
脈管形成の抑制に有用な組成物
【課題】組織における脈管形成を抑制する医薬組成物を提供する。
【解決手段】αvβ3拮抗物質として、αvβ3に対して免疫特異的な単クローン性抗体又はそのフラグメントを使用する。
【解決手段】αvβ3拮抗物質として、αvβ3に対して免疫特異的な単クローン性抗体又はそのフラグメントを使用する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、一般に医薬分野に関し、とりわけビトロネクチン(vitronectin)レセプター(αvβ3)の拮抗物質を用いて組織の脈管形成(angiogenesis)を抑制する方法および組成物に関する。
【背景技術】
【0002】
インテグリン(integrin)は、細胞外マトリックス蛋白に結合し、したがって、細胞−細胞および細胞−細胞外マトリックス相互作用(一般に粘着現象と呼ぶ)を仲介することが分かっている細胞性レセプター類である。多数のインテグリンおよびインテグリンと結合するそのリガンドが文献に記載されているが、多くのインテグリンの生物学的機能は不明のままである。インテグリンレセプターは、αおよびβサブユニットから形成される非共有結合異種二量体糖蛋白複合体という共通の構造特性をもつ蛋白類を構成する。
ビトロネクチンレセプター(ビトロネクチンに優先的に結合するその本来の特徴のためにこの名がある)は、αvβ1、αvβ3およびαvβ3と呼ばれる3種の異なるインテグリンと関係を有することが分かった(Horton,Int.J.Exp.Pathol.,71:741-759(1990))。αvβ1は、フィブロネクチン(fibronectin)およびビトロネクチンに結合する。αvβ3は、フィブリン、フィブリノーゲン、ラミニン、スロンボスポンジン(thrombospondin)、ビトロネクチン、ヴォン・ヴイレブランド因子(von Willebrand's factor)、オステオスポンチン(osteospontin)および骨唾液腺蛋白I(bone sialoprotein I)を含むきわめて多様なリガンドと結合する。αvβ5はビトロネクチンと結合する。これら3種のインテグリンが組織の細胞相互作用で果たす特定の細胞粘着の役割はなお研究中であるが、異なる生物学的機能を有する異なるインテグリンが存在することは明らかである。
【0003】
多数のインテグリンにとって、リガンドの重要な1つの認識部位は、アルギニン−グリシン−アスパラギン酸(RGD)のトリペプチド配列である。RGDは、ビトロネクチンレセプターインテグリンとして上記に認定されたリガンドの全てに見出される。このRGD認識部位は、RGD配列を含むポリペプチド(“ペプチド”)で模倣することができるが、このようなRGDペプチドはインテグリン機能の既知抑制物質である。RGDペプチドの配列および構造にしたがって特定のインテグリンを標的とするこの抑制特異性を変化させることができるということは、しかしながら言及に値する。
RGD認識部位の考察については以下の文献を参照されたい:Pierschbacherら、Nature,309:30-33(1984); Pierschbacherら、Proc.Natl.Acad.Sci.USA,81:5985-5988(1984)。種々のインテグリン特異性をもつ種々のRGDポリペプチドはまた下記の文献に記載された:Grantら、Cell,58:933-943(1989);Chereshら、Cell,58:945-953(1989);Aumailleyら、FEBS Letts.,291:50-54(1991);Pfaffら、J.Biol.Chem.,269:20233-20238(1994);さらに米国特許出願第4517686号、4578079号、4589881号、4614517号、4661111号、4792525号、4683291号、4879237号、4988621号、5041380号、および5061693号。
【0004】
脈管形成(angiogenesis)は、新たに発生した血管の組織内への増殖を伴う組織の血管新生(vascularization)の過程であり、新規血管新生(neo-vascularization)とも呼ばれる。この過程は、内皮細胞および平滑筋細胞の浸潤によって仲介される。この過程は3通りの態様のいずれか1つで進行すると考えられている:血管は予め存在する血管から成長する、新規な血管発生は前駆細胞から生じる(血管形成(vasculogenesis))、または存在する小血管の直径が拡大する(Bloodら、Bioch.Biophys.Acta,1023:89-118(1990))。血管内皮細胞は、少なくとも5つのRGD依存インテグリンを含むことが知られており、これらには、ビトロネクチンレセプター(αvβ3またはαvβ5)、コラーゲンI型およびIV型レセプター(α1β1)、ラミニンレセプター(α2β1)、フィブロネクチン/ラミニン/コラーゲンレセプター(α3β1)およびフィブロネクチンレセプター(α5β1)が含まれる(Davisら、J.Cell.Biochem.,51:206-218(1993))。平滑筋細胞は、少なくとも6つのRGD依存インテグリンを含むことが知られており、これらには、α5β1、αvβ3およびαvβ5が含まれる。
【0005】
脈管形成は、新生児の成長では重要な過程であるが、また創傷治癒および種々の臨床症状(組織の炎症、関節炎、腫瘍増殖、糖尿病性網膜症、網膜の新規血管新生による筋変性および同様な症状を含む)の病理発生においても重要である。
脈管形成に付随するこのような症状の発現は、脈管形成性疾患と呼ばれる(Folkmanら、Science,235:442-447(1987))。脈管形成は、一般に成人または成熟組織では見られないが、創傷治癒および黄体の増殖サイクルにおいては脈管形成が発生する。例えばモーセスらの論文を参照されたい(Mosesら、Science,248:1408-1410(1990))。
脈管形成の抑制は、腫瘍増殖を制限するために有用な治療であろうと提言された。脈管形成の抑制では、(1)例えばβFGF(線維芽細胞増殖因子)のような“脈管形成分子”の遊離の抑制、(2)例えば抗βFGFの使用による脈管形成分子の中和、(3)脈管形成刺激に対する内皮細胞の反応の抑制が提唱された。この後者の方法は注目を集め、フォークマンら(Folkmanら、Cancer Biology,3:89-96(1992))は、いくつかの内皮細胞反応抑制物質を記載したが、これにはコラゲナーゼ抑制物質、基底膜代謝回転抑制物質、止血性ステロイド、真菌由来脈管形成抑制物質、血小板因子4、スロンボスポンジン、関節炎薬剤(例えばD−ペニシラミンおよびチオリンゴ酸金)、ビタミンD3類似体、アルファーインターフェロン、および脈管形成の抑制に役立つ可能性があるものが含まれる。さらに、この他に提唱された脈管形成抑制物質については以下の文献を参照されたい:Bloodら、Bioch.Biophys.Acta.,1023:89-118(1990);Mosesら、Science,248:1408-1410(1990); Ingberら、Lab.Invest.,59:44-51(1988);並びに米国特許第5092885号、5112946号、5192744号および5202352号。前述の文献に記載された脈管形成抑制物質のいずれもαvβ3の抑制を標的としない。
【0006】
ビトロネクチンレセプターαvβ3を抑制するRGDを含むペプチドもまた報告された(Aumailleyら、FEBS Letts.,291:50-54(1991); Choiら、J.Vasc.Surg.,19:125-134(1994);Smithら、J.Biol.Chem.,265:12267-12271(1990);およびPfaffら、J.Biol.Chem.,269:20233-20238(1994))。脈管形成におけるインテグリンαvβ3の役割は、しかしながら、本発明以前には示唆されたこともなくまた明らかにされたこともなかった。
種々のインテグリンαまたはβサブユニットに免疫特異的単クローン性抗体を用いたインビトロでの細胞粘着抑制によって、αvβ3が、微小血管内皮細胞を含む様々な細胞型の細胞粘着に関与していることが示された(Davisら、J.Cell.Biol.,51:206-218(1993))。さらに、インビトロでRGDペプチドGRGDSの使用を報告したニコシアらの論文では、コラーゲンゲルで培養したラット大動脈からの“微小血管”の形成が抑制された(Nicosiaら、Am.J.Pathol.,138:829-833(1991))。しかしながら、コラーゲンゲル培養での“微小血管”形成のインビトロ抑制は、組織における脈管形成抑制のモデルではない。なぜならば、微小血管の構造が毛細血管芽と同じであるということも、またコラーゲンゲル培養における微小血管の形成が、もとのままの組織(例えば関節炎組織、腫瘍組織または脈管形成抑制が所望される病的組織)への新規血管の成長と同じであるということは示されていないからである。
したがって、本明細書で報告する研究以外に、本出願人らは、細胞粘着抑制物質を用いて脈管形成が組織で抑制できることを開示したものを全く知らない。特に、組織における脈管形成にαvβ3機能が必要とされ、またαvβ3拮抗物質が組織における脈管形成を抑制できるということはこれまで開示されたことはなかった。
【発明の開示】
【発明が解決しようとする課題】
【0007】
本発明は、組織における脈管形成はインテグリンαvβ3を必要とし、αvβ3の抑制物質は脈管形成を抑制することができることを明らかにする。本発明はまた、他のインテグリン(例えばαvβ5またはαvβ1)は、おそらくそれらは脈管形成の発生に必須ではないために、脈管形成を抑制しないことを明らかにする。
【0008】
したがって、本発明は、脈管形成抑制量のαvβ3拮抗物質を含む組成物を組織に投与することを含む、組織における脈管形成抑制方法を開示する。
処置されるべき組織は、脈管形成抑制が所望されるいずれの組織でもよく、例えば新規血管新生が発生しつつある病的組織である。代表的な組織は、炎症組織、固形腫瘍、転移、再狭窄を起こしている組織、および同様な組織を含む。
【課題を解決するための手段】
【0009】
本発明で使用されるαvβ3拮抗物質はαvβ3に結合することができ、さらにαvβ3が本来のリガンドに結合する能力を競合的に抑制することができる。
好ましくは、この拮抗物質は、他のインテグリンの間でαvβ3に対して特異性を示す。特に好ましい具体例では、このαvβ3拮抗物質は、フィブリノーゲンまたは他のRGD含有リガンドのαvβ3への結合を抑制するが、フィブロネクチンのαIIbβ3への結合を実質的に抑制しない。好ましいαvβ3拮抗物質は、ポリペプチドまたはαvβ3と免疫的に反応する単クローン性抗体もしくはその機能的フラグメントであろう。
【0010】
本開示の一部分を形成する図面は下記の通りである。
図1A−1Dは、正常皮膚および創傷治癒下の皮膚(肉芽形成組織と呼ぶ)におけるインテグリンサブユニットβ3およびβ1の組織分布を示す。β3およびβ1に対する抗体を用いた免疫組織化学的方法が、実施例3Aで述べるように実施された。図1Aおよび1Bは、それぞれ、正常皮膚および肉芽形成組織における抗β3の免疫活性を示す。図1Cおよび1Dは、それぞれ、正常皮膚および肉芽形成組織における抗β1の免疫活性を示す。
【0011】
図2A−2Dは、正常皮膚および創傷治癒下の皮膚(肉芽形成組織と呼ぶ)における、それぞれβ3およびβ1に結合するヴォン・ヴィレブランド因子およびラミニンリガンドの組織分布を示す。ヴォン・ヴィレブランド因子に対する抗体(抗vWF)およびラミニンに対する抗体(抗ラミニン)を用いた免疫組織化学的方法が、実施例3Bで述べるように実施された。図2Aおよび2Bは、それぞれ、正常皮膚および肉芽形成組織における抗vWFの免疫活性を示す。図2Cおよび2Dは、それぞれ、正常皮膚および肉芽形成組織における抗ラミニンの免疫活性を示す。
【0012】
図3A−3Dは、それぞれ膀胱癌、大腸癌、乳癌および肺癌の組織生検材料におけるビトロネクチンインテグリンレセプター、αvβ3の分布を示す。αvβ3に対するLM609抗体を用いた免疫組織化学的方法は、実施例3Cで述べるように実施された。
図4は、本発明の未処置10週齢ニワトリ胎児のCAMの典型的な顕微鏡写真を示す。その調製は実施例5Bで述べる。
【0013】
図5A−5Cは、本発明のCAM調製物のインテグリンβ1およびαvβ3の組織分布を示す。図5Aは、CSAT(抗β1抗体)を用いて免疫蛍光で免疫活性を検出した、未処置10週齢CAM調製物におけるβ1サブユニットの分布を示す。図5Bは、LM609(抗αvβ3抗体)を用いて免疫蛍光で免疫活性を検出した、未処置10週齢CAM調製物におけるαvβ3レセプターの分布を示す。図5Cは、LM609(抗αvβ3抗体)を用いて免疫蛍光で免疫活性を検出した、βFGF処置10週齢CAM調製物におけるαvβ3レセプターの分布を示す。
処置および結果は実施例5Cで述べる。
【0014】
図6は、実施例6Aで述べる未処置およびβFGF処置10週齢CAMのαvβ3およびβ1の相対的発現を測定した棒グラフを示す。平均蛍光濃度はY軸に、インテグリンプロフィルはX軸にプロットされている。
図7A−7Cは、それぞれ未処置、βFGF処置およびTNFα処置10週齢CAMの外観を示す。その方法および結果は実施例6Aに記載する。
【0015】
図8A−8Eは、実施例7A1)で述べるように、10日目CAMにおけるFGF誘発脈管形成に対する抗体局所処置の効果を示す。図8Aは、血管を欠く未処置CAM調製物を示す。図8Bは、以前に血管構造を欠いていた領域への、βFGF処置によって誘発された新しい血管構造の浸潤を示す。図8C,8Dおよび8Eは、それぞれβ1に対する抗体(抗β1;CSAT)、αvβ5に対する抗体(抗αvβ5;P3G2)およびαvβ3に対する抗体(抗αvβ3:LM609)の作用を示す。
【0016】
図9A−9Cは、実施例7D2)で述べるように、腫瘍によって誘発された脈管形成に対する合成ペプチド66203の静脈内注射の影響を示す。図9Aは、腫瘍誘発により生じた脈管形成に対するコントロールペプチドの静脈内処置の抑制効果の欠如を示す(コントロールペプチド腫瘍)。ペプチド66203の静脈内注射によるそのような脈管形成の抑制(環状RGD腫瘍)は、図9Bに示す。腫瘍処置領域に隣接する領域内の以前から存在する成熟した血管に対する、ペプチド66203の静脈内輸液後の抑制効果または細胞毒性の欠如は図9Cに示す(環状RGD隣接CAM)。
【0017】
図10A−10Cは、実施例7B1)で述べるように、増殖因子誘発脈管形成に対する単クローン性抗体の静脈内応用の効果を示す。図10Aは、抗体処置に曝されなかったβFGF誘発脈管形成を示す(コントロール)。図10Bに示すように、同じ調製物を抗αvβ5抗体P3G2で処置したとき脈管形成抑制は生じなかった。図10Cに示すように、抗αvβ3抗体LM609による処置で脈管形成抑制が生じた。
図11A−11Cは、実施例7Cに述べるように抗インテグリン抗体の局所適用後の胎児の脈管形成に対する効果を示す。それぞれ図11Aおよび11Bで示すように、脈管形成は、抗β1および抗αvβ5で6週齢CAMを処置しても抑制されなかった。対照的に、図11Cに示すように、抗αvβ3抗体LM609での処置は血管形成の抑制をもたらした。
図12は、実施例7D1)で述べるように、CAM調製物内の腫瘍に進入する血管数の測定を示す。このグラフでは、CSAT(抗β1)、LM609(抗αvβ3)、またはP3G2(抗αvβ5)のいずれかを局所的に用いて得られる血管の数がY軸に示されている。
図13A−13Dは、実施例9A1)aで述べるように、処置後7日の湿潤腫瘍重量と最初の腫瘍重量との間の比較を示す。各棒線は、各グループ当たり5−10個の腫瘍の平均±標準誤差を表す。腫瘍は、ヒトメラノーマ(M21−L)(図13A)、膵臓癌(Fg)(図13B)、肺癌(UCLAP−3)(図13C)および喉頭癌(HEp3)(図13D)CAM調製物に由来し、PBS、CSAT(抗β1)、またはLM609(抗αvβ3)で静脈内処置を施された。グラフでは、X軸に示したようにCSAT(抗β1)、LM609(抗αvβ3)またはPBSのいずれかを静脈内に用いて得られた腫瘍の重量がY軸に示されている。
【0018】
図14Aおよび14Bは、実施例9A1)aで述べるように、P3G2(抗αvβ5)(図14A)およびLM609(抗αvβ3)(図14B)で処置し、ヘマトキシリンおよびエオシンで染色した腫瘍の組織切片を示す。図14Aに示すように、コントロール抗体(P3G2)で処置した腫瘍は、腫瘍基質全体に存在する有糸分裂像(楔形)と同様に多数の血管(矢印)で示されるように、多くの生命活性を示す活発に分裂する腫瘍細胞を示した。対照的に、LM609(抗αvβ3)で処置した腫瘍では、あったとしてもごく僅かの生命活性を示す細胞または血管が検出されただけであった(図14B)。
図15A−15Eは、実施例9A1)bで述べるように、ペプチドで処置されたM21L腫瘍に対応し、下記の通りである。図15A、コントロール環状RADペプチド(69601);図15B、コントロール環状RGDペプチド(66203);図15C、同じ胎児から採取され環状RGDペプチド(66203)で処置された隣接CAM組織;さらに高倍率(×13)のコントロールRAD(69601)処置腫瘍(図15D)および環状RGDペプチド(66203)処置腫瘍(図15E)。図15Dは、RADコントロールペプチド(69601)
処置腫瘍の正常な血管を鮮明に表している。図15Eは、環状RGDペプチド(66203)処置腫瘍の破壊された血管(矢印)の例を示している。
【0019】
図16A−16Eは、実施例10で述べるウサギ眼球のインビボモデルアッセーにおける脈管形成拮抗物質による脈管形成の抑制を示す。図16Aおよび16Bは、βFGFおよびmAbP1F6(抗αvβ5)の存在下でのウサギの眼の脈管形成を示す。図16C、16Dおよび16Eは、βFGFおよびmAbLM609(抗αvβ3)の存在下でのウサギの眼の脈管形成の抑制を示す。
図17は、実施例11Aのマウス:ヒトキメラ−インビボマウスモデルの作製方法を示す工程図である。SCIDマウスの皮膚の一部分をヒト新生児包皮で置き換え、4週間治癒させた。移植が治癒した後、ヒト包皮にヒト腫瘍細胞を接種した。その後の4週間で、ヒトの皮膚からヒトの腫瘍内に増殖したヒト血管構造をもつヒトの腫瘍を含む測定可能な腫瘍が成長した。
図18は、それぞれ実施例12Aおよび12Bで述べるようにFACS分析で求めた、mAb処置およびペプチド処置CAMから得られ、アポプトーシスタグ(Apop Tag)で染色された単一細胞の%を示す。斜線棒および点刻棒は、それぞれアッセーの前24時間および48時間処置した胎児の細胞を表す。各々の棒線は3個の平均±標準誤差を表す。CAMは、mAbLM609(抗αvβ3)、CSAT(抗β1)またはPBSで実施例12A2)で述べるように処置された。CAMはまた、実施例12Bで述べるように、環状ペプチド69203(サイクロ−RGDfV、ペプチド203と表示)またはコントロール環状ペプチド69601(サイクロ−RADfV、ペプチド601と表示)で処置された。
【0020】
図19Aおよび19Bは、実施例12Cで述べるように、CSAT(抗β1)(図19A)またはLM609(抗αvβ3)(図19B)で処置した胎児のCAMの単一細胞懸濁をアポプトーシスタグおよび沃化プロピジウムで染色し、FACSで分析した結果をまとめたものを示す。Y軸は、アポプトーシスタグ染色の細胞数を表し、X軸は、沃化プロピジウム染色(DNA含有量)を表す。水平線は、アポプトーシスタグ染色の陰性ゲートを表す。左および右のパネルは、CSAT(抗β1)(図19A)およびLM609(抗αvβ3)(図19B)処置胎児のCAM細胞をそれぞれ示す。細胞サイクルの分析は、各状態当たり約8000例の分析によって実施した。
図20A−20Cは、実施例12Cで述べるように調製したCSAT(抗β1)処置胎児のCAM組織を表し、図20D−20Fは、LM609(抗αvβ3)処置胎児のCAM組織を表す。図20Aおよび20Dは、アポプトーシスタグで染色し、D.I.C.画像上に重ね合わせて蛍光(FITC)によって可視化させた組織を示す。図20Bおよび20Eは、mAbのLM609(抗αvβ3)で染色し蛍光(ローダミン)で可視化させた同じ組織を示す。図20Cおよび20Fは、アポプトーシスタグおよびLM609の両方で染色した同じ組織の合併させた画像を示し、ここで黄色の染色は共通の存在部位を表す。棒線は、左および右のパネルでそれぞれ15および50μmを表す。
【0021】
A.定義
アミノ酸残基:そのポリペプチド結合における化学的消化(加水分解)に際して形成されるアミノ酸。本明細書で述べるアミノ酸残基は、好ましくは“L”異性体形である。しかしながら、所望の機能特性が該ポリペプチドに保持されるかぎり、“D”異性体形残基をいずれのL−アミノ酸残基とも置換することができる。NH2は、ポリペプチドのアミノ末端に存在する遊離アミノ基を指す。COOHは、ポリペプチドのカルボキシ末端に存在する遊離カルボキシ基を指す。標準的なポリペプチド命名法(Biol.Chem.,243:3552-59(1969)に記載され、37CFR §1.822(b)(2)で採用された)に合わせ、アミノ酸残基の略語は下記の対応表に示す:
【0022】

【0023】
本明細書では、全てのアミノ酸残基配列は、その左と右の向きが、アミノ末端からカルボキシ末端の慣用的方向になっている式で表されているということは留意されねべならない。さらに、アミノ酸残基配列の始めと終わりのダッシュは、1つまたは2つ以上のアミノ酸残基のまた別の配列へのペプチド結合を示すことも留意されるべきである。
ポリペプチド:αアミノ基と隣接するアミノ酸残基のカルボキシ基との間でペプチド結合によって互いに連結された直線状のアミノ酸残基の列を指す。
ペプチド:本明細書で用いられているように、ポリペプチドとして互いに連結された50アミノ酸残基より大きくない直線状のアミノ酸残基の列を指す。
環状ペプチド:対応する直線状ペプチドに由来し、遊離のN−末端またはC−末端が存在せず、この場合、対応する直線状ペプチドのN−末端が、前記対応する直線状ペプチドのC−末端カルボキシレートとアミド結合を形成する。
蛋白:ポリペプチドの場合のように互いに連結された50アミノ酸残基より大きい直線状のアミノ酸残基の列を指す。
合成ペプチド:天然に生じる蛋白およびそのフラグメントにとらわれない、ペプチド結合で共に連結された化学的に生成されたアミノ酸残基の鎖を指す。
【0024】
B.一般的考察
一般に本発明は、脈管形成が特定のビトロネクチンレセプターαvβ3によって仲介され、さらにαvβ3機能の抑制は脈管形成を抑制するという発見に関連する。この発見は、脈管形成が種々の疾患の進行に果たす役割のゆえに重要である。脈管形成を抑制することによって、これら疾患に介入し、その症状を改善し、いくつかの場合にはこれら疾患を治癒させる。
新しい血管の増殖が、ある疾患に付随する病理の原因となるか、または遠因となる場合、脈管形成の抑制は、これらの疾患の有害な影響を軽減させるであろう。類リューマチ性関節炎、糖尿病性網膜症、炎症疾患、再狭窄などが例に含まれる。新しい血管の増殖が有害組織の支援に必要とされる場合、脈管形成の抑制は、該組織への血液供給を低下させ、したがって血液供給の要求で支えられている組織集団の減少に役立つ。その例には、腫瘍が厚さ数ミリを越えて増殖するために、さらに固形腫瘍の転移を成立させるために、新規血管新生が持続的な要求となる腫瘍の増殖が含まれる。
【0025】
一部には、この治療が脈管形成に対して高度に選択的で、他の生物学的過程に対してそのようなことはないという理由で、本発明の方法は効果的である。実施例で示すように、新しい血管の増殖のみが実質的なαvβ3を含み、したがって、この治療方法は成熟血管に副作用を与えない。さらに、αvβ3は正常な組織には広範囲に分布せず、むしろ新しい血管で選択的に見出され、したがって、この治療法は、新しい血管の増殖を選択的に標的とすることができる。
αvβ3のみを抑制することによって脈管形成を効果的に抑制することができるという発見は、潜在的に高い選択性をもち、したがって相対的に毒性が低い治療組成物の開発を可能にする。したがって、本発明は、1つまたは2つ以上のインテグリンを抑制する能力を有するペプチドを基本とする試薬の使用を開示するが、より選択的にαvβ3を抑制し、したがってαvβ3によって仲介される過程以外の他の生物学的過程を抑制するという副作用をもたない他の試薬をデザインすることが可能である。
例えば、本技術によって示されるように、αvβ3と免疫的に反応する高度に選択的な単クローン性抗体で、αvβ3機能の抑制について同様に選択的である抗体を調製することが可能である。さらに、本明細書で述べるように、αvβ3の抑制について選択的であるようにRGD含有ペプチドをデザインすることが可能である。
【0026】
本発明の発見以前には、αvβ3の生物学的機能と拮抗する試薬を用いて、脈管形成および脈管形成に付属する過程のどれもインビボで抑制することが可能であることは知られていなかった。
C.脈管形成抑制の方法
本発明は、組織の脈管形成の抑制、それによって脈管形成に依存する組織内の減少を抑制するための方法を提供する。一般に、本方法は、脈管形成抑制量のαvβ3拮抗物質を含む組成物を組織に投与することを含む。
上記で述べたように、脈管形成は、“発芽(sprouting)”、血管形成または血管拡大を含む組織の新規血管新生(この脈管形成過程の全てはαvβ3の発現によって仲介され、さらにαvβ3発現に依存する)を伴う種々の過程を含む。外傷治癒、黄体形成および胎児発生を除いて、大半の脈管形成過程は病的過程に付随し、したがって本治療方法の使用はこれら疾患に対して選択的で、有害な副作用をもたない。
【0027】
脈管形成が重要であると考えられている種々の疾患が存在する。これらは脈管形成性疾患と呼ばれ、炎症疾患(例えば免疫および非免疫性炎症、慢性関節性リューマチ並びに乾癬)、不適切なまたは時宜を得ない血管の侵襲(例えば糖尿病性網膜症、血管新生緑内障、再狭窄、アテローム性動脈硬化症プラークの毛細血管増殖および骨粗鬆症)および癌関連疾患(例えば固形腫瘍、固形腫瘍転移、血管線維腫、水晶体後線維増殖症、血管腫、カポジ肉腫、および腫瘍増殖を支援するために新規血管新生を必要とする同様な癌)を含む。
したがって、病的組織の脈管形成を抑制する方法は、これら疾患の症状を改善し、疾患によっては、これら疾患の治癒に寄与することができる。一具体例では、本発明は、組織内の脈管形成それ自体の抑制を意図する。組織内の脈管形成の程度、したがって本発明によって達成される抑制の程度は種々の方法で測定できる。これらの方法は、例えば、免疫組織化学によってαvβ3免疫陽性の非成熟および発生期の血管構造を検出する実施例で詳述される。
本明細書で述べるように、種々の組織または統合された組織を含む種々の器官(皮膚、筋肉、腸、結合組織、関節部、骨および脈管形成刺激に応じて血管が侵襲することができる同様な組織を含む)のいずれも、病的状態での脈管形成を支援することができる。
【0028】
したがって、関連する具体例では、処置される組織は炎症組織で、抑制される脈管形成は炎症組織の脈管形成であり、そこには炎症組織の新規血管新生が存在する。この種類では、本方法は、例えば慢性関節性リュウーマチの患者の関節炎組織、免疫もしくは非免疫性炎症組織、乾癬組織などの脈管形成の抑制を意図する。
多くの具体例で本発明にしたがって処置される対象は、望ましくは人間の患者であるが、本発明の原理は、本発明が、“患者”という用語に含まれることを意図する全ての哺乳類に関して有効であることを示唆していることは理解されよう。本明細書では、哺乳類という語が、脈管形成に付随する疾患の治療が所望される哺乳類のいずれの種も、特に農業用、家畜用哺乳類種も含むことは理解されよう。
別の関連する具体例では、処置される組織は、糖尿病性網膜症、黄斑の変性または血管新生緑内障の患者の網膜組織であり、抑制される脈管形成は網膜組織の脈管形成であり、そこには網膜組織の新規血管新生が存在する。
さらに別の関連する具体例では、処置される組織は、固形腫瘍、転移、皮膚癌、乳癌、血管腫または血管線維腫、および同様な癌をもつ患者の腫瘍組織であり、抑制される脈管形成は腫瘍組織の脈管形成で、そこには腫瘍組織の新規血管新生が存在する。本発明によって治療できる代表的な固形腫瘍組織は、肺、膵、乳房、結腸、喉頭、卵巣および同様な組織である。典型的な腫瘍組織の脈管形成およびその抑制は実施例で述べる。
【0029】
腫瘍組織の脈管形成の抑制は、新規血管新生が腫瘍増殖で果たす重要な役割のために特に好ましい具体例である。腫瘍組織で新規血管新生がない場合、この腫瘍組織は必要な栄養物を得ることができず増殖が鈍化し退縮し、ついには壊死を起こして腫瘍死をもたらす。
本発明は、換言すれば、本発明の方法にしたがって腫瘍の脈管形成を抑制することによって、腫瘍の新規血管新生を抑制する方法を提供する。同様に、本発明は、脈管形成を抑制する方法を実施することによって腫瘍の増殖を抑制する方法を提供する。
これらの方法はまた転移の形成に対して特に効果的である。その理由は、(1)転移の形成は、転移癌細胞が原発腫瘍を出ることができるように原発腫瘍の血管新生を必要とすること、および(2)二次的部位での転移の確立には転移癌の増殖を支援する新規血管新生が必要とされることである。
【0030】
関連する具体例では、本発明は、他の治療(例えば固形腫瘍に対抗し、さらに転移を制御する通常の化学療法)と結合させた方法を実施することを意図する。脈管形成抑制物質の投与は、典型的には化学療法の間またはそのあとで実施される。しかし、腫瘍組織が、腫瘍組織への血液供給と栄養の提供により脈管形成の回復を誘発することによって、脈管形成抑制物質の毒性攻撃に応答している場合は、一連の化学療法の後で時々脈管形成を抑制することは好ましい。さらに、固形腫瘍が除去された後、転移の予防として脈管形成抑制物質を投与することは好ましい。本方法が腫瘍の新規血管新生の抑制に適用される限りにおいて、本方法はまた、腫瘍組織の増殖抑制、腫瘍転移形成の抑制、成長腫瘍退縮に利用することができる。実施例では、本発明のαvβ3拮抗物質のただ1回の静脈投与後に、成長腫瘍の退縮が示される。
再狭窄は、経皮経管冠状動脈形成術の実施部位での平滑筋細胞(SMC)の移動と増殖の過程であるが、再狭窄は動脈形成術の成功を妨げる。再狭窄時のSMCの移動と増殖は、本発明で抑制可能な脈管形成の過程と考えられる。したがって、本発明はまた、動脈形成術処置の後で本方法にしたがって患者の脈管形成を抑制することによって再狭窄を抑制することを含む。再狭窄の抑制のためには、典型的にはαvβ3拮抗物質は、動脈形成術の処置の後約2日から約28日間、より典型的には前記処置後最初の約14日間投与される。
【0031】
組織の脈管形成を抑制するため、したがって脈管形成関連疾患の治療のために方法を実施するこの方法は、αvβ3の天然のリガンドとαvβ3が結合することを抑制することができるαvβ3拮抗物質の治療的に有効な量を含む組成物と、脈管形成が生じているか、または脈管形成が生じる危険性がある組織とを接触させることを含む。
αvβ3拮抗物質の投与のための用量範囲は、本明細書でさらに述べるように、拮抗物質の形態およびその効能に左右されるが、脈管形成および脈管形成によって仲介される症状が改善される所望の効果をもたらすために十分に多い量である。この投与量は副作用、例えば高粘性症候群、肺水腫、うっ血性心不全などを生じるほど多くてはいけない。一般には、用量は、年令、健康状態、性別、および患者の疾患の程度で変動し、当分野で技術を有する者が決定することが可能である。この用量はまた、いずれかの合併症の出現により個々の内科医が調節することもできる。
治療的に有効な量は、処置される組織の測定可能な脈管形成の抑制を生じるために十分なαvβ3拮抗物質の量、すなわち脈管形成抑制量である。脈管形成の抑制は、本明細書で述べるように免疫化学的に、または当業者に既知の他の方法でin situ で測定できる。
αvβ3拮抗物質が、αvβ3模倣物、RGD含有ペプチド、抗αvβ3単クローン性抗体またはそのフラグメントの形態を取ることができるかぎり、効能、したがって“治療的に有効な”量の表示は変動する可能性があることは理解されよう。
しかしながら、本アッセー方法で示されるように、本発明のαvβ3拮抗物質候補の効能を当業者は容易に評価できるであろう。
【0032】
αvβ3拮抗物質の効能は、CAMアッセーでの脈管形成の抑制、ウサギ眼球インビボアッセー、マウス:ヒトキメラインビボアッセーを含む種々の手段によって、さらに天然のリガンドのαvβ3への結合抑制(上記の方法は全て本明細書に記載されている)、および同様なアッセーによって測定できる。
好ましいαvβ3拮抗物質は、拮抗物質濃度0.5マイクロモル(um)未満、好ましくは0.1um未満、より好ましくは0.05umで溶液中で天然のリガンド(例えばフィブリノーゲンまたはビトロネクチン)のαvβ3への結合を実質的に抑制する能力を有する。“実質的に”とは、αvβ3拮抗物質の存在下における抑制によって、フィブリノーゲンの結合が少なくとも50%減少するのが観察されることを意味し、50%抑制は本明細書ではIC50値と呼ぶ。
【0033】
より好ましくはαvβ3拮抗物質は、他のインテグリンの中でαvβ3に対して選択性を示す。したがって、好ましいαvβ3拮抗物質は、実質的にαvβ3へのフィブリノーゲン結合を抑制するが、別のインテグリン(例えばαvβ1、αvβ5またはαIIbβ3)へのフィブリノーゲンの結合を実質的に抑制しない。特に、好ましいαvβ3拮抗物質は、αvβ3へのフィブリノーゲン結合抑制のIC50が別のインテグリンへのフィブリノーゲン結合抑制のIC50活性と比較して10倍から100倍低いものである。インテグリンへのフィブリノーゲン結合抑制のIC50活性を測定する典型的なアッセーは実施例で述べる。
単クローン性抗体形をとる本発明のαvβ3拮抗物質の治療的に有効な量は、典型的には、生理学的に容認できる組成物として投与されたとき、約0.01マイクログラム(ug)/ミリリットル(ml)から約100ug/ml、好ましくは約1ug/mlから約5ug/ml、通常は約5ug/mlの血中濃度を達成するために十分な量である。換言すれば、用量は、約0.1mg/kgから約300mg/kg、好ましくは約0.2mg/kgから約200mg/kg、最も好ましくは約0.5mg/kgから約20mg/kgで変動可能で、1日1回または2回以上、1日または数日投与される。
拮抗物質が単クローン性抗体のフラグメント形をとる場合、この量は、完全抗体の質量と比較したフラグメントの質量を基に容易に調整できる。好ましい血中モル濃度は、約2マイクロモル(uM)から約5ミリモル(mM)、好ましくは約100uMから1mMの抗体拮抗物質である。
【0034】
ポリペプチドまたは他の同様なサイズの小分子αvβ3模倣物の形をとる本発明のαvβ3拮抗物質の治療的に有効な量は、典型的には生理学的に容認できる組成物として投与されたとき、約0.1マイクログラム(ug)/ミリリットル(ml)から約200ug/ml、好ましくは約1ug/mlから約150ug/mlの血中濃度を達成するために十分な量である。1グラム分子当たり約500グラムの質量をもつポリペプチドを基にすると、好ましい血中モル濃度は、約2マイクロモル(uM)から約5ミリモル(mM)、好ましくは約100uMから1mMのポリペプチド拮抗物質である。換言すれば、体重当たりの用量は、約0.1mg/kgから約300mg/kg、好ましくは約0.2mg/kgから約200mg/kgで変動可能で、1日1回または2回以上、1日または数日投与される。
本発明の単クローン性抗体またはポリペプチドは、注射または時間をかけて徐々に輸液により非経口的に投与できる。体内の処置される組織は、典型的には全身的投与によってアクセス可能で、したがってもっとも頻繁には治療用組成物の静脈内投与によって処置されるが、目的の組織が標的分子を含む蓋然性が高い場合は他の組織および薬剤送達手段も含まれる。したがって、本発明の単クローン性抗体またはポリペプチドは、静脈内、腹腔内、筋肉内、皮下、体腔内、経皮的に投与でき、さらに蠕動的手段によって送達させることができる。
【0035】
本発明の単クローン性抗体またはポリペプチドを含む治療用組成物は、例えばユニットドースの注射によって通常の手段で注射される。“ユニットドース”という用語は、本発明の治療用組成物に関して用いられるときは、患者に対する単位投与量として適切な生理的に規定された単位を指す。各単位は、必要な希釈剤(すなわち担体または賦形剤)と結合させて所望の治療効果を生じるように計算された予め決定された活性物質の量を含む。
実施例に示すように好ましい具体例では、αvβ3拮抗物質は単回投与として静脈内に投与される。
【0036】
この組成物は、投与製剤に適合する態様で、さらに治療的に有効な量で投与される。投与される量およびタイミングは処置を受ける患者、患者の活性成分を利用する系、および治療効果の所望程度に左右される。投与に必要な活性成分の正確な量は医師の判定にしたがい、各個人に固有である。全身的に適用するために適切な投与量の範囲は本明細書に開示するが、しかしながら投与ルートに左右される。適切な投与様式もまた変更可能であるが、最初の投与が典型的となり1時間または1時間以上の間隔で注射または他の投与によるその後の反復投与が続く。また別には、インビボ治療のために特定された範囲の血中濃度を維持するために十分な持続的静脈輸液も含まれる。
本実施例で示すように、脈管形成抑制および腫瘍退縮は、拮抗剤との最初の接触後早くも7日で生じる。拮抗剤へのさらなる接触または延長接触は、好ましくは7日から6週間、より好ましくは約14から28日である。
【0037】
関連する具体例において、本実施例は、αvβ3の抑制とαvβ3を含む新規血管系の細胞のアポプトーシス誘発との間の関係を明らかにする。したがって、本発明はまた、組織内の新規血管系におけるアポプトーシスの抑制方法を含む。この方法は、全ての組織における脈管形成抑制に関して、さらにそのために記載された条件に関して実質的に本明細書で開示した通りに実施される。ただ1つの注目に値する違いは、効果のタイミングの違いである。これは、アポプトーシスは急激に、典型的には拮抗物質との接触後約48時間で現れ、一方、本明細書で述べるように、脈管形成の抑制および腫瘍の退縮はより緩徐に出現する。この違いは、投与時間および所望する効果に関して治療方法に影響を与える。新規血管構造のアポプトーシスのためには、典型的には投与は24時間から約4週間であるが、48時間から7日間が好ましい。
【0038】
D.治療用組成物
本発明は、本明細書で述べる治療方法の実施に有用な治療用組成物を含む。本発明の治療用組成物は、本明細書で述べるように、活性な成分として組成物中に溶解または懸濁させたαvβ3拮抗物質とともに、生理的に許容できる担体を含む。好ましい具体例では、治療用αvβ3拮抗物質組成物は、治療の目的で哺乳類または人間の患者に投与したとき抗原性をもたない。
本明細書で用いられるように、組成物、担体、希釈剤および試薬に関して用いるとき、“医薬的に許容できる”、“生理的に許容できる”という用語は、望ましくない生理的影響(例えば悪心、眩暈、胃の不快感など)を生じることなく哺乳類に投与することができることを示し、相互に用いられる。
【0039】
組成物中に溶解または懸濁させた活性成分を含む医薬組成物の調製は当分野ではよく理解されており、製剤によって限定する必要はない。典型的には、そのような組成物は、液体または懸濁物のいずれかとして注射可能なように調製されるが、使用前に液体となる溶液または懸濁物に適した固形形態もまた調製できる。
この調製物はまた乳濁物でもよい。
活性成分は、本明細書で述べる治療方法で使用するために適した量で、医薬的に許容でき、さらに活性成分と適合する賦形剤と混合される。適切な賦形剤は、例えば水、食塩水、デキストロース、グリセロール、またはエタノールなど、およびそれらの組み合わせである。さらに、所望する場合には、少量の補助物質、例えば湿潤剤もしくは乳化剤、pH緩衝剤など活性成分の効能を高める物質を含むことができる。
【0040】
本発明の治療組成物は、該組成物中の成分の医薬的に許容できる塩を含むことができる。医薬的に許容できる塩には酸付加塩(ポリペプチドの遊離アミノ基とで形成される)が含まれるが、前記酸付加塩は、例えば塩酸もしくは燐酸のような無機酸で形成されるか、または酢酸およびマンデリン酸などの有機酸で形成される。遊離カルボキシル基とで形成される塩はまた、無機塩基(例えば水酸化ナトリウム、カリウム、アンモニウム、カルシウムまたは第二鉄)で、さらに有機塩基(例えばイソプロピルアミン、トリメチルアミン、2−エチルアミノエタノール、ヒスチジン、プロカインなど)で誘導することができる。
環状ポリペプチドαvβ3拮抗物質の調製で用いる場合、特に好ましいものはTFAおよびHClの塩である。ペプチドの代表的な塩は実施例で述べる。
生理的に許容できる塩は当分野で周知である。典型的な液体担体は滅菌水溶液で、これは活性成分および水の他には物質を含まないか、または、生理的pH値で例えば燐酸ナトリウムのような緩衝液、生理的食塩水、もしくは両方(例えば燐酸緩衝食塩水)を含む。また別には、水性担体は、1つ以上の緩衝塩の他、例えば塩化ナトリウムおよびカリウムのような塩、デキストロース、ポリエチレングリコール並びに他の溶質を含むことができる。
液体組成物はまた、水とともにまたは水を除いた液相を含むことができる。付加されるそのような液相の代表例は、グリセリン、植物油(例えば綿実油)および水−油乳濁物である。
治療用組成物は、脈管形成抑制量の本発明のαvβ3拮抗物質を含み、典型的には、治療用組成物の総重量当たり少なくとも0.1重量%の量を含むように製剤化される。重量%は、組成物全体に対する抑制物質の重量比である。したがって、例えば、0.1重量%は完全な組成物100グラム当たり0.1グラムの抑制物質である。
【0041】
E.インテグリンαvβ3の拮抗物質
αvβ3拮抗物質は、組織での脈管形成を抑制するために本方法で用いられ、それらは、天然のαvβ3リガンドとの機能的な相互作用が干渉されるような態様でαvβ3と相互に作用する化合物を含む種々の形態をとることができる。典型的な拮抗物質は、αvβ3のリガンド結合部位に由来するαvβ3の類似体、αvβ3−リガンド結合反応に必要な構造領域を模倣するαvβ3もしくはαvβ3の天然リガンドのいずれかの模倣物、αvβ3に特異的な天然のリガンドの機能的結合ドメインに対応する(特にαvβ3の天然リガンドのRGD結合ドメインに対応する)配列を有するポリペプチド、およびαvβ3もしくは天然リガンドのいずれかと免疫反応を生じる抗体を含むが、本明細書で述べるようにこれらの全ては拮抗物質活性をもつ。
【0042】
1.ポリペプチド
ある具体例では、本発明は、ポリペプチド形をとるαvβ3拮抗物質を意図する。本明細書に述べるように、ポリペプチド(ペプチド)αvβ3拮抗物質は、αvβ3−リガンド相互作用に必要な領域にαvβ3の天然リガンドまたはαvβ3自体のいずれかの特徴的な配列を有し、さらにαvβ3拮抗物質活性を示す。好ましいαvβ3拮抗物質ペプチドはRGDトリペプチドを含み、さらにRGD含有領域内で天然のリガンドと配列として対応する。
好ましいRGD含有ポリペプチドは、αvβ3の天然リガンド(例えばフィブリノーゲン、ビトロネクチン、ヴォン・ヴィレブランド因子、ラミニン、スロンボスポンジンおよび同様な配列)のRGD含有領域のアミノ酸残基配列に対応する配列を有する。これらαvβ3リガンドの配列は周知である。したがって、αvβ3拮抗物質ペプチドは、天然リガンドのいずれかに由来することができるが、フィブリノーゲンおよびビトロネクチンが好ましい。
【0043】
特に好ましいαvβ3拮抗物質ペプチドは、前述のように他のインテグリンと比較したとき、その天然のリガンドとαvβ3との結合を優先的に抑制する。これらのαvβ3特異的ペプチドは、少なくともαvβ3に対する特異性が望ましくない副作用(例えば他のインテグリンの抑制)の発生を減少させるという理由から特に好ましい。αvβ3に対して選択性をもつ好ましいαvβ3拮抗物質ペプチドの識別は、典型的な結合抑制アッセー(例えば実施例で述べるようなELISAアッセー)で容易に明らかにできる。
ある具体例では、本発明のポリペプチドは、約100アミノ酸残基未満、好ましくは約60残基未満、より好ましくは約30残基未満を含む。ペプチドは直線でも環状でもよいが、特に好ましいペプチドは環状である。
好ましい環状および直線ペプチド並びにそれらの名称は実施例の表1に示す。
【0044】
本ポリペプチドは、それが必要な配列を含み、さらに例えば本明細書で述べるようなアッセーでαvβ3拮抗物質として機能することができる限り、αvβ3の天然のリガンドのアミノ酸残基配列と同一である必要はないことは理解されよう。
本ポリペプチドには、それがαvβ3拮抗物質である限り、本明細書にそのアミノ酸残基配列が示されているポリペプチドのいずれの類似体、フラグメントまたは化学的誘導体も含まれる。したがって、その使用に際して変化が一定の利点を提供する場合は、本ポリペプチドは、種々の変化、置換、挿入および欠失に付すことができる。この点に関して、本発明のαvβ3拮抗物質ポリペプチドは、列挙ペプチドの配列と同一であるよりはむしろ対応するものであり、この場合1つまたは2つ以上の変化があり、本明細書で規定されたアッセーの1つまたは2つ以上でαvβ3拮抗物質として機能する能力を保持する。
したがって、ポリペプチドは、ペプチド誘導体の種々の形態のいずれかをとることができ、これは、アミド、蛋白との共役物、環状化ペプチド、重合ペプチド、類似体、フラグメント、化学修飾ペプチドおよび同様な誘導体を含む。
【0045】
“類似体”という用語は、本明細書に具体的に示した配列と実質的に同一なアミノ酸残基配列をもついずれのポリペプチドも包含するが、このポリペプチドは、1つまたは2つ以上の残基が機能的に同様な残基と保存的に置換され、さらにそれは本明細書で述べるようにαvβ3拮抗物質活性を示す。保存的置換の例には、1個の非極性(疎水性)残基(例えばイソロイシン、バリン、ロイシンまたはメチオニン)の別のものとの置換、1個の極性(親水性)残基の別のものとの置換(例えばアルギニンとリジン間、グルタミンとアスパラギン間、グリシンとセリン間)、1つの塩基性残基(例えばリジン、アルギニンまたはヒスチジン)の別のものとの置換、または1つの酸性残基(例えばアスパラギン酸またはグルタミン酸)の別のものとの置換を含む。
“保存的置換”という言葉はまた、そのようなポリペプチドが必須の抑制活性を示すということを条件に、非誘導残基の代わりに化学的に誘導された残基の使用を包含する。
【0046】
“化学的に誘導された”とは、官能側鎖基の反応によって化学的に誘導された1つまたは2つ以上の残基を有するポリペプチドを指す。そのような誘導分子は、例えば、遊離アミノ基が、塩酸アミン、p−トルエンスルフォニル基、カルボベンゾキシ基、t−ブチルオキシカルボニル基、クロロアセチル基またはフォルミル基を形成するように誘導された分子を含む。遊離カルボキシ基は、塩、メチルおよびエチルエステルもしくはエステルの他の種類を形成するように誘導することができる。遊離ヒドロキシ基は、o−アシルまたはo−アルキル誘導体を形成するように誘導することができる。ヒスチジンのイミダゾル窒素は、N−im−ベンジルヒスチジンを形成するように誘導することができる。化学誘導体として含まれるものはまた、20種の標準アミノ酸の天然に生じる1つまたは2つ以上のアミノ酸誘導体を含むペプチドである。例えば、4−ヒドロキシプロリンはプロリンの代用にできる;5−ヒドロキシリジンはリジンの代用にできる;3−メチルヒスチジンはヒスチジンの代用にできる;ホモセリンはセリンの代用にできる;さらにオルニチンはリジンの代用にできる。本発明のポリペプチドはまた、必須の活性が維持される限り、1つまたは2つ以上の付加および/または欠失、または本明細書にその配列を示したポリペプチドの配列と関係がある残基をもついずれの配列も包含する。
“フラグメンド”という用語は、そのアミノ酸残基配列が本明細書に示されているポリペプチドのアミノ酸残基配列より短いアミノ酸残基配列をもつ一切のポリペプチドを指す。
【0047】
本発明のポリペプチドが、αvβ3の天然のリガンドの配列と同一でない配列を有する場合、それは、典型的には1つまたは2つ以上の保存的もしくは非保存的置換が行われたからで、通常は約30数%未満、好ましくは10数%未満のアミノ酸残基が置換される。付加残基はまた、“リンカー”を提供する目的のためにポリペプチドのいずれかの末端に付加できるが、このリンカーによって本発明のポリペプチドは都合よく標識もしくは固形マトリックス、または担体に固定することができる。
本発明のポリペプチドとともに用いることができる標識、固形マトリックスおよび担体は以下で述べる。
【0048】
アミノ酸残基リンカーは通常少なくとも1残基で、40またはそれより多い残基、もっと頻繁には1から10残基が可能であるが、αvβ3リガンドエピトープは形成しない。連結に用いられる典型的なアミノ酸残基はチロシン、システイン、リジン、グルタミン酸およびアスパラギン酸または同様なものである。さらに、本ポリペプチドは、別に特定されない場合は、末端−NH2のアシル化(例えばアセチル化またはチオグリコール酸のアミド化)によって、末端のカルボキシルアミド化(例えばアンモニア、メチルアミンとともに)および同様な末端の修飾によって修飾された配列のために、αvβ3リガンドの天然の配列とは異なっていてもよい。周知のように末端の修飾は、プロテイナーゼの消化による感受性を減少させるために有用で、したがって溶液中、特に生物学的液体(プロテアーゼが存在する可能性がある)中のポリペプチドの半減期を延長させるために役立つ。この点について、ポリペプチドの環状化はまた有用な末端修飾で、環状化によって形成される安定な構造のため、および本明細書で述べるようにそのような環状ペプチドについて見られる生物学的活性のゆえに特に好ましい。
【0049】
本発明のいずれのペプチドも、医薬的に許容できる塩の形態で用いることができる。本発明のペプチドと塩を形成することができる適切な酸は、無機酸、例えばトリフルオロ酢酸(TFA)、塩酸(HCl)、臭化水素酸、過塩素酸、硝酸、チオシアン酸、硫酸、燐酢酸、プロピオン酸、グリコール酸、乳酸、ピルビン酸、蓚酸、マロン酸、コハク酸、マレイン酸、フマル酸、アントラニル酸、桂皮酸、ナフタリンスルホン酸、スルファニル酸などを含む。HClおよびTFA塩は特に好ましい。
本発明のペプチドと塩を形成することができる適切な塩基は、無機塩基(例えば水酸化ナトリウム、水酸化アンモニウム、水酸化カリウムなど)、および有機塩基、例えばモノ−、ジ−、およびトリ−アルキル並びにアリールアミン(例えばトリエチルアミン、ジイソプロピルアミン、メチルアミン、ジメチルアミンなど)、並びに場合によっては置換されたエタノールアミン(例えばエタノールアミン、ジエタノールアミンなど)を含む。
【0050】
本発明のペプチド(本明細書ではまた本ポリペプチドと称される)は、前記ポリペプチド分野で習熟した者にとって周知の技術(組み換えDNA技術を含む)のいずれかにより合成できる。合成化学の技術(例えば固相メルリフィールド型合成)は、純度、抗原的特異性、望ましくない副産物を含まないこと、生成の容易さなどから好ましい。利用可能な多くの技術についての優れた要約は以下の文献で見出すことができる:Stewardら、“固相ペプチド合成(Solid Peptide Synthesis”、W.H.Freeman Co.,San Francisco,1969; Bodanszkyら、“ペプチド合成(Peptide Synthesis)”、John Wiley & Sons,第2版、1976; J.Meienhofer,“ホルモン蛋白およびペプチド(Hormonal Proteins and Peptides)”、第2巻、46ページ、アカデミックプレス(ニューヨーク)、1983; Merrifield,Adv.Enzymol.,32:221-96,1969; Fieldsら、Int.J.Peptide Protein Res.,35:161-214,1990; 並びに米国特許第4244946号(固相ペプチド合成に関して)およびSchroderら、“ペプチド(The Peptides)”、1巻、アカデミックプレス(ニューヨーク)、1965(古典的溶液合成に関して)。これらの文献の各々は参照により本明細書に含まれる。そのような合成で用いることができる適切な保護基は、上記の文献およびマッコーミーの論文(J.F.W.Mcomie,“有機化学の保護基(Protective Groups in Organic Chemistry)”、Plenum Press,ニューヨーク、1973(この文献は参照により本明細書に含まれる))に記載されている。
【0051】
一般に、意図される固相合成方法は、成長ペプチド鎖への1つまたは2つ以上のアミノ酸残基または適切に保護されたアミノ酸残基の連続的付加を含む。通常は、第一のアミノ酸残基のアミノ基またはカルボキシル基のいずれかが適切な、選択的に除去可能な保護基で保護されている。種々の、選択的に除去可能な保護基は、反応側鎖基をもつアミノ酸(例えばリジン)のために利用される。
具体例として固相合成を用いる場合、保護アミノ酸または誘導アミノ酸は、不活性な固形支持体にその未保護カルボキシルまたはアミノ基を介して附着されている。アミノ基またはカルボキシル基の保護基を続いて選択的に除去し、適切に保護された補完(アミノまたはカルボキシル)基をもつ前記配列の次のアミノ酸を混合し、固形支持体に既に付着した残基とアミド結合を形成させるために適切な条件下で反応させる。続いて、アミノまたはカルボキシル基の保護基をこの新しく付加したアミノ酸残基から除去し、(適切に保護された)次のアミノ酸をその後付加し、以下同様に行われる。最後に、所望のアミノ酸が適切な配列で連結され、一切の残余の末端保護基および側鎖基保護基(および固形支持体)は連続的にまたは同時に除去されて、最終的な直線状ポリペプチドが得られる。
【0052】
上記に記載したように具体例として調製した直線状ポリペプチドは、対応する環状ペプチドを形成させるために反応させることができる。ペプチドを環状化させるための典型的な方法は、ジンマーらの論文に記載されている(Zimmerら、“Peptides 1992”、393-394ページ、ESCOM Science Publishers,B.V.,1993)。典型的には、t−ブトキシカルボニル保護ペプチドのメチルエステルをメタノールに溶解させ、水酸化ナトリウム溶液を加え、混合物を20℃で反応させ、メチルエステル保護基を加水分解により除去する。溶媒を蒸発させた後、t−ブトキシカルボニル保護ペプチドを酸性化水溶液から酢酸エチルで抽出する。続いて、t−ブトキシカルボニル保護基をジオキサン補助溶媒中で穏やかな酸性条件下で除去する。このようにして得られた遊離アミノおよびカルボキシ末端をもつ非保護直線状ペプチドをその対応する環状ペプチドに変換させるために、この直線状ペプチドの希釈溶液をジクロロメタンとジメチルフォルムアミドの混合物中で、1−ヒドロキシベンゾトリアゾルとN−メチルモルフォリンの存在下でジシクロヘキシルカルボジイミドと反応させる。生じた環状ペプチドはクロマトグラフィーで精製される。
特に好ましい環状ペプチド合成方法は、ガーラスらの論文(Gurrathら、Eur.J. Biochem.,210:911-921(1992))および実施例に記載されている。本方法で使用するために特に好ましいペプチドは、c−(GrGDFV)(配列番号4)、c−(RGDfV)(配列番号5)、c−(RADfV)(配列番号6)、c−(RGDFv)(配列番号7)および直線状ペプチドYTAECKPQVTRGDVF(配列番号8)であり、ここで“c−”は環状ペプチドを指し、大文字はL−アミノ酸の一文字記号で、小文字はD−アミノ酸の一文字記号である。これらペプチドのアミノ酸残基配列はまた、配列番号4、5、6、7および8にそれぞれ示される。
【0053】
2.単クローン性抗体
一具体例で、本発明は、αvβ3と免疫的に反応し、本明細書で述べるようにαvβ3の天然のリガンドとαvβ3が結合するのを抑制する単クローン性抗体の形態をとるαvβ3拮抗物質を開示する。本発明はまた、この抗体を産生する細胞株、この細胞株を作製する方法、およびこの単クローン性抗体を生成する方法を開示する。
本発明の単クローン性抗体は、1)分離αvβ3と免疫的に反応し、2)フィブリノーゲンのαvβ3への結合を抑制する抗体分子を含む。優先的にαvβ3と結合する好ましい単クローン性抗体は、ハイブリドーマ細胞株ATCCHB9537が分泌するmAbLM609の免疫反応特性をもつ単クローン性抗体を含む。このハイブリドーマ細胞株ATCCHB9537は、ブダペスト条約の要請にしたがって、米国菌培養収集所(ATCC)(1301 Parklawn Drive,Rockville,MD,USA)に1987年9月15日に寄託された。
“抗体または抗体分子”という用語は、免疫グロブリン分子の集団および/または免疫グロブリンの免疫学的に活性な部分(すなわち抗体結合部位またはパラトープを含む分子)の集団を指す集合名詞として本明細書では用いられる。
“抗体結合部位”とは、抗原と特異的に結合する重鎖および軽鎖の可変並びに超可変領域から構成される抗体分子の構造部分である。
本発明で使用する典型的な抗体は、完全な免疫グロブリン分子、実質的に完全な免疫グロブリン分子および、パラトープを含む免疫グロブリン分子の部分(当分野でFab、Fab’、F(ab’)2およびF(v)として知られ、また抗体フラグメントと称される部分を含む)である。
【0054】
別の好ましい具体例では、本発明は、この発明の単クローン性抗体に由来するFabフラグメントを含む短縮免疫グロブリン分子を含む。Fcレセプターを欠くこのFabフラグメントは可溶性で、血清半減期に関して治療的利点をもち、さらに可溶性フラグメントを使用するという態様に関して診断的利点をもつ。可溶性Fabフラグメントの調製は、免疫学分野で一般に知られており、種々の方法で達成できる。
例えば、抗体のFabおよびF(ab’)2部分(フラグメント)は、周知の方法(例えば、Theofilopolous & Dixon,米国特許第4342566号を参照のこと)による実質的に完全な抗体のそれぞれパパインおよびペプシンによる蛋白分解反応によって調製される。Fab’抗体部分もまた周知であるが、2つの重鎖部分を連結するジスルフィド結合を例えばメルカプトエタノールで還元し、生じた蛋白メルカプタンを例えばヨードアセトアミドのような試薬でアルキル化してF(ab’)2部分から生成される。完全な免疫グロブリンを含む抗体が好ましく、本明細書で詳述するように用いられる。
【0055】
“単クローン性抗体”という語は、特定のエピトープと免疫的に反応することができるただ1種の抗体結合部位を含む抗体分子の集団を指す。したがって、単クローン性抗体は、それが免疫的に反応するいずれかのエピトープに対して典型的にはただ1つの結合親和性を示す。それゆえ、単クローン性抗体は、各々は異なるエピトープに対して免疫的に特異的である、複数の抗体結合部位をもつ抗体分子を含む可能性がある(例えば二特異性単クローン性抗体)。
単クローン性抗体は、典型的にはただ1種の抗体分子を分泌(産生)するハイブリドーマと呼ばれる単一の細胞クローンによって産生される抗体から成る。このハイブリドーマ細胞は、抗体産生細胞とミエローマまたは他の自己永続化細胞株とを融合させて形成される。そのような抗体の調製は、最初コーラーとミルシュタイン(Kohler & Milstein,Nature 256:495-497(1975))によって記載され、その内容は参照により本明細書に含まれる。また別の方法はゾーラによって記載された(Zola,“単クローン性抗体:術式の手引(Monoclonal Antibodies:A Manual of techniques)”CRC Press,Inc.(1987))。そのように調製したハイブリドーマの上清をαvβ3と免疫的に反応させ、さらに天然のリガンドへのαvβ3結合を抑制する抗体分子の存在についてスクリーニングすることができる。
【0056】
簡単に記せば、単クローン性抗体組成物を産生するハイブリドーマを生成するために、αvβ3源(例えば記載(Chereshら、J.Biol.Chem.,262:17703-17711(1987)にしたがってM21ヒトミエローマ細胞から分離したαvβ3)で強化免疫を施した哺乳類の脾臓から得たリンパ球とミエローマまたは他の自己永続化細胞株を融合させる。
ハイブリドーマを調製するために用いる細胞株はリンパ球と同じ種に由来することが好ましい。典型的には、マウス129GLX+株が好ましい哺乳類である。本発明で使用する適切なマウスミエローマには、ヒポキサンチン−アミノプテリン−チミジン−感受性(HAT)細胞株P3X63−Ag8.653、およびSp2/0−Ag14が含まれるが、これらは米国菌収集所(Rockville,MD)からそれぞれCRL1580およびCRL1581の名称で入手できる。
【0057】
典型的には、ポリエチレングリコール(PEG)1500を用いてミエローマ細胞と脾臓細胞を融合させる。融合ハイブリッドはHATに対する感受性によって選択される。本発明の単クローン性抗体を産生するハイブリドーマは、実施例で述べるように酵素結合免疫吸着アッセー(ELISA)を用いて識別される。
本発明の単クローン性抗体はまた、適切な特異性の抗体分子を分泌するハイブリドーマを含有する栄養培地を含む単クローン性ハイブリドーマの培養を開始することによって製造できる。培養は、ハイブリドーマが抗体分子を培養液中に分泌するために十分な時間、および条件下で維持される。続いて抗体含有培養液を採集する。その後、抗体分子を周知の技術でさらに分離する。
これらの組成物の調製のために有用な培地は当分野で周知であり、市販もされているが、合成培養液、系統内交配マウスなどを含む。代表的な合成培養液は、ダルベッコーの最少必須培地(Dulbecco's minimal essential medium)DMEM(Dulbeccoら、Virol.8:396(1959))で、4.5gm/lのグルコース、20mMグルタミンおよび20%ウシ胎児血清が補充される。
【0058】
単クローン性抗体、ハイブリドーマ細胞、またはハイブリドーマ細胞培養を製造する他の方法もまた周知である。例えば、以下の論文に記載された免疫学的全産物からの単クローン性抗体の分離方法を参照されたい:Sastryら、Proc.Natl.Acad.Sci.USA,86:5728-5732(1989); およびHuseら、Science,246:1275-1281(1989)。
また本発明に含まれるものは、本発明の単クローン性抗体を産生するハイブリドーマ細胞およびハイブリドーマ細胞を含む培養である。特に好ましいものは、単クローン性抗体mAbLM609を分泌するATCCHB9537と称されるハイブリドーマ細胞株である。mAbLM609はチェレッシュら(Chereshら、J.Biol.Chem.,262:17703-17711(1987)の記載のように調製されたが、その調製は実施例でもまた詳述する。
本発明は1具体例で、mAbLM609の免疫反応特性をもつ単クローン性抗体を含む。
【0059】
単クローン性抗体が本発明の単クローン性抗体と同じ(すなわち同等な)特異性(免疫反応特性)をもつか否かは、また、前者が予め選ばれた標的分子に後者が結合するのを妨げるか否かを調べることによって決定することが可能である。
固相に存在する標的分子の結合に関する標準的競合アッセーで本発明の単クローン性抗体によって結合減少が示されるように、もし被検単クローン性抗体が本発明の単クローン性抗体と競合するならば、その場合、2つの単クローン性抗体は、おそらく同じかまたは密接に関連するエピトープに結合するであろう。単クローン性抗体が本発明の単クローン性抗体の特異性をもつか否かを決定するまた別の方法は、通常反応性を有する標的分子と本発明の単クローン性抗体とを予め保温し、続いて被検単クローン性抗体を加え、被検単クローン性抗体が標的分子に結合する能力が抑制されるか否かを決定することである。被検単クローン性抗体が抑制されたならば、おそらく被検単クローン性抗体は、本発明の単クローン性抗体と同じか、または機能的に同等なエピトープ特異性をもつであろう。
単クローン性抗体が本発明の単クローン性抗体の特異性を有するか否かを決定するまた別の方法は、問題の抗体のCDR領域のアミノ酸残基配列を決定することである。そのCDR領域に同一または機能的に同等なアミノ酸残基配列もつ抗体分子は同じ結合特異性を有する。ポリペプチドの配列決定方法は当分野で周知である。
【0060】
抗体の免疫特異性、その標的分子結合能および、エピトープに対して抗体が示す付随する親和性は、この抗体が免疫的に反応するエピトープによって決定される。エピトープ特異性は、少なくとも部分的には、免疫グロブリンの重鎖の可変領域のアミノ酸残基配列によって、さらに部分的には軽鎖の可変領域アミノ酸残基配列によって決定される。
“結合特異性をもつ”という語を用いる場合、同等な単クローン性抗体が同じかまたは同様な免疫反応(結合)を示し、さらに予め選択された標的分子との結合に対して競合するということを示す。
ヒト化(humanized)単クローン性抗体は、特に人間に治療的に使用する場合はネズミの単クローン性抗体より特別な利点を提供する。具体的には、ヒトの抗体は“外来”抗原のように急激に血液循環から排除されず、さらに外来抗原および外来抗体と同じ態様で免疫系を活性化させない。“ヒト化”抗体の調製方法は当分野で一般に周知であり、本発明の抗体に容易に応用できる。
したがって、本発明は、一具体例において抗体の抗原結合能に実質的に干渉することなくヒトの免疫系成分を導入するために、移植によりヒト化させた本発明の単クローン性抗体を含む。
【0061】
3.αvβ3特異的模倣物(mimetics)
本発明は、“模倣物”と称されるポリペプチド、抗体および他の分子を含むαvβ3拮抗物質は、一般に本発明で用いることができることを示すが、この模倣物はαvβ3機能と干渉する能力を有する。特に好ましいものは、特にαvβ3の機能と干渉し、他のインテグリンの機能には干渉しない。
本明細書では、試薬がその必須の生物学的活性を保有している限り、種々の試薬が本方法で使用するために適切であることは理解されよう。これらの試薬は包括的に模倣物と称するが、それは、模倣物が、レセプターとリガンドの機能的相互作用に必要なαvβ3またはαvβ3リガンドのいずれかの結合ドメインを“模倣する”能力を保有し、したがって正常な機能に干渉(すなわち抑制)するからである。
【0062】
αvβ3模倣物は、抗体またはリガンド由来ペプチド以外で上記の特性を示すいずれの分子でもよい。それはペプチドの合成類似体でもよく、上記の結合ドメインの結合ポケットのような形状をした化合物または他の分子でもよい。
αvβ3模倣物のデザインは、当分野で既知の薬剤デザインのための様々な構造分析方法のいずれを用いても実施できる。これらの分析方法には、分子モデリング、二次元核磁気共鳴(2−DNMR)分析、X線結晶学、ペプチドのランダムスクリーニング、ペプチド類似体または他の化学ポリマーライブラリーおよび同様な薬剤デザイン方法が含まれる。
αvβ3拮抗物質は小さなポリペプチドまたは単クローン性抗体(αvβ3の選択的抑制という機能的特性を共有する2種の別々に異なる化学構造)がよいということを示す、本明細書で提示した広範囲の構造的証拠から見ると、本方法に有用なこのαvβ3拮抗物質の構造はあまり限定される必要はなく、本明細書で規定されるいずれのαvβ3模倣物も含むことができる。
【0063】
F.αvβ3の拮抗物質を識別する方法
本発明はまた、本方法にしたがって使用するαvβ3拮抗物質候補を識別する方法を開示する。これらのアッセー方法では、候補物質は、天然のリガンドへのαvβ3結合を抑制するその能力を調べられ、さらに、組織における脈管形成のその抑制能が調べられる。
第一のアッセーは、天然リガンドのαvβ3への直接結合の抑制を測定するが、好ましい具体例は実施例で詳細に記載される。このアッセーは典型的には、ELISAによって固相における分離αvβ3への天然リガンド(例えばフィブリノーゲン)の結合抑制の程度を測定する。
このアッセーはまた、αvβ3に特異性を示し、天然のリガンドが他のインテグリンに結合するのを抑制しない化合物を識別するためにも用いられる。この特異性アッセーは、ELISAアッセーを平行して行うことによって実施されるが、この場合、αvβ3および他のインテグリンは、天然のリガンドと結合するそれぞれの能力について、さらに、予め選択したリガンドと結合するインテグリンのそれぞれの能力を抑制する候補化合物について別々のアッセーチャンバーで同時にスクリーニングされる。好ましいスクリーニングアッセーの様式は実施例で述べる。
【0064】
第二のアッセーは、ニワトリの漿尿膜(CAM)での脈管形成を測定し、CAMアッセーと称される。このCAMアッセーは、他の研究者によって詳細に記載され、さらに腫瘍組織の脈管形成および新規血管新生の両方を測定するために用いられた(Ausprunkら、Am.J.Pathol.,79:597-618(1975); Ossonskiら、Cancer Res.,40:2300-2309(1980))。
CAMアッセーは、完全な組織の新規血管新生が生じ、実際のニワトリ胎児の血管がCAMまたはCAM上で成長する組織へと増殖していくので、十分に認知されたインビボの脈管形成のためのアッセーモデルである。
本明細書で示すように、CAMアッセーは、新規な血管増殖の量と程度の両方について新規血管新生の抑制を明示する。さらに、CAM上に移植されたいずれの組織(例えば腫瘍組織)の増殖も容易にモニターできる。最後に、このアッセーは、アッセー系の中に毒性に対する内部制御が存在するので特に有用である。ニワトリ胎児はいずれのテスト試薬にも曝すことができ、したがって、胎児の健康状態は毒性の指標である。
【0065】
第三のアッセーは、ウサギのインビボ眼球モデルで脈管形成を測定するが、これはウサギ眼球アッセーと称される。ウサギ眼球アッセーは、他の研究者によって詳細に記載され、さらに脈管形成抑制物質(例えばサリドマイド)の存在下で脈管形成および新規血管新生の両方を測定するために用いられた(D'Amatoら、Proc.Natl.Acad.Sci.,91:4082-4085(1994))。
新規血管新生過程(角膜の辺縁から角膜内へ増殖するウサギの血管によって実証される)が、眼球の本来透明な角膜を通して容易に見ることができるので、ウサギ眼球アッセーは、インビボ脈管形成の十分に認知されたアッセーモデルである。さらに、新規血管新生もしくは新規血管新生の退行の刺激または抑制の程度および量の両方が時間経過において容易にモニターできる。
【0066】
最後に、ウサギはいずれのテスト試薬にも曝すことができ、したがって、ウサギの健康状態がテスト試薬の毒性の指標である。
第四のアッセーは、マウス:ヒトキメラ−マウスモデルにおける脈管形成を測定し、キメラマウスアッセーと称される。このアッセーは、他の研究者によって詳細に記載され、さらに本明細書で、脈管形成、新規血管新生および腫瘍組織の退縮を測定するために詳述される(Yanら、J.Clin.Invest.,91:986-996(1993))。キメラマウスアッセーは、移植された皮膚移植片が正常なヒト皮膚組織と組織学的に類似し、完全組織の新規血管新生が生じ、この場合、実際のヒトの血管が移植されたヒトの皮膚から、移植されたヒト皮膚の表面のヒト腫瘍内に増殖していくので、インビボの脈管形成の有用なアッセーモデルである。ヒト移植片への新規血管新生の起始点は、ヒト特異的内皮細胞マーカーを用いて新規血管新生の免疫組織化学的染色によって明らかにすることができる。
【0067】
本明細書で示すように、キメラマウスアッセーは、新しい血管増殖の退行の量および程度の両方について新規血管新生の退行を明らかにする。さらに、移植皮膚上に移植されたいずれの組織(例えば腫瘍組織)の増殖に対する影響も容易にモニターできる。最後に、アッセー系の中に毒性に対する内部制御が存在するのでこのアッセーは有用である。キメラマウスはいずれのテスト試薬にも曝すことができ、したがって、マウスの健康状態は毒性の指標である。
【0068】
実施例
本発明に関する以下の実施例は説明のためであり、本発明を具体的に限定するものと解釈されるべきではない。さらにまた、当業者の視野内にある、現時点で明らかなまたは将来開発される種々の変形も、この後に続く本発明の請求の範囲内にあると考えられるべきである。
【0069】
1.合成ペプチドの調製
表1に示した直線状および環状ペプチドを、例えば以下の論文に記載された標準的固相合成技術を用いて合成した(Merrifield,Adv.Enzymol.,32:221-96(1969); G.B.Fields & R.L.Noble,Int.J.Peptide Protein Res.,35:161-214(1990))。
2グラム(g)のBOC-Gly-D-Arg-Gly-Asp-Phe-Val-OMe(配列番号1)を先ず60ミリリットル(ml)のメタノール(2Nの水酸化ナトリウム1.5mlが添加されている)に溶解して混合物を作製した。続いてこの混合物を20℃で3時間攪拌した。蒸発後、残留物を水にとり、希塩酸でpH3に酸性化し、エチル酢酸で抽出した。抽出物をNa2SO4上で乾燥させ、再度蒸発させて得られたBOC-Gly-D-Arg-Gly-Asp-Phe-Val-OH(配列番号2)をジオキサン中の2N塩酸20mlで、20℃2時間攪拌した。得られた混合物を蒸発させて、H-Gly-D-Arg-Gly-Asp-Phe-Val-OH(配列番号3)を得、続いて1800mlのジクロロメタンと200mlのジメチルホルムアミド(DMF)の混合物中に溶解し、その後0℃に冷却した。続いて、0.5gのジシクロヘキシルカルボジイミド(DCCI)、0.3gの1−ヒドロキシベンゾトリアゾル(HOBt)および0.23mlのN−メチルモルフォリンを攪拌しながら連続して加えた。
【0070】
得られた混合物を0℃でさらに24時間、20℃で48時間攪拌した。この溶液を濃縮し、混合ベッドイオン交換器で処理し塩を除去した。生じた樹脂をろ過で除去した後、清澄溶液を蒸発させ、残留物をクロマトグラフィーで精製してシクロ(-Gly-D-Arg-Gly-Asp-Phe-Val)(配列番号4)を回収した。表1に示した以下のペプチド(1文字コードのアミノ酸残基略語を用い、ペプチド番号により区別されている)が同様に得られた:シクロ(Arg-Gly-Asp-D-Phe-Val)(配列番号5);シクロ(Arg-Ala-Asp-D-Phe-Val)(配列番号6);シクロ(Arg-D-Ala-Asp-Phe-Val)(配列番号9);シクロ(Arg-Gly-Asp-Phe-D-Val)(配列番号7)。ペプチド62184の配列と同一の配列を有する66203と称するペプチドは、62184に存在するTFAの塩と異なりHCl塩を含むことだけが後者と異なっていた。実施例7(合成ペプチドが用いられている)で述べる脈管形成抑制アッセーでは、HClをもつこの66203ペプチドは、TFAをもつ同一のペプチドより脈管形成抑制でわずかに効果が高かった。
【0071】
【0072】
ペプチド62185の配列と同一の配列を有する69601と称するペプチドは、62184に存在するTFA塩ではなくHCl塩を含むことだけが後者と異なる。
環状ペプチドc−RADfV(69601)はフィブリノーゲンのインテグリンαvβ3への結合を抑制し、フィブリノーゲンのインテグリンαIIbβ3またはα5β1への結合は抑制しないことが示された(Pfaffら、J.Biol.Chem.,269:20233-20238(1994))。したがって、ペプチドc−RADfVはαvβ3に特異的である。
2.単クローン性抗体
ハイブリドーマATCC HB9537により分泌される単クローン性抗体LM609は、セファロース−ヒラマメレクチンビーズに吸着させた分離αvβ3を用いた免疫によって標準的ハイブリドーマ法で製造された。このαvβ3は、M21と称するヒトミエローマ細胞から分離し、抗体はチェレッシュらの記載にしたがって作製した(Chereshら、J.Biol.Chem.,262:17703-17711(1987))。M21細胞はモートン博士(Dr.D.L.Morton,University of California,ロサンゼルス、カリフォルニア)により提供され、2mMのL−グルタミン、50mg/mlの硫酸ジェンタマイシンおよび10%のウシ胎児血清を含むRPMI1640培養液で浮遊培養として増殖させた。
単クローン性抗体LM609は、特異的にαvβ3複合体と免疫的に反応し、αvサブユニットともβ3サブユニットともまたは他のインテグリンとも免疫的に反応しないことが示された。
【0073】
3.αvβ3発現の組織分布の特徴
A.抗インテグリンレセプター抗体による免疫蛍光
創傷治癒の間、血管の基底膜はいくつかの粘着蛋白(ヴォン・ヴィレブランド因子、フィブロネクチンおよびフィブリンを含む)を発現させる。さらに、粘着レセプターのうちのインテグリン類のいくつかは、培養平滑筋細胞および内皮細胞の表面で発現される(Cheresh、Proc.Natl.Acad.Sci.,USA,84:6471(1987);Janatら、J.Cell Physiol.,151:588(1992); Chengら、J.Cell Physiol.,139:275(1989)を参照のこと)。αvβ3(チェレッシュが記載したように(Proc.Natl.Acad.Sci.,USA,84:6471(1987))、ヴォン・ヴィレブランド因子、フィブリノーゲン(フィブリン)およびフィブロネクチンに対する内皮細胞レセプター)は、これらのインテグリンの中に存在する。このインテグリンは、内皮細胞の移動へと先導するカルシウム依存性シグナリング経路を開始させ、したがってリーベルシーら(Leavelseyら、J.Cell Biol.,121:163(1993))が記載したように、血管細胞生物学において基礎的な役割を果たすように思える。脈管形成中のαvβ3発現を調べるために、ヒトの創傷の肉芽形成組織または隣接正常皮膚を同意患者より採取し、1mlの燐酸緩衝食塩水で洗浄し、O.T.C.培地(Tissue Tek)に包埋した。この包埋組織を液体窒素中で約30から45秒瞬間冷凍した。クリオスタットミクロトーム上のこの冷凍ブロックから6ミクロンの厚さの切片を切り出し、続いてβ3インテグリン(αvβ3またはαIIbβ3)またはインテグリンのβ1亜類に特異的な抗体を用いて免疫ペルオキシダーゼ染色を施した。
【0074】
正常ヒト皮膚および創傷の肉芽形成組織の染色の結果は図1A−1Dに示す。
単クローン性抗体AP3およびLM534(それぞれβ3およびβ1インテグリンに対して誘導された)が、冷凍切片の免疫組織化学分析のために用いられた。4人の異なる供与者の組織を用いた実験で同一の結果が得られた。光学顕微鏡写真は300×の倍率で示されている。
αvβ3インテグリンは、肉芽形成組織の血管で豊富に発現されたが(図1B)、同じ供与者の正常皮膚の真皮および上皮では検出できなかった(図1A)。対照的に、β1インテグリンは、正常皮膚(図1C)および肉芽形成組織(図1D)の両方の血管および支質細胞で、さらにアダムスら(Adamsら、Cell,63:425(1991))の報告で先に示された通り上皮内の基底細胞で豊富に発現された。
B.抗リガンド抗体による免疫蛍光
それぞれβ3およびβ1インテグリン、ヴォン・ヴィレブランド因子およびラミニンに対するリガンドの存在について、上記のように調製したまた別の正常ヒト皮膚および肉芽形成組織の切片を調べた。ヴォン・ヴィレブランド因子は、正常ヒト皮膚(図2A)および肉芽形成組織(図2B)の血管に局在し、一方、ラミニンは、両組織の上皮基底膜の他、すべての血管に存在した(図2Cおよび2D)。
【0075】
C.癌組織の抗αvβ3抗体の分布
上記の分析に加えて、ヒトの患者から得た癌組織の生検材料もまたαvβ3の存在および分布について調べられた。インテグリンレセプター複合体、αvβ3に特異的な実施例2で調製された単クローン性抗体LM609で染色したという点を除き、組織は実施例1Aで述べたように調製された。さらに、代表的な腫瘍例をブリン固定液(Bulin Fixative)で8時間固定し、連続切片を切り出してH&E染色することによって、組織はまた顕微鏡による組織分析用に調製された。
膀胱癌、大腸癌、乳癌および肺癌組織の免疫ペルオキシダーゼ染色の結果は図3A−3Dに示す。αvβ3は、4種の癌生検材料に存在する血管にのみ豊富に発現し、これら組織に存在する他の細胞には全く存在しない。
ここに詳述した結果は、したがって、αvβ3インテグリンレセプターは、特定の組織型(すなわち肉芽形成組織、転移組織、および脈管形成が生じているその他の組織)で選択的に発現し、新しい血管の形成が停止している正常な組織では発現しないことを示している。これらの組織は、それゆえ、本発明の治療面についての理想的な標的を提供する。
【0076】
4.リガンド−レセプター結合アッセーにより検出されるαvβ3特異的合成ペプチドの同定
精製リガンド−レセプター結合アッセーで、αvβ3およびαIIbβ3レセプター結合活性と拮抗するそれらの能力を測定することによって、実施例1で調製された合成ペプチドをスクリーニングした。これら結合実験の方法は以下の論文に記載されているが(Barbasら、Proc.Natl.Acad.Sci.,USA,90:10003-10007(1993); Smithら、J.Biol.Chem.,265:11008-11013(1990); Pfaffら、J.Biol.Chem.,269:20233-20238(1994))、これらの内容は参照により本明細書に含まれる。
ここで述べるのは、レセプターが固形支持体に固定され、拮抗物質が可溶性であるリガンド−レセプター結合アッセーで拮抗物質を同定する方法である。さらに、リガンドが固形支持体に固定され、レセプターおよび拮抗物質が可溶性であるリガンド−レセプター結合アッセーも開示する。
簡単に記せば、選択した精製インテグリンを、ウェル当たり50ナノグラム(ng)の被覆濃度でタイターテック(Titertek)微量定量板(マイクロタイター)に別々に固定した。リガンド−レセプター結合アッセーで用いられたレセプターの精製は当分野で周知であり、当業者にとって通常の方法で容易に得ることができる。4℃で18時間保温した後、プレート上の非特異的結合部位をトリス緩衝食塩水中のウシ血清アルブミン(BSA)(10ミリグラム/ミリリットル(mg/ml))で封鎖した。抑制実験のためには、表1の種々の濃度の選択ペプチド(125I−ビトロネクチンまたは125I−フィブリノーゲンのインテグリンレセプター)をαvβ3およびαIIbβ3への結合を妨害する能力について調べた。これらのリガンドは、特定のインテグリン(αvβ3に対するビトロネクチンおよびαIIbβ3に対するフィブリノーゲン)について最適の結合を示したが、フィブリノーゲンのいずれかのレセプターへの結合を阻害する、ペプチドを用いた結合抑制実験は、レセプターのリガンドへの結合を1/2最大抑制するために必要なペプチド量のマイクロモル(uM)単位での正確な決定を可能にした。放射能標識リガンドは1nMの濃度で用いられ、結合は非標識合成ペプチドで別々に実施された。
【0077】
3時間保温した後、遊離リガンドを洗浄により除去し、結合リガンドをガンマ計測により検出した。別々に固定したαvβ3およびαIIbβ3レセプターへのレセプターおよび放射能標識フィブリノーゲンの結合を抑制するために表1に示した選択環状ペプチドを用いたアッセーで得られたデータは、データの点数間誤差が典型的には11%以下で高度に再現性があった。マイクロモル(uM)でのIC50(IC50uM)は2つのデータ点数の平均±標準偏差として表2に示したように表した。
【0078】
したがって、RGD含有またはRGD誘導環状ペプチド62181、62184、62185および62187(各々1個のD−アミノ酸残基をもつ)は、αIIbβ3レセプターへのフィブリノーゲン結合の抑制と比較したとき1/2最大抑制に必要なペプチド濃度はより小さいことで示されるように、αvβ3レセプターへの優先的なフィブリノーゲン結合抑制を示した。対照的に、その他のRGD含有またはRGD誘導環状ペプチド、62186、62175および62179は、αvβ3へのフィブリノーゲン結合阻害において有効でないか、またはαvβ3と比較してαIIbβ3へのフィブリノーゲン結合の優先的抑制を示した。これらの結果は、環状ペプチドRGDFV(ここでFはD−アミノ酸残基を示す)は、フィブリノーゲンのαvβ3インテグリンへの結合を特異的に抑制するが、αIIbβ3またはα5β1インテグリンへのそれは抑制しないというパフらの最近の報告(Pfaffら、J.Biol.Chem.,269:20233-20238(1994))と一致する。同様な結合抑制アッセーをRGDモチーフ(この配列はαvレセプターサブユニット、αIIbレセプターサブユニットまたはビトロネクチンリガンドアミノ酸残基配列に由来した)をもつ、または欠いている直線状ペプチドで実施した。直線状ペプチド、62880(VN由来アミノ酸残基35−49)、62411(αv由来アミノ酸残基676−687)、62503(αv由来アミノ酸残基655−667)および62502(αIIb由来アミノ酸残基配列296−306)の配列は表1に示す。これらペプチドの各々は、ビトロネクチン(VN)またはフィブリノーゲン(FG)のαIIbβ3またはαvβ3のいずれかへの結合抑制アッセーに別々に用いられた。各実験についての個々のアッセーのマイクロモルでのIC50のデータ(IC50uM)は表3に示す。
【0079】
【0080】
直線状ペプチドを用いた選択インテグリンレセプターへのリガンド結合抑制アッセーの結果は、αIIbβ3レセプターへのそれと比較して1/2最大抑制に必要なペプチド濃度がより小さいことによって示されるように、ペプチド62880のみが、FGまたはVNのαvβ3への1/2最大結合を抑制することに有効であることを示している。その他の直線状ペプチドのいずれもαvβ3へのリガンド結合を阻害する効果をもたなかった。ただし、ペプチド62502は、αIIbβ3へのVN結合の阻害に有効であった。
したがって、本明細書に記載するリガンド−レセプターアッセーは、本発明の実施でビトロネクチンレセプター(αvβ3)拮抗物質として使用されるように、特定のインテグリンレセプター(特にαvβ3)に対して選択的特異性を示す環状および直線状合成ペプチドの両方のスクリーニングに用いることができる。
【0081】
5.未処置ニワトリ漿尿膜(CAM)の特徴
A.CAMの調製
脈管形成は、正常な胎児の脈管形成が成熟血管の形成をもたらした後で、ニワトリの漿尿膜で誘発することができる。文献(Leibovichら、Nature,329:630(1987);Ausprunkら、Am.J.Pathol.,79:597(1975))に記載されたように、脈管形成は、特定のサイトカインまたは腫瘍フラグメントに反応して誘発されることが明らかにされた。CAMは、実施例6および7でそれぞれ述べるように、引き続き脈管形成および脈管形成抑制を誘発するためにニワトリ胎児から調製された。10日齢のニワトリ胎児はマックインタイヤー養鶏場(McIntyre Poultry,レークサイド、カリフォルニア)から入手し、60%の湿度で37℃で保温した。小さな工芸用ドリル(Dremel,Division of Emerson Electric Co.レーシーン、ウィスコンシン)を使って、直接卵の先端の気嚢上の卵殻に小孔を開けた。卵を光で検卵して予め調べた胎児血管の存在しない領域で卵の広い方の側面上に第二の孔をドリルで開けた。最初の孔に陰圧をかけ、その結果CAM(漿尿膜)を卵殻膜から引き剥がし、CAM上に偽の気嚢を作製した。小さな模型用輪状やすり(Dremel)を使って、1.0cm×1.0cmの真四角の窓を沈降CAMの上の卵殻に開けた。この小さな窓は下に横たわるCAMに直接アクセスすることを可能にした。
【0082】
得られたCAMの調製物を続いて以下の2通りのいずれかの時期で用いた:CAMへの追加の処置を行わない胎児発生6日(活発な新規血管新生が認められるステージ)、これは胎児の新規血管新生に対する影響を調べるために用いるモデルとなる;胎児発生10日、ここでは脈管形成は鎮静化している。後者の調製物は、したがって、実施例6で述べるようにサイトカイン処置または腫瘍との接触に反応して脈管形成再開を誘発させるために本発明で用いられた。
B.CAMの組織学
ニワトリ胎児CAMおよび/または、実施例8で述べるようにニワトリ胎児から切除したヒトの腫瘍の顕微鏡構造を調べるために、CAMおよび腫瘍を実施例3Aで述べたように冷凍切片用に調製した。免疫蛍光分析用に冷凍ブロックからクリオスタットミクロトームで6ミクロンの厚さの切片を切り出した。
【0083】
図4は、未処置10日齢CAMの血管のない領域の典型的な光学顕微鏡写真を示す。CAM系の脈管形成は、胎児発生のこのステージまでに鎮静化しており、この系は、隣接領域からその時点で一切血管が存在しないCAM領域へと、現存する血管から新規な血管構造の発生を刺激するために本発明で有用である
C.免疫蛍光で検出されるCAMのインテグリンプロフィル
CAM組織に存在するインテグリンレセプターの組織分布を見るために、腫瘍組織およびニワトリCAM組織の両方の6ミクロン(um)の冷凍切片をアセトンで30秒固定し、バックらの記載(Buckら、J.Cell Biol.,107:2351(1988))にしたがって、β1インテグリンサブユニットに特異的な単クローン性抗体、mAbCSAT(したがってコントロールとして使用)の10マイクログラム/ミリリットル(ug/ml)、または実施例2で調製したLM609で免疫蛍光染色を施した。一次染色の後、1:250希釈のヤギ抗マウスローダミン標識二次抗体(Tago)で染色し、一次免疫反応生成物を検出した。続いて、この切片をツァイス(Zeiss)の免疫蛍光化合物用顕微鏡で調べた。
【0084】
免疫蛍光分析の結果は、未処置10日ニワトリ胎児に存在する成熟血管はインテグリンβ1サブユニット(図5A)を発現することを示した。対照的に、図5Aに示した組織の連続切片では、LM609との免疫反応性は認められなかった(図5B)。したがって、LM609抗体によって検出されるインテグリンαvβ3は、10日齢の未処置ニワトリ胎児に存在する成熟血管では活発に発現されていなかった。CAMモデルおよび以下の実施例で示すように、血管が正常な胎児発生で新規の増殖を行っているか、またはサイトカインもしくは腫瘍によって誘発を受けているとき、血管はαvβ3を発現する。しかしながら活発な新規血管新生に続いてひとたび血管の成長が停止するや、αvβ3の発現は免疫蛍光分析によっては検出不能のレベルまで減少する。成熟血管における発現の欠如と較べて、脈管形成を経ている血管におけるこのようなαvβ3発現の調節は、CAMの脈管形成系を用いたモデル系として以下の実施例で示すように、脈管形成を制御しさらに抑制するために本発明の固有の能力を提供する。
【0085】
6.CAM脈管形成アッセー
A.増殖因子によって誘発される脈管形成
脈管形成は、実施例5Aで引用したようにサイトカインや増殖因子によって誘発されることが示された。ここで述べる実験では、実施例5で述べたCAM調製物での脈管形成は、この中で説明するようにCAM血管上に局所的に存在する増殖因子によって誘発された。
脈管形成は、ハンクス調整塩溶液(Hanks Balanced Salt Solution(HBSS))(GIBCO,グランドアイランド、ニューヨーク)または150ナノグラム/ミリリットル(ng/ml)の組み換え塩基性線維芽細胞増殖因子(βFGF)(Genzyme,ケンブリッジ、マサチューセッツ)を含むHBSSを飽和させた5ミリメートル(mm)×5mmのワットマンフィルターディスク(Whatman Filter Paper No.1)を10日齢のニワトリ胎児の血管のない領域に置くことによって誘発された。窓は後でテープを使って閉じた。他のアッセーでは、125ng/mlのβFGFも血管増殖を誘発するために効果があった。脈管形成は、72時間後光学顕微鏡でモニターした。CAMは瞬間冷凍し、6umの凍結切片をアセトンで固定して、10ug/mlの抗β1単クローン性抗体CSATまたはLM609のいずれかを用いて、実施例5Cで述べたように免疫蛍光によって染色した。
図5Cの免疫蛍光光学顕微鏡写真は、図5Bに示すように未処置のニワトリCAMでのαvβ3発現の欠如と対照的に、βFGF誘発脈管形成時にはニワトリCAM上に増強されたαvβ3が発現することを示す。αvβ3は、βFGF処置CAMの多くの血管で(75から80%)容易に検出できた。さらに、インテグリンβ1の発現は、未処置のCAMで認められる発現と変わりなく、β1はまた刺激血管で容易に検出可能であった。
【0086】
続いて、CAMクリオスタット切片のレーザー共焦点画像分析により、βFGF誘発脈管形成中のαvβ3とβ1インテグリンの相対的発現を定量した。続いてこの染色切片をツァイスのレーザー共焦点顕微鏡で調べた。25個のLM609染色血管および15個のCSAT染色血管(350から3500mm2の範囲にあり、平均サイズは〜1200mm2)を任意の視野から選び、レーザー共焦点画像分析による任意単位で単位面積当たりの各血管について平均ローダミン蛍光を測定した。データは血管の任意単位による平均蛍光量±標準誤差として表した。
【0087】
図6に表した結果は、ウィルコクソンの順位合計試験(Wilcoxon Rank Sum Test)(P<0.0001)で決定されたように、βFGF処置CAMではαvβ3染色は顕著に増強(4倍)されることを示した。一方、β1染色はβFGF処置の場合と顕著には異ならなかった。
さらに、このCAMアッセーを用いて、別の有効な脈管形成誘発物質である腫瘍壊死因子−α(TNFα)のβ1およびβ3インテグリンの発現における影響を調べた。βFGFまたはTNFαのいずれかをしみこませ、10日齢胎児のCAM上に静置したフィルターディスクは、72時間後に局所的脈管形成を促進させることが分かった。
これらの結果は、未処置(図7A)、βFGF処置(図7B)またはTNFα処置(図7C)CAMの光学顕微鏡写真で示す。血管は、βFGFおよびTNFα処置調製物では容易に識別できるが、未処置のCAMでは認められない。したがって、増殖因子/サイトカインの局所的適用は、隣接領域の成熟血管から最初は血管が存在しなかった領域へと脈管形成の誘導をもたらす。図5Cで示されたようにβFGFで誘発される血管および同時に発生するαvβ3発現から考えると、TNFαの処置は匹敵する活性をもたらす。
【0088】
これらの発見は、ヒトおよびニワトリの両方で、脈管形成中の血管は強化されたαvβ3発現を示すことを示唆する。これに一致して、培養内皮細胞上のαvβ3発現は、以下の論文に記載されたように(Janatら、J.Cell Physiol.,151:588(1992); Enensteinら、Exp.Cell Res.,203:499(1992);およびSwerlickら、J.Invest.Der.,99:715(1993))、種々のサイトカインによってインビトロで誘発することができる。
増殖因子誘発脈管形成の抗体およびペプチド抑制物質による影響は、実施例7Aおよび7Bに示される。
【0089】
B.胎児の脈管形成
胎児の新規血管系の自然状態での形成に対する脈管形成抑制物質の効果を調べるCAM調製物は、先に述べたように6日齢のニワトリ胎児であった。この発生段階では、血管は新規の増殖を行っており、したがってαvβ3が胎児の脈管形成に参画するか否かを決定するための有用な系を提供する。このCAM系は、アッセーを胎生10日目ではなく6日目に実施した点を除き上記のように調製された。本発明の抗体およびペプチドによる処置の胎児脈管形成に対する影響は実施例7Cに示す。
【0090】
C.腫瘍誘発脈管形成
腫瘍誘発脈管形成におけるαvβ3の役割を調べるために、先に増殖させ17日齢のニワトリ胎児のCAMから分離した種々のαvβ3陰性ヒトメラノーマおよび癌フラグメントを、ブルックスらの論文(Brooksら、J.Cell Biol.,122:1351(1993))および本明細書で述べるようにCAMアッセーに用いた。このフラグメントは、緩衝液の存在だけで強度の新規血管新生を誘発した。
脈管形成は、腫瘍フラグメントをCAM上に直接添加することによってCAM系で誘発された。ニワトリ胎児CAMの調製は、上記で述べた方法と同一であった。ディスクの代わりに、重量で50ミリグラム(mg)から55mgのヒトメラノーマ腫瘍M21L、ヒト肺癌腫瘍UCLAP−3、ヒト膵臓癌細胞株FG(Chereshら、cell,58:945-953(1989))、またはヒト喉頭癌細胞株HEp3(これら全てはαvβ3陰性腫瘍である)を、当初は血管が存在しない領域のCAM上に静置した。
【0091】
M21Lヒトメラノーマ細胞株、UCLAP−3ヒト肺癌細胞株、FG膵臓癌細胞株またはHEp3ヒト喉頭癌細胞株(全てαvβ3陰性)を用いて、ニワトリ胎児のCAM上に固形ヒト腫瘍を増殖させた。8×106のM21L、UCLAP−3およびFBまたは5×105のHEp3細胞の単一細胞懸濁を、全容積30マイクロリットル(ul)のHBSSとしてCAMに用いた。窓をテープで封じ、胎児を7日間保温して、ヒト腫瘍病巣を増殖させた。7日の終わりに(17日齢胎児)、腫瘍をCAMから切除し、周辺のCAM組織を切り取った。腫瘍を50mgから55mgの腫瘍フラグメントにスライスし、脈管形成アッセーまたは腫瘍増殖アッセーのいずれかに使用した。実施例6Aで述べたように、新たな10日齢のニワトリ胎児CAMの血管を欠いた領域にこの腫瘍フラグメントを静置した。
【0092】
実施例3Aで述べたように、αvβ3発現についてニワトリ胎児CAM上のインビボ増殖腫瘍をmAbLM609で染色した。腫瘍細胞の特異的染色は認められず、αvβ3発現の欠如を示唆した。
その後続いて、腫瘍誘発脈管形成に対する抗体およびペプチドの影響を測定するために、実施例7Dおよび7Eで述べるように処置した。腫瘍の退縮並びに脈管形成性血管および血管細胞のアポプトーシスに対する抗体およびペプチドの影響を測定するために、これらCAM腫瘍調製物をまた実施例8、9および12で述べるように処置した。
【0093】
7.CAMアッセーで測定された脈管形成の抑制
A.抑制物質の局所適用による増殖因子誘発脈管形成の抑制
1)単クローン性抗体による処置
αvβ3が脈管形成で活発な役割を果たすか否かを決定するために、βFGFまたはTNFαで飽和させたディスクをCAM上に静置し、続いて単クローン性抗体(mAbとも称する)、LM609(αvβ3に特異的)、CSAT(β1に特異的)またはP3G2(αvβ5に特異的)をこの調製物に添加した。
脈管形成は、βFGFで飽和させたフィルターディスクで10日齢胎児のCAMで誘発された。続いて、全容積25ulの滅菌HSBB中に25mgのmAbを含むHBSS50mlでこの円盤フィルターを0、24および48時間処理した。72時間で、CAMを採取し35mmのペトリ皿に入れ、1mlの燐酸緩衝食塩水で1度洗浄した。続いて、2人の観察者により二重盲検様式で、濾紙の裏側およびCAM組織をオリンパス立体顕微鏡で調べた。ディスクの直下のCAMの血管浸潤でCAMが>50%減少を示したとき、脈管形成抑制は顕著であると考えた。実験は、1条件につき6から7個の胎児を用いて、1抗体につき4回繰り返された。
【0094】
βFGF誘発脈管形成に対するmAb処置の影響は図8Aに示す。血管が存在しない未処置CAM調製物は、βFGF血管誘発(図8Bに示す)およびmAbによるCAMに対する影響(図8C−8E)との比較を提供するために図8Aに示した。mAbLM609で処理したCAMの約75%が、図8Eに示すように脈管形成の>50%抑制を示し、これらの多くは血管の浸潤を欠いているようであった。対照的に、緩衝コントロール(図8A)並びにmAbCSAT(図8C)およびP3G2(図8D)で処理したフィルターは、常に強度の血管新生を示した。
【0095】
同じ結果が脈管形成がTNFαで誘発されたとき得られた。血管を欠く領域近くの正常な血管の発達から出現する、以前から存在していた成熟血管に対するこれら同じ抗体の影響を調べるために、mAbで飽和させたフィルターディスクを、サイトカインを局所添加していない10日齢胎児のCAMの血管形成領域に静置した。立体顕微鏡下で可視化させて調べたとき、3種のmAbのいずれも以前から存在していた血管に影響を与えなかった。したがって、mAbLM609は新しい血管増殖のみを選択的に抑制し、隣接領域に存在する成熟血管には影響を与えなかった。同様な効果は、それぞれ実施例7A2)および7E2)で述べるように局所的にまたは静脈内に用いたとき、合成ペプチドでも認められた。
【0096】
2)合成ペプチドによる処置
増殖因子誘発脈管形成に対する環状および直線状ペプチドの影響を決定するために、また本発明の合成ペプチドを用いてCAMアッセーを実施した。実施例1のようにペプチドを調製し、80ugのペプチドが全容積25ulの滅菌HBSS中で提供された。このペプチド溶液を直ちにCAM調製物に適用し、続いて24時間、および48時間に再度適用した。72時間でこの濾紙および周辺CAM組織を切り出し、上記のように観察した。
【0097】
このアッセーの結果は、合成ペプチドが腫瘍誘発血管に静注された実施例7E2)で詳述する通り、図9A−9Cに示したものと同様であった。ここで、コントロールペプチド62186については、βFGF誘発血管は図9Aに示すように変化しないままであった。対照的に、環状RGDペプチド62814がフィルターに適用されたとき、血管の形成は抑制され、新しい血管構造を欠く領域が残された。この効果は、下記の実施例7E2)で述べるように図9Bで示される効果と外観上同様であった。さらに、静注されたペプチドについて図9Cでも示されるように、成熟血管が、増殖因子飽和フィルターが静置された場所から離れて存在する領域では、これら範囲外の血管に対して合成ペプチドの局所処理では効果は認められなかった。したがって、脈管形成に対するペプチドの抑制活性は、増殖因子によって誘発される脈管形成領域に限定され、以前から存在する近くの成熟血管には影響を与えず、周辺組織にはいかなる有害な細胞毒性ももたらさない。
実施例1で調製し、表1に示した他の合成ペプチドを用いて、同様なアッセーが実施される。
【0098】
B.抑制物質の静脈内適用による増殖因子誘発脈管形成の抑制
1)単クローン性抗体を用いた処置
CAM調製物に静脈内注射された単クローン性抗体の増殖因子誘発脈管形成に対する影響もまた、本発明で使用するために調べた。
静脈内注射のためのニワトリ胎児CAMの調製は、本質的には実施例7Aで述べた通りであるがいくつかの修飾を伴う。光で検卵する工程で際立つ血管を選び、それらの位置を示すために卵殻に印を付けた。卵殻にドリルで孔を開け、さらにCAMを落下させ、βFGF飽和濾紙を上記のようにCAMに静置した。窓を滅菌テープで封じ、胎児を孵卵器に戻した。24時間後、卵殻側面の先に選んだ際立つ血管上に直接注意深く第二の小孔を開けた。外側の卵殻を注意深く除去し、胎児膜は傷をつけずにおく。鉱物油(Perkin-Elmer Corp、ノーウォーク、コネチカット)の小滴で卵殻膜を透明にし、これによって血管を容易に見ることができる。精製滅菌mAbまたは合成ペプチド(後者は下記で詳述される)が、1胎児当たり全容積100ulの滅菌PBS中の200ugの投与量でただ1回30ゲージの注射針で血管に直接接種された。窓をテープで封じ、胎児を72時間まで保温した。フィルターディスクおよび周辺CAM組織を上記に述べたように分析した。
先にLM609が静脈内に接種されたCAM組織または腫瘍組織におけるLM609mAbの分布を知るために、本明細書および以下の実施例で示すように、HBSS中の2.5%BSAを用いて1時間室温で固定切片を遮断し、続いてヤギ抗マウスローダミン標識二次血清(Tago)の1:250希釈で染色した。その後、この切片をツァイスの免疫蛍光化合物顕微鏡で調べた。
【0099】
CAM調製物のβFGF誘発血管に対する静脈内抗体処理の結果は図10A−10Cに示す。図10Aに、βFGF処理の結果として誘発された脈管形成を示す。静脈内でmAbP3G2(抗αvβ5抗体)に曝されたとき、図10Bに示すようにβFGF誘発血管構造の状態に対して変化は認められなかった。対照的に、βFGF誘発脈管形成CAM調製物のLM609(抗αvβ3抗体)による処理は、図10Cに示すように新しい血管のフィルター領域への増殖を完全に抑制した。脈管形成に対するこの抑制効果は、したがってLM609抗αvβ3特異的抗体によるαvβ3レセプター活性の抑制から生じている。αvβ3の遮断はCAMのフィルター部位への新規血管構造形成を抑制しないので、αvβ5は、したがって新規血管の増殖のためのαvβ3と比較して必須ではない。
2)合成ペプチドによる処置
上記のCAM調製物の増殖因子誘発血管に、実施例1で調製した合成ペプチドを別々に静脈内注射する。血管の生存活性に対するこのペプチドの影響を同じように調べる。
【0100】
C.局所適用による胎児脈管形成の抑制
1)単クローン性抗体による処置
αvβ3が胎児の脈管形成に参画しているか否かを決定するために、CAMの血管の新規な成長に対するLM609の影響を6日齢の胎児(実施例5Aで述べるように活発な新規血管新生を特徴とするステージ)で調べた。CAMアッセーは、実施例6Cで述べたように調製し、続いてmAbで飽和させたディスクを6日齢の胎児のCAMにサイトカインの非存在下で局所的に適用した。3日後、CAMを切り出し写真を撮った。各実験は1群につき6個の胎児を含み、2回繰り返された。
抗体LM609(図11C)はこのような条件下で血管の増殖を防止したが、CSAT(図11A)またはP3G2(図11B)は防止しなかった。このことは、αvβ3は、脈管形成誘発のための添加増殖因子に依存しなかった胎児の新規血管新生において基本的な役割を果たすことを示唆している。
2)合成ペプチドによる処置
実施例1で調製した合成ペプチドを上記のように調製したCAM調製物に、実施例5A2)で述べたようにCAMへの局所的適用または血管への静脈内適用のいずれかで別々に加える。血管の生存活性に対するペプチドの影響を同じように調べる。
【0101】
D.局所適用による腫瘍誘発脈管形成の抑制
1)単クローン性抗体による処置
抗αvβ3拮抗物質、LM609並びにペプチド62181、62184、62185、62187および62880の胎児脈管形成への影響を調べた上記の脈管形成アッセーに加え、腫瘍誘発脈管形成におけるαvβ3の役割もまた調べられた。誘発物質として、αvβ3陰性ヒトM21−Lメラノーマフラグメント(それより前に増殖させ、17日齢のニワトリ胎児のCAMから分離されたもの)を用いた。このフラグメントは実施例6Cに述べたように調製した。
実施例7A1)で述べたように、25ulのHBSS中に25ugの濃度でmAbを腫瘍フラグメントに別々に局所適用し、続いて窓をテープで封じた。このmAbを同じ態様で24時間および48時間に再度加えた。72時間で、腫瘍および周辺CAM組織を実施例7A1)で述べたように調べた。
【0102】
実施例6Cで述べたように、腫瘍は、培養M21−L細胞(ヘルディング−ハーベルマンらの報告のようにインテグリンαvβ3を発現しない(Felding-Habermannら、J.Clin.Invest.,89:2018(1992))を10日齢のニワトリ胎児のCAMに移植することによって先ず誘導された。これらのαvβ3陰性フラグメントは、緩衝液単独で、またはmAbCSAT(抗β1)もしくはP3G2(抗αvβ5)の存在下で強度に新規血管新生を誘発した。対照的に、mAbLM609(抗αvβ3)は、殆どの血管が腫瘍塊および周辺CAMに浸潤するのを停止させた。
腫瘍誘発脈管形成に対するmAbの影響を定量するために、2人の観察者が、CAMの病巣面内の腫瘍に入り込む血管を立体顕微鏡下で二重盲検形式によって数えた。図12に示した各棒線は、2回の実験で各群12個のCAMから得た血管の平均数±標準誤差を示している。
【0103】
この定量分析は、ウィルコクソンの順位合計試験で求めたとき、緩衝液または他のmAb(P3G2もしくはCSAT)で処置した腫瘍と比較して、mAbLM609で処置した腫瘍に入る血管数は3倍減少することを明らかにした(P<0.0001)。M21−L腫瘍はαvβ3を発現しないという事実は、mAbLM609は、腫瘍細胞にではなく血管に直接影響を与えることによって脈管形成を抑制することを示唆している。これらの結果は、図3A−3Dに示した癌組織の生検材料におけるαvβ3の組織学的分布と一致するが、これらの図では、αvβ3の分布は腫瘍内の血管に限定され、腫瘍細胞自体には存在しなかつた。
【0104】
2)合成ペプチドによる処置
実施例1で調製した合成ペプチドが、上記のように腫瘍誘発脈管形成CAMアッセー系に局所的に適用される。血管の生存活性に対するこれらペプチドの影響が同じように調べられる。
【0105】
E.静脈内適用による腫瘍誘発脈管形成の抑制
1)単クローン性抗体による処置
実施例7D1)で述べるように調製した腫瘍誘発血管をまた、静脈内注射によりmAbで処置した。実施例7D1)で述べるように腫瘍をCAMに静置し、テープで窓を封じ、24時間後に200ugの精製mAbを先に述べたようにニワトリ胎児の血管に1回静脈内接種した。ニワトリ胎児をその後7日間保温した。
脈管形成の程度を続いて上記のように観察した。下記実施例8に述べるように、この期間経過後に腫瘍を切除してそれらの重量を調べ、腫瘍の増殖または退縮に対する抗体曝露の影響を決定した。
【0106】
2)合成ペプチドによる処置
CAMアッセー系の腫瘍誘発血管構造をペプチドに曝したときの影響もまた調べた。腫瘍−CAM調製物を上記のように用いたが、ただしこの場合、mAbの静脈内注射の代わりに、実施例1および実施例7A2)で述べたように調製した合成ペプチドが可視化された血管に別々に静脈内注射された。
HCl塩を含む環状ペプチド、66203およびコントロールペプチド62186を用いたCAMアッセーの結果は図9A−9Cに示す。図9Aで、コントロールペプチドによる処置は、腫瘍処理によって誘発され、CAMの本来は血管の存在しない領域に増殖した豊富で太い血管に影響を与えなかった。対照的に、環状RGDペプチド、66203(αvβ3に対する拮抗物質)がフィルターに適用されたとき、図9Bに示すように血管形成は抑制され、新しい血管構造の無い領域が残った。腫瘍を配置した場所の近くに位置する血管に対して有害な影響は全く存在しなかったことによって明らかなように、RGD含有ペプチドの抑制効果は特異的で局在的であった。
【0107】
したがって、図9Cでは、抑制ペプチドがCAMアッセー系に静脈内注射されたとき、腫瘍の配置場所の周辺ではあるが離れた領域のCAMに以前から存在する成熟血管に対しては影響は認められなかった。このような場所に以前から存在する血管は、これら血管内に流れている抑制物質によって影響を受けなかった。ただし、これら以前から存在する血管から腫瘍塊への新規な血管の形成は抑制された。したがって、実施例4のリガンド−レセプターアッセーでαvβ3の拮抗物質であることが以前に示された、66203および62184を含む合成ペプチドは、発達しつつある血管に限定される脈管形成を抑制するが、以前から存在する成熟した血管に対しては脈管形成を抑制しないことが明らかにされた。さらに、ペプチドの静脈内輸液は、図9Cの無傷の血管構造によって明らかなように、周辺領域に対して有害な細胞毒性を全くもたらさない。
同じようなアッセーが実施例1で調製し、表1に示す他の合成ペプチドを用いて実施される。
【0108】
8.CAMアッセーで測定したαvβ3拮抗物質による腫瘍組織の増殖抑制
実施例7D1)で述べたように増殖因子や腫瘍によって誘発される脈管形成に対する抗αvβ3拮抗物質の影響を視覚で評価することに加え、曝露した後の腫瘍塊への何らかの影響を測定することによってこの脈管形成への影響をまた測定した。この分析のために、腫瘍誘発脈管形成CAMアッセー系を実施例6Cおよび7Dで述べたように調製した。7日間の保温期間終了時に生じた腫瘍をCAMから切除し、一切の残余のCAM組織を除去して1mlの燐酸緩衝食塩水で洗浄し、各腫瘍の湿潤重量を求めた。
さらに、顕微鏡による組織分析用腫瘍の調製では、代表的な腫瘍例をブリン固定液で8時間固定し、パラフィンに包埋した。連続切片を切り出し、ヘマトキシリンおよびエオシン(H&E)で顕微鏡分析のために染色した(Gladsonら、J.Clin.Invest.,88:1924(1991))。切片の写真はオリンパス複合顕微鏡で250×で撮影した。
【0109】
A.局所適用
コントロール緩衝液(HBSS)、P3G2(抗αvβ5)またはLM609(抗αvβ3)の局所適用で得られた典型的なヒトメラノーマ腫瘍(M21L)の重量の結果は表4に示す。各処置につき多数の胎児の平均腫瘍重量(ミリグラム、mg)を調べ、表の下に示すように平均値の標準誤差と合わせて算出した。
【0110】
CAMアッセー系でαvβ3陰性ヒトメラノーマ腫瘍塊をLM609に曝露すると、未処置の平均腫瘍重量172mg±26は52mg±13に減少した。P3G2抗体はこの腫瘍塊に影響を与えなかった。したがって、αvβ3特異的LM609抗体の局所適用によるαvβ3レセプターの遮断は、前述の実施例に示したように脈管形成の抑制とともに腫瘍の退縮をもたらした。P3G2に曝露することによって生じた腫瘍塊の測定直径は、平均して約8mmから1cmであった。対照的に、LM609処置腫瘍は直径が平均して2から3mmであった。
これらの腫瘍の冷凍切片は、P3G2に曝された腫瘍については腫瘍細胞構造は無傷で、対照的にLM609に曝された腫瘍では組織化された細胞構造を欠いていることが明らかとなった。αvβ3レセプター活性は、したがってαvβ3発現新規血管構造の発達によって腫瘍塊に栄養を補給し続けるためにαvβ3陰性腫瘍にとって必須である。本発明のαvβ3拮抗物質によるαvβ3の遮断は、腫瘍への脈管形成の抑制をもたらし、最終的には腫瘍塊の減少をもたらす。
【0111】
B.静脈内適用
コントロール緩衝液(PBS、燐酸緩衝食塩水)、CSAT(抗β1)またはLM609(抗αvβ3)の静脈内適用によってもたらされた代表的な癌腫(UCLAP−3)の重量の結果は表5に示す。各処置につき多数の胎児の平均腫瘍重量を調べ、表の下に示すように平均値の標準誤差を合わせて算出した。
【0112】
CAMアッセー系におけるαvβ3陰性ヒト癌腫瘍塊のLM609への曝露は、未処置腫瘍の平均重量85mg±7を30mg±6に減少させた。CSAT抗体は腫瘍塊重量に顕著な影響を与えなかった。したがって、αvβ3特異的LM609抗体の静脈内適用によるαvβ3の遮断は、そのような処置が前述の実施例で示したように脈管形成抑制に加えて、上記でメラノーマ腫瘍塊の退縮をもたらしたように癌の退縮をもたらした。さらに、ヒトメラノーマ腫瘍の増殖は、LM609の静脈内注射によって同じように抑制された。
9.CAMアッセーで測定したαvβ3拮抗物質による腫瘍組織増殖の退縮
αvβ3拮抗物質の腫瘍増殖および生存に対する影響を調べるために、実施例5Aで述べたように、10日齢胎児のCAMにヒトメラノーマフラグメント並びに肺、膵および喉頭癌フラグメントを静置した。
【0113】
A.静脈内適用
1)単クローン性抗体による処置
a.LM609(抗αvβ3)およびCSAT(抗β1)による処置
αvβ3陰性ヒトメラノーマM21−L、膵癌FG、ヒト肺癌UCLAP−3またはヒト喉頭癌HEp3の癌フラグメントをCAMに移植後24時間して、胎児の静脈内にHBSのみ、またはmAbLM609(抗αvβ3)もしくはCSAT(抗β1)のいずれか(300ug/100ul)を一回注射した。腫瘍をさらに6日間増殖させた。保温期間の終了時に、腫瘍を注意深く切除し、周辺CAM組織を除去した。腫瘍切除は、別々の2人の研究者が容易に区別できる固形腫瘍塊を切除することによって実施した。これら腫瘍は識別が容易な辺縁を有し、したがって固形腫瘍塊とは容易に区別できる薄い半透明膜(CAM)は、腫瘍塊自体を傷つけることなく切除された。切除腫瘍の重量を測定し、形態学的および組織学的に調べた。
【0114】
図13に示すように、7日終了時の湿潤腫瘍重量を求め、処置前の最初の腫瘍重量と比較した。各棒線は1群につき5−10個の腫瘍の平均±SEを示す。調べた全ての腫瘍でコントロールと比較して、mAbLM609は顕著に(P<0.001)腫瘍増殖を抑制した。PBSまたはCSATで処置した腫瘍は全例で増殖した。対照的に、mAbLM609はこれら腫瘍の増殖を防止しただけでなく、殆どの事例で強度の退縮を誘発した。重要なことには、これらの腫瘍細胞はインテグリンαvβ3を発現せず、増殖抑制はこの抗体の新規血管構造に対する抗脈管形成作用によるものであり、腫瘍細胞に対する直接の作用ではないことを明示している。
b.LM609(抗αvβ3)およびP3G2(抗αvβ5)による処置
実施例5Aで述べたように、ヒトM21−Lメラノーマ腫瘍フラグメント(50mg)を10日齢胎児のCAMに移植した。24時間後、胎児の静脈内にPBSのみ、またはmAbLM609(抗αvβ3)もしくはP3G2(抗αvβ3)のいずれかを一回(300ug/100ul)注射した。実施例9A1)aで述べたように、腫瘍を増殖させ、ここで述べるように形態学的および組織学的に調べた。
mAbP3G2(抗αvβ5)またはLM609(抗αvβ3)で処置したM21−L腫瘍の代表例を形態学的に調べた。P3G2処置腫瘍は大きく(直径8mm)て良好な血管新生が認められたが、一方、mAbLM609で処置された抗体は、はるかに小さく(3mm)、検出可能な血管を欠いていた。
【0115】
組織学的切片を実施例9A1)aで述べたように、ヘマトキシリンおよびエオシンで染色した調製物でこれら腫瘍をさらに調べた。図14(上)に示すように、mAbP3G2(抗αvβ5)で処置した腫瘍は、腫瘍基質の多くの血管(矢印)の他に分裂像(楔形)でも示唆されるように活発に分裂する多数の生きた腫瘍細胞を示した。対照的に、mAbLM609(抗αvβ3)で処置した腫瘍では、元気な腫瘍細胞も血管もほとんど検出されなかった(図14下)。これらの結果は、インテグリンαvβ3の拮抗物質は腫瘍誘発脈管形成を抑制し、インビボでヒトの種々の腫瘍の増殖停止および退縮に導くことを示している。7日間の腫瘍増殖の後(17日齢胎児)で調べた胎児は、αvβ3拮抗物質で処置されたか否かにかかわらず、全体的な検査では正常のようであったということを指摘することは重要である。これらの発見は、このインテグリンの拮抗物質は発育胎児に毒性を持たないらしいということを示す。
【0116】
2)合成ペプチドによる処置
実施例5Aで述べたように、ヒトM21−Lメラノーマ腫瘍フラグメント(50mg)を10日齢胎児のCAMに移植した。24時間後に、胎児は、300ug/100ulのシクロ−RADfV(69601)またはシクロ−RGDfV(66203)のいずれかの静脈内注射を1回与えられた。合わせて72時間してから腫瘍を切除し、実施例9A1)で述べたように立体顕微鏡で形態学的および組織学的に調べた。
図15Aから15Eに示した写真は以下のように対応する:図15A、シクロRADfVペプチド(69601)で処置した2組のサンプル;図15B、シクロRGDfVペプチド(66203)で処置した2組のサンプル;図15C、シクロRGDfVペプチド(66203)で処置した同じ胎児から採取した隣接CAM組織;図15Dおよび15E、高倍率(13×)のペプチド処置腫瘍。図15Dは、コントロールペプチド(69601)で処置した腫瘍の正常な血管を示す。図15Eは、シクロ−RGDfVペプチド(66203)で処置した腫瘍の破壊された血管(矢印)の例を示す。
これらの結果は、ペプチド66203のみが血管形成を抑制し、さらに腫瘍に隣接するCAM組織の血管は影響されなかったことを示す。
【0117】
10.ウサギ眼球モデルインビボアッセーで測定したαvβ3拮抗物質による腫瘍組織の増殖退行
増殖因子誘発脈管形成に対する抗αvβ3拮抗物質の影響は、眼の角膜によって実証されるように本来透明な構造で観察することができる。新しい血管は角膜の辺縁(豊富な血液供給を有する)から角膜の中心部(本来血液供給をもたない)に向かって成長する。脈管形成刺激物質(例えばβFGF)は、角膜に適用されると角膜辺縁から新しい血管の成長を誘発する。脈管形成拮抗物質は、角膜に適用されると角膜の辺縁からの新しい血管の成長を抑制する。したがって、角膜は、角膜辺縁から丈夫なコラーゲンの詰まった角膜組織(これは容易に見ることができる)へと侵入する内皮細胞を介して脈管形成が進行する。ウサギ眼球モデルアッセーは、したがって化合物を直接眼の角膜に移植した後に続く脈管形成の刺激および抑制を直接観察するためのインビボモデルを提供する。
【0118】
A.ウサギ眼球インビボモデルアッセー
1)増殖因子によって誘発される脈管形成
脈管形成は、増殖因子βFGFを用いてウサギ眼球インビボモデルアッセーで誘発され、以下で詳述する。
a.増殖因子および単クローン性抗体を含むヒドロンペレットの調製
増殖因子およびmAbを含むヒドロンポリマーペレットを文献の記載にしたがって調製した(D'Amatoら、Proc.Natl.Acad.Sci.,91:4082-4085(1994))。個々のペレットは、βFGFを安定化させ、さらに周辺組織への緩徐な遊離を確保するためにスクラルファート(Carafet,Marion Merrell Dow Corporation)に結合させた増殖因子βFGF650ngを含んでいた。さらに、mAbLM609(抗αvβ3)またはmAbP1F6(抗αvβ5)のいずれか40μgをPBS中に含むヒドロンペレットを調製した。これらのペレットを、その表面に開けた2.5mmの中子を有する特別に調製したテフロンペッグの型に流し込んだ。約12μlの鋳造材料を各ペッグに入れ、滅菌フードで一晩重合させた。続いてペレットを紫外線照射で滅菌した。
【0119】
b.単クローン性抗体による処置
各実験は、一方の眼球にβFGFおよびLM609を含むペレットを、さらに他方の眼球にβFGFおよびマウスmAbP1F6(抗αvβ5)を含むペレットを与えた3匹のウサギから構成された。LM609(抗αvβ3)と他のmAbおよびPBSコントロールを比較するために対を形成する眼球試験を用いることは、被検mAb間での顕著な相違を明らかにすることができる厳密な試験のための手段を提供する。
P1F6mAbは、血管の内皮細胞の表面に見出されるが、おそらく脈管形成には必要ではないインテグリンαvβ5と免疫的に反応する。このmAbP1F6が脈管形成に必要か否かを決定するために、このmAbのみを含むペレットを調製し、下記に述べるよう調べて、このmAbは脈管形成を誘発しないことを確認した。
検査したmAbの全ては、周知の方法にしたがって蛋白AセファロースCL−4Bアフィニティーカラムクロマトグラフィーを用いて腹水から精製した。続いて溶出させた免疫グロブリンをPBSに対して透析し、デトキシ−ゲル(Detoxi-gel、Pierce Chemicals)で処理し内毒素を除去した。内毒素は強力な脈管形成刺激および炎症刺激物質であることが示された。mAbは、したがって、色素形成性カブトガニアメーバ様細胞溶解産物アッセー(Chromogenic Limulus Amebocyte Lysate Assay、Bio-Whittaker)を用いて内毒素の存在について検査し、検出可能な内毒素を含まないmAbのみをウサギ眼球モデルアッセーに用いた。
【0120】
βFGFおよびmAbLM609(抗αvβ3)またはP1F6(抗αvβ5)を含むヒドロンペレットをウサギの眼球に作製した角膜ポケットに挿入した。このヒドロンペレットはまたスクラルファートを含み、アッセー中にβFGFを安定化させる。個々のペレットは、ウサギの角膜の中央間質に外科的に作製した“ポケット”に移植された。この外科操作は、個々の角膜を写真により記録するためにカメラを搭載したビームスプリッターを備えたワイルドモデルM691手術用顕微鏡を用いて滅菌的術式で実施した。3mm×5mmの“ポケット”は、69ビーバーブレードを用いて角膜の厚さの半分まで3mmの切り目を入れて角膜間質に作製した。虹彩用へらを用いて間質は周辺で細かく切断され、辺縁から2mmの周辺端を用いてペレットを移植した。その後の14日間にβFGFおよびmAbは移植ペレットから周辺組織に拡散し、したがって角膜の辺縁から脈管形成が生じた。
【0121】
各処置の代表的な結果は図16Aから16Eに示す。出現する血管量を定量し、以下のように規定される時計時間の様式で記載する。時計が時間に分割されるのと同じ様式で、眼を12等分する。“血管の1時計時間”は、時計の1時間と同等な眼の領域を満たす血管の量を指す。βFGFのみを与えられた5匹のウサギは、新しい血管が角膜の辺縁から角膜の中央に向かって増殖している赤みがかった脈管形成を示したが、これは通常は血管をもたない。これらのウサギの1匹は、ペレットに対して1時計時間の血管だけしかもたなかった。βFGFおよびmAbLM609の両方を与えられた2匹のウサギには、検出可能な脈管形成が手術後14日まで全く存在しなかった。これらのウサギのうちの1匹には、14日までに3つの出血性出芽血管病巣があった。βFGFおよびmAbP3G2(抗αvβ5)を与えられたウサギのうちの2匹は強度の血管新生を示し、ここでは新しい血管は角膜の縁から角膜内に増殖した。これらのウサギのうち1匹には、ペレットに対して1から2時間の血管が存在しただけであった。ウサギ眼球モデルアッセーで実証されたように、mAbLM609(抗αvβ3)を与えられたウサギでは、増殖因子βFGFの存在下で正常な周辺血管に対しては脈管形成効果は認められなかった。対照的に、mAbP3G2(抗αvβ5)を与えられたウサギでは、増殖因子βFGFの存在下で正常な周辺血管に対して脈管形成が認められた。mAbLM609による角膜脈管形成の完全な抑制は、以前に報告されたいずれの抗脈管形成試薬よりも実質的に大きい。
【0122】
11.マウス:ヒトキメラアッセーで測定したαvβ3拮抗物質による腫瘍組織増殖のインビボ退行
マウス:ヒトキメラインビボモデルをSCIDマウスの皮膚の一部を新生児の包皮で置き換えることによって作製した(図17)。皮膚移植が定着した後、ヒトの包皮に癌細胞を接種した。測定可能な腫瘍が確立した後、mAbLM609(抗αvβ3)またはPBSのいずれかをマウス尾静脈に注射した。2−3週間してから、腫瘍を切り出し重量および組織を調べた。
A.マウス:ヒトキメラインビボアッセー
マウス:ヒトキメラインビボモデルは、実質的には文献にしたがって調製した(Yanら、J.Clin.Invest.,91:986-996(1993))。簡単に記せば、SCIDマウス(6−8週齢)から外科的に2cm2の領域の皮膚を切除し、ヒトの包皮で置き換えた。マウスを麻酔し、側面腹部領域の各々の側5cm2の毛を剃った。2か所の皮膚を筋膜まで完全に除去して2cm2の円形の移植床を調製した。ヒト新生児包皮由来の同じ大きさの完全な厚さのヒトの皮膚移植片をこの創傷床に静置しその場所に縫合した。移植片は皮膚に縫い付けたバンドエイドで被覆した。
【0123】
微細孔布テープも創傷を被覆するために用いた。
M21Lヒトメラノーマ細胞株またはMDA23.1乳癌細胞株(ATCCHTB26;mAbLM609を用いた組織切片の免疫反応性ではαvβ3陰性)を用いて、SCIDマウスのヒト皮膚移植片に固形ヒト腫瘍を形成させた。5×106M21−LまたはMDA23.1細胞の単一細胞浮遊液を経皮的にヒト皮膚移植片に注射した。続いてマウスを2から4週間観察して、測定可能なヒト腫瘍を増殖させた。
【0124】
B.静脈内適用
1)単クローン性抗体による処置
測定可能な腫瘍が増殖した後、M21L腫瘍細胞を注射したマウスに250μgのmAbLM609(抗αvβ3)またはPBSを、週に2度2から3週間尾静脈から静注した。この後、腫瘍を皮膚から切除し、周辺組織を除去した。表6の下に示したように各処置について数匹のマウスの平均腫瘍重量を調べた。
【0125】
【0126】
M21Lαvβ3陰性ヒト癌腫瘍塊をマウス:ヒトキメラアッセー系でLM609(抗αvβ3)に曝すことによって、PBS処置腫瘍の平均重量が198mgから113mgへと減少した。
mAbLM609(抗αvβ3)およびPBSで処置されたM21L腫瘍の代表例を形態学的に調べた。PBS処置腫瘍は大きく(直径8から10mm)血管新生は良好で、一方、mAbLM609(抗αvβ3)で処置した腫瘍ははるかに小さく(直径3から4mm)、検出可能な血管を欠いていた。
MDA23.1を注射した皮膚移植片に形成された腫瘍は検出および測定可能であった。成長腫瘍の形態学的検査によって、移植ヒト組織からMDA23.1腫瘍細胞へと新規血管新生が生じたことが明らかにされた。
【0127】
したがって、αvβ3特異的LM609抗体の静脈内適用によるαvβ3の遮断は、実施例9および10でそれぞれ述べたCAM系およびウサギ眼球モデル系と同じ態様でこのモデル系においても癌の退縮をもたらした。
12.CAMアッセーで測定される、インテグリンαvβ3の拮抗物質の存在下で細胞サイクルに入り、アポプトーシスが進行する血管細胞の刺激
脈管形成過程は、例えばβFGFおよびVEGFのようなサイトカインの血管細胞増殖を刺激する能力に依存する(Mignattiら、J.Cell.Biochem.,471:201(1991);Takeshitaら、J.Clin.Invest.,93:662(1994); およびKoyamaら、J.Cell.Physiol.,158:1(1994))。しかしながら、シグナリング現象は、これら血管細胞の成熟血管への分化を調節している可能性もあるようである。したがって、新規増殖または脈管形成が進行している血管細胞の増殖または分化と関係を有するシグナルとの干渉は、脈管形成の混乱をもたらすかもしれない。
【0128】
インテグリン連結(integrin ligation)現象は、アポプトーシスまたはインビトロのプログラムされた細胞死と同様に細胞増殖にも関与することが示された(Schwartz、Cancer Res.,51:1503(1993); Meredithら、Mol.Biol.Cell.,4:953(1993);Frischら、J.Cell Biol.,124:619(1994); およびRuoslahtiら、Cell,77:477(1994))。αvβ3拮抗物質の脈管形成に対する影響の綿密な調査によって、不連続で破壊された腫瘍付随血管の存在が明らかになった。したがって、血管連続性の欠如は血管細胞の選択的壊死またはアポプトーシスのためかもしれない。
この可能性を調べるために、増殖因子βFGFで脈管形成を誘発した後、本発明のmAbおよび環状ペプチドによる処置でCAMを調べた。
A.単クローン性抗体による処置
アポプトーシスは種々の方法で検出できるが、これらの方法は、DNAのフラグメント化を検出するために組織から分離したDNAの直接検査、さらに、フラグメント化DNAの遊離3’OH基を特異的に検出する抗体を用いた無傷の組織の3’OH基の検出を含む。
【0129】
1)DNAフラグメント化の分析
脈管形成は、実施例6Aで述べたように、10日齢胎児のCAMにβFGFで飽和させたフィルターディスクを静置することによって誘発された。LM609(抗αvβ3)によるCAMの免疫組織学的分析によって、βFGFによる脈管形成の開始後12から24時間で血管のαvβ3の発現は最高になることが明らかになった。したがって、βFGFによる刺激後24時間で、胎児の静脈内に100μlのPBSのみ、または300μgのmAbCSAT(抗β1)もしくはLM609(抗αvβ3)のいずれかを含むPBSを接種した。
DNAフラグメント化は、mAbLM609(抗αvβ3)、CSAT(抗β1)またはPBSの静脈内接種後24時間または48時間に、βFGF飽和フィルターディスク下のCAM組織を直接切除することによって検出した。切除CAM組織は滅菌PBSで3回洗浄し、細かくきざみ、0.25%の細菌性コラゲナーゼ(Worthington Biochemical; フリーホールド、ニュージャージー)に再浮遊させ、時々ボルテックスミキサーで攪拌しながら37℃で90分保温した。単一細胞浮遊液から等しい数のCAM細胞を用いてDNAを以前に報告されたように抽出した(Bissonetteら、Nature,359:552(1992))。簡単に記せば、等しい数のCAM細胞を10mMトリス−Cl(pH8.0)、0.5%(v/v)のトリトンX−100(シグマ、セントルイス、ミズーリー)中の10mMEDTAで溶解させた。細胞溶解物を4℃で15分16000×gで遠心し、可溶性フラグメント化DNAを完全なクロマチンペレットから分離させた。フラグメント化DNAを洗浄し沈澱させ、1.2%(w/v)アガーロースゲルで分析した。
【0130】
各処置から得た等しい数のCAM細胞から可溶性フラグメント化DNAを分離し、アガロースゲルで電気泳動的に分離し、臭化エチジウムで染色して可視化させた。処置後24時間で3種の異なる処置から得られたDNAフラグメント化の相対量には違いは検出されなかった。しかしながら、mAbLM609(抗αvβ3)による処置後48時間までに、mAbCSAT(抗β1)またはPBS単独で処置した胎児と比較したとき、DNAフラグメント化の顕著な増加が認められた。
2)細胞サイクルに入るための血管細胞の刺激
これらの過程におけるαvβ3の役割を実験的に調べるために、βFGFで処置したCAMまたは未処置のCAMから得た細胞を沃化プロピジウムで染色しさらにmAbLM609(抗αvβ3)で免疫的に反応させた。
【0131】
mAbLM609(抗αvβ3)、CSAT(抗β1)またはPBSで処置後24時間または48時間の胎児から分離したCAMを、上記のように細菌性コラゲナーゼで保温して単一細胞浮遊液にした。続いて、単一細胞を透過性にし、製造元(Oncor,ゲイサースバーグ、メリーランド)の指示にしたがいアポプトーシスタグ・インシトゥ・検出キット(Apop Tag Insitu Detection Kit)で染色した。アポプトーシスタグは、フラグメント化DNAの遊離3’OH基を特異的に検出する抗体である。そのような遊離3’OH基の検出は、アポプトーシス細胞の検出のための確立された方法である(Gavrieliら、L.Cell Biol.,119:493(1992)。
アポプトーシスタグ染色細胞を続いてPBS中の0.1%(v/v)トリトンX−100ですすぎ、0.5%(w/v)BSA、0.02%(w/v)アジ化ナトリウムおよびPBS中の200μg/mlのRNアーゼAを含むFACS緩衝液に再浮遊させた。細胞を1.5時間保温し、洗浄して蛍光活性化細胞選別によって分析した。細胞の蛍光は、ファックスキャンフローサイトメーター(FACScan flow cytometer)を用いて測定し、データは下記のように分析した。
【0132】
細胞の蛍光はファックスキャンフローサイトメーター(Becton Dickison、マウンテンビュー、カリフォルニア)で測定した。側面分散範囲(Side Scatter、SSC)および前方分散範囲(Forward scatter、FSC)は同時に決定し、全てのデータは、ファクスキャン研究用ソフト(Becton Dickison、マウンテンビュー、カリフォルニア)を備えたヒューレットパッカード(HP9000)コンピューターで収集した。データは、P.CリシスヴァージョンIソフト(P.C Lysis version I)(Becton Dickison、マウンテンビュー、カリフォルニア)で分析した。陰性コントロールゲートは、アポプトーシスタグキットの一次抗体を添加していない細胞浮遊液を用いて設定した。同一のゲート設定を両方の細胞集団に用い、異なる細胞処置につき約8000細胞を分析した。
mAb処置CAMから得られ、アポプトーシスタグで染色し、FACS分析で決定した単一細胞の%は図18に示す。斜線入り棒線は、分析の24時間前に処置した胎児の細胞を表す。点刻棒線は48時間前に処置した胎児の細胞を表す。
各棒線は、3つの複製の平均±標準誤差を表す。
図18に示すように、mAbLM609(抗αvβ3)で2日前に処置したCAMは、PBS単独またはCSAT(抗β1)のいずれかで処置したCAMと比較したときアポプトーシスタグ染色は3から4倍増加した。
【0133】
B.合成ペプチドによる処置
アポプトーシスに対する環状ペプチドの影響を決定するために、実施例6Aで述べたように、本発明の合成ペプチドを用いて増殖因子誘発脈管形成に関するCAMアッセーを実施した。ペプチド、シクロ−RGDfV(66203)およびシクロ−RADfV(69601)を実施例1で述べたように調製した。このペプチド溶液またはPBSをCAM調製物に300μg/mlの濃度で注射した。
24および48時間で濾紙および周辺CAM組織を切除してアポプトーシスタグで染色し、実施例12A2)で述べたようにアポプトーシスを検出した。
図18に示したように、ペプチド69203(スクロ−RGDfV)で2日前に処置したCAMは、PBS単独またはコントロールペプチド69601(シクロ−RADfV)のいずれかで処置したCAMと比較したとき、アポプトーシスタグ染色が3から4倍増加した。
C.アポプトーシスおよび細胞サイクルに対する単クローン性抗体による処置の影響
細胞サイクルに対する単クローン性抗体の影響を決定するために、沃化プロピジウムで染色することによって染色体DNAのコピー数について、さらにアポプトーシスタグで染色することによってアポプトーシスについて単一細胞浮遊液を調べた。
【0134】
mAbLM609(抗αvβ3)もしくはCSAT(抗β1)またはPBSで24時間または48時間前に処置したCAMの単一細胞浮遊液を、実施例12A1)で述べたように調製した。
アポプトーシスタグで細胞を染色するために、2.5%(w/v)BSAおよびPBS中の0.25%(w/v)アジ化ナトリウムを含む緩衝液で3回洗浄した。続いて細胞をPBS中の1%(w/v)パラホルムアルデヒドで15分固定し、その後上記のように3回洗浄した。非特異的結合を防止するために、単一細胞浮遊液をPBS中の5%(w/v)BSAで一晩4℃で処理した。続いて細胞を前述のように洗浄し、アポプトーシスタグで染色し、実施例12Aで述べたようにファックスキャンで細胞の蛍光を測定した。
各実験条件から得た細胞をPBS中の10μg/mlの沃化プロピジウム(シグマ、セントルイス、ミズーリー)で1時間染色し、PBSで2回洗浄し、アポプトーシスに典型的な核の特徴(クロマチン凝縮および分断化を含む)について調べた。アポプトーシス細胞の%を、少なくとも10から15個の任意に選んだ顕微鏡の視野から得た細胞の形態学的検査によって算定した。
【0135】
CSAT(抗β1)またはLM609(抗αvβ3)で処理した胎児のCAMの単一細胞浮遊液をアポプトーシスタグおよび沃化プロピジウムで染色し、FACSで分析した結果はまとめて図19に示す。Y軸はアポプトーシスタグ染色(アポプトーシス)を表し、X軸は沃化プロピジウム染色(DNA含有量)を表している。水平線は、アポプトーシスタグ染色の陰性ゲートを表す。左および右の写真はCSATおよびLM609処置胎児のCAM細胞をそれぞれ示す。細胞サイクル分析は、各条件につき約8000例の分析によって実施し、データは等高線図で表した。
DNA染料、沃化プロピジウムで染色した単一細胞サンプルによって、LM609(抗αvβ3)処置後48時間のCAM細胞の25−30%が核凝縮および/または分断化を示した。このような過程はアポプトーシスが進行している細胞の特徴である。これはCSAT(抗β1)処置CAMと対照的で、この場合90−95%の細胞が正常な核染色を示した。
【0136】
図19に示すように、LM609によるアポプトーシスの誘発と一致して、1コピー未満のDNAを含むピークに極めて多数の細胞が認められた(AO)。このピークは、後期のアポプトーシス細胞のフラグメント化DNAを表していることが以前に示された(Telfordら、Cytometry,13:137(1992))。さらに、これらのAO細胞は容易にアポプトーシスタグで染色され、アポプトーシス細胞を検出するこれら試薬の能力が確認された。しかしながら、AOで細胞が染色されることに加え、1コピーより多いDNAを含む極めて多くの細胞がアポプトーシスタグでもまた染色された(図19)。これらの結果は、LM609は、既に細胞サイクルに入った血管細胞でアポプトーシスを促進する能力を有することを明らかにしている。対照的に、細胞サイクルに入ったコントロールCAMから得られた細胞はごくわずかのアポプトーシスタグ染色を示し、コントロール処置CAMではアポプトーシス細胞が殆ど検出されなかったことと一致した。
【0137】
細胞サイクル(SおよびG2/M期)に入ったβFGF刺激CAMのこれらの細胞では、70%がLM609(抗αvβ3)で陽性染色を示した。これは、βFGF未処置CAMの細胞サイクルに入った細胞で認められた10%LM609染色に準えることができる。これらの発見は、βFGF刺激の後、αvβ3をもつ細胞の大半が活発な分裂を示していることを示唆する。
【0138】
これらの結果を合わせると、mAbLM609またはαvβ3の環状ペプチド拮抗物質の静脈内注射は、脈管形成抑制に続いてニワトリのCAM内でアポプトーシスを促進させることを示唆している。また、LM609との免疫反応性によってαvβ3の発現および、アポプトーシスタグとの免疫反応性によってアポプトーシスが進行している細胞について組織学的にCAMを調べた。LM609(抗αvβ3)、CSAT(抗β1)またはPBSで48時間前に処置した、実施例5Aで調製した胎児から切除したCAM切片を洗浄し、OTC(Baxter)に包埋し、液体窒素で瞬間冷凍した。CAM組織の6ミクロン切片を切り出し、アセトンで30秒固定し、使用まで−70℃で保存した。組織切片は、70%(v/v)エタノール中で簡単にすすぎ、続いてPBS中で3回洗浄して、染色用に調製した。次に、切片をPBS中の5%(v/v)BSAで2時間処理し、その後10μg/mlのmAbLM609で2時間保温した。続いて切片を洗浄し、1:50のローダミン共役ヤギ抗マウスIgG(Fisher Scientific,ピッツバーグ、ペンシルバニア)で2時間保温した。最後に、同じ切片を洗浄し、実施例12A2)で述べたようにアポプトーシスタグで染色した。染色組織をマウントし供焦点免疫蛍光顕微鏡で調べた。
【0139】
図20では、写真AからCはCSAT(抗β1)処置胎児のCAM組織を表し、写真DからFはLM609(抗αvβ3)処置胎児のCAM組織を表す。写真AおよびDは、アポプトーシスタグで染色し、D.I.C.画像に蛍光(FITC)を重焼きして可視化させた組織を示す。写真BおよびEはmAbLM609(抗αvβ3)で染色し、蛍光(ローダミン)で可視化させた同じ組織を示す。写真CおよびFは、アポプトーシスタグおよびLM609の両方で染色した同じ組織の結合画像を表し、黄色は同時存在を示す。棒線は、左および右の写真でそれぞれ15および50μmを表している。
図20(A−C)に示すように、CSATまたはPBSを静注した後アポプトーシスタグによる染色はきわめて僅かでさらに無秩序のようであるが、これは組織内に極めてわずかなレベルしかアポプトーシスが存在しないことを示唆している。対照的に、LM609または環状ペプチド203で先に処置された胎児のCAMは、アポプトーシスタグで濃く染色された多くの血管を示し、一方周辺の非血管細胞間ではごくわずかな反応性が認められただけである(図20D−E)。さらに、これらの組織の染色にアポプトーシスタグおよびLM609の両方を用いたとき(19Cおよび19F)、顕著な同時存在が、αvβ3拮抗物質で処置した胎児から得たCAMのこれらマーカー間でのみ認められた。これらの発見は、インビボで脈管形成を誘発した後、インテグリンαvβ3の抑制物質は選択的にαvβ3保有血管のアポプトーシスを促進することを明らかにした。
【0140】
脈管形成は、多くの分子的および細胞生物学的現象を含む複雑な過程であるが、一方、実証されたいくつかの流れは、血管細胞インテグリンαvβ3はこの過程で比較的後期における役割を果たすことを提唱している。第一に、免疫組織学的分析によって、血管細胞におけるαvβ3の発現はβFGFによる脈管形成の誘発後12−24時間で最大になることが明らかにされた。第二に、αvβ3の拮抗物質は多数の活性化物質によって誘発される脈管形成を妨害するが、このことは、このレセプターは、脈管形成へと続くおそらく一次的シグナリングに関する全ての現象から下流にある共通の経路に含まれることを示唆する。第三に、mAbLM609または環状ペプチド処置CAMは、これら脈管形成拮抗物質による処置後48時間まではDNAのほつれによって測定されるアポプトーシスで顕著な増加を示さなかった。最後に、αvβ3拮抗物質は、細胞サイクル進入が既に誘発された血管細胞のアポプトーシスを促進する。
【0141】
本明細書に提示した結果は、インテグリン連結現象がインビボで細胞の生存を調節することができるという最初の直接的証拠を提供する。したがって次のような仮説が提唱される:脈管形成がいったん開始すると、個々の血管細胞は分裂し脈管形成原に向かって移動し始め、その後、αvβ3連結は、分化と成熟血管形成へと進行する持続した細胞生存を可能にするシグナルを提供する。しかしながら、αvβ3連結が妨害されると、これら細胞はこの分子的指示を受け取ることができず、不履行によりアポプトーシスに陥る。この仮説はまた、分化が生じた後、成熟血管はもはやαvβ3シグナリングを生存のために必要とせず、したがってこのインテグリンに対して耐性をもつということを予想させるであろう。
最後に、本明細書に提示した結果は、インテグリンαvβ3の拮抗物質は、新形成または脈管形成を特徴とする他の疾患のための強力な治療方法を提供するであろう。第一に、αvβ3拮抗物質は、以前から存在する血管構造に影響を与えること無く新しく形成されつつある血管を破壊する。第二にこれらの拮抗物質は、ニワトリ胎児の生存活性に顕著な影響を与えず、それらの非毒性を示唆した。第三に、脈管形成は、脈管形成刺激にもかかわらず顕著に遮断された。最後に、αvβ3拮抗物質の全身的投与は、種々の組織学的に明瞭なヒト腫瘍の劇的な退縮をもたらした。
【0142】
したがって前述の実施例によって、インテグリンαvβ3は、種々の刺激による脈管形成において重要な役割をはたし、そのようなαvβ3は、新規脈管新生を特徴とする疾患に対する本発明のαvβ3拮抗物質の重要な治療標的であることが明らかになった。
前述の記載は、当業者が本発明を実施するために十分であろうと考えられる。本発明は、寄託された細胞株によって範囲が限定されるべきではない。なぜならば、寄託された具体例は、本発明の1つの特徴を単に詳述することを目的としており、機能的に同等ないずれの細胞株も本発明の範疇に入るからである。材料の寄託は、その最善の態様を含む本明細書中の記載が本発明の実施のためには不適当であるかもしれないことを承認するものではなく、また本明細書の記載に対して請求の範囲を制限することを意図したものでもない。実際、本明細書に記載したものに加え、当業者にとっては前述の記載から種々の修飾が明白であり、それらは添付の請求の範囲内に含まれるであろう。
【0143】
【図面の簡単な説明】
【0144】
【図1A】正常皮膚および創傷治癒下の皮膚(肉芽形成組織と呼ぶ)におけるインテグリンサブユニットβ3およびβ1の組織分布を示す。
【図1B】正常皮膚および創傷治癒下の皮膚(肉芽形成組織と呼ぶ)におけるインテグリンサブユニットβ3およびβ1の組織分布を示す。
【図1C】正常皮膚および創傷治癒下の皮膚(肉芽形成組織と呼ぶ)におけるインテグリンサブユニットβ3およびβ1の組織分布を示す。
【図1D】正常皮膚および創傷治癒下の皮膚(肉芽形成組織と呼ぶ)におけるインテグリンサブユニットβ3およびβ1の組織分布を示す。
【図2A】正常皮膚および創傷治癒下の皮膚(肉芽形成組織と呼ぶ)における、それぞれβ3およびβ1に結合するヴォン・ヴィレブランド因子およびラミニンリガンドの組織分布を示す。
【図2B】正常皮膚および創傷治癒下の皮膚(肉芽形成組織と呼ぶ)における、それぞれβ3およびβ1に結合するヴォン・ヴィレブランド因子およびラミニンリガンドの組織分布を示す。
【図2C】正常皮膚および創傷治癒下の皮膚(肉芽形成組織と呼ぶ)における、それぞれβ3およびβ1に結合するヴォン・ヴィレブランド因子およびラミニンリガンドの組織分布を示す。
【図2D】正常皮膚および創傷治癒下の皮膚(肉芽形成組織と呼ぶ)における、それぞれβ3およびβ1に結合するヴォン・ヴィレブランド因子およびラミニンリガンドの組織分布を示す。
【図3A】それぞれ膀胱癌、大腸癌、乳癌および肺癌の組織生検材料におけるビトロネクチンインテグリンレセプター、αvβ3の分布を示す。
【図3B】それぞれ膀胱癌、大腸癌、乳癌および肺癌の組織生検材料におけるビトロネクチンインテグリンレセプター、αvβ3の分布を示す。
【図3C】それぞれ膀胱癌、大腸癌、乳癌および肺癌の組織生検材料におけるビトロネクチンインテグリンレセプター、αvβ3の分布を示す。
【図3D】それぞれ膀胱癌、大腸癌、乳癌および肺癌の組織生検材料におけるビトロネクチンインテグリンレセプター、αvβ3の分布を示す。
【図4】本発明の未処置10週齢ニワトリ胎児のCAMの典型的な顕微鏡写真を示す。
【図5A】本発明のCAM調製物のインテグリンβ1およびαvβ3の組織分布を示す。
【図5B】本発明のCAM調製物のインテグリンβ1およびαvβ3の組織分布を示す。
【図5C】本発明のCAM調製物のインテグリンβ1およびαvβ3の組織分布を示す。
【図6】実施例6Aで述べる未処置およびβFGF処置10週齢CAMのαvβ3およびβ1の相対的発現を測定した棒グラフを示す。
【図7A】それぞれ未処置、βFGF処置およびTNFα処置10週齢CAMの外観を示す。
【図7B】それぞれ未処置、βFGF処置およびTNFα処置10週齢CAMの外観を示す。
【図7C】それぞれ未処置、βFGF処置およびTNFα処置10週齢CAMの外観を示す。
【図8A】実施例7A1)における、10日目CAMにおけるFGF誘発脈管形成に対する抗体局所処置の効果を示す。
【図8B】実施例7A1)における、10日目CAMにおけるFGF誘発脈管形成に対する抗体局所処置の効果を示す。
【図8C】実施例7A1)における、10日目CAMにおけるFGF誘発脈管形成に対する抗体局所処置の効果を示す。
【図8D】実施例7A1)における、10日目CAMにおけるFGF誘発脈管形成に対する抗体局所処置の効果を示す。
【図8E】実施例7A1)における、10日目CAMにおけるFGF誘発脈管形成に対する抗体局所処置の効果を示す。
【図9A】実施例7D2)における、腫瘍によって誘発された脈管形成に対する合成ペプチド66203の静脈内注射の影響を示す。
【図9B】実施例7D2)における、腫瘍によって誘発された脈管形成に対する合成ペプチド66203の静脈内注射の影響を示す。
【図9C】実施例7D2)における、腫瘍によって誘発された脈管形成に対する合成ペプチド66203の静脈内注射の影響を示す。
【図10A】実施例7B1)における、増殖因子誘発脈管形成に対する単クローン性抗体の静脈内応用の効果を示す。
【図10B】実施例7B1)における、増殖因子誘発脈管形成に対する単クローン性抗体の静脈内応用の効果を示す。
【図10C】実施例7B1)における、増殖因子誘発脈管形成に対する単クローン性抗体の静脈内応用の効果を示す。
【図11A】実施例7Cにおける、抗インテグリン抗体の局所適用後の胎児の脈管形成に対する効果を示す。
【図11B】実施例7Cにおける、抗インテグリン抗体の局所適用後の胎児の脈管形成に対する効果を示す。
【図11C】実施例7Cにおける、抗インテグリン抗体の局所適用後の胎児の脈管形成に対する効果を示す。
【図12】実施例7D1)における、CAM調製物内の腫瘍に進入する血管数の測定を示す。
【図13A】実施例9A1)aにおける、処置後7日の湿潤腫瘍重量と最初の腫瘍重量との間の比較を示す。
【図13B】実施例9A1)aにおける、処置後7日の湿潤腫瘍重量と最初の腫瘍重量との間の比較を示す。
【図13C】実施例9A1)aにおける、処置後7日の湿潤腫瘍重量と最初の腫瘍重量との間の比較を示す。
【図13D】実施例9A1)aにおける、処置後7日の湿潤腫瘍重量と最初の腫瘍重量との間の比較を示す。
【図14A】実施例9A1)aにおける、P3G2(抗αvβ5)(図14A)およびLM609(抗αvβ3)(図14B)で処置し、ヘマトキシリンおよびエオシンで染色した腫瘍の組織切片を示す。
【図14B】実施例9A1)aにおける、P3G2(抗αvβ5)(図14A)およびLM609(抗αvβ3)(図14B)で処置し、ヘマトキシリンおよびエオシンで染色した腫瘍の組織切片を示す。
【図15A】実施例9A1)bにおける、ペプチドで処置されたM21L腫瘍組織を示す。
【図15B】実施例9A1)bにおける、ペプチドで処置されたM21L腫瘍組織を示す。
【図15C】実施例9A1)bにおける、ペプチドで処置されたM21L腫瘍組織を示す。
【図15D】実施例9A1)bにおける、ペプチドで処置されたM21L腫瘍組織を示す。
【図15E】実施例9A1)bにおける、ペプチドで処置されたM21L腫瘍組織を示す。
【図16A】実施例10におけるウサギ眼球のインビボモデルアッセーにおける脈管形成拮抗物質による脈管形成の抑制を示す。
【図16B】実施例10におけるウサギ眼球のインビボモデルアッセーにおける脈管形成拮抗物質による脈管形成の抑制を示す。
【図16C】実施例10におけるウサギ眼球のインビボモデルアッセーにおける脈管形成拮抗物質による脈管形成の抑制を示す。
【図16D】実施例10におけるウサギ眼球のインビボモデルアッセーにおける脈管形成拮抗物質による脈管形成の抑制を示す。
【図16E】実施例10におけるウサギ眼球のインビボモデルアッセーにおける脈管形成拮抗物質による脈管形成の抑制を示す。
【図17】実施例11Aのマウス:ヒトキメラ−インビボマウスモデルの作製方法を示す工程図である。
【図18】FACS分析で求めた、mAb処置およびペプチド処置CAMから得られ、アポプトーシスタグ(Apop Tag)で染色された単一細胞の%を示す。
【図19】CSAT(抗β1)(図19A)またはLM609(抗αvβ3)(図19B)で処置した胎児のCAMの単一細胞懸濁をアポプトーシスタグおよび沃化プロピジウムで染色し、FACSで分析した結果をまとめたものを示す。
【図20A】処置胎児のCAM組織を表す。
【図20B】処置胎児のCAM組織を表す。
【図20C】処置胎児のCAM組織を表す。
【図20D】処置胎児のCAM組織を表す。
【図20E】処置胎児のCAM組織を表す。
【図20F】処置胎児のCAM組織を表す。
【技術分野】
【0001】
本発明は、一般に医薬分野に関し、とりわけビトロネクチン(vitronectin)レセプター(αvβ3)の拮抗物質を用いて組織の脈管形成(angiogenesis)を抑制する方法および組成物に関する。
【背景技術】
【0002】
インテグリン(integrin)は、細胞外マトリックス蛋白に結合し、したがって、細胞−細胞および細胞−細胞外マトリックス相互作用(一般に粘着現象と呼ぶ)を仲介することが分かっている細胞性レセプター類である。多数のインテグリンおよびインテグリンと結合するそのリガンドが文献に記載されているが、多くのインテグリンの生物学的機能は不明のままである。インテグリンレセプターは、αおよびβサブユニットから形成される非共有結合異種二量体糖蛋白複合体という共通の構造特性をもつ蛋白類を構成する。
ビトロネクチンレセプター(ビトロネクチンに優先的に結合するその本来の特徴のためにこの名がある)は、αvβ1、αvβ3およびαvβ3と呼ばれる3種の異なるインテグリンと関係を有することが分かった(Horton,Int.J.Exp.Pathol.,71:741-759(1990))。αvβ1は、フィブロネクチン(fibronectin)およびビトロネクチンに結合する。αvβ3は、フィブリン、フィブリノーゲン、ラミニン、スロンボスポンジン(thrombospondin)、ビトロネクチン、ヴォン・ヴイレブランド因子(von Willebrand's factor)、オステオスポンチン(osteospontin)および骨唾液腺蛋白I(bone sialoprotein I)を含むきわめて多様なリガンドと結合する。αvβ5はビトロネクチンと結合する。これら3種のインテグリンが組織の細胞相互作用で果たす特定の細胞粘着の役割はなお研究中であるが、異なる生物学的機能を有する異なるインテグリンが存在することは明らかである。
【0003】
多数のインテグリンにとって、リガンドの重要な1つの認識部位は、アルギニン−グリシン−アスパラギン酸(RGD)のトリペプチド配列である。RGDは、ビトロネクチンレセプターインテグリンとして上記に認定されたリガンドの全てに見出される。このRGD認識部位は、RGD配列を含むポリペプチド(“ペプチド”)で模倣することができるが、このようなRGDペプチドはインテグリン機能の既知抑制物質である。RGDペプチドの配列および構造にしたがって特定のインテグリンを標的とするこの抑制特異性を変化させることができるということは、しかしながら言及に値する。
RGD認識部位の考察については以下の文献を参照されたい:Pierschbacherら、Nature,309:30-33(1984); Pierschbacherら、Proc.Natl.Acad.Sci.USA,81:5985-5988(1984)。種々のインテグリン特異性をもつ種々のRGDポリペプチドはまた下記の文献に記載された:Grantら、Cell,58:933-943(1989);Chereshら、Cell,58:945-953(1989);Aumailleyら、FEBS Letts.,291:50-54(1991);Pfaffら、J.Biol.Chem.,269:20233-20238(1994);さらに米国特許出願第4517686号、4578079号、4589881号、4614517号、4661111号、4792525号、4683291号、4879237号、4988621号、5041380号、および5061693号。
【0004】
脈管形成(angiogenesis)は、新たに発生した血管の組織内への増殖を伴う組織の血管新生(vascularization)の過程であり、新規血管新生(neo-vascularization)とも呼ばれる。この過程は、内皮細胞および平滑筋細胞の浸潤によって仲介される。この過程は3通りの態様のいずれか1つで進行すると考えられている:血管は予め存在する血管から成長する、新規な血管発生は前駆細胞から生じる(血管形成(vasculogenesis))、または存在する小血管の直径が拡大する(Bloodら、Bioch.Biophys.Acta,1023:89-118(1990))。血管内皮細胞は、少なくとも5つのRGD依存インテグリンを含むことが知られており、これらには、ビトロネクチンレセプター(αvβ3またはαvβ5)、コラーゲンI型およびIV型レセプター(α1β1)、ラミニンレセプター(α2β1)、フィブロネクチン/ラミニン/コラーゲンレセプター(α3β1)およびフィブロネクチンレセプター(α5β1)が含まれる(Davisら、J.Cell.Biochem.,51:206-218(1993))。平滑筋細胞は、少なくとも6つのRGD依存インテグリンを含むことが知られており、これらには、α5β1、αvβ3およびαvβ5が含まれる。
【0005】
脈管形成は、新生児の成長では重要な過程であるが、また創傷治癒および種々の臨床症状(組織の炎症、関節炎、腫瘍増殖、糖尿病性網膜症、網膜の新規血管新生による筋変性および同様な症状を含む)の病理発生においても重要である。
脈管形成に付随するこのような症状の発現は、脈管形成性疾患と呼ばれる(Folkmanら、Science,235:442-447(1987))。脈管形成は、一般に成人または成熟組織では見られないが、創傷治癒および黄体の増殖サイクルにおいては脈管形成が発生する。例えばモーセスらの論文を参照されたい(Mosesら、Science,248:1408-1410(1990))。
脈管形成の抑制は、腫瘍増殖を制限するために有用な治療であろうと提言された。脈管形成の抑制では、(1)例えばβFGF(線維芽細胞増殖因子)のような“脈管形成分子”の遊離の抑制、(2)例えば抗βFGFの使用による脈管形成分子の中和、(3)脈管形成刺激に対する内皮細胞の反応の抑制が提唱された。この後者の方法は注目を集め、フォークマンら(Folkmanら、Cancer Biology,3:89-96(1992))は、いくつかの内皮細胞反応抑制物質を記載したが、これにはコラゲナーゼ抑制物質、基底膜代謝回転抑制物質、止血性ステロイド、真菌由来脈管形成抑制物質、血小板因子4、スロンボスポンジン、関節炎薬剤(例えばD−ペニシラミンおよびチオリンゴ酸金)、ビタミンD3類似体、アルファーインターフェロン、および脈管形成の抑制に役立つ可能性があるものが含まれる。さらに、この他に提唱された脈管形成抑制物質については以下の文献を参照されたい:Bloodら、Bioch.Biophys.Acta.,1023:89-118(1990);Mosesら、Science,248:1408-1410(1990); Ingberら、Lab.Invest.,59:44-51(1988);並びに米国特許第5092885号、5112946号、5192744号および5202352号。前述の文献に記載された脈管形成抑制物質のいずれもαvβ3の抑制を標的としない。
【0006】
ビトロネクチンレセプターαvβ3を抑制するRGDを含むペプチドもまた報告された(Aumailleyら、FEBS Letts.,291:50-54(1991); Choiら、J.Vasc.Surg.,19:125-134(1994);Smithら、J.Biol.Chem.,265:12267-12271(1990);およびPfaffら、J.Biol.Chem.,269:20233-20238(1994))。脈管形成におけるインテグリンαvβ3の役割は、しかしながら、本発明以前には示唆されたこともなくまた明らかにされたこともなかった。
種々のインテグリンαまたはβサブユニットに免疫特異的単クローン性抗体を用いたインビトロでの細胞粘着抑制によって、αvβ3が、微小血管内皮細胞を含む様々な細胞型の細胞粘着に関与していることが示された(Davisら、J.Cell.Biol.,51:206-218(1993))。さらに、インビトロでRGDペプチドGRGDSの使用を報告したニコシアらの論文では、コラーゲンゲルで培養したラット大動脈からの“微小血管”の形成が抑制された(Nicosiaら、Am.J.Pathol.,138:829-833(1991))。しかしながら、コラーゲンゲル培養での“微小血管”形成のインビトロ抑制は、組織における脈管形成抑制のモデルではない。なぜならば、微小血管の構造が毛細血管芽と同じであるということも、またコラーゲンゲル培養における微小血管の形成が、もとのままの組織(例えば関節炎組織、腫瘍組織または脈管形成抑制が所望される病的組織)への新規血管の成長と同じであるということは示されていないからである。
したがって、本明細書で報告する研究以外に、本出願人らは、細胞粘着抑制物質を用いて脈管形成が組織で抑制できることを開示したものを全く知らない。特に、組織における脈管形成にαvβ3機能が必要とされ、またαvβ3拮抗物質が組織における脈管形成を抑制できるということはこれまで開示されたことはなかった。
【発明の開示】
【発明が解決しようとする課題】
【0007】
本発明は、組織における脈管形成はインテグリンαvβ3を必要とし、αvβ3の抑制物質は脈管形成を抑制することができることを明らかにする。本発明はまた、他のインテグリン(例えばαvβ5またはαvβ1)は、おそらくそれらは脈管形成の発生に必須ではないために、脈管形成を抑制しないことを明らかにする。
【0008】
したがって、本発明は、脈管形成抑制量のαvβ3拮抗物質を含む組成物を組織に投与することを含む、組織における脈管形成抑制方法を開示する。
処置されるべき組織は、脈管形成抑制が所望されるいずれの組織でもよく、例えば新規血管新生が発生しつつある病的組織である。代表的な組織は、炎症組織、固形腫瘍、転移、再狭窄を起こしている組織、および同様な組織を含む。
【課題を解決するための手段】
【0009】
本発明で使用されるαvβ3拮抗物質はαvβ3に結合することができ、さらにαvβ3が本来のリガンドに結合する能力を競合的に抑制することができる。
好ましくは、この拮抗物質は、他のインテグリンの間でαvβ3に対して特異性を示す。特に好ましい具体例では、このαvβ3拮抗物質は、フィブリノーゲンまたは他のRGD含有リガンドのαvβ3への結合を抑制するが、フィブロネクチンのαIIbβ3への結合を実質的に抑制しない。好ましいαvβ3拮抗物質は、ポリペプチドまたはαvβ3と免疫的に反応する単クローン性抗体もしくはその機能的フラグメントであろう。
【0010】
本開示の一部分を形成する図面は下記の通りである。
図1A−1Dは、正常皮膚および創傷治癒下の皮膚(肉芽形成組織と呼ぶ)におけるインテグリンサブユニットβ3およびβ1の組織分布を示す。β3およびβ1に対する抗体を用いた免疫組織化学的方法が、実施例3Aで述べるように実施された。図1Aおよび1Bは、それぞれ、正常皮膚および肉芽形成組織における抗β3の免疫活性を示す。図1Cおよび1Dは、それぞれ、正常皮膚および肉芽形成組織における抗β1の免疫活性を示す。
【0011】
図2A−2Dは、正常皮膚および創傷治癒下の皮膚(肉芽形成組織と呼ぶ)における、それぞれβ3およびβ1に結合するヴォン・ヴィレブランド因子およびラミニンリガンドの組織分布を示す。ヴォン・ヴィレブランド因子に対する抗体(抗vWF)およびラミニンに対する抗体(抗ラミニン)を用いた免疫組織化学的方法が、実施例3Bで述べるように実施された。図2Aおよび2Bは、それぞれ、正常皮膚および肉芽形成組織における抗vWFの免疫活性を示す。図2Cおよび2Dは、それぞれ、正常皮膚および肉芽形成組織における抗ラミニンの免疫活性を示す。
【0012】
図3A−3Dは、それぞれ膀胱癌、大腸癌、乳癌および肺癌の組織生検材料におけるビトロネクチンインテグリンレセプター、αvβ3の分布を示す。αvβ3に対するLM609抗体を用いた免疫組織化学的方法は、実施例3Cで述べるように実施された。
図4は、本発明の未処置10週齢ニワトリ胎児のCAMの典型的な顕微鏡写真を示す。その調製は実施例5Bで述べる。
【0013】
図5A−5Cは、本発明のCAM調製物のインテグリンβ1およびαvβ3の組織分布を示す。図5Aは、CSAT(抗β1抗体)を用いて免疫蛍光で免疫活性を検出した、未処置10週齢CAM調製物におけるβ1サブユニットの分布を示す。図5Bは、LM609(抗αvβ3抗体)を用いて免疫蛍光で免疫活性を検出した、未処置10週齢CAM調製物におけるαvβ3レセプターの分布を示す。図5Cは、LM609(抗αvβ3抗体)を用いて免疫蛍光で免疫活性を検出した、βFGF処置10週齢CAM調製物におけるαvβ3レセプターの分布を示す。
処置および結果は実施例5Cで述べる。
【0014】
図6は、実施例6Aで述べる未処置およびβFGF処置10週齢CAMのαvβ3およびβ1の相対的発現を測定した棒グラフを示す。平均蛍光濃度はY軸に、インテグリンプロフィルはX軸にプロットされている。
図7A−7Cは、それぞれ未処置、βFGF処置およびTNFα処置10週齢CAMの外観を示す。その方法および結果は実施例6Aに記載する。
【0015】
図8A−8Eは、実施例7A1)で述べるように、10日目CAMにおけるFGF誘発脈管形成に対する抗体局所処置の効果を示す。図8Aは、血管を欠く未処置CAM調製物を示す。図8Bは、以前に血管構造を欠いていた領域への、βFGF処置によって誘発された新しい血管構造の浸潤を示す。図8C,8Dおよび8Eは、それぞれβ1に対する抗体(抗β1;CSAT)、αvβ5に対する抗体(抗αvβ5;P3G2)およびαvβ3に対する抗体(抗αvβ3:LM609)の作用を示す。
【0016】
図9A−9Cは、実施例7D2)で述べるように、腫瘍によって誘発された脈管形成に対する合成ペプチド66203の静脈内注射の影響を示す。図9Aは、腫瘍誘発により生じた脈管形成に対するコントロールペプチドの静脈内処置の抑制効果の欠如を示す(コントロールペプチド腫瘍)。ペプチド66203の静脈内注射によるそのような脈管形成の抑制(環状RGD腫瘍)は、図9Bに示す。腫瘍処置領域に隣接する領域内の以前から存在する成熟した血管に対する、ペプチド66203の静脈内輸液後の抑制効果または細胞毒性の欠如は図9Cに示す(環状RGD隣接CAM)。
【0017】
図10A−10Cは、実施例7B1)で述べるように、増殖因子誘発脈管形成に対する単クローン性抗体の静脈内応用の効果を示す。図10Aは、抗体処置に曝されなかったβFGF誘発脈管形成を示す(コントロール)。図10Bに示すように、同じ調製物を抗αvβ5抗体P3G2で処置したとき脈管形成抑制は生じなかった。図10Cに示すように、抗αvβ3抗体LM609による処置で脈管形成抑制が生じた。
図11A−11Cは、実施例7Cに述べるように抗インテグリン抗体の局所適用後の胎児の脈管形成に対する効果を示す。それぞれ図11Aおよび11Bで示すように、脈管形成は、抗β1および抗αvβ5で6週齢CAMを処置しても抑制されなかった。対照的に、図11Cに示すように、抗αvβ3抗体LM609での処置は血管形成の抑制をもたらした。
図12は、実施例7D1)で述べるように、CAM調製物内の腫瘍に進入する血管数の測定を示す。このグラフでは、CSAT(抗β1)、LM609(抗αvβ3)、またはP3G2(抗αvβ5)のいずれかを局所的に用いて得られる血管の数がY軸に示されている。
図13A−13Dは、実施例9A1)aで述べるように、処置後7日の湿潤腫瘍重量と最初の腫瘍重量との間の比較を示す。各棒線は、各グループ当たり5−10個の腫瘍の平均±標準誤差を表す。腫瘍は、ヒトメラノーマ(M21−L)(図13A)、膵臓癌(Fg)(図13B)、肺癌(UCLAP−3)(図13C)および喉頭癌(HEp3)(図13D)CAM調製物に由来し、PBS、CSAT(抗β1)、またはLM609(抗αvβ3)で静脈内処置を施された。グラフでは、X軸に示したようにCSAT(抗β1)、LM609(抗αvβ3)またはPBSのいずれかを静脈内に用いて得られた腫瘍の重量がY軸に示されている。
【0018】
図14Aおよび14Bは、実施例9A1)aで述べるように、P3G2(抗αvβ5)(図14A)およびLM609(抗αvβ3)(図14B)で処置し、ヘマトキシリンおよびエオシンで染色した腫瘍の組織切片を示す。図14Aに示すように、コントロール抗体(P3G2)で処置した腫瘍は、腫瘍基質全体に存在する有糸分裂像(楔形)と同様に多数の血管(矢印)で示されるように、多くの生命活性を示す活発に分裂する腫瘍細胞を示した。対照的に、LM609(抗αvβ3)で処置した腫瘍では、あったとしてもごく僅かの生命活性を示す細胞または血管が検出されただけであった(図14B)。
図15A−15Eは、実施例9A1)bで述べるように、ペプチドで処置されたM21L腫瘍に対応し、下記の通りである。図15A、コントロール環状RADペプチド(69601);図15B、コントロール環状RGDペプチド(66203);図15C、同じ胎児から採取され環状RGDペプチド(66203)で処置された隣接CAM組織;さらに高倍率(×13)のコントロールRAD(69601)処置腫瘍(図15D)および環状RGDペプチド(66203)処置腫瘍(図15E)。図15Dは、RADコントロールペプチド(69601)
処置腫瘍の正常な血管を鮮明に表している。図15Eは、環状RGDペプチド(66203)処置腫瘍の破壊された血管(矢印)の例を示している。
【0019】
図16A−16Eは、実施例10で述べるウサギ眼球のインビボモデルアッセーにおける脈管形成拮抗物質による脈管形成の抑制を示す。図16Aおよび16Bは、βFGFおよびmAbP1F6(抗αvβ5)の存在下でのウサギの眼の脈管形成を示す。図16C、16Dおよび16Eは、βFGFおよびmAbLM609(抗αvβ3)の存在下でのウサギの眼の脈管形成の抑制を示す。
図17は、実施例11Aのマウス:ヒトキメラ−インビボマウスモデルの作製方法を示す工程図である。SCIDマウスの皮膚の一部分をヒト新生児包皮で置き換え、4週間治癒させた。移植が治癒した後、ヒト包皮にヒト腫瘍細胞を接種した。その後の4週間で、ヒトの皮膚からヒトの腫瘍内に増殖したヒト血管構造をもつヒトの腫瘍を含む測定可能な腫瘍が成長した。
図18は、それぞれ実施例12Aおよび12Bで述べるようにFACS分析で求めた、mAb処置およびペプチド処置CAMから得られ、アポプトーシスタグ(Apop Tag)で染色された単一細胞の%を示す。斜線棒および点刻棒は、それぞれアッセーの前24時間および48時間処置した胎児の細胞を表す。各々の棒線は3個の平均±標準誤差を表す。CAMは、mAbLM609(抗αvβ3)、CSAT(抗β1)またはPBSで実施例12A2)で述べるように処置された。CAMはまた、実施例12Bで述べるように、環状ペプチド69203(サイクロ−RGDfV、ペプチド203と表示)またはコントロール環状ペプチド69601(サイクロ−RADfV、ペプチド601と表示)で処置された。
【0020】
図19Aおよび19Bは、実施例12Cで述べるように、CSAT(抗β1)(図19A)またはLM609(抗αvβ3)(図19B)で処置した胎児のCAMの単一細胞懸濁をアポプトーシスタグおよび沃化プロピジウムで染色し、FACSで分析した結果をまとめたものを示す。Y軸は、アポプトーシスタグ染色の細胞数を表し、X軸は、沃化プロピジウム染色(DNA含有量)を表す。水平線は、アポプトーシスタグ染色の陰性ゲートを表す。左および右のパネルは、CSAT(抗β1)(図19A)およびLM609(抗αvβ3)(図19B)処置胎児のCAM細胞をそれぞれ示す。細胞サイクルの分析は、各状態当たり約8000例の分析によって実施した。
図20A−20Cは、実施例12Cで述べるように調製したCSAT(抗β1)処置胎児のCAM組織を表し、図20D−20Fは、LM609(抗αvβ3)処置胎児のCAM組織を表す。図20Aおよび20Dは、アポプトーシスタグで染色し、D.I.C.画像上に重ね合わせて蛍光(FITC)によって可視化させた組織を示す。図20Bおよび20Eは、mAbのLM609(抗αvβ3)で染色し蛍光(ローダミン)で可視化させた同じ組織を示す。図20Cおよび20Fは、アポプトーシスタグおよびLM609の両方で染色した同じ組織の合併させた画像を示し、ここで黄色の染色は共通の存在部位を表す。棒線は、左および右のパネルでそれぞれ15および50μmを表す。
【0021】
A.定義
アミノ酸残基:そのポリペプチド結合における化学的消化(加水分解)に際して形成されるアミノ酸。本明細書で述べるアミノ酸残基は、好ましくは“L”異性体形である。しかしながら、所望の機能特性が該ポリペプチドに保持されるかぎり、“D”異性体形残基をいずれのL−アミノ酸残基とも置換することができる。NH2は、ポリペプチドのアミノ末端に存在する遊離アミノ基を指す。COOHは、ポリペプチドのカルボキシ末端に存在する遊離カルボキシ基を指す。標準的なポリペプチド命名法(Biol.Chem.,243:3552-59(1969)に記載され、37CFR §1.822(b)(2)で採用された)に合わせ、アミノ酸残基の略語は下記の対応表に示す:
【0022】
【0023】
本明細書では、全てのアミノ酸残基配列は、その左と右の向きが、アミノ末端からカルボキシ末端の慣用的方向になっている式で表されているということは留意されねべならない。さらに、アミノ酸残基配列の始めと終わりのダッシュは、1つまたは2つ以上のアミノ酸残基のまた別の配列へのペプチド結合を示すことも留意されるべきである。
ポリペプチド:αアミノ基と隣接するアミノ酸残基のカルボキシ基との間でペプチド結合によって互いに連結された直線状のアミノ酸残基の列を指す。
ペプチド:本明細書で用いられているように、ポリペプチドとして互いに連結された50アミノ酸残基より大きくない直線状のアミノ酸残基の列を指す。
環状ペプチド:対応する直線状ペプチドに由来し、遊離のN−末端またはC−末端が存在せず、この場合、対応する直線状ペプチドのN−末端が、前記対応する直線状ペプチドのC−末端カルボキシレートとアミド結合を形成する。
蛋白:ポリペプチドの場合のように互いに連結された50アミノ酸残基より大きい直線状のアミノ酸残基の列を指す。
合成ペプチド:天然に生じる蛋白およびそのフラグメントにとらわれない、ペプチド結合で共に連結された化学的に生成されたアミノ酸残基の鎖を指す。
【0024】
B.一般的考察
一般に本発明は、脈管形成が特定のビトロネクチンレセプターαvβ3によって仲介され、さらにαvβ3機能の抑制は脈管形成を抑制するという発見に関連する。この発見は、脈管形成が種々の疾患の進行に果たす役割のゆえに重要である。脈管形成を抑制することによって、これら疾患に介入し、その症状を改善し、いくつかの場合にはこれら疾患を治癒させる。
新しい血管の増殖が、ある疾患に付随する病理の原因となるか、または遠因となる場合、脈管形成の抑制は、これらの疾患の有害な影響を軽減させるであろう。類リューマチ性関節炎、糖尿病性網膜症、炎症疾患、再狭窄などが例に含まれる。新しい血管の増殖が有害組織の支援に必要とされる場合、脈管形成の抑制は、該組織への血液供給を低下させ、したがって血液供給の要求で支えられている組織集団の減少に役立つ。その例には、腫瘍が厚さ数ミリを越えて増殖するために、さらに固形腫瘍の転移を成立させるために、新規血管新生が持続的な要求となる腫瘍の増殖が含まれる。
【0025】
一部には、この治療が脈管形成に対して高度に選択的で、他の生物学的過程に対してそのようなことはないという理由で、本発明の方法は効果的である。実施例で示すように、新しい血管の増殖のみが実質的なαvβ3を含み、したがって、この治療方法は成熟血管に副作用を与えない。さらに、αvβ3は正常な組織には広範囲に分布せず、むしろ新しい血管で選択的に見出され、したがって、この治療法は、新しい血管の増殖を選択的に標的とすることができる。
αvβ3のみを抑制することによって脈管形成を効果的に抑制することができるという発見は、潜在的に高い選択性をもち、したがって相対的に毒性が低い治療組成物の開発を可能にする。したがって、本発明は、1つまたは2つ以上のインテグリンを抑制する能力を有するペプチドを基本とする試薬の使用を開示するが、より選択的にαvβ3を抑制し、したがってαvβ3によって仲介される過程以外の他の生物学的過程を抑制するという副作用をもたない他の試薬をデザインすることが可能である。
例えば、本技術によって示されるように、αvβ3と免疫的に反応する高度に選択的な単クローン性抗体で、αvβ3機能の抑制について同様に選択的である抗体を調製することが可能である。さらに、本明細書で述べるように、αvβ3の抑制について選択的であるようにRGD含有ペプチドをデザインすることが可能である。
【0026】
本発明の発見以前には、αvβ3の生物学的機能と拮抗する試薬を用いて、脈管形成および脈管形成に付属する過程のどれもインビボで抑制することが可能であることは知られていなかった。
C.脈管形成抑制の方法
本発明は、組織の脈管形成の抑制、それによって脈管形成に依存する組織内の減少を抑制するための方法を提供する。一般に、本方法は、脈管形成抑制量のαvβ3拮抗物質を含む組成物を組織に投与することを含む。
上記で述べたように、脈管形成は、“発芽(sprouting)”、血管形成または血管拡大を含む組織の新規血管新生(この脈管形成過程の全てはαvβ3の発現によって仲介され、さらにαvβ3発現に依存する)を伴う種々の過程を含む。外傷治癒、黄体形成および胎児発生を除いて、大半の脈管形成過程は病的過程に付随し、したがって本治療方法の使用はこれら疾患に対して選択的で、有害な副作用をもたない。
【0027】
脈管形成が重要であると考えられている種々の疾患が存在する。これらは脈管形成性疾患と呼ばれ、炎症疾患(例えば免疫および非免疫性炎症、慢性関節性リューマチ並びに乾癬)、不適切なまたは時宜を得ない血管の侵襲(例えば糖尿病性網膜症、血管新生緑内障、再狭窄、アテローム性動脈硬化症プラークの毛細血管増殖および骨粗鬆症)および癌関連疾患(例えば固形腫瘍、固形腫瘍転移、血管線維腫、水晶体後線維増殖症、血管腫、カポジ肉腫、および腫瘍増殖を支援するために新規血管新生を必要とする同様な癌)を含む。
したがって、病的組織の脈管形成を抑制する方法は、これら疾患の症状を改善し、疾患によっては、これら疾患の治癒に寄与することができる。一具体例では、本発明は、組織内の脈管形成それ自体の抑制を意図する。組織内の脈管形成の程度、したがって本発明によって達成される抑制の程度は種々の方法で測定できる。これらの方法は、例えば、免疫組織化学によってαvβ3免疫陽性の非成熟および発生期の血管構造を検出する実施例で詳述される。
本明細書で述べるように、種々の組織または統合された組織を含む種々の器官(皮膚、筋肉、腸、結合組織、関節部、骨および脈管形成刺激に応じて血管が侵襲することができる同様な組織を含む)のいずれも、病的状態での脈管形成を支援することができる。
【0028】
したがって、関連する具体例では、処置される組織は炎症組織で、抑制される脈管形成は炎症組織の脈管形成であり、そこには炎症組織の新規血管新生が存在する。この種類では、本方法は、例えば慢性関節性リュウーマチの患者の関節炎組織、免疫もしくは非免疫性炎症組織、乾癬組織などの脈管形成の抑制を意図する。
多くの具体例で本発明にしたがって処置される対象は、望ましくは人間の患者であるが、本発明の原理は、本発明が、“患者”という用語に含まれることを意図する全ての哺乳類に関して有効であることを示唆していることは理解されよう。本明細書では、哺乳類という語が、脈管形成に付随する疾患の治療が所望される哺乳類のいずれの種も、特に農業用、家畜用哺乳類種も含むことは理解されよう。
別の関連する具体例では、処置される組織は、糖尿病性網膜症、黄斑の変性または血管新生緑内障の患者の網膜組織であり、抑制される脈管形成は網膜組織の脈管形成であり、そこには網膜組織の新規血管新生が存在する。
さらに別の関連する具体例では、処置される組織は、固形腫瘍、転移、皮膚癌、乳癌、血管腫または血管線維腫、および同様な癌をもつ患者の腫瘍組織であり、抑制される脈管形成は腫瘍組織の脈管形成で、そこには腫瘍組織の新規血管新生が存在する。本発明によって治療できる代表的な固形腫瘍組織は、肺、膵、乳房、結腸、喉頭、卵巣および同様な組織である。典型的な腫瘍組織の脈管形成およびその抑制は実施例で述べる。
【0029】
腫瘍組織の脈管形成の抑制は、新規血管新生が腫瘍増殖で果たす重要な役割のために特に好ましい具体例である。腫瘍組織で新規血管新生がない場合、この腫瘍組織は必要な栄養物を得ることができず増殖が鈍化し退縮し、ついには壊死を起こして腫瘍死をもたらす。
本発明は、換言すれば、本発明の方法にしたがって腫瘍の脈管形成を抑制することによって、腫瘍の新規血管新生を抑制する方法を提供する。同様に、本発明は、脈管形成を抑制する方法を実施することによって腫瘍の増殖を抑制する方法を提供する。
これらの方法はまた転移の形成に対して特に効果的である。その理由は、(1)転移の形成は、転移癌細胞が原発腫瘍を出ることができるように原発腫瘍の血管新生を必要とすること、および(2)二次的部位での転移の確立には転移癌の増殖を支援する新規血管新生が必要とされることである。
【0030】
関連する具体例では、本発明は、他の治療(例えば固形腫瘍に対抗し、さらに転移を制御する通常の化学療法)と結合させた方法を実施することを意図する。脈管形成抑制物質の投与は、典型的には化学療法の間またはそのあとで実施される。しかし、腫瘍組織が、腫瘍組織への血液供給と栄養の提供により脈管形成の回復を誘発することによって、脈管形成抑制物質の毒性攻撃に応答している場合は、一連の化学療法の後で時々脈管形成を抑制することは好ましい。さらに、固形腫瘍が除去された後、転移の予防として脈管形成抑制物質を投与することは好ましい。本方法が腫瘍の新規血管新生の抑制に適用される限りにおいて、本方法はまた、腫瘍組織の増殖抑制、腫瘍転移形成の抑制、成長腫瘍退縮に利用することができる。実施例では、本発明のαvβ3拮抗物質のただ1回の静脈投与後に、成長腫瘍の退縮が示される。
再狭窄は、経皮経管冠状動脈形成術の実施部位での平滑筋細胞(SMC)の移動と増殖の過程であるが、再狭窄は動脈形成術の成功を妨げる。再狭窄時のSMCの移動と増殖は、本発明で抑制可能な脈管形成の過程と考えられる。したがって、本発明はまた、動脈形成術処置の後で本方法にしたがって患者の脈管形成を抑制することによって再狭窄を抑制することを含む。再狭窄の抑制のためには、典型的にはαvβ3拮抗物質は、動脈形成術の処置の後約2日から約28日間、より典型的には前記処置後最初の約14日間投与される。
【0031】
組織の脈管形成を抑制するため、したがって脈管形成関連疾患の治療のために方法を実施するこの方法は、αvβ3の天然のリガンドとαvβ3が結合することを抑制することができるαvβ3拮抗物質の治療的に有効な量を含む組成物と、脈管形成が生じているか、または脈管形成が生じる危険性がある組織とを接触させることを含む。
αvβ3拮抗物質の投与のための用量範囲は、本明細書でさらに述べるように、拮抗物質の形態およびその効能に左右されるが、脈管形成および脈管形成によって仲介される症状が改善される所望の効果をもたらすために十分に多い量である。この投与量は副作用、例えば高粘性症候群、肺水腫、うっ血性心不全などを生じるほど多くてはいけない。一般には、用量は、年令、健康状態、性別、および患者の疾患の程度で変動し、当分野で技術を有する者が決定することが可能である。この用量はまた、いずれかの合併症の出現により個々の内科医が調節することもできる。
治療的に有効な量は、処置される組織の測定可能な脈管形成の抑制を生じるために十分なαvβ3拮抗物質の量、すなわち脈管形成抑制量である。脈管形成の抑制は、本明細書で述べるように免疫化学的に、または当業者に既知の他の方法でin situ で測定できる。
αvβ3拮抗物質が、αvβ3模倣物、RGD含有ペプチド、抗αvβ3単クローン性抗体またはそのフラグメントの形態を取ることができるかぎり、効能、したがって“治療的に有効な”量の表示は変動する可能性があることは理解されよう。
しかしながら、本アッセー方法で示されるように、本発明のαvβ3拮抗物質候補の効能を当業者は容易に評価できるであろう。
【0032】
αvβ3拮抗物質の効能は、CAMアッセーでの脈管形成の抑制、ウサギ眼球インビボアッセー、マウス:ヒトキメラインビボアッセーを含む種々の手段によって、さらに天然のリガンドのαvβ3への結合抑制(上記の方法は全て本明細書に記載されている)、および同様なアッセーによって測定できる。
好ましいαvβ3拮抗物質は、拮抗物質濃度0.5マイクロモル(um)未満、好ましくは0.1um未満、より好ましくは0.05umで溶液中で天然のリガンド(例えばフィブリノーゲンまたはビトロネクチン)のαvβ3への結合を実質的に抑制する能力を有する。“実質的に”とは、αvβ3拮抗物質の存在下における抑制によって、フィブリノーゲンの結合が少なくとも50%減少するのが観察されることを意味し、50%抑制は本明細書ではIC50値と呼ぶ。
【0033】
より好ましくはαvβ3拮抗物質は、他のインテグリンの中でαvβ3に対して選択性を示す。したがって、好ましいαvβ3拮抗物質は、実質的にαvβ3へのフィブリノーゲン結合を抑制するが、別のインテグリン(例えばαvβ1、αvβ5またはαIIbβ3)へのフィブリノーゲンの結合を実質的に抑制しない。特に、好ましいαvβ3拮抗物質は、αvβ3へのフィブリノーゲン結合抑制のIC50が別のインテグリンへのフィブリノーゲン結合抑制のIC50活性と比較して10倍から100倍低いものである。インテグリンへのフィブリノーゲン結合抑制のIC50活性を測定する典型的なアッセーは実施例で述べる。
単クローン性抗体形をとる本発明のαvβ3拮抗物質の治療的に有効な量は、典型的には、生理学的に容認できる組成物として投与されたとき、約0.01マイクログラム(ug)/ミリリットル(ml)から約100ug/ml、好ましくは約1ug/mlから約5ug/ml、通常は約5ug/mlの血中濃度を達成するために十分な量である。換言すれば、用量は、約0.1mg/kgから約300mg/kg、好ましくは約0.2mg/kgから約200mg/kg、最も好ましくは約0.5mg/kgから約20mg/kgで変動可能で、1日1回または2回以上、1日または数日投与される。
拮抗物質が単クローン性抗体のフラグメント形をとる場合、この量は、完全抗体の質量と比較したフラグメントの質量を基に容易に調整できる。好ましい血中モル濃度は、約2マイクロモル(uM)から約5ミリモル(mM)、好ましくは約100uMから1mMの抗体拮抗物質である。
【0034】
ポリペプチドまたは他の同様なサイズの小分子αvβ3模倣物の形をとる本発明のαvβ3拮抗物質の治療的に有効な量は、典型的には生理学的に容認できる組成物として投与されたとき、約0.1マイクログラム(ug)/ミリリットル(ml)から約200ug/ml、好ましくは約1ug/mlから約150ug/mlの血中濃度を達成するために十分な量である。1グラム分子当たり約500グラムの質量をもつポリペプチドを基にすると、好ましい血中モル濃度は、約2マイクロモル(uM)から約5ミリモル(mM)、好ましくは約100uMから1mMのポリペプチド拮抗物質である。換言すれば、体重当たりの用量は、約0.1mg/kgから約300mg/kg、好ましくは約0.2mg/kgから約200mg/kgで変動可能で、1日1回または2回以上、1日または数日投与される。
本発明の単クローン性抗体またはポリペプチドは、注射または時間をかけて徐々に輸液により非経口的に投与できる。体内の処置される組織は、典型的には全身的投与によってアクセス可能で、したがってもっとも頻繁には治療用組成物の静脈内投与によって処置されるが、目的の組織が標的分子を含む蓋然性が高い場合は他の組織および薬剤送達手段も含まれる。したがって、本発明の単クローン性抗体またはポリペプチドは、静脈内、腹腔内、筋肉内、皮下、体腔内、経皮的に投与でき、さらに蠕動的手段によって送達させることができる。
【0035】
本発明の単クローン性抗体またはポリペプチドを含む治療用組成物は、例えばユニットドースの注射によって通常の手段で注射される。“ユニットドース”という用語は、本発明の治療用組成物に関して用いられるときは、患者に対する単位投与量として適切な生理的に規定された単位を指す。各単位は、必要な希釈剤(すなわち担体または賦形剤)と結合させて所望の治療効果を生じるように計算された予め決定された活性物質の量を含む。
実施例に示すように好ましい具体例では、αvβ3拮抗物質は単回投与として静脈内に投与される。
【0036】
この組成物は、投与製剤に適合する態様で、さらに治療的に有効な量で投与される。投与される量およびタイミングは処置を受ける患者、患者の活性成分を利用する系、および治療効果の所望程度に左右される。投与に必要な活性成分の正確な量は医師の判定にしたがい、各個人に固有である。全身的に適用するために適切な投与量の範囲は本明細書に開示するが、しかしながら投与ルートに左右される。適切な投与様式もまた変更可能であるが、最初の投与が典型的となり1時間または1時間以上の間隔で注射または他の投与によるその後の反復投与が続く。また別には、インビボ治療のために特定された範囲の血中濃度を維持するために十分な持続的静脈輸液も含まれる。
本実施例で示すように、脈管形成抑制および腫瘍退縮は、拮抗剤との最初の接触後早くも7日で生じる。拮抗剤へのさらなる接触または延長接触は、好ましくは7日から6週間、より好ましくは約14から28日である。
【0037】
関連する具体例において、本実施例は、αvβ3の抑制とαvβ3を含む新規血管系の細胞のアポプトーシス誘発との間の関係を明らかにする。したがって、本発明はまた、組織内の新規血管系におけるアポプトーシスの抑制方法を含む。この方法は、全ての組織における脈管形成抑制に関して、さらにそのために記載された条件に関して実質的に本明細書で開示した通りに実施される。ただ1つの注目に値する違いは、効果のタイミングの違いである。これは、アポプトーシスは急激に、典型的には拮抗物質との接触後約48時間で現れ、一方、本明細書で述べるように、脈管形成の抑制および腫瘍の退縮はより緩徐に出現する。この違いは、投与時間および所望する効果に関して治療方法に影響を与える。新規血管構造のアポプトーシスのためには、典型的には投与は24時間から約4週間であるが、48時間から7日間が好ましい。
【0038】
D.治療用組成物
本発明は、本明細書で述べる治療方法の実施に有用な治療用組成物を含む。本発明の治療用組成物は、本明細書で述べるように、活性な成分として組成物中に溶解または懸濁させたαvβ3拮抗物質とともに、生理的に許容できる担体を含む。好ましい具体例では、治療用αvβ3拮抗物質組成物は、治療の目的で哺乳類または人間の患者に投与したとき抗原性をもたない。
本明細書で用いられるように、組成物、担体、希釈剤および試薬に関して用いるとき、“医薬的に許容できる”、“生理的に許容できる”という用語は、望ましくない生理的影響(例えば悪心、眩暈、胃の不快感など)を生じることなく哺乳類に投与することができることを示し、相互に用いられる。
【0039】
組成物中に溶解または懸濁させた活性成分を含む医薬組成物の調製は当分野ではよく理解されており、製剤によって限定する必要はない。典型的には、そのような組成物は、液体または懸濁物のいずれかとして注射可能なように調製されるが、使用前に液体となる溶液または懸濁物に適した固形形態もまた調製できる。
この調製物はまた乳濁物でもよい。
活性成分は、本明細書で述べる治療方法で使用するために適した量で、医薬的に許容でき、さらに活性成分と適合する賦形剤と混合される。適切な賦形剤は、例えば水、食塩水、デキストロース、グリセロール、またはエタノールなど、およびそれらの組み合わせである。さらに、所望する場合には、少量の補助物質、例えば湿潤剤もしくは乳化剤、pH緩衝剤など活性成分の効能を高める物質を含むことができる。
【0040】
本発明の治療組成物は、該組成物中の成分の医薬的に許容できる塩を含むことができる。医薬的に許容できる塩には酸付加塩(ポリペプチドの遊離アミノ基とで形成される)が含まれるが、前記酸付加塩は、例えば塩酸もしくは燐酸のような無機酸で形成されるか、または酢酸およびマンデリン酸などの有機酸で形成される。遊離カルボキシル基とで形成される塩はまた、無機塩基(例えば水酸化ナトリウム、カリウム、アンモニウム、カルシウムまたは第二鉄)で、さらに有機塩基(例えばイソプロピルアミン、トリメチルアミン、2−エチルアミノエタノール、ヒスチジン、プロカインなど)で誘導することができる。
環状ポリペプチドαvβ3拮抗物質の調製で用いる場合、特に好ましいものはTFAおよびHClの塩である。ペプチドの代表的な塩は実施例で述べる。
生理的に許容できる塩は当分野で周知である。典型的な液体担体は滅菌水溶液で、これは活性成分および水の他には物質を含まないか、または、生理的pH値で例えば燐酸ナトリウムのような緩衝液、生理的食塩水、もしくは両方(例えば燐酸緩衝食塩水)を含む。また別には、水性担体は、1つ以上の緩衝塩の他、例えば塩化ナトリウムおよびカリウムのような塩、デキストロース、ポリエチレングリコール並びに他の溶質を含むことができる。
液体組成物はまた、水とともにまたは水を除いた液相を含むことができる。付加されるそのような液相の代表例は、グリセリン、植物油(例えば綿実油)および水−油乳濁物である。
治療用組成物は、脈管形成抑制量の本発明のαvβ3拮抗物質を含み、典型的には、治療用組成物の総重量当たり少なくとも0.1重量%の量を含むように製剤化される。重量%は、組成物全体に対する抑制物質の重量比である。したがって、例えば、0.1重量%は完全な組成物100グラム当たり0.1グラムの抑制物質である。
【0041】
E.インテグリンαvβ3の拮抗物質
αvβ3拮抗物質は、組織での脈管形成を抑制するために本方法で用いられ、それらは、天然のαvβ3リガンドとの機能的な相互作用が干渉されるような態様でαvβ3と相互に作用する化合物を含む種々の形態をとることができる。典型的な拮抗物質は、αvβ3のリガンド結合部位に由来するαvβ3の類似体、αvβ3−リガンド結合反応に必要な構造領域を模倣するαvβ3もしくはαvβ3の天然リガンドのいずれかの模倣物、αvβ3に特異的な天然のリガンドの機能的結合ドメインに対応する(特にαvβ3の天然リガンドのRGD結合ドメインに対応する)配列を有するポリペプチド、およびαvβ3もしくは天然リガンドのいずれかと免疫反応を生じる抗体を含むが、本明細書で述べるようにこれらの全ては拮抗物質活性をもつ。
【0042】
1.ポリペプチド
ある具体例では、本発明は、ポリペプチド形をとるαvβ3拮抗物質を意図する。本明細書に述べるように、ポリペプチド(ペプチド)αvβ3拮抗物質は、αvβ3−リガンド相互作用に必要な領域にαvβ3の天然リガンドまたはαvβ3自体のいずれかの特徴的な配列を有し、さらにαvβ3拮抗物質活性を示す。好ましいαvβ3拮抗物質ペプチドはRGDトリペプチドを含み、さらにRGD含有領域内で天然のリガンドと配列として対応する。
好ましいRGD含有ポリペプチドは、αvβ3の天然リガンド(例えばフィブリノーゲン、ビトロネクチン、ヴォン・ヴィレブランド因子、ラミニン、スロンボスポンジンおよび同様な配列)のRGD含有領域のアミノ酸残基配列に対応する配列を有する。これらαvβ3リガンドの配列は周知である。したがって、αvβ3拮抗物質ペプチドは、天然リガンドのいずれかに由来することができるが、フィブリノーゲンおよびビトロネクチンが好ましい。
【0043】
特に好ましいαvβ3拮抗物質ペプチドは、前述のように他のインテグリンと比較したとき、その天然のリガンドとαvβ3との結合を優先的に抑制する。これらのαvβ3特異的ペプチドは、少なくともαvβ3に対する特異性が望ましくない副作用(例えば他のインテグリンの抑制)の発生を減少させるという理由から特に好ましい。αvβ3に対して選択性をもつ好ましいαvβ3拮抗物質ペプチドの識別は、典型的な結合抑制アッセー(例えば実施例で述べるようなELISAアッセー)で容易に明らかにできる。
ある具体例では、本発明のポリペプチドは、約100アミノ酸残基未満、好ましくは約60残基未満、より好ましくは約30残基未満を含む。ペプチドは直線でも環状でもよいが、特に好ましいペプチドは環状である。
好ましい環状および直線ペプチド並びにそれらの名称は実施例の表1に示す。
【0044】
本ポリペプチドは、それが必要な配列を含み、さらに例えば本明細書で述べるようなアッセーでαvβ3拮抗物質として機能することができる限り、αvβ3の天然のリガンドのアミノ酸残基配列と同一である必要はないことは理解されよう。
本ポリペプチドには、それがαvβ3拮抗物質である限り、本明細書にそのアミノ酸残基配列が示されているポリペプチドのいずれの類似体、フラグメントまたは化学的誘導体も含まれる。したがって、その使用に際して変化が一定の利点を提供する場合は、本ポリペプチドは、種々の変化、置換、挿入および欠失に付すことができる。この点に関して、本発明のαvβ3拮抗物質ポリペプチドは、列挙ペプチドの配列と同一であるよりはむしろ対応するものであり、この場合1つまたは2つ以上の変化があり、本明細書で規定されたアッセーの1つまたは2つ以上でαvβ3拮抗物質として機能する能力を保持する。
したがって、ポリペプチドは、ペプチド誘導体の種々の形態のいずれかをとることができ、これは、アミド、蛋白との共役物、環状化ペプチド、重合ペプチド、類似体、フラグメント、化学修飾ペプチドおよび同様な誘導体を含む。
【0045】
“類似体”という用語は、本明細書に具体的に示した配列と実質的に同一なアミノ酸残基配列をもついずれのポリペプチドも包含するが、このポリペプチドは、1つまたは2つ以上の残基が機能的に同様な残基と保存的に置換され、さらにそれは本明細書で述べるようにαvβ3拮抗物質活性を示す。保存的置換の例には、1個の非極性(疎水性)残基(例えばイソロイシン、バリン、ロイシンまたはメチオニン)の別のものとの置換、1個の極性(親水性)残基の別のものとの置換(例えばアルギニンとリジン間、グルタミンとアスパラギン間、グリシンとセリン間)、1つの塩基性残基(例えばリジン、アルギニンまたはヒスチジン)の別のものとの置換、または1つの酸性残基(例えばアスパラギン酸またはグルタミン酸)の別のものとの置換を含む。
“保存的置換”という言葉はまた、そのようなポリペプチドが必須の抑制活性を示すということを条件に、非誘導残基の代わりに化学的に誘導された残基の使用を包含する。
【0046】
“化学的に誘導された”とは、官能側鎖基の反応によって化学的に誘導された1つまたは2つ以上の残基を有するポリペプチドを指す。そのような誘導分子は、例えば、遊離アミノ基が、塩酸アミン、p−トルエンスルフォニル基、カルボベンゾキシ基、t−ブチルオキシカルボニル基、クロロアセチル基またはフォルミル基を形成するように誘導された分子を含む。遊離カルボキシ基は、塩、メチルおよびエチルエステルもしくはエステルの他の種類を形成するように誘導することができる。遊離ヒドロキシ基は、o−アシルまたはo−アルキル誘導体を形成するように誘導することができる。ヒスチジンのイミダゾル窒素は、N−im−ベンジルヒスチジンを形成するように誘導することができる。化学誘導体として含まれるものはまた、20種の標準アミノ酸の天然に生じる1つまたは2つ以上のアミノ酸誘導体を含むペプチドである。例えば、4−ヒドロキシプロリンはプロリンの代用にできる;5−ヒドロキシリジンはリジンの代用にできる;3−メチルヒスチジンはヒスチジンの代用にできる;ホモセリンはセリンの代用にできる;さらにオルニチンはリジンの代用にできる。本発明のポリペプチドはまた、必須の活性が維持される限り、1つまたは2つ以上の付加および/または欠失、または本明細書にその配列を示したポリペプチドの配列と関係がある残基をもついずれの配列も包含する。
“フラグメンド”という用語は、そのアミノ酸残基配列が本明細書に示されているポリペプチドのアミノ酸残基配列より短いアミノ酸残基配列をもつ一切のポリペプチドを指す。
【0047】
本発明のポリペプチドが、αvβ3の天然のリガンドの配列と同一でない配列を有する場合、それは、典型的には1つまたは2つ以上の保存的もしくは非保存的置換が行われたからで、通常は約30数%未満、好ましくは10数%未満のアミノ酸残基が置換される。付加残基はまた、“リンカー”を提供する目的のためにポリペプチドのいずれかの末端に付加できるが、このリンカーによって本発明のポリペプチドは都合よく標識もしくは固形マトリックス、または担体に固定することができる。
本発明のポリペプチドとともに用いることができる標識、固形マトリックスおよび担体は以下で述べる。
【0048】
アミノ酸残基リンカーは通常少なくとも1残基で、40またはそれより多い残基、もっと頻繁には1から10残基が可能であるが、αvβ3リガンドエピトープは形成しない。連結に用いられる典型的なアミノ酸残基はチロシン、システイン、リジン、グルタミン酸およびアスパラギン酸または同様なものである。さらに、本ポリペプチドは、別に特定されない場合は、末端−NH2のアシル化(例えばアセチル化またはチオグリコール酸のアミド化)によって、末端のカルボキシルアミド化(例えばアンモニア、メチルアミンとともに)および同様な末端の修飾によって修飾された配列のために、αvβ3リガンドの天然の配列とは異なっていてもよい。周知のように末端の修飾は、プロテイナーゼの消化による感受性を減少させるために有用で、したがって溶液中、特に生物学的液体(プロテアーゼが存在する可能性がある)中のポリペプチドの半減期を延長させるために役立つ。この点について、ポリペプチドの環状化はまた有用な末端修飾で、環状化によって形成される安定な構造のため、および本明細書で述べるようにそのような環状ペプチドについて見られる生物学的活性のゆえに特に好ましい。
【0049】
本発明のいずれのペプチドも、医薬的に許容できる塩の形態で用いることができる。本発明のペプチドと塩を形成することができる適切な酸は、無機酸、例えばトリフルオロ酢酸(TFA)、塩酸(HCl)、臭化水素酸、過塩素酸、硝酸、チオシアン酸、硫酸、燐酢酸、プロピオン酸、グリコール酸、乳酸、ピルビン酸、蓚酸、マロン酸、コハク酸、マレイン酸、フマル酸、アントラニル酸、桂皮酸、ナフタリンスルホン酸、スルファニル酸などを含む。HClおよびTFA塩は特に好ましい。
本発明のペプチドと塩を形成することができる適切な塩基は、無機塩基(例えば水酸化ナトリウム、水酸化アンモニウム、水酸化カリウムなど)、および有機塩基、例えばモノ−、ジ−、およびトリ−アルキル並びにアリールアミン(例えばトリエチルアミン、ジイソプロピルアミン、メチルアミン、ジメチルアミンなど)、並びに場合によっては置換されたエタノールアミン(例えばエタノールアミン、ジエタノールアミンなど)を含む。
【0050】
本発明のペプチド(本明細書ではまた本ポリペプチドと称される)は、前記ポリペプチド分野で習熟した者にとって周知の技術(組み換えDNA技術を含む)のいずれかにより合成できる。合成化学の技術(例えば固相メルリフィールド型合成)は、純度、抗原的特異性、望ましくない副産物を含まないこと、生成の容易さなどから好ましい。利用可能な多くの技術についての優れた要約は以下の文献で見出すことができる:Stewardら、“固相ペプチド合成(Solid Peptide Synthesis”、W.H.Freeman Co.,San Francisco,1969; Bodanszkyら、“ペプチド合成(Peptide Synthesis)”、John Wiley & Sons,第2版、1976; J.Meienhofer,“ホルモン蛋白およびペプチド(Hormonal Proteins and Peptides)”、第2巻、46ページ、アカデミックプレス(ニューヨーク)、1983; Merrifield,Adv.Enzymol.,32:221-96,1969; Fieldsら、Int.J.Peptide Protein Res.,35:161-214,1990; 並びに米国特許第4244946号(固相ペプチド合成に関して)およびSchroderら、“ペプチド(The Peptides)”、1巻、アカデミックプレス(ニューヨーク)、1965(古典的溶液合成に関して)。これらの文献の各々は参照により本明細書に含まれる。そのような合成で用いることができる適切な保護基は、上記の文献およびマッコーミーの論文(J.F.W.Mcomie,“有機化学の保護基(Protective Groups in Organic Chemistry)”、Plenum Press,ニューヨーク、1973(この文献は参照により本明細書に含まれる))に記載されている。
【0051】
一般に、意図される固相合成方法は、成長ペプチド鎖への1つまたは2つ以上のアミノ酸残基または適切に保護されたアミノ酸残基の連続的付加を含む。通常は、第一のアミノ酸残基のアミノ基またはカルボキシル基のいずれかが適切な、選択的に除去可能な保護基で保護されている。種々の、選択的に除去可能な保護基は、反応側鎖基をもつアミノ酸(例えばリジン)のために利用される。
具体例として固相合成を用いる場合、保護アミノ酸または誘導アミノ酸は、不活性な固形支持体にその未保護カルボキシルまたはアミノ基を介して附着されている。アミノ基またはカルボキシル基の保護基を続いて選択的に除去し、適切に保護された補完(アミノまたはカルボキシル)基をもつ前記配列の次のアミノ酸を混合し、固形支持体に既に付着した残基とアミド結合を形成させるために適切な条件下で反応させる。続いて、アミノまたはカルボキシル基の保護基をこの新しく付加したアミノ酸残基から除去し、(適切に保護された)次のアミノ酸をその後付加し、以下同様に行われる。最後に、所望のアミノ酸が適切な配列で連結され、一切の残余の末端保護基および側鎖基保護基(および固形支持体)は連続的にまたは同時に除去されて、最終的な直線状ポリペプチドが得られる。
【0052】
上記に記載したように具体例として調製した直線状ポリペプチドは、対応する環状ペプチドを形成させるために反応させることができる。ペプチドを環状化させるための典型的な方法は、ジンマーらの論文に記載されている(Zimmerら、“Peptides 1992”、393-394ページ、ESCOM Science Publishers,B.V.,1993)。典型的には、t−ブトキシカルボニル保護ペプチドのメチルエステルをメタノールに溶解させ、水酸化ナトリウム溶液を加え、混合物を20℃で反応させ、メチルエステル保護基を加水分解により除去する。溶媒を蒸発させた後、t−ブトキシカルボニル保護ペプチドを酸性化水溶液から酢酸エチルで抽出する。続いて、t−ブトキシカルボニル保護基をジオキサン補助溶媒中で穏やかな酸性条件下で除去する。このようにして得られた遊離アミノおよびカルボキシ末端をもつ非保護直線状ペプチドをその対応する環状ペプチドに変換させるために、この直線状ペプチドの希釈溶液をジクロロメタンとジメチルフォルムアミドの混合物中で、1−ヒドロキシベンゾトリアゾルとN−メチルモルフォリンの存在下でジシクロヘキシルカルボジイミドと反応させる。生じた環状ペプチドはクロマトグラフィーで精製される。
特に好ましい環状ペプチド合成方法は、ガーラスらの論文(Gurrathら、Eur.J. Biochem.,210:911-921(1992))および実施例に記載されている。本方法で使用するために特に好ましいペプチドは、c−(GrGDFV)(配列番号4)、c−(RGDfV)(配列番号5)、c−(RADfV)(配列番号6)、c−(RGDFv)(配列番号7)および直線状ペプチドYTAECKPQVTRGDVF(配列番号8)であり、ここで“c−”は環状ペプチドを指し、大文字はL−アミノ酸の一文字記号で、小文字はD−アミノ酸の一文字記号である。これらペプチドのアミノ酸残基配列はまた、配列番号4、5、6、7および8にそれぞれ示される。
【0053】
2.単クローン性抗体
一具体例で、本発明は、αvβ3と免疫的に反応し、本明細書で述べるようにαvβ3の天然のリガンドとαvβ3が結合するのを抑制する単クローン性抗体の形態をとるαvβ3拮抗物質を開示する。本発明はまた、この抗体を産生する細胞株、この細胞株を作製する方法、およびこの単クローン性抗体を生成する方法を開示する。
本発明の単クローン性抗体は、1)分離αvβ3と免疫的に反応し、2)フィブリノーゲンのαvβ3への結合を抑制する抗体分子を含む。優先的にαvβ3と結合する好ましい単クローン性抗体は、ハイブリドーマ細胞株ATCCHB9537が分泌するmAbLM609の免疫反応特性をもつ単クローン性抗体を含む。このハイブリドーマ細胞株ATCCHB9537は、ブダペスト条約の要請にしたがって、米国菌培養収集所(ATCC)(1301 Parklawn Drive,Rockville,MD,USA)に1987年9月15日に寄託された。
“抗体または抗体分子”という用語は、免疫グロブリン分子の集団および/または免疫グロブリンの免疫学的に活性な部分(すなわち抗体結合部位またはパラトープを含む分子)の集団を指す集合名詞として本明細書では用いられる。
“抗体結合部位”とは、抗原と特異的に結合する重鎖および軽鎖の可変並びに超可変領域から構成される抗体分子の構造部分である。
本発明で使用する典型的な抗体は、完全な免疫グロブリン分子、実質的に完全な免疫グロブリン分子および、パラトープを含む免疫グロブリン分子の部分(当分野でFab、Fab’、F(ab’)2およびF(v)として知られ、また抗体フラグメントと称される部分を含む)である。
【0054】
別の好ましい具体例では、本発明は、この発明の単クローン性抗体に由来するFabフラグメントを含む短縮免疫グロブリン分子を含む。Fcレセプターを欠くこのFabフラグメントは可溶性で、血清半減期に関して治療的利点をもち、さらに可溶性フラグメントを使用するという態様に関して診断的利点をもつ。可溶性Fabフラグメントの調製は、免疫学分野で一般に知られており、種々の方法で達成できる。
例えば、抗体のFabおよびF(ab’)2部分(フラグメント)は、周知の方法(例えば、Theofilopolous & Dixon,米国特許第4342566号を参照のこと)による実質的に完全な抗体のそれぞれパパインおよびペプシンによる蛋白分解反応によって調製される。Fab’抗体部分もまた周知であるが、2つの重鎖部分を連結するジスルフィド結合を例えばメルカプトエタノールで還元し、生じた蛋白メルカプタンを例えばヨードアセトアミドのような試薬でアルキル化してF(ab’)2部分から生成される。完全な免疫グロブリンを含む抗体が好ましく、本明細書で詳述するように用いられる。
【0055】
“単クローン性抗体”という語は、特定のエピトープと免疫的に反応することができるただ1種の抗体結合部位を含む抗体分子の集団を指す。したがって、単クローン性抗体は、それが免疫的に反応するいずれかのエピトープに対して典型的にはただ1つの結合親和性を示す。それゆえ、単クローン性抗体は、各々は異なるエピトープに対して免疫的に特異的である、複数の抗体結合部位をもつ抗体分子を含む可能性がある(例えば二特異性単クローン性抗体)。
単クローン性抗体は、典型的にはただ1種の抗体分子を分泌(産生)するハイブリドーマと呼ばれる単一の細胞クローンによって産生される抗体から成る。このハイブリドーマ細胞は、抗体産生細胞とミエローマまたは他の自己永続化細胞株とを融合させて形成される。そのような抗体の調製は、最初コーラーとミルシュタイン(Kohler & Milstein,Nature 256:495-497(1975))によって記載され、その内容は参照により本明細書に含まれる。また別の方法はゾーラによって記載された(Zola,“単クローン性抗体:術式の手引(Monoclonal Antibodies:A Manual of techniques)”CRC Press,Inc.(1987))。そのように調製したハイブリドーマの上清をαvβ3と免疫的に反応させ、さらに天然のリガンドへのαvβ3結合を抑制する抗体分子の存在についてスクリーニングすることができる。
【0056】
簡単に記せば、単クローン性抗体組成物を産生するハイブリドーマを生成するために、αvβ3源(例えば記載(Chereshら、J.Biol.Chem.,262:17703-17711(1987)にしたがってM21ヒトミエローマ細胞から分離したαvβ3)で強化免疫を施した哺乳類の脾臓から得たリンパ球とミエローマまたは他の自己永続化細胞株を融合させる。
ハイブリドーマを調製するために用いる細胞株はリンパ球と同じ種に由来することが好ましい。典型的には、マウス129GLX+株が好ましい哺乳類である。本発明で使用する適切なマウスミエローマには、ヒポキサンチン−アミノプテリン−チミジン−感受性(HAT)細胞株P3X63−Ag8.653、およびSp2/0−Ag14が含まれるが、これらは米国菌収集所(Rockville,MD)からそれぞれCRL1580およびCRL1581の名称で入手できる。
【0057】
典型的には、ポリエチレングリコール(PEG)1500を用いてミエローマ細胞と脾臓細胞を融合させる。融合ハイブリッドはHATに対する感受性によって選択される。本発明の単クローン性抗体を産生するハイブリドーマは、実施例で述べるように酵素結合免疫吸着アッセー(ELISA)を用いて識別される。
本発明の単クローン性抗体はまた、適切な特異性の抗体分子を分泌するハイブリドーマを含有する栄養培地を含む単クローン性ハイブリドーマの培養を開始することによって製造できる。培養は、ハイブリドーマが抗体分子を培養液中に分泌するために十分な時間、および条件下で維持される。続いて抗体含有培養液を採集する。その後、抗体分子を周知の技術でさらに分離する。
これらの組成物の調製のために有用な培地は当分野で周知であり、市販もされているが、合成培養液、系統内交配マウスなどを含む。代表的な合成培養液は、ダルベッコーの最少必須培地(Dulbecco's minimal essential medium)DMEM(Dulbeccoら、Virol.8:396(1959))で、4.5gm/lのグルコース、20mMグルタミンおよび20%ウシ胎児血清が補充される。
【0058】
単クローン性抗体、ハイブリドーマ細胞、またはハイブリドーマ細胞培養を製造する他の方法もまた周知である。例えば、以下の論文に記載された免疫学的全産物からの単クローン性抗体の分離方法を参照されたい:Sastryら、Proc.Natl.Acad.Sci.USA,86:5728-5732(1989); およびHuseら、Science,246:1275-1281(1989)。
また本発明に含まれるものは、本発明の単クローン性抗体を産生するハイブリドーマ細胞およびハイブリドーマ細胞を含む培養である。特に好ましいものは、単クローン性抗体mAbLM609を分泌するATCCHB9537と称されるハイブリドーマ細胞株である。mAbLM609はチェレッシュら(Chereshら、J.Biol.Chem.,262:17703-17711(1987)の記載のように調製されたが、その調製は実施例でもまた詳述する。
本発明は1具体例で、mAbLM609の免疫反応特性をもつ単クローン性抗体を含む。
【0059】
単クローン性抗体が本発明の単クローン性抗体と同じ(すなわち同等な)特異性(免疫反応特性)をもつか否かは、また、前者が予め選ばれた標的分子に後者が結合するのを妨げるか否かを調べることによって決定することが可能である。
固相に存在する標的分子の結合に関する標準的競合アッセーで本発明の単クローン性抗体によって結合減少が示されるように、もし被検単クローン性抗体が本発明の単クローン性抗体と競合するならば、その場合、2つの単クローン性抗体は、おそらく同じかまたは密接に関連するエピトープに結合するであろう。単クローン性抗体が本発明の単クローン性抗体の特異性をもつか否かを決定するまた別の方法は、通常反応性を有する標的分子と本発明の単クローン性抗体とを予め保温し、続いて被検単クローン性抗体を加え、被検単クローン性抗体が標的分子に結合する能力が抑制されるか否かを決定することである。被検単クローン性抗体が抑制されたならば、おそらく被検単クローン性抗体は、本発明の単クローン性抗体と同じか、または機能的に同等なエピトープ特異性をもつであろう。
単クローン性抗体が本発明の単クローン性抗体の特異性を有するか否かを決定するまた別の方法は、問題の抗体のCDR領域のアミノ酸残基配列を決定することである。そのCDR領域に同一または機能的に同等なアミノ酸残基配列もつ抗体分子は同じ結合特異性を有する。ポリペプチドの配列決定方法は当分野で周知である。
【0060】
抗体の免疫特異性、その標的分子結合能および、エピトープに対して抗体が示す付随する親和性は、この抗体が免疫的に反応するエピトープによって決定される。エピトープ特異性は、少なくとも部分的には、免疫グロブリンの重鎖の可変領域のアミノ酸残基配列によって、さらに部分的には軽鎖の可変領域アミノ酸残基配列によって決定される。
“結合特異性をもつ”という語を用いる場合、同等な単クローン性抗体が同じかまたは同様な免疫反応(結合)を示し、さらに予め選択された標的分子との結合に対して競合するということを示す。
ヒト化(humanized)単クローン性抗体は、特に人間に治療的に使用する場合はネズミの単クローン性抗体より特別な利点を提供する。具体的には、ヒトの抗体は“外来”抗原のように急激に血液循環から排除されず、さらに外来抗原および外来抗体と同じ態様で免疫系を活性化させない。“ヒト化”抗体の調製方法は当分野で一般に周知であり、本発明の抗体に容易に応用できる。
したがって、本発明は、一具体例において抗体の抗原結合能に実質的に干渉することなくヒトの免疫系成分を導入するために、移植によりヒト化させた本発明の単クローン性抗体を含む。
【0061】
3.αvβ3特異的模倣物(mimetics)
本発明は、“模倣物”と称されるポリペプチド、抗体および他の分子を含むαvβ3拮抗物質は、一般に本発明で用いることができることを示すが、この模倣物はαvβ3機能と干渉する能力を有する。特に好ましいものは、特にαvβ3の機能と干渉し、他のインテグリンの機能には干渉しない。
本明細書では、試薬がその必須の生物学的活性を保有している限り、種々の試薬が本方法で使用するために適切であることは理解されよう。これらの試薬は包括的に模倣物と称するが、それは、模倣物が、レセプターとリガンドの機能的相互作用に必要なαvβ3またはαvβ3リガンドのいずれかの結合ドメインを“模倣する”能力を保有し、したがって正常な機能に干渉(すなわち抑制)するからである。
【0062】
αvβ3模倣物は、抗体またはリガンド由来ペプチド以外で上記の特性を示すいずれの分子でもよい。それはペプチドの合成類似体でもよく、上記の結合ドメインの結合ポケットのような形状をした化合物または他の分子でもよい。
αvβ3模倣物のデザインは、当分野で既知の薬剤デザインのための様々な構造分析方法のいずれを用いても実施できる。これらの分析方法には、分子モデリング、二次元核磁気共鳴(2−DNMR)分析、X線結晶学、ペプチドのランダムスクリーニング、ペプチド類似体または他の化学ポリマーライブラリーおよび同様な薬剤デザイン方法が含まれる。
αvβ3拮抗物質は小さなポリペプチドまたは単クローン性抗体(αvβ3の選択的抑制という機能的特性を共有する2種の別々に異なる化学構造)がよいということを示す、本明細書で提示した広範囲の構造的証拠から見ると、本方法に有用なこのαvβ3拮抗物質の構造はあまり限定される必要はなく、本明細書で規定されるいずれのαvβ3模倣物も含むことができる。
【0063】
F.αvβ3の拮抗物質を識別する方法
本発明はまた、本方法にしたがって使用するαvβ3拮抗物質候補を識別する方法を開示する。これらのアッセー方法では、候補物質は、天然のリガンドへのαvβ3結合を抑制するその能力を調べられ、さらに、組織における脈管形成のその抑制能が調べられる。
第一のアッセーは、天然リガンドのαvβ3への直接結合の抑制を測定するが、好ましい具体例は実施例で詳細に記載される。このアッセーは典型的には、ELISAによって固相における分離αvβ3への天然リガンド(例えばフィブリノーゲン)の結合抑制の程度を測定する。
このアッセーはまた、αvβ3に特異性を示し、天然のリガンドが他のインテグリンに結合するのを抑制しない化合物を識別するためにも用いられる。この特異性アッセーは、ELISAアッセーを平行して行うことによって実施されるが、この場合、αvβ3および他のインテグリンは、天然のリガンドと結合するそれぞれの能力について、さらに、予め選択したリガンドと結合するインテグリンのそれぞれの能力を抑制する候補化合物について別々のアッセーチャンバーで同時にスクリーニングされる。好ましいスクリーニングアッセーの様式は実施例で述べる。
【0064】
第二のアッセーは、ニワトリの漿尿膜(CAM)での脈管形成を測定し、CAMアッセーと称される。このCAMアッセーは、他の研究者によって詳細に記載され、さらに腫瘍組織の脈管形成および新規血管新生の両方を測定するために用いられた(Ausprunkら、Am.J.Pathol.,79:597-618(1975); Ossonskiら、Cancer Res.,40:2300-2309(1980))。
CAMアッセーは、完全な組織の新規血管新生が生じ、実際のニワトリ胎児の血管がCAMまたはCAM上で成長する組織へと増殖していくので、十分に認知されたインビボの脈管形成のためのアッセーモデルである。
本明細書で示すように、CAMアッセーは、新規な血管増殖の量と程度の両方について新規血管新生の抑制を明示する。さらに、CAM上に移植されたいずれの組織(例えば腫瘍組織)の増殖も容易にモニターできる。最後に、このアッセーは、アッセー系の中に毒性に対する内部制御が存在するので特に有用である。ニワトリ胎児はいずれのテスト試薬にも曝すことができ、したがって、胎児の健康状態は毒性の指標である。
【0065】
第三のアッセーは、ウサギのインビボ眼球モデルで脈管形成を測定するが、これはウサギ眼球アッセーと称される。ウサギ眼球アッセーは、他の研究者によって詳細に記載され、さらに脈管形成抑制物質(例えばサリドマイド)の存在下で脈管形成および新規血管新生の両方を測定するために用いられた(D'Amatoら、Proc.Natl.Acad.Sci.,91:4082-4085(1994))。
新規血管新生過程(角膜の辺縁から角膜内へ増殖するウサギの血管によって実証される)が、眼球の本来透明な角膜を通して容易に見ることができるので、ウサギ眼球アッセーは、インビボ脈管形成の十分に認知されたアッセーモデルである。さらに、新規血管新生もしくは新規血管新生の退行の刺激または抑制の程度および量の両方が時間経過において容易にモニターできる。
【0066】
最後に、ウサギはいずれのテスト試薬にも曝すことができ、したがって、ウサギの健康状態がテスト試薬の毒性の指標である。
第四のアッセーは、マウス:ヒトキメラ−マウスモデルにおける脈管形成を測定し、キメラマウスアッセーと称される。このアッセーは、他の研究者によって詳細に記載され、さらに本明細書で、脈管形成、新規血管新生および腫瘍組織の退縮を測定するために詳述される(Yanら、J.Clin.Invest.,91:986-996(1993))。キメラマウスアッセーは、移植された皮膚移植片が正常なヒト皮膚組織と組織学的に類似し、完全組織の新規血管新生が生じ、この場合、実際のヒトの血管が移植されたヒトの皮膚から、移植されたヒト皮膚の表面のヒト腫瘍内に増殖していくので、インビボの脈管形成の有用なアッセーモデルである。ヒト移植片への新規血管新生の起始点は、ヒト特異的内皮細胞マーカーを用いて新規血管新生の免疫組織化学的染色によって明らかにすることができる。
【0067】
本明細書で示すように、キメラマウスアッセーは、新しい血管増殖の退行の量および程度の両方について新規血管新生の退行を明らかにする。さらに、移植皮膚上に移植されたいずれの組織(例えば腫瘍組織)の増殖に対する影響も容易にモニターできる。最後に、アッセー系の中に毒性に対する内部制御が存在するのでこのアッセーは有用である。キメラマウスはいずれのテスト試薬にも曝すことができ、したがって、マウスの健康状態は毒性の指標である。
【0068】
実施例
本発明に関する以下の実施例は説明のためであり、本発明を具体的に限定するものと解釈されるべきではない。さらにまた、当業者の視野内にある、現時点で明らかなまたは将来開発される種々の変形も、この後に続く本発明の請求の範囲内にあると考えられるべきである。
【0069】
1.合成ペプチドの調製
表1に示した直線状および環状ペプチドを、例えば以下の論文に記載された標準的固相合成技術を用いて合成した(Merrifield,Adv.Enzymol.,32:221-96(1969); G.B.Fields & R.L.Noble,Int.J.Peptide Protein Res.,35:161-214(1990))。
2グラム(g)のBOC-Gly-D-Arg-Gly-Asp-Phe-Val-OMe(配列番号1)を先ず60ミリリットル(ml)のメタノール(2Nの水酸化ナトリウム1.5mlが添加されている)に溶解して混合物を作製した。続いてこの混合物を20℃で3時間攪拌した。蒸発後、残留物を水にとり、希塩酸でpH3に酸性化し、エチル酢酸で抽出した。抽出物をNa2SO4上で乾燥させ、再度蒸発させて得られたBOC-Gly-D-Arg-Gly-Asp-Phe-Val-OH(配列番号2)をジオキサン中の2N塩酸20mlで、20℃2時間攪拌した。得られた混合物を蒸発させて、H-Gly-D-Arg-Gly-Asp-Phe-Val-OH(配列番号3)を得、続いて1800mlのジクロロメタンと200mlのジメチルホルムアミド(DMF)の混合物中に溶解し、その後0℃に冷却した。続いて、0.5gのジシクロヘキシルカルボジイミド(DCCI)、0.3gの1−ヒドロキシベンゾトリアゾル(HOBt)および0.23mlのN−メチルモルフォリンを攪拌しながら連続して加えた。
【0070】
得られた混合物を0℃でさらに24時間、20℃で48時間攪拌した。この溶液を濃縮し、混合ベッドイオン交換器で処理し塩を除去した。生じた樹脂をろ過で除去した後、清澄溶液を蒸発させ、残留物をクロマトグラフィーで精製してシクロ(-Gly-D-Arg-Gly-Asp-Phe-Val)(配列番号4)を回収した。表1に示した以下のペプチド(1文字コードのアミノ酸残基略語を用い、ペプチド番号により区別されている)が同様に得られた:シクロ(Arg-Gly-Asp-D-Phe-Val)(配列番号5);シクロ(Arg-Ala-Asp-D-Phe-Val)(配列番号6);シクロ(Arg-D-Ala-Asp-Phe-Val)(配列番号9);シクロ(Arg-Gly-Asp-Phe-D-Val)(配列番号7)。ペプチド62184の配列と同一の配列を有する66203と称するペプチドは、62184に存在するTFAの塩と異なりHCl塩を含むことだけが後者と異なっていた。実施例7(合成ペプチドが用いられている)で述べる脈管形成抑制アッセーでは、HClをもつこの66203ペプチドは、TFAをもつ同一のペプチドより脈管形成抑制でわずかに効果が高かった。
【0071】
【0072】
ペプチド62185の配列と同一の配列を有する69601と称するペプチドは、62184に存在するTFA塩ではなくHCl塩を含むことだけが後者と異なる。
環状ペプチドc−RADfV(69601)はフィブリノーゲンのインテグリンαvβ3への結合を抑制し、フィブリノーゲンのインテグリンαIIbβ3またはα5β1への結合は抑制しないことが示された(Pfaffら、J.Biol.Chem.,269:20233-20238(1994))。したがって、ペプチドc−RADfVはαvβ3に特異的である。
2.単クローン性抗体
ハイブリドーマATCC HB9537により分泌される単クローン性抗体LM609は、セファロース−ヒラマメレクチンビーズに吸着させた分離αvβ3を用いた免疫によって標準的ハイブリドーマ法で製造された。このαvβ3は、M21と称するヒトミエローマ細胞から分離し、抗体はチェレッシュらの記載にしたがって作製した(Chereshら、J.Biol.Chem.,262:17703-17711(1987))。M21細胞はモートン博士(Dr.D.L.Morton,University of California,ロサンゼルス、カリフォルニア)により提供され、2mMのL−グルタミン、50mg/mlの硫酸ジェンタマイシンおよび10%のウシ胎児血清を含むRPMI1640培養液で浮遊培養として増殖させた。
単クローン性抗体LM609は、特異的にαvβ3複合体と免疫的に反応し、αvサブユニットともβ3サブユニットともまたは他のインテグリンとも免疫的に反応しないことが示された。
【0073】
3.αvβ3発現の組織分布の特徴
A.抗インテグリンレセプター抗体による免疫蛍光
創傷治癒の間、血管の基底膜はいくつかの粘着蛋白(ヴォン・ヴィレブランド因子、フィブロネクチンおよびフィブリンを含む)を発現させる。さらに、粘着レセプターのうちのインテグリン類のいくつかは、培養平滑筋細胞および内皮細胞の表面で発現される(Cheresh、Proc.Natl.Acad.Sci.,USA,84:6471(1987);Janatら、J.Cell Physiol.,151:588(1992); Chengら、J.Cell Physiol.,139:275(1989)を参照のこと)。αvβ3(チェレッシュが記載したように(Proc.Natl.Acad.Sci.,USA,84:6471(1987))、ヴォン・ヴィレブランド因子、フィブリノーゲン(フィブリン)およびフィブロネクチンに対する内皮細胞レセプター)は、これらのインテグリンの中に存在する。このインテグリンは、内皮細胞の移動へと先導するカルシウム依存性シグナリング経路を開始させ、したがってリーベルシーら(Leavelseyら、J.Cell Biol.,121:163(1993))が記載したように、血管細胞生物学において基礎的な役割を果たすように思える。脈管形成中のαvβ3発現を調べるために、ヒトの創傷の肉芽形成組織または隣接正常皮膚を同意患者より採取し、1mlの燐酸緩衝食塩水で洗浄し、O.T.C.培地(Tissue Tek)に包埋した。この包埋組織を液体窒素中で約30から45秒瞬間冷凍した。クリオスタットミクロトーム上のこの冷凍ブロックから6ミクロンの厚さの切片を切り出し、続いてβ3インテグリン(αvβ3またはαIIbβ3)またはインテグリンのβ1亜類に特異的な抗体を用いて免疫ペルオキシダーゼ染色を施した。
【0074】
正常ヒト皮膚および創傷の肉芽形成組織の染色の結果は図1A−1Dに示す。
単クローン性抗体AP3およびLM534(それぞれβ3およびβ1インテグリンに対して誘導された)が、冷凍切片の免疫組織化学分析のために用いられた。4人の異なる供与者の組織を用いた実験で同一の結果が得られた。光学顕微鏡写真は300×の倍率で示されている。
αvβ3インテグリンは、肉芽形成組織の血管で豊富に発現されたが(図1B)、同じ供与者の正常皮膚の真皮および上皮では検出できなかった(図1A)。対照的に、β1インテグリンは、正常皮膚(図1C)および肉芽形成組織(図1D)の両方の血管および支質細胞で、さらにアダムスら(Adamsら、Cell,63:425(1991))の報告で先に示された通り上皮内の基底細胞で豊富に発現された。
B.抗リガンド抗体による免疫蛍光
それぞれβ3およびβ1インテグリン、ヴォン・ヴィレブランド因子およびラミニンに対するリガンドの存在について、上記のように調製したまた別の正常ヒト皮膚および肉芽形成組織の切片を調べた。ヴォン・ヴィレブランド因子は、正常ヒト皮膚(図2A)および肉芽形成組織(図2B)の血管に局在し、一方、ラミニンは、両組織の上皮基底膜の他、すべての血管に存在した(図2Cおよび2D)。
【0075】
C.癌組織の抗αvβ3抗体の分布
上記の分析に加えて、ヒトの患者から得た癌組織の生検材料もまたαvβ3の存在および分布について調べられた。インテグリンレセプター複合体、αvβ3に特異的な実施例2で調製された単クローン性抗体LM609で染色したという点を除き、組織は実施例1Aで述べたように調製された。さらに、代表的な腫瘍例をブリン固定液(Bulin Fixative)で8時間固定し、連続切片を切り出してH&E染色することによって、組織はまた顕微鏡による組織分析用に調製された。
膀胱癌、大腸癌、乳癌および肺癌組織の免疫ペルオキシダーゼ染色の結果は図3A−3Dに示す。αvβ3は、4種の癌生検材料に存在する血管にのみ豊富に発現し、これら組織に存在する他の細胞には全く存在しない。
ここに詳述した結果は、したがって、αvβ3インテグリンレセプターは、特定の組織型(すなわち肉芽形成組織、転移組織、および脈管形成が生じているその他の組織)で選択的に発現し、新しい血管の形成が停止している正常な組織では発現しないことを示している。これらの組織は、それゆえ、本発明の治療面についての理想的な標的を提供する。
【0076】
4.リガンド−レセプター結合アッセーにより検出されるαvβ3特異的合成ペプチドの同定
精製リガンド−レセプター結合アッセーで、αvβ3およびαIIbβ3レセプター結合活性と拮抗するそれらの能力を測定することによって、実施例1で調製された合成ペプチドをスクリーニングした。これら結合実験の方法は以下の論文に記載されているが(Barbasら、Proc.Natl.Acad.Sci.,USA,90:10003-10007(1993); Smithら、J.Biol.Chem.,265:11008-11013(1990); Pfaffら、J.Biol.Chem.,269:20233-20238(1994))、これらの内容は参照により本明細書に含まれる。
ここで述べるのは、レセプターが固形支持体に固定され、拮抗物質が可溶性であるリガンド−レセプター結合アッセーで拮抗物質を同定する方法である。さらに、リガンドが固形支持体に固定され、レセプターおよび拮抗物質が可溶性であるリガンド−レセプター結合アッセーも開示する。
簡単に記せば、選択した精製インテグリンを、ウェル当たり50ナノグラム(ng)の被覆濃度でタイターテック(Titertek)微量定量板(マイクロタイター)に別々に固定した。リガンド−レセプター結合アッセーで用いられたレセプターの精製は当分野で周知であり、当業者にとって通常の方法で容易に得ることができる。4℃で18時間保温した後、プレート上の非特異的結合部位をトリス緩衝食塩水中のウシ血清アルブミン(BSA)(10ミリグラム/ミリリットル(mg/ml))で封鎖した。抑制実験のためには、表1の種々の濃度の選択ペプチド(125I−ビトロネクチンまたは125I−フィブリノーゲンのインテグリンレセプター)をαvβ3およびαIIbβ3への結合を妨害する能力について調べた。これらのリガンドは、特定のインテグリン(αvβ3に対するビトロネクチンおよびαIIbβ3に対するフィブリノーゲン)について最適の結合を示したが、フィブリノーゲンのいずれかのレセプターへの結合を阻害する、ペプチドを用いた結合抑制実験は、レセプターのリガンドへの結合を1/2最大抑制するために必要なペプチド量のマイクロモル(uM)単位での正確な決定を可能にした。放射能標識リガンドは1nMの濃度で用いられ、結合は非標識合成ペプチドで別々に実施された。
【0077】
3時間保温した後、遊離リガンドを洗浄により除去し、結合リガンドをガンマ計測により検出した。別々に固定したαvβ3およびαIIbβ3レセプターへのレセプターおよび放射能標識フィブリノーゲンの結合を抑制するために表1に示した選択環状ペプチドを用いたアッセーで得られたデータは、データの点数間誤差が典型的には11%以下で高度に再現性があった。マイクロモル(uM)でのIC50(IC50uM)は2つのデータ点数の平均±標準偏差として表2に示したように表した。
【0078】
したがって、RGD含有またはRGD誘導環状ペプチド62181、62184、62185および62187(各々1個のD−アミノ酸残基をもつ)は、αIIbβ3レセプターへのフィブリノーゲン結合の抑制と比較したとき1/2最大抑制に必要なペプチド濃度はより小さいことで示されるように、αvβ3レセプターへの優先的なフィブリノーゲン結合抑制を示した。対照的に、その他のRGD含有またはRGD誘導環状ペプチド、62186、62175および62179は、αvβ3へのフィブリノーゲン結合阻害において有効でないか、またはαvβ3と比較してαIIbβ3へのフィブリノーゲン結合の優先的抑制を示した。これらの結果は、環状ペプチドRGDFV(ここでFはD−アミノ酸残基を示す)は、フィブリノーゲンのαvβ3インテグリンへの結合を特異的に抑制するが、αIIbβ3またはα5β1インテグリンへのそれは抑制しないというパフらの最近の報告(Pfaffら、J.Biol.Chem.,269:20233-20238(1994))と一致する。同様な結合抑制アッセーをRGDモチーフ(この配列はαvレセプターサブユニット、αIIbレセプターサブユニットまたはビトロネクチンリガンドアミノ酸残基配列に由来した)をもつ、または欠いている直線状ペプチドで実施した。直線状ペプチド、62880(VN由来アミノ酸残基35−49)、62411(αv由来アミノ酸残基676−687)、62503(αv由来アミノ酸残基655−667)および62502(αIIb由来アミノ酸残基配列296−306)の配列は表1に示す。これらペプチドの各々は、ビトロネクチン(VN)またはフィブリノーゲン(FG)のαIIbβ3またはαvβ3のいずれかへの結合抑制アッセーに別々に用いられた。各実験についての個々のアッセーのマイクロモルでのIC50のデータ(IC50uM)は表3に示す。
【0079】
【0080】
直線状ペプチドを用いた選択インテグリンレセプターへのリガンド結合抑制アッセーの結果は、αIIbβ3レセプターへのそれと比較して1/2最大抑制に必要なペプチド濃度がより小さいことによって示されるように、ペプチド62880のみが、FGまたはVNのαvβ3への1/2最大結合を抑制することに有効であることを示している。その他の直線状ペプチドのいずれもαvβ3へのリガンド結合を阻害する効果をもたなかった。ただし、ペプチド62502は、αIIbβ3へのVN結合の阻害に有効であった。
したがって、本明細書に記載するリガンド−レセプターアッセーは、本発明の実施でビトロネクチンレセプター(αvβ3)拮抗物質として使用されるように、特定のインテグリンレセプター(特にαvβ3)に対して選択的特異性を示す環状および直線状合成ペプチドの両方のスクリーニングに用いることができる。
【0081】
5.未処置ニワトリ漿尿膜(CAM)の特徴
A.CAMの調製
脈管形成は、正常な胎児の脈管形成が成熟血管の形成をもたらした後で、ニワトリの漿尿膜で誘発することができる。文献(Leibovichら、Nature,329:630(1987);Ausprunkら、Am.J.Pathol.,79:597(1975))に記載されたように、脈管形成は、特定のサイトカインまたは腫瘍フラグメントに反応して誘発されることが明らかにされた。CAMは、実施例6および7でそれぞれ述べるように、引き続き脈管形成および脈管形成抑制を誘発するためにニワトリ胎児から調製された。10日齢のニワトリ胎児はマックインタイヤー養鶏場(McIntyre Poultry,レークサイド、カリフォルニア)から入手し、60%の湿度で37℃で保温した。小さな工芸用ドリル(Dremel,Division of Emerson Electric Co.レーシーン、ウィスコンシン)を使って、直接卵の先端の気嚢上の卵殻に小孔を開けた。卵を光で検卵して予め調べた胎児血管の存在しない領域で卵の広い方の側面上に第二の孔をドリルで開けた。最初の孔に陰圧をかけ、その結果CAM(漿尿膜)を卵殻膜から引き剥がし、CAM上に偽の気嚢を作製した。小さな模型用輪状やすり(Dremel)を使って、1.0cm×1.0cmの真四角の窓を沈降CAMの上の卵殻に開けた。この小さな窓は下に横たわるCAMに直接アクセスすることを可能にした。
【0082】
得られたCAMの調製物を続いて以下の2通りのいずれかの時期で用いた:CAMへの追加の処置を行わない胎児発生6日(活発な新規血管新生が認められるステージ)、これは胎児の新規血管新生に対する影響を調べるために用いるモデルとなる;胎児発生10日、ここでは脈管形成は鎮静化している。後者の調製物は、したがって、実施例6で述べるようにサイトカイン処置または腫瘍との接触に反応して脈管形成再開を誘発させるために本発明で用いられた。
B.CAMの組織学
ニワトリ胎児CAMおよび/または、実施例8で述べるようにニワトリ胎児から切除したヒトの腫瘍の顕微鏡構造を調べるために、CAMおよび腫瘍を実施例3Aで述べたように冷凍切片用に調製した。免疫蛍光分析用に冷凍ブロックからクリオスタットミクロトームで6ミクロンの厚さの切片を切り出した。
【0083】
図4は、未処置10日齢CAMの血管のない領域の典型的な光学顕微鏡写真を示す。CAM系の脈管形成は、胎児発生のこのステージまでに鎮静化しており、この系は、隣接領域からその時点で一切血管が存在しないCAM領域へと、現存する血管から新規な血管構造の発生を刺激するために本発明で有用である
C.免疫蛍光で検出されるCAMのインテグリンプロフィル
CAM組織に存在するインテグリンレセプターの組織分布を見るために、腫瘍組織およびニワトリCAM組織の両方の6ミクロン(um)の冷凍切片をアセトンで30秒固定し、バックらの記載(Buckら、J.Cell Biol.,107:2351(1988))にしたがって、β1インテグリンサブユニットに特異的な単クローン性抗体、mAbCSAT(したがってコントロールとして使用)の10マイクログラム/ミリリットル(ug/ml)、または実施例2で調製したLM609で免疫蛍光染色を施した。一次染色の後、1:250希釈のヤギ抗マウスローダミン標識二次抗体(Tago)で染色し、一次免疫反応生成物を検出した。続いて、この切片をツァイス(Zeiss)の免疫蛍光化合物用顕微鏡で調べた。
【0084】
免疫蛍光分析の結果は、未処置10日ニワトリ胎児に存在する成熟血管はインテグリンβ1サブユニット(図5A)を発現することを示した。対照的に、図5Aに示した組織の連続切片では、LM609との免疫反応性は認められなかった(図5B)。したがって、LM609抗体によって検出されるインテグリンαvβ3は、10日齢の未処置ニワトリ胎児に存在する成熟血管では活発に発現されていなかった。CAMモデルおよび以下の実施例で示すように、血管が正常な胎児発生で新規の増殖を行っているか、またはサイトカインもしくは腫瘍によって誘発を受けているとき、血管はαvβ3を発現する。しかしながら活発な新規血管新生に続いてひとたび血管の成長が停止するや、αvβ3の発現は免疫蛍光分析によっては検出不能のレベルまで減少する。成熟血管における発現の欠如と較べて、脈管形成を経ている血管におけるこのようなαvβ3発現の調節は、CAMの脈管形成系を用いたモデル系として以下の実施例で示すように、脈管形成を制御しさらに抑制するために本発明の固有の能力を提供する。
【0085】
6.CAM脈管形成アッセー
A.増殖因子によって誘発される脈管形成
脈管形成は、実施例5Aで引用したようにサイトカインや増殖因子によって誘発されることが示された。ここで述べる実験では、実施例5で述べたCAM調製物での脈管形成は、この中で説明するようにCAM血管上に局所的に存在する増殖因子によって誘発された。
脈管形成は、ハンクス調整塩溶液(Hanks Balanced Salt Solution(HBSS))(GIBCO,グランドアイランド、ニューヨーク)または150ナノグラム/ミリリットル(ng/ml)の組み換え塩基性線維芽細胞増殖因子(βFGF)(Genzyme,ケンブリッジ、マサチューセッツ)を含むHBSSを飽和させた5ミリメートル(mm)×5mmのワットマンフィルターディスク(Whatman Filter Paper No.1)を10日齢のニワトリ胎児の血管のない領域に置くことによって誘発された。窓は後でテープを使って閉じた。他のアッセーでは、125ng/mlのβFGFも血管増殖を誘発するために効果があった。脈管形成は、72時間後光学顕微鏡でモニターした。CAMは瞬間冷凍し、6umの凍結切片をアセトンで固定して、10ug/mlの抗β1単クローン性抗体CSATまたはLM609のいずれかを用いて、実施例5Cで述べたように免疫蛍光によって染色した。
図5Cの免疫蛍光光学顕微鏡写真は、図5Bに示すように未処置のニワトリCAMでのαvβ3発現の欠如と対照的に、βFGF誘発脈管形成時にはニワトリCAM上に増強されたαvβ3が発現することを示す。αvβ3は、βFGF処置CAMの多くの血管で(75から80%)容易に検出できた。さらに、インテグリンβ1の発現は、未処置のCAMで認められる発現と変わりなく、β1はまた刺激血管で容易に検出可能であった。
【0086】
続いて、CAMクリオスタット切片のレーザー共焦点画像分析により、βFGF誘発脈管形成中のαvβ3とβ1インテグリンの相対的発現を定量した。続いてこの染色切片をツァイスのレーザー共焦点顕微鏡で調べた。25個のLM609染色血管および15個のCSAT染色血管(350から3500mm2の範囲にあり、平均サイズは〜1200mm2)を任意の視野から選び、レーザー共焦点画像分析による任意単位で単位面積当たりの各血管について平均ローダミン蛍光を測定した。データは血管の任意単位による平均蛍光量±標準誤差として表した。
【0087】
図6に表した結果は、ウィルコクソンの順位合計試験(Wilcoxon Rank Sum Test)(P<0.0001)で決定されたように、βFGF処置CAMではαvβ3染色は顕著に増強(4倍)されることを示した。一方、β1染色はβFGF処置の場合と顕著には異ならなかった。
さらに、このCAMアッセーを用いて、別の有効な脈管形成誘発物質である腫瘍壊死因子−α(TNFα)のβ1およびβ3インテグリンの発現における影響を調べた。βFGFまたはTNFαのいずれかをしみこませ、10日齢胎児のCAM上に静置したフィルターディスクは、72時間後に局所的脈管形成を促進させることが分かった。
これらの結果は、未処置(図7A)、βFGF処置(図7B)またはTNFα処置(図7C)CAMの光学顕微鏡写真で示す。血管は、βFGFおよびTNFα処置調製物では容易に識別できるが、未処置のCAMでは認められない。したがって、増殖因子/サイトカインの局所的適用は、隣接領域の成熟血管から最初は血管が存在しなかった領域へと脈管形成の誘導をもたらす。図5Cで示されたようにβFGFで誘発される血管および同時に発生するαvβ3発現から考えると、TNFαの処置は匹敵する活性をもたらす。
【0088】
これらの発見は、ヒトおよびニワトリの両方で、脈管形成中の血管は強化されたαvβ3発現を示すことを示唆する。これに一致して、培養内皮細胞上のαvβ3発現は、以下の論文に記載されたように(Janatら、J.Cell Physiol.,151:588(1992); Enensteinら、Exp.Cell Res.,203:499(1992);およびSwerlickら、J.Invest.Der.,99:715(1993))、種々のサイトカインによってインビトロで誘発することができる。
増殖因子誘発脈管形成の抗体およびペプチド抑制物質による影響は、実施例7Aおよび7Bに示される。
【0089】
B.胎児の脈管形成
胎児の新規血管系の自然状態での形成に対する脈管形成抑制物質の効果を調べるCAM調製物は、先に述べたように6日齢のニワトリ胎児であった。この発生段階では、血管は新規の増殖を行っており、したがってαvβ3が胎児の脈管形成に参画するか否かを決定するための有用な系を提供する。このCAM系は、アッセーを胎生10日目ではなく6日目に実施した点を除き上記のように調製された。本発明の抗体およびペプチドによる処置の胎児脈管形成に対する影響は実施例7Cに示す。
【0090】
C.腫瘍誘発脈管形成
腫瘍誘発脈管形成におけるαvβ3の役割を調べるために、先に増殖させ17日齢のニワトリ胎児のCAMから分離した種々のαvβ3陰性ヒトメラノーマおよび癌フラグメントを、ブルックスらの論文(Brooksら、J.Cell Biol.,122:1351(1993))および本明細書で述べるようにCAMアッセーに用いた。このフラグメントは、緩衝液の存在だけで強度の新規血管新生を誘発した。
脈管形成は、腫瘍フラグメントをCAM上に直接添加することによってCAM系で誘発された。ニワトリ胎児CAMの調製は、上記で述べた方法と同一であった。ディスクの代わりに、重量で50ミリグラム(mg)から55mgのヒトメラノーマ腫瘍M21L、ヒト肺癌腫瘍UCLAP−3、ヒト膵臓癌細胞株FG(Chereshら、cell,58:945-953(1989))、またはヒト喉頭癌細胞株HEp3(これら全てはαvβ3陰性腫瘍である)を、当初は血管が存在しない領域のCAM上に静置した。
【0091】
M21Lヒトメラノーマ細胞株、UCLAP−3ヒト肺癌細胞株、FG膵臓癌細胞株またはHEp3ヒト喉頭癌細胞株(全てαvβ3陰性)を用いて、ニワトリ胎児のCAM上に固形ヒト腫瘍を増殖させた。8×106のM21L、UCLAP−3およびFBまたは5×105のHEp3細胞の単一細胞懸濁を、全容積30マイクロリットル(ul)のHBSSとしてCAMに用いた。窓をテープで封じ、胎児を7日間保温して、ヒト腫瘍病巣を増殖させた。7日の終わりに(17日齢胎児)、腫瘍をCAMから切除し、周辺のCAM組織を切り取った。腫瘍を50mgから55mgの腫瘍フラグメントにスライスし、脈管形成アッセーまたは腫瘍増殖アッセーのいずれかに使用した。実施例6Aで述べたように、新たな10日齢のニワトリ胎児CAMの血管を欠いた領域にこの腫瘍フラグメントを静置した。
【0092】
実施例3Aで述べたように、αvβ3発現についてニワトリ胎児CAM上のインビボ増殖腫瘍をmAbLM609で染色した。腫瘍細胞の特異的染色は認められず、αvβ3発現の欠如を示唆した。
その後続いて、腫瘍誘発脈管形成に対する抗体およびペプチドの影響を測定するために、実施例7Dおよび7Eで述べるように処置した。腫瘍の退縮並びに脈管形成性血管および血管細胞のアポプトーシスに対する抗体およびペプチドの影響を測定するために、これらCAM腫瘍調製物をまた実施例8、9および12で述べるように処置した。
【0093】
7.CAMアッセーで測定された脈管形成の抑制
A.抑制物質の局所適用による増殖因子誘発脈管形成の抑制
1)単クローン性抗体による処置
αvβ3が脈管形成で活発な役割を果たすか否かを決定するために、βFGFまたはTNFαで飽和させたディスクをCAM上に静置し、続いて単クローン性抗体(mAbとも称する)、LM609(αvβ3に特異的)、CSAT(β1に特異的)またはP3G2(αvβ5に特異的)をこの調製物に添加した。
脈管形成は、βFGFで飽和させたフィルターディスクで10日齢胎児のCAMで誘発された。続いて、全容積25ulの滅菌HSBB中に25mgのmAbを含むHBSS50mlでこの円盤フィルターを0、24および48時間処理した。72時間で、CAMを採取し35mmのペトリ皿に入れ、1mlの燐酸緩衝食塩水で1度洗浄した。続いて、2人の観察者により二重盲検様式で、濾紙の裏側およびCAM組織をオリンパス立体顕微鏡で調べた。ディスクの直下のCAMの血管浸潤でCAMが>50%減少を示したとき、脈管形成抑制は顕著であると考えた。実験は、1条件につき6から7個の胎児を用いて、1抗体につき4回繰り返された。
【0094】
βFGF誘発脈管形成に対するmAb処置の影響は図8Aに示す。血管が存在しない未処置CAM調製物は、βFGF血管誘発(図8Bに示す)およびmAbによるCAMに対する影響(図8C−8E)との比較を提供するために図8Aに示した。mAbLM609で処理したCAMの約75%が、図8Eに示すように脈管形成の>50%抑制を示し、これらの多くは血管の浸潤を欠いているようであった。対照的に、緩衝コントロール(図8A)並びにmAbCSAT(図8C)およびP3G2(図8D)で処理したフィルターは、常に強度の血管新生を示した。
【0095】
同じ結果が脈管形成がTNFαで誘発されたとき得られた。血管を欠く領域近くの正常な血管の発達から出現する、以前から存在していた成熟血管に対するこれら同じ抗体の影響を調べるために、mAbで飽和させたフィルターディスクを、サイトカインを局所添加していない10日齢胎児のCAMの血管形成領域に静置した。立体顕微鏡下で可視化させて調べたとき、3種のmAbのいずれも以前から存在していた血管に影響を与えなかった。したがって、mAbLM609は新しい血管増殖のみを選択的に抑制し、隣接領域に存在する成熟血管には影響を与えなかった。同様な効果は、それぞれ実施例7A2)および7E2)で述べるように局所的にまたは静脈内に用いたとき、合成ペプチドでも認められた。
【0096】
2)合成ペプチドによる処置
増殖因子誘発脈管形成に対する環状および直線状ペプチドの影響を決定するために、また本発明の合成ペプチドを用いてCAMアッセーを実施した。実施例1のようにペプチドを調製し、80ugのペプチドが全容積25ulの滅菌HBSS中で提供された。このペプチド溶液を直ちにCAM調製物に適用し、続いて24時間、および48時間に再度適用した。72時間でこの濾紙および周辺CAM組織を切り出し、上記のように観察した。
【0097】
このアッセーの結果は、合成ペプチドが腫瘍誘発血管に静注された実施例7E2)で詳述する通り、図9A−9Cに示したものと同様であった。ここで、コントロールペプチド62186については、βFGF誘発血管は図9Aに示すように変化しないままであった。対照的に、環状RGDペプチド62814がフィルターに適用されたとき、血管の形成は抑制され、新しい血管構造を欠く領域が残された。この効果は、下記の実施例7E2)で述べるように図9Bで示される効果と外観上同様であった。さらに、静注されたペプチドについて図9Cでも示されるように、成熟血管が、増殖因子飽和フィルターが静置された場所から離れて存在する領域では、これら範囲外の血管に対して合成ペプチドの局所処理では効果は認められなかった。したがって、脈管形成に対するペプチドの抑制活性は、増殖因子によって誘発される脈管形成領域に限定され、以前から存在する近くの成熟血管には影響を与えず、周辺組織にはいかなる有害な細胞毒性ももたらさない。
実施例1で調製し、表1に示した他の合成ペプチドを用いて、同様なアッセーが実施される。
【0098】
B.抑制物質の静脈内適用による増殖因子誘発脈管形成の抑制
1)単クローン性抗体を用いた処置
CAM調製物に静脈内注射された単クローン性抗体の増殖因子誘発脈管形成に対する影響もまた、本発明で使用するために調べた。
静脈内注射のためのニワトリ胎児CAMの調製は、本質的には実施例7Aで述べた通りであるがいくつかの修飾を伴う。光で検卵する工程で際立つ血管を選び、それらの位置を示すために卵殻に印を付けた。卵殻にドリルで孔を開け、さらにCAMを落下させ、βFGF飽和濾紙を上記のようにCAMに静置した。窓を滅菌テープで封じ、胎児を孵卵器に戻した。24時間後、卵殻側面の先に選んだ際立つ血管上に直接注意深く第二の小孔を開けた。外側の卵殻を注意深く除去し、胎児膜は傷をつけずにおく。鉱物油(Perkin-Elmer Corp、ノーウォーク、コネチカット)の小滴で卵殻膜を透明にし、これによって血管を容易に見ることができる。精製滅菌mAbまたは合成ペプチド(後者は下記で詳述される)が、1胎児当たり全容積100ulの滅菌PBS中の200ugの投与量でただ1回30ゲージの注射針で血管に直接接種された。窓をテープで封じ、胎児を72時間まで保温した。フィルターディスクおよび周辺CAM組織を上記に述べたように分析した。
先にLM609が静脈内に接種されたCAM組織または腫瘍組織におけるLM609mAbの分布を知るために、本明細書および以下の実施例で示すように、HBSS中の2.5%BSAを用いて1時間室温で固定切片を遮断し、続いてヤギ抗マウスローダミン標識二次血清(Tago)の1:250希釈で染色した。その後、この切片をツァイスの免疫蛍光化合物顕微鏡で調べた。
【0099】
CAM調製物のβFGF誘発血管に対する静脈内抗体処理の結果は図10A−10Cに示す。図10Aに、βFGF処理の結果として誘発された脈管形成を示す。静脈内でmAbP3G2(抗αvβ5抗体)に曝されたとき、図10Bに示すようにβFGF誘発血管構造の状態に対して変化は認められなかった。対照的に、βFGF誘発脈管形成CAM調製物のLM609(抗αvβ3抗体)による処理は、図10Cに示すように新しい血管のフィルター領域への増殖を完全に抑制した。脈管形成に対するこの抑制効果は、したがってLM609抗αvβ3特異的抗体によるαvβ3レセプター活性の抑制から生じている。αvβ3の遮断はCAMのフィルター部位への新規血管構造形成を抑制しないので、αvβ5は、したがって新規血管の増殖のためのαvβ3と比較して必須ではない。
2)合成ペプチドによる処置
上記のCAM調製物の増殖因子誘発血管に、実施例1で調製した合成ペプチドを別々に静脈内注射する。血管の生存活性に対するこのペプチドの影響を同じように調べる。
【0100】
C.局所適用による胎児脈管形成の抑制
1)単クローン性抗体による処置
αvβ3が胎児の脈管形成に参画しているか否かを決定するために、CAMの血管の新規な成長に対するLM609の影響を6日齢の胎児(実施例5Aで述べるように活発な新規血管新生を特徴とするステージ)で調べた。CAMアッセーは、実施例6Cで述べたように調製し、続いてmAbで飽和させたディスクを6日齢の胎児のCAMにサイトカインの非存在下で局所的に適用した。3日後、CAMを切り出し写真を撮った。各実験は1群につき6個の胎児を含み、2回繰り返された。
抗体LM609(図11C)はこのような条件下で血管の増殖を防止したが、CSAT(図11A)またはP3G2(図11B)は防止しなかった。このことは、αvβ3は、脈管形成誘発のための添加増殖因子に依存しなかった胎児の新規血管新生において基本的な役割を果たすことを示唆している。
2)合成ペプチドによる処置
実施例1で調製した合成ペプチドを上記のように調製したCAM調製物に、実施例5A2)で述べたようにCAMへの局所的適用または血管への静脈内適用のいずれかで別々に加える。血管の生存活性に対するペプチドの影響を同じように調べる。
【0101】
D.局所適用による腫瘍誘発脈管形成の抑制
1)単クローン性抗体による処置
抗αvβ3拮抗物質、LM609並びにペプチド62181、62184、62185、62187および62880の胎児脈管形成への影響を調べた上記の脈管形成アッセーに加え、腫瘍誘発脈管形成におけるαvβ3の役割もまた調べられた。誘発物質として、αvβ3陰性ヒトM21−Lメラノーマフラグメント(それより前に増殖させ、17日齢のニワトリ胎児のCAMから分離されたもの)を用いた。このフラグメントは実施例6Cに述べたように調製した。
実施例7A1)で述べたように、25ulのHBSS中に25ugの濃度でmAbを腫瘍フラグメントに別々に局所適用し、続いて窓をテープで封じた。このmAbを同じ態様で24時間および48時間に再度加えた。72時間で、腫瘍および周辺CAM組織を実施例7A1)で述べたように調べた。
【0102】
実施例6Cで述べたように、腫瘍は、培養M21−L細胞(ヘルディング−ハーベルマンらの報告のようにインテグリンαvβ3を発現しない(Felding-Habermannら、J.Clin.Invest.,89:2018(1992))を10日齢のニワトリ胎児のCAMに移植することによって先ず誘導された。これらのαvβ3陰性フラグメントは、緩衝液単独で、またはmAbCSAT(抗β1)もしくはP3G2(抗αvβ5)の存在下で強度に新規血管新生を誘発した。対照的に、mAbLM609(抗αvβ3)は、殆どの血管が腫瘍塊および周辺CAMに浸潤するのを停止させた。
腫瘍誘発脈管形成に対するmAbの影響を定量するために、2人の観察者が、CAMの病巣面内の腫瘍に入り込む血管を立体顕微鏡下で二重盲検形式によって数えた。図12に示した各棒線は、2回の実験で各群12個のCAMから得た血管の平均数±標準誤差を示している。
【0103】
この定量分析は、ウィルコクソンの順位合計試験で求めたとき、緩衝液または他のmAb(P3G2もしくはCSAT)で処置した腫瘍と比較して、mAbLM609で処置した腫瘍に入る血管数は3倍減少することを明らかにした(P<0.0001)。M21−L腫瘍はαvβ3を発現しないという事実は、mAbLM609は、腫瘍細胞にではなく血管に直接影響を与えることによって脈管形成を抑制することを示唆している。これらの結果は、図3A−3Dに示した癌組織の生検材料におけるαvβ3の組織学的分布と一致するが、これらの図では、αvβ3の分布は腫瘍内の血管に限定され、腫瘍細胞自体には存在しなかつた。
【0104】
2)合成ペプチドによる処置
実施例1で調製した合成ペプチドが、上記のように腫瘍誘発脈管形成CAMアッセー系に局所的に適用される。血管の生存活性に対するこれらペプチドの影響が同じように調べられる。
【0105】
E.静脈内適用による腫瘍誘発脈管形成の抑制
1)単クローン性抗体による処置
実施例7D1)で述べるように調製した腫瘍誘発血管をまた、静脈内注射によりmAbで処置した。実施例7D1)で述べるように腫瘍をCAMに静置し、テープで窓を封じ、24時間後に200ugの精製mAbを先に述べたようにニワトリ胎児の血管に1回静脈内接種した。ニワトリ胎児をその後7日間保温した。
脈管形成の程度を続いて上記のように観察した。下記実施例8に述べるように、この期間経過後に腫瘍を切除してそれらの重量を調べ、腫瘍の増殖または退縮に対する抗体曝露の影響を決定した。
【0106】
2)合成ペプチドによる処置
CAMアッセー系の腫瘍誘発血管構造をペプチドに曝したときの影響もまた調べた。腫瘍−CAM調製物を上記のように用いたが、ただしこの場合、mAbの静脈内注射の代わりに、実施例1および実施例7A2)で述べたように調製した合成ペプチドが可視化された血管に別々に静脈内注射された。
HCl塩を含む環状ペプチド、66203およびコントロールペプチド62186を用いたCAMアッセーの結果は図9A−9Cに示す。図9Aで、コントロールペプチドによる処置は、腫瘍処理によって誘発され、CAMの本来は血管の存在しない領域に増殖した豊富で太い血管に影響を与えなかった。対照的に、環状RGDペプチド、66203(αvβ3に対する拮抗物質)がフィルターに適用されたとき、図9Bに示すように血管形成は抑制され、新しい血管構造の無い領域が残った。腫瘍を配置した場所の近くに位置する血管に対して有害な影響は全く存在しなかったことによって明らかなように、RGD含有ペプチドの抑制効果は特異的で局在的であった。
【0107】
したがって、図9Cでは、抑制ペプチドがCAMアッセー系に静脈内注射されたとき、腫瘍の配置場所の周辺ではあるが離れた領域のCAMに以前から存在する成熟血管に対しては影響は認められなかった。このような場所に以前から存在する血管は、これら血管内に流れている抑制物質によって影響を受けなかった。ただし、これら以前から存在する血管から腫瘍塊への新規な血管の形成は抑制された。したがって、実施例4のリガンド−レセプターアッセーでαvβ3の拮抗物質であることが以前に示された、66203および62184を含む合成ペプチドは、発達しつつある血管に限定される脈管形成を抑制するが、以前から存在する成熟した血管に対しては脈管形成を抑制しないことが明らかにされた。さらに、ペプチドの静脈内輸液は、図9Cの無傷の血管構造によって明らかなように、周辺領域に対して有害な細胞毒性を全くもたらさない。
同じようなアッセーが実施例1で調製し、表1に示す他の合成ペプチドを用いて実施される。
【0108】
8.CAMアッセーで測定したαvβ3拮抗物質による腫瘍組織の増殖抑制
実施例7D1)で述べたように増殖因子や腫瘍によって誘発される脈管形成に対する抗αvβ3拮抗物質の影響を視覚で評価することに加え、曝露した後の腫瘍塊への何らかの影響を測定することによってこの脈管形成への影響をまた測定した。この分析のために、腫瘍誘発脈管形成CAMアッセー系を実施例6Cおよび7Dで述べたように調製した。7日間の保温期間終了時に生じた腫瘍をCAMから切除し、一切の残余のCAM組織を除去して1mlの燐酸緩衝食塩水で洗浄し、各腫瘍の湿潤重量を求めた。
さらに、顕微鏡による組織分析用腫瘍の調製では、代表的な腫瘍例をブリン固定液で8時間固定し、パラフィンに包埋した。連続切片を切り出し、ヘマトキシリンおよびエオシン(H&E)で顕微鏡分析のために染色した(Gladsonら、J.Clin.Invest.,88:1924(1991))。切片の写真はオリンパス複合顕微鏡で250×で撮影した。
【0109】
A.局所適用
コントロール緩衝液(HBSS)、P3G2(抗αvβ5)またはLM609(抗αvβ3)の局所適用で得られた典型的なヒトメラノーマ腫瘍(M21L)の重量の結果は表4に示す。各処置につき多数の胎児の平均腫瘍重量(ミリグラム、mg)を調べ、表の下に示すように平均値の標準誤差と合わせて算出した。
【0110】
CAMアッセー系でαvβ3陰性ヒトメラノーマ腫瘍塊をLM609に曝露すると、未処置の平均腫瘍重量172mg±26は52mg±13に減少した。P3G2抗体はこの腫瘍塊に影響を与えなかった。したがって、αvβ3特異的LM609抗体の局所適用によるαvβ3レセプターの遮断は、前述の実施例に示したように脈管形成の抑制とともに腫瘍の退縮をもたらした。P3G2に曝露することによって生じた腫瘍塊の測定直径は、平均して約8mmから1cmであった。対照的に、LM609処置腫瘍は直径が平均して2から3mmであった。
これらの腫瘍の冷凍切片は、P3G2に曝された腫瘍については腫瘍細胞構造は無傷で、対照的にLM609に曝された腫瘍では組織化された細胞構造を欠いていることが明らかとなった。αvβ3レセプター活性は、したがってαvβ3発現新規血管構造の発達によって腫瘍塊に栄養を補給し続けるためにαvβ3陰性腫瘍にとって必須である。本発明のαvβ3拮抗物質によるαvβ3の遮断は、腫瘍への脈管形成の抑制をもたらし、最終的には腫瘍塊の減少をもたらす。
【0111】
B.静脈内適用
コントロール緩衝液(PBS、燐酸緩衝食塩水)、CSAT(抗β1)またはLM609(抗αvβ3)の静脈内適用によってもたらされた代表的な癌腫(UCLAP−3)の重量の結果は表5に示す。各処置につき多数の胎児の平均腫瘍重量を調べ、表の下に示すように平均値の標準誤差を合わせて算出した。
【0112】
CAMアッセー系におけるαvβ3陰性ヒト癌腫瘍塊のLM609への曝露は、未処置腫瘍の平均重量85mg±7を30mg±6に減少させた。CSAT抗体は腫瘍塊重量に顕著な影響を与えなかった。したがって、αvβ3特異的LM609抗体の静脈内適用によるαvβ3の遮断は、そのような処置が前述の実施例で示したように脈管形成抑制に加えて、上記でメラノーマ腫瘍塊の退縮をもたらしたように癌の退縮をもたらした。さらに、ヒトメラノーマ腫瘍の増殖は、LM609の静脈内注射によって同じように抑制された。
9.CAMアッセーで測定したαvβ3拮抗物質による腫瘍組織増殖の退縮
αvβ3拮抗物質の腫瘍増殖および生存に対する影響を調べるために、実施例5Aで述べたように、10日齢胎児のCAMにヒトメラノーマフラグメント並びに肺、膵および喉頭癌フラグメントを静置した。
【0113】
A.静脈内適用
1)単クローン性抗体による処置
a.LM609(抗αvβ3)およびCSAT(抗β1)による処置
αvβ3陰性ヒトメラノーマM21−L、膵癌FG、ヒト肺癌UCLAP−3またはヒト喉頭癌HEp3の癌フラグメントをCAMに移植後24時間して、胎児の静脈内にHBSのみ、またはmAbLM609(抗αvβ3)もしくはCSAT(抗β1)のいずれか(300ug/100ul)を一回注射した。腫瘍をさらに6日間増殖させた。保温期間の終了時に、腫瘍を注意深く切除し、周辺CAM組織を除去した。腫瘍切除は、別々の2人の研究者が容易に区別できる固形腫瘍塊を切除することによって実施した。これら腫瘍は識別が容易な辺縁を有し、したがって固形腫瘍塊とは容易に区別できる薄い半透明膜(CAM)は、腫瘍塊自体を傷つけることなく切除された。切除腫瘍の重量を測定し、形態学的および組織学的に調べた。
【0114】
図13に示すように、7日終了時の湿潤腫瘍重量を求め、処置前の最初の腫瘍重量と比較した。各棒線は1群につき5−10個の腫瘍の平均±SEを示す。調べた全ての腫瘍でコントロールと比較して、mAbLM609は顕著に(P<0.001)腫瘍増殖を抑制した。PBSまたはCSATで処置した腫瘍は全例で増殖した。対照的に、mAbLM609はこれら腫瘍の増殖を防止しただけでなく、殆どの事例で強度の退縮を誘発した。重要なことには、これらの腫瘍細胞はインテグリンαvβ3を発現せず、増殖抑制はこの抗体の新規血管構造に対する抗脈管形成作用によるものであり、腫瘍細胞に対する直接の作用ではないことを明示している。
b.LM609(抗αvβ3)およびP3G2(抗αvβ5)による処置
実施例5Aで述べたように、ヒトM21−Lメラノーマ腫瘍フラグメント(50mg)を10日齢胎児のCAMに移植した。24時間後、胎児の静脈内にPBSのみ、またはmAbLM609(抗αvβ3)もしくはP3G2(抗αvβ3)のいずれかを一回(300ug/100ul)注射した。実施例9A1)aで述べたように、腫瘍を増殖させ、ここで述べるように形態学的および組織学的に調べた。
mAbP3G2(抗αvβ5)またはLM609(抗αvβ3)で処置したM21−L腫瘍の代表例を形態学的に調べた。P3G2処置腫瘍は大きく(直径8mm)て良好な血管新生が認められたが、一方、mAbLM609で処置された抗体は、はるかに小さく(3mm)、検出可能な血管を欠いていた。
【0115】
組織学的切片を実施例9A1)aで述べたように、ヘマトキシリンおよびエオシンで染色した調製物でこれら腫瘍をさらに調べた。図14(上)に示すように、mAbP3G2(抗αvβ5)で処置した腫瘍は、腫瘍基質の多くの血管(矢印)の他に分裂像(楔形)でも示唆されるように活発に分裂する多数の生きた腫瘍細胞を示した。対照的に、mAbLM609(抗αvβ3)で処置した腫瘍では、元気な腫瘍細胞も血管もほとんど検出されなかった(図14下)。これらの結果は、インテグリンαvβ3の拮抗物質は腫瘍誘発脈管形成を抑制し、インビボでヒトの種々の腫瘍の増殖停止および退縮に導くことを示している。7日間の腫瘍増殖の後(17日齢胎児)で調べた胎児は、αvβ3拮抗物質で処置されたか否かにかかわらず、全体的な検査では正常のようであったということを指摘することは重要である。これらの発見は、このインテグリンの拮抗物質は発育胎児に毒性を持たないらしいということを示す。
【0116】
2)合成ペプチドによる処置
実施例5Aで述べたように、ヒトM21−Lメラノーマ腫瘍フラグメント(50mg)を10日齢胎児のCAMに移植した。24時間後に、胎児は、300ug/100ulのシクロ−RADfV(69601)またはシクロ−RGDfV(66203)のいずれかの静脈内注射を1回与えられた。合わせて72時間してから腫瘍を切除し、実施例9A1)で述べたように立体顕微鏡で形態学的および組織学的に調べた。
図15Aから15Eに示した写真は以下のように対応する:図15A、シクロRADfVペプチド(69601)で処置した2組のサンプル;図15B、シクロRGDfVペプチド(66203)で処置した2組のサンプル;図15C、シクロRGDfVペプチド(66203)で処置した同じ胎児から採取した隣接CAM組織;図15Dおよび15E、高倍率(13×)のペプチド処置腫瘍。図15Dは、コントロールペプチド(69601)で処置した腫瘍の正常な血管を示す。図15Eは、シクロ−RGDfVペプチド(66203)で処置した腫瘍の破壊された血管(矢印)の例を示す。
これらの結果は、ペプチド66203のみが血管形成を抑制し、さらに腫瘍に隣接するCAM組織の血管は影響されなかったことを示す。
【0117】
10.ウサギ眼球モデルインビボアッセーで測定したαvβ3拮抗物質による腫瘍組織の増殖退行
増殖因子誘発脈管形成に対する抗αvβ3拮抗物質の影響は、眼の角膜によって実証されるように本来透明な構造で観察することができる。新しい血管は角膜の辺縁(豊富な血液供給を有する)から角膜の中心部(本来血液供給をもたない)に向かって成長する。脈管形成刺激物質(例えばβFGF)は、角膜に適用されると角膜辺縁から新しい血管の成長を誘発する。脈管形成拮抗物質は、角膜に適用されると角膜の辺縁からの新しい血管の成長を抑制する。したがって、角膜は、角膜辺縁から丈夫なコラーゲンの詰まった角膜組織(これは容易に見ることができる)へと侵入する内皮細胞を介して脈管形成が進行する。ウサギ眼球モデルアッセーは、したがって化合物を直接眼の角膜に移植した後に続く脈管形成の刺激および抑制を直接観察するためのインビボモデルを提供する。
【0118】
A.ウサギ眼球インビボモデルアッセー
1)増殖因子によって誘発される脈管形成
脈管形成は、増殖因子βFGFを用いてウサギ眼球インビボモデルアッセーで誘発され、以下で詳述する。
a.増殖因子および単クローン性抗体を含むヒドロンペレットの調製
増殖因子およびmAbを含むヒドロンポリマーペレットを文献の記載にしたがって調製した(D'Amatoら、Proc.Natl.Acad.Sci.,91:4082-4085(1994))。個々のペレットは、βFGFを安定化させ、さらに周辺組織への緩徐な遊離を確保するためにスクラルファート(Carafet,Marion Merrell Dow Corporation)に結合させた増殖因子βFGF650ngを含んでいた。さらに、mAbLM609(抗αvβ3)またはmAbP1F6(抗αvβ5)のいずれか40μgをPBS中に含むヒドロンペレットを調製した。これらのペレットを、その表面に開けた2.5mmの中子を有する特別に調製したテフロンペッグの型に流し込んだ。約12μlの鋳造材料を各ペッグに入れ、滅菌フードで一晩重合させた。続いてペレットを紫外線照射で滅菌した。
【0119】
b.単クローン性抗体による処置
各実験は、一方の眼球にβFGFおよびLM609を含むペレットを、さらに他方の眼球にβFGFおよびマウスmAbP1F6(抗αvβ5)を含むペレットを与えた3匹のウサギから構成された。LM609(抗αvβ3)と他のmAbおよびPBSコントロールを比較するために対を形成する眼球試験を用いることは、被検mAb間での顕著な相違を明らかにすることができる厳密な試験のための手段を提供する。
P1F6mAbは、血管の内皮細胞の表面に見出されるが、おそらく脈管形成には必要ではないインテグリンαvβ5と免疫的に反応する。このmAbP1F6が脈管形成に必要か否かを決定するために、このmAbのみを含むペレットを調製し、下記に述べるよう調べて、このmAbは脈管形成を誘発しないことを確認した。
検査したmAbの全ては、周知の方法にしたがって蛋白AセファロースCL−4Bアフィニティーカラムクロマトグラフィーを用いて腹水から精製した。続いて溶出させた免疫グロブリンをPBSに対して透析し、デトキシ−ゲル(Detoxi-gel、Pierce Chemicals)で処理し内毒素を除去した。内毒素は強力な脈管形成刺激および炎症刺激物質であることが示された。mAbは、したがって、色素形成性カブトガニアメーバ様細胞溶解産物アッセー(Chromogenic Limulus Amebocyte Lysate Assay、Bio-Whittaker)を用いて内毒素の存在について検査し、検出可能な内毒素を含まないmAbのみをウサギ眼球モデルアッセーに用いた。
【0120】
βFGFおよびmAbLM609(抗αvβ3)またはP1F6(抗αvβ5)を含むヒドロンペレットをウサギの眼球に作製した角膜ポケットに挿入した。このヒドロンペレットはまたスクラルファートを含み、アッセー中にβFGFを安定化させる。個々のペレットは、ウサギの角膜の中央間質に外科的に作製した“ポケット”に移植された。この外科操作は、個々の角膜を写真により記録するためにカメラを搭載したビームスプリッターを備えたワイルドモデルM691手術用顕微鏡を用いて滅菌的術式で実施した。3mm×5mmの“ポケット”は、69ビーバーブレードを用いて角膜の厚さの半分まで3mmの切り目を入れて角膜間質に作製した。虹彩用へらを用いて間質は周辺で細かく切断され、辺縁から2mmの周辺端を用いてペレットを移植した。その後の14日間にβFGFおよびmAbは移植ペレットから周辺組織に拡散し、したがって角膜の辺縁から脈管形成が生じた。
【0121】
各処置の代表的な結果は図16Aから16Eに示す。出現する血管量を定量し、以下のように規定される時計時間の様式で記載する。時計が時間に分割されるのと同じ様式で、眼を12等分する。“血管の1時計時間”は、時計の1時間と同等な眼の領域を満たす血管の量を指す。βFGFのみを与えられた5匹のウサギは、新しい血管が角膜の辺縁から角膜の中央に向かって増殖している赤みがかった脈管形成を示したが、これは通常は血管をもたない。これらのウサギの1匹は、ペレットに対して1時計時間の血管だけしかもたなかった。βFGFおよびmAbLM609の両方を与えられた2匹のウサギには、検出可能な脈管形成が手術後14日まで全く存在しなかった。これらのウサギのうちの1匹には、14日までに3つの出血性出芽血管病巣があった。βFGFおよびmAbP3G2(抗αvβ5)を与えられたウサギのうちの2匹は強度の血管新生を示し、ここでは新しい血管は角膜の縁から角膜内に増殖した。これらのウサギのうち1匹には、ペレットに対して1から2時間の血管が存在しただけであった。ウサギ眼球モデルアッセーで実証されたように、mAbLM609(抗αvβ3)を与えられたウサギでは、増殖因子βFGFの存在下で正常な周辺血管に対しては脈管形成効果は認められなかった。対照的に、mAbP3G2(抗αvβ5)を与えられたウサギでは、増殖因子βFGFの存在下で正常な周辺血管に対して脈管形成が認められた。mAbLM609による角膜脈管形成の完全な抑制は、以前に報告されたいずれの抗脈管形成試薬よりも実質的に大きい。
【0122】
11.マウス:ヒトキメラアッセーで測定したαvβ3拮抗物質による腫瘍組織増殖のインビボ退行
マウス:ヒトキメラインビボモデルをSCIDマウスの皮膚の一部を新生児の包皮で置き換えることによって作製した(図17)。皮膚移植が定着した後、ヒトの包皮に癌細胞を接種した。測定可能な腫瘍が確立した後、mAbLM609(抗αvβ3)またはPBSのいずれかをマウス尾静脈に注射した。2−3週間してから、腫瘍を切り出し重量および組織を調べた。
A.マウス:ヒトキメラインビボアッセー
マウス:ヒトキメラインビボモデルは、実質的には文献にしたがって調製した(Yanら、J.Clin.Invest.,91:986-996(1993))。簡単に記せば、SCIDマウス(6−8週齢)から外科的に2cm2の領域の皮膚を切除し、ヒトの包皮で置き換えた。マウスを麻酔し、側面腹部領域の各々の側5cm2の毛を剃った。2か所の皮膚を筋膜まで完全に除去して2cm2の円形の移植床を調製した。ヒト新生児包皮由来の同じ大きさの完全な厚さのヒトの皮膚移植片をこの創傷床に静置しその場所に縫合した。移植片は皮膚に縫い付けたバンドエイドで被覆した。
【0123】
微細孔布テープも創傷を被覆するために用いた。
M21Lヒトメラノーマ細胞株またはMDA23.1乳癌細胞株(ATCCHTB26;mAbLM609を用いた組織切片の免疫反応性ではαvβ3陰性)を用いて、SCIDマウスのヒト皮膚移植片に固形ヒト腫瘍を形成させた。5×106M21−LまたはMDA23.1細胞の単一細胞浮遊液を経皮的にヒト皮膚移植片に注射した。続いてマウスを2から4週間観察して、測定可能なヒト腫瘍を増殖させた。
【0124】
B.静脈内適用
1)単クローン性抗体による処置
測定可能な腫瘍が増殖した後、M21L腫瘍細胞を注射したマウスに250μgのmAbLM609(抗αvβ3)またはPBSを、週に2度2から3週間尾静脈から静注した。この後、腫瘍を皮膚から切除し、周辺組織を除去した。表6の下に示したように各処置について数匹のマウスの平均腫瘍重量を調べた。
【0125】
【0126】
M21Lαvβ3陰性ヒト癌腫瘍塊をマウス:ヒトキメラアッセー系でLM609(抗αvβ3)に曝すことによって、PBS処置腫瘍の平均重量が198mgから113mgへと減少した。
mAbLM609(抗αvβ3)およびPBSで処置されたM21L腫瘍の代表例を形態学的に調べた。PBS処置腫瘍は大きく(直径8から10mm)血管新生は良好で、一方、mAbLM609(抗αvβ3)で処置した腫瘍ははるかに小さく(直径3から4mm)、検出可能な血管を欠いていた。
MDA23.1を注射した皮膚移植片に形成された腫瘍は検出および測定可能であった。成長腫瘍の形態学的検査によって、移植ヒト組織からMDA23.1腫瘍細胞へと新規血管新生が生じたことが明らかにされた。
【0127】
したがって、αvβ3特異的LM609抗体の静脈内適用によるαvβ3の遮断は、実施例9および10でそれぞれ述べたCAM系およびウサギ眼球モデル系と同じ態様でこのモデル系においても癌の退縮をもたらした。
12.CAMアッセーで測定される、インテグリンαvβ3の拮抗物質の存在下で細胞サイクルに入り、アポプトーシスが進行する血管細胞の刺激
脈管形成過程は、例えばβFGFおよびVEGFのようなサイトカインの血管細胞増殖を刺激する能力に依存する(Mignattiら、J.Cell.Biochem.,471:201(1991);Takeshitaら、J.Clin.Invest.,93:662(1994); およびKoyamaら、J.Cell.Physiol.,158:1(1994))。しかしながら、シグナリング現象は、これら血管細胞の成熟血管への分化を調節している可能性もあるようである。したがって、新規増殖または脈管形成が進行している血管細胞の増殖または分化と関係を有するシグナルとの干渉は、脈管形成の混乱をもたらすかもしれない。
【0128】
インテグリン連結(integrin ligation)現象は、アポプトーシスまたはインビトロのプログラムされた細胞死と同様に細胞増殖にも関与することが示された(Schwartz、Cancer Res.,51:1503(1993); Meredithら、Mol.Biol.Cell.,4:953(1993);Frischら、J.Cell Biol.,124:619(1994); およびRuoslahtiら、Cell,77:477(1994))。αvβ3拮抗物質の脈管形成に対する影響の綿密な調査によって、不連続で破壊された腫瘍付随血管の存在が明らかになった。したがって、血管連続性の欠如は血管細胞の選択的壊死またはアポプトーシスのためかもしれない。
この可能性を調べるために、増殖因子βFGFで脈管形成を誘発した後、本発明のmAbおよび環状ペプチドによる処置でCAMを調べた。
A.単クローン性抗体による処置
アポプトーシスは種々の方法で検出できるが、これらの方法は、DNAのフラグメント化を検出するために組織から分離したDNAの直接検査、さらに、フラグメント化DNAの遊離3’OH基を特異的に検出する抗体を用いた無傷の組織の3’OH基の検出を含む。
【0129】
1)DNAフラグメント化の分析
脈管形成は、実施例6Aで述べたように、10日齢胎児のCAMにβFGFで飽和させたフィルターディスクを静置することによって誘発された。LM609(抗αvβ3)によるCAMの免疫組織学的分析によって、βFGFによる脈管形成の開始後12から24時間で血管のαvβ3の発現は最高になることが明らかになった。したがって、βFGFによる刺激後24時間で、胎児の静脈内に100μlのPBSのみ、または300μgのmAbCSAT(抗β1)もしくはLM609(抗αvβ3)のいずれかを含むPBSを接種した。
DNAフラグメント化は、mAbLM609(抗αvβ3)、CSAT(抗β1)またはPBSの静脈内接種後24時間または48時間に、βFGF飽和フィルターディスク下のCAM組織を直接切除することによって検出した。切除CAM組織は滅菌PBSで3回洗浄し、細かくきざみ、0.25%の細菌性コラゲナーゼ(Worthington Biochemical; フリーホールド、ニュージャージー)に再浮遊させ、時々ボルテックスミキサーで攪拌しながら37℃で90分保温した。単一細胞浮遊液から等しい数のCAM細胞を用いてDNAを以前に報告されたように抽出した(Bissonetteら、Nature,359:552(1992))。簡単に記せば、等しい数のCAM細胞を10mMトリス−Cl(pH8.0)、0.5%(v/v)のトリトンX−100(シグマ、セントルイス、ミズーリー)中の10mMEDTAで溶解させた。細胞溶解物を4℃で15分16000×gで遠心し、可溶性フラグメント化DNAを完全なクロマチンペレットから分離させた。フラグメント化DNAを洗浄し沈澱させ、1.2%(w/v)アガーロースゲルで分析した。
【0130】
各処置から得た等しい数のCAM細胞から可溶性フラグメント化DNAを分離し、アガロースゲルで電気泳動的に分離し、臭化エチジウムで染色して可視化させた。処置後24時間で3種の異なる処置から得られたDNAフラグメント化の相対量には違いは検出されなかった。しかしながら、mAbLM609(抗αvβ3)による処置後48時間までに、mAbCSAT(抗β1)またはPBS単独で処置した胎児と比較したとき、DNAフラグメント化の顕著な増加が認められた。
2)細胞サイクルに入るための血管細胞の刺激
これらの過程におけるαvβ3の役割を実験的に調べるために、βFGFで処置したCAMまたは未処置のCAMから得た細胞を沃化プロピジウムで染色しさらにmAbLM609(抗αvβ3)で免疫的に反応させた。
【0131】
mAbLM609(抗αvβ3)、CSAT(抗β1)またはPBSで処置後24時間または48時間の胎児から分離したCAMを、上記のように細菌性コラゲナーゼで保温して単一細胞浮遊液にした。続いて、単一細胞を透過性にし、製造元(Oncor,ゲイサースバーグ、メリーランド)の指示にしたがいアポプトーシスタグ・インシトゥ・検出キット(Apop Tag Insitu Detection Kit)で染色した。アポプトーシスタグは、フラグメント化DNAの遊離3’OH基を特異的に検出する抗体である。そのような遊離3’OH基の検出は、アポプトーシス細胞の検出のための確立された方法である(Gavrieliら、L.Cell Biol.,119:493(1992)。
アポプトーシスタグ染色細胞を続いてPBS中の0.1%(v/v)トリトンX−100ですすぎ、0.5%(w/v)BSA、0.02%(w/v)アジ化ナトリウムおよびPBS中の200μg/mlのRNアーゼAを含むFACS緩衝液に再浮遊させた。細胞を1.5時間保温し、洗浄して蛍光活性化細胞選別によって分析した。細胞の蛍光は、ファックスキャンフローサイトメーター(FACScan flow cytometer)を用いて測定し、データは下記のように分析した。
【0132】
細胞の蛍光はファックスキャンフローサイトメーター(Becton Dickison、マウンテンビュー、カリフォルニア)で測定した。側面分散範囲(Side Scatter、SSC)および前方分散範囲(Forward scatter、FSC)は同時に決定し、全てのデータは、ファクスキャン研究用ソフト(Becton Dickison、マウンテンビュー、カリフォルニア)を備えたヒューレットパッカード(HP9000)コンピューターで収集した。データは、P.CリシスヴァージョンIソフト(P.C Lysis version I)(Becton Dickison、マウンテンビュー、カリフォルニア)で分析した。陰性コントロールゲートは、アポプトーシスタグキットの一次抗体を添加していない細胞浮遊液を用いて設定した。同一のゲート設定を両方の細胞集団に用い、異なる細胞処置につき約8000細胞を分析した。
mAb処置CAMから得られ、アポプトーシスタグで染色し、FACS分析で決定した単一細胞の%は図18に示す。斜線入り棒線は、分析の24時間前に処置した胎児の細胞を表す。点刻棒線は48時間前に処置した胎児の細胞を表す。
各棒線は、3つの複製の平均±標準誤差を表す。
図18に示すように、mAbLM609(抗αvβ3)で2日前に処置したCAMは、PBS単独またはCSAT(抗β1)のいずれかで処置したCAMと比較したときアポプトーシスタグ染色は3から4倍増加した。
【0133】
B.合成ペプチドによる処置
アポプトーシスに対する環状ペプチドの影響を決定するために、実施例6Aで述べたように、本発明の合成ペプチドを用いて増殖因子誘発脈管形成に関するCAMアッセーを実施した。ペプチド、シクロ−RGDfV(66203)およびシクロ−RADfV(69601)を実施例1で述べたように調製した。このペプチド溶液またはPBSをCAM調製物に300μg/mlの濃度で注射した。
24および48時間で濾紙および周辺CAM組織を切除してアポプトーシスタグで染色し、実施例12A2)で述べたようにアポプトーシスを検出した。
図18に示したように、ペプチド69203(スクロ−RGDfV)で2日前に処置したCAMは、PBS単独またはコントロールペプチド69601(シクロ−RADfV)のいずれかで処置したCAMと比較したとき、アポプトーシスタグ染色が3から4倍増加した。
C.アポプトーシスおよび細胞サイクルに対する単クローン性抗体による処置の影響
細胞サイクルに対する単クローン性抗体の影響を決定するために、沃化プロピジウムで染色することによって染色体DNAのコピー数について、さらにアポプトーシスタグで染色することによってアポプトーシスについて単一細胞浮遊液を調べた。
【0134】
mAbLM609(抗αvβ3)もしくはCSAT(抗β1)またはPBSで24時間または48時間前に処置したCAMの単一細胞浮遊液を、実施例12A1)で述べたように調製した。
アポプトーシスタグで細胞を染色するために、2.5%(w/v)BSAおよびPBS中の0.25%(w/v)アジ化ナトリウムを含む緩衝液で3回洗浄した。続いて細胞をPBS中の1%(w/v)パラホルムアルデヒドで15分固定し、その後上記のように3回洗浄した。非特異的結合を防止するために、単一細胞浮遊液をPBS中の5%(w/v)BSAで一晩4℃で処理した。続いて細胞を前述のように洗浄し、アポプトーシスタグで染色し、実施例12Aで述べたようにファックスキャンで細胞の蛍光を測定した。
各実験条件から得た細胞をPBS中の10μg/mlの沃化プロピジウム(シグマ、セントルイス、ミズーリー)で1時間染色し、PBSで2回洗浄し、アポプトーシスに典型的な核の特徴(クロマチン凝縮および分断化を含む)について調べた。アポプトーシス細胞の%を、少なくとも10から15個の任意に選んだ顕微鏡の視野から得た細胞の形態学的検査によって算定した。
【0135】
CSAT(抗β1)またはLM609(抗αvβ3)で処理した胎児のCAMの単一細胞浮遊液をアポプトーシスタグおよび沃化プロピジウムで染色し、FACSで分析した結果はまとめて図19に示す。Y軸はアポプトーシスタグ染色(アポプトーシス)を表し、X軸は沃化プロピジウム染色(DNA含有量)を表している。水平線は、アポプトーシスタグ染色の陰性ゲートを表す。左および右の写真はCSATおよびLM609処置胎児のCAM細胞をそれぞれ示す。細胞サイクル分析は、各条件につき約8000例の分析によって実施し、データは等高線図で表した。
DNA染料、沃化プロピジウムで染色した単一細胞サンプルによって、LM609(抗αvβ3)処置後48時間のCAM細胞の25−30%が核凝縮および/または分断化を示した。このような過程はアポプトーシスが進行している細胞の特徴である。これはCSAT(抗β1)処置CAMと対照的で、この場合90−95%の細胞が正常な核染色を示した。
【0136】
図19に示すように、LM609によるアポプトーシスの誘発と一致して、1コピー未満のDNAを含むピークに極めて多数の細胞が認められた(AO)。このピークは、後期のアポプトーシス細胞のフラグメント化DNAを表していることが以前に示された(Telfordら、Cytometry,13:137(1992))。さらに、これらのAO細胞は容易にアポプトーシスタグで染色され、アポプトーシス細胞を検出するこれら試薬の能力が確認された。しかしながら、AOで細胞が染色されることに加え、1コピーより多いDNAを含む極めて多くの細胞がアポプトーシスタグでもまた染色された(図19)。これらの結果は、LM609は、既に細胞サイクルに入った血管細胞でアポプトーシスを促進する能力を有することを明らかにしている。対照的に、細胞サイクルに入ったコントロールCAMから得られた細胞はごくわずかのアポプトーシスタグ染色を示し、コントロール処置CAMではアポプトーシス細胞が殆ど検出されなかったことと一致した。
【0137】
細胞サイクル(SおよびG2/M期)に入ったβFGF刺激CAMのこれらの細胞では、70%がLM609(抗αvβ3)で陽性染色を示した。これは、βFGF未処置CAMの細胞サイクルに入った細胞で認められた10%LM609染色に準えることができる。これらの発見は、βFGF刺激の後、αvβ3をもつ細胞の大半が活発な分裂を示していることを示唆する。
【0138】
これらの結果を合わせると、mAbLM609またはαvβ3の環状ペプチド拮抗物質の静脈内注射は、脈管形成抑制に続いてニワトリのCAM内でアポプトーシスを促進させることを示唆している。また、LM609との免疫反応性によってαvβ3の発現および、アポプトーシスタグとの免疫反応性によってアポプトーシスが進行している細胞について組織学的にCAMを調べた。LM609(抗αvβ3)、CSAT(抗β1)またはPBSで48時間前に処置した、実施例5Aで調製した胎児から切除したCAM切片を洗浄し、OTC(Baxter)に包埋し、液体窒素で瞬間冷凍した。CAM組織の6ミクロン切片を切り出し、アセトンで30秒固定し、使用まで−70℃で保存した。組織切片は、70%(v/v)エタノール中で簡単にすすぎ、続いてPBS中で3回洗浄して、染色用に調製した。次に、切片をPBS中の5%(v/v)BSAで2時間処理し、その後10μg/mlのmAbLM609で2時間保温した。続いて切片を洗浄し、1:50のローダミン共役ヤギ抗マウスIgG(Fisher Scientific,ピッツバーグ、ペンシルバニア)で2時間保温した。最後に、同じ切片を洗浄し、実施例12A2)で述べたようにアポプトーシスタグで染色した。染色組織をマウントし供焦点免疫蛍光顕微鏡で調べた。
【0139】
図20では、写真AからCはCSAT(抗β1)処置胎児のCAM組織を表し、写真DからFはLM609(抗αvβ3)処置胎児のCAM組織を表す。写真AおよびDは、アポプトーシスタグで染色し、D.I.C.画像に蛍光(FITC)を重焼きして可視化させた組織を示す。写真BおよびEはmAbLM609(抗αvβ3)で染色し、蛍光(ローダミン)で可視化させた同じ組織を示す。写真CおよびFは、アポプトーシスタグおよびLM609の両方で染色した同じ組織の結合画像を表し、黄色は同時存在を示す。棒線は、左および右の写真でそれぞれ15および50μmを表している。
図20(A−C)に示すように、CSATまたはPBSを静注した後アポプトーシスタグによる染色はきわめて僅かでさらに無秩序のようであるが、これは組織内に極めてわずかなレベルしかアポプトーシスが存在しないことを示唆している。対照的に、LM609または環状ペプチド203で先に処置された胎児のCAMは、アポプトーシスタグで濃く染色された多くの血管を示し、一方周辺の非血管細胞間ではごくわずかな反応性が認められただけである(図20D−E)。さらに、これらの組織の染色にアポプトーシスタグおよびLM609の両方を用いたとき(19Cおよび19F)、顕著な同時存在が、αvβ3拮抗物質で処置した胎児から得たCAMのこれらマーカー間でのみ認められた。これらの発見は、インビボで脈管形成を誘発した後、インテグリンαvβ3の抑制物質は選択的にαvβ3保有血管のアポプトーシスを促進することを明らかにした。
【0140】
脈管形成は、多くの分子的および細胞生物学的現象を含む複雑な過程であるが、一方、実証されたいくつかの流れは、血管細胞インテグリンαvβ3はこの過程で比較的後期における役割を果たすことを提唱している。第一に、免疫組織学的分析によって、血管細胞におけるαvβ3の発現はβFGFによる脈管形成の誘発後12−24時間で最大になることが明らかにされた。第二に、αvβ3の拮抗物質は多数の活性化物質によって誘発される脈管形成を妨害するが、このことは、このレセプターは、脈管形成へと続くおそらく一次的シグナリングに関する全ての現象から下流にある共通の経路に含まれることを示唆する。第三に、mAbLM609または環状ペプチド処置CAMは、これら脈管形成拮抗物質による処置後48時間まではDNAのほつれによって測定されるアポプトーシスで顕著な増加を示さなかった。最後に、αvβ3拮抗物質は、細胞サイクル進入が既に誘発された血管細胞のアポプトーシスを促進する。
【0141】
本明細書に提示した結果は、インテグリン連結現象がインビボで細胞の生存を調節することができるという最初の直接的証拠を提供する。したがって次のような仮説が提唱される:脈管形成がいったん開始すると、個々の血管細胞は分裂し脈管形成原に向かって移動し始め、その後、αvβ3連結は、分化と成熟血管形成へと進行する持続した細胞生存を可能にするシグナルを提供する。しかしながら、αvβ3連結が妨害されると、これら細胞はこの分子的指示を受け取ることができず、不履行によりアポプトーシスに陥る。この仮説はまた、分化が生じた後、成熟血管はもはやαvβ3シグナリングを生存のために必要とせず、したがってこのインテグリンに対して耐性をもつということを予想させるであろう。
最後に、本明細書に提示した結果は、インテグリンαvβ3の拮抗物質は、新形成または脈管形成を特徴とする他の疾患のための強力な治療方法を提供するであろう。第一に、αvβ3拮抗物質は、以前から存在する血管構造に影響を与えること無く新しく形成されつつある血管を破壊する。第二にこれらの拮抗物質は、ニワトリ胎児の生存活性に顕著な影響を与えず、それらの非毒性を示唆した。第三に、脈管形成は、脈管形成刺激にもかかわらず顕著に遮断された。最後に、αvβ3拮抗物質の全身的投与は、種々の組織学的に明瞭なヒト腫瘍の劇的な退縮をもたらした。
【0142】
したがって前述の実施例によって、インテグリンαvβ3は、種々の刺激による脈管形成において重要な役割をはたし、そのようなαvβ3は、新規脈管新生を特徴とする疾患に対する本発明のαvβ3拮抗物質の重要な治療標的であることが明らかになった。
前述の記載は、当業者が本発明を実施するために十分であろうと考えられる。本発明は、寄託された細胞株によって範囲が限定されるべきではない。なぜならば、寄託された具体例は、本発明の1つの特徴を単に詳述することを目的としており、機能的に同等ないずれの細胞株も本発明の範疇に入るからである。材料の寄託は、その最善の態様を含む本明細書中の記載が本発明の実施のためには不適当であるかもしれないことを承認するものではなく、また本明細書の記載に対して請求の範囲を制限することを意図したものでもない。実際、本明細書に記載したものに加え、当業者にとっては前述の記載から種々の修飾が明白であり、それらは添付の請求の範囲内に含まれるであろう。
【0143】
【図面の簡単な説明】
【0144】
【図1A】正常皮膚および創傷治癒下の皮膚(肉芽形成組織と呼ぶ)におけるインテグリンサブユニットβ3およびβ1の組織分布を示す。
【図1B】正常皮膚および創傷治癒下の皮膚(肉芽形成組織と呼ぶ)におけるインテグリンサブユニットβ3およびβ1の組織分布を示す。
【図1C】正常皮膚および創傷治癒下の皮膚(肉芽形成組織と呼ぶ)におけるインテグリンサブユニットβ3およびβ1の組織分布を示す。
【図1D】正常皮膚および創傷治癒下の皮膚(肉芽形成組織と呼ぶ)におけるインテグリンサブユニットβ3およびβ1の組織分布を示す。
【図2A】正常皮膚および創傷治癒下の皮膚(肉芽形成組織と呼ぶ)における、それぞれβ3およびβ1に結合するヴォン・ヴィレブランド因子およびラミニンリガンドの組織分布を示す。
【図2B】正常皮膚および創傷治癒下の皮膚(肉芽形成組織と呼ぶ)における、それぞれβ3およびβ1に結合するヴォン・ヴィレブランド因子およびラミニンリガンドの組織分布を示す。
【図2C】正常皮膚および創傷治癒下の皮膚(肉芽形成組織と呼ぶ)における、それぞれβ3およびβ1に結合するヴォン・ヴィレブランド因子およびラミニンリガンドの組織分布を示す。
【図2D】正常皮膚および創傷治癒下の皮膚(肉芽形成組織と呼ぶ)における、それぞれβ3およびβ1に結合するヴォン・ヴィレブランド因子およびラミニンリガンドの組織分布を示す。
【図3A】それぞれ膀胱癌、大腸癌、乳癌および肺癌の組織生検材料におけるビトロネクチンインテグリンレセプター、αvβ3の分布を示す。
【図3B】それぞれ膀胱癌、大腸癌、乳癌および肺癌の組織生検材料におけるビトロネクチンインテグリンレセプター、αvβ3の分布を示す。
【図3C】それぞれ膀胱癌、大腸癌、乳癌および肺癌の組織生検材料におけるビトロネクチンインテグリンレセプター、αvβ3の分布を示す。
【図3D】それぞれ膀胱癌、大腸癌、乳癌および肺癌の組織生検材料におけるビトロネクチンインテグリンレセプター、αvβ3の分布を示す。
【図4】本発明の未処置10週齢ニワトリ胎児のCAMの典型的な顕微鏡写真を示す。
【図5A】本発明のCAM調製物のインテグリンβ1およびαvβ3の組織分布を示す。
【図5B】本発明のCAM調製物のインテグリンβ1およびαvβ3の組織分布を示す。
【図5C】本発明のCAM調製物のインテグリンβ1およびαvβ3の組織分布を示す。
【図6】実施例6Aで述べる未処置およびβFGF処置10週齢CAMのαvβ3およびβ1の相対的発現を測定した棒グラフを示す。
【図7A】それぞれ未処置、βFGF処置およびTNFα処置10週齢CAMの外観を示す。
【図7B】それぞれ未処置、βFGF処置およびTNFα処置10週齢CAMの外観を示す。
【図7C】それぞれ未処置、βFGF処置およびTNFα処置10週齢CAMの外観を示す。
【図8A】実施例7A1)における、10日目CAMにおけるFGF誘発脈管形成に対する抗体局所処置の効果を示す。
【図8B】実施例7A1)における、10日目CAMにおけるFGF誘発脈管形成に対する抗体局所処置の効果を示す。
【図8C】実施例7A1)における、10日目CAMにおけるFGF誘発脈管形成に対する抗体局所処置の効果を示す。
【図8D】実施例7A1)における、10日目CAMにおけるFGF誘発脈管形成に対する抗体局所処置の効果を示す。
【図8E】実施例7A1)における、10日目CAMにおけるFGF誘発脈管形成に対する抗体局所処置の効果を示す。
【図9A】実施例7D2)における、腫瘍によって誘発された脈管形成に対する合成ペプチド66203の静脈内注射の影響を示す。
【図9B】実施例7D2)における、腫瘍によって誘発された脈管形成に対する合成ペプチド66203の静脈内注射の影響を示す。
【図9C】実施例7D2)における、腫瘍によって誘発された脈管形成に対する合成ペプチド66203の静脈内注射の影響を示す。
【図10A】実施例7B1)における、増殖因子誘発脈管形成に対する単クローン性抗体の静脈内応用の効果を示す。
【図10B】実施例7B1)における、増殖因子誘発脈管形成に対する単クローン性抗体の静脈内応用の効果を示す。
【図10C】実施例7B1)における、増殖因子誘発脈管形成に対する単クローン性抗体の静脈内応用の効果を示す。
【図11A】実施例7Cにおける、抗インテグリン抗体の局所適用後の胎児の脈管形成に対する効果を示す。
【図11B】実施例7Cにおける、抗インテグリン抗体の局所適用後の胎児の脈管形成に対する効果を示す。
【図11C】実施例7Cにおける、抗インテグリン抗体の局所適用後の胎児の脈管形成に対する効果を示す。
【図12】実施例7D1)における、CAM調製物内の腫瘍に進入する血管数の測定を示す。
【図13A】実施例9A1)aにおける、処置後7日の湿潤腫瘍重量と最初の腫瘍重量との間の比較を示す。
【図13B】実施例9A1)aにおける、処置後7日の湿潤腫瘍重量と最初の腫瘍重量との間の比較を示す。
【図13C】実施例9A1)aにおける、処置後7日の湿潤腫瘍重量と最初の腫瘍重量との間の比較を示す。
【図13D】実施例9A1)aにおける、処置後7日の湿潤腫瘍重量と最初の腫瘍重量との間の比較を示す。
【図14A】実施例9A1)aにおける、P3G2(抗αvβ5)(図14A)およびLM609(抗αvβ3)(図14B)で処置し、ヘマトキシリンおよびエオシンで染色した腫瘍の組織切片を示す。
【図14B】実施例9A1)aにおける、P3G2(抗αvβ5)(図14A)およびLM609(抗αvβ3)(図14B)で処置し、ヘマトキシリンおよびエオシンで染色した腫瘍の組織切片を示す。
【図15A】実施例9A1)bにおける、ペプチドで処置されたM21L腫瘍組織を示す。
【図15B】実施例9A1)bにおける、ペプチドで処置されたM21L腫瘍組織を示す。
【図15C】実施例9A1)bにおける、ペプチドで処置されたM21L腫瘍組織を示す。
【図15D】実施例9A1)bにおける、ペプチドで処置されたM21L腫瘍組織を示す。
【図15E】実施例9A1)bにおける、ペプチドで処置されたM21L腫瘍組織を示す。
【図16A】実施例10におけるウサギ眼球のインビボモデルアッセーにおける脈管形成拮抗物質による脈管形成の抑制を示す。
【図16B】実施例10におけるウサギ眼球のインビボモデルアッセーにおける脈管形成拮抗物質による脈管形成の抑制を示す。
【図16C】実施例10におけるウサギ眼球のインビボモデルアッセーにおける脈管形成拮抗物質による脈管形成の抑制を示す。
【図16D】実施例10におけるウサギ眼球のインビボモデルアッセーにおける脈管形成拮抗物質による脈管形成の抑制を示す。
【図16E】実施例10におけるウサギ眼球のインビボモデルアッセーにおける脈管形成拮抗物質による脈管形成の抑制を示す。
【図17】実施例11Aのマウス:ヒトキメラ−インビボマウスモデルの作製方法を示す工程図である。
【図18】FACS分析で求めた、mAb処置およびペプチド処置CAMから得られ、アポプトーシスタグ(Apop Tag)で染色された単一細胞の%を示す。
【図19】CSAT(抗β1)(図19A)またはLM609(抗αvβ3)(図19B)で処置した胎児のCAMの単一細胞懸濁をアポプトーシスタグおよび沃化プロピジウムで染色し、FACSで分析した結果をまとめたものを示す。
【図20A】処置胎児のCAM組織を表す。
【図20B】処置胎児のCAM組織を表す。
【図20C】処置胎児のCAM組織を表す。
【図20D】処置胎児のCAM組織を表す。
【図20E】処置胎児のCAM組織を表す。
【図20F】処置胎児のCAM組織を表す。
【特許請求の範囲】
【請求項1】
脈管形成抑制量のαvβ3拮抗物質を含む、組織における脈管形成を抑制する医薬組成物であって、前記αvβ3拮抗物質が、αvβ3に対して免疫特異的な単クローン性抗体又はそのフラグメントであることを特徴とする医薬組成物。
【請求項2】
前記αvβ3拮抗物質が、αvβ3へのフィブリノーゲンの結合を抑制し、αIIbβ3またはαvβ1へのフィブリノーゲンの結合を実質的に抑制しない請求項1の医薬組成物。
【請求項3】
前記単クローン性抗体が、単クローン性抗体LM609(ATCCHB9537)の免疫反応特性を有する請求項1の医薬組成物。
【請求項4】
前記組織が、炎症を生じており、前記脈管形成が、炎症組織の脈管形成である請求項1〜3のいずれかに記載の医薬組成物。
【請求項5】
前記組織が、関節炎である請求項4に記載の医薬組成物。
【請求項6】
前記関節炎組織が、類リューマチ関節炎の哺乳類に存在する請求項5に記載の医薬組成物。
【請求項7】
前記脈管形成が、網膜脈管形成である、請求項1〜3のいずれかに記載の医薬組成物。
【請求項8】
前記組織が、血管腫である請求項1〜3のいずれかに記載の医薬組成物。
【請求項9】
前記組織が、固形腫瘍または固形腫瘍転移であり、前記脈管形成が、腫瘍の脈管形成である請求項1〜3のいずれかに記載の医薬組成物。
【請求項10】
前記脈管形成抑制量が、2μMから5mMである請求項1〜9のいずれかに記載の医薬組成物。
【請求項11】
化学療法と合わせて投与される請求項1〜10のいずれかに記載の医薬組成物。
【請求項12】
静脈内、経皮、関節滑液嚢内、筋肉内または経口投与の形態である請求項1〜11のいずれかに記載の医薬組成物。
【請求項13】
静脈内投与の形態である請求項1〜11のいずれかに記載の医薬組成物。
【請求項14】
脈管形成抑制量のαvβ3拮抗物質を含む、組織における腫瘍を退縮する医薬組成物であって、前記αvβ3拮抗物質が、αvβ3に対して免疫特異的な単クローン性抗体又はそのフラグメントであることを特徴とする医薬組成物。
【請求項15】
前記αvβ3拮抗物質が、αvβ3へのフィブリノーゲンの結合を抑制し、αIIbβ3またはαvβ1へのフィブリノーゲンの結合を実質的に抑制しない請求項14の医薬組成物。
【請求項16】
前記単クローン性抗体が、単クローン性抗体LM609(ATCCHB9537)の免疫反応特性を有する請求項14の医薬組成物。
【請求項17】
前記組織が、炎症を生じており、前記脈管形成が、炎症組織の脈管形成である請求項14〜16のいずれかに記載の医薬組成物。
【請求項18】
前記脈管形成抑制量が、2μMから5mMである請求項14〜16のいずれかに記載の医薬組成物。
【請求項19】
化学療法と合わせて投与される請求項14〜18のいずれかに記載の医薬組成物。
【請求項20】
静脈内、経皮、関節滑液嚢内、筋肉内または経口投与の形態である請求項14〜19のいずれかに記載の医薬組成物。
【請求項21】
静脈内投与の形態である請求項14〜19のいずれかに記載の医薬組成物。
【請求項22】
脈管形成抑制量のαvβ3拮抗物質を含む、組織における新規血管系のアポプトーシスを誘発するための医薬組成物であって、前記αvβ3拮抗物質が、αvβ3に対して免疫特異的な単クローン性抗体又はそのフラグメントであることを特徴とする医薬組成物。
【請求項23】
前記αvβ3拮抗物質が、αvβ3へのフィブリノーゲンの結合を抑制し、αIIbβ3またはαvβ1へのフィブリノーゲンの結合を実質的に抑制しない請求項22の医薬組成物。
【請求項24】
前記単クローン性抗体が、単クローン性抗体LM609(ATCCHB9537)の免疫反応特性を有する請求項22の医薬組成物。
【請求項25】
前記組織が、炎症を生じており、前記脈管形成が、炎症組織の脈管形成である請求項22〜24のいずれかに記載の医薬組成物。
【請求項26】
前記組織が、関節炎である請求項25に記載の医薬組成物。
【請求項27】
前記関節炎組織が、類リューマチ関節炎の哺乳類に存在する請求項26に記載の医薬組成物。
【請求項28】
前記脈管形成が、網膜脈管形成である、請求項22〜24のいずれかに記載の医薬組成物。
【請求項29】
前記組織が、血管腫である請求項22〜24のいずれかに記載の医薬組成物。
【請求項30】
前記組織が、固形腫瘍または固形腫瘍転移であり、前記脈管形成が、腫瘍の脈管形成である請求項22〜24のいずれかに記載の医薬組成物。
【請求項31】
前記脈管形成抑制量が、2μMから5mMである請求項22〜30のいずれかに記載の医薬組成物。
【請求項32】
化学療法と合わせて投与される請求項22〜31のいずれかに記載の医薬組成物。
【請求項33】
静脈内、経皮、関節滑液嚢内、筋肉内または経口投与の形態である請求項22〜32のいずれかに記載の医薬組成物。
【請求項34】
静脈内投与の形態である請求項22〜32のいずれかに記載の医薬組成物。
【請求項35】
関節炎、糖尿病性網膜症、黄斑変性、再狭窄、血管腫、又は肺、乳房又は結腸の固形腫瘍を治療するための医薬の製造におけるαvβ3拮抗物質の使用であって、前記医薬が、脈管形成抑制量のαvβ3拮抗物質を含み、前記αvβ3拮抗物質が、αvβ3に対して免疫特異的な単クローン性抗体又はそのフラグメントであることを特徴とする使用。
【請求項36】
前記医薬が、腫瘍の治療のためであり、前記脈管形成が、腫瘍脈管形成である請求項35に記載の使用。
【請求項37】
前記医薬が、関節炎の治療のためである請求項35に記載の使用。
【請求項38】
前記医薬が、類リューマチ関節炎の治療のためである請求項35に記載の使用。
【請求項39】
前記医薬が、糖尿病性網膜症の治療のためであり、前記脈管形成が、網膜脈管形成である請求項35に記載の使用。
【請求項40】
前記医薬が、黄斑変性の治療のためであり、前記脈管形成が、網膜脈管形成である請求項35に記載の使用。
【請求項41】
前記医薬が、血管腫の治療のためである請求項35に記載の使用。
【請求項42】
化学療法と合わせて投与される請求項35〜41のいずれかに記載の使用。
【請求項43】
前記医薬が、冠状動脈の血管形成術に続く再狭窄の危険性がある患者を治療するためである請求項35の使用。
【請求項44】
前記医薬が、1μg/mlから5μg/mlのαvβ3拮抗物質の血中濃度を達成するために投与される請求項35〜43のいずれかに記載の使用。
【請求項45】
前記医薬が、0.1mg/kgから300mg/kgの量のαvβ3拮抗物質が、1日1回または2回以上、1日または数日投与される請求項35〜43のいずれかに記載の使用。
【請求項46】
前記医薬が、静脈内、経皮、関節滑液嚢内、筋肉内または経口投与に適した形態である請求項35〜45のいずれかに記載の使用。
【請求項47】
前記医薬が、静脈内投与に適した形態である請求項35〜45のいずれかに記載の使用。
【請求項48】
前記αvβ3拮抗物質が、αvβ3へのフィブリノーゲンの結合を抑制し、αvβ3へのフィブリノーゲン結合抑制のIC50が、αIIbβ3またはαvβ1へのフィブリノーゲン結合抑制のIC50活性と比較して、少なくとも10倍である請求項35〜47のいずれかに記載の使用。
【請求項49】
前記単クローン性抗体又はそのフラグメントが、ヒト化されている請求項35〜48のいずれかに記載の使用。
【請求項50】
前記単クローン性抗体が、LM609(ATCC HB9537)である請求項35〜49いずれかに記載の使用。
【請求項51】
前記単クローン性抗体又はそのフラグメントが、単クローン性抗体LM609(ATCC HB9537)免疫反応特性を有する請求項35〜48のいずれかに記載の使用。
【請求項52】
前記医薬が、ヒト患者の治療のためである請求項35〜51のいずれかに記載の使用。
【請求項1】
脈管形成抑制量のαvβ3拮抗物質を含む、組織における脈管形成を抑制する医薬組成物であって、前記αvβ3拮抗物質が、αvβ3に対して免疫特異的な単クローン性抗体又はそのフラグメントであることを特徴とする医薬組成物。
【請求項2】
前記αvβ3拮抗物質が、αvβ3へのフィブリノーゲンの結合を抑制し、αIIbβ3またはαvβ1へのフィブリノーゲンの結合を実質的に抑制しない請求項1の医薬組成物。
【請求項3】
前記単クローン性抗体が、単クローン性抗体LM609(ATCCHB9537)の免疫反応特性を有する請求項1の医薬組成物。
【請求項4】
前記組織が、炎症を生じており、前記脈管形成が、炎症組織の脈管形成である請求項1〜3のいずれかに記載の医薬組成物。
【請求項5】
前記組織が、関節炎である請求項4に記載の医薬組成物。
【請求項6】
前記関節炎組織が、類リューマチ関節炎の哺乳類に存在する請求項5に記載の医薬組成物。
【請求項7】
前記脈管形成が、網膜脈管形成である、請求項1〜3のいずれかに記載の医薬組成物。
【請求項8】
前記組織が、血管腫である請求項1〜3のいずれかに記載の医薬組成物。
【請求項9】
前記組織が、固形腫瘍または固形腫瘍転移であり、前記脈管形成が、腫瘍の脈管形成である請求項1〜3のいずれかに記載の医薬組成物。
【請求項10】
前記脈管形成抑制量が、2μMから5mMである請求項1〜9のいずれかに記載の医薬組成物。
【請求項11】
化学療法と合わせて投与される請求項1〜10のいずれかに記載の医薬組成物。
【請求項12】
静脈内、経皮、関節滑液嚢内、筋肉内または経口投与の形態である請求項1〜11のいずれかに記載の医薬組成物。
【請求項13】
静脈内投与の形態である請求項1〜11のいずれかに記載の医薬組成物。
【請求項14】
脈管形成抑制量のαvβ3拮抗物質を含む、組織における腫瘍を退縮する医薬組成物であって、前記αvβ3拮抗物質が、αvβ3に対して免疫特異的な単クローン性抗体又はそのフラグメントであることを特徴とする医薬組成物。
【請求項15】
前記αvβ3拮抗物質が、αvβ3へのフィブリノーゲンの結合を抑制し、αIIbβ3またはαvβ1へのフィブリノーゲンの結合を実質的に抑制しない請求項14の医薬組成物。
【請求項16】
前記単クローン性抗体が、単クローン性抗体LM609(ATCCHB9537)の免疫反応特性を有する請求項14の医薬組成物。
【請求項17】
前記組織が、炎症を生じており、前記脈管形成が、炎症組織の脈管形成である請求項14〜16のいずれかに記載の医薬組成物。
【請求項18】
前記脈管形成抑制量が、2μMから5mMである請求項14〜16のいずれかに記載の医薬組成物。
【請求項19】
化学療法と合わせて投与される請求項14〜18のいずれかに記載の医薬組成物。
【請求項20】
静脈内、経皮、関節滑液嚢内、筋肉内または経口投与の形態である請求項14〜19のいずれかに記載の医薬組成物。
【請求項21】
静脈内投与の形態である請求項14〜19のいずれかに記載の医薬組成物。
【請求項22】
脈管形成抑制量のαvβ3拮抗物質を含む、組織における新規血管系のアポプトーシスを誘発するための医薬組成物であって、前記αvβ3拮抗物質が、αvβ3に対して免疫特異的な単クローン性抗体又はそのフラグメントであることを特徴とする医薬組成物。
【請求項23】
前記αvβ3拮抗物質が、αvβ3へのフィブリノーゲンの結合を抑制し、αIIbβ3またはαvβ1へのフィブリノーゲンの結合を実質的に抑制しない請求項22の医薬組成物。
【請求項24】
前記単クローン性抗体が、単クローン性抗体LM609(ATCCHB9537)の免疫反応特性を有する請求項22の医薬組成物。
【請求項25】
前記組織が、炎症を生じており、前記脈管形成が、炎症組織の脈管形成である請求項22〜24のいずれかに記載の医薬組成物。
【請求項26】
前記組織が、関節炎である請求項25に記載の医薬組成物。
【請求項27】
前記関節炎組織が、類リューマチ関節炎の哺乳類に存在する請求項26に記載の医薬組成物。
【請求項28】
前記脈管形成が、網膜脈管形成である、請求項22〜24のいずれかに記載の医薬組成物。
【請求項29】
前記組織が、血管腫である請求項22〜24のいずれかに記載の医薬組成物。
【請求項30】
前記組織が、固形腫瘍または固形腫瘍転移であり、前記脈管形成が、腫瘍の脈管形成である請求項22〜24のいずれかに記載の医薬組成物。
【請求項31】
前記脈管形成抑制量が、2μMから5mMである請求項22〜30のいずれかに記載の医薬組成物。
【請求項32】
化学療法と合わせて投与される請求項22〜31のいずれかに記載の医薬組成物。
【請求項33】
静脈内、経皮、関節滑液嚢内、筋肉内または経口投与の形態である請求項22〜32のいずれかに記載の医薬組成物。
【請求項34】
静脈内投与の形態である請求項22〜32のいずれかに記載の医薬組成物。
【請求項35】
関節炎、糖尿病性網膜症、黄斑変性、再狭窄、血管腫、又は肺、乳房又は結腸の固形腫瘍を治療するための医薬の製造におけるαvβ3拮抗物質の使用であって、前記医薬が、脈管形成抑制量のαvβ3拮抗物質を含み、前記αvβ3拮抗物質が、αvβ3に対して免疫特異的な単クローン性抗体又はそのフラグメントであることを特徴とする使用。
【請求項36】
前記医薬が、腫瘍の治療のためであり、前記脈管形成が、腫瘍脈管形成である請求項35に記載の使用。
【請求項37】
前記医薬が、関節炎の治療のためである請求項35に記載の使用。
【請求項38】
前記医薬が、類リューマチ関節炎の治療のためである請求項35に記載の使用。
【請求項39】
前記医薬が、糖尿病性網膜症の治療のためであり、前記脈管形成が、網膜脈管形成である請求項35に記載の使用。
【請求項40】
前記医薬が、黄斑変性の治療のためであり、前記脈管形成が、網膜脈管形成である請求項35に記載の使用。
【請求項41】
前記医薬が、血管腫の治療のためである請求項35に記載の使用。
【請求項42】
化学療法と合わせて投与される請求項35〜41のいずれかに記載の使用。
【請求項43】
前記医薬が、冠状動脈の血管形成術に続く再狭窄の危険性がある患者を治療するためである請求項35の使用。
【請求項44】
前記医薬が、1μg/mlから5μg/mlのαvβ3拮抗物質の血中濃度を達成するために投与される請求項35〜43のいずれかに記載の使用。
【請求項45】
前記医薬が、0.1mg/kgから300mg/kgの量のαvβ3拮抗物質が、1日1回または2回以上、1日または数日投与される請求項35〜43のいずれかに記載の使用。
【請求項46】
前記医薬が、静脈内、経皮、関節滑液嚢内、筋肉内または経口投与に適した形態である請求項35〜45のいずれかに記載の使用。
【請求項47】
前記医薬が、静脈内投与に適した形態である請求項35〜45のいずれかに記載の使用。
【請求項48】
前記αvβ3拮抗物質が、αvβ3へのフィブリノーゲンの結合を抑制し、αvβ3へのフィブリノーゲン結合抑制のIC50が、αIIbβ3またはαvβ1へのフィブリノーゲン結合抑制のIC50活性と比較して、少なくとも10倍である請求項35〜47のいずれかに記載の使用。
【請求項49】
前記単クローン性抗体又はそのフラグメントが、ヒト化されている請求項35〜48のいずれかに記載の使用。
【請求項50】
前記単クローン性抗体が、LM609(ATCC HB9537)である請求項35〜49いずれかに記載の使用。
【請求項51】
前記単クローン性抗体又はそのフラグメントが、単クローン性抗体LM609(ATCC HB9537)免疫反応特性を有する請求項35〜48のいずれかに記載の使用。
【請求項52】
前記医薬が、ヒト患者の治療のためである請求項35〜51のいずれかに記載の使用。
【図1A】


【図1B】

【図1C】

【図1D】

【図2A】

【図2B】

【図2C】

【図2D】
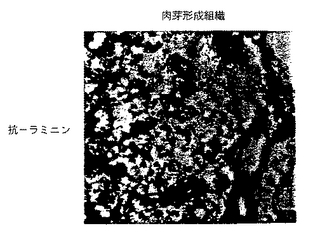
【図3A】

【図3B】

【図3C】

【図3D】

【図4】

【図5A】

【図5B】

【図5C】

【図6】

【図7A】

【図7B】

【図7C】
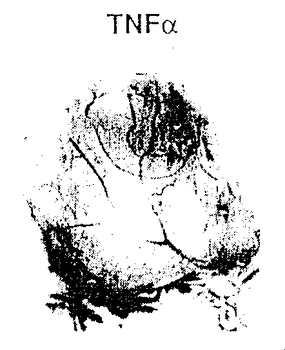
【図8A】

【図8B】

【図8C】

【図8D】

【図8E】

【図9A】
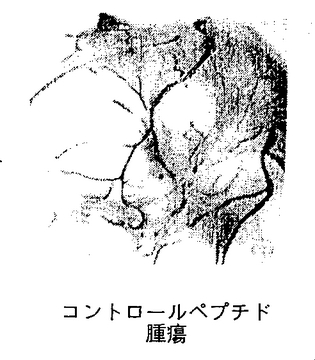
【図9B】

【図9C】

【図10A】

【図10B】
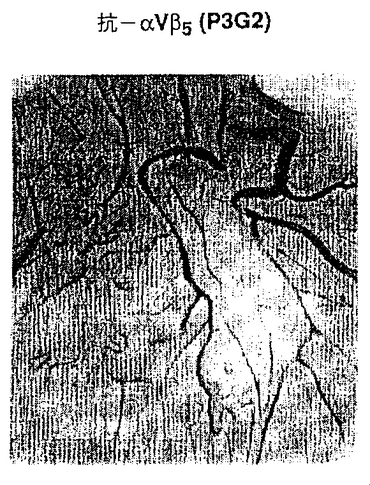
【図10C】

【図11A】

【図11B】

【図11C】
【図12】
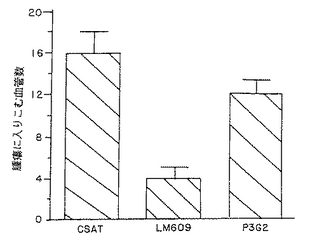
【図13A】

【図13B】
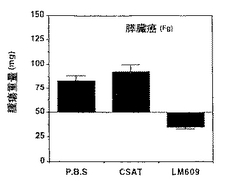
【図13C】

【図13D】
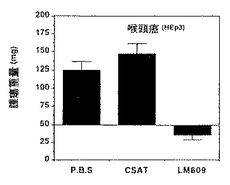
【図14A】
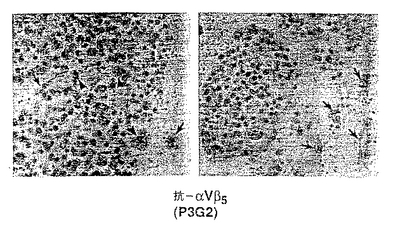
【図14B】

【図15A】
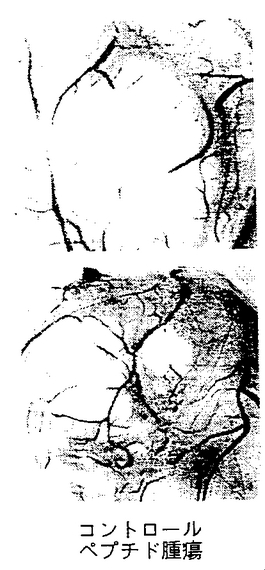
【図15B】

【図15C】
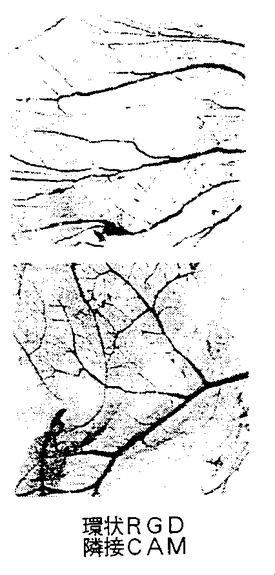
【図15D】

【図15E】

【図16A】

【図16B】

【図16C】

【図16D】

【図16E】

【図17】
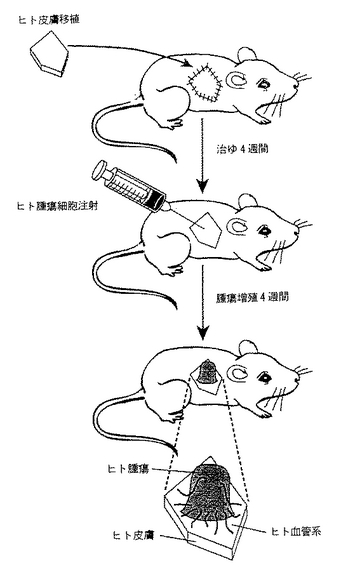
【図18】

【図19】
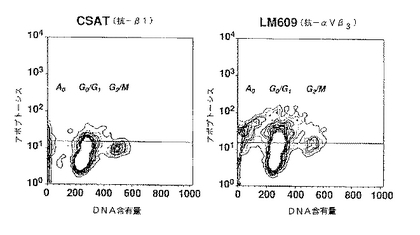
【図20A】
【図20B】

【図20C】

【図20D】

【図20E】

【図20F】

【図1B】
【図1C】
【図1D】
【図2A】
【図2B】
【図2C】
【図2D】
【図3A】
【図3B】
【図3C】
【図3D】
【図4】
【図5A】
【図5B】
【図5C】
【図6】
【図7A】
【図7B】
【図7C】
【図8A】
【図8B】
【図8C】
【図8D】
【図8E】
【図9A】
【図9B】
【図9C】
【図10A】
【図10B】
【図10C】
【図11A】
【図11B】
【図11C】
【図12】
【図13A】
【図13B】
【図13C】
【図13D】
【図14A】
【図14B】
【図15A】
【図15B】
【図15C】
【図15D】
【図15E】
【図16A】
【図16B】
【図16C】
【図16D】
【図16E】
【図17】
【図18】
【図19】
【図20A】
【図20B】
【図20C】
【図20D】
【図20E】
【図20F】
【公開番号】特開2006−273862(P2006−273862A)
【公開日】平成18年10月12日(2006.10.12)
【国際特許分類】
【出願番号】特願2006−142020(P2006−142020)
【出願日】平成18年5月22日(2006.5.22)
【分割の表示】特願平7−524675の分割
【原出願日】平成7年3月9日(1995.3.9)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.テフロン
【出願人】(593052785)ザ スクリップス リサーチ インスティテュート (91)
【Fターム(参考)】
【公開日】平成18年10月12日(2006.10.12)
【国際特許分類】
【出願日】平成18年5月22日(2006.5.22)
【分割の表示】特願平7−524675の分割
【原出願日】平成7年3月9日(1995.3.9)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.テフロン
【出願人】(593052785)ザ スクリップス リサーチ インスティテュート (91)
【Fターム(参考)】
[ Back to top ]