
脂質の生産のための遺伝的に改変された生物
本発明は、アシル−CoA:ステロールアシルトランスフェラーゼ/ステロールO−アシルトランスフェラーゼ(EC2.3.1.26)および/またはジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)および/またはレシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)および/またはアシルCoA−ワックスアルコールアシルトランスフェラーゼ(EC2.3.1.75)の活性が、対応する野生型生物と比較して低減るまたは排除されている、単離された、遺伝的に改変された非哺乳類生物、このような生物の使用方法、このような生物を作出するためのシャトルビヒクルおよびこのような生物の生産方法を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ある特定の酵素活性が低減および/または増強された、脂質、特に中性脂質の工業的生合成に有用な遺伝的に改変された非哺乳類生物、特に微生物に関する。本発明はさらに、このような生物の使用、このような生物を作出するためのシャトルビヒクルおよびこのような生物の生産方法に関する。
【背景技術】
【0002】
発明の背景および技術の現状
脂質は、もっぱらまたは部分的に、脂肪酸もしくはポリケチドのようなチオエステルのカルボアニオンに基づく縮合によって、および/またはプレノールもしくはステロールのようなイソプレン単位のカルボカチオンに基づく縮合によって生じる疎水性または両親媒性の小分子である。この群には、経済上極めて重要ないくつかの物質が含まれる。トリアシルグリセロール脂質群には例えば油脂類またはワックス類が含まれ、これらは例えば食品の成分としてまたは調理用に、石鹸、スキン製品、香料ならびに他のパーソナルケアおよび化粧製品の製造のために、塗料および他の木材処理製品の製造のために、電気工業における生分解性絶縁体として、生分解性作動液の製造のため、潤滑剤として、またはさらには従来のディーゼルの代替に使用可能なバイオディーゼルの主成分としてなど、極めて多様な状況で用いられる。イソプレノイド脂質スクアレンは、例えば、ワクチンまたは他の薬剤、栄養、化粧品ならびにOTC(over−the−counter)薬のアジュバントとして用いられる。スクアレンはまた、テルペン合成の構成ブロックとしても使用することができる。さらに、スクアレンは、生分解性潤滑剤として工業的に使用することができる。他の経済上重要な脂質としては、サポニン、ステロイドホルモン、ビタミンおよび原薬のような化合物の生産のための中枢的出発材料として用いられる、エルゴステロール、チモステロール、エピステロール、7−デヒドロコレステロールまたはラノステロールのようなステロールがある。
【0003】
脂質、特に中性脂質は、通常、細胞内の、脂質粒子として知られる特定の細胞内オルガネラに貯蔵される。これらの粒子は、ほんの少量のタンパク質が埋め込まれた極めて疎水性の高い単層からなる単純な構造を特徴とする。脂質は、加水分解がそれらの成分のリターンを代謝経路および/または異化経路に向けるまで、脂質粒子に貯蔵される。この脂質貯蔵物形成のプロセスは自然界で広く用いられ、あらゆるタイプの真核細胞が、脂肪体、脂質滴、油体またはオレオソームとも呼ぶことができる細胞内脂質粒子を含む。酵母サッカロミセス・セレビシエ(Saccharomyces cerevisiae)脂質滴は、細胞の総脂質含量の70%まで蓄積する。
【0004】
記載されている脂質の大部分は植物、動物または微生物などの天然源に由来するので、生きている細胞において脂質量を増やす、すなわち、脂質粒子中により多くの脂質を蓄積させる試みがなされた。この目的で、生物の代謝経路を改変することが知られている。特許文献1には、酵母におけるエルゴステロール代謝の遺伝子の過剰発現が細胞のエルゴステロール量の増大をもたらすとの記載がある。特許文献2には、ラノステロール−C14−デメチラーゼおよびHMG−CoA−レダクターゼ活性の増強に基づくチモステロールの生産方法が開示されている。特許文献3には、Δ22−デサチュラーゼ活性が低減され、HMG−CoAレダクターゼ、ラノステロールC14−デメチラーゼ、スクアレンエポキシダーゼおよびスクアレンシンセターゼ活性が増強されたトランスジェニック生物が記載されている。これらの生物は、ステロール脂質エルゴスタ−5,7−ジエノールの生産に使用することができる。従って、脂質生成生物を生物学的に改変することによって、生産される脂質の収量を著しく増大させることができる。
【0005】
しかしながら、得られる脂質化合物はほとんどの場合、不純で、混ざり合い、特に、その脂質がさらなる化学合成または改変の出発材料として用いられる場合には、分離および/または精製する必要がある。さらに、いずれの精製手順も、特に工業的規模で行われる場合には、高くつき、労力がかかり、環境を汚染する傾向がある。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】欧州特許第EP0486290A号
【特許文献2】国際公開第03/064652A号パンフレット
【特許文献3】国際公開第2004/083407A号パンフレット
【発明の概要】
【発明が解決しようとする課題】
【0007】
よって、本発明の基礎にある技術課題は、労力がかかり、コストがかかる精製手順を避けるために、より純度の高い形態で生物による脂質生産を可能とする手段および方法を提供することである。
【0008】
発明の概要および好ましい実施形態
この技術課題を解決するために、本発明は、請求項1に記載の生物を教示する。好ましい実施形態は、請求項1の従属項で明示する。その中の、「遺伝的に改変された」とは、現状の遺伝子工学の方法によってゲノムが改変された生物だけでなく、所望の遺伝子改変に従う突然変異体群から選択された生物(従来の突然変異誘発)も含むものとする。
【0009】
本発明は、特定の(中性)脂質がより純度の高い形態で脂質粒子に蓄積されるように生物を遺伝的に操作する、または生物学的に改変することができるという発見に基づく。本発明の生物は、生物によってより高い純度で生産させようとする脂質に応じて、所望でないステリルアシルエステルおよび/またはトリアシルグリセロールおよび/またはワックスエステルの合成を排除または低減するように改変される。中性脂質のより純度の高い形態での蓄積の他、驚くことに、潜在的に混入するステリルアシルエステルおよび/またはトリアシルグリセロールおよび/またはワックスエステルの低減または排除も所望の脂質のレベルおよび収量に増大をもたらす。これは、特定の理論に縛られるものではないが、脂質粒子内の貯蔵空間の増大および/または所望の脂質の合成のための基質の利用能の上昇によるものである可能性がある。
【0010】
「中性脂質」とは、荷電基を欠き、従って、二重膜に実質的な量では組み込まれ得ない脂質を意味する。この用語には、トリアシルグリセロール(TAG)、ステリルアシルエステル(SAE)およびワックスエステル(WE)が含まれる。スクアレンもこの種の「中性脂質」に属す。通常、脂質粒子の内容物は、組換え生物での所望の特定中性脂質の効率的生産に支障を来す種々の中性脂質の混合物である。この問題は、本発明により、生物の脂質体内で本来生成する中性脂質の少なくとも1つの種類をそこではもはや合成されないように生産者生物を改変することによって克服される。結果として、脂質粒子内に含まれる残りの中性脂質の組成はより純粋となり、従って、市販の商業目的での生産により魅力あるものとなる。
【0011】
一般に、より高い純度で合成させようとする中性脂質に応じて、または生合成において望ましくない混入物として除去しようとする中性脂質に応じて、請求項1に記載のいずれか1つの活性または種々の活性(2つ、3つまたは4つ)の組合せを用いることができる。よって、本発明の種々の実施形態が存在するが、以下、いくつかの特定の実施形態だけを詳細に記載する。
【0012】
よって、第一の実施形態では、本発明の生物は、ステリルアシルエステル(SAE)をもはや合成しないように改変される。ステリルアシルエステルは、ステロールと長鎖脂肪酸とのエステルである。一般に、SAEの合成は、細胞内でアシル−CoA:ステロールアシルトランスフェラーゼ/ステロールO−アシルトランスフェラーゼ(EC2.3.1.26)を含む酵素機構によって遂行される。触媒される反応は、ステロールと長鎖脂肪酸とのエステル化である。よって、好ましい実施形態では、本発明の生物は、アシル−CoA:ステロールアシルトランスフェラーゼ/ステロールO−アシルトランスフェラーゼ(EC2.3.1.26)の活性が、対応する非改変生物、好ましくは対応する野生型生物と比較して低減されるように改変される。
【0013】
酵母、特にS.セレビシエ(S. cerevisiae)では、ステリルアシルエステルを生じるステロールエステル化は、ステリルアシルトランスフェラーゼの2つのイソ酵素、すなわち、異なるステロール中間体に対して異なる特異的親和性を有するAre1pおよびAre2pによって遂行される。Are1pは、特に、ラノステロール、チモステロール、エルゴスタ−5,7−ジエノールなどのエルゴステロール前駆体と、7−デヒドロコレステロールなどの酵母にとって外来の物質とのエステル化をもたらす。Are2pは、酵母におけるエルゴステロール生合成経路の最終産物であるエルゴステロールのエステル化を優先的にもたらす。生物が酵母生物体、特にサッカロミセス(Saccharomyces)属のもの、好ましくはS.セレビシエである場合、Are1pまたはAre2pまたはその両者Are1pとAre2pの活性を、対応する非改変酵母生物、好ましくは対応する野生型生物と比較して低減または排除することができる。両酵素活性が排除される場合、細胞内でステロールのエステル化は起こらない。一方の活性のみを排除し、他方を維持することにより、所望により、ある特定のステロールをもたらす酵素活性がその生物において相応に維持または適合されていれば、細胞内のある特定のタイプのステリルアシルエステルを得ることが可能である。
【0014】
生物がステリルアシルエステルをもはや合成しないことを特徴とする本発明の第一の態様に関して、本発明の生物は、原則として、存在し得る任意の生物、植物生物、真菌生物または細菌生物であってよい。しかしながら、好ましくは、該生物は、その脂質粒子内にワックスエステルを本来貯蔵しないが、トリアシルグリセロールおよびステリルアシルエステルのような他の脂質だけを貯蔵する生物に由来する。よって、ステリルアシルエステルの合成のための生合成経路を排除することにより、このような生物はその脂質粒子内に本質的にトリアシルグリセロールのみを蓄積し、それにより、このような生物において、脂質粒子からかなり純粋な形態でトリアシルグリセロールを生産することが可能となる。それらの脂質粒子内にワックスエステルを本来蓄積しない生物の例としては、特に、大部分の動物、真菌細胞、および藻類とプランクトン細胞を除く大部分の植物細胞である。本発明の第一の態様に関する特に好ましい実施形態では、生物は真菌生物、好ましくは、ヤルロウィア(Yarrowia)、ロドトルラ(Rhodotorula)、リポミセス(Lipomyces)、カンジダ(Candida)、ロドスポリジウム(Rhodosporidium)、モルチエレラ(Mortierella)、ムコール(Mucor)、サッカロミセス(Saccharomyces)、ピキア(Pichia)、クルイベロミセス(Kluyveromyces)、アスペルギルス(Aspergillus)、ペニシリウム(Penicillium)およびディクチオステリウム(Dictyostelium)からなる群から選択される属の真菌生物である。ヤルロウィア・リポリチカ(Yarrowia lipolytica)、ロドトルラ・グルチニス(Rhodotorula glutinis)、リポミセス・スタルケイイ(Lipomyces starkeyi)、カンジダ・クルバタ(Candida curvata)、ロドスポリジウム・トルロイデス(Rhodosporidium tortuloides)、モルチエレラ・イサベリナ(Mortierella isabellina)、ムコール・ジャバニカス(Mucor javonicus)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、ピキア・パストリス(Pichia pastoris)、クルイベロミセス種(Kluyveromyces spec)、アスペルギルス種(Aspergillus spec)、ペニシリウム種(Penicillium spec)またはディクチオステリウム種(Dictyostelium spec)の真菌生物が特に好ましい。本発明の第一の態様の特に好ましい実施形態では、生物はS.セレビシエである。
【0015】
第二の実施形態では、本発明の生物は、ステリルアシルエステル(SAE)およびワックスエステル(WE)をもはや合成しないように改変される。
【0016】
一般に、WEの合成は、細胞内でアシルCoA−ワックスアルコールアシルトランスフェラーゼ(EC2.3.1.75)を含む酵素機構によって遂行される。よって、本発明の第二の態様の好ましい実施形態では、本発明の生物は、アシル−CoA:ステロールアシルトランスフェラーゼ/ステロールO−アシルトランスフェラーゼ(EC2.3.1.26)およびアシルCoA−ワックスアルコールアシルトランスフェラーゼ(EC2.3.1.75)の活性が、対応する非改変生物、好ましくは対応する野生型生物と比較し低減されるように改変される。
【0017】
本発明の第二の態様に関する本発明の生物は、ステリルアシルエステルおよびワックスエステルの合成が低減されている、好ましくは完全に排除されているので、脂質粒子内に主としてトリアシルグリセロールを蓄積する。従って、このような生物は、特に工業規模で、トリアシルグリセロールを純粋な形態で生産するのに好適である。
【0018】
本発明の第二の態様の生物は、好ましくは、ワックスエステルを本来生産し、それらを細胞の脂質粒子に蓄積する生物に由来する。ワックスエステルを生産し、貯蔵する生物は、例えば、表面の脱水を最小限にするために組織の疎水性被膜を設けるために用いられる植物および昆虫、藻類、プランクトン生物および細菌である。本発明の第二の態様の好ましい実施形態では、生物は細菌生物、好ましくは、マイコバクテリア(Mycobacterium)、ストレプトミセス(Streptomyces)、ロドコッカス(Rhodococcus)、ノカルジア(Nocardia)、バチルス(Bacillus)、コリネバクテリウム(Corynebacterium)、エシェリキア(Escherichia)または乳酸桿菌(Lactobacillus)属のものである。最も好ましくは、生物は大腸菌(Escherichia coli)である。
【0019】
さらに、このような本発明の生物では、その生物で本来生産されるトリアシルグリセロールを、またはその生物で本来生産される量で生産することが可能であるだけでなく、このような生物を該生物で本来生産される1以上の特定のトリアシルグリセロールの量を増大させるように遺伝的に改変すること、またはこのような生物をこのような生物によって本来生産されない1以上のトリアシルグリセロールの合成をもたらすように遺伝的に改変することも可能である。
【0020】
特に好ましい一実施形態では、本発明の生物によって蓄積されるトリアシルグリセロールは、グリセロールと少なくとも1つの不飽和脂肪酸とのエステルであり、より好ましくは、該不飽和脂肪酸はω−3脂肪酸である。原則として、該ω−3脂肪酸は存在し得る任意のω−3脂肪酸であり得るが、好ましくは、α−リノレン酸(ALA)、ステアリドン酸、エイコサテトラエン酸、エイコサペンタエン酸(EPA)、ドコサペンタエン酸(DPA)またはドコサヘキサエン酸(DHA)である。
【0021】
別の好ましい実施形態では、トリアシルグリセロール中の不飽和脂肪酸は、ω−6脂肪酸である。原則として、該ω−6脂肪酸は存在し得る任意のω−6脂肪酸であり得るが、好ましくは、リノール酸、γ−リノレン酸、エイコサジエン酸、ジホモ−γ−リノレン酸、アラキドン酸、ドコサジエン酸、アドレン酸、ドコサペンタエン酸またはカレンド酸である。
【0022】
別の好ましい実施形態では、トリアシルグリセロール中の不飽和脂肪酸は、ω−9脂肪酸である。原則として、ω−9脂肪酸は存在し得る任意のω−9脂肪酸であり得るが、好ましくは、オレイン酸、エイコセン酸、ミード酸、エルカ酸またはネルボン酸である。
【0023】
第一および第二の実施形態は、以下に記載されるようにさらに改変することができる。トリアシルグリセロールは一般に、ジアシルグリセロールと長鎖脂肪酸またはリン脂質のエステル化によって形成される。長鎖脂肪酸とのエステル化は、ジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)によって触媒される。リン脂質とのエステル化は、レシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)によって触媒される。特に好ましい実施形態では、本発明の第一または第二の態様の生物は、ジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)の活性および/またはレシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)の活性が、対応する非改変生物、好ましくは対応する野生型生物と比較して増強されていることを特徴とする。上述の2つの酵素の一方または両方の活性の増強は、生物の細胞内でのTAG合成の増大をもたらす。
【0024】
この他、生物がTGAに組み込まれるある特定の脂肪酸を生産可能となるように、またはそれらがこのような生物ですでに本来生産されている場合にはこのような脂肪酸の生産を増大させてその生物で生産されるTGAにおけるそれらの含量を増大させるように生物を遺伝的に改変することも可能である。例えば、生物で生産されるTGAにおいてエイコサペンタエン酸(EPA)、ドコサペンタエン酸(DPA)および/またはドコサヘキサン酸(DHA)などのω−3脂肪酸の量を増大させることが意図される場合には、Δ12脂肪酸デサチュラーゼ(EC1.14.19)、Δ15−デサチュラーゼ(EC1.14.19)、Δ6脂肪酸デサチュラーゼ(EC1.14.19.3)、脂肪エロンガーゼ、Δ5−デサチュラーゼ/Δ5−脂肪酸デサチュラーゼ(EC1.14.19)およびΔ4脂肪酸デサチュラーゼ(EC1.14.19)の酵素活性を増強することが有利であろう。よって、好ましい実施形態では、生物(organis)は、ジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)および/またはレシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)の活性と、Δ12脂肪酸デサチュラーゼ(EC1.14.19)、Δ15−デサチュラーゼ(EC1.14.19)、Δ6脂肪酸デサチュラーゼ(EC1.14.19.3)、脂肪酸エロンガーゼ、Δ5−デサチュラーゼ/Δ5−脂肪酸デサチュラーゼ(EC1.14.19)およびΔ4脂肪酸デサチュラーゼ(EC1.14.19)からなる群から選択される少なくとも1つの活性が、対応する非改変生物、好ましくは対応する野生型生物と比較して増強されていることを特徴とする。例えば、植物およびいくつかの真菌はω−3およびω−6脂肪酸を生産することができるが、例えばサッカロミセス・セレビシエは、ω−9脂肪酸のみ合成可能である。よって、本発明の生物がサッカロミセス・セレビシエである場合、ω−3脂肪酸および/またはω−6脂肪酸の生産に必要な酵素活性を細胞に導入することが望ましいと思われる。対応する遺伝子を酵母細胞に導入するためのアプローチは文献(例えば、国際公開第2004/101575号)に対応する遺伝子とともにすでに記載されている。特定の脂肪酸の、例えば酵母細胞での異種生合成は、例えば、Beaudoin et al. (Proc. Natl. Acad. Sci. USA 97(2000), 6421−6426)およびVeen et al. (Appl. Microbiol. Biotechnol. 63(2004), 635−646)に記載されている。
【0025】
本発明はまた、本発明の第一または第二の態様の生物を培養することを含む、トリアシルグリセロールの生産方法に関する。上記のように、これらの生物は、それらの脂質粒子内に本質的に純粋な形態で多量のトリアシルグリセロールを蓄積することができる。これらのTAGは、当技術分野で周知の方法に従って生物から単離することができる。例えば、細胞から脂質粒子を単離する方法およびこれらからその中に含まれている中性脂質を単離する方法は当技術分野で公知である。このような方法の例は、添付の実施例においてサッカロミセス・セレビシエ生物体に関して記載されている。
【0026】
本発明はまた、グリセロールと少なくとも1つの不飽和脂肪酸とのエステルであるトリアシルグリセロールの生産方法に関し、その方法は、
(i)阻害剤により、アシル−CoA:ステロールアシル−トランスフェラーゼ/ステロールO−アシルトランスフェラーゼ(EC2.3.1.26)および/またはアシルCoA−ワックスアルコールアシルトランスフェラーゼ(EC2.3.1.75)の活性を低減または排除すること;および
(ii)生物においてジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)および/またはレシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)の活性を、対応する非改変生物、好ましくは対応する野生型生物と比較して増強することを含む。この生物を培養し、TAGを単離することができる。
【0027】
生産されるトリアシルグリセロールに含まれる不飽和脂肪酸に関しては、好ましい実施形態では、ポリ不飽和脂肪酸の性質および増強される酵素活性の性質に関してすでに上記したものと同じことが当てはまる。特に好ましい実施形態では、ポリ不飽和脂肪酸はω−3脂肪酸であり、増強される酵素活性は、本発明の上記でω−3脂肪酸に関して詳しく記載されているものの少なくとも1つである。
【0028】
第三の実施形態では、本発明の生物は、トリアシルグリセロール(TAG)およびステリルアシルエステル(SAE)をもはや合成しないように改変される。上記のように、一般に、TAGの合成はジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)およびレシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)の酵素活性によって遂行される。よって、好ましい実施形態では、本発明の生物は、アシル−CoA:ステロールアシルトランスフェラーゼ/ステロールO−アシルトランスフェラーゼ(EC2.3.1.26)、ジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)およびレシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)の活性が、対応する非改変生物、好ましくは対応する野生型生物と比較して低減されるように改変される。
【0029】
好ましい実施形態では、本発明の第三の態様の生物は、ワックスエステルを本来生産および貯蔵する生物に由来する生物である。このような生物はすでに上記されている。好ましくは、生物はすでに上記されている細菌生物であり、最も好ましくは大腸菌である。TAGおよびSAEの合成が排除されているために、このような生物は主としてその脂質粒子内にワックスエステルを蓄積するのに好適である。特に好ましい実施形態では、このような生物はさらに、対応する非改変株、好ましくは対応する野生型株と比較してアシル−CoAワックスアルコールアシルトランスフェラーゼ(EC2.3.1.75)の活性に増強を示すことを特徴とする。これにより、生物において合成されるワックスエステルの量を増大されることが可能となる。生物の細胞内で所望の酵素の活性を増強するための手段および方法は当業者に公知であり、本明細書の以下に詳しく記載される。
【0030】
別の好ましい実施形態では、本発明の第三の態様の生物は、ワックスエステルを本来生産および貯蔵しない生物に由来する。このような生物は本発明の第一の態様に関して上記されている。好ましくは、このような生物は上記のような真菌生物、より好ましくはサッカロミセス属、最も好ましくはサッカロミセス・セレビシエ種である。TAGおよびSAEの合成が排除されているために、このような生物の脂質粒子は、以下に詳しく記載されるように、目的の脂質、例えば、スクアレンを蓄積するために使用することができる。
【0031】
第四の実施形態では、本発明の生物は、ワックスエステル(WE)、トリアシルグリセロール(TAG)およびステリルアシルエステル(SAE)をもはや合成しないように改変される。この態様では、生物は本質的にその脂質粒子内にいずれのWE、TAGおよびSEも貯蔵しない。このような生物は、その脂質粒子に、特に望まれる脂質を向けることができる生物を設計するのに特に有用である。例えば、このような生物に、後に脂質粒子に蓄積される別のタイプの脂質の合成を可能とする核酸分子を導入することができる。これにより、脂質粒子における本質的に純粋な目的脂質の合成を可能とする生物を提供することができる。
【0032】
好ましい実施形態では、本発明の第四の態様の生物は、アシル−CoA:ステロールアシルトランスフェラーゼ/ステロールO−アシルトランスフェラーゼ(EC2.3.1.26)、ジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)、レシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)およびアシルCoA−ワックスアルコールアシルトランスフェラーゼ(EC2.3.1.75)の活性が、対応する非改変生物、好ましくは対応する野生型生物と比較して低減されるように改変される。
【0033】
第五の実施形態では、本発明の生物は、細胞において脂質粒子内に蓄積される中性脂質を生産し、かつ、トリアシルグリセロール(TAG)をもはや合成しないように遺伝的に改変された、従って、該粒子において、その脂質粒子内に本来存在する該トリアシルグリセロールをもはや含まない生物である。上記のように一般に、TAGの合成は、例えば酵母においては、ジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)およびレシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)の酵素活性によって遂行される。よって、好ましい実施形態では、本発明の第五の態様の生物は、ジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)およびレシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)の活性が、対応する非改変生物、好ましくは対応する野生型生物と比較して低減されるように遺伝的に改変される。
【0034】
本発明の第五の態様の好ましい実施形態では、生物は、ワックスエステルを本来生産せず、その脂質粒子内に貯蔵しない生物に由来する。このような生物は上記されている。好ましくは、生物は上記のような真菌生物、より好ましくはサッカロミセス属、最も好ましくはサッカロミセス・セレビシエ種である。
【0035】
第六の実施形態では、本発明の生物は、細胞において脂質粒子内に蓄積される中性脂質を生産し、かつ、ワックスエステル(WE)およびトリアシルグリセロール(TAG)をもはや合成しないように遺伝的に改変された、従って、該粒子において、その脂質粒子内に本来存在するトリアシルグリセロールおよびワックスエステルをもはや含まない生物である。好ましい実施形態では、本発明の第六の態様の生物は、ジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)、レシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)およびアシルCoA−ワックスアルコールアシルトランスフェラーゼ(EC2.3.1.75)の活性が、対応する非改変生物、好ましくは対応する野生型生物と比較して低減されるように遺伝的に改変される。
【0036】
好ましい実施形態では、本発明の第五または第六の態様の生物、すなわち、トリアシルグリセロールをもはや合成しない、またはトリアシルグリセロールおよびワックスエステルをもはや合成しない生物は、その脂質粒子内にステロール脂質のアシルエステルを蓄積することができる。脂質粒子におけるステロール脂質のアシルエステルの蓄積は、その生物がこのようなエステルを本来合成するという事実によるものであり得、またはそれはステロール脂質のアシルエステルの生合成をもたらす対応する酵素をコードする遺伝子がそれらの細胞に導入されているという事実によるものであり得る。
【0037】
コレステロールおよびその誘導体などのステロール脂質は、グリセロリン脂質およびスフィンゴミエリンとともに膜脂質の重要な成分である。これらのステロイドは、同じ縮合4環コア構造も含むが、ホルモンおよびシグナル伝達分子として異なる生物学的役割を有する。C18ステロイドはエストロゲンファミリーを含み、C19ステロイドはテストステロンおよびアンドロステロンなどのアンドロゲンを含む。C21サブクラスは、プロゲストゲンならびに糖質コルチコイドおよび鉱質コルチコイドを含む。種々の形態のビタミンDを含むセコステロイドは、コア構造のB環の開裂を特徴とする。ステロールの他の例は胆汁酸およびそれらの結合体であり、これらは哺乳類ではコレステロールの酸化誘導体であり、肝臓で合成される。
【0038】
好ましい実施形態では、ステロール脂質はステロールまたはステロイドである。ステロイドは、一般に6−6−6−5様式で配置された4つの縮合環を有する炭素主鎖を特徴とするテルペノイド脂質である。ステロイドはこれらの環と結合している官能基およびこれらの環の酸化状態によって異なる。植物、動物および真菌には、何百もの異なるステロイドが見られる。全てのステロイドが細胞においてステロールラノステロール(動物および真菌)またはステロールシクロアルテノール(植物)のいずれかから製造される。両ステロールとも、トリテルペンスクアレンの環化に由来する。ステロイドには、エストロゲン、プロゲステロンおよびテストステロンが含まれる。
【0039】
ステロール(またはステロイドアルコール)は、A環の3位にヒドロキシル基を有するステロイドのサブグループである。それらはアセチル補酵素A(acetyl−conenzyme A)から合成される両親媒性脂質である。この分子全体は極めて平坦である。A環上のヒドロキシル基には極性がある。この脂肪鎖の残りの部分は非極性である。植物のステロールは植物ステロール(phytosterols)と呼ばれ、動物のステロールは動物ステロール(zoosterols)と呼ばれる。最も重要な動物ステロールはコレステロールおよびいくつかのステロイドホルモンであり、最も重要な植物ステロールはカンペステロール、シトステロールおよびスチグマステロールである。ステロールは、真核生物の生理に不可欠な役割を果たす。例えば、コレステロールは細胞膜の一部を形成し、その存在は細胞膜の流動性に影響を及ぼし、発生シグナル伝達における第二のメッセンジャーとして働く。植物ステロールはまた、ヒトの腸においてコレステロール吸収部位を遮断することも知られており、従って、ヒトにおいてコレステロールの低減を助ける。ヒトでは、ステロールは、例えば、日内周期、血液凝固などの重要なシグナルおよび代謝コミュニケーションを提供する働きをする。
【0040】
好ましい実施形態では、ステロールはコレステロール、コレステロール誘導体、7−デヒドロ−コレステロール、ラノステロール、ラノステロール誘導体、チモステロール、チモステロール誘導体、ラトステロール、ラトステロール誘導体、ククルビタシン、ククルバチン(cucurbatin)誘導体、エピステロール、エピステロール誘導体、テアステロン、テアステロン誘導体、カスタステロン、カスタステロン誘導体、チファステロール、チファステロール誘導体、カタステロン、カタステロン誘導体、シクロオイカレノール、シクロオイカレノール誘導体、シトステロール、シトステロール誘導体、イソフコステロール、イソフコステロール誘導体、フコステロール、フコステロール誘導体、ゴルゴステロール、ゴルゴステロール誘導体、エルゴステロール、エルゴステロール誘導体、スチグマステロールまたはスチグマステロール誘導体である。
【0041】
特に、7−デヒドロ−コレステロールのアシルエステルが蓄積される生物では、HMG−CoA−レダクターゼ(EC1.1.1.34)および/またはΔ24−レダクターゼおよび/またはラノステロールC14−デメチラーゼ/シトクロムP450 51(EC1.14.13.70)および/またはスクアレン−エポキシダーゼ/スクアレン−モノオキシゲナーゼ(EC1.14.99.7)の活性を、対応する野生型生物と比較して増強することができる。さらに、SAM:C−24ステロールメチルトランスフェラーゼおよび/またはC−22ステロールデサチュラーゼおよび/またはC−5ステロールデサチュラーゼの活性を、対応する野生型生物と比較して低減または排除することができる。エルゴステロールのアシルエステルが蓄積される生物では、HMG−CoA−レダクターゼ(EC1.1.1.34)および/またはラノステロールC14−デメチラーゼ/シトクロムP450 51(EC1.14.13.70)および/またはスクアレン−エポキシダーゼ/スクアレン−モノオキシゲナーゼ(EC1.14.99.7)の活性を、対応する野生型生物と比較して増強することができる。エピステロールのアシルエステルが蓄積される生物では、HMG−CoA−レダクターゼ(EC1.1.1.34)および/またはラノステロールC14−デメチラーゼ/シトクロムP450 51(EC1.14.13.70)および/またはスクアレン−エポキシダーゼ/スクアレン−モノオキシゲナーゼ(EC1.14.99.7)の活性を、対応する野生型生物と比較して増強することができ、かつ/またはΔ22−デサチュラーゼ/シトクロムP450 61(EC1.14.14.1)活性を、対応する野生型生物と比較して低減することができる。
【0042】
別の好ましい実施形態では、ステロイドはアンドロステロン、アンドロステロン誘導体、テストステロン、テストステロン誘導体、アンドロステンジオール、アンドロステンジオール誘導体、アンドロステンジオン、アンドロステンジオン誘導体、カルステロン、カルステロン誘導体、メタンドリオール、メタンドリオール誘導体、ボラステロン、ボラステロン誘導体、エピアンドロステロン、エピアンドロステロン誘導体、メスタノロン、メスタノロン誘導体、スタノロン、スタノロン誘導体、ステンボロン、ステンボロン誘導体、エピテストステロン、エピテストステロン誘導体、コルチゾール、コルチゾール誘導体、アルドステロン、アルドステロン誘導体、プレグネノロン、プレグネノロン誘導体、コルチゾン、コルチゾン誘導体、コルチコステロン、コルチコステロン誘導体、ノレシンドロン、ノレシンドロン誘導体、ウロコルチゾールまたはウロコルチゾール誘導体である。
【0043】
本発明の第五および第六の態様の好ましい実施形態では、ステロール脂質のアシルエステルを蓄積することができる生物は、個々のステロール脂質の合成に至る生合成経路を本来保有し、より好ましくは、個々のステロール脂質のより高い蓄積を得るために、対応する経路の酵素活性が対応する非改変生物と比較して増強されている生物である。
【0044】
好ましい実施形態では、本発明の態様1〜6の生物は、その脂質粒子内にプレノール脂質を蓄積することができる。プレノール脂質は、主としてメバロン酸(MVA)経路を介して生産される5炭素前駆体であるイソペンテニル二リン酸およびジメチルアリル二リン酸から合成される。プレノール脂質はイソプレノイド、キノンおよびヒドロキノン、ポリプレノール、ホパノイドおよび他のいくつかの希少なタイプを包含する。単純イソプレノイド(直鎖アルコール、二リン酸塩など)はC5単位の連続的付加によって形成され、これらのテルペン単位の数によって分類される。40を超える炭素を含む構造はポリテルペンとして知られる。カロテノイドは、抗酸化剤としておよびビタミンA前駆体として働く重要な単純イソプレノイドである。別の生物学的に重要なクラスの分子としては、非イソプレノイド起源のキノノイドコアと結合したイソプレノイドテールを含むキノンおよびヒドロキノンが挙げられる。ビタミンEおよびビタミンKならびにユビキノンがこのクラスの例である。細菌は、酸素と結合している末端のイソプレノイド単位が不飽和のままであるポリプレノール(バクトプレノールと呼ばれる)を合成するが、動物ポリプレノール(ドリコール)では、末端イソプレノイドは還元されている。
【0045】
好ましい実施形態では、脂質粒子内に蓄積されるプレノール脂質はイソプレノイドである。イソプレノイドとしては、例えば、C5イソプレノイド、C10イソプレノイド(モノテルペン)、C15イソプレノイド(セキテルペン)、C20イソプレノイド(ジテルペン)、C25イソプレノイド(セステルテルペン)、C30イソプレノイド(トリテルペン)、C40イソプレノイド(テトラテルペン)、ポリテルペンおよびレチノイドが挙げられる。
【0046】
特に好ましい実施形態では、イソプレノイドはトリテルペンである。トリテルペンは、6つのイソプレン単位からなり、基本分子式C30H48ー50を有する。この群は、例えば3S−スクアレン−2,3−エポキシド、スクアレン、プレスクアレン二リン酸、テトラヒマノール、α−アミリン、β−アミリン、ルペオール、酢酸ルペオール、タラキサステロール、アザジラクチンA、ネオクアッシン、クアッシンおよび3−アセチル−1−チグロイルアザジラクチニンを包含する。サメ肝油の成分である直鎖トリテルペンスクアレンは、2分子のピロリン酸ファルネシルの還元的結合から誘導される。その後、スクアレンは生合成的にプロセシングされ、全てのステロイドの構造前駆体であるラノステロールまたはシクロアルテノールのいずれかを生成する。
【0047】
特に好ましい実施形態では、脂質粒子内に蓄積されるプレノール脂質はスクアレンまたはスクアレン誘導体である。スクアレン誘導体は、スクアレンの場合には主鎖炭素原子に結合している水素原子の代わりに、主鎖炭素原子に結合している1以上の、特に、1〜10個または1〜4個の、付加的メチルまたはエチル基を含む。
【0048】
特に好ましい実施形態では、本発明の生物は好ましくは、本発明の第一、第三および第五の態様の態様に関して、その脂質粒子においてワックスエステルを生産および貯蔵しない生物、例えば、上記のような真菌生物、より好ましくはサッカロミセス属の真菌生物、最も好ましくは、サッカロミセス・セレビシエである。本発明の第一、第三および第五の態様に従い、トリアシルグリセロールおよび/またはステリルアシルエステルを合成しないように改変されたこのような生物は、その脂質粒子においてプレノール脂質、特に、スクアレンを合成および貯蔵することができ、それにより、脂質粒子内でこのようなプレノール脂質を本質的に純粋またはより高い純度の形態で生産することができる。よって、酵母生物においてTAGおよび/またはSAEの生合成のための合成経路を排除することにより、このような生物によって生産されるスクアレンの量および純度に関して生産を劇的に改善することがすでに可能である。
【0049】
このような生物におけるスクアレン生産のさらなる改善は、スクアレンの合成をもたらす酵素活性を増強し、かつ、スクアレンを他の化合物に変換する経路の酵素活性を低減するように生物の代謝を改変することによって達成することができる。
【0050】
スクアレン生合成経路の遺伝子は既知であり、例えば酵母S.セレビシエにおいてクローニングされている。主要なボトルネック酵素はHMG−CoA−レダクターゼ(HMG1)(Basson et al. (Mol. Cell. Biol. 8 (1988), 3793−3808)。よって、好ましい実施形態では、本発明の第一、第三または第五の態様の生物は脂質粒子内にスクアレンを蓄積することができ、かつ、HMG−CoA−レダクターゼ(EC1.1.1.34)の活性が対応する非改変生物、好ましくは対応する野生型生物と比較して増強されていることを特徴とする。HMG−CoA−レダクターゼ活性の増強は、当業者に周知の手段および方法によって達成することがき、それらは以下にもさらに詳細に記載される。好ましい実施形態では、HMG−CoA−レダクターゼの活性の増強は、その生物において、その酵素の触媒領域のみをコードし、膜結合ドメインはコードしないHMG−CoA−レダクターゼ遺伝子を発現させることによって達成される。このような変更は欧州特許第EP−A486290にすでに記載されている。この改変により、エルゴステロール生合成経路の中間体によるHMG−CoA−レダクターゼのフィードバック調節が回避される。別の好ましい実施形態では、本発明の生物においてHMG−CoA−レダクターゼをコードする遺伝子を、異種プロモーター、すなわち、HMG−CoA−レダクターゼ遺伝子にとって外来のプロモーターの制御下に置くが、特に、このプロモーターは、エルゴステロール生合成経路の中間体によってはその活性が調節されないプロモーターである。好適なプロモーターの例はADH1プロモーター、特に、おおよそ構成的発現を示すADH1「アベレージ」プロモーターである(Ruohonen et al., Journal of Biotechnology 39 (1995), 193−203)。
【0051】
酵母などのいくつかの生物では、スクアレンは他の化合物、特に、エルゴステロールに、また、エルゴステロール生合成経の中間体に変換され得る。ステロールエルゴステロール((22E)−エルゴスタ−5,7,22−トリエン−3−β−オール)の形成をもたらす化学反応および経路が麦角、酵母および糸状菌に見られる。これらの生物では、スクアレンはスクアレンモノオキシゲナーゼ/スクアレンエポキシダーゼ(EC1.14.99.7;スクアレンエポキシダーゼまたはERG1とも呼ばれる)によって(S)−2,3−エポキシスクアレンに変換され、これは次に2,3−オキシドスクアレン−ラノステロールシクライズ(cyclise)(EC5.4.99.7;ERG7)によってラノステロールに変換される。ラノステロールは次にシトクロムP450ラノステロール14a−デメチラーゼ(EC1.14.13.70;ERG11)によって、NADPHおよびO2を用いて、4,4−ジメチル−コレスタ−8,14,24−トリエノールに変換され、その後、C14ステロールレダクターゼ(EC1.3.1.70;ERG24)によって、NADPHを用いて、4,4−ジメチル−8,24−コレスタジエノールに変換される。この化合物は次にC−4ステロールメチルオキシダーゼ(EC1.14.13.72;ERG25)の作用によってさらに4−メチル−8,24−コレスタジエノールに変換される。この物質はC−3ステロールデヒドロゲナーゼ(EC1.1.1.170;ERG26)によって3−ケト−4−メチルチモステロールに変換される。次に、3−ケト−4−メチルチモステロールは、3−ケトステロールレダクターゼ(EC1.1.1.270;ERG27)によってチモステロールに変換される。チモステロールはそれ自体、次に、S−アデノシル−L−メチオニンとともに、SAM:C−24ステロールメチルトランスフェラーゼ(EC2.1.1.41;ERG6)の作用によってフェコステロールおよびS−アデノシル−ホモシステインに変換される。フェコステロールは次にC−8ステロールイソメラーゼ(ERG2)の作用によってエピステロールに変換され、その後、C−5ステロールデサチュラーゼ(EC1.14.21.6;ERG3)の作用によって、NADPHおよびO2を用いて、5,7,24(28)−エルゴスタトリエノールに変換される。この化合物はさらにC−22ステロールデサチュラーゼ(EC1.14.14;ERG5)の作用によって5,7,22,24(28)−エルゴスタテトラエノールに変換される。5,7,22,24(28)−エルゴスタテトラエノールは次にC−24ステロールレダクターゼ(EC1.3.1.71;ERG4)の作用によってエルゴステロールに変換される。本発明の生物が脂質粒子内にスクアレンを蓄積することが望まれる場合、また、この生物がエルゴステロール、またはスクアレンから始まるエルゴステロール経路の中間体を本来合成することができる生物に由来する場合には、その生物におけるスクアレンからエルゴステロールに至る経路の上述の酵素活性の1以上の活性を低減することが望ましいと思われる。原則として、上記エルゴステロール経路の上述の酵素のいずれか1つ、2以上または全てを低減することができる。好ましい実施形態では、スクアレンモノオキシゲナーゼ(EC1.14.99.7;スクアレンエポキシダーゼまたはERG1とも呼ばれる)の活性が低減されるが、一部のステロール合成は酵母細胞の生存に必要であるので完全には排除されない。また、エルゴステロールの生合成に関与する以下の酵素:SAM:C−24ステロールメチルトランスフェラーゼ(EC2.1.1.41)、C−22ステロールデサチュラーゼ(EC1.14.14)およびC−5ステロールデサチュラーゼ(EC1.14.21.6)の1以上の活性を低減することも可能である。このことは全てにおいて本発明の上述のいずれの実施形態にも当てはまる。
【0052】
本発明の一部として、しかしながら独立に、遺伝的に改変された生物に関して以下の特徴を適用する。
【0053】
本発明の範囲内で、酵母サッカロミセス・セレビシエにおける遺伝子SAK1および/またはHAP4の構成的過剰発現が脂質、特にスクアレンおよびステロールの生産性の著しい増大をもたらすことも判明した。遺伝子SAK1およびHAP4は呼吸−発酵フラックス分布に関連づけられる。
【0054】
遺伝子SAK1は、Snf1p/Snf4p複合体のリン酸化を担う上流セリン/トレオニンキナーゼをコードする。このリン酸化は、核に局在される活性なSnf1p/Snf4p複合体をもたらす。Snf1p/Snf4p複合体はタンパク質セリン/トレオニンキナーゼに属し、「ジオーキシーシフト(diauxic shift)」の際のグルコース抑制の軽減に中心的役割を果たす。このジオーキシーシフトとは、培地中のグルコースまたは他の発酵炭素源が消耗された際の発酵代謝から呼吸代謝への移行を表す。Snf1/Snf4複合体はそれぞれグルコース抑制および抑制解除を担うカスケードの階層を遙かに超える範囲である。ジオーキシーシフトの過程では、サッカロミセス・セレビシエの6000の遺伝子の約4分の1の発現レベルが有意に変化を受ける。これらの主要な変化は主として、発酵モードから呼吸モードへと移行される中枢代謝に携わる。発酵モードでは、酵母細胞はグルコースのような発酵可能な炭素源をエタノールと二酸化炭素へと代謝する。呼吸モードでは、生産されたエタノールが呼吸されて、細胞においてエネルギーを供給するアデノシン三リン酸(ATP)が得られる。ジオーキシーシフトの過程で、Snf1p/Snf4p複合体はリン酸化され、従って、Snf1−キナーゼSak1p、Tos1pおよびElm1pによって活性化される。活性化された複合体は核に局在され、そこでそれは2つの転写因子Mig1pおよびCat8pに影響を及ぼす。ジンクフィンガータンパク質Mig1pは転写レプレッサーであり、複合体としての多数のグルコース抑制遺伝子の特定のコンセンサス配列と結合するためにタンパク質Tup1pおよびCyc8pを動員し、それにより、このコンセンサス配列の下流の遺伝子の転写が抑制される。Cat8pは転写アクチベーターであり、培地中のグルコースが枯渇した際に少なくとも34の遺伝子の発現を誘導する。これらの遺伝子は主として、エタノールまたはアセテートのようなC2化合物の代謝に必要なグリオキシル酸シャントの遺伝子である。これに加え、クエン酸回路およびグリオキシル酸シャントの中間体の細胞内輸送を担う遺伝子もCat8pによって調節される。活性なSnf1p/Snf4p複合体はMig1pをリン酸化し、それによりこのタンパク質は不活性化され、核の外へ転流され、機能的ホモログであるCat8pおよびSip1pをリン酸化し、これがこれら2つのタンパク質の不活性をもたらす。従って、核に局在された活性なSnf1p/Snf4p複合体は、抑制カスケードのいくつかの転写因子に影響を及ぼすことから、グルコース抑制の軽減および発酵から呼吸への移行に大きく寄与する。よって、本発明の生物は、グルコースの存在下であってもより活性なSnf1p/Snf4p複合体を呈するようにさらに発達させることができる。この改変は、主要キナーゼとしてSnf1p/Snf4p複合体のリン酸化、従って、活性化を担うタンパク質Sak1pの転写脱調節によって達成される。
【0055】
Hap2/3/4/5タンパク質複合体は、多数のグルコース抑制遺伝子を調節する。これらは呼吸鎖およびクエン酸回路の主要な遺伝子である。この複合体はジオーキシーシフトの際にアクチベーターとしてこれらの遺伝子の転写を誘導し、呼吸−発酵バランスの呼吸方向へのシフトに強く寄与する。遺伝子HAP2、HAP3およびHAP5は構成的に発現される。本来、HAP4は、非発酵性炭素源での増殖の場合にのみ発現される。HAP4過剰発現変異株の培養の際に、この突然変異は増殖速度の増大、バイオマスおよび酢酸の生産、ならびにグリセロールおよびエタノールの生成の低下をもたらすのが見られた。Hap2/3/4/5タンパク質複合体は、呼吸能、ミトコンドリアの生合成およびクエン酸回路の炭素フラックスを高めることによって呼吸系を拡張する。
【0056】
本発明の範囲内での別の観察として、遺伝子FLD1の欠失が、酵母、特に酵母サッカロミセス・セレビシエのような生物において脂質生産、特に、スクアレン生産の予期できない著しい増大をもたらすというものがある。遺伝子FLD1は、脂質滴の形態、数および大きさに関与するタンパク質セイピンをコードする。FLD1の過剰発現は脂質粒子の融合をもたらし、その結果、酵母細胞において著しく拡大した、しかしながら数は少ない脂質粒子が形成されることが報告されている。野生型酵母株では、スクアレンはステロール生合成経路の中間体であり、細胞に蓄積されない。酵母において、欧州特許第EP−A486290号に記載されるように酵素の触媒部分のみをコードし、膜結合ドメインはコードしないHMG−CoA−レダクターゼ遺伝子を発現させることによって達成されるHMG−CoA−レダクターゼ活性の増強は、細胞においてスクアレンの蓄積をもたらす。この蓄積は、エルゴステロール生合成経路の中間体によるHMG−CoA−レダクターゼのフィードバック調節の回避によるものである。本発明の枠組みの範囲で、遺伝子FLD1の欠失は、HMG−CoA−レダクターゼ活性の増強を示す酵母株において、脂質生産、特にスクアレン生産の予期できない著しい増大をもたらすことが認められた。
【0057】
SAK1、HAP4およびFLD1に関する上記の発見は、上記のような本発明の6つの態様のいずれにおいても、さらなる改変および実施形態をもたらす。しかしながら、これらの発見は、上記の改変を含まないが、個々の非改変生物によって典型的に合成されるあらゆる脂質を合成する能力を有する生物にも独立した価値を持つ。これに関して、非改変生物は、野生型の他、対応する野生型生物と比較して高いHMG−CoA−レダクターゼ(EC1.1.1.34)活性を有する生物を含む。
【0058】
よって、本発明はまた、遺伝子HAP4および/またはSAK1が、転写において脱調節され、かつ/または過剰発現され、かつ/また遺伝子FLD1が抑制もしくは不活性化もしくは欠失された(全て、対応する野生型生物と比較して)、単離された遺伝的に改変された非哺乳類生物を含む。具体的には、遺伝子HAP4およびSAK1の一方または双方を(所望により)ADH1プロモーターのような構成的に活性なプロモーターの制御下に置くことができる。この変形形態のさらなる実施形態では、遺伝子REG1を、対応する野生型生物と比較して抑制または不活性化または欠失させる。この他、本発明に関してこれまでに述べたことも全て、同様にこの独立した態様に当てはまり、これまでに開示された特徴はいずれも、同様にこの独立した態様に組み込むことができる。
【0059】
本発明はまた、第二の脂質を生産する方法に関し、その方法では、本発明の生物を用い、培養し、その生物から第二の脂質を単離する。例えば、プレノール脂質を生産するための本発明の方法は、その脂質粒子内にプレノール脂質を蓄積することができる、上記のような本発明の第三または第四の態様の生物を培養することを含む。蓄積されるプレノール脂質に関しては、好ましい実施形態では、本発明の第三および第四の態様の生物に関して上記したものと同じことが当てはまる。対応する生物において低減または増強される酵素活性についても同じことが言える。別の例として、ステリルアシルエステルを生産する方法は、本発明の第五または第六の態様による生物を培養することを含む。
【0060】
上記のようなタンパク質または酵素の活性における、対応する非改変生物と比較しての低減は、好ましくは少なくとも50%、より好ましくは少なくとも75%、いっそうより好ましくは少なくとも80%、特に好ましくは少なくとも90%、最も好ましくは100%である。100%の低減は、該生物に存在する該酵素のタンパク質または酵素活性が存在しないことを意味する。上記のようなタンパク質または酵素の活性における、対応する非改変生物と比較しての増強は、好ましくは少なくとも10%、より好ましくは少なくとも50%、いっそうより好ましくは少なくとも200%、特に好ましくは少なくとも1000%である。「増強する」とは、また、非改変生物が測定可能なこのような活性を含まない場合の、酵素またはタンパク質の酵素活性(測定できる場合はもちろん)の存在も含む。「活性が増強する」とは、さらに、タンパク質が酵素ではないあらゆる場合において(HAP4など)、タンパク質の量の増加を包含する。そして、上記の増強のパラメーターは同様に量にも当てはまる。酵素活性は、所定量の抽出物に、その抽出物から生成物への反応を触媒する所定量の酵素および所望によりさらに必要な反応成分を加えること、および所定時間内に合成された生成物の量を測定することによって測定することができる。上述の酵素の活性の具体的な測定方法は、例えば、国際公開第03/064650A1号に開示されている。その文献に記載されていない上述の酵素の活性の測定も同様に実施される。
【0061】
細胞において、ある酵素の活性を低減する方法は、例えば、その酵素をコードする遺伝子の遺伝子発現を低減または排除することであり、例えば、対応する遺伝子の上流により弱いプロモーターを付与することによるか、またはその遺伝子および/もしくは関連のプロモーターを完全もしくは部分的に欠失させ、かつ/もしくは生物の細胞内に酵素阻害剤を加えて翻訳される酵素を阻害する、かつ/または細胞にsiRNAを組み込んで活性な転写物の量を低減する、かつ/またはその遺伝子の突然変異によって活性の低い変異体を生成することによる。
【0062】
遺伝子発現の低減とは、個々の酵素をコードするヌクレオチド配列の遺伝子発現のレベルが、対応する非改変生物、好ましくは対応する野生型生物における該ヌクレオチド配列の遺伝子発現レベルと比較して低減されることを意味する。活性に関して上記に示したものと同じレベルの低減を適用する。遺伝子発現レベルを低減する手段および方法は、例えば、合成される対応するmRNAもしくはタンパク質の量の決定、または個々のタンパク質の酵素活性の決定を含む。好ましい実施形態では、遺伝子発現のレベルは、例えばノーザンブロットにおいてmRNAの量を測定することによって決定される。別の好ましい実施形態では、遺伝子発現のレベルは、例えばウエスタンブロットにおいて合成される対応するタンパク質の量を測定することにより、または対応する酵素活性の量を決定することによって決定される。遺伝子発現の低減はまた、着目する遺伝子を機能しないようにすることによって達成することもできる。遺伝子を機能しないようにするための1つの可能性のある手段が遺伝子破壊である。
【0063】
酵素の活性を増強する方法は、その酵素をコードする遺伝子(異種または同種)が構成的に活性な(同種または異種)プロモーターの制御下に配置されるように、かつ/またはその酵素をコードする(異種または同種)遺伝子のコピー数が増え、かつ/もしくは突然変異によって活性が増強されるように転写の脱調節が達成されるような細胞の形質転換を含む。
【0064】
酵素活性の増強または低減を果たすための具体例を実施例に示す。しかしながら、当業者ならば、本明細書で特に開示する必要なく、当技術分野で周知の他の方法を適用することもできよう。
【0065】
本発明で用いる酵素として好適な遺伝子配列を以下に開示するが、同じ酵素活性を有する他の遺伝子配列も同様に使用可能である。特定の遺伝子配列またはそれによりコードされるタンパク質配列には本発明の範囲内の関連のある構造的特徴はないが、同じEC(Enzyme Commision)番号に分類されている。
【0066】
アシル−CoA:ステロールアシルトランスフェラーゼ/ステロールO−アシルトランスフェラーゼ(EC2.3.1.26)の遺伝子配列は、NC_001135.4、NC_001147.6、NM_005891、NM_144784、NM_153728を含む。
【0067】
ジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)の遺伝子配列は、NC_001147.5、XM_002478787、NM_123089、XM_002378082、NM_032564、NM_001012345、NM_010046、XM_002146497を含む。
【0068】
レシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)の遺伝子配列は、NC_001147.6、NM_008490、NM_001162568、NM_000229、NM_001005715、NM_017024、NM_001082190を含む。
【0069】
アシルCoA−ワックスアルコールアシルトランスフェラーゼ(EC2.3.1.75)の遺伝子配列は、NM_123089、NM_177448を含む。
【0070】
HMG−Co0A−レダクターゼの遺伝子配列は、NC_001145、NM_106299、NC_003421.2、NC_009784.1、NC_003028.3、NC_007308.3、および図5の配列(末端切断型tHMG1)を含む。
【0071】
C−24ステロールメチルトランスフェラーゼの遺伝子配列は、NC_001145、NC_000911.1、NC_003423.3、XM_505173、XM_716615を含む。
【0072】
C−22ステロールデサチュラーゼの遺伝子配列は、NC_003424.3、NC_009046.1、NC_001145.2、XM_500188、XM_711840を含む。
【0073】
C−5ステロールデサチュラーゼの遺伝子配列は、NC_001144、S46162、NG_009446、NM_053642、NM_001035356、XM_503090、XM_708519を含む。
【0074】
HAP4の遺伝子配列は、NC_001143.7、XM_448596、XM_001645329を含む。
【0075】
SAK1の遺伝子配列は、NC_001137.2、XM_502591、XM_448319、XM_453478、NM_208704を含む。
【0076】
REG1の遺伝子配列は、NC_001136.8、XM_500990、XM_448729、XM_455276を含む。
【0077】
FLD1の遺伝子配列は、NC_001144.4、NM_210286、XM_001647166、XM_449778を含む。
【0078】
7−デヒドロコレステロールレダクターゼの遺伝子配列は、NM_103926、NM_001360、NM_007856、NM_203904、NM_001014927、NM_201330、NM_022389、NM_001131727、NM_001087087、XM_001497598、XM_001174160、XM_001099101、BM490402、CA753545を含む。
【0079】
24−デヒドロコレステロールレダクターゼの遺伝子配列は、NM_014762、NM_001016800、NM_001094456、NM_001008645、NM_001103276、NM_001080148、NM_053272、NM_00103128、XM_001488247、AB125202、XM_001153751を含む。
【0080】
ラノステロールステロール14−デメチラーゼの遺伝子配列は、NC_001140.5、XM_500518、EF059165、XM_445876、XM_454109を含む。
【0081】
スクアレンモノオキシゲナーゼの遺伝子配列は、NC_001139.8、M64994、XM_503994、XM_706801、XM_455763を含む。
【0082】
好適なプロモーター(異種発現される個々の異なる酵素のプロモーターは同じまたは異なる場合がある)の遺伝子配列は、NC_001142、NC_001139、NC_001147、NC_001139、NC_001148、NC_001135、NC_001136を含む。
【0083】
一般に、本発明の生物(上記の態様のそれに関わらず)存在し得る任意の非ヒト、好ましくは非哺乳類生物であり得る。好適な生物の例は、特許請求の範囲に示されている。
【図面の簡単な説明】
【0084】
【図1】酵母サッカロミセス・セレビシエの構築された変異株由来の全脂質抽出物の薄層クロマトグラフィー。
【図2】酵母サッカロミセス・セレビシエにおけるスクアレンの生合成を模式的に示す。
【図3】酵母サッカロミセス・セレビシエにおけるトリアシルグリセロールの生合成を模式的に示す。
【図4】酵母サッカロミセス・セレビシエにおけるステリルアシルエステルの生合成を模式的に示す。
【図5】末端切断型HMG CoA−レダクターゼ、tHMG1の配列を示す。
【発明を実施するための形態】
【0085】
以下の実施例は単に本発明を説明するためのものである。
【0086】
実施例では、以下の材料および方法を用いた。
【0087】
1.制限切断
プラスミド(1〜10μg)の制限酵素処理は30μlバッチで行った。このために、24μlのH2O中にDNAを取り、3μlの対応するバッファー、1μlのRSA(ウシ血清アルブミン)および2μlの酵素と混合した。酵素濃度は、DNA量に応じて1単位/μlまたは5単位/μlとした。場合によっては、バッチにさらに1μlのRNアーゼを加えてtRNAを分解した。この制限バッチを37℃で2時間インキュベートした。制限酵素処理はミニゲルで管理した。
【0088】
2.ゲル電気泳動
ゲル電気泳動はミニゲルまたはワイドミニゲル装置で行った。ミニゲル(約20ml、8バッグ)およびワイドミニゲル(50ml、15または30バッグ)はTAE中1%のアガロースからなった。移動バッファーとして1*TAEを用いた。サンプル(10μl)と3μlの停止液を混合し、適用した。HindIIIで切断したI−DNAを標品として用いた(23.1kb;9.4kb;6.6kb;4.4kb;2.3kb;2.0kb;0.6kbにバンド)。分離のために、45〜60分間、80Vの電圧をかけた。その後、ゲルを臭化エチジウム溶液で染色し、ビデオ撮影システムINTASを用いてUV光下で保持、またはオレンジ色のフィルターを用いて写真撮影した。
【0089】
3.ゲル溶出
所望の断片を、ゲル溶出を用いて単離した。この制限調製物をミニゲルのいくつかのバッグに適用し、分離した。[λ]−HindIIIおよび「サクリファイス・トレース(sacrifice trace)」のみを臭化エチジウム溶液で染色し、UV光下で観察し、所望の断片を標識した。結果として、残りのバッグでは臭化エチジウムおよびUV光によってDNAを損傷させずにすんだ。染色されたゲル片と非染色ゲル片をアラインすることにより、非染色ゲル片から所望の断片を、その標識に基づいて切り出すことができた。単離する断片を含むこのアガロース片を透析管に入れ、少量のTAEバッファーとともに気泡が入らないように密閉し、BioRad−ミニゲル装置に入れた。移動バッファーは1*TAEからなり、40分間、電圧を100Vとした。次に、流動極性を2分間変更して透析管に付着しているDNAを遊離させた。透析管の、DNA断片を含むバッファーを反応槽に移し、エタノール沈殿法を行った。このために、DNA溶液に、一部(1/10)量の3M酢酸ナトリウム、tRNA(1μl/50μl溶液)および2.5倍の容量の氷冷96%エタノールを加えた。このバッチを−20℃で30分間インキュベートした後、4℃で30分間、12,000rpmにて遠心分離した。DNAペレットを乾燥させ、10〜50μlのH2O(DNA量による)に取った。
【0090】
4.クレノウ処理
DNA断片の突出末端をクレノウ処理により作出し、「平滑末端」を得た。DNA1μg当たり、以下のバッチをピペットで一緒に吸い取った。この場合、夾雑物がクレノウ−ポリメラーゼを阻害しないように、DNAはエタノール沈殿法からのものとするべきである。インキュベーションは37℃で30分間、その後、70℃でさらに5分間行い、反応を停止させた。DNAはこのバッチからエタノール沈殿法によって得、10μlのH2Oに取った。
【0091】
5.ライゲーション
連結させるDNA断片を合わせた。最終量13.1μlには、ベクター−インサート比1:5で約0.5μgのDNAを含んだ。このサンプルを70℃で45秒間インキュベートし、室温まで冷却し(約3分)、その後、氷上で10分間インキュベートした。次に、ライゲーションバッファー:2.6μlの500mmol TrisHCl、pH7.5および1.3μlの100mmol MgCl2を加え、それらを氷上でさらに10分間インキュベートした。1μlの500mmol DTTおよび1μlの10mmol ATPを加えた後、1μlのリガーゼ(1単位/μl)を氷上でさらに10分間加えた。この処理は全体的に、隣接するDNA末端が再び分離しないようできる限り振盪させないように行うべきである。ライゲーションは4℃で一晩行った。
【0092】
6.大腸菌の形質転換
成分大腸菌(E. coli)NM522細胞をライゲーション調製物のDNAで形質転換させた。陽性対照として、バッチに50ngのpScL3プラスミドを供給し、ヌル対照として、バッチにDNAを供給しなかった。各形質転換調製物について、100μlの8%PEG溶液、10μlのDNAおよび200μlのコンピテント細胞(大腸菌NM522)をピペットで卓上遠沈管に入れた。これらのバッチを氷上に30分間置き、断続的に振盪した。その後、42℃で1分間熱ショックを行った。再生のため、1mlのLB培地を細胞に加え、37℃にて90分間シェーカー上でインキュベートした。各100μlの未希釈バッチ、1:10希釈液および1:100希釈液をLB+アンピシリンプレートに平らに拡げ、37℃で一晩インキュベートした。
【0093】
7.大腸菌からのプラスミド単離(Miniprep)
大腸菌コロニーを卓上遠沈管の1.5mlのLB+アンピシリン培地中で、37℃および120rpmにて一晩培養した。翌日、細胞を5000rpm、4℃で5分間遠心分離し、ペレットを50μlのTEバッファーに取った。各バッチを100μlの0.2N NaOH、1%SDS溶液と混合し、氷上に5分間置いた(細胞の溶解)。次に、400μlの酢酸Na/NaCl溶液(230μlのH2O、130μlの3M酢酸ナトリウム、および40μlの5M NaCl)を加え、バッチを混合し、氷上にさらに15分間置いた(タンパク質の沈殿)。11,000rpmで15分間遠心分離した後、プラスミドDNAを含む上清をエッペンドルフ管に移した。上清が完全に透明でなければ、もう一度遠心分離を行った。この上清を360μlの氷冷イソプロパノールと混合し、−20℃で30分間インキュベートした(DNAの沈殿)。DNAを遠心分離し(15分、12,000rpm、4℃)、上清を廃棄し、ペレットを100μlの氷冷96%エタノールで洗浄し、−20℃で15分間インキュベートし、再び遠心分離を行った(15分、12,000rpm、4℃)。ペレットをスピードバキュームで乾燥させた後、100μlのH2Oに取った。制限分析により、プラスミドDNAの特性決定を行った。このために、各バッチ10μlを制限酵素で処理し、ワイドミニゲルでのゲル電気泳動により分離した(上記参照)。
【0094】
8.大腸菌でのプラスミド単離(Maxiprep)
より多量のプラスミドDNAを単離するために、maxiprep法を行った。100mlのLB+アンピシリン培地を含む2つのプランジャーに、単離されるプラスミドを担持するコロニーまたは100μlの凍結培養物を接種し、37℃、120rpmで一晩インキュベートする。翌日、培養物(200ml)をGSAビーカーに移し、10分間4000rpm(2600*g)で遠心分離した。細胞ペレットを6mlのTEバッファーに再懸濁させた。細胞壁を消化するために、1.2mlのリゾチーム溶液(20mg/mlTEバッファー)を加え、室温で10分間インキュベートした。次に、12mlの0.2N NaOH、1%SDS溶液を用いて細胞溶解を行い、室温でさらに5分間インキュベートした。9mlの冷却した3M酢酸ナトリウム溶液(pH4.8)を加え、15分間氷上でインキュベートすることによってタンパク質を沈殿させた。遠心分離(GSA:13,000rpm(27,500*g)、20分、4℃)の後、DNAを含む上清を新しいGSAビーカーに移し、15mlの氷冷イソプロパノールを用い、−20℃で30分間インキュベートしてDNAを沈殿させた。このDNAペレットを5mlの氷冷エタノールで洗浄し、風乾した(約30〜60分)。その後、これを1mlのH2Oに再懸濁させた。このプラスミドを制限分析により調査した。濃度はミニゲルに希釈液を加えることによって求めた。塩含量を引き下げるために、30〜60分微量透析を行った(孔径0.025μm)。
【0095】
9.酵母の形質転換
酵母の形質転換のため、サッカロミセス・セレビシエ(S. cerevisiae)AH22株の前培養物を調製した。20mlのYE培地を含むプランジャーに100μlの凍結培養物を接種し、28℃、120rpmで一晩インキュベートした。主培養は、10μl、20μlまたは50μlの前培養物を接種した100mlのYE培地を含むプランジャーにて同じ条件下で行った。
【0096】
9.1 コンピテント細胞の作製
翌日、トーマチャンバー(Thoma chamber)を用いてプランジャーを計数し、3〜5*107細胞/mlを保有するプランジャーを用いてこの手順を続けた。遠心分離(GSA:5000rpm(4000*g)、10分)により細胞を採取した。細胞ペレットを10mlのTEバッファーに再懸濁させ、2本の卓上遠沈管に分けた(各5ml)。細胞を6000rpmで3分間遠心分離し、各5mlのTEバッファーで2回洗浄した。次に、細胞ペレットを109細胞につき330μlの酢酸リチウムバッファーに取り、無菌の50mlエルレンマイヤーフラスコに移し、28℃で1時間振盪させた。結果として、これらの細胞は形質転換能があった。
【0097】
9.2 形質転換
各形質転換調製物について、15μlのニシン精子DNA(10mg/ml)、10μlの形質転換されるDNA(約0.5μg)および330μlの成分細胞をピペットで卓上遠沈管に入れ、28℃で30分間インキュベートした(振盪させない)。次に、700μlの50%PEG6000を加え、振盪させずに28℃でさらに1時間インキュベートした。その後、42℃で5分間熱ショックを行った。100μlのこの懸濁液を選択培地(YNB、Difco)に播種し、ロイシン原栄養株を選択した。G418耐性の選択の場合には、熱ショック後に細胞の再生を行う(9.3再生段階を参照)。
【0098】
9.3 再生段階
選択マーカーはG418耐性であるので、細胞は耐性遺伝子の発現に時間を要した。形質転換調製物を4mlのYE培地と混合し、シェーカー(120rpm)にて28℃で一晩インキュベートした。翌日、細胞を遠心分離し(6,000rpm、3分)、1mlのYE培地に取り、100μlまたは200μlをYE+G418プレートに平らに拡げた。これらのプレートを28℃で数日間インキュベートした。
【0099】
10.PCRの反応条件
ポリメラーゼ連鎖反応の反応条件は個々の場合ごとに至適化しなければならず、いずれのバッチにも必ずしも有効というわけではない。従って、とりわけ、DNAの使用量、塩濃度および融解温度は変動し得る。課題の本発明者らの処方物では、エッペンドルフキャップ中に、サーモサイクラーで用いるのに好適な以下の物質:PCR調製物として総容量50μlとなるように水に溶解させた5μlのスーパーバッファー、8μlのdNTP(各0.625μM)、5’−プライマー、3’−プライマーおよび0.2μgのマトリックスDNAを合わせるのが有利であることが分かり、2μl(〜0.1U)のSuper Taqポリメラーゼに加えた。このバッチを短時間で遠心分離し、1滴のオイルでカバーした。増幅には37〜40回のサイクルを選択した。
【0100】
11.S.セレビシエからの脂質粒子の単離
酵母細胞を28℃で72時間、250rpmで往復振盪しながら、50mlのWMVIII最小培地中で増殖させた。細胞を遠心分離により採取し、Leber et al.(Leber R,Zinser E,Zellnig G,Paltauf F,Daum G. Characterization of lipid particles of the yeast,Saccharomyces cerevisiae. Yeast. 1994 Nov;10(11):1421−1428)に従って脂質粒子を単離および精製した。ステロール分析では、脂質粒子は、GCにより総ステロールを定量するために室温、30%メタノール性KOH中で16時間鹸化するか、またはクロロホルム/メタノール(4:1)でステロールを直接抽出し、遊離ステロールとエステル化ステロールを識別するためのTLCによって、もしくは遊離ステロールの定量のためのGCによって分析するかのいずれかを行った。
【0101】
12.スクアレンおよびステロールの分析
細胞全体の脂質および脂質粒子中の総脂質を定量するために、GC分析の前にサンプルを鹸化した。125OD600の細胞を0.5N HCl中、100℃で20分間処理し、室温まで冷却した。その後、3gのKOH、およびピロガロール(2g/l)を含む12.5mlのメタノールを加えた。鹸化のために、混合物を水浴中、70℃で2時間インキュベートした。加水分解されたエステルをn−ヘキサンで抽出した。非鹸化画分を2mlのn−ヘキサンに再懸濁させた。スクアレンおよびステロールを、スクアレンおよびコレステロールを内部標準としてGCにより定量した。スクアレンおよびステロールを、150から250℃までプログラムされたキャピラリーカラム(25m×0.25mm×0.25μm[膜厚];Chrompack CPSil5)を備えたHewlett−Packard 5890ガスクロマトグラフで分離した。温度は最初2分間150℃とした後、15℃/分で最終温度250℃まで昇温し、この温度で20分間保持した。直線的速度を30cm/秒とし、ヘリウムをキャリアガスとして用い、注入はスプリットレスモードで行った。注入容量は1μlとした。各ピークの面積を計算し、細胞乾重1gに対して換算した。各サンプルを2回測定した。同定にはエルゴステロールおよびスクアレンの標品を用いた。
【0102】
中性脂質を抽出し、Sorger and Daum(J.Bacteriol.184(2002),519−524)の方法で定量した。詳細には、中性脂質の定量では、抽出物をシリカゲル60プレートに適用し、クロマトグラムを、軽質石油−ジエチルエーテル−酢酸(25:25:1、vol/vol/vol)の溶媒系を用いて3分の1の距離まで展開させた。その後、プレートを軽く乾燥させ、軽質石油−ジエチルエーテル(49:1、vol/vol)の溶媒系を用いてプレートの上部までさらに展開させた。中性脂質は、TLCチャンバー内で薄層プレートをヨウ素蒸気で染色することにより可視化した。定量は比重走査によって行った。
【0103】
スクアレンおよびステロール分析のための酵母サッカロミセス・セレビシエ株の標準的な培養手順は次の通りとした。
前培養:100ml振盪フラスコ中の20mlのWMVIII培地に20μlの対応するグリセロール保存液に接種し、30℃、150rpmで48時間培養した。
主培養:バッフル付き250ml振盪フラスコ中の50mlのWMVIII培地に1%の前培養物を接種し、30℃、150rpmで72時間培養した。
【0104】
実施例1
S.セレビシエAH22ura3における遺伝子ARE1およびARE2の欠失
ベクターpUG6(Gueldener U,Heck S,Fiedler T,Beinhauer JD and Hegemann JH (1996).A new efficient gene disruption cassette for repeated use in budding yeast. Nucleic Acids Res 24 2519−24)を用いて、遺伝子ARE1およびARE2を欠失させた。
【0105】
両遺伝子の欠失は、同じ方法によって連続的に行った。まず、ARE1を欠失させ、その後、ARE2を欠失させた。プラスミドを調製した後、pUG6の断片をPCRによって増幅し、loxP−kanMX−loxPからなるツールを得た。ARE1 respec.ARE2コード配列の5’および3’配列をpUG6ベクターのloxP領域と融合させるためのプライマーを構築した。
【0106】
得られたPCR産物は、KanR遺伝子、loxP部位およびS.セレビシエAH22ura3における組込み形質転換のためのARE1 respec.ARE2相同領域からなる。酵母における相同組換えが、標的配列の欠失をもたらす。
【0107】
G418に対する耐性を陽性クローンの選択に用いた。この酵母株においてARE1 respec.ARE2コード領域は欠失されていた。さらなる遺伝子を欠失させるための株を作製するために、この株からG418耐性を除去した。この目的で、この株をpSH47で形質転換させた(Guldner et al., 1996)。このベクターは、loxP部位に隣接するKanR遺伝子を除去するためのcre−リコンビナーゼを有する。
【0108】
pSH47を定着させるため、この株を5−FOA(5−フルオロオロト酸)(1g/L)寒天プレート上で対抗選択した。得られた株は遺伝子ARE1およびARE2の双方を欠失している。
【0109】
実施例2
S.セレビシエAH22ura3における遺伝子DGA1およびLRO1の欠失
ベクターpUG6(Guldner et al.,1996)を用いて、遺伝子DGA1およびLRO1を欠失させた。
【0110】
両遺伝子の欠失は、同じ方法によって連続的に行った。まず、DGA1を欠失させ、その後、LRO1を欠失させた。プラスミドを調製した後、pUG6の断片をPCRによって増幅し、loxP−kanMX−loxPからなるツールを得た。
【0111】
DGA1 respec.LRO1コード配列の5’および3’配列をpUG6ベクターのloxP領域と融合させるためのプライマーを構築した。
【0112】
得られたPCR産物は、KanR遺伝子、loxP部位およびS.セレビシエAH22ura3における組込み形質転換のためのDGA1 respec.LRO1相同領域からなる。酵母における相同組換えが、標的配列の欠失をもたらす。
【0113】
G418に対する耐性を陽性クローンの選択に用いた。この酵母株においてDGA1 respec.LRO1コード領域は欠失されていた。さらなる遺伝子を欠失させるための株を作製するために、この株からG418耐性を除去した。この目的で、この株をpSH47で形質転換させた(Guldner et al., 1996)。このベクターは、loxP部位に隣接するKanR遺伝子を除去するためのcre−リコンビナーゼを有する。
【0114】
pSH47を定着させるため、この株を5−FOA(5−フルオロオロト酸)(1g/L)寒天プレート上で対抗選択した。
【0115】
得られた株は遺伝子DGA1およびLRO1の双方を欠失している。
【0116】
実施例3
S.セレビシエAH22ura3are1are2における遺伝子DGA1およびLRO1の欠失
ベクターpUG6(Guldner et al., 1996)を用いて、遺伝子DGA1およびLRO1を欠失させた。
【0117】
両遺伝子の欠失は、同じ方法によって連続的に行った。まず、DGA1を欠失させ、その後、LRO1を欠失させた。プラスミドを調製した後、pUG6の断片をPCRによって増幅し、loxP−kanMX−loxPからなるツールを得た。DGA1 respec.LRO1コード配列の5’および3’配列をpUG6ベクターのloxP領域と融合させるためのプライマーを構築した。
【0118】
得られたPCR産物は、KanR遺伝子、loxP部位およびS.セレビシエAH22ura3における組込み形質転換のためのDGA1 respec.LRO1相同領域からなる。酵母における相同組換えが、標的配列の欠失をもたらす。
【0119】
G418に対する耐性を陽性クローンの選択に用いた。この酵母株においてDGA1 respec.LRO1コード領域は欠失されていた。さらなる遺伝子を欠失させるための株を作製するために、この株からG418耐性を除去した。この目的で、この株をpSH47で形質転換させた(Guldner et al., 1996)。このベクターは、loxP部位に隣接するKanR遺伝子を除去するためのcre−リコンビナーゼを有する。
【0120】
pSH47を定着させるため、この株を5−FOA(5−フルオロオロト酸)(1g/L)寒天プレート上で対抗選択した。
【0121】
得られた株は遺伝子ARE1、ARE2、DGA1およびLRO1の4つを欠失している。
【0122】
実施例4
エピソームプラスミドを用いた、実施例1〜3から得られた酵母株および参照株としてのAH22ura3におけるt−HMG1の発現
tHMGのDNA配列(Basson et al. (Mol. Cell. Biol. 8 (1988), 3793−3808))を、標準的な方法を用いてサッカロミセス・セレビシエS288C(Mortimer and Johnston (Genetics 113 (1986), 35−43))のゲノムDNAからPCRにより増幅した。この場合に用いるプライマーはDNAオリゴマーtHMG−5’およびtHMG−3’である。得られたDNA断片を、クレノウ処理後にクローニングベクターpUC19(Yanisch−Perron et al. (1985): Improved M13 phage cloning vectors and host strains: nucleotide sequences of the M13mp18 and pUC19 vectors. In: Gene. Bd. 33, S. 103−119)に導入し、ベクターpUC19−tHMGを得た。プラスミドを単離し、pUC19−tHMGをエンドヌクレアーゼEcoRIおよびBamHIで制限処理した後、得られた断片を、これもまたEcoRIおよびBamHIで処理した酵母発現ベクターpPT2b(Lang and Looman (Appl. Microbiol. Biotechnol. 44(1995),147−156))に導入した。作出されたプラスミドpPT2b−tHMGは、末端切断型ADH1−プロモーター(Bennetzen and Hall (Yeast 7(1982),475−477))およびTRP1−ターミネーター(Tschumper G, Carbon J. Sequence of a yeast DNA fragment containing a chromosomal replicator and the TRP1 gene. Gene. 1980 Jul;10(2):157−166)を含み、その間にtHMG−DNA断片が見られる。ベクターpPT2b−tHMGからエンドヌクレアーゼEcoRVおよびNruIによってDNA切片を単離した。このDNA切片は、いわゆる中程度の長さのADH1−プロモーター、tHMG遺伝子およびTRP1−ターミネーターを含む。このDNA切片を、エンドヌクレアーゼSphIおよびDNAポリメラーゼで処理した酵母ベクターYEp13(Fischhoff et al. (Gene 27 (1984), 239−251))に導入した。
【0123】
S.セレビシエAH22URA3are1are2、AH22URA3dga1lro1、AH22URA3are1are2dga1lro1およびAH22URA3をYEpH2で形質転換した。参照として、プラスミドYEp13を用いた。
【0124】
実施例5
染色体組込みプラスミドYDpUHK3を用いた、実施例1〜3から得られた酵母株および参照株としてのAH22ura3におけるt−HMG1の染色体組込みおよび過剰発現
ベクターYEpH2をエンドヌクレアーゼEcoRVおよびNruIで処理した。このようにして、以下の領域:テトラサイクリン耐性遺伝子由来の転写活性化領域(Sidhu and Bollon (10 (1990) 157−166))、中程度の長さのADH1−プロモーター、tHMGおよびTRP1−ターミネーター(発現カセット)を有するDNA断片を作出した。このDNA断片を、StuIで処理したベクターYDpU(Berben et al. 1991 Berben G., Dumont J., Gilliquet V, Bolle P−A. and Hilger F. (1991): ”The YDp plasmids: a uniform set of vectors bearing versatile gene disruption cassettes for” Saccharomyces cerevisiae”. ”Yeast 7, 475−477)に導入した。作出されたベクターYDpUH2/12をエンドヌクレアーゼSmaIで処理し、カナマイシン耐性をコードするDNA配列と連結する(Webster, T. D., Dickson, R. C. (1983) Direct selection of Saccharomyces cerevisiae resistant to the antiobiotic G418 following transformation with a DNA vector carrying the kanamycin−resistance gene of Tn903. Gene 26: 243−252)。作出された構築物(YDpUHK3)をEcoRVで処理した。酵母株サッカロミセス・セレビシエAH22をこの構築物で形質転換させた。この例の場合と同様に、酵母を線状化ベクターで形質転換すると、染色体のURA3遺伝子座にベクター全体が組み込まれる。組み込まれたベクターに由来する、発現カセット(大腸菌起源、E.coli−アンピシリン耐性遺伝子、TEF−プロモーターおよびカナマイシン耐性遺伝子)の部分ではない領域を除去するために、形質転換酵母を、ウラシル栄養要求酵母を増長するFOA選択(Boeke et al. (Methods in Enzymology 154 (1987), 164−175))による選択圧に曝した。選択でいうところのウラシル栄養要求株はAH22tH3ura8の名称を持ち、URA3遺伝子に染色体組込みとしてのtHMG1発現カセットを有する。
【0125】
表1に示される酵母株を、それらのスクアレン生産性/含量について評価した。この目的で、これらの株を30℃、150rpmで振盪しながら72時間、WMVIII培地で培養した。細胞を0.5M煮沸HCl中で破砕した後、脂質を20mlのn−ヘキサンで2回抽出し、GC/MSで分析/定量した(詳細は第12項を参照)。以下のデータが得られた(表1)。
【0126】
【表1】

【0127】
実施例6
S.セレビシエAH22tH3ura8are1are2における遺伝子FLD1の欠失
ベクターpUG6(Guldner et al., 1996)を用いて、遺伝子FLD1を欠失させた。
【0128】
プラスミドを調製した後、pUG6の断片をPCRにより増幅し、loxP−kanMX−loxPからなるツールを得た。FLD1コード配列の5’および3’配列をpUG6ベクターのloxP領域と融合させるためのプライマーを構築した。
【0129】
得られたPCR産物は、KanR遺伝子、loxP部位およびS.セレビシエAH22tH3ura8are1are2における組込み形質転換のためのFLD1相同領域からなる。酵母における相同組換えが、標的配列の欠失をもたらす。
【0130】
G418に対する耐性を陽性クローンの選択に用いた。この酵母株においてFLD1コード領域は欠失されていた。さらなる遺伝子を欠失させるための株を作製するために、この株からG418耐性を除去した。この目的で、この株をpSH47で形質転換させた(Guldner et al., 1996)。このベクターは、loxP部位に隣接するKanR遺伝子を除去するためのcre−リコンビナーゼを有する。
【0131】
pSH47を定着させるため、この株を5−FOA(5−フルオロオロト酸)(1g/L)寒天プレート上で対抗選択した。
【0132】
得られた株は遺伝子ARE1、ARE2およびFLD1の3つを欠失しており、AH22tH3ura8are1are2fld1と呼ばれる。
【0133】
表2に示される酵母株を、それらのスクアレン生産性/含量について評価した。この目的で、これらの株を30℃、150rpmで振盪しながら72時間、WMVIII培地で培養した。細胞を0.5M煮沸HCl中で破砕した後、脂質を20mlのn−ヘキサンで2回抽出し、GC/MSで分析/定量した(詳細は第12項を参照)。以下のデータが得られた(表2)。
【0134】
【表2】
【0135】
実施例7
実施例1〜5から得られた酵母株における遺伝子SAK1およびHAP4のエピソーム過剰発現
遺伝子SAK1およびHAP4のエピソーム過剰発現のため、これらの遺伝子を発現ベクターpFlat1およびpFlat3にそれぞれクローニングした。この目的で、これら2つの遺伝子をS.セレビシエS288c株由来の染色体DNAから、5’末端にNotI制限部位を、3’末端にXhoI制限部位を導入するプライマーを用いて増幅した。得られたPCR断片、ならびにベクターpFlat1およびpFlat3を制限エンドヌクレアーゼXhoIおよびNotIで制限処理した。制限酵素で処理したPCR断片および線状化ベクターを連結し、ベクターpFlat1−SAK1およびpFlat3−HAP4を得た。これらのベクターはそれぞれ、SAK1およびHAP4の強い構成的発現をもたらすために、構成型のADH1プロモーターとTRP1ターミネーターの隣接した1コピーの遺伝子SAK1およびHAP4を有する。
【0136】
プラスミドpFlat3を構築するために、プラスミドYEp24をSphIで切断し、ADH1プロモーターとTRP1ターミネーターを含み、その間にプラスミドpUC19の多重クローニング部位を有する900bpのSphI断片を、プラスミドpPT2Bから挿入した。この多重クローニング部位を、NotIおよびXhoIの制限部位を含むポリリンカーを挿入することにより拡大した。選択のためのURA3遺伝子を有する、得られたプラスミドpFlat1をNcoI制限処理によって線状化し、クレノウポリメラーゼで平滑末端とし、酵母LEU2遺伝子を含むYDpLの、平滑末端化されたBamHI断片を組み込んだ。得られたベクターをpFlat3とした。
【0137】
プラスミドpFlat1−SAK1およびpFlat3−HAP4、また、対照としてのエンプティープラスミドpFlat1およびpFlat3を用い、実施例1〜6から得られた酵母株を形質転換した。
【0138】
表3に示される酵母株を、それらのスクアレン生産性/含量について評価した。この目的で、これらの株を30℃、150rpmで振盪しながら72時間、WMVIII培地で培養した。細胞を0.5M煮沸HCl中で破砕した後、脂質を20mlのn−ヘキサンで2回抽出し、GC/MSで分析/定量した(詳細は第12項を参照)。以下のデータが得られた(表3)。
【0139】
【表3】
【0140】
実施例8
構築された株の中性脂質組成を薄層クロマトグラフィーにより評価した。全脂質抽出および薄層クロマトグラフィーは第12項に従って行った。図1は、野生型株AH22ura3、脱調節型HMG−CoAレダクターゼを有するAH22tH3ura8株、および2つの二重欠失株AH22tH3ura8Δare1Δare2とAH22tH3ura8Δdga1Δlro1の全脂質/中性脂質組成を示す。図1は、脱調節型HMG−CoAレダクターゼを発現し、高い量のスクアレンを生産するレーン3〜8の株に比べ、野生型株(AH22ura3、レーン1および2)は極めて低量のスクアレンしか生産しないことを示唆する。ステリルエステルの形成を担う酵素をコードする遺伝子(are1、are2)およびトリアシルグリセロールの形成を担う酵素をコードする遺伝子(dga1、lro1)の欠失は、対応する株においてこれらの成分の完全な欠失をもたらす(レーン5〜8に黒い四角で示す)。これらの脂質成分をスクアレン、オレイン酸コレステリル、トリオレイン酸、オレイン酸およびエルゴステロールの標品によって同定した(示されていない)。
【技術分野】
【0001】
本発明は、ある特定の酵素活性が低減および/または増強された、脂質、特に中性脂質の工業的生合成に有用な遺伝的に改変された非哺乳類生物、特に微生物に関する。本発明はさらに、このような生物の使用、このような生物を作出するためのシャトルビヒクルおよびこのような生物の生産方法に関する。
【背景技術】
【0002】
発明の背景および技術の現状
脂質は、もっぱらまたは部分的に、脂肪酸もしくはポリケチドのようなチオエステルのカルボアニオンに基づく縮合によって、および/またはプレノールもしくはステロールのようなイソプレン単位のカルボカチオンに基づく縮合によって生じる疎水性または両親媒性の小分子である。この群には、経済上極めて重要ないくつかの物質が含まれる。トリアシルグリセロール脂質群には例えば油脂類またはワックス類が含まれ、これらは例えば食品の成分としてまたは調理用に、石鹸、スキン製品、香料ならびに他のパーソナルケアおよび化粧製品の製造のために、塗料および他の木材処理製品の製造のために、電気工業における生分解性絶縁体として、生分解性作動液の製造のため、潤滑剤として、またはさらには従来のディーゼルの代替に使用可能なバイオディーゼルの主成分としてなど、極めて多様な状況で用いられる。イソプレノイド脂質スクアレンは、例えば、ワクチンまたは他の薬剤、栄養、化粧品ならびにOTC(over−the−counter)薬のアジュバントとして用いられる。スクアレンはまた、テルペン合成の構成ブロックとしても使用することができる。さらに、スクアレンは、生分解性潤滑剤として工業的に使用することができる。他の経済上重要な脂質としては、サポニン、ステロイドホルモン、ビタミンおよび原薬のような化合物の生産のための中枢的出発材料として用いられる、エルゴステロール、チモステロール、エピステロール、7−デヒドロコレステロールまたはラノステロールのようなステロールがある。
【0003】
脂質、特に中性脂質は、通常、細胞内の、脂質粒子として知られる特定の細胞内オルガネラに貯蔵される。これらの粒子は、ほんの少量のタンパク質が埋め込まれた極めて疎水性の高い単層からなる単純な構造を特徴とする。脂質は、加水分解がそれらの成分のリターンを代謝経路および/または異化経路に向けるまで、脂質粒子に貯蔵される。この脂質貯蔵物形成のプロセスは自然界で広く用いられ、あらゆるタイプの真核細胞が、脂肪体、脂質滴、油体またはオレオソームとも呼ぶことができる細胞内脂質粒子を含む。酵母サッカロミセス・セレビシエ(Saccharomyces cerevisiae)脂質滴は、細胞の総脂質含量の70%まで蓄積する。
【0004】
記載されている脂質の大部分は植物、動物または微生物などの天然源に由来するので、生きている細胞において脂質量を増やす、すなわち、脂質粒子中により多くの脂質を蓄積させる試みがなされた。この目的で、生物の代謝経路を改変することが知られている。特許文献1には、酵母におけるエルゴステロール代謝の遺伝子の過剰発現が細胞のエルゴステロール量の増大をもたらすとの記載がある。特許文献2には、ラノステロール−C14−デメチラーゼおよびHMG−CoA−レダクターゼ活性の増強に基づくチモステロールの生産方法が開示されている。特許文献3には、Δ22−デサチュラーゼ活性が低減され、HMG−CoAレダクターゼ、ラノステロールC14−デメチラーゼ、スクアレンエポキシダーゼおよびスクアレンシンセターゼ活性が増強されたトランスジェニック生物が記載されている。これらの生物は、ステロール脂質エルゴスタ−5,7−ジエノールの生産に使用することができる。従って、脂質生成生物を生物学的に改変することによって、生産される脂質の収量を著しく増大させることができる。
【0005】
しかしながら、得られる脂質化合物はほとんどの場合、不純で、混ざり合い、特に、その脂質がさらなる化学合成または改変の出発材料として用いられる場合には、分離および/または精製する必要がある。さらに、いずれの精製手順も、特に工業的規模で行われる場合には、高くつき、労力がかかり、環境を汚染する傾向がある。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】欧州特許第EP0486290A号
【特許文献2】国際公開第03/064652A号パンフレット
【特許文献3】国際公開第2004/083407A号パンフレット
【発明の概要】
【発明が解決しようとする課題】
【0007】
よって、本発明の基礎にある技術課題は、労力がかかり、コストがかかる精製手順を避けるために、より純度の高い形態で生物による脂質生産を可能とする手段および方法を提供することである。
【0008】
発明の概要および好ましい実施形態
この技術課題を解決するために、本発明は、請求項1に記載の生物を教示する。好ましい実施形態は、請求項1の従属項で明示する。その中の、「遺伝的に改変された」とは、現状の遺伝子工学の方法によってゲノムが改変された生物だけでなく、所望の遺伝子改変に従う突然変異体群から選択された生物(従来の突然変異誘発)も含むものとする。
【0009】
本発明は、特定の(中性)脂質がより純度の高い形態で脂質粒子に蓄積されるように生物を遺伝的に操作する、または生物学的に改変することができるという発見に基づく。本発明の生物は、生物によってより高い純度で生産させようとする脂質に応じて、所望でないステリルアシルエステルおよび/またはトリアシルグリセロールおよび/またはワックスエステルの合成を排除または低減するように改変される。中性脂質のより純度の高い形態での蓄積の他、驚くことに、潜在的に混入するステリルアシルエステルおよび/またはトリアシルグリセロールおよび/またはワックスエステルの低減または排除も所望の脂質のレベルおよび収量に増大をもたらす。これは、特定の理論に縛られるものではないが、脂質粒子内の貯蔵空間の増大および/または所望の脂質の合成のための基質の利用能の上昇によるものである可能性がある。
【0010】
「中性脂質」とは、荷電基を欠き、従って、二重膜に実質的な量では組み込まれ得ない脂質を意味する。この用語には、トリアシルグリセロール(TAG)、ステリルアシルエステル(SAE)およびワックスエステル(WE)が含まれる。スクアレンもこの種の「中性脂質」に属す。通常、脂質粒子の内容物は、組換え生物での所望の特定中性脂質の効率的生産に支障を来す種々の中性脂質の混合物である。この問題は、本発明により、生物の脂質体内で本来生成する中性脂質の少なくとも1つの種類をそこではもはや合成されないように生産者生物を改変することによって克服される。結果として、脂質粒子内に含まれる残りの中性脂質の組成はより純粋となり、従って、市販の商業目的での生産により魅力あるものとなる。
【0011】
一般に、より高い純度で合成させようとする中性脂質に応じて、または生合成において望ましくない混入物として除去しようとする中性脂質に応じて、請求項1に記載のいずれか1つの活性または種々の活性(2つ、3つまたは4つ)の組合せを用いることができる。よって、本発明の種々の実施形態が存在するが、以下、いくつかの特定の実施形態だけを詳細に記載する。
【0012】
よって、第一の実施形態では、本発明の生物は、ステリルアシルエステル(SAE)をもはや合成しないように改変される。ステリルアシルエステルは、ステロールと長鎖脂肪酸とのエステルである。一般に、SAEの合成は、細胞内でアシル−CoA:ステロールアシルトランスフェラーゼ/ステロールO−アシルトランスフェラーゼ(EC2.3.1.26)を含む酵素機構によって遂行される。触媒される反応は、ステロールと長鎖脂肪酸とのエステル化である。よって、好ましい実施形態では、本発明の生物は、アシル−CoA:ステロールアシルトランスフェラーゼ/ステロールO−アシルトランスフェラーゼ(EC2.3.1.26)の活性が、対応する非改変生物、好ましくは対応する野生型生物と比較して低減されるように改変される。
【0013】
酵母、特にS.セレビシエ(S. cerevisiae)では、ステリルアシルエステルを生じるステロールエステル化は、ステリルアシルトランスフェラーゼの2つのイソ酵素、すなわち、異なるステロール中間体に対して異なる特異的親和性を有するAre1pおよびAre2pによって遂行される。Are1pは、特に、ラノステロール、チモステロール、エルゴスタ−5,7−ジエノールなどのエルゴステロール前駆体と、7−デヒドロコレステロールなどの酵母にとって外来の物質とのエステル化をもたらす。Are2pは、酵母におけるエルゴステロール生合成経路の最終産物であるエルゴステロールのエステル化を優先的にもたらす。生物が酵母生物体、特にサッカロミセス(Saccharomyces)属のもの、好ましくはS.セレビシエである場合、Are1pまたはAre2pまたはその両者Are1pとAre2pの活性を、対応する非改変酵母生物、好ましくは対応する野生型生物と比較して低減または排除することができる。両酵素活性が排除される場合、細胞内でステロールのエステル化は起こらない。一方の活性のみを排除し、他方を維持することにより、所望により、ある特定のステロールをもたらす酵素活性がその生物において相応に維持または適合されていれば、細胞内のある特定のタイプのステリルアシルエステルを得ることが可能である。
【0014】
生物がステリルアシルエステルをもはや合成しないことを特徴とする本発明の第一の態様に関して、本発明の生物は、原則として、存在し得る任意の生物、植物生物、真菌生物または細菌生物であってよい。しかしながら、好ましくは、該生物は、その脂質粒子内にワックスエステルを本来貯蔵しないが、トリアシルグリセロールおよびステリルアシルエステルのような他の脂質だけを貯蔵する生物に由来する。よって、ステリルアシルエステルの合成のための生合成経路を排除することにより、このような生物はその脂質粒子内に本質的にトリアシルグリセロールのみを蓄積し、それにより、このような生物において、脂質粒子からかなり純粋な形態でトリアシルグリセロールを生産することが可能となる。それらの脂質粒子内にワックスエステルを本来蓄積しない生物の例としては、特に、大部分の動物、真菌細胞、および藻類とプランクトン細胞を除く大部分の植物細胞である。本発明の第一の態様に関する特に好ましい実施形態では、生物は真菌生物、好ましくは、ヤルロウィア(Yarrowia)、ロドトルラ(Rhodotorula)、リポミセス(Lipomyces)、カンジダ(Candida)、ロドスポリジウム(Rhodosporidium)、モルチエレラ(Mortierella)、ムコール(Mucor)、サッカロミセス(Saccharomyces)、ピキア(Pichia)、クルイベロミセス(Kluyveromyces)、アスペルギルス(Aspergillus)、ペニシリウム(Penicillium)およびディクチオステリウム(Dictyostelium)からなる群から選択される属の真菌生物である。ヤルロウィア・リポリチカ(Yarrowia lipolytica)、ロドトルラ・グルチニス(Rhodotorula glutinis)、リポミセス・スタルケイイ(Lipomyces starkeyi)、カンジダ・クルバタ(Candida curvata)、ロドスポリジウム・トルロイデス(Rhodosporidium tortuloides)、モルチエレラ・イサベリナ(Mortierella isabellina)、ムコール・ジャバニカス(Mucor javonicus)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、ピキア・パストリス(Pichia pastoris)、クルイベロミセス種(Kluyveromyces spec)、アスペルギルス種(Aspergillus spec)、ペニシリウム種(Penicillium spec)またはディクチオステリウム種(Dictyostelium spec)の真菌生物が特に好ましい。本発明の第一の態様の特に好ましい実施形態では、生物はS.セレビシエである。
【0015】
第二の実施形態では、本発明の生物は、ステリルアシルエステル(SAE)およびワックスエステル(WE)をもはや合成しないように改変される。
【0016】
一般に、WEの合成は、細胞内でアシルCoA−ワックスアルコールアシルトランスフェラーゼ(EC2.3.1.75)を含む酵素機構によって遂行される。よって、本発明の第二の態様の好ましい実施形態では、本発明の生物は、アシル−CoA:ステロールアシルトランスフェラーゼ/ステロールO−アシルトランスフェラーゼ(EC2.3.1.26)およびアシルCoA−ワックスアルコールアシルトランスフェラーゼ(EC2.3.1.75)の活性が、対応する非改変生物、好ましくは対応する野生型生物と比較し低減されるように改変される。
【0017】
本発明の第二の態様に関する本発明の生物は、ステリルアシルエステルおよびワックスエステルの合成が低減されている、好ましくは完全に排除されているので、脂質粒子内に主としてトリアシルグリセロールを蓄積する。従って、このような生物は、特に工業規模で、トリアシルグリセロールを純粋な形態で生産するのに好適である。
【0018】
本発明の第二の態様の生物は、好ましくは、ワックスエステルを本来生産し、それらを細胞の脂質粒子に蓄積する生物に由来する。ワックスエステルを生産し、貯蔵する生物は、例えば、表面の脱水を最小限にするために組織の疎水性被膜を設けるために用いられる植物および昆虫、藻類、プランクトン生物および細菌である。本発明の第二の態様の好ましい実施形態では、生物は細菌生物、好ましくは、マイコバクテリア(Mycobacterium)、ストレプトミセス(Streptomyces)、ロドコッカス(Rhodococcus)、ノカルジア(Nocardia)、バチルス(Bacillus)、コリネバクテリウム(Corynebacterium)、エシェリキア(Escherichia)または乳酸桿菌(Lactobacillus)属のものである。最も好ましくは、生物は大腸菌(Escherichia coli)である。
【0019】
さらに、このような本発明の生物では、その生物で本来生産されるトリアシルグリセロールを、またはその生物で本来生産される量で生産することが可能であるだけでなく、このような生物を該生物で本来生産される1以上の特定のトリアシルグリセロールの量を増大させるように遺伝的に改変すること、またはこのような生物をこのような生物によって本来生産されない1以上のトリアシルグリセロールの合成をもたらすように遺伝的に改変することも可能である。
【0020】
特に好ましい一実施形態では、本発明の生物によって蓄積されるトリアシルグリセロールは、グリセロールと少なくとも1つの不飽和脂肪酸とのエステルであり、より好ましくは、該不飽和脂肪酸はω−3脂肪酸である。原則として、該ω−3脂肪酸は存在し得る任意のω−3脂肪酸であり得るが、好ましくは、α−リノレン酸(ALA)、ステアリドン酸、エイコサテトラエン酸、エイコサペンタエン酸(EPA)、ドコサペンタエン酸(DPA)またはドコサヘキサエン酸(DHA)である。
【0021】
別の好ましい実施形態では、トリアシルグリセロール中の不飽和脂肪酸は、ω−6脂肪酸である。原則として、該ω−6脂肪酸は存在し得る任意のω−6脂肪酸であり得るが、好ましくは、リノール酸、γ−リノレン酸、エイコサジエン酸、ジホモ−γ−リノレン酸、アラキドン酸、ドコサジエン酸、アドレン酸、ドコサペンタエン酸またはカレンド酸である。
【0022】
別の好ましい実施形態では、トリアシルグリセロール中の不飽和脂肪酸は、ω−9脂肪酸である。原則として、ω−9脂肪酸は存在し得る任意のω−9脂肪酸であり得るが、好ましくは、オレイン酸、エイコセン酸、ミード酸、エルカ酸またはネルボン酸である。
【0023】
第一および第二の実施形態は、以下に記載されるようにさらに改変することができる。トリアシルグリセロールは一般に、ジアシルグリセロールと長鎖脂肪酸またはリン脂質のエステル化によって形成される。長鎖脂肪酸とのエステル化は、ジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)によって触媒される。リン脂質とのエステル化は、レシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)によって触媒される。特に好ましい実施形態では、本発明の第一または第二の態様の生物は、ジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)の活性および/またはレシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)の活性が、対応する非改変生物、好ましくは対応する野生型生物と比較して増強されていることを特徴とする。上述の2つの酵素の一方または両方の活性の増強は、生物の細胞内でのTAG合成の増大をもたらす。
【0024】
この他、生物がTGAに組み込まれるある特定の脂肪酸を生産可能となるように、またはそれらがこのような生物ですでに本来生産されている場合にはこのような脂肪酸の生産を増大させてその生物で生産されるTGAにおけるそれらの含量を増大させるように生物を遺伝的に改変することも可能である。例えば、生物で生産されるTGAにおいてエイコサペンタエン酸(EPA)、ドコサペンタエン酸(DPA)および/またはドコサヘキサン酸(DHA)などのω−3脂肪酸の量を増大させることが意図される場合には、Δ12脂肪酸デサチュラーゼ(EC1.14.19)、Δ15−デサチュラーゼ(EC1.14.19)、Δ6脂肪酸デサチュラーゼ(EC1.14.19.3)、脂肪エロンガーゼ、Δ5−デサチュラーゼ/Δ5−脂肪酸デサチュラーゼ(EC1.14.19)およびΔ4脂肪酸デサチュラーゼ(EC1.14.19)の酵素活性を増強することが有利であろう。よって、好ましい実施形態では、生物(organis)は、ジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)および/またはレシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)の活性と、Δ12脂肪酸デサチュラーゼ(EC1.14.19)、Δ15−デサチュラーゼ(EC1.14.19)、Δ6脂肪酸デサチュラーゼ(EC1.14.19.3)、脂肪酸エロンガーゼ、Δ5−デサチュラーゼ/Δ5−脂肪酸デサチュラーゼ(EC1.14.19)およびΔ4脂肪酸デサチュラーゼ(EC1.14.19)からなる群から選択される少なくとも1つの活性が、対応する非改変生物、好ましくは対応する野生型生物と比較して増強されていることを特徴とする。例えば、植物およびいくつかの真菌はω−3およびω−6脂肪酸を生産することができるが、例えばサッカロミセス・セレビシエは、ω−9脂肪酸のみ合成可能である。よって、本発明の生物がサッカロミセス・セレビシエである場合、ω−3脂肪酸および/またはω−6脂肪酸の生産に必要な酵素活性を細胞に導入することが望ましいと思われる。対応する遺伝子を酵母細胞に導入するためのアプローチは文献(例えば、国際公開第2004/101575号)に対応する遺伝子とともにすでに記載されている。特定の脂肪酸の、例えば酵母細胞での異種生合成は、例えば、Beaudoin et al. (Proc. Natl. Acad. Sci. USA 97(2000), 6421−6426)およびVeen et al. (Appl. Microbiol. Biotechnol. 63(2004), 635−646)に記載されている。
【0025】
本発明はまた、本発明の第一または第二の態様の生物を培養することを含む、トリアシルグリセロールの生産方法に関する。上記のように、これらの生物は、それらの脂質粒子内に本質的に純粋な形態で多量のトリアシルグリセロールを蓄積することができる。これらのTAGは、当技術分野で周知の方法に従って生物から単離することができる。例えば、細胞から脂質粒子を単離する方法およびこれらからその中に含まれている中性脂質を単離する方法は当技術分野で公知である。このような方法の例は、添付の実施例においてサッカロミセス・セレビシエ生物体に関して記載されている。
【0026】
本発明はまた、グリセロールと少なくとも1つの不飽和脂肪酸とのエステルであるトリアシルグリセロールの生産方法に関し、その方法は、
(i)阻害剤により、アシル−CoA:ステロールアシル−トランスフェラーゼ/ステロールO−アシルトランスフェラーゼ(EC2.3.1.26)および/またはアシルCoA−ワックスアルコールアシルトランスフェラーゼ(EC2.3.1.75)の活性を低減または排除すること;および
(ii)生物においてジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)および/またはレシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)の活性を、対応する非改変生物、好ましくは対応する野生型生物と比較して増強することを含む。この生物を培養し、TAGを単離することができる。
【0027】
生産されるトリアシルグリセロールに含まれる不飽和脂肪酸に関しては、好ましい実施形態では、ポリ不飽和脂肪酸の性質および増強される酵素活性の性質に関してすでに上記したものと同じことが当てはまる。特に好ましい実施形態では、ポリ不飽和脂肪酸はω−3脂肪酸であり、増強される酵素活性は、本発明の上記でω−3脂肪酸に関して詳しく記載されているものの少なくとも1つである。
【0028】
第三の実施形態では、本発明の生物は、トリアシルグリセロール(TAG)およびステリルアシルエステル(SAE)をもはや合成しないように改変される。上記のように、一般に、TAGの合成はジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)およびレシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)の酵素活性によって遂行される。よって、好ましい実施形態では、本発明の生物は、アシル−CoA:ステロールアシルトランスフェラーゼ/ステロールO−アシルトランスフェラーゼ(EC2.3.1.26)、ジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)およびレシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)の活性が、対応する非改変生物、好ましくは対応する野生型生物と比較して低減されるように改変される。
【0029】
好ましい実施形態では、本発明の第三の態様の生物は、ワックスエステルを本来生産および貯蔵する生物に由来する生物である。このような生物はすでに上記されている。好ましくは、生物はすでに上記されている細菌生物であり、最も好ましくは大腸菌である。TAGおよびSAEの合成が排除されているために、このような生物は主としてその脂質粒子内にワックスエステルを蓄積するのに好適である。特に好ましい実施形態では、このような生物はさらに、対応する非改変株、好ましくは対応する野生型株と比較してアシル−CoAワックスアルコールアシルトランスフェラーゼ(EC2.3.1.75)の活性に増強を示すことを特徴とする。これにより、生物において合成されるワックスエステルの量を増大されることが可能となる。生物の細胞内で所望の酵素の活性を増強するための手段および方法は当業者に公知であり、本明細書の以下に詳しく記載される。
【0030】
別の好ましい実施形態では、本発明の第三の態様の生物は、ワックスエステルを本来生産および貯蔵しない生物に由来する。このような生物は本発明の第一の態様に関して上記されている。好ましくは、このような生物は上記のような真菌生物、より好ましくはサッカロミセス属、最も好ましくはサッカロミセス・セレビシエ種である。TAGおよびSAEの合成が排除されているために、このような生物の脂質粒子は、以下に詳しく記載されるように、目的の脂質、例えば、スクアレンを蓄積するために使用することができる。
【0031】
第四の実施形態では、本発明の生物は、ワックスエステル(WE)、トリアシルグリセロール(TAG)およびステリルアシルエステル(SAE)をもはや合成しないように改変される。この態様では、生物は本質的にその脂質粒子内にいずれのWE、TAGおよびSEも貯蔵しない。このような生物は、その脂質粒子に、特に望まれる脂質を向けることができる生物を設計するのに特に有用である。例えば、このような生物に、後に脂質粒子に蓄積される別のタイプの脂質の合成を可能とする核酸分子を導入することができる。これにより、脂質粒子における本質的に純粋な目的脂質の合成を可能とする生物を提供することができる。
【0032】
好ましい実施形態では、本発明の第四の態様の生物は、アシル−CoA:ステロールアシルトランスフェラーゼ/ステロールO−アシルトランスフェラーゼ(EC2.3.1.26)、ジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)、レシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)およびアシルCoA−ワックスアルコールアシルトランスフェラーゼ(EC2.3.1.75)の活性が、対応する非改変生物、好ましくは対応する野生型生物と比較して低減されるように改変される。
【0033】
第五の実施形態では、本発明の生物は、細胞において脂質粒子内に蓄積される中性脂質を生産し、かつ、トリアシルグリセロール(TAG)をもはや合成しないように遺伝的に改変された、従って、該粒子において、その脂質粒子内に本来存在する該トリアシルグリセロールをもはや含まない生物である。上記のように一般に、TAGの合成は、例えば酵母においては、ジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)およびレシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)の酵素活性によって遂行される。よって、好ましい実施形態では、本発明の第五の態様の生物は、ジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)およびレシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)の活性が、対応する非改変生物、好ましくは対応する野生型生物と比較して低減されるように遺伝的に改変される。
【0034】
本発明の第五の態様の好ましい実施形態では、生物は、ワックスエステルを本来生産せず、その脂質粒子内に貯蔵しない生物に由来する。このような生物は上記されている。好ましくは、生物は上記のような真菌生物、より好ましくはサッカロミセス属、最も好ましくはサッカロミセス・セレビシエ種である。
【0035】
第六の実施形態では、本発明の生物は、細胞において脂質粒子内に蓄積される中性脂質を生産し、かつ、ワックスエステル(WE)およびトリアシルグリセロール(TAG)をもはや合成しないように遺伝的に改変された、従って、該粒子において、その脂質粒子内に本来存在するトリアシルグリセロールおよびワックスエステルをもはや含まない生物である。好ましい実施形態では、本発明の第六の態様の生物は、ジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)、レシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)およびアシルCoA−ワックスアルコールアシルトランスフェラーゼ(EC2.3.1.75)の活性が、対応する非改変生物、好ましくは対応する野生型生物と比較して低減されるように遺伝的に改変される。
【0036】
好ましい実施形態では、本発明の第五または第六の態様の生物、すなわち、トリアシルグリセロールをもはや合成しない、またはトリアシルグリセロールおよびワックスエステルをもはや合成しない生物は、その脂質粒子内にステロール脂質のアシルエステルを蓄積することができる。脂質粒子におけるステロール脂質のアシルエステルの蓄積は、その生物がこのようなエステルを本来合成するという事実によるものであり得、またはそれはステロール脂質のアシルエステルの生合成をもたらす対応する酵素をコードする遺伝子がそれらの細胞に導入されているという事実によるものであり得る。
【0037】
コレステロールおよびその誘導体などのステロール脂質は、グリセロリン脂質およびスフィンゴミエリンとともに膜脂質の重要な成分である。これらのステロイドは、同じ縮合4環コア構造も含むが、ホルモンおよびシグナル伝達分子として異なる生物学的役割を有する。C18ステロイドはエストロゲンファミリーを含み、C19ステロイドはテストステロンおよびアンドロステロンなどのアンドロゲンを含む。C21サブクラスは、プロゲストゲンならびに糖質コルチコイドおよび鉱質コルチコイドを含む。種々の形態のビタミンDを含むセコステロイドは、コア構造のB環の開裂を特徴とする。ステロールの他の例は胆汁酸およびそれらの結合体であり、これらは哺乳類ではコレステロールの酸化誘導体であり、肝臓で合成される。
【0038】
好ましい実施形態では、ステロール脂質はステロールまたはステロイドである。ステロイドは、一般に6−6−6−5様式で配置された4つの縮合環を有する炭素主鎖を特徴とするテルペノイド脂質である。ステロイドはこれらの環と結合している官能基およびこれらの環の酸化状態によって異なる。植物、動物および真菌には、何百もの異なるステロイドが見られる。全てのステロイドが細胞においてステロールラノステロール(動物および真菌)またはステロールシクロアルテノール(植物)のいずれかから製造される。両ステロールとも、トリテルペンスクアレンの環化に由来する。ステロイドには、エストロゲン、プロゲステロンおよびテストステロンが含まれる。
【0039】
ステロール(またはステロイドアルコール)は、A環の3位にヒドロキシル基を有するステロイドのサブグループである。それらはアセチル補酵素A(acetyl−conenzyme A)から合成される両親媒性脂質である。この分子全体は極めて平坦である。A環上のヒドロキシル基には極性がある。この脂肪鎖の残りの部分は非極性である。植物のステロールは植物ステロール(phytosterols)と呼ばれ、動物のステロールは動物ステロール(zoosterols)と呼ばれる。最も重要な動物ステロールはコレステロールおよびいくつかのステロイドホルモンであり、最も重要な植物ステロールはカンペステロール、シトステロールおよびスチグマステロールである。ステロールは、真核生物の生理に不可欠な役割を果たす。例えば、コレステロールは細胞膜の一部を形成し、その存在は細胞膜の流動性に影響を及ぼし、発生シグナル伝達における第二のメッセンジャーとして働く。植物ステロールはまた、ヒトの腸においてコレステロール吸収部位を遮断することも知られており、従って、ヒトにおいてコレステロールの低減を助ける。ヒトでは、ステロールは、例えば、日内周期、血液凝固などの重要なシグナルおよび代謝コミュニケーションを提供する働きをする。
【0040】
好ましい実施形態では、ステロールはコレステロール、コレステロール誘導体、7−デヒドロ−コレステロール、ラノステロール、ラノステロール誘導体、チモステロール、チモステロール誘導体、ラトステロール、ラトステロール誘導体、ククルビタシン、ククルバチン(cucurbatin)誘導体、エピステロール、エピステロール誘導体、テアステロン、テアステロン誘導体、カスタステロン、カスタステロン誘導体、チファステロール、チファステロール誘導体、カタステロン、カタステロン誘導体、シクロオイカレノール、シクロオイカレノール誘導体、シトステロール、シトステロール誘導体、イソフコステロール、イソフコステロール誘導体、フコステロール、フコステロール誘導体、ゴルゴステロール、ゴルゴステロール誘導体、エルゴステロール、エルゴステロール誘導体、スチグマステロールまたはスチグマステロール誘導体である。
【0041】
特に、7−デヒドロ−コレステロールのアシルエステルが蓄積される生物では、HMG−CoA−レダクターゼ(EC1.1.1.34)および/またはΔ24−レダクターゼおよび/またはラノステロールC14−デメチラーゼ/シトクロムP450 51(EC1.14.13.70)および/またはスクアレン−エポキシダーゼ/スクアレン−モノオキシゲナーゼ(EC1.14.99.7)の活性を、対応する野生型生物と比較して増強することができる。さらに、SAM:C−24ステロールメチルトランスフェラーゼおよび/またはC−22ステロールデサチュラーゼおよび/またはC−5ステロールデサチュラーゼの活性を、対応する野生型生物と比較して低減または排除することができる。エルゴステロールのアシルエステルが蓄積される生物では、HMG−CoA−レダクターゼ(EC1.1.1.34)および/またはラノステロールC14−デメチラーゼ/シトクロムP450 51(EC1.14.13.70)および/またはスクアレン−エポキシダーゼ/スクアレン−モノオキシゲナーゼ(EC1.14.99.7)の活性を、対応する野生型生物と比較して増強することができる。エピステロールのアシルエステルが蓄積される生物では、HMG−CoA−レダクターゼ(EC1.1.1.34)および/またはラノステロールC14−デメチラーゼ/シトクロムP450 51(EC1.14.13.70)および/またはスクアレン−エポキシダーゼ/スクアレン−モノオキシゲナーゼ(EC1.14.99.7)の活性を、対応する野生型生物と比較して増強することができ、かつ/またはΔ22−デサチュラーゼ/シトクロムP450 61(EC1.14.14.1)活性を、対応する野生型生物と比較して低減することができる。
【0042】
別の好ましい実施形態では、ステロイドはアンドロステロン、アンドロステロン誘導体、テストステロン、テストステロン誘導体、アンドロステンジオール、アンドロステンジオール誘導体、アンドロステンジオン、アンドロステンジオン誘導体、カルステロン、カルステロン誘導体、メタンドリオール、メタンドリオール誘導体、ボラステロン、ボラステロン誘導体、エピアンドロステロン、エピアンドロステロン誘導体、メスタノロン、メスタノロン誘導体、スタノロン、スタノロン誘導体、ステンボロン、ステンボロン誘導体、エピテストステロン、エピテストステロン誘導体、コルチゾール、コルチゾール誘導体、アルドステロン、アルドステロン誘導体、プレグネノロン、プレグネノロン誘導体、コルチゾン、コルチゾン誘導体、コルチコステロン、コルチコステロン誘導体、ノレシンドロン、ノレシンドロン誘導体、ウロコルチゾールまたはウロコルチゾール誘導体である。
【0043】
本発明の第五および第六の態様の好ましい実施形態では、ステロール脂質のアシルエステルを蓄積することができる生物は、個々のステロール脂質の合成に至る生合成経路を本来保有し、より好ましくは、個々のステロール脂質のより高い蓄積を得るために、対応する経路の酵素活性が対応する非改変生物と比較して増強されている生物である。
【0044】
好ましい実施形態では、本発明の態様1〜6の生物は、その脂質粒子内にプレノール脂質を蓄積することができる。プレノール脂質は、主としてメバロン酸(MVA)経路を介して生産される5炭素前駆体であるイソペンテニル二リン酸およびジメチルアリル二リン酸から合成される。プレノール脂質はイソプレノイド、キノンおよびヒドロキノン、ポリプレノール、ホパノイドおよび他のいくつかの希少なタイプを包含する。単純イソプレノイド(直鎖アルコール、二リン酸塩など)はC5単位の連続的付加によって形成され、これらのテルペン単位の数によって分類される。40を超える炭素を含む構造はポリテルペンとして知られる。カロテノイドは、抗酸化剤としておよびビタミンA前駆体として働く重要な単純イソプレノイドである。別の生物学的に重要なクラスの分子としては、非イソプレノイド起源のキノノイドコアと結合したイソプレノイドテールを含むキノンおよびヒドロキノンが挙げられる。ビタミンEおよびビタミンKならびにユビキノンがこのクラスの例である。細菌は、酸素と結合している末端のイソプレノイド単位が不飽和のままであるポリプレノール(バクトプレノールと呼ばれる)を合成するが、動物ポリプレノール(ドリコール)では、末端イソプレノイドは還元されている。
【0045】
好ましい実施形態では、脂質粒子内に蓄積されるプレノール脂質はイソプレノイドである。イソプレノイドとしては、例えば、C5イソプレノイド、C10イソプレノイド(モノテルペン)、C15イソプレノイド(セキテルペン)、C20イソプレノイド(ジテルペン)、C25イソプレノイド(セステルテルペン)、C30イソプレノイド(トリテルペン)、C40イソプレノイド(テトラテルペン)、ポリテルペンおよびレチノイドが挙げられる。
【0046】
特に好ましい実施形態では、イソプレノイドはトリテルペンである。トリテルペンは、6つのイソプレン単位からなり、基本分子式C30H48ー50を有する。この群は、例えば3S−スクアレン−2,3−エポキシド、スクアレン、プレスクアレン二リン酸、テトラヒマノール、α−アミリン、β−アミリン、ルペオール、酢酸ルペオール、タラキサステロール、アザジラクチンA、ネオクアッシン、クアッシンおよび3−アセチル−1−チグロイルアザジラクチニンを包含する。サメ肝油の成分である直鎖トリテルペンスクアレンは、2分子のピロリン酸ファルネシルの還元的結合から誘導される。その後、スクアレンは生合成的にプロセシングされ、全てのステロイドの構造前駆体であるラノステロールまたはシクロアルテノールのいずれかを生成する。
【0047】
特に好ましい実施形態では、脂質粒子内に蓄積されるプレノール脂質はスクアレンまたはスクアレン誘導体である。スクアレン誘導体は、スクアレンの場合には主鎖炭素原子に結合している水素原子の代わりに、主鎖炭素原子に結合している1以上の、特に、1〜10個または1〜4個の、付加的メチルまたはエチル基を含む。
【0048】
特に好ましい実施形態では、本発明の生物は好ましくは、本発明の第一、第三および第五の態様の態様に関して、その脂質粒子においてワックスエステルを生産および貯蔵しない生物、例えば、上記のような真菌生物、より好ましくはサッカロミセス属の真菌生物、最も好ましくは、サッカロミセス・セレビシエである。本発明の第一、第三および第五の態様に従い、トリアシルグリセロールおよび/またはステリルアシルエステルを合成しないように改変されたこのような生物は、その脂質粒子においてプレノール脂質、特に、スクアレンを合成および貯蔵することができ、それにより、脂質粒子内でこのようなプレノール脂質を本質的に純粋またはより高い純度の形態で生産することができる。よって、酵母生物においてTAGおよび/またはSAEの生合成のための合成経路を排除することにより、このような生物によって生産されるスクアレンの量および純度に関して生産を劇的に改善することがすでに可能である。
【0049】
このような生物におけるスクアレン生産のさらなる改善は、スクアレンの合成をもたらす酵素活性を増強し、かつ、スクアレンを他の化合物に変換する経路の酵素活性を低減するように生物の代謝を改変することによって達成することができる。
【0050】
スクアレン生合成経路の遺伝子は既知であり、例えば酵母S.セレビシエにおいてクローニングされている。主要なボトルネック酵素はHMG−CoA−レダクターゼ(HMG1)(Basson et al. (Mol. Cell. Biol. 8 (1988), 3793−3808)。よって、好ましい実施形態では、本発明の第一、第三または第五の態様の生物は脂質粒子内にスクアレンを蓄積することができ、かつ、HMG−CoA−レダクターゼ(EC1.1.1.34)の活性が対応する非改変生物、好ましくは対応する野生型生物と比較して増強されていることを特徴とする。HMG−CoA−レダクターゼ活性の増強は、当業者に周知の手段および方法によって達成することがき、それらは以下にもさらに詳細に記載される。好ましい実施形態では、HMG−CoA−レダクターゼの活性の増強は、その生物において、その酵素の触媒領域のみをコードし、膜結合ドメインはコードしないHMG−CoA−レダクターゼ遺伝子を発現させることによって達成される。このような変更は欧州特許第EP−A486290にすでに記載されている。この改変により、エルゴステロール生合成経路の中間体によるHMG−CoA−レダクターゼのフィードバック調節が回避される。別の好ましい実施形態では、本発明の生物においてHMG−CoA−レダクターゼをコードする遺伝子を、異種プロモーター、すなわち、HMG−CoA−レダクターゼ遺伝子にとって外来のプロモーターの制御下に置くが、特に、このプロモーターは、エルゴステロール生合成経路の中間体によってはその活性が調節されないプロモーターである。好適なプロモーターの例はADH1プロモーター、特に、おおよそ構成的発現を示すADH1「アベレージ」プロモーターである(Ruohonen et al., Journal of Biotechnology 39 (1995), 193−203)。
【0051】
酵母などのいくつかの生物では、スクアレンは他の化合物、特に、エルゴステロールに、また、エルゴステロール生合成経の中間体に変換され得る。ステロールエルゴステロール((22E)−エルゴスタ−5,7,22−トリエン−3−β−オール)の形成をもたらす化学反応および経路が麦角、酵母および糸状菌に見られる。これらの生物では、スクアレンはスクアレンモノオキシゲナーゼ/スクアレンエポキシダーゼ(EC1.14.99.7;スクアレンエポキシダーゼまたはERG1とも呼ばれる)によって(S)−2,3−エポキシスクアレンに変換され、これは次に2,3−オキシドスクアレン−ラノステロールシクライズ(cyclise)(EC5.4.99.7;ERG7)によってラノステロールに変換される。ラノステロールは次にシトクロムP450ラノステロール14a−デメチラーゼ(EC1.14.13.70;ERG11)によって、NADPHおよびO2を用いて、4,4−ジメチル−コレスタ−8,14,24−トリエノールに変換され、その後、C14ステロールレダクターゼ(EC1.3.1.70;ERG24)によって、NADPHを用いて、4,4−ジメチル−8,24−コレスタジエノールに変換される。この化合物は次にC−4ステロールメチルオキシダーゼ(EC1.14.13.72;ERG25)の作用によってさらに4−メチル−8,24−コレスタジエノールに変換される。この物質はC−3ステロールデヒドロゲナーゼ(EC1.1.1.170;ERG26)によって3−ケト−4−メチルチモステロールに変換される。次に、3−ケト−4−メチルチモステロールは、3−ケトステロールレダクターゼ(EC1.1.1.270;ERG27)によってチモステロールに変換される。チモステロールはそれ自体、次に、S−アデノシル−L−メチオニンとともに、SAM:C−24ステロールメチルトランスフェラーゼ(EC2.1.1.41;ERG6)の作用によってフェコステロールおよびS−アデノシル−ホモシステインに変換される。フェコステロールは次にC−8ステロールイソメラーゼ(ERG2)の作用によってエピステロールに変換され、その後、C−5ステロールデサチュラーゼ(EC1.14.21.6;ERG3)の作用によって、NADPHおよびO2を用いて、5,7,24(28)−エルゴスタトリエノールに変換される。この化合物はさらにC−22ステロールデサチュラーゼ(EC1.14.14;ERG5)の作用によって5,7,22,24(28)−エルゴスタテトラエノールに変換される。5,7,22,24(28)−エルゴスタテトラエノールは次にC−24ステロールレダクターゼ(EC1.3.1.71;ERG4)の作用によってエルゴステロールに変換される。本発明の生物が脂質粒子内にスクアレンを蓄積することが望まれる場合、また、この生物がエルゴステロール、またはスクアレンから始まるエルゴステロール経路の中間体を本来合成することができる生物に由来する場合には、その生物におけるスクアレンからエルゴステロールに至る経路の上述の酵素活性の1以上の活性を低減することが望ましいと思われる。原則として、上記エルゴステロール経路の上述の酵素のいずれか1つ、2以上または全てを低減することができる。好ましい実施形態では、スクアレンモノオキシゲナーゼ(EC1.14.99.7;スクアレンエポキシダーゼまたはERG1とも呼ばれる)の活性が低減されるが、一部のステロール合成は酵母細胞の生存に必要であるので完全には排除されない。また、エルゴステロールの生合成に関与する以下の酵素:SAM:C−24ステロールメチルトランスフェラーゼ(EC2.1.1.41)、C−22ステロールデサチュラーゼ(EC1.14.14)およびC−5ステロールデサチュラーゼ(EC1.14.21.6)の1以上の活性を低減することも可能である。このことは全てにおいて本発明の上述のいずれの実施形態にも当てはまる。
【0052】
本発明の一部として、しかしながら独立に、遺伝的に改変された生物に関して以下の特徴を適用する。
【0053】
本発明の範囲内で、酵母サッカロミセス・セレビシエにおける遺伝子SAK1および/またはHAP4の構成的過剰発現が脂質、特にスクアレンおよびステロールの生産性の著しい増大をもたらすことも判明した。遺伝子SAK1およびHAP4は呼吸−発酵フラックス分布に関連づけられる。
【0054】
遺伝子SAK1は、Snf1p/Snf4p複合体のリン酸化を担う上流セリン/トレオニンキナーゼをコードする。このリン酸化は、核に局在される活性なSnf1p/Snf4p複合体をもたらす。Snf1p/Snf4p複合体はタンパク質セリン/トレオニンキナーゼに属し、「ジオーキシーシフト(diauxic shift)」の際のグルコース抑制の軽減に中心的役割を果たす。このジオーキシーシフトとは、培地中のグルコースまたは他の発酵炭素源が消耗された際の発酵代謝から呼吸代謝への移行を表す。Snf1/Snf4複合体はそれぞれグルコース抑制および抑制解除を担うカスケードの階層を遙かに超える範囲である。ジオーキシーシフトの過程では、サッカロミセス・セレビシエの6000の遺伝子の約4分の1の発現レベルが有意に変化を受ける。これらの主要な変化は主として、発酵モードから呼吸モードへと移行される中枢代謝に携わる。発酵モードでは、酵母細胞はグルコースのような発酵可能な炭素源をエタノールと二酸化炭素へと代謝する。呼吸モードでは、生産されたエタノールが呼吸されて、細胞においてエネルギーを供給するアデノシン三リン酸(ATP)が得られる。ジオーキシーシフトの過程で、Snf1p/Snf4p複合体はリン酸化され、従って、Snf1−キナーゼSak1p、Tos1pおよびElm1pによって活性化される。活性化された複合体は核に局在され、そこでそれは2つの転写因子Mig1pおよびCat8pに影響を及ぼす。ジンクフィンガータンパク質Mig1pは転写レプレッサーであり、複合体としての多数のグルコース抑制遺伝子の特定のコンセンサス配列と結合するためにタンパク質Tup1pおよびCyc8pを動員し、それにより、このコンセンサス配列の下流の遺伝子の転写が抑制される。Cat8pは転写アクチベーターであり、培地中のグルコースが枯渇した際に少なくとも34の遺伝子の発現を誘導する。これらの遺伝子は主として、エタノールまたはアセテートのようなC2化合物の代謝に必要なグリオキシル酸シャントの遺伝子である。これに加え、クエン酸回路およびグリオキシル酸シャントの中間体の細胞内輸送を担う遺伝子もCat8pによって調節される。活性なSnf1p/Snf4p複合体はMig1pをリン酸化し、それによりこのタンパク質は不活性化され、核の外へ転流され、機能的ホモログであるCat8pおよびSip1pをリン酸化し、これがこれら2つのタンパク質の不活性をもたらす。従って、核に局在された活性なSnf1p/Snf4p複合体は、抑制カスケードのいくつかの転写因子に影響を及ぼすことから、グルコース抑制の軽減および発酵から呼吸への移行に大きく寄与する。よって、本発明の生物は、グルコースの存在下であってもより活性なSnf1p/Snf4p複合体を呈するようにさらに発達させることができる。この改変は、主要キナーゼとしてSnf1p/Snf4p複合体のリン酸化、従って、活性化を担うタンパク質Sak1pの転写脱調節によって達成される。
【0055】
Hap2/3/4/5タンパク質複合体は、多数のグルコース抑制遺伝子を調節する。これらは呼吸鎖およびクエン酸回路の主要な遺伝子である。この複合体はジオーキシーシフトの際にアクチベーターとしてこれらの遺伝子の転写を誘導し、呼吸−発酵バランスの呼吸方向へのシフトに強く寄与する。遺伝子HAP2、HAP3およびHAP5は構成的に発現される。本来、HAP4は、非発酵性炭素源での増殖の場合にのみ発現される。HAP4過剰発現変異株の培養の際に、この突然変異は増殖速度の増大、バイオマスおよび酢酸の生産、ならびにグリセロールおよびエタノールの生成の低下をもたらすのが見られた。Hap2/3/4/5タンパク質複合体は、呼吸能、ミトコンドリアの生合成およびクエン酸回路の炭素フラックスを高めることによって呼吸系を拡張する。
【0056】
本発明の範囲内での別の観察として、遺伝子FLD1の欠失が、酵母、特に酵母サッカロミセス・セレビシエのような生物において脂質生産、特に、スクアレン生産の予期できない著しい増大をもたらすというものがある。遺伝子FLD1は、脂質滴の形態、数および大きさに関与するタンパク質セイピンをコードする。FLD1の過剰発現は脂質粒子の融合をもたらし、その結果、酵母細胞において著しく拡大した、しかしながら数は少ない脂質粒子が形成されることが報告されている。野生型酵母株では、スクアレンはステロール生合成経路の中間体であり、細胞に蓄積されない。酵母において、欧州特許第EP−A486290号に記載されるように酵素の触媒部分のみをコードし、膜結合ドメインはコードしないHMG−CoA−レダクターゼ遺伝子を発現させることによって達成されるHMG−CoA−レダクターゼ活性の増強は、細胞においてスクアレンの蓄積をもたらす。この蓄積は、エルゴステロール生合成経路の中間体によるHMG−CoA−レダクターゼのフィードバック調節の回避によるものである。本発明の枠組みの範囲で、遺伝子FLD1の欠失は、HMG−CoA−レダクターゼ活性の増強を示す酵母株において、脂質生産、特にスクアレン生産の予期できない著しい増大をもたらすことが認められた。
【0057】
SAK1、HAP4およびFLD1に関する上記の発見は、上記のような本発明の6つの態様のいずれにおいても、さらなる改変および実施形態をもたらす。しかしながら、これらの発見は、上記の改変を含まないが、個々の非改変生物によって典型的に合成されるあらゆる脂質を合成する能力を有する生物にも独立した価値を持つ。これに関して、非改変生物は、野生型の他、対応する野生型生物と比較して高いHMG−CoA−レダクターゼ(EC1.1.1.34)活性を有する生物を含む。
【0058】
よって、本発明はまた、遺伝子HAP4および/またはSAK1が、転写において脱調節され、かつ/または過剰発現され、かつ/また遺伝子FLD1が抑制もしくは不活性化もしくは欠失された(全て、対応する野生型生物と比較して)、単離された遺伝的に改変された非哺乳類生物を含む。具体的には、遺伝子HAP4およびSAK1の一方または双方を(所望により)ADH1プロモーターのような構成的に活性なプロモーターの制御下に置くことができる。この変形形態のさらなる実施形態では、遺伝子REG1を、対応する野生型生物と比較して抑制または不活性化または欠失させる。この他、本発明に関してこれまでに述べたことも全て、同様にこの独立した態様に当てはまり、これまでに開示された特徴はいずれも、同様にこの独立した態様に組み込むことができる。
【0059】
本発明はまた、第二の脂質を生産する方法に関し、その方法では、本発明の生物を用い、培養し、その生物から第二の脂質を単離する。例えば、プレノール脂質を生産するための本発明の方法は、その脂質粒子内にプレノール脂質を蓄積することができる、上記のような本発明の第三または第四の態様の生物を培養することを含む。蓄積されるプレノール脂質に関しては、好ましい実施形態では、本発明の第三および第四の態様の生物に関して上記したものと同じことが当てはまる。対応する生物において低減または増強される酵素活性についても同じことが言える。別の例として、ステリルアシルエステルを生産する方法は、本発明の第五または第六の態様による生物を培養することを含む。
【0060】
上記のようなタンパク質または酵素の活性における、対応する非改変生物と比較しての低減は、好ましくは少なくとも50%、より好ましくは少なくとも75%、いっそうより好ましくは少なくとも80%、特に好ましくは少なくとも90%、最も好ましくは100%である。100%の低減は、該生物に存在する該酵素のタンパク質または酵素活性が存在しないことを意味する。上記のようなタンパク質または酵素の活性における、対応する非改変生物と比較しての増強は、好ましくは少なくとも10%、より好ましくは少なくとも50%、いっそうより好ましくは少なくとも200%、特に好ましくは少なくとも1000%である。「増強する」とは、また、非改変生物が測定可能なこのような活性を含まない場合の、酵素またはタンパク質の酵素活性(測定できる場合はもちろん)の存在も含む。「活性が増強する」とは、さらに、タンパク質が酵素ではないあらゆる場合において(HAP4など)、タンパク質の量の増加を包含する。そして、上記の増強のパラメーターは同様に量にも当てはまる。酵素活性は、所定量の抽出物に、その抽出物から生成物への反応を触媒する所定量の酵素および所望によりさらに必要な反応成分を加えること、および所定時間内に合成された生成物の量を測定することによって測定することができる。上述の酵素の活性の具体的な測定方法は、例えば、国際公開第03/064650A1号に開示されている。その文献に記載されていない上述の酵素の活性の測定も同様に実施される。
【0061】
細胞において、ある酵素の活性を低減する方法は、例えば、その酵素をコードする遺伝子の遺伝子発現を低減または排除することであり、例えば、対応する遺伝子の上流により弱いプロモーターを付与することによるか、またはその遺伝子および/もしくは関連のプロモーターを完全もしくは部分的に欠失させ、かつ/もしくは生物の細胞内に酵素阻害剤を加えて翻訳される酵素を阻害する、かつ/または細胞にsiRNAを組み込んで活性な転写物の量を低減する、かつ/またはその遺伝子の突然変異によって活性の低い変異体を生成することによる。
【0062】
遺伝子発現の低減とは、個々の酵素をコードするヌクレオチド配列の遺伝子発現のレベルが、対応する非改変生物、好ましくは対応する野生型生物における該ヌクレオチド配列の遺伝子発現レベルと比較して低減されることを意味する。活性に関して上記に示したものと同じレベルの低減を適用する。遺伝子発現レベルを低減する手段および方法は、例えば、合成される対応するmRNAもしくはタンパク質の量の決定、または個々のタンパク質の酵素活性の決定を含む。好ましい実施形態では、遺伝子発現のレベルは、例えばノーザンブロットにおいてmRNAの量を測定することによって決定される。別の好ましい実施形態では、遺伝子発現のレベルは、例えばウエスタンブロットにおいて合成される対応するタンパク質の量を測定することにより、または対応する酵素活性の量を決定することによって決定される。遺伝子発現の低減はまた、着目する遺伝子を機能しないようにすることによって達成することもできる。遺伝子を機能しないようにするための1つの可能性のある手段が遺伝子破壊である。
【0063】
酵素の活性を増強する方法は、その酵素をコードする遺伝子(異種または同種)が構成的に活性な(同種または異種)プロモーターの制御下に配置されるように、かつ/またはその酵素をコードする(異種または同種)遺伝子のコピー数が増え、かつ/もしくは突然変異によって活性が増強されるように転写の脱調節が達成されるような細胞の形質転換を含む。
【0064】
酵素活性の増強または低減を果たすための具体例を実施例に示す。しかしながら、当業者ならば、本明細書で特に開示する必要なく、当技術分野で周知の他の方法を適用することもできよう。
【0065】
本発明で用いる酵素として好適な遺伝子配列を以下に開示するが、同じ酵素活性を有する他の遺伝子配列も同様に使用可能である。特定の遺伝子配列またはそれによりコードされるタンパク質配列には本発明の範囲内の関連のある構造的特徴はないが、同じEC(Enzyme Commision)番号に分類されている。
【0066】
アシル−CoA:ステロールアシルトランスフェラーゼ/ステロールO−アシルトランスフェラーゼ(EC2.3.1.26)の遺伝子配列は、NC_001135.4、NC_001147.6、NM_005891、NM_144784、NM_153728を含む。
【0067】
ジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)の遺伝子配列は、NC_001147.5、XM_002478787、NM_123089、XM_002378082、NM_032564、NM_001012345、NM_010046、XM_002146497を含む。
【0068】
レシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)の遺伝子配列は、NC_001147.6、NM_008490、NM_001162568、NM_000229、NM_001005715、NM_017024、NM_001082190を含む。
【0069】
アシルCoA−ワックスアルコールアシルトランスフェラーゼ(EC2.3.1.75)の遺伝子配列は、NM_123089、NM_177448を含む。
【0070】
HMG−Co0A−レダクターゼの遺伝子配列は、NC_001145、NM_106299、NC_003421.2、NC_009784.1、NC_003028.3、NC_007308.3、および図5の配列(末端切断型tHMG1)を含む。
【0071】
C−24ステロールメチルトランスフェラーゼの遺伝子配列は、NC_001145、NC_000911.1、NC_003423.3、XM_505173、XM_716615を含む。
【0072】
C−22ステロールデサチュラーゼの遺伝子配列は、NC_003424.3、NC_009046.1、NC_001145.2、XM_500188、XM_711840を含む。
【0073】
C−5ステロールデサチュラーゼの遺伝子配列は、NC_001144、S46162、NG_009446、NM_053642、NM_001035356、XM_503090、XM_708519を含む。
【0074】
HAP4の遺伝子配列は、NC_001143.7、XM_448596、XM_001645329を含む。
【0075】
SAK1の遺伝子配列は、NC_001137.2、XM_502591、XM_448319、XM_453478、NM_208704を含む。
【0076】
REG1の遺伝子配列は、NC_001136.8、XM_500990、XM_448729、XM_455276を含む。
【0077】
FLD1の遺伝子配列は、NC_001144.4、NM_210286、XM_001647166、XM_449778を含む。
【0078】
7−デヒドロコレステロールレダクターゼの遺伝子配列は、NM_103926、NM_001360、NM_007856、NM_203904、NM_001014927、NM_201330、NM_022389、NM_001131727、NM_001087087、XM_001497598、XM_001174160、XM_001099101、BM490402、CA753545を含む。
【0079】
24−デヒドロコレステロールレダクターゼの遺伝子配列は、NM_014762、NM_001016800、NM_001094456、NM_001008645、NM_001103276、NM_001080148、NM_053272、NM_00103128、XM_001488247、AB125202、XM_001153751を含む。
【0080】
ラノステロールステロール14−デメチラーゼの遺伝子配列は、NC_001140.5、XM_500518、EF059165、XM_445876、XM_454109を含む。
【0081】
スクアレンモノオキシゲナーゼの遺伝子配列は、NC_001139.8、M64994、XM_503994、XM_706801、XM_455763を含む。
【0082】
好適なプロモーター(異種発現される個々の異なる酵素のプロモーターは同じまたは異なる場合がある)の遺伝子配列は、NC_001142、NC_001139、NC_001147、NC_001139、NC_001148、NC_001135、NC_001136を含む。
【0083】
一般に、本発明の生物(上記の態様のそれに関わらず)存在し得る任意の非ヒト、好ましくは非哺乳類生物であり得る。好適な生物の例は、特許請求の範囲に示されている。
【図面の簡単な説明】
【0084】
【図1】酵母サッカロミセス・セレビシエの構築された変異株由来の全脂質抽出物の薄層クロマトグラフィー。
【図2】酵母サッカロミセス・セレビシエにおけるスクアレンの生合成を模式的に示す。
【図3】酵母サッカロミセス・セレビシエにおけるトリアシルグリセロールの生合成を模式的に示す。
【図4】酵母サッカロミセス・セレビシエにおけるステリルアシルエステルの生合成を模式的に示す。
【図5】末端切断型HMG CoA−レダクターゼ、tHMG1の配列を示す。
【発明を実施するための形態】
【0085】
以下の実施例は単に本発明を説明するためのものである。
【0086】
実施例では、以下の材料および方法を用いた。
【0087】
1.制限切断
プラスミド(1〜10μg)の制限酵素処理は30μlバッチで行った。このために、24μlのH2O中にDNAを取り、3μlの対応するバッファー、1μlのRSA(ウシ血清アルブミン)および2μlの酵素と混合した。酵素濃度は、DNA量に応じて1単位/μlまたは5単位/μlとした。場合によっては、バッチにさらに1μlのRNアーゼを加えてtRNAを分解した。この制限バッチを37℃で2時間インキュベートした。制限酵素処理はミニゲルで管理した。
【0088】
2.ゲル電気泳動
ゲル電気泳動はミニゲルまたはワイドミニゲル装置で行った。ミニゲル(約20ml、8バッグ)およびワイドミニゲル(50ml、15または30バッグ)はTAE中1%のアガロースからなった。移動バッファーとして1*TAEを用いた。サンプル(10μl)と3μlの停止液を混合し、適用した。HindIIIで切断したI−DNAを標品として用いた(23.1kb;9.4kb;6.6kb;4.4kb;2.3kb;2.0kb;0.6kbにバンド)。分離のために、45〜60分間、80Vの電圧をかけた。その後、ゲルを臭化エチジウム溶液で染色し、ビデオ撮影システムINTASを用いてUV光下で保持、またはオレンジ色のフィルターを用いて写真撮影した。
【0089】
3.ゲル溶出
所望の断片を、ゲル溶出を用いて単離した。この制限調製物をミニゲルのいくつかのバッグに適用し、分離した。[λ]−HindIIIおよび「サクリファイス・トレース(sacrifice trace)」のみを臭化エチジウム溶液で染色し、UV光下で観察し、所望の断片を標識した。結果として、残りのバッグでは臭化エチジウムおよびUV光によってDNAを損傷させずにすんだ。染色されたゲル片と非染色ゲル片をアラインすることにより、非染色ゲル片から所望の断片を、その標識に基づいて切り出すことができた。単離する断片を含むこのアガロース片を透析管に入れ、少量のTAEバッファーとともに気泡が入らないように密閉し、BioRad−ミニゲル装置に入れた。移動バッファーは1*TAEからなり、40分間、電圧を100Vとした。次に、流動極性を2分間変更して透析管に付着しているDNAを遊離させた。透析管の、DNA断片を含むバッファーを反応槽に移し、エタノール沈殿法を行った。このために、DNA溶液に、一部(1/10)量の3M酢酸ナトリウム、tRNA(1μl/50μl溶液)および2.5倍の容量の氷冷96%エタノールを加えた。このバッチを−20℃で30分間インキュベートした後、4℃で30分間、12,000rpmにて遠心分離した。DNAペレットを乾燥させ、10〜50μlのH2O(DNA量による)に取った。
【0090】
4.クレノウ処理
DNA断片の突出末端をクレノウ処理により作出し、「平滑末端」を得た。DNA1μg当たり、以下のバッチをピペットで一緒に吸い取った。この場合、夾雑物がクレノウ−ポリメラーゼを阻害しないように、DNAはエタノール沈殿法からのものとするべきである。インキュベーションは37℃で30分間、その後、70℃でさらに5分間行い、反応を停止させた。DNAはこのバッチからエタノール沈殿法によって得、10μlのH2Oに取った。
【0091】
5.ライゲーション
連結させるDNA断片を合わせた。最終量13.1μlには、ベクター−インサート比1:5で約0.5μgのDNAを含んだ。このサンプルを70℃で45秒間インキュベートし、室温まで冷却し(約3分)、その後、氷上で10分間インキュベートした。次に、ライゲーションバッファー:2.6μlの500mmol TrisHCl、pH7.5および1.3μlの100mmol MgCl2を加え、それらを氷上でさらに10分間インキュベートした。1μlの500mmol DTTおよび1μlの10mmol ATPを加えた後、1μlのリガーゼ(1単位/μl)を氷上でさらに10分間加えた。この処理は全体的に、隣接するDNA末端が再び分離しないようできる限り振盪させないように行うべきである。ライゲーションは4℃で一晩行った。
【0092】
6.大腸菌の形質転換
成分大腸菌(E. coli)NM522細胞をライゲーション調製物のDNAで形質転換させた。陽性対照として、バッチに50ngのpScL3プラスミドを供給し、ヌル対照として、バッチにDNAを供給しなかった。各形質転換調製物について、100μlの8%PEG溶液、10μlのDNAおよび200μlのコンピテント細胞(大腸菌NM522)をピペットで卓上遠沈管に入れた。これらのバッチを氷上に30分間置き、断続的に振盪した。その後、42℃で1分間熱ショックを行った。再生のため、1mlのLB培地を細胞に加え、37℃にて90分間シェーカー上でインキュベートした。各100μlの未希釈バッチ、1:10希釈液および1:100希釈液をLB+アンピシリンプレートに平らに拡げ、37℃で一晩インキュベートした。
【0093】
7.大腸菌からのプラスミド単離(Miniprep)
大腸菌コロニーを卓上遠沈管の1.5mlのLB+アンピシリン培地中で、37℃および120rpmにて一晩培養した。翌日、細胞を5000rpm、4℃で5分間遠心分離し、ペレットを50μlのTEバッファーに取った。各バッチを100μlの0.2N NaOH、1%SDS溶液と混合し、氷上に5分間置いた(細胞の溶解)。次に、400μlの酢酸Na/NaCl溶液(230μlのH2O、130μlの3M酢酸ナトリウム、および40μlの5M NaCl)を加え、バッチを混合し、氷上にさらに15分間置いた(タンパク質の沈殿)。11,000rpmで15分間遠心分離した後、プラスミドDNAを含む上清をエッペンドルフ管に移した。上清が完全に透明でなければ、もう一度遠心分離を行った。この上清を360μlの氷冷イソプロパノールと混合し、−20℃で30分間インキュベートした(DNAの沈殿)。DNAを遠心分離し(15分、12,000rpm、4℃)、上清を廃棄し、ペレットを100μlの氷冷96%エタノールで洗浄し、−20℃で15分間インキュベートし、再び遠心分離を行った(15分、12,000rpm、4℃)。ペレットをスピードバキュームで乾燥させた後、100μlのH2Oに取った。制限分析により、プラスミドDNAの特性決定を行った。このために、各バッチ10μlを制限酵素で処理し、ワイドミニゲルでのゲル電気泳動により分離した(上記参照)。
【0094】
8.大腸菌でのプラスミド単離(Maxiprep)
より多量のプラスミドDNAを単離するために、maxiprep法を行った。100mlのLB+アンピシリン培地を含む2つのプランジャーに、単離されるプラスミドを担持するコロニーまたは100μlの凍結培養物を接種し、37℃、120rpmで一晩インキュベートする。翌日、培養物(200ml)をGSAビーカーに移し、10分間4000rpm(2600*g)で遠心分離した。細胞ペレットを6mlのTEバッファーに再懸濁させた。細胞壁を消化するために、1.2mlのリゾチーム溶液(20mg/mlTEバッファー)を加え、室温で10分間インキュベートした。次に、12mlの0.2N NaOH、1%SDS溶液を用いて細胞溶解を行い、室温でさらに5分間インキュベートした。9mlの冷却した3M酢酸ナトリウム溶液(pH4.8)を加え、15分間氷上でインキュベートすることによってタンパク質を沈殿させた。遠心分離(GSA:13,000rpm(27,500*g)、20分、4℃)の後、DNAを含む上清を新しいGSAビーカーに移し、15mlの氷冷イソプロパノールを用い、−20℃で30分間インキュベートしてDNAを沈殿させた。このDNAペレットを5mlの氷冷エタノールで洗浄し、風乾した(約30〜60分)。その後、これを1mlのH2Oに再懸濁させた。このプラスミドを制限分析により調査した。濃度はミニゲルに希釈液を加えることによって求めた。塩含量を引き下げるために、30〜60分微量透析を行った(孔径0.025μm)。
【0095】
9.酵母の形質転換
酵母の形質転換のため、サッカロミセス・セレビシエ(S. cerevisiae)AH22株の前培養物を調製した。20mlのYE培地を含むプランジャーに100μlの凍結培養物を接種し、28℃、120rpmで一晩インキュベートした。主培養は、10μl、20μlまたは50μlの前培養物を接種した100mlのYE培地を含むプランジャーにて同じ条件下で行った。
【0096】
9.1 コンピテント細胞の作製
翌日、トーマチャンバー(Thoma chamber)を用いてプランジャーを計数し、3〜5*107細胞/mlを保有するプランジャーを用いてこの手順を続けた。遠心分離(GSA:5000rpm(4000*g)、10分)により細胞を採取した。細胞ペレットを10mlのTEバッファーに再懸濁させ、2本の卓上遠沈管に分けた(各5ml)。細胞を6000rpmで3分間遠心分離し、各5mlのTEバッファーで2回洗浄した。次に、細胞ペレットを109細胞につき330μlの酢酸リチウムバッファーに取り、無菌の50mlエルレンマイヤーフラスコに移し、28℃で1時間振盪させた。結果として、これらの細胞は形質転換能があった。
【0097】
9.2 形質転換
各形質転換調製物について、15μlのニシン精子DNA(10mg/ml)、10μlの形質転換されるDNA(約0.5μg)および330μlの成分細胞をピペットで卓上遠沈管に入れ、28℃で30分間インキュベートした(振盪させない)。次に、700μlの50%PEG6000を加え、振盪させずに28℃でさらに1時間インキュベートした。その後、42℃で5分間熱ショックを行った。100μlのこの懸濁液を選択培地(YNB、Difco)に播種し、ロイシン原栄養株を選択した。G418耐性の選択の場合には、熱ショック後に細胞の再生を行う(9.3再生段階を参照)。
【0098】
9.3 再生段階
選択マーカーはG418耐性であるので、細胞は耐性遺伝子の発現に時間を要した。形質転換調製物を4mlのYE培地と混合し、シェーカー(120rpm)にて28℃で一晩インキュベートした。翌日、細胞を遠心分離し(6,000rpm、3分)、1mlのYE培地に取り、100μlまたは200μlをYE+G418プレートに平らに拡げた。これらのプレートを28℃で数日間インキュベートした。
【0099】
10.PCRの反応条件
ポリメラーゼ連鎖反応の反応条件は個々の場合ごとに至適化しなければならず、いずれのバッチにも必ずしも有効というわけではない。従って、とりわけ、DNAの使用量、塩濃度および融解温度は変動し得る。課題の本発明者らの処方物では、エッペンドルフキャップ中に、サーモサイクラーで用いるのに好適な以下の物質:PCR調製物として総容量50μlとなるように水に溶解させた5μlのスーパーバッファー、8μlのdNTP(各0.625μM)、5’−プライマー、3’−プライマーおよび0.2μgのマトリックスDNAを合わせるのが有利であることが分かり、2μl(〜0.1U)のSuper Taqポリメラーゼに加えた。このバッチを短時間で遠心分離し、1滴のオイルでカバーした。増幅には37〜40回のサイクルを選択した。
【0100】
11.S.セレビシエからの脂質粒子の単離
酵母細胞を28℃で72時間、250rpmで往復振盪しながら、50mlのWMVIII最小培地中で増殖させた。細胞を遠心分離により採取し、Leber et al.(Leber R,Zinser E,Zellnig G,Paltauf F,Daum G. Characterization of lipid particles of the yeast,Saccharomyces cerevisiae. Yeast. 1994 Nov;10(11):1421−1428)に従って脂質粒子を単離および精製した。ステロール分析では、脂質粒子は、GCにより総ステロールを定量するために室温、30%メタノール性KOH中で16時間鹸化するか、またはクロロホルム/メタノール(4:1)でステロールを直接抽出し、遊離ステロールとエステル化ステロールを識別するためのTLCによって、もしくは遊離ステロールの定量のためのGCによって分析するかのいずれかを行った。
【0101】
12.スクアレンおよびステロールの分析
細胞全体の脂質および脂質粒子中の総脂質を定量するために、GC分析の前にサンプルを鹸化した。125OD600の細胞を0.5N HCl中、100℃で20分間処理し、室温まで冷却した。その後、3gのKOH、およびピロガロール(2g/l)を含む12.5mlのメタノールを加えた。鹸化のために、混合物を水浴中、70℃で2時間インキュベートした。加水分解されたエステルをn−ヘキサンで抽出した。非鹸化画分を2mlのn−ヘキサンに再懸濁させた。スクアレンおよびステロールを、スクアレンおよびコレステロールを内部標準としてGCにより定量した。スクアレンおよびステロールを、150から250℃までプログラムされたキャピラリーカラム(25m×0.25mm×0.25μm[膜厚];Chrompack CPSil5)を備えたHewlett−Packard 5890ガスクロマトグラフで分離した。温度は最初2分間150℃とした後、15℃/分で最終温度250℃まで昇温し、この温度で20分間保持した。直線的速度を30cm/秒とし、ヘリウムをキャリアガスとして用い、注入はスプリットレスモードで行った。注入容量は1μlとした。各ピークの面積を計算し、細胞乾重1gに対して換算した。各サンプルを2回測定した。同定にはエルゴステロールおよびスクアレンの標品を用いた。
【0102】
中性脂質を抽出し、Sorger and Daum(J.Bacteriol.184(2002),519−524)の方法で定量した。詳細には、中性脂質の定量では、抽出物をシリカゲル60プレートに適用し、クロマトグラムを、軽質石油−ジエチルエーテル−酢酸(25:25:1、vol/vol/vol)の溶媒系を用いて3分の1の距離まで展開させた。その後、プレートを軽く乾燥させ、軽質石油−ジエチルエーテル(49:1、vol/vol)の溶媒系を用いてプレートの上部までさらに展開させた。中性脂質は、TLCチャンバー内で薄層プレートをヨウ素蒸気で染色することにより可視化した。定量は比重走査によって行った。
【0103】
スクアレンおよびステロール分析のための酵母サッカロミセス・セレビシエ株の標準的な培養手順は次の通りとした。
前培養:100ml振盪フラスコ中の20mlのWMVIII培地に20μlの対応するグリセロール保存液に接種し、30℃、150rpmで48時間培養した。
主培養:バッフル付き250ml振盪フラスコ中の50mlのWMVIII培地に1%の前培養物を接種し、30℃、150rpmで72時間培養した。
【0104】
実施例1
S.セレビシエAH22ura3における遺伝子ARE1およびARE2の欠失
ベクターpUG6(Gueldener U,Heck S,Fiedler T,Beinhauer JD and Hegemann JH (1996).A new efficient gene disruption cassette for repeated use in budding yeast. Nucleic Acids Res 24 2519−24)を用いて、遺伝子ARE1およびARE2を欠失させた。
【0105】
両遺伝子の欠失は、同じ方法によって連続的に行った。まず、ARE1を欠失させ、その後、ARE2を欠失させた。プラスミドを調製した後、pUG6の断片をPCRによって増幅し、loxP−kanMX−loxPからなるツールを得た。ARE1 respec.ARE2コード配列の5’および3’配列をpUG6ベクターのloxP領域と融合させるためのプライマーを構築した。
【0106】
得られたPCR産物は、KanR遺伝子、loxP部位およびS.セレビシエAH22ura3における組込み形質転換のためのARE1 respec.ARE2相同領域からなる。酵母における相同組換えが、標的配列の欠失をもたらす。
【0107】
G418に対する耐性を陽性クローンの選択に用いた。この酵母株においてARE1 respec.ARE2コード領域は欠失されていた。さらなる遺伝子を欠失させるための株を作製するために、この株からG418耐性を除去した。この目的で、この株をpSH47で形質転換させた(Guldner et al., 1996)。このベクターは、loxP部位に隣接するKanR遺伝子を除去するためのcre−リコンビナーゼを有する。
【0108】
pSH47を定着させるため、この株を5−FOA(5−フルオロオロト酸)(1g/L)寒天プレート上で対抗選択した。得られた株は遺伝子ARE1およびARE2の双方を欠失している。
【0109】
実施例2
S.セレビシエAH22ura3における遺伝子DGA1およびLRO1の欠失
ベクターpUG6(Guldner et al.,1996)を用いて、遺伝子DGA1およびLRO1を欠失させた。
【0110】
両遺伝子の欠失は、同じ方法によって連続的に行った。まず、DGA1を欠失させ、その後、LRO1を欠失させた。プラスミドを調製した後、pUG6の断片をPCRによって増幅し、loxP−kanMX−loxPからなるツールを得た。
【0111】
DGA1 respec.LRO1コード配列の5’および3’配列をpUG6ベクターのloxP領域と融合させるためのプライマーを構築した。
【0112】
得られたPCR産物は、KanR遺伝子、loxP部位およびS.セレビシエAH22ura3における組込み形質転換のためのDGA1 respec.LRO1相同領域からなる。酵母における相同組換えが、標的配列の欠失をもたらす。
【0113】
G418に対する耐性を陽性クローンの選択に用いた。この酵母株においてDGA1 respec.LRO1コード領域は欠失されていた。さらなる遺伝子を欠失させるための株を作製するために、この株からG418耐性を除去した。この目的で、この株をpSH47で形質転換させた(Guldner et al., 1996)。このベクターは、loxP部位に隣接するKanR遺伝子を除去するためのcre−リコンビナーゼを有する。
【0114】
pSH47を定着させるため、この株を5−FOA(5−フルオロオロト酸)(1g/L)寒天プレート上で対抗選択した。
【0115】
得られた株は遺伝子DGA1およびLRO1の双方を欠失している。
【0116】
実施例3
S.セレビシエAH22ura3are1are2における遺伝子DGA1およびLRO1の欠失
ベクターpUG6(Guldner et al., 1996)を用いて、遺伝子DGA1およびLRO1を欠失させた。
【0117】
両遺伝子の欠失は、同じ方法によって連続的に行った。まず、DGA1を欠失させ、その後、LRO1を欠失させた。プラスミドを調製した後、pUG6の断片をPCRによって増幅し、loxP−kanMX−loxPからなるツールを得た。DGA1 respec.LRO1コード配列の5’および3’配列をpUG6ベクターのloxP領域と融合させるためのプライマーを構築した。
【0118】
得られたPCR産物は、KanR遺伝子、loxP部位およびS.セレビシエAH22ura3における組込み形質転換のためのDGA1 respec.LRO1相同領域からなる。酵母における相同組換えが、標的配列の欠失をもたらす。
【0119】
G418に対する耐性を陽性クローンの選択に用いた。この酵母株においてDGA1 respec.LRO1コード領域は欠失されていた。さらなる遺伝子を欠失させるための株を作製するために、この株からG418耐性を除去した。この目的で、この株をpSH47で形質転換させた(Guldner et al., 1996)。このベクターは、loxP部位に隣接するKanR遺伝子を除去するためのcre−リコンビナーゼを有する。
【0120】
pSH47を定着させるため、この株を5−FOA(5−フルオロオロト酸)(1g/L)寒天プレート上で対抗選択した。
【0121】
得られた株は遺伝子ARE1、ARE2、DGA1およびLRO1の4つを欠失している。
【0122】
実施例4
エピソームプラスミドを用いた、実施例1〜3から得られた酵母株および参照株としてのAH22ura3におけるt−HMG1の発現
tHMGのDNA配列(Basson et al. (Mol. Cell. Biol. 8 (1988), 3793−3808))を、標準的な方法を用いてサッカロミセス・セレビシエS288C(Mortimer and Johnston (Genetics 113 (1986), 35−43))のゲノムDNAからPCRにより増幅した。この場合に用いるプライマーはDNAオリゴマーtHMG−5’およびtHMG−3’である。得られたDNA断片を、クレノウ処理後にクローニングベクターpUC19(Yanisch−Perron et al. (1985): Improved M13 phage cloning vectors and host strains: nucleotide sequences of the M13mp18 and pUC19 vectors. In: Gene. Bd. 33, S. 103−119)に導入し、ベクターpUC19−tHMGを得た。プラスミドを単離し、pUC19−tHMGをエンドヌクレアーゼEcoRIおよびBamHIで制限処理した後、得られた断片を、これもまたEcoRIおよびBamHIで処理した酵母発現ベクターpPT2b(Lang and Looman (Appl. Microbiol. Biotechnol. 44(1995),147−156))に導入した。作出されたプラスミドpPT2b−tHMGは、末端切断型ADH1−プロモーター(Bennetzen and Hall (Yeast 7(1982),475−477))およびTRP1−ターミネーター(Tschumper G, Carbon J. Sequence of a yeast DNA fragment containing a chromosomal replicator and the TRP1 gene. Gene. 1980 Jul;10(2):157−166)を含み、その間にtHMG−DNA断片が見られる。ベクターpPT2b−tHMGからエンドヌクレアーゼEcoRVおよびNruIによってDNA切片を単離した。このDNA切片は、いわゆる中程度の長さのADH1−プロモーター、tHMG遺伝子およびTRP1−ターミネーターを含む。このDNA切片を、エンドヌクレアーゼSphIおよびDNAポリメラーゼで処理した酵母ベクターYEp13(Fischhoff et al. (Gene 27 (1984), 239−251))に導入した。
【0123】
S.セレビシエAH22URA3are1are2、AH22URA3dga1lro1、AH22URA3are1are2dga1lro1およびAH22URA3をYEpH2で形質転換した。参照として、プラスミドYEp13を用いた。
【0124】
実施例5
染色体組込みプラスミドYDpUHK3を用いた、実施例1〜3から得られた酵母株および参照株としてのAH22ura3におけるt−HMG1の染色体組込みおよび過剰発現
ベクターYEpH2をエンドヌクレアーゼEcoRVおよびNruIで処理した。このようにして、以下の領域:テトラサイクリン耐性遺伝子由来の転写活性化領域(Sidhu and Bollon (10 (1990) 157−166))、中程度の長さのADH1−プロモーター、tHMGおよびTRP1−ターミネーター(発現カセット)を有するDNA断片を作出した。このDNA断片を、StuIで処理したベクターYDpU(Berben et al. 1991 Berben G., Dumont J., Gilliquet V, Bolle P−A. and Hilger F. (1991): ”The YDp plasmids: a uniform set of vectors bearing versatile gene disruption cassettes for” Saccharomyces cerevisiae”. ”Yeast 7, 475−477)に導入した。作出されたベクターYDpUH2/12をエンドヌクレアーゼSmaIで処理し、カナマイシン耐性をコードするDNA配列と連結する(Webster, T. D., Dickson, R. C. (1983) Direct selection of Saccharomyces cerevisiae resistant to the antiobiotic G418 following transformation with a DNA vector carrying the kanamycin−resistance gene of Tn903. Gene 26: 243−252)。作出された構築物(YDpUHK3)をEcoRVで処理した。酵母株サッカロミセス・セレビシエAH22をこの構築物で形質転換させた。この例の場合と同様に、酵母を線状化ベクターで形質転換すると、染色体のURA3遺伝子座にベクター全体が組み込まれる。組み込まれたベクターに由来する、発現カセット(大腸菌起源、E.coli−アンピシリン耐性遺伝子、TEF−プロモーターおよびカナマイシン耐性遺伝子)の部分ではない領域を除去するために、形質転換酵母を、ウラシル栄養要求酵母を増長するFOA選択(Boeke et al. (Methods in Enzymology 154 (1987), 164−175))による選択圧に曝した。選択でいうところのウラシル栄養要求株はAH22tH3ura8の名称を持ち、URA3遺伝子に染色体組込みとしてのtHMG1発現カセットを有する。
【0125】
表1に示される酵母株を、それらのスクアレン生産性/含量について評価した。この目的で、これらの株を30℃、150rpmで振盪しながら72時間、WMVIII培地で培養した。細胞を0.5M煮沸HCl中で破砕した後、脂質を20mlのn−ヘキサンで2回抽出し、GC/MSで分析/定量した(詳細は第12項を参照)。以下のデータが得られた(表1)。
【0126】
【表1】
【0127】
実施例6
S.セレビシエAH22tH3ura8are1are2における遺伝子FLD1の欠失
ベクターpUG6(Guldner et al., 1996)を用いて、遺伝子FLD1を欠失させた。
【0128】
プラスミドを調製した後、pUG6の断片をPCRにより増幅し、loxP−kanMX−loxPからなるツールを得た。FLD1コード配列の5’および3’配列をpUG6ベクターのloxP領域と融合させるためのプライマーを構築した。
【0129】
得られたPCR産物は、KanR遺伝子、loxP部位およびS.セレビシエAH22tH3ura8are1are2における組込み形質転換のためのFLD1相同領域からなる。酵母における相同組換えが、標的配列の欠失をもたらす。
【0130】
G418に対する耐性を陽性クローンの選択に用いた。この酵母株においてFLD1コード領域は欠失されていた。さらなる遺伝子を欠失させるための株を作製するために、この株からG418耐性を除去した。この目的で、この株をpSH47で形質転換させた(Guldner et al., 1996)。このベクターは、loxP部位に隣接するKanR遺伝子を除去するためのcre−リコンビナーゼを有する。
【0131】
pSH47を定着させるため、この株を5−FOA(5−フルオロオロト酸)(1g/L)寒天プレート上で対抗選択した。
【0132】
得られた株は遺伝子ARE1、ARE2およびFLD1の3つを欠失しており、AH22tH3ura8are1are2fld1と呼ばれる。
【0133】
表2に示される酵母株を、それらのスクアレン生産性/含量について評価した。この目的で、これらの株を30℃、150rpmで振盪しながら72時間、WMVIII培地で培養した。細胞を0.5M煮沸HCl中で破砕した後、脂質を20mlのn−ヘキサンで2回抽出し、GC/MSで分析/定量した(詳細は第12項を参照)。以下のデータが得られた(表2)。
【0134】
【表2】
【0135】
実施例7
実施例1〜5から得られた酵母株における遺伝子SAK1およびHAP4のエピソーム過剰発現
遺伝子SAK1およびHAP4のエピソーム過剰発現のため、これらの遺伝子を発現ベクターpFlat1およびpFlat3にそれぞれクローニングした。この目的で、これら2つの遺伝子をS.セレビシエS288c株由来の染色体DNAから、5’末端にNotI制限部位を、3’末端にXhoI制限部位を導入するプライマーを用いて増幅した。得られたPCR断片、ならびにベクターpFlat1およびpFlat3を制限エンドヌクレアーゼXhoIおよびNotIで制限処理した。制限酵素で処理したPCR断片および線状化ベクターを連結し、ベクターpFlat1−SAK1およびpFlat3−HAP4を得た。これらのベクターはそれぞれ、SAK1およびHAP4の強い構成的発現をもたらすために、構成型のADH1プロモーターとTRP1ターミネーターの隣接した1コピーの遺伝子SAK1およびHAP4を有する。
【0136】
プラスミドpFlat3を構築するために、プラスミドYEp24をSphIで切断し、ADH1プロモーターとTRP1ターミネーターを含み、その間にプラスミドpUC19の多重クローニング部位を有する900bpのSphI断片を、プラスミドpPT2Bから挿入した。この多重クローニング部位を、NotIおよびXhoIの制限部位を含むポリリンカーを挿入することにより拡大した。選択のためのURA3遺伝子を有する、得られたプラスミドpFlat1をNcoI制限処理によって線状化し、クレノウポリメラーゼで平滑末端とし、酵母LEU2遺伝子を含むYDpLの、平滑末端化されたBamHI断片を組み込んだ。得られたベクターをpFlat3とした。
【0137】
プラスミドpFlat1−SAK1およびpFlat3−HAP4、また、対照としてのエンプティープラスミドpFlat1およびpFlat3を用い、実施例1〜6から得られた酵母株を形質転換した。
【0138】
表3に示される酵母株を、それらのスクアレン生産性/含量について評価した。この目的で、これらの株を30℃、150rpmで振盪しながら72時間、WMVIII培地で培養した。細胞を0.5M煮沸HCl中で破砕した後、脂質を20mlのn−ヘキサンで2回抽出し、GC/MSで分析/定量した(詳細は第12項を参照)。以下のデータが得られた(表3)。
【0139】
【表3】
【0140】
実施例8
構築された株の中性脂質組成を薄層クロマトグラフィーにより評価した。全脂質抽出および薄層クロマトグラフィーは第12項に従って行った。図1は、野生型株AH22ura3、脱調節型HMG−CoAレダクターゼを有するAH22tH3ura8株、および2つの二重欠失株AH22tH3ura8Δare1Δare2とAH22tH3ura8Δdga1Δlro1の全脂質/中性脂質組成を示す。図1は、脱調節型HMG−CoAレダクターゼを発現し、高い量のスクアレンを生産するレーン3〜8の株に比べ、野生型株(AH22ura3、レーン1および2)は極めて低量のスクアレンしか生産しないことを示唆する。ステリルエステルの形成を担う酵素をコードする遺伝子(are1、are2)およびトリアシルグリセロールの形成を担う酵素をコードする遺伝子(dga1、lro1)の欠失は、対応する株においてこれらの成分の完全な欠失をもたらす(レーン5〜8に黒い四角で示す)。これらの脂質成分をスクアレン、オレイン酸コレステリル、トリオレイン酸、オレイン酸およびエルゴステロールの標品によって同定した(示されていない)。
【特許請求の範囲】
【請求項1】
アシル−CoA:ステロールアシルトランスフェラーゼ/ステロールO−アシルトランスフェラーゼ(EC2.3.1.26)および/またはジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)および/またはレシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)および/またはアシル−CoA:長鎖アルコールO−アシルトランスフェラーゼ(EC2.3.1.75)の活性が対応する野生型生物と比較して低減または排除されており、HMG−CoA−レダクターゼ(EC1.1.1.34)の活性が、対応する野生型生物と比較して増強されている、単離された、遺伝的に改変された非哺乳類生物。
【請求項2】
対応する野生型生物の脂質粒子に蓄積される少なくとも第一の脂質、好ましくは、トリアシルグリセロールおよび/またはステリルアシルエステルおよび/またはワックスエステルがもはや合成されず、
第一の脂質とは異なる第二の脂質が遺伝的に改変された生物の脂質粒子に蓄積され、
該第二の脂質が好ましくは中性脂質であり、より好ましくはプレノール脂質からなる群から選択され、特にイソプレノイドであり、いっそうより好ましくはトリテルペンであり、最も好ましくは、スクアレン、スクアレン誘導体(ここで、スクアレン誘導体は、スクアレンの主鎖炭素原子に結合した水素原子の代わりに主鎖炭素原子に結合した1以上の、特に1〜10または1〜4個の付加的メチルまたはエチル基を含む)およびステロール脂質のアシルエステル、好ましくはステロールまたはステロイドから選択される、
請求項1に記載の生物。
【請求項3】
SAM:C−24ステロールメチルトランスフェラーゼ(EC2.1.1.41)、C−22ステロールデサチュラーゼ(EC1.14.14.−)およびC−5ステロールデサチュラーゼ(EC1.14.21.6)からなる群から選択される活性のうち1つまたは2つまたは全てが、対応する野生型生物と比較して低減または排除されている、請求項1または2に記載の生物。
【請求項4】
スクアレンモノオキシゲナーゼ(EC1.14.99.7)、ステロール14−デメチラーゼ(EC1.14.13.70)および7−デヒドロコレステロールレダクターゼ(EC1.3.1.21)からなる群から選択される活性のうち1つまたは2つまたは全てが、対応する野生型生物と比較して増強されている、請求項1〜3のいずれか一項に記載の生物。
【請求項5】
請求項1に記載の改変された活性の代わりにまたはそれに加えて、HAP4pおよび/またはSAK1pの活性および/または量が増強され、かつ/またはREG1pおよび/またはFLD1pの活性および/または量が低減または排除されている、請求項1〜4のいずれか一項に記載の生物。
【請求項6】
ステロールがコレステロール、コレステロール誘導体、7−デヒドロ−コレステロール、ラノステロール、ラノステロール誘導体、チモステロール、チモステロール誘導体、ラトステロール、ラトステロール誘導体、ククルビタシン、ククルバチン(cucurbatin)誘導体、エピステロール、エピステロール誘導体、テアステロン、テアステロン誘導体、カスタステロン、カスタステロン誘導体、チファステロール、チファステロール誘導体、カタステロン、カタステロン誘導体、シクロオイカレノール、シクロオイカレノール誘導体、シトステロール、シトステロール誘導体、イソフコステロール、イソフコステロール誘導体、フコステロール、フコステロール誘導体、ゴルゴステロール、ゴルゴステロール誘導体、エルゴステロール、エルゴステロール誘導体、スチグマステロールまたはスチグマステロール誘導体であるか、または
ステロイドが、アンドロステロン、アンドロステロン誘導体、テストステロン、テストステロン誘導体、アンドロステンジオール、アンドロステンジオール誘導体、アンドロステンジオン、アンドロステンジオン誘導体、カルステロン、カルステロン誘導体、メタンドリオール、メタンドリオール誘導体、ボラステロン、ボラステロン誘導体、エピアンドロステロン、エピアンドロステロン誘導体、メスタノロン、メスタノロン誘導体、スタノロン、スタノロン誘導体、ステンボロン、ステンボロン誘導体、エピテストステロン、エピテストステロン誘導体、コルチゾール、コルチゾール誘導体、アルドステロン、アルドステロン誘導体、プレグネノロン、プレグネノロン誘導体、コルチゾン、コルチゾン誘導体、コルチコステロン、コルチコステロン誘導体、ノレシンドロン、ノレシンドロン誘導体、ウロコルチゾールまたはウロコルチゾール誘導体である、
請求項1〜5のいずれか一項に記載の生物。
【請求項7】
生物が原核生物、好ましくは細菌であり、より好ましくはマイコバクテリア属、ストレプトミセス属、ロドコッカス属、ノカルジア属、バチルス属、大腸菌(Escherichia coli)、コリネバクテリウム属、アセトバクター属、アシネトバクター属、シュードモナス属、乳酸菌種またはストレプトミセス種からなる群から選択されるか、または
生物が真核生物、好ましくは真菌、植物、藻類または昆虫細胞であり、より好ましくはヤルロウィア・リポリチカ(Yarrowia lipolytica)、ロドトルラ・グルチニス(Rhodotorula glutinis)、リポミセス・スタルケイイ(Lipomyces starkeyi)、カンジダ・クルバタ(Candida curvata)、ロドスポリジウム・トルロイデス(Rhodosporidium tortuloides)、モルチエレラ・イサベリナ(Mortierella isabellina)、ムコール・ジャバニカス(Mucor javonicus)、サッカロミセス(Saccharomyces)、ピキア・パストリス(Pichia pastoris)、クルイベロミセス種(Kluyveromyces spec)、リゾープス属(Rhizopus)、フサリウム属(Fusarium)、フシジウム属(Fusidium)、ジベレラ属(Gibberella)、モルチエレラ属(Mortierella)、トリコデルマ属(Trichoderma)、アスペルギルス種(Aspergillus spec)、ペニシリウム種(Penicillium spec)またはディクチオステリウム種(Dictyostelium spec)、特に、サッカロミセス属、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、サッカロミセス・デルブルエキイ(Saccharomyces delbruckii)、サッカロミセス・イタリカス(Saccharomyces italicus)、サッカロミセス・エリプソイデウス(Saccharomyces ellipsoideus)、サッカロミセス・フェルメンタティ(Saccharomyces fermentati)、サッカロミセス・クルイベリ(Saccharomyces kluyveri)、サッカロミセス・クルセイ(Saccharomyces krusei)、サッカロミセス・ラクティス(Saccharomyces lactis)、サッカロミセス・マルキシアヌス(Saccharomyces marxianus)、サッカロミセス・ミクロエリプソイデス(Saccharomyces microellipsoides)、サッカロミセス・モンタヌス(Saccharomyces montanus)、サッカロミセス・ノルベンシス(Saccharomyces norbensis)、サッカロミセス・オレアセウス(Saccharomyces oleaceus)、サッカロミセス・パラドキサス(Saccharomyces paradoxus)、サッカロミセス・パストリアヌス(Saccharomyces pastorianus)、サッカロミセス・プレトリエンシス(Saccharomyces pretoriensis)、サッカロミセス・ロゼイ(Saccharomyces rosei)、サッカロミセス・ロウキシイ(Saccharomyces rouxii)、サッカロミセス・ウバルム(Saccharomyces uvarum)、サッカロミセス・ルドウィギイ(Saccharomycodes ludwigii)、またはクルイベロミセス・ラクティス(K. lactis)、クルイベロミセス・マルキシアヌス・バー・マルキシアヌス(K. marxianus var. marxianus)、クルイベロミセス・サーモトレランス(K. thermotolerans)などのクルイベロミセス属、カンジダ・ウチリス(Candida utilis)、カンジダ・トロピカリス(Candida tropicalis)、カンジダ・アルビカンス(Candida albicans)、カンジダ・リポリチカ(Candida lipolytica)およびカンジダ・ベルサチリス(Candida versatilis)などのカンジダ属、ピキア・スティピディス(Pichia stipidis)、ピキア・パストリス(Piachia pastoris)およびピキア・ソルビトフィラ(Pichia sorbitophila)などのピキア属、クリプトコッカス属(Cryptococcus)、デバリオミセス属(Debaromyces)、ハンセヌラ属(Hansenula)、サッカロミコプシス属(Saccharomycecopsis)、サッカロミコデス属(Saccharomycodes)、シゾサッカロミセス属(Schizosaccharomyces)、ウィッケルハミア属(Wickerhamia)、デバリオミセス属(Debayomyces)、ハンセニアスポラ属(Hanseniaspora)、クロエケラ属(Kloeckera)、チゴサッカロミセス属(Zygosaccharomyces)、オガタエア属(Ogataea)、クライシア属(Kuraishia)、コマガタエラ(Komagataella)、メトシュニコウィア属(Metschnikowia)、ウィリオプシス属(Williopsis)、ナカザワエア属(Nakazawaea)、クリプトコッカス属(Cryptococcus)、トルラスポラ属(Torulaspora)、ブルレラ属(Bullera)、ロドトルラ属(Rhodotorula)、ウィロプシス属(Willopsis)およびスポロボロミセス属の酵母からなる群、またはラッカセイ、ナタネ、カノーラ、ヒマワリ(unflower)、アフロール(afflor)、ケシ、マスタード、アサ、トウゴマ(Rizinus)、オリーブ、アマランス、メキシカンチーア種子、ゴマ、キンセンカ、ザクロ、サンカップ、モウズイカ、アザミ、ノバラ、ヘーゼルナッツ、アーモンド、マカダミア、アボカド、ゲッケイジュ、カボチャ、アマ、ダイズ、ピスタチオ、ルリチザ、ココナッツ、クルミ、トウモロコシ、コムギ、ライムギ、エンバク、ライコムギ、コメ、オオムギ、ワタ、キャッサバ、コショウ、マリーゴールド、ナス科植物、ジャガイモ、ヤシ、タバコ、ナス、ビジア属(Vicia)、エンドウ、アルファルファ、コーヒー、ココア、茶、ヤナギ、牧草、フィスコミトレラ・オデル・セラトドン(physcomitrella oder ceratodon)からなる群、またはクリプチスタ(Cryptista)、緑色鞭藻類(Chloromonadophyceae)、黄緑藻類(Xanthophycea)、海産渦鞭毛藻類(Crypthecodinium)、黄金色植物(Chrysophyta)、珪藻類(Bacillariophyta)、褐藻類(Phaeophyta)、紅藻類(Rhodophyta)、緑藻類(Chlorophyta)、ハプト植物、クリプチスタ(Cryptista)、ユーグレナ動物門(Euglenozoa)、渦鞭毛動物門(Dinozoa)、クロララクニオン植物門(Chlorarachniophyta)からなる群、またはヨトウガ(Spodoptera frugiperda)、キンウワバ(Trichoplusia ni)、ヨトウガ(Mamestra brassicae)、ショウジョウバエ(Drosophila)由来の細胞からなる群から選択される、請求項1〜6のいずれか一項に記載の生物。
【請求項8】
生物を培養すること、およびその生物から脂質を単離することを含む、脂質、好ましくは第二の脂質の生産のための、請求項1〜7のいずれか一項に記載の生物の使用。
【請求項9】
ワクチンのための、生分解性潤滑剤としての、化粧品および/または医薬品中のアジュバントとしての、第二の脂質とは異なる化粧的にまたは薬学的に有効な物質を含むアジュバント処方物の調製における、請求項8に従って得られた第二の脂質(ここで、第二の脂質は化粧的にまたは薬学的に有効な物質と混合され、投与、好ましくは局所投与または経口投与用にガレン製剤的に調製される)の使用。
【請求項10】
アシル−CoA:ステロールアシルトランスフェラーゼ/ステロールO−アシルトランスフェラーゼ(EC2.3.1.26)および/またはジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)および/またはレシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)および/またはスクアレンモノオキシゲナーゼ(EC1.14.99.7)および/またはステロール14−デメチラーゼ(EC1.14.13.70)および/または7−デヒドロコレステロールレダクターゼ(EC1.3.1.21)および/またはHAP4pおよび/またはSAK1pのうちの1つまたはいくつかの活性を有するタンパク質をコードする1つの核酸またはいくつかの同じもしくは異なる核酸を含み、この核酸が少なくとも1つの、好ましくは構成的に活性なプロモーターの制御下にある、核酸構築物、特に、プラスミドまたは組込み発現カセット。
【請求項11】
HMG−CoA−レダクターゼ(EC1.1.1.34)活性を有するタンパク質をコードする核酸がさらに含まれ、該タンパク質が好ましくは構成的に活性なプロモーターの制御下にある、請求項10に記載の核酸構築物。
【請求項12】
原種生物が形質転換され、該原種生物が、SAM:C−24ステロールメチルトランスフェラーゼ(EC2.1.1.41)、C−22ステロールデサチュラーゼおよび/またはC−5ステロールデサチュラーゼ(EC1.14.21.6)および/またはアシル−CoA:ステロールアシルトランスフェラーゼ/ステロールO−アシルトランスフェラーゼ(EC2.3.1.26)および/またはジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)および/またはレシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)および/またはアシル−CoA:長鎖アルコールO−アシルトランスフェラーゼ(EC2.3.1.75)および/またはREG1pおよび/またはFLD1pからなる群から選択される活性の1つまたは2つまたは全てが、対応する野生型生物と比較して低減または排除されている野生型生物または原種生物である、請求項1〜7のいずれか一項に記載の生物を生産するための請求項10または11に記載の核酸構築物の使用。
【請求項1】
アシル−CoA:ステロールアシルトランスフェラーゼ/ステロールO−アシルトランスフェラーゼ(EC2.3.1.26)および/またはジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)および/またはレシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)および/またはアシル−CoA:長鎖アルコールO−アシルトランスフェラーゼ(EC2.3.1.75)の活性が対応する野生型生物と比較して低減または排除されており、HMG−CoA−レダクターゼ(EC1.1.1.34)の活性が、対応する野生型生物と比較して増強されている、単離された、遺伝的に改変された非哺乳類生物。
【請求項2】
対応する野生型生物の脂質粒子に蓄積される少なくとも第一の脂質、好ましくは、トリアシルグリセロールおよび/またはステリルアシルエステルおよび/またはワックスエステルがもはや合成されず、
第一の脂質とは異なる第二の脂質が遺伝的に改変された生物の脂質粒子に蓄積され、
該第二の脂質が好ましくは中性脂質であり、より好ましくはプレノール脂質からなる群から選択され、特にイソプレノイドであり、いっそうより好ましくはトリテルペンであり、最も好ましくは、スクアレン、スクアレン誘導体(ここで、スクアレン誘導体は、スクアレンの主鎖炭素原子に結合した水素原子の代わりに主鎖炭素原子に結合した1以上の、特に1〜10または1〜4個の付加的メチルまたはエチル基を含む)およびステロール脂質のアシルエステル、好ましくはステロールまたはステロイドから選択される、
請求項1に記載の生物。
【請求項3】
SAM:C−24ステロールメチルトランスフェラーゼ(EC2.1.1.41)、C−22ステロールデサチュラーゼ(EC1.14.14.−)およびC−5ステロールデサチュラーゼ(EC1.14.21.6)からなる群から選択される活性のうち1つまたは2つまたは全てが、対応する野生型生物と比較して低減または排除されている、請求項1または2に記載の生物。
【請求項4】
スクアレンモノオキシゲナーゼ(EC1.14.99.7)、ステロール14−デメチラーゼ(EC1.14.13.70)および7−デヒドロコレステロールレダクターゼ(EC1.3.1.21)からなる群から選択される活性のうち1つまたは2つまたは全てが、対応する野生型生物と比較して増強されている、請求項1〜3のいずれか一項に記載の生物。
【請求項5】
請求項1に記載の改変された活性の代わりにまたはそれに加えて、HAP4pおよび/またはSAK1pの活性および/または量が増強され、かつ/またはREG1pおよび/またはFLD1pの活性および/または量が低減または排除されている、請求項1〜4のいずれか一項に記載の生物。
【請求項6】
ステロールがコレステロール、コレステロール誘導体、7−デヒドロ−コレステロール、ラノステロール、ラノステロール誘導体、チモステロール、チモステロール誘導体、ラトステロール、ラトステロール誘導体、ククルビタシン、ククルバチン(cucurbatin)誘導体、エピステロール、エピステロール誘導体、テアステロン、テアステロン誘導体、カスタステロン、カスタステロン誘導体、チファステロール、チファステロール誘導体、カタステロン、カタステロン誘導体、シクロオイカレノール、シクロオイカレノール誘導体、シトステロール、シトステロール誘導体、イソフコステロール、イソフコステロール誘導体、フコステロール、フコステロール誘導体、ゴルゴステロール、ゴルゴステロール誘導体、エルゴステロール、エルゴステロール誘導体、スチグマステロールまたはスチグマステロール誘導体であるか、または
ステロイドが、アンドロステロン、アンドロステロン誘導体、テストステロン、テストステロン誘導体、アンドロステンジオール、アンドロステンジオール誘導体、アンドロステンジオン、アンドロステンジオン誘導体、カルステロン、カルステロン誘導体、メタンドリオール、メタンドリオール誘導体、ボラステロン、ボラステロン誘導体、エピアンドロステロン、エピアンドロステロン誘導体、メスタノロン、メスタノロン誘導体、スタノロン、スタノロン誘導体、ステンボロン、ステンボロン誘導体、エピテストステロン、エピテストステロン誘導体、コルチゾール、コルチゾール誘導体、アルドステロン、アルドステロン誘導体、プレグネノロン、プレグネノロン誘導体、コルチゾン、コルチゾン誘導体、コルチコステロン、コルチコステロン誘導体、ノレシンドロン、ノレシンドロン誘導体、ウロコルチゾールまたはウロコルチゾール誘導体である、
請求項1〜5のいずれか一項に記載の生物。
【請求項7】
生物が原核生物、好ましくは細菌であり、より好ましくはマイコバクテリア属、ストレプトミセス属、ロドコッカス属、ノカルジア属、バチルス属、大腸菌(Escherichia coli)、コリネバクテリウム属、アセトバクター属、アシネトバクター属、シュードモナス属、乳酸菌種またはストレプトミセス種からなる群から選択されるか、または
生物が真核生物、好ましくは真菌、植物、藻類または昆虫細胞であり、より好ましくはヤルロウィア・リポリチカ(Yarrowia lipolytica)、ロドトルラ・グルチニス(Rhodotorula glutinis)、リポミセス・スタルケイイ(Lipomyces starkeyi)、カンジダ・クルバタ(Candida curvata)、ロドスポリジウム・トルロイデス(Rhodosporidium tortuloides)、モルチエレラ・イサベリナ(Mortierella isabellina)、ムコール・ジャバニカス(Mucor javonicus)、サッカロミセス(Saccharomyces)、ピキア・パストリス(Pichia pastoris)、クルイベロミセス種(Kluyveromyces spec)、リゾープス属(Rhizopus)、フサリウム属(Fusarium)、フシジウム属(Fusidium)、ジベレラ属(Gibberella)、モルチエレラ属(Mortierella)、トリコデルマ属(Trichoderma)、アスペルギルス種(Aspergillus spec)、ペニシリウム種(Penicillium spec)またはディクチオステリウム種(Dictyostelium spec)、特に、サッカロミセス属、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、サッカロミセス・デルブルエキイ(Saccharomyces delbruckii)、サッカロミセス・イタリカス(Saccharomyces italicus)、サッカロミセス・エリプソイデウス(Saccharomyces ellipsoideus)、サッカロミセス・フェルメンタティ(Saccharomyces fermentati)、サッカロミセス・クルイベリ(Saccharomyces kluyveri)、サッカロミセス・クルセイ(Saccharomyces krusei)、サッカロミセス・ラクティス(Saccharomyces lactis)、サッカロミセス・マルキシアヌス(Saccharomyces marxianus)、サッカロミセス・ミクロエリプソイデス(Saccharomyces microellipsoides)、サッカロミセス・モンタヌス(Saccharomyces montanus)、サッカロミセス・ノルベンシス(Saccharomyces norbensis)、サッカロミセス・オレアセウス(Saccharomyces oleaceus)、サッカロミセス・パラドキサス(Saccharomyces paradoxus)、サッカロミセス・パストリアヌス(Saccharomyces pastorianus)、サッカロミセス・プレトリエンシス(Saccharomyces pretoriensis)、サッカロミセス・ロゼイ(Saccharomyces rosei)、サッカロミセス・ロウキシイ(Saccharomyces rouxii)、サッカロミセス・ウバルム(Saccharomyces uvarum)、サッカロミセス・ルドウィギイ(Saccharomycodes ludwigii)、またはクルイベロミセス・ラクティス(K. lactis)、クルイベロミセス・マルキシアヌス・バー・マルキシアヌス(K. marxianus var. marxianus)、クルイベロミセス・サーモトレランス(K. thermotolerans)などのクルイベロミセス属、カンジダ・ウチリス(Candida utilis)、カンジダ・トロピカリス(Candida tropicalis)、カンジダ・アルビカンス(Candida albicans)、カンジダ・リポリチカ(Candida lipolytica)およびカンジダ・ベルサチリス(Candida versatilis)などのカンジダ属、ピキア・スティピディス(Pichia stipidis)、ピキア・パストリス(Piachia pastoris)およびピキア・ソルビトフィラ(Pichia sorbitophila)などのピキア属、クリプトコッカス属(Cryptococcus)、デバリオミセス属(Debaromyces)、ハンセヌラ属(Hansenula)、サッカロミコプシス属(Saccharomycecopsis)、サッカロミコデス属(Saccharomycodes)、シゾサッカロミセス属(Schizosaccharomyces)、ウィッケルハミア属(Wickerhamia)、デバリオミセス属(Debayomyces)、ハンセニアスポラ属(Hanseniaspora)、クロエケラ属(Kloeckera)、チゴサッカロミセス属(Zygosaccharomyces)、オガタエア属(Ogataea)、クライシア属(Kuraishia)、コマガタエラ(Komagataella)、メトシュニコウィア属(Metschnikowia)、ウィリオプシス属(Williopsis)、ナカザワエア属(Nakazawaea)、クリプトコッカス属(Cryptococcus)、トルラスポラ属(Torulaspora)、ブルレラ属(Bullera)、ロドトルラ属(Rhodotorula)、ウィロプシス属(Willopsis)およびスポロボロミセス属の酵母からなる群、またはラッカセイ、ナタネ、カノーラ、ヒマワリ(unflower)、アフロール(afflor)、ケシ、マスタード、アサ、トウゴマ(Rizinus)、オリーブ、アマランス、メキシカンチーア種子、ゴマ、キンセンカ、ザクロ、サンカップ、モウズイカ、アザミ、ノバラ、ヘーゼルナッツ、アーモンド、マカダミア、アボカド、ゲッケイジュ、カボチャ、アマ、ダイズ、ピスタチオ、ルリチザ、ココナッツ、クルミ、トウモロコシ、コムギ、ライムギ、エンバク、ライコムギ、コメ、オオムギ、ワタ、キャッサバ、コショウ、マリーゴールド、ナス科植物、ジャガイモ、ヤシ、タバコ、ナス、ビジア属(Vicia)、エンドウ、アルファルファ、コーヒー、ココア、茶、ヤナギ、牧草、フィスコミトレラ・オデル・セラトドン(physcomitrella oder ceratodon)からなる群、またはクリプチスタ(Cryptista)、緑色鞭藻類(Chloromonadophyceae)、黄緑藻類(Xanthophycea)、海産渦鞭毛藻類(Crypthecodinium)、黄金色植物(Chrysophyta)、珪藻類(Bacillariophyta)、褐藻類(Phaeophyta)、紅藻類(Rhodophyta)、緑藻類(Chlorophyta)、ハプト植物、クリプチスタ(Cryptista)、ユーグレナ動物門(Euglenozoa)、渦鞭毛動物門(Dinozoa)、クロララクニオン植物門(Chlorarachniophyta)からなる群、またはヨトウガ(Spodoptera frugiperda)、キンウワバ(Trichoplusia ni)、ヨトウガ(Mamestra brassicae)、ショウジョウバエ(Drosophila)由来の細胞からなる群から選択される、請求項1〜6のいずれか一項に記載の生物。
【請求項8】
生物を培養すること、およびその生物から脂質を単離することを含む、脂質、好ましくは第二の脂質の生産のための、請求項1〜7のいずれか一項に記載の生物の使用。
【請求項9】
ワクチンのための、生分解性潤滑剤としての、化粧品および/または医薬品中のアジュバントとしての、第二の脂質とは異なる化粧的にまたは薬学的に有効な物質を含むアジュバント処方物の調製における、請求項8に従って得られた第二の脂質(ここで、第二の脂質は化粧的にまたは薬学的に有効な物質と混合され、投与、好ましくは局所投与または経口投与用にガレン製剤的に調製される)の使用。
【請求項10】
アシル−CoA:ステロールアシルトランスフェラーゼ/ステロールO−アシルトランスフェラーゼ(EC2.3.1.26)および/またはジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)および/またはレシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)および/またはスクアレンモノオキシゲナーゼ(EC1.14.99.7)および/またはステロール14−デメチラーゼ(EC1.14.13.70)および/または7−デヒドロコレステロールレダクターゼ(EC1.3.1.21)および/またはHAP4pおよび/またはSAK1pのうちの1つまたはいくつかの活性を有するタンパク質をコードする1つの核酸またはいくつかの同じもしくは異なる核酸を含み、この核酸が少なくとも1つの、好ましくは構成的に活性なプロモーターの制御下にある、核酸構築物、特に、プラスミドまたは組込み発現カセット。
【請求項11】
HMG−CoA−レダクターゼ(EC1.1.1.34)活性を有するタンパク質をコードする核酸がさらに含まれ、該タンパク質が好ましくは構成的に活性なプロモーターの制御下にある、請求項10に記載の核酸構築物。
【請求項12】
原種生物が形質転換され、該原種生物が、SAM:C−24ステロールメチルトランスフェラーゼ(EC2.1.1.41)、C−22ステロールデサチュラーゼおよび/またはC−5ステロールデサチュラーゼ(EC1.14.21.6)および/またはアシル−CoA:ステロールアシルトランスフェラーゼ/ステロールO−アシルトランスフェラーゼ(EC2.3.1.26)および/またはジアシルグリセロールアシルトランスフェラーゼ/ジアシルグリセロールO−アシルトランスフェラーゼ(EC2.3.1.20)および/またはレシチンコレステロールアシルトランスフェラーゼ/リン脂質:ジアシルグリセロールアシルトランスフェラーゼ(EC2.3.1.158)および/またはアシル−CoA:長鎖アルコールO−アシルトランスフェラーゼ(EC2.3.1.75)および/またはREG1pおよび/またはFLD1pからなる群から選択される活性の1つまたは2つまたは全てが、対応する野生型生物と比較して低減または排除されている野生型生物または原種生物である、請求項1〜7のいずれか一項に記載の生物を生産するための請求項10または11に記載の核酸構築物の使用。
【図5】


【図1】
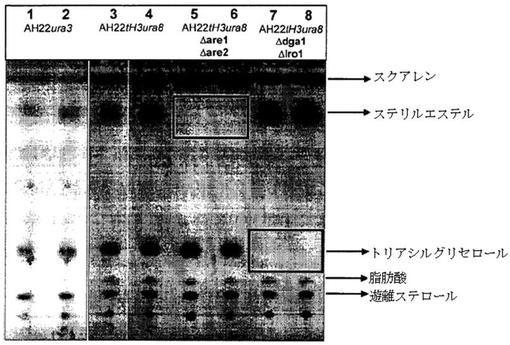
【図2】
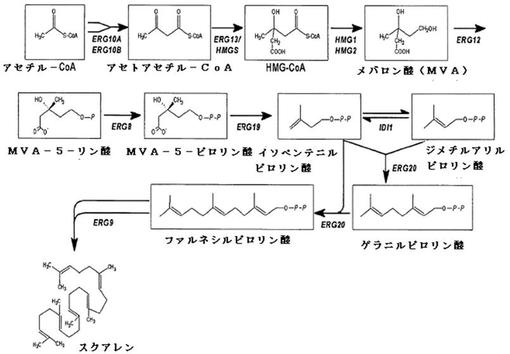
【図3】
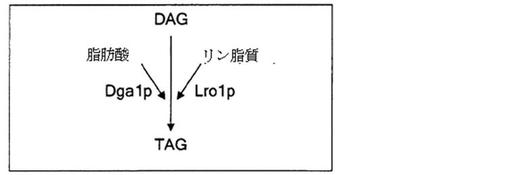
【図4】
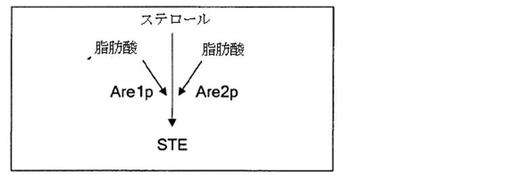
【図1】
【図2】
【図3】
【図4】
【公表番号】特表2013−502903(P2013−502903A)
【公表日】平成25年1月31日(2013.1.31)
【国際特許分類】
【出願番号】特願2012−525904(P2012−525904)
【出願日】平成22年8月4日(2010.8.4)
【国際出願番号】PCT/EP2010/004925
【国際公開番号】WO2011/023298
【国際公開日】平成23年3月3日(2011.3.3)
【出願人】(511282575)オーガノバランス ゲーエムベーハー (2)
【Fターム(参考)】
【公表日】平成25年1月31日(2013.1.31)
【国際特許分類】
【出願日】平成22年8月4日(2010.8.4)
【国際出願番号】PCT/EP2010/004925
【国際公開番号】WO2011/023298
【国際公開日】平成23年3月3日(2011.3.3)
【出願人】(511282575)オーガノバランス ゲーエムベーハー (2)
【Fターム(参考)】
[ Back to top ]