
脱メチル化および細胞の再プログラミングを促進するためのシチジンデアミナーゼ関連薬剤の使用
哺乳動物細胞における脱メチル化を調節するための方法、組成物およびキットが提供される。候補薬剤を、哺乳動物細胞におけるゲノムDNA脱メチル化の調節における活性についてスクリーニングするための方法、組成物およびキットも提供される。これらの方法、組成物およびキットは、人工多能性幹細胞(iPS)および体細胞をin vitroで作製することにおいて、および、癌およびゲノム刷り込みにおける欠損から生じる障害を含めたヒトの障害を治療するために使用される。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願の相互参照
米国特許法第119条(e)に準じて、本出願は、2010年12月18日出願の米国仮特許出願第61/284,519号の出願日について優先権を主張する;その開示は、参照により本明細書に組み込まれる。
【0002】
政府の権利
本発明は、国立保健研究機構(National Institutes of Health)から授与されたAG009521およびAG024987の下、政府支援で行われた。政府は、本発明において一定の権利を有する。
【0003】
本発明は、哺乳動物細胞におけるゲノムDNA脱メチル化を誘導するための方法および組成物、ならびに、候補薬剤を、哺乳動物細胞におけるゲノムDNA脱メチル化の調節における活性についてスクリーニングするための方法および組成物に関する。
【背景技術】
【0004】
いくつかの因子を導入することによって体細胞の核を多能性に、または別の細胞系譜の体細胞に再プログラミングすることにより、再生医療の分野における主要なブレイクスルーである、患者に特異的な人工多能性細胞(iPS)および患者に特異的な体細胞の生成が可能になった。しかし、これらのプロセスには時間がかかり(2〜3週間)、非同期的であり、頻度が低く(<0.1%)(例えば、Takahashi、K.ら(2007)Cell 131:861〜72;Takahashi、K.&Yamanaka、S.(2006)Cell 126:663〜76;Wernig、M.ら(2007)Nature 448:318〜24;Wernig、M.ら(2008)Nat Biotechnolを参照されたい)、DNA脱メチル化がネックになっている(Mikkelsen、T.S.ら(2008)Nature 454、49〜55)。したがって、哺乳動物細胞におけるDNA脱メチル化を調節する機構を解明すること、および脱メチル化を促進する薬剤を同定することは、臨床および研究の興味の対象である。本発明は、これらの問題を解決するものである。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Rai、K.ら(2008)「DNA demethylation in Zebrafish Involves the Coupling of a Deaminase、a Glycosylase、and Gadd45」Cell 135:1201〜1212(ゼブラフィッシュにおけるDNA脱メチル化の基礎をなす機構についての記載を見いだすことができる。)
【発明の概要】
【0006】
哺乳動物細胞における脱メチル化を調節するための方法、組成物およびキットが提供される。これらの方法、組成物およびキットは、細胞運命の再プログラミングを導くことにおいて、例えば、体細胞から人工多能性幹細胞(iPS)を作製することにおいて、および体細胞を、別の細胞運命に向け直すことにおいて使用される。体細胞は、例えば、メチル化の欠損から生じる、またはそれによって悪化するヒトの障害、例えば、癌および異常なゲノム刷り込みに関連する障害の治療において使用するために、in vitroで、およびin vivoで作製することができる。候補薬剤を、哺乳動物細胞におけるゲノムDNA脱メチル化の調節における活性についてスクリーニングするための方法、組成物およびキットも提供される。
【0007】
本発明の一態様では、哺乳動物細胞におけるゲノムDNAメチル化の量を減少させるための方法であって、対象とする遺伝子(複数可)の発現を制御するプロモーター領域におけるヌクレオチドのメチル化を減少させることを含む方法が提供される。方法は、最初の哺乳動物細胞を、有効量の、シチジンデアミナーゼ(CD)活性を促進する薬剤と接触させるステップを含み、例えば、哺乳動物細胞は、脱メチル化を許容する体細胞を含めた体細胞である。
【0008】
一部の実施形態では、CD活性を促進する薬剤は、活性化誘導性シチジンデアミナーゼ(AID)ポリペプチドまたはAIDポリペプチドをコードする核酸である。一部の実施形態では、CD活性を促進する薬剤は、アポリポタンパク質B RNA編集触媒成分(APOBEC)ポリペプチドまたはAPOBECポリペプチドをコードする核酸である。一部の実施形態では、細胞を表5のポリペプチドとも接触させる。ある特定の実施形態では、表5のポリペプチドは、メチル化シトシンのヒドロキシル化メチルシトシンへの変換を促進する薬剤、例えば、tetタンパク質、例えば、tet1またはtet2である。一部の実施形態では、接触させるステップをin vitroで実施する。
【0009】
一部の実施形態では、最初の哺乳動物細胞は、体細胞、例えば、脱メチル化を許容する体細胞である。一部のそのような実施形態では、作製される細胞は、人工多能性幹細胞(iPS)である。一部の実施形態では、方法は、体細胞を、iPS細胞への運命を促進する1種または複数種の因子と接触させるステップをさらに含む。一部の実施形態では、作製される細胞は、出発細胞とは異なる系列の体細胞である。一部のそのような実施形態では、方法は、体細胞を、所望の体細胞運命を促進する1種または複数種の因子と接触させるステップをさらに含む。
【0010】
一部の実施形態では、最初の細胞は、多能性幹細胞、例えば、脱メチル化を許容する多能性幹細胞である。一部のそのような実施形態では、多能性幹細胞から作製される細胞は、体細胞である。一部のそのような実施形態では、方法は、多能性幹細胞を、所望の体細胞運命を促進する1種または複数種の因子と接触させるステップをさらに含む。
【0011】
一部の実施形態では、接触させるステップは、in vivoで、ゲノムDNA脱メチル化療法を必要とする対象において実施される。一部のそのような実施形態では、最初の細胞は、腫瘍細胞、例えば、脱メチル化を許容する腫瘍細胞であり、対象は、癌に罹患している対象である。他のそのような実施形態では、最初の細胞は、形質転換されていない体細胞、例えば、脱メチル化を許容する体細胞である。
【0012】
本発明の一態様では、候補薬剤を、細胞においてゲノムDNA脱メチル化活性を調節する活性についてスクリーニングする方法が提供される。そのような方法では、細胞の第1の集団を、有効量の、シチジンデアミナーゼ(CD)活性を促進する薬剤とin vitroで接触させる。次いで、この集団の亜集団を候補薬剤と接触させ、一方、第2の集団、すなわち、対照集団は、候補薬剤と接触させない。次いで、候補薬剤と接触させた亜集団の特性を、候補薬剤と接触させなかった細胞の亜集団の特性と比較し、第1の亜集団と第2の亜集団との間の細胞の特性の違いにより、候補薬剤が細胞におけるゲノムDNA脱メチル化活性を調節することが示される。
【0013】
一部の実施形態では、CD活性を促進する薬剤は、AIDポリペプチドまたはAIDポリペプチドをコードする核酸である。一部の実施形態では、第1の集団の細胞は、腫瘍細胞、すなわち腫瘍由来の細胞である。ある特定の実施形態では、腫瘍細胞におけるゲノムDNA脱メチル化を調節する候補薬剤は、癌における腫瘍成長を調節する薬剤である。
【0014】
一部の実施形態では、第1の集団の細胞は、体細胞、またはES細胞と体細胞から作製された異核共存体である。一部の実施形態では、体細胞のDNAのゲノムDNA脱メチル化を調節する候補薬剤は、体細胞のiPS細胞化の誘導を調節する薬剤である。
【0015】
本発明の一態様では、シチジンデアミナーゼのDNA脱メチル化活性の調節における活性があるタンパク質を同定するための方法が提供される。そのような方法では、細胞の集団を、シチジンデアミナーゼをコードする配列を含む核酸と接触させ、シチジンデアミナーゼを細胞の粗タンパク質抽出物から沈殿させ、免疫沈降物を質量分析に供し、ここで、質量分析によって同定された1種または複数種のタンパク質は、シチジンデアミナーゼの脱メチル化活性に欠かせないものである。一部の実施形態では、シチジンデアミナーゼは、AIDまたはAPOBECである。一部の実施形態では、同定されるタンパク質は、表5のタンパク質である。
【0016】
本発明は、以下の詳細な説明を付属図と併せて読むと最もよく理解される。本特許または出願のファイルは、カラーで製作された少なくとも1つの図面を含有する。カラーの図面(複数可)を伴う本特許または特許出願公開のコピーは、要請し、必要な料金を支払えば、特許庁により提供されるであろう。一般的な実施に従って、図のさまざまな特徴は一定の縮尺ではないことが強調される。それどころか、さまざまな特徴の次元は、明瞭にするために任意に拡大または縮小されている。図面には、以下の図が含まれる。
【図面の簡単な説明】
【0017】
【図1】マウスES細胞×ヒト線維芽細胞の異核共存体の単離および特徴付けを示す図である。a、異核共存体の融合スキームである。GFP+マウスES細胞(mES)をDsRed+ヒト初代線維芽細胞(hFb)と2:1の比率で共培養し、次いで、ポリエチレングリコール(PEG)を使用して融合した。b、GFP+mES、DsRed+hFb、および融合の2日後に選別したGFP+DsRed+異核共存体(第1の選別異核共存体)のFACSプロファイルである。異核共存体集団をさらに再選別して(第2の選別異核共存体)、集団の約80%の濃縮(濃縮)にし、分析した。異核共存体を単離するために使用した選別ゲートが示されている。c、融合の2日後に2回選別し、サイトスピンしたGFP+とDsRed+の異核共存体の代表的な画像が示されている。異核共存体の核は、別個のままであり、融合していない。Hoechst33342(青色)は核を示し、示されている異核共存体は3つの明るいマウスの核および1つの均一に染色されたヒトの核を有する。スケールバー=50μm。d、GFP+DsRed+異核共存体を融合の2日後に2回選別し、サイトスピンし、Ki−67(青色)について染色して細胞分裂を評価した。GFP蛍光によって示される異核共存体は、Ki−67(青色)について陰性である2つの別個の核(矢印)、それとは対照的に、Ki−67について陽性に染色された単核細胞(矢じり)を有する。スケールバー=50μm。e、融合の1日後、2日後および3日後に選別し、サイトスピンした異核共存体を、Ki−67染色に基づいてスコア化し、98(±2)%の異核共存体が非分裂性であった(平均±標準誤差、p<0.05)。f、GFP−ES細胞を使用して生成した異核共存体(補足的な方法を参照されたい)を、融合の1日後にヒト線維芽細胞マーカーであるThy1.1を使用して濃縮し、サイトスピンし、Hoechst33258を使用してBrdU(緑色)および核(青色)について染色した。示されている異核共存体(矢印)は、3つの均一に染色されたヒトの核および1つの明るい点状のマウスの核を有し、BrdUについて陰性であった。対照的に、示されたヒト単核細胞(矢じり)は、BrdUについて陽性に染色される。スケールバー=50μm。g、融合の1日後、2日後および3日後に濃縮し、サイトスピンした異核共存体を、核およびBrdUの染色に基づいてスコア化した。異核共存体の94(±3)%においてDNA複製は起こらなかった(平均±標準誤差、p<0.05)。
【図2】単一細胞レベルでの異核共存体におけるヒト線維芽細胞の多能性遺伝子の発現の時間経過を示す図である。a、Oct4、NanogおよびGAPDHに対するヒト特異的なプライマーを、0日目の融合していない共培養物、および融合の1日後、2日後および3日後に単離した異核共存体(mESXhFb)のRT−PCRのために使用した。hOct4およびhNanogはどちらも、異核共存体試料において上方制御され、異核共存体におけるヒト線維芽細胞核の再プログラミングの急速な開始が示されている。b、リアルタイムPCRを使用して、融合の1日後、2日後および3日後に単離した異核共存体におけるOct4(灰色)およびNanog(黒色)の上方制御を、ヒト特異的なプライマーを使用して評価した(平均±標準偏差)。融合していない共培養物は、0日目の対照としての機能を果たし、hOct4およびhNanogの発現を、hGAPDHの発現に対して正規化した。0日目ならびに1日目、2日目および3日目に、hOct4の遺伝子発現とhNanogの遺伝子発現の間に統計的有意差が観察された(*は、p<0.03を示す)。示されているデータは、3つの独立した融合実験からのものである。c、異核共存体集団における再プログラミングの効率を評価するための単一の異核共存体のネステッドPCR。融合の3日後に異核共存体を80%まで濃縮し、単一細胞として単離した。示されている通り、単一の異核共存体に対して、GAPDH(G)、Oct4(O)およびNanog(N)に対するヒト特異的なプライマーを使用して直接逆転写およびネステッドPCRを同時に行った。単回の融合実験で分析された12個の異核共存体が示されている。2つの追加的な融合実験で分析された10個の異核共存体および31個の異核共存体が図7に示されている。d、Oct4のみを発現している異核共存体ならびにOct4とNanogの両方を発現している異核共存体の割合は70±13%であり、これにより、高い割合の異核共存体が多能性への再プログラミングを開始することが示されている。示されているデータは、3つの独立した融合実験の概要である(平均±標準偏差)。
【図3】異核共存体における、ヒト線維芽細胞の多能性遺伝子のプロモーターにおけるDNA脱メチル化の時間経過を示す図である。a、異核共存体におけるヒトOct4プロモーターおよびヒトNanogプロモーターのメチル化状態のバイサルファイト配列決定分析。異核共存体におけるヒトOct4プロモーターおよびヒトNanogプロモーターはどちらも、融合の1日後、2日後および3日後に、共培養した対照と比較して急速かつ進行性のDNA脱メチル化を示す。白い丸は非メチル化CpGジヌクレオチドを示し、黒い丸はメチル化CpGジヌクレオチドを示す。b、融合後の、異核共存体におけるヒトOct4プロモーターのパーセント脱メチル化により、脱メチル化が3日目に80%まで進行性に増加したことが示されている。c、融合後の、異核共存体におけるヒトNanogプロモーターのパーセント脱メチル化により、脱メチル化が3日目に56%まで進行性に増加したことが示されている。各時点で、2から3の独立した実験において少なくとも10クローンを分析した;代表的な10クローンが示されている。
【図4】異核共存体において、ヒト線維芽細胞の多能性への再プログラミングを開始するためのAID依存性のDNA脱メチル化の必要性を示す図である。a、リアルタイムPCRによって評価した、siRNA処理に供した異核共存体におけるAIDおよびヒト多能性遺伝子の発現。si−1、si−2、si−3およびsi−4は、AIDを対象とする別個のsiRNAである。融合の2日後に単離した異核共存体を、si−3およびsi−4で処理し、3日目に単離した異核共存体を、si−1およびsi−2で処理した。マウスAID転写物およびヒトAID転写物の総レベルを、変性プライマーのセットを使用して評価し、一方、hOct4およびhNanogに対してヒト特異的なプライマーを使用した。遺伝子発現を、AIDの発現についてはGAPDH(変性プライマー)に対して、ヒトOct4の発現およびヒトNanogの発現についてはhGAPDHに対して内部で正規化した。次いで、試料を、対応する、対照siRNAで処理した2日目の試料または3日目の試料に対して正規化し、代表的なsi対照(100%)が示されている。si−3、si−4、si−1、およびsi−2で処理した異核共存体におけるAIDの発現は、対照と比較して低下した。4種のsiRNA全てによるAIDのノックダウンにより、多能性遺伝子であるhOct4およびhNanogの発現が遮断された。b、融合の2日目および3日目に、si−3/si−4およびsi−1/si−2によってAIDをノックダウンした際のヒトOct4プロモーターおよびヒトNanogプロモーターは、それぞれ、メチル化されたままであり、これは、脱メチル化が阻害されたことを示し、異核共存体におけるDNA脱メチル化および核の再プログラミングにおけるAIDの役割を裏づけている。c、si−3/si−4、およびsi−1/si−2によってAIDをノックダウンした際のヒトOct4プロモーターおよびヒトNanogプロモーターでのパーセント脱メチル化により、それらのそれぞれの、si対照で処理した対照試料の2日目および3日目と比較して、脱メチル化が遮断されたことが示された。d、ヒトAIDは、線維芽細胞における重度にメチル化されたヒトNanog遺伝子およびヒトOct4遺伝子のプロモーターに動員される。mESおよびhFbにおいて、抗AID抗体を用いてクロマチン免疫沈降を行った。バックグラウンドのIgGシグナルと比較したAIDの占有率が示されている(平均±標準誤差)。hFbならびに陽性対照としてmES、CμおよびCdx2において、それぞれ有意なAIDの結合が検出された(p<0.02)。hOct4およびhNanogのメチル化プロモーターへのAIDの結合は、ヒト線維芽細胞において有意であった(p<0.02)が、マウスES細胞における非メチル化Oct4プロモーターおよび非メチル化Nanogプロモーターへの有意な結合は観察されなかった。e、異核共存体における多能性への再プログラミングにおけるAID依存性のDNA脱メチル化のモデル。デアミナーゼAIDと一緒に作用し得る哺乳動物のDNAデメチラーゼ複合体(X、Y、およびZ)の他の推定上の構成成分は、まだ同定されていない。
【図5】異核共存体の生成、siRNAノックダウン、hAID過剰発現、およびレスキュー実験のためのスキームを示す図である。
【図6】BrdU実験のための異核共存体のThy1.1濃縮を示す図である。a、ヒト線維芽細胞に対するThy1.1の特異性。a(パネル上部)に示されているように、GFP+マウスES細胞はヒト特異的なThy1.1抗体に結合しないが、dsRed+ヒト線維芽細胞の100%がThy1.1(パネル下部)に結合し、これは、Thy1.1をヒト線維芽細胞に対する細胞表面マーカーとして特異的に使用することができることを示している。b、Thy1.1抗体を使用した異核共存体の濃縮。ビオチン化したThy1.1を使用して、PEG処理したmES共培養物およびhFb共培養物を標識し、ストレプトアビジン磁気ビーズを使用してヒト線維芽細胞および異核共存体(mESX hFb)について濃縮した。b(左下)に示されているように、濃縮前にはPEG処理したmES共培養物およびFb共培養物に0.1〜1%の異核共存体が存在するが、磁気ビーズ濃縮後には、Thy1.1陽性の異核共存体および線維芽細胞は、それぞれ10%および80%であった。
【図7】異核共存体はDNA複製を行わないことを示す図である。GFP−(非GFP)のES細胞およびヒト線維芽細胞を使用して生成した異核共存体を、融合の1日後に、ヒト線維芽細胞マーカーであるThy1.1を使用して濃縮し(補足的な方法を参照されたい)、サイトスピンし、BrdU(緑色の)および核(青色)についてHoechst33258を用いて染色した。示されている異核共存体(矢印)は、3つの均一に染色されたヒト核および1つの明るい点状のマウスの核を有し、BrdUについては陰性であり、これは、異核共存体においてDNA複製がないことを示している。対照的に、示されている単核細胞(矢じり)は、BrdUについて陽性に染色される。磁性ストレプトアビジンビーズは、免疫蛍光による画像において小さな黒い丸として細胞の表面上に見ることができる。スケールバー=50ミクロン。
【図8】種間の異核共存体における多能性遺伝子の活性化を試験するためのヒト特異的なプライマーの検証についての図である。a、Oct4およびNanogに対して設計したヒト特異的なプライマーは、ヒトES細胞(hES)由来のヒト転写物を選択的に増幅するが、マウスES細胞(mES)由来のOct4転写物およびNanog転写物は検出しない。b、ヒトOct4プロモーターおよびヒトNanogプロモーターのメチル化状態を評価するために設計したヒトおよびバイサルファイトに特異的なプライマーにより、hFbからのみ産物が増幅され、mESからは増幅されなかった。c、単一の異核共存体におけるヒトGAPDH、Oct4転写物およびNanog転写物の発現を評価するために設計したネステッドPCRのプライマーの種特異性。ヒトGAPDHプライマーを用いるとhES細胞とヒト初代線維芽細胞(hFb)の両方で産物が検出されるが、mESでは産物は検出されない、ヒトOct4転写物およびヒトNanog転写物は、hES細胞のみで検出され、hFbまたはmESでは産物は検出されない。
【図9】共培養した対照における、ある期間にわたるヒト多能性遺伝子の発現。0日目および3日目の、マウスES細胞およびhFbの融合していない共培養物を、多能性遺伝子であるOct4およびNanogに対するヒト特異的なプライマーを使用してリアルタイムPCRによって分析した。遺伝子発現の倍率変化を得るために、遺伝子発現をhGAPDHおよび0日目の対照に対して正規化した。データにより、hOct4およびhNanogの発現が0日目から3日目まで変化しないままであることが示されている。
【図10】異核共存体におけるヒト多能性遺伝子の活性化および発現を示すグラフである。多能性遺伝子であるEssrb、TDGF1、Sox2ならびに細胞周期制御因子であるKlf4およびc−mycに対するヒト特異的なプライマーを、0日目の融合していない共培養物および融合の2日後に単離した異核共存体(mESX hFb)のリアルタイムPCRのために使用した。遺伝子発現の倍率変化を得るために、遺伝子発現をhGAPDHおよび0日目の対照に対して正規化した。異核共存体の試料においてEssrb、TDGF1およびc−mycが誘導された。ヒト線維芽細胞においてすでに発現されているSox2はそれ以上増加せず、Essrbと相互に交換することができることが公知である(Feng、B.ら(2009)Nat Cell Biol 11、197〜203)Klf4もそれ以上増加しない。
【図11】単一の異核共存体における再プログラミングの効率を示す図である。図2cにおいて、単一の融合実験から得られた12個の異核共存体からの結果が示されている。本図には、2つの追加的な独立した融合実験から得られた異核共存体における多能性遺伝子の発現が示されている(それぞれn=10および31)。3つの独立した実験の結果の概要および統計値は図2dに示されている。
【図12】相対的なAID mRNAの発現を示す図である。AIDの転写レベルを、B−リンパ球細胞系(Ramos)、マウス胚性幹細胞(mES)およびヒト線維芽細胞(hFb)においてリアルタイムPCRによって評価した。転写レベルをGAPDHに対して正規化する。Ramos、mES、およびhFbにおけるAIDの発現の比率は、およそ100:15:5である。
【図13】AID標的配列を対象とするsiRNA:ヒトおよびマウスについてのアラインメントを示す図である。ヒト(配列番号105〜配列番号114)およびマウス(配列番号115〜配列番号124)のAID mRNAにおけるsiRNA標的配列およびそれらの対応する標的、ならびにAID転写物に沿ったそれらの相対的な位置が示されている。標的配列の上にミスマッチ(*)が示されている。
【図14】マウスES細胞およびヒト線維芽細胞におけるAIDノックダウンの効力を示すグラフである。siRNAトランスフェクションの3日後にマウスES細胞およびヒト線維芽細胞におけるAIDのノックダウンをリアルタイムPCRによって評価した。AID si−1、AID si−2、AID si−3およびAID si−4は、AIDを対象とする別個のsiRNAであり(図13に示されている配列)、それらにより、マウスES細胞におけるAIDの転写(GAPDHに対して正規化した)が、それぞれ、トランスフェクションの3日後に、対照siRNAと比較して81(±13)%、79(±12)%、70(±8)%、および99(±0.1)%低下した。ヒト線維芽細胞では、AID mRNAのレベルは、siRNA1、siRNA2、siRNA3およびsiRNA4によって、それぞれ46(±11)%、72(±23)%、99(±0.1)%および99(±0.1)%低下した。
【図15】マウスES細胞におけるAIDタンパク質およびノックダウンの検出を示す図である。a、マウスES細胞全体の溶解物2mgからの免疫沈降後のAIDタンパク質の検出。1%のインプット(20μg)をローディングした。b、si−1でトランスフェクトした3日後の濃縮されたマウスES細胞溶解物(170μg)におけるAIDタンパク質レベルの検出。α−チューブリンは、ローディング対照として示されている。c、α−チューブリンに対して正規化されたAIDタンパク質レベルの定量化。si−1で処理したES細胞におけるAIDタンパク質レベルは、si対照試料と比較して12%まで低下した。
【図16】異核共存体におけるヒトOct4プロモーターおよびヒトNanogプロモーターについて収集されたバイサルファイト配列データを示す図である。合計330クローンについて配列決定した。
【図17】ヒトAIDの過剰発現によって異核共存体における再プログラミングの開始は加速されないことを示す図である。融合の1日後の異核共存体の線維芽細胞における、ヒトAID(hAID)の一過性の過剰発現を伴う、またはそれを伴わない、ヒトOct4プロモーターおよびヒトNanogプロモーターのメチル化状態についてのバイサルファイト配列決定分析(図5)。a、hAIDレベルをリアルタイムPCRによって評価し、2つの別々の融合実験において、1日目の異核共存体において、それぞれ2倍および4倍に上方制御されることが見いだされた。b、d、ヒトOct4プロモーターおよびヒトNanogプロモーターのDNA脱メチル化の程度は、hAIDが過剰発現されても増加しない。2つの独立した融合実験について同様の結果が得られた。白い丸は、非メチル化CpGジヌクレオチドを示し、黒い丸は、メチル化CpGジヌクレオチドを示す。2つの独立した融合実験において少なくとも10クローンを分析した;代表的な10クローンが示されている。c、e、融合の1日後に、hAIDの一過性の過剰発現を伴う、またはそれを伴わない、異核共存体におけるヒトOct4プロモーターおよびヒトNanogプロモーターにおいて観察されたパーセント脱メチル化。Oct4プロモーターおよびNanogプロモーターにおけるDNA脱メチル化は、hAIDが過剰発現していても増加しない。
【図18】ヒトAIDの過剰発現により、異核共存体におけるAIDが一過性にノックダウンされている間、再プログラミングの開始がレスキューされることを示す図である。si−RNAターゲティングAID(si−1)でトランスフェクトした異核共存体において過剰発現しているヒトAID(hAID)によってレスキュー実験を行った(図5参照)。a、hAIDレベルを、リアルタイムPCRによって評価し、2つの別々の融合実験において、2日目の異核共存体において、それぞれ2.5倍および4倍に上方制御されることが見いだされた。b、ヒト特異的なプライマーを使用したリアルタイムPCRによって評価された通り、hAIDの過剰発現により、2日目の異核共存体において、対照と比較して多能性遺伝子であるhOct4の発現が部分的にレスキューされ、hNanog遺伝子発現が完全にレスキューされる。発現レベルを、同じ2日目の試料においてhGAPDHに対して正規化し、次いで0日目の対照に対して正規化する。c、d、融合の2日後に単離した異核共存体を、ヒトOct4プロモーターおよびヒトNanogプロモーターのメチル化状態についてバイサルファイト配列決定分析に供した。どちらのプロモーターも脱メチル化を示し、これは、AIDの下方制御によって引き起こされる再プログラミングの遮断は、hAIDの過剰発現により、Nanogプロモーターの脱メチル化の完全なレスキューおよびOct4プロモーターの脱メチル化の部分的なレスキューを伴って克服される。Oct4プロモーターの脱メチル化は、72%の対照レベルと比較すると、8%脱メチル化(si−1)から22%脱メチル化(hAID+si−1)までレスキューされる。Nanogプロモーターの脱メチル化は、31%脱メチル化(si−1)から51%脱メチル化(hAID+si−1)までレスキューされる。47%の対照レベルと比較すると、完全なレスキューが観察される。
【図19】バイサルファイト特異的配列決定およびChIPアッセイにおいて調査したCpG密度を示すヒトOct4プロモーター(配列番号125)およびヒトNanogプロモーター(配列番号126)のマップである。バイサルファイト特異的なアンプリコンの配列が示されている。ヒトOct4プロモーターおよびヒトNanogプロモーターにおけるCpG部位が肉太活字で示されている。ChIPプライマーの適用範囲の領域(リアルタイムPCRのアンプリコン)が示されている。ATG開始コドンからの距離が示されている。
【発明を実施するための形態】
【0018】
本方法および組成物について記載する前に、本発明は記載されている特定の方法または組成物に限定されず、それ自体は当然変動することが理解されるべきである。本発明の範囲は、添付の特許請求の範囲によってのみ限定されるので、本明細書において使用される用語法は、特定の実施形態を記載するためだけのものであり、限定的なものではないことも理解される。
【0019】
さまざまな値が提供される場合、文脈によりそうでないことが明確に規定されない限り、下限の単位の10分の1まで、その範囲の上限と下限の間に入る値のそれぞれも、明確に開示されていると理解される。任意の明示された値、または明示された範囲の間に入る値および任意の他の明示された値、またはその明示された範囲の間に入る値のより小さな範囲のそれぞれが本発明の範囲内に包含される。これらのより小さな範囲の上限と下限は、それぞれ独立に、その範囲に含めることができる、またはその範囲から排除することができ、そのより小さな範囲に限界のいずれかが含まれる、どちらの限界も含まれない、またはどちらの限界も含まれる各範囲も、本発明に包含され、明示された範囲内の任意の明確に排除される限界に支配される。明示された範囲が限界の一方または両方を含む場合、それらの含まれる限界のいずれか、またはその両方を除く範囲も、本発明に包含される。
【0020】
別段の定義のない限り、本明細書において使用される全ての技術用語および科学用語は、本発明が属する分野の当業者に一般に理解されているものと同じ意味を有する。本明細書に記載の方法および材料と類似した、またはそれと等しい任意の方法および材料を本発明の実施または試験において使用することができるが、一部の潜在的な好ましい方法および材料がここに記載されている。本明細書で言及されている全ての刊行物が、引用されている刊行物に関連して方法および/または材料を開示し記載するために、参照により本明細書に組み込まれる。矛盾がある範囲では、組み込まれた刊行物の開示のいずれにも本開示が取って代わることが理解される。
【0021】
本明細書および添付の特許請求の範囲において使用される、単数形「a(1つの)」、「an(1つの)」、および「the(その)」は、文脈によりそうでないことが明確に規定されない限り、複数の指示対象を包含することに留意しなければならない。したがって、例えば、「細胞」への言及は、複数のそのような細胞を包含し、「ペプチド」への言及は、1種または複数種のペプチドおよびその等価物、例えば、当業者に公知のポリペプチドなどへの言及を包含する。
【0022】
本明細書で考察されている刊行物は、単に、本出願の出願日より前のそれらの開示について提供されている。本明細書における全ては、本発明が、先行発明の理由でそのような刊行物に先立つ権限がないことの容認であると解釈されるべきではない。さらに、提供されている刊行物の日付は、それぞれ独立に確認する必要があり得る実際の刊行物の日付とは異なる可能性がある。
【0023】
定義
哺乳動物細胞における脱メチル化を調節するための方法、組成物およびキットが提供される。候補薬剤を、哺乳動物細胞におけるゲノムDNA脱メチル化活性のレベルの調節における活性についてスクリーニングするための方法、組成物およびキットも提供される。これらの方法、組成物およびキットは、人工多能性幹細胞(iPS)および体細胞をin vitroで作製することにおいて、および、癌およびゲノム刷り込みにおける欠損から生じる障害を含めたヒトの障害をin vivoで治療するために使用される。これらの、または他の本発明の目的、利点および特徴は、以下に十分に記載されている組成物および方法の詳細を読めば、当業者には明白になるであろう。
【0024】
「DNAメチル化」または単に「メチル化」は、DNAにメチル基が付加されることを意味する。DNAにメチル基が付加される反応は、酵素のDNAメチルトランスフェラーゼ(DNMT)によって触媒される。脊椎動物では、DNAメチル化は、一般には、ヌクレオチドシトシンにおいて、通常CpG部位(シトシン−リン酸−グアニン部位;すなわち、DNA配列においてシトシンのすぐ次にグアニンが続く場所)で起こる。これにより、シトシンが、本明細書では互換的に「5−メチルシトシン」「5−meC」、および「メチル化シトシン」と称される5−メチルシトシンに変換される。付加されたメチル基により、シトシンの構造が、その塩基対合の性質が変化することなく変化する。「GCリッチ」領域(すなわち、約65%のCG残基でできている)であるCpG配列およびCpGアイランドのメチル化の程度は、多くの場合、遺伝子の転写活性に関連し、高度にメチル化されたCpGアイランドを含有するプロモーターは一般にはサイレントであり、非メチル化CpGアイランドおよびあまりメチル化されていないCpGアイランドを含有するプロモーターは、一般には活性である。
【0025】
「DNA脱メチル化」または単に「脱メチル化」は、CpG配列がメチル化CpG配列から非メチル化CpG配列に変換されることを意味する。
【0026】
「DNA脱メチル化を許容する細胞」または「脱メチル化を許容する細胞」は、そのCpG配列をメチル化CpG配列から非メチル化CpG配列に変換することができる細胞を意味する。細胞に5−meCpGリッチヌクレオチド配列を含むDNAベクターを提供すること、および、ベクターによって供給されたヌクレオチド配列を、例えば、バイサルファイト配列決定またはメチラーゼ感受性制限エンドヌクレアーゼ消化によって回収し分析してそのヌクレオチド配列のCpG配列が脱メチル化されているかどうかを決定して、細胞において活性化誘導性シチジンデアミナーゼ(AID)(GenBank受託番号NM_020661)をコードするcDNAを過剰発現させることによって細胞が脱メチル化を許容するかどうかを決定することができる。
【0027】
「シチジンデアミナーゼ活性」または「CD活性」は、リボース環(シチジン)またはデオキシリボース環(デオキシシチジン)に付着しているシトシンヌクレオシドまたは5−メチルシトシンヌクレオシドからアミン基が除去される酵素経路の活性を意味する。シトシンからアミン基が除去されることにより、ヌクレオシドがウラシルに変換され、一方、5−メチルシトシンからアミン基が除去されることにより、ヌクレオシドがチミンに変更される。例えば、以下の略図を参照されたい:
【0028】
【化1】

【0029】
「多能性幹細胞」または「多能性細胞」は、生物体において全種類の細胞に分化する能力を有する細胞を意味する。多能性細胞は、奇形腫を形成し得、生きている生物体において外胚葉組織、中胚葉組織、または内胚葉組織の一因となっている。多能性幹細胞の例は、胚性幹(ES)細胞、胚性生殖幹(EG)細胞、および人工多能性幹(iPS)細胞である。
【0030】
「胚性幹細胞」または「ES細胞」は、a)自己再生することができ、b)生物体において分化して全種類の細胞を産生することができ、かつc)発生中の生物体の胞胚の内部細胞塊に由来する細胞を意味する。ES細胞は、生物体において全種類の細胞に分化する能力を維持させながら、長期にわたって培養することができる。培養物中で、ES細胞は、一般には、大きな核細胞質比、明確な境界および目立った核を有する平らなコロニーとして成長する。さらに、ES細胞は、SSEA−3、SSEA−4、TRA−1−60、TRA−1−81、およびアルカリホスファターゼを発現するが、SSEA−1は発現しない。ES細胞を生成し、特徴づける方法の例は、例えば、その開示が参照により本明細書に組み込まれる米国特許第7,029,913号、米国特許第5,843,780号、および米国特許第6,200,806号において見ることができる。
【0031】
「胚性生殖幹細胞」、「胚性生殖細胞」または「EG細胞」は、a)自己再生することができ、b)生物体において分化して全種類の細胞を産生することができ、かつc)生殖細胞および生殖細胞前駆体、例えば、始原生殖細胞に由来する、すなわち、精子および卵子になる細胞を意味する。胚性生殖細胞(EG細胞)は、上記の胚性幹細胞と同様の性質を有すると考えられている。EG細胞を生成し、特徴づける方法の例は、例えば、その開示が参照により本明細書に組み込まれる、米国特許第7,153,684号;Matsui、Yら、(1992)Cell 70:841;Shamblott、M.ら(2001)Proc.Natl.Acad.Sci.USA 98:113;Shamblott、M.ら(1998)Proc.Natl.Acad.Sci.USA、95:13726;およびKoshimizu、U.ら(1996)Development、122:1235において見ることができる。
【0032】
「人工多能性幹細胞」または「iPS細胞」は、a)自己再生することができ、b)生物体において分化して全種類の細胞を産生することができ、かつc)体細胞に由来する細胞を意味する。iPS細胞は、ES細胞様の形態を有し、大きな核細胞質比、明確な境界および目立った核を有する平らなコロニーとして成長する。さらに、iPS細胞は、これらに限定されないが、アルカリホスファターゼ、SSEA3、SSEA4、Sox2、Oct3/4、Nanog、TRA160、TRA181、TDGF 1、Dnmt3b、FoxD3、GDF3、Cyp26a1、TERT、およびzfp42を含めた当業者に公知の1種または複数種の重要な多能性マーカーを発現する。iPS細胞を生成し、特徴づける方法の例は、例えば、その開示が参照により本明細書に組み込まれる、出願番号:US20090047263、US20090068742、US20090191159、US20090227032、US20090246875、およびUS20090304646において見ることができる。
【0033】
「体細胞」は、実験的な操作をしなければ、普通は生物体において全種類の細胞を生じさせない、生物体における任意の細胞を意味する。言い換えれば、体細胞は、自然には体の3つの胚葉、すなわち、外胚葉、中胚葉および内胚葉の全ての細胞を生成しない、十分に分化した細胞である。例えば、体細胞は、神経細胞と神経前駆細胞の両方を含み、神経前駆細胞は、中枢神経系の全部または一部の細胞型を自然に生じさせることができ得るが、中胚葉系列または内胚葉系列の細胞を生じさせることはできない。
【0034】
「再プログラミング因子」は、細胞に作用して転写を変化させ、それによって細胞を多能性に再プログラミングする生物活性因子の1種または複数種、すなわち、混合物を意味する。細胞に再プログラミング因子を提供する、すなわち細胞を再プログラミング因子と接触させる本発明の方法では、これらの再プログラミング因子は、個々に、または単一の組成物として、すなわち、再プログラミング因子を予備混合した組成物として細胞に提供することができる。因子は、同じモル比で、または異なるモル比で提供することができる。因子は、本発明の細胞を培養する過程において、一回または多数回提供することができる。
【0035】
「再プログラミングの効率」は、細胞のin vitro培養物の、別の細胞型の細胞を生じるように再プログラムされる能力を意味する。薬剤、例えば、シチジンデアミナーゼ活性を促進する薬剤の存在下で増強された再プログラミング効率を示す細胞は、その薬剤と接触させた場合に、その薬剤と接触させなかった細胞と比較して、別の細胞型の細胞を生じる増強された能力を示す。増強されるとは、細胞培養物が、薬剤と接触させなかった細胞培養物の能力の少なくとも50%、約100%、約200%、約300%、約400%、約600%、約1000%、約2000%、少なくとも約5000%の、新しい細胞型を生じさせる能力を有することを意味する。言い換えれば、細胞培養物により、薬剤と接触させていない細胞の集団によって産生される新しい細胞型の細胞の約1.5倍、約2倍、約3倍、約4倍、約6倍、約10倍、約20倍、約30倍、約50倍、約100倍、約200倍を超える新しい細胞型の細胞が産生される。本出願の一部の実施形態では、再プログラミングの効率を増強する薬剤は、所望の細胞運命を獲得する間に活性になることが当技術分野で公知であるプロモーターにおけるDNAメチル化の量を、例えば、薬剤がない場合に観察されるDNAメチル化の量と比較して1.5分の1以下、すなわち、約1.5分の1、約2分の1、約3分の1、約4分の1、約6分の1、または約10分の1またはそれ未満に減少させる薬剤である。本出願の一部の実施形態では、再プログラミングの効率を増強する薬剤は、所望の細胞運命を獲得する間に活性になることが当技術分野で公知であるプロモーターによって調節される遺伝子の転写量を、例えば、薬剤がない場合に観察される転写量と比較して約1.5倍以上、すなわち、約1.5倍、約2倍、約3倍、約4倍、約6倍、または約10倍またはそれ以上増加させる薬剤である。
【0036】
「プロモーター配列」は、細胞においてRNAポリメラーゼに結合し、下流(3’方向)のコード配列の転写を開始させることができるDNA調節領域である。本発明を明確にする目的において、プロモーター配列は、その3’末端に転写開始部位が結合し、バックグラウンドを超えて検出可能なレベルで転写を開始するために必要な最小数の塩基またはエレメントを含むように上流(5’方向)に伸長している。プロモーター配列内に、転写開始部位、ならびにRNAポリメラーゼの結合に関与するタンパク質結合ドメインが見いだされる。真核生物のプロモーターは、多くの場合、常にではないが、「TATA」ボックスおよび「CAT」ボックスを含有する。誘導性プロモーターを含めた種々のプロモーターを使用して、本発明の種々のベクターを駆動することができる。プロモーター配列からの転写活性は、プロモーターがメチル化されている程度によって調節することができる。
【0037】
「治療」、「治療すること」などの用語は、本明細書では、一般に、所望の薬理的効果および/または生理的効果を得ることを意味するように使用される。効果は、疾患またはその症状を完全にまたは部分的に予防するという点で予防的であってよく、かつ/または、疾患および/または疾患に起因する有害作用を部分的または完全に治癒させるという点で治療的であってよい。「治療」は、本明細書で使用される場合、哺乳動物における疾患の任意の治療を包含し、(a)疾患の素因があり得るが、今のところはまだそれを有すると診断されていない対象において疾患が生じることを予防すること;(b)疾患を抑制すること、すなわち、その発生を制止すること;または(c)疾患を軽減すること、すなわち、疾患の退縮を引き起こすことを含む。治療剤は、疾患または害が発症する前、その間、またはその後に投与することができる。進行中の疾患を治療し、その治療により患者の望ましくない臨床症状を安定させる、または低減することが特に興味深い。そのような治療は、患部組織の機能が完全に喪失する前に行うことが望ましい。本主題の療法は、疾患の症候性の段階、およびある場合では疾患の症候性の段階後に施されることが望ましい。
【0038】
「個体」、「対象」、「宿主」および「患者」という用語は、本明細書では互換的に使用され、診断、治療、または療法が望まれる任意の哺乳動物の対象、特に、ヒトを指す。
【0039】
シチジンデアミナーゼ(CD)活性を促進する薬剤
哺乳動物細胞におけるメチル化の量を調節するための方法、組成物およびキットが提供される。本発明の一態様では、哺乳動物細胞におけるゲノムDNAメチル化の量を、細胞を、シチジンデアミナーゼ活性を促進する1種または複数種の薬剤と接触させることによって減少させる。上記の通り、シチジンデアミナーゼ(CD)活性は、DNAまたはRNAにおけるシトシンまたは5−メチルシトシンからアミノ基を除去する酵素活性である。本出願において使用されるシチジンデアミナーゼ活性を促進する薬剤の例は、シチジンデアミナーゼのAID/APOBECクラスのポリペプチドおよび断片、ならびにこれらのポリペプチドおよび断片をコードする核酸である。
【0040】
活性化誘導性シチジンデアミナーゼは、AID、AICDA、ARP2、CDA2、またはHIGM2とも称され、適応性の体液性免疫系におけるその役割について最もよく知られているシチジンデアミナーゼであり、免疫グロブリン遺伝子座のDNAにおけるシトシン残基を脱アミノ化して、抗体遺伝子の多様化を強化する(免疫グロブリンV遺伝子の体細胞超変異および遺伝子変換ならびにIgC遺伝子のスイッチ組換え)。「AID遺伝子産物」、「AIDポリペプチド」、「AIDペプチド」、および「AIDタンパク質」という用語は、本明細書では互換的に使用され、ネイティブな配列のAIDポリペプチド、AIDポリペプチド変異体、AIDポリペプチド断片およびキメラAIDポリペプチドを指す。AIDポリペプチドおよびそれをコードする核酸のネイティブな配列は、GenBank受託番号NM_020661(配列番号1、配列番号2)において見ることができる。
【0041】
アポリポタンパク質B RNA編集触媒成分タンパク質は、APOBECタンパク質とも称され、シチジンを脱アミノ化するタンパク質のファミリーである。「APOBEC遺伝子産物」、「APOBECポリペプチド」、「APOBECペプチド」、および「APOBECタンパク質」という用語は、本明細書では互換的に使用され、APOBECポリペプチド、APOBECポリペプチド変異体、APOBECポリペプチド断片およびキメラのAPOBECポリペプチドのネイティブな配列を指す。APOBECファミリーの始祖メンバーであるAPOBEC1は、シトシン6666を脱アミノ化してウラシルにし、それによって中途終止コドンを創出し、短縮されたアポリポタンパク質Bポリペプチド鎖の組織特異的な産生を強化することによってアポリポタンパク質B RNAを編集する複合体の触媒成分である。APOBEC1ポリペプチドおよびそれをコードする核酸のネイティブなヒト配列は、GenBank受託番号NM_001644(配列番号3、配列番号:4)において見ることができる。APOBEC3ファミリーのメンバー(APOBEC3F、APOBEC3GおよびAPOBEC3H)は、レトロウイルスの感染を制限する先天性免疫経路において、レトロウイルスの第1の鎖cDNA複製中間体においてシトシンを脱アミノ化すること、またはウイルスのゲノムに致死的な超変異を生じさせることによって役割を果たし;APOBEC3F(KA6、ARP8、MGC74891、およびBK150C2.4.mRNAとしても公知である)のネイティブなヒト配列は、GenBank受託番号NM_145298.5(アイソフォームa)(配列番号5、配列番号6)およびNM_001006666.1(アイソフォームb)(配列番号7、配列番号8)において見ることができ;APOBEC3G(ARP9、CEM15、MDS019、FLJ12740、bK150C2.7およびdJ494G10.1としても公知である)のネイティブなヒト配列は、GenBank受託番号NM_021822.2(配列番号9、配列番号10)において見ることができ;APOBEC3H(ARP10としても公知である)のネイティブなヒト配列は、GenBank受託番号NM_001166003.1(アイソフォーム1)(配列番号11、配列番号12)、NM_181773.3(アイソフォーム2)(配列番号13、配列番号14)NM_001166002.1(アイソフォーム3)(配列番号15、配列番号16)、NM_001166004.1(アイソフォーム4)(配列番号17、配列番号18)において見ることができる。シチジンデアミナーゼのAPOBECファミリーの他のメンバーとしては、そのネイティブなヒト配列が、GenBank受託番号NM_006789(配列番号19、配列番号20)において見ることができるAPOBEC2(ARP1およびARCD1としても公知である);そのネイティブなヒト配列が、GenBank受託番号NM_145699.3(配列番号21、配列番号22)において見ることができるAPOBEC3(Phorbolin1、ARP3、PHRBN、およびbK150C2.1としても公知である);そのネイティブなヒト配列が、GenBank受託番号NM_004900(配列番号23、配列番号24)において見ることができるAPOBEC3B(ARP4、ARCD3、PHRBNL、APOBEC1L、FLJ21201、bK150C2.2およびDJ742C19.2としても公知である);そのネイティブなヒト配列が、GenBank受託番号NM_014508.2(配列番号25、配列番号26)において見ることができるAPOBEC3C(PBI、ARP5、ARDC2、ARDC4、APOBEC1L、MGC19485、およびbK150C2.3としても公知である);およびそのネイティブなヒト配列が、GenBank受託番号NM_152426.3(配列番号27,配列番号28)において見ることができるAPOBEC3D(ARP6、APOBEC3E、およびAPOBEC3DEとしても公知である)が挙げられる。
【0042】
シチジンデアミナーゼのAID/APOBECクラスおよびタンパク質のこのクラスの間で保存されているドメインに関するより多くの情報は、その開示が参照により本明細書に組み込まれる、Conticello、S.G.ら(2005)Molecular Biologyおよび Evolution 22(2)367〜377において見ることができる。
【0043】
一部の実施形態では、CD活性、したがって、ゲノムDNA脱メチル化を促進する薬剤は、AIDポリペプチドである。AIDポリペプチドは、シチジンの脱アミノ化を促進するAID配列を含むポリペプチドである。AIDポリペプチドは、AIDの完全なポリペプチド配列またはシチジンデアミナーゼ活性を有するAIDの断片、例えば、C末端の10アミノ酸を抜いた全長のポリペプチド(Barretoら(2003)Mol.Cell 12(2):501〜8)に対して50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、98%または100%の配列同一性を有するポリペプチドを含んでよい。そのような断片は、当業者が当技術分野で周知の一般的な生化学的技法および遺伝学的技法を使用して容易に同定可能である。全長のAIDまたはそのシチジンデアミナーゼ活性ドメインのポリペプチド配列に対して50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、98%または100%の配列同一性を有するポリペプチドをコードする核酸、およびこれらの核酸を含むベクターも、本発明に包含される。
【0044】
一部の実施形態では、CD活性、したがって、ゲノムDNA脱メチル化を促進する薬剤は、APOBECポリペプチドである。APOBECポリペプチドは、シチジンの脱アミノ化を促進するAPOBEC配列を含むポリペプチドである。APOBECポリペプチドは、APOBECの完全なポリペプチド配列またはシチジンデアミナーゼ活性を有するAPOBECの断片に対して50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、98%または100%の配列同一性を有するポリペプチドを含んでよい。そのような断片は、当業者が当技術分野で周知の一般的な生化学的技法および遺伝学的技法を使用して容易に同定可能である。全長のAPOBECポリペプチドまたはそれらのシチジンデアミナーゼ活性ドメインのいずれかのポリペプチド配列に対して50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、98%または100%の配列同一性を有するポリペプチドをコードする核酸、およびこれらの核酸を含むベクターも、本発明に包含される。
【0045】
上記の通り、本発明において使用するために適した薬剤は、シチジンデアミナーゼタンパク質のAID/APOBECクラスのポリペプチドおよび断片、ならびにこれらのポリペプチドおよび断片をコードする核酸を含む。一部の実施形態では、CD活性を促進する1種または複数種の薬剤は、核内作用性の非組み込み型ポリペプチドである。言い換えれば、対象細胞を、CD活性を促進するポリペプチド(「CD活性ポリペプチド」)と接触させ、核内で作用させる。非組み込み型とは、ポリペプチドが対象細胞、すなわち脱メチル化活性を促進することが望まれる細胞のゲノム内に組み込まれないことを意味する。
【0046】
CD活性ポリペプチドの細胞膜を渡る輸送を促進するために、CD活性ポリペプチド配列をポリペプチド浸透性ドメインに融合することができる。いくつもの浸透性ドメインが当技術分野で公知であり、ペプチド、ペプチド模倣薬、および非ペプチド担体を含めた本発明の核内作用性の非組み込み型ポリペプチドにおいて使用することができる。例えば、浸透性ペプチドは、ペネトラチンと称されるDrosophila melanogasterの転写因子であるAntennapaediaの第3のアルファヘリックス由来であってよい。別の例として、浸透性ペプチドは、例えば、天然に存在するtatタンパク質のアミノ酸49〜57を含み得るHIV−1tat基本領域アミノ酸配列を含む。他の浸透性ドメインとしては、ポリアルギニンモチーフ、例えば、HIV−1revタンパク質のアミノ酸34〜56の領域、ノナアルギニン、オクタアルギニンなどが挙げられる。(例えば、移行ペプチドおよびペプトイドの教示に関して参照により本明細書に明確に組み込まれる、Futakiら(2003)Curr Protein Pept Sci.2003 Apr;4(2):87〜96;およびWenderら(2000)Proc.Natl.Acad.Sci.U.S.A、2000年11月21日;97(24):13003〜8;米国特許出願公開第20030220334号;同第20030083256号;同第20030032593号;および同第20030022831号を参照されたい)。ノナアルギニン(R9)配列は、特徴づけられているより効率的なPTDの1つである(Wenderら2000;Uemuraら2002)。
【0047】
CD活性ポリペプチドは、当技術分野で公知の従来の方法を使用して、in vitro合成によって調製することができる。種々の市販の合成装置、例えば、Applied Biosystems、Inc.、Beckmanなどによる自動合成機が利用可能である。合成機を使用することによって、天然に存在するアミノ酸を非天然アミノ酸で置換することができる。特定の配列および調製様式は、利便性、経済的側面、必要な純度などによって決定される。シチジンデアミナーゼ活性ポリペプチドを無細胞系において調製する他の方法としては、例えば、参照により本明細書に組み込まれる、米国特許出願第61/271,000号において教示されている方法が挙げられる。
【0048】
CD活性ポリペプチドは、組換え合成の従来の方法に従って単離し、精製することもできる。発現宿主の溶解物を調製し、その溶解物を、HPLC、排除クロマトグラフィー、ゲル電気泳動、アフィニティークロマトグラフィー、または他の精製技法を使用して精製する。ほとんどの場合、使用される組成物は、産物の調製およびその精製の方法に関連する汚染物質に関して、少なくとも20重量%、通常、少なくとも約75重量%、好ましくは少なくとも約95重量%、および治療のためには、通常少なくとも約99.5重量%の所望の産物を含む。通常、百分率は、総タンパク質量に基づく。CD活性ポリペプチドは、組換えによって、直接だけでなく、異種ポリペプチド、例えば、成熟タンパク質またはポリペプチドのN末端に特異的な切断部位を有するポリペプチドとの融合ポリペプチドとしても作製することができる。発現ベクターは、通常、選択マーカーとも称される選択遺伝子を含有する、この遺伝子は、選択培地中で成長させる形質転換された宿主細胞の生存または成長に必要なタンパク質をコードする。
【0049】
当技術分野において公知の方法によって精製した後、標準のタンパク質形質導入方法によってCD活性ポリペプチドを対象細胞に提供する。ある場合では、タンパク質形質導入方法は、細胞を担体および少なくとも1つの精製したCD活性ポリペプチドを含有する組成物と接触させることを含む。適切な担体およびそれらを使用するための方法の例としては、これらに限定されないが、米国特許第6,841,535号に記載のChariot(商標)(Active Motif、Inc.、Carlsbad、Calif.);Bioport(商標)(Gene Therapy Systems、Inc.、San Diego、Calif.)、GenomeONE(Cosmo Bio Co.、Ltd.、Tokyo、Japan)、およびProteoJuice(登録商標)(Novagen、Madison、Wis.)などの市販の試薬、または例えば、米国特許出願第10/138,593号に記載のナノ粒子タンパク質形質導入試薬が挙げられる。
【0050】
他の実施形態では、CD活性を促進する1種または複数種の薬剤は、CD活性ポリペプチドをコードする核酸である。CD活性ポリペプチドをコードする核酸を対象細胞に提供するために使用されるベクターは、一般には、核酸の発現を駆動するため、すなわち、転写を活性化するために適したプロモーターを含む。これは、遍在的に作用するプロモーター、例えば、CMV−β−アクチンプロモーター、または誘導性プロモーター、例えば特定の細胞集団において活性なプロモーターまたはテトラサイクリンなどの薬物が存在することに反応するプロモーターなどを含んでよい。転写活性化とは、標的細胞において転写が基礎レベルを少なくとも約10倍、少なくとも約100倍、通常少なくとも約1000倍超えて増加することを意味する。さらに、核酸を提供するために使用するベクターは、後で除去しなければならない遺伝子を含んでよく、その際は、例えば、Cre/Loxなどのリコンビナーゼ系、または、例えば、TK、bcl−xsなどのヘルペスウイルスなどの選択毒性を可能にする遺伝子を含めることによって該遺伝子が破壊されて発現される細胞を用いる。
【0051】
CD活性ポリペプチドをコードする核酸は、対象細胞に直接提供することができる。言い換えれば、細胞を、CD活性ポリペプチドをコードする核酸を含むベクターと、ベクターが細胞に取り込まれるように接触させる。電気穿孔、塩化カルシウムトランスフェクション、およびリポフェクションなどの、細胞を核酸ベクターと接触させるための方法は当技術分野で周知である。このように核酸を送達するベクターは、通常、エピソームとして、例えば、プラスミドまたはミニサークルDNAとして維持される。
【0052】
あるいは、核酸は、ウイルスによって対象細胞に提供することができる。言い換えれば、細胞を、CD活性ポリペプチドをコードする核酸を含むウイルス粒子と接触させる。そのような方法には、レトロウイルス、例えば、レンチウイルスが特に適している。一般に使用されるレトロウイルスベクターは「不完全である」、すなわち、増殖性感染のために必要なウイルスタンパク質を産生することができない。もっと正確に言えば、ベクターの複製には、パッケージング細胞系において成長させることが必要である。対象とする核酸を含むウイルス粒子を生成するために、核酸を含むレトロウイルスの核酸をパッケージング細胞系によってウイルスカプシド中にパッケージする。異なるパッケージング細胞系からは、カプシド中に組み入れられる異なる外被タンパク質がもたらされ、この外被タンパク質により、ウイルス粒子の細胞に対する特異性が決定される。外被タンパク質は、狭宿主性、広宿主性および異種栄養性の少なくとも3種類のものである。狭宿主性外被タンパク質、例えば、MMLVを用いてパッケージしたレトロウイルスは、大部分のマウスおよびラット細胞型に感染することができ、これは、BOSC23(Pearら(1993)P.N.A.S.90:8392〜8396)などの狭宿主性パッケージング細胞系を使用することによって生成される。広宿主性外被タンパク質、例えば、4070A(Danosら、上記)を担持するレトロウイルスは、ヒト、イヌおよびマウスを含めた大部分の哺乳動物細胞型に感染することができ、これは、PA12(Millerら(1985)Mol.Cell.Biol.5:431〜437);PA317(Millerら(1986)Mol.Cell.Biol.6:2895〜2902);GRIP(Danosら(1988)PNAS 85:6460〜6464)などの広宿主性パッケージング細胞系を使用することによって生成される。異種栄養性外被タンパク質、例えば、AKR envを用いてパッケージしたレトロウイルスは、マウスの細胞以外の大部分の哺乳動物細胞型に感染することができる。適切なパッケージング細胞系を使用して、対象細胞がパッケージされたウイルス粒子の標的になることを確実にすることができる。シチジンデアミナーゼ活性を促進するポリペプチドをコードする核酸を含むレトロウイルスベクターをパッケージング細胞系に導入する方法およびパッケージング系によって生成されるウイルス粒子を採取する方法は当技術分野で周知である。
【0053】
本発明の方法では、哺乳動物細胞におけるゲノムDNAメチル化の量を、有効量の、CD活性を促進する1種または複数種の薬剤と接触させることによって減少させる。細胞におけるゲノムDNAメチル化を減少させるために十分な薬剤の量は、細胞におけるCD活性を促進するために十分な薬剤の量、すなわち、細胞におけるシトシンおよび5−メチルシトシンからのアミノ基の除去を促進するために十分な量である。この量は、シトシンヌクレオシドまたは5−メチルシトシンヌクレオシドの、それぞれウラシルヌクレオシドまたはチミンヌクレオシドへの変換を測定する当技術分野で公知のいくつものアッセイによって経験的に決定することができ、ここで、細胞におけるゲノムDNAメチル化の量を減少させるための薬剤の有効量は、5%以上、すなわち、5%、10%、20%、40%、60%、80%、または100%のシトシンまたは5−メチルシトシンの、ウラシルまたはチミンへの変換を誘導する量である。例えば、dCのdU(デオキシウラシル)への脱アミノ化の程度は、その開示が参照により本明細書に組み込まれるBransteitter、R.ら((2003)PNAS 100(7):4102〜4107)に記載のウラシルDNAグリコシラーゼ(UDG)および脱プリン部位エンドヌクレアーゼ(APE)を使用することによってアッセイすることができる。そのようなアッセイでは、5’末端を32Pで標識したDNA基質またはRNA基質(例えば、100nM)を、シチジンデアミナーゼ活性を促進する薬剤と一緒にインキュベートする。次いで、相補DNA鎖を基質とアニーリングさせ、その後、UDGおよびAPEと一緒にインキュベートする。インキュベートした後、反応を終了させ、反応生成物を変性ポリアクリルアミドゲル電気泳動(PAGE)によって分離し、ホスフォイメージングによって可視化し、ここで、標識された末端からシチジンまでの長さに対応する短い放射性生成物が存在することにより、シチジンにおけるCD活性を反映する、元のシチジンにおける切れ目が示される。別の例として、dCの脱アミノ化は、同様にBransteitter、R.ら、上記に記載されているプライマー伸長のジデオキシヌクレオチドによる終結を使用することによって検出することができる。そのようなアッセイでは、CD活性を促進する薬剤と反応するDNA基質またはRNA基質を3倍過剰の18−mer32P−標識プライマーとアニーリングさせ、3種のdNTPと、それに加えて2’,3’−ジデオキシアデノシン(ddA)三リン酸または2’,3’−ジデオキシグアノシン(ddG)三リン酸のいずれかの存在下でT7シーケナーゼを使用することによってプライマーを伸長する。基質−伸長プライマー複合体を熱変性させ、分離した鎖を相補DNA鎖とアニーリングさせ、上記のUDGおよびAPEと一緒にインキュベートする。反応生成物を変性PAGEによって分離し、ホスフォイメージングによって可視化し、ここで、脱アミノ化効率は、ddAミックスを用いた伸長反応から、C鋳型における、およびC鋳型を通り越した組み込まれたバンドの強度と比較したC/U鋳型の逆のバンドの強度の比として算出する。効率は、ddGミックスを用いた伸長反応からも、鋳型Cを通り越した組み込まれたバンドの強度の、C鋳型における、およびC鋳型を通り越した組み込まれたバンドの強度に対する比として算出することができる。このように、CD活性を促進する薬剤を同定することができ、有効量の、CD活性を促進する薬剤を経験的に決定することができる。
【0054】
ゲノムDNAメチル化の量を減少させるために十分である薬剤の有効量は、その薬剤で処理した後のDNAメチル化の程度をアッセイすることによっても決定することができる。細胞におけるゲノムDNA脱メチル化の量を減少させる薬剤の有効量は、DNA配列内のメチル化CpG配列の数の、1.5分の1以上への低下、すなわち、1.5分の1、2分の1、3分の1、4分の1、5分の1、10分の1、または20分の1またはそれ以下への低下を誘導する量である。CpG配列のメチル化の状態をアッセイするためのいくつかの方法、例えば、制限エンドヌクレアーゼ消化およびバイサルファイト配列決定が当技術分野で周知である。制限エンドヌクレアーゼ消化では、5−メチルシトシンを含有するCpG配列(例えば、CmeCGG)を、制限酵素Hpallによる切断に対する5−メチルシトシンを含有する配列の抵抗性によって、非メチル化シトシンを含有するCpG配列(CCGG)と区別することができる。対照的に、メチル化CpG配列および非メチル化CpG配列は、制限酵素Msplによって同等によく消化される。このことに基づいて、ゲノムDNAを、a)CpG配列の位置およびb)これらの配列が非メチル化(すなわち、Hpall制限に対して感受性が高い)配列であるか、またはメチル化(すなわち、Hpall制限に対して抵抗性である)配列であるかを決定するために、別々の反応においてMsplおよびHpallを用いた制限エンドヌクレアーゼ消化に供することができる。バイサルファイト配列決定では、DNAをバイサルファイトで処理することにより、シトシン残基がウラシルに変換されるが、5−メチルシトシン残基は影響されないままである;したがって、バイサルファイト処理により、DNA配列に、個々のシトシン残基のメチル化状態に応じた特異的な変化が導入され、これにより、DNAのセグメントのメチル化状態に関する単一のヌクレオチド分解情報がもたらされる。そのメチル化の状態についてアッセイすることができるゲノムDNAの領域の例としては、OCT4、NANOG、RB1、CDKN2AINK4A、CDKN2AARF、CDH1、CDH13、TIMP3、VHL、MLH1、MGMT、BRCA1、GSTP1、SMARCA3、RASSF1A、SOCS1、ESR1、DAPK1のプロモーター領域が挙げられる。アッセイすることができるゲノムDNAの他の領域は、その開示が参照により本明細書に組み込まれる、Costello、J.F.ら(2000)Nature Genet.25:132〜138、Song、F.ら、(2005)PNAS 102:3336〜3341、およびRobertson、K.D.(2005)Nature Review Genetics 6:597〜610に記載されている。
【0055】
細胞におけるゲノムDNAメチル化の量を減少させるために十分である薬剤の有効量は、細胞におけるメチル化感受性遺伝子の発現の変化をアッセイすることによって決定することもできる。メチル化感受性遺伝子は、発現レベルがそれらのプロモーターのメチル化の状態に対して感受性である遺伝子である。
【0056】
いくつかの遺伝子のプロモーターにおけるCpG配列のメチル化の増加は、メチル化感受性遺伝子プロモーターの転写活性の低下およびメチル化感受性遺伝子の発現の低下を伴う可能性があり、一方、これらの遺伝子のプロモーターにおけるCpG配列の脱メチル化は、メチル化感受性遺伝子プロモーターの転写活性の増加およびこれらの遺伝子の発現の増加を伴う可能性がある。有効量の、メチル化感受性遺伝子プロモーターの脱メチル化を促進する薬剤により、その遺伝子の発現の少なくとも約2倍の増加が誘導される。細胞とCD活性を促進する薬剤を接触させた後の遺伝子発現レベルの変化は、細胞を薬剤と接触させる前、およびその後に、遺伝子のRNAレベルおよび/またはタンパク質レベルを、例えば、RT−PCR、ノーザンブロットハイブリダイゼーション、ウエスタンブロットハイブリダイゼーションまたはELISAによって測定することによってアッセイすることができる。メチル化感受性遺伝子は当技術分野で周知であり、それらとしては、例えば、前の段落において列挙されているものなどの遺伝子が挙げられる。
【0057】
細胞
本発明の方法において使用するために適した細胞は、ヒト、霊長類、家畜動物、農場動物、および動物園の動物、実験動物または愛玩動物、例えば、イヌ、ネコ、ウシ、ウマ、ヒツジ、ブタ、ヤギ、ウサギ、ラット、マウスなどを含めた任意の哺乳動物細胞であってよい。細胞におけるゲノムDNA脱メチル化活性の量の増加が示された本発明の複数の態様では、細胞は、脱メチル化を許容する細胞であることが好ましい。脱メチル化を許容する細胞は、それらのCpG配列がメチル化CpG配列から非メチル化CpG配列に変換され得る細胞である。細胞が脱メチル化を許容するかどうかは、細胞において活性化誘導性シチジンデアミナーゼ(AID)をコードするcDNAを過剰発現させ、細胞にCpGリッチDNAを保有するベクターを提供し、外因的に供給されたCpGリッチDNAを回収し、例えば、バイサルファイト配列決定またはメチラーゼ特異的制限エンドヌクレアーゼ消化によって、メチル化の程度について分析することによって決定することができる。
【0058】
方法において使用するための対象とする細胞が脱メチル化を許容しない(not permissive to demetylation)、すなわち、脱メチル化を許容しない(demetylation−impermissive)細胞であると決定された場合では、脱メチル化を許容しない細胞を、有効量の、メチル化シトシンのヒドロキシル化メチルシトシンへの変換を促進する1種または複数種の薬剤、G:Tミスマッチ特異的な修復活性を促進する1種または複数種の薬剤、および/または成長停止およびDNA損傷誘導性の45(GADD45)活性を促進する1種または複数種の薬剤と接触させることによって、脱メチル化を許容する細胞になるように誘導することができる。
【0059】
メチル化シトシンのヒドロキシル化メチルシトシンへの変換を促進する薬剤は、脱アミノ化のためにメチル化核酸を刺激する。メチル化シトシンのヒドロキシル化メチルシトシンへの変換を促進する薬剤の例は、tetタンパク質のポリペプチドおよび断片、すなわち、tet1(Genbank受託番号:NM030625.2;配列番号29および配列番号30)、およびtet2(Genbank受託番号:NM_001127208.1配列番号31および配列番号32(アイソフォームa);およびGenbank受託番号:NM_017628.3、配列番号33および配列番号34(アイソフォームb))、ならびにこれらのポリペプチドをコードする核酸である。
【0060】
G:Tミスマッチ特異的な修復活性を促進する薬剤は、G/Tミスマッチからのチミン部分の除去およびこれらのチミン部分のシトシン部分との交換を促進する薬剤である。G:Tミスマッチ特異的な修復活性を促進する薬剤の例は、メチル結合ドメインタンパク質(メチル−CpG結合ドメインポリペプチドとしても公知である)およびタンパク質チミンDNAグリコシラーゼ(TDG)のポリペプチドおよび断片、ならびにこれらのポリペプチドをコードする核酸である。
【0061】
メチル結合ドメインタンパク質は、メチル−CpG結合ドメインのそれぞれにおける存在によって関連づけられる核タンパク質である。タンパク質のこのクラスには:MECP2、MBD1、MBD2、MBD3、およびMBD4の5つのメンバーがある。特に興味深いのは、メチルCpG部位でのDNA修復において機能することができる、細菌のDNA修復酵素とのタンパク質配列の類似性を有するメンバー、例えば、MBD4である。細胞を、脱メチル化に許容的になるように誘導することにおいて使用されるMBD4ポリペプチドおよびそれらをコードする核酸は、GenBank受託番号NM_003925.1(配列番号35および配列番号36)においてその配列を見ることができるMED1としても公知のヒトMBD4のアミノ酸配列と少なくとも70%同一であるアミノ酸配列を含むポリペプチドである。
【0062】
チミンDNAグリコシラーゼ(TDG)タンパク質は、5−メチルシトシンおよびシトシンの自発的な脱アミノ化によって引き起こされる遺伝子の突然変異に対する細胞防御において、G/Tミスマッチからチミン部分を除去し、グアニンとの誤対合からウラシル部分および5−ブロモウラシル部分を除去することによって中心的な役割を果たす酵素である。細胞を、脱メチル化に許容的になるように誘導することにおいて使用されるTDGポリペプチドおよびそれらをコードする核酸は、GenBank受託番号NM_003211.4(配列番号37および配列番号38)においてその配列を見ることができるヒトTDGのアミノ酸配列と少なくとも70%同一であるアミノ酸配列を含むポリペプチドである。
【0063】
成長停止およびDNA損傷誘導性の45(GADD45)タンパク質は、ストレス性の成長停止の状態およびDNA損傷剤処理の後にレベルが増加するタンパク質である。細胞を、脱メチル化に許容的になるように誘導することにおいて使用されるGADD45ポリペプチドおよびそれらをコードする核酸は、ヒトGADD45α(GenBank受託番号NM_001924.2(配列番号39および配列番号40)、GADD45β(GenBank受託番号NM_015675.2(配列番号41および配列番号42)、またはGADD45γ(GenBank受託番号NM_006705.3(配列番号43および配列番号44)のアミノ酸配列と少なくとも70%同一であるアミノ酸配列を含むポリペプチドである。
【0064】
G:Tミスマッチ特異的な修復活性を促進する薬剤(複数可)およびGADD45活性を促進する薬剤(複数可)は、上記のCD活性を促進する薬剤を提供するための方法によって、ポリペプチドまたはそれらのポリペプチドをコードする核酸として提供することができる。細胞は、シチジンデアミナーゼ活性を促進する1種または複数種の薬剤と接触させることと並行して、上記の方法によって脱メチル化に許容的になるように誘導することができる。あるいは、まず、細胞を脱メチル化に許容的にし、次いでCD活性を促進する1種または複数種の薬剤と接触させることができる。
【0065】
in vitroにおける方法および使用
本発明の一部の方法では、細胞を、CD活性を促進する1種または複数種の薬剤とin vitroで接触させる。これらの実施形態において対象とする、脱メチル化を許容する哺乳動物細胞、および脱メチル化を許容するように誘導することができる哺乳動物細胞としては、多能性幹細胞、例えば、ES細胞、iPS細胞、胚性生殖細胞;体細胞、例えば、線維芽細胞、造血細胞、ニューロン、筋肉細胞、骨細胞、血管内皮細胞、腸細胞など、およびそれらの系列が限られた前駆体および前駆物質;ならびに、当技術分野において周知であり、下記の実施例に記載されている、2種類以上の細胞の融合物である異核共存体が挙げられる。細胞は、樹立細胞系由来であってよい、または、初代細胞であってよく、「初代細胞」、「初代細胞系」、および「初代培養物」は、本明細書では、対象に由来し、in vitroにおいて培養の限られた数の継代、すなわち、分裂にわたって成長させることができる細胞および細胞培養物を指すために互換的に使用される。例えば、初代培養物は、0回、1回、2回、4回、5回、10回、または15回、しかし危機段階を経るには満たない回数継代することができる培養物である。一般には、本発明の初代細胞系は、in vitroにおいて10回の継代未満維持される。
【0066】
対象細胞は、新生児、若年者または成人由来であってよい新鮮細胞または凍結細胞から、および皮膚、筋肉、骨髄、末梢血、臍帯血、脾臓、肝臓、膵臓、肺、腸、胃、および他の分化した組織を含めた組織から単離することができる。組織は、生きているドナーからの生検またはアフェレーシスによって得ることができる、または死亡したもしくは瀕死のドナーから死亡の約48時間以内に得ることができる、または新鮮に凍結させた組織、死亡の約12時間以内に凍結させ、約−20℃未満、通常およそ液体窒素の温度(−190℃)で無制限に維持した組織から得ることができる。組織から細胞を単離するために、分散または懸濁のために適切な溶液を使用することができる。そのような溶液は、一般に、低濃度、一般に5〜25mMの許容できる緩衝液と併せて、ウシ胎仔血清または他の天然に存在する因子を補充することが都合よい、平衡塩類溶液、例えば、通常の生理食塩水、PBS、ハンクス平衡塩類溶液などである。都合のよい緩衝液としては、HEPES、リン酸緩衝液、乳酸緩衝液などが挙げられる。
【0067】
in vitroでシチジンデアミナーゼ活性を促進する1種または複数種の薬剤と接触させる細胞は、薬剤(複数可)の存在下で約30分間〜約24時間、例えば、1時間、1.5時間、2時間、2.5時間、3時間、3.5時間、4時間、5時間、6時間、7時間、8時間、12時間、16時間、18時間、20時間、または約30分間から約24時間までの任意の他の時間インキュベートすることができ、これを約毎日〜約4日ごと、例えば、1.5日ごと、2日ごと、3日ごと、または約毎日から約4日ごとまでの任意の他の頻度で繰り返すことができる。薬剤(複数可)は、1回または複数回、例えば、1回、2回、3回、または4回以上対象細胞に提供することができ、各接触事象の後、細胞を薬剤(複数可)と一緒にいくらかの時間、例えば、16〜24時間インキュベートしておき、その後、培地を新鮮な培地と交換し、細胞をさらに培養する。
【0068】
本発明の一部の方法では、CD活性を促進する薬剤と接触させる、脱メチル化を許容する細胞は、脱メチル化を許容する体細胞である。これらの方法のいくつかでは、脱メチル化を許容する体細胞を、異なる細胞系譜の体細胞になるように再プログラムする。言い換えれば、本発明の方法を使用して、ある系列の体細胞から別の系列の体細胞への変換を促進することができる。異なる系列の体細胞は、当技術分野で周知のマーカーおよび形態によって容易に同定可能である。
【0069】
脱メチル化を許容する体細胞を、CD活性を促進する薬剤と接触させる一部の方法では、脱メチル化を許容する体細胞を、人工多能性幹(iPS)細胞になるように再プログラムする。言い換えれば、作製される細胞は、iPS細胞である。上記の通り、iPS細胞は、ES細胞様の形態(例えば、大きな核細胞質比、明確な境界および目立った核を有する平らなコロニーとして成長する)が、体細胞に由来する多能性幹細胞である。
【0070】
本発明の一部の方法では、CD活性を促進する薬剤と接触させる脱メチル化を許容する細胞は、多能性幹細胞、例えば、胚性幹(ES)細胞、胚性生殖(EG)細胞、または人工多能性幹(iPS)細胞である。これらの方法では、脱メチル化を許容する多能性幹細胞を、体細胞になるように再プログラムする。言い換えれば、本発明の方法を使用して、多能性幹細胞の体細胞へのプログラミングを促進することができる。体細胞の例としては、外胚葉系(例えば、神経および線維芽細胞)、中胚葉系(例えば、心筋細胞)、または内胚葉(例えば、膵臓細胞)系由来の任意の分化細胞が挙げられる。体細胞は、1種または複数種の:膵ベータ細胞、神経幹細胞、神経(例えば、ドーパミン作動性神経)、乏突起膠細胞、乏突起膠細胞前駆細胞、肝細胞、肝幹細胞、星状膠細胞、ミオサイト、造血細胞、心筋細胞などであってよい。上に示されている通り、多能性幹細胞に由来する体細胞は、最終分化細胞であってよい、またはそれらは、特定の系列の細胞を生じることができてよい。例えば、多能性細胞は、種々の多分化能細胞型、例えば、神経幹細胞、心臓幹細胞、または肝幹細胞に分化させることができる。次いで、幹細胞は、新しい細胞型にさらに分化させることができ、例えば、神経幹細胞は、神経に分化させることができ;心臓幹細胞は、心筋細胞に分化させることができ;肝幹細胞は、肝細胞に分化させることができる。そのような方法によって作製される体細胞は、上記の通り、当技術分野で周知の特定の細胞系列のマーカーおよび形態によって、そのようなものとして容易に同定可能である。
【0071】
脱メチル化を許容する細胞の他の細胞型への再プログラミングを促進するために、脱メチル化を許容する細胞を、細胞の再プログラミングを促進する1種または複数種の薬剤と接触させる追加的なステップを実施することができる。このステップは、脱メチル化を許容する細胞を、CD活性を促進する薬剤と接触させる前、脱メチル化を許容する細胞を、CD活性を促進する薬剤と接触させると同時に、または脱メチル化を許容する細胞を、CD活性を促進する薬剤と接触させた後で実行することができる。細胞の再プログラミングを促進する薬剤は、ポリペプチド、核酸薬剤、または小分子薬剤であってよい。このステップにおいて提供することができる薬剤の例としては、これらに限定されないが、GSK−3阻害剤、例えば、CHIR99021など(Li、W.ら(2009)Stem Cells、Epub Oct.16 2009,);HDAC阻害剤、例えば、バルプロ酸など(その開示が参照により本明細書に組み込まれるHuangfu、D.(2008)Nature Biotechnol 26(7)795〜797;およびUS20090191159に記載);ヒストンメチルトランスフェラーゼ阻害剤、例えば、G9aヒストンメチルトランスフェラーゼ阻害剤、例えば、BIX−01294、など(Shi、Yら(2008)Cell Stem Cell 3(5):568〜574);ジヒドロピリジン受容体のアゴニスト、例えば、BayK8644、など(Shi、Yら(2008)Cell Stem Cell 3(5):568〜574);およびTGFβシグナル伝達の阻害剤、例えば、RepSoxなど(lchida、JK.ら(2009)Cell Stem Cell 5(5):491〜503)が挙げられる。このステップにおいて提供することができる薬剤の他の例としては、再プログラミング因子が挙げられる。上記の通り、再プログラミング因子は、細胞に作用して転写を変化させ、それによって細胞を新しい細胞運命に再プログラミングする生物活性因子である。
【0072】
ある細胞系譜の体細胞の、別の細胞系譜の体細胞への再プログラミングを促進する薬剤の多数の例が当技術分野で公知であり、そのいずれも、本発明において使用することができる。これらとしては、例えば、筋肉に特異的な性質を色素細胞、神経細胞、脂肪細胞、肝細胞および線維芽細胞内に誘導する再プログラミング因子MYOD(筋原性因子1;GenBank受託番号NM_002478.4およびNP_002469.2)、例えば、Weintraub、H.W.らProc.Natl.Acad.Sci USA 86:5434〜5438;Davis、R.L.ら(1987)Cell 51 :987〜1000;Schafer、B.W.ら(1990)Nature 344:454〜8)を参照されたい;NEUROG3(neurogenin3、NGN3;GenBank受託番号NM_020999.2およびNP_066279.2)、PDX1(膵臓および十二指腸ホメオボックス1;GenBank受託番号.NM_000209.3およびNP_000200.1)および組み合わせて、in vivoで膵臓の外分泌細胞を機能性β細胞に効率的に変換することができるMafA(v−maf筋腱膜線維肉腫癌遺伝子ホモログA;GenBank受託番号NM_201589.2およびNP_963883.2)、例えば、Zhou、Q.ら(2008)Nature 455:627〜32)を参照されたい;およびB細胞において単独で、または線維芽細胞においてPu.1(脾フォーカス形成ウイルス(SFFV)プロウイルス組み込み癌遺伝子、SPI1;GenBank受託番号NM_001080547.1、NP_001074016.1、NM_003120.2およびNP_003111.2)と組み合わせてマクロファージ特性を誘導する、C/EBPa(CCAAT/エンハンサー結合性タンパク質、アルファ;GenBank受託番号NM004364.3およびNP_004355.2)、例えば、Bussmann、L.H.ら(2009)Cell Stem Cell 5:554〜66;Feng、R.ら(2008)Proc Natl Acad Sci USA 105:6057〜62;Xie、H.ら(2004)Cell 117:663〜76)を参照されたい、が挙げられる。他の薬剤としては、IL2受容体(IL受容体2AおよびIL受容体2B;GenBank受託番号NM_000417.2、NP_000408.1、NM_000878.2およびNP_000869.1)およびGM−CSF受容体(コロニー刺激因子2受容体、アルファ(CSF2RA)およびコロニー刺激因子2受容体、ベータ(CSF2RB);GenBank受託番号NM_001161529.1、NP_001155001.1、NM_000395.2、およびNP_000386.1)、委任リンパ球前駆細胞において骨髄の変換を誘導する、例えば、Kondo、M.ら(2000)Nature 407:383〜6)を参照されたい、が挙げられる。上に列挙されているGenBank受託番号に記載の薬剤、ならびにこれらのポリペプチドをコードする核酸のアミノ酸配列と少なくとも70%、75%、80%、85%、90%、91%、92%、95%、97%、99%、または100%同一であるアミノ酸配列を含むポリペプチドは、本発明の方法において、ある脱メチル化を許容する細胞系譜の体細胞の、別の細胞系譜の体細胞への再プログラミングを促進する薬剤として使用される。
【0073】
体細胞のiPS細胞への再プログラミングを促進する薬剤の多数の例が当技術分野で公知であり、そのいずれも、本発明において使用することができ、例えば、その開示が参照により本明細書に組み込まれる、米国特許出願第20090047263号、US20090068742、US20090191159、US20090227032、US20090246875、およびUS20090304646を参照されたい。これらとしては、例えば、再プログラミング因子Oct3/4、(POUクラス5ホメオボックス1(POU5F1);GenBank受託番号NP_002692およびNM_002701);Sox2(性決定領域Yボックス2タンパク質;GenBank受託番号NP_003097およびNM_003106):Klf4(クルッペル様因子4;GenBank受託番号NP_004226およびNM_004235);c−Myc(骨髄細胞腫症ウイルス癌遺伝子ホモログ;GenBank受託番号NP_002458およびNM_002467);Nanog(Nanogホメオボックス;GenBank受託番号NP_079141およびNM_024865);およびLin−28(C.elegansのLin−28ホモログ;GenBank受託番号NP_078950およびNM_024674)が挙げられる。上に列挙されているGenBank受託番号に記載の薬剤のアミノ酸配列と少なくとも70%、75%、80%、85%、90%、91%、92%、95%、97%、99%、または100%同一であるアミノ酸配列を含むポリペプチド、ならびにこれらのポリペプチドをコードする核酸は、本発明の方法において、脱メチル化を許容する体細胞の、iPS細胞への再プログラミングを促進する薬剤として使用される。
【0074】
多能性幹細胞の、体細胞への再プログラミングを促進する薬剤の多数の例は当技術分野で公知であり、そのいずれも、本発明において使用することができる。例えば、神経幹細胞は、多能性細胞を、NOG(noggin;GenBank受託番号NM_005450.4およびNP_005441.1)または他の骨形成タンパク質アンタゴニスト(Itsyksonら、(2005)、Mol、Cell Neurosci.、30(1):24〜36)の存在下で浮遊凝集体として培養することによって、または多能性細胞を浮遊液中で、増殖因子、例えば、FGF−2(線維芽細胞増殖因子2、塩基性線維芽細胞増殖因子(bFGF)としても公知である;GenBank受託番号NM_002006.4およびNP_001997.5)、例えば、Zhangら、(2001)、Nat.Biotech.(19):1129〜1133を参照されたい、の存在下で培養して凝集体を形成することによって生成することができる。ある場合では、FGF−2を含有する無血清培地中で凝集体を培養する。別の例では、FGF−2を含む無血清培地の存在下で多能性細胞をマウス間質細胞系、例えばPA6と共培養する。さらに別の例では、FGF−2を含有する無血清培地に多能性細胞を直接移して分化を直接誘導する。
【0075】
多能性細胞に由来する神経幹は、神経、乏突起膠細胞、または星状膠細胞に分化させることができる。多くの場合、神経幹細胞を生成するために用いる条件は、神経、乏突起膠細胞、または星状膠細胞を生成するためにも使用することができる。例えば、ドーパミン作動性ニューロンへの分化を促進するために、多能性細胞またはそれに由来する神経幹細胞を、無血清条件下でPA6マウス間質細胞系と共培養することができる。例えば、Kawasakiら、(2000)Neuron、28(1):3140を参照されたい。他の方法も記載されており、例えば、Pompら、(2005)、Stem Cells 23(7):923〜30;米国特許第6,395,546号、例えば、Leeら、(2000)、Nature Biotechnol.、18:675〜679を参照されたい。多能性細胞またはそれに由来する神経幹細胞の乏突起膠細胞への分化は、例えば、多能性細胞または神経幹細胞を間質細胞と共培養することによって(例えば、Hermannら(2004)、J Cell Sci.117(Pt19):4411〜22を参照されたい)、または多能性細胞または神経幹細胞を、インターロイキン(IL)−6受容体(GenBank受託番号NM_000565.2およびNP_000556.1)、またはその誘導体がIL−6サイトカイン(GenBank受託番号NM_000600.3およびNP_000591.1)またはその誘導体に連結している融合タンパク質の存在下で培養することによって促進することができる。乏突起膠細胞は、当技術分野で公知の他の方法によって多能性細胞から生成することもできる。例えば、Kangら、(2007)Stem Cells 25、419〜424を参照されたい。星状膠細胞は、多能性細胞またはそれに由来する神経幹細胞から、例えば、bFGFおよびEGF(上皮増殖因子;GenBank受託番号NM_001963.3およびNP_001954.2)を伴う神経形成培地の存在下で多能性細胞または神経幹細胞を培養することによって作製することもできる。例えば、Brustleら、(1999)、Science、285:754〜756を参照されたい。
【0076】
多能性細胞は、当技術分野で公知の方法によって膵ベータ細胞に分化させることができる。例えば、Lumelskyら、(2001)Science、292:1389〜1394;Assadyら、(2001)、Diabetes、50:1691〜1697;D’Amourら、(2006)、Nat.Biotechnol.、24:1392〜1401.;D’Amourら、(2005)、Nat.Biotechnol.23:1534〜1541。方法は、多能性細胞を、アクチビンA(インヒビン、ベータA(INHBA);GenBank受託番号NM_002192.2およびNP_002183.1)を補充した無血清培地中で培養し、その後、オールトランスレチノイン酸を補充した無血清培地の存在下で培養し、その後、bFGFおよびニコチンアミドを補充した無血清培地の存在下で培養することを含んでよい。例えば、Jiangら、(2007)、Cell Res.、4:333〜444。他の例では、方法は、多能性細胞を、無血清培地、アクチビンA、およびWntタンパク質(例えば、GenBank受託番号NM_005430、NM_003391、NM_004185、NM_030753、NM_033131、NM_030761、NM_003392、NM_032642、NM_006522、NM_004625、NM_058238、NM_058244、NM_003393、NM_003395、NM_003396、NM_025216、NM_003394、Wnt−11NM_004626、およびNM_016087)の存在下で、約0.5日から約6日間まで、例えば、約0.5日、1日、2日間、3日間、4日間、5日間、6日間培養し;その後、約0.1%から約2%まで、例えば、0.2%のFBSおよびアクチビンAの存在下で約1日から約4日間まで、例えば、約1日、2日間、3日間、または4日間培養し;その後、2%のFBS、FGF10(線維芽細胞増殖因子10、GenBank受託番号NM_004465.1およびNP_004456.1)、KAAD−シクロパミン(ケト−N−アミノエチルアミノカプロイルジヒドロシンナモイルシクロパミン)およびレチノイン酸の存在下で約1日から約5日間まで、例えば、1日、2日間、3日間、4日間、または5日間培養し、;その後、1%のB27、ガンマセレクターゼ阻害剤およびエクステンジン−4(extendin−4)と一緒に約1日から約4日間、例えば、1日、2日間、3日間、または4日間培養し;最後に、1%のB27、エクステンジン−4、IGF−1、およびHGFの存在下で約1日から約4日間、例えば、1日、2日間、3日間、または4日間培養することを含む。
【0077】
多能性細胞から肝細胞または肝幹細胞を分化させることができる。例えば、多能性細胞を酪酸ナトリウムの存在下で培養することにより、肝細胞を生成することができる。例えば、Rambhatlaら、(2003)、Cell Transplant 12:1〜11を参照されたい。別の例では、肝細胞は、多能性細胞を、血清を含まない培地中、アクチビンAの存在下で培養し、その後、細胞をFGF4(線維芽細胞増殖因子−4;GenBank受託番号NM_002007.2およびNP_001998.1)およびBMP2(骨形成タンパク質−2;GenBank受託番号NM_001200.2およびNP_001191.1)中で培養することによって作製することができる。例えば、Caiら、(2007)Hepatology 45(5):1229〜39。例示的な実施形態では、多能性細胞を、アクチビンAの存在下で約2日間から約6日間まで、例えば、約2日間、約3日間、約4日間、約5日間、または約6日間培養し、次いで、多能性細胞を、HGF(肝細胞増殖因子;GenBank受託番号NM_010427.4およびNP_034557.3)の存在下で約5日間から約10日間まで、例えば、約5日間、約6日間、約7日間、約8日間、約9日間、または約10日間培養することによって肝細胞または肝幹細胞に分化させる。
【0078】
多能性細胞は、心筋細胞に分化させることもできる。骨形成タンパク質(BMP)シグナル伝達を阻害することにより、心筋細胞(または心筋細胞)の生成をもたらすことができる。例えば、Yuasaら、(2005)、Nat.Biotechnol.、23(5):607〜11を参照されたい。したがって、例示的な実施形態では、多能性細胞をNOG(noggin)の存在下で約2日間から約6日間まで、例えば、約2日間、約3日間、約4日間、約5日間、または約6日間培養し、その後、胚葉体を形成させ、胚葉体を約1週間から約4週間、例えば、約1週間、約2週間、約3週間、または約4週間培養する。他の例では、心筋細胞は、多能性細胞を、LIF(白血病抑制因子;GenBank受託番号NM_002309.3およびNP_002300.1)の存在下で培養することによって、または多能性細胞を当技術分野で公知の他の方法に供してES細胞から心筋細胞を生成することによって生成することができる。例えば、Baderら、(2000)、Circ.Res.、86:787〜794、Kehatら、(2001)、J.Clin.Invest.、108:407〜414;Mummeryら、(2003)、Circulation、107:2733〜2740。
【0079】
多能性細胞から他の細胞種を生成する方法の例としては:(1)脂肪細胞を生成するために、多能性細胞を、レチノイン酸、LIF、甲状腺ホルモンおよびインスリンの存在下で培養すること(例えば、Daniら、(1997)、J.Cell Sci.、110:1279〜1285);(2)軟骨細胞を生成するために、多能性細胞を、BMP2またはBMP4(GenBank受託番号NM_001202.3、NP_001193.2、NM_130850.2、NP_570911.2、NM_130851.2、およびNP_570912.2)の存在下で培養すること(例えば、Kramerら、(2000)、Mech.Dev.、92:193〜205);(3)多能性細胞を、平滑筋を生成するための条件下で培養すること(例えば、Yamashitaら、(2000)、Nature、408:92〜96);(4)ケラチノサイトを生成するために、多能性細胞を、ベータ−1インテグリン(GenBank受託番号NM_002211.3およびNP_002202.2)の存在下で培養すること(例えば、Baguttiら、(1996)、Dev.Biol.、179:184〜196);(5)マクロファージを生成するために、多能性細胞を、IL3(インターロイキン−3;GenBank受託番号NM_000588.3およびNP_000579.2)およびCSF1(コロニー刺激因子、マクロファージ;GenBank受託番号NM_000757.4、NP_000748.3)の存在下で培養すること(例えば、Lieschkeおよび Dunn(1995)、Exp.Hemat.、23:328〜334);(6)肥満細胞を生成するために、多能性細胞を、IL−3およびSCF(造血幹細胞因子、kitリガンドとしても公知の幹細胞因子;GenBank受託番号NM_000899.3およびNP_000890.1)の存在下で培養すること(例えば、Tsaiら、(2000)、Proc.Natl.Acad.Sci.USA、97:9186〜9190);(7)メラニン形成細胞を生成するために、多能性細胞を、デキサメタゾンおよびSCFの存在下で培養すること(例えば、Yamaneら、(1999)、Dev.Dyn.、216:450〜458);(8)骨芽細胞を生成するために、多能性細胞を、デキサメタゾン、レチノイン酸、アスコルビン酸、ベータ−グリセロリン酸の存在下で胎児のマウス骨芽細胞と共培養すること(例えば、Butteryら、(2001)、Tissue Eng.、7:89〜99);(9)骨芽細胞を生成するために、多能性細胞を、骨形成性の因子の存在下で培養すること(例えば、Sottileら、(2003)、Cloning Stem Cells、5:149〜155);(10)骨格筋細胞を生成するために、多能性細胞においてインスリン様増殖因子−2を過剰発現させ、その細胞をジメチルスルホキシドの存在下で培養すること(例えば、Prelleら、(2000)、Biochem.Biophys.Res.Commun.、277:631〜638を参照されたい);(11)多能性細胞を、白色血液細胞を生成するための条件に供すること;または、(12)造血前駆細胞を生成するために、多能性細胞を、BMP4ならびに:SCF、FLT3(fms関連チロシンキナーゼ3;GenBank受託番号NM_004119.2およびNP_004110.2)、IL−3、IL−6(インターロイキン6;GenBank受託番号M_000600.3およびNP_000591.1)、およびCSF3(コロニー刺激因子、顆粒球;GenBank受託番号NM_000759.2およびNP_000750.1)の1つまたは複数の存在下で培養すること(例えば、Chadwickら、(2003)、Blood、102:906〜915を参照されたい)が挙げられる。
【0080】
上に列挙されているGenBank受託番号に記載の薬剤のアミノ酸配列と少なくとも70%、75%、80%、85%、90%、91%、92%、95%、97%、99%、または100%同一であるアミノ酸配列を含むポリペプチド、ならびにこれらのポリペプチドをコードする核酸は、本発明の方法において、多能性細胞の体細胞への再プログラミングを促進する薬剤として使用される。
【0081】
細胞の再プログラミングを促進する薬剤は、これらに限定されないが、CD活性を促進する薬剤について上記されている方法を含めた当技術分野で周知の方法によって、脱メチル化を許容する細胞に提供することができる。薬剤は、個別に、または単一の組成物として、すなわち、薬剤の予備混合した組成物として提供することができる。薬剤は、同時に、または違う時間に逐次的に対象細胞に加えることができる。一部の実施形態では、少なくとも2種の薬剤のセット、例えば、Oct3/4ポリペプチドおよびSox2ポリペプチドを提供する。一部の実施形態では、3種の薬剤のセット、例えば、Oct3/4ポリペプチド、Sox2ポリペプチド、およびKlf4ポリペプチドを提供する。一部の実施形態では、4種の薬剤のセット、例えば、Oct3/4ポリペプチド、Sox2ポリペプチド、Klf4ポリペプチド、およびc−Mycポリペプチドを提供する。CD活性を促進する薬剤(複数可)と同様に、薬剤(複数可)は、対象細胞に1回または複数回提供することができ、各接触事象の後、細胞を薬剤と一緒にいくらかの時間、例えば、16〜24時間インキュベートしておき、その後、培地を新鮮な培地と交換し、細胞をさらに培養する。
【0082】
脱メチル化を許容する細胞を、CD活性を促進する薬剤(複数可)と接触させた後、所望の細胞の成長を促進するために、接触させた細胞を培養する。細胞を培養して上記のiPS細胞または体細胞の特定の型の成長を促進するための方法、上記の体細胞の特定の型のiPS細胞クローンまたはクローンを単離するための方法、およびそれらの細胞クローンの細胞を培養して上記のiPS細胞または体細胞の特定の型の成長を促進する方法は当技術分野で周知であり、そのいずれも、本発明において、再プログラムされた脱メチル化を許容する細胞から所望の細胞を成長させ、単離し、再培養するために使用することができる。
【0083】
細胞を、CD活性を促進する薬剤(複数可)と接触させることによって脱メチル化を許容する細胞培養物の細胞におけるゲノムDNAメチル化の量を減少させることにより、脱メチル化を許容する細胞の、所望の細胞型への再プログラミングの効率が、CD活性を促進する薬剤が存在しない場合に観察される効率と比較して増加する。言い換えれば、体細胞および細胞培養物は、再プログラミングを促進することが当技術分野で公知の因子の存在下でCD活性を促進する1種または複数種の薬剤と接触させた場合に、CD活性を促進する1種または複数種の薬剤と接触させなかった細胞と比較して所望の細胞型を生じさせるための能力が増強されることが実証されている。増強されるとは、体細胞培養物が、CD活性を促進する薬剤と接触させなかった細胞の集団の所望の細胞型を生じさせる能力の少なくとも約50%、約100%、約200%、約300%、約400%、約600%、約1000%、少なくとも約2000%の、所望の細胞型を生じさせる能力を有することを意味する。言い換えれば、脱メチル化を許容する細胞の培養物により、CD活性を促進する1種または複数種の薬剤と接触させていない脱メチル化を許容する細胞の集団によって産生される所望の細胞型の細胞数の約1.5倍、約2倍、約3倍、約4倍、約6倍、約10倍、約20倍、約30倍、約50倍、約100倍、約200倍が産生される。再プログラミングの効率は、所望の細胞型を獲得すると脱メチル化された状態になることが当技術分野で公知のプロモーターにおけるメチル化の量をアッセイすることによって決定することができる。そのような場合では、それらのプロモーターにおけるメチル化の量が、CD活性を促進する薬剤が存在しない場合に観察されたメチル化の量の約1.5分の1、約2分の1、約3分の1、約4分の1、約6分の1、約10分の1未満である場合に、CD活性を促進する薬剤が存在することに起因する再プログラミングの効率の増強が観察される。その代わりにまたはそれに加えて、再プログラミングの効率は、所望の細胞系を獲得するとより高度に発現されることが当技術分野で公知の遺伝子の発現レベルをアッセイすることによって決定することができる。そのような場合では、これらの遺伝子の発現レベルが、CD活性を促進する薬剤が存在しない場合に観察される発現レベルよりも約1.5倍、約2倍、約3倍、約4倍、約6倍、約10倍大きい場合に、CD活性を促進する薬剤が存在することに起因する再プログラミングの効率の増強が観察される。
【0084】
上記のin vitroにおける方法によって再プログラムされた脱メチル化を許容する細胞から得られる細胞は、疾患(例えば、遺伝的欠陥)を治療するための療法として使用することができる。特に、上記の方法によって脱メチル化を許容する体細胞から得られる体細胞および上記の方法によって多能性幹細胞体細胞から得られる体細胞は、広範囲の疾患または障害に罹患している対象に、例えば、レシピレントにおいて分化している細胞または分化した細胞を再構成する、または補うために移入することができる。同様に、脱メチル化を許容する体細胞から得られる人工多能性幹細胞を、広範囲の疾患または障害に罹患している対象に移入することができる、または、それらをin vitroで種々の細胞系譜の体細胞に分化させ、次いで広範囲の疾患または障害に罹患している対象に移入することができる。これらに限定されないが、多能性細胞を分化させる方法を含めた、多能性細胞をより特殊化した細胞型に分化させる方法は多数あり、これらを、上記の通り、幹細胞、特にES細胞を、体細胞になるように再プログラムするために使用することができる。
【0085】
療法は、疾患の原因を治療することを対象としてよい;あるいは、療法は、疾患または状態の影響を治療するものであってよい。例えば、由来細胞は、対象の傷害部位に、またはその近くに移入することができる;または細胞は、細胞が傷害部位に移動する、またはそこに向かうことができるように対象に導入することができる。移入された細胞は、損傷を受けた、または傷害を受けた細胞と有利に置き換え、対象の全体的な状態の改善を可能にすることができる。ある場合には、移入された細胞により、組織再生または修復が刺激され得る。
【0086】
ある場合では、由来細胞または由来細胞の亜集団は、対象に移入する前に精製または単離することができる。ある場合では、所望の細胞型に特異的な1種または複数種のモノクローナル抗体を細胞集団と一緒にインキュベートし、結合した細胞を単離する。他の場合では、細胞の所望の亜集団に細胞型に特異的なプロモーターの制御下にあるレポーター遺伝子を発現させ、次いで、それを用いて由来細胞またはその亜集団を精製または単離する。
【0087】
ある場合では、種々の目的で、例えば、機能喪失突然変異を有する遺伝子を置き換えるため、マーカー遺伝子を提供するためなどに、遺伝子を、対象に移入する前に脱メチル化を許容する細胞またはそれに由来する細胞に導入することができる。あるいは、アンチセンスmRNAまたはリボザイムを発現させるベクターを導入し、それによって望ましくない遺伝子の発現を遮断する。遺伝子療法の他の方法は、薬物耐性遺伝子を導入して、正常な前駆細胞が利点を有することを可能にし、選択圧、例えば、多数の薬物耐性遺伝子(MDR)、またはbcl−2などの抗アポトーシス遺伝子に供することである。種々の当技術分野で公知の技法、例えば、上記の通り、電気穿孔、カルシウム沈降DNA(calcium precipitated DNA)、融合、トランスフェクション、リポフェクション、感染などを用いて、標的細胞に核酸を導入することができる。DNAを導入する特定の様式は、本発明の実施には重大ではない。
【0088】
遺伝子改変された脱メチル化を許容する細胞またはそれに由来する細胞を有することを立証するために、種々の技法を使用することができる。細胞のゲノムは制限されている可能性があり、増幅して、または増幅せずに使用する。ポリメラーゼ連鎖反応;ゲル電気泳動;制限分析;サザンブロット、ノーザンブロット、およびウエスタンブロット;配列決定などは、全て使用することができる。種々の条件下で細胞を成長させて、細胞が、導入されたDNAを発現する能力を維持しながら骨髄系列の全てに成熟化可能なことを確実にすることができる。in vitroおよびin vivoにおける種々の試験を使用して、確実に細胞の多能性の能力(capability)が維持されるようにすることができる。
【0089】
対象への治療の施行数は変動し得る。誘導された細胞および/または分化した細胞の対象への導入は、一回の事象であってよいが、ある特定の状況では、そのような治療によって引き出される改善は限られた期間にわたるものであり、持続的な一連の反復治療が必要になる可能性がある。他の場合では、効果が観察される前に細胞を多数回投与することが必要になり得る。正確なプロトコールは、疾患または状態、疾患の病期および治療されている個々の対象のパラメータに左右される。
【0090】
細胞は、以下の経路のいずれかによって対象に導入することができる:非経口的経路、静脈内経路、動脈内経路、筋肉内経路、皮下経路、経皮的経路、気管内経路、腹腔内経路、または脊髄液内経路。
【0091】
神経疾患または神経障害に罹患している対象には、特に、本発明の方法によって得られる細胞を利用する療法が有益であり得る。いくつかの手法では、神経性の状態、例えば、アルツハイマー病、パーキンソン病、多発性硬化症、脳梗塞、脊髄損傷、または他の中枢神経系障害を治療するために、神経幹細胞または神経系細胞を傷害部位に移植することができる。例えば、Morizaneら、(2008)、Cell Tissue Res.、331(1):323〜326;Couttsおよび Keirstead(2008)、Exp.Neurol.、209(2):368〜377;Goswamiおよび Rao(2007)、Drugs、10(10):713〜719を参照されたい。パーキンソン病を治療するために、ドーパミン作用性ニューロンをパーキンソン病の対象の線条体に移植することができる。多発性硬化症を治療するために、乏突起膠細胞または乏突起膠細胞の前駆体をMSに罹患している対象に移入することができる。本発明の方法によって得られる細胞は、脳および脊髄を修復するために、病変へのそれらの移動を標的にすることができる手掛かりに反応するように遺伝子工学で操作することもできる。例えば、Chenら、(2007)、Stem Cell Rev.、3(4):280〜288。
【0092】
神経性障害以外の疾患も、本発明の方法によって生成される細胞を利用する療法によって治療することができる。虚血性心筋症、伝導疾患、および先天性欠損などの変性心疾患には、心筋細胞またはそれらの前駆物質を移植することが有効であり得る。例えば、Janssensら、(2006)、Lancet、367:113〜121を参照されたい。
【0093】
膵島細胞(またはランゲルハンス島の初代細胞)を糖尿病(例えば、糖尿病、1型)に罹患している対象に移植することができる。例えば、Burnsら、(2006)Curr.Stem Cell Res.Ther.、2:255〜266を参照されたい。一部の実施形態では、本発明の方法によって得られる膵ベータ細胞を糖尿病(例えば、1型糖尿病)に罹患している対象に移植することができる。
【0094】
他の例では、本発明の方法によって得られる肝細胞または肝幹細胞を、肝疾患、例えば、肝炎、硬変症、または肝不全に罹患している対象に移植する。
【0095】
本発明の方法によって得られる造血細胞または造血幹細胞(HSC)を血液の癌、または他の血液障害または免疫障害に罹患している対象に移植することができる。造血細胞またはHSCによって治療される可能性がある血液の癌の例としては:急性リンパ芽球性白血病、急性骨髄芽球性白血病、慢性骨髄性白血病(CML)、ホジキン病、多発性骨髄腫、および非ホジキンリンパ腫が挙げられる。多くの場合、そのような疾患に罹患している対象は、急速に分裂している血液細胞を死滅させるために放射線治療および/または化学療法薬による治療を受けなければならない。本発明の方法によって得られるHSCをこれらの対象に導入することは、細胞の枯渇した貯蔵を再充填させるのに役立ち得る。
【0096】
ある場合では、本発明の方法によって得られる造血細胞またはHSCは、癌と直接戦うためにも使用することができる。例えば、同種異系のHSCの移植は、腎癌の治療において見込みがあることが示されている。例えば、Childsら、(2000)、N.Engl.J.M編、343:750〜758を参照されたい。一部の実施形態では、本発明の方法によって得られる同種異系のHSC、または自己のHSCでさえ、腎癌または他の癌を治療するために対象に導入することができる。
【0097】
本発明の方法によって得られる造血細胞またはHSCは、血液細胞以外の細胞または組織、例えば、筋肉、血管、または骨を生成または修復するためにも対象に導入することができる。そのような治療は、多数の障害に対して有用であり得る。
【0098】
ある場合では、本発明の方法によって得られる細胞を免疫無防備状態の動物、例えば、SCIDマウスに移入し、分化させる。移植された細胞は、分化細胞型と腫瘍細胞の混合物を形成し得る。対象とする特異的な分化細胞型は、系列特異的なマーカーを使用することによって、例えば、蛍光活性化細胞選別(FACS)または他の選別方法、例えば、磁気活性化細胞選別(MACS)によって、腫瘍細胞から離して選択し、精製することができる。次いで、疾患または状態を治療するために分化細胞を対象(例えば、自己の対象、HLAが一致した対象)に移植することができる。疾患または状態は、造血障害、内分泌欠乏、神経変性障害、脱毛、または本明細書に記載の他の疾患または状態であってよい。
【0099】
本発明の方法によって得られる細胞は、任意の生理的に許容される培地に投与することができる。本発明の方法によって得られる細胞は、単独で、または、例えば、それらが移植された組織におけるそれらの成長および/または組織化を支持するために、適切な基質またはマトリックスと一緒に提供することができる。通常、少なくとも1×105個、好ましくは1×106個以上の細胞を投与する。細胞は、注射、カテーテルなどによって導入することができる。細胞は、液体窒素の温度で凍結させ、長期間貯蔵することができ、解凍して使用することができる。凍結させる場合、細胞は通常10%のDMSO、50%のFCS、40%のRPMI1640培地中で貯蔵する。解凍したら、前駆細胞の増殖および分化に関連する増殖因子および/または間質細胞を使用することによって細胞を増やす。
【0100】
in vivoにおける方法および使用
一部の実施形態では、例えば、ゲノムDNA脱メチル化療法を必要とする対象において、脱メチル化を許容する細胞を、CD活性を促進する1種または複数種の薬剤とin vivoで接触させる。
【0101】
ポリペプチド、小分子および核酸を対象に投与するためのいくつもの当技術分野において周知の方法のいずれかによって、細胞を、CD活性を促進する薬剤(複数可)とin vivoで接触させることができる。薬剤は、種々の製剤に組み込むことができる。より詳細には、薬剤は、適切な薬学的に許容される担体または希釈剤と組み合わせることによって、医薬組成物に処方することができ、また、錠剤、カプセル剤、散剤、顆粒剤、軟膏剤、液剤、坐剤、注射剤、吸入剤、ゲル剤、ミクロスフェア、およびエアロゾル剤などの固体の形態、半固体の形態、液体の形態または気体の形態の調製物に処方することができる。そのようなものとして、シチジンデアミナーゼ活性を促進する薬剤(複数可)の投与は、経口投与、頬側投与、直腸投与、非経口投与、腹腔内投与、皮内投与、経皮投与、気管内(intracheal)投与などを含めた種々のやり方で実現することができる。活性薬剤は、投与後に全身にわたってよい、または局部投与、壁内投与を使用すること、もしくは埋め込み部位で活性用量を保持するために作用する移植片を使用することによって局所的にすることができる。活性薬剤は、即時活性用に処方することができる、または、持続的放出用に処方することができる。
【0102】
いくつかの状態、特に、中枢神経系の状態のために、薬剤を、血液脳関門(BBB)を渡るように処方することが必要であり得る。血液脳関門(BBB)を通って薬物送達するための1つの戦略は、マンニトールもしくはロイコトリエンなどの浸透性の手段によって、またはブラジキニンなどの血管作動性物質を使用することによって生化学的に、BBBを破壊することを伴う。特異的な薬剤を脳腫瘍にターゲティングするためにBBB開口を使用する可能性も1つの選択肢である。組成物を血管内注射によって投与する場合、BBB破壊剤は本発明の治療用組成物と同時投与することができる。BBBを通過させる他の戦略は、カベオリン1に媒介されるトランスサイトーシス、グルコース担体およびアミノ酸担体などの担体に媒介される輸送体、インスリンまたはトランスフェリンの受容体媒介性トランスサイトーソイス、およびp−糖タンパク質などの活性な流出輸送体を含めた内在性の輸送系の使用を伴ってよい。活性な輸送部分を本発明において使用するための治療用化合物とコンジュゲートして、血管の内皮壁を渡る輸送を容易にすることもできる。あるいは、BBBの反対側への治療用薬剤の薬物送達は、局所送達、例えば、くも膜下腔内送達、例えば、オンマイヤーレザバー(Ommaya reservoir)を通る(例えば、参照により本明細書に組み込まれる、米国特許第5,222,982号および同第5,385,582号を参照されたい);ボーラス注射、例えば、シリンジによって、例えば、硝子体内または頭蓋内に;連続な注入、例えば、対流を伴うカニューレ挿入によって(例えば、参照により本明細書に組み込まれる米国特許出願第20070254842号を参照されたい);または、薬剤が可逆的に添付されているデバイスを埋め込むこと(例えば、参照により本明細書に組み込まれる、米国特許出願第20080081064号および同第20090196903号を参照されたい)によってよい。
【0103】
投与するべきCD活性を促進する薬剤(複数可)の有効量または有効用量を算出することは当業者の技術の範囲内であり、当業者にとっては常套的である。言うまでもなく、投与するべき最終的な量は、投与経路および治療される障害または状態の性質に左右される。
【0104】
医薬に含めるために、CD活性を促進する薬剤(複数可)を商業的な市販の供給源から得ることができる。一般的な提案として、用量当たりの非経口的に投与する化合物の薬学的有効量の総量は、用量反応曲線によって測定することができる範囲内になる。
【0105】
治療的投与のために使用されるCD活性を促進する薬剤(複数可)は、滅菌しなければならない。滅菌は、滅菌濾過膜(例えば、0.2μmの膜)を通して濾過することによって容易に実現される。治療用組成物は、一般に、滅菌アクセスポートを有する容器、例えば、静脈内溶液バッグまたは皮下注射針で穴をあけることができる止め栓を有するバイアル内に置く。CD活性を促進する薬剤(複数可)は、普通は、単位用量容器または多回用量容器、例えば、密封したアンプルまたはバイアル内で、水溶液として、または再構成するための凍結乾燥製剤として貯蔵する。凍結乾燥製剤の例として、10mLのバイアルを5mlの滅菌濾過した化合物1%(w/v)水溶液で満たし、生じた混合物を凍結乾燥する。輸液は、凍結乾燥した化合物を、注射用静菌水を用いて再構成することによって調製する。
【0106】
医薬組成物は、所望の製剤に応じて、動物またはヒトに投与するための医薬組成物を処方するために一般に使用される、ビヒクルと定義される、薬学的に許容できる無毒性の担体または希釈剤を含んでよい。希釈剤は、組み合わせの生物活性に影響を及ぼさないように選択する。そのような希釈剤の例は、蒸留水、緩衝水、生理食塩水、PBS、リンゲル液、ブドウ糖溶液、およびハンクス液である。さらに、医薬組成物または製剤は、他の担体、アジュバント、または非毒性、非治療的、非免疫原性の安定剤、賦形剤なども含んでよい。組成物は、生理的条件に近づけるための追加的な物質、例えば、pH調整剤および緩衝剤、毒性調整剤、湿潤剤および界面活性剤などをも含んでよい。
【0107】
組成物は、例えば抗酸化剤などの任意の種々の安定化剤も含んでよい。医薬組成物がポリペプチドを含む場合、ポリペプチドは、ポリペプチドのin vivoにおける安定性を増強する、またはそうでなければその薬理学的性質を増強する(例えば、ポリペプチドの半減期を増加させる、その毒性を低下させる、溶解性または取り込みを増強する)種々の周知の化合物と複合させることができる。そのような修飾剤または複合剤の例としては、硫酸、グルコン酸、クエン酸およびリン酸が挙げられる。組成物のポリペプチドは、それらのin vivo属性を増強する分子と複合させることもできる。そのような分子としては、例えば、炭水化物、ポリアミン、アミノ酸、他のペプチド、イオン(例えば、ナトリウム、カリウム、カルシウム、マグネシウム、マンガン)、および脂質が挙げられる。
【0108】
投与の種々の型に適している製剤に関する別の手引きは、Remington’s Pharmaceutical Sciences、Mace Publishing Company、Philadelphia、Pa.、第17版(1985)において見ることができる。薬物送達ための方法についての簡単な総説については、Langer、Science 249:1527〜1533(1990)を参照されたい。
【0109】
医薬組成物は、予防的な治療および/または治療的な治療のために投与することができる。活性成分の毒性および治療効果は、細胞培養物および/または実験動物において標準の薬学的手順に従って決定することができ、例えば、LD50(集団の50%に対して致死的な用量)およびED50(集団の50%に対して治療的に有効な用量)を決定することを含む。毒性効果と治療効果との間の用量比は治療指数であり、LD50/ED50比として表すことができる。大きな治療的指標を示す化合物が好ましい。
【0110】
細胞培養試験および/または動物試験から得られたデータを、ヒトに対してさまざまな投与量を処方することにおいて使用することができる。活性成分の投与量は、一般には、ED50を含むさまざまな循環濃度の範囲内に並び、毒性は低い。投与量は、使用する剤形および利用する投与経路に応じてこの範囲内で変動してよい。
【0111】
医薬組成物を処方するために使用する構成成分は、高純度のものであることが好ましく、潜在的に有害な汚染物質を実質的に含まない(例えば、少なくともNational Food(NF)グレード、一般に、少なくとも分析グレード、より一般には少なくとも医薬グレード)。さらに、in vivoで使用するための組成物は、通常滅菌される。所与の化合物を使用する前に合成しなければならない限りでは、生じた生成物は、一般には、合成プロセスまたは精製プロセスの間に存在する可能性がある任意の潜在的に毒性の薬剤、特に、任意の内毒素を実質的に含まない。非経口投与するための組成物も滅菌され、実質的に等張性であり、GMP条件下で製造される。
【0112】
特定の患者に与えるべき有効量の治療用組成物は、種々の因子に左右され、そのいくつかは患者によって異なる。有能な臨床医は、必要に応じて、疾患状態の進行を停止または逆転させるために、患者に投与するための有効量の治療剤を決定することができる。LD50動物データ、および薬剤に関して入手可能な他の情報を利用して、臨床医は、投与経路に応じて、個人に対する最大の安全用量を決定することができる。例えば、治療用組成物が投与される流体が大きいことを考慮すると、静脈内投与する用量は、くも膜下腔内投与する用量よりも高くてよい。同様に、体から急速に取り除かれる組成物は、治療的な濃度を維持するために、高用量で、または反復用量で投与することができる。有能な臨床医は、通常の技術を利用して、常套的な臨床試験の過程で特定の治療薬の投与量を最適化することができる。
【0113】
本方法を用いて治療することができる哺乳動物種としては、イヌおよびネコ;ウマ;ウシ;ヒツジなど、ならびに霊長類、特に、ヒトが挙げられる。動物モデル、特に小さな哺乳動物、例えば、マウス、ウサギなどを実験的調査のために使用することができる。他の使用としては、活性な脱メチル化シグナル伝達の存在下で特異的な効果を調査することが望ましい調査が挙げられる。
【0114】
本発明の方法は、併用療法においても使用される。例えば、いくつもの薬剤、例えば、化学療法剤、キナーゼ阻害剤、アンジオスタチン、エンドスタチン、VEGF阻害剤などが癌の治療において有用であり得る。本発明のCD活性を促進する薬剤(複数可)とこれらの他の薬剤を組み合わせて使用することは、個々の薬物について必要な投与量が低減すること、および異なる薬物の相補的な効果という利点を有し得る。
【0115】
上記の通り、本発明は、ヒト患者などの哺乳動物の治療において、ゲノムDNA脱メチル化療法を必要とする対象に使用される。そのような対象の例は、プロモーターが高メチル化されていることによって異常にサイレンシングされた遺伝子に関連する状態に罹患している対象である。そのような状態を特徴とする疾患に罹患している患者には、係属中の特許請求された発明治療プロトコールが大いに有益である。
【0116】
そのような状態の1つの例は癌である。いくつもの遺伝子、すなわち、メチル化感受性遺伝子が、癌では異常に高メチル化され、サイレンシングされていることが公知である。これらとしては、細胞周期制御に関与する遺伝子(例えば、RB1、CDKN2AINK4A、CDKN2AARF)、腫瘍細胞の浸潤に関与する遺伝子(例えば、CDH1、CDH13、TIMP3、VHL)、DNA修復に関与する遺伝子(例えば、MLH1、MGMT、BRCA1、GSTP1)、クロマチンリモデリングに関与する遺伝子(例えばSMARCA3)、細胞シグナル伝達に関与する遺伝子(例えば、RASSF1A、SOCS1)、転写に関与する遺伝子(例えば、ESR1)、およびアポトーシスに関与する遺伝子(例えば、DAPK1)が挙げられる。したがって、本発明の方法および組成物は、癌、例えば、神経膠腫、髄芽腫、結腸癌、結腸直腸癌、乳癌、または白血病に罹患している対象における腫瘍成長および癌の進行を阻害することにおいて使用される。「癌」という用語は、一般には、哺乳動物における、細胞の成長/増殖が調節されていないことを特徴とする生理的状態を指す。癌の例としては、これらに限定されないが、細胞腫、リンパ腫、芽細胞種、および白血病が挙げられる。癌のより詳細な例としては、これらに限定されないが、慢性リンパ球性白血病(CLL)、非小細胞癌(NSCLC)を含めた肺癌、乳癌、卵巣癌、子宮頸部癌、子宮内膜癌、前立腺癌、結腸直腸癌、腸のカルチノイド、膀胱癌、胃癌、膵臓癌、肝癌(肝細胞癌)、肝芽腫、食道癌、肺腺癌、中皮腫、滑膜肉腫、骨肉腫、頭頸部扁平上皮癌、若年性鼻咽腔血管線維腫、脂肪肉腫、甲状腺癌、黒色腫、基底細胞癌(BCC)、髄芽腫およびデスモイドが挙げられる。特定の癌と上記の対象とする遺伝子のメチル化状態との間の相関は、その開示が参照により本明細書に組み込まれるRobertson、K.D.(2005)Nature Review Genetics 6:597〜610において見ることができる。
【0117】
腫瘍成長および癌の進行を阻害するためのCD活性を促進する薬剤(複数可)の有効量は、上述のメチル化感受性遺伝子の1種または複数種の発現を、in vitroおよびin vivoで、例えば2倍以上増加させ、かつ/またはin vitroにおける癌細胞の増殖速度またはin vivoにおける腫瘍の増殖阻害に測定可能な低下をもたらす量である。例えば、好ましい成長阻害剤は、腫瘍成長を、適切な対照と比較して少なくとも約5%、少なくとも約10%、少なくとも約20%、好ましくは約20%〜約50%、さらに好ましくは、50%超(例えば、約50%〜約100%)阻害し、対照は、一般には、試験されているシチジンデアミナーゼ活性を促進する薬剤(複数可)で処理していない癌細胞である。薬剤を体重1kg当たり約1μgから約100mgまでで投与することにより、腫瘍サイズまたは細胞増殖が抗体を最初に投与してから約5日〜3ヶ月以内、好ましくは約5〜30日以内に低下した場合、その薬剤はin vivoにおいて成長阻害性である。特定の態様では、腫瘍サイズは、療法を開始した時のそのサイズと比較して低下する。
【0118】
本発明の方法によって治療することができる、プロモーターが高メチル化されていることによって異常にサイレンシングされた遺伝子に関連する状態の別の例は、異常なゲノム刷り込みに関連する状態である。ゲノム刷り込みでは、特定の遺伝子は、由来する親(parent of origin)に特異的な様式で発現している。ゲノム刷り込みは、刷り込まれた遺伝子が、母親から受け継がれた対立遺伝子からのみ発現されるか、または父親から受け継がれた対立遺伝子からのみ発現される、古典的なメンデル遺伝と無関係の遺伝プロセスである。ゲノム刷り込みは、遺伝子の配列を変化させることなく単一対立遺伝子性遺伝子発現を実現するためにメチル化およびヒストン修飾を必要とする。これらの後成的な目印は、生殖系列において確立され、生物体の全ての体細胞全体を通して維持される。
【0119】
本発明の方法によって治療可能な、異常なゲノム刷り込みに関連するいくつもの状態が同定されている。例えば、胎児および出生後の過成長、器官の肥大、腫瘍リスクの増加、および顔面の異常を特徴とするベックウィズ−ヴィーデマン症候群では、IGF2/H19刷り込み制御領域1における母系対立遺伝子の新規のメチル化が観察される。精神遅滞、肥満、低身長、および行動上の問題を特徴とするプラダー・ウィリー症候群では、PWS遺伝子の父系対立遺伝子の新規のメチル化が観察される。腎臓の副甲状腺ホルモン抵抗性を特徴とする偽性副甲状腺機能低下症1B型では、NESP55の母系対立遺伝子の新規のメチル化が観察される。本発明の方法は、これらの遺伝子座における脱メチル化を促進し、それによって適切な遺伝子発現を回復すること使用される。
【0120】
本発明の方法によって治療することができる、プロモーターが高メチル化されていることによって異常にサイレンシングされた遺伝子に関連する状態の別の例は、反復不安定性(repeat instability)疾患に関連する状態である。これらの疾患では、反復配列が拡大することにより、それらの配列の近くの遺伝子の発現に影響を及ぼす異常なメチル化が生じる。本発明の方法によって治療可能な反復不安定性に関連するいくつもの状態が同定されている。例えば、精神遅滞、巨睾丸症、および自閉症的行動を特徴とする脆弱X症候群では、FMRI遺伝子の5’UTRのCGG反復が拡大することにより、5’UTR配列が新規にメチル化され、FMRI遺伝子が異常にサイレンシングされる。別の例として、肢および顔面の筋肉の衰弱および消耗、筋緊張症、ならびに白内障を特徴とする筋緊張性ジストロフィー(DM1)では、DMPK遺伝子のUTR内のCTG反復が拡大することにより、拡大したCTG反復の近くのCpGアイランドが新規にメチル化され、それによって今度はSIX5遺伝子が攪乱され、サイレンシングされる。本発明の方法は、これらの遺伝子座における脱メチル化を促進し、それによって適切な遺伝子発現を回復することに使用される。
【0121】
スクリーニング方法。
本明細書に記載の方法は、候補薬剤を、脱メチル化を調節する活性についてスクリーニングするための有用なシステムを提供する。そのために、CD活性を促進する薬剤が、脱メチル化を増強することに対する強力な効果を有することが示されている。CD活性を阻害する薬剤を、脱メチル化が起こっている細胞を含む細胞培養物系に加えることにより、この脱メチル化活性が強力に抑制され、したがって、Oct4およびNanogなどのメチル化感受性遺伝子の転写活性プロモーターの量が低下する。この脱メチル化活性の抑制ならびにその後の、これらのプロモーターにおけるメチル化の増加および転写活性のサイレンシングは、早ければ、脱メチル化している細胞を、CD活性を阻害する薬剤と接触させた1日後に観察され得、3日目までにこれらのメチル化感受性遺伝子がほぼ完全にサイレンシングされる。
【0122】
生物学的に活性な薬剤を求めるスクリーニングアッセイでは、細胞、通常細胞培養物を、CD活性を促進する薬剤の存在下で、対象とする薬剤と接触させ、候補薬剤の効果を、アウトプットパラメータ、例えばメチル化CpG配列の量、メチル化感受性遺伝子の発現などを上記の方法によってモニターすることによって評価する。
【0123】
パラメータは、細胞の数量化できる構成成分、特に、望ましくはハイスループットシステムにおいて正確に測定することができる構成成分である。パラメータは、細胞表面決定基、受容体、タンパク質、またはそのコンフォメーションの修飾または翻訳後修飾、脂質、炭水化物、有機分子または無機分子、核酸、例えば、mRNA、DNAなどを含めた任意の細胞構成成分または細胞産物、またはそのような細胞構成成分に由来する部分、またはそれらの組み合わせであってよい。大部分のパラメータにより定量的な読み取りがもたらされるが、ある場合には、半定量的な結果または定性的な結果が許容される。読み取りは、単一の決定値を含んでよい、または、平均値、中央値もしくは分散などを含んでよい。特徴的に、多数の同じアッセイから、各パラメータについてさまざまなパラメータの読み取り値が得られる。変動性が予測され、試験パラメータのセットのそれぞれについてのさまざまな値は、単一の値をもたらすために使用される一般的な統計学的方法を用いた標準の統計学的な方法を使用して得られる。
【0124】
例えば、薬剤を、脱メチル化活性を促進する活性について、例えば、候補薬剤を、CD活性を促進する薬剤の存在下で細胞培養物に加えることによってスクリーニングすることができる。観察されるメチル化の量が減少すること、例えば、メチル化感受性遺伝子のプロモーターまたは外因的に供給された5−meCpGリッチ核酸の5−メチルシトシンの数が、候補薬剤が存在しない培養物において観察されるものに対して、例えば、1.5分の1、2分の1、3分の1またはそれ以下に減少することにより、候補薬剤が脱メチル化を促進する薬剤であったことが示される。そのような実施形態では、細胞は、脱メチル化を許容する細胞であってよい、または、脱メチル化を許容しない細胞であってよい。
【0125】
あるいは、例えば、候補薬剤を、CD活性を促進する薬剤の存在下で細胞培養物に加えることによって、脱メチル化活性を抑制する活性について薬剤をスクリーニングすることができる。観察されるメチル化の量、例えば、メチル化感受性遺伝子のプロモーターまたは外因的に供給された5−meCpGリッチ核酸の5−メチルシトシンの数が、候補薬剤が存在しない培養物において観察されるものと比較して、全く減少しないこと、またはほんの少量しか減少しないことにより、候補薬剤が脱メチル化を抑制する薬剤であったことが示される。そのような実施形態では、培養細胞は、脱メチル化を許容する細胞である。
【0126】
スクリーニングの対象とされる候補薬剤としては、多数の化学的クラス、主として有機金属分子を含み得る有機分子、無機分子、遺伝子配列などを包含する既知化合物および未知化合物が挙げられる。本発明の重要な態様は、毒性試験などを含めた、候補薬物の評価である。
【0127】
候補薬剤は、構造的な相互作用、特に水素結合に必要な官能基を含む有機分子を含み、一般には少なくともアミン基、カルボニル基、ヒドロキシル基またはカルボキシル基を含み、しばしば、官能性化学基の少なくとも2つを含む。候補薬剤は、多くの場合、上記の官能基の1つまたは複数で置換された環式炭素または複素環式構造および/または芳香族または多芳香族の構造を含む。候補薬剤は、ペプチド、ポリヌクレオチド、糖類、脂肪酸、ステロイド、プリン、ピリミジン、誘導体、構造的な類似体またはそれらの組み合わせを含めた生体分子の中にも見いだされる。薬理活性のある薬物、遺伝的活性のある分子などが包含される。対象とする化合物としては、化学療法剤、ホルモンまたはホルモンアンタゴニストなどが挙げられる。本発明に適した医薬品の例は、「The Pharmacological Basis of Therapeutics」Goodmanおよび Gilman、McGraw−Hill、New York、N.Y.、(1996)、第9版に記載されているものである。毒素、生物戦剤および化学戦剤(biological and chemical warfare agent)も包含される。例えば、Somani、S.M.(編)「Chemical Warfare Agents」Academic Press、New York、1992)を参照されたい。
【0128】
スクリーニングの対象とされる候補薬剤としては、核酸、例えば、siRNA、shRNA、アンチセンス分子、またはmiRNAをコードする核酸、またはポリペプチドをコードする核酸も挙げられる。核酸を標的細胞に移入するために有用な多くのベクターが利用可能である。ベクターは、エピソームとして、例えば、プラスミド、ミニサークルDNA、サイトメガロウイルス、アデノウイルスなどのウイルス由来のベクターとして維持することができる、またはベクターは、相同組換えまたはランダムな組み込みによって標的細胞のゲノムに組み込むことができる。例えば、MMLV、HIV−1、ALVなどのレトロウイルス由来のベクター。ベクターは、対象細胞に直接提供することができる。言い換えれば、多能性細胞を、対象とする核酸を含むベクターと、ベクターが細胞に取り込まれるように接触させる。
【0129】
電気穿孔、塩化カルシウムトランスフェクション、およびリポフェクションなどの、細胞を核酸ベクターと接触させるための方法は当技術分野で周知である。あるいは、対象とする核酸は、ウイルスによって対象細胞に提供することができる。言い換えれば、多能性細胞を、対象とする核酸を含むウイルス粒子と接触させる。本発明の方法には、レトロウイルス、例えば、レンチウイルスが特に適している。一般に使用されるレトロウイルスベクターは、「不完全である」、すなわち、増殖性感染のために必要なウイルスタンパク質を産生することができない。もっと正確に言えば、ベクターの複製には、パッケージング細胞系において成長させることが必要である。対象とする核酸を含むウイルス粒子を生成するために、核酸を含むレトロウイルスの核酸をパッケージング細胞系によってウイルスカプシド中にパッケージする。異なるパッケージング細胞系からは、カプシド中に組み入れられる異なる外被タンパク質がもたらされ、この外被タンパク質により、ウイルス粒子の細胞に対する特異性が決定される。外被タンパク質は、狭宿主性、広宿主性および異種栄養性の少なくとも3種類のものである。狭宿主性外被タンパク質、例えば、MMLVを用いてパッケージしたレトロウイルスは、大部分のマウスおよびラット細胞型に感染することができ、これは、BOSC23などの狭宿主性パッケージング細胞系を使用することによって生成される(Pearら(1993)P.N.A.S.90:8392〜8396)。広宿主性外被タンパク質、例えば、4070A(Danosら、上記)を担持するレトロウイルスは、ヒト、イヌおよびマウスを含めた大部分の哺乳動物細胞型に感染することができ、これは、PA12(Millerら(1985)Mol.Cell.Biol.5:431〜437);PA317(Millerら(1986)Mol.Cell.Biol.6:2895〜2902);GRIP(Danosら(1988)PNAS 85:6460〜6464)などの広宿主性パッケージング細胞系を使用することによって生成される。異種栄養性外被タンパク質、例えば、AKR envを用いてパッケージしたレトロウイルスは、マウスの細胞以外の大部分の哺乳動物細胞型に感染することができる。適切なパッケージング細胞系を使用して、対象CD33+分化体細胞がパッケージされたウイルス粒子の標的になることを確実にすることができる。再プログラミング因子をコードする核酸を含むレトロウイルスベクターをパッケージング細胞系に導入する方法、およびパッケージング系によって生成されるウイルス粒子を採取する方法は当技術分野で周知である。
【0130】
対象とする核酸を対象細胞に提供するために使用するベクターは、一般には、対象とする核酸の発現を駆動する、すなわち、転写を活性化するために適したプロモーターを含む。これは、遍在的に作用するプロモーター、例えば、CMV−b−アクチンプロモーター、または誘導性プロモーター、例えば特定の細胞集団において活性なプロモーターまたはテトラサイクリンなどの薬物が存在することに反応するプロモーターなどを含んでよい。転写活性化とは、標的細胞において転写が基礎レベルを少なくとも約10倍、少なくとも約100倍、通常少なくとも約1000倍超えて増加することを意味する。さらに、対象細胞に再プログラミング因子を提供するために使用するベクターは、後で除去しなければならない遺伝子を含んでよく、その際は、例えば、Cre/Loxなどのリコンビナーゼ系、または、例えば、TK、bcl−xsなどのヘルペスウイルスなどの選択毒性を可能にする遺伝子を含めることによって該遺伝子が破壊されて発現される細胞を用いる。
【0131】
スクリーニングの対象とされる候補薬剤としては、ポリペプチドも挙げられる。そのようなポリペプチドは、場合によって、生成物の溶解性を増加させるポリペプチドドメインと融合することができる。ドメインは、定義済みのプロテアーゼ切断部位、例えば、TEVプロテアーゼによって切断されるTEV配列を通じてポリペプチドに連結することができる。リンカーは、1つまたは複数の可動性の配列、例えば、1個から10個までのグリシン残基も含んでよい。一部の実施形態では、融合タンパク質の切断は、生成物の溶解性を維持する緩衝液中、例えば、0.5Mから2Mまでの尿素の存在下、溶解性を増加させるポリペプチドおよび/またはポリヌクレオチドの存在下などで行う。対象とするドメインとしては、エンドソーム溶解性(endosomolytic)ドメイン、例えば、インフルエンザHAドメイン;および作製に役立つ他のポリペプチド、例えば、IF2ドメイン、GSTドメイン、GRPEドメインなどが挙げられる。
【0132】
候補ポリペプチド薬剤を、凝集のシグナル伝達を細胞内で阻害するその能力についてアッセイする場合、ポリペプチドは、ポリペプチド浸透性ドメインと融合した対象とするポリペプチド配列を含んでよい。いくつもの浸透性ドメインが当技術分野で公知であり、ペプチド、ペプチド模倣薬、および非ペプチド担体を含めた本発明の非組み込み型のポリペプチドにおいて使用することができる。例えば、浸透性ペプチドは、ペネトラチンと称され、アミノ酸配列RQIKIWFQNRRMKWKKを含む、Drosophila melanogasterの転写因子であるAntennapaediaの第3のアルファヘリックス由来であってよい。別の例として、浸透性ペプチドは、例えば、天然に存在するtatタンパク質のアミノ酸49〜57を含み得るHIV−1tat基本領域アミノ酸配列を含む。他の浸透性ドメインとしては、ポリアルギニンモチーフ、例えば、HIV−1revタンパク質のアミノ酸34〜56の領域、ノナアルギニン、オクタアルギニンなどが挙げられる。(例えば、移行ペプチドおよびペプトイドの教示に関して参照により本明細書に明確に組み込まれる、Futakiら(2003)Curr Protein Pept Sci.2003年4月;4(2):87〜96;およびWenderら(2000)Proc.Natl.Acad.Sci.U.S.A 2000年11月21日;97(24):13003〜8;米国特許出願公開第20030220334号;同第20030083256号;同第20030032593号;および同第20030022831号を参照されたい)。ノナアルギニン(R9)配列は、特徴づけられているより効率的なPTDの1つである(Wenderら2000;Uemuraら2002)。
【0133】
候補ポリペプチド薬剤を、凝集のシグナル伝達を細胞外で阻害するその能力についてアッセイする場合、ポリペプチドは、安定性が改善されるように処方することができる。例えば、ペプチドをペグ化することができ、ポリエチレンオキシ基により、血流中の寿命が増強される。追加的な機能性をもたらすため、例えば、in vivoにおける安定性を増加させるために、ポリペプチドを別のポリペプチドと融合することができる。一般に、そのような融合パートナーは、安定な血漿タンパク質であり、それにより、例えば、融合物として存在するとポリペプチドのin vivo血漿内半減期が延長され、具体的には、そのような安定な血漿タンパク質は免疫グロブリン定常ドメインである。安定な血漿タンパク質が通常多量体の形態、例えば、免疫グロブリンまたはリポタンパク質で見いだされ、その中の同じポリペプチドまたは異なるポリペプチド鎖が、通常ジスルフィド結合および/または非共有結合して集合した多鎖ポリペプチドを形成しているほとんどの場合、ポリペプチドを含有する本明細書の融合物も、安定な血漿タンパク質前駆物質と実質的に同じ構造を有する多量体として作製し、使用する。これらの多量体が含むポリペプチド薬剤は同種のものである、または、これらの多量体は2種以上のポリペプチド薬剤を含有してよい。
【0134】
候補ポリペプチド薬剤は、原核細胞によって又は真核生物から産生することができ、ほどくこと、例えば、熱変性、DTT低下などによってさらに加工し、当技術分野で公知の方法を使用してさらに再び折りたたむことができる。一次配列を変化させない対象とする修飾としては、ポリペプチドの化学的な誘導体化、例えば、アシル化、アセチル化、カルボキシル化、アミド化などが挙げられる。グリコシル化の修飾、例えば、ポリペプチドのグリコシル化パターンを、その合成およびプロセシングの間に、またはさらなるプロセシングステップにおいて修飾することによって;例えば、ポリペプチドを、哺乳動物のグリコシル化酵素または脱グリコシル酵素などのグリコシル化に影響を及ぼす酵素に曝露させることによって行われる修飾も包含される。リン酸化されたアミノ酸残基、例えば、ホスホチロシン、ホスホセリン、またはホスホスレオニンを有する配列も包含される。ポリペプチドは、それらのタンパク質分解に対する耐性を改善するため、または溶解特性を最適化するため、またはそれらを治療剤としてより適切にするために、通常の分子生物学的技法および合成化学を使用して修飾することができる。そのようなポリペプチドの類似体としては、天然に存在するL−アミノ酸以外の残基、例えば、D−アミノ酸または天然に存在しない合成のアミノ酸を含有するものが挙げられる。アミノ酸残基の一部または全部をD−アミノ酸で置換することができる。
【0135】
候補ポリペプチド薬剤は、当技術分野で公知の従来の方法を使用して、in vitro合成によって調製することができる。種々の市販の合成装置、例えば、Applied Biosystems、Inc.、Beckmanなどによる自動合成機が利用可能である。合成機を使用することによって、天然に存在するアミノ酸を非天然アミノ酸で置換することができる。特定の配列および調製様式は、利便性、経済的側面、必要な純度などによって決定される。あるいは、候補ポリペプチド薬剤は、組換え合成の従来の方法に従って単離し、精製することができる。発現宿主の溶解物を調製し、溶解物を、HPLC、排除クロマトグラフィー、ゲル電気泳動、アフィニティークロマトグラフィー、または他の精製技法を使用して精製することができる。ほとんどの場合、使用される組成物は、産物の調製およびその精製の方法に関連する汚染物質に関して、少なくとも20重量%、通常、少なくとも約75重量%、好ましくは少なくとも約95重量%、および治療のためには、通常少なくとも約99.5重量%の所望の産物を含む。通常、百分率は、総タンパク質量に基づく。
【0136】
ある場合では、スクリーニングされる候補ポリペプチド薬剤は抗体である。「抗体」または「抗体部分」という用語は、エピトープに適合し、それを認識する特定の形状を有する任意のポリペプチド鎖含有分子構造を含むものとし、ここで、1つまたは複数の非共有結合性の相互作用により、分子構造とエピトープとの間の複合体が安定化する。所与の構造の特異的または選択的な適合およびその特異的なエピトープは、時には、「鍵と鍵穴」適合と称される。典型的な抗体分子は、免疫グロブリンであり、全ての供給源、例えば、ヒト、げっ歯類、ウサギ、雌ウシ、ヒツジ、ブタ、イヌ、他の哺乳動物、ニワトリ、他のトリなど由来の全種類の免疫グロブリン、IgG、IgM、IgA、IgE、IgDなどが、「抗体」であるとみなされる。本発明において利用される抗体は、ポリクローナル抗体またはモノクローナル抗体のいずれかである。抗体は、一般には、細胞が培養されている培地中に提供される。
【0137】
候補薬剤を含めた化合物は、合成化合物または天然化合物のライブラリーを含めた多種多様な供給源から得られる。例えば、ランダム化されたオリゴヌクレオチドおよびオリゴペプチドの発現を含めた、生体分子を含めた多種多様な有機化合物のランダムな合成および指向性合成のための多数の手段が利用可能である。あるいは、細菌の抽出物、真菌の抽出物、植物の抽出物および動物の抽出物の形態の天然化合物のライブラリーが利用可能である、または容易に作製される。さらに、天然の、または合成的に作製したライブラリーおよび化合物は、従来の化学的手段、物理的手段および生化学的手段によって容易に修飾され、また、それを使用して、組み合わせのライブラリーを作製することができる。公知の薬理薬剤(pharmacological agent)を、例えば、アシル化、アルキル化、エステル化、アミド化(amidification)などの直接的またはランダムな化学修飾に供して構造類似体を作製することができる。
【0138】
候補薬剤は、通常、薬剤を欠く細胞と併せて、少なくとも1つ、通常複数の細胞試料に加えることによって、その生物活性についてスクリーニングする。薬剤に反応したパラメータの変化を測定し、結果を、参照培養物と比較すること、例えば、薬剤の存在下と非存在下で比較すること、他の薬剤を用いて得られた参照培養物と比較することなどによって評価する。
【0139】
薬剤は、溶液に都合よく加えられる、または培養物中の細胞の培地に対して容易に可溶性の形態である。薬剤は、フロースルーシステムで、断続的または連続的な流れとして加えることができる、あるいは、化合物のボーラスを、単独でまたは増加的に、別の静的な溶液に加える。フロースルーシステムでは、2つの流体を使用し、一方は生理的に中性の溶液であり、他方は加えた試験化合物と同じ溶液である。第1の流体を、細胞を通過させ、その後、第2の流体を通過させる。単一溶液法では、試験化合物のボーラスを細胞の周囲の培地の体積に加える。培地の構成成分の全体的な濃度は、ボーラスを加えることによって、またはフロースルー法における2つの溶液間で有意に変化するべきではない。
【0140】
さまざまな濃度に対する示差的な反応を得るために、異なる薬剤の濃度を用いて複数のアッセイを平行して実行することができる。当技術分野で公知の通り、薬剤の有効濃度の決定には、一般には1:10、または他のlog尺度の、希釈によって生じるさまざまな濃度を用いる。必要であれば、第2の一連の希釈を用いて濃度をさらに細かく区分することができる。一般には、これらの濃度の1つは陰性対照としての機能を果たす、すなわち、ゼロ濃度または薬剤の検出レベルを下回る濃度、または表現型における検出可能な変化を生じさせない薬剤の濃度またはそれを下回る濃度である。
【0141】
選択マーカーの存在を数量化するために種々の方法を利用することができる。例えば、DNAメチル化の状態を、例えば、特定のCpG配列において測定するために、クロマチン免疫沈降(ChIP)を実施して内在性DNAを単離することができ、次いで、それを制限エンドヌクレアーゼHpallで消化して脱メチル化の程度を決定することができる、またはバイサルファイト配列決定を行うことができる。存在する分子の量を測定するために、例えば、メチル化感受性遺伝子の発現を測定する場合、都合のよい方法は、分子を、蛍光性、発光性、放射性、酵素的に活性などであり得る検出可能部分、特に、高い親和性を有するパラメータへの結合に特異的な分子で標識することである。蛍光部分は、実質的にあらゆる生体分子、構造、または細胞型を標識するために容易に利用可能である。免疫蛍光部分は、特定のタンパク質だけでなく、特異的なコンフォメーション、切断産物、またはリン酸化のような部位修飾にも結合するように誘導することができる。個々のペプチドおよびタンパク質は、自己蛍光するように、例えば、それらを細胞内で緑色蛍光タンパク質キメラとして発現させることによって遺伝子工学で操作することができる(概説について、Jonesら(1999)Trends Biotechnol.17(12):477〜81を参照されたい)。
【0142】
例えば上記のものなどのスクリーニングは、特定の生物系における脱メチル化の調節における活性を有する薬剤を同定するために合わせてあつらえることができる。例えば、腫瘍細胞において、細胞周期、腫瘍−細胞浸潤、DNA修復、クロマチンリモデリング、細胞シグナル伝達、転写およびアポトーシスを調節する遺伝子などのメチル化感受性遺伝子のプロモーターの脱メチル化を促進する薬剤は、これらの遺伝子の脱メチル化を促進すること、したがって、腫瘍においてこれらの遺伝子を発現させることに使用し、それによって、癌細胞の増殖および腫瘍成長を阻止することができる。別の例として、体細胞またはES細胞と体細胞の異核共存体における多能性遺伝子であるOct4およびNanogなどのメチル化感受性遺伝子のプロモーターにおける脱メチル化を促進する薬剤は、iPS細胞を作製するための公知の方法において多能性に関連する遺伝子の脱メチル化を促進することに使用することができる。いくつかのそのような場合、例えば、体細胞では、これらの方法は、スクリーニングする目的で、iPS表現型をさらに促進するために、細胞に再プログラミング因子を提供するステップを含んでよい。
【0143】
キットを提供することができ、キットは、本明細書に記載のように、CD活性を促進する1種または複数種の薬剤および細胞を、脱メチル化を許容するように誘導するための試薬を含む。対象とする組み合わせは、1種または複数種のAIDポリペプチドもしくはAPOBECポリペプチド、またはそれらのペプチドをコードする核酸を含むベクターおよび再プログラミングを促進する1種または複数種の薬剤を含んでよい。キットは、対象の細胞におけるDNAメチル化の状態を決定するために適した試薬をさらに含んでよい。キットは、チューブ、緩衝液など、および使用説明書も含んでよい。
【実施例】
【0144】
以下の実施例は、本発明をどのように行い、使用するかについての完全な開示および記載を当業者に提供するために提示され、出願人らが彼らの発明とみなすものの範囲を限定するものではなく、また、以下の実験が、実施した全ての実験または唯一の実験であることを示すものでもない。使用される数字(例えば、量、温度など)に関して正確さを確実にするための努力がなされているが、いくらかの実験的な誤差および偏差が考慮されるべきである。別段の指定のない限り、部分は重量による部分であり、分子量は重量平均分子量であり、温度は摂氏度であり、圧力は、大気または大気付近の圧力である。
【実施例1】
【0145】
核の多能性への再プログラミングに必須である新規の初期制御因子を同定するために、我々は、体細胞の分化した状態を維持することに固有の原理の解明において有用であることが立証された異核共存体の我々の以前の経験を活用した。特に、これらの以前の我々の研究およびその他の研究により、ヒト細胞の「最終分化した」状態は固定されておらず、変化させることができ、他の分化した状態に典型的な以前はサイレントであった遺伝子の発現が誘導されることが示された(Blau、H.M.ら(1983)Cell 32、1171〜801;Baron、M.H.&Maniatis、T.(1986)Cell 46、591〜602;Wright、W.E.(1984)Exp Cell Res 151、55〜69;Spear、B.T.&Tilghman、S.M.(1990)Mol Cell Biol 10、5047〜54;Chiu、CP.&Blau、H.M.R(1984)Cell 37、879〜87)。我々は、(1)再プログラミングは、全てのES細胞因子の存在下で起こり、(2)再プログラミングの開始は、融合と同時に開始され、(3)再プログラミングは、融合した非分裂細胞において評価され、かつ(4)種差により融合した細胞型の転写物が区別されるので、異核共存体を用いて、機構を解明し、多能性への再プログラミングの開始において役割を有する新規の遺伝子を同定することができると推論した。
【0146】
材料および方法
フローサイトメトリーによる異核共存体の生成および単離。GFP+マウスES細胞およびDsRed+ヒト胎児の肺の初代線維芽細胞を、以前に記載されている通りレトロウイルス構築物を用いた形質導入によって生成し(Palermo、A.ら(2009)Faseb J)、融合して非分裂性の、多核異核共存体を形成した。細胞をまず、ES培地中で12時間共培養し、次いで、PEG1500(Roche)を用いて37℃で2分間処理し、その後DMEMで4回連続的に洗浄した。ES培地を洗浄した後に交換し、その後12時間ごとに交換した。GFP+/DsRed+異核共存体をフローサイトメトリー(FACSVantage SE、BD)によって2回選別し、遺伝子発現およびメチル化について解析した。
【0147】
免疫蛍光法。異核共存体を2.5%v/vのヤギ血清および1mMのEDTAを伴うPBS中で2回選別し、900rpmで5分間サイトスピンした。サイトスピンしたGFP+/DsRed+異核共存体をHoechst33342で染色し、画像化した。抗体染色のために、サイトスピンした細胞を固定し、透過処理し、PBS中20%のFBSを使用してブロッキングした。細胞を、一次抗体マウス抗Ki−67(Dako Denmark A/S)と一緒に、ブロッキング緩衝液中1:100希釈で1時間インキュベートし、PBS中で3回すすぎ、次いで、ヤギ抗マウスCascade blue二次抗体(Millipore)と一緒に1:500希釈で30分間インキュベートし、3回すすぎ、Fluoromount−Gを用いて封入し、画像化した。画像を、落射蛍光顕微鏡(Axioplan2;Carl Zeiss MicroImaging、Inc.)、Fluar20×/0.75対物レンズまたは40×/0.90対物レンズ、およびデジタルカメラ(ORCA−ER C4742−95;Hamamatsu Photonics)を使用して取得した。取得するために使用したソフトウェアはOpenLab4.0.2(Improvision)であった。
【0148】
PEG誘導性融合の3時間後にBrdUをmES共培養物およびhFb共培養物に加えた。標識化および抗体染色を、BrdU Labeling and Detection Kit I(Roche)を使用して行った。
【0149】
遺伝子発現の解析。RNAを、ES細胞、線維芽細胞、および融合後またはRNeasy microキット(Qiagen)を使用してsiRNA処理した後、違う時間に2回選別した異核共存体から調製した。各試料について全RNAを、Superscript First−Strand Synthesis System for RT−PCR(Invitrogen)を使用して逆転写した。逆転写された材料をGo GreenTaq DNAポリメラーゼ(Promega)を使用してPCRに供した。Oct4、NanogおよびGAPDHの発現を解析するためのヒト特異的なプライマーを設計した。siRNA処理実験においてAIDおよびGAPDHに対して使用したプライマーにより、ヒト転写物とマウス転写物の両方を増幅して異核共存体におけるAIDおよびGAPDHの総レベルを評価した。RT−PCRおよび定量的PCRのために使用するヒト特異的なプライマーは、:hOct4 F 5’−TCGAGAACCGAGTGAGAGGC−3’(配列番号45)、R−5’−CACACTCGGACCACATCCTTC−3’(配列番号46);hNanog F 5’−CCAACATCCTGAACCTCAGCTAC−3’(配列番号47)、R 5’−GCCTTCTGCGTCACACCATT−3’(配列番号48);hGAPDH F 5’−TGTCCCCACTGCCAACGTGTCA−3’(配列番号49)、R 5’−AGCGTCAAAGGTGGAGGAGTGGGT−3’(配列番号50)である。siRNA処理後のノックダウンを評価するための非種特異的なプライマー配列は、以下の通りである:GAPDH F 5’−ACCACAGTCCATGCCATCAC−3’(配列番号51)、R 5’−TCCACCACCCTGTTGCTGTA−3’(配列番号52);AID F 5’−AAAATGTCCGCTGGGCTAAG−3’(配列番号53)、R 5’−AGGTCCCAGTCCGAGATGTAG−3’(配列番号54)。
【0150】
リアルタイムPCR。リアルタイムPCRを、ABI 7900HT リアルタイムPCRシステムを使用して、Sybr Green PCRミックス(Applied Biosystems)を用いて行った。試料を94℃で2分間、40×(94℃で20秒間、58℃で45秒間)でサイクルにかけた。
【0151】
【表1】
【0152】
単一細胞RT−PCR。単一の異核共存体をFACS(FACSVantage SE、BD)によって直接選別して、9μl一定分量のRT−PCR溶解緩衝液を含有するPCRチューブに入れた。緩衝液の構成成分は、市販のRT−PCR緩衝液(Superscript One−Step RT−PCR Kit Reaction Buffer、Invitrogen)、RNアーゼ阻害剤(Protector RNase Inhibitor、Roche)および0.15%のIGEPAL界面活性剤(Sigma)を含んだ。短パルスで急速回転させた後、PCR−チューブをすぐに衝撃凍結し、その後の分析のために−80℃で貯蔵した。
【0153】
二段階マルチプレックスネステッド単一細胞RT−PCRのために、細胞溶解物を、まず、SuperScript One−Step RT−PCR Kit(Invitrogen)を使用して、Oct4、NanogおよびGAPDHに対するヒトおよび遺伝子に特異的なプライマー対を用いて逆転写した(表2、外部プライマー;図5b)。簡単に述べると、同じPCR細胞−溶解チューブに、遺伝子特異的なプライマー対およびRNアーゼ阻害剤を含有するRT−PCR反応混合物を加えることによって、RT−PCRを行った。第1ラウンドPCRおよび第2ラウンドPCRならびにより大きな特異性を確実にするネステッドRT−PCR用のイントロンスパニングプライマーセットを設計し、使用することによって、ゲノム産物を排除した。第1のステップでは、55℃で30分間逆転写反応を行い、その後94℃で2分間のステップを行った。その後に、以下の通りPCR増幅を30サイクル行った:94℃で30秒間;58℃で30秒;68℃で30秒間。最終的なPCRステップにおいて、反応産物を68℃で3分間インキュベートした。完了した反応産物を4℃で貯蔵した。
【0154】
PCRプロトコールの第2のステップにおいて、第1のステップからの完了したRT−PCR反応産物を水で1:1希釈した。これらの反応産物の1パーセントを、PCRの第2ラウンドのために複製して新しい反応チューブに移し、PCRの第2ラウンドを遺伝子のそれぞれについて別々に、より大きな特異性のためにネステッド遺伝子特異的な内部プライマーを使用して、総反応容積20μlで行った(Platinum Taq Super−Mix HF、Invitrogen)。以下の通りPCR増幅を30サイクル行った:94℃で30秒間;58℃で30秒間;68℃で30秒間。最終的なPCRステップにおいて、反応産物を68℃で3分間インキュベートした。完了した反応産物を4℃で貯蔵した。次いで、第2ラウンドのPCR産物を反応容積の5分の1および1.4%アガロースゲルを用いてゲル電気泳動に供した。
【0155】
【表2】
【0156】
DNAメチル化解析。FACSで選別した異核共存体(細胞2,000〜10,000個)を20μlのPBS中に収集した。DNeasy Tissue Kit(Qiagen)を使用してDNAを抽出した。Epitect Bisulfite Kit(Qiagen)を使用してバイサルファイト処理を行った。ヒトおよびバイサルファイトに特異的なプライマーを使用して、ヒトOct4プロモーターおよびヒトNanogプロモーターの領域についてのネステッドPCRを行った(表3)。試料を、94℃で2分間、30×(94℃で20秒間、61℃で30秒間、68℃で30秒間)の第1の、ネステッドPCRのサイクルにかけた。バイサルファイトに特異的なPCR増幅の第2ラウンドからのPCR産物を、以前に記載されている通りクローニングし、配列決定した(Zhang、F.ら(2007)Proc Natl Acad Sci USA 104、4395〜400)。
【0157】
【表3】
【0158】
siRNAトランスフェクション。siRNAトランスフェクションのために、ES細胞および初代線維芽細胞を、トランスフェクトする前日に50〜60%集密に播いた。silmporter(Millipore)を使用してsiRNA(Dharmacon)をトランスフェクトした。
【0159】
クロマチン免疫沈降。クロマチン免疫沈降を、表4で提供されるプライマーを使用して、Dahlおよび Collas((2008)Nat Protoc 3、1032〜45)によって以前に記載されている通り行った。ChIPデータは、入力DNAに対して正規化されたものとして提示され、エラーバーは標準誤差(sem)を示す。
【0160】
【表4】
【0161】
統計分析。データは、平均±標準誤差として示されている。スチューデントのt検定を用いた群間の比較は両側分布を仮定している。
【0162】
異核共存体のThy1.1(CD90)濃縮。PEGで処理したGFP−(非GFP)mESとDsRed+hFbの共培養物をトリプシン処理し、FACS緩衝液3mLに再懸濁させた。細胞を、ビオチンマウス抗ヒトCD90(BD Pharmingen)と一緒に、1:5000希釈で、室温で30分間インキュベートした。細胞を1回洗浄し、FACS緩衝液3mLに再懸濁させ、10μlのDynabeads Biotin Binder(Invitrogen)と一緒に室温で30分間インキュベートした。ビーズを磁気分離によって除去し、2回洗浄し、濃縮された異核共存体をサイトスピンした。
【0163】
免疫沈降およびウエスタンブロット。マウスES細胞を、IP緩衝液(20mMのトリス、pH7.5、1mMのDTT、0.5mMのEDTA、350mMのNaCI、10%(vol/vol)のグリセロール、10μMのZnCIに溶解させた。細胞全体の溶解物を室温で30分間予め清澄化(pre−clear)し、その後、AIDをプルダウンした。簡単に述べると、細胞溶解物、次いでAIDを、ウサギポリクローナルAID抗体と架橋結合したProtein A Plus Agarose beads(Pierce)を使用してプルダウンした。細胞溶解物2mgから免疫沈降を行った。
【0164】
mESにおけるAIDタンパク質ノックダウンを可視化するために、si対照 またはsi−1でトランスフェクトした3日後に細胞溶解物を回収した。これらの試料中のAIDの検出を、細胞全体の溶解物170μgから、抗マウス−AID(L7E7、Cell Signaling、1:500希釈)を用いて行った。膜を取り除き、抗マウスα−チューブリン(Sigma、1:20,000希釈)を用いてローディング対照を探索した。AIDの免疫沈降を同じL7E7抗体を用いて検出した。
【0165】
結果
種間の異核共存体を作製するために、GFPレポーター遺伝子を形質導入したマウス胚性幹細胞(mES)を、DsRedレポーター遺伝子を形質導入した初代ヒト線維芽細胞(hFb)と共培養し、ポリエチレングリコール(PEG)を使用して融合した(図1a;図5のスキーム)。融合GFP+DsRed+異核共存体は、FACSによって容易に選別され(図1b)、蛍光顕微鏡を用いて同定され、Hoechst33342またはHoechst33258を用いて可視化すると明瞭に染色されたヒトおよびマウスの核を含有した(それぞれ図1cおよび図1f)。PEG融合の効率は低いので(0.6〜1.0%)、GFP+DsRed+異核共存体を2回選別し、80%の純度まで濃縮した(図1b)。増殖している細胞にのみ存在する核タンパク質であるKi−67に対する抗体を使用して、融合後にアッセイした3日間にわたって、異核共存体の98(±2)%において細胞分裂が起こらなかったことを決定した(図1d、e)。さらに、同じ期間にわたって、異核共存体の94(±4)%においてBrdU標識は検出されず、これは、DNA複製が起こらなかったことを示している(図1f、g;図6;図7)。遺伝子の投与量および各細胞型が寄与するタンパク質の割合により、体細胞における核の再プログラミングの方向が決定されるので、多能性の状態への再プログラミングを支持するために、ES細胞が線維芽細胞に数でまさる(2:1)ようにインプット細胞の比率を歪めた。
【0166】
ヒト線維芽細胞においてES細胞特異的な遺伝子が誘導されたかどうかを決定するために、ヒトOct4およびヒトNanogの誘導を、種特異的プライマーを使用して、遍在するGAPDHと比較してアッセイした(図8)。融合の1日後、2日後および3日後に選別した異核共存体から単離したmRNAを半定量RT−PCRおよびリアルタイムPCRによって評価した(図2a、b)。使用した0日目の対照は、(a)単独のヒト線維芽細胞;(b)mESとhFbのPEG前、融合していない共培養物;または(c)PEGおよび融合の効果に対する対照としてPEGで処理したヒト線維芽細胞のいずれかであった。上記の0日目の対照は全て同様の結果を生じた。異核共存体におけるヒトOct4転写物およびヒトNanog転写物の両方の誘導は、早ければ融合の1日後にはっきり見えたが(図2a、b)、対照でははっきり見えず(図9)、これは、2つの重要なヒト多能性遺伝子の発現の開始は、異核共存体において急速であることを示している。1日目までに、同じ試料におけるヒトOct4およびヒトNanogの発現(GAPDHに対して正規化された)は、融合していない共培養した対照(0日目)に対して5倍に増加し、2日目および3日目に10倍で持続された(図2b)。ヒト特異的プライマーを用いて、Oct4およびNanogに加えて、他の重要な多能性遺伝子が誘導されたかどうかを、リアルタイムPCRを使用して決定した。ES細胞の自己再生を維持するために不可欠であり、Oct4およびNanogの標的になることが示されているEssrb(Bhattacharya、B.ら(2004)Blood 103、2956〜64)およびTDGF1(Bhattacharya、B.ら(2004)Blood 103、2956〜64)(クリプト(cripto))は、融合の2日後の異核共存体において、それぞれ3倍および2.5倍に上方制御されることが見いだされた(図10)。ヒト線維芽細胞においてすでに発現されているSox2およびそのプロモーターは、マウス線維芽細胞における所見と一致して、融合前に広範囲にわたって脱メチル化されており;その発現は、融合後に増加しなかった。機能的に、Essrbと相互に交換することができるKlf4(Feng、B.ら(2009)Nat Cell Biol 11、197〜203)の発現は、融合の2日後の異核共存体において変化しなかった(図10)。
【0167】
融合後のヒト線維芽細胞における核の再プログラミングの効率を評価するために、単一のFACSで選別した異核共存体を3種のヒト転写物、Oct4、Nanog、およびGAPDH(対照)について、いずれの場合にも2セットのヒト特異的なプライマーを用いて、ネステッドRT−PCRによって解析した(図2c)。マウスES細胞(対照)ではヒト遺伝子産物は全く検出されず、ヒト線維芽細胞(対照)ではヒトGAPDHのみが検出された(図8)。対照的に、3つの独立した融合実験から融合の3日後に単回のFACSで選別した異核共存体の70%でヒトOct4とヒトNanogの両方が発現され(図2c、d;図11)、これは、高い割合の異核共存体で多能性への再プログラミングが開始されたことを示している。このことは、例えば、Takahashi、K.ら(2007)Cell 131、861〜72;Takahashi、K.&Yamanaka、S.(2006)Cell 126、663〜76;Wernig、M.ら(2007)Nature 448、318〜24;and Wernig、M.ら(2008)Nat Biotechnolにおいて観察された、2〜3週間の集団全体のiPS細胞(<0.1%)における、Oct4およびNanogの発現の遅く非能率的な誘導と著しい対照をなす。
【0168】
DNA脱メチル化は、線維芽細胞のiPS細胞への再プログラミングの主要な限定ステップであることが示されているので、異核共存体におけるヒトOct4プロモーターおよびヒトNanogプロモーターの脱メチル化の時間経過および程度を対照と比較して解析した。融合の1日後、2日後および3日後に異核共存体からDNAを単離し、バイサルファイト変換に供した。ヒトOct4プロモーターおよびヒトNanogプロモーターを、ヒトおよびバイサルファイトに特異的なプライマーを用いてPCRによって増幅し(表3、図8)、産物をクローニングし、配列決定した。ヒトOct4プロモーターおよびヒトNanogプロモーターにおけるDNA脱メチル化が明白であり、3日を通して増加した(図3a)。対照的に、βグロビンHS2遺伝子座は、全体を通してメチル化されたままであり、これは、DNA脱メチル化が特異的であったことを示している。ヒトOct4プロモーターおよびヒトNanogプロモーターにおける脱メチル化CpG部位の時間経過および進行性の蓄積(図3b、c)は、リアルタイムPCRを用いて同じ3日間にわたって観察された転写物の蓄積の進行性の増加(図2b)に匹敵した。特に、ヒト体細胞における多能性遺伝子のプロモーターの脱メチル化および活性化は、Ki−67またはBrdUによって標識されていない場合に起こる(図1e、g);したがって、脱メチル化は活性であり、細胞分裂およびDNA複製と無関係である。
【0169】
哺乳動物の多能性の生殖細胞において検出され(Morgan、H.D.ら(2004)J Biol Chem 279、52353〜60)、ゼブラフィッシュにおける受精後の活性なDNA脱メチル化に関係づけられる(Rai、K.ら(2008)Cell 135、1201〜12)ので、AIDの発現について、リアルタイムPCRを用いてマウスES細胞およびヒト線維芽細胞をアッセイした。体細胞におけるAIDの発現は、一般にBリンパ球に限られると考えられているが、ヒト線維芽細胞ならびにマウスES細胞において、B−リンパ球細胞系であるRamosと比較して非常にレベルが低下してはいるが(それぞれ5%および15%)、AID mRNAが検出された(図12)。これらの細胞におけるAIDの役割を調査するために、マウスおよびヒトAID mRNAのレベルを、AIDコード領域内の異なる配列に対する3種の別個の非オーバーラップsiRNA、および、オフターゲットの効果を除外し、結果がAIDに特異的であることを確実にするために、AIDの非コード3’UTRに特異的な第4のsiRNAをトランスフェクトすることによって一過性にノックダウンした(図13)。AIDコード領域と50%同一である第5のsiRNAを対照(si対照)として使用した。ノックダウンの程度およびタイミングを、siRNA−1、siRNA−2、siRNA−3およびsiRNA−4により、AIDの転写が、トランスフェクションの3日後に対照siRNAと比較してそれぞれ81(±13)%、79(±12)%、70(±8)%、および99(±0.1)%低下した対照マウスES細胞においてまず確認した(図14、上部)。免疫沈降、その後にウエスタンブロットを使用するとマウスES細胞においてAIDタンパク質が検出され、同様に濃縮されたES細胞溶解物全体においてもAIDタンパク質が検出された(図15)。ES細胞溶解物におけるsiRNA1によるAIDノックダウンを検証し、対照タンパク質レベルが88%低下したことはmRNAが81%低下したこととよく相関した(図15)。ヒト線維芽細胞では、AIDの転写は、siRNA1、siRNA2、siRNA3およびsiRNA4により、それぞれ46(±11)%、72(±23)%、99(±0.1)%および99(±0.1)%低下した(図14、下部)。これらのデータは、ES細胞および線維芽細胞の両方において、AIDが存在し、4つの別個のsiRNAによって効率的に低下させることができることを示している。
【0170】
AIDノックダウンに供した異核共存体における再プログラミングの開始を評価するために、GAPDHと比較したOct4およびNanogの発現をリアルタイムPCRによって評価した。異核共存体実験のために、融合の24時間前にマウスES細胞とヒト線維芽細胞の両方にsiRNAをトランスフェクトした(スキームについては図5を参照されたい)。異核共存体において、リアルタイムPCRによって持続性のAIDのノックダウンが検出された。siRNA1およびsiRNA2を使用すると、AIDは、融合の3日後に、対照siRNAでトランスフェクトした異核共存体と比較して77(±6)%および35(±2)%低下した(図4a)。siRNA3および4siRNAにより、異核共存体においてより強力なノックダウンが引き起こされ、AIDは、融合の2日後に、siRNA対照と比較して96(±1)%、および89(±3)%低下した(図4a)。著しく、Oct4の発現は、siRNA1およびsiRNA2を使用すると、融合の3日後に、対照siRNAと比較して0.9(±0.6)%および9(±2)%まで低下した(図4a)。同様に、siRNA1およびsiRNA2を使用すると、Nanogの発現は、融合の3日後に、siRNA対照と比較して1.5(±0.4)%および1.5(±0.1)%まで著しく低下した(図4a)。siRNA3およびsiRNA4の存在下で、Oct4の発現は、融合の2日後に、siRNA対照と比較して8(±2)%および4(±3)%まで低下し、一方、Nanogの発現は19(±12)%および7(±4)%まで低下した。ここで使用した4種のsiRNA全ては、Oct4およびNanogの発現を少なくとも80%遮断することにおいて同様の効果を有した。これらの知見は、35%ノックダウンにより、96%ノックダウンに匹敵する多能性遺伝子の誘導の阻害が導かれたので、AIDの効果は非常に投与量感受性であることを示している。まとめると、これらのデータは、ここで使用したAIDに対する4種のsiRNA全てが、Oct4およびNanogの活性化を少なくとも80%遮断することにおいて同様に強力な効果を有したことを示している。
【0171】
プロモーターの脱メチル化に対するAIDの効果を評価するために、異核共存体におけるヒトOct4プロモーターおよびヒトNanogプロモーターのCpGのメチル化状態をアッセイした。siRNA1およびsiRNA2を使用してAIDノックダウンに供した3日目の異核共存体では、ヒトOct4プロモーターにおけるCpG脱メチル化の程度は、対照の82%と比較してそれぞれ26%および6%まで低下した(図4b、c)。Nanogプロモーターについて、CpGの脱メチル化は、対照の53%脱メチル化と比較して、それぞれ24%および25%まで低下した(図4b、c)。siRNA3およびsiRNA4を使用すると、Oct4プロモーターにおけるCpGの脱メチル化の程度は、2日目の対照試料における72%と比較してそれぞれ18%および8%まで低下し、一方、Nanogプロモーターについては、CpGの脱メチル化の程度は、対照における48%と比較してそれぞれ3%および<1%まで低下した(図4b、c)。siRNAノックダウン実験の全てについてのバイサルファイト配列決定データの概要が図16に示されている。AIDがノックダウンされるとOct4プロモーターおよびNanogプロモーターの脱メチル化が低下することに対応して、Oct4転写物およびNanog転写物の誘導が、2日目および3日目に、対照と比較して少なくとも80%低下した(図4a)。これらのデータは、プロモーターの脱メチル化が、これらの2つの多能性遺伝子の発現に欠かせないこと、およびAIDが、体細胞の再プログラミングにおいて、哺乳動物のDNAが脱メチル化されるために必要であることを示している。
【0172】
再プログラミングを開始するためのAIDの必要性をさらに調査するために、異核共存体において、siRNAノックダウンによって引き起こされるDNA脱メチル化の遮断をレスキューするその能力を試験した。AIDレベルを増加させることによってsiRNAノックダウンが克服されるかどうかを試験するために、マウスES細胞において、siRNAをトランスフェクトする前にhAIDを一過性に過剰発現させた(図5のスキームを参照されたい)。2つの別々の実験において、hAIDを、AID siRNAが存在しない異核共存体における対照と比較して2倍または4倍過剰発現させた場合、融合の1日後にプロモーターの脱メチル化または再プログラミングは加速されなかった(図17)。これは、おそらく、異核共存体では起こるまでに少なくとも1日が必要になり得る、ヒトOct4プロモーターの脱メチル化のカイネティクス、または再プログラミングを加速するためにhAIDと協調して働く追加的な因子がないことに起因し得る。しかし、siRNA−1による一過性のノックダウンを受けている異核共存体において、すなわち、siRNAの存在下でhAIDを過剰発現させると、Nanogプロモーターの脱メチル化が完全にレスキューされ、遺伝子が発現され、また、Oct4プロモーターの脱メチル化が部分的にレスキューされ、遺伝子が発現された(図18)。これらのデータは、追加されたhAIDは機能性であり、siRNAのどんな非特異的な効果も除外することを示しており、さらに、多能性への再プログラミングの開始時のDNA脱メチル化におけるAIDの特異的かつ不可欠な役割を確認している。
【0173】
ヒトOct4プロモーターおよびヒトNanogプロモーターのDNA脱メチル化におけるAIDの役割をさらに検証するために、抗AID抗体を用いてクロマチン免疫沈降(ChIP)実験を行うことにより、AIDがそれらのプロモーター領域に特異的に結合するかどうかを試験した。ChIP実験において評価したプロモーター領域は、バイサルファイト配列決定によってCpGの脱メチル化について分析された(図4d;図19)Oct4プロモーター領域およびNanogプロモーター領域の範囲内になるように設計した。ヒト線維芽細胞では、ChIP解析により、ヒトOct4プロモーター(6倍)およびヒトNanog(8倍)プロモーターの両方に対するAIDの有意な結合が示された(図4d)。したがって、AIDは、線維芽細胞において、再プログラミングの間に脱メチル化を受けるヒトOct4およびヒトNanogの重度にメチル化されたプロモーター領域に結合する。対照として、IgM定常領域(Cμ)のプロモーターへのAIDの結合は、予測通り有意であったが(Okazaki、I.M.ら(2002)Nature 416、340〜5)、線維芽細胞において発現されるThy1.1への結合は観察されなかった。
【0174】
線維芽細胞と対照的に、マウスのOct4およびNanogのプロモーター領域では、おそらく、これらのプロモーターが発現され、脱メチル化されているので、ES細胞におけるAIDタンパク質は高レベルであるにもかかわらず、AIDの結合は観察されなかった(図4d)。対照として、未分化のES細胞では発現されていない遺伝子であるCdx2のプロモーターにおいてAIDの結合が検出されたが、以前報告された通り、p53プロモーターでは検出されなかった。まとめると、これらの所見により、DNA脱メチル化におけるAIDの直接関与、なならびに多能性への再プログラミングの開始を導くヒトのNanogおよびOct4の持続的発現についての強力な裏付けがもたらされる。
【0175】
考察
DNA脱メチル化は、遺伝子サイレンシングを克服し、時間的にかつ空間的に制御された哺乳動物の遺伝子の発現を誘導するために不可欠であるが、何年も取り組まれているにもかかわらず、コンセンサスな哺乳動物のDNAデメチラーゼは未だ同定されていない。5メチル−シトシンDNAグリコシラーゼによるDNA脱メチル化の証拠が、植物体において示されているが(Gong、Z.ら(2002)Cell 111、803〜14;Choi、Y.ら(2002)Cell 110、33〜42)、チミンDNAグリコシラーゼ(TDG)またはメチル−CpG−結合性ドメインタンパク質4(Mbd4)などの哺乳動物の相同体では比較に適した機能が示されていない(Cortazar、D.ら(2007)DNA Repair(Amst)6、489〜504;Millar、C.B.ら(2002)Science 297、403〜5)。
【0176】
AIDは、B細胞における抗体多様性の生成、RNAの編集および抗ウイルス(性)の反応における役割が確立されているシトシンデアミナーゼのファミリーに属する(AID、サブグループApobec1、Apobec2およびApobec3)(Conticello、S.G.ら(2007)Adv Immunol 94、37〜73)。AIDおよびApobec1はどちらも前駆体生殖細胞、卵母細胞および初期胚において発現され、in vitroで頑強な5−メチルシトシンデアミナーゼ活性を有し(Morgan、H.D.ら(2004)J Biol Chem 279、52353〜60)、塩基除去DNA修復(BER)経路によって修復されるT−Gミスマッチをもたらし、理論的に複製を伴わないDNA脱メチル化を導き得る。最近、ゼブラフィッシュの胚において、AIDは、シトシンの脱アミノ化に影響を及ぼし、Mbd4による塩基除去を導く、Mbd4およびGadd45aとの三者間タンパク質複合体のメンバーであると意味づけられた(Rai、K.ら(2008)Cell 135、1201〜12)。第3の構成成分であるGadd45aは酵素活性を欠き、アフリカツメガエルの卵母細胞における修復に媒介されるDNA脱メチル化および遺伝子活性化におけるその役割については、まだ議論が続いている(Barreto、G.ら(20070 Nature 445、671〜5;Jin、S.G.ら(2008)PLoS Genet 4、e1000013)。
【0177】
本明細書において提供されるデータにより、AIDを活性な哺乳動物細胞におけるDNA脱メチル化に関連づける証拠がもたらされ、AID依存性のDNA脱メチル化が、ヒト線維芽細胞における多能性の誘導に必要な早期の後成的な変化であることが実証されている。異核共存体においてAIDをノックダウンすることにより、線維芽細胞核におけるヒトOct4プロモーターおよびヒトNanogプロモーターのDNA脱メチル化が妨げられた。これと一貫して、ヒト体性線維芽細胞において、AID依存性のDNA脱メチル化が低下すると、これらの多能性因子の発現および核の多能性への再プログラミングの開始が阻害され、これにより、AIDが、再プログラミングの開始に決定的な調節因子であることの強力な証拠がもたらされる。AIDの低下の阻害性の効果は、hAIDの過剰発現によりレスキューされ、Nanogに対しては完全なレスキューが観察され、Oct4に対しては部分的なレスキューが観察された。さらに、ヒト線維芽細胞において、サイレントなメチル化されたOct4プロモーターおよびNanogプロモーターにおいてAIDの結合が観察されたが、マウスES細胞における活性な、非メチル化Oct4プロモーターおよび非メチル化Nanogプロモーターでは観察されず、これは、DNA脱メチル化におけるAIDの特異的な役割を実証している。
【0178】
本発明で実現された高効率の異核共存体における再プログラミングにより、iPS細胞の生成を導く安定な再プログラミングの第1の公知のマーカーであるOct4およびNanogを含めた5つの多能性遺伝子の誘導に欠かせない調節因子を発見することが可能になった。ここで、異核共存体プラットフォームを、(a)AIDと一緒に働いて活性なDNA脱メチル化を媒介する可能性がある哺乳動物のDNA脱メチル化複合体(グリコシラーゼおよび他のDNA修復酵素)の他の構成成分を解明するため(図4e)、(b)細胞融合後にすぐに発現されるヒト遺伝子についてスクリーニングすることによって核の再プログラミングの追加的な調節因子についての不偏の検索を行うために活用することができる。今後の研究により、AIDの単独での、またはこれらの他の分子と併せての発現によりiPS細胞の生成が加速されるかどうかが示されるであろう。
【実施例2】
【0179】
AIDの潜在性相互作用物質を同定し、哺乳動物のDNA脱メチル化を調整する機能性分子プレーヤーについて理解するために、質量分析を使用した。以下のAID構築物を使用した:1)pHAGE−STEMCCAレンチウイルスベクターにクローニングした、タンパク質のN末端に2つのタンデムなFLAGタグを含有するヒトAID、および2)pHAGE−STEMCCAレンチウイルスベクターにクローニングした、タンパク質のC末端も2つのタンデムなFLAGタグを含有するヒトAID。その後、これらの構築物を含有するウイルスを使用してマウス胚性幹細胞(CGR8)に感染させ、Flag−ヒトAIDを過剰発現している安定な細胞系を選択した。対照として、2×FLAGタグのみを含有するレンチウイルスベクターを使用した。
【0180】
AIDを発現している安定なES細胞系、および対照の2×Flagを、FLAGタグに対する抗体を使用してAIDタンパク質を免疫沈降させるために、細胞質抽出物および核抽出物に分画した。生じた複合体を質量分析に供した。分析において、AIDは最も豊富なタンパク質であることが見いだされ、AIDに随伴するいくつもの独特のタンパク質が同定された(表5)。
【0181】
【表5−1】
【0182】
【表5−2】
【0183】
【表5−3】
【0184】
【表5−4】
【0185】
【表5−5】
【0186】
上記では、ただ単に本発明の原理が例示されている。当業者は本明細書においてはっきりとは記載されていない、または示されていないが、本発明の原理を具体化し、その主旨および範囲内に含まれる種々の取り合わせを考案することができることが理解されよう。さらに、本明細書において列挙されている全ての例および条件付きの言葉は、主に、読者が本発明の原理および当技術分野を推進するために出願人らが提供する概念を理解するために役立つことが意図され、そのような具体的に列挙された実施例および条件に限定されないと解釈されるべきである。さらに、本明細書において、本発明の原理、態様および実施形態ならびにその特定の実施例が列挙されている全ての記載は、それらの構造的等価物と機能的等価物の両方を包含するものとする。さらに、そのような等価物は、現在公知の等価物と今後開発される等価物、すなわち、構造にかかわらず、同じ機能を実施する、開発される任意の要素の両方を包含するものとする。本発明の範囲は、したがって、本明細書に示され、記載されている例示的な実施形態に限定されるものではない。もっと正確に言うと、本発明の範囲および主旨は、添付の特許請求の範囲によって具体化される。
【図1−1】
【図1−2】
【図2−1】
【図2−2】
【技術分野】
【0001】
関連出願の相互参照
米国特許法第119条(e)に準じて、本出願は、2010年12月18日出願の米国仮特許出願第61/284,519号の出願日について優先権を主張する;その開示は、参照により本明細書に組み込まれる。
【0002】
政府の権利
本発明は、国立保健研究機構(National Institutes of Health)から授与されたAG009521およびAG024987の下、政府支援で行われた。政府は、本発明において一定の権利を有する。
【0003】
本発明は、哺乳動物細胞におけるゲノムDNA脱メチル化を誘導するための方法および組成物、ならびに、候補薬剤を、哺乳動物細胞におけるゲノムDNA脱メチル化の調節における活性についてスクリーニングするための方法および組成物に関する。
【背景技術】
【0004】
いくつかの因子を導入することによって体細胞の核を多能性に、または別の細胞系譜の体細胞に再プログラミングすることにより、再生医療の分野における主要なブレイクスルーである、患者に特異的な人工多能性細胞(iPS)および患者に特異的な体細胞の生成が可能になった。しかし、これらのプロセスには時間がかかり(2〜3週間)、非同期的であり、頻度が低く(<0.1%)(例えば、Takahashi、K.ら(2007)Cell 131:861〜72;Takahashi、K.&Yamanaka、S.(2006)Cell 126:663〜76;Wernig、M.ら(2007)Nature 448:318〜24;Wernig、M.ら(2008)Nat Biotechnolを参照されたい)、DNA脱メチル化がネックになっている(Mikkelsen、T.S.ら(2008)Nature 454、49〜55)。したがって、哺乳動物細胞におけるDNA脱メチル化を調節する機構を解明すること、および脱メチル化を促進する薬剤を同定することは、臨床および研究の興味の対象である。本発明は、これらの問題を解決するものである。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Rai、K.ら(2008)「DNA demethylation in Zebrafish Involves the Coupling of a Deaminase、a Glycosylase、and Gadd45」Cell 135:1201〜1212(ゼブラフィッシュにおけるDNA脱メチル化の基礎をなす機構についての記載を見いだすことができる。)
【発明の概要】
【0006】
哺乳動物細胞における脱メチル化を調節するための方法、組成物およびキットが提供される。これらの方法、組成物およびキットは、細胞運命の再プログラミングを導くことにおいて、例えば、体細胞から人工多能性幹細胞(iPS)を作製することにおいて、および体細胞を、別の細胞運命に向け直すことにおいて使用される。体細胞は、例えば、メチル化の欠損から生じる、またはそれによって悪化するヒトの障害、例えば、癌および異常なゲノム刷り込みに関連する障害の治療において使用するために、in vitroで、およびin vivoで作製することができる。候補薬剤を、哺乳動物細胞におけるゲノムDNA脱メチル化の調節における活性についてスクリーニングするための方法、組成物およびキットも提供される。
【0007】
本発明の一態様では、哺乳動物細胞におけるゲノムDNAメチル化の量を減少させるための方法であって、対象とする遺伝子(複数可)の発現を制御するプロモーター領域におけるヌクレオチドのメチル化を減少させることを含む方法が提供される。方法は、最初の哺乳動物細胞を、有効量の、シチジンデアミナーゼ(CD)活性を促進する薬剤と接触させるステップを含み、例えば、哺乳動物細胞は、脱メチル化を許容する体細胞を含めた体細胞である。
【0008】
一部の実施形態では、CD活性を促進する薬剤は、活性化誘導性シチジンデアミナーゼ(AID)ポリペプチドまたはAIDポリペプチドをコードする核酸である。一部の実施形態では、CD活性を促進する薬剤は、アポリポタンパク質B RNA編集触媒成分(APOBEC)ポリペプチドまたはAPOBECポリペプチドをコードする核酸である。一部の実施形態では、細胞を表5のポリペプチドとも接触させる。ある特定の実施形態では、表5のポリペプチドは、メチル化シトシンのヒドロキシル化メチルシトシンへの変換を促進する薬剤、例えば、tetタンパク質、例えば、tet1またはtet2である。一部の実施形態では、接触させるステップをin vitroで実施する。
【0009】
一部の実施形態では、最初の哺乳動物細胞は、体細胞、例えば、脱メチル化を許容する体細胞である。一部のそのような実施形態では、作製される細胞は、人工多能性幹細胞(iPS)である。一部の実施形態では、方法は、体細胞を、iPS細胞への運命を促進する1種または複数種の因子と接触させるステップをさらに含む。一部の実施形態では、作製される細胞は、出発細胞とは異なる系列の体細胞である。一部のそのような実施形態では、方法は、体細胞を、所望の体細胞運命を促進する1種または複数種の因子と接触させるステップをさらに含む。
【0010】
一部の実施形態では、最初の細胞は、多能性幹細胞、例えば、脱メチル化を許容する多能性幹細胞である。一部のそのような実施形態では、多能性幹細胞から作製される細胞は、体細胞である。一部のそのような実施形態では、方法は、多能性幹細胞を、所望の体細胞運命を促進する1種または複数種の因子と接触させるステップをさらに含む。
【0011】
一部の実施形態では、接触させるステップは、in vivoで、ゲノムDNA脱メチル化療法を必要とする対象において実施される。一部のそのような実施形態では、最初の細胞は、腫瘍細胞、例えば、脱メチル化を許容する腫瘍細胞であり、対象は、癌に罹患している対象である。他のそのような実施形態では、最初の細胞は、形質転換されていない体細胞、例えば、脱メチル化を許容する体細胞である。
【0012】
本発明の一態様では、候補薬剤を、細胞においてゲノムDNA脱メチル化活性を調節する活性についてスクリーニングする方法が提供される。そのような方法では、細胞の第1の集団を、有効量の、シチジンデアミナーゼ(CD)活性を促進する薬剤とin vitroで接触させる。次いで、この集団の亜集団を候補薬剤と接触させ、一方、第2の集団、すなわち、対照集団は、候補薬剤と接触させない。次いで、候補薬剤と接触させた亜集団の特性を、候補薬剤と接触させなかった細胞の亜集団の特性と比較し、第1の亜集団と第2の亜集団との間の細胞の特性の違いにより、候補薬剤が細胞におけるゲノムDNA脱メチル化活性を調節することが示される。
【0013】
一部の実施形態では、CD活性を促進する薬剤は、AIDポリペプチドまたはAIDポリペプチドをコードする核酸である。一部の実施形態では、第1の集団の細胞は、腫瘍細胞、すなわち腫瘍由来の細胞である。ある特定の実施形態では、腫瘍細胞におけるゲノムDNA脱メチル化を調節する候補薬剤は、癌における腫瘍成長を調節する薬剤である。
【0014】
一部の実施形態では、第1の集団の細胞は、体細胞、またはES細胞と体細胞から作製された異核共存体である。一部の実施形態では、体細胞のDNAのゲノムDNA脱メチル化を調節する候補薬剤は、体細胞のiPS細胞化の誘導を調節する薬剤である。
【0015】
本発明の一態様では、シチジンデアミナーゼのDNA脱メチル化活性の調節における活性があるタンパク質を同定するための方法が提供される。そのような方法では、細胞の集団を、シチジンデアミナーゼをコードする配列を含む核酸と接触させ、シチジンデアミナーゼを細胞の粗タンパク質抽出物から沈殿させ、免疫沈降物を質量分析に供し、ここで、質量分析によって同定された1種または複数種のタンパク質は、シチジンデアミナーゼの脱メチル化活性に欠かせないものである。一部の実施形態では、シチジンデアミナーゼは、AIDまたはAPOBECである。一部の実施形態では、同定されるタンパク質は、表5のタンパク質である。
【0016】
本発明は、以下の詳細な説明を付属図と併せて読むと最もよく理解される。本特許または出願のファイルは、カラーで製作された少なくとも1つの図面を含有する。カラーの図面(複数可)を伴う本特許または特許出願公開のコピーは、要請し、必要な料金を支払えば、特許庁により提供されるであろう。一般的な実施に従って、図のさまざまな特徴は一定の縮尺ではないことが強調される。それどころか、さまざまな特徴の次元は、明瞭にするために任意に拡大または縮小されている。図面には、以下の図が含まれる。
【図面の簡単な説明】
【0017】
【図1】マウスES細胞×ヒト線維芽細胞の異核共存体の単離および特徴付けを示す図である。a、異核共存体の融合スキームである。GFP+マウスES細胞(mES)をDsRed+ヒト初代線維芽細胞(hFb)と2:1の比率で共培養し、次いで、ポリエチレングリコール(PEG)を使用して融合した。b、GFP+mES、DsRed+hFb、および融合の2日後に選別したGFP+DsRed+異核共存体(第1の選別異核共存体)のFACSプロファイルである。異核共存体集団をさらに再選別して(第2の選別異核共存体)、集団の約80%の濃縮(濃縮)にし、分析した。異核共存体を単離するために使用した選別ゲートが示されている。c、融合の2日後に2回選別し、サイトスピンしたGFP+とDsRed+の異核共存体の代表的な画像が示されている。異核共存体の核は、別個のままであり、融合していない。Hoechst33342(青色)は核を示し、示されている異核共存体は3つの明るいマウスの核および1つの均一に染色されたヒトの核を有する。スケールバー=50μm。d、GFP+DsRed+異核共存体を融合の2日後に2回選別し、サイトスピンし、Ki−67(青色)について染色して細胞分裂を評価した。GFP蛍光によって示される異核共存体は、Ki−67(青色)について陰性である2つの別個の核(矢印)、それとは対照的に、Ki−67について陽性に染色された単核細胞(矢じり)を有する。スケールバー=50μm。e、融合の1日後、2日後および3日後に選別し、サイトスピンした異核共存体を、Ki−67染色に基づいてスコア化し、98(±2)%の異核共存体が非分裂性であった(平均±標準誤差、p<0.05)。f、GFP−ES細胞を使用して生成した異核共存体(補足的な方法を参照されたい)を、融合の1日後にヒト線維芽細胞マーカーであるThy1.1を使用して濃縮し、サイトスピンし、Hoechst33258を使用してBrdU(緑色)および核(青色)について染色した。示されている異核共存体(矢印)は、3つの均一に染色されたヒトの核および1つの明るい点状のマウスの核を有し、BrdUについて陰性であった。対照的に、示されたヒト単核細胞(矢じり)は、BrdUについて陽性に染色される。スケールバー=50μm。g、融合の1日後、2日後および3日後に濃縮し、サイトスピンした異核共存体を、核およびBrdUの染色に基づいてスコア化した。異核共存体の94(±3)%においてDNA複製は起こらなかった(平均±標準誤差、p<0.05)。
【図2】単一細胞レベルでの異核共存体におけるヒト線維芽細胞の多能性遺伝子の発現の時間経過を示す図である。a、Oct4、NanogおよびGAPDHに対するヒト特異的なプライマーを、0日目の融合していない共培養物、および融合の1日後、2日後および3日後に単離した異核共存体(mESXhFb)のRT−PCRのために使用した。hOct4およびhNanogはどちらも、異核共存体試料において上方制御され、異核共存体におけるヒト線維芽細胞核の再プログラミングの急速な開始が示されている。b、リアルタイムPCRを使用して、融合の1日後、2日後および3日後に単離した異核共存体におけるOct4(灰色)およびNanog(黒色)の上方制御を、ヒト特異的なプライマーを使用して評価した(平均±標準偏差)。融合していない共培養物は、0日目の対照としての機能を果たし、hOct4およびhNanogの発現を、hGAPDHの発現に対して正規化した。0日目ならびに1日目、2日目および3日目に、hOct4の遺伝子発現とhNanogの遺伝子発現の間に統計的有意差が観察された(*は、p<0.03を示す)。示されているデータは、3つの独立した融合実験からのものである。c、異核共存体集団における再プログラミングの効率を評価するための単一の異核共存体のネステッドPCR。融合の3日後に異核共存体を80%まで濃縮し、単一細胞として単離した。示されている通り、単一の異核共存体に対して、GAPDH(G)、Oct4(O)およびNanog(N)に対するヒト特異的なプライマーを使用して直接逆転写およびネステッドPCRを同時に行った。単回の融合実験で分析された12個の異核共存体が示されている。2つの追加的な融合実験で分析された10個の異核共存体および31個の異核共存体が図7に示されている。d、Oct4のみを発現している異核共存体ならびにOct4とNanogの両方を発現している異核共存体の割合は70±13%であり、これにより、高い割合の異核共存体が多能性への再プログラミングを開始することが示されている。示されているデータは、3つの独立した融合実験の概要である(平均±標準偏差)。
【図3】異核共存体における、ヒト線維芽細胞の多能性遺伝子のプロモーターにおけるDNA脱メチル化の時間経過を示す図である。a、異核共存体におけるヒトOct4プロモーターおよびヒトNanogプロモーターのメチル化状態のバイサルファイト配列決定分析。異核共存体におけるヒトOct4プロモーターおよびヒトNanogプロモーターはどちらも、融合の1日後、2日後および3日後に、共培養した対照と比較して急速かつ進行性のDNA脱メチル化を示す。白い丸は非メチル化CpGジヌクレオチドを示し、黒い丸はメチル化CpGジヌクレオチドを示す。b、融合後の、異核共存体におけるヒトOct4プロモーターのパーセント脱メチル化により、脱メチル化が3日目に80%まで進行性に増加したことが示されている。c、融合後の、異核共存体におけるヒトNanogプロモーターのパーセント脱メチル化により、脱メチル化が3日目に56%まで進行性に増加したことが示されている。各時点で、2から3の独立した実験において少なくとも10クローンを分析した;代表的な10クローンが示されている。
【図4】異核共存体において、ヒト線維芽細胞の多能性への再プログラミングを開始するためのAID依存性のDNA脱メチル化の必要性を示す図である。a、リアルタイムPCRによって評価した、siRNA処理に供した異核共存体におけるAIDおよびヒト多能性遺伝子の発現。si−1、si−2、si−3およびsi−4は、AIDを対象とする別個のsiRNAである。融合の2日後に単離した異核共存体を、si−3およびsi−4で処理し、3日目に単離した異核共存体を、si−1およびsi−2で処理した。マウスAID転写物およびヒトAID転写物の総レベルを、変性プライマーのセットを使用して評価し、一方、hOct4およびhNanogに対してヒト特異的なプライマーを使用した。遺伝子発現を、AIDの発現についてはGAPDH(変性プライマー)に対して、ヒトOct4の発現およびヒトNanogの発現についてはhGAPDHに対して内部で正規化した。次いで、試料を、対応する、対照siRNAで処理した2日目の試料または3日目の試料に対して正規化し、代表的なsi対照(100%)が示されている。si−3、si−4、si−1、およびsi−2で処理した異核共存体におけるAIDの発現は、対照と比較して低下した。4種のsiRNA全てによるAIDのノックダウンにより、多能性遺伝子であるhOct4およびhNanogの発現が遮断された。b、融合の2日目および3日目に、si−3/si−4およびsi−1/si−2によってAIDをノックダウンした際のヒトOct4プロモーターおよびヒトNanogプロモーターは、それぞれ、メチル化されたままであり、これは、脱メチル化が阻害されたことを示し、異核共存体におけるDNA脱メチル化および核の再プログラミングにおけるAIDの役割を裏づけている。c、si−3/si−4、およびsi−1/si−2によってAIDをノックダウンした際のヒトOct4プロモーターおよびヒトNanogプロモーターでのパーセント脱メチル化により、それらのそれぞれの、si対照で処理した対照試料の2日目および3日目と比較して、脱メチル化が遮断されたことが示された。d、ヒトAIDは、線維芽細胞における重度にメチル化されたヒトNanog遺伝子およびヒトOct4遺伝子のプロモーターに動員される。mESおよびhFbにおいて、抗AID抗体を用いてクロマチン免疫沈降を行った。バックグラウンドのIgGシグナルと比較したAIDの占有率が示されている(平均±標準誤差)。hFbならびに陽性対照としてmES、CμおよびCdx2において、それぞれ有意なAIDの結合が検出された(p<0.02)。hOct4およびhNanogのメチル化プロモーターへのAIDの結合は、ヒト線維芽細胞において有意であった(p<0.02)が、マウスES細胞における非メチル化Oct4プロモーターおよび非メチル化Nanogプロモーターへの有意な結合は観察されなかった。e、異核共存体における多能性への再プログラミングにおけるAID依存性のDNA脱メチル化のモデル。デアミナーゼAIDと一緒に作用し得る哺乳動物のDNAデメチラーゼ複合体(X、Y、およびZ)の他の推定上の構成成分は、まだ同定されていない。
【図5】異核共存体の生成、siRNAノックダウン、hAID過剰発現、およびレスキュー実験のためのスキームを示す図である。
【図6】BrdU実験のための異核共存体のThy1.1濃縮を示す図である。a、ヒト線維芽細胞に対するThy1.1の特異性。a(パネル上部)に示されているように、GFP+マウスES細胞はヒト特異的なThy1.1抗体に結合しないが、dsRed+ヒト線維芽細胞の100%がThy1.1(パネル下部)に結合し、これは、Thy1.1をヒト線維芽細胞に対する細胞表面マーカーとして特異的に使用することができることを示している。b、Thy1.1抗体を使用した異核共存体の濃縮。ビオチン化したThy1.1を使用して、PEG処理したmES共培養物およびhFb共培養物を標識し、ストレプトアビジン磁気ビーズを使用してヒト線維芽細胞および異核共存体(mESX hFb)について濃縮した。b(左下)に示されているように、濃縮前にはPEG処理したmES共培養物およびFb共培養物に0.1〜1%の異核共存体が存在するが、磁気ビーズ濃縮後には、Thy1.1陽性の異核共存体および線維芽細胞は、それぞれ10%および80%であった。
【図7】異核共存体はDNA複製を行わないことを示す図である。GFP−(非GFP)のES細胞およびヒト線維芽細胞を使用して生成した異核共存体を、融合の1日後に、ヒト線維芽細胞マーカーであるThy1.1を使用して濃縮し(補足的な方法を参照されたい)、サイトスピンし、BrdU(緑色の)および核(青色)についてHoechst33258を用いて染色した。示されている異核共存体(矢印)は、3つの均一に染色されたヒト核および1つの明るい点状のマウスの核を有し、BrdUについては陰性であり、これは、異核共存体においてDNA複製がないことを示している。対照的に、示されている単核細胞(矢じり)は、BrdUについて陽性に染色される。磁性ストレプトアビジンビーズは、免疫蛍光による画像において小さな黒い丸として細胞の表面上に見ることができる。スケールバー=50ミクロン。
【図8】種間の異核共存体における多能性遺伝子の活性化を試験するためのヒト特異的なプライマーの検証についての図である。a、Oct4およびNanogに対して設計したヒト特異的なプライマーは、ヒトES細胞(hES)由来のヒト転写物を選択的に増幅するが、マウスES細胞(mES)由来のOct4転写物およびNanog転写物は検出しない。b、ヒトOct4プロモーターおよびヒトNanogプロモーターのメチル化状態を評価するために設計したヒトおよびバイサルファイトに特異的なプライマーにより、hFbからのみ産物が増幅され、mESからは増幅されなかった。c、単一の異核共存体におけるヒトGAPDH、Oct4転写物およびNanog転写物の発現を評価するために設計したネステッドPCRのプライマーの種特異性。ヒトGAPDHプライマーを用いるとhES細胞とヒト初代線維芽細胞(hFb)の両方で産物が検出されるが、mESでは産物は検出されない、ヒトOct4転写物およびヒトNanog転写物は、hES細胞のみで検出され、hFbまたはmESでは産物は検出されない。
【図9】共培養した対照における、ある期間にわたるヒト多能性遺伝子の発現。0日目および3日目の、マウスES細胞およびhFbの融合していない共培養物を、多能性遺伝子であるOct4およびNanogに対するヒト特異的なプライマーを使用してリアルタイムPCRによって分析した。遺伝子発現の倍率変化を得るために、遺伝子発現をhGAPDHおよび0日目の対照に対して正規化した。データにより、hOct4およびhNanogの発現が0日目から3日目まで変化しないままであることが示されている。
【図10】異核共存体におけるヒト多能性遺伝子の活性化および発現を示すグラフである。多能性遺伝子であるEssrb、TDGF1、Sox2ならびに細胞周期制御因子であるKlf4およびc−mycに対するヒト特異的なプライマーを、0日目の融合していない共培養物および融合の2日後に単離した異核共存体(mESX hFb)のリアルタイムPCRのために使用した。遺伝子発現の倍率変化を得るために、遺伝子発現をhGAPDHおよび0日目の対照に対して正規化した。異核共存体の試料においてEssrb、TDGF1およびc−mycが誘導された。ヒト線維芽細胞においてすでに発現されているSox2はそれ以上増加せず、Essrbと相互に交換することができることが公知である(Feng、B.ら(2009)Nat Cell Biol 11、197〜203)Klf4もそれ以上増加しない。
【図11】単一の異核共存体における再プログラミングの効率を示す図である。図2cにおいて、単一の融合実験から得られた12個の異核共存体からの結果が示されている。本図には、2つの追加的な独立した融合実験から得られた異核共存体における多能性遺伝子の発現が示されている(それぞれn=10および31)。3つの独立した実験の結果の概要および統計値は図2dに示されている。
【図12】相対的なAID mRNAの発現を示す図である。AIDの転写レベルを、B−リンパ球細胞系(Ramos)、マウス胚性幹細胞(mES)およびヒト線維芽細胞(hFb)においてリアルタイムPCRによって評価した。転写レベルをGAPDHに対して正規化する。Ramos、mES、およびhFbにおけるAIDの発現の比率は、およそ100:15:5である。
【図13】AID標的配列を対象とするsiRNA:ヒトおよびマウスについてのアラインメントを示す図である。ヒト(配列番号105〜配列番号114)およびマウス(配列番号115〜配列番号124)のAID mRNAにおけるsiRNA標的配列およびそれらの対応する標的、ならびにAID転写物に沿ったそれらの相対的な位置が示されている。標的配列の上にミスマッチ(*)が示されている。
【図14】マウスES細胞およびヒト線維芽細胞におけるAIDノックダウンの効力を示すグラフである。siRNAトランスフェクションの3日後にマウスES細胞およびヒト線維芽細胞におけるAIDのノックダウンをリアルタイムPCRによって評価した。AID si−1、AID si−2、AID si−3およびAID si−4は、AIDを対象とする別個のsiRNAであり(図13に示されている配列)、それらにより、マウスES細胞におけるAIDの転写(GAPDHに対して正規化した)が、それぞれ、トランスフェクションの3日後に、対照siRNAと比較して81(±13)%、79(±12)%、70(±8)%、および99(±0.1)%低下した。ヒト線維芽細胞では、AID mRNAのレベルは、siRNA1、siRNA2、siRNA3およびsiRNA4によって、それぞれ46(±11)%、72(±23)%、99(±0.1)%および99(±0.1)%低下した。
【図15】マウスES細胞におけるAIDタンパク質およびノックダウンの検出を示す図である。a、マウスES細胞全体の溶解物2mgからの免疫沈降後のAIDタンパク質の検出。1%のインプット(20μg)をローディングした。b、si−1でトランスフェクトした3日後の濃縮されたマウスES細胞溶解物(170μg)におけるAIDタンパク質レベルの検出。α−チューブリンは、ローディング対照として示されている。c、α−チューブリンに対して正規化されたAIDタンパク質レベルの定量化。si−1で処理したES細胞におけるAIDタンパク質レベルは、si対照試料と比較して12%まで低下した。
【図16】異核共存体におけるヒトOct4プロモーターおよびヒトNanogプロモーターについて収集されたバイサルファイト配列データを示す図である。合計330クローンについて配列決定した。
【図17】ヒトAIDの過剰発現によって異核共存体における再プログラミングの開始は加速されないことを示す図である。融合の1日後の異核共存体の線維芽細胞における、ヒトAID(hAID)の一過性の過剰発現を伴う、またはそれを伴わない、ヒトOct4プロモーターおよびヒトNanogプロモーターのメチル化状態についてのバイサルファイト配列決定分析(図5)。a、hAIDレベルをリアルタイムPCRによって評価し、2つの別々の融合実験において、1日目の異核共存体において、それぞれ2倍および4倍に上方制御されることが見いだされた。b、d、ヒトOct4プロモーターおよびヒトNanogプロモーターのDNA脱メチル化の程度は、hAIDが過剰発現されても増加しない。2つの独立した融合実験について同様の結果が得られた。白い丸は、非メチル化CpGジヌクレオチドを示し、黒い丸は、メチル化CpGジヌクレオチドを示す。2つの独立した融合実験において少なくとも10クローンを分析した;代表的な10クローンが示されている。c、e、融合の1日後に、hAIDの一過性の過剰発現を伴う、またはそれを伴わない、異核共存体におけるヒトOct4プロモーターおよびヒトNanogプロモーターにおいて観察されたパーセント脱メチル化。Oct4プロモーターおよびNanogプロモーターにおけるDNA脱メチル化は、hAIDが過剰発現していても増加しない。
【図18】ヒトAIDの過剰発現により、異核共存体におけるAIDが一過性にノックダウンされている間、再プログラミングの開始がレスキューされることを示す図である。si−RNAターゲティングAID(si−1)でトランスフェクトした異核共存体において過剰発現しているヒトAID(hAID)によってレスキュー実験を行った(図5参照)。a、hAIDレベルを、リアルタイムPCRによって評価し、2つの別々の融合実験において、2日目の異核共存体において、それぞれ2.5倍および4倍に上方制御されることが見いだされた。b、ヒト特異的なプライマーを使用したリアルタイムPCRによって評価された通り、hAIDの過剰発現により、2日目の異核共存体において、対照と比較して多能性遺伝子であるhOct4の発現が部分的にレスキューされ、hNanog遺伝子発現が完全にレスキューされる。発現レベルを、同じ2日目の試料においてhGAPDHに対して正規化し、次いで0日目の対照に対して正規化する。c、d、融合の2日後に単離した異核共存体を、ヒトOct4プロモーターおよびヒトNanogプロモーターのメチル化状態についてバイサルファイト配列決定分析に供した。どちらのプロモーターも脱メチル化を示し、これは、AIDの下方制御によって引き起こされる再プログラミングの遮断は、hAIDの過剰発現により、Nanogプロモーターの脱メチル化の完全なレスキューおよびOct4プロモーターの脱メチル化の部分的なレスキューを伴って克服される。Oct4プロモーターの脱メチル化は、72%の対照レベルと比較すると、8%脱メチル化(si−1)から22%脱メチル化(hAID+si−1)までレスキューされる。Nanogプロモーターの脱メチル化は、31%脱メチル化(si−1)から51%脱メチル化(hAID+si−1)までレスキューされる。47%の対照レベルと比較すると、完全なレスキューが観察される。
【図19】バイサルファイト特異的配列決定およびChIPアッセイにおいて調査したCpG密度を示すヒトOct4プロモーター(配列番号125)およびヒトNanogプロモーター(配列番号126)のマップである。バイサルファイト特異的なアンプリコンの配列が示されている。ヒトOct4プロモーターおよびヒトNanogプロモーターにおけるCpG部位が肉太活字で示されている。ChIPプライマーの適用範囲の領域(リアルタイムPCRのアンプリコン)が示されている。ATG開始コドンからの距離が示されている。
【発明を実施するための形態】
【0018】
本方法および組成物について記載する前に、本発明は記載されている特定の方法または組成物に限定されず、それ自体は当然変動することが理解されるべきである。本発明の範囲は、添付の特許請求の範囲によってのみ限定されるので、本明細書において使用される用語法は、特定の実施形態を記載するためだけのものであり、限定的なものではないことも理解される。
【0019】
さまざまな値が提供される場合、文脈によりそうでないことが明確に規定されない限り、下限の単位の10分の1まで、その範囲の上限と下限の間に入る値のそれぞれも、明確に開示されていると理解される。任意の明示された値、または明示された範囲の間に入る値および任意の他の明示された値、またはその明示された範囲の間に入る値のより小さな範囲のそれぞれが本発明の範囲内に包含される。これらのより小さな範囲の上限と下限は、それぞれ独立に、その範囲に含めることができる、またはその範囲から排除することができ、そのより小さな範囲に限界のいずれかが含まれる、どちらの限界も含まれない、またはどちらの限界も含まれる各範囲も、本発明に包含され、明示された範囲内の任意の明確に排除される限界に支配される。明示された範囲が限界の一方または両方を含む場合、それらの含まれる限界のいずれか、またはその両方を除く範囲も、本発明に包含される。
【0020】
別段の定義のない限り、本明細書において使用される全ての技術用語および科学用語は、本発明が属する分野の当業者に一般に理解されているものと同じ意味を有する。本明細書に記載の方法および材料と類似した、またはそれと等しい任意の方法および材料を本発明の実施または試験において使用することができるが、一部の潜在的な好ましい方法および材料がここに記載されている。本明細書で言及されている全ての刊行物が、引用されている刊行物に関連して方法および/または材料を開示し記載するために、参照により本明細書に組み込まれる。矛盾がある範囲では、組み込まれた刊行物の開示のいずれにも本開示が取って代わることが理解される。
【0021】
本明細書および添付の特許請求の範囲において使用される、単数形「a(1つの)」、「an(1つの)」、および「the(その)」は、文脈によりそうでないことが明確に規定されない限り、複数の指示対象を包含することに留意しなければならない。したがって、例えば、「細胞」への言及は、複数のそのような細胞を包含し、「ペプチド」への言及は、1種または複数種のペプチドおよびその等価物、例えば、当業者に公知のポリペプチドなどへの言及を包含する。
【0022】
本明細書で考察されている刊行物は、単に、本出願の出願日より前のそれらの開示について提供されている。本明細書における全ては、本発明が、先行発明の理由でそのような刊行物に先立つ権限がないことの容認であると解釈されるべきではない。さらに、提供されている刊行物の日付は、それぞれ独立に確認する必要があり得る実際の刊行物の日付とは異なる可能性がある。
【0023】
定義
哺乳動物細胞における脱メチル化を調節するための方法、組成物およびキットが提供される。候補薬剤を、哺乳動物細胞におけるゲノムDNA脱メチル化活性のレベルの調節における活性についてスクリーニングするための方法、組成物およびキットも提供される。これらの方法、組成物およびキットは、人工多能性幹細胞(iPS)および体細胞をin vitroで作製することにおいて、および、癌およびゲノム刷り込みにおける欠損から生じる障害を含めたヒトの障害をin vivoで治療するために使用される。これらの、または他の本発明の目的、利点および特徴は、以下に十分に記載されている組成物および方法の詳細を読めば、当業者には明白になるであろう。
【0024】
「DNAメチル化」または単に「メチル化」は、DNAにメチル基が付加されることを意味する。DNAにメチル基が付加される反応は、酵素のDNAメチルトランスフェラーゼ(DNMT)によって触媒される。脊椎動物では、DNAメチル化は、一般には、ヌクレオチドシトシンにおいて、通常CpG部位(シトシン−リン酸−グアニン部位;すなわち、DNA配列においてシトシンのすぐ次にグアニンが続く場所)で起こる。これにより、シトシンが、本明細書では互換的に「5−メチルシトシン」「5−meC」、および「メチル化シトシン」と称される5−メチルシトシンに変換される。付加されたメチル基により、シトシンの構造が、その塩基対合の性質が変化することなく変化する。「GCリッチ」領域(すなわち、約65%のCG残基でできている)であるCpG配列およびCpGアイランドのメチル化の程度は、多くの場合、遺伝子の転写活性に関連し、高度にメチル化されたCpGアイランドを含有するプロモーターは一般にはサイレントであり、非メチル化CpGアイランドおよびあまりメチル化されていないCpGアイランドを含有するプロモーターは、一般には活性である。
【0025】
「DNA脱メチル化」または単に「脱メチル化」は、CpG配列がメチル化CpG配列から非メチル化CpG配列に変換されることを意味する。
【0026】
「DNA脱メチル化を許容する細胞」または「脱メチル化を許容する細胞」は、そのCpG配列をメチル化CpG配列から非メチル化CpG配列に変換することができる細胞を意味する。細胞に5−meCpGリッチヌクレオチド配列を含むDNAベクターを提供すること、および、ベクターによって供給されたヌクレオチド配列を、例えば、バイサルファイト配列決定またはメチラーゼ感受性制限エンドヌクレアーゼ消化によって回収し分析してそのヌクレオチド配列のCpG配列が脱メチル化されているかどうかを決定して、細胞において活性化誘導性シチジンデアミナーゼ(AID)(GenBank受託番号NM_020661)をコードするcDNAを過剰発現させることによって細胞が脱メチル化を許容するかどうかを決定することができる。
【0027】
「シチジンデアミナーゼ活性」または「CD活性」は、リボース環(シチジン)またはデオキシリボース環(デオキシシチジン)に付着しているシトシンヌクレオシドまたは5−メチルシトシンヌクレオシドからアミン基が除去される酵素経路の活性を意味する。シトシンからアミン基が除去されることにより、ヌクレオシドがウラシルに変換され、一方、5−メチルシトシンからアミン基が除去されることにより、ヌクレオシドがチミンに変更される。例えば、以下の略図を参照されたい:
【0028】
【化1】
【0029】
「多能性幹細胞」または「多能性細胞」は、生物体において全種類の細胞に分化する能力を有する細胞を意味する。多能性細胞は、奇形腫を形成し得、生きている生物体において外胚葉組織、中胚葉組織、または内胚葉組織の一因となっている。多能性幹細胞の例は、胚性幹(ES)細胞、胚性生殖幹(EG)細胞、および人工多能性幹(iPS)細胞である。
【0030】
「胚性幹細胞」または「ES細胞」は、a)自己再生することができ、b)生物体において分化して全種類の細胞を産生することができ、かつc)発生中の生物体の胞胚の内部細胞塊に由来する細胞を意味する。ES細胞は、生物体において全種類の細胞に分化する能力を維持させながら、長期にわたって培養することができる。培養物中で、ES細胞は、一般には、大きな核細胞質比、明確な境界および目立った核を有する平らなコロニーとして成長する。さらに、ES細胞は、SSEA−3、SSEA−4、TRA−1−60、TRA−1−81、およびアルカリホスファターゼを発現するが、SSEA−1は発現しない。ES細胞を生成し、特徴づける方法の例は、例えば、その開示が参照により本明細書に組み込まれる米国特許第7,029,913号、米国特許第5,843,780号、および米国特許第6,200,806号において見ることができる。
【0031】
「胚性生殖幹細胞」、「胚性生殖細胞」または「EG細胞」は、a)自己再生することができ、b)生物体において分化して全種類の細胞を産生することができ、かつc)生殖細胞および生殖細胞前駆体、例えば、始原生殖細胞に由来する、すなわち、精子および卵子になる細胞を意味する。胚性生殖細胞(EG細胞)は、上記の胚性幹細胞と同様の性質を有すると考えられている。EG細胞を生成し、特徴づける方法の例は、例えば、その開示が参照により本明細書に組み込まれる、米国特許第7,153,684号;Matsui、Yら、(1992)Cell 70:841;Shamblott、M.ら(2001)Proc.Natl.Acad.Sci.USA 98:113;Shamblott、M.ら(1998)Proc.Natl.Acad.Sci.USA、95:13726;およびKoshimizu、U.ら(1996)Development、122:1235において見ることができる。
【0032】
「人工多能性幹細胞」または「iPS細胞」は、a)自己再生することができ、b)生物体において分化して全種類の細胞を産生することができ、かつc)体細胞に由来する細胞を意味する。iPS細胞は、ES細胞様の形態を有し、大きな核細胞質比、明確な境界および目立った核を有する平らなコロニーとして成長する。さらに、iPS細胞は、これらに限定されないが、アルカリホスファターゼ、SSEA3、SSEA4、Sox2、Oct3/4、Nanog、TRA160、TRA181、TDGF 1、Dnmt3b、FoxD3、GDF3、Cyp26a1、TERT、およびzfp42を含めた当業者に公知の1種または複数種の重要な多能性マーカーを発現する。iPS細胞を生成し、特徴づける方法の例は、例えば、その開示が参照により本明細書に組み込まれる、出願番号:US20090047263、US20090068742、US20090191159、US20090227032、US20090246875、およびUS20090304646において見ることができる。
【0033】
「体細胞」は、実験的な操作をしなければ、普通は生物体において全種類の細胞を生じさせない、生物体における任意の細胞を意味する。言い換えれば、体細胞は、自然には体の3つの胚葉、すなわち、外胚葉、中胚葉および内胚葉の全ての細胞を生成しない、十分に分化した細胞である。例えば、体細胞は、神経細胞と神経前駆細胞の両方を含み、神経前駆細胞は、中枢神経系の全部または一部の細胞型を自然に生じさせることができ得るが、中胚葉系列または内胚葉系列の細胞を生じさせることはできない。
【0034】
「再プログラミング因子」は、細胞に作用して転写を変化させ、それによって細胞を多能性に再プログラミングする生物活性因子の1種または複数種、すなわち、混合物を意味する。細胞に再プログラミング因子を提供する、すなわち細胞を再プログラミング因子と接触させる本発明の方法では、これらの再プログラミング因子は、個々に、または単一の組成物として、すなわち、再プログラミング因子を予備混合した組成物として細胞に提供することができる。因子は、同じモル比で、または異なるモル比で提供することができる。因子は、本発明の細胞を培養する過程において、一回または多数回提供することができる。
【0035】
「再プログラミングの効率」は、細胞のin vitro培養物の、別の細胞型の細胞を生じるように再プログラムされる能力を意味する。薬剤、例えば、シチジンデアミナーゼ活性を促進する薬剤の存在下で増強された再プログラミング効率を示す細胞は、その薬剤と接触させた場合に、その薬剤と接触させなかった細胞と比較して、別の細胞型の細胞を生じる増強された能力を示す。増強されるとは、細胞培養物が、薬剤と接触させなかった細胞培養物の能力の少なくとも50%、約100%、約200%、約300%、約400%、約600%、約1000%、約2000%、少なくとも約5000%の、新しい細胞型を生じさせる能力を有することを意味する。言い換えれば、細胞培養物により、薬剤と接触させていない細胞の集団によって産生される新しい細胞型の細胞の約1.5倍、約2倍、約3倍、約4倍、約6倍、約10倍、約20倍、約30倍、約50倍、約100倍、約200倍を超える新しい細胞型の細胞が産生される。本出願の一部の実施形態では、再プログラミングの効率を増強する薬剤は、所望の細胞運命を獲得する間に活性になることが当技術分野で公知であるプロモーターにおけるDNAメチル化の量を、例えば、薬剤がない場合に観察されるDNAメチル化の量と比較して1.5分の1以下、すなわち、約1.5分の1、約2分の1、約3分の1、約4分の1、約6分の1、または約10分の1またはそれ未満に減少させる薬剤である。本出願の一部の実施形態では、再プログラミングの効率を増強する薬剤は、所望の細胞運命を獲得する間に活性になることが当技術分野で公知であるプロモーターによって調節される遺伝子の転写量を、例えば、薬剤がない場合に観察される転写量と比較して約1.5倍以上、すなわち、約1.5倍、約2倍、約3倍、約4倍、約6倍、または約10倍またはそれ以上増加させる薬剤である。
【0036】
「プロモーター配列」は、細胞においてRNAポリメラーゼに結合し、下流(3’方向)のコード配列の転写を開始させることができるDNA調節領域である。本発明を明確にする目的において、プロモーター配列は、その3’末端に転写開始部位が結合し、バックグラウンドを超えて検出可能なレベルで転写を開始するために必要な最小数の塩基またはエレメントを含むように上流(5’方向)に伸長している。プロモーター配列内に、転写開始部位、ならびにRNAポリメラーゼの結合に関与するタンパク質結合ドメインが見いだされる。真核生物のプロモーターは、多くの場合、常にではないが、「TATA」ボックスおよび「CAT」ボックスを含有する。誘導性プロモーターを含めた種々のプロモーターを使用して、本発明の種々のベクターを駆動することができる。プロモーター配列からの転写活性は、プロモーターがメチル化されている程度によって調節することができる。
【0037】
「治療」、「治療すること」などの用語は、本明細書では、一般に、所望の薬理的効果および/または生理的効果を得ることを意味するように使用される。効果は、疾患またはその症状を完全にまたは部分的に予防するという点で予防的であってよく、かつ/または、疾患および/または疾患に起因する有害作用を部分的または完全に治癒させるという点で治療的であってよい。「治療」は、本明細書で使用される場合、哺乳動物における疾患の任意の治療を包含し、(a)疾患の素因があり得るが、今のところはまだそれを有すると診断されていない対象において疾患が生じることを予防すること;(b)疾患を抑制すること、すなわち、その発生を制止すること;または(c)疾患を軽減すること、すなわち、疾患の退縮を引き起こすことを含む。治療剤は、疾患または害が発症する前、その間、またはその後に投与することができる。進行中の疾患を治療し、その治療により患者の望ましくない臨床症状を安定させる、または低減することが特に興味深い。そのような治療は、患部組織の機能が完全に喪失する前に行うことが望ましい。本主題の療法は、疾患の症候性の段階、およびある場合では疾患の症候性の段階後に施されることが望ましい。
【0038】
「個体」、「対象」、「宿主」および「患者」という用語は、本明細書では互換的に使用され、診断、治療、または療法が望まれる任意の哺乳動物の対象、特に、ヒトを指す。
【0039】
シチジンデアミナーゼ(CD)活性を促進する薬剤
哺乳動物細胞におけるメチル化の量を調節するための方法、組成物およびキットが提供される。本発明の一態様では、哺乳動物細胞におけるゲノムDNAメチル化の量を、細胞を、シチジンデアミナーゼ活性を促進する1種または複数種の薬剤と接触させることによって減少させる。上記の通り、シチジンデアミナーゼ(CD)活性は、DNAまたはRNAにおけるシトシンまたは5−メチルシトシンからアミノ基を除去する酵素活性である。本出願において使用されるシチジンデアミナーゼ活性を促進する薬剤の例は、シチジンデアミナーゼのAID/APOBECクラスのポリペプチドおよび断片、ならびにこれらのポリペプチドおよび断片をコードする核酸である。
【0040】
活性化誘導性シチジンデアミナーゼは、AID、AICDA、ARP2、CDA2、またはHIGM2とも称され、適応性の体液性免疫系におけるその役割について最もよく知られているシチジンデアミナーゼであり、免疫グロブリン遺伝子座のDNAにおけるシトシン残基を脱アミノ化して、抗体遺伝子の多様化を強化する(免疫グロブリンV遺伝子の体細胞超変異および遺伝子変換ならびにIgC遺伝子のスイッチ組換え)。「AID遺伝子産物」、「AIDポリペプチド」、「AIDペプチド」、および「AIDタンパク質」という用語は、本明細書では互換的に使用され、ネイティブな配列のAIDポリペプチド、AIDポリペプチド変異体、AIDポリペプチド断片およびキメラAIDポリペプチドを指す。AIDポリペプチドおよびそれをコードする核酸のネイティブな配列は、GenBank受託番号NM_020661(配列番号1、配列番号2)において見ることができる。
【0041】
アポリポタンパク質B RNA編集触媒成分タンパク質は、APOBECタンパク質とも称され、シチジンを脱アミノ化するタンパク質のファミリーである。「APOBEC遺伝子産物」、「APOBECポリペプチド」、「APOBECペプチド」、および「APOBECタンパク質」という用語は、本明細書では互換的に使用され、APOBECポリペプチド、APOBECポリペプチド変異体、APOBECポリペプチド断片およびキメラのAPOBECポリペプチドのネイティブな配列を指す。APOBECファミリーの始祖メンバーであるAPOBEC1は、シトシン6666を脱アミノ化してウラシルにし、それによって中途終止コドンを創出し、短縮されたアポリポタンパク質Bポリペプチド鎖の組織特異的な産生を強化することによってアポリポタンパク質B RNAを編集する複合体の触媒成分である。APOBEC1ポリペプチドおよびそれをコードする核酸のネイティブなヒト配列は、GenBank受託番号NM_001644(配列番号3、配列番号:4)において見ることができる。APOBEC3ファミリーのメンバー(APOBEC3F、APOBEC3GおよびAPOBEC3H)は、レトロウイルスの感染を制限する先天性免疫経路において、レトロウイルスの第1の鎖cDNA複製中間体においてシトシンを脱アミノ化すること、またはウイルスのゲノムに致死的な超変異を生じさせることによって役割を果たし;APOBEC3F(KA6、ARP8、MGC74891、およびBK150C2.4.mRNAとしても公知である)のネイティブなヒト配列は、GenBank受託番号NM_145298.5(アイソフォームa)(配列番号5、配列番号6)およびNM_001006666.1(アイソフォームb)(配列番号7、配列番号8)において見ることができ;APOBEC3G(ARP9、CEM15、MDS019、FLJ12740、bK150C2.7およびdJ494G10.1としても公知である)のネイティブなヒト配列は、GenBank受託番号NM_021822.2(配列番号9、配列番号10)において見ることができ;APOBEC3H(ARP10としても公知である)のネイティブなヒト配列は、GenBank受託番号NM_001166003.1(アイソフォーム1)(配列番号11、配列番号12)、NM_181773.3(アイソフォーム2)(配列番号13、配列番号14)NM_001166002.1(アイソフォーム3)(配列番号15、配列番号16)、NM_001166004.1(アイソフォーム4)(配列番号17、配列番号18)において見ることができる。シチジンデアミナーゼのAPOBECファミリーの他のメンバーとしては、そのネイティブなヒト配列が、GenBank受託番号NM_006789(配列番号19、配列番号20)において見ることができるAPOBEC2(ARP1およびARCD1としても公知である);そのネイティブなヒト配列が、GenBank受託番号NM_145699.3(配列番号21、配列番号22)において見ることができるAPOBEC3(Phorbolin1、ARP3、PHRBN、およびbK150C2.1としても公知である);そのネイティブなヒト配列が、GenBank受託番号NM_004900(配列番号23、配列番号24)において見ることができるAPOBEC3B(ARP4、ARCD3、PHRBNL、APOBEC1L、FLJ21201、bK150C2.2およびDJ742C19.2としても公知である);そのネイティブなヒト配列が、GenBank受託番号NM_014508.2(配列番号25、配列番号26)において見ることができるAPOBEC3C(PBI、ARP5、ARDC2、ARDC4、APOBEC1L、MGC19485、およびbK150C2.3としても公知である);およびそのネイティブなヒト配列が、GenBank受託番号NM_152426.3(配列番号27,配列番号28)において見ることができるAPOBEC3D(ARP6、APOBEC3E、およびAPOBEC3DEとしても公知である)が挙げられる。
【0042】
シチジンデアミナーゼのAID/APOBECクラスおよびタンパク質のこのクラスの間で保存されているドメインに関するより多くの情報は、その開示が参照により本明細書に組み込まれる、Conticello、S.G.ら(2005)Molecular Biologyおよび Evolution 22(2)367〜377において見ることができる。
【0043】
一部の実施形態では、CD活性、したがって、ゲノムDNA脱メチル化を促進する薬剤は、AIDポリペプチドである。AIDポリペプチドは、シチジンの脱アミノ化を促進するAID配列を含むポリペプチドである。AIDポリペプチドは、AIDの完全なポリペプチド配列またはシチジンデアミナーゼ活性を有するAIDの断片、例えば、C末端の10アミノ酸を抜いた全長のポリペプチド(Barretoら(2003)Mol.Cell 12(2):501〜8)に対して50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、98%または100%の配列同一性を有するポリペプチドを含んでよい。そのような断片は、当業者が当技術分野で周知の一般的な生化学的技法および遺伝学的技法を使用して容易に同定可能である。全長のAIDまたはそのシチジンデアミナーゼ活性ドメインのポリペプチド配列に対して50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、98%または100%の配列同一性を有するポリペプチドをコードする核酸、およびこれらの核酸を含むベクターも、本発明に包含される。
【0044】
一部の実施形態では、CD活性、したがって、ゲノムDNA脱メチル化を促進する薬剤は、APOBECポリペプチドである。APOBECポリペプチドは、シチジンの脱アミノ化を促進するAPOBEC配列を含むポリペプチドである。APOBECポリペプチドは、APOBECの完全なポリペプチド配列またはシチジンデアミナーゼ活性を有するAPOBECの断片に対して50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、98%または100%の配列同一性を有するポリペプチドを含んでよい。そのような断片は、当業者が当技術分野で周知の一般的な生化学的技法および遺伝学的技法を使用して容易に同定可能である。全長のAPOBECポリペプチドまたはそれらのシチジンデアミナーゼ活性ドメインのいずれかのポリペプチド配列に対して50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、98%または100%の配列同一性を有するポリペプチドをコードする核酸、およびこれらの核酸を含むベクターも、本発明に包含される。
【0045】
上記の通り、本発明において使用するために適した薬剤は、シチジンデアミナーゼタンパク質のAID/APOBECクラスのポリペプチドおよび断片、ならびにこれらのポリペプチドおよび断片をコードする核酸を含む。一部の実施形態では、CD活性を促進する1種または複数種の薬剤は、核内作用性の非組み込み型ポリペプチドである。言い換えれば、対象細胞を、CD活性を促進するポリペプチド(「CD活性ポリペプチド」)と接触させ、核内で作用させる。非組み込み型とは、ポリペプチドが対象細胞、すなわち脱メチル化活性を促進することが望まれる細胞のゲノム内に組み込まれないことを意味する。
【0046】
CD活性ポリペプチドの細胞膜を渡る輸送を促進するために、CD活性ポリペプチド配列をポリペプチド浸透性ドメインに融合することができる。いくつもの浸透性ドメインが当技術分野で公知であり、ペプチド、ペプチド模倣薬、および非ペプチド担体を含めた本発明の核内作用性の非組み込み型ポリペプチドにおいて使用することができる。例えば、浸透性ペプチドは、ペネトラチンと称されるDrosophila melanogasterの転写因子であるAntennapaediaの第3のアルファヘリックス由来であってよい。別の例として、浸透性ペプチドは、例えば、天然に存在するtatタンパク質のアミノ酸49〜57を含み得るHIV−1tat基本領域アミノ酸配列を含む。他の浸透性ドメインとしては、ポリアルギニンモチーフ、例えば、HIV−1revタンパク質のアミノ酸34〜56の領域、ノナアルギニン、オクタアルギニンなどが挙げられる。(例えば、移行ペプチドおよびペプトイドの教示に関して参照により本明細書に明確に組み込まれる、Futakiら(2003)Curr Protein Pept Sci.2003 Apr;4(2):87〜96;およびWenderら(2000)Proc.Natl.Acad.Sci.U.S.A、2000年11月21日;97(24):13003〜8;米国特許出願公開第20030220334号;同第20030083256号;同第20030032593号;および同第20030022831号を参照されたい)。ノナアルギニン(R9)配列は、特徴づけられているより効率的なPTDの1つである(Wenderら2000;Uemuraら2002)。
【0047】
CD活性ポリペプチドは、当技術分野で公知の従来の方法を使用して、in vitro合成によって調製することができる。種々の市販の合成装置、例えば、Applied Biosystems、Inc.、Beckmanなどによる自動合成機が利用可能である。合成機を使用することによって、天然に存在するアミノ酸を非天然アミノ酸で置換することができる。特定の配列および調製様式は、利便性、経済的側面、必要な純度などによって決定される。シチジンデアミナーゼ活性ポリペプチドを無細胞系において調製する他の方法としては、例えば、参照により本明細書に組み込まれる、米国特許出願第61/271,000号において教示されている方法が挙げられる。
【0048】
CD活性ポリペプチドは、組換え合成の従来の方法に従って単離し、精製することもできる。発現宿主の溶解物を調製し、その溶解物を、HPLC、排除クロマトグラフィー、ゲル電気泳動、アフィニティークロマトグラフィー、または他の精製技法を使用して精製する。ほとんどの場合、使用される組成物は、産物の調製およびその精製の方法に関連する汚染物質に関して、少なくとも20重量%、通常、少なくとも約75重量%、好ましくは少なくとも約95重量%、および治療のためには、通常少なくとも約99.5重量%の所望の産物を含む。通常、百分率は、総タンパク質量に基づく。CD活性ポリペプチドは、組換えによって、直接だけでなく、異種ポリペプチド、例えば、成熟タンパク質またはポリペプチドのN末端に特異的な切断部位を有するポリペプチドとの融合ポリペプチドとしても作製することができる。発現ベクターは、通常、選択マーカーとも称される選択遺伝子を含有する、この遺伝子は、選択培地中で成長させる形質転換された宿主細胞の生存または成長に必要なタンパク質をコードする。
【0049】
当技術分野において公知の方法によって精製した後、標準のタンパク質形質導入方法によってCD活性ポリペプチドを対象細胞に提供する。ある場合では、タンパク質形質導入方法は、細胞を担体および少なくとも1つの精製したCD活性ポリペプチドを含有する組成物と接触させることを含む。適切な担体およびそれらを使用するための方法の例としては、これらに限定されないが、米国特許第6,841,535号に記載のChariot(商標)(Active Motif、Inc.、Carlsbad、Calif.);Bioport(商標)(Gene Therapy Systems、Inc.、San Diego、Calif.)、GenomeONE(Cosmo Bio Co.、Ltd.、Tokyo、Japan)、およびProteoJuice(登録商標)(Novagen、Madison、Wis.)などの市販の試薬、または例えば、米国特許出願第10/138,593号に記載のナノ粒子タンパク質形質導入試薬が挙げられる。
【0050】
他の実施形態では、CD活性を促進する1種または複数種の薬剤は、CD活性ポリペプチドをコードする核酸である。CD活性ポリペプチドをコードする核酸を対象細胞に提供するために使用されるベクターは、一般には、核酸の発現を駆動するため、すなわち、転写を活性化するために適したプロモーターを含む。これは、遍在的に作用するプロモーター、例えば、CMV−β−アクチンプロモーター、または誘導性プロモーター、例えば特定の細胞集団において活性なプロモーターまたはテトラサイクリンなどの薬物が存在することに反応するプロモーターなどを含んでよい。転写活性化とは、標的細胞において転写が基礎レベルを少なくとも約10倍、少なくとも約100倍、通常少なくとも約1000倍超えて増加することを意味する。さらに、核酸を提供するために使用するベクターは、後で除去しなければならない遺伝子を含んでよく、その際は、例えば、Cre/Loxなどのリコンビナーゼ系、または、例えば、TK、bcl−xsなどのヘルペスウイルスなどの選択毒性を可能にする遺伝子を含めることによって該遺伝子が破壊されて発現される細胞を用いる。
【0051】
CD活性ポリペプチドをコードする核酸は、対象細胞に直接提供することができる。言い換えれば、細胞を、CD活性ポリペプチドをコードする核酸を含むベクターと、ベクターが細胞に取り込まれるように接触させる。電気穿孔、塩化カルシウムトランスフェクション、およびリポフェクションなどの、細胞を核酸ベクターと接触させるための方法は当技術分野で周知である。このように核酸を送達するベクターは、通常、エピソームとして、例えば、プラスミドまたはミニサークルDNAとして維持される。
【0052】
あるいは、核酸は、ウイルスによって対象細胞に提供することができる。言い換えれば、細胞を、CD活性ポリペプチドをコードする核酸を含むウイルス粒子と接触させる。そのような方法には、レトロウイルス、例えば、レンチウイルスが特に適している。一般に使用されるレトロウイルスベクターは「不完全である」、すなわち、増殖性感染のために必要なウイルスタンパク質を産生することができない。もっと正確に言えば、ベクターの複製には、パッケージング細胞系において成長させることが必要である。対象とする核酸を含むウイルス粒子を生成するために、核酸を含むレトロウイルスの核酸をパッケージング細胞系によってウイルスカプシド中にパッケージする。異なるパッケージング細胞系からは、カプシド中に組み入れられる異なる外被タンパク質がもたらされ、この外被タンパク質により、ウイルス粒子の細胞に対する特異性が決定される。外被タンパク質は、狭宿主性、広宿主性および異種栄養性の少なくとも3種類のものである。狭宿主性外被タンパク質、例えば、MMLVを用いてパッケージしたレトロウイルスは、大部分のマウスおよびラット細胞型に感染することができ、これは、BOSC23(Pearら(1993)P.N.A.S.90:8392〜8396)などの狭宿主性パッケージング細胞系を使用することによって生成される。広宿主性外被タンパク質、例えば、4070A(Danosら、上記)を担持するレトロウイルスは、ヒト、イヌおよびマウスを含めた大部分の哺乳動物細胞型に感染することができ、これは、PA12(Millerら(1985)Mol.Cell.Biol.5:431〜437);PA317(Millerら(1986)Mol.Cell.Biol.6:2895〜2902);GRIP(Danosら(1988)PNAS 85:6460〜6464)などの広宿主性パッケージング細胞系を使用することによって生成される。異種栄養性外被タンパク質、例えば、AKR envを用いてパッケージしたレトロウイルスは、マウスの細胞以外の大部分の哺乳動物細胞型に感染することができる。適切なパッケージング細胞系を使用して、対象細胞がパッケージされたウイルス粒子の標的になることを確実にすることができる。シチジンデアミナーゼ活性を促進するポリペプチドをコードする核酸を含むレトロウイルスベクターをパッケージング細胞系に導入する方法およびパッケージング系によって生成されるウイルス粒子を採取する方法は当技術分野で周知である。
【0053】
本発明の方法では、哺乳動物細胞におけるゲノムDNAメチル化の量を、有効量の、CD活性を促進する1種または複数種の薬剤と接触させることによって減少させる。細胞におけるゲノムDNAメチル化を減少させるために十分な薬剤の量は、細胞におけるCD活性を促進するために十分な薬剤の量、すなわち、細胞におけるシトシンおよび5−メチルシトシンからのアミノ基の除去を促進するために十分な量である。この量は、シトシンヌクレオシドまたは5−メチルシトシンヌクレオシドの、それぞれウラシルヌクレオシドまたはチミンヌクレオシドへの変換を測定する当技術分野で公知のいくつものアッセイによって経験的に決定することができ、ここで、細胞におけるゲノムDNAメチル化の量を減少させるための薬剤の有効量は、5%以上、すなわち、5%、10%、20%、40%、60%、80%、または100%のシトシンまたは5−メチルシトシンの、ウラシルまたはチミンへの変換を誘導する量である。例えば、dCのdU(デオキシウラシル)への脱アミノ化の程度は、その開示が参照により本明細書に組み込まれるBransteitter、R.ら((2003)PNAS 100(7):4102〜4107)に記載のウラシルDNAグリコシラーゼ(UDG)および脱プリン部位エンドヌクレアーゼ(APE)を使用することによってアッセイすることができる。そのようなアッセイでは、5’末端を32Pで標識したDNA基質またはRNA基質(例えば、100nM)を、シチジンデアミナーゼ活性を促進する薬剤と一緒にインキュベートする。次いで、相補DNA鎖を基質とアニーリングさせ、その後、UDGおよびAPEと一緒にインキュベートする。インキュベートした後、反応を終了させ、反応生成物を変性ポリアクリルアミドゲル電気泳動(PAGE)によって分離し、ホスフォイメージングによって可視化し、ここで、標識された末端からシチジンまでの長さに対応する短い放射性生成物が存在することにより、シチジンにおけるCD活性を反映する、元のシチジンにおける切れ目が示される。別の例として、dCの脱アミノ化は、同様にBransteitter、R.ら、上記に記載されているプライマー伸長のジデオキシヌクレオチドによる終結を使用することによって検出することができる。そのようなアッセイでは、CD活性を促進する薬剤と反応するDNA基質またはRNA基質を3倍過剰の18−mer32P−標識プライマーとアニーリングさせ、3種のdNTPと、それに加えて2’,3’−ジデオキシアデノシン(ddA)三リン酸または2’,3’−ジデオキシグアノシン(ddG)三リン酸のいずれかの存在下でT7シーケナーゼを使用することによってプライマーを伸長する。基質−伸長プライマー複合体を熱変性させ、分離した鎖を相補DNA鎖とアニーリングさせ、上記のUDGおよびAPEと一緒にインキュベートする。反応生成物を変性PAGEによって分離し、ホスフォイメージングによって可視化し、ここで、脱アミノ化効率は、ddAミックスを用いた伸長反応から、C鋳型における、およびC鋳型を通り越した組み込まれたバンドの強度と比較したC/U鋳型の逆のバンドの強度の比として算出する。効率は、ddGミックスを用いた伸長反応からも、鋳型Cを通り越した組み込まれたバンドの強度の、C鋳型における、およびC鋳型を通り越した組み込まれたバンドの強度に対する比として算出することができる。このように、CD活性を促進する薬剤を同定することができ、有効量の、CD活性を促進する薬剤を経験的に決定することができる。
【0054】
ゲノムDNAメチル化の量を減少させるために十分である薬剤の有効量は、その薬剤で処理した後のDNAメチル化の程度をアッセイすることによっても決定することができる。細胞におけるゲノムDNA脱メチル化の量を減少させる薬剤の有効量は、DNA配列内のメチル化CpG配列の数の、1.5分の1以上への低下、すなわち、1.5分の1、2分の1、3分の1、4分の1、5分の1、10分の1、または20分の1またはそれ以下への低下を誘導する量である。CpG配列のメチル化の状態をアッセイするためのいくつかの方法、例えば、制限エンドヌクレアーゼ消化およびバイサルファイト配列決定が当技術分野で周知である。制限エンドヌクレアーゼ消化では、5−メチルシトシンを含有するCpG配列(例えば、CmeCGG)を、制限酵素Hpallによる切断に対する5−メチルシトシンを含有する配列の抵抗性によって、非メチル化シトシンを含有するCpG配列(CCGG)と区別することができる。対照的に、メチル化CpG配列および非メチル化CpG配列は、制限酵素Msplによって同等によく消化される。このことに基づいて、ゲノムDNAを、a)CpG配列の位置およびb)これらの配列が非メチル化(すなわち、Hpall制限に対して感受性が高い)配列であるか、またはメチル化(すなわち、Hpall制限に対して抵抗性である)配列であるかを決定するために、別々の反応においてMsplおよびHpallを用いた制限エンドヌクレアーゼ消化に供することができる。バイサルファイト配列決定では、DNAをバイサルファイトで処理することにより、シトシン残基がウラシルに変換されるが、5−メチルシトシン残基は影響されないままである;したがって、バイサルファイト処理により、DNA配列に、個々のシトシン残基のメチル化状態に応じた特異的な変化が導入され、これにより、DNAのセグメントのメチル化状態に関する単一のヌクレオチド分解情報がもたらされる。そのメチル化の状態についてアッセイすることができるゲノムDNAの領域の例としては、OCT4、NANOG、RB1、CDKN2AINK4A、CDKN2AARF、CDH1、CDH13、TIMP3、VHL、MLH1、MGMT、BRCA1、GSTP1、SMARCA3、RASSF1A、SOCS1、ESR1、DAPK1のプロモーター領域が挙げられる。アッセイすることができるゲノムDNAの他の領域は、その開示が参照により本明細書に組み込まれる、Costello、J.F.ら(2000)Nature Genet.25:132〜138、Song、F.ら、(2005)PNAS 102:3336〜3341、およびRobertson、K.D.(2005)Nature Review Genetics 6:597〜610に記載されている。
【0055】
細胞におけるゲノムDNAメチル化の量を減少させるために十分である薬剤の有効量は、細胞におけるメチル化感受性遺伝子の発現の変化をアッセイすることによって決定することもできる。メチル化感受性遺伝子は、発現レベルがそれらのプロモーターのメチル化の状態に対して感受性である遺伝子である。
【0056】
いくつかの遺伝子のプロモーターにおけるCpG配列のメチル化の増加は、メチル化感受性遺伝子プロモーターの転写活性の低下およびメチル化感受性遺伝子の発現の低下を伴う可能性があり、一方、これらの遺伝子のプロモーターにおけるCpG配列の脱メチル化は、メチル化感受性遺伝子プロモーターの転写活性の増加およびこれらの遺伝子の発現の増加を伴う可能性がある。有効量の、メチル化感受性遺伝子プロモーターの脱メチル化を促進する薬剤により、その遺伝子の発現の少なくとも約2倍の増加が誘導される。細胞とCD活性を促進する薬剤を接触させた後の遺伝子発現レベルの変化は、細胞を薬剤と接触させる前、およびその後に、遺伝子のRNAレベルおよび/またはタンパク質レベルを、例えば、RT−PCR、ノーザンブロットハイブリダイゼーション、ウエスタンブロットハイブリダイゼーションまたはELISAによって測定することによってアッセイすることができる。メチル化感受性遺伝子は当技術分野で周知であり、それらとしては、例えば、前の段落において列挙されているものなどの遺伝子が挙げられる。
【0057】
細胞
本発明の方法において使用するために適した細胞は、ヒト、霊長類、家畜動物、農場動物、および動物園の動物、実験動物または愛玩動物、例えば、イヌ、ネコ、ウシ、ウマ、ヒツジ、ブタ、ヤギ、ウサギ、ラット、マウスなどを含めた任意の哺乳動物細胞であってよい。細胞におけるゲノムDNA脱メチル化活性の量の増加が示された本発明の複数の態様では、細胞は、脱メチル化を許容する細胞であることが好ましい。脱メチル化を許容する細胞は、それらのCpG配列がメチル化CpG配列から非メチル化CpG配列に変換され得る細胞である。細胞が脱メチル化を許容するかどうかは、細胞において活性化誘導性シチジンデアミナーゼ(AID)をコードするcDNAを過剰発現させ、細胞にCpGリッチDNAを保有するベクターを提供し、外因的に供給されたCpGリッチDNAを回収し、例えば、バイサルファイト配列決定またはメチラーゼ特異的制限エンドヌクレアーゼ消化によって、メチル化の程度について分析することによって決定することができる。
【0058】
方法において使用するための対象とする細胞が脱メチル化を許容しない(not permissive to demetylation)、すなわち、脱メチル化を許容しない(demetylation−impermissive)細胞であると決定された場合では、脱メチル化を許容しない細胞を、有効量の、メチル化シトシンのヒドロキシル化メチルシトシンへの変換を促進する1種または複数種の薬剤、G:Tミスマッチ特異的な修復活性を促進する1種または複数種の薬剤、および/または成長停止およびDNA損傷誘導性の45(GADD45)活性を促進する1種または複数種の薬剤と接触させることによって、脱メチル化を許容する細胞になるように誘導することができる。
【0059】
メチル化シトシンのヒドロキシル化メチルシトシンへの変換を促進する薬剤は、脱アミノ化のためにメチル化核酸を刺激する。メチル化シトシンのヒドロキシル化メチルシトシンへの変換を促進する薬剤の例は、tetタンパク質のポリペプチドおよび断片、すなわち、tet1(Genbank受託番号:NM030625.2;配列番号29および配列番号30)、およびtet2(Genbank受託番号:NM_001127208.1配列番号31および配列番号32(アイソフォームa);およびGenbank受託番号:NM_017628.3、配列番号33および配列番号34(アイソフォームb))、ならびにこれらのポリペプチドをコードする核酸である。
【0060】
G:Tミスマッチ特異的な修復活性を促進する薬剤は、G/Tミスマッチからのチミン部分の除去およびこれらのチミン部分のシトシン部分との交換を促進する薬剤である。G:Tミスマッチ特異的な修復活性を促進する薬剤の例は、メチル結合ドメインタンパク質(メチル−CpG結合ドメインポリペプチドとしても公知である)およびタンパク質チミンDNAグリコシラーゼ(TDG)のポリペプチドおよび断片、ならびにこれらのポリペプチドをコードする核酸である。
【0061】
メチル結合ドメインタンパク質は、メチル−CpG結合ドメインのそれぞれにおける存在によって関連づけられる核タンパク質である。タンパク質のこのクラスには:MECP2、MBD1、MBD2、MBD3、およびMBD4の5つのメンバーがある。特に興味深いのは、メチルCpG部位でのDNA修復において機能することができる、細菌のDNA修復酵素とのタンパク質配列の類似性を有するメンバー、例えば、MBD4である。細胞を、脱メチル化に許容的になるように誘導することにおいて使用されるMBD4ポリペプチドおよびそれらをコードする核酸は、GenBank受託番号NM_003925.1(配列番号35および配列番号36)においてその配列を見ることができるMED1としても公知のヒトMBD4のアミノ酸配列と少なくとも70%同一であるアミノ酸配列を含むポリペプチドである。
【0062】
チミンDNAグリコシラーゼ(TDG)タンパク質は、5−メチルシトシンおよびシトシンの自発的な脱アミノ化によって引き起こされる遺伝子の突然変異に対する細胞防御において、G/Tミスマッチからチミン部分を除去し、グアニンとの誤対合からウラシル部分および5−ブロモウラシル部分を除去することによって中心的な役割を果たす酵素である。細胞を、脱メチル化に許容的になるように誘導することにおいて使用されるTDGポリペプチドおよびそれらをコードする核酸は、GenBank受託番号NM_003211.4(配列番号37および配列番号38)においてその配列を見ることができるヒトTDGのアミノ酸配列と少なくとも70%同一であるアミノ酸配列を含むポリペプチドである。
【0063】
成長停止およびDNA損傷誘導性の45(GADD45)タンパク質は、ストレス性の成長停止の状態およびDNA損傷剤処理の後にレベルが増加するタンパク質である。細胞を、脱メチル化に許容的になるように誘導することにおいて使用されるGADD45ポリペプチドおよびそれらをコードする核酸は、ヒトGADD45α(GenBank受託番号NM_001924.2(配列番号39および配列番号40)、GADD45β(GenBank受託番号NM_015675.2(配列番号41および配列番号42)、またはGADD45γ(GenBank受託番号NM_006705.3(配列番号43および配列番号44)のアミノ酸配列と少なくとも70%同一であるアミノ酸配列を含むポリペプチドである。
【0064】
G:Tミスマッチ特異的な修復活性を促進する薬剤(複数可)およびGADD45活性を促進する薬剤(複数可)は、上記のCD活性を促進する薬剤を提供するための方法によって、ポリペプチドまたはそれらのポリペプチドをコードする核酸として提供することができる。細胞は、シチジンデアミナーゼ活性を促進する1種または複数種の薬剤と接触させることと並行して、上記の方法によって脱メチル化に許容的になるように誘導することができる。あるいは、まず、細胞を脱メチル化に許容的にし、次いでCD活性を促進する1種または複数種の薬剤と接触させることができる。
【0065】
in vitroにおける方法および使用
本発明の一部の方法では、細胞を、CD活性を促進する1種または複数種の薬剤とin vitroで接触させる。これらの実施形態において対象とする、脱メチル化を許容する哺乳動物細胞、および脱メチル化を許容するように誘導することができる哺乳動物細胞としては、多能性幹細胞、例えば、ES細胞、iPS細胞、胚性生殖細胞;体細胞、例えば、線維芽細胞、造血細胞、ニューロン、筋肉細胞、骨細胞、血管内皮細胞、腸細胞など、およびそれらの系列が限られた前駆体および前駆物質;ならびに、当技術分野において周知であり、下記の実施例に記載されている、2種類以上の細胞の融合物である異核共存体が挙げられる。細胞は、樹立細胞系由来であってよい、または、初代細胞であってよく、「初代細胞」、「初代細胞系」、および「初代培養物」は、本明細書では、対象に由来し、in vitroにおいて培養の限られた数の継代、すなわち、分裂にわたって成長させることができる細胞および細胞培養物を指すために互換的に使用される。例えば、初代培養物は、0回、1回、2回、4回、5回、10回、または15回、しかし危機段階を経るには満たない回数継代することができる培養物である。一般には、本発明の初代細胞系は、in vitroにおいて10回の継代未満維持される。
【0066】
対象細胞は、新生児、若年者または成人由来であってよい新鮮細胞または凍結細胞から、および皮膚、筋肉、骨髄、末梢血、臍帯血、脾臓、肝臓、膵臓、肺、腸、胃、および他の分化した組織を含めた組織から単離することができる。組織は、生きているドナーからの生検またはアフェレーシスによって得ることができる、または死亡したもしくは瀕死のドナーから死亡の約48時間以内に得ることができる、または新鮮に凍結させた組織、死亡の約12時間以内に凍結させ、約−20℃未満、通常およそ液体窒素の温度(−190℃)で無制限に維持した組織から得ることができる。組織から細胞を単離するために、分散または懸濁のために適切な溶液を使用することができる。そのような溶液は、一般に、低濃度、一般に5〜25mMの許容できる緩衝液と併せて、ウシ胎仔血清または他の天然に存在する因子を補充することが都合よい、平衡塩類溶液、例えば、通常の生理食塩水、PBS、ハンクス平衡塩類溶液などである。都合のよい緩衝液としては、HEPES、リン酸緩衝液、乳酸緩衝液などが挙げられる。
【0067】
in vitroでシチジンデアミナーゼ活性を促進する1種または複数種の薬剤と接触させる細胞は、薬剤(複数可)の存在下で約30分間〜約24時間、例えば、1時間、1.5時間、2時間、2.5時間、3時間、3.5時間、4時間、5時間、6時間、7時間、8時間、12時間、16時間、18時間、20時間、または約30分間から約24時間までの任意の他の時間インキュベートすることができ、これを約毎日〜約4日ごと、例えば、1.5日ごと、2日ごと、3日ごと、または約毎日から約4日ごとまでの任意の他の頻度で繰り返すことができる。薬剤(複数可)は、1回または複数回、例えば、1回、2回、3回、または4回以上対象細胞に提供することができ、各接触事象の後、細胞を薬剤(複数可)と一緒にいくらかの時間、例えば、16〜24時間インキュベートしておき、その後、培地を新鮮な培地と交換し、細胞をさらに培養する。
【0068】
本発明の一部の方法では、CD活性を促進する薬剤と接触させる、脱メチル化を許容する細胞は、脱メチル化を許容する体細胞である。これらの方法のいくつかでは、脱メチル化を許容する体細胞を、異なる細胞系譜の体細胞になるように再プログラムする。言い換えれば、本発明の方法を使用して、ある系列の体細胞から別の系列の体細胞への変換を促進することができる。異なる系列の体細胞は、当技術分野で周知のマーカーおよび形態によって容易に同定可能である。
【0069】
脱メチル化を許容する体細胞を、CD活性を促進する薬剤と接触させる一部の方法では、脱メチル化を許容する体細胞を、人工多能性幹(iPS)細胞になるように再プログラムする。言い換えれば、作製される細胞は、iPS細胞である。上記の通り、iPS細胞は、ES細胞様の形態(例えば、大きな核細胞質比、明確な境界および目立った核を有する平らなコロニーとして成長する)が、体細胞に由来する多能性幹細胞である。
【0070】
本発明の一部の方法では、CD活性を促進する薬剤と接触させる脱メチル化を許容する細胞は、多能性幹細胞、例えば、胚性幹(ES)細胞、胚性生殖(EG)細胞、または人工多能性幹(iPS)細胞である。これらの方法では、脱メチル化を許容する多能性幹細胞を、体細胞になるように再プログラムする。言い換えれば、本発明の方法を使用して、多能性幹細胞の体細胞へのプログラミングを促進することができる。体細胞の例としては、外胚葉系(例えば、神経および線維芽細胞)、中胚葉系(例えば、心筋細胞)、または内胚葉(例えば、膵臓細胞)系由来の任意の分化細胞が挙げられる。体細胞は、1種または複数種の:膵ベータ細胞、神経幹細胞、神経(例えば、ドーパミン作動性神経)、乏突起膠細胞、乏突起膠細胞前駆細胞、肝細胞、肝幹細胞、星状膠細胞、ミオサイト、造血細胞、心筋細胞などであってよい。上に示されている通り、多能性幹細胞に由来する体細胞は、最終分化細胞であってよい、またはそれらは、特定の系列の細胞を生じることができてよい。例えば、多能性細胞は、種々の多分化能細胞型、例えば、神経幹細胞、心臓幹細胞、または肝幹細胞に分化させることができる。次いで、幹細胞は、新しい細胞型にさらに分化させることができ、例えば、神経幹細胞は、神経に分化させることができ;心臓幹細胞は、心筋細胞に分化させることができ;肝幹細胞は、肝細胞に分化させることができる。そのような方法によって作製される体細胞は、上記の通り、当技術分野で周知の特定の細胞系列のマーカーおよび形態によって、そのようなものとして容易に同定可能である。
【0071】
脱メチル化を許容する細胞の他の細胞型への再プログラミングを促進するために、脱メチル化を許容する細胞を、細胞の再プログラミングを促進する1種または複数種の薬剤と接触させる追加的なステップを実施することができる。このステップは、脱メチル化を許容する細胞を、CD活性を促進する薬剤と接触させる前、脱メチル化を許容する細胞を、CD活性を促進する薬剤と接触させると同時に、または脱メチル化を許容する細胞を、CD活性を促進する薬剤と接触させた後で実行することができる。細胞の再プログラミングを促進する薬剤は、ポリペプチド、核酸薬剤、または小分子薬剤であってよい。このステップにおいて提供することができる薬剤の例としては、これらに限定されないが、GSK−3阻害剤、例えば、CHIR99021など(Li、W.ら(2009)Stem Cells、Epub Oct.16 2009,);HDAC阻害剤、例えば、バルプロ酸など(その開示が参照により本明細書に組み込まれるHuangfu、D.(2008)Nature Biotechnol 26(7)795〜797;およびUS20090191159に記載);ヒストンメチルトランスフェラーゼ阻害剤、例えば、G9aヒストンメチルトランスフェラーゼ阻害剤、例えば、BIX−01294、など(Shi、Yら(2008)Cell Stem Cell 3(5):568〜574);ジヒドロピリジン受容体のアゴニスト、例えば、BayK8644、など(Shi、Yら(2008)Cell Stem Cell 3(5):568〜574);およびTGFβシグナル伝達の阻害剤、例えば、RepSoxなど(lchida、JK.ら(2009)Cell Stem Cell 5(5):491〜503)が挙げられる。このステップにおいて提供することができる薬剤の他の例としては、再プログラミング因子が挙げられる。上記の通り、再プログラミング因子は、細胞に作用して転写を変化させ、それによって細胞を新しい細胞運命に再プログラミングする生物活性因子である。
【0072】
ある細胞系譜の体細胞の、別の細胞系譜の体細胞への再プログラミングを促進する薬剤の多数の例が当技術分野で公知であり、そのいずれも、本発明において使用することができる。これらとしては、例えば、筋肉に特異的な性質を色素細胞、神経細胞、脂肪細胞、肝細胞および線維芽細胞内に誘導する再プログラミング因子MYOD(筋原性因子1;GenBank受託番号NM_002478.4およびNP_002469.2)、例えば、Weintraub、H.W.らProc.Natl.Acad.Sci USA 86:5434〜5438;Davis、R.L.ら(1987)Cell 51 :987〜1000;Schafer、B.W.ら(1990)Nature 344:454〜8)を参照されたい;NEUROG3(neurogenin3、NGN3;GenBank受託番号NM_020999.2およびNP_066279.2)、PDX1(膵臓および十二指腸ホメオボックス1;GenBank受託番号.NM_000209.3およびNP_000200.1)および組み合わせて、in vivoで膵臓の外分泌細胞を機能性β細胞に効率的に変換することができるMafA(v−maf筋腱膜線維肉腫癌遺伝子ホモログA;GenBank受託番号NM_201589.2およびNP_963883.2)、例えば、Zhou、Q.ら(2008)Nature 455:627〜32)を参照されたい;およびB細胞において単独で、または線維芽細胞においてPu.1(脾フォーカス形成ウイルス(SFFV)プロウイルス組み込み癌遺伝子、SPI1;GenBank受託番号NM_001080547.1、NP_001074016.1、NM_003120.2およびNP_003111.2)と組み合わせてマクロファージ特性を誘導する、C/EBPa(CCAAT/エンハンサー結合性タンパク質、アルファ;GenBank受託番号NM004364.3およびNP_004355.2)、例えば、Bussmann、L.H.ら(2009)Cell Stem Cell 5:554〜66;Feng、R.ら(2008)Proc Natl Acad Sci USA 105:6057〜62;Xie、H.ら(2004)Cell 117:663〜76)を参照されたい、が挙げられる。他の薬剤としては、IL2受容体(IL受容体2AおよびIL受容体2B;GenBank受託番号NM_000417.2、NP_000408.1、NM_000878.2およびNP_000869.1)およびGM−CSF受容体(コロニー刺激因子2受容体、アルファ(CSF2RA)およびコロニー刺激因子2受容体、ベータ(CSF2RB);GenBank受託番号NM_001161529.1、NP_001155001.1、NM_000395.2、およびNP_000386.1)、委任リンパ球前駆細胞において骨髄の変換を誘導する、例えば、Kondo、M.ら(2000)Nature 407:383〜6)を参照されたい、が挙げられる。上に列挙されているGenBank受託番号に記載の薬剤、ならびにこれらのポリペプチドをコードする核酸のアミノ酸配列と少なくとも70%、75%、80%、85%、90%、91%、92%、95%、97%、99%、または100%同一であるアミノ酸配列を含むポリペプチドは、本発明の方法において、ある脱メチル化を許容する細胞系譜の体細胞の、別の細胞系譜の体細胞への再プログラミングを促進する薬剤として使用される。
【0073】
体細胞のiPS細胞への再プログラミングを促進する薬剤の多数の例が当技術分野で公知であり、そのいずれも、本発明において使用することができ、例えば、その開示が参照により本明細書に組み込まれる、米国特許出願第20090047263号、US20090068742、US20090191159、US20090227032、US20090246875、およびUS20090304646を参照されたい。これらとしては、例えば、再プログラミング因子Oct3/4、(POUクラス5ホメオボックス1(POU5F1);GenBank受託番号NP_002692およびNM_002701);Sox2(性決定領域Yボックス2タンパク質;GenBank受託番号NP_003097およびNM_003106):Klf4(クルッペル様因子4;GenBank受託番号NP_004226およびNM_004235);c−Myc(骨髄細胞腫症ウイルス癌遺伝子ホモログ;GenBank受託番号NP_002458およびNM_002467);Nanog(Nanogホメオボックス;GenBank受託番号NP_079141およびNM_024865);およびLin−28(C.elegansのLin−28ホモログ;GenBank受託番号NP_078950およびNM_024674)が挙げられる。上に列挙されているGenBank受託番号に記載の薬剤のアミノ酸配列と少なくとも70%、75%、80%、85%、90%、91%、92%、95%、97%、99%、または100%同一であるアミノ酸配列を含むポリペプチド、ならびにこれらのポリペプチドをコードする核酸は、本発明の方法において、脱メチル化を許容する体細胞の、iPS細胞への再プログラミングを促進する薬剤として使用される。
【0074】
多能性幹細胞の、体細胞への再プログラミングを促進する薬剤の多数の例は当技術分野で公知であり、そのいずれも、本発明において使用することができる。例えば、神経幹細胞は、多能性細胞を、NOG(noggin;GenBank受託番号NM_005450.4およびNP_005441.1)または他の骨形成タンパク質アンタゴニスト(Itsyksonら、(2005)、Mol、Cell Neurosci.、30(1):24〜36)の存在下で浮遊凝集体として培養することによって、または多能性細胞を浮遊液中で、増殖因子、例えば、FGF−2(線維芽細胞増殖因子2、塩基性線維芽細胞増殖因子(bFGF)としても公知である;GenBank受託番号NM_002006.4およびNP_001997.5)、例えば、Zhangら、(2001)、Nat.Biotech.(19):1129〜1133を参照されたい、の存在下で培養して凝集体を形成することによって生成することができる。ある場合では、FGF−2を含有する無血清培地中で凝集体を培養する。別の例では、FGF−2を含む無血清培地の存在下で多能性細胞をマウス間質細胞系、例えばPA6と共培養する。さらに別の例では、FGF−2を含有する無血清培地に多能性細胞を直接移して分化を直接誘導する。
【0075】
多能性細胞に由来する神経幹は、神経、乏突起膠細胞、または星状膠細胞に分化させることができる。多くの場合、神経幹細胞を生成するために用いる条件は、神経、乏突起膠細胞、または星状膠細胞を生成するためにも使用することができる。例えば、ドーパミン作動性ニューロンへの分化を促進するために、多能性細胞またはそれに由来する神経幹細胞を、無血清条件下でPA6マウス間質細胞系と共培養することができる。例えば、Kawasakiら、(2000)Neuron、28(1):3140を参照されたい。他の方法も記載されており、例えば、Pompら、(2005)、Stem Cells 23(7):923〜30;米国特許第6,395,546号、例えば、Leeら、(2000)、Nature Biotechnol.、18:675〜679を参照されたい。多能性細胞またはそれに由来する神経幹細胞の乏突起膠細胞への分化は、例えば、多能性細胞または神経幹細胞を間質細胞と共培養することによって(例えば、Hermannら(2004)、J Cell Sci.117(Pt19):4411〜22を参照されたい)、または多能性細胞または神経幹細胞を、インターロイキン(IL)−6受容体(GenBank受託番号NM_000565.2およびNP_000556.1)、またはその誘導体がIL−6サイトカイン(GenBank受託番号NM_000600.3およびNP_000591.1)またはその誘導体に連結している融合タンパク質の存在下で培養することによって促進することができる。乏突起膠細胞は、当技術分野で公知の他の方法によって多能性細胞から生成することもできる。例えば、Kangら、(2007)Stem Cells 25、419〜424を参照されたい。星状膠細胞は、多能性細胞またはそれに由来する神経幹細胞から、例えば、bFGFおよびEGF(上皮増殖因子;GenBank受託番号NM_001963.3およびNP_001954.2)を伴う神経形成培地の存在下で多能性細胞または神経幹細胞を培養することによって作製することもできる。例えば、Brustleら、(1999)、Science、285:754〜756を参照されたい。
【0076】
多能性細胞は、当技術分野で公知の方法によって膵ベータ細胞に分化させることができる。例えば、Lumelskyら、(2001)Science、292:1389〜1394;Assadyら、(2001)、Diabetes、50:1691〜1697;D’Amourら、(2006)、Nat.Biotechnol.、24:1392〜1401.;D’Amourら、(2005)、Nat.Biotechnol.23:1534〜1541。方法は、多能性細胞を、アクチビンA(インヒビン、ベータA(INHBA);GenBank受託番号NM_002192.2およびNP_002183.1)を補充した無血清培地中で培養し、その後、オールトランスレチノイン酸を補充した無血清培地の存在下で培養し、その後、bFGFおよびニコチンアミドを補充した無血清培地の存在下で培養することを含んでよい。例えば、Jiangら、(2007)、Cell Res.、4:333〜444。他の例では、方法は、多能性細胞を、無血清培地、アクチビンA、およびWntタンパク質(例えば、GenBank受託番号NM_005430、NM_003391、NM_004185、NM_030753、NM_033131、NM_030761、NM_003392、NM_032642、NM_006522、NM_004625、NM_058238、NM_058244、NM_003393、NM_003395、NM_003396、NM_025216、NM_003394、Wnt−11NM_004626、およびNM_016087)の存在下で、約0.5日から約6日間まで、例えば、約0.5日、1日、2日間、3日間、4日間、5日間、6日間培養し;その後、約0.1%から約2%まで、例えば、0.2%のFBSおよびアクチビンAの存在下で約1日から約4日間まで、例えば、約1日、2日間、3日間、または4日間培養し;その後、2%のFBS、FGF10(線維芽細胞増殖因子10、GenBank受託番号NM_004465.1およびNP_004456.1)、KAAD−シクロパミン(ケト−N−アミノエチルアミノカプロイルジヒドロシンナモイルシクロパミン)およびレチノイン酸の存在下で約1日から約5日間まで、例えば、1日、2日間、3日間、4日間、または5日間培養し、;その後、1%のB27、ガンマセレクターゼ阻害剤およびエクステンジン−4(extendin−4)と一緒に約1日から約4日間、例えば、1日、2日間、3日間、または4日間培養し;最後に、1%のB27、エクステンジン−4、IGF−1、およびHGFの存在下で約1日から約4日間、例えば、1日、2日間、3日間、または4日間培養することを含む。
【0077】
多能性細胞から肝細胞または肝幹細胞を分化させることができる。例えば、多能性細胞を酪酸ナトリウムの存在下で培養することにより、肝細胞を生成することができる。例えば、Rambhatlaら、(2003)、Cell Transplant 12:1〜11を参照されたい。別の例では、肝細胞は、多能性細胞を、血清を含まない培地中、アクチビンAの存在下で培養し、その後、細胞をFGF4(線維芽細胞増殖因子−4;GenBank受託番号NM_002007.2およびNP_001998.1)およびBMP2(骨形成タンパク質−2;GenBank受託番号NM_001200.2およびNP_001191.1)中で培養することによって作製することができる。例えば、Caiら、(2007)Hepatology 45(5):1229〜39。例示的な実施形態では、多能性細胞を、アクチビンAの存在下で約2日間から約6日間まで、例えば、約2日間、約3日間、約4日間、約5日間、または約6日間培養し、次いで、多能性細胞を、HGF(肝細胞増殖因子;GenBank受託番号NM_010427.4およびNP_034557.3)の存在下で約5日間から約10日間まで、例えば、約5日間、約6日間、約7日間、約8日間、約9日間、または約10日間培養することによって肝細胞または肝幹細胞に分化させる。
【0078】
多能性細胞は、心筋細胞に分化させることもできる。骨形成タンパク質(BMP)シグナル伝達を阻害することにより、心筋細胞(または心筋細胞)の生成をもたらすことができる。例えば、Yuasaら、(2005)、Nat.Biotechnol.、23(5):607〜11を参照されたい。したがって、例示的な実施形態では、多能性細胞をNOG(noggin)の存在下で約2日間から約6日間まで、例えば、約2日間、約3日間、約4日間、約5日間、または約6日間培養し、その後、胚葉体を形成させ、胚葉体を約1週間から約4週間、例えば、約1週間、約2週間、約3週間、または約4週間培養する。他の例では、心筋細胞は、多能性細胞を、LIF(白血病抑制因子;GenBank受託番号NM_002309.3およびNP_002300.1)の存在下で培養することによって、または多能性細胞を当技術分野で公知の他の方法に供してES細胞から心筋細胞を生成することによって生成することができる。例えば、Baderら、(2000)、Circ.Res.、86:787〜794、Kehatら、(2001)、J.Clin.Invest.、108:407〜414;Mummeryら、(2003)、Circulation、107:2733〜2740。
【0079】
多能性細胞から他の細胞種を生成する方法の例としては:(1)脂肪細胞を生成するために、多能性細胞を、レチノイン酸、LIF、甲状腺ホルモンおよびインスリンの存在下で培養すること(例えば、Daniら、(1997)、J.Cell Sci.、110:1279〜1285);(2)軟骨細胞を生成するために、多能性細胞を、BMP2またはBMP4(GenBank受託番号NM_001202.3、NP_001193.2、NM_130850.2、NP_570911.2、NM_130851.2、およびNP_570912.2)の存在下で培養すること(例えば、Kramerら、(2000)、Mech.Dev.、92:193〜205);(3)多能性細胞を、平滑筋を生成するための条件下で培養すること(例えば、Yamashitaら、(2000)、Nature、408:92〜96);(4)ケラチノサイトを生成するために、多能性細胞を、ベータ−1インテグリン(GenBank受託番号NM_002211.3およびNP_002202.2)の存在下で培養すること(例えば、Baguttiら、(1996)、Dev.Biol.、179:184〜196);(5)マクロファージを生成するために、多能性細胞を、IL3(インターロイキン−3;GenBank受託番号NM_000588.3およびNP_000579.2)およびCSF1(コロニー刺激因子、マクロファージ;GenBank受託番号NM_000757.4、NP_000748.3)の存在下で培養すること(例えば、Lieschkeおよび Dunn(1995)、Exp.Hemat.、23:328〜334);(6)肥満細胞を生成するために、多能性細胞を、IL−3およびSCF(造血幹細胞因子、kitリガンドとしても公知の幹細胞因子;GenBank受託番号NM_000899.3およびNP_000890.1)の存在下で培養すること(例えば、Tsaiら、(2000)、Proc.Natl.Acad.Sci.USA、97:9186〜9190);(7)メラニン形成細胞を生成するために、多能性細胞を、デキサメタゾンおよびSCFの存在下で培養すること(例えば、Yamaneら、(1999)、Dev.Dyn.、216:450〜458);(8)骨芽細胞を生成するために、多能性細胞を、デキサメタゾン、レチノイン酸、アスコルビン酸、ベータ−グリセロリン酸の存在下で胎児のマウス骨芽細胞と共培養すること(例えば、Butteryら、(2001)、Tissue Eng.、7:89〜99);(9)骨芽細胞を生成するために、多能性細胞を、骨形成性の因子の存在下で培養すること(例えば、Sottileら、(2003)、Cloning Stem Cells、5:149〜155);(10)骨格筋細胞を生成するために、多能性細胞においてインスリン様増殖因子−2を過剰発現させ、その細胞をジメチルスルホキシドの存在下で培養すること(例えば、Prelleら、(2000)、Biochem.Biophys.Res.Commun.、277:631〜638を参照されたい);(11)多能性細胞を、白色血液細胞を生成するための条件に供すること;または、(12)造血前駆細胞を生成するために、多能性細胞を、BMP4ならびに:SCF、FLT3(fms関連チロシンキナーゼ3;GenBank受託番号NM_004119.2およびNP_004110.2)、IL−3、IL−6(インターロイキン6;GenBank受託番号M_000600.3およびNP_000591.1)、およびCSF3(コロニー刺激因子、顆粒球;GenBank受託番号NM_000759.2およびNP_000750.1)の1つまたは複数の存在下で培養すること(例えば、Chadwickら、(2003)、Blood、102:906〜915を参照されたい)が挙げられる。
【0080】
上に列挙されているGenBank受託番号に記載の薬剤のアミノ酸配列と少なくとも70%、75%、80%、85%、90%、91%、92%、95%、97%、99%、または100%同一であるアミノ酸配列を含むポリペプチド、ならびにこれらのポリペプチドをコードする核酸は、本発明の方法において、多能性細胞の体細胞への再プログラミングを促進する薬剤として使用される。
【0081】
細胞の再プログラミングを促進する薬剤は、これらに限定されないが、CD活性を促進する薬剤について上記されている方法を含めた当技術分野で周知の方法によって、脱メチル化を許容する細胞に提供することができる。薬剤は、個別に、または単一の組成物として、すなわち、薬剤の予備混合した組成物として提供することができる。薬剤は、同時に、または違う時間に逐次的に対象細胞に加えることができる。一部の実施形態では、少なくとも2種の薬剤のセット、例えば、Oct3/4ポリペプチドおよびSox2ポリペプチドを提供する。一部の実施形態では、3種の薬剤のセット、例えば、Oct3/4ポリペプチド、Sox2ポリペプチド、およびKlf4ポリペプチドを提供する。一部の実施形態では、4種の薬剤のセット、例えば、Oct3/4ポリペプチド、Sox2ポリペプチド、Klf4ポリペプチド、およびc−Mycポリペプチドを提供する。CD活性を促進する薬剤(複数可)と同様に、薬剤(複数可)は、対象細胞に1回または複数回提供することができ、各接触事象の後、細胞を薬剤と一緒にいくらかの時間、例えば、16〜24時間インキュベートしておき、その後、培地を新鮮な培地と交換し、細胞をさらに培養する。
【0082】
脱メチル化を許容する細胞を、CD活性を促進する薬剤(複数可)と接触させた後、所望の細胞の成長を促進するために、接触させた細胞を培養する。細胞を培養して上記のiPS細胞または体細胞の特定の型の成長を促進するための方法、上記の体細胞の特定の型のiPS細胞クローンまたはクローンを単離するための方法、およびそれらの細胞クローンの細胞を培養して上記のiPS細胞または体細胞の特定の型の成長を促進する方法は当技術分野で周知であり、そのいずれも、本発明において、再プログラムされた脱メチル化を許容する細胞から所望の細胞を成長させ、単離し、再培養するために使用することができる。
【0083】
細胞を、CD活性を促進する薬剤(複数可)と接触させることによって脱メチル化を許容する細胞培養物の細胞におけるゲノムDNAメチル化の量を減少させることにより、脱メチル化を許容する細胞の、所望の細胞型への再プログラミングの効率が、CD活性を促進する薬剤が存在しない場合に観察される効率と比較して増加する。言い換えれば、体細胞および細胞培養物は、再プログラミングを促進することが当技術分野で公知の因子の存在下でCD活性を促進する1種または複数種の薬剤と接触させた場合に、CD活性を促進する1種または複数種の薬剤と接触させなかった細胞と比較して所望の細胞型を生じさせるための能力が増強されることが実証されている。増強されるとは、体細胞培養物が、CD活性を促進する薬剤と接触させなかった細胞の集団の所望の細胞型を生じさせる能力の少なくとも約50%、約100%、約200%、約300%、約400%、約600%、約1000%、少なくとも約2000%の、所望の細胞型を生じさせる能力を有することを意味する。言い換えれば、脱メチル化を許容する細胞の培養物により、CD活性を促進する1種または複数種の薬剤と接触させていない脱メチル化を許容する細胞の集団によって産生される所望の細胞型の細胞数の約1.5倍、約2倍、約3倍、約4倍、約6倍、約10倍、約20倍、約30倍、約50倍、約100倍、約200倍が産生される。再プログラミングの効率は、所望の細胞型を獲得すると脱メチル化された状態になることが当技術分野で公知のプロモーターにおけるメチル化の量をアッセイすることによって決定することができる。そのような場合では、それらのプロモーターにおけるメチル化の量が、CD活性を促進する薬剤が存在しない場合に観察されたメチル化の量の約1.5分の1、約2分の1、約3分の1、約4分の1、約6分の1、約10分の1未満である場合に、CD活性を促進する薬剤が存在することに起因する再プログラミングの効率の増強が観察される。その代わりにまたはそれに加えて、再プログラミングの効率は、所望の細胞系を獲得するとより高度に発現されることが当技術分野で公知の遺伝子の発現レベルをアッセイすることによって決定することができる。そのような場合では、これらの遺伝子の発現レベルが、CD活性を促進する薬剤が存在しない場合に観察される発現レベルよりも約1.5倍、約2倍、約3倍、約4倍、約6倍、約10倍大きい場合に、CD活性を促進する薬剤が存在することに起因する再プログラミングの効率の増強が観察される。
【0084】
上記のin vitroにおける方法によって再プログラムされた脱メチル化を許容する細胞から得られる細胞は、疾患(例えば、遺伝的欠陥)を治療するための療法として使用することができる。特に、上記の方法によって脱メチル化を許容する体細胞から得られる体細胞および上記の方法によって多能性幹細胞体細胞から得られる体細胞は、広範囲の疾患または障害に罹患している対象に、例えば、レシピレントにおいて分化している細胞または分化した細胞を再構成する、または補うために移入することができる。同様に、脱メチル化を許容する体細胞から得られる人工多能性幹細胞を、広範囲の疾患または障害に罹患している対象に移入することができる、または、それらをin vitroで種々の細胞系譜の体細胞に分化させ、次いで広範囲の疾患または障害に罹患している対象に移入することができる。これらに限定されないが、多能性細胞を分化させる方法を含めた、多能性細胞をより特殊化した細胞型に分化させる方法は多数あり、これらを、上記の通り、幹細胞、特にES細胞を、体細胞になるように再プログラムするために使用することができる。
【0085】
療法は、疾患の原因を治療することを対象としてよい;あるいは、療法は、疾患または状態の影響を治療するものであってよい。例えば、由来細胞は、対象の傷害部位に、またはその近くに移入することができる;または細胞は、細胞が傷害部位に移動する、またはそこに向かうことができるように対象に導入することができる。移入された細胞は、損傷を受けた、または傷害を受けた細胞と有利に置き換え、対象の全体的な状態の改善を可能にすることができる。ある場合には、移入された細胞により、組織再生または修復が刺激され得る。
【0086】
ある場合では、由来細胞または由来細胞の亜集団は、対象に移入する前に精製または単離することができる。ある場合では、所望の細胞型に特異的な1種または複数種のモノクローナル抗体を細胞集団と一緒にインキュベートし、結合した細胞を単離する。他の場合では、細胞の所望の亜集団に細胞型に特異的なプロモーターの制御下にあるレポーター遺伝子を発現させ、次いで、それを用いて由来細胞またはその亜集団を精製または単離する。
【0087】
ある場合では、種々の目的で、例えば、機能喪失突然変異を有する遺伝子を置き換えるため、マーカー遺伝子を提供するためなどに、遺伝子を、対象に移入する前に脱メチル化を許容する細胞またはそれに由来する細胞に導入することができる。あるいは、アンチセンスmRNAまたはリボザイムを発現させるベクターを導入し、それによって望ましくない遺伝子の発現を遮断する。遺伝子療法の他の方法は、薬物耐性遺伝子を導入して、正常な前駆細胞が利点を有することを可能にし、選択圧、例えば、多数の薬物耐性遺伝子(MDR)、またはbcl−2などの抗アポトーシス遺伝子に供することである。種々の当技術分野で公知の技法、例えば、上記の通り、電気穿孔、カルシウム沈降DNA(calcium precipitated DNA)、融合、トランスフェクション、リポフェクション、感染などを用いて、標的細胞に核酸を導入することができる。DNAを導入する特定の様式は、本発明の実施には重大ではない。
【0088】
遺伝子改変された脱メチル化を許容する細胞またはそれに由来する細胞を有することを立証するために、種々の技法を使用することができる。細胞のゲノムは制限されている可能性があり、増幅して、または増幅せずに使用する。ポリメラーゼ連鎖反応;ゲル電気泳動;制限分析;サザンブロット、ノーザンブロット、およびウエスタンブロット;配列決定などは、全て使用することができる。種々の条件下で細胞を成長させて、細胞が、導入されたDNAを発現する能力を維持しながら骨髄系列の全てに成熟化可能なことを確実にすることができる。in vitroおよびin vivoにおける種々の試験を使用して、確実に細胞の多能性の能力(capability)が維持されるようにすることができる。
【0089】
対象への治療の施行数は変動し得る。誘導された細胞および/または分化した細胞の対象への導入は、一回の事象であってよいが、ある特定の状況では、そのような治療によって引き出される改善は限られた期間にわたるものであり、持続的な一連の反復治療が必要になる可能性がある。他の場合では、効果が観察される前に細胞を多数回投与することが必要になり得る。正確なプロトコールは、疾患または状態、疾患の病期および治療されている個々の対象のパラメータに左右される。
【0090】
細胞は、以下の経路のいずれかによって対象に導入することができる:非経口的経路、静脈内経路、動脈内経路、筋肉内経路、皮下経路、経皮的経路、気管内経路、腹腔内経路、または脊髄液内経路。
【0091】
神経疾患または神経障害に罹患している対象には、特に、本発明の方法によって得られる細胞を利用する療法が有益であり得る。いくつかの手法では、神経性の状態、例えば、アルツハイマー病、パーキンソン病、多発性硬化症、脳梗塞、脊髄損傷、または他の中枢神経系障害を治療するために、神経幹細胞または神経系細胞を傷害部位に移植することができる。例えば、Morizaneら、(2008)、Cell Tissue Res.、331(1):323〜326;Couttsおよび Keirstead(2008)、Exp.Neurol.、209(2):368〜377;Goswamiおよび Rao(2007)、Drugs、10(10):713〜719を参照されたい。パーキンソン病を治療するために、ドーパミン作用性ニューロンをパーキンソン病の対象の線条体に移植することができる。多発性硬化症を治療するために、乏突起膠細胞または乏突起膠細胞の前駆体をMSに罹患している対象に移入することができる。本発明の方法によって得られる細胞は、脳および脊髄を修復するために、病変へのそれらの移動を標的にすることができる手掛かりに反応するように遺伝子工学で操作することもできる。例えば、Chenら、(2007)、Stem Cell Rev.、3(4):280〜288。
【0092】
神経性障害以外の疾患も、本発明の方法によって生成される細胞を利用する療法によって治療することができる。虚血性心筋症、伝導疾患、および先天性欠損などの変性心疾患には、心筋細胞またはそれらの前駆物質を移植することが有効であり得る。例えば、Janssensら、(2006)、Lancet、367:113〜121を参照されたい。
【0093】
膵島細胞(またはランゲルハンス島の初代細胞)を糖尿病(例えば、糖尿病、1型)に罹患している対象に移植することができる。例えば、Burnsら、(2006)Curr.Stem Cell Res.Ther.、2:255〜266を参照されたい。一部の実施形態では、本発明の方法によって得られる膵ベータ細胞を糖尿病(例えば、1型糖尿病)に罹患している対象に移植することができる。
【0094】
他の例では、本発明の方法によって得られる肝細胞または肝幹細胞を、肝疾患、例えば、肝炎、硬変症、または肝不全に罹患している対象に移植する。
【0095】
本発明の方法によって得られる造血細胞または造血幹細胞(HSC)を血液の癌、または他の血液障害または免疫障害に罹患している対象に移植することができる。造血細胞またはHSCによって治療される可能性がある血液の癌の例としては:急性リンパ芽球性白血病、急性骨髄芽球性白血病、慢性骨髄性白血病(CML)、ホジキン病、多発性骨髄腫、および非ホジキンリンパ腫が挙げられる。多くの場合、そのような疾患に罹患している対象は、急速に分裂している血液細胞を死滅させるために放射線治療および/または化学療法薬による治療を受けなければならない。本発明の方法によって得られるHSCをこれらの対象に導入することは、細胞の枯渇した貯蔵を再充填させるのに役立ち得る。
【0096】
ある場合では、本発明の方法によって得られる造血細胞またはHSCは、癌と直接戦うためにも使用することができる。例えば、同種異系のHSCの移植は、腎癌の治療において見込みがあることが示されている。例えば、Childsら、(2000)、N.Engl.J.M編、343:750〜758を参照されたい。一部の実施形態では、本発明の方法によって得られる同種異系のHSC、または自己のHSCでさえ、腎癌または他の癌を治療するために対象に導入することができる。
【0097】
本発明の方法によって得られる造血細胞またはHSCは、血液細胞以外の細胞または組織、例えば、筋肉、血管、または骨を生成または修復するためにも対象に導入することができる。そのような治療は、多数の障害に対して有用であり得る。
【0098】
ある場合では、本発明の方法によって得られる細胞を免疫無防備状態の動物、例えば、SCIDマウスに移入し、分化させる。移植された細胞は、分化細胞型と腫瘍細胞の混合物を形成し得る。対象とする特異的な分化細胞型は、系列特異的なマーカーを使用することによって、例えば、蛍光活性化細胞選別(FACS)または他の選別方法、例えば、磁気活性化細胞選別(MACS)によって、腫瘍細胞から離して選択し、精製することができる。次いで、疾患または状態を治療するために分化細胞を対象(例えば、自己の対象、HLAが一致した対象)に移植することができる。疾患または状態は、造血障害、内分泌欠乏、神経変性障害、脱毛、または本明細書に記載の他の疾患または状態であってよい。
【0099】
本発明の方法によって得られる細胞は、任意の生理的に許容される培地に投与することができる。本発明の方法によって得られる細胞は、単独で、または、例えば、それらが移植された組織におけるそれらの成長および/または組織化を支持するために、適切な基質またはマトリックスと一緒に提供することができる。通常、少なくとも1×105個、好ましくは1×106個以上の細胞を投与する。細胞は、注射、カテーテルなどによって導入することができる。細胞は、液体窒素の温度で凍結させ、長期間貯蔵することができ、解凍して使用することができる。凍結させる場合、細胞は通常10%のDMSO、50%のFCS、40%のRPMI1640培地中で貯蔵する。解凍したら、前駆細胞の増殖および分化に関連する増殖因子および/または間質細胞を使用することによって細胞を増やす。
【0100】
in vivoにおける方法および使用
一部の実施形態では、例えば、ゲノムDNA脱メチル化療法を必要とする対象において、脱メチル化を許容する細胞を、CD活性を促進する1種または複数種の薬剤とin vivoで接触させる。
【0101】
ポリペプチド、小分子および核酸を対象に投与するためのいくつもの当技術分野において周知の方法のいずれかによって、細胞を、CD活性を促進する薬剤(複数可)とin vivoで接触させることができる。薬剤は、種々の製剤に組み込むことができる。より詳細には、薬剤は、適切な薬学的に許容される担体または希釈剤と組み合わせることによって、医薬組成物に処方することができ、また、錠剤、カプセル剤、散剤、顆粒剤、軟膏剤、液剤、坐剤、注射剤、吸入剤、ゲル剤、ミクロスフェア、およびエアロゾル剤などの固体の形態、半固体の形態、液体の形態または気体の形態の調製物に処方することができる。そのようなものとして、シチジンデアミナーゼ活性を促進する薬剤(複数可)の投与は、経口投与、頬側投与、直腸投与、非経口投与、腹腔内投与、皮内投与、経皮投与、気管内(intracheal)投与などを含めた種々のやり方で実現することができる。活性薬剤は、投与後に全身にわたってよい、または局部投与、壁内投与を使用すること、もしくは埋め込み部位で活性用量を保持するために作用する移植片を使用することによって局所的にすることができる。活性薬剤は、即時活性用に処方することができる、または、持続的放出用に処方することができる。
【0102】
いくつかの状態、特に、中枢神経系の状態のために、薬剤を、血液脳関門(BBB)を渡るように処方することが必要であり得る。血液脳関門(BBB)を通って薬物送達するための1つの戦略は、マンニトールもしくはロイコトリエンなどの浸透性の手段によって、またはブラジキニンなどの血管作動性物質を使用することによって生化学的に、BBBを破壊することを伴う。特異的な薬剤を脳腫瘍にターゲティングするためにBBB開口を使用する可能性も1つの選択肢である。組成物を血管内注射によって投与する場合、BBB破壊剤は本発明の治療用組成物と同時投与することができる。BBBを通過させる他の戦略は、カベオリン1に媒介されるトランスサイトーシス、グルコース担体およびアミノ酸担体などの担体に媒介される輸送体、インスリンまたはトランスフェリンの受容体媒介性トランスサイトーソイス、およびp−糖タンパク質などの活性な流出輸送体を含めた内在性の輸送系の使用を伴ってよい。活性な輸送部分を本発明において使用するための治療用化合物とコンジュゲートして、血管の内皮壁を渡る輸送を容易にすることもできる。あるいは、BBBの反対側への治療用薬剤の薬物送達は、局所送達、例えば、くも膜下腔内送達、例えば、オンマイヤーレザバー(Ommaya reservoir)を通る(例えば、参照により本明細書に組み込まれる、米国特許第5,222,982号および同第5,385,582号を参照されたい);ボーラス注射、例えば、シリンジによって、例えば、硝子体内または頭蓋内に;連続な注入、例えば、対流を伴うカニューレ挿入によって(例えば、参照により本明細書に組み込まれる米国特許出願第20070254842号を参照されたい);または、薬剤が可逆的に添付されているデバイスを埋め込むこと(例えば、参照により本明細書に組み込まれる、米国特許出願第20080081064号および同第20090196903号を参照されたい)によってよい。
【0103】
投与するべきCD活性を促進する薬剤(複数可)の有効量または有効用量を算出することは当業者の技術の範囲内であり、当業者にとっては常套的である。言うまでもなく、投与するべき最終的な量は、投与経路および治療される障害または状態の性質に左右される。
【0104】
医薬に含めるために、CD活性を促進する薬剤(複数可)を商業的な市販の供給源から得ることができる。一般的な提案として、用量当たりの非経口的に投与する化合物の薬学的有効量の総量は、用量反応曲線によって測定することができる範囲内になる。
【0105】
治療的投与のために使用されるCD活性を促進する薬剤(複数可)は、滅菌しなければならない。滅菌は、滅菌濾過膜(例えば、0.2μmの膜)を通して濾過することによって容易に実現される。治療用組成物は、一般に、滅菌アクセスポートを有する容器、例えば、静脈内溶液バッグまたは皮下注射針で穴をあけることができる止め栓を有するバイアル内に置く。CD活性を促進する薬剤(複数可)は、普通は、単位用量容器または多回用量容器、例えば、密封したアンプルまたはバイアル内で、水溶液として、または再構成するための凍結乾燥製剤として貯蔵する。凍結乾燥製剤の例として、10mLのバイアルを5mlの滅菌濾過した化合物1%(w/v)水溶液で満たし、生じた混合物を凍結乾燥する。輸液は、凍結乾燥した化合物を、注射用静菌水を用いて再構成することによって調製する。
【0106】
医薬組成物は、所望の製剤に応じて、動物またはヒトに投与するための医薬組成物を処方するために一般に使用される、ビヒクルと定義される、薬学的に許容できる無毒性の担体または希釈剤を含んでよい。希釈剤は、組み合わせの生物活性に影響を及ぼさないように選択する。そのような希釈剤の例は、蒸留水、緩衝水、生理食塩水、PBS、リンゲル液、ブドウ糖溶液、およびハンクス液である。さらに、医薬組成物または製剤は、他の担体、アジュバント、または非毒性、非治療的、非免疫原性の安定剤、賦形剤なども含んでよい。組成物は、生理的条件に近づけるための追加的な物質、例えば、pH調整剤および緩衝剤、毒性調整剤、湿潤剤および界面活性剤などをも含んでよい。
【0107】
組成物は、例えば抗酸化剤などの任意の種々の安定化剤も含んでよい。医薬組成物がポリペプチドを含む場合、ポリペプチドは、ポリペプチドのin vivoにおける安定性を増強する、またはそうでなければその薬理学的性質を増強する(例えば、ポリペプチドの半減期を増加させる、その毒性を低下させる、溶解性または取り込みを増強する)種々の周知の化合物と複合させることができる。そのような修飾剤または複合剤の例としては、硫酸、グルコン酸、クエン酸およびリン酸が挙げられる。組成物のポリペプチドは、それらのin vivo属性を増強する分子と複合させることもできる。そのような分子としては、例えば、炭水化物、ポリアミン、アミノ酸、他のペプチド、イオン(例えば、ナトリウム、カリウム、カルシウム、マグネシウム、マンガン)、および脂質が挙げられる。
【0108】
投与の種々の型に適している製剤に関する別の手引きは、Remington’s Pharmaceutical Sciences、Mace Publishing Company、Philadelphia、Pa.、第17版(1985)において見ることができる。薬物送達ための方法についての簡単な総説については、Langer、Science 249:1527〜1533(1990)を参照されたい。
【0109】
医薬組成物は、予防的な治療および/または治療的な治療のために投与することができる。活性成分の毒性および治療効果は、細胞培養物および/または実験動物において標準の薬学的手順に従って決定することができ、例えば、LD50(集団の50%に対して致死的な用量)およびED50(集団の50%に対して治療的に有効な用量)を決定することを含む。毒性効果と治療効果との間の用量比は治療指数であり、LD50/ED50比として表すことができる。大きな治療的指標を示す化合物が好ましい。
【0110】
細胞培養試験および/または動物試験から得られたデータを、ヒトに対してさまざまな投与量を処方することにおいて使用することができる。活性成分の投与量は、一般には、ED50を含むさまざまな循環濃度の範囲内に並び、毒性は低い。投与量は、使用する剤形および利用する投与経路に応じてこの範囲内で変動してよい。
【0111】
医薬組成物を処方するために使用する構成成分は、高純度のものであることが好ましく、潜在的に有害な汚染物質を実質的に含まない(例えば、少なくともNational Food(NF)グレード、一般に、少なくとも分析グレード、より一般には少なくとも医薬グレード)。さらに、in vivoで使用するための組成物は、通常滅菌される。所与の化合物を使用する前に合成しなければならない限りでは、生じた生成物は、一般には、合成プロセスまたは精製プロセスの間に存在する可能性がある任意の潜在的に毒性の薬剤、特に、任意の内毒素を実質的に含まない。非経口投与するための組成物も滅菌され、実質的に等張性であり、GMP条件下で製造される。
【0112】
特定の患者に与えるべき有効量の治療用組成物は、種々の因子に左右され、そのいくつかは患者によって異なる。有能な臨床医は、必要に応じて、疾患状態の進行を停止または逆転させるために、患者に投与するための有効量の治療剤を決定することができる。LD50動物データ、および薬剤に関して入手可能な他の情報を利用して、臨床医は、投与経路に応じて、個人に対する最大の安全用量を決定することができる。例えば、治療用組成物が投与される流体が大きいことを考慮すると、静脈内投与する用量は、くも膜下腔内投与する用量よりも高くてよい。同様に、体から急速に取り除かれる組成物は、治療的な濃度を維持するために、高用量で、または反復用量で投与することができる。有能な臨床医は、通常の技術を利用して、常套的な臨床試験の過程で特定の治療薬の投与量を最適化することができる。
【0113】
本方法を用いて治療することができる哺乳動物種としては、イヌおよびネコ;ウマ;ウシ;ヒツジなど、ならびに霊長類、特に、ヒトが挙げられる。動物モデル、特に小さな哺乳動物、例えば、マウス、ウサギなどを実験的調査のために使用することができる。他の使用としては、活性な脱メチル化シグナル伝達の存在下で特異的な効果を調査することが望ましい調査が挙げられる。
【0114】
本発明の方法は、併用療法においても使用される。例えば、いくつもの薬剤、例えば、化学療法剤、キナーゼ阻害剤、アンジオスタチン、エンドスタチン、VEGF阻害剤などが癌の治療において有用であり得る。本発明のCD活性を促進する薬剤(複数可)とこれらの他の薬剤を組み合わせて使用することは、個々の薬物について必要な投与量が低減すること、および異なる薬物の相補的な効果という利点を有し得る。
【0115】
上記の通り、本発明は、ヒト患者などの哺乳動物の治療において、ゲノムDNA脱メチル化療法を必要とする対象に使用される。そのような対象の例は、プロモーターが高メチル化されていることによって異常にサイレンシングされた遺伝子に関連する状態に罹患している対象である。そのような状態を特徴とする疾患に罹患している患者には、係属中の特許請求された発明治療プロトコールが大いに有益である。
【0116】
そのような状態の1つの例は癌である。いくつもの遺伝子、すなわち、メチル化感受性遺伝子が、癌では異常に高メチル化され、サイレンシングされていることが公知である。これらとしては、細胞周期制御に関与する遺伝子(例えば、RB1、CDKN2AINK4A、CDKN2AARF)、腫瘍細胞の浸潤に関与する遺伝子(例えば、CDH1、CDH13、TIMP3、VHL)、DNA修復に関与する遺伝子(例えば、MLH1、MGMT、BRCA1、GSTP1)、クロマチンリモデリングに関与する遺伝子(例えばSMARCA3)、細胞シグナル伝達に関与する遺伝子(例えば、RASSF1A、SOCS1)、転写に関与する遺伝子(例えば、ESR1)、およびアポトーシスに関与する遺伝子(例えば、DAPK1)が挙げられる。したがって、本発明の方法および組成物は、癌、例えば、神経膠腫、髄芽腫、結腸癌、結腸直腸癌、乳癌、または白血病に罹患している対象における腫瘍成長および癌の進行を阻害することにおいて使用される。「癌」という用語は、一般には、哺乳動物における、細胞の成長/増殖が調節されていないことを特徴とする生理的状態を指す。癌の例としては、これらに限定されないが、細胞腫、リンパ腫、芽細胞種、および白血病が挙げられる。癌のより詳細な例としては、これらに限定されないが、慢性リンパ球性白血病(CLL)、非小細胞癌(NSCLC)を含めた肺癌、乳癌、卵巣癌、子宮頸部癌、子宮内膜癌、前立腺癌、結腸直腸癌、腸のカルチノイド、膀胱癌、胃癌、膵臓癌、肝癌(肝細胞癌)、肝芽腫、食道癌、肺腺癌、中皮腫、滑膜肉腫、骨肉腫、頭頸部扁平上皮癌、若年性鼻咽腔血管線維腫、脂肪肉腫、甲状腺癌、黒色腫、基底細胞癌(BCC)、髄芽腫およびデスモイドが挙げられる。特定の癌と上記の対象とする遺伝子のメチル化状態との間の相関は、その開示が参照により本明細書に組み込まれるRobertson、K.D.(2005)Nature Review Genetics 6:597〜610において見ることができる。
【0117】
腫瘍成長および癌の進行を阻害するためのCD活性を促進する薬剤(複数可)の有効量は、上述のメチル化感受性遺伝子の1種または複数種の発現を、in vitroおよびin vivoで、例えば2倍以上増加させ、かつ/またはin vitroにおける癌細胞の増殖速度またはin vivoにおける腫瘍の増殖阻害に測定可能な低下をもたらす量である。例えば、好ましい成長阻害剤は、腫瘍成長を、適切な対照と比較して少なくとも約5%、少なくとも約10%、少なくとも約20%、好ましくは約20%〜約50%、さらに好ましくは、50%超(例えば、約50%〜約100%)阻害し、対照は、一般には、試験されているシチジンデアミナーゼ活性を促進する薬剤(複数可)で処理していない癌細胞である。薬剤を体重1kg当たり約1μgから約100mgまでで投与することにより、腫瘍サイズまたは細胞増殖が抗体を最初に投与してから約5日〜3ヶ月以内、好ましくは約5〜30日以内に低下した場合、その薬剤はin vivoにおいて成長阻害性である。特定の態様では、腫瘍サイズは、療法を開始した時のそのサイズと比較して低下する。
【0118】
本発明の方法によって治療することができる、プロモーターが高メチル化されていることによって異常にサイレンシングされた遺伝子に関連する状態の別の例は、異常なゲノム刷り込みに関連する状態である。ゲノム刷り込みでは、特定の遺伝子は、由来する親(parent of origin)に特異的な様式で発現している。ゲノム刷り込みは、刷り込まれた遺伝子が、母親から受け継がれた対立遺伝子からのみ発現されるか、または父親から受け継がれた対立遺伝子からのみ発現される、古典的なメンデル遺伝と無関係の遺伝プロセスである。ゲノム刷り込みは、遺伝子の配列を変化させることなく単一対立遺伝子性遺伝子発現を実現するためにメチル化およびヒストン修飾を必要とする。これらの後成的な目印は、生殖系列において確立され、生物体の全ての体細胞全体を通して維持される。
【0119】
本発明の方法によって治療可能な、異常なゲノム刷り込みに関連するいくつもの状態が同定されている。例えば、胎児および出生後の過成長、器官の肥大、腫瘍リスクの増加、および顔面の異常を特徴とするベックウィズ−ヴィーデマン症候群では、IGF2/H19刷り込み制御領域1における母系対立遺伝子の新規のメチル化が観察される。精神遅滞、肥満、低身長、および行動上の問題を特徴とするプラダー・ウィリー症候群では、PWS遺伝子の父系対立遺伝子の新規のメチル化が観察される。腎臓の副甲状腺ホルモン抵抗性を特徴とする偽性副甲状腺機能低下症1B型では、NESP55の母系対立遺伝子の新規のメチル化が観察される。本発明の方法は、これらの遺伝子座における脱メチル化を促進し、それによって適切な遺伝子発現を回復すること使用される。
【0120】
本発明の方法によって治療することができる、プロモーターが高メチル化されていることによって異常にサイレンシングされた遺伝子に関連する状態の別の例は、反復不安定性(repeat instability)疾患に関連する状態である。これらの疾患では、反復配列が拡大することにより、それらの配列の近くの遺伝子の発現に影響を及ぼす異常なメチル化が生じる。本発明の方法によって治療可能な反復不安定性に関連するいくつもの状態が同定されている。例えば、精神遅滞、巨睾丸症、および自閉症的行動を特徴とする脆弱X症候群では、FMRI遺伝子の5’UTRのCGG反復が拡大することにより、5’UTR配列が新規にメチル化され、FMRI遺伝子が異常にサイレンシングされる。別の例として、肢および顔面の筋肉の衰弱および消耗、筋緊張症、ならびに白内障を特徴とする筋緊張性ジストロフィー(DM1)では、DMPK遺伝子のUTR内のCTG反復が拡大することにより、拡大したCTG反復の近くのCpGアイランドが新規にメチル化され、それによって今度はSIX5遺伝子が攪乱され、サイレンシングされる。本発明の方法は、これらの遺伝子座における脱メチル化を促進し、それによって適切な遺伝子発現を回復することに使用される。
【0121】
スクリーニング方法。
本明細書に記載の方法は、候補薬剤を、脱メチル化を調節する活性についてスクリーニングするための有用なシステムを提供する。そのために、CD活性を促進する薬剤が、脱メチル化を増強することに対する強力な効果を有することが示されている。CD活性を阻害する薬剤を、脱メチル化が起こっている細胞を含む細胞培養物系に加えることにより、この脱メチル化活性が強力に抑制され、したがって、Oct4およびNanogなどのメチル化感受性遺伝子の転写活性プロモーターの量が低下する。この脱メチル化活性の抑制ならびにその後の、これらのプロモーターにおけるメチル化の増加および転写活性のサイレンシングは、早ければ、脱メチル化している細胞を、CD活性を阻害する薬剤と接触させた1日後に観察され得、3日目までにこれらのメチル化感受性遺伝子がほぼ完全にサイレンシングされる。
【0122】
生物学的に活性な薬剤を求めるスクリーニングアッセイでは、細胞、通常細胞培養物を、CD活性を促進する薬剤の存在下で、対象とする薬剤と接触させ、候補薬剤の効果を、アウトプットパラメータ、例えばメチル化CpG配列の量、メチル化感受性遺伝子の発現などを上記の方法によってモニターすることによって評価する。
【0123】
パラメータは、細胞の数量化できる構成成分、特に、望ましくはハイスループットシステムにおいて正確に測定することができる構成成分である。パラメータは、細胞表面決定基、受容体、タンパク質、またはそのコンフォメーションの修飾または翻訳後修飾、脂質、炭水化物、有機分子または無機分子、核酸、例えば、mRNA、DNAなどを含めた任意の細胞構成成分または細胞産物、またはそのような細胞構成成分に由来する部分、またはそれらの組み合わせであってよい。大部分のパラメータにより定量的な読み取りがもたらされるが、ある場合には、半定量的な結果または定性的な結果が許容される。読み取りは、単一の決定値を含んでよい、または、平均値、中央値もしくは分散などを含んでよい。特徴的に、多数の同じアッセイから、各パラメータについてさまざまなパラメータの読み取り値が得られる。変動性が予測され、試験パラメータのセットのそれぞれについてのさまざまな値は、単一の値をもたらすために使用される一般的な統計学的方法を用いた標準の統計学的な方法を使用して得られる。
【0124】
例えば、薬剤を、脱メチル化活性を促進する活性について、例えば、候補薬剤を、CD活性を促進する薬剤の存在下で細胞培養物に加えることによってスクリーニングすることができる。観察されるメチル化の量が減少すること、例えば、メチル化感受性遺伝子のプロモーターまたは外因的に供給された5−meCpGリッチ核酸の5−メチルシトシンの数が、候補薬剤が存在しない培養物において観察されるものに対して、例えば、1.5分の1、2分の1、3分の1またはそれ以下に減少することにより、候補薬剤が脱メチル化を促進する薬剤であったことが示される。そのような実施形態では、細胞は、脱メチル化を許容する細胞であってよい、または、脱メチル化を許容しない細胞であってよい。
【0125】
あるいは、例えば、候補薬剤を、CD活性を促進する薬剤の存在下で細胞培養物に加えることによって、脱メチル化活性を抑制する活性について薬剤をスクリーニングすることができる。観察されるメチル化の量、例えば、メチル化感受性遺伝子のプロモーターまたは外因的に供給された5−meCpGリッチ核酸の5−メチルシトシンの数が、候補薬剤が存在しない培養物において観察されるものと比較して、全く減少しないこと、またはほんの少量しか減少しないことにより、候補薬剤が脱メチル化を抑制する薬剤であったことが示される。そのような実施形態では、培養細胞は、脱メチル化を許容する細胞である。
【0126】
スクリーニングの対象とされる候補薬剤としては、多数の化学的クラス、主として有機金属分子を含み得る有機分子、無機分子、遺伝子配列などを包含する既知化合物および未知化合物が挙げられる。本発明の重要な態様は、毒性試験などを含めた、候補薬物の評価である。
【0127】
候補薬剤は、構造的な相互作用、特に水素結合に必要な官能基を含む有機分子を含み、一般には少なくともアミン基、カルボニル基、ヒドロキシル基またはカルボキシル基を含み、しばしば、官能性化学基の少なくとも2つを含む。候補薬剤は、多くの場合、上記の官能基の1つまたは複数で置換された環式炭素または複素環式構造および/または芳香族または多芳香族の構造を含む。候補薬剤は、ペプチド、ポリヌクレオチド、糖類、脂肪酸、ステロイド、プリン、ピリミジン、誘導体、構造的な類似体またはそれらの組み合わせを含めた生体分子の中にも見いだされる。薬理活性のある薬物、遺伝的活性のある分子などが包含される。対象とする化合物としては、化学療法剤、ホルモンまたはホルモンアンタゴニストなどが挙げられる。本発明に適した医薬品の例は、「The Pharmacological Basis of Therapeutics」Goodmanおよび Gilman、McGraw−Hill、New York、N.Y.、(1996)、第9版に記載されているものである。毒素、生物戦剤および化学戦剤(biological and chemical warfare agent)も包含される。例えば、Somani、S.M.(編)「Chemical Warfare Agents」Academic Press、New York、1992)を参照されたい。
【0128】
スクリーニングの対象とされる候補薬剤としては、核酸、例えば、siRNA、shRNA、アンチセンス分子、またはmiRNAをコードする核酸、またはポリペプチドをコードする核酸も挙げられる。核酸を標的細胞に移入するために有用な多くのベクターが利用可能である。ベクターは、エピソームとして、例えば、プラスミド、ミニサークルDNA、サイトメガロウイルス、アデノウイルスなどのウイルス由来のベクターとして維持することができる、またはベクターは、相同組換えまたはランダムな組み込みによって標的細胞のゲノムに組み込むことができる。例えば、MMLV、HIV−1、ALVなどのレトロウイルス由来のベクター。ベクターは、対象細胞に直接提供することができる。言い換えれば、多能性細胞を、対象とする核酸を含むベクターと、ベクターが細胞に取り込まれるように接触させる。
【0129】
電気穿孔、塩化カルシウムトランスフェクション、およびリポフェクションなどの、細胞を核酸ベクターと接触させるための方法は当技術分野で周知である。あるいは、対象とする核酸は、ウイルスによって対象細胞に提供することができる。言い換えれば、多能性細胞を、対象とする核酸を含むウイルス粒子と接触させる。本発明の方法には、レトロウイルス、例えば、レンチウイルスが特に適している。一般に使用されるレトロウイルスベクターは、「不完全である」、すなわち、増殖性感染のために必要なウイルスタンパク質を産生することができない。もっと正確に言えば、ベクターの複製には、パッケージング細胞系において成長させることが必要である。対象とする核酸を含むウイルス粒子を生成するために、核酸を含むレトロウイルスの核酸をパッケージング細胞系によってウイルスカプシド中にパッケージする。異なるパッケージング細胞系からは、カプシド中に組み入れられる異なる外被タンパク質がもたらされ、この外被タンパク質により、ウイルス粒子の細胞に対する特異性が決定される。外被タンパク質は、狭宿主性、広宿主性および異種栄養性の少なくとも3種類のものである。狭宿主性外被タンパク質、例えば、MMLVを用いてパッケージしたレトロウイルスは、大部分のマウスおよびラット細胞型に感染することができ、これは、BOSC23などの狭宿主性パッケージング細胞系を使用することによって生成される(Pearら(1993)P.N.A.S.90:8392〜8396)。広宿主性外被タンパク質、例えば、4070A(Danosら、上記)を担持するレトロウイルスは、ヒト、イヌおよびマウスを含めた大部分の哺乳動物細胞型に感染することができ、これは、PA12(Millerら(1985)Mol.Cell.Biol.5:431〜437);PA317(Millerら(1986)Mol.Cell.Biol.6:2895〜2902);GRIP(Danosら(1988)PNAS 85:6460〜6464)などの広宿主性パッケージング細胞系を使用することによって生成される。異種栄養性外被タンパク質、例えば、AKR envを用いてパッケージしたレトロウイルスは、マウスの細胞以外の大部分の哺乳動物細胞型に感染することができる。適切なパッケージング細胞系を使用して、対象CD33+分化体細胞がパッケージされたウイルス粒子の標的になることを確実にすることができる。再プログラミング因子をコードする核酸を含むレトロウイルスベクターをパッケージング細胞系に導入する方法、およびパッケージング系によって生成されるウイルス粒子を採取する方法は当技術分野で周知である。
【0130】
対象とする核酸を対象細胞に提供するために使用するベクターは、一般には、対象とする核酸の発現を駆動する、すなわち、転写を活性化するために適したプロモーターを含む。これは、遍在的に作用するプロモーター、例えば、CMV−b−アクチンプロモーター、または誘導性プロモーター、例えば特定の細胞集団において活性なプロモーターまたはテトラサイクリンなどの薬物が存在することに反応するプロモーターなどを含んでよい。転写活性化とは、標的細胞において転写が基礎レベルを少なくとも約10倍、少なくとも約100倍、通常少なくとも約1000倍超えて増加することを意味する。さらに、対象細胞に再プログラミング因子を提供するために使用するベクターは、後で除去しなければならない遺伝子を含んでよく、その際は、例えば、Cre/Loxなどのリコンビナーゼ系、または、例えば、TK、bcl−xsなどのヘルペスウイルスなどの選択毒性を可能にする遺伝子を含めることによって該遺伝子が破壊されて発現される細胞を用いる。
【0131】
スクリーニングの対象とされる候補薬剤としては、ポリペプチドも挙げられる。そのようなポリペプチドは、場合によって、生成物の溶解性を増加させるポリペプチドドメインと融合することができる。ドメインは、定義済みのプロテアーゼ切断部位、例えば、TEVプロテアーゼによって切断されるTEV配列を通じてポリペプチドに連結することができる。リンカーは、1つまたは複数の可動性の配列、例えば、1個から10個までのグリシン残基も含んでよい。一部の実施形態では、融合タンパク質の切断は、生成物の溶解性を維持する緩衝液中、例えば、0.5Mから2Mまでの尿素の存在下、溶解性を増加させるポリペプチドおよび/またはポリヌクレオチドの存在下などで行う。対象とするドメインとしては、エンドソーム溶解性(endosomolytic)ドメイン、例えば、インフルエンザHAドメイン;および作製に役立つ他のポリペプチド、例えば、IF2ドメイン、GSTドメイン、GRPEドメインなどが挙げられる。
【0132】
候補ポリペプチド薬剤を、凝集のシグナル伝達を細胞内で阻害するその能力についてアッセイする場合、ポリペプチドは、ポリペプチド浸透性ドメインと融合した対象とするポリペプチド配列を含んでよい。いくつもの浸透性ドメインが当技術分野で公知であり、ペプチド、ペプチド模倣薬、および非ペプチド担体を含めた本発明の非組み込み型のポリペプチドにおいて使用することができる。例えば、浸透性ペプチドは、ペネトラチンと称され、アミノ酸配列RQIKIWFQNRRMKWKKを含む、Drosophila melanogasterの転写因子であるAntennapaediaの第3のアルファヘリックス由来であってよい。別の例として、浸透性ペプチドは、例えば、天然に存在するtatタンパク質のアミノ酸49〜57を含み得るHIV−1tat基本領域アミノ酸配列を含む。他の浸透性ドメインとしては、ポリアルギニンモチーフ、例えば、HIV−1revタンパク質のアミノ酸34〜56の領域、ノナアルギニン、オクタアルギニンなどが挙げられる。(例えば、移行ペプチドおよびペプトイドの教示に関して参照により本明細書に明確に組み込まれる、Futakiら(2003)Curr Protein Pept Sci.2003年4月;4(2):87〜96;およびWenderら(2000)Proc.Natl.Acad.Sci.U.S.A 2000年11月21日;97(24):13003〜8;米国特許出願公開第20030220334号;同第20030083256号;同第20030032593号;および同第20030022831号を参照されたい)。ノナアルギニン(R9)配列は、特徴づけられているより効率的なPTDの1つである(Wenderら2000;Uemuraら2002)。
【0133】
候補ポリペプチド薬剤を、凝集のシグナル伝達を細胞外で阻害するその能力についてアッセイする場合、ポリペプチドは、安定性が改善されるように処方することができる。例えば、ペプチドをペグ化することができ、ポリエチレンオキシ基により、血流中の寿命が増強される。追加的な機能性をもたらすため、例えば、in vivoにおける安定性を増加させるために、ポリペプチドを別のポリペプチドと融合することができる。一般に、そのような融合パートナーは、安定な血漿タンパク質であり、それにより、例えば、融合物として存在するとポリペプチドのin vivo血漿内半減期が延長され、具体的には、そのような安定な血漿タンパク質は免疫グロブリン定常ドメインである。安定な血漿タンパク質が通常多量体の形態、例えば、免疫グロブリンまたはリポタンパク質で見いだされ、その中の同じポリペプチドまたは異なるポリペプチド鎖が、通常ジスルフィド結合および/または非共有結合して集合した多鎖ポリペプチドを形成しているほとんどの場合、ポリペプチドを含有する本明細書の融合物も、安定な血漿タンパク質前駆物質と実質的に同じ構造を有する多量体として作製し、使用する。これらの多量体が含むポリペプチド薬剤は同種のものである、または、これらの多量体は2種以上のポリペプチド薬剤を含有してよい。
【0134】
候補ポリペプチド薬剤は、原核細胞によって又は真核生物から産生することができ、ほどくこと、例えば、熱変性、DTT低下などによってさらに加工し、当技術分野で公知の方法を使用してさらに再び折りたたむことができる。一次配列を変化させない対象とする修飾としては、ポリペプチドの化学的な誘導体化、例えば、アシル化、アセチル化、カルボキシル化、アミド化などが挙げられる。グリコシル化の修飾、例えば、ポリペプチドのグリコシル化パターンを、その合成およびプロセシングの間に、またはさらなるプロセシングステップにおいて修飾することによって;例えば、ポリペプチドを、哺乳動物のグリコシル化酵素または脱グリコシル酵素などのグリコシル化に影響を及ぼす酵素に曝露させることによって行われる修飾も包含される。リン酸化されたアミノ酸残基、例えば、ホスホチロシン、ホスホセリン、またはホスホスレオニンを有する配列も包含される。ポリペプチドは、それらのタンパク質分解に対する耐性を改善するため、または溶解特性を最適化するため、またはそれらを治療剤としてより適切にするために、通常の分子生物学的技法および合成化学を使用して修飾することができる。そのようなポリペプチドの類似体としては、天然に存在するL−アミノ酸以外の残基、例えば、D−アミノ酸または天然に存在しない合成のアミノ酸を含有するものが挙げられる。アミノ酸残基の一部または全部をD−アミノ酸で置換することができる。
【0135】
候補ポリペプチド薬剤は、当技術分野で公知の従来の方法を使用して、in vitro合成によって調製することができる。種々の市販の合成装置、例えば、Applied Biosystems、Inc.、Beckmanなどによる自動合成機が利用可能である。合成機を使用することによって、天然に存在するアミノ酸を非天然アミノ酸で置換することができる。特定の配列および調製様式は、利便性、経済的側面、必要な純度などによって決定される。あるいは、候補ポリペプチド薬剤は、組換え合成の従来の方法に従って単離し、精製することができる。発現宿主の溶解物を調製し、溶解物を、HPLC、排除クロマトグラフィー、ゲル電気泳動、アフィニティークロマトグラフィー、または他の精製技法を使用して精製することができる。ほとんどの場合、使用される組成物は、産物の調製およびその精製の方法に関連する汚染物質に関して、少なくとも20重量%、通常、少なくとも約75重量%、好ましくは少なくとも約95重量%、および治療のためには、通常少なくとも約99.5重量%の所望の産物を含む。通常、百分率は、総タンパク質量に基づく。
【0136】
ある場合では、スクリーニングされる候補ポリペプチド薬剤は抗体である。「抗体」または「抗体部分」という用語は、エピトープに適合し、それを認識する特定の形状を有する任意のポリペプチド鎖含有分子構造を含むものとし、ここで、1つまたは複数の非共有結合性の相互作用により、分子構造とエピトープとの間の複合体が安定化する。所与の構造の特異的または選択的な適合およびその特異的なエピトープは、時には、「鍵と鍵穴」適合と称される。典型的な抗体分子は、免疫グロブリンであり、全ての供給源、例えば、ヒト、げっ歯類、ウサギ、雌ウシ、ヒツジ、ブタ、イヌ、他の哺乳動物、ニワトリ、他のトリなど由来の全種類の免疫グロブリン、IgG、IgM、IgA、IgE、IgDなどが、「抗体」であるとみなされる。本発明において利用される抗体は、ポリクローナル抗体またはモノクローナル抗体のいずれかである。抗体は、一般には、細胞が培養されている培地中に提供される。
【0137】
候補薬剤を含めた化合物は、合成化合物または天然化合物のライブラリーを含めた多種多様な供給源から得られる。例えば、ランダム化されたオリゴヌクレオチドおよびオリゴペプチドの発現を含めた、生体分子を含めた多種多様な有機化合物のランダムな合成および指向性合成のための多数の手段が利用可能である。あるいは、細菌の抽出物、真菌の抽出物、植物の抽出物および動物の抽出物の形態の天然化合物のライブラリーが利用可能である、または容易に作製される。さらに、天然の、または合成的に作製したライブラリーおよび化合物は、従来の化学的手段、物理的手段および生化学的手段によって容易に修飾され、また、それを使用して、組み合わせのライブラリーを作製することができる。公知の薬理薬剤(pharmacological agent)を、例えば、アシル化、アルキル化、エステル化、アミド化(amidification)などの直接的またはランダムな化学修飾に供して構造類似体を作製することができる。
【0138】
候補薬剤は、通常、薬剤を欠く細胞と併せて、少なくとも1つ、通常複数の細胞試料に加えることによって、その生物活性についてスクリーニングする。薬剤に反応したパラメータの変化を測定し、結果を、参照培養物と比較すること、例えば、薬剤の存在下と非存在下で比較すること、他の薬剤を用いて得られた参照培養物と比較することなどによって評価する。
【0139】
薬剤は、溶液に都合よく加えられる、または培養物中の細胞の培地に対して容易に可溶性の形態である。薬剤は、フロースルーシステムで、断続的または連続的な流れとして加えることができる、あるいは、化合物のボーラスを、単独でまたは増加的に、別の静的な溶液に加える。フロースルーシステムでは、2つの流体を使用し、一方は生理的に中性の溶液であり、他方は加えた試験化合物と同じ溶液である。第1の流体を、細胞を通過させ、その後、第2の流体を通過させる。単一溶液法では、試験化合物のボーラスを細胞の周囲の培地の体積に加える。培地の構成成分の全体的な濃度は、ボーラスを加えることによって、またはフロースルー法における2つの溶液間で有意に変化するべきではない。
【0140】
さまざまな濃度に対する示差的な反応を得るために、異なる薬剤の濃度を用いて複数のアッセイを平行して実行することができる。当技術分野で公知の通り、薬剤の有効濃度の決定には、一般には1:10、または他のlog尺度の、希釈によって生じるさまざまな濃度を用いる。必要であれば、第2の一連の希釈を用いて濃度をさらに細かく区分することができる。一般には、これらの濃度の1つは陰性対照としての機能を果たす、すなわち、ゼロ濃度または薬剤の検出レベルを下回る濃度、または表現型における検出可能な変化を生じさせない薬剤の濃度またはそれを下回る濃度である。
【0141】
選択マーカーの存在を数量化するために種々の方法を利用することができる。例えば、DNAメチル化の状態を、例えば、特定のCpG配列において測定するために、クロマチン免疫沈降(ChIP)を実施して内在性DNAを単離することができ、次いで、それを制限エンドヌクレアーゼHpallで消化して脱メチル化の程度を決定することができる、またはバイサルファイト配列決定を行うことができる。存在する分子の量を測定するために、例えば、メチル化感受性遺伝子の発現を測定する場合、都合のよい方法は、分子を、蛍光性、発光性、放射性、酵素的に活性などであり得る検出可能部分、特に、高い親和性を有するパラメータへの結合に特異的な分子で標識することである。蛍光部分は、実質的にあらゆる生体分子、構造、または細胞型を標識するために容易に利用可能である。免疫蛍光部分は、特定のタンパク質だけでなく、特異的なコンフォメーション、切断産物、またはリン酸化のような部位修飾にも結合するように誘導することができる。個々のペプチドおよびタンパク質は、自己蛍光するように、例えば、それらを細胞内で緑色蛍光タンパク質キメラとして発現させることによって遺伝子工学で操作することができる(概説について、Jonesら(1999)Trends Biotechnol.17(12):477〜81を参照されたい)。
【0142】
例えば上記のものなどのスクリーニングは、特定の生物系における脱メチル化の調節における活性を有する薬剤を同定するために合わせてあつらえることができる。例えば、腫瘍細胞において、細胞周期、腫瘍−細胞浸潤、DNA修復、クロマチンリモデリング、細胞シグナル伝達、転写およびアポトーシスを調節する遺伝子などのメチル化感受性遺伝子のプロモーターの脱メチル化を促進する薬剤は、これらの遺伝子の脱メチル化を促進すること、したがって、腫瘍においてこれらの遺伝子を発現させることに使用し、それによって、癌細胞の増殖および腫瘍成長を阻止することができる。別の例として、体細胞またはES細胞と体細胞の異核共存体における多能性遺伝子であるOct4およびNanogなどのメチル化感受性遺伝子のプロモーターにおける脱メチル化を促進する薬剤は、iPS細胞を作製するための公知の方法において多能性に関連する遺伝子の脱メチル化を促進することに使用することができる。いくつかのそのような場合、例えば、体細胞では、これらの方法は、スクリーニングする目的で、iPS表現型をさらに促進するために、細胞に再プログラミング因子を提供するステップを含んでよい。
【0143】
キットを提供することができ、キットは、本明細書に記載のように、CD活性を促進する1種または複数種の薬剤および細胞を、脱メチル化を許容するように誘導するための試薬を含む。対象とする組み合わせは、1種または複数種のAIDポリペプチドもしくはAPOBECポリペプチド、またはそれらのペプチドをコードする核酸を含むベクターおよび再プログラミングを促進する1種または複数種の薬剤を含んでよい。キットは、対象の細胞におけるDNAメチル化の状態を決定するために適した試薬をさらに含んでよい。キットは、チューブ、緩衝液など、および使用説明書も含んでよい。
【実施例】
【0144】
以下の実施例は、本発明をどのように行い、使用するかについての完全な開示および記載を当業者に提供するために提示され、出願人らが彼らの発明とみなすものの範囲を限定するものではなく、また、以下の実験が、実施した全ての実験または唯一の実験であることを示すものでもない。使用される数字(例えば、量、温度など)に関して正確さを確実にするための努力がなされているが、いくらかの実験的な誤差および偏差が考慮されるべきである。別段の指定のない限り、部分は重量による部分であり、分子量は重量平均分子量であり、温度は摂氏度であり、圧力は、大気または大気付近の圧力である。
【実施例1】
【0145】
核の多能性への再プログラミングに必須である新規の初期制御因子を同定するために、我々は、体細胞の分化した状態を維持することに固有の原理の解明において有用であることが立証された異核共存体の我々の以前の経験を活用した。特に、これらの以前の我々の研究およびその他の研究により、ヒト細胞の「最終分化した」状態は固定されておらず、変化させることができ、他の分化した状態に典型的な以前はサイレントであった遺伝子の発現が誘導されることが示された(Blau、H.M.ら(1983)Cell 32、1171〜801;Baron、M.H.&Maniatis、T.(1986)Cell 46、591〜602;Wright、W.E.(1984)Exp Cell Res 151、55〜69;Spear、B.T.&Tilghman、S.M.(1990)Mol Cell Biol 10、5047〜54;Chiu、CP.&Blau、H.M.R(1984)Cell 37、879〜87)。我々は、(1)再プログラミングは、全てのES細胞因子の存在下で起こり、(2)再プログラミングの開始は、融合と同時に開始され、(3)再プログラミングは、融合した非分裂細胞において評価され、かつ(4)種差により融合した細胞型の転写物が区別されるので、異核共存体を用いて、機構を解明し、多能性への再プログラミングの開始において役割を有する新規の遺伝子を同定することができると推論した。
【0146】
材料および方法
フローサイトメトリーによる異核共存体の生成および単離。GFP+マウスES細胞およびDsRed+ヒト胎児の肺の初代線維芽細胞を、以前に記載されている通りレトロウイルス構築物を用いた形質導入によって生成し(Palermo、A.ら(2009)Faseb J)、融合して非分裂性の、多核異核共存体を形成した。細胞をまず、ES培地中で12時間共培養し、次いで、PEG1500(Roche)を用いて37℃で2分間処理し、その後DMEMで4回連続的に洗浄した。ES培地を洗浄した後に交換し、その後12時間ごとに交換した。GFP+/DsRed+異核共存体をフローサイトメトリー(FACSVantage SE、BD)によって2回選別し、遺伝子発現およびメチル化について解析した。
【0147】
免疫蛍光法。異核共存体を2.5%v/vのヤギ血清および1mMのEDTAを伴うPBS中で2回選別し、900rpmで5分間サイトスピンした。サイトスピンしたGFP+/DsRed+異核共存体をHoechst33342で染色し、画像化した。抗体染色のために、サイトスピンした細胞を固定し、透過処理し、PBS中20%のFBSを使用してブロッキングした。細胞を、一次抗体マウス抗Ki−67(Dako Denmark A/S)と一緒に、ブロッキング緩衝液中1:100希釈で1時間インキュベートし、PBS中で3回すすぎ、次いで、ヤギ抗マウスCascade blue二次抗体(Millipore)と一緒に1:500希釈で30分間インキュベートし、3回すすぎ、Fluoromount−Gを用いて封入し、画像化した。画像を、落射蛍光顕微鏡(Axioplan2;Carl Zeiss MicroImaging、Inc.)、Fluar20×/0.75対物レンズまたは40×/0.90対物レンズ、およびデジタルカメラ(ORCA−ER C4742−95;Hamamatsu Photonics)を使用して取得した。取得するために使用したソフトウェアはOpenLab4.0.2(Improvision)であった。
【0148】
PEG誘導性融合の3時間後にBrdUをmES共培養物およびhFb共培養物に加えた。標識化および抗体染色を、BrdU Labeling and Detection Kit I(Roche)を使用して行った。
【0149】
遺伝子発現の解析。RNAを、ES細胞、線維芽細胞、および融合後またはRNeasy microキット(Qiagen)を使用してsiRNA処理した後、違う時間に2回選別した異核共存体から調製した。各試料について全RNAを、Superscript First−Strand Synthesis System for RT−PCR(Invitrogen)を使用して逆転写した。逆転写された材料をGo GreenTaq DNAポリメラーゼ(Promega)を使用してPCRに供した。Oct4、NanogおよびGAPDHの発現を解析するためのヒト特異的なプライマーを設計した。siRNA処理実験においてAIDおよびGAPDHに対して使用したプライマーにより、ヒト転写物とマウス転写物の両方を増幅して異核共存体におけるAIDおよびGAPDHの総レベルを評価した。RT−PCRおよび定量的PCRのために使用するヒト特異的なプライマーは、:hOct4 F 5’−TCGAGAACCGAGTGAGAGGC−3’(配列番号45)、R−5’−CACACTCGGACCACATCCTTC−3’(配列番号46);hNanog F 5’−CCAACATCCTGAACCTCAGCTAC−3’(配列番号47)、R 5’−GCCTTCTGCGTCACACCATT−3’(配列番号48);hGAPDH F 5’−TGTCCCCACTGCCAACGTGTCA−3’(配列番号49)、R 5’−AGCGTCAAAGGTGGAGGAGTGGGT−3’(配列番号50)である。siRNA処理後のノックダウンを評価するための非種特異的なプライマー配列は、以下の通りである:GAPDH F 5’−ACCACAGTCCATGCCATCAC−3’(配列番号51)、R 5’−TCCACCACCCTGTTGCTGTA−3’(配列番号52);AID F 5’−AAAATGTCCGCTGGGCTAAG−3’(配列番号53)、R 5’−AGGTCCCAGTCCGAGATGTAG−3’(配列番号54)。
【0150】
リアルタイムPCR。リアルタイムPCRを、ABI 7900HT リアルタイムPCRシステムを使用して、Sybr Green PCRミックス(Applied Biosystems)を用いて行った。試料を94℃で2分間、40×(94℃で20秒間、58℃で45秒間)でサイクルにかけた。
【0151】
【表1】
【0152】
単一細胞RT−PCR。単一の異核共存体をFACS(FACSVantage SE、BD)によって直接選別して、9μl一定分量のRT−PCR溶解緩衝液を含有するPCRチューブに入れた。緩衝液の構成成分は、市販のRT−PCR緩衝液(Superscript One−Step RT−PCR Kit Reaction Buffer、Invitrogen)、RNアーゼ阻害剤(Protector RNase Inhibitor、Roche)および0.15%のIGEPAL界面活性剤(Sigma)を含んだ。短パルスで急速回転させた後、PCR−チューブをすぐに衝撃凍結し、その後の分析のために−80℃で貯蔵した。
【0153】
二段階マルチプレックスネステッド単一細胞RT−PCRのために、細胞溶解物を、まず、SuperScript One−Step RT−PCR Kit(Invitrogen)を使用して、Oct4、NanogおよびGAPDHに対するヒトおよび遺伝子に特異的なプライマー対を用いて逆転写した(表2、外部プライマー;図5b)。簡単に述べると、同じPCR細胞−溶解チューブに、遺伝子特異的なプライマー対およびRNアーゼ阻害剤を含有するRT−PCR反応混合物を加えることによって、RT−PCRを行った。第1ラウンドPCRおよび第2ラウンドPCRならびにより大きな特異性を確実にするネステッドRT−PCR用のイントロンスパニングプライマーセットを設計し、使用することによって、ゲノム産物を排除した。第1のステップでは、55℃で30分間逆転写反応を行い、その後94℃で2分間のステップを行った。その後に、以下の通りPCR増幅を30サイクル行った:94℃で30秒間;58℃で30秒;68℃で30秒間。最終的なPCRステップにおいて、反応産物を68℃で3分間インキュベートした。完了した反応産物を4℃で貯蔵した。
【0154】
PCRプロトコールの第2のステップにおいて、第1のステップからの完了したRT−PCR反応産物を水で1:1希釈した。これらの反応産物の1パーセントを、PCRの第2ラウンドのために複製して新しい反応チューブに移し、PCRの第2ラウンドを遺伝子のそれぞれについて別々に、より大きな特異性のためにネステッド遺伝子特異的な内部プライマーを使用して、総反応容積20μlで行った(Platinum Taq Super−Mix HF、Invitrogen)。以下の通りPCR増幅を30サイクル行った:94℃で30秒間;58℃で30秒間;68℃で30秒間。最終的なPCRステップにおいて、反応産物を68℃で3分間インキュベートした。完了した反応産物を4℃で貯蔵した。次いで、第2ラウンドのPCR産物を反応容積の5分の1および1.4%アガロースゲルを用いてゲル電気泳動に供した。
【0155】
【表2】
【0156】
DNAメチル化解析。FACSで選別した異核共存体(細胞2,000〜10,000個)を20μlのPBS中に収集した。DNeasy Tissue Kit(Qiagen)を使用してDNAを抽出した。Epitect Bisulfite Kit(Qiagen)を使用してバイサルファイト処理を行った。ヒトおよびバイサルファイトに特異的なプライマーを使用して、ヒトOct4プロモーターおよびヒトNanogプロモーターの領域についてのネステッドPCRを行った(表3)。試料を、94℃で2分間、30×(94℃で20秒間、61℃で30秒間、68℃で30秒間)の第1の、ネステッドPCRのサイクルにかけた。バイサルファイトに特異的なPCR増幅の第2ラウンドからのPCR産物を、以前に記載されている通りクローニングし、配列決定した(Zhang、F.ら(2007)Proc Natl Acad Sci USA 104、4395〜400)。
【0157】
【表3】
【0158】
siRNAトランスフェクション。siRNAトランスフェクションのために、ES細胞および初代線維芽細胞を、トランスフェクトする前日に50〜60%集密に播いた。silmporter(Millipore)を使用してsiRNA(Dharmacon)をトランスフェクトした。
【0159】
クロマチン免疫沈降。クロマチン免疫沈降を、表4で提供されるプライマーを使用して、Dahlおよび Collas((2008)Nat Protoc 3、1032〜45)によって以前に記載されている通り行った。ChIPデータは、入力DNAに対して正規化されたものとして提示され、エラーバーは標準誤差(sem)を示す。
【0160】
【表4】
【0161】
統計分析。データは、平均±標準誤差として示されている。スチューデントのt検定を用いた群間の比較は両側分布を仮定している。
【0162】
異核共存体のThy1.1(CD90)濃縮。PEGで処理したGFP−(非GFP)mESとDsRed+hFbの共培養物をトリプシン処理し、FACS緩衝液3mLに再懸濁させた。細胞を、ビオチンマウス抗ヒトCD90(BD Pharmingen)と一緒に、1:5000希釈で、室温で30分間インキュベートした。細胞を1回洗浄し、FACS緩衝液3mLに再懸濁させ、10μlのDynabeads Biotin Binder(Invitrogen)と一緒に室温で30分間インキュベートした。ビーズを磁気分離によって除去し、2回洗浄し、濃縮された異核共存体をサイトスピンした。
【0163】
免疫沈降およびウエスタンブロット。マウスES細胞を、IP緩衝液(20mMのトリス、pH7.5、1mMのDTT、0.5mMのEDTA、350mMのNaCI、10%(vol/vol)のグリセロール、10μMのZnCIに溶解させた。細胞全体の溶解物を室温で30分間予め清澄化(pre−clear)し、その後、AIDをプルダウンした。簡単に述べると、細胞溶解物、次いでAIDを、ウサギポリクローナルAID抗体と架橋結合したProtein A Plus Agarose beads(Pierce)を使用してプルダウンした。細胞溶解物2mgから免疫沈降を行った。
【0164】
mESにおけるAIDタンパク質ノックダウンを可視化するために、si対照 またはsi−1でトランスフェクトした3日後に細胞溶解物を回収した。これらの試料中のAIDの検出を、細胞全体の溶解物170μgから、抗マウス−AID(L7E7、Cell Signaling、1:500希釈)を用いて行った。膜を取り除き、抗マウスα−チューブリン(Sigma、1:20,000希釈)を用いてローディング対照を探索した。AIDの免疫沈降を同じL7E7抗体を用いて検出した。
【0165】
結果
種間の異核共存体を作製するために、GFPレポーター遺伝子を形質導入したマウス胚性幹細胞(mES)を、DsRedレポーター遺伝子を形質導入した初代ヒト線維芽細胞(hFb)と共培養し、ポリエチレングリコール(PEG)を使用して融合した(図1a;図5のスキーム)。融合GFP+DsRed+異核共存体は、FACSによって容易に選別され(図1b)、蛍光顕微鏡を用いて同定され、Hoechst33342またはHoechst33258を用いて可視化すると明瞭に染色されたヒトおよびマウスの核を含有した(それぞれ図1cおよび図1f)。PEG融合の効率は低いので(0.6〜1.0%)、GFP+DsRed+異核共存体を2回選別し、80%の純度まで濃縮した(図1b)。増殖している細胞にのみ存在する核タンパク質であるKi−67に対する抗体を使用して、融合後にアッセイした3日間にわたって、異核共存体の98(±2)%において細胞分裂が起こらなかったことを決定した(図1d、e)。さらに、同じ期間にわたって、異核共存体の94(±4)%においてBrdU標識は検出されず、これは、DNA複製が起こらなかったことを示している(図1f、g;図6;図7)。遺伝子の投与量および各細胞型が寄与するタンパク質の割合により、体細胞における核の再プログラミングの方向が決定されるので、多能性の状態への再プログラミングを支持するために、ES細胞が線維芽細胞に数でまさる(2:1)ようにインプット細胞の比率を歪めた。
【0166】
ヒト線維芽細胞においてES細胞特異的な遺伝子が誘導されたかどうかを決定するために、ヒトOct4およびヒトNanogの誘導を、種特異的プライマーを使用して、遍在するGAPDHと比較してアッセイした(図8)。融合の1日後、2日後および3日後に選別した異核共存体から単離したmRNAを半定量RT−PCRおよびリアルタイムPCRによって評価した(図2a、b)。使用した0日目の対照は、(a)単独のヒト線維芽細胞;(b)mESとhFbのPEG前、融合していない共培養物;または(c)PEGおよび融合の効果に対する対照としてPEGで処理したヒト線維芽細胞のいずれかであった。上記の0日目の対照は全て同様の結果を生じた。異核共存体におけるヒトOct4転写物およびヒトNanog転写物の両方の誘導は、早ければ融合の1日後にはっきり見えたが(図2a、b)、対照でははっきり見えず(図9)、これは、2つの重要なヒト多能性遺伝子の発現の開始は、異核共存体において急速であることを示している。1日目までに、同じ試料におけるヒトOct4およびヒトNanogの発現(GAPDHに対して正規化された)は、融合していない共培養した対照(0日目)に対して5倍に増加し、2日目および3日目に10倍で持続された(図2b)。ヒト特異的プライマーを用いて、Oct4およびNanogに加えて、他の重要な多能性遺伝子が誘導されたかどうかを、リアルタイムPCRを使用して決定した。ES細胞の自己再生を維持するために不可欠であり、Oct4およびNanogの標的になることが示されているEssrb(Bhattacharya、B.ら(2004)Blood 103、2956〜64)およびTDGF1(Bhattacharya、B.ら(2004)Blood 103、2956〜64)(クリプト(cripto))は、融合の2日後の異核共存体において、それぞれ3倍および2.5倍に上方制御されることが見いだされた(図10)。ヒト線維芽細胞においてすでに発現されているSox2およびそのプロモーターは、マウス線維芽細胞における所見と一致して、融合前に広範囲にわたって脱メチル化されており;その発現は、融合後に増加しなかった。機能的に、Essrbと相互に交換することができるKlf4(Feng、B.ら(2009)Nat Cell Biol 11、197〜203)の発現は、融合の2日後の異核共存体において変化しなかった(図10)。
【0167】
融合後のヒト線維芽細胞における核の再プログラミングの効率を評価するために、単一のFACSで選別した異核共存体を3種のヒト転写物、Oct4、Nanog、およびGAPDH(対照)について、いずれの場合にも2セットのヒト特異的なプライマーを用いて、ネステッドRT−PCRによって解析した(図2c)。マウスES細胞(対照)ではヒト遺伝子産物は全く検出されず、ヒト線維芽細胞(対照)ではヒトGAPDHのみが検出された(図8)。対照的に、3つの独立した融合実験から融合の3日後に単回のFACSで選別した異核共存体の70%でヒトOct4とヒトNanogの両方が発現され(図2c、d;図11)、これは、高い割合の異核共存体で多能性への再プログラミングが開始されたことを示している。このことは、例えば、Takahashi、K.ら(2007)Cell 131、861〜72;Takahashi、K.&Yamanaka、S.(2006)Cell 126、663〜76;Wernig、M.ら(2007)Nature 448、318〜24;and Wernig、M.ら(2008)Nat Biotechnolにおいて観察された、2〜3週間の集団全体のiPS細胞(<0.1%)における、Oct4およびNanogの発現の遅く非能率的な誘導と著しい対照をなす。
【0168】
DNA脱メチル化は、線維芽細胞のiPS細胞への再プログラミングの主要な限定ステップであることが示されているので、異核共存体におけるヒトOct4プロモーターおよびヒトNanogプロモーターの脱メチル化の時間経過および程度を対照と比較して解析した。融合の1日後、2日後および3日後に異核共存体からDNAを単離し、バイサルファイト変換に供した。ヒトOct4プロモーターおよびヒトNanogプロモーターを、ヒトおよびバイサルファイトに特異的なプライマーを用いてPCRによって増幅し(表3、図8)、産物をクローニングし、配列決定した。ヒトOct4プロモーターおよびヒトNanogプロモーターにおけるDNA脱メチル化が明白であり、3日を通して増加した(図3a)。対照的に、βグロビンHS2遺伝子座は、全体を通してメチル化されたままであり、これは、DNA脱メチル化が特異的であったことを示している。ヒトOct4プロモーターおよびヒトNanogプロモーターにおける脱メチル化CpG部位の時間経過および進行性の蓄積(図3b、c)は、リアルタイムPCRを用いて同じ3日間にわたって観察された転写物の蓄積の進行性の増加(図2b)に匹敵した。特に、ヒト体細胞における多能性遺伝子のプロモーターの脱メチル化および活性化は、Ki−67またはBrdUによって標識されていない場合に起こる(図1e、g);したがって、脱メチル化は活性であり、細胞分裂およびDNA複製と無関係である。
【0169】
哺乳動物の多能性の生殖細胞において検出され(Morgan、H.D.ら(2004)J Biol Chem 279、52353〜60)、ゼブラフィッシュにおける受精後の活性なDNA脱メチル化に関係づけられる(Rai、K.ら(2008)Cell 135、1201〜12)ので、AIDの発現について、リアルタイムPCRを用いてマウスES細胞およびヒト線維芽細胞をアッセイした。体細胞におけるAIDの発現は、一般にBリンパ球に限られると考えられているが、ヒト線維芽細胞ならびにマウスES細胞において、B−リンパ球細胞系であるRamosと比較して非常にレベルが低下してはいるが(それぞれ5%および15%)、AID mRNAが検出された(図12)。これらの細胞におけるAIDの役割を調査するために、マウスおよびヒトAID mRNAのレベルを、AIDコード領域内の異なる配列に対する3種の別個の非オーバーラップsiRNA、および、オフターゲットの効果を除外し、結果がAIDに特異的であることを確実にするために、AIDの非コード3’UTRに特異的な第4のsiRNAをトランスフェクトすることによって一過性にノックダウンした(図13)。AIDコード領域と50%同一である第5のsiRNAを対照(si対照)として使用した。ノックダウンの程度およびタイミングを、siRNA−1、siRNA−2、siRNA−3およびsiRNA−4により、AIDの転写が、トランスフェクションの3日後に対照siRNAと比較してそれぞれ81(±13)%、79(±12)%、70(±8)%、および99(±0.1)%低下した対照マウスES細胞においてまず確認した(図14、上部)。免疫沈降、その後にウエスタンブロットを使用するとマウスES細胞においてAIDタンパク質が検出され、同様に濃縮されたES細胞溶解物全体においてもAIDタンパク質が検出された(図15)。ES細胞溶解物におけるsiRNA1によるAIDノックダウンを検証し、対照タンパク質レベルが88%低下したことはmRNAが81%低下したこととよく相関した(図15)。ヒト線維芽細胞では、AIDの転写は、siRNA1、siRNA2、siRNA3およびsiRNA4により、それぞれ46(±11)%、72(±23)%、99(±0.1)%および99(±0.1)%低下した(図14、下部)。これらのデータは、ES細胞および線維芽細胞の両方において、AIDが存在し、4つの別個のsiRNAによって効率的に低下させることができることを示している。
【0170】
AIDノックダウンに供した異核共存体における再プログラミングの開始を評価するために、GAPDHと比較したOct4およびNanogの発現をリアルタイムPCRによって評価した。異核共存体実験のために、融合の24時間前にマウスES細胞とヒト線維芽細胞の両方にsiRNAをトランスフェクトした(スキームについては図5を参照されたい)。異核共存体において、リアルタイムPCRによって持続性のAIDのノックダウンが検出された。siRNA1およびsiRNA2を使用すると、AIDは、融合の3日後に、対照siRNAでトランスフェクトした異核共存体と比較して77(±6)%および35(±2)%低下した(図4a)。siRNA3および4siRNAにより、異核共存体においてより強力なノックダウンが引き起こされ、AIDは、融合の2日後に、siRNA対照と比較して96(±1)%、および89(±3)%低下した(図4a)。著しく、Oct4の発現は、siRNA1およびsiRNA2を使用すると、融合の3日後に、対照siRNAと比較して0.9(±0.6)%および9(±2)%まで低下した(図4a)。同様に、siRNA1およびsiRNA2を使用すると、Nanogの発現は、融合の3日後に、siRNA対照と比較して1.5(±0.4)%および1.5(±0.1)%まで著しく低下した(図4a)。siRNA3およびsiRNA4の存在下で、Oct4の発現は、融合の2日後に、siRNA対照と比較して8(±2)%および4(±3)%まで低下し、一方、Nanogの発現は19(±12)%および7(±4)%まで低下した。ここで使用した4種のsiRNA全ては、Oct4およびNanogの発現を少なくとも80%遮断することにおいて同様の効果を有した。これらの知見は、35%ノックダウンにより、96%ノックダウンに匹敵する多能性遺伝子の誘導の阻害が導かれたので、AIDの効果は非常に投与量感受性であることを示している。まとめると、これらのデータは、ここで使用したAIDに対する4種のsiRNA全てが、Oct4およびNanogの活性化を少なくとも80%遮断することにおいて同様に強力な効果を有したことを示している。
【0171】
プロモーターの脱メチル化に対するAIDの効果を評価するために、異核共存体におけるヒトOct4プロモーターおよびヒトNanogプロモーターのCpGのメチル化状態をアッセイした。siRNA1およびsiRNA2を使用してAIDノックダウンに供した3日目の異核共存体では、ヒトOct4プロモーターにおけるCpG脱メチル化の程度は、対照の82%と比較してそれぞれ26%および6%まで低下した(図4b、c)。Nanogプロモーターについて、CpGの脱メチル化は、対照の53%脱メチル化と比較して、それぞれ24%および25%まで低下した(図4b、c)。siRNA3およびsiRNA4を使用すると、Oct4プロモーターにおけるCpGの脱メチル化の程度は、2日目の対照試料における72%と比較してそれぞれ18%および8%まで低下し、一方、Nanogプロモーターについては、CpGの脱メチル化の程度は、対照における48%と比較してそれぞれ3%および<1%まで低下した(図4b、c)。siRNAノックダウン実験の全てについてのバイサルファイト配列決定データの概要が図16に示されている。AIDがノックダウンされるとOct4プロモーターおよびNanogプロモーターの脱メチル化が低下することに対応して、Oct4転写物およびNanog転写物の誘導が、2日目および3日目に、対照と比較して少なくとも80%低下した(図4a)。これらのデータは、プロモーターの脱メチル化が、これらの2つの多能性遺伝子の発現に欠かせないこと、およびAIDが、体細胞の再プログラミングにおいて、哺乳動物のDNAが脱メチル化されるために必要であることを示している。
【0172】
再プログラミングを開始するためのAIDの必要性をさらに調査するために、異核共存体において、siRNAノックダウンによって引き起こされるDNA脱メチル化の遮断をレスキューするその能力を試験した。AIDレベルを増加させることによってsiRNAノックダウンが克服されるかどうかを試験するために、マウスES細胞において、siRNAをトランスフェクトする前にhAIDを一過性に過剰発現させた(図5のスキームを参照されたい)。2つの別々の実験において、hAIDを、AID siRNAが存在しない異核共存体における対照と比較して2倍または4倍過剰発現させた場合、融合の1日後にプロモーターの脱メチル化または再プログラミングは加速されなかった(図17)。これは、おそらく、異核共存体では起こるまでに少なくとも1日が必要になり得る、ヒトOct4プロモーターの脱メチル化のカイネティクス、または再プログラミングを加速するためにhAIDと協調して働く追加的な因子がないことに起因し得る。しかし、siRNA−1による一過性のノックダウンを受けている異核共存体において、すなわち、siRNAの存在下でhAIDを過剰発現させると、Nanogプロモーターの脱メチル化が完全にレスキューされ、遺伝子が発現され、また、Oct4プロモーターの脱メチル化が部分的にレスキューされ、遺伝子が発現された(図18)。これらのデータは、追加されたhAIDは機能性であり、siRNAのどんな非特異的な効果も除外することを示しており、さらに、多能性への再プログラミングの開始時のDNA脱メチル化におけるAIDの特異的かつ不可欠な役割を確認している。
【0173】
ヒトOct4プロモーターおよびヒトNanogプロモーターのDNA脱メチル化におけるAIDの役割をさらに検証するために、抗AID抗体を用いてクロマチン免疫沈降(ChIP)実験を行うことにより、AIDがそれらのプロモーター領域に特異的に結合するかどうかを試験した。ChIP実験において評価したプロモーター領域は、バイサルファイト配列決定によってCpGの脱メチル化について分析された(図4d;図19)Oct4プロモーター領域およびNanogプロモーター領域の範囲内になるように設計した。ヒト線維芽細胞では、ChIP解析により、ヒトOct4プロモーター(6倍)およびヒトNanog(8倍)プロモーターの両方に対するAIDの有意な結合が示された(図4d)。したがって、AIDは、線維芽細胞において、再プログラミングの間に脱メチル化を受けるヒトOct4およびヒトNanogの重度にメチル化されたプロモーター領域に結合する。対照として、IgM定常領域(Cμ)のプロモーターへのAIDの結合は、予測通り有意であったが(Okazaki、I.M.ら(2002)Nature 416、340〜5)、線維芽細胞において発現されるThy1.1への結合は観察されなかった。
【0174】
線維芽細胞と対照的に、マウスのOct4およびNanogのプロモーター領域では、おそらく、これらのプロモーターが発現され、脱メチル化されているので、ES細胞におけるAIDタンパク質は高レベルであるにもかかわらず、AIDの結合は観察されなかった(図4d)。対照として、未分化のES細胞では発現されていない遺伝子であるCdx2のプロモーターにおいてAIDの結合が検出されたが、以前報告された通り、p53プロモーターでは検出されなかった。まとめると、これらの所見により、DNA脱メチル化におけるAIDの直接関与、なならびに多能性への再プログラミングの開始を導くヒトのNanogおよびOct4の持続的発現についての強力な裏付けがもたらされる。
【0175】
考察
DNA脱メチル化は、遺伝子サイレンシングを克服し、時間的にかつ空間的に制御された哺乳動物の遺伝子の発現を誘導するために不可欠であるが、何年も取り組まれているにもかかわらず、コンセンサスな哺乳動物のDNAデメチラーゼは未だ同定されていない。5メチル−シトシンDNAグリコシラーゼによるDNA脱メチル化の証拠が、植物体において示されているが(Gong、Z.ら(2002)Cell 111、803〜14;Choi、Y.ら(2002)Cell 110、33〜42)、チミンDNAグリコシラーゼ(TDG)またはメチル−CpG−結合性ドメインタンパク質4(Mbd4)などの哺乳動物の相同体では比較に適した機能が示されていない(Cortazar、D.ら(2007)DNA Repair(Amst)6、489〜504;Millar、C.B.ら(2002)Science 297、403〜5)。
【0176】
AIDは、B細胞における抗体多様性の生成、RNAの編集および抗ウイルス(性)の反応における役割が確立されているシトシンデアミナーゼのファミリーに属する(AID、サブグループApobec1、Apobec2およびApobec3)(Conticello、S.G.ら(2007)Adv Immunol 94、37〜73)。AIDおよびApobec1はどちらも前駆体生殖細胞、卵母細胞および初期胚において発現され、in vitroで頑強な5−メチルシトシンデアミナーゼ活性を有し(Morgan、H.D.ら(2004)J Biol Chem 279、52353〜60)、塩基除去DNA修復(BER)経路によって修復されるT−Gミスマッチをもたらし、理論的に複製を伴わないDNA脱メチル化を導き得る。最近、ゼブラフィッシュの胚において、AIDは、シトシンの脱アミノ化に影響を及ぼし、Mbd4による塩基除去を導く、Mbd4およびGadd45aとの三者間タンパク質複合体のメンバーであると意味づけられた(Rai、K.ら(2008)Cell 135、1201〜12)。第3の構成成分であるGadd45aは酵素活性を欠き、アフリカツメガエルの卵母細胞における修復に媒介されるDNA脱メチル化および遺伝子活性化におけるその役割については、まだ議論が続いている(Barreto、G.ら(20070 Nature 445、671〜5;Jin、S.G.ら(2008)PLoS Genet 4、e1000013)。
【0177】
本明細書において提供されるデータにより、AIDを活性な哺乳動物細胞におけるDNA脱メチル化に関連づける証拠がもたらされ、AID依存性のDNA脱メチル化が、ヒト線維芽細胞における多能性の誘導に必要な早期の後成的な変化であることが実証されている。異核共存体においてAIDをノックダウンすることにより、線維芽細胞核におけるヒトOct4プロモーターおよびヒトNanogプロモーターのDNA脱メチル化が妨げられた。これと一貫して、ヒト体性線維芽細胞において、AID依存性のDNA脱メチル化が低下すると、これらの多能性因子の発現および核の多能性への再プログラミングの開始が阻害され、これにより、AIDが、再プログラミングの開始に決定的な調節因子であることの強力な証拠がもたらされる。AIDの低下の阻害性の効果は、hAIDの過剰発現によりレスキューされ、Nanogに対しては完全なレスキューが観察され、Oct4に対しては部分的なレスキューが観察された。さらに、ヒト線維芽細胞において、サイレントなメチル化されたOct4プロモーターおよびNanogプロモーターにおいてAIDの結合が観察されたが、マウスES細胞における活性な、非メチル化Oct4プロモーターおよび非メチル化Nanogプロモーターでは観察されず、これは、DNA脱メチル化におけるAIDの特異的な役割を実証している。
【0178】
本発明で実現された高効率の異核共存体における再プログラミングにより、iPS細胞の生成を導く安定な再プログラミングの第1の公知のマーカーであるOct4およびNanogを含めた5つの多能性遺伝子の誘導に欠かせない調節因子を発見することが可能になった。ここで、異核共存体プラットフォームを、(a)AIDと一緒に働いて活性なDNA脱メチル化を媒介する可能性がある哺乳動物のDNA脱メチル化複合体(グリコシラーゼおよび他のDNA修復酵素)の他の構成成分を解明するため(図4e)、(b)細胞融合後にすぐに発現されるヒト遺伝子についてスクリーニングすることによって核の再プログラミングの追加的な調節因子についての不偏の検索を行うために活用することができる。今後の研究により、AIDの単独での、またはこれらの他の分子と併せての発現によりiPS細胞の生成が加速されるかどうかが示されるであろう。
【実施例2】
【0179】
AIDの潜在性相互作用物質を同定し、哺乳動物のDNA脱メチル化を調整する機能性分子プレーヤーについて理解するために、質量分析を使用した。以下のAID構築物を使用した:1)pHAGE−STEMCCAレンチウイルスベクターにクローニングした、タンパク質のN末端に2つのタンデムなFLAGタグを含有するヒトAID、および2)pHAGE−STEMCCAレンチウイルスベクターにクローニングした、タンパク質のC末端も2つのタンデムなFLAGタグを含有するヒトAID。その後、これらの構築物を含有するウイルスを使用してマウス胚性幹細胞(CGR8)に感染させ、Flag−ヒトAIDを過剰発現している安定な細胞系を選択した。対照として、2×FLAGタグのみを含有するレンチウイルスベクターを使用した。
【0180】
AIDを発現している安定なES細胞系、および対照の2×Flagを、FLAGタグに対する抗体を使用してAIDタンパク質を免疫沈降させるために、細胞質抽出物および核抽出物に分画した。生じた複合体を質量分析に供した。分析において、AIDは最も豊富なタンパク質であることが見いだされ、AIDに随伴するいくつもの独特のタンパク質が同定された(表5)。
【0181】
【表5−1】
【0182】
【表5−2】
【0183】
【表5−3】
【0184】
【表5−4】
【0185】
【表5−5】
【0186】
上記では、ただ単に本発明の原理が例示されている。当業者は本明細書においてはっきりとは記載されていない、または示されていないが、本発明の原理を具体化し、その主旨および範囲内に含まれる種々の取り合わせを考案することができることが理解されよう。さらに、本明細書において列挙されている全ての例および条件付きの言葉は、主に、読者が本発明の原理および当技術分野を推進するために出願人らが提供する概念を理解するために役立つことが意図され、そのような具体的に列挙された実施例および条件に限定されないと解釈されるべきである。さらに、本明細書において、本発明の原理、態様および実施形態ならびにその特定の実施例が列挙されている全ての記載は、それらの構造的等価物と機能的等価物の両方を包含するものとする。さらに、そのような等価物は、現在公知の等価物と今後開発される等価物、すなわち、構造にかかわらず、同じ機能を実施する、開発される任意の要素の両方を包含するものとする。本発明の範囲は、したがって、本明細書に示され、記載されている例示的な実施形態に限定されるものではない。もっと正確に言うと、本発明の範囲および主旨は、添付の特許請求の範囲によって具体化される。
【図1−1】
【図1−2】
【図2−1】
【図2−2】
【特許請求の範囲】
【請求項1】
哺乳動物細胞におけるゲノムDNAメチル化の量を減少させる方法であって、
前記哺乳動物細胞を、ゲノムDNAメチル化が減少するような条件下で、有効量の、シチジンデアミナーゼ(CD)活性を促進する1種または複数種の薬剤と接触させるステップを含む方法。
【請求項2】
CD活性を促進する1種または複数種の薬剤が活性化誘導性シチジンデアミナーゼ(AID)ポリペプチドまたはAIDポリペプチドをコードする核酸である、請求項1に記載の方法。
【請求項3】
CD活性を促進する1種または複数種の薬剤がアポリポタンパク質B RNA編集触媒成分(APOBEC)ポリペプチドまたはAPOBECペプチドをコードする核酸である、請求項1に記載の方法。
【請求項4】
前記哺乳動物細胞を、選択されるタンパク質と接触させるステップをさらに含み、CD活性を促進する1種または複数種の薬剤がtetタンパク質をさらに含む、請求項1に記載の方法。
【請求項5】
前記接触させるステップが、in vitroで実施される、請求項1に記載の方法。
【請求項6】
前記接触させる哺乳動物細胞が、脱メチル化を許容する体細胞である、請求項1に記載の方法。
【請求項7】
接触させる哺乳動物細胞が、前記接触させるステップの後に人工多能性幹(iPS)細胞になる、請求項6に記載の方法。
【請求項8】
前記脱メチル化を許容する体細胞を、iPS細胞への運命を促進する1種または複数種の因子と接触させるステップをさらに含む、請求項7に記載の方法。
【請求項9】
接触させる哺乳動物細胞が、脱メチル化を許容する体細胞の細胞系譜とは異なる細胞系譜の体細胞になる、請求項6に記載の方法。
【請求項10】
前記脱メチル化を許容する体細胞を、所望の体細胞運命を促進する1種または複数種の因子と接触させるステップをさらに含む、請求項9に記載の方法。
【請求項11】
接触させる哺乳動物細胞が多能性幹細胞である、請求項1に記載の方法。
【請求項12】
多能性幹細胞が、胚性幹(ES)細胞、胚性生殖幹(EG)細胞、および人工多能性幹(iPS)細胞からなる群から選択される、請求項11に記載の方法。
【請求項13】
作製される細胞が体細胞である、請求項11に記載の方法。
【請求項14】
前記多能性細胞を、所望の体細胞運命を促進する1種または複数種の因子と接触させるステップをさらに含む、請求項13に記載の方法。
【請求項15】
前記接触させるステップが、ゲノムDNA脱メチル化療法を必要とする対象において実施される、請求項1に記載の方法。
【請求項16】
前記細胞が癌細胞であり、前記対象が、癌に罹患している対象である、請求項15に記載の方法。
【請求項17】
候補薬剤を、細胞におけるゲノムDNA脱メチル化活性の調節における活性についてスクリーニングする方法であって、
細胞の集団を、有効量の、シチジンデアミナーゼ(CD)活性を促進する薬剤と接触させるステップと、
候補薬剤と接触させた細胞を、シチジンデアミナーゼ活性を促進する薬剤と接触させたが候補薬剤と接触させていない細胞の集団と比較するステップであって、
第1の亜集団と第2の亜集団との間の特性の違いにより、候補薬剤がゲノムDNA脱メチル化活性を調節することが示されるステップと
を含む方法。
【請求項18】
CD活性を促進する薬剤が、AIDペプチドまたはAIDペプチドをコードする核酸である、請求項17に記載の方法。
【請求項19】
前記第1の細胞の集団が腫瘍細胞である、請求項17に記載の方法。
【請求項20】
腫瘍細胞におけるゲノムの脱メチル化を調節する候補薬剤が、癌における腫瘍成長を調節する薬剤である、請求項19に記載の方法。
【請求項21】
前記第1の細胞の集団が、体細胞またはES細胞と体細胞から作製された異核共存体である、請求項17に記載の方法。
【請求項22】
体細胞におけるゲノムの脱メチル化を調節する候補薬剤が、体細胞の多能性の誘導を調節する薬剤である、請求項21に記載の方法。
【請求項23】
シチジンデアミナーゼのDNA脱メチル化活性の調節における活性を有するタンパク質を同定するための方法であって、
細胞の集団を、シチジンデアミナーゼをコードする配列を含む核酸と接触させるステップと、
前記細胞の粗タンパク質抽出物からシチジンデアミナーゼを免疫沈降させるステップと、
前記免疫沈降物を質量分析に供するステップとを含み、
質量分析によって同定された1種または複数種のタンパク質が、前記シチジンデアミナーゼの脱メチル化活性に欠かせないものである、方法。
【請求項1】
哺乳動物細胞におけるゲノムDNAメチル化の量を減少させる方法であって、
前記哺乳動物細胞を、ゲノムDNAメチル化が減少するような条件下で、有効量の、シチジンデアミナーゼ(CD)活性を促進する1種または複数種の薬剤と接触させるステップを含む方法。
【請求項2】
CD活性を促進する1種または複数種の薬剤が活性化誘導性シチジンデアミナーゼ(AID)ポリペプチドまたはAIDポリペプチドをコードする核酸である、請求項1に記載の方法。
【請求項3】
CD活性を促進する1種または複数種の薬剤がアポリポタンパク質B RNA編集触媒成分(APOBEC)ポリペプチドまたはAPOBECペプチドをコードする核酸である、請求項1に記載の方法。
【請求項4】
前記哺乳動物細胞を、選択されるタンパク質と接触させるステップをさらに含み、CD活性を促進する1種または複数種の薬剤がtetタンパク質をさらに含む、請求項1に記載の方法。
【請求項5】
前記接触させるステップが、in vitroで実施される、請求項1に記載の方法。
【請求項6】
前記接触させる哺乳動物細胞が、脱メチル化を許容する体細胞である、請求項1に記載の方法。
【請求項7】
接触させる哺乳動物細胞が、前記接触させるステップの後に人工多能性幹(iPS)細胞になる、請求項6に記載の方法。
【請求項8】
前記脱メチル化を許容する体細胞を、iPS細胞への運命を促進する1種または複数種の因子と接触させるステップをさらに含む、請求項7に記載の方法。
【請求項9】
接触させる哺乳動物細胞が、脱メチル化を許容する体細胞の細胞系譜とは異なる細胞系譜の体細胞になる、請求項6に記載の方法。
【請求項10】
前記脱メチル化を許容する体細胞を、所望の体細胞運命を促進する1種または複数種の因子と接触させるステップをさらに含む、請求項9に記載の方法。
【請求項11】
接触させる哺乳動物細胞が多能性幹細胞である、請求項1に記載の方法。
【請求項12】
多能性幹細胞が、胚性幹(ES)細胞、胚性生殖幹(EG)細胞、および人工多能性幹(iPS)細胞からなる群から選択される、請求項11に記載の方法。
【請求項13】
作製される細胞が体細胞である、請求項11に記載の方法。
【請求項14】
前記多能性細胞を、所望の体細胞運命を促進する1種または複数種の因子と接触させるステップをさらに含む、請求項13に記載の方法。
【請求項15】
前記接触させるステップが、ゲノムDNA脱メチル化療法を必要とする対象において実施される、請求項1に記載の方法。
【請求項16】
前記細胞が癌細胞であり、前記対象が、癌に罹患している対象である、請求項15に記載の方法。
【請求項17】
候補薬剤を、細胞におけるゲノムDNA脱メチル化活性の調節における活性についてスクリーニングする方法であって、
細胞の集団を、有効量の、シチジンデアミナーゼ(CD)活性を促進する薬剤と接触させるステップと、
候補薬剤と接触させた細胞を、シチジンデアミナーゼ活性を促進する薬剤と接触させたが候補薬剤と接触させていない細胞の集団と比較するステップであって、
第1の亜集団と第2の亜集団との間の特性の違いにより、候補薬剤がゲノムDNA脱メチル化活性を調節することが示されるステップと
を含む方法。
【請求項18】
CD活性を促進する薬剤が、AIDペプチドまたはAIDペプチドをコードする核酸である、請求項17に記載の方法。
【請求項19】
前記第1の細胞の集団が腫瘍細胞である、請求項17に記載の方法。
【請求項20】
腫瘍細胞におけるゲノムの脱メチル化を調節する候補薬剤が、癌における腫瘍成長を調節する薬剤である、請求項19に記載の方法。
【請求項21】
前記第1の細胞の集団が、体細胞またはES細胞と体細胞から作製された異核共存体である、請求項17に記載の方法。
【請求項22】
体細胞におけるゲノムの脱メチル化を調節する候補薬剤が、体細胞の多能性の誘導を調節する薬剤である、請求項21に記載の方法。
【請求項23】
シチジンデアミナーゼのDNA脱メチル化活性の調節における活性を有するタンパク質を同定するための方法であって、
細胞の集団を、シチジンデアミナーゼをコードする配列を含む核酸と接触させるステップと、
前記細胞の粗タンパク質抽出物からシチジンデアミナーゼを免疫沈降させるステップと、
前記免疫沈降物を質量分析に供するステップとを含み、
質量分析によって同定された1種または複数種のタンパク質が、前記シチジンデアミナーゼの脱メチル化活性に欠かせないものである、方法。
【図3】

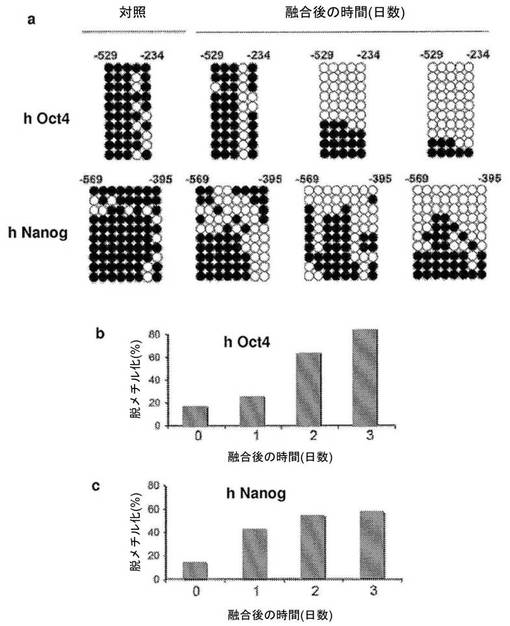
【図4−1】
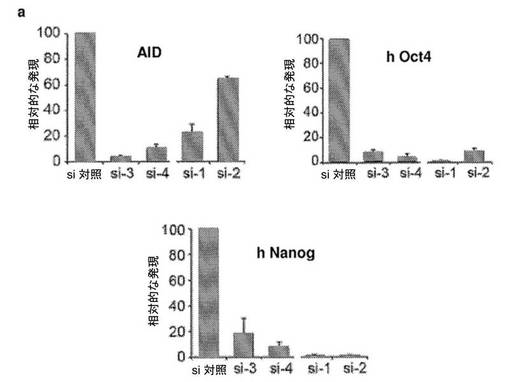
【図4−2】
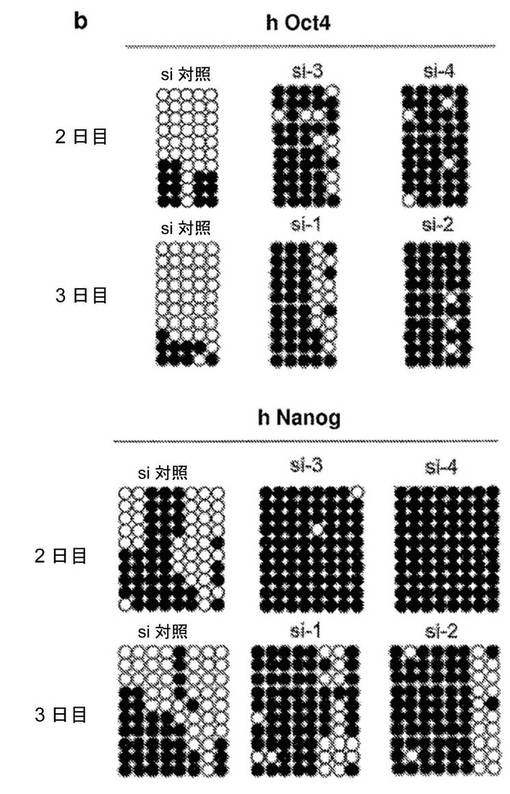
【図4−3】
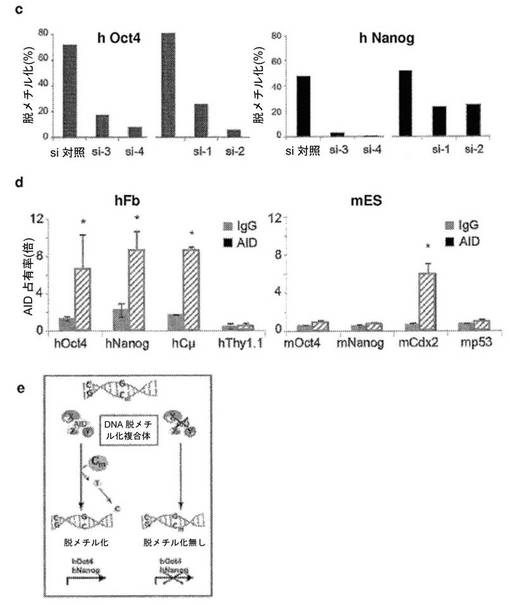
【図5】
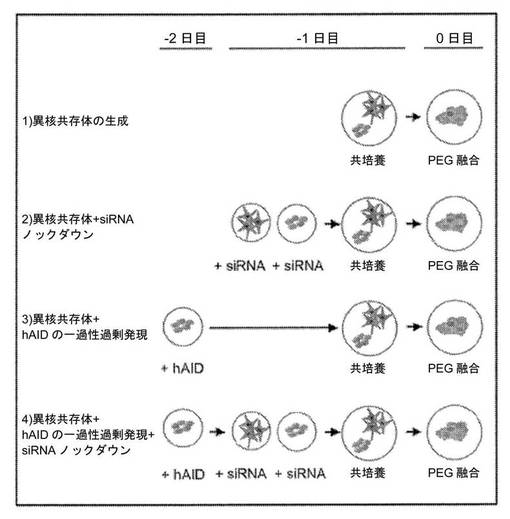
【図6−1】
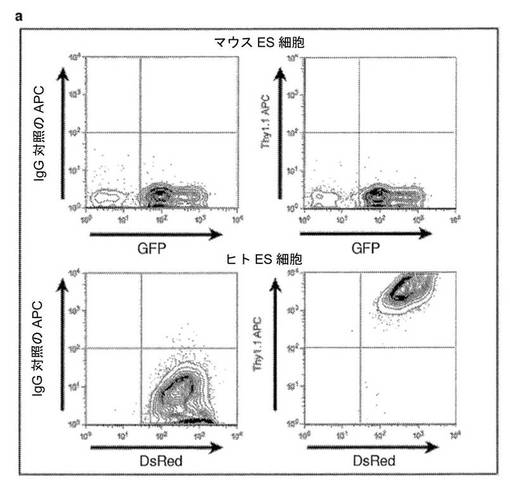
【図6−2】
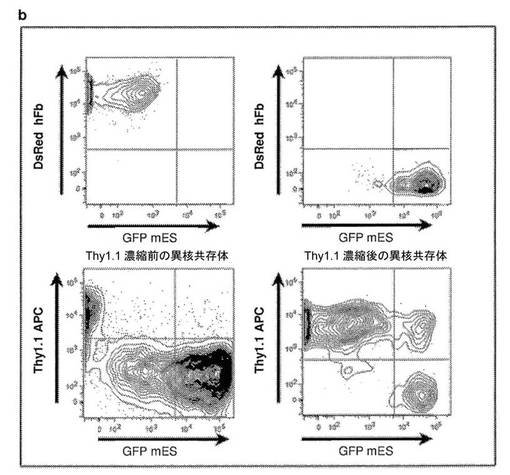
【図7】

【図8】
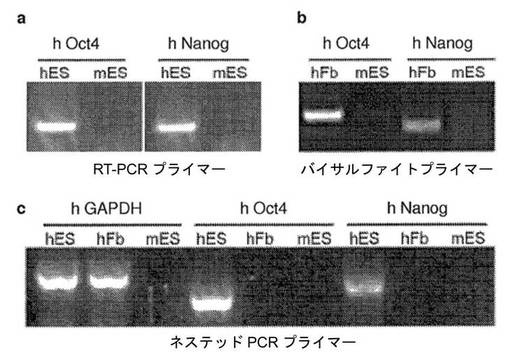
【図9】
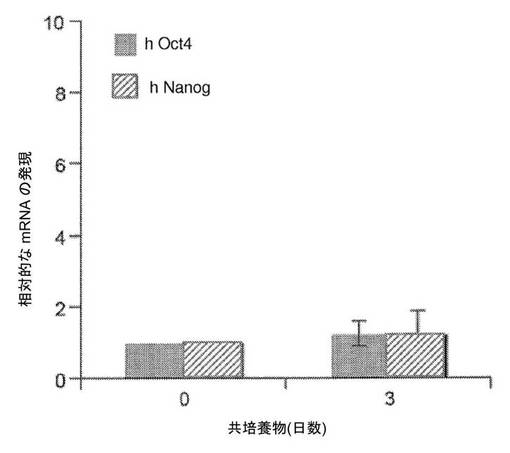
【図10】
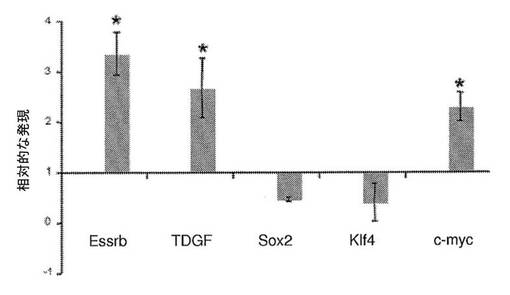
【図11】
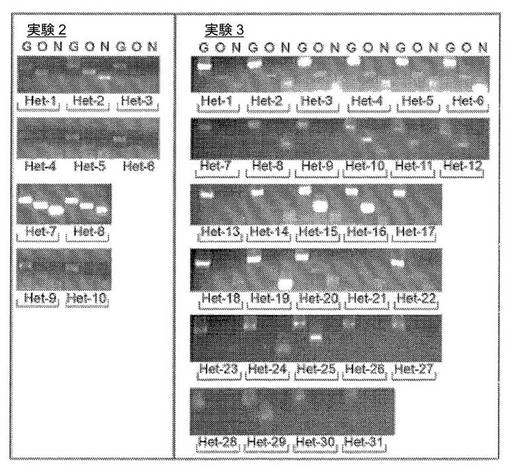
【図12】
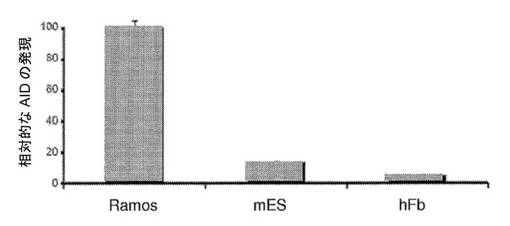
【図13】
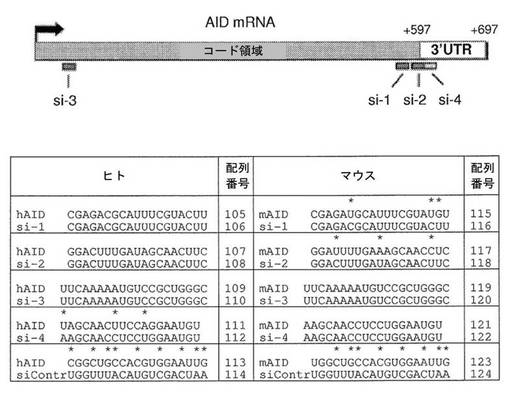
【図14】
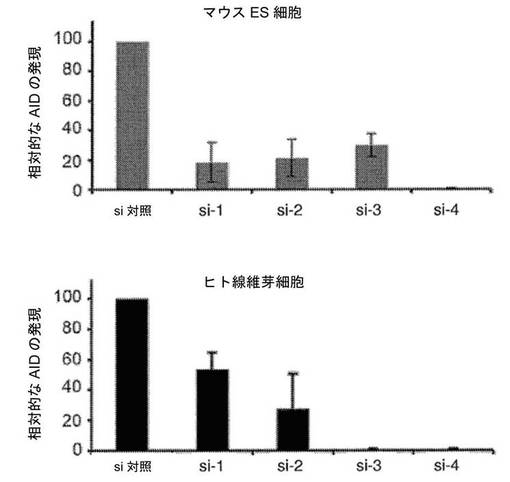
【図15】
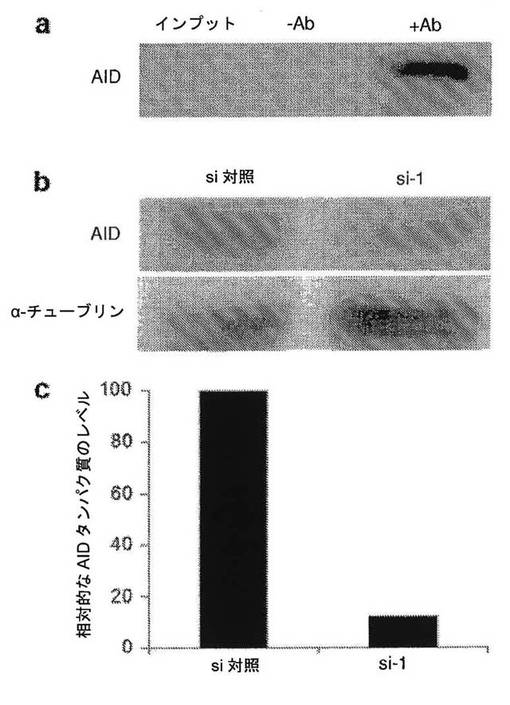
【図16−1】
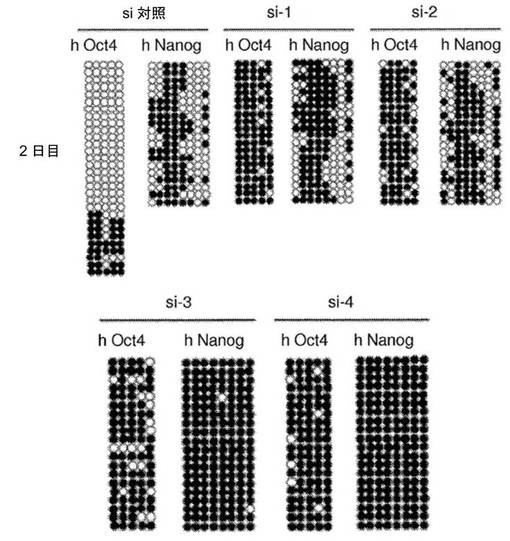
【図16−2】
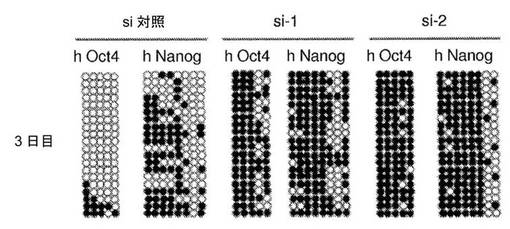
【図17】
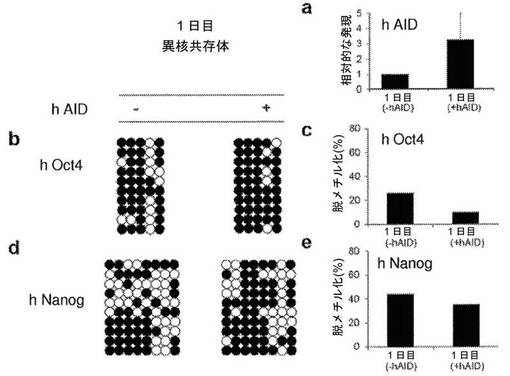
【図18−1】
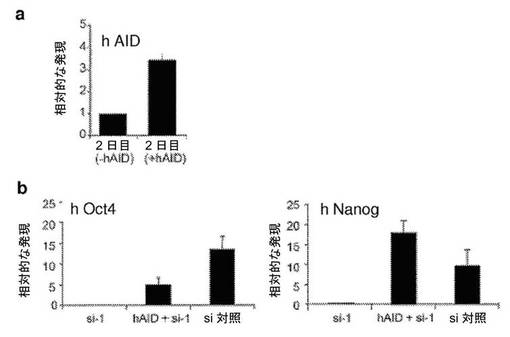
【図18−2】
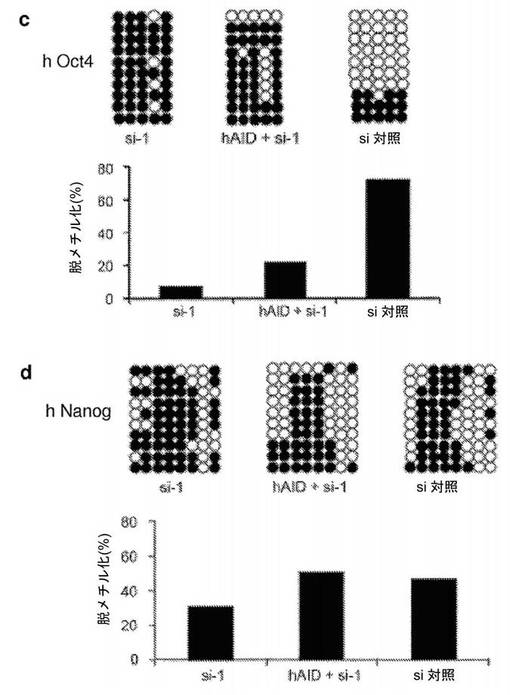
【図19】
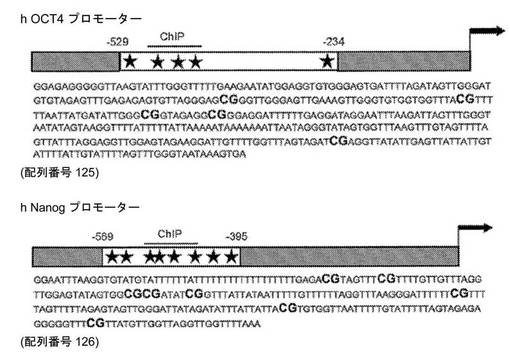
【図4−1】
【図4−2】
【図4−3】
【図5】
【図6−1】
【図6−2】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16−1】
【図16−2】
【図17】
【図18−1】
【図18−2】
【図19】
【公表番号】特表2013−514779(P2013−514779A)
【公表日】平成25年5月2日(2013.5.2)
【国際特許分類】
【出願番号】特願2012−544877(P2012−544877)
【出願日】平成22年12月17日(2010.12.17)
【国際出願番号】PCT/US2010/060976
【国際公開番号】WO2011/075627
【国際公開日】平成23年6月23日(2011.6.23)
【出願人】(503115205)ボード オブ トラスティーズ オブ ザ レランド スタンフォード ジュニア ユニバーシティ (69)
【Fターム(参考)】
【公表日】平成25年5月2日(2013.5.2)
【国際特許分類】
【出願日】平成22年12月17日(2010.12.17)
【国際出願番号】PCT/US2010/060976
【国際公開番号】WO2011/075627
【国際公開日】平成23年6月23日(2011.6.23)
【出願人】(503115205)ボード オブ トラスティーズ オブ ザ レランド スタンフォード ジュニア ユニバーシティ (69)
【Fターム(参考)】
[ Back to top ]