレート上に播種し、37℃で1日インキュベートした(80〜90%コンフルエント)。感染当日に細胞を数回すすいでから、ウイルス上清(新鮮BEGMで1:3希釈したもの)(8μg/mlポリブレンを含有するもの)を37℃で6時間接種した。細胞を数回すすいでから、新鮮培地と共にさらに2日間インキュベートした。細胞を4%パラホルムアルデヒドで30分間固定し、顕微鏡で調べた。図59参照。結果を表2に要約する。
【0334】
(表2)
【0335】
本明細書に提示する結果は、GFPレンチウイルスに感染したNHBEが6時間後に明らかな形態変化を示さなかったことを証明している。6時間の1回感染により、フローサイトメトリーで評価して50%を超えるGFP陽性細胞が生じた。SAECの感染効率も50%を超えているようだった。本明細書に提示する結果は、所望する任意の遺伝子を使って遺伝子改変肺臓細胞を生成させ得ることを証明している。
【0336】
マウス肺臓への気道上皮細胞の注入
哺乳動物の肺臓中に気道上皮細胞を注入することの実現可能性を決定するために、次の実験を計画した。簡単に述べると、5μmマイクロスフェアとPBSの1:1混合物100μlをC57BL/6Jマウス、雌、約10週齢(Jackson Lab)に、気管を通して注入した。マウスは注入後数分間生き続けた。肺臓を直ちに収集し、ホルマリンで固定した。5μmパラフィン包埋切片をH&Eで染色した。この試験は、気道への点滴注入によって送達された細胞サイズの粒子が哺乳動物の肺臓内に検出されるかどうかを決定するために、最初の実現可能性試験として行われた。マイクロスフェアが注入されたマウス肺臓のH&E像からの結果は、かなりの数のマイクロスフェアがマウス肺臓の全ての葉に存在することを証明した。図60参照。これらの結果は、気管アプローチを使った注入を細胞点滴注入に使用できることを証明している。
【0337】
次の一組の実験には、GFP標識細胞を外植されたマウス肺臓に注入することを含めた。簡単に述べると、細胞をGFPレトロウイルスに6時間感染させた。C57BL/6Jマウスを安楽死させ、気管を露出し、近位端を縫合糸で結紮した。100μlのGFPレトロウイルス感染SAECを、マウスの肺臓に、気管を通して注入した(続いて、細胞を押し込むために200μlの空気を注入した)。気管の遠位端を結紮した。肺臓を外植し、直ちに10%ホルマリンで固定した。5μmパラフィン包埋切片を脱パラフィンし、再水和し、抗原回復(10mMクエン酸緩衝液pH6を使用)を行い、透過処理し(Triton Xを使用)、ブロッキングした。ウサギポリクローナル抗GFP抗体(Abcam製)を切片に適用し、次にAlexa Fluor 555コンジュゲートヤギ抗ウサギIgGを適用した。スライドをDAPIで対比染色した。外植された肺臓にGFP陽性細胞が見いだされることが観察された。図61参照。本明細書に提示する結果は、肺臓内への細胞の注入の成功を証明すると共に、導入遺伝子(GFP)を形質導入されたヒト上皮細胞が肺上皮に付着したことを証明している。
【0338】
マウス肺臓へのGFP標識細胞の注入の実現可能性を決定するために、次の一組の実験を計画した。簡単に述べると、マウス(C57BL/6J、雌、約10週齢)を麻酔し、気管を露出した。100μlのGFPレトロウイルス感染ヒト気道上皮細胞を気管を通して肺臓に注入した(続いて200μlの空気を注入した)。図62参照。マウスを6時間〜3日間回復させた。収集当日に、血液を除去するために肺動脈を通して肺臓をPBSで灌流し、次に気管を通して10%ホルマリンで灌流し、切り離した。収集した肺臓をホルマリン中でさらに4時間固定した。5μmパラフィン包埋切片を、本明細書の他の項で論じるように、GFPについて染色した。GFP陽性ヒト気道上皮細胞(NHBEとSAECの両方)が、気道内への点滴注入後、何日間も、マウス肺臓に見いだされることが観察された。図63参照。これは、ヒト上皮細胞(これは培養下で成長させたものであり、関心対象の導入遺伝子を形質導入されたものである)が、レシピエント哺乳動物の肺臓中に送達され、レシピエント肺臓に付着して、生き残り、増殖できることを示している。
【0339】
実施例7:脱細胞化人工肺臓の埋植
生きているラットレシピエントに脱細胞化人工肺臓を埋植することの実現可能性を示すために、この実験を計画した。先の実施例に従って脱細胞化人工ラット肺臓を調製した。成体雄ラットを、ケタミンとキシラジンの腹腔内注射で麻酔した。次に、ラットの気管に挿管し、麻酔を維持するためにフォーレンと混合した100%酸素で換気した。滅菌条件下で、胸骨正中切開によって胸郭を開いた。肋骨を両側に後退させて、正常に膨らんでいる肺臓と拍動している心臓を暴露した。全身ヘパリン化後に、未変性左肺臓をまとめて(in toto)切除した。次に、手術用顕微鏡下で、脱細胞化人工肺臓を、レシピエントの肺動脈、肺静脈および左主気管支に、10-0縫合糸を使って吻合した。
【0340】
肺動脈および肺静脈上の血管鉗子を取り除いた後、血管は人工肺臓を正常に灌流することがわかった。また、埋植された人工肺臓は容易に換気され、固有の未変性右肺と同様に拡張および収縮を繰り返した。図64に、換気サイクル中の拡張時と収縮時における、埋植された人工肺臓の写真を示す。したがって、本明細書に記載の技法を使って作製された人工肺臓は、哺乳動物に埋植可能であると共に、器官全体を通る灌流および気道の容易な換気を可能にするという観点から機能的である。埋植期間の全体にわたって、埋植された人工肺臓からの出血または空気漏出の証拠はなかった。
【0341】
本明細書で言及するどの特許、特許出願および刊行物の開示も、参照によりそのまま、本明細書に組み入れられる。
【0342】
本発明を具体的態様について開示したが、当業者が本発明の真の要旨および範囲から逸脱することなく本発明の他の態様および変形を考案し得ることは、明らかである。本願の特許請求の範囲は、そのような態様および等価な変形を全て包含すると解釈されるものとする。
【背景技術】
【0001】
発明の背景
毎年400,000人のアメリカ人が肺臓疾患で死亡する。さらに憂慮されることに、他の主要疾患カテゴリについては死亡率が減少しつつあるのに(心臓疾患、がんおよび脳卒中)、肺臓疾患による死亡率は増加しつつある。嚢胞性線維症、気腫/COPD、および特発性肺線維症を含むいくつかの肺臓疾患については、肺臓移植(lung transplantation)が今なお唯一の根治的処置である。しかし、肺臓移植後の患者生存率は、5年時点で50%、10年時点で24%しかない[Mondrinos et al., 2008, Tissue Eng 14:361-8(非特許文献1)]。したがって、移植に使用することができる人工(engineered)肺臓組織の開発が、強く求められている。人工肺臓組織の利点の一つは、患者自身の細胞を使って組織を成長させることができ、そうすることによって、現在の肺臓移植で要求されるような強い免疫抑制の必要を回避することができるという点である。免疫抑制は移植された器官の拒絶を防ぐために必要であるが、感染、悪性疾患、腎機能障害、心血管障害、および神経障害を含む広範な問題につながり得る[Pietra et al., 2000, J Clin Invest 106:1003-10(非特許文献2);Christie et al., 2009, J Heart Lung Transplant 28:1031-49(非特許文献3)]。
【0002】
組織工学は、細胞学的、分子学的、技術的および医学的進歩を組み合わせて埋植(implantation)に適した代用組織を作出しようとする成長分野である。前途有望な研究が、血管、膀胱、心臓弁、および心臓組織を含むさまざまな組織について行われている[Nichols et al., 2008, Proc Am Thor Soc 5:723-30(非特許文献4);Satchell et al., 2004, J Am Soc Nephrol 15:566-74(非特許文献5);Atala et al., 2006, Lancet 367:1241-6(非特許文献6);Orfanos et al., 2004, Intensive Care Med 30:1702-14(非特許文献7)]。しかし、肺臓は、実験室で工学的に操作(engineer)することが困難な組織である。呼吸の機械的圧力に耐えることができ、内皮、上皮および間葉細胞の成長を支持することができ、しかも、極めて異なっているが密接に近接している2つのコンパートメント間のガス交換のための手段を提供する複雑なマトリックスを、肺臓は必要とする。
【0003】
臨床現場における潜在的な患者使用の他に、人工肺臓組織は、肺の生物学および生理学の広範囲にわたる種々の重要な局面を研究するために、実験室で使用することもできる。インビトロ三次元肺臓培養モデルはごくわずかしかない[Vandenbroucke et al., 2008, Ann N Y Acad Sci 1123:134-4(非特許文献8)]。そのうえ、肺の内皮および上皮細胞は他の多くの細胞タイプよりも実験室で培養することが困難であり[Malda et al., 2004, Biomaterials 25:5773-80(非特許文献9);Reichenspurner, 2005, J Heart Lung Transplant 24:119-30(非特許文献10)]、肺の前駆細胞および幹細胞生物学の分野は進展が比較的遅かったBlaisdell et al., 2009, Stem Cells 27:2263-70(非特許文献11);Muratore et al., 2008, J Surg Res 155(2):225-30(非特許文献12)]。したがって、当技術分野では、未変性(native)肺環境の重要な特質を再現するインビトロ肺臓組織の開発が必要とされている。本発明は当技術分野におけるこの必要を満たすものである。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】Mondrinos et al., 2008, Tissue Eng 14:361-8
【非特許文献2】Pietra et al., 2000, J Clin Invest 106:1003-10
【非特許文献3】Christie et al., 2009, J Heart Lung Transplant 28:1031-49
【非特許文献4】Nichols et al., 2008, Proc Am Thor Soc 5:723-30
【非特許文献5】Satchell et al., 2004, J Am Soc Nephrol 15:566-74
【非特許文献6】Atala et al., 2006, Lancet 367:1241-6
【非特許文献7】Orfanos et al., 2004, Intensive Care Med 30:1702-14
【非特許文献8】Vandenbroucke et al., 2008, Ann N Y Acad Sci 1123:134-4
【非特許文献9】Malda et al., 2004, Biomaterials 25:5773-80
【非特許文献10】Reichenspurner, 2005, J Heart Lung Transplant 24:119-30
【非特許文献11】Blaisdell et al., 2009, Stem Cells 27:2263-70
【非特許文献12】Muratore et al., 2008, J Surg Res 155(2):225-30
【発明の概要】
【0005】
発明の概略
本発明は、細胞成長を支持する能力を有する脱細胞化組織を提供する。好ましくは、脱細胞化組織は、脱細胞化前の対応する天然組織の特徴を示す。より好ましくは、組織は肺臓である。
【0006】
ある態様では、脱細胞化組織が、他の点では同一な脱細胞化前の組織の形態と実質的に類似する形態を示す。
【0007】
別の態様において、請求項1の脱細胞化組織は、対応する天然組織の細胞外マトリックスを保っており、当該細胞外マトリックスは、実質的にインタクト(intact)な外面を含む。
【0008】
別の態様では、免疫原性マーカーが脱細胞化組織から実質的に除去されている。
【0009】
ある態様では、脱細胞化組織が、対応する天然組織の機械的性質と実質的に類似する機械的性質を示す。
【0010】
本発明は、三次元スキャフォールド(scaffold)と細胞の集団とを含む組成物を提供する。好ましくは、組成物は、肺臓細胞の分化状態を支持し維持する能力を有する。
【0011】
ある態様では、三次元スキャフォールドが脱細胞化組織である。
【0012】
別の態様では、組成物が、インタクトな気道樹および血管網を示す。
【0013】
別の態様では、細胞の集団が幹細胞を含む。
【0014】
別の態様では、細胞の集団が上皮細胞および内皮細胞を含む。
【0015】
別の態様では、細胞が遺伝子改変されている。ある態様では、細胞がCFTR遺伝子を発現するように遺伝子改変されている。
【0016】
ある態様では、組成物が、肺胞上皮細胞の分化状態を支持し維持する能力を有する。
【0017】
別の態様では、スキャフォールドが、フィブロネクチン、ラミニン、コラーゲン、糖タンパク質、トロンボスポンジン、エラスチン、フィブリリン、ムコ多糖、糖脂質、ヘパリン硫酸、コンドロイチン硫酸、ケラチン硫酸、グリコサミノグリカン、ヒアルロン酸、プロテオグリカン、ビトロネクチン、ポリ-D-リジン、多糖、およびそれらの組合せからなる群より選択される生体適合性材料を含む。
【0018】
ある態様では、細胞が、分枝形態形成の誘導に関連する遺伝子発現を示す。
【0019】
別の態様では、組成物が肺臓組織の特徴を含む。いくつかの例では、特徴が、分枝形態形成、遠位肺臓上皮細胞分化、上皮成長、血管発生、およびそれらの組合せからなる群より選択される。
【0020】
本発明は、肺臓細胞の分化状態を支持し維持する能力を有する人工三次元組織を製作する方法を提供する。本方法は、播種済スキャフォールドを作製するために脱細胞化スキャフォールドに細胞の集団を播種する工程を含む。
【0021】
本発明は、肺臓組織の健康状態を調整する試験作用因子の能力に関して試験作用因子をスクリーニングするためのインビトロ法を提供する。本方法は、試験作用因子を人工三次元肺臓組織モデルに接触させる工程と、試験作用因子がモデルに及ぼす効果を測定する工程とを含む。モデルに加えられる変更はいずれも、その試験作用因子が肺臓組織の健康状態を調整できることのしるしである。
【0022】
ある態様では、試験作用因子が、化学剤、医薬、ペプチド、核酸、および放射線からなる群より選択される。
【0023】
別の態様では、試験作用因子が治療剤のための送達媒体である。
【0024】
ある態様において、本方法は、細胞数、面積、体積、形状、形態、マーカー発現または染色体断片化に対する試験作用因子の効果を決定する工程を含む。
【0025】
別の態様において、本方法は、肺臓組織モデルに対して所望の効果を持つ作用因子を選択する工程を含む。
【0026】
本発明は、哺乳動物における肺臓欠陥を軽減または処置する方法を提供する。本方法は、肺臓細胞の分化状態を支持し維持する能力を有する三次元構築物を含む組成物の治療有効量を哺乳動物に投与し、それによってその哺乳動物における肺臓欠陥を軽減または処置する工程を含む。
【0027】
本発明は、細胞成長を支持する能力を有する脱細胞化組織を含む埋植可能な組成物を提供する。好ましくは、脱細胞化組織は、脱細胞化前の対応する天然組織の特徴を示す。
【0028】
ある態様において、埋植可能な組成物は細胞の集団を含む。好ましくは、埋植可能な組成物は、肺臓組織の分化状態を支持し維持する能力を有する。
【図面の簡単な説明】
【0029】
本発明を例証するために、本発明の一定の態様を図面に図示する。ただし、本発明は、図面に図示する態様の厳密な配置および道具に限定されるわけではない。
【0030】
【図1A】図1A〜1Dを含む図1は、未変性肺臓および脱細胞化肺臓のH&E染色および定量的DNAアッセイを図示する一連の画像である。細胞材料は除去されたが、それでもスキャフォールドのアーキテクチャは保たれたことを、図1は証明している。DNAは未変性レベルの約1.2%まで除去された。*はp<0.01を示す。
【図1B】図1A〜1Dを含む図1は、未変性肺臓および脱細胞化肺臓のH&E染色および定量的DNAアッセイを図示する一連の画像である。細胞材料は除去されたが、それでもスキャフォールドのアーキテクチャは保たれたことを、図1は証明している。DNAは未変性レベルの約1.2%まで除去された。*はp<0.01を示す。
【図1C】図1A〜1Dを含む図1は、未変性肺臓および脱細胞化肺臓のH&E染色および定量的DNAアッセイを図示する一連の画像である。細胞材料は除去されたが、それでもスキャフォールドのアーキテクチャは保たれたことを、図1は証明している。DNAは未変性レベルの約1.2%まで除去された。*はp<0.01を示す。
【図1D】図1A〜1Dを含む図1は、未変性肺臓および脱細胞化肺臓のH&E染色および定量的DNAアッセイを図示する一連の画像である。細胞材料は除去されたが、それでもスキャフォールドのアーキテクチャは保たれたことを、図1は証明している。DNAは未変性レベルの約1.2%まで除去された。*はp<0.01を示す。図1Dは脱細胞化肺臓の画像である。
【図2】図2Aおよび図2Bを含む図2は、脱細胞化スキャフォールド中の残存DNAに関する染色を図示する一連の画像である。DNAがDAPIで染色されている。比較が可能なように、画像は同じ時間、露出した。
【図3】図3は、MHCクラスIおよびII抗原に関するウェスタンブロットであり、脱細胞化スキャフォールドにおけるMHCクラスIまたはII抗原の欠如を証明している。
【図4】図4Aおよび4Bを含む図4は、未変性肺臓および脱細胞化肺臓におけるコラーゲン染色を図示する一連の画像である。コラーゲンIは緑色に染色され、コラーゲンIVは赤色に染色され、核はDAPIで青色に対比染色される。コラーゲンIは大血管の近くに見いだされ、コラーゲンIVは実質全体に分布している。未変性肺臓では、実質中の赤血球が自己蛍光ゆえに緑色に見えることに注意されたい。
【図5】図5Aおよび5Bを含む図5は、未変性肺臓および脱細胞化肺臓の走査型EMを図示する一連の画像である。肺胞中隔はインタクトである。スケールバーは、左側のパネルでは100μm、右側のパネルでは20μmである。
【図6】図6A〜6Cを含む図6は、未変性肺臓および脱細胞化肺臓の透過型EMを図示する一連の画像である。脱細胞化灌流圧を30mmHg未満に維持すると、肺胞基底膜が保たれる。Cは毛細管を示し、Aは肺胞を示し、Sは肺胞中隔を示す。スケールバーは全てのパネルにおいて2μmである。
【図7】図7は、脱細胞化肺臓の透過型EMを図示する画像であり、保存された毛細管を証明している。脱細胞化のための灌流圧は20mmHg未満だった。Cは毛細管を示し、Aは肺胞を示す。肺胞および毛細管の寸法は、脱細胞化マトリックスの圧縮ゆえに、インビボにある時よりも小さく見える。スケールバーは、上側のパネルでは2μm、左下のパネルでは1μm、右下のパネルでは500nmである。
【図8】脱細胞化スキャフォールドによる5μmマイクロスフェアの保持を図示するグラフである。マイクロスフェアアッセイは、脱細胞化時の低い灌流圧(<30mmHg)によって、気道コンパートメントにおける5μm粒子の95%の保持が可能になることを証明している。*は、未変性物との比較で、p<0.05を示す。
【図9】図9Aおよび9Bを含む図9は、未変性肺臓および脱細胞化肺臓の脈管構造のマイクロCTの一連の画像である。全体として、58μmの解像度でイメージングした場合、脱細胞化スキャフォールドは未変性物と類似しているようである。
【図10】図10Aおよび10Bを含む図10は、未変性肺臓および脱細胞化肺臓の脈管構造の高解像度マイクロCTを図示する一連の画像である。これらのスキャンの解像度は6.5μmである。
【図11】図11は、機械的試験プロトコールを図示する図解である。簡単に述べると、肺臓組織の条片を上側のプレートに取り付け、次にそれを下げて、組織を下側のプレートに取り付ける。組織を20%歪みまで周期的に伸張した後、破損するまで伸張する。
【図12】図12A〜12Cを含む図12は、未変性肺臓および脱細胞化肺臓のコラーゲン染色およびコラーゲン含量を図示する一連の画像である。Massonのトリクローム染色により、未変性肺臓および脱細胞化肺臓の両方に、波状の暗青色線維が明らかになる。定量的アッセイは、未変性肺臓および脱細胞化肺臓ではコラーゲンが保存されるが、ドデシル硫酸ナトリウム(SDS)を使った脱細胞化ではコラーゲンが失われることを証明している。*はp<0.01を示す。
【図13】図13A〜13Cを含む図13は、未変性肺臓および脱細胞化肺臓に関するエラスチン組織化学(Verhoff-van Geison)を図示する一連の画像である。図13は、未変性肺臓および脱細胞化肺臓の両方において、波状の暗いエラスチン線維を図示している。脱細胞化肺臓では、未変性物と比較して、エラスチンがいくらか保存されることが、定量的アッセイによって証明される。*はp<0.01を示す。
【図14】図14A〜14Cを含む図14は、未変性肺臓および脱細胞化肺臓に関するGAG組織化学(アルシアンブルー)を図示する一連の画像である。未変性肺臓では青色のGAG染色が図示されるが、脱細胞化肺臓にはそれらが存在しない。脱細胞化肺臓では、未変性肺臓と比較して、硫酸化GAGが失われることが、定量的アッセイによって証明される。*はp<0.01を示す。
【図15】図15は、未変性肺臓および脱細胞化肺臓の応力-歪み曲線を図示する画像である。SDSは、ドデシル硫酸ナトリウムで処理した肺臓を示す。
【図16】図16は、未変性、脱細胞化およびSDS脱細胞化肺臓の極限引張強さを図示する図表である。SDSはドデシル硫酸ナトリウムを使って脱細胞化した肺臓を示す。*は、未変性物との比較で、p<0.01を示す。
【図17】図17Aおよび17Bを含む図17は、インビトロ肺臓培養に使用されるバイオリアクタの概略図を図示する一連の画像である。
【図18】図18Aおよび18Bを含む図18は、インビトロ肺臓培養時の肺動脈圧および気管圧を図示する一連の画像である。灌流量は約5ml/分である。
【図19】図19A〜19Cを含む図19は、肺臓アーキテクチャ(architecture)および気道上皮に対する空気による換気と液体による換気の効果を対比して図示する一連の画像である。空気換気は3日間の培養後に気道の拡張と気道上皮の破壊を引き起こす。
【図20】図20Aおよび20Bを含む図20は、未変性肺臓培養時の細胞アポトーシスおよび細胞数に対する血管灌流および圧力の効果を図示する一連の画像である。未変性物との比較で、*はp<0.01を示し、#はp<0.05を示す。
【図21】図21A〜21Dを含む図21は、未変性肺臓および灌流培養肺臓におけるCCSPおよびSPC発現の比較を図示する一連の画像である。CCSPおよびSPCは赤く染色され、核はDAPIで青く対比染色される。
【図22】図22Aおよび22Bを含む図22は、未変性肺臓および灌流培養肺臓におけるPECAM発現の比較を図示する一連の画像である。灌流肺臓培養(30mmHg)ではPECAM発現が依然として認められる。PECAMは赤く染色され、核はDAPIで青く対比染色される。
【図23】図23Aおよび23Bを含む図23は、未変性肺臓培養時の細胞アポトーシスおよび細胞数に対する換気の効果を図示する一連の画像である。未変性物との比較で、*はp<0.01を示し、#はp<0.05を示す。
【図24】図24A〜24Cを含む図24は、未変性肺臓および換気培養肺臓におけるアポトーシス核を図示する一連の画像である。単一接続による換気は、未変性肺臓または気道「ループ」による換気と比較して、はるかに高い比率のアポトーシス核をもたらした。アポトーシス核はTUNELによって褐色に染色され、正常な核は緑色に対比染色される。
【図25−1】図25A〜25Jを含む図25は、未変性肺臓および7日間培養した肺臓における肺胞構造を図示する一連の画像である。細胞形態、肺胞構造、および中隔アーキテクチャは、未変性肺臓と培養換気肺臓の間で、類似しているようである。
【図25−2】図25A〜25Jを含む図25は、未変性肺臓および7日間培養した肺臓における肺胞構造を図示する一連の画像である。細胞形態、肺胞構造、および中隔アーキテクチャは、未変性肺臓と培養換気肺臓の間で、類似しているようである。図25C〜25Jは、7日間のインビトロ換気肺臓培養後の肺細胞分化の維持を図示している。
【図26】図26は、換気が肺脈管構造の受動的灌流を可能にすることを証明する画像である。マイクロスフェアは、もっぱらインビトロ培養時の肺臓の換気運動のみによって、血管および毛細管中に見いだされる。
【図27】図27Aおよび27Bを含む図27は、脱細胞化スキャフォールド上で培養した不死化上皮細胞株MLE-12のH&E染色を図示する一連の画像である。
【図28A】図28A〜図28Fを含む図28は、単離された新生児肺細胞の肺マーカーパネルのフローサイトメトリー染色を図示する一連の画像である。青色または緑色の曲線はアイソタイプ対照染色であり、赤色は表示した抗原である。
【図28B】図28A〜図28Fを含む図28は、単離された新生児肺細胞の肺マーカーパネルのフローサイトメトリー染色を図示する一連の画像である。青色または緑色の曲線はアイソタイプ対照染色であり、赤色は表示した抗原である。
【図28C】図28A〜図28Fを含む図28は、単離された新生児肺細胞の肺マーカーパネルのフローサイトメトリー染色を図示する一連の画像である。青色または緑色の曲線はアイソタイプ対照染色であり、赤色は表示した抗原である。
【図28D】図28A〜図28Fを含む図28は、単離された新生児肺細胞の肺マーカーパネルのフローサイトメトリー染色を図示する一連の画像である。青色または緑色の曲線はアイソタイプ対照染色であり、赤色は表示した抗原である。
【図28E】図28A〜図28Fを含む図28は、単離された新生児肺細胞の肺マーカーパネルのフローサイトメトリー染色を図示する一連の画像である。青色または緑色の曲線はアイソタイプ対照染色であり、赤色は表示した抗原である。
【図28F】図28A〜図28Fを含む図28は、単離された新生児肺細胞の肺マーカーパネルのフローサイトメトリー染色を図示する一連の画像である。青色または緑色の曲線はアイソタイプ対照染色であり、赤色は表示した抗原である。
【図29】図29は、培養8日時点の人工肺臓のH&E染色を図示する画像である。ここでは条件が上皮細胞成長に最適化されている。
【図30】図30Aおよび30Bを含む図30は、培養4日および8日時点の人工肺臓のPCNA染色を図示する一連の画像である。増殖性核はPCNAに関して褐色に染まり、陰性核はヘマトキシリンで対比染色される。
【図31】図31Aおよび31Bを含む図31は、培養4日および8日時点の人工肺臓のTUNEL染色を図示する一連の画像である。陽性核は褐色であり、陰性核はメチルグリーンで対比染色される。
【図32】図32A〜32Bを含む図32は、4日時点の未変性肺臓および人工肺臓のクララ細胞分泌タンパク質(CCSP)染色を図示する一連の画像である。CCSPは赤色に染色され、核はDAPIで青色に対比染色される。
【図33】図33A〜33Cを含む図33は、4日および8日時点の未変性肺臓および人工肺臓のサーファクタントタンパク質C染色を図示する一連の画像である。SPCは赤色に染色され、核はDAPIで青色に対比染色される。
【図34】図34A〜34Cを含む図34は、4日時点の未変性肺臓および人工肺臓のアクアポリン5染色を図示する一連の画像である。AQPは赤色に染色され、核はDAPIで青色に対比染色される。
【図35】図35Aおよび35Bを含む図35は、人工肺臓組織におけるSPCおよびCCSPに関する二重染色を図示する一連の画像である。SPCは緑色に染色され、CCSPは赤色に染色され、核はDAPIで青色に対比染色される。SPC-CCSP二重陽性細胞は黄色に見える。
【図36】図36A〜36Cを含む図36は、未変性肺臓および人工肺臓における基底細胞に関するサイトケラチン14染色を図示する一連の画像である。サイトケラチンは赤色に染まり、核はDAPIで青色に対比染色される。
【図37】図37は、人工肺臓におけるサイトケラチン14およびCCSPに関する二重染色を図示する画像である。サイトケラチン14は赤色に染色され、CCSPは緑色に染色され、核はDAPIで青色に対比染色される。
【図38】図38Aおよび38Bを含む図38は、未変性肺臓および人工肺臓のα-アクチン染色を図示する一連の画像である。α-アクチンは緑色に染色され、核はDAPIで青色に対比染色される。
【図39】図39A〜39Fを含む図39は、上皮発生に対する培地組成の効果を図示する一連の画像である。BGJb培地中で培養すると、上皮構造は、SPC顆粒の頂端部発現を起こし、CCSP発現の喪失を伴う。DMEM培地では、細胞がSPCとCCSPの発現をどちらも保ち、SPC発現は散在性に細胞質発現である。
【図40】図40は、人工上皮組織におけるサーファクタント発現を図示する画像である。「Lad」はタンパク質ラダーであり;表示のバンドは20および25kDaであり;「Nat」は未変性肺臓組織であり;「Vent」はDMEM培地で換気した人工肺臓組織であり;「Perf」はDMEM培地で灌流した人工肺臓組織であり;「DMEM」はDMEM培地中の静置培養人工肺臓であり;「BGJb」はBGJb培地中の静置培養人工肺臓であり;「ALI」は空気で換気した人工肺臓であり;「Decell」は脱細胞化スキャフォールドである。
【図41】図41A〜41Cを含む図41は、人工肺臓組織における上皮発生に対する空気による換気の効果を図示する一連の画像である。AQP発現は、実質細胞(これはSPCについても陽性である(図41B))中に認められると共に(左上)、立方上皮細胞にも時折強い発現が認められる(右上)。立方上皮のCCSP発現も認められる(図41C)。
【図42】図42Aおよび42Bを含む図42は、未変性肺臓および人工肺臓における繊毛上皮を図示する一連の画像である。人工肺臓については繊毛細胞が赤色で強調されている。
【図43】図43Aおよび43Bを含む図43は、人工肺臓培養に対する灌流および換気の効果を図示する一連の画像である。
【図44】図44A〜44Dを含む図44は、人工肺臓培養における細胞増殖およびアポトーシスに対する灌流および換気の効果を図示する一連の画像である。
【図45】図45Aおよび45Bを含む図45は、人工肺臓組織におけるCCSP発現に対する灌流および換気の効果を図示する一連の画像である。CCSPは赤色に染色され、核はDAPIで青色に対比染色される。
【図46】図46A〜46Bを含む図46は、人工肺臓組織におけるSPC発現に対する灌流および換気の効果を図示する一連の画像である。SPCは赤色に染色され、核はDAPIで青色に対比染色される。
【図47】図47は、ラット肺臓微小血管内皮細胞を播種したフィブロネクチン被覆脱細胞化スキャフォールドのH&E染色を図示する画像である。
【図48】図48Aおよび48Bを含む図48は、灌流人工肺臓内皮と換気人工肺臓内皮のH&E染色を対比して図示する一連の画像である。
【図49】図49Aおよび49Bを含む図49は、灌流人工肺臓内皮と換気人工肺臓内皮のTUNEL染色を対比して図示する一連の画像である。換気だけで培養されたECは、灌流肺臓よりも、実質的に多くアポトーシスを起こす。アポトーシス核はTUNELによって褐色に染まり、陰性核はメチルグリーンで対比染色される。
【図50】図50は、人工肺臓組織における内皮細胞間の密着結合形成の画像による証明である。内皮細胞には星印を付けてあり、それらは広範な細胞間結合によって分離されている。スケールバーは500nmである。
【図51】図51Aおよび51Bを含む図51は、未変性肺臓および人工肺臓におけるVE-カドヘリンの発現を図示する一連の画像である。VE-カドヘリンは赤色に染色され、核はDAPIで青色に対比染色される。
【図52】図52は、2メガダルトンFITC標識デキストランに対する、内皮細胞のみを播種した人工肺臓の透過性を図示する図表である。*は、脱細胞化スキャフォールドとの比較で、p<0.05を示す。
【図53】図53は、人工組織の極限引張強さを図示する図表である。未変性肺臓および脱細胞化肺臓の強さも示している。
【図54】図54A〜54Cを含む図54は、人工内皮組織の成長に対する培地の影響を図示する一連の画像である。人工灌流内皮を表示の培地タイプ中で培養した。H&E組織像を左側のパネルに示す。一方、右側のパネルは褐色(TUNELによる)のアポトーシス核を示し、正常な核はメチルグリーンで対比染色される。
【図55】図55は、CHAPS緩衝液中で4〜8時間インキュベートすることによって調製された脱細胞化気管が、コラーゲンマトリックスを維持し、大半の細胞の、組織からの除去を示したことを証明する一連の画像である。
【図56】図56は、脱細胞化気管が未変性気管に見られる3タイプのCOLの全てを含有していたことを証明する一連の画像である。
【図57】図57は、脱細胞化気管がNHBEの接着および成長を支持したことを証明する一連の画像である。
【図58】図58は、脱細胞化気管がSAECの接着および成長を支持したことを証明する一連の画像である。
【図59】図59は、GFPレンチウイルスに感染したNHBEが6時間後に明白な形態変化を示さなかったことを証明する一連の画像である。
【図60】図60は、気道への点滴注入による送達後に、かなりの数のマイクロスフェアが、マウス肺臓の全ての葉に存在していたことを証明する一連の画像である。
【図61】図61は、肺臓への細胞の注入の成功と、導入遺伝子(GFP)を形質導入されたヒト上皮細胞が肺臓上皮に接着したこととを証明する一連の画像である。
【図62】図62は、トリプシン処理前に見られる注入に使用したGFP細胞を図示する一連の画像である。
【図63A】図63A〜63Cを含む図63は、GFP陽性ヒト気道上皮細胞(NHBEとSAECの両方)が、気道への点滴注入後、何日間も、マウス肺臓に見いだされたことを証明する一連の画像である。
【図63B】図63A〜63Cを含む図63は、GFP陽性ヒト気道上皮細胞(NHBEとSAECの両方)が、気道への点滴注入後、何日間も、マウス肺臓に見いだされたことを証明する一連の画像である。
【図63C】図63A〜63Cを含む図63は、GFP陽性ヒト気道上皮細胞(NHBEとSAECの両方)が、気道への点滴注入後、何日間も、マウス肺臓に見いだされたことを証明する一連の画像である。
【図64】図64Aおよび64Bを含む図64は、換気サイクル中の拡張時および収縮時における、埋植された人工肺臓を示す一連の画像である。
【発明を実施するための形態】
【0031】
発明の詳細な説明
本発明は人工肺臓組織を提供する。本発明は、一つには、天然肺臓組織の特徴を示すように三次元肺臓組織を生成させることができるという発見に基づいている。
【0032】
ある態様では、人工肺臓組織が脱細胞化未変性肺臓組織から誘導される。脱細胞化組織は細胞およびDNAを実質的に欠いている。好ましくは、脱細胞化組織が免疫原性分子も欠いている。より好ましくは、脱細胞化組織は、細胞の付着と増殖にとって大切であるいくつかの重要な細胞外マトリックス分子を保っている。
【0033】
本発明は組織を脱細胞化する方法を包含する。この脱細胞化方法は、肺臓の細胞外マトリックスの重要な局面を保つと共に、肺臓の細胞外マトリックスに対するいかなる損傷も最小限に抑えつつ、組織から細胞材料および核材料を除去する工程を含む。ある態様では、脱細胞化プロセスが、組織から抗原性分子を除去し、それによって組織を非免疫原性にする工程も含む。ある態様では、本発明の脱細胞化プロセスが、細胞培養と完全に適合すると同時に障壁機能を提供する脱細胞化スキャフォールドを生成させる工程を含む。好ましくは、脱細胞化スキャフォールドは、インタクトな気道樹および血管網を持つ肺臓スキャフォールドである。
【0034】
本発明はバイオリアクタも包含する。好ましくは、バイオリアクタは、任意の三次元組織のインビトロ培養を支持する能力を有する。ある態様では、バイオリアクタが、陰圧によって肺臓を換気する能力を有するだけでなく、生理的速度および圧力で血管灌流および換気を提供する能力も持つ。バイオリアクタは、例えば脈管構造を通した培地の灌流、気道を出入りする培地または空気の動き、陰(ならびに陽)圧による肺臓の換気などを可能にする。
【0035】
本発明の肺臓組織のインビトロ三次元モデルは、肺臓発生生物学を調べるのに役立つ。また、本モデルは、その他、例えば創薬、毒性試験、疾患病理学などにも役立つ。
【0036】
本発明は、肺臓組織をインビトロで生成させることができるという発見にも関係する。インビトロモデルは、宿主循環と密接に連係された導管上皮から構成される肺胞形成単位を連想させる構造の形成を再現する。したがって本発明は、再生医学の一形態として、血管化された肺組織を生成させるための方法および組成物を提供する。
【0037】
本発明は、哺乳動物、好ましくはヒトにおける肺臓欠陥を軽減または処置する方法も提供する。本方法は、それを必要とする哺乳動物に、本発明の三次元構築物を含む組成物の治療有効量を投与し、それによってその哺乳動物における肺臓欠陥を軽減または処置する工程を含む。
【0038】
定義
別段の定義がない限り、本明細書において使用する技術用語および科学用語は全て、一般に、本発明が属する技術分野の当業者が通常理解しているものと同じ意味を持つ。一般に、本明細書において使用する用語体系、ならびに細胞培養、分子遺伝学、有機化学、ならびに核酸化学およびハイブリダイゼーションにおける実験手法は、当技術分野において周知であり、通常使用されているものである。
【0039】
核酸とペプチドの合成には標準的な技法を使用する。これらの技法および手法は、一般に、本明細書の至るところに記載する当技術分野における常法や種々の一般的参考文献(例えばSambrookおよびRussell(2001)「Molecular Cloning, A Laboratory Approach」(Cold Spring Harbor Laboratory Press、ニューヨーク州コールドスプリングハーバー)ならびにAusubel et al.(2002)「Current Protocols in Molecular Biology」(John Wiley & Sons、ニューヨーク州ニューヨーク))に従って行われる。
【0040】
冠詞「a」および「an」(「ある」「1つの」)は、本明細書では、1つの、または1つより多くの(すなわち少なくとも1つの)、その冠詞の文法上の対象を指すために使用される。例えば、「ある要素」とは、1つの要素または1つより多くの要素を意味する。
【0041】
「約」という用語は当業者には理解され、それが使用される文脈に基づいて、ある程度は変動するであろう。
【0042】
「前駆体細胞」、「前駆細胞」、および「幹細胞」という用語は、当技術分野では互換的に使用され、本明細書において使用される場合は、無限回にわたって有糸分裂して自分自身を再生するか、所望の細胞タイプへと分化するであろう子孫細胞を生み出す能力を、潜在的に有する、多能性の、または分化拘束されていない(lineage-uncommitted)前駆細胞を指す。多能性幹細胞とは対照的に、分化拘束された(lineage-committed)前駆細胞は、一般に、表現型が互いに異なる多数の細胞タイプを生じる能力は持たないと考えられる。その代わりに、前駆細胞は、1つか、場合によっては2つの、分化拘束された細胞タイプを生じる。
【0043】
本明細書において使用する「脱分化」という用語は、細胞が、専門性の低い状態に戻ることを指す。脱分化後、そのような細胞は、再プログラミング前に可能であったものと比較して、より多くの、または異なる細胞タイプへと、分化する能力を有するであろう。逆分化(すなわち脱分化)のプロセスは分化よりも複雑である可能性が高く、より原始的になるように細胞を「再プログラムする」ことが必要である。
【0044】
本明細書にいう「スキャフォールド」は、細胞の接着と増殖に適した表面を提供する生体適合性材料を含む構造を指す。スキャフォールドは、さらに、機械的安定性および機械的支持も提供し得る。スキャフォールドは、三次元形状に影響を及ぼすようにまたは三次元形状の境界を定めるように特定の形状または形態をとるか、増殖細胞の集団が呈する形態をとり得る。そのような形状または形態には、薄膜(例えば第3の次元よりも実質的に大きな2つの次元を持つ形態)、リボン、ひも、シート、平らな円板、円柱、球、三次元無定形の形状などがあるが、これらに限定されるわけではない。
【0045】
本明細書にいう「生体適合性」とは、哺乳動物に埋植された時にその哺乳動物において有害な応答を惹起しない、任意の材料を指す。生体適合性材料は、個体中に導入された場合に、その個体にとって毒性または有害でなく、その哺乳動物におけるその材料の免疫学的拒絶を誘導することもない。
【0046】
本明細書にいう「自家」とは、後にその材料が再導入されることになる個体と同じ個体に由来する、生物学的材料を指す。
【0047】
本明細書にいう「同種異系」とは、その材料が導入されることになる個体と同じ種の遺伝学的に異なる個体に由来する、生物学的材料を指す。
【0048】
本明細書において、「グラフト(graft)」とは、典型的には欠陥を置き換え、矯正し、または他の方法で克服するために個体に埋植される、細胞、組織または器官を指す。グラフトはさらにスキャフォールドを含んでもよい。組織または器官は、同じ個体から派生する細胞からなることができ、このグラフトを、本明細書では、以下の交換可能な用語で表す:「オートグラフト(autograft)」、「自家トランスプラント(autologous transplant)」、「自家インプラント(autologous implant)」および「自家グラフト」。同じ種の遺伝学的に異なる個体に由来する細胞を含むグラフトを、本明細書では、以下の交換可能な用語で表す:「アログラフト(allograft)」、「同種異系トランスプラント」、「同種異系インプラント」および「同種異系グラフト」。ある個体の一卵性双生児である兄妹から得られるグラフトを、本明細書では、「アイソグラフト(isograft)」、「同種同系トランスプラント」、「同種同系インプラント」または「同種同系グラフト」という。「キセノグラフト(xenograft)」、「異種トランスプラント」または「異種インプラント」とは、ある個体から、種が異なる別の個体へのグラフトを指す。
【0049】
本明細書において使用する「組織グラフティング(tissue grafting)」および「組織再建(tissue reconstructing)」という用語は、どちらも、肺臓欠陥または軟部組織欠陥などの組織欠陥を処置または軽減するために、グラフトを個体に埋植することを指す。
【0050】
本明細書において、疾患、欠陥、障害または状態を「軽減する」とは、その疾患、欠陥、障害または状態の1つまたは複数の症状の重症度を低減することを意味する。
【0051】
本明細書にいう「処置する」とは、ある疾患、欠陥、障害、または有害な状態などの症状を患者が経験する頻度を低減することを意味する。
【0052】
本明細書にいう「治療有効量」は、その組成物の投与を受ける個体に有益な効果を与えるのに十分であるような、本発明の組成物の量である。
【0053】
本明細書において使用する「成長培地」という用語は、細胞の成長を促進する培養培地を指すものとする。成長培地は、一般に、動物血清を含有するであろう。場合によっては、成長培地が動物血清を含有しないこともあるだろう。
【0054】
「分化培地」は、本明細書では、完全には分化していない幹細胞、胎児肺細胞または他のそのような前駆細胞が、その培地中でインキュベートされた時に、分化細胞の特徴の一部または全部を持つ細胞へと発生するように添加物を含むまたは添加物を欠く、細胞成長培地を指すために使用される。
【0055】
本明細書において使用する「成長因子産物」という用語は、細胞に対して成長効果、増殖効果、分化効果、または栄養効果を有する、タンパク質、ペプチド、マイトジェン、または他の分子を指す。成長因子には、線維芽細胞成長因子(FGF)、塩基性線維芽細胞成長因子(bFGF)、酸性線維芽細胞成長因子(aFGF)、上皮成長因子(EGF)、インスリン様成長因子I(IGF-T)、インスリン様成長因子II(IGF-II)、血小板由来成長因子(PDGF)、血管内皮細胞成長因子(VEGF)、アクチビンA、骨形態形成タンパク質(BMP)、インスリン、成長ホルモン、エリスロポエチン、トロンボポエチン、インターロイキン3(IL-3)、インターロイキン6(IL-6)、インターロイキン7(IL-7)、マクロファージコロニー刺激因子、c-kitリガンド/幹細胞因子、オステオプロテゲリンリガンド、インスリン、神経成長因子、毛様体神経栄養因子、サイトカイン、ケモカイン、モルフォゲン、中和抗体、他のタンパク質、および小分子などが含まれるが、これらに限定されるわけではない。好ましくは、FGFは、FGF2、FGF7、FGF10、およびその任意の組合せから選択される群から選択される。
【0056】
「単離された細胞」とは、組織または哺乳動物においてその単離された細胞に天然に付随している他の構成要素および/または細胞から分離された細胞を指す。
【0057】
本明細書にいう「胎児肺細胞」(FPC)は、胚の肺臓組織から単離された細胞を指す。FPCの混合集団は、上皮、間葉および内皮細胞を含み得るが、これらに限るわけではない。
【0058】
本明細書にいう「上皮細胞」とは、身体の外面を形成し、器官、腔および粘膜表面を裏打ちする細胞を意味する。
【0059】
本明細書にいう「内皮細胞」とは、血管およびリンパ管ならびに他のさまざまな体腔を裏打ちする細胞を意味する。
【0060】
本明細書にいう「実質的に精製された」細胞は、本質的に他の細胞タイプを含まない細胞である。したがって、実質的に精製された細胞とは、その天然状態ではそれに通常付随している他の細胞タイプから精製された細胞を指す。
【0061】
「拡大性(expandability)」は、本明細書では、増殖して、例えば数を拡大するか、細胞の集団の場合であれば、集団倍加を起こすという、細胞の能力を指すために使用される。
【0062】
「肺臓特異的」という用語は、身体の他の組織と比較して肺臓において優勢に発現する核酸分子またはポリペプチドを指す。ある好ましい態様では、「肺臓特異的」核酸分子またはポリペプチドが、身体の他のどの組織よりも5倍高いレベルで発現する。より好ましい態様では、「肺臓特異的」核酸分子またはポリペプチドが、身体の他のどの組織よりも10倍高いレベルで発現し、より好ましくは、身体の他のどの組織よりも少なくとも15倍、20倍、25倍、50倍または100倍高いレベルで発現する。核酸分子レベルは、ノーザンブロットハイブリダイゼーションなどの核酸ハイブリダイゼーションまたは定量PCRによって測定することができる。ポリペプチドレベルは、タンパク質レベルを正確に測定することが知られている任意の方法、例えばウェスタンブロット分析などによって測定することができる。
【0063】
「増殖」は、本明細書では、類似する形態の、特に細胞の、複製または増倍を指すために使用される。すなわち増殖は、より多数の細胞の産生を包含し、例えば、細胞の数を単純に数え上げること、3H-チミジンの細胞への取り込みを測定することなどによって測定することができる。
【0064】
本明細書にいう「組織工学」とは、組織代用(tissue replacement)または組織再建に使用するためにエクスビボで組織を生成させるプロセスを指す。組織工学は「再生医学」の一例であり、これは、細胞、遺伝子または他の生物学的構成単位の組込みと、生物工学的材料および技術とによる、組織および器官の修復または代用への取り組みを包含する。
【0065】
本明細書にいう「内在性」とは、ある生物、細胞または系に由来する、またはそれらの内部で産生された、任意の材料を指す。
【0066】
「外因性」とは、ある生物、細胞、または系に導入された、またはそれらの外で産生された、任意の材料を指す。
【0067】
「コードする」とは、所定の配列のヌクレオチド(すなわちrRNA、tRNAおよびmRNA)または所定の配列のアミノ酸とその結果生じる生物学的性質とを有する他のポリマーおよび高分子を生物学的プロセスにおいて合成するための鋳型として役立つという、遺伝子、cDNA、またはmRNAなどのポリヌクレオチドにおけるヌクレオチドの特異的配列が持つ固有の性質を指す。したがって、ある遺伝子に対応するmRNAの転写および翻訳が、細胞内または他の生物学的系内で、あるタンパク質を産生するのであれば、その遺伝子はそのタンパク質をコードしている。コード鎖(mRNA配列と同一のヌクレオチド配列を持ち、配列表に通常記載されるもの)と、遺伝子またはcDNAの転写のための鋳型として使用される非コード鎖は、どちらも、タンパク質、またはその遺伝子もしくはcDNAの他の産物をコードしているということができる。
【0068】
別段の指定がない限り、「アミノ酸配列をコードするヌクレオチド配列」には、互いの縮重型であって同じアミノ酸配列をコードする、全てのヌクレオチド配列が含まれる。タンパク質およびRNAをコードするヌクレオチド配列はイントロンを含み得る。
【0069】
「単離された核酸」とは、天然の状態でそれに隣接している配列から分離されている核酸セグメントまたは核酸フラグメント、すなわち通常はそのフラグメントに近接している配列(すなわち、それが天然に存在するゲノム中でそのフラグメントに近接している配列)から取り出されたDNAフラグメントを指す。この用語は、その核酸に天然に付随する他の構成要素、すなわち、細胞内でそれに天然に付随しているRNAまたはDNAまたはタンパク質から実質的に精製された核酸にも適用される。したがってこの用語には、例えば、ベクターに組み込まれているか、自律的に複製するプラスミドもしくはウイルスに組み込まれているか、または原生生物もしくは真核生物のゲノムDNAに組み込まれている組換えDNA、または他の配列から独立して別個の分子(すなわちcDNAまたはPCRもしくは制限酵素消化によって作り出されるゲノムフラグメントもしくはcDNAフラグメント)として存在する組換えDNAが含まれる。また、追加のポリペプチド配列をコードするハイブリッド遺伝子の一部である組換えDNAも含まれる。
【0070】
本発明については、よく見られる核酸塩基に対して、以下の略号を使用する。「A」はアデノシンを指し、「C」はシトシンを指し、「G」はグアノシンを指し、「T」はチミジンを指し、「U」はウリジンを指す。
【0071】
本明細書において使用する「転写制御下にある」または「作動的に連結された」という表現は、そのプロモーターがそのポリヌクレオチドに対して正しい位置と向きにあって、RNAポリメラーゼの開始およびポリヌクレオチドの発現を制御することを意味する。
【0072】
本明細書において使用する「プロモーター/調節配列」という用語は、そのプロモーター/制御配列に機能的に連結される遺伝子産物の発現にとって必要な核酸配列を意味する。いくつかの例では、この配列はコアプロモーター配列であることができ、他の例では、この配列はエンハンサー配列および遺伝子産物の発現に必要な他の調節要素も含むことができる。プロモーター/調節配列は、例えば、遺伝子産物を組織特異的に発現させるものであることができる。
【0073】
「構成的」プロモーターは、ある遺伝子産物をコードまたは指定するポリヌクレオチドと機能的に連結された場合に、ある細胞において、その細胞の大半のまたは全ての生理的条件下で、その遺伝子産物を産生させる、ヌクレオチド配列である。
【0074】
「誘導性」プロモーターは、ある遺伝子産物をコードまたは指定するポリヌクレオチドと機能的に連結された場合に、ある細胞において、実質上、そのプロモーターに対応する誘導因子がその細胞中に存在する場合にのみ、その遺伝子産物を産生させる、ヌクレオチド配列である。
【0075】
本明細書において使用する「組織」という用語には、例えば骨、神経組織、腱および靱帯を含む線維性結合組織、軟骨、硬膜、心膜、筋、肺臓、心臓弁、静脈および動脈ならびに他の脈管構造、真皮、脂肪組織、または腺組織が含まれるが、これらに限定されるわけではない。
【0076】
「組織特異的」プロモーターは、ある遺伝子産物をコードまたは指定するポリヌクレオチドと機能的に連結された場合に、ある細胞において、実質上、その細胞がそのプロモーターに対応する組織タイプの細胞である場合にのみ、その遺伝子産物を産生させる、ヌクレオチド配列である。
【0077】
「ベクター」は、単離された核酸を含み、その単離された核酸を細胞の内部に送達するために使用することができる、組成物である。限定するわけではないが、線状ポリヌクレオチド、イオン性または両親媒性化合物と会合したポリヌクレオチド、プラスミド、およびウイルスなど、数多くのベクターが当技術分野において公知である。したがって「ベクター」という用語には、自律的に複製するプラスミドまたはウイルスが含まれる。また、この用語は、細胞への核酸の導入を容易にする非プラスミドおよび非ウイルス化合物、例えばポリリジン化合物、リポソームなどを包含すると解釈すべきでもある。ウイルスベクターの例には、アデノウイルスベクター、アデノ随伴ウイルスベクター、レトロウイルスベクターなどがあるが、これらに限定されるわけではない。
【0078】
「発現ベクター」とは、発現させようとするヌクレオチド配列に作動的に連結された発現制御配列を含む組換えポリヌクレオチドを含むベクターを指す。発現ベクターは、発現にとって十分なシス作用要素を含み、発現のための他の要素は、宿主細胞によって供給されるか、インビトロ発現系で供給され得る。発現ベクターには、当技術分野において公知であるもの、例えばコスミド、プラスミド(すなわち裸のプラスミドまたはリポソームに含有されたプラスミド)およびウイルスなどであって、組換えポリヌクレオチドが組み込まれているものが、全て含まれる。
【0079】
本明細書において使用する「対象」および「患者」という用語は、互換的に使用される。本明細書にいう対象は、好ましくは、哺乳動物、例えば非霊長類(例えばウシ、ブタ、ウマ、ネコ、イヌ、ラットなど)および霊長類(例えばサルおよびヒト)、最も好ましくはヒトである。
【0080】
説明
本発明は、人工三次元肺組織、およびその三次元肺組織を製作する方法を提供する。好ましくは、肺組織は肺臓組織である。ある態様において、人工肺組織は、天然肺組織によって例示される分枝形態形成を示す。したがって本発明は、天然肺組織を模倣するインビトロモデルを提供する。インビトロ三次元肺組織モデルは、例えば創薬、毒性試験、疾患病理学などに役立つ。
【0081】
本発明は、細胞材料は除去するが細胞外マトリックスの重要な構成要素は保つ技法を使って肺臓組織を脱細胞化するのに有用な手法の発見に基づく。脱細胞化肺臓マトリックスの開発は、組織工学的応用のためのスキャフォールドとして重要である。したがって本発明は、組織または器官を実質的に脱細胞化するための方法を包含する。好ましくは、本方法は、組織または器官が、移植時に、レシピエントの免疫系によって拒絶されないように、組織または器官の免疫原性を著しく低減または排除する。本方法は、ドナーから組織を取り出し、組織または器官の細胞の実質上全てを除去するために組織を加工する工程を含む。本方法はさらに、レシピエントに埋植するために、幹細胞や胎児細胞など(ただしこれらに限るわけではない)の細胞を播種することによって、脱細胞化スキャフォールドに再集合させる工程を含む。好ましくは、脱細胞化スキャフォールドに、非免疫原性細胞を播種する。ある態様では、脱細胞化スキャフォールドに、意図するレシピエントにとって自家である細胞を播種する。処理し、置換しようとする組織のタイプに依存して、播種済インプラントをレシピエントに埋植した時に適当な組織が形成されるように、当技術分野において公知であるか今後公知になる異なる幹細胞が選択される。
【0082】
いくつかの例では、人工三次元肺組織が、組織上で培養された細胞を含む。本発明の脱細胞化組織上での培養には、任意の適切な細胞を使用することができる。いくつかの例では、肺臓組織を再生させるために、幹細胞を脱細胞化組織上で培養する。いくつかの例では、胎児または新生児肺細胞(NPC)を脱細胞化組織上で培養する。いくつかの例では、上皮細胞、間葉細胞、および内皮細胞(ただしこれらに限るわけではない)を含むNPCの混合集団を使用する。
【0083】
播種後に、スキャフォールド上の細胞は、任意で、拡大培地または分化培地に供されるか、組織特異的成長因子の存在下で培養される。次に、その組成物を、それを必要とする対象に埋植する。対象は哺乳動物であることができるが、好ましくはヒトであり、成長および埋植のための細胞の供給源は任意の哺乳動物、好ましくはヒトである。埋植された組成物はインビボでさらなる細胞成長を支持するので、組織再建をもたらす。したがって、本発明は、組織グラフティング療法のための人工三次元肺組織の使用を提供する。
【0084】
本発明は、インビボでの肺組織の生成も包含する。好ましくは、血管化した肺組織を、インビボで生成させる。ある局面では、インビボ肺組織形成を容易にするために、哺乳動物に、脱細胞化組織との関連において、胎児肺細胞が投与される。
【0085】
本発明では、脱細胞化組織に、新生児肺細胞または成人肺細胞などの適切な細胞を播種することができ、その結果生じる組成物は、血管化した三次元肺組織モデルとして、前臨床インビトロ薬理試験、生理学的試験および科学的試験に使用できることが証明される。また、脱細胞化組織には、新生児肺細胞または自家肺細胞などの適切な細胞を播種することができ、その結果生じる組成物は、インビボでの組織再建に使用することができる。
【0086】
本発明の組成物および方法には無数の有用な応用がある。本組成物は、個体における組織欠陥を軽減または処置するための治療方法に使用することができる。本組成物は治療化合物を同定するためにインビトロまたはインビボで使用することもでき、したがって治療能力を有し得る。
【0087】
脱細胞化
本発明は、当技術分野において公知の組織工学的技法を上回る進歩を提供する。具体的には、本発明は、脱細胞化組織(好ましくは哺乳動物から得られる脱細胞化天然組織)を出発源として使用して、人工組織スキャフォールドを製作する方法を提供する。
【0088】
脱細胞化プロセスは化学的方法に依拠する。ある局面では、脱細胞化に使用される化学溶液(脱細胞化溶液ともいう)は一般に、少なくとも高張溶液、界面活性剤、およびキレート剤を含む。好ましくは、高張溶液は高張塩化ナトリウム溶液である。好ましくは、界面活性剤はCHAPSなどの両性イオン性界面活性剤である。好ましくは、キレート剤はEDTAである。
【0089】
ある態様において、脱細胞化溶液は、細胞との浸透圧適合性のための緩衝液(例えばPBS)を含むことができる。いくつかの例では、脱細胞化溶液が、1つまたは複数のコラゲナーゼ、1つまたは複数のディスパーゼ、1つまたは複数のDNアーゼ、またはトリプシンなどのプロテアーゼなどの酵素(ただしこれらに限定されるわけではない)も含むことができる。いくつかの例では、脱細胞化溶液が、上記に加えて、または上記の代わりに、1つまたは複数の酵素の阻害剤(例えばプロテアーゼ阻害剤、ヌクレアーゼ阻害剤、および/またはコラゲナーゼ阻害剤)を含むことができる。
【0090】
ある態様において、組織を脱細胞化するための本発明の方法は、組織を脱細胞化溶液で灌流する工程を含む。脱細胞化溶液を組織に灌流させる圧力は、望ましい圧力に調節することができる。好ましくは、脱細胞化溶液を組織に約30mmHg未満の灌流圧で灌流させる。より好ましくは、脱細胞化溶液を組織に約20mmHg未満の圧力で灌流させる。
【0091】
ある態様では、細胞除去を達成するために脱細胞化溶液を肺臓組織の気道中に導入することができる。
【0092】
ある態様では、本発明の脱細胞化組織が、本質的に、血管樹のECM構成要素を含む、その組織の全てまたは大半の領域の細胞外マトリックス(ECM)構成要素からなる。ECM構成要素は、以下の物質をどれでも含むことができ、または以下の物質の全てを含むことができる:基底膜などの所定の構造として組織化された状態を保つことができる、フィブロネクチン、フィブリリン、ラミニン、エラスチン、コラーゲンファミリーのメンバー(例えばコラーゲンI、III、およびIV)、グリコサミノグリカン、礎質(ground substance)、細網線維およびトロンボスポンジン。脱細胞化の成功は、標準的な組織染色手法を使って組織切片中に検出可能な筋フィラメント、内皮細胞、平滑筋細胞、上皮細胞および核が存在しないことと定義される。そうでない場合もあるが、好ましくは、残留細胞片も脱細胞化組織から除去された。
【0093】
ある態様では、天然組織の脱細胞化プロセスが、組織の未変性三次元構造を保存する。すなわち、ECM構成要素を含む組織の形態およびアーキテクチャは、脱細胞化のプロセス中およびプロセス後に維持される。ECMの形態およびアーキテクチャは、視覚的および/または組織学的に調べることができる。例えば、固形臓器(solid organ)の外面上または器官もしくは組織の脈管構造内の基底膜は、脱細胞化によって除去されたり、著しく損傷されたりすべきでない。また、ECMの原線維は、脱細胞化されていない器官または臓器のそれと類似しているか、著しくは変化していないべきである。
【0094】
ある態様では、例えば脱細胞化組織を保存するために、または再細胞化に備えて脱細胞化組織を調製するために、かつ/または再細胞化プロセス中の細胞を補助もしくは刺激するために、1つまたは複数の化合物を脱細胞化組織中または脱細胞化組織上に適用することができる。そのような化合物には、1つまたは複数の成長因子(例えばVEGF、DKK-1、FGF、BMP-1、BMP-4、SDF-1、IGF、およびHGF)、免疫調整剤(例えばサイトカイン、グルココルチコイド、IL2Rアンタゴニスト、ロイコトリエンアンタゴニスト)、および/または凝固カスケードを修飾する因子(例えばアスピリン、ヘパリン結合タンパク質、およびヘパリン)などがあるが、これらに限定されるわけではない。また、脱細胞化された器官または組織をさらに、例えば照射(例えばUV、γ線)処理して、脱細胞化組織上または脱細胞化組織中に残っているあらゆるタイプの微生物を減少させ、またはその存在を排除することもできる。
【0095】
脱細胞化組織を生成させるための本発明の脱細胞化溶液の使用は、脈管構造を含む基礎をなすECMと元の組織の他の総体的な形態上の特質とをインタクトな状態で残しつつ、組織の細胞を破壊するための制御された正確な方法を提供する。したがって、脱細胞化されたスキャフォールドは、適当な細胞による播種に適している。このプロセスがインビトロで行われる場合、播種された組織は、代用組織として、レシピエントへの埋植に適している。脱細胞化組織そのものに加えて、本発明は、そのようなスキャフォールドから構築される人工組織の製造方法も包含する。
【0096】
本発明は、組織工学において使用するための組織スキャフォールドを作製するのに適した方法を提供する。組織の供給源は限定されないが、例示的な態様では、組織が、比較的大きな動物、または(関心対象の組織に関して)ヒトと類似する解剖学的形態を持つと認識されている動物、例えばブタ、ウシ、ウマ、サル、または類人猿に由来する。いくつかの態様では、組織の供給源がヒトであり、その使用により、当該スキャフォールドに基づく人工組織の拒絶の可能性を低減することができる。好ましい態様において、本方法は、肺胞中隔が保存された肺胞アーキテクチャなど、組織のインタクトな血管構造を残す。本明細書において使用する「インタクト」という用語は、ある要素がその要素の元の機能を相当程度果たす能力を有するような状態を指す。
【0097】
ある態様では、脱細胞化肺臓が、正常肺臓マトリックスのいくつかの重要な特徴を保っている。例えば、脱細胞化肺臓は、コラーゲン、エラスチン、フィブロネクチン、およびプロテオグリカンの少なくとも1つまたは複数を含む。
【0098】
脱細胞化組織は、主要組織適合遺伝子複合体(MHC)クラスIまたはII抗原を保っていないので、組織は、レシピエントに投与された時に、有害な免疫応答を引き起こさない。
【0099】
脱細胞化組織は正常な未変性肺臓の機械的性質を保っている。脱細胞化組織は正常な未変性肺臓の障壁機能の一部も保っている。
【0100】
バイオリアクタ
本発明は、組織を脱細胞化しかつ/または再細胞化するためのシステム(例えばバイオリアクタ)を提供する。バイオリアクタは、細胞生存性、細胞の分化状態、および肺臓形態の維持を可能にする。脱細胞化スキャフォールドは、適切な細胞源と共にバイオリアクタ中で培養すると、肺の内皮、上皮、および間葉細胞を含む広範な細胞タイプの接着および増殖を支持することができる。本発明のバイオリアクタには、生体(vivo)環境の重要な特質が組み込まれている。本バイオリアクタは、脱細胞化および/または再細胞化プロセスを最適化するための変更が可能なように設計された。ある態様では、バイオリアクタが、使用者の指定した流量かつ哺乳動物の生理的流量レベルおよび生理的圧力レベル内で、脈管構造を通して培地を灌流させる能力を有する。別の態様では、バイオリアクタが、気管を通して空気または培地で組織(例えば肺臓)を換気する能力を有する。好ましくは、正常な生理学的条件と合致するように陰圧換気法が使用されるが、陽圧を使った換気を行うこともできる。さらに別の態様において、バイオリアクタは、組織の血管コンパートメントと気道コンパートメントを異なる培地タイプで浸すことを可能にする能力を有する。別の態様において、バイオリアクタは、培養培地へのガス交換を可能にすると同時に、換気に関する所望の要件を満たす。別の態様では、バイオリアクタが、圧力測定(例えば肺動脈圧および気管圧の測定)を可能にするためのポート(port)を持つ。好ましくは、圧力は正常な生理学的値の範囲内にある。別の態様では、バイオリアクタが、定期的な培地交換を可能にする手段を持つ。
【0101】
本発明のバイオリアクタは、一般に、組織にカニューレを挿入するための少なくとも1つのカニューレ挿入装置、カニューレを通して培地を灌流させるための灌流器具、および器官または組織の滅菌環境を維持するための手段(例えば封じ込めシステム)を含む。カニューレ挿入装置は、一般に、組織の血管、導管、および/または腔中に導入するための適当なサイズの中空チューブを含む。典型的には、組織中の1つまたは複数の血管、導管、および/または腔にカニューレが挿入される。灌流器具は、液体(例えば細胞破壊培地)のための保持容器および1つまたは複数のカニューレを通して器官越しに液体を移動させるための機構(例えばポンプ、空気圧、重力)を含むことができる。脱細胞化および/または再細胞化中の組織の滅菌性は、本明細書の他の項で論じる方法を使って維持することができる。
【0102】
バイオリアクタは、本明細書に記載するとおり、組織を脱細胞化および再細胞化するために使用することができる。本プロセスは、一定の灌流特徴(例えば圧力、体積、フローパターン、温度、ガス、pH)、機械的力(例えば心室壁運動および心室壁応力)、および電気刺激(例えばペーシング)について監視することができる。灌流の有効性は流出液および組織切片において評価することができる。灌流体積、フローパターン、温度、O2およびCO2分圧、ならびにpHは、標準的な方法を使って監視することができる。
【0103】
バイオリアクタおよび/または組織を監視するために、センサを使用することができる。ソノミクロメトリー、微小検圧法、および/またはコンダクタンス測定を使って、圧力-体積を取得することができる。例えば、センサを使って、カニューレが挿入された器官または組織を通って移動する液体の圧力;系内の周囲温度および/または器官もしくは組織の温度;カニューレが挿入された器官または組織を通って移動する液体のpHおよび/または流量;および/または再細胞化組織の生物学的活性を監視することができる。そのような特質を監視するためのセンサを持つことに加えて、組織を脱細胞化および/または再細胞化するためのシステムは、そのような特質を維持または調節するための手段も含むことができる。そのような特質を維持または調節するための手段は、温度計、サーモスタット、電極、圧力センサ、オーバーフロー弁、液体の流量を変化させるための弁、溶液のpHを変化させるために使用される溶液への流体接続を開け閉めするための弁、風船、外部ペースメーカー、および/またはコンプライアンスチャンバなどの構成要素を含むことができる。安定した状態(例えば温度)の確保に役立つように、チャンバ、リザーバ、およびチューブには水ジャケットを装着することができる。
【0104】
バイオリアクタは、細胞の生存および分化を支持するために、肺臓組織に十分な栄養素供給および機械的刺激を与える能力を有する。バイオリアクタは、インビトロ肺臓組織培養および人工肺臓組織培養に使用することができる。好ましくは、本発明の脱細胞化肺臓スキャフォールドを使って人工肺臓組織を培養するために、バイオリアクタを使用することができる。
【0105】
肺臓組織の真の3次元セグメントをインビトロ培養する能力を有するバイオリアクタの開発は、臨床的に有用な人工肺臓組織の開発における重要な一工程である。例えば、人工肺臓をレシピエントに埋植する前に、人工肺臓組織の成長および成熟をバイオリアクタ中で行うことにより、最終的な埋植肺臓組織のインビボでの機能性を強化することができる。また、肺の生物学、生理学および発生の研究を支援するために、インビトロ肺臓培養のためのバイオリアクタを使用することもできる。すなわち、肺臓内皮および上皮細胞の相互作用による肺胞-毛細管障壁の形成を、本発明の人工肺臓組織およびバイオリアクタを使って研究することができる。当業者は、現在使用されているさまざまな動物モデルよりもよく制御された環境で、肺臓の挙動を研究することが可能になるだろう。人工肺臓組織およびバイオリアクタは、時間も費用もかかるヒト治験または動物治験に進む前に、ヒト組織および動物組織における薬理学的な試験および調査を行うために使用することもできる。
【0106】
組成物
本発明の組成物は、人工肺臓組織を含む。好ましくは、人工肺臓組織は、以下に挙げる性質の任意の1つまたは複数を示す:1)開存性の灌流された脈管構造と、換気することができる開存性の気道樹とが存在する、脈管構造および気道;2)人工肺臓が、レシピエントの生理学的必要を支えるために、気道と血管コンパートメントとの間で十分な気体を交換する能力を有し、最も好ましくは、肺静脈中の酸素分圧が少なくとも50mmHgであるような、ガス交換;3)人工組織が、必要とされる全ての動き(特に呼吸運動および血管灌流)にも外科的埋植中の操作にも耐え得るほど十分に強いような、力学的性質;4)レシピエントに埋植された時に人工肺臓組織が免疫応答を惹起しないような、免疫原性。
【0107】
本発明の組成物および方法は、任意の適切な細胞を使って実施することができる。好ましくは、適切な細胞または細胞群は再生性であり、本発明の脱細胞化組織を再細胞化するために使用することができる。再生性細胞の一例には、幹細胞、胚性幹細胞、成人幹細胞、臍帯血細胞、組織由来の幹細胞または前駆細胞、骨髄由来の幹細胞または前駆細胞、血液由来の幹細胞または前駆細胞、間葉系幹細胞(MSC)、骨格筋由来の細胞、複能性成人前駆細胞(MAPC)、胎児肺細胞、分化肺上皮細胞、肺前駆細胞、血管前駆細胞、分化血管細胞などがあるが、これらに限定されるわけではない。使用することができる他の再生性細胞には、骨髄由来の幹細胞、例えば骨髄単核球(BM-MNC)、内皮または血管幹細胞または前駆細胞、および末梢血由来の幹細胞、例えば内皮前駆細胞(EPC)などがある。
【0108】
好ましくは、適切な細胞は、哺乳動物、より好ましくは霊長類、さらに好ましくはヒトから単離される。本発明の方法において有用な細胞は、本明細書において(例えば実施例の項で)論じる方法を使って、または当技術分野で公知の任意の方法によって、単離される。単離に続いて、その適切な細胞を、培養培地中で培養する。
【0109】
限定ではない一例として、新生児肺細胞(NPC)を細胞の培養に関して詳述する。しかし、適切な細胞に合わせて培養条件を変更できることは、当業者にはわかるだろう。肺細胞の成長を支持する培地製剤には、イーグル最小必須培地、ADC-1、LPM(ウシ血清アルブミン非含有)、F10(HAM)、F12(HAM)、DCCM1、DCCM2、RPMI1640、BGJ培地(フィトン-ジャクソンの改良を含むものおよび含まないもの)、イーグル基礎培地(アールの塩類ベース(salt base)が添加されたBME)、ダルベッコ変法イーグル培地(血清を含まないDMEM)、ヤマネ(Yamane)、IMEM-20、グラスゴー変法イーグル培地(GMEM)、リーボビッツL-15培地、マッコイの5A培地、M199培地(アールの塩類ベースを含むM199E)、M199培地(ハンクスの塩類ベースを含むM199H)、イーグル最小必須培地(アールの塩類ベースを含むMEM-E)、イーグル最小必須培地(ハンクスの塩類ベースを含むMEM-H)およびイーグル最小必須培地(非必須アミノ酸を含むMEM-NAA)などがあるが、これらに限定されるわけではない。
【0110】
本発明の方法に役立つ培地のさらなる限定でない例は、ウシまたは他の種の胎児血清を、少なくとも1%〜約30%、好ましくは少なくとも約5%〜15%、最も好ましくは約10%の濃度で含有し得る。ウシまたは他の種の胎児抽出物は、約1%〜30%、好ましくは少なくとも約5%〜15%、最も好ましくは約10%の濃度で存在することができる。
【0111】
典型的には、NPC培養培地は、基礎培地、血清および抗生物質/抗真菌物質を含む。好ましい基礎培地の一つは、DMEM/F12(1:1)である。好ましい血清はウシ胎児血清(FBS)であるが、ウマ血清またはヒト血清を含む他の血清も使用することができる。好ましくは、NPCの成長を支持するために、20%までのFBSが上記の培地に加えられるであろう。しかし、NPC成長にとって必要なFBS中の成長因子、サイトカインおよびホルモンが同定され、それらが成長培地に適当な濃度で与えられるのであれば、合成培地を使用することもできる。さらに、追加の構成要素を培養培地に加え得ることも、理解される。そのような構成要素には、抗生物質、抗真菌物質、アルブミン、成長因子、アミノ酸、および細胞の培養に関して当技術分野で公知である他の構成要素などが含まれるが、これらに限定されるわけではない。培地に加えることができる抗生物質には、ペニシリンおよびストレプトマイシンがあるが、これらに限定されるわけではない。培養培地中のペニシリンの濃度は1mlあたり約10〜約200単位である。培養培地中のストレプトマイシンの濃度は約10〜約200μg/mlである。しかし本発明は、決して、NPCの培養に関して、どの一培地にも限定されると解釈されるべきではない。むしろ、組織培養において肺細胞を支持する能力を有する任意の培地を使用することができる。
【0112】
また、NPC培養培地には、少なくとも1つの成長因子を補足することもできる。好ましくは、成長因子は線維芽細胞成長因子(FGF)である。例えば、FGF10、FGF7、FGF2の任意の組合せを、NPC培養培地に補足することができる。FGF7の好ましい濃度は、約0.1〜100ng/ml(およびその中間にある任意の整数)であり、より好ましくは、濃度は約10ng/mlである。FGF10の好ましい濃度は約1〜200ng/ml(およびその中間にある任意の整数)であり、より好ましくは、濃度は約25ng/mlである。FGF2の好ましい濃度は約1〜200ng/ml(およびその中間にある任意の整数)であり、より好ましくは、濃度は約25ng/mlである。
【0113】
単離に続いて、NPCは、培養器具中の培養培地において、一定の期間、または細胞がコンフルエントに達するまで、培養した後、細胞を別の培養器具に移すことができる。最初のプレーティングに続いて、細胞を約6日間培養維持することにより、継代第0代(P0)の集団を得ることができる。細胞は、無限回にわたって継代することができ、各継代は、細胞を約6〜7日間培養することを含み、その間、細胞倍加時間は約3〜約5日の範囲に及び得る。培養器具は、細胞をインビトロで培養するために一般に使用される任意の培養器具であることができる。
【0114】
NPCは、ある期間にわたって、または細胞が一定レベルのコンフルエンスに達するまで、FGFを補足した培養培地中で培養することができる。好ましくは、コンフルエンスのレベルは70%より高い。より好ましくはコンフルエンスのレベルは、90%より高い。期間は、インビトロでの細胞の培養に適した任意の期間であることができる。NPC培養培地は、NPCの培養中はいつでも取り替えることができる。好ましくは、3〜4日ごとに培養培地を取り替える。次に、NPCを培養器具から収集し、その後、直ちにそれらを使用するか、低温保存して、後に使用するまで貯蔵しておくことができる。NPCはトリプシン処理、EDTA処理、または培養器具から細胞を収集するために使用される他の任意の手法によって、収集することができる。
【0115】
本明細書において記載するNPCは、常法に従って低温保存することができる。好ましくは、約100万〜1000万個の細胞を、10%DMSOを含有する培養培地に入れて、液体N2の気相中で低温保存する。凍結細胞は、37℃の浴中で旋回させることによって融解し、新鮮な成長培地中で再懸濁し、上述のように拡大することができる。
【0116】
本発明は、スキャフォールドに「播種(seed)」する細胞も提供する。NPCはスキャフォールド上で培養することができる。細胞は、細胞を分化培地中で培養することにより、インビトロで分化することもできる。あるいは、細胞が哺乳動物内で組織と接触するか、細胞が、組織から放出される物質(例えば成長因子、酵素、またはホルモン)による影響を受けるように、組織の十分近くにある場合には、細胞はインビボで分化することもできる。言い換えると、マトリックスのNPCは、組織からのシグナルを受け取ることにより、肺臓などの組織と接触することができる。そのようなシグナリングは、例えば、NPCの表面上またはNPCに由来する細胞の表面上にある受容体が、哺乳動物内で組織によって放出された成長因子、酵素、またはホルモンなどの分子を結合し、その分子からのシグナルを伝達する場合に、起こるだろう。これらの作用物質は分化を誘導するので、NPCは、それらが置かれた組織中の分化細胞によって通常発現されるものと同じタンパク質の一部を、そして場合によっては(全てではないとしても)大半を、発現するようになる。
【0117】
上記の代わりに、または上記に加えて、マトリックスのNPCは、その細胞の環境に、ある物質(例えば成長因子、酵素、ホルモン、または他のシグナリング分子)を加えることによって、分化するように誘導することができる。例えば、ある物質を、本発明の生物学的足場に加えることができる。
【0118】
NPCとそれに付随する細胞マトリックスは最終的に完全に分化した状態になることができ、状況によっては(例えば組織学的に成熟した完全な組織を再現するために細胞を使用する場合には)それが望ましい場合もあるが、処置を成功させるには、投与された細胞の全てが完全に分化する必要はなく、細胞マトリックスのNPCは、哺乳動物を処置するのに十分な点まで分化するだけでよい。その点には、マトリックスを患者に投与する前に到達するか、マトリックスを患者に投与した後に到達することができる。
【0119】
分化は、マトリックスの細胞が、埋植部位にある成熟細胞と基本的に同じ表現型を発現する場合に起こる。例えば、本発明の定義を目的とする場合、肺臓中に埋植された細胞マトリックスのNPCは、それが肺臓(例えば肺胞上皮細胞)によって発現されるタンパク質と本質的に同じタンパク質を発現する場合に、分化しているという。肺臓マーカーに対する抗体は市販されているか、そうでなくても、容易に取得することができる。
【0120】
分化細胞は、その外部形態によって、およびそれらが他の細胞と形成する接続によって、同定することもできる。例えば、肺臓細胞に分化する細胞は、細気管支に似た複雑な形態を発生することができる。例えば本発明は、三次元スキャフォールド上でNPCを培養することにより、成熟した肺臓細胞の特徴を示すことができるという、新規な発見に基づいている。
【0121】
器官または組織を生成させるために脱細胞化器官中および脱細胞化器官上に導入される細胞の数は、器官(例えば器官の種類、器官のサイズおよび重量)または組織にも、再生性細胞のタイプおよび発生段階にも依存する。異なるタイプの細胞は、その細胞が到達するであろう集団密度に関して、異なる傾向を持ち得る。同様に、異なる器官または組織は異なる密度で細胞化され得る。例えば、脱細胞化器官または組織には、少なくとも約1,000(例えば少なくとも10,000、100,000、1,000,000、10,000,000、または100,000,000)個の再生性細胞を播種することができ、あるいは、脱細胞化器官または組織は、そこに付着した約1,000細胞/mg組織(湿重量、すなわち脱細胞化前)〜約10,000,000細胞/mg組織(湿重量)を持つことができる。
【0122】
細胞は、1つまたは複数の場所への注入によって、脱細胞化器官または組織に導入することができる。また、2タイプ以上の細胞(すなわち細胞のカクテル)を脱細胞化器官または組織に導入することができる。例えば、細胞のカクテルを脱細胞化器官または組織中の複数の場所に注入するか、異なる細胞タイプを脱細胞化器官または組織の異なる部分に注入することができる。注入の代わりに、または注入に加えて、再生性細胞または細胞のカクテルは、カニューレを挿入した脱細胞化器官または組織への灌流によって導入することができる。例えば、細胞は灌流培地を使って脱細胞化器官中に灌流することができ、次にその灌流培地を拡大培地および/または分化培地に変えて、再生性細胞の成長および/または分化を誘導することができる。肺臓組織の場合、細胞は、気管を通して気道コンパートメントに導入するか、肺動脈もしくは肺静脈を通して血管コンパートメントに導入するか、またはその両方に導入することができる。
【0123】
再細胞化中、器官または組織は、再生性細胞の少なくとも一部が脱細胞化器官または組織内および脱細胞化器官または組織上で増倍しかつ/または分化することができるような条件下に維持される。それらの条件には、適当な温度および/または圧力、電気的および/または機械的活動、力、適当な量のO2および/またはCO2、適当な量の湿気、および滅菌またはほぼ滅菌状態が含まれるが、これらに限定されるわけではない。再細胞化中、脱細胞化器官または組織およびそこに付着した細胞は、適切な環境に維持される。例えば細胞は、栄養補給(例えば栄養素および/またはグルコースなどの炭素源)、外因性のホルモンもしくは成長因子、および/または特定のpHを必要とし得る。
【0124】
細胞は、脱細胞化器官または組織にとって同種異系(例えばヒト細胞を播種されたヒト脱細胞化器官または組織)であるか、再生性細胞は脱細胞化器官または組織にとって異種(例えばヒト細胞を播種されたブタ脱細胞化器官または組織)であることができる。
【0125】
いくつかの例では、本明細書に記載する方法によって生成させる器官または組織が、患者に移植されるものである。これらの場合、脱細胞化器官または組織を再細胞化するために使用される細胞は、再生性細胞が患者にとって自家細胞になるように、患者から得ることができる。患者の細胞は、例えば、当技術分野において公知の方法を使って、血液、骨髄、組織、または器官から、人生の異なる段階で(例えば、出生前または周産期に、思春期中に、または成人として)得ることができる。あるいは、脱細胞化器官または組織を再細胞化するために使用される細胞は、患者にとって同種同系の細胞(すなわち、一卵性双生児の一方から得られるもの)であるか、細胞は、例えば患者の親類から得られる、または患者とは親類関係にないHLA適合個体から得られる、ヒトリンパ球抗原(HLA)適合細胞であるか、または細胞は、例えば非HLA適合ドナーから得られる、患者にとって同種異系の細胞であることができる。
【0126】
細胞の供給源(例えば自家であるかないか)とは関係なく、脱細胞化固形臓器は、患者にとって、自家、同種異系または異種であることができる。
【0127】
一定の例では、脱細胞化組織が、インビボで(例えば組織が個体中に移植された後で)、細胞によって再細胞化され得る。インビボ再細胞化は、例えば本明細書に記載する細胞のいずれかを使って、上述のように行うことができる(例えば注入および/または灌流)。上記に代えて、または上記に加えて、内在性細胞による脱細胞化器官または組織のインビボ播種も、自然に起こるか、または再細胞化組織に送達される因子によって媒介され得る。
【0128】
遺伝子改変
本発明は、哺乳動物における肺臓細胞治療を容易にするために本発明の脱細胞化組織を使用することができるという発見に関する。
【0129】
別の態様において、脱細胞化肺臓組織は、肺上皮細胞などの所望の肺臓細胞を培養するために使用することができる。細胞は、遺伝子改変されていてもされていなくても、気腫、閉塞性細気管支炎、および嚢胞性線維症を含む(ただしこれらに限定されるわけではない)肺臓疾患を処置するために使用することができる。例えば、本発明の脱細胞化組織は、ヒト肺気道上皮細胞を培養するための基質として使用することができる。次に、培養ヒト気道上皮細胞を、例えば気管点滴注入、吸入または注入などによって、レシピエントに送達することができる。培養下に拡大されるそのような細胞は、レシピエントにおける治療を達成するために使用することができる。脱細胞化肺臓組織(例えば気管)は、組織培養プラスチック製品などの典型的な細胞培養環境では成長させることが通常は極めて困難な肺上皮細胞を培養し拡大するための傑出したプラットフォームになる。
【0130】
遺伝子治療との関連では、レシピエントの肺臓への細胞の送達に先だって、脱細胞化組織上で培養される細胞を、関心対象の遺伝子で処理することができる。いくつかの例では、そのような細胞ベースの遺伝子送達が、アデノウイルス遺伝子送達ベクターの吸入などといった肺臓への他の遺伝子送達手段に対して、著しい利点を示すことができる。宿主への細胞ベースの遺伝子送達が持つこの優位性は、吸入された遺伝子ベクターが典型的には、粘液層および宿主免疫系によって課される障壁により、低い細胞形質導入効率をもたらすという知見に由来する。細胞中に前もって挿入された治療遺伝子の送達では、レシピエント肺臓細胞への遺伝子治療ベクターの貫入に付随する問題が回避される。
【0131】
本発明の脱細胞化肺臓組織は、標準的な細胞培養プラスチック製品での培養と比較して、上皮細胞などの肺臓細胞を高度に生存可能かつ分化した状態で成長させるための便利で効率のよい手段を提供する。そして、脱細胞化マトリックス上での肺上皮細胞などの肺細胞の拡大は、細胞治療にとって有効であるような十分に多数の細胞を与える。また、脱細胞化マトリックス上での肺臓上皮細胞の拡大は、インビトロで培養細胞を遺伝子治療ベクターで処理するためのプラットフォームにもなる。選ばれた遺伝子をインビトロでトランスフェクトされた細胞は、次に、任意で、関心対象の導入遺伝子を発現する細胞だけを選択するために精製し、次いで、そのような細胞治療を必要とするレシピエントに導入することができる。そのようなアプローチは、嚢胞性線維症などの遺伝的肺臓疾患の処置において、とりわけ価値があるだろう。
【0132】
ある態様において、本発明は、嚢胞性線維症を処置する方法を提供する。本方法は、上皮細胞などの関心対象の細胞に、正常型のCFTR遺伝子(この遺伝子の突然変異型が嚢胞性線維症の原因遺伝子である)をトランスフェクトする工程を含む。そのようなトランスフェクト細胞を、気管への点滴注入、吸入、または他の導入手段によって患者に送達すれば、これらの患者への遺伝子ベクターの送達に付随してきた著しい困難が軽減される。こうして、嚢胞性線維症における有効な細胞治療および遺伝子送達を実現することができる。しかし本発明はCFTR遺伝子をトランスフェクトした細胞による嚢胞性線維症の処置だけに限定されるべきではない。むしろ本発明は、肺臓細胞に関連する任意の疾患または障害の処置を包含する。
【0133】
したがって本発明は、本発明の脱細胞化組織上で培養された遺伝子改変細胞(例えば肺細胞)の使用を提供する。遺伝子改変は、例えば外因性遺伝子(「導入遺伝子」)の発現をもたらすか、内在性遺伝子の発現の変化をもたらし得る。そのような遺伝子改変は治療上の利益を持ち得る。あるいは、遺伝子改変は、例えば本発明の組成物を個体に埋植した後に、そのように修飾された細胞を追跡しまたは同定するための手段にもなり得る。細胞の追跡には、移植された遺伝子改変細胞の移動、同化および生存を追跡することが含まれ得る。遺伝子改変は少なくとも第2の遺伝子を含むこともできる。第2の遺伝子は、例えば、選択可能な抗生物質耐性遺伝子または他の選択可能マーカーをコードし得る。
【0134】
細胞を追跡するのに有用なタンパク質には、緑色蛍光タンパク質(GFP)、他の蛍光タンパク質のいずれか(例えば強化緑色、シアン、黄色、青色および赤色蛍光タンパク質;Clontech、カリフォルニア州パロアルト)または他のタグタンパク質(例えばLacZ、FLAG-tag、Myc、His6など)などがあるが、これらに限定されるわけではない。
【0135】
細胞の遺伝子改変の目的が生物学的活性物質の産生にある場合、その物質は一般に、所与の障害の処置にとって有用なものであるだろう。例えば、細胞が骨または軟部組織形成に関連する一定の成長因子産物を分泌するように、細胞を遺伝子改変することが望ましい場合があるだろう。組織修復に関係する他の内在性細胞タイプの成長を誘導するための成長因子産物も有用である。例えば、内在性毛細管および/または微小血管内皮細胞を刺激するための成長因子は、軟部組織欠陥の修復において、特に大体積の欠陥に、役立ち得る。
【0136】
本発明の細胞は、細胞の培養にとって有益な分子、例えば栄養因子、成長因子、サイトカインなどを産生するように外因性遺伝物質を細胞中に導入することにより、遺伝子改変することができる。また、そのような分子を産生するように細胞を遺伝子改変することにより、細胞は、それを必要とする哺乳動物中に移植された時に、哺乳動物に対して追加の治療効果を与えることができる。例えば遺伝子改変細胞は、哺乳動物中の移植部位に近接する細胞にとって有益な分子を分泌することができる。
【0137】
肺細胞は、当業者に知られる任意の方法を使って遺伝子改変することができる。例えばSambrook et al.(2001)「Molecular Cloning: A Laboratory Manual」(Cold Spring Harbor Laboratory Press、ニューヨーク州コールドスプリングハーバー)およびAusubel et al.編(1997)「Current Protocols in Molecular Biology」(John Wiley & Sons、ニューヨーク州ニューヨーク)を参照されたい。例えば肺細胞を、導入遺伝子を含む核酸が含まれている発現ベクターに暴露して、導入遺伝子が細胞内で発現されるのに適当な条件下で核酸が細胞内に導入されるようにする。導入遺伝子は一般に、適切なプロモーターに機能的に連結されたポリヌクレオチドを含む発現カセットである。ポリヌクレオチドはタンパク質をコードするか、生物学的に活性なRNA(例えばアンチセンスRNAまたはリボザイム)をコードすることができる。したがって、例えばポリヌクレオチドは、毒素に対する耐性を付与する遺伝子、ホルモン(例えばペプチド成長ホルモン、ホルモン放出因子、性ホルモン、副腎皮質刺激ホルモン、サイトカイン(例えばインターフェリン(interferin)、インターロイキン、リンフォカイン)など)、細胞表面結合型細胞内シグナリング部分(例えば細胞接着分子、ホルモン受容体など)、所与の系譜の分化を促進する因子(例えば骨形態形成タンパク質(BMP))などをコードすることができる。
【0138】
発現カセット内では、コードポリヌクレオチドが適切なプロモーターに機能的に連結される。適切なプロモーターの例には、原核生物プロモーターおよびウイルスプロモーター(例えばレトロウイルスITR、LTR、前初期ウイルスプロモーター(IEp)、例えばヘルペスウイルスIEp(例えばICP4-IEpおよびICP0-IEEp)、サイトメガロウイルス(CMV)IEp、および他のウイルスプロモーター、例えばラウス肉腫ウイルス(RSV)プロモーター、およびマウス白血病ウイルス(MLV)プロモーター)が含まれる。他の適切なプロモーターは、真核生物プロモーター、例えばエンハンサー(例えばウサギβ-グロビン調節要素)、構成的に活性なプロモーター(例えばβ-アクチンプロモーターなど)、シグナル特異的プロモーター(例えばRU486に応答するプロモーターなどの誘導性プロモーター)、組織特異的プロモーターである。事前に定められた細胞との関連で遺伝子発現を駆動するのに適したプロモーターを選択することは、当技術分野の技能で十分に可能である。発現カセットは、2つ以上のコードポリヌクレオチドを含むことができ、所望どおりに他の要素(例えばポリアデニル化配列、膜挿入シグナルまたは分泌リーダーをコードする配列、リボソームエントリー配列、転写調節要素(例えばエンハンサー、サイレンサーなど)など)を含むことができる。
【0139】
導入遺伝子を含有する発現カセットは、細胞に導入遺伝子を送達するのに適した遺伝子ベクター中に組み込まれるべきである。所望する最終用途に応じて、任意のそれらベクターを、細胞を遺伝子改変するために、そのように使用することができる(例えばプラスミド、裸のDNA、アデノウイルス、アデノ随伴ウイルス、ヘルペスウイルス、レンチウイルス、パピローマウイルス、レトロウイルスなどのウイルスなど)。そのようなベクター内に所望の発現カセットを構築する任意の方法を使用することができ、それらの多くは当技術分野において周知である(例えば直接クローニング、相同組換えなど)。ベクターの選択が、細胞中にベクターを導入するために使用される一般に当技術分野において公知である方法(例えばプロトプラスト融合による方法、リン酸カルシウム沈殿法、遺伝子銃、エレクトロポレーション、DEAEデキストランまたは脂質担体によるトランスフェクション、ウイルスベクターによる感染など)を、おおむね決めることになる。
【0140】
ポリメラーゼ連鎖反応(PCR)、リガーゼ連鎖反応(LCR)、および他のDNAまたはRNAポリメラーゼ媒介技法を含むインビトロ増幅法を当業者に実行させるのに足りる技法の例が、Sambrook et al.「MOLECULAR CLONING: A LABORATORY MANUAL」第1〜3巻(第3版、Cold Spring Harbor Press、NY 2001)に見いだされる。
【0141】
いったんあるタンパク質に関して核酸をクローニングしたら、当業者は、さまざまな肺臓細胞中で組換え遺伝子を発現させることができる。当業者は、所望の導入遺伝子を発現させるために利用することができる数多くの発現系を知っていると予期される。
【0142】
本発明は、天然肺臓組織を模倣する人工三次元組織を提供する。天然肺臓組織を模倣する複合材料およびスキャフォールドを作り出す能力は、先行技術の方法より高度な組織および組織集合体の修復および再生を可能にし、先行技術の方法を使って達成できるものよりも、より正確な組織学的構造および機能を示す。例えば、人工肺臓組織は、出芽構造および伸長する管状構造を示す細胞を含む。そのうえ、細胞は、形態形成および肺臓上皮分化に関与する遺伝子を発現する。形態形成および肺臓上皮分化に関与する遺伝子には、遠位上皮マーカー遺伝子SpCおよびSpB、間葉由来モルフォゲンFGF10、FGFr2、および血管内皮成長因子A(VEGF)などがあるが、これらに限定されるわけではない。
【0143】
投与
本発明では、インビトロ環境とインビボ環境の両方での人工組織の使用が考えられる。したがって本発明は、研究目的および治療目的もしくは医学的/獣医学的目的での人工組織の使用を提供する。研究環境では、この技術に関して、数多くの実用的応用が存在する。そのような応用の一例は、エクスビボがんモデル(例えばさまざまなアブレーション技法(例えば放射線処置、化学療法処置、またはそれらの組合せを含む)の有効性を実験室で調べ、そうすることによって処置方法を最適化するために病気の患者を使用することを避けるためのものなど)における人工組織の使用である。例えば、新しく摘出した肺臓をバイオリアクタに取り付け、その肺臓を処理して組織をアブレートすることができる。インビボ使用の別の例は、組織工学のための使用である。
【0144】
本発明の人工組織はインビボでも役立つ。数ある用途のなかでも、対象(本明細書においては「患者」と互換的に使用され、ヒトと動物の両方を包含するものとする)のインビボ処置法を挙げることができる。一般に、一定の態様に関して、対象の処置方法は、対象中または対象の表面に本発明の人工組織を埋植する工程を含み、組織の埋植は対象における検出可能な変化をもたらす。検出可能な変化は、自然の知覚を使ってまたは人造の装置を使って検出することができる任意の変化であることができる。本発明では任意のタイプの処置(例えば疾患または障害の治療的処置、皮膚の斑点の美容的処置など)が想定されるが、多くの態様では、処置が、対象の疾患、障害、または他の苦痛の原因の治療的処置である。したがって検出可能な変化は、対象を冒す疾患または障害の少なくとも1つの臨床症状における変化(好ましくは改善)の検出であることができる。例示的なインビボ治療法には、腫瘍処置後の器官の再生、医学的装置を埋植するための手術部位の調製、皮膚グラフティング、および組織または器官(疾患または障害によって損傷を受けたか破壊されたものなど)の一部または全部の代用が含まれる。例示的な器官または組織には、以下に挙げるものが含まれる:心臓、肺臓、肝臓、腎臓、膀胱、脳、耳、目、または皮膚。対象はヒトまたは動物であることができるという事実を考慮すると、本発明は医学的応用と獣医学的応用の両方を持つ。
【0145】
ある態様において、本方法は、処理される組織の細胞を殺し、組織スキャフォールドを作り出すために、組織を本発明の脱細胞化方法に暴露する工程を含む。本方法はさらに、組織スキャフォールドに細胞を播種する工程、および播種された細胞を組織スキャフォールドの中または上で増殖させる工程を含むことができる。増殖により、健常で機能的な細胞を含有する再生組織が作製される。
【0146】
本発明は、人工肺臓組織を、それを必要とする哺乳動物中に埋植することによって、患者を処置する方法も提供する。いくつかの例では、人工肺臓組織が、適切な細胞、例えばNPCを含む。しかし本発明は、いかなる特定タイプの細胞にも限定されるべきでない。埋植後、グラフトされた細胞は、それに内在性組織の特徴を発生させる環境キューに応答することができる。好ましくは、細胞は、導管構造を形成する分化遠位上皮細胞(proSpC発現性)から構成される組織型(histiotypic)肺胞様構造を形成する。このように、埋植された細胞は、それを周囲の組織に似せる特徴を発生させるであろう。これらの方法を使って、生物学的足場は組織を増強することができ、本発明の生物学的足場は、組織工学に使用することができ、従来のいかなる組織工学的背景でも使用することができる。
【0147】
したがって本発明は組織再生への応用を包含する。組織再生治療アプローチの目的は、高密度の修復コンピテント細胞(または局所環境による影響を受けたときにコンピテントになることができる細胞)を、初期の創傷力学と最終的な新組織生成の両方を最適化するような形で、欠陥部位に送達することである。本発明の組成物は、個体における肺臓組織欠陥を軽減または処置するのにとりわけ有用である。有利なことに、本発明の組成物は、改善された肺臓組織再生をもたらす。具体的には、本発明の組成物の結果として、組織再生が、より迅速に達成される。
【0148】
有利なことに、本発明の組成物および方法は、先行技術の方法に対する改善を表す。好ましくは、肺臓欠陥を処置するために使用される組成物は、本明細書の他の項で説明するように、NPCを含み、より好ましくは、3次元培養物を生成するようにスキャフォールド上に播種されてインビトロで培養されたNPCを含む。
【0149】
創薬のためのモデル
本発明は、肺臓の疾患または障害に関して、試験化合物の治療活性の評価を可能にするのに適したインビトロ法を提供する。好ましくは、本方法は、人工三次元肺臓組織の使用を含む。
【0150】
本方法は、脱細胞化組織を使って開発されたモデルに基づく。いくつかの例では、脱細胞化組織に適切な細胞を播種することができる。いくつかの例では、上皮、間葉、および内皮細胞を含有するNPCの混合集団を使って、三次元人工肺臓組織を生成させる。例えば、NPCを三次元脱細胞化肺臓組織内に置く。こうして、本モデルには、成長および近接細胞との細胞間コミュニケーションに対するNPCの影響が組み入れられる。三次元肺臓組織は天然肺臓組織を模倣し、例えば人工肺臓組織は天然肺臓組織によって例示される分枝形態形成を示す。
【0151】
本モデルは肺臓組織の病変に対して薬物を試験するのに役立つ。また、本モデルは、肺臓組織の病変に対する、特定の治療剤送達媒体の効果を調べて、例えば異なる送達系によって投与された同じ薬剤の効果を比較するために使用するか、または単に送達媒体そのもの(例えばウイルスベクター)が肺臓の病変に影響を及ぼす能力を有するかどうかを評価するために使用することができる。
【0152】
ある態様において、本発明は、肺臓組織の健康状態を調整する試験作用因子の能力について、試験作用因子をスクリーニングするためのインビトロ法を提供する。本方法は、試験作用因子を人工三次元肺臓組織モデルと接触させる工程、および試験作用因子が肺臓組織モデルに対して持つ効果を測定する工程を含む。試験作用因子の存在下でモデルに加わる変化はいずれも、その試験作用因子が肺臓組織の健康状態を調整できることのしるしである。
【0153】
別の態様において、本発明は、ある試験作用因子が肺臓組織に対して持つ効果を観察するためのインビトロ法であって、以下の工程を含む方法を提供する:
a)正常肺臓組織をモデル化することを意図した少なくとも1つの三次元肺臓組織モデルを用意する工程;
b)試験作用因子を肺臓組織モデルと接触させる工程;および
c)試験作用因子が肺臓組織モデルに対して持つ効果を観察する工程。
【0154】
本組織モデルは、スキャフォールド(例えばコラーゲンマトリックス)上の三次元的細胞アレイと、少なくとも1つの試験細胞とを含む構築物である。本方法は、肺臓組織の病変に対する試験作用因子の効果を観察する工程を含む。しかし、本方法は、肺臓組織上の個々の細胞タイプに対する試験作用因子の効果を観察する工程をさらに含んでもよい。
【0155】
試験作用因子は、化学剤(例えば毒素)、医薬、ペプチド、タンパク質(例えば抗体、サイトカイン、酵素など)および核酸(タンパク質、アンチセンス剤(すなわち、ターゲット細胞タイプ中で発現するターゲットRNAに相補的な配列を含む核酸、例えばRNAiまたはsiRNA)、リボザイムなどの治療剤をコードし得る、遺伝子医薬品および導入された遺伝子を含む)を含む、任意の作用因子であることができる。上記に加えて、または上記に代えて、試験作用因子は、放射線(例えば電離放射線、UV光または熱)などの物理的作用因子であってもよく、これらは単独で試験するか、化学剤および他の作用因子と組合せて試験することができる。
【0156】
本モデルは送達媒体を試験するためにも使用することができる。これらは、従来の医薬製剤から遺伝子送達媒体まで、任意の形態をとり得る。例えば本モデルは、2つまたはそれ以上の異なる送達系(例えばデポー製剤と放出制御製剤)によって投与された同じ薬剤の治療効果に対する効果を比較するために使用することができる。ある特定媒体がそれだけで肺臓組織に対して効果を持つかどうかを調べるためにも使用することができる。遺伝子ベースの治療薬の使用が増加するにつれて、考え得る種々の送達系に付随する安全性の問題が、ますます重要になる。したがって本発明のモデルは、例えば裸のDNAまたはRNAなどの核酸治療薬のための送達系、ウイルスベクター(例えばレトロウイルスベクターまたはアデノウイルスベクター)、リポソームなどの性質を調べるために使用することができる。したがって試験作用因子は、任意の治療剤が付随しているまたは付随していない、任意の適当なタイプの送達媒体であることができる。
【0157】
試験作用因子は、試験されるモデルに、任意の適切な手段を使って加えることができる。例えば、試験作用因子をモデルの表面上に滴下して、拡散または他の方法でモデルに進入させるか、試験作用因子を栄養培地に加え、コラーゲンゲルを通して拡散させることができる。本モデルは、電離放射線、UV光または熱などの物理的作用因子の効果を、単独で、または化学剤と組み合わせて(例えば光線力学的治療において)、試験するのにも適している。
【0158】
試験作用因子がモデルに対して持つ効果の観察は、さまざまな方法を使って達成することができる。例えば、ある特定の作用因子は、アポトーシスに入るように細胞を誘導するかもしれない。細胞における検出可能な変化には、細胞の面積、体積、形状、形態、マーカー発現(例えば細胞表面マーカー発現)または他の適切な特徴、例えば染色体断片化が含まれ得る。細胞増殖に対する試験作用因子の効果を観察するために細胞数を監視することもでき、これは、存在する特定細胞タイプの数を計数することなどによって直接的に分析するか、特定細胞塊のサイズを測定することなどによって間接的に分析することができる。これらは、例えば適切な蛍光細胞染色などを使って、インタクトなモデル上で直接的または間接的に観察することができる。これは、生きているモデルを連続分析するための、生体染色色素または遺伝子的に導入された蛍光マーカー(例えば緑色蛍光タンパク質)による細胞の事前標識(pre-labeling)によって行うか、固定化と、ヨウ化プロピジウムまたは蛍光標識抗体などの蛍光物質による事後標識(post-labeling)とによって行うことができる。あるいは、特定のmRNA種の発現について試験するために、通常の組織学的方法、例えば適切な細胞ターゲットに対する抗体を使用する免疫組織化学、またはインサイチューハイブリダイゼーションによって、モデルを処理することもできる。さらにまた、コンピュータシステムおよびソフトウェアを使って細胞をさまざまな時点でイメージングし、例えば細胞の密度、場所および/または形態などの任意の変化を検出することによって、これを自動/ロボット的または半自動的に行うこともできる。特に、共焦点レーザー走査型顕微鏡法は、インタクトなモデルの三次元解析を可能にする。したがって、通常は従来の二次元培養中の細胞に対してのみ可能である細胞挙動の定量的解析を、インタクトな三次元肺臓組織モデルに直接応用することが可能である。このようにして、細胞の増殖、アポトーシス、壊死、移動およびマトリックス浸潤、その他の定量的連続分析が、従来の二次元細胞培養と生きている動物モデルの間の間隙を埋める三次元肺臓組織モデルで得られる。
【0159】
実験例
以下に実験例を挙げて本発明をさらに詳しく説明する。これらの実施例は、例示のために記載するに過ぎず、別段の指定がない限り、限定を意図するものではない。したがって本発明は、決して、以下の実施例に限定されると解釈されるべきではなく、本発明は、本明細書が提供する教示の結果として明白になる、ありとあらゆる変形を包含すると解釈すべきである。
【実施例】
【0160】
実施例1:ラット肺臓の脱細胞化および脱細胞化スキャフォールドの形態学的特徴付け
脱細胞化器官は、組織工学スキャフォールドとしての使用に、いくつかの利点をもたらす。ある局面において、脱細胞化スキャフォールドは、肺臓の場合であれば血管系および気道網を含む組織機能のための適当な三次元構成を持つ。また、細胞外マトリックス(ECM)構成要素は、種を超えて広く保存されているので、異種埋植時に脱細胞化スキャフォールドが免疫応答を誘導する可能性は低い[Bernard et al., 1983, Biochemistry 1983;22:5213-23]。別の局面において、未変性ECMは、細胞の付着、伝播、成長および分化にとって、最適な基質を提供する。
【0161】
本発明の脱細胞化プロセスの目標は、肺臓のECMの重要な局面を保ちかつ肺臓のECMに対するいかなる損傷も最小限に抑えつつ、細胞材料および核材料を除去することである。本明細書に提示する結果は、未変性肺臓組織を脱細胞化して、細胞構成要素および抗原性分子を除去し、しかも重要な細胞外マトリックス分子は保つことが可能であることを証明している。ある局面において、本発明の脱細胞化プロセスの目標は、細胞培養と完全に適合すると同時に障壁機能を提供する脱細胞化肺臓スキャフォールドを生成させることである。また、脱細胞化肺臓スキャフォールドは、インタクトな気道樹および血管網を持つことが望ましい。
【0162】
この研究では化学的な脱細胞化法を使用した。この研究で使用した化学薬品には、塩化ナトリウム、CHAPS、およびEDTAが含まれる。高張塩化ナトリウム溶液は、細胞を効率よく溶解することができるが、組織からの細胞構成要素の除去を補助することはない。CHAPSは両性イオン性界面活性剤であり、これは細胞材料の効率の良い可溶化を可能にし、それゆえに細胞材料の効率の良い除去を可能にする。EDTAは、ECMへの細胞付着の破壊を助ける重要な二価イオン(すなわちCa2+)を結合するキレート剤である。また、溶液は高いアルカリ性を持ち、それが、細胞質細胞構成要素と、可溶化しなければマトリックスを詰まらせるGAGとを、可溶化するのに役立つ[Gilbert et al., 2008 J Surg Res 152(l):135-9]]。
【0163】
これらの実験で使用した材料および方法を以下に説明する。
【0164】
材料および方法
器官収集
肺臓を若年成体(3ヶ月齢)雄フィッシャー344ラットから収集した。動物実験の作業は全て、イェール大学施設内動物実験委員会(Yale University Institutional Animal Care and Use Committee)からの承認を受けて行われた。動物をペントバルビタールナトリウム(Sigma、40mg/kg)の腹腔内注射によって麻酔した。麻酔の誘導後に、肋骨縁の直下で横切開によって腹部に入った。横隔膜を穿刺し、胸郭を切って、肺臓を露出させた。50U/mlヘパリン(Sigma)を含有するPBSで右心室を通して肺臓を灌流した。灌流が完了した後、心臓、肺臓および気管を切り離し、ひとまとめにして取り出した。
【0165】
バイオリアクタ構成要素
バイオリアクタ構成要素はCole-Parmer(イリノイ州バーノンヒルズ)から入手した。シリコーン栓と500mlのガラス製広口瓶とがバイオリアクタの基礎をなした。灌流ループおよび空気換気を含む、肺臓への必要な接続が可能になるように、シリコーン栓を通して、サイズL/S14およびL/S16のPharMedチューブ(オハイオ州ウェストレーク)を挿入した。圧力は、灌流ポンプと肺動脈への接続との間のTruWave圧変換器(Edwards Lifesciences、カリフォルニア州アービン)を使って監視した。灌流はMasterflex L/S可変速ローラーポンプ(Masterflex、イリノイ州バーノンヒルズ)を使って達成した。
【0166】
脱細胞化プロセス
脱細胞化に使用した流体は、PBS中の8mM CHAPS、1M NaCl、25mM EDTAだった。化学薬品は全てSigmaから入手し、PBSはGibcoから入手した。バイオリアクタを脱細胞化流体で満たし、バイオリアクタを37℃に保たれたインキュベータに移した。灌流圧は肺動脈幹への流入時に監視し、30または20mmHg未満に維持した。脱細胞化流体は以下の時点で新鮮な流体と交換した:30分、1時間、2時間、4時間、6時間。大半の条件について、4時間後または6時間後に脱細胞化を停止した。
【0167】
DNAアッセイ
Quant-iT PicoGreen dsDNAアッセイキット(Invitrogen、オレゴン州ユージーン)を、製造者の説明書に従って使用することにより、組織のDNA含量を定量した。簡単に述べると、組織試料を重量測定し、凍結乾燥し、TE緩衝液で希釈し、Quant-iT PicoGreen試薬と混合した。蛍光を485nmで励起して535nmで測定し、標準曲線を使ってDNA含量を定量した。未変性試料と脱細胞化試料の両方について、少なくとも4つの試料を測定した。
【0168】
ウェスタンブロット
ウェスタンブロッティング用の組織を、プロテアーゼ阻害剤(Sigma)を添加した冷RIPA緩衝液(Boston Bioproducts)中で消化し、15,000rpmで30秒間ホモジナイズした。4℃で1時間インキュベートした後、14,000gで25分間の遠心分離によって、不溶性粒子を除去した。タンパク質濃度をBradfordアッセイ[Bradford, 1976, Anal Biochem 72:248-54]によって定量した後、Laemmli還元緩衝液(Boston Bioproducts)中、65℃で25分間、煮沸した。試料を分析まで-80℃で保存した。25〜30μgのタンパク質を使って、さまざまなパーセンテージのポリアクリルアミドゲルで、試料を泳動した。電気泳動後に、タンパク質をニトロセルロース膜に転写した。膜をTBS中ですすいだ後、0.05%Tween-20を含むTBS(TBS-T)中の5%脱脂粉乳(NFDM)または3%ウシ血清アルブミン中で、1時間ブロッキングした。一次抗体を、TBS-T中の2%NFDMまたは3%BSA中で、一晩適用した。二次抗体は、ロバまたはヤギにおいて産生させたものを、Santa Cruzから入手し、1:2000の希釈度で室温において1時間適用した。Supersignal West Picoから得た基質(これを5分間適用してから、フィルム現像した)を使ってタンパク質を検出した。
【0169】
免疫蛍光
組織ブロックを3.7%ホルムアルデヒド(Sigma)中で4時間固定した後、70%エタノールに移し、パラフィンに包埋した。イェール大学組織学中核施設(Yale University Histology core facility)によって薄い(5μm)切片が調製された。組織切片をキシレン中で脱パラフィンし、エタノール勾配によって再水和し、緩衝液(PBS+0.2%Triton-X)中で15分間すすいだ。抗原回復(antigen retrieval)を0.01Mクエン酸、pH6.0中、70℃で20分間行った。室温まで冷却した後、切片を緩衝液中ですすぎ、次にPBS中、5%ウシ血清アルブミン(BSA)および0.75%グリシンで、室温において1時間ブロッキングした。一次抗体を、ブロッキング緩衝液中、適当な濃度で、4℃において一晩適用した。スライドを緩衝液中で3回すすいだ後、二次抗体をブロッキング緩衝液中、1:500希釈で、室温において1時間適用した。二次抗体は、Invitrogenから入手したAlexFluor555ロバ抗ヤギまたは抗ウサギ抗体およびAlexaFluor488ニワトリ抗ウサギ抗体とした。DAPI含有封入剤(Vector Labs)を使ってスライドを封入し、Zeiss Axiovert 200M倒立蛍光顕微鏡を使って画像を取得した。
【0170】
走査型電子顕微鏡法
0.1Mカコジル酸緩衝液(EMD Biosciences、ニュージャージー州ギブスタウン)中の2%グルタルアルデヒドおよび2.5%パラホルムアルデヒドを使って試料を室温で2時間固定した後、カコジル酸緩衝液中ですすぎ、薄切し、エタノール勾配によって脱水した。試料をヘキサメチルジシラザン中で10分間さらに脱水し、一晩乾燥した後、金でスパッタコーティングし、イェール大学地質学および地球物理学施設(Yale University Geology and Geophysics facility)においてJOEL JXA-8600を使って分析した。
【0171】
透過型電子顕微鏡法
PBS中の4%パラホルムアルデヒドを使って試料を固定した後、0.1Mカコジル酸ナトリウム緩衝固定剤(pH7.4)中の2%グルタルアルデヒドおよび2.5%パラホルムアルデヒドに、室温で2時間入れておいた。試料を0.1Mカコジル酸ナトリウム緩衝液中で3回すすぎ、1%四酸化オスミウム中で1時間、後固定し、次に、マレイン酸緩衝液(pH5.2)中の2%酢酸ウランにおいて、さらに1時間、一括して染色した。次に、試料をすすぎ、段階的エタノール系列によって脱水し、エポン樹脂を浸透させ、60℃で一晩ベーキングした。硬化したブロックを、Leica UltraCut UCTを使って切断し、ニッケルグリッド上に60nm切片を集め、2%酢酸ウランおよびクエン酸鉛を使って染色した。80kVのFEI Tencai Biotwin TEMで試料を観察した。Morada CCDデジタルカメラを使用し、iTEM(オリンパス)ソフトウェアを使って、画像を撮影した。
【0172】
マイクロスフェア保持
脱細胞化肺臓または未変性肺臓を、本明細書の他の項で説明するように、カニューレに取り付け、その肺臓を、気管を通して、5μmのマイクロスフェアを含有するPBSで膨らませた。次に、脈管構造を、PBS各10mlのすすぎ液で3回フラッシングした。マイクロスフェアをdH2O中で2回洗浄して、細片を除去し、溶解しなければ未変性肺臓の読みに影響を及ぼすであろう細胞を全て溶解した。4.9μm〜5.1μmの粒子を測定するように設定したCoulter計数器を使って、各試料中のマイクロスフェア濃度を定量し、マイクロスフェア注入前に測定したベースラインの読みと比較した。
【0173】
マイクロCTイメージング
未変性肺臓または脱細胞化肺臓を10%中性緩衝ホルマリン(Sigma)中で固定し、気道または脈管構造を通して造影剤を注入した。造影剤はPBS中の20%ビスマスおよび5%ゼラチン(Sigma)とした。造影剤の注入後、肺臓を氷浴で冷却してゼラチンを重合させた。
【0174】
肺臓全体に関して、有効検出器ピクセルサイズ0.029mmに設定したマイクロCTイメージングシステム(GE eXplore Locus SP、GE Healthcare)で肺脈管構造をイメージングした。マイクロCTは、ピークX線管電圧60kV、管電流80mA、1600ミリ秒/フレーム、22検出器ビニングモデル、720ビュー、および0.5°刻み/ビューで作動させた。1つの葉(右上葉)の高解像度イメージングのために、試料をコンピュータ制御された回転ステージ上に配置し、垂直軸の周りを、0.4°の回転間隔で、360°スキャンした。管はピーク値80kVおよび80mAで作動させる。1×1に設定した検出器ビニングモデルおよび0.0065mmの解像度で、各ビューごとの露出時間は、典型的には3000ミリ秒とした。どちらの取得も、肺臓または1つの葉を貫く連続的な軸方向VFFフォーマット画像のセットをもたらした。
【0175】
Microview Software(GE Healthcare)を使用することにより、生データを補正し、ボクセル寸法58μm×58μm×58μmで再構築して、肺臓中の血管樹全体を可視化した。高品質の血管樹(1つの葉)については、ボクセル寸法を6.5μm×6.5μm×6.5μmに設定した。このソフトウェアは、生データから最大値投影像を再構築するためにも使用した。
【0176】
多平面再構成、空間フィルタリング、およびボリュームレンダリング技法により、データセットを、横断面、矢状断面、前頭断面、ハイブリッド断面、および3Dフォーマットで見ることを可能にした。対象抽出および関心領域測定には二値画像を使用した。三次元ボリューム画像は、改変Feldkampフィルタ補正逆投影アルゴリズムを使用することにより、斜視図から再構築される。しかしこの系では、ラット肺臓全体(視野、約3.0cm)を、58μmという小さな典型的立方ボクセル寸法を持つ画像で、調べることができる。各ボクセルの不透明度は16ビットグレースケール値によって表される。
【0177】
実験の結果を以下に説明する。
【0178】
脱細胞化方法
本発明に提示する結果は、インタクトな齧歯類肺臓の完全な葉から細胞材料を除去する脱細胞化方法を証明している。1M NaCl、8mM CHAPSおよび25mM EDTAを使った脱細胞化は、細胞材料を除去するのに最適であり、しかもコラーゲンまたはエラスチン線維を除去せず(組織像に基づく)、マトリックスの構造的完全性も損傷しないらしい(機械的試験に基づく)ことが観察された。これに比して、SDSを含有する溶液による脱細胞化は、マトリックスの機械的強度を損なうことが見いだされた。他の条件は、細胞材料を効率よく除去しないか、マトリックスの完全性の著しい低下を引き起こすことが見いだされた。
【0179】
組織学的分析
組織学的検査を使って数多くの脱細胞化肺臓スキャフォールドを特徴付けた。核およびDNAに関するH&E染色およびDAPI染色によれば、脱細胞化肺臓は、インタクトな細胞をただ一つも示さなかった。時折、巻き戻されたDNAまたは細胞抗原が観察されたが、インタクトな細胞は観察されなかった。図1は、未変性肺臓および脱細胞化肺臓のH&E染色を示し、一方、図2は、残存DNAに関するDAPI染色を示している。標準的組織切片では肺胞中隔がインタクトに見え、大きい気道および血管もそうであったという事実に基づいて、肺構造の保存も観察された。
【0180】
DNA含量
細胞材料の完全な除去は、いくつかの理由で重要である。第1に、スキャフォールドを組織工学的応用に使用するつもりである場合は、スキャフォールドに新しい細胞源を播種する前に、全ての細胞がスキャフォールドから除去されることを確実にしなければならない。人工組織がインビボ応用に使用される場合は、いかなる残存細胞材料も、再播種済スキャフォールドの評価を複雑にするだけでなく、免疫合併症を引き起こすだろう[Conconi et al., 2005, Transpl Int 18:727-34;Macchiarini et al., 2008, Lancet 372(9655):2023-30;Alexander et al., 2009, Cell Transplant 18:255-9]。そのため、本発明のスキャフォールドは、MHCクラスIおよびII抗原がどちらも脱細胞化スキャフォールド中に存在しないことが確認された。第2に、細胞外マトリックスの肺臓力学への寄与を個別に評価するために、全ての細胞構成要素を除去すべきである。末梢肺臓力学に寄与し得る2クラスの構成要素は、細胞材料と細胞外マトリックスである。細胞外マトリックスはさらに、主としてコラーゲン、エラスチン、およびプロテオグリカンに分割することができる[Cavalcante et al., 2005, J Appl Physiol 98:672-9;Dunsmore et al., 1996, Am J Physiol 270:L3-27;Ito et al., 2005, J Appl Physiol 98:503-11;Suki et al., 2005, J Appl Physiol 98:1892-9]。脱細胞化スキャフォールドからの細胞構成要素の除去を確実にすることにより、スキャフォールドの機械的性質を評価することができる。
【0181】
細胞材料の除去を証拠付けるために、定量的DNAアッセイを行った。未変性肺臓と比較して脱細胞化スキャフォールドではDNA含量の激しい減少が観察された(図1C)。脱細胞化スキャフォールドは未変性肺臓中に見いだされるDNAの約1.2%を含有し、それは乾燥重量1mgあたり1.83±0.29ngのDNAに相当した。これに比して、未変性肺臓の場合は、38.7±5.8ng/mgである。残存DNAを最小限に抑えるためにスキャフォールドの十分な洗浄を一般に使用することができるが、図2においてDAPI染色が巻き戻されたDNAの小さなクラスターを示していることから証明されるように、全てのDNAの完全な除去は難しく、少量のDNAが残った。DNA含量の激しい低下は細胞除去を示し、組織学的所見と共に、生存可能な細胞材料の全てがスキャフォールドから欠けていることを確認した。
【0182】
脱細胞化スキャフォールドでは、DNAのほぼ99%が除去されていることが証明された。少量のDNAがマトリックス中に残るが、それは、図2に示すようにDNAの細長い鎖として存在する。DAPI染色によれば、核構造におけるこの残存DNAの組織化は観察されなかった。
【0183】
未変性肺臓と比較して98.8%のDNAが除去され、残留DNA濃度は組織1mg(乾燥重量)あたり1.83ngのDNAであることが観察された。これは、他の脱細胞化心臓組織について他の研究者らが観察した16.6ng/mgという残存DNAレベルと比較して優れており[Ott et al., 2008, Nat Med 14:213-21]、このレベルが、本研究および他の研究[Gilbert et al., 2008 J Surg Res 152(l):135-9]の場合のように乾燥重量ではなく、湿重量に規格化されていることを考えると、とりわけそうである。しかし、観察される残存DNAのレベルは、皮膚グラフトに使用される市販のおよび実験室で作製されたECMスキャフォールド(一部のスキャフォールドは1.13ng/mgもの残存DNAを持っていたものの[Gilbert et al., 2008 J Surg Res 152(l):135-9]、大半のスキャフォールドは乾燥重量1mgあたり0.2ng未満のDNAを示す)に見られるものよりも高い。
【0184】
免疫原性
脱細胞化スキャフォールドの免疫原性を、主要組織適合遺伝子複合体(MHC)クラスIおよびII抗原に関する染色によって特徴付けた。MHCクラスIおよびIIタンパク質は、抗原-特異的免疫応答において重要な膜糖タンパク質である。MHCクラスI抗原は全ての有核細胞において発現し、MHCクラスII抗原は免疫系の特殊化した細胞上に見いだされる。MHCクラスI抗原は生物が「自己」を「非自己」と識別することを可能にするので、将来的に動物モデル中に人工肺臓組織を埋植した際の免疫問題を回避するには、脱細胞化スキャフォールドから除去することが重要である。図3に、MHCクラスIおよびII抗原ならびにβ-アクチンに関するウェスタンブロッティングの結果を図示する。免疫ブロッティングによってMHCクラスIおよびII抗原の両方の完全な喪失が観察されたことから、脱細胞化スキャフォールドは、組織工学的応用に使用しても、著しい免疫応答を惹起しないと予想されることが確認された。β-アクチンも失われたことは、細胞材料の非存在と合致する。宿主に埋植された場合にスキャフォールドが免疫応答を惹起する可能性は低いと考えられる。
【0185】
細胞外マトリックスの特徴付け
コラーゲン:コラーゲンは、肺臓の最も重要な構造的構成要素であり、組織の総合的な機械的強度を、主に担っている。免疫蛍光を使って、図4に示すように、未変性肺臓および脱細胞化肺臓におけるコラーゲンIおよびIVの分布を特徴付けた。コラーゲンIおよびIVはどちらも脱細胞化マトリックスによって保持されており、コラーゲンIは主により大きな気道および脈管構造付近に認められ、コラーゲンIVは実質全体に認められる。類似する染色パターンが、未変性肺臓にも脱細胞化肺臓にも認められた。これらのコラーゲンサブタイプがそれぞれ解剖学上適切な場所に保存されることにより、人工肺臓組織の開発時に、細胞タイプの選択的沈着が可能になるだろう。
【0186】
脱細胞化スキャフォールドの走査型EM評価
走査型電子顕微鏡法(SEM)を使って、脱細胞化肺臓スキャフォールドの微細構造を評価した。図5に、細胞が除去され、しかも肺胞アーキテクチャが全体的に維持されていることを証明するサンプル画像を示す。脱細胞化肺臓中の肺胞はわずかにしぼんでいるように見えたが、これは固定の人為的結果である。未変性肺臓は、固定剤で肺臓を膨らませることによって固定したが、脱細胞化肺臓は、加圧時に肺胞コンパートメント内に固定剤流体を含有することができないので、しぼんだ外観の肺臓を与える。しかし、肺胞アーキテクチャは概して類似しており、肺胞中隔が保存されている。これらの結果は、組織学的研究による知見と共に、全体的な肺の気道アーキテクチャおよび肺胞中隔を含む肺胞構造が、脱細胞化スキャフォールドにおいてインタクトであったことを示している。
【0187】
スキャフォールド超微細構造に対する灌流圧の影響
走査型EM研究に加えて、透過型EM(TEM)も使って、毛細管-肺胞基底膜を調べた。インタクトな毛細管網の存在は、脱細胞化スキャフォールドが、肺胞腔への高分子の移行に対抗することを可能にし、人工肺臓組織における毛細管内皮の成長にとって適切な基質にもなるので、これは、脱細胞化スキャフォールドの決定的に重要な特質である。
【0188】
図7Aおよび7Bに、未変性肺臓と、血管灌流圧の制御なしで脱細胞化した肺臓のTEM像を図示する。このような条件下では、肺胞基底膜は時折、同定することができず、毛細管を見いだすことができなかった。いかなる特定の理論にも束縛されることは望まないが、基底膜および超微細構造の損傷は、脱細胞化プロセス中の灌流圧を最小限に抑え、脱細胞化を開始する前に脈管構造を最大限に血管拡張しておくことによって減少させることができると考えられる。脱細胞化流体は生理的流量未満の流量で脈管構造に灌流させたが、大量の細胞溶解ならびに脈管構造における細胞タンパク質およびDNAの蓄積ゆえに、血管灌流圧は脱細胞化中に超生理的になり得る。そこで、肺動脈圧を注意深く監視し、この圧力が約20〜30mmHg未満に厳密に保たれるように、脱細胞化バイオリアクタおよび灌流量を調整した。血管拡張剤ニトロプルシドナトリウムを利用して、初期灌流圧を最低限に抑えた。
【0189】
図6Cに、圧力を約30mmHg未満に保って脱細胞化したスキャフォールドのTEM像を示す。これらの条件下では、インタクトな連続的肺胞基底膜が観察された。コラーゲン線維および他のマトリックス構成要素は、肺胞中隔内に保たれている。しかし、肺胞を取り囲んで豊富に存在するはずの透明な毛細管構造の存在は一切認められない。
【0190】
脱細胞化スキャフォールドにおける毛細管構造の保持
齧歯類の肺血管系における典型的な圧力は15mmHg未満であり[Lee et al., 1999, Cell 99:301-12]、これは上記の研究で利用した30mmHgよりもかなり低い。灌流圧を下げるために灌流の流量を低下させ、血管拡張剤を使用したにもかかわらず、脱細胞化灌流圧を30mmHg未満に維持することは困難だった。しかし、脱細胞化プロトコールをわずかに変更すれば脱細胞化時の灌流を約20mmHg未満の圧力で行うことが可能になることが、発見された。重要なことに、これは毛細管構造の保持を可能にした。この変更には、脈管構造を通して脱細胞化流体の灌流を始める前に、気道コンパートメントを脱細胞化流体で洗浄することが含まれた。その結果、血管灌流圧が、特に脱細胞化プロセスの開始時において、有意に低下した。図8に示すように、この技法は、脱細胞化スキャフォールドにおける毛細管構造の保持を可能にした。毛細管の保持は、脱細胞化肺臓スキャフォールドの作出における重要な進展であると考えられる。
【0191】
スキャフォールドはインタクトな気道樹および血管網を保持するべきである。マイクロCTイメージングに加えて走査型および透過型電子顕微鏡法を使用することにより、全体として、スキャフォールドは脱細胞化プロセス後に極めてよく保存されていることが証明された。走査型EMは、所定の組織学的検査と共に、スキャフォールドが総体的にインタクトであり、大きな欠陥を持たない(すなわち肺胞および肺胞中隔はインタクトに見える)ことを証明した。透過型EMは、肺胞基底膜がよく保存されること、および少なくとも一部の毛細管が保たれることを証明した。マイクロCTイメージングは、脈管構造が、下は直径100μmの血管まで、インタクトであることを証明した。
【0192】
透過性評価
肺臓がインビボで機能するには、肺胞および間質腔への大量の血液損失を回避するために、肺臓が、開存性で漏出しにくい連続した脈管構造を持たなければならない。5μmマイクロスフェアを気道コンパートメント中に保持して脈管構造へのこれらの高分子の輸送を許さない脱細胞化肺臓スキャフォールドの能力を評価した。脈管構造中に保たれる必要があるであろう血液の主要構成要素である赤血球のサイズを模倣するために、5μm粒子を使用した。したがって、気道から脈管構造への5μm粒子の漏出を、脱細胞化膜を横切るそのような粒子の移動に著しい指向性はないという仮定の下で評価した。
【0193】
未変性肺臓、灌流圧を制御せずに(一定の灌流流量で)脱細胞化した肺臓、および血管拡張後に制御された灌流圧(30mmHg未満)で脱細胞化した肺臓の透過性を決定した。結果を図8に示す。この結果は、TEM所見を、より大規模に確認するものだった。高い(制御されない)灌流圧による脱細胞化が39%の漏出につながるのに対し、低圧脱細胞化の場合は5.7%、未変性肺臓の場合は2.1%だった。
【0194】
マイクロCTイメージング
マイクロCTイメージングを使って、脱細胞化肺臓スキャフォールドの気道および血管コンパートメントの開存性を評価した。この技法により、肺臓スキャフォールドの三次元像を得ることが可能になり、気道および血管コンパートメントの開存度の同定が容易になる。
【0195】
図9に58μmの解像度で脈管構造の画像を示す。この解像度で、大血管はインタクトであることが示され(図9の最上段のパネル)、未変性試料および脱細胞化試料が概して類似していることが、下段および中段のパネルに示されている。脈管構造の高解像度画像(6.8μm)を図10に示す。ここでは血管が三次元投影像(最大値投影像)として示されている。これらの画像では、わずかな血管漏出が、脱細胞化スキャフォールドの一部の領域に示される曇りとして、同定された。
【0196】
脱細胞化マトリックスの決定的に重要な特質は未変性三次元構造の保存である。脱細胞化スキャフォールドの構造がどの程度保存されているかを評価するために、走査型および透過型EM、マイクロCT、およびマイクロスフェア透過性アッセイの組合せを使用した。脱細胞化肺臓の超微細構造特徴をSEMで調べたところ、肺胞アーキテクチャおよび肺胞中隔の維持が証明された。透過型EMにより、完全にインタクトな肺胞基底膜ならびにコラーゲンおよびエラスチン線維が証明された。これらのEM所見は、肺臓マトリックスを脱細胞化する他の研究であって、そのような構造が保たれるもの[Lwebuga-Mukasa et al., 1986, Exp Cell Res 162:423-35]と合致している。脱細胞化時の血管灌流圧を厳密に制御することにより、本明細書に提示する結果は、毛細管の保持を証明している。マイクロCTイメージングにより、控えめに見積もっても下は直径100μmの血管までの血管網の保持が証明され、それより小さな血管もかなりの数がインタクトである。
【0197】
実施例2:脱細胞化肺臓組織の機械的完全性に対する細胞外マトリックス構成要素の寄与
スキャフォールドの機械的性質に焦点を絞って脱細胞化スキャフォールドの組成をさらに詳しく評価するために、以下の実験を計画した。いかなる特定の理論にも束縛されることは望まないが、脱細胞化肺臓スキャフォールドは、主にコラーゲンおよびエラスチンからの寄与によって、未変性肺臓の顕著な機械的特質を保っていると考えられる。本明細書に提示する結果は、細胞の寄与には依存しない肺臓力学を研究するためのプラットフォームとしての脱細胞化肺臓組織の有用性を証明している。
【0198】
本明細書に提示する結果は、コラーゲン含量が保たれ、エラスチン含量が未変性レベルの約40%のレベルで保たれ、一方、グリコサミノグリカンは、大部分が、脱細胞化スキャフォールドから失われることを証明している。
【0199】
以下にこれらの実験で使用した材料および方法を説明する。
【0200】
材料および方法
器官収集および脱細胞化
本明細書の他の項で説明したように肺臓組織を収集し、脱細胞化した。
【0201】
組織学的分析
組織学的検査を使って数多くの脱細胞化肺臓スキャフォールドを特徴付け、細胞材料の除去を確認した。組織を固定し、パラフィン包埋し、薄切りした。分析は、標準的なヘマトキシリンおよびエオシン染色(H&E)、コラーゲンに関するMassonのトリクローム染色、エラスチンに関するVerhoff van Gieson染色、およびプロテオグリカンに関するアルシアンブルー染色、ならびに4'6-ジアミジノ-2-フェニルインドール(DAPI)を用いるDNAの染色によって行った。
【0202】
コラーゲンアッセイ
改変されたGrantの方法[Grant 1964, 1964, J Clin Pathol 17:685-6]を使ってOH-プロリンを検出する比色アッセイで、コラーゲンを定量した。肺臓試料を凍結乾燥し、重量測定した後、パパイン(140μg/ml)中、60℃で一晩インキュベートした(Sigma)。パパイン消化した試料を6N HCl中、115℃で18時間インキュベートし、中和し、クロラミンTで酸化し、p-ジメチルアミノベンズアルデヒドと反応させた。吸光度を波長550nmで測定し、1:10(w/w)というヒドロキシプロリン対コラーゲンの比を使って、組織のコラーゲン含量を算出した。未変性試料および脱細胞化試料について少なくとも4つの試料を測定した。
【0203】
エラスチンアッセイ
Fastinエラスチン・アッセイ・キット(Biocolor、北アイルランド・ベルファスト)を使ってエラスチンを定量した。肺臓試料をまず凍結乾燥し、重量を測定してから、Foronjy et al.[Foronjy et al., 2008, Am J Physiol Lung Cell Mol Physiol 294:L1149-57]に記載の方法に従って、エラスチンを抽出した。試料を0.25Mシュウ酸と共に100℃でインキュベートした後、10,000gで遠心分離し、上清を取り置いた。5回の抽出から得られた上清をプールし、6回目の抽出から得られた上清も、組織中にもうエラスチンが残っていないことを確かめるために測定した。10,000分子量カットオフフィルタ(Millipore)を使ってシュウ酸を除去した後、dH2Oに再懸濁し、Fastinエラスチンキットを使って、製造者の説明書に従って分析した。未変性試料および脱細胞化試料について、少なくとも4つの試料を測定した。
【0204】
硫酸化グリコサミノグリカンアッセイ
Blyscan GAGアッセイキットを使って、コンドロイチン、デルマタン、ヘパランおよびケラタン硫酸を含む硫酸化グリコサミノグリカン(sGAG)を定量した。パパイン消化した試料(上記コラーゲンアッセイについて説明したように調製したもの)を、製造者の説明書に従ってアッセイした。簡単に述べると、硫酸化GAGを1,9-ジメチル-メチレンブルー色素で標識し、吸光度を650nmで測定した。
【0205】
機械的試験
10Nロードセルを装着したInstron 5848を使って未変性肺臓および脱細胞化肺臓試料を分析した。既知寸法の組織切片を20%歪みまで10サイクルにわたって周期的にプレ伸張して弾性的性質を調べた後、破損するまで伸張して、極限引張強さ(UTS)を評価した。試験プロトコールの図解については図11を参照されたい。組織寸法を使って、工学応力および工学歪みを力と距離から算出した。
【0206】
実験の結果を以下に説明する。
【0207】
コラーゲンおよびエラスチン含量
図12Cに示すように、脱細胞化スキャフォールド中のコラーゲン含量は、未変性肺臓と識別できなかった。コラーゲンは肺臓の機械的強度に重要な役割を果たすので、コラーゲンのこの保存は重要である。コラーゲン含量は、図12に示すMassonのトリクロームによる組織化学的染色でも維持されていた。同様に図12Cに示すように、適切でないことがわかった脱細胞化法の一つであるSDSで脱細胞化したスキャフォールドでは、コラーゲン含量が減少した。コラーゲンのこの喪失は、SDS脱細胞化スキャフォールドにおける低下した機械的完全性と相関すると考えられる。
【0208】
定量的アッセイでも組織染色でも証明されるとおり(図13)、脱細胞化スキャフォールドでは、エラスチン含量も(減少はするが)保存される。エラスチン線維は肺臓に弾性を与え、吸入後の肺臓の弛緩(したがって呼気)において重要な役割を果たす組織の自然反跳にとって、決定的に重要である。脱細胞化プロセス中にこれらの線維が保持されることは、決定的に重要である。というのも、それは、肺細胞集団をスキャフォールドに再播種しようとする努力において、肺臓スキャフォールドが適切に換気されることを可能にするからである。スキャフォールドは未変性エラスチン含量の60%を失ったが、残ったエラスチンは、本明細書の他の項で論じる機械的試験結果からわかるように、肺臓に弾性機能を与えるのに十分だった。
【0209】
全体として、これらの重要なECM構成要素の保持は、スキャフォールドが生理的レベルの機械適応力を経験することを可能にした。これは、重要なことである。というのも、さまざまな発生および細胞分化プロセスが機械的刺激に依拠するからである。また、ECMは、マトリックスへの細胞付着を補助するのに決定的に重要であり、これらの未変性ECM構成要素の保持は細胞の付着と伝播を容易にし、したがって生体工学的(bioengineered)肺臓組織の開発を容易にする。
【0210】
プロテオグリカン含量
プロテオグリカンは、1つまたは複数のグリコサミノグリカン(GAG)鎖に連結したコアタンパク質からなる。大半のGAGは硫酸化されており、それが定量的アッセイによるそれらの検出を可能にする。その結果を図14に示す。脱細胞化スキャフォールドのGAG含量は、未変性肺臓より有意に低かった(未変性肺臓レベルの約6%)。プロテオグリカンは、細胞表面上または細胞外マトリックス内に見いだされ[Ferdous et al., 2007, Tissue Engineering 13:1893-904]、それらの除去は、一つには、細胞結合型GAGの除去によるものである。しかし、ECM内に見いだされるGAGも、脱細胞化溶液によって可溶化され得る。図14は、プロテオグリカンに関するアルシアンブルー組織染色であり、これは、脱細胞化肺臓スキャフォールド中に残っているGAGの量が未変性肺臓と比較して減少したことを示しており、定量的アッセイの結果が確認された。
【0211】
機械的特徴付け
末梢肺臓条片の機械的試験を使って、未変性肺臓試料および脱細胞化肺臓試料の両方の準静的力学を評価した。応力-歪み曲線の弾性域は、未変性試料と脱細胞化試料がどちらも履歴挙動を表したことを示している。履歴現象は肺臓が粘弾性材料であることを証明しており、拡大曲線と弛緩曲線の間の相違は、弛緩中に回復されないエネルギーを表す。また、図15に示すように、試料はクリープを起こさなかった。仮に肺臓組織がクリープを起こすとすると、拡張後に元の位置に収縮することはなく、それゆえに肺臓は決して完全には収縮せず、ガス交換が損なわれるであろう。この適当な弾性肺臓挙動の保存は、肺臓スキャフォールドにとって重要である。なぜなら、肺弾性の喪失は、いくつかの疾患状態、とりわけ肺気腫に見られるからである[Gelb et al., 2002, Chest 121:715-21]。
【0212】
極限引張強さ(UTS)は破損時の試料上の応力であり、材料の強さの尺度である。図16に示すように、脱細胞化試料のUTSは、未変性試料のそれと識別できなかった。しかし、ドデシル硫酸ナトリウム(SDS)を含有する緩衝液中で試料を脱細胞化した場合は、UTSの減少によって証明されるように、機械的完全性が損なわれた。SDSはコラーゲンを分解して、組織の断片化および腫脹を引き起こすことができ[Bodnar et al., 1986, Thorac Cardiovasc Surg. 34(2):82-5;Gilbert et al., 2006, Biomaterials 27:3675-83]、組織の伸展性(extensibility)を増加させることも示されている[Mirsadraee et al., 2006, Tissue Eng 12:763-73]。SDSはイオン性が高い両親媒性界面活性剤であり、その疎水領域はタンパク質と相互作用することができ、一方、親水性部分は、特に負に帯電している場合には、水と結合して組織腫脹を引き起こす[Bodnar et al., 1986, Thorac Cardiovasc Surg. 34(2):82-5]。他の研究ではSDS処理によるUTSの減少が常に見られるわけではなかったが[Mirsadraee et al., 2006, Tissue Eng 12:763-73]、これは、組織の相違によるものだろう。Mirsadraee et al.は心臓周囲組織を研究したが、これは肺臓よりはるかに密に充填されたコラーゲン線維を含有している。肺臓では、組織の幾何学ゆえに、コラーゲン線維が高度に分散されており、SDSが誘発する腫脹は、定量的コラーゲンアッセイで見られるように、はるかに容易に、コラーゲンの除去につながり得る。
【0213】
本明細書に提示する結果は、脱細胞化スキャフォールドが、関連するインビボでの生理的力に耐え得ることを証明している。
【0214】
本明細書に提示する結果により、コラーゲンとエラスチンはどちらも機能的レベルで保存されることが確認された。これらの知見により、肺臓力学への主な寄与は、コラーゲンとエラスチンによってなされ、細胞構成成分またはプロテオグリカンによるものではないことが確認された。本明細書に提示する結果により、未変性肺臓の特徴を示す脱細胞化肺臓スキャフォールド(これらの特徴は、肺臓スキャフォールドを、組織工学的応用のための前途有望な基質にすると共に、詳細なマトリックス力学ならびに肺臓の生物学、発生および生理学を研究するためのプラットフォームにする)の作製が証明された。
【0215】
実施例3:3次元肺臓組織のインビトロ培養のためのバイオリアクタの設計と検証
バイオリアクタを使って3次元肺臓組織をインビトロで培養することができる。そのようなバイオリアクタの開発は、人工肺臓組織の成長に関する研究を行うのに有益であるばかりでなく、肺生物学の研究にも有益であるだろう。成人肺臓細胞の長期インビトロ培養を可能にするシステムは、今のところない。
【0216】
肺臓組織全体をインビトロ培養するためのバイオリアクタを設計するために、以下の実験を行った。細胞の生存および分化を支持するために肺臓組織に十分な栄養素供給および機械的刺激を与え得ることを目指して、一連の設計制約を満たすように、バイオリアクタを設計した。細胞の生存性および分化状態の維持によって証明される肺臓組織の全葉のインビトロ培養をバイオリアクタが支持できるかどうかを評価するために、実験を設計した。バイオリアクタを評価するための過程では、バイオリアクタにおける肺臓の生存に対する灌流および換気の効果を評価した。本明細書に提示する結果は、バイオリアクタがインビトロ肺臓組織培養に使用することができ、それゆえに肺臓組織のエンジニアリングに応用可能であることを証明している。
【0217】
これらの実験で使用した材料および方法を以下に説明する。
【0218】
材料および方法
全肺臓培養
肺臓を若年成体(3ヶ月齢)雄フィッシャー344ラットから収集した。動物実験の作業は全て、イェール大学施設内動物実験委員会からの承認を受けて行われた。動物をペントバルビタールナトリウム(Sigma、40mg/kg)の腹腔内注射によって麻酔した。麻酔の誘導後に、胸部および腹部にエタノールを噴霧し、肋骨縁の直下で横切開を施し、腹腔に入った。横隔膜を穿刺し、肺臓に触れないように注意しながら肋骨を後退させた。下大静脈を切断し、50U/mlヘパリン(Sigma)および1μg/mlニトロプルシドナトリウム(Sigma)を含有する20〜30mlのPBSで右心室を通して肺臓を灌流した。次に気管を切り離し、できるだけ高く切った。心臓および肺臓への残りの接続を全て切り離して、心臓、肺臓および気管をひとまとめにして動物から取り出せるようにした。
【0219】
カニューレ装着
器官の除去後に、気管と、心臓の右側を通して肺動脈幹とに、カニューレを接続した。心尖を円刃刀で切り落とし、直角カニューレを右心室を通して肺動脈幹へと挿入した。このカニューレにシリンジを取り付け、漏出を伴わない適正なカニューレ設置と肺臓の十分な灌流とを確かめるために、5〜10mlのヘパリン化食塩水を注入した。次に、このカニューレを心臓に縫合糸で固定した。別の真っ直ぐな返し付(barb-end)カニューレを気管に挿入し、縫合糸で固定した。次に、肺臓をバイオリアクタに接続し、本明細書の他の項で説明するプロトコールに従って、脱細胞化した。
【0220】
カニューレを、右心室を通して肺動脈と、気管とに取り付け、肺臓をバイオリアクタに接続した。PBS中の2%アンホテリシン、ペニシリンおよびストレプトマイシンで気道を洗浄した後、PBSで2回洗浄し、次にバイオリアクタを培地で満たして、培養を開始した。血管灌流および換気を実験条件の指図どおりに行った。
【0221】
バイオリアクタの構成要素と組み立て
バイオリアクタの構成要素は、別段の注記がない限り、Cole-Parmer(イリノイ州バーノンヒルズ)から入手した。シリコーン栓と500mlのガラス製広口瓶とがバイオリアクタの基礎をなした。灌流ループ、気管接続、空気換気、および培地交換ポートを含む肺臓への必要な接続が可能になるように、シリコーン栓を通して、サイズL/S14およびL/S16のPharMedチューブ(オハイオ州ウェストレーク)を挿入した。圧力は、灌流ポンプと肺動脈への接続との間のTruWave圧変換器(Edwards Lifesciences、カリフォルニア州アービン)を使って監視した。灌流はMasterflex L/S可変速ローラーポンプ(Masterflex、イリノイ州バーノンヒルズ)を使って達成した。換気は、多チャンネルプログラマブルシリンジポンプ(Cole Parmer)を使って行い、吸気と呼気をそれぞれ10mlの体積で30秒間かけて行った。バイオリアクタの略図を図17に示す。
【0222】
組織学的検査および免疫蛍光法
所望の培養期間後に、肺臓を固定し、パラフィン包埋し、薄切りした。所定の組織学的検査(H&E)を、アクアポリン5(I型上皮)、サーファクタントタンパク質C(II型上皮)、CCSP(クララ細胞)、およびPECAM-1(内皮)に関する免疫蛍光法と共に行った。切片をキシレン中で脱パラフィンし、再水和し、0.2%Triton-Xを含むPBS(緩衝液)と共に15分間インキュベートした。PBS中の0.02Mクエン酸を使って、75〜85℃で20分間、抗原回復を行った後、切片を緩衝液中ですすいだ。ブロッキングは、PBS+1%ウシ血清アルブミンおよび0.75%グリシンを使って室温で1時間行った。一次抗体を緩衝液ですすぎ落とし、二次抗体を1:500の希釈度で室温において1時間適用した。二次抗体はInvitrogenから入手した(AlexaFluor555またはAlexaFluor488x)。画像はZeiss Axiovert 200M倒立蛍光顕微鏡を使って取得した。
【0223】
細胞増殖は、増殖性細胞核抗原(PCNA)に関する染色によって評価し(Zymed、カリフォルニア州サンフランシスコ)、アポトーシス核を、末端デオキシヌクレオチジルトランスフェラーゼdUTPニック末端ラベリング(TUNEL)染色(Calbiochem、カリフォルニア州サンディエゴ)で検出した。どちらのアッセイも製造者の説明書に従って行った。
【0224】
マイクロスフェア換気アッセイ
バイオリアクタにおける肺臓の換気が、培地の移動を誘発して脈管構造を灌流するのに十分であるかどうかを決定するために、5μmポリスチレンマイクロスフェア(SPI Supplies、ペンシルバニア州ウェストチェスター)を使って簡単なアッセイを開発した。肺臓を、本明細書の他の項で説明するように、バイオリアクタに接続し、換気したが、灌流はしなかった。バイオリアクタチャンバを、1000万個のマイクロスフェアを含有する100mlの培地(培地1mlあたり10万個のマイクロスフェア)で満たした。培養を換気だけで3時間続けた。次に肺臓を固定し、パラフィン包埋し、薄切し、所定の組織学的検査(H&E)を使って分析した。
【0225】
実験の結果を以下に説明する。
【0226】
バイオリアクタ設計要件
バイオリアクタには、齧歯類インビボ環境の重要な特質を組み入れると共に、所望する条件に応じて使用者がいくつかの重要なパラメータを調整できるようにも設計した。設計目標は次のとおりである。
・システムは、使用者が指定した、生理的レベルの範囲内にある速度で、脈管構造を通して培地を灌流させる能力をもたなければならない。
・システムは、空気または培地を使って気管を通して肺臓を換気する能力をもたなければならない。正常な生理的条件と合致するように、陰圧換気が好ましく、肺臓を絶えず換気できることが好ましい。
・バイオリアクタは、好ましくは、肺臓の血管コンパートメントと気道コンパートメントとを異なる培地タイプで浸すことを可能にするべきである。
・バイオリアクタは、換気に関する上記の要件を満たすと同時に、培養培地へのガス交換も可能としなければならない。
・バイオリアクタは、肺動脈および気管圧の圧力測定を可能にするポートを持たなければならない。圧力は、理想的には、正常な生理学的値の範囲内にあるべきであり、肺動脈圧は15〜30mmHg未満である[Li et al., 2004, Proc Natl Acad Sci USA 101:11488-93]。
・バイオリアクタは、定期的な培地交換を可能にする手段を持たなければならない。
・バイオリアクタは、標準的な組織培養インキュベータの物理的境界内に収まり得るように、小さくかつ自給式でなければならない。
・全てのバイオリアクタ構成要素は、安価かつ入手が容易でなければならない。
・バイオリアクタおよび全ての構成要素は(好ましくはオートクレーブによる)滅菌が、可能でなければならない。
【0227】
上記の基準を満たすバイオリアクタを設計し、構築した。そのバイオリアクタの図解を図17に示す。
【0228】
バイオリアクタ灌流システム
肺臓への灌流は、バイオリアクタ本体から肺動脈へと培地を循環させるローラーポンプによって行った。使用者は灌流量を指定することができる。心臓の右心室を通した肺動脈幹へのカニューレの接続が容易になるように、ラットの心臓を肺臓に取り付けたままにしておく。ただし肺静脈は灌流ループに直接的には接続しなかった。むしろ肺静脈は心臓の左側からメインバイオリアクタリザーバに直接的に排液した。肺臓の静脈排出路はメインバイオリアクタ中に直接出る。
【0229】
肺臓を通る灌流量は使用者の指定に合わせて設定することができる。成体ラットにおける生理的流量は40〜80ml/分であるが、人工組織培養の場合、典型的には、流量はこの値よりはるかに低い。成体ラットの場合、酸素化されるには、全血液量が肺臓を通過しなければならないのに対し、人工組織培養中は、肺細胞の成長を支持するのに十分な培地を灌流するだけでよい。したがって、人工的培養(engineered culture)時の灌流量は、正常な生理学的シャントゆえに肺臓の血流量が心拍出量の8〜10%しかないラット胎児におけるそれに近い[Hislop et al., 2000, Ped Resp Rev 1:321-7]。圧力プロファイルは、肺血管圧を低下させるために使用することができるニトロプルシドナトリウムなどの血管拡張剤を使って、ある程度制御することができる。典型的には、灌流圧は、肺動脈系に見られる最大値である約30mmHg未満に保たれる[Li et al., 2004, Proc Natl Acad Sci USA 101:11488-93]。
【0230】
バイオリアクタ換気システム
バイオリアクタは陽圧換気と陰圧換気がどちらも可能だった。インビボでは、呼吸は通常、陰圧換気によって達成される。横隔膜が収縮し、胸郭が拡大して、胸腔内に陰圧を生じ、それが、この圧力不均衡を緩和するべく肺臓内に空気を流入させる。吸気後に、呼吸筋が弛緩し、肺臓は受動的にしぼむ。
【0231】
陰圧換気はバイオリアクタにおける換気の主要モードである。肺臓を取り巻く陰圧を達成するために、バイオリアクタのメインチャンバは完全に気密でなければならない。これは、全ての空気口および圧力監視口を閉じることによって達成される。次に、シリンジポンプを使って、バイオリアクタ本体から所定の体積の空気を抜いて、陰圧を生じさせる。この圧力を緩和させるための唯一の経路は、別個のリザーバに接続されている気管を通して培地(または空気)が肺臓に流入する経路である。次に、バイオリアクタ本体中に空気を押し戻す方向に、シリンジポンプを逆転させる。これはチャンバー内での陰圧の強化を逆戻りさせ、培地(または空気)は気管リザーバに向かって逆流する。この時、肺臓は受動的にしぼむ。
【0232】
気管カニューレは一方向弁を利用する:
図17に図示するように、気管への接続には、Y字連結器と、バイオリアクタ本体に向かって開く一方向弁が必要である。このタイプの接続は、流体の気道コンパートメントからの漏出ゆえに必要になる。吸気時は、ある体積の培地が肺臓に入る。しかし、この培地の一部は肺胞膜を横切って間質腔または脈管構造へと漏出する。それゆえ、吸気時に肺臓に進入した培地の全てが、呼気時に気管リザーバに戻ることができるわけではない。図17に示す設計には、吸気時には全ての培地が肺臓に進入することを許すという特徴が組み込まれている。しかし呼気時には、培地は肺臓から、またはバイオリアクタ本体から一方向弁を通して、気管リザーバに戻ることができる。
【0233】
バイオリアクタは、シリンジポンプを気管カニューレまたは気管リザーバに直接接続することによって、陽圧換気を利用することもできる。
【0234】
気管入口の改造
バイオリアクタに関して、換気時に、肺臓気道コンパートメントに十分な新鮮培地が供給されないことが観察された。いかなる特定の理論にも束縛されることは望まないが、これは、主として、気管と別個の気管リザーバとの間のチューブ内に含まれる培地の体積ゆえに、同じ培地が気管を出入りして、気管に進入する新鮮培地が不十分になっているからであったと考えられる。気道培地流ループ中の「死腔」が、呼吸時に新鮮培地が肺臓組織に到達することを妨げた。そのため、図17に概説するように、換気時に培地が異なる経路を辿って肺臓に出入りするように、バイオリアクタを改造した。この改造により、各吸息と共に気管に入る培地の大半は気管リザーバから直接供給される(したがって気管から出てくる培地と比較して「新鮮」である)ことになった。
【0235】
バイオリアクタ培養中の酸素供給:
十分な酸素含量があることを保証するために、肺臓培養中のバイオリアクタにおける組織培養培地の酸素含量を測定した。特に、陰圧換気(この間、バイオリアクタ本体は気密であり、酸素進入のための唯一の入口は気管リザーバを介したものである)中に、十分な酸素送達があることを保証する必要がある。酸素分圧は、培養の間、著しくは低下せず、正常組織培養培地におけるレベルと同じ6.0〜7.0mg/Lを保つことがわかった。これらのレベルは、80〜100mmHgという正常な生理的レベルを上回っている(6〜7mg/Lは分圧137〜159mmHgに相当する)。
【0236】
バイオリアクタ圧力プロファイル
圧力が予想限度内または生理学的限度内にあることを保証するために、バイオリアクタ中で培養されている人工肺臓組織の気管および肺動脈における圧力プロファイルを測定した。図18に代表的プロファイルを示す。灌流圧は、典型的には、約2〜30mmHgに保たれた。記載の例では、ベースライン灌流圧が10〜17mmHgで変動した。しかし、陰圧換気の効果がこのプロファイルに重ね合わされて、陰圧「吸息」時には灌流圧が0〜7mmHgまで低下した。この効果は生理学的に見られ、肺脈管構造の圧力は息の吸い込みに伴って低下する。バイオリアクタでは、肺静脈がメインチャンバに直接排液し、このメインチャンバは、「胸腔」としても機能し、肺臓を換気するために陰圧を生じさせた場所である。これが、バイオリアクタから灌流脈管構造への陰圧の伝達を増加させる役割を果たした。
【0237】
灌流圧プロファイルから、最大負「胸腔」圧は約-12mmHgであり、生理学的値とほぼ一致した。それゆえに、吸気中は、この陰圧が気道に加えられ、流体(または空気)を気管リザーバから肺臓へと動かす。この圧力は、気道樹を上がるにつれて徐々に減少し、気管の入口では-3mmHgだった。注目すべきことに、気管への入口における圧力は、大気圧に拘束されるので、生理的には本質的にゼロだった。しかし、バイオリアクタでは、気管と気管リザーバの間の長さ(ここで圧力が0に達する)により、この圧力が吸気中はわずかに負のままだった。
【0238】
培地および酸素要求量
以下の結果は、バイオリアクタ中で培養される齧歯類肺臓に要求される培地および空気の体積の決定に役立たせることを目的とした一連の計算を示す。
【0239】
組織培養比較:
インビトロ組織培養時は、500万個の細胞に、3日ごとに12mlの培地を供給することが一般的である。成体齧歯類肺臓が1億個の細胞を含有すると仮定すると、これは、3日ごとに多くて240mlという培地要求量に相当する。しかしこれは過剰評価であるだろう。というのも、組織培養中の細胞が一般に活発に複製するのに対して、インタクトな齧歯類肺臓中の多く細胞は休止状態にあり、それゆえに培地要求量が低いからである。
【0240】
グルコース消費要求量:
灌流ラット肺臓のグルコース消費量は、1時間あたり乾燥重量1グラムあたり43μmolであることが証明されている[Kerr et al., 1979, Am J Physiol 236:E229-33]。成体ラットの肺臓は約150〜250mgの乾燥重量を持ち[Inokawa et al., 2006, Ann Thorac Surg 82:1219-25]、組織培養培地は、典型的には、5.5mmol/Lのグルコースを含有する。それゆえに、成体ラットの肺臓は、そのグルコース消費要求量を供給するために、1日あたり28〜47mlの組織培養培地を要求するだろう。
【0241】
酸素要求量:
肺動脈内皮細胞は、1分あたり細胞100万個あたり6nmolの酸素を消費し[Xu et al., 2007, Proc Natl Acad Sci USA 104:1342-7]、一方、ラットII型上皮細胞は1分あたり1.25nmolを消費する[Dobbs et al., 1980, Biochim Biophys Acta 618:510-23]。成体ラット肺臓には1億個の細胞があり、肺臓中の全ての細胞が高い方の速度で酸素を消費すると仮定すると、ラット肺臓は1日あたり多くて26mgの酸素を要求するだろう。組織培養培地は、1リットルあたり約6mgの酸素を含有し、バイオリアクタは約300mlの培地を含有する。したがって培地は、新鮮培地の交換1回(3日ごと)につき1.8mgの酸素を供給することができる。さらに、酸素はバイオリアクタ中の空気にも含まれる。バイオリアクタ本体には約200mlの空気がある。培養器中の空気は約20%のO2を含有し、これは海面位、37℃で、空気1リットルあたり約260mgの酸素に相当する。したがってバイオリアクタ中の空気は約52mgの酸素を含有する。
【0242】
本発明のバイオリアクタは、上記の計算に基づいて、培養齧歯類肺臓の培地および酸素要求量を供給する。通例、バイオリアクタには合計240mlの培地を供給することができ(バイオリアクタ本体に180mlおよび気管リザーバに60ml)、バイオリアクタ中の空気は毎日交換することができる。これらの条件は、培養肺臓に必要以上の栄養素および酸素を提供するのに足りると考えられる。
【0243】
全肺臓培養
肺臓バイオリアクタの設計を検証し最適化するために、全未変性齧歯類肺臓のインビトロ培養を使用した。肺臓をバイオリアクタ中で最長7日間培養した。バイオリアクタは細胞の生存および分化ならびに肺臓形態を維持するのに十分な栄養素供給および機械的刺激を提供することが証明された。
【0244】
細胞生存、肺臓形態および細胞分化状態の維持に対するバイオリアクタ条件の効果を調べるためにも、バイオリアクタにおける未変性肺臓の培養を使用した。まず最初に、肺臓形態に対する空気換気と液体(培地)換気の効果を比較した。次に、細胞生存に対する換気技法および栄養素送達の効果を評価した。細胞の生存および分化に対する血管灌流圧の効果も評価した。7日間の培養中に細胞分化を維持するバイオリアクタの能力も評価した。
【0245】
全体的肺臓形態に対する空気換気と培地換気の効果:
バイオリアクタ中で培養される肺臓を培地または大気(約20% O2)で換気する効果を評価した。培地による換気は、改善された栄養素送達(これは、バイオリアクタでは、大きな気道に供給する灌流された気管支循環がないので、より一層重要になり得る)を提供するだろうから、改善された細胞生存をもたらすだろうと考えられる。しかし、空気による換気は、成体肺臓が条件づけられている条件であり、肺上皮は気液界面の存在下で培養されることが多く、それはラット胎児肺臓における適当な肺発生を可能にすることが示されている[Funkhouser et al., 1976, Biochem Biophys Res Comm 70:630-7]。そこで、培地による換気が、気液界面の欠如による上皮分化状態の喪失をもたらすかどうかを調べるための実験も計画した。
【0246】
3日間の培養後に、培地で換気した肺臓と空気で換気した肺臓との間に、有意な相違が認められた。図19に示すように、培地換気は未変性肺臓と同じように見えたが、空気換気肺臓は著しく拡張した気道を示し、気道中の細胞片が明らかだった(図19C)。そのうえ、図19Cの中央のパネルは、空気換気肺臓の気管支上皮および細気管支上皮が完全になくなっていることを示し、この所見は、肺臓全体にわたって一貫していた。さらにまた、図19Cの右側のパネルに示すように、拡張した末梢気腔も明白だった。
【0247】
(空気による換気に加えて)脈管構造に培地を灌流させても気道上皮は剥皮され、換気を行わずに脈管構造に培地を灌流させた場合は気道上皮がインタクトであることも観察された。これは、気道上皮の喪失が十分な培地の欠如によるものではなく、空気換気の効果に関係することを示唆している。気管支循環はどの培養でも灌流されないことが観察された。
【0248】
上皮細胞は、その生理的位置と同じく気液界面で培養されることが多い。気液界面(ALI)は上皮分化を誘導するために利用されることが多く[Gruenert et al., 1995, Am J Physiol 268:L347-60;Wong et al., 2009, J Clin Invest 1 19:336-48;Hosokawa et al., 2007, Connect Tissue Res 48:9-18]、気液界面の欠如は絨毛形成の減少につながる可能性がある[Ostrowski et al., 1995, Exp Lung Res 21:957-70;Yeh et al., 2007, Laryngoscope 117:1439-44]。さらにまた、気液界面は、インビトロ培養時に、II型上皮によるサーファクタントの分泌が維持されることを可能にする[Mason et al., 2002, Am J Physiol Lung Cell Mol Physiol 282:L249-58]。したがって、ALIの非存在下では、細胞分化状態の相違が生じるだろうと予想された。しかし、7日まで行った培養物について図25に示すように、培地で換気した肺臓におけるクララ細胞分泌タンパク質(CCSP)、サーファクタントタンパク質C(SPC)、またはアクアポリン(AQP)の発現パターンには、有意な変化が観察されなかった。
【0249】
細胞生存に対する灌流の効果:
インビトロ肺臓培養を灌流だけで支持することができるか、そしてもしそうなら、どの灌流圧が最適であるかを決定するために、バイオリアクタ中の培養未変性肺臓における細胞生存および細胞分化に対する血管灌流の効果を調べた。これらの実験を複雑にするのは、肺臓の外植後に血管透過性が急速に増加するという事実だった。10mmHgの圧力を使った単離肺臓の灌流は、10分以内に肺臓水腫を引き起こし得る[Wierup et al., 2005, J Heart Lung Transplant 24:379-85]。外植から5〜10分以内に血管漏出が観察され、小粒子(半径28nm)の3〜4%が肺胞-毛細管膜を横切って漏出した。広範な肺微小血管漏出は、遠位毛細管および静脈構造への培地送達を減少させるか、無くしてしまうことさえあり得る。したがって、流れを遠方まで送達し、遠位毛細管を開存させておくには、約1〜10mmHgという生理的レベルより高い血管灌流圧が要求されるだろう[Li et al., 2004, Proc Natl Acad Sci USA 101:11488-93]。
【0250】
バイオリアクタにおける3日間の未変性肺臓培養時の細胞生存に対する血管灌流圧の効果を調べた。図20に示すように、最高30mmHgの高い灌流圧は、10または20mmHgの圧力と比較して、少ないアポトーシス細胞と、高い細胞密度とをもたらした。しかし、灌流圧とは無関係に、血管灌流による細胞分化の維持は不十分だった。実質的に低いCCSPレベルおよびSPCレベルが観察され(図21)、アクアポリン発現はほとんど全く存在しなかった。図22に示すように、PECAM発現は、脈管構造の大きな血管では観察されたが、毛細管では、減少した発現が観察された。これらの実験により、灌流だけでは、十分な細胞生存または細胞分化を維持するのに十分でないことが証明された。
【0251】
気道コンパートメントにおける培地流路が細胞生存に及ぼす効果:
培地による換気は肺臓形態および細胞分化の維持を可能にしたが、換気培養肺臓では、未変性肺臓と比較して、有意に高いアポトーシス細胞率が観察された(図23および28参照)。これは、新鮮培地の送達が不十分であったためと考えられるので、換気中の気道コンパートメントへの新鮮培地の送達を増加させるためにバイオリアクタを改造する実験を計画した。図17に示し、本明細書の他の項で説明するように、バイオリアクタ本体を気管リザーバに接続する1本のラインがある。このチューブの長さは約40〜45cmであり、3〜3.5mlの培地を含有する。換気中は、陰圧吸気時に約2.5〜3.0mlの培地が肺臓に引き込まれ、同じ体積の培地がチューブを通って気管広口瓶に戻る。それゆえに、各「吸息」時に肺臓に進入する培地のうち、この約2.5〜3.0mlは新鮮ではなく、ただ単にチューブから肺臓へと戻るにすぎない。したがって、肺臓への実際の培地送達は、新鮮な培地を使った換気によって送達されるであろうものよりはるかに少ない。
【0252】
バイオリアクタ本体と気管リザーバとの間に第2の接続が付加されるように、バイオリアクタの設計を改造した。一方向逆止弁を使って、一方の接続を吸気中の培地送達に使用し、他方の接続を呼気中の培地還流に使用した。この改造により、「再利用される」培地が各換気サイクルにつき約2.5〜3.0mlからわずか約0.75mlに減少し、それゆえに新鮮培地の送達量が著しく増加した。
【0253】
このバイオリアクタ改造の効果を図24に示す。この図では、追加の呼吸ラインが細胞の生存を改善することが示された。アポトーシス細胞のパーセンテージは、1本のラインで換気した場合の21.5%(図27の「換気のみ」)から、「ループ」換気では3.9%まで低下した。これに対し、未変性肺臓では0.5%である。
【0254】
「ループ」換気は「再利用される」培地の量を減らすことによって肺臓への培地の送達を増加させるが、血管還流の追加によって培地送達を増加させることもできる。灌流は、換気と共同して、細胞アポトーシスを、単一ラインのみの場合の21.5%から7.9%まで低下させた(図23)。「ループ」換気への改造は単一ライン換気と比較して総細胞数をわずかに増加させたが(図23B)、これは有意な差ではなかった。単一ライン換気でも「ループ」換気でも、未変性肺臓と比較すると細胞数は減少したが、統計的に有意ではなかった。
【0255】
本明細書に提示する結果は、「ループ」換気への改造または補足的血液灌流を使って十分な新鮮培地が肺臓に送達されるのであれば、換気だけでも、3日間の培養中はバイオリアクタにおける未変性肺臓組織の生存が可能になることを証明している。ループ換気は最善の総合的結果を示し、細胞のアポトーシスを最小限に抑えると同時に、培養肺臓組織における細胞数を最大にする。
【0256】
細胞形態、細胞分化および肺胞構造:
バイオリアクタの設計をさらに詳しく検証するために、未変性肺臓の7日間培養を行った。これらの培養では、本明細書の他の項で説明する「ループ」改造を施した、培地による換気を利用し、いかなる血管灌流も行わなかった。血管灌流は利用しなかったが、今後の研究では、「ループ」換気を使った長期換気培養への灌流の付加を探究するかもしれない。
【0257】
肺臓を細胞増殖、アポトーシスおよび細胞分化について、アクアポリン5(I型上皮)、サーファクタントタンパク質C(II型上皮)、クララ細胞分泌タンパク質(クララ細胞)、およびPECAM-1(内皮)に関する染色により、組織学的検査で評価した。図29Aおよび29Bに示すように、全体的な肺アーキテクチャは肺胞構造を含めて保存された。低倍率画像は培地呼吸に関して図19に示したものと識別できなかった。さらにまた、図29C〜29Jに示すように、細胞マーカーの発現パターンは、未変性肺臓と実質的に異ならなかった。
【0258】
換気は単独で肺臓の脈管構造の受動的灌流を可能にする:
換気は単独で、内皮を含むいくつかの重要な肺臓細胞タイプの細胞生存と細胞表現型の維持とを、7日間まで可能にできることが証明された。しかしこれは、脈管構造を通した能動的灌流が存在しない状態でのことであり、それが最初は意外であった。灌流の欠如が内皮の生存または分化に影響を及ぼさない理由を調べるために、5μmマイクロスフェアを使って、脈管構造への流体の移動に換気が及ぼす影響を調べる実験を行った。換気によって誘発される物理的動きは、バイオリアクタ中の培地に向かって開いている脈管構造に入ったり出たりする培地の受動的移動を引き起こすのに十分であったと考えられる。この実験では、5μmマイクロスフェアを含有する培地で満たしたバイオリアクタ中で肺臓を3時間換気した。もし肺臓の脈管構造内にマイクロスフェアが観察されたら、それは、換気によって受動的灌流が誘導されたことを示す。図26に示すように、マイクロスフェアは大血管にも一部の毛細管にも見いだされた。これらの結果は、灌流はそれだけで、脈管構造における培地の移動を誘発するのに十分であり、したがって灌流の欠如にもかかわらず内皮の維持を可能にすることを示している。
【0259】
脈管構造における培地のこの移動は、換気による肺臓の物理的運動の結果である。このような大きな粒子を脈管構造中に移動させるには、拡散だけでは不十分であるだろう。拡散による脈管構造中へのマイクロスフェアの予想される移動は、フィックの第二法則を使って概算することができる。
【0260】
細胞の生存および分化の支持
本明細書に提示する結果は、血管灌流だけでは、細胞生存や、II型上皮によるサーファクタント産生を含む細胞分化を支持するには、十分でないことを証明した。しかし、培地による陰圧換気は、広範な細胞生存(未変性レベルの95.1%まで)を支持し、上皮および内皮の分化を維持するのに十分であった。
【0261】
本実施例の実験の全体的目的は、人工肺臓組織の今後の培養にこのバイオリアクタを使用するために、全齧歯類肺臓をインビトロで長期間にわたって培養する能力を有するバイオリアクタの根拠ある設計を証明することだった。上記の結果は、人工肺臓培養における使用に適した設計の成功を証明している。
【0262】
実施例4:人工肺臓組織における上皮発生
本明細書に提示する結果は、脱細胞化スキャフォールドが細胞毒性を持たず、上皮、内皮、および間葉細胞を含む広範な肺細胞タイプの接着と増殖を支持することを証明している。いくつかの例では、肺臓組織のエンジニアリングに、幹細胞を使用することができる。人工肺臓組織が有用であるためには、それを脈管構造および気道に接続することができなければならない。気道は肺胞とつながっていなくてはならず、血管接続は肺胞を取り巻く緻密な毛細管網に通じなくてはならない。
【0263】
本明細書に提示する結果は、人工肺臓組織の発生が、培地タイプ、灌流、換気、および気液界面の存在を含む重要なバイオリアクタ条件によって左右されることを証明している。いくつかの例では、培養培地による換気(すなわち「液体換気」)が、気道構造および上皮細胞の分化を助ける。さらにまた、気液界面における人工組織の小片の静置培養は、組織成長に対して有意な効果を示し、バイオリアクタでは、空気による換気が細胞の分化および上皮構造の発生に影響を及ぼすことを示した。
【0264】
これらの実験で使用した材料および方法を以下に説明する。
【0265】
材料および方法
スキャフォールド調製
成体Fischer 344ラットの肺臓を収集し、本明細書の他の項で説明するように脱細胞化した。脱細胞化操作に続いて、スキャフォールドを滅菌水(10回交換)中ですすぎ、次に、PBS中の10%ペニシリン/ストレプトマイシンにおいて少なくとも12時間すすいだ。後の培養では、DNA残存物の除去を助けるために、10%FBS中でもすすいだ。次に、完全な灌流および呼吸システムを取り付けた新しい滅菌バイオリアクタに、肺臓を移した。次に肺臓をPBSで2回すすぎ、培養に使用する培地で1回すすいだ。
【0266】
新生児細胞単離
肺臓を本明細書の他の項で論じるように新生児(約7日齢)ラットから単離した。次に肺臓を70%エタノール中で10秒間すすぎ、ダルベッコ変法イーグル培地(DMEM、Gibco)中で2回すすいだ後、滅菌乾燥ペトリ皿に移した。肺臓を円刃刀で5分間切り刻んでから、エラスターゼ消化用の円錐管に移した。DNアーゼ、コラゲナーゼおよびエラスターゼはWorthington Biochemical(ニュージャージー州レークウッド)から入手した。エラスターゼ消化は、DMEM中の4U/mlエラスターゼを100U/ml DNアーゼと共に使用し、撹拌しながら室温で20分間行った。次に、組織塊を70μmナイロンフィルタで濾過し、DMEMですすいだ。消化されなかった塊を新しいチューブに移し、Ca2+およびMg2+を含む1:1 DMEM:PBS中の1mg/mlコラゲナーゼ溶液において、撹拌しながら室温で1時間、コラゲナーゼ消化した。コラゲナーゼ消化組織を再び70μmフィルタで濾過し、消化されなかった断片をシリンジプランジャーを使って物理的に破砕した。残った組織をDMEMですすぎ、70μmフィルタで濾過した。コラゲナーゼ消化およびエラスターゼ消化から得た細胞を合わせてから、DMEM中で3回洗浄し、培養に使用する培地中で1回洗浄した。トリパンブルー色素排除法を使って細胞の生存性を評価した後、本明細書の他の項で説明するように、細胞を脱細胞化スキャフォールドに播種した。
【0267】
新生児細胞播種
肺細胞単離と脱細胞化スキャフォールドの調製後に、単離された細胞を、培養に使用する培地に懸濁した。気道コンパートメントの播種には、バイオリアクタあたり15mlの細胞懸濁液を気管リザーバに注入し、陰圧換気で肺臓の気道コンパートメント中に細胞を移動させることによって、細胞を播種した。脈管構造の播種には、バイオリアクタあたり3mlの細胞懸濁液を肺動脈中に注入した。灌流も換気も行わずに細胞を終夜付着させた後、実験条件に応じて、灌流および/または換気を開始した。
【0268】
人工組織培養
播種後、肺臓を一晩静置培養してから、灌流または換気を開始した。灌流および換気は、実験条件に応じて変更した。培養培地は週に2回入れ換えた。換気条件については、バイオリアクタ中の空気を手作業で交換できるように毎日1回、短時間停止した時間を除けば、肺臓を絶えず換気した。人工組織の断片の培養については、一晩播種した後、スキャフォールドをバイオリアクタから取り出し、滅菌剪刀を使って小さい(1〜3mm)断片に切断した。その断片を培養用ペトリ皿に移し、表示がある場合には、後に、気液界面培養のための0.4μmフィルタインサートを含むペトリ皿に移した。
【0269】
フローサイトメトリー
細胞単離後に、細胞を緩衝液(2mM EDTAおよび0.5%ウシ血清アルブミンを含むPBS)中ですすいだ。細胞内抗原を染色するために、細胞を室温において1%ホルムアルデヒドで15分間固定し、次にPBS中の0.2%Triton-Xで透過処理した。一次抗体を、1:100の希釈度で、室温にて30分間、緩衝液中で適用した。緩衝液中で3回すすいだ後、二次抗体を1:100の希釈度で室温にて20分間適用した。細胞は、イェール大学医学部細胞選別施設(Yale School of Medicine Cell Sorting Facility)のBecton-Dickinson FACSCalibur装置で分析した。
【0270】
実験の結果を以下に説明する。
【0271】
スキャフォールドには細胞毒性はない
まず最初に、腫瘍由来肺臓上皮細胞株MLE-12を、脱細胞化肺臓スキャフォールドにおける予備培養実験に使用した。これらの実験は、スキャフォールドに細胞毒性がないことを証明するため、および脱細胞化スキャフォールドを利用する培養のためのバイオリアクタシステムの一階妥当性(first-order validity)を証明するために行った。MLE-12細胞株は、脈管構造を通した培地の灌流を伴うバイオリアクタにおける脱細胞化スキャフォールド上での10日までの培養期間中、ロバストな細胞成長を示すことが観察された。組織像を図27に示す。細胞は、3日の時点では極めて原始的な肺胞構造を形成するようだったが、その後は著しく増殖し、7日までに無制御な細胞成長を示す。腫瘍由来の細胞株であるから、これは予想される帰結である。これらの実験は、バイオリアクタおよび脱細胞化肺臓スキャフォールドの検証における予備段階であり、これにより、新鮮単離新生児肺細胞を使った以後の実験が正当化された。
【0272】
新生児肺細胞の収集
齧歯類肺臓上皮はインビトロで培養することが難しいので、また新生児ラットの肺細胞は若く比較的可塑性が高いので[Massaro et al., 1985, J Clin Invest 76:1297-305;Meyrick et al., 1982, Am Rev Respir Dis 125:468-73]、肺細胞タイプの不均質な混合物を表す多数の細胞を単離できることを含むいくつかの理由から、新生児肺細胞を選択した。
【0273】
細胞数、生存性、およびフローサイトメトリーに基づく細胞タイプの分布に基づいて、細胞単離の条件を最適化した。使用した主なマーカーは、サーファクタントタンパク質C(SPC;II型肺細胞)、アクアポリン5(AQP;I型肺細胞)、クララ細胞分泌タンパク質(CCSP;クララ細胞)、および血小板内皮細胞接着分子1(PECAM-1;内皮細胞)である。細胞単離の条件は、総合的な細胞数および生存性を最適化する反復実験の結果として選択した。酵素およびインキュベーション条件の選択は、総細胞数、トリパンブルー色素排除法およびフローサイトメトリー分析によって評価される細胞の収量および生存性に基づいて最適化した。
【0274】
「最適化された」単離レジメンの下で得られた試料肺臓単離のフローサイトメトリーデータを図28に示す。典型的な単離では、細胞の5〜10%がCCSP陽性であり、細胞の40〜60%がSPC陽性であり、細胞の2〜8%がAQP陽性であり、細胞の1〜2%がサイトケラチン14陽性であり、細胞の10〜30%がPECAM-1陽性であり、細胞の5〜10%がα-アクチン陽性である。サイトスピン調製物を使って、CCSPおよびSPCに関する染色を確認した。これらのパーセンテージの大半は予想される範囲内にあるが、I型肺細胞(AQP陽性)については、未変性肺臓におけるその割合だけに基づけば、もっと高い収量が予想されるだろう。しかし、I型肺細胞は非常にもろく、それらの多くは細胞収集プロセスに耐え抜く可能性が低い。ひと腹の子(7〜12匹)からの総細胞収量は細胞約1億個であり、生存率は75〜85%だった。
【0275】
人工肺臓培養のためのバイオリアクタ条件の予備的確認
主に、組織学的検査による細胞密度、生存性および形態、ならびにいくらかのタンパク質発現評価に基づいて、さまざまな変量および適切な条件を探究するために、実験を計画した。評価した条件を以下に簡単に述べる。
【0276】
培地選択:基礎培地と血清濃度をどちらも変化させて、いくつかの培地組成を評価した。BGJb(無血清)、10%FBSを含むDMEM、15%FBSを含むEGM-2、およびEGM-2+15%FBSとBGJbの3対1混合物を含むいくつかの培地条件を使って、上皮再集合(repopulation)、主にII型上皮を観察した。組織学的検査および免疫蛍光染色によれば、培地タイプBGJbおよびDMEM+10%FBSは、総合的上皮成長にとって優れた条件をもたらした。その結果、以後の実験にはこれらの培地を使用した。ただし本発明はいかなる具体的培地にも限定されるべきでない。これは、所望する増殖および分化を促進する任意の培地を使用することができるからである。
【0277】
灌流および換気:予備実験では灌流と換気の両方を使用した。例えば、栄養素を供給するために培地を2〜5ml/分の流速で灌流した。1回の吸息による1日1回の換気の効果も評価した。しかし、その結果得られる人工肺臓培養に有意差は検出されなかった。1日1回の換気では明確な利益が観察されなかったにもかかわらず、この最低レベルの換気は、より生理的な培養環境を与えるであろうと推定された。それゆえに、以降の実験の大部分では、培養物を灌流し、1回の吸息により、1日1回、換気した。
【0278】
脱細胞化スキャフォールドは上皮、内皮および間葉細胞の成長を支持する
以下の実験は、発生工学的(development engineered)肺臓組織にとっての、脱細胞化スキャフォールド、肺臓バイオリアクタ、および単離新生児肺細胞集団の妥当性を証明するために計画した。これらの人工組織の培養に使用した詳細な条件を表1に示す。表には、これらの条件が人工組織の成長に及ぼす効果を評価するために具体的に調べた条件も記載する。ただし本発明はこれらの条件に限定されるわけではない。むしろ、本発明には、適用可能な条件であれば、その条件がバイオリアクタとの関連で人工肺臓の生成を促進する限り、いずれも包含される。
【0279】
(表1)人工肺臓培養のためのバイオリアクタ条件
【0280】
上皮細胞再集合の証明
本明細書に提示する結果は、脱細胞化肺臓スキャフォールドへの上皮細胞の接着および増殖を証明する。これらの実験には、DMEM+10%FBS培地、2ml/分での脈管構造の灌流、1日1回の換気、非被覆(uncoated)脱細胞化スキャフォールド、および無選別新生児肺細胞集団(本明細書の他の項で説明する「最適化された」条件)を使用した。
【0281】
図29に、人工肺臓組織のH&E染色を示す。4日時点で、著しい数の、組織化され立方上皮で裏打ちされた発生中の上皮構造が観察され、8日時点では、観察されるそのような構造が少なくなって、多くの細胞がより成熟した表現型をとった。4日時点では、多くの細胞が増殖性であったが、8日時点では、観察される増殖性細胞は少なくなった(図30)。4日時点でも、8日時点でも、著しい数のアポトーシス細胞は観察されなかった(図31)。
【0282】
免疫蛍光法を使って、I型肺細胞にはアクアポリン5を、II型肺細胞にはサーファクタントタンパク質Cを、そしてクララ細胞にはクララ細胞分泌タンパク質を使用して、重要な上皮細胞マーカーの発現を証拠立てた。図33に示すように、培養物ではII型上皮細胞が一般に優勢であり、後期の時点では特にそうだった。クララ細胞は4日時点で最も高密度に観察され、8日時点では少なくなった(図32)。4日時点ではアクアポリンに関する染色も観察されたが、それより遅い時点では、観察されるアクアポリン染色が著しく減少した(図34)。
【0283】
I型上皮に関するアクアポリン染色は、培養4日の時点では、形状が立方形である細胞を表すことが観察された。これは、機能している肺臓において肺胞を裏打ちする細胞としてのそれらの通常の扁平な形態とは相容れない。さらにまた、これらの立方形細胞は、図36および図37に見られるように、しばしばSPCまたはCCSPのいずれかについても陽性に染色された。したがって、4日時点でアクアポリンを発現する細胞が、平坦な形態と他のマーカーを伴わないアクアポリンの発現とによって示唆されるであろう成熟したI型上皮である可能性は低い。
【0284】
I型肺細胞はインビボでは、肺胞上皮の局所常在前駆体細胞であるII型上皮細胞に由来する。発生中の未変性肺臓では、胎児呼吸運動がなければI型肺細胞が最終分化状態を獲得せず[Inanlou et al., 2005, Dev Dyn 233:772-82]、組織像およびTEMでは形状が立方形のままである[Inanlou et al., 2005, Histol Histopathol 20:1261-6]。
【0285】
I型肺細胞分化の欠如は、定期的に換気されないこれらの培養物では、意外なことではなかった。アクアポリンを発現する細胞は、おそらく、I型上皮に完全には分化していなかった局所前駆体細胞から生じるのだろう。したがって脱細胞化スキャフォールドは肺上皮の付着および増殖を支持することができる。II型上皮、ならびにクララ細胞、および完全に分化したI型上皮細胞の前駆体である可能性が高い細胞のロバストな成長が観察された。本発明者らは、これらの所見を、脈管構造を通した培地灌流下に、呼吸運動を時折行うだけで観察している。
【0286】
上皮前駆細胞再集合
脱細胞化スキャフォールドにおける2タイプの肺上皮前駆細胞の成長が観察された。CCSPとSPCについて二重に陽性である細胞は、気管支肺胞幹細胞と呼ばれる局所前駆細胞であると報告されており、これはクララ細胞にもII型肺細胞にも分化することができ、気管支肺胞管接合部に見いだされる[Lane et al., 2007, Regenerative Medicine 2:407-15;Kim et al., 2005, Cell 121:823-35]。図35に、これらの細胞の予想される生理学的位置である末端細気管支の外観と合致する構造中に見いだされる、そのような二重陽性細胞を示す。
【0287】
基底細胞は肺気道の局所幹細胞であり、それらは円柱上皮の下に存在して、近位気道の上皮にとって再生性細胞源として役立つ。図36は、基底細胞マーカーであるサイトケラチン14について陽性である細胞を示している。比較のために、未変性肺臓を図36Aに示す。さらにまた、サイトケラチン14とCCSPに関する二重染色を図37に示す。図37は、クララ細胞が気道を裏打ちし、基底細胞がそれらの下に横たわっていることも示しており、これらは、その正常な解剖学的位置に合致している。これらの細胞は、時には、未変性肺臓におけるそれらの位置(図36B)と合致して、より大きい気道構造の下に見いだされるが、大きな気道には関係しないと思われるクラスター中にも見いだされる(図36C)。
【0288】
上皮細胞および内皮細胞の成長に加えて、間葉細胞も、脱細胞化肺臓スキャフォールドに再集合することができる。図38は、平滑筋および筋線維芽細胞を染色するα-アクチンの免疫蛍光染色を示す。これらの人工組織は、上皮成長にとって有利であることが示された同じ条件下で培養された。間葉細胞が、未変性肺臓におけるそれらの位置と合致して、発生中の上皮構造の下および間に位置することも見いだされた。したがって、本明細書に提示する結果は、脱細胞化スキャフォールドが間葉細胞の成長にとっても適切な基質であることと、新生児肺細胞の集団には生きた間葉細胞が含まれていたことを証明している。
【0289】
培地組成が上皮分化に及ぼす効果
培地タイプは、細胞の成長と分化に、したがって人工肺臓組織の発生に、著しい影響を持つことができる。これらの相違の一部をより詳しく調べるために、上皮分化に関して無血清培地(BGJb)と血清含有培地(DMEM+10%FBS)を使って人工肺臓培養の成長を比較した。これらの実験では、まず最初に、細胞をDMEM+10%FBS中のスキャフォールド上に播種し、この培地中で2日間培養した後、BGJb(無血清)に4日間かけて徐々に移行し、最後に純粋なBGJb培地中で2日間培養した。無血清培地への移行は、サーファクタントの発現に対する実質的な影響を引き起こした。無血清培地は、DMEM+10%FBSと比較して、より多くのサーファクタント(SPC)の頂端部発現につながることが観察された(図39Cおよび39D)。これは、無血清培地(BGJb)では、ウェスタンブロット上で、サーファクタントの発現が有意に増加することと一致する(図40;「DMEM」および「BGJb」と記したレーンを比較のこと)。さらにまた、サーファクタントの形態が、未変性肺臓と、はるかによく合致していた(BGJb培地では大半のサーファクタントが21kDaプロSPC形として認められた)。
【0290】
さらにまた、無血清培地への移行は、図43Fおよび43Eに記載の免疫蛍光によって認められるCCSP発現の現象につながる。どちらの培地タイプでも、散在性のCCSP発現が発生中の上皮構造の管腔に観察された。これらの培養は小さい組織切片で行われたので、この表現型は、灌流または換気の欠如によるものであったと考えられる。
【0291】
バイオリアクタにおける肺臓発生に気液界面が及ぼす効果
気液界面を作り出すために、人工肺臓をまず、培地による換気下で4日間培養して、細胞を付着させ増殖させた。培養の最後の4日間は換気を培地から濾過大気に切り換えた。
【0292】
空気による換気は、気道上皮に重度の損傷を引き起こすと共に、一部の肺胞壁に破壊的な変化を引き起こした(図19)。そこで、空気換気によって未変性肺臓に引き起こされる損傷を減少させる努力として、空気換気に使用する一回換気量を、液体または空気呼吸に関する約2.0〜2.5mlという先の値から、約50%減少させた。
【0293】
バイオリアクタにおける空気換気の存在に起因する細胞付着または細胞形態の変化は観察されなかった。しかし、バイオリアクタで培養された人工肺臓における空気による換気は、I型上皮の分化マーカーであるアクアポリンの発現の誘導につながることが観察された。これは、典型的にはサーファクタントタンパク質C(II型上皮を示す)についてのみ染色される実質の細胞でも、発生中の上皮構造における立方形細胞の偶発的染色でも認められた。観察された染色パターンは、アクアポリン発現細胞の大半がSPCも発現する可能性が極めて高いことを示している。図42Bからわかるように、実質中の細胞の事実上全てがSPCを発現し、一方、実質細胞のサブセットはAQPを発現する(図41A)。
【0294】
これらの知見は、気液界面がII型上皮からI型上皮への分化を誘導していることを、強く示唆している。II型上皮は、I型上皮にとって公知の局所前駆細胞であるので、この分化は意外なことではない[Adamson et al., 1974, Lab Invest 30:35-42;Fehrenbach, 2001, Respir Res 2:33-46]。そのうえ、図40に示すように、空気換気人工肺臓では、サーファクタントタンパク質Cの発現量の減少が観察された(レーン「ALI」を、培地で換気(「Vent」)および灌流(「Perf」)した肺と比較されたい)。これは、II型上皮からI型上皮への分化とも合致する。さらにまた、図42に示すように、絨毛上皮細胞の数の増加も観察された。これは空気界面を導入した結果である可能性が高い。気道上皮をインビトロで培養すると、液体から空気界面への細胞の移行が絨毛発現を誘導する[You et al., 2002, Am J Physiol Lung Cell Mol Physiol 283:L1315-21;Davidson et al., 2004, J Cyst Fibros 3 Suppl 2:59-62]。そのうえ、気液界面の欠如は絨毛形成の減少につながり得る[Ostrowski et al., 1995, Exp Lung Res 21 :957-70;Yeh et al., 2007, Laryngoscope 117:1439-44]。したがっていくつかの例では、空気界面が、インビトロでの肺臓組織の再生に、重要な影響を持つ。
【0295】
灌流および換気が人工肺臓組織における上皮構造の発生に及ぼす効果
灌流および換気は、バイオリアクタにおける肺臓組織の培養に著しい影響を及ぼすことができる。換気は、I型およびII型肺細胞の分化を含む肺臓上皮発生にも著しい影響を持つ[Inanlou et al., 2005, Histol Histopathol 20:1261-6;Inanlou et al., 2005, Dev Dyn 233:772-82;Inanlou et al., 2005, Dev Dyn 232:43-54]。そこで、バイオリアクタにおける8日間の培養中に灌流および換気が人工肺臓発生に及ぼす効果を比較した。これらの実験では、条件を、他の項で論じる換気実験時に利用したものと同じ条件とし、DMEM+10%FBSの培養培地、非被覆スキャフォールド、および無選別新生児肺細胞集団を使用した。ただし、培養物は、脈管構造を通して2ml/分で灌流するか、培地を使って1吸息/分で連続的に換気した。
【0296】
図50に示すように、上皮構造を形成する細胞の多くが、CCSPに関して陽性に染色される。換気した培養物ではCCSPに関する染色が観察されたが、細胞はより平坦な形態を獲得した。これは、換気が、肺胞上皮(I型またはII型肺細胞)に向かって分化するようにCCSP発現細胞を誘導していることを示すのだろう。
【0297】
換気が存在しない場合は、図43のH&E染色で目に見えるエオシン好性の物質、したがってタンパク質性である可能性が高い物質で、発生中の上皮構造の管腔が満たされた。この物質は、図45に見られるように、CCSPについても陽性に染色された。このCCSPの蓄積は、これらの上皮構造を裏打ちするクララ細胞がCCSPを産生していることを示す。さらにまた、換気によるこの物質の除去は、気道樹が依然としてインタクトであって流体を通すことを示し、これら発生中の上皮構造が気道樹の一部であって、マトリックス内でランダムには増殖しないことも示唆する。
【0298】
図46に示すように、灌流と換気は、SPCの発現に対して、有意な効果を持たなかった。細胞の大半は、換気条件下でも、灌流条件下でも、SPCについて陽性だった。
【0299】
実施例5:人工肺臓組織における内皮発生
脱細胞化スキャフォールドが人工肺臓内皮の成長を支持することができるかどうかを決定するために、また、いくつかの具体的因子が人工内皮の発生に及ぼす効果を、血管コンパートメントと気道コンパートメントの間の機能的な内皮障壁の形成に影響を及ぼすそれらの因子の能力に焦点を絞って評価するために、以下の実験を計画した。
【0300】
これらの実験で使用した材料および方法を以下に説明する。
【0301】
材料および方法
スキャフォールドの調製
脱細胞化スキャフォールドは、本明細書の他の項で説明するように調製した。
【0302】
新生児細胞の単離と播種
新生児ラット肺細胞は、本明細書の他の項で説明するように単離した。細胞を、本明細書の他の項で説明するように、スキャフォールドに播種した。
【0303】
内皮細胞培養
ラット肺臓微小血管内皮細胞をVEC Technologies(ニューヨーク州レンセリア)から入手し、フィブロネクチン被覆(約1μg/cm2、Gibco)組織培養器上、10%FBSと追加成長因子(supplemental growth factors)(VEC Technologies)を含むMCDB-131完全培地中で成長させた。
【0304】
人工内皮培養のための最適化された条件
スキャフォールドを1mgのフィブロネクチン(Gibco)で被覆し、37℃のPBS 60ml中、脈管構造を通して灌流した後、PBSおよび培地ですすいだ。培養0日目と培養2日目または3日目の2回、各回800万〜1000万個のラット肺臓微小血管ECを播種した(2回の播種のそれぞれについて、肺臓1つあたり2つのT150培養フラスコを使用した)。0.25%トリプシン(Gibco)を使って細胞を組織培養プレートからトリプシン処理し、細胞塊を除去するために40μmフィルタで濾過し、約3mlの培地に入れた単回ボーラス注射として、肺動脈に注入した。細胞を1時間接着させた後、脈管構造を通して約1.5ml/分で灌流を開始した。1〜2時間後に、7〜10日の培養期間の残りの部分は、灌流量を3ml/分に増加させた。培地は3〜4日ごとに換えた。
【0305】
免疫蛍光法
本明細書の他の項で説明するように、組織試料を調製し、染色した。
【0306】
透過型電子顕微鏡法(TEM)
本明細書の他の項で説明するように、試料を調製し、分析した。
【0307】
微粒子保持
巨大高分子程度のサイズを持つ小さな粒子に対する全ラット肺臓の透過性を評価するために、アッセイを開発した。このアッセイでは、気道-血管障壁を横切って起こるFITC標識デキストラン溶液の漏出を定量した。2,000,000Daの分子量を持つFITC標識デキストランをSigma(ミズーリ州セントルイス)から入手した。未変性肺臓および0.025%トリプシンで2分間処理した未変性肺臓の透過性を測定することによってアッセイの検証を行った。肺臓をヘパリン化PBSで灌流し、通常のバイオリアクタカニューレに接続した。ベースライン洗浄液試料を得た後、トリプシン処理肺臓は、PBS中の0.025%トリプシン10mlで灌流し、室温で2分間留まらせてから、10mlのPBSですすいだ。FITC標識デキストラン溶液(1mg/ml)を肺動脈に注入し、次に20mlのPBSでフラッシングした。次に、2つの洗浄液試料を直ちに、気管から連続して採取した。蛍光プレートリーダーを使って蛍光を測定し、データを標準曲線に当てはめた。脱細胞化肺臓または人工肺臓で行う場合は、マイクロスフェアアッセイ(3.2.9章参照)の場合と同様に、このアッセイを気道によって行った。したがって、FITC-デキストランを気道に注入し、脈管構造をPBSでフラッシングした。
【0308】
実験の結果を以下に説明する。
【0309】
細胞供給源:市販されているラット肺臓微小血管EC源を使用した。これらの細胞を、フィブロネクチン被覆したスキャフォールド中に播種して、EC特異的培地の存在下で培養すると、図47に示すように、内皮細胞の実質的成長が観察された。
【0310】
内皮スクリーニング実験の要約:本明細書の他の項で論じるスクリーニング実験により、人工内皮培養に適合する一組の条件を同定することができた。帰結は、主に、細胞生存性に関する組織学的検査と、免疫蛍光法でのPECAMの発現とによって評価した。これらのパイロット試験の結果、個別の条件が人工肺臓内皮に及ぼす影響を体系的に評価することができるように、スキャフォールド内部での内皮細胞成長を可能にする一組の条件が得られた。
【0311】
人工内皮の培養に適していると同定された条件は、次のとおりだった:精製されインビトロで拡大されたラット肺臓微小血管EC集団の使用;フィブロネクチン被覆スキャフォールド;およびEC特異的培地(10%FBSおよび追加成長因子を含むMCDB-131)の使用。
【0312】
FITC-デキストラン透過性アッセイの検証
巨大高分子程度のサイズを持つ小さな粒子に対する全ラット肺臓の透過性を評価するためのアッセイを開発した。このアッセイでは、気道-血管障壁を横切って起こるFITC標識デキストラン溶液の漏出を定量した。このアッセイは、培養の過程で繰り返し使用することができ、細胞培養に害を及ぼさない材料を使用し、全肺臓の透過性の測定をもたらす。さらにまた、アッセイを固定直前に行えば、FITC-デキストランを、抗FITC抗体を使って組織切片上で同定することができるだろう。
【0313】
FITC標識デキストランは2,000,000Daの分子量を持つ。単分散デキストランの場合、ストークス-アインシュタイン半径(nm)は、rs=0.0488(MW)0.437によって分子量と関連づけられる[Venturoli et al., 2005, Am J Physiol Renal Physiol 288:F605-13;Oliver et al., 1992, J Am Soc Nephrol 3:214-28]。2MDaデキストランの場合は、これにより、27.7nmという半径が得られる。未変性肺臓と、脈管構造を希釈トリプシンで短時間灌流することによって「漏出しやすく」した未変性肺臓の透過性を評価することによって、このアッセイを検証した。FITC標識デキストラン溶液を肺動脈に注入した後、食塩水でフラッシングした。次に、2つの洗浄液試料を直ちに、気管から連続して採取した。図48に示すように、肺臓の透過性は、基底膜への内皮付着が破壊されることにより、予想どおり、トリプシン処理によって増加する。しかし、未変性肺臓でも、このアッセイでは測定可能な漏出を与えた。この血管透過性の劣化は、動物の屠殺とデキストラン注入との間の遅延、ならびに肺臓組織の取扱の結果である。
【0314】
脱細胞化肺臓または人工肺臓で行う場合は、マイクロスフェアアッセイの場合と同様に、このアッセイを気道によって行った。したがって、FITC-デキストランを気道に注入し、脈管構造を食塩水でフラッシングした。血管コンパートメントに移動したデキストランを漏出として測定した。脱細胞化肺臓または人工肺臓では、組織が流体に対して高い透過性を持ち、気道洗浄後に還流試料を得ることができないので、アッセイをこの方法で行った。したがって、デキストランを単回ボーラス洗浄液として気道に注入し、脈管構造をフラッシングして、気道-血管障壁を横切って起こるFITC-デキストランの流出を測定した。
【0315】
人工肺臓内皮に対する還流と換気の効果
具体的条件が人工内皮組織の発生に及ぼす効果を、機能的内皮障壁の形成に焦点を絞って評価するために、以下の実験を計画した。灌流しながら人工肺臓内皮を培養することの効果と換気しながら人工肺臓内皮を培養することの効果を比較した。換気と灌流の両方を、内皮細胞の生存および増殖に関して評価し、透過型EMを使って細胞間結合の形成についても評価した。
【0316】
図48の組織像に示されるように、灌流は人工肺臓内皮の成長を実質的に改善することが観察された。また、図49に示すように、またH&E組織像におけるそれらの外観不良と合致して、換気では、より多くのアポトーシス細胞が観察された。
【0317】
また、灌流培養物と換気培養物を、細胞間結合の存在について、透過型EMおよびVE-カドヘリン染色を使って分析した。内皮細胞観の密着結合は、隣接する細胞を互いに堅く連結し、よってこれらの傍細胞間隙間での流体の動きを阻害するので、障壁機能の重要な手段である[Majno et al., 1961, J Biophys Biochem Cytol. 11:571-605]。これらの細胞結合が弱いか存在しない場合は、脈管構造外への流体の漏出が起こって、それが肺水腫を引き起こし得る[Orfanos et al., 2004, Intensive Care Med 30:1702-14;Maniatis et al., 2008, Vascular Pharmacology 49:119-33]。
【0318】
図50に示すように、TEMを使って、灌流人工組織における細胞間結合を観察した。全ての細胞が密着結合形成を示したわけではなかった。
【0319】
VE-カドヘリンに関する免疫蛍光法を使って細胞間結合形成を評価した。図51に示すように、灌流人工肺臓内皮には、VE-カドヘリンに関してロバストな染色が見いだされた。換気組織は、組織像およびTEMでのそれらの外観不良ゆえに、VE-カドヘリンについて染色しなかった。
【0320】
人工肺臓内皮の障壁機能の評価
血管床と気腔の間に機能的障壁を形成する人工肺臓内皮の能力を評価するために以下の実験を計画した。これは、肺胞への流体漏出を減少させ、よってガス交換を可能にするために重要であり、人工肺臓組織にとっての目標の重要な構成要素である。肺臓内皮によって提供される障壁機能を評価するために、透過性アッセイを使って、気腔から脈管構造への小さい(55nm)FITC-デキストラン粒子の移動を測定した。このアッセイは、本明細書の他の項で説明するように開発され、検証された。
【0321】
このアッセイでは、FITC-デキストランを気道コンパートメントに注入し、脈管構造を食塩水でフラッシングすることにより、肺胞-血管障壁を横切って漏出した量を測定した。脱細胞化スキャフォールドの場合、そのような小粒子に対する障壁機能は本質的に存在せず、事実上全て(98.4%)のデキストランが肺胞から脈管構造中に移動して、血管すすぎ液と共に回収された。これに対し、未変性肺臓は、同様に処理した場合に、12.9%の漏出を示す(図52)。
【0322】
灌流された人工肺臓内皮組織では、気道コンパートメントにおける30%までのデキストランの保持が、7〜10日の培養期間後に証明された。換気内皮組織は87%の透過性を示した。
【0323】
灌流人工肺臓内皮に関するこれらの知見は、特に蛍光免疫法によるロバストなVE-カドヘリン発現(図51)およびTEMによる細胞間結合形成という知見と合わせて考えると、人工内皮組織における機能的内皮障壁の形成が起こったことを示している。
【0324】
図63は人工組織の極限引張強さを図示する図表である。未変性肺臓と脱細胞化肺臓の強さも示されている。この図は、人工組織の極限引張応力が未変性肺臓とも脱細胞化マトリックスとも同等であることを示している。したがって、組織工学的肺臓の機械的性質は再細胞化プロセス後も維持される。
【0325】
実施例6:肺臓細胞治療
本明細書に提示する結果は、哺乳動物における肺臓細胞治療を達成するための脱細胞化肺臓組織の使用を証明する。一般に、工程には、気管の脱細胞化、脱細胞化気管組織内の細胞外マトリックス構成要素の検出、脱細胞化気管マトリックスにおけるヒト気管支(大気道)および小気道肺上皮細胞の培養、所望の遺伝子によるヒト肺上皮細胞の遺伝子治療、および点滴注入による肺臓へのヒト肺上皮細胞の点滴注入が含まれ、レシピエント肺臓における細胞の付着および生存が確認される。
【0326】
気管の脱細胞化
ブタ気管を収集し、血液を除去するためにPBS中ですすいだ。断片を切断し、10%中性緩衝ホルマリンで固定し、パラフィンに包埋し、5mm切片に切断した。残りを5つの環に切断し、CHAPS緩衝液(pH13.5)中で撹拌しながら、2時間、4時間および8時間の時点でCHAPS緩衝液を交換して、2〜24時間インキュベートした。表示した時点において、組織をCHAPS緩衝液から取り出し、PBSですすいだ。断片を切断し、10%ホルマリンで固定し、脱細胞化を確認するための組織学的分析用に加工した。
【0327】
CHAPS緩衝液中で4〜8時間インキュベートして調製した脱細胞化気管は、コラーゲンマトリックスを維持しており、細胞の大半が組織から除去されている(軟骨層を除く)ことが観察された。図55参照。以下の実験は、撹拌しながら6時間のCHAPSインキュベーションを行うことによる脱細胞化気管の調製に基づいた。
【0328】
脱細胞化の前後に気管中の細胞外マトリックスを検出するために、これらの実験を計画した。簡単に述べると、組織の断片を未変性気管および脱細胞化気管から切り取り、10%ホルマリンで固定した。5μmパラフィン包埋切片をH&EおよびMassonのトリクロームで染色した。パラフィン包埋切片を、コラーゲン(COL)、フィブロネクチン(FN)およびラミニン(LN)を含む細胞外マトリックスタンパク質についても免疫染色した。切片を脱パラフィンし、再水和し、抗原回復(プロテイナーゼKを使用)を行い、ブロッキングした。自家製のウサギ抗ヒトFN、COL I、COL III、COL IV、COL VまたはLN抗体を切片に適用し、次にAlexa Fluor 546コンジュゲートヤギ抗ウサギIgGを適用した。スライドをDAPIで対比染色した。未変性気管は、COL I、COL III、COL Vについて陽性に染色されるが、他のECMタンパク質については陽性に染色されないことが観察された。脱細胞化気管は、未変性気管に見られる3タイプのCOLを全て含有していた。図56参照。これらの結果は、脱細胞化気管組織が、インビトロで気道上皮細胞の接着、成長および分化を支持することを示唆している。
【0329】
脱細胞化気管組織上での気道上皮細胞の成長
脱細胞化組織上で細胞を成長させるために、次の一組の実験を計画した。簡単に述べると、ヒト気管支/気管上皮細胞(NHBE)を気管支上皮成長培地(BEGM)中で培養した。脱細胞化気管(6時間CHAPS緩衝液インキュベーション)を、滅菌PBSで十分に(少なくとも24時間は)すすいだ。軟骨層(ならびに外膜層)をはがして、後続の細胞播種のために気管粘膜および粘膜下層を残した。組織を約5×5cm2のサイズに切断し、6ウェルプレート中のトランスウェルインサート(孔径0.4μm)上に、上皮面を上向きにして置いた。
【0330】
細胞を脱細胞化気管組織に次のように播種した。50μlの細胞(密度3.5×106細胞/ml)を脱細胞化気管組織の上皮面に加え、37℃で30分間インキュベートした。次に、新鮮な培養培地をトランスウェルインサートの上側と下側の両方に加え、細胞をさらにインキュベートした。表示した時間で、組織をトランスウェルから取り出し、10%ホルマリンで固定した。パラフィン包埋した5μm切片をH&Eで染色した。脱細胞化気管はNHBEの接着と成長を支持することが観察された。図57参照。
【0331】
導入遺伝子でトランスフェクトされたヒト小気道上皮細胞(SAEC)を脱細胞化組織上で成長させるために、次の一組の実験を計画した。簡単に述べると、ヒトSAECに、GFPレトロウイルスを6時間トランスフェクトした。細胞を6ウェルプレート中の脱細胞化気管組織に次のように播種した。50μlの細胞(密度5×106細胞/ml)を脱細胞化気管組織の上皮面(約0.5×1.0cm2)に加え、37℃で30分間インキュベートした。次に、新鮮な培地を加え、細胞をさらにインキュベートした。脱細胞化気管はSAECの接着と成長も支持することが観察された。図58参照。さらにまた、脱細胞化気管は、関心対象の遺伝子がトランスフェクトされたヒト肺上皮細胞を培養することの原理証明として、GFPがトランスフェクトされたSAECの接着および成長も支持した。
【0332】
気道上皮細胞の遺伝子治療
肺臓細胞の遺伝子治療に脱細胞化組織を使用することの実現可能性を決定するために、次の一組の実験を計画した。遺伝子改変は次のように行った。Phoenixパッケージング細胞株を使ってEGFP(強化GFP)レトロウイルス上清を調製した。EGFP DNAをLZRSpBMNベクターに挿入した。NHBEおよびSAECを80%コンフルエントを超えるまで成長させた。感染当日に細胞を数回すすぎ、ウイルス上清(8μg/mlポリブレンを含有するもの)を37℃で6時間接種した。細胞を数回すすぎ、新鮮培地中で一晩インキュベートした。次に、フローサイトメトリーを使ってGFPについて分析した。感染細胞はGFPについて陽性に染色されることが観察された。例えばNHBE細胞は非感染細胞と比較してGFPについて18.5%の陽性染色を示した。SAEC細胞は、非感染細胞と比較してGFPについて16%の陽性染色を示した。本明細書に提示する結果は、レトロウイルスを使ってこの培養系でヒト肺上皮細胞に関心対象の遺伝子を感染させ得ることを証明している。しかし、導入遺伝子を送達するための他の手段も本発明には包含される。例えば、レンチウイルス系を使用することの実現可能性を調べるために、次の一組の実験を計画した。
【0333】
GFPレンチウイルスによる感染は次のように行った。EGFPレンチウイルス上清を無血清培地中の293T細胞で調製した。CMVプロモーターを持つpSicoRベクターにGFP DNAを挿入した。NHBEを1×105細胞/ウェルの密度で6ウェルプレート上に播種し、37℃で1日インキュベートした(80〜90%コンフルエント)。感染当日に細胞を数回すすいでから、ウイルス上清(新鮮BEGMで1:3希釈したもの)(8μg/mlポリブレンを含有するもの)を37℃で6時間接種した。細胞を数回すすいでから、新鮮培地と共にさらに2日間インキュベートした。細胞を4%パラホルムアルデヒドで30分間固定し、顕微鏡で調べた。図59参照。結果を表2に要約する。
【0334】
(表2)
【0335】
本明細書に提示する結果は、GFPレンチウイルスに感染したNHBEが6時間後に明らかな形態変化を示さなかったことを証明している。6時間の1回感染により、フローサイトメトリーで評価して50%を超えるGFP陽性細胞が生じた。SAECの感染効率も50%を超えているようだった。本明細書に提示する結果は、所望する任意の遺伝子を使って遺伝子改変肺臓細胞を生成させ得ることを証明している。
【0336】
マウス肺臓への気道上皮細胞の注入
哺乳動物の肺臓中に気道上皮細胞を注入することの実現可能性を決定するために、次の実験を計画した。簡単に述べると、5μmマイクロスフェアとPBSの1:1混合物100μlをC57BL/6Jマウス、雌、約10週齢(Jackson Lab)に、気管を通して注入した。マウスは注入後数分間生き続けた。肺臓を直ちに収集し、ホルマリンで固定した。5μmパラフィン包埋切片をH&Eで染色した。この試験は、気道への点滴注入によって送達された細胞サイズの粒子が哺乳動物の肺臓内に検出されるかどうかを決定するために、最初の実現可能性試験として行われた。マイクロスフェアが注入されたマウス肺臓のH&E像からの結果は、かなりの数のマイクロスフェアがマウス肺臓の全ての葉に存在することを証明した。図60参照。これらの結果は、気管アプローチを使った注入を細胞点滴注入に使用できることを証明している。
【0337】
次の一組の実験には、GFP標識細胞を外植されたマウス肺臓に注入することを含めた。簡単に述べると、細胞をGFPレトロウイルスに6時間感染させた。C57BL/6Jマウスを安楽死させ、気管を露出し、近位端を縫合糸で結紮した。100μlのGFPレトロウイルス感染SAECを、マウスの肺臓に、気管を通して注入した(続いて、細胞を押し込むために200μlの空気を注入した)。気管の遠位端を結紮した。肺臓を外植し、直ちに10%ホルマリンで固定した。5μmパラフィン包埋切片を脱パラフィンし、再水和し、抗原回復(10mMクエン酸緩衝液pH6を使用)を行い、透過処理し(Triton Xを使用)、ブロッキングした。ウサギポリクローナル抗GFP抗体(Abcam製)を切片に適用し、次にAlexa Fluor 555コンジュゲートヤギ抗ウサギIgGを適用した。スライドをDAPIで対比染色した。外植された肺臓にGFP陽性細胞が見いだされることが観察された。図61参照。本明細書に提示する結果は、肺臓内への細胞の注入の成功を証明すると共に、導入遺伝子(GFP)を形質導入されたヒト上皮細胞が肺上皮に付着したことを証明している。
【0338】
マウス肺臓へのGFP標識細胞の注入の実現可能性を決定するために、次の一組の実験を計画した。簡単に述べると、マウス(C57BL/6J、雌、約10週齢)を麻酔し、気管を露出した。100μlのGFPレトロウイルス感染ヒト気道上皮細胞を気管を通して肺臓に注入した(続いて200μlの空気を注入した)。図62参照。マウスを6時間〜3日間回復させた。収集当日に、血液を除去するために肺動脈を通して肺臓をPBSで灌流し、次に気管を通して10%ホルマリンで灌流し、切り離した。収集した肺臓をホルマリン中でさらに4時間固定した。5μmパラフィン包埋切片を、本明細書の他の項で論じるように、GFPについて染色した。GFP陽性ヒト気道上皮細胞(NHBEとSAECの両方)が、気道内への点滴注入後、何日間も、マウス肺臓に見いだされることが観察された。図63参照。これは、ヒト上皮細胞(これは培養下で成長させたものであり、関心対象の導入遺伝子を形質導入されたものである)が、レシピエント哺乳動物の肺臓中に送達され、レシピエント肺臓に付着して、生き残り、増殖できることを示している。
【0339】
実施例7:脱細胞化人工肺臓の埋植
生きているラットレシピエントに脱細胞化人工肺臓を埋植することの実現可能性を示すために、この実験を計画した。先の実施例に従って脱細胞化人工ラット肺臓を調製した。成体雄ラットを、ケタミンとキシラジンの腹腔内注射で麻酔した。次に、ラットの気管に挿管し、麻酔を維持するためにフォーレンと混合した100%酸素で換気した。滅菌条件下で、胸骨正中切開によって胸郭を開いた。肋骨を両側に後退させて、正常に膨らんでいる肺臓と拍動している心臓を暴露した。全身ヘパリン化後に、未変性左肺臓をまとめて(in toto)切除した。次に、手術用顕微鏡下で、脱細胞化人工肺臓を、レシピエントの肺動脈、肺静脈および左主気管支に、10-0縫合糸を使って吻合した。
【0340】
肺動脈および肺静脈上の血管鉗子を取り除いた後、血管は人工肺臓を正常に灌流することがわかった。また、埋植された人工肺臓は容易に換気され、固有の未変性右肺と同様に拡張および収縮を繰り返した。図64に、換気サイクル中の拡張時と収縮時における、埋植された人工肺臓の写真を示す。したがって、本明細書に記載の技法を使って作製された人工肺臓は、哺乳動物に埋植可能であると共に、器官全体を通る灌流および気道の容易な換気を可能にするという観点から機能的である。埋植期間の全体にわたって、埋植された人工肺臓からの出血または空気漏出の証拠はなかった。
【0341】
本明細書で言及するどの特許、特許出願および刊行物の開示も、参照によりそのまま、本明細書に組み入れられる。
【0342】
本発明を具体的態様について開示したが、当業者が本発明の真の要旨および範囲から逸脱することなく本発明の他の態様および変形を考案し得ることは、明らかである。本願の特許請求の範囲は、そのような態様および等価な変形を全て包含すると解釈されるものとする。
【特許請求の範囲】
【請求項1】
細胞成長を支持する能力を有する脱細胞化組織であって、脱細胞化前の対応する天然組織の特徴を示す、脱細胞化組織。
【請求項2】
肺臓である、請求項1記載の脱細胞化組織。
【請求項3】
他の点では同一な脱細胞化前の組織の形態と実質的に類似する形態を示す、請求項1記載の脱細胞化組織。
【請求項4】
対応する天然組織の細胞外マトリックスを保持する脱細胞化組織であって、該細胞外マトリックスが、実質的にインタクトな外面を含む、請求項1記載の脱細胞化組織。
【請求項5】
免疫原性マーカーが実質的に除去されている、請求項1記載の脱細胞化組織。
【請求項6】
対応する天然組織の機械的性質と実質的に類似する機械的性質を示す、請求項1記載の脱細胞化組織。
【請求項7】
肺臓細胞の分化状態を支持し維持する能力を有する、三次元スキャフォールドと細胞の集団とを含む組成物。
【請求項8】
三次元スキャフォールドが脱細胞化組織である、請求項7記載の組成物。
【請求項9】
インタクトな気道樹および血管網を示す、請求項7記載の組成物。
【請求項10】
前記集団が幹細胞を含む、請求項7記載の組成物。
【請求項11】
前記集団が上皮細胞および内皮細胞を含む、請求項7記載の組成物。
【請求項12】
前記細胞が遺伝子改変されている、請求項7記載の組成物。
【請求項13】
肺胞上皮細胞の分化状態を支持し維持する能力を有する、請求項7記載の組成物。
【請求項14】
スキャフォールドが、フィブロネクチン、ラミニン、コラーゲン、糖タンパク質、トロンボスポンジン、エラスチン、フィブリリン、ムコ多糖、糖脂質、ヘパリン硫酸、コンドロイチン硫酸、ケラチン硫酸、グリコサミノグリカン、ヒアルロン酸、プロテオグリカン、ビトロネクチン、ポリ-D-リジン、多糖、およびそれらの組合せからなる群より選択される生体適合性材料を含む、請求項7記載の組成物。
【請求項15】
分枝形態形成の誘導に関連する遺伝子発現を示す細胞を含む、請求項7記載の組成物。
【請求項16】
前記遺伝子がCFTRである、請求項7記載の組成物。
【請求項17】
分枝形態形成、遠位肺臓上皮細胞分化、上皮成長、血管発生、およびそれらの組合せからなる群より選択される肺臓組織の特徴を含む、請求項7記載の組成物。
【請求項18】
播種済スキャフォールドを作製するために脱細胞化スキャフォールドに細胞の集団を播種する工程を含む、肺臓細胞の分化状態を支持し維持する能力を有する人工三次元組織を製作する方法。
【請求項19】
脱細胞化スキャフォールドがインタクトな気道樹および血管網を示す、請求項18記載の方法。
【請求項20】
前記集団が幹細胞を含む、請求項18記載の方法。
【請求項21】
前記集団が上皮細胞および内皮細胞を含む、請求項18記載の方法。
【請求項22】
前記細胞が遺伝子改変されている、請求項18記載の方法。
【請求項23】
脱細胞化スキャフォールドが肺胞上皮細胞の分化状態を支持し維持する能力を有する、請求項18記載の方法。
【請求項24】
スキャフォールドが、フィブロネクチン、ラミニン、コラーゲン、糖タンパク質、トロンボスポンジン、エラスチン、フィブリリン、ムコ多糖、糖脂質、ヘパリン硫酸、コンドロイチン硫酸、ケラチン硫酸、グリコサミノグリカン、ヒアルロン酸、プロテオグリカン、ビトロネクチン、ポリ-D-リジン、多糖、およびそれらの組合せからなる群より選択される生体適合性材料を含む、請求項18記載の方法。
【請求項25】
分枝形態形成の誘導に関連する遺伝子発現を示す細胞を含む、請求項18記載の方法。
【請求項26】
前記遺伝子がCFTRである、請求項18記載の方法。
【請求項27】
人工三次元組織が、分枝形態形成、遠位肺臓上皮細胞分化、上皮成長、血管発生、およびそれらの組合せからなる群より選択される肺臓組織の特徴を示す、請求項18記載の方法。
【請求項28】
試験作用因子を人工三次元肺臓組織モデルに接触させる工程と、該試験作用因子が該モデルに及ぼす効果を測定する工程とを含む、肺臓組織の健康状態を調整する試験作用因子の能力に関して試験作用因子をスクリーニングするためのインビトロ方法であって、該モデルに加えられるいずれの変更も、試験作用因子が肺臓組織の健康状態を調整できることを示す、インビトロ方法。
【請求項29】
人工三次元組織が脱細胞化スキャフォールドに由来する、請求項28記載の方法。
【請求項30】
試験作用因子が、化学剤、医薬、ペプチド、核酸、および放射線からなる群より選択される、請求項28記載の方法。
【請求項31】
試験作用因子が治療剤のための送達媒体である、請求項28記載の方法。
【請求項32】
細胞数、面積、体積、形状、形態、マーカー発現、または染色体断片化に対する試験作用因子の効果を決定する工程を含む、請求項28記載の方法。
【請求項33】
肺臓組織モデルに対して所望の効果を持つ作用因子を選択する工程をさらに含む、請求項28記載の方法。
【請求項34】
肺臓細胞の分化状態を支持し維持する能力を有する三次元構築物を含む組成物の治療有効量を哺乳動物に投与し、それによって哺乳動物における肺臓欠陥を軽減または処置する工程を含む、哺乳動物における肺臓欠陥を軽減または処置する方法。
【請求項35】
細胞成長を支持する能力を有する脱細胞化組織を含む埋植可能な組成物であって、脱細胞化組織が脱細胞化前の対応する天然組織の特徴を示す、組成物。
【請求項36】
細胞の集団を含み、肺臓細胞の分化状態を支持し維持する能力を有する、請求項35記載の組成物。
【請求項37】
前記集団が幹細胞を含む、請求項36記載の組成物。
【請求項38】
前記集団が上皮細胞および内皮細胞を含む、請求項36記載の組成物。
【請求項39】
前記細胞が遺伝子改変されている、請求項36記載の組成物。
【請求項1】
細胞成長を支持する能力を有する脱細胞化組織であって、脱細胞化前の対応する天然組織の特徴を示す、脱細胞化組織。
【請求項2】
肺臓である、請求項1記載の脱細胞化組織。
【請求項3】
他の点では同一な脱細胞化前の組織の形態と実質的に類似する形態を示す、請求項1記載の脱細胞化組織。
【請求項4】
対応する天然組織の細胞外マトリックスを保持する脱細胞化組織であって、該細胞外マトリックスが、実質的にインタクトな外面を含む、請求項1記載の脱細胞化組織。
【請求項5】
免疫原性マーカーが実質的に除去されている、請求項1記載の脱細胞化組織。
【請求項6】
対応する天然組織の機械的性質と実質的に類似する機械的性質を示す、請求項1記載の脱細胞化組織。
【請求項7】
肺臓細胞の分化状態を支持し維持する能力を有する、三次元スキャフォールドと細胞の集団とを含む組成物。
【請求項8】
三次元スキャフォールドが脱細胞化組織である、請求項7記載の組成物。
【請求項9】
インタクトな気道樹および血管網を示す、請求項7記載の組成物。
【請求項10】
前記集団が幹細胞を含む、請求項7記載の組成物。
【請求項11】
前記集団が上皮細胞および内皮細胞を含む、請求項7記載の組成物。
【請求項12】
前記細胞が遺伝子改変されている、請求項7記載の組成物。
【請求項13】
肺胞上皮細胞の分化状態を支持し維持する能力を有する、請求項7記載の組成物。
【請求項14】
スキャフォールドが、フィブロネクチン、ラミニン、コラーゲン、糖タンパク質、トロンボスポンジン、エラスチン、フィブリリン、ムコ多糖、糖脂質、ヘパリン硫酸、コンドロイチン硫酸、ケラチン硫酸、グリコサミノグリカン、ヒアルロン酸、プロテオグリカン、ビトロネクチン、ポリ-D-リジン、多糖、およびそれらの組合せからなる群より選択される生体適合性材料を含む、請求項7記載の組成物。
【請求項15】
分枝形態形成の誘導に関連する遺伝子発現を示す細胞を含む、請求項7記載の組成物。
【請求項16】
前記遺伝子がCFTRである、請求項7記載の組成物。
【請求項17】
分枝形態形成、遠位肺臓上皮細胞分化、上皮成長、血管発生、およびそれらの組合せからなる群より選択される肺臓組織の特徴を含む、請求項7記載の組成物。
【請求項18】
播種済スキャフォールドを作製するために脱細胞化スキャフォールドに細胞の集団を播種する工程を含む、肺臓細胞の分化状態を支持し維持する能力を有する人工三次元組織を製作する方法。
【請求項19】
脱細胞化スキャフォールドがインタクトな気道樹および血管網を示す、請求項18記載の方法。
【請求項20】
前記集団が幹細胞を含む、請求項18記載の方法。
【請求項21】
前記集団が上皮細胞および内皮細胞を含む、請求項18記載の方法。
【請求項22】
前記細胞が遺伝子改変されている、請求項18記載の方法。
【請求項23】
脱細胞化スキャフォールドが肺胞上皮細胞の分化状態を支持し維持する能力を有する、請求項18記載の方法。
【請求項24】
スキャフォールドが、フィブロネクチン、ラミニン、コラーゲン、糖タンパク質、トロンボスポンジン、エラスチン、フィブリリン、ムコ多糖、糖脂質、ヘパリン硫酸、コンドロイチン硫酸、ケラチン硫酸、グリコサミノグリカン、ヒアルロン酸、プロテオグリカン、ビトロネクチン、ポリ-D-リジン、多糖、およびそれらの組合せからなる群より選択される生体適合性材料を含む、請求項18記載の方法。
【請求項25】
分枝形態形成の誘導に関連する遺伝子発現を示す細胞を含む、請求項18記載の方法。
【請求項26】
前記遺伝子がCFTRである、請求項18記載の方法。
【請求項27】
人工三次元組織が、分枝形態形成、遠位肺臓上皮細胞分化、上皮成長、血管発生、およびそれらの組合せからなる群より選択される肺臓組織の特徴を示す、請求項18記載の方法。
【請求項28】
試験作用因子を人工三次元肺臓組織モデルに接触させる工程と、該試験作用因子が該モデルに及ぼす効果を測定する工程とを含む、肺臓組織の健康状態を調整する試験作用因子の能力に関して試験作用因子をスクリーニングするためのインビトロ方法であって、該モデルに加えられるいずれの変更も、試験作用因子が肺臓組織の健康状態を調整できることを示す、インビトロ方法。
【請求項29】
人工三次元組織が脱細胞化スキャフォールドに由来する、請求項28記載の方法。
【請求項30】
試験作用因子が、化学剤、医薬、ペプチド、核酸、および放射線からなる群より選択される、請求項28記載の方法。
【請求項31】
試験作用因子が治療剤のための送達媒体である、請求項28記載の方法。
【請求項32】
細胞数、面積、体積、形状、形態、マーカー発現、または染色体断片化に対する試験作用因子の効果を決定する工程を含む、請求項28記載の方法。
【請求項33】
肺臓組織モデルに対して所望の効果を持つ作用因子を選択する工程をさらに含む、請求項28記載の方法。
【請求項34】
肺臓細胞の分化状態を支持し維持する能力を有する三次元構築物を含む組成物の治療有効量を哺乳動物に投与し、それによって哺乳動物における肺臓欠陥を軽減または処置する工程を含む、哺乳動物における肺臓欠陥を軽減または処置する方法。
【請求項35】
細胞成長を支持する能力を有する脱細胞化組織を含む埋植可能な組成物であって、脱細胞化組織が脱細胞化前の対応する天然組織の特徴を示す、組成物。
【請求項36】
細胞の集団を含み、肺臓細胞の分化状態を支持し維持する能力を有する、請求項35記載の組成物。
【請求項37】
前記集団が幹細胞を含む、請求項36記載の組成物。
【請求項38】
前記集団が上皮細胞および内皮細胞を含む、請求項36記載の組成物。
【請求項39】
前記細胞が遺伝子改変されている、請求項36記載の組成物。
【図1A】


【図1B】

【図1C】

【図1D】
【図2】

【図3】
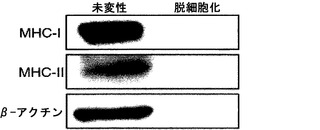
【図4】
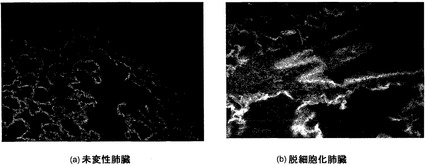
【図5】

【図6】
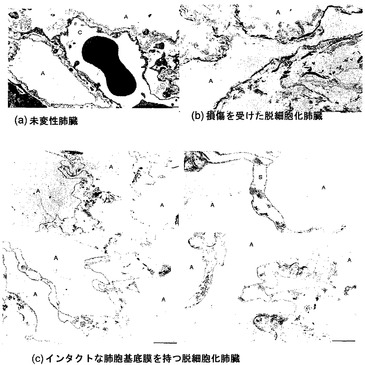
【図7】
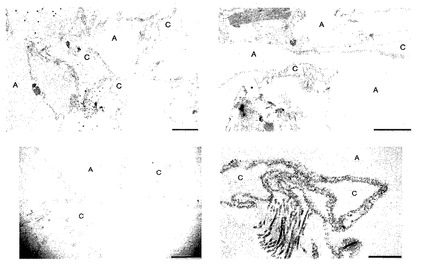
【図8】
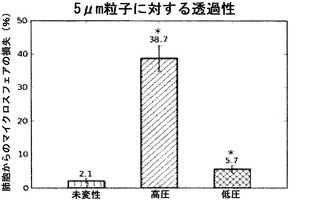
【図9】
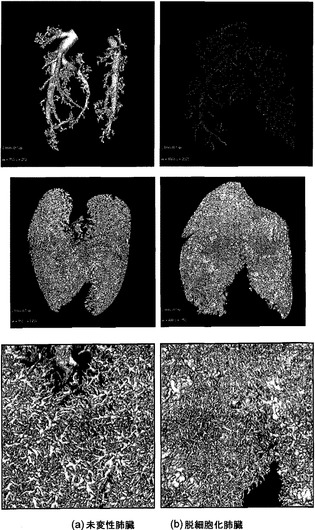
【図10】
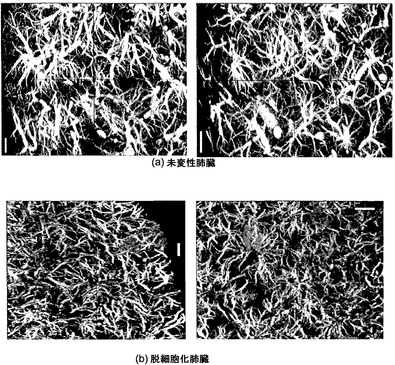
【図11】
【図12】
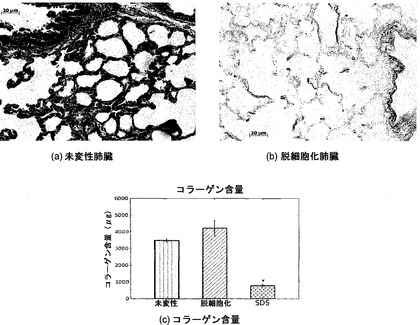
【図13】
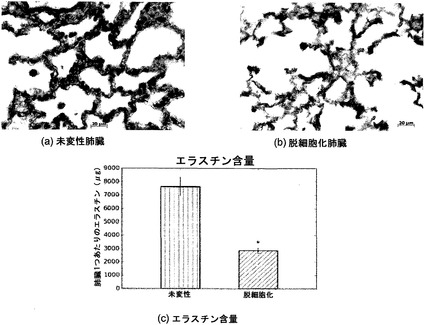
【図14】
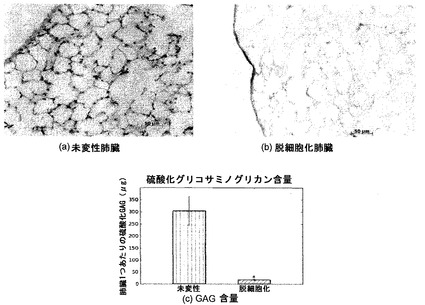
【図15】
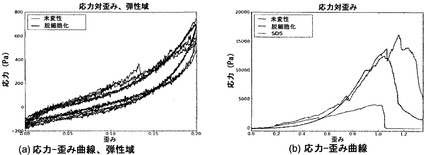
【図16】
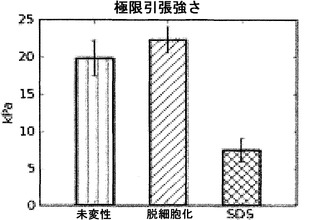
【図17】
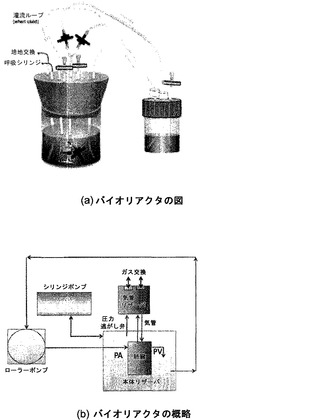
【図18】
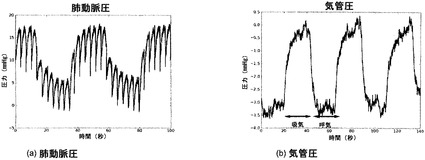
【図19】
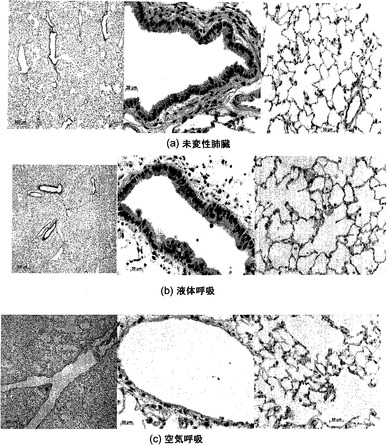
【図20】
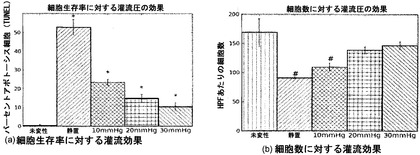
【図21】

【図22】

【図23】
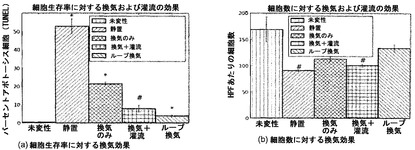
【図24】

【図25−1】
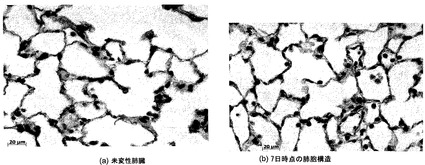
【図25−2】

【図26】
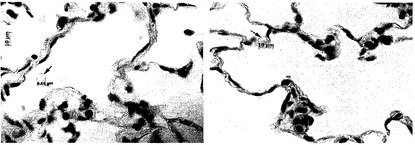
【図27】
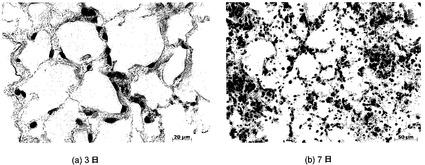
【図28A】
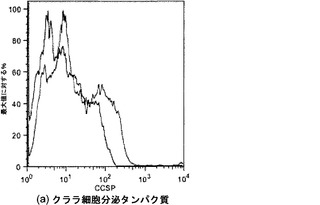
【図28B】
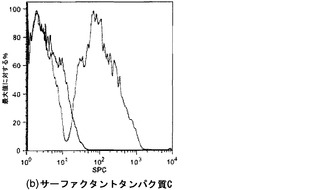
【図28C】
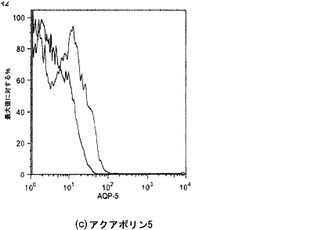
【図28D】
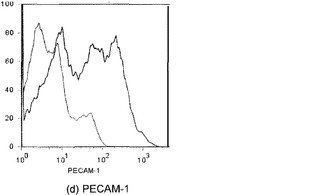
【図28E】
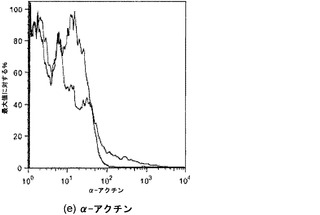
【図28F】
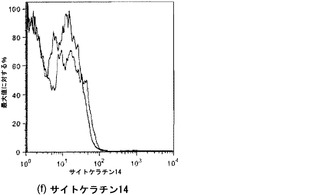
【図29】
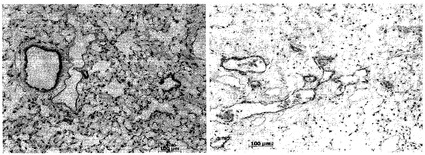
【図30】
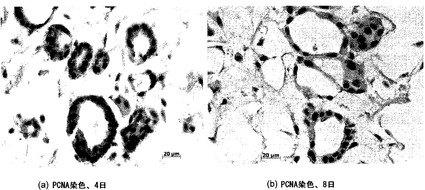
【図31】
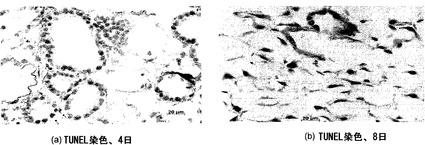
【図32】

【図33】

【図34】

【図35】

【図36】

【図37】

【図38】
【図39】

【図40】

【図41】

【図42】
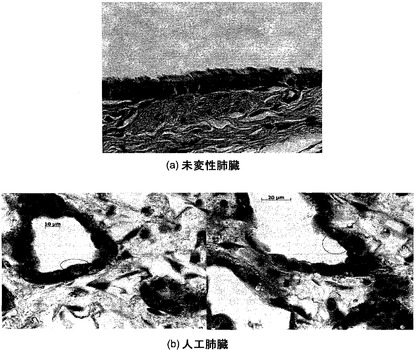
【図43】
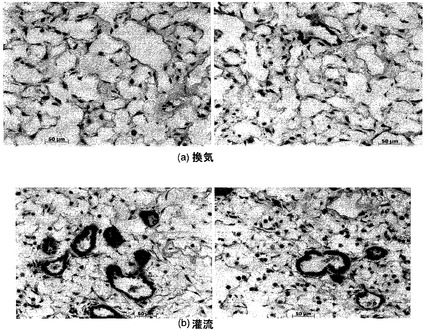
【図44】
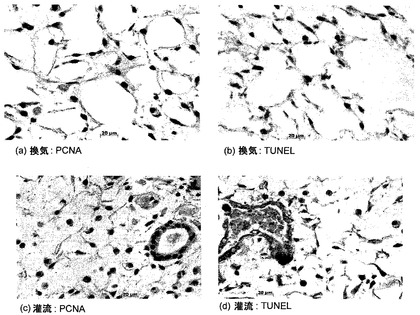
【図45】

【図46】

【図47】

【図48】
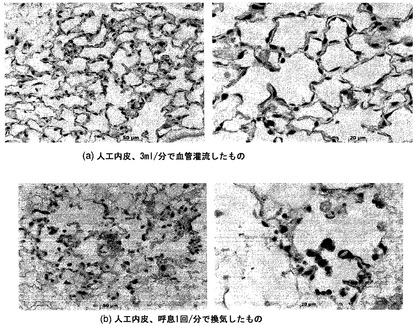
【図49】
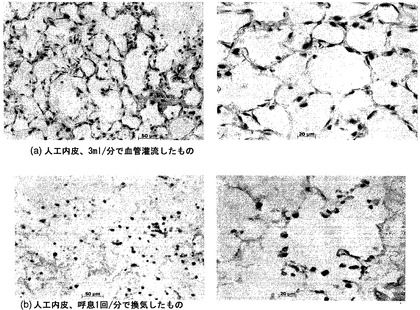
【図50】
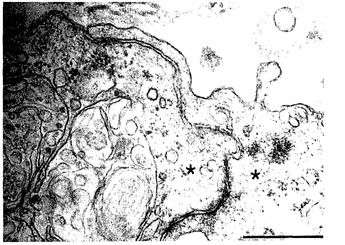
【図51】

【図52】
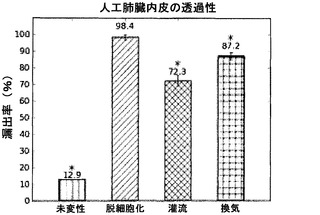
【図53】
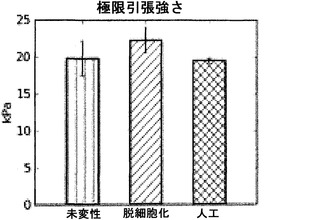
【図54】
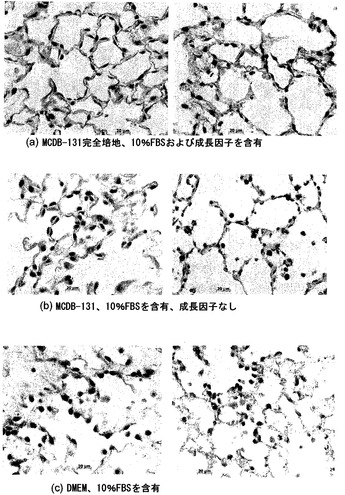
【図55】
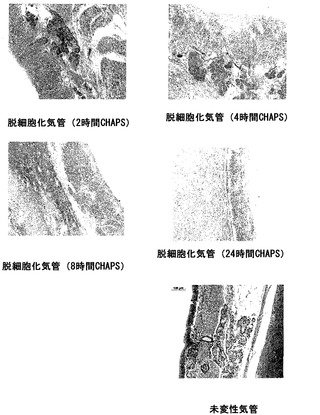
【図56】

【図57】
【図58】
【図59】

【図60】
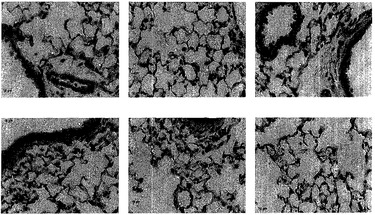
【図61】

【図62】

【図63A】

【図63B】

【図63C】

【図64】
【図1B】
【図1C】
【図1D】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25−1】
【図25−2】
【図26】
【図27】
【図28A】
【図28B】
【図28C】
【図28D】
【図28E】
【図28F】
【図29】
【図30】
【図31】
【図32】
【図33】
【図34】
【図35】
【図36】
【図37】
【図38】
【図39】
【図40】
【図41】
【図42】
【図43】
【図44】
【図45】
【図46】
【図47】
【図48】
【図49】
【図50】
【図51】
【図52】
【図53】
【図54】
【図55】
【図56】
【図57】
【図58】
【図59】
【図60】
【図61】
【図62】
【図63A】
【図63B】
【図63C】
【図64】
【公表番号】特表2012−516699(P2012−516699A)
【公表日】平成24年7月26日(2012.7.26)
【国際特許分類】
【出願番号】特願2011−549260(P2011−549260)
【出願日】平成22年2月4日(2010.2.4)
【国際出願番号】PCT/US2010/023213
【国際公開番号】WO2010/091188
【国際公開日】平成22年8月12日(2010.8.12)
【出願人】(503469393)イエール ユニバーシティ (11)
【Fターム(参考)】
【公表日】平成24年7月26日(2012.7.26)
【国際特許分類】
【出願日】平成22年2月4日(2010.2.4)
【国際出願番号】PCT/US2010/023213
【国際公開番号】WO2010/091188
【国際公開日】平成22年8月12日(2010.8.12)
【出願人】(503469393)イエール ユニバーシティ (11)
【Fターム(参考)】
[ Back to top ]