
胚、卵母細胞、および幹細胞の撮像および評価
1つ以上の胚もしくは多能性細胞の発達の可能性および/または1つ以上の胚もしくは多能性細胞における染色体異常の存在を判定するための方法、組成物、およびキットを提供する。これらの方法、組成物、およびキットは、ヒトにおける不妊の治療に最も有用である、生体外での胚および卵母細胞の識別に用途を見出す。

【発明の詳細な説明】
【技術分野】
【0001】
関連出願の相互参照
本出願は、2010年5月7日に出願された米国仮特許出願第61/332,651号および2009年8月22日に出願された米国仮特許出願第61/236,085号に対して優先権を主張し、それらの双方は、参照によりそれらの全体が本明細書に組み込まれる。
【0002】
本発明は、生物学的および臨床試験の分野、特にヒトおよび動物の双方からの接合体/胚、卵母細胞、および幹細胞の撮像および評価に関する。
【背景技術】
【0003】
不妊は、妊娠可能年齢の夫婦の10〜15%に影響を及ぼす一般的な健康問題である。2006年には、米国だけで、約140,000回の生体外受精(IVF)が実施された(cdc.gov/art)。これは、変わりやすくまた明確に定義されていない場合が多い、着床および満期児までの発達の可能性を有する胚を毎年100万個以上培養する結果となった。平均して30%の出生は、多胎妊娠をもたらしたが、IVF後の1回当りの出生率は、わずか29%であった(cdc.gov/art)。多胎妊娠は、流産、早産、および低出産率等の、母親と胎児の双方に有害転帰になる。IVF失敗の潜在的な原因は多様であるが、1978年にIVFが導入されて以来、主な課題の1つは、移植に最も適し、満期妊娠をもたらす可能性が最もある胚を識別することである。
【0004】
ヒトの胚の生物学の研究は困難なままであり、研究費の対象外であることが多いため、基本的な胚の発達の分野における理解は限られている。したがって、胚の発達の現在の知識の大半は、モデル生物の研究に由来する。しかしながら、異なる種からの胚は、類似する発達期を経るが、タイミングは種により異なる。これらの相違、およびその他の多くにより、1つの種から他の種に対して直接推定することが不適切となる(Taft,R.E.(2008)Theriogenology 69(1):10−16)。ヒトの発達の一般的な経路、ならびに基本的な根底にある分子決定要因は、ヒトの胚の発達に固有である。例えば、マウスにおいて、胚の転写は、受精の約12時間後に活性化され、同時に第1卵割され、一方、ヒトにおいて、胚遺伝子活性化(EGA)は、およそ8細胞期である、3日目に生じる(Bell,C.E.,et al.(2008)Mol.Hum.Reprod.14:691−701、Braude,P.,et al.(1988)Nature 332:459−461、Hamatani,T.et al.(2004)Proc.Natl.Acad.Sci.101:10326−10331、Dobson,T.et al.(2004)Human Molecular Genetics 13(14):1461−1470)。加えて、初期のヒトの発達で調節される遺伝子は、固有である(Dobson,T.et al.(2004)Human Molecular Genetics13(14):1461−1470)。さらに、マウス等の他の種において、生体外で培養された胚の85%超は、哺乳類の発達において最初の主要な目印の1つである胚盤胞期に達し、一方で、培養されたヒトの胚は、約30〜50%の平均胚盤胞形成率を有し、断片化および発達停止等のモザイク現象および異常表現型の発生率が高い(Rienzi,L.et al.(2005)Reprod.Biomed.Online 10:669−681、Alikani,M.,et al.(2005) Mol.Hum.Reprod.11:335−344、Keltz,M.D.,et al.(2006)Fertil.Steril.86:321−324、French,D.B.,et al.(2009)Fertil.Steril.)。そのような相違にもかかわらず、着床前の胚の発達の研究の多くは、モデル生物に由来し、ヒトの胚の発達に関連付けることが困難である(Zernicka−Goetz,M.(2002)Development 129:815−829、Wang,Q.,et al.(2004)Dev Cell.6:133−144、Bell,C.E.,et al.(2008)Mol.Hum.Reprod.14:691−701、Zernicka−Goetz,M.(2006)Curr.Opin.Genet.Dev.16:406−412、Mtango,N.R.,et al.(2008)Int.Rev.Cell.Mol.Biol.268:223−290)。
【0005】
従来、IVFの臨床において、ヒトの胚の生存能は、均一な大きさの単核卵割球の存在、および細胞断片化の程度等の、単純な形態学的観測により評価されてきた(Rijinders PM,Jansen CAM.(1998)Hum Reprod 13:2869−73、Milki AA,et al.(2002)Fertil Steril 77:1191−5)。最近では、胚の長期培養(5日目の胚盤胞期まで)および着床前の遺伝子診断(PGD)を介した染色体の状態の分析等の付加的な方法も、胚の質を評価するために使用されている(Milki A,et al.(2000)Fertil Steril 73:126−9、Fragouli E,(2009)Fertil Steril Jun 21[EPub ahead of print]、El−Toukhy T,et al.(2009)Hum Reprod 6:20、Vanneste E,et al.(2009)Nat Med 15:577−83)。しかしながら、これらの方法には、培養期間を延長し、胚の整合性を乱すという潜在的なリスクも存在する(Manipalviratn S,et al.(2009)Fertil Steril 91:305−15、Mastenbroek S,et al.(2007)N Engl J Med.357:9−17)。
【0006】
最近、微速度撮像が初期の胚の発達を観測するのに有用なツールであり得ることが示されている。いくつかの方法は、卵細胞質内精子注入(ICSI)後、ヒトの胚の発達を監視するために微速度撮像を使用してきた(Nagy et al.(1994)Human Reproduction.9(9):1743−1748、Payne et al.(1997)Human Reproduction.12:532−541)。極体突出および前核形成が分析され、3日目の良好な形態と関連付けられた。しかしながら、パラメータは、胚盤胞形成または妊娠の転帰と相関しなかった。他の方法は、ヒトの胚の生存能を予測するための指標として第1分割の開始に目を向けてきた(Fenwick,et al.(2002)Human Reproduction,17:407−412、Lundin,et al.(2001)Human Reproduction 16:2652−2657)。しかしながら、これらの方法は、細胞質分裂の持続時間、または初期分割間の時間間隔の重要性を認識しない。
【0007】
他の方法は、初期の胚の発達中、細胞分割のタイミングおよび程度を測定するために微速度撮像を使用してきた(WO/2007/144001)。しかしながら、これらの方法は、ウシの胚の微速度撮像のための基本的で一般的な方法しか開示せず、これは、発達の可能性、形態学的挙動、分子および後成的プログラム、ならびに移植を取り巻くタイミングおよびパラメータに関して、実質的にヒトの胚と異なる。例えば、ウシの胚は、ヒトの胚と比較して、着床するのに実質的により長くかかる(それぞれ、30日および9日)。(Taft,(2008)Theriogenology 69(1):10−16。さらに、ヒトの胚の生存能を予測し得る具体的な撮像パラメータ、または時間間隔は、開示されていない。
【0008】
最近では、受精後の最初の24時間の間、ヒトの胚の発達を観測するために、微速度撮像が使用されている(Lemmen et al.(2008)Reproductive BioMedicine Online 17(3):385−391)。最初の分割後の核の同調性は、妊娠の転帰と相関することが分かった。しかしながら、この研究は、初期の第1分割が重要な予測パラメータではないと結論付けられ、これは、前の研究と矛盾する(Fenwick,et al.(2002)Human Reproduction 17:407−412、Lundin,et al.(2001)Human Reproduction 16:2652−2657)。
【0009】
最後に、胚の分子プログラムまたは染色体構成との相関によって撮像パラメータを検証した研究はない。よって、ヒトの胚の評価方法は、いくつかの概念に欠け、本方法によって改善され得、これは、微速度顕微鏡検査、画像分析、ならびに撮像パラメータの分子プロファイルと染色体構成との相関の新規適用を含む。本発明はこれらの問題に対処する。
【発明の概要】
【0010】
1つ以上の胚もしくは多能性細胞における1つ以上の胚もしくは多能性細胞の発達の可能性を判定するための方法、組成物、およびキットを提供する。これらの方法、組成物、およびキットは、良好な発達の可能性、すなわち、胚盤胞に発達する能力または潜在的可能性を有する、生体外における胚および卵母細胞を識別するのに使用され、よって、これはヒト等における不妊を治療する方法に有用である。
【0011】
本発明の一部の態様において、胚または多能性細胞の発達の可能性を判定するための方法を提供する。そのような態様において、胚または多能性細胞の1つ以上の細胞パラメータは、細胞パラメータ測定値を導き出すように測定される。次いで、細胞パラメータは、胚または多能性細胞の発達の可能性の判定を提供するために利用され、この判定は、臨床的行動方針を導くために使用され得る。一部の実施形態において、細胞パラメータは、微速度顕微鏡検査により測定可能である形態学的事象である。一部の実施形態において、例えば、胚がアッセイされる時、1つ以上の細胞パラメータは、細胞質分裂事象、例えば、細胞質分裂1の持続時間、細胞質分裂1と細胞質分裂2との間の時間間隔、および細胞質分裂2と細胞質分裂3との間の時間間隔である。ある実施形態において、細胞周期1の持続時間も細胞パラメータとして利用される。一部の実施形態において、細胞パラメータ測定値は、それを基準の胚からの比較可能な細胞パラメータ値と比較し、この比較の結果を使用して、胚の発達の可能性の判定を提供することにより利用される。一部の実施形態において、胚はヒトの胚である。一部の実施形態において、細胞パラメータは、遺伝子発現測定値を導き出すように測定される遺伝子発現レベルである。一部の実施形態において、遺伝子発現測定値は、それを1つの基準の多能性細胞もしくは胚もしくはそれらからの1つ以上の細胞の遺伝子発現測定値と比較することにより利用され、ここで、この比較の結果は、多能性細胞もしくは胚の発達の可能性の判定を提供するように利用される。一部の実施形態において、胚はヒトの胚である。
【0012】
本発明の一部の態様において、群における他の胚または多能性細胞に対して、胚または多様性細胞をそれらの発達の可能性についてランク付けするための方法を提供する。そのような実施形態において、群における胚または多能性細胞の1つ以上の細胞パラメータは、胚または多能性細胞のそれぞれについての細胞パラメータ測定値を導き出すように測定される。次いで、細胞パラメータ測定値は、群における胚または多能性細胞のそれぞれの発達の可能性を判定するために相互に利用され、この判定は、臨床的行動方針を導くために使用され得る。一部の実施形態において、細胞パラメータは、微速度顕微鏡検査により測定可能である形態学的事象である。一部の実施形態において、例えば、胚がランク付けされる時、1つ以上の細胞パラメータは、細胞質分裂事象、例えば、細胞質分裂1の持続時間、細胞質分裂1と細胞質分裂2との間の時間間隔、および細胞質分裂2と細胞質分裂3との間の時間間隔である。ある実施形態において、細胞周期1の持続時間も測定される。一部の実施形態において、細胞パラメータは、1つ以上の遺伝子の発現レベルである。一部の実施形態において、1つ以上の細胞パラメータ測定値は、群における胚または多能性細胞のそれぞれからの細胞パラメータ測定値を相互に比較して、相互に対して、胚または多能性細胞の発達の可能性を判定することにより利用される。一部の実施形態において、1つ以上の細胞パラメータ測定値は、各細胞パラメータ測定値を基準の胚または多能性細胞からの細胞パラメータ測定値と比較して、各胚または多能性細胞についての発達の可能性を判定すること、およびこれらの発達の可能性を比較して、相互に対して、胚または多能性細胞の発達の可能性を判定することにより利用される。
【0013】
本発明の一部の態様において、生殖補助(IVF)を受ける女性に移植するための、良好な発達の可能性を有する胚を提供するための方法を提供する。そのような態様において、1つ以上の胚は、胚の発達に十分な条件下で培養される。次いで、1つ以上の細胞パラメータは、1つ以上の胚において、細胞パラメータ測定値を導き出すように測定される。次いで、細胞パラメータ測定値は、1つ以上の胚の発達の可能性の判定を提供するために利用される。次いで、良好な発達の可能性を示す1つ以上の胚は、女性に移植される。
【図面の簡単な説明】
【0014】
本発明は、付随の図面と併せて以下の詳述を読む場合に最もよく理解される。一般的な実践において、図面の様々な特徴は、原寸に比例しないことが強調される。一方、様々な特徴の大きさは、明確にするために、任意に拡大または縮小される。図面に含まれるのは、以下の図である。
【図1】胚を評価するために使用されるプロセスを示すフローチャートである。
【図2】6日間にわたる細胞分裂および分割を示す一連の写真である。画像は、1日目から6日目までラベルが付けられている。スケールバーは50μmを表す。
【図3】1細胞胚(接合体)から胚盤胞への発達に成功した比率を示す棒グラフである。4つの個別の実験に亘って、微速度顕微鏡検査を介して、5〜6日目まで、合計100個の胚を観測した。各示された時期(胚盤胞、8細胞、4〜7細胞、2〜3細胞、および1細胞)に達した細胞の比率を示す。
【図4】示された時間の間追跡された、一連の3つの異なる胚である(上段、中段、および下段)。
【図5】本評価に使用される期間の間の時間経過を示す図であり、第1細胞質分裂の持続時間、第1と第2の分割との間の時間(細胞質分裂1の分離と細胞質分裂2の開始との間の時間間隔として測定される)、および第2と第3の有糸分裂との間の時間(細胞質分裂2の開始と細胞質分裂3の開始との間の時間間隔として測定される)を含む。
【図6】3つの事象の測定値を示す3D点グラフであり、第1細胞質分裂の持続時間、第1と第2の細胞分割の間の時間間隔(細胞質分裂1の分離と細胞質分裂2の開始との間の時間間隔として測定される)、および第2の細胞分割と第3の細胞分割との間の時間(細胞質分裂2の開始と細胞質分裂3の開始との間の時間間隔として測定される)を含む。胚盤胞期に達する胚(丸で示される)は、3Dグラフ上で共に密集していることが示され、一方、胚盤胞に達する前に停止する胚(Xで示される)は、全体に散らばっている。
【図7】3つの動的形態学的パラメータを使用して胚盤胞形成を予測するための受信者動作特性(ROC)曲線を示すグラフである。
【図8】6つの停止した1〜2細胞胚および5つの正常な1〜2細胞胚からの52の遺伝子の遺伝子発現レベルを示すレーダーグラフである。正常な胚と異常な胚との間の発現レベルの相違は、マンホイットニー検定により判定されると、黄色でハイライトされ、星印で示されるこれらの遺伝子において、統計的に有意であった。
【図9】停止した2細胞胚および正常な2細胞胚における、異なる遺伝子の発現レベルを示す棒グラフである。選択されたいくつかの停止した2細胞胚の微速度画像を上部に示す。
【図10】停止した4細胞胚および正常な4細胞胚における、図9に示される同じ遺伝子の比較を示す棒グラフである。選択されたいくつかの停止した4細胞胚の微速度画像を上部に示す。
【図11】4つの明確なパターンを有する遺伝子発現パターン(ESSP)を示す一連の棒グラフである。胚の遺伝子活性化(2日目)前および3日目の典型的な発現前の早期移植の時間を示す。
【図12】異なる期の単一卵割球からの遺伝子の遺伝子発現を示す。(A)異なる細胞期でプロットされた単一の卵割球からのCTNNB1およびCDX2の2つの遺伝子の遺伝子発現であり、例えば、2細胞、3細胞、桑実胚、および胚盤胞の異なる期におけるこれらの遺伝子発現レベルにおける変化を示す。(B)接合体プログラムから発現した遺伝子と比較した、母性プログラムで発現した遺伝子を表す棒グラフで表された遺伝子発現特性。
【図13】胚の生存能を評価するために、微速度画像分析および相関分子分析を使用するためのモデル図である。
【図14】生体外での卵母細胞成熟中の3つの発達期を示す一連の写真である。
【図15】生体外での卵母細胞成熟後の胚の発達のプロセスを示す一連の写真である。
【図16】卵母細胞を評価するために使用されるプロセスを示すフローチャートである。
【図17】幹細胞および多能性幹細胞を評価するために使用されるプロセスを示すフローチャートである。
【図18】ニューロンロゼットに分化する、誘発された多能性幹細胞のプロセスを示す一連の写真である。
【図19】発現レベルについてアッセイされた遺伝子が分類され得る分類表であり、分類毎に遺伝子の数を含む。
【図20】141の正常に発達した単一胚および単一卵割球の遺伝子発現分析中に識別された、4つの胚期特異的パターン(ESSP)、および遺伝子をそれらの分類のうちのそれぞれ1つに分類した表である。
【図21】胚盤胞形成を予測するための撮像パラメータの能力を示す自動化画像分析を示す。(A)単一胚についての追跡アルゴリズムの結果を示す。(B)分析された14個一組の胚を示す。(C)細胞質分裂の持続時間、および第1と第2の有糸分裂の間の時間についての手動画像分析の自動化分析との比較を示す。(D)良好な胚盤胞形態の不良な胚盤胞形態との比較を示す。
【図22】本発明による暗視野顕微鏡の概略図であり、左側の差し込みは、レーザー加工暗視野パッチ設定を示す。
【図23】インキュベータ内に設置するため、およびコンピュータに接続するために支持台上に載置された、図22に図示される3つの並んだ顕微鏡の写真である。図23Aは顕微鏡を示し、図23Bはインキュベータ内の顕微鏡を示す。
【図24】本研究で使用された画像捕捉ソフトウェアのスクリーンショットであり、3つのチャネルから撮像された胚を示す。
【図25A−D】実験2のステーション2からの選択された微速度画像を示す4枚の一連の写真である。図25Aおよび25Bは、培地を交換する前に捉えた画像であり、図25Cおよび25Dは、培地を交換した後に捉えた画像である。
【図26A−B】正常な胚および異常な胚の細胞活性を示すグラフ(左)および細胞写真(右)である。図26Aおよび図26Bは、合わせて、3日目に、胚が類似する形態を有するが、それらの細胞活性プロットは、大きく異なり、それらのうちの1つのみが胚盤胞に発達することを示す。
【図27A−B】マイクロウェルを備えた、特注のペトリ皿の図である。図27Aは、寸法とともに皿の図を示し、図27Bは、マイクロウェルの3D表示を示す。
【図28A−B】事前画像登録をした、および事前画像登録をしない細胞活性を示すグラフである。図28Aおよび28Bは、併せると、登録が結果を整理し、胚の移動および回転によるスパイクを取り除くことを示す。
【図29A−B】正常な胚および異常な胚の細胞活性を示すグラフ(左)および細胞写真(右)である。図29Aおよび図29Bは、併せて、3日目に、胚が類似する形態を有するが、それらの細胞活性プロットは大きく異なり、それらのうちの1つのみが胚盤胞に発達することを示す。
【図30】胚の発達中の連続する組の画像間における画素強度の差を示すグラフである。これは、胚の生存能を評価するために単独で使用されるか、またはどのくらいの粒子(予測された胚モデル)が使用されるべきかを判定することにより、粒子フィルタ等の他のアルゴリズムを改善するための方式として使用され得る。
【図31A−G】種々の細胞期の2D追跡の結果を示す7枚の一連の写真である。フレーム番号で示される細胞の進行は、各画像組と関連する:フレーム15(図31A)、45(B)、48(C)、189(D)、190(E)、196(F)および234(G)。下段は、重ね合わせたシミュレーション画像を示す。輪郭は、可視細胞膜であり、白い点線は、遮蔽された膜である。画像フレームは、5分毎に捉えられ、数画像のみが示される。
【図32A−B】3D細胞追跡の2つの成功例を示す一連の写真および図である。胚の各写真の図は、それぞれ、フレーム314およびフレーム228においてモデルの側面からの視点を示すフレーム314およびフレーム228を除き、3Dモデルの上からの視点を示す。画像フレームは、5分毎に捉えられた。
【図33】1細胞〜2細胞分割についての粒子フィルタ結果の図表示である。データ点は、細胞中心の3D位置である。1細胞モデル、2細胞モデル、3細胞モデル、および4細胞モデルについての点を示す。上段は、予測後の粒子、そして下段は、再サンプリング後の粒子を示す。
【図34A−B】14個一組の胚における自動化対手動の画像分析の比較を示すグラフである。図34Aは、第1細胞質分裂の持続時間についての比較を示し、図34Bは、第1と第2の有糸分裂の間の時間についての比較を示す。
【図35】胚をモデル化し、ある形態パラメータを測定するために画像分析がどのように使用されるかを示すフローチャートである。
【発明を実施するための形態】
【0015】
本発明の方法および組成物を説明する前に、本発明は、説明される特定の方法または組成物に限定されず、そのようなものとして、当然のことながら、変動し得ることを理解する。本発明の範囲は、付属の請求項によってのみ限定されるため、本明細書に使用される用語は、単に特定の実施形態を説明する目的のためであり、限定することが意図されないことも理解する。
【0016】
値の範囲が提供される場合、その範囲の上限と下限の間の各介在値も、文脈に明確に指示されない限り、下限の単位の10分の1まで、具体的に開示されることを理解する。規定範囲にあるいかなる規定値または介在値と、その規定範囲にある他のいかなる規定値または介在値との間の各小範囲は、本発明内に包含される。これらの小範囲の上限および下限は、独立して範囲に含まれるか、または除外され、小範囲にいずれかが含まれる、いずれも含まれない、および双方が含まれる各範囲も、本発明内に包含され、規定範囲において特に除外されるいかなる限度も対象となる。規定範囲が1つ、または双方の限度を含む場合において、これらの含まれた限度のいずれか、または双方を除外する範囲も本発明に含まれる。
【0017】
特に定義されない限り、本明細書に使用される全ての技術および科学的用語は、本発明が属する技術の当業者に一般に理解されるのと同様の意味を持つ。本明細書に説明されるものと類似、または同等のあらゆる方法および材料は、本発明の実践または試験において使用され得るが、一部の可能性のある、および好ましい方法ならびに材料をここで説明する。本明細書に言及される全ての刊行物は、引用される刊行物に関連する方法および/または材料を公開および説明するために参照により本明細書に組み込まれる。組み込まれた刊行物のあらゆる開示と矛盾がある点では、本開示が優先することを理解する。
【0018】
本明細書および付属の請求項に使用される、単数形「a」、「an」、および「the」は、文脈に明確に示されない限り、複数形の指示物を含むことに留意するべきである。よって、例えば、「細胞(a cell)」との言及は、複数のそのような細胞を含み、「ペプチド」との言及は、当業者に公知である、1つ以上のペプチドおよびその等価物、例えば、ポリペプチドへの言及を含み、他も同様である。
【0019】
本明細書において論じられる刊行物は、単に、本出願の出願日前の開示であるために、提供される。本明細書におけるいかなる記載も、本発明が、先の発明の長所によって、そのような刊行物に先行する権利を与えられていないと認めるものとして解釈されるべきではない。さらに、提供された刊行物の日付は、それぞれに確認される必要がある実際の公表日とは異なっている場合がある。
【0020】
定義
1つ以上の胚もしくは多能性細胞の発達の可能性および/または1つ以上の胚もしくは多能性細胞における染色体異常の存在を判定するための方法、組成物、およびキットを提供する。これらの方法、組成物、およびキットは、ヒトにおける不妊の治療に最も有用である生体外における胚および卵母細胞を識別するのに使用される。本発明のこれらの、および他の目的、利点、および特色は、以下にさらに詳しく説明する、本発明の方法および組成物の詳細を読むことにより当業者に明らかになるだろう。
【0021】
「発達の可能性」および「発達能力」という用語は、本明細書において、健康な胚または多能性細胞の成長もしくは発達する能力または潜在的可能性を指すように使用される。
【0022】
「胚」という用語は、本明細書において、2つの半数体配偶子細胞、例えば、未受精の二次卵母細胞および精子細胞が合体して、二倍体全能性細胞、例えば、受精卵を形成する時に形成される接合体と、直後の細胞分割、すなわち、胚分割から桑実胚、すなわち、16細胞期および胚盤胞期(分化した栄養外胚葉および内細胞塊を有する)までを生じる胚の双方を指すように使用される。
【0023】
「多能性細胞」という用語は、本明細書において、生物において、複数の細胞型に分化する能力を有するあらゆる細胞を意味するように使用される。多能性細胞の例としては、幹細胞卵母細胞、および1細胞胚(すなわち、接合体)が挙げられる。
【0024】
「幹細胞」という用語は、本明細書において、(a)自己再生する能力を有する、および(b)多様な分化細胞型を生じる可能性を有する細胞、または細胞集団を指すように使用される。しばしば、幹細胞は、複数の細胞の系列を生じる可能性を有する。本明細書に使用される、幹細胞は、全能性幹細胞、例えば、生物の胚および胚外組織の全てを生じる受精卵母細胞;多能性幹細胞、例えば、生物の胚組織の全て、すなわち、内胚葉、中胚葉、および外胚葉系列を生じる胚幹(ES)細胞、胚生殖(EG)細胞、または誘発された多能性幹(iPS)細胞;多能性幹細胞、例えば、生物の胚組織のうちの少なくとも2つ、すなわち、内胚葉、中胚葉、および外胚葉系列のうちの少なくとも2つを生じる間葉系幹細胞であるか、または特定の組織の複数の分化細胞型を生じる組織特異的幹細胞であり得る。組織特異的幹細胞は、特定の組織の細胞を生じる組織特異的胚細胞、ならびに成人の組織に属し、その組織の細胞、例えば、中枢神経系の細胞の全てを生じる神経幹細胞、骨格筋を生じる衛星細胞、および造血系の細胞の全てを生じる造血幹細胞を生じる体性幹細胞を含む。
【0025】
「卵母細胞」という用語は、本明細書において、未受精の女性生殖細胞または生殖体を指すように使用される。本出願の卵母細胞は、一次卵母細胞または二次卵母細胞であり得、一次卵母細胞である場合、それらは、減数分裂Iを経るように位置付けられるか、またはそれを経て、二次卵母細胞である場合、減数分裂IIを経るように位置付けられるか、またはそれを経る。
【0026】
「減数分裂」とは、生殖体の産生をもたらす細胞周期事象を意味する。最初の減数分裂細胞周期または減数分裂Iにおいて、細胞の染色体は複製され、2つの娘細胞に分配される。次いで、これらの娘細胞は、第2の減数分裂細胞周期または減数分裂IIで分割し、これは、DNA合成を伴わず、半数の染色体を有する生殖体をもたらす。
【0027】
「胚胞」期とは、減数分裂Iの細胞周期の前期I、すなわち核材料の第1分割前と相関する一次卵母細胞の成熟期を意味する。この期の卵母細胞は、「胚胞卵母細胞」とも呼ばれ、特徴的に大きな核のため、胚胞と呼ばれる。生体外で培養された正常なヒトの卵母細胞において、胚胞は、成熟開始約6〜24時間後に生じる。
【0028】
「中期I」期とは、減数分裂Iの細胞周期の中期Iと相関する一次卵母細胞の成熟期を意味する。胚胞卵母細胞との比較において、中期Iの卵母細胞は、大きい、明確に定義された核を持たない。生体外で培養された正常なヒトの卵母細胞において、中期Iは、成熟開始約12〜36時間後に生じる。
【0029】
「中期II」期とは、減数分裂IIの細胞周期の中期IIと相関する二次卵母細胞の成熟期を意味する。中期IIは、第1極体の突出により区別される。生体外で培養された正常なヒトの卵母細胞において、中期IIは、成熟開始約24〜48時間後に生じる。
【0030】
「有糸分裂細胞周期」とは、細胞の染色体の複製、ならびにこれらの染色体および細胞の細胞質物質の2つの娘細胞への分割をもたらす事象を意味する。有糸分裂細胞周期は、静止期と有糸分裂の2つの期に分けられる。静止期において、細胞は成長し、そのDNAを複製する。有糸分裂において、細胞は、細胞分割を開始し、完了するが、最初にその核材料を分配し、その後、その細胞質材料とその分配した核材料(細胞質分裂)を2つの個別の細胞に分割する。
【0031】
「第1の有糸分裂細胞周期」または「細胞周期1」とは、受精から第1細胞質分裂事象、すなわち、受精卵母細胞の2つの娘細胞への分割の完了までの時間間隔を意味する。卵母細胞が生体外受精である場合、ヒト絨毛性ゴナドトロピン(HCG)の注入(一般に、卵母細胞回収前に投与される)と第1細胞質分裂事象の完了との間の時間間隔が、代理時間間隔として使用され得る。
【0032】
「第2の有糸分裂細胞周期」または「細胞周期2」とは、胚で観測される第2細胞周期事象を意味し、有糸分裂による受精卵母細胞からの娘細胞の産生と、有糸分裂によるこれらの娘細胞(「先行娘細胞」または娘細胞A)のうちの1つからの第1の一組の孫娘細胞の産生との間の時間間隔である。細胞周期2が完了すると、胚は3つの細胞から成る。言い換えると、細胞周期2は、2細胞を含有する胚と3細胞を含有する胚との間の時間として視覚的に識別することができる。
【0033】
「第3の有糸分裂細胞周期」または「細胞周期3」とは、胚で観測される第3細胞周期事象を意味し、通常、有糸分裂による受精卵母細胞からの娘細胞の産生から有糸分裂による第2娘細胞(「遅れ娘細胞」または娘細胞B)からの第2の一組の孫娘細胞の産生までの時間間隔である。細胞周期3が完了すると、胚は4つの細胞から成る。言い換えると、細胞周期3は、3細胞を含有する胚と4細胞を含有する胚との間の時間として視覚的に識別することができる。
【0034】
「第1分割事象」とは、第1分割、すなわち、卵母細胞の2つの娘細胞への分割、すなわち、細胞周期1を意味する。第1の分割事象が完了すると、胚は2つの細胞から成る。
【0035】
「第2分割事象」とは、第2の一組の分割、すなわち、先行娘細胞の2つの孫娘細胞への分割、および遅れ娘細胞の2つの孫娘細胞への分割を意味する。言い換えると、第2の分割事象は、細胞周期2および細胞周期3の双方から成る。第2の分割が完了すると、胚は4つの細胞から成る。
【0036】
「第3分割事象」とは、第3の一組の分割、すなわち、全ての孫娘細胞の分割を意味する。第3分割事象が完了すると、胚は、通常、8つの細胞から成る。
【0037】
「細胞質分裂」または「細胞分割」とは、細胞が細胞分割を受ける有糸分裂の時期を意味する。言い換えると、細胞の分配された核材料およびその細胞質材料が分割されて、2つの娘細胞を産生する有糸分裂期である。細胞質分裂の期間は、細胞膜の圧縮(「分裂溝」)が最初に観測された時から、その圧縮事象の分離、すなわち、2つの娘細胞の生成の間の期間または時間枠として識別可能である。分裂溝の開始は、細胞膜の湾曲が凸面(外側に向かって丸い)から凹面(くぼみまたはへこみを伴う内側への屈曲)に変化する点として視覚的に識別され得る。これは、2つの分裂溝を示す白色矢印により、図4の上部パネルに図示される。細胞伸長の開始は、細胞質分裂の開始を標識するためにも使用され得、この場合、細胞質分裂の期間は、細胞伸長の開始と細胞分割の分離との間の期間として定義される。
【0038】
「第1細胞質分裂」または「細胞質分裂1」とは、受精後最初の細胞分割事象、すなわち、受精卵母細胞が分割して、2つの娘細胞を産生することを意味する。第1細胞質分裂は、一般に、受精約1日後に生じる。
【0039】
「第2細胞質分裂」または「細胞質分裂2」とは、第2細胞分割事象が胚において観測される、すなわち、受精卵母細胞の娘細胞(「先行娘細胞」または娘A)が第1の一組の2つの孫娘細胞に分割することを意味する。
【0040】
「第3細胞質分裂」または「細胞質分裂3」とは、第3細胞分割事象が胚において観測される、すなわち、受精卵母細胞の他の娘細胞(「遅れ娘細胞」または娘B)が第2の一組の2つの孫娘細胞に分割することを意味する。
【0041】
「位置合わせマーカー(fiduciary marker)」または「位置合わせマーカー(fiducial marker)」という用語は、参照点または測定点として使用するための、産生された画像に現れる撮像システムの視野域で使用される目標物である。撮像対象物の中または上に設置される何かか、または光学装置のレチクルの印もしくは一組の印のいずれかであり得る。
【0042】
「マイクロウェル」という用語は、細胞規模で寸法決定された、好ましくは、単一真核細胞を収容するために提供される容器を指す。
【0043】
目的の多能性細胞および胚
本発明の方法において、1つ以上の胚または多能性細胞は、胚または多能性細胞の1つ以上の細胞パラメータを測定し、これらの測定値を利用して、胚または多能性細胞の発達の可能性を判定することにより、その発達の可能性について評価される。よって、得られた情報は、例えば、生体外受精胚を移植するか、培養した1つの細胞または複数の細胞を移植するか、という臨床的決定を導くために使用され得る。
【0044】
本発明の方法により評価され得る胚の例としては、1細胞胚(接合体とも称される)、2細胞胚、3細胞胚、4細胞胚、5細胞胚、6細胞胚、8細胞胚等、通常、最大16細胞胚までが挙げられ、それらのいずれも、あらゆる都合のよい様式、例えば、生体内で成熟した卵母細胞、または生体外で成熟した卵母細胞に由来し得る。
【0045】
本発明の方法により評価され得る多能性細胞の例としては、全能性幹細胞、例えば、一次卵母細胞および二次卵母細胞等の卵母細胞、多能性幹細胞、例えば、ES細胞、EG細胞、iPS細胞等、多能性細胞、例えば、間葉系幹細胞、ならびに組織特異的幹細胞が挙げられる。これらは、あらゆる発育期から、例えば、胚、新生児、若年、または成人、ならびにいずれかの性別、すなわちXXまたはXYであり得る。
【0046】
胚および多能性細胞は、あらゆる生物、例えば、あらゆる哺乳類種、例えば、ヒト、霊長類、ウマ、ウシ、ブタ、イヌ、ネコ等に由来し得る。好ましくは、それらはヒトに由来する。それらは、予め凍結され得、例えば、胚は、1細胞期で凍結保存され、その後解凍されるか、または凍結および解凍された卵母細胞および幹細胞であり得る。代替的に、それらは、新たに調製された、例えば、生体外受精技法により卵母細胞から新たに調製された胚、生体外成熟技法を通して新たに採取および/もしくは新たに成熟させた、または生体外で生殖細胞に分化され、卵母細胞に成熟させた多能性幹細胞に由来する卵母細胞、当該分野において公知の方法による解離および組織の培養により新たに調製された幹細胞等であり得る。それらは、例えば、胚については、生体外受精の分野で使用される場合等の条件下で(例えば、米国特許第6,610,543号、米国特許第6,130,086号、米国特許第5,837,543号を参照し、それらの開示は、参照により本明細書に組み込まれる)、卵母細胞においては、卵母細胞の成熟を促進するために当該分野において使用される場合等の条件下で(例えば、米国特許第5,882,928号、および米国特許第6,281,013号を参照し、それらの開示は、参照により本明細書に組み込まれる)、幹細胞においては、増殖を促進するために当該分野において使用される場合等の条件下で(例えば、米国特許第6,777,233号、米国特許第7037892号、米国特許第7,029,913号、米国特許第5,843,780号、および米国特許第6,200,806号、米国特許出願第2009/0047263号、米国特許出願第2009/0068742号を参照し、それらの開示は、参照により本明細書に組み込まれる)評価される試料の生存、成長、および/または発達を促進するために当該分野において公知のあらゆる都合のよい条件下で培養され得る。しばしば、胚/多能性細胞は、評価される特定の胚/多能性細胞の必要性に合わせた、血清もしくは血清代替物、アミノ酸、および成長因子を添加されたノックアウトDMEM、DMEM−F12またはイスコフ改変ダルベッコ培地等の市販の培地で培養される。
【0047】
微速度撮像分析
一部の実施形態において、胚/多能性細胞は、微速度撮像により細胞パラメータを測定することにより評価される。胚/多能性細胞は、標準の培養皿で培養され得る。代替的に、胚/多能性細胞は、特注の培養皿、例えば、本明細書に記載される光学品質のマイクロウェルを備える特注の培養皿で培養され得る。そのような特注の培養皿において、各マイクロウェルは、単一の胚/多能性細胞を保持し、各マイクロウェルの底面は、細胞の有糸分裂プロセスを追跡するのに十分な解像度を備えた単一の小型顕微鏡により、単一皿内の胚の全群が同時に撮像できるように光学品質仕上げである。マイクロウェルの全群は、培養皿の同一の培地液滴を共有し、培地液滴を安定させるためにマイクロウェルの周囲に位置付けられた外壁、ならびにマイクロウェルの近くに設置される位置合わせマーカーも含むことができる。表面の疎水性は、プラズマエッチングまたは他の処理により調節することができ、培地で満たされた時にマイクロウェル中で気泡が形成されるのを防止する。標準の培養皿または特注の培養皿が利用されるかどうかにかかわらず、培養中、1つ以上の発達胚は、同一の培養培地で培養される、例えば、1皿当り1〜30個の胚が培養され得る。
【0048】
画像は、経時的に取得され、その後、1つ以上の細胞パラメータの測定値を導き出すように分析される。微速度撮像は、デジタル画像保存および分析用に装備されるあらゆるコンピュータ制御顕微鏡で、例えば、加熱台およびインキュベーションチャンバを装備した倒立顕微鏡、または従来のインキュベータの内部に収まる特注の小型顕微鏡アレイで実施され得る。並んだ小型顕微鏡は、同一インキュベータ中の複数の試料皿を同時に培養することができ、連続画像捕捉間の最小時間間隔に対して制限がない複数のチャネルを収容するように拡張可能である。複数の顕微鏡の使用は、試料を移動する必要性を排除し、これは、システムの正確性および全体的なシステムの信頼度を改善する。インキュベータ中の個々の顕微鏡は、部分的に、または完全に隔離することができ、各培養皿に独自の制御された環境を提供する。これは、他の試料の環境を乱すことなく、皿の撮像ステーションへの、およびそこからの移動を可能にする。
【0049】
微速度撮像用の撮像システムは、明視野照明、暗視野照明、位相差、ホフマン変調コントラスト、微分干渉コントラスト、または蛍光を利用することができる。一部の実施形態において、暗視野照明は、後続の特色抽出および画像分析のために、強調した画像コントラストを提供するために使用され得る。加えて、赤色または近赤外線源は、光毒性を低減し、細胞膜と細胞の内側部分との間のコントラスト比を改善するために使用され得る。
【0050】
取得された画像は、ライブビデオと同様に継続して、または微速度撮影と同様に断続してのいずれかで保存され得、ここで、対象物は、静止画で繰り返し撮像される。好ましくは、画像間の時間間隔は、以下に説明される有意な形態学的事象を捕捉するために、1〜30分の間であるべきである。代替的な実施形態において、画像間の時間間隔は、細胞活性量により変動し得る。例えば、活性期間中、画像は、数秒毎または数分毎に撮られ、一方、不活性期間中、画像は、10分もしくは15分毎、またはそれ以上毎に撮られ得る。捕捉した画像に対するリアルタイム画像分析は、いつ、およびどのように時間間隔が変動するかを検知するために使用される。我々の方法において、試料が受ける総光量は、5日間の撮像において、約24分の連続低線量光曝露に相当すると推定される。微速度撮像システムの光強度は、低電力のLED(例えば、通常の100Wのハロゲン球電球と比較して1WのLEDを使用)、および高感度のカメラセンサにより、生殖補助顕微鏡で通常使用される光強度より有意に低い。よって、微速度撮像システムを使用する胚が受ける総光エネルギー量は、IVF臨床での日常的取り扱い中に受けるエネルギー量と同程度か、またはそれ未満である。加えて、曝露時間は、胚/多能性細胞への総光曝露量を低減するために有意に短縮され得る。画像当り0.5秒の光曝露で、5分毎に捕捉された画像の2日間の撮像において、総低線量光曝露量は、5分未満である。
【0051】
画像取得後、画像は抽出され、異なる細胞パラメータ、例えば、細胞の大きさ、透明帯の厚さ、断片化の程度、細胞分割によって生じる娘細胞の対称性、最初のいくつかの有糸分裂間の時間間隔、および細胞質分裂の持続時間について分析される。
【0052】
微速度撮像により測定され得る細胞パラメータは、一般に、形態学的事象である。例えば、胚の評価において、微速度撮像は、細胞質分裂事象、例えば、細胞質分裂1、細胞質分裂2、細胞質分裂3、または細胞質分裂4の持続時間を測定するために使用され得、ここで、細胞質分裂事象の持続時間は、分裂溝(細胞質分裂の開始)の最初の観測と分裂溝の2つの娘細胞への分離(すなわち、2つの娘細胞の産生)との間の時間間隔として定義される。別の目的のパラメータは、細胞周期事象、例えば、細胞周期1、細胞周期2、細胞周期3、または細胞周期4の持続時間であり、ここで、細胞周期事象の持続時間は、細胞の産生(細胞周期1においては、卵子の受精、後の細胞周期においては、細胞質分裂の分離時)とその細胞からの2つの娘細胞の産生との間の時間間隔として定義される。微速度撮像により測定され得る他の目的の細胞パラメータは、これらの細胞事象により定義される時間間隔、例えば、(a)細胞質分裂1の開始と細胞質分裂2の開始との間の間隔、細胞質分裂1の分離と細胞質分裂2の分離との間の間隔、細胞質分裂1の開始と細胞質分裂2の分離との間の間隔、もしくは細胞質分裂1の分離と細胞質分裂2の開始との間の間隔のうちのいずれか1つとして定義可能である、細胞質分裂1と細胞質分裂2との間の時間間隔、または(b)細胞質分裂2の開始と細胞質分裂3の開始との間の間隔、もしくは細胞質分裂2の分離と細胞質分裂3の分離との間の間隔、もしくは細胞質分裂2の開始と細胞質分裂3の分離との間の間隔、もしくは細胞質分裂2の分離と細胞質分裂3の開始との間の間隔のうちのいずれか1つとして定義可能である、細胞質分裂2と細胞質分裂3との間の時間間隔を含む。
【0053】
生体外受精の目的において、生体外環境に対する培養条件の欠点による胚の損失を低減するため、および培養中に生じ得る後成的エラーに関連する有害転帰の可能性を低減するために、胚は、発達の初期に、例えば、2日目、または3日目までに、すなわち、8細胞期に至るまでに子宮に移植されることが有利であると考えられる(Katari et al.(2009)Hum Mol Genet.18(20):3769−78、Sepulveda et al.(2009)Fertil Steril.91(5):1765−70)。したがって、より長期の分析、例えば、約36時間、約54時間、約60時間、約72時間、約84時間、約96時間、またはそれ以上も、本方法によって意図されるが、細胞パラメータの測定は、受精から2日以内に行われることが好ましい。
【0054】
微速度撮像により評価され得る成熟卵母細胞の細胞パラメータの例としては、卵母細胞膜の形態の変化、例えば、透明帯からの分離の速度および程度;卵母細胞核の形態の変化、例えば、胚胞崩壊(GVBD)の開始、完了、および速度;細胞質および核における顆粒移動の速度および方向;卵母細胞および第1極体の細胞質分裂、ならびに第1極体の突出の移動および/または持続時間が挙げられるが、これらに限定されない。他のパラメータは、成熟二次卵母細胞、および第2極体の細胞質分裂の持続時間を含む。
【0055】
微速度撮像により評価され得る幹細胞または幹細胞集団の細胞パラメータの例としては、細胞質分裂事象の持続時間、細胞質分裂事象間の時間、細胞質分裂事象前およびその間の幹細胞の大きさおよび形状、細胞質分裂事象により産生された娘細胞の数、分裂溝の空間的配位、観測された非対称分割の速度および/または数(すなわち、1つの娘細胞が幹細胞を維持し、一方、他は分化する)、観測された対称分割の速度および/または数(すなわち、双方の娘細胞が幹細胞として留まるか、双方が分化するかのいずれかである)、および細胞質分裂事象の分離と幹細胞が分化し始める時との間の時間間隔が挙げられるが、これらに限定されない。
【0056】
パラメータは、手動で測定されるか、または自動的に、例えば、画像分析ソフトウェアにより測定され得る。画像分析ソフトウェアが利用される場合、例えば、仮定される胚/多能性細胞モデルの分布を生成する、単純な光学モデルに基づき画像をシュミレーションする、およびこれらのシミュレーションを観測した画像データと比較する、遂次モンテカルロ法に基づく確率的モデル推定技法を利用する画像分析アルゴリズムが使用され得る。そのような確率的モデルによる推定が利用される場合、細胞は、あらゆる適切な形状として、例えば、2D空間における楕円の集まり、3D空間における楕円体の集まり等としてモデル化され得る。遮蔽および深度の不明確さに対処するために、方法は、予想された物理的挙動に対応する幾何学的制約を強化することができる。確実性を改善させるために、画像は、1つ以上の焦点面で捕捉され得る。
【0057】
遺伝子発現分析
一部の実施形態において、胚または多能性細胞は、遺伝子発現を測定することにより評価され得る。そのような実施形態において、細胞パラメータは、遺伝子発現レベル、または遺伝子発現プロファイルである。1つ以上の発現を判定すること、すなわち、発現プロファイルまたは発現評価を得ることは、1つ以上の目的の遺伝子の核酸転写、例えば、mRNAを測定することにより、または1つ以上の目的の遺伝子の発現産物である、1つ以上の異なるタンパク質/ポリペプチドのレベル、例えば、プロテオーム発現プロファイルを測定することにより行われる。言い換えると、「発現プロファイル」および「発現評価」という用語は、RNAレベルまたはタンパク質レベルの遺伝子発現プロファイルを含むように広範に使用される。
【0058】
一部の実施形態において、遺伝子の発現は、核酸発現プロファイルを得ることにより評価され得、ここで、試料中の1つ以上の核酸の量またはレベル、例えば、1つ以上の目的の遺伝子の核酸転写が判定される。これらの実施形態において、発現プロファイルを生成するためにアッセイされる試料は、核酸試料である。核酸試料は、評価される胚または細胞の目的の遺伝子の発現情報を含む複数の別個の核酸、または別個の核酸の集団を含む。核酸は、試料が得られる宿主細胞または組織の発現情報を保有する限り、RNAまたはDNA核酸、例えば、mRNA、cRNA、cDNA等を含み得る。試料は、当該分野において公知である多くの異なる方式で、例えば、細胞からのmRNAの単離により調製され(ここで、単離されたmRNAはそのまま使用される)、差次的発現分野において公知であるように、cDNA、cRNA等を調製するために増幅、利用され得る。試料は、標準のプロトコルを使用して、単一細胞、例えば、目的の多能性細胞の培養物の多能性細胞、もしくは目的の胚からの単一細胞(卵割球)から、またはいくつかの細胞、例えば、多能性細胞の培養物の画分、または目的の胚の2、3、もしくは4つ以上の卵割球から調製され得る。
【0059】
発現プロファイルは、あらゆる都合のよいプロトコルを使用して、最初の核酸試料から生成され得る。差次的遺伝子発現分析の分野において利用されるもののような、発現プロファイルを生成する種々の異なる手段が公知であるが、発現プロファイルを生成するための代表的な都合のよいプロトコルの種類の1つは、アレイベースの遺伝子発現プロファイル生成プロトコルである。そのような用途は、生成されるプロファイルにおいて、アッセイされる/プロファイルされる遺伝子のそれぞれの「プローブ」核酸を示す核酸が利用されるハイブリダイゼーションアッセイである。これらのアッセイにおいて、標的核酸の試料がアッセイされる最初の核酸試料から最初に調製され、ここで、調製は、標識、例えば、シグナル産生システムの構成要素で標的核酸を標識することを含み得る。標的核酸試料調製後、ハイブリダイゼーション条件下で試料をアレイと接触させ、それによって、複合体がアレイ表面に付着するプローブ配列に相補的である標的核酸間で形成される。その後、ハイブリッド形成された複合体の存在は、定性的または定量的のいずれかで検出される。
【0060】
本発明の方法で利用される発現プロファイルを生成するために実践され得る具体的なハイブリダイゼーション技術は、米国特許第5,143,854号、第5,288,644号、第5,324,633号、第5,432,049号、第5,470,710号、第5,492,806号、第5,503,980号、第5,510,270号、第5,525,464号、第5,547,839号、第5,580,732号、第5,661,028号、第5,800,992号(それらの開示は、参照により本明細書に組み込まれる)、ならびに国際公開第WO95/21265号、第WO96/31622号、第WO97/10365号、第WO97/27317号、欧州特許第373 203号、および第EP785 280号に記載される技術を含む。これらの方法において、発現がアッセイされる表現型決定遺伝子のそれぞれについてのプローブを含む複数の「プローブ」核酸が、上述のように標的核酸と接触させられる。接触は、ハイブリダイゼーション条件下、例えば、厳密なハイブリダイゼーション条件下で行われ、その後、未結合核酸は除去される。本明細書に使用される、「厳密なアッセイ条件」という用語は、所望の特異性を提供するのに十分に相補的でない結合構成要素間での結合対の形成にあまり適していないが、アッセイにおいて所望の特異性レベルを提供するのに十分に相補的である核酸の結合対、例えば、表面結合および溶液相を産生するのに適している条件を指す。厳密なアッセイ条件は、ハイブリダイゼーションおよび洗浄条件の双方の総和または組み合わせ(全体性)である。
【0061】
得られたハイブリッド形成された核酸のパターンは、プローブされた遺伝子のそれぞれの発現に関する情報を提供し、ここで、発現情報は、遺伝子が発現するかどうか、および通常どんなレベルで発現するかに関してであり、発現データ、すなわち、発現プロファイル(例えば、トランスクリプトソームの形態で)は、定性的および定量的の双方であり得る。
【0062】
代替的に、試料中の1つ以上の核酸のレベルを定量化するための非アレイベースの方法が利用され得、増幅プロトコルに基づくもの、例えば、定量的PCR、逆転写PCR(RT−PCR)、リアルタイムPCR等を含む、ポリメラーゼ連鎖反応(PCR)ベースのアッセイを含む。
【0063】
一部の実施形態において、遺伝子の発現は、プロテオーム発現プロファイルを得ることにより評価され得、ここで、例えば、目的の遺伝子によりコードされるタンパク質/ポリペプチド等の、試料中の1つ以上のタンパク質/ポリペプチドの量またはレベルが判定される。これらの実施形態において、方法において利用される発現プロファイルを生成するためにアッセイされる試料は、タンパク質の試料である。発現プロファイルがプロテオーム発現プロファイル、すなわち、試料中の1つ以上のタンパク質レベルのプロファイルである場合、タンパク質レベルを評価するためのあらゆる都合のよいプロトコルが利用され得、アッセイされた試料中の1つ以上のタンパク質のレベルが判定される。
【0064】
タンパク質レベルについてアッセイする種々の異なる手段は、当該分野において公知であるが、タンパク質レベルをアッセイするための代表的な都合のよいプロトコルの種類は、ELISAである。ELISAおよびELISAベースのアッセイにおいて、目的のタンパク質に特異的な1つ以上の抗体は、選択された固体表面上、好ましくは、ポリスチレンマイクロタイタープレートのウェル等の、タンパク質親和性を示す表面に固定され得る。不完全に吸着された材料を除去するために洗浄した後、アッセイプレートウェルは、ウシ血清アルブミン(BSA)、カゼイン、または粉乳の溶液等の、試験試料に関して抗原性的に中性であることが知られている非特異的「ブロッキング」タンパク質でコーティングされる。これは、固定化表面上の非特異的吸着部位のブロッキングを可能にし、これによって、抗原の表面への非特異的結合により生じるバックグラウンドを低減する。未結合のブロッキングタンパク質を除去するために洗浄した後、免疫複合体(抗原/抗体)形成をもたらす条件下で、固定化表面を試験される試料と接触させる。そのような条件は、リン酸緩衝生理食塩水(PBS)/TweenまたはPBS/Triton−X100中にBSAまたはウシガンマグロブリン(BGG)を希釈する等、希釈液で試料を希釈することを含み、これは、非特異的バックグラウンドの低減も補助する傾向にあり、(他の温度も使用され得るが)およそ約25°〜27℃の温度で約2〜4時間、試料をインキュベートすることを可能にする。インキュベーション後、非免疫複合材料を除去するために、抗血清接触面を洗浄する。例示的な洗浄手順は、PBS/Tween、PBS/Triton−X100、またはホウ酸緩衝液等の溶液で洗浄することを含む。その後、結合免疫複合体を第1の抗体と異なる標的に特異性を有する第2の抗体に曝し、第2の抗体の結合を検出することにより、免疫複合体形成の発生および量が判定される。特定の実施形態において、第2の抗体は、関連酵素、例えば、ウレアーゼ、ペルオキシダーゼ、またはアルカリホスファターゼを有し、これは、適切な発色基質を用いてインキュベートすることにより、有色の沈殿物を生成する。例えば、ウレアーゼまたはペルオキシダーゼ抱合抗ヒトIgGは、免疫複合体形成の発達に好ましい時間および条件下で(例えば、PBS/Tween等のPBS含有溶液中で、室温で2時間インキュベートする)利用され得る。第2の抗体を用いたそのようなインキュベーション、および未結合材料を除去するための洗浄の後、例えば、ウレアーゼ標識の場合においては、尿素およびブロモクレゾールパープル、またはペルオキシダーゼ標識の場合においては、2,2’−アジノ−ジ−(3−エチル−ベンズチアゾリン)−6−スルホン酸(ABTS)およびH2O2等の、発色基質を用いてインキュベーションすることにより、標識の量を定量化する。その後、定量化は、例えば、可視スペクトル分光光度計を使用して、色の生成の程度を測定することにより達成される。
【0065】
先行フォーマットは、最初に試料をアッセイプレートに結合することにより変更され得る。次いで、一次抗体を、アッセイプレートを用いてインキュベートした後、一次抗体に特異性を有する標識された第2の抗体を使用して、結合した一次抗体を検出する。
【0066】
抗体または複数の抗体が固定される固体基質は、広範な種々の材料から、および広範な種々の形状、例えば、マイクロタイタープレート、マイクロビーズ、計深棒、樹脂粒子等に作製され得る。基質は、シグナル対ノイズ比を最大にするように、バックグラウンド結合を最小にするように、ならびに分離および費用を軽減するように選択され得る。洗浄は、基質が使用されるのに最も適した手段、例えば、リザーバからビーズもしくは計深棒を除去する、マイクロタイタープレート等のリザーバを空にする、もしくは希釈する、またはビーズ、粒子、クロマトグラフカラム、もしくはフィルタを洗浄溶液または溶媒ですすぐことにより達成され得る。
【0067】
代替的に、試料中の1つ以上のタンパク質のレベルを測定するために、非ELISAベースの方法が利用され得る。代表的な例としては、質量分析、プロテオームアレイ、xMAP(商標)微小球体技術、フローサイトメトリー、ウエスタンブロット、および免疫組織化学が挙げられるが、これらに限定されない。
【0068】
得られたデータは、プローブされた遺伝子のそれぞれの発現に関する情報を提供し、発現情報は、遺伝子が発現するかどうか、および通常どんなレベルで発現するかに関してであり、また発現データは、定性的および定量的の双方であり得る。
【0069】
発現プロファイルの生成において、一部の実施形態において、試料は、少なくとも1つの遺伝子/タンパク質、時折、複数の遺伝子/タンパク質についての発現データを含む発現プロファイルを生成するためにアッセイされ、ここで、複数とは、少なくとも2つの異なる遺伝子/タンパク質、多くの場合、少なくとも約3つ、通常少なくとも約10以上、およびより一般には、50以上、もしくは100以上等の少なくとも約15以上の異なる遺伝子/タンパク質を意味する。
【0070】
最も広い意味において、発現の評価は、定性的または定量的であり得る。そのようなものとして、検出が定性的である場合、本方法は、核酸または発現産物などの標的検体がアッセイされる試料中に存在するかどうかの測定または評価、例えば鑑定を提供する。また他の実施形態において、本方法は、標的検体がアッセイされる試料中に存在するかどうかの定量的検出、すなわち、例えば、アッセイされる試料中の核酸またはタンパク質などの標的検体の実際の量もしくは相対的存在量の評価または鑑定を提供する。そのような実施形態において、定量的検出は絶対的であり得、もしその方法が試料中の例えば標的核酸もしくはタンパク質の2つ以上の異なる検体を検出する方法である場合、相対的であり得る。そのようなものとして、例えば、試料中の核酸またはタンパク質の標的検体を定量化する場面において使用される時、「定量化」という用語は、絶対的または相対的定量化を指すことができる。絶対的定量化は、1つ以上の対照検体の既知濃度を含むことにより、および既知の対照検体を用いて標的検体の検出レベルを参照する、すなわち、標準化することにより(例えば、標準曲線の生成を介して)達成され得る。代替的に、相対的定量化は、2つ以上の異なる標的検体間の検出レベルまたは量を比較して、例えば、相互に対比して、2つ以上の異なる検体のそれぞれの相対的定量化を提供することにより達成され得る。
【0071】
発現レベルが接合体の発達の可能性を予測する遺伝子の例としては、コフィリン(NM_005507)、DIAPH1(NM_001079812、NM_005219)、ECT2(NM_018098)、MYLC2/MYL5(NM_002477)、DGCR8(NM_022720)、Dicer/DICER1(NM_030621、NM_177438)、TARBP2(NM_004178、NM_134323、NM_134324)、CPEB1(NM_001079533、NM_001079534、NM_001079535、NM_030594)、Symplekin/SYMPK(NM_004819)、YBX2(NM_015982)、ZAR1(NM_175619)、CTNNB1(NM_001098209、NM_001098210、NM_001098210、NM_001904)、DNMT3B(NM_006892、NM_175848、NM_175849、NM_175850)、TERT(NM_198253、NM_198255)、YY1(NM_003403)、IFGR2/IFNGR2(NM_005534)、BTF3(NM_001037637、NM_001207)、およびNELF(NM_001130969、NM_001130970、NM_001130971、NM_015537)が挙げられる。発現レベルが胚の発達の可能性を予測する細胞パラメータとして機能する他の遺伝子を図8に提供する。遺伝子発現レベル測定値を導き出すことにおいて、発現レベルは、評価された後、標準対照に対して標準化されることが多く、標準対照とは例えばGAPDHもしくはRPLPOという発達を通して一定であることが知られている遺伝子、またはその時間点での発現が知られている遺伝子、の試料中の発現レベルである。
【0072】
遺伝子発現レベルは、単一細胞、例えば、目的の胚からの卵割球、もしくは単離した卵母細胞、もしくは幹細胞の培養物から単離した細胞等から判定され得るか、またはそれらは、胚、例えば、目的の全胚までを含む、2、3、または4個以上の目的の胚の卵割球、もしくは幹細胞の全培養物までを含む、幹細胞の培養物からの複数の細胞等から決定され得る。
【0073】
他の態様において、本発明は、単一細胞で遺伝子型の特定および遺伝子発現分析を同時に実施するためのプロトコルを含む。胚において、これは、単一細胞が胚から取り出され、そのDNAが核型異常または特定の疾患遺伝子について試験される手順である、着床前遺伝子診断(PGD)を改善するために使用され得る。我々の方法は、遺伝的および遺伝子発現分析を同時に行うことを可能にする。方法は、以下のステップを含む:(1)単一細胞を少量の培地または緩衝液中に収集する、(2)遺伝子型特定及び遺伝子発現分析用プライマーの混合物を使用して、一工程の逆転写およびポリメラーゼ連鎖反応(PCR)増幅を実施する、(3)18サイクル未満のPCR後の増幅したcDNAのアリコートを収集して、増幅の直線性を保存する、(4)cDNAアリコートを使用して、定量的リアルタイムPCR等の標準技法を用いて遺伝子発現分析を実施する、(5)残りの試料を使用して、2回目のPCRを実施し、遺伝子型特定の目的のために、遺伝子情報をさらに増幅する、および(6)ゲル電気泳動等の標準技法を使用して、遺伝子型を特定する。
【0074】
画像および/または遺伝子発現分析から発達の可能性を判定
細胞パラメータ測定値が得られたら、測定値を利用して、胚/多能性細胞の発達の可能性を判定する。上述の通り、「発達の可能性」および「発達能力」という用語は、多能性細胞もしくは組織の成長する、もしくは発達する能力または潜在的可能性を指す。例えば、卵母細胞または胚において、発達の可能性は、その卵母細胞もしくは胚の健康な胚盤胞に成長する、または発達する能力もしくは潜在的可能性であり得る。別の例として、幹細胞の場合、発達の可能性は、1つ以上の目的の細胞、例えば、神経、筋肉、B−もしくはT細胞等に成長もしくは発達する能力または潜在的可能性である。一部の実施形態において、卵母細胞または胚の発達の可能性とは、その卵母細胞もしくは胚が健康な胚盤胞に発達する;子宮への移植が成功する;妊娠期間を経る;および/もしくは生きて生まれる、能力または潜在的可能性である。一部の実施形態において、多能性細胞の発達の可能性とは、その多能性細胞の1つ以上の目的の細胞、例えば、神経、筋肉、B−もしくはT細胞等に発達する、および/または生体内において目的の組織に寄与する、能力もしくは潜在的可能性である。
【0075】
「良好な発達の可能性」とは、胚/多能性細胞が統計的に所望の通りに発達する可能性を意味する、すなわち、所望の通りに、55%、60%、70%、80%、90%、95%以上の確率、例えば、100%の発達の確率を有する。言い換えると、良好な発達の可能性の判定を導き出すために使用される細胞パラメータ測定値を示す100のうちの55、100のうちの60、100うちの70、100のうちの80、100のうちの90、100のうちの95、または100のうちの100の胚もしくは多能性細胞が、実際、所望通りに発達を続ける。逆に、「不良な発達の可能性」とは、胚/多能性細胞が統計的に所望の通りに発達する可能性がないことを意味する、すなわち、所望の通りに、50%、40%、30%、20%、10%、5%以下の確率、例えば、0%の発達の確率を有する。言い換えると、不良な発達の可能性の判定を導き出すために使用される細胞パラメータ測定値を示す100のうちのわずか50、100のうちの40、100のうちの30、100のうちの20、100のうちの10、または100のうちの5以下の胚もしくは多能性細胞が、実際、所望の通りに、発達を続ける。本明細書に使用される、「正常」または「健康な」胚および多能性細胞とは、良好な発達の可能性を示し、一方、「異常な」胚および多能性細胞は、不良な発達の可能性を示す。
【0076】
一部の実施形態において、細胞パラメータ測定値は、胚/多能性細胞の発達の可能性を判定するために直接使用される。言い換えると、測定値自体の絶対値は、発達の可能性を判定するのに十分である。細胞パラメータを測定するために微速度撮像を使用する実施形態におけるこの例としては、次の(a)約0〜30分、例えば、約6〜20分、平均して約12〜14分続く細胞質分裂1、(b)約20〜27時間、例えば、約25〜27時間続く細胞周期1、(c)約8〜15時間、例えば、約9〜13時間、約11+/−2.1時間の平均値の細胞質分裂1の分離と細胞質分裂2の開始との間の時間間隔、(d)約0〜5時間、例えば、約0〜3時間、約1+/−1.6時間の平均時間の細胞質分裂2の開始と細胞質分裂3の開始との間の時間間隔が挙げられるがこれらに限定されず、それらのうちのいずれかが、単独または組み合わせで、ヒトの胚における良好な発達の可能性を示す。直接測定値の例としては、(a)約30分以上、例えば、約32、35、40、45、50、55、もしくは60分以上続く細胞質分裂1、(b)約27時間以上、例えば、28、29、もしくは30時間以上続く細胞周期1、(c)15時間以上、例えば、約16、17、18、19、もしくは20時間以上、または8時間未満、例えば、7、5、4、もしくは3時間以下続く、細胞質分裂1の分離と細胞質分裂2の開始との間の時間間隔、(d)6、7、8、9、もしくは10時間以上である、細胞質分裂2の開始と細胞質分裂3の開始との間の時間間隔が挙げられるが、これらに限定されず、それらのうちのいずれかが、単独または組合せで、ヒトの胚において不良な発達の可能性を示す。
【0077】
一部の実施形態において、細胞パラメータ測定値は、それを基準もしくは対照の胚/多能性細胞からの細胞パラメータ測定値と比較し、この比較の結果を使用して、胚/多能性細胞の発達の可能性の判定を提供することにより利用される。本明細書に使用される、「基準」および「対照」という用語は、所与の胚/多能性細胞の細胞パラメータ測定値を解釈し、それに対して発達の可能性の判定を行うために使用される、規格化された胚または細胞を意味する。基準または対照は、所望の表現型、例えば、良好な発達の可能性を有することが知られている胚/多能性細胞であり得、したがって、陽性基準または対照の胚/多能性細胞であり得る。代替的に、基準/対照の胚/多能性細胞は、所望の表現型を有さないことが知られている胚/多能性細胞であり得、したがって、陰性基準/対照の胚/多能性細胞であり得る。
【0078】
ある実施形態において、得られた細胞パラメータ測定値を、単一の基準/対照の胚/多能性細胞からの比較可能な細胞パラメータ測定値と比較し、アッセイされる胚/細胞の表現型に関する情報を得る。また他の実施形態において、得られた細胞パラメータ測定値を、2つ以上の異なる基準/胚もしくは多能性細胞からの比較可能な細胞パラメータ測定値と比較し、アッセイした胚/細胞の表現型に関するより詳細な情報を得る。例えば、アッセイされる胚もしくは多能性細胞から得られた細胞パラメータ測定値を、陽性および陰性の胚もしくは多能性細胞の双方と比較し、胚/細胞が目的の表現型を有するかに関する確実な情報を得る。
【0079】
例として、正常なヒトの胚における、すなわち、良好な発達の可能性を有する細胞質分裂1は、約0〜30分、より一般には、約6〜20分、平均して約12〜14分、すなわち、約1、2、3、4、もしくは5分、より一般には、約6、7、8、9、10、11、12、13、14、15、16、17、18、19、もしくは20分、一部の場合においては、21、22、23、24、25、26、27、28、29、もしくは最大約30分である。正常な基準の胚において観測されるものと比較して、評価される胚の細胞質分裂1が完了する時間が長ければ、不良な発達の可能性を示す。第2の例として、正常な胚の細胞周期1、すなわち、受精から細胞質分裂1完了までの時間は、通常、約20〜27時間、より一般には、約25〜27時間、すなわち、約15、16、17、18、もしくは19時間、より一般には、約20、21、22、23、もしくは24時間、より一般には、約25、26、もしくは27時間で完了する。正常な基準の胚において観測されるものと比較して、評価される胚における長い細胞周期1は、不良な発達の可能性を示す。第3の例として、正常なヒトの胚における細胞質分裂1の分離および細胞質分裂2の開始は、約8〜15時間、より頻繁には、約9〜13時間であって、約11+/−2.1時間の平均値であり、すなわち、6、7、もしくは8時間、より一般には、約9、10、11、12、13、14、もしくは最大約15時間である。正常な基準の胚において観測されるものと比較して、評価される胚における長い、または短い細胞周期2は、不良な発達の可能性を示す。第4の例として、正常なヒトの胚における細胞質分裂2の開始と細胞質分裂3の開始との間の時間間隔、すなわち、第2と第3の有糸分裂の同時発生は、一般に、約0〜5時間、より一般には、約0、1、2、もしくは3時間、平均時間は約1+/−1.6時間であり、正常な基準の胚において観測されるものと比較して、評価される胚における細胞質分裂2と細胞質分裂3の完了との間の長い間隔は、不良な発達の可能性を示す。最後に、発達の可能性を評価するためのパラメータとして遺伝子発現レベルを使用する時に、この実施形態がどのように適用されるかの例として、コフィリン、DIAPH1、ECT2、MYLC2、DGCR8、ダイサー、TARBP2、CPEB1、シンプレキン、YBX2、ZAR1、CTNNB1、DNMT3B、TERT、YY1、IFGR2、BTF3、および/またはNELFの低発現レベル、すなわち、正常な基準の2細胞胚において観測されるものと比較して、評価される2細胞胚において、1.5倍、2倍、3倍、4倍、5倍、10倍、20倍、50倍、または100倍低い発現は、不良な発達の可能性示し、一方、正常な基準の2細胞胚において観測されるものと同等もしくはそれより大きい発現は、良好な発達の可能性を示す。他の例は、例えば、評価される胚/多能性細胞と一緒に1つ以上の基準の胚または多能性細胞を観測することによる、経験的データに由来する。あらゆる基準の胚/多能性細胞、例えば、良好な発達の可能性を有する正常な基準試料、または不良な発達の可能性を有する異常な基準試料が利用され得る。一部の場合において、2つ以上の基準試料が利用され得る、例えば、正常な基準試料および異常な基準試料の双方が使用され得る。
【0080】
一部の実施形態において、微速度顕微鏡検査および発現プロファイリングの双方ではなく、微速度顕微鏡検査または発現プロファイリングにより導き出される細胞パラメータ測定値を使用することが望ましい場合がある。他の実施形態において、微速度顕微鏡検査により導き出される細胞パラメータ測定値および発現プロファイリングにより導き出される細胞パラメータ測定値を使用することが望ましい場合がある。
【0081】
上述の通り、胚もしくは多能性細胞の発達の可能性を判定するために、1つ以上のパラメータが測定され、利用され得る。一部の実施形態において、単一パラメータの測定値は、発達の可能性の判定を導き出すのに十分であり得る。一部の実施形態において、2つ以上のパラメータ、例えば、2つの細胞パラメータ、3つの細胞パラメータ、または4つ以上の細胞パラメータの測定値を利用することが望ましい場合がある。
【0082】
ある実施形態において、複数のパラメータは、より高い感度および特異性を提供し得るため、複数のパラメータをアッセイすることが望ましい場合がある。感度とは、そうであると正しく識別される実際の陽性物の比率を意味する。これは、数学的に、
【0083】
【表1】
として表され得る。よって、「陽性物」が、良好な発達の可能性を有する、すなわち、胚盤胞に発達する胚であり、「陰性物」が、不良な発達の可能性を有する、すなわち、胚盤胞に発達しない胚である方法において、100%の感度は、試験がそのように胚盤胞に発達する全ての胚を認識することを意味する。一部の実施形態において、アッセイの感度は、約70%、80%、90%、95%、98%以上、例えば、100%であり得る。特異性とは、そうであると正しく識別される陰性の比率を意味する。これは、数学的に、
【表2】
として表され得る。よって、陽性物が、良好な発達の可能性を有する、すなわち、胚盤胞に発達する胚であり、「陰性物」が、不良な発達の可能性を有する、すなわち、胚盤胞に発達しない胚である方法において、100%の特異性は、試験がそのように胚盤胞に発達しない、すなわち、胚盤胞期前に停止する、全ての胚を認識することを意味する。一部の実施形態において、アッセイの特異性は、約70%、80%、90%、95%、98%以上、例えば、100%であり得る。
【0084】
以下の実施例のセクションおよび図7に示す通り、3つのパラメータの使用は、94%の感度および93%の特異性を提供し、カットオフ点は、胚盤胞分布の標準偏差の3倍である。言い換えると、本発明の方法は、胚盤胞に発達するであろう胚の数を94%の確率(感度)で、および胚盤胞期前に停止するであろう胚の数を93%の確率(特異性)で正確に識別することができる。加えて、指定の平均値および/またはカットオフ点は、これらの値を計算するために使用されるデータセットならびに具体的な用途により修正され得る。
【0085】
一部の実施形態において、胚または多能性細胞の評価は、本発明の胚/多能性細胞、例えば、「発達の可能性の評価」、「染色体異常の評価」等の、当業者の評価を含む書面報告書を作成することを含む。よって、本発明の方法は、そのような評価の結果を提供する報告書を作成する、または出力するステップをさらに含む場合があり、この報告書は、電子媒体(例えば、コンピュータモニタ上に電子表示)の形態で、または有形媒体(例えば、用紙に印刷された報告書もしくは他の有形媒体)の形式で提供され得る。
【0086】
本明細書に記載される、「報告書」とは、本発明の方法により導き出された評価に関する目的の情報を提供する報告要素を含む、電子または有形文書である。本発明の報告書は、完全に、または部分的に電子的に作成され得る。本発明の報告書は、少なくとも本発明の胚または多能性細胞の発達の可能性の評価、染色体異常の存在の確率の評価等を含む。本発明の報告書は、1)試験施設に関する情報、2)業務提供者の情報、3)対象のデータ、4)試料データ、5)評価がどのように導き出されたかに関する情報を提供する詳細な評価報告セクション、例えば、a)取得した細胞パラメータ測定値、b)もしあれば、利用された基準値、および6)他の特色のうちの1つ以上をさらに含む場合がある。
【0087】
報告書は、試験施設についての情報を含む場合があり、この情報は、試料収集および/またはデータ生成が行われた、病院、クリニック、または研究所に関連する。試料収集は、試料がどのように生成されたか、例えば、対象からどのように採取されたか、および/またはどのように培養されたか等を含む場合がある。データ生成は、画像がどのように取得されたか、または遺伝子発現プロファイルがどのように分析されたかを含む場合がある。この情報は、例えば、試験施設の名称および所在地、アッセイを実施した、および/または入力データを入力した研究技術者の身元、アッセイが実施された、および/または分析された日付および時間、試料および/または結果データが保存される場所、アッセイに使用された試薬(例えば、キット等)のロット番号等に関する1つ以上の詳細を含む場合がある。この情報の報告欄は、一般に、ユーザにより提供される情報を使用して作成され得る。
【0088】
報告書は、業務提供者についての情報を含む場合があり、これは、ユーザが居る医療機関の外部、または医療機関の内部に位置し得る。そのようは情報の例としては、業務提供者の名称および所在地、調査者の氏名、ならびに必要な場合、または所望する場合、試料の調製および/もしくはデータ生成を行った個人の氏名を含む場合がある。この情報の報告欄は、一般に、ユーザにより入力されたデータを使用して作成され得、これは、予め書かれた選択項目の中から(例えば、ドロップダウンメニューを使用して)選択され得る。報告書における他の業務提供者の情報は、結果および/または解釈的報告についての技術情報の連絡先を含む場合がある。
【0089】
報告書は、対象データセクションを含み得、卵母細胞または多能性細胞が採取された対象の病歴、患者の年齢、生体外受精周期の特徴(例えば、受精率、3日目の卵胞刺激ホルモン(FSH)レベル)、およびいつ卵母細胞が採取されたか、接合体/胚コホートパラメータ(例えば、胚の総数)を含む。この対象データは、胚の評価を改善するために、および/または移植する胚の最適数の判定を補助するために統合され得る。報告書は、対象(例えば、氏名、対象の生年月日(DOB)、性別、郵送先住所および/または自宅住所、医療記録番号(MRN)、医療機関の部屋番号および/またはベッド番号)、保険情報等)、発達の可能性の評価を指示した対象の医師、または他の医療専門家の氏名、ならびに指示した医師と異なる場合、対象のケアに対して責任がある職員医師(例えば、かかりつけの医師)の氏名を識別する情報等の管理上の対象のデータ(つまり、発達の可能性の評価に必須ではないデータ)も含む場合がある。
【0090】
報告書は、試料データセクションを含む場合があり、これは、試料の種類(胚または多能性細胞、および多能性細胞の種類)、どのように試料が取り扱われたか(例えば、保存温度、調製プロトコル)、ならびに収集された日付および時間等の、評価において分析された生物学的試料についての情報を提供し得る。この情報の報告欄は、一般に、ユーザにより入力されたデータを使用して作成され得、これらの一部は、予め書かれた選択項目(例えば、ドロップダウンメニューを使用して)として提供され得る。
【0091】
報告書は、評価報告セクションを含む場合があり、これは、本明細書に記載される、評価/判定がどのように導き出されたかに関する情報を含み得る。解釈上の報告書は、例えば、評価される胚または多能性細胞の微速度画像、および/または遺伝子発現結果を含み得る。報告書の評価部分は、任意に、推奨セクションも含み得る。例えば、結果が胚の良好な発達の可能性を示す場合、当該分野において推奨されるように、推奨は、不妊治療の間に子宮に移植される限られた胚の数についての推奨を含み得る。
【0092】
報告書は、付加的要素または変更要素を含み得ることも容易に理解されるだろう。例えば、電子である場合、報告書は、報告書の選択された要素についてのより詳細な情報を提供する内部または外部データベースを提示するハイパーリンクを含有し得る。例えば、報告書の患者のデータ要素は、電子的患者記録へのハイパーリンク、またはそのような患者記録にアクセスするためのサイトを含み得、この患者記録は、機密データベースに保持される。この後者の実施形態は、病院内システム、またはクリニック内環境において興味あるものであり得る。電子形式である場合、報告書は、例えば、コンピュータメモリ、zip、CD、DVD等のコンピュータ可読媒体等の、適切な物理的媒体上に記録される。
【0093】
報告書は、上の要素の全て、または一部を含み得るが、但し、報告書は、一般に、少なくとも、ユーザにより要求された分析(例えば、発達の可能性の評価)を提供するのに十分な要素を含むものとすることは、容易に理解されるだろう。
【0094】
実用性
上述の通り、本発明の方法は、それらの発達の可能性を判定するために、胚または多能性細胞を評価するために使用され得る。この発達の可能性の判定は、臨床的判断および/または行動を導くために使用され得る。例えば、妊娠率を増加させるために、臨床医は、複数の胚を患者に移植する場合が多く、母親と胎児の双方に対して健康上のリスクを引き起こす多胎妊娠をもたらす可能性がある。本発明の方法から得られた結果を使用することにより、移植され、胎児に発達する胚の発達の可能性は、移植前に判定され、リスクを最小にしながら、満期妊娠の成功の確率を最大にするように、実施者がいくつの胚を移植するかを判断することができる。
【0095】
本発明の方法に従ってなされた評価は、胚または多能性細胞の群において、それらの発達の可能性について胚または多能性細胞をランク付けすることにも用途を見出すことができる。例えば、一部の場合において、複数の胚が、胚盤胞に発達できる可能性がある、すなわち、良好な発達の可能性を有する。しかしながら、一部の胚は、他より胚盤胞期または高品質の胚盤胞に達する可能性がある、すなわち、他の胚より良好な発達の可能性を有する。そのような場合において、本発明の方法は、群内で胚をランク付けするために使用され得る。そのような方法において、各胚/多能性細胞についての細胞パラメータ測定値を導き出すように、各胚/多能性細胞についての1つ以上の細胞パラメータが測定される。次いで、相互に対して胚または多能性細胞の発達の可能性を判定するために、胚または多能性細胞のそれぞれからの1つ以上の細胞パラメータ測定値が利用される。一部の実施形態において、それらを相互に直接比較して、胚または多能性細胞の発達の可能性を判定することにより、胚または多能性細胞のそれぞれからの細胞パラメータ測定値が利用される。一部の実施形態において、細胞パラメータ測定値を基準の胚/多能性細胞からの細胞パラメータ測定値と比較して、各胚/多能性細胞の発達の可能性を判定し、その後、各胚/多能性細胞の判定された発達の可能性を比較して、相互に対して胚または多能性細胞の発達の可能性を判定することにより、胚または多能性細胞のそれぞれからの細胞パラメータ測定値が利用される。このようにして、例えば、複数の接合体/胚を評価する実施者は、最良質の胚のみ、すなわち、最良の発達の可能性を有するものを選択して、リスクを最小にしながら、満期妊娠の成功の確率を最大にするように移植することができる。
【0096】
本発明の方法に従ってなされる評価は、生体外で成熟させた卵母細胞、および生体外で培養された幹細胞の発達の可能性を判定することにも用途を見出すことができる。本発明の方法によって得られた卵母細胞の発達の可能性についての情報は、受精させる卵母細胞の実施者の選択を導くことができ、それらの卵母細胞から胚盤胞を得る高い成功確率をもたらす。同様に、幹細胞の発達の可能性についての情報は、例えば、それを必要とする対象の組織を生体内で再構成または交換する処置に使用するための幹細胞を実施者が選択するために情報を与えることができる。
【0097】
試薬、デバイス、およびキット
上述の方法のうちの1つ以上を実践するための試薬、デバイス、およびそのキットを提供する。本発明の試薬、デバイス、およびそのキットは、非常に多様である。目的の試薬およびデバイスは、前述の細胞パラメータのいずれかを測定する方法に関する上述のものを含み、ここで、そのような試薬は、実施される特定の測定プロトコルに応じて、培養プレート、培養培地、顕微鏡、撮像ソフトウェア、撮像分析ソフトウェア、核酸プライマーのアレイ、核酸プローブ、抗体、シグナル産生系試薬等を含み得る。例えば、試薬は、上述の通り、コフィリン、DIAPH1、ECT2、MYLC2/MYL5、DGCR8、ダイサー/DICER1、TARBP2、CPEB1、シンプレキン/SYMPK、YBX2、ZAR1、CTNNB1、DNMT3B、TERT、YY1、IFGR2/IFNGR2、BTF3、およびNELFの遺伝子のうちの1つ以上に特異的であるPCRプライマーを含み得る。試薬の他の例としては、目的の遺伝子、またはこれらの目的の遺伝子によりコードされるタンパク質に対する抗体のうちの1つ以上に特異的であるアレイが挙げられる。
【0098】
上の構成要素に加え、本発明のキットは、本発明の方法を実践するための説明書をさらに含む。これらの説明書は、種々な形態で本発明のキットに提示され得、それらのうちの1つ以上は、キットに提示され得る。これらの説明書が提示され得る一形態は、適切な媒体または基材上に印刷された情報として、例えば情報が印刷された用紙(単数または複数)等として、キットのパッケージ内、添付文書内等に提示され得る。また別の手段は、情報が記録されたコンピュータ可読媒体、例えば、ディスケット、CD等である。提示され得るまた別の手段は、移動サイトの情報にアクセスするためにインターネットを介して使用され得るウェブサイトアドレスである。あらゆる都合のよい手段がキットに提示され得る。
【0099】
顕微鏡アレイを備える自動細胞撮像
【0100】
上述の方法の一部は、微速度撮像を介して胚および幹細胞の発達を観測する機能を必要とする。これは、標準のインキュベータの内部に収まる小型の多チャネル顕微鏡アレイから成るシステムを使用して達成され得る。これは、物理的に皿を移動する必要がなく、複数の試料を迅速かつ同時に撮像することを可能にする。図20に示される例示的な原型の1つは、暗視野照明を備える3チャネル顕微鏡から成るが、他の型の照明も使用され得る。「3チャネル」とは、3つの個別の培養皿を同時に撮像する3つの独立した顕微鏡を有することを意味する。ステッパモーターは、3D画像スタックを焦点に合わせる、または取得するために、焦点位置を調節するために使用される。我々はそれでヒトの胚を観測したが、白色光LEDは、照明に使用され、赤色または近赤外線(IR)LEDを使用することにより、細胞膜と細胞の内側部分との間のコントラスト比を改善させることができる。この改善されたコントラスト比は、手動および自動の双方の画像分析を補助することができる。加えて、赤外域へ移動することにより、試料に対する光毒性を低減することができる。画像は、他の型のカメラが使用され得るが、低費用の高解像度のウェブカメラによって捕捉される。
【0101】
図22に示す、上述の原型システムの各顕微鏡は、1〜30までの胚を含有し得る培養皿を撮像するために使用される。顕微鏡は、LEDにより生成されるあらゆる熱を分散させる補助をするヒートシンクに接続された白色光LEDからの光を収集し、これは、短い曝露時間としては非常に小さい。光は、直接光を防ぐための従来の暗視野パッチを通り、集光レンズを通って、培養され、研究される胚を保有する培養皿である「ペトリ皿」と標識された試料上へ達する。培養皿は、胚の順序を維持し、皿がインキュベータへ、およびインキュベータから運ばれる間にそれらが移動するのを防止する補助をするウェルを有し得る。ウェルは、胚が同じ培地液滴を共有できるのに十分に緊密に配置され得る。次いで、分散した光は、顕微鏡の対物レンズを通り、その後、色消しダブレットを通り、CMOSセンサ上へ通過する。CMOSセンサは、デジタルカメラとして機能し、上述の画像分析および追跡のためにコンピュータに接続される。
【0102】
この設計は、非常に多くのチャネルおよび異なる照明技法を提供するために容易に拡張可能であり、試料を供給するための流体デバイスを収容するように変更することができる。加えて、本設計は、フィードバック制御システムと統合することができ、ここで、温度、CO2(対照pHに対して)、および培地等の培養条件がフィードバックおよび撮像データに基づき、リアルタイムで最適化される。このシステムは、ヒトの胚の発達の微速度ビデオを取得するために使用され、これは、生体外受精(IVF)処置における胚の生存能を判定する実用性を有する。他の用途は、幹細胞療法、薬物スクリーニング、および組織工学を含む。
【0103】
デバイスの一実施形態において、照明は、アルミニウムヒートシンンク上に載置され、BuckPuck電流制御ドライバにより電力を供給されたLuxeon白色発光ダイオード(LED)により提供される。LEDからの光は、視準レンズを通過する。視準された光は、その後、図22に示される特注のレーザ加工パッチストップを通過し、非球面集光レンズを使用して中空の光錐に焦点が合わせられる。試料を直接透過する光は、対物レンズによって排除され、一方、試料によって分散される光は、収集される。視界を増大するためにより小さい倍率が使用されるか、または解像度を増加するためにより大きい倍率が使用され得るが、一実施形態において、20X倍率のOlympus対物レンズが使用される。次いで、収集された光は、色消しダブレットレンズ(すなわち、鏡筒)を通過して、色収差および球面収差の作用を低減する。代替的に、撮像対物レンズから収集された光は、鏡筒の代わりとして機能する、対向する方向に向けられた別の対物レンズを通過し得る。一構成において、撮像対物レンズは10X対物レンズであり得る一方、鏡筒対物レンズは4X対物レンズであり得る。得られた画像は、2メガ画素解像度(1600×1200画素)を備えたCMOSセンサにより捕捉される。異なる型のセンサおよび解像度も使用され得る。
【0104】
図23Aは、3つの同一顕微鏡を備える、多チャネル顕微鏡アレイの写真を示す。全ての光学構成要素は、鏡筒に載置される。アレイシステムの操作において、ペトリ皿は、手動の2軸傾斜台上に載置されるアクリルプラットフォーム上に装填され、これは、光学軸に対する画像面の調節を可能にする。これらの台は、顕微鏡の底部に固定され、最初に位置合わせをした後、動かない。LED、コリメータレンズ、パッチストップ、および集光レンズから成る照明モジュールは、位置付け、および照明光の焦点合わせのために、手動のxyz台上に載置される。対物レンズ、色消しレンズ、およびCMOSセンサから成る撮像モジュールも、視界の位置付け、および対物レンズの焦点合わせのために、手動のxyz台上に載置される。撮像モジュールの3つ全ては、移動台に取り付けられ、単一レバーアームにより支持され、これは、ステッパモーターを使用して作動する。これは、コンピュータ制御による焦点合わせおよび画像スタックの自動捕捉を可能にする。他の方法の自動焦点合わせならびに操作が使用され得る。
【0105】
顕微鏡アレイは、図23Bに示されるように、標準のインキュベータ内部に設置された。CMOS画像センサは、USB接続を介してインキュベータの内部に位置する単一ハブに接続され、これは、他の通信および電源線とともに外部PCにつながる。全ての電気ケーブルは、シリコーン接着剤で密封されたゴム製ストッパーの中心を通ってインキュベータを退出する。
【0106】
上述の顕微鏡アレイは、初期のヒトの胚の発達および接合体から胚盤胞期までの確認された成長の微速度画像を記録するために使用された。4つの異なる実験は、合計242個の胚を監視した。この群のうち、100個が最大5または6日目まで撮像され、他は、遺伝子発現分析のために種々の時間点で撮像ステーションから取り出された。画像捕捉ソフトウェアおよび撮像された胚のスクリーンショットを図24に示す。画像は、5分毎に捕捉され、画像当りおよそ1秒の低光曝露であった。試料が受けた総光量は、連続曝露の24分に相当し、取り扱い中、IVFクリニックで経験する総レベルと類似する。画像当り1秒の持続時間の光曝露が低減され得る。ヒトの胚で作業する前に、我々は、胚盤胞形成率および遺伝子発現パターンの双方が撮像プロセスによって影響を受けないことを確実にするために、マウスの着床前の胚で広範な対照実験を実施した。
【0107】
図25および26は、微速度配列からの選択した画像を示す。画像は、1日目、2.5日目、4日目、および5.5日目について示される。図25に示される配列において、9つの胚のうちの3つが胚盤胞に発達し、図26に示される配列において、12の胚のうち5つが胚盤胞に発達した。胚は3日目に培地交換を受けたため、写真視野中のそれらの位置は、移動しているが、個々の胚は、経時的に追跡された。遂次的培地の使用は、胚の発達の期特異的必要条件を満たす必要がある。培地交換中、胚を数分間撮像ステーションから取り出し、新しいペトリ皿に移動した。培地交換中の各胚の同一性の追跡を維持するために、1つの皿から他の皿への試料の移動をビデオテープ録画して、胚が混同されなかったことを確認した。このプロセスは、遺伝子発現分析のために試料の収集中にも使用された。胚の同一性を追跡する課題は、特定の順序で胚を配置するのを補助するためのウェルを使用することにより回避され得る。
【0108】
マイクロウェルを備えたペトリ皿
【0109】
異なるステーション間でペトリ皿を移動する場合、胚が時折移動し、それによって胚の同一性の追跡の維持を困難にする。これは、微速度撮像が1つのステーション上で実施され、胚が胚の選択および移動のために第2ステーションに後に移動される場合、問題となる。一方法は、個々のペトリ皿で胚を培養することである。しかしながら、これは、各胚が独自の培地滴液を有することが必要となる。通常のIVF処置において、患者の胚の全てを同じペトリ皿上で、かつ同じ培地滴液中で培養することが一般に望ましい。この問題に対処するために、我々は、マイクロウェルを備えた特注のペトリ皿を設計した。これは、胚が移動するのを防ぎ、インキュベータまたは撮像ステーションに、またはそこから移動される時に、ペトリ皿上でのそれらの配置を維持する。加えて、ウェルは、それらが同じ培地液滴を共有し、全てが同じ顕微鏡によって同時に表示され得るように十分に小さく、緊密に配置される。各マイクロウェルの底面は、光学品質の仕上げである。図27Aは、一実施形態の寸法を伴う図を示す。この型において、25のマイクロウェルが1.7×1.7mmの視界内に緊密に配置されている。図27Bは、3D表示のマイクロウェルを示し、これは、皿の表面内に約100ミクロン陥凹する。文字、数字、および他の印を含む位置合わせマーカーが、識別を補助するために皿の上に含まれている。
【0110】
本明細書に引用される全ての参考文献は、参照によりそれらの全体が組み込まれる。
【実施例】
【0111】
以下の実施例は、本発明をどのように行い、使用するかの開示、および説明を当業者に提供するために提示され、発明者が、彼らの発明と見なすものの範囲を制限することを意図せず、またそれらは、以下の実験が全て、または唯一の実施された実験であることを表すことを意図しない。使用された数字(例えば、量、温度等)に関する正確性を確実にする試みがなされたが、一部の実験エラーおよび逸脱が考慮されるべきである。特に記載のない限り、部は重量部であり、分子量は重量平均分子量であり、温度は摂氏温度であり、圧力は大気圧または大気圧付近である。
【0112】
試料源
【0113】
本研究に使用された全ての胚は、複数の胎生学者によって多年度にわたり収集され、受精され、凍結保存された。我々の研究における患者当りの胚の平均数は3つであり、所定のIVFセンターで出会った全年齢群を含んだ。特に、これらの実験に使用された胚の全ては、IVF生成され(ICSIとは対照的に)、そのため、胚は、相対的に正常な機能(少なくとも卵丘、帯、および卵細胞膜を透過し、前核を形成するそれらの能力に関して)を有する精子に由来した。刺激プロトコルは、標準のロング・リュープロン・プロトコルであった(cdc.gov/art)。過剰なヒトの胚の凍結保存は、室温(22+2℃)で25分間、凍結用培地(1.5Mの1,2プロパンジオール+0.2Mのスクロース)にそれらを設置することにより達成された。その後、緩慢凍結プロトコル(−1℃/分〜−6.5℃;5分間保持;播種;5分間保持、−0.5℃/分〜−80℃;液体窒素中に浸す)を使用して、胚を凍結した。委員。いかなる保護された健康情報も胚と関連し得ない。
【0114】
凍結保存された大量の胚の組を検証し、次の観測を行った:1)目印に関して、胚は、2細胞への分裂(2日目初期に生じる)、RNA分解の開始(1〜3日目に生じる)、4および8細胞への分裂(それぞれ、2日目後期と3日目後期に生じる)、胚ゲノムの活性(8細胞期の3日目)、および桑実胚および胚盤胞の形成(それぞれ、4日目と5日目に生じる)を含む、正常な胚の発達を示すタイミングを示した。2)胚は、臨床設定で得られる胚に特有である胚盤胞期に達する能力を示した。これは、発達するであろう、および発達しないであろうこれらの「選別」が、1細胞期の凍結保存前に行われないため(3日目または胚盤胞期の発達の後期に凍結保存された胚に特有であるように)、おそらく、胚が2PN期で凍結保存され、IVFクリニックで遭遇した多くの胚を表したという事実による。よって、我々のデータは、これらの胚が通常のIVFクリニックで観測されるものと比較して、類似する胚盤胞形成率を示したことを確認する。3)以前の研究は、新鮮な胚と比較した時、2PN期で凍結される胚が発達、着床、臨床妊娠、および出産において類似する可能性を示すことを示した。他の研究も、凍結卵母細胞について類似する結果を示し、ヒトの胚の発達の最も初期の事象は、凍結保存後、適切な時系列を維持することを示唆する。4)我々は、受精時間または解凍時間に依存しないパラメータに焦点をあてた。我々が測定した最初のパラメータ(第1細胞質分裂の持続時間)は、持続時間が短く(ca10〜15分)、本研究において、受精時間に依存しない(最終結果にかかわらず、全ての胚において、独立して測定され得る)。さらに、全ての後続のパラメータは、この初期の測定点に対して測定され、胚盤胞への発達に成功した胚と発達に失敗した胚との間で比較される。5)最後に、3PNである新鮮な(未凍結)胚は、新鮮な正常な胚と同じ時間枠に沿って発達することが知られていることに留意し、我々は、我々がStanford IVFクリニックより得た新鮮な3PNの胚において、パラメータを比較し、それらが我々の凍結保存された胚、または公表された報告のものと異ならないことを観測した。
【0115】
実験計画
【0116】
4つの実験組において、我々は、242個の前核期の胚の発達(それぞれ、61、80、64、および37個)を追跡した。各実験組において、ヒトの接合体を1日目に解凍し、複数のプレート上で、少数群で培養した。各プレートは、独立して、別の撮像ステーション上で暗視野下、微速度顕微鏡検査で観測された。約24時間間隔で、1プレートの胚を撮像システムから取り出し、高処理リアルタイム定量PCR遺伝子発現分析のために、単一胚、または単一細胞(卵割球)として収集した。各プレートは、通常、採取時に予想された発達期に達した胚(「正常」と称される)と、初期の発達期で停止した、もしくは遅延した、または大規模に断片化したもの(「異常」と称される)との混合物を含有した。胚は、単一のインタクトな胚として分析されるか、または単一の卵割球に解離された後、遺伝子特異的RNA増幅されるかのいずれかであった。胚盤胞形成を監視するために、胚の一部(242個のうち100個)を5または6日目まで撮像した。
【0117】
ヒトの胚培養および顕微鏡検査
【0118】
液体窒素貯蔵タンクからクリオバイアルを取り出すことによって、ヒトの胚を解凍し、それらを室温に設置した。バイアルが解凍したら、開封し、胚を切開顕微鏡下で視覚化した。次いで、バイアルの内容物を3003培養皿の底に注いだ。胚を液滴中に置き、各胚の生存を評価し、記録した。室温で、1.0Mの1,2プロパンジオール+0.2Mのスクロースを含有する3037培養皿に5分間、その後、0.5Mの1,2プロパンジオール+0.2Mのスクロースに5分間、そして0.0Mの1,2プロパンジオール+0.2Mのスクロースに5分間、胚を移動した。続いて、油下、マイクロドロップを使用して、1日目と3日目の間に10%のQuinn’s Advantage血清タンパク質代替物(SPS;CooperSurgical)を添加したQuinn’s Advantage分裂培地(CooperSurgical)、そして3日目後に10%のSPSを含むQuinn’s Advantage胚盤胞培地(CooperSurgical)で胚を培養した。全ての実験は、第1実験中の2つのステーションを除き、同じ種類の分裂培地を使用し、これは、Global培地(LifeGlobal,Guilford,CT)を使用した。この少数集団(12個の胚)において、胚は、わずかに低い胚盤胞形成率(12のうち3つ、または25%)を示したが、我々の予測パラメータの感度および特異性は、双方とも100%であった。
【0119】
複数の試料の同時分析に対応する、ならびに異なるプラットフォーム全体のデータの一貫性を検証するために、複数のシステムで微速度撮像を実施した。システムは、7つの個々の顕微鏡から成る:(1)Tokai Hit加熱台、白色光Luxeon LED、および暗視野照明用のアパーチャを装備した、改良されたOlympus IX−70/71顕微鏡2台、(2)加熱台、白色光Luxeon LED、およびホフマン変調コントラスト照明(注意:これらのシステムは、暗視野照明がパラメータの測定に好ましいと決定された後、4つの実験のうちの最初の間にのみ使用された)を装備した、改良されたOlympus CKX−40/41顕微鏡2台、および(3)白色光Luxeon LEDおよび暗視野照明用のアパーチャを装備した、インキュベータの内部に収まる特注の3チャネル小型顕微鏡アレイ。これらの異なるシステム上で培養された胚の間の発達挙動、胚盤胞形成率、または遺伝子発現プロファイルにおいて有意な相違は観測されず、実際、胚盤胞予測における我々のパラメータは、複数のシステムおよび実験全体において一貫していた。
【0120】
全てのシステムの光強度は、低電力のLED(通常の100Wハロゲン電球と比較して)、および高感度のカメラセンサにより、生殖補助顕微鏡で通常使用される光より有意に低い。光学電力計を使用して、我々は、473nmの波長の通常の生殖補助顕微鏡(Olympus IX−71ホフマン変調コントラスト)が、倍率により、ほぼ7〜10mWの範囲であると判定し、一方、我々の撮像システムの電力は、同じ波長で0.2〜0.3mWの間であると測定された。画像は、最大5または6日目までの間、5分毎に1秒の曝露時間で捕捉され、約24分の連続光曝露をもたらす。0.3mWの電力で、これは、通常の生殖補助顕微鏡下、ほぼ1分の曝露に相当する。
【0121】
相関する撮像、および遺伝子発現実験中の各胚の同一性を追跡するために、我々は、立体顕微鏡上にビデオカメラを取り付け、培地交換および試料収集中の試料の移動プロセスを録画した。我々は、マウス着床前の胚(n=56)、およびヒトの胚の少数集団(n=22)を用いた対照実験を実施し、撮像された胚と対照の胚との間の胚盤胞形成率に有意な相違を(p=0.96)観測しなかった。
【0122】
高処理qRT−PCR分析
【0123】
単一の胚または単一の卵割球qRT−PCR分析において、胚を最初に酸性タイロード液で処理し、透明帯を除去した。単一の卵割球を収集するために、厳密にピペット操作しながら、5〜20分間、37℃でHEPES(CooperSurgical)を含むQuinn’s Advantage Ca2+Mg2+を含まない培地中で胚をインキュベートした。試料を10μlの反応緩衝液中に直接収集し、後続の一工程逆転写/前置増幅反応を前述のように実施した。逆転写および前置増幅反応中、プールした20X ABI assay−on−demand qRT−PCRプライマーおよびプローブミックス(Applied Biosystems)を遺伝子特異的プライマーとして使用した。 前述のように、ABI assay−on−demand qRT−PCRプローブを使用して、Fluidigm Biomark 96.96 Dynamic Arrayを用いて、高処理qRT−PCR反応を実施した。全ての試料を3つまたは4つのテクニカルレプリケート(technical replicate)技術的な複写物に装填した。qRT−PCRデータ分析をqBasePlus (Biogazelle)、Microsoft Excel、および特注のソフトウェアを用いて実施した。データ品質不良(例えば、不良なPCR増幅曲線)、または評価された胚において一貫して低発現〜発現なしであること、のいずれかであれば、特定の遺伝子をデータ分析から排除した。卵割球期の分析において、使用した母性転写パネルは、DAZL、GDF3、IFITM1、STELLAR、SYCP3、VASA、GDF9、PDCD5、ZAR1、およびZP1を含み、一方、胚遺伝子パネルは、ATF7IP、CCNA1、EIF1AX、EIF4A3、H2AFZ、HSP70.1、JARID1B、LSM3、PABPC1、およびSERTAD1を含む。基準遺伝子であるGAPDHおよびRPLP0に対する、ならびに遺伝子平均に対する各遺伝子の発現レベルは、geNormおよびΔΔCt法を使用して計算された。GAPDHおよびRPLP0は、経験的に、GAPDHについては1.18および46%、ならびにRPLP0については1.18および34%の遺伝子安定値および変動係数に基づき、本研究の基準遺伝子として選択された。これらは、我々が試験した10のハウススキーピング遺伝子の中で最も安定しており、十分通常の異種試料の組の範囲内であった。第2に、我々は、予想通り、単一卵割球において、RPLP0およびGAPDHの転写量が、ヒトの発達の最初の3日間のEGA前の新規転写の不在下、1細胞期と8細胞期との間の分割当り約1Ct値減少し、各細胞が各卵割毎にmRNAのプールの約半分を受け継ぐという予測と一致することを観測した。第3に、我々は、EGA開始後、単一の卵割球におけるこれらの基準遺伝子の発現レベルは、8細胞と桑実胚期との間、安定した状態を保ったことに留意した。インタクトな胚のレベルで、RPLP0およびGAPDHの双方のCt値は、発達全体を通して桑実胚期までほとんど一定を保ち、後の胚盤胞期でわずかに増加するが、おそらく存在する多数の卵割球の転写レベルの増加によるものである。この研究で実施された遺伝子発現分析の大半は、桑実胚期前の発達期に焦点をあてたが、その時、基準遺伝子の発現レベルは非常に安定していた。
【0124】
自動化細胞追跡
【0125】
我々の細胞追跡アルゴリズムは、遂次モンテカルロ法に基づく確率的フレームワークを使用し、これは、コンピュータビジョンの分野において、粒子フィルタと呼ばれることが多い。粒子フィルタは、経時的に状態、対照、および測定の3つの主要変数の伝搬を追跡する。状態変数は、胚のモデルであり、楕円の集まりとして表される。対照変数は、状態変数を変換する入力であり、我々の細胞伝搬およびビジョンモデルから成る。測定変数は、状態の観測であり、微速度顕微鏡によって取得された我々の画像から成る。各時間ステップでの現状態の我々の推定は、事後確率分布を用いて表され、これは、粒子と呼ばれる一組の重み付き試料で近似される。我々は、粒子および胚モデルの用語を交換可能に使用し、ここで、粒子は所与の時間の胚モデルの仮定の1つである。初期化後、粒子フィルタは、予測、測定、および更新の3つのステップを繰り返し適用する。
【0126】
予測: 細胞は2D空間で楕円として表され、各細胞は、配向および重複インデックスを有する。重複インデックスは、細胞の相対的高さを指定する。一般に、我々が予測したい2種類の挙動があり、それは、細胞運動と細胞分割である。細胞運動において、我々の対照入力は、粒子を捕え、位置、配向、ならびに主軸および短軸の長さを含む、各細胞の各パラメータを無作為に撹乱する。撹乱は、比較的小さい分散(初期値の5%)の正規分布から無作為に試料サンプリングされる。細胞分割において、我々は以下のアプローチを使用する。所与の時間点で、各粒子において、我々は、細胞のうちの1つが分割するという50%の確率を割り当てる。この値は、経験的に選択され、現在の構成を良好に網羅しながら、広範な可能な細胞分割に及ぶ。分割が予測される場合、分割細胞が無作為に選択される。細胞が分割することが選択される時に、我々は、楕円の主軸に沿って対称分割を適用し、同じ大きさおよび形状の2つの娘細胞を産生する。次いで、我々は、娘細胞の各値を無作為に撹乱する。最後に、我々は、残りの細胞に対するそれらの集合的重複を維持しながら、2つの娘細胞の重複インデックスを無作為に選択する。
【0127】
対照入力を適用した後、我々は、各粒子をシミュレーション画像に変換する。これは、重複インデックスを使用して、各細胞の楕円形状をシミュレーション画像に投影することにより達成される。対応する画素値は、1のバイナリー値に設定され、観測された画像データと比較可能な膜厚を作製するために拡大される。胚は、部分的に透明であり、焦点外の光が収集されるため、胚の底部の細胞膜は、時々しか可視でない。したがって、遮蔽された細胞膜は、10%の確率で加えられる。実践において、我々は、これらの遮蔽された膜点が、正確な形状モデル化に重要であるが、それらが可視縁部に類似しないようにそれらを十分に散在させることが重要であることを発見した。
【0128】
測定:我々が仮定モデルの分布を生成したら、対応するシミュレーション画像を実際の顕微鏡画像と比較する。顕微鏡画像は、主曲率に基づいた方法の後、閾値処理を使用して、細胞膜のバイナリー画像を作製するために前処理される。比較の正確性は、対称切断型チャンバ距離を使用して評価され、これは、次いで、重量または可能性を各粒子に割り当てるために使用される。
【0129】
更新:重量が割り当てられた後、次の反復のために新しい粒子の組を作製するために、これらの重量に比例して粒子を選択する。これは、最も高確率の領域の粒子分布に焦点をあてる。低確率の粒子は破棄され、一方、高確率の粒子は乗算される。粒子の再サンプリングは、低分散法を使用して実施される。
【0130】
胚がモデル化されたら、我々は、本文で説明する、細胞質分裂の持続時間、および有糸分裂間の時間等の動的撮像パラメータを抽出することができる。我々の細胞追跡ソフトウェアは、事前にMatlabに実装され、計算時間は、粒子の数により各画像において数秒から30秒の範囲であった。我々の現在のソフトウェアのバージョンは、Cで実装され、計算時間は、粒子の数により1〜5秒の範囲である。
【0131】
実施例1
胚の発達の可能性を判定するための撮像分析
【0132】
方法
接合体とも称される凍結された1細胞のヒトの胚を解凍し、培養物中に設置し、IVF処置で使用されるような条件下で培養した。上でより詳細に記載するように、これらの胚は、2PN期で凍結され、よって、無差別に凍結保存されたため、通常の生体外受精(IVF)集団を表すと思われる。これは、新規周期中、最高品質であると認知されたものの移動後の発達の後期で通常凍結保存される胚と対称的である。一部の実験において、胚は、標準の培養皿に設置された。他の実験において、胚は、光学品質のマイクロウェルを備える特注の培養皿で培養された。
【0133】
皿当り通常1〜30個の成長胚は、デジタル画像保存および分析用に装備されたコンピュータ制御顕微鏡を用いて、微速度撮像により個別に追跡された。一部の場合において、微速度撮像は、加熱台およびインキュベーションチャンバを装備した倒立顕微鏡を用いて実施された。他の場合において、微速度撮像は、従来のインキュベータの内部に収まる特注の小型顕微鏡アレイを用いて実施され、これは、同じインキュベータ内の複数の試料皿を同時に培養することを可能にし、連続した画像捕捉間の最小時間間隔に対して制限がない複数のチャネルを収容するように拡張可能である。複数の顕微鏡の使用は、試料を移動する必要性を排除し、これは、システムの正確性および全体的なシステムの信頼度を改善した。撮像システムは暗視野を使用し、他の照明が十分であったであろうことが留意されたが、これは、後続の特色抽出および画像分析のために強化された画像コントラストを提供した。インキュベータ中の個々の顕微鏡は、相互に隔離することができ、各培養皿に独自の制御された環境を提供する。これは、他の試料の環境を乱すことなく、皿の撮像ステーションへの、およびそこからの移動を可能にした。
【0134】
微速度画像は、次の細胞パラメータのうちの少なくとも1つの測定を含む、後続の細胞形態の分析のために収集された:第1細胞質分裂の持続時間、第1と第2の細胞分割との間の時間間隔、および第2と第3の細胞分割との間の時間間隔。図に示される画像は、最大5または6日間、5分毎に1秒の曝露時間で撮られた。以下により詳細に説明されるように、第1細胞質分裂は、一般に、受精1日後に生じ、約14分の間続く。第1および第2の細胞分割は、一般に、平均約11時間で分離される。第2および第3の細胞分割は、一般に、平均約1時間で分離される。よって、撮像は、受精後約36時間(数時間の加減)続く期間にわたった。
【0135】
結果
培養における健康なヒトの着床前の胚の発達時系列は、微速度撮像により6日間にわたり記録された(図2)。正常なヒトの接合体は、2日目の初期に第1分裂を受けることが観測された。続いて、胚は、4日目に桑実胚に密集する前に、それぞれ、2日目と3日目の後期に4細胞および8細胞に分裂する。第1の形態学的に明らかな細胞分化は、胚盤胞形成中の5及び6日目に観測され、その時、全能性卵割球は、胎盤のような胚外構造をもたらす栄養外胚葉細胞、または生体内で胎児に、そして生体外で多能性胚幹細胞に発達する、内細胞塊のいずれかに分化する。
【0136】
我々は、次に、4つの独立した実験組で242個の普通に受精させた胚の発達を追跡し、5または6日目まで培養された試料の中で、正常な胚および停止した胚の分布を記録した。242個の胚のうち、100個は、5または6日目まで培養され、胚盤胞形成率は、33%〜53%の間であることが観測され、通常のIVFクリニックでの胚盤胞形成率と類似した(図3)。残りの胚は、異なる発達期で停止し、最も一般的には、2細胞と8細胞期との間であり、異常と定義された(図3)胚盤胞期への胚の発達の成功を予測する定量的撮像パラメータを識別するために、我々は、卵割球の大きさ、透明帯の厚さ、断片化の程度、第1細胞周期の長さ、最初のいくつかの有糸分裂間の時間間隔、および第1細胞質分裂の持続時間を含む、微速度ビデオからいくつかのパラメータを抽出し、分析した。発達的に正常な胚と異常な胚の双方のビデオ画像分析中、我々は、多くの停止した胚が第1細胞分割中、異常な細胞質分裂を受けたことを観測した。正常な胚は、速やかに制御された様式で、分裂溝の出現から娘細胞の完全分離まで14.3+/−6.0分の狭い時間枠で細胞質分裂を完了した。これは図4の上部に示される。逆に、異常な胚は、一般に、2つの異常な細胞質分裂表現型のうちの1つを示した。軽度の表現型において、細胞質分裂の形態およびメカニズムは、正常のように見えたが、プロセスを完了するのに必要とした時間は長く、さらに数分から1時間の範囲であった(図4)。時折、わずかに延長した細胞質分裂を受けた胚でも、胚盤胞に発達した。より重度の表現型において、細胞質分裂の形態およびメカニズムは撹乱された。例えば、図4の下部パネルの例に示されるように、胚は、片側だけ分裂溝を形成し、最終的により小さい構成要素に断片化する前に、数時間、異常な一連の膜ラフリング事象を受けた。そのような挙動の他の変形が観測された。加えて、これらのより重度の表現型を示す異常な胚は断片化され、胚の断片化が後に異常な胚発達をもたらす異常な細胞質分裂の副産物の可能性がある直接的な証拠を提供する。
【0137】
我々の撮像結果のより詳細な分析は、正常な胚が胚の遺伝子活性(EGA)が始まる前の初期の分割中、細胞質分裂および有糸分裂において厳密なタイミングに従うことを示し、胚の発達の可能性が、受け継がれた母性プログラムにより予め決定されることを示唆する。特に、我々は、厳密に制御された初期の胚の細胞周期において、3つの一次的な間隔:(1)第1細胞質分裂の持続時間、(2)第1と第2との間の有糸分裂の時間間隔、および(3)第2と第3との間の有糸分裂の同時発生に留意した。これらの3つの時間間隔と形態変化との間の関係を図5に示す。正常な胚において、我々は、パラメータがそれぞれ約14.3+/−6.0分、11.1+/−2.1時間、および1.0+/−1.6時間であると測定した(ここで、平均+/−標準偏差として与えられる)。
【0138】
我々は、単一細胞期で開始する3PN(三倍体)の少数組(n=10)の新鮮な(凍結保存されていない)胚の撮像を実施した。3PNの胚は、正常な新鮮な胚から少なくとも最初の3つの細胞周期と同様の目印事象の時系列をたどることが示されている。これらの胚は、撮像システムを検証するために(しかし、技術的理由のため、胚盤胞まで遂行されなかった)、我々の主要な実験前に撮像された。この新鮮な胚の組のうち、胚の3つが我々の凍結保存された2PNの胚と類似する事象の時系列をたどり、細胞質分裂の持続時間は、15〜30分の範囲であり、第1と第2の有糸分裂の間の時間は、9.6〜13.8時間の範囲であり、第2と第3の有糸分裂との間の時間は、0.3〜1.0時間の範囲であった。しかしながら、胚の7つにおいて、我々は、3つの分裂溝の同時出現、わずかに延長した細胞質分裂、および最終的な3つの娘細胞への分離を特徴とする固有の細胞質分裂表現型を観測した(図4)。これらの胚は、15〜70分の範囲の細胞質分裂の持続時間(分裂溝の開始から3つの娘細胞への完全分離までの間の時間を特徴とする)、8.7〜12.7時間の範囲の第1と第2の有糸分裂(3細胞〜4細胞)の間の時間、および0.3〜2.6時間の範囲の第2と第3の有糸分裂(4細胞〜5細胞)との間の時間を有する。この観測は、異常な胚によって示される多種多様な細胞質分裂表現型とともに、我々の凍結保存された胚が凍結保存プロセスによって発達的に遅れたわけではなく、2つの卵割球に分裂する新鮮な接合体と類似して挙動することを示唆する。
【0139】
0〜33分の間の第1細胞質分裂、7.8〜14.3時間の間の第1と第2の有糸分裂の間の時間、および0〜5.8時間の間の第2と第3の有糸分裂との間の時間を有することにより、それぞれ94%および93%の感度および特異性で、胚盤胞期に達した胚を予測することができた(図6)。逆に、これらの枠のうちの1つ以上外の値を示した胚は、停止することが予測された。胚盤胞の発達に成功した全ての正常な胚は、3つ全てのパラメータにおいて、類似する値を示した。逆に、異常な胚は、それらが間隔を完了するのに費やした時間の長さにおいて、かなりの量のばらつきを示した(図6)。我々は、(1)第1細胞質分裂を完了する期間が正常より長いと、不良な発達の可能性を示す、(2)第1と第2の細胞分割との間の間隔が正常より長い、もしくは短いと、不良な発達の可能性を示す、および(3)第2と第3の細胞分割との間の間隔が正常より長いと、不良な発達の可能性を示すことを観測した。よって、これらのパラメータは、胚が胚盤胞形成に進む能力、および胚盤胞の質を予測した。
【0140】
最後に、我々は、各パラメータは自動的に胚の発達の可能性を予測したが、3つ全てのパラメータの使用は、双方が90%を超えた感度および特異性を提供し、標準偏差の3倍のカットオフ点であったことに留意した。これらのパラメータの受信者操作特性(ROC)曲線を図7に示す。この図の曲線は、種々の標準偏差カットオフの真の陽性率(感度)対偽の陽性率(1−特異性)を示す。このROCを導き出すために、以下の数字を使用した:真の陽性の数=34(胚盤胞に達することを正確に予測された);真の陰性の数=54(停止することを正確に予測された);偽の陽性の数=4(胚盤胞に達することを不正確に予測された);偽の陰性の数=2(停止することを不正確に予測された)。
【0141】
考察
我々の分析は、最初の3つの卵割の間の有糸分裂および細胞質分裂において、厳密なタイミングに従う胚は、胚盤胞期に発達すること、拡張内細胞塊(ICM)を伴う高品質の胚盤胞を形成することの双方の可能性が非常に高いことを示す。動的形態パラメータは、IVF処置中、移植または凍結保存に最適な胚を選択するために使用され得る。これらのパラメータは、異なる質の胚盤胞を区別するためにも使用され得、群内の相対的な胚の発達の可能性のランク付けを可能にする。IVFクリニックにおける標準的な実践は、8細胞期(3日目)で移植することである。胚盤胞移植は、3日目の移植と比較して、最大2倍の着床率を有するため、一部のクリニックは、胚を胚盤胞期(5日目)まで培養することを選択する。しかしながら、多くのクリニックは、後成的障害のリスクが増加するため、延長した培養を避けている。予測的撮像パラメータは、4細胞期(2日目)まで、および胚遺伝子活性前の胚の生存能を予測するために使用され得る。これは、通常実践されるより丸1日早く、胚がそれらの分子プログラムの著しい変化を受ける前の胚の移植または凍結保存を可能にする。これは、一番最適な胚がPGDまたは他の種類の分析用に選択されることも可能にする。
【0142】
実施例2
遺伝子発現分析を通した撮像パラメータの検証、および発達の可能性を判定するための遺伝子発分析の使用
【0143】
方法
接合体とも称される凍結された1細胞のヒトの胚を解凍し、培養物中に設置し、IVF処置で使用されるような条件下で培養した。一部の実験において、胚を標準の培養皿に設置した。他の実験において、胚は、光学品質のマイクロウェルを備える特注の培養皿で培養された。
【0144】
胚を培養および撮像システムから取り出し、遺伝子発現分析用に、単一胚または単一細胞(卵割球)のいずれかとして収集した。各プレートは、通常、胚の混合物を含有し、一部は採取時に予想された発達期に達し、他は初期の発達期で停止したか、大規模に断片化した。採取時に予想された発達期に達した胚は、「正常」と分類され、一方、停止した胚は、「異常」とみなされた。例えば、試料収集のために、プレートの胚を2日目後期に撮像ステーションから取り出した時に、4細胞期およびそれ以降に達したいずれかの胚は正常と識別され、一方、4細胞期に達しなかった胚は停止と標識されたであろう。これらの停止した胚は、それらが停止した発達期により分類されるため、2日目後期で2つの卵割球のみの胚は、停止した2細胞胚として分析されたであろう。試料収集時に形態的に死滅し、多孔質のように見える胚(例えば、卵割球の退化)を排除するよう注意を払った。生存しているように見える胚(正常および停止の双方において)のみを遺伝子発現分析に使用した。しかしながら、収集時中に正常に見えた胚が、それらが後期に成長可能であっても、最終的に停止する可能性もあり得る。これらの分類のそれぞれを代表する胚の遺伝子発現分析は、定量的RT−PCR(qRT−PCR)により実施された。約24時間間隔で、高処理qRT−PCR遺伝子発現分析用に、胚を個々の撮像システムから収集し、最大96の遺伝子の多重反応を96の試料に対してアッセイした。遺伝子発現分析をFluidigm Biomark Systemを用いて実施し、これは、ナノリットル量で、最大9216の同時TaqMan assay−based qRT−PCR反応を実行できる。
【0145】
結果
形態事象の根底にあり得る分子メカニズムを解明するために、我々は、相関した遺伝子発現プロファイリングを実施した。ハウスキーピング遺伝子、生殖細胞マーカー、母性因子、EGAマーカー、栄養膜マーカー、内細胞塊マーカー、多能性マーカー、後成的制御因子、転写因子、ホルモン受容体、およびその他を含む、異なる分類に属する96の異なる遺伝子の発現レベルを試料毎にアッセイした(図19の表1)。わずかに異なるが重複する2つの組の遺伝子を2つの異なる実験組でアッセイし、ヒトの胚の運命についての固有の遺伝子診断の組を提供する。固有の遺伝子組は、モデル生物からの胚、またはヒトの胚性幹細胞に関するデータ、ならびに我々の未公開マイクロアレイデータから収集された。ヒトの着床前の胚におけるこれらの遺伝子組の発現状態は、この研究において始めて明らかにされる。
【0146】
基準遺伝子のGAPDHおよびRPLPOに対する、ならびに遺伝子平均に対する各遺伝子の発現値は、geNorm(El−Toukhy T,et al.(2009)Hum Reprod)およびAACt(Vanneste E,et al.(2009)Nat Med15:577−83)法を使用して計算された。遺伝子安定値および変動係数は、GAPDHにおいて1.18および46%であり、RPLPOにおいて1.18および34%であり、我々が試験を行った10のハウスキーピング遺伝子の中で最も安定しており、十分に通常の異種試料組の範囲内であった。単一の卵割球において、予想通り、RPLPOおよびGAPDHの転写量は、1細胞期と8細胞期との間の分割当りおよそ1Ct値減少し、これは、卵割の二分作用ならびに最初の3日のヒトの発達中のEGAの欠損による。単一の卵割球におけるこれらの基準遺伝子の発現レベルは、8細胞から桑実胚期との間、安定した状態を保った。全胚レベルで、RPLPOおよびGAPDHの双方のCt値は、桑実胚期までの発達全体にわたり非常に一定であった。RPLPOおよびGAPDHの発現レベルは、胚盤胞において有意に増加し、これは、存在する卵割球の数の増加による可能性が高い。これら変動は、RPLPOおよびGAPDHならびに基準遺伝子の有効性に影響を及ぼさなかった。この研究で実施された遺伝子発現分析の大半は、桑実胚期前の発達期に焦点をあてており、その時、基準遺伝子の発現レベルは非常に安定していた。
【0147】
正常な胚と異常な胚との間の差次的遺伝子発現 図8は、対数目盛上のレーダーグラフにプロットした、6つの異常な1〜2細胞胚、および5つの正常な1〜2細胞胚からの52の遺伝子の平均発現レベルを示す。停止した胚は、一般に、正常な胚と比較して、mRNA量の減少を示し、細胞質分裂、RNAプロセシング、およびmiRNA生合成を促進した遺伝子が最も強く影響を受けた。星印で強調された遺伝子は、マンホイットニー検定により判定された、正常な胚と異常な胚との間に、統計的に有意な相違(p<0.05)を示す。これらの18の遺伝子は、コフィリン、DIAPH1、ECT2、MYLC2、DGCR8、ダイサー、TARBP2、CPEB1、シンプレキン、YBX2、ZAR1、CTNNB1、DNMT3B、TERT、YY1、IFGR2、BTF3およびNELFである。各遺伝子は、図に示される群に属し、それは、細胞質分裂:コフィリン、DIAPH1、ECT2、およびMYCL2;miRNA生合成:DGCR8、ダイサー、およびTARBP2;RNAプロセシング:YBX2;母性因子:ZAR1;ハウスキーピング:CTNNB1;多能性:DNMT3B、TERT、およびYY1;受容体:IGFR2;ならびに転写因子:BTF3およびNELFである。大半の場合、これらの遺伝子の発現は、停止した1および2細胞胚より正常な1および2細胞胚で高かった。
【0148】
興味深いことに、特定の遺伝子分類は、他より異常な胚でより影響を受けた。例えば、異常な胚において、ハウスキーピング遺伝子、ホルモン受容体、および母性因子の大半は、遺伝子発現において大きく変更されず、一方、細胞質分裂およびmiRNA生合成に関与する多くの遺伝子は、有意に減少した発現を示した。さらに、影響を受けた遺伝子の中で、一部の遺伝子が他より正常な胚と異常なとの間で非常に大きな相違を示した。例えば、DGCR8、ダイサー、およびTARBP2等の、miRNA生合成経路に関与する遺伝子は、異常な胚において、高度に減少した発現レベルを示した。特に、最も強く影響を受けた2つの遺伝子であるCPEB1およびシンプレキンは、転写のポリ(A)テールの長さを操作することにより母系mRNA保存および再活性化を調節する同じ分子メカニズムに属した(Bettegowda,A.et al.(2007)Front.Biosci.12:3713−3726)。これらのデータは、胚の異常性は、胚のmRNA調節プログラムの欠損と相関することを示唆する。
【0149】
細胞質分裂 の遺伝子発現プロファイルとの相関 遺伝子発現分析を主要な細胞質分裂構成要素をコードした遺伝子を用いて実施した。各胚の同一性は、立体顕微鏡上にカメラを取り付け、培地交換および試料収集中の試料移動のプロセスをビデオ撮影することにより追跡された。異常な胚の遺伝子発現プロファイルを評価する時に、我々は、主要な細胞質分裂構成要素において、異常な細胞質分裂と低遺伝子発現レベルとの間に強力な相関関係を観測した。興味深いことに、異常な胚の遺伝子発現プロファイルは、それらの異常な形態学的表現型と同じくらい多様であり、可変であった。
【0150】
細胞質分裂遺伝子発現は、正常な2細胞胚と異常な2細胞胚との関係(図9)、および正常と異常な4細胞胚との関係(図10)では変動したことが判明した。図9および10は、正常な2細胞のヒトの胚において(図9)、および正常な4細胞胚(図10)において最も高度に発現した遺伝子の相対的発現を示し、異なる細胞質分裂表現型と相関した。図9に示すように、第1細胞質分裂中に異常な膜ラフリングを示した停止した2細胞胚は、試験した全ての細胞質分裂調節遺伝子において有意に減少した発現レベルを有した。図9で相違を示した遺伝子は、アニリン、コフィリン、DIAPH1、DIAPH2、DNM2、ECT2、MKLP2、MYCL2、およびRhoAである。正常な発現レベルは、右側の棒に示され、各遺伝子において高いことが分かる。図9のグラフ上の写真において、異常な2細胞胚を示し、スケールバーは50μmを表す。図10は、片側のみの細胞質分裂溝を有する異常な細胞質分裂を受けた停止した4細胞胚からの結果を示し、第1分割中の非常に延長した細胞質分裂は、細胞質分裂制御因子であるアニリンおよびECT2において減少した発現を示した。図10のスケールバーも50μmを表す。
【0151】
胚期特異的遺伝子発現パターン 図11は、141個の正常に発達した単一胚および単一卵割球の遺伝子発現分析中に識別された、4つの胚期特定パターン(ESSP)を示す。4つのESSPのうちのそれぞれ1つに該当する遺伝子を表2に列挙する(図20)。図11のプロットは、類似する発現パターンに基づく分類、およびそれらの発現値の平均化(基準遺伝子に対して)により作製された。ESSPの相対的発現レベルは、類似する発現パターンを有する遺伝子の発現レベルを平均化することによって計算された。遺伝子発現レベルは、異なる細胞期、すなわち、1c=1細胞、M=桑実胚、B=胚盤胞に対してプロットされる。図11において、4つのESSPのそれぞれにおける遺伝子の相対的発現は、1細胞(1c)から桑実胚および胚盤胞までの発現の関数として示される。ESSP1は母性的遺伝を示し、ESSP2は遺伝子転写活性を示し、ESSP3は後期活性を示し、ESSP4は持続的転写を示す。ESSP2に示されるように、IVFクリニックにおける移植点は3日目に生じ、その時、胚は、胚遺伝子活性により著しい発達変化を受けている。微速度画像データは、胚の発達の可能性が4細胞期までに識別され得、それによって2日目、およびこの遺伝子活性前の早期胚の移植を可能にする。この早期移植は、IVF処置の成功率を改善するのに有用である。
【0152】
表2(図20)は、識別された4つのESSPのそれぞれに属する遺伝子を列挙する。各遺伝子の相対的遺伝子発現レベルは、基準遺伝子(GAPDHおよびRPLPO)に対して、および遺伝子平均に対して計算された。胚の発達の時系列に対する各遺伝子の発現パターンは、次の4つのESSPのうちの1つに従った:ESSPパターン(1)初期:始めは高く、ゆっくり低下し、胚盤胞前に止まる遺伝子;ESSPパターン(2)中期:4細胞期後に始まる遺伝子;ESSPパターン(3)後期:桑実胚または胚盤胞で始まる遺伝子;およびESSPパターン(4)一定:比較的一定した発現値を有する遺伝子。
【0153】
ESSP1は、母性的に受け継がれた遺伝子のパターンを示す。これらの転写は 接合体期で高い発現レベルで開始し、続いて、胚が胚盤胞に発達するにつれて低下した。これらの転写の半減期は、約21時間であった。GDF9およびZAR1等の他のモデル生物、ならびに生殖細胞(卵母細胞)特異的遺伝子であるVASAおよびDAZLからの旧知の母性因子は、このカテゴリーに分類された。ESSP2は胚活性化遺伝子を含み、これは、4細胞期後、最初に胚において転写された。このカテゴリーにおける一部の遺伝子は、2つの活性化の波を示すように見え、第1のより小さい波は、5〜6細胞期のときであり、第2のより大きい波は8細胞期のときである。EIF1AX31およびJARID1 B32等の他のモデル生物からの既知のEGA遺伝子は、このカテゴリーに分類された。ESSP3は、胚盤胞期まで発現しなかった遅い活性化遺伝子から成っており、栄養膜マーカーGCM1を含む。ESSP4は、発達全体を通して、基準遺伝子に対して安定した発現を維持した持続的転写を含有した。これらの遺伝子の半減期は193時間であり、ESSP1より約9倍長かった。このカテゴリーは、ハウスキーピング遺伝子、転写因子、後成的制御因子、ホルモン受容体およびその他の混合物を含んだ。これらの遺伝子発現の4パターンは、61個の単一の正常な胚および卵割球の試料を使用した別の実験組でも確認された。
【0154】
第1分割中に異常な細胞質分裂および有糸分裂挙動を示す異常な胚は、特に胚性RNA操作に関与する遺伝子において、高度に不安定な遺伝子発現プロファイルと相関した。よって、これらの方法論を混合して、着床前の胚の生存能を予測するために使用され得る方法を提供することができる。結果は、異常な胚が、RNAプロセシングおよびmiRNA生合成における欠損プログラムと共に一生を開始し、母性mRNAの過剰な分解を生じることを示唆する。そのような未制御のRNA分解の確率的性質は、転写の不作為な破壊をもたらし、異常な胚において観測される広範な異常表現型を生じる。減少したレベルのmiRNAは、制御された母性RNA分解において欠損を生じ、異なる期で発達停止をもたらす。
【0155】
個々の卵割球分析 ヒトの着床前の胚において、分子分化がいつ始まるかを評価するために、異なる発達期の17個の胚から採取した単一卵割球のCDX2の発現レベルを分析した。図12Aは、2細胞から胚盤胞までの発達期の関数として、CTBBN1(暗色の棒)およびCDX2(薄色の棒)の2つの遺伝子の相対的発現レベルを示す。見て分かるように、CDX2は、4細胞期前の胚からの一部の単一卵割球において散発的に低レベルで発現した(図12A)。しかしながら、6細胞期以降、どの胚も、有意なレベルでCDX2を発現した少なくとも1つの卵割球を含有した。同じく図12Aに示されるハウスキーピング遺伝子のCTNNB1の発現レベルは、同じ胚からの卵割球の間で一定しており、CDX2の異種発現パターンがqPCR人工産物ではないことを示した。独立した実験からのデータは、類似する観測を示す。これらの結果は、ヒトの着床前の胚における分子分化が早ければ4細胞期の直後に生じる可能性があることを示す。
【0156】
興味深いことに、単一卵割球の遺伝子発現プロファイルの検査は、異なる発達期に対応する遺伝子発現特性を有する卵割球を含有した胚を明らかにした。いずれかの所与の時間でのいずれかの所与の胚の遺伝子発現プロファイルは、母性mRNA分解およびEGAの総和に相当する。初期の発達年齢の若い卵割球は、通常、多量の母性転写と、少量の接合体遺伝子とを含有し、その反対が、より進んだ発達年齢の老いた卵割球において当てはまる。この実験において、母性プログラムは、10のESSP1マーカー(母性転写)の平均発現値として定義され、胚プログラムは、10のESSP2マーカー(胚転写)の平均発現値により定義される。使用された母性転写パネルは、DAZL、GDF3、IFITM1、STELLAR、SYCP3、VASA、GDF9、PDCD5、ZAR1、およびZP1を含み、一方、胚遺伝子パネルは、ATF7IP、CCNA1、EIF1 AX、EIF4A3、H2AFZ、HSP70.1、JARID1 B、LSM3、PABPC1、およびSERTAD1を含む。この特定の8細胞胚から収集に成功した6つの卵割球のうち、3つの卵割球が正常な3細胞胚試料からの卵割球に類似する遺伝子発現特性を示し、一方、他の3つの卵割球は、正常な8細胞胚試料からの卵割球に類似した(図12B)。この観測の最も可能性のある説明は、胚内の細胞の亜集団の停止である。この部分的な停止表現型は、我々が試験した試料の中の別の9細胞胚および2つの桑実胚でも観測された。それらの正常な姉妹細胞と同じ時間量を培養で費やした停止した卵割球において、母性転写レベルが高いままであったという事実は、母性RNAの分解が、単に時間とともに生じる自発的なプロセスではなく、ミクロRNA(miRNA)等の特定のRNA分解メカニズムの機能を必要とする可能性が高いことを示す。これらのデータは、母性mRNA分解が哺乳類の胚形成中の保存された発達事象であり、正常な胚の発達に必要であるというさらなる証拠も提供する(Bettegowda,A.,et al.(2008)Reprod.Fertil.Dev.20:45−53)。加えて、これらのデータは、胚における個別の卵割球が自律的であり、相互に独立して発達できることを示唆する。さらに、これらの結果は、試験される細胞のmRNAのレベル(遺伝子発現レベルを示す)について試験するために、本明細書に記載される遺伝子発現レベル試験を使用することができることを示し、ここで、RNAは、母性プログラムの一部であることが知られている遺伝子であり、胚発達の後期におけるそのような発現レベルの持続性は、異常転帰の可能性と相関するか、または胚プログラムの一部であり、ここで、経時的な不在は、異常な転帰の可能性を示す。本明細書で検証された母性プログラム遺伝子は、ZAR1、PDCD5、NLRP5、H5F1、GDF9、およびBNC2である。他の母性作用遺伝子も知られており、使用することができる。
【0157】
胚遺伝子活性化 本方法は、少なくとも一部、異常な発達的に停止した胚が、EGA(胚遺伝子活性化)が生じる前の最初の3分割中、異常な細胞質分裂および有糸分裂タイミングを頻繁に示すという見解に基づく。これは、胚の発達の運命が、母性遺伝によって大きく判定され、200134のHARDYらにより実施されたヒトの着床前の発達の数学的モデルに顕著に従う見解である。さらに、細胞質分裂および有糸分裂の異常性は、MIRNA生合成および母性MRNAマスキング、保存および再活性化を調節する遺伝子の母性転写の減少したレベルと強く相関する。MIRNAは、生物の発達および分化を含む、多様な生物学的プロセスにおけるMRNA分解を促進することにより翻訳を調節する(BLAKAJ,A.&LIN,H.(2008)J.BIOL.CHEM.283:9505−9508、STEFANI,G.&SLACK,F.J.(2008)NAT.REV.MOL.CELL BIOL.9:219−230)。モデル生物からの増加した証拠は、MIRNAが初期の胚における母性転写分解の主要な制御因子であり得ることを示す(BETTEGOWDA,A.,ET AL.(2008)REPROD.FERTIL.DEV.20:45−53)。よって、MIRNA生合成における欠損は、異常な胚発達をもたらす可能性がある。一方、母性MRNAを適切に操作できないことも不良な胚形成をもたらす。哺乳類の卵母細胞は、母親の出産前に初期の胚成長を支持するのに必要な母性RNA転写物の大規模なプールを合成する。これらの転写物は、それらが受精後再活性化されるまで、圧縮され、長期間の間保存される。この母性RNA操作プログラムの欠損は、母性転写物の量および質に影響を及ぼす可能性があり、よって、発達が成功する可能性を危うくする。
【0158】
胚生存能を評価するためのモデル 図13は、相関する撮像および分子分析に基づくヒトの胚の発達のモデルを示す。胚盤胞への発達の成功を予測するための重要な短時間、および胚発達の図を含む、接合体から胚盤胞までの発達の時系列を示す。図の主要な分子データは、ヒトの胚が母親から受け継いだ独特の卵母細胞RNAの組により一生を開始することを示す。このRNAの組が、卵子の特異的RNA操作プログラムによって維持され、適切にパッケージされる。受精後、卵母細胞から胚への変移が始まる時に、卵子に特異的な一部の母性RNA(ESSP1;胚期特異的パターン1)の分解が分解されなければならない。並行して、発達が続くと、他のRNAが各卵割球に理想的に等しく分配される(ESSP4)。分解の成功とRNAの分配は、細胞自律的様式で、胚ゲノム活性化(EGA)、およびESSP2の遺伝子の転写に達する。特に、卵割中、胚性卵割球は、独立して、停止、または進行し得る。胚における細胞自律性発達の転帰は、個々の卵割球が停止または進行し得ることであり、8細胞胚が桑実胚期およびそれ以降に進行すると、胚盤胞の質は、停止した、または8細胞以降に進行した細胞の数によって影響を受ける。撮像データは、成功または失敗を予測する重要な発達期間があることを示す:第1細胞質分裂、第2卵割、および第2および第3の卵割の同時発生。これらのパラメータは、前述の細胞追跡アルゴリズムおよびソフトウェアを使用して、自動的に測定され得る。記載されるシステムおよび方法は、主要な撮像予測装置を用いて胚の転帰を診断するために使用され得、発達初期(EGA前)により少ない胚の移植を可能にする。
【0159】
実施例3
卵母細胞成熟および後続の胚の発達の撮像
【0160】
結果
現在のIVF処置の主な制限の1つは、卵母細胞の質および利用可能度である。例えば、現在のIVFプロトコルは、小さい周期プールから卵母細胞を動員し、受精に少数の卵母細胞(例えば、1〜20)を提供する。さらに、IVF処置中、ホルモン刺激後に回収された約20%の卵母細胞が未成熟として分類され、通常、現在の培養条件下において、胚の発達の可能性が低いため破棄される。
【0161】
卵母細胞プールを増加する一方法は、生体外成熟を通してである。図14は、胚胞、中期I、および中期IIを含む、生体外成熟中の3つの発達期を示す。胚胞および中期I期は未成熟卵母細胞として分類され、一方、中期IIは第1極体の存在のため、成熟として分類され、これは、生体外成熟開始24〜48時間後に生じる。図15は、生体外で成熟された卵母細胞の胚の発達を示す。
【0162】
卵母細胞プールを増加する別の方法は、一次および二次プールから卵母細胞を動員し、最大数千の卵母細胞を提供する。この処置において、休止状態の卵胞を卵巣から動員し、生体外でプログラムし、正常な染色体構成、後成的状態、RNA発現、および形態を有する卵母細胞を産生する。他の態様において、卵母細胞は、生体外で生殖細胞に分化され、ヒトの卵母細胞に成熟させた多能性幹細胞に由来し得る。
【0163】
図14に図示する、生体外卵母細胞の成熟プロセスは、本発明の方法における測定値および分析用の細胞パラメータを定義するために使用され得るいくつかの細胞変化によって特徴付けられる。これらは、例えば、卵母細胞膜の形態の変化、例えば、透明帯からの分離の速度および程度;卵母細胞核の形態の変化、例えば、胚胞崩壊(GVBD)の開始、完了、および速度;細胞質および核における顆粒移動の速度および方向;ならびに第1極体の移動および/または突出を含む。
【0164】
実施例4
幹細胞分化の撮像
【0165】
結果
微速度画像分析は、生存能、発達の可能性、および幹細胞、誘発された多能性幹細胞(iPSC)およびヒト胚性幹細胞(hESC)等の、他の細胞種の転帰を評価するためにも使用され得る。幹細胞の発達の可能性は、微速度画像分析を使用して、細胞発達および分化中の形態変化を測定することにより評価され得る(図17)。次いで、分化した細胞は、生体内移植または他の使用のために、分析および選択され得る。細胞質分裂の持続時間、有糸分裂事象間の時間、細胞の大きさおよび形状、細胞の数、細胞の運動、分割パターン、分化、非対称分割(ここで、1つの娘細胞が幹細胞を維持し、一方、他は分化する)、対称分割(ここで、双方の娘細胞が幹細胞として留まるか、または双方が分化するかのいずれか)、および運命特定(幹細胞がいつ分化するかを正確に判定する)等の、幹細胞のいくつかのパラメータが、微速度画像データから抽出され、分析され得る。
【0166】
幹細胞療法の基本式は、未分化幹細胞が生体外で培養され、特定の細胞種に分化され、続いて、損傷した組織および/または臓器の再生のために受給者に移植され得ることである。微速度画像分析は、成熟組織への組み込みが可能である非腫瘍原性の分化した後代を形成する幹細胞を識別するために、高処理の非侵襲性デバイスとして使用され得る。潜在的な用途は、アルツハイマー病およびパーキンソン病等の神経障害、血管系障害、および心臓病、関節炎等の筋肉および骨格障害、自己免疫疾患、および癌の治療、ならびに標的および新規薬物療法を評価することによる創薬を含む。
【0167】
ヒトにおいて、損傷した組織は、一般に、体の幹細胞からの連続動員および分化により置換される。しかしながら、体の再生するための能力は加齢とともに低下する。この一例は、括約筋欠損による尿失禁である。筋線維の数および神経密度は年齢とともに減少するため、加齢は、括約筋欠損の主な原因の一つであると考えられている。失禁を有する患者を治療するために、iPSCは、分化した平滑筋細胞を産生するために、膣壁組織から培養された線維芽細胞に由来し得る。次いで、これらの分化した細胞は、生体内に移植され得る。移植前に、多分化能、分化、メチル化、および腫瘍形成能に関してiPSCを特徴付けるために、微速度画像分析が使用され得る。他の用途は、パーキンソン病の患者の皮膚細胞に由来し、移植用にニューロンに分化されたiPSCの微速度撮像を含む(図18)。
【0168】
実施例5
【0169】
自動化分析による撮像パラメータの検証
【0170】
我々の微速度画像データによって明らかなように、ヒトの胚の発達は、コホート内の胚間で高度に変動するプロセスであり、胚は、細胞分割中に広範な挙動を示し得る。よって、高度に異常な細胞質分裂(図4)の持続時間等のある発達事象の手動特徴付けは、解釈に左右され得る。我々の撮像パラメータおよび胚盤胞形成を系統的に予測する機能を検証するために、我々は、最大4細胞期までの細胞分割の自動化追跡用のアルゴリズムを開発した。我々の追跡アルゴリズムは、遂次モンテカルロ法に基づく確率モデル推定技法を利用する。この技法は、仮定された胚モデルの分布を生成し、単純な光学モデルに基づき画像をシミュレーションし、これらのシミュレーションを観測された画像データと比較することにより機能する(図21a)。
【0171】
胚は、位置、配向、および重複インデックスを伴う楕円の集まりとして(細胞の相対的高さを表すため)モデル化された。これらのモデルを用いて、細胞質分裂の持続時間および有糸分裂間の時間を抽出することができる。細胞質分裂は、通常、最初の細胞質分裂溝の出現(ここで、双極陥凹が分裂軸に沿って形成される)から娘細胞の完全分離までにより定義される。我々は、1細胞から2細胞分割前の細胞伸長の持続時間として、細胞質分裂を近似することにより問題を簡素化した。軸長の相違が15%を超えた場合(経験的に選択される)、細胞が伸長したと判断される。有糸分裂間の時間は、各モデルの細胞数を数えることにより容易に抽出される。
【0172】
我々は、14個一組のヒトの胚(図21b)に対して我々のアルゴリズムを試験し、自動化測定値を手動画像分析と比較した(図21c、図21d)。このデータ組において、14個の胚のうちの8つが良好な形態で胚盤胞期に達した(図21e上部)。自動化測定値は、手動測定値と密接に一致し、8つの胚全てが、胚盤胞に達するように正確に予測された。14個の胚のうちの2つが不良な形態で胚盤胞に達した(不良な質の内細胞塊、図21e下部)。これらの胚において、手動評価は、1つが胚盤胞に達し、1つが停止するであろうと示され、一方、自動化評価は、双方が停止するであろうと予測された。最後に、14個の胚のうちの4つが胚盤胞期前に停止し、双方の方法により、停止することが正確に予測された。
【0173】
粒子フィルタフレームワーク
【0174】
粒子フィルタは、モンテカルロシミュレーションに基づくモデル推定技法である。それは、仮定モデルの分布を生成し、これらのモデルを観測されたデータと比較することによって未知または「隠れ」モデルを推定するために使用される。任意運動力学および測定値の不確実性に対応するその能力によって、それは細胞分割の追跡に対して理想的な候補となる。
【0175】
粒子フィルタは、経時的に、状態x、対照u、および測定値zの3つの主要変数の伝搬を追跡する。状態変数xは、我々が推定したい胚のモデルであり、楕円(2Dにおいて)または楕円体(3Dにおいて)の集まりとして表される。対照変数uは、状態変数を変換する入力であり、我々の細胞伝搬および分割モデルから成る。測定値変数zは、状態の観測であり、微速度顕微鏡によって取得された我々の画像から成る。これらのパラメータは、以下のセクションでより詳細に説明される。
【0176】
各時間ステップtにおける現在の状態xの推定は、事後確率分布で表される。この事後は、時折、信念とも称され、現在の状態xt、所与の全ての過去の画像測定値z1:t、および過去の対照u1:tの条件付確率として定義される。
【数1】
【0177】
粒子フィルタは、事後を一組の重み付き試料、または粒子によって近似し、
【数2】
として示される。
【0178】
ここで、Mは、粒子の数である。粒子および胚モデルという用語は、本明細書において交換可能に使用される。よって、単一の粒子xt[m](ここで、1<=m<=M)は、時間tにおける胚モデルの仮定の1つである。
【0179】
初期化後、粒子フィルタは、3ステップを繰り返し適用する。第1ステップは予測であり、ここで、各粒子は、対照入力
【数3】
を使用して伝搬される。
【0180】
得られた粒子の組は、事前確率の近似である。第2ステップは測定値更新であり、ここで、各粒子は、現在の測定値の確率に対応する重要な重さを割り当てられる。
【数4】
【0181】
重み付き粒子の組は、事後bel(xt)の近似である。
【0182】
粒子フィルタの主要な構成要素は、第3ステップに入り、ここで、粒子の組がそれらの重さによって再サンプリングされる。この再サンプリングステップは、最高確率の領域における粒子分布に焦点をあてる。
【0183】
細胞表示
【0184】
細胞は、2D空間において楕円として表示される。各細胞は、デカルト座標において、主軸、短軸、および2次元位置を有し、等式:
【数5】
により与えられる。
【0185】
各楕円は、向かう方向θ(yaw)も有し、これは、楕円にx−y平面で回転することを可能にする。楕円は、ほぼ常に相互に重複するため、我々は、重複インデックスhも示し、これは、重複の順序(または細胞の相対的高さ)を指定する。時間tにおける各胚モデルのパラメータは、したがって、
【数6】
として与えられる。
【0186】
ここで、Nは、そのモデルにおける細胞の数である。
【0187】
細胞の撹乱および分割
【0188】
粒子フィルタの第1ステップは予測であり、ここで、各粒子は、対照入力を使用して伝搬される。我々の用途において、我々がモデル化したい挙動が2種類ある。第1の挙動の種類は細胞運動を含み、これは、翻訳、yaw角周囲の回転、および主軸および短軸の長さの変化を含む。第2の挙動の種類は細胞分裂であり、ここで、細胞は2つの新しい細胞に分裂する。
【0189】
細胞運動をモデル化するために、我々の対照入力は粒子をとり、各細胞の各値を無作為に撹乱する:x0i、y0i、ai、bi、θi。撹乱は、比較的小さい分散(通常、初期値の%に設定される)の正規分布から無作為にサンプリングされる。
【0190】
細胞分割をモデル化するために、我々は以下のアプローチを使用する。所与の時間点で、各粒子において、我々は、細胞のうちの1つが分割するという50%の確率を割り当てる。この値は、経験的に選択され、現在の構成を良好に網羅しながら、広範な可能な細胞分割に及ぶ。分割が予測される場合、分割細胞は無作為に選択される。より複雑なモデルには、粒子中の細胞の数、およびそれらの分割パターンの履歴等のさらなる要因を考慮に入れ、実際のデータから観測された挙動に基づき、モデルを作製することが可能である。
【0191】
細胞が分割することが選択される時、楕円の主軸に沿った対称分割が適用され、同じ大きさおよび形状の2つの娘細胞が産生される。次いで、娘細胞の各値は、無作為に撹乱される。撹乱は、正規分布から、しかし、新しい細胞形状における大きな変動に対処するためにより大きな分散(初期値の10%)で再度サンプリングされる。最後に、残りの細胞に対してそれらの集合的重複を維持しながら、2つの娘細胞の重複インデックスを無作為に選択する。
【0192】
画像シミュレーション
【0193】
対照入力を各粒子に適用した後、粒子表示は、実際の画像と比較され得るシミュレーション画像に変換されなければならない。正確な画像シミュレーションは、難しい作業であり得、光線追跡技法および光学モデルの使用を必要とする場合が多い。現実的な画像のシミュレーションを試みるより、本発明の方法は、画像において容易に識別可能である特色をシミュレーションすることに焦点をあてる。具体的に、細胞膜の画像をシミュレーションする。
【0194】
考慮に入れられなければならない物理的観測が2つある。第1に、顕微鏡は、胚を通して単一面に焦点が合わせられるが、視野深度は、かなり大きく、焦点外の光は、ほぼ胚全体から収集される。第2に、胚は部分的に透明であり、これは、胚の下の細胞膜が胚の上の細胞を通して見られる時がある(いつもではない)ことを意味する。
【0195】
これらの物理的観測を念頭において、ここで、画像シミュレーションモデルを説明する。各細胞において、その対応する楕円形状は、重複インデックスhを使用してシミュレーション画像上に投影される。対応する画素値は、1のバイナリー値に設定され、観測された画像データと比較可能な膜厚を作製するために拡大される。重複インデックスhは、細胞が相互の上に重なり合う順序を指定する。遮蔽された細胞膜は、時々のみ可視的であるため、遮蔽点が検出される場合、それらは、低確率(通常、約10%)でシミュレーション画像に置かれる。実践において、これらの遮蔽された膜点は、正確な形状モデル化に必要であるが、それらが可視縁部に類似しないようにそれらを十分に散在させることが重要である。
【0196】
画像の前処理
【0197】
測定値変数zをここで説明する。本発明の方法のゴールは、シミュレーション画像と比較するために、顕微鏡画像による細胞膜のバイナリー画像を抽出することである。これらの膜は、高曲率および高コントラストを示すが、強度または色に基づく閾値化技法を使用して、容易に抽出されない。したがって、主曲率に基づく検出器が利用される。この方法はヘッセ作用素:
【数7】
を使用する。
【0198】
ここで、Ixx、Ixy、およびIyyは、画素位置sおよびガウススケールσで評価された二次偏導関数である。2x2ヘッセ行列の固有値は、主曲率についての情報を提供し、一方、固有値のサインは、「くぼみ」を「うね」43と区別する。明るいピークまたはうねを検出するために、各画素における主曲率が
【数8】
として計算される。
【0199】
ここで、λ2は、最小固有値である。変動する厚さの膜を検出するために、スケールの範囲に対してヘッセ作用素(すなわちσmin<=σ<=σmax)を適用し、この範囲に対する最大曲率を抽出する。最後に、ヘッセ画像を閾値化し、抽出した細胞膜のバイナリー画像を作製する。閾値レベルは、通常、ヘッセにおいて画素値の標準偏差の2倍に設定される。
【0200】
粒子の重さ
【0201】
「粒子フィルタフレームワーク」という表題のセクションで説明したように、粒子フィルタの第2の主要なステップは、測定値の更新であり、ここで、粒子は、特定モデルを考慮した、現在の測定値の確率に対応する重要な重さを割り当てられる。我々の場合、重要な重さは、上述の前処理された顕微鏡画像を、これも上述のシミュレーション画像と比較することにより判定される。
【0202】
この問題は、以前に検証され、ここで、標準化した相互情報を使用してシミュレーション画像を実際の画像と比較することにより、粒子フィルタの重さを計算した。このアプローチは、占有格子マッチングの考えに類似し、これは、双方が占有される(値1)か、双方が占有されない(値0)のいずれかである画素位置を検索する。これらの方法は、シミュレーション画像および実際の画像の形状は類似するが、わずかに不整合である時に問題がある。代わりに、説明される方法は、面取り距離に基づく尤度関数を使用し、これは、1つの点集合から別の点集合までの最も近い距離の平均値を測定する。それぞれ、シミュレーション画像と実際の画像において非ゼロ画素に対応する点A(大きさmの実数の組)およびB(大きさnの実数の組)の二組が定義される。点集合AからBの前方面取り距離は、
【数9】
として得られる。
【0203】
後方面取り距離も同様に定義される。本方法は、対称面取り距離を利用し、これは、いかに良くシミュレーション画像が実際の画像と一致したか、ならびにいかに良く実際の画像がシミュレーション画像と一致したかの基準を提供する。
【数10】
【0204】
実践において、個々の距離の測定値は、ノイズの影響を削減するために切り捨てられる。計算時間を削減するために、距離は、画像の距離変換の画素位置を調べることにより判定される。
【0205】
面取り距離は、推定モデルを考慮した、我々のデータ測定値の尤度基準として使用される。つまり、時間tにおける、所与の画像測定値ztおよび粒子モデルxt[m]において、粒子の重要な重さ(particle importance weight)は、
【数11】
として得られる。
【0206】
定数λは、通常1に設定され、尤度分布の「平坦性」を制御するように変動され得る。
【0207】
粒子再サンプリングおよび動的割り付け
【0208】
粒子フィルタの第3の主要なステップは再サンプリングであり、ここで、粒子は、それらの重さに比例して選択され、新しい組の粒子を作製する。低確率の粒子は破棄され、一方、高確率の粒子は乗算される。再サンプリングの効率的なアルゴリズムの開発に対して多くの従来の研究があった。本方法は、変動が少ないアプローチを使用する。
【0209】
粒子フィルタの重要な課題は、粒子の数の選択である。最も簡単な選択は、固定値、例えば、M=1000を使用することである。その後、各時間ステップにおいて、一組Mの粒子を同じ大きさの別の組に変換する。用途の内容において、細胞が不活性である、または単にわずかに大きさおよび位置を変更する、比較的長い期間が存在し得る。この観測の利点は、細胞活性量によって粒子の数を動的に割り付けることにより、処理負荷を低減するために利用される。つまり、細胞が活性であり、分割している時、我々は、粒子の数を増加し、細胞が不活性である時、我々は、粒子の数を削減する。
【0210】
細胞活性の程度を測定するために、新しい画像(顕微鏡によって取得)と前の画像との間の画素強度の2乗和の差(SSD)を計算する。ノイズを削減するために、最初に画像をガウスフィルタで平滑化し、SSD値を因果的移動平均で経時的に平滑化する。次いで、この値に比例して粒子の数を動的に調節し、100<M<1000の範囲内に収まるように切り捨てる。図30は、粒子の数がどのように1細胞期から4細胞期に分割する胚に割り当てられ得るかを示すグラフである。この方法は、事前画像登録は実施されなかったため、単に画像における「活性」量の基準を提供するだけであり、細胞分割と胚運動(翻訳および/または回転)を区別しないことに留意するべきである。この状況において(粒子の数を判定する)、粒子の数はいずれかの事象においても増加するはずであるため、これは許容される。実践において、我々は、最も可能性のある胚モデルの細胞の数に基づき、粒子の数も調節する。つまり、より多くの細胞が画像に存在すると考えられる時に、より多くの粒子が生成される。
【0211】
2次元追跡の限界
【0212】
上述の2D細胞追跡アルゴリズムは、胚の細胞の数、ならびにそれらの2D形状を判定するのに有用である。しかしながら、根底にある物理的表示がないという事実により制限される。これは、胚の生存能を評価するための、自動的な細胞分割の追跡に重要である場合も、重要でない場合もある。例えば、細胞質分裂の持続時間、および細胞分割間の時間等の特定のパラメータは、2D細胞追跡アルゴリズムを使用して測定され得る。次のセクションにおいて、我々は、我々の2Dモデルを3Dに拡張する。2D画像から3D形状を推定することにより生じる遮蔽および深度の曖昧さに対処するために、幾何制約および細胞容量の保存に対する制約を適用する。
【0213】
細胞表示および3次元追跡
【0214】
このセクションは、細胞分割の3D追跡用のアルゴリズムを説明する。2Dアルゴリズムからのステップの多くがこのアルゴリズムに持ち越されるが、主要な例外がいくつかある。3Dの使用に新しい細胞表示がある。ここで、細胞は3D空間で楕円体として表され、等式:
【数12】
により得られる。
【0215】
各楕円体も、向かう方向θ、ピッチψ、および回転αを有する。よって、時間tにおける胚モデルの表示は、
【数13】
として得られる。
【0216】
この改訂されたモデルの重要な作用の1つは、2D画像から3D形状を推測することに関連する曖昧さが存在し得るということである。例えば、形状が球状である細胞は、より長い主軸およびより大きいピッチ回転を有する細胞と類似する外見を有するであろう。後に示すが、区別するために十分な情報が利用可能になるまで(例えば、細胞分割等の事象から)、粒子分布は、これらの複数の仮定を維持するため、これは大きな問題ではない。
【0217】
楕円体は、剛体と考えられる、つまり、変形は明示的にモデル化されない。しかしながら、我々は、隣接する楕円体間のわずかな重複を可能にし、これらの重複する領域において、我々は、細胞が相互に対して平坦化されると想定する。それは胚において一般的に観測されるため、これは重要な考察であり、我々は、以下のセクションでそれを説明する。
【0218】
細胞撹乱および分割
【0219】
我々の3D細胞分割および撹乱モデルは、セクション4の「細胞撹乱および分割」のモデルと類似するが、主要な例外がいくつかある。3D形状の推定は、容量の保存を強化するために使用され得る。これは、細胞が任意に、特にz方向に大きく成長することを防止する。容量保存は、2つの状況において適用される。第1に、細胞撹乱において、軸aおよびbは変動し、cは、容量がその個々の細胞のために保存されるように計算される。第2に、細胞分割において、以下の制約が適用される。
【数14】
【0220】
ここで、下付き文字pは親細胞を示し、下付き文字d1およびd2は2つの娘細胞を示す。実践において、我々は、胚の総容量を原容量のプラス/マイナス5%の間で変動させることにより、これらの制約のわずかな違反を可能にする。これは、初期の容量推定における潜在的な不正確性を相殺するために使用される。
【0221】
細胞が3Dで分割することが選択される場合、その分割は、以下の方式でモデル化される。第1に、選択した単一細胞において、楕円の長軸に沿った分割が適用され、これは、構成によりa、b、またはcのいずれかであり得る。娘細胞は、大きさが等しく、均一に間隔がとられるように初期化され、親細胞の回転が考慮に入れられる。次いで、これらのパラメータは、広範の可能な構成を網羅するように撹乱され、ここでも初期値の10%に設定された変数の正規分布を使用する。
【0222】
幾何制約
【0223】
遮蔽および深度の曖昧さの問題は、容量の保存を通して部分的に軽減される。しかしながら、隣接する楕円体の空間的関係に関する制約も必要とされる。第1の制約は、細胞は、半径の20%を超える重複が禁止されることである。許容される量で重複する細胞において、それらが相互に対して平坦化したという想定がなされる。説明される粒子モデルは、画像シミュレーション中、楕円体を交差する内側点を無視することにより、この現象を表す。これは、経験的に動機づけされ、物理的に観測された挙動とよく相関する。
【0224】
第2の制約は、細胞を近接に保つことが課されることである。この制約は、ヒトの胚の物理的挙動に直接関係し、ここで、細胞は、透明帯と呼ばれる膜により制約される。帯は、球殻としてモデル化され、境界条件を課すためにそれを使用する。帯の半径は、1細胞胚の半径より30%大きく設定される。
【0225】
これらの制約は以下の通り強化される。所与の時間における各粒子において、上述のように、新しい粒子を生成するために無作為な対照入力が適用される。いずれかの物理的制約が犯された場合、新しい粒子は廃棄され、新しい無作為な対照が適用される。一定数の試みの後、満足する新しい粒子が生成されない場合、その粒子は廃棄される。
【0226】
画像シミュレーション
【0227】
実施例で使用される暗視野照明の利点は、細胞膜が細胞内部より多く光を分散することである。この作用は、細胞膜が光学軸(z軸)に平行である位置で最も顕著である。したがって、画像をシミュレーションするために、これらの位置が我々の3Dモデルにおいて検索され、これは、それらの回転のため楕円体の赤道に位置する必要はない。可視縁部および遮蔽縁部に関しても、上述のように、同じ規則に従う。
【0228】
2Dでの細胞追跡実施例
【0229】
この実施例は、自動化細胞顕微鏡に関し、細胞分割の2D追跡のために上述のアルゴリズムを使用する。このモデルは、画像中の細胞の数、ならびに細胞膜の2D輪郭を追跡するために設計された。第1ステップは画像の取得であり、これは、画像シミュレーションおよび画像前処理等の以下のセクションを動機づける。この実施例の微速度画像列は、10X対物レンズを備えた特注のOlympus IX−50倒立顕微鏡を用いて取得された。顕微鏡は、暗視野照明に改変され、ここで、中空の光錐は、光源と集光レンズとの間に円形アパーチャを設置することにより、試料に焦点を合わせる。対物レンズは、試料によって分散された光を収集し、直接透過した光を排除し、暗い背景上に明るい画像を産生する。暗視野照明の利点は、細胞膜が細胞内部より多く光を分散する傾向にあり、それによって、それらのコントラストを増強する。顕微鏡は、加熱台および特注のインキュベーションチャンバが装備されており、最大5または6日間にわたる胚の培養を可能にする。画像は、IX−50のサイドポート上に載置されたOlympus SLRデジタルカメラにより、5分間隔で捕捉された。
【0230】
胚の撮像は、それらが接合体、またはほぼ球形状の受精卵である時に開始された。粒子の組を初期化するために、セクション6の「画像の前処理」に記載するように、閾値ヘッセを計算し、最小2乗を使用してそれに円を適合させる。次いで、等分布からサンプリングした、無作為配向の円として全ての粒子を初期化する。
【0231】
図31は、1細胞期から4細胞期までの細胞分割を追跡するための2Dアルゴリズムの結果を示す。結果は、部分的に遮蔽される底に面する細胞においてさえ、細胞膜がアルゴリズムによりうまく抽出されることを示す。大半の粒子フィルタ用途において、「単一」の最適モデルは、粒子分布からの重み付き状態パラメータの和として表される場合が多いことに留意するべきである。しかしながら、本明細書に提示される結果において、高確率の粒子が表示される。
【0232】
3Dでの細胞追跡実施例
【0233】
図32は、1細胞期から4細胞期までの追跡のための上述の3Dアルゴリズムの成功した2つの用途を示す。図33は、粒子が1細胞から2細胞への分割中にどのように分散されるかの例を示す図である(図32に示される最初の実施例と対応する)。このプロットは、各細胞の中心の3D位置を示す。細胞が分割し始めると、予測は、どの娘細胞が他の上に重なり合うかに関する曖昧さを示すが、これは数フレーム内に解決される。
【0234】
予測パラメータの抽出
【0235】
前述の方法を使用して、胚がモデル化されたら、特定のパラメータがモデルから抽出され得る。通常、最適なモデルまたは最も高確率のモデルが使用される。これらのパラメータは、例えば、第1細胞質分裂の持続時間、第1と第2の細胞分割との間の時間、および第2と第3の細胞分割との間の時間を含む。細胞質分裂の持続時間は、細胞のモデルが2つの細胞に分裂する前にどのくらい長く伸長したかを測定することにより近似され得る。伸長は、楕円の主軸対短軸の比率を調べることにより測定され得る。モデルから抽出され得る他のパラメータは、受精と第1細胞分割との間の時間、細胞および分割プロセスの形状および対称性、分割の角度、断片化等を含む。パラメータは、2D細胞追跡アルゴリズムまたは3D細胞追跡アルゴリズムのいずれかを使用して抽出され得る。
【0236】
細胞質分裂は、最初の細胞質分裂溝の出現から娘細胞の完全な分離までによって定義される。我々の胚モデルは、変形不可能な楕円から成るため、細胞質分裂溝の出現の識別は困難な課題である。一方法は、楕円の変形を可能にするが、これはより複雑な追跡問題をもたらす。別の方法は、前処理顕微鏡画像の曲率の変化を探すことであるが、これは、胚のモデルから直接予測パラメータを測定しようとする我々の目的を無にする。よって、我々は、第1細胞質分裂の持続時間を、1細胞から2細胞分割前までの細胞伸長の持続時間として近似することにより問題を簡潔化する。伸長は、楕円の短軸bに対する主軸aの比率を計算することにより定量化される。細胞は、
【数15】
である場合、伸長したと判断される。
【0237】
この15%の値は、経験的に選択され、この特定のデータセットにおいて良く機能するが、他の値も使用され得る。胚モデルが2細胞に分割したら、我々は、1細胞モデルの伸長の持続時間を計算することにより、第1細胞質分裂の近似持続時間を抽出することができる。
【0238】
原理上、有糸分裂事象間の時間を測定することは容易である。例えば、第1と第2の有糸分裂の間の時間は、2細胞モデルと3細胞モデルとの間の時間として測定され得る。しかしながら、一部の場合において、胚は、独特な、無作為な挙動を示す場合がある。これは、例えば、1細胞から2細胞、2細胞から見かけ3もしくは4細胞に進み、そして2細胞に戻ることを含む。説明されるアルゴリズムは、この種の挙動を追跡することができるが、有糸分裂事象間の時間間隔の判定に対する課題を提起する。
【0239】
この挙動に対処するための一方法は、以下の通りである:2細胞と3細胞のモデルとの間の時間を測定する代わりに(第1と第2の有糸分裂の間の時間を見出すため)、これは、2細胞モデルが最も高確率の画像フレームの数を単純に数えることにより近似され得る。これは、一部の場合においてよく機能するが、有糸分裂事象間の真の時間を必ず表すとは限らない。細胞の数に基づきモデルに対する制限を強化することにより、これらの事象に対処することができる。つまり、各反復で分布から最適なモデル、または最も高確率のモデルを選択する場合、モデルの細胞の数は、常に同じままであるか、または増加するが、決して減少しないことが必要とされる。この制約を強化した後、有糸分裂事象間の時間を計算することは容易である。この制約は、ごくわずかな変動を示し得る追跡結果をフィルタにかけるのにも有用であり、これは、モデルが、例えば1細胞と2細胞のモデルの間を前後に切り替わる時に時折生じる。
【0240】
予測パラメータを抽出するための方法
【0241】
図35は、上述の方法を要約したフローチャートを示す。フローチャートは、(これは複数の胚または他の種類の細胞および幹細胞にも適用され得るが)単一の胚がどのように分析され得るかを示す。第1ステップにおいて、胚の画像が微速度顕微鏡を用いて取得される(「測定値」)。この画像は、ファイルに保存され、後の時間点で再度開くことができる。必要はないが、画像は、一般に、ある特色を強調するために前処理される。可能な胚構造のモデルが予測され、画像はそれらのモデルからシミュレーションされる(「予測」)。シミュレーション画像は、前述の細胞膜の画像、または前処理前の顕微鏡画像をより正確に表す画像を含み得る。次いで、モデルは、前処理された顕微鏡画像と比較される(「比較」)。この比較を使用して、最良の予測が保たれ、一方、不良な予測は廃棄される。次いで、得られた予測の組は、次の画像の予測を改善するために使用される。複数の連続画像においてこのプロセスを実施した後、例えば、細胞質分裂の持続時間、および有糸分裂事象間の時間等の、形態学的パラメータを最良のモデルから直接測定することが可能である。これらのパラメータは、前述の胚の生存能を評価するために使用され得る。
【0242】
実施例7
【0243】
細胞活性の自動化分析
【0244】
上述の方法は、顕微鏡を介して細胞の発達を追跡する機能を必要とする。胚において、複数の胚を追跡することが望ましく、これは同一の皿で一緒に培養される。本明細書で使用される分析方法は、画像が定期的に撮影されることも必要である(例えば、胚においては、1〜5日にわたり1〜30分毎、異なる時間間隔が幹細胞等の他の細胞型において使用され得る)。したがって、自動的に胚の発達を追跡する撮像方法が考案された。
【0245】
微速度顕微鏡検査において、細胞を制御された条件下で成長させ、運動性(環境内での運動)、増殖(成長および分割)、および形態変化(大きさおよび形状)等のプロセスを監視するために、長時間にわたり撮像した。実験期間および生成された膨大な画像データにより、細胞分割の持続時間および細胞分割間の時間等のパラメータの抽出は、面倒な仕事であり得る。これは、複数の試料が同時に撮像される高処理用途において、特にそうである。よって、所望の情報を自動的に抽出することができる画像分析ソフトウェアが必要である。
【0246】
胚の生存能を評価する一方法は、画像中の「細胞活性」量を測定することである。これは、単純に、遂次画像の組を選び、それらの画素値を比較することにより達成され得る。より具体的には、各新しい画像における細胞活性量を測定するために、全ての重複する画素iに関して、I’と示される新しい画像とI’と示される前の画像との間の画素強度の2乗和の差(SSD)を計算する
【数16】
【0247】
ノイズを削減するために、最初に画像をガウスフィルタで平滑化する。図28は、単一の胚の1日目〜3日目までの細胞活性のプロットを示す。示すように、ヒトの胚における1細胞から2細胞への分割、2細胞から4細胞への分割、4細胞から8細胞への分割に対応する鋭いピークが存在する。ピークの幅は、細胞分割の持続時間を表す。
【0248】
このアプローチの限界の1つは、SSD計量が画像中の活性量のみを測定し、胚運動(移動または回転)等の事象が細胞分割と非常に同じに見えることである。この問題に対する解決策の1つは、SSDを計算する前に画像登録を実施することである。画像登録は、それらを同じ座標系に整合するために、2つの画像間の幾何学的関係を見出すプロセスであり、種々の異なる技法を使用して達成され得る。例えば、レーベンバーグマルカート反復非線形ルーチンの変形を使用することができ、これは、重複する画素強度におけるSSDを最小にすることにより画像を表示する。LMアルゴリズムは、3×3のホモグラフィ行列:
【数17】
を使用して画素位置を変換する。
【0249】
ここで、目的画素位置x’およびy’は、
【数18】
として標準化される。
【0250】
よって、
【数19】
である。
【0251】
ホモグラフィ行列は、種々の画像変換に適用することができ、本用途における妥当な選択は、剛性体(ユークリッド)変換であろう。これは、翻訳および面内回転(カメラ軸に沿って)の胚の画像を整合するだろう。しかしながら、わずかに一般化し、アフィン変換を使用することが可能であり、これは、画像の歪みを可能にする。この一般化は、測定されようとするシグナルにより望ましい場合も、望ましくない場合もある。運動等式は、よって、
【数20】
となる。
【0252】
LMアルゴリズムは、最初に、連鎖法則:
【数21】
を使用して、未知の運動パラメータhkに関するeの偏導関数を計算する。
【0253】
アフィン運動パラメータにおいて、これらの偏導関数は、
【数22】
となる。
【0254】
次に、これらの偏導関数を使用して、LMアルゴリズムは、各画素からの寄与を加えることにより、近似ヘッセ行列A(大きさ6×6の実数の組において)および重み付き勾配ベクトル(大きさ6x1の実数の組において)を計算する。
【数23】
【0255】
最後に、運動パラメータは、増分運動を加えることにより更新され得る。
【数24】
【0256】
ここで、定数λは、運動更新の刻み幅を調節し、Iは単位行列である。
【0257】
アルゴリズムの各反復で、最初の画像は更新された運動推定値によって歪められ、重複する領域の画素強度のSSDを計算することにより第2の画像と比較される。本用途は、連続画像間の胚の運動が非常に小さいと想定し、したがって、少数で一定数の反復のみが行われる。図28Bは、各画像の組において行われた画像登録なし(28A)および画像登録あり(28B)の細胞活性のプロットを示す。レーベンバーグマルカートルーチンの誤差関数はSSDであるため、単に各表示の残余誤差をプロットする。図29は、正常および異常な胚の発達についての細胞の活性のプロットを比較する。胚生学者が通常形態を評価するであろう点である3日目に、胚は同じように見え、双方とも潜在的に生存可能であると考えられる。しかしながら、胚のうちの1つは、通常の一連の細胞分割を受け、一方、他方は、1細胞胚から複数の細胞および断片に分裂するため、それらの細胞活性プロットは、大幅に異なる。予想通り、正常な活性プロットを有する胚は、最終的に、5.5日目までに胚盤胞に達する。
【0258】
画素強度のSSDを計算する前に、他の種類の画像登録が使用され得る。これは、例えば、相互相関、標準化した相互相関、相互位相相関、相互情報、特徴検出および追跡、スケール不変特徴変換(SIFT)、オプティカルフロー、および勾配降下を含む。特徴およびコントラスト強調等の表示前の画像の前処理は、望ましい場合も、望ましくない場合もある。
【0259】
胚の生存能を評価するためのモデル
【0260】
図13は、相関する撮像および分子分析に基づくヒトの胚の発達のモデルを示す。胚盤胞への発達の成功を予測するための重要な短時間、および胚の発達の図を含む、接合体から胚盤胞までの発達の時系列を示す。図の主要な分子データは、ヒトの胚が母親から受け継がれた独特の卵母細胞RNAの組により一生を開始することを示す。このRNAの組が、卵子の特異的RNA操作プログラムによって維持され、適切にパッケージされる。受精後、卵母細胞から胚への変移が始まる時に、卵子に特異的な一部の母性RNA(ESSP1;胚期特異的パターン1)の分解が分解されなければならない。並行して、発達が続くと、他のRNAが各卵割球に理想的に等しく分配される(ESSP4)。分解の成功とRNAの分配は、細胞自律的様式で、胚ゲノム活性化(EGA)、およびESSP2の遺伝子の転写に達する。特に、卵割中、胚性卵割球は、独立して、停止、または進行し得る。胚における細胞自律性発達の転帰は、個々の卵割球が停止または進行し得ることであり、8細胞胚が桑実胚期およびそれ以降に進行すると、胚盤胞の質は、停止した、または8細胞以降に進行した細胞の数によって影響を受ける。撮像データは、成功または失敗を予測する重要な発達期間があることを示す:第1細胞質分裂、第2卵割、および第2および第3の卵割の同時発生。これらのパラメータは、前述の細胞追跡アルゴリズムおよびソフトウェアを使用して、自動的に測定され得る。記載されるシステムおよび方法は、主要な撮像予測装置を用いて胚の転帰を診断するために使用され得、発達初期(EGA前)により少ない胚の移植を可能にする。自動対手動画像分析の比較
【0261】
図34は、一組14の胚における手動画像分析に対する自動化画像分析の比較を示す。胚1から10(プロットに標識される)は、異なる形態を有した胚盤胞期に達した。胚11から14は停止し、胚盤胞に達しなかった。図34Aは、第1細胞質分裂の持続時間の測定の比較を示し、図34Bは、第1と第2の有糸分裂の間の時間の測定の比較を示す。示されるように、2つの方法は、一般に、良好な一致を示す。第1細胞質分裂の持続時間におけるわずかな矛盾が予想されるが、それらは、我々の自動化分析が、前述の通り、伸長を測定することにより近似を行うという事実に起因し得るためである。いくつかの場合において、細胞質分裂の持続時間ならびに第1と第2の有糸分裂の間の時間の双方において、自動化分析と手動分析との間により大きな不一致がある。これは、異常な胚のいくつかに生じ、手動で特徴付けること、ならびに自動的に追跡することの双方が困難である異常な挙動に起因する。この群の胚において、最初の2つの基準(第1細胞質分裂の持続時間、および第1と第2の有糸分裂の間の時間)のみを使用することにより、自動化アルゴリズムは、ゼロの偽陽性を有する。これは、偽陽性が回避されなければならないIVF処置において、非常に重要であろう。手動画像分析は、偽陰性(胚9)が1つであり、一方、自動化アルゴリズムは、偽陰性(胚9および10)が2つであった。しかしながら、胚9および10の双方は技術的に胚盤胞期に達したが、これらは他の胚盤胞と比較して不良な形態を示し、移植の候補にはあまり最適ではないだろう。手動画像分析において、胚14は、これらの2つの基準に基づきおそらく偽陽性であり、第3のパラメータの第2と第3の有糸分裂との間の持続時間は、真の陰性を与える必要がある。しかしながら、自動化アルゴリズムは、最初の2つの基準のみを使用して、正しい予測を行う。これらの結果は、我々の自動化アルゴリズムが胚盤胞対非胚盤胞を適切に予測し、ならびに異なる質間の胚盤胞を区別できることを示す。よって、複数の胚が良好な発達の可能性を有することが判定される時、IVF処置中、移植のための上位1つまたは2つの胚を選択するために、それらの相対的な質のランク付けを計算することが可能である。
【0262】
前述されるものは、単に本発明の原則を解説するにすぎない。本明細書に明白に説明もしくは表されてはいないが、当業者は、本発明の原則を具体化し、その精神と範囲内に含まれるものである様々な構造を考案し得ることが理解されるであろう。さらに、本明細書に記載される全ての実施例および条件言語は、主に、読者が、本発明の原則、ならびに本発明者らによって貢献された当技術分野を推進するための概念を理解することを補助することを意図しており、かかる具体的に記載される実施例および条件に限定されることはない解釈されたい。さらに、原則、態様、および本発明の実施形態、ならびにその特異的実施例を記載する本明細書の全ての陳述は、その構造的および機能的同等物の双方を包含することを意図している。加えて、かかる同等物が、現在既知の同等物および将来的に開発される同等物の双方を含むことを意図している、すなわち、構造にかかわらず、同一機能を実施する開発されるいずれの要素をもである。したがって、本発明の範囲は、本明細書で表され、説明される例示的実施形態に限定されることを意図していない。正確には、本発明の範囲および精神は、添付の特許請求の範囲によって具体化される。
【技術分野】
【0001】
関連出願の相互参照
本出願は、2010年5月7日に出願された米国仮特許出願第61/332,651号および2009年8月22日に出願された米国仮特許出願第61/236,085号に対して優先権を主張し、それらの双方は、参照によりそれらの全体が本明細書に組み込まれる。
【0002】
本発明は、生物学的および臨床試験の分野、特にヒトおよび動物の双方からの接合体/胚、卵母細胞、および幹細胞の撮像および評価に関する。
【背景技術】
【0003】
不妊は、妊娠可能年齢の夫婦の10〜15%に影響を及ぼす一般的な健康問題である。2006年には、米国だけで、約140,000回の生体外受精(IVF)が実施された(cdc.gov/art)。これは、変わりやすくまた明確に定義されていない場合が多い、着床および満期児までの発達の可能性を有する胚を毎年100万個以上培養する結果となった。平均して30%の出生は、多胎妊娠をもたらしたが、IVF後の1回当りの出生率は、わずか29%であった(cdc.gov/art)。多胎妊娠は、流産、早産、および低出産率等の、母親と胎児の双方に有害転帰になる。IVF失敗の潜在的な原因は多様であるが、1978年にIVFが導入されて以来、主な課題の1つは、移植に最も適し、満期妊娠をもたらす可能性が最もある胚を識別することである。
【0004】
ヒトの胚の生物学の研究は困難なままであり、研究費の対象外であることが多いため、基本的な胚の発達の分野における理解は限られている。したがって、胚の発達の現在の知識の大半は、モデル生物の研究に由来する。しかしながら、異なる種からの胚は、類似する発達期を経るが、タイミングは種により異なる。これらの相違、およびその他の多くにより、1つの種から他の種に対して直接推定することが不適切となる(Taft,R.E.(2008)Theriogenology 69(1):10−16)。ヒトの発達の一般的な経路、ならびに基本的な根底にある分子決定要因は、ヒトの胚の発達に固有である。例えば、マウスにおいて、胚の転写は、受精の約12時間後に活性化され、同時に第1卵割され、一方、ヒトにおいて、胚遺伝子活性化(EGA)は、およそ8細胞期である、3日目に生じる(Bell,C.E.,et al.(2008)Mol.Hum.Reprod.14:691−701、Braude,P.,et al.(1988)Nature 332:459−461、Hamatani,T.et al.(2004)Proc.Natl.Acad.Sci.101:10326−10331、Dobson,T.et al.(2004)Human Molecular Genetics 13(14):1461−1470)。加えて、初期のヒトの発達で調節される遺伝子は、固有である(Dobson,T.et al.(2004)Human Molecular Genetics13(14):1461−1470)。さらに、マウス等の他の種において、生体外で培養された胚の85%超は、哺乳類の発達において最初の主要な目印の1つである胚盤胞期に達し、一方で、培養されたヒトの胚は、約30〜50%の平均胚盤胞形成率を有し、断片化および発達停止等のモザイク現象および異常表現型の発生率が高い(Rienzi,L.et al.(2005)Reprod.Biomed.Online 10:669−681、Alikani,M.,et al.(2005) Mol.Hum.Reprod.11:335−344、Keltz,M.D.,et al.(2006)Fertil.Steril.86:321−324、French,D.B.,et al.(2009)Fertil.Steril.)。そのような相違にもかかわらず、着床前の胚の発達の研究の多くは、モデル生物に由来し、ヒトの胚の発達に関連付けることが困難である(Zernicka−Goetz,M.(2002)Development 129:815−829、Wang,Q.,et al.(2004)Dev Cell.6:133−144、Bell,C.E.,et al.(2008)Mol.Hum.Reprod.14:691−701、Zernicka−Goetz,M.(2006)Curr.Opin.Genet.Dev.16:406−412、Mtango,N.R.,et al.(2008)Int.Rev.Cell.Mol.Biol.268:223−290)。
【0005】
従来、IVFの臨床において、ヒトの胚の生存能は、均一な大きさの単核卵割球の存在、および細胞断片化の程度等の、単純な形態学的観測により評価されてきた(Rijinders PM,Jansen CAM.(1998)Hum Reprod 13:2869−73、Milki AA,et al.(2002)Fertil Steril 77:1191−5)。最近では、胚の長期培養(5日目の胚盤胞期まで)および着床前の遺伝子診断(PGD)を介した染色体の状態の分析等の付加的な方法も、胚の質を評価するために使用されている(Milki A,et al.(2000)Fertil Steril 73:126−9、Fragouli E,(2009)Fertil Steril Jun 21[EPub ahead of print]、El−Toukhy T,et al.(2009)Hum Reprod 6:20、Vanneste E,et al.(2009)Nat Med 15:577−83)。しかしながら、これらの方法には、培養期間を延長し、胚の整合性を乱すという潜在的なリスクも存在する(Manipalviratn S,et al.(2009)Fertil Steril 91:305−15、Mastenbroek S,et al.(2007)N Engl J Med.357:9−17)。
【0006】
最近、微速度撮像が初期の胚の発達を観測するのに有用なツールであり得ることが示されている。いくつかの方法は、卵細胞質内精子注入(ICSI)後、ヒトの胚の発達を監視するために微速度撮像を使用してきた(Nagy et al.(1994)Human Reproduction.9(9):1743−1748、Payne et al.(1997)Human Reproduction.12:532−541)。極体突出および前核形成が分析され、3日目の良好な形態と関連付けられた。しかしながら、パラメータは、胚盤胞形成または妊娠の転帰と相関しなかった。他の方法は、ヒトの胚の生存能を予測するための指標として第1分割の開始に目を向けてきた(Fenwick,et al.(2002)Human Reproduction,17:407−412、Lundin,et al.(2001)Human Reproduction 16:2652−2657)。しかしながら、これらの方法は、細胞質分裂の持続時間、または初期分割間の時間間隔の重要性を認識しない。
【0007】
他の方法は、初期の胚の発達中、細胞分割のタイミングおよび程度を測定するために微速度撮像を使用してきた(WO/2007/144001)。しかしながら、これらの方法は、ウシの胚の微速度撮像のための基本的で一般的な方法しか開示せず、これは、発達の可能性、形態学的挙動、分子および後成的プログラム、ならびに移植を取り巻くタイミングおよびパラメータに関して、実質的にヒトの胚と異なる。例えば、ウシの胚は、ヒトの胚と比較して、着床するのに実質的により長くかかる(それぞれ、30日および9日)。(Taft,(2008)Theriogenology 69(1):10−16。さらに、ヒトの胚の生存能を予測し得る具体的な撮像パラメータ、または時間間隔は、開示されていない。
【0008】
最近では、受精後の最初の24時間の間、ヒトの胚の発達を観測するために、微速度撮像が使用されている(Lemmen et al.(2008)Reproductive BioMedicine Online 17(3):385−391)。最初の分割後の核の同調性は、妊娠の転帰と相関することが分かった。しかしながら、この研究は、初期の第1分割が重要な予測パラメータではないと結論付けられ、これは、前の研究と矛盾する(Fenwick,et al.(2002)Human Reproduction 17:407−412、Lundin,et al.(2001)Human Reproduction 16:2652−2657)。
【0009】
最後に、胚の分子プログラムまたは染色体構成との相関によって撮像パラメータを検証した研究はない。よって、ヒトの胚の評価方法は、いくつかの概念に欠け、本方法によって改善され得、これは、微速度顕微鏡検査、画像分析、ならびに撮像パラメータの分子プロファイルと染色体構成との相関の新規適用を含む。本発明はこれらの問題に対処する。
【発明の概要】
【0010】
1つ以上の胚もしくは多能性細胞における1つ以上の胚もしくは多能性細胞の発達の可能性を判定するための方法、組成物、およびキットを提供する。これらの方法、組成物、およびキットは、良好な発達の可能性、すなわち、胚盤胞に発達する能力または潜在的可能性を有する、生体外における胚および卵母細胞を識別するのに使用され、よって、これはヒト等における不妊を治療する方法に有用である。
【0011】
本発明の一部の態様において、胚または多能性細胞の発達の可能性を判定するための方法を提供する。そのような態様において、胚または多能性細胞の1つ以上の細胞パラメータは、細胞パラメータ測定値を導き出すように測定される。次いで、細胞パラメータは、胚または多能性細胞の発達の可能性の判定を提供するために利用され、この判定は、臨床的行動方針を導くために使用され得る。一部の実施形態において、細胞パラメータは、微速度顕微鏡検査により測定可能である形態学的事象である。一部の実施形態において、例えば、胚がアッセイされる時、1つ以上の細胞パラメータは、細胞質分裂事象、例えば、細胞質分裂1の持続時間、細胞質分裂1と細胞質分裂2との間の時間間隔、および細胞質分裂2と細胞質分裂3との間の時間間隔である。ある実施形態において、細胞周期1の持続時間も細胞パラメータとして利用される。一部の実施形態において、細胞パラメータ測定値は、それを基準の胚からの比較可能な細胞パラメータ値と比較し、この比較の結果を使用して、胚の発達の可能性の判定を提供することにより利用される。一部の実施形態において、胚はヒトの胚である。一部の実施形態において、細胞パラメータは、遺伝子発現測定値を導き出すように測定される遺伝子発現レベルである。一部の実施形態において、遺伝子発現測定値は、それを1つの基準の多能性細胞もしくは胚もしくはそれらからの1つ以上の細胞の遺伝子発現測定値と比較することにより利用され、ここで、この比較の結果は、多能性細胞もしくは胚の発達の可能性の判定を提供するように利用される。一部の実施形態において、胚はヒトの胚である。
【0012】
本発明の一部の態様において、群における他の胚または多能性細胞に対して、胚または多様性細胞をそれらの発達の可能性についてランク付けするための方法を提供する。そのような実施形態において、群における胚または多能性細胞の1つ以上の細胞パラメータは、胚または多能性細胞のそれぞれについての細胞パラメータ測定値を導き出すように測定される。次いで、細胞パラメータ測定値は、群における胚または多能性細胞のそれぞれの発達の可能性を判定するために相互に利用され、この判定は、臨床的行動方針を導くために使用され得る。一部の実施形態において、細胞パラメータは、微速度顕微鏡検査により測定可能である形態学的事象である。一部の実施形態において、例えば、胚がランク付けされる時、1つ以上の細胞パラメータは、細胞質分裂事象、例えば、細胞質分裂1の持続時間、細胞質分裂1と細胞質分裂2との間の時間間隔、および細胞質分裂2と細胞質分裂3との間の時間間隔である。ある実施形態において、細胞周期1の持続時間も測定される。一部の実施形態において、細胞パラメータは、1つ以上の遺伝子の発現レベルである。一部の実施形態において、1つ以上の細胞パラメータ測定値は、群における胚または多能性細胞のそれぞれからの細胞パラメータ測定値を相互に比較して、相互に対して、胚または多能性細胞の発達の可能性を判定することにより利用される。一部の実施形態において、1つ以上の細胞パラメータ測定値は、各細胞パラメータ測定値を基準の胚または多能性細胞からの細胞パラメータ測定値と比較して、各胚または多能性細胞についての発達の可能性を判定すること、およびこれらの発達の可能性を比較して、相互に対して、胚または多能性細胞の発達の可能性を判定することにより利用される。
【0013】
本発明の一部の態様において、生殖補助(IVF)を受ける女性に移植するための、良好な発達の可能性を有する胚を提供するための方法を提供する。そのような態様において、1つ以上の胚は、胚の発達に十分な条件下で培養される。次いで、1つ以上の細胞パラメータは、1つ以上の胚において、細胞パラメータ測定値を導き出すように測定される。次いで、細胞パラメータ測定値は、1つ以上の胚の発達の可能性の判定を提供するために利用される。次いで、良好な発達の可能性を示す1つ以上の胚は、女性に移植される。
【図面の簡単な説明】
【0014】
本発明は、付随の図面と併せて以下の詳述を読む場合に最もよく理解される。一般的な実践において、図面の様々な特徴は、原寸に比例しないことが強調される。一方、様々な特徴の大きさは、明確にするために、任意に拡大または縮小される。図面に含まれるのは、以下の図である。
【図1】胚を評価するために使用されるプロセスを示すフローチャートである。
【図2】6日間にわたる細胞分裂および分割を示す一連の写真である。画像は、1日目から6日目までラベルが付けられている。スケールバーは50μmを表す。
【図3】1細胞胚(接合体)から胚盤胞への発達に成功した比率を示す棒グラフである。4つの個別の実験に亘って、微速度顕微鏡検査を介して、5〜6日目まで、合計100個の胚を観測した。各示された時期(胚盤胞、8細胞、4〜7細胞、2〜3細胞、および1細胞)に達した細胞の比率を示す。
【図4】示された時間の間追跡された、一連の3つの異なる胚である(上段、中段、および下段)。
【図5】本評価に使用される期間の間の時間経過を示す図であり、第1細胞質分裂の持続時間、第1と第2の分割との間の時間(細胞質分裂1の分離と細胞質分裂2の開始との間の時間間隔として測定される)、および第2と第3の有糸分裂との間の時間(細胞質分裂2の開始と細胞質分裂3の開始との間の時間間隔として測定される)を含む。
【図6】3つの事象の測定値を示す3D点グラフであり、第1細胞質分裂の持続時間、第1と第2の細胞分割の間の時間間隔(細胞質分裂1の分離と細胞質分裂2の開始との間の時間間隔として測定される)、および第2の細胞分割と第3の細胞分割との間の時間(細胞質分裂2の開始と細胞質分裂3の開始との間の時間間隔として測定される)を含む。胚盤胞期に達する胚(丸で示される)は、3Dグラフ上で共に密集していることが示され、一方、胚盤胞に達する前に停止する胚(Xで示される)は、全体に散らばっている。
【図7】3つの動的形態学的パラメータを使用して胚盤胞形成を予測するための受信者動作特性(ROC)曲線を示すグラフである。
【図8】6つの停止した1〜2細胞胚および5つの正常な1〜2細胞胚からの52の遺伝子の遺伝子発現レベルを示すレーダーグラフである。正常な胚と異常な胚との間の発現レベルの相違は、マンホイットニー検定により判定されると、黄色でハイライトされ、星印で示されるこれらの遺伝子において、統計的に有意であった。
【図9】停止した2細胞胚および正常な2細胞胚における、異なる遺伝子の発現レベルを示す棒グラフである。選択されたいくつかの停止した2細胞胚の微速度画像を上部に示す。
【図10】停止した4細胞胚および正常な4細胞胚における、図9に示される同じ遺伝子の比較を示す棒グラフである。選択されたいくつかの停止した4細胞胚の微速度画像を上部に示す。
【図11】4つの明確なパターンを有する遺伝子発現パターン(ESSP)を示す一連の棒グラフである。胚の遺伝子活性化(2日目)前および3日目の典型的な発現前の早期移植の時間を示す。
【図12】異なる期の単一卵割球からの遺伝子の遺伝子発現を示す。(A)異なる細胞期でプロットされた単一の卵割球からのCTNNB1およびCDX2の2つの遺伝子の遺伝子発現であり、例えば、2細胞、3細胞、桑実胚、および胚盤胞の異なる期におけるこれらの遺伝子発現レベルにおける変化を示す。(B)接合体プログラムから発現した遺伝子と比較した、母性プログラムで発現した遺伝子を表す棒グラフで表された遺伝子発現特性。
【図13】胚の生存能を評価するために、微速度画像分析および相関分子分析を使用するためのモデル図である。
【図14】生体外での卵母細胞成熟中の3つの発達期を示す一連の写真である。
【図15】生体外での卵母細胞成熟後の胚の発達のプロセスを示す一連の写真である。
【図16】卵母細胞を評価するために使用されるプロセスを示すフローチャートである。
【図17】幹細胞および多能性幹細胞を評価するために使用されるプロセスを示すフローチャートである。
【図18】ニューロンロゼットに分化する、誘発された多能性幹細胞のプロセスを示す一連の写真である。
【図19】発現レベルについてアッセイされた遺伝子が分類され得る分類表であり、分類毎に遺伝子の数を含む。
【図20】141の正常に発達した単一胚および単一卵割球の遺伝子発現分析中に識別された、4つの胚期特異的パターン(ESSP)、および遺伝子をそれらの分類のうちのそれぞれ1つに分類した表である。
【図21】胚盤胞形成を予測するための撮像パラメータの能力を示す自動化画像分析を示す。(A)単一胚についての追跡アルゴリズムの結果を示す。(B)分析された14個一組の胚を示す。(C)細胞質分裂の持続時間、および第1と第2の有糸分裂の間の時間についての手動画像分析の自動化分析との比較を示す。(D)良好な胚盤胞形態の不良な胚盤胞形態との比較を示す。
【図22】本発明による暗視野顕微鏡の概略図であり、左側の差し込みは、レーザー加工暗視野パッチ設定を示す。
【図23】インキュベータ内に設置するため、およびコンピュータに接続するために支持台上に載置された、図22に図示される3つの並んだ顕微鏡の写真である。図23Aは顕微鏡を示し、図23Bはインキュベータ内の顕微鏡を示す。
【図24】本研究で使用された画像捕捉ソフトウェアのスクリーンショットであり、3つのチャネルから撮像された胚を示す。
【図25A−D】実験2のステーション2からの選択された微速度画像を示す4枚の一連の写真である。図25Aおよび25Bは、培地を交換する前に捉えた画像であり、図25Cおよび25Dは、培地を交換した後に捉えた画像である。
【図26A−B】正常な胚および異常な胚の細胞活性を示すグラフ(左)および細胞写真(右)である。図26Aおよび図26Bは、合わせて、3日目に、胚が類似する形態を有するが、それらの細胞活性プロットは、大きく異なり、それらのうちの1つのみが胚盤胞に発達することを示す。
【図27A−B】マイクロウェルを備えた、特注のペトリ皿の図である。図27Aは、寸法とともに皿の図を示し、図27Bは、マイクロウェルの3D表示を示す。
【図28A−B】事前画像登録をした、および事前画像登録をしない細胞活性を示すグラフである。図28Aおよび28Bは、併せると、登録が結果を整理し、胚の移動および回転によるスパイクを取り除くことを示す。
【図29A−B】正常な胚および異常な胚の細胞活性を示すグラフ(左)および細胞写真(右)である。図29Aおよび図29Bは、併せて、3日目に、胚が類似する形態を有するが、それらの細胞活性プロットは大きく異なり、それらのうちの1つのみが胚盤胞に発達することを示す。
【図30】胚の発達中の連続する組の画像間における画素強度の差を示すグラフである。これは、胚の生存能を評価するために単独で使用されるか、またはどのくらいの粒子(予測された胚モデル)が使用されるべきかを判定することにより、粒子フィルタ等の他のアルゴリズムを改善するための方式として使用され得る。
【図31A−G】種々の細胞期の2D追跡の結果を示す7枚の一連の写真である。フレーム番号で示される細胞の進行は、各画像組と関連する:フレーム15(図31A)、45(B)、48(C)、189(D)、190(E)、196(F)および234(G)。下段は、重ね合わせたシミュレーション画像を示す。輪郭は、可視細胞膜であり、白い点線は、遮蔽された膜である。画像フレームは、5分毎に捉えられ、数画像のみが示される。
【図32A−B】3D細胞追跡の2つの成功例を示す一連の写真および図である。胚の各写真の図は、それぞれ、フレーム314およびフレーム228においてモデルの側面からの視点を示すフレーム314およびフレーム228を除き、3Dモデルの上からの視点を示す。画像フレームは、5分毎に捉えられた。
【図33】1細胞〜2細胞分割についての粒子フィルタ結果の図表示である。データ点は、細胞中心の3D位置である。1細胞モデル、2細胞モデル、3細胞モデル、および4細胞モデルについての点を示す。上段は、予測後の粒子、そして下段は、再サンプリング後の粒子を示す。
【図34A−B】14個一組の胚における自動化対手動の画像分析の比較を示すグラフである。図34Aは、第1細胞質分裂の持続時間についての比較を示し、図34Bは、第1と第2の有糸分裂の間の時間についての比較を示す。
【図35】胚をモデル化し、ある形態パラメータを測定するために画像分析がどのように使用されるかを示すフローチャートである。
【発明を実施するための形態】
【0015】
本発明の方法および組成物を説明する前に、本発明は、説明される特定の方法または組成物に限定されず、そのようなものとして、当然のことながら、変動し得ることを理解する。本発明の範囲は、付属の請求項によってのみ限定されるため、本明細書に使用される用語は、単に特定の実施形態を説明する目的のためであり、限定することが意図されないことも理解する。
【0016】
値の範囲が提供される場合、その範囲の上限と下限の間の各介在値も、文脈に明確に指示されない限り、下限の単位の10分の1まで、具体的に開示されることを理解する。規定範囲にあるいかなる規定値または介在値と、その規定範囲にある他のいかなる規定値または介在値との間の各小範囲は、本発明内に包含される。これらの小範囲の上限および下限は、独立して範囲に含まれるか、または除外され、小範囲にいずれかが含まれる、いずれも含まれない、および双方が含まれる各範囲も、本発明内に包含され、規定範囲において特に除外されるいかなる限度も対象となる。規定範囲が1つ、または双方の限度を含む場合において、これらの含まれた限度のいずれか、または双方を除外する範囲も本発明に含まれる。
【0017】
特に定義されない限り、本明細書に使用される全ての技術および科学的用語は、本発明が属する技術の当業者に一般に理解されるのと同様の意味を持つ。本明細書に説明されるものと類似、または同等のあらゆる方法および材料は、本発明の実践または試験において使用され得るが、一部の可能性のある、および好ましい方法ならびに材料をここで説明する。本明細書に言及される全ての刊行物は、引用される刊行物に関連する方法および/または材料を公開および説明するために参照により本明細書に組み込まれる。組み込まれた刊行物のあらゆる開示と矛盾がある点では、本開示が優先することを理解する。
【0018】
本明細書および付属の請求項に使用される、単数形「a」、「an」、および「the」は、文脈に明確に示されない限り、複数形の指示物を含むことに留意するべきである。よって、例えば、「細胞(a cell)」との言及は、複数のそのような細胞を含み、「ペプチド」との言及は、当業者に公知である、1つ以上のペプチドおよびその等価物、例えば、ポリペプチドへの言及を含み、他も同様である。
【0019】
本明細書において論じられる刊行物は、単に、本出願の出願日前の開示であるために、提供される。本明細書におけるいかなる記載も、本発明が、先の発明の長所によって、そのような刊行物に先行する権利を与えられていないと認めるものとして解釈されるべきではない。さらに、提供された刊行物の日付は、それぞれに確認される必要がある実際の公表日とは異なっている場合がある。
【0020】
定義
1つ以上の胚もしくは多能性細胞の発達の可能性および/または1つ以上の胚もしくは多能性細胞における染色体異常の存在を判定するための方法、組成物、およびキットを提供する。これらの方法、組成物、およびキットは、ヒトにおける不妊の治療に最も有用である生体外における胚および卵母細胞を識別するのに使用される。本発明のこれらの、および他の目的、利点、および特色は、以下にさらに詳しく説明する、本発明の方法および組成物の詳細を読むことにより当業者に明らかになるだろう。
【0021】
「発達の可能性」および「発達能力」という用語は、本明細書において、健康な胚または多能性細胞の成長もしくは発達する能力または潜在的可能性を指すように使用される。
【0022】
「胚」という用語は、本明細書において、2つの半数体配偶子細胞、例えば、未受精の二次卵母細胞および精子細胞が合体して、二倍体全能性細胞、例えば、受精卵を形成する時に形成される接合体と、直後の細胞分割、すなわち、胚分割から桑実胚、すなわち、16細胞期および胚盤胞期(分化した栄養外胚葉および内細胞塊を有する)までを生じる胚の双方を指すように使用される。
【0023】
「多能性細胞」という用語は、本明細書において、生物において、複数の細胞型に分化する能力を有するあらゆる細胞を意味するように使用される。多能性細胞の例としては、幹細胞卵母細胞、および1細胞胚(すなわち、接合体)が挙げられる。
【0024】
「幹細胞」という用語は、本明細書において、(a)自己再生する能力を有する、および(b)多様な分化細胞型を生じる可能性を有する細胞、または細胞集団を指すように使用される。しばしば、幹細胞は、複数の細胞の系列を生じる可能性を有する。本明細書に使用される、幹細胞は、全能性幹細胞、例えば、生物の胚および胚外組織の全てを生じる受精卵母細胞;多能性幹細胞、例えば、生物の胚組織の全て、すなわち、内胚葉、中胚葉、および外胚葉系列を生じる胚幹(ES)細胞、胚生殖(EG)細胞、または誘発された多能性幹(iPS)細胞;多能性幹細胞、例えば、生物の胚組織のうちの少なくとも2つ、すなわち、内胚葉、中胚葉、および外胚葉系列のうちの少なくとも2つを生じる間葉系幹細胞であるか、または特定の組織の複数の分化細胞型を生じる組織特異的幹細胞であり得る。組織特異的幹細胞は、特定の組織の細胞を生じる組織特異的胚細胞、ならびに成人の組織に属し、その組織の細胞、例えば、中枢神経系の細胞の全てを生じる神経幹細胞、骨格筋を生じる衛星細胞、および造血系の細胞の全てを生じる造血幹細胞を生じる体性幹細胞を含む。
【0025】
「卵母細胞」という用語は、本明細書において、未受精の女性生殖細胞または生殖体を指すように使用される。本出願の卵母細胞は、一次卵母細胞または二次卵母細胞であり得、一次卵母細胞である場合、それらは、減数分裂Iを経るように位置付けられるか、またはそれを経て、二次卵母細胞である場合、減数分裂IIを経るように位置付けられるか、またはそれを経る。
【0026】
「減数分裂」とは、生殖体の産生をもたらす細胞周期事象を意味する。最初の減数分裂細胞周期または減数分裂Iにおいて、細胞の染色体は複製され、2つの娘細胞に分配される。次いで、これらの娘細胞は、第2の減数分裂細胞周期または減数分裂IIで分割し、これは、DNA合成を伴わず、半数の染色体を有する生殖体をもたらす。
【0027】
「胚胞」期とは、減数分裂Iの細胞周期の前期I、すなわち核材料の第1分割前と相関する一次卵母細胞の成熟期を意味する。この期の卵母細胞は、「胚胞卵母細胞」とも呼ばれ、特徴的に大きな核のため、胚胞と呼ばれる。生体外で培養された正常なヒトの卵母細胞において、胚胞は、成熟開始約6〜24時間後に生じる。
【0028】
「中期I」期とは、減数分裂Iの細胞周期の中期Iと相関する一次卵母細胞の成熟期を意味する。胚胞卵母細胞との比較において、中期Iの卵母細胞は、大きい、明確に定義された核を持たない。生体外で培養された正常なヒトの卵母細胞において、中期Iは、成熟開始約12〜36時間後に生じる。
【0029】
「中期II」期とは、減数分裂IIの細胞周期の中期IIと相関する二次卵母細胞の成熟期を意味する。中期IIは、第1極体の突出により区別される。生体外で培養された正常なヒトの卵母細胞において、中期IIは、成熟開始約24〜48時間後に生じる。
【0030】
「有糸分裂細胞周期」とは、細胞の染色体の複製、ならびにこれらの染色体および細胞の細胞質物質の2つの娘細胞への分割をもたらす事象を意味する。有糸分裂細胞周期は、静止期と有糸分裂の2つの期に分けられる。静止期において、細胞は成長し、そのDNAを複製する。有糸分裂において、細胞は、細胞分割を開始し、完了するが、最初にその核材料を分配し、その後、その細胞質材料とその分配した核材料(細胞質分裂)を2つの個別の細胞に分割する。
【0031】
「第1の有糸分裂細胞周期」または「細胞周期1」とは、受精から第1細胞質分裂事象、すなわち、受精卵母細胞の2つの娘細胞への分割の完了までの時間間隔を意味する。卵母細胞が生体外受精である場合、ヒト絨毛性ゴナドトロピン(HCG)の注入(一般に、卵母細胞回収前に投与される)と第1細胞質分裂事象の完了との間の時間間隔が、代理時間間隔として使用され得る。
【0032】
「第2の有糸分裂細胞周期」または「細胞周期2」とは、胚で観測される第2細胞周期事象を意味し、有糸分裂による受精卵母細胞からの娘細胞の産生と、有糸分裂によるこれらの娘細胞(「先行娘細胞」または娘細胞A)のうちの1つからの第1の一組の孫娘細胞の産生との間の時間間隔である。細胞周期2が完了すると、胚は3つの細胞から成る。言い換えると、細胞周期2は、2細胞を含有する胚と3細胞を含有する胚との間の時間として視覚的に識別することができる。
【0033】
「第3の有糸分裂細胞周期」または「細胞周期3」とは、胚で観測される第3細胞周期事象を意味し、通常、有糸分裂による受精卵母細胞からの娘細胞の産生から有糸分裂による第2娘細胞(「遅れ娘細胞」または娘細胞B)からの第2の一組の孫娘細胞の産生までの時間間隔である。細胞周期3が完了すると、胚は4つの細胞から成る。言い換えると、細胞周期3は、3細胞を含有する胚と4細胞を含有する胚との間の時間として視覚的に識別することができる。
【0034】
「第1分割事象」とは、第1分割、すなわち、卵母細胞の2つの娘細胞への分割、すなわち、細胞周期1を意味する。第1の分割事象が完了すると、胚は2つの細胞から成る。
【0035】
「第2分割事象」とは、第2の一組の分割、すなわち、先行娘細胞の2つの孫娘細胞への分割、および遅れ娘細胞の2つの孫娘細胞への分割を意味する。言い換えると、第2の分割事象は、細胞周期2および細胞周期3の双方から成る。第2の分割が完了すると、胚は4つの細胞から成る。
【0036】
「第3分割事象」とは、第3の一組の分割、すなわち、全ての孫娘細胞の分割を意味する。第3分割事象が完了すると、胚は、通常、8つの細胞から成る。
【0037】
「細胞質分裂」または「細胞分割」とは、細胞が細胞分割を受ける有糸分裂の時期を意味する。言い換えると、細胞の分配された核材料およびその細胞質材料が分割されて、2つの娘細胞を産生する有糸分裂期である。細胞質分裂の期間は、細胞膜の圧縮(「分裂溝」)が最初に観測された時から、その圧縮事象の分離、すなわち、2つの娘細胞の生成の間の期間または時間枠として識別可能である。分裂溝の開始は、細胞膜の湾曲が凸面(外側に向かって丸い)から凹面(くぼみまたはへこみを伴う内側への屈曲)に変化する点として視覚的に識別され得る。これは、2つの分裂溝を示す白色矢印により、図4の上部パネルに図示される。細胞伸長の開始は、細胞質分裂の開始を標識するためにも使用され得、この場合、細胞質分裂の期間は、細胞伸長の開始と細胞分割の分離との間の期間として定義される。
【0038】
「第1細胞質分裂」または「細胞質分裂1」とは、受精後最初の細胞分割事象、すなわち、受精卵母細胞が分割して、2つの娘細胞を産生することを意味する。第1細胞質分裂は、一般に、受精約1日後に生じる。
【0039】
「第2細胞質分裂」または「細胞質分裂2」とは、第2細胞分割事象が胚において観測される、すなわち、受精卵母細胞の娘細胞(「先行娘細胞」または娘A)が第1の一組の2つの孫娘細胞に分割することを意味する。
【0040】
「第3細胞質分裂」または「細胞質分裂3」とは、第3細胞分割事象が胚において観測される、すなわち、受精卵母細胞の他の娘細胞(「遅れ娘細胞」または娘B)が第2の一組の2つの孫娘細胞に分割することを意味する。
【0041】
「位置合わせマーカー(fiduciary marker)」または「位置合わせマーカー(fiducial marker)」という用語は、参照点または測定点として使用するための、産生された画像に現れる撮像システムの視野域で使用される目標物である。撮像対象物の中または上に設置される何かか、または光学装置のレチクルの印もしくは一組の印のいずれかであり得る。
【0042】
「マイクロウェル」という用語は、細胞規模で寸法決定された、好ましくは、単一真核細胞を収容するために提供される容器を指す。
【0043】
目的の多能性細胞および胚
本発明の方法において、1つ以上の胚または多能性細胞は、胚または多能性細胞の1つ以上の細胞パラメータを測定し、これらの測定値を利用して、胚または多能性細胞の発達の可能性を判定することにより、その発達の可能性について評価される。よって、得られた情報は、例えば、生体外受精胚を移植するか、培養した1つの細胞または複数の細胞を移植するか、という臨床的決定を導くために使用され得る。
【0044】
本発明の方法により評価され得る胚の例としては、1細胞胚(接合体とも称される)、2細胞胚、3細胞胚、4細胞胚、5細胞胚、6細胞胚、8細胞胚等、通常、最大16細胞胚までが挙げられ、それらのいずれも、あらゆる都合のよい様式、例えば、生体内で成熟した卵母細胞、または生体外で成熟した卵母細胞に由来し得る。
【0045】
本発明の方法により評価され得る多能性細胞の例としては、全能性幹細胞、例えば、一次卵母細胞および二次卵母細胞等の卵母細胞、多能性幹細胞、例えば、ES細胞、EG細胞、iPS細胞等、多能性細胞、例えば、間葉系幹細胞、ならびに組織特異的幹細胞が挙げられる。これらは、あらゆる発育期から、例えば、胚、新生児、若年、または成人、ならびにいずれかの性別、すなわちXXまたはXYであり得る。
【0046】
胚および多能性細胞は、あらゆる生物、例えば、あらゆる哺乳類種、例えば、ヒト、霊長類、ウマ、ウシ、ブタ、イヌ、ネコ等に由来し得る。好ましくは、それらはヒトに由来する。それらは、予め凍結され得、例えば、胚は、1細胞期で凍結保存され、その後解凍されるか、または凍結および解凍された卵母細胞および幹細胞であり得る。代替的に、それらは、新たに調製された、例えば、生体外受精技法により卵母細胞から新たに調製された胚、生体外成熟技法を通して新たに採取および/もしくは新たに成熟させた、または生体外で生殖細胞に分化され、卵母細胞に成熟させた多能性幹細胞に由来する卵母細胞、当該分野において公知の方法による解離および組織の培養により新たに調製された幹細胞等であり得る。それらは、例えば、胚については、生体外受精の分野で使用される場合等の条件下で(例えば、米国特許第6,610,543号、米国特許第6,130,086号、米国特許第5,837,543号を参照し、それらの開示は、参照により本明細書に組み込まれる)、卵母細胞においては、卵母細胞の成熟を促進するために当該分野において使用される場合等の条件下で(例えば、米国特許第5,882,928号、および米国特許第6,281,013号を参照し、それらの開示は、参照により本明細書に組み込まれる)、幹細胞においては、増殖を促進するために当該分野において使用される場合等の条件下で(例えば、米国特許第6,777,233号、米国特許第7037892号、米国特許第7,029,913号、米国特許第5,843,780号、および米国特許第6,200,806号、米国特許出願第2009/0047263号、米国特許出願第2009/0068742号を参照し、それらの開示は、参照により本明細書に組み込まれる)評価される試料の生存、成長、および/または発達を促進するために当該分野において公知のあらゆる都合のよい条件下で培養され得る。しばしば、胚/多能性細胞は、評価される特定の胚/多能性細胞の必要性に合わせた、血清もしくは血清代替物、アミノ酸、および成長因子を添加されたノックアウトDMEM、DMEM−F12またはイスコフ改変ダルベッコ培地等の市販の培地で培養される。
【0047】
微速度撮像分析
一部の実施形態において、胚/多能性細胞は、微速度撮像により細胞パラメータを測定することにより評価される。胚/多能性細胞は、標準の培養皿で培養され得る。代替的に、胚/多能性細胞は、特注の培養皿、例えば、本明細書に記載される光学品質のマイクロウェルを備える特注の培養皿で培養され得る。そのような特注の培養皿において、各マイクロウェルは、単一の胚/多能性細胞を保持し、各マイクロウェルの底面は、細胞の有糸分裂プロセスを追跡するのに十分な解像度を備えた単一の小型顕微鏡により、単一皿内の胚の全群が同時に撮像できるように光学品質仕上げである。マイクロウェルの全群は、培養皿の同一の培地液滴を共有し、培地液滴を安定させるためにマイクロウェルの周囲に位置付けられた外壁、ならびにマイクロウェルの近くに設置される位置合わせマーカーも含むことができる。表面の疎水性は、プラズマエッチングまたは他の処理により調節することができ、培地で満たされた時にマイクロウェル中で気泡が形成されるのを防止する。標準の培養皿または特注の培養皿が利用されるかどうかにかかわらず、培養中、1つ以上の発達胚は、同一の培養培地で培養される、例えば、1皿当り1〜30個の胚が培養され得る。
【0048】
画像は、経時的に取得され、その後、1つ以上の細胞パラメータの測定値を導き出すように分析される。微速度撮像は、デジタル画像保存および分析用に装備されるあらゆるコンピュータ制御顕微鏡で、例えば、加熱台およびインキュベーションチャンバを装備した倒立顕微鏡、または従来のインキュベータの内部に収まる特注の小型顕微鏡アレイで実施され得る。並んだ小型顕微鏡は、同一インキュベータ中の複数の試料皿を同時に培養することができ、連続画像捕捉間の最小時間間隔に対して制限がない複数のチャネルを収容するように拡張可能である。複数の顕微鏡の使用は、試料を移動する必要性を排除し、これは、システムの正確性および全体的なシステムの信頼度を改善する。インキュベータ中の個々の顕微鏡は、部分的に、または完全に隔離することができ、各培養皿に独自の制御された環境を提供する。これは、他の試料の環境を乱すことなく、皿の撮像ステーションへの、およびそこからの移動を可能にする。
【0049】
微速度撮像用の撮像システムは、明視野照明、暗視野照明、位相差、ホフマン変調コントラスト、微分干渉コントラスト、または蛍光を利用することができる。一部の実施形態において、暗視野照明は、後続の特色抽出および画像分析のために、強調した画像コントラストを提供するために使用され得る。加えて、赤色または近赤外線源は、光毒性を低減し、細胞膜と細胞の内側部分との間のコントラスト比を改善するために使用され得る。
【0050】
取得された画像は、ライブビデオと同様に継続して、または微速度撮影と同様に断続してのいずれかで保存され得、ここで、対象物は、静止画で繰り返し撮像される。好ましくは、画像間の時間間隔は、以下に説明される有意な形態学的事象を捕捉するために、1〜30分の間であるべきである。代替的な実施形態において、画像間の時間間隔は、細胞活性量により変動し得る。例えば、活性期間中、画像は、数秒毎または数分毎に撮られ、一方、不活性期間中、画像は、10分もしくは15分毎、またはそれ以上毎に撮られ得る。捕捉した画像に対するリアルタイム画像分析は、いつ、およびどのように時間間隔が変動するかを検知するために使用される。我々の方法において、試料が受ける総光量は、5日間の撮像において、約24分の連続低線量光曝露に相当すると推定される。微速度撮像システムの光強度は、低電力のLED(例えば、通常の100Wのハロゲン球電球と比較して1WのLEDを使用)、および高感度のカメラセンサにより、生殖補助顕微鏡で通常使用される光強度より有意に低い。よって、微速度撮像システムを使用する胚が受ける総光エネルギー量は、IVF臨床での日常的取り扱い中に受けるエネルギー量と同程度か、またはそれ未満である。加えて、曝露時間は、胚/多能性細胞への総光曝露量を低減するために有意に短縮され得る。画像当り0.5秒の光曝露で、5分毎に捕捉された画像の2日間の撮像において、総低線量光曝露量は、5分未満である。
【0051】
画像取得後、画像は抽出され、異なる細胞パラメータ、例えば、細胞の大きさ、透明帯の厚さ、断片化の程度、細胞分割によって生じる娘細胞の対称性、最初のいくつかの有糸分裂間の時間間隔、および細胞質分裂の持続時間について分析される。
【0052】
微速度撮像により測定され得る細胞パラメータは、一般に、形態学的事象である。例えば、胚の評価において、微速度撮像は、細胞質分裂事象、例えば、細胞質分裂1、細胞質分裂2、細胞質分裂3、または細胞質分裂4の持続時間を測定するために使用され得、ここで、細胞質分裂事象の持続時間は、分裂溝(細胞質分裂の開始)の最初の観測と分裂溝の2つの娘細胞への分離(すなわち、2つの娘細胞の産生)との間の時間間隔として定義される。別の目的のパラメータは、細胞周期事象、例えば、細胞周期1、細胞周期2、細胞周期3、または細胞周期4の持続時間であり、ここで、細胞周期事象の持続時間は、細胞の産生(細胞周期1においては、卵子の受精、後の細胞周期においては、細胞質分裂の分離時)とその細胞からの2つの娘細胞の産生との間の時間間隔として定義される。微速度撮像により測定され得る他の目的の細胞パラメータは、これらの細胞事象により定義される時間間隔、例えば、(a)細胞質分裂1の開始と細胞質分裂2の開始との間の間隔、細胞質分裂1の分離と細胞質分裂2の分離との間の間隔、細胞質分裂1の開始と細胞質分裂2の分離との間の間隔、もしくは細胞質分裂1の分離と細胞質分裂2の開始との間の間隔のうちのいずれか1つとして定義可能である、細胞質分裂1と細胞質分裂2との間の時間間隔、または(b)細胞質分裂2の開始と細胞質分裂3の開始との間の間隔、もしくは細胞質分裂2の分離と細胞質分裂3の分離との間の間隔、もしくは細胞質分裂2の開始と細胞質分裂3の分離との間の間隔、もしくは細胞質分裂2の分離と細胞質分裂3の開始との間の間隔のうちのいずれか1つとして定義可能である、細胞質分裂2と細胞質分裂3との間の時間間隔を含む。
【0053】
生体外受精の目的において、生体外環境に対する培養条件の欠点による胚の損失を低減するため、および培養中に生じ得る後成的エラーに関連する有害転帰の可能性を低減するために、胚は、発達の初期に、例えば、2日目、または3日目までに、すなわち、8細胞期に至るまでに子宮に移植されることが有利であると考えられる(Katari et al.(2009)Hum Mol Genet.18(20):3769−78、Sepulveda et al.(2009)Fertil Steril.91(5):1765−70)。したがって、より長期の分析、例えば、約36時間、約54時間、約60時間、約72時間、約84時間、約96時間、またはそれ以上も、本方法によって意図されるが、細胞パラメータの測定は、受精から2日以内に行われることが好ましい。
【0054】
微速度撮像により評価され得る成熟卵母細胞の細胞パラメータの例としては、卵母細胞膜の形態の変化、例えば、透明帯からの分離の速度および程度;卵母細胞核の形態の変化、例えば、胚胞崩壊(GVBD)の開始、完了、および速度;細胞質および核における顆粒移動の速度および方向;卵母細胞および第1極体の細胞質分裂、ならびに第1極体の突出の移動および/または持続時間が挙げられるが、これらに限定されない。他のパラメータは、成熟二次卵母細胞、および第2極体の細胞質分裂の持続時間を含む。
【0055】
微速度撮像により評価され得る幹細胞または幹細胞集団の細胞パラメータの例としては、細胞質分裂事象の持続時間、細胞質分裂事象間の時間、細胞質分裂事象前およびその間の幹細胞の大きさおよび形状、細胞質分裂事象により産生された娘細胞の数、分裂溝の空間的配位、観測された非対称分割の速度および/または数(すなわち、1つの娘細胞が幹細胞を維持し、一方、他は分化する)、観測された対称分割の速度および/または数(すなわち、双方の娘細胞が幹細胞として留まるか、双方が分化するかのいずれかである)、および細胞質分裂事象の分離と幹細胞が分化し始める時との間の時間間隔が挙げられるが、これらに限定されない。
【0056】
パラメータは、手動で測定されるか、または自動的に、例えば、画像分析ソフトウェアにより測定され得る。画像分析ソフトウェアが利用される場合、例えば、仮定される胚/多能性細胞モデルの分布を生成する、単純な光学モデルに基づき画像をシュミレーションする、およびこれらのシミュレーションを観測した画像データと比較する、遂次モンテカルロ法に基づく確率的モデル推定技法を利用する画像分析アルゴリズムが使用され得る。そのような確率的モデルによる推定が利用される場合、細胞は、あらゆる適切な形状として、例えば、2D空間における楕円の集まり、3D空間における楕円体の集まり等としてモデル化され得る。遮蔽および深度の不明確さに対処するために、方法は、予想された物理的挙動に対応する幾何学的制約を強化することができる。確実性を改善させるために、画像は、1つ以上の焦点面で捕捉され得る。
【0057】
遺伝子発現分析
一部の実施形態において、胚または多能性細胞は、遺伝子発現を測定することにより評価され得る。そのような実施形態において、細胞パラメータは、遺伝子発現レベル、または遺伝子発現プロファイルである。1つ以上の発現を判定すること、すなわち、発現プロファイルまたは発現評価を得ることは、1つ以上の目的の遺伝子の核酸転写、例えば、mRNAを測定することにより、または1つ以上の目的の遺伝子の発現産物である、1つ以上の異なるタンパク質/ポリペプチドのレベル、例えば、プロテオーム発現プロファイルを測定することにより行われる。言い換えると、「発現プロファイル」および「発現評価」という用語は、RNAレベルまたはタンパク質レベルの遺伝子発現プロファイルを含むように広範に使用される。
【0058】
一部の実施形態において、遺伝子の発現は、核酸発現プロファイルを得ることにより評価され得、ここで、試料中の1つ以上の核酸の量またはレベル、例えば、1つ以上の目的の遺伝子の核酸転写が判定される。これらの実施形態において、発現プロファイルを生成するためにアッセイされる試料は、核酸試料である。核酸試料は、評価される胚または細胞の目的の遺伝子の発現情報を含む複数の別個の核酸、または別個の核酸の集団を含む。核酸は、試料が得られる宿主細胞または組織の発現情報を保有する限り、RNAまたはDNA核酸、例えば、mRNA、cRNA、cDNA等を含み得る。試料は、当該分野において公知である多くの異なる方式で、例えば、細胞からのmRNAの単離により調製され(ここで、単離されたmRNAはそのまま使用される)、差次的発現分野において公知であるように、cDNA、cRNA等を調製するために増幅、利用され得る。試料は、標準のプロトコルを使用して、単一細胞、例えば、目的の多能性細胞の培養物の多能性細胞、もしくは目的の胚からの単一細胞(卵割球)から、またはいくつかの細胞、例えば、多能性細胞の培養物の画分、または目的の胚の2、3、もしくは4つ以上の卵割球から調製され得る。
【0059】
発現プロファイルは、あらゆる都合のよいプロトコルを使用して、最初の核酸試料から生成され得る。差次的遺伝子発現分析の分野において利用されるもののような、発現プロファイルを生成する種々の異なる手段が公知であるが、発現プロファイルを生成するための代表的な都合のよいプロトコルの種類の1つは、アレイベースの遺伝子発現プロファイル生成プロトコルである。そのような用途は、生成されるプロファイルにおいて、アッセイされる/プロファイルされる遺伝子のそれぞれの「プローブ」核酸を示す核酸が利用されるハイブリダイゼーションアッセイである。これらのアッセイにおいて、標的核酸の試料がアッセイされる最初の核酸試料から最初に調製され、ここで、調製は、標識、例えば、シグナル産生システムの構成要素で標的核酸を標識することを含み得る。標的核酸試料調製後、ハイブリダイゼーション条件下で試料をアレイと接触させ、それによって、複合体がアレイ表面に付着するプローブ配列に相補的である標的核酸間で形成される。その後、ハイブリッド形成された複合体の存在は、定性的または定量的のいずれかで検出される。
【0060】
本発明の方法で利用される発現プロファイルを生成するために実践され得る具体的なハイブリダイゼーション技術は、米国特許第5,143,854号、第5,288,644号、第5,324,633号、第5,432,049号、第5,470,710号、第5,492,806号、第5,503,980号、第5,510,270号、第5,525,464号、第5,547,839号、第5,580,732号、第5,661,028号、第5,800,992号(それらの開示は、参照により本明細書に組み込まれる)、ならびに国際公開第WO95/21265号、第WO96/31622号、第WO97/10365号、第WO97/27317号、欧州特許第373 203号、および第EP785 280号に記載される技術を含む。これらの方法において、発現がアッセイされる表現型決定遺伝子のそれぞれについてのプローブを含む複数の「プローブ」核酸が、上述のように標的核酸と接触させられる。接触は、ハイブリダイゼーション条件下、例えば、厳密なハイブリダイゼーション条件下で行われ、その後、未結合核酸は除去される。本明細書に使用される、「厳密なアッセイ条件」という用語は、所望の特異性を提供するのに十分に相補的でない結合構成要素間での結合対の形成にあまり適していないが、アッセイにおいて所望の特異性レベルを提供するのに十分に相補的である核酸の結合対、例えば、表面結合および溶液相を産生するのに適している条件を指す。厳密なアッセイ条件は、ハイブリダイゼーションおよび洗浄条件の双方の総和または組み合わせ(全体性)である。
【0061】
得られたハイブリッド形成された核酸のパターンは、プローブされた遺伝子のそれぞれの発現に関する情報を提供し、ここで、発現情報は、遺伝子が発現するかどうか、および通常どんなレベルで発現するかに関してであり、発現データ、すなわち、発現プロファイル(例えば、トランスクリプトソームの形態で)は、定性的および定量的の双方であり得る。
【0062】
代替的に、試料中の1つ以上の核酸のレベルを定量化するための非アレイベースの方法が利用され得、増幅プロトコルに基づくもの、例えば、定量的PCR、逆転写PCR(RT−PCR)、リアルタイムPCR等を含む、ポリメラーゼ連鎖反応(PCR)ベースのアッセイを含む。
【0063】
一部の実施形態において、遺伝子の発現は、プロテオーム発現プロファイルを得ることにより評価され得、ここで、例えば、目的の遺伝子によりコードされるタンパク質/ポリペプチド等の、試料中の1つ以上のタンパク質/ポリペプチドの量またはレベルが判定される。これらの実施形態において、方法において利用される発現プロファイルを生成するためにアッセイされる試料は、タンパク質の試料である。発現プロファイルがプロテオーム発現プロファイル、すなわち、試料中の1つ以上のタンパク質レベルのプロファイルである場合、タンパク質レベルを評価するためのあらゆる都合のよいプロトコルが利用され得、アッセイされた試料中の1つ以上のタンパク質のレベルが判定される。
【0064】
タンパク質レベルについてアッセイする種々の異なる手段は、当該分野において公知であるが、タンパク質レベルをアッセイするための代表的な都合のよいプロトコルの種類は、ELISAである。ELISAおよびELISAベースのアッセイにおいて、目的のタンパク質に特異的な1つ以上の抗体は、選択された固体表面上、好ましくは、ポリスチレンマイクロタイタープレートのウェル等の、タンパク質親和性を示す表面に固定され得る。不完全に吸着された材料を除去するために洗浄した後、アッセイプレートウェルは、ウシ血清アルブミン(BSA)、カゼイン、または粉乳の溶液等の、試験試料に関して抗原性的に中性であることが知られている非特異的「ブロッキング」タンパク質でコーティングされる。これは、固定化表面上の非特異的吸着部位のブロッキングを可能にし、これによって、抗原の表面への非特異的結合により生じるバックグラウンドを低減する。未結合のブロッキングタンパク質を除去するために洗浄した後、免疫複合体(抗原/抗体)形成をもたらす条件下で、固定化表面を試験される試料と接触させる。そのような条件は、リン酸緩衝生理食塩水(PBS)/TweenまたはPBS/Triton−X100中にBSAまたはウシガンマグロブリン(BGG)を希釈する等、希釈液で試料を希釈することを含み、これは、非特異的バックグラウンドの低減も補助する傾向にあり、(他の温度も使用され得るが)およそ約25°〜27℃の温度で約2〜4時間、試料をインキュベートすることを可能にする。インキュベーション後、非免疫複合材料を除去するために、抗血清接触面を洗浄する。例示的な洗浄手順は、PBS/Tween、PBS/Triton−X100、またはホウ酸緩衝液等の溶液で洗浄することを含む。その後、結合免疫複合体を第1の抗体と異なる標的に特異性を有する第2の抗体に曝し、第2の抗体の結合を検出することにより、免疫複合体形成の発生および量が判定される。特定の実施形態において、第2の抗体は、関連酵素、例えば、ウレアーゼ、ペルオキシダーゼ、またはアルカリホスファターゼを有し、これは、適切な発色基質を用いてインキュベートすることにより、有色の沈殿物を生成する。例えば、ウレアーゼまたはペルオキシダーゼ抱合抗ヒトIgGは、免疫複合体形成の発達に好ましい時間および条件下で(例えば、PBS/Tween等のPBS含有溶液中で、室温で2時間インキュベートする)利用され得る。第2の抗体を用いたそのようなインキュベーション、および未結合材料を除去するための洗浄の後、例えば、ウレアーゼ標識の場合においては、尿素およびブロモクレゾールパープル、またはペルオキシダーゼ標識の場合においては、2,2’−アジノ−ジ−(3−エチル−ベンズチアゾリン)−6−スルホン酸(ABTS)およびH2O2等の、発色基質を用いてインキュベーションすることにより、標識の量を定量化する。その後、定量化は、例えば、可視スペクトル分光光度計を使用して、色の生成の程度を測定することにより達成される。
【0065】
先行フォーマットは、最初に試料をアッセイプレートに結合することにより変更され得る。次いで、一次抗体を、アッセイプレートを用いてインキュベートした後、一次抗体に特異性を有する標識された第2の抗体を使用して、結合した一次抗体を検出する。
【0066】
抗体または複数の抗体が固定される固体基質は、広範な種々の材料から、および広範な種々の形状、例えば、マイクロタイタープレート、マイクロビーズ、計深棒、樹脂粒子等に作製され得る。基質は、シグナル対ノイズ比を最大にするように、バックグラウンド結合を最小にするように、ならびに分離および費用を軽減するように選択され得る。洗浄は、基質が使用されるのに最も適した手段、例えば、リザーバからビーズもしくは計深棒を除去する、マイクロタイタープレート等のリザーバを空にする、もしくは希釈する、またはビーズ、粒子、クロマトグラフカラム、もしくはフィルタを洗浄溶液または溶媒ですすぐことにより達成され得る。
【0067】
代替的に、試料中の1つ以上のタンパク質のレベルを測定するために、非ELISAベースの方法が利用され得る。代表的な例としては、質量分析、プロテオームアレイ、xMAP(商標)微小球体技術、フローサイトメトリー、ウエスタンブロット、および免疫組織化学が挙げられるが、これらに限定されない。
【0068】
得られたデータは、プローブされた遺伝子のそれぞれの発現に関する情報を提供し、発現情報は、遺伝子が発現するかどうか、および通常どんなレベルで発現するかに関してであり、また発現データは、定性的および定量的の双方であり得る。
【0069】
発現プロファイルの生成において、一部の実施形態において、試料は、少なくとも1つの遺伝子/タンパク質、時折、複数の遺伝子/タンパク質についての発現データを含む発現プロファイルを生成するためにアッセイされ、ここで、複数とは、少なくとも2つの異なる遺伝子/タンパク質、多くの場合、少なくとも約3つ、通常少なくとも約10以上、およびより一般には、50以上、もしくは100以上等の少なくとも約15以上の異なる遺伝子/タンパク質を意味する。
【0070】
最も広い意味において、発現の評価は、定性的または定量的であり得る。そのようなものとして、検出が定性的である場合、本方法は、核酸または発現産物などの標的検体がアッセイされる試料中に存在するかどうかの測定または評価、例えば鑑定を提供する。また他の実施形態において、本方法は、標的検体がアッセイされる試料中に存在するかどうかの定量的検出、すなわち、例えば、アッセイされる試料中の核酸またはタンパク質などの標的検体の実際の量もしくは相対的存在量の評価または鑑定を提供する。そのような実施形態において、定量的検出は絶対的であり得、もしその方法が試料中の例えば標的核酸もしくはタンパク質の2つ以上の異なる検体を検出する方法である場合、相対的であり得る。そのようなものとして、例えば、試料中の核酸またはタンパク質の標的検体を定量化する場面において使用される時、「定量化」という用語は、絶対的または相対的定量化を指すことができる。絶対的定量化は、1つ以上の対照検体の既知濃度を含むことにより、および既知の対照検体を用いて標的検体の検出レベルを参照する、すなわち、標準化することにより(例えば、標準曲線の生成を介して)達成され得る。代替的に、相対的定量化は、2つ以上の異なる標的検体間の検出レベルまたは量を比較して、例えば、相互に対比して、2つ以上の異なる検体のそれぞれの相対的定量化を提供することにより達成され得る。
【0071】
発現レベルが接合体の発達の可能性を予測する遺伝子の例としては、コフィリン(NM_005507)、DIAPH1(NM_001079812、NM_005219)、ECT2(NM_018098)、MYLC2/MYL5(NM_002477)、DGCR8(NM_022720)、Dicer/DICER1(NM_030621、NM_177438)、TARBP2(NM_004178、NM_134323、NM_134324)、CPEB1(NM_001079533、NM_001079534、NM_001079535、NM_030594)、Symplekin/SYMPK(NM_004819)、YBX2(NM_015982)、ZAR1(NM_175619)、CTNNB1(NM_001098209、NM_001098210、NM_001098210、NM_001904)、DNMT3B(NM_006892、NM_175848、NM_175849、NM_175850)、TERT(NM_198253、NM_198255)、YY1(NM_003403)、IFGR2/IFNGR2(NM_005534)、BTF3(NM_001037637、NM_001207)、およびNELF(NM_001130969、NM_001130970、NM_001130971、NM_015537)が挙げられる。発現レベルが胚の発達の可能性を予測する細胞パラメータとして機能する他の遺伝子を図8に提供する。遺伝子発現レベル測定値を導き出すことにおいて、発現レベルは、評価された後、標準対照に対して標準化されることが多く、標準対照とは例えばGAPDHもしくはRPLPOという発達を通して一定であることが知られている遺伝子、またはその時間点での発現が知られている遺伝子、の試料中の発現レベルである。
【0072】
遺伝子発現レベルは、単一細胞、例えば、目的の胚からの卵割球、もしくは単離した卵母細胞、もしくは幹細胞の培養物から単離した細胞等から判定され得るか、またはそれらは、胚、例えば、目的の全胚までを含む、2、3、または4個以上の目的の胚の卵割球、もしくは幹細胞の全培養物までを含む、幹細胞の培養物からの複数の細胞等から決定され得る。
【0073】
他の態様において、本発明は、単一細胞で遺伝子型の特定および遺伝子発現分析を同時に実施するためのプロトコルを含む。胚において、これは、単一細胞が胚から取り出され、そのDNAが核型異常または特定の疾患遺伝子について試験される手順である、着床前遺伝子診断(PGD)を改善するために使用され得る。我々の方法は、遺伝的および遺伝子発現分析を同時に行うことを可能にする。方法は、以下のステップを含む:(1)単一細胞を少量の培地または緩衝液中に収集する、(2)遺伝子型特定及び遺伝子発現分析用プライマーの混合物を使用して、一工程の逆転写およびポリメラーゼ連鎖反応(PCR)増幅を実施する、(3)18サイクル未満のPCR後の増幅したcDNAのアリコートを収集して、増幅の直線性を保存する、(4)cDNAアリコートを使用して、定量的リアルタイムPCR等の標準技法を用いて遺伝子発現分析を実施する、(5)残りの試料を使用して、2回目のPCRを実施し、遺伝子型特定の目的のために、遺伝子情報をさらに増幅する、および(6)ゲル電気泳動等の標準技法を使用して、遺伝子型を特定する。
【0074】
画像および/または遺伝子発現分析から発達の可能性を判定
細胞パラメータ測定値が得られたら、測定値を利用して、胚/多能性細胞の発達の可能性を判定する。上述の通り、「発達の可能性」および「発達能力」という用語は、多能性細胞もしくは組織の成長する、もしくは発達する能力または潜在的可能性を指す。例えば、卵母細胞または胚において、発達の可能性は、その卵母細胞もしくは胚の健康な胚盤胞に成長する、または発達する能力もしくは潜在的可能性であり得る。別の例として、幹細胞の場合、発達の可能性は、1つ以上の目的の細胞、例えば、神経、筋肉、B−もしくはT細胞等に成長もしくは発達する能力または潜在的可能性である。一部の実施形態において、卵母細胞または胚の発達の可能性とは、その卵母細胞もしくは胚が健康な胚盤胞に発達する;子宮への移植が成功する;妊娠期間を経る;および/もしくは生きて生まれる、能力または潜在的可能性である。一部の実施形態において、多能性細胞の発達の可能性とは、その多能性細胞の1つ以上の目的の細胞、例えば、神経、筋肉、B−もしくはT細胞等に発達する、および/または生体内において目的の組織に寄与する、能力もしくは潜在的可能性である。
【0075】
「良好な発達の可能性」とは、胚/多能性細胞が統計的に所望の通りに発達する可能性を意味する、すなわち、所望の通りに、55%、60%、70%、80%、90%、95%以上の確率、例えば、100%の発達の確率を有する。言い換えると、良好な発達の可能性の判定を導き出すために使用される細胞パラメータ測定値を示す100のうちの55、100のうちの60、100うちの70、100のうちの80、100のうちの90、100のうちの95、または100のうちの100の胚もしくは多能性細胞が、実際、所望通りに発達を続ける。逆に、「不良な発達の可能性」とは、胚/多能性細胞が統計的に所望の通りに発達する可能性がないことを意味する、すなわち、所望の通りに、50%、40%、30%、20%、10%、5%以下の確率、例えば、0%の発達の確率を有する。言い換えると、不良な発達の可能性の判定を導き出すために使用される細胞パラメータ測定値を示す100のうちのわずか50、100のうちの40、100のうちの30、100のうちの20、100のうちの10、または100のうちの5以下の胚もしくは多能性細胞が、実際、所望の通りに、発達を続ける。本明細書に使用される、「正常」または「健康な」胚および多能性細胞とは、良好な発達の可能性を示し、一方、「異常な」胚および多能性細胞は、不良な発達の可能性を示す。
【0076】
一部の実施形態において、細胞パラメータ測定値は、胚/多能性細胞の発達の可能性を判定するために直接使用される。言い換えると、測定値自体の絶対値は、発達の可能性を判定するのに十分である。細胞パラメータを測定するために微速度撮像を使用する実施形態におけるこの例としては、次の(a)約0〜30分、例えば、約6〜20分、平均して約12〜14分続く細胞質分裂1、(b)約20〜27時間、例えば、約25〜27時間続く細胞周期1、(c)約8〜15時間、例えば、約9〜13時間、約11+/−2.1時間の平均値の細胞質分裂1の分離と細胞質分裂2の開始との間の時間間隔、(d)約0〜5時間、例えば、約0〜3時間、約1+/−1.6時間の平均時間の細胞質分裂2の開始と細胞質分裂3の開始との間の時間間隔が挙げられるがこれらに限定されず、それらのうちのいずれかが、単独または組み合わせで、ヒトの胚における良好な発達の可能性を示す。直接測定値の例としては、(a)約30分以上、例えば、約32、35、40、45、50、55、もしくは60分以上続く細胞質分裂1、(b)約27時間以上、例えば、28、29、もしくは30時間以上続く細胞周期1、(c)15時間以上、例えば、約16、17、18、19、もしくは20時間以上、または8時間未満、例えば、7、5、4、もしくは3時間以下続く、細胞質分裂1の分離と細胞質分裂2の開始との間の時間間隔、(d)6、7、8、9、もしくは10時間以上である、細胞質分裂2の開始と細胞質分裂3の開始との間の時間間隔が挙げられるが、これらに限定されず、それらのうちのいずれかが、単独または組合せで、ヒトの胚において不良な発達の可能性を示す。
【0077】
一部の実施形態において、細胞パラメータ測定値は、それを基準もしくは対照の胚/多能性細胞からの細胞パラメータ測定値と比較し、この比較の結果を使用して、胚/多能性細胞の発達の可能性の判定を提供することにより利用される。本明細書に使用される、「基準」および「対照」という用語は、所与の胚/多能性細胞の細胞パラメータ測定値を解釈し、それに対して発達の可能性の判定を行うために使用される、規格化された胚または細胞を意味する。基準または対照は、所望の表現型、例えば、良好な発達の可能性を有することが知られている胚/多能性細胞であり得、したがって、陽性基準または対照の胚/多能性細胞であり得る。代替的に、基準/対照の胚/多能性細胞は、所望の表現型を有さないことが知られている胚/多能性細胞であり得、したがって、陰性基準/対照の胚/多能性細胞であり得る。
【0078】
ある実施形態において、得られた細胞パラメータ測定値を、単一の基準/対照の胚/多能性細胞からの比較可能な細胞パラメータ測定値と比較し、アッセイされる胚/細胞の表現型に関する情報を得る。また他の実施形態において、得られた細胞パラメータ測定値を、2つ以上の異なる基準/胚もしくは多能性細胞からの比較可能な細胞パラメータ測定値と比較し、アッセイした胚/細胞の表現型に関するより詳細な情報を得る。例えば、アッセイされる胚もしくは多能性細胞から得られた細胞パラメータ測定値を、陽性および陰性の胚もしくは多能性細胞の双方と比較し、胚/細胞が目的の表現型を有するかに関する確実な情報を得る。
【0079】
例として、正常なヒトの胚における、すなわち、良好な発達の可能性を有する細胞質分裂1は、約0〜30分、より一般には、約6〜20分、平均して約12〜14分、すなわち、約1、2、3、4、もしくは5分、より一般には、約6、7、8、9、10、11、12、13、14、15、16、17、18、19、もしくは20分、一部の場合においては、21、22、23、24、25、26、27、28、29、もしくは最大約30分である。正常な基準の胚において観測されるものと比較して、評価される胚の細胞質分裂1が完了する時間が長ければ、不良な発達の可能性を示す。第2の例として、正常な胚の細胞周期1、すなわち、受精から細胞質分裂1完了までの時間は、通常、約20〜27時間、より一般には、約25〜27時間、すなわち、約15、16、17、18、もしくは19時間、より一般には、約20、21、22、23、もしくは24時間、より一般には、約25、26、もしくは27時間で完了する。正常な基準の胚において観測されるものと比較して、評価される胚における長い細胞周期1は、不良な発達の可能性を示す。第3の例として、正常なヒトの胚における細胞質分裂1の分離および細胞質分裂2の開始は、約8〜15時間、より頻繁には、約9〜13時間であって、約11+/−2.1時間の平均値であり、すなわち、6、7、もしくは8時間、より一般には、約9、10、11、12、13、14、もしくは最大約15時間である。正常な基準の胚において観測されるものと比較して、評価される胚における長い、または短い細胞周期2は、不良な発達の可能性を示す。第4の例として、正常なヒトの胚における細胞質分裂2の開始と細胞質分裂3の開始との間の時間間隔、すなわち、第2と第3の有糸分裂の同時発生は、一般に、約0〜5時間、より一般には、約0、1、2、もしくは3時間、平均時間は約1+/−1.6時間であり、正常な基準の胚において観測されるものと比較して、評価される胚における細胞質分裂2と細胞質分裂3の完了との間の長い間隔は、不良な発達の可能性を示す。最後に、発達の可能性を評価するためのパラメータとして遺伝子発現レベルを使用する時に、この実施形態がどのように適用されるかの例として、コフィリン、DIAPH1、ECT2、MYLC2、DGCR8、ダイサー、TARBP2、CPEB1、シンプレキン、YBX2、ZAR1、CTNNB1、DNMT3B、TERT、YY1、IFGR2、BTF3、および/またはNELFの低発現レベル、すなわち、正常な基準の2細胞胚において観測されるものと比較して、評価される2細胞胚において、1.5倍、2倍、3倍、4倍、5倍、10倍、20倍、50倍、または100倍低い発現は、不良な発達の可能性示し、一方、正常な基準の2細胞胚において観測されるものと同等もしくはそれより大きい発現は、良好な発達の可能性を示す。他の例は、例えば、評価される胚/多能性細胞と一緒に1つ以上の基準の胚または多能性細胞を観測することによる、経験的データに由来する。あらゆる基準の胚/多能性細胞、例えば、良好な発達の可能性を有する正常な基準試料、または不良な発達の可能性を有する異常な基準試料が利用され得る。一部の場合において、2つ以上の基準試料が利用され得る、例えば、正常な基準試料および異常な基準試料の双方が使用され得る。
【0080】
一部の実施形態において、微速度顕微鏡検査および発現プロファイリングの双方ではなく、微速度顕微鏡検査または発現プロファイリングにより導き出される細胞パラメータ測定値を使用することが望ましい場合がある。他の実施形態において、微速度顕微鏡検査により導き出される細胞パラメータ測定値および発現プロファイリングにより導き出される細胞パラメータ測定値を使用することが望ましい場合がある。
【0081】
上述の通り、胚もしくは多能性細胞の発達の可能性を判定するために、1つ以上のパラメータが測定され、利用され得る。一部の実施形態において、単一パラメータの測定値は、発達の可能性の判定を導き出すのに十分であり得る。一部の実施形態において、2つ以上のパラメータ、例えば、2つの細胞パラメータ、3つの細胞パラメータ、または4つ以上の細胞パラメータの測定値を利用することが望ましい場合がある。
【0082】
ある実施形態において、複数のパラメータは、より高い感度および特異性を提供し得るため、複数のパラメータをアッセイすることが望ましい場合がある。感度とは、そうであると正しく識別される実際の陽性物の比率を意味する。これは、数学的に、
【0083】
【表1】
として表され得る。よって、「陽性物」が、良好な発達の可能性を有する、すなわち、胚盤胞に発達する胚であり、「陰性物」が、不良な発達の可能性を有する、すなわち、胚盤胞に発達しない胚である方法において、100%の感度は、試験がそのように胚盤胞に発達する全ての胚を認識することを意味する。一部の実施形態において、アッセイの感度は、約70%、80%、90%、95%、98%以上、例えば、100%であり得る。特異性とは、そうであると正しく識別される陰性の比率を意味する。これは、数学的に、
【表2】
として表され得る。よって、陽性物が、良好な発達の可能性を有する、すなわち、胚盤胞に発達する胚であり、「陰性物」が、不良な発達の可能性を有する、すなわち、胚盤胞に発達しない胚である方法において、100%の特異性は、試験がそのように胚盤胞に発達しない、すなわち、胚盤胞期前に停止する、全ての胚を認識することを意味する。一部の実施形態において、アッセイの特異性は、約70%、80%、90%、95%、98%以上、例えば、100%であり得る。
【0084】
以下の実施例のセクションおよび図7に示す通り、3つのパラメータの使用は、94%の感度および93%の特異性を提供し、カットオフ点は、胚盤胞分布の標準偏差の3倍である。言い換えると、本発明の方法は、胚盤胞に発達するであろう胚の数を94%の確率(感度)で、および胚盤胞期前に停止するであろう胚の数を93%の確率(特異性)で正確に識別することができる。加えて、指定の平均値および/またはカットオフ点は、これらの値を計算するために使用されるデータセットならびに具体的な用途により修正され得る。
【0085】
一部の実施形態において、胚または多能性細胞の評価は、本発明の胚/多能性細胞、例えば、「発達の可能性の評価」、「染色体異常の評価」等の、当業者の評価を含む書面報告書を作成することを含む。よって、本発明の方法は、そのような評価の結果を提供する報告書を作成する、または出力するステップをさらに含む場合があり、この報告書は、電子媒体(例えば、コンピュータモニタ上に電子表示)の形態で、または有形媒体(例えば、用紙に印刷された報告書もしくは他の有形媒体)の形式で提供され得る。
【0086】
本明細書に記載される、「報告書」とは、本発明の方法により導き出された評価に関する目的の情報を提供する報告要素を含む、電子または有形文書である。本発明の報告書は、完全に、または部分的に電子的に作成され得る。本発明の報告書は、少なくとも本発明の胚または多能性細胞の発達の可能性の評価、染色体異常の存在の確率の評価等を含む。本発明の報告書は、1)試験施設に関する情報、2)業務提供者の情報、3)対象のデータ、4)試料データ、5)評価がどのように導き出されたかに関する情報を提供する詳細な評価報告セクション、例えば、a)取得した細胞パラメータ測定値、b)もしあれば、利用された基準値、および6)他の特色のうちの1つ以上をさらに含む場合がある。
【0087】
報告書は、試験施設についての情報を含む場合があり、この情報は、試料収集および/またはデータ生成が行われた、病院、クリニック、または研究所に関連する。試料収集は、試料がどのように生成されたか、例えば、対象からどのように採取されたか、および/またはどのように培養されたか等を含む場合がある。データ生成は、画像がどのように取得されたか、または遺伝子発現プロファイルがどのように分析されたかを含む場合がある。この情報は、例えば、試験施設の名称および所在地、アッセイを実施した、および/または入力データを入力した研究技術者の身元、アッセイが実施された、および/または分析された日付および時間、試料および/または結果データが保存される場所、アッセイに使用された試薬(例えば、キット等)のロット番号等に関する1つ以上の詳細を含む場合がある。この情報の報告欄は、一般に、ユーザにより提供される情報を使用して作成され得る。
【0088】
報告書は、業務提供者についての情報を含む場合があり、これは、ユーザが居る医療機関の外部、または医療機関の内部に位置し得る。そのようは情報の例としては、業務提供者の名称および所在地、調査者の氏名、ならびに必要な場合、または所望する場合、試料の調製および/もしくはデータ生成を行った個人の氏名を含む場合がある。この情報の報告欄は、一般に、ユーザにより入力されたデータを使用して作成され得、これは、予め書かれた選択項目の中から(例えば、ドロップダウンメニューを使用して)選択され得る。報告書における他の業務提供者の情報は、結果および/または解釈的報告についての技術情報の連絡先を含む場合がある。
【0089】
報告書は、対象データセクションを含み得、卵母細胞または多能性細胞が採取された対象の病歴、患者の年齢、生体外受精周期の特徴(例えば、受精率、3日目の卵胞刺激ホルモン(FSH)レベル)、およびいつ卵母細胞が採取されたか、接合体/胚コホートパラメータ(例えば、胚の総数)を含む。この対象データは、胚の評価を改善するために、および/または移植する胚の最適数の判定を補助するために統合され得る。報告書は、対象(例えば、氏名、対象の生年月日(DOB)、性別、郵送先住所および/または自宅住所、医療記録番号(MRN)、医療機関の部屋番号および/またはベッド番号)、保険情報等)、発達の可能性の評価を指示した対象の医師、または他の医療専門家の氏名、ならびに指示した医師と異なる場合、対象のケアに対して責任がある職員医師(例えば、かかりつけの医師)の氏名を識別する情報等の管理上の対象のデータ(つまり、発達の可能性の評価に必須ではないデータ)も含む場合がある。
【0090】
報告書は、試料データセクションを含む場合があり、これは、試料の種類(胚または多能性細胞、および多能性細胞の種類)、どのように試料が取り扱われたか(例えば、保存温度、調製プロトコル)、ならびに収集された日付および時間等の、評価において分析された生物学的試料についての情報を提供し得る。この情報の報告欄は、一般に、ユーザにより入力されたデータを使用して作成され得、これらの一部は、予め書かれた選択項目(例えば、ドロップダウンメニューを使用して)として提供され得る。
【0091】
報告書は、評価報告セクションを含む場合があり、これは、本明細書に記載される、評価/判定がどのように導き出されたかに関する情報を含み得る。解釈上の報告書は、例えば、評価される胚または多能性細胞の微速度画像、および/または遺伝子発現結果を含み得る。報告書の評価部分は、任意に、推奨セクションも含み得る。例えば、結果が胚の良好な発達の可能性を示す場合、当該分野において推奨されるように、推奨は、不妊治療の間に子宮に移植される限られた胚の数についての推奨を含み得る。
【0092】
報告書は、付加的要素または変更要素を含み得ることも容易に理解されるだろう。例えば、電子である場合、報告書は、報告書の選択された要素についてのより詳細な情報を提供する内部または外部データベースを提示するハイパーリンクを含有し得る。例えば、報告書の患者のデータ要素は、電子的患者記録へのハイパーリンク、またはそのような患者記録にアクセスするためのサイトを含み得、この患者記録は、機密データベースに保持される。この後者の実施形態は、病院内システム、またはクリニック内環境において興味あるものであり得る。電子形式である場合、報告書は、例えば、コンピュータメモリ、zip、CD、DVD等のコンピュータ可読媒体等の、適切な物理的媒体上に記録される。
【0093】
報告書は、上の要素の全て、または一部を含み得るが、但し、報告書は、一般に、少なくとも、ユーザにより要求された分析(例えば、発達の可能性の評価)を提供するのに十分な要素を含むものとすることは、容易に理解されるだろう。
【0094】
実用性
上述の通り、本発明の方法は、それらの発達の可能性を判定するために、胚または多能性細胞を評価するために使用され得る。この発達の可能性の判定は、臨床的判断および/または行動を導くために使用され得る。例えば、妊娠率を増加させるために、臨床医は、複数の胚を患者に移植する場合が多く、母親と胎児の双方に対して健康上のリスクを引き起こす多胎妊娠をもたらす可能性がある。本発明の方法から得られた結果を使用することにより、移植され、胎児に発達する胚の発達の可能性は、移植前に判定され、リスクを最小にしながら、満期妊娠の成功の確率を最大にするように、実施者がいくつの胚を移植するかを判断することができる。
【0095】
本発明の方法に従ってなされた評価は、胚または多能性細胞の群において、それらの発達の可能性について胚または多能性細胞をランク付けすることにも用途を見出すことができる。例えば、一部の場合において、複数の胚が、胚盤胞に発達できる可能性がある、すなわち、良好な発達の可能性を有する。しかしながら、一部の胚は、他より胚盤胞期または高品質の胚盤胞に達する可能性がある、すなわち、他の胚より良好な発達の可能性を有する。そのような場合において、本発明の方法は、群内で胚をランク付けするために使用され得る。そのような方法において、各胚/多能性細胞についての細胞パラメータ測定値を導き出すように、各胚/多能性細胞についての1つ以上の細胞パラメータが測定される。次いで、相互に対して胚または多能性細胞の発達の可能性を判定するために、胚または多能性細胞のそれぞれからの1つ以上の細胞パラメータ測定値が利用される。一部の実施形態において、それらを相互に直接比較して、胚または多能性細胞の発達の可能性を判定することにより、胚または多能性細胞のそれぞれからの細胞パラメータ測定値が利用される。一部の実施形態において、細胞パラメータ測定値を基準の胚/多能性細胞からの細胞パラメータ測定値と比較して、各胚/多能性細胞の発達の可能性を判定し、その後、各胚/多能性細胞の判定された発達の可能性を比較して、相互に対して胚または多能性細胞の発達の可能性を判定することにより、胚または多能性細胞のそれぞれからの細胞パラメータ測定値が利用される。このようにして、例えば、複数の接合体/胚を評価する実施者は、最良質の胚のみ、すなわち、最良の発達の可能性を有するものを選択して、リスクを最小にしながら、満期妊娠の成功の確率を最大にするように移植することができる。
【0096】
本発明の方法に従ってなされる評価は、生体外で成熟させた卵母細胞、および生体外で培養された幹細胞の発達の可能性を判定することにも用途を見出すことができる。本発明の方法によって得られた卵母細胞の発達の可能性についての情報は、受精させる卵母細胞の実施者の選択を導くことができ、それらの卵母細胞から胚盤胞を得る高い成功確率をもたらす。同様に、幹細胞の発達の可能性についての情報は、例えば、それを必要とする対象の組織を生体内で再構成または交換する処置に使用するための幹細胞を実施者が選択するために情報を与えることができる。
【0097】
試薬、デバイス、およびキット
上述の方法のうちの1つ以上を実践するための試薬、デバイス、およびそのキットを提供する。本発明の試薬、デバイス、およびそのキットは、非常に多様である。目的の試薬およびデバイスは、前述の細胞パラメータのいずれかを測定する方法に関する上述のものを含み、ここで、そのような試薬は、実施される特定の測定プロトコルに応じて、培養プレート、培養培地、顕微鏡、撮像ソフトウェア、撮像分析ソフトウェア、核酸プライマーのアレイ、核酸プローブ、抗体、シグナル産生系試薬等を含み得る。例えば、試薬は、上述の通り、コフィリン、DIAPH1、ECT2、MYLC2/MYL5、DGCR8、ダイサー/DICER1、TARBP2、CPEB1、シンプレキン/SYMPK、YBX2、ZAR1、CTNNB1、DNMT3B、TERT、YY1、IFGR2/IFNGR2、BTF3、およびNELFの遺伝子のうちの1つ以上に特異的であるPCRプライマーを含み得る。試薬の他の例としては、目的の遺伝子、またはこれらの目的の遺伝子によりコードされるタンパク質に対する抗体のうちの1つ以上に特異的であるアレイが挙げられる。
【0098】
上の構成要素に加え、本発明のキットは、本発明の方法を実践するための説明書をさらに含む。これらの説明書は、種々な形態で本発明のキットに提示され得、それらのうちの1つ以上は、キットに提示され得る。これらの説明書が提示され得る一形態は、適切な媒体または基材上に印刷された情報として、例えば情報が印刷された用紙(単数または複数)等として、キットのパッケージ内、添付文書内等に提示され得る。また別の手段は、情報が記録されたコンピュータ可読媒体、例えば、ディスケット、CD等である。提示され得るまた別の手段は、移動サイトの情報にアクセスするためにインターネットを介して使用され得るウェブサイトアドレスである。あらゆる都合のよい手段がキットに提示され得る。
【0099】
顕微鏡アレイを備える自動細胞撮像
【0100】
上述の方法の一部は、微速度撮像を介して胚および幹細胞の発達を観測する機能を必要とする。これは、標準のインキュベータの内部に収まる小型の多チャネル顕微鏡アレイから成るシステムを使用して達成され得る。これは、物理的に皿を移動する必要がなく、複数の試料を迅速かつ同時に撮像することを可能にする。図20に示される例示的な原型の1つは、暗視野照明を備える3チャネル顕微鏡から成るが、他の型の照明も使用され得る。「3チャネル」とは、3つの個別の培養皿を同時に撮像する3つの独立した顕微鏡を有することを意味する。ステッパモーターは、3D画像スタックを焦点に合わせる、または取得するために、焦点位置を調節するために使用される。我々はそれでヒトの胚を観測したが、白色光LEDは、照明に使用され、赤色または近赤外線(IR)LEDを使用することにより、細胞膜と細胞の内側部分との間のコントラスト比を改善させることができる。この改善されたコントラスト比は、手動および自動の双方の画像分析を補助することができる。加えて、赤外域へ移動することにより、試料に対する光毒性を低減することができる。画像は、他の型のカメラが使用され得るが、低費用の高解像度のウェブカメラによって捕捉される。
【0101】
図22に示す、上述の原型システムの各顕微鏡は、1〜30までの胚を含有し得る培養皿を撮像するために使用される。顕微鏡は、LEDにより生成されるあらゆる熱を分散させる補助をするヒートシンクに接続された白色光LEDからの光を収集し、これは、短い曝露時間としては非常に小さい。光は、直接光を防ぐための従来の暗視野パッチを通り、集光レンズを通って、培養され、研究される胚を保有する培養皿である「ペトリ皿」と標識された試料上へ達する。培養皿は、胚の順序を維持し、皿がインキュベータへ、およびインキュベータから運ばれる間にそれらが移動するのを防止する補助をするウェルを有し得る。ウェルは、胚が同じ培地液滴を共有できるのに十分に緊密に配置され得る。次いで、分散した光は、顕微鏡の対物レンズを通り、その後、色消しダブレットを通り、CMOSセンサ上へ通過する。CMOSセンサは、デジタルカメラとして機能し、上述の画像分析および追跡のためにコンピュータに接続される。
【0102】
この設計は、非常に多くのチャネルおよび異なる照明技法を提供するために容易に拡張可能であり、試料を供給するための流体デバイスを収容するように変更することができる。加えて、本設計は、フィードバック制御システムと統合することができ、ここで、温度、CO2(対照pHに対して)、および培地等の培養条件がフィードバックおよび撮像データに基づき、リアルタイムで最適化される。このシステムは、ヒトの胚の発達の微速度ビデオを取得するために使用され、これは、生体外受精(IVF)処置における胚の生存能を判定する実用性を有する。他の用途は、幹細胞療法、薬物スクリーニング、および組織工学を含む。
【0103】
デバイスの一実施形態において、照明は、アルミニウムヒートシンンク上に載置され、BuckPuck電流制御ドライバにより電力を供給されたLuxeon白色発光ダイオード(LED)により提供される。LEDからの光は、視準レンズを通過する。視準された光は、その後、図22に示される特注のレーザ加工パッチストップを通過し、非球面集光レンズを使用して中空の光錐に焦点が合わせられる。試料を直接透過する光は、対物レンズによって排除され、一方、試料によって分散される光は、収集される。視界を増大するためにより小さい倍率が使用されるか、または解像度を増加するためにより大きい倍率が使用され得るが、一実施形態において、20X倍率のOlympus対物レンズが使用される。次いで、収集された光は、色消しダブレットレンズ(すなわち、鏡筒)を通過して、色収差および球面収差の作用を低減する。代替的に、撮像対物レンズから収集された光は、鏡筒の代わりとして機能する、対向する方向に向けられた別の対物レンズを通過し得る。一構成において、撮像対物レンズは10X対物レンズであり得る一方、鏡筒対物レンズは4X対物レンズであり得る。得られた画像は、2メガ画素解像度(1600×1200画素)を備えたCMOSセンサにより捕捉される。異なる型のセンサおよび解像度も使用され得る。
【0104】
図23Aは、3つの同一顕微鏡を備える、多チャネル顕微鏡アレイの写真を示す。全ての光学構成要素は、鏡筒に載置される。アレイシステムの操作において、ペトリ皿は、手動の2軸傾斜台上に載置されるアクリルプラットフォーム上に装填され、これは、光学軸に対する画像面の調節を可能にする。これらの台は、顕微鏡の底部に固定され、最初に位置合わせをした後、動かない。LED、コリメータレンズ、パッチストップ、および集光レンズから成る照明モジュールは、位置付け、および照明光の焦点合わせのために、手動のxyz台上に載置される。対物レンズ、色消しレンズ、およびCMOSセンサから成る撮像モジュールも、視界の位置付け、および対物レンズの焦点合わせのために、手動のxyz台上に載置される。撮像モジュールの3つ全ては、移動台に取り付けられ、単一レバーアームにより支持され、これは、ステッパモーターを使用して作動する。これは、コンピュータ制御による焦点合わせおよび画像スタックの自動捕捉を可能にする。他の方法の自動焦点合わせならびに操作が使用され得る。
【0105】
顕微鏡アレイは、図23Bに示されるように、標準のインキュベータ内部に設置された。CMOS画像センサは、USB接続を介してインキュベータの内部に位置する単一ハブに接続され、これは、他の通信および電源線とともに外部PCにつながる。全ての電気ケーブルは、シリコーン接着剤で密封されたゴム製ストッパーの中心を通ってインキュベータを退出する。
【0106】
上述の顕微鏡アレイは、初期のヒトの胚の発達および接合体から胚盤胞期までの確認された成長の微速度画像を記録するために使用された。4つの異なる実験は、合計242個の胚を監視した。この群のうち、100個が最大5または6日目まで撮像され、他は、遺伝子発現分析のために種々の時間点で撮像ステーションから取り出された。画像捕捉ソフトウェアおよび撮像された胚のスクリーンショットを図24に示す。画像は、5分毎に捕捉され、画像当りおよそ1秒の低光曝露であった。試料が受けた総光量は、連続曝露の24分に相当し、取り扱い中、IVFクリニックで経験する総レベルと類似する。画像当り1秒の持続時間の光曝露が低減され得る。ヒトの胚で作業する前に、我々は、胚盤胞形成率および遺伝子発現パターンの双方が撮像プロセスによって影響を受けないことを確実にするために、マウスの着床前の胚で広範な対照実験を実施した。
【0107】
図25および26は、微速度配列からの選択した画像を示す。画像は、1日目、2.5日目、4日目、および5.5日目について示される。図25に示される配列において、9つの胚のうちの3つが胚盤胞に発達し、図26に示される配列において、12の胚のうち5つが胚盤胞に発達した。胚は3日目に培地交換を受けたため、写真視野中のそれらの位置は、移動しているが、個々の胚は、経時的に追跡された。遂次的培地の使用は、胚の発達の期特異的必要条件を満たす必要がある。培地交換中、胚を数分間撮像ステーションから取り出し、新しいペトリ皿に移動した。培地交換中の各胚の同一性の追跡を維持するために、1つの皿から他の皿への試料の移動をビデオテープ録画して、胚が混同されなかったことを確認した。このプロセスは、遺伝子発現分析のために試料の収集中にも使用された。胚の同一性を追跡する課題は、特定の順序で胚を配置するのを補助するためのウェルを使用することにより回避され得る。
【0108】
マイクロウェルを備えたペトリ皿
【0109】
異なるステーション間でペトリ皿を移動する場合、胚が時折移動し、それによって胚の同一性の追跡の維持を困難にする。これは、微速度撮像が1つのステーション上で実施され、胚が胚の選択および移動のために第2ステーションに後に移動される場合、問題となる。一方法は、個々のペトリ皿で胚を培養することである。しかしながら、これは、各胚が独自の培地滴液を有することが必要となる。通常のIVF処置において、患者の胚の全てを同じペトリ皿上で、かつ同じ培地滴液中で培養することが一般に望ましい。この問題に対処するために、我々は、マイクロウェルを備えた特注のペトリ皿を設計した。これは、胚が移動するのを防ぎ、インキュベータまたは撮像ステーションに、またはそこから移動される時に、ペトリ皿上でのそれらの配置を維持する。加えて、ウェルは、それらが同じ培地液滴を共有し、全てが同じ顕微鏡によって同時に表示され得るように十分に小さく、緊密に配置される。各マイクロウェルの底面は、光学品質の仕上げである。図27Aは、一実施形態の寸法を伴う図を示す。この型において、25のマイクロウェルが1.7×1.7mmの視界内に緊密に配置されている。図27Bは、3D表示のマイクロウェルを示し、これは、皿の表面内に約100ミクロン陥凹する。文字、数字、および他の印を含む位置合わせマーカーが、識別を補助するために皿の上に含まれている。
【0110】
本明細書に引用される全ての参考文献は、参照によりそれらの全体が組み込まれる。
【実施例】
【0111】
以下の実施例は、本発明をどのように行い、使用するかの開示、および説明を当業者に提供するために提示され、発明者が、彼らの発明と見なすものの範囲を制限することを意図せず、またそれらは、以下の実験が全て、または唯一の実施された実験であることを表すことを意図しない。使用された数字(例えば、量、温度等)に関する正確性を確実にする試みがなされたが、一部の実験エラーおよび逸脱が考慮されるべきである。特に記載のない限り、部は重量部であり、分子量は重量平均分子量であり、温度は摂氏温度であり、圧力は大気圧または大気圧付近である。
【0112】
試料源
【0113】
本研究に使用された全ての胚は、複数の胎生学者によって多年度にわたり収集され、受精され、凍結保存された。我々の研究における患者当りの胚の平均数は3つであり、所定のIVFセンターで出会った全年齢群を含んだ。特に、これらの実験に使用された胚の全ては、IVF生成され(ICSIとは対照的に)、そのため、胚は、相対的に正常な機能(少なくとも卵丘、帯、および卵細胞膜を透過し、前核を形成するそれらの能力に関して)を有する精子に由来した。刺激プロトコルは、標準のロング・リュープロン・プロトコルであった(cdc.gov/art)。過剰なヒトの胚の凍結保存は、室温(22+2℃)で25分間、凍結用培地(1.5Mの1,2プロパンジオール+0.2Mのスクロース)にそれらを設置することにより達成された。その後、緩慢凍結プロトコル(−1℃/分〜−6.5℃;5分間保持;播種;5分間保持、−0.5℃/分〜−80℃;液体窒素中に浸す)を使用して、胚を凍結した。委員。いかなる保護された健康情報も胚と関連し得ない。
【0114】
凍結保存された大量の胚の組を検証し、次の観測を行った:1)目印に関して、胚は、2細胞への分裂(2日目初期に生じる)、RNA分解の開始(1〜3日目に生じる)、4および8細胞への分裂(それぞれ、2日目後期と3日目後期に生じる)、胚ゲノムの活性(8細胞期の3日目)、および桑実胚および胚盤胞の形成(それぞれ、4日目と5日目に生じる)を含む、正常な胚の発達を示すタイミングを示した。2)胚は、臨床設定で得られる胚に特有である胚盤胞期に達する能力を示した。これは、発達するであろう、および発達しないであろうこれらの「選別」が、1細胞期の凍結保存前に行われないため(3日目または胚盤胞期の発達の後期に凍結保存された胚に特有であるように)、おそらく、胚が2PN期で凍結保存され、IVFクリニックで遭遇した多くの胚を表したという事実による。よって、我々のデータは、これらの胚が通常のIVFクリニックで観測されるものと比較して、類似する胚盤胞形成率を示したことを確認する。3)以前の研究は、新鮮な胚と比較した時、2PN期で凍結される胚が発達、着床、臨床妊娠、および出産において類似する可能性を示すことを示した。他の研究も、凍結卵母細胞について類似する結果を示し、ヒトの胚の発達の最も初期の事象は、凍結保存後、適切な時系列を維持することを示唆する。4)我々は、受精時間または解凍時間に依存しないパラメータに焦点をあてた。我々が測定した最初のパラメータ(第1細胞質分裂の持続時間)は、持続時間が短く(ca10〜15分)、本研究において、受精時間に依存しない(最終結果にかかわらず、全ての胚において、独立して測定され得る)。さらに、全ての後続のパラメータは、この初期の測定点に対して測定され、胚盤胞への発達に成功した胚と発達に失敗した胚との間で比較される。5)最後に、3PNである新鮮な(未凍結)胚は、新鮮な正常な胚と同じ時間枠に沿って発達することが知られていることに留意し、我々は、我々がStanford IVFクリニックより得た新鮮な3PNの胚において、パラメータを比較し、それらが我々の凍結保存された胚、または公表された報告のものと異ならないことを観測した。
【0115】
実験計画
【0116】
4つの実験組において、我々は、242個の前核期の胚の発達(それぞれ、61、80、64、および37個)を追跡した。各実験組において、ヒトの接合体を1日目に解凍し、複数のプレート上で、少数群で培養した。各プレートは、独立して、別の撮像ステーション上で暗視野下、微速度顕微鏡検査で観測された。約24時間間隔で、1プレートの胚を撮像システムから取り出し、高処理リアルタイム定量PCR遺伝子発現分析のために、単一胚、または単一細胞(卵割球)として収集した。各プレートは、通常、採取時に予想された発達期に達した胚(「正常」と称される)と、初期の発達期で停止した、もしくは遅延した、または大規模に断片化したもの(「異常」と称される)との混合物を含有した。胚は、単一のインタクトな胚として分析されるか、または単一の卵割球に解離された後、遺伝子特異的RNA増幅されるかのいずれかであった。胚盤胞形成を監視するために、胚の一部(242個のうち100個)を5または6日目まで撮像した。
【0117】
ヒトの胚培養および顕微鏡検査
【0118】
液体窒素貯蔵タンクからクリオバイアルを取り出すことによって、ヒトの胚を解凍し、それらを室温に設置した。バイアルが解凍したら、開封し、胚を切開顕微鏡下で視覚化した。次いで、バイアルの内容物を3003培養皿の底に注いだ。胚を液滴中に置き、各胚の生存を評価し、記録した。室温で、1.0Mの1,2プロパンジオール+0.2Mのスクロースを含有する3037培養皿に5分間、その後、0.5Mの1,2プロパンジオール+0.2Mのスクロースに5分間、そして0.0Mの1,2プロパンジオール+0.2Mのスクロースに5分間、胚を移動した。続いて、油下、マイクロドロップを使用して、1日目と3日目の間に10%のQuinn’s Advantage血清タンパク質代替物(SPS;CooperSurgical)を添加したQuinn’s Advantage分裂培地(CooperSurgical)、そして3日目後に10%のSPSを含むQuinn’s Advantage胚盤胞培地(CooperSurgical)で胚を培養した。全ての実験は、第1実験中の2つのステーションを除き、同じ種類の分裂培地を使用し、これは、Global培地(LifeGlobal,Guilford,CT)を使用した。この少数集団(12個の胚)において、胚は、わずかに低い胚盤胞形成率(12のうち3つ、または25%)を示したが、我々の予測パラメータの感度および特異性は、双方とも100%であった。
【0119】
複数の試料の同時分析に対応する、ならびに異なるプラットフォーム全体のデータの一貫性を検証するために、複数のシステムで微速度撮像を実施した。システムは、7つの個々の顕微鏡から成る:(1)Tokai Hit加熱台、白色光Luxeon LED、および暗視野照明用のアパーチャを装備した、改良されたOlympus IX−70/71顕微鏡2台、(2)加熱台、白色光Luxeon LED、およびホフマン変調コントラスト照明(注意:これらのシステムは、暗視野照明がパラメータの測定に好ましいと決定された後、4つの実験のうちの最初の間にのみ使用された)を装備した、改良されたOlympus CKX−40/41顕微鏡2台、および(3)白色光Luxeon LEDおよび暗視野照明用のアパーチャを装備した、インキュベータの内部に収まる特注の3チャネル小型顕微鏡アレイ。これらの異なるシステム上で培養された胚の間の発達挙動、胚盤胞形成率、または遺伝子発現プロファイルにおいて有意な相違は観測されず、実際、胚盤胞予測における我々のパラメータは、複数のシステムおよび実験全体において一貫していた。
【0120】
全てのシステムの光強度は、低電力のLED(通常の100Wハロゲン電球と比較して)、および高感度のカメラセンサにより、生殖補助顕微鏡で通常使用される光より有意に低い。光学電力計を使用して、我々は、473nmの波長の通常の生殖補助顕微鏡(Olympus IX−71ホフマン変調コントラスト)が、倍率により、ほぼ7〜10mWの範囲であると判定し、一方、我々の撮像システムの電力は、同じ波長で0.2〜0.3mWの間であると測定された。画像は、最大5または6日目までの間、5分毎に1秒の曝露時間で捕捉され、約24分の連続光曝露をもたらす。0.3mWの電力で、これは、通常の生殖補助顕微鏡下、ほぼ1分の曝露に相当する。
【0121】
相関する撮像、および遺伝子発現実験中の各胚の同一性を追跡するために、我々は、立体顕微鏡上にビデオカメラを取り付け、培地交換および試料収集中の試料の移動プロセスを録画した。我々は、マウス着床前の胚(n=56)、およびヒトの胚の少数集団(n=22)を用いた対照実験を実施し、撮像された胚と対照の胚との間の胚盤胞形成率に有意な相違を(p=0.96)観測しなかった。
【0122】
高処理qRT−PCR分析
【0123】
単一の胚または単一の卵割球qRT−PCR分析において、胚を最初に酸性タイロード液で処理し、透明帯を除去した。単一の卵割球を収集するために、厳密にピペット操作しながら、5〜20分間、37℃でHEPES(CooperSurgical)を含むQuinn’s Advantage Ca2+Mg2+を含まない培地中で胚をインキュベートした。試料を10μlの反応緩衝液中に直接収集し、後続の一工程逆転写/前置増幅反応を前述のように実施した。逆転写および前置増幅反応中、プールした20X ABI assay−on−demand qRT−PCRプライマーおよびプローブミックス(Applied Biosystems)を遺伝子特異的プライマーとして使用した。 前述のように、ABI assay−on−demand qRT−PCRプローブを使用して、Fluidigm Biomark 96.96 Dynamic Arrayを用いて、高処理qRT−PCR反応を実施した。全ての試料を3つまたは4つのテクニカルレプリケート(technical replicate)技術的な複写物に装填した。qRT−PCRデータ分析をqBasePlus (Biogazelle)、Microsoft Excel、および特注のソフトウェアを用いて実施した。データ品質不良(例えば、不良なPCR増幅曲線)、または評価された胚において一貫して低発現〜発現なしであること、のいずれかであれば、特定の遺伝子をデータ分析から排除した。卵割球期の分析において、使用した母性転写パネルは、DAZL、GDF3、IFITM1、STELLAR、SYCP3、VASA、GDF9、PDCD5、ZAR1、およびZP1を含み、一方、胚遺伝子パネルは、ATF7IP、CCNA1、EIF1AX、EIF4A3、H2AFZ、HSP70.1、JARID1B、LSM3、PABPC1、およびSERTAD1を含む。基準遺伝子であるGAPDHおよびRPLP0に対する、ならびに遺伝子平均に対する各遺伝子の発現レベルは、geNormおよびΔΔCt法を使用して計算された。GAPDHおよびRPLP0は、経験的に、GAPDHについては1.18および46%、ならびにRPLP0については1.18および34%の遺伝子安定値および変動係数に基づき、本研究の基準遺伝子として選択された。これらは、我々が試験した10のハウススキーピング遺伝子の中で最も安定しており、十分通常の異種試料の組の範囲内であった。第2に、我々は、予想通り、単一卵割球において、RPLP0およびGAPDHの転写量が、ヒトの発達の最初の3日間のEGA前の新規転写の不在下、1細胞期と8細胞期との間の分割当り約1Ct値減少し、各細胞が各卵割毎にmRNAのプールの約半分を受け継ぐという予測と一致することを観測した。第3に、我々は、EGA開始後、単一の卵割球におけるこれらの基準遺伝子の発現レベルは、8細胞と桑実胚期との間、安定した状態を保ったことに留意した。インタクトな胚のレベルで、RPLP0およびGAPDHの双方のCt値は、発達全体を通して桑実胚期までほとんど一定を保ち、後の胚盤胞期でわずかに増加するが、おそらく存在する多数の卵割球の転写レベルの増加によるものである。この研究で実施された遺伝子発現分析の大半は、桑実胚期前の発達期に焦点をあてたが、その時、基準遺伝子の発現レベルは非常に安定していた。
【0124】
自動化細胞追跡
【0125】
我々の細胞追跡アルゴリズムは、遂次モンテカルロ法に基づく確率的フレームワークを使用し、これは、コンピュータビジョンの分野において、粒子フィルタと呼ばれることが多い。粒子フィルタは、経時的に状態、対照、および測定の3つの主要変数の伝搬を追跡する。状態変数は、胚のモデルであり、楕円の集まりとして表される。対照変数は、状態変数を変換する入力であり、我々の細胞伝搬およびビジョンモデルから成る。測定変数は、状態の観測であり、微速度顕微鏡によって取得された我々の画像から成る。各時間ステップでの現状態の我々の推定は、事後確率分布を用いて表され、これは、粒子と呼ばれる一組の重み付き試料で近似される。我々は、粒子および胚モデルの用語を交換可能に使用し、ここで、粒子は所与の時間の胚モデルの仮定の1つである。初期化後、粒子フィルタは、予測、測定、および更新の3つのステップを繰り返し適用する。
【0126】
予測: 細胞は2D空間で楕円として表され、各細胞は、配向および重複インデックスを有する。重複インデックスは、細胞の相対的高さを指定する。一般に、我々が予測したい2種類の挙動があり、それは、細胞運動と細胞分割である。細胞運動において、我々の対照入力は、粒子を捕え、位置、配向、ならびに主軸および短軸の長さを含む、各細胞の各パラメータを無作為に撹乱する。撹乱は、比較的小さい分散(初期値の5%)の正規分布から無作為に試料サンプリングされる。細胞分割において、我々は以下のアプローチを使用する。所与の時間点で、各粒子において、我々は、細胞のうちの1つが分割するという50%の確率を割り当てる。この値は、経験的に選択され、現在の構成を良好に網羅しながら、広範な可能な細胞分割に及ぶ。分割が予測される場合、分割細胞が無作為に選択される。細胞が分割することが選択される時に、我々は、楕円の主軸に沿って対称分割を適用し、同じ大きさおよび形状の2つの娘細胞を産生する。次いで、我々は、娘細胞の各値を無作為に撹乱する。最後に、我々は、残りの細胞に対するそれらの集合的重複を維持しながら、2つの娘細胞の重複インデックスを無作為に選択する。
【0127】
対照入力を適用した後、我々は、各粒子をシミュレーション画像に変換する。これは、重複インデックスを使用して、各細胞の楕円形状をシミュレーション画像に投影することにより達成される。対応する画素値は、1のバイナリー値に設定され、観測された画像データと比較可能な膜厚を作製するために拡大される。胚は、部分的に透明であり、焦点外の光が収集されるため、胚の底部の細胞膜は、時々しか可視でない。したがって、遮蔽された細胞膜は、10%の確率で加えられる。実践において、我々は、これらの遮蔽された膜点が、正確な形状モデル化に重要であるが、それらが可視縁部に類似しないようにそれらを十分に散在させることが重要であることを発見した。
【0128】
測定:我々が仮定モデルの分布を生成したら、対応するシミュレーション画像を実際の顕微鏡画像と比較する。顕微鏡画像は、主曲率に基づいた方法の後、閾値処理を使用して、細胞膜のバイナリー画像を作製するために前処理される。比較の正確性は、対称切断型チャンバ距離を使用して評価され、これは、次いで、重量または可能性を各粒子に割り当てるために使用される。
【0129】
更新:重量が割り当てられた後、次の反復のために新しい粒子の組を作製するために、これらの重量に比例して粒子を選択する。これは、最も高確率の領域の粒子分布に焦点をあてる。低確率の粒子は破棄され、一方、高確率の粒子は乗算される。粒子の再サンプリングは、低分散法を使用して実施される。
【0130】
胚がモデル化されたら、我々は、本文で説明する、細胞質分裂の持続時間、および有糸分裂間の時間等の動的撮像パラメータを抽出することができる。我々の細胞追跡ソフトウェアは、事前にMatlabに実装され、計算時間は、粒子の数により各画像において数秒から30秒の範囲であった。我々の現在のソフトウェアのバージョンは、Cで実装され、計算時間は、粒子の数により1〜5秒の範囲である。
【0131】
実施例1
胚の発達の可能性を判定するための撮像分析
【0132】
方法
接合体とも称される凍結された1細胞のヒトの胚を解凍し、培養物中に設置し、IVF処置で使用されるような条件下で培養した。上でより詳細に記載するように、これらの胚は、2PN期で凍結され、よって、無差別に凍結保存されたため、通常の生体外受精(IVF)集団を表すと思われる。これは、新規周期中、最高品質であると認知されたものの移動後の発達の後期で通常凍結保存される胚と対称的である。一部の実験において、胚は、標準の培養皿に設置された。他の実験において、胚は、光学品質のマイクロウェルを備える特注の培養皿で培養された。
【0133】
皿当り通常1〜30個の成長胚は、デジタル画像保存および分析用に装備されたコンピュータ制御顕微鏡を用いて、微速度撮像により個別に追跡された。一部の場合において、微速度撮像は、加熱台およびインキュベーションチャンバを装備した倒立顕微鏡を用いて実施された。他の場合において、微速度撮像は、従来のインキュベータの内部に収まる特注の小型顕微鏡アレイを用いて実施され、これは、同じインキュベータ内の複数の試料皿を同時に培養することを可能にし、連続した画像捕捉間の最小時間間隔に対して制限がない複数のチャネルを収容するように拡張可能である。複数の顕微鏡の使用は、試料を移動する必要性を排除し、これは、システムの正確性および全体的なシステムの信頼度を改善した。撮像システムは暗視野を使用し、他の照明が十分であったであろうことが留意されたが、これは、後続の特色抽出および画像分析のために強化された画像コントラストを提供した。インキュベータ中の個々の顕微鏡は、相互に隔離することができ、各培養皿に独自の制御された環境を提供する。これは、他の試料の環境を乱すことなく、皿の撮像ステーションへの、およびそこからの移動を可能にした。
【0134】
微速度画像は、次の細胞パラメータのうちの少なくとも1つの測定を含む、後続の細胞形態の分析のために収集された:第1細胞質分裂の持続時間、第1と第2の細胞分割との間の時間間隔、および第2と第3の細胞分割との間の時間間隔。図に示される画像は、最大5または6日間、5分毎に1秒の曝露時間で撮られた。以下により詳細に説明されるように、第1細胞質分裂は、一般に、受精1日後に生じ、約14分の間続く。第1および第2の細胞分割は、一般に、平均約11時間で分離される。第2および第3の細胞分割は、一般に、平均約1時間で分離される。よって、撮像は、受精後約36時間(数時間の加減)続く期間にわたった。
【0135】
結果
培養における健康なヒトの着床前の胚の発達時系列は、微速度撮像により6日間にわたり記録された(図2)。正常なヒトの接合体は、2日目の初期に第1分裂を受けることが観測された。続いて、胚は、4日目に桑実胚に密集する前に、それぞれ、2日目と3日目の後期に4細胞および8細胞に分裂する。第1の形態学的に明らかな細胞分化は、胚盤胞形成中の5及び6日目に観測され、その時、全能性卵割球は、胎盤のような胚外構造をもたらす栄養外胚葉細胞、または生体内で胎児に、そして生体外で多能性胚幹細胞に発達する、内細胞塊のいずれかに分化する。
【0136】
我々は、次に、4つの独立した実験組で242個の普通に受精させた胚の発達を追跡し、5または6日目まで培養された試料の中で、正常な胚および停止した胚の分布を記録した。242個の胚のうち、100個は、5または6日目まで培養され、胚盤胞形成率は、33%〜53%の間であることが観測され、通常のIVFクリニックでの胚盤胞形成率と類似した(図3)。残りの胚は、異なる発達期で停止し、最も一般的には、2細胞と8細胞期との間であり、異常と定義された(図3)胚盤胞期への胚の発達の成功を予測する定量的撮像パラメータを識別するために、我々は、卵割球の大きさ、透明帯の厚さ、断片化の程度、第1細胞周期の長さ、最初のいくつかの有糸分裂間の時間間隔、および第1細胞質分裂の持続時間を含む、微速度ビデオからいくつかのパラメータを抽出し、分析した。発達的に正常な胚と異常な胚の双方のビデオ画像分析中、我々は、多くの停止した胚が第1細胞分割中、異常な細胞質分裂を受けたことを観測した。正常な胚は、速やかに制御された様式で、分裂溝の出現から娘細胞の完全分離まで14.3+/−6.0分の狭い時間枠で細胞質分裂を完了した。これは図4の上部に示される。逆に、異常な胚は、一般に、2つの異常な細胞質分裂表現型のうちの1つを示した。軽度の表現型において、細胞質分裂の形態およびメカニズムは、正常のように見えたが、プロセスを完了するのに必要とした時間は長く、さらに数分から1時間の範囲であった(図4)。時折、わずかに延長した細胞質分裂を受けた胚でも、胚盤胞に発達した。より重度の表現型において、細胞質分裂の形態およびメカニズムは撹乱された。例えば、図4の下部パネルの例に示されるように、胚は、片側だけ分裂溝を形成し、最終的により小さい構成要素に断片化する前に、数時間、異常な一連の膜ラフリング事象を受けた。そのような挙動の他の変形が観測された。加えて、これらのより重度の表現型を示す異常な胚は断片化され、胚の断片化が後に異常な胚発達をもたらす異常な細胞質分裂の副産物の可能性がある直接的な証拠を提供する。
【0137】
我々の撮像結果のより詳細な分析は、正常な胚が胚の遺伝子活性(EGA)が始まる前の初期の分割中、細胞質分裂および有糸分裂において厳密なタイミングに従うことを示し、胚の発達の可能性が、受け継がれた母性プログラムにより予め決定されることを示唆する。特に、我々は、厳密に制御された初期の胚の細胞周期において、3つの一次的な間隔:(1)第1細胞質分裂の持続時間、(2)第1と第2との間の有糸分裂の時間間隔、および(3)第2と第3との間の有糸分裂の同時発生に留意した。これらの3つの時間間隔と形態変化との間の関係を図5に示す。正常な胚において、我々は、パラメータがそれぞれ約14.3+/−6.0分、11.1+/−2.1時間、および1.0+/−1.6時間であると測定した(ここで、平均+/−標準偏差として与えられる)。
【0138】
我々は、単一細胞期で開始する3PN(三倍体)の少数組(n=10)の新鮮な(凍結保存されていない)胚の撮像を実施した。3PNの胚は、正常な新鮮な胚から少なくとも最初の3つの細胞周期と同様の目印事象の時系列をたどることが示されている。これらの胚は、撮像システムを検証するために(しかし、技術的理由のため、胚盤胞まで遂行されなかった)、我々の主要な実験前に撮像された。この新鮮な胚の組のうち、胚の3つが我々の凍結保存された2PNの胚と類似する事象の時系列をたどり、細胞質分裂の持続時間は、15〜30分の範囲であり、第1と第2の有糸分裂の間の時間は、9.6〜13.8時間の範囲であり、第2と第3の有糸分裂との間の時間は、0.3〜1.0時間の範囲であった。しかしながら、胚の7つにおいて、我々は、3つの分裂溝の同時出現、わずかに延長した細胞質分裂、および最終的な3つの娘細胞への分離を特徴とする固有の細胞質分裂表現型を観測した(図4)。これらの胚は、15〜70分の範囲の細胞質分裂の持続時間(分裂溝の開始から3つの娘細胞への完全分離までの間の時間を特徴とする)、8.7〜12.7時間の範囲の第1と第2の有糸分裂(3細胞〜4細胞)の間の時間、および0.3〜2.6時間の範囲の第2と第3の有糸分裂(4細胞〜5細胞)との間の時間を有する。この観測は、異常な胚によって示される多種多様な細胞質分裂表現型とともに、我々の凍結保存された胚が凍結保存プロセスによって発達的に遅れたわけではなく、2つの卵割球に分裂する新鮮な接合体と類似して挙動することを示唆する。
【0139】
0〜33分の間の第1細胞質分裂、7.8〜14.3時間の間の第1と第2の有糸分裂の間の時間、および0〜5.8時間の間の第2と第3の有糸分裂との間の時間を有することにより、それぞれ94%および93%の感度および特異性で、胚盤胞期に達した胚を予測することができた(図6)。逆に、これらの枠のうちの1つ以上外の値を示した胚は、停止することが予測された。胚盤胞の発達に成功した全ての正常な胚は、3つ全てのパラメータにおいて、類似する値を示した。逆に、異常な胚は、それらが間隔を完了するのに費やした時間の長さにおいて、かなりの量のばらつきを示した(図6)。我々は、(1)第1細胞質分裂を完了する期間が正常より長いと、不良な発達の可能性を示す、(2)第1と第2の細胞分割との間の間隔が正常より長い、もしくは短いと、不良な発達の可能性を示す、および(3)第2と第3の細胞分割との間の間隔が正常より長いと、不良な発達の可能性を示すことを観測した。よって、これらのパラメータは、胚が胚盤胞形成に進む能力、および胚盤胞の質を予測した。
【0140】
最後に、我々は、各パラメータは自動的に胚の発達の可能性を予測したが、3つ全てのパラメータの使用は、双方が90%を超えた感度および特異性を提供し、標準偏差の3倍のカットオフ点であったことに留意した。これらのパラメータの受信者操作特性(ROC)曲線を図7に示す。この図の曲線は、種々の標準偏差カットオフの真の陽性率(感度)対偽の陽性率(1−特異性)を示す。このROCを導き出すために、以下の数字を使用した:真の陽性の数=34(胚盤胞に達することを正確に予測された);真の陰性の数=54(停止することを正確に予測された);偽の陽性の数=4(胚盤胞に達することを不正確に予測された);偽の陰性の数=2(停止することを不正確に予測された)。
【0141】
考察
我々の分析は、最初の3つの卵割の間の有糸分裂および細胞質分裂において、厳密なタイミングに従う胚は、胚盤胞期に発達すること、拡張内細胞塊(ICM)を伴う高品質の胚盤胞を形成することの双方の可能性が非常に高いことを示す。動的形態パラメータは、IVF処置中、移植または凍結保存に最適な胚を選択するために使用され得る。これらのパラメータは、異なる質の胚盤胞を区別するためにも使用され得、群内の相対的な胚の発達の可能性のランク付けを可能にする。IVFクリニックにおける標準的な実践は、8細胞期(3日目)で移植することである。胚盤胞移植は、3日目の移植と比較して、最大2倍の着床率を有するため、一部のクリニックは、胚を胚盤胞期(5日目)まで培養することを選択する。しかしながら、多くのクリニックは、後成的障害のリスクが増加するため、延長した培養を避けている。予測的撮像パラメータは、4細胞期(2日目)まで、および胚遺伝子活性前の胚の生存能を予測するために使用され得る。これは、通常実践されるより丸1日早く、胚がそれらの分子プログラムの著しい変化を受ける前の胚の移植または凍結保存を可能にする。これは、一番最適な胚がPGDまたは他の種類の分析用に選択されることも可能にする。
【0142】
実施例2
遺伝子発現分析を通した撮像パラメータの検証、および発達の可能性を判定するための遺伝子発分析の使用
【0143】
方法
接合体とも称される凍結された1細胞のヒトの胚を解凍し、培養物中に設置し、IVF処置で使用されるような条件下で培養した。一部の実験において、胚を標準の培養皿に設置した。他の実験において、胚は、光学品質のマイクロウェルを備える特注の培養皿で培養された。
【0144】
胚を培養および撮像システムから取り出し、遺伝子発現分析用に、単一胚または単一細胞(卵割球)のいずれかとして収集した。各プレートは、通常、胚の混合物を含有し、一部は採取時に予想された発達期に達し、他は初期の発達期で停止したか、大規模に断片化した。採取時に予想された発達期に達した胚は、「正常」と分類され、一方、停止した胚は、「異常」とみなされた。例えば、試料収集のために、プレートの胚を2日目後期に撮像ステーションから取り出した時に、4細胞期およびそれ以降に達したいずれかの胚は正常と識別され、一方、4細胞期に達しなかった胚は停止と標識されたであろう。これらの停止した胚は、それらが停止した発達期により分類されるため、2日目後期で2つの卵割球のみの胚は、停止した2細胞胚として分析されたであろう。試料収集時に形態的に死滅し、多孔質のように見える胚(例えば、卵割球の退化)を排除するよう注意を払った。生存しているように見える胚(正常および停止の双方において)のみを遺伝子発現分析に使用した。しかしながら、収集時中に正常に見えた胚が、それらが後期に成長可能であっても、最終的に停止する可能性もあり得る。これらの分類のそれぞれを代表する胚の遺伝子発現分析は、定量的RT−PCR(qRT−PCR)により実施された。約24時間間隔で、高処理qRT−PCR遺伝子発現分析用に、胚を個々の撮像システムから収集し、最大96の遺伝子の多重反応を96の試料に対してアッセイした。遺伝子発現分析をFluidigm Biomark Systemを用いて実施し、これは、ナノリットル量で、最大9216の同時TaqMan assay−based qRT−PCR反応を実行できる。
【0145】
結果
形態事象の根底にあり得る分子メカニズムを解明するために、我々は、相関した遺伝子発現プロファイリングを実施した。ハウスキーピング遺伝子、生殖細胞マーカー、母性因子、EGAマーカー、栄養膜マーカー、内細胞塊マーカー、多能性マーカー、後成的制御因子、転写因子、ホルモン受容体、およびその他を含む、異なる分類に属する96の異なる遺伝子の発現レベルを試料毎にアッセイした(図19の表1)。わずかに異なるが重複する2つの組の遺伝子を2つの異なる実験組でアッセイし、ヒトの胚の運命についての固有の遺伝子診断の組を提供する。固有の遺伝子組は、モデル生物からの胚、またはヒトの胚性幹細胞に関するデータ、ならびに我々の未公開マイクロアレイデータから収集された。ヒトの着床前の胚におけるこれらの遺伝子組の発現状態は、この研究において始めて明らかにされる。
【0146】
基準遺伝子のGAPDHおよびRPLPOに対する、ならびに遺伝子平均に対する各遺伝子の発現値は、geNorm(El−Toukhy T,et al.(2009)Hum Reprod)およびAACt(Vanneste E,et al.(2009)Nat Med15:577−83)法を使用して計算された。遺伝子安定値および変動係数は、GAPDHにおいて1.18および46%であり、RPLPOにおいて1.18および34%であり、我々が試験を行った10のハウスキーピング遺伝子の中で最も安定しており、十分に通常の異種試料組の範囲内であった。単一の卵割球において、予想通り、RPLPOおよびGAPDHの転写量は、1細胞期と8細胞期との間の分割当りおよそ1Ct値減少し、これは、卵割の二分作用ならびに最初の3日のヒトの発達中のEGAの欠損による。単一の卵割球におけるこれらの基準遺伝子の発現レベルは、8細胞から桑実胚期との間、安定した状態を保った。全胚レベルで、RPLPOおよびGAPDHの双方のCt値は、桑実胚期までの発達全体にわたり非常に一定であった。RPLPOおよびGAPDHの発現レベルは、胚盤胞において有意に増加し、これは、存在する卵割球の数の増加による可能性が高い。これら変動は、RPLPOおよびGAPDHならびに基準遺伝子の有効性に影響を及ぼさなかった。この研究で実施された遺伝子発現分析の大半は、桑実胚期前の発達期に焦点をあてており、その時、基準遺伝子の発現レベルは非常に安定していた。
【0147】
正常な胚と異常な胚との間の差次的遺伝子発現 図8は、対数目盛上のレーダーグラフにプロットした、6つの異常な1〜2細胞胚、および5つの正常な1〜2細胞胚からの52の遺伝子の平均発現レベルを示す。停止した胚は、一般に、正常な胚と比較して、mRNA量の減少を示し、細胞質分裂、RNAプロセシング、およびmiRNA生合成を促進した遺伝子が最も強く影響を受けた。星印で強調された遺伝子は、マンホイットニー検定により判定された、正常な胚と異常な胚との間に、統計的に有意な相違(p<0.05)を示す。これらの18の遺伝子は、コフィリン、DIAPH1、ECT2、MYLC2、DGCR8、ダイサー、TARBP2、CPEB1、シンプレキン、YBX2、ZAR1、CTNNB1、DNMT3B、TERT、YY1、IFGR2、BTF3およびNELFである。各遺伝子は、図に示される群に属し、それは、細胞質分裂:コフィリン、DIAPH1、ECT2、およびMYCL2;miRNA生合成:DGCR8、ダイサー、およびTARBP2;RNAプロセシング:YBX2;母性因子:ZAR1;ハウスキーピング:CTNNB1;多能性:DNMT3B、TERT、およびYY1;受容体:IGFR2;ならびに転写因子:BTF3およびNELFである。大半の場合、これらの遺伝子の発現は、停止した1および2細胞胚より正常な1および2細胞胚で高かった。
【0148】
興味深いことに、特定の遺伝子分類は、他より異常な胚でより影響を受けた。例えば、異常な胚において、ハウスキーピング遺伝子、ホルモン受容体、および母性因子の大半は、遺伝子発現において大きく変更されず、一方、細胞質分裂およびmiRNA生合成に関与する多くの遺伝子は、有意に減少した発現を示した。さらに、影響を受けた遺伝子の中で、一部の遺伝子が他より正常な胚と異常なとの間で非常に大きな相違を示した。例えば、DGCR8、ダイサー、およびTARBP2等の、miRNA生合成経路に関与する遺伝子は、異常な胚において、高度に減少した発現レベルを示した。特に、最も強く影響を受けた2つの遺伝子であるCPEB1およびシンプレキンは、転写のポリ(A)テールの長さを操作することにより母系mRNA保存および再活性化を調節する同じ分子メカニズムに属した(Bettegowda,A.et al.(2007)Front.Biosci.12:3713−3726)。これらのデータは、胚の異常性は、胚のmRNA調節プログラムの欠損と相関することを示唆する。
【0149】
細胞質分裂 の遺伝子発現プロファイルとの相関 遺伝子発現分析を主要な細胞質分裂構成要素をコードした遺伝子を用いて実施した。各胚の同一性は、立体顕微鏡上にカメラを取り付け、培地交換および試料収集中の試料移動のプロセスをビデオ撮影することにより追跡された。異常な胚の遺伝子発現プロファイルを評価する時に、我々は、主要な細胞質分裂構成要素において、異常な細胞質分裂と低遺伝子発現レベルとの間に強力な相関関係を観測した。興味深いことに、異常な胚の遺伝子発現プロファイルは、それらの異常な形態学的表現型と同じくらい多様であり、可変であった。
【0150】
細胞質分裂遺伝子発現は、正常な2細胞胚と異常な2細胞胚との関係(図9)、および正常と異常な4細胞胚との関係(図10)では変動したことが判明した。図9および10は、正常な2細胞のヒトの胚において(図9)、および正常な4細胞胚(図10)において最も高度に発現した遺伝子の相対的発現を示し、異なる細胞質分裂表現型と相関した。図9に示すように、第1細胞質分裂中に異常な膜ラフリングを示した停止した2細胞胚は、試験した全ての細胞質分裂調節遺伝子において有意に減少した発現レベルを有した。図9で相違を示した遺伝子は、アニリン、コフィリン、DIAPH1、DIAPH2、DNM2、ECT2、MKLP2、MYCL2、およびRhoAである。正常な発現レベルは、右側の棒に示され、各遺伝子において高いことが分かる。図9のグラフ上の写真において、異常な2細胞胚を示し、スケールバーは50μmを表す。図10は、片側のみの細胞質分裂溝を有する異常な細胞質分裂を受けた停止した4細胞胚からの結果を示し、第1分割中の非常に延長した細胞質分裂は、細胞質分裂制御因子であるアニリンおよびECT2において減少した発現を示した。図10のスケールバーも50μmを表す。
【0151】
胚期特異的遺伝子発現パターン 図11は、141個の正常に発達した単一胚および単一卵割球の遺伝子発現分析中に識別された、4つの胚期特定パターン(ESSP)を示す。4つのESSPのうちのそれぞれ1つに該当する遺伝子を表2に列挙する(図20)。図11のプロットは、類似する発現パターンに基づく分類、およびそれらの発現値の平均化(基準遺伝子に対して)により作製された。ESSPの相対的発現レベルは、類似する発現パターンを有する遺伝子の発現レベルを平均化することによって計算された。遺伝子発現レベルは、異なる細胞期、すなわち、1c=1細胞、M=桑実胚、B=胚盤胞に対してプロットされる。図11において、4つのESSPのそれぞれにおける遺伝子の相対的発現は、1細胞(1c)から桑実胚および胚盤胞までの発現の関数として示される。ESSP1は母性的遺伝を示し、ESSP2は遺伝子転写活性を示し、ESSP3は後期活性を示し、ESSP4は持続的転写を示す。ESSP2に示されるように、IVFクリニックにおける移植点は3日目に生じ、その時、胚は、胚遺伝子活性により著しい発達変化を受けている。微速度画像データは、胚の発達の可能性が4細胞期までに識別され得、それによって2日目、およびこの遺伝子活性前の早期胚の移植を可能にする。この早期移植は、IVF処置の成功率を改善するのに有用である。
【0152】
表2(図20)は、識別された4つのESSPのそれぞれに属する遺伝子を列挙する。各遺伝子の相対的遺伝子発現レベルは、基準遺伝子(GAPDHおよびRPLPO)に対して、および遺伝子平均に対して計算された。胚の発達の時系列に対する各遺伝子の発現パターンは、次の4つのESSPのうちの1つに従った:ESSPパターン(1)初期:始めは高く、ゆっくり低下し、胚盤胞前に止まる遺伝子;ESSPパターン(2)中期:4細胞期後に始まる遺伝子;ESSPパターン(3)後期:桑実胚または胚盤胞で始まる遺伝子;およびESSPパターン(4)一定:比較的一定した発現値を有する遺伝子。
【0153】
ESSP1は、母性的に受け継がれた遺伝子のパターンを示す。これらの転写は 接合体期で高い発現レベルで開始し、続いて、胚が胚盤胞に発達するにつれて低下した。これらの転写の半減期は、約21時間であった。GDF9およびZAR1等の他のモデル生物、ならびに生殖細胞(卵母細胞)特異的遺伝子であるVASAおよびDAZLからの旧知の母性因子は、このカテゴリーに分類された。ESSP2は胚活性化遺伝子を含み、これは、4細胞期後、最初に胚において転写された。このカテゴリーにおける一部の遺伝子は、2つの活性化の波を示すように見え、第1のより小さい波は、5〜6細胞期のときであり、第2のより大きい波は8細胞期のときである。EIF1AX31およびJARID1 B32等の他のモデル生物からの既知のEGA遺伝子は、このカテゴリーに分類された。ESSP3は、胚盤胞期まで発現しなかった遅い活性化遺伝子から成っており、栄養膜マーカーGCM1を含む。ESSP4は、発達全体を通して、基準遺伝子に対して安定した発現を維持した持続的転写を含有した。これらの遺伝子の半減期は193時間であり、ESSP1より約9倍長かった。このカテゴリーは、ハウスキーピング遺伝子、転写因子、後成的制御因子、ホルモン受容体およびその他の混合物を含んだ。これらの遺伝子発現の4パターンは、61個の単一の正常な胚および卵割球の試料を使用した別の実験組でも確認された。
【0154】
第1分割中に異常な細胞質分裂および有糸分裂挙動を示す異常な胚は、特に胚性RNA操作に関与する遺伝子において、高度に不安定な遺伝子発現プロファイルと相関した。よって、これらの方法論を混合して、着床前の胚の生存能を予測するために使用され得る方法を提供することができる。結果は、異常な胚が、RNAプロセシングおよびmiRNA生合成における欠損プログラムと共に一生を開始し、母性mRNAの過剰な分解を生じることを示唆する。そのような未制御のRNA分解の確率的性質は、転写の不作為な破壊をもたらし、異常な胚において観測される広範な異常表現型を生じる。減少したレベルのmiRNAは、制御された母性RNA分解において欠損を生じ、異なる期で発達停止をもたらす。
【0155】
個々の卵割球分析 ヒトの着床前の胚において、分子分化がいつ始まるかを評価するために、異なる発達期の17個の胚から採取した単一卵割球のCDX2の発現レベルを分析した。図12Aは、2細胞から胚盤胞までの発達期の関数として、CTBBN1(暗色の棒)およびCDX2(薄色の棒)の2つの遺伝子の相対的発現レベルを示す。見て分かるように、CDX2は、4細胞期前の胚からの一部の単一卵割球において散発的に低レベルで発現した(図12A)。しかしながら、6細胞期以降、どの胚も、有意なレベルでCDX2を発現した少なくとも1つの卵割球を含有した。同じく図12Aに示されるハウスキーピング遺伝子のCTNNB1の発現レベルは、同じ胚からの卵割球の間で一定しており、CDX2の異種発現パターンがqPCR人工産物ではないことを示した。独立した実験からのデータは、類似する観測を示す。これらの結果は、ヒトの着床前の胚における分子分化が早ければ4細胞期の直後に生じる可能性があることを示す。
【0156】
興味深いことに、単一卵割球の遺伝子発現プロファイルの検査は、異なる発達期に対応する遺伝子発現特性を有する卵割球を含有した胚を明らかにした。いずれかの所与の時間でのいずれかの所与の胚の遺伝子発現プロファイルは、母性mRNA分解およびEGAの総和に相当する。初期の発達年齢の若い卵割球は、通常、多量の母性転写と、少量の接合体遺伝子とを含有し、その反対が、より進んだ発達年齢の老いた卵割球において当てはまる。この実験において、母性プログラムは、10のESSP1マーカー(母性転写)の平均発現値として定義され、胚プログラムは、10のESSP2マーカー(胚転写)の平均発現値により定義される。使用された母性転写パネルは、DAZL、GDF3、IFITM1、STELLAR、SYCP3、VASA、GDF9、PDCD5、ZAR1、およびZP1を含み、一方、胚遺伝子パネルは、ATF7IP、CCNA1、EIF1 AX、EIF4A3、H2AFZ、HSP70.1、JARID1 B、LSM3、PABPC1、およびSERTAD1を含む。この特定の8細胞胚から収集に成功した6つの卵割球のうち、3つの卵割球が正常な3細胞胚試料からの卵割球に類似する遺伝子発現特性を示し、一方、他の3つの卵割球は、正常な8細胞胚試料からの卵割球に類似した(図12B)。この観測の最も可能性のある説明は、胚内の細胞の亜集団の停止である。この部分的な停止表現型は、我々が試験した試料の中の別の9細胞胚および2つの桑実胚でも観測された。それらの正常な姉妹細胞と同じ時間量を培養で費やした停止した卵割球において、母性転写レベルが高いままであったという事実は、母性RNAの分解が、単に時間とともに生じる自発的なプロセスではなく、ミクロRNA(miRNA)等の特定のRNA分解メカニズムの機能を必要とする可能性が高いことを示す。これらのデータは、母性mRNA分解が哺乳類の胚形成中の保存された発達事象であり、正常な胚の発達に必要であるというさらなる証拠も提供する(Bettegowda,A.,et al.(2008)Reprod.Fertil.Dev.20:45−53)。加えて、これらのデータは、胚における個別の卵割球が自律的であり、相互に独立して発達できることを示唆する。さらに、これらの結果は、試験される細胞のmRNAのレベル(遺伝子発現レベルを示す)について試験するために、本明細書に記載される遺伝子発現レベル試験を使用することができることを示し、ここで、RNAは、母性プログラムの一部であることが知られている遺伝子であり、胚発達の後期におけるそのような発現レベルの持続性は、異常転帰の可能性と相関するか、または胚プログラムの一部であり、ここで、経時的な不在は、異常な転帰の可能性を示す。本明細書で検証された母性プログラム遺伝子は、ZAR1、PDCD5、NLRP5、H5F1、GDF9、およびBNC2である。他の母性作用遺伝子も知られており、使用することができる。
【0157】
胚遺伝子活性化 本方法は、少なくとも一部、異常な発達的に停止した胚が、EGA(胚遺伝子活性化)が生じる前の最初の3分割中、異常な細胞質分裂および有糸分裂タイミングを頻繁に示すという見解に基づく。これは、胚の発達の運命が、母性遺伝によって大きく判定され、200134のHARDYらにより実施されたヒトの着床前の発達の数学的モデルに顕著に従う見解である。さらに、細胞質分裂および有糸分裂の異常性は、MIRNA生合成および母性MRNAマスキング、保存および再活性化を調節する遺伝子の母性転写の減少したレベルと強く相関する。MIRNAは、生物の発達および分化を含む、多様な生物学的プロセスにおけるMRNA分解を促進することにより翻訳を調節する(BLAKAJ,A.&LIN,H.(2008)J.BIOL.CHEM.283:9505−9508、STEFANI,G.&SLACK,F.J.(2008)NAT.REV.MOL.CELL BIOL.9:219−230)。モデル生物からの増加した証拠は、MIRNAが初期の胚における母性転写分解の主要な制御因子であり得ることを示す(BETTEGOWDA,A.,ET AL.(2008)REPROD.FERTIL.DEV.20:45−53)。よって、MIRNA生合成における欠損は、異常な胚発達をもたらす可能性がある。一方、母性MRNAを適切に操作できないことも不良な胚形成をもたらす。哺乳類の卵母細胞は、母親の出産前に初期の胚成長を支持するのに必要な母性RNA転写物の大規模なプールを合成する。これらの転写物は、それらが受精後再活性化されるまで、圧縮され、長期間の間保存される。この母性RNA操作プログラムの欠損は、母性転写物の量および質に影響を及ぼす可能性があり、よって、発達が成功する可能性を危うくする。
【0158】
胚生存能を評価するためのモデル 図13は、相関する撮像および分子分析に基づくヒトの胚の発達のモデルを示す。胚盤胞への発達の成功を予測するための重要な短時間、および胚発達の図を含む、接合体から胚盤胞までの発達の時系列を示す。図の主要な分子データは、ヒトの胚が母親から受け継いだ独特の卵母細胞RNAの組により一生を開始することを示す。このRNAの組が、卵子の特異的RNA操作プログラムによって維持され、適切にパッケージされる。受精後、卵母細胞から胚への変移が始まる時に、卵子に特異的な一部の母性RNA(ESSP1;胚期特異的パターン1)の分解が分解されなければならない。並行して、発達が続くと、他のRNAが各卵割球に理想的に等しく分配される(ESSP4)。分解の成功とRNAの分配は、細胞自律的様式で、胚ゲノム活性化(EGA)、およびESSP2の遺伝子の転写に達する。特に、卵割中、胚性卵割球は、独立して、停止、または進行し得る。胚における細胞自律性発達の転帰は、個々の卵割球が停止または進行し得ることであり、8細胞胚が桑実胚期およびそれ以降に進行すると、胚盤胞の質は、停止した、または8細胞以降に進行した細胞の数によって影響を受ける。撮像データは、成功または失敗を予測する重要な発達期間があることを示す:第1細胞質分裂、第2卵割、および第2および第3の卵割の同時発生。これらのパラメータは、前述の細胞追跡アルゴリズムおよびソフトウェアを使用して、自動的に測定され得る。記載されるシステムおよび方法は、主要な撮像予測装置を用いて胚の転帰を診断するために使用され得、発達初期(EGA前)により少ない胚の移植を可能にする。
【0159】
実施例3
卵母細胞成熟および後続の胚の発達の撮像
【0160】
結果
現在のIVF処置の主な制限の1つは、卵母細胞の質および利用可能度である。例えば、現在のIVFプロトコルは、小さい周期プールから卵母細胞を動員し、受精に少数の卵母細胞(例えば、1〜20)を提供する。さらに、IVF処置中、ホルモン刺激後に回収された約20%の卵母細胞が未成熟として分類され、通常、現在の培養条件下において、胚の発達の可能性が低いため破棄される。
【0161】
卵母細胞プールを増加する一方法は、生体外成熟を通してである。図14は、胚胞、中期I、および中期IIを含む、生体外成熟中の3つの発達期を示す。胚胞および中期I期は未成熟卵母細胞として分類され、一方、中期IIは第1極体の存在のため、成熟として分類され、これは、生体外成熟開始24〜48時間後に生じる。図15は、生体外で成熟された卵母細胞の胚の発達を示す。
【0162】
卵母細胞プールを増加する別の方法は、一次および二次プールから卵母細胞を動員し、最大数千の卵母細胞を提供する。この処置において、休止状態の卵胞を卵巣から動員し、生体外でプログラムし、正常な染色体構成、後成的状態、RNA発現、および形態を有する卵母細胞を産生する。他の態様において、卵母細胞は、生体外で生殖細胞に分化され、ヒトの卵母細胞に成熟させた多能性幹細胞に由来し得る。
【0163】
図14に図示する、生体外卵母細胞の成熟プロセスは、本発明の方法における測定値および分析用の細胞パラメータを定義するために使用され得るいくつかの細胞変化によって特徴付けられる。これらは、例えば、卵母細胞膜の形態の変化、例えば、透明帯からの分離の速度および程度;卵母細胞核の形態の変化、例えば、胚胞崩壊(GVBD)の開始、完了、および速度;細胞質および核における顆粒移動の速度および方向;ならびに第1極体の移動および/または突出を含む。
【0164】
実施例4
幹細胞分化の撮像
【0165】
結果
微速度画像分析は、生存能、発達の可能性、および幹細胞、誘発された多能性幹細胞(iPSC)およびヒト胚性幹細胞(hESC)等の、他の細胞種の転帰を評価するためにも使用され得る。幹細胞の発達の可能性は、微速度画像分析を使用して、細胞発達および分化中の形態変化を測定することにより評価され得る(図17)。次いで、分化した細胞は、生体内移植または他の使用のために、分析および選択され得る。細胞質分裂の持続時間、有糸分裂事象間の時間、細胞の大きさおよび形状、細胞の数、細胞の運動、分割パターン、分化、非対称分割(ここで、1つの娘細胞が幹細胞を維持し、一方、他は分化する)、対称分割(ここで、双方の娘細胞が幹細胞として留まるか、または双方が分化するかのいずれか)、および運命特定(幹細胞がいつ分化するかを正確に判定する)等の、幹細胞のいくつかのパラメータが、微速度画像データから抽出され、分析され得る。
【0166】
幹細胞療法の基本式は、未分化幹細胞が生体外で培養され、特定の細胞種に分化され、続いて、損傷した組織および/または臓器の再生のために受給者に移植され得ることである。微速度画像分析は、成熟組織への組み込みが可能である非腫瘍原性の分化した後代を形成する幹細胞を識別するために、高処理の非侵襲性デバイスとして使用され得る。潜在的な用途は、アルツハイマー病およびパーキンソン病等の神経障害、血管系障害、および心臓病、関節炎等の筋肉および骨格障害、自己免疫疾患、および癌の治療、ならびに標的および新規薬物療法を評価することによる創薬を含む。
【0167】
ヒトにおいて、損傷した組織は、一般に、体の幹細胞からの連続動員および分化により置換される。しかしながら、体の再生するための能力は加齢とともに低下する。この一例は、括約筋欠損による尿失禁である。筋線維の数および神経密度は年齢とともに減少するため、加齢は、括約筋欠損の主な原因の一つであると考えられている。失禁を有する患者を治療するために、iPSCは、分化した平滑筋細胞を産生するために、膣壁組織から培養された線維芽細胞に由来し得る。次いで、これらの分化した細胞は、生体内に移植され得る。移植前に、多分化能、分化、メチル化、および腫瘍形成能に関してiPSCを特徴付けるために、微速度画像分析が使用され得る。他の用途は、パーキンソン病の患者の皮膚細胞に由来し、移植用にニューロンに分化されたiPSCの微速度撮像を含む(図18)。
【0168】
実施例5
【0169】
自動化分析による撮像パラメータの検証
【0170】
我々の微速度画像データによって明らかなように、ヒトの胚の発達は、コホート内の胚間で高度に変動するプロセスであり、胚は、細胞分割中に広範な挙動を示し得る。よって、高度に異常な細胞質分裂(図4)の持続時間等のある発達事象の手動特徴付けは、解釈に左右され得る。我々の撮像パラメータおよび胚盤胞形成を系統的に予測する機能を検証するために、我々は、最大4細胞期までの細胞分割の自動化追跡用のアルゴリズムを開発した。我々の追跡アルゴリズムは、遂次モンテカルロ法に基づく確率モデル推定技法を利用する。この技法は、仮定された胚モデルの分布を生成し、単純な光学モデルに基づき画像をシミュレーションし、これらのシミュレーションを観測された画像データと比較することにより機能する(図21a)。
【0171】
胚は、位置、配向、および重複インデックスを伴う楕円の集まりとして(細胞の相対的高さを表すため)モデル化された。これらのモデルを用いて、細胞質分裂の持続時間および有糸分裂間の時間を抽出することができる。細胞質分裂は、通常、最初の細胞質分裂溝の出現(ここで、双極陥凹が分裂軸に沿って形成される)から娘細胞の完全分離までにより定義される。我々は、1細胞から2細胞分割前の細胞伸長の持続時間として、細胞質分裂を近似することにより問題を簡素化した。軸長の相違が15%を超えた場合(経験的に選択される)、細胞が伸長したと判断される。有糸分裂間の時間は、各モデルの細胞数を数えることにより容易に抽出される。
【0172】
我々は、14個一組のヒトの胚(図21b)に対して我々のアルゴリズムを試験し、自動化測定値を手動画像分析と比較した(図21c、図21d)。このデータ組において、14個の胚のうちの8つが良好な形態で胚盤胞期に達した(図21e上部)。自動化測定値は、手動測定値と密接に一致し、8つの胚全てが、胚盤胞に達するように正確に予測された。14個の胚のうちの2つが不良な形態で胚盤胞に達した(不良な質の内細胞塊、図21e下部)。これらの胚において、手動評価は、1つが胚盤胞に達し、1つが停止するであろうと示され、一方、自動化評価は、双方が停止するであろうと予測された。最後に、14個の胚のうちの4つが胚盤胞期前に停止し、双方の方法により、停止することが正確に予測された。
【0173】
粒子フィルタフレームワーク
【0174】
粒子フィルタは、モンテカルロシミュレーションに基づくモデル推定技法である。それは、仮定モデルの分布を生成し、これらのモデルを観測されたデータと比較することによって未知または「隠れ」モデルを推定するために使用される。任意運動力学および測定値の不確実性に対応するその能力によって、それは細胞分割の追跡に対して理想的な候補となる。
【0175】
粒子フィルタは、経時的に、状態x、対照u、および測定値zの3つの主要変数の伝搬を追跡する。状態変数xは、我々が推定したい胚のモデルであり、楕円(2Dにおいて)または楕円体(3Dにおいて)の集まりとして表される。対照変数uは、状態変数を変換する入力であり、我々の細胞伝搬および分割モデルから成る。測定値変数zは、状態の観測であり、微速度顕微鏡によって取得された我々の画像から成る。これらのパラメータは、以下のセクションでより詳細に説明される。
【0176】
各時間ステップtにおける現在の状態xの推定は、事後確率分布で表される。この事後は、時折、信念とも称され、現在の状態xt、所与の全ての過去の画像測定値z1:t、および過去の対照u1:tの条件付確率として定義される。
【数1】
【0177】
粒子フィルタは、事後を一組の重み付き試料、または粒子によって近似し、
【数2】
として示される。
【0178】
ここで、Mは、粒子の数である。粒子および胚モデルという用語は、本明細書において交換可能に使用される。よって、単一の粒子xt[m](ここで、1<=m<=M)は、時間tにおける胚モデルの仮定の1つである。
【0179】
初期化後、粒子フィルタは、3ステップを繰り返し適用する。第1ステップは予測であり、ここで、各粒子は、対照入力
【数3】
を使用して伝搬される。
【0180】
得られた粒子の組は、事前確率の近似である。第2ステップは測定値更新であり、ここで、各粒子は、現在の測定値の確率に対応する重要な重さを割り当てられる。
【数4】
【0181】
重み付き粒子の組は、事後bel(xt)の近似である。
【0182】
粒子フィルタの主要な構成要素は、第3ステップに入り、ここで、粒子の組がそれらの重さによって再サンプリングされる。この再サンプリングステップは、最高確率の領域における粒子分布に焦点をあてる。
【0183】
細胞表示
【0184】
細胞は、2D空間において楕円として表示される。各細胞は、デカルト座標において、主軸、短軸、および2次元位置を有し、等式:
【数5】
により与えられる。
【0185】
各楕円は、向かう方向θ(yaw)も有し、これは、楕円にx−y平面で回転することを可能にする。楕円は、ほぼ常に相互に重複するため、我々は、重複インデックスhも示し、これは、重複の順序(または細胞の相対的高さ)を指定する。時間tにおける各胚モデルのパラメータは、したがって、
【数6】
として与えられる。
【0186】
ここで、Nは、そのモデルにおける細胞の数である。
【0187】
細胞の撹乱および分割
【0188】
粒子フィルタの第1ステップは予測であり、ここで、各粒子は、対照入力を使用して伝搬される。我々の用途において、我々がモデル化したい挙動が2種類ある。第1の挙動の種類は細胞運動を含み、これは、翻訳、yaw角周囲の回転、および主軸および短軸の長さの変化を含む。第2の挙動の種類は細胞分裂であり、ここで、細胞は2つの新しい細胞に分裂する。
【0189】
細胞運動をモデル化するために、我々の対照入力は粒子をとり、各細胞の各値を無作為に撹乱する:x0i、y0i、ai、bi、θi。撹乱は、比較的小さい分散(通常、初期値の%に設定される)の正規分布から無作為にサンプリングされる。
【0190】
細胞分割をモデル化するために、我々は以下のアプローチを使用する。所与の時間点で、各粒子において、我々は、細胞のうちの1つが分割するという50%の確率を割り当てる。この値は、経験的に選択され、現在の構成を良好に網羅しながら、広範な可能な細胞分割に及ぶ。分割が予測される場合、分割細胞は無作為に選択される。より複雑なモデルには、粒子中の細胞の数、およびそれらの分割パターンの履歴等のさらなる要因を考慮に入れ、実際のデータから観測された挙動に基づき、モデルを作製することが可能である。
【0191】
細胞が分割することが選択される時、楕円の主軸に沿った対称分割が適用され、同じ大きさおよび形状の2つの娘細胞が産生される。次いで、娘細胞の各値は、無作為に撹乱される。撹乱は、正規分布から、しかし、新しい細胞形状における大きな変動に対処するためにより大きな分散(初期値の10%)で再度サンプリングされる。最後に、残りの細胞に対してそれらの集合的重複を維持しながら、2つの娘細胞の重複インデックスを無作為に選択する。
【0192】
画像シミュレーション
【0193】
対照入力を各粒子に適用した後、粒子表示は、実際の画像と比較され得るシミュレーション画像に変換されなければならない。正確な画像シミュレーションは、難しい作業であり得、光線追跡技法および光学モデルの使用を必要とする場合が多い。現実的な画像のシミュレーションを試みるより、本発明の方法は、画像において容易に識別可能である特色をシミュレーションすることに焦点をあてる。具体的に、細胞膜の画像をシミュレーションする。
【0194】
考慮に入れられなければならない物理的観測が2つある。第1に、顕微鏡は、胚を通して単一面に焦点が合わせられるが、視野深度は、かなり大きく、焦点外の光は、ほぼ胚全体から収集される。第2に、胚は部分的に透明であり、これは、胚の下の細胞膜が胚の上の細胞を通して見られる時がある(いつもではない)ことを意味する。
【0195】
これらの物理的観測を念頭において、ここで、画像シミュレーションモデルを説明する。各細胞において、その対応する楕円形状は、重複インデックスhを使用してシミュレーション画像上に投影される。対応する画素値は、1のバイナリー値に設定され、観測された画像データと比較可能な膜厚を作製するために拡大される。重複インデックスhは、細胞が相互の上に重なり合う順序を指定する。遮蔽された細胞膜は、時々のみ可視的であるため、遮蔽点が検出される場合、それらは、低確率(通常、約10%)でシミュレーション画像に置かれる。実践において、これらの遮蔽された膜点は、正確な形状モデル化に必要であるが、それらが可視縁部に類似しないようにそれらを十分に散在させることが重要である。
【0196】
画像の前処理
【0197】
測定値変数zをここで説明する。本発明の方法のゴールは、シミュレーション画像と比較するために、顕微鏡画像による細胞膜のバイナリー画像を抽出することである。これらの膜は、高曲率および高コントラストを示すが、強度または色に基づく閾値化技法を使用して、容易に抽出されない。したがって、主曲率に基づく検出器が利用される。この方法はヘッセ作用素:
【数7】
を使用する。
【0198】
ここで、Ixx、Ixy、およびIyyは、画素位置sおよびガウススケールσで評価された二次偏導関数である。2x2ヘッセ行列の固有値は、主曲率についての情報を提供し、一方、固有値のサインは、「くぼみ」を「うね」43と区別する。明るいピークまたはうねを検出するために、各画素における主曲率が
【数8】
として計算される。
【0199】
ここで、λ2は、最小固有値である。変動する厚さの膜を検出するために、スケールの範囲に対してヘッセ作用素(すなわちσmin<=σ<=σmax)を適用し、この範囲に対する最大曲率を抽出する。最後に、ヘッセ画像を閾値化し、抽出した細胞膜のバイナリー画像を作製する。閾値レベルは、通常、ヘッセにおいて画素値の標準偏差の2倍に設定される。
【0200】
粒子の重さ
【0201】
「粒子フィルタフレームワーク」という表題のセクションで説明したように、粒子フィルタの第2の主要なステップは、測定値の更新であり、ここで、粒子は、特定モデルを考慮した、現在の測定値の確率に対応する重要な重さを割り当てられる。我々の場合、重要な重さは、上述の前処理された顕微鏡画像を、これも上述のシミュレーション画像と比較することにより判定される。
【0202】
この問題は、以前に検証され、ここで、標準化した相互情報を使用してシミュレーション画像を実際の画像と比較することにより、粒子フィルタの重さを計算した。このアプローチは、占有格子マッチングの考えに類似し、これは、双方が占有される(値1)か、双方が占有されない(値0)のいずれかである画素位置を検索する。これらの方法は、シミュレーション画像および実際の画像の形状は類似するが、わずかに不整合である時に問題がある。代わりに、説明される方法は、面取り距離に基づく尤度関数を使用し、これは、1つの点集合から別の点集合までの最も近い距離の平均値を測定する。それぞれ、シミュレーション画像と実際の画像において非ゼロ画素に対応する点A(大きさmの実数の組)およびB(大きさnの実数の組)の二組が定義される。点集合AからBの前方面取り距離は、
【数9】
として得られる。
【0203】
後方面取り距離も同様に定義される。本方法は、対称面取り距離を利用し、これは、いかに良くシミュレーション画像が実際の画像と一致したか、ならびにいかに良く実際の画像がシミュレーション画像と一致したかの基準を提供する。
【数10】
【0204】
実践において、個々の距離の測定値は、ノイズの影響を削減するために切り捨てられる。計算時間を削減するために、距離は、画像の距離変換の画素位置を調べることにより判定される。
【0205】
面取り距離は、推定モデルを考慮した、我々のデータ測定値の尤度基準として使用される。つまり、時間tにおける、所与の画像測定値ztおよび粒子モデルxt[m]において、粒子の重要な重さ(particle importance weight)は、
【数11】
として得られる。
【0206】
定数λは、通常1に設定され、尤度分布の「平坦性」を制御するように変動され得る。
【0207】
粒子再サンプリングおよび動的割り付け
【0208】
粒子フィルタの第3の主要なステップは再サンプリングであり、ここで、粒子は、それらの重さに比例して選択され、新しい組の粒子を作製する。低確率の粒子は破棄され、一方、高確率の粒子は乗算される。再サンプリングの効率的なアルゴリズムの開発に対して多くの従来の研究があった。本方法は、変動が少ないアプローチを使用する。
【0209】
粒子フィルタの重要な課題は、粒子の数の選択である。最も簡単な選択は、固定値、例えば、M=1000を使用することである。その後、各時間ステップにおいて、一組Mの粒子を同じ大きさの別の組に変換する。用途の内容において、細胞が不活性である、または単にわずかに大きさおよび位置を変更する、比較的長い期間が存在し得る。この観測の利点は、細胞活性量によって粒子の数を動的に割り付けることにより、処理負荷を低減するために利用される。つまり、細胞が活性であり、分割している時、我々は、粒子の数を増加し、細胞が不活性である時、我々は、粒子の数を削減する。
【0210】
細胞活性の程度を測定するために、新しい画像(顕微鏡によって取得)と前の画像との間の画素強度の2乗和の差(SSD)を計算する。ノイズを削減するために、最初に画像をガウスフィルタで平滑化し、SSD値を因果的移動平均で経時的に平滑化する。次いで、この値に比例して粒子の数を動的に調節し、100<M<1000の範囲内に収まるように切り捨てる。図30は、粒子の数がどのように1細胞期から4細胞期に分割する胚に割り当てられ得るかを示すグラフである。この方法は、事前画像登録は実施されなかったため、単に画像における「活性」量の基準を提供するだけであり、細胞分割と胚運動(翻訳および/または回転)を区別しないことに留意するべきである。この状況において(粒子の数を判定する)、粒子の数はいずれかの事象においても増加するはずであるため、これは許容される。実践において、我々は、最も可能性のある胚モデルの細胞の数に基づき、粒子の数も調節する。つまり、より多くの細胞が画像に存在すると考えられる時に、より多くの粒子が生成される。
【0211】
2次元追跡の限界
【0212】
上述の2D細胞追跡アルゴリズムは、胚の細胞の数、ならびにそれらの2D形状を判定するのに有用である。しかしながら、根底にある物理的表示がないという事実により制限される。これは、胚の生存能を評価するための、自動的な細胞分割の追跡に重要である場合も、重要でない場合もある。例えば、細胞質分裂の持続時間、および細胞分割間の時間等の特定のパラメータは、2D細胞追跡アルゴリズムを使用して測定され得る。次のセクションにおいて、我々は、我々の2Dモデルを3Dに拡張する。2D画像から3D形状を推定することにより生じる遮蔽および深度の曖昧さに対処するために、幾何制約および細胞容量の保存に対する制約を適用する。
【0213】
細胞表示および3次元追跡
【0214】
このセクションは、細胞分割の3D追跡用のアルゴリズムを説明する。2Dアルゴリズムからのステップの多くがこのアルゴリズムに持ち越されるが、主要な例外がいくつかある。3Dの使用に新しい細胞表示がある。ここで、細胞は3D空間で楕円体として表され、等式:
【数12】
により得られる。
【0215】
各楕円体も、向かう方向θ、ピッチψ、および回転αを有する。よって、時間tにおける胚モデルの表示は、
【数13】
として得られる。
【0216】
この改訂されたモデルの重要な作用の1つは、2D画像から3D形状を推測することに関連する曖昧さが存在し得るということである。例えば、形状が球状である細胞は、より長い主軸およびより大きいピッチ回転を有する細胞と類似する外見を有するであろう。後に示すが、区別するために十分な情報が利用可能になるまで(例えば、細胞分割等の事象から)、粒子分布は、これらの複数の仮定を維持するため、これは大きな問題ではない。
【0217】
楕円体は、剛体と考えられる、つまり、変形は明示的にモデル化されない。しかしながら、我々は、隣接する楕円体間のわずかな重複を可能にし、これらの重複する領域において、我々は、細胞が相互に対して平坦化されると想定する。それは胚において一般的に観測されるため、これは重要な考察であり、我々は、以下のセクションでそれを説明する。
【0218】
細胞撹乱および分割
【0219】
我々の3D細胞分割および撹乱モデルは、セクション4の「細胞撹乱および分割」のモデルと類似するが、主要な例外がいくつかある。3D形状の推定は、容量の保存を強化するために使用され得る。これは、細胞が任意に、特にz方向に大きく成長することを防止する。容量保存は、2つの状況において適用される。第1に、細胞撹乱において、軸aおよびbは変動し、cは、容量がその個々の細胞のために保存されるように計算される。第2に、細胞分割において、以下の制約が適用される。
【数14】
【0220】
ここで、下付き文字pは親細胞を示し、下付き文字d1およびd2は2つの娘細胞を示す。実践において、我々は、胚の総容量を原容量のプラス/マイナス5%の間で変動させることにより、これらの制約のわずかな違反を可能にする。これは、初期の容量推定における潜在的な不正確性を相殺するために使用される。
【0221】
細胞が3Dで分割することが選択される場合、その分割は、以下の方式でモデル化される。第1に、選択した単一細胞において、楕円の長軸に沿った分割が適用され、これは、構成によりa、b、またはcのいずれかであり得る。娘細胞は、大きさが等しく、均一に間隔がとられるように初期化され、親細胞の回転が考慮に入れられる。次いで、これらのパラメータは、広範の可能な構成を網羅するように撹乱され、ここでも初期値の10%に設定された変数の正規分布を使用する。
【0222】
幾何制約
【0223】
遮蔽および深度の曖昧さの問題は、容量の保存を通して部分的に軽減される。しかしながら、隣接する楕円体の空間的関係に関する制約も必要とされる。第1の制約は、細胞は、半径の20%を超える重複が禁止されることである。許容される量で重複する細胞において、それらが相互に対して平坦化したという想定がなされる。説明される粒子モデルは、画像シミュレーション中、楕円体を交差する内側点を無視することにより、この現象を表す。これは、経験的に動機づけされ、物理的に観測された挙動とよく相関する。
【0224】
第2の制約は、細胞を近接に保つことが課されることである。この制約は、ヒトの胚の物理的挙動に直接関係し、ここで、細胞は、透明帯と呼ばれる膜により制約される。帯は、球殻としてモデル化され、境界条件を課すためにそれを使用する。帯の半径は、1細胞胚の半径より30%大きく設定される。
【0225】
これらの制約は以下の通り強化される。所与の時間における各粒子において、上述のように、新しい粒子を生成するために無作為な対照入力が適用される。いずれかの物理的制約が犯された場合、新しい粒子は廃棄され、新しい無作為な対照が適用される。一定数の試みの後、満足する新しい粒子が生成されない場合、その粒子は廃棄される。
【0226】
画像シミュレーション
【0227】
実施例で使用される暗視野照明の利点は、細胞膜が細胞内部より多く光を分散することである。この作用は、細胞膜が光学軸(z軸)に平行である位置で最も顕著である。したがって、画像をシミュレーションするために、これらの位置が我々の3Dモデルにおいて検索され、これは、それらの回転のため楕円体の赤道に位置する必要はない。可視縁部および遮蔽縁部に関しても、上述のように、同じ規則に従う。
【0228】
2Dでの細胞追跡実施例
【0229】
この実施例は、自動化細胞顕微鏡に関し、細胞分割の2D追跡のために上述のアルゴリズムを使用する。このモデルは、画像中の細胞の数、ならびに細胞膜の2D輪郭を追跡するために設計された。第1ステップは画像の取得であり、これは、画像シミュレーションおよび画像前処理等の以下のセクションを動機づける。この実施例の微速度画像列は、10X対物レンズを備えた特注のOlympus IX−50倒立顕微鏡を用いて取得された。顕微鏡は、暗視野照明に改変され、ここで、中空の光錐は、光源と集光レンズとの間に円形アパーチャを設置することにより、試料に焦点を合わせる。対物レンズは、試料によって分散された光を収集し、直接透過した光を排除し、暗い背景上に明るい画像を産生する。暗視野照明の利点は、細胞膜が細胞内部より多く光を分散する傾向にあり、それによって、それらのコントラストを増強する。顕微鏡は、加熱台および特注のインキュベーションチャンバが装備されており、最大5または6日間にわたる胚の培養を可能にする。画像は、IX−50のサイドポート上に載置されたOlympus SLRデジタルカメラにより、5分間隔で捕捉された。
【0230】
胚の撮像は、それらが接合体、またはほぼ球形状の受精卵である時に開始された。粒子の組を初期化するために、セクション6の「画像の前処理」に記載するように、閾値ヘッセを計算し、最小2乗を使用してそれに円を適合させる。次いで、等分布からサンプリングした、無作為配向の円として全ての粒子を初期化する。
【0231】
図31は、1細胞期から4細胞期までの細胞分割を追跡するための2Dアルゴリズムの結果を示す。結果は、部分的に遮蔽される底に面する細胞においてさえ、細胞膜がアルゴリズムによりうまく抽出されることを示す。大半の粒子フィルタ用途において、「単一」の最適モデルは、粒子分布からの重み付き状態パラメータの和として表される場合が多いことに留意するべきである。しかしながら、本明細書に提示される結果において、高確率の粒子が表示される。
【0232】
3Dでの細胞追跡実施例
【0233】
図32は、1細胞期から4細胞期までの追跡のための上述の3Dアルゴリズムの成功した2つの用途を示す。図33は、粒子が1細胞から2細胞への分割中にどのように分散されるかの例を示す図である(図32に示される最初の実施例と対応する)。このプロットは、各細胞の中心の3D位置を示す。細胞が分割し始めると、予測は、どの娘細胞が他の上に重なり合うかに関する曖昧さを示すが、これは数フレーム内に解決される。
【0234】
予測パラメータの抽出
【0235】
前述の方法を使用して、胚がモデル化されたら、特定のパラメータがモデルから抽出され得る。通常、最適なモデルまたは最も高確率のモデルが使用される。これらのパラメータは、例えば、第1細胞質分裂の持続時間、第1と第2の細胞分割との間の時間、および第2と第3の細胞分割との間の時間を含む。細胞質分裂の持続時間は、細胞のモデルが2つの細胞に分裂する前にどのくらい長く伸長したかを測定することにより近似され得る。伸長は、楕円の主軸対短軸の比率を調べることにより測定され得る。モデルから抽出され得る他のパラメータは、受精と第1細胞分割との間の時間、細胞および分割プロセスの形状および対称性、分割の角度、断片化等を含む。パラメータは、2D細胞追跡アルゴリズムまたは3D細胞追跡アルゴリズムのいずれかを使用して抽出され得る。
【0236】
細胞質分裂は、最初の細胞質分裂溝の出現から娘細胞の完全な分離までによって定義される。我々の胚モデルは、変形不可能な楕円から成るため、細胞質分裂溝の出現の識別は困難な課題である。一方法は、楕円の変形を可能にするが、これはより複雑な追跡問題をもたらす。別の方法は、前処理顕微鏡画像の曲率の変化を探すことであるが、これは、胚のモデルから直接予測パラメータを測定しようとする我々の目的を無にする。よって、我々は、第1細胞質分裂の持続時間を、1細胞から2細胞分割前までの細胞伸長の持続時間として近似することにより問題を簡潔化する。伸長は、楕円の短軸bに対する主軸aの比率を計算することにより定量化される。細胞は、
【数15】
である場合、伸長したと判断される。
【0237】
この15%の値は、経験的に選択され、この特定のデータセットにおいて良く機能するが、他の値も使用され得る。胚モデルが2細胞に分割したら、我々は、1細胞モデルの伸長の持続時間を計算することにより、第1細胞質分裂の近似持続時間を抽出することができる。
【0238】
原理上、有糸分裂事象間の時間を測定することは容易である。例えば、第1と第2の有糸分裂の間の時間は、2細胞モデルと3細胞モデルとの間の時間として測定され得る。しかしながら、一部の場合において、胚は、独特な、無作為な挙動を示す場合がある。これは、例えば、1細胞から2細胞、2細胞から見かけ3もしくは4細胞に進み、そして2細胞に戻ることを含む。説明されるアルゴリズムは、この種の挙動を追跡することができるが、有糸分裂事象間の時間間隔の判定に対する課題を提起する。
【0239】
この挙動に対処するための一方法は、以下の通りである:2細胞と3細胞のモデルとの間の時間を測定する代わりに(第1と第2の有糸分裂の間の時間を見出すため)、これは、2細胞モデルが最も高確率の画像フレームの数を単純に数えることにより近似され得る。これは、一部の場合においてよく機能するが、有糸分裂事象間の真の時間を必ず表すとは限らない。細胞の数に基づきモデルに対する制限を強化することにより、これらの事象に対処することができる。つまり、各反復で分布から最適なモデル、または最も高確率のモデルを選択する場合、モデルの細胞の数は、常に同じままであるか、または増加するが、決して減少しないことが必要とされる。この制約を強化した後、有糸分裂事象間の時間を計算することは容易である。この制約は、ごくわずかな変動を示し得る追跡結果をフィルタにかけるのにも有用であり、これは、モデルが、例えば1細胞と2細胞のモデルの間を前後に切り替わる時に時折生じる。
【0240】
予測パラメータを抽出するための方法
【0241】
図35は、上述の方法を要約したフローチャートを示す。フローチャートは、(これは複数の胚または他の種類の細胞および幹細胞にも適用され得るが)単一の胚がどのように分析され得るかを示す。第1ステップにおいて、胚の画像が微速度顕微鏡を用いて取得される(「測定値」)。この画像は、ファイルに保存され、後の時間点で再度開くことができる。必要はないが、画像は、一般に、ある特色を強調するために前処理される。可能な胚構造のモデルが予測され、画像はそれらのモデルからシミュレーションされる(「予測」)。シミュレーション画像は、前述の細胞膜の画像、または前処理前の顕微鏡画像をより正確に表す画像を含み得る。次いで、モデルは、前処理された顕微鏡画像と比較される(「比較」)。この比較を使用して、最良の予測が保たれ、一方、不良な予測は廃棄される。次いで、得られた予測の組は、次の画像の予測を改善するために使用される。複数の連続画像においてこのプロセスを実施した後、例えば、細胞質分裂の持続時間、および有糸分裂事象間の時間等の、形態学的パラメータを最良のモデルから直接測定することが可能である。これらのパラメータは、前述の胚の生存能を評価するために使用され得る。
【0242】
実施例7
【0243】
細胞活性の自動化分析
【0244】
上述の方法は、顕微鏡を介して細胞の発達を追跡する機能を必要とする。胚において、複数の胚を追跡することが望ましく、これは同一の皿で一緒に培養される。本明細書で使用される分析方法は、画像が定期的に撮影されることも必要である(例えば、胚においては、1〜5日にわたり1〜30分毎、異なる時間間隔が幹細胞等の他の細胞型において使用され得る)。したがって、自動的に胚の発達を追跡する撮像方法が考案された。
【0245】
微速度顕微鏡検査において、細胞を制御された条件下で成長させ、運動性(環境内での運動)、増殖(成長および分割)、および形態変化(大きさおよび形状)等のプロセスを監視するために、長時間にわたり撮像した。実験期間および生成された膨大な画像データにより、細胞分割の持続時間および細胞分割間の時間等のパラメータの抽出は、面倒な仕事であり得る。これは、複数の試料が同時に撮像される高処理用途において、特にそうである。よって、所望の情報を自動的に抽出することができる画像分析ソフトウェアが必要である。
【0246】
胚の生存能を評価する一方法は、画像中の「細胞活性」量を測定することである。これは、単純に、遂次画像の組を選び、それらの画素値を比較することにより達成され得る。より具体的には、各新しい画像における細胞活性量を測定するために、全ての重複する画素iに関して、I’と示される新しい画像とI’と示される前の画像との間の画素強度の2乗和の差(SSD)を計算する
【数16】
【0247】
ノイズを削減するために、最初に画像をガウスフィルタで平滑化する。図28は、単一の胚の1日目〜3日目までの細胞活性のプロットを示す。示すように、ヒトの胚における1細胞から2細胞への分割、2細胞から4細胞への分割、4細胞から8細胞への分割に対応する鋭いピークが存在する。ピークの幅は、細胞分割の持続時間を表す。
【0248】
このアプローチの限界の1つは、SSD計量が画像中の活性量のみを測定し、胚運動(移動または回転)等の事象が細胞分割と非常に同じに見えることである。この問題に対する解決策の1つは、SSDを計算する前に画像登録を実施することである。画像登録は、それらを同じ座標系に整合するために、2つの画像間の幾何学的関係を見出すプロセスであり、種々の異なる技法を使用して達成され得る。例えば、レーベンバーグマルカート反復非線形ルーチンの変形を使用することができ、これは、重複する画素強度におけるSSDを最小にすることにより画像を表示する。LMアルゴリズムは、3×3のホモグラフィ行列:
【数17】
を使用して画素位置を変換する。
【0249】
ここで、目的画素位置x’およびy’は、
【数18】
として標準化される。
【0250】
よって、
【数19】
である。
【0251】
ホモグラフィ行列は、種々の画像変換に適用することができ、本用途における妥当な選択は、剛性体(ユークリッド)変換であろう。これは、翻訳および面内回転(カメラ軸に沿って)の胚の画像を整合するだろう。しかしながら、わずかに一般化し、アフィン変換を使用することが可能であり、これは、画像の歪みを可能にする。この一般化は、測定されようとするシグナルにより望ましい場合も、望ましくない場合もある。運動等式は、よって、
【数20】
となる。
【0252】
LMアルゴリズムは、最初に、連鎖法則:
【数21】
を使用して、未知の運動パラメータhkに関するeの偏導関数を計算する。
【0253】
アフィン運動パラメータにおいて、これらの偏導関数は、
【数22】
となる。
【0254】
次に、これらの偏導関数を使用して、LMアルゴリズムは、各画素からの寄与を加えることにより、近似ヘッセ行列A(大きさ6×6の実数の組において)および重み付き勾配ベクトル(大きさ6x1の実数の組において)を計算する。
【数23】
【0255】
最後に、運動パラメータは、増分運動を加えることにより更新され得る。
【数24】
【0256】
ここで、定数λは、運動更新の刻み幅を調節し、Iは単位行列である。
【0257】
アルゴリズムの各反復で、最初の画像は更新された運動推定値によって歪められ、重複する領域の画素強度のSSDを計算することにより第2の画像と比較される。本用途は、連続画像間の胚の運動が非常に小さいと想定し、したがって、少数で一定数の反復のみが行われる。図28Bは、各画像の組において行われた画像登録なし(28A)および画像登録あり(28B)の細胞活性のプロットを示す。レーベンバーグマルカートルーチンの誤差関数はSSDであるため、単に各表示の残余誤差をプロットする。図29は、正常および異常な胚の発達についての細胞の活性のプロットを比較する。胚生学者が通常形態を評価するであろう点である3日目に、胚は同じように見え、双方とも潜在的に生存可能であると考えられる。しかしながら、胚のうちの1つは、通常の一連の細胞分割を受け、一方、他方は、1細胞胚から複数の細胞および断片に分裂するため、それらの細胞活性プロットは、大幅に異なる。予想通り、正常な活性プロットを有する胚は、最終的に、5.5日目までに胚盤胞に達する。
【0258】
画素強度のSSDを計算する前に、他の種類の画像登録が使用され得る。これは、例えば、相互相関、標準化した相互相関、相互位相相関、相互情報、特徴検出および追跡、スケール不変特徴変換(SIFT)、オプティカルフロー、および勾配降下を含む。特徴およびコントラスト強調等の表示前の画像の前処理は、望ましい場合も、望ましくない場合もある。
【0259】
胚の生存能を評価するためのモデル
【0260】
図13は、相関する撮像および分子分析に基づくヒトの胚の発達のモデルを示す。胚盤胞への発達の成功を予測するための重要な短時間、および胚の発達の図を含む、接合体から胚盤胞までの発達の時系列を示す。図の主要な分子データは、ヒトの胚が母親から受け継がれた独特の卵母細胞RNAの組により一生を開始することを示す。このRNAの組が、卵子の特異的RNA操作プログラムによって維持され、適切にパッケージされる。受精後、卵母細胞から胚への変移が始まる時に、卵子に特異的な一部の母性RNA(ESSP1;胚期特異的パターン1)の分解が分解されなければならない。並行して、発達が続くと、他のRNAが各卵割球に理想的に等しく分配される(ESSP4)。分解の成功とRNAの分配は、細胞自律的様式で、胚ゲノム活性化(EGA)、およびESSP2の遺伝子の転写に達する。特に、卵割中、胚性卵割球は、独立して、停止、または進行し得る。胚における細胞自律性発達の転帰は、個々の卵割球が停止または進行し得ることであり、8細胞胚が桑実胚期およびそれ以降に進行すると、胚盤胞の質は、停止した、または8細胞以降に進行した細胞の数によって影響を受ける。撮像データは、成功または失敗を予測する重要な発達期間があることを示す:第1細胞質分裂、第2卵割、および第2および第3の卵割の同時発生。これらのパラメータは、前述の細胞追跡アルゴリズムおよびソフトウェアを使用して、自動的に測定され得る。記載されるシステムおよび方法は、主要な撮像予測装置を用いて胚の転帰を診断するために使用され得、発達初期(EGA前)により少ない胚の移植を可能にする。自動対手動画像分析の比較
【0261】
図34は、一組14の胚における手動画像分析に対する自動化画像分析の比較を示す。胚1から10(プロットに標識される)は、異なる形態を有した胚盤胞期に達した。胚11から14は停止し、胚盤胞に達しなかった。図34Aは、第1細胞質分裂の持続時間の測定の比較を示し、図34Bは、第1と第2の有糸分裂の間の時間の測定の比較を示す。示されるように、2つの方法は、一般に、良好な一致を示す。第1細胞質分裂の持続時間におけるわずかな矛盾が予想されるが、それらは、我々の自動化分析が、前述の通り、伸長を測定することにより近似を行うという事実に起因し得るためである。いくつかの場合において、細胞質分裂の持続時間ならびに第1と第2の有糸分裂の間の時間の双方において、自動化分析と手動分析との間により大きな不一致がある。これは、異常な胚のいくつかに生じ、手動で特徴付けること、ならびに自動的に追跡することの双方が困難である異常な挙動に起因する。この群の胚において、最初の2つの基準(第1細胞質分裂の持続時間、および第1と第2の有糸分裂の間の時間)のみを使用することにより、自動化アルゴリズムは、ゼロの偽陽性を有する。これは、偽陽性が回避されなければならないIVF処置において、非常に重要であろう。手動画像分析は、偽陰性(胚9)が1つであり、一方、自動化アルゴリズムは、偽陰性(胚9および10)が2つであった。しかしながら、胚9および10の双方は技術的に胚盤胞期に達したが、これらは他の胚盤胞と比較して不良な形態を示し、移植の候補にはあまり最適ではないだろう。手動画像分析において、胚14は、これらの2つの基準に基づきおそらく偽陽性であり、第3のパラメータの第2と第3の有糸分裂との間の持続時間は、真の陰性を与える必要がある。しかしながら、自動化アルゴリズムは、最初の2つの基準のみを使用して、正しい予測を行う。これらの結果は、我々の自動化アルゴリズムが胚盤胞対非胚盤胞を適切に予測し、ならびに異なる質間の胚盤胞を区別できることを示す。よって、複数の胚が良好な発達の可能性を有することが判定される時、IVF処置中、移植のための上位1つまたは2つの胚を選択するために、それらの相対的な質のランク付けを計算することが可能である。
【0262】
前述されるものは、単に本発明の原則を解説するにすぎない。本明細書に明白に説明もしくは表されてはいないが、当業者は、本発明の原則を具体化し、その精神と範囲内に含まれるものである様々な構造を考案し得ることが理解されるであろう。さらに、本明細書に記載される全ての実施例および条件言語は、主に、読者が、本発明の原則、ならびに本発明者らによって貢献された当技術分野を推進するための概念を理解することを補助することを意図しており、かかる具体的に記載される実施例および条件に限定されることはない解釈されたい。さらに、原則、態様、および本発明の実施形態、ならびにその特異的実施例を記載する本明細書の全ての陳述は、その構造的および機能的同等物の双方を包含することを意図している。加えて、かかる同等物が、現在既知の同等物および将来的に開発される同等物の双方を含むことを意図している、すなわち、構造にかかわらず、同一機能を実施する開発されるいずれの要素をもである。したがって、本発明の範囲は、本明細書で表され、説明される例示的実施形態に限定されることを意図していない。正確には、本発明の範囲および精神は、添付の特許請求の範囲によって具体化される。
【特許請求の範囲】
【請求項1】
ヒトの胚または多能性細胞の発達の可能性を判定するための方法であって、
(a)前記ヒトの胚または多能性細胞の1つ以上の細胞パラメータを測定して、細胞パラメータ測定値を導き出すことと、
(b)前記細胞パラメータ測定値を利用して、前記胚または多能性細胞の発達の可能性の判定を提供することと、を含む、方法。
【請求項2】
前記細胞パラメータは、微速度顕微鏡検査により測定可能である、請求項1に記載の方法。
【請求項3】
胚の発達の可能性が判定され、前記1つ以上の細胞パラメータが、
(i)細胞質分裂事象の持続時間、
(ii)細胞質分裂1と細胞質分裂2との間の時間間隔、および
(iii)細胞質分裂2と細胞質分裂3との間の時間間隔から成る群から選択される、請求項2に記載の方法。
【請求項4】
前記ヒトの胚の良好な発達の可能性は、
(i)約0分〜約30分の細胞質分裂1の持続時間、
(ii)約8〜15時間の細胞質分裂1の分離と細胞質分裂2の開始との間の時間間隔、および/または
(iii)約0〜5時間の細胞質分裂2の開始と細胞質分裂3の開始との間の時間間隔により示される、請求項3に記載の方法。
【請求項5】
前記測定するステップは、細胞周期1の持続時間を測定することをさらに含む、請求項4に記載の方法。
【請求項6】
前記ヒトの胚の良好な発達の可能性は、約20〜27時間の細胞周期1の持続時間により示される、請求項5に記載の方法。
【請求項7】
前記利用するステップは、前記細胞パラメータ測定値を基準のヒトの胚からの細胞パラメータ測定値と比較し、前記比較の結果を利用して、前記ヒトの胚の発達の可能性の判定を提供することを含む、請求項3に記載の方法。
【請求項8】
前記ヒトの胚の不良な発達の可能性は、
(a)前記ヒトの胚の細胞質分裂1が、前記基準のヒトの胚より長い、
(b)前記ヒトの胚の細胞質分裂1の分離と細胞質分裂2の開始との間の時間間隔が、前記基準のヒトの胚より長い、または短い、
(c)前記ヒトの胚の細胞質分裂2の開始と細胞質分裂3の開始との間の時間間隔が、前記基準のヒトの胚より長いことにより示される、請求項7に記載の方法。
【請求項9】
前記測定は、細胞周期1の持続時間を測定することをさらに含む、請求項8に記載の方法。
【請求項10】
前記ヒトの胚の不良な発達の可能性は、前記胚の細胞周期1が前記基準のヒトの胚より長いことにより示される、請求項9に記載の方法。
【請求項11】
前記1つ以上の細胞パラメータは、遺伝子発現測定値を導き出すために測定される遺伝子発現レベルである、請求項1に記載の方法。
【請求項12】
胚の発達の可能性が判定され、前記1つ以上の遺伝子は、コフィリン、DIAPH1、ECT2、MYLC2/MYL5、DGCR8、ダイサー/DICER1、TARBP2、CPEB1、シンプレキン/SYMPK、YBX2、ZAR1、CTNNB1、DNMT3B、TERT、YY1、IFGR2/IFNGR2、BTF3、およびNELFから成る群から選択される、請求項11に記載の方法。
【請求項13】
前記利用するステップは、前記遺伝子発現測定値を基準の胚または多能性細胞からの遺伝子発現測定値と比較することと、前記比較の結果を使用して、前記ヒトの胚または多能性細胞の発達の可能性の判定を提供することと、を含む、請求項11に記載の方法。
【請求項14】
胚の発達の可能性が判定され、前記基準の胚に対して、前記胚におけるコフィリン、DIAPH1、ECT2、MYLC2/MYL5、DGCR8、ダイサー/DICER1、TARBP2、CPEB1、シンプレキン/SYMPK、YBX2、ZAR1、CTNNB1、DNMT3B、TERT、YY1、IFGR2/IFNGR2、BTF3、およびNEFから成る群から選択される1つ以上の遺伝子のより低い発現レベルは、不良な発達の可能性を示す、請求項13に記載の方法。
【請求項15】
相互に対してヒトの胚または多能性細胞をランク付けするための方法であって、
(a)各ヒトの胚または多能性細胞について少なくとも1つの細胞パラメータを測定して、各胚または多能性細胞の細胞パラメータ測定値を導き出すステップと、
(b)前記胚または多能性細胞のそれぞれからの前記少なくとも1つの細胞パラメータ測定値を利用して、相互に対して前記ヒトの胚または多能性細胞をランク付けするステップと、を含む、方法。
【請求項16】
前記少なくとも1つの細胞パラメータは、微速度顕微鏡検査により測定可能である、請求項15に記載の方法。
【請求項17】
胚がランク付けされ、前記少なくとも1つの細胞パラメータは、
(i)細胞質分裂事象の持続時間、
(ii)細胞質分裂1と細胞質分裂2との間の時間間隔、および
(iii)細胞質分裂2と細胞質分裂3との間の時間間隔から成る群から選択される、請求項16に記載の方法。
【請求項18】
前記測定ステップは、細胞周期1の持続時間を測定することをさらに含む、請求項17に記載の方法。
【請求項19】
前記1つ以上の細胞パラメータは、1つ以上の遺伝子の発現レベルである、請求項1に記載の方法。
【請求項20】
前記1つ以上の遺伝子は、コフィリン、DIAPH1、ECT2、MYLC2/MYL5、DGCR8、ダイサー/DICER1、TARBP2、CPEB1、シンプレキン/SYMPK、YBX2、ZAR1、CTNNB1、DNMT3B、TERT、YY1、IFGR2/IFNGR2、BTF3、およびNELFから成る群から選択される、請求項19に記載の方法。
【請求項21】
前記利用するステップは、前記胚または多能性細胞のそれぞれからの前記細胞パラメータ測定値を相互に比較して、相互に比較した胚または多能性細胞の発達の可能性を判定することにより達成される、請求項15に記載の方法。
【請求項22】
前記利用するステップは、前記胚または多能性細胞のそれぞれからの前記細胞パラメータ測定値を基準の胚または多能性細胞からの細胞パラメータ測定値と比較して、各胚または多能性細胞の発達の可能性を判定することにより、および各胚または多能性細胞の発達の可能性を比較して、相互に比較して胚または多能性細胞の発達の可能性を判定することにより達成される、請求項15に記載の方法。
【請求項23】
良好な発達の可能性を有するヒトの胚を女性に提供する方法であって、
(a)胚の発達に十分な条件下で、1つ以上の胚を培養するステップと、
(b)前記1つ以上の胚において1つ以上の細胞パラメータを測定して、細胞パラメータ測定値を導き出すステップと、
(c)前記細胞パラメータ測定値を利用して、前記1つ以上の胚の発達の可能性の判定を提供するステップと、
(d)良好な発達の可能性を示す前記1つ以上の胚を、それを必要とする女性に移すステップと、を含む、方法。
【請求項24】
前記胚は、生体外で卵母細胞を受精させることにより産生される、請求項23に記載の方法。
【請求項25】
前記卵母細胞は、生体外で成熟する、請求項23に記載の方法。
【請求項26】
前記卵母細胞は、生体内で成熟する、請求項23に記載の方法。
【請求項27】
生体外で成熟した卵母細胞は、成長因子を添加される、請求項25に記載の方法。
【請求項28】
培養中の細胞を自動撮像するための装置であって、
(a)インキュベータの内部に設置された複数の顕微鏡と、
(b)ステージ上に固定された培養皿に焦点を合わせるように配置された各顕微鏡と、
(c)前記培養皿中の細胞を照射するための光源を備えた各顕微鏡と、
(d)撮像カメラを備えた各顕微鏡と、
(e)1つ以上のカメラからの画像を保存し、経時的に連続画像を分析するためにプログラムされたコンピュータと、を備える、装置。
【請求項29】
前記光源は、暗視野照明を提供する、請求項28に記載の装置。
【請求項30】
前記光源は、赤色または近赤外線である、請求項28に記載の装置。
【請求項31】
細胞培養のための装置であって、
(a)マイクロウェルを備えたペトリ皿と、
(b)光学品質仕上げの底面を有する各マイクロウェルと、
(c)単一媒体滴液を共有できるように一緒に緊密に配置されるマイクロウェルと、を備える、装置。
【請求項32】
外壁が、媒体滴液を保持するために、マイクロウェルの周囲に位置付けられている、請求項31に記載の装置。
【請求項33】
位置合わせマーカーが、マイクロウェルの近くに設置される、請求項31に記載の装置。
【請求項34】
胚細胞または幹細胞を成長させる発達の可能性を判定するためのシステムであって、
(a)顕微鏡と、
(b)前記顕微鏡から画像を取得するための撮像カメラと、
(c)前記カメラからの画像を保存するためのコンピュータと、
(d)(i)細胞質分裂の持続時間、(ii)有糸分裂事象間の時間、のうちの少なくとも1つを測定するために、前記細胞のモデルを作製するためのソフトウェアと、を備える、システム。
【請求項35】
分裂細胞がピクセルを有する一連のデジタル画像としてサンプリングされる、培養中の可動な分裂細胞を顕微鏡的に撮像するための自動化方法であって、
(a)前記一連のピクセルを計算された形状として表すステップと、
(b)前記計算された形状の値を乱すことにより、細胞運動および分裂による前記一連のピクセルにおける変化を予測するステップと、
(c)予測された形状から一連のシミュレーション画像を生成するステップと、
(d)シミュレーション画像を実際の画像と比較するステップと、
(e)予測された形状の正確性を判定するために比較を使用するステップと、を含む、自動化方法。
【請求項36】
画像シミュレーションは、1つ以上の細胞膜の輪郭をシミュレーションすることを含む、請求項35に記載の方法。
【請求項37】
前記多能性細胞は卵母細胞であり、前記1つ以上の細胞パラメータは、
(i)極体を産生する細胞質分裂事象の持続時間、および
(ii)減数分裂細胞周期間の時間間隔から成る群より選択される、請求項2に記載の方法。
【請求項1】
ヒトの胚または多能性細胞の発達の可能性を判定するための方法であって、
(a)前記ヒトの胚または多能性細胞の1つ以上の細胞パラメータを測定して、細胞パラメータ測定値を導き出すことと、
(b)前記細胞パラメータ測定値を利用して、前記胚または多能性細胞の発達の可能性の判定を提供することと、を含む、方法。
【請求項2】
前記細胞パラメータは、微速度顕微鏡検査により測定可能である、請求項1に記載の方法。
【請求項3】
胚の発達の可能性が判定され、前記1つ以上の細胞パラメータが、
(i)細胞質分裂事象の持続時間、
(ii)細胞質分裂1と細胞質分裂2との間の時間間隔、および
(iii)細胞質分裂2と細胞質分裂3との間の時間間隔から成る群から選択される、請求項2に記載の方法。
【請求項4】
前記ヒトの胚の良好な発達の可能性は、
(i)約0分〜約30分の細胞質分裂1の持続時間、
(ii)約8〜15時間の細胞質分裂1の分離と細胞質分裂2の開始との間の時間間隔、および/または
(iii)約0〜5時間の細胞質分裂2の開始と細胞質分裂3の開始との間の時間間隔により示される、請求項3に記載の方法。
【請求項5】
前記測定するステップは、細胞周期1の持続時間を測定することをさらに含む、請求項4に記載の方法。
【請求項6】
前記ヒトの胚の良好な発達の可能性は、約20〜27時間の細胞周期1の持続時間により示される、請求項5に記載の方法。
【請求項7】
前記利用するステップは、前記細胞パラメータ測定値を基準のヒトの胚からの細胞パラメータ測定値と比較し、前記比較の結果を利用して、前記ヒトの胚の発達の可能性の判定を提供することを含む、請求項3に記載の方法。
【請求項8】
前記ヒトの胚の不良な発達の可能性は、
(a)前記ヒトの胚の細胞質分裂1が、前記基準のヒトの胚より長い、
(b)前記ヒトの胚の細胞質分裂1の分離と細胞質分裂2の開始との間の時間間隔が、前記基準のヒトの胚より長い、または短い、
(c)前記ヒトの胚の細胞質分裂2の開始と細胞質分裂3の開始との間の時間間隔が、前記基準のヒトの胚より長いことにより示される、請求項7に記載の方法。
【請求項9】
前記測定は、細胞周期1の持続時間を測定することをさらに含む、請求項8に記載の方法。
【請求項10】
前記ヒトの胚の不良な発達の可能性は、前記胚の細胞周期1が前記基準のヒトの胚より長いことにより示される、請求項9に記載の方法。
【請求項11】
前記1つ以上の細胞パラメータは、遺伝子発現測定値を導き出すために測定される遺伝子発現レベルである、請求項1に記載の方法。
【請求項12】
胚の発達の可能性が判定され、前記1つ以上の遺伝子は、コフィリン、DIAPH1、ECT2、MYLC2/MYL5、DGCR8、ダイサー/DICER1、TARBP2、CPEB1、シンプレキン/SYMPK、YBX2、ZAR1、CTNNB1、DNMT3B、TERT、YY1、IFGR2/IFNGR2、BTF3、およびNELFから成る群から選択される、請求項11に記載の方法。
【請求項13】
前記利用するステップは、前記遺伝子発現測定値を基準の胚または多能性細胞からの遺伝子発現測定値と比較することと、前記比較の結果を使用して、前記ヒトの胚または多能性細胞の発達の可能性の判定を提供することと、を含む、請求項11に記載の方法。
【請求項14】
胚の発達の可能性が判定され、前記基準の胚に対して、前記胚におけるコフィリン、DIAPH1、ECT2、MYLC2/MYL5、DGCR8、ダイサー/DICER1、TARBP2、CPEB1、シンプレキン/SYMPK、YBX2、ZAR1、CTNNB1、DNMT3B、TERT、YY1、IFGR2/IFNGR2、BTF3、およびNEFから成る群から選択される1つ以上の遺伝子のより低い発現レベルは、不良な発達の可能性を示す、請求項13に記載の方法。
【請求項15】
相互に対してヒトの胚または多能性細胞をランク付けするための方法であって、
(a)各ヒトの胚または多能性細胞について少なくとも1つの細胞パラメータを測定して、各胚または多能性細胞の細胞パラメータ測定値を導き出すステップと、
(b)前記胚または多能性細胞のそれぞれからの前記少なくとも1つの細胞パラメータ測定値を利用して、相互に対して前記ヒトの胚または多能性細胞をランク付けするステップと、を含む、方法。
【請求項16】
前記少なくとも1つの細胞パラメータは、微速度顕微鏡検査により測定可能である、請求項15に記載の方法。
【請求項17】
胚がランク付けされ、前記少なくとも1つの細胞パラメータは、
(i)細胞質分裂事象の持続時間、
(ii)細胞質分裂1と細胞質分裂2との間の時間間隔、および
(iii)細胞質分裂2と細胞質分裂3との間の時間間隔から成る群から選択される、請求項16に記載の方法。
【請求項18】
前記測定ステップは、細胞周期1の持続時間を測定することをさらに含む、請求項17に記載の方法。
【請求項19】
前記1つ以上の細胞パラメータは、1つ以上の遺伝子の発現レベルである、請求項1に記載の方法。
【請求項20】
前記1つ以上の遺伝子は、コフィリン、DIAPH1、ECT2、MYLC2/MYL5、DGCR8、ダイサー/DICER1、TARBP2、CPEB1、シンプレキン/SYMPK、YBX2、ZAR1、CTNNB1、DNMT3B、TERT、YY1、IFGR2/IFNGR2、BTF3、およびNELFから成る群から選択される、請求項19に記載の方法。
【請求項21】
前記利用するステップは、前記胚または多能性細胞のそれぞれからの前記細胞パラメータ測定値を相互に比較して、相互に比較した胚または多能性細胞の発達の可能性を判定することにより達成される、請求項15に記載の方法。
【請求項22】
前記利用するステップは、前記胚または多能性細胞のそれぞれからの前記細胞パラメータ測定値を基準の胚または多能性細胞からの細胞パラメータ測定値と比較して、各胚または多能性細胞の発達の可能性を判定することにより、および各胚または多能性細胞の発達の可能性を比較して、相互に比較して胚または多能性細胞の発達の可能性を判定することにより達成される、請求項15に記載の方法。
【請求項23】
良好な発達の可能性を有するヒトの胚を女性に提供する方法であって、
(a)胚の発達に十分な条件下で、1つ以上の胚を培養するステップと、
(b)前記1つ以上の胚において1つ以上の細胞パラメータを測定して、細胞パラメータ測定値を導き出すステップと、
(c)前記細胞パラメータ測定値を利用して、前記1つ以上の胚の発達の可能性の判定を提供するステップと、
(d)良好な発達の可能性を示す前記1つ以上の胚を、それを必要とする女性に移すステップと、を含む、方法。
【請求項24】
前記胚は、生体外で卵母細胞を受精させることにより産生される、請求項23に記載の方法。
【請求項25】
前記卵母細胞は、生体外で成熟する、請求項23に記載の方法。
【請求項26】
前記卵母細胞は、生体内で成熟する、請求項23に記載の方法。
【請求項27】
生体外で成熟した卵母細胞は、成長因子を添加される、請求項25に記載の方法。
【請求項28】
培養中の細胞を自動撮像するための装置であって、
(a)インキュベータの内部に設置された複数の顕微鏡と、
(b)ステージ上に固定された培養皿に焦点を合わせるように配置された各顕微鏡と、
(c)前記培養皿中の細胞を照射するための光源を備えた各顕微鏡と、
(d)撮像カメラを備えた各顕微鏡と、
(e)1つ以上のカメラからの画像を保存し、経時的に連続画像を分析するためにプログラムされたコンピュータと、を備える、装置。
【請求項29】
前記光源は、暗視野照明を提供する、請求項28に記載の装置。
【請求項30】
前記光源は、赤色または近赤外線である、請求項28に記載の装置。
【請求項31】
細胞培養のための装置であって、
(a)マイクロウェルを備えたペトリ皿と、
(b)光学品質仕上げの底面を有する各マイクロウェルと、
(c)単一媒体滴液を共有できるように一緒に緊密に配置されるマイクロウェルと、を備える、装置。
【請求項32】
外壁が、媒体滴液を保持するために、マイクロウェルの周囲に位置付けられている、請求項31に記載の装置。
【請求項33】
位置合わせマーカーが、マイクロウェルの近くに設置される、請求項31に記載の装置。
【請求項34】
胚細胞または幹細胞を成長させる発達の可能性を判定するためのシステムであって、
(a)顕微鏡と、
(b)前記顕微鏡から画像を取得するための撮像カメラと、
(c)前記カメラからの画像を保存するためのコンピュータと、
(d)(i)細胞質分裂の持続時間、(ii)有糸分裂事象間の時間、のうちの少なくとも1つを測定するために、前記細胞のモデルを作製するためのソフトウェアと、を備える、システム。
【請求項35】
分裂細胞がピクセルを有する一連のデジタル画像としてサンプリングされる、培養中の可動な分裂細胞を顕微鏡的に撮像するための自動化方法であって、
(a)前記一連のピクセルを計算された形状として表すステップと、
(b)前記計算された形状の値を乱すことにより、細胞運動および分裂による前記一連のピクセルにおける変化を予測するステップと、
(c)予測された形状から一連のシミュレーション画像を生成するステップと、
(d)シミュレーション画像を実際の画像と比較するステップと、
(e)予測された形状の正確性を判定するために比較を使用するステップと、を含む、自動化方法。
【請求項36】
画像シミュレーションは、1つ以上の細胞膜の輪郭をシミュレーションすることを含む、請求項35に記載の方法。
【請求項37】
前記多能性細胞は卵母細胞であり、前記1つ以上の細胞パラメータは、
(i)極体を産生する細胞質分裂事象の持続時間、および
(ii)減数分裂細胞周期間の時間間隔から成る群より選択される、請求項2に記載の方法。
【図4】

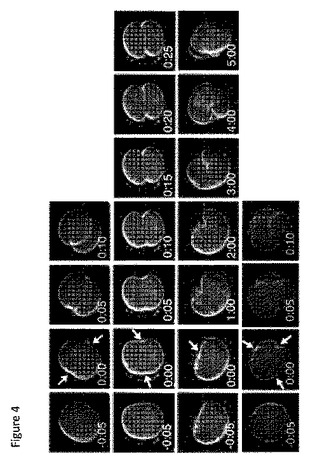
【図15A】

【図15B】

【図15C】

【図15D】

【図15E】

【図15F】

【図18a】

【図18b】

【図18c】

【図18d】

【図18e】


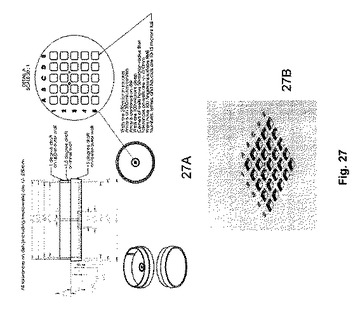
【図1】
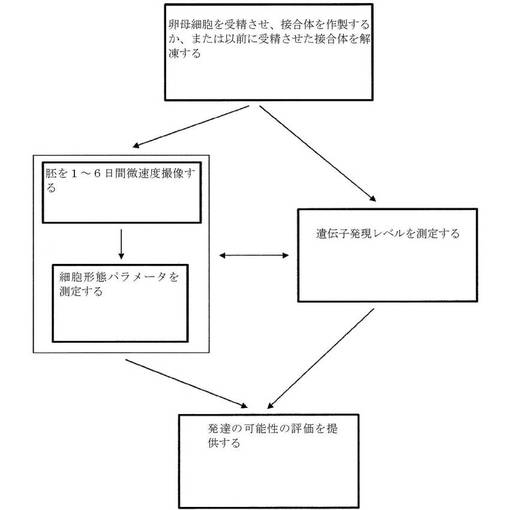
【図2】
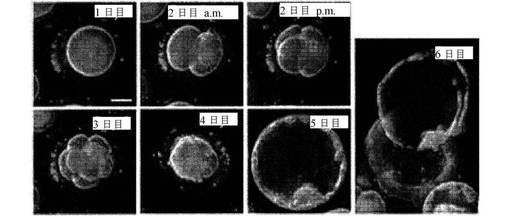
【図3】
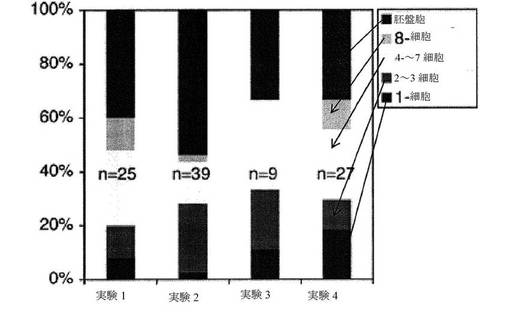
【図5】
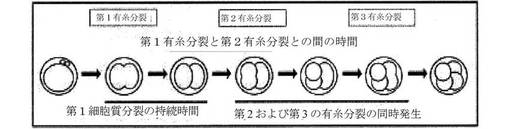
【図6】

【図7】
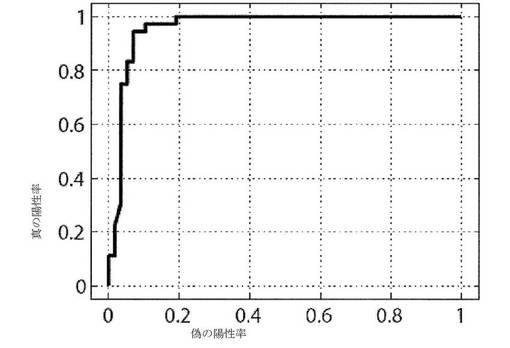
【図8】
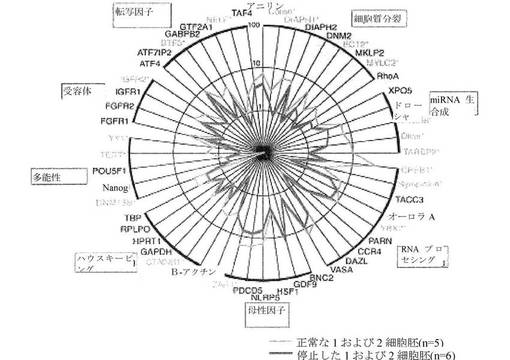
【図9】
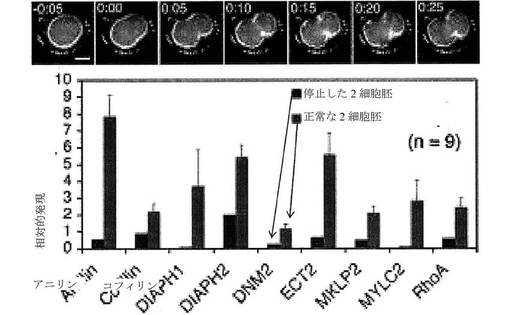
【図10】
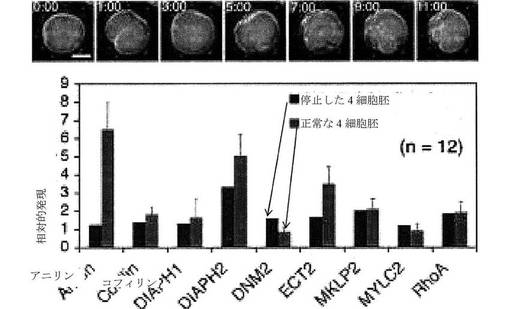
【図11】
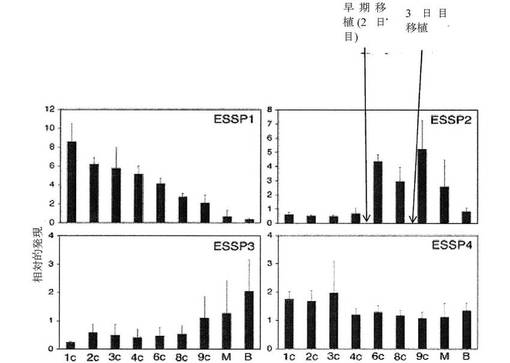
【図12】

【図13】
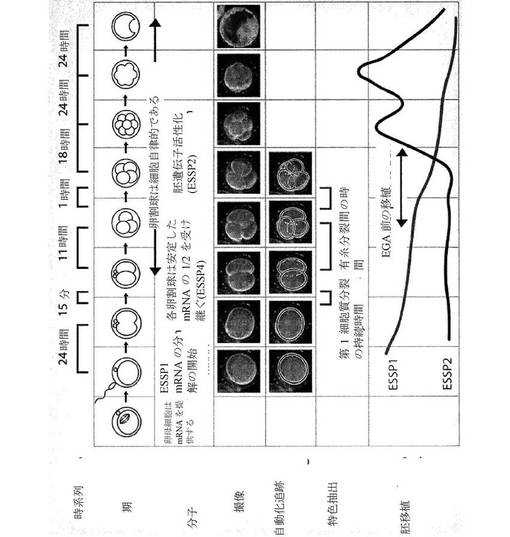
【図14】
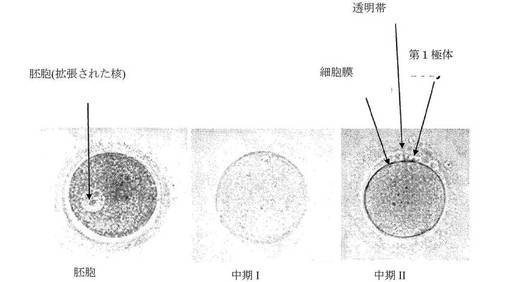
【図16】

【図17】
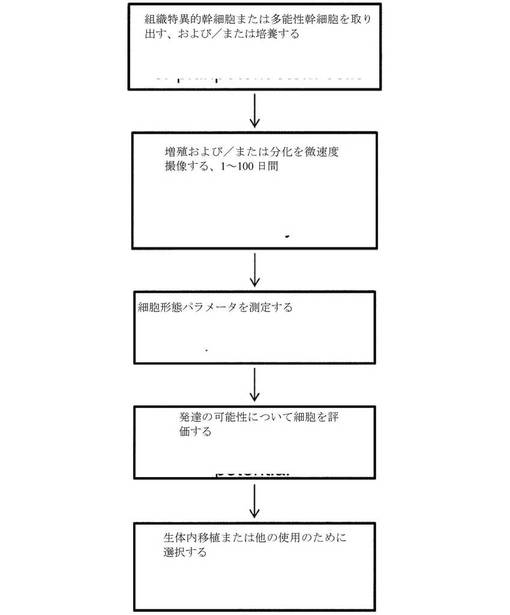
【図19】

【図20】

【図21】

【図22】
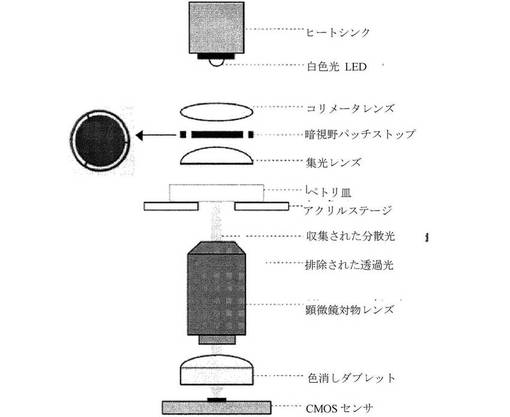
【図24】
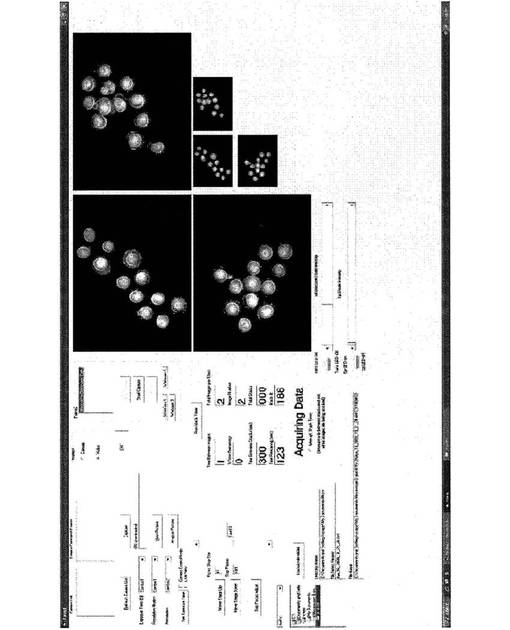
【図25A−D】

【図26A−B】

【図27A−B】
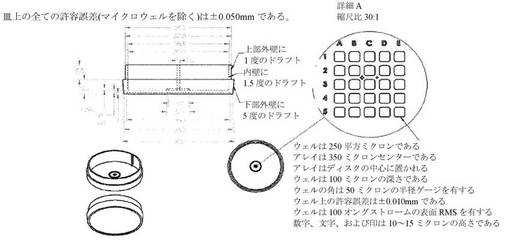
【図28A−B】
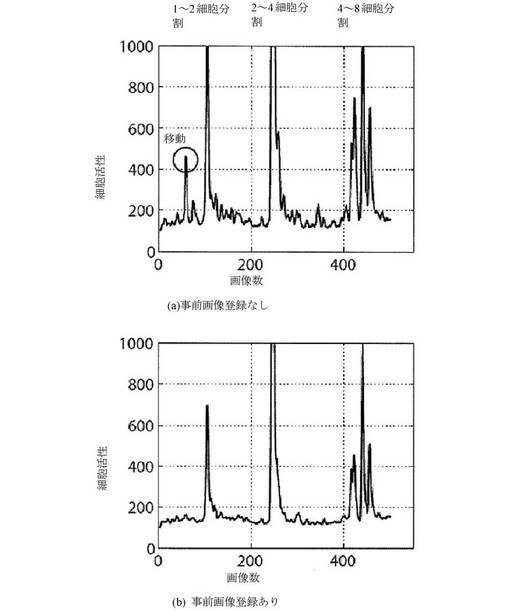
【図29A−B】
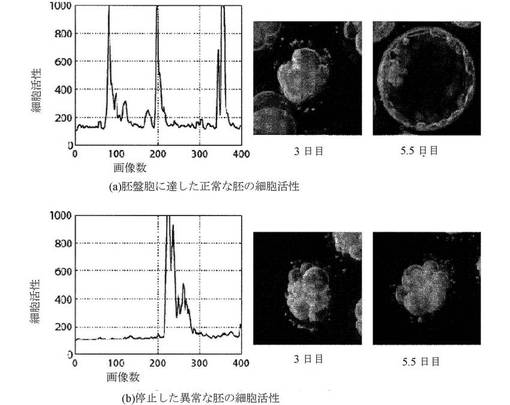
【図30】

【図31A−G】

【図32A−B】
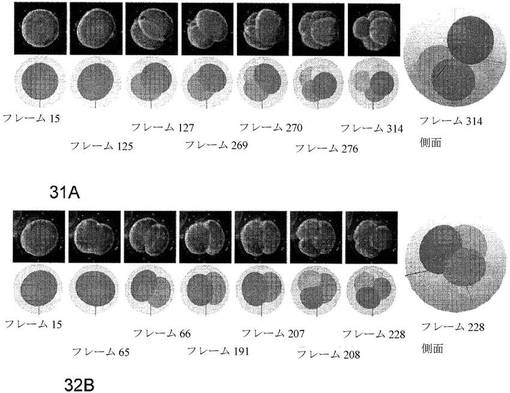
【図33】

【図34A−B】

【図35】

【図15A】
【図15B】
【図15C】
【図15D】
【図15E】
【図15F】
【図18a】
【図18b】
【図18c】
【図18d】
【図18e】
【図1】
【図2】
【図3】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図16】
【図17】
【図19】
【図20】
【図21】
【図22】
【図24】
【図25A−D】
【図26A−B】
【図27A−B】
【図28A−B】
【図29A−B】
【図30】
【図31A−G】
【図32A−B】
【図33】
【図34A−B】
【図35】
【公表番号】特表2013−502233(P2013−502233A)
【公表日】平成25年1月24日(2013.1.24)
【国際特許分類】
【出願番号】特願2012−526882(P2012−526882)
【出願日】平成22年8月23日(2010.8.23)
【国際出願番号】PCT/US2010/046343
【国際公開番号】WO2011/025736
【国際公開日】平成23年3月3日(2011.3.3)
【出願人】(500429147)ザ ボード オブ トラスティーズ オブ ザ リーランド スタンフォード ジュニア ユニバーシティ (13)
【Fターム(参考)】
【公表日】平成25年1月24日(2013.1.24)
【国際特許分類】
【出願日】平成22年8月23日(2010.8.23)
【国際出願番号】PCT/US2010/046343
【国際公開番号】WO2011/025736
【国際公開日】平成23年3月3日(2011.3.3)
【出願人】(500429147)ザ ボード オブ トラスティーズ オブ ザ リーランド スタンフォード ジュニア ユニバーシティ (13)
【Fターム(参考)】
[ Back to top ]