
胚性幹細胞の分化を誘導する方法及びその用途
本発明は、胚性幹細胞の分化した神経細胞への分化を誘導する方法を提供する。本発明は、さらに、分化した神経細胞、及び分化した神経細胞を含む細胞の集団を生じさせる方法を提供する。さらに、本発明は、被験者において脊髄を再増殖させる方法、及び治療の必要がある被験者において神経組織の変性を治療する方法を提供する。本発明は、さらに、神経前駆細胞、分化した神経細胞、及び同用途を提供する。また、分化した神経細胞を含むトランスジェニック非ヒト動物も提供される。本発明は、さらに、分化した神経細胞の集団を単離する方法に向けられる。最後に、本発明は、ニューロンの変性に関連した症状の治療における使用のための薬剤の同定方法を提供する。
【発明の詳細な説明】
【発明の開示】
【0001】
発明の背景
数百もの異なるニューロンの種類が脊椎の中枢神経系(CNS)の発生の間に生成され、ニューロン回路の形成に必須の細胞の多様性を確立する。特定の種類又はクラスのCNSニューロンの選択的な変性は、多くの神経学上の障害の根源である。この理解は、神経変性性障害を治療するという見地から、ニューロンの補給可能な源として作用するかもしれない前駆細胞の集団を規定することにおいて興味を生じさせてきた。系統的様式においてニューロンの分化の特定の経路に沿ったそのような前駆細胞を支配すること(directing)は難しいことが証明されたが、単に、CNSニューロンのほとんどのクラスを生じさせる通常の発生の経路がほとんど定義されていないままであるからではない。
【0002】
前駆細胞の神経学上の潜在性の研究は、3つの主要なクラスに焦点が合わされてきた:(1)胚又は成人の神経組織由来の神経前駆細胞(Alvarez-Buylla et al.,A unified hypothesis on the lineage of neural stem cells.Nat.Rev.Neurosci.,2:287-93,2001;Gage,F.H.,Mammalian neural stem cells.Science,287:1433-38,2000;Temple,S.,The development of neural stem cells.Nature,414:112-17,2001;Uchida et al.,Direct isolation of human central nervous system stem cells.Proc.Natl.Acad.Sci.USA,97:14720-725,2000);(2)他の組織及び器官由来の非神経前駆細胞(Brazelton et al.,From Marrow to brain:expression of neuronal phenotypes in adult mice.Science,290:1775-79,2000;Mezey et al.,Turning blood into brain:cells bearing neuronal antigens generated in vitro from bone marrow.Science,290:1779-82,2000;Terada et al.,Bone marrow cells adopt the phenotype of other cells by spontaneous fusion.Nature,416:545-48,2002);及び(3)胚性幹(ES)細胞(Bain et al.,Embryonic stem cells express neuronal properties in vitro.Dev.Biol.,168:342-57,1995;Reubinoff et al.,Neural progenitors from human embryonic stem cells.Nat.Biotechnol.,19:1134-40,2001;Schuldiner et al.,Induced neuronal differentiation of human embryonic stem cells.Brain Res.,913:201-05,2001;Zhang et al.,In vitro differentiation of transplantable neural precursors from human embryonic stem cells.Nat.Biotechnol.,19:1129-33,2001;Rathjen et al.,Directed differentiation of pluripotent cells to neural lineages:homogenous formation and differentiation of a neurectderm population.Development,129:2649-61,2002)。ES細胞は、ニューロン及び神経細胞の両方を生成し、そして幾つかの例においては、中脳ドーパミン作用性のニューロンを含む、特定のクラスのニューロンに特徴的な細胞種マーカーを発現する能力を有する(Kawasaki et al.,Induction of midbrain dopaminergic neurons from ES cells by stromal cell-derived inducing activity.Neuron,28:31-40,2000;Lee et al.,Efficient generation of midbrain and hindbrain neurons from mouse embryonic stem cells.Nat.Biotechnol.,18:675-79,2000)。これらの利点にも拘わらず、しかしながら、本発明の前までは、ES細胞が容易に特定のニューロン細胞種を生成することができることも、それらが神経発生の正常なプログラムを再現できることも、知られていなかった。
【0003】
脊髄の運動ニューロンは、ニューロン特異化(specification)の関連経路の多くが定義された一つのCNSニューロンサブタイプを代表する(Jessell et al.,Neuronal specification in the spinal cord:inductive signals and transcriptional codes.Nat.Rev.Genet.,1:20-29,2000;Lee et al.,Transcriptional networks regulating neuronal identity in the developing spinal cord.Nat.Neurosci.,4補遺:1183-91,2001)。脊髄の運動ニューロンの生成は、幾つかの発生段階を含むことが明白である。最初に、外胚葉細胞が、吻側(rostral)神経特性−BMP,FGF,及びWntシグナリングの制御を通して達成されたプロセスを獲得する(Munoz-Sanjuan et al.,Neural induction,the default model and embryonic stem cells.Nat.Rev.Neurosci.,3:271-80,2002;Wilson et al.,Neural induction:toward a unifying mechanism.Nat.Neurosci.,3:4補遺:1161-68,2001)。これらの吻側神経前駆細胞はレチノイン酸(RA)を含む尾部側化シグナリングに応答して脊髄の位置的アイデンティティーを獲得する(Blumberg et al.,An essential role for retinoic signaling in anteroposterior neural patterning.Development,124:373-79,1997;Durston et al.,Retinoids and related signals in early development of the vertebrate central nerve system.Curr.Top.Dev.Biol.,40:111-75,1998;Muhr et al.,Convergent inductive signals specify midbrain,hindbrain,and spinal cord identity in gastrula stage chick embryos.Neuron,23:689-702,1999)。次に、脊髄の前駆細胞は、ソニックヘッジホッグ蛋白質(SHh)の腹側化アクションに対する応答として運動ニューロン前駆細胞のアイデンティティーを獲得する(Briscoe et al.,Specification of neuronal fates in the ventral neural tube.Curr.Opin.Neurobiol.,11:43-49,2001)。
【0004】
SHhシグナリングによる運動ニューロン前駆細胞のアイデンティティーの特異化は、ホメオドメイン(HD)及び基本的なヘリックス−ループ−ヘリックス(bHLH)転写因子の発現の背腹側パターンの確立を通して媒介する(Briscoe et al.,前掲)。相対的に高いレベルのSHhシグナリングにおいては、pMNドメインと呼ばれる別の前駆ドメインが確立される;このドメイン内では、細胞は介在ニューロンよりも運動ニューロンの生成に委ねられるらしい(Briscoe et al.,A homeodomain protein code specifies progenitor cell identity and neuronal fate in the ventral neural tube.Cell,101:435-45,2000)。
【0005】
MNドメイン内の前駆細胞は、2つのHD蛋白質(Pax6及びNkx6.1)及びbHLH蛋白質(Olig2)の発現により特徴付けされる(Ericson et al.,Pax controls progenitor cell identity and neuronal fate in response to graded SHh signaling.Cell,90:169-80,1997;Sander et al.,Ventral neural patterning by Nkx homeobox genes:Nkx6.1 controls somatic motor neuron and ventral interneuron fates.Genes Dev.,14:2134-39,2000;Vallstedt et al.,Different levels of repressor activity assign redundant and specific roles to Nkx6 genes in motor neuron and interneuron specification.Neuron,31:743-55,2001;Novitch et al.,Coordination regulation of motor neuron subtype identity and pan-neuronal properties by the bHLH repressor Olig2 and neurogenin2 in the coordinate induction of pan-neuronal and subtype-specific properties of motoneurons.Neuron,31:757-71,2001)。これらの3つの転写因子の各々は、脊髄の運動ニューロンのアイデンティティーの明確化において本質的な役割を担う(Ericson et al.,Pax6 controls progenitor cell identity and neuronal fate in response to graded SHh signaling.Cell,90:169-80,1997;Sander et al.,Different levels of repressor activity assign redundant and specific roles to Nkx6 genes in motor neuron and interneuron specification.Neuron,31:743-55,2001;Novitch et al.,Coordinate regulation of motor neuron subtype identity anf pan-neuronal properties by the bHLH repressor Olig2.Neuron,31:773-89,2001;Mizuguchi et al.,Combinatorial roles of olig2 and neurogenin2 in the coordinated induction of pan-neuronal and subtype-specific properties of motoneurons.Neuron,31:757-71,2001;Zhou et al.,The bHLH transcription factors OLIG2 and OLIG1 couple neuronal and glial subtype specification.Cell,109:61-73,2002;Lu et al.,Common developmental requirement for Olig function indicates a motor neuron/oligodendrocyte connection.Cell,109:75-86,2002)。さらに、それらの組み合わされた活性が細胞サイクルを離れて(out of)運動ニューロン前駆体を操作し(drive)(Novitch et al.,Coordinate regulation of motor neuron subtype identity and pan-neuronal properties by the bHLH repressor Olig2.Neuron,31:773-89,2001;Mizuguchi et al.,Combinatorial roles of olig2 and neurogenin2 in the coordinated induction of pan-neuronal and subtype-specific properties of motoneurons.Neuron,31:757-71,2001)、そして有糸分裂後の運動ニューロンのアイデンティティーを強化する(consolidate)下流の転写因子(特に、HD蛋白質,HB9)の発現を指示する(Pfaff et al.,Requirement for LIM homeobox gene Isl1 in motor neuron generation reveals a motor neuron-dependent step in interneuron differentiation.Cell,84:309-20,1996;Arber et al.,Requirement for the homeobox gene Hb9 in the consolidation of motor neuron identity.Neuron,23:659-764,199;Thaler et al.,Active suppression of interneuron programs within developing motor neurons revealed by analysis of homeodomain factor HB9.Neuron,23:675-87.1999)。
【0006】
上記の発見は、神経発生の通常の経路における洞察が、合理的な様式にて前駆細胞、例えばES細胞の異種セットを、特定のCNSニューロンサブタイプへ向けるために適用できることを示唆する。ES細胞は運動ニューロンの分子特性の幾つかを伴う細胞を生成することが報告された(Renoncourt et al.,Neurons derived in vitro from ES cells express homeoproteins characteristic of motoneurons and interneurons.Mech.Dev.,79:185-97,1998)。しかしながら、本発明の前には、これらのニューロンの生成の経路も、それらのインビボ発生能力も、十分に探索されていなかった。
【0007】
発明の概要
発明者らは、神経管の吻尾間の(rostrocaudal)軸索及び背腹間の軸索に沿って機能し、且つインビボにおいて運動ニューロンの運命を特定する細胞外シグナリング因子が、インビトロにおいて、マウスES細胞の分化を機能性脊髄運運動ニューロンへ向けるために利用され得るか否かを試験した。本明細書において開示されるとおり、発明者らは、神経化されたES細胞のRAへの暴露が脊髄の前駆細胞の分化を誘導すること、及びヘッジホッグ(Hh)シグナリングの活性化が運動ニューロンを生成するためにこれらの細胞を指示することを確定した。発明者らは、Hhシグナリングにより誘発された運動ニューロンの分化の固有の分子経路がインビボにおいて運動ニューロンの生成に関与する経路を模倣することも示した。GFP−に基づく運動ニューロンマーキング法の使用は、ES−細胞−由来の運動ニューロンの純粋な集団の特性決定を許容した。そのような運動ニューロンは、インビボにおいて胚性脊髄を再増殖させ(repopulating)、軸索を末梢へ伸長させ、そして骨格筋との接触の部位においてシナプス分化を受けることができる。即ち、神経発生の通常の経路を合理的様式においてくつがえす(subverted)ことにより、ES細胞をCNSニューロンの特定のサブクラスへ向かわせることができる。
【0008】
従って、本発明は、胚性幹細胞を、分化した神経細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子及び背側化又は腹側化胚性シグナリング因子に接触させることにより、胚性幹細胞の分化した神経細胞への分化を誘導する方法を提供する。また、この方法により生産した、分化した神経細胞も提供される。
【0009】
本発明は、分化した神経細胞を生産する方法も提供し、(a)胚性幹細胞の培養物を得るか又は生じさせ;(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させ;そして(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させることによる。また、この方法により生産された、分化した神経細胞を含む細胞集団も提供される。
【0010】
本発明は、被験者において脊髄を再増殖させる(repopulation)方法も提供し、(a)胚性幹細胞の培養物を得るか又は生じさせ;(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させ;(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させ;そして(e)分化した神経細胞を被験者の脊髄に移植することによる。
【0011】
本発明は、治療の必要な被験者において神経組織の変性を治療する方法にも向けられ、(a)胚性幹細胞の培養物を得るか又は生じさせ;(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させ;(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させ;そして(e)神経組織の変性を治療するのに有効な量にて分化した神経細胞を被験者に移植することによる。
【0012】
さらに、本発明は、(a)胚性幹細胞の培養物を得るか又は生じさせ;(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;そして(c)任意に、神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させることにより生じさせた神経前駆細胞を提供する。また、神経の発生、機能、及び死を分析すること、又はシナプスの分化を監視することにおける、これらの神経前駆細胞の用途も提供される。
【0013】
本発明は、さらに、(a)胚性幹細胞の培養物を得るか又は生じさせ;(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させ;そして(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させることにより生じさせた神経前駆細胞を提供する。また、これらの分化した神経細胞を含む非ヒトトランスジェニック動物及び神経の発生、機能、及び死を分析すること、又はシナプスの分化を監視することにおける、これらの分化した神経細胞の用途も提供される。
【0014】
本発明は、分化した神経細胞の集団を単離及び又は精製する方法にも向けられ、(a)増強された緑色蛍光蛋白質(eGFP)を発現する胚性幹細胞の培養物を得るか又は生じさせ;(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;(c)神経前駆細胞を、eGFPを発現する分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させ;(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させ;(e)分化した神経細胞においてeGFPの発現を検出し;そして(f)eGFPを発現する分化した神経細胞を単離することによる。
【0015】
最後に、本発明は、神経の変性に関連した症状を治療することにおける使用のための薬剤を同定する方法も提供され、(a)胚性幹細胞の培養物を得るか又は生じさせ;(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量のレチノイン酸に接触させ;(c)神経前駆細胞においてヘッジホッグシグナリング経路を活性化することによりニューロンを生じさせるが、但し、ニューロンの幾つか又は全てが変性しており;(d)変性したニューロンを候補の薬剤と接触させ;そして(e)当該薬剤が幾つか又は全ての変性したニューロンの再生を増強するか否かを決定することによる。
【0016】
本発明の追加の側面は、続く説明から明らかになる。
【0017】
発明の詳細な説明
発生の間、特定の方向特性又は位置特性を幹細胞に授与するシグナリング因子の制御を含む一連の発生段階を通して、神経細胞が胚性幹細胞から生成される。最初に、外胚葉の細胞が吻側又は尾部側の神経特性を獲得し、そして吻側化(rostralizing)及び/又は尾部側化(caudalizing)の胚性シグナリング因子の制御を通して、吻側の又は尾部側の前駆細胞に分化すると言える。その後、神経前駆細胞はさらに分化して、背側化(dorsalizing)及び/又は腹側化(ventralizing)胚性シグナリング因子の作用に応答して、前駆細胞のサブタイプのアイデンティティーを獲得するか、又は完全に分化した神経細胞になる。
【0018】
例えば、運動ニューロンは様々なシグナリング因子に応答してES細胞から発生すると言える。最初に、BMP,FGF,及びWntシグナリングの制御により、外胚葉細胞が吻側神経特性を獲得する。結果の吻側の神経前駆細胞は、次に、レチノイン酸(RA)を含む尾部側のシグナルに応答して、脊髄の位置に関するアイデンティティーを獲得する。SHhシグナリングによる運動ニューロン前駆体のアイデンティティーの明確化は、ホメオドメイン(HD)及び基本的ヘリックス−ループ−ヘリックス(bHLH)転写因子の発現の背腹パターンの確立を通して介在される。相対的に高いレベルのSHhシグナリングにおいては、pMNドメインと呼ばれる別の前駆体ドメインが確立される。このドメイン内では、細胞が介在ニューロンよりも運動ニューロンの生成にかかわるらしい。pMNドメイン内の前駆細胞は、2つのHD蛋白質、Pax6及びNkx6.1、及びbHLH蛋白質、Olig2の発現により特徴付けられる。これらの3つの転写因子の各々は、脊髄の運動ニューロンアイデンティティーの明確化において必須の役割を有する。さらに、それらの組み合わされた活性が細胞サイクルを離れて(out of)運動ニューロン前駆細胞を操作し(drive)、そして有糸分裂後の運動ニューロンのアイデンティティーを強化する下流の転写因子(特に、HD蛋白質、HB9)の発現を指示する。
【0019】
脊髄の運動ニューロンは運動性挙動の制御において必須の役割を演じるし、それらの変性は多くの神経障害の基礎となる。運動ニューロンの生成に関与する誘導性シグナル及び転写因子が同定されたことから、正常な発生への洞察を用いて、合理的な様式にて幹細胞を運動ニューロンの運命に向けさせ得ることを示唆する。発明者らは、本明細書において、マウスの胚性幹(ES)細胞が外部からのシグナリング因子により誘導されることにより、最初に脊髄前駆細胞に分化でき、次に脊髄の運動ニューロンに分化できることを明らかにする。ES−細胞由来の神経前駆体からの運動ニューロンの生成の分子経路は、インビボにおいて配備されるそれを再現する。胚性脊髄へ導入されたES−細胞由来の運動ニューロンは、腹側角を増殖させ(populate)、軸索を末梢に伸ばし、そして標的の骨格筋との接触部位においてシナプス分化を受ける。即ち、通常の神経発生の経路に関与する誘導性シグナルは、ES細胞がCNSニューロンの特定のクラスを形成することを指示できる。
【0020】
従って、本発明は、分化した神経細胞への胚性幹細胞の分化を誘導する方法、及びこの方法により生成された分化した神経細胞を提供する。本明細書にて使用される用語「胚性幹細胞の分化を誘導する」は、胚性幹細胞を活性化するか、開始するか、又は刺激することにより分化を受けること−発生の間に細胞が構造上並びに機能上特化されることになる細胞プロセスを意味する。胚性幹(ES)細胞は、培養された細胞であり、胚盤胞のステージの胚の多能性内部細胞隗由来であり、無期限に複製することができる。一般に、ES細胞は他の細胞に分化する能力を有する(即ち、それらは多能性);即ち、それらは新規な細胞の連続する源として機能するかもしれない。本発明の胚性幹細胞はあらゆる細胞から得て良いが、好ましくは哺乳類(例えば、ヒト、家畜動物、又は市販の動物)から得る。本発明の一つの態様において、胚性幹細胞はマウスの胚性幹細胞である。別の好ましい態様において、胚性幹細胞はヒトから得られる。
【0021】
さらに本明細書において使用される「分化した神経細胞」は、中枢神経系(CNS)又は末梢神経系(PNS)の一部分化した細胞又は完全に分化した細胞であり、そして、限定ではないが、完全に分化したガングリオン細胞、グリア(又は神経グリア)細胞(例えば、星状細胞、乏突起神経膠細胞、又はシュワン細胞)、顆粒細胞、ニューロン細胞(又はニューロン)、及び星形細胞、並びにそれらのあらゆる神経前駆細胞を含む。前駆細胞は、発生及び分化の間に、一連の細胞分割により別の細胞系列を生じる親細胞である。神経前駆細胞は、例えば、最後にはCNS又はPNSの完全に分化した神経細胞に発生する細胞系列に委ねられる(committed);しかしながら、そのような神経前駆細胞は神経細胞の特定の種類、又はサブクラスに供されたことがまだない。最初に、神経前駆細胞は吻側の特性を獲得し(例えば、吻側神経前駆細胞)、続いて位置的アイデンティティーを獲得してよい(例えば、小脳前駆細胞、大脳前駆細胞、又は脊髄前駆細胞)。そのような一部分化した神経前駆細胞は、特定の種類の神経細胞に分化して(例えば、星状細胞、ガングリオン細胞、顆粒細胞、ニューロン、乏突起神経膠細胞、シュワン細胞、又は星形細胞の前駆細胞)、そして以後は、完全に分化した前駆細胞(例えば星状細胞、ガングリオン細胞、顆粒細胞、ニューロン(背根ガングリオン(DRG)ニューロン、運動ニューロン、末梢ニューロン、脊髄ニューロン、又は腹側介在ニューロン)、乏突起神経膠細胞、シュワン細胞、又は星形細胞)を生じるように委ねられるようになる。従って、本発明の一部分化した神経細胞は、神経のアイデンティティーを有し、方向アイデンティティー又は位置アイデンティティーを獲得した細胞であるか、又は特定のクラスの神経細胞に発生するが完全に分化した神経細胞には発生しないことを委ねられた細胞であってよい。
【0022】
さらに、本明細書にて使用される「神経細胞」又は「ニューロン」は、核及び周囲の細胞質;幾つかの短い放射突起(processes)(星状細胞);及び一つの長い突起(軸索)、小枝様のブランチで終わるもの(末端星状細胞(telodendrons))、及びそのコースに沿って伸びるブランチを有してよいもの(側枝(collaterals))を含む、細胞ボディー(周核体)から一般にはなる神経系の、伝導性(conducting)細胞又は神経細胞である。ニューロンの例は、限定ではないが、背根ガングリオン(DRG)のニューロン、運動ニューロン、末梢ニューロン、知覚ニューロン、脊髄のニューロン、及び原側介在ニューロンを含み、それら全てはコリン作動性、ドーパミン作動性、又はセロトニン作動性であってよい。
【0023】
ES細胞の一部分化したか又は完全に分化した神経細胞への分化は、公知の細胞又は分子の手法、及び本明細書にて開示されたアッセイ及び方法により検出してよい。本発明の一つの態様において、分化した神経細胞は、運動ニューロンである(例えば、有糸分裂後の運動ニューロン又は脊髄の運動ニューロン)。本発明の別の態様において、分化した神経細胞は遺伝子を標識され、本明細書にて記載されたように、増強された緑色蛍光蛋白質(eGFP)を発現する。eGFP遺伝子マーカーは、分化した神経細胞の集団を単離及び/又は精製するための方法において、又は脊髄の制御を監視するための方法において特に有用かもしれず、以下に記載されるとおりである。
【0024】
本発明の方法は、胚性幹細胞を、吻側化又は尾部側化の胚性シグナリング因子、及び背側化又は腹側化の敗勢シグナリング因子に、分化した神経細胞を精製するのに有効な量にて接触させることを含む。本明細書にて使用される「因子」は、蛋白質、ポリペプチド、ペプチド、核酸(DNA又はRNAを含む)、抗体、Fab断片、F(ab’)2断片、分子、化合物、抗生物質、薬剤、及びそれらのあらゆる組み合わせを含む。Fab断片は抗体の単価の抗原結合断片であり、パパイン消化により生成される。F(ab’)2断片は抗体の二価の抗原結合断片であり、ペプシン消化により生成される。
【0025】
さらに本明細書にて使用される「胚性シグナリング因子」は、発生及び分化の推移の間に、胚の中で細胞へ情報を運ぶ因子である。そのような情報は、例えば、胚の中の細胞の方向又は位置に関してよく、そして情報を受けたそれらの細胞によりたどられる分化の経路を決定してよい。本発明の胚性シグナリング因子は、例えば、形態学上の因子又は神経因子であってよい。本発明における使用のための胚性シグナリング因子の例は、限定ではないが、レチノイン酸(RA)、骨形態蛋白質(BMPs)(例えば、BMP1からBMP7,BMP2及びBMP4を含む)、繊維芽成長因子(FGFs)(例えば、アルファFGF,FGF8,FGF10,HBGF1,塩基性FGF,ベータFGF,及びHBGF)、ヘッジホッグ蛋白質(例えば、ソニックヘッジホッグ(SHh)及びインディアンヘッジホッグ(IHh)、ヘッジホッグシグナリングのアゴニスト(例えば、Hh−Ag1.3)、及びヘッジホッグシグナリング経路の他の活性化因子を含む。
【0026】
さらに、本明細書にて使用される「吻側化胚性シグナリング因子」は、対象又は被験者の頭部又は上位の方向に誘導するか、寄与する(contribute)か、又は発生する因子を意味する。対照的に、「尾部側化胚性シグナリング因子」は、対象又は被験者の尻、尾又は下位の方向に誘導するか、寄与する(contribute)か、又は発生する因子を意味する。吻側アイデンティティーの割り当てに寄与する胚性シグナリング因子は、ES細胞の神経前駆細胞への分化をもたらすと言える。本発明における使用のための吻側化及び尾部側化胚性シグナリング因子の例は、限定ではないが、RA,BMP,FGF,及びWntを含む。RAは発生の間に吻側アイデンティティーを割り当てる役割を担うことが知られている。最近、Wntシグナリングも吻側アイデンティティーを割り当てる役割を担うことが示された(Nordstrom et al.,Progressive induction of caudal neural character be graded Wnt signaling.Nat.Neurosci.,5(6):525-32,2002)。即ち、本発明の一つの態様において、吻側化胚性シグナリング因子はRA又はWntである。発明の別の態様においては、尾部側化胚性シグナリング因子がRA又はWntである。
【0027】
本明細書にて使用される「背側化胚性シグナリング因子」は、対象又は被験者の背中又は後位の方向に誘導するか、寄与する(contribute)か、又は発生する因子を意味する。対照的に、本明細書にて使用される「腹側化胚性シグナリング因子」は、対象又は被験者の前部又は前方の方向に誘導するか、寄与する(contribute)か、又は発生する因子を意味する。背側アイデンティティーの割り当てに寄与する胚性シグナリング因子は、ES細胞の神経前駆細胞への分化又は完全に分化した神経細胞への分化をもたらすと言える。本発明における使用のための背側化及び腹側化胚性シグナリング因子の例は、限定ではないが、BMP,Wnt,ヘッジホッグ蛋白質、ヘッジホッグシグナリングのアゴニスト、及びヘッジホッグシグナリング経路の他の活性化因子を含む。SHhシグナリングは、例えば、運動ニューロン前駆細胞のアイデンティティーを脊髄前駆細胞に提供することが知られている。
【0028】
本発明の一つの態様において、背側化胚性シグナリング因子は、BMP又はWntである。本発明の別の態様において、腹側化胚性シグナリング因子は、ヘッジホッグシグナリング経路の他の活性化因子である。本明細書にて使用される「ヘッジホッグシグナリング」は、ヘッジホッグファミリーの蛋白質(例えば、ソニックヘッジホッグ(SHh)及びインディアンヘッジホッグ(IHh))のあらゆるメンバーの生化学シグナリング、特に発生及び分化の間に胚中の細胞の方向及び位置に関係するシグナリングを意味する。ヘッジホッグシグナリング経路の「活性化因子」は、ヘッジホッグシグナリングを作用させるか又は生じさせる因子、及びヘッジホッグシグナリング経路においてヘッジホッグシグナリングを誘導するか、開始させるか、又は刺激する因子を含む。例示として、ヘッジホッグシグナリングの活性化因子はヘッジホッグ蛋白質(例えば、SHh、IHh)又はヘッジホッグシグナリングのアゴニスト(例えば、Hh−Ag1.3)であってよい。本明細書にて使用されるヘッジホッグシグナリングの「アゴニスト」は、細胞中のヘッジホッグシグナリングが開始するか、刺激されるか、又は誘導されるように、天然に生じる物質により通常は刺激される細胞受容体に親和性を有し、且つ当該受容体において生理活性を刺激する因子である。
【0029】
胚性シグナリング因子、BMP,FGF,Wnt,SHh,及びIHhは蛋白質である。本明細書にて使用される「BMP」はBMP蛋白質及びBMPアナログの両方を含み;「FGF」はFGF蛋白質及びFGFアナログの両方を含み;「Wnt」はWnt蛋白質及びWntアナログの両方を含み;「SHh」はSHh蛋白質及びSHhアナログの両方を含み;「IHh」はIHh蛋白質及びIHhアナログの両方を含む。他に示さない限り、「蛋白質」は、蛋白質、蛋白質ドメイン、ポリペプチド又はペプチドを含む。本明細書にて使用される特定の蛋白質の「アナログ」は、当該蛋白質の生物活性を有し、且つ当該蛋白質と60%又はそれより高い(好ましくは、70%又はそれより高い)のアミノ酸配列相同性を有する、当該蛋白質の機能性バリアントである。「アナログ」は、相同な三次元コンフォメーションを有する蛋白質のバリアントを含む。さらに本明細書にて使用される用語「生物活性」は、発生及び分化の間のBMP,FGF,Wnt,SHh,又はIHh蛋白質のシグナリング活性を意味し、以下に記載される。
【0030】
さらに、本明細書にて使用される「保存的置換」は、置換されたアミノ酸残基に機能上均等なアミノ酸置換であり、それらが極性又は立体の配置を有するか、又はそれらが置換された残基と同じクラスに属するためである(例えば、疎水性、酸性又は塩基性)。本明細書にて使用される用語「保存的置換」は、本発明の蛋白質が発生及び分化の間にシグナリングを作用させる能力に対して取るに足らない(inconsequential)効果を有する置換を含む。
【0031】
本明細書にて論じられるとおり、BMP,FGF,Wnt,SHh,又はIHhは蛋白質である。対照的に、RA及びビタミンAはモルフォゲンであると信じられるアルデヒド分子である。RAは容易に利用可能であり;例えばシグマケミカル社(セントルイス、ミズーリ)から得てよい。本発明の他の因子は、合成によるか、組換えによるか、又は天然の細胞から単離してよい。好ましくは、蛋白質を組換えにより生産し、慣用の技術及び当該蛋白質をコードするcDNAを用いる。SHh及びIHh(並びにHh−Ag1.3)もキュリス社(ケンブリッジ、マサチューセッツ)から得てよい。
【0032】
本発明の方法は、胚性幹細胞を、吻側化及び/又は尾部側化胚性シグナリング因子及び背側化及び/又は腹側化胚性シグナリング因子に接触させることを含む。胚性シグナリング因子は、分化した神経細胞を生じさせるのに有効な量にて提供される。これらの量は、本明細書にて開示された公知の手法及び方法に基づいて、当業者により容易に決定される。本発明の好ましい態様においては、胚性幹細胞を、背側化及び/又は腹側化胚性シグナリング因子に接触させる前に、吻側化及び/又は尾部側化胚性シグナリング因子に接触させる。
【0033】
発明者らは、神経栄養性因子の存在下で培養されたニューロンが生存して且つ突起を念入りに作ることを証明した。従って、別の態様において、本発明の方法は、さらに、背側化又は腹側化胚性シグナリング因子との接触後に、少なくとも一つの神経栄養性因子に胚性幹細胞を接触させる工程を含む。本明細書にて使用される「神経栄養性因子」は、神経組織の栄養又はメンテナンスに関与する因子である。神経栄養性因子は、委ねられた神経前駆細胞の発生及び分化であるか、又はそれらは分化した神経細胞の成長及び生存を誘導するかもしれない。古典的な例の神経栄養性因子は、NGF(神経成長因子)である。本発明における使用のための神経栄養性因子の他の例は、限定ではないが、GDNF,NGF,NT3,CNTF,及びBDNFを含む。これらの因子はR&Dシステムズ社(ミネアポリス、ミネソタ)から得てよい。本発明の神経栄養性因子は、CNS又はPNSの完全に分化した神経細胞(例えば、ニューロン)を生産するのに有効な量で提供される。この量は、本明細書にて開示された公知の手法及び方法に基づいて、当業者により容易に決定してよい。
【0034】
本発明の方法においては、胚性幹(ES)細胞を、被験者においてインビトロ又はインビボにて有効な量の胚性シグナリング因子に接触させてよい。胚性シグナリング因子を細胞内へ導入することにより当該因子をES細胞に接触させてよい。接触をインビトロにて行う場合、因子を直接培養培地に加えてよく、本明細書に記載されているとおりである。或いは、因子を被験者に導入するか(例えば、因子を被験者の細胞に導入することにより)、又は因子を被験者に投与することにより、被験者内でインビボにてESに接触させてよい。被験者は胚性動物又は発生した動物であってよいが、好ましくは哺乳類(例えば、ヒト、家畜、又は市販の動物)である。より好ましくは、被験者はヒトである。胚性シグナリング因子をインビボにてES細胞に接触させる場合、被験者は好ましくは胚である。しかしながら、ES細胞を完全に成長したヒト又は動物被験者に移植すること、及び胚性シグナリング因子を次にヒトに移植することにより、ES細胞の分化した神経細胞への被験者内でのインビボにての分化を作用させることは、本発明の範囲内である。
【0035】
本発明の因子は、インビトロ又はインビボの何れかにおいて被験者の中で、蛋白質、核酸及び他の薬剤の導入及び投与に使用される公知の技術により、ES細胞と接触させてよく、例えば注射及び輸液(transfusion)である。標的のES細胞を被験者の特定の部分に局在化する場合は、注射によるか又は幾つかの他の手段により(例えば、因子を血液又は他の体液に導入することにより)、ES細胞へ直接導入することが望まれるかもしれない。
【0036】
胚性シグナリング因子が蛋白質又は他の分子である場合、本明細書に開示された慣用の技術及び方法により、直接ES細胞に導入してよい。さらに、蛋白質の胚性シグナリング因子は、蛋白質阻害剤の発現を許容する様式にて、因子をコードする核酸をES細胞に導入することにより、間接にES細胞に導入してよい。因子は、限定ではないが、エレクトロポレーション、DEAEデキストラントランスフェクション、リン酸カルシウムトランスフェクション、モノカチオンリポソーム融合、ポリカチオンリポソーム融合、プロトプラスト融合、インビボの電場の創製、DNA−コートされたマイクロ発射砲撃(microprojectile bombardment)、組換え体複製欠損ウイルスを用いた注入、相同組換え、インビボ遺伝子治療、エクスビボ遺伝子治療、ウイルスベクター、及び裸のDNAの移行(transfer)、又はそれらのあらゆる組み合わせを含む、慣用の当業界公知の手法を用いて、インビトロ又はインビボにて、ES細胞に導入してよい。遺伝子治療に適した組換えウイルスベクターは、限定ではないが、レトロウイルス、HSV,アデノウイルス、アデノ随伴ウイルス、セミリキフォレストウイルス、サイトメガロウイルス、及びワクシニアウイルスのようなウイルスのゲノム由来のベクターを含む。使用される核酸の量は、分化した神経細胞を生じさせるのに有効な蛋白質因子の量を発現するのに十分な量である。これらの量は、当業者により容易に決定してよい。蛋白質の胚性シグナリング因子をコードする核酸を適当なES細胞中へ慣用の手法を用いてインビトロにて導入することにより、ES細胞中の蛋白質因子の発現を達成してよい。蛋白質の胚性シグナリング因子を発現するES細胞は、次に、インビボにおいて分化した神経細胞を生じさせるために被験者に導入されてよい。
【0037】
本発明の方法によれば、胚性シグナリング因子は、限定ではないが、経口投与、非経口投与、及び経皮投与を含む公知の手法によりヒト又は動物被験者に投与してよい。好ましくは、因子を頭蓋内、脊髄内(intraspinal)、脳髄内、又は皮下注射により、非経口により投与する。本発明の因子は、ES細胞と胚性シグナリング因子の間のインビボの接触をもたらすための上記の方法の何れかに従い、被験者に投与してよい。
【0038】
経口投与のためには、胚性シグナリング因子製剤をカプセル、タブレット、粉末、顆粒としてか、又はサスペンジョンとして与えてよい。製剤は、慣用の付加物、例えば、ラクトース、マニトール、コーンスターチ、又はポテトスターチを有してよい。製剤は、結合剤、例えば結晶セルロース、セルロース誘導体、アカシア、コーンスターチ、又はゼラチンと共に与えてもよい。さらに、製剤は、崩壊剤、例えば、コーンスターチ、ポテトスターチ、又はカルボキシメチルセルロースナトリウムと共に与えてもよい。製剤は、二塩基カルシウムリン酸無水物又はスターチグリコール酸ナトリウムと共に与えてもよい。最後に、製剤は、潤滑剤、例えば、タルク又はステアリン酸マグネシウムと共に与えてもよい。
【0039】
非経口投与のためには(即ち、消化管以外の経路を通した注入による投与)、胚性シグナリング因子を、好ましくは被験者の血液と等張の滅菌水溶液と化合してよい。そのような製剤は、固形活性成分を、生理学上融和性の物質、例えば塩化ナトリウム、グリシン等を含み、且つ生理条件に融和性の緩衝化されたpHを有する水の中に溶解することにより、調製してよい。製剤は、ユニット又は複数用量コンテナー、例えばシールされたアンプル又はバイアル内にて与えてよい。製剤は、限定ではないが、筋膜上(epifascial)、関節内(intracapsular)、頭蓋内、皮膚内、髄腔内、筋肉内、眼窩内(intraorbital)、腹腔内、脊髄内(intraspinal)、胸骨内、血管内、静脈内、柔組織、皮下又は舌下を含むあらゆる注入様式によりデリバリーしてよい。
【0040】
経皮投与のためには、胚性シグナリング因子を、皮膚の因子に対する浸透性を増加させ、そして因子が皮膚を通して血流に浸透することを可能にさせる皮膚浸透増強剤、例えば、プロピレングリコール、ポリエチレングリコール、イソプロパノール、エタノール、オレイン酸、N−メチルピロリドン等と組み合わせてよい。因子/増強剤組成物は、さらに、ポリマー物質、例えば、エチルセルロース、ヒドロキシプロピルセルロース、エチレン/ビニルアセテート、ポリビニルピロリドン等と組み合わせることにより、上記組成物をゲル形態にて提供してもよく、溶剤、例えば塩化メチレンに溶解することにより、所望の粘度まで蒸発させ、そして次にベーキング物質に塗布することによりパッチを提供してよい。
【0041】
本発明は、ES細胞の分化した神経細胞(脊髄運動ニューロンを含む)への分化を誘導し、そして、そうして生成された神経細胞を、増強された緑色蛍光蛋白質(eGFP)を遺伝子マーカーとして使用して精製及び単離する方法を提供する。本明細書に記載されたインビトロES細胞分化誘導方法は、ニューロンの源、又は被験者への移植物のために利用可能なCNS又はPNSの他の神経細胞を提供する。即ち、この方法は、神経組織の変性に関連した症状を治療することにおける使用のために、神経細胞を生成するのに特に有用である。
【0042】
本明細書にて使用される用語「神経組織」は、本発明の分化した神経細胞及びそれらの前駆細胞(progenitors)を含む、神経系の組織を意味する。本明細書にて使用される神経組織が低下したか又は劣った機能活性形態に変化する、神経組織の劣化の状態を意味する。ES細胞の(例えば、脊髄運動ニューロンへの)分化を誘導することにより、本明細書に記載された方法は、分化した神経細胞の生成及び続く移植の必要がある被験者へのその移植を通して、被験者内の様々な損傷した及び/又は変性した神経組織を再増殖させる(repopulating)こと(例えば、脊髄の再増殖)において有用であると信じられる。
【0043】
従って、本発明は、神経組織変性の治療の必要のある被験者において神経組織変性を治療する方法を提供し、上記の方法に従い、ES細胞の分化した神経細胞への分化を誘導し、そして分化した神経細胞を被験者に移植し、それにより神経組織の変性を治療することを含む。例示として、本発明の方法は、以下の工程:(a)胚性幹細胞の培養物を得るか又は生じさせ;(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の、吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の、背側化又は腹側化胚性シグナリング因子に接触させ;(d)任意に、神経細胞のサブクラスを、少なくとも一つの神経栄養因子と接触させ;そして(e)神経組織の変性を治療するのに有効な量にて、分化した神経細胞を被験者に移植することを含む。発明の一つの態様において、被験者は胚である。発明の別の態様において、被験者はヒトである。好ましくは、被験者は神経組織の変性を有する。
【0044】
神経組織の変性は、中枢神経系(CNS)又は末梢神経系(PNS)において生じてよく、そして限定ではないが、初期の神経病の症状(例えば、神経変性疾患)、CNS及びPNS外傷(trauma)及び損傷、及び非神経の機能不全の後天性の二次作用(例えば、変性事象、病原性事象、又は外傷性事象に対して二次的な神経の損失を含む、様々な異常、症状、及び因子に起因するか又は関連するかもしれない。CNS外傷の例は、限定ではないが、しみ(blunt)の外傷、低酸素症、及び侵入性外傷を含む。非神経の機能不全の後天性の二次作用の例は、限定ではないが、脳性麻痺、先天的水頭症、筋ジストロフィー、発作(stroke)、及び血管性痴呆(vascular dementia)、並びに以下:脳出血、発生障害(例えば、脳の欠損、例えば先天的水頭症、又は脊髄の欠損、例えば脊髄披裂(spina bifida))、糖尿病性脳障害、過敏性脳障害、頭蓋内動脈瘤、貧血、腎不全、クモ膜下出血、脳及び脊髄に対する外傷、化学治療剤及び抗ウイルス剤のような治療剤による治療、脳及び脊髄の血管損傷、及び神経組織が変性しやすい他の疾患又は症状の何れかによりもたらされる神経変性を含む。
【0045】
本発明の一つの態様において、神経組織の変性はPNS中の末梢神経障害である。本明細書にて使用される用語「末梢神経障害」は、感覚の損失、筋肉の弱化、筋肉の萎縮、深部反射の低下の症候群、及び/又は血管運動神経の兆候を意味する。末梢神経障害を有する被験者においては、ミエリン鞘(シュワン細胞)が主に冒されているかもしれないか、又は軸索が主に冒されているかもしれない。末梢神経障害は、単一の神経(単一神経障害(mononeuropathy))、別のエリアの2つ又はそれより多い神経(複合性単一神経障害(multiple neuropathy))又は多数の神経を同時に(多発性神経障害(polyneuropathy))冒すかもしれない。
【0046】
本明細書に開示された方法により治療されるかもしれない末梢神経障害の例は、限定ではないが、急性又は慢性の炎症性の多発性神経障害に関連する末梢神経障害、筋委縮性側索硬化症(ALS)、コラーゲン血管障害(例えば、多発性動脈瘤紅斑、慢性関節リウマチ、シェーグレン症候群、又は全身性紅斑性踉狼瘡)、ジフテリア、ギラン−バレー症候群、遺伝性末梢神経障害(例えば、シャルコー−マリー−ツース病(Charcot-Marie-Tooth disease)(タイプI,タイプII及び全てのタイプを含む)、遺伝性の運動ニューロン障害及び知覚ニューロン障害(タイプI,II及びIII及び腓骨筋肉萎縮)、遺伝性圧脆弱性神経障害(HNPP)、感染性疾患(例えば、後天性免役不全症候群(AIDS))、ライム病(例えば、Borrelia burgdorferiによる感染)、微生物の侵入(例えば、ハンセン病−神経の外傷後の神経障害の全世界での主な原因)、大脳白質萎縮症、代謝性の疾患又は障害(例えば、アミロイド症、真性糖尿病、甲状腺機能低下症、ポルフィリン症、類肉腫症、又は尿毒症)、神経繊維腫症、栄養不足、準新生物形成(paraneoplastic)疾患、腓骨神経麻痺、ポリオ、ポルフィリン症、ポリオ後症候群、プロテウス症候群、圧迫麻痺(pressure paralysis)(例えば、手根管症候群)、進行性延髄麻痺、橈骨神経麻痺、脊髄筋肉萎縮(SMA)、毒性薬剤(例えば、バルビツール、一酸化炭素、クロロブタノール、ダプソン、エメチン、重金属、ヘキソバルビツール、鉛、ニトロフラントニン、オルソジニトロフェナル、フェニトイン、ピリドキシン、スルフォナミド、トリオルトクレシルリン酸、ビンカのアルカロイド、多くの溶剤、他の工業性の毒、及び特定のAIDS薬(ジダノシン及びザルシタビンを含む)、外傷(神経の外傷−末梢神経障害の全世界的な主な原因)、尺骨麻痺(Beers and Berkow,編纂、The Merck Manual of Diagnosis and Therapy,第17版(Whitehouse Station,NJ:Merck Research Laboratories,1999)第183章)。本発明の好ましい態様において、末梢神経障害はALS又はSMAである。
【0047】
本発明の別の態様において、神経組織の変性は神経変性疾患である。本明細書に記載された方法により処置してよい神経変性疾患の例は、限定ではないが、アルツハイマー病、筋委縮性側索硬化症(ルーゲーリック病)、ビンスワンガー病、ハンチントンコレラ、多発性硬化症、重症筋無力症、パーキンソン病、及びピック病(Pick's disease)を含む。脱髄の症状に関連した神経組織変性を治療するために本明細書に記載された方法を使用することも、本発明の範囲内である。脱髄症の例は、限定ではないが、急性散在性脳脊髄炎(ADEM)、急性横断性脊髄炎、急性ウイルス性脳炎、副腎白質萎縮症(ALD)、副腎脊髄神経障害(adrenomyeloneuropathy)、AIDS−液胞脊髄障害、HTLV−付随性脊髄障害、レーベル遺伝子視神経症、多発性硬化症(MS)、進行性の多病巣性白質脳症(PML)、準急性硬化症汎脳炎、及び熱帯性痙攣性不全対麻痺を含む。
【0048】
本発明の分化した神経細胞は、当業界において公知の標準手法、並びに本明細書に記載された方法により、治療の必要がある被験者に移植してよい。例示として、胚のボディー(多能性ES細胞に由来してよい)を適当な胚性シグナリング因子(例えば、RA及びHh−Ag1.3)により誘導することにより、分化した神経細胞を生成してよい。誘導から適当な時間の後に(例えば、誘導の3−4日後)、胚のボディー(EBs)を移植のために調製し(例えば、部分的に粉砕された)、そして次に被験者に移植してよい(例えば、チキン、HHステージ15−17の脊髄へ)。移植された組織を順応させる(accommodate)ため、移植の前に被験者に吸引による外傷を与えてよい(suction-lesioned)。約1/2−1のEB(又はEB均等物)を、次に、被験者の吻側頸部、尾部側頸部、胸部又は腰部領域において2−5の体節にわたるセグメントへ移植してよい。
【0049】
本発明の一つの態様において、分化した神経細胞は被験者の脊髄に移植されて、それにより被験者の脊髄を再増殖させ、そして神経組織の変性はALS又はSMAに関連した神経障害である。移植されたニューロンが脊髄の運動ニューロンである場合、吻側化及び尾部側化胚性シグナリング因子は、好ましくはレチノイン酸であり、そして背側化又は腹側化胚性シグナリング因子は、好ましくはヘッジホッグシグナリング経路の活性化因子である。
【0050】
本発明の方法において、分化した神経細胞は、神経組織の変性を治療するのに有効な量において、治療の必要のある被験者に移植される。本明細書にて使用される用語「神経組織の変性を治療するのに有効」は、神経組織の変性の臨床上の障害又は兆候を改善するか又は最小にするのに有効なことを意味する。例えば、神経組織の変性が末梢神経障害である場合、末梢神経障害の臨床上の障害又は兆候は、血管運動神経の兆候を緩和すること、深部反射を増加させること、筋肉萎縮を減少させること、感覚の機能を回復させること、及び筋肉を強化することにより、改善するか又は最小化してよい。治療の必要な被験者において神経組織の変性を治療するのに有効な分化した神経組織細胞の量は、各ケースの特定の因子に依存して変更され、神経組織変性の種類、神経組織変性のステージ、被験者の体重、被験者の症状の重度、分化した神経細胞の種類及び移植の方法を含む。この量は、当業者により、臨床試験及び本明細書に開示された方法を含む公知の手法に基づいて、容易に決定してよい。
【0051】
分化した神経細胞へのES細胞の分化を誘導する上記の方法に関して、本発明は、さらに、分化した神経細胞を生成する方法を提供するが、(a)ES細胞の培養物を得るか又は生じさせ;(b)ES細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させ;そして(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させることを含む。本発明は、細胞の集団も提供し、この方法により生成した、分化した神経細胞を含む。一つの態様において、細胞の全て又は一部がeGFPを発現する。本発明の分化した神経細胞が脊髄の運動ニューロンである場合、吻側化及び/又は尾部側化胚性シグナリング因子は、好ましくはレチノイン酸であり、そして背側化又は腹側化胚性シグナリング因子は、好ましくはヘッジホッグシグナリング経路の活性化因子である。
【0052】
本発明の方法において、工程(b)−(d)の何れかは、インビトロか、又は被験者中でインビボにて実施してよい。インビトロの工程の後に、細胞を被験者に移植するが、残りの工程はインビボにて実施される。従って、本発明の方法は、さらに、神経前駆細胞又は分化した神経細胞を被験者に移植する工程を含む。例えば、ES細胞の培養物を、インビトロにおいて吻側化及び/又は尾部側化胚性シグナリング因子に接触させることにより、神経前駆細胞を生成してよい。そうして生成された神経前駆細胞を次に被験者に移植してよいが、工程(c)及び(d)はインビボにて実施する。
【0053】
別の態様においては、ES細胞の培養物を、インビトロにおいて吻側化及び/又は尾部側化胚性シグナリング因子に接触させることにより、神経前駆細胞を生成してよい。神経前駆細胞を、次に、背側化又は腹側化胚性シグナリング因子にインビトロにて接触させることにより、分化した神経細胞を生成してよい。以後、分化した神経細胞を被験者に移植してよく、工程(d)をインビボにて実施する。
【0054】
さらなる別法においては、ES細胞の培養物を、インビトロにおいて吻側化及び/又は尾部側化胚性シグナリング因子に接触させることにより、神経前駆細胞を生成し;神経前駆細胞を、背側化又は腹側化胚性シグナリング因子にインビトロにて接触させることにより、分化した神経細胞を生成し;そして、任意に、分化した神経細胞を、インビトロにおいて少なくとも一つの神経栄養因子と接触させてよい。分化した神経細胞を、次に、被験者に移植してよい。本発明の一つの態様において、ニューロンを被験者の脊髄に移植する。
【0055】
本発明は、さらに、被験者の脊髄を再増殖させる方法を提供するが、(a)ES細胞の培養物を得るか又は生じさせ;(b)ES細胞の培養物を、神経前駆細胞を生じさせるのに有効な量にて、吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量にて、背側化又は腹側化胚性シグナリング因子に接触させ;(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させ;そして(e)分化した神経細胞を、被験者の脊髄に移植することを含む。分化した神経細胞は、脊髄を再増殖させるのに有効な量にて被験者の脊髄に移植される。この量は、公知の手法及び本明細書に開示された方法に基づいて、当業者により容易に決定してよい。
【0056】
本発明の好ましい態様によれば、吻側化及び/又は尾部側化胚性シグナリング因子は、レチノイン酸であり、そして背側化又は腹側化胚性シグナリング因子は、ヘッジホッグシグナリング経路の活性化因子である。本発明のこの方法は、神経組織の変性、特に末梢神経障害の被験者を治療するのに有用かもしれない。従って、本発明の別の態様において、被験者は末梢神経障害を有する。好ましくは、末梢神経障害はALS又はSMAである。本発明のさらに別の態様において、移植された、分化した神経細胞は、eGFPを発現し、それにより、脊髄再増殖の監視を容易にさせる。
【0057】
本発明は、さらに、工程:(a)ES細胞の培養物を得るか又は生じさせ;(b)ES細胞の培養物を、神経前駆細胞を生じさせるのに有効な量にて、吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;そして(c)任意に、神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量にて、背側化又は腹側化胚性シグナリング因子に接触させることを含む方法により生産される神経前駆細胞を提供する。また、工程:(a)ES細胞の培養物を得るか又は生じさせ;(b)ES細胞の培養物を、神経前駆細胞を生じさせるのに有効な量にて、吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量にて、背側化又は腹側化胚性シグナリング因子に接触させ;そして(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させることを含む方法により生産される神経前駆細胞も提供する。発明の一つの態様において、分化した神経細胞はeGFPを発現する。
【0058】
CNSニューロンの特定のクラスの選択的変性は多くの神経障害の基になるから、ニューロンの成長、生存、及び活性の研究は、相変わらず優先事項である。不幸なことに、しかしながら、生きているニューロンは、そのような研究において容易に利用可能ではない。そのため、本発明は研究される神経細胞の可能性を秘めた無制限な源を提供するため、神経科学及び神経学の分野の研究者に特に重要になる。従って、本発明は、特定の研究エリアにおいて上記の前駆細胞及び分化した神経細胞の用途も提供する。
【0059】
本発明の神経前駆細胞及び分化した神経細胞は、ニューロン(特に、運動ニューロン)の発生、機能及び死の分析−運動ニューロン疾患の完全な理解に必須の研究において有用になる。さらに、本発明の神経前駆細胞及び分化した神経細胞は、標的筋肉との接触のブイにおいてシナプスの分化を監視するのに有用になる。最後に、本発明の神経前駆細胞及び分化した神経細胞は、ALS又はSMA患者において検出された変異の一つを運ぶ変性した運動ニューロンと正常な健康な運動ニューロンの直接の比較を容易にする(例えば、スーパーオキシドジスムターゼ(SOD)又は生存運動ニューロン蛋白質(SMN)遺伝子内の変異)。そのような比較のために、よく知られた技術及び本明細書に記載された方法を用いて、健康な神経細胞及び疾患の神経細胞の両方を生産してよい。あるいは、SOD又はSMN変異を有する細胞を、ALS又はSMAを有する生きた患者又は死んだ患者から単離してよい。
【0060】
本明細書に記載されるとおり、発明者らは、増強された緑色蛍光蛋白質(eGFP)発現のおかげで生きた運動ニューロンが同定できたES細胞系を構築した。運動ニューロン特異的トランス遺伝子発現を授与するマウスHB9遺伝子の9−kb 5’領域の制御下でeGFPのcDNAを発現するトランスジェニックマウス系列を生じさせた。トランスジェニックファウンダーマウスをHB9とeGFPの発現の比較によりスクリーニングした。一つのマウス系列、mHB9−Gfp1bは高レベルのeGFPをE9.5−P10マウスの脊髄運動にの細胞ボディーにおいて発現し、内生HB9のパターンと平行なパターンにおいてであった。さらに、eGFPの発現が運動ニューロンの軸索及び樹状突起において高レベルで検出された。ES細胞系(HBG3)はmHB9−Gfp1bトランスジェニックマウス由来であった;これらの細胞は生殖系(germ line)を通してeGfp遺伝子を伝搬して、類似の運動ニューロン選択的パターンにおいてeGFPの発現を指示した。
【0061】
従って、本発明は、ES細胞がeGFPを発現する、本発明の方法においての使用のための胚性幹(ES)細胞を含むトランスジェニック非ヒト動物系をさらに提供する。非ヒト動物は適当な動物(例えば、ネコ、家畜、イヌ、ウマ、ヒヤギ、齧歯類、及びヒツジ)であってよいが、それは好ましくは齧歯類である。より好ましくは、非ヒト動物はラット又はマウスである。本発明のトランスジェニック非ヒト動物は、遺伝子操作されたトランスジェニック動物のための様々な技術により製造してよく、当業界公知のものを含む。発明の一つの態様において、トランスジェニック非ヒト動物のゲノムは、増強された緑色蛍光蛋白質(eGFP)遺伝子を含む。このトランスジェニック動物から確立された動物系列は、その胚の中に、eGFPを発現するES細胞を含む。
【0062】
本明細書にて使用される用語「トランスジェニック非ヒト動物」は、そのゲノムがトランス遺伝子の導入により変更された、実験操作により製造された、遺伝子操作された非ヒト動物を意味する。さらに本明細書にて使用される用語「トランス遺伝子」は、実験操作により動物のゲノムに導入された核酸分子(例えば、DNA、遺伝子、又はそれらの断片)を意味し、導入された遺伝子は当該動物にとって内生ではないか、又は動物にとって内生の遺伝子の修飾形態か又は変異形態である。内生遺伝子の修飾形態又は変異形態は、ヒトの介入を通して製造してよい(例えば、点変異の導入、フレームシフトの導入、内生遺伝子の部分又は断片の欠失、選択可能なマーカー遺伝子の挿入、停止コドンの挿入等による)。
【0063】
トランスジェニック非ヒト動物は、ヒトの介入を含む幾つかの方法により製造してよく、限定ではないが、非ヒト動物の胚生幹細胞、新たに受精した卵、又は初期胚へのトランス遺伝子の導入;非ヒト動物の体細胞及び/又は生殖細胞の染色体へのトランス遺伝子の組込み;及び本明細書に記載されたあらゆる方法を含む。本発明の好ましい態様において、トランスジェニック非ヒト動物は、トランス遺伝子のES細胞への導入により製造される。
【0064】
例示として、本発明のトランスジェニックマウス系は、マウスHB9遺伝子の5’上流領域(Arber et al.,Requirement for the homeobox gene HB9 in the consolidation of motor neuron identity.Neuron,23:659-764,1999)、続く5’スプライス基質(Choi et al.,A generic intron increases gene expression in transgenic mice.Mol.Cell Biol.,11:3070-74,1991)、eGFP遺伝子、及びウシ成長ホルモンポリアデニレーションシグナルを含む断片(例えば、約9kB)を使用する構築物の前核注入により確立してよい。ES細胞系を、次に、異型接合胚盤胞から遊動してよく、以前に記載されたとおりである(Abbondanzo et al.,Derivation of embryonic stem cell lines.Methods Enzymol.,225:803-23,1993)。遺伝子系列は、マウス胚盤胞を発生させるその能力及び生殖系の遺伝(germ transmission)に関するその能力に基づいて選択してよい。胚生幹細胞は、次に、本明細書に記載された方法における使用のために、トランスジェニック動物から単離してよい。
【0065】
本発明の一つの態様において、トランスジェニック動物は、スーパーオキシドジスムターゼ(SOD)又は生存運動ニューロン蛋白質(SMN)遺伝子が変異した(例えば、SOD−1−G93S変異)か又は選択的に不活性化されることによりその内生SOD又はSMN遺伝子内に破壊をもたらす、ゲノムを有する。本明細書にて使用される「破壊」は、機能するSOD又はSMN蛋白質の正常な発現を妨害する、SOD又はSMN遺伝子内の変異(即ち、遺伝物質内の永遠の遺伝性の変化)を意味する(例えば、それが変異SOD又はSMN蛋白質をもたらすか;それが正常な量のSOD又はSMN蛋白質の発現を妨害するか;又はそれがSOD又はSMN蛋白質の発現を妨害する)。破壊の例は、限定ではないが、点変異、フレームシフト変異の導入、内生遺伝子の一部又は断片の欠失、選択可能なマーカー遺伝子の挿入、及び停止コドンの挿入を含む。本明細書にて使用される「変異体」は、野生型遺伝子(又はその遺伝子産物)に比較して、その配列内(又はその機能特性)に少なくとも一つの修飾を呈する遺伝子(又はその遺伝子産物)を意味する。対照的に、用語「野生型」は、その天然源(例えば、天然集団)中でもっとも頻繁に見いだされる、特定の遺伝子(又はその遺伝子産物)に関して特徴的な遺伝子型(又は表現型)を意味する。野生型動物は、例えば、SOD又はSMN蛋白質を発現する。
【0066】
本発明のトランスジェニック非ヒト動物における選択的不活性化は、様々な方法により達成してよく、そして異型接合破壊(一つのSOD対立遺伝子又は一つのSMN対立遺伝子が破壊されて、結果のトランスジェニック動物が変異に関して異型接合である)又は同型接合破壊(SOD又はSMNの対立遺伝子両方が破壊されて、結果のトランスジェニック動物が変異に関して同型接合である)の何れかをもたらしてよい。本発明の一つの態様において、トランスジェニック動物の内生SOD又はSMN遺伝子は、SOD又はSMN遺伝子産物に共通の領域をコードする核酸配列との相同組換えにより破壊される。例示として、相同組換えによる破壊は、SOD又はSMN遺伝子内のノックアウト変異、特に少なくとも一つの欠失がSOD又はSMN遺伝子の少なくとも一つのエクソン内に導入されたノックアウト変異を生じてよい。さらに、SOD又はSMN遺伝子内の破壊が、異種選択可能マーカー遺伝子の内生SOD又はSMN遺伝子への挿入をもたらしてよい。
【0067】
eGFP遺伝子を発現し、そしてSOD又はSMN遺伝子内にノックアウト変異を有するトランスジェニック非ヒト動物の系列を確立する一つの方法は、以下の工程を含んでよい:(a)SOD又はSMN標的ベクターを生成し;(b)標的ベクターを非ヒト動物のレシピエント細胞に導入し;(c)処理されたレシピエント細胞を非ヒト動物の胚盤胞に導入することにより、処理された胚盤胞を生成し;(d)処理された胚盤胞を偽妊娠非ヒト動物に導入し;(e)移植された胚盤胞を予定日まで発生させ;(f)そのゲノムがノックアウト破壊をその内生SOD又はSMN遺伝子内に含むトランスジェニック非ヒト動物を同定し;(g)トランスジェニック非ヒト動物を繁殖させることにより、野生型に比較して低下したSOD又はSMN蛋白質の発現を呈するトランスジェニック非ヒト動物を得て;そして(h)eGFPを発現し、そして野生型に比較して低下したSOD又はSMN蛋白質の発現を呈するトランスジェニック非ヒト動物を得るために、当該トランスジェニック非ヒト動物を、そのゲノムがeGFP遺伝子を含むトランスジェニック非ヒト動物と交配させることを含む。eGFPを発現するトランスジェニック非ヒト動物は、上記の方法に従い得てよい。SOD又はSMN蛋白質を過剰に発現するトランスジェニック非ヒト動物を提供することも、本発明の範囲内である。
【0068】
本発明のeGFP発現トランスジェニック動物系は、ES細胞の源及び移植、キメラ、及び二重標識組換え実験のための組織を提供する。本発明のトランスジェニック動物系は、運動ニューロン変性の遺伝モデルも提供する。例えば、トランスジェニック動物系由来の動物のゲノムがそのSOD遺伝子内に変異を有する場合、それは運動ニューロン変性、特にALSを治療するのに有用かもしれない薬剤をスクリーニングするための薬学モデルとして使用してよい。このモデルは緩慢な変性の開始を呈し得るはずであるが、その結果は劇的であると予測される。さらに、トランスジェニック動物のゲノムがそのSMN遺伝子内に変異を有する場合、それは運動ニューロン変性、特にSMAを治療するのに有用かもしれない薬剤をスクリーニングするための薬学モデルとして使用してよい。このモデルは素早い変性の開始を呈すると予測される。
【0069】
本発明のトランスジェニック非ヒト動物由来のES細胞は、本明細書に記載された方法において使用してよく、インビトロ又はインビボの分化、及び被験者への再導入を含む。さらに、eGFPを発現するトランスジェニック動物系由来のES細胞及びSOD又はSMN遺伝子内に変異を有するトランスジェニック動物系由来のES細胞を交配してよい。変異体EPA細胞の挙動を、次に、細胞及び/又は組織がeGFPを発現する、発生した動物又はキメラ胚の顕微鏡観察により追跡する。
【0070】
本発明は、さらに、分化した神経細胞の純粋な集団を単離する方法、及び/又は分化した神経細胞の集団を精製する方法を提供し、工程:(a)増強された緑色蛍光蛋白質(eGFP)を発現するES細胞の培養物を得るか又は生じさせ;(b)ES細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の、吻側化及び/又は尾部側化胚性シグナリング因子に接触させるが、但し、神経前駆細胞の幾つか又は全てもeGFPを発現し;(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の、背側化又は腹側化胚性シグナリング因子に接触させるが、但し、分化した神経細胞もeGFPを発現し;(d)任意に、分化した神経細胞のサブクラスを、少なくとも一つの神経栄養因子と接触させ;(e)分化した神経細胞の中でeGFPの発現を検出し;そして(f)eGFPを発現する分化した神経細胞を単離することを含む。発明の一つの態様において、分化した神経細胞は、運動ニューロンである。eGFPを発現するES細胞は、上記に通りにして本発明のトランスジェニック動物から得てよく、あるいは本明細書に開示された他の方法に従い作成してよい。
【0071】
本発明の方法によると、eGFPの発現は、インビトロ又はインビボの何れかのアッセイにより分化した神経細胞中で検出してよい。本明細書にて使用された「発現」は、少なくとも一つのmRNA転写物中のeGFP遺伝子の転写、又は少なくとも一つのmRNAのeGFP蛋白質への翻訳を意味する。分化した神経細胞は、eGFP蛋白質、eGFPのcDNA、又はeGFPのmRNAをアッセイすることによる、eGFP発現をアッセイしてよい。eGFPの適切な形態は、本明細書に開示された特定の技術に基づいて明らかとなる。
【0072】
分化した神経細胞は、eGFP発現に関してアッセイしてよく、そしてeGFP発現は当業界で公知のアッセイ及び方法を用いて、分化した神経細胞内で検出してよい。eGFPは培養物中及びインビボにおいて細胞を標識するための非侵入性マーカーを提供するから、eGFPの発現は、好ましくは、イメージング技術、特にブライトフィールド、相、及び蛍光イメージング技術を用いて、分化した神経細胞中で検出され、本明細書に開示されるとおりである。高レベルのeGFPを発現する分化した神経細胞は、次に、以下に記載される、それらのeGFP蛍光及びフォワードライトスキャンに基づいて、分類することにより(例えば、Beckman-Coulter Altraフローサイトメトリーを用いた、FACSソーティング)、細胞懸濁液から単離してよい。
【0073】
他の方法も、本発明の分化した神経細胞内でeGFP発現を検出するのに使用してよい。そのような検出方法の例は、限定ではないが、ハイブリダイゼーション分析、イメージング技術、免疫学上の技術、免疫沈殿、放射検出、ウエスタンブロット分析、及び本明細書に開示されたあらゆる追加のアッセイ又は検出方法を含む。例えば、eGFP蛋白質又はeGFP核酸と反応性の薬剤を用いて、eGFP発現に関して分化した神経細胞をアッセイしてよい。本明細書にて使用される「反応性」は、eGFPに親和性を有するか、eGFPに結合するか、又はeGFPに対して向けられる薬剤を意味する。さらに本明細書にて使用される「薬剤」は、蛋白質、ポリペプチド、ペプチド、核酸(DNA又はRNAを含む)、抗体、Fab断片、F(ab’)2断片、分子、化合物、抗生物質、薬品(drug)、及びそれらのあらゆる組み合わせを含む。本発明の一つの態様において、eGFPと反応性の薬剤は、抗体である(例えば、αGFP(Molecular Probes,Inc.,Eugene,OR))。
【0074】
分化した神経細胞内でのeGFP発現の検出の後に、細胞中のeGFP発現の程度を、もし必要なら様々な定量アッセイのうちの一つを用いて、測定するか又は定量してよい。そのようなアッセイは当業者に知られており、eGFP蛋白質の量を測定するための、免疫組織化学、免疫細胞学、フローサイトメトリー、質量分光分析、ウエスタンブロット分析、又はELISAを含んでよい。
【0075】
本発明は、さらに、ニューロン(例えば、運動ニューロン)の変性に関連した症状を治療することにおける使用のための薬剤を同定する方法を提供する。本明細書にて使用される「ニューロンの変性」は、ニューロンが劣ったか又は低い機能活性の形態に変化するニューロンの変性の症状を意味する。ニューロンの変性に関連した症状の例は、本明細書に記載された、末梢神経障害、脱ミエリン症状、及び根本的な神経学の症状(例えば、神経変性疾患)、CNS及びPNS外傷及び障害、及び非神経機能不全の後天性二次作用(例えば、変性性、病理性、又は外傷性の自己に対して二次的な神経損失)を含む。
【0076】
本発明の方法は、(a)胚性幹細胞の培養物を得るか又は生じさせ;(b)神経前駆細胞を生成するのに有効なレチノイン酸の量に、当該胚性幹細胞の培養物を接触させ;(c)神経前駆細胞中のヘッジホッグシグナリング経路を活性化することにより、神経の幾つか又は全てが変性したニューロンを生成し;(d)変性したニューロンを候補薬剤に接触させ;そして(e)薬剤が変性したニューロンの幾つか又は全ての再生を増強するか否か決定する工程を含む。本明細書にて使用される「再生を増強する」は、変性していたニューロン(神経突起及びミエリン鞘)の部分的又は完全な成長(又は再成長)を増大させるか(augment)、改善するか、又は増加させる(increase)ことを意味する。ニューロンの再生は、中枢神経系及び末梢神経系の両方のニューロンにおいて生じてよい。本発明の一つの態様において、上記薬剤は、変性した運動ニューロンの再生を増強する。
【0077】
本発明の方法においては、本明細書に記載された、因子(又は薬剤)と細胞の間の接触をもたらすあらゆる方法、及び誘導及び投与のあらゆる方法により、変性したニューロンを候補の薬剤と接触させてよい。ニューロンの再生、及び増強された再生は、公知の手法により測定するか又は検出してよく、ミエリン−特異的且つ軸索特異的な蛋白質のためのウエスタンブロッティング、形態計測を伴う電子顕微鏡、及び当業者に知られているあらゆる方法、分子手法、及びアッセイを含む。さらに、ミエリンの成長を、g−比、軸索:ミエリン結合の完全性の一つの測定法を用いてアッセイしてよい。g−比は軸索とミエリンの全直径割る軸索の直径として定義される。この比は、何れかの一定のサイズの軸索に関しての相対的なミエリンの信頼できる測定値を提供する(Bieri et al.,Abnormal nerve conduction studies in mice expressing a mutant form of the POU transcription factor,SCIP.J.Neurosci.Res.,50:821-28,1997)。多数の研究が証明したことは、0.6のg−比がほとんどの繊維には標準であるということである(Waxman and Bennett,Relative conduction velocities of small melinated and nonmyelinated fibers in the central nervous system.Nature New Biol.,238:217,1972)。
【0078】
本発明の一つの態様において、変性したニューロン内のゲノミックDNAはSOD又はSMN遺伝子内に変異を有する。そのような変性したニューロンは、例えば、上記のとおり、そのゲノムが変異したSOD又はSMN遺伝子を含む本発明のトランスジェニック動物から単離してよい。発明の一つの態様において、変性したニューロンは、増強された緑色蛍光蛋白質(eGFP)を発現する。そのような変性したニューロンは、上記のとおり、そのゲノムがeGFP遺伝子を含む本発明のトランスジェニック動物から単離してよい。SOD及びeGFP遺伝子の両方を発現するか、又はSMN及びeGFP遺伝子の両方を発現する変性したニューロンは、SOD又はSMN変異を有するES細胞を、eGFP遺伝子を含むES細胞と交配することにより生成してよい。
【0079】
本発明は以下の実施例において記載されるが、発明の理解を助けるために記載されるのであり、以後の請求の範囲において定義される発明の範囲を、如何なる意味においても、限定するものとして解釈されるべきではない。
【0080】
実施例
実施例1−ES−細胞培養
野生型(MM13又はW9.5)又はHB9::GFPトランスジェニックマウス由来の(HBG3)ES細胞を、ES−細胞培地(4500mg/lのグルコース及び2250mg/lの重炭酸ナトリウムと共に、1x非必須アミノ酸、1xヌクレオシド(全てがSpecialty Media,Lavellette,ニュージャージーから)、0.1mMの2−メルカプトエタノール(Sigma Chemical Co.,セントルイス、ミズーリ)を付加したダルベッコ修飾イーグル培地(DMEM))、2mM L−グルタミン(Gibco)、1xペニシリン/ストレプトマイシン(Gibco BRL,ガイセルスベルグ、メリーランド)、15%のFBS(HyClone,ロウガン、ユタ)、及び1000μ/mlのLIF(Chemicon,テメキュラ、カリフォルニア)の中で初期マウス胚繊維芽細胞の供給層上で生育させた。ES−細胞のコロニーをプレートから2日後に一部解離させ(dissociated)、そしてDF培地(グルコース(4500mg/l,Sigma)、2mM L−グルタミン(Gibco)、1xペニシリン/ストレプトマイシン(Gibco)、0.1mMの2−メルカプトエタノール(Sigma)、1xインスリン−トランスフェリン−セレニウムサプリメント(Gibco)、20nMのプロゲステロン(Sigma)、及び60μMのプトレシン(Sigma)を付加されたDMEM/F12をESK10培地(LIFとFBSを除いたが、10%のノックアウト血清補充(Gibco)を追加したES細胞培地)と1:1混合した)からなるDFK5培地の中で培養した。培地を2日後に、レチノイン酸(RA)(100nMから2μM)(Sigma)、ソニックヘッジホッグ蛋白質(SHh−N;300nM)(Curis,Inc.,ケンブリッジ、マサチューセッツ)、ヘッジホッグアゴニストHh−Ag1.3(1−1000nM)(Curis,Inc.)、又はヘッジホッグブロッキング抗体(5EI,30μg/ml)を付加した新鮮な培地と交換した。胚のボディー(EBs)をさらに2−5日間培養した。幾つかの実験のために、ES細胞をストロマPA6細胞系(Riken Cell Bank,日本)の単層上で、DFK5培地単独か、又は記載されたとおりに(Kawasaki et al.,Induction of midbrain dopaminergic neurons fom ES cells by stromal cell-derived inducing activity.Neuron,28:31-40,2000)、2μMのRAを追加して生育させた。
【0081】
幾つかのHBG3 ES細胞由来のEBsを誘導の4日後にパパイン分離システム(Worthington Biochemical Corp.,フリーホールド、ニュージャージー)を用いて分離し、内生eGFP発現に基づいてFACS分類し、マトリゲル(BD)をコートされたペトリ皿又はテラサキウエル上にプレートし、そして5%のウマ血清(Gibco)、1xのB−27サプリメント(Gibco)、1xのインスリン−トランスフェリン−セレンサプリメント(Gibco)、20nMのプロゲステロン(Sigma)、及び60μMのプトレシン(Sigma)を付加されたF12培地(Specialty Media)内で培養した。神経栄養生因子(GDNF,NT3,CNTF,及びBDNF)(10ng/ml;R&D Systems,Inc.,ミネアポリス、ミネソタ)の混合物を、選択された実験に含ませた。
【0082】
実施例2−免役細胞化学
胚ボディー(EBs)を4%パラホルムアルデヒドにより固定化し、そしてアクリオスタット上で切片化した。免役染色のため、以下の抗体を使用した:ウサギαHB9,αOlig2,αNkx2.2,αLhx3,αDbx1(Pierani et al.,Control of interneuron fate in the developing spinal cord by the progenitor homeodomain protein Dbx1.Neuroon,29(2):367-84,2001),αIrx3,αeGFP(1:2000,Molecular Probes,Inc.,ユージーン、オレゴン),αSox1(1:500)(Pevny et al.,Generation of purified neural precursors from embryonic stem cells by lineage selection.Curr.Biol.,8:971-74,1998),αOtx2(1:1000)(Mallamacu et al.,OTX2 homeoprotein in the developing central nervous system and migratory cells of the olfactory area.Mech.Dev.,58:165-78,1996),αHoxC5(Liu et al.,Assigning the positional identity of spinal motor neurons.Rostrocaudal patterning of Hox-c expression by FGFs,Gdf11,and retinoids.Neuron,32:997-1012,2001),及びαcHX10;マウスαPax7,αPax6,αHB9,αLim1,αNkx6.1,αNkx2.2,αEn1,αLim2(マウス特異的),αNCAM(マウス特異的),αSC1(チキン特異的),αNeuN(1:500,Chemicon),αTuj1(1:2000)(BabCo,バークレー、カリフォルニア),αSyb(マウス特異的;1:1000)(Synaptic Systems),αSyn(1:1000,Synaptic Systems),及びα神経繊維;モルモットα−Isl1,抗−Olig2,抗−Hoxc6(Liu et al.,Assigning the positional identity of spinal motor neurons.Rostrocaudal patterning of Hox-c expression by FGFs,Gdf11,and retinoids.Neuron,32:997-1012,2001);ヤギ−αChAT(1:200)(Chemicon),αVAChT(マウス特異的;1:1000)(Chemicon)(Novitch et al.,Coordinate regulation of motor neuron subtype identity and pan-neuronal properties by the bHLH repressor Olig2.Neuron,31:773-89,2001,特定しない限り)。
【0083】
実施例3−eGFPトランスジェニックマウス系の生成
マウスHB9遺伝子の5’上流領域(Arber et al.,Requirement for the homeobox gene Hb9 in the consolidation of motor neuron identity.Neuron,23:659-764,1999)、続く5’スプライス基質(Choi et al.,A generic intron increases gene expression in transgenic mice.Mol.Cell Biol.,11:3070-74:1991)、eGFP遺伝子、及びウシ成長ホルモンポリアデニル化シグナルを含む約9kbの断片を用いた構築物の前核注入により、トランスジェニックマウス系を確立した。HB9の約9kBのNotI断片は、E9.5からP10を試験したトランスジェニック胚内でeGFP発現が運動ニューロンを発生させるよう指令するのに十分である。一つのトランスジェニック系において(mHB9−Gfp1b)、eGFP発現のパターンは内生HB9の発現のプロフィールにぴったりと一致した。ES−細胞系を、前に記載されたとおりに(Abbondanzo et al.,Derivation of embryonic stem cell lines.Methods Enzymol.,225:803-23,1993)、mHB9−Gfp1b異型接合胚盤胞から誘導した。一つの選択された系(HBG3)について、マウス胚盤胞を発生させるように統合するその能力及び生殖系列の移行(germline transmission)のためのその能力に関して検査した。
【0084】
実施例4−HBG3−細胞−由来の運動ニューロンのFACS単離
HBG3 ES−細胞の胚ボディーをパパイン分離系(Worthington Biochemical Corp.)を用いて分離した。高いレベルのeGFPを発現する細胞を、Beckman-Counter Altraフローサイトメトリーを用いて、プロット中に示されたゲートを用いてそれらのeGFP蛍光及びフォーワード光散乱に基づいて、細胞懸濁液から分類した。一般に、入力した細胞の20−30%は高いレベルのeGFPを発現した。上記分析は、このeGFP−高(Hi)細胞群が>95%純粋な集団の運動ニューロンを構成要素としたことを確証した。典型的なFACS分類の収量はEBあたり3000−5000のGFP−高細胞である。
【0085】
実施例5−HBG3 ES−細胞−由来の運動ニューロンのインビボ移植
HBG3 ES細胞由来のEBsを2μMのRA及び1μMのHh−Ag1.3により誘導した。誘導の3−4日後に、EBsを一部粉砕して、チキンのHHステージ15−17の脊髄に移植し、吸引損傷して移植された組織を適応させた。約1/2−1のEB均等物が、チキン胚脊髄の吻側頸部、尾部側頸部、胸部、又は腰部領域において2から5の体節にわたるセグメントに移植した。幾つかの胚を移植の3日後に回収し、固定化し、ビブラトーム上で切片化し、そしてeGFPに対する抗体及び細胞−又は種−特異的抗体により染色した。他の胚(胸部脊髄への移植物を受ける)は7日目に回収した。それらの胸郭を切開し、ローダミン−コンジュゲートされたα−ブンガロトキシン(2μg/ml,Molecular Probe,Inc.)及びGFP,Syb,Syn,又はVAChTに対する抗体によりホウルマウント(wholemount)として染色した。
【0086】
以下に記載されるとおり、発明者らの実験結果は、レチノイン酸が胚ボディー(EBs)中の神経前駆細胞上に尾部側化効果を有すること、運動ニューロン前駆体特異化が尾部側化EBsにおいてはヘッジホッグ依存性であること、eGFP−標識されたES−由来のニューロンは生成及び単離に成功するかもしれないこと、そしてeGFP−標識されたES−細胞由来の運動ニューロンは胚の脊髄内で分化することを示す。
【0087】
RAは胚ボディー中で神経前駆体を尾部側化する
マウスES細胞が運動ニューロンを生じさせる能力を検査し始めるため、発明者らは、凝集培養において2日間マウスES細胞を生育させたが、胚ボディー(EBs)の形成をもたらす方法であった(Bain et al.,Embryonic stem cells express neuronal propertiesin vitro.Dev.Biol..168:342-57,1995)。最初に約1000の細胞からなったEBsは、さらに1−7日間、追加の因子の存在又は不在下で、懸濁培養にて保持した。
【0088】
制御条件下では、2−3日間生育させたEBsは、全神経前駆体マーカーSox1発現する細胞を、あるとしても僅かしか含まなかった(図1)(Pevny et al.,Generation of purified neural precursors from embryonic stem cells by lineage selection.Curr.Biol.,8:971-74,1998;Wood et al.,Comparative expression of the mouse Sox1,Sox2 and Sox3 genes from pre-gastrulation to early somite stages.Mech.Dev.,86:19-201,1999)。同様に、5日目に検査されたEBsは、神経核マーカーNeuNの発現により評価されるニューロン及びMAb TuJ1により認識される神経βチューブリンアイソフォームを、あったとしても僅かしか含まなかった(図1B及び示さないデータ)。対照的に、EBsのRA(100nMから2μM)への2−3日の暴露は、多数のSox1+細胞の存在をもたらし;5日間で、多数のNeuN+,TuJ1+有糸分裂後のニューロンが検出された(図1B及び示さないデータ)。即ち、マウスES細胞由来のEBsはニューロンを生じさせるように誘導することができる。Bain et al.,Embryonic stem cells express neuronal properties in vitro.Dev.Biol.,168:342-57,1995;及びXiam et al.,Peering into early neurogenesis with embryonic stem cells.Trends in Neurosci.,24:685-86,2001も参照。
【0089】
発明者らは、次に、両者の同時発現が初期中脳の位置アイデンティティーの指標である、Otx2とEn1の発現をアッセイすることにより(Davis et al.,Expression pattern of the homeobox-containing genes En-1 and En-2 and the proto-oncogene int-1 diverge during mouse development.Genes Dev.,2:1736-44,1988;Mallamaci et al.,OTX2 homeoprotein in the developing central nervous system and migratory cells of the olfactory area.Mech.Dev.,58:165-78,1996)、そして、脊髄の位置アイデンティティーマーカーであるHox蛋白質Hoxc5,Hoxc6及びHoxc8の発現をアッセイすることにより(Belting et al.,Multiple phases of expression and regulation of mouse Hoxc8 during early embryogenesis.J.Exp.Zool.,282:196-222,1998;Liu et al.,Assigning the pisitional identity of spinal motor neurons.Rostrocaudal patterning of hox-c expression by FGFs,Gdf11,and retinoids.Neuron,32:997-1012,2001)、RA−暴露されたEBs内に形成された神経前駆細胞の吻側位置のアイデンティティーを決定した。3日間RAに暴露されたEBs中の細胞はOtx2とEn1の発現を欠き、そしてHoxc5及びHoxc6を発現したが、Hoxc8は発現しなかった(図1B及び示さなかったデータ)。神経Hox−c発現のこのプロフィールは、吻側頸部脊髄の位置的特性を伴う細胞の指標である(Belting et al.,Multiple phases of expression and regulation of mouse Hoxc8 during early embryogenesis.J.Exp.Zool.,282:196-222,1998;Liu et al.,Assigning the pisitional identity of spinal motor neurons.Rostrocaudal patterning of hox-c expression by FGFs,Gdf11,and retinoids.Neuron,32:997-1012,2001)。
【0090】
ES細胞のRAへの暴露は神経の分化及び脊髄位置的マーカーの発現の両方を促進させるから、このプロトコルはRAのES−由来の神経細胞に対する尾部側化作用の直接の試験を可能にさせない。よって、発明者らは、RA−非依存性経路を通してES細胞を神経化する手段を探した。これを達成するため、発明者らは、神経誘導性シグナルの源としてPA6ストロマ細胞系を使用した(Kawasaki et al.,Induction of midbrain dopaminergic neurons from ES cells to PA6 cell activity for 5 days induced activity.Neuron,28:31-40,2000)。マウスES細胞のPA6細胞活性への5日間の暴露は、莫大な数のSox1+細胞を誘導し、それらの多くはOtx2及びEn1を同時に発現した;対照的に、Hoxc5又はHoxc6を発現した細胞はなかった(図1C及び示さなかったデータ)。さらに、6日間で、多数のOtx2+,TuJ1+ニューロンが検出された(図1C)。ES細胞のPA6細胞活性及びRA(2μM)への暴露は、なおも多数のSox1+を誘導した;しかしながら、これらの細胞はOtx2及びEn1の発現を欠いたし、多くが今はHoxc5及びHoxc6を発現した(図1C及び示さなかったデータ)。合わせると、これらの発見は、インビボにおけるように(Muhr et al.,Convergent inductive signals specify midbrain,hindbrain,and spinal cord identity in gastrula stage chick embryos.Neuron,23:689-702,1999)、初期中脳と同様な位置的特性の神経化ES細胞が尾部側化されることにより、RAへの暴露に際して脊髄と同様な位置的特性になり得ることを示す。
【0091】
運動ニューロン前駆体特異化は尾部側化胚ボディーにおいてHh−依存性である
RAに暴露されたEBs内に存在した、脊髄の位置的特性を有する神経前駆体が運動ニューロン前駆体に分化できるか否かを検査するため、発明者らは、運動ニューロン前駆体アイデンティティーの特異化において連続的な工程を規定するHD及びbHLHの転写因子の発現を監視した(図2A)。神経前駆細胞のPax7,Irx3,Dbx1,Pax6,Nkx6.1,Nkx2.2,及びOlig2の詳述された別個のセットが、神経管の背−腹軸に沿って配置された(Briscoe et al.,Specification of neuronal fates in the ventral axis of the neural tube.Curr.Opin.Neurobiol.,11:43-49,2001;図2B)。脊髄の運動ニューロン前駆体がpMNドメイン中に発見された;それらはPax6,Nkx6.1及びOlig2を発現し、そしてPax7,Irx3,Dbx1及びNkx2.2を排除した(図2B及び示さなかったデータ)。RAに3日間暴露されたEBsは、Pax7,Pax6,Irx3及びDbx1を発現する細胞を多数含み、そしてNkx6.1,Olig2又はNkx2.2を発現する細胞はあったとしてもほとんど含まなかった(図2C及び2D及び示さなかったデータ)。転写因子の発現のこのプロフィールは、介在ニューロンを生じる背側及び中間の脊髄内に位置する前駆細胞の特徴である(図2B)(Lee et al.,The specification of dorsal cell fates in the vertebrate central nervous system.Annu.Rev.Neurosci.,22:261-94,1999)。
【0092】
運動ニューロン前駆体の誘導は、SHh活性に依存する(Briscoe et al.,Specification of neuronal fates in the ventral axis of the neural tube.Curr.Opin.Neurobiol.,11:43-49,2001)。よって、発明者らは、SHhシグナリングがRAに暴露されたEBsにおいて前駆細胞転写因子発現のプロフィールを変化させるか否かを検査した。SHhシグナリング経路を達成するため、発明者らは、SHhシグナリングの特異的小分子アゴニストであるHh−Ag1.3(Frank-Kamenetsky et al.,Chemical genetics of Hh signaling:Identification and characterization of Smoothened agonists and antagonists.Submitted,2002)又は組換えSHh−N蛋白質(Roelink et al.,Floor plate and motor neuron induction by different concentration of the amino-terminal cleavage product of sonic hedgehog antoproteolysis.Cell,81:445-55,1995)を使用し、本質的に同一の結果を得た。
【0093】
RA処理されたEBsの10nMのHh−Ag1.3への3日間の暴露は、転写因子発現のプロフィールに顕著な変化をもたらした。Pax7発現は完全に抑圧され、Pax6+及びIrx3+細胞の数は増加し、Dbx1+細胞の数は減少し、そして僅かなNkx6.1+及びOlig2+細胞が検出された(図2C及び2D及び示さなかったデータ)。RA処理されたEBsの1μMのHh−Ag1.3への暴露は、Dbx1発現を事実上は除去し、Irx3+及びPax6+細胞の数を顕著に減少させ、そして大きく増加した数のNkx6.1+及びOlig2+細胞を誘導した(図2C及び2D)。Hh−Ag1.3に暴露されたEBs内のOlig2+細胞の多くがBrdUを取込んだことから(図10)、それらは循環する前駆体であることが示された。この濃度のHh−Ag1.3において、僅かなNkx2.2+細胞も検出された(図2C及び2D)が、HNF3β+フロアープレート細胞は検出されなかった(データは示さず)。Hhシグナリングに応答性のEBs内の前駆細胞転写因子マーカーの発現のプロフィールの背側から腹側へのシフトは、神経管前駆体の挙動によく似ている(Ericson et al.,Two critical periods of Sonic Hedgehog signaling required for the specification of motor neuron identity.Cell,87:661-73,1996;Ericson et al.,Pax6 controls progenitor cell identity and neuronal fate in response to graded SHh signaling.Cell,90:169-80,1997;Ericson et al.,A homeodomain protein code specifies progenitor cell identity and neuronal fate in the ventral neural tube.Cell,101:435-45,2000)。特に、高レベルのSHhシグナリングは、運動ニューロン前駆体のHD及びbHLH蛋白質発現の特徴のプロフィールを発現した多数の細胞を生じさせた。
【0094】
運動ニューロンがRA−及びHh−暴露されたEBs内に存在する尾部側化され且つ腹側化された前駆細胞から生じたか否かを決定するため、発明者らは、体細胞の運動ニューロンにより選択的且つ不変に発現されるHD蛋白質、HB9の発現を分析した(Pfaff et al.,Requirement for LIM homeobox gene Isl1 in motor neuron generation reveals a motor neuron-dependent step in interneuron differentiation.Cell,84:309-20,1996;Arber et al.,Requirement for the homeobox gene Hb9 in the consolidation of motor neuron identity.Neuron,659-764,1999;Thaler et al.,Active suppression of interneuron programs within developing motor neuron revealed by analysis of homeodomain factor HB9.Neuron,23:675-87,1999)(図3A)。5日間単独で生育されたRAに暴露されたEBsは僅かなHB9+ニューロンを確かに含んだ(図3B;7±2 HB9+ニューロン/切片)。これらのニューロンがHh−依存性様式にて生じたのか否かを試験するため、発明者らは、機能をブロックする抗−Hh抗体の存在下でRA−暴露EBsを生育させた(mAb 5E1 Ig,30μg/ml)(Ericson et al.,Two critical periods of Sonic Hedgehog signalng required for the specification of motor neuron identity.Cell,87:661-73,1996)。抗−Hh存在下で生育させたRAに暴露されたEBsはHB9+ニューロンを含まなかったことから(図3B)、Hhの体内の源は、RAに暴露されたEBsに見いだされる僅かな運動ニューロンの分化のために必要とされる。
【0095】
発明者らは、運動ニューロンの生成がRAに暴露されたEBs内のHhシグナリングのレベルを評価することにより増強できるか否かも検査した。RA処理されたEBsのSHh(300nM)又はHh−Ag1.3(1μM)への暴露は、生じた運動ニューロンの数を顕著に増加させ;典型的には、これらのEBs中の20−30%の細胞がHB9を発現した(約5000のHB9+ニューロン;509±57のHB9+ニューロン/切片;約10−15の切片/EB)(図3B及び示さなかったデータ)。これらのHB9+細胞はNeuNとTuJ1を同時発現し(図3C及び3D)、そしてBrdUを取込まなかった(図10)ことからそれらは有糸分裂後のニューロンであることが示される。HB9+ニューロンは、RA暴露なしでHh−Ag1.3(1μM)により5日間処理されたEBsにおいて生じなかった(図3)。即ち、EBs内の運動ニューロンの生成はRAの尾部側化作用及びHhシグナルの腹側化作用の両方に依存する。
【0096】
有糸分裂後の運動ニューロンの分子特性をより詳細に査定するため、発明者らは、脊髄の運動ニューロンのサブセット:Isl1,Lhx3及びLim1により発現されるLIM HD蛋白質の発現を検査した(Tsuchida et al.,Topographic organization of embryonic motor neurons defined by expression of LIM homeobox genes.Cell,79:957-70,1994;Sharma et al.,LIM homeodomain factors Lhx3 and Lhx4 assign subtype identities for motor neurons.Cell,95:817-28,1998;Kania et al.,Coordinate roles for LIM homeobox genes in directing the dorsoventral trajectory of motor axons in the vertebrate limb.Cell,102:161-73,2000)(図3A)。3つの全てのLIM HD蛋白質はRA及びHhに暴露されたEBsにおいてHB9+運動ニューロンにより発現された。70%を超えるHB9+運動ニューロンがIsl1とLhx3を同時発現した(図3E及び3F)が、はるかに少ないフラクション(<5%)がLim1を同時発現した(図3G)。HB9+ニューロンは何れも後脳(hindbrain)の内臓の(visceral)運動ニューロンのマーカーPhox2bを同時発現しなかった(Pattyn et al.,Control of hindbrain motor neuron differentiation by the homeobox gene Phox2b.Development,127:1349-58,2000)。側面の(lateral)運動柱(LMC)ニューロンのプロフィールの指標である、HB9とLim1を同時発現する運動ニューロンの低い比率(Kania et al.,Coordinate roles for LIM homeobox genes in directing the dorsoventral trajectory of motor axons in the vertebrate limb.Cell,102:161-73,2000)は、運動ニューロンの吻側頸部のアイデンティティーと一致しており、Hox−c蛋白質発現のプロフィールにより明らかになった(Belting et al.,Multiple phases of expression and regulation of mouse Hoxc8 during early embryogenesis.J.Exp.Zool.,282:196-222,1998;Liu et al.,Assigning the positional identity of spinal motor neurons.Rostrocaudal patterning of Hox-c expression by FGFs,Gdf11,and retinoids.Neuron,32:997-1012,2001)。
【0097】
SHhは特別な(graded)様式にて作用するから、腹側の介在ニューロンは運動ニューロンの生成に十分な濃度よりほんの僅か低い濃度にて誘導される(Briscoe and Ericson,Specification of neuronal fates in the ventral tube.Curr.Opin.Neurobiol.,11(1):43-49,2001)。よって、発明者らは、腹側の介在ニューロンも1μMのHh−Ag1.3に暴露されたRA処理したEBsにおいて生じるか否かを検査した。Lhx3+,Chx10+V2介在ニューロン及びLim1/2+(V0及びV1同様)介在ニューロンがこれらのEBs内で検出された(図3G及び3H及び示されなかったデータ)。即ち、初期神経組織を用いたように(Briscoe and Ericson,Specification of neuronal fates in the ventral tube.Curr.Opin.Neurobiol.,11(1):43-49,2001)、EBs中の細胞が暴露されたHhシグナリングのレベルにおける小さなバリエーションが、腹側の介在ニューロン及び脊髄の運動ニューロンの両方の生成をもたらす。
【0098】
eGFP標識されたES細胞由来の運動ニューロンが生成されて単離されるかもしれない
RA及びHhシグナリングによりEBs内で誘導された腹側の神経の不均一性は、発明者らが、ES細胞由来の運動ニューロンを同定し、精製し、そして操作する方法を開発することを促した。よって、発明者らは、増強された緑色蛍光蛋白質(eGFP)の発現により同定可能な運動ニューロンを生じさせ得るES細胞系をデザインした。運動ニューロン特異的トランス遺伝子発現を授与するマウスHB9遺伝子の9−kbの5’領域の制御下でeGFPのcDNAを発現する、トランスジェニックマウス系を生成した(図4A)(Arber et al.,Requirement for the homeobox gene HB9 in the consolidation of motor neuron identity.Neuron,23:659-764,1999)。HB9とeGFPの発現のパターンを比較することにより、トランスジェニックファウンダーマウスをスクリーニングした。一つのマウス系列、mHB9−Gfp1bが高いレベルのeGFPをE9.5−P10マウスにて脊髄運動ニューロンの細胞ボディー内で、内生HB9と平行したパターンで発現したことがわかった(図4B−4G)。さらに、eGFP発現が高レベルで運動ニューロンの軸索及び樹状突起内に検出された(図4C及び示されなかったデータ)。ES細胞系(HBG3)をmHB9−Gfp1bトランスジェニックマウスから誘導した;これらの細胞は、生殖系を通してeGFP遺伝子を遺伝し、そして類似の運動ニューロン選択パターンにてeGFPの発現を指示した(データは示さず)。
【0099】
発明者らは、HBG3 ES細胞が、RA及びHh暴露の条件下で、他のマウスES細胞によく似た様式にて運動ニューロンに分化したことを発見した。Hh−Ag1.3なしでRA中に5日間生育させたHBG3 ES細胞から生じさせたEBsは、僅かなHB9+運動ニューロンを含み、これらの全てはそれらの身体及び神経突起中でeGFPを同時発現した(図5A)。eGFP+/HB9+運動ニューロンの数はHh−Ag1.3への暴露に際して増加し、濃度依存様式であった(図5B及び5C)。典型的には、eGFP+/HB9+運動ニューロンは、RA及び1μMのHh−Ag1.3へ暴露されたEBs中に存在する細胞の全数の20−30%を占めた(1000;図5C)。RA存在下におけるEBsのHh−Ag1.3(1μM)への暴露は、eGFP+/HB9+運動ニューロンを生じさせなかった(−1000;図5C)。即ち、HBG3 EB細胞は、RA及びHhシグナリング経路の活性化を通して十分な様式にてeGFP+/HB9+運動ニューロンを生じさせるために誘導することもできる。
【0100】
発明者らは、HBG3 ES細胞由来の運動ニューロンが胚由来の運動ニューロンの特徴であるインビトロの分化の局面を経験するか否かを試験するためにeGFP発現を使用した。RA−及びHh−暴露されたEBs(図5D)を単一の細胞又は小さな凝集体に分離し、そして神経栄養因子の存在下でマトリゲル基質上にプレートした。ES細胞由来の運動ニューロンは接着して長い突起を延ばし、eGFP発現により可視化できた(図5R−5F)。これらのES細胞由来の運動ニューロンの形態上の特徴はmHB9−Gfp1bマウスの胚脊髄から単離されたeGFP標識運動ニューロンの特徴と似ていた(図6J及び6K及び示さなかったデータ)。さらに、生じさせたeGFP+/HB9+運動ニューロンは、インビトロにてコリンアセチルトランスフラーゼを発現した(図5G及び5H)ことから、それらのコリン作用性神経伝達物質の状態の指標である。
【0101】
発明者らは、初期胚運動ニューロンとの直接の比較において、HBG3 ES細胞由来の運動ニューロンの神経栄養性因子依存性も検査した。発明者らは、RA及びHhに暴露されたHBG3−由来のEBsを単一細胞懸濁液へ分離し、そして蛍光活性化細胞分類(FACS)を用いて高いレベルのeGFPを発現する細胞を単離した(図6A)。平行して、発明者らは、E10.5 mHB9−Gfp1b胚の吻側頸部脊髄から初期胚運動ニューロンをFACSにより単離した(データは示さず)。何れかの源から、高レベルのeGFPを発現したニューロンの>98%はHBも発現し(図6B及び6C)、そしてこれらのニューロンは長い突起を延ばした(図6D、及び6E及び示さなかったデータ)。HBG3 ES細胞から生じさせた(図6F−6I及び6L)か又はE10.5 mHB9−Gfp1b胚から単離した(図6J−6L)精製されたeGFP+/HB9+運動ニューロンの生存は、神経栄養性因子に対する類似の依存性を呈した(例えば、Bloch-Gallego et al.,Survival in vitro of motoneurons identified or purified by novel antibody-based methods is selectively enhanced by muscle-derived factors.Development,111:221-32,1991;及びCamu et al.,Purification of embryonic rat motoneurons by panning on a monoclonal antibody to the low-affinity NGF receptor.J.Neurosci.Methods,44:59-70,1992を参照)。
【0102】
ES細胞由来のeGFP+運動ニューロンは胚脊髄内で分化する
HBG3 ES細胞由来の運動ニューロンのインビトロ生存及び分化は、発明者らが、インビボにて脊髄へ再導入された場合にこれらのニューロンの挙動を検査することを促した。RA及びHh−Ag1.3暴露の3−4日後にEBsを単離して(有糸分裂後の運動ニューロンが最初に出現したとき)、小凝集体に分散させ、そしてHHステージ15−17のチキン胚の脊髄の将来の吻側(又は尾部側)頸部、胸部又は腰部レベルに移植した(図7A)が、当該ステージは内生運動ニューロンの分化が始まるステージである(Hollyday et al.,An antoradiographic study of the formation of the lateral motor column in the chick embryo.Brain Res.,132:197-208,1977)。操作された胚をさらに3−7日間、HHステージ27−36まで発生させた。
【0103】
HHステージ27において操作された胚の脊髄の分析は、多数のeGFP+/HB9+運動ニューロンの存在を明らかにした(図7B−7J及び7O−7Q)。EBグラフトの最初の背腹の置き換えは制御されなかったが、発明者らは、3−7日後に検査したときに、チキンの脊髄内のマウスの運動ニューロン及び介在ニューロンの位置において著しい分離を検出した(図7E)。ほとんど一定して、eGFP+/HB9+運動ニューロンが内生運動ニューロンのそれに特徴的な腹側両面の位置内に見いだされたが、EB由来のLim2+介在ニューロン(齧歯類特異的抗体により認識される)は脊髄の背腹軸に沿って散在した(図7E)。莫大なeGFP+/HB9+運動ニューロンが、グラフティングのセグメントレベルの除去にも拘わらず、Lhx3を同時発現し、そしてLim1の発現を欠いた(図7F及び示さなかったデータ)。即ち、インビトロにおいて観察されたeGFP+/HB9+のES細胞の運動ニューロンはインビボグラフティングにより変化しなかったことが明白である。
【0104】
Olig2+運動ニューロン前駆体がグラフト化されたEBs内に存在したことから、インサイチュにて末期の分化を受けたグラフト化前駆細胞由来のチキンの脊髄の中に見いだされた幾つか又は全部のeGFP+運動ニューロンであり得る。従って、発明者らは、インビトロにおいてHBG3細胞から生じさせた有糸分裂後運動ニューロンがインビボにおいて脊髄の中で生存することもできるか否かを試験した。eGFP+/HB9+のES細胞由来の運動ニューロンを分離したEBsからFACS分類により単離して、次に、チキンの脊髄にグラフト化した。3日後、eGFP+/HB9+運動ニューロンが脊髄の腹側領域内に見いだされた(図7C及び7D)。即ち、有糸分裂後のES細胞由来の運動ニューロンはインビボにおいて生存も分化もできる。
【0105】
特定の幹細胞集団が体細胞との融合を受けることが報告された(Terada et al.,Bone marrow cells adopt the phenotype of other cells by spontaneous cell fusion.Nature,416:542-45,2002;Ying et al.,Changing potency by spontaneous cell fusion.Nature,545-48,2002)ことから、発明者らが、HBG3 ES細胞由来のeGFP+/HB9+運動ニューロンがチキンの脊髄のニューロンと体細胞ハイブリッドを形成したかもしれない可能性を試験することを促した。発明者らは、eGFP+/HB9+運動ニューロン細胞ボディーがマウスオリジンのニューロン表面蛋白質、例えば齧歯類特異的抗体により認識される−NCAM(図7G)を発現したが、チキンオリジンの表面蛋白質、例えばチキン特異的抗体により認識されるSCI(図7I)は発現しなかったことを発見した。同様に、グラフト化された運動ニューロンの軸索はマウスの表面蛋白質を発現したが、チキンの表面蛋白質は発現しなかった(図7H及び7I)。これらの発見は、体細胞融合が起こることに対する直接の証拠を提供する。
【0106】
追加の対照として、発明者らは、インビトロにおけるRAへの暴露によりニューロン化及び尾部側化されたが、抗Hh抗体(5E1;30μg/ml)の注入により腹側化を妨げられたHBG3 EBsのグラフトの運命を検査した。多数のマウスNCAM+細胞及びLim2+介在ニューロンが見いだされたが、eGFP+/HB9+運動ニューロンは検出されなかった(図7K−7N)。即ち、RAに暴露されたEBs内に存在する細胞は、宿主チキン組織により提供されたシグナルへの暴露に際してインビボの運動ニューロンの運命には向けられなかった。
【0107】
さらに、発明者らは、HBG3 ES細胞由来の運動ニューロンが脊髄の外側に伸びるか否かを検査した。多数のeGFP+軸索が腹側の根(roots)に検出された(図7B,7C,7D及び7O)。HHステージ27の胚のeGFP+運動軸索の末梢突起のパターンの分析は、HBG3 ES細胞由来の運動ニューロンが吻側頸部レベルにてグラフト化されて、軸索筋肉組織に突起したこと(データは示さず)、胸部レベルでグラフト化された運動ニューロンが軸索及び体壁筋肉組織の両方に対して突起したこと(図7O及び7P)、そして四肢のレベル(尾部側頸部又は腰部脊髄)でグラフト化された運動ニューロンが四肢の背側及び腹側の二等分に軸索を突起させたことを明らかにした(図7Q及び示さなかったデータ)。即ち、ES細胞由来の運動ニューロンは、脊髄の異なるセグメントレベルを集団化し(populate)、そしてインビボにて宿主体細胞運動ニューロンにより選択された主要な末梢軸索経路に沿って突起することが可能である。
【0108】
発明者らは、eGFP+運動ニューロンの軸索が筋肉標的に到達することも発見した。例えば、eGFP+運動軸索の広範囲な内部への成長及び分枝化が、HHステージ30−35において肋間筋肉中に検出された(図8A)。発明者らは、これらのeGFP+運動軸索が筋肉接触の部位において末端シナプスの分化の側面を呈するか否かを検査した。特に、発明者らは、神経の末端分化の3つのマーカー:齧歯類特異的抗体により認識されるシナプトブレビン(Syb);齧歯類特異的抗体により認識される小胞性(vesicular)AChトランスポーター(VAChT);及び齧歯類及びチキンの両方の蛋白質を検出する抗体により認識されるシナプトタグミン(Syn)を分析した。筋肉と接触した部位において、eGFP+運動軸索ブランチが前シナプス分化(specializations)のサインを示したことが、Syb,Syn及びVAChTの局所(focal)発現により明らかにされた(図8B−8E)。
【0109】
発明者らは、次に、eGFP+運動軸索の前シナプス末端分化がACh受容体の分配を分析することにより筋肉表面膜の後シナプス分化と整列化された(aligned)か否かを検査した。eGFP+運動軸索の末端分化の多くが筋肉表面上でACh受容体の局所クラスターと整列化されたことが、ローダミン−α−ブンガロトキシン(Rh−α−BTX)標識により定義された(図8F)。即ち、ES細胞由来の運動ニューロンの軸索は、インビボにおいて標的の骨格筋によりシナプスを形成することが明白であった。
【0110】
上で詳述されたとおり、発明者らは、インビボにおいて特定のニューロンのサブタイプへの神経前駆細胞の変換に関与する、細胞外誘導性シグナル及び転写因子の描写が、CNSニューロンの特定のクラスへのES細胞の分化に対する、発生学に基づく合理的なアプローチを可能にさせる。発明者らの発見は、高い効率で脊髄運動ニューロンを生じさせることが可能なこと、及びES細胞からの運動ニューロンの生成の経路がインビボにおいて運動ニューロンの生成の工程を再現することを示す。ES細胞由来の運動ニューロンは、インビボにて腹側脊髄を再増殖し、軸索を末梢に伸長し、そして筋肉標的によりシナプスを形成する。これらの研究は、正常な発生シグナリングカスケード、特に細胞外誘導性シグナルの調節へ洞察を適用する可能性を確立することにより、CNSニューロンの特定のクラスへのES細胞の分化を指示する。ES細胞由来の運動ニューロンが標的筋肉細胞に神経を分布させる能力は、脊髄の損傷及び運動ニューロンの変性疾患の哺乳類モデルにおいて運動機能を回復させるための、そのようなニューロンの使用の系統的評価の可能性を提供する。
【0111】
ES細胞からの運動ニューロン生成の経路
インビボの運動ニューロンの運命の特異化は、3つの連続する工程:(a)外胚葉細胞の初期神経化;(b)神経細胞の二次的尾部側化;及び(c)尾部側化された神経細胞の腹側化にあると考えることができる(Jessell et al.,Neuronal specification in the spinal cord:inductive signals and transcriptional codes.Nat.Rev.Genet.,1:20-29,2000;Wilson et al.,Neural induction:toward a unifying mechanism.Nat.Neurosci.,補遺4:1161-1168,2001)。本明細書中で論じられた、発明者らの結果は、ES細胞の運動ニューロンへの分化の経路が神経細胞の尾部側化及び腹側化に関与する通常のプログラムに似ている証拠を提供する。
【0112】
それらの最終的な位置的アイデンティティーにも拘わらず、神経プレート細胞は最初に吻側特性を有するらしい(Munoz-Sanjuan et al.,Neural induction:the default model and embryonic stem cells.Nat.Rev.Neurosci.3:271-80,2002)。脊髄の特性の次の負担(imposition)は、軸に近い中胚葉により提供されるRA−媒介性シグナルを含むらしい(Muhr et al.,Convergent inductive signals specify midbrain,hindbrain,and spinal cord identity in gastrula stage chick embryos.Neuron,23:689-702,1999;Wilson et al.,Neural induction:toward a unifying mechanism.Nat.Neurosci.,補遺4:1161-68,2001)。特に、RAは中脳又は吻側後脳アイデンティティーを犠牲にして脊髄の特性を特定することが示された(Muhr et al.,Convergent inductive signals specify midbrain,hindbrain,and spinal cord identity in gastrula stage chick embryos.Neuron,23:689-702,1999)。発明者らは、PA6細胞活性への暴露により神経化されたES細胞が中脳の位置的特性を発現する(Kawasaki et al.,Induction of midbrain dopaminergic neurons from ES cells to PA6 cell activity for 5 days induced activity.Neuron,28:31-40,2000も参照)が、RAへの暴露に際して頸部脊髄位置アイデンティティーに変換できることを発見した。この研究において記載された運動ニューロン生成におけるRA暴露のための要求は、中脳のドーパミン作用性神経分化のRAが媒介する阻害により補われる(Kawasaki et al.,Induction of midbrain dopaminergic neurons from ES cells to PA6 cell activity for 5 days induced activity.Neuron,28:31-40,2000)。合わせて、これらの発見は、神経管の吻側尾部側の軸をパターン化することにおけるその通常の役割に平行した様式にて、神経化されたEBs内でRAが脊髄の位置的アイデンティティーを促進させるという見解を支持する。
【0113】
EBs中でRAにより誘導された脊髄細胞が何故吻側頸部の位置的アイデンティティーを有するのであろうか?これに対するもっともらしい理由は、一般的脊髄前駆体のアイデンティティーの特異化の後の時点で、脊髄細胞の吻側尾部側の位置的アイデンティティーが進行中のRAシグナリングにより影響され続けるという発見から来る(Liu et al.,Assigning the positional identity of spinal motor neurons.Rostrocaudal patterning of Hox-c expression by FGFs,Gdf11,and retinoids.Neuron,32:997-1012,2001)。特に、RAが媒介するシグナリングは、胸部及び腰部の位置的アイデンティティーの犠牲として吻側頸部脊髄の位置的特性の獲得を促進する(Liu et al.,Assigning the positional identity of spinal motor neurons.Rostrocaudal patterning of Hox-c expression by FGFs,Gdf11,and retinoids.Neuron,32:997-1012,2001)。即ち、神経化されたES細胞のRAへの暴露は、最初の脊髄の位置的特性の創発(emergence)、及びこれらの細胞の吻側頸部アイデンティティーへの限定の両方の説明となるらしい。
【0114】
脊髄の前駆体アイデンティティーを獲得したES細胞も、運動ニューロン前駆体特異化の通常の経路をたどるらしい。これに関する証拠の一つのラインは、Hhシグナリングに対するそれらの細胞の感受性である。インビボにおけるような(Chiang et al.,Cyclopia and defective axial patterning in mice lacking Sonic Hedgehog gene function.Nature,383,407-13,1996)、ES細胞由来の脊髄前駆体からの運動ニューロンの生成は、完全にHhシグナリングに依存する。即ち、追加されたHhの不在下での僅かな運動ニューロンの生成は、EBs内の低レベルの内生Hh活性の発現によりもたらされるらしく、おそらくは、IHh又はSHhにより媒介される(Byrd et al.,Hedgehog is required for murine yolk sac angiogenesis.Development,129:361-72,2002;Dyer et al.,Indian hedgehog activates hematopoiesis and vasculogenesis and can respecify prospective neurectodermal cell fate in the mouse embryo.Development,128:1717-30,2001)。逆に、EBs内でHhシグナリングのレベルを上昇させると、運動ニューロン生成を顕著に増強する。
【0115】
証拠の第2のラインは、Hhシグナリングにより導き出される前駆体転写因子の発現のプロフィールにより提供される。Hhシグナリングの不在下では、EBs内の脊髄前駆体は、背及び中間の神経前駆体アイデンティティーを呈する。Hhシグナリングの活性化は初期脊髄前駆細胞の応答を厳密に模倣する濃度依存性様式において背側前駆体決定基の発現を失わせる(Briscoe et al.,Specification of neuronal fates in the ventral axis of the neural tube.Curr.Opin.Neurobiol.,11:43-49,2001)。さらに、高レベルのHhシグナリング活性はNkx6.1とOlig2の発現を誘導し−当該2つの前駆細胞転写因子はインビボにおいては深く運動ニューロン生成にリンクする(Sander et al.,Ventral neural patterning by Nkx homeobox genes:Nkx6.1 controls somatic motor neuron and ventral interneuron fates.Genes Dev.,14:2134-39,2000;Vallstedt et al.,Different levels of repressor activity assign redundant and specific roles to Nkx6 genes in motor neuron and interneuron specification.Neuron,31:743-55,2001;Novitch et al.,Coordinate regulation of motor neuron subtype identity and pan-neuronal properties by the bHLH repressor Olig2.Neuron,31:773-89,2001;Mizuguchi et al.,Combinatorial roles of olig2 and neurogenin2 in the coordinated induction of pan-neuronal and subtype-specific properties of motoneurons.Neuron,31:757-71,2001)。合わせると、これらの発見は、神経化ES細胞が初期神経プレート細胞の挙動によく似た様式で吻側及び背腹側パターン化シグナルの両方に応答し、最初に脊髄前駆細胞を生じ、そして次に運動ニューロン前駆体を生じることを示す。
【0116】
発明者らの研究は、ES細胞からの神経分化の初期の相に関与する工程を扱ってこなかった。インビボでは、神経の誘導が阻害性(blockade)BMPシグナリングに依存するらしく(Munoz-Sanjuan et al.,Neural induction,the default model and embryonic stem cells.Nat.Rev.Neurosci.,3:271-80,2002)、そして少なくともチキンにおいてはFGFsへの暴露に依存するらしい(Streit et al.,Initiation of neural induction by FGF signaling before gastrulation.Nature,406:74-78,2000;Wilson et al.,An early requirement for FGF signalling in the acquisition of neural cell fate in the chick embryo.Curr.Biol.,10421-429,2000;Wilson et al.,The status of Wnt signalling regulates neural and epidermal fates in the chick embryo.Nature,411:325-30,2001)。最近の研究は、ES細胞中の神経誘導におけるBMPシグナリングの阻害のための役割を示唆している(Gratsch et al.,Noggin and chordin have distinct activities in promoting lineage commitment of mouse embryonic stem (ES) cells.Dev.Biol.,245:83-94,2002;Tropepe et al.,Direct neural fate specification from embryonic stem cells:a primitive mammalian neural stem cell stage acquired through a default mechanism.Neuron,30:65-78,2001)。しかしながら、ES細胞における神経特異化の初期の工程がインビボにおいて神経誘導の経路に如何にして順応するかを決定するには、追加の研究が必要である。
【0117】
インビボにおけるES細胞由来の運動ニューロンの分化
幾つかの観察は、ES細胞由来の運動ニューロンも、それらの胚の類似物(counterpart)に似た様式にて挙動することを示す。最初に、eGFP+運動ニューロンが典型的には吻側脊髄に制限され、しばしば内生運動ニューロンの特徴である腹側面(ventrolateral)位置における。対照的に、ES細胞由来の脊髄介在ニューロンは脊髄の背腹軸のいたるところに散在する。これらの観察は、ES細胞由来の運動ニューロン及び介在ニューロンが宿主脊髄へのそれらの誘導後に分離の活動的なプロセスを受けることを示唆する。別の可能性は、腹側脊髄を増殖させられない運動ニューロンが、死ぬか又はeGFP発現を失うことである。このあとの可能性に対する一つの意見は、Nkx6又はMNR2蛋白質の発現の欠如(misexpression)に応答して背側脊髄中で生じた運動ニューロンが正規の場所ではない背側の位置において生存及び分化しないことである(Tanabe et al.,Specification of motor neuron identity by the MNR2 homeodomain protein.Cell,95:67-80,1998;Briscoe et al.,A homeodomain protein code specifies progenitor cell identity and neuronal fate in the ventral neural tube.Cell,101:435-45,2000)。
【0118】
第2に、ES細胞由来の運動ニューロンは腹側の根(ventral root)を通して脊髄から軸索を突出させ、そして体細胞の運動ニューロンにより選択される(taken)主要な末梢経路を選択する。ES細胞由来の運動ニューロンの軸索の四肢(limb)への突起は注目に値するが、これらのニューロンのうち極めて僅かなものしか、四肢−突起LMCニューロンに適したLIM HDコードを発現しないからである(Tsuchida et al.,Topographic organization of embryonic motor neurons defined by expression of LIM homeobox genes.Cell,79:957-70,1994)。運動ニューロンのLIM HDコードと末梢軸索の軌道の間の類似のミスマッチが2つの他の実験コンテクストにおいて発見された。Lhx3の過剰発現は、脊髄の四肢レベルにおいて運動ニューロンに対してMMC分子アイデンティティーを授与する;しかし、これらの運動ニューロンの多くは四肢へ軸索を突出させる(Sharma et al.,Genetic and epigenetic mechanisms contribute to motor neuron pathfinding.Nature,406:515-19,2000)。さらに、脊髄の四肢レベルに位置した胸部レベルの運動ニューロンはなおも四肢に突出することができる(O'Braien et al.,Development and survival of thoracic motorneurons and hindlimb musculture following transplantation of the thoracic neural tube to the lumbar region in the chick embryo:anatomical aspects.J.Neurobiol.,21:313-40,1990)。即ち、ES細胞由来の運動ニューロンの軸索の四肢への突起は、運動ニューロンが末梢経路の変更されたか又は制限された選択に対向する(face)ような環境においてLIM HDコードが無効にされ(overridden)得るという事実を反映するらしい。
【0119】
第3に、標的筋肉へ到着すると、ES細胞由来の運動ニューロンの軸索は、前シナプス分化の多数のサイン、注目すべきはコリン作用性神経伝達物質の特性の発現を呈する。さらに、これらの前シナプス特異化が、後シナプス筋膜上のACh受容体の局所クラスターとの一列整列(in alignment with)において見いだされることから、ES細胞由来の運動ニューロンと骨格筋の間に形成されるシナプス接触が機能することが示唆される。合わせると、これらのインビボの研究は、脊髄中の新たに生じた運動ニューロンがその骨格筋標的に神経を分布させることを通して、ES細胞由来の運動ニューロンが通常の発生プログラムにて連続工程をうまく通り抜けることができることを示す。
【0120】
最後に、誘導性シグナルがES細胞を運動ニューロンに変換できることによる効果は、ES細胞が暴露される吻側尾部側及び背側腹側パターン化シグナルのアイデンティティーと濃度の系統的なバリエーションを通して、CNSニューロンの他の予め定義されたクラスを生じさせるための一般的な戦略を示唆する。ES細胞を遺伝的に操作する要求無しに、単に細胞外因子の使用を通して、細胞の運命を指示する可能性(例えば、Kyba et al.,HoxB4 confers definitive lymphoid-myeloid engraftment potential on embryonic stem cell and yolk sac hematopoietic progenitors.Cell,109:29-37,2002)は、この戦略のヒトES細胞及び神経前駆細胞の他のクラスへの直接の拡張を可能にするかもしれない。
【0121】
前記の発明は明確化及び理解のためにいくらか詳細に説明されたが、開示を読んで、当業者は、形式及び細部の様々な変化が特許請求の範囲の発明の真の範囲を逸脱することなくなされ得ることを認識する。
【図面の簡単な説明】
【0122】
【図1】図1は、神経化された胚性細胞においてレチノイン酸が尾部側の位置のアイデンティティーを誘導することを示す。(A)脊髄の運動ニューロンアイデンティティーの明確化の連続する工程。神経誘導性シグナルは初期(primitive)外胚葉細胞を神経の運命に変換する。神経誘導の初期の相の間は、前駆細胞が腹側の(anterior)アイデンティティーを発現する。レチノイン酸(RA)を含む外来性のシグナルは、吻側の神経プレート細胞を進行性のより尾部側のアイデンティティーへ変換する。尾部側の神経前駆体はソニックヘッジホッグ(SHh)シグナリングの影響下では運動ニューロンに変換される。(B)RA(2μM)の存在又は不在下で生育された胚のボディー(EBs)内の全神経(pan-neural)及び吻側尾部側(rostrocaudal)のマーカーの発現。初期神経プレートマーカーSox1の発現が、RA存在下で3日間培養されたEBsにおいて増加した。otx2発現は、RA不在下で3日間生育させたEBsにおいて存在したが、RA存在下では消失した。Hoxc6はRAへ5日間暴露されたEBsにおいて発現した。NeuN発現により特徴付けされた多数の有糸分裂後のニューロンがRA存在下でEBsを5日間生育させた時に、観察された。(C)PA6細胞上で6日間培養されたES細胞はニューロンマーカー、Sox1を発現した。多くのSox1+細胞がEn1を同時発現したが、Hoxc6を発現しなかった。多くのTuJ1+ニューロンがOtx2を同時発現した。RA(2μM)のPA6細胞上で生育させたES細胞への添加はEn1及びOtx2発現の消滅、及び前駆細胞及びTuJ1+有糸分裂後のニューロン内でのHoxc6の発現をもたらした。これらニューロンの多くはHB9を発現したため、運動ニューロンであった(図9参照)。
【図2】図2は、胚ボディー(EBs)内の神経前駆細胞のヘッジホッグ−依存性腹側化を示す。(A)脊髄の運動ニューロン生成のSHh−活性化転写経路の模式的概要。運動ニューロンの生成を阻害する蛋白質はlrx3及びNkx2.2である。残りの蛋白質は運動ニューロンの生成に必要である。(B)E9.5マウス胚の尾部側の神経管内のHD及びbHLH転写因子の発現のパターン。運動ニューロンを生じる前駆体ドメインを、Olig2,Nkx6.1の同時発現及び低レベルのPax6により特徴付けした。(C)RA(2μM)単独又は10nM又は1μMのHh−Ag1.3共存下の何れかでの、3日間生育させたES−細胞由来のEBsにおける転写因子発現のプロフィール。(D)RA及び異なる濃度のHh−Ag1.3の存在下でのEBs内の転写因子発現の定量化。描写されたのは、各条件下でアッセイされた8つのEBsからの、セクションあたりの、平均±s.e.m.の値である。
【図3】図3は、胚ボディー(EBs)内の運動ニューロン及び介在ニューロン転写因子の発現を描写する。(A)HB9及びLhx3の同時発現は新たに生成された運動ニューロンを定義した。Lhx3は単独でV2介在ニューロンにより発現された。HB9とIsl1の同時発現は運動ニューロンのサブセットを定義した。幾つかの側面(lateral)運動柱(LMC)ニューロンはIsl1不在下で高いレベルのHB9を発現した;対照的に、他のLMCニューロンはIsl1を発現したが、極めて低レベルでHB9を発現したにすぎなかった。Isl1は幾つかの背面側の介在ニューロンによっても発現された。HB9とLin1の同時発現は、側面のLMCニューロンを定義した。多数の介在ニューロンがLim1も発現した。(B)Hh−Ag1.3(1μM)の存在下であるがRA不在下で5日間生育させたEBsはHB9+ニューロンを含まなかった。RA(2μM)の存在下であるがHh−Ag1.3不在下で5日間生育させたEBsはHB9+ニューロンを少量含んだ。RA及び抗−Hh抗体(mAb 5E1;30μg/ml)の存在下で5日間生育させたEBsは、HB9+ニューロンを含まなかった。RA及び組換えSHh−N(300nM)存在下で5日間生育させたEBsは、多くのHB9+ニューロンを含んだ。(C)RA及びHh−Ag1.3(1μM)存在下で5日間生育させたEBsのニューロン内のHB9及びNeuNの同時発現。(D)RA存在下で5日間生育させて、さらに2日間培養されたEBsの分離により得られたTuJ1+によりの核内のHB9の局在。HB9発現を欠いた他のニューロンも存在した。(E)RA及びHh−Ag1.3(1μM)存在下で5日間生育させたEBs内の運動ニューロン内のHB9とLhx3の同時発現。Lhx3のみを発現した細胞はV2介在ニューロンであった。(F)RA及びHh−Ag1.3(1μM)存在下で5日間生育させたEBs内の運動ニューロン内のHB9とIsl1の同時発現。(G)RA及びHh−Ag1.3(1μM)存在下で5日間生育させたEBs内の少数のニューロン内でHB9とLim1が同時発現された。Lim1のみを発現した細胞は介在ニューロンであった。(H)RA及びHh−Ag1.3(1μM)存在下で5日間生育させたEBs内のV2介在ニューロン内のChx10とLhx3の同時発現。
【図4】図4は、トランスジェニックマウスの胚の脊髄内のeGFP−標識された運動ニューロンを示す。(A)mHB9−Gfp1bトランスジェニックマウスの系の生成のためのコンストラクト。(B)E10.5 mHB9−Gfp1bマウス胚における増強された緑色蛍光蛋白質(eGFP)発現のパターン。eGFPは、腹側の脊髄及び脊髄から伸びた運動神経において高度に発現された。(C)eGFP発現が、E10.5mHB9−Gfp1b胚の腹側角内の運動ニューロン細胞ボディー及び樹上突起中に検出された。eGFP発現は、腹根により脊髄を興奮させる運動ニューロン軸索内でも検出された。このラインにおいては、E10.5において、後ではなく、極めて低い発現が背根のガングリオンのニューロン(運動ニューロン内のそれより<20倍)及び腹側の介在ニューロン(図4C及び4E中の矢印)において検出された。(D)最初の有糸分裂後の運動ニューロンが生成されたとき(E9.5)の、mHB9−Gfp1b胚の前肢レベルの脊髄内のHB9及びeGFPの発現のパターン。(E−G)E10.5 mHB9−Gfp1b胚の胸部脊髄内のHB9及びeGFPの発現のパターン。eGFPは運動ニューロン内でHB9と共に発現された。空洞ゾーンの近くに位置する新たな有糸分裂後の運動ニューロンは、脊髄の側面領域内の成熟運動ニューロンよりも低いレベルのeGFPを発現したことに注目されたい。
【図5】図5は、HBG ES細胞にインビトロで由来するeGFP−標識された運動ニューロンを描写する。(A)RA存在下で5日間生育させた胚ボディー(EBs)内のHB9+,Isl1+運動ニューロンは、細胞ボディー及び軸索内でeGFPを発現した。(B)eGFP+運動ニューロンの数はHh−Ag1.3(1μM)への暴露に際して増加した。(C)RA及び濃度を増加させたHh−Ag1.3(0から22μM)の存在下で5日間生育させたEBs内のeGFP+運動ニューロンの定量化。2.2μMのHh−Ag1.3において、EBs中の22.5%の細胞がeGFPを発現した。高い濃度(10μM)においては、hh−Ag1.3はEBsにおける細胞の分化に有害となった。「−1000」と記されたデータ点は、1μMのHh−Ag1.3存在下であるがRA不在下で生育させたEBsの培養物を意味する。(D)RA及びHh−Ag1.3(1μM)存在下で5日間生育させたEBs内でのeGFP発現を示す、明視野及び蛍光イメージ。(E)RA及びHh−Ag1.3(1μM)存在下で5日間生育させ、そしてさらに2日間マトリゲル基質上にプレートされた胚ボディー内でのeGFP発現を示す相及び蛍光イメージ。多数のeGFP+運動軸索がこれらの凝集物から出現した。(F)運動ニューロンから伸びた神経をeGFPが標識することを示す相及び蛍光イメージ。(G,H)7日間インビトロにおいて生育させたEBs−細胞由来の運動ニューロン内のHB9,eGFP,及びコリンアセチルトランスフェラーゼ(ChAT)の同時発現。HB9及びeGFP発現を欠いたChAT+ニューロンはおそらく介在ニューロンであった。
【図6】図6は、eGFP+HBG3 ES−細胞由来の運動ニューロンの単離を示す。(A)Hh−Ag1.3(1μM)単独、RA(1μM)単独、又は当該2つの因子の組み合わせの存在下で分化した、HBG3 ES細胞のFACS分析(細胞密度プロット)。高いレベルのeGFPを発現する運動ニューロンを分離するために使用されたゲートは、ダッシュの長方形により記した。(B,C)FACS分離により単離された、精製されたES−細胞運動ニューロンは高いレベルのeGFPを発現した。(D,E)24時間マトリゲル上で生育させた場合の、eGFP+ニューロン及び伸びた神経突起。(F,G)神経栄養因子(NT3,BDNF,CNTF,及びGDNF)の存在下で培養された、FACS分離されたES−細胞由来の運動ニューロンは、インビトロにおいて3日間生存し、そして突起(processes)を念入りに作る(elaborate)(F)。生存した運動ニューロンはeGFPの発現を維持した(G)。(H,I)神経栄養因子の不在下で培養したほとんどのFACS分離されたES−細胞由来の運動ニューロンはインビトロにおいて3日以内に死んだ。(J,K)FACS分離された初期吻側頸部運動ニューロンは、ES−細胞由来の運動ニューロンのそれと同じ様式で挙動した。神経栄養因子の不在下では、ニューロンは生存してeGFP+突起を念入りに作った。(L)神経栄養因子の存在下(NT)又は不在下(対照)での培養物中の3日後の初期頸部運動ニューロン(cMN)及びES−細胞由来の運動ニューロン(ES−MN)生存の定量化(プレートされた細胞のパーセンテージ)。平均±s.e.m.;6ウエルをスコアした。
【図7】図7は、インビボにおける移植されたES−細胞由来の運動ニューロンの完成(integration)を描写する。(A)HBG3 ES−細胞由来の運動ニューロン富裕化EBsの、ハンブルガー−ハミルトン(HH)ステージの15匹のチキン脊髄への移植を示す模式的ダイアグラム。レシピエントのチキンの胚の脊髄を吸引により損傷させ、そして約1/2−1の部分的に粉砕されたEBをガラス製のマイクロピペットを用いて確実なスペースに移植した。(B)HHステージ27の宿主のチキン胚の胸部及び腰部の脊髄内のeGFP+運動ニューロン(運動ニューロン富裕化されたEBsをHHステージ27において移植した)の位置を示す明視野/蛍光イメージ(腹側の視野)。脊髄から出現するeGFP+軸索に注目。(C,D)FACS−分離された運動ニューロンの(HHステージ16の胚に移植)ステージ27における胸部脊髄内の位置。横断面は、腹側の脊髄中のeGFP+運動ニューロンを明らかにした(D)。(E−J)ステージ15における運動ニューロン富裕化EBsの片側のみの移植後の、吻側頸部レベルのHHステージ27のチキンの脊髄の横断面図。運動ニューロンは脊髄の腹側角において主に見いだされ、マウス特異的Lim2抗体により標識された移植された介在ニューロンから分離される(E)。多数のES−細胞由来の運動ニューロンがeGFPとLhx3を同時発現した(F)。移植されたマウスES−細胞由来の運動ニューロン(G)及び軸索(矢印、H)を齧歯類特異的NCAM抗体で標識したが、それらはチキン運動ニューロンマーカー蛋白質、SCIを発現しなかった(I,J)。eGFP−,NCAM+の軸索は床板(floor plate)を横切ったが、脊髄から突き出なかった(矢印、G,H)。(K−N)HHステージ16において尾部側化されたEBsの移植物(RA(2μM)及び抗−Hh抗体(5E1,30μg/ml)の存在下で生育させた)を受けた、HHステージ27における胸部の脊髄の横断面図。非マウス由来の運動ニューロンが、eGFP発現(K)によるか又はマウス特異的HB9抗体(L)の何れかにより、検出された。対照的に、多数のマウス由来のNCAM+(M)及びLim2+(N)ニューロンが存在した。(O−Q)ステージ16−17における運動ニューロン富裕化EBsの移植物を受けた、胸部(O,P)、又は腰部(Q)レベルにおけるHHステージ27のチキン脊髄の横断面図。eGFP+運動ニューロン細胞ボディーが腹側脊髄において濃縮された。グラフト化された組織内の幾つかの異所性のeGFP+運動ニューロンは脊髄のルーメン内にトラップされたままであった。eGFP+軸索は脊髄を興奮させたが、主に、腹根を通してであり、そして軸索(O−Q)、ボディーの壁(O,P)及び背側及び腹側の肢(Q)を供給する全ての主要な神経ブランチに沿って突き出た。全ての末梢の軸索の経路は神経繊維(NF)発現により検出された。前ガングリオン運動軸索により占められた腹側正中に並んだ神経内のeGFP+軸索の不在に注目。
【図8】図8は、インビボにおけるES−細胞由来の運動ニューロンのシナプス分化を示す。(A)HHステージ35のチキン胚胸郭のホールマウント(wholemount)調製。ES−細胞由来のeGFP+軸索は肋間筋を脈状にしなかった(innervated)。(B,C)神経の部位のeGFP+軸索の末端のシナプトブレビン(Syb)と小胞性のACh輸送体(VAChT)が筋肉と接触する。抗−Syb及びVAChT抗体はマウスの蛋白質を認識したが、チキンの蛋白質を認識しなかった。(D)肋間筋を供給する5つの神経ブランチ内の神経繊維(NF)とeGFPの発現。幾つかのeGFP+軸索はNF発現を欠いたことに注目されたい。α−ブンガロトキシン(αBTX)発現により定義されたとおり、eGFP+軸索の末端はACh受容体クラスターと同時に生じた(coincided)。(E)eGFP+運動軸索におけるシナプトタグミン発現とαBTX標識の一致(coincidence)。(F)eGFP+運動軸索におけるSyb発現とαBTX標識の一致。
【図9】図9は、PA6細胞上で生育した尾部側化されたES細胞は運動ニューロンに分化することを示す。(A,B)10日間PA6細胞上で生育したES細胞はHB9+上でニューロンに分化しなかった(A)。2μMのRAの2日目の培養培地への添加は、多数のHB9+上でニューロンの生成、並びにOlig2−発現前駆体の生成をもたらした(B)。(C)2μMのRA存在下で10日間PA6細胞上で生育した運動ニューロン含有ES細胞コロニーの定量。平均±s.d.;3ウエル。
【図10】図10は、ES内の運動ニューロン前駆体がBrdUを取込むことを示す。(A−C)固定化前の1時間のBrdU標識(20μg/ml)は、RA及びHh−Ag1.3存在下で3日間生育したES内の多数のBrdU+,Olig2+前駆体を明らかにした。5日間生育したES内のHB9+運動ニューロンはBrdUを取り込まなかった(B)が、少数のBrdU+,Olig2+は存在した(C)。
【発明の開示】
【0001】
発明の背景
数百もの異なるニューロンの種類が脊椎の中枢神経系(CNS)の発生の間に生成され、ニューロン回路の形成に必須の細胞の多様性を確立する。特定の種類又はクラスのCNSニューロンの選択的な変性は、多くの神経学上の障害の根源である。この理解は、神経変性性障害を治療するという見地から、ニューロンの補給可能な源として作用するかもしれない前駆細胞の集団を規定することにおいて興味を生じさせてきた。系統的様式においてニューロンの分化の特定の経路に沿ったそのような前駆細胞を支配すること(directing)は難しいことが証明されたが、単に、CNSニューロンのほとんどのクラスを生じさせる通常の発生の経路がほとんど定義されていないままであるからではない。
【0002】
前駆細胞の神経学上の潜在性の研究は、3つの主要なクラスに焦点が合わされてきた:(1)胚又は成人の神経組織由来の神経前駆細胞(Alvarez-Buylla et al.,A unified hypothesis on the lineage of neural stem cells.Nat.Rev.Neurosci.,2:287-93,2001;Gage,F.H.,Mammalian neural stem cells.Science,287:1433-38,2000;Temple,S.,The development of neural stem cells.Nature,414:112-17,2001;Uchida et al.,Direct isolation of human central nervous system stem cells.Proc.Natl.Acad.Sci.USA,97:14720-725,2000);(2)他の組織及び器官由来の非神経前駆細胞(Brazelton et al.,From Marrow to brain:expression of neuronal phenotypes in adult mice.Science,290:1775-79,2000;Mezey et al.,Turning blood into brain:cells bearing neuronal antigens generated in vitro from bone marrow.Science,290:1779-82,2000;Terada et al.,Bone marrow cells adopt the phenotype of other cells by spontaneous fusion.Nature,416:545-48,2002);及び(3)胚性幹(ES)細胞(Bain et al.,Embryonic stem cells express neuronal properties in vitro.Dev.Biol.,168:342-57,1995;Reubinoff et al.,Neural progenitors from human embryonic stem cells.Nat.Biotechnol.,19:1134-40,2001;Schuldiner et al.,Induced neuronal differentiation of human embryonic stem cells.Brain Res.,913:201-05,2001;Zhang et al.,In vitro differentiation of transplantable neural precursors from human embryonic stem cells.Nat.Biotechnol.,19:1129-33,2001;Rathjen et al.,Directed differentiation of pluripotent cells to neural lineages:homogenous formation and differentiation of a neurectderm population.Development,129:2649-61,2002)。ES細胞は、ニューロン及び神経細胞の両方を生成し、そして幾つかの例においては、中脳ドーパミン作用性のニューロンを含む、特定のクラスのニューロンに特徴的な細胞種マーカーを発現する能力を有する(Kawasaki et al.,Induction of midbrain dopaminergic neurons from ES cells by stromal cell-derived inducing activity.Neuron,28:31-40,2000;Lee et al.,Efficient generation of midbrain and hindbrain neurons from mouse embryonic stem cells.Nat.Biotechnol.,18:675-79,2000)。これらの利点にも拘わらず、しかしながら、本発明の前までは、ES細胞が容易に特定のニューロン細胞種を生成することができることも、それらが神経発生の正常なプログラムを再現できることも、知られていなかった。
【0003】
脊髄の運動ニューロンは、ニューロン特異化(specification)の関連経路の多くが定義された一つのCNSニューロンサブタイプを代表する(Jessell et al.,Neuronal specification in the spinal cord:inductive signals and transcriptional codes.Nat.Rev.Genet.,1:20-29,2000;Lee et al.,Transcriptional networks regulating neuronal identity in the developing spinal cord.Nat.Neurosci.,4補遺:1183-91,2001)。脊髄の運動ニューロンの生成は、幾つかの発生段階を含むことが明白である。最初に、外胚葉細胞が、吻側(rostral)神経特性−BMP,FGF,及びWntシグナリングの制御を通して達成されたプロセスを獲得する(Munoz-Sanjuan et al.,Neural induction,the default model and embryonic stem cells.Nat.Rev.Neurosci.,3:271-80,2002;Wilson et al.,Neural induction:toward a unifying mechanism.Nat.Neurosci.,3:4補遺:1161-68,2001)。これらの吻側神経前駆細胞はレチノイン酸(RA)を含む尾部側化シグナリングに応答して脊髄の位置的アイデンティティーを獲得する(Blumberg et al.,An essential role for retinoic signaling in anteroposterior neural patterning.Development,124:373-79,1997;Durston et al.,Retinoids and related signals in early development of the vertebrate central nerve system.Curr.Top.Dev.Biol.,40:111-75,1998;Muhr et al.,Convergent inductive signals specify midbrain,hindbrain,and spinal cord identity in gastrula stage chick embryos.Neuron,23:689-702,1999)。次に、脊髄の前駆細胞は、ソニックヘッジホッグ蛋白質(SHh)の腹側化アクションに対する応答として運動ニューロン前駆細胞のアイデンティティーを獲得する(Briscoe et al.,Specification of neuronal fates in the ventral neural tube.Curr.Opin.Neurobiol.,11:43-49,2001)。
【0004】
SHhシグナリングによる運動ニューロン前駆細胞のアイデンティティーの特異化は、ホメオドメイン(HD)及び基本的なヘリックス−ループ−ヘリックス(bHLH)転写因子の発現の背腹側パターンの確立を通して媒介する(Briscoe et al.,前掲)。相対的に高いレベルのSHhシグナリングにおいては、pMNドメインと呼ばれる別の前駆ドメインが確立される;このドメイン内では、細胞は介在ニューロンよりも運動ニューロンの生成に委ねられるらしい(Briscoe et al.,A homeodomain protein code specifies progenitor cell identity and neuronal fate in the ventral neural tube.Cell,101:435-45,2000)。
【0005】
MNドメイン内の前駆細胞は、2つのHD蛋白質(Pax6及びNkx6.1)及びbHLH蛋白質(Olig2)の発現により特徴付けされる(Ericson et al.,Pax controls progenitor cell identity and neuronal fate in response to graded SHh signaling.Cell,90:169-80,1997;Sander et al.,Ventral neural patterning by Nkx homeobox genes:Nkx6.1 controls somatic motor neuron and ventral interneuron fates.Genes Dev.,14:2134-39,2000;Vallstedt et al.,Different levels of repressor activity assign redundant and specific roles to Nkx6 genes in motor neuron and interneuron specification.Neuron,31:743-55,2001;Novitch et al.,Coordination regulation of motor neuron subtype identity and pan-neuronal properties by the bHLH repressor Olig2 and neurogenin2 in the coordinate induction of pan-neuronal and subtype-specific properties of motoneurons.Neuron,31:757-71,2001)。これらの3つの転写因子の各々は、脊髄の運動ニューロンのアイデンティティーの明確化において本質的な役割を担う(Ericson et al.,Pax6 controls progenitor cell identity and neuronal fate in response to graded SHh signaling.Cell,90:169-80,1997;Sander et al.,Different levels of repressor activity assign redundant and specific roles to Nkx6 genes in motor neuron and interneuron specification.Neuron,31:743-55,2001;Novitch et al.,Coordinate regulation of motor neuron subtype identity anf pan-neuronal properties by the bHLH repressor Olig2.Neuron,31:773-89,2001;Mizuguchi et al.,Combinatorial roles of olig2 and neurogenin2 in the coordinated induction of pan-neuronal and subtype-specific properties of motoneurons.Neuron,31:757-71,2001;Zhou et al.,The bHLH transcription factors OLIG2 and OLIG1 couple neuronal and glial subtype specification.Cell,109:61-73,2002;Lu et al.,Common developmental requirement for Olig function indicates a motor neuron/oligodendrocyte connection.Cell,109:75-86,2002)。さらに、それらの組み合わされた活性が細胞サイクルを離れて(out of)運動ニューロン前駆体を操作し(drive)(Novitch et al.,Coordinate regulation of motor neuron subtype identity and pan-neuronal properties by the bHLH repressor Olig2.Neuron,31:773-89,2001;Mizuguchi et al.,Combinatorial roles of olig2 and neurogenin2 in the coordinated induction of pan-neuronal and subtype-specific properties of motoneurons.Neuron,31:757-71,2001)、そして有糸分裂後の運動ニューロンのアイデンティティーを強化する(consolidate)下流の転写因子(特に、HD蛋白質,HB9)の発現を指示する(Pfaff et al.,Requirement for LIM homeobox gene Isl1 in motor neuron generation reveals a motor neuron-dependent step in interneuron differentiation.Cell,84:309-20,1996;Arber et al.,Requirement for the homeobox gene Hb9 in the consolidation of motor neuron identity.Neuron,23:659-764,199;Thaler et al.,Active suppression of interneuron programs within developing motor neurons revealed by analysis of homeodomain factor HB9.Neuron,23:675-87.1999)。
【0006】
上記の発見は、神経発生の通常の経路における洞察が、合理的な様式にて前駆細胞、例えばES細胞の異種セットを、特定のCNSニューロンサブタイプへ向けるために適用できることを示唆する。ES細胞は運動ニューロンの分子特性の幾つかを伴う細胞を生成することが報告された(Renoncourt et al.,Neurons derived in vitro from ES cells express homeoproteins characteristic of motoneurons and interneurons.Mech.Dev.,79:185-97,1998)。しかしながら、本発明の前には、これらのニューロンの生成の経路も、それらのインビボ発生能力も、十分に探索されていなかった。
【0007】
発明の概要
発明者らは、神経管の吻尾間の(rostrocaudal)軸索及び背腹間の軸索に沿って機能し、且つインビボにおいて運動ニューロンの運命を特定する細胞外シグナリング因子が、インビトロにおいて、マウスES細胞の分化を機能性脊髄運運動ニューロンへ向けるために利用され得るか否かを試験した。本明細書において開示されるとおり、発明者らは、神経化されたES細胞のRAへの暴露が脊髄の前駆細胞の分化を誘導すること、及びヘッジホッグ(Hh)シグナリングの活性化が運動ニューロンを生成するためにこれらの細胞を指示することを確定した。発明者らは、Hhシグナリングにより誘発された運動ニューロンの分化の固有の分子経路がインビボにおいて運動ニューロンの生成に関与する経路を模倣することも示した。GFP−に基づく運動ニューロンマーキング法の使用は、ES−細胞−由来の運動ニューロンの純粋な集団の特性決定を許容した。そのような運動ニューロンは、インビボにおいて胚性脊髄を再増殖させ(repopulating)、軸索を末梢へ伸長させ、そして骨格筋との接触の部位においてシナプス分化を受けることができる。即ち、神経発生の通常の経路を合理的様式においてくつがえす(subverted)ことにより、ES細胞をCNSニューロンの特定のサブクラスへ向かわせることができる。
【0008】
従って、本発明は、胚性幹細胞を、分化した神経細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子及び背側化又は腹側化胚性シグナリング因子に接触させることにより、胚性幹細胞の分化した神経細胞への分化を誘導する方法を提供する。また、この方法により生産した、分化した神経細胞も提供される。
【0009】
本発明は、分化した神経細胞を生産する方法も提供し、(a)胚性幹細胞の培養物を得るか又は生じさせ;(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させ;そして(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させることによる。また、この方法により生産された、分化した神経細胞を含む細胞集団も提供される。
【0010】
本発明は、被験者において脊髄を再増殖させる(repopulation)方法も提供し、(a)胚性幹細胞の培養物を得るか又は生じさせ;(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させ;(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させ;そして(e)分化した神経細胞を被験者の脊髄に移植することによる。
【0011】
本発明は、治療の必要な被験者において神経組織の変性を治療する方法にも向けられ、(a)胚性幹細胞の培養物を得るか又は生じさせ;(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させ;(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させ;そして(e)神経組織の変性を治療するのに有効な量にて分化した神経細胞を被験者に移植することによる。
【0012】
さらに、本発明は、(a)胚性幹細胞の培養物を得るか又は生じさせ;(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;そして(c)任意に、神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させることにより生じさせた神経前駆細胞を提供する。また、神経の発生、機能、及び死を分析すること、又はシナプスの分化を監視することにおける、これらの神経前駆細胞の用途も提供される。
【0013】
本発明は、さらに、(a)胚性幹細胞の培養物を得るか又は生じさせ;(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させ;そして(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させることにより生じさせた神経前駆細胞を提供する。また、これらの分化した神経細胞を含む非ヒトトランスジェニック動物及び神経の発生、機能、及び死を分析すること、又はシナプスの分化を監視することにおける、これらの分化した神経細胞の用途も提供される。
【0014】
本発明は、分化した神経細胞の集団を単離及び又は精製する方法にも向けられ、(a)増強された緑色蛍光蛋白質(eGFP)を発現する胚性幹細胞の培養物を得るか又は生じさせ;(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;(c)神経前駆細胞を、eGFPを発現する分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させ;(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させ;(e)分化した神経細胞においてeGFPの発現を検出し;そして(f)eGFPを発現する分化した神経細胞を単離することによる。
【0015】
最後に、本発明は、神経の変性に関連した症状を治療することにおける使用のための薬剤を同定する方法も提供され、(a)胚性幹細胞の培養物を得るか又は生じさせ;(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量のレチノイン酸に接触させ;(c)神経前駆細胞においてヘッジホッグシグナリング経路を活性化することによりニューロンを生じさせるが、但し、ニューロンの幾つか又は全てが変性しており;(d)変性したニューロンを候補の薬剤と接触させ;そして(e)当該薬剤が幾つか又は全ての変性したニューロンの再生を増強するか否かを決定することによる。
【0016】
本発明の追加の側面は、続く説明から明らかになる。
【0017】
発明の詳細な説明
発生の間、特定の方向特性又は位置特性を幹細胞に授与するシグナリング因子の制御を含む一連の発生段階を通して、神経細胞が胚性幹細胞から生成される。最初に、外胚葉の細胞が吻側又は尾部側の神経特性を獲得し、そして吻側化(rostralizing)及び/又は尾部側化(caudalizing)の胚性シグナリング因子の制御を通して、吻側の又は尾部側の前駆細胞に分化すると言える。その後、神経前駆細胞はさらに分化して、背側化(dorsalizing)及び/又は腹側化(ventralizing)胚性シグナリング因子の作用に応答して、前駆細胞のサブタイプのアイデンティティーを獲得するか、又は完全に分化した神経細胞になる。
【0018】
例えば、運動ニューロンは様々なシグナリング因子に応答してES細胞から発生すると言える。最初に、BMP,FGF,及びWntシグナリングの制御により、外胚葉細胞が吻側神経特性を獲得する。結果の吻側の神経前駆細胞は、次に、レチノイン酸(RA)を含む尾部側のシグナルに応答して、脊髄の位置に関するアイデンティティーを獲得する。SHhシグナリングによる運動ニューロン前駆体のアイデンティティーの明確化は、ホメオドメイン(HD)及び基本的ヘリックス−ループ−ヘリックス(bHLH)転写因子の発現の背腹パターンの確立を通して介在される。相対的に高いレベルのSHhシグナリングにおいては、pMNドメインと呼ばれる別の前駆体ドメインが確立される。このドメイン内では、細胞が介在ニューロンよりも運動ニューロンの生成にかかわるらしい。pMNドメイン内の前駆細胞は、2つのHD蛋白質、Pax6及びNkx6.1、及びbHLH蛋白質、Olig2の発現により特徴付けられる。これらの3つの転写因子の各々は、脊髄の運動ニューロンアイデンティティーの明確化において必須の役割を有する。さらに、それらの組み合わされた活性が細胞サイクルを離れて(out of)運動ニューロン前駆細胞を操作し(drive)、そして有糸分裂後の運動ニューロンのアイデンティティーを強化する下流の転写因子(特に、HD蛋白質、HB9)の発現を指示する。
【0019】
脊髄の運動ニューロンは運動性挙動の制御において必須の役割を演じるし、それらの変性は多くの神経障害の基礎となる。運動ニューロンの生成に関与する誘導性シグナル及び転写因子が同定されたことから、正常な発生への洞察を用いて、合理的な様式にて幹細胞を運動ニューロンの運命に向けさせ得ることを示唆する。発明者らは、本明細書において、マウスの胚性幹(ES)細胞が外部からのシグナリング因子により誘導されることにより、最初に脊髄前駆細胞に分化でき、次に脊髄の運動ニューロンに分化できることを明らかにする。ES−細胞由来の神経前駆体からの運動ニューロンの生成の分子経路は、インビボにおいて配備されるそれを再現する。胚性脊髄へ導入されたES−細胞由来の運動ニューロンは、腹側角を増殖させ(populate)、軸索を末梢に伸ばし、そして標的の骨格筋との接触部位においてシナプス分化を受ける。即ち、通常の神経発生の経路に関与する誘導性シグナルは、ES細胞がCNSニューロンの特定のクラスを形成することを指示できる。
【0020】
従って、本発明は、分化した神経細胞への胚性幹細胞の分化を誘導する方法、及びこの方法により生成された分化した神経細胞を提供する。本明細書にて使用される用語「胚性幹細胞の分化を誘導する」は、胚性幹細胞を活性化するか、開始するか、又は刺激することにより分化を受けること−発生の間に細胞が構造上並びに機能上特化されることになる細胞プロセスを意味する。胚性幹(ES)細胞は、培養された細胞であり、胚盤胞のステージの胚の多能性内部細胞隗由来であり、無期限に複製することができる。一般に、ES細胞は他の細胞に分化する能力を有する(即ち、それらは多能性);即ち、それらは新規な細胞の連続する源として機能するかもしれない。本発明の胚性幹細胞はあらゆる細胞から得て良いが、好ましくは哺乳類(例えば、ヒト、家畜動物、又は市販の動物)から得る。本発明の一つの態様において、胚性幹細胞はマウスの胚性幹細胞である。別の好ましい態様において、胚性幹細胞はヒトから得られる。
【0021】
さらに本明細書において使用される「分化した神経細胞」は、中枢神経系(CNS)又は末梢神経系(PNS)の一部分化した細胞又は完全に分化した細胞であり、そして、限定ではないが、完全に分化したガングリオン細胞、グリア(又は神経グリア)細胞(例えば、星状細胞、乏突起神経膠細胞、又はシュワン細胞)、顆粒細胞、ニューロン細胞(又はニューロン)、及び星形細胞、並びにそれらのあらゆる神経前駆細胞を含む。前駆細胞は、発生及び分化の間に、一連の細胞分割により別の細胞系列を生じる親細胞である。神経前駆細胞は、例えば、最後にはCNS又はPNSの完全に分化した神経細胞に発生する細胞系列に委ねられる(committed);しかしながら、そのような神経前駆細胞は神経細胞の特定の種類、又はサブクラスに供されたことがまだない。最初に、神経前駆細胞は吻側の特性を獲得し(例えば、吻側神経前駆細胞)、続いて位置的アイデンティティーを獲得してよい(例えば、小脳前駆細胞、大脳前駆細胞、又は脊髄前駆細胞)。そのような一部分化した神経前駆細胞は、特定の種類の神経細胞に分化して(例えば、星状細胞、ガングリオン細胞、顆粒細胞、ニューロン、乏突起神経膠細胞、シュワン細胞、又は星形細胞の前駆細胞)、そして以後は、完全に分化した前駆細胞(例えば星状細胞、ガングリオン細胞、顆粒細胞、ニューロン(背根ガングリオン(DRG)ニューロン、運動ニューロン、末梢ニューロン、脊髄ニューロン、又は腹側介在ニューロン)、乏突起神経膠細胞、シュワン細胞、又は星形細胞)を生じるように委ねられるようになる。従って、本発明の一部分化した神経細胞は、神経のアイデンティティーを有し、方向アイデンティティー又は位置アイデンティティーを獲得した細胞であるか、又は特定のクラスの神経細胞に発生するが完全に分化した神経細胞には発生しないことを委ねられた細胞であってよい。
【0022】
さらに、本明細書にて使用される「神経細胞」又は「ニューロン」は、核及び周囲の細胞質;幾つかの短い放射突起(processes)(星状細胞);及び一つの長い突起(軸索)、小枝様のブランチで終わるもの(末端星状細胞(telodendrons))、及びそのコースに沿って伸びるブランチを有してよいもの(側枝(collaterals))を含む、細胞ボディー(周核体)から一般にはなる神経系の、伝導性(conducting)細胞又は神経細胞である。ニューロンの例は、限定ではないが、背根ガングリオン(DRG)のニューロン、運動ニューロン、末梢ニューロン、知覚ニューロン、脊髄のニューロン、及び原側介在ニューロンを含み、それら全てはコリン作動性、ドーパミン作動性、又はセロトニン作動性であってよい。
【0023】
ES細胞の一部分化したか又は完全に分化した神経細胞への分化は、公知の細胞又は分子の手法、及び本明細書にて開示されたアッセイ及び方法により検出してよい。本発明の一つの態様において、分化した神経細胞は、運動ニューロンである(例えば、有糸分裂後の運動ニューロン又は脊髄の運動ニューロン)。本発明の別の態様において、分化した神経細胞は遺伝子を標識され、本明細書にて記載されたように、増強された緑色蛍光蛋白質(eGFP)を発現する。eGFP遺伝子マーカーは、分化した神経細胞の集団を単離及び/又は精製するための方法において、又は脊髄の制御を監視するための方法において特に有用かもしれず、以下に記載されるとおりである。
【0024】
本発明の方法は、胚性幹細胞を、吻側化又は尾部側化の胚性シグナリング因子、及び背側化又は腹側化の敗勢シグナリング因子に、分化した神経細胞を精製するのに有効な量にて接触させることを含む。本明細書にて使用される「因子」は、蛋白質、ポリペプチド、ペプチド、核酸(DNA又はRNAを含む)、抗体、Fab断片、F(ab’)2断片、分子、化合物、抗生物質、薬剤、及びそれらのあらゆる組み合わせを含む。Fab断片は抗体の単価の抗原結合断片であり、パパイン消化により生成される。F(ab’)2断片は抗体の二価の抗原結合断片であり、ペプシン消化により生成される。
【0025】
さらに本明細書にて使用される「胚性シグナリング因子」は、発生及び分化の推移の間に、胚の中で細胞へ情報を運ぶ因子である。そのような情報は、例えば、胚の中の細胞の方向又は位置に関してよく、そして情報を受けたそれらの細胞によりたどられる分化の経路を決定してよい。本発明の胚性シグナリング因子は、例えば、形態学上の因子又は神経因子であってよい。本発明における使用のための胚性シグナリング因子の例は、限定ではないが、レチノイン酸(RA)、骨形態蛋白質(BMPs)(例えば、BMP1からBMP7,BMP2及びBMP4を含む)、繊維芽成長因子(FGFs)(例えば、アルファFGF,FGF8,FGF10,HBGF1,塩基性FGF,ベータFGF,及びHBGF)、ヘッジホッグ蛋白質(例えば、ソニックヘッジホッグ(SHh)及びインディアンヘッジホッグ(IHh)、ヘッジホッグシグナリングのアゴニスト(例えば、Hh−Ag1.3)、及びヘッジホッグシグナリング経路の他の活性化因子を含む。
【0026】
さらに、本明細書にて使用される「吻側化胚性シグナリング因子」は、対象又は被験者の頭部又は上位の方向に誘導するか、寄与する(contribute)か、又は発生する因子を意味する。対照的に、「尾部側化胚性シグナリング因子」は、対象又は被験者の尻、尾又は下位の方向に誘導するか、寄与する(contribute)か、又は発生する因子を意味する。吻側アイデンティティーの割り当てに寄与する胚性シグナリング因子は、ES細胞の神経前駆細胞への分化をもたらすと言える。本発明における使用のための吻側化及び尾部側化胚性シグナリング因子の例は、限定ではないが、RA,BMP,FGF,及びWntを含む。RAは発生の間に吻側アイデンティティーを割り当てる役割を担うことが知られている。最近、Wntシグナリングも吻側アイデンティティーを割り当てる役割を担うことが示された(Nordstrom et al.,Progressive induction of caudal neural character be graded Wnt signaling.Nat.Neurosci.,5(6):525-32,2002)。即ち、本発明の一つの態様において、吻側化胚性シグナリング因子はRA又はWntである。発明の別の態様においては、尾部側化胚性シグナリング因子がRA又はWntである。
【0027】
本明細書にて使用される「背側化胚性シグナリング因子」は、対象又は被験者の背中又は後位の方向に誘導するか、寄与する(contribute)か、又は発生する因子を意味する。対照的に、本明細書にて使用される「腹側化胚性シグナリング因子」は、対象又は被験者の前部又は前方の方向に誘導するか、寄与する(contribute)か、又は発生する因子を意味する。背側アイデンティティーの割り当てに寄与する胚性シグナリング因子は、ES細胞の神経前駆細胞への分化又は完全に分化した神経細胞への分化をもたらすと言える。本発明における使用のための背側化及び腹側化胚性シグナリング因子の例は、限定ではないが、BMP,Wnt,ヘッジホッグ蛋白質、ヘッジホッグシグナリングのアゴニスト、及びヘッジホッグシグナリング経路の他の活性化因子を含む。SHhシグナリングは、例えば、運動ニューロン前駆細胞のアイデンティティーを脊髄前駆細胞に提供することが知られている。
【0028】
本発明の一つの態様において、背側化胚性シグナリング因子は、BMP又はWntである。本発明の別の態様において、腹側化胚性シグナリング因子は、ヘッジホッグシグナリング経路の他の活性化因子である。本明細書にて使用される「ヘッジホッグシグナリング」は、ヘッジホッグファミリーの蛋白質(例えば、ソニックヘッジホッグ(SHh)及びインディアンヘッジホッグ(IHh))のあらゆるメンバーの生化学シグナリング、特に発生及び分化の間に胚中の細胞の方向及び位置に関係するシグナリングを意味する。ヘッジホッグシグナリング経路の「活性化因子」は、ヘッジホッグシグナリングを作用させるか又は生じさせる因子、及びヘッジホッグシグナリング経路においてヘッジホッグシグナリングを誘導するか、開始させるか、又は刺激する因子を含む。例示として、ヘッジホッグシグナリングの活性化因子はヘッジホッグ蛋白質(例えば、SHh、IHh)又はヘッジホッグシグナリングのアゴニスト(例えば、Hh−Ag1.3)であってよい。本明細書にて使用されるヘッジホッグシグナリングの「アゴニスト」は、細胞中のヘッジホッグシグナリングが開始するか、刺激されるか、又は誘導されるように、天然に生じる物質により通常は刺激される細胞受容体に親和性を有し、且つ当該受容体において生理活性を刺激する因子である。
【0029】
胚性シグナリング因子、BMP,FGF,Wnt,SHh,及びIHhは蛋白質である。本明細書にて使用される「BMP」はBMP蛋白質及びBMPアナログの両方を含み;「FGF」はFGF蛋白質及びFGFアナログの両方を含み;「Wnt」はWnt蛋白質及びWntアナログの両方を含み;「SHh」はSHh蛋白質及びSHhアナログの両方を含み;「IHh」はIHh蛋白質及びIHhアナログの両方を含む。他に示さない限り、「蛋白質」は、蛋白質、蛋白質ドメイン、ポリペプチド又はペプチドを含む。本明細書にて使用される特定の蛋白質の「アナログ」は、当該蛋白質の生物活性を有し、且つ当該蛋白質と60%又はそれより高い(好ましくは、70%又はそれより高い)のアミノ酸配列相同性を有する、当該蛋白質の機能性バリアントである。「アナログ」は、相同な三次元コンフォメーションを有する蛋白質のバリアントを含む。さらに本明細書にて使用される用語「生物活性」は、発生及び分化の間のBMP,FGF,Wnt,SHh,又はIHh蛋白質のシグナリング活性を意味し、以下に記載される。
【0030】
さらに、本明細書にて使用される「保存的置換」は、置換されたアミノ酸残基に機能上均等なアミノ酸置換であり、それらが極性又は立体の配置を有するか、又はそれらが置換された残基と同じクラスに属するためである(例えば、疎水性、酸性又は塩基性)。本明細書にて使用される用語「保存的置換」は、本発明の蛋白質が発生及び分化の間にシグナリングを作用させる能力に対して取るに足らない(inconsequential)効果を有する置換を含む。
【0031】
本明細書にて論じられるとおり、BMP,FGF,Wnt,SHh,又はIHhは蛋白質である。対照的に、RA及びビタミンAはモルフォゲンであると信じられるアルデヒド分子である。RAは容易に利用可能であり;例えばシグマケミカル社(セントルイス、ミズーリ)から得てよい。本発明の他の因子は、合成によるか、組換えによるか、又は天然の細胞から単離してよい。好ましくは、蛋白質を組換えにより生産し、慣用の技術及び当該蛋白質をコードするcDNAを用いる。SHh及びIHh(並びにHh−Ag1.3)もキュリス社(ケンブリッジ、マサチューセッツ)から得てよい。
【0032】
本発明の方法は、胚性幹細胞を、吻側化及び/又は尾部側化胚性シグナリング因子及び背側化及び/又は腹側化胚性シグナリング因子に接触させることを含む。胚性シグナリング因子は、分化した神経細胞を生じさせるのに有効な量にて提供される。これらの量は、本明細書にて開示された公知の手法及び方法に基づいて、当業者により容易に決定される。本発明の好ましい態様においては、胚性幹細胞を、背側化及び/又は腹側化胚性シグナリング因子に接触させる前に、吻側化及び/又は尾部側化胚性シグナリング因子に接触させる。
【0033】
発明者らは、神経栄養性因子の存在下で培養されたニューロンが生存して且つ突起を念入りに作ることを証明した。従って、別の態様において、本発明の方法は、さらに、背側化又は腹側化胚性シグナリング因子との接触後に、少なくとも一つの神経栄養性因子に胚性幹細胞を接触させる工程を含む。本明細書にて使用される「神経栄養性因子」は、神経組織の栄養又はメンテナンスに関与する因子である。神経栄養性因子は、委ねられた神経前駆細胞の発生及び分化であるか、又はそれらは分化した神経細胞の成長及び生存を誘導するかもしれない。古典的な例の神経栄養性因子は、NGF(神経成長因子)である。本発明における使用のための神経栄養性因子の他の例は、限定ではないが、GDNF,NGF,NT3,CNTF,及びBDNFを含む。これらの因子はR&Dシステムズ社(ミネアポリス、ミネソタ)から得てよい。本発明の神経栄養性因子は、CNS又はPNSの完全に分化した神経細胞(例えば、ニューロン)を生産するのに有効な量で提供される。この量は、本明細書にて開示された公知の手法及び方法に基づいて、当業者により容易に決定してよい。
【0034】
本発明の方法においては、胚性幹(ES)細胞を、被験者においてインビトロ又はインビボにて有効な量の胚性シグナリング因子に接触させてよい。胚性シグナリング因子を細胞内へ導入することにより当該因子をES細胞に接触させてよい。接触をインビトロにて行う場合、因子を直接培養培地に加えてよく、本明細書に記載されているとおりである。或いは、因子を被験者に導入するか(例えば、因子を被験者の細胞に導入することにより)、又は因子を被験者に投与することにより、被験者内でインビボにてESに接触させてよい。被験者は胚性動物又は発生した動物であってよいが、好ましくは哺乳類(例えば、ヒト、家畜、又は市販の動物)である。より好ましくは、被験者はヒトである。胚性シグナリング因子をインビボにてES細胞に接触させる場合、被験者は好ましくは胚である。しかしながら、ES細胞を完全に成長したヒト又は動物被験者に移植すること、及び胚性シグナリング因子を次にヒトに移植することにより、ES細胞の分化した神経細胞への被験者内でのインビボにての分化を作用させることは、本発明の範囲内である。
【0035】
本発明の因子は、インビトロ又はインビボの何れかにおいて被験者の中で、蛋白質、核酸及び他の薬剤の導入及び投与に使用される公知の技術により、ES細胞と接触させてよく、例えば注射及び輸液(transfusion)である。標的のES細胞を被験者の特定の部分に局在化する場合は、注射によるか又は幾つかの他の手段により(例えば、因子を血液又は他の体液に導入することにより)、ES細胞へ直接導入することが望まれるかもしれない。
【0036】
胚性シグナリング因子が蛋白質又は他の分子である場合、本明細書に開示された慣用の技術及び方法により、直接ES細胞に導入してよい。さらに、蛋白質の胚性シグナリング因子は、蛋白質阻害剤の発現を許容する様式にて、因子をコードする核酸をES細胞に導入することにより、間接にES細胞に導入してよい。因子は、限定ではないが、エレクトロポレーション、DEAEデキストラントランスフェクション、リン酸カルシウムトランスフェクション、モノカチオンリポソーム融合、ポリカチオンリポソーム融合、プロトプラスト融合、インビボの電場の創製、DNA−コートされたマイクロ発射砲撃(microprojectile bombardment)、組換え体複製欠損ウイルスを用いた注入、相同組換え、インビボ遺伝子治療、エクスビボ遺伝子治療、ウイルスベクター、及び裸のDNAの移行(transfer)、又はそれらのあらゆる組み合わせを含む、慣用の当業界公知の手法を用いて、インビトロ又はインビボにて、ES細胞に導入してよい。遺伝子治療に適した組換えウイルスベクターは、限定ではないが、レトロウイルス、HSV,アデノウイルス、アデノ随伴ウイルス、セミリキフォレストウイルス、サイトメガロウイルス、及びワクシニアウイルスのようなウイルスのゲノム由来のベクターを含む。使用される核酸の量は、分化した神経細胞を生じさせるのに有効な蛋白質因子の量を発現するのに十分な量である。これらの量は、当業者により容易に決定してよい。蛋白質の胚性シグナリング因子をコードする核酸を適当なES細胞中へ慣用の手法を用いてインビトロにて導入することにより、ES細胞中の蛋白質因子の発現を達成してよい。蛋白質の胚性シグナリング因子を発現するES細胞は、次に、インビボにおいて分化した神経細胞を生じさせるために被験者に導入されてよい。
【0037】
本発明の方法によれば、胚性シグナリング因子は、限定ではないが、経口投与、非経口投与、及び経皮投与を含む公知の手法によりヒト又は動物被験者に投与してよい。好ましくは、因子を頭蓋内、脊髄内(intraspinal)、脳髄内、又は皮下注射により、非経口により投与する。本発明の因子は、ES細胞と胚性シグナリング因子の間のインビボの接触をもたらすための上記の方法の何れかに従い、被験者に投与してよい。
【0038】
経口投与のためには、胚性シグナリング因子製剤をカプセル、タブレット、粉末、顆粒としてか、又はサスペンジョンとして与えてよい。製剤は、慣用の付加物、例えば、ラクトース、マニトール、コーンスターチ、又はポテトスターチを有してよい。製剤は、結合剤、例えば結晶セルロース、セルロース誘導体、アカシア、コーンスターチ、又はゼラチンと共に与えてもよい。さらに、製剤は、崩壊剤、例えば、コーンスターチ、ポテトスターチ、又はカルボキシメチルセルロースナトリウムと共に与えてもよい。製剤は、二塩基カルシウムリン酸無水物又はスターチグリコール酸ナトリウムと共に与えてもよい。最後に、製剤は、潤滑剤、例えば、タルク又はステアリン酸マグネシウムと共に与えてもよい。
【0039】
非経口投与のためには(即ち、消化管以外の経路を通した注入による投与)、胚性シグナリング因子を、好ましくは被験者の血液と等張の滅菌水溶液と化合してよい。そのような製剤は、固形活性成分を、生理学上融和性の物質、例えば塩化ナトリウム、グリシン等を含み、且つ生理条件に融和性の緩衝化されたpHを有する水の中に溶解することにより、調製してよい。製剤は、ユニット又は複数用量コンテナー、例えばシールされたアンプル又はバイアル内にて与えてよい。製剤は、限定ではないが、筋膜上(epifascial)、関節内(intracapsular)、頭蓋内、皮膚内、髄腔内、筋肉内、眼窩内(intraorbital)、腹腔内、脊髄内(intraspinal)、胸骨内、血管内、静脈内、柔組織、皮下又は舌下を含むあらゆる注入様式によりデリバリーしてよい。
【0040】
経皮投与のためには、胚性シグナリング因子を、皮膚の因子に対する浸透性を増加させ、そして因子が皮膚を通して血流に浸透することを可能にさせる皮膚浸透増強剤、例えば、プロピレングリコール、ポリエチレングリコール、イソプロパノール、エタノール、オレイン酸、N−メチルピロリドン等と組み合わせてよい。因子/増強剤組成物は、さらに、ポリマー物質、例えば、エチルセルロース、ヒドロキシプロピルセルロース、エチレン/ビニルアセテート、ポリビニルピロリドン等と組み合わせることにより、上記組成物をゲル形態にて提供してもよく、溶剤、例えば塩化メチレンに溶解することにより、所望の粘度まで蒸発させ、そして次にベーキング物質に塗布することによりパッチを提供してよい。
【0041】
本発明は、ES細胞の分化した神経細胞(脊髄運動ニューロンを含む)への分化を誘導し、そして、そうして生成された神経細胞を、増強された緑色蛍光蛋白質(eGFP)を遺伝子マーカーとして使用して精製及び単離する方法を提供する。本明細書に記載されたインビトロES細胞分化誘導方法は、ニューロンの源、又は被験者への移植物のために利用可能なCNS又はPNSの他の神経細胞を提供する。即ち、この方法は、神経組織の変性に関連した症状を治療することにおける使用のために、神経細胞を生成するのに特に有用である。
【0042】
本明細書にて使用される用語「神経組織」は、本発明の分化した神経細胞及びそれらの前駆細胞(progenitors)を含む、神経系の組織を意味する。本明細書にて使用される神経組織が低下したか又は劣った機能活性形態に変化する、神経組織の劣化の状態を意味する。ES細胞の(例えば、脊髄運動ニューロンへの)分化を誘導することにより、本明細書に記載された方法は、分化した神経細胞の生成及び続く移植の必要がある被験者へのその移植を通して、被験者内の様々な損傷した及び/又は変性した神経組織を再増殖させる(repopulating)こと(例えば、脊髄の再増殖)において有用であると信じられる。
【0043】
従って、本発明は、神経組織変性の治療の必要のある被験者において神経組織変性を治療する方法を提供し、上記の方法に従い、ES細胞の分化した神経細胞への分化を誘導し、そして分化した神経細胞を被験者に移植し、それにより神経組織の変性を治療することを含む。例示として、本発明の方法は、以下の工程:(a)胚性幹細胞の培養物を得るか又は生じさせ;(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の、吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の、背側化又は腹側化胚性シグナリング因子に接触させ;(d)任意に、神経細胞のサブクラスを、少なくとも一つの神経栄養因子と接触させ;そして(e)神経組織の変性を治療するのに有効な量にて、分化した神経細胞を被験者に移植することを含む。発明の一つの態様において、被験者は胚である。発明の別の態様において、被験者はヒトである。好ましくは、被験者は神経組織の変性を有する。
【0044】
神経組織の変性は、中枢神経系(CNS)又は末梢神経系(PNS)において生じてよく、そして限定ではないが、初期の神経病の症状(例えば、神経変性疾患)、CNS及びPNS外傷(trauma)及び損傷、及び非神経の機能不全の後天性の二次作用(例えば、変性事象、病原性事象、又は外傷性事象に対して二次的な神経の損失を含む、様々な異常、症状、及び因子に起因するか又は関連するかもしれない。CNS外傷の例は、限定ではないが、しみ(blunt)の外傷、低酸素症、及び侵入性外傷を含む。非神経の機能不全の後天性の二次作用の例は、限定ではないが、脳性麻痺、先天的水頭症、筋ジストロフィー、発作(stroke)、及び血管性痴呆(vascular dementia)、並びに以下:脳出血、発生障害(例えば、脳の欠損、例えば先天的水頭症、又は脊髄の欠損、例えば脊髄披裂(spina bifida))、糖尿病性脳障害、過敏性脳障害、頭蓋内動脈瘤、貧血、腎不全、クモ膜下出血、脳及び脊髄に対する外傷、化学治療剤及び抗ウイルス剤のような治療剤による治療、脳及び脊髄の血管損傷、及び神経組織が変性しやすい他の疾患又は症状の何れかによりもたらされる神経変性を含む。
【0045】
本発明の一つの態様において、神経組織の変性はPNS中の末梢神経障害である。本明細書にて使用される用語「末梢神経障害」は、感覚の損失、筋肉の弱化、筋肉の萎縮、深部反射の低下の症候群、及び/又は血管運動神経の兆候を意味する。末梢神経障害を有する被験者においては、ミエリン鞘(シュワン細胞)が主に冒されているかもしれないか、又は軸索が主に冒されているかもしれない。末梢神経障害は、単一の神経(単一神経障害(mononeuropathy))、別のエリアの2つ又はそれより多い神経(複合性単一神経障害(multiple neuropathy))又は多数の神経を同時に(多発性神経障害(polyneuropathy))冒すかもしれない。
【0046】
本明細書に開示された方法により治療されるかもしれない末梢神経障害の例は、限定ではないが、急性又は慢性の炎症性の多発性神経障害に関連する末梢神経障害、筋委縮性側索硬化症(ALS)、コラーゲン血管障害(例えば、多発性動脈瘤紅斑、慢性関節リウマチ、シェーグレン症候群、又は全身性紅斑性踉狼瘡)、ジフテリア、ギラン−バレー症候群、遺伝性末梢神経障害(例えば、シャルコー−マリー−ツース病(Charcot-Marie-Tooth disease)(タイプI,タイプII及び全てのタイプを含む)、遺伝性の運動ニューロン障害及び知覚ニューロン障害(タイプI,II及びIII及び腓骨筋肉萎縮)、遺伝性圧脆弱性神経障害(HNPP)、感染性疾患(例えば、後天性免役不全症候群(AIDS))、ライム病(例えば、Borrelia burgdorferiによる感染)、微生物の侵入(例えば、ハンセン病−神経の外傷後の神経障害の全世界での主な原因)、大脳白質萎縮症、代謝性の疾患又は障害(例えば、アミロイド症、真性糖尿病、甲状腺機能低下症、ポルフィリン症、類肉腫症、又は尿毒症)、神経繊維腫症、栄養不足、準新生物形成(paraneoplastic)疾患、腓骨神経麻痺、ポリオ、ポルフィリン症、ポリオ後症候群、プロテウス症候群、圧迫麻痺(pressure paralysis)(例えば、手根管症候群)、進行性延髄麻痺、橈骨神経麻痺、脊髄筋肉萎縮(SMA)、毒性薬剤(例えば、バルビツール、一酸化炭素、クロロブタノール、ダプソン、エメチン、重金属、ヘキソバルビツール、鉛、ニトロフラントニン、オルソジニトロフェナル、フェニトイン、ピリドキシン、スルフォナミド、トリオルトクレシルリン酸、ビンカのアルカロイド、多くの溶剤、他の工業性の毒、及び特定のAIDS薬(ジダノシン及びザルシタビンを含む)、外傷(神経の外傷−末梢神経障害の全世界的な主な原因)、尺骨麻痺(Beers and Berkow,編纂、The Merck Manual of Diagnosis and Therapy,第17版(Whitehouse Station,NJ:Merck Research Laboratories,1999)第183章)。本発明の好ましい態様において、末梢神経障害はALS又はSMAである。
【0047】
本発明の別の態様において、神経組織の変性は神経変性疾患である。本明細書に記載された方法により処置してよい神経変性疾患の例は、限定ではないが、アルツハイマー病、筋委縮性側索硬化症(ルーゲーリック病)、ビンスワンガー病、ハンチントンコレラ、多発性硬化症、重症筋無力症、パーキンソン病、及びピック病(Pick's disease)を含む。脱髄の症状に関連した神経組織変性を治療するために本明細書に記載された方法を使用することも、本発明の範囲内である。脱髄症の例は、限定ではないが、急性散在性脳脊髄炎(ADEM)、急性横断性脊髄炎、急性ウイルス性脳炎、副腎白質萎縮症(ALD)、副腎脊髄神経障害(adrenomyeloneuropathy)、AIDS−液胞脊髄障害、HTLV−付随性脊髄障害、レーベル遺伝子視神経症、多発性硬化症(MS)、進行性の多病巣性白質脳症(PML)、準急性硬化症汎脳炎、及び熱帯性痙攣性不全対麻痺を含む。
【0048】
本発明の分化した神経細胞は、当業界において公知の標準手法、並びに本明細書に記載された方法により、治療の必要がある被験者に移植してよい。例示として、胚のボディー(多能性ES細胞に由来してよい)を適当な胚性シグナリング因子(例えば、RA及びHh−Ag1.3)により誘導することにより、分化した神経細胞を生成してよい。誘導から適当な時間の後に(例えば、誘導の3−4日後)、胚のボディー(EBs)を移植のために調製し(例えば、部分的に粉砕された)、そして次に被験者に移植してよい(例えば、チキン、HHステージ15−17の脊髄へ)。移植された組織を順応させる(accommodate)ため、移植の前に被験者に吸引による外傷を与えてよい(suction-lesioned)。約1/2−1のEB(又はEB均等物)を、次に、被験者の吻側頸部、尾部側頸部、胸部又は腰部領域において2−5の体節にわたるセグメントへ移植してよい。
【0049】
本発明の一つの態様において、分化した神経細胞は被験者の脊髄に移植されて、それにより被験者の脊髄を再増殖させ、そして神経組織の変性はALS又はSMAに関連した神経障害である。移植されたニューロンが脊髄の運動ニューロンである場合、吻側化及び尾部側化胚性シグナリング因子は、好ましくはレチノイン酸であり、そして背側化又は腹側化胚性シグナリング因子は、好ましくはヘッジホッグシグナリング経路の活性化因子である。
【0050】
本発明の方法において、分化した神経細胞は、神経組織の変性を治療するのに有効な量において、治療の必要のある被験者に移植される。本明細書にて使用される用語「神経組織の変性を治療するのに有効」は、神経組織の変性の臨床上の障害又は兆候を改善するか又は最小にするのに有効なことを意味する。例えば、神経組織の変性が末梢神経障害である場合、末梢神経障害の臨床上の障害又は兆候は、血管運動神経の兆候を緩和すること、深部反射を増加させること、筋肉萎縮を減少させること、感覚の機能を回復させること、及び筋肉を強化することにより、改善するか又は最小化してよい。治療の必要な被験者において神経組織の変性を治療するのに有効な分化した神経組織細胞の量は、各ケースの特定の因子に依存して変更され、神経組織変性の種類、神経組織変性のステージ、被験者の体重、被験者の症状の重度、分化した神経細胞の種類及び移植の方法を含む。この量は、当業者により、臨床試験及び本明細書に開示された方法を含む公知の手法に基づいて、容易に決定してよい。
【0051】
分化した神経細胞へのES細胞の分化を誘導する上記の方法に関して、本発明は、さらに、分化した神経細胞を生成する方法を提供するが、(a)ES細胞の培養物を得るか又は生じさせ;(b)ES細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させ;そして(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させることを含む。本発明は、細胞の集団も提供し、この方法により生成した、分化した神経細胞を含む。一つの態様において、細胞の全て又は一部がeGFPを発現する。本発明の分化した神経細胞が脊髄の運動ニューロンである場合、吻側化及び/又は尾部側化胚性シグナリング因子は、好ましくはレチノイン酸であり、そして背側化又は腹側化胚性シグナリング因子は、好ましくはヘッジホッグシグナリング経路の活性化因子である。
【0052】
本発明の方法において、工程(b)−(d)の何れかは、インビトロか、又は被験者中でインビボにて実施してよい。インビトロの工程の後に、細胞を被験者に移植するが、残りの工程はインビボにて実施される。従って、本発明の方法は、さらに、神経前駆細胞又は分化した神経細胞を被験者に移植する工程を含む。例えば、ES細胞の培養物を、インビトロにおいて吻側化及び/又は尾部側化胚性シグナリング因子に接触させることにより、神経前駆細胞を生成してよい。そうして生成された神経前駆細胞を次に被験者に移植してよいが、工程(c)及び(d)はインビボにて実施する。
【0053】
別の態様においては、ES細胞の培養物を、インビトロにおいて吻側化及び/又は尾部側化胚性シグナリング因子に接触させることにより、神経前駆細胞を生成してよい。神経前駆細胞を、次に、背側化又は腹側化胚性シグナリング因子にインビトロにて接触させることにより、分化した神経細胞を生成してよい。以後、分化した神経細胞を被験者に移植してよく、工程(d)をインビボにて実施する。
【0054】
さらなる別法においては、ES細胞の培養物を、インビトロにおいて吻側化及び/又は尾部側化胚性シグナリング因子に接触させることにより、神経前駆細胞を生成し;神経前駆細胞を、背側化又は腹側化胚性シグナリング因子にインビトロにて接触させることにより、分化した神経細胞を生成し;そして、任意に、分化した神経細胞を、インビトロにおいて少なくとも一つの神経栄養因子と接触させてよい。分化した神経細胞を、次に、被験者に移植してよい。本発明の一つの態様において、ニューロンを被験者の脊髄に移植する。
【0055】
本発明は、さらに、被験者の脊髄を再増殖させる方法を提供するが、(a)ES細胞の培養物を得るか又は生じさせ;(b)ES細胞の培養物を、神経前駆細胞を生じさせるのに有効な量にて、吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量にて、背側化又は腹側化胚性シグナリング因子に接触させ;(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させ;そして(e)分化した神経細胞を、被験者の脊髄に移植することを含む。分化した神経細胞は、脊髄を再増殖させるのに有効な量にて被験者の脊髄に移植される。この量は、公知の手法及び本明細書に開示された方法に基づいて、当業者により容易に決定してよい。
【0056】
本発明の好ましい態様によれば、吻側化及び/又は尾部側化胚性シグナリング因子は、レチノイン酸であり、そして背側化又は腹側化胚性シグナリング因子は、ヘッジホッグシグナリング経路の活性化因子である。本発明のこの方法は、神経組織の変性、特に末梢神経障害の被験者を治療するのに有用かもしれない。従って、本発明の別の態様において、被験者は末梢神経障害を有する。好ましくは、末梢神経障害はALS又はSMAである。本発明のさらに別の態様において、移植された、分化した神経細胞は、eGFPを発現し、それにより、脊髄再増殖の監視を容易にさせる。
【0057】
本発明は、さらに、工程:(a)ES細胞の培養物を得るか又は生じさせ;(b)ES細胞の培養物を、神経前駆細胞を生じさせるのに有効な量にて、吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;そして(c)任意に、神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量にて、背側化又は腹側化胚性シグナリング因子に接触させることを含む方法により生産される神経前駆細胞を提供する。また、工程:(a)ES細胞の培養物を得るか又は生じさせ;(b)ES細胞の培養物を、神経前駆細胞を生じさせるのに有効な量にて、吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量にて、背側化又は腹側化胚性シグナリング因子に接触させ;そして(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させることを含む方法により生産される神経前駆細胞も提供する。発明の一つの態様において、分化した神経細胞はeGFPを発現する。
【0058】
CNSニューロンの特定のクラスの選択的変性は多くの神経障害の基になるから、ニューロンの成長、生存、及び活性の研究は、相変わらず優先事項である。不幸なことに、しかしながら、生きているニューロンは、そのような研究において容易に利用可能ではない。そのため、本発明は研究される神経細胞の可能性を秘めた無制限な源を提供するため、神経科学及び神経学の分野の研究者に特に重要になる。従って、本発明は、特定の研究エリアにおいて上記の前駆細胞及び分化した神経細胞の用途も提供する。
【0059】
本発明の神経前駆細胞及び分化した神経細胞は、ニューロン(特に、運動ニューロン)の発生、機能及び死の分析−運動ニューロン疾患の完全な理解に必須の研究において有用になる。さらに、本発明の神経前駆細胞及び分化した神経細胞は、標的筋肉との接触のブイにおいてシナプスの分化を監視するのに有用になる。最後に、本発明の神経前駆細胞及び分化した神経細胞は、ALS又はSMA患者において検出された変異の一つを運ぶ変性した運動ニューロンと正常な健康な運動ニューロンの直接の比較を容易にする(例えば、スーパーオキシドジスムターゼ(SOD)又は生存運動ニューロン蛋白質(SMN)遺伝子内の変異)。そのような比較のために、よく知られた技術及び本明細書に記載された方法を用いて、健康な神経細胞及び疾患の神経細胞の両方を生産してよい。あるいは、SOD又はSMN変異を有する細胞を、ALS又はSMAを有する生きた患者又は死んだ患者から単離してよい。
【0060】
本明細書に記載されるとおり、発明者らは、増強された緑色蛍光蛋白質(eGFP)発現のおかげで生きた運動ニューロンが同定できたES細胞系を構築した。運動ニューロン特異的トランス遺伝子発現を授与するマウスHB9遺伝子の9−kb 5’領域の制御下でeGFPのcDNAを発現するトランスジェニックマウス系列を生じさせた。トランスジェニックファウンダーマウスをHB9とeGFPの発現の比較によりスクリーニングした。一つのマウス系列、mHB9−Gfp1bは高レベルのeGFPをE9.5−P10マウスの脊髄運動にの細胞ボディーにおいて発現し、内生HB9のパターンと平行なパターンにおいてであった。さらに、eGFPの発現が運動ニューロンの軸索及び樹状突起において高レベルで検出された。ES細胞系(HBG3)はmHB9−Gfp1bトランスジェニックマウス由来であった;これらの細胞は生殖系(germ line)を通してeGfp遺伝子を伝搬して、類似の運動ニューロン選択的パターンにおいてeGFPの発現を指示した。
【0061】
従って、本発明は、ES細胞がeGFPを発現する、本発明の方法においての使用のための胚性幹(ES)細胞を含むトランスジェニック非ヒト動物系をさらに提供する。非ヒト動物は適当な動物(例えば、ネコ、家畜、イヌ、ウマ、ヒヤギ、齧歯類、及びヒツジ)であってよいが、それは好ましくは齧歯類である。より好ましくは、非ヒト動物はラット又はマウスである。本発明のトランスジェニック非ヒト動物は、遺伝子操作されたトランスジェニック動物のための様々な技術により製造してよく、当業界公知のものを含む。発明の一つの態様において、トランスジェニック非ヒト動物のゲノムは、増強された緑色蛍光蛋白質(eGFP)遺伝子を含む。このトランスジェニック動物から確立された動物系列は、その胚の中に、eGFPを発現するES細胞を含む。
【0062】
本明細書にて使用される用語「トランスジェニック非ヒト動物」は、そのゲノムがトランス遺伝子の導入により変更された、実験操作により製造された、遺伝子操作された非ヒト動物を意味する。さらに本明細書にて使用される用語「トランス遺伝子」は、実験操作により動物のゲノムに導入された核酸分子(例えば、DNA、遺伝子、又はそれらの断片)を意味し、導入された遺伝子は当該動物にとって内生ではないか、又は動物にとって内生の遺伝子の修飾形態か又は変異形態である。内生遺伝子の修飾形態又は変異形態は、ヒトの介入を通して製造してよい(例えば、点変異の導入、フレームシフトの導入、内生遺伝子の部分又は断片の欠失、選択可能なマーカー遺伝子の挿入、停止コドンの挿入等による)。
【0063】
トランスジェニック非ヒト動物は、ヒトの介入を含む幾つかの方法により製造してよく、限定ではないが、非ヒト動物の胚生幹細胞、新たに受精した卵、又は初期胚へのトランス遺伝子の導入;非ヒト動物の体細胞及び/又は生殖細胞の染色体へのトランス遺伝子の組込み;及び本明細書に記載されたあらゆる方法を含む。本発明の好ましい態様において、トランスジェニック非ヒト動物は、トランス遺伝子のES細胞への導入により製造される。
【0064】
例示として、本発明のトランスジェニックマウス系は、マウスHB9遺伝子の5’上流領域(Arber et al.,Requirement for the homeobox gene HB9 in the consolidation of motor neuron identity.Neuron,23:659-764,1999)、続く5’スプライス基質(Choi et al.,A generic intron increases gene expression in transgenic mice.Mol.Cell Biol.,11:3070-74,1991)、eGFP遺伝子、及びウシ成長ホルモンポリアデニレーションシグナルを含む断片(例えば、約9kB)を使用する構築物の前核注入により確立してよい。ES細胞系を、次に、異型接合胚盤胞から遊動してよく、以前に記載されたとおりである(Abbondanzo et al.,Derivation of embryonic stem cell lines.Methods Enzymol.,225:803-23,1993)。遺伝子系列は、マウス胚盤胞を発生させるその能力及び生殖系の遺伝(germ transmission)に関するその能力に基づいて選択してよい。胚生幹細胞は、次に、本明細書に記載された方法における使用のために、トランスジェニック動物から単離してよい。
【0065】
本発明の一つの態様において、トランスジェニック動物は、スーパーオキシドジスムターゼ(SOD)又は生存運動ニューロン蛋白質(SMN)遺伝子が変異した(例えば、SOD−1−G93S変異)か又は選択的に不活性化されることによりその内生SOD又はSMN遺伝子内に破壊をもたらす、ゲノムを有する。本明細書にて使用される「破壊」は、機能するSOD又はSMN蛋白質の正常な発現を妨害する、SOD又はSMN遺伝子内の変異(即ち、遺伝物質内の永遠の遺伝性の変化)を意味する(例えば、それが変異SOD又はSMN蛋白質をもたらすか;それが正常な量のSOD又はSMN蛋白質の発現を妨害するか;又はそれがSOD又はSMN蛋白質の発現を妨害する)。破壊の例は、限定ではないが、点変異、フレームシフト変異の導入、内生遺伝子の一部又は断片の欠失、選択可能なマーカー遺伝子の挿入、及び停止コドンの挿入を含む。本明細書にて使用される「変異体」は、野生型遺伝子(又はその遺伝子産物)に比較して、その配列内(又はその機能特性)に少なくとも一つの修飾を呈する遺伝子(又はその遺伝子産物)を意味する。対照的に、用語「野生型」は、その天然源(例えば、天然集団)中でもっとも頻繁に見いだされる、特定の遺伝子(又はその遺伝子産物)に関して特徴的な遺伝子型(又は表現型)を意味する。野生型動物は、例えば、SOD又はSMN蛋白質を発現する。
【0066】
本発明のトランスジェニック非ヒト動物における選択的不活性化は、様々な方法により達成してよく、そして異型接合破壊(一つのSOD対立遺伝子又は一つのSMN対立遺伝子が破壊されて、結果のトランスジェニック動物が変異に関して異型接合である)又は同型接合破壊(SOD又はSMNの対立遺伝子両方が破壊されて、結果のトランスジェニック動物が変異に関して同型接合である)の何れかをもたらしてよい。本発明の一つの態様において、トランスジェニック動物の内生SOD又はSMN遺伝子は、SOD又はSMN遺伝子産物に共通の領域をコードする核酸配列との相同組換えにより破壊される。例示として、相同組換えによる破壊は、SOD又はSMN遺伝子内のノックアウト変異、特に少なくとも一つの欠失がSOD又はSMN遺伝子の少なくとも一つのエクソン内に導入されたノックアウト変異を生じてよい。さらに、SOD又はSMN遺伝子内の破壊が、異種選択可能マーカー遺伝子の内生SOD又はSMN遺伝子への挿入をもたらしてよい。
【0067】
eGFP遺伝子を発現し、そしてSOD又はSMN遺伝子内にノックアウト変異を有するトランスジェニック非ヒト動物の系列を確立する一つの方法は、以下の工程を含んでよい:(a)SOD又はSMN標的ベクターを生成し;(b)標的ベクターを非ヒト動物のレシピエント細胞に導入し;(c)処理されたレシピエント細胞を非ヒト動物の胚盤胞に導入することにより、処理された胚盤胞を生成し;(d)処理された胚盤胞を偽妊娠非ヒト動物に導入し;(e)移植された胚盤胞を予定日まで発生させ;(f)そのゲノムがノックアウト破壊をその内生SOD又はSMN遺伝子内に含むトランスジェニック非ヒト動物を同定し;(g)トランスジェニック非ヒト動物を繁殖させることにより、野生型に比較して低下したSOD又はSMN蛋白質の発現を呈するトランスジェニック非ヒト動物を得て;そして(h)eGFPを発現し、そして野生型に比較して低下したSOD又はSMN蛋白質の発現を呈するトランスジェニック非ヒト動物を得るために、当該トランスジェニック非ヒト動物を、そのゲノムがeGFP遺伝子を含むトランスジェニック非ヒト動物と交配させることを含む。eGFPを発現するトランスジェニック非ヒト動物は、上記の方法に従い得てよい。SOD又はSMN蛋白質を過剰に発現するトランスジェニック非ヒト動物を提供することも、本発明の範囲内である。
【0068】
本発明のeGFP発現トランスジェニック動物系は、ES細胞の源及び移植、キメラ、及び二重標識組換え実験のための組織を提供する。本発明のトランスジェニック動物系は、運動ニューロン変性の遺伝モデルも提供する。例えば、トランスジェニック動物系由来の動物のゲノムがそのSOD遺伝子内に変異を有する場合、それは運動ニューロン変性、特にALSを治療するのに有用かもしれない薬剤をスクリーニングするための薬学モデルとして使用してよい。このモデルは緩慢な変性の開始を呈し得るはずであるが、その結果は劇的であると予測される。さらに、トランスジェニック動物のゲノムがそのSMN遺伝子内に変異を有する場合、それは運動ニューロン変性、特にSMAを治療するのに有用かもしれない薬剤をスクリーニングするための薬学モデルとして使用してよい。このモデルは素早い変性の開始を呈すると予測される。
【0069】
本発明のトランスジェニック非ヒト動物由来のES細胞は、本明細書に記載された方法において使用してよく、インビトロ又はインビボの分化、及び被験者への再導入を含む。さらに、eGFPを発現するトランスジェニック動物系由来のES細胞及びSOD又はSMN遺伝子内に変異を有するトランスジェニック動物系由来のES細胞を交配してよい。変異体EPA細胞の挙動を、次に、細胞及び/又は組織がeGFPを発現する、発生した動物又はキメラ胚の顕微鏡観察により追跡する。
【0070】
本発明は、さらに、分化した神経細胞の純粋な集団を単離する方法、及び/又は分化した神経細胞の集団を精製する方法を提供し、工程:(a)増強された緑色蛍光蛋白質(eGFP)を発現するES細胞の培養物を得るか又は生じさせ;(b)ES細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の、吻側化及び/又は尾部側化胚性シグナリング因子に接触させるが、但し、神経前駆細胞の幾つか又は全てもeGFPを発現し;(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の、背側化又は腹側化胚性シグナリング因子に接触させるが、但し、分化した神経細胞もeGFPを発現し;(d)任意に、分化した神経細胞のサブクラスを、少なくとも一つの神経栄養因子と接触させ;(e)分化した神経細胞の中でeGFPの発現を検出し;そして(f)eGFPを発現する分化した神経細胞を単離することを含む。発明の一つの態様において、分化した神経細胞は、運動ニューロンである。eGFPを発現するES細胞は、上記に通りにして本発明のトランスジェニック動物から得てよく、あるいは本明細書に開示された他の方法に従い作成してよい。
【0071】
本発明の方法によると、eGFPの発現は、インビトロ又はインビボの何れかのアッセイにより分化した神経細胞中で検出してよい。本明細書にて使用された「発現」は、少なくとも一つのmRNA転写物中のeGFP遺伝子の転写、又は少なくとも一つのmRNAのeGFP蛋白質への翻訳を意味する。分化した神経細胞は、eGFP蛋白質、eGFPのcDNA、又はeGFPのmRNAをアッセイすることによる、eGFP発現をアッセイしてよい。eGFPの適切な形態は、本明細書に開示された特定の技術に基づいて明らかとなる。
【0072】
分化した神経細胞は、eGFP発現に関してアッセイしてよく、そしてeGFP発現は当業界で公知のアッセイ及び方法を用いて、分化した神経細胞内で検出してよい。eGFPは培養物中及びインビボにおいて細胞を標識するための非侵入性マーカーを提供するから、eGFPの発現は、好ましくは、イメージング技術、特にブライトフィールド、相、及び蛍光イメージング技術を用いて、分化した神経細胞中で検出され、本明細書に開示されるとおりである。高レベルのeGFPを発現する分化した神経細胞は、次に、以下に記載される、それらのeGFP蛍光及びフォワードライトスキャンに基づいて、分類することにより(例えば、Beckman-Coulter Altraフローサイトメトリーを用いた、FACSソーティング)、細胞懸濁液から単離してよい。
【0073】
他の方法も、本発明の分化した神経細胞内でeGFP発現を検出するのに使用してよい。そのような検出方法の例は、限定ではないが、ハイブリダイゼーション分析、イメージング技術、免疫学上の技術、免疫沈殿、放射検出、ウエスタンブロット分析、及び本明細書に開示されたあらゆる追加のアッセイ又は検出方法を含む。例えば、eGFP蛋白質又はeGFP核酸と反応性の薬剤を用いて、eGFP発現に関して分化した神経細胞をアッセイしてよい。本明細書にて使用される「反応性」は、eGFPに親和性を有するか、eGFPに結合するか、又はeGFPに対して向けられる薬剤を意味する。さらに本明細書にて使用される「薬剤」は、蛋白質、ポリペプチド、ペプチド、核酸(DNA又はRNAを含む)、抗体、Fab断片、F(ab’)2断片、分子、化合物、抗生物質、薬品(drug)、及びそれらのあらゆる組み合わせを含む。本発明の一つの態様において、eGFPと反応性の薬剤は、抗体である(例えば、αGFP(Molecular Probes,Inc.,Eugene,OR))。
【0074】
分化した神経細胞内でのeGFP発現の検出の後に、細胞中のeGFP発現の程度を、もし必要なら様々な定量アッセイのうちの一つを用いて、測定するか又は定量してよい。そのようなアッセイは当業者に知られており、eGFP蛋白質の量を測定するための、免疫組織化学、免疫細胞学、フローサイトメトリー、質量分光分析、ウエスタンブロット分析、又はELISAを含んでよい。
【0075】
本発明は、さらに、ニューロン(例えば、運動ニューロン)の変性に関連した症状を治療することにおける使用のための薬剤を同定する方法を提供する。本明細書にて使用される「ニューロンの変性」は、ニューロンが劣ったか又は低い機能活性の形態に変化するニューロンの変性の症状を意味する。ニューロンの変性に関連した症状の例は、本明細書に記載された、末梢神経障害、脱ミエリン症状、及び根本的な神経学の症状(例えば、神経変性疾患)、CNS及びPNS外傷及び障害、及び非神経機能不全の後天性二次作用(例えば、変性性、病理性、又は外傷性の自己に対して二次的な神経損失)を含む。
【0076】
本発明の方法は、(a)胚性幹細胞の培養物を得るか又は生じさせ;(b)神経前駆細胞を生成するのに有効なレチノイン酸の量に、当該胚性幹細胞の培養物を接触させ;(c)神経前駆細胞中のヘッジホッグシグナリング経路を活性化することにより、神経の幾つか又は全てが変性したニューロンを生成し;(d)変性したニューロンを候補薬剤に接触させ;そして(e)薬剤が変性したニューロンの幾つか又は全ての再生を増強するか否か決定する工程を含む。本明細書にて使用される「再生を増強する」は、変性していたニューロン(神経突起及びミエリン鞘)の部分的又は完全な成長(又は再成長)を増大させるか(augment)、改善するか、又は増加させる(increase)ことを意味する。ニューロンの再生は、中枢神経系及び末梢神経系の両方のニューロンにおいて生じてよい。本発明の一つの態様において、上記薬剤は、変性した運動ニューロンの再生を増強する。
【0077】
本発明の方法においては、本明細書に記載された、因子(又は薬剤)と細胞の間の接触をもたらすあらゆる方法、及び誘導及び投与のあらゆる方法により、変性したニューロンを候補の薬剤と接触させてよい。ニューロンの再生、及び増強された再生は、公知の手法により測定するか又は検出してよく、ミエリン−特異的且つ軸索特異的な蛋白質のためのウエスタンブロッティング、形態計測を伴う電子顕微鏡、及び当業者に知られているあらゆる方法、分子手法、及びアッセイを含む。さらに、ミエリンの成長を、g−比、軸索:ミエリン結合の完全性の一つの測定法を用いてアッセイしてよい。g−比は軸索とミエリンの全直径割る軸索の直径として定義される。この比は、何れかの一定のサイズの軸索に関しての相対的なミエリンの信頼できる測定値を提供する(Bieri et al.,Abnormal nerve conduction studies in mice expressing a mutant form of the POU transcription factor,SCIP.J.Neurosci.Res.,50:821-28,1997)。多数の研究が証明したことは、0.6のg−比がほとんどの繊維には標準であるということである(Waxman and Bennett,Relative conduction velocities of small melinated and nonmyelinated fibers in the central nervous system.Nature New Biol.,238:217,1972)。
【0078】
本発明の一つの態様において、変性したニューロン内のゲノミックDNAはSOD又はSMN遺伝子内に変異を有する。そのような変性したニューロンは、例えば、上記のとおり、そのゲノムが変異したSOD又はSMN遺伝子を含む本発明のトランスジェニック動物から単離してよい。発明の一つの態様において、変性したニューロンは、増強された緑色蛍光蛋白質(eGFP)を発現する。そのような変性したニューロンは、上記のとおり、そのゲノムがeGFP遺伝子を含む本発明のトランスジェニック動物から単離してよい。SOD及びeGFP遺伝子の両方を発現するか、又はSMN及びeGFP遺伝子の両方を発現する変性したニューロンは、SOD又はSMN変異を有するES細胞を、eGFP遺伝子を含むES細胞と交配することにより生成してよい。
【0079】
本発明は以下の実施例において記載されるが、発明の理解を助けるために記載されるのであり、以後の請求の範囲において定義される発明の範囲を、如何なる意味においても、限定するものとして解釈されるべきではない。
【0080】
実施例
実施例1−ES−細胞培養
野生型(MM13又はW9.5)又はHB9::GFPトランスジェニックマウス由来の(HBG3)ES細胞を、ES−細胞培地(4500mg/lのグルコース及び2250mg/lの重炭酸ナトリウムと共に、1x非必須アミノ酸、1xヌクレオシド(全てがSpecialty Media,Lavellette,ニュージャージーから)、0.1mMの2−メルカプトエタノール(Sigma Chemical Co.,セントルイス、ミズーリ)を付加したダルベッコ修飾イーグル培地(DMEM))、2mM L−グルタミン(Gibco)、1xペニシリン/ストレプトマイシン(Gibco BRL,ガイセルスベルグ、メリーランド)、15%のFBS(HyClone,ロウガン、ユタ)、及び1000μ/mlのLIF(Chemicon,テメキュラ、カリフォルニア)の中で初期マウス胚繊維芽細胞の供給層上で生育させた。ES−細胞のコロニーをプレートから2日後に一部解離させ(dissociated)、そしてDF培地(グルコース(4500mg/l,Sigma)、2mM L−グルタミン(Gibco)、1xペニシリン/ストレプトマイシン(Gibco)、0.1mMの2−メルカプトエタノール(Sigma)、1xインスリン−トランスフェリン−セレニウムサプリメント(Gibco)、20nMのプロゲステロン(Sigma)、及び60μMのプトレシン(Sigma)を付加されたDMEM/F12をESK10培地(LIFとFBSを除いたが、10%のノックアウト血清補充(Gibco)を追加したES細胞培地)と1:1混合した)からなるDFK5培地の中で培養した。培地を2日後に、レチノイン酸(RA)(100nMから2μM)(Sigma)、ソニックヘッジホッグ蛋白質(SHh−N;300nM)(Curis,Inc.,ケンブリッジ、マサチューセッツ)、ヘッジホッグアゴニストHh−Ag1.3(1−1000nM)(Curis,Inc.)、又はヘッジホッグブロッキング抗体(5EI,30μg/ml)を付加した新鮮な培地と交換した。胚のボディー(EBs)をさらに2−5日間培養した。幾つかの実験のために、ES細胞をストロマPA6細胞系(Riken Cell Bank,日本)の単層上で、DFK5培地単独か、又は記載されたとおりに(Kawasaki et al.,Induction of midbrain dopaminergic neurons fom ES cells by stromal cell-derived inducing activity.Neuron,28:31-40,2000)、2μMのRAを追加して生育させた。
【0081】
幾つかのHBG3 ES細胞由来のEBsを誘導の4日後にパパイン分離システム(Worthington Biochemical Corp.,フリーホールド、ニュージャージー)を用いて分離し、内生eGFP発現に基づいてFACS分類し、マトリゲル(BD)をコートされたペトリ皿又はテラサキウエル上にプレートし、そして5%のウマ血清(Gibco)、1xのB−27サプリメント(Gibco)、1xのインスリン−トランスフェリン−セレンサプリメント(Gibco)、20nMのプロゲステロン(Sigma)、及び60μMのプトレシン(Sigma)を付加されたF12培地(Specialty Media)内で培養した。神経栄養生因子(GDNF,NT3,CNTF,及びBDNF)(10ng/ml;R&D Systems,Inc.,ミネアポリス、ミネソタ)の混合物を、選択された実験に含ませた。
【0082】
実施例2−免役細胞化学
胚ボディー(EBs)を4%パラホルムアルデヒドにより固定化し、そしてアクリオスタット上で切片化した。免役染色のため、以下の抗体を使用した:ウサギαHB9,αOlig2,αNkx2.2,αLhx3,αDbx1(Pierani et al.,Control of interneuron fate in the developing spinal cord by the progenitor homeodomain protein Dbx1.Neuroon,29(2):367-84,2001),αIrx3,αeGFP(1:2000,Molecular Probes,Inc.,ユージーン、オレゴン),αSox1(1:500)(Pevny et al.,Generation of purified neural precursors from embryonic stem cells by lineage selection.Curr.Biol.,8:971-74,1998),αOtx2(1:1000)(Mallamacu et al.,OTX2 homeoprotein in the developing central nervous system and migratory cells of the olfactory area.Mech.Dev.,58:165-78,1996),αHoxC5(Liu et al.,Assigning the positional identity of spinal motor neurons.Rostrocaudal patterning of Hox-c expression by FGFs,Gdf11,and retinoids.Neuron,32:997-1012,2001),及びαcHX10;マウスαPax7,αPax6,αHB9,αLim1,αNkx6.1,αNkx2.2,αEn1,αLim2(マウス特異的),αNCAM(マウス特異的),αSC1(チキン特異的),αNeuN(1:500,Chemicon),αTuj1(1:2000)(BabCo,バークレー、カリフォルニア),αSyb(マウス特異的;1:1000)(Synaptic Systems),αSyn(1:1000,Synaptic Systems),及びα神経繊維;モルモットα−Isl1,抗−Olig2,抗−Hoxc6(Liu et al.,Assigning the positional identity of spinal motor neurons.Rostrocaudal patterning of Hox-c expression by FGFs,Gdf11,and retinoids.Neuron,32:997-1012,2001);ヤギ−αChAT(1:200)(Chemicon),αVAChT(マウス特異的;1:1000)(Chemicon)(Novitch et al.,Coordinate regulation of motor neuron subtype identity and pan-neuronal properties by the bHLH repressor Olig2.Neuron,31:773-89,2001,特定しない限り)。
【0083】
実施例3−eGFPトランスジェニックマウス系の生成
マウスHB9遺伝子の5’上流領域(Arber et al.,Requirement for the homeobox gene Hb9 in the consolidation of motor neuron identity.Neuron,23:659-764,1999)、続く5’スプライス基質(Choi et al.,A generic intron increases gene expression in transgenic mice.Mol.Cell Biol.,11:3070-74:1991)、eGFP遺伝子、及びウシ成長ホルモンポリアデニル化シグナルを含む約9kbの断片を用いた構築物の前核注入により、トランスジェニックマウス系を確立した。HB9の約9kBのNotI断片は、E9.5からP10を試験したトランスジェニック胚内でeGFP発現が運動ニューロンを発生させるよう指令するのに十分である。一つのトランスジェニック系において(mHB9−Gfp1b)、eGFP発現のパターンは内生HB9の発現のプロフィールにぴったりと一致した。ES−細胞系を、前に記載されたとおりに(Abbondanzo et al.,Derivation of embryonic stem cell lines.Methods Enzymol.,225:803-23,1993)、mHB9−Gfp1b異型接合胚盤胞から誘導した。一つの選択された系(HBG3)について、マウス胚盤胞を発生させるように統合するその能力及び生殖系列の移行(germline transmission)のためのその能力に関して検査した。
【0084】
実施例4−HBG3−細胞−由来の運動ニューロンのFACS単離
HBG3 ES−細胞の胚ボディーをパパイン分離系(Worthington Biochemical Corp.)を用いて分離した。高いレベルのeGFPを発現する細胞を、Beckman-Counter Altraフローサイトメトリーを用いて、プロット中に示されたゲートを用いてそれらのeGFP蛍光及びフォーワード光散乱に基づいて、細胞懸濁液から分類した。一般に、入力した細胞の20−30%は高いレベルのeGFPを発現した。上記分析は、このeGFP−高(Hi)細胞群が>95%純粋な集団の運動ニューロンを構成要素としたことを確証した。典型的なFACS分類の収量はEBあたり3000−5000のGFP−高細胞である。
【0085】
実施例5−HBG3 ES−細胞−由来の運動ニューロンのインビボ移植
HBG3 ES細胞由来のEBsを2μMのRA及び1μMのHh−Ag1.3により誘導した。誘導の3−4日後に、EBsを一部粉砕して、チキンのHHステージ15−17の脊髄に移植し、吸引損傷して移植された組織を適応させた。約1/2−1のEB均等物が、チキン胚脊髄の吻側頸部、尾部側頸部、胸部、又は腰部領域において2から5の体節にわたるセグメントに移植した。幾つかの胚を移植の3日後に回収し、固定化し、ビブラトーム上で切片化し、そしてeGFPに対する抗体及び細胞−又は種−特異的抗体により染色した。他の胚(胸部脊髄への移植物を受ける)は7日目に回収した。それらの胸郭を切開し、ローダミン−コンジュゲートされたα−ブンガロトキシン(2μg/ml,Molecular Probe,Inc.)及びGFP,Syb,Syn,又はVAChTに対する抗体によりホウルマウント(wholemount)として染色した。
【0086】
以下に記載されるとおり、発明者らの実験結果は、レチノイン酸が胚ボディー(EBs)中の神経前駆細胞上に尾部側化効果を有すること、運動ニューロン前駆体特異化が尾部側化EBsにおいてはヘッジホッグ依存性であること、eGFP−標識されたES−由来のニューロンは生成及び単離に成功するかもしれないこと、そしてeGFP−標識されたES−細胞由来の運動ニューロンは胚の脊髄内で分化することを示す。
【0087】
RAは胚ボディー中で神経前駆体を尾部側化する
マウスES細胞が運動ニューロンを生じさせる能力を検査し始めるため、発明者らは、凝集培養において2日間マウスES細胞を生育させたが、胚ボディー(EBs)の形成をもたらす方法であった(Bain et al.,Embryonic stem cells express neuronal propertiesin vitro.Dev.Biol..168:342-57,1995)。最初に約1000の細胞からなったEBsは、さらに1−7日間、追加の因子の存在又は不在下で、懸濁培養にて保持した。
【0088】
制御条件下では、2−3日間生育させたEBsは、全神経前駆体マーカーSox1発現する細胞を、あるとしても僅かしか含まなかった(図1)(Pevny et al.,Generation of purified neural precursors from embryonic stem cells by lineage selection.Curr.Biol.,8:971-74,1998;Wood et al.,Comparative expression of the mouse Sox1,Sox2 and Sox3 genes from pre-gastrulation to early somite stages.Mech.Dev.,86:19-201,1999)。同様に、5日目に検査されたEBsは、神経核マーカーNeuNの発現により評価されるニューロン及びMAb TuJ1により認識される神経βチューブリンアイソフォームを、あったとしても僅かしか含まなかった(図1B及び示さないデータ)。対照的に、EBsのRA(100nMから2μM)への2−3日の暴露は、多数のSox1+細胞の存在をもたらし;5日間で、多数のNeuN+,TuJ1+有糸分裂後のニューロンが検出された(図1B及び示さないデータ)。即ち、マウスES細胞由来のEBsはニューロンを生じさせるように誘導することができる。Bain et al.,Embryonic stem cells express neuronal properties in vitro.Dev.Biol.,168:342-57,1995;及びXiam et al.,Peering into early neurogenesis with embryonic stem cells.Trends in Neurosci.,24:685-86,2001も参照。
【0089】
発明者らは、次に、両者の同時発現が初期中脳の位置アイデンティティーの指標である、Otx2とEn1の発現をアッセイすることにより(Davis et al.,Expression pattern of the homeobox-containing genes En-1 and En-2 and the proto-oncogene int-1 diverge during mouse development.Genes Dev.,2:1736-44,1988;Mallamaci et al.,OTX2 homeoprotein in the developing central nervous system and migratory cells of the olfactory area.Mech.Dev.,58:165-78,1996)、そして、脊髄の位置アイデンティティーマーカーであるHox蛋白質Hoxc5,Hoxc6及びHoxc8の発現をアッセイすることにより(Belting et al.,Multiple phases of expression and regulation of mouse Hoxc8 during early embryogenesis.J.Exp.Zool.,282:196-222,1998;Liu et al.,Assigning the pisitional identity of spinal motor neurons.Rostrocaudal patterning of hox-c expression by FGFs,Gdf11,and retinoids.Neuron,32:997-1012,2001)、RA−暴露されたEBs内に形成された神経前駆細胞の吻側位置のアイデンティティーを決定した。3日間RAに暴露されたEBs中の細胞はOtx2とEn1の発現を欠き、そしてHoxc5及びHoxc6を発現したが、Hoxc8は発現しなかった(図1B及び示さなかったデータ)。神経Hox−c発現のこのプロフィールは、吻側頸部脊髄の位置的特性を伴う細胞の指標である(Belting et al.,Multiple phases of expression and regulation of mouse Hoxc8 during early embryogenesis.J.Exp.Zool.,282:196-222,1998;Liu et al.,Assigning the pisitional identity of spinal motor neurons.Rostrocaudal patterning of hox-c expression by FGFs,Gdf11,and retinoids.Neuron,32:997-1012,2001)。
【0090】
ES細胞のRAへの暴露は神経の分化及び脊髄位置的マーカーの発現の両方を促進させるから、このプロトコルはRAのES−由来の神経細胞に対する尾部側化作用の直接の試験を可能にさせない。よって、発明者らは、RA−非依存性経路を通してES細胞を神経化する手段を探した。これを達成するため、発明者らは、神経誘導性シグナルの源としてPA6ストロマ細胞系を使用した(Kawasaki et al.,Induction of midbrain dopaminergic neurons from ES cells to PA6 cell activity for 5 days induced activity.Neuron,28:31-40,2000)。マウスES細胞のPA6細胞活性への5日間の暴露は、莫大な数のSox1+細胞を誘導し、それらの多くはOtx2及びEn1を同時に発現した;対照的に、Hoxc5又はHoxc6を発現した細胞はなかった(図1C及び示さなかったデータ)。さらに、6日間で、多数のOtx2+,TuJ1+ニューロンが検出された(図1C)。ES細胞のPA6細胞活性及びRA(2μM)への暴露は、なおも多数のSox1+を誘導した;しかしながら、これらの細胞はOtx2及びEn1の発現を欠いたし、多くが今はHoxc5及びHoxc6を発現した(図1C及び示さなかったデータ)。合わせると、これらの発見は、インビボにおけるように(Muhr et al.,Convergent inductive signals specify midbrain,hindbrain,and spinal cord identity in gastrula stage chick embryos.Neuron,23:689-702,1999)、初期中脳と同様な位置的特性の神経化ES細胞が尾部側化されることにより、RAへの暴露に際して脊髄と同様な位置的特性になり得ることを示す。
【0091】
運動ニューロン前駆体特異化は尾部側化胚ボディーにおいてHh−依存性である
RAに暴露されたEBs内に存在した、脊髄の位置的特性を有する神経前駆体が運動ニューロン前駆体に分化できるか否かを検査するため、発明者らは、運動ニューロン前駆体アイデンティティーの特異化において連続的な工程を規定するHD及びbHLHの転写因子の発現を監視した(図2A)。神経前駆細胞のPax7,Irx3,Dbx1,Pax6,Nkx6.1,Nkx2.2,及びOlig2の詳述された別個のセットが、神経管の背−腹軸に沿って配置された(Briscoe et al.,Specification of neuronal fates in the ventral axis of the neural tube.Curr.Opin.Neurobiol.,11:43-49,2001;図2B)。脊髄の運動ニューロン前駆体がpMNドメイン中に発見された;それらはPax6,Nkx6.1及びOlig2を発現し、そしてPax7,Irx3,Dbx1及びNkx2.2を排除した(図2B及び示さなかったデータ)。RAに3日間暴露されたEBsは、Pax7,Pax6,Irx3及びDbx1を発現する細胞を多数含み、そしてNkx6.1,Olig2又はNkx2.2を発現する細胞はあったとしてもほとんど含まなかった(図2C及び2D及び示さなかったデータ)。転写因子の発現のこのプロフィールは、介在ニューロンを生じる背側及び中間の脊髄内に位置する前駆細胞の特徴である(図2B)(Lee et al.,The specification of dorsal cell fates in the vertebrate central nervous system.Annu.Rev.Neurosci.,22:261-94,1999)。
【0092】
運動ニューロン前駆体の誘導は、SHh活性に依存する(Briscoe et al.,Specification of neuronal fates in the ventral axis of the neural tube.Curr.Opin.Neurobiol.,11:43-49,2001)。よって、発明者らは、SHhシグナリングがRAに暴露されたEBsにおいて前駆細胞転写因子発現のプロフィールを変化させるか否かを検査した。SHhシグナリング経路を達成するため、発明者らは、SHhシグナリングの特異的小分子アゴニストであるHh−Ag1.3(Frank-Kamenetsky et al.,Chemical genetics of Hh signaling:Identification and characterization of Smoothened agonists and antagonists.Submitted,2002)又は組換えSHh−N蛋白質(Roelink et al.,Floor plate and motor neuron induction by different concentration of the amino-terminal cleavage product of sonic hedgehog antoproteolysis.Cell,81:445-55,1995)を使用し、本質的に同一の結果を得た。
【0093】
RA処理されたEBsの10nMのHh−Ag1.3への3日間の暴露は、転写因子発現のプロフィールに顕著な変化をもたらした。Pax7発現は完全に抑圧され、Pax6+及びIrx3+細胞の数は増加し、Dbx1+細胞の数は減少し、そして僅かなNkx6.1+及びOlig2+細胞が検出された(図2C及び2D及び示さなかったデータ)。RA処理されたEBsの1μMのHh−Ag1.3への暴露は、Dbx1発現を事実上は除去し、Irx3+及びPax6+細胞の数を顕著に減少させ、そして大きく増加した数のNkx6.1+及びOlig2+細胞を誘導した(図2C及び2D)。Hh−Ag1.3に暴露されたEBs内のOlig2+細胞の多くがBrdUを取込んだことから(図10)、それらは循環する前駆体であることが示された。この濃度のHh−Ag1.3において、僅かなNkx2.2+細胞も検出された(図2C及び2D)が、HNF3β+フロアープレート細胞は検出されなかった(データは示さず)。Hhシグナリングに応答性のEBs内の前駆細胞転写因子マーカーの発現のプロフィールの背側から腹側へのシフトは、神経管前駆体の挙動によく似ている(Ericson et al.,Two critical periods of Sonic Hedgehog signaling required for the specification of motor neuron identity.Cell,87:661-73,1996;Ericson et al.,Pax6 controls progenitor cell identity and neuronal fate in response to graded SHh signaling.Cell,90:169-80,1997;Ericson et al.,A homeodomain protein code specifies progenitor cell identity and neuronal fate in the ventral neural tube.Cell,101:435-45,2000)。特に、高レベルのSHhシグナリングは、運動ニューロン前駆体のHD及びbHLH蛋白質発現の特徴のプロフィールを発現した多数の細胞を生じさせた。
【0094】
運動ニューロンがRA−及びHh−暴露されたEBs内に存在する尾部側化され且つ腹側化された前駆細胞から生じたか否かを決定するため、発明者らは、体細胞の運動ニューロンにより選択的且つ不変に発現されるHD蛋白質、HB9の発現を分析した(Pfaff et al.,Requirement for LIM homeobox gene Isl1 in motor neuron generation reveals a motor neuron-dependent step in interneuron differentiation.Cell,84:309-20,1996;Arber et al.,Requirement for the homeobox gene Hb9 in the consolidation of motor neuron identity.Neuron,659-764,1999;Thaler et al.,Active suppression of interneuron programs within developing motor neuron revealed by analysis of homeodomain factor HB9.Neuron,23:675-87,1999)(図3A)。5日間単独で生育されたRAに暴露されたEBsは僅かなHB9+ニューロンを確かに含んだ(図3B;7±2 HB9+ニューロン/切片)。これらのニューロンがHh−依存性様式にて生じたのか否かを試験するため、発明者らは、機能をブロックする抗−Hh抗体の存在下でRA−暴露EBsを生育させた(mAb 5E1 Ig,30μg/ml)(Ericson et al.,Two critical periods of Sonic Hedgehog signalng required for the specification of motor neuron identity.Cell,87:661-73,1996)。抗−Hh存在下で生育させたRAに暴露されたEBsはHB9+ニューロンを含まなかったことから(図3B)、Hhの体内の源は、RAに暴露されたEBsに見いだされる僅かな運動ニューロンの分化のために必要とされる。
【0095】
発明者らは、運動ニューロンの生成がRAに暴露されたEBs内のHhシグナリングのレベルを評価することにより増強できるか否かも検査した。RA処理されたEBsのSHh(300nM)又はHh−Ag1.3(1μM)への暴露は、生じた運動ニューロンの数を顕著に増加させ;典型的には、これらのEBs中の20−30%の細胞がHB9を発現した(約5000のHB9+ニューロン;509±57のHB9+ニューロン/切片;約10−15の切片/EB)(図3B及び示さなかったデータ)。これらのHB9+細胞はNeuNとTuJ1を同時発現し(図3C及び3D)、そしてBrdUを取込まなかった(図10)ことからそれらは有糸分裂後のニューロンであることが示される。HB9+ニューロンは、RA暴露なしでHh−Ag1.3(1μM)により5日間処理されたEBsにおいて生じなかった(図3)。即ち、EBs内の運動ニューロンの生成はRAの尾部側化作用及びHhシグナルの腹側化作用の両方に依存する。
【0096】
有糸分裂後の運動ニューロンの分子特性をより詳細に査定するため、発明者らは、脊髄の運動ニューロンのサブセット:Isl1,Lhx3及びLim1により発現されるLIM HD蛋白質の発現を検査した(Tsuchida et al.,Topographic organization of embryonic motor neurons defined by expression of LIM homeobox genes.Cell,79:957-70,1994;Sharma et al.,LIM homeodomain factors Lhx3 and Lhx4 assign subtype identities for motor neurons.Cell,95:817-28,1998;Kania et al.,Coordinate roles for LIM homeobox genes in directing the dorsoventral trajectory of motor axons in the vertebrate limb.Cell,102:161-73,2000)(図3A)。3つの全てのLIM HD蛋白質はRA及びHhに暴露されたEBsにおいてHB9+運動ニューロンにより発現された。70%を超えるHB9+運動ニューロンがIsl1とLhx3を同時発現した(図3E及び3F)が、はるかに少ないフラクション(<5%)がLim1を同時発現した(図3G)。HB9+ニューロンは何れも後脳(hindbrain)の内臓の(visceral)運動ニューロンのマーカーPhox2bを同時発現しなかった(Pattyn et al.,Control of hindbrain motor neuron differentiation by the homeobox gene Phox2b.Development,127:1349-58,2000)。側面の(lateral)運動柱(LMC)ニューロンのプロフィールの指標である、HB9とLim1を同時発現する運動ニューロンの低い比率(Kania et al.,Coordinate roles for LIM homeobox genes in directing the dorsoventral trajectory of motor axons in the vertebrate limb.Cell,102:161-73,2000)は、運動ニューロンの吻側頸部のアイデンティティーと一致しており、Hox−c蛋白質発現のプロフィールにより明らかになった(Belting et al.,Multiple phases of expression and regulation of mouse Hoxc8 during early embryogenesis.J.Exp.Zool.,282:196-222,1998;Liu et al.,Assigning the positional identity of spinal motor neurons.Rostrocaudal patterning of Hox-c expression by FGFs,Gdf11,and retinoids.Neuron,32:997-1012,2001)。
【0097】
SHhは特別な(graded)様式にて作用するから、腹側の介在ニューロンは運動ニューロンの生成に十分な濃度よりほんの僅か低い濃度にて誘導される(Briscoe and Ericson,Specification of neuronal fates in the ventral tube.Curr.Opin.Neurobiol.,11(1):43-49,2001)。よって、発明者らは、腹側の介在ニューロンも1μMのHh−Ag1.3に暴露されたRA処理したEBsにおいて生じるか否かを検査した。Lhx3+,Chx10+V2介在ニューロン及びLim1/2+(V0及びV1同様)介在ニューロンがこれらのEBs内で検出された(図3G及び3H及び示されなかったデータ)。即ち、初期神経組織を用いたように(Briscoe and Ericson,Specification of neuronal fates in the ventral tube.Curr.Opin.Neurobiol.,11(1):43-49,2001)、EBs中の細胞が暴露されたHhシグナリングのレベルにおける小さなバリエーションが、腹側の介在ニューロン及び脊髄の運動ニューロンの両方の生成をもたらす。
【0098】
eGFP標識されたES細胞由来の運動ニューロンが生成されて単離されるかもしれない
RA及びHhシグナリングによりEBs内で誘導された腹側の神経の不均一性は、発明者らが、ES細胞由来の運動ニューロンを同定し、精製し、そして操作する方法を開発することを促した。よって、発明者らは、増強された緑色蛍光蛋白質(eGFP)の発現により同定可能な運動ニューロンを生じさせ得るES細胞系をデザインした。運動ニューロン特異的トランス遺伝子発現を授与するマウスHB9遺伝子の9−kbの5’領域の制御下でeGFPのcDNAを発現する、トランスジェニックマウス系を生成した(図4A)(Arber et al.,Requirement for the homeobox gene HB9 in the consolidation of motor neuron identity.Neuron,23:659-764,1999)。HB9とeGFPの発現のパターンを比較することにより、トランスジェニックファウンダーマウスをスクリーニングした。一つのマウス系列、mHB9−Gfp1bが高いレベルのeGFPをE9.5−P10マウスにて脊髄運動ニューロンの細胞ボディー内で、内生HB9と平行したパターンで発現したことがわかった(図4B−4G)。さらに、eGFP発現が高レベルで運動ニューロンの軸索及び樹状突起内に検出された(図4C及び示されなかったデータ)。ES細胞系(HBG3)をmHB9−Gfp1bトランスジェニックマウスから誘導した;これらの細胞は、生殖系を通してeGFP遺伝子を遺伝し、そして類似の運動ニューロン選択パターンにてeGFPの発現を指示した(データは示さず)。
【0099】
発明者らは、HBG3 ES細胞が、RA及びHh暴露の条件下で、他のマウスES細胞によく似た様式にて運動ニューロンに分化したことを発見した。Hh−Ag1.3なしでRA中に5日間生育させたHBG3 ES細胞から生じさせたEBsは、僅かなHB9+運動ニューロンを含み、これらの全てはそれらの身体及び神経突起中でeGFPを同時発現した(図5A)。eGFP+/HB9+運動ニューロンの数はHh−Ag1.3への暴露に際して増加し、濃度依存様式であった(図5B及び5C)。典型的には、eGFP+/HB9+運動ニューロンは、RA及び1μMのHh−Ag1.3へ暴露されたEBs中に存在する細胞の全数の20−30%を占めた(1000;図5C)。RA存在下におけるEBsのHh−Ag1.3(1μM)への暴露は、eGFP+/HB9+運動ニューロンを生じさせなかった(−1000;図5C)。即ち、HBG3 EB細胞は、RA及びHhシグナリング経路の活性化を通して十分な様式にてeGFP+/HB9+運動ニューロンを生じさせるために誘導することもできる。
【0100】
発明者らは、HBG3 ES細胞由来の運動ニューロンが胚由来の運動ニューロンの特徴であるインビトロの分化の局面を経験するか否かを試験するためにeGFP発現を使用した。RA−及びHh−暴露されたEBs(図5D)を単一の細胞又は小さな凝集体に分離し、そして神経栄養因子の存在下でマトリゲル基質上にプレートした。ES細胞由来の運動ニューロンは接着して長い突起を延ばし、eGFP発現により可視化できた(図5R−5F)。これらのES細胞由来の運動ニューロンの形態上の特徴はmHB9−Gfp1bマウスの胚脊髄から単離されたeGFP標識運動ニューロンの特徴と似ていた(図6J及び6K及び示さなかったデータ)。さらに、生じさせたeGFP+/HB9+運動ニューロンは、インビトロにてコリンアセチルトランスフラーゼを発現した(図5G及び5H)ことから、それらのコリン作用性神経伝達物質の状態の指標である。
【0101】
発明者らは、初期胚運動ニューロンとの直接の比較において、HBG3 ES細胞由来の運動ニューロンの神経栄養性因子依存性も検査した。発明者らは、RA及びHhに暴露されたHBG3−由来のEBsを単一細胞懸濁液へ分離し、そして蛍光活性化細胞分類(FACS)を用いて高いレベルのeGFPを発現する細胞を単離した(図6A)。平行して、発明者らは、E10.5 mHB9−Gfp1b胚の吻側頸部脊髄から初期胚運動ニューロンをFACSにより単離した(データは示さず)。何れかの源から、高レベルのeGFPを発現したニューロンの>98%はHBも発現し(図6B及び6C)、そしてこれらのニューロンは長い突起を延ばした(図6D、及び6E及び示さなかったデータ)。HBG3 ES細胞から生じさせた(図6F−6I及び6L)か又はE10.5 mHB9−Gfp1b胚から単離した(図6J−6L)精製されたeGFP+/HB9+運動ニューロンの生存は、神経栄養性因子に対する類似の依存性を呈した(例えば、Bloch-Gallego et al.,Survival in vitro of motoneurons identified or purified by novel antibody-based methods is selectively enhanced by muscle-derived factors.Development,111:221-32,1991;及びCamu et al.,Purification of embryonic rat motoneurons by panning on a monoclonal antibody to the low-affinity NGF receptor.J.Neurosci.Methods,44:59-70,1992を参照)。
【0102】
ES細胞由来のeGFP+運動ニューロンは胚脊髄内で分化する
HBG3 ES細胞由来の運動ニューロンのインビトロ生存及び分化は、発明者らが、インビボにて脊髄へ再導入された場合にこれらのニューロンの挙動を検査することを促した。RA及びHh−Ag1.3暴露の3−4日後にEBsを単離して(有糸分裂後の運動ニューロンが最初に出現したとき)、小凝集体に分散させ、そしてHHステージ15−17のチキン胚の脊髄の将来の吻側(又は尾部側)頸部、胸部又は腰部レベルに移植した(図7A)が、当該ステージは内生運動ニューロンの分化が始まるステージである(Hollyday et al.,An antoradiographic study of the formation of the lateral motor column in the chick embryo.Brain Res.,132:197-208,1977)。操作された胚をさらに3−7日間、HHステージ27−36まで発生させた。
【0103】
HHステージ27において操作された胚の脊髄の分析は、多数のeGFP+/HB9+運動ニューロンの存在を明らかにした(図7B−7J及び7O−7Q)。EBグラフトの最初の背腹の置き換えは制御されなかったが、発明者らは、3−7日後に検査したときに、チキンの脊髄内のマウスの運動ニューロン及び介在ニューロンの位置において著しい分離を検出した(図7E)。ほとんど一定して、eGFP+/HB9+運動ニューロンが内生運動ニューロンのそれに特徴的な腹側両面の位置内に見いだされたが、EB由来のLim2+介在ニューロン(齧歯類特異的抗体により認識される)は脊髄の背腹軸に沿って散在した(図7E)。莫大なeGFP+/HB9+運動ニューロンが、グラフティングのセグメントレベルの除去にも拘わらず、Lhx3を同時発現し、そしてLim1の発現を欠いた(図7F及び示さなかったデータ)。即ち、インビトロにおいて観察されたeGFP+/HB9+のES細胞の運動ニューロンはインビボグラフティングにより変化しなかったことが明白である。
【0104】
Olig2+運動ニューロン前駆体がグラフト化されたEBs内に存在したことから、インサイチュにて末期の分化を受けたグラフト化前駆細胞由来のチキンの脊髄の中に見いだされた幾つか又は全部のeGFP+運動ニューロンであり得る。従って、発明者らは、インビトロにおいてHBG3細胞から生じさせた有糸分裂後運動ニューロンがインビボにおいて脊髄の中で生存することもできるか否かを試験した。eGFP+/HB9+のES細胞由来の運動ニューロンを分離したEBsからFACS分類により単離して、次に、チキンの脊髄にグラフト化した。3日後、eGFP+/HB9+運動ニューロンが脊髄の腹側領域内に見いだされた(図7C及び7D)。即ち、有糸分裂後のES細胞由来の運動ニューロンはインビボにおいて生存も分化もできる。
【0105】
特定の幹細胞集団が体細胞との融合を受けることが報告された(Terada et al.,Bone marrow cells adopt the phenotype of other cells by spontaneous cell fusion.Nature,416:542-45,2002;Ying et al.,Changing potency by spontaneous cell fusion.Nature,545-48,2002)ことから、発明者らが、HBG3 ES細胞由来のeGFP+/HB9+運動ニューロンがチキンの脊髄のニューロンと体細胞ハイブリッドを形成したかもしれない可能性を試験することを促した。発明者らは、eGFP+/HB9+運動ニューロン細胞ボディーがマウスオリジンのニューロン表面蛋白質、例えば齧歯類特異的抗体により認識される−NCAM(図7G)を発現したが、チキンオリジンの表面蛋白質、例えばチキン特異的抗体により認識されるSCI(図7I)は発現しなかったことを発見した。同様に、グラフト化された運動ニューロンの軸索はマウスの表面蛋白質を発現したが、チキンの表面蛋白質は発現しなかった(図7H及び7I)。これらの発見は、体細胞融合が起こることに対する直接の証拠を提供する。
【0106】
追加の対照として、発明者らは、インビトロにおけるRAへの暴露によりニューロン化及び尾部側化されたが、抗Hh抗体(5E1;30μg/ml)の注入により腹側化を妨げられたHBG3 EBsのグラフトの運命を検査した。多数のマウスNCAM+細胞及びLim2+介在ニューロンが見いだされたが、eGFP+/HB9+運動ニューロンは検出されなかった(図7K−7N)。即ち、RAに暴露されたEBs内に存在する細胞は、宿主チキン組織により提供されたシグナルへの暴露に際してインビボの運動ニューロンの運命には向けられなかった。
【0107】
さらに、発明者らは、HBG3 ES細胞由来の運動ニューロンが脊髄の外側に伸びるか否かを検査した。多数のeGFP+軸索が腹側の根(roots)に検出された(図7B,7C,7D及び7O)。HHステージ27の胚のeGFP+運動軸索の末梢突起のパターンの分析は、HBG3 ES細胞由来の運動ニューロンが吻側頸部レベルにてグラフト化されて、軸索筋肉組織に突起したこと(データは示さず)、胸部レベルでグラフト化された運動ニューロンが軸索及び体壁筋肉組織の両方に対して突起したこと(図7O及び7P)、そして四肢のレベル(尾部側頸部又は腰部脊髄)でグラフト化された運動ニューロンが四肢の背側及び腹側の二等分に軸索を突起させたことを明らかにした(図7Q及び示さなかったデータ)。即ち、ES細胞由来の運動ニューロンは、脊髄の異なるセグメントレベルを集団化し(populate)、そしてインビボにて宿主体細胞運動ニューロンにより選択された主要な末梢軸索経路に沿って突起することが可能である。
【0108】
発明者らは、eGFP+運動ニューロンの軸索が筋肉標的に到達することも発見した。例えば、eGFP+運動軸索の広範囲な内部への成長及び分枝化が、HHステージ30−35において肋間筋肉中に検出された(図8A)。発明者らは、これらのeGFP+運動軸索が筋肉接触の部位において末端シナプスの分化の側面を呈するか否かを検査した。特に、発明者らは、神経の末端分化の3つのマーカー:齧歯類特異的抗体により認識されるシナプトブレビン(Syb);齧歯類特異的抗体により認識される小胞性(vesicular)AChトランスポーター(VAChT);及び齧歯類及びチキンの両方の蛋白質を検出する抗体により認識されるシナプトタグミン(Syn)を分析した。筋肉と接触した部位において、eGFP+運動軸索ブランチが前シナプス分化(specializations)のサインを示したことが、Syb,Syn及びVAChTの局所(focal)発現により明らかにされた(図8B−8E)。
【0109】
発明者らは、次に、eGFP+運動軸索の前シナプス末端分化がACh受容体の分配を分析することにより筋肉表面膜の後シナプス分化と整列化された(aligned)か否かを検査した。eGFP+運動軸索の末端分化の多くが筋肉表面上でACh受容体の局所クラスターと整列化されたことが、ローダミン−α−ブンガロトキシン(Rh−α−BTX)標識により定義された(図8F)。即ち、ES細胞由来の運動ニューロンの軸索は、インビボにおいて標的の骨格筋によりシナプスを形成することが明白であった。
【0110】
上で詳述されたとおり、発明者らは、インビボにおいて特定のニューロンのサブタイプへの神経前駆細胞の変換に関与する、細胞外誘導性シグナル及び転写因子の描写が、CNSニューロンの特定のクラスへのES細胞の分化に対する、発生学に基づく合理的なアプローチを可能にさせる。発明者らの発見は、高い効率で脊髄運動ニューロンを生じさせることが可能なこと、及びES細胞からの運動ニューロンの生成の経路がインビボにおいて運動ニューロンの生成の工程を再現することを示す。ES細胞由来の運動ニューロンは、インビボにて腹側脊髄を再増殖し、軸索を末梢に伸長し、そして筋肉標的によりシナプスを形成する。これらの研究は、正常な発生シグナリングカスケード、特に細胞外誘導性シグナルの調節へ洞察を適用する可能性を確立することにより、CNSニューロンの特定のクラスへのES細胞の分化を指示する。ES細胞由来の運動ニューロンが標的筋肉細胞に神経を分布させる能力は、脊髄の損傷及び運動ニューロンの変性疾患の哺乳類モデルにおいて運動機能を回復させるための、そのようなニューロンの使用の系統的評価の可能性を提供する。
【0111】
ES細胞からの運動ニューロン生成の経路
インビボの運動ニューロンの運命の特異化は、3つの連続する工程:(a)外胚葉細胞の初期神経化;(b)神経細胞の二次的尾部側化;及び(c)尾部側化された神経細胞の腹側化にあると考えることができる(Jessell et al.,Neuronal specification in the spinal cord:inductive signals and transcriptional codes.Nat.Rev.Genet.,1:20-29,2000;Wilson et al.,Neural induction:toward a unifying mechanism.Nat.Neurosci.,補遺4:1161-1168,2001)。本明細書中で論じられた、発明者らの結果は、ES細胞の運動ニューロンへの分化の経路が神経細胞の尾部側化及び腹側化に関与する通常のプログラムに似ている証拠を提供する。
【0112】
それらの最終的な位置的アイデンティティーにも拘わらず、神経プレート細胞は最初に吻側特性を有するらしい(Munoz-Sanjuan et al.,Neural induction:the default model and embryonic stem cells.Nat.Rev.Neurosci.3:271-80,2002)。脊髄の特性の次の負担(imposition)は、軸に近い中胚葉により提供されるRA−媒介性シグナルを含むらしい(Muhr et al.,Convergent inductive signals specify midbrain,hindbrain,and spinal cord identity in gastrula stage chick embryos.Neuron,23:689-702,1999;Wilson et al.,Neural induction:toward a unifying mechanism.Nat.Neurosci.,補遺4:1161-68,2001)。特に、RAは中脳又は吻側後脳アイデンティティーを犠牲にして脊髄の特性を特定することが示された(Muhr et al.,Convergent inductive signals specify midbrain,hindbrain,and spinal cord identity in gastrula stage chick embryos.Neuron,23:689-702,1999)。発明者らは、PA6細胞活性への暴露により神経化されたES細胞が中脳の位置的特性を発現する(Kawasaki et al.,Induction of midbrain dopaminergic neurons from ES cells to PA6 cell activity for 5 days induced activity.Neuron,28:31-40,2000も参照)が、RAへの暴露に際して頸部脊髄位置アイデンティティーに変換できることを発見した。この研究において記載された運動ニューロン生成におけるRA暴露のための要求は、中脳のドーパミン作用性神経分化のRAが媒介する阻害により補われる(Kawasaki et al.,Induction of midbrain dopaminergic neurons from ES cells to PA6 cell activity for 5 days induced activity.Neuron,28:31-40,2000)。合わせて、これらの発見は、神経管の吻側尾部側の軸をパターン化することにおけるその通常の役割に平行した様式にて、神経化されたEBs内でRAが脊髄の位置的アイデンティティーを促進させるという見解を支持する。
【0113】
EBs中でRAにより誘導された脊髄細胞が何故吻側頸部の位置的アイデンティティーを有するのであろうか?これに対するもっともらしい理由は、一般的脊髄前駆体のアイデンティティーの特異化の後の時点で、脊髄細胞の吻側尾部側の位置的アイデンティティーが進行中のRAシグナリングにより影響され続けるという発見から来る(Liu et al.,Assigning the positional identity of spinal motor neurons.Rostrocaudal patterning of Hox-c expression by FGFs,Gdf11,and retinoids.Neuron,32:997-1012,2001)。特に、RAが媒介するシグナリングは、胸部及び腰部の位置的アイデンティティーの犠牲として吻側頸部脊髄の位置的特性の獲得を促進する(Liu et al.,Assigning the positional identity of spinal motor neurons.Rostrocaudal patterning of Hox-c expression by FGFs,Gdf11,and retinoids.Neuron,32:997-1012,2001)。即ち、神経化されたES細胞のRAへの暴露は、最初の脊髄の位置的特性の創発(emergence)、及びこれらの細胞の吻側頸部アイデンティティーへの限定の両方の説明となるらしい。
【0114】
脊髄の前駆体アイデンティティーを獲得したES細胞も、運動ニューロン前駆体特異化の通常の経路をたどるらしい。これに関する証拠の一つのラインは、Hhシグナリングに対するそれらの細胞の感受性である。インビボにおけるような(Chiang et al.,Cyclopia and defective axial patterning in mice lacking Sonic Hedgehog gene function.Nature,383,407-13,1996)、ES細胞由来の脊髄前駆体からの運動ニューロンの生成は、完全にHhシグナリングに依存する。即ち、追加されたHhの不在下での僅かな運動ニューロンの生成は、EBs内の低レベルの内生Hh活性の発現によりもたらされるらしく、おそらくは、IHh又はSHhにより媒介される(Byrd et al.,Hedgehog is required for murine yolk sac angiogenesis.Development,129:361-72,2002;Dyer et al.,Indian hedgehog activates hematopoiesis and vasculogenesis and can respecify prospective neurectodermal cell fate in the mouse embryo.Development,128:1717-30,2001)。逆に、EBs内でHhシグナリングのレベルを上昇させると、運動ニューロン生成を顕著に増強する。
【0115】
証拠の第2のラインは、Hhシグナリングにより導き出される前駆体転写因子の発現のプロフィールにより提供される。Hhシグナリングの不在下では、EBs内の脊髄前駆体は、背及び中間の神経前駆体アイデンティティーを呈する。Hhシグナリングの活性化は初期脊髄前駆細胞の応答を厳密に模倣する濃度依存性様式において背側前駆体決定基の発現を失わせる(Briscoe et al.,Specification of neuronal fates in the ventral axis of the neural tube.Curr.Opin.Neurobiol.,11:43-49,2001)。さらに、高レベルのHhシグナリング活性はNkx6.1とOlig2の発現を誘導し−当該2つの前駆細胞転写因子はインビボにおいては深く運動ニューロン生成にリンクする(Sander et al.,Ventral neural patterning by Nkx homeobox genes:Nkx6.1 controls somatic motor neuron and ventral interneuron fates.Genes Dev.,14:2134-39,2000;Vallstedt et al.,Different levels of repressor activity assign redundant and specific roles to Nkx6 genes in motor neuron and interneuron specification.Neuron,31:743-55,2001;Novitch et al.,Coordinate regulation of motor neuron subtype identity and pan-neuronal properties by the bHLH repressor Olig2.Neuron,31:773-89,2001;Mizuguchi et al.,Combinatorial roles of olig2 and neurogenin2 in the coordinated induction of pan-neuronal and subtype-specific properties of motoneurons.Neuron,31:757-71,2001)。合わせると、これらの発見は、神経化ES細胞が初期神経プレート細胞の挙動によく似た様式で吻側及び背腹側パターン化シグナルの両方に応答し、最初に脊髄前駆細胞を生じ、そして次に運動ニューロン前駆体を生じることを示す。
【0116】
発明者らの研究は、ES細胞からの神経分化の初期の相に関与する工程を扱ってこなかった。インビボでは、神経の誘導が阻害性(blockade)BMPシグナリングに依存するらしく(Munoz-Sanjuan et al.,Neural induction,the default model and embryonic stem cells.Nat.Rev.Neurosci.,3:271-80,2002)、そして少なくともチキンにおいてはFGFsへの暴露に依存するらしい(Streit et al.,Initiation of neural induction by FGF signaling before gastrulation.Nature,406:74-78,2000;Wilson et al.,An early requirement for FGF signalling in the acquisition of neural cell fate in the chick embryo.Curr.Biol.,10421-429,2000;Wilson et al.,The status of Wnt signalling regulates neural and epidermal fates in the chick embryo.Nature,411:325-30,2001)。最近の研究は、ES細胞中の神経誘導におけるBMPシグナリングの阻害のための役割を示唆している(Gratsch et al.,Noggin and chordin have distinct activities in promoting lineage commitment of mouse embryonic stem (ES) cells.Dev.Biol.,245:83-94,2002;Tropepe et al.,Direct neural fate specification from embryonic stem cells:a primitive mammalian neural stem cell stage acquired through a default mechanism.Neuron,30:65-78,2001)。しかしながら、ES細胞における神経特異化の初期の工程がインビボにおいて神経誘導の経路に如何にして順応するかを決定するには、追加の研究が必要である。
【0117】
インビボにおけるES細胞由来の運動ニューロンの分化
幾つかの観察は、ES細胞由来の運動ニューロンも、それらの胚の類似物(counterpart)に似た様式にて挙動することを示す。最初に、eGFP+運動ニューロンが典型的には吻側脊髄に制限され、しばしば内生運動ニューロンの特徴である腹側面(ventrolateral)位置における。対照的に、ES細胞由来の脊髄介在ニューロンは脊髄の背腹軸のいたるところに散在する。これらの観察は、ES細胞由来の運動ニューロン及び介在ニューロンが宿主脊髄へのそれらの誘導後に分離の活動的なプロセスを受けることを示唆する。別の可能性は、腹側脊髄を増殖させられない運動ニューロンが、死ぬか又はeGFP発現を失うことである。このあとの可能性に対する一つの意見は、Nkx6又はMNR2蛋白質の発現の欠如(misexpression)に応答して背側脊髄中で生じた運動ニューロンが正規の場所ではない背側の位置において生存及び分化しないことである(Tanabe et al.,Specification of motor neuron identity by the MNR2 homeodomain protein.Cell,95:67-80,1998;Briscoe et al.,A homeodomain protein code specifies progenitor cell identity and neuronal fate in the ventral neural tube.Cell,101:435-45,2000)。
【0118】
第2に、ES細胞由来の運動ニューロンは腹側の根(ventral root)を通して脊髄から軸索を突出させ、そして体細胞の運動ニューロンにより選択される(taken)主要な末梢経路を選択する。ES細胞由来の運動ニューロンの軸索の四肢(limb)への突起は注目に値するが、これらのニューロンのうち極めて僅かなものしか、四肢−突起LMCニューロンに適したLIM HDコードを発現しないからである(Tsuchida et al.,Topographic organization of embryonic motor neurons defined by expression of LIM homeobox genes.Cell,79:957-70,1994)。運動ニューロンのLIM HDコードと末梢軸索の軌道の間の類似のミスマッチが2つの他の実験コンテクストにおいて発見された。Lhx3の過剰発現は、脊髄の四肢レベルにおいて運動ニューロンに対してMMC分子アイデンティティーを授与する;しかし、これらの運動ニューロンの多くは四肢へ軸索を突出させる(Sharma et al.,Genetic and epigenetic mechanisms contribute to motor neuron pathfinding.Nature,406:515-19,2000)。さらに、脊髄の四肢レベルに位置した胸部レベルの運動ニューロンはなおも四肢に突出することができる(O'Braien et al.,Development and survival of thoracic motorneurons and hindlimb musculture following transplantation of the thoracic neural tube to the lumbar region in the chick embryo:anatomical aspects.J.Neurobiol.,21:313-40,1990)。即ち、ES細胞由来の運動ニューロンの軸索の四肢への突起は、運動ニューロンが末梢経路の変更されたか又は制限された選択に対向する(face)ような環境においてLIM HDコードが無効にされ(overridden)得るという事実を反映するらしい。
【0119】
第3に、標的筋肉へ到着すると、ES細胞由来の運動ニューロンの軸索は、前シナプス分化の多数のサイン、注目すべきはコリン作用性神経伝達物質の特性の発現を呈する。さらに、これらの前シナプス特異化が、後シナプス筋膜上のACh受容体の局所クラスターとの一列整列(in alignment with)において見いだされることから、ES細胞由来の運動ニューロンと骨格筋の間に形成されるシナプス接触が機能することが示唆される。合わせると、これらのインビボの研究は、脊髄中の新たに生じた運動ニューロンがその骨格筋標的に神経を分布させることを通して、ES細胞由来の運動ニューロンが通常の発生プログラムにて連続工程をうまく通り抜けることができることを示す。
【0120】
最後に、誘導性シグナルがES細胞を運動ニューロンに変換できることによる効果は、ES細胞が暴露される吻側尾部側及び背側腹側パターン化シグナルのアイデンティティーと濃度の系統的なバリエーションを通して、CNSニューロンの他の予め定義されたクラスを生じさせるための一般的な戦略を示唆する。ES細胞を遺伝的に操作する要求無しに、単に細胞外因子の使用を通して、細胞の運命を指示する可能性(例えば、Kyba et al.,HoxB4 confers definitive lymphoid-myeloid engraftment potential on embryonic stem cell and yolk sac hematopoietic progenitors.Cell,109:29-37,2002)は、この戦略のヒトES細胞及び神経前駆細胞の他のクラスへの直接の拡張を可能にするかもしれない。
【0121】
前記の発明は明確化及び理解のためにいくらか詳細に説明されたが、開示を読んで、当業者は、形式及び細部の様々な変化が特許請求の範囲の発明の真の範囲を逸脱することなくなされ得ることを認識する。
【図面の簡単な説明】
【0122】
【図1】図1は、神経化された胚性細胞においてレチノイン酸が尾部側の位置のアイデンティティーを誘導することを示す。(A)脊髄の運動ニューロンアイデンティティーの明確化の連続する工程。神経誘導性シグナルは初期(primitive)外胚葉細胞を神経の運命に変換する。神経誘導の初期の相の間は、前駆細胞が腹側の(anterior)アイデンティティーを発現する。レチノイン酸(RA)を含む外来性のシグナルは、吻側の神経プレート細胞を進行性のより尾部側のアイデンティティーへ変換する。尾部側の神経前駆体はソニックヘッジホッグ(SHh)シグナリングの影響下では運動ニューロンに変換される。(B)RA(2μM)の存在又は不在下で生育された胚のボディー(EBs)内の全神経(pan-neural)及び吻側尾部側(rostrocaudal)のマーカーの発現。初期神経プレートマーカーSox1の発現が、RA存在下で3日間培養されたEBsにおいて増加した。otx2発現は、RA不在下で3日間生育させたEBsにおいて存在したが、RA存在下では消失した。Hoxc6はRAへ5日間暴露されたEBsにおいて発現した。NeuN発現により特徴付けされた多数の有糸分裂後のニューロンがRA存在下でEBsを5日間生育させた時に、観察された。(C)PA6細胞上で6日間培養されたES細胞はニューロンマーカー、Sox1を発現した。多くのSox1+細胞がEn1を同時発現したが、Hoxc6を発現しなかった。多くのTuJ1+ニューロンがOtx2を同時発現した。RA(2μM)のPA6細胞上で生育させたES細胞への添加はEn1及びOtx2発現の消滅、及び前駆細胞及びTuJ1+有糸分裂後のニューロン内でのHoxc6の発現をもたらした。これらニューロンの多くはHB9を発現したため、運動ニューロンであった(図9参照)。
【図2】図2は、胚ボディー(EBs)内の神経前駆細胞のヘッジホッグ−依存性腹側化を示す。(A)脊髄の運動ニューロン生成のSHh−活性化転写経路の模式的概要。運動ニューロンの生成を阻害する蛋白質はlrx3及びNkx2.2である。残りの蛋白質は運動ニューロンの生成に必要である。(B)E9.5マウス胚の尾部側の神経管内のHD及びbHLH転写因子の発現のパターン。運動ニューロンを生じる前駆体ドメインを、Olig2,Nkx6.1の同時発現及び低レベルのPax6により特徴付けした。(C)RA(2μM)単独又は10nM又は1μMのHh−Ag1.3共存下の何れかでの、3日間生育させたES−細胞由来のEBsにおける転写因子発現のプロフィール。(D)RA及び異なる濃度のHh−Ag1.3の存在下でのEBs内の転写因子発現の定量化。描写されたのは、各条件下でアッセイされた8つのEBsからの、セクションあたりの、平均±s.e.m.の値である。
【図3】図3は、胚ボディー(EBs)内の運動ニューロン及び介在ニューロン転写因子の発現を描写する。(A)HB9及びLhx3の同時発現は新たに生成された運動ニューロンを定義した。Lhx3は単独でV2介在ニューロンにより発現された。HB9とIsl1の同時発現は運動ニューロンのサブセットを定義した。幾つかの側面(lateral)運動柱(LMC)ニューロンはIsl1不在下で高いレベルのHB9を発現した;対照的に、他のLMCニューロンはIsl1を発現したが、極めて低レベルでHB9を発現したにすぎなかった。Isl1は幾つかの背面側の介在ニューロンによっても発現された。HB9とLin1の同時発現は、側面のLMCニューロンを定義した。多数の介在ニューロンがLim1も発現した。(B)Hh−Ag1.3(1μM)の存在下であるがRA不在下で5日間生育させたEBsはHB9+ニューロンを含まなかった。RA(2μM)の存在下であるがHh−Ag1.3不在下で5日間生育させたEBsはHB9+ニューロンを少量含んだ。RA及び抗−Hh抗体(mAb 5E1;30μg/ml)の存在下で5日間生育させたEBsは、HB9+ニューロンを含まなかった。RA及び組換えSHh−N(300nM)存在下で5日間生育させたEBsは、多くのHB9+ニューロンを含んだ。(C)RA及びHh−Ag1.3(1μM)存在下で5日間生育させたEBsのニューロン内のHB9及びNeuNの同時発現。(D)RA存在下で5日間生育させて、さらに2日間培養されたEBsの分離により得られたTuJ1+によりの核内のHB9の局在。HB9発現を欠いた他のニューロンも存在した。(E)RA及びHh−Ag1.3(1μM)存在下で5日間生育させたEBs内の運動ニューロン内のHB9とLhx3の同時発現。Lhx3のみを発現した細胞はV2介在ニューロンであった。(F)RA及びHh−Ag1.3(1μM)存在下で5日間生育させたEBs内の運動ニューロン内のHB9とIsl1の同時発現。(G)RA及びHh−Ag1.3(1μM)存在下で5日間生育させたEBs内の少数のニューロン内でHB9とLim1が同時発現された。Lim1のみを発現した細胞は介在ニューロンであった。(H)RA及びHh−Ag1.3(1μM)存在下で5日間生育させたEBs内のV2介在ニューロン内のChx10とLhx3の同時発現。
【図4】図4は、トランスジェニックマウスの胚の脊髄内のeGFP−標識された運動ニューロンを示す。(A)mHB9−Gfp1bトランスジェニックマウスの系の生成のためのコンストラクト。(B)E10.5 mHB9−Gfp1bマウス胚における増強された緑色蛍光蛋白質(eGFP)発現のパターン。eGFPは、腹側の脊髄及び脊髄から伸びた運動神経において高度に発現された。(C)eGFP発現が、E10.5mHB9−Gfp1b胚の腹側角内の運動ニューロン細胞ボディー及び樹上突起中に検出された。eGFP発現は、腹根により脊髄を興奮させる運動ニューロン軸索内でも検出された。このラインにおいては、E10.5において、後ではなく、極めて低い発現が背根のガングリオンのニューロン(運動ニューロン内のそれより<20倍)及び腹側の介在ニューロン(図4C及び4E中の矢印)において検出された。(D)最初の有糸分裂後の運動ニューロンが生成されたとき(E9.5)の、mHB9−Gfp1b胚の前肢レベルの脊髄内のHB9及びeGFPの発現のパターン。(E−G)E10.5 mHB9−Gfp1b胚の胸部脊髄内のHB9及びeGFPの発現のパターン。eGFPは運動ニューロン内でHB9と共に発現された。空洞ゾーンの近くに位置する新たな有糸分裂後の運動ニューロンは、脊髄の側面領域内の成熟運動ニューロンよりも低いレベルのeGFPを発現したことに注目されたい。
【図5】図5は、HBG ES細胞にインビトロで由来するeGFP−標識された運動ニューロンを描写する。(A)RA存在下で5日間生育させた胚ボディー(EBs)内のHB9+,Isl1+運動ニューロンは、細胞ボディー及び軸索内でeGFPを発現した。(B)eGFP+運動ニューロンの数はHh−Ag1.3(1μM)への暴露に際して増加した。(C)RA及び濃度を増加させたHh−Ag1.3(0から22μM)の存在下で5日間生育させたEBs内のeGFP+運動ニューロンの定量化。2.2μMのHh−Ag1.3において、EBs中の22.5%の細胞がeGFPを発現した。高い濃度(10μM)においては、hh−Ag1.3はEBsにおける細胞の分化に有害となった。「−1000」と記されたデータ点は、1μMのHh−Ag1.3存在下であるがRA不在下で生育させたEBsの培養物を意味する。(D)RA及びHh−Ag1.3(1μM)存在下で5日間生育させたEBs内でのeGFP発現を示す、明視野及び蛍光イメージ。(E)RA及びHh−Ag1.3(1μM)存在下で5日間生育させ、そしてさらに2日間マトリゲル基質上にプレートされた胚ボディー内でのeGFP発現を示す相及び蛍光イメージ。多数のeGFP+運動軸索がこれらの凝集物から出現した。(F)運動ニューロンから伸びた神経をeGFPが標識することを示す相及び蛍光イメージ。(G,H)7日間インビトロにおいて生育させたEBs−細胞由来の運動ニューロン内のHB9,eGFP,及びコリンアセチルトランスフェラーゼ(ChAT)の同時発現。HB9及びeGFP発現を欠いたChAT+ニューロンはおそらく介在ニューロンであった。
【図6】図6は、eGFP+HBG3 ES−細胞由来の運動ニューロンの単離を示す。(A)Hh−Ag1.3(1μM)単独、RA(1μM)単独、又は当該2つの因子の組み合わせの存在下で分化した、HBG3 ES細胞のFACS分析(細胞密度プロット)。高いレベルのeGFPを発現する運動ニューロンを分離するために使用されたゲートは、ダッシュの長方形により記した。(B,C)FACS分離により単離された、精製されたES−細胞運動ニューロンは高いレベルのeGFPを発現した。(D,E)24時間マトリゲル上で生育させた場合の、eGFP+ニューロン及び伸びた神経突起。(F,G)神経栄養因子(NT3,BDNF,CNTF,及びGDNF)の存在下で培養された、FACS分離されたES−細胞由来の運動ニューロンは、インビトロにおいて3日間生存し、そして突起(processes)を念入りに作る(elaborate)(F)。生存した運動ニューロンはeGFPの発現を維持した(G)。(H,I)神経栄養因子の不在下で培養したほとんどのFACS分離されたES−細胞由来の運動ニューロンはインビトロにおいて3日以内に死んだ。(J,K)FACS分離された初期吻側頸部運動ニューロンは、ES−細胞由来の運動ニューロンのそれと同じ様式で挙動した。神経栄養因子の不在下では、ニューロンは生存してeGFP+突起を念入りに作った。(L)神経栄養因子の存在下(NT)又は不在下(対照)での培養物中の3日後の初期頸部運動ニューロン(cMN)及びES−細胞由来の運動ニューロン(ES−MN)生存の定量化(プレートされた細胞のパーセンテージ)。平均±s.e.m.;6ウエルをスコアした。
【図7】図7は、インビボにおける移植されたES−細胞由来の運動ニューロンの完成(integration)を描写する。(A)HBG3 ES−細胞由来の運動ニューロン富裕化EBsの、ハンブルガー−ハミルトン(HH)ステージの15匹のチキン脊髄への移植を示す模式的ダイアグラム。レシピエントのチキンの胚の脊髄を吸引により損傷させ、そして約1/2−1の部分的に粉砕されたEBをガラス製のマイクロピペットを用いて確実なスペースに移植した。(B)HHステージ27の宿主のチキン胚の胸部及び腰部の脊髄内のeGFP+運動ニューロン(運動ニューロン富裕化されたEBsをHHステージ27において移植した)の位置を示す明視野/蛍光イメージ(腹側の視野)。脊髄から出現するeGFP+軸索に注目。(C,D)FACS−分離された運動ニューロンの(HHステージ16の胚に移植)ステージ27における胸部脊髄内の位置。横断面は、腹側の脊髄中のeGFP+運動ニューロンを明らかにした(D)。(E−J)ステージ15における運動ニューロン富裕化EBsの片側のみの移植後の、吻側頸部レベルのHHステージ27のチキンの脊髄の横断面図。運動ニューロンは脊髄の腹側角において主に見いだされ、マウス特異的Lim2抗体により標識された移植された介在ニューロンから分離される(E)。多数のES−細胞由来の運動ニューロンがeGFPとLhx3を同時発現した(F)。移植されたマウスES−細胞由来の運動ニューロン(G)及び軸索(矢印、H)を齧歯類特異的NCAM抗体で標識したが、それらはチキン運動ニューロンマーカー蛋白質、SCIを発現しなかった(I,J)。eGFP−,NCAM+の軸索は床板(floor plate)を横切ったが、脊髄から突き出なかった(矢印、G,H)。(K−N)HHステージ16において尾部側化されたEBsの移植物(RA(2μM)及び抗−Hh抗体(5E1,30μg/ml)の存在下で生育させた)を受けた、HHステージ27における胸部の脊髄の横断面図。非マウス由来の運動ニューロンが、eGFP発現(K)によるか又はマウス特異的HB9抗体(L)の何れかにより、検出された。対照的に、多数のマウス由来のNCAM+(M)及びLim2+(N)ニューロンが存在した。(O−Q)ステージ16−17における運動ニューロン富裕化EBsの移植物を受けた、胸部(O,P)、又は腰部(Q)レベルにおけるHHステージ27のチキン脊髄の横断面図。eGFP+運動ニューロン細胞ボディーが腹側脊髄において濃縮された。グラフト化された組織内の幾つかの異所性のeGFP+運動ニューロンは脊髄のルーメン内にトラップされたままであった。eGFP+軸索は脊髄を興奮させたが、主に、腹根を通してであり、そして軸索(O−Q)、ボディーの壁(O,P)及び背側及び腹側の肢(Q)を供給する全ての主要な神経ブランチに沿って突き出た。全ての末梢の軸索の経路は神経繊維(NF)発現により検出された。前ガングリオン運動軸索により占められた腹側正中に並んだ神経内のeGFP+軸索の不在に注目。
【図8】図8は、インビボにおけるES−細胞由来の運動ニューロンのシナプス分化を示す。(A)HHステージ35のチキン胚胸郭のホールマウント(wholemount)調製。ES−細胞由来のeGFP+軸索は肋間筋を脈状にしなかった(innervated)。(B,C)神経の部位のeGFP+軸索の末端のシナプトブレビン(Syb)と小胞性のACh輸送体(VAChT)が筋肉と接触する。抗−Syb及びVAChT抗体はマウスの蛋白質を認識したが、チキンの蛋白質を認識しなかった。(D)肋間筋を供給する5つの神経ブランチ内の神経繊維(NF)とeGFPの発現。幾つかのeGFP+軸索はNF発現を欠いたことに注目されたい。α−ブンガロトキシン(αBTX)発現により定義されたとおり、eGFP+軸索の末端はACh受容体クラスターと同時に生じた(coincided)。(E)eGFP+運動軸索におけるシナプトタグミン発現とαBTX標識の一致(coincidence)。(F)eGFP+運動軸索におけるSyb発現とαBTX標識の一致。
【図9】図9は、PA6細胞上で生育した尾部側化されたES細胞は運動ニューロンに分化することを示す。(A,B)10日間PA6細胞上で生育したES細胞はHB9+上でニューロンに分化しなかった(A)。2μMのRAの2日目の培養培地への添加は、多数のHB9+上でニューロンの生成、並びにOlig2−発現前駆体の生成をもたらした(B)。(C)2μMのRA存在下で10日間PA6細胞上で生育した運動ニューロン含有ES細胞コロニーの定量。平均±s.d.;3ウエル。
【図10】図10は、ES内の運動ニューロン前駆体がBrdUを取込むことを示す。(A−C)固定化前の1時間のBrdU標識(20μg/ml)は、RA及びHh−Ag1.3存在下で3日間生育したES内の多数のBrdU+,Olig2+前駆体を明らかにした。5日間生育したES内のHB9+運動ニューロンはBrdUを取り込まなかった(B)が、少数のBrdU+,Olig2+は存在した(C)。
【特許請求の範囲】
【請求項1】
胚性幹細胞を、分化した神経細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子及び背側化又は腹側化胚性シグナリング因子に接触させることを含む、胚性幹細胞の分化した神経細胞への分化を誘導する方法。
【請求項2】
胚性幹細胞がマウスの胚性幹細胞である、請求項1記載の方法。
【請求項3】
胚性幹細胞がヒトの胚性幹細胞である、請求項1記載の方法。
【請求項4】
吻側化胚性シグナリング因子がレチノイン酸又はWntである、請求項1記載の方法。
【請求項5】
尾部側化胚性シグナリング因子がレチノイン酸又はWntである、請求項1記載の方法。
【請求項6】
背側化胚性シグナリング因子がBMP又はWntである、請求項1記載の方法。
【請求項7】
腹側化胚性シグナリング因子がヘッジホッグシグナリング経路の活性化因子である、請求項1記載の方法。
【請求項8】
ヘッジホッグシグナリング経路の活性化因子がヘッジホッグ蛋白質又はヘッジホッグシグナリングのアゴニストである、請求項1記載の方法。
【請求項9】
ヘッジホッグシグナリング経路の活性化因子がSHh又はHh−Ag1.3である、請求項1記載の方法。
【請求項10】
背側化又は腹側化胚性シグナリング因子に接触させる前に、胚性幹細胞を吻側化及び/又は尾部側化胚性シグナリング因子に接触させる、請求項1記載の方法。
【請求項11】
背側化又は腹側化胚性シグナリング因子に接触させた後に、胚性幹細胞を少なくとも一つの神経栄養因子と接触させることをさらに含む、請求項10記載の方法。
【請求項12】
分化した神経細胞が、星状細胞、ニューロン、乏突起神経膠細胞、又はシュワン細胞からなる群から選択される、請求項1記載の方法。
【請求項13】
分化した神経細胞が運動ニューロンである、請求項1記載の方法。
【請求項14】
運動ニューロンが脊髄運動ニューロンである、請求項1記載の方法。
【請求項15】
分化した神経細胞が増強された緑色蛍光蛋白質(eGFP)を発現する、請求項1記載の方法。
【請求項16】
接触をインビボにて被験者において作用させる、請求項1記載の方法。
【請求項17】
胚性シグナリング因子を被験者に投与することにより、接触をインビボにて被験者において作用させる、請求項16記載の方法。
【請求項18】
接触をインビトロにて作用させる、請求項1記載の方法。
【請求項19】
分化した神経細胞を被験者に移植する工程をさらに含む、請求項18記載の方法。
【請求項20】
分化した神経細胞を被験者の脊髄に移植する、請求項19記載の方法。
【請求項21】
被験者が胚である、請求項19記載の方法。
【請求項22】
被験者がヒトである、請求項19記載の方法。
【請求項23】
被験者が神経組織の変性を有する、請求項19記載の方法。
【請求項24】
神経組織の変性が末梢神経障害又は神経変性疾患である、請求項23記載の方法。
【請求項25】
末梢神経障害が、筋委縮性側索硬化症(ALS)、神経の外傷、準新生物形成(paraneoplastic)疾患、ポリオ、ポリオ後症候群、進行性延髄麻痺、及び脊髄筋肉萎縮(SMA)からなる群から選択される症状に関連する、請求項24記載の方法。
【請求項26】
神経変性疾患が、アルツハイマー病、筋委縮性側索硬化症(ルーゲーリック病)、ビンスワンガー病、ハンチントンコレラ、多発性硬化症、重症筋無力症、パーキンソン病、及びピック病(Pick's disease)からなる群から選択される、請求項24記載の方法。
【請求項27】
請求項1記載の方法により生じた、分化した神経細胞。
【請求項28】
分化した神経細胞が増強された緑色蛍光蛋白質(eGFP)を発現する、請求項27記載の分化した神経細胞。
【請求項29】
分化した神経細胞を生産する方法であって、
(a)胚性幹細胞の培養物を得るか又は生じさせ;
(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;
(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させ;そして
(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させる
工程を含む方法。
【請求項30】
胚性幹細胞がマウスの胚性幹細胞である、請求項29記載の方法。
【請求項31】
胚性幹細胞がヒトの胚性幹細胞である、請求項29記載の方法。
【請求項32】
吻側化胚性シグナリング因子がレチノイン酸又はWntである、請求項29記載の方法。
【請求項33】
尾部側化胚性シグナリング因子がレチノイン酸又はWntである、請求項29記載の方法。
【請求項34】
背側化胚性シグナリング因子がBMP又はWntである、請求項29記載の方法。
【請求項35】
腹側化胚性シグナリング因子がヘッジホッグシグナリング経路の活性化因子である、請求項29記載の方法。
【請求項36】
ヘッジホッグシグナリング経路の活性化因子がヘッジホッグ蛋白質又はヘッジホッグシグナリングのアゴニストである、請求項35記載の方法。
【請求項37】
ヘッジホッグシグナリング経路の活性化因子がSHh又はHh−Ag1.3である、請求項35記載の方法。
【請求項38】
神経前駆細胞が、DRGニューロンの神経前駆細胞、運動ニューロンの神経前駆細胞、末梢ニューロンの神経前駆細胞、脊髄ニューロンの神経前駆細胞、及び腹側介在ニューロンの神経前駆細胞からなる群から選択される、請求項29記載の方法。
【請求項39】
分化した神経細胞が、星状細胞、ニューロン、乏突起神経膠細胞、又はシュワン細胞からなる群から選択される、請求項29記載の方法。
【請求項40】
分化した神経細胞が運動ニューロンである、請求項39記載の方法。
【請求項41】
分化した神経細胞が有糸分裂後である、請求項40記載の方法。
【請求項42】
運動ニューロンが脊髄運動ニューロンである、請求項40記載の方法。
【請求項43】
分化した神経細胞が増強された緑色蛍光蛋白質(eGFP)を発現する、請求項29記載の方法。
【請求項44】
工程(b)−(d)をインビボにて被験者において実施する、請求項29記載の方法。
【請求項45】
工程(b)、(c)、及び(d)をインビトロにて実施する、請求項29記載の方法。
【請求項46】
分化した神経細胞を被験者に移植する工程をさらに含む、請求項45記載の方法。
【請求項47】
分化した神経細胞を被験者の脊髄に移植する、請求項46記載の方法。
【請求項48】
神経前駆細胞を被験者に移植すること及び分化した神経細胞を被験者に移植することからなる群から選択される追加の工程をさらに含む、請求項45記載の方法。
【請求項49】
被験者が胚である、請求項48記載の方法。
【請求項50】
被験者がヒトである、請求項48記載の方法。
【請求項51】
被験者が神経組織の変性を有する、請求項48記載の方法。
【請求項52】
神経組織の変性が末梢神経障害又は神経変性疾患である、請求項51記載の方法。
【請求項53】
末梢神経障害が、筋委縮性側索硬化症(ALS)、神経の外傷、準新生物形成(paraneoplastic)疾患、ポリオ、ポリオ後症候群、進行性延髄麻痺、及び脊髄筋肉萎縮(SMA)からなる群から選択される症状に関連する、請求項52記載の方法。
【請求項54】
神経変性疾患が、アルツハイマー病、筋委縮性側索硬化症(ルーゲーリック病)、ビンスワンガー病、ハンチントンコレラ、多発性硬化症、重症筋無力症、パーキンソン病、及びピック病(Pick's disease)からなる群から選択される、請求項52記載の方法。
【請求項55】
請求項29記載の方法により生じた分化した神経細胞を含む、細胞の集団。
【請求項56】
分化した神経細胞が増強された緑色蛍光蛋白質(eGFP)を発現する、請求項55記載の分化した神経細胞。
【請求項57】
被験者において脊髄を再増殖させる方法であって、
(a)胚性幹細胞の培養物を得るか又は生じさせ;
(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;
(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させ;
(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させ;そして
(e)分化した神経細胞を被験者の脊髄に移植する
工程を含む方法。
【請求項58】
胚性幹細胞がマウスの胚性幹細胞である、請求項57記載の方法。
【請求項59】
胚性幹細胞がヒトの胚性幹細胞である、請求項57記載の方法。
【請求項60】
尾部側化胚性シグナリング因子がレチノイン酸である、請求項57記載の方法。
【請求項61】
腹側化胚性シグナリング因子がヘッジホッグシグナリング経路の活性化因子である、請求項57記載の方法。
【請求項62】
ヘッジホッグシグナリング経路の活性化因子がヘッジホッグ蛋白質又はヘッジホッグシグナリングのアゴニストである、請求項61記載の方法。
【請求項63】
ヘッジホッグシグナリング経路の活性化因子がSHh又はHh−Ag1.3である、請求項61記載の方法。
【請求項64】
神経前駆細胞が、DRGニューロンの神経前駆細胞、運動ニューロンの神経前駆細胞、末梢ニューロンの神経前駆細胞、脊髄ニューロンの神経前駆細胞、及び腹側介在ニューロンの神経前駆細胞からなる群から選択される、請求項57記載の方法。
【請求項65】
分化した神経細胞が、星状細胞、ニューロン、乏突起神経膠細胞、又はシュワン細胞からなる群から選択される、請求項57記載の方法。
【請求項66】
分化した神経細胞が運動ニューロンである、請求項65記載の方法。
【請求項67】
分化した神経細胞が有糸分裂後である、請求項66記載の方法。
【請求項68】
運動ニューロンが脊髄運動ニューロンである、請求項66記載の方法。
【請求項69】
分化した神経細胞が増強された緑色蛍光蛋白質(eGFP)を発現する、請求項57記載の方法。
【請求項70】
被験者が胚である、請求項57記載の方法。
【請求項71】
被験者がヒトである、請求項57記載の方法。
【請求項72】
被験者が神経組織の変性を有する、請求項57記載の方法。
【請求項73】
神経組織の変性が末梢神経障害又は神経変性疾患である、請求項72記載の方法。
【請求項74】
末梢神経障害が、筋委縮性側索硬化症(ALS)、神経の外傷、準新生物形成(paraneoplastic)疾患、ポリオ、ポリオ後症候群、進行性延髄麻痺、及び脊髄筋肉萎縮(SMA)からなる群から選択される症状に関連する、請求項73記載の方法。
【請求項75】
末梢神経障害がALS又はSMAである、請求項74記載の方法。
【請求項76】
治療の必要な被験者において神経組織の変性を治療する方法であって、
(a)胚性幹細胞の培養物を得るか又は生じさせ;
(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;
(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させ;
(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させ;そして
(e)神経組織の変性を治療するのに有効な量にて分化した神経細胞を被験者に移植する
工程を含む方法。
【請求項77】
神経組織の変性が末梢神経障害又は神経変性疾患である、請求項76記載の方法。
【請求項78】
末梢神経障害が、筋委縮性側索硬化症(ALS)、神経の外傷、準新生物形成(paraneoplastic)疾患、ポリオ、ポリオ後症候群、進行性延髄麻痺、及び脊髄筋肉萎縮(SMA)からなる群から選択される症状に関連する、請求項77記載の方法。
【請求項79】
神経変性疾患が、アルツハイマー病、筋委縮性側索硬化症(ルーゲーリック病)、ビンスワンガー病、ハンチントンコレラ、多発性硬化症、重症筋無力症、パーキンソン病、及びピック病(Pick's disease)からなる群から選択される、請求項77記載の方法。
【請求項80】
胚性幹細胞がマウスの胚性幹細胞である、請求項76記載の方法。
【請求項81】
胚性幹細胞がヒトの胚性幹細胞である、請求項76記載の方法。
【請求項82】
吻側化胚性シグナリング因子がレチノイン酸又はWntである、請求項76記載の方法。
【請求項83】
尾部側化胚性シグナリング因子がレチノイン酸又はWntである、請求項76記載の方法。
【請求項84】
背側化胚性シグナリング因子がBMP又はWntである、請求項76記載の方法。
【請求項85】
腹側化胚性シグナリング因子がヘッジホッグシグナリング経路の活性化因子である、請求項76記載の方法。
【請求項86】
ヘッジホッグシグナリング経路の活性化因子がヘッジホッグ蛋白質又はヘッジホッグシグナリングのアゴニストである、請求項85記載の方法。
【請求項87】
ヘッジホッグシグナリング経路の活性化因子がSHh又はHh−Ag1.3である、請求項85記載の方法。
【請求項88】
神経前駆細胞が、DRGニューロンの神経前駆細胞、運動ニューロンの神経前駆細胞、末梢ニューロンの神経前駆細胞、脊髄ニューロンの神経前駆細胞、及び腹側介在ニューロンの神経前駆細胞からなる群から選択される、請求項76記載の方法。
【請求項89】
分化した神経細胞が、星状細胞、ニューロン、乏突起神経膠細胞、又はシュワン細胞からなる群から選択される、請求項76記載の方法。
【請求項90】
分化した神経細胞が運動ニューロンである、請求項89記載の方法。
【請求項91】
分化した神経細胞が有糸分裂後である、請求項90記載の方法。
【請求項92】
運動ニューロンが脊髄運動ニューロンである、請求項90記載の方法。
【請求項93】
被験者が胚である、請求項76記載の方法。
【請求項94】
被験者がヒトである、請求項76記載の方法。
【請求項95】
工程:
(a)胚性幹細胞の培養物を得るか又は生じさせ;
(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;そして
(c)任意に、神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させること
により生じさせた神経前駆細胞。
【請求項96】
増強された緑色蛍光蛋白質(eGFP)を発現する、請求項95記載の神経前駆細胞。
【請求項97】
神経の発生、機能、及び死を分析すること、又はシナプスの分化を監視することにおける、請求項95記載の神経前駆細胞の使用。
【請求項98】
工程:
(a)胚性幹細胞の培養物を得るか又は生じさせ;
(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;
(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させ;そして
(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させること
により生じさせた、分化した神経細胞。
【請求項99】
増強された緑色蛍光蛋白質(eGFP)を発現する、請求項98記載の分化した神経細胞。
【請求項100】
神経の発生、機能、及び死を分析すること、又はシナプスの分化を監視することにおける、請求項95記載の分化した神経細胞の使用。
【請求項101】
増強された緑色蛍光蛋白質(eGFP)を発現する請求項98記載の胚性幹細胞を含む、トランスジェニック非ヒト動物系。
【請求項101】
分化した神経細胞の集団を単離及び又は精製する方法であって、
(a)増強された緑色蛍光蛋白質(eGFP)を発現する胚性幹細胞の培養物を得るか又は生じさせ;
(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;
(c)神経前駆細胞を、eGFPを発現する分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させ;
(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させ;
(e)分化した神経細胞においてeGFPの発現を検出し;そして
(f)eGFPを発現する分化した神経細胞を単離する
工程を含む方法。
【請求項103】
神経の変性に関連した症状を治療することにおける使用のための薬剤を同定する方法であって、
(a)胚性幹細胞の培養物を得るか又は生じさせ;
(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量のレチノイン酸に接触させ;
(c)神経前駆細胞においてヘッジホッグシグナリング経路を活性化することによりニューロンを生じさせるが、但し、ニューロンの幾つか又は全てが変性しており;
(d)変性したニューロンを候補の薬剤と接触させ;そして
(e)当該薬剤が幾つか又は全ての変性したニューロンの再生を増強するか否かを決定する
工程を含む方法。
【請求項104】
変性した神経ニューロン中のゲノミックDNAがSOD又はSMN遺伝子中に変異を有する、請求項103記載の方法。
【請求項105】
変性した神経ニューロンが増強された緑色蛍光蛋白質(eGFP)を発現する、請求項98記載の方法。
【請求項1】
胚性幹細胞を、分化した神経細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子及び背側化又は腹側化胚性シグナリング因子に接触させることを含む、胚性幹細胞の分化した神経細胞への分化を誘導する方法。
【請求項2】
胚性幹細胞がマウスの胚性幹細胞である、請求項1記載の方法。
【請求項3】
胚性幹細胞がヒトの胚性幹細胞である、請求項1記載の方法。
【請求項4】
吻側化胚性シグナリング因子がレチノイン酸又はWntである、請求項1記載の方法。
【請求項5】
尾部側化胚性シグナリング因子がレチノイン酸又はWntである、請求項1記載の方法。
【請求項6】
背側化胚性シグナリング因子がBMP又はWntである、請求項1記載の方法。
【請求項7】
腹側化胚性シグナリング因子がヘッジホッグシグナリング経路の活性化因子である、請求項1記載の方法。
【請求項8】
ヘッジホッグシグナリング経路の活性化因子がヘッジホッグ蛋白質又はヘッジホッグシグナリングのアゴニストである、請求項1記載の方法。
【請求項9】
ヘッジホッグシグナリング経路の活性化因子がSHh又はHh−Ag1.3である、請求項1記載の方法。
【請求項10】
背側化又は腹側化胚性シグナリング因子に接触させる前に、胚性幹細胞を吻側化及び/又は尾部側化胚性シグナリング因子に接触させる、請求項1記載の方法。
【請求項11】
背側化又は腹側化胚性シグナリング因子に接触させた後に、胚性幹細胞を少なくとも一つの神経栄養因子と接触させることをさらに含む、請求項10記載の方法。
【請求項12】
分化した神経細胞が、星状細胞、ニューロン、乏突起神経膠細胞、又はシュワン細胞からなる群から選択される、請求項1記載の方法。
【請求項13】
分化した神経細胞が運動ニューロンである、請求項1記載の方法。
【請求項14】
運動ニューロンが脊髄運動ニューロンである、請求項1記載の方法。
【請求項15】
分化した神経細胞が増強された緑色蛍光蛋白質(eGFP)を発現する、請求項1記載の方法。
【請求項16】
接触をインビボにて被験者において作用させる、請求項1記載の方法。
【請求項17】
胚性シグナリング因子を被験者に投与することにより、接触をインビボにて被験者において作用させる、請求項16記載の方法。
【請求項18】
接触をインビトロにて作用させる、請求項1記載の方法。
【請求項19】
分化した神経細胞を被験者に移植する工程をさらに含む、請求項18記載の方法。
【請求項20】
分化した神経細胞を被験者の脊髄に移植する、請求項19記載の方法。
【請求項21】
被験者が胚である、請求項19記載の方法。
【請求項22】
被験者がヒトである、請求項19記載の方法。
【請求項23】
被験者が神経組織の変性を有する、請求項19記載の方法。
【請求項24】
神経組織の変性が末梢神経障害又は神経変性疾患である、請求項23記載の方法。
【請求項25】
末梢神経障害が、筋委縮性側索硬化症(ALS)、神経の外傷、準新生物形成(paraneoplastic)疾患、ポリオ、ポリオ後症候群、進行性延髄麻痺、及び脊髄筋肉萎縮(SMA)からなる群から選択される症状に関連する、請求項24記載の方法。
【請求項26】
神経変性疾患が、アルツハイマー病、筋委縮性側索硬化症(ルーゲーリック病)、ビンスワンガー病、ハンチントンコレラ、多発性硬化症、重症筋無力症、パーキンソン病、及びピック病(Pick's disease)からなる群から選択される、請求項24記載の方法。
【請求項27】
請求項1記載の方法により生じた、分化した神経細胞。
【請求項28】
分化した神経細胞が増強された緑色蛍光蛋白質(eGFP)を発現する、請求項27記載の分化した神経細胞。
【請求項29】
分化した神経細胞を生産する方法であって、
(a)胚性幹細胞の培養物を得るか又は生じさせ;
(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;
(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させ;そして
(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させる
工程を含む方法。
【請求項30】
胚性幹細胞がマウスの胚性幹細胞である、請求項29記載の方法。
【請求項31】
胚性幹細胞がヒトの胚性幹細胞である、請求項29記載の方法。
【請求項32】
吻側化胚性シグナリング因子がレチノイン酸又はWntである、請求項29記載の方法。
【請求項33】
尾部側化胚性シグナリング因子がレチノイン酸又はWntである、請求項29記載の方法。
【請求項34】
背側化胚性シグナリング因子がBMP又はWntである、請求項29記載の方法。
【請求項35】
腹側化胚性シグナリング因子がヘッジホッグシグナリング経路の活性化因子である、請求項29記載の方法。
【請求項36】
ヘッジホッグシグナリング経路の活性化因子がヘッジホッグ蛋白質又はヘッジホッグシグナリングのアゴニストである、請求項35記載の方法。
【請求項37】
ヘッジホッグシグナリング経路の活性化因子がSHh又はHh−Ag1.3である、請求項35記載の方法。
【請求項38】
神経前駆細胞が、DRGニューロンの神経前駆細胞、運動ニューロンの神経前駆細胞、末梢ニューロンの神経前駆細胞、脊髄ニューロンの神経前駆細胞、及び腹側介在ニューロンの神経前駆細胞からなる群から選択される、請求項29記載の方法。
【請求項39】
分化した神経細胞が、星状細胞、ニューロン、乏突起神経膠細胞、又はシュワン細胞からなる群から選択される、請求項29記載の方法。
【請求項40】
分化した神経細胞が運動ニューロンである、請求項39記載の方法。
【請求項41】
分化した神経細胞が有糸分裂後である、請求項40記載の方法。
【請求項42】
運動ニューロンが脊髄運動ニューロンである、請求項40記載の方法。
【請求項43】
分化した神経細胞が増強された緑色蛍光蛋白質(eGFP)を発現する、請求項29記載の方法。
【請求項44】
工程(b)−(d)をインビボにて被験者において実施する、請求項29記載の方法。
【請求項45】
工程(b)、(c)、及び(d)をインビトロにて実施する、請求項29記載の方法。
【請求項46】
分化した神経細胞を被験者に移植する工程をさらに含む、請求項45記載の方法。
【請求項47】
分化した神経細胞を被験者の脊髄に移植する、請求項46記載の方法。
【請求項48】
神経前駆細胞を被験者に移植すること及び分化した神経細胞を被験者に移植することからなる群から選択される追加の工程をさらに含む、請求項45記載の方法。
【請求項49】
被験者が胚である、請求項48記載の方法。
【請求項50】
被験者がヒトである、請求項48記載の方法。
【請求項51】
被験者が神経組織の変性を有する、請求項48記載の方法。
【請求項52】
神経組織の変性が末梢神経障害又は神経変性疾患である、請求項51記載の方法。
【請求項53】
末梢神経障害が、筋委縮性側索硬化症(ALS)、神経の外傷、準新生物形成(paraneoplastic)疾患、ポリオ、ポリオ後症候群、進行性延髄麻痺、及び脊髄筋肉萎縮(SMA)からなる群から選択される症状に関連する、請求項52記載の方法。
【請求項54】
神経変性疾患が、アルツハイマー病、筋委縮性側索硬化症(ルーゲーリック病)、ビンスワンガー病、ハンチントンコレラ、多発性硬化症、重症筋無力症、パーキンソン病、及びピック病(Pick's disease)からなる群から選択される、請求項52記載の方法。
【請求項55】
請求項29記載の方法により生じた分化した神経細胞を含む、細胞の集団。
【請求項56】
分化した神経細胞が増強された緑色蛍光蛋白質(eGFP)を発現する、請求項55記載の分化した神経細胞。
【請求項57】
被験者において脊髄を再増殖させる方法であって、
(a)胚性幹細胞の培養物を得るか又は生じさせ;
(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;
(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させ;
(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させ;そして
(e)分化した神経細胞を被験者の脊髄に移植する
工程を含む方法。
【請求項58】
胚性幹細胞がマウスの胚性幹細胞である、請求項57記載の方法。
【請求項59】
胚性幹細胞がヒトの胚性幹細胞である、請求項57記載の方法。
【請求項60】
尾部側化胚性シグナリング因子がレチノイン酸である、請求項57記載の方法。
【請求項61】
腹側化胚性シグナリング因子がヘッジホッグシグナリング経路の活性化因子である、請求項57記載の方法。
【請求項62】
ヘッジホッグシグナリング経路の活性化因子がヘッジホッグ蛋白質又はヘッジホッグシグナリングのアゴニストである、請求項61記載の方法。
【請求項63】
ヘッジホッグシグナリング経路の活性化因子がSHh又はHh−Ag1.3である、請求項61記載の方法。
【請求項64】
神経前駆細胞が、DRGニューロンの神経前駆細胞、運動ニューロンの神経前駆細胞、末梢ニューロンの神経前駆細胞、脊髄ニューロンの神経前駆細胞、及び腹側介在ニューロンの神経前駆細胞からなる群から選択される、請求項57記載の方法。
【請求項65】
分化した神経細胞が、星状細胞、ニューロン、乏突起神経膠細胞、又はシュワン細胞からなる群から選択される、請求項57記載の方法。
【請求項66】
分化した神経細胞が運動ニューロンである、請求項65記載の方法。
【請求項67】
分化した神経細胞が有糸分裂後である、請求項66記載の方法。
【請求項68】
運動ニューロンが脊髄運動ニューロンである、請求項66記載の方法。
【請求項69】
分化した神経細胞が増強された緑色蛍光蛋白質(eGFP)を発現する、請求項57記載の方法。
【請求項70】
被験者が胚である、請求項57記載の方法。
【請求項71】
被験者がヒトである、請求項57記載の方法。
【請求項72】
被験者が神経組織の変性を有する、請求項57記載の方法。
【請求項73】
神経組織の変性が末梢神経障害又は神経変性疾患である、請求項72記載の方法。
【請求項74】
末梢神経障害が、筋委縮性側索硬化症(ALS)、神経の外傷、準新生物形成(paraneoplastic)疾患、ポリオ、ポリオ後症候群、進行性延髄麻痺、及び脊髄筋肉萎縮(SMA)からなる群から選択される症状に関連する、請求項73記載の方法。
【請求項75】
末梢神経障害がALS又はSMAである、請求項74記載の方法。
【請求項76】
治療の必要な被験者において神経組織の変性を治療する方法であって、
(a)胚性幹細胞の培養物を得るか又は生じさせ;
(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;
(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させ;
(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させ;そして
(e)神経組織の変性を治療するのに有効な量にて分化した神経細胞を被験者に移植する
工程を含む方法。
【請求項77】
神経組織の変性が末梢神経障害又は神経変性疾患である、請求項76記載の方法。
【請求項78】
末梢神経障害が、筋委縮性側索硬化症(ALS)、神経の外傷、準新生物形成(paraneoplastic)疾患、ポリオ、ポリオ後症候群、進行性延髄麻痺、及び脊髄筋肉萎縮(SMA)からなる群から選択される症状に関連する、請求項77記載の方法。
【請求項79】
神経変性疾患が、アルツハイマー病、筋委縮性側索硬化症(ルーゲーリック病)、ビンスワンガー病、ハンチントンコレラ、多発性硬化症、重症筋無力症、パーキンソン病、及びピック病(Pick's disease)からなる群から選択される、請求項77記載の方法。
【請求項80】
胚性幹細胞がマウスの胚性幹細胞である、請求項76記載の方法。
【請求項81】
胚性幹細胞がヒトの胚性幹細胞である、請求項76記載の方法。
【請求項82】
吻側化胚性シグナリング因子がレチノイン酸又はWntである、請求項76記載の方法。
【請求項83】
尾部側化胚性シグナリング因子がレチノイン酸又はWntである、請求項76記載の方法。
【請求項84】
背側化胚性シグナリング因子がBMP又はWntである、請求項76記載の方法。
【請求項85】
腹側化胚性シグナリング因子がヘッジホッグシグナリング経路の活性化因子である、請求項76記載の方法。
【請求項86】
ヘッジホッグシグナリング経路の活性化因子がヘッジホッグ蛋白質又はヘッジホッグシグナリングのアゴニストである、請求項85記載の方法。
【請求項87】
ヘッジホッグシグナリング経路の活性化因子がSHh又はHh−Ag1.3である、請求項85記載の方法。
【請求項88】
神経前駆細胞が、DRGニューロンの神経前駆細胞、運動ニューロンの神経前駆細胞、末梢ニューロンの神経前駆細胞、脊髄ニューロンの神経前駆細胞、及び腹側介在ニューロンの神経前駆細胞からなる群から選択される、請求項76記載の方法。
【請求項89】
分化した神経細胞が、星状細胞、ニューロン、乏突起神経膠細胞、又はシュワン細胞からなる群から選択される、請求項76記載の方法。
【請求項90】
分化した神経細胞が運動ニューロンである、請求項89記載の方法。
【請求項91】
分化した神経細胞が有糸分裂後である、請求項90記載の方法。
【請求項92】
運動ニューロンが脊髄運動ニューロンである、請求項90記載の方法。
【請求項93】
被験者が胚である、請求項76記載の方法。
【請求項94】
被験者がヒトである、請求項76記載の方法。
【請求項95】
工程:
(a)胚性幹細胞の培養物を得るか又は生じさせ;
(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;そして
(c)任意に、神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させること
により生じさせた神経前駆細胞。
【請求項96】
増強された緑色蛍光蛋白質(eGFP)を発現する、請求項95記載の神経前駆細胞。
【請求項97】
神経の発生、機能、及び死を分析すること、又はシナプスの分化を監視することにおける、請求項95記載の神経前駆細胞の使用。
【請求項98】
工程:
(a)胚性幹細胞の培養物を得るか又は生じさせ;
(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;
(c)神経前駆細胞を、分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させ;そして
(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させること
により生じさせた、分化した神経細胞。
【請求項99】
増強された緑色蛍光蛋白質(eGFP)を発現する、請求項98記載の分化した神経細胞。
【請求項100】
神経の発生、機能、及び死を分析すること、又はシナプスの分化を監視することにおける、請求項95記載の分化した神経細胞の使用。
【請求項101】
増強された緑色蛍光蛋白質(eGFP)を発現する請求項98記載の胚性幹細胞を含む、トランスジェニック非ヒト動物系。
【請求項101】
分化した神経細胞の集団を単離及び又は精製する方法であって、
(a)増強された緑色蛍光蛋白質(eGFP)を発現する胚性幹細胞の培養物を得るか又は生じさせ;
(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量の吻側化及び/又は尾部側化胚性シグナリング因子に接触させ;
(c)神経前駆細胞を、eGFPを発現する分化した神経細胞を生じさせるのに有効な量の背側化又は腹側化胚性シグナリング因子に接触させ;
(d)任意に、分化した神経細胞を、少なくとも一つの神経栄養因子と接触させ;
(e)分化した神経細胞においてeGFPの発現を検出し;そして
(f)eGFPを発現する分化した神経細胞を単離する
工程を含む方法。
【請求項103】
神経の変性に関連した症状を治療することにおける使用のための薬剤を同定する方法であって、
(a)胚性幹細胞の培養物を得るか又は生じさせ;
(b)胚性幹細胞の培養物を、神経前駆細胞を生じさせるのに有効な量のレチノイン酸に接触させ;
(c)神経前駆細胞においてヘッジホッグシグナリング経路を活性化することによりニューロンを生じさせるが、但し、ニューロンの幾つか又は全てが変性しており;
(d)変性したニューロンを候補の薬剤と接触させ;そして
(e)当該薬剤が幾つか又は全ての変性したニューロンの再生を増強するか否かを決定する
工程を含む方法。
【請求項104】
変性した神経ニューロン中のゲノミックDNAがSOD又はSMN遺伝子中に変異を有する、請求項103記載の方法。
【請求項105】
変性した神経ニューロンが増強された緑色蛍光蛋白質(eGFP)を発現する、請求項98記載の方法。
【図1】


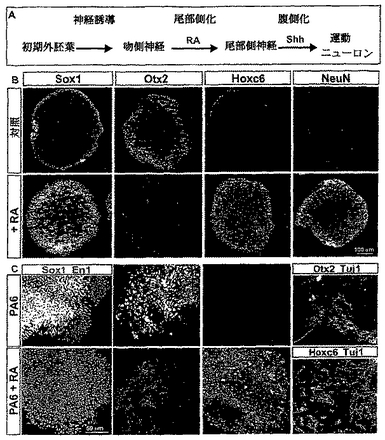
【図2】
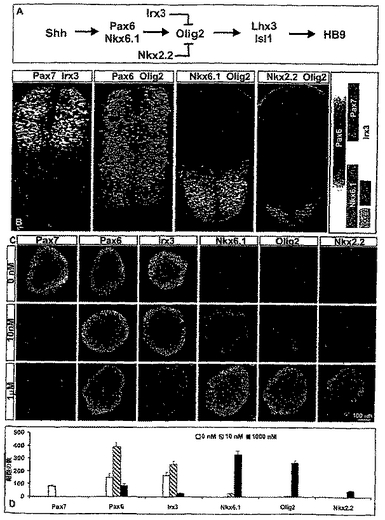
【図3】
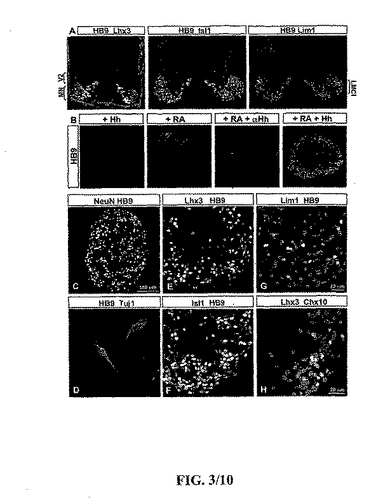
【図4】
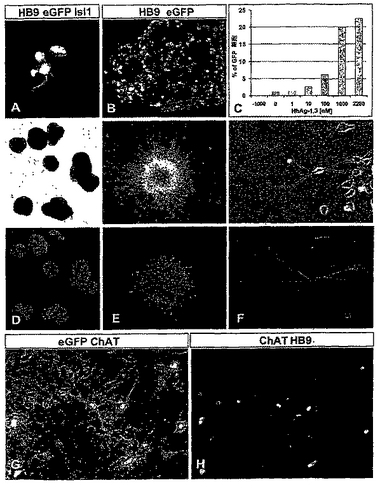
【図5】
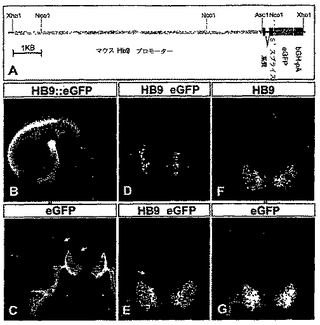
【図6】
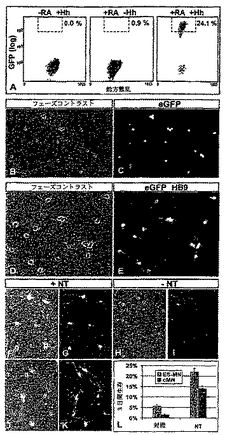
【図7】
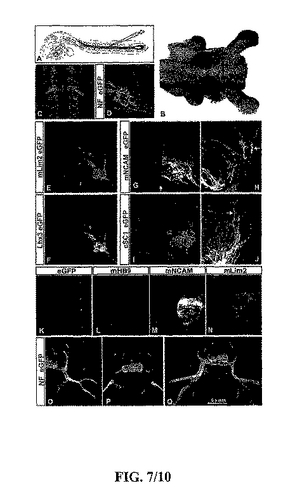
【図8】
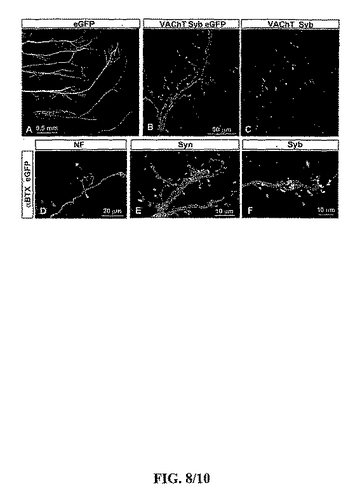
【図9】
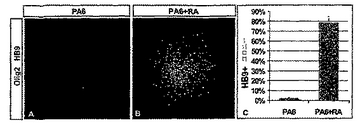
【図10】
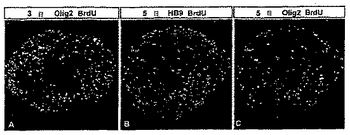
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公表番号】特表2006−517084(P2006−517084A)
【公表日】平成18年7月20日(2006.7.20)
【国際特許分類】
【出願番号】特願2004−521501(P2004−521501)
【出願日】平成15年6月26日(2003.6.26)
【国際出願番号】PCT/US2003/020399
【国際公開番号】WO2004/007665
【国際公開日】平成16年1月22日(2004.1.22)
【出願人】(505019998)トラスティーズ・オブ・コロンビア・ユニバーシティ・イン・ザ・シティ・オブ・ニューヨーク (7)
【Fターム(参考)】
【公表日】平成18年7月20日(2006.7.20)
【国際特許分類】
【出願日】平成15年6月26日(2003.6.26)
【国際出願番号】PCT/US2003/020399
【国際公開番号】WO2004/007665
【国際公開日】平成16年1月22日(2004.1.22)
【出願人】(505019998)トラスティーズ・オブ・コロンビア・ユニバーシティ・イン・ザ・シティ・オブ・ニューヨーク (7)
【Fターム(参考)】
[ Back to top ]