
脊椎動物Serrate遺伝子のヌクレオチドおよびタンパク質配列およびそれらに基づく方法
【課題】脊椎動物のSerrate遺伝子のヌクレオチド配列、およびそれらのコード化されたタンパク質のアミノ酸配列、さらには、それらの誘導体(例えば、断片)および類似体を提供する。
【解決手段】精製された脊椎動物のSerrateタンパク質。一つの実施形態において、本発明は、機能的に活性な、すなわち全長(野性型)Serrateタンパク質に関連する1以上の公知の機能活性を示すことができる、本発明の脊椎動物Serrate誘導体および類似体に関する。
【解決手段】精製された脊椎動物のSerrateタンパク質。一つの実施形態において、本発明は、機能的に活性な、すなわち全長(野性型)Serrateタンパク質に関連する1以上の公知の機能活性を示すことができる、本発明の脊椎動物Serrate誘導体および類似体に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、補助金番号 GM 29093 およびNS 26084のもとに米国厚生省(Department of Health and HumanServices)から授与された政府援助金によって部分的に成された。米国政府は本発明に一定の権利を有する。
1.序論
本発明は脊椎動物Serrate遺伝子およびそれらによってコードされるタンパク質産物、ならびに上記タンパク質産物の誘導体および類似体に関する。脊椎動物Serrateタンパク質、誘導体および抗体の作製もまた提供される。本発明はさらに治療用組成物、ならびに診断および治療方法に関する。
【背景技術】
【0002】
2.発明の背景
ショウジョウバエにおける遺伝子分析は、複雑な発生経路を細かく調べ、そして相互作用している遺伝子座を同定するのに極めて有用であった。しかし、遺伝子の相互作用の下に横たわるプロセスの正確な性質を理解するには、該遺伝子のタンパク質産物の知識が必要である。
【0003】
発生学的、遺伝子的および分子的証拠は、ショウジョウバエにおける外胚葉分化の初期段階は細胞の相互作用に依存することを示している(Doe およびGoodman,1985,Dev.Biol.111:206-219; TechnauおよびCampos-Ortega,1986,Dev.Biol.195:445-454;Vassinら,1985,J.Neurogenet.2:291-308;de la Conchaら,1988,Genetics 118:499-508; Xuら,1990,Genes Dev.4:464-475;Artavanis-Tsakonas,1988,Trends Genet.4:95-100)。突然変異分析は、外胚葉細胞の表皮経路と神経経路の間の選択に影響を及ぼす、接合体的に(zygotically)作用する遺伝子の小グループ(いわゆる神経形成遺伝子座)を明らかにしている(Poulson,1937,Proc.Natl.Acad.Sci.23:133-137;Lehmannら,1983,Wilhelm Roux's Arch.Dev.Biol.192:62-74; Jurgensら,1984,WilhelmRoux's Arch.Dev.Biol.193:283-295; Wieschausら,1984,Wilhelm Roux's Arch.Dev.Biol.193:296-307;Nusslein-Volhard ら,1984,Wilhelm Roux's Arch.Dev.Biol.193:267-282)。接合体性神経形成遺伝子座[Notch(N)、Delta(Dl)、mastermind(mam)、EnhancerofSplit(E(spl)、neuralized(neu)、およびbig brain(bib)]のいずれか1つにおけるヌル突然変異は、腹側および側面の表皮構造を犠牲にして神経系の肥大をもたらす。この効果は、表皮前駆細胞を誤って神経経路に入れたためであり、そしてこのことは、神経形成遺伝子機能は神経形成領域内の細胞を神経になる運命から上皮になる運命に方向転換させるために必要であることを暗示している。Serrateは、Notch遺伝子座と相互作用することが可能な遺伝子ユニットとして同定された(Xuら,1990,GenesDev.4:464-475)。これらの遺伝子的および発生的考察は、以下の仮説に導いた。すなわち、神経形成遺伝子座のタンパク質産物は、適切な表皮発生に必要な細胞相互作用機構の構成要素として機能する、という仮説である(Artavanis-Tsakonas,S.,1988,TrendsGenet.4:95-100)。
【0004】
突然変異分析はまた、神経形成遺伝子の作用は多面発現的であって、胚発生のみに限定されないことを明らかにしている。例えば、個眼、剛毛、羽根の形成(これらもまた細胞相互作用に依存することが知られている)は神経形成突然変異によって影響される(Morganら,1925,Bibliogr.Genet.2:1-226; Welsons,1956,Dros.Inf.Serv.30:157-158;Preissら,1988,EMBO J.7:3917-3927; ShellenbargerおよびMohler,1978,Dev.Biol.62:432-446;TechnauおよびCampos-Ortega,1986,Wilhelm Roux's Dev.Biol.195:445-454; Tomlson およびReady,1987,Dev.Biol.120:366-376;CaganおよびReady,1989,Genes Dev.3:1099-1112)。
【0005】
配列分析(Whartonら,1985,Cell43:567-581; Kidd およびYoung,1986,Mol.Cell.Biol.6:3094-3108; Vassinら,1987,EMBO J.6:3431-3440;Kopczynskiら,1988,Genes Dev.2:1723-1735)は、神経形成遺伝子座の2つ、すなわちNotchおよびDeltaは、膜を一回貫く膜貫通タンパク質をコードしているように思われることを示した。Notch遺伝子は、Notch/lin-12リピートと称する、3個のシステインが豊富なリピートを後に従えた36個の表皮増殖因子(EGF)に似たタンデムなリピートを含む大きいN末端細胞外ドメインを有する約300kdのタンパク質(本発明者らはこのタンパク質を示すために「Notch」という用語を用いる)をコードしている(Whartonら,1985,Cell43:567-581; Kidd およびYoung,1986,Mol.Cell.Biol.6:3094-3108; Yochemら,1988,Nature335:547-550)。Delta は、細胞外ドメイン内に9個のEGFに似たリピートを有する約100 kdのタンパク質(本発明者らはDLZM、すなわち顕著な接合体および母体の転写物のタンパク質産物を示すために「Delta」という用語を用いる;Kopczynskiら,1988,Genes Dev.2:1723-1735)をコードしている(Vassinら,1987,EMBO J.6:3431-3440;Kopczynskiら,1988,Genes Dev.2:1723-1735)。分子的研究は以下の示唆をもたらした。すなわち、Notch およびDelta は、初期の発生的決定に関与する細胞コミュニケーション作用機構の生化学的に相互作用する要素を構成する、という示唆である(Fehonら,1990,Cell61:523-534)。
【0006】
EGFに似たモチーフは、血液凝固カスケードに関与するタンパク質を含む種々のタンパク質に見いだされている(FurieおよびFurie,1988,Cell 53:505-518)。特に、このモチーフは血液凝固因子IXおよびX等の細胞外タンパク質(Reesら,1988,EMBOJ.7:2053-2061; FurieおよびFurie,1988,Cell 53:505-518)、他のショウジョウバエ遺伝子(Knustら,1987,EMBOJ.761-766; Rothbergら,1988,Cell 55:1047-1059)、および幾つかの細胞表面受容体タンパク質、例えばトロンボモジュリン(Suzukiら,1987,EMBOJ.6:1891-1897)およびLDL受容体(Sudhofら,1985,Science 228:815-822)に見いだされている。タンパク質結合部位がトロンボモジュリンおよびウロキナーゼのEGFリピートドメインにマップされた(Kurosawaら,J.Biol.Chem.263:5993-5996; Appella ら,1987,J.Biol.Chem.262:4437-4440)。ショウジョウバエSerrate遺伝子はすでにクローン化され、特徴付けされている(1993年6月24日付けPCT公開 WO 93/12141)。しかし、脊椎動物Serrate遺伝子を得ようとする試みにもかかわらず、本発明がなされるまで、いかなる脊椎動物Serrate遺伝子も入手不可能であった。
【0007】
上記文献の引用は、それらの文献が本発明の先行技術であると承認したものと解釈して
はならない。
【発明の概要】
【課題を解決するための手段】
【0008】
3.発明の概略
本発明は脊椎動物Serrate遺伝子(ヒトSerrate遺伝子および他の種の関連する遺伝子)のヌクレオチド配列、およびそれらの遺伝子によってコードされるタンパク質のアミノ酸配列、ならびにそれらの誘導体(例えば断片)および類似体に関する。前記のヌクレオチド配列とハイブリダイゼーション可能な、またはそれに相補的な核酸もまた提供される。特定の実施態様において、前記Serrateタンパク質はヒトタンパク質である。
【0009】
本発明は、機能的に活性な、すなわち全長(野性型)Serrateタンパク質に関連する1以上の公知の機能活性を示すことができる、本発明の脊椎動物Serrate誘導体および類似体に関する。このような機能活性は抗原性〔抗Serrate抗体に結合する(またはSerrateと結合を競合する)能力〕、免疫原性(Serrateに結合する抗体を生成する能力)、Notchまたは他のトポリズミック(toporythmic)なタンパク質またはその断片に結合する(またはSerrateと結合を競合する)能力(「接着性」("adhesiveness"))、Serrate受容体に結合する(またはSerrateと結合を競合する)能力を含むが、これらだけに限定されない。本明細書に使用する「トポリズミックなタンパク質」とは、Notch、Delta、Serrate、EnhancerofSplit およびDeltexのタンパク質産物、ならびに、例えばハイブリダイズするそれらの遺伝子配列の能力、またはDelta、SerrateまたはNotchとの相同性、または表現型の相互作用を示す遺伝子の能力によって同定可能な、この相互に作用する遺伝子ファミリーの他のメンバーのタンパク質産物を言う。
【0010】
本発明はさらに、細胞内ドメイン、細胞外ドメイン、膜貫通ドメイン、膜結合領域、またはSerrateタンパク質の1以上のEGFに似た(相同の)リピートまたはこれらの任意の組合せを含むがこれらだけに限定されない、Serrateタンパク質の1以上のドメインを含む脊椎動物Serrateの断片(およびそれらの誘導体および類似体)に関する。
【0011】
脊椎動物Serrateに対する抗体、その誘導体および類似体がさらに提供される。
【0012】
脊椎動物Serrateタンパク質、その誘導体および類似体の例えば組換え手段による作製方法もまた提供される。
【0013】
本発明はまた、脊椎動物Serrateタンパク質および核酸に基づく治療および診断方法な
らびに組成物に関する。本発明は、本発明の治療用化合物を投与することによる細胞の運
命、すなわち分化、の障害に対する治療を提供する。このような治療用化合物(本明細書
では「治療剤」と呼ぶ)は、脊椎動物Serrateタンパク質ならびにそれらの類似体および
誘導体(断片を含む);それらに対する抗体;脊椎動物Serrateタンパク質、類似体また
は誘導体をコードする核酸;および脊椎動物Serrateアンチセンス核酸を含む。好ましい
実施態様において、本発明の治療剤は癌状態を治療するため、または前腫瘍状態あるいは
非悪性腫瘍状態から腫瘍状態または悪性腫瘍状態への進行を阻止するために投与される。
他の特定の実施態様において、本発明の治療剤は神経系の障害を治療するため、または組
織の再生及び修復を促進するために投与される。
【0014】
1つの実施態様において、Notchおよび/またはSerrate機能に拮抗する、またはこれらを抑制する治療剤(以後「アンタゴニスト治療剤」と称する)が治療的効果のために投与される。別の実施態様においては、Notchおよび/またはSerrate機能を促進する治療剤(以後「アゴニスト治療剤」と称する)が治療的効果のために投与される。
【0015】
Notchおよび/またはSerrateタンパク質の異常な、または望ましくないレベルの発現、
または活性、または局在化を伴う細胞の運命に関する疾患、特に過剰増殖性(例えば癌)
または過少増殖性疾患は、後により詳しく記述するようにそれらのレベルを検出すること
により診断することができる。
【0016】
好ましい側面において、本発明の治療剤は、Notchタンパク質またはその断片への結合
を媒介する脊椎動物Serrateの少なくとも断片(本明細書では「接着性断片」と称する)
からなるタンパク質である。
本発明は、例えば以下の項目を提供する。
(項目1)
精製された脊椎動物のSerrateタンパク質。
(項目2)
ヒトのタンパク質である項目1に記載のタンパク質。
(項目3)
哺乳動物のタンパク質である項目1に記載のタンパク質。
(項目4)
配列番号2のアミノ酸番号30〜1218に実質的に記載されているアミノ酸配列を含
有する項目2に記載のタンパク質。
(項目5)
配列番号4のアミノ酸番号1〜1257に実質的に記載されているアミノ酸配列を含有
する項目2に記載のタンパク質。
(項目6)
ATCCに寄託されて受託番号68876号を指定されたプラスミドSerFLまたは該プラスミド中のSerrate配列にハイブリダイズ可能な核酸によってコードされる精製されたヒトタンパク質。
(項目7)
ATCCに寄託されて受託番号97068号を指定されたプラスミドpBS39によってコードされる
項目2に記載のタンパク質。
(項目8)
ATCCに寄託されて受託番号 号を指定されたプラスミドpBS15によってコードされるSerrateアミノ酸配列を含有する項目2に記載のタンパク質。
(項目9)
ATCCに寄託されて受託番号 号を指定されたプラスミドpBS3-2によってコードされるSerrateアミノ酸配列を含有する項目2に記載のタンパク質。
(項目10)
項目1のタンパク質の精製された断片であって、Serrateタンパク質の1つ以上の機
能活性を示すことが可能である前記断片。
(項目11)
項目2のタンパク質の精製された断片であって、ヒトまたはキイロショウジョウバエ(D.melanogaster)のSerrateタンパク質の1つ以上の機能活性を示すことが可能である前記断片。
(項目12)
ヒトのSerrateタンパク質に対する抗体と結合可能である項目2または7に記載のタンパク質の精製された断片。
(項目13)
項目10に記載の断片を含有する分子。
(項目14)
脊椎動物のSerrateタンパク質の精製された断片であって、前記タンパク質の細胞外ドメイン、DSLドメイン、表皮増殖因子様のリピートドメイン、システインに富むドメイン、膜貫通ドメインおよび細胞内ドメインからなる群から選ばれるドメインを含有する前記断片。
(項目15)
脊椎動物のSerrateタンパク質の精製された断片であって、前記タンパク質のDSLドメインを含有する前記断片。
(項目16)
脊椎動物のSerrateタンパク質の精製された断片であって、前記タンパク質の表皮増殖因子に相同性のリピートを含有する前記断片。
(項目17)
前記Serrateタンパク質がヒトSerrateタンパク質である項目14に記載の断片。
(項目18)
Notchタンパク質またはDeltaタンパク質に相同性の領域を含有し、かつ少なくとも10個のアミノ酸からなる脊椎動物のSerrateタンパク質の精製した断片。
(項目19)
Serrateタンパク質ではない第2のタンパク質のアミノ酸配列に共有結合によって融合している少なくとも10個のアミノ酸からなる脊椎動物のSerrateタンパク質の断片を含有するキメラタンパク質。
(項目20)
Serrateタンパク質の前記断片が抗Serrate抗体によって結合され得る断片である項目19に記載のキメラタンパク質。
(項目21)
前記Serrateタンパク質がヒトのタンパク質である項目19に記載のキメラタンパク質。
(項目22)
Serrateタンパク質の1つ以上の機能活性を示すことが可能である項目19に記載のキメラタンパク質。
(項目23)
脊椎動物のSerrateタンパク質の精製した断片であって、(a)抗Serrate抗体によって結合可能であり、(b)前記タンパク質の膜貫通および細胞内ドメインを欠失しており、かつ(c)前記Serrateタンパク質の少なくとも10個のアミノ酸からなる前記断片。
(項目24)
脊椎動物のSerrateタンパク質の精製した断片であって、(a)抗Serrate抗体によって結合可能であり、(b)前記タンパク質の細胞外ドメインを欠失しており、かつ(c)前記Serrateタンパク質の少なくとも10個のアミノ酸からなる前記断片。
(項目25)
Notchタンパク質に結合可能な脊椎動物Serrateタンパク質の精製した断片。
(項目26)
前記Serrateタンパク質の表皮増殖因子様リピートを欠失している項目25に記載の断片。
(項目27)
前記Serrateタンパク質がヒトSerrateタンパク質である項目23、24、25または26に記載の断片。
(項目28)
配列番号2または配列番号4の断片である項目29に記載の断片。
(項目29)
項目25に記載の断片を含有する分子。
(項目30)
項目1に記載のSerrateタンパク質と結合可能であり、かつショウジョウバエ(Drosophila)のSerrateタンパク質とは結合しない抗体。
(項目31)
項目2に記載のSerrateタンパク質と結合可能であり、かつショウジョウバエ(Drosophila)のSerrateタンパク質とは結合しない抗体。
(項目32)
モノクローナルである項目30に記載の抗体。
(項目33)
項目32に記載の抗体の断片を含有する分子であって、該断片が脊椎動物のSerrate
タンパク質と結合可能である前記分子。
(項目34)
脊椎動物のSerrateタンパク質をコードするヌクレオチド配列を含有する単離した核酸。
(項目35)
DNAである項目34に記載の核酸。
(項目36)
項目34に記載のヌクレオチド配列に完全に相補的なヌクレオチド配列を含有する単
離した核酸。
(項目37)
項目2に記載のSerrateタンパク質をコードするヌクレオチド配列を含有する単離し
た核酸。
(項目38)
ATCCに寄託して受託番号97068号を指定されたプラスミドpBS39に含有されるSerrateコード配列を含有する単離した核酸。
(項目39)
ATCCに寄託して受託番号68876号を指定されたプラスミドSerFLおよび該プラスミド中のSerrate配列にハイブリダイズ可能な単離したヒト核酸。
(項目40)
ATCCに寄託して受託番号 号を指定されたプラスミドpBS3-2に含有されるSerrateコード配列を含有する単離した核酸。
(項目41)
ATCCに寄託して受託番号 号を指定されたプラスミドpBS15に含有されるSerrateコード配列を含有する単離した核酸。
(項目42)
タンパク質をコードするヌクレオチド配列を含有する単離した核酸であって、前記タン
パク質が配列番号4のアミノ酸番号1〜1257を含有する前記核酸。
(項目43)
少なくとも8個のヌクレオチドからなる脊椎動物のSerrate遺伝子の断片を含有する単離された核酸。
(項目44)
項目14、15、16または25に記載の断片をコードするヌクレオチド配列を含有する単離した核酸。
(項目45)
前記断片がヒトSerrateタンパク質の断片である項目44に記載の核酸。
(項目46)
項目12に記載の断片をコードするヌクレオチド配列を含有する単離された核酸。
(項目47)
タンパク質をコードするヌクレオチド配列を含有する単離した核酸であって、前記タンパク質が配列番号2のアミノ酸番号30〜1218を含有する前記核酸。
(項目48)
項目21に記載のタンパク質をコードするヌクレオチド配列を含有する単離された核酸。
(項目49)
項目34、37または43に記載の核酸を含有する組換え細胞。
(項目50)
項目38、40または41に記載の核酸を含有する組換え細胞。
(項目51)
項目34または37に記載の核酸を含有する組換え細胞を増殖させて、コードされた前記Serrateタンパク質が前記細胞によって発現されるようにする工程と、前記発現されたSerrateタンパク質を回収する工程とを包含するSerrateタンパク質の生成方法。
(項目52)
項目38、40または41に記載の核酸を含有する組換え細胞を増殖させて、コードされたSerrateタンパク質が前記細胞によって発現されるようにする工程と、前記発現されたSerrateタンパク質を回収する工程とを包含するSerrateタンパク質の生成方法。
(項目53)
項目45に記載の核酸を含有する組換え細胞を増殖させて、コードされたタンパク質が前記細胞によって発現されるようにする工程と、前記発現されたタンパク質を回収する工程とを包含するタンパク質の生成方法。
(項目54)
Serrateタンパク質の断片を含有するタンパク質の生成方法であって、項目46に記載の核酸を含有する組換え細胞を増殖させて、コードされたタンパク質が前記細胞によって発現されるようにする工程と、前記発現されたタンパク質を回収する工程とを包含する前記方法。
(項目55)
項目51に記載の方法による生産物。
(項目56)
項目52に記載の方法による生産物。
(項目57)
項目53に記載の方法による生産物。
(項目58)
項目54に記載の方法による生産物。
(項目59)
治療上有効な量の脊椎動物Serrateタンパク質と、製剤上許容される担体とを含有する医薬組成物。
(項目60)
前記Serrateタンパク質がヒトSerrateタンパク質である項目59に記載の組成物。
(項目61)
治療上有効な量の項目14、15、16または25に記載の断片と、製剤上許容される担体とを含有する医薬組成物。
(項目62)
治療上有効な量の項目12に記載の断片と、製剤上許容される担体とを含有する医薬組成物。
(項目63)
脊椎動物Serrateタンパク質の断片を含有する分子を治療上有効な量で含有する医薬組成物であって、その誘導体または類似体が、Notchタンパク質への結合能、またはNotchタンパク質の表皮増殖因子様リピート11および12を含有する分子への結合能により特徴付けられる前記組成物。
(項目64)
治療上有効な量の項目34、36または37に記載の核酸と、製剤上許容される担体とを含有する医薬組成物。
(項目65)
治療上有効な量の項目44に記載の核酸と、製剤上許容される担体とを含有する医薬組成物。
(項目66)
治療上有効な量の項目46に記載の核酸と、製剤上許容される担体とを含有する医薬組成物。
(項目67)
治療上有効な量の項目30に記載の抗体と、製剤上許容される担体とを含有する医薬組成物。
(項目68)
治療上有効な量の項目30に記載の抗体の結合ドメインを含有する前記抗体の断片または誘導体と、製剤上許容される担体とを含有する医薬組成物。
(項目69)
被験者の疾病または障害を治療または予防する方法であって、かかる治療または予防が望まれる被験者に、Notchタンパク質に結合可能な脊椎動物Serrateタンパク質またはその誘導体を治療上有効な量で投与することを含んでなる前記方法。
(項目70)
前記疾病または障害が、類似の非悪性腫瘍サンプルにおけるNotch活性または発現と比べて、高いNotch活性、またはNotchタンパク質または抗Notch抗体によって結合されるNotch誘導体の高い発現によって特徴付けられる悪性腫瘍である項目69に記載の方法。
(項目71)
前記疾病または障害が、子宮頸癌、乳癌、結腸癌、黒色腫、精上皮腫および肺癌からなる群から選ばれる項目69に記載の方法。
(項目72)
前記被験者がヒトである項目69に記載の方法。
(項目73)
前記Serrateタンパク質がヒトSerrateタンパク質である項目69に記載の方法。
(項目74)
被験者の疾病または障害を治療または予防する方法であって、かかる治療または予防が望まれる被験者に、治療上有効な量の分子を投与すること含んでなり、前記分子が、(a)10個のヌクレオチドを含有し、(b)脊椎動物のSerrate遺伝子に特異的なRNA転写の少なくとも10ヌクレオチド部分に完全に相補的な配列を含有し、かつ(c)前記RNA転写物にハイブリダイズ可能なオリゴヌクレオチドである前記方法。
(項目75)
被験者の疾病または障害を治療または予防する方法であって、かかる治療または予防が望まれる被験者に、治療上有効な量の項目34、37または46に記載の核酸を投与することを含んでなる前記方法。
(項目76)
被験者の疾病または障害を治療または予防する方法であって、かかる治療または予防が望まれる被験者に、治療上有効な量の項目32に記載の抗体を投与することを含んでなる前記方法。
(項目77)
前記疾病または障害が中枢神経系の疾病または障害である項目73に記載の方法。
(項目78)
10個のヌクレオチドを含有し、かつ脊椎動物のSerrate遺伝子に特異的なRNA転写物の少なくとも10ヌクレオチド部分に完全に相補的な配列を含有する単離されたオリゴヌクレオチドであって、前記RNA転写物にハイブリダイズ可能である前記オリゴヌクレオチド。
(項目79)
項目78に記載のオリゴヌクレオチドと、製剤上許容される担体とを含有する医薬組成物。
(項目80)
Serrateタンパク質をコードする核酸配列の細胞における発現の抑制方法であって、前記細胞に、有効量の項目78に記載のオリゴヌクレオチドを供給することを含んでなる前記方法。
(項目81)
患者における異常レベルのNotch-Serrateタンパク質の結合活性によって特徴付けられる疾病または障害の診断方法であって、前記患者由来のサンプル中のNotchタンパク質の、脊椎動物Serrateタンパク質への結合能を測定することを含んでなり、正常な個体由来の類似サンプルで見られる結合能と比べて、前記Notchタンパク質の、前記Serrateタンパク質への結合能の増加または減少が、前記患者における前記疾病または障害の存在を示す前記方法。
(項目82)
患者由来のサンプルにおける脊椎動物Serrateタンパク質のレベルを測定することを含んでなる、異常レベルのSerrateタンパク質によって特徴付けられる前記患者における疾病または障害の診断方法であって、正常な個体由来の類似サンプルで見られる前記Serrateタンパク質のレベルと比べて、前記Serrateタンパク質のレベルの増加および減少が、前記患者における前記疾病または障害の存在を示す前記方法。
【0017】
3.1.定義
本明細書においては、何ら強調なしに遺伝子の名前によって示されるタンパク質産物に
対し、遺伝子の名前を強調したもの、つまりイタリック体にしたものまたは下線を引いた
ものは遺伝子を表すものとする。例えば、「Serrate」はSerrate遺伝子を示すものとし、
これに対し「Serrate」はSerrate遺伝子のタンパク質産物を示すものとする。
【図面の簡単な説明】
【0018】
【図1A】Human Serrate-1(Human Jagged-1(HJ1)としても知られている)のヌクレオチド配列(配列番号1)およびタンパク質配列(配列番号2)。
【図1B】Human Serrate-1(Human Jagged-1(HJ1)としても知られている)のヌクレオチド配列(配列番号1)およびタンパク質配列(配列番号2)。
【図1C】Human Serrate-1(Human Jagged-1(HJ1)としても知られている)のヌクレオチド配列(配列番号1)およびタンパク質配列(配列番号2)。
【図1D】Human Serrate-1(Human Jagged-1(HJ1)としても知られている)のヌクレオチド配列(配列番号1)およびタンパク質配列(配列番号2)。
【図1E】Human Serrate-1(Human Jagged-1(HJ1)としても知られている)のヌクレオチド配列(配列番号1)およびタンパク質配列(配列番号2)。
【図1F】Human Serrate-1(Human Jagged-1(HJ1)としても知られている)のヌクレオチド配列(配列番号1)およびタンパク質配列(配列番号2)。
【図1G】Human Serrate-1(Human Jagged-1(HJ1)としても知られている)のヌクレオチド配列(配列番号1)およびタンパク質配列(配列番号2)。
【図2A】ヒト胎児脳cDNAライブラリーより単離したクローンpBS15 およびpBS3-2の配列を組み合わせてコンピュータを用いて作製したHuman Serrate-2(HumanJagged-2(HJ2)としても知られている)の「完全な」ヌクレオチド配列(配列番号3)およびアミノ酸配列(配列番号4)。シグナル配列とSDLドメインの間のHumanSerrate-2 の部分をコードする領域には、約120 ヌクレオチドの欠失がある。
【図2B】ヒト胎児脳cDNAライブラリーより単離したクローンpBS15 およびpBS3-2の配列を組み合わせてコンピュータを用いて作製したHuman Serrate-2(HumanJagged-2(HJ2)としても知られている)の「完全な」ヌクレオチド配列(配列番号3)およびアミノ酸配列(配列番号4)。シグナル配列とSDLドメインの間のHumanSerrate-2 の部分をコードする領域には、約120 ヌクレオチドの欠失がある。
【図2C】ヒト胎児脳cDNAライブラリーより単離したクローンpBS15 およびpBS3-2の配列を組み合わせてコンピュータを用いて作製したHuman Serrate-2(HumanJagged-2(HJ2)としても知られている)の「完全な」ヌクレオチド配列(配列番号3)およびアミノ酸配列(配列番号4)。シグナル配列とSDLドメインの間のHumanSerrate-2 の部分をコードする領域には、約120 ヌクレオチドの欠失がある。
【図2D】ヒト胎児脳cDNAライブラリーより単離したクローンpBS15 およびpBS3-2の配列を組み合わせてコンピュータを用いて作製したHuman Serrate-2(HumanJagged-2(HJ2)としても知られている)の「完全な」ヌクレオチド配列(配列番号3)およびアミノ酸配列(配列番号4)。シグナル配列とSDLドメインの間のHumanSerrate-2 の部分をコードする領域には、約120 ヌクレオチドの欠失がある。
【図2E】ヒト胎児脳cDNAライブラリーより単離したクローンpBS15 およびpBS3-2の配列を組み合わせてコンピュータを用いて作製したHuman Serrate-2(HumanJagged-2(HJ2)としても知られている)の「完全な」ヌクレオチド配列(配列番号3)およびアミノ酸配列(配列番号4)。シグナル配列とSDLドメインの間のHumanSerrate-2 の部分をコードする領域には、約120 ヌクレオチドの欠失がある。
【図2F】ヒト胎児脳cDNAライブラリーより単離したクローンpBS15 およびpBS3-2の配列を組み合わせてコンピュータを用いて作製したHuman Serrate-2(HumanJagged-2(HJ2)としても知られている)の「完全な」ヌクレオチド配列(配列番号3)およびアミノ酸配列(配列番号4)。シグナル配列とSDLドメインの間のHumanSerrate-2 の部分をコードする領域には、約120 ヌクレオチドの欠失がある。
【図2G】ヒト胎児脳cDNAライブラリーより単離したクローンpBS15 およびpBS3-2の配列を組み合わせてコンピュータを用いて作製したHuman Serrate-2(HumanJagged-2(HJ2)としても知られている)の「完全な」ヌクレオチド配列(配列番号3)およびアミノ酸配列(配列番号4)。シグナル配列とSDLドメインの間のHumanSerrate-2 の部分をコードする領域には、約120 ヌクレオチドの欠失がある。
【図3A】ニワトリSerrate(C-Serrate)cDNAのヌクレオチド配列(配列番号5)。
【図3B】ニワトリSerrate(C-Serrate)cDNAのヌクレオチド配列(配列番号5)。
【図4A】C-Serrate(シグナル配列のアミノ末端を欠く)のアミノ酸配列(配列番号6)。シグナル配列(成熟タンパク質の予測されるアミノ末端を示す)に続く推定上の開裂部位を矢印で示す;DSLドメインを星印で示す;EGFに似たリピート(ELR)に破線を施してある;ELRと膜貫通ドメインの間のシステインが豊富な領域を矢印の間に示す;そしてシグナル膜貫通ドメイン(アミノ酸1042と1066の間)を太字で示す。
【図4B】C-Serrate(シグナル配列のアミノ末端を欠く)のアミノ酸配列(配列番号6)。シグナル配列(成熟タンパク質の予測されるアミノ末端を示す)に続く推定上の開裂部位を矢印で示す;DSLドメインを星印で示す;EGFに似たリピート(ELR)に破線を施してある;ELRと膜貫通ドメインの間のシステインが豊富な領域を矢印の間に示す;そしてシグナル膜貫通ドメイン(アミノ酸1042と1066の間)を太字で示す。
【図5】キイロショウジョウバエDelta(配列番号7)およびSerrate(配列番号8)ならびにC-Serrate(配列番号6)のアミノ末端配列のアライメント。示された領域はシグナル配列の末端からDSLドメインの末端に伸びている。DSLドメインが示されている。3つのタンパク質全てにおいて同一のアミノ酸を四角で囲んである。
【図6】C-Serrateと比較したショウジョウバエDeltaおよびショウジョウバエSerrateのドメイン構造を示す図。C-SerrateおよびショウジョウバエSerrateにのみ存在する、EGFリピートのすぐ下流にある第2のシステインに富む領域は、示されていない。疎水性領域を黒で示す;DSLドメインに碁盤模様を付し、そしてEGFに似たリピートに平行線の陰影を付してある。
【発明を実施するための形態】
【0019】
5.発明の詳細な説明
本発明は脊椎動物Serrate遺伝子のヌクレオチド配列、およびそれらによってコードさ
れるタンパク質のアミノ酸配列に関する。本発明はさらに脊椎動物Serrateタンパク質の断片および他の誘導体、および類似体に関する。このような断片または誘導体をコードする核酸もまた本発明の範囲内にある。本発明は多数の異なる種の脊椎動物Serrate遺伝子、およびそれらによってコードされるタンパク質を提供する。本発明のSerrate遺伝子は、ヒトSerrateおよび脊椎動物種における関連する遺伝子(相同体)を含む。特定の実施態様において、Serrate遺伝子およびタンパク質は哺乳動物由来である。本発明の好ましい実施態様において、Serrateタンパク質はヒトタンパク質である。最も好ましい実施態様において、Serrateタンパク質はHumanSerrate-1 またはHuman Serrate-2 である。上記タンパク質および誘導体の例えば組換え法による作製が提供される。
【0020】
本発明は機能的に活性な、すなわち全長(野性型)Serrateタンパク質に関連する1以上の公知の機能活性を示すことができる、本発明の脊椎動物Serrate誘導体および類似体に関する。このような機能活性は抗原性〔抗Serrate抗体に結合する(またはSerrateと結合を競合する)能力〕、免疫原性(Serrateに結合する抗体を生成する能力)、Notchまたは他のトポリズミックなタンパク質またはその断片に結合する(またはSerrateと結合を競合する)能力(「接着性」)、Serrate受容体に結合する(またはSerrateと結合を競合する)能力を含むが、これらだけに限定されない。本明細書に使用する「トポリズミックなタンパク質」とは、Notch、Delta、Serrate、Enhancerof SplitおよびDeltexのタンパク質産物、ならびに、例えばハイブリダイズするそれらの遺伝子配列の能力、またはDelta、Serrateまたは Notchとの相同性、または表現型の相互作用を示す遺伝子の能力によって同定可能な、この相互に作用する遺伝子ファミリーの他のメンバーのタンパク質産物を言う。
【0021】
本発明はさらに、細胞内ドメイン、細胞外ドメイン、膜貫通ドメイン、膜関連領域、ま
たはSerrateタンパク質の1以上のEGFに似た(相同の)リピート、またはこれらの任
意の組合せを含むがこれらだけに限定されない、Serrateタンパク質の1以上のドメイン
を含む脊椎動物Serrateの断片(およびそれらの誘導体および類似体)に関する。
【0022】
Serrateに対する抗体、その誘導体および類似体がさらに提供される。
【0023】
後に示すように、Serrateは発生および他の生理的プロセスにおいて、特にNotchへのリガンドとして重大な役割を果たす。この役割は、細胞の運命(分化)の決定に関与している。特に、Serrateは中枢神経系における細胞の運命の決定において主要な役割を果たすと思われる。分化および他の生理的プロセスの研究および操作において、本発明の核酸およびアミノ酸配列およびそれらに対する抗体を、ヒトおよび他の種のSerratemRNAおよびタンパク質の検出および定量化のために用いて、それらの発現を研究し、Serrateおよび断片および他の誘導体および類似体を作製することができる。本発明はまた、Serrateタンパク質および核酸配列に基づく治療および診断方法、ならびに組成物に関する。本発明は、本発明の治療用化合物を投与することによる細胞の運命(すなわち分化)の疾患に対する治療を提供する。このような治療用化合物(本明細書では「治療剤」と呼ぶ)は、脊椎動物Serrateタンパク質ならびにそれらの類似体および誘導体(断片を含む);それらに対する抗体;脊椎動物Serrateタンパク質、類似体または誘導体をコードする核酸;および脊椎動物Serrateアンチセンス核酸を含む。好ましい実施態様において、本発明の治療剤は癌状態を治療するため、または前腫瘍状態あるいは非悪性腫瘍状態から腫瘍状態または悪性腫瘍状態への進行を阻止するために投与される。他の特定の実施態様において、本発明の治療剤は神経系疾患を治療するため、または組織の再生及び補修を促進するために投与される。
【0024】
1つの実施態様において、Notchおよび/またはSerrate機能に拮抗する、またはこれらを抑制する治療剤(以後「アンタゴニスト治療剤」と称する)が治療的効果のために投与される。別の実施態様においては、Notchおよび/またはSerrate機能を促進する治療剤(以後「アゴニスト治療剤」と称する)が治療的効果のために投与される。
【0025】
Notchおよび/またはSerrateタンパク質の異常な、または望ましくないレベルの発現、
または活性、または局在化を伴う細胞の運命に関する疾患、特に過剰増殖性(例えば癌)
または過少増殖性疾患は、後により詳しく記述するようにそれらのレベルを検出すること
により診断することができる。
【0026】
好ましい側面において、本発明の治療剤は、Notchタンパク質またはその断片への結合
を媒介する脊椎動物Serrateの少なくとも断片(本明細書では「接着性断片」と称する)
からなるタンパク質である。
【0027】
本発明は、なかんずくマウスSerrate相同体のクローニング(第6節)、アフリカツメガエル(カエル)Serrate相同体のクローニング(第7節)、ニワトリSerrate相同体のクローニング(第8節)、およびヒトSerrate相同体であるHuman Serrate-1(HJ1)およびHumanSerrate-2(HJ2)のクローニング(第9節)を開示する後述の実施例によって例証される。
【0028】
開示を明確にするため、限定するためではないが、本発明の詳細な記述を以下の分節に分ける。
【0029】
5.1.Serrate遺伝子の単離
本発明は、脊椎動物Serrate核酸のヌクレオチド配列に関する。特定の実施態様において、脊椎動物Serrate核酸は図1(配列番号1)、図2(配列番号3)、図3(配列番号6)に示されるcDNA配列又はそれらのコード領域、又は脊椎動物Serrateタンパク質をコードする核酸(例えば、配列番号2、4又は6の配列を有する)を含む。
【0030】
本発明は、脊椎動物Serrate配列の少なくとも8個のヌクレオチド(すなわち、ハイブリダイズ可能な部分)からなる核酸を提供する。他の実施態様において、核酸は脊椎動物Serrate配列の少なくとも10個の(連続した)ヌクレオチド、25個のヌクレオチド、50個のヌクレオチド、100個のヌクレオチド、150個のヌクレオチドまたは200個のヌクレオチド、または全長脊椎動物Serrateコード配列からなる。本発明はまた、上記の配列とハイブリダイズ可能な、またはそれらに相補的な核酸に関する。特定の側面において、Serrate遺伝子の少なくとも10、25、50、100または200個のヌクレオチドまたは全コード領域に相補的な配列を含む核酸が提供される。
【0031】
特定の実施態様において、低ストリンジェンシー条件下で脊椎動物Serrate核酸(例えば配列番号1の配列を有する)または脊椎動物Serrate誘導体をコードする核酸とハイブリダイズ可能な核酸が提供される。非限定的な例としては、そのような低ストリンジェンシー条件を用いる方法は以下の通りである(ShiloおよびWeinberg,1981,Proc.Natl.Acad.Sci.USA78:6789-6792 も参照されたい):DNAを含有するフィルターを、35% ホルムアミド、5X SSC、50 mM Tris-HCl(pH7.5)、5mM EDTA、0.1% PVP、0.1% Ficoll、1% BSAおよび500 μg/ml変性サケ精子DNAを含有する溶液中で40℃で6 時間前処理する。同じ溶液に以下の改変を加えて、その中でハイブリダイゼーションを実施する:0.02% PVP、0.02% Ficoll、0.2% BSA、100 μg/ml サケ精子DNA、10%(wt/vol)硫酸デキストラン、および5-20 X106 cpmの32P標識プローブを用いる。ハイブリダイゼーション混合物中でフィルターを40℃で18〜20時間インキュベートし、次に2XSSC、25 mM Tris-HCl(pH7.4)、5 mM EDTAおよび0.1% SDSを含有する溶液を用いて55℃で1.5 時間洗浄する。洗浄溶液を新鮮な溶液と交換し、さらに60℃で1.5時間インキュベートする。フィルターを乾燥状態でブロットし、オートラジオグラフィーのため露光する。必要であれば、65〜68℃でフィルターに三回目の洗浄を施し、フィルムに再度露光する。使用できる他の低ストリンジェンシー条件は当分野で周知である(例えば、交差種(cross-species)ハイブリダイゼーションに用いられる条件)。
【0032】
別の特定の実施態様において、高ストリンジェンシー条件下で脊椎動物Serrate核酸とハイブリダイズ可能な核酸が提供される。非限定的な例としては、そのような高ストリンジェンシー条件を用いる方法は以下の通りである:DNAを含有するフィルターのプレハイブリダイゼーションを65℃で8時間から一晩、6X SSC、50 mM Tris-HCl(pH 7.5)、5 mM EDTA、0.02% PVP、0.02% Ficoll、0.02% BSAおよび500 μg/ml 変性サケ精子DNAからなる緩衝液中で実施する。100 μg/ml 変性サケ精子DNAおよび5-20 X 106cpmの32P標識プローブを含有するプレハイブリダイゼーション混合物中で、フィルターを65℃で48時間ハイブリダイズさせる。フィルターの洗浄は、2X SSC、0.01%PVP、0.01% Ficollおよび0.01% BSA を含有する溶液を用いて37℃で1 時間実施する。次に、オートラジオグラフィーの前に、0.1X SSCを用いて50℃で45分間洗浄する。使用できる他の高ストリンジェンシー条件は当分野で周知である。
【0033】
脊椎動物Serrateタンパク質の断片および誘導体をコードする核酸(第5.6節参照)および脊椎動物Serrateアンチセンス核酸(第5.11節参照)がさらに提供される。容易に明らかなように、本明細書に用いる「Serrateタンパク質の断片または部分をコードする核酸」という表現は、Serrateタンパク質の記述された断片または部分のみをコードする核酸をさすと解釈されるべきであって、連続した配列としてのSerrateタンパク質の他の隣接部分をささないものとする。
【0034】
他のトポリズミックタンパク質との相同領域を含む脊椎動物Serrate核酸の断片もまた提供される。他の種のSerrateタンパク質のDSL領域(ショウジョウバエDeltaおよびSerrateとの相同領域)もまた提供される。DeltaとSerrateの間の保存された領域(配列番号8のSerrateアミノ酸63〜73、124〜134、149〜158、195〜206、214〜219および250〜259、またはDSLドメインによって表される領域、等)をコードする核酸もまた提供される。
【0035】
限定するためではなく特定の例として提示される、脊椎動物Serrate遺伝子クローニン
グの特定の実施態様は以下の通りである。
【0036】
発現クローニング(当分野で良く知られている技法)のため、当分野で公知の方法によ
り発現ライブラリーを構築する。例えば、mRNA(例:ヒトの)を単離し、cDNAを
作製して、宿主細胞によって発現されうるように発現ベクター(例:バクテリオファージ
誘導体)に連結し、次にこのベクターを宿主細胞に導入する。次に、種々のスクリーニン
グアッセイを用いて発現されたSerrate産物を選択することができる。1つの実施態様に
おいて、選択のために抗Serrate抗体を用いることができる。
【0037】
別の好ましい側面においては、選択に先立って、ゲノムまたはcDNAライブラリー中の所望の配列を増幅するためにPCRが用いられる。公知のSerrate配列を表すオリゴヌクレオチドプライマーをPCRプライマーとして用いることができる。好ましい側面において、オリゴヌクレオチドプライマーは、SerrateとDeltaの間で強い相同性を有するSerrate保存セグメント(segment)の少なくとも一部をコードする。合成オリゴヌクレオチドをプライマーとして用いて、興味のある源(RNAまたはDNA)、好ましくはcDNAライブラリーからPCRによって配列を増幅することができる。PCRは、例えばPerkin-ElmerCetusサーマルサイクラーおよびTaq ポリメラーゼ(Gene AmpTM)を用いて実施することができる。増幅されるDNAは、任意の真核生物種由来のmRNAまたはcDNAまたはゲノムDNAを含みうる。PCR反応に用いるため、数個の異なる縮重プライマーの合成を選択することができる。PCR反応を開始させる(priming)ために用いるハイブリダイゼーション条件のストリンジェンシーを変えて、公知のSerrateヌクレオチド配列と単離される核酸相同体との間により大きい又はより小さい度合いの核酸配列類似性をもたらすことも可能である。交差種ハイブリダイゼーションのためには、低ストリンジェンシー条件が好ましい。同種ハイブリダイゼーションのためには、中程度にストリンジェントな条件が好ましい。Serrate相同体の1セグメントを上首尾に増幅した後、このセグメントをクローン化し、配列決定し、そして完全なcDNAまたはゲノムクローンを単離するためのプローブとして用いることができる。次に、このクローンが、後に記述する遺伝子の完全なヌクレオチド配列の決定、遺伝子発現の分析、および機能分析のための遺伝子タンパク質産物の産生を可能とする。このようにして、Serrateタンパク質をコードするさらなる遺伝子を同定することができる。このような方法を、後述の種々の実施例の節において例として提示する。
【0038】
上記の方法は、脊椎動物Serrateのクローンを得る方法についての下記の一般的説明を
制限するものではない。
【0039】
脊椎動物細胞はいずれも可能性としてSerrate遺伝子の分子クローニングのための核酸
供給源となることができる。Serrateをコードする核酸配列はヒト、ブタ、ウシ、ネコ、
トリ、ウマ、イヌ、ならびにさらなる霊長類の供給源、等から単離することができる。例
えば、本発明者らはマウス、アフリカツメガエルおよびヒトの適切な大きさの断片を、c
DNAライブラリーおよびショウジョウバエSerrateプライマーを用いて、PCRにより
増幅した。DNAはクローン化されたDNA(例:DNA「ライブラリー」)から当分野
で公知の標準的方法により、化学合成、cDNAクローニング、または所望の細胞から精
製したゲノムDNAあるいはその断片のクローニングによって得ることができる。(例え
ば、Sambrookら,1989,MolecularCloning,A Laboratory Manual,第2版,Cold Sprin
g HarborLaboratory Press,Cold Spring Harbor,New York; Glover,D.M.(編),1985
,DNACloning: A Practical Approach,MRL Press,Ltd.,Oxford,U.K.,Vol.I,II
参照)。ゲノムDNAから得たクローンは、コード領域に加えて調節およびイントロンD
NA領域を含んでいる可能性がある。cDNAから得たクローンは、エキソン配列のみを
含む。供給源が何であれ、遺伝子の増殖のためには、これを適切なベクター中に分子的に
クローン化しなければならない。
【0040】
ゲノムDNA由来の遺伝子の分子クローニングにおいては、DNA断片が生成され、そ
のうちのあるものが所望の遺伝子をコードしている。種々の制限酵素を用いてDNAを特
定の部位で開裂することができる。または、マンガンの存在下でDNAse を用いてDNAを
断片化することができる。または、例えば超音波処理により、DNAを物理的に剪断する
ことができる。次に、アガロースおよびポリアクリルアミドゲル電気泳動およびカラムク
ロマトグラフィーを含むがこれらだけに限定されない標準的技法により、直鎖状DNA断
片を大きさによって分離することができる。
【0041】
ひとたびDNA断片を生成したならば、所望の遺伝子を含有する特定のDNA断片の同
定を種々の方法で実施することができる。例えば、(任意の種の)Serrate遺伝子または
その特定のRNA、またはその断片(例えば細胞外ドメイン、第5.6 節参照)が入手可能
で、精製および標識することができる場合、生成されたDNA断片を標識化プローブとの
核酸ハイブリダイゼーションによりスクリーニングすることができる(Benton,W.およびD
avis,R.,1977,Science196:180; Grunstein,M.およびHogness,D.,1975,Proc.Na
tl.Acad.Sci.USA72:3961)。プローブに実質的相同性を有するDNA断片はハイブリ
ダイズする。また、制限酵素消化、および公知の制限地図(もしそのようなものがあるな
らば)による断片サイズの期待されるサイズとの比較によって適切な断片を同定すること
も可能である。さらに、遺伝子の特性に基づいて選択を実施することができる。または、
遺伝子の発現産物の物理的、化学的または免疫学的特性に基づくアッセイによって該遺伝
子の存在を検出することができる。例えば、Serrateについて公知な電気泳動における移
動、等電点電気泳動挙動、加水分解消化地図、受容体結合活性、in vitro凝集活性(「接
着性」)または抗原特性に類似した、または同一のものを有するタンパク質を産生するc
DNAクローン、または適切なmRNAをハイブリッド選択(hybrid-select)するDNA
クローンを選択することができる。Serrateに対する抗体が使用可能な場合は、ELIS
A(酵素結合イムノソルベントアッセイ)型手順を用いて、標識化抗体を推定上のSerrat
e合成クローンと結合させることによりSerrateタンパク質を同定することができる。
【0042】
Serrate遺伝子は、核酸ハイブリダイゼーションによるmRNA選択とそれに続くin vitro翻訳によって同定することも可能である。この方法では、断片を用いてハイブリダイゼーションにより相補的mRNAを単離する。このようなDNA断片は、入手可能な、別の種(例:ヒト、ニワトリ)の精製SerrateDNAを表してもよい。単離されたmRNAの単離された産物のinvitro翻訳産物の免疫沈降分析または機能分析(例えば、in vitroでの凝集能;受容体との結合;後記参照)は、mRNAを同定し、そしてその結果、所望の配列を含む相補的DNA断片を同定する。さらに、Serrateタンパク質に対して特異的に誘導された固定化抗体への細胞から単離されたポリソームの吸着により、特定のmRNAを選択することができる。(吸着されたポリソームから)選択されたmRNAを鋳型として用いて、放射性標識化SerratecDNAを合成することができる。次に、放射性標識化mRNAまたはcDNAをプローブとして用いて、他のゲノムDNA断片の中からSerrateDNA断片を同定することができる。
【0043】
SerrateゲノムDNAの単離に代わるものとしては、公知の配列から遺伝子配列それ自
体を化学的に合成すること、またはSerrateタンパク質をコードするmRNAに対するc
DNAを作製すること、が含まれるが、これらだけに限定されない。例えば、Serrate遺
伝子のcDNAクローニングのためのRNAを、Serrateを発現する細胞から単離するこ
とができる。他の方法も可能であり、それらは本発明の範囲内にある。
【0044】
次に、同定され単離された遺伝子を適切なクローニングベクターに挿入することができる。当分野で公知の多数のベクター-宿主系が使用できる。使用できるベクターはプラスミドまたは改変されたウイルスを含むがそれらだけに限定されない。また、ベクター系は用いられる宿主細胞と適合性のものでなければならない。そのようなベクターは、ラムダ誘導体等のバクテリオファージ、PBR322またはpUCプラスミド誘導体等のプラスミドを含むがそれらだけに限定されない。クローニングベクターへの挿入は、例えばDNA断片を相補的付着末端を有するクローニングベクターに連結することにより達成できる。しかし、DNAを断片化するのに用いられた相補的制限部位がクローニングベクターに存在しない場合は、DNA分子の末端を酵素的に改変することができる。または、ヌクレオチド配列(リンカー)をDNA末端に連結することにより所望の任意の部位を作製することができる。このような連結されたリンカーは、制限エンドヌクレアーゼ認識配列をコードする特定の化学的に合成されたオリゴヌクレオチドを含むことができる。別の方法では、開裂したベクターおよびSerrate遺伝子をホモポリマー的テーリング(tailing)により改変することができる。遺伝子配列の多数のコピーが生成されるように、組換え分子を形質転換、トランスフェクション、感染、エレクトロポレーション、等により宿主細胞に導入することができる。
【0045】
別の方法においては、所望の遺伝子を「ショットガン」法により適切なクローニングベ
クターに挿入した後に同定および単離することができる。クローニングベクターに挿入す
る前に、例えばサイズ分画による所望の遺伝子の富化が実施できる。
【0046】
特定の実施態様において、単離されたSerrate遺伝子、cDNA、または合成DNA配
列を組み込んだ組換えDNA分子を用いた宿主細胞の形質転換は、遺伝子の多数コピーの
生成を可能とする。したがって、形質転換体を増殖させ、該形質転換体から組換えDNA
分子を単離し、そして必要であれば単離された組換えDNAから挿入遺伝子を回収するこ
とにより、遺伝子を大量に得ることができる。
【0047】
本発明により提供されるSerrate配列は、天然のSerrateタンパク質に見いだされるアミノ酸配列と実質的に同一のアミノ酸配列をコードする核酸配列、および機能的に等価のアミノ酸を有するコードされたアミノ酸配列を包含する。これらすべてはSerrate誘導体に関する第5.6節(後述)に記述されている。
【0048】
5.2.Serrate遺伝子の発現
脊椎動物Serrateタンパク質またはその機能的に活性な断片または他の誘導体(第5.6節参照)をコードするヌクレオチド配列を適切な発現ベクター、すなわち、挿入されたタンパク質コード配列の転写および翻訳に必要な要素を含有するベクターに挿入することができる。必要な転写および翻訳シグナルは、天然の脊椎動物Serrate遺伝子および/またはそのフランキング領域によって供給することもできる。種々の宿主−ベクター系を用いてタンパク質コード配列を発現することができる。これらは、ウイルス(例:ワクシニアウイルス、アデノウイルス、等)によって感染させた哺乳動物細胞系;ウイルス(例:バキュロウイルス)によって感染させた昆虫細胞系;酵母ベクターを含有する酵母等の微生物;またはバクテリオファージ、DNA、プラスミドDNAまたはコスミドDNAによって形質転換した細菌を含むが、それらだけに限定されない。ベクターの発現要素は、その強度および特異性において異なる。使用する宿主−ベクター系により、多数の適切な転写および翻訳要素のうち任意のものを用いることができる。特定の実施態様において、Serrate遺伝子の接着性部分が発現される。別の特定の実施態様において、HumanSerrate遺伝子またはヒトSerrate遺伝子の機能的に活性な部分をコードする配列(Human Serrate-1(HJ1)またはHumanSerrate-2(HJ2)、等)が発現される。さらに別の実施態様において、細胞外ドメインを含むSerrateの断片、または他の誘導体、またはSerrateの類似体が発現される。
【0049】
DNA断片のベクターへの挿入のために以前に記述された任意の方法を用いて、適切な転写/翻訳制御シグナルおよびタンパク質コード配列からなるキメラ遺伝子を含有する発現ベクターを構築することができる。これらの方法は、in vitro組換えDNAおよび合成技法およびin vivo組換え体(遺伝子組換え)を含みうる。Serrateタンパク質またはペプチド断片をコードする核酸配列の発現は、組換えDNA分子によって形質転換された宿主中で該Serrateタンパク質またはペプチドが発現されるように、第2の核酸配列によって調節することができる。例えば、当分野で公知の任意のプロモーター/エンハンサー要素によってSerrateタンパク質の発現を制御することができる。トポリズミックな遺伝子発現を制御するために使用できるプロモーターは以下のものを含むがそれらだけに限定されない。すなわち、SV40初期プロモーター領域(BernoistおよびChambon,1981,Naure 290:304-310)、ラウス肉腫ウイルスの3'長末端リピートに含まれるプロモーター(Yamamotoら,1980,Cell22:787-797)、ヘルペスチミジンキナーゼプロモーター(Wagnerら,1981,Proc.Natl.Acad.Sci.USA78:1441-1445)、メタロチオネイン遺伝子の調節配列(Brinster ら,1982,Nature 296:39-42)、β-ラクタマーゼプロモーター(Villa-Kamaroffら,1978,Proc.Natl.Acad.Sci.USA 75:3727-3731)等の原核細胞発現ベクター、またはtac プロモーター(DeBoer ら,1983,Proc.Natl.Acad.Sci.USA80:21-25); Scientific American,1980,242:74-94「組換え細菌由来の有用なタンパク質」をも参照; ノパリンシンセターゼプロモーター領域を含む植物発現ベクター(Herrera-Estrellaら,Nature303:209-213)またはカリフラワーモザイクウイルス 35S RNAプロモーター(Gardnerら,1981,Nucl.Acids Res.9:2871)、および光合成酵素リブロースビスリン酸カルボキシラーゼのプロモーター(Herrera-Estrellaら,1984,Nature310:115-120);酵母または他の真菌由来のプロモーター要素、例えばGal 4プロモーター、ADC(アルコールデヒドロゲナーゼ)プロモーター、PGK(ホスホグリセロールキナーゼ)プロモーター、アルカリホスファターゼプロモーター、および組織特異性を示し、トランスジェニック動物に使用されてきた下記の動物転写制御領域:膵臓腺房細胞において活性なエラスターゼI遺伝子制御領域(Swiftら,1984,Cell38:639-646; Ornitzら,1986,Cold Spring Harbor Symp.Quant.Biol.50:399-409;MacDonald,1987,Hepatology 7:425-515); 膵臓ベータ細胞において活性なインスリン遺伝子制御領域(Hanahan,1985,Nature315:115-122)、リンパ系細胞において活性な免疫グロブリン遺伝子制御領域(Grosschedlら,1984,Cell 38:647-658;Adamesら,1985,Nature 318:533-538; Alexanderら,1987,Mol.Cell.Biol.7:1436-1444)、精巣、乳房、リンパ系およびマスト細胞において活性なマウス乳腺癌ウイルス制御領域(Lederら,1986,Cell45:485-495)、肝臓において活性なアルブミン遺伝子制御領域(Pinkertら,1987,Genes and Devel.1:268-276)、肝臓において活性なαフェトプロテイン遺伝子制御領域(Krumlaufら,1985,Mol.Cell.Biol.5:1639-1648;Hammerら,1987,Science 235:53-58)、肝臓において活性なα 1-アンチトリプシン遺伝子制御領域(Kelseyら,1987,Genesand Devel.1:161-171)、骨髄様細胞において活性なβグロビン遺伝子制御領域(Mogramら,1985,Nature 315:338-340;Kolliasら,1986,Cell 46:89-94)、脳の稀突起膠細胞において活性なミエリン塩基性タンパク質遺伝子制御領域(Readheadら,1987,Cell48:703-712)、骨格筋において活性なミオシンL鎖2遺伝子制御領域(Sani,1985,Nature 314:283-286)および視床下部において活性なゴナドトロピン放出ホルモン遺伝子制御領域(Masonら,1986,Science234:1372-1378)である。
【0050】
Serrate遺伝子挿入配列を有する発現ベクターは、3つの一般的アプローチによって同
定することができる。すなわち、(a)核酸ハイブリダイゼーション、(b)「マーカー
」遺伝子機能の存在または不在、および(c)挿入配列の発現、である。第1のアプロー
チでは、挿入されたトポリズミックな遺伝子に相同な配列を含むプローブを用いた核酸ハ
イブリダイゼーションによって、発現ベクターに挿入された外来遺伝子の存在を検出する
ことができる。第2のアプローチでは、ベクターへの外来遺伝子の挿入によってもたらさ
れる特定の「マーカー」遺伝子機能(例えば、チミジンキナーゼ活性、抗生物質耐性、形
質転換表現型、バキュロウイルスにおける閉鎖体の形成、等)の存在または不在に基づい
て、組換えベクター/宿主系を同定および選択することができる。例えば、Serrate遺伝
子がベクターのマーカー遺伝子配列内に挿入されると、Serrate挿入配列を有する組換え
体はマーカー遺伝子機能の不在によって同定することができる。第3のアプローチでは、
組換え発現ベクターは、組換え体によって発現された外来遺伝子産物をアッセイすること
により同定することができる。このようなアッセイは、in vitroアッセイ系において、例
えばSerrate遺伝子産物の物理的または機能的特性(例えば、Notchとの凝集(結合)、受容体との結合、抗体との結合)に基づくことができる。
【0051】
特定の組換えDNA分子がひとたび同定され単離されたならば、当分野で公知の幾つか
の方法を用いてそれを増殖させることができる。ひとたび適切な宿主系および増殖条件が
確立されたならば、組換え発現ベクターを大量に増殖させ、調製することができる。前に
説明したように、使用できる発現ベクターは下記のベクターまたはその誘導体を含むが、
それらだけに限定されない。すなわち、2〜3名を挙げるならば、ワクシニアウイルスま
たはアデノウイルス等のヒトまたは動物ウイルス;バキュロウイルス等の昆虫ウイルス;
酵母ベクター;バクテリオファージベクター(例えばラムダ)、およびプラスミドおよび
コスミドDNAベクターである。
【0052】
さらに、挿入配列の発現を変調する、または所望の特定の様式で遺伝子産物を改変しプ
ロセシングする、宿主細胞株を選択することができる。特定のプロモーターからの発現は
、特定の誘導物質(inducer)の存在下で増大させることができる。したがって、遺伝子工
学的に作製されたSerrateタンパク質の発現を制御することができる。さらに、異なる宿
主細胞は、タンパク質の翻訳および翻訳後プロセシングおよび修飾(例えば、シグナル配
列等のグリコシル化、開裂)のための特徴的で特異的な作用機構を有する。適切な細胞系
または宿主系を選択して、発現された外来タンパク質の所望の修飾およびプロセシングを
確実にすることができる。例えば、細菌系における発現を用いて非グリコシル化コアタン
パク質産物を産生することができる。酵母における発現は、グリコシル化産物を産生する
。哺乳動物細胞における発現を用いて、異種哺乳動物トポリズミックタンパク質の「天然
の」グリコシル化を確実にすることができる。さらに、異なるベクター/宿主発現系は加
水分解開裂等のプロセシング反応に異なる程度の影響を及ぼしうる。
【0053】
他の特定の実施態様において、Serrateタンパク質、断片、類似体、または誘導体を融
合またはキメラタンパク質産物〔ペプチド結合を介して(異なるタンパク質の)異種タン
パク質配列に結合した上記タンパク質、断片、類似体、または誘導体を含む〕として発現
させることができる。このようなキメラ産物は、所望のアミノ酸配列をコードする適切な
核酸配列を当分野で公知の方法により適切なコードフレーム内で相互に連結し、そして該
キメラ産物を当分野で周知の方法により発現させることにより、作製することができる。
または、そのようなキメラ産物は例えばペプチド合成機を用いて、タンパク質合成技法に
より作製することができる。
【0054】
cDNA配列およびゲノム配列の両者ともクローン化し、発現させることが可能である
。
【0055】
5.3.Serrate遺伝子産物の同定および精製
特定の側面において、本発明は脊椎動物Serrate(好ましくはヒトSerrate相同体)、および抗原決定基を含む(すなわち、抗体によって認識されうる)、または機能的に活性なその断片および誘導体のアミノ酸配列、ならびにそれらをコードする核酸配列を提供する。本明細書に用いる「機能的に活性な」物質とは、全長(野性型)Serrateタンパク質に関連する1以上の公知の機能的活性〔例えば、Notchまたはその一部との結合、他の任意のSerrateリガンドとの結合、抗原性(抗Serrate抗体との結合)、等〕を示す物質をいう。
【0056】
特定の実施態様において、本発明は少なくとも6アミノ酸、10アミノ酸、25アミノ酸、50アミノ酸、または少なくとも75アミノ酸からなる脊椎動物Serrateタンパク質の断片を提供する。他の実施態様において、本発明のタンパク質はSerrateタンパク質の細胞外ドメイン、DSLドメイン、表皮増殖因子に似たリピート(ELR)ドメイン、ELRの1つまたは任意の組合せ、システインに富む領域、膜貫通ドメイン、または細胞内(細胞質)ドメイン、またはNotchに結合する部分、または上記のものの任意の組合せを含む、または本質的にそれ(ら)から成る。脊椎動物Serrateタンパク質の上記領域の幾つか又は全部を欠く断片、または断片を含むタンパク質もまた提供される。上記のタンパク質および断片をコードする核酸が提供される。
【0057】
脊椎動物Serrate遺伝子配列を発現する組換え体がひとたび同定されたならば、遺伝子
産物を分析することができる。これは、該産物を放射性標識し、次にゲル電気泳動、イム
ノアッセイ、等により分析することを含む、産物の物理的または機能的特性に基づくアッ
セイによって達成することができる。
【0058】
Serrateタンパク質がひとたび同定されたならば、クロマトグラフィー(例えば、イオ
ン交換、アフィニティーおよびサイジングカラムクロマトグラフィー)、遠心、示差溶解
性を含む標準的方法、またはタンパク質精製のための他の標準的技法により、該タンパク
質を単離し、精製することができる。機能的特性は、任意の適切なアッセイを用いて評価
することができる(第5.7 節参照)。
【0059】
または、組換え体によって産生されたSerrateタンパク質がひとたび同定されたならば
、組換え体に含有されているキメラ遺伝子の核酸配列から上記タンパク質のアミノ酸配列
を推定することができる。その結果、当分野で公知の標準的な化学的方法により該タンパ
ク質を合成することができる(例えば、Hunkapiller,M.ら,1984,Nature 310:105-111
参照)。
【0060】
本発明の特定の実施態様において、組換えDNA技法により産生されたものであれ、化
学合成法により作製されたものであれ、そのようなSerrateタンパク質は一次アミノ酸配
列として図1、2または3(それぞれ配列番号2、4または6)に実質的に示すアミノ酸
配列の全部または一部を含むタンパク質、ならびにそれらの断片および他の誘導体および
類似体を包含するが、それらだけに限定されない。
【0061】
5.4.Serrate遺伝子およびタンパク質の構造
Serrate遺伝子およびタンパク質の構造を、当分野で公知の種々の方法によって分析す
ることができる。
5.4.1.遺伝子分析
脊椎動物Serrate遺伝子に対応するクローン化DNAまたはcDNAを、サザンハイブリダイゼーション(Southern,E.M.,1975,J.Mol.Biol.98:503-517)、ノーザンハイブリダイゼーション(例えば、Freemanら,1983,Proc.Natl.Acad.Sci.USA80:4094-4098参照)、制限エンドヌクレアーゼマッピング(Maniatis,T.,1982,Molecular Cloning,ALaboratory Manual,Cold Spring Harbor,N.Y.)およびDNA配列分析を含むがこれらだけに限定されない方法によって分析することができる。ポリメラーゼチェーンリアクション(PCR;米国特許第4,683,202、4,683,195および4,889,818号;Gyllensteinら,1988,Proc.Natl.Acad.Sci.USA 85:7652-7656; Ochmanら,1988,Genetics120:621-623; Loh ら,1989,Science 243:217-220)およびその後のSerrate特異的プローブを用いたサザンハイブリダイゼーションは、種々の細胞型由来のDNA中におけるSerrate遺伝子の検出を可能とする。PCR以外の増幅方法もよく知られており、それらも採用することができる。1つの実施態様において、サザンハイブリダイゼーションを用いてSerrateの遺伝子的連結を確認することができる。ノーザンハイブリダイゼーション分析を用いてSerrate遺伝子の発現を確認することができる。種々の発生または活性状態にある種々の細胞型をSerrate発現に関して試験することができる。そのような技法およびそれらの結果の例を後出の第6節に記述する。サザンおよびノーザンハイブリダイゼーションの両方についてのハイブリダイゼーション条件のストリンジェンシーは、用いられた特定のSerrateプローブに対して所望の程度の関連性を有する核酸の検出を確実にするように操作することができる。
【0062】
制限エンドヌクレアーゼマッピングを用いてSerrate遺伝子の遺伝子構造を大まかに決
定することができる。特定の実施態様において、制限酵素による開裂を用いて後出の図2
に示す制限地図を引き出すことができる。制限エンドヌクレアーゼ開裂によって引き出さ
れる制限地図は、DNA配列分析によって確認することができる。
【0063】
DNA配列分析は、マクサム・ギルバート法(1980,Meth.Enzyml.65:499-560)、サンガージデオキシ法(Sanger,F.ら,1977,Proc.Natl.Acad.Sci.USA74:5463)、T7 DNAポリメラーゼの使用(TaborおよびRichardson,米国特許第4,795,699号)または自動DNAシークエネーター(例えば、AppliedBiosystems,Foster City,CA)の使用を含むが、それらだけに限定されない当分野で公知の任意の技法により実施することができる。代表的Serrate遺伝子のcDNA配列は図1および2に実質的に示す配列を含み、そして後出の第9節に記述されている。
【0064】
5.4.2.タンパク質分析
Serrateタンパク質のアミノ酸配列は、DNA配列からの推定により、または例えば自
動アミノ酸シークエンサーを用いた上記タンパク質の直接配列決定により、引き出すこと
ができる。代表的Serrateタンパク質のアミノ酸配列は図1に実質的に示す配列を含み、
そして、アミノ酸番号30〜1219によって示される代表的成熟タンパク質と共に後出の第9節に詳述されている。
【0065】
Serrateタンパク質配列を親水性分析(Hopp,T.およびWoods,K.,1981,Proc.Natl.
Acad.Sci.USA78:3824)によってさらに特徴付けることができる。親水性プロフィール
を用いて、Serrateタンパク質の疎水性および親水性領域、およびそのような領域をコー
ドする遺伝子配列の対応する領域を同定することができる。
【0066】
第2に、構造分析(Chou,P.およびFasman,G.,1974,Biochemistry13:222)を実施し
て、特異的二次構造を取るSerrateの領域を同定することができる。
【0067】
操作、翻訳、および二次構造の予測、ならびにオープンリーディングフレームの予測お
よびプロッティングもまた、当分野で利用可能なコンピュータソフトウエアプログラムを
用いて達成することができる。
【0068】
構造分析の他の方法もまた採用することができる。これらは、X線結晶学(Engstom,A.
,1974,Biochem.Exp.Biol.11:7-13)およびコンピュータモデリング(Fletterick,R.
およびZoller,M.(編),1986,「コンピュータグラフィックおよび分子モデリング」
,CurrentCommunications in Molecular Biology,Cold Spring Harbor Laboratory,Co
ld SpringHarbor,New York)を含むが、これらだけに限定されない。
5.5.Serrateタンパク質およびその誘導体に対する抗体の生成
本発明によれば、脊椎動物Serrateタンパク質、その断片または他の誘導体、またはその類似体を免疫原として用いて、そのような免疫原を認識する抗体を生成することができる。そのような抗体は、ポリクローナル、モノクローナル、キメラ、一本鎖、Fab断片およびFab 発現ライブラリーを含むが、これらだけに限定されない。特定の実施態様において、ヒトSerrateに対する抗体が産生される。別の実施態様において、Serrateの細胞外ドメインに対する抗体が産生される。別の実施態様において、Serrateの細胞内ドメインに対する抗体が産生される。
【0069】
Serrateタンパク質または誘導体または類似体に対するポリクローナル抗体の産生のた
めに、当分野で公知の種々の方法が使用できる。特定の実施態様において、図1に示す配
列またはそのサブ配列によってコードされるSerrateタンパク質のエピトープに対するウ
サギポリクローナル抗体を得ることができる。抗体の産生のため、天然のSerrateタンパ
ク質、またはその合成物、またはその誘導体(例:断片)を注射することにより、ウサギ
、マウス、ラット、等を含むがこれらだけに限定されない種々の宿主動物を免疫感作する
ことができる。宿主の種によって、免疫応答を増大させるため、種々のアジュバントを用
いることができる。これらのアジュバントには以下のものが含まれるが、それらだけに限
定されない。すなわち、フロイントアジュバント(完全および不完全)、水酸化アルミニ
ウム等のミネラルゲル、リゾレシチン、プルロニックポリオール等の界面活性剤、ポリア
ニオン、ペプチド、オイルエマルジョン、スカシ貝ヘモシアニン、ジニトロフェノール、
およびBCG(カルメット−ゲラン杆菌)およびコリネバクテリウム・パルバム(Coryne
bacteriumparvum)等の可能性として有用なヒトアジュバントである。
【0070】
脊椎動物Serrateタンパク質配列またはその類似体に向けられたモノクローナル抗体の調製のため、培養されている連続的継代細胞系による抗体分子の産生を提供する任意の技法を用いることができる。例えば、KohlerおよびMilsteinによって最初に開発されたハイブリドーマ技法(Nature,1975,256:495-497)、ならびにトリオーマ(trioma)技法、ヒトB細胞ハイブリドーマ技法(Kozborら,1983,ImmunologyToday,4:72)およびヒトモノクローナル抗体を産生するEBV−ハイブリドーマ技法(Coleら,1985,Monoclonal Antibodiesand Cancer Therapy,Alan R.Liss,Inc.,pp.77-96)である。本発明の付加的実施態様においては、最近の技術(PCT/US90/02545)を用いて、無菌動物においてモノクローナル抗体を産生することができる。本発明によれば、ヒト抗体が使用可能であり、そしてこれはヒトハイブリドーマ(Coteら,1983,Proc.Natl.Acad.Sci.USA80:2026-2030)を用いて、またはEBVウイルスを用いてin vitroでヒトB細胞を形質転換して(Coleら,1985,MonoclonalAntibodies and Cancer Therapy,Alan R.Liss,pp.77-96)得ることができる。実際、本発明によれば、Serrateに特異的なマウス抗体分子由来の遺伝子を適切な生物活性を有するヒト抗体分子由来の遺伝子と共にスプライシングすることにより「キメラ」抗体(Morrisonら,1984,Proc.Natl.Acad.Sci.USA81:6851-6855; Neubergerら,1984,Nature,312:604-608; Takedaら,1985,Nature,314:452-454)を産生するために開発された技法を用いることができる。そのような抗体は本発明の範囲内にある。
【0071】
本発明によれば、一本鎖抗体(米国特許第4,946,778号)を産生するために記述された
技法を、Serrate特異的一本鎖抗体の産生のために適合させることができる。本発明の付
加的実施態様は、Fab発現ライブラリー(Huseら,1989,Science 246:1275-1281)の構築
のために記述された技法を用いて、Serrateタンパク質、誘導体または類似体に対し所望
の特異性を有するモノクローナルFab 断片の迅速で容易な同定を可能とする。
【0072】
分子のイディオタイプを含有する抗体断片を公知の技法により生成することができる。
例えば、そのような断片は抗体分子のペプシン消化により生成されるF(ab')2断片;F(ab'
)2断片のジスルフィド架橋を還元することにより生成されるFab'断片、および抗体分子を
パパインおよび還元剤で処理することにより生成されるFab 断片を含むが、これらだけに
限定されない。
【0073】
抗体の産生において、所望の抗体のスクリーニングは、当分野で公知の技法、例えばELISA(酵素結合イムノソルベントアッセイ)により達成することができる。例えば、Serrateタンパク質の特定ドメインを認識する抗体を選択するため、そのようなドメインを有するSerrate断片と結合する産物について、生成されたハイブリドーマをアッセイすることができる。脊椎動物(例:ヒト)Serrateに特異的な抗体を選択するためには、脊椎動物Serrateとの陽性結合およびショウジョウバエSerrateとの結合の欠如に基づいて選択することができる。別の実施態様においては、ヒトSerrateとの結合、および他の種のSerrateとの不結合によって選択することができる。
【0074】
上記の抗体は、本発明のタンパク質配列(例えば、後出の第5.7 節参照)の局在化と活
性に関する当分野で公知の方法に、例えば診断法等においてそれらのタンパク質のイメー
ジイングのため、適当な生理的サンプル中のそれらのレベルを測定するため、に用いるこ
とができる。
【0075】
Serrateタンパク質のあるドメインに特異的な抗体もまた提供される。特定の実施態様において、SerrateのNotch結合断片に結合する抗体が提供される。
【0076】
本発明の別の実施態様(後出参照)において、抗Serrate抗体および結合ドメインを有
するその断片は治療剤である。
5.6.Serrateタンパク質、誘導体および類似体
本発明はさらに脊椎動物Serrateタンパク質、およびSerrateタンパク質の誘導体(断片を含むがそれだけに限定されない)および類似体に関する。脊椎動物Serrateタンパク質誘導体およびタンパク質類似体をコードする核酸もまた提供される。1つの実施態様において、Serrateタンパク質は後出の第5.1節に記述される脊椎動物Serrate核酸によってコードされる。特定の側面において、上記タンパク質、誘導体または類似体はカエル、マウス、ラット、ブタ、ウシ、イヌ、サル、またはヒトSerrateタンパク質である。
【0077】
脊椎動物Serrateに関連する誘導体および類似体の作製および使用は本発明の範囲内にある。特定の実施態様において、上記誘導体または類似体は機能的に活性である、すなわち全長、野性型Serrateタンパク質に関連する1以上の機能的活性を示すことができる。1例としては、所望の免疫原性または抗原性を有するそのような誘導体または類似体は、例えばイムノアッセイにおいて、免疫感作のため、Serrate活性の抑制のため、等に用いることができる。所望のSerrate特性(例えば、Notchまたは他のトポリズミックなタンパク質との結合、細胞表面受容体との結合)を保持する、またはこれを抑制する、そのような分子は、それぞれそのような特性およびその生理的相関物の誘導物質またはインヒビターとして用いることができる。特定の実施態様は、抗Serrate抗体によって結合されうるが、Notchタンパク質または他のトポリズミックなタンパク質と結合しないSerrate断片に関する。Serrateの誘導体または類似体は、第5.7節に記載のアッセイを含むがそれらだけに限定されない当分野で公知の方法によって、所望の活性について試験することができる。
【0078】
特に、Serrate誘導体は、機能的に等価な分子をもたらす置換、付加または欠失によりSerrate配列を変更することによって作製することができる。ヌクレオチドコード配列の縮重により、Serrate遺伝子として実質的に同一のアミノ酸配列をコードする他のDNA配列が本発明の実施において使用できる。これらのDNA配列には以下のものが含まれるが、それらだけに限定されない。すなわち、配列内で機能的に等価なアミノ酸残基をコードする異なるコドンとの置換によって変更された、その結果表に現れない変化(silentchange)を生じているSerrate遺伝子の全部または一部を含むヌクレオチド配列である。同様に、本発明のSerrate誘導体は、一次アミノ酸配列として変更された配列(配列内で機能的に等価なアミノ酸残基が置換され、表に現れない変化を生じている)を含むSerrateタンパク質のアミノ酸配列の全部または一部を含むものを包含するが、それらだけに限定されない。例えば、配列内の1以上のアミノ酸残基を、機能的等価物として作用する類似の極性を有する別のアミノ酸に置換することが可能で、これは表に現れない変更をもたらす。配列内のアミノ酸の置換物(Substitute)は、当該アミノ酸が属するクラスの他のメンバーから選択することができる。例えば、非極性(疎水性)アミノ酸はアラニン、ロイシン、イソロイシン、バリン、プロリン、フェニルアラニン、トリプトファンおよびメチオニンを含む。極性中性アミノ酸は、グリシン、セリン、トレオニン、システイン、チロシン、アスパラギン、およびグルタミンを含む。正に荷電した(塩基性)アミノ酸は、アルギニン、リシンおよびヒスチジンを含む。負に荷電した(酸性)アミノ酸は、アスパラギン酸およびグルタミン酸を含む。
【0079】
本発明の特定の実施態様において、Serrateタンパク質の少なくとも10個の(連続した)アミノ酸からなる脊椎動物Serrateタンパク質の断片からなる、またはそれを含むタンパク質が提供される。他の実施態様において、上記断片はSerrateタンパク質の少なくとも20個または50個のアミノ酸からなる。特定の実施態様において、そのような断片は35、100または200アミノ酸より小さい。脊椎動物Serrateの誘導体または類似体は、脊椎動物Serrateまたはその断片に実質的に相同な(例えば、同じ大きさのアミノ酸配列にわたって少なくとも30%同一)ペプチド、またはそれをコードする核酸が脊椎動物Serrateのコード配列とハイブリダイズ可能であるペプチドを含むが、それらだけに限定されない。
【0080】
本発明のSerrate誘導体および類似体は当分野で公知の種々の方法により作製すること
ができる。それらの作製をもたらす操作は、遺伝子またはタンパク質レベルで起こりうる
。例えば、クローン化Serrate遺伝子配列を当分野で公知の多数の戦略のうち任意のもの
により改変することができる(Maniatis,T.,1990,Molecular Cloning,A Laboratory M
anual,第2版,ColdSprings Harbor Laboratory,Cold Spring Harbor,New York)。上
記配列を制限エンドヌクレアーゼを用いて適切な部位で開裂し、次に所望であればさらに
酵素的に改変し、そして単離してin vitroで連結することができる。Serrateの誘導体ま
たは類似体をコードする遺伝子の作製においては、改変された遺伝子が、所望のSerrate
活性がコードされている遺伝子領域において、翻訳停止シグナルによって中断されること
なく、Serrateと同じ翻訳リーディングフレーム内に確実に留まるように注意しなければ
ならない。
【0081】
さらに、Serrateをコードする核酸配列をinvitroまたはin vivoで突然変異させて、翻訳開始および/または停止配列を創出および/または破壊し、またはコード領域に変異を創出し、および/または新たな制限エンドヌクレアーゼ部位を形成し、または既存のそのような部位を破壊し、さらなるinvitro改変を容易にすることができる。当分野で公知の突然変異誘発のための任意の技法を用いることができる。これらの技法は、in vitro部位特異的突然変異誘発(Hutchinson,C.ら,1978, J. Biol. Chem. 253:6551)、TAB(登録商標)リンカー(Pharmacia)の使用、等を含むが、それらだけに限定されない。
【0082】
Serrate 配列の操作をタンパク質レベルで実施することもできる。本発明の範囲内に含
まれるものは、例えば、グリコシル化、アセチル化、リン酸化、アミド化、既知の保護/
遮断基による誘導体化、タンパク質分解切断、抗体分子または他の細胞リガンドへの連結
、その他によって、翻訳中もしくは翻訳後に異なる修飾を受けたSerrate タンパク質断片
またはその他の誘導体もしくは類似体である。既知の手法による多数の化学的修飾のどれ
を実施してもよく、限定するわけではないが、臭化シアン、トリプシン、キモトリプシン
、パパイン、V8プロテアーゼ、NaBH4による特異的化学切断;アセチル化、ホルミル
化、酸化、還元;ツニカマイシンの存在下での代謝合成;その他が含まれる。
【0083】
その他に、Serrateの類似体および誘導体を化学的に合成することもできる。例えば、ペプチド合成装置の使用によって、所望のドメイン(第5.6.1節参照)を含んでいる、またはinvitroで所望の凝集活性を仲介する、あるいは受容体に結合するSerrate タンパク質の一部分に相当するペプチドを合成することができる。さらに、所望ならば、Serrate配列中に、置換または付加によって非古典的アミノ酸または化学的アミノ酸類似体を導入することもできる。非古典的アミノ酸として、限定するわけではないが、通常のアミノ酸のD−異性体、α−アミノイソ酪酸、4−アミノ酪酸、ヒドロキシプロリン、サルコシン、シトルリン、システイン酸、t−ブチルグリシン、t−ブチルアラニン、フェニルグリシン、シクロヘキシルアラニン、β−アラニン、修飾アミノ酸(β−メチルアミノ酸、Cα−メチルアミノ酸、およびNα−メチルアミノ酸など)が含まれる。
【0084】
特定の態様において、このSerrate 誘導体は、アミノ−またはカルボキシル末端の位置で別のタンパク質のアミノ酸配列にペプチド結合を介して結合した脊椎動物のSerrateタンパク質または(好ましくは少なくともSerrate タンパク質のドメインまたはモチーフ、あるいは少なくともSerrate タンパク質の10アミノ酸からなる)それらの断片を含む、キメラ、または融合タンパク質である。一態様において、こうしたキメラタンパク質は、このタンパク質をコードする(別のタンパク質をコードする配列にフレーム内連結したSerrateコード配列を含む)核酸の組み換え発現によって製造される。こうしたキメラ産物は、当業界で既知の方法によって、固有のコードフレーム内に所望のアミノ酸配列をコードする適当な核酸配列同士を連結させ、当業界で通常知られた方法によってキメラ産物を発現させることによって製造することができる。あるいはまた、例えばペプチド合成装置の使用によるタンパク質合成技術によって、こうしたキメラ産物を製造してもよい。特定の態様において、キメラタンパク質が発現されて、その細胞によって成熟Serrateタンパク質に転換されるように、異種シグナル配列を有する成熟脊椎動物Serrate タンパク質をコードするキメラ核酸を発現させる。限定するわけではないが、また別の例において、Serrateおよび別のトポリスミック(toporythmic)遺伝子、例えばDeltaの両方をコードする部分を含む組み換え分子を本発明にしたがって構築することができる。こうした組み換え分子中にコードされたタンパク質はSerrate およびDeltaの両方が組み合わされた特性を示し、そしてアゴニストおよびアンタゴニストを含む新規な生物学的活性プロフィールを表現する。また、Serrate およびDelta の一次配列はコンピュータシミュレーションを使用するこの分子の三次構造の予測に使用することもできる(Hoppand Woods,1981,Proc.Natl.Acad.Sci.U.S.A.783824-3828);Serrate /Deltaキメラ組み換え遺伝子は三次構造および生物学的機能間の相関関係を参照して設計することができるはずである。同様に、任意の異種タンパク質をコードする配列に融合した脊椎動物Serrateの一部分を含むキメラ遺伝子を構築することもできる。ある特定の態様は、少なくとも10アミノ酸の脊椎動物Serrate断片を含むキメラタンパク質に関係するものである。
【0085】
別の特定の態様において、Serrate 誘導体は別のトポリスミックタンパク質と相同の領
域を含むSerrate断片である。第1のタンパク質のある領域を、第2のタンパク質中のこ
の領域に含まれるアミノ酸の数と等しい数のアミノ酸を有するある配列に比較したとき、
この領域のアミノ酸配列が、30%以上同一であるかまたは75%以上同一もしくは保存的変
化に属する場合、ここではこれを“相同性”とみなすこととする。例えば、こうしたSerr
ate 断片として、Delta、またはDSLドメインもしくはこれらの部分に相同な1以上の領
域を含有するものがある。
【0086】
誘導体および類似体のその他の特定の態様を下記の亜節および下記の実施例の節に記載
する。
5.6.1.Serrate タンパク質の1以上のドメインを含有するSerrate誘導体
特定の1態様において、本発明は、脊椎動物Serrate 誘導体または類似体、特に、限定
するわけではないが、細胞外ドメイン、DSL ドメイン、ELR ドメイン、システインに富ん
だドメイン、膜貫通ドメイン、細胞内ドメイン、膜結合領域、およびSerrate タンパク質
の1以上のEGF-様リピート単位(ELR)、または前記の任意の組合せを含む、Serrateタ
ンパク質の1以上のドメインを含むか、またはこれらからなる脊椎動物Serrate 断片およ
び誘導体に関する。ヒトおよびヒナドリSerrate タンパク質に関する特定の例について、
それぞれ実施例第9および8節においてこうしたドメインが同定される。
【0087】
特定の1態様において、脊椎動物Serrate の特定の断片を含む分子は、相当するSerrate タンパク質中でショウジョウバエ(Drosophila)Serrate および/またはDeltaタンパク質の特定の断片に最も相同な断片を含むものである。特定の1態様において、こうした分子は配列番号10、12、または18に相同なアミノ酸配列を含むかまたはこれで構成されている。別法として、Serrate相同体のドメインを含む断片を第5.3.2節に記載したようなタンパク質分析法によって同定することができる。
5.6.2.トポリスミックタンパク質ドメインへの結合を仲介するSerrate誘導体
本発明はまた、トポリスミックタンパク質への結合を仲介する(そしてここではこれら
を“接着性”と称する)脊椎動物Serrate 断片、およびこうした断片の類似体または誘導
体、ならびにこれらをコードする核酸配列を提供する。
【0088】
特定の1態様において、Serrate の接着性断片は、ショウジョウバエSerrate 配列のほぼ第85-283番目または第79-282番目のアミノ酸に最も相同なSerrateの部分を含むものである(PCT 公報、国際公開公報第93/12141号、1993年6月24日付、参照)。
【0089】
特別の1態様において、Serrate タンパク質の接着性断片はDSL ドメインまたはその部分を含んでいる。Notch への結合を仲介するDSL ドメイン内のサブ断片を、欠失性変異体を発現する構築物の分析によって同定することができる。
【0090】
トポリスミックタンパク質(好ましくはNotch)への結合能力は、こうしたトポリスミックタンパク質を発現する細胞およびSerrate またはSerrate 誘導体を発現する細胞を使用したinvitroでの凝集アッセイによって、証明することができる(第5.7節参照)。すなわち、あるSerrate 断片のNotch タンパク質への結合能力は、第1の細胞の表面に発現されたSerrate断片の、第2の細胞の表面に発現されたNotch タンパク質への結合能力で証明することができる。
【0091】
こうしたアッセイにおいて使用するための、トポリスミックタンパク質またはその接着
性ドメインをコードする核酸配列は、霊長類供給源と同様にヒト、ブタ、ウシ、ネコ、ト
リ、ウマ、イヌ、または昆虫、ならびに既知のトポリスミック遺伝子の相同体が同定され
る他のどんな種からも単離することができる。
5.7.Serrateタンパク質、誘導体および類似体のアッセイ
脊椎動物Serrateタンパク質、誘導体および類似体の機能的活性を各種の方法によって
アッセイすることができる。
【0092】
例えば、抗Serrate抗体に結合する、またはこの結合について野性型Serrate と競合する能力をアッセイする場合の1態様において、当業界で既知の各種のイムノアッセイを使用することができ、これらには、限定するわけではないが、ラジオイムノアッセイ、ELISA(酵素結合イムノソルベントアッセイ)、“サンドイッチ”イムノアッセイ、免疫放射線アッセイ、ゲル拡散沈降反応、免疫拡散アッセイ、(例えば金コロイド、酵素または放射性同位元素標識を使用した)insitu イムノアッセイ、ウェスタンブロット、沈降反応、凝集アッセイ(例えばゲル凝集アッセイ、血球凝集アッセイ)、補体結合アッセイ、免疫蛍光アッセイ、タンパク質Aアッセイ、免疫電気泳動アッセイその他などの技術を使用する競合および非競合アッセイシステムが含まれる。1態様において、抗体の結合を一次抗体上の標識を検出することによって検出する。別の態様においては、一次抗体に対する二次抗体または試薬の結合を検出することによって、一次抗体を検出する。さらに別の態様においては、二次抗体を標識する。イムノアッセイにおける結合を検出するための多くの手段が当業界で知られており、これらは本発明の範囲内に含まれる。
【0093】
トポリスミックタンパク質、例えばNotch への結合を仲介する能力をアッセイする別の態様において、1993年6月24日付のPCT 公報、国際公開公報第93/12141号に記載されているものなどのinvitro凝集アッセイを実施することができる(Fehon ら、1990,Cell 61:523-534;Rebay ら、1991,Cell67:687-699をも参照)。
【0094】
Serrate に対する受容体を同定する別の態様において、受容体の結合を例えば当業界で
周知の手段によってアッセイすることができる。別の態様において、Serrate 受容体を発
現する細胞に対するSerrateの結合の生理学的相関性(シグナル形質導入)をアッセイす
ることができる。
【0095】
別の態様における、昆虫またはその他のモデルシステムにおいて、野性型脊椎動物Serrate の誘導体または類似体であるSerrate 変異体の表現型の影響を調べるための遺伝学的研究を実施することができる。
【0096】
その他の方法は当業者が知ることができるであろうし、それらは本発明の範囲内に含まれる。
【0097】
5.8.治療上の用途
本発明は、本発明の治療用化合物の投与による細胞死または分化の障害の治療を提供する。こうした治療用化合物(ここでは、“治療剤(Therapeutics)”と称する)には以下のものが含まれる;脊椎動物Serrate タンパク質ならびにその類似体および誘導体(断片を含む)(例えばここに上記したもの);(ここに上記したような)それらに対する抗体;脊椎動物Serrateタンパク質、類似体または誘導体(例えばここに上記したもの)をコードする核酸;およびSerrate アンチセンス核酸。上述したように、本発明のアンタゴニスト治療剤は脊椎動物Serrateの機能および/またはNotch の機能(Serrate はNotch の1リガンドであるから)に対してアンタゴニスト作用または阻害をする治療剤である。こうしたアンタゴニスト治療剤は(例えばショウジョウバエにおける)遺伝学的アッセイも利用することができるが、最も好ましくは既知の通常のinvitroアッセイ、例えばSerrate の別のタンパク質(例えばNotch タンパク質)への結合を阻害する、または好ましくはin vitroでまたは細胞培養物中でアッセイされる任意の既知のNotchまたはSerrate の機能を阻害する能力に基づくものを使用することによって、同定される。好ましい態様において、アンタゴニスト治療剤はNotch への結合を仲介するSerrateの断片などの機能的に活性な断片を含むタンパク質またはその誘導体、もしくはそれらに対する抗体である。別の特定の態様において、こうしたアンタゴニスト治療剤はNotchに結合するSerrate の断片を含む分子を発現することができる核酸、またはSerrate アンチセンス核酸(本文の第5.11節参照)である。特定の治療剤の効果および罹病した組織の治療にとってその投与が必要かどうかを判定するために、好ましくは下記のような適当なinvitroまたはin vivo アッセイを利用すべきであることを留意すべきである。なぜならば、組織の罹病の進行経過によってアンタゴニストまたはアゴニスト治療剤のどちらが必要とされるかが決まるからである。
【0098】
さらに、投与の様式、例えばSerrate タンパク質または誘導体を可溶性形状で投与する
か、またはSerrateタンパク質の細胞内組み換え発現のためのこれをコードする核酸を介
して投与するか、はそれがアゴニストまたはアンタゴニストのどちらで作用するかに影響
を与える。
【0099】
本発明の別の態様において、相同的組み換えによるSerrate の不活性化を促進するため
に、アンタゴニスト治療剤として、脊椎動物Serrate遺伝子の一部分を含有する核酸を使
用する(Kollerand Smithies,1989,Proc.Natl.Acad.Sci.USA 86:8932-8935;Zijlstraら
、1989,Nature342:435-438)。
【0100】
上記のような本発明のアゴニスト治療剤はSerrate の機能を促進する。こうしたアゴニ
スト治療剤として、限定するわけではないが、Serrate への結合を仲介するNotch の一部
分を含むタンパク質および誘導体、ならびにこれらをコードする核酸(これらがコードし
ている産物をinvivo で発現するように投与することができる)が含まれる。
【0101】
本発明の治療剤のその他の記述および供給源は本文の第5.1節から第5.7節に見ら
れる。
【0102】
所望のSerrate特性、例えばNotch への結合、細胞内リガンドへの結合、を保持、あるいは阻害する分子は、こうした特性およびその生理学的相関物のそれぞれ誘発剤または阻害剤として治療のために使用することができる。特定の1態様において、Notchに結合する脊椎動物Serrate の一部分の配列を含有するペプチド(例えば第10−50番目または第15−25番目のアミノ酸;特に約10,15,20または25アミノ酸)をNotch機能にアンタゴニスト作用させるために使用する。特定の1態様において、Notch の発現の増加に関連するヒトまたはその他の悪性腫瘍(例えば、子宮頚ガン、結腸ガン、乳ガン、扁平腺ガン(下記参照))の治療または予防のためにこうしたアンタゴニスト治療剤を使用する。限定するわけではないが、下記の実施例に記載するアッセイを含む、当業界で既知の操作法によって、Serrateの誘導体または類似体の所望の活性を試験することができる。例えば、欠失変異体を発現させ、いくつかの方法(例えばinvitro 細胞凝集アッセイ、相互作用トラップシステム)の中のいずれかによって発現産物のNotchへの結合をアッセイすることによって、Notch EGF-リピート単位(ELR)11および12に結合し、DSLドメインよりも小さな脊椎動物Serrate 断片を含む分子を得て、選択することができる。これらの方法のいくつかを下記の実施例の節に記載する。特定の1態様において、所望の活性を有するペプチドを選択するために、ペプチドライブラリーをスクリーニングすることができる。こうしたスクリーニングは、例えばNotchまたはNotch ELR 11および12反復単位を含有する分子に対する結合に関するアッセイによって実施することができる。
【0103】
本発明のアゴニストおよびアンタゴニスト治療剤は細胞死の障害に対する治療上の有用
性を有する。アゴニスト治療剤は以下の障害において治療用に(予防上も含んで)投与される;(1)Notch またはSerrate 機能の欠落または(正常または所望の数値に対する)減少が関与する疾病または障害、例えば、Notch またはSerrateタンパク質が欠失、遺伝的欠損、生物学的に不活性または過少活性、もしくは過少発現である患者;および(2)in vitro(またはin vivo)アッセイ(下記参照)がSerrateアゴニストの投与の有用性を示す疾病または障害。Notch またはSerrate 機能の欠落または減少は、例えば患者の組織のサンプルを(例えば生検組織から)取り、発現されたNotchまたはSerrate タンパク質のタンパク質量、構造および/または活性についてこれをin vitroでアッセイすることによって、容易に検出することができる。当業界で標準的な多くの方法をここで使用することができる。これらとして限定するわけではないが、NotchまたはSerrate タンパク質の検出および/または可視化のためのイムノアッセイ(例えばウェスタンブロット、ドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動による免疫沈降、免疫細胞化学、その他)、および/または、NotchまたはSerrate mRNAをそれぞれ検出および/または可視化することによってNotch またはSerrate の発現を検出するためのハイブリダイゼーションアッセイ(例えばノーザンアッセイ、ドットブロットinsitu ハイブリダイゼーション、その他)が含まれる。
【0104】
ある特定のアゴニスト治療剤またはアンタゴニスト治療剤の投与が必要とされるかどうかを判定するために使用することができるin vitroアッセイには、患者の組織サンプルを培養して増殖させて、治療剤に暴露するかまたは別の方法でこれを投与し、組織サンプルに対するその治療剤の効果を観察する、invitro細胞培養アッセイが含まれる。患者が悪性腫瘍を有している1態様において、この悪性腫瘍からの細胞サンプルをプレートにとり、培養して増殖させて、その後この細胞を治療剤に暴露する。悪性腫瘍細胞の生存または増殖を(例えば終末分化を促進することによって)阻害する治療剤をinvivo の治療用に選択する。こうした生存および/または増殖を評価するために、当業界で標準的な多くのアッセイを使用することができる。;例えば、細胞の増殖を、3H-チミジンの取り込みの測定、細胞数の直接の計測、プロトオンコジーン(例えばfoc,myc)などの既知の遺伝子の転写活性または細胞周期マーカーの変化の検出によってアッセイすることができる。;細胞の生存度はトリパンブルー染色によって評価することができ、分化は形態学その他における変化を基礎として視覚的に評価することができる。特定の場合には、悪性腫瘍細胞培養物を(1)アゴニスト治療剤、および(2)アンタゴニスト治療剤に別々に暴露する。アッセイの結果によってどちらの型の治療剤が治療効果を有するかが示される。
【0105】
別の1態様において、それぞれ高または低増殖性障害を有する、または有すると推定さ
れる組織からの患者の細胞サンプルに対して所望の効果、細胞増殖の阻害または促進を示
す治療剤が有用なものとして示される。こうした高または低増殖性障害には、限定するわ
けではないが、下記の第5.8.1節から第5.8.3節に記載されたものが含まれる。
【0106】
もう一つの特定の実施態様において、神経損傷または神経系変性障害の治療に使用する
ための治療薬が提示されているが、該治療薬は、in vitroでの神経再生の促進/罹患した
患者タイプの神経細胞からの神経突起の伸展を呈する。
【0107】
更に、NotchまたはSerrate優位活性化表現型(「機能獲得」突然変異)が関与することが判明または知られている疾患または障害において、本発明の拮抗治療薬の投与もまた提示されている。NotchまたはSerrate優位陰性表現型(「機能損失」突然変異)が関与することが判明または知られている疾患または障害において、本発明の作用治療薬の投与が提示されている。hsp70熱ショックプロモーターならびに眼特異的プロモーターの下で一連のドロゾフィラNotch欠失突然変異体を異所的に発現することによって、Notchタンパク質の種々の構造ドメインの機能をinvivoで調べた(Rebay ら.,1993,Cell 74:319-329 を参照されたい)。2つのクラスの優位表現型、すなわち、Notch機能損失突然変異を示すもの、およびNotch機能獲得突然変異を示すものが観測された。ほとんどの細胞外配列が欠損したタンパク質の過剰発現により優位「活性化」表現型を生じ、一方、ほとんどの細胞内配列が欠損したタンパク質の過剰発現により優位「陰性」表現型を生じた。これらの結果から次のことを示唆された。すなわち、Notchは受容体(この受容体の細胞外ドメインはリガンド結合を媒介する)として機能し、その結果、細胞質ドメインは発生シグナルを伝達することが示唆された。本発明者らは、SerrateがNotchELR 11および12に結合することを明らかにした(PCT公開第WO93/12141号を参照されたい)。
【0108】
種々の特定の実施態様において、患者の障害に関与する細胞タイプの代表的細胞を用い
てin vitroアッセイを行い、治療薬がこうした細胞タイプに所望の効果を示すかを決定することができる。
【0109】
もう1つの実施態様において、患者の組織サンプルの細胞で、前腫瘍性であるとの疑いのあるものを、同様に、プレーティングまたはinvitroで増殖させて、治療薬に晒す。より正常な(すなわち、前腫瘍状態、腫瘍状態、悪性状態、または形質転換表現型の徴候がより弱い)細胞表現型を生じる治療薬を、治療用に選択する。当該技術分野の多くのアッセイ標準を使用して、前腫瘍状態、腫瘍状態、または形質転換もしくは悪性表現型が存在するかを評価することができる。例えば、形質転換表現型に関連した特徴(invivoでの腫瘍形成能に関連した一連のin vitroの特徴)としては、より丸形の細胞形態、より弱い基質結合、接触抑制の欠損、固定依存性の欠損、プラスミノーゲンアクチベータなどのプロテアーゼの放出、糖輸送の増大、血清必要量の減少、胎児性抗原の発現、250,000ダルトンの表面タンパク質の消失などが挙げられる(Luriaら.,1978,General Virology,第3版.,John Wiley & Sons,New York pp.436-446を参照されたい)。
【0110】
他の特定の実施態様において、治療される特定の患者に由来する細胞サンプルを使用するのではなく、本発明に従って、次のような細胞系統を使用して、上述のin vitroアッセイを行うことができる。すなわち、該細胞系統が、治療もしくは予防が望まれる悪性障害、腫瘍性障害、もしくは前腫瘍性障害に由来するか、またはそれらに関連した特徴を呈するか、あるいは作用対象となることが望まれる神経細胞タイプもしくは他の細胞タイプに由来する場合である。
【0111】
拮抗治療薬は、(1)NotchまたはSerrate機能のレベルの増大(正常または所望のレベルと比較して)が関与する疾患または障害において、例えば、NotchまたはSerrateタンパク質が過剰発現されるか、または過剰活性である疾患または障害において、および(2)invitro(またはin vivo)アッセイからSerrate拮抗薬の投与の効用があることが示唆される疾患または障害において、治療のために(予防のために)投与される。NotchまたはSerrate機能のレベルの増大は、上述したような方法によりタンパク質および/またはRNAを定量することによって、容易に検出することができる。患者の組織サンプルの細胞、または適切な細胞系統もしくは細胞型を用いたinvitroアッセイを、治療薬の効用を決定するために上述したように実施することができる。
5.8.1.悪性疾患
拮抗治療薬または作用治療薬を介在させたときの効能について上述したように試験する
ことができるとともに、治療薬の効用の徴候の観察に基づいて治療することができる悪性
または前腫瘍状態としては、以下の5.8.1および5.9.1節に記載のものが挙げられるが、これらに限定されるものではない。
【0112】
in vitro(および/またはinvivo)で試験することができるとともに、適切なアッセ
イの結果を観察し、これに基づいて本発明に従った治療が行えるタイプの細胞を含む悪性
腫瘍および関連障害としては、表1に列挙したものが挙げられるが、これらに限定される
ものではない(こうした障害の総説については、Fishman ら.,1985,Medicine,第2版.
,J.B.LippincottCo.,Philadelphia を参照されたい)。
【0113】
【表1】

特定の実施態様において、上皮組織(例えば、頸部、食道、および肺の上皮組織)の悪性疾患または異常増殖変化(例えば、化成および異形成)を治療または予防することができる。
【0114】
結腸および頸部の悪性腫瘍では、それらの非悪性組織と比べると、ヒトNotch の発現が増大する(1994年4月14日公開のPCT公開番号第WO94/07474号を参照されたい。この全体を参考として本明細書に組み入れる)。従って、特定の実施態様において、有効量の拮抗治療薬(例えば、Notch機能と拮抗するSerrate誘導体)を投与することによって、結腸または頸部の悪性疾患または前悪性疾患の変化を治療または予防することができる。結腸癌および頸部癌の中でNotch発現の増大が見られるということは、多数のより癌性かつ高増殖性の状態が、アップレギュレートされたNotchを呈することを示唆する。従って、特定の実施態様において、Notch機能と拮抗する拮抗治療薬を投与することによって、種々の癌、例えば、乳癌、扁平上皮腺癌、セミノーマ、および肺癌、ならびにそれらの中で生じた前悪性疾患の変化、更に他の高増殖性障害を治療または予防することができる。
【0115】
5.8.2.神経系障害
拮抗治療薬または作用治療薬を介在させたときの効能について上述したように試験する
ことができるとともに、治療薬の効用の徴候の観察に基づいて治療することができる細胞
型を含む神経系障害としては、軸索の切断、ニューロンの縮小もしくは退化、または髄鞘
脱落のいずれかを生じる神経系損傷および疾患または障害が挙げられるが、これらに限定
されるものではない。本発明にしたがって患者(ヒトおよびヒト以外の哺乳類の患者を含
む)の治療が可能な神経系病変としては、中枢神経系(脊髄、脳を含む)または末梢神経
系の以下の病変が挙げられるが、これらに限定されるものではない:
(i)外傷性病変(物理的損傷により生じるかまたは手術に伴う病変、例えば、神経系の
一部を切断する病変または圧迫性損傷が含まれる);
(ii)虚血性病変(神経系の一部で酸素が欠乏することにより神経損傷を生じるかまたは
死亡するもので、脳梗塞もしくは虚血、または脊髄梗塞もしくは虚血が含まれる);
(iii)悪性病変(神経系関連悪性疾患または非神経系組織由来の悪性疾患のいずれかで
ある悪性組織によって、神経系の一部が破壊または損傷を受けたもの);
(iv)感染性病変(感染の結果として、例えば、膿瘍によって、あるいはヒト免疫不全ウ
イルス、帯状庖疹ウイルス、もしくは単純庖疹ウイルスの感染と関連して、またはライム
病、結核、梅毒と関連して、神経系の一部が破壊または損傷を受けたもの);
(v)変性性病変(変性過程の結果として神経系の一部が破壊または損傷を受けたもので
、パーキンソン病、アルツハイマー病、ハンティングトン舞踏病、または筋萎縮性側索硬
化症が含まれるが、これらに限定されるものではない);
(vi)栄養性疾患または障害に関連した病変(栄養障害または代謝異常によって神経系の
一部が破壊または損傷を受けたもので、ビタミンB12欠乏症、葉酸欠乏症、ウェルニッケ
病、たばこ性‐アルコール性弱視、マルキアファーヴァ‐ビニャミ病(脳梁の原発性変性
)及びアルコール性小脳変性が含まれるが、これらに限定されるものではない);
(vii)全身性疾患に関連した神経系病変(糖尿病(糖尿病性ニューロパシー、ベル麻痺
)、全身性紅斑性狼瘡、癌腫、またはサルコイドーシスが含まれるが、これらに限定され
るものではない);
(viii)毒性物質、例えば、アルコール、鉛、または特定の神経毒によって引き起こされ
る病変;及び
(ix)脱髄病変(脱髄疾患によって神経系の一部が破壊または損傷を受けたもので、多発
性硬化症、ヒト免疫不全ウイルス関連ミエロパシーまたは種々の病因、進行性多病巣白質
脳症、および橋中央ミエリン溶解が含まれるが、これらに限定されるものではない)。
【0116】
ニューロンの生存または分化を促進する生物活性を試験することによって、神経系疾患
の治療に有用な本発明に係る治療薬を選択することが可能である(5.8 節を参照されたい
)。例えば、以下の効果のいずれかを誘発する治療薬が本発明に有用であるが、これらに
限定されるものではない:
(i)培養中のニューロンの生存時間の増大;
(ii)培養中またはinvivoでのニューロンの発芽の増大;
(iii)培養中またはinvivoでのニューロン関連分子(例えば、運動ニューロンに関するコリンアセチルトランスフェラーゼまたはアセチルコリンエステラーゼ)の生成の増大;
(iv)invivoでのニューロン機能不全の徴候の増大。
【0117】
こうした効果は、当該技術分野で公知のいかなる方法によっても測定することが可能である。好ましい実施態様において、ただし、これに限定されるものではないが、ニューロンの生存の増大は、Arakawaら(1990,J.Neurosci.10:3507-3515)により報告された方法により測定可能であり;ニューロンの発芽の増大は、Pestronkら(1980,Exp.Neurol.70:65-82)またはBrownら(1981,Ann.Rev.Neurosci.4:17-42)により報告された方法により測定可能であり;ニューロン関連分子の生成の増大は、測定される分子にもよるが、バイオアッセイ、酵素アッセイ、抗体結合、ノーザンブロットアッセイなどにより測定可能であり;運動ニューロンの機能不全は、運動ニューロン疾患の物理的症状発現、例えば衰弱、運動ニューロン伝導速度、または機能障害を評価することにより測定可能である。
【0118】
特定の実施態様において、本発明に従って治療可能な運動ニューロン疾患としては、運
動ニューロンおよび神経系の他のコンポーネントに影響を与える恐れのある梗塞、感染、
毒との接触、外傷、外科的損傷、変性性疾患、または悪性疾患、ならびにニューロンに選
択的に影響を及ぼす疾患、例えば筋萎縮性側索硬化症が挙げられるが、これらに限定され
るものではなく、更に、進行性棘筋萎縮症、進行性球麻痺、原発性側索硬化症、小児およ
び若年筋萎縮症、小児期進行性球麻痺(ファツィオ‐ロンド症候群)、ポリオおよびポリ
オ後症候群、遺伝性運動感覚ニューロパシー(シャルコー‐マリー‐トゥース病)も含ま
れるが、これらに限定されるものではない。
【0119】
5.8.3.組織修復および再生
本発明のもう1つの実施態様において、組織再生および修復を促進するために、本発明
の治療薬が使用される(例えば、良性異常増殖疾患の治療のために使用されるが、これに
限定されるものではない)。特定の実施態様は、肝硬変(瘢痕が正常な肝臓再生過程を阻
害する状態)の治療、ケロイド(過形成性瘢痕)形成(瘢痕過程により正常の更新が妨害
されて皮膚の美観が損なわれた状態)、乾癬(皮膚の過剰増殖および固有細胞運命決定の
遅延を特徴とするよく見かける皮膚の状態)、および禿頭症(末端分化毛包(Notchに富
んだ組織)が正常に機能しない状態)の治療に関連する。もう1つの実施態様において、
本発明の治療薬を使用して、内耳の感覚上皮の変性性疾患または外傷性疾患の治療を行う
。
【0120】
5.9.予防的使用
5.9.1.悪性疾患
本発明の治療薬は、腫瘍状態または悪性状態への進行を防止するために投与できる。これには表1に列挙した疾患が対象となるが、これらに限定されるものではない。こうした投与が行えるのは、上述したアッセイにおいて治療薬がこうした疾患の治療または防止に有効であることが示された場合である。こうした予防的使用が利用できるのは、上述したような腫瘍または癌へ進化することが知られているか、またはそうした進化の疑いのある状態のとき、具体的には、過形成、化生、または特に異形成が関与した非腫瘍性細胞成長が起こったときである(こうした異常成長状態の総説については、Robbins及びAngell,1976,Basic Pathology,第2版.,W.B.Saunders Co.,Philadelphia,pp.68-79を参照されたい)。過形成とは、組織または器官の中の細胞数の増加が関係する調節された細胞増殖の形態のことであり、構造または機能上の有意な変化は見られない。1つだけ例を挙げると、子宮内膜過形成は、しばしば子宮内膜癌へ進行する。化生とは、成人細胞(すなわち十分に分化した細胞)の1つのタイプが成人細胞のもう1つのタイプと置き換わる調節された細胞成長の形態のことである。化生は、上皮組織細胞または結合組織細胞で起こる可能性がある。非定型性化生には、いくらか障害を有する化生性上皮が含まれる。異形成とは、しばしば癌の前兆となるものであり、主に上皮に見うけられる;これは非腫瘍性細胞成長の中で最も障害を有する形態であり、それぞれの細胞の均一性および細胞の構造上の配列が失われる。異形成細胞はしばしば異常に大きくかつ強く染色された核を有し、多形性を呈する。異形成は、慢性的刺激または炎症が起こるの場合に特徴的に生じるもので、頸部、呼吸経路、口腔、および胆嚢によく見うけられる。
【0121】
過形成、化生、または異形成を特徴とする異常細胞成長が存在する代わりに、またはその成長が存在することに加えて、患者の細胞サンプルがin vivoで呈するか、またはin vitroで呈する形質転換表現型または悪性表現型のうちの1つ以上の特徴が存在することは、本発明の治療薬を予防/治療のための投与することが望ましいことを示唆する。先に述べたように、形質転換表現型のこうした特徴としては、形態の変化、より弱い基質結合、接触抑制の欠損、固定依存性の欠損、プロテアーゼの放出、糖輸送の増大、血清必要量の減少、胎児性抗原の発現、250,000ダルトンの細胞表面タンパク質の消失などが挙げられる(形質転換表現型または悪性表現型に関連した特徴については、同上.,pp.84-90をも参照されたい)。
【0122】
特定の実施態様において、白斑症、上皮の外観上良性の過形成または異形成疾患、また
はボーエン病、insitu癌は、予防的介入が望ましいと考えられる前腫瘍性疾患である。
【0123】
もう1つの実施態様において、線維嚢胞病(嚢胞性過形成、乳房異形成、特に腺疾患(
良性上皮過形成))は、予防的介入が望ましいと考えられる。
【0124】
他の実施態様において、悪性疾患に対する以下の素因のうちの1つ以上を呈する患者を、有効量の治療薬の投与により治療する。その要因とは次の通りである:悪性疾患に関連した染色体転座(例えば、慢性骨髄性白血病に対するフィラデルフィア染色体、濾胞性リンパ腫に対するt(14;18)など)、家族性ポリープ症またはガードナー症候群(結腸癌の恐れのある前兆)、良性単一クローン性高ガンマグロブリン血症(多発性骨髄腫の恐れのある前兆)、およびメンデル型(遺伝的)継承様式を示す癌または前癌性病(例えば、結腸の家族性ポリープ症、ガードナー症候群、遺伝性外骨腫症、多内分泌腺腫症、アミロイド生成を伴う髄甲状腺癌および褐色細胞腫、ポッツ‐ジェガース症候群、フォン・レックリングハウゼンの神経線維腫病、網膜芽細胞腫、頸動脈小体腫瘍、皮膚黒色癌、眼内性黒色癌、色素性乾皮症、毛細管拡張性運動失調症、チェディアック‐東症候群、白子症、ファンコーニ再生不良性貧血、並びにブルーム症候群;Robbins及び Angell,1976,Basic Pathology,第2版.,W.B.Saunders Co.,Philadelphia,pp.112-113を参照されたい)の者と1親等の関係。
【0125】
もう1つの特定の実施態様において、本発明の拮抗治療薬をヒト患者に投与して、乳癌
、結腸癌、または子宮頚癌への進行を防ぐ。
5.9.2.他の障害
他の実施態様において、本発明の治療薬を投与して、5.8.2.節に記載の神経系障害また
は5.8.3.節に記載の他の障害(例えば、肝硬変、乾癬、ケロイド、禿頭症)を防ぐことが
できる。
【0126】
5.10.治療または予防の効用の実証
本発明の治療薬をinvivoで試験して、所望の治療活性または予防活性を調べることが
できる。例えば、こうした化合物を好適な動物モデル系で試験してからヒトにおいて試験
することができる。好適な動物としては、ラット、マウス、ニワトリ、ウシ、サル、ウサ
ギなどが挙げられるが、これらに限定されるものではない。in vivo試験に対しては、ヒ
トに投与する前に、当該技術分野で公知のいかなる動物モデル系を使用してもよい。
【0127】
5.11.Serrate発現のアンチセンス調節
本発明は、脊椎動物Serrateまたはその一部をコードする遺伝子またはcDNAに対してアンチセンスである少なくとも6個または少なくとも10個のヌクレオチドの核酸の治療または予防への使用を提供する。本明細書中で使用される「アンチセンス」とは、ある種の配列相補性によって脊椎動物SerrateRNA(好ましくはmRNA)の一部とハイブリッドを形成することが可能な核酸を意味する。こうしたアンチセンス核酸は、本発明の拮抗治療薬としての効用を有し、セクション5.8およびそのサブセクションに既に記載したような障害の治療または予防に使用することができる。
【0128】
本発明のアンチセンス核酸は、細胞に直接投与することができるか、または外来導入配列の転写によって細胞内で生成させることができる、二本鎖または一本鎖のRNAもしくはDNAまたはそれらの修飾体もしくは誘導体であるオリゴヌクレオチドであってもよい。
【0129】
特定の実施態様において、本発明により提供されるSerrateアンチセンス核酸は、腫瘍または他の障害(ただし、これらの腫瘍タイプまたは障害の細胞がSerrate遺伝子またはNotch遺伝子を発現することを(in vitroまたはin vivoで)実証することができるもの)の治療に使用することができる。これはRNAまたはタンパク質を検出することにより実証が可能である。
【0130】
本発明は更に、以下のセクション5.12に記載されているように、医薬品として許容しうる担体中に有効量の本発明のSerrateアンチセンス核酸を含む医薬用組成物を提供する。また、本発明の医薬用組成物の投与を含む、障害(例えば、セクション5.8および5.9に記載の障害)の治療および予防の方法が提供される。
【0131】
もう1つの実施態様において、本発明は、原核細胞または真核細胞中でのSerrate核酸配列の発現を抑制する方法に関する。この方法には、本発明のアンチセンス脊椎動物Serrate核酸を含む有効量の組成物を細胞に供給することが含まれる。
【0132】
Serrateアンチセンス核酸およびそれらの使用については、以下で詳細に説明する。
5.11.1.脊椎動物Serrateアンチセンス核酸
脊椎動物Serrateアンチセンス核酸は、少なくとも6個のヌクレオチド、好ましくはオリゴヌクレオチド(オリゴヌクレオチドの数の好ましい範囲は10個〜約50個である)から成る。特定の態様において、オリゴヌクレオチドは、Serrate遺伝子に対してアンチセンスである少なくとも10個のヌクレオチド、少なくとも15個のヌクレオチド、少なくとも100個のヌクレオチド、または少なくとも200個のヌクレオチドを含有する。オリゴヌクレオチドは、一本鎖または二本鎖の、DNAもしくはRNA、またはそれらのキメラ混合物もしくは誘導体もしくは修飾体であってもよい。オリゴヌクレオチドの塩基部分、糖部分、またはホスフェート主鎖を修飾してもよい。オリゴヌクレオチドには、ペプチドなどの他の追加基、あるいは細胞膜(例えば、Letsingeret al.,1989,Proc.Natl.Acad.Sci.U.S.A.86:6553-6556; Lemaitre et al.,1987,Proc.Natl.Acad.Sci.84:648-652;1988年12月15日公開のPCT公開第WO88/09810号を参照されたい)もしくは血液脳関門(例えば、1988年4月25日公開のPCT公開第WO89/10134号を参照されたい)を通過する輸送の促進剤、ハイブリダイゼーションに起因する開裂剤(例えば、Krolet al.,1988,BioTechniques 6:958-976を参照されたい)、またはインターカレート剤(例えば、Zon,1988,Pharm.Res.5:539-549を参照されたい)が含まれていてもよい。
【0133】
本発明の好ましい態様において、脊椎動物Serrateアンチセンスオリゴヌクレオチド(好ましくは一本鎖DNA)が提供される。最も好ましい態様において、こうしたオリゴヌクレオチドには、Serrate(最も好ましくはヒトSerrate同族体)のSH3結合ドメインまたはNotch結合ドメインをコードする配列に対してアンチセンスである配列が含まれる。オリゴヌクレオチドの構造の任意の位置を、当該技術分野で一般的に知られた置換基を用いて修飾してもよい。
【0134】
Serrateアンチセンスオリゴヌクレオチドには、5-フロオロウラシル、5-ブロモウラシル、5-クロロウラシル、5-ヨードウラシル、ヒポキサンチン、キサンチン、4-アセチルシトシン、5-(カルボキシヒドロキシメチル)ウラシル、5-カルボキシメチルアミノメチル-2-チオウリジン、5-カルボキシメチルアミノメチルウラシル、ジヒドロウラシル、β-D-ガラクトシルキューオシン、イノシン、N6-イソペンテニルアデニン、1-メチルグアニン、1-メチルイノシン、2,2-ジメチルグアニン、2-メチルアデニン、2-メチルグアニン、3-メチルシトシン、5-メチルシトシン、N6-アデニン、7-メチルグアニン、5-メチルアミノメチルウラシル、5-メトキシアミノメチル-2-チオウラシル、β-D-マンノシルキューオシン、5'-メトキシカルボキシメチルウラシル、5-メトキシウラシル、2-メチルチオ-N6-イソペンテニルアデニン、ウラシル-5-オキシ酢酸(v)、ワイブトキソシン、プソイドウラシル、キューオシン、2-チオシトシン、5-メチル-2-チオウラシル、2-チオウラシル、4-チオウラシル、5-メチルウラシル、ウラシル-5-オキシ酢酸メチルエステル、ウラシル-5-オキシ酢酸(v)、5-メチル-2-チオウラシル、3-(3-アミノ-3-N-2-カルボキシプロピル)ウラシル、(acp3)w、および2,6-ジアミノプリンから選ばれる少なくとも1つの修飾された塩基部分が含まれていてもよい。
【0135】
別の実施態様において、オリゴヌクレオチドは、アラビノース、2-フルオロアラビノー
ス、キシルロース、およびヘキソースから成る群より選ばれる少なくとも1つの修飾され
た糖部分を含むが、糖部分がこれらに限定されるものではない。
【0136】
更に別の実施態様において、オリゴヌクレオチドは、ホスホロチオエート、ホスホロジ
チオエート、ホスホラミドチオエート、ホスホラミデート、ホスホルジアミデート、メチ
ルホスホネート、アルキルホスホトリエステル、およびこれらのホルムアセタールまたは
類似体から成る群より選ばれる少なくとも1つの修飾されたホスフェート主鎖を含む。
【0137】
更に別の実施態様において、オリゴヌクレオチドはα-アノマーオリゴヌクレオチドで
ある。α-アノマーオリゴヌクレオチドは、通常のβ-単位とは異なり鎖が互いに平行に走
る相補RNAと共に特異的な二本鎖ハイブリッドを形成する(Gautier et al.,1987,Nu
cl.Acids Res.15:6625-6641)。
【0138】
オリゴヌクレオチドは他の分子(例えば、ペプチド、ハイブリダイゼーションに起因す
る架橋剤、輸送剤、ハイブリダイゼーションに起因する開裂剤など)とコンジュゲートし
てもよい。
【0139】
本発明のオリゴヌクレオチドは、当該技術分野で公知の標準的な方法で、例えば、DN
A自動合成装置(例えば、Biosearch、Applied Biosystemsなどから市販されている装置
)を用いて合成することが可能である。例として、ホスホロチオエートオリゴヌクレオチ
ドは、Steinらの方法で合成可能であり(1988,Nucl.AcidsRes.16:3209)、メチルホ
スホネートオリゴヌクレオチドは、調節された細孔のガラスポリマー担体を使用して調製
可能である(Sarinet al.,1988,Proc.Natl.Acad.Sci.U.S.A.85:7448-7451)。
【0140】
特定の実施態様において、Serrateアンチセンスオリゴヌクレオチドは、触媒RNAすなわちリボザイムを含む(1990年10月4日公開のPCT国際公開第WO90/11364;Sarver et al.,1990,Science247:1222-1225号を参照されたい)。別の実施態様において、オリゴヌクレオチドは、2'-O-メチルリボヌクレオチド(Inoue et al.,1987,Nucl.AcidsRes.15:6131-6148)またはキメラRNA‐DNA類似体(Inoue et al.,1987,FEBS Lett.215:327-330)である。
【0141】
別の実施態様において、本発明のSerrateアンチセンス核酸が、外来配列の転写によって細胞内で生成される。例えば、ベクターをin vivoで導入し、細胞がこのベクターを捕らえ、細胞内でこのベクターまたはその一部を転写し、本発明のアンチセンス核酸(RNA)を生成することができる。こうしたベクターには、Serrateアンチセンス核酸をコードする配列が含まれている。このベクターは、転写されて所望のアンチセンスRNAを生成するかぎり、エピソームの状態を保持することができるか、または染色体と一体化することができる。これらのベクターは、当該技術分野で標準的な組換えDNA技法によって得ることができる。ベクターは、哺乳類細胞中での複製および発現のために使用されるプラスミド、ウイルス、または当該技術分野で公知の他のものであってもよい。SerrateアンチセンスRNAをコードする配列の発現を、当該技術分野で公知の任意のプロモーターを用いて行い、哺乳動物好ましくはヒトの細胞において機能させることができる。これらのプロモーターは誘導性または構成的であってもよい。こうしたプロモーターとしては、SV40初期プロモーター領域(Bernoistand Chambon,1981,Nature 290:304-310)、ラウス肉腫ウイルスの3'-長末端反復配列に含まれるプロモーター(Yamamotoet al.,1980,Cell 22:787-797)、ヘルペスチミジンキナーゼプロモーター(Wagner et al.,1981,Proc.Natl.Acad.Sci.U.S.A.78:1441-1445)、メタロチオネイン遺伝子の調節配列(Brinsteret al.,1982,Nature 296:39-42)が挙げられるが、これらに限定されるものではない。
【0142】
本発明のアンチセンス核酸には、脊椎動物Serrate遺伝子、好ましくはヒトSerrate遺伝子に対して特異的なRNA転写産物の少なくとも一部と相補的な配列が含まれる。しかしながら、絶対的な相補性が好ましいものではあるが、それが必要というわけではない。本明細書中に記載されている「RNAの少なくとも一部と相補的な」配列とは、RNAとハイブリッドを形成して安定な二本鎖を形成するのに十分な相補性を有する配列を意味する。二本鎖Serrateアンチセンス核酸の場合は、二本鎖DNAのうちの一本の鎖を試験するか、または三本鎖の形成を調べてもよい。ハイブリッド形成能は、アンチセンス核酸の相補性の度合いと長さの両方に依存する。一般的には、ハイブリッド形成する核酸が長いほど、それに含まれるより多くの塩基がSerrateRNAとの不一致を起こすが、依然として安定な二本鎖(または三本鎖。ただし場合による)を形成する。当業者は、標準的な手順を使用してハイブリッド化複合体の融点を測定することにより、不一致の許容範囲を確認することができる。
5.11.2.脊椎動物Serrateアンチセンス核酸の治療上の効用
脊椎動物Serrateアンチセンス核酸を使用して、SerrateまたはNotchを発現することが分かっている細胞タイプの悪性疾患または他の障害を治療(または防止)することができる。特定の実施態様において、悪性疾患とは、頸部癌、乳癌、もしくは結腸癌、または扁平上皮腺癌のことである。こうした発現を試験することができる悪性細胞、腫瘍性細胞、および前腫瘍性細胞としては、セクション5.8.1および5.9.1で既に記載した細胞が挙げられるが、これらに限定されるものではない。好ましい実施態様において、一本鎖DNAアンチセンスSerrateオリゴヌクレオチドが使用される。
【0143】
SerrateまたはNotchRNAを発現する悪性(特に腫瘍)細胞タイプは、当該技術分野で公知の種々の方法によって同定することができる。こうした方法としては、SerrateまたはNotch特異性核酸とのハイブリダイゼーション(例えば、ノーザンハイブリダイゼーション、ドットブロットハイブリダイゼーション、in situ ハイブリダイゼーション)、該細胞タイプに由来するRNAがin vitroでNotchまたはSerrateに翻訳される能力の観測、イムノアッセイなどが挙げられるが、これらに限定されるものではない。好ましい実施態様において、例えば、免疫細胞化学によるか、またはinsituハイブリダイゼーションによって、治療前にNotchまたはSerrate発現に関して患者の原発性腫瘍組織を調べる。
【0144】
医薬品として許容しうる担体中に有効量の脊椎動物Serrateアンチセンス核酸を含む本
発明の医薬用組成物は、NotchまたはSerrate RNAまたはタンパク質を発現するタイプ
の悪性疾患を患った患者に投与することができる。
【0145】
特定の障害または状態の治療に有効なSerrateアンチセンス核酸の量は、障害または状
態の特性に依存するが、標準的な臨床技術により決定することができる。可能ならば、治
療する腫瘍タイプのアンチセンス細胞毒性を、in vitroで、次に有用な動物モデル系で測
定してから、ヒトにおいて試験および使用することが望ましい。
【0146】
特定の実施態様において、脊椎動物Serrateアンチセンス核酸を含む医薬用組成物は、
リポソーム、ミクロ粒子、またはマイクロカプセルを介して投与される。本発明の種々の
実施態様において、こうした組成物を使用してSerrateアンチセンス核酸を持続的に放出
することが有用である。特定の実施態様において、抗体を介して特定の識別可能な腫瘍抗
原を標的とするリポソームを利用することが望ましい(Leonetti et al.,1990,Proc.N
atl.Acad.Sci.U.S.A.87:2448-2451;Renneisen et al.,1990,J.Biol.Chem.265:
16337-16342)。
5.12.治療/予防のための投与および組成物
本発明は、有効量の本発明の治療薬を被検者に投与することによって治療(および予防
)を行う方法を提供する。好ましい態様において、治療薬は実質的に精製されたものであ
る。被検者は好ましくは動物であり、ウシ、ブタ、ニワトリなどの動物が含まれ、好まし
くは哺乳動物、最も好ましくはヒトであるが、これらに限定されるものではない。
【0147】
種々のデリバリーシステムが公知であり、これらは本発明の治療薬の投与に使用できる
。例えば、リポソーム、ミクロ粒子、マイクロカプセル中への封入、組換え細胞による発
現、受容体媒介エンドサイトーシス(例えば、Wu and Wu,1987,J.Biol.Chem.262:44
29-4432を参照されたい)、治療薬である核酸をレトロウイルスベクターまたは他のベク
ターの一部として構築することなどが使用できる。導入方法としては、皮内、筋肉内、腹
腔内、静脈内、皮下、鼻腔内、硬膜外、口腔を介した経路が挙げられるが、これらに限定
されるものではない。この化合物は、任意の便利な経路で投与してもよい。例えば、注入
またはボーラス注射によって、表皮または粘膜皮膚の内面(例えば、口腔粘膜、直腸粘膜
、および腸粘膜など)を介した吸収によって投与してもよい。更に、この化合物を他の生
物活性薬剤と共に投与してもよい。投与は、全身的でも局所的でもよい。また、脳室内注
射および髄腔内注射などの任意の好適な経路で、本発明の医薬用組成物を中枢神経系に導
入することが望ましい。脳室内注射は、例えば、レザバー(オムヤマ・レザバーなど)に
取り付けられた脳室内カテーテルを用いて容易に行える。また、肺への投与は、例えば、
吸入器または噴霧器を使用し、エアゾール剤と共に処方することによって実施することが
できる。
【0148】
特定の実施態様において、治療が必要な領域に本発明の医薬用組成物を局所的に投与す
ることが望ましい。こうした局所的投与は、例えば、外科手術中の局所注入、局所施用(
例えば、外科手術後の創傷用包帯と併用して)によって、注射によって、カテーテルを用
いて、座薬を用いて、または移植片(該移植片は、多孔性、非多孔性、または膠状の材料
から成るもので、例えば、シアラスチック(sialastic)膜などの膜または繊維などである
)を用いて行うことができるが、これらに限定されるものではない。1実施態様において
、悪性腫瘍または腫瘍性組織もしくは前腫瘍性組織の部位(すなわち形成部位)に直接注
射を行うことによって、投与することができる。
【0149】
もう1つの実施態様において、治療薬をベシクル中に入れて、特にリポソームに入れて
送達することができる(Langer,Science 249:1527-1533(1990); Treat et al.,in Lipo
somes in theTherapy of Infectious Disease and Cancer,Lopez-Berestein and Fidle
r(eds.),Liss,NewYork,pp.353-365(1989); Lopez-Berestein,ibid.,pp.317-327
を参照されたい;一般的な内容についてもこれらの文献を参照されたい)。
【0150】
更に別の実施態様において、制御放出システムで治療薬を送達することができる。1実施態様では、ポンプを使用してもよい(Langer,supra; Sefton,CRC Crit.Ref.Biomed.Eng.14:201(1987);Buchwald et al.,Surgery 88:507(1980); Saudek et al.,N.Engl.J.Med.321:574(1989)を参照されたい)。別の実施態様において、ポリマー材料を使用することができる(MedicalApplications of Controlled Release,Langer and Wise(eds.),CRC Pres.,Boca Raton,Florida(1974);Controlled Drug Bioavailability,Drug Product Design and Performance,Smolen andBall(eds.),Wiley,New York(1984); Ranger and Peppas,J.Macromol.Sci.Rev.Macromol.Chem.23:61(1983)を参照されたい;更にLevyet al.,Science 228;190(1985); During et al.,Ann.Neurol.25:351(1989); Howard etal.,J.Neurosurg.71:105(1989)をも参照されたい)。更に別の実施態様において、放出制御システムを治療標的(すなわち、脳)の近傍に配置することができる。この場合、全身用量のわずか一部が必要となるだけである(Goodson,inMedical Applications of Controlled Release,supra,vol.2,pp.115-138(1984)を参照されたい)。
【0151】
その他の放出制御システムについては、Langerの総説(Science 249:1527-1533(1990)の中で説明されている。
【0152】
治療薬が核酸であり、この核酸がタンパク質治療薬をコードする場合の特定の実施態様において、この核酸を次のようにしてin vivoで投与し、そのコードされたタンパク質の発現を促進することができる。すなわち、この核酸を適切な核酸発現ベクターの一部として構築して投与し、細胞内に取り込ませることによって、例えば、レトロウイルスベクターを用いて(米国特許第4,980,286号を参照されたい)、または直接注射することによって、またはミクロ粒子衝撃(例えば、遺伝子ガン;Dupont製Biolistic)によって、または脂質、細胞表面受容体、もしくは移入剤を塗布することによって、または核に入り込むことが知られているホメオボックス様ペプチド(例えば、Joliotet al.,1991,Proc.Natl.Acad.Sci.USA 88:1864-1868を参照されたい)に結合させて投与することなどによって実施することができる。この他に、核酸治療薬を細胞内に導入し、宿主細胞DNA内に取り込んで相同的組換えにより発現させることができる。
【0153】
個々の障害の治療または予防に関連した特定の実施態様において、以下の形態の投与を
行うのが好ましい:
また、本発明は、医薬組成物も提供する。かかる組成物は、治療上有効な量の治療薬及
び製剤学的に許容できる担体を含む。特定の態様において、「製剤学的に許容できる」と
いう用語は、連邦政府又は州政府の統制機関によって認可された、ということ又は動物、
より特定的にはヒトにおける使用のための米国薬局方もしくは他の一般的に認可された薬
局方に記載された、ということを意味する。
【0154】
「担体」という用語は、治療薬とともに投与される希釈剤、アジュバント、賦形剤又はビヒクルをいう。そのような製剤学的担体は、水及び落花生由、大豆油、鉱油、ごま油などのような、石油、動物、植物、又は合成源由来のものを含むオイル等の無菌液でもよい。医薬組成物が静脈内投与される場合、好ましい担体は水である。また、生理食塩水並びに水性ブドウ糖(dextrose)及びグリセロール溶液も、特に注射溶液用の液状担体として用いることができる。適当な製剤学的賦形剤としては、デンプン、グルコース、ラクトース、スクロース、ゼラチン、麦芽、米、小麦、白亜(chalk)、シリカゲル、ステアリン酸ナトリウム、グリセリンモノステアラート、タルク、塩化ナトリウム、脱脂粉乳、グリセロール、プロピレン、グリコール、水、エタノール等が挙げられる。組成物には、所望により少量の加湿もしくは乳化剤又はpH緩衝剤を含ませることもできる。これらの組成物は、溶液、懸濁液、エマルジョン、錠剤、丸剤、カプセル剤、散剤、徐放性剤等の剤形を取り得る。組成物は、伝統的な結合剤及びトリグリセリド等の担体を用いて坐剤として処方することができる。経口製剤としては、製剤学的グレードのマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、ナトリウムサッカリン、セルロース、炭酸マグネシウム等の標準的な担体を挙げることができる。適当な製剤学的担体の例は、E.W.Martinによる「レミントンの薬学(Remington'sPharmaceutical Sciences)」に記載されている。このような組成物には、治療上有効な量の治療薬が、好ましくは患者に適した投与剤形を提供するような適量の担体とともに精製された状態で含まれる。その処方は投与様式に合わせるのがよい。
【0155】
好ましい態様においては、組成物はヒトへの静脈内投与に適合した医薬組成物としてルーチン的な手順で処方される。静脈内投与用の組成物は、典型的には無菌等張水性緩衝液に溶かした溶液である。必要ならば、組成物に可溶化剤及び注射部位の痛みを取り除くためのリドカイン等の局所麻酔剤をも含ませることができる。一般的に、処方成分は、別々に、あるいは例えば凍結乾燥粉末として、又は活性剤の量を示すアンプルもしくはサッシェ等の気密性密封容器中に入れた水分のない濃縮液として単位剤形(unit dosage form)中に混合して提供される。組成物を輸液によって投与する場合、組成物は無菌の製剤学的グレードの水又は生理食塩水を含む輸液ボトルを用いて調剤され得る。組成物を注射によって投与する場合、処方成分を投与前に混合し得るように、注射用の無菌水又は生理食塩水のアンプルを提供することができる。
【0156】
本発明の治療薬は、中性の状態又は塩の状態で処方され得る。製剤学的に許容できる塩
としては、塩酸、リン酸、酢酸、シュウ酸、酒石酸等から誘導される塩のような遊離のア
ミノ基により形成された塩、及び水酸化ナトリウム、水酸化カリウム、水酸化アンモニウ
ム、水酸化カルシウム、水酸化第二鉄、イソプロピルアミン、トリエチルアミン、2-エチ
ルアミノエタノール、ヒスチジン、プロカイン等から誘導される塩のような遊離のカルボ
キシル基により形成された塩が挙げられる。
【0157】
特定の障害又は病態の治療に有効である本発明の治療薬の量は、その障害又は病態の性質に依存し、標準的な臨床技術によって決定することができる。さらに、最適用量範囲を確認するのを助けるためにin vitroアッセイを任意に用い得る。また、その処方に用いられる厳密な用量は投与経路、疾患又は障害の重篤度にも依存し、開業医の判断及び各患者の情況によって決定される。しかしながら、静脈内投与に適した用量範囲は、一般的に、体重1kg当たり活性化合物約20〜500μg である。鼻腔内投与に適した用量範囲は、約0.01pg/kg(体重)〜1mg/kg(体重)である。有効な用量はin vitro又は動物モデル試験系から得られた用量−応答曲線から推定し得る。
【0158】
坐剤には、一般的に、0.5〜10重量%の範囲の活性成分が含まれ、経口製剤には、好ましくは10〜95%の活性成分が含まれる。
【0159】
また、本発明は、本発明の医薬組成物の1以上の成分で充填された1以上の容器を含む
医薬パック又はキットも提供する。このような容器には、医薬品又は生物学的製品の製造
、使用又は販売を規制する政府機関によって定められたフォームの注意書き(この注意書
きは、前記機関によるヒトへの投与のための製造、使用又は販売の認可を示すものである
)を任意に添付することができる。
【0160】
5.13.診断的効用
脊椎動物Serrateタンパク質、類似体、誘導体及びその部分配列(subsequences)、脊椎動物Serrate 核酸(及びそれと相補的な配列)、抗脊椎動物Serrate抗体は、診断における用途を有する。そのような分子は、Serrate 発現に影響する種々の病態、疾患及び障害を検出し、予知し、診断し、もしくは監視するために、又はその治療を監視するために、イムノアッセイ等のアッセイに用いることができる。特に、そのようなイムノアッセイは、患者から得られたサンプルを免疫特異的結合が生じ得るような条件下で抗Serrate抗体と接触させ、その抗体による全特異的結合の量を検出又は測定することを含む方法によって行うことができる。特定の態様においては、好ましくは抗Notch 抗体との結合との関連における組織区分(tissuesections)でのそのような抗体の結合を用いて、疾患状態におけるNotch 及び/又はSerrate の異常な局在化又は異常なレベルのNotch-Serrate共同局在化を検出することができる。特定の態様においては、Serrate に対する抗体を用いて、患者の組織又は血清サンプルにおけるSerrate の存在(そのSerrateの異常なレベルは疾患症状の指標になる。)をアッセイすることができる。内因性Notch タンパク質におけるSerrate 結合能力の異常なレベル、又は内因性Serrateタンパク質におけるNotch(又は他のSerrate リガンド)に対する結合能力の異常なレベルは、細胞運命(cell fate)の障害(例えば、癌等)の指標となり得る。「異常なレベル」とは、体の一部分由来の類似のサンプル又はその障害を有さない被験者由来の類似のサンプル中に存在するレベル又は該レベルを代表する標準レベルに比べて高い又は低いレベルを意味する。
【0161】
使用可能なイムノアッセイとしては、少しだけ名を挙げると、ウェスタンブロット、ラ
ジオイムノアッセイ、ELISA(酵素結合イムノソルベントアッセイ)、「サンドイッチ」
イムノアッセイ、免疫沈降アッセイ、沈降反応、ゲル拡散沈降反応、免疫拡散アッセイ、
凝集アッセイ、補体固定アッセイ、免疫放射線アッセイ、蛍光イムノアッセイ、タンパク
質Aイムノアッセイ等の技術を用いる競合及び非競合アッセイ系が含まれるがこれらに限
定されるものではない。
【0162】
また、脊椎動物Serrate 遺伝子、相補配列を含む関連のある核酸配列及び部分配列、並
びにその他のトポリズミック(toporythmic)遺伝子配列も、ハイブリダイゼーションアッ
セイに使用することができる。脊椎動物Serrate核酸配列又は少なくとも約8のヌクレオ
チドを含むその部分配列は、ハイブリダイゼーションプローブとして使用することができ
る。ハイブリダイゼーションアッセイを用いて上記したようなSerrate 発現及び/又は活
性における異常な変化と関連する病態、障害又は疾患状態を検出し、予知し、診断し、又
は監視することができる。特に、そのようなハイブリダイゼーションアッセイは、核酸を
含むサンプルをSerrateDNA又はRNAにハイブリダイズ可能な核酸プローブとハイブ
リダイゼーションが生じ得るような条件下で接触させ、結果として生じるハイブリダイゼ
ーションを検出又は測定することを含む方法によって行われる。
【0163】
さらに、SerrateはNotch に結合するので、脊椎動物Serrate 又はその結合部分を用いて、例えば、結腸及び子宮頸癌のようなNotch 発現の増加を示す悪性腫瘍のスクリーニングにおけるサンプル中のNotchの存在及び/又は量をアッセイすることができる。
【実施例】
【0164】
6.マウスSerrate相同体の単離及び特性付け
M-Serrate-1と称するマウスSerrate 相同体を下記のようにして単離した:
マウスSerrate -1遺伝子
組織起源:10.5日齢マウス胎仔RNA
単離法:
a)上記のRNAに対するcDNAをランダムプライミング
b)下記を用いた上記cDNAのPCR
増幅条件:供給バッファー50μl にcDNA 50ng、プライマー各1μg、dNTP's 0.2mM
、Taq 1.8U(Perkin-Elmer)を入れた液、94℃/30秒、45℃/2分、72℃/1分で40サイクル、各サイクルを2秒延長。
【0165】
両末端で配列決定された 1.8kbの断片が得られ、C-Serrate -1に相当するものと同定された。
【0166】
M-Serrate-1の部分DNA配列:
発現パターン:発現パターンは、発達中の中枢神経系、末梢神経系、肢、腎臓、水晶体、及び脈管系における発現を含む、C-Serrate -1(ニワトリSerrate)(下記の第11節参照)で観察されたものと同様であると決定された。
7.ツメガエルSerrate 相同体の単離及び特性付け
ツメガエル Serrate-1と称するツメガエルSerrate 相同体を下記のようにして単離した:
ツメガエルSerrate -1遺伝子
組織起源:神経胚期の胚RNA
単離法:
a)上記のRNAに対するcDNAをランダムプライミング
b)下記を用いたPCR
増幅条件:供給バッファー50μl にcDNA 50ng 、プライマー各1μg 、dNTP's 0.2mM、Taq 1.8U(Perkin-Elmer)を入れた液、94℃/30秒、45℃/2分、72℃/1分で40サイクル、各サイクルを2秒延長。
【0167】
部分的に配列決定されてC-Serrate -1との関係が確認された〜700bp の断片が得られた。
8.ニワトリ(CHICK)Serrate 相同体の単離及び特性付け
本実施例では、ニワトリSerrate 相同体C-Serrate 、並びに2つのニワトリNotch相同体C-Notch -1及びC-Notch -2の断片のクローニング及び配列を、初期胚形成中のそれらの発現パターンとともに報告する。C-Serrateの転写のパターンは胚の多くの領域におけるC-Notch -1の転写のパターンと共通点があり、これは、C-Notch-1 が、ショウジョウバエのNotchのように、Serrate の受容体であることを示唆するものである。特に、Notch 及びSerrate は、発達中の中枢及び抹消神経系の神経形成領域において発現する。
【0168】
本発明者らのデータは、Notch の公知のリガンドであるSerrate が節足動物からコルダ
ートまで保存されていることを示している。共通する発現パターンは、Notch とのその機
能的な関係の保存を示唆するものであり、ニワトリの成長、特にその中枢神経系の発達は
、いくつかの特異的な位置でのC-Notch-1 とSerrate の相互作用が関与しているということを示唆する。
材料及び方法
胚
白レグホンニワトリの卵をパークファーム大学(University Park Farm)から入手し、38
℃でインキュベートした。胚をHamburger 及びHamilton(1951,J.Exp.Zool.88:49-92)にしたがって段階づけした(staged)。
Notchのニワトリ相同体のクローニング
ニワトリNotch1及びNotch2遺伝子の約1000塩基対PCR断片を、縮重プライマー及びLardelli及びLendahl(1993,Exp.Cell.Res.204:364-372)に概説されたPCR条件を用いて、耳外植片RNA(下記参照)から増幅した。そのPCR断片をBluescriptKS-にサブクローニングし、配列決定してDIGアンチセンスRNAプローブの作成用の鋳型として用いた(RNA転写キット,Stratagene; DIG RNA標識混合物,BoehringerMannheim)。
ショウジョウバエSerrate のニワトリ相同体のクローニング
耳外植片を段階8から段階13の胚から切断した。各耳外植片は、2つの耳杯、後脳及び咽頭の間にある短い区画、並びにそれに結びついた頭外胚葉及び間葉から構成されていた。RNAを標準プロトコール(Sambrooket al.,1989,in Molecular Cloning: A Laboratory Manual,2nd ed.,Cold SpringHarbor Laboratory Press,コールドスプリングハーバー(Cold Spring Harbor)、ニューヨーク)を一部変更したものを用いて抽出し、ポリA+mRNAをPolyATtractmRNA単離システム(Promega)を用いて全RNAから単離した。第1のcDNA鎖をSuperScriptPreamplificationシステム(Gibco)を用いて合成した。
【0169】
PCR及び縮重プライマーを用いて、耳外植片cDNA由来のショウジョウバエ遺伝子
Serrate に相同性のあるニワトリ遺伝子の断片を増幅した。プライマーは、ハエのDelta
タンパク質及びSerrateタンパク質の両方で見出されるペプチドモチーフを認識するよう
に設計した:
1)プライマー1,5-CGI(T/C)TITGC(T/C)TIAA(G/A)(G/C)AITA(C/T)CA-3'(配列番号17)は、ハエDeltaタンパク質及びSerrate タンパク質のアミノ末端に位置するモチーフRLCLK(E/H)YQ(配列番号18)に相当する。
【0170】
2)プライマー2,5'-TCIATGCAIGTICCICC(A/G)TT-3'(配列番号11)は、いくつかのE
GF様リピートに見出されるモチーフNGGTCID(配列番号12)に相当する。PCR条件は
下記のとおりである:94℃1分間、45℃ 1.5分間及び72℃2分間の35サイクル;その後72℃10分間の最終伸長工程。長さ約 900塩基対のPCR産物を精製し、BluescriptKS-(Stratagene)にサブクローニングし、そのDNA配列を部分的に決定してそれがほぼSerrate 相同体であることを確認した。次いで、それを用いて2つのcDNAライブラリー:
1)段階8〜13の耳外植片をランダムプライミングしたcDNAライブラリー
2)段階17のニワトリ脊髄をオリゴdTプライミングしたcDNAライブラリー
をスクリーニングすることによってより大きなcDNAクローンを回収した。共通しているcDNAを単離し、その遺伝子の全コーディング領域をいっしょにほとんどカバーする2つ(9及び3A.1という)をBluescript KS-にサブクローニングした。DNA配列を、2本鎖Nested Deletion キット(Pharmacia)、及びシークエナーゼ酵素(米国BiochemicalCorporation)を用いるサンガージデオキシチェーンターミネーション法を用いて作製したネスト欠失シリーズ(nested deletion series)から決定した。配列を一列に並べてGenewarks2.3 及びIntelligenetics を用いて分析した。相同性調査はプログラムSharq を用いて行った。
【0171】
オープンリーディングフレームの5’最末端(the most 5’end)を得るために、Lardell
iら(1994,Mechanismsof Development 46:123-136)の方法を用いる他の多くのライブラ
リー(cDNA及びゲノムライブラリー)のスクリーニングを含む、多くの他のPCRに
基づく方法を用いた。
insitu ハイブリダイゼーション
遺伝子転写のパターンをDIG標識RNAプローブ、並びに:
1)高度にストリンジェントな全載(high-stringencywholemount)in situ ハイブリダイ
ゼーションプロトコール、及び
2)Strahleら(1994,Trends in Genet.10:7)のプロトコールに基づいたクリオスタッ
ト区画上でのinsitu ハイブリダイゼーションを用いるin situ ハイブリダイゼーションによって決定した。
結果
脊椎動物胚中のニワトリSerrate の可能性のある役割を見抜くために、Notch とSerrate の機能的結合がショウジョウバエ中で生じているということから、そのSerrate発現とニワトリNotch の発現との関係を調べた。2つのニワトリNotch 相同体を下記のようにして得た。
C-Notch-1及びC-Notch -2は、それぞれ、齧歯類Notch -1及びNotch -2遺伝子の明白な相対物である
2日齢ニワトリ胚から調製したcDNA及び公知の齧歯類Notch 相同体と共通の保存領域に基づいた縮重プライマーを用いて、PCRによってニワトリ中のNotch 相同体を調査した。この方法において、C-Notch-1及びC-Notch-2と呼ばれている2つの別個の遺伝子のそれぞれ長さ約1000ヌクレオチド断片を得た。それらの断片は第3のNotch/lin12 リピートから最後の5つほどのEGF様リピートを含むものにまで伸長する。EGF様リピートは、非常に多くのタンパク質(その大部分は、他の点ではNotchとは関係がない)に存在する。しかしながら、3つのNotch/lin12 リピートは、Notch 遺伝子ファミリーに特有のものであり、その公知のメンバー全てで見出される。C-Notch-1は、齧歯類Notch1(Weinmasterら,1991,Development 113:199-205)と高度のアミノ酸同一性を示し、齧歯類Notch1と概ね似たようなドメインで発現する(下記参照)。齧歯類Notch遺伝子の中で、C-Notch -2はNotch 2(Weinmasterら,1992,Development116:931-941)とほとんど類似しているようである。
【0172】
in situ ハイブリダイゼーションによって初期胚におけるC-Notch-1の発現パターンを調べた。C-Notch-1は、1〜2日齢ニワトリ胚で、神経管、前原体節期中葉胚、腎形成性中葉胚(予期中葉胚(prospectivemesoderm))、鼻プラコード、耳プラコード/小嚢、水晶体プラコード、上鰓プラコード、心臓における脈管系の内皮内層(endothelial lining)、及び肢芽の外胚葉性頂堤(AER)を含む、多くの明瞭に定義されたドメインにおいて発現した。これらの部位は、報告されている齧歯類における同等の段階でのNotch1発現の部位と適合する(表2)。
【0173】
発現データとともに配列データを取り上げると、C-Notch -1は齧歯類Notch1のニワト
リオルソローグ(ortholog)、又はその非常に類似した類縁物のいずれかであると結論付け
られる。
【0174】
【表2】
C-Serrate はショウジョウバエSerrateの相同体であり、Notchファミリーに属する受容
体のリガンドの候補をコードする
ショウジョウバエにおいては、Notchに対する2つのリガンドが知られており、関連する2つの遺伝子Delta 及びSerrate によりコードされている。これらの遺伝子に対応するアミノ酸配列は、Notchに対するinvitro結合に必要でありかつ十分な領域、DSL モチ−フを含む5'末端において相同である。Sarrate のニワトリ相同体の断片を単離するために、DSL モチ−フのいずれかの側の配列を認識するように設計されたPCR及び縮重プライマーを使用した(材料及び方法を参照)。900塩基対のPCR 断片を回収し、ライブラリーをスクリーニングするのに使用し、オーバーラップcDNAクローンを単離することができた。これらのcDNAクローンのDNA配列は、いくつかの5'塩基のみを欠く、3582ヌクレオチドの殆ど完全な単一のオープンリーディングフレームを示すものであった。ショウジョウバエDelta及びSerrate のアミノ酸配列との比較により、ニワトリSerrate タンパク質のシグナル配列の部分をコードするコード配列の部分のみが失われていることが示された。
【0175】
前記ヌクレオチド配列(配列番号5)(図3)の翻訳から、1230アミノ酸のタンパク質(配列番号6)(図4)が予測される。ヒドロパシープロットにより、トランスメンブランドメインに特徴的な単一の疎水性領域があることが明らかにされている(Kyteand Doolittle,1982,J.Mol.Biol.157:105-132)。さらに、前記タンパク質は、その細胞外ドメインにタンデム配列で結合している16個のEFG-様リピートを有する。ニワトリ配列を、D.melanogasterDelta 及びSerrate の配列と比較することにより、前記クローンがSerrate のニワトリ相同体をコードしていることが示されている(図5、図6)。ショウジョウバエSerrateが14個のEFG-様リピートを有し、リピート4、6及び10に大きな挿入物を有しているのに対し、前記ニワトリ相同体は2つの余分なEFG-様リピートを有し、10番目のリピート中に16アミノ酸の小さい挿入物を1つ有するだけである。両方のタンパク質は、EFG-様リピートとトランスメンブランドメインとの間に2番目のシステインリッチ領域を有し、この領域のシステインの間隔は2つのタンパク質において殆ど同じである(ショウジョウバエSerrateのCX2CXCX6CX4CX15CX5CX7CX4CX5CとC-SerrateのCX2CXCX6CX4CX9CX5CX7CX4CX5Cとを比較せよ)。C-Serrate の細胞内ドメインは、ショウジョウバエ Delta及びSerrate のいずれの細胞内ドメインとも有意な相同性を有していない。
C-Serrate は中枢神経系、頭プラコード、腎中胚葉、脈管系、及び体肢芽間葉において発
現される
ステージ4〜ステージ21の早期の胚形成の間に、約12時間の間隔で、全載標本中においてin situ ハイブリダイゼーションを行い、C-Serrateの発現を調べた。後期の段階については凍結切片についてinsitu ハイブリダイゼーションを行うことにより調べた。
【0176】
全載物に見られるように、C-Serrate の早期の発現の主要な部位は、中枢神経系、頭プ
ラコード、腎中胚葉、脈管系、及び体肢芽間葉の5つの表題の下にグループ分けすること
ができる。
中枢神経系
C-Serrate の最初の検出可能な発現は、段階6(0体節/24時間)における中枢神経系において、神経板の背面部分内で見られた。段階10(9〜10体節/35.5時間)までに、間脳予定域において強い縞状の発現が見られた。菱脳及び脊髄予定域において別の弱い染色が見られた。
【0177】
ステージ13において、神経管にいくつかの発現のパッチが見られた。間脳においては、
神経分節D2に対応すると見える、強い三角形の縞状の発現が見られた。前部中脳底床に2つのパッチ(正中線の各側に1つずつ)が見られ、中脳背部において拡散した染色が見られた。菱脳及び吻側脊髄においては、正中線の各側に2つの縦方向の発現の縞が見られ、1つは神経管の背部縁に沿ったものであり、2番目のものはより腹側のものであり、底板に隣接するものであった。両者は(ラット)Notch1発現のドメイン内に位置していた。腹側の縞の前部端は中脳/菱脳境界にあった。背側の縞は背側中脳における発現に連続していた。前部脊髄においては、発現はより小さなスポット状となり、縞はC-Serrateを発現する離れ離れに分散した細胞に置き変わった。
【0178】
ステージ17(58時間)においては、間脳及び中脳においては発現は変化しなかった。菱
脳及び脊髄においては、別の2つの縦の縞があった。1つは背腹方向軸に沿った中央のも
のであり、第2のものはより広いより腹側の縞であった。これらの縞の前部端は菱脳の小
片2の前部境界に一致していた。菱脳における4つの縦方向の縞は全て胚の脊髄に続いて
おり、その後部端に向かって減少していた。これらの発現の縞は少なくともステージ31(E
7)以下で維持されていた。ステージ21(84時間)までに大脳半球に別の発現が見られ、視蓋中の細胞における入り交じった分布で強い発現が見られた。
【0179】
頭プラコード
C-Serrate が、全ての頭プラコード、即ち水晶体プラコード、鼻プラコード、耳プラコ
ード/耳胞及び上鰓プラコードに発現され、三叉神経プラコード(形態学的には十分に確
定されていない)に対応し得る、耳プラコード前部に対する頭蓋外胚葉パッチに発現され
ることは驚くべきことである。
【0180】
水晶体プラコードにおいては、ステージ11において既に発現がみられ、急速に非常に強くなり、少なくともステージ21まで維持された。鼻プラコードにおいては発現はより弱く、ステージ13から検出されたに過ぎなかった。発現はやはり少なくともステージ21まで維持された。
【0181】
耳プラコードについても同様に、発現はステージ10において見えるようになり、ステージ11の初期(12〜14体節、42.5時間)まで強かった。奇妙なことに、耳発現ドメイン、即ちそこでは前記遺伝子が発現されなかったプラコードの前腹側領域中に「孔」が見られた。さらに、プラコードが陥入して耳胞を形成するに従って、前側部及び後内側分裂極において最も強い発現が見られた。さらに後になって、耳胞が内耳の膜迷路に変化するに従って、C-Serrate発現は知覚パッチに限定されるようになった。
【0182】
鰓上発現は、ステージ13/14 において、第1及び第2の鰓裂の背側の縁周辺の外胚葉で
強い染色として見られた。これは鰓裂の内層の深部と、鰓嚢の内皮内層とにおける遺伝子
の発現を伴っており、ここでこれらの2つの上皮は互いに隣接している。
【0183】
最後に、ステージ11において、大きくて強力であるが、一時的なパッチ発現が、耳原基に対してちょうど前部腹側の頭蓋外胚葉において見られた。その位置から、これは三叉神経プラコードの領域であるか、あるいはそれを含むものであると考えられた。
腎中胚葉
発生中の中腎細管中のステージ10及びより段階の進んだ胚(ステージ17〜21)からの中間中胚葉の細胞において発現が検出された。
体肢芽
C-Serrate mRNAは、発生中の体肢芽の遠位端における間葉のパッチに局在していた
。これは体肢の成長における役割を示唆している可能性がある。
その他の部位
発現は、尾芽、尿膜柄においても見られ、後期においてはおそらくその他の組織におい
ても見られるであろう。
C-Serrate 発現の全ての主要な部位はC-Notch-1発現のドメイン内にある
C-SerrateのDSLドメイン及び隣接するN-末端領域が保存されていることは、それがNotchファミリーに属する受容体についてのリガンドとして機能することを示唆している。従って、C-Serrate発現がNotch遺伝子の発現を伴う部位が見出されるであろうと予測された。そのような部位においては、Serrate-Notchシグナリングにより細胞が連絡を取り合っていることを示すものとして、2つの遺伝子のオーバーラップした、あるいは連続的な発現が起こり得る。実際にどのようなオーバーラップが起こるのかを発見するために、インキュベーション8日(E8)までのステージの範囲にわたって、insitu ハイブリダイゼーションにより示される、C-Serrate の発現パターンをC-Notch-1のものと比較した。C-Serrate発現の観察された部位の全ては実際に、C-Notch-1 の発現ドメイン内あるいは非常に近い位置にあった(表III)。
【0184】
【表3】
昆虫の神経発生におけるNotch 及びその相手の重要性により、前記相同遺伝子が脊椎動
物CNS の発生に関与しているかどうかを知ることは特に重要であった。C-SerrateはCNS
で発現され、その発現パターンはNotch相同体のものとの顕著な相関を示すものである。
【0185】
C-Notch-1及びC-Serrate アンチセンスRNAプローブとハイブリダイズしたニワトリ
6日胎児の脊髄の横断切片について分析した。C-Notch-1は先に記載したように管腔領域
全体に発現され、この領域内ではSerrateが強く発現された2つの小さいパッチが存在し
た。
考察
ショウジョウバエの発生においては、Notch遺伝子の産物を介した細胞間シグナリングが、分化した細胞型の詳細なパターンを特定する最終的な細胞の予定運命決定において主要な役割を果している。このシグナリング経路においてはNotchタンパク質がトランスメンブラン受容体として同定されており、この経路は神経発生におけるその役割において最もよく知られているものである。即ち、NotchあるいはNotchを介するシグナル伝達に必要なその他の遺伝子の任意のセットにおける機能欠失突然変異が神経外胚葉における細胞の予定運命を変え、表皮性のはずであった細胞を代わりに神経性のものにするはずである。しかしNotch-依存性シグナリングは、神経組織におけるのと同様に非神経組織においても重要である。これは例えば、卵子発生、筋発生、マルピギー管の形成、及び消化管において、そして網膜、末梢小感覚体及び中枢神経系の発生において、分化のモードの選択を制御する。これらの場合の殆どにおいて、Notchにより伝達されるシグナルは側方抑制を媒介するようであり、側方抑制とは特定の経路で分化するようになった細胞、例えば神経芽細胞が、そのごく近傍にあるものが同様な行動をとることを阻害するという細胞の相互作用の一種である。これにより隣接する細胞は異なる経路で行動するようにされ、異なる細胞種のきめこまかいパターンを形成する。
【0186】
しかし、これはNotchを介して伝達されるシグナルの唯一の機能ではないとするのに十分な理由がある。2種類のNotchの直接のリガンドが同定されている。これらは、Delta及びSerrate 遺伝子の産物である。両者は、Notch自身と同様に、その細胞外ドメインにおいてEFG-様リピートのタンデム配列を有するトランスメンブランタンパク質をコードしている。Delta及びSerrate タンパク質の両方は、細胞付着アッセイにおいてNotchに結合することが示されており、in vitroにおいてNotchと相互作用するのに必要にして十分なモチーフ、いわゆるEBDあるいはDSLドメインを含むアミノ末端において相同性を有する大きな領域を共有している。このような生化学的類似性にもかかわらず、それらはかなり異なる発生上の機能を有しているようである。Serrateはハエにおいて多くの部位に発現されるが、肩の盤、翅の盤、及び平均根盤においてのみ必要とされるようである。Serrate の機能が突然変異により欠損された場合はこれらの構造体は成長できなくなる。翅盤の研究により、それが特異的にSerrateに依存する翅縁であることが示されている。Serrate を欠いていると、この重要なシグナリング領域及び成長中心は形成できず、Serrate が翅盤の腹側部分でGAL4-UASプロモーターのもとに異所的に発現されると、異所性の翅縁組織が誘導され、異所性の増殖を起こす。Notchは翅縁においてSerrateの受容体であるようであり、これはNotch のいくつかの突然変異対立遺伝子が、翅縁発生の類似した阻害をおこし、二種の遺伝子の効果において対立遺伝子特異的相互作用が見られることによる。
【0187】
本明細書においては、ショウジョウバエ遺伝子Serrate の相同体の同定及び完全長配列、及びラット/マウスNotch1及びNotch2のニワトリ相同体の同定及び部分配列を記載するものである。
【0188】
ニワトリSerratecDNA内には、その細胞外ドメインに16個のEFG リピートを有する大きいトランスメンブランタンパク質をコードすると予測される単一のオープンリーディングフレームが存在する。それは非常によく保存されたDSLモチーフを有しており、それがNotchと直接相互作用するはずであることを示唆している。ニワトリSerrate の細胞内ドメインは、ショウジョウバエDelta 及びSerrateの細胞内ドメインを含む現在のデータベースのいずれとも相同性を示さない。しかし、ニワトリ及びヒトSerrate の細胞内ドメイン(12節を参照)は殆ど同一であることが指摘される。
【0189】
in situ ハイブリダイゼーションにより、初期胚発生の間においてC-Notch-1及びC-Serrateの位置的分布を調べた。C-Notch-1及びC-Serrateは、それらが同時に発現されるいくつかの領域(CNS、耳、鰓領域、水晶体、心臓、鼻プラコード及び中腎)を含む発現のダイナミックで複雑なパターンを示す。このオーバーラップした発現と、C-SerrateがNotch結合ドメインに非常によく保存された領域を有するという知見とを合わせて考えると、この受容体/リガンド相互作用がショウジョウバエから脊椎動物まで保存されてきたことが示唆されている。
【0190】
ショウジョウバエにおいてはNotch受容体はかなり広範に分布しており、そのリガンドは、オーバーラップしているがより限定的なドメインにおいて見られる。ニワトリにおいても同様な状況が見られる。
【0191】
ハエNotchは、ショウジョウバエの発生の多くの段階において必要である。特に中枢神経系及び末梢感覚器官の発生における側方抑制におけるその役割が最もよく研究された例である。しかし、Notchは多機能受容体であり、種々のシグナリング分子(Delta及びSerrateを含む)と相互反応でき、側方抑制の枠組内には容易には適合しない発生過程においても相互反応できる。Delta が側方抑制におけるシグナリング分子であることを示す証拠はあるが、Serrateが側方抑制に関与していることを示唆するデータはない。そうではなく、Serrate は幼生の背側成虫盤、即ち肩、平均根及び翅盤の発生に必要であるようである。最後のものはこれらの過程のもっともよく研究されたものであるが、これにおいては、Serrate及びNotchは、翅の発生の全体としての有機的結合に必要な構造、背腹側翅縁の発生に重要である。
【0192】
C-Serrateが重要な機能を有していることは、特にそのNotch-結合ドメインのその配列が保存されていることから推論できる。C-Serrate について本明細書に記載した発現パターンは以下の情報を与えるものである。第一に、Serrate遺伝子はC-Notch-1が発現される部位あるいは隣接する部位において発現されるので(おそらくはその他のNotch相同体とともに)、C-SerrateはC-Notch-1(あるいは同様な発現パターンを有するその他のニワトリNotch相同体)に結合することによりその作用を発揮する可能性が極めて高い。第二に、発生中の腎臓、脈管系及び体肢芽における発現は中胚葉及び外胚葉間の誘導的シグナリングにおける関与を反映しているものであり得、前記のシグナリングはこれらの器官のすべてのものの発生において重要な部分を演ずるものである。例えば体肢芽においては、C-Serrateは遠位中胚葉において発現され、C-Notch-1は重畳する先端外胚葉隆起において発現され、この維持は下部の中胚葉からのシグナルに依存することが知られている。頭プラコードにおいては、同様な役割が可能であるが、誘導的シグナリングについての証拠はより弱いものであり、C-Serrateは同様に、プラコード上皮内の細胞間の連絡、例えばプラコード細胞の分化の特定のモードを制御することに関与している可能性がある。
【0193】
何がCNS 内のC-Serrateの発現の奇妙に限定されたドメイン内でのその機能であろうか?一つの可能性は、これがオリゴ樹状細胞の生産の制御に関与しているということであり、これは神経管の頭尾軸に沿って伸びる組織の狭いバンドから始まると報告されている。
【0194】
9.ヒトSerrate相同体の単離と特徴付け
ヒトSerrate 配列のクローンを以下に記載するようにして得た。
【0195】
ポリメラーゼ連鎖反応(PCR)を使用してヒト胎盤cDNAライブラリーからDNAを増幅した。この反応で使用した縮重オリゴヌクレオチドプライマーは、ショウジョウバエSerrate及びショウジョウバエDelta の間の相同性の高いアミノ末端領域に基づいて設計した(図5参照)。この相同性の高い領域は5'“DSL”ドメインを含んでおり、これはDelta及びSerrate のNotch-結合部分をコードすると考えられる。2種類のPCR 産物、350 bp断片と1.2 kb断片を単離して使用した。これらのPCR断片は32P により標識し、17〜18週齢の胎児から作製された市販のヒト胎児脳cDNΛライブラリー(以前からStratageneより入手可能)をスクリーニングするのに使用した。ここでこのcDNAをλ-ZapベクターのEcoRI部位に挿入した。
【0196】
前記1.2 kb断片は、106クローン中の1つのクローンにハイブリダイズし、スクリーニングした。単離されたファージλクローンを製造元の使用説明書に従ってプラスミドに変換することによりλDNAからこの断片を回収し、ベクターBluescriptKS-(Stratagene)のEcoRI 部位中の挿入物としてSerrate-相同cDNAを得た。このプラスミドは“pBS39”と命名し、このcDNAクローンに対応する遺伝子をヒトSerrate-1と称する(ヒトJagged-1(“HJ1”)としても知られる)。この単離されたcDNAは6464ヌクレオチド長で、完全なオープンリーディングフレーム並びに5'及び3'非翻訳領域を含んでいた(図1)。Sequenase(登録商標)配列決定システム(U.S.BiochemicalCorp.)を用いて、5及び6%Sequagelアクリルアミド配列決定ゲル上で配列決定を行った。
【0197】
前記350 bp断片は、約1.1及び3.1 kb長のcDNA挿入物を含む2種類のクローンとハイブリダイズした。これらの挿入物を含むプラスミド構築物はそれぞれpBS14 及びpBS15 と称する。それぞれのクローンを単離し、そのそれぞれの挿入物をλcDNAから回収し、上記のように配列決定した。pBS14挿入物のヌクレオチド配列は、pBS15 cDNA挿入物内に内部的に含まれている配列の1.1 kb部分に同一であり、従ってこのクローンについてはそれ以上の特徴付けは行わなかった。3.1kb pBS15挿入物の配列は、挿入物の5'末端の20ヌクレオチドを除く全てにわたるオープンリーディングフレームをコードしていた。この予測されたオープンリーディングフレームのアミノ末端残基に位置するメチオニンはpBS39中のヒトSerrate-1(HJ-1)cDNAクローンによりコードされた開始メチオニンと相同であった。pBS15 のcDNA挿入物をコードする遺伝子はヒトSerrate-2と命名し、これはヒトJagged-2(“HJ2”)としても知られる。
【0198】
前記pBS15(HJ2)3.1kb挿入物を次に32P で標識し、別のヒト胎児脳ライブラリー(Clontechからのもの)をスクリーニングするのに使用した。このライブラリーは、25〜26週齢胎児から作製されたcDNAをλgt11のEcoRI部位にクローニングしたものである。このスクリーニングにより3種類の可能性のある陽性クローンが同定された。cDNAを単離するために、液体溶解物からλgt11DNAを調製し、DEAEカラム上で精製した。精製されたDNAを次いでEcoRIで切断し、cDNA挿入物を単離し、Bluescript KS-のEcoRI 部位中にサブクローニングした。前記のcDNAを含むBluescript構築物をpBS3-15、pBS3-2及びpBS3-20と命名した。これらのcDNAクローンのうちの2種類、pBS3-2及びpBS3-20はpBS15 と部分的にオーバーラップする配列を含んでおり、さらに特徴付けした。pBS3-2は、pBS15cDNA挿入物のヌクレオチド1210からポリアデニル化シグナルの直後にわたる3.2 kb挿入物を有していた。pBS3-20 の2.6 kb挿入物について制限地図を作成し、部分的に配列決定して3'及び5'末端を決定した。この分析によれば、pBS3-20挿入物はpBS3-2cDNA挿入物内に完全に含まれる核酸配列を有していることが示され、従ってpBS3-20 はそれ以上特徴付けしなかった。pBS3-15 の挿入物はBluescriptベクター断片夾雑物として測定した。
【0199】
コンピューター上で作成された「完全」ヒトSerrate-2(HJ2)cDNA(配列番号3)の推定アミノ酸配列(配列番号4)の、pBS39からのヒトSerrate-1(HJ1)の推定アミノ酸配列(配列番号2)とのアライメントにより、pBS15(HJ2)挿入物によりコードされた領域内で、仮想的シグナル配列とDSLドメインの開始部との間にあるフレームシフトを生じる約120 塩基のギャップが判明した(図2)。pBS15 挿入物のギャップ中の失われたヌクレオチドは配列番号3のヌクレオチド240 及び241 の間に位置すべきものである。この失われた領域はおそらく、Stratageneライブラリー構築の際のクローニングの人工的産物により生じたものと思われる。
【0200】
アンカーPCR、RACE及びTakaraの延長PCR技術を使用したHJ2 の5'末端をクローン化する試みは成功しなかった。しかし、HJ2 の5'末端を含む可能性のある3種類のヒトゲノムクローンがヒトゲノムコスミドライブラリーのスクリーニングから得られた。このライブラリーは、pWE15ベクターのBamHI 部位中に導入した単一のXhoI部位中に30 kb 断片をクローニングしたものである(未改変ベクターはStratageneより入手可能である)。このコスミドライブラリーを、pBS15(HJ2)の5'末端から増幅されたPCR断片によりスクリーニングし、3種類の陽性コスミドクローンを単離した。2種類の異なるプライマーのセットを使用し、コスミドクローンをテンプレートとして使用して、pBS15の5'末端に対応するDNAを増幅したところ、両方のセットにより単一のバンドが生成され、これをサブクローニングしたが、いずれもPCR人工産物を含んでいることが判明した。コスミドクローンの部分はPCRなしに直接サブクローニングし、pBS15 において失われているDNAの120 ヌクレオチド部分を含むコスミドクローンの部分を得た。
【0201】
ヒトSerrate-1 相同体(HJ1)をコードするpBS39 cDNA挿入物を配列決定したところ、その遺伝子産物の完全なコード配列を含んでいた。そのヌクレオチド(配列番号1)及びタンパク質(配列番号2)の配列を図2に示す。ヒトSerrate-1(HJ1)のヌクレオチド配列をMacVectorソフトウェア(International Biotechnology Inc.,New Haven,CT)を使用して翻訳した。コード領域は配列番号1のヌクレオチド番号371-4024からなる。DNAStar(Madison,WI)のProteanタンパク質分析ソフトウェアプログラムを使用して単一のペプチド及びトランスメンブラン領域を予測した(疎水性に基づいた)。シグナルペプチドは、配列番号2のアミノ酸14-29(配列番号1のヌクレオチド番号410-457によりコードされる)からなると予測され、これにより成熟タンパク質のアミノ末端はアミノ酸番号30のGly から開始すると予測された。トランスメンブランドメインは、配列番号2のアミノ酸番号1068-1089であると予測され、配列番号1のヌクレオチド番号3572-3637によりコードされる。ショウジョウバエDelta 及びSerrate と相同性を有する領域であり、Notch(特にNotch ELR 11及び12)との結合を媒介すると予測されるコンセンサス(DSL)ドメインは、配列番号2のアミノ酸185-229にわたり、配列番号1のヌクレオチド番号923-1057によりコードされる。アミノ酸配列中の上皮成長因子様(ELR)リピートは目視同定した。以下の15の(完全長)ELR及び3の部分ELRが同定された。
従って、全ELRドメインはアミノ酸番号234-896(配列番号1のヌクレオチド番号1070-3058 によりコードされる)である。細胞外ドメインは、配列番号2のアミノ酸番号1-1067であると予測され、配列番号1のヌクレオチド番号371-3571によりコードされる(成熟タンパク質中のアミノ酸番号30-1067;配列番号1のヌクレオチド番号458-3571によりコードされる)。細胞内(細胞質)ドメインは、配列番号2のアミノ酸番号1090-1218であると予測され、配列番号1のヌクレオチド番号3638-4024 によりコードされる。
【0202】
特定の種類のヒト組織中におけるHJ1 の発現は、放射性標識pBS39 でClontech Human Multiple Tissue Northern Blotをプローブすることにより確認した。前記プローブは約6.6kbの単一のバンドとハイブリダイズし、心臓、脳、胎盤、肺、骨格筋、膵臓、肝臓及び腎臓を含むアッセイした全ての組織において発現された。HJ1 が成人骨格筋及び心筋において発現されたという観察は特に興味深いものであり、これは成人の筋肉繊維は細胞外マトリックスの層により完全に包囲されており、従ってこれらの細胞におけるHJ1の役割が直接の細胞間伝達にあるということはありそうにないからである。
【0203】
コンピュータ上で作成した「完全」(内部欠失を含む)ヒトSerrate-2(HJ2)cDNAヌクレオチド配列(配列番号3)及びアミノ酸配列(配列番号4)を図2に示す。ヌクレオチド配列はMacVectorソフトウェア(International Biotechnology Inc.,New Haven,CT)を使用して翻訳した。コード領域は配列番号3のヌクレオチド番号332-4102からなる。DNAStar(Madison,WI)のProteanタンパク質分析ソフトウェアプログラムを使用してシグナルペプチド及びトランスメンブラン領域を予測した(疎水性に基づいた)。トランスメンブランドメインは、配列番号4のアミノ酸番号912-933であると予測され、配列番号3のヌクレオチド番号3065-3130 によりコードされる。ショウジョウバエDelta 及びSerrate と相同性を有する領域であり、Notch(特にNotchELR 11及び12)との結合を媒介すると予測されるコンセンサス(DSL)ドメインは、配列番号4のアミノ酸26-70にわたり、配列番号3のヌクレオチド番号407-541によりコードされる。アミノ酸配列中の上皮成長因子様(ELR)リピートは目視同定した。以下の15の(完全長)ELR及び3の部分ELRが同定された。
従って、全ELRドメインはアミノ酸番号75-735(配列番号3のヌクレオチド番号554-2536によりコードされる)である。細胞外ドメインは、配列番号4のアミノ酸番号1-912であると予測され、配列番号3のヌクレオチド番号332-3064によりコードされる。細胞内(細胞質)ドメインは、配列番号4のアミノ酸番号934-1257であると予測され、配列番号3のヌクレオチド番号3131-4102によりコードされる。
【0204】
ヒトSerrate-1(HJ1)と同様に、コンピュータ上で作成した「完全」(内部欠失を有する)ヒトSerrate-2(HJ2)cDNA(配列番号3)は、16の完全なEGFリピート及び2つの中断されたEGF リピート、並びに仮想的Notchリガンド中にのみ見られたDSL ドメインとして知られる診断的には陰性のEFG リピートを含むタンパク質をコードする。コンピュータで作成された「完全」ヒトSerrate-2(HJ2)のオープンリーディングフレームは約1400アミノ酸長であり、HJ1及びラットSerrate相同体Jaggedのカルボキシ末端よりも約182 アミノ酸だけ長い。完全HJ2 及びHJ1 の間にはタンパク質のアミノ末端部分において有意な相同性が存在するが、この相同性はHJ1のアミノ酸番号1029付近の仮想的トランスメンブランドメインの直前で失われる。長いCOOH-末端テイルの存在はHJ2 の何らかの別の機能あるいは調節の可能性を意味するものであることから、この結果は、特に興味深いものである。
【0205】
「完全」(内部欠失を有する)ヒトSerrate-2(HJ2)cDNA(配列番号3)配列は、pBS15及びpBS3-2の配列オーバーラップ中に存在するAccI、DraIIIあるいはBamHI のための唯一の制限部位を利用して構築することができ、これらの酵素はそれぞれヌクレオチド1431、2648及び2802においてpBS15挿入物を切断する。
【0206】
特定の種類のヒト組織中におけるHJ2 の発現を、放射性標識クローンpBS15 でClontech Human Multiple Tissue NorthernBlotをプローブすることにより確認した。このプローブは約5.2 kbの単一のバンドとハイブリダイズし、心臓、脳、胎盤、肺、骨格筋及び膵臓において発現されたが、肝臓及び腎臓においては存在しないかあるいは殆ど検出できなかった。上述したHJ1発現の場合と同様に、HJ2 のpBS15 挿入物成分が成人の骨格筋及び心筋において発現されたという観察は特に興味深いものであり、これは成人筋肉繊維は細胞外マトリックスの層により完全に包囲されており、従ってこれらの細胞におけるHJ2の役割が直接の細胞間伝達にあるということことはありそうにないからである。
【0207】
発現構築物を単離されたクローンを使用して作製した。クローンをそのベクターからEcoRI 制限断片として切り出し、発現ベクターのEcoRI 制限部位中にサブクローニングした。これにより正しいリーディングフレーム中のサブクローンからのヒトSerrateタンパク質産物の発現が可能となった。この方法を使用して、サイトメガロウイルスプロモーターの制御下での発現用の発現ベクター中にpBS39 のHJ1cDNA挿入物をクローニングした発現構築物を作製し、3T3 及びHAKAT ヒトケラチノサイト系の両方においてHJ1を発現させた。
【0208】
10.微生物の寄託
完全長ヒトSerrate-1(HJ1)をコードするEcoRI断片を含むプラスミドpBS39 は、American Type Culture Collection,1201 Parklawn Drive,Rockville,Maryland20852に、特許手続上の微生物の寄託の国際的承認に関するブダペスト条約の規定に基づき、1995年2月28日に寄託し、受託番号97068 を得た。
【0209】
BluescriptKS-のEcoRI部位にクローニングしたヒトSerrate-2(HJ2)のアミノ末端をコードする3.1 kb EcoRI断片を含むプラスミドpBS15 は、AmericanType CultureCollection,1201 Parklawn Drive,Rockville,Maryland 20852に、特許手続上の微生物の寄託の国際的承認に関するブダペスト条約の規定に基づき、1995年3月5日に寄託し、受託番号 を得た。
【0210】
BluescriptKS-のEcoRI部位にクローニングしたヒトSerrate-2(HJ2)のカルボキシ末端をコードする3.2 kb EcoRI断片を含むプラスミドpBS3-2は、AmericanType Culture Collection,1201 Parklawn Drive,Rockville,Maryland 20852に、特許手続上の微生物の寄託の国際的承認に関するブダペスト条約の規定に基づき、1995年3月5日に寄託し、受託番号 を得た。
【0211】
本発明は、寄託された微生物あるいは本明細書に記載した特定の態様によりその範囲を限定されるものではない。実際に、これまでの記載及び添付の図面から、本明細書に記載したものに加えて本発明の種々の改変が当業者に明らかとなるであろう。そのような改変は添付の請求の範囲の範囲内に含まれるものである。
【0212】
本明細書に引用した種々の文献は、引用によりその全体を本明細書の一部とするものである。
【0213】
【技術分野】
【0001】
本発明は、補助金番号 GM 29093 およびNS 26084のもとに米国厚生省(Department of Health and HumanServices)から授与された政府援助金によって部分的に成された。米国政府は本発明に一定の権利を有する。
1.序論
本発明は脊椎動物Serrate遺伝子およびそれらによってコードされるタンパク質産物、ならびに上記タンパク質産物の誘導体および類似体に関する。脊椎動物Serrateタンパク質、誘導体および抗体の作製もまた提供される。本発明はさらに治療用組成物、ならびに診断および治療方法に関する。
【背景技術】
【0002】
2.発明の背景
ショウジョウバエにおける遺伝子分析は、複雑な発生経路を細かく調べ、そして相互作用している遺伝子座を同定するのに極めて有用であった。しかし、遺伝子の相互作用の下に横たわるプロセスの正確な性質を理解するには、該遺伝子のタンパク質産物の知識が必要である。
【0003】
発生学的、遺伝子的および分子的証拠は、ショウジョウバエにおける外胚葉分化の初期段階は細胞の相互作用に依存することを示している(Doe およびGoodman,1985,Dev.Biol.111:206-219; TechnauおよびCampos-Ortega,1986,Dev.Biol.195:445-454;Vassinら,1985,J.Neurogenet.2:291-308;de la Conchaら,1988,Genetics 118:499-508; Xuら,1990,Genes Dev.4:464-475;Artavanis-Tsakonas,1988,Trends Genet.4:95-100)。突然変異分析は、外胚葉細胞の表皮経路と神経経路の間の選択に影響を及ぼす、接合体的に(zygotically)作用する遺伝子の小グループ(いわゆる神経形成遺伝子座)を明らかにしている(Poulson,1937,Proc.Natl.Acad.Sci.23:133-137;Lehmannら,1983,Wilhelm Roux's Arch.Dev.Biol.192:62-74; Jurgensら,1984,WilhelmRoux's Arch.Dev.Biol.193:283-295; Wieschausら,1984,Wilhelm Roux's Arch.Dev.Biol.193:296-307;Nusslein-Volhard ら,1984,Wilhelm Roux's Arch.Dev.Biol.193:267-282)。接合体性神経形成遺伝子座[Notch(N)、Delta(Dl)、mastermind(mam)、EnhancerofSplit(E(spl)、neuralized(neu)、およびbig brain(bib)]のいずれか1つにおけるヌル突然変異は、腹側および側面の表皮構造を犠牲にして神経系の肥大をもたらす。この効果は、表皮前駆細胞を誤って神経経路に入れたためであり、そしてこのことは、神経形成遺伝子機能は神経形成領域内の細胞を神経になる運命から上皮になる運命に方向転換させるために必要であることを暗示している。Serrateは、Notch遺伝子座と相互作用することが可能な遺伝子ユニットとして同定された(Xuら,1990,GenesDev.4:464-475)。これらの遺伝子的および発生的考察は、以下の仮説に導いた。すなわち、神経形成遺伝子座のタンパク質産物は、適切な表皮発生に必要な細胞相互作用機構の構成要素として機能する、という仮説である(Artavanis-Tsakonas,S.,1988,TrendsGenet.4:95-100)。
【0004】
突然変異分析はまた、神経形成遺伝子の作用は多面発現的であって、胚発生のみに限定されないことを明らかにしている。例えば、個眼、剛毛、羽根の形成(これらもまた細胞相互作用に依存することが知られている)は神経形成突然変異によって影響される(Morganら,1925,Bibliogr.Genet.2:1-226; Welsons,1956,Dros.Inf.Serv.30:157-158;Preissら,1988,EMBO J.7:3917-3927; ShellenbargerおよびMohler,1978,Dev.Biol.62:432-446;TechnauおよびCampos-Ortega,1986,Wilhelm Roux's Dev.Biol.195:445-454; Tomlson およびReady,1987,Dev.Biol.120:366-376;CaganおよびReady,1989,Genes Dev.3:1099-1112)。
【0005】
配列分析(Whartonら,1985,Cell43:567-581; Kidd およびYoung,1986,Mol.Cell.Biol.6:3094-3108; Vassinら,1987,EMBO J.6:3431-3440;Kopczynskiら,1988,Genes Dev.2:1723-1735)は、神経形成遺伝子座の2つ、すなわちNotchおよびDeltaは、膜を一回貫く膜貫通タンパク質をコードしているように思われることを示した。Notch遺伝子は、Notch/lin-12リピートと称する、3個のシステインが豊富なリピートを後に従えた36個の表皮増殖因子(EGF)に似たタンデムなリピートを含む大きいN末端細胞外ドメインを有する約300kdのタンパク質(本発明者らはこのタンパク質を示すために「Notch」という用語を用いる)をコードしている(Whartonら,1985,Cell43:567-581; Kidd およびYoung,1986,Mol.Cell.Biol.6:3094-3108; Yochemら,1988,Nature335:547-550)。Delta は、細胞外ドメイン内に9個のEGFに似たリピートを有する約100 kdのタンパク質(本発明者らはDLZM、すなわち顕著な接合体および母体の転写物のタンパク質産物を示すために「Delta」という用語を用いる;Kopczynskiら,1988,Genes Dev.2:1723-1735)をコードしている(Vassinら,1987,EMBO J.6:3431-3440;Kopczynskiら,1988,Genes Dev.2:1723-1735)。分子的研究は以下の示唆をもたらした。すなわち、Notch およびDelta は、初期の発生的決定に関与する細胞コミュニケーション作用機構の生化学的に相互作用する要素を構成する、という示唆である(Fehonら,1990,Cell61:523-534)。
【0006】
EGFに似たモチーフは、血液凝固カスケードに関与するタンパク質を含む種々のタンパク質に見いだされている(FurieおよびFurie,1988,Cell 53:505-518)。特に、このモチーフは血液凝固因子IXおよびX等の細胞外タンパク質(Reesら,1988,EMBOJ.7:2053-2061; FurieおよびFurie,1988,Cell 53:505-518)、他のショウジョウバエ遺伝子(Knustら,1987,EMBOJ.761-766; Rothbergら,1988,Cell 55:1047-1059)、および幾つかの細胞表面受容体タンパク質、例えばトロンボモジュリン(Suzukiら,1987,EMBOJ.6:1891-1897)およびLDL受容体(Sudhofら,1985,Science 228:815-822)に見いだされている。タンパク質結合部位がトロンボモジュリンおよびウロキナーゼのEGFリピートドメインにマップされた(Kurosawaら,J.Biol.Chem.263:5993-5996; Appella ら,1987,J.Biol.Chem.262:4437-4440)。ショウジョウバエSerrate遺伝子はすでにクローン化され、特徴付けされている(1993年6月24日付けPCT公開 WO 93/12141)。しかし、脊椎動物Serrate遺伝子を得ようとする試みにもかかわらず、本発明がなされるまで、いかなる脊椎動物Serrate遺伝子も入手不可能であった。
【0007】
上記文献の引用は、それらの文献が本発明の先行技術であると承認したものと解釈して
はならない。
【発明の概要】
【課題を解決するための手段】
【0008】
3.発明の概略
本発明は脊椎動物Serrate遺伝子(ヒトSerrate遺伝子および他の種の関連する遺伝子)のヌクレオチド配列、およびそれらの遺伝子によってコードされるタンパク質のアミノ酸配列、ならびにそれらの誘導体(例えば断片)および類似体に関する。前記のヌクレオチド配列とハイブリダイゼーション可能な、またはそれに相補的な核酸もまた提供される。特定の実施態様において、前記Serrateタンパク質はヒトタンパク質である。
【0009】
本発明は、機能的に活性な、すなわち全長(野性型)Serrateタンパク質に関連する1以上の公知の機能活性を示すことができる、本発明の脊椎動物Serrate誘導体および類似体に関する。このような機能活性は抗原性〔抗Serrate抗体に結合する(またはSerrateと結合を競合する)能力〕、免疫原性(Serrateに結合する抗体を生成する能力)、Notchまたは他のトポリズミック(toporythmic)なタンパク質またはその断片に結合する(またはSerrateと結合を競合する)能力(「接着性」("adhesiveness"))、Serrate受容体に結合する(またはSerrateと結合を競合する)能力を含むが、これらだけに限定されない。本明細書に使用する「トポリズミックなタンパク質」とは、Notch、Delta、Serrate、EnhancerofSplit およびDeltexのタンパク質産物、ならびに、例えばハイブリダイズするそれらの遺伝子配列の能力、またはDelta、SerrateまたはNotchとの相同性、または表現型の相互作用を示す遺伝子の能力によって同定可能な、この相互に作用する遺伝子ファミリーの他のメンバーのタンパク質産物を言う。
【0010】
本発明はさらに、細胞内ドメイン、細胞外ドメイン、膜貫通ドメイン、膜結合領域、またはSerrateタンパク質の1以上のEGFに似た(相同の)リピートまたはこれらの任意の組合せを含むがこれらだけに限定されない、Serrateタンパク質の1以上のドメインを含む脊椎動物Serrateの断片(およびそれらの誘導体および類似体)に関する。
【0011】
脊椎動物Serrateに対する抗体、その誘導体および類似体がさらに提供される。
【0012】
脊椎動物Serrateタンパク質、その誘導体および類似体の例えば組換え手段による作製方法もまた提供される。
【0013】
本発明はまた、脊椎動物Serrateタンパク質および核酸に基づく治療および診断方法な
らびに組成物に関する。本発明は、本発明の治療用化合物を投与することによる細胞の運
命、すなわち分化、の障害に対する治療を提供する。このような治療用化合物(本明細書
では「治療剤」と呼ぶ)は、脊椎動物Serrateタンパク質ならびにそれらの類似体および
誘導体(断片を含む);それらに対する抗体;脊椎動物Serrateタンパク質、類似体また
は誘導体をコードする核酸;および脊椎動物Serrateアンチセンス核酸を含む。好ましい
実施態様において、本発明の治療剤は癌状態を治療するため、または前腫瘍状態あるいは
非悪性腫瘍状態から腫瘍状態または悪性腫瘍状態への進行を阻止するために投与される。
他の特定の実施態様において、本発明の治療剤は神経系の障害を治療するため、または組
織の再生及び修復を促進するために投与される。
【0014】
1つの実施態様において、Notchおよび/またはSerrate機能に拮抗する、またはこれらを抑制する治療剤(以後「アンタゴニスト治療剤」と称する)が治療的効果のために投与される。別の実施態様においては、Notchおよび/またはSerrate機能を促進する治療剤(以後「アゴニスト治療剤」と称する)が治療的効果のために投与される。
【0015】
Notchおよび/またはSerrateタンパク質の異常な、または望ましくないレベルの発現、
または活性、または局在化を伴う細胞の運命に関する疾患、特に過剰増殖性(例えば癌)
または過少増殖性疾患は、後により詳しく記述するようにそれらのレベルを検出すること
により診断することができる。
【0016】
好ましい側面において、本発明の治療剤は、Notchタンパク質またはその断片への結合
を媒介する脊椎動物Serrateの少なくとも断片(本明細書では「接着性断片」と称する)
からなるタンパク質である。
本発明は、例えば以下の項目を提供する。
(項目1)
精製された脊椎動物のSerrateタンパク質。
(項目2)
ヒトのタンパク質である項目1に記載のタンパク質。
(項目3)
哺乳動物のタンパク質である項目1に記載のタンパク質。
(項目4)
配列番号2のアミノ酸番号30〜1218に実質的に記載されているアミノ酸配列を含
有する項目2に記載のタンパク質。
(項目5)
配列番号4のアミノ酸番号1〜1257に実質的に記載されているアミノ酸配列を含有
する項目2に記載のタンパク質。
(項目6)
ATCCに寄託されて受託番号68876号を指定されたプラスミドSerFLまたは該プラスミド中のSerrate配列にハイブリダイズ可能な核酸によってコードされる精製されたヒトタンパク質。
(項目7)
ATCCに寄託されて受託番号97068号を指定されたプラスミドpBS39によってコードされる
項目2に記載のタンパク質。
(項目8)
ATCCに寄託されて受託番号 号を指定されたプラスミドpBS15によってコードされるSerrateアミノ酸配列を含有する項目2に記載のタンパク質。
(項目9)
ATCCに寄託されて受託番号 号を指定されたプラスミドpBS3-2によってコードされるSerrateアミノ酸配列を含有する項目2に記載のタンパク質。
(項目10)
項目1のタンパク質の精製された断片であって、Serrateタンパク質の1つ以上の機
能活性を示すことが可能である前記断片。
(項目11)
項目2のタンパク質の精製された断片であって、ヒトまたはキイロショウジョウバエ(D.melanogaster)のSerrateタンパク質の1つ以上の機能活性を示すことが可能である前記断片。
(項目12)
ヒトのSerrateタンパク質に対する抗体と結合可能である項目2または7に記載のタンパク質の精製された断片。
(項目13)
項目10に記載の断片を含有する分子。
(項目14)
脊椎動物のSerrateタンパク質の精製された断片であって、前記タンパク質の細胞外ドメイン、DSLドメイン、表皮増殖因子様のリピートドメイン、システインに富むドメイン、膜貫通ドメインおよび細胞内ドメインからなる群から選ばれるドメインを含有する前記断片。
(項目15)
脊椎動物のSerrateタンパク質の精製された断片であって、前記タンパク質のDSLドメインを含有する前記断片。
(項目16)
脊椎動物のSerrateタンパク質の精製された断片であって、前記タンパク質の表皮増殖因子に相同性のリピートを含有する前記断片。
(項目17)
前記Serrateタンパク質がヒトSerrateタンパク質である項目14に記載の断片。
(項目18)
Notchタンパク質またはDeltaタンパク質に相同性の領域を含有し、かつ少なくとも10個のアミノ酸からなる脊椎動物のSerrateタンパク質の精製した断片。
(項目19)
Serrateタンパク質ではない第2のタンパク質のアミノ酸配列に共有結合によって融合している少なくとも10個のアミノ酸からなる脊椎動物のSerrateタンパク質の断片を含有するキメラタンパク質。
(項目20)
Serrateタンパク質の前記断片が抗Serrate抗体によって結合され得る断片である項目19に記載のキメラタンパク質。
(項目21)
前記Serrateタンパク質がヒトのタンパク質である項目19に記載のキメラタンパク質。
(項目22)
Serrateタンパク質の1つ以上の機能活性を示すことが可能である項目19に記載のキメラタンパク質。
(項目23)
脊椎動物のSerrateタンパク質の精製した断片であって、(a)抗Serrate抗体によって結合可能であり、(b)前記タンパク質の膜貫通および細胞内ドメインを欠失しており、かつ(c)前記Serrateタンパク質の少なくとも10個のアミノ酸からなる前記断片。
(項目24)
脊椎動物のSerrateタンパク質の精製した断片であって、(a)抗Serrate抗体によって結合可能であり、(b)前記タンパク質の細胞外ドメインを欠失しており、かつ(c)前記Serrateタンパク質の少なくとも10個のアミノ酸からなる前記断片。
(項目25)
Notchタンパク質に結合可能な脊椎動物Serrateタンパク質の精製した断片。
(項目26)
前記Serrateタンパク質の表皮増殖因子様リピートを欠失している項目25に記載の断片。
(項目27)
前記Serrateタンパク質がヒトSerrateタンパク質である項目23、24、25または26に記載の断片。
(項目28)
配列番号2または配列番号4の断片である項目29に記載の断片。
(項目29)
項目25に記載の断片を含有する分子。
(項目30)
項目1に記載のSerrateタンパク質と結合可能であり、かつショウジョウバエ(Drosophila)のSerrateタンパク質とは結合しない抗体。
(項目31)
項目2に記載のSerrateタンパク質と結合可能であり、かつショウジョウバエ(Drosophila)のSerrateタンパク質とは結合しない抗体。
(項目32)
モノクローナルである項目30に記載の抗体。
(項目33)
項目32に記載の抗体の断片を含有する分子であって、該断片が脊椎動物のSerrate
タンパク質と結合可能である前記分子。
(項目34)
脊椎動物のSerrateタンパク質をコードするヌクレオチド配列を含有する単離した核酸。
(項目35)
DNAである項目34に記載の核酸。
(項目36)
項目34に記載のヌクレオチド配列に完全に相補的なヌクレオチド配列を含有する単
離した核酸。
(項目37)
項目2に記載のSerrateタンパク質をコードするヌクレオチド配列を含有する単離し
た核酸。
(項目38)
ATCCに寄託して受託番号97068号を指定されたプラスミドpBS39に含有されるSerrateコード配列を含有する単離した核酸。
(項目39)
ATCCに寄託して受託番号68876号を指定されたプラスミドSerFLおよび該プラスミド中のSerrate配列にハイブリダイズ可能な単離したヒト核酸。
(項目40)
ATCCに寄託して受託番号 号を指定されたプラスミドpBS3-2に含有されるSerrateコード配列を含有する単離した核酸。
(項目41)
ATCCに寄託して受託番号 号を指定されたプラスミドpBS15に含有されるSerrateコード配列を含有する単離した核酸。
(項目42)
タンパク質をコードするヌクレオチド配列を含有する単離した核酸であって、前記タン
パク質が配列番号4のアミノ酸番号1〜1257を含有する前記核酸。
(項目43)
少なくとも8個のヌクレオチドからなる脊椎動物のSerrate遺伝子の断片を含有する単離された核酸。
(項目44)
項目14、15、16または25に記載の断片をコードするヌクレオチド配列を含有する単離した核酸。
(項目45)
前記断片がヒトSerrateタンパク質の断片である項目44に記載の核酸。
(項目46)
項目12に記載の断片をコードするヌクレオチド配列を含有する単離された核酸。
(項目47)
タンパク質をコードするヌクレオチド配列を含有する単離した核酸であって、前記タンパク質が配列番号2のアミノ酸番号30〜1218を含有する前記核酸。
(項目48)
項目21に記載のタンパク質をコードするヌクレオチド配列を含有する単離された核酸。
(項目49)
項目34、37または43に記載の核酸を含有する組換え細胞。
(項目50)
項目38、40または41に記載の核酸を含有する組換え細胞。
(項目51)
項目34または37に記載の核酸を含有する組換え細胞を増殖させて、コードされた前記Serrateタンパク質が前記細胞によって発現されるようにする工程と、前記発現されたSerrateタンパク質を回収する工程とを包含するSerrateタンパク質の生成方法。
(項目52)
項目38、40または41に記載の核酸を含有する組換え細胞を増殖させて、コードされたSerrateタンパク質が前記細胞によって発現されるようにする工程と、前記発現されたSerrateタンパク質を回収する工程とを包含するSerrateタンパク質の生成方法。
(項目53)
項目45に記載の核酸を含有する組換え細胞を増殖させて、コードされたタンパク質が前記細胞によって発現されるようにする工程と、前記発現されたタンパク質を回収する工程とを包含するタンパク質の生成方法。
(項目54)
Serrateタンパク質の断片を含有するタンパク質の生成方法であって、項目46に記載の核酸を含有する組換え細胞を増殖させて、コードされたタンパク質が前記細胞によって発現されるようにする工程と、前記発現されたタンパク質を回収する工程とを包含する前記方法。
(項目55)
項目51に記載の方法による生産物。
(項目56)
項目52に記載の方法による生産物。
(項目57)
項目53に記載の方法による生産物。
(項目58)
項目54に記載の方法による生産物。
(項目59)
治療上有効な量の脊椎動物Serrateタンパク質と、製剤上許容される担体とを含有する医薬組成物。
(項目60)
前記Serrateタンパク質がヒトSerrateタンパク質である項目59に記載の組成物。
(項目61)
治療上有効な量の項目14、15、16または25に記載の断片と、製剤上許容される担体とを含有する医薬組成物。
(項目62)
治療上有効な量の項目12に記載の断片と、製剤上許容される担体とを含有する医薬組成物。
(項目63)
脊椎動物Serrateタンパク質の断片を含有する分子を治療上有効な量で含有する医薬組成物であって、その誘導体または類似体が、Notchタンパク質への結合能、またはNotchタンパク質の表皮増殖因子様リピート11および12を含有する分子への結合能により特徴付けられる前記組成物。
(項目64)
治療上有効な量の項目34、36または37に記載の核酸と、製剤上許容される担体とを含有する医薬組成物。
(項目65)
治療上有効な量の項目44に記載の核酸と、製剤上許容される担体とを含有する医薬組成物。
(項目66)
治療上有効な量の項目46に記載の核酸と、製剤上許容される担体とを含有する医薬組成物。
(項目67)
治療上有効な量の項目30に記載の抗体と、製剤上許容される担体とを含有する医薬組成物。
(項目68)
治療上有効な量の項目30に記載の抗体の結合ドメインを含有する前記抗体の断片または誘導体と、製剤上許容される担体とを含有する医薬組成物。
(項目69)
被験者の疾病または障害を治療または予防する方法であって、かかる治療または予防が望まれる被験者に、Notchタンパク質に結合可能な脊椎動物Serrateタンパク質またはその誘導体を治療上有効な量で投与することを含んでなる前記方法。
(項目70)
前記疾病または障害が、類似の非悪性腫瘍サンプルにおけるNotch活性または発現と比べて、高いNotch活性、またはNotchタンパク質または抗Notch抗体によって結合されるNotch誘導体の高い発現によって特徴付けられる悪性腫瘍である項目69に記載の方法。
(項目71)
前記疾病または障害が、子宮頸癌、乳癌、結腸癌、黒色腫、精上皮腫および肺癌からなる群から選ばれる項目69に記載の方法。
(項目72)
前記被験者がヒトである項目69に記載の方法。
(項目73)
前記Serrateタンパク質がヒトSerrateタンパク質である項目69に記載の方法。
(項目74)
被験者の疾病または障害を治療または予防する方法であって、かかる治療または予防が望まれる被験者に、治療上有効な量の分子を投与すること含んでなり、前記分子が、(a)10個のヌクレオチドを含有し、(b)脊椎動物のSerrate遺伝子に特異的なRNA転写の少なくとも10ヌクレオチド部分に完全に相補的な配列を含有し、かつ(c)前記RNA転写物にハイブリダイズ可能なオリゴヌクレオチドである前記方法。
(項目75)
被験者の疾病または障害を治療または予防する方法であって、かかる治療または予防が望まれる被験者に、治療上有効な量の項目34、37または46に記載の核酸を投与することを含んでなる前記方法。
(項目76)
被験者の疾病または障害を治療または予防する方法であって、かかる治療または予防が望まれる被験者に、治療上有効な量の項目32に記載の抗体を投与することを含んでなる前記方法。
(項目77)
前記疾病または障害が中枢神経系の疾病または障害である項目73に記載の方法。
(項目78)
10個のヌクレオチドを含有し、かつ脊椎動物のSerrate遺伝子に特異的なRNA転写物の少なくとも10ヌクレオチド部分に完全に相補的な配列を含有する単離されたオリゴヌクレオチドであって、前記RNA転写物にハイブリダイズ可能である前記オリゴヌクレオチド。
(項目79)
項目78に記載のオリゴヌクレオチドと、製剤上許容される担体とを含有する医薬組成物。
(項目80)
Serrateタンパク質をコードする核酸配列の細胞における発現の抑制方法であって、前記細胞に、有効量の項目78に記載のオリゴヌクレオチドを供給することを含んでなる前記方法。
(項目81)
患者における異常レベルのNotch-Serrateタンパク質の結合活性によって特徴付けられる疾病または障害の診断方法であって、前記患者由来のサンプル中のNotchタンパク質の、脊椎動物Serrateタンパク質への結合能を測定することを含んでなり、正常な個体由来の類似サンプルで見られる結合能と比べて、前記Notchタンパク質の、前記Serrateタンパク質への結合能の増加または減少が、前記患者における前記疾病または障害の存在を示す前記方法。
(項目82)
患者由来のサンプルにおける脊椎動物Serrateタンパク質のレベルを測定することを含んでなる、異常レベルのSerrateタンパク質によって特徴付けられる前記患者における疾病または障害の診断方法であって、正常な個体由来の類似サンプルで見られる前記Serrateタンパク質のレベルと比べて、前記Serrateタンパク質のレベルの増加および減少が、前記患者における前記疾病または障害の存在を示す前記方法。
【0017】
3.1.定義
本明細書においては、何ら強調なしに遺伝子の名前によって示されるタンパク質産物に
対し、遺伝子の名前を強調したもの、つまりイタリック体にしたものまたは下線を引いた
ものは遺伝子を表すものとする。例えば、「Serrate」はSerrate遺伝子を示すものとし、
これに対し「Serrate」はSerrate遺伝子のタンパク質産物を示すものとする。
【図面の簡単な説明】
【0018】
【図1A】Human Serrate-1(Human Jagged-1(HJ1)としても知られている)のヌクレオチド配列(配列番号1)およびタンパク質配列(配列番号2)。
【図1B】Human Serrate-1(Human Jagged-1(HJ1)としても知られている)のヌクレオチド配列(配列番号1)およびタンパク質配列(配列番号2)。
【図1C】Human Serrate-1(Human Jagged-1(HJ1)としても知られている)のヌクレオチド配列(配列番号1)およびタンパク質配列(配列番号2)。
【図1D】Human Serrate-1(Human Jagged-1(HJ1)としても知られている)のヌクレオチド配列(配列番号1)およびタンパク質配列(配列番号2)。
【図1E】Human Serrate-1(Human Jagged-1(HJ1)としても知られている)のヌクレオチド配列(配列番号1)およびタンパク質配列(配列番号2)。
【図1F】Human Serrate-1(Human Jagged-1(HJ1)としても知られている)のヌクレオチド配列(配列番号1)およびタンパク質配列(配列番号2)。
【図1G】Human Serrate-1(Human Jagged-1(HJ1)としても知られている)のヌクレオチド配列(配列番号1)およびタンパク質配列(配列番号2)。
【図2A】ヒト胎児脳cDNAライブラリーより単離したクローンpBS15 およびpBS3-2の配列を組み合わせてコンピュータを用いて作製したHuman Serrate-2(HumanJagged-2(HJ2)としても知られている)の「完全な」ヌクレオチド配列(配列番号3)およびアミノ酸配列(配列番号4)。シグナル配列とSDLドメインの間のHumanSerrate-2 の部分をコードする領域には、約120 ヌクレオチドの欠失がある。
【図2B】ヒト胎児脳cDNAライブラリーより単離したクローンpBS15 およびpBS3-2の配列を組み合わせてコンピュータを用いて作製したHuman Serrate-2(HumanJagged-2(HJ2)としても知られている)の「完全な」ヌクレオチド配列(配列番号3)およびアミノ酸配列(配列番号4)。シグナル配列とSDLドメインの間のHumanSerrate-2 の部分をコードする領域には、約120 ヌクレオチドの欠失がある。
【図2C】ヒト胎児脳cDNAライブラリーより単離したクローンpBS15 およびpBS3-2の配列を組み合わせてコンピュータを用いて作製したHuman Serrate-2(HumanJagged-2(HJ2)としても知られている)の「完全な」ヌクレオチド配列(配列番号3)およびアミノ酸配列(配列番号4)。シグナル配列とSDLドメインの間のHumanSerrate-2 の部分をコードする領域には、約120 ヌクレオチドの欠失がある。
【図2D】ヒト胎児脳cDNAライブラリーより単離したクローンpBS15 およびpBS3-2の配列を組み合わせてコンピュータを用いて作製したHuman Serrate-2(HumanJagged-2(HJ2)としても知られている)の「完全な」ヌクレオチド配列(配列番号3)およびアミノ酸配列(配列番号4)。シグナル配列とSDLドメインの間のHumanSerrate-2 の部分をコードする領域には、約120 ヌクレオチドの欠失がある。
【図2E】ヒト胎児脳cDNAライブラリーより単離したクローンpBS15 およびpBS3-2の配列を組み合わせてコンピュータを用いて作製したHuman Serrate-2(HumanJagged-2(HJ2)としても知られている)の「完全な」ヌクレオチド配列(配列番号3)およびアミノ酸配列(配列番号4)。シグナル配列とSDLドメインの間のHumanSerrate-2 の部分をコードする領域には、約120 ヌクレオチドの欠失がある。
【図2F】ヒト胎児脳cDNAライブラリーより単離したクローンpBS15 およびpBS3-2の配列を組み合わせてコンピュータを用いて作製したHuman Serrate-2(HumanJagged-2(HJ2)としても知られている)の「完全な」ヌクレオチド配列(配列番号3)およびアミノ酸配列(配列番号4)。シグナル配列とSDLドメインの間のHumanSerrate-2 の部分をコードする領域には、約120 ヌクレオチドの欠失がある。
【図2G】ヒト胎児脳cDNAライブラリーより単離したクローンpBS15 およびpBS3-2の配列を組み合わせてコンピュータを用いて作製したHuman Serrate-2(HumanJagged-2(HJ2)としても知られている)の「完全な」ヌクレオチド配列(配列番号3)およびアミノ酸配列(配列番号4)。シグナル配列とSDLドメインの間のHumanSerrate-2 の部分をコードする領域には、約120 ヌクレオチドの欠失がある。
【図3A】ニワトリSerrate(C-Serrate)cDNAのヌクレオチド配列(配列番号5)。
【図3B】ニワトリSerrate(C-Serrate)cDNAのヌクレオチド配列(配列番号5)。
【図4A】C-Serrate(シグナル配列のアミノ末端を欠く)のアミノ酸配列(配列番号6)。シグナル配列(成熟タンパク質の予測されるアミノ末端を示す)に続く推定上の開裂部位を矢印で示す;DSLドメインを星印で示す;EGFに似たリピート(ELR)に破線を施してある;ELRと膜貫通ドメインの間のシステインが豊富な領域を矢印の間に示す;そしてシグナル膜貫通ドメイン(アミノ酸1042と1066の間)を太字で示す。
【図4B】C-Serrate(シグナル配列のアミノ末端を欠く)のアミノ酸配列(配列番号6)。シグナル配列(成熟タンパク質の予測されるアミノ末端を示す)に続く推定上の開裂部位を矢印で示す;DSLドメインを星印で示す;EGFに似たリピート(ELR)に破線を施してある;ELRと膜貫通ドメインの間のシステインが豊富な領域を矢印の間に示す;そしてシグナル膜貫通ドメイン(アミノ酸1042と1066の間)を太字で示す。
【図5】キイロショウジョウバエDelta(配列番号7)およびSerrate(配列番号8)ならびにC-Serrate(配列番号6)のアミノ末端配列のアライメント。示された領域はシグナル配列の末端からDSLドメインの末端に伸びている。DSLドメインが示されている。3つのタンパク質全てにおいて同一のアミノ酸を四角で囲んである。
【図6】C-Serrateと比較したショウジョウバエDeltaおよびショウジョウバエSerrateのドメイン構造を示す図。C-SerrateおよびショウジョウバエSerrateにのみ存在する、EGFリピートのすぐ下流にある第2のシステインに富む領域は、示されていない。疎水性領域を黒で示す;DSLドメインに碁盤模様を付し、そしてEGFに似たリピートに平行線の陰影を付してある。
【発明を実施するための形態】
【0019】
5.発明の詳細な説明
本発明は脊椎動物Serrate遺伝子のヌクレオチド配列、およびそれらによってコードさ
れるタンパク質のアミノ酸配列に関する。本発明はさらに脊椎動物Serrateタンパク質の断片および他の誘導体、および類似体に関する。このような断片または誘導体をコードする核酸もまた本発明の範囲内にある。本発明は多数の異なる種の脊椎動物Serrate遺伝子、およびそれらによってコードされるタンパク質を提供する。本発明のSerrate遺伝子は、ヒトSerrateおよび脊椎動物種における関連する遺伝子(相同体)を含む。特定の実施態様において、Serrate遺伝子およびタンパク質は哺乳動物由来である。本発明の好ましい実施態様において、Serrateタンパク質はヒトタンパク質である。最も好ましい実施態様において、Serrateタンパク質はHumanSerrate-1 またはHuman Serrate-2 である。上記タンパク質および誘導体の例えば組換え法による作製が提供される。
【0020】
本発明は機能的に活性な、すなわち全長(野性型)Serrateタンパク質に関連する1以上の公知の機能活性を示すことができる、本発明の脊椎動物Serrate誘導体および類似体に関する。このような機能活性は抗原性〔抗Serrate抗体に結合する(またはSerrateと結合を競合する)能力〕、免疫原性(Serrateに結合する抗体を生成する能力)、Notchまたは他のトポリズミックなタンパク質またはその断片に結合する(またはSerrateと結合を競合する)能力(「接着性」)、Serrate受容体に結合する(またはSerrateと結合を競合する)能力を含むが、これらだけに限定されない。本明細書に使用する「トポリズミックなタンパク質」とは、Notch、Delta、Serrate、Enhancerof SplitおよびDeltexのタンパク質産物、ならびに、例えばハイブリダイズするそれらの遺伝子配列の能力、またはDelta、Serrateまたは Notchとの相同性、または表現型の相互作用を示す遺伝子の能力によって同定可能な、この相互に作用する遺伝子ファミリーの他のメンバーのタンパク質産物を言う。
【0021】
本発明はさらに、細胞内ドメイン、細胞外ドメイン、膜貫通ドメイン、膜関連領域、ま
たはSerrateタンパク質の1以上のEGFに似た(相同の)リピート、またはこれらの任
意の組合せを含むがこれらだけに限定されない、Serrateタンパク質の1以上のドメイン
を含む脊椎動物Serrateの断片(およびそれらの誘導体および類似体)に関する。
【0022】
Serrateに対する抗体、その誘導体および類似体がさらに提供される。
【0023】
後に示すように、Serrateは発生および他の生理的プロセスにおいて、特にNotchへのリガンドとして重大な役割を果たす。この役割は、細胞の運命(分化)の決定に関与している。特に、Serrateは中枢神経系における細胞の運命の決定において主要な役割を果たすと思われる。分化および他の生理的プロセスの研究および操作において、本発明の核酸およびアミノ酸配列およびそれらに対する抗体を、ヒトおよび他の種のSerratemRNAおよびタンパク質の検出および定量化のために用いて、それらの発現を研究し、Serrateおよび断片および他の誘導体および類似体を作製することができる。本発明はまた、Serrateタンパク質および核酸配列に基づく治療および診断方法、ならびに組成物に関する。本発明は、本発明の治療用化合物を投与することによる細胞の運命(すなわち分化)の疾患に対する治療を提供する。このような治療用化合物(本明細書では「治療剤」と呼ぶ)は、脊椎動物Serrateタンパク質ならびにそれらの類似体および誘導体(断片を含む);それらに対する抗体;脊椎動物Serrateタンパク質、類似体または誘導体をコードする核酸;および脊椎動物Serrateアンチセンス核酸を含む。好ましい実施態様において、本発明の治療剤は癌状態を治療するため、または前腫瘍状態あるいは非悪性腫瘍状態から腫瘍状態または悪性腫瘍状態への進行を阻止するために投与される。他の特定の実施態様において、本発明の治療剤は神経系疾患を治療するため、または組織の再生及び補修を促進するために投与される。
【0024】
1つの実施態様において、Notchおよび/またはSerrate機能に拮抗する、またはこれらを抑制する治療剤(以後「アンタゴニスト治療剤」と称する)が治療的効果のために投与される。別の実施態様においては、Notchおよび/またはSerrate機能を促進する治療剤(以後「アゴニスト治療剤」と称する)が治療的効果のために投与される。
【0025】
Notchおよび/またはSerrateタンパク質の異常な、または望ましくないレベルの発現、
または活性、または局在化を伴う細胞の運命に関する疾患、特に過剰増殖性(例えば癌)
または過少増殖性疾患は、後により詳しく記述するようにそれらのレベルを検出すること
により診断することができる。
【0026】
好ましい側面において、本発明の治療剤は、Notchタンパク質またはその断片への結合
を媒介する脊椎動物Serrateの少なくとも断片(本明細書では「接着性断片」と称する)
からなるタンパク質である。
【0027】
本発明は、なかんずくマウスSerrate相同体のクローニング(第6節)、アフリカツメガエル(カエル)Serrate相同体のクローニング(第7節)、ニワトリSerrate相同体のクローニング(第8節)、およびヒトSerrate相同体であるHuman Serrate-1(HJ1)およびHumanSerrate-2(HJ2)のクローニング(第9節)を開示する後述の実施例によって例証される。
【0028】
開示を明確にするため、限定するためではないが、本発明の詳細な記述を以下の分節に分ける。
【0029】
5.1.Serrate遺伝子の単離
本発明は、脊椎動物Serrate核酸のヌクレオチド配列に関する。特定の実施態様において、脊椎動物Serrate核酸は図1(配列番号1)、図2(配列番号3)、図3(配列番号6)に示されるcDNA配列又はそれらのコード領域、又は脊椎動物Serrateタンパク質をコードする核酸(例えば、配列番号2、4又は6の配列を有する)を含む。
【0030】
本発明は、脊椎動物Serrate配列の少なくとも8個のヌクレオチド(すなわち、ハイブリダイズ可能な部分)からなる核酸を提供する。他の実施態様において、核酸は脊椎動物Serrate配列の少なくとも10個の(連続した)ヌクレオチド、25個のヌクレオチド、50個のヌクレオチド、100個のヌクレオチド、150個のヌクレオチドまたは200個のヌクレオチド、または全長脊椎動物Serrateコード配列からなる。本発明はまた、上記の配列とハイブリダイズ可能な、またはそれらに相補的な核酸に関する。特定の側面において、Serrate遺伝子の少なくとも10、25、50、100または200個のヌクレオチドまたは全コード領域に相補的な配列を含む核酸が提供される。
【0031】
特定の実施態様において、低ストリンジェンシー条件下で脊椎動物Serrate核酸(例えば配列番号1の配列を有する)または脊椎動物Serrate誘導体をコードする核酸とハイブリダイズ可能な核酸が提供される。非限定的な例としては、そのような低ストリンジェンシー条件を用いる方法は以下の通りである(ShiloおよびWeinberg,1981,Proc.Natl.Acad.Sci.USA78:6789-6792 も参照されたい):DNAを含有するフィルターを、35% ホルムアミド、5X SSC、50 mM Tris-HCl(pH7.5)、5mM EDTA、0.1% PVP、0.1% Ficoll、1% BSAおよび500 μg/ml変性サケ精子DNAを含有する溶液中で40℃で6 時間前処理する。同じ溶液に以下の改変を加えて、その中でハイブリダイゼーションを実施する:0.02% PVP、0.02% Ficoll、0.2% BSA、100 μg/ml サケ精子DNA、10%(wt/vol)硫酸デキストラン、および5-20 X106 cpmの32P標識プローブを用いる。ハイブリダイゼーション混合物中でフィルターを40℃で18〜20時間インキュベートし、次に2XSSC、25 mM Tris-HCl(pH7.4)、5 mM EDTAおよび0.1% SDSを含有する溶液を用いて55℃で1.5 時間洗浄する。洗浄溶液を新鮮な溶液と交換し、さらに60℃で1.5時間インキュベートする。フィルターを乾燥状態でブロットし、オートラジオグラフィーのため露光する。必要であれば、65〜68℃でフィルターに三回目の洗浄を施し、フィルムに再度露光する。使用できる他の低ストリンジェンシー条件は当分野で周知である(例えば、交差種(cross-species)ハイブリダイゼーションに用いられる条件)。
【0032】
別の特定の実施態様において、高ストリンジェンシー条件下で脊椎動物Serrate核酸とハイブリダイズ可能な核酸が提供される。非限定的な例としては、そのような高ストリンジェンシー条件を用いる方法は以下の通りである:DNAを含有するフィルターのプレハイブリダイゼーションを65℃で8時間から一晩、6X SSC、50 mM Tris-HCl(pH 7.5)、5 mM EDTA、0.02% PVP、0.02% Ficoll、0.02% BSAおよび500 μg/ml 変性サケ精子DNAからなる緩衝液中で実施する。100 μg/ml 変性サケ精子DNAおよび5-20 X 106cpmの32P標識プローブを含有するプレハイブリダイゼーション混合物中で、フィルターを65℃で48時間ハイブリダイズさせる。フィルターの洗浄は、2X SSC、0.01%PVP、0.01% Ficollおよび0.01% BSA を含有する溶液を用いて37℃で1 時間実施する。次に、オートラジオグラフィーの前に、0.1X SSCを用いて50℃で45分間洗浄する。使用できる他の高ストリンジェンシー条件は当分野で周知である。
【0033】
脊椎動物Serrateタンパク質の断片および誘導体をコードする核酸(第5.6節参照)および脊椎動物Serrateアンチセンス核酸(第5.11節参照)がさらに提供される。容易に明らかなように、本明細書に用いる「Serrateタンパク質の断片または部分をコードする核酸」という表現は、Serrateタンパク質の記述された断片または部分のみをコードする核酸をさすと解釈されるべきであって、連続した配列としてのSerrateタンパク質の他の隣接部分をささないものとする。
【0034】
他のトポリズミックタンパク質との相同領域を含む脊椎動物Serrate核酸の断片もまた提供される。他の種のSerrateタンパク質のDSL領域(ショウジョウバエDeltaおよびSerrateとの相同領域)もまた提供される。DeltaとSerrateの間の保存された領域(配列番号8のSerrateアミノ酸63〜73、124〜134、149〜158、195〜206、214〜219および250〜259、またはDSLドメインによって表される領域、等)をコードする核酸もまた提供される。
【0035】
限定するためではなく特定の例として提示される、脊椎動物Serrate遺伝子クローニン
グの特定の実施態様は以下の通りである。
【0036】
発現クローニング(当分野で良く知られている技法)のため、当分野で公知の方法によ
り発現ライブラリーを構築する。例えば、mRNA(例:ヒトの)を単離し、cDNAを
作製して、宿主細胞によって発現されうるように発現ベクター(例:バクテリオファージ
誘導体)に連結し、次にこのベクターを宿主細胞に導入する。次に、種々のスクリーニン
グアッセイを用いて発現されたSerrate産物を選択することができる。1つの実施態様に
おいて、選択のために抗Serrate抗体を用いることができる。
【0037】
別の好ましい側面においては、選択に先立って、ゲノムまたはcDNAライブラリー中の所望の配列を増幅するためにPCRが用いられる。公知のSerrate配列を表すオリゴヌクレオチドプライマーをPCRプライマーとして用いることができる。好ましい側面において、オリゴヌクレオチドプライマーは、SerrateとDeltaの間で強い相同性を有するSerrate保存セグメント(segment)の少なくとも一部をコードする。合成オリゴヌクレオチドをプライマーとして用いて、興味のある源(RNAまたはDNA)、好ましくはcDNAライブラリーからPCRによって配列を増幅することができる。PCRは、例えばPerkin-ElmerCetusサーマルサイクラーおよびTaq ポリメラーゼ(Gene AmpTM)を用いて実施することができる。増幅されるDNAは、任意の真核生物種由来のmRNAまたはcDNAまたはゲノムDNAを含みうる。PCR反応に用いるため、数個の異なる縮重プライマーの合成を選択することができる。PCR反応を開始させる(priming)ために用いるハイブリダイゼーション条件のストリンジェンシーを変えて、公知のSerrateヌクレオチド配列と単離される核酸相同体との間により大きい又はより小さい度合いの核酸配列類似性をもたらすことも可能である。交差種ハイブリダイゼーションのためには、低ストリンジェンシー条件が好ましい。同種ハイブリダイゼーションのためには、中程度にストリンジェントな条件が好ましい。Serrate相同体の1セグメントを上首尾に増幅した後、このセグメントをクローン化し、配列決定し、そして完全なcDNAまたはゲノムクローンを単離するためのプローブとして用いることができる。次に、このクローンが、後に記述する遺伝子の完全なヌクレオチド配列の決定、遺伝子発現の分析、および機能分析のための遺伝子タンパク質産物の産生を可能とする。このようにして、Serrateタンパク質をコードするさらなる遺伝子を同定することができる。このような方法を、後述の種々の実施例の節において例として提示する。
【0038】
上記の方法は、脊椎動物Serrateのクローンを得る方法についての下記の一般的説明を
制限するものではない。
【0039】
脊椎動物細胞はいずれも可能性としてSerrate遺伝子の分子クローニングのための核酸
供給源となることができる。Serrateをコードする核酸配列はヒト、ブタ、ウシ、ネコ、
トリ、ウマ、イヌ、ならびにさらなる霊長類の供給源、等から単離することができる。例
えば、本発明者らはマウス、アフリカツメガエルおよびヒトの適切な大きさの断片を、c
DNAライブラリーおよびショウジョウバエSerrateプライマーを用いて、PCRにより
増幅した。DNAはクローン化されたDNA(例:DNA「ライブラリー」)から当分野
で公知の標準的方法により、化学合成、cDNAクローニング、または所望の細胞から精
製したゲノムDNAあるいはその断片のクローニングによって得ることができる。(例え
ば、Sambrookら,1989,MolecularCloning,A Laboratory Manual,第2版,Cold Sprin
g HarborLaboratory Press,Cold Spring Harbor,New York; Glover,D.M.(編),1985
,DNACloning: A Practical Approach,MRL Press,Ltd.,Oxford,U.K.,Vol.I,II
参照)。ゲノムDNAから得たクローンは、コード領域に加えて調節およびイントロンD
NA領域を含んでいる可能性がある。cDNAから得たクローンは、エキソン配列のみを
含む。供給源が何であれ、遺伝子の増殖のためには、これを適切なベクター中に分子的に
クローン化しなければならない。
【0040】
ゲノムDNA由来の遺伝子の分子クローニングにおいては、DNA断片が生成され、そ
のうちのあるものが所望の遺伝子をコードしている。種々の制限酵素を用いてDNAを特
定の部位で開裂することができる。または、マンガンの存在下でDNAse を用いてDNAを
断片化することができる。または、例えば超音波処理により、DNAを物理的に剪断する
ことができる。次に、アガロースおよびポリアクリルアミドゲル電気泳動およびカラムク
ロマトグラフィーを含むがこれらだけに限定されない標準的技法により、直鎖状DNA断
片を大きさによって分離することができる。
【0041】
ひとたびDNA断片を生成したならば、所望の遺伝子を含有する特定のDNA断片の同
定を種々の方法で実施することができる。例えば、(任意の種の)Serrate遺伝子または
その特定のRNA、またはその断片(例えば細胞外ドメイン、第5.6 節参照)が入手可能
で、精製および標識することができる場合、生成されたDNA断片を標識化プローブとの
核酸ハイブリダイゼーションによりスクリーニングすることができる(Benton,W.およびD
avis,R.,1977,Science196:180; Grunstein,M.およびHogness,D.,1975,Proc.Na
tl.Acad.Sci.USA72:3961)。プローブに実質的相同性を有するDNA断片はハイブリ
ダイズする。また、制限酵素消化、および公知の制限地図(もしそのようなものがあるな
らば)による断片サイズの期待されるサイズとの比較によって適切な断片を同定すること
も可能である。さらに、遺伝子の特性に基づいて選択を実施することができる。または、
遺伝子の発現産物の物理的、化学的または免疫学的特性に基づくアッセイによって該遺伝
子の存在を検出することができる。例えば、Serrateについて公知な電気泳動における移
動、等電点電気泳動挙動、加水分解消化地図、受容体結合活性、in vitro凝集活性(「接
着性」)または抗原特性に類似した、または同一のものを有するタンパク質を産生するc
DNAクローン、または適切なmRNAをハイブリッド選択(hybrid-select)するDNA
クローンを選択することができる。Serrateに対する抗体が使用可能な場合は、ELIS
A(酵素結合イムノソルベントアッセイ)型手順を用いて、標識化抗体を推定上のSerrat
e合成クローンと結合させることによりSerrateタンパク質を同定することができる。
【0042】
Serrate遺伝子は、核酸ハイブリダイゼーションによるmRNA選択とそれに続くin vitro翻訳によって同定することも可能である。この方法では、断片を用いてハイブリダイゼーションにより相補的mRNAを単離する。このようなDNA断片は、入手可能な、別の種(例:ヒト、ニワトリ)の精製SerrateDNAを表してもよい。単離されたmRNAの単離された産物のinvitro翻訳産物の免疫沈降分析または機能分析(例えば、in vitroでの凝集能;受容体との結合;後記参照)は、mRNAを同定し、そしてその結果、所望の配列を含む相補的DNA断片を同定する。さらに、Serrateタンパク質に対して特異的に誘導された固定化抗体への細胞から単離されたポリソームの吸着により、特定のmRNAを選択することができる。(吸着されたポリソームから)選択されたmRNAを鋳型として用いて、放射性標識化SerratecDNAを合成することができる。次に、放射性標識化mRNAまたはcDNAをプローブとして用いて、他のゲノムDNA断片の中からSerrateDNA断片を同定することができる。
【0043】
SerrateゲノムDNAの単離に代わるものとしては、公知の配列から遺伝子配列それ自
体を化学的に合成すること、またはSerrateタンパク質をコードするmRNAに対するc
DNAを作製すること、が含まれるが、これらだけに限定されない。例えば、Serrate遺
伝子のcDNAクローニングのためのRNAを、Serrateを発現する細胞から単離するこ
とができる。他の方法も可能であり、それらは本発明の範囲内にある。
【0044】
次に、同定され単離された遺伝子を適切なクローニングベクターに挿入することができる。当分野で公知の多数のベクター-宿主系が使用できる。使用できるベクターはプラスミドまたは改変されたウイルスを含むがそれらだけに限定されない。また、ベクター系は用いられる宿主細胞と適合性のものでなければならない。そのようなベクターは、ラムダ誘導体等のバクテリオファージ、PBR322またはpUCプラスミド誘導体等のプラスミドを含むがそれらだけに限定されない。クローニングベクターへの挿入は、例えばDNA断片を相補的付着末端を有するクローニングベクターに連結することにより達成できる。しかし、DNAを断片化するのに用いられた相補的制限部位がクローニングベクターに存在しない場合は、DNA分子の末端を酵素的に改変することができる。または、ヌクレオチド配列(リンカー)をDNA末端に連結することにより所望の任意の部位を作製することができる。このような連結されたリンカーは、制限エンドヌクレアーゼ認識配列をコードする特定の化学的に合成されたオリゴヌクレオチドを含むことができる。別の方法では、開裂したベクターおよびSerrate遺伝子をホモポリマー的テーリング(tailing)により改変することができる。遺伝子配列の多数のコピーが生成されるように、組換え分子を形質転換、トランスフェクション、感染、エレクトロポレーション、等により宿主細胞に導入することができる。
【0045】
別の方法においては、所望の遺伝子を「ショットガン」法により適切なクローニングベ
クターに挿入した後に同定および単離することができる。クローニングベクターに挿入す
る前に、例えばサイズ分画による所望の遺伝子の富化が実施できる。
【0046】
特定の実施態様において、単離されたSerrate遺伝子、cDNA、または合成DNA配
列を組み込んだ組換えDNA分子を用いた宿主細胞の形質転換は、遺伝子の多数コピーの
生成を可能とする。したがって、形質転換体を増殖させ、該形質転換体から組換えDNA
分子を単離し、そして必要であれば単離された組換えDNAから挿入遺伝子を回収するこ
とにより、遺伝子を大量に得ることができる。
【0047】
本発明により提供されるSerrate配列は、天然のSerrateタンパク質に見いだされるアミノ酸配列と実質的に同一のアミノ酸配列をコードする核酸配列、および機能的に等価のアミノ酸を有するコードされたアミノ酸配列を包含する。これらすべてはSerrate誘導体に関する第5.6節(後述)に記述されている。
【0048】
5.2.Serrate遺伝子の発現
脊椎動物Serrateタンパク質またはその機能的に活性な断片または他の誘導体(第5.6節参照)をコードするヌクレオチド配列を適切な発現ベクター、すなわち、挿入されたタンパク質コード配列の転写および翻訳に必要な要素を含有するベクターに挿入することができる。必要な転写および翻訳シグナルは、天然の脊椎動物Serrate遺伝子および/またはそのフランキング領域によって供給することもできる。種々の宿主−ベクター系を用いてタンパク質コード配列を発現することができる。これらは、ウイルス(例:ワクシニアウイルス、アデノウイルス、等)によって感染させた哺乳動物細胞系;ウイルス(例:バキュロウイルス)によって感染させた昆虫細胞系;酵母ベクターを含有する酵母等の微生物;またはバクテリオファージ、DNA、プラスミドDNAまたはコスミドDNAによって形質転換した細菌を含むが、それらだけに限定されない。ベクターの発現要素は、その強度および特異性において異なる。使用する宿主−ベクター系により、多数の適切な転写および翻訳要素のうち任意のものを用いることができる。特定の実施態様において、Serrate遺伝子の接着性部分が発現される。別の特定の実施態様において、HumanSerrate遺伝子またはヒトSerrate遺伝子の機能的に活性な部分をコードする配列(Human Serrate-1(HJ1)またはHumanSerrate-2(HJ2)、等)が発現される。さらに別の実施態様において、細胞外ドメインを含むSerrateの断片、または他の誘導体、またはSerrateの類似体が発現される。
【0049】
DNA断片のベクターへの挿入のために以前に記述された任意の方法を用いて、適切な転写/翻訳制御シグナルおよびタンパク質コード配列からなるキメラ遺伝子を含有する発現ベクターを構築することができる。これらの方法は、in vitro組換えDNAおよび合成技法およびin vivo組換え体(遺伝子組換え)を含みうる。Serrateタンパク質またはペプチド断片をコードする核酸配列の発現は、組換えDNA分子によって形質転換された宿主中で該Serrateタンパク質またはペプチドが発現されるように、第2の核酸配列によって調節することができる。例えば、当分野で公知の任意のプロモーター/エンハンサー要素によってSerrateタンパク質の発現を制御することができる。トポリズミックな遺伝子発現を制御するために使用できるプロモーターは以下のものを含むがそれらだけに限定されない。すなわち、SV40初期プロモーター領域(BernoistおよびChambon,1981,Naure 290:304-310)、ラウス肉腫ウイルスの3'長末端リピートに含まれるプロモーター(Yamamotoら,1980,Cell22:787-797)、ヘルペスチミジンキナーゼプロモーター(Wagnerら,1981,Proc.Natl.Acad.Sci.USA78:1441-1445)、メタロチオネイン遺伝子の調節配列(Brinster ら,1982,Nature 296:39-42)、β-ラクタマーゼプロモーター(Villa-Kamaroffら,1978,Proc.Natl.Acad.Sci.USA 75:3727-3731)等の原核細胞発現ベクター、またはtac プロモーター(DeBoer ら,1983,Proc.Natl.Acad.Sci.USA80:21-25); Scientific American,1980,242:74-94「組換え細菌由来の有用なタンパク質」をも参照; ノパリンシンセターゼプロモーター領域を含む植物発現ベクター(Herrera-Estrellaら,Nature303:209-213)またはカリフラワーモザイクウイルス 35S RNAプロモーター(Gardnerら,1981,Nucl.Acids Res.9:2871)、および光合成酵素リブロースビスリン酸カルボキシラーゼのプロモーター(Herrera-Estrellaら,1984,Nature310:115-120);酵母または他の真菌由来のプロモーター要素、例えばGal 4プロモーター、ADC(アルコールデヒドロゲナーゼ)プロモーター、PGK(ホスホグリセロールキナーゼ)プロモーター、アルカリホスファターゼプロモーター、および組織特異性を示し、トランスジェニック動物に使用されてきた下記の動物転写制御領域:膵臓腺房細胞において活性なエラスターゼI遺伝子制御領域(Swiftら,1984,Cell38:639-646; Ornitzら,1986,Cold Spring Harbor Symp.Quant.Biol.50:399-409;MacDonald,1987,Hepatology 7:425-515); 膵臓ベータ細胞において活性なインスリン遺伝子制御領域(Hanahan,1985,Nature315:115-122)、リンパ系細胞において活性な免疫グロブリン遺伝子制御領域(Grosschedlら,1984,Cell 38:647-658;Adamesら,1985,Nature 318:533-538; Alexanderら,1987,Mol.Cell.Biol.7:1436-1444)、精巣、乳房、リンパ系およびマスト細胞において活性なマウス乳腺癌ウイルス制御領域(Lederら,1986,Cell45:485-495)、肝臓において活性なアルブミン遺伝子制御領域(Pinkertら,1987,Genes and Devel.1:268-276)、肝臓において活性なαフェトプロテイン遺伝子制御領域(Krumlaufら,1985,Mol.Cell.Biol.5:1639-1648;Hammerら,1987,Science 235:53-58)、肝臓において活性なα 1-アンチトリプシン遺伝子制御領域(Kelseyら,1987,Genesand Devel.1:161-171)、骨髄様細胞において活性なβグロビン遺伝子制御領域(Mogramら,1985,Nature 315:338-340;Kolliasら,1986,Cell 46:89-94)、脳の稀突起膠細胞において活性なミエリン塩基性タンパク質遺伝子制御領域(Readheadら,1987,Cell48:703-712)、骨格筋において活性なミオシンL鎖2遺伝子制御領域(Sani,1985,Nature 314:283-286)および視床下部において活性なゴナドトロピン放出ホルモン遺伝子制御領域(Masonら,1986,Science234:1372-1378)である。
【0050】
Serrate遺伝子挿入配列を有する発現ベクターは、3つの一般的アプローチによって同
定することができる。すなわち、(a)核酸ハイブリダイゼーション、(b)「マーカー
」遺伝子機能の存在または不在、および(c)挿入配列の発現、である。第1のアプロー
チでは、挿入されたトポリズミックな遺伝子に相同な配列を含むプローブを用いた核酸ハ
イブリダイゼーションによって、発現ベクターに挿入された外来遺伝子の存在を検出する
ことができる。第2のアプローチでは、ベクターへの外来遺伝子の挿入によってもたらさ
れる特定の「マーカー」遺伝子機能(例えば、チミジンキナーゼ活性、抗生物質耐性、形
質転換表現型、バキュロウイルスにおける閉鎖体の形成、等)の存在または不在に基づい
て、組換えベクター/宿主系を同定および選択することができる。例えば、Serrate遺伝
子がベクターのマーカー遺伝子配列内に挿入されると、Serrate挿入配列を有する組換え
体はマーカー遺伝子機能の不在によって同定することができる。第3のアプローチでは、
組換え発現ベクターは、組換え体によって発現された外来遺伝子産物をアッセイすること
により同定することができる。このようなアッセイは、in vitroアッセイ系において、例
えばSerrate遺伝子産物の物理的または機能的特性(例えば、Notchとの凝集(結合)、受容体との結合、抗体との結合)に基づくことができる。
【0051】
特定の組換えDNA分子がひとたび同定され単離されたならば、当分野で公知の幾つか
の方法を用いてそれを増殖させることができる。ひとたび適切な宿主系および増殖条件が
確立されたならば、組換え発現ベクターを大量に増殖させ、調製することができる。前に
説明したように、使用できる発現ベクターは下記のベクターまたはその誘導体を含むが、
それらだけに限定されない。すなわち、2〜3名を挙げるならば、ワクシニアウイルスま
たはアデノウイルス等のヒトまたは動物ウイルス;バキュロウイルス等の昆虫ウイルス;
酵母ベクター;バクテリオファージベクター(例えばラムダ)、およびプラスミドおよび
コスミドDNAベクターである。
【0052】
さらに、挿入配列の発現を変調する、または所望の特定の様式で遺伝子産物を改変しプ
ロセシングする、宿主細胞株を選択することができる。特定のプロモーターからの発現は
、特定の誘導物質(inducer)の存在下で増大させることができる。したがって、遺伝子工
学的に作製されたSerrateタンパク質の発現を制御することができる。さらに、異なる宿
主細胞は、タンパク質の翻訳および翻訳後プロセシングおよび修飾(例えば、シグナル配
列等のグリコシル化、開裂)のための特徴的で特異的な作用機構を有する。適切な細胞系
または宿主系を選択して、発現された外来タンパク質の所望の修飾およびプロセシングを
確実にすることができる。例えば、細菌系における発現を用いて非グリコシル化コアタン
パク質産物を産生することができる。酵母における発現は、グリコシル化産物を産生する
。哺乳動物細胞における発現を用いて、異種哺乳動物トポリズミックタンパク質の「天然
の」グリコシル化を確実にすることができる。さらに、異なるベクター/宿主発現系は加
水分解開裂等のプロセシング反応に異なる程度の影響を及ぼしうる。
【0053】
他の特定の実施態様において、Serrateタンパク質、断片、類似体、または誘導体を融
合またはキメラタンパク質産物〔ペプチド結合を介して(異なるタンパク質の)異種タン
パク質配列に結合した上記タンパク質、断片、類似体、または誘導体を含む〕として発現
させることができる。このようなキメラ産物は、所望のアミノ酸配列をコードする適切な
核酸配列を当分野で公知の方法により適切なコードフレーム内で相互に連結し、そして該
キメラ産物を当分野で周知の方法により発現させることにより、作製することができる。
または、そのようなキメラ産物は例えばペプチド合成機を用いて、タンパク質合成技法に
より作製することができる。
【0054】
cDNA配列およびゲノム配列の両者ともクローン化し、発現させることが可能である
。
【0055】
5.3.Serrate遺伝子産物の同定および精製
特定の側面において、本発明は脊椎動物Serrate(好ましくはヒトSerrate相同体)、および抗原決定基を含む(すなわち、抗体によって認識されうる)、または機能的に活性なその断片および誘導体のアミノ酸配列、ならびにそれらをコードする核酸配列を提供する。本明細書に用いる「機能的に活性な」物質とは、全長(野性型)Serrateタンパク質に関連する1以上の公知の機能的活性〔例えば、Notchまたはその一部との結合、他の任意のSerrateリガンドとの結合、抗原性(抗Serrate抗体との結合)、等〕を示す物質をいう。
【0056】
特定の実施態様において、本発明は少なくとも6アミノ酸、10アミノ酸、25アミノ酸、50アミノ酸、または少なくとも75アミノ酸からなる脊椎動物Serrateタンパク質の断片を提供する。他の実施態様において、本発明のタンパク質はSerrateタンパク質の細胞外ドメイン、DSLドメイン、表皮増殖因子に似たリピート(ELR)ドメイン、ELRの1つまたは任意の組合せ、システインに富む領域、膜貫通ドメイン、または細胞内(細胞質)ドメイン、またはNotchに結合する部分、または上記のものの任意の組合せを含む、または本質的にそれ(ら)から成る。脊椎動物Serrateタンパク質の上記領域の幾つか又は全部を欠く断片、または断片を含むタンパク質もまた提供される。上記のタンパク質および断片をコードする核酸が提供される。
【0057】
脊椎動物Serrate遺伝子配列を発現する組換え体がひとたび同定されたならば、遺伝子
産物を分析することができる。これは、該産物を放射性標識し、次にゲル電気泳動、イム
ノアッセイ、等により分析することを含む、産物の物理的または機能的特性に基づくアッ
セイによって達成することができる。
【0058】
Serrateタンパク質がひとたび同定されたならば、クロマトグラフィー(例えば、イオ
ン交換、アフィニティーおよびサイジングカラムクロマトグラフィー)、遠心、示差溶解
性を含む標準的方法、またはタンパク質精製のための他の標準的技法により、該タンパク
質を単離し、精製することができる。機能的特性は、任意の適切なアッセイを用いて評価
することができる(第5.7 節参照)。
【0059】
または、組換え体によって産生されたSerrateタンパク質がひとたび同定されたならば
、組換え体に含有されているキメラ遺伝子の核酸配列から上記タンパク質のアミノ酸配列
を推定することができる。その結果、当分野で公知の標準的な化学的方法により該タンパ
ク質を合成することができる(例えば、Hunkapiller,M.ら,1984,Nature 310:105-111
参照)。
【0060】
本発明の特定の実施態様において、組換えDNA技法により産生されたものであれ、化
学合成法により作製されたものであれ、そのようなSerrateタンパク質は一次アミノ酸配
列として図1、2または3(それぞれ配列番号2、4または6)に実質的に示すアミノ酸
配列の全部または一部を含むタンパク質、ならびにそれらの断片および他の誘導体および
類似体を包含するが、それらだけに限定されない。
【0061】
5.4.Serrate遺伝子およびタンパク質の構造
Serrate遺伝子およびタンパク質の構造を、当分野で公知の種々の方法によって分析す
ることができる。
5.4.1.遺伝子分析
脊椎動物Serrate遺伝子に対応するクローン化DNAまたはcDNAを、サザンハイブリダイゼーション(Southern,E.M.,1975,J.Mol.Biol.98:503-517)、ノーザンハイブリダイゼーション(例えば、Freemanら,1983,Proc.Natl.Acad.Sci.USA80:4094-4098参照)、制限エンドヌクレアーゼマッピング(Maniatis,T.,1982,Molecular Cloning,ALaboratory Manual,Cold Spring Harbor,N.Y.)およびDNA配列分析を含むがこれらだけに限定されない方法によって分析することができる。ポリメラーゼチェーンリアクション(PCR;米国特許第4,683,202、4,683,195および4,889,818号;Gyllensteinら,1988,Proc.Natl.Acad.Sci.USA 85:7652-7656; Ochmanら,1988,Genetics120:621-623; Loh ら,1989,Science 243:217-220)およびその後のSerrate特異的プローブを用いたサザンハイブリダイゼーションは、種々の細胞型由来のDNA中におけるSerrate遺伝子の検出を可能とする。PCR以外の増幅方法もよく知られており、それらも採用することができる。1つの実施態様において、サザンハイブリダイゼーションを用いてSerrateの遺伝子的連結を確認することができる。ノーザンハイブリダイゼーション分析を用いてSerrate遺伝子の発現を確認することができる。種々の発生または活性状態にある種々の細胞型をSerrate発現に関して試験することができる。そのような技法およびそれらの結果の例を後出の第6節に記述する。サザンおよびノーザンハイブリダイゼーションの両方についてのハイブリダイゼーション条件のストリンジェンシーは、用いられた特定のSerrateプローブに対して所望の程度の関連性を有する核酸の検出を確実にするように操作することができる。
【0062】
制限エンドヌクレアーゼマッピングを用いてSerrate遺伝子の遺伝子構造を大まかに決
定することができる。特定の実施態様において、制限酵素による開裂を用いて後出の図2
に示す制限地図を引き出すことができる。制限エンドヌクレアーゼ開裂によって引き出さ
れる制限地図は、DNA配列分析によって確認することができる。
【0063】
DNA配列分析は、マクサム・ギルバート法(1980,Meth.Enzyml.65:499-560)、サンガージデオキシ法(Sanger,F.ら,1977,Proc.Natl.Acad.Sci.USA74:5463)、T7 DNAポリメラーゼの使用(TaborおよびRichardson,米国特許第4,795,699号)または自動DNAシークエネーター(例えば、AppliedBiosystems,Foster City,CA)の使用を含むが、それらだけに限定されない当分野で公知の任意の技法により実施することができる。代表的Serrate遺伝子のcDNA配列は図1および2に実質的に示す配列を含み、そして後出の第9節に記述されている。
【0064】
5.4.2.タンパク質分析
Serrateタンパク質のアミノ酸配列は、DNA配列からの推定により、または例えば自
動アミノ酸シークエンサーを用いた上記タンパク質の直接配列決定により、引き出すこと
ができる。代表的Serrateタンパク質のアミノ酸配列は図1に実質的に示す配列を含み、
そして、アミノ酸番号30〜1219によって示される代表的成熟タンパク質と共に後出の第9節に詳述されている。
【0065】
Serrateタンパク質配列を親水性分析(Hopp,T.およびWoods,K.,1981,Proc.Natl.
Acad.Sci.USA78:3824)によってさらに特徴付けることができる。親水性プロフィール
を用いて、Serrateタンパク質の疎水性および親水性領域、およびそのような領域をコー
ドする遺伝子配列の対応する領域を同定することができる。
【0066】
第2に、構造分析(Chou,P.およびFasman,G.,1974,Biochemistry13:222)を実施し
て、特異的二次構造を取るSerrateの領域を同定することができる。
【0067】
操作、翻訳、および二次構造の予測、ならびにオープンリーディングフレームの予測お
よびプロッティングもまた、当分野で利用可能なコンピュータソフトウエアプログラムを
用いて達成することができる。
【0068】
構造分析の他の方法もまた採用することができる。これらは、X線結晶学(Engstom,A.
,1974,Biochem.Exp.Biol.11:7-13)およびコンピュータモデリング(Fletterick,R.
およびZoller,M.(編),1986,「コンピュータグラフィックおよび分子モデリング」
,CurrentCommunications in Molecular Biology,Cold Spring Harbor Laboratory,Co
ld SpringHarbor,New York)を含むが、これらだけに限定されない。
5.5.Serrateタンパク質およびその誘導体に対する抗体の生成
本発明によれば、脊椎動物Serrateタンパク質、その断片または他の誘導体、またはその類似体を免疫原として用いて、そのような免疫原を認識する抗体を生成することができる。そのような抗体は、ポリクローナル、モノクローナル、キメラ、一本鎖、Fab断片およびFab 発現ライブラリーを含むが、これらだけに限定されない。特定の実施態様において、ヒトSerrateに対する抗体が産生される。別の実施態様において、Serrateの細胞外ドメインに対する抗体が産生される。別の実施態様において、Serrateの細胞内ドメインに対する抗体が産生される。
【0069】
Serrateタンパク質または誘導体または類似体に対するポリクローナル抗体の産生のた
めに、当分野で公知の種々の方法が使用できる。特定の実施態様において、図1に示す配
列またはそのサブ配列によってコードされるSerrateタンパク質のエピトープに対するウ
サギポリクローナル抗体を得ることができる。抗体の産生のため、天然のSerrateタンパ
ク質、またはその合成物、またはその誘導体(例:断片)を注射することにより、ウサギ
、マウス、ラット、等を含むがこれらだけに限定されない種々の宿主動物を免疫感作する
ことができる。宿主の種によって、免疫応答を増大させるため、種々のアジュバントを用
いることができる。これらのアジュバントには以下のものが含まれるが、それらだけに限
定されない。すなわち、フロイントアジュバント(完全および不完全)、水酸化アルミニ
ウム等のミネラルゲル、リゾレシチン、プルロニックポリオール等の界面活性剤、ポリア
ニオン、ペプチド、オイルエマルジョン、スカシ貝ヘモシアニン、ジニトロフェノール、
およびBCG(カルメット−ゲラン杆菌)およびコリネバクテリウム・パルバム(Coryne
bacteriumparvum)等の可能性として有用なヒトアジュバントである。
【0070】
脊椎動物Serrateタンパク質配列またはその類似体に向けられたモノクローナル抗体の調製のため、培養されている連続的継代細胞系による抗体分子の産生を提供する任意の技法を用いることができる。例えば、KohlerおよびMilsteinによって最初に開発されたハイブリドーマ技法(Nature,1975,256:495-497)、ならびにトリオーマ(trioma)技法、ヒトB細胞ハイブリドーマ技法(Kozborら,1983,ImmunologyToday,4:72)およびヒトモノクローナル抗体を産生するEBV−ハイブリドーマ技法(Coleら,1985,Monoclonal Antibodiesand Cancer Therapy,Alan R.Liss,Inc.,pp.77-96)である。本発明の付加的実施態様においては、最近の技術(PCT/US90/02545)を用いて、無菌動物においてモノクローナル抗体を産生することができる。本発明によれば、ヒト抗体が使用可能であり、そしてこれはヒトハイブリドーマ(Coteら,1983,Proc.Natl.Acad.Sci.USA80:2026-2030)を用いて、またはEBVウイルスを用いてin vitroでヒトB細胞を形質転換して(Coleら,1985,MonoclonalAntibodies and Cancer Therapy,Alan R.Liss,pp.77-96)得ることができる。実際、本発明によれば、Serrateに特異的なマウス抗体分子由来の遺伝子を適切な生物活性を有するヒト抗体分子由来の遺伝子と共にスプライシングすることにより「キメラ」抗体(Morrisonら,1984,Proc.Natl.Acad.Sci.USA81:6851-6855; Neubergerら,1984,Nature,312:604-608; Takedaら,1985,Nature,314:452-454)を産生するために開発された技法を用いることができる。そのような抗体は本発明の範囲内にある。
【0071】
本発明によれば、一本鎖抗体(米国特許第4,946,778号)を産生するために記述された
技法を、Serrate特異的一本鎖抗体の産生のために適合させることができる。本発明の付
加的実施態様は、Fab発現ライブラリー(Huseら,1989,Science 246:1275-1281)の構築
のために記述された技法を用いて、Serrateタンパク質、誘導体または類似体に対し所望
の特異性を有するモノクローナルFab 断片の迅速で容易な同定を可能とする。
【0072】
分子のイディオタイプを含有する抗体断片を公知の技法により生成することができる。
例えば、そのような断片は抗体分子のペプシン消化により生成されるF(ab')2断片;F(ab'
)2断片のジスルフィド架橋を還元することにより生成されるFab'断片、および抗体分子を
パパインおよび還元剤で処理することにより生成されるFab 断片を含むが、これらだけに
限定されない。
【0073】
抗体の産生において、所望の抗体のスクリーニングは、当分野で公知の技法、例えばELISA(酵素結合イムノソルベントアッセイ)により達成することができる。例えば、Serrateタンパク質の特定ドメインを認識する抗体を選択するため、そのようなドメインを有するSerrate断片と結合する産物について、生成されたハイブリドーマをアッセイすることができる。脊椎動物(例:ヒト)Serrateに特異的な抗体を選択するためには、脊椎動物Serrateとの陽性結合およびショウジョウバエSerrateとの結合の欠如に基づいて選択することができる。別の実施態様においては、ヒトSerrateとの結合、および他の種のSerrateとの不結合によって選択することができる。
【0074】
上記の抗体は、本発明のタンパク質配列(例えば、後出の第5.7 節参照)の局在化と活
性に関する当分野で公知の方法に、例えば診断法等においてそれらのタンパク質のイメー
ジイングのため、適当な生理的サンプル中のそれらのレベルを測定するため、に用いるこ
とができる。
【0075】
Serrateタンパク質のあるドメインに特異的な抗体もまた提供される。特定の実施態様において、SerrateのNotch結合断片に結合する抗体が提供される。
【0076】
本発明の別の実施態様(後出参照)において、抗Serrate抗体および結合ドメインを有
するその断片は治療剤である。
5.6.Serrateタンパク質、誘導体および類似体
本発明はさらに脊椎動物Serrateタンパク質、およびSerrateタンパク質の誘導体(断片を含むがそれだけに限定されない)および類似体に関する。脊椎動物Serrateタンパク質誘導体およびタンパク質類似体をコードする核酸もまた提供される。1つの実施態様において、Serrateタンパク質は後出の第5.1節に記述される脊椎動物Serrate核酸によってコードされる。特定の側面において、上記タンパク質、誘導体または類似体はカエル、マウス、ラット、ブタ、ウシ、イヌ、サル、またはヒトSerrateタンパク質である。
【0077】
脊椎動物Serrateに関連する誘導体および類似体の作製および使用は本発明の範囲内にある。特定の実施態様において、上記誘導体または類似体は機能的に活性である、すなわち全長、野性型Serrateタンパク質に関連する1以上の機能的活性を示すことができる。1例としては、所望の免疫原性または抗原性を有するそのような誘導体または類似体は、例えばイムノアッセイにおいて、免疫感作のため、Serrate活性の抑制のため、等に用いることができる。所望のSerrate特性(例えば、Notchまたは他のトポリズミックなタンパク質との結合、細胞表面受容体との結合)を保持する、またはこれを抑制する、そのような分子は、それぞれそのような特性およびその生理的相関物の誘導物質またはインヒビターとして用いることができる。特定の実施態様は、抗Serrate抗体によって結合されうるが、Notchタンパク質または他のトポリズミックなタンパク質と結合しないSerrate断片に関する。Serrateの誘導体または類似体は、第5.7節に記載のアッセイを含むがそれらだけに限定されない当分野で公知の方法によって、所望の活性について試験することができる。
【0078】
特に、Serrate誘導体は、機能的に等価な分子をもたらす置換、付加または欠失によりSerrate配列を変更することによって作製することができる。ヌクレオチドコード配列の縮重により、Serrate遺伝子として実質的に同一のアミノ酸配列をコードする他のDNA配列が本発明の実施において使用できる。これらのDNA配列には以下のものが含まれるが、それらだけに限定されない。すなわち、配列内で機能的に等価なアミノ酸残基をコードする異なるコドンとの置換によって変更された、その結果表に現れない変化(silentchange)を生じているSerrate遺伝子の全部または一部を含むヌクレオチド配列である。同様に、本発明のSerrate誘導体は、一次アミノ酸配列として変更された配列(配列内で機能的に等価なアミノ酸残基が置換され、表に現れない変化を生じている)を含むSerrateタンパク質のアミノ酸配列の全部または一部を含むものを包含するが、それらだけに限定されない。例えば、配列内の1以上のアミノ酸残基を、機能的等価物として作用する類似の極性を有する別のアミノ酸に置換することが可能で、これは表に現れない変更をもたらす。配列内のアミノ酸の置換物(Substitute)は、当該アミノ酸が属するクラスの他のメンバーから選択することができる。例えば、非極性(疎水性)アミノ酸はアラニン、ロイシン、イソロイシン、バリン、プロリン、フェニルアラニン、トリプトファンおよびメチオニンを含む。極性中性アミノ酸は、グリシン、セリン、トレオニン、システイン、チロシン、アスパラギン、およびグルタミンを含む。正に荷電した(塩基性)アミノ酸は、アルギニン、リシンおよびヒスチジンを含む。負に荷電した(酸性)アミノ酸は、アスパラギン酸およびグルタミン酸を含む。
【0079】
本発明の特定の実施態様において、Serrateタンパク質の少なくとも10個の(連続した)アミノ酸からなる脊椎動物Serrateタンパク質の断片からなる、またはそれを含むタンパク質が提供される。他の実施態様において、上記断片はSerrateタンパク質の少なくとも20個または50個のアミノ酸からなる。特定の実施態様において、そのような断片は35、100または200アミノ酸より小さい。脊椎動物Serrateの誘導体または類似体は、脊椎動物Serrateまたはその断片に実質的に相同な(例えば、同じ大きさのアミノ酸配列にわたって少なくとも30%同一)ペプチド、またはそれをコードする核酸が脊椎動物Serrateのコード配列とハイブリダイズ可能であるペプチドを含むが、それらだけに限定されない。
【0080】
本発明のSerrate誘導体および類似体は当分野で公知の種々の方法により作製すること
ができる。それらの作製をもたらす操作は、遺伝子またはタンパク質レベルで起こりうる
。例えば、クローン化Serrate遺伝子配列を当分野で公知の多数の戦略のうち任意のもの
により改変することができる(Maniatis,T.,1990,Molecular Cloning,A Laboratory M
anual,第2版,ColdSprings Harbor Laboratory,Cold Spring Harbor,New York)。上
記配列を制限エンドヌクレアーゼを用いて適切な部位で開裂し、次に所望であればさらに
酵素的に改変し、そして単離してin vitroで連結することができる。Serrateの誘導体ま
たは類似体をコードする遺伝子の作製においては、改変された遺伝子が、所望のSerrate
活性がコードされている遺伝子領域において、翻訳停止シグナルによって中断されること
なく、Serrateと同じ翻訳リーディングフレーム内に確実に留まるように注意しなければ
ならない。
【0081】
さらに、Serrateをコードする核酸配列をinvitroまたはin vivoで突然変異させて、翻訳開始および/または停止配列を創出および/または破壊し、またはコード領域に変異を創出し、および/または新たな制限エンドヌクレアーゼ部位を形成し、または既存のそのような部位を破壊し、さらなるinvitro改変を容易にすることができる。当分野で公知の突然変異誘発のための任意の技法を用いることができる。これらの技法は、in vitro部位特異的突然変異誘発(Hutchinson,C.ら,1978, J. Biol. Chem. 253:6551)、TAB(登録商標)リンカー(Pharmacia)の使用、等を含むが、それらだけに限定されない。
【0082】
Serrate 配列の操作をタンパク質レベルで実施することもできる。本発明の範囲内に含
まれるものは、例えば、グリコシル化、アセチル化、リン酸化、アミド化、既知の保護/
遮断基による誘導体化、タンパク質分解切断、抗体分子または他の細胞リガンドへの連結
、その他によって、翻訳中もしくは翻訳後に異なる修飾を受けたSerrate タンパク質断片
またはその他の誘導体もしくは類似体である。既知の手法による多数の化学的修飾のどれ
を実施してもよく、限定するわけではないが、臭化シアン、トリプシン、キモトリプシン
、パパイン、V8プロテアーゼ、NaBH4による特異的化学切断;アセチル化、ホルミル
化、酸化、還元;ツニカマイシンの存在下での代謝合成;その他が含まれる。
【0083】
その他に、Serrateの類似体および誘導体を化学的に合成することもできる。例えば、ペプチド合成装置の使用によって、所望のドメイン(第5.6.1節参照)を含んでいる、またはinvitroで所望の凝集活性を仲介する、あるいは受容体に結合するSerrate タンパク質の一部分に相当するペプチドを合成することができる。さらに、所望ならば、Serrate配列中に、置換または付加によって非古典的アミノ酸または化学的アミノ酸類似体を導入することもできる。非古典的アミノ酸として、限定するわけではないが、通常のアミノ酸のD−異性体、α−アミノイソ酪酸、4−アミノ酪酸、ヒドロキシプロリン、サルコシン、シトルリン、システイン酸、t−ブチルグリシン、t−ブチルアラニン、フェニルグリシン、シクロヘキシルアラニン、β−アラニン、修飾アミノ酸(β−メチルアミノ酸、Cα−メチルアミノ酸、およびNα−メチルアミノ酸など)が含まれる。
【0084】
特定の態様において、このSerrate 誘導体は、アミノ−またはカルボキシル末端の位置で別のタンパク質のアミノ酸配列にペプチド結合を介して結合した脊椎動物のSerrateタンパク質または(好ましくは少なくともSerrate タンパク質のドメインまたはモチーフ、あるいは少なくともSerrate タンパク質の10アミノ酸からなる)それらの断片を含む、キメラ、または融合タンパク質である。一態様において、こうしたキメラタンパク質は、このタンパク質をコードする(別のタンパク質をコードする配列にフレーム内連結したSerrateコード配列を含む)核酸の組み換え発現によって製造される。こうしたキメラ産物は、当業界で既知の方法によって、固有のコードフレーム内に所望のアミノ酸配列をコードする適当な核酸配列同士を連結させ、当業界で通常知られた方法によってキメラ産物を発現させることによって製造することができる。あるいはまた、例えばペプチド合成装置の使用によるタンパク質合成技術によって、こうしたキメラ産物を製造してもよい。特定の態様において、キメラタンパク質が発現されて、その細胞によって成熟Serrateタンパク質に転換されるように、異種シグナル配列を有する成熟脊椎動物Serrate タンパク質をコードするキメラ核酸を発現させる。限定するわけではないが、また別の例において、Serrateおよび別のトポリスミック(toporythmic)遺伝子、例えばDeltaの両方をコードする部分を含む組み換え分子を本発明にしたがって構築することができる。こうした組み換え分子中にコードされたタンパク質はSerrate およびDeltaの両方が組み合わされた特性を示し、そしてアゴニストおよびアンタゴニストを含む新規な生物学的活性プロフィールを表現する。また、Serrate およびDelta の一次配列はコンピュータシミュレーションを使用するこの分子の三次構造の予測に使用することもできる(Hoppand Woods,1981,Proc.Natl.Acad.Sci.U.S.A.783824-3828);Serrate /Deltaキメラ組み換え遺伝子は三次構造および生物学的機能間の相関関係を参照して設計することができるはずである。同様に、任意の異種タンパク質をコードする配列に融合した脊椎動物Serrateの一部分を含むキメラ遺伝子を構築することもできる。ある特定の態様は、少なくとも10アミノ酸の脊椎動物Serrate断片を含むキメラタンパク質に関係するものである。
【0085】
別の特定の態様において、Serrate 誘導体は別のトポリスミックタンパク質と相同の領
域を含むSerrate断片である。第1のタンパク質のある領域を、第2のタンパク質中のこ
の領域に含まれるアミノ酸の数と等しい数のアミノ酸を有するある配列に比較したとき、
この領域のアミノ酸配列が、30%以上同一であるかまたは75%以上同一もしくは保存的変
化に属する場合、ここではこれを“相同性”とみなすこととする。例えば、こうしたSerr
ate 断片として、Delta、またはDSLドメインもしくはこれらの部分に相同な1以上の領
域を含有するものがある。
【0086】
誘導体および類似体のその他の特定の態様を下記の亜節および下記の実施例の節に記載
する。
5.6.1.Serrate タンパク質の1以上のドメインを含有するSerrate誘導体
特定の1態様において、本発明は、脊椎動物Serrate 誘導体または類似体、特に、限定
するわけではないが、細胞外ドメイン、DSL ドメイン、ELR ドメイン、システインに富ん
だドメイン、膜貫通ドメイン、細胞内ドメイン、膜結合領域、およびSerrate タンパク質
の1以上のEGF-様リピート単位(ELR)、または前記の任意の組合せを含む、Serrateタ
ンパク質の1以上のドメインを含むか、またはこれらからなる脊椎動物Serrate 断片およ
び誘導体に関する。ヒトおよびヒナドリSerrate タンパク質に関する特定の例について、
それぞれ実施例第9および8節においてこうしたドメインが同定される。
【0087】
特定の1態様において、脊椎動物Serrate の特定の断片を含む分子は、相当するSerrate タンパク質中でショウジョウバエ(Drosophila)Serrate および/またはDeltaタンパク質の特定の断片に最も相同な断片を含むものである。特定の1態様において、こうした分子は配列番号10、12、または18に相同なアミノ酸配列を含むかまたはこれで構成されている。別法として、Serrate相同体のドメインを含む断片を第5.3.2節に記載したようなタンパク質分析法によって同定することができる。
5.6.2.トポリスミックタンパク質ドメインへの結合を仲介するSerrate誘導体
本発明はまた、トポリスミックタンパク質への結合を仲介する(そしてここではこれら
を“接着性”と称する)脊椎動物Serrate 断片、およびこうした断片の類似体または誘導
体、ならびにこれらをコードする核酸配列を提供する。
【0088】
特定の1態様において、Serrate の接着性断片は、ショウジョウバエSerrate 配列のほぼ第85-283番目または第79-282番目のアミノ酸に最も相同なSerrateの部分を含むものである(PCT 公報、国際公開公報第93/12141号、1993年6月24日付、参照)。
【0089】
特別の1態様において、Serrate タンパク質の接着性断片はDSL ドメインまたはその部分を含んでいる。Notch への結合を仲介するDSL ドメイン内のサブ断片を、欠失性変異体を発現する構築物の分析によって同定することができる。
【0090】
トポリスミックタンパク質(好ましくはNotch)への結合能力は、こうしたトポリスミックタンパク質を発現する細胞およびSerrate またはSerrate 誘導体を発現する細胞を使用したinvitroでの凝集アッセイによって、証明することができる(第5.7節参照)。すなわち、あるSerrate 断片のNotch タンパク質への結合能力は、第1の細胞の表面に発現されたSerrate断片の、第2の細胞の表面に発現されたNotch タンパク質への結合能力で証明することができる。
【0091】
こうしたアッセイにおいて使用するための、トポリスミックタンパク質またはその接着
性ドメインをコードする核酸配列は、霊長類供給源と同様にヒト、ブタ、ウシ、ネコ、ト
リ、ウマ、イヌ、または昆虫、ならびに既知のトポリスミック遺伝子の相同体が同定され
る他のどんな種からも単離することができる。
5.7.Serrateタンパク質、誘導体および類似体のアッセイ
脊椎動物Serrateタンパク質、誘導体および類似体の機能的活性を各種の方法によって
アッセイすることができる。
【0092】
例えば、抗Serrate抗体に結合する、またはこの結合について野性型Serrate と競合する能力をアッセイする場合の1態様において、当業界で既知の各種のイムノアッセイを使用することができ、これらには、限定するわけではないが、ラジオイムノアッセイ、ELISA(酵素結合イムノソルベントアッセイ)、“サンドイッチ”イムノアッセイ、免疫放射線アッセイ、ゲル拡散沈降反応、免疫拡散アッセイ、(例えば金コロイド、酵素または放射性同位元素標識を使用した)insitu イムノアッセイ、ウェスタンブロット、沈降反応、凝集アッセイ(例えばゲル凝集アッセイ、血球凝集アッセイ)、補体結合アッセイ、免疫蛍光アッセイ、タンパク質Aアッセイ、免疫電気泳動アッセイその他などの技術を使用する競合および非競合アッセイシステムが含まれる。1態様において、抗体の結合を一次抗体上の標識を検出することによって検出する。別の態様においては、一次抗体に対する二次抗体または試薬の結合を検出することによって、一次抗体を検出する。さらに別の態様においては、二次抗体を標識する。イムノアッセイにおける結合を検出するための多くの手段が当業界で知られており、これらは本発明の範囲内に含まれる。
【0093】
トポリスミックタンパク質、例えばNotch への結合を仲介する能力をアッセイする別の態様において、1993年6月24日付のPCT 公報、国際公開公報第93/12141号に記載されているものなどのinvitro凝集アッセイを実施することができる(Fehon ら、1990,Cell 61:523-534;Rebay ら、1991,Cell67:687-699をも参照)。
【0094】
Serrate に対する受容体を同定する別の態様において、受容体の結合を例えば当業界で
周知の手段によってアッセイすることができる。別の態様において、Serrate 受容体を発
現する細胞に対するSerrateの結合の生理学的相関性(シグナル形質導入)をアッセイす
ることができる。
【0095】
別の態様における、昆虫またはその他のモデルシステムにおいて、野性型脊椎動物Serrate の誘導体または類似体であるSerrate 変異体の表現型の影響を調べるための遺伝学的研究を実施することができる。
【0096】
その他の方法は当業者が知ることができるであろうし、それらは本発明の範囲内に含まれる。
【0097】
5.8.治療上の用途
本発明は、本発明の治療用化合物の投与による細胞死または分化の障害の治療を提供する。こうした治療用化合物(ここでは、“治療剤(Therapeutics)”と称する)には以下のものが含まれる;脊椎動物Serrate タンパク質ならびにその類似体および誘導体(断片を含む)(例えばここに上記したもの);(ここに上記したような)それらに対する抗体;脊椎動物Serrateタンパク質、類似体または誘導体(例えばここに上記したもの)をコードする核酸;およびSerrate アンチセンス核酸。上述したように、本発明のアンタゴニスト治療剤は脊椎動物Serrateの機能および/またはNotch の機能(Serrate はNotch の1リガンドであるから)に対してアンタゴニスト作用または阻害をする治療剤である。こうしたアンタゴニスト治療剤は(例えばショウジョウバエにおける)遺伝学的アッセイも利用することができるが、最も好ましくは既知の通常のinvitroアッセイ、例えばSerrate の別のタンパク質(例えばNotch タンパク質)への結合を阻害する、または好ましくはin vitroでまたは細胞培養物中でアッセイされる任意の既知のNotchまたはSerrate の機能を阻害する能力に基づくものを使用することによって、同定される。好ましい態様において、アンタゴニスト治療剤はNotch への結合を仲介するSerrateの断片などの機能的に活性な断片を含むタンパク質またはその誘導体、もしくはそれらに対する抗体である。別の特定の態様において、こうしたアンタゴニスト治療剤はNotchに結合するSerrate の断片を含む分子を発現することができる核酸、またはSerrate アンチセンス核酸(本文の第5.11節参照)である。特定の治療剤の効果および罹病した組織の治療にとってその投与が必要かどうかを判定するために、好ましくは下記のような適当なinvitroまたはin vivo アッセイを利用すべきであることを留意すべきである。なぜならば、組織の罹病の進行経過によってアンタゴニストまたはアゴニスト治療剤のどちらが必要とされるかが決まるからである。
【0098】
さらに、投与の様式、例えばSerrate タンパク質または誘導体を可溶性形状で投与する
か、またはSerrateタンパク質の細胞内組み換え発現のためのこれをコードする核酸を介
して投与するか、はそれがアゴニストまたはアンタゴニストのどちらで作用するかに影響
を与える。
【0099】
本発明の別の態様において、相同的組み換えによるSerrate の不活性化を促進するため
に、アンタゴニスト治療剤として、脊椎動物Serrate遺伝子の一部分を含有する核酸を使
用する(Kollerand Smithies,1989,Proc.Natl.Acad.Sci.USA 86:8932-8935;Zijlstraら
、1989,Nature342:435-438)。
【0100】
上記のような本発明のアゴニスト治療剤はSerrate の機能を促進する。こうしたアゴニ
スト治療剤として、限定するわけではないが、Serrate への結合を仲介するNotch の一部
分を含むタンパク質および誘導体、ならびにこれらをコードする核酸(これらがコードし
ている産物をinvivo で発現するように投与することができる)が含まれる。
【0101】
本発明の治療剤のその他の記述および供給源は本文の第5.1節から第5.7節に見ら
れる。
【0102】
所望のSerrate特性、例えばNotch への結合、細胞内リガンドへの結合、を保持、あるいは阻害する分子は、こうした特性およびその生理学的相関物のそれぞれ誘発剤または阻害剤として治療のために使用することができる。特定の1態様において、Notchに結合する脊椎動物Serrate の一部分の配列を含有するペプチド(例えば第10−50番目または第15−25番目のアミノ酸;特に約10,15,20または25アミノ酸)をNotch機能にアンタゴニスト作用させるために使用する。特定の1態様において、Notch の発現の増加に関連するヒトまたはその他の悪性腫瘍(例えば、子宮頚ガン、結腸ガン、乳ガン、扁平腺ガン(下記参照))の治療または予防のためにこうしたアンタゴニスト治療剤を使用する。限定するわけではないが、下記の実施例に記載するアッセイを含む、当業界で既知の操作法によって、Serrateの誘導体または類似体の所望の活性を試験することができる。例えば、欠失変異体を発現させ、いくつかの方法(例えばinvitro 細胞凝集アッセイ、相互作用トラップシステム)の中のいずれかによって発現産物のNotchへの結合をアッセイすることによって、Notch EGF-リピート単位(ELR)11および12に結合し、DSLドメインよりも小さな脊椎動物Serrate 断片を含む分子を得て、選択することができる。これらの方法のいくつかを下記の実施例の節に記載する。特定の1態様において、所望の活性を有するペプチドを選択するために、ペプチドライブラリーをスクリーニングすることができる。こうしたスクリーニングは、例えばNotchまたはNotch ELR 11および12反復単位を含有する分子に対する結合に関するアッセイによって実施することができる。
【0103】
本発明のアゴニストおよびアンタゴニスト治療剤は細胞死の障害に対する治療上の有用
性を有する。アゴニスト治療剤は以下の障害において治療用に(予防上も含んで)投与される;(1)Notch またはSerrate 機能の欠落または(正常または所望の数値に対する)減少が関与する疾病または障害、例えば、Notch またはSerrateタンパク質が欠失、遺伝的欠損、生物学的に不活性または過少活性、もしくは過少発現である患者;および(2)in vitro(またはin vivo)アッセイ(下記参照)がSerrateアゴニストの投与の有用性を示す疾病または障害。Notch またはSerrate 機能の欠落または減少は、例えば患者の組織のサンプルを(例えば生検組織から)取り、発現されたNotchまたはSerrate タンパク質のタンパク質量、構造および/または活性についてこれをin vitroでアッセイすることによって、容易に検出することができる。当業界で標準的な多くの方法をここで使用することができる。これらとして限定するわけではないが、NotchまたはSerrate タンパク質の検出および/または可視化のためのイムノアッセイ(例えばウェスタンブロット、ドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動による免疫沈降、免疫細胞化学、その他)、および/または、NotchまたはSerrate mRNAをそれぞれ検出および/または可視化することによってNotch またはSerrate の発現を検出するためのハイブリダイゼーションアッセイ(例えばノーザンアッセイ、ドットブロットinsitu ハイブリダイゼーション、その他)が含まれる。
【0104】
ある特定のアゴニスト治療剤またはアンタゴニスト治療剤の投与が必要とされるかどうかを判定するために使用することができるin vitroアッセイには、患者の組織サンプルを培養して増殖させて、治療剤に暴露するかまたは別の方法でこれを投与し、組織サンプルに対するその治療剤の効果を観察する、invitro細胞培養アッセイが含まれる。患者が悪性腫瘍を有している1態様において、この悪性腫瘍からの細胞サンプルをプレートにとり、培養して増殖させて、その後この細胞を治療剤に暴露する。悪性腫瘍細胞の生存または増殖を(例えば終末分化を促進することによって)阻害する治療剤をinvivo の治療用に選択する。こうした生存および/または増殖を評価するために、当業界で標準的な多くのアッセイを使用することができる。;例えば、細胞の増殖を、3H-チミジンの取り込みの測定、細胞数の直接の計測、プロトオンコジーン(例えばfoc,myc)などの既知の遺伝子の転写活性または細胞周期マーカーの変化の検出によってアッセイすることができる。;細胞の生存度はトリパンブルー染色によって評価することができ、分化は形態学その他における変化を基礎として視覚的に評価することができる。特定の場合には、悪性腫瘍細胞培養物を(1)アゴニスト治療剤、および(2)アンタゴニスト治療剤に別々に暴露する。アッセイの結果によってどちらの型の治療剤が治療効果を有するかが示される。
【0105】
別の1態様において、それぞれ高または低増殖性障害を有する、または有すると推定さ
れる組織からの患者の細胞サンプルに対して所望の効果、細胞増殖の阻害または促進を示
す治療剤が有用なものとして示される。こうした高または低増殖性障害には、限定するわ
けではないが、下記の第5.8.1節から第5.8.3節に記載されたものが含まれる。
【0106】
もう一つの特定の実施態様において、神経損傷または神経系変性障害の治療に使用する
ための治療薬が提示されているが、該治療薬は、in vitroでの神経再生の促進/罹患した
患者タイプの神経細胞からの神経突起の伸展を呈する。
【0107】
更に、NotchまたはSerrate優位活性化表現型(「機能獲得」突然変異)が関与することが判明または知られている疾患または障害において、本発明の拮抗治療薬の投与もまた提示されている。NotchまたはSerrate優位陰性表現型(「機能損失」突然変異)が関与することが判明または知られている疾患または障害において、本発明の作用治療薬の投与が提示されている。hsp70熱ショックプロモーターならびに眼特異的プロモーターの下で一連のドロゾフィラNotch欠失突然変異体を異所的に発現することによって、Notchタンパク質の種々の構造ドメインの機能をinvivoで調べた(Rebay ら.,1993,Cell 74:319-329 を参照されたい)。2つのクラスの優位表現型、すなわち、Notch機能損失突然変異を示すもの、およびNotch機能獲得突然変異を示すものが観測された。ほとんどの細胞外配列が欠損したタンパク質の過剰発現により優位「活性化」表現型を生じ、一方、ほとんどの細胞内配列が欠損したタンパク質の過剰発現により優位「陰性」表現型を生じた。これらの結果から次のことを示唆された。すなわち、Notchは受容体(この受容体の細胞外ドメインはリガンド結合を媒介する)として機能し、その結果、細胞質ドメインは発生シグナルを伝達することが示唆された。本発明者らは、SerrateがNotchELR 11および12に結合することを明らかにした(PCT公開第WO93/12141号を参照されたい)。
【0108】
種々の特定の実施態様において、患者の障害に関与する細胞タイプの代表的細胞を用い
てin vitroアッセイを行い、治療薬がこうした細胞タイプに所望の効果を示すかを決定することができる。
【0109】
もう1つの実施態様において、患者の組織サンプルの細胞で、前腫瘍性であるとの疑いのあるものを、同様に、プレーティングまたはinvitroで増殖させて、治療薬に晒す。より正常な(すなわち、前腫瘍状態、腫瘍状態、悪性状態、または形質転換表現型の徴候がより弱い)細胞表現型を生じる治療薬を、治療用に選択する。当該技術分野の多くのアッセイ標準を使用して、前腫瘍状態、腫瘍状態、または形質転換もしくは悪性表現型が存在するかを評価することができる。例えば、形質転換表現型に関連した特徴(invivoでの腫瘍形成能に関連した一連のin vitroの特徴)としては、より丸形の細胞形態、より弱い基質結合、接触抑制の欠損、固定依存性の欠損、プラスミノーゲンアクチベータなどのプロテアーゼの放出、糖輸送の増大、血清必要量の減少、胎児性抗原の発現、250,000ダルトンの表面タンパク質の消失などが挙げられる(Luriaら.,1978,General Virology,第3版.,John Wiley & Sons,New York pp.436-446を参照されたい)。
【0110】
他の特定の実施態様において、治療される特定の患者に由来する細胞サンプルを使用するのではなく、本発明に従って、次のような細胞系統を使用して、上述のin vitroアッセイを行うことができる。すなわち、該細胞系統が、治療もしくは予防が望まれる悪性障害、腫瘍性障害、もしくは前腫瘍性障害に由来するか、またはそれらに関連した特徴を呈するか、あるいは作用対象となることが望まれる神経細胞タイプもしくは他の細胞タイプに由来する場合である。
【0111】
拮抗治療薬は、(1)NotchまたはSerrate機能のレベルの増大(正常または所望のレベルと比較して)が関与する疾患または障害において、例えば、NotchまたはSerrateタンパク質が過剰発現されるか、または過剰活性である疾患または障害において、および(2)invitro(またはin vivo)アッセイからSerrate拮抗薬の投与の効用があることが示唆される疾患または障害において、治療のために(予防のために)投与される。NotchまたはSerrate機能のレベルの増大は、上述したような方法によりタンパク質および/またはRNAを定量することによって、容易に検出することができる。患者の組織サンプルの細胞、または適切な細胞系統もしくは細胞型を用いたinvitroアッセイを、治療薬の効用を決定するために上述したように実施することができる。
5.8.1.悪性疾患
拮抗治療薬または作用治療薬を介在させたときの効能について上述したように試験する
ことができるとともに、治療薬の効用の徴候の観察に基づいて治療することができる悪性
または前腫瘍状態としては、以下の5.8.1および5.9.1節に記載のものが挙げられるが、これらに限定されるものではない。
【0112】
in vitro(および/またはinvivo)で試験することができるとともに、適切なアッセ
イの結果を観察し、これに基づいて本発明に従った治療が行えるタイプの細胞を含む悪性
腫瘍および関連障害としては、表1に列挙したものが挙げられるが、これらに限定される
ものではない(こうした障害の総説については、Fishman ら.,1985,Medicine,第2版.
,J.B.LippincottCo.,Philadelphia を参照されたい)。
【0113】
【表1】
特定の実施態様において、上皮組織(例えば、頸部、食道、および肺の上皮組織)の悪性疾患または異常増殖変化(例えば、化成および異形成)を治療または予防することができる。
【0114】
結腸および頸部の悪性腫瘍では、それらの非悪性組織と比べると、ヒトNotch の発現が増大する(1994年4月14日公開のPCT公開番号第WO94/07474号を参照されたい。この全体を参考として本明細書に組み入れる)。従って、特定の実施態様において、有効量の拮抗治療薬(例えば、Notch機能と拮抗するSerrate誘導体)を投与することによって、結腸または頸部の悪性疾患または前悪性疾患の変化を治療または予防することができる。結腸癌および頸部癌の中でNotch発現の増大が見られるということは、多数のより癌性かつ高増殖性の状態が、アップレギュレートされたNotchを呈することを示唆する。従って、特定の実施態様において、Notch機能と拮抗する拮抗治療薬を投与することによって、種々の癌、例えば、乳癌、扁平上皮腺癌、セミノーマ、および肺癌、ならびにそれらの中で生じた前悪性疾患の変化、更に他の高増殖性障害を治療または予防することができる。
【0115】
5.8.2.神経系障害
拮抗治療薬または作用治療薬を介在させたときの効能について上述したように試験する
ことができるとともに、治療薬の効用の徴候の観察に基づいて治療することができる細胞
型を含む神経系障害としては、軸索の切断、ニューロンの縮小もしくは退化、または髄鞘
脱落のいずれかを生じる神経系損傷および疾患または障害が挙げられるが、これらに限定
されるものではない。本発明にしたがって患者(ヒトおよびヒト以外の哺乳類の患者を含
む)の治療が可能な神経系病変としては、中枢神経系(脊髄、脳を含む)または末梢神経
系の以下の病変が挙げられるが、これらに限定されるものではない:
(i)外傷性病変(物理的損傷により生じるかまたは手術に伴う病変、例えば、神経系の
一部を切断する病変または圧迫性損傷が含まれる);
(ii)虚血性病変(神経系の一部で酸素が欠乏することにより神経損傷を生じるかまたは
死亡するもので、脳梗塞もしくは虚血、または脊髄梗塞もしくは虚血が含まれる);
(iii)悪性病変(神経系関連悪性疾患または非神経系組織由来の悪性疾患のいずれかで
ある悪性組織によって、神経系の一部が破壊または損傷を受けたもの);
(iv)感染性病変(感染の結果として、例えば、膿瘍によって、あるいはヒト免疫不全ウ
イルス、帯状庖疹ウイルス、もしくは単純庖疹ウイルスの感染と関連して、またはライム
病、結核、梅毒と関連して、神経系の一部が破壊または損傷を受けたもの);
(v)変性性病変(変性過程の結果として神経系の一部が破壊または損傷を受けたもので
、パーキンソン病、アルツハイマー病、ハンティングトン舞踏病、または筋萎縮性側索硬
化症が含まれるが、これらに限定されるものではない);
(vi)栄養性疾患または障害に関連した病変(栄養障害または代謝異常によって神経系の
一部が破壊または損傷を受けたもので、ビタミンB12欠乏症、葉酸欠乏症、ウェルニッケ
病、たばこ性‐アルコール性弱視、マルキアファーヴァ‐ビニャミ病(脳梁の原発性変性
)及びアルコール性小脳変性が含まれるが、これらに限定されるものではない);
(vii)全身性疾患に関連した神経系病変(糖尿病(糖尿病性ニューロパシー、ベル麻痺
)、全身性紅斑性狼瘡、癌腫、またはサルコイドーシスが含まれるが、これらに限定され
るものではない);
(viii)毒性物質、例えば、アルコール、鉛、または特定の神経毒によって引き起こされ
る病変;及び
(ix)脱髄病変(脱髄疾患によって神経系の一部が破壊または損傷を受けたもので、多発
性硬化症、ヒト免疫不全ウイルス関連ミエロパシーまたは種々の病因、進行性多病巣白質
脳症、および橋中央ミエリン溶解が含まれるが、これらに限定されるものではない)。
【0116】
ニューロンの生存または分化を促進する生物活性を試験することによって、神経系疾患
の治療に有用な本発明に係る治療薬を選択することが可能である(5.8 節を参照されたい
)。例えば、以下の効果のいずれかを誘発する治療薬が本発明に有用であるが、これらに
限定されるものではない:
(i)培養中のニューロンの生存時間の増大;
(ii)培養中またはinvivoでのニューロンの発芽の増大;
(iii)培養中またはinvivoでのニューロン関連分子(例えば、運動ニューロンに関するコリンアセチルトランスフェラーゼまたはアセチルコリンエステラーゼ)の生成の増大;
(iv)invivoでのニューロン機能不全の徴候の増大。
【0117】
こうした効果は、当該技術分野で公知のいかなる方法によっても測定することが可能である。好ましい実施態様において、ただし、これに限定されるものではないが、ニューロンの生存の増大は、Arakawaら(1990,J.Neurosci.10:3507-3515)により報告された方法により測定可能であり;ニューロンの発芽の増大は、Pestronkら(1980,Exp.Neurol.70:65-82)またはBrownら(1981,Ann.Rev.Neurosci.4:17-42)により報告された方法により測定可能であり;ニューロン関連分子の生成の増大は、測定される分子にもよるが、バイオアッセイ、酵素アッセイ、抗体結合、ノーザンブロットアッセイなどにより測定可能であり;運動ニューロンの機能不全は、運動ニューロン疾患の物理的症状発現、例えば衰弱、運動ニューロン伝導速度、または機能障害を評価することにより測定可能である。
【0118】
特定の実施態様において、本発明に従って治療可能な運動ニューロン疾患としては、運
動ニューロンおよび神経系の他のコンポーネントに影響を与える恐れのある梗塞、感染、
毒との接触、外傷、外科的損傷、変性性疾患、または悪性疾患、ならびにニューロンに選
択的に影響を及ぼす疾患、例えば筋萎縮性側索硬化症が挙げられるが、これらに限定され
るものではなく、更に、進行性棘筋萎縮症、進行性球麻痺、原発性側索硬化症、小児およ
び若年筋萎縮症、小児期進行性球麻痺(ファツィオ‐ロンド症候群)、ポリオおよびポリ
オ後症候群、遺伝性運動感覚ニューロパシー(シャルコー‐マリー‐トゥース病)も含ま
れるが、これらに限定されるものではない。
【0119】
5.8.3.組織修復および再生
本発明のもう1つの実施態様において、組織再生および修復を促進するために、本発明
の治療薬が使用される(例えば、良性異常増殖疾患の治療のために使用されるが、これに
限定されるものではない)。特定の実施態様は、肝硬変(瘢痕が正常な肝臓再生過程を阻
害する状態)の治療、ケロイド(過形成性瘢痕)形成(瘢痕過程により正常の更新が妨害
されて皮膚の美観が損なわれた状態)、乾癬(皮膚の過剰増殖および固有細胞運命決定の
遅延を特徴とするよく見かける皮膚の状態)、および禿頭症(末端分化毛包(Notchに富
んだ組織)が正常に機能しない状態)の治療に関連する。もう1つの実施態様において、
本発明の治療薬を使用して、内耳の感覚上皮の変性性疾患または外傷性疾患の治療を行う
。
【0120】
5.9.予防的使用
5.9.1.悪性疾患
本発明の治療薬は、腫瘍状態または悪性状態への進行を防止するために投与できる。これには表1に列挙した疾患が対象となるが、これらに限定されるものではない。こうした投与が行えるのは、上述したアッセイにおいて治療薬がこうした疾患の治療または防止に有効であることが示された場合である。こうした予防的使用が利用できるのは、上述したような腫瘍または癌へ進化することが知られているか、またはそうした進化の疑いのある状態のとき、具体的には、過形成、化生、または特に異形成が関与した非腫瘍性細胞成長が起こったときである(こうした異常成長状態の総説については、Robbins及びAngell,1976,Basic Pathology,第2版.,W.B.Saunders Co.,Philadelphia,pp.68-79を参照されたい)。過形成とは、組織または器官の中の細胞数の増加が関係する調節された細胞増殖の形態のことであり、構造または機能上の有意な変化は見られない。1つだけ例を挙げると、子宮内膜過形成は、しばしば子宮内膜癌へ進行する。化生とは、成人細胞(すなわち十分に分化した細胞)の1つのタイプが成人細胞のもう1つのタイプと置き換わる調節された細胞成長の形態のことである。化生は、上皮組織細胞または結合組織細胞で起こる可能性がある。非定型性化生には、いくらか障害を有する化生性上皮が含まれる。異形成とは、しばしば癌の前兆となるものであり、主に上皮に見うけられる;これは非腫瘍性細胞成長の中で最も障害を有する形態であり、それぞれの細胞の均一性および細胞の構造上の配列が失われる。異形成細胞はしばしば異常に大きくかつ強く染色された核を有し、多形性を呈する。異形成は、慢性的刺激または炎症が起こるの場合に特徴的に生じるもので、頸部、呼吸経路、口腔、および胆嚢によく見うけられる。
【0121】
過形成、化生、または異形成を特徴とする異常細胞成長が存在する代わりに、またはその成長が存在することに加えて、患者の細胞サンプルがin vivoで呈するか、またはin vitroで呈する形質転換表現型または悪性表現型のうちの1つ以上の特徴が存在することは、本発明の治療薬を予防/治療のための投与することが望ましいことを示唆する。先に述べたように、形質転換表現型のこうした特徴としては、形態の変化、より弱い基質結合、接触抑制の欠損、固定依存性の欠損、プロテアーゼの放出、糖輸送の増大、血清必要量の減少、胎児性抗原の発現、250,000ダルトンの細胞表面タンパク質の消失などが挙げられる(形質転換表現型または悪性表現型に関連した特徴については、同上.,pp.84-90をも参照されたい)。
【0122】
特定の実施態様において、白斑症、上皮の外観上良性の過形成または異形成疾患、また
はボーエン病、insitu癌は、予防的介入が望ましいと考えられる前腫瘍性疾患である。
【0123】
もう1つの実施態様において、線維嚢胞病(嚢胞性過形成、乳房異形成、特に腺疾患(
良性上皮過形成))は、予防的介入が望ましいと考えられる。
【0124】
他の実施態様において、悪性疾患に対する以下の素因のうちの1つ以上を呈する患者を、有効量の治療薬の投与により治療する。その要因とは次の通りである:悪性疾患に関連した染色体転座(例えば、慢性骨髄性白血病に対するフィラデルフィア染色体、濾胞性リンパ腫に対するt(14;18)など)、家族性ポリープ症またはガードナー症候群(結腸癌の恐れのある前兆)、良性単一クローン性高ガンマグロブリン血症(多発性骨髄腫の恐れのある前兆)、およびメンデル型(遺伝的)継承様式を示す癌または前癌性病(例えば、結腸の家族性ポリープ症、ガードナー症候群、遺伝性外骨腫症、多内分泌腺腫症、アミロイド生成を伴う髄甲状腺癌および褐色細胞腫、ポッツ‐ジェガース症候群、フォン・レックリングハウゼンの神経線維腫病、網膜芽細胞腫、頸動脈小体腫瘍、皮膚黒色癌、眼内性黒色癌、色素性乾皮症、毛細管拡張性運動失調症、チェディアック‐東症候群、白子症、ファンコーニ再生不良性貧血、並びにブルーム症候群;Robbins及び Angell,1976,Basic Pathology,第2版.,W.B.Saunders Co.,Philadelphia,pp.112-113を参照されたい)の者と1親等の関係。
【0125】
もう1つの特定の実施態様において、本発明の拮抗治療薬をヒト患者に投与して、乳癌
、結腸癌、または子宮頚癌への進行を防ぐ。
5.9.2.他の障害
他の実施態様において、本発明の治療薬を投与して、5.8.2.節に記載の神経系障害また
は5.8.3.節に記載の他の障害(例えば、肝硬変、乾癬、ケロイド、禿頭症)を防ぐことが
できる。
【0126】
5.10.治療または予防の効用の実証
本発明の治療薬をinvivoで試験して、所望の治療活性または予防活性を調べることが
できる。例えば、こうした化合物を好適な動物モデル系で試験してからヒトにおいて試験
することができる。好適な動物としては、ラット、マウス、ニワトリ、ウシ、サル、ウサ
ギなどが挙げられるが、これらに限定されるものではない。in vivo試験に対しては、ヒ
トに投与する前に、当該技術分野で公知のいかなる動物モデル系を使用してもよい。
【0127】
5.11.Serrate発現のアンチセンス調節
本発明は、脊椎動物Serrateまたはその一部をコードする遺伝子またはcDNAに対してアンチセンスである少なくとも6個または少なくとも10個のヌクレオチドの核酸の治療または予防への使用を提供する。本明細書中で使用される「アンチセンス」とは、ある種の配列相補性によって脊椎動物SerrateRNA(好ましくはmRNA)の一部とハイブリッドを形成することが可能な核酸を意味する。こうしたアンチセンス核酸は、本発明の拮抗治療薬としての効用を有し、セクション5.8およびそのサブセクションに既に記載したような障害の治療または予防に使用することができる。
【0128】
本発明のアンチセンス核酸は、細胞に直接投与することができるか、または外来導入配列の転写によって細胞内で生成させることができる、二本鎖または一本鎖のRNAもしくはDNAまたはそれらの修飾体もしくは誘導体であるオリゴヌクレオチドであってもよい。
【0129】
特定の実施態様において、本発明により提供されるSerrateアンチセンス核酸は、腫瘍または他の障害(ただし、これらの腫瘍タイプまたは障害の細胞がSerrate遺伝子またはNotch遺伝子を発現することを(in vitroまたはin vivoで)実証することができるもの)の治療に使用することができる。これはRNAまたはタンパク質を検出することにより実証が可能である。
【0130】
本発明は更に、以下のセクション5.12に記載されているように、医薬品として許容しうる担体中に有効量の本発明のSerrateアンチセンス核酸を含む医薬用組成物を提供する。また、本発明の医薬用組成物の投与を含む、障害(例えば、セクション5.8および5.9に記載の障害)の治療および予防の方法が提供される。
【0131】
もう1つの実施態様において、本発明は、原核細胞または真核細胞中でのSerrate核酸配列の発現を抑制する方法に関する。この方法には、本発明のアンチセンス脊椎動物Serrate核酸を含む有効量の組成物を細胞に供給することが含まれる。
【0132】
Serrateアンチセンス核酸およびそれらの使用については、以下で詳細に説明する。
5.11.1.脊椎動物Serrateアンチセンス核酸
脊椎動物Serrateアンチセンス核酸は、少なくとも6個のヌクレオチド、好ましくはオリゴヌクレオチド(オリゴヌクレオチドの数の好ましい範囲は10個〜約50個である)から成る。特定の態様において、オリゴヌクレオチドは、Serrate遺伝子に対してアンチセンスである少なくとも10個のヌクレオチド、少なくとも15個のヌクレオチド、少なくとも100個のヌクレオチド、または少なくとも200個のヌクレオチドを含有する。オリゴヌクレオチドは、一本鎖または二本鎖の、DNAもしくはRNA、またはそれらのキメラ混合物もしくは誘導体もしくは修飾体であってもよい。オリゴヌクレオチドの塩基部分、糖部分、またはホスフェート主鎖を修飾してもよい。オリゴヌクレオチドには、ペプチドなどの他の追加基、あるいは細胞膜(例えば、Letsingeret al.,1989,Proc.Natl.Acad.Sci.U.S.A.86:6553-6556; Lemaitre et al.,1987,Proc.Natl.Acad.Sci.84:648-652;1988年12月15日公開のPCT公開第WO88/09810号を参照されたい)もしくは血液脳関門(例えば、1988年4月25日公開のPCT公開第WO89/10134号を参照されたい)を通過する輸送の促進剤、ハイブリダイゼーションに起因する開裂剤(例えば、Krolet al.,1988,BioTechniques 6:958-976を参照されたい)、またはインターカレート剤(例えば、Zon,1988,Pharm.Res.5:539-549を参照されたい)が含まれていてもよい。
【0133】
本発明の好ましい態様において、脊椎動物Serrateアンチセンスオリゴヌクレオチド(好ましくは一本鎖DNA)が提供される。最も好ましい態様において、こうしたオリゴヌクレオチドには、Serrate(最も好ましくはヒトSerrate同族体)のSH3結合ドメインまたはNotch結合ドメインをコードする配列に対してアンチセンスである配列が含まれる。オリゴヌクレオチドの構造の任意の位置を、当該技術分野で一般的に知られた置換基を用いて修飾してもよい。
【0134】
Serrateアンチセンスオリゴヌクレオチドには、5-フロオロウラシル、5-ブロモウラシル、5-クロロウラシル、5-ヨードウラシル、ヒポキサンチン、キサンチン、4-アセチルシトシン、5-(カルボキシヒドロキシメチル)ウラシル、5-カルボキシメチルアミノメチル-2-チオウリジン、5-カルボキシメチルアミノメチルウラシル、ジヒドロウラシル、β-D-ガラクトシルキューオシン、イノシン、N6-イソペンテニルアデニン、1-メチルグアニン、1-メチルイノシン、2,2-ジメチルグアニン、2-メチルアデニン、2-メチルグアニン、3-メチルシトシン、5-メチルシトシン、N6-アデニン、7-メチルグアニン、5-メチルアミノメチルウラシル、5-メトキシアミノメチル-2-チオウラシル、β-D-マンノシルキューオシン、5'-メトキシカルボキシメチルウラシル、5-メトキシウラシル、2-メチルチオ-N6-イソペンテニルアデニン、ウラシル-5-オキシ酢酸(v)、ワイブトキソシン、プソイドウラシル、キューオシン、2-チオシトシン、5-メチル-2-チオウラシル、2-チオウラシル、4-チオウラシル、5-メチルウラシル、ウラシル-5-オキシ酢酸メチルエステル、ウラシル-5-オキシ酢酸(v)、5-メチル-2-チオウラシル、3-(3-アミノ-3-N-2-カルボキシプロピル)ウラシル、(acp3)w、および2,6-ジアミノプリンから選ばれる少なくとも1つの修飾された塩基部分が含まれていてもよい。
【0135】
別の実施態様において、オリゴヌクレオチドは、アラビノース、2-フルオロアラビノー
ス、キシルロース、およびヘキソースから成る群より選ばれる少なくとも1つの修飾され
た糖部分を含むが、糖部分がこれらに限定されるものではない。
【0136】
更に別の実施態様において、オリゴヌクレオチドは、ホスホロチオエート、ホスホロジ
チオエート、ホスホラミドチオエート、ホスホラミデート、ホスホルジアミデート、メチ
ルホスホネート、アルキルホスホトリエステル、およびこれらのホルムアセタールまたは
類似体から成る群より選ばれる少なくとも1つの修飾されたホスフェート主鎖を含む。
【0137】
更に別の実施態様において、オリゴヌクレオチドはα-アノマーオリゴヌクレオチドで
ある。α-アノマーオリゴヌクレオチドは、通常のβ-単位とは異なり鎖が互いに平行に走
る相補RNAと共に特異的な二本鎖ハイブリッドを形成する(Gautier et al.,1987,Nu
cl.Acids Res.15:6625-6641)。
【0138】
オリゴヌクレオチドは他の分子(例えば、ペプチド、ハイブリダイゼーションに起因す
る架橋剤、輸送剤、ハイブリダイゼーションに起因する開裂剤など)とコンジュゲートし
てもよい。
【0139】
本発明のオリゴヌクレオチドは、当該技術分野で公知の標準的な方法で、例えば、DN
A自動合成装置(例えば、Biosearch、Applied Biosystemsなどから市販されている装置
)を用いて合成することが可能である。例として、ホスホロチオエートオリゴヌクレオチ
ドは、Steinらの方法で合成可能であり(1988,Nucl.AcidsRes.16:3209)、メチルホ
スホネートオリゴヌクレオチドは、調節された細孔のガラスポリマー担体を使用して調製
可能である(Sarinet al.,1988,Proc.Natl.Acad.Sci.U.S.A.85:7448-7451)。
【0140】
特定の実施態様において、Serrateアンチセンスオリゴヌクレオチドは、触媒RNAすなわちリボザイムを含む(1990年10月4日公開のPCT国際公開第WO90/11364;Sarver et al.,1990,Science247:1222-1225号を参照されたい)。別の実施態様において、オリゴヌクレオチドは、2'-O-メチルリボヌクレオチド(Inoue et al.,1987,Nucl.AcidsRes.15:6131-6148)またはキメラRNA‐DNA類似体(Inoue et al.,1987,FEBS Lett.215:327-330)である。
【0141】
別の実施態様において、本発明のSerrateアンチセンス核酸が、外来配列の転写によって細胞内で生成される。例えば、ベクターをin vivoで導入し、細胞がこのベクターを捕らえ、細胞内でこのベクターまたはその一部を転写し、本発明のアンチセンス核酸(RNA)を生成することができる。こうしたベクターには、Serrateアンチセンス核酸をコードする配列が含まれている。このベクターは、転写されて所望のアンチセンスRNAを生成するかぎり、エピソームの状態を保持することができるか、または染色体と一体化することができる。これらのベクターは、当該技術分野で標準的な組換えDNA技法によって得ることができる。ベクターは、哺乳類細胞中での複製および発現のために使用されるプラスミド、ウイルス、または当該技術分野で公知の他のものであってもよい。SerrateアンチセンスRNAをコードする配列の発現を、当該技術分野で公知の任意のプロモーターを用いて行い、哺乳動物好ましくはヒトの細胞において機能させることができる。これらのプロモーターは誘導性または構成的であってもよい。こうしたプロモーターとしては、SV40初期プロモーター領域(Bernoistand Chambon,1981,Nature 290:304-310)、ラウス肉腫ウイルスの3'-長末端反復配列に含まれるプロモーター(Yamamotoet al.,1980,Cell 22:787-797)、ヘルペスチミジンキナーゼプロモーター(Wagner et al.,1981,Proc.Natl.Acad.Sci.U.S.A.78:1441-1445)、メタロチオネイン遺伝子の調節配列(Brinsteret al.,1982,Nature 296:39-42)が挙げられるが、これらに限定されるものではない。
【0142】
本発明のアンチセンス核酸には、脊椎動物Serrate遺伝子、好ましくはヒトSerrate遺伝子に対して特異的なRNA転写産物の少なくとも一部と相補的な配列が含まれる。しかしながら、絶対的な相補性が好ましいものではあるが、それが必要というわけではない。本明細書中に記載されている「RNAの少なくとも一部と相補的な」配列とは、RNAとハイブリッドを形成して安定な二本鎖を形成するのに十分な相補性を有する配列を意味する。二本鎖Serrateアンチセンス核酸の場合は、二本鎖DNAのうちの一本の鎖を試験するか、または三本鎖の形成を調べてもよい。ハイブリッド形成能は、アンチセンス核酸の相補性の度合いと長さの両方に依存する。一般的には、ハイブリッド形成する核酸が長いほど、それに含まれるより多くの塩基がSerrateRNAとの不一致を起こすが、依然として安定な二本鎖(または三本鎖。ただし場合による)を形成する。当業者は、標準的な手順を使用してハイブリッド化複合体の融点を測定することにより、不一致の許容範囲を確認することができる。
5.11.2.脊椎動物Serrateアンチセンス核酸の治療上の効用
脊椎動物Serrateアンチセンス核酸を使用して、SerrateまたはNotchを発現することが分かっている細胞タイプの悪性疾患または他の障害を治療(または防止)することができる。特定の実施態様において、悪性疾患とは、頸部癌、乳癌、もしくは結腸癌、または扁平上皮腺癌のことである。こうした発現を試験することができる悪性細胞、腫瘍性細胞、および前腫瘍性細胞としては、セクション5.8.1および5.9.1で既に記載した細胞が挙げられるが、これらに限定されるものではない。好ましい実施態様において、一本鎖DNAアンチセンスSerrateオリゴヌクレオチドが使用される。
【0143】
SerrateまたはNotchRNAを発現する悪性(特に腫瘍)細胞タイプは、当該技術分野で公知の種々の方法によって同定することができる。こうした方法としては、SerrateまたはNotch特異性核酸とのハイブリダイゼーション(例えば、ノーザンハイブリダイゼーション、ドットブロットハイブリダイゼーション、in situ ハイブリダイゼーション)、該細胞タイプに由来するRNAがin vitroでNotchまたはSerrateに翻訳される能力の観測、イムノアッセイなどが挙げられるが、これらに限定されるものではない。好ましい実施態様において、例えば、免疫細胞化学によるか、またはinsituハイブリダイゼーションによって、治療前にNotchまたはSerrate発現に関して患者の原発性腫瘍組織を調べる。
【0144】
医薬品として許容しうる担体中に有効量の脊椎動物Serrateアンチセンス核酸を含む本
発明の医薬用組成物は、NotchまたはSerrate RNAまたはタンパク質を発現するタイプ
の悪性疾患を患った患者に投与することができる。
【0145】
特定の障害または状態の治療に有効なSerrateアンチセンス核酸の量は、障害または状
態の特性に依存するが、標準的な臨床技術により決定することができる。可能ならば、治
療する腫瘍タイプのアンチセンス細胞毒性を、in vitroで、次に有用な動物モデル系で測
定してから、ヒトにおいて試験および使用することが望ましい。
【0146】
特定の実施態様において、脊椎動物Serrateアンチセンス核酸を含む医薬用組成物は、
リポソーム、ミクロ粒子、またはマイクロカプセルを介して投与される。本発明の種々の
実施態様において、こうした組成物を使用してSerrateアンチセンス核酸を持続的に放出
することが有用である。特定の実施態様において、抗体を介して特定の識別可能な腫瘍抗
原を標的とするリポソームを利用することが望ましい(Leonetti et al.,1990,Proc.N
atl.Acad.Sci.U.S.A.87:2448-2451;Renneisen et al.,1990,J.Biol.Chem.265:
16337-16342)。
5.12.治療/予防のための投与および組成物
本発明は、有効量の本発明の治療薬を被検者に投与することによって治療(および予防
)を行う方法を提供する。好ましい態様において、治療薬は実質的に精製されたものであ
る。被検者は好ましくは動物であり、ウシ、ブタ、ニワトリなどの動物が含まれ、好まし
くは哺乳動物、最も好ましくはヒトであるが、これらに限定されるものではない。
【0147】
種々のデリバリーシステムが公知であり、これらは本発明の治療薬の投与に使用できる
。例えば、リポソーム、ミクロ粒子、マイクロカプセル中への封入、組換え細胞による発
現、受容体媒介エンドサイトーシス(例えば、Wu and Wu,1987,J.Biol.Chem.262:44
29-4432を参照されたい)、治療薬である核酸をレトロウイルスベクターまたは他のベク
ターの一部として構築することなどが使用できる。導入方法としては、皮内、筋肉内、腹
腔内、静脈内、皮下、鼻腔内、硬膜外、口腔を介した経路が挙げられるが、これらに限定
されるものではない。この化合物は、任意の便利な経路で投与してもよい。例えば、注入
またはボーラス注射によって、表皮または粘膜皮膚の内面(例えば、口腔粘膜、直腸粘膜
、および腸粘膜など)を介した吸収によって投与してもよい。更に、この化合物を他の生
物活性薬剤と共に投与してもよい。投与は、全身的でも局所的でもよい。また、脳室内注
射および髄腔内注射などの任意の好適な経路で、本発明の医薬用組成物を中枢神経系に導
入することが望ましい。脳室内注射は、例えば、レザバー(オムヤマ・レザバーなど)に
取り付けられた脳室内カテーテルを用いて容易に行える。また、肺への投与は、例えば、
吸入器または噴霧器を使用し、エアゾール剤と共に処方することによって実施することが
できる。
【0148】
特定の実施態様において、治療が必要な領域に本発明の医薬用組成物を局所的に投与す
ることが望ましい。こうした局所的投与は、例えば、外科手術中の局所注入、局所施用(
例えば、外科手術後の創傷用包帯と併用して)によって、注射によって、カテーテルを用
いて、座薬を用いて、または移植片(該移植片は、多孔性、非多孔性、または膠状の材料
から成るもので、例えば、シアラスチック(sialastic)膜などの膜または繊維などである
)を用いて行うことができるが、これらに限定されるものではない。1実施態様において
、悪性腫瘍または腫瘍性組織もしくは前腫瘍性組織の部位(すなわち形成部位)に直接注
射を行うことによって、投与することができる。
【0149】
もう1つの実施態様において、治療薬をベシクル中に入れて、特にリポソームに入れて
送達することができる(Langer,Science 249:1527-1533(1990); Treat et al.,in Lipo
somes in theTherapy of Infectious Disease and Cancer,Lopez-Berestein and Fidle
r(eds.),Liss,NewYork,pp.353-365(1989); Lopez-Berestein,ibid.,pp.317-327
を参照されたい;一般的な内容についてもこれらの文献を参照されたい)。
【0150】
更に別の実施態様において、制御放出システムで治療薬を送達することができる。1実施態様では、ポンプを使用してもよい(Langer,supra; Sefton,CRC Crit.Ref.Biomed.Eng.14:201(1987);Buchwald et al.,Surgery 88:507(1980); Saudek et al.,N.Engl.J.Med.321:574(1989)を参照されたい)。別の実施態様において、ポリマー材料を使用することができる(MedicalApplications of Controlled Release,Langer and Wise(eds.),CRC Pres.,Boca Raton,Florida(1974);Controlled Drug Bioavailability,Drug Product Design and Performance,Smolen andBall(eds.),Wiley,New York(1984); Ranger and Peppas,J.Macromol.Sci.Rev.Macromol.Chem.23:61(1983)を参照されたい;更にLevyet al.,Science 228;190(1985); During et al.,Ann.Neurol.25:351(1989); Howard etal.,J.Neurosurg.71:105(1989)をも参照されたい)。更に別の実施態様において、放出制御システムを治療標的(すなわち、脳)の近傍に配置することができる。この場合、全身用量のわずか一部が必要となるだけである(Goodson,inMedical Applications of Controlled Release,supra,vol.2,pp.115-138(1984)を参照されたい)。
【0151】
その他の放出制御システムについては、Langerの総説(Science 249:1527-1533(1990)の中で説明されている。
【0152】
治療薬が核酸であり、この核酸がタンパク質治療薬をコードする場合の特定の実施態様において、この核酸を次のようにしてin vivoで投与し、そのコードされたタンパク質の発現を促進することができる。すなわち、この核酸を適切な核酸発現ベクターの一部として構築して投与し、細胞内に取り込ませることによって、例えば、レトロウイルスベクターを用いて(米国特許第4,980,286号を参照されたい)、または直接注射することによって、またはミクロ粒子衝撃(例えば、遺伝子ガン;Dupont製Biolistic)によって、または脂質、細胞表面受容体、もしくは移入剤を塗布することによって、または核に入り込むことが知られているホメオボックス様ペプチド(例えば、Joliotet al.,1991,Proc.Natl.Acad.Sci.USA 88:1864-1868を参照されたい)に結合させて投与することなどによって実施することができる。この他に、核酸治療薬を細胞内に導入し、宿主細胞DNA内に取り込んで相同的組換えにより発現させることができる。
【0153】
個々の障害の治療または予防に関連した特定の実施態様において、以下の形態の投与を
行うのが好ましい:
また、本発明は、医薬組成物も提供する。かかる組成物は、治療上有効な量の治療薬及
び製剤学的に許容できる担体を含む。特定の態様において、「製剤学的に許容できる」と
いう用語は、連邦政府又は州政府の統制機関によって認可された、ということ又は動物、
より特定的にはヒトにおける使用のための米国薬局方もしくは他の一般的に認可された薬
局方に記載された、ということを意味する。
【0154】
「担体」という用語は、治療薬とともに投与される希釈剤、アジュバント、賦形剤又はビヒクルをいう。そのような製剤学的担体は、水及び落花生由、大豆油、鉱油、ごま油などのような、石油、動物、植物、又は合成源由来のものを含むオイル等の無菌液でもよい。医薬組成物が静脈内投与される場合、好ましい担体は水である。また、生理食塩水並びに水性ブドウ糖(dextrose)及びグリセロール溶液も、特に注射溶液用の液状担体として用いることができる。適当な製剤学的賦形剤としては、デンプン、グルコース、ラクトース、スクロース、ゼラチン、麦芽、米、小麦、白亜(chalk)、シリカゲル、ステアリン酸ナトリウム、グリセリンモノステアラート、タルク、塩化ナトリウム、脱脂粉乳、グリセロール、プロピレン、グリコール、水、エタノール等が挙げられる。組成物には、所望により少量の加湿もしくは乳化剤又はpH緩衝剤を含ませることもできる。これらの組成物は、溶液、懸濁液、エマルジョン、錠剤、丸剤、カプセル剤、散剤、徐放性剤等の剤形を取り得る。組成物は、伝統的な結合剤及びトリグリセリド等の担体を用いて坐剤として処方することができる。経口製剤としては、製剤学的グレードのマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、ナトリウムサッカリン、セルロース、炭酸マグネシウム等の標準的な担体を挙げることができる。適当な製剤学的担体の例は、E.W.Martinによる「レミントンの薬学(Remington'sPharmaceutical Sciences)」に記載されている。このような組成物には、治療上有効な量の治療薬が、好ましくは患者に適した投与剤形を提供するような適量の担体とともに精製された状態で含まれる。その処方は投与様式に合わせるのがよい。
【0155】
好ましい態様においては、組成物はヒトへの静脈内投与に適合した医薬組成物としてルーチン的な手順で処方される。静脈内投与用の組成物は、典型的には無菌等張水性緩衝液に溶かした溶液である。必要ならば、組成物に可溶化剤及び注射部位の痛みを取り除くためのリドカイン等の局所麻酔剤をも含ませることができる。一般的に、処方成分は、別々に、あるいは例えば凍結乾燥粉末として、又は活性剤の量を示すアンプルもしくはサッシェ等の気密性密封容器中に入れた水分のない濃縮液として単位剤形(unit dosage form)中に混合して提供される。組成物を輸液によって投与する場合、組成物は無菌の製剤学的グレードの水又は生理食塩水を含む輸液ボトルを用いて調剤され得る。組成物を注射によって投与する場合、処方成分を投与前に混合し得るように、注射用の無菌水又は生理食塩水のアンプルを提供することができる。
【0156】
本発明の治療薬は、中性の状態又は塩の状態で処方され得る。製剤学的に許容できる塩
としては、塩酸、リン酸、酢酸、シュウ酸、酒石酸等から誘導される塩のような遊離のア
ミノ基により形成された塩、及び水酸化ナトリウム、水酸化カリウム、水酸化アンモニウ
ム、水酸化カルシウム、水酸化第二鉄、イソプロピルアミン、トリエチルアミン、2-エチ
ルアミノエタノール、ヒスチジン、プロカイン等から誘導される塩のような遊離のカルボ
キシル基により形成された塩が挙げられる。
【0157】
特定の障害又は病態の治療に有効である本発明の治療薬の量は、その障害又は病態の性質に依存し、標準的な臨床技術によって決定することができる。さらに、最適用量範囲を確認するのを助けるためにin vitroアッセイを任意に用い得る。また、その処方に用いられる厳密な用量は投与経路、疾患又は障害の重篤度にも依存し、開業医の判断及び各患者の情況によって決定される。しかしながら、静脈内投与に適した用量範囲は、一般的に、体重1kg当たり活性化合物約20〜500μg である。鼻腔内投与に適した用量範囲は、約0.01pg/kg(体重)〜1mg/kg(体重)である。有効な用量はin vitro又は動物モデル試験系から得られた用量−応答曲線から推定し得る。
【0158】
坐剤には、一般的に、0.5〜10重量%の範囲の活性成分が含まれ、経口製剤には、好ましくは10〜95%の活性成分が含まれる。
【0159】
また、本発明は、本発明の医薬組成物の1以上の成分で充填された1以上の容器を含む
医薬パック又はキットも提供する。このような容器には、医薬品又は生物学的製品の製造
、使用又は販売を規制する政府機関によって定められたフォームの注意書き(この注意書
きは、前記機関によるヒトへの投与のための製造、使用又は販売の認可を示すものである
)を任意に添付することができる。
【0160】
5.13.診断的効用
脊椎動物Serrateタンパク質、類似体、誘導体及びその部分配列(subsequences)、脊椎動物Serrate 核酸(及びそれと相補的な配列)、抗脊椎動物Serrate抗体は、診断における用途を有する。そのような分子は、Serrate 発現に影響する種々の病態、疾患及び障害を検出し、予知し、診断し、もしくは監視するために、又はその治療を監視するために、イムノアッセイ等のアッセイに用いることができる。特に、そのようなイムノアッセイは、患者から得られたサンプルを免疫特異的結合が生じ得るような条件下で抗Serrate抗体と接触させ、その抗体による全特異的結合の量を検出又は測定することを含む方法によって行うことができる。特定の態様においては、好ましくは抗Notch 抗体との結合との関連における組織区分(tissuesections)でのそのような抗体の結合を用いて、疾患状態におけるNotch 及び/又はSerrate の異常な局在化又は異常なレベルのNotch-Serrate共同局在化を検出することができる。特定の態様においては、Serrate に対する抗体を用いて、患者の組織又は血清サンプルにおけるSerrate の存在(そのSerrateの異常なレベルは疾患症状の指標になる。)をアッセイすることができる。内因性Notch タンパク質におけるSerrate 結合能力の異常なレベル、又は内因性Serrateタンパク質におけるNotch(又は他のSerrate リガンド)に対する結合能力の異常なレベルは、細胞運命(cell fate)の障害(例えば、癌等)の指標となり得る。「異常なレベル」とは、体の一部分由来の類似のサンプル又はその障害を有さない被験者由来の類似のサンプル中に存在するレベル又は該レベルを代表する標準レベルに比べて高い又は低いレベルを意味する。
【0161】
使用可能なイムノアッセイとしては、少しだけ名を挙げると、ウェスタンブロット、ラ
ジオイムノアッセイ、ELISA(酵素結合イムノソルベントアッセイ)、「サンドイッチ」
イムノアッセイ、免疫沈降アッセイ、沈降反応、ゲル拡散沈降反応、免疫拡散アッセイ、
凝集アッセイ、補体固定アッセイ、免疫放射線アッセイ、蛍光イムノアッセイ、タンパク
質Aイムノアッセイ等の技術を用いる競合及び非競合アッセイ系が含まれるがこれらに限
定されるものではない。
【0162】
また、脊椎動物Serrate 遺伝子、相補配列を含む関連のある核酸配列及び部分配列、並
びにその他のトポリズミック(toporythmic)遺伝子配列も、ハイブリダイゼーションアッ
セイに使用することができる。脊椎動物Serrate核酸配列又は少なくとも約8のヌクレオ
チドを含むその部分配列は、ハイブリダイゼーションプローブとして使用することができ
る。ハイブリダイゼーションアッセイを用いて上記したようなSerrate 発現及び/又は活
性における異常な変化と関連する病態、障害又は疾患状態を検出し、予知し、診断し、又
は監視することができる。特に、そのようなハイブリダイゼーションアッセイは、核酸を
含むサンプルをSerrateDNA又はRNAにハイブリダイズ可能な核酸プローブとハイブ
リダイゼーションが生じ得るような条件下で接触させ、結果として生じるハイブリダイゼ
ーションを検出又は測定することを含む方法によって行われる。
【0163】
さらに、SerrateはNotch に結合するので、脊椎動物Serrate 又はその結合部分を用いて、例えば、結腸及び子宮頸癌のようなNotch 発現の増加を示す悪性腫瘍のスクリーニングにおけるサンプル中のNotchの存在及び/又は量をアッセイすることができる。
【実施例】
【0164】
6.マウスSerrate相同体の単離及び特性付け
M-Serrate-1と称するマウスSerrate 相同体を下記のようにして単離した:
マウスSerrate -1遺伝子
組織起源:10.5日齢マウス胎仔RNA
単離法:
a)上記のRNAに対するcDNAをランダムプライミング
b)下記を用いた上記cDNAのPCR
増幅条件:供給バッファー50μl にcDNA 50ng、プライマー各1μg、dNTP's 0.2mM
、Taq 1.8U(Perkin-Elmer)を入れた液、94℃/30秒、45℃/2分、72℃/1分で40サイクル、各サイクルを2秒延長。
【0165】
両末端で配列決定された 1.8kbの断片が得られ、C-Serrate -1に相当するものと同定された。
【0166】
M-Serrate-1の部分DNA配列:
発現パターン:発現パターンは、発達中の中枢神経系、末梢神経系、肢、腎臓、水晶体、及び脈管系における発現を含む、C-Serrate -1(ニワトリSerrate)(下記の第11節参照)で観察されたものと同様であると決定された。
7.ツメガエルSerrate 相同体の単離及び特性付け
ツメガエル Serrate-1と称するツメガエルSerrate 相同体を下記のようにして単離した:
ツメガエルSerrate -1遺伝子
組織起源:神経胚期の胚RNA
単離法:
a)上記のRNAに対するcDNAをランダムプライミング
b)下記を用いたPCR
増幅条件:供給バッファー50μl にcDNA 50ng 、プライマー各1μg 、dNTP's 0.2mM、Taq 1.8U(Perkin-Elmer)を入れた液、94℃/30秒、45℃/2分、72℃/1分で40サイクル、各サイクルを2秒延長。
【0167】
部分的に配列決定されてC-Serrate -1との関係が確認された〜700bp の断片が得られた。
8.ニワトリ(CHICK)Serrate 相同体の単離及び特性付け
本実施例では、ニワトリSerrate 相同体C-Serrate 、並びに2つのニワトリNotch相同体C-Notch -1及びC-Notch -2の断片のクローニング及び配列を、初期胚形成中のそれらの発現パターンとともに報告する。C-Serrateの転写のパターンは胚の多くの領域におけるC-Notch -1の転写のパターンと共通点があり、これは、C-Notch-1 が、ショウジョウバエのNotchのように、Serrate の受容体であることを示唆するものである。特に、Notch 及びSerrate は、発達中の中枢及び抹消神経系の神経形成領域において発現する。
【0168】
本発明者らのデータは、Notch の公知のリガンドであるSerrate が節足動物からコルダ
ートまで保存されていることを示している。共通する発現パターンは、Notch とのその機
能的な関係の保存を示唆するものであり、ニワトリの成長、特にその中枢神経系の発達は
、いくつかの特異的な位置でのC-Notch-1 とSerrate の相互作用が関与しているということを示唆する。
材料及び方法
胚
白レグホンニワトリの卵をパークファーム大学(University Park Farm)から入手し、38
℃でインキュベートした。胚をHamburger 及びHamilton(1951,J.Exp.Zool.88:49-92)にしたがって段階づけした(staged)。
Notchのニワトリ相同体のクローニング
ニワトリNotch1及びNotch2遺伝子の約1000塩基対PCR断片を、縮重プライマー及びLardelli及びLendahl(1993,Exp.Cell.Res.204:364-372)に概説されたPCR条件を用いて、耳外植片RNA(下記参照)から増幅した。そのPCR断片をBluescriptKS-にサブクローニングし、配列決定してDIGアンチセンスRNAプローブの作成用の鋳型として用いた(RNA転写キット,Stratagene; DIG RNA標識混合物,BoehringerMannheim)。
ショウジョウバエSerrate のニワトリ相同体のクローニング
耳外植片を段階8から段階13の胚から切断した。各耳外植片は、2つの耳杯、後脳及び咽頭の間にある短い区画、並びにそれに結びついた頭外胚葉及び間葉から構成されていた。RNAを標準プロトコール(Sambrooket al.,1989,in Molecular Cloning: A Laboratory Manual,2nd ed.,Cold SpringHarbor Laboratory Press,コールドスプリングハーバー(Cold Spring Harbor)、ニューヨーク)を一部変更したものを用いて抽出し、ポリA+mRNAをPolyATtractmRNA単離システム(Promega)を用いて全RNAから単離した。第1のcDNA鎖をSuperScriptPreamplificationシステム(Gibco)を用いて合成した。
【0169】
PCR及び縮重プライマーを用いて、耳外植片cDNA由来のショウジョウバエ遺伝子
Serrate に相同性のあるニワトリ遺伝子の断片を増幅した。プライマーは、ハエのDelta
タンパク質及びSerrateタンパク質の両方で見出されるペプチドモチーフを認識するよう
に設計した:
1)プライマー1,5-CGI(T/C)TITGC(T/C)TIAA(G/A)(G/C)AITA(C/T)CA-3'(配列番号17)は、ハエDeltaタンパク質及びSerrate タンパク質のアミノ末端に位置するモチーフRLCLK(E/H)YQ(配列番号18)に相当する。
【0170】
2)プライマー2,5'-TCIATGCAIGTICCICC(A/G)TT-3'(配列番号11)は、いくつかのE
GF様リピートに見出されるモチーフNGGTCID(配列番号12)に相当する。PCR条件は
下記のとおりである:94℃1分間、45℃ 1.5分間及び72℃2分間の35サイクル;その後72℃10分間の最終伸長工程。長さ約 900塩基対のPCR産物を精製し、BluescriptKS-(Stratagene)にサブクローニングし、そのDNA配列を部分的に決定してそれがほぼSerrate 相同体であることを確認した。次いで、それを用いて2つのcDNAライブラリー:
1)段階8〜13の耳外植片をランダムプライミングしたcDNAライブラリー
2)段階17のニワトリ脊髄をオリゴdTプライミングしたcDNAライブラリー
をスクリーニングすることによってより大きなcDNAクローンを回収した。共通しているcDNAを単離し、その遺伝子の全コーディング領域をいっしょにほとんどカバーする2つ(9及び3A.1という)をBluescript KS-にサブクローニングした。DNA配列を、2本鎖Nested Deletion キット(Pharmacia)、及びシークエナーゼ酵素(米国BiochemicalCorporation)を用いるサンガージデオキシチェーンターミネーション法を用いて作製したネスト欠失シリーズ(nested deletion series)から決定した。配列を一列に並べてGenewarks2.3 及びIntelligenetics を用いて分析した。相同性調査はプログラムSharq を用いて行った。
【0171】
オープンリーディングフレームの5’最末端(the most 5’end)を得るために、Lardell
iら(1994,Mechanismsof Development 46:123-136)の方法を用いる他の多くのライブラ
リー(cDNA及びゲノムライブラリー)のスクリーニングを含む、多くの他のPCRに
基づく方法を用いた。
insitu ハイブリダイゼーション
遺伝子転写のパターンをDIG標識RNAプローブ、並びに:
1)高度にストリンジェントな全載(high-stringencywholemount)in situ ハイブリダイ
ゼーションプロトコール、及び
2)Strahleら(1994,Trends in Genet.10:7)のプロトコールに基づいたクリオスタッ
ト区画上でのinsitu ハイブリダイゼーションを用いるin situ ハイブリダイゼーションによって決定した。
結果
脊椎動物胚中のニワトリSerrate の可能性のある役割を見抜くために、Notch とSerrate の機能的結合がショウジョウバエ中で生じているということから、そのSerrate発現とニワトリNotch の発現との関係を調べた。2つのニワトリNotch 相同体を下記のようにして得た。
C-Notch-1及びC-Notch -2は、それぞれ、齧歯類Notch -1及びNotch -2遺伝子の明白な相対物である
2日齢ニワトリ胚から調製したcDNA及び公知の齧歯類Notch 相同体と共通の保存領域に基づいた縮重プライマーを用いて、PCRによってニワトリ中のNotch 相同体を調査した。この方法において、C-Notch-1及びC-Notch-2と呼ばれている2つの別個の遺伝子のそれぞれ長さ約1000ヌクレオチド断片を得た。それらの断片は第3のNotch/lin12 リピートから最後の5つほどのEGF様リピートを含むものにまで伸長する。EGF様リピートは、非常に多くのタンパク質(その大部分は、他の点ではNotchとは関係がない)に存在する。しかしながら、3つのNotch/lin12 リピートは、Notch 遺伝子ファミリーに特有のものであり、その公知のメンバー全てで見出される。C-Notch-1は、齧歯類Notch1(Weinmasterら,1991,Development 113:199-205)と高度のアミノ酸同一性を示し、齧歯類Notch1と概ね似たようなドメインで発現する(下記参照)。齧歯類Notch遺伝子の中で、C-Notch -2はNotch 2(Weinmasterら,1992,Development116:931-941)とほとんど類似しているようである。
【0172】
in situ ハイブリダイゼーションによって初期胚におけるC-Notch-1の発現パターンを調べた。C-Notch-1は、1〜2日齢ニワトリ胚で、神経管、前原体節期中葉胚、腎形成性中葉胚(予期中葉胚(prospectivemesoderm))、鼻プラコード、耳プラコード/小嚢、水晶体プラコード、上鰓プラコード、心臓における脈管系の内皮内層(endothelial lining)、及び肢芽の外胚葉性頂堤(AER)を含む、多くの明瞭に定義されたドメインにおいて発現した。これらの部位は、報告されている齧歯類における同等の段階でのNotch1発現の部位と適合する(表2)。
【0173】
発現データとともに配列データを取り上げると、C-Notch -1は齧歯類Notch1のニワト
リオルソローグ(ortholog)、又はその非常に類似した類縁物のいずれかであると結論付け
られる。
【0174】
【表2】
C-Serrate はショウジョウバエSerrateの相同体であり、Notchファミリーに属する受容
体のリガンドの候補をコードする
ショウジョウバエにおいては、Notchに対する2つのリガンドが知られており、関連する2つの遺伝子Delta 及びSerrate によりコードされている。これらの遺伝子に対応するアミノ酸配列は、Notchに対するinvitro結合に必要でありかつ十分な領域、DSL モチ−フを含む5'末端において相同である。Sarrate のニワトリ相同体の断片を単離するために、DSL モチ−フのいずれかの側の配列を認識するように設計されたPCR及び縮重プライマーを使用した(材料及び方法を参照)。900塩基対のPCR 断片を回収し、ライブラリーをスクリーニングするのに使用し、オーバーラップcDNAクローンを単離することができた。これらのcDNAクローンのDNA配列は、いくつかの5'塩基のみを欠く、3582ヌクレオチドの殆ど完全な単一のオープンリーディングフレームを示すものであった。ショウジョウバエDelta及びSerrate のアミノ酸配列との比較により、ニワトリSerrate タンパク質のシグナル配列の部分をコードするコード配列の部分のみが失われていることが示された。
【0175】
前記ヌクレオチド配列(配列番号5)(図3)の翻訳から、1230アミノ酸のタンパク質(配列番号6)(図4)が予測される。ヒドロパシープロットにより、トランスメンブランドメインに特徴的な単一の疎水性領域があることが明らかにされている(Kyteand Doolittle,1982,J.Mol.Biol.157:105-132)。さらに、前記タンパク質は、その細胞外ドメインにタンデム配列で結合している16個のEFG-様リピートを有する。ニワトリ配列を、D.melanogasterDelta 及びSerrate の配列と比較することにより、前記クローンがSerrate のニワトリ相同体をコードしていることが示されている(図5、図6)。ショウジョウバエSerrateが14個のEFG-様リピートを有し、リピート4、6及び10に大きな挿入物を有しているのに対し、前記ニワトリ相同体は2つの余分なEFG-様リピートを有し、10番目のリピート中に16アミノ酸の小さい挿入物を1つ有するだけである。両方のタンパク質は、EFG-様リピートとトランスメンブランドメインとの間に2番目のシステインリッチ領域を有し、この領域のシステインの間隔は2つのタンパク質において殆ど同じである(ショウジョウバエSerrateのCX2CXCX6CX4CX15CX5CX7CX4CX5CとC-SerrateのCX2CXCX6CX4CX9CX5CX7CX4CX5Cとを比較せよ)。C-Serrate の細胞内ドメインは、ショウジョウバエ Delta及びSerrate のいずれの細胞内ドメインとも有意な相同性を有していない。
C-Serrate は中枢神経系、頭プラコード、腎中胚葉、脈管系、及び体肢芽間葉において発
現される
ステージ4〜ステージ21の早期の胚形成の間に、約12時間の間隔で、全載標本中においてin situ ハイブリダイゼーションを行い、C-Serrateの発現を調べた。後期の段階については凍結切片についてinsitu ハイブリダイゼーションを行うことにより調べた。
【0176】
全載物に見られるように、C-Serrate の早期の発現の主要な部位は、中枢神経系、頭プ
ラコード、腎中胚葉、脈管系、及び体肢芽間葉の5つの表題の下にグループ分けすること
ができる。
中枢神経系
C-Serrate の最初の検出可能な発現は、段階6(0体節/24時間)における中枢神経系において、神経板の背面部分内で見られた。段階10(9〜10体節/35.5時間)までに、間脳予定域において強い縞状の発現が見られた。菱脳及び脊髄予定域において別の弱い染色が見られた。
【0177】
ステージ13において、神経管にいくつかの発現のパッチが見られた。間脳においては、
神経分節D2に対応すると見える、強い三角形の縞状の発現が見られた。前部中脳底床に2つのパッチ(正中線の各側に1つずつ)が見られ、中脳背部において拡散した染色が見られた。菱脳及び吻側脊髄においては、正中線の各側に2つの縦方向の発現の縞が見られ、1つは神経管の背部縁に沿ったものであり、2番目のものはより腹側のものであり、底板に隣接するものであった。両者は(ラット)Notch1発現のドメイン内に位置していた。腹側の縞の前部端は中脳/菱脳境界にあった。背側の縞は背側中脳における発現に連続していた。前部脊髄においては、発現はより小さなスポット状となり、縞はC-Serrateを発現する離れ離れに分散した細胞に置き変わった。
【0178】
ステージ17(58時間)においては、間脳及び中脳においては発現は変化しなかった。菱
脳及び脊髄においては、別の2つの縦の縞があった。1つは背腹方向軸に沿った中央のも
のであり、第2のものはより広いより腹側の縞であった。これらの縞の前部端は菱脳の小
片2の前部境界に一致していた。菱脳における4つの縦方向の縞は全て胚の脊髄に続いて
おり、その後部端に向かって減少していた。これらの発現の縞は少なくともステージ31(E
7)以下で維持されていた。ステージ21(84時間)までに大脳半球に別の発現が見られ、視蓋中の細胞における入り交じった分布で強い発現が見られた。
【0179】
頭プラコード
C-Serrate が、全ての頭プラコード、即ち水晶体プラコード、鼻プラコード、耳プラコ
ード/耳胞及び上鰓プラコードに発現され、三叉神経プラコード(形態学的には十分に確
定されていない)に対応し得る、耳プラコード前部に対する頭蓋外胚葉パッチに発現され
ることは驚くべきことである。
【0180】
水晶体プラコードにおいては、ステージ11において既に発現がみられ、急速に非常に強くなり、少なくともステージ21まで維持された。鼻プラコードにおいては発現はより弱く、ステージ13から検出されたに過ぎなかった。発現はやはり少なくともステージ21まで維持された。
【0181】
耳プラコードについても同様に、発現はステージ10において見えるようになり、ステージ11の初期(12〜14体節、42.5時間)まで強かった。奇妙なことに、耳発現ドメイン、即ちそこでは前記遺伝子が発現されなかったプラコードの前腹側領域中に「孔」が見られた。さらに、プラコードが陥入して耳胞を形成するに従って、前側部及び後内側分裂極において最も強い発現が見られた。さらに後になって、耳胞が内耳の膜迷路に変化するに従って、C-Serrate発現は知覚パッチに限定されるようになった。
【0182】
鰓上発現は、ステージ13/14 において、第1及び第2の鰓裂の背側の縁周辺の外胚葉で
強い染色として見られた。これは鰓裂の内層の深部と、鰓嚢の内皮内層とにおける遺伝子
の発現を伴っており、ここでこれらの2つの上皮は互いに隣接している。
【0183】
最後に、ステージ11において、大きくて強力であるが、一時的なパッチ発現が、耳原基に対してちょうど前部腹側の頭蓋外胚葉において見られた。その位置から、これは三叉神経プラコードの領域であるか、あるいはそれを含むものであると考えられた。
腎中胚葉
発生中の中腎細管中のステージ10及びより段階の進んだ胚(ステージ17〜21)からの中間中胚葉の細胞において発現が検出された。
体肢芽
C-Serrate mRNAは、発生中の体肢芽の遠位端における間葉のパッチに局在していた
。これは体肢の成長における役割を示唆している可能性がある。
その他の部位
発現は、尾芽、尿膜柄においても見られ、後期においてはおそらくその他の組織におい
ても見られるであろう。
C-Serrate 発現の全ての主要な部位はC-Notch-1発現のドメイン内にある
C-SerrateのDSLドメイン及び隣接するN-末端領域が保存されていることは、それがNotchファミリーに属する受容体についてのリガンドとして機能することを示唆している。従って、C-Serrate発現がNotch遺伝子の発現を伴う部位が見出されるであろうと予測された。そのような部位においては、Serrate-Notchシグナリングにより細胞が連絡を取り合っていることを示すものとして、2つの遺伝子のオーバーラップした、あるいは連続的な発現が起こり得る。実際にどのようなオーバーラップが起こるのかを発見するために、インキュベーション8日(E8)までのステージの範囲にわたって、insitu ハイブリダイゼーションにより示される、C-Serrate の発現パターンをC-Notch-1のものと比較した。C-Serrate発現の観察された部位の全ては実際に、C-Notch-1 の発現ドメイン内あるいは非常に近い位置にあった(表III)。
【0184】
【表3】
昆虫の神経発生におけるNotch 及びその相手の重要性により、前記相同遺伝子が脊椎動
物CNS の発生に関与しているかどうかを知ることは特に重要であった。C-SerrateはCNS
で発現され、その発現パターンはNotch相同体のものとの顕著な相関を示すものである。
【0185】
C-Notch-1及びC-Serrate アンチセンスRNAプローブとハイブリダイズしたニワトリ
6日胎児の脊髄の横断切片について分析した。C-Notch-1は先に記載したように管腔領域
全体に発現され、この領域内ではSerrateが強く発現された2つの小さいパッチが存在し
た。
考察
ショウジョウバエの発生においては、Notch遺伝子の産物を介した細胞間シグナリングが、分化した細胞型の詳細なパターンを特定する最終的な細胞の予定運命決定において主要な役割を果している。このシグナリング経路においてはNotchタンパク質がトランスメンブラン受容体として同定されており、この経路は神経発生におけるその役割において最もよく知られているものである。即ち、NotchあるいはNotchを介するシグナル伝達に必要なその他の遺伝子の任意のセットにおける機能欠失突然変異が神経外胚葉における細胞の予定運命を変え、表皮性のはずであった細胞を代わりに神経性のものにするはずである。しかしNotch-依存性シグナリングは、神経組織におけるのと同様に非神経組織においても重要である。これは例えば、卵子発生、筋発生、マルピギー管の形成、及び消化管において、そして網膜、末梢小感覚体及び中枢神経系の発生において、分化のモードの選択を制御する。これらの場合の殆どにおいて、Notchにより伝達されるシグナルは側方抑制を媒介するようであり、側方抑制とは特定の経路で分化するようになった細胞、例えば神経芽細胞が、そのごく近傍にあるものが同様な行動をとることを阻害するという細胞の相互作用の一種である。これにより隣接する細胞は異なる経路で行動するようにされ、異なる細胞種のきめこまかいパターンを形成する。
【0186】
しかし、これはNotchを介して伝達されるシグナルの唯一の機能ではないとするのに十分な理由がある。2種類のNotchの直接のリガンドが同定されている。これらは、Delta及びSerrate 遺伝子の産物である。両者は、Notch自身と同様に、その細胞外ドメインにおいてEFG-様リピートのタンデム配列を有するトランスメンブランタンパク質をコードしている。Delta及びSerrate タンパク質の両方は、細胞付着アッセイにおいてNotchに結合することが示されており、in vitroにおいてNotchと相互作用するのに必要にして十分なモチーフ、いわゆるEBDあるいはDSLドメインを含むアミノ末端において相同性を有する大きな領域を共有している。このような生化学的類似性にもかかわらず、それらはかなり異なる発生上の機能を有しているようである。Serrateはハエにおいて多くの部位に発現されるが、肩の盤、翅の盤、及び平均根盤においてのみ必要とされるようである。Serrate の機能が突然変異により欠損された場合はこれらの構造体は成長できなくなる。翅盤の研究により、それが特異的にSerrateに依存する翅縁であることが示されている。Serrate を欠いていると、この重要なシグナリング領域及び成長中心は形成できず、Serrate が翅盤の腹側部分でGAL4-UASプロモーターのもとに異所的に発現されると、異所性の翅縁組織が誘導され、異所性の増殖を起こす。Notchは翅縁においてSerrateの受容体であるようであり、これはNotch のいくつかの突然変異対立遺伝子が、翅縁発生の類似した阻害をおこし、二種の遺伝子の効果において対立遺伝子特異的相互作用が見られることによる。
【0187】
本明細書においては、ショウジョウバエ遺伝子Serrate の相同体の同定及び完全長配列、及びラット/マウスNotch1及びNotch2のニワトリ相同体の同定及び部分配列を記載するものである。
【0188】
ニワトリSerratecDNA内には、その細胞外ドメインに16個のEFG リピートを有する大きいトランスメンブランタンパク質をコードすると予測される単一のオープンリーディングフレームが存在する。それは非常によく保存されたDSLモチーフを有しており、それがNotchと直接相互作用するはずであることを示唆している。ニワトリSerrate の細胞内ドメインは、ショウジョウバエDelta 及びSerrateの細胞内ドメインを含む現在のデータベースのいずれとも相同性を示さない。しかし、ニワトリ及びヒトSerrate の細胞内ドメイン(12節を参照)は殆ど同一であることが指摘される。
【0189】
in situ ハイブリダイゼーションにより、初期胚発生の間においてC-Notch-1及びC-Serrateの位置的分布を調べた。C-Notch-1及びC-Serrateは、それらが同時に発現されるいくつかの領域(CNS、耳、鰓領域、水晶体、心臓、鼻プラコード及び中腎)を含む発現のダイナミックで複雑なパターンを示す。このオーバーラップした発現と、C-SerrateがNotch結合ドメインに非常によく保存された領域を有するという知見とを合わせて考えると、この受容体/リガンド相互作用がショウジョウバエから脊椎動物まで保存されてきたことが示唆されている。
【0190】
ショウジョウバエにおいてはNotch受容体はかなり広範に分布しており、そのリガンドは、オーバーラップしているがより限定的なドメインにおいて見られる。ニワトリにおいても同様な状況が見られる。
【0191】
ハエNotchは、ショウジョウバエの発生の多くの段階において必要である。特に中枢神経系及び末梢感覚器官の発生における側方抑制におけるその役割が最もよく研究された例である。しかし、Notchは多機能受容体であり、種々のシグナリング分子(Delta及びSerrateを含む)と相互反応でき、側方抑制の枠組内には容易には適合しない発生過程においても相互反応できる。Delta が側方抑制におけるシグナリング分子であることを示す証拠はあるが、Serrateが側方抑制に関与していることを示唆するデータはない。そうではなく、Serrate は幼生の背側成虫盤、即ち肩、平均根及び翅盤の発生に必要であるようである。最後のものはこれらの過程のもっともよく研究されたものであるが、これにおいては、Serrate及びNotchは、翅の発生の全体としての有機的結合に必要な構造、背腹側翅縁の発生に重要である。
【0192】
C-Serrateが重要な機能を有していることは、特にそのNotch-結合ドメインのその配列が保存されていることから推論できる。C-Serrate について本明細書に記載した発現パターンは以下の情報を与えるものである。第一に、Serrate遺伝子はC-Notch-1が発現される部位あるいは隣接する部位において発現されるので(おそらくはその他のNotch相同体とともに)、C-SerrateはC-Notch-1(あるいは同様な発現パターンを有するその他のニワトリNotch相同体)に結合することによりその作用を発揮する可能性が極めて高い。第二に、発生中の腎臓、脈管系及び体肢芽における発現は中胚葉及び外胚葉間の誘導的シグナリングにおける関与を反映しているものであり得、前記のシグナリングはこれらの器官のすべてのものの発生において重要な部分を演ずるものである。例えば体肢芽においては、C-Serrateは遠位中胚葉において発現され、C-Notch-1は重畳する先端外胚葉隆起において発現され、この維持は下部の中胚葉からのシグナルに依存することが知られている。頭プラコードにおいては、同様な役割が可能であるが、誘導的シグナリングについての証拠はより弱いものであり、C-Serrateは同様に、プラコード上皮内の細胞間の連絡、例えばプラコード細胞の分化の特定のモードを制御することに関与している可能性がある。
【0193】
何がCNS 内のC-Serrateの発現の奇妙に限定されたドメイン内でのその機能であろうか?一つの可能性は、これがオリゴ樹状細胞の生産の制御に関与しているということであり、これは神経管の頭尾軸に沿って伸びる組織の狭いバンドから始まると報告されている。
【0194】
9.ヒトSerrate相同体の単離と特徴付け
ヒトSerrate 配列のクローンを以下に記載するようにして得た。
【0195】
ポリメラーゼ連鎖反応(PCR)を使用してヒト胎盤cDNAライブラリーからDNAを増幅した。この反応で使用した縮重オリゴヌクレオチドプライマーは、ショウジョウバエSerrate及びショウジョウバエDelta の間の相同性の高いアミノ末端領域に基づいて設計した(図5参照)。この相同性の高い領域は5'“DSL”ドメインを含んでおり、これはDelta及びSerrate のNotch-結合部分をコードすると考えられる。2種類のPCR 産物、350 bp断片と1.2 kb断片を単離して使用した。これらのPCR断片は32P により標識し、17〜18週齢の胎児から作製された市販のヒト胎児脳cDNΛライブラリー(以前からStratageneより入手可能)をスクリーニングするのに使用した。ここでこのcDNAをλ-ZapベクターのEcoRI部位に挿入した。
【0196】
前記1.2 kb断片は、106クローン中の1つのクローンにハイブリダイズし、スクリーニングした。単離されたファージλクローンを製造元の使用説明書に従ってプラスミドに変換することによりλDNAからこの断片を回収し、ベクターBluescriptKS-(Stratagene)のEcoRI 部位中の挿入物としてSerrate-相同cDNAを得た。このプラスミドは“pBS39”と命名し、このcDNAクローンに対応する遺伝子をヒトSerrate-1と称する(ヒトJagged-1(“HJ1”)としても知られる)。この単離されたcDNAは6464ヌクレオチド長で、完全なオープンリーディングフレーム並びに5'及び3'非翻訳領域を含んでいた(図1)。Sequenase(登録商標)配列決定システム(U.S.BiochemicalCorp.)を用いて、5及び6%Sequagelアクリルアミド配列決定ゲル上で配列決定を行った。
【0197】
前記350 bp断片は、約1.1及び3.1 kb長のcDNA挿入物を含む2種類のクローンとハイブリダイズした。これらの挿入物を含むプラスミド構築物はそれぞれpBS14 及びpBS15 と称する。それぞれのクローンを単離し、そのそれぞれの挿入物をλcDNAから回収し、上記のように配列決定した。pBS14挿入物のヌクレオチド配列は、pBS15 cDNA挿入物内に内部的に含まれている配列の1.1 kb部分に同一であり、従ってこのクローンについてはそれ以上の特徴付けは行わなかった。3.1kb pBS15挿入物の配列は、挿入物の5'末端の20ヌクレオチドを除く全てにわたるオープンリーディングフレームをコードしていた。この予測されたオープンリーディングフレームのアミノ末端残基に位置するメチオニンはpBS39中のヒトSerrate-1(HJ-1)cDNAクローンによりコードされた開始メチオニンと相同であった。pBS15 のcDNA挿入物をコードする遺伝子はヒトSerrate-2と命名し、これはヒトJagged-2(“HJ2”)としても知られる。
【0198】
前記pBS15(HJ2)3.1kb挿入物を次に32P で標識し、別のヒト胎児脳ライブラリー(Clontechからのもの)をスクリーニングするのに使用した。このライブラリーは、25〜26週齢胎児から作製されたcDNAをλgt11のEcoRI部位にクローニングしたものである。このスクリーニングにより3種類の可能性のある陽性クローンが同定された。cDNAを単離するために、液体溶解物からλgt11DNAを調製し、DEAEカラム上で精製した。精製されたDNAを次いでEcoRIで切断し、cDNA挿入物を単離し、Bluescript KS-のEcoRI 部位中にサブクローニングした。前記のcDNAを含むBluescript構築物をpBS3-15、pBS3-2及びpBS3-20と命名した。これらのcDNAクローンのうちの2種類、pBS3-2及びpBS3-20はpBS15 と部分的にオーバーラップする配列を含んでおり、さらに特徴付けした。pBS3-2は、pBS15cDNA挿入物のヌクレオチド1210からポリアデニル化シグナルの直後にわたる3.2 kb挿入物を有していた。pBS3-20 の2.6 kb挿入物について制限地図を作成し、部分的に配列決定して3'及び5'末端を決定した。この分析によれば、pBS3-20挿入物はpBS3-2cDNA挿入物内に完全に含まれる核酸配列を有していることが示され、従ってpBS3-20 はそれ以上特徴付けしなかった。pBS3-15 の挿入物はBluescriptベクター断片夾雑物として測定した。
【0199】
コンピューター上で作成された「完全」ヒトSerrate-2(HJ2)cDNA(配列番号3)の推定アミノ酸配列(配列番号4)の、pBS39からのヒトSerrate-1(HJ1)の推定アミノ酸配列(配列番号2)とのアライメントにより、pBS15(HJ2)挿入物によりコードされた領域内で、仮想的シグナル配列とDSLドメインの開始部との間にあるフレームシフトを生じる約120 塩基のギャップが判明した(図2)。pBS15 挿入物のギャップ中の失われたヌクレオチドは配列番号3のヌクレオチド240 及び241 の間に位置すべきものである。この失われた領域はおそらく、Stratageneライブラリー構築の際のクローニングの人工的産物により生じたものと思われる。
【0200】
アンカーPCR、RACE及びTakaraの延長PCR技術を使用したHJ2 の5'末端をクローン化する試みは成功しなかった。しかし、HJ2 の5'末端を含む可能性のある3種類のヒトゲノムクローンがヒトゲノムコスミドライブラリーのスクリーニングから得られた。このライブラリーは、pWE15ベクターのBamHI 部位中に導入した単一のXhoI部位中に30 kb 断片をクローニングしたものである(未改変ベクターはStratageneより入手可能である)。このコスミドライブラリーを、pBS15(HJ2)の5'末端から増幅されたPCR断片によりスクリーニングし、3種類の陽性コスミドクローンを単離した。2種類の異なるプライマーのセットを使用し、コスミドクローンをテンプレートとして使用して、pBS15の5'末端に対応するDNAを増幅したところ、両方のセットにより単一のバンドが生成され、これをサブクローニングしたが、いずれもPCR人工産物を含んでいることが判明した。コスミドクローンの部分はPCRなしに直接サブクローニングし、pBS15 において失われているDNAの120 ヌクレオチド部分を含むコスミドクローンの部分を得た。
【0201】
ヒトSerrate-1 相同体(HJ1)をコードするpBS39 cDNA挿入物を配列決定したところ、その遺伝子産物の完全なコード配列を含んでいた。そのヌクレオチド(配列番号1)及びタンパク質(配列番号2)の配列を図2に示す。ヒトSerrate-1(HJ1)のヌクレオチド配列をMacVectorソフトウェア(International Biotechnology Inc.,New Haven,CT)を使用して翻訳した。コード領域は配列番号1のヌクレオチド番号371-4024からなる。DNAStar(Madison,WI)のProteanタンパク質分析ソフトウェアプログラムを使用して単一のペプチド及びトランスメンブラン領域を予測した(疎水性に基づいた)。シグナルペプチドは、配列番号2のアミノ酸14-29(配列番号1のヌクレオチド番号410-457によりコードされる)からなると予測され、これにより成熟タンパク質のアミノ末端はアミノ酸番号30のGly から開始すると予測された。トランスメンブランドメインは、配列番号2のアミノ酸番号1068-1089であると予測され、配列番号1のヌクレオチド番号3572-3637によりコードされる。ショウジョウバエDelta 及びSerrate と相同性を有する領域であり、Notch(特にNotch ELR 11及び12)との結合を媒介すると予測されるコンセンサス(DSL)ドメインは、配列番号2のアミノ酸185-229にわたり、配列番号1のヌクレオチド番号923-1057によりコードされる。アミノ酸配列中の上皮成長因子様(ELR)リピートは目視同定した。以下の15の(完全長)ELR及び3の部分ELRが同定された。
従って、全ELRドメインはアミノ酸番号234-896(配列番号1のヌクレオチド番号1070-3058 によりコードされる)である。細胞外ドメインは、配列番号2のアミノ酸番号1-1067であると予測され、配列番号1のヌクレオチド番号371-3571によりコードされる(成熟タンパク質中のアミノ酸番号30-1067;配列番号1のヌクレオチド番号458-3571によりコードされる)。細胞内(細胞質)ドメインは、配列番号2のアミノ酸番号1090-1218であると予測され、配列番号1のヌクレオチド番号3638-4024 によりコードされる。
【0202】
特定の種類のヒト組織中におけるHJ1 の発現は、放射性標識pBS39 でClontech Human Multiple Tissue Northern Blotをプローブすることにより確認した。前記プローブは約6.6kbの単一のバンドとハイブリダイズし、心臓、脳、胎盤、肺、骨格筋、膵臓、肝臓及び腎臓を含むアッセイした全ての組織において発現された。HJ1 が成人骨格筋及び心筋において発現されたという観察は特に興味深いものであり、これは成人の筋肉繊維は細胞外マトリックスの層により完全に包囲されており、従ってこれらの細胞におけるHJ1の役割が直接の細胞間伝達にあるということはありそうにないからである。
【0203】
コンピュータ上で作成した「完全」(内部欠失を含む)ヒトSerrate-2(HJ2)cDNAヌクレオチド配列(配列番号3)及びアミノ酸配列(配列番号4)を図2に示す。ヌクレオチド配列はMacVectorソフトウェア(International Biotechnology Inc.,New Haven,CT)を使用して翻訳した。コード領域は配列番号3のヌクレオチド番号332-4102からなる。DNAStar(Madison,WI)のProteanタンパク質分析ソフトウェアプログラムを使用してシグナルペプチド及びトランスメンブラン領域を予測した(疎水性に基づいた)。トランスメンブランドメインは、配列番号4のアミノ酸番号912-933であると予測され、配列番号3のヌクレオチド番号3065-3130 によりコードされる。ショウジョウバエDelta 及びSerrate と相同性を有する領域であり、Notch(特にNotchELR 11及び12)との結合を媒介すると予測されるコンセンサス(DSL)ドメインは、配列番号4のアミノ酸26-70にわたり、配列番号3のヌクレオチド番号407-541によりコードされる。アミノ酸配列中の上皮成長因子様(ELR)リピートは目視同定した。以下の15の(完全長)ELR及び3の部分ELRが同定された。
従って、全ELRドメインはアミノ酸番号75-735(配列番号3のヌクレオチド番号554-2536によりコードされる)である。細胞外ドメインは、配列番号4のアミノ酸番号1-912であると予測され、配列番号3のヌクレオチド番号332-3064によりコードされる。細胞内(細胞質)ドメインは、配列番号4のアミノ酸番号934-1257であると予測され、配列番号3のヌクレオチド番号3131-4102によりコードされる。
【0204】
ヒトSerrate-1(HJ1)と同様に、コンピュータ上で作成した「完全」(内部欠失を有する)ヒトSerrate-2(HJ2)cDNA(配列番号3)は、16の完全なEGFリピート及び2つの中断されたEGF リピート、並びに仮想的Notchリガンド中にのみ見られたDSL ドメインとして知られる診断的には陰性のEFG リピートを含むタンパク質をコードする。コンピュータで作成された「完全」ヒトSerrate-2(HJ2)のオープンリーディングフレームは約1400アミノ酸長であり、HJ1及びラットSerrate相同体Jaggedのカルボキシ末端よりも約182 アミノ酸だけ長い。完全HJ2 及びHJ1 の間にはタンパク質のアミノ末端部分において有意な相同性が存在するが、この相同性はHJ1のアミノ酸番号1029付近の仮想的トランスメンブランドメインの直前で失われる。長いCOOH-末端テイルの存在はHJ2 の何らかの別の機能あるいは調節の可能性を意味するものであることから、この結果は、特に興味深いものである。
【0205】
「完全」(内部欠失を有する)ヒトSerrate-2(HJ2)cDNA(配列番号3)配列は、pBS15及びpBS3-2の配列オーバーラップ中に存在するAccI、DraIIIあるいはBamHI のための唯一の制限部位を利用して構築することができ、これらの酵素はそれぞれヌクレオチド1431、2648及び2802においてpBS15挿入物を切断する。
【0206】
特定の種類のヒト組織中におけるHJ2 の発現を、放射性標識クローンpBS15 でClontech Human Multiple Tissue NorthernBlotをプローブすることにより確認した。このプローブは約5.2 kbの単一のバンドとハイブリダイズし、心臓、脳、胎盤、肺、骨格筋及び膵臓において発現されたが、肝臓及び腎臓においては存在しないかあるいは殆ど検出できなかった。上述したHJ1発現の場合と同様に、HJ2 のpBS15 挿入物成分が成人の骨格筋及び心筋において発現されたという観察は特に興味深いものであり、これは成人筋肉繊維は細胞外マトリックスの層により完全に包囲されており、従ってこれらの細胞におけるHJ2の役割が直接の細胞間伝達にあるということことはありそうにないからである。
【0207】
発現構築物を単離されたクローンを使用して作製した。クローンをそのベクターからEcoRI 制限断片として切り出し、発現ベクターのEcoRI 制限部位中にサブクローニングした。これにより正しいリーディングフレーム中のサブクローンからのヒトSerrateタンパク質産物の発現が可能となった。この方法を使用して、サイトメガロウイルスプロモーターの制御下での発現用の発現ベクター中にpBS39 のHJ1cDNA挿入物をクローニングした発現構築物を作製し、3T3 及びHAKAT ヒトケラチノサイト系の両方においてHJ1を発現させた。
【0208】
10.微生物の寄託
完全長ヒトSerrate-1(HJ1)をコードするEcoRI断片を含むプラスミドpBS39 は、American Type Culture Collection,1201 Parklawn Drive,Rockville,Maryland20852に、特許手続上の微生物の寄託の国際的承認に関するブダペスト条約の規定に基づき、1995年2月28日に寄託し、受託番号97068 を得た。
【0209】
BluescriptKS-のEcoRI部位にクローニングしたヒトSerrate-2(HJ2)のアミノ末端をコードする3.1 kb EcoRI断片を含むプラスミドpBS15 は、AmericanType CultureCollection,1201 Parklawn Drive,Rockville,Maryland 20852に、特許手続上の微生物の寄託の国際的承認に関するブダペスト条約の規定に基づき、1995年3月5日に寄託し、受託番号 を得た。
【0210】
BluescriptKS-のEcoRI部位にクローニングしたヒトSerrate-2(HJ2)のカルボキシ末端をコードする3.2 kb EcoRI断片を含むプラスミドpBS3-2は、AmericanType Culture Collection,1201 Parklawn Drive,Rockville,Maryland 20852に、特許手続上の微生物の寄託の国際的承認に関するブダペスト条約の規定に基づき、1995年3月5日に寄託し、受託番号 を得た。
【0211】
本発明は、寄託された微生物あるいは本明細書に記載した特定の態様によりその範囲を限定されるものではない。実際に、これまでの記載及び添付の図面から、本明細書に記載したものに加えて本発明の種々の改変が当業者に明らかとなるであろう。そのような改変は添付の請求の範囲の範囲内に含まれるものである。
【0212】
本明細書に引用した種々の文献は、引用によりその全体を本明細書の一部とするものである。
【0213】
【特許請求の範囲】
【請求項1】
明細書に記載の発明。
【請求項1】
明細書に記載の発明。
【図1A】


【図1B】

【図1C】

【図1D】

【図1E】

【図1F】

【図1G】

【図2A】

【図2B】

【図2C】

【図2D】

【図2E】

【図2F】

【図2G】

【図3A】

【図3B】

【図4A】

【図4B】

【図5】

【図6】
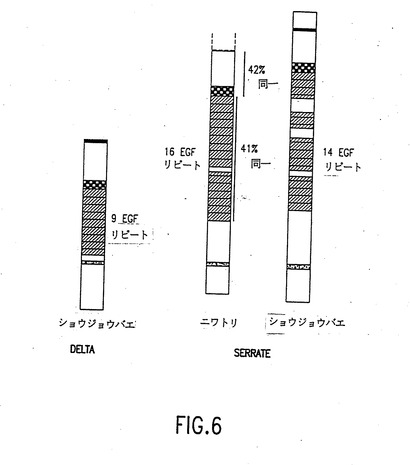
【図1B】
【図1C】
【図1D】
【図1E】
【図1F】
【図1G】
【図2A】
【図2B】
【図2C】
【図2D】
【図2E】
【図2F】
【図2G】
【図3A】
【図3B】
【図4A】
【図4B】
【図5】
【図6】
【公開番号】特開2010−273685(P2010−273685A)
【公開日】平成22年12月9日(2010.12.9)
【国際特許分類】
【出願番号】特願2010−166570(P2010−166570)
【出願日】平成22年7月23日(2010.7.23)
【分割の表示】特願2007−260391(P2007−260391)の分割
【原出願日】平成8年3月7日(1996.3.7)
【出願人】(392019352)イェール ユニバーシティー (38)
【氏名又は名称原語表記】YALE UNIVERSITY
【出願人】(592210108)インペリアル キャンサー リサーチ テクノロジー リミテッド (5)
【Fターム(参考)】
【公開日】平成22年12月9日(2010.12.9)
【国際特許分類】
【出願日】平成22年7月23日(2010.7.23)
【分割の表示】特願2007−260391(P2007−260391)の分割
【原出願日】平成8年3月7日(1996.3.7)
【出願人】(392019352)イェール ユニバーシティー (38)
【氏名又は名称原語表記】YALE UNIVERSITY
【出願人】(592210108)インペリアル キャンサー リサーチ テクノロジー リミテッド (5)
【Fターム(参考)】
[ Back to top ]