
腫瘍の治療に使用できるPTENの細胞外形態の同定
配列番号1を含んでいる単離されたヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long), その断片およびアナログ, それをコードしている核酸及びそれを含んでいる組成物が提供される。PTEN-long, その断片およびアナログを用いて、固形腫瘍における血管形成を阻害する、固形腫瘍を治療する、および固形腫瘍の成長を阻害する方法が提供される。
【発明の詳細な説明】
【発明の説明】
【0001】
本出願は、2009年2月17日に出願された米国仮出願番号61/207,974の優先権を主張するものであり、その内容は本明細書中に参照により援用される。
【0002】
本出願に開示された研究は、国立癌研究所(National Cancer Institute)からの助成金番号CA082783による政府のサポートによりなされた。従って、米国政府は、本発明に特定の権利を有する。
【0003】
本出願の全体において、様々な文献が括弧内の著者名及び西暦年により参照される。これら文献の完全なる書誌的事項は、本明細書の「参照文献」に記載される。これら公開の開示の全体は、本明細書中に引用することにより本出願に援用され、当該発明が属する技術の状況をより完全に記載する。
【0004】
[背景]
PTEN 腫瘍抑制因子(WO98/34624を参照されたい、この公報は本明細書中にその全体が参照によって援用される)は、重要な二次メッセンジャー ホスファチジルイノシトール 3,4,5-三リン酸を脱リン酸化する細胞質ホスファターゼである(Maehama and Dixon 1998)。この活性は、抗-アポトーシス経路, 細胞周期の進行および細胞代謝の増加を含むAktのPIP3活性化により開始される多くの腫瘍形成性のシグナルをダウンレギュレートする(Sulis and Parsons 2003)。癌におけるPTENの役割は、多くの異なる腫瘍タイプにおいて遺伝的な又は機能的な頻繁な欠損から明らかである(Bonneau and Longy 2000)。最初にグリア癌(glial cancers)において欠失していることが発見され、以後は前立腺, 乳房, 子宮内膜, メラノサイト, 腎臓 および 肺の腫瘍形成と関係づけられてきた。また、PTENの生殖系列の変異は、Cowden's 症候群などの遺伝性の癌素因症候群(cancer predisposition syndromes)とも関連した(Eng 2003)。PTEN欠損のマウスモデルは、多くの異なる組織タイプにおいてヘテロ接合性マウスおよび組織特異的ノックアウトの双方で腫瘍抑制因子としての役割を再現した(Di Cristofano, Pesce et al. 1998; Kwabi-Addo, Giri et al. 2001; Petrocelli and Slingerland 2001; You, Castrillon et al. 2002; Fraser, Zhu et al. 2004)。
【0005】
PTENタンパク質は、N末端の二重特異性ホスファターゼドメイン, およびC末端のC2 リン脂質結合ドメイン, 続いて調節に重要(リン酸化部位が内部に見いだされるので)な定まった形を持たない尾部(unstructured tail)を含む(Lee, Yang et al. 1999; Vazquez, Ramaswamy et al. 2000; Torres and Pulido 2001; Vazquez, Grossman et al. 2001)。PTEN タンパク質は、大抵は細胞質性である。しかしながら、核にPTENが存在するとの証拠が増加しており、その局在化はNEDD4-1によるタンパク質のモノユビキチン化により制御される(Baker 2007; Wang, Trotman et al. 2007)。
【0006】
5'UTRのリボソームスキャニングは、開始コドン AUGでの翻訳開始に先行する。リボソームが適切な開始コドンを決定する実際の手段の理解はいまだに不完全であるが、mRNA自身および開始前複合体(pre-initiation complex)がどこでそのスキャニングを緩徐にし、翻訳を開始させるかを指令する配列の双方の特定の特性が存在する。古典的な「コザック配列」CCACCATGG(下線で示したATGが開始コドンである)は、開始に関して最も好都合な配列であることが示されている(Kozak 1991)。また、mRNAの二次構造は、おそらくヘリカーゼを必要とし、リーディングの前に二次構造を融解する開始前複合体のスキャニングを事実上緩徐化することにより開始を促進する(Kozak 1990)。
【0007】
特定の転写物において、翻訳開始は、非-AUGコドンから生じえる。通常、これはほんの僅かなパーセンテージの転写物から翻訳されたトータルタンパク質を含み、結果はN-端で変化する混合種のタンパク質である。Kozakは、非-AUG コドンからの翻訳の開始の効率を明らかし、GUG および CUGの双方がインビトロで翻訳を開始する能力があるが、効率的が非常に低いことを見出した(Kozak 1989)。さらなる研究によって、メチオニンの利用可能性によりいまだに不明確である機構をかいして翻訳開始の乱雑性(promiscuity)を変化させえることが示され、おそらくeIF2(43S 開始前複合体のコンポーネント)の栄養分感受性キナーゼ(nutrient sensitive kinase)によるリン酸化が関与することが示された(Hershey 1991; Hann 1994)。
【0008】
幾つかのタンパク質は、代替の開始コドンから翻訳されることが示された。転写因子 c-mycは代替の上流 CUG 開始コドンを有し、翻訳された場合に14 アミノ酸がタンパク質のN-末端に付加される(Hann and Eisenman 1984)。この代替のアイソフォームが、選択的にバーキットリンパ腫において破壊されたことが示された(Hann, King et al. 1988)。組織培養において、メチオニンが低濃度である場合、長い形態のmycが高い細胞密度で優先的に転写される(Hann, Sloan-Brown et al. 1992)。さらなる研究によって、長い形態のc-mycは成長阻害性であり、古典的なc-myc タンパク質と異なるセットの転写標的を有することが明らかとされた(Hann, Dixit et al. 1994). (Florkiewicz and Sommer 1989) (Prats, Kaghad et al. 1989)。
【0009】
さらに、タンパク質の実際の細胞内局在が代替の開始コドンにより決定できることが知られている。マウス 癌原遺伝子 int-2の場合、上流 CUG コドンからの代替の開始は核局在化をコードするが、AUG コドンは分泌性経路に局在化するためのシグナルペプチドをコードする(Acland, Dixon et al. 1990)。類似する現象はヒト FGF3において記載され、AUGから翻訳されたタンパク質は分泌経路に向かうが、上流 CUGから翻訳されたタンパク質は核に局在する(Kiefer, Acland et al. 1994)。さらにまた、幾つかの真核生物のタンパク質(例えば、TEF-1 および PRPS-3)においては、タンパク質は完全にCUGコドンから開始される(Taira, Iizasa et al. 1990; Xiao, Davidson et al. 1991)。
【0010】
分泌に向かうタンパク質は、シグナルペプチドと称される疎水性アミノ酸のストレッチ(a stretch )により小胞体を標的とする(Blobel, Walter et al. 1979)。通常、タンパク質のN-端に認められるシグナルペプチドは、翻訳の際にシグナル識別粒子 (SRP)に結合し、リボソームの停止を生じ、粗面小胞体にトランスロケーションし、ここでSRPレセプターと結合する。一旦リボソームがドッキングしたら、SRP-SRP レセプター複合体が放出され、翻訳がSec61トランスロコンを通りERの内腔を通り再開される。次に、シグナルペプチドが可溶性タンパク質のケースにおいては切断され、Secトランスロコンからタンパク質が放出される。膜を貫通するタンパク質の場合、膜貫通ヘリックスは、ER トランスロケーションのためのシグナルペプチドとして貢献する。これらのタンパク質は、ゴルジにおけるグリコシル化により広範に修飾され、分泌ベシクル中で形質膜にシャトルされる(Alberts 2002)。
【0011】
癌において重要であることが示された幾つかの分泌タンパク質が存在する。例えば、Wnt シグナル伝達経路は、肺癌において変更されることが示された。Wntは、Frizzled レセプターのファミリーに対する分泌型リガンドである。FrizzledのWnt 活性化により乱れを生じ、β-カテニン デグラデーション 複合体(APCを含む)が解離を生じ、これによりβ-カテニンのレベルが上昇し、そして核にトランスロケーションし、核でTCF 転写因子と相互作用し、トランス活性化(transactivate)が許容される。遺伝性および散発性(sporadic)の双方の結腸癌でAPCにおける変異を不活性化すること、またβ-カテニンにおける変異を活性化することが詳細に言及されている。さらに、幾つかの細胞外のリガンドアンタゴニスト(例えば、SFRP および Wnt-5a)は、Wntと同じFrizzled レセプターと競合する。双方との腫瘍抑制因子であることが示された; SFRP ノックアウトマウスは、リンパ様腫瘍を発生し、Wnt-5aのエピジェネティック サイレンシングがメラノーマにおいて検出された。
【0012】
PTENの全ての公開された報告は、このタンパク質が細胞質または核の何れかに局在することを指摘する。幾つかの細胞外タンパク質(glypicans および syndecans)は、分泌経路に関与する幾つかのタンパク質(reticulocalbinおよびcalumenin)のようにPTENと結合することが見出された。PTENが分泌経路に進入して、係る相互作用が許容されることが想定された。実際、本出願で開示されたとおり、新規の異なって翻訳されるタンパク質が存在し、このタンパク質はPTEN-longと命名され、N末端シグナルペプチドを含み、細胞外に分泌される。
【0013】
[発明の概要]
配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいる単離されたヒトのホスファターゼおよびtensin ホモログ ロング ポリペプチド(PTEN-long)または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログ,またはその各々のバリアント(variant)。
【0014】
(i) 配列番号1の残基 1〜173,または(ii) 配列番号5の残基 1〜173を含んでいるそのアナログ,または(iii) 配列番号1の残基 22〜516を含むポリペプチド。
【0015】
配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22 〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを含む薬学的組成物。
【0016】
対象における固形腫瘍(solid tumor)を治療するための方法であって、対象における固形腫瘍を治療するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【0017】
対象における固形腫瘍の成長を阻害するための方法であって、対象における固形腫瘍の成長を阻害するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【0018】
対象の固形腫瘍における血管形成を阻害するための方法であって、対象の固形腫瘍における血管形成を阻害するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【0019】
対象の固形腫瘍における血管の血管上皮細胞(vascular epithelial cell)のアポトーシスを誘導するための方法であって、対象の固形腫瘍における血管の血管上皮細胞のアポトーシスを誘導するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【0020】
対象における固形腫瘍を治療するための方法であって、配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5を含んでいるそのアナログをコードする特定量の発現ベクターを、PTEN-long, PTEN-long 断片,又はそのアナログを前記固形腫瘍の細胞中で対象における固形腫瘍を治療するため有効な量で発現するように前記対象に投与することを含む方法。
【0021】
対象における固形腫瘍の治療に使用するための、対象における固形腫瘍の成長を阻害するための、対象の固形腫瘍における血管上皮細胞のアポトーシスを誘導するための、または対象の固形腫瘍における血管形成を阻害するための、配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long)または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログ。
【0022】
それぞれPTEN-long, PTEN-long 断片,又はそのアナログの血漿半減期を増加させる非-ペプチド因子に結合させた、配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログを含む組成物。
【0023】
配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long)をコードする,または配列番号1の残基 22〜516を含んでいるその断片をコードする,または配列番号5を含んでいるそのアナログをコードする単離された核酸。
【0024】
配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログをコードしている核酸を含む発現ベクター。
【0025】
配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログを発現する能力のある形質転換された細胞であって、前記細胞はそのゲノムにPTEN-longまたはそのアナログをコードする組換えDNAが統合される形質転換された細胞。
【0026】
配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログを発現する能力のある宿主細胞であって、PTEN-longまたはそのアナログをコードするプラスミドを含む宿主細胞。
【0027】
それぞれPTEN-long, PTEN-long 断片,又はそのアナログの血漿半減期を増加させる非-ペプチド因子に結合させた配列番号1を含んでいるPTEN-long,または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5を含んでいるそのアナログを作るため、(1) 配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5を含んでいるそのアナログ, および(2) PTEN-long又はそのアナログの血漿半減期を増加させる非-ペプチド因子を混合(admixing)することを含む方法。
【0028】
配列番号2または配列番号6の核酸配列または配列番号 2または配列番号6の核酸配列の相補物と,または配列番号2 配列番号6とストリンジェントな条件下で特異的にハイブリダイズするその相補物の少なくとも20ヌクレオチド断片からなる単離された核酸分子。
【0029】
単離された抗体,又はその断片であって、(1) 配列番号1のアミノ酸残基 1-173を含むPTEN-ロング ポリペプチドまたは(2) 配列番号5を含んでいるそのアナログまたは(3) 配列番号3で表される配列を有しているアミノ酸または(4) 配列番号1の残基 22〜516を含んでいるPTEN-longの断片と結合する抗体又はその断片。
【0030】
単離された抗体又はその断片であって、前記抗体又はその断片はPTEN-longのコンホメーションエピトープと結合し、前記PTEN-longは配列番号1の残基 1-173で表される配列を含むが、しかし、前記抗体はPTENとは結合しない。
【0031】
単離された抗体又はその断片であって、前記抗体又はその断片はPTEN-longのエピトープと結合し、前記PTEN-longは配列番号1の残基 1-173で表される配列を含むが、しかし、前記抗体はPTENとは結合しない。
【0032】
配列番号1,または配列番号5のペプチド断片であって、抗-腫瘍, 抗-血管形成性(anti-angiogenic),または抗-アポトーシス活性を有する断片。
【図面の簡単な説明】
【0033】
【図1】PTEN-long 構築物の概略図。内因性の開始部位または代替の開始部位の何れかからPTENの発現を駆動するため作出された組み合わせを示している発現構築物。標準(Canonical)のPTENは黒色で示され、UTRにおける翻訳領域は灰色で示される。
【図2】ヒト PTEN mRNAの概略図。PTEN mRNAは、示された標準のATG開始コドンとインフレームの上流の173 アミノ酸をコードする。翻訳は、標準のATGから上流でヌクレオチド -519のCTGで開始される。拡大されたものは、配列番号1の残基 1-173に示される。
【図3】PTEN オルソログのN-端のアラインメント。示した種のPTEN タンパク質配列をVector NTI (Invitrogen)でBLOSUM62 スコア マトリックスを用いて整列させた。ヒト および マウスの双方の拡張したN末端配列(アステリクス)は、標準のAUG開始コドンから-519 (H. sapiens) および -520 (M. musculus)でのCUG 代替開始コドン用いて公開されたmRNAからORFinder (NCBI)を用いて翻訳された。ヒト (NM_000314) および マウス (NM_008960)のmRNA 配列。ミツバチ(Apis mellifera)の配列は、ベイラー医科大学のミツバチゲノムプロジェクトから得られた。線虫(Caenorhabditis elegans)のPTEN (Daf-18)のタンパク質配列を、Wormbaseからダウンロードした。ウシ(Bos Taurus)(XM_613125)およびチンパンジー(Pan troglodytes)(XP_521544)を、NCBIからダウンロードした。
【図4】PTEN-longの存在の証拠 A) 異なる細胞株の二つの異なるPTEN抗体での調査。MCF10A および HEK293は、PTENに関して野生型である。BT549およびHCC1937はPTEN ヌルであり、ZR-75-1はPTENに変異(L136)を有する;B) PTENおよびPTEN-longの双方を認識するPTENに対するモノクローナル抗体での異なる細胞株のさらなる調査;C) Wt ES 細胞は、大量のPTEN-longを発現する。PTEN-longは、これらの細胞において安定的なPTEN shRNA発現に感受性であり、PTEN ノックアウト細胞においては全く存在しない。多くのpAktレベルは、PTENのレベルに逆にしたがう;D) PTEN siRNAは、HEK293細胞においてPTENおよびPTEN-long双方のノックダウンを生じる。E) PTENヌルPC3細胞におけるプラスミドの外因性の発現。PTENorfは、単に開始コドン AUGからのORFをコードする(レーン 2)。ATR (ATR = 代替の翻訳領域)の付加によって、PTEN-longを弱く翻訳できる(レーン 3)。上流の開始部位のATGへの変異は、タンパク質の相補物(complement)を完全にPTEN-longにシフトした(レーン 5 および 6)。ATG 開始コドンのATAへの変異は、55kDaバンドを無効にする(レーン 4 および 6)。E) 抗体は5'ATRによりコードされたアミノ酸に対するものであり、HEK293同様PTENorfまたは5'ATR (ATG/ATG)をコードしているプラスミドの何れかを過剰発現しているPTENヌル U87細胞株からの双方の細胞溶解産物に使用された。PTEN-longは、5'ATRを過剰発現している細胞にのみ認められた。U87 細胞で観察されたバックグラウンドのバンドは、ブロットの下部に存在する。
【図5】シグナルペプチド予測。PTEN 5'UTR 配列を翻訳し、SignalIP3.0に入力した。真核生物シグナルペプチドのためのHidden markovモデルを予測のため使用した。N-領域は、シグナルペプチドの陽性に荷電したN末端配列を示す。H-領域は、シグナルペプチドの疎水性コアである。C-領域は、通常疎水性コアのヘリックスを破壊するプロリンによりマークされる穏やかに極性の領域である。切断可能性は、タンパク質がERの内腔に放出されることを許容するシグナルペプチドを放出する切断部位の予測である。(Dalbey and Heijne, 2002)。切断がポジション 21で発生することが予測される。
【図6】コンカナバリンAプルダウン。HEK293細胞を溶解し、コンカナバリン セファロースを使用してグリコシル化タンパク質をプルダウンした。溶出物をSDS-PAGEで分離し、PTEN (6H2.1)で免疫ブロットした。PTEN-longの濃縮は、プルダウン 対 インプットにより観察できる。長いPTENのバンドの濃縮に注意されたい。PTENは、複数の潜在的なO-グリコシル化部位を有するが、しかし、僅か一つのN-グリコシル化部位を有する。HEK293 細胞中でPTEN相補物の一部がグリコシル化されるかどうかを決定するプルダウンアッセイにレクチン コンカナバリン-A(糖部分と結合する)を我々は使用した。我々はPTENの混合物を精製でき、PTEN-longは凡そ50%であった(正常なPTENに対してPTEN-longの多大なる濃縮)。この事項は、PTEN-longがグリコシル化されること、またPTENの細胞質の55kDa形態がグリコシル化されるか又はPTEN-longが細胞外で切断されるかの何れかを示す。
【図7】PTEN および PTEN-βは、ヘパランと結合する。マウス肝抽出物を1ml HiTrap ヘパランセファロース (Amersham) カラムに通過させた。そのカラムを500mM NaClで洗浄し、タンパク質を連続的に1M NaClのカラム容量で溶出した。フラクションをPTEN モノクローナル抗体を用いてPTENに関してSDS-PAGEで分析した。PTENは、以前に高度に陰性に荷電した種に親和性を有することが示され、PTENの特性により高度に陰イオン性のPtdIns(3,4,5)P3を好むことが導かれる(Das, Dixon et al. 2003)。ヘパランは最も陰性に荷電した生物学的分子(biological molecules)の一つであるので、我々はヘパランが実際にPTENの細胞外基質への結合を媒介すると仮定した。マウスの肝臓からのタンパク質抽出物を用いて、我々はPTENがヘパランと高親和性に結合することを発見した。さらにまた、ヘパリン アガロース カラムから1M NaClを用いたPTENの連続的な溶出によって、PTEN-longも溶出された。
【図8】プロテアーゼプロテクションアッセイ。HEK293細胞をプロテイナーゼ Kの濃度を増加させて再懸濁した。終濃度 0.2%でTritonを、最高濃度のプロテイナーゼ Kを含んでいる反応に加えた。反応をPMSFで停止し、細胞溶解産物をlaemlli 緩衝剤で作った。溶解産物を8% ポリアクリルアミドゲル上でSDS-PAGEで分離し、PTEN (6H2.1), AKT, E-カドヘリンに関して免疫ブロットした。PTENの免疫ブロットの大きなバンドが、PTEN-longと指定された。これらのデータは、E-カドヘリン および PTEN-longが大部分は細胞表面にあることを示す。
【図9】馴化培地(conditioned media)のヘパリン親和性精製からのPTEN および PTEN-longの高塩溶出(High salt elution)。馴化培地からの親和性精製後、PTEN および PTEN-longはHiTrap ヘパリン (Amersham) カラムから溶出できる。PTENの尾部に対するモノクローナル抗体(上記)および5'ATRにおいて翻訳されたアミノ酸に特異的な抗体の双方は、凡そ55kDaの質量のタンパク質バンドを認識する。
【図10】ヒト血清からのPTENの精製。AB 血液からのヒト血清をプロテインA/Gで抗体のプレクリア(precleared)をしてヘパリンセファロースに供試した。溶出物をSDS-PAGEで分離し、PTENに関して又は重鎖混入のための対照に対する二次単独で免疫ブロットした。
【図11】図 11A - 11C。 PTEN-longの抗-血管形成活性。(A) PTEN-longは、発生している網膜中の血管および毛細管のサブセットに発現する。この発現パターンは、PTENの標準形(canonical form)とはっきりと対照を成し、これらの領域に生じる血管退行(vascular regression)の誘導におけるPTEN-longの役割と矛盾しない。この血管退行のプロセスが全網膜の溶解産物のウエスタンブロットのとおり高酸素により 最上部右(B)、また低酸素条件下で内皮細胞におけるPTEN-longの欠損(C)により誘導される場合、この関係はPTEN-longの顕著なアップレギュレーションにより強められる。これらの所見は、糖尿病性網膜症, 同様に高増殖性の血管障害(hyper-proliferative vascular disorders)などの抗-血管新生治療としてのPTEN-longの有用性を示す。矢印は、CD34 および PTEN-long 陽性の組織(血管)を示す。
【図12】PTEN-longのアポトーシス促進活性。指摘のとおり、アポトーシスは、精製 PTEN-longで24 hrs処理されたMCF-10A乳房上皮細胞において誘導された。カスパーゼ 3 切断は、アポトーシス活性の指標(indicative)であった。
【図13】PTEN-longでのマウスの処理。PTEN-longまたは空ベクター対照のいずれかで十日処理し、キャリパー測定で測定した腫瘍サイズのグラフ。293 細胞を、pcDNA3.1 His V5 ベクターにおけるATG/ATG PTEN-longでトランスフェクトした。トランスフェクションの48 hr 後、細胞質ライセート(Cytoplasmic lysates)を作出し、V5-抗体ビーズを通過させ、V5 ペプチドで溶出した。V5-ビーズ精製溶出物のウエスタンブロットを下に示す。PTEN-longが腫瘍を治療するため使用できるだろうとの最初の観察。異種移植片を、スキッドマウスの乳房の脂肪パッドに注射したU87 グリオブラストーマ 細胞 (百万)を用いて樹立した。治療を移植の凡そ二週後に開始した。
【図14】PTEN-longでのマウスの治療の結果。グラフは、14日間の対照およびPTEN-longの注射で処理したマウスの生存率(surviving fraction)を示す(日数)。
【図15】PTEN (5'UTRを欠損しているPTENorf), PTEN-long, およびG からRへの変異をアミノ酸305に有するPTEN-long(PTENorfにおけるG129R変異に匹敵する)の示した構築物を、293 細胞にトランスフェクトした。これらの細胞からの精製タンパク質を用いて、PTEN-longが活性なホスファターゼであること、またPTEN-long G305R 変異(これはPTENにおけるG129Rである)がホスファターゼ活性を低下させたことが示された。
【図16】ホスファターゼ活性は、PTEN-long 活性に必須であり、PTEN-long(G305R) 変異の実験で示された。PTENの文献に基づいて、C2 ドメインの内側に作った短縮体(truncations)はタンパク質を不安定化することが知られている。また、PTENの結晶構造に基づいて、C2 ドメイン および ホスファターゼドメインの間の相互作用がホスファターゼ活性に重要であると信じられている。従って、C-端でのPTEN-long 活性のための最小ドメインは、C2 ドメインを必要とするが、尾部を必要としない。N-端で、予測された切断部位は、アミノ酸 21である。従って、タンパク質の機能領域は、この領域内にある。この点に関して、U87 腫瘍をPTENと又はPTEN-longと対比して処理した場合、有意な効果はPTEN治療では観察されず、PTEN-long 治療でのみ観察されることに注意することが重要である。
【図17】His および V5 タグを有するpcDNA3.1 発現ベクター中のATG/ATG-PTEN longでトランスフェクトされた293 細胞からのPTEN-longの精製。Ni+ カラム 溶出後、溶出液をゲル濾過カラムで分離した。OD280を青線で示す。PTEN-longは、フラクション 7-18に濃縮された。本実験の収量は、凡そ1 mgであった。矢印は、PTEN-long および 移動性が変化したPTEN-long産物(altered migrating PTEN-long products)を示す。
【図18】図 18A - 18B。 情報(readout)として細胞死を用いたLNCaP 前立腺癌細胞とPTEN-long 精製 タンパク質との用量反応(タンパク質はARVYSにより精製された)。1xは、0.33 マイクログラム / mlと等しい。細胞を成長因子なしの培地で処理した。24 hr後、細胞を血清フリー培地で洗浄し、Laemmliサンプル緩衝剤で溶解した。示したタンパク質に関してウエスタンブロットを行った。(B) PTEN-long, PTEN-long(G305R)またはMock対照で処理したU87 グリオブラストーマ細胞は、PARPの切断およびセリン 473でのpAKT シグナルのダウンレギュレーションにより示されるようなアポトーシスの誘導を示す。これらのデータによって、PTEN-longがアポトーシスを誘導し、PI3K/AKT シグナル伝達を減少できることがさらに確認される。
【図19】図 19A - 19C。 AKTA 精製 PTEN-long タンパク質は、ノギスとxenogen動物生体イメージングシステムと連動させたルシフェラーゼレポーターを用いることとの双方により測定されたとおり、五日間にわたり腫瘍サイズを減少できた。マウスは、~0.05mg PTEN-long/日を五日間与えられた。異種移植片は、FUW-ルシフェラーゼ-neoで感染させたためルシフェラーゼを発現する乳房の脂肪パッドに注射された百万の U87 グリオブラストーマ細胞で樹立された。マウスは、Xenogen Imaging Systemでのイメージングの10 分前に腹腔内にルシフェリンを注射された。(A) 治療前(左パネル)および五日治療時(右パネル)の4 マウスに関するルシフェラーゼ測定。(B) 治療前と5 日治療間のキャリパー測定(cm2)。(C) パネルにイメージされXenogenシステムにより検出されたフォトン(Photons)。同齢集団の四マウスに関する標準誤差が示される。0日から5日で検出されたフォトンに関するスチューデントのt-検定。
【図20】独立した実験において、U87 腫瘍をPTEN (orf-403 アミノ酸; n=5), PTEN-long (G305R; n=5)または野生型 PTEN-long (n=4)での処理前に1.5 cm2に成長させた。処理の5 日後、平均のルミネセンス変化(average change luminescence)は、PTEN-long 処理マウスで有意な減少を示すが、PTENまたはPTEN-long(G305R)処理したコホートに関して減少しないことを示す。ルミネセンスの減少は、腫瘍サイズの減少と相関する。これらのデータによって、PTEN-longは機能するために5'ATR および ホスファターゼ 活性を必要とすることが実証された。
【図21】図 21A - 21C。 PTEN-long-処理したU87 異種移植片の分析によって、アポトーシスの活性化およびPI3K シグナル伝達の阻害が実証された。腫瘍を上記のとおり5 日処理した。(A) PTEN-longの野生型 および G305Rで処理した腫瘍を、示した処理の5 日後に収穫し、ウエスタン分析のため溶解した。PTEN-longに関する野生型タンパク質は、FOXO および AKTのリン酸化が減少し、カスパーゼ-3切断を活性化することができた。(B) 示したとおり、5 日処理した代表的な腫瘍を、ホルマリンで固定し、パラフィン包埋した。切片を切断されたカスパーゼ-3(アポトーシスのマーカー)を検出する抗体で染色した。PTEN-long 処理細胞は、アポトーシス細胞のパーセントの有意な増加を有する(P=0.0419, スチューデントのt-検定)。(C) 切断されたカスパーゼ-3 染色の代表的な像。
【図22】図 22A - 22B。 5 日PTEN-longで処理した同じ腫瘍において、血管の数は非常に減少した。PTEN-longの野生型およびG305Rで処理した腫瘍を、示した処理の5 日後に収穫し、ホルマリンで固定し、パラフィン包埋した。切片をCD31(血管に沿ってならぶ内皮細胞のマーカー)を検出する抗体で染色した。(A) PTEN-long 処理細胞は、血管数/視野(40x 対物)の有意な減少を有した(P=0.007579, スチューデントのt-検定)。(B) CD31染色の代表的なイメージ。
【図23】図 23A - 23B。 U87異種移植片は六群(n=3/群)で樹立され、PTEN-longを筋肉内(IM), 腹腔内(IP), 腫瘍内(IT), 皮下 (SC), 静脈内(IV)注射で四 日治療した。(A) キャリパーにより測定された0日〜4日の腫瘍サイズ(CM2)の変化の平均。(B) xenogen イメージングからの代表的な像を右に示す。このデータから、全ての注射法が無処置のマウスと比較して腫瘍成長に影響すること、またIMで処理した同齢集団のみが退行量の有意な減少を示したことを我々は結論づけることができる。
【図24】図24A - 24C。 異種移植実験を乳房, 脳, および 前立腺に由来する6細胞株で行った。上部は、四つの乳癌細胞株で記された変化である。(A, B, およびD)。示した処置日にキャリパーで測定した腫瘍表面領域(cm2)のグラフ。(C) ヘマトクリット-1143 細胞における変化は、僅かに処置の24 時間後に認められた。全ての四つの細胞株において、処置後に腫瘍サイズが明らかに減少する。
【図25】PTEN-longは細胞と結合する。PTEN-long タンパク質をU87 細胞培地に氷上で 10 分間加え、固定し、PTEN-longを認識する抗体でPTEN-longを染色した。
【図26】Milesアッセイ: 血管透過性の誘導はPTEN-longにより阻害された。この阻害は、精製タンパク質をPTEN抗体(6H2.1)とプレインキュベーションすることにより逆転させることができる。PTEN-longは、VEGFによる血管透過誘導を阻害できる。この誘導は、PTEN-longをPTENに対する抗体とプレインキュベーションすることにより回復できたが、対照IgGではできなかった。
【0034】
[発明の詳細な記載]
配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいる単離されたヒトのホスファターゼおよびtensin ホモログ ロング ポリペプチド(PTEN-long)または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログ,またはその各々のバリアント。
【0035】
(i) 配列番号1の残基 1〜173,または(ii) 配列番号5の残基 1〜173を含んでいるそのアナログ,または(iii) 配列番号1の残基 22〜516を含むポリペプチド。
【0036】
配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22 〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを含む薬学的組成物。
【0037】
対象における固形腫瘍を治療するための方法であって、対象における固形腫瘍を治療するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含むヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【0038】
対象における固形腫瘍の成長を阻害するための方法であって、対象における固形腫瘍の成長を阻害するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【0039】
対象の固形腫瘍における血管形成を阻害するための方法であって、対象の固形腫瘍における血管形成を阻害するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【0040】
対象の固形腫瘍における血管の血管上皮細胞(vascular epithelial cell)のアポトーシスを誘導するための方法であって、対象の固形腫瘍における血管の血管上皮細胞のアポトーシスを誘導するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【0041】
本出願に記載された方法の態様において、腫瘍は、癌性腫瘍(cancerous tumor)である。本出願に記載された方法の態様において、癌性腫瘍は、対象のグリア細胞, 前立腺, 卵巣, 子宮, 子宮内膜, 乳房, メラノサイト, 腎臓, 肺, 結腸, 頭部(head), 頸部(neck),または膵臓の腫瘍である。
【0042】
本出願に記載された方法の態様において、癌性腫瘍は、PTENにより若しくはPI3K経路により活性化される又はPTEN-陰性である。
【0043】
本出願に記載された方法の態様において、PTEN-long,またはそのアナログ,またはPTEN-long 断片は、固形腫瘍への直接的な導入により対象に投与される。本出願に記載された方法の態様において、PTEN-long,またはそのアナログ,またはPTEN-long 断片は、固形腫瘍に注射される。本出願に記載された方法の態様において、PTEN-long,またはそのアナログ,またはPTEN-long 断片は、直接的にカテーテルで固形腫瘍に導入される。本出願に記載された方法の態様において、PTEN-long,またはそのアナログ,またはPTEN-long 断片は、固形腫瘍に供給する血管への直接的な導入により対象に投与される。本出願に記載された方法の態様において、PTEN-long,またはそのアナログ,またはPTEN-long 断片は、固形腫瘍に供給する血管に注射される。本出願に記載された方法の態様において、PTEN-long,またはそのアナログ,またはPTEN-long 断片は、直接的にカテーテルで固形腫瘍に供給する血管に導入される。本出願に記載された方法の態様において、PTEN-long,またはそのアナログ,またはPTEN-long 断片は、対象の静脈内に投与される。本出願に記載された方法の態様において、PTEN-long,またはそのアナログ,またはPTEN-long 断片は、対象の皮下に投与される。
【0044】
対象における固形腫瘍を治療するための方法であって、配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5を含んでいるそのアナログをコードする特定量の発現ベクターを、PTEN-long, PTEN-long 断片,又はそのアナログを前記固形腫瘍の細胞中で対象における固形腫瘍を治療するため有効な量で発現するように前記対象に投与することを含む方法。
【0045】
対象における固形腫瘍の治療に使用するための、対象における固形腫瘍の成長を阻害するための、対象の固形腫瘍における血管上皮細胞のアポトーシスを誘導するための、または対象の固形腫瘍における血管形成を阻害するための、配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long)または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログ。
【0046】
本出願に記載された使用のための化合物の態様において、腫瘍は、癌性腫瘍(cancerous tumor)である。本出願に記載された方法の態様において、癌性腫瘍は、グリア, 前立腺, 乳房, 子宮内膜, メラノサイト, 腎臓または肺癌の腫瘍である。
【0047】
本出願に記載された使用のための化合物の態様において、癌性腫瘍は、PTENにより若しくはPI3K経路により活性化される又はPTEN-陰性である。
【0048】
それぞれPTEN-long, PTEN-long 断片,又はそのアナログの血漿半減期を増加させる非-ペプチド因子に結合させた、配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログを含む組成物。
【0049】
本出願に記載された組成物の態様において、さらに薬学的担体を含んでいる。本出願において記載された組成物の態様において、血漿半減期を増加させる非-ペプチド因子は、ポリエチレングリコール(PEG)である。記載された組成物の態様において、PEGは、PTEN-long, PTEN-long 断片, またはそのアナログのC-末端またはN-末端に結合される。記載された組成物の態様において、血漿半減期を増加させる非-ペプチド因子は、9-フルオレニルメチル クロロホルマート (Fmoc)または(7-スルホ)-9-フルオレニルメトキシカルボニルである。
【0050】
配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long)をコードする,または配列番号1の残基 22〜516を含んでいるその断片をコードする,または配列番号5を含んでいるそのアナログをコードする単離された核酸。
【0051】
一態様において、核酸は、配列番号2または配列番号6で表される配列を有している連続的なヌクレオチドを含む。一態様において、核酸は、配列番号2または配列番号6で表される配列の連続的なヌクレオチド残基503〜2243を含む。一態様において、核酸は、配列番号3または配列番号7で表される配列を有している連続的なヌクレオチドを含む。一態様において、核酸は、配列番号3または配列番号7で表される配列の連続的なヌクレオチド残基503〜2243を含む。一態様において、核酸はRNAである。一態様において、核酸はDNAである。一態様において、核酸はcDNAである。
【0052】
配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログをコードしている核酸を含む発現ベクター。
【0053】
配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログを発現する能力のある形質転換された細胞であって、前記細胞はそのゲノムにPTEN-longまたはそのアナログをコードする組換えDNAが統合される形質転換された細胞。
【0054】
配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログを発現する能力のある宿主細胞であって、PTEN-longまたはそのアナログをコードするプラスミドを含む宿主細胞。
【0055】
一態様において、宿主細胞は、細菌細胞である。一態様において、宿主細胞は、哺乳類細胞である。
【0056】
それぞれPTEN-long, PTEN-long 断片,又はそのアナログの血漿半減期を増加させる非-ペプチド因子に結合させた配列番号1を含んでいるPTEN-long,または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5を含んでいるそのアナログを作るため、(1) 配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5を含んでいるそのアナログ, および(2) PTEN-long又はそのアナログの血漿半減期を増加させる非-ペプチド因子を混合(admixing)することを含む方法。
【0057】
配列番号2または配列番号6の核酸配列の相補物と,または配列番号2 配列番号6とストリンジェントな条件下で特異的にハイブリダイズする配列番号2または配列番号6の核酸配列または配列番号 2または配列番号6の核酸配列の相補物の少なくとも20ヌクレオチドの断片からなる単離された核酸分子。
【0058】
単離された抗体,又はその断片であって、(1) 配列番号1のアミノ酸残基 1-173を含むPTEN-ロング ポリペプチドまたは(2) 配列番号5を含んでいるそのアナログまたは(3) 配列番号3で表される配列を有しているアミノ酸または(4) 配列番号1の残基 22〜516を含んでいるPTEN-longの断片と結合する抗体又はその断片。
【0059】
一態様において、前記抗体はモノクローナル抗体である。一態様において、前記抗体は、抗体断片である。一態様において、前記抗体断片は、Fab, Fab', F(ab')2, またはFv 断片である。一態様において、前記抗体断片は、単鎖抗体(single-chain antibody)である。一態様において、前記抗体はヒト化抗体である。
【0060】
単離された抗体又はその断片であって、前記抗体又はその断片はPTEN-longのコンホメーションエピトープと結合し、前記PTEN-longは配列番号1の残基 1〜173で表される配列を含むが、しかし、前記抗体はPTENとは結合しない。
【0061】
一態様において、前記抗体は、配列番号1の残基 153〜173の何れかで構成されるエピトープと結合する。
【0062】
単離された抗体又はその断片であって、前記抗体又はその断片はPTEN-longのエピトープと結合し、前記PTEN-longは配列番号1の残基 1〜173で表される配列を含むが、しかし、前記抗体はPTENとは結合しない。
【0063】
一態様において、前記抗体は、配列番号1の残基 153〜173(または、その部分)と結合する。
【0064】
配列番号1,または配列番号5のペプチド断片であって、抗-腫瘍, 抗-血管形成性,または抗-アポトーシス活性を有する断片。一態様において、前記ペプチドは、5〜10, 10〜20, 20〜30, または30〜40 アミノ酸を含む。一態様において、前記ペプチドは、配列番号1または配列番号5のアミノ酸 1〜173を含む。一態様において、前記ペプチドは、配列番号1の連続的なアミノ酸残基 22〜516を含む。一態様において、前記ペプチドは、配列番号1または配列番号5の連続的なアミノ酸残基 174〜576を含まない。
【0065】
本出願に使用される物質の「予防上有効な(prophylactically effective)」量は、前記物質が投与される対象における所定の病理学的なコンディションの開始を阻止する又は遅延させるための効果的な量である。
【0066】
本出願に使用される物質の「治療上有効な(therapeutically effective)」量は、前記物質が投与される対象が患っている所定の病理学的なコンディションの症状または原因(cause)を治療する, 寛解させる又は低下させるための効果的な量である。
【0067】
一態様において、治療上または予防上有効な量は、約 1 mg の因子/対象〜約 1 g の因子/対象/投薬(dosing)である。別の態様において、治療上または予防上有効な量は、約 10 mg の因子/対象〜500 mg の因子/対象である。さらなる態様において、治療上または予防上有効な量は、約 50 mg の因子/対象〜200 mg の因子/対象である。更なる一態様において、治療上または予防上有効な量は、約 100 mg の因子/対象である。なお更なる態様において、治療上または予防上有効な量は、50 mgの因子/対象, 100 mgの因子/対象, 150 mgの因子/対象, 200 mgの因子/対象, 250 mgの因子/対象, 300 mgの因子/対象, 400 mgの因子/対象および 500 mgの因子/対象から選択される。
【0068】
因子(agent)を「投与すること」は、当業者に既知の様々な方法および送達システムの何れかを用いて成立させる(effected)又は行うことができる。投与することは、例えば、静脈内, 経口, 経鼻, 腹腔内, 脳脊髄液を介して, 移植片を介して, 経粘膜, 経皮, 筋肉内, 血管内, 動脈内, 冠内, 心筋内 または皮下であってもよい。
【0069】
「PTEN-long アナログ(PTEN-long analogue)」の用語は、一または二以上の修飾されたアミノ酸を有するが、配列番号1と少なくとも 90%の配列類似性(sequence similarity)を保持し、以下に記載されたインビボ研究により測定される腫瘍成長の阻害の活性を保持する又は改善するポリペプチドを包含する。PTEN自身(即ち、配列番号1の残基 174-576のみで規定されるポリペプチド)は、明示的に除外される。
【0070】
「PTEN-long バリアント(PTEN-long variant)」の用語は、一または二以上の付加的なアミノ酸(典型的には、PTEN-longのN-末端またはC-末端の何れか、または双方に5未満の付加的なアミノ酸を有している)を有し、以下に記載されたインビボ研究により測定される腫瘍成長の阻害活性を保持する又は改善するポリペプチドを包含する。
【0071】
「PTEN-long アナログのバリアント(PTEN-long analogue variant)」の用語は、一または二以上の付加的なアミノ酸〔典型的には、PTEN-longアナログ(即ち、配列番号5)のN-末端またはC-末端の何れか、または双方に5未満の付加的なアミノ酸を有している〕を有し、以下に記載されたインビボ研究により測定される腫瘍成長の阻害活性を保持する又は改善するポリペプチドを包含する。
【0072】
PTEN-long アナログを参照している本出願の全ての態様は、必要な変更を加えて、PTEN-long バリアントに適用可能である。
【0073】
PTEN-longは、別にPTEN-beta, PTEN-β, PTEN-Sと参照される。
【0074】
PTEN-long, PTEN-long アナログ, PTEN-long バリアント, PTEN-long アナログのバリアント, 又はその各々の組み合わせのための注射可能な薬物配送システムは、溶液、懸濁液、ゲル、ミクロスフェアおよびポリマー性の注射可能物質(polymeric injectables)を含み、また溶解性変更薬剤(solubility-altering agents)(例えば、エタノール、プロピレン・グリコールおよびスクロース)およびポリマー〔例えば、ポリカプリルアクトン(polycaprylactones)およびPLGA's〕などの賦形剤を含むことができる。インプラント可能なシステム(Implantable systems)は、ロッド(rods)およびディスク(discs)を含み、また非限定の例のPLGAおよびポリカプリルアクトンなどの賦形剤を含有することができる。
【0075】
本発明の組成物および化合物のための経口の送達システムは、錠剤およびカプセル剤を含む。これらは、結合剤〔例えば、ヒドロキシプロピルメチルセルロース、ポリビニールピロリドン、他のセルロース系材料(cellulosic materials)およびスターチ〕、希釈剤〔例えば、ラクトースおよび他の糖類、スターチ、リン酸二カルシウム(dicalcium phosphate)およびセルロース系材料〕、崩壊剤(例えば、スターチポリマーおよびセルロース系材料)および潤滑剤(例えば、ステアレートおよびタルク)などの賦形剤を含有してもよい。
【0076】
本発明の組成物および化合物のための経粘膜的な送達システムは、パッチ(patches)、錠剤、座剤、ペッサリー(pessaries)、ゲルおよびクリームを含み、溶解剤および増強剤(例えば、プロピレン・グリコール、胆汁酸塩およびアミノ酸)などの賦形剤、並びに他のビヒクル(例えば、ポリエチレン・グリコール、脂肪酸エステルおよび誘導体、並びにヒドロキシプロピルメチルセルロースおよびヒアルロン酸などの親水性ポリマー)を含有することができる。
【0077】
本発明の組成物および化合物のための経皮的な送達システムは、例えば、水溶性および非水溶性のゲル、クリーム、複合乳剤(multiple emulsions)、ミクロ乳剤(microemulsions)、リポソーム、軟膏、水溶性および非水溶性の溶液、ローション(lotions)、エアロゾル(aerosols)、ハイドロカーボン基材および粉末(hydrocarbon bases and powders)を含み、且つそれは溶解剤(solubilizers)、浸透増強剤(permeation enhancers)(例えば、脂肪酸、脂肪酸エステル、脂肪アルコールおよびアミノ酸)、および親水性のポリマー(例えば、ポリカルボフィルおよびポリビニールピロリドン)などの賦形剤を含有することが可能である。一態様において、薬学的に許容される担体は、リポソームまたは経皮的な増強剤(transdermal enhancer)である。
【0078】
本発明の組成物および化合物の再構成可能(reconstitutable)な送達システムのための溶液、懸濁液、粉剤には、懸濁剤〔例えば、ガム、ザンタン(zanthans)、セルロース(cellulosics)および糖〕、保湿剤(例えば、ソルビトール)、溶解剤(例えば、エタノール、水、PEGおよびプロピレングリコール)、界面活性剤(例えば、ラウリル硫酸ナトリウム、Spans、Tweens、およびセチルピリジン)、保存剤および抗酸化剤〔例えば、パラベン(parabens)、ビタミンEおよびC、およびアスコルビン酸〕、抗固化剤(anti-caking agents)、コーティング剤、およびキレート化剤(例えば、EDTA)などのビヒクルが含まれる。
【0079】
本出願に使用される「5'ATR」は、以下の実験のセクションに記載される5'代替の翻訳領域である。
【0080】
本出願に使用される、「薬学的な担体」は、動物またはヒトに対して本発明(instant)の化合物または組成物を送達するための薬学的に許容される溶媒, 懸濁剤またはビヒクルである。担体は、液体, エアロゾル, ゲルまたは固形であってもよく、計画された投与の様式を考慮して選択される。
【0081】
「非-ペプチド因子(Non-peptide agent)」は、限定されることなく、グリコマー(glycomer), ポリマー, 小分子〔即ち、1000未満の分子量を有している炭化水素に基づく分子(hydrocarbon-based molecule)または有機分子〕, 脂質, リポソームを含む任意の化学物質(chemical entity)を意味する。非-ペプチド因子の例には、PEG, Fmoc およびFMSが含まれるが、これらに限定されない。
【0082】
本出願に使用される「固形腫瘍(Solid Tumor)」は、癌性(cancerous)の及び非癌性の固形腫瘍を含む。癌性の固形腫瘍には、限定されることなく、胆道癌; グリオブラストーマ および髄芽腫を含む脳癌; 乳癌; 子宮頚癌(cervical cancer); 絨毛癌; 結腸癌; 子宮内膜癌; 食道癌; 胃癌; ボーエン病およびパジェット病を含む上皮内の新生物; 肝臓癌; 肺癌; ホジキン病およびリンパ球性リンパ腫を含むリンパ腫; 神経芽細胞腫; 扁平上皮癌を含む口腔癌; 上皮細胞, 間質性細胞, 胚細胞および間充織細胞から発生するものを含む卵巣癌; 膵臓癌; 前立腺癌; 結腸直腸癌; 平滑筋肉腫, 横紋筋肉腫, 脂肪肉腫, 線維肉腫および骨肉腫を含む肉腫; メラノーマ, カポジ肉腫, 基底細胞癌(basocellular cancer)および扁平上皮細胞癌を含む皮膚癌; 胚腫瘍(セミノーマ, 非-セミノーマ[奇形腫, 絨毛癌]), 間質腫瘍(stromal tumors)および胚細胞腫瘍を含む精巣癌; 甲状腺腺癌(thyroid adenocarcinoma)および髄様癌(medullar carcinoma)を含む甲状腺癌; および腺癌およびウィルムス腫瘍を含む腎臓癌が含まれるが、非固形組織の腫瘍、例えば、急性のリンパ性および骨髄性の白血病; 多発性骨髄腫; AIDS関連白血病および成人T細胞白血病リンパ腫(adult T-cell leukemia lymphoma)を含む白血病および他の血液学的な新生物は除外される。
【0083】
配列表の配列番号1は、全長のPTEN-long タンパク質配列である。配列番号1のアミノ酸残基1〜173は、5' ATRのmRNA配列(以下の配列番号2の記載を参照されたい)によりコードされるPTEN-longの新規の残基を表す。PTENの標準の開始メチオニンは、配列番号1のアミノ酸残基 174である。
【0084】
配列番号2は、全長 PTEN-long mRNA配列である。PTEN-longの5' ATR配列は、配列番号2のヌクレオチド 513(非標準のCUG開始コドン)で開始し、配列番号2のヌクレオチド 1031で終了する。5' ATRは、配列番号2のヌクレオチド 1032での標準のAUG開始コドンで始まり、配列番号2のヌクレオチド 2243で終わるPTENオープンリーディングフレームとインフレームである。従って、PTEN-longのオープンリーディングフレームは、配列番号2のヌクレオチド 513から配列番号2のヌクレオチド 2243に伸び、PTENのN-端への173アミノ酸の付加が導びかれる。このタンパク質は、PTEN-longと参照される。
【0085】
配列番号3は、全長 PTEN-long mRNA配列に対応するcDNAである。
【0086】
配列番号4は、配列番号1のアミノ酸残基 153〜173を含んでいるPTEN-long タンパク質配列のペプチドである。このユニークなペプチドは、PTENに認められないPTEN-longに由来するユニークなエピトープを表す。
【0087】
配列番号5は、全長 PTEN-long アナログのタンパク質配列である。PTEN-longのアミノ酸残基 1は、前記アナログにおいてLeuからMetへと変化してタンパク質収量が増加する。
【0088】
配列番号6は、修飾された全長 PTEN-long アナログのmRNA配列である。PTEN-longアナログの5' ATR配列は、配列番号6のヌクレオチド 513(操作されたAUG開始コドン)で開始し、配列番号6のヌクレオチド 1031で終了する。
【0089】
配列番号7は、修飾された全長 PTEN-longアナログのmRNA配列に対応する修飾されたcDNAである。
【0090】
範囲が明細書中で与えられる場合、前記範囲はその範囲の全ての整数とその範囲内の0.1単位, および任意の細分範囲(sub-range)を含むことが理解される。例えば、77〜90%の範囲は、77.0%, 77.1%, 77.2%, 77.3%, 77.4%, 77.5%, 77.6%, 77.7%, 77.8%, 77.9%, 80.0%, 80.1%, 80.2%, 80.3%, 80.4%, 80.5%, 80.6%, 80.7%, 80.8%, 80.9%, および90.0%, 同様に80%〜81.5%の範囲などを含む。
【0091】
本出願に記載された様々な要素の全ての組み合わせは、本発明の範囲内である。
【0092】
本発明は以下の例を参照することに更に理解される。 当業者は、詳細に説明される特定の実験が、本願の特許請求の範囲においてより完全に記載される本発明の単なる例であることを容易に理解する。
【0093】
[例]
PTEN 腫瘍抑制因子は、癌において最も一般的に変更される遺伝子の一つであり、ホスファチジルイノシトール3,4,5-三リン酸の脂質ホスファターゼとして機能し、これが次にホスファチジルイノシトール 3-キナーゼ(PI3K) および Aktからの腫瘍形成性のシグナル伝達を抑制する。PTEN mRNAを検査することによって、5' 非翻訳領域 (UTR)が770bpについてPTENのオープンリーディングフレーム (ORF)とインフレームであることが明らかとされた。このUTR ORF内で、標準のAUG開始コドンの513塩基対上流での弱いコザック配列内に代替のCUG開始コドンが存在する。標準のPTEN ORFの発現により凡そ55kDaに移動するタンパク質が発現するが、5'UTRを含んでいるPTEN cDNAの発現によりPTEN-longと称される70kDaの第二のタンパク質を産生することができる。開始部位の変異は、55kDa PTENが標準の開始コドンでの翻訳から産生されるが、他方でPTEN-longは上流の代替の開始部位から開始されることを指摘する。異なる PTEN 抗体での免疫ブロットによって、複数の細胞株にPTEN-longが内因性(endogenous)に存在することが実証された。マウスES細胞におけるノックダウンおよびノックアウト研究によって、この大きなタンパク質が実際にPTENであることが確認された。付加されたN末端タンパク質配列がシグナルペプチドおよび切断部位をコードしており、PTEN-longが分泌経路に進入することを指摘している。PTEN-longは優先的にレクチンコンカナバリンAと結合し、グリコシル化されることが示される。さらにまた、PTEN-longは、PTENに対する抗体やヘパラン硫酸の双方を用いる親和性精製により馴化培地(conditioned media)から精製できる。また、PTEN-longは、インビボでのプロテアーゼプロテクションアッセイにおけるデグラデーションに感受性であるが、正常なPTENは感受性ではなく、PTEN-longが細胞膜の外側に局在することを示している。
【0094】
試薬, 細胞株および抗体 - プロテイナーゼ K および コンカナバリン- AをSigma (St. Louis, MO)から購入した。ヘパリン セファロース および HiTrapヘパリンHPカラムをAmersham (Piscataway, NJ)から購入した。PTENに対する抗体をCell Signaling (Danvers MA) およびCascade (Winchester MA)から購入した。Akt 抗体をCell Signaling (Danvers MA)から購入し、E カドヘリン抗体をUpstate Millipore (Billerica, Ma)から購入した。PTENの新規の翻訳において認められたエピトープ PRHQQLLPSLSSFFFSHRLPD (配列番号3)に対して産生されたポリクローナル親和性精製抗体については、Zymed Laboratories (South San Fransisco, CA)が行った。二次抗体をPierce (Rockford, IL)から購入した。HEK293, ZR-75-1, SKBR-3, MDAMB-361, BT549, および PC3をATCC (Manassas, VA)から得て、提供されたガイドラインにしたがい成長させた。
【0095】
プラスミドおよび構築物 ― pCEP4-PTEN(PTENの完全なオープンリーディングフレームおよび5'-非翻訳領域をコードしている)は、前に報告されたとおり、PTEN cDNA(NCBIにU90351として寄託された)をpCEP4(Invitrogen)のNotI部位にクローニングすることにより作出された(Li, Simpson et al. 1998)。5'UTRは、このプラスミドにおいてクローニングのため使用されるオリジナルのNotI 制限部位の上流の配列をコードしているアダプタをライゲーションすることによりさらに拡張された。前記アダプタは、標準の開始部位の-513に局在する第一の可能な代替のCTG開始コドンの10 塩基対上流までコードした。また、推定上の代替の開始部位をATGに変異させたアダプタを使用して、長い形態が効率的に発現されるだろう第二セットの発現構築物を作出した。さらに、標準の開始コドンのATAへの突然変異誘発も行い、トータルで4つの異なる構築物が生じた(図 5.1)。これらの4つのバリエーション(同様に、オリジナルのPTENのオープンリーディングフレーム)を、安定な発現のためMSCV(Clontech, Mountainview, CA)レトロウイルスベクターに感染によりサブクローンした。
【0096】
プロテアーゼプロテクションアッセイ ― HEK293 細胞をトリプシンなしで氷冷PBSに収集し、5x105の細胞のアリクウォット(aliquots)を0.5ug/ml〜10ug/mlでプロテイナーゼ Kの濃度を増加させて30 分間インキュベーションした。Triton 0.1%での対照を、示したタンパク質を分解するプロテイナーゼ Kの能力を検証するため含めた。反応を5mM PMSFで停止した。細胞を2x Laemmliサンプル緩衝剤(125nM Tris pH 6.8, 20% グリセロール, 0.05% ブロモフェノールブルー, 4% SDS, 10% 2-メルカプトエタノール)で溶解し、PTEN, Akt および E カドヘリンに関して免疫ブロットした。
【0097】
マウスの肝臓からのPTEN精製 ― C57BL6 マウスの肝臓を液体窒素中で急速凍結(snap frozen)し、粉砕し、TNN緩衝剤(50mM Tris pH 7.4, 150mM NaCl, 0.5% NP-40, 5mM EDTA, 3% グリセロール, 1mM DTT, 1x 哺乳類のプロテアーゼカクテルインヒビター [Sigma])に再懸濁した。懸濁液を乳鉢でホモジナイズし、40,000RPMで4 度で1時間遠心分離した。上清を連続的に0.45 ミクロンおよび0.22ミクロンのフィルターで濾過した。セファクリル 200 サイズ排除カラム(Amersham)をTNNで事前に平衡化し、サンプルを0.3ml/hrの速度でアプライし、続いて緩衝液をアプライした。2mlのフラクションを収集し、低分子量のサンプルをプールし、事前に平衡化したHiTrap Hepain HPカラム(Amersham)にアプライした。カラムを三カラム容量のTNNで洗浄し、タンパク質を0.3M, 0.5M および 1M NaCl TNN溶液の段階的な 3x カラム容量で溶出した。0.5mlの増加分のフラクションを収集し、PTENに関して免疫ブロットした。
【0098】
培地からのPTENのヘパリン精製 ― HEK293細胞を15cmディッシュ中で10%FBS DMEMでコンフルエントまで成長させた。細胞をFBSなしの15mlのDMEMで一晩インキュベーションした。20 プレートから培地を収集し、0.45 ミクロンのフィルターを通して濾過した。1mlのヘパリン HP カラムを、AktaPrime (AmershamBioscience)を用いて4ml/minの流速で4℃でDMEMで平衡化した。馴化培地を、カラムに1ml/minで通して通過させた。カラムを10容量のBC200 (200mM Nacl, 50mMT Tris pH7.4, 1mM EDTA, 0.2% Triton X-100)で洗浄した。タンパク質を5mlの1M NaClで1ml/minで1mlのフラクションに溶出した。各フラクションのタンパク質濃度を280nmのODで決定した。各フラクションの半分を、トリクロロ酢酸を20%で沈殿させ、冷アセトンで洗浄し、減圧下で乾燥させた。タンパク質を20 ulのLaemmli溶解緩衝剤に再構成し、PTENおよびPTEN-longに対する抗体を用いて免疫ブロットした。
【0099】
血清からのPTEN精製 ― AB血漿からのヒト血清をSigmaから得た。1mlの血清を、0.45ミクロンのフィルターを通して濾過し、プロテインA/G アガロースを用いて1 時間インキュベーションして抗体をプレクリアした。ヘパリン-アガロースを、セファロースの対照と共に一晩プレクリアした血清でインキュベーションし、次の日にBC150 (150mM NaCl, 25mM Tris pH7.4, 1% NP-40, 0.25% Na デオキシコール酸, 1mM EDTA)で洗浄した。タンパク質をlaemmliサンプル緩衝剤で溶出し、PTENに関して又は重鎖の混入に関して二次のみで免疫ブロットした。
【0100】
コンカナバリンA プルダウン ― HEK293細胞をサブコンフルエンスでBC500 (500mM NaCl, 20mM Tris pH 7.4, 1% Triton X-100, 1mM MnCl2, 1mM CaCl2, 1x プロテアーゼインヒビターカクテル)で溶解した。細胞溶解産物を、遠心分離し、濾過した。プルダウンを、20 マイクロリッターのコンカナバリンAセファロース(Sigma)で1 時間、4℃で行った。樹脂を、BC500で洗浄し、タンパク質をLaemmliサンプル緩衝剤で溶出した。
【0101】
結果
PTEN mRNAは上流に代替の開始部位を有する
NCBIに寄託されたPTEN mRNA(Li and Sun 1997; Steck, Pershouse et al. 1997)は、広範囲にわたる5'UTRを含む。5'UTR領域中の凡そ770bpの連続配列(contiguous sequence)は、開始コドンとインフレームである。メチオニンは、この領域でコードされない; しかしながら、標準の開始コドンから-519で始まる幾つかの代替の開始CUGコドンが存在する。この配列の翻訳から、scansite (scansite.mit.edu) および prosite (www.ebi.ac.uk/ppsearch)にしたがって同定可能なドメインは明らかとされなかった。この全体領域の翻訳によって、173アミノ酸がPTENに加えられ、その分子量を凡そ70 キロダルトンに増加させるだろう(図 2 および 配列番号1)。
【0102】
他のPTENオルソログのアラインメントによって、ヒト UTRの翻訳配列が様々な種のPTENのオープンリーディングフレームに認められることが明らかとされた。チンパンジー, ウシ, ミツバチおよび線虫は、全てヒト 5'UTRの翻訳産物と相同なタンパク質配列を含む(図 3)。さらにまた、ヒト 5'UTR および マウス PTEN 5'UTRのアラインメントは、広範囲にわたりヌクレオチド相同性を示した(示さず)。マウス 5'UTRは522 塩基対にわたり標準の開始コドンとインフレームで翻訳され、ヒト 5'UTRの翻訳と比較した場合に高度に相同なタンパク質配列であることが明らかとされた(図 5.3)。5'UTRの相同性や他の種の翻訳されたタンパク質にヒト 5'UTRから由来するアミノ酸配列が実際に存在することは、この配列が進化上の重要であることを示している。
【0103】
PTEN mRNAは、代替の上流部位から翻訳を開始できる。PTEN ORFの過剰発現は、55kDaの単一のタンパク質バンドを生じた。5'UTRを含めることによって、約70kDaの二番目のより大きいタンパク質バンドを生じた。PTEN免疫ブロットにおけるより大きなタンパク質バンドは、幾つかの細胞株にも内因性に存在し、異なるモノクローナル抗体により検出可能であった(図 4)。この大きいタンパク質バンドは、マウスの野生型 ES 細胞に存在し、PTENノックアウトマウスのES細胞には存在せず、PTEN shRNAを安定に発現しているマウス ES クローンにおいて減少した(図 4)。また、HEK293細胞においてヒト PTEN タンパク質のsiRNAを用いたノックダウンによって、70 kDaタンパク質のノックダウンを生じる。
【0104】
PTENヌル PC3 細胞株においてPTENのORFをコードしているプラスミドの発現は、55kDaタンパク質の産生を生じる。5'UTRもコードするプラスミドが過剰発現された場合、70kDaタンパク質が産生された。CTGからATGへの上流の推定上の開始コドンの変異(図 1, 「ATG/ATG」)は、優勢に免疫ブロットパターンを70 kDaの形態にシフトした(図 4)。また、55kDaのバンドは、突然変異誘発によりATG開始コドンに由来することが確認された(図 4 「CTG/ATA」)。
【0105】
従って、5' UTRは、長い形態のPTENの翻訳を開始するため十分であった。従って、5' UTRはAlternately Translated Regionのため5'ATRと命名され、検出された大きいタンパク質は「PTEN-long」と命名された。
【0106】
親和性精製したポリクローナル抗体は、5'ATRから翻訳されたアミノ酸に対し産生され、過剰発現研究において組換え型のPTEN-longの産生、またHEK293 細胞における内因性の形態を確認するため使用された(図 4)。HEK293 細胞の全細胞溶解産物の免疫ブロットおよびPC3 細胞における過剰発現研究から、複数の形態のPTEN-longが認められ、潜在的な翻訳後修飾, 記載されていないスプライス形態(undocumented splice forms)または5'ATRにおける代替の開始コドンのいずれかを指摘している。
【0107】
PTEN-longはN末端シグナルペプチドをコードする
PTEN-longのN末端配列は、膜貫通配列またはシグナルペプチドのいずれかの指標であろうアラニンの長いストレッチ(a long stretch)を含む。SignalIP 3.0を用いた翻訳配列の分析によって、高い確率(>95%)で前記配列がシグナルペプチドを含むことが予測された(図 5)。シグナルペプチドは、特徴的に塩基性アミノ酸と続く疎水性のストレッチとから構成される。推定上の疎水性の膜貫通ヘリックスは、プロリンにより破壊され、やや極性の配列が続く。また、配列が切断されることも予測され、このタンパク質がERの内腔に放出されることを指摘している。
【0108】
PTENはグリコシル化される
分泌された細胞外のタンパク質の特徴の一つは、ゴルジ体における複合糖部分(complex sugar moieties)の付加である(グリコシル化として知られるプロセス)。糖はN-グリコシル化を介して共通配列 N-X-S/T (Xはプロリンではない)においてアスパラギンに付加できる(Gupta and Brunak 2002); またセリン, スレオニンおよびチロシンのヒドロキシル基はO-グリコシル化の標的となりえる(Julenius, Molgaard et al. 2005)。PTENは、複数のO-グリコシル化部位を有するが、しかし、僅か一つのN-グリコシル化部位を有する。HEK293 細胞中でPTEN相補物(PTEN complement)の部分がグリコシル化されたかどうかを決定するプルダウンアッセイにレクチン コンカナバリン-A(糖部分と結合する)を使用した。凡そ50%のPTEN-longのPTENの混合物をこれらの細胞から精製した(図 6)。この事項は、PTEN-longがグリコシル化されること、またPTENの細胞質の55kDa形態がグリコシル化されるか又はPTEN-longが細胞外で切断されることを示す。
【0109】
PTEN-longはヘパランと結合し、細胞表面に認められる
PTENは幾つかのプロテオグリカン(例えば、syndecans および glypicans)と結合し、膜の外側のリーフレット(leaflet)に付着することが認められる。これらのプロテオグリカンは、最もヘパラン化した細胞外分子のうちの二つである(Blero, Zhang et al. 2005)。PTENは以前に高度に陰性に荷電した種に高い親和性を有することが示され、PTENの特性により高度に陰イオン性のPIP3を好むことが導かれる(Das, Dixon et al. 2003)。ヘパランは最も陰性に荷電した生物学的分子(biological molecules)の一つであるので、ヘパランがPTENの細胞外基質への結合を媒介することが可能であった。マウスの肝臓からのタンパク質抽出物を用いて、PTENがヘパランと高親和性に結合することが発見された。さらにまた、ヘパリン アガロース カラムから1M NaClを用いたPTENの連続的な溶出によって、PTEN-longも溶出された(図7)。
【0110】
細胞膜の外面に突き出たPTEN-longは、プロテアーゼのデグラデーションに感受性である。プロテアーゼプロテクションアッセイにおいて、生細胞をプロテアーゼとインキュベーションし、細胞外のタンパク質のみが分解された(というのも、脂質膜はプロテアーゼを不透過性であり、全ての細胞内タンパク質の防御に貢献するからである)。HEK293細胞を付着培養からPBSで穏やかに撹拌して除去し、プロテイナーゼ Kの濃度を増加させて懸濁した。反応をPMSFで停止し、細胞をlaemlli 緩衝剤で溶解した。PTEN-longは、既知の細胞外タンパク質であるE-カドヘリンと共にプロテイナーゼ Kでの処理に感受性を示した(図 8)。他方でPTENは中等度のプロテアーゼ感受性を示し、55kDa種のいくつかも細胞外(グリコシル化されるので)であるか又は幾らかの細胞溶解がアッセイの間に発生し、これにより細胞質のPTENがプロテイナーゼ Kに暴露されることを示している。プロテイナーゼ Kに暴露された場合にPTENが分解されることを証明するため、膜を透過するtritonでの対照を含めた。これがPTEN本来(PTEN proper)であるか又は55kDaに移動し、PTEN抗体のC末端エピトープを保持するPTEN-longの切断形態であるかどうかはまだ分からない。このデータは、PTEN-longが細胞表面にあることを指摘している。
【0111】
可溶性のPTEN-longは培地に分泌される
タンパク質の部分が可溶性であり、細胞環境に放出される可能性。
【0112】
ヘパリンセファロースを、HEK293細胞で馴化した血清フリー培地からPTEN-longを親和性精製するため使用した。カラムの溶出によって、培地中に50kDaの分子量に移動するPTENの存在が明らかとされた(図 9)。PTEN-long 特異的な抗体での免疫ブロットは同じ50kDa種を明らかとし、このタンパク質が代替の開始部位から翻訳される配列およびPTEN モノクローナル抗体のC末端エピトープの配列を保持することを示している。この事項は、55kDaであることが観察されたPTENの部分が実際に上流の開始コドンに由来する切断された翻訳産物であることを暗示する。
【0113】
培地へのPTENの分泌は、さらにATG/ATG 構築物でトランスフェクトされたHEK293細胞においてPTEN-longを過剰発現することにより確認された。これらの細胞を使用して、血清フリーの馴化培地で一晩産生させた。PTEN モノクローナル抗体 6H2.1を使用して、1mlの培地からPTENを免疫沈降した。大きいPTENバンドが、より低い55kDaバンドと共に培地から良好に免疫沈降された。前記タンパク質が過剰発現されたので、前記タンパク質の適切なプロセシングはおそらく生じず、完全なサイズの70kDa PTENの分泌が生じる。
【0114】
PTENはヒト血清中に認められる。
【0115】
生理的な分泌物の最良の供給源の一つは血清である。ヘパリンセファロースを使用してヒト血清からPTENを親和性精製した。ヒト血清を遠心分離し、濾過して粒子性の物質を除去した。次にBC150に1:5に希釈し、頻繁にプロテインA/GでプレクリアしてIgGを除去した。血清を少量のヘパリンセファロースでバッチインキュベーションした。ヘパリン セファロースをlaemmli 緩衝剤で溶出し、溶出液をPTENに関してブロットし、また重鎖の混入を排除するため二次抗体単独でブロットした。PTEN および PTEN-longは、双方ヒト血清に認められた(図 10)。
【0116】
PTEN-longの抗-血管形成活性
PTEN-longの抗-血管形成の役目を以下に示す:
(1) PTEN-longは通常はマウスの発生している網膜において弱く発現するが、新生児の発生の間で退行(involution)/細胞死を経験している血管に高レベルの発現が認められる(図 11);
(2) PTEN-longは、腫瘍中のアポトーシスしている血管に認められる。さらにまた、PTEN-longで処理された上皮細胞(トランスフェクトされた細胞から部分的に精製された)は、細胞の遊走を阻害し、アポトーシスを誘導した。(図12)。精製したPTEN-longは、カスパーゼ-3 切断により測定されたとおり培養においてU87, HUVEC 内皮細胞, または293細胞のアポトーシスの活性化と関連する細胞死も誘導できる。
【0117】
PTEN-longのインビボでの抗-腫瘍および抗-血管形成活性
図 13は、PTEN-longでのマウスの処理を示す、 (A) マウス (n=5)にグリオブラストーマ細胞株U87が注射され、異種移植片が乳房の脂肪パッドの2部位(左および右)に形成された。腫瘍移植後、一つの腫瘍にPTEN-longを直接注射し、反対側の腫瘍には注射されなかった(w/ PTEN-long)。また、5 マウスの対照セットは、空ベクターでトランスフェクトされた細胞から由来するmockの精製タンパク質の調製物を注射された(空ベクター)。再び、反対側の腫瘍は注射されなかった(w/空ベクター)。マウスを1-11日および13-14日に処理した。最大の直径(cm)を、示した日にノギスで測定した。腫瘍容量が >1 cmに達した時、マウスを屠殺した。(B) タンパク質をPTEN-long発現ベクターを293細胞にトランスフェクションすることにより調製し、V5親和性樹脂を用いて部分的に精製し、V5 ペプチドで溶出した。図14は、14日間の対照注射で処理したマウスの生存率を示す(日数)。
【0118】
網膜染色
p7のマウス網膜中のPTEN-longおよび血管の染色によって、PTEN-longがこのポイントでマウスの網膜の血管発生において最初に退行(regress)する硝子体管(hyaloid vessels)を選択的に染色することが明らかとされた。PTEN-longに対する抗体は、エピトープ: N-PRHQQLLPSLSSFFFSHRLPD-C (配列番号3)に対するものである。管(Vessel)の染色をBS1-レクチンで行った。
【0119】
精製
PTEN-longの精製のための一つの方法において、293 細胞をATG/ATG PTEN-longでトランスフェクトし、細胞溶解産物をNi+ アフィニティーカラムに通過させた。PTEN-longを、AKTA PurifierでNi+ カラムでイミダゾール溶出緩衝剤を用いて一貫して(consistently)精製した。
【0120】
腫瘍退行
PTEN (orf 403 アミノ酸)またはPTEN-longのいずれかで注射前にトランスフェクトされたU87 細胞の異種移植片。注射後の7 日間、PTENを過剰発現している同齢集団(n= 4のうち4)と比較してPTEN-longを過剰発現している同齢集団において乳房血管の減少が認められた。これはPTEN-longが腫瘍環境に影響しえることを示唆する。
【0121】
考察
細胞溶解産物や組織からのPTEN免疫ブロットにおいて二番目のより大きいタンパク質バンドを定期的に観察した。より大きなバンドがPTENであることが確認される証拠には、次の事項が含まれる:
大きなタンパク質バンドは、異なるPTENモノクローナル抗体により検出された;
細胞がPTENに対するsiRNAで処理される又はPTEN座位がマウスにおいてノックアウトされる場合、大きなタンパク質は存在しない。PTENの5'UTRは、PTENの古典的な開始コドンと700 塩基対以上にわたりインフレームであることが観察された。さらに、PTENの522 塩基対上流のCUGコドンが存在し、翻訳される場合にPTEN免疫ブロットにおける大きなタンパク質のバンドのサイズを説明することができた。強いコザック配列とは関連しないが、-1 シトシン および +1 グアノシン配列が保持される。翻訳され、PTEN ORFに付加される場合、凡そ70kDaのタンパク質が産生されるはずであり、その分子量は観察された大きなPTENバンドの分子量である。
【0122】
この配列の翻訳は、実際のコード配列内の幾つかのPTENオルソログに既に存在する。マウスの組織溶解産物において類似するバンドが観察されたので、マウス 5'UTRも点検した。マウスの5'UTR ヌクレオチド配列は、ヒトの5'UTRと高度に相同的であり、同様に開始コドンとインフレームである。二つの潜在的な代替の開始コドンが-522 および -516に存在し、これらの部位からのこの配列の翻訳はヒト配列と90%+ 相同的なアミノ酸配列を示す。この推定上のタンパク質の保存は著しく、この配列の進化上の重要性が示された。この配列をよりよく記載するため、PTENの5'ATRまたは代替の翻訳領域と改名して翻訳の潜在性を記載した。
【0123】
PTENのオープンリーディングフレームを5'ATRと共にクローン化してプラスミドを構築し、この組換え型のPTENの発現を標準の403のアミノ酸を産生するオープンリーディングフレーム単独と比較した。5'ATRを含めることにより二番目の大きなPTENタンパク質バンドが生じ、このバンドは55kDaに移動する単一のバンドを作るPTENのまさに標準のORFを含んでいる発現プラスミドと比べて約70kDaに移動する。大きなタンパク質は、翻訳されたトータルのタンパク質の僅か小部分に勘定される; しかしながら、推定上の開始部位のATGへの変異によって、タンパク質比は主に大きな形態にシフトした。
【0124】
5'ATRのタンパク質配列の保存は、進化のアーティファクト以上であることを示す。N-末端は脂肪族アミノ酸のストレッチを含み、これが膜貫通配列であることが予測された。PrositeおよびSignal 3.0IPの使用によって、PTEN-longのN-末端がシグナルペプチドであり、それに直接続くプロテアーゼ切断部位を有することが予測された。
【0125】
PTEN-longが細胞の細胞外表面に局在するかどうかをインビボでのプロテアーゼプロテクションアッセイを使用して検査した。PTEN-longは細胞外のプロテアーゼの量を増加させることにより進行性のデグラデーションを示したが、PTENは示さなかった。この結果は、少なくとも幾らかのPTEN-longが細胞外にあり、少なくとも部分的に細胞膜の外側のリーフレットに付着することを示している。この結果は最も魅力的な結果であり、細胞膜の外側のリーフレットにおける活性リピッドホスファターゼの意味が提起された。外膜結合性のプロテオグリカンの二つのファミリー(glypicans および syndecans)は、以前にPTENタンパク質複合体中に同定された。
【0126】
細胞表面におけるPTENの存在は、可溶性の分泌されるPTENの可能性を排除しない。Syndecans および glypicansは、最も多量にヘパラン化したプロテオグリカンのうちの二つである。ヘパランは高度に陰性に荷電したグリコサミノグリカンであり、またPTENは陰イオンへの親和性を有することが示されており、高度に陰性に荷電したPIP3を基質として選択することを部分的に説明している。マウス肝臓からのPTEN精製を最適化する実験によって、PTEN および PTEN-longの双方がヘパリン セファロース カラムを用いて精製できることが明らかとされた。さらにまた、約50kDaのタンパク質を、HEK293 細胞で馴化した血清フリー培地からヘパリンセファロースカラムを用いて精製した。精製されたタンパク質は、PTENに特異的なモノクローナル抗体およびPTEN-longに存在するユニークなアミノ酸残基に対するポリクローナル抗体により認識された。前に、PTEN-long抗体は、70kDa付近のタンパク質バンドのみを認識した。双方の抗体が一つのバンドを認識できたとの観察によって、タンパク分解プロセシングがおそらく発生すること、免疫ブロットで観察されたタンパク質が双方の抗体のエピトープを保持するPTENの断片であることが示される。また、PTEN および PTEN-longの双方は、ヒト血清からヘパリン親和性精製を用いて精製できた。
【0127】
過去10年の文献によって、PTENの配列の想定(assuming)が蓄積されてきた。代替の部位から翻訳され、外側のリーフレットや細胞外間隙の双方に分泌されるPTENの新規の形態が存在するとの証拠を本出願は提供する。
【0128】
インビボの結果は、PTEN-longがヒト血清に通常存在する新規の抗-腫瘍化合物であり、抗-血管形成やアポトーシス促進特性を有することを示す。
【0129】
[参照文献]
Acland, P., M. Dixon, et al. (1990). "Subcellular fate of the int-2 oncoprotein is determined by choice of initiation codon." Nature 343(6259): 662-5.
Alberts, B. (2002). Molecular biology of the cell. New York, Garland Science.
Baker, S. J. (2007). "PTEN enters the nuclear age." Cell 128(1): 25-8.
Blero, D., J. Zhang, et al. (2005). "Phosphatidylinositol 3,4,5-trisphosphate modulation in SHIP2-deficient mouse embryonic fibroblasts." Febs J 272(10): 2512-22.
Blobel, G., P. Walter, et al. (1979). "Translocation of proteins across membranes: the signal hypothesis and beyond." Symp Soc Exp Biol 33: 9-36.
Bonneau, D. and M. Longy (2000). "Mutations of the human PTEN gene." Hum Mutat 16(2): 109-22.
Di Cristofano, A., B. Pesce, et al. (1998). "Pten is essential for embryonic development and tumour suppression." Nat Genet 19(4): 348-55.
Eng, C. (2003). "PTEN: one gene, many syndromes." Hum Mutat 22(3): 183-98.
Florkiewicz, R. Z. and A. Sommer (1989). "Human basic fibroblast growth factor gene encodes four polypeptides: three initiate translation from non-AUG codons." Proc Natl Acad Sci U S A 86(11): 3978-81.
Fraser, M. M., X. Zhu, et al. (2004). "Pten loss causes hypertrophy and increased proliferation of astrocytes in vivo." Cancer Res 64(21): 7773-9.
Gupta, R. and S. Brunak (2002). "Prediction of glycosylation across the human proteome and the correlation to protein function." Pac Symp Biocomput: 310-22.Hann, S. R. (1994). "Regulation and function of non-AUG-initiated proto-oncogenes." Biochimie 76(9): 880-6.
Hann, S. R., M. Dixit, et al. (1994). "The alternatively initiated c-Myc proteins differentially regulate transcription through a noncanonical DNA-binding site." Genes Dev 8(20): 2441-52.
Hann, S. R. and R. N. Eisenman (1984). "Proteins encoded by the human c-myc oncogene: differential expression in neoplastic cells." Mol Cell Biol 4(11): 2486-97.
Hann, S. R., M. W. King, et al. (1988). "A non-AUG translational initiation in c-myc exon 1 generates an N-terminally distinct protein whose synthesis is disrupted in Burkitt's lymphomas." Cell 52(2): 185-95.
Hann, S. R., K. Sloan-Brown, et al. (1992). "Translational activation of the non-AUG-initiated c-myc 1 protein at high cell densities due to methionine deprivation." Genes Dev 6(7): 1229-40.
Hershey, J. W. (1991). "Translational control in mammalian cells." Annu Rev Biochem 60: 717-55.
Julenius, K., A. Molgaard, et al. (2005). "Prediction, conservation analysis, and structural characterization of mammalian mucin-type O-glycosylation sites." Glycobiology 15(2): 153-64.
Kiefer, P., P. Acland, et al. (1994). "Competition between nuclear localization and secretory signals determines the subcellular fate of a single CUG-initiated form of FGF3." Embo J 13(17): 4126-36.
Kozak, M. (1989). "Context effects and inefficient initiation at non-AUG codons in eucaryotic cell-free translation systems." Mol Cell Biol 9(11): 5073-80.
Kozak, M. (1990). "Downstream secondary structure facilitates recognition of initiator codons by eukaryotic ribosomes." Proc Natl Acad Sci U S A 87(21): 8301-5.
Kozak, M. (1991). "An analysis of vertebrate mRNA sequences: intimations of translational control." J Cell Biol 115(4): 887-903.
Kwabi-Addo, B., D. Giri, et al. (2001). "Haploinsufficiency of the Pten tumor suppressor gene promotes prostate cancer progression." Proc Natl Acad Sci U S A 98(20): 11563-8.
Lee, J. O., H. Yang, et al. (1999). "Crystal structure of the PTEN tumor suppressor: implications for its phosphoinositide phosphatase activity and membrane association." Cell 99(3): 323-34.
Li, J., L. Simpson, et al. (1998). "The PTEN/MMAC1 tumor suppressor induces cell death that is rescued by the AKT/protein kinase B oncogene." Cancer Res 58(24): 5667-72.
Maehama, T. and J. E. Dixon (1998). "The tumor suppressor, PTEN/MMAC1, dephosphorylates the lipid second messenger, phosphatidylinositol 3,4,5-trisphosphate." J Biol Chem 273(22): 13375-8.
Petrocelli, T. and J. M. Slingerland (2001). "PTEN deficiency: a role in mammary carcinogenesis." Breast Cancer Res 3(6): 356-60.
Prats, H., M. Kaghad, et al. (1989). "High molecular mass forms of basic fibroblast growth factor are initiated by alternative CUG codons." Proc Natl Acad Sci U S A 86(6): 1836-40.
Sulis, M. L. and R. Parsons (2003). "PTEN: from pathology to biology." Trends Cell Biol 13(9): 478-83.
Taira, M., T. Iizasa, et al. (1990). "A human testis-specific mRNA for phosphoribosylpyrophosphate synthetase that initiates from a non-AUG codon." J Biol Chem 265(27): 16491-7.
Torres, J. and R. Pulido (2001). "The tumor suppressor PTEN is phosphorylated by the protein kinase CK2 at its C terminus. Implications for PTEN stability to proteasome-mediated degradation." J Biol Chem 276(2): 993-8.
Vazquez, F., S. R. Grossman, et al. (2001). "Phosphorylation of the PTEN tail acts as an inhibitory switch by preventing its recruitment into a protein complex." J Biol Chem 276(52): 48627-30.
Vazquez, F., S. Ramaswamy, et al. (2000). "Phosphorylation of the PTEN tail regulates protein stability and function." Mol Cell Biol 20(14): 5010-8.
Wang, X., L. C. Trotman, et al. (2007). "NEDD4-1 is a proto-oncogenic ubiquitin ligase for PTEN." Cell 128(1): 129-39.
Xiao, J. H., I. Davidson, et al. (1991). "Cloning, expression, and transcriptional properties of the human enhancer factor TEF-1." Cell 65(4): 551-68.
You, M. J., D. H. Castrillon, et al. (2002). "Genetic analysis of Pten and Ink4a/Arf interactions in the suppression of tumorigenesis in mice." Proc Natl Acad Sci U S A 99(3): 1455-60
【発明の説明】
【0001】
本出願は、2009年2月17日に出願された米国仮出願番号61/207,974の優先権を主張するものであり、その内容は本明細書中に参照により援用される。
【0002】
本出願に開示された研究は、国立癌研究所(National Cancer Institute)からの助成金番号CA082783による政府のサポートによりなされた。従って、米国政府は、本発明に特定の権利を有する。
【0003】
本出願の全体において、様々な文献が括弧内の著者名及び西暦年により参照される。これら文献の完全なる書誌的事項は、本明細書の「参照文献」に記載される。これら公開の開示の全体は、本明細書中に引用することにより本出願に援用され、当該発明が属する技術の状況をより完全に記載する。
【0004】
[背景]
PTEN 腫瘍抑制因子(WO98/34624を参照されたい、この公報は本明細書中にその全体が参照によって援用される)は、重要な二次メッセンジャー ホスファチジルイノシトール 3,4,5-三リン酸を脱リン酸化する細胞質ホスファターゼである(Maehama and Dixon 1998)。この活性は、抗-アポトーシス経路, 細胞周期の進行および細胞代謝の増加を含むAktのPIP3活性化により開始される多くの腫瘍形成性のシグナルをダウンレギュレートする(Sulis and Parsons 2003)。癌におけるPTENの役割は、多くの異なる腫瘍タイプにおいて遺伝的な又は機能的な頻繁な欠損から明らかである(Bonneau and Longy 2000)。最初にグリア癌(glial cancers)において欠失していることが発見され、以後は前立腺, 乳房, 子宮内膜, メラノサイト, 腎臓 および 肺の腫瘍形成と関係づけられてきた。また、PTENの生殖系列の変異は、Cowden's 症候群などの遺伝性の癌素因症候群(cancer predisposition syndromes)とも関連した(Eng 2003)。PTEN欠損のマウスモデルは、多くの異なる組織タイプにおいてヘテロ接合性マウスおよび組織特異的ノックアウトの双方で腫瘍抑制因子としての役割を再現した(Di Cristofano, Pesce et al. 1998; Kwabi-Addo, Giri et al. 2001; Petrocelli and Slingerland 2001; You, Castrillon et al. 2002; Fraser, Zhu et al. 2004)。
【0005】
PTENタンパク質は、N末端の二重特異性ホスファターゼドメイン, およびC末端のC2 リン脂質結合ドメイン, 続いて調節に重要(リン酸化部位が内部に見いだされるので)な定まった形を持たない尾部(unstructured tail)を含む(Lee, Yang et al. 1999; Vazquez, Ramaswamy et al. 2000; Torres and Pulido 2001; Vazquez, Grossman et al. 2001)。PTEN タンパク質は、大抵は細胞質性である。しかしながら、核にPTENが存在するとの証拠が増加しており、その局在化はNEDD4-1によるタンパク質のモノユビキチン化により制御される(Baker 2007; Wang, Trotman et al. 2007)。
【0006】
5'UTRのリボソームスキャニングは、開始コドン AUGでの翻訳開始に先行する。リボソームが適切な開始コドンを決定する実際の手段の理解はいまだに不完全であるが、mRNA自身および開始前複合体(pre-initiation complex)がどこでそのスキャニングを緩徐にし、翻訳を開始させるかを指令する配列の双方の特定の特性が存在する。古典的な「コザック配列」CCACCATGG(下線で示したATGが開始コドンである)は、開始に関して最も好都合な配列であることが示されている(Kozak 1991)。また、mRNAの二次構造は、おそらくヘリカーゼを必要とし、リーディングの前に二次構造を融解する開始前複合体のスキャニングを事実上緩徐化することにより開始を促進する(Kozak 1990)。
【0007】
特定の転写物において、翻訳開始は、非-AUGコドンから生じえる。通常、これはほんの僅かなパーセンテージの転写物から翻訳されたトータルタンパク質を含み、結果はN-端で変化する混合種のタンパク質である。Kozakは、非-AUG コドンからの翻訳の開始の効率を明らかし、GUG および CUGの双方がインビトロで翻訳を開始する能力があるが、効率的が非常に低いことを見出した(Kozak 1989)。さらなる研究によって、メチオニンの利用可能性によりいまだに不明確である機構をかいして翻訳開始の乱雑性(promiscuity)を変化させえることが示され、おそらくeIF2(43S 開始前複合体のコンポーネント)の栄養分感受性キナーゼ(nutrient sensitive kinase)によるリン酸化が関与することが示された(Hershey 1991; Hann 1994)。
【0008】
幾つかのタンパク質は、代替の開始コドンから翻訳されることが示された。転写因子 c-mycは代替の上流 CUG 開始コドンを有し、翻訳された場合に14 アミノ酸がタンパク質のN-末端に付加される(Hann and Eisenman 1984)。この代替のアイソフォームが、選択的にバーキットリンパ腫において破壊されたことが示された(Hann, King et al. 1988)。組織培養において、メチオニンが低濃度である場合、長い形態のmycが高い細胞密度で優先的に転写される(Hann, Sloan-Brown et al. 1992)。さらなる研究によって、長い形態のc-mycは成長阻害性であり、古典的なc-myc タンパク質と異なるセットの転写標的を有することが明らかとされた(Hann, Dixit et al. 1994). (Florkiewicz and Sommer 1989) (Prats, Kaghad et al. 1989)。
【0009】
さらに、タンパク質の実際の細胞内局在が代替の開始コドンにより決定できることが知られている。マウス 癌原遺伝子 int-2の場合、上流 CUG コドンからの代替の開始は核局在化をコードするが、AUG コドンは分泌性経路に局在化するためのシグナルペプチドをコードする(Acland, Dixon et al. 1990)。類似する現象はヒト FGF3において記載され、AUGから翻訳されたタンパク質は分泌経路に向かうが、上流 CUGから翻訳されたタンパク質は核に局在する(Kiefer, Acland et al. 1994)。さらにまた、幾つかの真核生物のタンパク質(例えば、TEF-1 および PRPS-3)においては、タンパク質は完全にCUGコドンから開始される(Taira, Iizasa et al. 1990; Xiao, Davidson et al. 1991)。
【0010】
分泌に向かうタンパク質は、シグナルペプチドと称される疎水性アミノ酸のストレッチ(a stretch )により小胞体を標的とする(Blobel, Walter et al. 1979)。通常、タンパク質のN-端に認められるシグナルペプチドは、翻訳の際にシグナル識別粒子 (SRP)に結合し、リボソームの停止を生じ、粗面小胞体にトランスロケーションし、ここでSRPレセプターと結合する。一旦リボソームがドッキングしたら、SRP-SRP レセプター複合体が放出され、翻訳がSec61トランスロコンを通りERの内腔を通り再開される。次に、シグナルペプチドが可溶性タンパク質のケースにおいては切断され、Secトランスロコンからタンパク質が放出される。膜を貫通するタンパク質の場合、膜貫通ヘリックスは、ER トランスロケーションのためのシグナルペプチドとして貢献する。これらのタンパク質は、ゴルジにおけるグリコシル化により広範に修飾され、分泌ベシクル中で形質膜にシャトルされる(Alberts 2002)。
【0011】
癌において重要であることが示された幾つかの分泌タンパク質が存在する。例えば、Wnt シグナル伝達経路は、肺癌において変更されることが示された。Wntは、Frizzled レセプターのファミリーに対する分泌型リガンドである。FrizzledのWnt 活性化により乱れを生じ、β-カテニン デグラデーション 複合体(APCを含む)が解離を生じ、これによりβ-カテニンのレベルが上昇し、そして核にトランスロケーションし、核でTCF 転写因子と相互作用し、トランス活性化(transactivate)が許容される。遺伝性および散発性(sporadic)の双方の結腸癌でAPCにおける変異を不活性化すること、またβ-カテニンにおける変異を活性化することが詳細に言及されている。さらに、幾つかの細胞外のリガンドアンタゴニスト(例えば、SFRP および Wnt-5a)は、Wntと同じFrizzled レセプターと競合する。双方との腫瘍抑制因子であることが示された; SFRP ノックアウトマウスは、リンパ様腫瘍を発生し、Wnt-5aのエピジェネティック サイレンシングがメラノーマにおいて検出された。
【0012】
PTENの全ての公開された報告は、このタンパク質が細胞質または核の何れかに局在することを指摘する。幾つかの細胞外タンパク質(glypicans および syndecans)は、分泌経路に関与する幾つかのタンパク質(reticulocalbinおよびcalumenin)のようにPTENと結合することが見出された。PTENが分泌経路に進入して、係る相互作用が許容されることが想定された。実際、本出願で開示されたとおり、新規の異なって翻訳されるタンパク質が存在し、このタンパク質はPTEN-longと命名され、N末端シグナルペプチドを含み、細胞外に分泌される。
【0013】
[発明の概要]
配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいる単離されたヒトのホスファターゼおよびtensin ホモログ ロング ポリペプチド(PTEN-long)または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログ,またはその各々のバリアント(variant)。
【0014】
(i) 配列番号1の残基 1〜173,または(ii) 配列番号5の残基 1〜173を含んでいるそのアナログ,または(iii) 配列番号1の残基 22〜516を含むポリペプチド。
【0015】
配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22 〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを含む薬学的組成物。
【0016】
対象における固形腫瘍(solid tumor)を治療するための方法であって、対象における固形腫瘍を治療するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【0017】
対象における固形腫瘍の成長を阻害するための方法であって、対象における固形腫瘍の成長を阻害するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【0018】
対象の固形腫瘍における血管形成を阻害するための方法であって、対象の固形腫瘍における血管形成を阻害するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【0019】
対象の固形腫瘍における血管の血管上皮細胞(vascular epithelial cell)のアポトーシスを誘導するための方法であって、対象の固形腫瘍における血管の血管上皮細胞のアポトーシスを誘導するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【0020】
対象における固形腫瘍を治療するための方法であって、配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5を含んでいるそのアナログをコードする特定量の発現ベクターを、PTEN-long, PTEN-long 断片,又はそのアナログを前記固形腫瘍の細胞中で対象における固形腫瘍を治療するため有効な量で発現するように前記対象に投与することを含む方法。
【0021】
対象における固形腫瘍の治療に使用するための、対象における固形腫瘍の成長を阻害するための、対象の固形腫瘍における血管上皮細胞のアポトーシスを誘導するための、または対象の固形腫瘍における血管形成を阻害するための、配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long)または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログ。
【0022】
それぞれPTEN-long, PTEN-long 断片,又はそのアナログの血漿半減期を増加させる非-ペプチド因子に結合させた、配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログを含む組成物。
【0023】
配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long)をコードする,または配列番号1の残基 22〜516を含んでいるその断片をコードする,または配列番号5を含んでいるそのアナログをコードする単離された核酸。
【0024】
配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログをコードしている核酸を含む発現ベクター。
【0025】
配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログを発現する能力のある形質転換された細胞であって、前記細胞はそのゲノムにPTEN-longまたはそのアナログをコードする組換えDNAが統合される形質転換された細胞。
【0026】
配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログを発現する能力のある宿主細胞であって、PTEN-longまたはそのアナログをコードするプラスミドを含む宿主細胞。
【0027】
それぞれPTEN-long, PTEN-long 断片,又はそのアナログの血漿半減期を増加させる非-ペプチド因子に結合させた配列番号1を含んでいるPTEN-long,または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5を含んでいるそのアナログを作るため、(1) 配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5を含んでいるそのアナログ, および(2) PTEN-long又はそのアナログの血漿半減期を増加させる非-ペプチド因子を混合(admixing)することを含む方法。
【0028】
配列番号2または配列番号6の核酸配列または配列番号 2または配列番号6の核酸配列の相補物と,または配列番号2 配列番号6とストリンジェントな条件下で特異的にハイブリダイズするその相補物の少なくとも20ヌクレオチド断片からなる単離された核酸分子。
【0029】
単離された抗体,又はその断片であって、(1) 配列番号1のアミノ酸残基 1-173を含むPTEN-ロング ポリペプチドまたは(2) 配列番号5を含んでいるそのアナログまたは(3) 配列番号3で表される配列を有しているアミノ酸または(4) 配列番号1の残基 22〜516を含んでいるPTEN-longの断片と結合する抗体又はその断片。
【0030】
単離された抗体又はその断片であって、前記抗体又はその断片はPTEN-longのコンホメーションエピトープと結合し、前記PTEN-longは配列番号1の残基 1-173で表される配列を含むが、しかし、前記抗体はPTENとは結合しない。
【0031】
単離された抗体又はその断片であって、前記抗体又はその断片はPTEN-longのエピトープと結合し、前記PTEN-longは配列番号1の残基 1-173で表される配列を含むが、しかし、前記抗体はPTENとは結合しない。
【0032】
配列番号1,または配列番号5のペプチド断片であって、抗-腫瘍, 抗-血管形成性(anti-angiogenic),または抗-アポトーシス活性を有する断片。
【図面の簡単な説明】
【0033】
【図1】PTEN-long 構築物の概略図。内因性の開始部位または代替の開始部位の何れかからPTENの発現を駆動するため作出された組み合わせを示している発現構築物。標準(Canonical)のPTENは黒色で示され、UTRにおける翻訳領域は灰色で示される。
【図2】ヒト PTEN mRNAの概略図。PTEN mRNAは、示された標準のATG開始コドンとインフレームの上流の173 アミノ酸をコードする。翻訳は、標準のATGから上流でヌクレオチド -519のCTGで開始される。拡大されたものは、配列番号1の残基 1-173に示される。
【図3】PTEN オルソログのN-端のアラインメント。示した種のPTEN タンパク質配列をVector NTI (Invitrogen)でBLOSUM62 スコア マトリックスを用いて整列させた。ヒト および マウスの双方の拡張したN末端配列(アステリクス)は、標準のAUG開始コドンから-519 (H. sapiens) および -520 (M. musculus)でのCUG 代替開始コドン用いて公開されたmRNAからORFinder (NCBI)を用いて翻訳された。ヒト (NM_000314) および マウス (NM_008960)のmRNA 配列。ミツバチ(Apis mellifera)の配列は、ベイラー医科大学のミツバチゲノムプロジェクトから得られた。線虫(Caenorhabditis elegans)のPTEN (Daf-18)のタンパク質配列を、Wormbaseからダウンロードした。ウシ(Bos Taurus)(XM_613125)およびチンパンジー(Pan troglodytes)(XP_521544)を、NCBIからダウンロードした。
【図4】PTEN-longの存在の証拠 A) 異なる細胞株の二つの異なるPTEN抗体での調査。MCF10A および HEK293は、PTENに関して野生型である。BT549およびHCC1937はPTEN ヌルであり、ZR-75-1はPTENに変異(L136)を有する;B) PTENおよびPTEN-longの双方を認識するPTENに対するモノクローナル抗体での異なる細胞株のさらなる調査;C) Wt ES 細胞は、大量のPTEN-longを発現する。PTEN-longは、これらの細胞において安定的なPTEN shRNA発現に感受性であり、PTEN ノックアウト細胞においては全く存在しない。多くのpAktレベルは、PTENのレベルに逆にしたがう;D) PTEN siRNAは、HEK293細胞においてPTENおよびPTEN-long双方のノックダウンを生じる。E) PTENヌルPC3細胞におけるプラスミドの外因性の発現。PTENorfは、単に開始コドン AUGからのORFをコードする(レーン 2)。ATR (ATR = 代替の翻訳領域)の付加によって、PTEN-longを弱く翻訳できる(レーン 3)。上流の開始部位のATGへの変異は、タンパク質の相補物(complement)を完全にPTEN-longにシフトした(レーン 5 および 6)。ATG 開始コドンのATAへの変異は、55kDaバンドを無効にする(レーン 4 および 6)。E) 抗体は5'ATRによりコードされたアミノ酸に対するものであり、HEK293同様PTENorfまたは5'ATR (ATG/ATG)をコードしているプラスミドの何れかを過剰発現しているPTENヌル U87細胞株からの双方の細胞溶解産物に使用された。PTEN-longは、5'ATRを過剰発現している細胞にのみ認められた。U87 細胞で観察されたバックグラウンドのバンドは、ブロットの下部に存在する。
【図5】シグナルペプチド予測。PTEN 5'UTR 配列を翻訳し、SignalIP3.0に入力した。真核生物シグナルペプチドのためのHidden markovモデルを予測のため使用した。N-領域は、シグナルペプチドの陽性に荷電したN末端配列を示す。H-領域は、シグナルペプチドの疎水性コアである。C-領域は、通常疎水性コアのヘリックスを破壊するプロリンによりマークされる穏やかに極性の領域である。切断可能性は、タンパク質がERの内腔に放出されることを許容するシグナルペプチドを放出する切断部位の予測である。(Dalbey and Heijne, 2002)。切断がポジション 21で発生することが予測される。
【図6】コンカナバリンAプルダウン。HEK293細胞を溶解し、コンカナバリン セファロースを使用してグリコシル化タンパク質をプルダウンした。溶出物をSDS-PAGEで分離し、PTEN (6H2.1)で免疫ブロットした。PTEN-longの濃縮は、プルダウン 対 インプットにより観察できる。長いPTENのバンドの濃縮に注意されたい。PTENは、複数の潜在的なO-グリコシル化部位を有するが、しかし、僅か一つのN-グリコシル化部位を有する。HEK293 細胞中でPTEN相補物の一部がグリコシル化されるかどうかを決定するプルダウンアッセイにレクチン コンカナバリン-A(糖部分と結合する)を我々は使用した。我々はPTENの混合物を精製でき、PTEN-longは凡そ50%であった(正常なPTENに対してPTEN-longの多大なる濃縮)。この事項は、PTEN-longがグリコシル化されること、またPTENの細胞質の55kDa形態がグリコシル化されるか又はPTEN-longが細胞外で切断されるかの何れかを示す。
【図7】PTEN および PTEN-βは、ヘパランと結合する。マウス肝抽出物を1ml HiTrap ヘパランセファロース (Amersham) カラムに通過させた。そのカラムを500mM NaClで洗浄し、タンパク質を連続的に1M NaClのカラム容量で溶出した。フラクションをPTEN モノクローナル抗体を用いてPTENに関してSDS-PAGEで分析した。PTENは、以前に高度に陰性に荷電した種に親和性を有することが示され、PTENの特性により高度に陰イオン性のPtdIns(3,4,5)P3を好むことが導かれる(Das, Dixon et al. 2003)。ヘパランは最も陰性に荷電した生物学的分子(biological molecules)の一つであるので、我々はヘパランが実際にPTENの細胞外基質への結合を媒介すると仮定した。マウスの肝臓からのタンパク質抽出物を用いて、我々はPTENがヘパランと高親和性に結合することを発見した。さらにまた、ヘパリン アガロース カラムから1M NaClを用いたPTENの連続的な溶出によって、PTEN-longも溶出された。
【図8】プロテアーゼプロテクションアッセイ。HEK293細胞をプロテイナーゼ Kの濃度を増加させて再懸濁した。終濃度 0.2%でTritonを、最高濃度のプロテイナーゼ Kを含んでいる反応に加えた。反応をPMSFで停止し、細胞溶解産物をlaemlli 緩衝剤で作った。溶解産物を8% ポリアクリルアミドゲル上でSDS-PAGEで分離し、PTEN (6H2.1), AKT, E-カドヘリンに関して免疫ブロットした。PTENの免疫ブロットの大きなバンドが、PTEN-longと指定された。これらのデータは、E-カドヘリン および PTEN-longが大部分は細胞表面にあることを示す。
【図9】馴化培地(conditioned media)のヘパリン親和性精製からのPTEN および PTEN-longの高塩溶出(High salt elution)。馴化培地からの親和性精製後、PTEN および PTEN-longはHiTrap ヘパリン (Amersham) カラムから溶出できる。PTENの尾部に対するモノクローナル抗体(上記)および5'ATRにおいて翻訳されたアミノ酸に特異的な抗体の双方は、凡そ55kDaの質量のタンパク質バンドを認識する。
【図10】ヒト血清からのPTENの精製。AB 血液からのヒト血清をプロテインA/Gで抗体のプレクリア(precleared)をしてヘパリンセファロースに供試した。溶出物をSDS-PAGEで分離し、PTENに関して又は重鎖混入のための対照に対する二次単独で免疫ブロットした。
【図11】図 11A - 11C。 PTEN-longの抗-血管形成活性。(A) PTEN-longは、発生している網膜中の血管および毛細管のサブセットに発現する。この発現パターンは、PTENの標準形(canonical form)とはっきりと対照を成し、これらの領域に生じる血管退行(vascular regression)の誘導におけるPTEN-longの役割と矛盾しない。この血管退行のプロセスが全網膜の溶解産物のウエスタンブロットのとおり高酸素により 最上部右(B)、また低酸素条件下で内皮細胞におけるPTEN-longの欠損(C)により誘導される場合、この関係はPTEN-longの顕著なアップレギュレーションにより強められる。これらの所見は、糖尿病性網膜症, 同様に高増殖性の血管障害(hyper-proliferative vascular disorders)などの抗-血管新生治療としてのPTEN-longの有用性を示す。矢印は、CD34 および PTEN-long 陽性の組織(血管)を示す。
【図12】PTEN-longのアポトーシス促進活性。指摘のとおり、アポトーシスは、精製 PTEN-longで24 hrs処理されたMCF-10A乳房上皮細胞において誘導された。カスパーゼ 3 切断は、アポトーシス活性の指標(indicative)であった。
【図13】PTEN-longでのマウスの処理。PTEN-longまたは空ベクター対照のいずれかで十日処理し、キャリパー測定で測定した腫瘍サイズのグラフ。293 細胞を、pcDNA3.1 His V5 ベクターにおけるATG/ATG PTEN-longでトランスフェクトした。トランスフェクションの48 hr 後、細胞質ライセート(Cytoplasmic lysates)を作出し、V5-抗体ビーズを通過させ、V5 ペプチドで溶出した。V5-ビーズ精製溶出物のウエスタンブロットを下に示す。PTEN-longが腫瘍を治療するため使用できるだろうとの最初の観察。異種移植片を、スキッドマウスの乳房の脂肪パッドに注射したU87 グリオブラストーマ 細胞 (百万)を用いて樹立した。治療を移植の凡そ二週後に開始した。
【図14】PTEN-longでのマウスの治療の結果。グラフは、14日間の対照およびPTEN-longの注射で処理したマウスの生存率(surviving fraction)を示す(日数)。
【図15】PTEN (5'UTRを欠損しているPTENorf), PTEN-long, およびG からRへの変異をアミノ酸305に有するPTEN-long(PTENorfにおけるG129R変異に匹敵する)の示した構築物を、293 細胞にトランスフェクトした。これらの細胞からの精製タンパク質を用いて、PTEN-longが活性なホスファターゼであること、またPTEN-long G305R 変異(これはPTENにおけるG129Rである)がホスファターゼ活性を低下させたことが示された。
【図16】ホスファターゼ活性は、PTEN-long 活性に必須であり、PTEN-long(G305R) 変異の実験で示された。PTENの文献に基づいて、C2 ドメインの内側に作った短縮体(truncations)はタンパク質を不安定化することが知られている。また、PTENの結晶構造に基づいて、C2 ドメイン および ホスファターゼドメインの間の相互作用がホスファターゼ活性に重要であると信じられている。従って、C-端でのPTEN-long 活性のための最小ドメインは、C2 ドメインを必要とするが、尾部を必要としない。N-端で、予測された切断部位は、アミノ酸 21である。従って、タンパク質の機能領域は、この領域内にある。この点に関して、U87 腫瘍をPTENと又はPTEN-longと対比して処理した場合、有意な効果はPTEN治療では観察されず、PTEN-long 治療でのみ観察されることに注意することが重要である。
【図17】His および V5 タグを有するpcDNA3.1 発現ベクター中のATG/ATG-PTEN longでトランスフェクトされた293 細胞からのPTEN-longの精製。Ni+ カラム 溶出後、溶出液をゲル濾過カラムで分離した。OD280を青線で示す。PTEN-longは、フラクション 7-18に濃縮された。本実験の収量は、凡そ1 mgであった。矢印は、PTEN-long および 移動性が変化したPTEN-long産物(altered migrating PTEN-long products)を示す。
【図18】図 18A - 18B。 情報(readout)として細胞死を用いたLNCaP 前立腺癌細胞とPTEN-long 精製 タンパク質との用量反応(タンパク質はARVYSにより精製された)。1xは、0.33 マイクログラム / mlと等しい。細胞を成長因子なしの培地で処理した。24 hr後、細胞を血清フリー培地で洗浄し、Laemmliサンプル緩衝剤で溶解した。示したタンパク質に関してウエスタンブロットを行った。(B) PTEN-long, PTEN-long(G305R)またはMock対照で処理したU87 グリオブラストーマ細胞は、PARPの切断およびセリン 473でのpAKT シグナルのダウンレギュレーションにより示されるようなアポトーシスの誘導を示す。これらのデータによって、PTEN-longがアポトーシスを誘導し、PI3K/AKT シグナル伝達を減少できることがさらに確認される。
【図19】図 19A - 19C。 AKTA 精製 PTEN-long タンパク質は、ノギスとxenogen動物生体イメージングシステムと連動させたルシフェラーゼレポーターを用いることとの双方により測定されたとおり、五日間にわたり腫瘍サイズを減少できた。マウスは、~0.05mg PTEN-long/日を五日間与えられた。異種移植片は、FUW-ルシフェラーゼ-neoで感染させたためルシフェラーゼを発現する乳房の脂肪パッドに注射された百万の U87 グリオブラストーマ細胞で樹立された。マウスは、Xenogen Imaging Systemでのイメージングの10 分前に腹腔内にルシフェリンを注射された。(A) 治療前(左パネル)および五日治療時(右パネル)の4 マウスに関するルシフェラーゼ測定。(B) 治療前と5 日治療間のキャリパー測定(cm2)。(C) パネルにイメージされXenogenシステムにより検出されたフォトン(Photons)。同齢集団の四マウスに関する標準誤差が示される。0日から5日で検出されたフォトンに関するスチューデントのt-検定。
【図20】独立した実験において、U87 腫瘍をPTEN (orf-403 アミノ酸; n=5), PTEN-long (G305R; n=5)または野生型 PTEN-long (n=4)での処理前に1.5 cm2に成長させた。処理の5 日後、平均のルミネセンス変化(average change luminescence)は、PTEN-long 処理マウスで有意な減少を示すが、PTENまたはPTEN-long(G305R)処理したコホートに関して減少しないことを示す。ルミネセンスの減少は、腫瘍サイズの減少と相関する。これらのデータによって、PTEN-longは機能するために5'ATR および ホスファターゼ 活性を必要とすることが実証された。
【図21】図 21A - 21C。 PTEN-long-処理したU87 異種移植片の分析によって、アポトーシスの活性化およびPI3K シグナル伝達の阻害が実証された。腫瘍を上記のとおり5 日処理した。(A) PTEN-longの野生型 および G305Rで処理した腫瘍を、示した処理の5 日後に収穫し、ウエスタン分析のため溶解した。PTEN-longに関する野生型タンパク質は、FOXO および AKTのリン酸化が減少し、カスパーゼ-3切断を活性化することができた。(B) 示したとおり、5 日処理した代表的な腫瘍を、ホルマリンで固定し、パラフィン包埋した。切片を切断されたカスパーゼ-3(アポトーシスのマーカー)を検出する抗体で染色した。PTEN-long 処理細胞は、アポトーシス細胞のパーセントの有意な増加を有する(P=0.0419, スチューデントのt-検定)。(C) 切断されたカスパーゼ-3 染色の代表的な像。
【図22】図 22A - 22B。 5 日PTEN-longで処理した同じ腫瘍において、血管の数は非常に減少した。PTEN-longの野生型およびG305Rで処理した腫瘍を、示した処理の5 日後に収穫し、ホルマリンで固定し、パラフィン包埋した。切片をCD31(血管に沿ってならぶ内皮細胞のマーカー)を検出する抗体で染色した。(A) PTEN-long 処理細胞は、血管数/視野(40x 対物)の有意な減少を有した(P=0.007579, スチューデントのt-検定)。(B) CD31染色の代表的なイメージ。
【図23】図 23A - 23B。 U87異種移植片は六群(n=3/群)で樹立され、PTEN-longを筋肉内(IM), 腹腔内(IP), 腫瘍内(IT), 皮下 (SC), 静脈内(IV)注射で四 日治療した。(A) キャリパーにより測定された0日〜4日の腫瘍サイズ(CM2)の変化の平均。(B) xenogen イメージングからの代表的な像を右に示す。このデータから、全ての注射法が無処置のマウスと比較して腫瘍成長に影響すること、またIMで処理した同齢集団のみが退行量の有意な減少を示したことを我々は結論づけることができる。
【図24】図24A - 24C。 異種移植実験を乳房, 脳, および 前立腺に由来する6細胞株で行った。上部は、四つの乳癌細胞株で記された変化である。(A, B, およびD)。示した処置日にキャリパーで測定した腫瘍表面領域(cm2)のグラフ。(C) ヘマトクリット-1143 細胞における変化は、僅かに処置の24 時間後に認められた。全ての四つの細胞株において、処置後に腫瘍サイズが明らかに減少する。
【図25】PTEN-longは細胞と結合する。PTEN-long タンパク質をU87 細胞培地に氷上で 10 分間加え、固定し、PTEN-longを認識する抗体でPTEN-longを染色した。
【図26】Milesアッセイ: 血管透過性の誘導はPTEN-longにより阻害された。この阻害は、精製タンパク質をPTEN抗体(6H2.1)とプレインキュベーションすることにより逆転させることができる。PTEN-longは、VEGFによる血管透過誘導を阻害できる。この誘導は、PTEN-longをPTENに対する抗体とプレインキュベーションすることにより回復できたが、対照IgGではできなかった。
【0034】
[発明の詳細な記載]
配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいる単離されたヒトのホスファターゼおよびtensin ホモログ ロング ポリペプチド(PTEN-long)または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログ,またはその各々のバリアント。
【0035】
(i) 配列番号1の残基 1〜173,または(ii) 配列番号5の残基 1〜173を含んでいるそのアナログ,または(iii) 配列番号1の残基 22〜516を含むポリペプチド。
【0036】
配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22 〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを含む薬学的組成物。
【0037】
対象における固形腫瘍を治療するための方法であって、対象における固形腫瘍を治療するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含むヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【0038】
対象における固形腫瘍の成長を阻害するための方法であって、対象における固形腫瘍の成長を阻害するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【0039】
対象の固形腫瘍における血管形成を阻害するための方法であって、対象の固形腫瘍における血管形成を阻害するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【0040】
対象の固形腫瘍における血管の血管上皮細胞(vascular epithelial cell)のアポトーシスを誘導するための方法であって、対象の固形腫瘍における血管の血管上皮細胞のアポトーシスを誘導するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【0041】
本出願に記載された方法の態様において、腫瘍は、癌性腫瘍(cancerous tumor)である。本出願に記載された方法の態様において、癌性腫瘍は、対象のグリア細胞, 前立腺, 卵巣, 子宮, 子宮内膜, 乳房, メラノサイト, 腎臓, 肺, 結腸, 頭部(head), 頸部(neck),または膵臓の腫瘍である。
【0042】
本出願に記載された方法の態様において、癌性腫瘍は、PTENにより若しくはPI3K経路により活性化される又はPTEN-陰性である。
【0043】
本出願に記載された方法の態様において、PTEN-long,またはそのアナログ,またはPTEN-long 断片は、固形腫瘍への直接的な導入により対象に投与される。本出願に記載された方法の態様において、PTEN-long,またはそのアナログ,またはPTEN-long 断片は、固形腫瘍に注射される。本出願に記載された方法の態様において、PTEN-long,またはそのアナログ,またはPTEN-long 断片は、直接的にカテーテルで固形腫瘍に導入される。本出願に記載された方法の態様において、PTEN-long,またはそのアナログ,またはPTEN-long 断片は、固形腫瘍に供給する血管への直接的な導入により対象に投与される。本出願に記載された方法の態様において、PTEN-long,またはそのアナログ,またはPTEN-long 断片は、固形腫瘍に供給する血管に注射される。本出願に記載された方法の態様において、PTEN-long,またはそのアナログ,またはPTEN-long 断片は、直接的にカテーテルで固形腫瘍に供給する血管に導入される。本出願に記載された方法の態様において、PTEN-long,またはそのアナログ,またはPTEN-long 断片は、対象の静脈内に投与される。本出願に記載された方法の態様において、PTEN-long,またはそのアナログ,またはPTEN-long 断片は、対象の皮下に投与される。
【0044】
対象における固形腫瘍を治療するための方法であって、配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5を含んでいるそのアナログをコードする特定量の発現ベクターを、PTEN-long, PTEN-long 断片,又はそのアナログを前記固形腫瘍の細胞中で対象における固形腫瘍を治療するため有効な量で発現するように前記対象に投与することを含む方法。
【0045】
対象における固形腫瘍の治療に使用するための、対象における固形腫瘍の成長を阻害するための、対象の固形腫瘍における血管上皮細胞のアポトーシスを誘導するための、または対象の固形腫瘍における血管形成を阻害するための、配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long)または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログ。
【0046】
本出願に記載された使用のための化合物の態様において、腫瘍は、癌性腫瘍(cancerous tumor)である。本出願に記載された方法の態様において、癌性腫瘍は、グリア, 前立腺, 乳房, 子宮内膜, メラノサイト, 腎臓または肺癌の腫瘍である。
【0047】
本出願に記載された使用のための化合物の態様において、癌性腫瘍は、PTENにより若しくはPI3K経路により活性化される又はPTEN-陰性である。
【0048】
それぞれPTEN-long, PTEN-long 断片,又はそのアナログの血漿半減期を増加させる非-ペプチド因子に結合させた、配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログを含む組成物。
【0049】
本出願に記載された組成物の態様において、さらに薬学的担体を含んでいる。本出願において記載された組成物の態様において、血漿半減期を増加させる非-ペプチド因子は、ポリエチレングリコール(PEG)である。記載された組成物の態様において、PEGは、PTEN-long, PTEN-long 断片, またはそのアナログのC-末端またはN-末端に結合される。記載された組成物の態様において、血漿半減期を増加させる非-ペプチド因子は、9-フルオレニルメチル クロロホルマート (Fmoc)または(7-スルホ)-9-フルオレニルメトキシカルボニルである。
【0050】
配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long)をコードする,または配列番号1の残基 22〜516を含んでいるその断片をコードする,または配列番号5を含んでいるそのアナログをコードする単離された核酸。
【0051】
一態様において、核酸は、配列番号2または配列番号6で表される配列を有している連続的なヌクレオチドを含む。一態様において、核酸は、配列番号2または配列番号6で表される配列の連続的なヌクレオチド残基503〜2243を含む。一態様において、核酸は、配列番号3または配列番号7で表される配列を有している連続的なヌクレオチドを含む。一態様において、核酸は、配列番号3または配列番号7で表される配列の連続的なヌクレオチド残基503〜2243を含む。一態様において、核酸はRNAである。一態様において、核酸はDNAである。一態様において、核酸はcDNAである。
【0052】
配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログをコードしている核酸を含む発現ベクター。
【0053】
配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログを発現する能力のある形質転換された細胞であって、前記細胞はそのゲノムにPTEN-longまたはそのアナログをコードする組換えDNAが統合される形質転換された細胞。
【0054】
配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログを発現する能力のある宿主細胞であって、PTEN-longまたはそのアナログをコードするプラスミドを含む宿主細胞。
【0055】
一態様において、宿主細胞は、細菌細胞である。一態様において、宿主細胞は、哺乳類細胞である。
【0056】
それぞれPTEN-long, PTEN-long 断片,又はそのアナログの血漿半減期を増加させる非-ペプチド因子に結合させた配列番号1を含んでいるPTEN-long,または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5を含んでいるそのアナログを作るため、(1) 配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5を含んでいるそのアナログ, および(2) PTEN-long又はそのアナログの血漿半減期を増加させる非-ペプチド因子を混合(admixing)することを含む方法。
【0057】
配列番号2または配列番号6の核酸配列の相補物と,または配列番号2 配列番号6とストリンジェントな条件下で特異的にハイブリダイズする配列番号2または配列番号6の核酸配列または配列番号 2または配列番号6の核酸配列の相補物の少なくとも20ヌクレオチドの断片からなる単離された核酸分子。
【0058】
単離された抗体,又はその断片であって、(1) 配列番号1のアミノ酸残基 1-173を含むPTEN-ロング ポリペプチドまたは(2) 配列番号5を含んでいるそのアナログまたは(3) 配列番号3で表される配列を有しているアミノ酸または(4) 配列番号1の残基 22〜516を含んでいるPTEN-longの断片と結合する抗体又はその断片。
【0059】
一態様において、前記抗体はモノクローナル抗体である。一態様において、前記抗体は、抗体断片である。一態様において、前記抗体断片は、Fab, Fab', F(ab')2, またはFv 断片である。一態様において、前記抗体断片は、単鎖抗体(single-chain antibody)である。一態様において、前記抗体はヒト化抗体である。
【0060】
単離された抗体又はその断片であって、前記抗体又はその断片はPTEN-longのコンホメーションエピトープと結合し、前記PTEN-longは配列番号1の残基 1〜173で表される配列を含むが、しかし、前記抗体はPTENとは結合しない。
【0061】
一態様において、前記抗体は、配列番号1の残基 153〜173の何れかで構成されるエピトープと結合する。
【0062】
単離された抗体又はその断片であって、前記抗体又はその断片はPTEN-longのエピトープと結合し、前記PTEN-longは配列番号1の残基 1〜173で表される配列を含むが、しかし、前記抗体はPTENとは結合しない。
【0063】
一態様において、前記抗体は、配列番号1の残基 153〜173(または、その部分)と結合する。
【0064】
配列番号1,または配列番号5のペプチド断片であって、抗-腫瘍, 抗-血管形成性,または抗-アポトーシス活性を有する断片。一態様において、前記ペプチドは、5〜10, 10〜20, 20〜30, または30〜40 アミノ酸を含む。一態様において、前記ペプチドは、配列番号1または配列番号5のアミノ酸 1〜173を含む。一態様において、前記ペプチドは、配列番号1の連続的なアミノ酸残基 22〜516を含む。一態様において、前記ペプチドは、配列番号1または配列番号5の連続的なアミノ酸残基 174〜576を含まない。
【0065】
本出願に使用される物質の「予防上有効な(prophylactically effective)」量は、前記物質が投与される対象における所定の病理学的なコンディションの開始を阻止する又は遅延させるための効果的な量である。
【0066】
本出願に使用される物質の「治療上有効な(therapeutically effective)」量は、前記物質が投与される対象が患っている所定の病理学的なコンディションの症状または原因(cause)を治療する, 寛解させる又は低下させるための効果的な量である。
【0067】
一態様において、治療上または予防上有効な量は、約 1 mg の因子/対象〜約 1 g の因子/対象/投薬(dosing)である。別の態様において、治療上または予防上有効な量は、約 10 mg の因子/対象〜500 mg の因子/対象である。さらなる態様において、治療上または予防上有効な量は、約 50 mg の因子/対象〜200 mg の因子/対象である。更なる一態様において、治療上または予防上有効な量は、約 100 mg の因子/対象である。なお更なる態様において、治療上または予防上有効な量は、50 mgの因子/対象, 100 mgの因子/対象, 150 mgの因子/対象, 200 mgの因子/対象, 250 mgの因子/対象, 300 mgの因子/対象, 400 mgの因子/対象および 500 mgの因子/対象から選択される。
【0068】
因子(agent)を「投与すること」は、当業者に既知の様々な方法および送達システムの何れかを用いて成立させる(effected)又は行うことができる。投与することは、例えば、静脈内, 経口, 経鼻, 腹腔内, 脳脊髄液を介して, 移植片を介して, 経粘膜, 経皮, 筋肉内, 血管内, 動脈内, 冠内, 心筋内 または皮下であってもよい。
【0069】
「PTEN-long アナログ(PTEN-long analogue)」の用語は、一または二以上の修飾されたアミノ酸を有するが、配列番号1と少なくとも 90%の配列類似性(sequence similarity)を保持し、以下に記載されたインビボ研究により測定される腫瘍成長の阻害の活性を保持する又は改善するポリペプチドを包含する。PTEN自身(即ち、配列番号1の残基 174-576のみで規定されるポリペプチド)は、明示的に除外される。
【0070】
「PTEN-long バリアント(PTEN-long variant)」の用語は、一または二以上の付加的なアミノ酸(典型的には、PTEN-longのN-末端またはC-末端の何れか、または双方に5未満の付加的なアミノ酸を有している)を有し、以下に記載されたインビボ研究により測定される腫瘍成長の阻害活性を保持する又は改善するポリペプチドを包含する。
【0071】
「PTEN-long アナログのバリアント(PTEN-long analogue variant)」の用語は、一または二以上の付加的なアミノ酸〔典型的には、PTEN-longアナログ(即ち、配列番号5)のN-末端またはC-末端の何れか、または双方に5未満の付加的なアミノ酸を有している〕を有し、以下に記載されたインビボ研究により測定される腫瘍成長の阻害活性を保持する又は改善するポリペプチドを包含する。
【0072】
PTEN-long アナログを参照している本出願の全ての態様は、必要な変更を加えて、PTEN-long バリアントに適用可能である。
【0073】
PTEN-longは、別にPTEN-beta, PTEN-β, PTEN-Sと参照される。
【0074】
PTEN-long, PTEN-long アナログ, PTEN-long バリアント, PTEN-long アナログのバリアント, 又はその各々の組み合わせのための注射可能な薬物配送システムは、溶液、懸濁液、ゲル、ミクロスフェアおよびポリマー性の注射可能物質(polymeric injectables)を含み、また溶解性変更薬剤(solubility-altering agents)(例えば、エタノール、プロピレン・グリコールおよびスクロース)およびポリマー〔例えば、ポリカプリルアクトン(polycaprylactones)およびPLGA's〕などの賦形剤を含むことができる。インプラント可能なシステム(Implantable systems)は、ロッド(rods)およびディスク(discs)を含み、また非限定の例のPLGAおよびポリカプリルアクトンなどの賦形剤を含有することができる。
【0075】
本発明の組成物および化合物のための経口の送達システムは、錠剤およびカプセル剤を含む。これらは、結合剤〔例えば、ヒドロキシプロピルメチルセルロース、ポリビニールピロリドン、他のセルロース系材料(cellulosic materials)およびスターチ〕、希釈剤〔例えば、ラクトースおよび他の糖類、スターチ、リン酸二カルシウム(dicalcium phosphate)およびセルロース系材料〕、崩壊剤(例えば、スターチポリマーおよびセルロース系材料)および潤滑剤(例えば、ステアレートおよびタルク)などの賦形剤を含有してもよい。
【0076】
本発明の組成物および化合物のための経粘膜的な送達システムは、パッチ(patches)、錠剤、座剤、ペッサリー(pessaries)、ゲルおよびクリームを含み、溶解剤および増強剤(例えば、プロピレン・グリコール、胆汁酸塩およびアミノ酸)などの賦形剤、並びに他のビヒクル(例えば、ポリエチレン・グリコール、脂肪酸エステルおよび誘導体、並びにヒドロキシプロピルメチルセルロースおよびヒアルロン酸などの親水性ポリマー)を含有することができる。
【0077】
本発明の組成物および化合物のための経皮的な送達システムは、例えば、水溶性および非水溶性のゲル、クリーム、複合乳剤(multiple emulsions)、ミクロ乳剤(microemulsions)、リポソーム、軟膏、水溶性および非水溶性の溶液、ローション(lotions)、エアロゾル(aerosols)、ハイドロカーボン基材および粉末(hydrocarbon bases and powders)を含み、且つそれは溶解剤(solubilizers)、浸透増強剤(permeation enhancers)(例えば、脂肪酸、脂肪酸エステル、脂肪アルコールおよびアミノ酸)、および親水性のポリマー(例えば、ポリカルボフィルおよびポリビニールピロリドン)などの賦形剤を含有することが可能である。一態様において、薬学的に許容される担体は、リポソームまたは経皮的な増強剤(transdermal enhancer)である。
【0078】
本発明の組成物および化合物の再構成可能(reconstitutable)な送達システムのための溶液、懸濁液、粉剤には、懸濁剤〔例えば、ガム、ザンタン(zanthans)、セルロース(cellulosics)および糖〕、保湿剤(例えば、ソルビトール)、溶解剤(例えば、エタノール、水、PEGおよびプロピレングリコール)、界面活性剤(例えば、ラウリル硫酸ナトリウム、Spans、Tweens、およびセチルピリジン)、保存剤および抗酸化剤〔例えば、パラベン(parabens)、ビタミンEおよびC、およびアスコルビン酸〕、抗固化剤(anti-caking agents)、コーティング剤、およびキレート化剤(例えば、EDTA)などのビヒクルが含まれる。
【0079】
本出願に使用される「5'ATR」は、以下の実験のセクションに記載される5'代替の翻訳領域である。
【0080】
本出願に使用される、「薬学的な担体」は、動物またはヒトに対して本発明(instant)の化合物または組成物を送達するための薬学的に許容される溶媒, 懸濁剤またはビヒクルである。担体は、液体, エアロゾル, ゲルまたは固形であってもよく、計画された投与の様式を考慮して選択される。
【0081】
「非-ペプチド因子(Non-peptide agent)」は、限定されることなく、グリコマー(glycomer), ポリマー, 小分子〔即ち、1000未満の分子量を有している炭化水素に基づく分子(hydrocarbon-based molecule)または有機分子〕, 脂質, リポソームを含む任意の化学物質(chemical entity)を意味する。非-ペプチド因子の例には、PEG, Fmoc およびFMSが含まれるが、これらに限定されない。
【0082】
本出願に使用される「固形腫瘍(Solid Tumor)」は、癌性(cancerous)の及び非癌性の固形腫瘍を含む。癌性の固形腫瘍には、限定されることなく、胆道癌; グリオブラストーマ および髄芽腫を含む脳癌; 乳癌; 子宮頚癌(cervical cancer); 絨毛癌; 結腸癌; 子宮内膜癌; 食道癌; 胃癌; ボーエン病およびパジェット病を含む上皮内の新生物; 肝臓癌; 肺癌; ホジキン病およびリンパ球性リンパ腫を含むリンパ腫; 神経芽細胞腫; 扁平上皮癌を含む口腔癌; 上皮細胞, 間質性細胞, 胚細胞および間充織細胞から発生するものを含む卵巣癌; 膵臓癌; 前立腺癌; 結腸直腸癌; 平滑筋肉腫, 横紋筋肉腫, 脂肪肉腫, 線維肉腫および骨肉腫を含む肉腫; メラノーマ, カポジ肉腫, 基底細胞癌(basocellular cancer)および扁平上皮細胞癌を含む皮膚癌; 胚腫瘍(セミノーマ, 非-セミノーマ[奇形腫, 絨毛癌]), 間質腫瘍(stromal tumors)および胚細胞腫瘍を含む精巣癌; 甲状腺腺癌(thyroid adenocarcinoma)および髄様癌(medullar carcinoma)を含む甲状腺癌; および腺癌およびウィルムス腫瘍を含む腎臓癌が含まれるが、非固形組織の腫瘍、例えば、急性のリンパ性および骨髄性の白血病; 多発性骨髄腫; AIDS関連白血病および成人T細胞白血病リンパ腫(adult T-cell leukemia lymphoma)を含む白血病および他の血液学的な新生物は除外される。
【0083】
配列表の配列番号1は、全長のPTEN-long タンパク質配列である。配列番号1のアミノ酸残基1〜173は、5' ATRのmRNA配列(以下の配列番号2の記載を参照されたい)によりコードされるPTEN-longの新規の残基を表す。PTENの標準の開始メチオニンは、配列番号1のアミノ酸残基 174である。
【0084】
配列番号2は、全長 PTEN-long mRNA配列である。PTEN-longの5' ATR配列は、配列番号2のヌクレオチド 513(非標準のCUG開始コドン)で開始し、配列番号2のヌクレオチド 1031で終了する。5' ATRは、配列番号2のヌクレオチド 1032での標準のAUG開始コドンで始まり、配列番号2のヌクレオチド 2243で終わるPTENオープンリーディングフレームとインフレームである。従って、PTEN-longのオープンリーディングフレームは、配列番号2のヌクレオチド 513から配列番号2のヌクレオチド 2243に伸び、PTENのN-端への173アミノ酸の付加が導びかれる。このタンパク質は、PTEN-longと参照される。
【0085】
配列番号3は、全長 PTEN-long mRNA配列に対応するcDNAである。
【0086】
配列番号4は、配列番号1のアミノ酸残基 153〜173を含んでいるPTEN-long タンパク質配列のペプチドである。このユニークなペプチドは、PTENに認められないPTEN-longに由来するユニークなエピトープを表す。
【0087】
配列番号5は、全長 PTEN-long アナログのタンパク質配列である。PTEN-longのアミノ酸残基 1は、前記アナログにおいてLeuからMetへと変化してタンパク質収量が増加する。
【0088】
配列番号6は、修飾された全長 PTEN-long アナログのmRNA配列である。PTEN-longアナログの5' ATR配列は、配列番号6のヌクレオチド 513(操作されたAUG開始コドン)で開始し、配列番号6のヌクレオチド 1031で終了する。
【0089】
配列番号7は、修飾された全長 PTEN-longアナログのmRNA配列に対応する修飾されたcDNAである。
【0090】
範囲が明細書中で与えられる場合、前記範囲はその範囲の全ての整数とその範囲内の0.1単位, および任意の細分範囲(sub-range)を含むことが理解される。例えば、77〜90%の範囲は、77.0%, 77.1%, 77.2%, 77.3%, 77.4%, 77.5%, 77.6%, 77.7%, 77.8%, 77.9%, 80.0%, 80.1%, 80.2%, 80.3%, 80.4%, 80.5%, 80.6%, 80.7%, 80.8%, 80.9%, および90.0%, 同様に80%〜81.5%の範囲などを含む。
【0091】
本出願に記載された様々な要素の全ての組み合わせは、本発明の範囲内である。
【0092】
本発明は以下の例を参照することに更に理解される。 当業者は、詳細に説明される特定の実験が、本願の特許請求の範囲においてより完全に記載される本発明の単なる例であることを容易に理解する。
【0093】
[例]
PTEN 腫瘍抑制因子は、癌において最も一般的に変更される遺伝子の一つであり、ホスファチジルイノシトール3,4,5-三リン酸の脂質ホスファターゼとして機能し、これが次にホスファチジルイノシトール 3-キナーゼ(PI3K) および Aktからの腫瘍形成性のシグナル伝達を抑制する。PTEN mRNAを検査することによって、5' 非翻訳領域 (UTR)が770bpについてPTENのオープンリーディングフレーム (ORF)とインフレームであることが明らかとされた。このUTR ORF内で、標準のAUG開始コドンの513塩基対上流での弱いコザック配列内に代替のCUG開始コドンが存在する。標準のPTEN ORFの発現により凡そ55kDaに移動するタンパク質が発現するが、5'UTRを含んでいるPTEN cDNAの発現によりPTEN-longと称される70kDaの第二のタンパク質を産生することができる。開始部位の変異は、55kDa PTENが標準の開始コドンでの翻訳から産生されるが、他方でPTEN-longは上流の代替の開始部位から開始されることを指摘する。異なる PTEN 抗体での免疫ブロットによって、複数の細胞株にPTEN-longが内因性(endogenous)に存在することが実証された。マウスES細胞におけるノックダウンおよびノックアウト研究によって、この大きなタンパク質が実際にPTENであることが確認された。付加されたN末端タンパク質配列がシグナルペプチドおよび切断部位をコードしており、PTEN-longが分泌経路に進入することを指摘している。PTEN-longは優先的にレクチンコンカナバリンAと結合し、グリコシル化されることが示される。さらにまた、PTEN-longは、PTENに対する抗体やヘパラン硫酸の双方を用いる親和性精製により馴化培地(conditioned media)から精製できる。また、PTEN-longは、インビボでのプロテアーゼプロテクションアッセイにおけるデグラデーションに感受性であるが、正常なPTENは感受性ではなく、PTEN-longが細胞膜の外側に局在することを示している。
【0094】
試薬, 細胞株および抗体 - プロテイナーゼ K および コンカナバリン- AをSigma (St. Louis, MO)から購入した。ヘパリン セファロース および HiTrapヘパリンHPカラムをAmersham (Piscataway, NJ)から購入した。PTENに対する抗体をCell Signaling (Danvers MA) およびCascade (Winchester MA)から購入した。Akt 抗体をCell Signaling (Danvers MA)から購入し、E カドヘリン抗体をUpstate Millipore (Billerica, Ma)から購入した。PTENの新規の翻訳において認められたエピトープ PRHQQLLPSLSSFFFSHRLPD (配列番号3)に対して産生されたポリクローナル親和性精製抗体については、Zymed Laboratories (South San Fransisco, CA)が行った。二次抗体をPierce (Rockford, IL)から購入した。HEK293, ZR-75-1, SKBR-3, MDAMB-361, BT549, および PC3をATCC (Manassas, VA)から得て、提供されたガイドラインにしたがい成長させた。
【0095】
プラスミドおよび構築物 ― pCEP4-PTEN(PTENの完全なオープンリーディングフレームおよび5'-非翻訳領域をコードしている)は、前に報告されたとおり、PTEN cDNA(NCBIにU90351として寄託された)をpCEP4(Invitrogen)のNotI部位にクローニングすることにより作出された(Li, Simpson et al. 1998)。5'UTRは、このプラスミドにおいてクローニングのため使用されるオリジナルのNotI 制限部位の上流の配列をコードしているアダプタをライゲーションすることによりさらに拡張された。前記アダプタは、標準の開始部位の-513に局在する第一の可能な代替のCTG開始コドンの10 塩基対上流までコードした。また、推定上の代替の開始部位をATGに変異させたアダプタを使用して、長い形態が効率的に発現されるだろう第二セットの発現構築物を作出した。さらに、標準の開始コドンのATAへの突然変異誘発も行い、トータルで4つの異なる構築物が生じた(図 5.1)。これらの4つのバリエーション(同様に、オリジナルのPTENのオープンリーディングフレーム)を、安定な発現のためMSCV(Clontech, Mountainview, CA)レトロウイルスベクターに感染によりサブクローンした。
【0096】
プロテアーゼプロテクションアッセイ ― HEK293 細胞をトリプシンなしで氷冷PBSに収集し、5x105の細胞のアリクウォット(aliquots)を0.5ug/ml〜10ug/mlでプロテイナーゼ Kの濃度を増加させて30 分間インキュベーションした。Triton 0.1%での対照を、示したタンパク質を分解するプロテイナーゼ Kの能力を検証するため含めた。反応を5mM PMSFで停止した。細胞を2x Laemmliサンプル緩衝剤(125nM Tris pH 6.8, 20% グリセロール, 0.05% ブロモフェノールブルー, 4% SDS, 10% 2-メルカプトエタノール)で溶解し、PTEN, Akt および E カドヘリンに関して免疫ブロットした。
【0097】
マウスの肝臓からのPTEN精製 ― C57BL6 マウスの肝臓を液体窒素中で急速凍結(snap frozen)し、粉砕し、TNN緩衝剤(50mM Tris pH 7.4, 150mM NaCl, 0.5% NP-40, 5mM EDTA, 3% グリセロール, 1mM DTT, 1x 哺乳類のプロテアーゼカクテルインヒビター [Sigma])に再懸濁した。懸濁液を乳鉢でホモジナイズし、40,000RPMで4 度で1時間遠心分離した。上清を連続的に0.45 ミクロンおよび0.22ミクロンのフィルターで濾過した。セファクリル 200 サイズ排除カラム(Amersham)をTNNで事前に平衡化し、サンプルを0.3ml/hrの速度でアプライし、続いて緩衝液をアプライした。2mlのフラクションを収集し、低分子量のサンプルをプールし、事前に平衡化したHiTrap Hepain HPカラム(Amersham)にアプライした。カラムを三カラム容量のTNNで洗浄し、タンパク質を0.3M, 0.5M および 1M NaCl TNN溶液の段階的な 3x カラム容量で溶出した。0.5mlの増加分のフラクションを収集し、PTENに関して免疫ブロットした。
【0098】
培地からのPTENのヘパリン精製 ― HEK293細胞を15cmディッシュ中で10%FBS DMEMでコンフルエントまで成長させた。細胞をFBSなしの15mlのDMEMで一晩インキュベーションした。20 プレートから培地を収集し、0.45 ミクロンのフィルターを通して濾過した。1mlのヘパリン HP カラムを、AktaPrime (AmershamBioscience)を用いて4ml/minの流速で4℃でDMEMで平衡化した。馴化培地を、カラムに1ml/minで通して通過させた。カラムを10容量のBC200 (200mM Nacl, 50mMT Tris pH7.4, 1mM EDTA, 0.2% Triton X-100)で洗浄した。タンパク質を5mlの1M NaClで1ml/minで1mlのフラクションに溶出した。各フラクションのタンパク質濃度を280nmのODで決定した。各フラクションの半分を、トリクロロ酢酸を20%で沈殿させ、冷アセトンで洗浄し、減圧下で乾燥させた。タンパク質を20 ulのLaemmli溶解緩衝剤に再構成し、PTENおよびPTEN-longに対する抗体を用いて免疫ブロットした。
【0099】
血清からのPTEN精製 ― AB血漿からのヒト血清をSigmaから得た。1mlの血清を、0.45ミクロンのフィルターを通して濾過し、プロテインA/G アガロースを用いて1 時間インキュベーションして抗体をプレクリアした。ヘパリン-アガロースを、セファロースの対照と共に一晩プレクリアした血清でインキュベーションし、次の日にBC150 (150mM NaCl, 25mM Tris pH7.4, 1% NP-40, 0.25% Na デオキシコール酸, 1mM EDTA)で洗浄した。タンパク質をlaemmliサンプル緩衝剤で溶出し、PTENに関して又は重鎖の混入に関して二次のみで免疫ブロットした。
【0100】
コンカナバリンA プルダウン ― HEK293細胞をサブコンフルエンスでBC500 (500mM NaCl, 20mM Tris pH 7.4, 1% Triton X-100, 1mM MnCl2, 1mM CaCl2, 1x プロテアーゼインヒビターカクテル)で溶解した。細胞溶解産物を、遠心分離し、濾過した。プルダウンを、20 マイクロリッターのコンカナバリンAセファロース(Sigma)で1 時間、4℃で行った。樹脂を、BC500で洗浄し、タンパク質をLaemmliサンプル緩衝剤で溶出した。
【0101】
結果
PTEN mRNAは上流に代替の開始部位を有する
NCBIに寄託されたPTEN mRNA(Li and Sun 1997; Steck, Pershouse et al. 1997)は、広範囲にわたる5'UTRを含む。5'UTR領域中の凡そ770bpの連続配列(contiguous sequence)は、開始コドンとインフレームである。メチオニンは、この領域でコードされない; しかしながら、標準の開始コドンから-519で始まる幾つかの代替の開始CUGコドンが存在する。この配列の翻訳から、scansite (scansite.mit.edu) および prosite (www.ebi.ac.uk/ppsearch)にしたがって同定可能なドメインは明らかとされなかった。この全体領域の翻訳によって、173アミノ酸がPTENに加えられ、その分子量を凡そ70 キロダルトンに増加させるだろう(図 2 および 配列番号1)。
【0102】
他のPTENオルソログのアラインメントによって、ヒト UTRの翻訳配列が様々な種のPTENのオープンリーディングフレームに認められることが明らかとされた。チンパンジー, ウシ, ミツバチおよび線虫は、全てヒト 5'UTRの翻訳産物と相同なタンパク質配列を含む(図 3)。さらにまた、ヒト 5'UTR および マウス PTEN 5'UTRのアラインメントは、広範囲にわたりヌクレオチド相同性を示した(示さず)。マウス 5'UTRは522 塩基対にわたり標準の開始コドンとインフレームで翻訳され、ヒト 5'UTRの翻訳と比較した場合に高度に相同なタンパク質配列であることが明らかとされた(図 5.3)。5'UTRの相同性や他の種の翻訳されたタンパク質にヒト 5'UTRから由来するアミノ酸配列が実際に存在することは、この配列が進化上の重要であることを示している。
【0103】
PTEN mRNAは、代替の上流部位から翻訳を開始できる。PTEN ORFの過剰発現は、55kDaの単一のタンパク質バンドを生じた。5'UTRを含めることによって、約70kDaの二番目のより大きいタンパク質バンドを生じた。PTEN免疫ブロットにおけるより大きなタンパク質バンドは、幾つかの細胞株にも内因性に存在し、異なるモノクローナル抗体により検出可能であった(図 4)。この大きいタンパク質バンドは、マウスの野生型 ES 細胞に存在し、PTENノックアウトマウスのES細胞には存在せず、PTEN shRNAを安定に発現しているマウス ES クローンにおいて減少した(図 4)。また、HEK293細胞においてヒト PTEN タンパク質のsiRNAを用いたノックダウンによって、70 kDaタンパク質のノックダウンを生じる。
【0104】
PTENヌル PC3 細胞株においてPTENのORFをコードしているプラスミドの発現は、55kDaタンパク質の産生を生じる。5'UTRもコードするプラスミドが過剰発現された場合、70kDaタンパク質が産生された。CTGからATGへの上流の推定上の開始コドンの変異(図 1, 「ATG/ATG」)は、優勢に免疫ブロットパターンを70 kDaの形態にシフトした(図 4)。また、55kDaのバンドは、突然変異誘発によりATG開始コドンに由来することが確認された(図 4 「CTG/ATA」)。
【0105】
従って、5' UTRは、長い形態のPTENの翻訳を開始するため十分であった。従って、5' UTRはAlternately Translated Regionのため5'ATRと命名され、検出された大きいタンパク質は「PTEN-long」と命名された。
【0106】
親和性精製したポリクローナル抗体は、5'ATRから翻訳されたアミノ酸に対し産生され、過剰発現研究において組換え型のPTEN-longの産生、またHEK293 細胞における内因性の形態を確認するため使用された(図 4)。HEK293 細胞の全細胞溶解産物の免疫ブロットおよびPC3 細胞における過剰発現研究から、複数の形態のPTEN-longが認められ、潜在的な翻訳後修飾, 記載されていないスプライス形態(undocumented splice forms)または5'ATRにおける代替の開始コドンのいずれかを指摘している。
【0107】
PTEN-longはN末端シグナルペプチドをコードする
PTEN-longのN末端配列は、膜貫通配列またはシグナルペプチドのいずれかの指標であろうアラニンの長いストレッチ(a long stretch)を含む。SignalIP 3.0を用いた翻訳配列の分析によって、高い確率(>95%)で前記配列がシグナルペプチドを含むことが予測された(図 5)。シグナルペプチドは、特徴的に塩基性アミノ酸と続く疎水性のストレッチとから構成される。推定上の疎水性の膜貫通ヘリックスは、プロリンにより破壊され、やや極性の配列が続く。また、配列が切断されることも予測され、このタンパク質がERの内腔に放出されることを指摘している。
【0108】
PTENはグリコシル化される
分泌された細胞外のタンパク質の特徴の一つは、ゴルジ体における複合糖部分(complex sugar moieties)の付加である(グリコシル化として知られるプロセス)。糖はN-グリコシル化を介して共通配列 N-X-S/T (Xはプロリンではない)においてアスパラギンに付加できる(Gupta and Brunak 2002); またセリン, スレオニンおよびチロシンのヒドロキシル基はO-グリコシル化の標的となりえる(Julenius, Molgaard et al. 2005)。PTENは、複数のO-グリコシル化部位を有するが、しかし、僅か一つのN-グリコシル化部位を有する。HEK293 細胞中でPTEN相補物(PTEN complement)の部分がグリコシル化されたかどうかを決定するプルダウンアッセイにレクチン コンカナバリン-A(糖部分と結合する)を使用した。凡そ50%のPTEN-longのPTENの混合物をこれらの細胞から精製した(図 6)。この事項は、PTEN-longがグリコシル化されること、またPTENの細胞質の55kDa形態がグリコシル化されるか又はPTEN-longが細胞外で切断されることを示す。
【0109】
PTEN-longはヘパランと結合し、細胞表面に認められる
PTENは幾つかのプロテオグリカン(例えば、syndecans および glypicans)と結合し、膜の外側のリーフレット(leaflet)に付着することが認められる。これらのプロテオグリカンは、最もヘパラン化した細胞外分子のうちの二つである(Blero, Zhang et al. 2005)。PTENは以前に高度に陰性に荷電した種に高い親和性を有することが示され、PTENの特性により高度に陰イオン性のPIP3を好むことが導かれる(Das, Dixon et al. 2003)。ヘパランは最も陰性に荷電した生物学的分子(biological molecules)の一つであるので、ヘパランがPTENの細胞外基質への結合を媒介することが可能であった。マウスの肝臓からのタンパク質抽出物を用いて、PTENがヘパランと高親和性に結合することが発見された。さらにまた、ヘパリン アガロース カラムから1M NaClを用いたPTENの連続的な溶出によって、PTEN-longも溶出された(図7)。
【0110】
細胞膜の外面に突き出たPTEN-longは、プロテアーゼのデグラデーションに感受性である。プロテアーゼプロテクションアッセイにおいて、生細胞をプロテアーゼとインキュベーションし、細胞外のタンパク質のみが分解された(というのも、脂質膜はプロテアーゼを不透過性であり、全ての細胞内タンパク質の防御に貢献するからである)。HEK293細胞を付着培養からPBSで穏やかに撹拌して除去し、プロテイナーゼ Kの濃度を増加させて懸濁した。反応をPMSFで停止し、細胞をlaemlli 緩衝剤で溶解した。PTEN-longは、既知の細胞外タンパク質であるE-カドヘリンと共にプロテイナーゼ Kでの処理に感受性を示した(図 8)。他方でPTENは中等度のプロテアーゼ感受性を示し、55kDa種のいくつかも細胞外(グリコシル化されるので)であるか又は幾らかの細胞溶解がアッセイの間に発生し、これにより細胞質のPTENがプロテイナーゼ Kに暴露されることを示している。プロテイナーゼ Kに暴露された場合にPTENが分解されることを証明するため、膜を透過するtritonでの対照を含めた。これがPTEN本来(PTEN proper)であるか又は55kDaに移動し、PTEN抗体のC末端エピトープを保持するPTEN-longの切断形態であるかどうかはまだ分からない。このデータは、PTEN-longが細胞表面にあることを指摘している。
【0111】
可溶性のPTEN-longは培地に分泌される
タンパク質の部分が可溶性であり、細胞環境に放出される可能性。
【0112】
ヘパリンセファロースを、HEK293細胞で馴化した血清フリー培地からPTEN-longを親和性精製するため使用した。カラムの溶出によって、培地中に50kDaの分子量に移動するPTENの存在が明らかとされた(図 9)。PTEN-long 特異的な抗体での免疫ブロットは同じ50kDa種を明らかとし、このタンパク質が代替の開始部位から翻訳される配列およびPTEN モノクローナル抗体のC末端エピトープの配列を保持することを示している。この事項は、55kDaであることが観察されたPTENの部分が実際に上流の開始コドンに由来する切断された翻訳産物であることを暗示する。
【0113】
培地へのPTENの分泌は、さらにATG/ATG 構築物でトランスフェクトされたHEK293細胞においてPTEN-longを過剰発現することにより確認された。これらの細胞を使用して、血清フリーの馴化培地で一晩産生させた。PTEN モノクローナル抗体 6H2.1を使用して、1mlの培地からPTENを免疫沈降した。大きいPTENバンドが、より低い55kDaバンドと共に培地から良好に免疫沈降された。前記タンパク質が過剰発現されたので、前記タンパク質の適切なプロセシングはおそらく生じず、完全なサイズの70kDa PTENの分泌が生じる。
【0114】
PTENはヒト血清中に認められる。
【0115】
生理的な分泌物の最良の供給源の一つは血清である。ヘパリンセファロースを使用してヒト血清からPTENを親和性精製した。ヒト血清を遠心分離し、濾過して粒子性の物質を除去した。次にBC150に1:5に希釈し、頻繁にプロテインA/GでプレクリアしてIgGを除去した。血清を少量のヘパリンセファロースでバッチインキュベーションした。ヘパリン セファロースをlaemmli 緩衝剤で溶出し、溶出液をPTENに関してブロットし、また重鎖の混入を排除するため二次抗体単独でブロットした。PTEN および PTEN-longは、双方ヒト血清に認められた(図 10)。
【0116】
PTEN-longの抗-血管形成活性
PTEN-longの抗-血管形成の役目を以下に示す:
(1) PTEN-longは通常はマウスの発生している網膜において弱く発現するが、新生児の発生の間で退行(involution)/細胞死を経験している血管に高レベルの発現が認められる(図 11);
(2) PTEN-longは、腫瘍中のアポトーシスしている血管に認められる。さらにまた、PTEN-longで処理された上皮細胞(トランスフェクトされた細胞から部分的に精製された)は、細胞の遊走を阻害し、アポトーシスを誘導した。(図12)。精製したPTEN-longは、カスパーゼ-3 切断により測定されたとおり培養においてU87, HUVEC 内皮細胞, または293細胞のアポトーシスの活性化と関連する細胞死も誘導できる。
【0117】
PTEN-longのインビボでの抗-腫瘍および抗-血管形成活性
図 13は、PTEN-longでのマウスの処理を示す、 (A) マウス (n=5)にグリオブラストーマ細胞株U87が注射され、異種移植片が乳房の脂肪パッドの2部位(左および右)に形成された。腫瘍移植後、一つの腫瘍にPTEN-longを直接注射し、反対側の腫瘍には注射されなかった(w/ PTEN-long)。また、5 マウスの対照セットは、空ベクターでトランスフェクトされた細胞から由来するmockの精製タンパク質の調製物を注射された(空ベクター)。再び、反対側の腫瘍は注射されなかった(w/空ベクター)。マウスを1-11日および13-14日に処理した。最大の直径(cm)を、示した日にノギスで測定した。腫瘍容量が >1 cmに達した時、マウスを屠殺した。(B) タンパク質をPTEN-long発現ベクターを293細胞にトランスフェクションすることにより調製し、V5親和性樹脂を用いて部分的に精製し、V5 ペプチドで溶出した。図14は、14日間の対照注射で処理したマウスの生存率を示す(日数)。
【0118】
網膜染色
p7のマウス網膜中のPTEN-longおよび血管の染色によって、PTEN-longがこのポイントでマウスの網膜の血管発生において最初に退行(regress)する硝子体管(hyaloid vessels)を選択的に染色することが明らかとされた。PTEN-longに対する抗体は、エピトープ: N-PRHQQLLPSLSSFFFSHRLPD-C (配列番号3)に対するものである。管(Vessel)の染色をBS1-レクチンで行った。
【0119】
精製
PTEN-longの精製のための一つの方法において、293 細胞をATG/ATG PTEN-longでトランスフェクトし、細胞溶解産物をNi+ アフィニティーカラムに通過させた。PTEN-longを、AKTA PurifierでNi+ カラムでイミダゾール溶出緩衝剤を用いて一貫して(consistently)精製した。
【0120】
腫瘍退行
PTEN (orf 403 アミノ酸)またはPTEN-longのいずれかで注射前にトランスフェクトされたU87 細胞の異種移植片。注射後の7 日間、PTENを過剰発現している同齢集団(n= 4のうち4)と比較してPTEN-longを過剰発現している同齢集団において乳房血管の減少が認められた。これはPTEN-longが腫瘍環境に影響しえることを示唆する。
【0121】
考察
細胞溶解産物や組織からのPTEN免疫ブロットにおいて二番目のより大きいタンパク質バンドを定期的に観察した。より大きなバンドがPTENであることが確認される証拠には、次の事項が含まれる:
大きなタンパク質バンドは、異なるPTENモノクローナル抗体により検出された;
細胞がPTENに対するsiRNAで処理される又はPTEN座位がマウスにおいてノックアウトされる場合、大きなタンパク質は存在しない。PTENの5'UTRは、PTENの古典的な開始コドンと700 塩基対以上にわたりインフレームであることが観察された。さらに、PTENの522 塩基対上流のCUGコドンが存在し、翻訳される場合にPTEN免疫ブロットにおける大きなタンパク質のバンドのサイズを説明することができた。強いコザック配列とは関連しないが、-1 シトシン および +1 グアノシン配列が保持される。翻訳され、PTEN ORFに付加される場合、凡そ70kDaのタンパク質が産生されるはずであり、その分子量は観察された大きなPTENバンドの分子量である。
【0122】
この配列の翻訳は、実際のコード配列内の幾つかのPTENオルソログに既に存在する。マウスの組織溶解産物において類似するバンドが観察されたので、マウス 5'UTRも点検した。マウスの5'UTR ヌクレオチド配列は、ヒトの5'UTRと高度に相同的であり、同様に開始コドンとインフレームである。二つの潜在的な代替の開始コドンが-522 および -516に存在し、これらの部位からのこの配列の翻訳はヒト配列と90%+ 相同的なアミノ酸配列を示す。この推定上のタンパク質の保存は著しく、この配列の進化上の重要性が示された。この配列をよりよく記載するため、PTENの5'ATRまたは代替の翻訳領域と改名して翻訳の潜在性を記載した。
【0123】
PTENのオープンリーディングフレームを5'ATRと共にクローン化してプラスミドを構築し、この組換え型のPTENの発現を標準の403のアミノ酸を産生するオープンリーディングフレーム単独と比較した。5'ATRを含めることにより二番目の大きなPTENタンパク質バンドが生じ、このバンドは55kDaに移動する単一のバンドを作るPTENのまさに標準のORFを含んでいる発現プラスミドと比べて約70kDaに移動する。大きなタンパク質は、翻訳されたトータルのタンパク質の僅か小部分に勘定される; しかしながら、推定上の開始部位のATGへの変異によって、タンパク質比は主に大きな形態にシフトした。
【0124】
5'ATRのタンパク質配列の保存は、進化のアーティファクト以上であることを示す。N-末端は脂肪族アミノ酸のストレッチを含み、これが膜貫通配列であることが予測された。PrositeおよびSignal 3.0IPの使用によって、PTEN-longのN-末端がシグナルペプチドであり、それに直接続くプロテアーゼ切断部位を有することが予測された。
【0125】
PTEN-longが細胞の細胞外表面に局在するかどうかをインビボでのプロテアーゼプロテクションアッセイを使用して検査した。PTEN-longは細胞外のプロテアーゼの量を増加させることにより進行性のデグラデーションを示したが、PTENは示さなかった。この結果は、少なくとも幾らかのPTEN-longが細胞外にあり、少なくとも部分的に細胞膜の外側のリーフレットに付着することを示している。この結果は最も魅力的な結果であり、細胞膜の外側のリーフレットにおける活性リピッドホスファターゼの意味が提起された。外膜結合性のプロテオグリカンの二つのファミリー(glypicans および syndecans)は、以前にPTENタンパク質複合体中に同定された。
【0126】
細胞表面におけるPTENの存在は、可溶性の分泌されるPTENの可能性を排除しない。Syndecans および glypicansは、最も多量にヘパラン化したプロテオグリカンのうちの二つである。ヘパランは高度に陰性に荷電したグリコサミノグリカンであり、またPTENは陰イオンへの親和性を有することが示されており、高度に陰性に荷電したPIP3を基質として選択することを部分的に説明している。マウス肝臓からのPTEN精製を最適化する実験によって、PTEN および PTEN-longの双方がヘパリン セファロース カラムを用いて精製できることが明らかとされた。さらにまた、約50kDaのタンパク質を、HEK293 細胞で馴化した血清フリー培地からヘパリンセファロースカラムを用いて精製した。精製されたタンパク質は、PTENに特異的なモノクローナル抗体およびPTEN-longに存在するユニークなアミノ酸残基に対するポリクローナル抗体により認識された。前に、PTEN-long抗体は、70kDa付近のタンパク質バンドのみを認識した。双方の抗体が一つのバンドを認識できたとの観察によって、タンパク分解プロセシングがおそらく発生すること、免疫ブロットで観察されたタンパク質が双方の抗体のエピトープを保持するPTENの断片であることが示される。また、PTEN および PTEN-longの双方は、ヒト血清からヘパリン親和性精製を用いて精製できた。
【0127】
過去10年の文献によって、PTENの配列の想定(assuming)が蓄積されてきた。代替の部位から翻訳され、外側のリーフレットや細胞外間隙の双方に分泌されるPTENの新規の形態が存在するとの証拠を本出願は提供する。
【0128】
インビボの結果は、PTEN-longがヒト血清に通常存在する新規の抗-腫瘍化合物であり、抗-血管形成やアポトーシス促進特性を有することを示す。
【0129】
[参照文献]
Acland, P., M. Dixon, et al. (1990). "Subcellular fate of the int-2 oncoprotein is determined by choice of initiation codon." Nature 343(6259): 662-5.
Alberts, B. (2002). Molecular biology of the cell. New York, Garland Science.
Baker, S. J. (2007). "PTEN enters the nuclear age." Cell 128(1): 25-8.
Blero, D., J. Zhang, et al. (2005). "Phosphatidylinositol 3,4,5-trisphosphate modulation in SHIP2-deficient mouse embryonic fibroblasts." Febs J 272(10): 2512-22.
Blobel, G., P. Walter, et al. (1979). "Translocation of proteins across membranes: the signal hypothesis and beyond." Symp Soc Exp Biol 33: 9-36.
Bonneau, D. and M. Longy (2000). "Mutations of the human PTEN gene." Hum Mutat 16(2): 109-22.
Di Cristofano, A., B. Pesce, et al. (1998). "Pten is essential for embryonic development and tumour suppression." Nat Genet 19(4): 348-55.
Eng, C. (2003). "PTEN: one gene, many syndromes." Hum Mutat 22(3): 183-98.
Florkiewicz, R. Z. and A. Sommer (1989). "Human basic fibroblast growth factor gene encodes four polypeptides: three initiate translation from non-AUG codons." Proc Natl Acad Sci U S A 86(11): 3978-81.
Fraser, M. M., X. Zhu, et al. (2004). "Pten loss causes hypertrophy and increased proliferation of astrocytes in vivo." Cancer Res 64(21): 7773-9.
Gupta, R. and S. Brunak (2002). "Prediction of glycosylation across the human proteome and the correlation to protein function." Pac Symp Biocomput: 310-22.Hann, S. R. (1994). "Regulation and function of non-AUG-initiated proto-oncogenes." Biochimie 76(9): 880-6.
Hann, S. R., M. Dixit, et al. (1994). "The alternatively initiated c-Myc proteins differentially regulate transcription through a noncanonical DNA-binding site." Genes Dev 8(20): 2441-52.
Hann, S. R. and R. N. Eisenman (1984). "Proteins encoded by the human c-myc oncogene: differential expression in neoplastic cells." Mol Cell Biol 4(11): 2486-97.
Hann, S. R., M. W. King, et al. (1988). "A non-AUG translational initiation in c-myc exon 1 generates an N-terminally distinct protein whose synthesis is disrupted in Burkitt's lymphomas." Cell 52(2): 185-95.
Hann, S. R., K. Sloan-Brown, et al. (1992). "Translational activation of the non-AUG-initiated c-myc 1 protein at high cell densities due to methionine deprivation." Genes Dev 6(7): 1229-40.
Hershey, J. W. (1991). "Translational control in mammalian cells." Annu Rev Biochem 60: 717-55.
Julenius, K., A. Molgaard, et al. (2005). "Prediction, conservation analysis, and structural characterization of mammalian mucin-type O-glycosylation sites." Glycobiology 15(2): 153-64.
Kiefer, P., P. Acland, et al. (1994). "Competition between nuclear localization and secretory signals determines the subcellular fate of a single CUG-initiated form of FGF3." Embo J 13(17): 4126-36.
Kozak, M. (1989). "Context effects and inefficient initiation at non-AUG codons in eucaryotic cell-free translation systems." Mol Cell Biol 9(11): 5073-80.
Kozak, M. (1990). "Downstream secondary structure facilitates recognition of initiator codons by eukaryotic ribosomes." Proc Natl Acad Sci U S A 87(21): 8301-5.
Kozak, M. (1991). "An analysis of vertebrate mRNA sequences: intimations of translational control." J Cell Biol 115(4): 887-903.
Kwabi-Addo, B., D. Giri, et al. (2001). "Haploinsufficiency of the Pten tumor suppressor gene promotes prostate cancer progression." Proc Natl Acad Sci U S A 98(20): 11563-8.
Lee, J. O., H. Yang, et al. (1999). "Crystal structure of the PTEN tumor suppressor: implications for its phosphoinositide phosphatase activity and membrane association." Cell 99(3): 323-34.
Li, J., L. Simpson, et al. (1998). "The PTEN/MMAC1 tumor suppressor induces cell death that is rescued by the AKT/protein kinase B oncogene." Cancer Res 58(24): 5667-72.
Maehama, T. and J. E. Dixon (1998). "The tumor suppressor, PTEN/MMAC1, dephosphorylates the lipid second messenger, phosphatidylinositol 3,4,5-trisphosphate." J Biol Chem 273(22): 13375-8.
Petrocelli, T. and J. M. Slingerland (2001). "PTEN deficiency: a role in mammary carcinogenesis." Breast Cancer Res 3(6): 356-60.
Prats, H., M. Kaghad, et al. (1989). "High molecular mass forms of basic fibroblast growth factor are initiated by alternative CUG codons." Proc Natl Acad Sci U S A 86(6): 1836-40.
Sulis, M. L. and R. Parsons (2003). "PTEN: from pathology to biology." Trends Cell Biol 13(9): 478-83.
Taira, M., T. Iizasa, et al. (1990). "A human testis-specific mRNA for phosphoribosylpyrophosphate synthetase that initiates from a non-AUG codon." J Biol Chem 265(27): 16491-7.
Torres, J. and R. Pulido (2001). "The tumor suppressor PTEN is phosphorylated by the protein kinase CK2 at its C terminus. Implications for PTEN stability to proteasome-mediated degradation." J Biol Chem 276(2): 993-8.
Vazquez, F., S. R. Grossman, et al. (2001). "Phosphorylation of the PTEN tail acts as an inhibitory switch by preventing its recruitment into a protein complex." J Biol Chem 276(52): 48627-30.
Vazquez, F., S. Ramaswamy, et al. (2000). "Phosphorylation of the PTEN tail regulates protein stability and function." Mol Cell Biol 20(14): 5010-8.
Wang, X., L. C. Trotman, et al. (2007). "NEDD4-1 is a proto-oncogenic ubiquitin ligase for PTEN." Cell 128(1): 129-39.
Xiao, J. H., I. Davidson, et al. (1991). "Cloning, expression, and transcriptional properties of the human enhancer factor TEF-1." Cell 65(4): 551-68.
You, M. J., D. H. Castrillon, et al. (2002). "Genetic analysis of Pten and Ink4a/Arf interactions in the suppression of tumorigenesis in mice." Proc Natl Acad Sci U S A 99(3): 1455-60
【特許請求の範囲】
【請求項1】
配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいる単離されたヒトのホスファターゼおよびtensin ホモログ ロング ポリペプチド(PTEN-long)または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログ,またはその各々のバリアント。
【請求項2】
(i) 配列番号1の残基 1〜173,または(ii) 配列番号5の残基 1〜173を含んでいるそのアナログ,または(iii) 配列番号1の残基 22〜516を含むポリペプチド。
【請求項3】
配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22 〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを含む薬学的組成物。
【請求項4】
対象における固形腫瘍を治療するための方法であって、前記対象における固形腫瘍を治療するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【請求項5】
対象における固形腫瘍の成長を阻害するための方法であって、前記対象における固形腫瘍の成長を阻害するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【請求項6】
対象の固形腫瘍における血管形成を阻害するための方法であって、前記対象の固形腫瘍における血管形成を阻害するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【請求項7】
対象の固形腫瘍における血管の血管上皮細胞(vascular epithelial cell)のアポトーシスを誘導するための方法であって、前記対象の固形腫瘍における血管の血管上皮細胞のアポトーシスを誘導するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【請求項8】
請求項4, 5, 6 または7に記載の方法であって、前記腫瘍は癌性腫瘍である方法。
【請求項9】
請求項8に記載の方法であって、前記癌性腫瘍は、対象のグリア細胞, 前立腺, 卵巣, 子宮, 子宮内膜, 乳房, メラノサイト, 腎臓, 肺, 結腸, 頭部(head), 頸部(neck),または膵臓の腫瘍である方法。
【請求項10】
請求項8に記載の方法であって、前記癌性腫瘍は、PTENにより又はPI3K経路により活性化される又はPTEN-陰性である方法。
【請求項11】
請求項4〜10の何れか一項に記載の方法であって、前記PTEN-long,またはそのアナログ,またはPTEN-long 断片は、前記固形腫瘍への直接的な導入により前記対象に投与される方法。
【請求項12】
請求項11に記載の方法であって、前記PTEN-long,またはそのアナログ,またはPTEN-long 断片は、前記固形腫瘍に注射される方法。
【請求項13】
請求項11に記載の方法であって、前記PTEN-long,またはそのアナログ,またはPTEN-long 断片は、直接的にカテーテルで前記固形腫瘍に導入される方法。
【請求項14】
請求項4〜10の何れか一項に記載の方法であって、前記PTEN-long,またはそのアナログ,またはPTEN-long 断片は、前記固形腫瘍に供給する血管への直接的な導入により前記対象に投与される方法。
【請求項15】
請求項14に記載の方法であって、前記PTEN-long,またはそのアナログ,またはPTEN-long 断片は、前記固形腫瘍へ供給する血管に注射される方法。
【請求項16】
請求項14に記載の方法であって、前記PTEN-long,またはそのアナログ,またはPTEN-long 断片は、直接的にカテーテルで前記固形腫瘍へ供給する血管に導入される方法。
【請求項17】
請求項4〜10の何れか一項に記載の方法であって、前記PTEN-long,またはそのアナログ,またはPTEN-long 断片は、前記対象の静脈内に投与される方法。
【請求項18】
請求項4〜10の何れか一項に記載の方法であって、前記PTEN-long,またはそのアナログ,またはPTEN-long 断片は、前記対象の皮下に投与される方法。
【請求項19】
対象における固形腫瘍を治療するための方法であって、配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5を含んでいるそのアナログをコードする特定量の発現ベクターを、PTEN-long, PTEN-long 断片,又はそのアナログを前記固形腫瘍の細胞中で前記対象における固形腫瘍を治療するための有効な量で発現するように前記対象に投与することを含む方法。
【請求項20】
対象における固形腫瘍の治療に使用するための、対象における固形腫瘍の成長を阻害するための、対象の固形腫瘍における血管上皮細胞のアポトーシスを誘導するための、または対象の固形腫瘍における血管形成を阻害するためのの、配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long)または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログ。
【請求項21】
前記腫瘍は癌性腫瘍である請求項20に記載のPTEN-long又はその断片又はそのアナログ。
【請求項22】
前記癌性腫瘍はグリア, 前立腺, 乳房, 子宮内膜, メラノサイト, 腎臓または肺癌の腫瘍である、請求項21に記載のPTEN-long又はその断片又はそのアナログ。
【請求項23】
前記癌性腫瘍はPTENによりPI3K経路により活性化される又はPTEN-陰性である、請求項21に記載のPTEN-long又はその断片又はそのアナログ。
【請求項24】
それぞれPTEN-long, PTEN-long 断片,又はそのアナログの血漿半減期を増加させる非-ペプチド因子に結合させた、配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログを含む組成物。
【請求項25】
薬学的な担体をさらに含む、請求項24に記載の組成物。
【請求項26】
血漿半減期を増加させる非-ペプチド因子はポリエチレングリコール(PEG)である、請求項24に記載の組成物。
【請求項27】
前記PEGはPTEN-long, PTEN-long 断片, またはそのアナログのC-末端またはN-末端に結合された、請求項26に記載の組成物。
【請求項28】
前記血漿半減期を増加させる非-ペプチド因子は9-フルオレニルメチル クロロホルマート (Fmoc)または(7-スルホ)-9-フルオレニルメトキシカルボニルである、請求項24に記載の組成物。
【請求項29】
配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long)をコードする,または配列番号1の残基 22〜516を含んでいるその断片をコードする,または配列番号5を含んでいるそのアナログをコードする単離された核酸。
【請求項30】
配列番号2または配列番号6で表される配列を有している連続的なヌクレオチドを含む、請求項29に記載の核酸。
【請求項31】
配列番号2または配列番号6で表される配列の連続的なヌクレオチド残基503〜2243を含む、請求項30に記載の核酸。
【請求項32】
配列番号3または配列番号7で表される配列を有している連続的なヌクレオチドを含む、請求項29に記載の核酸。
【請求項33】
配列番号3または配列番号7で表される配列の連続的なヌクレオチド残基503〜2243を含む、請求項32に記載の核酸。
【請求項34】
前記核酸がRNAである、請求項29に記載の核酸。
【請求項35】
前記核酸がDNAである、請求項33に記載の核酸。
【請求項36】
前記核酸がcDNAである、請求項35に記載の核酸。
【請求項37】
配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログをコードしている核酸を含む発現ベクター。
【請求項38】
配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログを発現する能力のある形質転換された細胞であって、前記細胞はそのゲノムにPTEN-longまたはそのアナログをコードする組換えDNAが統合された形質転換された細胞。
【請求項39】
配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログを発現する能力のある宿主細胞であって、PTEN-longまたはそのアナログをコードするプラスミドを含む宿主細胞。
【請求項40】
前記宿主細胞は細菌細胞である、請求項39に記載の宿主細胞。
【請求項41】
前記宿主細胞は哺乳類細胞である、請求項39に記載の宿主細胞。
【請求項42】
それぞれPTEN-long, PTEN-long 断片,又はそのアナログの血漿半減期を増加させる非-ペプチド因子に結合させた配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5を含んでいるそのアナログを作るため、(1) 配列番号1を含んでいるPTEN-long,または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5を含んでいるそのアナログ, および(2) PTEN-long又はそのアナログの血漿半減期を増加させる非-ペプチド因子を混合(admixing)することを含む方法。
【請求項43】
配列番号 2または配列番号6の核酸配列の相補物と,または配列番号2 配列番号6とストリンジェントな条件下で特異的にハイブリダイズする、配列番号2または配列番号6の核酸配列又はその相補物の少なくとも20ヌクレオチドの断片からなる単離された核酸分子。
【請求項44】
(1) 配列番号1のアミノ酸残基 1〜173を含むPTEN-ロング ポリペプチドまたは(2) 配列番号5を含んでいるそのアナログまたは(3) 配列番号3で表される配列を有しているアミノ酸または(4) 配列番号1の残基 22〜516を含んでいるPTEN-longの断片と結合する単離された抗体又はその断片。
【請求項45】
前記抗体がモノクローナル抗体である、請求項44に記載の単離された抗体。
【請求項46】
前記抗体が抗体断片である、請求項44に記載の単離された抗体。
【請求項47】
Fab, Fab', F(ab')2, またはFv断片である、請求項46に記載の抗体断片。
【請求項48】
前記抗体断片が単鎖抗体である、請求項47に記載の抗体。
【請求項49】
ヒト化抗体である、請求項44に記載の抗体。
【請求項50】
PTEN-longのコンホメーションエピトープと結合し、前記PTEN-longは配列番号1の残基 1〜173で表される配列を含むが、しかし、前記抗体はPTENと結合しない単離された抗体又はその断片。
【請求項51】
配列番号1の残基 153〜173の何れかで構成されるエピトープと結合する、請求項50に記載の抗体。
【請求項52】
PTEN-longのエピトープと結合し、前記PTEN-longは配列番号1の残基 1〜173で表される配列を含むが、しかし、前記抗体はPTENと結合しない単離された抗体又はその断片。
【請求項53】
配列番号1の残基 153〜173又はその部分と結合する、請求項52に記載の抗体。
【請求項54】
抗-腫瘍, 抗-血管形成性,または抗-アポトーシス活性を有する、配列番号1,または配列番号5のペプチド断片。
【請求項55】
5〜10, 10〜20, 20〜30, または30〜40 アミノ酸を含む、請求項54に記載のペプチド。
【請求項56】
配列番号1または配列番号5のアミノ酸 1〜173を含む、請求項54に記載のペプチド。
【請求項57】
配列番号1の連続的なアミノ酸残基 22〜516を含む、請求項54に記載のペプチド。
【請求項58】
配列番号1または配列番号5の連続的なアミノ酸残基 174〜576を含まない、請求項54または57に記載のペプチド。
【請求項1】
配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいる単離されたヒトのホスファターゼおよびtensin ホモログ ロング ポリペプチド(PTEN-long)または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログ,またはその各々のバリアント。
【請求項2】
(i) 配列番号1の残基 1〜173,または(ii) 配列番号5の残基 1〜173を含んでいるそのアナログ,または(iii) 配列番号1の残基 22〜516を含むポリペプチド。
【請求項3】
配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22 〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを含む薬学的組成物。
【請求項4】
対象における固形腫瘍を治療するための方法であって、前記対象における固形腫瘍を治療するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【請求項5】
対象における固形腫瘍の成長を阻害するための方法であって、前記対象における固形腫瘍の成長を阻害するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【請求項6】
対象の固形腫瘍における血管形成を阻害するための方法であって、前記対象の固形腫瘍における血管形成を阻害するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【請求項7】
対象の固形腫瘍における血管の血管上皮細胞(vascular epithelial cell)のアポトーシスを誘導するための方法であって、前記対象の固形腫瘍における血管の血管上皮細胞のアポトーシスを誘導するため有効な量の配列番号1で表される配列を有している連続的なアミノ酸残基を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5で表される配列を有している連続的なアミノ酸残基を含んでいるそのアナログを前記対象に投与することを含む方法。
【請求項8】
請求項4, 5, 6 または7に記載の方法であって、前記腫瘍は癌性腫瘍である方法。
【請求項9】
請求項8に記載の方法であって、前記癌性腫瘍は、対象のグリア細胞, 前立腺, 卵巣, 子宮, 子宮内膜, 乳房, メラノサイト, 腎臓, 肺, 結腸, 頭部(head), 頸部(neck),または膵臓の腫瘍である方法。
【請求項10】
請求項8に記載の方法であって、前記癌性腫瘍は、PTENにより又はPI3K経路により活性化される又はPTEN-陰性である方法。
【請求項11】
請求項4〜10の何れか一項に記載の方法であって、前記PTEN-long,またはそのアナログ,またはPTEN-long 断片は、前記固形腫瘍への直接的な導入により前記対象に投与される方法。
【請求項12】
請求項11に記載の方法であって、前記PTEN-long,またはそのアナログ,またはPTEN-long 断片は、前記固形腫瘍に注射される方法。
【請求項13】
請求項11に記載の方法であって、前記PTEN-long,またはそのアナログ,またはPTEN-long 断片は、直接的にカテーテルで前記固形腫瘍に導入される方法。
【請求項14】
請求項4〜10の何れか一項に記載の方法であって、前記PTEN-long,またはそのアナログ,またはPTEN-long 断片は、前記固形腫瘍に供給する血管への直接的な導入により前記対象に投与される方法。
【請求項15】
請求項14に記載の方法であって、前記PTEN-long,またはそのアナログ,またはPTEN-long 断片は、前記固形腫瘍へ供給する血管に注射される方法。
【請求項16】
請求項14に記載の方法であって、前記PTEN-long,またはそのアナログ,またはPTEN-long 断片は、直接的にカテーテルで前記固形腫瘍へ供給する血管に導入される方法。
【請求項17】
請求項4〜10の何れか一項に記載の方法であって、前記PTEN-long,またはそのアナログ,またはPTEN-long 断片は、前記対象の静脈内に投与される方法。
【請求項18】
請求項4〜10の何れか一項に記載の方法であって、前記PTEN-long,またはそのアナログ,またはPTEN-long 断片は、前記対象の皮下に投与される方法。
【請求項19】
対象における固形腫瘍を治療するための方法であって、配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5を含んでいるそのアナログをコードする特定量の発現ベクターを、PTEN-long, PTEN-long 断片,又はそのアナログを前記固形腫瘍の細胞中で前記対象における固形腫瘍を治療するための有効な量で発現するように前記対象に投与することを含む方法。
【請求項20】
対象における固形腫瘍の治療に使用するための、対象における固形腫瘍の成長を阻害するための、対象の固形腫瘍における血管上皮細胞のアポトーシスを誘導するための、または対象の固形腫瘍における血管形成を阻害するためのの、配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long)または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログ。
【請求項21】
前記腫瘍は癌性腫瘍である請求項20に記載のPTEN-long又はその断片又はそのアナログ。
【請求項22】
前記癌性腫瘍はグリア, 前立腺, 乳房, 子宮内膜, メラノサイト, 腎臓または肺癌の腫瘍である、請求項21に記載のPTEN-long又はその断片又はそのアナログ。
【請求項23】
前記癌性腫瘍はPTENによりPI3K経路により活性化される又はPTEN-陰性である、請求項21に記載のPTEN-long又はその断片又はそのアナログ。
【請求項24】
それぞれPTEN-long, PTEN-long 断片,又はそのアナログの血漿半減期を増加させる非-ペプチド因子に結合させた、配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログを含む組成物。
【請求項25】
薬学的な担体をさらに含む、請求項24に記載の組成物。
【請求項26】
血漿半減期を増加させる非-ペプチド因子はポリエチレングリコール(PEG)である、請求項24に記載の組成物。
【請求項27】
前記PEGはPTEN-long, PTEN-long 断片, またはそのアナログのC-末端またはN-末端に結合された、請求項26に記載の組成物。
【請求項28】
前記血漿半減期を増加させる非-ペプチド因子は9-フルオレニルメチル クロロホルマート (Fmoc)または(7-スルホ)-9-フルオレニルメトキシカルボニルである、請求項24に記載の組成物。
【請求項29】
配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long)をコードする,または配列番号1の残基 22〜516を含んでいるその断片をコードする,または配列番号5を含んでいるそのアナログをコードする単離された核酸。
【請求項30】
配列番号2または配列番号6で表される配列を有している連続的なヌクレオチドを含む、請求項29に記載の核酸。
【請求項31】
配列番号2または配列番号6で表される配列の連続的なヌクレオチド残基503〜2243を含む、請求項30に記載の核酸。
【請求項32】
配列番号3または配列番号7で表される配列を有している連続的なヌクレオチドを含む、請求項29に記載の核酸。
【請求項33】
配列番号3または配列番号7で表される配列の連続的なヌクレオチド残基503〜2243を含む、請求項32に記載の核酸。
【請求項34】
前記核酸がRNAである、請求項29に記載の核酸。
【請求項35】
前記核酸がDNAである、請求項33に記載の核酸。
【請求項36】
前記核酸がcDNAである、請求項35に記載の核酸。
【請求項37】
配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログをコードしている核酸を含む発現ベクター。
【請求項38】
配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログを発現する能力のある形質転換された細胞であって、前記細胞はそのゲノムにPTEN-longまたはそのアナログをコードする組換えDNAが統合された形質転換された細胞。
【請求項39】
配列番号1を含んでいるヒト ホスファターゼ および tensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,又は配列番号5を含んでいるそのアナログを発現する能力のある宿主細胞であって、PTEN-longまたはそのアナログをコードするプラスミドを含む宿主細胞。
【請求項40】
前記宿主細胞は細菌細胞である、請求項39に記載の宿主細胞。
【請求項41】
前記宿主細胞は哺乳類細胞である、請求項39に記載の宿主細胞。
【請求項42】
それぞれPTEN-long, PTEN-long 断片,又はそのアナログの血漿半減期を増加させる非-ペプチド因子に結合させた配列番号1を含んでいるヒト ホスファターゼおよびtensin ホモログ ロング ポリペプチド (PTEN-long),または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5を含んでいるそのアナログを作るため、(1) 配列番号1を含んでいるPTEN-long,または配列番号1の残基 22〜516を含んでいるその断片,または配列番号5を含んでいるそのアナログ, および(2) PTEN-long又はそのアナログの血漿半減期を増加させる非-ペプチド因子を混合(admixing)することを含む方法。
【請求項43】
配列番号 2または配列番号6の核酸配列の相補物と,または配列番号2 配列番号6とストリンジェントな条件下で特異的にハイブリダイズする、配列番号2または配列番号6の核酸配列又はその相補物の少なくとも20ヌクレオチドの断片からなる単離された核酸分子。
【請求項44】
(1) 配列番号1のアミノ酸残基 1〜173を含むPTEN-ロング ポリペプチドまたは(2) 配列番号5を含んでいるそのアナログまたは(3) 配列番号3で表される配列を有しているアミノ酸または(4) 配列番号1の残基 22〜516を含んでいるPTEN-longの断片と結合する単離された抗体又はその断片。
【請求項45】
前記抗体がモノクローナル抗体である、請求項44に記載の単離された抗体。
【請求項46】
前記抗体が抗体断片である、請求項44に記載の単離された抗体。
【請求項47】
Fab, Fab', F(ab')2, またはFv断片である、請求項46に記載の抗体断片。
【請求項48】
前記抗体断片が単鎖抗体である、請求項47に記載の抗体。
【請求項49】
ヒト化抗体である、請求項44に記載の抗体。
【請求項50】
PTEN-longのコンホメーションエピトープと結合し、前記PTEN-longは配列番号1の残基 1〜173で表される配列を含むが、しかし、前記抗体はPTENと結合しない単離された抗体又はその断片。
【請求項51】
配列番号1の残基 153〜173の何れかで構成されるエピトープと結合する、請求項50に記載の抗体。
【請求項52】
PTEN-longのエピトープと結合し、前記PTEN-longは配列番号1の残基 1〜173で表される配列を含むが、しかし、前記抗体はPTENと結合しない単離された抗体又はその断片。
【請求項53】
配列番号1の残基 153〜173又はその部分と結合する、請求項52に記載の抗体。
【請求項54】
抗-腫瘍, 抗-血管形成性,または抗-アポトーシス活性を有する、配列番号1,または配列番号5のペプチド断片。
【請求項55】
5〜10, 10〜20, 20〜30, または30〜40 アミノ酸を含む、請求項54に記載のペプチド。
【請求項56】
配列番号1または配列番号5のアミノ酸 1〜173を含む、請求項54に記載のペプチド。
【請求項57】
配列番号1の連続的なアミノ酸残基 22〜516を含む、請求項54に記載のペプチド。
【請求項58】
配列番号1または配列番号5の連続的なアミノ酸残基 174〜576を含まない、請求項54または57に記載のペプチド。
【図1】



【図2】
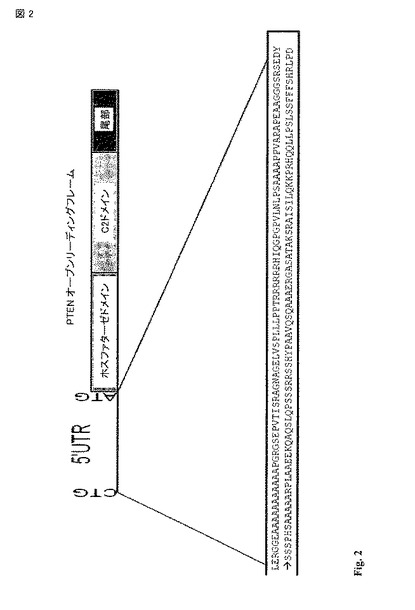
【図3】

【図4】
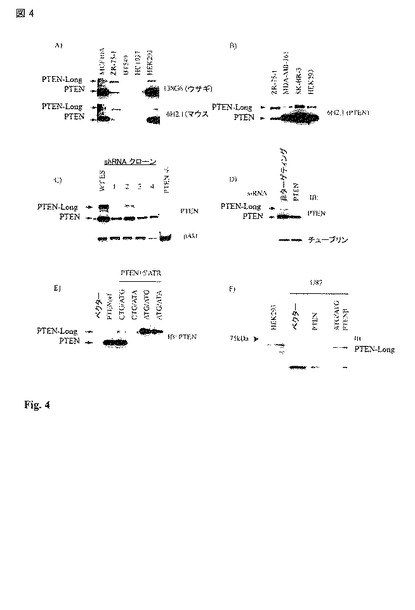
【図5】
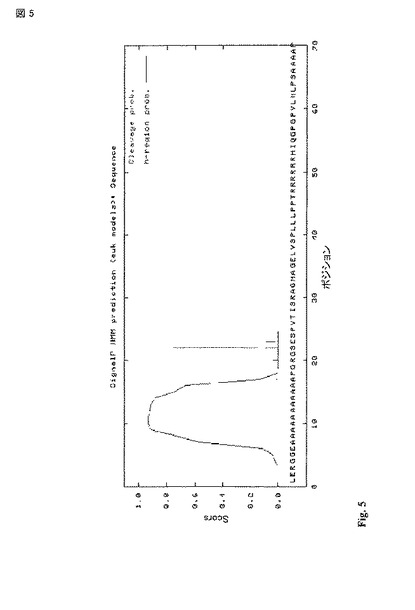
【図6】
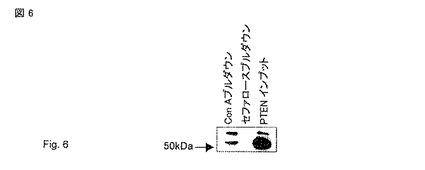
【図7】
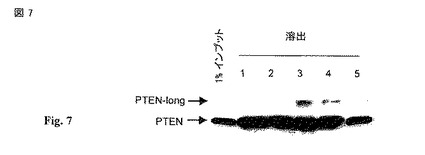
【図8】
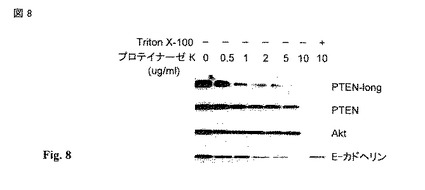
【図9】
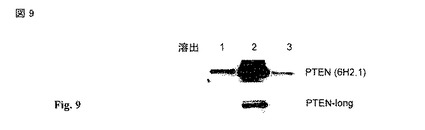
【図10】
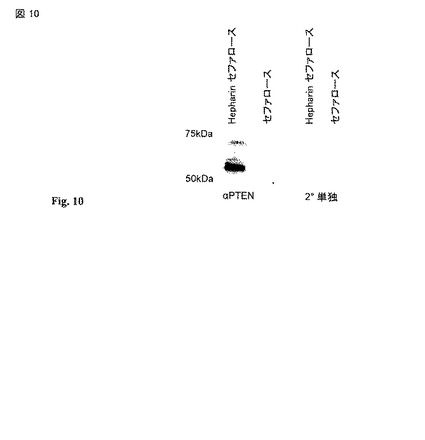
【図11】
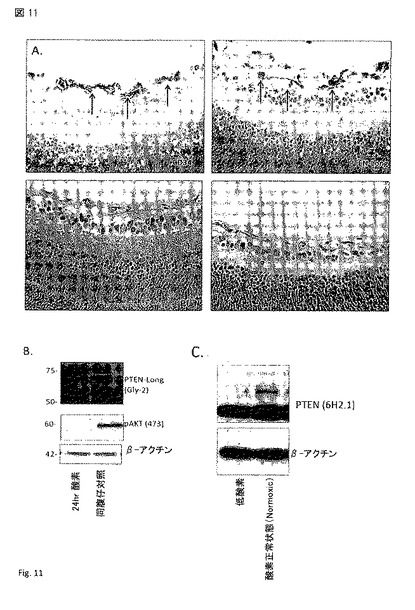
【図12】

【図13】
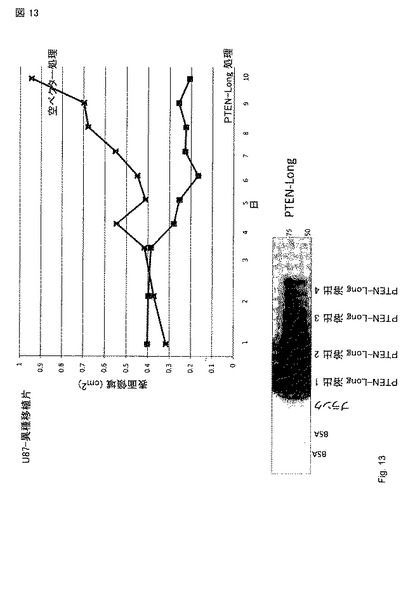
【図14】
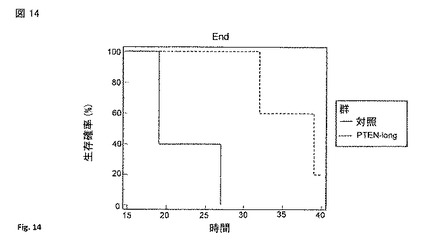
【図15】
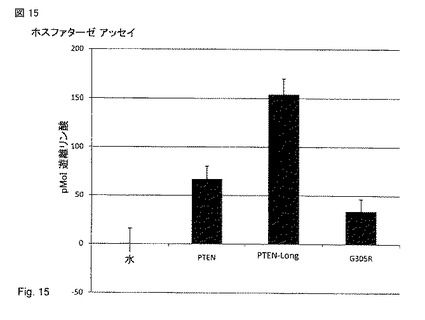
【図16】
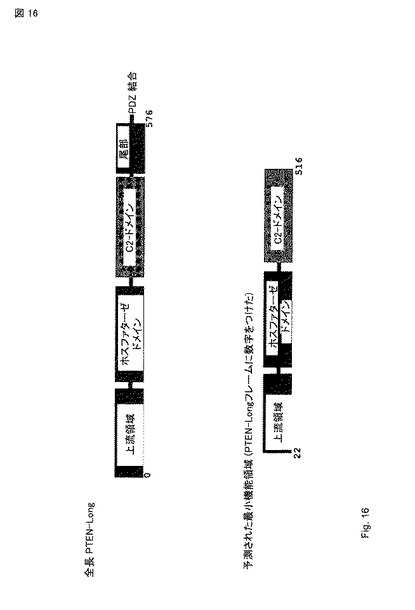
【図17】
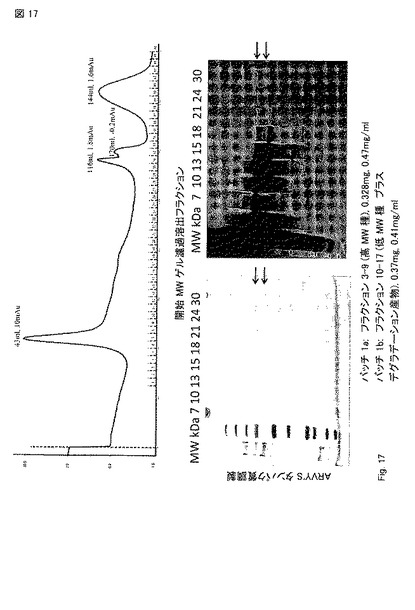
【図18】
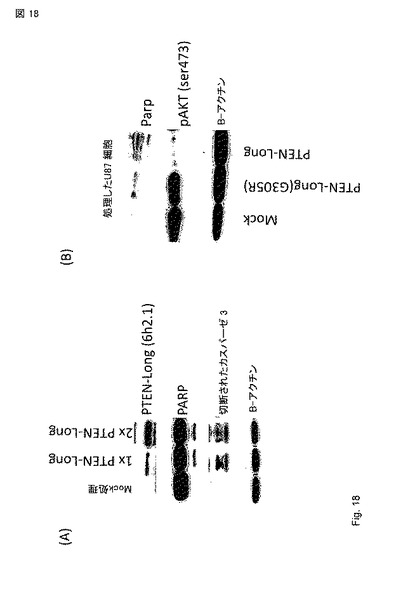
【図19】
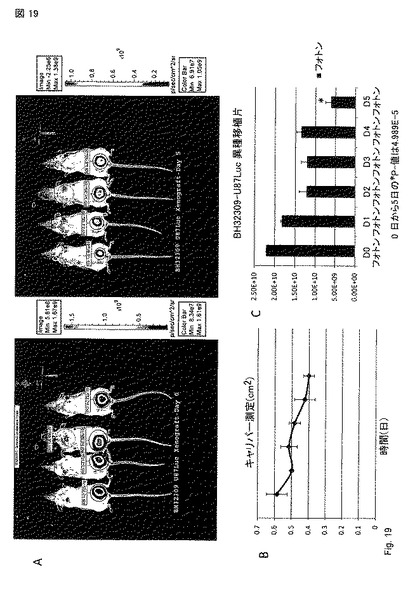
【図20】

【図21】
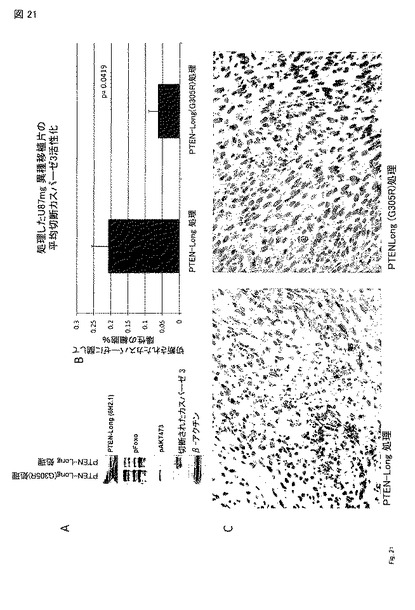
【図22】
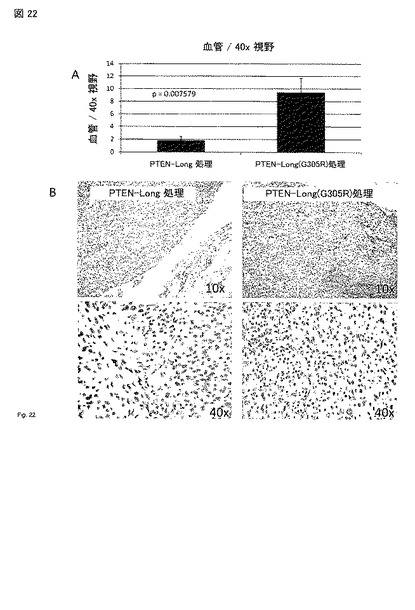
【図23】
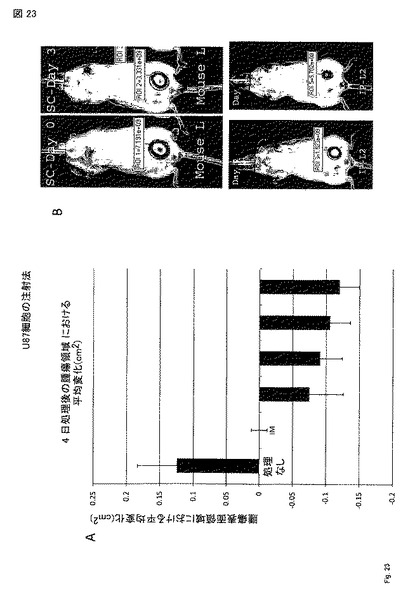
【図24】
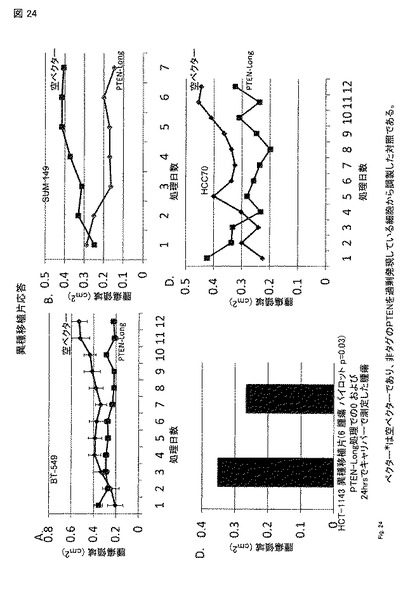
【図25】
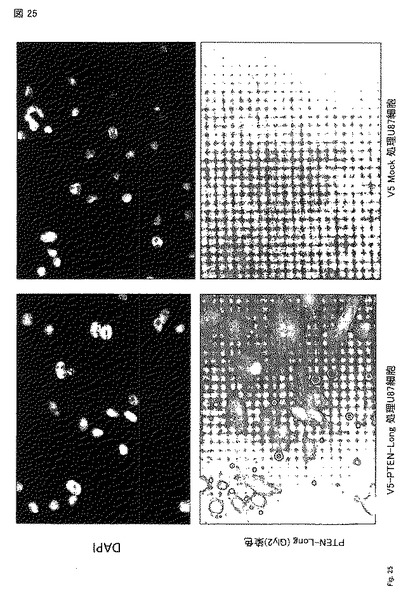
【図26】
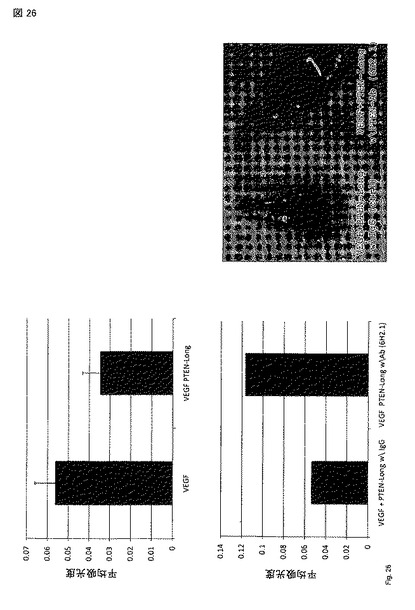
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【公表番号】特表2012−517827(P2012−517827A)
【公表日】平成24年8月9日(2012.8.9)
【国際特許分類】
【出願番号】特願2011−551062(P2011−551062)
【出願日】平成22年2月17日(2010.2.17)
【国際出願番号】PCT/US2010/000469
【国際公開番号】WO2010/096173
【国際公開日】平成22年8月26日(2010.8.26)
【出願人】(592104782)ザ・トラスティーズ・オブ・コランビア・ユニバーシティー・イン・ザ・シティー・オブ・ニューヨーク (21)
【氏名又は名称原語表記】THE TRUSTEES OF COLUMBIA UNIVERSITY IN THE CITY OF NEW YORK
【Fターム(参考)】
【公表日】平成24年8月9日(2012.8.9)
【国際特許分類】
【出願日】平成22年2月17日(2010.2.17)
【国際出願番号】PCT/US2010/000469
【国際公開番号】WO2010/096173
【国際公開日】平成22年8月26日(2010.8.26)
【出願人】(592104782)ザ・トラスティーズ・オブ・コランビア・ユニバーシティー・イン・ザ・シティー・オブ・ニューヨーク (21)
【氏名又は名称原語表記】THE TRUSTEES OF COLUMBIA UNIVERSITY IN THE CITY OF NEW YORK
【Fターム(参考)】
[ Back to top ]