
脂肪酸の低下した飽和レベルを伴う種子および油
本発明は、「無飽和」カノーラ油を提供する。本発明は、そのような油を作製するために使用することができる種子も提供する。これらの種子を作製する植物も、本発明に含まれる。本発明は更に、植物に最適化されたΔ9デサチュラーゼ遺伝子を提供する。
【発明の詳細な説明】
【背景技術】
【0001】
関連出願の相互参照
本願は、2004年10月8日に出願された米国特許仮出願第60/617,532号の優先権を請求するものである。
【0002】
発明の背景
食物脂肪摂取の主な給源として次第に、植物由来の油が、動物由来の油および脂肪と置き換わっている。しかしほとんどの先進国における飽和脂肪摂取は、総カロリー消費の約15%〜20%を維持している。より健康な生活スタイルを促進する努力において、米国農務省(USDA)は最近、飽和脂肪は一日のカロリー摂取の10%未満とすることを推奨している。消費者の注意を喚起するために、USDAにより公布された最新の表示指針は、「低飽和」表示するためには14gサービング当たり1.0g未満であること、および「無飽和」表示するためには14gサービング当たり0.5g未満であることを現在求めている。このことは、「低飽和」および「無飽和」表示を付けるためには、各々、植物油の飽和脂肪酸含量は、7%および3.5%未満であることを必要とすることを意味する。これらの指針の発布以降、「低飽和」油を求める消費者が急増している。今のところこれには、主にカノーラ油が当てはまり、ヒマワリ油およびベニバナ油がはるかに低い程度当てはまる。
【0003】
植物または動物のいずれが起源であるかに関わらず、油の特徴は、炭素原子および水素原子の数に加え、脂肪酸鎖を含む二重結合の数および位置により主に決定される。植物由来の油のほとんどは、変動量のパルミチン酸(16:0)、ステアリン酸(18:0)、オレイン酸(18:1)、リノール酸(18:2)およびリノレン酸(18:3)の脂肪酸から構成される。パルミチン酸およびステアリン酸の炭素鎖は、水素原子により飽和され、従って二重結合を有さないので、それらは従来「飽和された」と称され;これらは、可能な限り最大数の水素原子を含む。しかしオレイン酸、リノール酸およびリノレン酸は、各々1、2および3個の二重結合を有する18-炭素の脂肪酸鎖である。オレイン酸は典型的には、一価不飽和脂肪酸と考えられるのに対し、リノール酸およびリノレン酸は、多価不飽和脂肪酸と考えられる。米国農務省は、「無飽和体(no saturate)または無飽和(no sat)」製品を、(脂肪酸の総量と比較して)3.5質量%未満の合計飽和脂肪酸を有する製品と規定している。
【0004】
不飽和脂肪(一価不飽和および多価不飽和)は有益であるが(特に適度に消費される場合)、飽和脂肪およびトランス脂肪は有益ではない。飽和脂肪およびトランス脂肪は、血中LDLコレステロールレベルを上昇する。同じく食事性コレステロールは、LDLコレステロールを上昇し、LDLが上昇しなくとも心臓疾患の原因となることがある。従って健康な食事の一部として、飽和脂肪、トランス脂肪、およびコレステロールの少ない食品を選択することが推奨される。
【0005】
最近の研究から、主要食事性脂肪成分としての一価不飽和体(monounsaturate)、特に高レベルのオレイン酸の健康上の価値が確立されている。このような食事は、高飽和脂肪酸食の結果である動脈硬化症の発生を減少させると考えられる。従って、高含量の一価不飽和体を有する食用植物油が必要である。種子の変異誘発を使用し、4%を超えない飽和脂肪酸含量を有するナタネ油が作製されている(PCT国際特許出願公開番号 国際公開公報第91/15578号)。
【0006】
1985年に世界で供給された食用油の13%以上が一般に、ナタネまたはマスタードとして公知の油糧種子作物アブラナ(Brassica)種から作製された。アブラナは、ダイズおよびヤシに続く、3番目に重要な食用油の供給源である。アブラナは、比較的低温で発芽および生育することができるので、これは、温度の低い農業地域において栽培することができ、更にはより温暖な地帯における冬期作物として使用される、数少ない商業的に重要な食用油糧種子作物のひとつでもある。更に全般的に植物油、特にナタネ油は、合成油または鉱油/ナフテン系油のものと同等の性能を提供する可能性を有し、生分解性である点でも非常に望ましい利点があるので、工業的用途でのこれらの使用が検討されることがかなり増えてきている。
【0007】
カノーラ油は、全ての植物油の中で最低レベルの飽和脂肪酸を有する。「カノーラ」は、種子の総脂肪酸含量を元にエルカ酸(C22:1)含量が最大でも2質量%であり(好ましくは最大でも0.5質量%であり、最も好ましくは本質的に0質量%である)、ならびに圧搾後、脱脂された(油非含有)粗挽き粉1g当たり30マイクロモル未満を含有する風乾された粗挽き粉を作製する、ナタネ(アブラナ)を意味する。これらの種類のナタネは、この種の従来の亜種と比べ、それらの食用適性が際だっている。
【0008】
植物油の修飾は、化学的に実行されてもよい。この方法を使用し、約3%未満の飽和脂肪酸を含むサラダ油/料理用油が得られており(米国特許第4,948,811号);この油は、化学反応によるか、または飽和脂質の物理的分離により、形成され得る。望ましい特徴の油を実現するための「遺伝子操作」の使用が全般的に言及されている(第3段、58行など参照)。しかし特定の油糧種子植物が、望ましい特徴の植物油を提供するために修飾される方法の詳細は明らかではない。
【0009】
典型的には、植物油の脂肪酸組成物は、従来の育種技術により修飾される。これらの技術は、現存する生殖質を、脂肪酸組成に影響を及ぼす天然の変異の供給源として使用する。このような変異は、適当なスクリーニングの使用により明らかにされ、かつ選択され、引き続きの育種と組み合わせられる。例えばそのような方法は、ナタネ油中の長鎖脂肪酸エルカ酸エステルの量を減少させ(Stefansson, B. R., High and Low Erucic Acid Rapeseed Oils, Kramer J. K. G. et al., eds;Academic Press, ニューヨーク;pp. 144-161(1983))、およびコーン油中の一価不飽和脂肪酸オレイン酸エステルの量を増加する(米国特許出願第07/554,526号)ために使用されている。
【0010】
最近、変異原を使用し、そこから所望の特性を選択するために使用可能な変異のプールを増加することが試みられている。しかし変異原は一般に、既に存在する遺伝子の失活または修飾により作用し、特定の機能の喪失または低下を生じる。従って変異誘発による新規特徴の導入は、既に存在するいくつかの形質の喪失に左右されることが多い。加えて、変異原による所望の目的の達成は、一般に不確かである。わずかな型の植物油中の修飾された脂肪酸組成のみが、この方法を用いて実現される。このような脂肪酸組成に影響を及ぼす「作出された」変異の一例は、ナタネ油中の多価不飽和脂肪酸、特にリノール酸エステルおよびリノレン酸エステルの減少、それに伴う一価不飽和脂肪酸オレイン酸エステルの増加である(AuId, M., et al., (1992) Crop Sci. 印刷中)。別のものは、ナタネ油中の飽和脂肪酸の減少である(PCT国際公開公報第91/15578号)。しかし、種子油合成の生化学は複雑であり、十分に理解されておらず;ナタネ油において観察される脂肪酸組成の変化に寄与するいくつかの機構が存在する(PCT国際公開公報第91/15578号)。そのような変化に影響を及ぼすための変異誘発の使用は、本質的にランダムであり、非特異的である。
【0011】
遺伝子操作を使用し脂肪酸組成を修飾する可能性は、理論的には、特定の望ましい遺伝子の正確に制御された導入、更には特定の望ましくない遺伝子または遺伝子産物の失活をもたらす。従って既に存在する遺伝子とは完全に独立した新規形質が植物に導入されるか、または予め選択された遺伝子が失活もしくは修飾される。しかし脂肪酸組成を修飾するために遺伝子操作を効果的に使用するためのひとつの根拠となるものは、脂肪酸の合成およびプロセシングを調節する植物細胞の作業時の機構の理論的に正確なモデルである。
【0012】
米国特許第6,495,738号(同じく国際公開公報第99/50430号参照)は、コーン油およびタバコ種子中の飽和脂肪酸レベルは、植物細胞内での真菌パルミチン酸エステル-CoAΔ9デサチュラーゼの発現により変更することができることを開示している。これらのタンパク質は優先的に、その分子のCoA部分からの2個の水素原子の除去、および9番目と10番目の炭素原子の間への二重結合の追加により、パルミチン酸エステル-CoA分子を酵素的に不飽和化し、その結果パルミトレイル-CoA(16:1Δ9)を生成する可能性が高い。パルミトレイル-CoAは最終的には、種子油に取り込まれ、その結果該油の総飽和体(saturate)レベルを低下する。コーン油の総飽和体脂肪酸レベルは、平均約13.9%であるが、これは先に考察した現在の表示指針には合致しない。更にトウモロコシは典型的には、ダイズ、カノーラ、ヒマワリなどと比べ、油糧作物とは見なされていない。事実トウモロコシから生成および抽出された油は、デンプン抽出において使用される湿式圧搾プロセスの副産物と見なされている。そのために、コーン油の飽和レベルを修飾することについて関心はほとんど無い。
【0013】
油糧種子においては、脂肪酸合成は主に色素体で生じること、および新たに合成された脂肪酸は色素体から細胞質へ輸送されることが、前提条件とされている。細胞質において、これらは、トリグリセリドの構築に使用され、これは小胞体(endoreticular)膜において生じる。
【0014】
脂肪酸合成の主要生成物は、パルミチン酸エステル(16:0)であり、これはステアリン酸エステル(18:0)へ効率的に伸長するように見える。依然色素体において、この飽和脂肪酸は次にΔ9デサチュラーゼとして公知の酵素により不飽和化され、1個または複数の炭素-炭素二重結合を導入する。具体的には、ステアリン酸エステルは、色素体Δ9デサチュラーゼ酵素により迅速に不飽和化され、オレイン酸エステル(18:1)を生じることができる。実際パルミチン酸エステルは、色素体Δ9デサチュラーゼにより、パルミトレイン酸エステル(16:1)にも不飽和化されるが、この脂肪酸は、ほとんどの植物油において極微量(0〜0.2%)であるように見える。
【0015】
従って、色素体における脂肪酸合成の主要生成物は、パルミチン酸エステル、ステアリン酸エステル、およびオレイン酸エステルである。ほとんどの油において、飽和脂肪酸ははるかに少ない割合で存在するので、オレイン酸エステルが、合成される主要脂肪酸である。
【0016】
引き続きの細胞質中の色素体の外側での植物脂肪酸の不飽和化は、オレイン酸エステルに限定されるように見え、これはリノール酸エステル(18:2)およびリノレン酸エステル(18:3)へ不飽和化され得る。加えて植物に応じて、オレイン酸エステルは更に、伸長(20:1、22:1、および/または24:1に対する)によるか、または官能基の付加により、修飾され得る。その後これらの脂肪酸は、飽和脂肪酸パルミチン酸エステルおよびステアリン酸エステルと共に、トリグリセリドへ構築され得る。
【0017】
植物のΔ9デサチュラーゼ酵素は、可溶性である。これは、色素体ストロマ内に位置し、および基質として、主にステアリル-ACPであるACPへエステル化された新たに合成された脂肪酸を使用する。これは、小胞体膜(ER、またはミクロソーム)に位置し、基質としてCo-Aへエステル化された脂肪酸を使用し、ならびに飽和脂肪酸パルミチン酸エステルおよびステアリン酸エステルの両方を不飽和化する、酵母Δ9デサチュラーゼ酵素とは対照的である。米国特許第5,723,595号および同第6,706,950号は、植物デサチュラーゼに関する。
【0018】
酵母Δ9デサチュラーゼ遺伝子は、出芽酵母(Saccharomyces cerevisiae)から単離され、クローニングされ、配列決定されている(Stukey, J.E. et al., J. Biol. Chem. 264:16537-16544(1989);Stukey, J.E. et al., J. Biol. Chem. 265:20144-20149(1990))。この遺伝子は、それが明らかに過剰発現される条件下で、出芽酵母株を形質転換するためにも使用されており、これはトリグリセリドの染色のためにナイルレッドを使用する蛍光顕微鏡により決定されるように、形質転換された酵母細胞において、増大した貯蔵脂質の蓄積を生じる(米国特許第5,057,419号)。この脂肪酸組成物は、特徴決定されていない。この参考文献は、単離された酵母Δ9デサチュラーゼ遺伝子からの情報を使用し、最初に他のデサチュラーゼ遺伝子を酵母から、または他の生物から単離し、次にこれらの遺伝子を酵母または植物へ条件下で再導入する、「一般的」考察を含んでいる。これは生成された油およびその脂肪酸組成の修飾のための高発現につながることは推測される。
【0019】
引き続き、酵母Δ9デサチュラーゼ遺伝子は、タバコ葉組織へ導入され(Polashcok, J. et al., FASEB J. 5:A1157(1991))、およびこの組織において明らかに発現されたことが報告された。更にこの遺伝子は、トマトにおいて発現された。Wang et al., J. Agric Food Chem., 44:3399-3402 (1996);および、C. Wang et al., Phytochemistry, 58:227-232(2001)参照のこと。タバコおよびトマトの両方においてある種の不飽和体の若干の増加および一部の飽和体の若干の減少が報告されたが、タバコおよびトマトは明らかに油糧作物ではない。この酵母遺伝子は、セイヨウアブラナ(Brassica napus)へも導入された(米国特許第5,777,201号参照)。パルミチン酸エステルおよびステアリン酸エステル(飽和)の減少ならびにパルミトレイン酸エステルおよびオレイン酸エステル(不飽和)の増加が報告されたが(前記特許の実施例7の表1aおよび1b参照)、この言及は、下記「詳細な説明」の項の始めにより詳細に考察されている。国際公開公報第00/11012号および米国特許第6,825,335号は、植物において発現するための合成酵母デサチュラーゼ遺伝子に関連しており、ここでこの遺伝子は、デサチュラーゼドメインおよびcyt b5ドメインを含む。これらの参考文献の「背景」の項は、脂肪酸合成を詳細に説明している。
【0020】
植物油が食事性または工業用かどうかの性能特性は、その脂肪酸プロファイルにより、すなわち油中に存在する脂肪酸の種類、ならびに各種類の相対量および絶対量により、実質的に決定される。脂肪酸プロファイルと性能特性の間のいくつかの関係はわかっているが、多くは不明のままである。それにもかからわず、植物油に存在する不飽和の種類および量は、食事性および工業用の両方の用途に関連している。
【0021】
標準のカノーラ油は、約8〜12%リノレン酸を含有し、これは酸化的、従って香味、安定性に関して、ダイズ油と同様の範疇に入る。カノーラ油の酸化安定度は、不飽和の量を減少させるための水素化、抗酸化剤の添加、およびより良い酸化安定度を有する1種または複数の油の配合など、多くの方法で改善することができる。例えばカノーラ油とヒマワリのような低リノレン酸油との配合は、18:3レベルを低下し、その結果油の安定性を向上する。しかしこれらの処理は、必然的に油の価格を上昇し、その他の複雑さを有し得る。例えば、水素化は、飽和脂肪酸のレベルおよびトランス不飽和量の両方を増加する傾向があり、これらは両方とも食事用途には望ましくない。
【0022】
高オレイン酸油は使用可能であるが、しかし、そのような精選油の追加費用の可能性に加え、非常に高レベルのオレイン酸のために育種された作物からの植物油は、高レベルの多不飽和脂肪酸、特にリノール酸および/またはリノレン酸をかなり保持するので、これらは工業用途は満たさないことを証明することができる。そのような油は、料理用油としての使用を含む食事用途には依然極めて有用であるが、工業用途において見出されたより厳密な条件下では、不適切な酸化安定度を有する。例え抗酸化剤の添加が、工業用途に必要な酸化安定度のレベルをこれらの油にもたらすのに十分ではないとしても;これは恐らく、これらの油において見出される、極めて高い易酸化性を伴うリノレン酸のレベルに起因していると考えられる。
【0023】
酸化安定度は、熱および圧力の条件下および化学副産物の存在下で、滑沢剤の寿命を延長するために、工業用途に重要である。このような用途においても同じく、リノレン酸、およびより少ない程度にリノール酸は、酸化安定度の不良が原因となっている。
【0024】
従って、農学的に生育でき、および食事用途において使用するためにこれが適格であるのに十分なレベルの酸化安定度を有する種子油を作製し、ならびに加えて工業用途における使用を見出すために、単独で十分に安定であるか、または抗酸化剤に十分に反応性であるような、様々なセイヨウアブラナを得ることは望ましいと考えられる。
【0025】
欧州特許出願第EP 323753号、米国特許第5,840,946号、および米国特許第5,638,637号は、オレイン酸含量80〜90%(総脂肪酸含量についての質量%)および2%を超えないエルカ酸を有するナタネ油を開示している。オレイン酸含量を改善するためには、変異誘発が使用された。米国特許第5,840,946号の「特許請求の範囲」は更に、その油は、2%を超えないエルカ酸含量、および3.5%未満のα-リノレン酸含量、ならびに7%を超えないステアリン酸およびパルミチン酸の形の飽和脂肪酸含量も有することを特定している。これらの特許は、変異誘発に続く選択に関連している。
【0026】
米国特許第5,387,758号;同第5,434,283号;および、同第5,545,821号は、2〜4%のステアリン酸およびパルミチン酸の組合せ(質量%)、ならびに約2質量%を超えないエルカ酸含量を有するナタネに関する。変異誘発を使用し、ステアリン酸およびパルミチン酸の含量を低下する。
【0027】
国際公開公報第92/03919号、および米国特許第5,668,299号;同第5,861,187号;および、同第6,084,157号は、変更された脂肪酸プロファイルを有するナタネ種子、植物、および油に関する。そのようなプロファイルのいくつかが言及されており、それらの全ては、パルミチン酸含量約7%〜約12%、リノール含量約14%〜約20%、ステアリン酸含量約0.8%〜約1.1%、およびα-リノレン酸含量約7%〜約9%、更にはある範囲のFDA飽和体(FDA saturate)と組合せた、最大エルカ酸含量約2%を予想している。これらの特許は、飽和脂肪酸および「FDA飽和体」を、ラウリン酸(C12:0)、ミリスチン酸(C14:0)、パルミチン酸(C16:0)、およびステアリン酸(C18:0)の合計と定義している。
【0028】
国際公開公報第93/06714号、および米国特許第6,270,828号;同第6,562,397号;同第6,680,396号;および、同第6,689,409号は、低下したグルコシノレート(従って低下したイオウ)を伴い、更にはα-リノレン酸含量が約2%〜約7%であるカノーラ油および種子に関する。
【0029】
米国特許第6,169,190号は、オレイン系脂肪酸含量が約71〜77%およびリノレン酸含量が約3%未満であるカノーラ種子由来の油に関する。34〜55の間のオレイン:リノレン比も、特許請求されている。
【0030】
米国特許第6,063,947号および同第5,850,026号は、カノーラ種子から得られた油、それに関連したカノーラ植物、およびその油の製造法を特許請求しており、ここでこの油は、約80%よりも多い(約86〜89%)オレイン酸含量、約2%〜約6%のリノール酸含量、2.5%未満(約1〜2%)のα-リノレン酸含量、および約2%未満のエルカ酸含量(加水分解後)を有する。これらの特許は、ミクロソームオレイン酸デサチュラーゼ(オレイン酸をリノール酸へ転換するΔ-12デサチュラーゼ)およびミクロソームリノール酸デサチュラーゼ(リノール酸をα-リノレン酸へ転換するΔ-15デサチュラーゼ)遺伝子発現の種子特異的阻害に関する。
【0031】
米国特許第5,952,544号は、炭素15と16の間の反応を触媒する、植物色素体またはミクロソームΔ-15脂肪酸デサチュラーゼ酵素の断片を特許請求している。
【0032】
米国特許第4,627,192号および同第4,743,402号は、約80〜94%(それらの総脂肪酸含量に対して)のオレイン酸含量およびリノール対オレイン比約0.09未満を有するヒマワリ種子およびヒマワリ油に関する。これらのヒマワリ植物は、伝統的育種技術により得られた。
【0033】
国際公開公報第2003002751号は、植物の油表現型を変更するためのキナーゼ遺伝子などの使用に関する。
【0034】
既に有益な油糧種子作物の脂肪酸プロファイルに有意に(および望ましく)作用する、特に植物および油のその他の局面に有害に作用することなく、飽和脂肪のレベルを低下する、Δ9デサチュラーゼ遺伝子の能力は、予測不可能である。
【発明の開示】
【0035】
発明の簡単な概要
本発明の目的は、「無飽和」カノーラ油を提供することである。本発明は、一部、ある植物種子の飽和脂肪酸を減少させる方法にも関する。これらの結果は驚くべきことに、カノーラ(アブラナ)におけるΔ9デサチュラーゼ遺伝子の使用により実現された。この技術は、本明細書において開示されたように、他の植物にも適用することができる。本発明に含まれるのは、そのような油および種子を生成することが可能である植物、好ましくはカノーラである。本発明は、該植物から種子および油も提供し、ここでこれらの油は、これまで達成されていないような、特に有利な特徴および脂肪酸プロファイルを有する。本発明はなお更に、植物に最適化されたΔ9デサチュラーゼ遺伝子を提供する。一部の好ましい態様において、好ましい植物は、本発明のΔ9デサチュラーゼ遺伝子の少なくとも2種のコピーを含む。このような植物により生成される種子は驚くべきことに、遺伝子サイレンシングの作用を示さないが、更に驚くことに総飽和体レベルは低下する。
【0036】
配列の簡単な説明
SEQ ID NO:1は、本明細書において使用される、植物に最適化されたΔ9デサチュラーゼ遺伝子のオープンリーディングフレームの核酸配列を示す。
SEQ ID NO:2は、コザック配列およびBamHIクローニング部位(残基1-10)に続くSEQ ID NO:1のORFと、ORFの末端の翻訳終結因子(残基1379-1381)の配列を示す。
SEQ ID NO:3は、Δ9遺伝子の増幅に使用されたΔ9フォワードBプライマーの核酸配列を示す。
SEQ ID NO:4は、Δ9遺伝子の増幅に使用されたΔ9リバースBプライマーの核酸配列を示す。
SEQ ID NO:5は、SEQ ID NO:1によりコードされたアミノ酸配列を示す。
【0037】
発明の詳細な説明
本発明は、「無飽和」カノーラ油を提供する。本発明は一部、ある植物種子の飽和脂肪酸を減少させる方法にも関する。これらの結果は驚くべきことに、植物、好ましくは油糧植物、更により好ましくはカノーラ(アブラナ)中の飽和脂肪酸の「無飽和」レベルを驚く程生じるが、Δ9デサチュラーゼ遺伝子の使用により実現された。本発明は、そのような植物を含み、ならびに該植物由来の種子および油も提供し、ここでこれらの油は、これまで達成されていないような、特に有利な特性および脂肪酸プロファイルを有する。
【0038】
アスペルギルス・ニジュランス(Aspergillus nidulans)ミクロソームΔ9-CoAデサチュラーゼ遺伝子が本明細書において例示されている。このΔ9デサチュラーゼは、膜結合型酵素であり、16:0-CoAおよび18:0-CoAの16:1-CoAおよび18:1-CoAへの反応(Δ9位置での二重結合の付加)を触媒する。本発明は更に、他の飽和体、例えばC20:0、C22:0およびC24:0のレベルも、非常に驚きに値しかつ有利に減少したが、C16:1およびC18:1不飽和体(unsaturate)は増加した(C18:2およびC18:3はほとんどまたは全く増加せず、場合によってはこれらの比較的安定していない多価不飽和体(polyunsaturate)は減少する)点が驚きに値する。従来は、既に望ましい(しかし最適ではない)脂肪酸プロファイルを有するアブラナおよび他の「優れた」油糧種子植物において、これが良好な酵素であるかどうか(この遺伝子は十分に発現されるかどうかを含む)は不明であった。(例えばトウモロコシの飽和体においてわずかに10%の減少が得られた。)。
【0039】
「発明の背景」の項において言及したように、様々な生物および植物の複雑な脂肪酸プロファイルおよび代謝経路、ならびにそれらの異なる生理的細胞機構を考えると、例えこの遺伝子および酵素がアブラナにおいてある作用を有したとしても、それらの作用は有益であると予想することはできないと考えられる。「発明の背景」の項においてにおいて考察したように、望ましい脂肪酸の1型または複数型の増加は、他の望ましい脂肪酸の減少、望ましくない脂肪酸の増加、および農学的ペナルティ(すなわち、修飾された植物に対する他のあからさまな有害作用)を生じることが多い。本発明は、莢サイズ、種子収量、種子サイズ、油収量などの他の価値のある農学的特徴に対応する有害作用を伴わずに実践することができることも驚くべきことであった。単独のトランスジェニック挿入断片に関して、植物ホモ接合体の有害作用は存在しなかった。一部のデュアルホモ接合体スタック(ふたつのトランスジェニック事象の交差により生じた)は、莢数および結実の減少を示したが;その原因は未だ不明である。しかし表27は、非トランスジェニック対照に類似した種子収量を有するスタック(すなわち、明らかに増加したコピー数事象)、更には「無飽和」組成も含んでいる。
【0040】
予想外のことに、デサチュラーゼ間の差異、更には酵母、真菌、植物、および動物のΔ9デサチュラーゼ間の差異のために、別の理由も生じる。デサチュラーゼの差異は、ひとつには、様々なデサチュラーゼの給源生物の細胞構造の差異に起因し得る。米国特許第5,777,201号の酵母デサチュラーゼは、「発明の背景」の項で考察されている。これは、本明細書において例示されているアスペルギルスデサチュラーゼよりも長い(510個アミノ酸対455個アミノ酸)。加えて約400個のアミノ酸全体の同一性はわずかに約52%である(BLASTおよびBestFitの両方により決定、Smith-Watermanプログラム;両方ともEMBOSSで実行)。その特許の実施例7の表1aおよび1bは、酵母デサチュラーゼを用いて達成される飽和体の減少は、カノーラにおいて例証されたアスペルギルスデサチュラーゼにより本発明に従い達成されるものよりもはるかに弱いことを示している。酵母デサチュラーゼの比較的弱い性能に関する可能性のある説明であり得る様々な要因が存在する。例えばそのタンパク質は、植物において本来不安定であるかもしれない(しかし本デサチュラーゼは、カノーラにおいて極めて明らかに非常に安定している)。これらは、触媒効率、基質親和性、補因子親和性などの、その他の酵素特性も異なり得る。
【0041】
米国特許第5,723,595号および同第6,706,950号の紅花デサチュラーゼと比較すると、この紅花デサチュラーゼ(396個アミノ酸)は、本明細書で例示されているアスペルギルスデサチュラーゼ(455個アミノ酸)よりもより短い。紅花デサチュラーゼは、色素体においても見つかっているが、本アスペルギルスデサチュラーゼは、ER/ミクロソーム/細胞質コンパートメントにおいて見つかっている。更に、紅花デサチュラーゼは、色素体において見出されるアシル-ACP基質を使用するが、アスペルギルスデサチュラーゼは、細胞質コンパートメントにおいて見出されるアシル-CoA基質を使用する。従って本発明に関して、アシル-CoA基質のプールの実質的部分は、アスペルギルスデサチュラーゼに使用可能であるかどうかは不明である。
【0042】
従って、本Δ9デサチュラーゼは、それらから特に油の改善された食品品質について優れた特性を有するカノーラ植物、種子、および油を得ることができることがわかったことは大きな驚きであった。非常に驚くべきことであるが、飽和脂肪酸の60%を超える減少が、アラビドプシス(Arabidopsis)において実現され、および飽和脂肪酸の43%を超える減少がカノーラにおいて実現された。再度、このことがいずれかの適当な植物の最良の脂肪酸プロファイルのひとつを既に生じた植物において実現されたことに注目することは重要である。本発明は、以下により詳細に示されかつ説明されるように、驚きに値しかつ有利な脂肪酸プロファイルおよび比を実現するためにも使用された。ステアリン酸は飽和脂肪酸であると考えられるが、これはコレステロール低下作用を有することがわかっている。従って比較的高いレベルのステアリン酸は、有益である。同様に比較的高いレベルのアラキドン酸は、望ましいものである。本明細書のデータに示されたように、本発明の種子由来の油は、これら2種の脂肪酸の有利なプロファイルを、例えば望ましいレベルのバクセン酸と共に有する。同じく、有利なレベルのこれらの脂肪酸および/または総飽和体は、本発明の業務用品質の植物(例えば矮性植物とは対照的)における望ましい高さ、収量、および他の有益な特徴と組合せて存在することができることも本明細書において示される。本発明のこのような植物に関する例証的データも本明細書において示されている。
【0043】
本発明は、例証されたデサチュラーゼに限定されないことは注意すべきである。様々なデサチュラーゼおよびΔ9デサチュラーゼは、GENBANKから入手可能であり、ならびにこれらの酵素の配列の相違点を観察および比較するための配列アラインメントは、標準の手法を用いて行うことができる。本明細書に例証されたものに類似した酵素は、本発明に従い使用することができる。
【0044】
例えば本アスペルギルスデサチュラーゼは、ふたつのドメインを有する。第一のドメイン(その分子のアミノ-末端側のおよそ2/3)は、デサチュラーゼドメインであり、第二のドメイン(分子のC-末端側のほぼ1/3)は、シトクロムb5ドメインである。例えばSEQ ID NO:5の残基62-279は、例えば、脂肪酸デサチュラーゼgnl|CDD|25523 pfam00487の残基4-233と整列化することができる。SEQ ID NO:5の残基332-407は、gnl|CDD|22935 pfam00173の残基1-74(シトクロムb5ドメイン)と整列化することができる。SEQ ID NO:5の残基17-305は、脂肪酸デサチュラーゼgnl|CDD|11113 COG1398(OLE1)の脂質代謝ドメインの残基3-288と整列化することができる。SEQ ID NO:5の残基301-449は、gnl|CDD|14396 COG5274のCYB5(脂質代謝に関与したシトクロムb)の残基11-163と整列化することができる。シトクロムb5を欠いているSEQ ID NO:5のデサチュラーゼドメインは機能することができ、これは植物の色素体デサチュラーゼの一般的構造であるからである。細胞質コンパートメントにおいて見出されるアシル-CoA基質を使用する、公開された推定されたミクロソームのマツのデサチュラーゼ(LOCUS AF438199)も存在し、これはcytb5ドメインを欠いている。アスペルギルスシトクロムb5ドメインを、別の生物のそれと、例え植物細胞質デサチュラーゼ由来のものであっても、交換することも可能である。これらのドメイン、またはこれらのドメインのいずれかもしくは両方をコードしているセグメントは、以下により詳細に説明されるように、本発明の分子を定義するために、プローブとして使用することができる。
【0045】
従って、本発明に有用な遺伝子およびタンパク質は、特別に例証された完全長配列のみではなく、これらの配列の部分、セグメントおよび/または断片(完全長分子と比べ内部および/または末端の欠失を含む)、それらの変種、変異体、キメラおよび融合体も含む。本発明において使用されるタンパク質は、本明細書に特別に例示されたタンパク質の特徴的酵素活性を維持する限りは、置換されたアミノ酸を有することができる。「変種」遺伝子は、例示されたタンパク質と同じタンパク質または同等の機能を有する同等のタンパク質をコードしているヌクレオチド配列を有する。「変種タンパク質」および「同等なタンパク質」という用語は、例示されたタンパク質と同じまたは本質的に同じ生物学的/機能的活性を有するタンパク質を意味する。本明細書において使用される「同等な」配列という表現は、機能を改善またはこれに有害に作用しないようなアミノ酸の置換、欠失、付加、または挿入を有する配列を意味する。機能を保持している断片も、この定義に含まれる。例示されたタンパク質の対応する断片と同じまたは同様の機能を保持する断片および他の同等物は、本発明の範囲内である。アミノ酸の置換または付加のような変化は、タンパク質のプロテアーゼ安定性の増加(または減少)のような様々な目的で行うことができる(タンパク質の機能を物質的に/実質的に減少させることはない)。
【0046】
遺伝子の変動は、例えば点変異を作出する常法を用い容易に構築することができる。加えて米国特許第5,605,793号は、例えば、ランダム断片化後に、DNA再構築体を使用することによる追加の分子多様性を作出する方法を開示している。変種遺伝子を使用し、変種タンパク質を作出することができ;組換え宿主を使用し、変種タンパク質を生成することができる。これらの「遺伝子シャッフリング」技術を用い、本明細書に例示された配列の(例えば)5、10、15、20、25、30、35、40、45、50、55、60個のいずれかの近接残基(アミノ酸またはヌクレオチド)を含む、同等の遺伝子およびタンパク質を構築することができる。
【0047】
完全長遺伝子の断片は、市販のエキソヌクレアーゼまたはエンドヌクレアーゼを使用し、常法に従い作製することができる。例えばBal31などの酵素または部位特異的突然変異誘発を使用し、これらの遺伝子の末端から、系統的にヌクレオチドを切断することができる。同じく、活性断片をコードしている遺伝子を、様々な制限酵素を用いて得ることができる。プロテアーゼは、これらのタンパク質の活性断片を直接得るために使用することができる。
【0048】
本タンパク質は、短縮型でありかつ機能活性を維持することは、本明細書に開示された本発明の範囲内である。「短縮型タンパク質」とは、タンパク質の一部が切断され、切断後も依然酵素活性を示すことを意味する。更に効果的に切断されたタンパク質は、該タンパク質をコードしているDNA塩基が、制限エンドヌクレアーゼによる消化、または当業者が使用可能な他の技術のいずれかを通じて除去される分子生物学技術を用い作製することができる。短縮後、該タンパク質は、大腸菌(Escherichia coli)、バキュロウイルス、植物-ベースのウイルスシステム、酵母などの異種システムにおいて発現され、その後本明細書に説明されたような昆虫アッセイ法に配置し、活性を決定することができる。短縮型タンパク質は、全体の完全長配列よりも短いものを有するにもかかわらず、それらが機能活性を保持するように、うまく作製することができることは当技術分野において周知である。バシラス・スリンギエンシス(B.t.)毒素を、短縮(コア毒素)型において使用することができることは当技術分野において周知である。例えば、Adangらの論文(Gene 36:289-300 (1985), 「Characterized full-length and truncated plasmid clones of the crystal protein of Bacillus thuringiensis subsp kurstaki HD-73 and their toxicity to Manduca sexta」)を参照のこと。幼虫ホルモンエステラーゼを含む、殺虫活性を保持する短縮型タンパク質の別の例も存在する(米国特許第5,674,485号、The Regents of the University of California)。本明細書に使用されるように、「毒素」という用語は、機能的活性のある短縮型を含むことも意味する。
【0049】
本発明において使用するためのタンパク質および遺伝子は、例えばオリゴヌクレオチドプローブを用いて、定義、同定および/または入手することができる。これらのプローブは、適当な標識により検出可能であることができるか、または国際公開公報第93/16094号に開示されたような、固有の蛍光を生じることができる、検出可能なヌクレオチド配列である。これらのプローブ(および本発明のポリヌクレオチド)は、DNA、RNA、またはPNAであってよい。本発明の合成プローブ(およびポリヌクレオチド)は、アデニン(A)、シトシン(C)、グアニン(G)、チミン(T)、およびウラシル(U;RNA分子について)に加え、イノシン(4種の塩基全てと対形成が可能である中性の塩基;時には、合成プローブにおいて4種の塩基全ての混合物の代わりに使用される)も有することができる。従って、本明細書において合成した、縮重オリゴヌクレオチドが言及され、および「N」または「n」が総称的に使用される場合、「N」または「n」は、G、A、T、C、またはイノシンであることができる。本明細書において使用される多義性(ambiguity)暗号は、本願の出願の慣例として標準のIUPAC命名法に従う(例えばRはAまたはGを意味し、YはCまたはTを意味するなど)。
【0050】
当技術分野において周知であるように、プローブ分子が核酸試料とハイブリダイズする場合、これはプローブおよび試料は、実質的に相同/類似/同一であると当然推定することができる。好ましくは、例えばKeller, G.H.、M.M. Manakの著書(DNA Probes, Stockton Press, ニューヨーク, NY, pp. 169-170(1987))に説明されたような、当技術分野において周知の技術により、ポリヌクレオチドのハイブリダイゼーションが最初に行われ、それに低、中、高ストリンジェンシー条件下での洗浄が続く。例えば本明細書に記されたように、低ストリンジェンシー条件は、2xSSC(標準クエン酸ナトリウム、塩化ナトリウム緩衝液)/0.1%SDS(ドデシル硫酸ナトリウム)による室温で15分間の最初の洗浄により実現することができる。典型的には2回の洗浄が行われる。次により高いストリンジェンシーは、塩濃度を低下することによるか、および/または温度を上昇することにより実現することができる。例えば先に説明された洗浄には、0.1xSSC/0.1%SDSによる各15分間室温での2回の洗浄、それに続く0.1xSSC/0.1%SDSによる各30分間55℃での洗浄を続けることができる。これらの温度を、本明細書に記しかつ当業者に公知である他のハイブリダイゼーションおよび洗浄プロトコールと共に使用することができる(例えばSSPEを、SSCの代わりの塩として使用することができる)。2xSSC/0.1%SDSは、2OxSSC 50mlおよび10%SDS 5mlを水445mlに添加することにより調製することができる。2OxSSCは、NaCl(175.3g/0.150M)、クエン酸ナトリウム(88.2g/0.015M)、および水を一緒にし、10N NaOHによりpHを7.0に調節し、その後容量を1Lに調節することにより調製することができる。10%SDSは、SDS 10gをオートクレーブにかけた水50mlに溶解し、その後100mlに希釈することにより調製することができる。
【0051】
プローブの検出は、ハイブリダイゼーションが維持されるかどうかを公知の様式で決定する手段を提供する。このようなプローブ分析は、本発明の毒素-コードしている遺伝子を同定する迅速な方法を提供する。本発明においてプローブとして使用されるヌクレオチドセグメントは、DNA合成装置および標準手法を用い合成することができる。これらのヌクレオチド配列は、本発明の遺伝子を増幅するためのPCRプライマーとしても使用することができる。
【0052】
所定のポリヌクレオチドとのハイブリダイゼーションは、本発明のタンパク質および遺伝子の同定、発見および/または定義に使用することができる技術である。本明細書において使用されるハイブリダイゼーションの「ストリンジェント」条件は、本明細書に説明された条件と、同じまたはほぼ同じ程度のハイブリダイゼーション特異性を実現する条件を意味する。32P標識した遺伝子特異的プローブによるサザンブロットの際に固定されたDNAのハイブリダイゼーションは、標準法により行うことができる(例えばManiatis, T., E.F. Fritsch, J. Sambrook [1982] Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, コールドスプリングハーバー, NY参照)。一般にハイブリダイゼーションおよび引き続きの洗浄は、標的配列の検出を可能にする条件下で行われる。2本鎖DNA遺伝子プローブに関して、ハイブリダイゼーションは、6xSSPE、5xデンハルト液、0.1%SDS、0.1mg/ml変性DNAの中において、DNAハイブリッドの融解温度(Tm)より20〜25℃低い温度で一晩実行することができる。融解温度は、下記式で説明される(Beltz, G. A., K.A. Jacobs, T.H. Eickbush, P.T. Cherbas, and F.C. Kafatos [1983] Methods of Enzymology, R. Wu, L. Grossman and K. Moldave [eds.] Academic Press, ニューヨーク, 100:266-285)。
Tm=81.5℃+16.6 Log[Na+]+0.41(%G+C)−0.61(%ホルムアミド)−600/塩基対の中の二重鎖の長さ
洗浄は典型的には、以下のように実行される。
1)1xSSPE、0.1%SDS中室温で15分間を2回(低ストリンジェンシー洗浄)
2)0.2xSSPE、0.1%SDS中Tm-20℃で15分間を1回(中ストリンジェンシー洗浄)
【0053】
オリゴヌクレオチドプローブに関して、ハイブリダイゼーションは、6xSSPE、5xデンハルト液、0.1%SDS、0.1mg/ml変性DNAの中において、ハイブリッドの融解温度(Tm)より10〜20℃低い温度で一晩実行することができる。オリゴヌクレオチドプローブのTmは、下記式により決定された。
Tm(℃)=2(T/A塩基対数)+4(G/C塩基対数) (Suggs, S.V., T. Miyake, E.H. Kawashime, M.J. Johnson, K. Itakura, and R.B. Wallace [1981] ICN-UCLA Symp. Dev. Biol. Using Purified Genes, D.D. Brown [ed.], Academic Press, ニューヨーク, 23:683-693)。
【0054】
洗浄は、以下のように実行することができる。
1)1xSSPE、0.1%SDS中室温で15分間を2回(低ストリンジェンシー洗浄)
2)1xSSPE、0.1%SDS中ハイブリダイゼーション温度で15分間を1回(中ストリンジェンシー洗浄)
【0055】
一般に、ストリンジェンシーを変更するために、塩および/または温度を変更することができる。長さが>70またはそのような塩基の標識されたDNA断片では、下記の条件が使用される。
低:1または2xSSPE、室温
低:1または2xSSPE、42℃
中:0.2xまたは1xSSPE、65℃
高:0.1xSSPE、65℃。
【0056】
二重鎖形成および安定性は、ハイブリッドの2本鎖の間の実質的相補性に左右され、および前述のように、ある程度のミスマッチは許容され得る。従って本発明のプローブ配列は、説明された配列の変異(単独および複数の両方)、欠失、挿入、およびそれらの組合せを含み、ここで該変異、挿入および欠失は、関心のある標的ポリヌクレオチドとの安定したハイブリッドの形成を可能にする。変異、挿入、および欠失は、多くの様式で所定のポリヌクレオチド配列を作出することができ、およびこれらの方法は、当業者に公知である。他の方法は、今後公知となると思われる。
【0057】
遺伝暗号の縮重/縮退のために、多種多様なDNA配列が、本明細書において開示されたアミノ酸配列をコードすることができる。これは、同じ、または本質的に同じ酵素をコードしている代替のDNA配列を作成するために、訓練を受けた当業者の技術内である。これらの変種DNA配列は、本発明の範囲内である。
【0058】
本発明は、例えば以下を含む:
1)野生型生物から得たタンパク質;
2)変異から生じた変種;
3)保存的アミノ酸置換を生じることによりデザインされた変種;および
4)本TCタンパク質をコードしている複数の異なる配列のランダム断片化および再構築により作出された変種(DNAシャッフリング)。例えば米国特許第5,605,793号参照。
【0059】
本タンパク質をコードしているDNA配列は、野生型配列、変異配列、または予め決定されたタンパク質を発現するようにデザインされた合成配列であることができる。例えばポリアデニル化シグナルを避け、および植物に好ましいコドンを使用することにより、植物において高度に発現するようにデザインされたDNA配列は、特に有用である。
【0060】
ある種のタンパク質および遺伝子が、本明細書において特別に例示されている。これらのタンパク質および遺伝子は単なる例証であるので、本発明は例証されたタンパク質と同じまたは同様の機能性を有する変種または同等のタンパク質(および、それらの同等物をコードしているヌクレオチド配列)の使用を含むことは、容易に明らかであるはずである。同等のタンパク質は、例証された酵素(またはそれらの活性断片)とのアミノ酸類似性(および/または相同性)を有すると思われる。好ましい本発明のポリヌクレオチドおよびタンパク質は、より狭い同一性および/または類似性範囲に関して定義することができる。例えば、酵素タンパク質の同一性および/または類似性は、本明細書に例示されたまたは示唆された配列と比較して40%、41%、42%、43%、44%、45%、46%、47%、48%、49%、50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、または99%であることができる。先に列記されたいずれかの数字を使用し、上限および下限を定義することができる。例えば、本発明のタンパク質は、例えば例示されたタンパク質と、50〜90%の同一性を有すると定義することができる。
【0061】
特に指定しない限りは、本明細書で使用されるふたつの核酸の%配列同一性および/または類似性は、KarlinおよびAltschulの論文(Proc. Natl. Acad. Sci. USA 90:5873-5877(1993))において改変された、KarlinおよびAltschulのアルゴリズム(Proc. Natl. Acad. Sci. USA 87:2264-2268(1990))を用いて決定される。このようなアルゴリズムは、Altschulらの論文(J. MoL. Biol. 215:402-410(1990))のNBLASTおよびXBLASTプログラムに組み込まれる。BLASTヌクレオチド検索は、NBLASTプログラムにより、スコア=100、ワードレングス=12で実行される。ギャップ付きBLASTは、Altschulら(Nucl. Acids Res. 25:3389-3402(1997))に説明されたように使用することができる。BLASTおよびギャップ付きBLASTプログラムを使用する場合、代表的プログラム(NBLASTおよびXBLAST)のデフォルトのパラメータが使用される。NCBI/NIHのウェブサイトを参照のこと。
【0062】
比較目的でギャップ付きアラインメントを得るために、Vector NTI Suite 8(InforMax, Inc., ノースベセスタ, MD, U.S.A.)のAlignX関数を、デフォルトのパラメータで使用することができる。典型的にはこれらは、ギャップオープニングペナルティ15、ギャップイクステンションペナルティ6.66、およびギャップセパレーションペナルティ範囲8であると考えられる。2種またはそれよりも多い配列を整列化し、この方法で、または当技術分野において周知の他の技術を用い比較することができる。このようなアラインメントを分析することにより、本ポリペプチドの比較的保存された領域および保存されない領域を同定することができる。これは、例えば、1個または複数のアミノ酸残基の修飾または置換によるポリペプチド配列の変化が、許容されると予想され得るかどうかの評価に有用である。
【0063】
アミノ酸の相同性/類似性/同一性は、典型的には(しかし必ずしもではない)、その活性を説明するタンパク質、または活性に最終的に寄与する三次元形態の決定に関与したタンパク質の領域において、最高である。これに関して、ある種のアミノ酸置換は許容でき、および耐容できると予想することができる。例えばこれらの置換は、活性に重要でないタンパク質の領域内であることができる。タンパク質の結晶構造の分析、およびソフトウェアによるタンパク質構造のモデリングは、実際タンパク質の特性を変更および/または機能を増大するために、修飾することができるタンパク質の領域を同定する(部位特異的突然変異、シャッフリングなどを使用する)ために使用することができる。
【0064】
タンパク質の様々な特性および三次元特徴も、タンパク質の活性/機能に有害に作用することなく、変更することができる。保存的アミノ酸置換は、許容され/分子の三次元形態に有害に作用しないと予想され得る。アミノ酸は、以下のクラスに入れることができる:非極性、非帯電極性、塩基性および酸性。それによりひとつのクラスのアミノ酸が、同じ型の別のアミノ酸と交換される保存的置換は、その置換がその化合物の生物学的活性に有害でない限りは、本発明の範囲内に収まる。以下のリストは、各クラスに属するアミノ酸の例を提供する。

【0065】
場合によっては、非保存的置換も行うことができる。重要な要因は、これらの置換は、タンパク質の機能活性/生物学的活性/酵素活性を有意に損なってはならないことである。
【0066】
植物における異種遺伝子の高度な発現を得るために、例えば、該遺伝子が植物細胞においてより効率的に発現されるように、これらを再操作することが好ましいことがある。配列は一般に、植物における最適化された発現のためにデザインすることができるか、またはこれらは植物の特定の型における最適化された発現のためにデザインすることができる。カノーラは、該植物中での異種遺伝子の発現レベルを増大するように形質転換する前に、異種遺伝子を再デザインすることが好ましいそのような植物のひとつである。従って真菌タンパク質をコードしている遺伝子のデザインにおける追加の工程は、例えば、異なる型の生物における最適な発現のための異種遺伝子の再操作である。植物発現について最適化される合成遺伝子の作製に関する指針は、例えば、米国特許第5,380,831号に見出されることができる。植物における発現について最適化された配列は、本明細書において、SEQ ID NO:1(これはSEQ ID NO:5に示された例示されたタンパク質をコードしている)として例示されている。
【0067】
本明細書で使用される「単離された」ポリヌクレオチドおよび/またはタンパク質、ならびに「精製された」タンパク質の言及は、それらが天然では一緒に見られる他の分子と会合しない場合、これらの分子を意味する。従って「単離された」および/または「精製された」の言及は、本明細書に説明されたような「人手(hand of man)」の関与を示している。例えば、発現のために植物へ入れられた本発明の真菌ポリヌクレオチド(または「遺伝子」)は、「単離されたポリヌクレオチド」である。同様に本発明のタンパク質は、植物により作製された場合、「単離されたタンパク質」である。
【0068】
「組換え」分子は、組換えられた分子を意味する。核酸分子に関して使用される場合、この用語は、分子生物学的技術により一緒に結びつけられる核酸配列で構成される分子を意味する。「組換え」という用語は、タンパク質またはポリペプチドに関して使用される場合には、1種または複数の組換え核酸分子を使用し作製されるタンパク質分子を意味する。
【0069】
「異種」という用語は、核酸配列について使用される場合は、天然には結びつけられない核酸配列、または天然とは異なる位置で結びつけられる核酸配列に、ライゲーションされるか、またはライゲーションするように操作される、ヌクレオチド配列を意味する。従って「異種」という用語は、核酸分子が、遺伝子操作を用い、すなわち人的介入により、操作されることを意味する。従って本発明の遺伝子は、異種プロモーター(または、転写調節領域が、関心のある配列へ機能的に連結されている場合には、関心のあるヌクレオチド配列の転写を媒介もしくは変調することが可能なヌクレオチド配列を意味する「転写調節領域」)に機能的に連結することができる。好ましい異種プロモーターは、植物プロモーターであることができる。プロモーターおよび/または転写調節領域および関心のある配列は、これらの配列が、転写調節領域により媒介または変調される関心のある配列の転写を可能にするように機能的に連結されている場合には、「機能的に連結されている」。一部の態様において、機能的に連結されるために、転写調節領域は、関心のある配列と同一鎖上に位置することができる。転写調節領域は、一部の態様において、関心のある配列の5'側に位置することができる。そのような態様において、転写調節領域は、関心のある配列のすぐ5'側であることができるか、またはこれらの領域の間に配列が介在することができる。転写調節領域と関心のある配列の機能的連結には、転写調節領域に結合される適当な分子(例えば、トランスジェニックアクチベータータンパク質)が必要であり、従って本発明は、インビトロまたはインビボのいずれかにおいて、そのような分子が提供される態様を包含している。
【0070】
本発明に使用するためのタンパク質を得る多くの方法が存在する。例えば、本明細書において開示されたタンパク質に対する抗体を使用し、混合物から他のタンパク質を同定および単離することができる。特に抗体は、最も一定でありかつ他のタンパク質から最も異なるようなタンパク質の部分に対し生じると考えられる。その後これらの抗体を使用し、免疫沈降、酵素結合免疫吸着アッセイ法(ELISA)、またはイムノブロッティングにより、特徴的活性を伴う同等のタンパク質を特別に同定することができる。本明細書において開示されたタンパク質または同等のタンパク質、またはこれらのタンパク質の断片に対する抗体は、標準の手法を用い、容易に調製することができる。このような抗体は、本発明の局面である。
【0071】
本明細書において言及されたかまたは示唆されたいずれかの単離体に「由来する」または「から入手可能」なタンパク質は、そのタンパク質(または同様のタンパク質)を例示された単離体または他の給源、例えば他の真菌株もしくは細菌株、または植物(例えば、このタンパク質を作製するように操作された植物)から得ることができることを意味する。「に由来する」は、この言外の意味も有し、ならびに例えばポリヌクレオチドが植物における発現のために修飾されているような、所定の型の真菌または細菌から入手可能なポリヌクレオチド(およびタンパク質)を含む。当業者は、真菌の遺伝子およびタンパク質の説明が与えられたならば、植物を、そのタンパク質を産生するように操作することができることを容易に認識すると思われる。抗体調製物、核酸プローブ(例えばDNAおよびRNA)などは、本明細書において開示されたポリヌクレオチドおよび/またはアミノ酸配列を用いて調製することができ、ならびに他の(天然の)給源からの他のタンパク質遺伝子のスクリーニングおよび回収に使用することができる。
【0072】
本発明の油は、高度の酸化安定度を維持するが、より低いレベルの飽和脂肪酸およびより高いレベルの不飽和脂肪酸を含む。好ましい本発明の油は、総飽和脂肪酸含量3.5%未満、オレイン酸含量少なくとも75%(および好ましくは驚くべきことに80%未満)、ならびに多価不飽和脂肪酸含量20%未満(およびより好ましくは15%未満、更により好ましくは10%未満、および更により好ましくは9%未満)を有する。本発明は、総脂肪酸含量の2.5%を超えない(および好ましくはそれ未満)総飽和脂肪酸含量(C:14、C:16、C:18、C:20、C:22、およびC:24)を有する、好ましくは前述の範囲のオレイン酸を伴う、カノーラ種子を実現するためにも使用することができる。油の不安定性に寄与する18:2および18:3のレベルは、増加しないか、または好ましくは低下する(食品用途のために)。これらの特定の脂肪酸のいずれか、それらの組合せのいずれか、および特にC18多価不飽和体の一方または両方についての範囲のエンドポイントは、本明細書に示された図および表のいずれかから得ることができる。
【0073】
本発明は、総飽和体が3.5%未満の精製油/脱臭油を生じる、農学的に精選されたカノーラ種子を提供するために使用することができる。これらの植物に由来した油は、様々な最終製品の配合に使用することができるか、またはこれらは「無飽和」(または「低飽和」)製品のための独立型(stand-alone)フライ用油として使用することができる。
【0074】
特に記さない限りは、カノーラ種子の所定の集合の飽和脂肪酸含量は、油が、種子の粉砕により種子から除去され、およびメタノールおよび水酸化ナトリウムとの反応後、脂肪酸メチルエステルとして抽出されるような標準の手法により決定することができる。次に得られるエステルを、脂肪酸含量について、不飽和度および鎖長を元にした分離を可能にするキャピラリーカラムを使用する気液クロマトグラフィーにより分析される。この分析手法は、例えば、J. K. Daunらの論文(J. Amer. Oil Chem. Soc. 60:1751-1754(1983))に説明されている。
【0075】
カノーラ種子の脂肪酸組成は、「ハーフ種子」分析、「単独/全種子」分析、または「バルク種子」分析のいずれかについて以下に説明されたように決定された。「ハーフ種子」分析について、胚由来の子葉組織の一部を除去し、分析し;その後残りの種子を保存し、所望の場合には発芽させた。ハーフ種子技術は、安定して遺伝的に制御された脂肪酸変異の選択(およびその後の育種および交雑)の点で若干信頼性に欠けるが、本発明は、好ましい遺伝子が導入され、かつ安定した系統の作出に使用されたことを明らかにしている。特徴的でない変異とは異なり、遺伝子は、植物に導入されおよび安定して維持されることは当技術分野において周知である。従って、本明細書に説明された分析は、本遺伝子の利用性および指定された特徴を有するカノーラ油を実現することができることを明らかにしている。
【0076】
脂肪酸の「無飽和体(No saturate)」(すなわち、「無飽和」)レベルは、市販のNexera 710(カノーラ)生殖質由来のトランスジェニック系統からの種子において達成された。無飽和レベルは、合わせた飽和体3.5%未満として定義される。加えて、低下された飽和体が、Westarカノーラ系統、および同じ形質転換構築体を伴う他のCrucifer(アラビドプシス)の両方において認められた。注目すべきことに、単独の種子中の飽和体レベルは、一部の種子について2.6〜2.7%に低下した。本発明は、2.5%またはそれ未満の総飽和体を伴う種子を作製するためにも使用することができる。これは、油の加工は、総飽和「スコア」を約0.5〜1%追加するので重要であり、このことは加工された油製品は、依然標準の試験手法を用い測定できるFDAにより規定された「無飽和」レベルに達することを意味する。この許容レベルを有することは、単に試験装置のあるレベルの夾雑(より高い飽和種子により)を可能にする(特にプラント操作者が、種子バッチの識別の維持に余り習熟していない場合)のみではなく、圃場での成長条件のあるレベルの変動(より多くの飽和体を作製する傾向がある高温など)および隣接圃場の改良されないカノーラからの花粉の漂流(drifting)による交雑受粉(これは望ましい遺伝子を希釈する)も可能にする。
【0077】
米国食品医薬品局は、「飽和脂肪」を、21 CFR 101.9(c)(2)(i)において、「二重結合を含まない全ての脂肪酸の合計として定義された、サービング中の飽和脂肪のグラム数の表記」と規定している。特に指定しない限り、これは本明細書において「総飽和体」および「総飽和脂肪」について使用される定義である。食品のサービングは、21 CFR 101.9(c)(2)(i)において、その食品がサービング中に総脂肪の0.5g未満を含有する場合には、「飽和脂肪なし」と考えられる。「総脂肪」は、21 CFR 101.9(c)(2)において、「総脂質脂肪酸として定義されおよびトリグリセリドとして表された、サービング中の総脂肪のグラム数の表記」と規定される。様々な種類の食品に関する「サービングサイズ」が、21 CFR 101.12(b)において規定されており、これは、油のサービングは大さじ1杯または15mlと規定している。本明細書において使用されるように、これは14gを意味すると理解される。従って「無飽和」カノーラ油(または、飽和脂肪を含まないカノーラ油)は、本明細書において、サービング中の総飽和脂肪0.5g未満を含有するカノーラ油と定義される(カノーラ油14gは、脂肪14gを含む)。別の表現をすると、「無飽和」カノーラ油は、総飽和体3.57%未満を含有する(総飽和体0.5gを、総脂肪14gで除算)。特に指定しない限りは、本明細書の脂肪酸のパーセントは全て、その脂肪酸が成分である油の質量%である。
【0078】
本明細書に記されたように、本発明は驚くべきことに、カノーラ種子から油を得るために使用することができ、ここで該油は、総飽和体3.57%未満を含有する。油は、前段において言及したような、当技術分野において周知の手法を用い、本種子から得ることができ、およびこの油は、本明細書に例証された技術を含む周知の技術を用い、含量についてアッセイすることができる。別に特定しない限りは、ハーフ種子油データおよび圃場油(field oil)データを作製するために使用された分析は、塩基触媒されたトランスエステル化反応(AOCS Ce 2-66、代替法)を使用した。このプロトコールは、本明細書に説明されたケン化/酸エステル化プロトコールに類似しているが、ただこのケン化/酸エステル化プロトコールは、総脂質を測定し、その大半は塩基触媒されたトランスエステル化反応により検出されたトリアシルグリセリド由来の同じ脂肪酸であることが異なる。
【0079】
市販のNexera 710生殖質において、酸化不安定度に寄与している18:3脂肪酸のレベルは、比較的普遍であった。従って本発明は、単に低級飽和脂肪を伴う植物、種子、および油を提供するのみではなく、非常に驚くべきことに他の有益な特性を維持している植物、種子、および油も提供する。すなわち本発明の植物および遺伝子は、驚くべきことに、植物の他の有利な特徴に有害に作用することなく、使用することができる。
【0080】
好ましい態様において、本発明は、複数種の本発明のΔ9デサチュラーゼ遺伝子の発現されたコピーを含む植物を提供する。本明細書に示された結果は、この遺伝子の多コピーの発現は、驚くべきことにカノーラ植物の脂肪酸プロファイルを改善した(飽和脂肪レベルは大きく低下した)ことを示している。これは、当技術分野は、同じ遺伝子の多コピーの発現に関してこれまで予測不可能であったので部分的に驚くべきことである。「遺伝子サイレンシング」は、異種遺伝子の多コピー(例えばゲノム中の異なる位置に挿入された)の使用に対して示す公知の現象のひとつである。同じく複数の形質転換事象を得ることを試みることは理想的ではない。従って複数種(2、3、4種など)のΔ9デサチュラーゼ事象を含む植物を作出する動機は存在しなかった。そのような植物が実際に改善された特徴を有することも期待されななかった。
【0081】
アブラナ科(Cruciferous)植物のふたつの例が、本明細書において特に例示されている:セイヨウアブラナ(Brassica napus)(カノーラ)およびアラビドプシス(Arabidopsis)。しかし当技術分野において公知であるように、他のアブラナ種および他のアブラナ科を、例えば、カノーラなどにおける品種改良および望ましい形質の開発に使用することができる。従って本発明で使用することができるその他のそのような植物は、カブ(Brassica rapa)、カラシナ(Brassica juncea)、ブラッシカ・カリナタ(Brassica carinata)、クロガラシ(Brassica nigra)、キャベツ(Brassica oleracea)、ダイコン(Raphanus sativus)、およびシロガラシ(Sinapis alba)を含む。ダイズ、ダイズ植物、およびダイズ油も、本発明に従う改善について試験することができる。
【0082】
好ましい態様において、本発明のΔ9デサチュラーゼ遺伝子は、植物発現のために最適化される。従って本発明は、植物に最適化されたΔ9デサチュラーゼ遺伝子も提供する。本明細書において例示された最適化は、好ましいコドンおよびコザック翻訳開始領域の導入、ならびに望ましくない配列の除去を含んだ。この遺伝子は、β-ファセオリンプロモーター(強力な双子葉種子貯蔵タンパク質プロモーター)により駆動される。
【0083】
その発現が油合成と同時発生するためのプロモーター(例えばACP、エロンガーゼ)を使用し、発現はタンパク質貯蔵よりもより早期に生じるので、更に飽和体を減少させることができる。(タバコ構築体がnos 3'UTRを使用する前、およびトウモロコシ構築体が構成的メイズ(maize)ユビキチン-1プロモーターおよびnos 3'UTRを使用する前)。ビシリン、レクチン、クルシフェリン、グリシン、およびコングリシニンプロモーター、US20030005485 A1に開示された植物種子プロモーター、US20030159173 A1のエロンガーゼプロモーター、および米国特許第5,767,363号のACPプロモーターを含む、他の双子葉種子プロモーターを、本発明に従い使用することができる。同じく例えば、US6100450A(種子特異性、胚において発現、8段8行);US20030159173A1(0044段の種子特異性プロモーター;例はUSP、ホルデニン、ACP、ナピン、FatB3、およびFatB4である);WO9218634A1(緒言は、他の特許の1〜7頁の種子特異性プロモーターを考察している);WO0116340A1(7頁13行は、「種子特異的」プロモーターの定義を提供し、これは典型的には、他の組織においては5%未満で発現する;10頁19〜29行は、アルブミン、グロブリン、ビシリンおよびレグミン様タンパク質などの、種子貯蔵タンパク質、非貯蔵オレオシン、ACP、サチュラーゼ、デサチュラーゼ、エロンガーゼのような脂肪酸代謝に関連したプロモーターを考察している);WO2003014347A2(プロモーター定義は23〜25頁:好ましくは種子特異性が2倍より大きい);US20030233677A1(0033段は、「種子プロモーター」の例を提供する[ナピン、ACCase、2Sアルブミン、ファセオリン、オレオシン、ゼイン、グルテリン、デンプン合成酵素、デンプン分枝酵素]);WO2003092361A2(15頁は、「プロモーター」の定義を提供する;17頁の冒頭は、ゼイン、7S貯蔵タンパク質、ブラジルナッツタンパク質、phe非含有タンパク質、アルブミン、β-コングリシン、11S、α-ホルドチオニン(hordothionin)、アルセリン、レクチン、グルテニンを含む、プロモーター例および参照特許を提供する(貯蔵タンパク質のみ));US20030148300 A1(請求項8参照、ナピンプロモーター、ファセオリンプロモーター、ダイズトリプシンインヒビタープロモーター、ACPプロモーター、ステアロイル-ACPデサチュラーゼプロモーター、ダイズ7Sプロモーター、オレオシンプロモーター、コングリシンプロモーター、オレオシンプロモーター、胚発生期に豊富な(embryogenesis-abundant)タンパク質プロモーター、胚グロブリンプロモーター、アルセリン5、ナピンプロモーター、および酸性キチナーゼプロモーター);米国特許第5,777,201号(6段、30〜50行、構成的プロモーター、種子-および/または発生上で調節されたプロモーター、例えば植物脂肪酸脂質生合成遺伝子[ACP、アシルトランスフェラーゼ、デサチュラーゼ、脂質転移タンパク質]、または種子プロモーター[ナピン、クルシフェリン、コングリシニン、レクチン]、または誘導性プロモーター[光、熱、創傷インデューサー])も参照のこと。
【0084】
高等植物の色素体は、遺伝子操作の魅力的標的である。葉緑体(色素体のひとつの型)形質転換が実現され、利点がある。例えば米国特許第5,932,479号;同第6,004,782号;および同第6,642,053号参照。同じく米国特許第5,693,507号および同第6,680,426号参照。葉緑体ゲノムの形質転換の利点は:形質転換された葉緑体は、母系遺伝のみであり、従って花粉による他の植物へ異系交配により伝達されないので、潜在的に環境に安全であること;外来遺伝子の高コピー数を実現する可能性があること;および、高度にエネルギー依存型である葉緑体へのタンパク質の輸入が減少されるので、植物エネルギーコストを削減すること;が挙げられる。
【0085】
植物色素体(葉緑体、アミロプラスト、エライオプラスト、エチオプラスト、クロモプラスト 有色体など)は、光合成に加え、アミノ酸、複合糖質、脂肪酸、および色素などの多くの産業上重要な化合物の生成に寄与している主要な生合成中心である。色素体は、原色素体として公知の共通の前駆体に由来し;従って、所定の植物種中の色素体は全て、同じ遺伝的内容物を有する。
【0086】
ほとんどの植物の色素体は、母系遺伝である。結果的に、核において発現された異種遺伝子とは異なり、色素体において発現された異種遺伝子は、花粉により拡散しない(disseminate)。従って植物色素体へ導入された形質は、野生型類縁に伝達されないと考えられる。このことは、これらの形質は野生型類縁へは伝達されないので、除草剤耐性などの、自然条件または化学条件に対する忍容性または抵抗性に関する植物の遺伝子操作に利点をもたらす。
【0087】
高等植物の色素体ゲノム(プラストーム)は、葉細胞1個当たり1,900〜50,000コピーで存在する、120〜160kbの環状二本鎖DNA分子である(Palmer, 1991)。一般に植物細胞は、小型の120〜160kbの環状ゲノムを500〜10,000コピーを含み、その各分子は、大きい(約25kb)逆方向反復を有する。従って、植物細胞を、関心のある特定の遺伝子の最大20,000コピーを含むように操作することは可能であり;これは潜在的に、非常に高レベルの外来遺伝子発現を生じることができる。
【0088】
本発明の油は、工業用途に加え、様々な食品用途に適用可能であり、特にそれらに合わせて作製することができる。本発明は、料理用油を除き、それ自身、ポテトチップなどの「無飽和」製品を含む(米国特許第6,689,409号参照、これはジャガイモおよびカノーラ油を含有するフライ食品組成物を特許請求しているが;しかし本発明は、米国特許第6,689,409号に開示された組成物を改善するために使用することができる。)。
【0089】
本発明の植物は、特徴および形質の様々な望ましい組合せを実現するために、他の植物と交雑することができる。より更なる改善は、本植物の交雑、公知の育種技術の使用、ならびに追加のもしくはその他の有益な形質および特徴を有する他のカノーラ系統のような生殖質の他の有利な給源により行うことができる。別の例は、色素体Δ9デサチュラーゼを有する系統との交雑である。
【0090】
従って本発明を使用し、変動する環境条件下で、業務用品質油中の、総飽和脂肪酸3.5%未満(および、育種家種子における種子油中の総飽和脂肪酸3%未満)を実現することができる。これは、貯蔵タンパク質の品質および量を低下することなく、カノーラ粗挽き粉中の消化が悪い繊維の増加を伴わずに、および1エーカー当たりの種子収量(または他の望ましい農学的形質)に負の影響を及ぼさずに、達成され得る。
【0091】
以下に、本明細書において使用される脂肪酸の一般名を、それらの炭素原子および二重結合の数と共に列記する。飽和脂肪は、0個の二重結合を有する。
【0092】
(表1)
*=Δ9位の二重結合
**=Δ-11位の二重結合
【0093】
本明細書において言及されたかもしくは引用された全ての特許、特許出願、仮出願、および刊行物は、それらの全体が、本明細書の系統的技術と矛盾しない程度に、本明細書に参照として組入れられている。
【0094】
特に明記または暗示しない限りは、本明細書において使用される「1つの(a)、1つの(an)、およびその(the)」という用語は、「少なくともひとつ」を表す。
【0095】
下記実施例は、本発明を実践する手法を例証するものである。これらの実施例は、限定として構築されるものではない。全てのパーセントは質量%であり、および溶媒混合物の割合は、特に指定しない限りは容量%である。
【0096】
実施例1−Δ9デサチュラーゼ遺伝子再構築
植物-好ましい翻訳コドンへのアスペルギルス・ニジュランス配列の変化、独自の制限酵素部位の導入、および望ましくない配列および一部の二次構造の除去の組合せにより、本発明のΔ9デサチュラーゼ遺伝子を、植物発現のために再デザインした。再デザインされた遺伝子は、Operon, Inc.で合成された。このポリヌクレオチドのオープンリーディングフレームの配列は、ここでSEQ ID NO:1として示した。コザック配列およびBamHIクローニング部位(キャップ)が先立つORFの配列、加えてORFの末端の翻訳ターミネーター(キャップ)は、SEQ ID NO:2に記した。
【0097】
実施例2−Δ9デサチュラーゼ植物形質転換ベクターの構築
BamHI-BstEII遺伝子断片を、Pvβ-ファセオリンプロモーターとPvβ-ファセオリン3' UTR(pPhas-UTR)の間で、ベクターにクローニングした。この構築体は、pOILと称した。プロモーター-遺伝子-UTR断片を、NotIによる消化によりpOILから切り出し、平滑末端化し、ベクターpOEA1の平滑Pmel部位へクローニングした。最終のベクターを、pPD9-OEA1と称した。
【0098】
実施例3−pPD9-OEA1による植物形質転換
プラスミドベクターpPD9-OEA1を、アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)[C58GV3101株(C58ClRifR) pMP90 (GmR), Koncz and Schell、 Mol. Gen. Genet (1986)]へ形質転換した。その後Δ9-デサチュラーゼ植物を、アグロバクテリウム・ツメファシエンスが媒介した植物の形質転換により得た。
【0099】
アラビドプシスを、当技術分野において周知の手法である「浸潤法」により形質転換した。植物は、自殖し、乾燥した種子を、FAME(脂肪酸メチルエステル)分析のために収穫した。
【0100】
カノーラの形質転換に使用したプロトコールは、Katavicの論文[Katavic, Campbell, L., Friesen, L., Palmer, D., Keller, W., and Taylor, D.C. (1996), 「Agrobacterium- mediated genetic transformation of selected high erucic acid B. napus cultivars」、第4回Canadian Plant Tissue Culture and Genetic Engineering Conference, サスカトーン, SK, 1996年6月1-4日]に説明されたものであり、DAS's Nexera系統のために改変された。胚軸切片を、セイヨウアブラナ(B. napus)、cv WestarまたはNexera 710の6-日齢の実生から単離し、ならびにカルス誘導培地(callus initiation medium)において培養し、その後形質転換した。形質転換の日に、胚軸を、Δ9-デサチュラーゼ遺伝子を含むプラスミドDNAの断片が細胞染色体に組み込まれるような、形質遺伝子を伴うプラスミドpPD9-OEA1を含むアグロバクテリウム培養物と共に同時インキュベーションした。共培養期間後、胚軸を、選択剤としてアンモニウムグルフォシネートを含むカルス誘導培地へ移した。健全な耐性カルス組織を得、約12〜16週間新鮮な選択培地へ繰り返して移した。植物を、再生し、Convironsグロースチャンバーへ移した。植物を自殖し、種子を得た。トランスジェニック植物が不稔である場合、これらは改良していないNexera 710系統の花粉で交雑した。乾燥種子を、FAME分析のために収穫した。
【0101】
実施例4−カノーラ事象選別法、およびカノーラの結果のまとめ
特に記さない限りは、下記手法を用い、Nex710/Δ9カノーラ種子を得、そのデータは、引き続きの実施例に提示した。
【0102】
一般に、発育、選択、または選別事象に関して以下の4種の主要な工程が存在する。最初のトランスジェニック事象の選別、T1およびT2種子の選別、T1またはT2植物の選別、ならびに圃場性能による事象の選別。形質転換されたカルスは、最初にT0植物へ再生された。
【0103】
最初の選別に関し、107推定トランスジェニックを、作物栽培学によりおよびサザンブロットにより(単純または複雑、すなわち3コピーよりも多いコピー)スクリーニングした。1事象につき複数の種子をスクリーニングした。T1種子飽和体を決定し、C16:1/C16:0比を決定し(触媒効率を暗示する)、ならびに生化学的表現型の分離を決定した。これらのデータおよび種子利用度(時期、量)を元に、種子の限定されたサブセットを先に進めた。
【0104】
T1およびT2種子の選別(少数の事象は、1世代により進行される)に関して、ハーフ種子分析を行い、可能性のあるホモ接合体を同定した。このデータを元に、分離集団からハーフ種子を、温室生育のために選択した。
【0105】
次の主要な工程は、T1またはT2植物の選別である(少数の事象は、1世代により進行される)。サザンを、導入遺伝子の組込みの完全性を決定するために行った。接合状態も、インベーダーアッセイ法(Third Wave Technologies, Inc.)により決定した。これらのデータを用い、圃場試験のための種子を選択した。
【0106】
T1温室試験のために、1事象につき30個の種子に、ハーフ種子分析を施した。Nex710カノーラを、市販品チェックとして使用した。事象内の全ての個体は、接合状態をサンプリングし、Δ9およびPATに関して対立遺伝子コピー数を決定した。この後、PCRおよびインドールアセトアミド加水分解酵素(IAAH;負のスコア化またはスクリーニングが可能なマーカー)分析を行った。この事象内の全ての個体は、葉を着色し(leaf painted)、PAT分離比を決定した。次にこれらの個体に、サザン分析を行い、「無飽和」プロファイル、正のIAAH、ならびにPATおよびΔ9のホモ接合性に取り組んだ。
【0107】
次に、これらはT2分析に使用した(前述の各植物由来の100個の種子を、ハーフ種子分析した)。インベーダーを使用し、コピー数を証明し、かつこの事象は、D9およびPATについて分離したかどうかを調べた。葉の着色を使用し、この系統はPATについて分離するかどうかを調べた。10個の種子バルクの脂肪酸データを、ハーフ種子データ、インベーダー結果、LP、およびIAAH-陽性を元に植物から収集した。
【0108】
第四の主要工程は、圃場性能による選別事象であった(T2またはT3種子を植え付け、T2またはT3植物、その後T3またはT4種子を分析した)。トランスジェニック事象の広範なサンプリングを行った。農学的、サザン、および接合状態を分析した。バッチ種子油分析も行った。これらのデータを元に、遺伝子量を増加する交雑に関する事象を選択した。
【0109】
以下の選択判定基準を用い、系統を圃場試験に進めた。23事象(6レパートリー/エントリー(reps/entry))からの224系統(ヌルを含む)を、3つの場所で、2つ組の苗床(in replicated nurseries)で評価した。6のT3事象および17のT2事象を、植え付けた。圃場へ移す系統/事象の選択は、各植物由来の種子の挿入数およびハーフ種子分析、それに続く10種子バルクの脂肪酸分析を元にした。選択された系統の総飽和割合の範囲は、ほぼ3.3〜4.5%の範囲であった。以下のT3事象を、更なる発育のために選択した。
【0110】
(表2A)
【0111】
以下のT2事象は、更なる発育のために選択した。
【0112】
(表2B)
【0113】
圃場試験は、以下のように行った。農学的評価を、引き続きの実施例において考察したように行い、農学的ペナルティが、Δ9(D9)に関連していることを確認した。更なるコピー数の証明のために、15のインベーダー葉組織試料を、最も有望なT3系統(プラス同胞-ヌル)の28種から収集し、系統がPATから分離しているかどうかを決定した。D9系統を、総飽和3.5%未満を有することを元に選択した。
【0114】
10種のサザン組織試料を、3種の最も有望なT2系統から採取し、これを総飽和3.5%未満を有することを元に選択した。全ての組織-サンプリングした植物を、自家受粉した。脂肪酸分析は、自殖した植物由来の10種の種子バルク(455試料)、およびOP条(rows)由来のバルク種子試料1g(1445)を元に行った。
【0115】
「最良の」T4事象は、以下であった。
【0116】
(表3A)
【0117】
「最良の」T3事象は、以下であった。
【0118】
(表3B)
【0119】
一般に、Δ9に関連した大きい農学的ペナルティは観察されず、および一部の系統は、種子質量の約10%の増加を示した。農学的結果は、下記実施例18においてより詳細に考察している。個々の脂肪酸成分の分布に関する全般的観察は、下記実施例11-13において考察している。事象218-11.30、36-11.19、および69-11.19に関する平均TSATデータ、TSATの変化率(%)、およびある脂肪酸成分の変化率(%)は、表4-7に示した。一般に、同胞-ヌルおよび野生型と比較して、TSATの〜30%から〜40%の減少が観察された。しかし更なる改良が、以下により詳細に考察されており、例えば更に交雑を行うことができる。
【0120】
(表4)3場所にわたる事象218-11.30からの全系統に関する平均TSAT
【0121】
(表5)C16:0、C16:1、C18:0、およびC18:1対ヌルおよび野生型対照Nex 710のTSAT変化率(%)
【0122】
(表6)C16:0、C16:1、C18:0およびC18:1対ヌルおよび野生型対照 Nex 710のTSAT変化率(%)
【0123】
(表7)C16:0、C16:1、C18:0およびC18:1対ヌルおよび野生型対照 Nex 710のTSAT変化率(%)
【0124】
実施例5−植物の分子特徴
葉試料は、DNA分析のために採取し、PCRおよびサザン分析により導入遺伝子の存在を証明し、ならびに場合によってはELISAによりPATタンパク質の発現を確認した。
【0125】
5A. Δ9デサチュラーゼPCR分析のプロトコール:
以下の2種のプライマーを使用した。
Δ9フォワードB:
Δ9リバースB:
このプライマー対は、Δ9遺伝子の増幅後に、〜1380bp断片を生じた。
【0126】
このスクリーニングにおいて、MJ Tetradサーマルサイクラーにより、以下のサイクリングプロトコールを使用した。
1. 94℃、2分間
2. 94℃、1分間
3. 50℃、2分間
4. 72℃、3分間+5秒間/伸長サイクル
5. 工程2-4の25回の反復
6. 分析の準備ができるまで、または少なくとも2分間、4℃
【0127】
5B. サザン分析のための植物ゲノムDNAの抽出プロトコール:
Qiagenから入手したDNeasy Plant Maxi Kitを使用した。そのブックレットのプロトコールを、溶離部分に以下の変更を行い使用した。緩衝液AEは、DNA等級水(Fisher No. BP561-1)で1:10希釈した。予め65℃に温めた希釈したAE緩衝液0.75mlを用い、2回の溶離を行った。DNAは、イソプロパノールで沈殿し、70%エタノールで洗浄した。DNAペレットを、100μlの1X TE緩衝液中に再懸濁した。DNA濃度を定量した。DNA 6μgをアリコートとし、最終容積を40μlに調節した。試料を、-20℃で保存した。
【0128】
5C. FAME分析(メタノール性H2SO4による種子からの直接のFAME合成):
FAME分析のためのプロトコールは、以下である。
【0129】
GC Specs
ガスクロマトグラフィー:デュアル注入口およびデュアルフレームイオン化検出器を備えた、Hewlett-Packard 6890
データシステム:HP Chemstation、Leap Technologies, カルボロ, NC 27510, PAL System
カラム:J&Wキャピラリーカラム、DB-23、6OMx内径0.25mm、マイクロフィルム厚0.15、最大操作温度250℃。カタログ番号:122-2361
温度プロファイル:平衡時間:1分間。開始温度:50℃。開始時間:3分間。上昇率:40℃/分。最終温度:240℃。最終時間:7.25分間。
【0130】
アラビドプシスのFAME手法:
15:0標準100μl(50μg)を、清潔な16x125mmガラスチューブに添加した(内部標準ストック液:2:1クロロホルム:イソプロパノール中500μg/mlのC15:0 TAG)。蒸発している水浴中、窒素下で55℃で標準を乾燥する。
乾燥した時点で、2%DMPを伴う1Nメタノール性H2SO4 2mlを(100mlについて:メタノール95,22ml、H2SO4 2.772ml、DMP=2,2-ジメトキシプロパン2ml)添加する。チューブを85℃に加熱する。
清潔な16x125mmガラスチューブ中で〜5mgのアラビドプシス種子を秤量し、正確な質量を記録する。
リパーゼを破壊するために、温メタノール性H2SO4 を、標準と共に種子の入ったチューブに添加し、85℃で15分間インキュベーションする。
バイアルを〜50℃に冷却し、その後種子をミニグラインダー中ガラス製乳棒で粉砕する。
試料をチューブに戻し、少なくとも1時間窒素下85℃でインキュベーションする。バイアルを氷上で冷却する。最初に0.9%NaCl 0.5ml、その後ヘキサン中の17:0標準250μl(0.1335mg/mlメチルエステルストック液)を添加する。激しく攪拌し、1000gで5分間遠心する。
パスツールピペットで、100〜200μlを、コニカルインサート(conical insert)(0.5ml)を伴う1.5mlバイアルへ移す。
5μlをGCへ注入する。
【0131】
カノーラに関するFAME手法:
15:0標準100μl(50μg)を、清潔な16x125mmガラスチューブに添加した(内部標準ストック液:2:1クロロホルム:イソプロパノール中500μg/mlのC15:0 TAG)。蒸発している水浴中、窒素下で55℃で標準を乾燥する。
乾燥した時点で、2%DMPを伴う1Nメタノール性H2SO4 2ml(100mlについて:メタノール95.22ml、H2SO4 2.772ml、DMP=2,2-ジメトキシプロパン2ml)を添加する。チューブを85℃に加熱する。
清潔なチューブ中で1個のカノーラ種子を秤量し、正確な質量を記録する。
リパーゼを破壊するために、種子材料を温メタノール性H2SO4と共に、標準の入ったチューブに添加し、85℃で15分間インキュベーションする。
バイアルを〜50℃に冷却し、その後種子をガラス製乳棒で粉砕する。
少なくとも1時間N2下85℃でインキュベーションする。
バイアルを氷上で冷却する。最初に0.9%NaCl 0.5ml、その後ヘキサン中の17:0標準250μl(0.1335mg/mlメチルエステルストック液)を添加する。激しく攪拌し、1000gで5分間遠心する。
パスツールピペットで、100〜200μlを、コニカルインサート(0.5ml)を伴う1.5mlバイアルへ移す。
5μlをGCへ注入する。
【0132】
実施例6−アラビドプシスの結果
最初の結果は、図1に図示し、これは飽和脂肪酸の60%よりも大きい減少が達成されたことを示している。同じく16:0(4.4%)よりもより多くの16:1(例えば5.9%)が達成された。
【0133】
Δ9-CoA-デサチュラーゼ法による無飽和油
FAME分析
約18個の追加の形質転換体に由来するT2種子を分析した。このデータ(表8および図2参照)は、最大60〜70%の「飽和体」の減少を示している。これは、示された最初のデータ(図1参照)よりも、更に強力な飽和脂肪酸の減少である。T2世代は、依然分離されることに注目することは大事であり;従って次の世代でより良い成績の(performing)系統が期待される。この点は、形質が固定されおよび系統が導入遺伝子についてホモ接合体であるまで、T1、T2、T3、および本明細書の別所に記された他の最初の世代(カノーラ系統を含む)全てについてあてはまる。(形質が固定される安定した系統および植物が作出され、および次の実施例で説明される。)。
【0134】
(表8)
値は、単独の試料調製物およびGC試行に由来する(平均ではない)。
TF-21は、野生型(非形質転換)植物として挙動し;恐らく説明は、遺伝子サイレンシングまたは非トランスジェニック回避(グルフォシネート系除草剤による不適切な選択)を含む。
「?」は、GCクロマトグラムのピークの正体は疑問があるか、または不明であることを示す。
【0135】
実施例7−Westarデータ
実施例5Cに説明されたものに類似したプロトコールを、周知の「Westar」カノーラ由来のカノーラ系統に適用した。図3に図示したように、示された飽和脂肪は、43%よりも多く減少され、24:0が含まれた場合には、50%の減少が実現された。
【0136】
実施例8−Nexera 710データの例
本明細書の別所に説明されたものに類似したプロトコールを、周知の「Nexera 710」市販の精選カノーラ由来のカノーラ系統に適用した。総飽和体は、本明細書に説明し、以下に特定した方法を用い計算した。
【0137】
総飽和は、16:0+18:0+20:0+22:0+24:0脂肪酸の和に由来する。単独の種子中の一部の注目すべき飽和体レベルは、表9に示した。油プロファイルは、mol%値として表されている。mol%値は、各特定の脂肪酸の式量を計算に組込んでいる。これは、その脂肪酸種の式量で除算された、所定の脂肪酸種の質量(ピーク面積、または%脂肪酸の直接計算に使用される同じ値)を使用する。
【0138】
(表9)
【0139】
最低の総飽和体を伴う種子に由来するプロファイルを、分析した(種子113a 11.19#4、113a 11.19 #8、5 11.19 #6および5 11.19#8)。未修飾のNexera 710生殖質値は、同じFAME分析の試行から得た。植物は、Convironsグロースチャンバーにおいて生育し、そのため実際のmol%値は、圃場で生育した種子とは異なる。一般に、トランスジェニック植物は、16:0、18:0および20:0の減少、ならびに16:1の増加を示す。18:0レベルは一般に、Nexera 710の平均1.4%(上限2.08%、下限0.81%)から、選択トランスジェニック材料の0.1%平均(上限0.6%、下限0%)までに収まる。同じく16:0レベルは、平均4.6%(上限5.12%〜下限4%)から、3.0%(上限3.41%〜下限2.63%)に収まる。同様に、20:0レベルは、Nexera 710の0.5%平均から、選択されたトランスジェニックの0%まで下落する。16:1レベルは、Nexera 710において検出不能であり、トランスジェニックにおいて平均2.3%(上限2.71%〜下限1.72%)まで増加する。平均18:3レベルは、小さいトランスジェニック集団においてわずかに増加したが、値の範囲は、未修飾のNexera 710試料と重複した。これらの結果は、以下のようにまとめられる。
【0140】
(表10)
【0141】
実施例9-更なるカノーラデータ
本明細書の別所に説明されたものに類似したプロトコールを、周知の「Nexera 710」市販の精選カノーラに由来する追加のカノーラ系統に適用した。以下に示したように、総飽和体および脂肪酸の個々の種類の質量%を、本明細書に考察された方法を用いて計算した。
【0142】
図4A-Cおよび5A-Cは、各々、事象36-11.19および218-11.30からの代表的結果を示し、これは本発明の実践により得ることができる減少した飽和脂肪レベルを明らかにしている。本発明に従い更なる操作を行うことにより、ここに例示された飽和脂肪レベルは、更に低下することができる。このデータは全て、示されたように自殖したトランスジェニックカノーラ植物から得た。
【0143】
まとめると、事象36-11.19に関して、温室生育した植物からのT2ハーフ種子分析は、総飽和体を2.57%と低くした。温室生育した植物からのT3全種子に関する総飽和体は、3.66%と低かった。結果は、図4Aに図示している(数値データは、図4Bおよび4Cである)。事象218-11.30温室生育した植物に関して、T2ハーフ種子分析は、総飽和体が2.71%と低いことを明らかにした。T3全種子は、3.37%と低い総飽和体を有した。結果は、図5Aに図示している(数値データは、図5Bおよび5Cである)。参考のために、NATREONは、圃場条件下で、平均して総飽和体6.5%を有した。
【0144】
本発明の更なる改善することにより(自殖、他の優れた系統との交雑、デサチュラーゼ遺伝子コピー数の増加[追加の形質転換、更なる育種交雑などによる]、デサチュラーゼ発現時期の変化、および変異などの追加ラウンド)、総飽和脂肪レベルの更に大きいレベルの減少が実現され得る。
【0145】
実施例10−更なるカノーラデータの分析−総飽和脂肪の%減少
実施例9に示されたデータは、本発明の様々な局面を例示するために、様々な計算において使用することができる。例えば、総飽和脂肪の%減少は、所定の植物の総飽和体の、対照系統の総飽和体による最初の除算、それに続く100%からの減算により計算することができる。本発明によりもたらされるこのような低下の例は、以下に例示される。結果は、最も近い自然数(非小数)に丸めた概数であることができる。
【0146】
(表11)
【0147】
グラフ、図、表または本明細書に考察された他のものに示された数値は全て、本発明の境界および限界を規定するためのエンドポイントとして使用することができる。同様に、先に示されたものおよびより詳細に以下に考察されるもののような、これらの数値を使用する計算は、本発明の境界および限界を規定するために使用することができる。表12-14は、事象218-11.30、36-11.19、および69-11.19を伴う系統に関する、更なる代表的結果および計算を示している。
【0148】
(表12)交雑に関する事象218-11.30 HS選択
【0149】
(表13)交雑に関する事象36-11.19 HS選択
【0150】
(表14)交雑に関する事象69-11.19 HS選択
【0151】
実施例11−更なるカノーラデータの分析−詳細な脂肪酸プロファイル
再度、図4A-Cおよび5A-Cは、事象218-11.30および36-11.19を有する様々な植物の脂肪酸プロファイルを示す代表的結果を示している。一般に、これらの結果は、16:0および18:0レベルが大きく低下されるのみではなく(対応する不飽和体レベルの結果としての増加を伴う)、20:0、22:0、および24:0レベルも有利かつ驚くべきかつ予想外のことに、低下したことを示している。場合によっては、18:2および18:3レベルも低下することができ、これは改良された油の酸化安定性を増強する。更に、先に示唆された比のいずれか(例えば16:0-16:1、18:0-18:1、および例えば18:0-[20:0+22:0+24:0])を使用し、本発明を実践する上での有利な結果を規定することができる。総C20:0+C22:0+C24:0の合計した減少率も、驚くべきことに本発明により実現された。従って本発明は、本明細書に例示されたような、有利かつ改善された脂肪酸プロファイルを有する植物を提供する。本発明に従い更なる改良を作製することにより、より良い飽和体の減少、「無飽和」の増加、およびより良い比を実現することができる。
【0152】
例えば、以下の図5Cおよび図4Bのデータを使用し、様々な計算を、本発明の達成を例示するために使用することができる。図5Cおよび図4Bからの試料データは、下記表に示されている。各指定された脂肪酸の量は、各事象について、括弧内には関連する対照植物について記されている。
【0153】
(表15)
【0154】
218-11.30事象に注目すると、C20:0、C22:0、およびC24:0成分による飽和体への総寄与(contribution)は、対照において1.08%であるが、これらの成分は有利なことに本発明のカノーラ系統においては0.26%に減少している。このことは、これらの飽和体における1/4を超える減少を示している。同様に、各成分は、独立しているとみなすことができる。同じく218-11.30事象に注目すると、C20:0成分は、対照/野生型において0.62%であるが、これは本発明の植物系統において約1/5.6に減少される(0.11%へ低下)。C22:0成分は、約1/2.7に減少され:d-9デサチュラーゼ植物系統においては0.12%であり、これはデサチュラーゼ遺伝子を欠いている対照系統の0.32%よりも低い。C24:0は、0.14%から0.03%へと約1/4.7に減少される。
【0155】
事象36(または36-11.19)に関して、ふたつの系統が、C20:0、C22:0、およびC24:0含量を、各々0.32、0.15、および0.04、ならびに0.30、0.14、および0.07を有する。各々0.64、0.42、および0.21を有する対照と比べ、これらの系統は、約半分のC20:0、および少なくとも約1/3のC22:0およびC24:0の減少を有する。先に言及された第一の系統は実際に、C24:0の1/5を超える減少を示している。
【0156】
これらの好ましい脂肪酸成分のいずれかについて、本発明の他の系統のいずれかについて、非常に多くの同様の計算を行うことができることは、直ぐに明らかになると思われる。これらの例示は、限定として構築されるものではなく、そのような新規の低下および比は、本発明を定義するために使用することができる。
【0157】
実施例12−次世代の更なるハーフ種子データ
更なるハーフ種子FAME分析を、表16に示す。この図は、T3世代における2.64%、およびT4世代における2.66%と低い総飽和を示している。表17は、各系統に存在するD-9デサチュラーゼ遺伝子のコピー数を示している(表16の試料IDおよび表17のID行を参照のこと)。コピー数の作用は、実施例14および19においてより詳細に考察されている。
【0158】
(表16)
【0159】
(表17)
【0160】
実施例13−実施例12のハーフ種子データの更なる分析、ならびに脂肪酸シフト、不飽和体増加、および飽和体減少を示す更なるデータ
T3圃場試験からのハーフ種子データを、脂肪酸含量の「総飽和体」データに対する様々な比較を図示するために、プロットした。図6Aおよび6Bは、ヌル(非機能的挿入断片を伴う事象)および野生型対照(形質転換していない系統)と比べ、トランスジェニック事象におけるC16:0の減少およびC16:1の増加を明らかに示している。図6Cおよび6Dは、ヌルおよび野生型対照と比べ、トランスジェニック事象におけるC18:0の減少およびC18:1の増加を明らかに示している。図6Eおよび6Fは、ヌルおよび野生型対照と比べ、トランスジェニック事象におけるC20:0およびC22:0の各々減少を明らかに示している。
【0161】
同様の結果が、実施例4の考察で得られたデータを用い、棒グラフにおいても示されている。図6Gおよび6Hは、ヌルおよび野生型対照と比べ、トランスジェニック事象におけるC16:0のシフトおよび減少、ならびにC16:1のシフトおよび増加を明らかに示している。図6Iおよび6Jは、ヌルおよび野生型対照と比べ、トランスジェニック事象におけるC18:0のシフトおよび減少、ならびにC18:1のシフトおよび増加を明らかに示している。図6Kおよび6Lは、C18:2およびC18:3について同様の棒グラフを示している。
【0162】
先に考察されたプロットおよび図6Mも、既に良好であるNex 710系統と比べ、総飽和体の非常に驚くべき減少を明らかに図示している。図6Nは、1000個の種子の分布を示している。
【0163】
実施例14−複数のΔ9デサチュラーゼ遺伝子による飽和脂肪レベルの減少
温室の増加および圃場試験からの結果は、総飽和体の減少とアスペルギルスΔ9デサチュラーゼコピー数の間に相関があることを示唆している。
【0164】
14A:事象選別および導入遺伝子コピー数の作用のFAME分析プロトコール
試料調製
1. 個々の種子の質量を得、プラスチック製母プレート上に配置する。
2. 各ウェルに2, 1/8"ボールを加える。
3. 母プレートを液体ハンドラーHamiltonで取り上げ:IS(C11:0)および代理物(C15:0 FAEE)と共にヘプタン400μLを添加し、その後ナトリウムメトキシド(0.5N)100μLを添加する。
4. キャップを、ストリップキャップに嵌め、5分間Genoにより粉砕する(1x500回)。
5. ストリップキャップを再配置する。
6. プレートの蓋をゴム製マット上に置く(余分のシーリング)。蓋を、黒色絶縁(electrical)テープで巻き、母プレートの底を外し、バイアルを露出する。
7. プレートを、ボルテックス/ヒーター上に15分間、37℃/60rpmで配置する(ボルテックスの孔には、砂が詰められている)。
8. プレートをx3500rpmで2分間遠心する。
9. 底蓋を戻し、蓋/ストリップキャップを取り外す。上層350μlを、Hamiltonを用いて、娘プレートへ移す。
10. その後抽出プレートへ、ヘプタン400μLを、ISおよび代理物と共に添加する。
11. 工程5から10を更に2回繰り返す(合計3回移す)。
12. 最後に移した後、抽出プレートをHamilton上に維持する。抽出物50μlを、C11と共にヘプタン450μlを含有するアルミニウムブロック中に搭載されたガラスインサートに移す。
13. GCへ注入する。
【0165】
GC/FID分析
GCパラメータ:
1試料につき、1μl注入、スプリットレス
カラム-
DB23、15m、内径0.25mm、およびフィルム厚0.15μm
GCパラメータ:
オーブン温度プログラム−
70℃で2.15分間維持(スプリットレス)
70℃〜150℃、25℃/分
150℃〜180℃、5℃/分
180℃〜220℃、25℃/分
220℃で2分間維持。
注入温度−230℃
検出温度−240℃
メイクアップガス−窒素 25mL/分
FID燃料−空気 400mL/分、水素 40mL/分
フロント注入器:掃流時間:1分間、掃流:35ml/分
バック注入器:掃流時間:2.15分間、掃流:35ml/分
フロント投入圧:1.0mL/分−定常流
バック投入圧:1.0mL/分−定常流
試行時間: 14.95分間
流量−ヘリウム定常流 1mL/分
【0166】
捕捉配列
96試料を、ライナー上での構造(build)が最小になるように、フロントカラムおよびバックカラム間で分離する。試料の一覧を、注入される試料の種類に応じて、5種の注入法と共に作製している。最初の注入された5試料は常に以下である。
1. マトリックス
2. マトリックス
3. 標準1
4. カノーラ陽性対照
5. 試薬ブランク
6-27. カノーラ試料
33. 標準2
34-54. カノーラ試料
55. 標準3
各リストは、16試料の3事象を含む(48試料)。
標準は、以下のように分布したFAME合計25ppmを含む。
【0167】
(表18)
【0168】
14B:おおよそのコピー数に対する予備サザンブロット分析
これらの目的に使用したDNA調製プロトコールは、以下に示した。DNA約6μgを、HindIIIで消化し、消化したDNAを、0.75%アガロースゲル上を流した。正帯電したナイロンメンブレン上へのブロッティングおよびハイブリダイゼーションは、典型的プロトコールに従った(Maniatis, Roche Applied Science, Inc.)。プローブは、アスペルギルスΔ9デサチュラーゼ遺伝子に由来したDIG標識した(Roche Applied Science, Inc.のキット)PCR産物からなった。洗浄は、2xSSC/0.1%SDSにより室温で5分間を2回、その後65℃の0.1xSSC/0.1%SDSで2回行った。ハイブリダイズされたバンドは、DIG-ルミネセント検出キットにより、製造業者(Roche Applied Science, Inc.)の指示に従い可視化した。ハイブリダイズしているバンドを計測し、トランスジェニック試料は最初に、1〜3本のバンドを示した場合は、「単純」、または3本よりも多いバンドを示した場合は、「複雑」と説明した。
【0169】
14C:トランスジェニック事象における導入遺伝子コピー数および飽和脂肪酸の減少の比較
下記定義を、表に適用した。
【0170】
【0171】
(表19)
【0172】
「飽和体の減少の比」は一般に同じトランスジェニック事象の種子を使用したので、これを事象のランク付けのために使用した。この直接比較は、様々な時点で組織培養により生じた植物と成長した植物の間の変動を低下することを補助している。
【0173】
前記データは、みかけの遺伝子量作用を示し;導入遺伝子のより多いコピーは、より多くの有効な飽和脂肪酸減少を引き起こす傾向がある。例えば、先に示された57種の「非対照」植物が存在する。これらの57種の植物は、19種の植物の3群に分けることができる。植物の一番上のセット(飽和体の最良の減少および最低レベルの飽和体を示している)は、19種の「複雑」事象(デサチュラーゼ遺伝子の3個よりも多いコピー)の11種を有する。このセットは、「単純」または「恐らく単純」と特徴付けられた19事象のうちの8種を含んだ。19種の植物の中間のセットは、「複雑」事象を19種中わずかに6種有した(19種中13種の「単純」または「恐らく単純」事象)。なお更に、19種の植物の第三のセット(飽和体の比較的最低の減少を示す)は、わずかに3種の複雑事象を含み、19事象中の16種は、「単純」または「恐らく単純」であった。従って、3個よりも多いデサチュラーゼコピーを有する細胞を伴う植物は、わずかに1個のこの遺伝子のコピーを有する細胞を伴う植物よりも、飽和体をより良く減少することを示しているように見える。
【0174】
実施例15−オレイン酸およびバクセン酸の分離
バクセン酸は、Δ-11位に二重結合を伴うC18:1である。バクセン酸は、色素体の外側でのΔ9 C16:1の伸長により形成される。本明細書において考察した他の分析法は、オレイン酸およびバクセン酸のピークを一緒に「オレイン酸」と記される単独の組成の割合とするので、その追跡は重要である。すなわち、特に記されない限りは、これらふたつは分離されなかった。バクセン酸の貢献を減算することにより、オレイン酸の貢献割合が、好ましくかつ有利なことに(および驚くべきことに)、80%未満に維持されるが、依然総飽和体の減少(全体の「無飽和」または「低飽和」レベルに対し)は達成されることが、明らかにされる。以下のふたつの実施例において示したように、これを説明するために、2種類の分析を使用した。
【0175】
実施例16−カノーラΔ9種子抽出およびバクセン酸SOPに関する分析
図7Aおよび7Bは、下記プロトコールを使用し得られたデータを図示している。
【0176】
試料調製(前述のものと同じ)
1. 個々の種子の質量を得、プラスチック製母プレート上に配置する。
2. 各ウェルに2, 1/8"ボールを加える。
3. 母プレートを液体ハンドラーHamiltonで取り上げ:IS(C11:0)および代理物(C15:0 FAEE)と共にヘプタン400μLを添加し、その後ナトリウムメトキシド(0.5N)100μLを添加する。
4. キャップを、ストリップキャップに嵌め、5分間Genoにより粉砕する(1x500回)。
5. ストリップキャップを再配置する。
6. プレートの蓋をゴム製マット上に置く(余分のシーリング)。蓋を、黒色絶縁テープで巻き、母プレートの底を外し、バイアルを露出する。
7. プレートを、ボルテックス/ヒーター上に15分間、37℃/60rpmで配置する(ボルテックスの孔には、砂が詰められている)。
8. プレートをx3500rpmで2分間遠心する。
9. 底蓋を戻し、蓋/ストリップキャップを取り外す。上層350μlを、Hamiltonを用いて、娘プレートへ移す。
10. その後抽出プレートへ、ヘプタン400μLを、ISおよび代理物と共に添加する。
11. 工程5から10を更に2回繰り返す(合計3回移す)。
12. 最後に移した後、抽出プレートをHamilton上に維持する。抽出物50μlを、C11と共にヘプタン450μlを含有するアルミニウムブロック中に搭載されたガラスインサートに移す。
13. GCへ注入する。
【0177】
GC/FID分析(FAMEプロファイルとは異なる)
GCパラメータ:
1試料につき、1μl注入、スプリットレス
カラム-
SGEのBPX 70、15m、内径0.25mm、およびフィルム厚0.25μm
GCパラメータ:
オーブン温度プログラム−
70℃で2.15分間維持(スプリットレス)
70℃〜140℃、25℃/分
140℃で14分間維持
140℃〜180℃、10℃/分
180℃で3分間維持
180℃〜220℃、25℃/分
220℃で3分間維持。
注入温度−230℃
検出温度−240℃
メイクアップガス−窒素 25mL/分
FID燃料−空気 400mL/分、水素 40mL/分
フロント注入器:掃流時間:1分間、掃流:35ml/分
バック注入器:掃流時間:2.15分間、掃流:35ml/分
フロント投入圧:1.0mL/分−定常流
バック投入圧:1.0mL/分−定常流
試行時間: 30.55分間
流量−ヘリウム定常流 1mL/分
【0178】
捕捉配列
96試料を、ライナー上での構造が最小になるように、フロントカラムおよびバックカラム間で分離する。試料の一覧を、注入される試料の種類に応じて、5種の注入法と共に作製している。最初の注入された5試料は常に以下である。
1. マトリックス
2. マトリックス
3. 標準1
4. 試薬ブランク
5-32. カノーラ試料
33. 標準2
34-54. カノーラ試料
55. 標準3
【0179】
各リストは、8試料の6事象を含む(1種子=1試料)(48試料)。
【0180】
標準は、以下のように分布したFAME合計200ppmを含む。
【0181】
(表20)
【0182】
実施例17−ガスクロマトグラフィー/質量分析/飛行型を使用するバクセン酸寄与の分析
表21は、下記のプロトコールから得られたデータを示している。表21において、T4について、脂質割合(%)は低下されず;これは、トランスジェニック系統において40.8%で維持された(対照系統と同じ)。
【0183】
(表21)
*:処理法により検出限界未満である場合、デフォルトは0であった。
捕捉および処理は、HP Chemserverパッケージを使用
【0184】
機器の説明
Lecoから入手した飛行時間型質量分析装置Pegasus IIIは、ガスクロマトグラフィーHP 6890と連結した。
CTC Analytics technologyから入手したCombi Pal自動注入器を、10μlシリンジと共に、HP 6890に装備した。
【0185】
GC法
GCパラメータ:
1試料につき、1〜3μl注入、スプリットレス
カラム-
SolGel Wax、30m、内径0.25mm、およびフィルム厚0.25μm
GCパラメータ:
オーブン温度プログラム−
70℃〜175℃で25℃/分(スプリットレス)
175℃で25分間維持
175℃〜230℃、50℃/分
230℃で3分間維持
注入温度−230℃
検出温度−300℃
バック注入器:掃流時間:30秒、掃流:20ml/分
バック投入圧:2mL/分−定常流
試行時間: 33.3分間
流量−ヘリウム定常流 2mL/分
【0186】
質量分析器
質量選択:
50〜600amuから収集した質量
フィラメントバイアス:-70V
イオン給源:225℃
検出器:
検出電圧:1600V
捕捉速度:10スペクトル/秒
溶媒は100秒遅れ
【0187】
フラグメンテーション
ChromaTOFソフトウェアは、より良い分離および解釈のために、フラグメンテーションをコンパイルし、および同時移動するピークを解析する。
【0188】
脂肪酸の同定は、同じ条件で注入された標準液(下記参照)を元に、保持時間およびフラグメンテーション合致、ならびに/または先に説明されたソフトウェアに含まれたNIST/EPA/NIHデータベースとの良好な合致により行う。
【0189】
cis 11-オクタデセン酸メチルエステルとしても公知であるバクセン酸メチルエステルは、Sigmaから得たバクセン酸(CAS:506-17-2)から作製された標準物を試行することにより、カノーラ種子抽出物から同定された。保持時間は、1207秒であり、標準品と853の合致および先に説明された文献と814の合致を生じる。
【0190】
注入された標準品の組成
【0191】
(表22)
【0192】
バクセン酸メチルエステル法:
メチル化された生成物を、この酸100mgのメチル化後に得た。
−100mgを、30mlのガラスバイアル中のMeOHCl 0.5N(Supelco)の5mlに溶解。
−窒素下70℃で1時間加熱。
−室温に冷却後、0.9%NaClを含有する水5mlを添加。
−3回連続したヘキサン(15ml)抽出により、ヘキサン中にエステルを分配。
−この有機層を、HNaCO3を2.5%含有する水15mlと混合することにより、酸残渣を中和。
−有機層を、N2下で蒸発させ、室温で油状の透明な液体を得、これはバクセン酸メチルエステルに対応している。
【0193】
実施例18−優れた農学的形質を維持しながらの「無飽和」の達成
様々な圃場で、農学的測定を行い、トランスジェニックと対照を比較した。結論は、全般的に、このトランスジェニック植物は、Nex 710対照と、同様に挙動し、および類似した形質を示した(はるかに改善された飽和体レベル以外)。従って本導入遺伝子は、植物の健康に一貫して負の作用は有さない。形質および等級化システムのまとめを、表23に示している。様々な判定基準を、様々な場所の圃場で使用した。
【0194】
表24-27は、いくつかの代表的結果を数値で表している。下記の略号を、これらの図において使用した。
DTF=開花までの日数 EOF=開花終了までの日数
HT=高さ SC=不稔性の数
DTM=成熟までの日数 LSV=晩期活力
LOD=倒伏 SD WT=種子質量
【0195】
表24は、事象#218-11.30からの系統の農学的データを示している。表25は、事象#36-11.19からの系統の農学的データを示している。表26は、事象69-11.19からの系統の農学的データを示している。これらのことは、本発明、すなわち「無飽和」カノーラの実現は、驚くべきことに、他の重要な農学的形質への有害な作用を伴わずに実践することができることを明らかにしている。
【0196】
(表23)
【0197】
(表24)事象#218-11.30からの最良の性能の系統の農学的データのまとめ
【0198】
(表25)事象#36-11.19からの最良の性能の系統の農学的データのまとめ
【0199】
(表26)事象#69-11.19からの最良の性能の系統の農学的データのまとめ
【0200】
実施例19−遺伝子量の作用;Δ9デサチュラーゼ遺伝子の多コピーの挿入による飽和体の更なる減少
本実施例は更に、Δ9デサチュラーゼ遺伝子の「スタック」している多コピーは、驚くべき遺伝子量の作用を有し、このことは、カノーラのような油糧種子植物中の飽和体の更なる低下にも使用することができることを示している。以下は、特に記さない限りは、FAME手法を用いて測定された温室データである。
【0201】
表27は、未変性のNex 710、シングル事象(218、36、69)、ならびにインベーダーアッセイ法に基づき選択されたF2植物を辿るF3種子パッケージの各々由来のF3 10種子バルク脂肪酸データを示す。(表27の最後の2列は、Nex 710対照と比較した、各植物に関するおおよその平均総飽和体、および総飽和体の平均減少の概数を示す)。その世代で、サザンデータは使用しなかった。従って飽和体発現およびインベーダーアッセイ法によって、推定されたスタックおよび推定された親同胞に加えヌルの選択を行い、F3植物の生育およびサザンを使用する再試験により確認した。
【0202】
例えば、統計学的には分析されないが、41種の「スタック」系統からの試料は、平均総飽和体3.5%を有した。21種の「シングル」系統は、平均総飽和3.84%を有した。
【0203】
表28は、コピー数および接合状態を確認するために植直しした推定されたF3スタック、推定された親同胞、およびヌルのまとめを含んでいる。TDN0400141、TDN0400142、TDN0400145、TDN0400155、TDN0400158、およびTDN0400160と命名された系統は、ホモ接合体スタックであると推定された。系統TDN0400189、TDN0400143、TDN0400197、TDN0400167、およびTDN0400184は、スタック外の親同胞であると推定された。系統TDN0400198、TDN0400199は、これらのスタックの自殖されたヌルとして進めた。TDN0400202、TDN0400204、およびTDN0400208は、シングル事象である。この材料は、現在も圃場にあるか、または最近収穫された。従って圃場データは、まだ入手できていない。
【0204】
図8および9は、F3植物からのDNAの2個のゲル泳動の写真である。サザン分析についての単離したDNAと同様の問題点にこの工程で遭遇し、そのため提供されたこれらの植物の9種全てがゲルに出現したわけではない。これらのゲルを元に、系統TDN0400141、TDN0400142、および恐らくTDN0400158(わずかに2種の植物)は、ホモ接合体スタックである。TDN0400145は、依然事象36について分離しており、およびTDN0400155は、依然事象69について分離している。系統TDN0400160は、バンドの奇数分離を有するように見え、これは全て3種のシングル事象に分離していることを示している。TDN0400160について、経過観察を行った。この交雑からの植物#8は、2.6%と低い飽和レベルを有し、これは高い接合状態のトリプルスタックと一致している。追加の分子分析は、このことを確認している。親同胞であると推定された系統の全てが、これらのゲル中に出現したわけではない(表29の事象カラムにおいて強調された系統TDN0400184、TDN0400189、およびTDN0400197、以下に考察)。単独の植物脂肪酸データを元に、TDN0400184由来の9種の植物(同胞でない(sibbed-out)シングル事象218であると推定される)は、推定されたスタックと同じくらい低い飽和レベルを有した。すなわち、このデータの傾向は、スタックは、「同胞でない」事象と比べ、飽和体の減少を一貫して示す(2種のトランスジェニック事象の交雑から回復された単純トランスジェニック事象)。
【0205】
表29は、Nex 710、シングル事象、および各単独のF4植物からの10種の種子バルク脂肪酸データを含む。各推定されたスタック、ヌル、シングル事象などの9種の植物は、再生し、分子分析のために組織試料とし、および脂肪酸分析のための種子セットで維持した。
【0206】
(表27)
【0207】
(表28)
【0208】
(表29)
【図面の簡単な説明】
【0209】
【図1】アラビドプシスにおいて、飽和脂肪酸の60%を超える減少が実現されたことを示している。このグラフは、単独のアラビドプシス事象に関するT2およびT3種子データをまとめている。
【図2】18種の追加の形質転換体由来のT2アラビドプシス種子における、最大60〜70%の「飽和体」の減少を示している。このグラフに図示されたデータは、表8に示された数値データおよびより早期の数値データと組み合わせられた。
【図3】Westarカノーラにおいて、飽和脂肪が、43%よりも多く減少したこと(および、24:0が含まれる場合は、50%減少が達成されたこと)を示している。
【図4A】対照と比較した、事象36-11.19を含む様々なカノーラ植物由来の種子の総飽和を比較する棒グラフを示す。
【図4B】棒グラフにより図示された数値データを表す。
【図4C】棒グラフにより図示された数値データを表す。
【図5A】対照と比較した、事象218-11.30を含む様々なカノーラ植物由来の種子の総飽和を比較する棒グラフを示す。
【図5B】棒グラフにより図示された数値データを表す。
【図5C】棒グラフにより図示された数値データを表す。
【図6】T3圃場試験由来のハーフ種子データを示す。図6Aおよび6Bは、ヌル対照(導入遺伝子が植物から分離された事象)および野生型対照(非形質転換系統)と比較したトランスジェニック事象におけるC16:0の減少およびC16:1の増加を明らかに示している。図6Cおよび6Dは、ヌル対照および野生型対照と比較したトランスジェニック事象におけるC18:0の減少およびC18:1の増加を明らかに示している。図6Eおよび6Fは、ヌル対照および野生型対照と比較したトランスジェニック事象におけるC20:0およびC22:0の各々の減少を明らかに示している。図6Gおよび6Hは、ヌル対照および野生型対照と比較したトランスジェニック事象におけるC16:0のシフトおよび減少、ならびにC16:1のシフトおよび増加を明らかに示している。図6Iおよび6Jは、ヌル対照および野生型対照と比較したトランスジェニック事象におけるC18:0のシフトおよび減少、ならびにC18:1のシフトおよび増加を明らかに示している。図6Kおよび6Lは、C18:2およびC18:3の同様の棒グラフを示している。図6Mは更に、既存の非常に良好なNex 710系統と比べた、総飽和の減少を図示している。図6Nは、1000個の種子についての分布を示している。
【図7】実施例16のプロトコールを使用して得たデータを示している。
【図8】実施例19において考察した、F3植物由来のDNAによる2個のゲル泳動の図である。
【図9】実施例19において考察した、F3植物由来のDNAによる2個のゲル泳動の図である。
【背景技術】
【0001】
関連出願の相互参照
本願は、2004年10月8日に出願された米国特許仮出願第60/617,532号の優先権を請求するものである。
【0002】
発明の背景
食物脂肪摂取の主な給源として次第に、植物由来の油が、動物由来の油および脂肪と置き換わっている。しかしほとんどの先進国における飽和脂肪摂取は、総カロリー消費の約15%〜20%を維持している。より健康な生活スタイルを促進する努力において、米国農務省(USDA)は最近、飽和脂肪は一日のカロリー摂取の10%未満とすることを推奨している。消費者の注意を喚起するために、USDAにより公布された最新の表示指針は、「低飽和」表示するためには14gサービング当たり1.0g未満であること、および「無飽和」表示するためには14gサービング当たり0.5g未満であることを現在求めている。このことは、「低飽和」および「無飽和」表示を付けるためには、各々、植物油の飽和脂肪酸含量は、7%および3.5%未満であることを必要とすることを意味する。これらの指針の発布以降、「低飽和」油を求める消費者が急増している。今のところこれには、主にカノーラ油が当てはまり、ヒマワリ油およびベニバナ油がはるかに低い程度当てはまる。
【0003】
植物または動物のいずれが起源であるかに関わらず、油の特徴は、炭素原子および水素原子の数に加え、脂肪酸鎖を含む二重結合の数および位置により主に決定される。植物由来の油のほとんどは、変動量のパルミチン酸(16:0)、ステアリン酸(18:0)、オレイン酸(18:1)、リノール酸(18:2)およびリノレン酸(18:3)の脂肪酸から構成される。パルミチン酸およびステアリン酸の炭素鎖は、水素原子により飽和され、従って二重結合を有さないので、それらは従来「飽和された」と称され;これらは、可能な限り最大数の水素原子を含む。しかしオレイン酸、リノール酸およびリノレン酸は、各々1、2および3個の二重結合を有する18-炭素の脂肪酸鎖である。オレイン酸は典型的には、一価不飽和脂肪酸と考えられるのに対し、リノール酸およびリノレン酸は、多価不飽和脂肪酸と考えられる。米国農務省は、「無飽和体(no saturate)または無飽和(no sat)」製品を、(脂肪酸の総量と比較して)3.5質量%未満の合計飽和脂肪酸を有する製品と規定している。
【0004】
不飽和脂肪(一価不飽和および多価不飽和)は有益であるが(特に適度に消費される場合)、飽和脂肪およびトランス脂肪は有益ではない。飽和脂肪およびトランス脂肪は、血中LDLコレステロールレベルを上昇する。同じく食事性コレステロールは、LDLコレステロールを上昇し、LDLが上昇しなくとも心臓疾患の原因となることがある。従って健康な食事の一部として、飽和脂肪、トランス脂肪、およびコレステロールの少ない食品を選択することが推奨される。
【0005】
最近の研究から、主要食事性脂肪成分としての一価不飽和体(monounsaturate)、特に高レベルのオレイン酸の健康上の価値が確立されている。このような食事は、高飽和脂肪酸食の結果である動脈硬化症の発生を減少させると考えられる。従って、高含量の一価不飽和体を有する食用植物油が必要である。種子の変異誘発を使用し、4%を超えない飽和脂肪酸含量を有するナタネ油が作製されている(PCT国際特許出願公開番号 国際公開公報第91/15578号)。
【0006】
1985年に世界で供給された食用油の13%以上が一般に、ナタネまたはマスタードとして公知の油糧種子作物アブラナ(Brassica)種から作製された。アブラナは、ダイズおよびヤシに続く、3番目に重要な食用油の供給源である。アブラナは、比較的低温で発芽および生育することができるので、これは、温度の低い農業地域において栽培することができ、更にはより温暖な地帯における冬期作物として使用される、数少ない商業的に重要な食用油糧種子作物のひとつでもある。更に全般的に植物油、特にナタネ油は、合成油または鉱油/ナフテン系油のものと同等の性能を提供する可能性を有し、生分解性である点でも非常に望ましい利点があるので、工業的用途でのこれらの使用が検討されることがかなり増えてきている。
【0007】
カノーラ油は、全ての植物油の中で最低レベルの飽和脂肪酸を有する。「カノーラ」は、種子の総脂肪酸含量を元にエルカ酸(C22:1)含量が最大でも2質量%であり(好ましくは最大でも0.5質量%であり、最も好ましくは本質的に0質量%である)、ならびに圧搾後、脱脂された(油非含有)粗挽き粉1g当たり30マイクロモル未満を含有する風乾された粗挽き粉を作製する、ナタネ(アブラナ)を意味する。これらの種類のナタネは、この種の従来の亜種と比べ、それらの食用適性が際だっている。
【0008】
植物油の修飾は、化学的に実行されてもよい。この方法を使用し、約3%未満の飽和脂肪酸を含むサラダ油/料理用油が得られており(米国特許第4,948,811号);この油は、化学反応によるか、または飽和脂質の物理的分離により、形成され得る。望ましい特徴の油を実現するための「遺伝子操作」の使用が全般的に言及されている(第3段、58行など参照)。しかし特定の油糧種子植物が、望ましい特徴の植物油を提供するために修飾される方法の詳細は明らかではない。
【0009】
典型的には、植物油の脂肪酸組成物は、従来の育種技術により修飾される。これらの技術は、現存する生殖質を、脂肪酸組成に影響を及ぼす天然の変異の供給源として使用する。このような変異は、適当なスクリーニングの使用により明らかにされ、かつ選択され、引き続きの育種と組み合わせられる。例えばそのような方法は、ナタネ油中の長鎖脂肪酸エルカ酸エステルの量を減少させ(Stefansson, B. R., High and Low Erucic Acid Rapeseed Oils, Kramer J. K. G. et al., eds;Academic Press, ニューヨーク;pp. 144-161(1983))、およびコーン油中の一価不飽和脂肪酸オレイン酸エステルの量を増加する(米国特許出願第07/554,526号)ために使用されている。
【0010】
最近、変異原を使用し、そこから所望の特性を選択するために使用可能な変異のプールを増加することが試みられている。しかし変異原は一般に、既に存在する遺伝子の失活または修飾により作用し、特定の機能の喪失または低下を生じる。従って変異誘発による新規特徴の導入は、既に存在するいくつかの形質の喪失に左右されることが多い。加えて、変異原による所望の目的の達成は、一般に不確かである。わずかな型の植物油中の修飾された脂肪酸組成のみが、この方法を用いて実現される。このような脂肪酸組成に影響を及ぼす「作出された」変異の一例は、ナタネ油中の多価不飽和脂肪酸、特にリノール酸エステルおよびリノレン酸エステルの減少、それに伴う一価不飽和脂肪酸オレイン酸エステルの増加である(AuId, M., et al., (1992) Crop Sci. 印刷中)。別のものは、ナタネ油中の飽和脂肪酸の減少である(PCT国際公開公報第91/15578号)。しかし、種子油合成の生化学は複雑であり、十分に理解されておらず;ナタネ油において観察される脂肪酸組成の変化に寄与するいくつかの機構が存在する(PCT国際公開公報第91/15578号)。そのような変化に影響を及ぼすための変異誘発の使用は、本質的にランダムであり、非特異的である。
【0011】
遺伝子操作を使用し脂肪酸組成を修飾する可能性は、理論的には、特定の望ましい遺伝子の正確に制御された導入、更には特定の望ましくない遺伝子または遺伝子産物の失活をもたらす。従って既に存在する遺伝子とは完全に独立した新規形質が植物に導入されるか、または予め選択された遺伝子が失活もしくは修飾される。しかし脂肪酸組成を修飾するために遺伝子操作を効果的に使用するためのひとつの根拠となるものは、脂肪酸の合成およびプロセシングを調節する植物細胞の作業時の機構の理論的に正確なモデルである。
【0012】
米国特許第6,495,738号(同じく国際公開公報第99/50430号参照)は、コーン油およびタバコ種子中の飽和脂肪酸レベルは、植物細胞内での真菌パルミチン酸エステル-CoAΔ9デサチュラーゼの発現により変更することができることを開示している。これらのタンパク質は優先的に、その分子のCoA部分からの2個の水素原子の除去、および9番目と10番目の炭素原子の間への二重結合の追加により、パルミチン酸エステル-CoA分子を酵素的に不飽和化し、その結果パルミトレイル-CoA(16:1Δ9)を生成する可能性が高い。パルミトレイル-CoAは最終的には、種子油に取り込まれ、その結果該油の総飽和体(saturate)レベルを低下する。コーン油の総飽和体脂肪酸レベルは、平均約13.9%であるが、これは先に考察した現在の表示指針には合致しない。更にトウモロコシは典型的には、ダイズ、カノーラ、ヒマワリなどと比べ、油糧作物とは見なされていない。事実トウモロコシから生成および抽出された油は、デンプン抽出において使用される湿式圧搾プロセスの副産物と見なされている。そのために、コーン油の飽和レベルを修飾することについて関心はほとんど無い。
【0013】
油糧種子においては、脂肪酸合成は主に色素体で生じること、および新たに合成された脂肪酸は色素体から細胞質へ輸送されることが、前提条件とされている。細胞質において、これらは、トリグリセリドの構築に使用され、これは小胞体(endoreticular)膜において生じる。
【0014】
脂肪酸合成の主要生成物は、パルミチン酸エステル(16:0)であり、これはステアリン酸エステル(18:0)へ効率的に伸長するように見える。依然色素体において、この飽和脂肪酸は次にΔ9デサチュラーゼとして公知の酵素により不飽和化され、1個または複数の炭素-炭素二重結合を導入する。具体的には、ステアリン酸エステルは、色素体Δ9デサチュラーゼ酵素により迅速に不飽和化され、オレイン酸エステル(18:1)を生じることができる。実際パルミチン酸エステルは、色素体Δ9デサチュラーゼにより、パルミトレイン酸エステル(16:1)にも不飽和化されるが、この脂肪酸は、ほとんどの植物油において極微量(0〜0.2%)であるように見える。
【0015】
従って、色素体における脂肪酸合成の主要生成物は、パルミチン酸エステル、ステアリン酸エステル、およびオレイン酸エステルである。ほとんどの油において、飽和脂肪酸ははるかに少ない割合で存在するので、オレイン酸エステルが、合成される主要脂肪酸である。
【0016】
引き続きの細胞質中の色素体の外側での植物脂肪酸の不飽和化は、オレイン酸エステルに限定されるように見え、これはリノール酸エステル(18:2)およびリノレン酸エステル(18:3)へ不飽和化され得る。加えて植物に応じて、オレイン酸エステルは更に、伸長(20:1、22:1、および/または24:1に対する)によるか、または官能基の付加により、修飾され得る。その後これらの脂肪酸は、飽和脂肪酸パルミチン酸エステルおよびステアリン酸エステルと共に、トリグリセリドへ構築され得る。
【0017】
植物のΔ9デサチュラーゼ酵素は、可溶性である。これは、色素体ストロマ内に位置し、および基質として、主にステアリル-ACPであるACPへエステル化された新たに合成された脂肪酸を使用する。これは、小胞体膜(ER、またはミクロソーム)に位置し、基質としてCo-Aへエステル化された脂肪酸を使用し、ならびに飽和脂肪酸パルミチン酸エステルおよびステアリン酸エステルの両方を不飽和化する、酵母Δ9デサチュラーゼ酵素とは対照的である。米国特許第5,723,595号および同第6,706,950号は、植物デサチュラーゼに関する。
【0018】
酵母Δ9デサチュラーゼ遺伝子は、出芽酵母(Saccharomyces cerevisiae)から単離され、クローニングされ、配列決定されている(Stukey, J.E. et al., J. Biol. Chem. 264:16537-16544(1989);Stukey, J.E. et al., J. Biol. Chem. 265:20144-20149(1990))。この遺伝子は、それが明らかに過剰発現される条件下で、出芽酵母株を形質転換するためにも使用されており、これはトリグリセリドの染色のためにナイルレッドを使用する蛍光顕微鏡により決定されるように、形質転換された酵母細胞において、増大した貯蔵脂質の蓄積を生じる(米国特許第5,057,419号)。この脂肪酸組成物は、特徴決定されていない。この参考文献は、単離された酵母Δ9デサチュラーゼ遺伝子からの情報を使用し、最初に他のデサチュラーゼ遺伝子を酵母から、または他の生物から単離し、次にこれらの遺伝子を酵母または植物へ条件下で再導入する、「一般的」考察を含んでいる。これは生成された油およびその脂肪酸組成の修飾のための高発現につながることは推測される。
【0019】
引き続き、酵母Δ9デサチュラーゼ遺伝子は、タバコ葉組織へ導入され(Polashcok, J. et al., FASEB J. 5:A1157(1991))、およびこの組織において明らかに発現されたことが報告された。更にこの遺伝子は、トマトにおいて発現された。Wang et al., J. Agric Food Chem., 44:3399-3402 (1996);および、C. Wang et al., Phytochemistry, 58:227-232(2001)参照のこと。タバコおよびトマトの両方においてある種の不飽和体の若干の増加および一部の飽和体の若干の減少が報告されたが、タバコおよびトマトは明らかに油糧作物ではない。この酵母遺伝子は、セイヨウアブラナ(Brassica napus)へも導入された(米国特許第5,777,201号参照)。パルミチン酸エステルおよびステアリン酸エステル(飽和)の減少ならびにパルミトレイン酸エステルおよびオレイン酸エステル(不飽和)の増加が報告されたが(前記特許の実施例7の表1aおよび1b参照)、この言及は、下記「詳細な説明」の項の始めにより詳細に考察されている。国際公開公報第00/11012号および米国特許第6,825,335号は、植物において発現するための合成酵母デサチュラーゼ遺伝子に関連しており、ここでこの遺伝子は、デサチュラーゼドメインおよびcyt b5ドメインを含む。これらの参考文献の「背景」の項は、脂肪酸合成を詳細に説明している。
【0020】
植物油が食事性または工業用かどうかの性能特性は、その脂肪酸プロファイルにより、すなわち油中に存在する脂肪酸の種類、ならびに各種類の相対量および絶対量により、実質的に決定される。脂肪酸プロファイルと性能特性の間のいくつかの関係はわかっているが、多くは不明のままである。それにもかからわず、植物油に存在する不飽和の種類および量は、食事性および工業用の両方の用途に関連している。
【0021】
標準のカノーラ油は、約8〜12%リノレン酸を含有し、これは酸化的、従って香味、安定性に関して、ダイズ油と同様の範疇に入る。カノーラ油の酸化安定度は、不飽和の量を減少させるための水素化、抗酸化剤の添加、およびより良い酸化安定度を有する1種または複数の油の配合など、多くの方法で改善することができる。例えばカノーラ油とヒマワリのような低リノレン酸油との配合は、18:3レベルを低下し、その結果油の安定性を向上する。しかしこれらの処理は、必然的に油の価格を上昇し、その他の複雑さを有し得る。例えば、水素化は、飽和脂肪酸のレベルおよびトランス不飽和量の両方を増加する傾向があり、これらは両方とも食事用途には望ましくない。
【0022】
高オレイン酸油は使用可能であるが、しかし、そのような精選油の追加費用の可能性に加え、非常に高レベルのオレイン酸のために育種された作物からの植物油は、高レベルの多不飽和脂肪酸、特にリノール酸および/またはリノレン酸をかなり保持するので、これらは工業用途は満たさないことを証明することができる。そのような油は、料理用油としての使用を含む食事用途には依然極めて有用であるが、工業用途において見出されたより厳密な条件下では、不適切な酸化安定度を有する。例え抗酸化剤の添加が、工業用途に必要な酸化安定度のレベルをこれらの油にもたらすのに十分ではないとしても;これは恐らく、これらの油において見出される、極めて高い易酸化性を伴うリノレン酸のレベルに起因していると考えられる。
【0023】
酸化安定度は、熱および圧力の条件下および化学副産物の存在下で、滑沢剤の寿命を延長するために、工業用途に重要である。このような用途においても同じく、リノレン酸、およびより少ない程度にリノール酸は、酸化安定度の不良が原因となっている。
【0024】
従って、農学的に生育でき、および食事用途において使用するためにこれが適格であるのに十分なレベルの酸化安定度を有する種子油を作製し、ならびに加えて工業用途における使用を見出すために、単独で十分に安定であるか、または抗酸化剤に十分に反応性であるような、様々なセイヨウアブラナを得ることは望ましいと考えられる。
【0025】
欧州特許出願第EP 323753号、米国特許第5,840,946号、および米国特許第5,638,637号は、オレイン酸含量80〜90%(総脂肪酸含量についての質量%)および2%を超えないエルカ酸を有するナタネ油を開示している。オレイン酸含量を改善するためには、変異誘発が使用された。米国特許第5,840,946号の「特許請求の範囲」は更に、その油は、2%を超えないエルカ酸含量、および3.5%未満のα-リノレン酸含量、ならびに7%を超えないステアリン酸およびパルミチン酸の形の飽和脂肪酸含量も有することを特定している。これらの特許は、変異誘発に続く選択に関連している。
【0026】
米国特許第5,387,758号;同第5,434,283号;および、同第5,545,821号は、2〜4%のステアリン酸およびパルミチン酸の組合せ(質量%)、ならびに約2質量%を超えないエルカ酸含量を有するナタネに関する。変異誘発を使用し、ステアリン酸およびパルミチン酸の含量を低下する。
【0027】
国際公開公報第92/03919号、および米国特許第5,668,299号;同第5,861,187号;および、同第6,084,157号は、変更された脂肪酸プロファイルを有するナタネ種子、植物、および油に関する。そのようなプロファイルのいくつかが言及されており、それらの全ては、パルミチン酸含量約7%〜約12%、リノール含量約14%〜約20%、ステアリン酸含量約0.8%〜約1.1%、およびα-リノレン酸含量約7%〜約9%、更にはある範囲のFDA飽和体(FDA saturate)と組合せた、最大エルカ酸含量約2%を予想している。これらの特許は、飽和脂肪酸および「FDA飽和体」を、ラウリン酸(C12:0)、ミリスチン酸(C14:0)、パルミチン酸(C16:0)、およびステアリン酸(C18:0)の合計と定義している。
【0028】
国際公開公報第93/06714号、および米国特許第6,270,828号;同第6,562,397号;同第6,680,396号;および、同第6,689,409号は、低下したグルコシノレート(従って低下したイオウ)を伴い、更にはα-リノレン酸含量が約2%〜約7%であるカノーラ油および種子に関する。
【0029】
米国特許第6,169,190号は、オレイン系脂肪酸含量が約71〜77%およびリノレン酸含量が約3%未満であるカノーラ種子由来の油に関する。34〜55の間のオレイン:リノレン比も、特許請求されている。
【0030】
米国特許第6,063,947号および同第5,850,026号は、カノーラ種子から得られた油、それに関連したカノーラ植物、およびその油の製造法を特許請求しており、ここでこの油は、約80%よりも多い(約86〜89%)オレイン酸含量、約2%〜約6%のリノール酸含量、2.5%未満(約1〜2%)のα-リノレン酸含量、および約2%未満のエルカ酸含量(加水分解後)を有する。これらの特許は、ミクロソームオレイン酸デサチュラーゼ(オレイン酸をリノール酸へ転換するΔ-12デサチュラーゼ)およびミクロソームリノール酸デサチュラーゼ(リノール酸をα-リノレン酸へ転換するΔ-15デサチュラーゼ)遺伝子発現の種子特異的阻害に関する。
【0031】
米国特許第5,952,544号は、炭素15と16の間の反応を触媒する、植物色素体またはミクロソームΔ-15脂肪酸デサチュラーゼ酵素の断片を特許請求している。
【0032】
米国特許第4,627,192号および同第4,743,402号は、約80〜94%(それらの総脂肪酸含量に対して)のオレイン酸含量およびリノール対オレイン比約0.09未満を有するヒマワリ種子およびヒマワリ油に関する。これらのヒマワリ植物は、伝統的育種技術により得られた。
【0033】
国際公開公報第2003002751号は、植物の油表現型を変更するためのキナーゼ遺伝子などの使用に関する。
【0034】
既に有益な油糧種子作物の脂肪酸プロファイルに有意に(および望ましく)作用する、特に植物および油のその他の局面に有害に作用することなく、飽和脂肪のレベルを低下する、Δ9デサチュラーゼ遺伝子の能力は、予測不可能である。
【発明の開示】
【0035】
発明の簡単な概要
本発明の目的は、「無飽和」カノーラ油を提供することである。本発明は、一部、ある植物種子の飽和脂肪酸を減少させる方法にも関する。これらの結果は驚くべきことに、カノーラ(アブラナ)におけるΔ9デサチュラーゼ遺伝子の使用により実現された。この技術は、本明細書において開示されたように、他の植物にも適用することができる。本発明に含まれるのは、そのような油および種子を生成することが可能である植物、好ましくはカノーラである。本発明は、該植物から種子および油も提供し、ここでこれらの油は、これまで達成されていないような、特に有利な特徴および脂肪酸プロファイルを有する。本発明はなお更に、植物に最適化されたΔ9デサチュラーゼ遺伝子を提供する。一部の好ましい態様において、好ましい植物は、本発明のΔ9デサチュラーゼ遺伝子の少なくとも2種のコピーを含む。このような植物により生成される種子は驚くべきことに、遺伝子サイレンシングの作用を示さないが、更に驚くことに総飽和体レベルは低下する。
【0036】
配列の簡単な説明
SEQ ID NO:1は、本明細書において使用される、植物に最適化されたΔ9デサチュラーゼ遺伝子のオープンリーディングフレームの核酸配列を示す。
SEQ ID NO:2は、コザック配列およびBamHIクローニング部位(残基1-10)に続くSEQ ID NO:1のORFと、ORFの末端の翻訳終結因子(残基1379-1381)の配列を示す。
SEQ ID NO:3は、Δ9遺伝子の増幅に使用されたΔ9フォワードBプライマーの核酸配列を示す。
SEQ ID NO:4は、Δ9遺伝子の増幅に使用されたΔ9リバースBプライマーの核酸配列を示す。
SEQ ID NO:5は、SEQ ID NO:1によりコードされたアミノ酸配列を示す。
【0037】
発明の詳細な説明
本発明は、「無飽和」カノーラ油を提供する。本発明は一部、ある植物種子の飽和脂肪酸を減少させる方法にも関する。これらの結果は驚くべきことに、植物、好ましくは油糧植物、更により好ましくはカノーラ(アブラナ)中の飽和脂肪酸の「無飽和」レベルを驚く程生じるが、Δ9デサチュラーゼ遺伝子の使用により実現された。本発明は、そのような植物を含み、ならびに該植物由来の種子および油も提供し、ここでこれらの油は、これまで達成されていないような、特に有利な特性および脂肪酸プロファイルを有する。
【0038】
アスペルギルス・ニジュランス(Aspergillus nidulans)ミクロソームΔ9-CoAデサチュラーゼ遺伝子が本明細書において例示されている。このΔ9デサチュラーゼは、膜結合型酵素であり、16:0-CoAおよび18:0-CoAの16:1-CoAおよび18:1-CoAへの反応(Δ9位置での二重結合の付加)を触媒する。本発明は更に、他の飽和体、例えばC20:0、C22:0およびC24:0のレベルも、非常に驚きに値しかつ有利に減少したが、C16:1およびC18:1不飽和体(unsaturate)は増加した(C18:2およびC18:3はほとんどまたは全く増加せず、場合によってはこれらの比較的安定していない多価不飽和体(polyunsaturate)は減少する)点が驚きに値する。従来は、既に望ましい(しかし最適ではない)脂肪酸プロファイルを有するアブラナおよび他の「優れた」油糧種子植物において、これが良好な酵素であるかどうか(この遺伝子は十分に発現されるかどうかを含む)は不明であった。(例えばトウモロコシの飽和体においてわずかに10%の減少が得られた。)。
【0039】
「発明の背景」の項において言及したように、様々な生物および植物の複雑な脂肪酸プロファイルおよび代謝経路、ならびにそれらの異なる生理的細胞機構を考えると、例えこの遺伝子および酵素がアブラナにおいてある作用を有したとしても、それらの作用は有益であると予想することはできないと考えられる。「発明の背景」の項においてにおいて考察したように、望ましい脂肪酸の1型または複数型の増加は、他の望ましい脂肪酸の減少、望ましくない脂肪酸の増加、および農学的ペナルティ(すなわち、修飾された植物に対する他のあからさまな有害作用)を生じることが多い。本発明は、莢サイズ、種子収量、種子サイズ、油収量などの他の価値のある農学的特徴に対応する有害作用を伴わずに実践することができることも驚くべきことであった。単独のトランスジェニック挿入断片に関して、植物ホモ接合体の有害作用は存在しなかった。一部のデュアルホモ接合体スタック(ふたつのトランスジェニック事象の交差により生じた)は、莢数および結実の減少を示したが;その原因は未だ不明である。しかし表27は、非トランスジェニック対照に類似した種子収量を有するスタック(すなわち、明らかに増加したコピー数事象)、更には「無飽和」組成も含んでいる。
【0040】
予想外のことに、デサチュラーゼ間の差異、更には酵母、真菌、植物、および動物のΔ9デサチュラーゼ間の差異のために、別の理由も生じる。デサチュラーゼの差異は、ひとつには、様々なデサチュラーゼの給源生物の細胞構造の差異に起因し得る。米国特許第5,777,201号の酵母デサチュラーゼは、「発明の背景」の項で考察されている。これは、本明細書において例示されているアスペルギルスデサチュラーゼよりも長い(510個アミノ酸対455個アミノ酸)。加えて約400個のアミノ酸全体の同一性はわずかに約52%である(BLASTおよびBestFitの両方により決定、Smith-Watermanプログラム;両方ともEMBOSSで実行)。その特許の実施例7の表1aおよび1bは、酵母デサチュラーゼを用いて達成される飽和体の減少は、カノーラにおいて例証されたアスペルギルスデサチュラーゼにより本発明に従い達成されるものよりもはるかに弱いことを示している。酵母デサチュラーゼの比較的弱い性能に関する可能性のある説明であり得る様々な要因が存在する。例えばそのタンパク質は、植物において本来不安定であるかもしれない(しかし本デサチュラーゼは、カノーラにおいて極めて明らかに非常に安定している)。これらは、触媒効率、基質親和性、補因子親和性などの、その他の酵素特性も異なり得る。
【0041】
米国特許第5,723,595号および同第6,706,950号の紅花デサチュラーゼと比較すると、この紅花デサチュラーゼ(396個アミノ酸)は、本明細書で例示されているアスペルギルスデサチュラーゼ(455個アミノ酸)よりもより短い。紅花デサチュラーゼは、色素体においても見つかっているが、本アスペルギルスデサチュラーゼは、ER/ミクロソーム/細胞質コンパートメントにおいて見つかっている。更に、紅花デサチュラーゼは、色素体において見出されるアシル-ACP基質を使用するが、アスペルギルスデサチュラーゼは、細胞質コンパートメントにおいて見出されるアシル-CoA基質を使用する。従って本発明に関して、アシル-CoA基質のプールの実質的部分は、アスペルギルスデサチュラーゼに使用可能であるかどうかは不明である。
【0042】
従って、本Δ9デサチュラーゼは、それらから特に油の改善された食品品質について優れた特性を有するカノーラ植物、種子、および油を得ることができることがわかったことは大きな驚きであった。非常に驚くべきことであるが、飽和脂肪酸の60%を超える減少が、アラビドプシス(Arabidopsis)において実現され、および飽和脂肪酸の43%を超える減少がカノーラにおいて実現された。再度、このことがいずれかの適当な植物の最良の脂肪酸プロファイルのひとつを既に生じた植物において実現されたことに注目することは重要である。本発明は、以下により詳細に示されかつ説明されるように、驚きに値しかつ有利な脂肪酸プロファイルおよび比を実現するためにも使用された。ステアリン酸は飽和脂肪酸であると考えられるが、これはコレステロール低下作用を有することがわかっている。従って比較的高いレベルのステアリン酸は、有益である。同様に比較的高いレベルのアラキドン酸は、望ましいものである。本明細書のデータに示されたように、本発明の種子由来の油は、これら2種の脂肪酸の有利なプロファイルを、例えば望ましいレベルのバクセン酸と共に有する。同じく、有利なレベルのこれらの脂肪酸および/または総飽和体は、本発明の業務用品質の植物(例えば矮性植物とは対照的)における望ましい高さ、収量、および他の有益な特徴と組合せて存在することができることも本明細書において示される。本発明のこのような植物に関する例証的データも本明細書において示されている。
【0043】
本発明は、例証されたデサチュラーゼに限定されないことは注意すべきである。様々なデサチュラーゼおよびΔ9デサチュラーゼは、GENBANKから入手可能であり、ならびにこれらの酵素の配列の相違点を観察および比較するための配列アラインメントは、標準の手法を用いて行うことができる。本明細書に例証されたものに類似した酵素は、本発明に従い使用することができる。
【0044】
例えば本アスペルギルスデサチュラーゼは、ふたつのドメインを有する。第一のドメイン(その分子のアミノ-末端側のおよそ2/3)は、デサチュラーゼドメインであり、第二のドメイン(分子のC-末端側のほぼ1/3)は、シトクロムb5ドメインである。例えばSEQ ID NO:5の残基62-279は、例えば、脂肪酸デサチュラーゼgnl|CDD|25523 pfam00487の残基4-233と整列化することができる。SEQ ID NO:5の残基332-407は、gnl|CDD|22935 pfam00173の残基1-74(シトクロムb5ドメイン)と整列化することができる。SEQ ID NO:5の残基17-305は、脂肪酸デサチュラーゼgnl|CDD|11113 COG1398(OLE1)の脂質代謝ドメインの残基3-288と整列化することができる。SEQ ID NO:5の残基301-449は、gnl|CDD|14396 COG5274のCYB5(脂質代謝に関与したシトクロムb)の残基11-163と整列化することができる。シトクロムb5を欠いているSEQ ID NO:5のデサチュラーゼドメインは機能することができ、これは植物の色素体デサチュラーゼの一般的構造であるからである。細胞質コンパートメントにおいて見出されるアシル-CoA基質を使用する、公開された推定されたミクロソームのマツのデサチュラーゼ(LOCUS AF438199)も存在し、これはcytb5ドメインを欠いている。アスペルギルスシトクロムb5ドメインを、別の生物のそれと、例え植物細胞質デサチュラーゼ由来のものであっても、交換することも可能である。これらのドメイン、またはこれらのドメインのいずれかもしくは両方をコードしているセグメントは、以下により詳細に説明されるように、本発明の分子を定義するために、プローブとして使用することができる。
【0045】
従って、本発明に有用な遺伝子およびタンパク質は、特別に例証された完全長配列のみではなく、これらの配列の部分、セグメントおよび/または断片(完全長分子と比べ内部および/または末端の欠失を含む)、それらの変種、変異体、キメラおよび融合体も含む。本発明において使用されるタンパク質は、本明細書に特別に例示されたタンパク質の特徴的酵素活性を維持する限りは、置換されたアミノ酸を有することができる。「変種」遺伝子は、例示されたタンパク質と同じタンパク質または同等の機能を有する同等のタンパク質をコードしているヌクレオチド配列を有する。「変種タンパク質」および「同等なタンパク質」という用語は、例示されたタンパク質と同じまたは本質的に同じ生物学的/機能的活性を有するタンパク質を意味する。本明細書において使用される「同等な」配列という表現は、機能を改善またはこれに有害に作用しないようなアミノ酸の置換、欠失、付加、または挿入を有する配列を意味する。機能を保持している断片も、この定義に含まれる。例示されたタンパク質の対応する断片と同じまたは同様の機能を保持する断片および他の同等物は、本発明の範囲内である。アミノ酸の置換または付加のような変化は、タンパク質のプロテアーゼ安定性の増加(または減少)のような様々な目的で行うことができる(タンパク質の機能を物質的に/実質的に減少させることはない)。
【0046】
遺伝子の変動は、例えば点変異を作出する常法を用い容易に構築することができる。加えて米国特許第5,605,793号は、例えば、ランダム断片化後に、DNA再構築体を使用することによる追加の分子多様性を作出する方法を開示している。変種遺伝子を使用し、変種タンパク質を作出することができ;組換え宿主を使用し、変種タンパク質を生成することができる。これらの「遺伝子シャッフリング」技術を用い、本明細書に例示された配列の(例えば)5、10、15、20、25、30、35、40、45、50、55、60個のいずれかの近接残基(アミノ酸またはヌクレオチド)を含む、同等の遺伝子およびタンパク質を構築することができる。
【0047】
完全長遺伝子の断片は、市販のエキソヌクレアーゼまたはエンドヌクレアーゼを使用し、常法に従い作製することができる。例えばBal31などの酵素または部位特異的突然変異誘発を使用し、これらの遺伝子の末端から、系統的にヌクレオチドを切断することができる。同じく、活性断片をコードしている遺伝子を、様々な制限酵素を用いて得ることができる。プロテアーゼは、これらのタンパク質の活性断片を直接得るために使用することができる。
【0048】
本タンパク質は、短縮型でありかつ機能活性を維持することは、本明細書に開示された本発明の範囲内である。「短縮型タンパク質」とは、タンパク質の一部が切断され、切断後も依然酵素活性を示すことを意味する。更に効果的に切断されたタンパク質は、該タンパク質をコードしているDNA塩基が、制限エンドヌクレアーゼによる消化、または当業者が使用可能な他の技術のいずれかを通じて除去される分子生物学技術を用い作製することができる。短縮後、該タンパク質は、大腸菌(Escherichia coli)、バキュロウイルス、植物-ベースのウイルスシステム、酵母などの異種システムにおいて発現され、その後本明細書に説明されたような昆虫アッセイ法に配置し、活性を決定することができる。短縮型タンパク質は、全体の完全長配列よりも短いものを有するにもかかわらず、それらが機能活性を保持するように、うまく作製することができることは当技術分野において周知である。バシラス・スリンギエンシス(B.t.)毒素を、短縮(コア毒素)型において使用することができることは当技術分野において周知である。例えば、Adangらの論文(Gene 36:289-300 (1985), 「Characterized full-length and truncated plasmid clones of the crystal protein of Bacillus thuringiensis subsp kurstaki HD-73 and their toxicity to Manduca sexta」)を参照のこと。幼虫ホルモンエステラーゼを含む、殺虫活性を保持する短縮型タンパク質の別の例も存在する(米国特許第5,674,485号、The Regents of the University of California)。本明細書に使用されるように、「毒素」という用語は、機能的活性のある短縮型を含むことも意味する。
【0049】
本発明において使用するためのタンパク質および遺伝子は、例えばオリゴヌクレオチドプローブを用いて、定義、同定および/または入手することができる。これらのプローブは、適当な標識により検出可能であることができるか、または国際公開公報第93/16094号に開示されたような、固有の蛍光を生じることができる、検出可能なヌクレオチド配列である。これらのプローブ(および本発明のポリヌクレオチド)は、DNA、RNA、またはPNAであってよい。本発明の合成プローブ(およびポリヌクレオチド)は、アデニン(A)、シトシン(C)、グアニン(G)、チミン(T)、およびウラシル(U;RNA分子について)に加え、イノシン(4種の塩基全てと対形成が可能である中性の塩基;時には、合成プローブにおいて4種の塩基全ての混合物の代わりに使用される)も有することができる。従って、本明細書において合成した、縮重オリゴヌクレオチドが言及され、および「N」または「n」が総称的に使用される場合、「N」または「n」は、G、A、T、C、またはイノシンであることができる。本明細書において使用される多義性(ambiguity)暗号は、本願の出願の慣例として標準のIUPAC命名法に従う(例えばRはAまたはGを意味し、YはCまたはTを意味するなど)。
【0050】
当技術分野において周知であるように、プローブ分子が核酸試料とハイブリダイズする場合、これはプローブおよび試料は、実質的に相同/類似/同一であると当然推定することができる。好ましくは、例えばKeller, G.H.、M.M. Manakの著書(DNA Probes, Stockton Press, ニューヨーク, NY, pp. 169-170(1987))に説明されたような、当技術分野において周知の技術により、ポリヌクレオチドのハイブリダイゼーションが最初に行われ、それに低、中、高ストリンジェンシー条件下での洗浄が続く。例えば本明細書に記されたように、低ストリンジェンシー条件は、2xSSC(標準クエン酸ナトリウム、塩化ナトリウム緩衝液)/0.1%SDS(ドデシル硫酸ナトリウム)による室温で15分間の最初の洗浄により実現することができる。典型的には2回の洗浄が行われる。次により高いストリンジェンシーは、塩濃度を低下することによるか、および/または温度を上昇することにより実現することができる。例えば先に説明された洗浄には、0.1xSSC/0.1%SDSによる各15分間室温での2回の洗浄、それに続く0.1xSSC/0.1%SDSによる各30分間55℃での洗浄を続けることができる。これらの温度を、本明細書に記しかつ当業者に公知である他のハイブリダイゼーションおよび洗浄プロトコールと共に使用することができる(例えばSSPEを、SSCの代わりの塩として使用することができる)。2xSSC/0.1%SDSは、2OxSSC 50mlおよび10%SDS 5mlを水445mlに添加することにより調製することができる。2OxSSCは、NaCl(175.3g/0.150M)、クエン酸ナトリウム(88.2g/0.015M)、および水を一緒にし、10N NaOHによりpHを7.0に調節し、その後容量を1Lに調節することにより調製することができる。10%SDSは、SDS 10gをオートクレーブにかけた水50mlに溶解し、その後100mlに希釈することにより調製することができる。
【0051】
プローブの検出は、ハイブリダイゼーションが維持されるかどうかを公知の様式で決定する手段を提供する。このようなプローブ分析は、本発明の毒素-コードしている遺伝子を同定する迅速な方法を提供する。本発明においてプローブとして使用されるヌクレオチドセグメントは、DNA合成装置および標準手法を用い合成することができる。これらのヌクレオチド配列は、本発明の遺伝子を増幅するためのPCRプライマーとしても使用することができる。
【0052】
所定のポリヌクレオチドとのハイブリダイゼーションは、本発明のタンパク質および遺伝子の同定、発見および/または定義に使用することができる技術である。本明細書において使用されるハイブリダイゼーションの「ストリンジェント」条件は、本明細書に説明された条件と、同じまたはほぼ同じ程度のハイブリダイゼーション特異性を実現する条件を意味する。32P標識した遺伝子特異的プローブによるサザンブロットの際に固定されたDNAのハイブリダイゼーションは、標準法により行うことができる(例えばManiatis, T., E.F. Fritsch, J. Sambrook [1982] Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, コールドスプリングハーバー, NY参照)。一般にハイブリダイゼーションおよび引き続きの洗浄は、標的配列の検出を可能にする条件下で行われる。2本鎖DNA遺伝子プローブに関して、ハイブリダイゼーションは、6xSSPE、5xデンハルト液、0.1%SDS、0.1mg/ml変性DNAの中において、DNAハイブリッドの融解温度(Tm)より20〜25℃低い温度で一晩実行することができる。融解温度は、下記式で説明される(Beltz, G. A., K.A. Jacobs, T.H. Eickbush, P.T. Cherbas, and F.C. Kafatos [1983] Methods of Enzymology, R. Wu, L. Grossman and K. Moldave [eds.] Academic Press, ニューヨーク, 100:266-285)。
Tm=81.5℃+16.6 Log[Na+]+0.41(%G+C)−0.61(%ホルムアミド)−600/塩基対の中の二重鎖の長さ
洗浄は典型的には、以下のように実行される。
1)1xSSPE、0.1%SDS中室温で15分間を2回(低ストリンジェンシー洗浄)
2)0.2xSSPE、0.1%SDS中Tm-20℃で15分間を1回(中ストリンジェンシー洗浄)
【0053】
オリゴヌクレオチドプローブに関して、ハイブリダイゼーションは、6xSSPE、5xデンハルト液、0.1%SDS、0.1mg/ml変性DNAの中において、ハイブリッドの融解温度(Tm)より10〜20℃低い温度で一晩実行することができる。オリゴヌクレオチドプローブのTmは、下記式により決定された。
Tm(℃)=2(T/A塩基対数)+4(G/C塩基対数) (Suggs, S.V., T. Miyake, E.H. Kawashime, M.J. Johnson, K. Itakura, and R.B. Wallace [1981] ICN-UCLA Symp. Dev. Biol. Using Purified Genes, D.D. Brown [ed.], Academic Press, ニューヨーク, 23:683-693)。
【0054】
洗浄は、以下のように実行することができる。
1)1xSSPE、0.1%SDS中室温で15分間を2回(低ストリンジェンシー洗浄)
2)1xSSPE、0.1%SDS中ハイブリダイゼーション温度で15分間を1回(中ストリンジェンシー洗浄)
【0055】
一般に、ストリンジェンシーを変更するために、塩および/または温度を変更することができる。長さが>70またはそのような塩基の標識されたDNA断片では、下記の条件が使用される。
低:1または2xSSPE、室温
低:1または2xSSPE、42℃
中:0.2xまたは1xSSPE、65℃
高:0.1xSSPE、65℃。
【0056】
二重鎖形成および安定性は、ハイブリッドの2本鎖の間の実質的相補性に左右され、および前述のように、ある程度のミスマッチは許容され得る。従って本発明のプローブ配列は、説明された配列の変異(単独および複数の両方)、欠失、挿入、およびそれらの組合せを含み、ここで該変異、挿入および欠失は、関心のある標的ポリヌクレオチドとの安定したハイブリッドの形成を可能にする。変異、挿入、および欠失は、多くの様式で所定のポリヌクレオチド配列を作出することができ、およびこれらの方法は、当業者に公知である。他の方法は、今後公知となると思われる。
【0057】
遺伝暗号の縮重/縮退のために、多種多様なDNA配列が、本明細書において開示されたアミノ酸配列をコードすることができる。これは、同じ、または本質的に同じ酵素をコードしている代替のDNA配列を作成するために、訓練を受けた当業者の技術内である。これらの変種DNA配列は、本発明の範囲内である。
【0058】
本発明は、例えば以下を含む:
1)野生型生物から得たタンパク質;
2)変異から生じた変種;
3)保存的アミノ酸置換を生じることによりデザインされた変種;および
4)本TCタンパク質をコードしている複数の異なる配列のランダム断片化および再構築により作出された変種(DNAシャッフリング)。例えば米国特許第5,605,793号参照。
【0059】
本タンパク質をコードしているDNA配列は、野生型配列、変異配列、または予め決定されたタンパク質を発現するようにデザインされた合成配列であることができる。例えばポリアデニル化シグナルを避け、および植物に好ましいコドンを使用することにより、植物において高度に発現するようにデザインされたDNA配列は、特に有用である。
【0060】
ある種のタンパク質および遺伝子が、本明細書において特別に例示されている。これらのタンパク質および遺伝子は単なる例証であるので、本発明は例証されたタンパク質と同じまたは同様の機能性を有する変種または同等のタンパク質(および、それらの同等物をコードしているヌクレオチド配列)の使用を含むことは、容易に明らかであるはずである。同等のタンパク質は、例証された酵素(またはそれらの活性断片)とのアミノ酸類似性(および/または相同性)を有すると思われる。好ましい本発明のポリヌクレオチドおよびタンパク質は、より狭い同一性および/または類似性範囲に関して定義することができる。例えば、酵素タンパク質の同一性および/または類似性は、本明細書に例示されたまたは示唆された配列と比較して40%、41%、42%、43%、44%、45%、46%、47%、48%、49%、50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、または99%であることができる。先に列記されたいずれかの数字を使用し、上限および下限を定義することができる。例えば、本発明のタンパク質は、例えば例示されたタンパク質と、50〜90%の同一性を有すると定義することができる。
【0061】
特に指定しない限りは、本明細書で使用されるふたつの核酸の%配列同一性および/または類似性は、KarlinおよびAltschulの論文(Proc. Natl. Acad. Sci. USA 90:5873-5877(1993))において改変された、KarlinおよびAltschulのアルゴリズム(Proc. Natl. Acad. Sci. USA 87:2264-2268(1990))を用いて決定される。このようなアルゴリズムは、Altschulらの論文(J. MoL. Biol. 215:402-410(1990))のNBLASTおよびXBLASTプログラムに組み込まれる。BLASTヌクレオチド検索は、NBLASTプログラムにより、スコア=100、ワードレングス=12で実行される。ギャップ付きBLASTは、Altschulら(Nucl. Acids Res. 25:3389-3402(1997))に説明されたように使用することができる。BLASTおよびギャップ付きBLASTプログラムを使用する場合、代表的プログラム(NBLASTおよびXBLAST)のデフォルトのパラメータが使用される。NCBI/NIHのウェブサイトを参照のこと。
【0062】
比較目的でギャップ付きアラインメントを得るために、Vector NTI Suite 8(InforMax, Inc., ノースベセスタ, MD, U.S.A.)のAlignX関数を、デフォルトのパラメータで使用することができる。典型的にはこれらは、ギャップオープニングペナルティ15、ギャップイクステンションペナルティ6.66、およびギャップセパレーションペナルティ範囲8であると考えられる。2種またはそれよりも多い配列を整列化し、この方法で、または当技術分野において周知の他の技術を用い比較することができる。このようなアラインメントを分析することにより、本ポリペプチドの比較的保存された領域および保存されない領域を同定することができる。これは、例えば、1個または複数のアミノ酸残基の修飾または置換によるポリペプチド配列の変化が、許容されると予想され得るかどうかの評価に有用である。
【0063】
アミノ酸の相同性/類似性/同一性は、典型的には(しかし必ずしもではない)、その活性を説明するタンパク質、または活性に最終的に寄与する三次元形態の決定に関与したタンパク質の領域において、最高である。これに関して、ある種のアミノ酸置換は許容でき、および耐容できると予想することができる。例えばこれらの置換は、活性に重要でないタンパク質の領域内であることができる。タンパク質の結晶構造の分析、およびソフトウェアによるタンパク質構造のモデリングは、実際タンパク質の特性を変更および/または機能を増大するために、修飾することができるタンパク質の領域を同定する(部位特異的突然変異、シャッフリングなどを使用する)ために使用することができる。
【0064】
タンパク質の様々な特性および三次元特徴も、タンパク質の活性/機能に有害に作用することなく、変更することができる。保存的アミノ酸置換は、許容され/分子の三次元形態に有害に作用しないと予想され得る。アミノ酸は、以下のクラスに入れることができる:非極性、非帯電極性、塩基性および酸性。それによりひとつのクラスのアミノ酸が、同じ型の別のアミノ酸と交換される保存的置換は、その置換がその化合物の生物学的活性に有害でない限りは、本発明の範囲内に収まる。以下のリストは、各クラスに属するアミノ酸の例を提供する。
【0065】
場合によっては、非保存的置換も行うことができる。重要な要因は、これらの置換は、タンパク質の機能活性/生物学的活性/酵素活性を有意に損なってはならないことである。
【0066】
植物における異種遺伝子の高度な発現を得るために、例えば、該遺伝子が植物細胞においてより効率的に発現されるように、これらを再操作することが好ましいことがある。配列は一般に、植物における最適化された発現のためにデザインすることができるか、またはこれらは植物の特定の型における最適化された発現のためにデザインすることができる。カノーラは、該植物中での異種遺伝子の発現レベルを増大するように形質転換する前に、異種遺伝子を再デザインすることが好ましいそのような植物のひとつである。従って真菌タンパク質をコードしている遺伝子のデザインにおける追加の工程は、例えば、異なる型の生物における最適な発現のための異種遺伝子の再操作である。植物発現について最適化される合成遺伝子の作製に関する指針は、例えば、米国特許第5,380,831号に見出されることができる。植物における発現について最適化された配列は、本明細書において、SEQ ID NO:1(これはSEQ ID NO:5に示された例示されたタンパク質をコードしている)として例示されている。
【0067】
本明細書で使用される「単離された」ポリヌクレオチドおよび/またはタンパク質、ならびに「精製された」タンパク質の言及は、それらが天然では一緒に見られる他の分子と会合しない場合、これらの分子を意味する。従って「単離された」および/または「精製された」の言及は、本明細書に説明されたような「人手(hand of man)」の関与を示している。例えば、発現のために植物へ入れられた本発明の真菌ポリヌクレオチド(または「遺伝子」)は、「単離されたポリヌクレオチド」である。同様に本発明のタンパク質は、植物により作製された場合、「単離されたタンパク質」である。
【0068】
「組換え」分子は、組換えられた分子を意味する。核酸分子に関して使用される場合、この用語は、分子生物学的技術により一緒に結びつけられる核酸配列で構成される分子を意味する。「組換え」という用語は、タンパク質またはポリペプチドに関して使用される場合には、1種または複数の組換え核酸分子を使用し作製されるタンパク質分子を意味する。
【0069】
「異種」という用語は、核酸配列について使用される場合は、天然には結びつけられない核酸配列、または天然とは異なる位置で結びつけられる核酸配列に、ライゲーションされるか、またはライゲーションするように操作される、ヌクレオチド配列を意味する。従って「異種」という用語は、核酸分子が、遺伝子操作を用い、すなわち人的介入により、操作されることを意味する。従って本発明の遺伝子は、異種プロモーター(または、転写調節領域が、関心のある配列へ機能的に連結されている場合には、関心のあるヌクレオチド配列の転写を媒介もしくは変調することが可能なヌクレオチド配列を意味する「転写調節領域」)に機能的に連結することができる。好ましい異種プロモーターは、植物プロモーターであることができる。プロモーターおよび/または転写調節領域および関心のある配列は、これらの配列が、転写調節領域により媒介または変調される関心のある配列の転写を可能にするように機能的に連結されている場合には、「機能的に連結されている」。一部の態様において、機能的に連結されるために、転写調節領域は、関心のある配列と同一鎖上に位置することができる。転写調節領域は、一部の態様において、関心のある配列の5'側に位置することができる。そのような態様において、転写調節領域は、関心のある配列のすぐ5'側であることができるか、またはこれらの領域の間に配列が介在することができる。転写調節領域と関心のある配列の機能的連結には、転写調節領域に結合される適当な分子(例えば、トランスジェニックアクチベータータンパク質)が必要であり、従って本発明は、インビトロまたはインビボのいずれかにおいて、そのような分子が提供される態様を包含している。
【0070】
本発明に使用するためのタンパク質を得る多くの方法が存在する。例えば、本明細書において開示されたタンパク質に対する抗体を使用し、混合物から他のタンパク質を同定および単離することができる。特に抗体は、最も一定でありかつ他のタンパク質から最も異なるようなタンパク質の部分に対し生じると考えられる。その後これらの抗体を使用し、免疫沈降、酵素結合免疫吸着アッセイ法(ELISA)、またはイムノブロッティングにより、特徴的活性を伴う同等のタンパク質を特別に同定することができる。本明細書において開示されたタンパク質または同等のタンパク質、またはこれらのタンパク質の断片に対する抗体は、標準の手法を用い、容易に調製することができる。このような抗体は、本発明の局面である。
【0071】
本明細書において言及されたかまたは示唆されたいずれかの単離体に「由来する」または「から入手可能」なタンパク質は、そのタンパク質(または同様のタンパク質)を例示された単離体または他の給源、例えば他の真菌株もしくは細菌株、または植物(例えば、このタンパク質を作製するように操作された植物)から得ることができることを意味する。「に由来する」は、この言外の意味も有し、ならびに例えばポリヌクレオチドが植物における発現のために修飾されているような、所定の型の真菌または細菌から入手可能なポリヌクレオチド(およびタンパク質)を含む。当業者は、真菌の遺伝子およびタンパク質の説明が与えられたならば、植物を、そのタンパク質を産生するように操作することができることを容易に認識すると思われる。抗体調製物、核酸プローブ(例えばDNAおよびRNA)などは、本明細書において開示されたポリヌクレオチドおよび/またはアミノ酸配列を用いて調製することができ、ならびに他の(天然の)給源からの他のタンパク質遺伝子のスクリーニングおよび回収に使用することができる。
【0072】
本発明の油は、高度の酸化安定度を維持するが、より低いレベルの飽和脂肪酸およびより高いレベルの不飽和脂肪酸を含む。好ましい本発明の油は、総飽和脂肪酸含量3.5%未満、オレイン酸含量少なくとも75%(および好ましくは驚くべきことに80%未満)、ならびに多価不飽和脂肪酸含量20%未満(およびより好ましくは15%未満、更により好ましくは10%未満、および更により好ましくは9%未満)を有する。本発明は、総脂肪酸含量の2.5%を超えない(および好ましくはそれ未満)総飽和脂肪酸含量(C:14、C:16、C:18、C:20、C:22、およびC:24)を有する、好ましくは前述の範囲のオレイン酸を伴う、カノーラ種子を実現するためにも使用することができる。油の不安定性に寄与する18:2および18:3のレベルは、増加しないか、または好ましくは低下する(食品用途のために)。これらの特定の脂肪酸のいずれか、それらの組合せのいずれか、および特にC18多価不飽和体の一方または両方についての範囲のエンドポイントは、本明細書に示された図および表のいずれかから得ることができる。
【0073】
本発明は、総飽和体が3.5%未満の精製油/脱臭油を生じる、農学的に精選されたカノーラ種子を提供するために使用することができる。これらの植物に由来した油は、様々な最終製品の配合に使用することができるか、またはこれらは「無飽和」(または「低飽和」)製品のための独立型(stand-alone)フライ用油として使用することができる。
【0074】
特に記さない限りは、カノーラ種子の所定の集合の飽和脂肪酸含量は、油が、種子の粉砕により種子から除去され、およびメタノールおよび水酸化ナトリウムとの反応後、脂肪酸メチルエステルとして抽出されるような標準の手法により決定することができる。次に得られるエステルを、脂肪酸含量について、不飽和度および鎖長を元にした分離を可能にするキャピラリーカラムを使用する気液クロマトグラフィーにより分析される。この分析手法は、例えば、J. K. Daunらの論文(J. Amer. Oil Chem. Soc. 60:1751-1754(1983))に説明されている。
【0075】
カノーラ種子の脂肪酸組成は、「ハーフ種子」分析、「単独/全種子」分析、または「バルク種子」分析のいずれかについて以下に説明されたように決定された。「ハーフ種子」分析について、胚由来の子葉組織の一部を除去し、分析し;その後残りの種子を保存し、所望の場合には発芽させた。ハーフ種子技術は、安定して遺伝的に制御された脂肪酸変異の選択(およびその後の育種および交雑)の点で若干信頼性に欠けるが、本発明は、好ましい遺伝子が導入され、かつ安定した系統の作出に使用されたことを明らかにしている。特徴的でない変異とは異なり、遺伝子は、植物に導入されおよび安定して維持されることは当技術分野において周知である。従って、本明細書に説明された分析は、本遺伝子の利用性および指定された特徴を有するカノーラ油を実現することができることを明らかにしている。
【0076】
脂肪酸の「無飽和体(No saturate)」(すなわち、「無飽和」)レベルは、市販のNexera 710(カノーラ)生殖質由来のトランスジェニック系統からの種子において達成された。無飽和レベルは、合わせた飽和体3.5%未満として定義される。加えて、低下された飽和体が、Westarカノーラ系統、および同じ形質転換構築体を伴う他のCrucifer(アラビドプシス)の両方において認められた。注目すべきことに、単独の種子中の飽和体レベルは、一部の種子について2.6〜2.7%に低下した。本発明は、2.5%またはそれ未満の総飽和体を伴う種子を作製するためにも使用することができる。これは、油の加工は、総飽和「スコア」を約0.5〜1%追加するので重要であり、このことは加工された油製品は、依然標準の試験手法を用い測定できるFDAにより規定された「無飽和」レベルに達することを意味する。この許容レベルを有することは、単に試験装置のあるレベルの夾雑(より高い飽和種子により)を可能にする(特にプラント操作者が、種子バッチの識別の維持に余り習熟していない場合)のみではなく、圃場での成長条件のあるレベルの変動(より多くの飽和体を作製する傾向がある高温など)および隣接圃場の改良されないカノーラからの花粉の漂流(drifting)による交雑受粉(これは望ましい遺伝子を希釈する)も可能にする。
【0077】
米国食品医薬品局は、「飽和脂肪」を、21 CFR 101.9(c)(2)(i)において、「二重結合を含まない全ての脂肪酸の合計として定義された、サービング中の飽和脂肪のグラム数の表記」と規定している。特に指定しない限り、これは本明細書において「総飽和体」および「総飽和脂肪」について使用される定義である。食品のサービングは、21 CFR 101.9(c)(2)(i)において、その食品がサービング中に総脂肪の0.5g未満を含有する場合には、「飽和脂肪なし」と考えられる。「総脂肪」は、21 CFR 101.9(c)(2)において、「総脂質脂肪酸として定義されおよびトリグリセリドとして表された、サービング中の総脂肪のグラム数の表記」と規定される。様々な種類の食品に関する「サービングサイズ」が、21 CFR 101.12(b)において規定されており、これは、油のサービングは大さじ1杯または15mlと規定している。本明細書において使用されるように、これは14gを意味すると理解される。従って「無飽和」カノーラ油(または、飽和脂肪を含まないカノーラ油)は、本明細書において、サービング中の総飽和脂肪0.5g未満を含有するカノーラ油と定義される(カノーラ油14gは、脂肪14gを含む)。別の表現をすると、「無飽和」カノーラ油は、総飽和体3.57%未満を含有する(総飽和体0.5gを、総脂肪14gで除算)。特に指定しない限りは、本明細書の脂肪酸のパーセントは全て、その脂肪酸が成分である油の質量%である。
【0078】
本明細書に記されたように、本発明は驚くべきことに、カノーラ種子から油を得るために使用することができ、ここで該油は、総飽和体3.57%未満を含有する。油は、前段において言及したような、当技術分野において周知の手法を用い、本種子から得ることができ、およびこの油は、本明細書に例証された技術を含む周知の技術を用い、含量についてアッセイすることができる。別に特定しない限りは、ハーフ種子油データおよび圃場油(field oil)データを作製するために使用された分析は、塩基触媒されたトランスエステル化反応(AOCS Ce 2-66、代替法)を使用した。このプロトコールは、本明細書に説明されたケン化/酸エステル化プロトコールに類似しているが、ただこのケン化/酸エステル化プロトコールは、総脂質を測定し、その大半は塩基触媒されたトランスエステル化反応により検出されたトリアシルグリセリド由来の同じ脂肪酸であることが異なる。
【0079】
市販のNexera 710生殖質において、酸化不安定度に寄与している18:3脂肪酸のレベルは、比較的普遍であった。従って本発明は、単に低級飽和脂肪を伴う植物、種子、および油を提供するのみではなく、非常に驚くべきことに他の有益な特性を維持している植物、種子、および油も提供する。すなわち本発明の植物および遺伝子は、驚くべきことに、植物の他の有利な特徴に有害に作用することなく、使用することができる。
【0080】
好ましい態様において、本発明は、複数種の本発明のΔ9デサチュラーゼ遺伝子の発現されたコピーを含む植物を提供する。本明細書に示された結果は、この遺伝子の多コピーの発現は、驚くべきことにカノーラ植物の脂肪酸プロファイルを改善した(飽和脂肪レベルは大きく低下した)ことを示している。これは、当技術分野は、同じ遺伝子の多コピーの発現に関してこれまで予測不可能であったので部分的に驚くべきことである。「遺伝子サイレンシング」は、異種遺伝子の多コピー(例えばゲノム中の異なる位置に挿入された)の使用に対して示す公知の現象のひとつである。同じく複数の形質転換事象を得ることを試みることは理想的ではない。従って複数種(2、3、4種など)のΔ9デサチュラーゼ事象を含む植物を作出する動機は存在しなかった。そのような植物が実際に改善された特徴を有することも期待されななかった。
【0081】
アブラナ科(Cruciferous)植物のふたつの例が、本明細書において特に例示されている:セイヨウアブラナ(Brassica napus)(カノーラ)およびアラビドプシス(Arabidopsis)。しかし当技術分野において公知であるように、他のアブラナ種および他のアブラナ科を、例えば、カノーラなどにおける品種改良および望ましい形質の開発に使用することができる。従って本発明で使用することができるその他のそのような植物は、カブ(Brassica rapa)、カラシナ(Brassica juncea)、ブラッシカ・カリナタ(Brassica carinata)、クロガラシ(Brassica nigra)、キャベツ(Brassica oleracea)、ダイコン(Raphanus sativus)、およびシロガラシ(Sinapis alba)を含む。ダイズ、ダイズ植物、およびダイズ油も、本発明に従う改善について試験することができる。
【0082】
好ましい態様において、本発明のΔ9デサチュラーゼ遺伝子は、植物発現のために最適化される。従って本発明は、植物に最適化されたΔ9デサチュラーゼ遺伝子も提供する。本明細書において例示された最適化は、好ましいコドンおよびコザック翻訳開始領域の導入、ならびに望ましくない配列の除去を含んだ。この遺伝子は、β-ファセオリンプロモーター(強力な双子葉種子貯蔵タンパク質プロモーター)により駆動される。
【0083】
その発現が油合成と同時発生するためのプロモーター(例えばACP、エロンガーゼ)を使用し、発現はタンパク質貯蔵よりもより早期に生じるので、更に飽和体を減少させることができる。(タバコ構築体がnos 3'UTRを使用する前、およびトウモロコシ構築体が構成的メイズ(maize)ユビキチン-1プロモーターおよびnos 3'UTRを使用する前)。ビシリン、レクチン、クルシフェリン、グリシン、およびコングリシニンプロモーター、US20030005485 A1に開示された植物種子プロモーター、US20030159173 A1のエロンガーゼプロモーター、および米国特許第5,767,363号のACPプロモーターを含む、他の双子葉種子プロモーターを、本発明に従い使用することができる。同じく例えば、US6100450A(種子特異性、胚において発現、8段8行);US20030159173A1(0044段の種子特異性プロモーター;例はUSP、ホルデニン、ACP、ナピン、FatB3、およびFatB4である);WO9218634A1(緒言は、他の特許の1〜7頁の種子特異性プロモーターを考察している);WO0116340A1(7頁13行は、「種子特異的」プロモーターの定義を提供し、これは典型的には、他の組織においては5%未満で発現する;10頁19〜29行は、アルブミン、グロブリン、ビシリンおよびレグミン様タンパク質などの、種子貯蔵タンパク質、非貯蔵オレオシン、ACP、サチュラーゼ、デサチュラーゼ、エロンガーゼのような脂肪酸代謝に関連したプロモーターを考察している);WO2003014347A2(プロモーター定義は23〜25頁:好ましくは種子特異性が2倍より大きい);US20030233677A1(0033段は、「種子プロモーター」の例を提供する[ナピン、ACCase、2Sアルブミン、ファセオリン、オレオシン、ゼイン、グルテリン、デンプン合成酵素、デンプン分枝酵素]);WO2003092361A2(15頁は、「プロモーター」の定義を提供する;17頁の冒頭は、ゼイン、7S貯蔵タンパク質、ブラジルナッツタンパク質、phe非含有タンパク質、アルブミン、β-コングリシン、11S、α-ホルドチオニン(hordothionin)、アルセリン、レクチン、グルテニンを含む、プロモーター例および参照特許を提供する(貯蔵タンパク質のみ));US20030148300 A1(請求項8参照、ナピンプロモーター、ファセオリンプロモーター、ダイズトリプシンインヒビタープロモーター、ACPプロモーター、ステアロイル-ACPデサチュラーゼプロモーター、ダイズ7Sプロモーター、オレオシンプロモーター、コングリシンプロモーター、オレオシンプロモーター、胚発生期に豊富な(embryogenesis-abundant)タンパク質プロモーター、胚グロブリンプロモーター、アルセリン5、ナピンプロモーター、および酸性キチナーゼプロモーター);米国特許第5,777,201号(6段、30〜50行、構成的プロモーター、種子-および/または発生上で調節されたプロモーター、例えば植物脂肪酸脂質生合成遺伝子[ACP、アシルトランスフェラーゼ、デサチュラーゼ、脂質転移タンパク質]、または種子プロモーター[ナピン、クルシフェリン、コングリシニン、レクチン]、または誘導性プロモーター[光、熱、創傷インデューサー])も参照のこと。
【0084】
高等植物の色素体は、遺伝子操作の魅力的標的である。葉緑体(色素体のひとつの型)形質転換が実現され、利点がある。例えば米国特許第5,932,479号;同第6,004,782号;および同第6,642,053号参照。同じく米国特許第5,693,507号および同第6,680,426号参照。葉緑体ゲノムの形質転換の利点は:形質転換された葉緑体は、母系遺伝のみであり、従って花粉による他の植物へ異系交配により伝達されないので、潜在的に環境に安全であること;外来遺伝子の高コピー数を実現する可能性があること;および、高度にエネルギー依存型である葉緑体へのタンパク質の輸入が減少されるので、植物エネルギーコストを削減すること;が挙げられる。
【0085】
植物色素体(葉緑体、アミロプラスト、エライオプラスト、エチオプラスト、クロモプラスト 有色体など)は、光合成に加え、アミノ酸、複合糖質、脂肪酸、および色素などの多くの産業上重要な化合物の生成に寄与している主要な生合成中心である。色素体は、原色素体として公知の共通の前駆体に由来し;従って、所定の植物種中の色素体は全て、同じ遺伝的内容物を有する。
【0086】
ほとんどの植物の色素体は、母系遺伝である。結果的に、核において発現された異種遺伝子とは異なり、色素体において発現された異種遺伝子は、花粉により拡散しない(disseminate)。従って植物色素体へ導入された形質は、野生型類縁に伝達されないと考えられる。このことは、これらの形質は野生型類縁へは伝達されないので、除草剤耐性などの、自然条件または化学条件に対する忍容性または抵抗性に関する植物の遺伝子操作に利点をもたらす。
【0087】
高等植物の色素体ゲノム(プラストーム)は、葉細胞1個当たり1,900〜50,000コピーで存在する、120〜160kbの環状二本鎖DNA分子である(Palmer, 1991)。一般に植物細胞は、小型の120〜160kbの環状ゲノムを500〜10,000コピーを含み、その各分子は、大きい(約25kb)逆方向反復を有する。従って、植物細胞を、関心のある特定の遺伝子の最大20,000コピーを含むように操作することは可能であり;これは潜在的に、非常に高レベルの外来遺伝子発現を生じることができる。
【0088】
本発明の油は、工業用途に加え、様々な食品用途に適用可能であり、特にそれらに合わせて作製することができる。本発明は、料理用油を除き、それ自身、ポテトチップなどの「無飽和」製品を含む(米国特許第6,689,409号参照、これはジャガイモおよびカノーラ油を含有するフライ食品組成物を特許請求しているが;しかし本発明は、米国特許第6,689,409号に開示された組成物を改善するために使用することができる。)。
【0089】
本発明の植物は、特徴および形質の様々な望ましい組合せを実現するために、他の植物と交雑することができる。より更なる改善は、本植物の交雑、公知の育種技術の使用、ならびに追加のもしくはその他の有益な形質および特徴を有する他のカノーラ系統のような生殖質の他の有利な給源により行うことができる。別の例は、色素体Δ9デサチュラーゼを有する系統との交雑である。
【0090】
従って本発明を使用し、変動する環境条件下で、業務用品質油中の、総飽和脂肪酸3.5%未満(および、育種家種子における種子油中の総飽和脂肪酸3%未満)を実現することができる。これは、貯蔵タンパク質の品質および量を低下することなく、カノーラ粗挽き粉中の消化が悪い繊維の増加を伴わずに、および1エーカー当たりの種子収量(または他の望ましい農学的形質)に負の影響を及ぼさずに、達成され得る。
【0091】
以下に、本明細書において使用される脂肪酸の一般名を、それらの炭素原子および二重結合の数と共に列記する。飽和脂肪は、0個の二重結合を有する。
【0092】
(表1)
*=Δ9位の二重結合
**=Δ-11位の二重結合
【0093】
本明細書において言及されたかもしくは引用された全ての特許、特許出願、仮出願、および刊行物は、それらの全体が、本明細書の系統的技術と矛盾しない程度に、本明細書に参照として組入れられている。
【0094】
特に明記または暗示しない限りは、本明細書において使用される「1つの(a)、1つの(an)、およびその(the)」という用語は、「少なくともひとつ」を表す。
【0095】
下記実施例は、本発明を実践する手法を例証するものである。これらの実施例は、限定として構築されるものではない。全てのパーセントは質量%であり、および溶媒混合物の割合は、特に指定しない限りは容量%である。
【0096】
実施例1−Δ9デサチュラーゼ遺伝子再構築
植物-好ましい翻訳コドンへのアスペルギルス・ニジュランス配列の変化、独自の制限酵素部位の導入、および望ましくない配列および一部の二次構造の除去の組合せにより、本発明のΔ9デサチュラーゼ遺伝子を、植物発現のために再デザインした。再デザインされた遺伝子は、Operon, Inc.で合成された。このポリヌクレオチドのオープンリーディングフレームの配列は、ここでSEQ ID NO:1として示した。コザック配列およびBamHIクローニング部位(キャップ)が先立つORFの配列、加えてORFの末端の翻訳ターミネーター(キャップ)は、SEQ ID NO:2に記した。
【0097】
実施例2−Δ9デサチュラーゼ植物形質転換ベクターの構築
BamHI-BstEII遺伝子断片を、Pvβ-ファセオリンプロモーターとPvβ-ファセオリン3' UTR(pPhas-UTR)の間で、ベクターにクローニングした。この構築体は、pOILと称した。プロモーター-遺伝子-UTR断片を、NotIによる消化によりpOILから切り出し、平滑末端化し、ベクターpOEA1の平滑Pmel部位へクローニングした。最終のベクターを、pPD9-OEA1と称した。
【0098】
実施例3−pPD9-OEA1による植物形質転換
プラスミドベクターpPD9-OEA1を、アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)[C58GV3101株(C58ClRifR) pMP90 (GmR), Koncz and Schell、 Mol. Gen. Genet (1986)]へ形質転換した。その後Δ9-デサチュラーゼ植物を、アグロバクテリウム・ツメファシエンスが媒介した植物の形質転換により得た。
【0099】
アラビドプシスを、当技術分野において周知の手法である「浸潤法」により形質転換した。植物は、自殖し、乾燥した種子を、FAME(脂肪酸メチルエステル)分析のために収穫した。
【0100】
カノーラの形質転換に使用したプロトコールは、Katavicの論文[Katavic, Campbell, L., Friesen, L., Palmer, D., Keller, W., and Taylor, D.C. (1996), 「Agrobacterium- mediated genetic transformation of selected high erucic acid B. napus cultivars」、第4回Canadian Plant Tissue Culture and Genetic Engineering Conference, サスカトーン, SK, 1996年6月1-4日]に説明されたものであり、DAS's Nexera系統のために改変された。胚軸切片を、セイヨウアブラナ(B. napus)、cv WestarまたはNexera 710の6-日齢の実生から単離し、ならびにカルス誘導培地(callus initiation medium)において培養し、その後形質転換した。形質転換の日に、胚軸を、Δ9-デサチュラーゼ遺伝子を含むプラスミドDNAの断片が細胞染色体に組み込まれるような、形質遺伝子を伴うプラスミドpPD9-OEA1を含むアグロバクテリウム培養物と共に同時インキュベーションした。共培養期間後、胚軸を、選択剤としてアンモニウムグルフォシネートを含むカルス誘導培地へ移した。健全な耐性カルス組織を得、約12〜16週間新鮮な選択培地へ繰り返して移した。植物を、再生し、Convironsグロースチャンバーへ移した。植物を自殖し、種子を得た。トランスジェニック植物が不稔である場合、これらは改良していないNexera 710系統の花粉で交雑した。乾燥種子を、FAME分析のために収穫した。
【0101】
実施例4−カノーラ事象選別法、およびカノーラの結果のまとめ
特に記さない限りは、下記手法を用い、Nex710/Δ9カノーラ種子を得、そのデータは、引き続きの実施例に提示した。
【0102】
一般に、発育、選択、または選別事象に関して以下の4種の主要な工程が存在する。最初のトランスジェニック事象の選別、T1およびT2種子の選別、T1またはT2植物の選別、ならびに圃場性能による事象の選別。形質転換されたカルスは、最初にT0植物へ再生された。
【0103】
最初の選別に関し、107推定トランスジェニックを、作物栽培学によりおよびサザンブロットにより(単純または複雑、すなわち3コピーよりも多いコピー)スクリーニングした。1事象につき複数の種子をスクリーニングした。T1種子飽和体を決定し、C16:1/C16:0比を決定し(触媒効率を暗示する)、ならびに生化学的表現型の分離を決定した。これらのデータおよび種子利用度(時期、量)を元に、種子の限定されたサブセットを先に進めた。
【0104】
T1およびT2種子の選別(少数の事象は、1世代により進行される)に関して、ハーフ種子分析を行い、可能性のあるホモ接合体を同定した。このデータを元に、分離集団からハーフ種子を、温室生育のために選択した。
【0105】
次の主要な工程は、T1またはT2植物の選別である(少数の事象は、1世代により進行される)。サザンを、導入遺伝子の組込みの完全性を決定するために行った。接合状態も、インベーダーアッセイ法(Third Wave Technologies, Inc.)により決定した。これらのデータを用い、圃場試験のための種子を選択した。
【0106】
T1温室試験のために、1事象につき30個の種子に、ハーフ種子分析を施した。Nex710カノーラを、市販品チェックとして使用した。事象内の全ての個体は、接合状態をサンプリングし、Δ9およびPATに関して対立遺伝子コピー数を決定した。この後、PCRおよびインドールアセトアミド加水分解酵素(IAAH;負のスコア化またはスクリーニングが可能なマーカー)分析を行った。この事象内の全ての個体は、葉を着色し(leaf painted)、PAT分離比を決定した。次にこれらの個体に、サザン分析を行い、「無飽和」プロファイル、正のIAAH、ならびにPATおよびΔ9のホモ接合性に取り組んだ。
【0107】
次に、これらはT2分析に使用した(前述の各植物由来の100個の種子を、ハーフ種子分析した)。インベーダーを使用し、コピー数を証明し、かつこの事象は、D9およびPATについて分離したかどうかを調べた。葉の着色を使用し、この系統はPATについて分離するかどうかを調べた。10個の種子バルクの脂肪酸データを、ハーフ種子データ、インベーダー結果、LP、およびIAAH-陽性を元に植物から収集した。
【0108】
第四の主要工程は、圃場性能による選別事象であった(T2またはT3種子を植え付け、T2またはT3植物、その後T3またはT4種子を分析した)。トランスジェニック事象の広範なサンプリングを行った。農学的、サザン、および接合状態を分析した。バッチ種子油分析も行った。これらのデータを元に、遺伝子量を増加する交雑に関する事象を選択した。
【0109】
以下の選択判定基準を用い、系統を圃場試験に進めた。23事象(6レパートリー/エントリー(reps/entry))からの224系統(ヌルを含む)を、3つの場所で、2つ組の苗床(in replicated nurseries)で評価した。6のT3事象および17のT2事象を、植え付けた。圃場へ移す系統/事象の選択は、各植物由来の種子の挿入数およびハーフ種子分析、それに続く10種子バルクの脂肪酸分析を元にした。選択された系統の総飽和割合の範囲は、ほぼ3.3〜4.5%の範囲であった。以下のT3事象を、更なる発育のために選択した。
【0110】
(表2A)
【0111】
以下のT2事象は、更なる発育のために選択した。
【0112】
(表2B)
【0113】
圃場試験は、以下のように行った。農学的評価を、引き続きの実施例において考察したように行い、農学的ペナルティが、Δ9(D9)に関連していることを確認した。更なるコピー数の証明のために、15のインベーダー葉組織試料を、最も有望なT3系統(プラス同胞-ヌル)の28種から収集し、系統がPATから分離しているかどうかを決定した。D9系統を、総飽和3.5%未満を有することを元に選択した。
【0114】
10種のサザン組織試料を、3種の最も有望なT2系統から採取し、これを総飽和3.5%未満を有することを元に選択した。全ての組織-サンプリングした植物を、自家受粉した。脂肪酸分析は、自殖した植物由来の10種の種子バルク(455試料)、およびOP条(rows)由来のバルク種子試料1g(1445)を元に行った。
【0115】
「最良の」T4事象は、以下であった。
【0116】
(表3A)
【0117】
「最良の」T3事象は、以下であった。
【0118】
(表3B)
【0119】
一般に、Δ9に関連した大きい農学的ペナルティは観察されず、および一部の系統は、種子質量の約10%の増加を示した。農学的結果は、下記実施例18においてより詳細に考察している。個々の脂肪酸成分の分布に関する全般的観察は、下記実施例11-13において考察している。事象218-11.30、36-11.19、および69-11.19に関する平均TSATデータ、TSATの変化率(%)、およびある脂肪酸成分の変化率(%)は、表4-7に示した。一般に、同胞-ヌルおよび野生型と比較して、TSATの〜30%から〜40%の減少が観察された。しかし更なる改良が、以下により詳細に考察されており、例えば更に交雑を行うことができる。
【0120】
(表4)3場所にわたる事象218-11.30からの全系統に関する平均TSAT
【0121】
(表5)C16:0、C16:1、C18:0、およびC18:1対ヌルおよび野生型対照Nex 710のTSAT変化率(%)
【0122】
(表6)C16:0、C16:1、C18:0およびC18:1対ヌルおよび野生型対照 Nex 710のTSAT変化率(%)
【0123】
(表7)C16:0、C16:1、C18:0およびC18:1対ヌルおよび野生型対照 Nex 710のTSAT変化率(%)
【0124】
実施例5−植物の分子特徴
葉試料は、DNA分析のために採取し、PCRおよびサザン分析により導入遺伝子の存在を証明し、ならびに場合によってはELISAによりPATタンパク質の発現を確認した。
【0125】
5A. Δ9デサチュラーゼPCR分析のプロトコール:
以下の2種のプライマーを使用した。
Δ9フォワードB:
Δ9リバースB:
このプライマー対は、Δ9遺伝子の増幅後に、〜1380bp断片を生じた。
【0126】
このスクリーニングにおいて、MJ Tetradサーマルサイクラーにより、以下のサイクリングプロトコールを使用した。
1. 94℃、2分間
2. 94℃、1分間
3. 50℃、2分間
4. 72℃、3分間+5秒間/伸長サイクル
5. 工程2-4の25回の反復
6. 分析の準備ができるまで、または少なくとも2分間、4℃
【0127】
5B. サザン分析のための植物ゲノムDNAの抽出プロトコール:
Qiagenから入手したDNeasy Plant Maxi Kitを使用した。そのブックレットのプロトコールを、溶離部分に以下の変更を行い使用した。緩衝液AEは、DNA等級水(Fisher No. BP561-1)で1:10希釈した。予め65℃に温めた希釈したAE緩衝液0.75mlを用い、2回の溶離を行った。DNAは、イソプロパノールで沈殿し、70%エタノールで洗浄した。DNAペレットを、100μlの1X TE緩衝液中に再懸濁した。DNA濃度を定量した。DNA 6μgをアリコートとし、最終容積を40μlに調節した。試料を、-20℃で保存した。
【0128】
5C. FAME分析(メタノール性H2SO4による種子からの直接のFAME合成):
FAME分析のためのプロトコールは、以下である。
【0129】
GC Specs
ガスクロマトグラフィー:デュアル注入口およびデュアルフレームイオン化検出器を備えた、Hewlett-Packard 6890
データシステム:HP Chemstation、Leap Technologies, カルボロ, NC 27510, PAL System
カラム:J&Wキャピラリーカラム、DB-23、6OMx内径0.25mm、マイクロフィルム厚0.15、最大操作温度250℃。カタログ番号:122-2361
温度プロファイル:平衡時間:1分間。開始温度:50℃。開始時間:3分間。上昇率:40℃/分。最終温度:240℃。最終時間:7.25分間。
【0130】
アラビドプシスのFAME手法:
15:0標準100μl(50μg)を、清潔な16x125mmガラスチューブに添加した(内部標準ストック液:2:1クロロホルム:イソプロパノール中500μg/mlのC15:0 TAG)。蒸発している水浴中、窒素下で55℃で標準を乾燥する。
乾燥した時点で、2%DMPを伴う1Nメタノール性H2SO4 2mlを(100mlについて:メタノール95,22ml、H2SO4 2.772ml、DMP=2,2-ジメトキシプロパン2ml)添加する。チューブを85℃に加熱する。
清潔な16x125mmガラスチューブ中で〜5mgのアラビドプシス種子を秤量し、正確な質量を記録する。
リパーゼを破壊するために、温メタノール性H2SO4 を、標準と共に種子の入ったチューブに添加し、85℃で15分間インキュベーションする。
バイアルを〜50℃に冷却し、その後種子をミニグラインダー中ガラス製乳棒で粉砕する。
試料をチューブに戻し、少なくとも1時間窒素下85℃でインキュベーションする。バイアルを氷上で冷却する。最初に0.9%NaCl 0.5ml、その後ヘキサン中の17:0標準250μl(0.1335mg/mlメチルエステルストック液)を添加する。激しく攪拌し、1000gで5分間遠心する。
パスツールピペットで、100〜200μlを、コニカルインサート(conical insert)(0.5ml)を伴う1.5mlバイアルへ移す。
5μlをGCへ注入する。
【0131】
カノーラに関するFAME手法:
15:0標準100μl(50μg)を、清潔な16x125mmガラスチューブに添加した(内部標準ストック液:2:1クロロホルム:イソプロパノール中500μg/mlのC15:0 TAG)。蒸発している水浴中、窒素下で55℃で標準を乾燥する。
乾燥した時点で、2%DMPを伴う1Nメタノール性H2SO4 2ml(100mlについて:メタノール95.22ml、H2SO4 2.772ml、DMP=2,2-ジメトキシプロパン2ml)を添加する。チューブを85℃に加熱する。
清潔なチューブ中で1個のカノーラ種子を秤量し、正確な質量を記録する。
リパーゼを破壊するために、種子材料を温メタノール性H2SO4と共に、標準の入ったチューブに添加し、85℃で15分間インキュベーションする。
バイアルを〜50℃に冷却し、その後種子をガラス製乳棒で粉砕する。
少なくとも1時間N2下85℃でインキュベーションする。
バイアルを氷上で冷却する。最初に0.9%NaCl 0.5ml、その後ヘキサン中の17:0標準250μl(0.1335mg/mlメチルエステルストック液)を添加する。激しく攪拌し、1000gで5分間遠心する。
パスツールピペットで、100〜200μlを、コニカルインサート(0.5ml)を伴う1.5mlバイアルへ移す。
5μlをGCへ注入する。
【0132】
実施例6−アラビドプシスの結果
最初の結果は、図1に図示し、これは飽和脂肪酸の60%よりも大きい減少が達成されたことを示している。同じく16:0(4.4%)よりもより多くの16:1(例えば5.9%)が達成された。
【0133】
Δ9-CoA-デサチュラーゼ法による無飽和油
FAME分析
約18個の追加の形質転換体に由来するT2種子を分析した。このデータ(表8および図2参照)は、最大60〜70%の「飽和体」の減少を示している。これは、示された最初のデータ(図1参照)よりも、更に強力な飽和脂肪酸の減少である。T2世代は、依然分離されることに注目することは大事であり;従って次の世代でより良い成績の(performing)系統が期待される。この点は、形質が固定されおよび系統が導入遺伝子についてホモ接合体であるまで、T1、T2、T3、および本明細書の別所に記された他の最初の世代(カノーラ系統を含む)全てについてあてはまる。(形質が固定される安定した系統および植物が作出され、および次の実施例で説明される。)。
【0134】
(表8)
値は、単独の試料調製物およびGC試行に由来する(平均ではない)。
TF-21は、野生型(非形質転換)植物として挙動し;恐らく説明は、遺伝子サイレンシングまたは非トランスジェニック回避(グルフォシネート系除草剤による不適切な選択)を含む。
「?」は、GCクロマトグラムのピークの正体は疑問があるか、または不明であることを示す。
【0135】
実施例7−Westarデータ
実施例5Cに説明されたものに類似したプロトコールを、周知の「Westar」カノーラ由来のカノーラ系統に適用した。図3に図示したように、示された飽和脂肪は、43%よりも多く減少され、24:0が含まれた場合には、50%の減少が実現された。
【0136】
実施例8−Nexera 710データの例
本明細書の別所に説明されたものに類似したプロトコールを、周知の「Nexera 710」市販の精選カノーラ由来のカノーラ系統に適用した。総飽和体は、本明細書に説明し、以下に特定した方法を用い計算した。
【0137】
総飽和は、16:0+18:0+20:0+22:0+24:0脂肪酸の和に由来する。単独の種子中の一部の注目すべき飽和体レベルは、表9に示した。油プロファイルは、mol%値として表されている。mol%値は、各特定の脂肪酸の式量を計算に組込んでいる。これは、その脂肪酸種の式量で除算された、所定の脂肪酸種の質量(ピーク面積、または%脂肪酸の直接計算に使用される同じ値)を使用する。
【0138】
(表9)
【0139】
最低の総飽和体を伴う種子に由来するプロファイルを、分析した(種子113a 11.19#4、113a 11.19 #8、5 11.19 #6および5 11.19#8)。未修飾のNexera 710生殖質値は、同じFAME分析の試行から得た。植物は、Convironsグロースチャンバーにおいて生育し、そのため実際のmol%値は、圃場で生育した種子とは異なる。一般に、トランスジェニック植物は、16:0、18:0および20:0の減少、ならびに16:1の増加を示す。18:0レベルは一般に、Nexera 710の平均1.4%(上限2.08%、下限0.81%)から、選択トランスジェニック材料の0.1%平均(上限0.6%、下限0%)までに収まる。同じく16:0レベルは、平均4.6%(上限5.12%〜下限4%)から、3.0%(上限3.41%〜下限2.63%)に収まる。同様に、20:0レベルは、Nexera 710の0.5%平均から、選択されたトランスジェニックの0%まで下落する。16:1レベルは、Nexera 710において検出不能であり、トランスジェニックにおいて平均2.3%(上限2.71%〜下限1.72%)まで増加する。平均18:3レベルは、小さいトランスジェニック集団においてわずかに増加したが、値の範囲は、未修飾のNexera 710試料と重複した。これらの結果は、以下のようにまとめられる。
【0140】
(表10)
【0141】
実施例9-更なるカノーラデータ
本明細書の別所に説明されたものに類似したプロトコールを、周知の「Nexera 710」市販の精選カノーラに由来する追加のカノーラ系統に適用した。以下に示したように、総飽和体および脂肪酸の個々の種類の質量%を、本明細書に考察された方法を用いて計算した。
【0142】
図4A-Cおよび5A-Cは、各々、事象36-11.19および218-11.30からの代表的結果を示し、これは本発明の実践により得ることができる減少した飽和脂肪レベルを明らかにしている。本発明に従い更なる操作を行うことにより、ここに例示された飽和脂肪レベルは、更に低下することができる。このデータは全て、示されたように自殖したトランスジェニックカノーラ植物から得た。
【0143】
まとめると、事象36-11.19に関して、温室生育した植物からのT2ハーフ種子分析は、総飽和体を2.57%と低くした。温室生育した植物からのT3全種子に関する総飽和体は、3.66%と低かった。結果は、図4Aに図示している(数値データは、図4Bおよび4Cである)。事象218-11.30温室生育した植物に関して、T2ハーフ種子分析は、総飽和体が2.71%と低いことを明らかにした。T3全種子は、3.37%と低い総飽和体を有した。結果は、図5Aに図示している(数値データは、図5Bおよび5Cである)。参考のために、NATREONは、圃場条件下で、平均して総飽和体6.5%を有した。
【0144】
本発明の更なる改善することにより(自殖、他の優れた系統との交雑、デサチュラーゼ遺伝子コピー数の増加[追加の形質転換、更なる育種交雑などによる]、デサチュラーゼ発現時期の変化、および変異などの追加ラウンド)、総飽和脂肪レベルの更に大きいレベルの減少が実現され得る。
【0145】
実施例10−更なるカノーラデータの分析−総飽和脂肪の%減少
実施例9に示されたデータは、本発明の様々な局面を例示するために、様々な計算において使用することができる。例えば、総飽和脂肪の%減少は、所定の植物の総飽和体の、対照系統の総飽和体による最初の除算、それに続く100%からの減算により計算することができる。本発明によりもたらされるこのような低下の例は、以下に例示される。結果は、最も近い自然数(非小数)に丸めた概数であることができる。
【0146】
(表11)
【0147】
グラフ、図、表または本明細書に考察された他のものに示された数値は全て、本発明の境界および限界を規定するためのエンドポイントとして使用することができる。同様に、先に示されたものおよびより詳細に以下に考察されるもののような、これらの数値を使用する計算は、本発明の境界および限界を規定するために使用することができる。表12-14は、事象218-11.30、36-11.19、および69-11.19を伴う系統に関する、更なる代表的結果および計算を示している。
【0148】
(表12)交雑に関する事象218-11.30 HS選択
【0149】
(表13)交雑に関する事象36-11.19 HS選択
【0150】
(表14)交雑に関する事象69-11.19 HS選択
【0151】
実施例11−更なるカノーラデータの分析−詳細な脂肪酸プロファイル
再度、図4A-Cおよび5A-Cは、事象218-11.30および36-11.19を有する様々な植物の脂肪酸プロファイルを示す代表的結果を示している。一般に、これらの結果は、16:0および18:0レベルが大きく低下されるのみではなく(対応する不飽和体レベルの結果としての増加を伴う)、20:0、22:0、および24:0レベルも有利かつ驚くべきかつ予想外のことに、低下したことを示している。場合によっては、18:2および18:3レベルも低下することができ、これは改良された油の酸化安定性を増強する。更に、先に示唆された比のいずれか(例えば16:0-16:1、18:0-18:1、および例えば18:0-[20:0+22:0+24:0])を使用し、本発明を実践する上での有利な結果を規定することができる。総C20:0+C22:0+C24:0の合計した減少率も、驚くべきことに本発明により実現された。従って本発明は、本明細書に例示されたような、有利かつ改善された脂肪酸プロファイルを有する植物を提供する。本発明に従い更なる改良を作製することにより、より良い飽和体の減少、「無飽和」の増加、およびより良い比を実現することができる。
【0152】
例えば、以下の図5Cおよび図4Bのデータを使用し、様々な計算を、本発明の達成を例示するために使用することができる。図5Cおよび図4Bからの試料データは、下記表に示されている。各指定された脂肪酸の量は、各事象について、括弧内には関連する対照植物について記されている。
【0153】
(表15)
【0154】
218-11.30事象に注目すると、C20:0、C22:0、およびC24:0成分による飽和体への総寄与(contribution)は、対照において1.08%であるが、これらの成分は有利なことに本発明のカノーラ系統においては0.26%に減少している。このことは、これらの飽和体における1/4を超える減少を示している。同様に、各成分は、独立しているとみなすことができる。同じく218-11.30事象に注目すると、C20:0成分は、対照/野生型において0.62%であるが、これは本発明の植物系統において約1/5.6に減少される(0.11%へ低下)。C22:0成分は、約1/2.7に減少され:d-9デサチュラーゼ植物系統においては0.12%であり、これはデサチュラーゼ遺伝子を欠いている対照系統の0.32%よりも低い。C24:0は、0.14%から0.03%へと約1/4.7に減少される。
【0155】
事象36(または36-11.19)に関して、ふたつの系統が、C20:0、C22:0、およびC24:0含量を、各々0.32、0.15、および0.04、ならびに0.30、0.14、および0.07を有する。各々0.64、0.42、および0.21を有する対照と比べ、これらの系統は、約半分のC20:0、および少なくとも約1/3のC22:0およびC24:0の減少を有する。先に言及された第一の系統は実際に、C24:0の1/5を超える減少を示している。
【0156】
これらの好ましい脂肪酸成分のいずれかについて、本発明の他の系統のいずれかについて、非常に多くの同様の計算を行うことができることは、直ぐに明らかになると思われる。これらの例示は、限定として構築されるものではなく、そのような新規の低下および比は、本発明を定義するために使用することができる。
【0157】
実施例12−次世代の更なるハーフ種子データ
更なるハーフ種子FAME分析を、表16に示す。この図は、T3世代における2.64%、およびT4世代における2.66%と低い総飽和を示している。表17は、各系統に存在するD-9デサチュラーゼ遺伝子のコピー数を示している(表16の試料IDおよび表17のID行を参照のこと)。コピー数の作用は、実施例14および19においてより詳細に考察されている。
【0158】
(表16)
【0159】
(表17)
【0160】
実施例13−実施例12のハーフ種子データの更なる分析、ならびに脂肪酸シフト、不飽和体増加、および飽和体減少を示す更なるデータ
T3圃場試験からのハーフ種子データを、脂肪酸含量の「総飽和体」データに対する様々な比較を図示するために、プロットした。図6Aおよび6Bは、ヌル(非機能的挿入断片を伴う事象)および野生型対照(形質転換していない系統)と比べ、トランスジェニック事象におけるC16:0の減少およびC16:1の増加を明らかに示している。図6Cおよび6Dは、ヌルおよび野生型対照と比べ、トランスジェニック事象におけるC18:0の減少およびC18:1の増加を明らかに示している。図6Eおよび6Fは、ヌルおよび野生型対照と比べ、トランスジェニック事象におけるC20:0およびC22:0の各々減少を明らかに示している。
【0161】
同様の結果が、実施例4の考察で得られたデータを用い、棒グラフにおいても示されている。図6Gおよび6Hは、ヌルおよび野生型対照と比べ、トランスジェニック事象におけるC16:0のシフトおよび減少、ならびにC16:1のシフトおよび増加を明らかに示している。図6Iおよび6Jは、ヌルおよび野生型対照と比べ、トランスジェニック事象におけるC18:0のシフトおよび減少、ならびにC18:1のシフトおよび増加を明らかに示している。図6Kおよび6Lは、C18:2およびC18:3について同様の棒グラフを示している。
【0162】
先に考察されたプロットおよび図6Mも、既に良好であるNex 710系統と比べ、総飽和体の非常に驚くべき減少を明らかに図示している。図6Nは、1000個の種子の分布を示している。
【0163】
実施例14−複数のΔ9デサチュラーゼ遺伝子による飽和脂肪レベルの減少
温室の増加および圃場試験からの結果は、総飽和体の減少とアスペルギルスΔ9デサチュラーゼコピー数の間に相関があることを示唆している。
【0164】
14A:事象選別および導入遺伝子コピー数の作用のFAME分析プロトコール
試料調製
1. 個々の種子の質量を得、プラスチック製母プレート上に配置する。
2. 各ウェルに2, 1/8"ボールを加える。
3. 母プレートを液体ハンドラーHamiltonで取り上げ:IS(C11:0)および代理物(C15:0 FAEE)と共にヘプタン400μLを添加し、その後ナトリウムメトキシド(0.5N)100μLを添加する。
4. キャップを、ストリップキャップに嵌め、5分間Genoにより粉砕する(1x500回)。
5. ストリップキャップを再配置する。
6. プレートの蓋をゴム製マット上に置く(余分のシーリング)。蓋を、黒色絶縁(electrical)テープで巻き、母プレートの底を外し、バイアルを露出する。
7. プレートを、ボルテックス/ヒーター上に15分間、37℃/60rpmで配置する(ボルテックスの孔には、砂が詰められている)。
8. プレートをx3500rpmで2分間遠心する。
9. 底蓋を戻し、蓋/ストリップキャップを取り外す。上層350μlを、Hamiltonを用いて、娘プレートへ移す。
10. その後抽出プレートへ、ヘプタン400μLを、ISおよび代理物と共に添加する。
11. 工程5から10を更に2回繰り返す(合計3回移す)。
12. 最後に移した後、抽出プレートをHamilton上に維持する。抽出物50μlを、C11と共にヘプタン450μlを含有するアルミニウムブロック中に搭載されたガラスインサートに移す。
13. GCへ注入する。
【0165】
GC/FID分析
GCパラメータ:
1試料につき、1μl注入、スプリットレス
カラム-
DB23、15m、内径0.25mm、およびフィルム厚0.15μm
GCパラメータ:
オーブン温度プログラム−
70℃で2.15分間維持(スプリットレス)
70℃〜150℃、25℃/分
150℃〜180℃、5℃/分
180℃〜220℃、25℃/分
220℃で2分間維持。
注入温度−230℃
検出温度−240℃
メイクアップガス−窒素 25mL/分
FID燃料−空気 400mL/分、水素 40mL/分
フロント注入器:掃流時間:1分間、掃流:35ml/分
バック注入器:掃流時間:2.15分間、掃流:35ml/分
フロント投入圧:1.0mL/分−定常流
バック投入圧:1.0mL/分−定常流
試行時間: 14.95分間
流量−ヘリウム定常流 1mL/分
【0166】
捕捉配列
96試料を、ライナー上での構造(build)が最小になるように、フロントカラムおよびバックカラム間で分離する。試料の一覧を、注入される試料の種類に応じて、5種の注入法と共に作製している。最初の注入された5試料は常に以下である。
1. マトリックス
2. マトリックス
3. 標準1
4. カノーラ陽性対照
5. 試薬ブランク
6-27. カノーラ試料
33. 標準2
34-54. カノーラ試料
55. 標準3
各リストは、16試料の3事象を含む(48試料)。
標準は、以下のように分布したFAME合計25ppmを含む。
【0167】
(表18)
【0168】
14B:おおよそのコピー数に対する予備サザンブロット分析
これらの目的に使用したDNA調製プロトコールは、以下に示した。DNA約6μgを、HindIIIで消化し、消化したDNAを、0.75%アガロースゲル上を流した。正帯電したナイロンメンブレン上へのブロッティングおよびハイブリダイゼーションは、典型的プロトコールに従った(Maniatis, Roche Applied Science, Inc.)。プローブは、アスペルギルスΔ9デサチュラーゼ遺伝子に由来したDIG標識した(Roche Applied Science, Inc.のキット)PCR産物からなった。洗浄は、2xSSC/0.1%SDSにより室温で5分間を2回、その後65℃の0.1xSSC/0.1%SDSで2回行った。ハイブリダイズされたバンドは、DIG-ルミネセント検出キットにより、製造業者(Roche Applied Science, Inc.)の指示に従い可視化した。ハイブリダイズしているバンドを計測し、トランスジェニック試料は最初に、1〜3本のバンドを示した場合は、「単純」、または3本よりも多いバンドを示した場合は、「複雑」と説明した。
【0169】
14C:トランスジェニック事象における導入遺伝子コピー数および飽和脂肪酸の減少の比較
下記定義を、表に適用した。
【0170】
【0171】
(表19)
【0172】
「飽和体の減少の比」は一般に同じトランスジェニック事象の種子を使用したので、これを事象のランク付けのために使用した。この直接比較は、様々な時点で組織培養により生じた植物と成長した植物の間の変動を低下することを補助している。
【0173】
前記データは、みかけの遺伝子量作用を示し;導入遺伝子のより多いコピーは、より多くの有効な飽和脂肪酸減少を引き起こす傾向がある。例えば、先に示された57種の「非対照」植物が存在する。これらの57種の植物は、19種の植物の3群に分けることができる。植物の一番上のセット(飽和体の最良の減少および最低レベルの飽和体を示している)は、19種の「複雑」事象(デサチュラーゼ遺伝子の3個よりも多いコピー)の11種を有する。このセットは、「単純」または「恐らく単純」と特徴付けられた19事象のうちの8種を含んだ。19種の植物の中間のセットは、「複雑」事象を19種中わずかに6種有した(19種中13種の「単純」または「恐らく単純」事象)。なお更に、19種の植物の第三のセット(飽和体の比較的最低の減少を示す)は、わずかに3種の複雑事象を含み、19事象中の16種は、「単純」または「恐らく単純」であった。従って、3個よりも多いデサチュラーゼコピーを有する細胞を伴う植物は、わずかに1個のこの遺伝子のコピーを有する細胞を伴う植物よりも、飽和体をより良く減少することを示しているように見える。
【0174】
実施例15−オレイン酸およびバクセン酸の分離
バクセン酸は、Δ-11位に二重結合を伴うC18:1である。バクセン酸は、色素体の外側でのΔ9 C16:1の伸長により形成される。本明細書において考察した他の分析法は、オレイン酸およびバクセン酸のピークを一緒に「オレイン酸」と記される単独の組成の割合とするので、その追跡は重要である。すなわち、特に記されない限りは、これらふたつは分離されなかった。バクセン酸の貢献を減算することにより、オレイン酸の貢献割合が、好ましくかつ有利なことに(および驚くべきことに)、80%未満に維持されるが、依然総飽和体の減少(全体の「無飽和」または「低飽和」レベルに対し)は達成されることが、明らかにされる。以下のふたつの実施例において示したように、これを説明するために、2種類の分析を使用した。
【0175】
実施例16−カノーラΔ9種子抽出およびバクセン酸SOPに関する分析
図7Aおよび7Bは、下記プロトコールを使用し得られたデータを図示している。
【0176】
試料調製(前述のものと同じ)
1. 個々の種子の質量を得、プラスチック製母プレート上に配置する。
2. 各ウェルに2, 1/8"ボールを加える。
3. 母プレートを液体ハンドラーHamiltonで取り上げ:IS(C11:0)および代理物(C15:0 FAEE)と共にヘプタン400μLを添加し、その後ナトリウムメトキシド(0.5N)100μLを添加する。
4. キャップを、ストリップキャップに嵌め、5分間Genoにより粉砕する(1x500回)。
5. ストリップキャップを再配置する。
6. プレートの蓋をゴム製マット上に置く(余分のシーリング)。蓋を、黒色絶縁テープで巻き、母プレートの底を外し、バイアルを露出する。
7. プレートを、ボルテックス/ヒーター上に15分間、37℃/60rpmで配置する(ボルテックスの孔には、砂が詰められている)。
8. プレートをx3500rpmで2分間遠心する。
9. 底蓋を戻し、蓋/ストリップキャップを取り外す。上層350μlを、Hamiltonを用いて、娘プレートへ移す。
10. その後抽出プレートへ、ヘプタン400μLを、ISおよび代理物と共に添加する。
11. 工程5から10を更に2回繰り返す(合計3回移す)。
12. 最後に移した後、抽出プレートをHamilton上に維持する。抽出物50μlを、C11と共にヘプタン450μlを含有するアルミニウムブロック中に搭載されたガラスインサートに移す。
13. GCへ注入する。
【0177】
GC/FID分析(FAMEプロファイルとは異なる)
GCパラメータ:
1試料につき、1μl注入、スプリットレス
カラム-
SGEのBPX 70、15m、内径0.25mm、およびフィルム厚0.25μm
GCパラメータ:
オーブン温度プログラム−
70℃で2.15分間維持(スプリットレス)
70℃〜140℃、25℃/分
140℃で14分間維持
140℃〜180℃、10℃/分
180℃で3分間維持
180℃〜220℃、25℃/分
220℃で3分間維持。
注入温度−230℃
検出温度−240℃
メイクアップガス−窒素 25mL/分
FID燃料−空気 400mL/分、水素 40mL/分
フロント注入器:掃流時間:1分間、掃流:35ml/分
バック注入器:掃流時間:2.15分間、掃流:35ml/分
フロント投入圧:1.0mL/分−定常流
バック投入圧:1.0mL/分−定常流
試行時間: 30.55分間
流量−ヘリウム定常流 1mL/分
【0178】
捕捉配列
96試料を、ライナー上での構造が最小になるように、フロントカラムおよびバックカラム間で分離する。試料の一覧を、注入される試料の種類に応じて、5種の注入法と共に作製している。最初の注入された5試料は常に以下である。
1. マトリックス
2. マトリックス
3. 標準1
4. 試薬ブランク
5-32. カノーラ試料
33. 標準2
34-54. カノーラ試料
55. 標準3
【0179】
各リストは、8試料の6事象を含む(1種子=1試料)(48試料)。
【0180】
標準は、以下のように分布したFAME合計200ppmを含む。
【0181】
(表20)
【0182】
実施例17−ガスクロマトグラフィー/質量分析/飛行型を使用するバクセン酸寄与の分析
表21は、下記のプロトコールから得られたデータを示している。表21において、T4について、脂質割合(%)は低下されず;これは、トランスジェニック系統において40.8%で維持された(対照系統と同じ)。
【0183】
(表21)
*:処理法により検出限界未満である場合、デフォルトは0であった。
捕捉および処理は、HP Chemserverパッケージを使用
【0184】
機器の説明
Lecoから入手した飛行時間型質量分析装置Pegasus IIIは、ガスクロマトグラフィーHP 6890と連結した。
CTC Analytics technologyから入手したCombi Pal自動注入器を、10μlシリンジと共に、HP 6890に装備した。
【0185】
GC法
GCパラメータ:
1試料につき、1〜3μl注入、スプリットレス
カラム-
SolGel Wax、30m、内径0.25mm、およびフィルム厚0.25μm
GCパラメータ:
オーブン温度プログラム−
70℃〜175℃で25℃/分(スプリットレス)
175℃で25分間維持
175℃〜230℃、50℃/分
230℃で3分間維持
注入温度−230℃
検出温度−300℃
バック注入器:掃流時間:30秒、掃流:20ml/分
バック投入圧:2mL/分−定常流
試行時間: 33.3分間
流量−ヘリウム定常流 2mL/分
【0186】
質量分析器
質量選択:
50〜600amuから収集した質量
フィラメントバイアス:-70V
イオン給源:225℃
検出器:
検出電圧:1600V
捕捉速度:10スペクトル/秒
溶媒は100秒遅れ
【0187】
フラグメンテーション
ChromaTOFソフトウェアは、より良い分離および解釈のために、フラグメンテーションをコンパイルし、および同時移動するピークを解析する。
【0188】
脂肪酸の同定は、同じ条件で注入された標準液(下記参照)を元に、保持時間およびフラグメンテーション合致、ならびに/または先に説明されたソフトウェアに含まれたNIST/EPA/NIHデータベースとの良好な合致により行う。
【0189】
cis 11-オクタデセン酸メチルエステルとしても公知であるバクセン酸メチルエステルは、Sigmaから得たバクセン酸(CAS:506-17-2)から作製された標準物を試行することにより、カノーラ種子抽出物から同定された。保持時間は、1207秒であり、標準品と853の合致および先に説明された文献と814の合致を生じる。
【0190】
注入された標準品の組成
【0191】
(表22)
【0192】
バクセン酸メチルエステル法:
メチル化された生成物を、この酸100mgのメチル化後に得た。
−100mgを、30mlのガラスバイアル中のMeOHCl 0.5N(Supelco)の5mlに溶解。
−窒素下70℃で1時間加熱。
−室温に冷却後、0.9%NaClを含有する水5mlを添加。
−3回連続したヘキサン(15ml)抽出により、ヘキサン中にエステルを分配。
−この有機層を、HNaCO3を2.5%含有する水15mlと混合することにより、酸残渣を中和。
−有機層を、N2下で蒸発させ、室温で油状の透明な液体を得、これはバクセン酸メチルエステルに対応している。
【0193】
実施例18−優れた農学的形質を維持しながらの「無飽和」の達成
様々な圃場で、農学的測定を行い、トランスジェニックと対照を比較した。結論は、全般的に、このトランスジェニック植物は、Nex 710対照と、同様に挙動し、および類似した形質を示した(はるかに改善された飽和体レベル以外)。従って本導入遺伝子は、植物の健康に一貫して負の作用は有さない。形質および等級化システムのまとめを、表23に示している。様々な判定基準を、様々な場所の圃場で使用した。
【0194】
表24-27は、いくつかの代表的結果を数値で表している。下記の略号を、これらの図において使用した。
DTF=開花までの日数 EOF=開花終了までの日数
HT=高さ SC=不稔性の数
DTM=成熟までの日数 LSV=晩期活力
LOD=倒伏 SD WT=種子質量
【0195】
表24は、事象#218-11.30からの系統の農学的データを示している。表25は、事象#36-11.19からの系統の農学的データを示している。表26は、事象69-11.19からの系統の農学的データを示している。これらのことは、本発明、すなわち「無飽和」カノーラの実現は、驚くべきことに、他の重要な農学的形質への有害な作用を伴わずに実践することができることを明らかにしている。
【0196】
(表23)
【0197】
(表24)事象#218-11.30からの最良の性能の系統の農学的データのまとめ
【0198】
(表25)事象#36-11.19からの最良の性能の系統の農学的データのまとめ
【0199】
(表26)事象#69-11.19からの最良の性能の系統の農学的データのまとめ
【0200】
実施例19−遺伝子量の作用;Δ9デサチュラーゼ遺伝子の多コピーの挿入による飽和体の更なる減少
本実施例は更に、Δ9デサチュラーゼ遺伝子の「スタック」している多コピーは、驚くべき遺伝子量の作用を有し、このことは、カノーラのような油糧種子植物中の飽和体の更なる低下にも使用することができることを示している。以下は、特に記さない限りは、FAME手法を用いて測定された温室データである。
【0201】
表27は、未変性のNex 710、シングル事象(218、36、69)、ならびにインベーダーアッセイ法に基づき選択されたF2植物を辿るF3種子パッケージの各々由来のF3 10種子バルク脂肪酸データを示す。(表27の最後の2列は、Nex 710対照と比較した、各植物に関するおおよその平均総飽和体、および総飽和体の平均減少の概数を示す)。その世代で、サザンデータは使用しなかった。従って飽和体発現およびインベーダーアッセイ法によって、推定されたスタックおよび推定された親同胞に加えヌルの選択を行い、F3植物の生育およびサザンを使用する再試験により確認した。
【0202】
例えば、統計学的には分析されないが、41種の「スタック」系統からの試料は、平均総飽和体3.5%を有した。21種の「シングル」系統は、平均総飽和3.84%を有した。
【0203】
表28は、コピー数および接合状態を確認するために植直しした推定されたF3スタック、推定された親同胞、およびヌルのまとめを含んでいる。TDN0400141、TDN0400142、TDN0400145、TDN0400155、TDN0400158、およびTDN0400160と命名された系統は、ホモ接合体スタックであると推定された。系統TDN0400189、TDN0400143、TDN0400197、TDN0400167、およびTDN0400184は、スタック外の親同胞であると推定された。系統TDN0400198、TDN0400199は、これらのスタックの自殖されたヌルとして進めた。TDN0400202、TDN0400204、およびTDN0400208は、シングル事象である。この材料は、現在も圃場にあるか、または最近収穫された。従って圃場データは、まだ入手できていない。
【0204】
図8および9は、F3植物からのDNAの2個のゲル泳動の写真である。サザン分析についての単離したDNAと同様の問題点にこの工程で遭遇し、そのため提供されたこれらの植物の9種全てがゲルに出現したわけではない。これらのゲルを元に、系統TDN0400141、TDN0400142、および恐らくTDN0400158(わずかに2種の植物)は、ホモ接合体スタックである。TDN0400145は、依然事象36について分離しており、およびTDN0400155は、依然事象69について分離している。系統TDN0400160は、バンドの奇数分離を有するように見え、これは全て3種のシングル事象に分離していることを示している。TDN0400160について、経過観察を行った。この交雑からの植物#8は、2.6%と低い飽和レベルを有し、これは高い接合状態のトリプルスタックと一致している。追加の分子分析は、このことを確認している。親同胞であると推定された系統の全てが、これらのゲル中に出現したわけではない(表29の事象カラムにおいて強調された系統TDN0400184、TDN0400189、およびTDN0400197、以下に考察)。単独の植物脂肪酸データを元に、TDN0400184由来の9種の植物(同胞でない(sibbed-out)シングル事象218であると推定される)は、推定されたスタックと同じくらい低い飽和レベルを有した。すなわち、このデータの傾向は、スタックは、「同胞でない」事象と比べ、飽和体の減少を一貫して示す(2種のトランスジェニック事象の交雑から回復された単純トランスジェニック事象)。
【0205】
表29は、Nex 710、シングル事象、および各単独のF4植物からの10種の種子バルク脂肪酸データを含む。各推定されたスタック、ヌル、シングル事象などの9種の植物は、再生し、分子分析のために組織試料とし、および脂肪酸分析のための種子セットで維持した。
【0206】
(表27)
【0207】
(表28)
【0208】
(表29)
【図面の簡単な説明】
【0209】
【図1】アラビドプシスにおいて、飽和脂肪酸の60%を超える減少が実現されたことを示している。このグラフは、単独のアラビドプシス事象に関するT2およびT3種子データをまとめている。
【図2】18種の追加の形質転換体由来のT2アラビドプシス種子における、最大60〜70%の「飽和体」の減少を示している。このグラフに図示されたデータは、表8に示された数値データおよびより早期の数値データと組み合わせられた。
【図3】Westarカノーラにおいて、飽和脂肪が、43%よりも多く減少したこと(および、24:0が含まれる場合は、50%減少が達成されたこと)を示している。
【図4A】対照と比較した、事象36-11.19を含む様々なカノーラ植物由来の種子の総飽和を比較する棒グラフを示す。
【図4B】棒グラフにより図示された数値データを表す。
【図4C】棒グラフにより図示された数値データを表す。
【図5A】対照と比較した、事象218-11.30を含む様々なカノーラ植物由来の種子の総飽和を比較する棒グラフを示す。
【図5B】棒グラフにより図示された数値データを表す。
【図5C】棒グラフにより図示された数値データを表す。
【図6】T3圃場試験由来のハーフ種子データを示す。図6Aおよび6Bは、ヌル対照(導入遺伝子が植物から分離された事象)および野生型対照(非形質転換系統)と比較したトランスジェニック事象におけるC16:0の減少およびC16:1の増加を明らかに示している。図6Cおよび6Dは、ヌル対照および野生型対照と比較したトランスジェニック事象におけるC18:0の減少およびC18:1の増加を明らかに示している。図6Eおよび6Fは、ヌル対照および野生型対照と比較したトランスジェニック事象におけるC20:0およびC22:0の各々の減少を明らかに示している。図6Gおよび6Hは、ヌル対照および野生型対照と比較したトランスジェニック事象におけるC16:0のシフトおよび減少、ならびにC16:1のシフトおよび増加を明らかに示している。図6Iおよび6Jは、ヌル対照および野生型対照と比較したトランスジェニック事象におけるC18:0のシフトおよび減少、ならびにC18:1のシフトおよび増加を明らかに示している。図6Kおよび6Lは、C18:2およびC18:3の同様の棒グラフを示している。図6Mは更に、既存の非常に良好なNex 710系統と比べた、総飽和の減少を図示している。図6Nは、1000個の種子についての分布を示している。
【図7】実施例16のプロトコールを使用して得たデータを示している。
【図8】実施例19において考察した、F3植物由来のDNAによる2個のゲル泳動の図である。
【図9】実施例19において考察した、F3植物由来のDNAによる2個のゲル泳動の図である。
【特許請求の範囲】
【請求項1】
3.5%未満の総飽和体(saturate)および80%未満のオレイン酸を含有する油性画分を有する種子を生成する、カノーラ植物。
【請求項2】
前記油性画分が、70%〜78%のオレイン酸を含有する、請求項1記載の植物。
【請求項3】
前記油性画分が、3%を超えないリノレン酸を含有する、請求項1記載の植物。
【請求項4】
前記油性画分が、70%〜78%のオレイン酸および3.5%を超えないリノレン酸を含有する、請求項1記載の植物。
【請求項5】
請求項1記載のカノーラ植物から生じた、種子。
【請求項6】
3.5%未満の総飽和体および80%未満のオレイン酸を含有する、カノーラ油。
【請求項7】
ジャガイモ材料および、請求項6記載のカノーラ油を含有する、フライ食品組成物。
【請求項8】
前記油性画分が、2.7%を超えない総飽和体を有する、請求項1記載のカノーラ植物。
【請求項9】
2.7%を超えない総飽和体を含有する油性画分を有する、請求項5記載のカノーラ種子。
【請求項10】
2.7%を超えない総飽和体を含有する油性画分を有する、請求項6記載のカノーラ油。
【請求項11】
ジャガイモ材料を、請求項6記載のカノーラ油で揚げる工程を含む、フライ食品組成物の製造法。
【請求項12】
Δ9デサチュラーゼタンパク質をコードしている、カノーラ植物のゲノムに安定して組み込まれた、少なくとも1種のポリヌクレオチドを含む、カノーラ植物であって、SEQ ID NO:5のタンパク質をコードしている核酸分子の全相補体が、洗浄後に該ポリヌクレオチドとのハイブリダイゼーションを維持し、ここで該洗浄条件が、2xSSC(Standard Saline Citrate)および0.1%SDS(Sodium Dodecyl Sulfate)による室温で15分間である、植物。
【請求項13】
前記核酸分子が、SEQ ID NO:1を含む、請求項12記載の植物。
【請求項14】
前記洗浄条件が、0.1xSSCおよび0.1%SDSによる、室温で15分間である、請求項12記載の植物。
【請求項15】
前記洗浄条件が、0.1xSSCおよび0.1%SDSによる、55℃で30分間である、請求項12記載の植物。
【請求項16】
前記ゲノムが、2種の前記ポリヌクレオチドを含む、請求項12記載の植物。
【請求項17】
前記ゲノムが、3種の前記ポリヌクレオチドを含む、請求項12記載の植物。
【請求項18】
前記ポリヌクレオチドが、種子特異的プロモーターへ機能的に連結されている、請求項12記載の植物。
【請求項19】
請求項5記載の種子により生育された植物。
【請求項20】
トランスジェニックカノーラ植物の少なくとも1種の種子の油性画分中の飽和脂肪を、対応する野生型カノーラ植物の種子の野生型油性画分と比較して、減少させる方法であって、Δ9デサチュラーゼタンパク質をコードしているポリヌクレオチドを発現するカノーラ植物を作出することを含み、ここで該タンパク質をコードしているヌクレオチド分子が、SEQ ID NO:1の分子とハイブリダイズする、方法。
【請求項21】
油性画分が、80%未満のオレイン酸を含有する、請求項20記載の方法。
【請求項22】
前記タンパク質が、前記対応する野生型カノーラ植物と比較して、前記トランスジェニックカノーラ植物の前記種子の油性画分中のパルミチン酸(16:0)の減少、ベヘン酸(22:0)の減少、およびパルミトレイン酸(16:1)の増加を引き起こす、請求項20記載の方法。
【請求項23】
前記飽和脂肪が、少なくとも60%減少される、請求項20記載の方法。
【請求項24】
SEQ ID NO:1に示されたヌクレオチド配列を含む、ポリヌクレオチド。
【請求項25】
前記植物が、高さが少なくとも100cmであり、平均種子質量が3mgを上回る、請求項1記載の植物。
【請求項1】
3.5%未満の総飽和体(saturate)および80%未満のオレイン酸を含有する油性画分を有する種子を生成する、カノーラ植物。
【請求項2】
前記油性画分が、70%〜78%のオレイン酸を含有する、請求項1記載の植物。
【請求項3】
前記油性画分が、3%を超えないリノレン酸を含有する、請求項1記載の植物。
【請求項4】
前記油性画分が、70%〜78%のオレイン酸および3.5%を超えないリノレン酸を含有する、請求項1記載の植物。
【請求項5】
請求項1記載のカノーラ植物から生じた、種子。
【請求項6】
3.5%未満の総飽和体および80%未満のオレイン酸を含有する、カノーラ油。
【請求項7】
ジャガイモ材料および、請求項6記載のカノーラ油を含有する、フライ食品組成物。
【請求項8】
前記油性画分が、2.7%を超えない総飽和体を有する、請求項1記載のカノーラ植物。
【請求項9】
2.7%を超えない総飽和体を含有する油性画分を有する、請求項5記載のカノーラ種子。
【請求項10】
2.7%を超えない総飽和体を含有する油性画分を有する、請求項6記載のカノーラ油。
【請求項11】
ジャガイモ材料を、請求項6記載のカノーラ油で揚げる工程を含む、フライ食品組成物の製造法。
【請求項12】
Δ9デサチュラーゼタンパク質をコードしている、カノーラ植物のゲノムに安定して組み込まれた、少なくとも1種のポリヌクレオチドを含む、カノーラ植物であって、SEQ ID NO:5のタンパク質をコードしている核酸分子の全相補体が、洗浄後に該ポリヌクレオチドとのハイブリダイゼーションを維持し、ここで該洗浄条件が、2xSSC(Standard Saline Citrate)および0.1%SDS(Sodium Dodecyl Sulfate)による室温で15分間である、植物。
【請求項13】
前記核酸分子が、SEQ ID NO:1を含む、請求項12記載の植物。
【請求項14】
前記洗浄条件が、0.1xSSCおよび0.1%SDSによる、室温で15分間である、請求項12記載の植物。
【請求項15】
前記洗浄条件が、0.1xSSCおよび0.1%SDSによる、55℃で30分間である、請求項12記載の植物。
【請求項16】
前記ゲノムが、2種の前記ポリヌクレオチドを含む、請求項12記載の植物。
【請求項17】
前記ゲノムが、3種の前記ポリヌクレオチドを含む、請求項12記載の植物。
【請求項18】
前記ポリヌクレオチドが、種子特異的プロモーターへ機能的に連結されている、請求項12記載の植物。
【請求項19】
請求項5記載の種子により生育された植物。
【請求項20】
トランスジェニックカノーラ植物の少なくとも1種の種子の油性画分中の飽和脂肪を、対応する野生型カノーラ植物の種子の野生型油性画分と比較して、減少させる方法であって、Δ9デサチュラーゼタンパク質をコードしているポリヌクレオチドを発現するカノーラ植物を作出することを含み、ここで該タンパク質をコードしているヌクレオチド分子が、SEQ ID NO:1の分子とハイブリダイズする、方法。
【請求項21】
油性画分が、80%未満のオレイン酸を含有する、請求項20記載の方法。
【請求項22】
前記タンパク質が、前記対応する野生型カノーラ植物と比較して、前記トランスジェニックカノーラ植物の前記種子の油性画分中のパルミチン酸(16:0)の減少、ベヘン酸(22:0)の減少、およびパルミトレイン酸(16:1)の増加を引き起こす、請求項20記載の方法。
【請求項23】
前記飽和脂肪が、少なくとも60%減少される、請求項20記載の方法。
【請求項24】
SEQ ID NO:1に示されたヌクレオチド配列を含む、ポリヌクレオチド。
【請求項25】
前記植物が、高さが少なくとも100cmであり、平均種子質量が3mgを上回る、請求項1記載の植物。
【図1】


【図2】
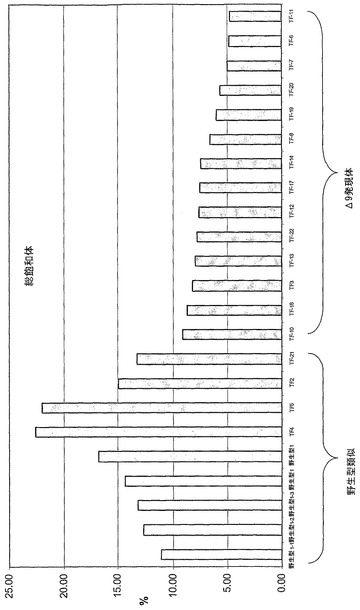
【図3】
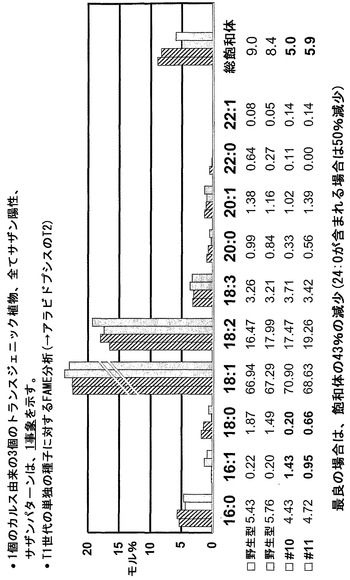
【図4A】

【図4B】

【図4C】
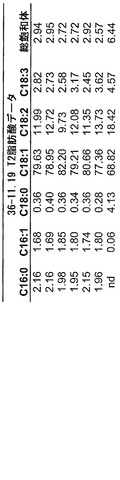
【図5A】

【図5B】
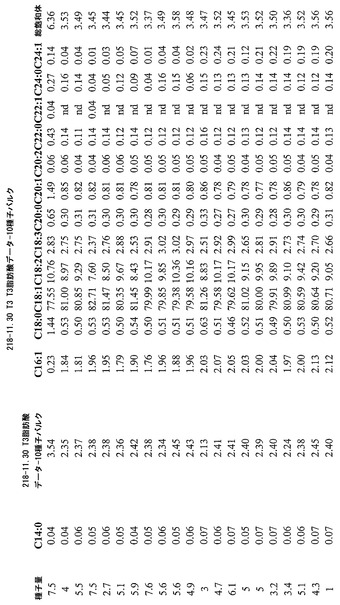
【図5C】
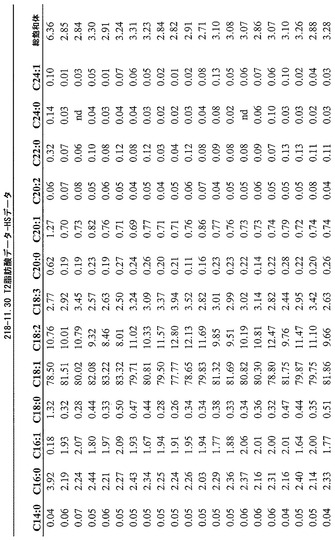
【図6A】
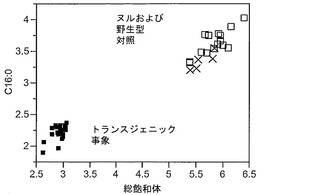
【図6B】
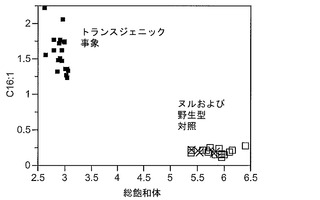
【図6C】
【図6D】
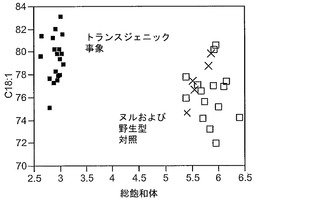
【図6E】
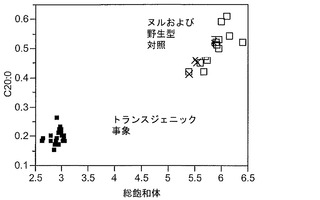
【図6F】
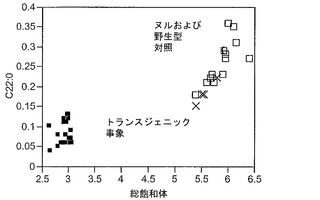
【図6G】
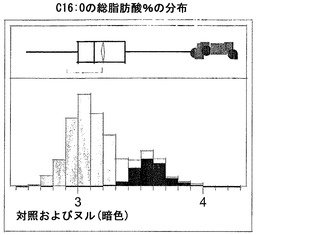
【図6H】
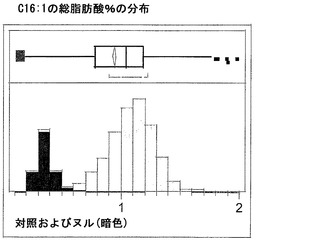
【図6I】
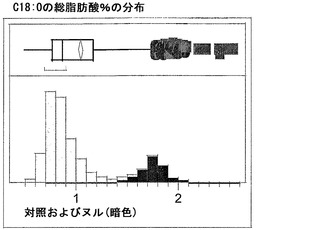
【図6J】
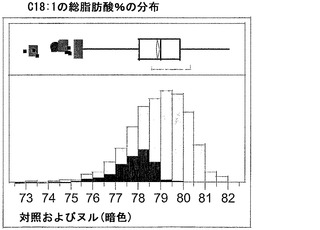
【図6K】
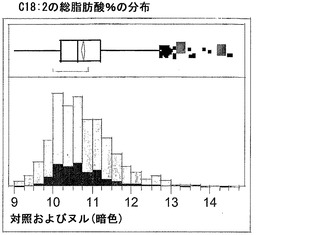
【図6L】
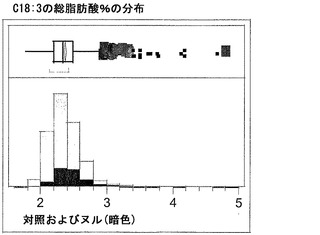
【図6M】
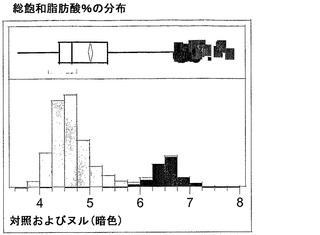
【図6N】
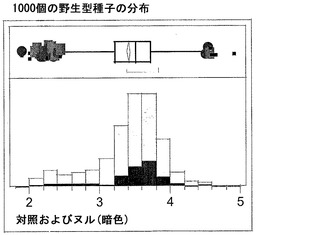
【図7】
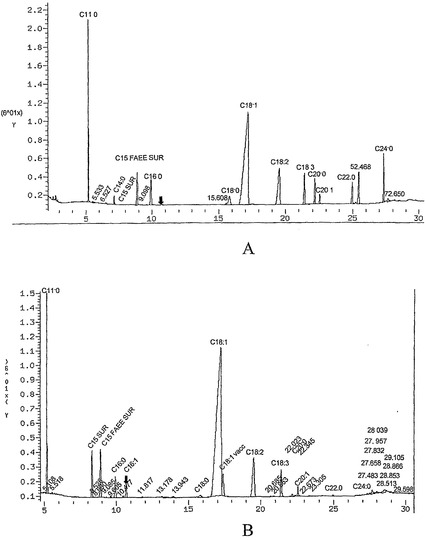
【図8】

【図9】
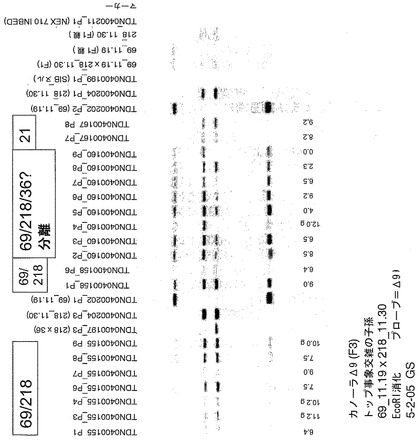
【図2】
【図3】
【図4A】
【図4B】
【図4C】
【図5A】
【図5B】
【図5C】
【図6A】
【図6B】
【図6C】
【図6D】
【図6E】
【図6F】
【図6G】
【図6H】
【図6I】
【図6J】
【図6K】
【図6L】
【図6M】
【図6N】
【図7】
【図8】
【図9】
【公表番号】特表2008−515433(P2008−515433A)
【公表日】平成20年5月15日(2008.5.15)
【国際特許分類】
【出願番号】特願2007−535825(P2007−535825)
【出願日】平成17年10月7日(2005.10.7)
【国際出願番号】PCT/US2005/036052
【国際公開番号】WO2006/042049
【国際公開日】平成18年4月20日(2006.4.20)
【出願人】(505412443)ダウ アグロサイエンス リミテッド ライアビリティー カンパニー (10)
【Fターム(参考)】
【公表日】平成20年5月15日(2008.5.15)
【国際特許分類】
【出願日】平成17年10月7日(2005.10.7)
【国際出願番号】PCT/US2005/036052
【国際公開番号】WO2006/042049
【国際公開日】平成18年4月20日(2006.4.20)
【出願人】(505412443)ダウ アグロサイエンス リミテッド ライアビリティー カンパニー (10)
【Fターム(参考)】
[ Back to top ]