
腫瘍性疾患の治療的処置にて用いるための成体幹細胞由来の微細小胞(MV)
本願発明は腫瘍の治療的処置の分野に関する。本願発明者らは成体幹細胞より由来の微細小胞が、腫瘍性疾患を患っている患者に投与された場合に、著しい抗腫瘍作用を発揮することを見出した。好ましい微細小胞は、骨髄間充織幹細胞、糸球体間充織幹細胞または非楕円形肝臓幹細胞より誘導される。
【発明の詳細な説明】
【技術分野】
【0001】
本願発明は腫瘍性疾患の治療的処置に関する。
【背景技術】
【0002】
造血幹細胞移植は、充実性腫瘍の患者にて腫瘍阻害作用を発揮し、ならびに転移性の乳癌、腎臓癌、卵巣癌、前立腺癌および膵臓癌にて抗腫瘍作用を発揮することが知られている。
【0003】
ヒト骨髄間充織幹細胞(BM−MSC)は多種の器官の修復および組織の修復に寄与することが明らかにされ、実証研究はMSCの移植が心臓、肝臓および腎臓を含むいくつかの器官の機能性および構造的回復に有益な効果のある可能性があることを示唆した。
【0004】
幹細胞の微小環境は、増殖を阻害し、分化を促進するシグナルを提供することで発癌を防止するのに不可欠な役割を発揮するようである。
【0005】
しかしながら、幹細胞の治療指標における使用は、かかる幹細胞療法の腫瘍形成能の危険性が報告されていることを考えれば、それほど望ましいものではない(Amariglio Nら、Donor-derived brain tumor following neural stem cell transplantation in an ataxia telangiectasia patient. PLoS Med. 2009 Feb 17;6(2))。
【0006】
細胞由来の微細小胞(MV)は、特徴的な抗原を発現する細胞によって放出される小胞であって、その細胞から膜および細胞質成分を生成して担持し、細胞通信の新たな機構であると記載されている。最近になって、本願発明者らはヒト内皮前駆細胞(EPC)、BM−MSCおよび肝臓幹細胞から由来の微細小胞が、標的となる分化した細胞を内皮細胞、尿細管上皮細胞および肝細胞として再プログラム化し(Deregibus MCら、Endothelial progenitor cell derived microvesicles activate an angiogenic program in endothelial cells by a horizontal transfer of mRNA. Blood. 2007 Oct 1;110(7):2440-8;Bruno Sら、Mesenchymal stem cell-derived microvesicles protect against acute tubular injury. J Am Soc Nephrol. 2009 May;20(5):1053-67;Herrera MBら、Human liver stem cell-derived microvesicles accelerate hepatic regeneration in hepatectomized rats. J Cell Mol Med. 2009 Jul 24)、組織の再生および修復に貢献し得る、遺伝物質(mRNA)の移動のためのビヒクルとして供し得ることを明らかにした。
【発明の概要】
【発明が解決しようとする課題】
【0007】
国際特許出願WO2009/087361は、誘導物質を未分化細胞の第一集団に適用することで分化細胞を産生し、ついで微細小胞をその分化細胞より単離することを開示する。この点については、WO2009/087361の30頁の第1段落のすべてを参照のこと。未分化細胞の第1集団は、例えば、骨髄間充織幹細胞である。しかし、WO2009/087361は未分化細胞より微細小胞を直接得ることを開示していない。
【課題を解決するための手段】
【0008】
本願発明者らは、この度、成体肝細胞、好ましくは骨髄または糸球体間充識幹細胞より、あるいは非楕円形肝臓幹細胞より由来の微細小胞(MV)が、インビトロおよびインビボの両方で顕著な抗腫瘍活性を示し、それにより癌の治療的処置のための対応するすべての幹細胞にわたって別途利点を示しうることを見出した。本願発明の成体幹細胞より由来の微細小胞の抗腫瘍活性は、該微細小胞の、種々のヒト癌細胞株の増殖およびアポトーシスに対する効果、ならびにその毛細血管様構造の形成に対する効果を測定することによりインビボにて明らかにされた。微細小胞の抗腫瘍活性もまた、MV−処置の腫瘍増殖に対する効果を測定することでインビボのマウス実験にて確認された。
【0009】
成体幹細胞より由来の微細小胞の抗腫瘍活性の観察は先行技術では全く予期せぬものである。間充織幹細胞(MSC)の調製物は、実際、いくつかの組織にて再生作用を発揮することが知られている。例えば、骨髄由来のMSCは、可溶性細胞外マトリックス糖蛋白、サイトカインおよび成長因子を含む、多くの栄養性分子を分泌することにより、自然と、血液生成を支持することが知られている。その上、内皮幹細胞より由来の微細小胞は、WO2009/050742より、インビトロおよびインビボの両方で、血管形成およびアポトーシスに対する耐性を促進することが分かっている。WO2009/057165では、幹細胞より由来の微細小胞は、損傷した組織または器官の内皮および上皮再生を誘発することが明らかにされた。
【0010】
WO2009/105044にて、間充織幹細胞より由来の微細小胞は、多数かつ多種の疾患の治療に用いるのに適すると推論している。癌の処置について、実験的証拠はなく、理論的説明もなく、具体的な方法も提供されていない。
【0011】
かくして、本願発明の第1の態様は、腫瘍性疾患の治療的処置に用いるための成体幹細胞より由来の微細小胞である。
【0012】
これに関連して、微細小胞は、それらを投与する同一患者より採取される成体幹細胞に由来する必要はないことを理解すべきである。むしろ、違う対象に由来して調製され、医薬の形態にて維持され、次にその必要とする患者に投与されてもよい。これはいわゆる同種アプローチである。
【0013】
好ましい実施態様において、成体幹細胞はヒト間充織幹細胞またはヒト肝臓幹細胞である。好ましいヒト肝臓幹細胞は、WO2006/126219に開示される間充織および胚幹細胞マーカーの両方を発現するヒト非楕円形肝臓幹細胞(HLSC)である。この細胞株は、成体幹細胞が、肝細胞マーカーを発現し、成熟肝細胞、インスリン産生細胞、骨形成原細胞および上皮細胞への分化能を有する、好ましくはアルブミン、α−フェト蛋白、CK18、CD44、CD29、CD73、CD146、CD105、CD90からなる群より選択されるマーカーを発現し、好ましくはCD133、CD117、CK19、CD34、チトクロムP450からなる群より選択されるマーカーを発現しない、成体組織より単離される、非楕円形ヒト肝臓多能性前駆細胞株である点で特に特徴付けられる。
【0014】
もう一つ別の好ましい実施態様において、ヒト間充織幹細胞は、ヒト成体骨髄(BM−MSC)より誘導される。もう一つ別の好ましい実施態様にて、ヒト間充織幹細胞は、欧州特許出願番号第08425708.8に開示されるように、ヒト成体脱糸球体嚢(Gl−MSC)より誘導される。これらの細胞はさらには、該細胞がCD133陰性、CD146陽性およびCD34陰性であり、有足細胞、内皮細胞およびメサンギウム細胞への分可能を有し、好ましくはまたCD24、Pax−2、CD31、CD29、CD44、CD73、CD90、CD105、CD166、ナノグ、ムサシ、ビメンチン、ネスチンからなる群より選択されるマーカーを発現し、好ましくはα−SMA、Oct−4、CD45、サイトケラチン、CD80、CD86、CD40からなる群より選択されるマーカーを発現しない点で特徴付けられる。
【0015】
本願発明の一の実施態様によれば、腫瘍性疾患は、肝臓腫瘍(例、肝癌)、上皮腫瘍(例、カポシ肉腫)、乳房腫瘍(例、乳腺腺癌)、肺腫瘍、前立腺腫瘍、胃腫瘍、結腸腫瘍および卵巣腫瘍からなる群より選択される。
【0016】
本願発明のもう一つ別の態様は、上記した腫瘍性疾患の治療的処置のための医薬の製造にて、上記の成体幹細胞より誘導される微細小胞を用いることである。
【0017】
一の実施態様にて、該治療的処置は、一または複数の細胞傷害性または細胞増殖抑制性剤を投与することを含む。適当な細胞傷害性および細胞増殖抑制性剤として、例えば、パクリタセル(Paclitaxel)、レナリドミド(Lenalidomide)、ポマリドミド
(Pomalidomide)、エピルビシン(Epirubicin)、5FU、スニチニブ(Sunitinib)、ラ−パチニブ(La-patinib)、カネルチニブ(Canertinib)、シクロホスファミド
(cyclophosphamide)、ドキソルビシン(doxorubicin)、レナリドミデン/デキサメタゾン(Lenalidomiden/Dexamethason)、ポ−マリドミド(Po-malidomide)/デキサメタゾン、カルボプラチン(Carboplatin)、ラパマイシン(Rapamycin)、ミトキサントロン(mitoxantron)、オキサリプラチン(oxaliplatin)、ドキセタセル(docetaxel)、ビノレルビン(vinorelbin)、ビンクリスチン(vincristine)およびその組み合わせが挙げられる。ドキソルビシンおよび/またはビンクリスチンと、成体幹細胞より由来のMVとの組み合わせの投与が極めて好ましい。というのも、かかる組み合わせは相乗作用を発揮することが示されたからである(図13を参照)。
【0018】
微細小胞は、その必要とする患者に、局所または全身投与のいずれかで投与される。局所および全身投与の両方に適する医薬剤形は、例えば、注射可能な剤形である。一例として、微細小胞が、充実性腫瘍で局所的な腫瘍内(i.t.)注射にて、あるいは充実性腫瘍の場合と、転移癌の場合の両方で静脈内注射または注入にて投与される。投与されるべきMVの適当な用量は複数の因子に依存するが、一般に、受容者の体重1kg当たり0.1−200μg、好ましくは受容者の体重1kg当たり1−150μg、さらにより好ましくは受容者の体重1kg当たり3−120μgで構成される。
【0019】
本願明細書にて用いられる「成体幹細胞より由来の微細小胞(MV)」なる語は、少なくとも部分的に成体幹細胞より誘導される、膜粒子に言及する。同様に、「成体幹細胞」なる語は、増殖能(自己再生能)および分化能(塑性能)を有し、それによってライフサイクルの終わりを迎える特化された細胞系統の成熟細胞に取って代わる、未分化または一部未分化の細胞を包含する。本願明細書にて用いられる「成体幹細胞」なる語は、無制限の自己再生能および多能性塑性を有する肝細胞、および多能性塑性を有し、ある場合には制限された自己再生能を有する前駆細胞の両方を包含する。好ましい実施態様において、「成体幹細胞」は、少なくとも2種の、より好ましくは少なくとも3種の異なる型の特化された、十分に分化された成熟細胞への分化能を有する、多能性または複数の能力の塑性を有することを意味する。
【0020】
本願明細書の文脈の中で、「成体幹細胞」なる語は、胚盤胞の内細胞塊より単離される「胚幹細胞」とは異なり、意図的に、成体組織より単離される幹細胞を意味する。成体幹細胞はまた、「体性幹細胞」としても知られている。
【0021】
本願明細書の文脈の中で、「成体幹細胞より由来の微細小胞(MV)」なる表現は、意図的に、微細小胞が未分化の幹細胞より直接誘導されることを意味する。
【0022】
本願明細書の文脈の中で、「直接」なる語は、微細小胞を得る前にいずれの分化工程がなされることも、生じることもなく、微細小胞が未分化の成体幹細胞より誘導されることを意味する。
【0023】
本願発明にて使用される成体幹細胞より誘導される微細小胞は、一般に、形状が球体であり、100nm〜5μm、より典型的には0.2〜1μmの範囲の直径を有する。仮に粒子の形状が球体でない場合、上記した値は該粒子の最大の寸法に言及するものである。
【0024】
本願発明にて使用される微細小胞をもたらす幹細胞は、本願明細書の実験のセクションにて記載されるように単離されてもよい。次に、微細小胞(MV)は、単離された幹細胞の上澄から、例えば本願明細書の実験のセクションに開示される超遠心分離により得られてもよい。ついで、その単離されたMVは、1または複数の抗凍結剤を配合した懸濁液中、超低温、例、−80℃で凍らせることで使用するまで貯蔵されてもよい。適当な抗凍結剤は、例えば、ジメチルスルホキシド(DMSO)およびグリセロールである。DMSOの細胞懸濁液の1容量%濃度での使用は、細胞の良好な保存および患者に再び注入された場合の毒性作用の制限を保証する。抗凍結剤として言及されてもよい他の物質は、いわゆる、密なバリアを細胞表面に形成し、細胞内脱水を減らすように作用する高分子量物質である。
【0025】
本願発明のさらなる目的および利点は、単に例示として用いられる、以下の実施例からさらに明らかになるであろう。実施例において、以下の図面が参照される。
【図面の簡単な説明】
【0026】
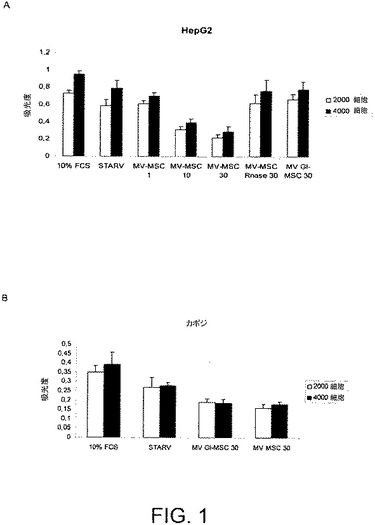
【図1】図1AおよびBは、HepG2細胞(図1A)およびKS細胞(図1B)を異なる量の微細小胞(MV)と一緒にインキュベートすることで、ビヒクル単独でインキュベートされた対照となる細胞と比べて、増殖が顕著に阻害されることを示す。HepG2およびKS細胞の増殖は、異なる量のBM−MSCからのMV(1、10および30μg/ml)、またはRNaseで前処理されている、もしくはされていないGl−MSCからのMVと一緒にインキュベーションした48時間後に、BrdU取り込みアッセイで評価された。結果は3回の実験の平均値±SDで表される。
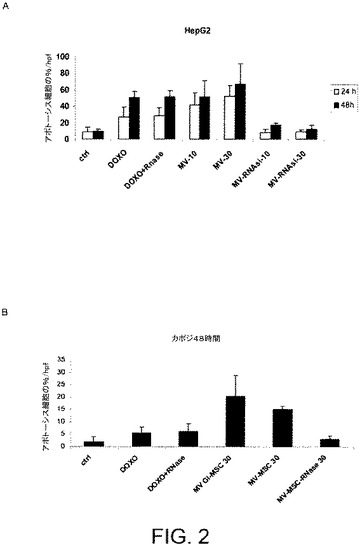
【図2】図2AおよびBは、HepG2およびKS細胞とMVとのインキュベーションが、ビヒクル単独でインキュベートされた対照と、同様にドキソルビシン刺激と比べて、アポトーシスを顕著に促進することを示す。HepG2およびKS細胞のアポトーシスは、BM−MSC(RNaseで前処理されているか、されていない)またはGl−MSCからの異なる量のMVと一緒にインキュベーションした24時間および/または48時間後のアポトーシス細胞の割合としてTunelアッセイで評価された。結果は3回の平均値±SDで表される。
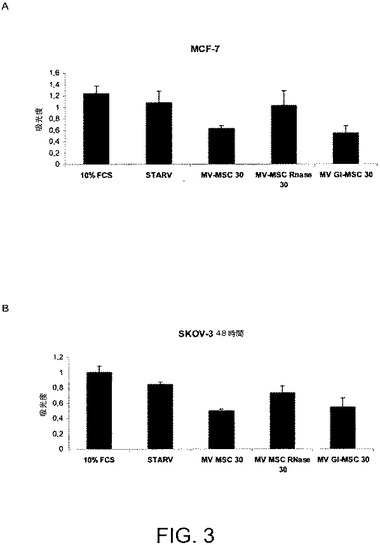
【図3】図3Aおよび3Bは、MCF−7細胞(図3A)およびSKOV−3細胞(図3B)を、BM−MSCからの30μg/mlのMVと一緒に48時間インキュベートすることで、ビヒクル単独でインキュベートされた対照細胞と比べて増殖が顕著に阻害されることを示す。MCF−7およびSKOV−3細胞の増殖をBM−MSCからの、またはGl−MSCからの30μg/mlのMVと一緒にインキュベートした48時間後にBrdU取り込みアッセイにより評価した。結果は3回の実験の平均値±SDで表される。
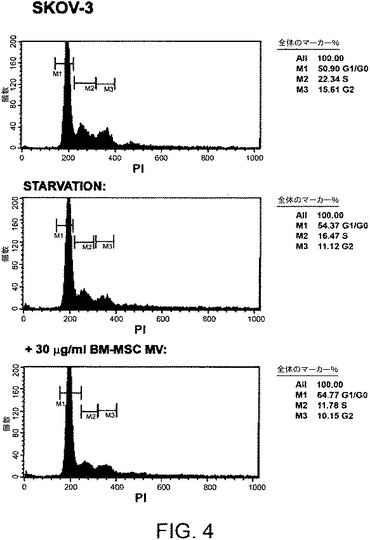
【図4】BM−MSCからのMVが、特にSKOV−3細胞のG0/G1フェーズにて細胞の増加を誘発することを示す。DNA含量を、10%FCSと共に、FCSを除き(飢餓)、30μg/mlのMVの存在下で24時間培養したSKOV−3細胞にて測定した。MVの存在下、G0/G1フェーズにて細胞数の増加が観察される。
【図5】図5Aおよび5BはMVのインビトロでの血管形成における作用を評価するのに実施された実験の結果を示す。HUVECおよびTECのマトリゲル内での毛細血管様構造を形成する能力について評価した。
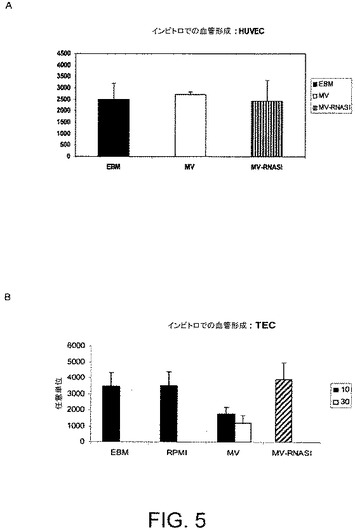
【図6】MVのインビトロでのアポトーシスにおける作用を評価するのに実施された実験の結果を示す。HUVECおよびTECのアポトーシスを、異なる量のMVと一緒にインキュベートした48時間後のアポトーシス細胞の割合としてTunelアッセイで評価した。
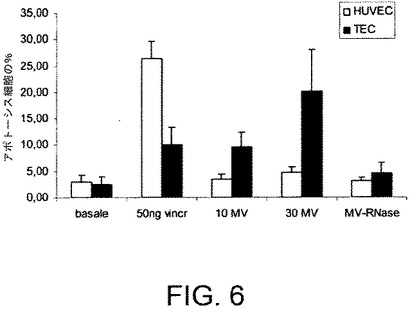
【図7】MVのTECの増殖に対する作用を評価するのに実施された実験の結果を示す。TECの増殖は、RNaseで前処理されているか、またはされていないBM−MSCからの異なる量のMV(10および30μg/ml)と一緒にインキュベートした48時間後にBrdU取り込みアッセイで評価した。結果は2回の実験の平均値±SDとして表される。
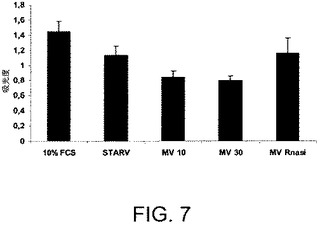
【図8】HepG2異種移植片のSCIDマウスに腫瘍内投与された、RNaseで処理されているか、またはされていないMVのインビボでの抗腫瘍活性を示す。腫瘍塊を毎週該移植片の2つの直交する寸法をキャリパーを用いて測定することにより決定した。結果は腫瘍塊の増大の割合として表され:第1処置(HepG2注射の1週間後)の腫瘍塊を慣例により100%値として固定する。
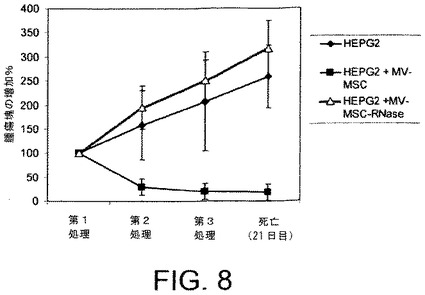
【図9】BM−MSCから由来のMVがインビボにて腫瘍増殖を減少させることを示す。A)BM−MSCから由来のMVで処置されている(右図)または処理されていない(左図)HepG2腫瘍の代表例;B)BM−MSCからのMVで処置されている(右図)または処理されていない(左図)切除されたHepG2腫瘍の代表例;C)BM−MSCからのMVで処置されている(右図)または処理されていない(左図)HepG2腫瘍のヘマトキシリンおよびエオシン染色。
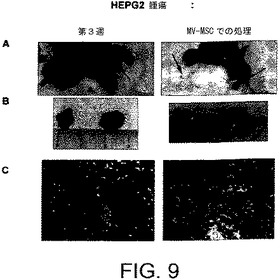
【図10】MVのRNaseを用いる前処理がBM−MSCから由来のMVの抗腫瘍活性を無効にすることを示す。A)MV−RNaseで処置されている切除されたHepG2腫瘍の代表例;B)MV−RNaseで処置されているHepG2腫瘍のヘマトキシリンおよびエオシン染色。
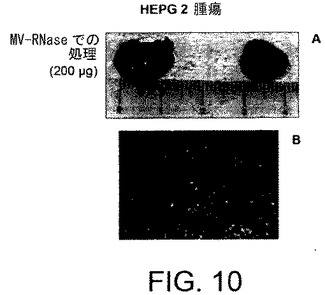
【図11】HepG2上のBrdUをベースとする増殖アッセイの結果を示すグラフである。HepG2をDMEM単独中に、あるいは異なる量のHLSCより由来のMVを補足したDEMEにて培養した。3日後、HepG2増殖がBrdU取り込みアッセイを用いて定量された。
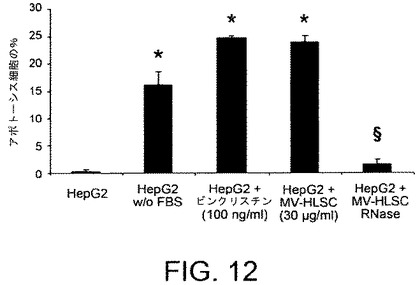
【図12】HepG2細胞上のアポトーシスアッセイの結果を示すグラフである。HepG2細胞は、DMEM単独中で、または10%FCSを含むDMEM中で、ビンクリスチン(100ng/ml)またはMV−HLSC(30μg/ml)またはMV−HLSC(RNaseで前処理されている、30μg/ml)を補足して培養された。24時間後に分析を行った。
【図13】基底条件下のHepG2細胞、あるいはビトロネクチン、ドキソルビシン、MV−HLSCまたはビトロネクチン+MV−HLSCおよびドキソルビシン+MV−HLSCで処理したHepG2細胞でのアポトーシスアッセイの結果を示すグラフである。
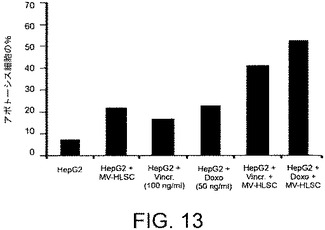
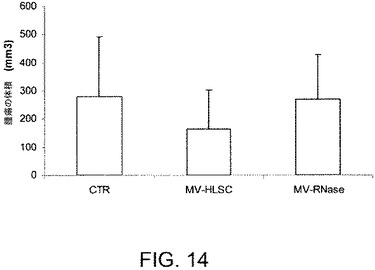
【図14】MV−HLSC(n=3)、ビヒクル(n=2)またはRNase処理のMV−HLSC(n=3)でi.t.処理された後で、マウスを殺した際の、回収されたHepG2腫瘍の体積を測定することで得られたデータを示すグラフである。腫瘍の体積は該移植片の2つの直交する寸法をキャリパーを用いて毎週測定することにより決定された。
【図15】HLSC−MV処理によるインビボでの腫瘍増殖の阻害および腫瘍内アポトーシスの誘発を示す顕微鏡写真を図示する。A)4週間後に回収されたHepG2腫瘍のアポトーシス、PCNAおよびヘマトキシリン&エオシン染色を示す代表的顕微鏡写真;B)MV−処理されたマウスから回収されたHepG2腫瘍のアポトーシス、PCNAおよびヘマトキシリン&エオシン染色を示す代表的顕微鏡写真;C)MV−RNAase処理されたマウスから回収されたHepG2腫瘍のアポトーシス、PCNAおよびヘマトキシリン&エオシン染色を示す代表的顕微鏡写真。
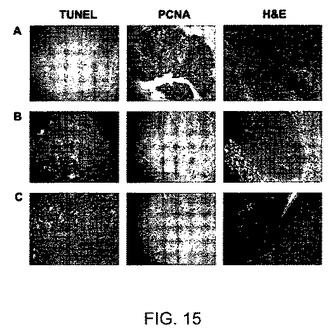
【図16】MCF−7細胞を異なる濃度のHLSC−MVと共にインキュベートすることにより実施されたインビトロでの増殖アッセイの結果を示すグラフである。MCF−7細胞の増殖は、2、10、15および30μg/mlのHLSC細胞から由来のMVと共に48時間インキュベートした後に、BrdU取り込みアッセイにより評価された。該実験を3回重複して行った。P<0.05。
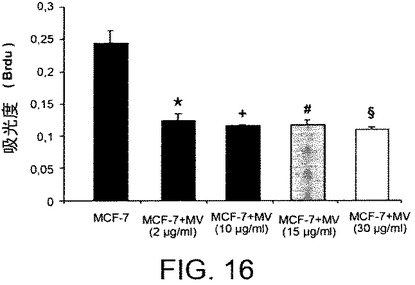
【図17】カポジ細胞(KS)を異なる濃度のHLSC−MVと共にインキュベートすることにより実施されたインビトロでの増殖アッセイの結果を示すグラフである。カポジ細胞の増殖は、2、10、15および30μg/mlのHLSC細胞から由来のMVと共に48時間インキュベートした後に、BrdU取り込みアッセイにより評価された。該実験を3回重複して行った。P<0.05。
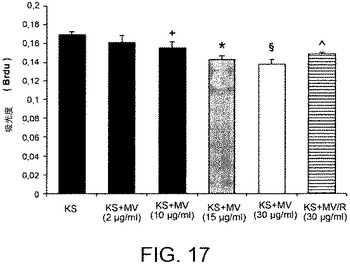
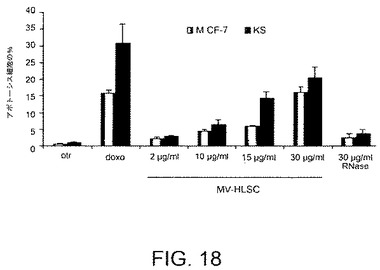
【図18】MCF−7細胞およびカポジ細胞をHLSC−MVと共にインキュベートすることにより実施されたインビトロでのアポトーシスアッセイの結果を示すグラフである。アポトーシスは異なる量のMV(2、10、15および30μl/ml)および30μg/mlのRNase処理されたMVと共にインキュベートされた48時間後のアポトーシス細胞の割合としてTUNELアッセイにより評価された。アポトーシス誘発の陽性対照としてドキソルビシンを用いた。陰性対照にて、MCF−7細胞およびカポジ細胞をビヒクル単独で処理した。該実験を3回重複して行った。P<0.05。
【発明を実施するための形態】
【0027】
1.間充織幹細胞(MSC)からの微細小胞(MV)
【0028】
1.1 MSCの単離および特徴付け
【0029】
骨髄細胞をフィコール(Ficoll)グラジエント(密度:1022g/ml;Sigma-Aldrich、St. Louis、MO)上に積層させ、1500rpmで30分間遠心分離に付した。単核細胞を間充織幹細胞基底培地(MSCBM、Lonza)の存在下で培養させた。5日間培養した後、培地を変換した。単離された細胞を拡張させるために、付着した単層を、第1継代について15日目に、その後の継代については7日毎に37℃で5分間トリプシン処理に付して分離した。細胞を10000細胞/cm2の密度で播き、継代6より後の細胞は使用されなかった。
【0030】
糸球体からのMSC集団(Gl−MSC)が、Bruno Sら、Isolation and characterization of resident mesenchymal stem cells in human glomeruli. Stem Cells Dev. 2009;18:867-880に記載されるように、手術で摘出された腎臓の皮質の正常な部分より得られた。皮質の切開した後で、糸球体の懸濁液を標準的な確立された方法を用いて集めた:一連の等級別のメッシュ(60および120メッシュ)に通した後、最上部にある120メッシュのシーブで糸球体を集めた。次に、糸球体を自然沈降(室温で10分間)により円錐管の底部で集め、ボーマン嚢の臓側板を、10mlのピペットを用いて吸引/排出を数回行うことで機械的に、かつコラゲナーゼI(Sigma, St.Louis, MO)を用いて2分間消化することで酵素的に除去した。ついで、細胞およびボーマン嚢を取り出すために、糸球体を自然沈降により円錐管の底部に集め、それをフィブロネクチンを被覆したT25フラスコ(Falcon、BD Bioscience、Two Oak Park、Bedford、MA)に移した。糸球体を間充織幹細胞基礎培地(MSCBM、Lonza)の存在下で培養した。細胞を放置し、継代前に集密とさせた:継代4になるまでの継代間の間隔は変動し(3−7日)、それからは約7日で確立された。
【0031】
各継代の際に、細胞を計数し、細胞蛍光分析および免疫蛍光分析により免疫表現型について分析した。細胞蛍光分析は、次の抗体、すべてのフィコエリトリン(PE)またはフルオレセインイソチオシアネート(FITC)接合体:抗−CD105、−CD29、−CD31、−CD146、−CD44、−CD90(Dakocytomation、Copenhagen、Denmark);−CD73、−CD34、−CD45、−CD80、−CD86、−CD166、HLA−I(Becton Dickinson Biosciences Pharmingen、San Jose、CA);−CD133(Miltenyi Biotec、Auburn);KDR(R&D Systems、Abington、U.K.);−HLA−II(Chemicon International Temecula、CA)、−CD40(Immunotech、Beckman Coulter)、−CD154(Serotec、Raleigh、NC USA)モノクローナル抗体を用いて実施された。マウスIgG同形対照はDakocytomationからのものであった。インキュベーションはすべて、0.1%ウシ血清アルブミンおよび0.1%アジ化ナトリウムを含有する、100μlのリン酸緩衝セイライン(PBS)中、4℃で行われた。各サンプルについて、10000個の細胞をFACSCalibur血球計算器(BD Biosciences Pharmingen)で分析した。ゲーティングは陰性対照に基づいて構成され、行われたすべての分析にて補償対照が含められた。セルクエストソフトウェア(Cell Quest software)(BD Biosciences Pharmingen)を用いて各実験からゲート化された集団についての集団割合および数が生成された。
【0032】
間接免疫蛍光を、チャンバースライド(Nalgen Nunc International、Rochester、NY、USA)上で培養され、2%シュークロースを含有し、必要ならば、ヘペス−トリトンX100緩衝剤(Sigma、St. Louis、MO)で透過処理された、4%パラホルムアルデヒドに固定された、MSCについて行った。次の抗体:マウスモノクローナル抗ビネンチン(Sigma)およびウサギポリクローナル抗−フォン・ヴィレブランド因子(Dakocytomation)を用いた。一次抗体を省略したもの、または非免疫ウサギまたはマウスIgGと置き換えられたものが、適当であるならば、対照として使用された。Alexa Fluor488抗−ウサギおよび 抗−マウスTexas Red(Molecular Probes、Leiden、The Netherlands)が二次抗体として使用された。共焦点顕微鏡分析が、Zeiss LSM 5 Pascal Model Confocal Microscope(Carl Zeiss International、Germany)を用いて実施された。ヘキスト33258染料(Sigma)が核染色のために添加された。
【0033】
BM−MSCおよびGL−MSC調製物は、CD45、CD14およびCD34などの造血マーカーを発現しなかった。該調製物は副刺激性分子(CD80、CD86およびCD40)も内皮マーカー(CD31、フォン・ヴィレブランド因子、KDR)も発現しなかった。異なる培養継代での細胞調製物はすべて典型的なMSCマーカー:CD105、CD73、CD44、CD90、CD166およびCD146を発現した。それらはまたHLAクラスIも発現した。
【0034】
MSCの脂質生成、骨形成および軟骨形成の分化能は、Pittenger MF、Martin BJ、Mesenchymal and their potential as cardiac therapeutics. Circ. Res 2004;95:9-20に記載されるように測定された。簡単には、MSCをAdipogenic Medium(Lonza)で3週間培養した。分化を評価するのに、細胞を4%パラホルムアルデヒド中室温で20分間固定し、メタノール(Sigma)中0.5%のOil Red O(Sigma)を用いて室温で20分間染色した。
【0035】
骨形成分化は、MSCをOsteogenic Medium(Lonza)で培養することにより評価された。培地を週に二回3週間変えた。分化を評価するのに、細胞を4%パラホルムアルデヒド中20分間固定し、Alizarin Red(pH4.1)(Lonza)を用いて室温で20分間染色した。
【0036】
軟骨形成の分化では、2.5x105のMSCを15mlの円錐形ポリプロピレン管(Falcon BD Bioscience)中150gで5分間遠心分離に付し、DMEMで2回洗浄した。該ペレットを10ng/mlのTransforming Growth Factor β3(Lonza)を補足した軟骨培地(Lonza)にて培養した。該培地を28日間で3日毎に変えた。ペレットを4%パラホルムアルデヒド中に一夜固定し、パラフィン包埋切片を0.1%サフラニンO(Sigma)を用いてグリコサミノグリカンについて、および1%アルシャンブルーを用いて硫酸化プロテオグリカンについて染色した。
【0037】
1.2 MSCから由来のMVの単離および特徴付け
【0038】
FCSを除去し、0.5%のBSA(Sigma)を補足したRPMIにて培養された、上記のように得られたBM−MSCまたはGL−MSCの上澄から微細小胞(MV)を得た。2000gで20分間遠心分離に付して残骸を除去した後、無細胞上澄を100000gで超遠心分離器(Beckman Coulter Optima L-90K)を用いて4℃で1時間遠心分離に付し、25mMのN−2−ヒドロキシエチルピペラジン−N’−2−エタンスルホン酸(HEPES)(Sigma)を含有する無血清培地199にて洗浄し、同条件下で第2の超遠心分離に供した。MVのエンドトキシン汚染物を、Limulus試験により、製造業者の使用説明書(Charles River Laboratories、Inc.、Wilmington、MA、USA)に従って排除し、MVを−80℃で貯蔵した。
【0039】
選択される実験においては、MVを1U/mlのRNase(Ambion Inc.、Austin、TX、USA)を用いて37℃で1時間処理し、10U/mlのRNase阻害剤(Ambion Inc.)を添加することで反応を停止させ、超遠心分離によりMVを洗浄した。
【0040】
1.3 MSCから由来のMVを用いて行われるインビトロ実験
【0041】
癌細胞株 ヒト肝臓癌細胞株(HepG2)、ヒト乳腺腺癌細胞株(MCF−7)およびヒト卵巣癌細胞株(SKOV−3)を、10%ウシ胎仔血清(FCS、Euroclone)、100U/mlのペニシリン、100mg/mlのストレプトマイシンおよび1%のグルタミン(すべて、Sigmaより入手)を含有する低グルコースDMEM(Euroclone)中に培養し、5%CO2、37℃の加湿雰囲気下でインキュベーターに維持した。カポジ肉腫細胞(KS細胞)の初代培養を、免疫抑制療法下にある、腎臓同種移植を受けた患者の皮膚病変より得、10%のFCS、100μg/mlのペニシリンおよび100μg/mlのストレプトマイシンを補足したRPMI1640培地にて培養した。
【0042】
腫瘍内皮細胞(TEC):単離および培養 MACSシステム(Miltenyi Biotec、Auburn、CA、USA)を用いる磁気細胞分類により、磁気ビーズに結合した抗−CD105抗体を用いて、明細胞型腎細胞癌の試料よりTECを単離した。TEC細胞株を確立し、上皮細胞増殖因子(10ng/ml)、ヒドロコルチゾン(1mg/ml)、ウシ脳抽出物(すべて、Lonzaより入手)および10%のFCSを補足した内皮基底完全培地(EBM)中に培養を維持した。TECは形態学、vWF抗原、CD105、CD146および血管内皮カドヘリンに対して陽性染色を示し、サイトケラチンおよびデスミン(Bussolati Bら、Altered angiogenesis and survival in endothelial cells derived from renal carcinoma. FASEB J 2003;17:1159-1161)に対して陰性染色を示すことにより、内皮細胞として特徴付けられる。
【0043】
ヒト臍帯静脈内皮細胞(HUVEC):単離および培養 ヒト臍帯静脈内皮細胞(HUVEC)を、以前に記載されるように(Bussolati Bら、Vascular endothelial growth factor receptor-1 modulates vascular endothelial growth factor-mediated angiogenesis via nitric oxide. Am J Pathol. 2001Sep;159(3):993-1008)、臍帯静脈より得、EBMおよび10%FCSに維持された。実験は第2および第3継代のHUVECで行われた。
【0044】
細胞増殖 HepG2、MCF−7、SKOV−3、KS細胞またはHUVECを、2000または4000細胞/ウェルで、FCSを除く、RNaseで前処理されている、または前処理されていない異なる濃度の微細小胞を有するDMEM(Sigma)中の96−ウェルに播種した。DNA合成が、48時間培養した後、5−ブロモ−2’−デオキシ−ウリジン(BrdU)の細胞DNAへの取り込みとして検出された。ついで、細胞を0.5Mのエタノール/HClで固定し、ヌクレアーゼと共にインキュベートし、該DNAを消化した。DNAへのBrdUの取り込みを抗−BrdUペルオキシダーゼ接合のmAbを用いて検出し、可溶性発色基質(Roche Applied Science、Mannheim、Germany)で可視化した。光学密度を405nmの下でELISAを用いて測定した。
【0045】
細胞周期分析 ヒト癌細胞株を、30μg/mlの異なるMVの調製物を用いて24時間刺激し、トリプシンにより分離し、冷80%エタノール中に固定した。細胞を−20℃に少なくとも24時間維持し、次にPBSで洗浄した。ついで、該細胞を室温でヨウ化プロピジウム(50μg/ml)(Sigma)と一緒に1時間インキュベートし、RNase(200μg/ml)(Sigma)および0.5%のNonidetP40(Sigma)含有の溶液中で該DNAを染色した。各サンプルについて、50000個の細胞をFACSCalibur血球計算器(BD Biosciences Pharmingen)で分析した。
【0046】
アポトーシスアッセイ HepG2、MCF−7、SKOV−3、KS細胞、HUVECまたはTECが、8000個の細胞/ウェルで、10%FCSを含み、ドキソルビシン(100ng/ml、Sigma)または異なる濃度(10および30μg/ml)のRNaseで前処理されている、またはされていないMVの存在下で、DMEM(Sigma)中に配置された96−ウェルに播種された。アポトーシスをTUNELアッセイ(ApopTag Oncor、Gaithersburg、MD、USA)により評価した。処理して24または48時間経過した後、該細胞をPBSで洗浄し、1%パラホルムアルデヒド(pH7.4)中に4℃で15分間固定させ、PBSにて2回洗浄し、ついで予め冷却したエタノール−酢酸(2:1)に−20℃で5分間ポスト固定させた。試料を酵素末端デオキシヌクレオチジルトランスフェラーゼ(TdT)で処理した。ついで、該細胞をフルオレセインとの加熱した抗ジゴキシゲニン接合体で処理し、室温で30分間インキュベートした。試料を1μg/mlのヨウ化プロピジウム含有の培地に取り込み、該細胞を免疫蛍光により分析した。結果は赤色蛍光発光性細胞(全細胞)に対する緑色蛍光発光性細胞(アポトーシスを起こした細胞)の割合として表される。
【0047】
インビトロでの血管形成 24ウェルのプレートを成長因子を減少させるマトリゲル(BD Biosciences)を用いて4℃で被覆し、加湿雰囲気下、37℃、5%CO2で30分間インキュベートさせた。HUVECまたはTECを、マトリゲル被覆のウェルの5%FCSを含むRPMIまたはEBM中に、5x104個細胞/ウェルの密度で、異なる濃度の、RNaseで処理されているか、処理されていないMVの存在または不在の下で播種させた。インキュベーションから6時間経過した後、該細胞をニコン倒立顕微鏡(Nikon)で観察し、実験結果を記録した。結果は、管の長さの平均として表され、MicroImage分析システム(Cast Imaging)で測定され、任意単位にて表され、3回の異なる実験からの2つの重複したウェルにて20xの倍率で5個所の異なるフィールドにて評価された。
【0048】
統計分析 異なる実験操作からのデータはすべて平均値±SDとして表される。統計分析は、必要に応じて、Newmann−Keulsのマルチ比較試験を備えたANOVAで行われた。
【0049】
1.4 インビトロ結果
【0050】
1.4.1 BM−MSCおよびGL−MSCより由来のMVの腫瘍細胞株に対するインビトロ生物学的効果
ヒトBM−MSCから由来のMVの抗腫瘍活性を、HepG2、MCF−7、SKOV−3およびKS細胞株に対してその増殖を阻害し、そのアポトーシスを誘発する能力を測定することによりインビトロにて評価した。
【0051】
図1はHepG2細胞(図1A)およびKS細胞(図1B)を異なる量のMVと共に48時間インキュベートすると、ビヒクル単独でインキュベートされた対照細胞と比べて増殖が顕著に阻害されることを示す。
【0052】
図2はHepG2細胞(図2A)およびKS細胞(図2B)をMVと共に24および48時間インキュベートすると、ビヒクル単独でインキュベートされた対照細胞と比べて、ドキソルビシン刺激と同様に、アポトーシスが顕著に促進されることを示す。
【0053】
MVによりシャトルされるRNAの完全な分解を誘発するようにMVをRNaseと一緒にインキュベートした場合、MVにより惹起されるHepG2およびKS細胞に対するアンチ増殖およびプロアポトーシス作用は小さくなった(図1および2)。MVのRNase処理そのものはドキソルビシンにより誘発される癌細胞株のアポトーシスを干渉しなかった(図2)。
【0054】
反対に、MCF−7細胞およびSKOV−3細胞と30μg/mlのBM−MSCからのMVとの48時間に及ぶインキュベーションは、ビヒクル単独でインキュベートされた対照細胞と比べて増殖を顕著に阻害した(図3Aおよび3B)が、アポトーシスを促進しなかった。これら2種の腫瘍細胞株にて、本願発明者らはまた、細胞がG0/G1フェーズにある割合を、細胞がSおよびG2フェーズにある割合と比較して評価するために、細胞周期をヨウ化プロピジウム染色技法を用いて検討した。本願発明者らは、BrdUの取り込みで観察される増殖の阻害を説明しうる、BM−MSCからのMVがG0/G1フェーズにある細胞、特にSKOV−3細胞(図4)の増大を誘発することを観察した。
【0055】
本願発明者らはまた、Gl−MSCから由来のMVの腫瘍細胞株の増殖およびアポトーシスに対する作用を試験した。Gl−MSCはHepG2細胞株の増殖およびアポトーシスに影響を与えなかった。反対に、Gl−MSCから由来のMVはKS細胞の増殖を阻害し、そのアポトーシスを誘発した(図1および2)。その上、MCF−7およびSKOV−3細胞と30μg/mlのGl−MSCから由来のMVとの48時間に及ぶインキュベーションは、ビヒクル単独でインキュベートした対照の細胞と比べて増殖を阻害した(図3)が、アポトーシスを促進しなかった。
【0056】
1.4.2 ヒト線維芽細胞から由来のMV
【0057】
ヒト線維芽細胞から由来のMVは種々の癌細胞株の増殖を阻害せず、そのアポトーシスを誘発しなかった(図示せず)。
【0058】
1.4.3 BM−MSCから由来のMVの内皮細胞に対するインビトロでの効果
【0059】
本願発明者らはまた、BM−MSCから由来のMVのHUVECおよび腫瘍内皮細胞(TEC)の増殖、アポトーシスおよび毛細血管様形成に対するインビトロ作用を検討した。
【0060】
MV処理はHUVECの増殖に影響を及ぼさず(図示せず)、その毛細血管様形成能にも影響しなかった(図5A)。加えて、HUVECの種々の量のMVとの48時間にわたるインキュベーションはアポトーシスを誘発しなかった(図6)。
【0061】
反対に、TECの種々の量のMVとの48時間にわたるインキュベーションは、ビヒクル単独でインキュベートした対照となる細胞と比べて、増殖を顕著に阻害し(図7)、アポトーシスを顕著に促進した(図6)。TECの増殖は、RNaseで前処理されている、またはされていないBM−MSCからの種々の量(10および30μg/ml)のMVと共にインキュベートして48時間経過した後にBrdUの取り込みアッセイにより評価した。図7の結果は2回の実験の平均値±SDとして表される。
【0062】
MVをRNaseと共にインキュベートした場合、MVにより惹起されるTECに対するアポトーシス作用は著しく減少した。マトリゲルに播種されたTECの種々の量のMVとのインキュベーションは、TECの毛細血管様構造をインビトロにて形成する能力を著しく阻害した。MVのRNaseでの前処理は、MVの細管形成に対する阻害作用を失効させた(図5B)。
【0063】
1.5 MSCより由来のMVを用いるインビボ実験
【0064】
腫瘍形成 3x106個のHepG2細胞を集め、SCIDマウス(Charles River、Jackson Laboratories、Bar Harbor、ME)に皮下移植した。トリプシン−EDTAを用いて収穫した培養細胞をPBSで洗浄し、マイクロ血球計算器のチャンバーにて計数し、100μlのDMEMおよび100μlのマトリゲルマトリックス(Becton Dickinson)に再び懸濁させた。細胞を氷冷させ、1mlのシリンジを用いる26−ゲージ針を介してSCIDマウスの左背中に皮下注射した。動物を活動および健康状態について毎日モニター観察し、体重測定および腫瘍塊の測定を3日毎に行った。腫瘍塊は移植片の2つの直交する直径をキャリパー計測により測定し、式:1/2axb2(式中、aは長軸であり、bは短軸である)を用いて計算した(Hou Jら、Experimental therapy of hepatoma with artemisin and its derivatives:in vitro and in vivo activity、chemosensitization and mechanism of action. Clin Cancer Research. 2008;14:5519-5530))。1週間後、移植された腫瘍が約15mm3の体積に達した時に、本願発明者らは、週に一度のMVの腫瘍内注射を開始した。最初の処理は100μgのMV(RNaseで処理された、またはされていない)で、最大容量が20μlであり;その後の腫瘍内注射は50μgのMV(RNaseで処理された、またはされていない)で、最大容量が20μlであった。対照となるマウスには、発明者らは同じ容量のビヒクルを単独で腫瘍内注射した。マウスをランダムに3つの処理群:a)MVを腫瘍内注射した群(n=8);b)RNaseで処理されたMVを腫瘍内注射した群(n=8);およびc)同じ容量のビヒクルを単独で注射した対照群(n=5)に分けた。マトリゲル注射から3週間後に、マウスを殺し、腫瘍を回収し、組織構造について加工処理した。
【0065】
1.6 インビボ結果
【0066】
1.6.1 BM−MSCから由来のMVのHepg2腫瘍増殖に対するインビボでの生物学的作用
【0067】
腫瘍形成および増殖がSCIDマウスにてBM−MSCより由来のMVにより阻害される。BM−MSCより由来のMVのインビボにおける腫瘍形成および増殖に対する効果を測定するために、SCIDマウスに、マトリゲルの存在下にてHepG2を皮下注射した。注射した1週間後に、腫瘍の体積が約15mm3であったならば、発明者らは週に一度のMV(RNaseで処理された、またはされていない)の腫瘍内注射を、20μlの最大容量にて開始した。最初の処理は100μgのMVであり;その後の腫瘍内注射は50μgのMVであった。対照となるマウスには、発明者らは20μlのビヒクルを単独で腫瘍内注射した。
【0068】
マトリゲル注射から3週間経過した後、すべての腫瘍を回収し、分析した。HepG2異種移植片実験にて、MVの腫瘍内注射は腫瘍増殖に対して阻害剤作用を示した(図8)。腫瘍の大きさおよび体積は、MVで処理されたSCIDマウスにて有意に小さく(図9AおよびB)、組織学的分析はMVで処理されたHepG2腫瘍にて壊死の部分を示した(図9C)。RNaseで前処理されたMVを注射された腫瘍は、対照となる腫瘍と大きさおよび組織構造にて差異がなかった(図10AおよびB)。
【0069】
2. 肝臓幹細胞から由来の微細小胞(MV)
【0070】
2.1 成体ヒト肝臓幹細胞(HLSC)の単離および特徴付け
【0071】
HLSCは、L−グルタミン(5mM)、ヘペス(12mM、pH7.4)、ペニシリン(50IU/ml)、ストレプトマイシン(50μg/ml)(すべて、Sigma、St. Louisより入手)、FCS(10%)を補足した、最小必須培地/内皮細胞基底培地−1(α−MEM/EBM)(3:1)(Gibco/Cambrex)にて培養された、Cambrex Bio Science Verviers S.p.r.l.(Verviers、Belgium)より入手の、低温保存された正常なヒト肝細胞より単離された。拡張した細胞をT−75フラスコに移し、それらが集密に達した時に分析された。
【0072】
肝癌細胞株HepG2を10%ウシ胎仔血清(FBS)含有のダルベッコ修飾イーグル培地(DMEM)で培養させた。
【0073】
2.2 HLSCから由来のMVの単離
【0074】
MVを2%ウシ胎仔血清(FBS)補足のMEM−アルファにて培養されたHLSCの上澄より得た。血清無しで一夜インキュベートされた細胞の生存能をトリパンブルーの排除により検出した。2000gで20分間遠心分離に付し、残骸を取り除いた後、無細胞上澄を100000g(Beckman Coulter Optima L-90K超遠心分離器)で4℃で1時間遠心分離に付し、25mMのN−2−ヒドロキシエチルピペラジン−N’−2−エタンスルホン酸(HEPES)(Sigma)含有の無血清培地199で洗浄し、同じ条件下で第2の超遠心分離に供した。インビトロおよびインビボにて、蛍光顕微鏡法またはFACS分析によりMVを追跡するのに、幹細胞由来のMVが赤色蛍光脂肪族発色団を脂質二重層中に挿入するPKH26色素(Sigma)で標識された。標識後、MVを洗浄し、10000gで1時間4℃で超遠心分離に供した。MVペレットを培地199に懸濁させ、蛋白含量をブラッドフォード法(BioRad、Hercules、CA)により定量した。MVの汚染エンドトキシンをリムルス試験により製造者の説明書(Charles River Laboratories、Inc.、Wilmington、MA)に従って排除し、MVを−80℃で貯蔵した。ヨウ化プロピジウムで染色した後のMV懸濁液上でなされた形態学的分析はアポトーシス体の存在を明らかにしなかった。
【0075】
選択された実験にて、HLSCからのMVは1U/mlのRNase(Ambion Inc.、Austin、TX)で1時間37℃で処理され、10U/mlのRNase阻害剤(Ambion Inc.)を添加することで反応を停止させ、MVを超遠心分離で洗浄した。RNase処理の効果を、TRIZOL試薬(Invitrogen、Carlsbad、CA)を用いてRNA抽出を行った後に、抽出された全RNAを分光光度計分析により評価した(未処理:1.3±0.2μg RNA/mgの蛋白MV;RNase処理:<0.2μg RNA/mgの蛋白MV)。加えて、RNAase処理および処理されていないMVより抽出されたRNAをオリゴdTでなされるレトロトランスクリプションにより標識し、0.6%アガロース上で分析し、RNase処理によるRNAの完全な分解を示した。対照として、MVを1U/mlのDNase(Ambion Inc.)で1時間37℃で処理した。
【0076】
2.3 HLSCより単離されたMVを用いて実施されたインビトロ実験
【0077】
増殖アッセイ DNA分析は、製造者の仕様説明書に従って酵素免役吸着測定キット(Chemicon、Temecula、CA)を用いる5−ブロモ−2−デオキシウリジン(BrdU)の細胞DNAへの取り込みとして検出された。簡単には、洗浄した後、細胞を10モル/lのBrdUと一緒に6〜10時間、37℃で、5%CO2にて、加湿雰囲気下でインキュベートさせ、0.5モル/Lのエタノール/HClで固定し、ヌクレアーゼと一緒にインキュベートし、DNAを消化した。DNA中に取り込まれたBrdUを、抗BrdUペルオキシダーゼ接合モノクローナル抗体を用いて検出し、可溶性発色基質で可視化した。光学密度を酵素免疫吸着測定リーダーを用いて405nmで測定した。
【0078】
アポトーシスアッセイ アポトーシスを末端dUTPニックエンド標識アッセイ(ApoTag;Oncor、Gaithersburg、MD)を用いて評価した。細胞(8x103/ウェル)を96ウェルプレートで培養し、リン酸緩衝セイライン(PBS)に懸濁させ、PBS中の1%パラホルムアルデヒド(pH7.4)にて15分間、4℃で、つづいて予め冷却したエタノール/酢酸(2:1)で5分間−20℃で固定させた。細胞を末端デオキシヌクレオチドトランスフェラーゼ酵素で処理し、加湿チャンバーにて1時間37℃でインキュベートし、ついで加温したフルオレセインイソチオシアネート接合のアンチジゴキシゲニンで30分間室温で処理した。洗浄した後、試料を1g/mlのヨウ化プロピジウム含有の培地に固定し、細胞を免疫蛍光により分析した。
【0079】
統計分析 種々の実験操作のデータはすべて平均±SDとして表される。統計分析は、必要に応じて、ニューマン−クールズの多重比較試験のANOVAにより実施された。
【0080】
2.4 インビトロ結果
【0081】
2.4.1 HLSCより由来のMVのHepG2肝癌細胞株の増殖に対する効果
【0082】
本願発明者らはHLSCより由来のMVのHepG2肝癌細胞株の増殖に対する効果を評価した。簡単には、HepG2細胞を種々の量の(10、20および30μg/ml)のHLSC由来のMVそのものと共にインキュベートするか、またはRNaseで3日間処理した。インキュベーションの終わりに、HepG2培養物を計数するか、0.5Mのエタノール/HClに固定し、ヌクレアーゼと一緒にインキュベートし、そのDNAを消化した。該DNAに取り込まれたBrdUを抗BrdUペルオキシダーゼ接合モノクローナル抗体を用いて検出し、可溶性発色基質で可視化した。光学密度をELISAリーダーを用いて405nmで測定した。図11に示されるように、HLSCより由来のMVはHepG2増殖を顕著に阻害しうる。このことはまたRNase処理のMVにも適用する。
【0083】
2.4.2 HLSCより由来のMVのHepG2肝癌細胞株のアポトーシスに対する効果
【0084】
HLSC誘導のMVのHepG2のアポトーシス誘発能を評価した。簡単には、HepG2を8000細胞/ウェルの密度で96−ウェルプレートの10%FCSを補足したDMEMに播種し、FCSの不在下で、癌の化学療法にて使用される2種の有糸分裂阻害剤である、ビンクリスチン(100ng/ml)またはドキソルビシン(50ng/ml)で処理することにより、あるいはMV処理(30μg/ml)で培養してアポトーシスを誘発した。対照として、MVをまた1U/mlのRNase18(Ambion、Austin、TX)で37℃1時間処理し、癌細胞増殖の阻害への寄与がMVによって癌細胞にデリバリーされたmRNAの水平移動に依存するかどうかを評価した。アポトーシスをTUNELアッセイ分析を用いて24および72時間で評価した。図12に示されるように、HLSCより由来のMVは、ビンクリスチンにより誘発されるアポトーシスと比べて、HepG2のアポトーシスを誘発することが可能であった。反対に、RNase処理はアポトーシスを誘発しなかった。加えて、HepG2のビンクリスチン+MV−HLSCまたはドキソルビシン+MV−HLSCでの処理は図13に示されるように相加作用をもたらす。
【0085】
2.5 HLSCより単離されるMVで行われるインビボ実験
【0086】
細胞培養 ヒト肝癌細胞HepG2を、10%ウシ胎仔血清、100μg/mlのペニシリンおよび100μg/mlのストレプトマイシンを補足したDEMEで培養し、5%CO2、37℃の加湿雰囲気のインキュベーター中に維持した。
【0087】
ヒト肝臓幹細胞(HLSC)を、10%ウシ胎仔血清、100μg/mlのペニシリンおよび100μg/mlのストレプトマイシンを補足した、α−MEM/EBM(3:1)にて培養した。EBMはhEGF(ヒト上皮成長因子)、ヒドロコルチゾン、GA(ゲンタマイシン)、BBE(ウシ脳抽出物)で復元された。
【0088】
微細小胞(MV)のHLSCからの単離 MVを2%ウシ胎仔血清を補足したα−MEM培地にて培養したHLSCの上澄より得た。2000gで20分間遠心分離に付して残骸を除去した後、無細胞上澄を100000gで1時間4℃で遠心分離に付し、25mMのN−2−ヒドロキシエチルピペラジン−N’−2−エタンスルホン酸(HEPES)を含有する血清不含培地199で洗浄し、同じ条件下で第2の超遠心分離に供した。MVペレットを0.1%のDMSOの培地199に懸濁させ、蛋白含量をブラッドフォード法を用いて定量した。
【0089】
実験の設計 雄の4ないし5月齢のSCIDをCharles River laboratoriesより得た。すべてのマウスを無菌状態の檻に入れ、1週間保持して順応させた。0日目に、マトリゲル基底膜マトリックスを含む血清不含DMEMに再び懸濁させた3x106のHepG2腫瘍細胞を1:1の割合で2回注射して投与した。細胞懸濁液を合計0.2mlの容量でSCIDマウスの左鼠径部に注射した。すべてのマウスを無作為に3つの処理群:20μlの腫瘍内(i.t.)MV注射(n=3);20μlのi.t.PBS注射(n=2)および20μlのi.t.MV−RNase処理の注射(n=3)に分けた。7日目に、腫瘍細胞移植後の処理を開始した。腫瘍は7日目より触診可能となり;0.1%のDMSOを補足したM199に懸濁させた、50または100μgのMVを腫瘍移植の7、12、14および18日後に注射した。腫瘍の体積が約15mm3になった時に治療を開始した。動物を活性および健康状態について毎日モニター観察し、体重測定および腫瘍塊の測定を3日毎に行った。
【0090】
腫瘍をキャリパーで測定した。腫瘍塊を、キャリパーにより移植片の2つの直交する直径を測定し、式:1/2a x b2(式中、aは長軸であり、bは短軸である)を用いて計算した。28日目に動物を殺し、さらなる分析のために腫瘍を集めた。
【0091】
形態学研究 腫瘍を10%緩衝化中性ホルマリンに固定し、慣用的に加工処理し、パラフィンに包埋し、5μmの薄片にし、顕微鏡試験のためにH&Eで染色した。増殖を検出するための免疫組織化学を抗PCNAモノクローナル抗体を用いて行った。切片を抗マウスHRP二次抗体で遮断して標識化した(1:300希釈)。一次抗体を省略したもの、または非免疫マウスIgGと置き換えられたものが、対照として使用された。アポトーシスをTUNELによりパラフィン包埋組織の切片にて評価した。10個の連続しない切片をアポトーシスの陽性腫瘍細胞について630の倍率で計数した。ヘキスト33258色素を核染色のために添加した。
【0092】
統計分析 種々の実験操作のデータはすべて平均±SDとして表される。統計分析は、必要に応じて、ニューマン−クールズの多重比較試験のANOVAにより実施された。
【0093】
2.6 インビボ結果
【0094】
2.6.1 腫瘍の成長および増殖がSCIDマウスの肝癌異種移植片実験にてHLSCから由来のMVによって阻害された
【0095】
HLSCから由来のMVのインビボでの腫瘍増殖に対する効果を測定するために、SCIDマウスに、ヒト肝細胞癌の細胞株HepG2を皮下移植した。HepG2注射の1および2週間後で、腫瘍の体積が約15mm3であった際に、マウスにMV(50または100μg)を最大20μlの容量で腫瘍内注射して処理した。対照となるマウスにおいては、腫瘍に20μlのビヒクルを単独で注射した。HepG2の注射から3および4週間後に、すべての腫瘍を回収し、分析した。この異種移植片実験にて、MVの腫瘍内注射(図14)は腫瘍成長に対して阻害剤の効果を示した。加えて、組織学的分析はMVで処理された腫瘍にて壊死の領域を示し(図15B)、抗増殖作用がPCNA染色を用いて観察された(図15B)。
【0096】
2.6.2 HLSCより由来のMVによるSCIDマウスにおける肝癌異種移植片実験でのアポトーシスの誘発
【0097】
腫瘍内アポトーシスの効果を測定するために、MVで処理された腫瘍からのパラフィン切片をTUNELで分析した。MV処理はビヒクル単独で処理された腫瘍と比べて腫瘍を誘発した(図15A)。
【0098】
2.7 HLSCから由来のMVの、MCF−7乳腺腺癌およぼカポジ肉腫(KS)細胞に対するインビトロでの生物学的効果
【0099】
1.7.1 材料および方法
【0100】
細胞培養 ヒト非楕円形肝臓幹細胞(HLSC)を、10%ウシ胎仔血清、100μg/mlのペニシリンおよび100μg/mlのストレプトマイシンを補足した、α−MEM/EBM(3:1)にて培養した。EBMはhEGF(ヒト上皮成長因子)、ヒドロコルチゾン、GA(ゲンタマイシン)、BBE(ウシ脳抽出物)で復元された。
【0101】
MCF−7乳腺腺癌細胞株をAmerican Type Culture Collection(Manassas、VA)より入手し、10%FCS、100μg/mlのペニシリンおよび100μg/mlのストレプトマイシンを補足した、DMEMにて培養し、インキュベーター中、5%CO2の加湿雰囲気下、37℃で維持した。
【0102】
カポジ肉腫細胞(KS細胞)の初代培養を、免疫抑制療法を受けている腎同種移植片を有する患者の皮膚病変から入手し、10%FCS、100μg/mlのペニシリンおよび100μg/mlのストレプトマイシンを補足した、RPMI1640培地にて培養した。
【0103】
MVの単離 2%ウシ胎仔血清(FBS)を補足した、MEM−アルファにて18時間培養したHLSCの上澄からMVを得た。選択された実験にて、MVをFBSの不在下で集めた。2%のFBSおよび血清無しで一夜インキュベートされた細胞の生存能をトリパンブルーの排除により検出した(90%より大きい、データは示さず)。2000gで20分間遠心分離に付し、残骸を取り除いた後、無細胞の上澄を100000g(Beckman Coulter Optima L-90K超遠心分離器)で1時間4℃で遠心分離に付し、25mMのN−2−ヒドロキシエチルピペラジン−N'−2−エタンスルホン酸(HEPES)含有の無血清培地199で洗浄し、同じ条件下で第2の超遠心分離に供した。MV細胞を培地199に懸濁させ、蛋白含量をブラッドフォード法で測定した。MVを−80℃で貯蔵した。ヨウ化プロピジウムで染色した後のMV懸濁液上でなされた形態学的分析はアポトーシス体の存在を明らかにしなかった。
【0104】
RNase処理 選択された実験にて、HLSCからのMVを1U/mlのRNaseで1時間37℃で処理した。10U/mlのRNase阻害剤を添加することで反応を停止させ、MVを超遠心分離により洗浄した。
【0105】
細胞増殖 HLSCから由来のMVが種々の腫瘍に対してその抗腫瘍活性を発揮するかどうかを調べるために、MCF−7乳腺腺癌細胞およびカポジ肉腫細胞から由来の細胞株を、各々、8000細胞/ウェルの密度で、種々の濃度のMV(2;10;15;および30μg/mlおよび30μg/mlのRNase処理のMV)を含む、96−ウェルプレートのDMEMおよびRPMIに播種した。DNA合成を、48時間培養した後の、5−ブロモ−2’−デオキシ−ウリジン(BrdU)の細胞DNAへの取り込みとして検出した。次に、細胞を0.5Mのエタノール/HClで固定し、ヌクレアーゼと一緒にインキュベートし、DNAを消化させた。該DNAに取り込まれたBrdUを、抗−BrdUペルオキシダーゼ接合mAbを用いて検出し、可溶性発色基質を用いて可視化した。光学密度は405nmでELISAリーダーを用いて測定された。
【0106】
アポトーシスアッセイ MCF−7およびKS細胞を8000細胞/ウェルの密度で96−ウェルの10%FCSを含み、ドキソルビシン(100ng/ml)または種々の濃度のMV(2;10;15および30μg/mlおよび30μg/mlのRNase処理されたMV)の存在する低グルコースのDMEMに播種した。アポトーシスをTUNELアッセイを用いて評価した。
【0107】
2/7/2 結果:HLSCより由来のMVはMCF−7細胞およびKS細胞のインビトロでの増殖を阻害した
【0108】
MCF−7乳腺腺癌細胞およびカポジ肉腫細胞と、2、10、15および30μg/mlのHLSC−6Bから由来のMV(図16および17)との48時間に及ぶインキュベーションは、ビヒクル単独でインキュベートされた対照細胞と比べて、細胞増殖を顕著に阻害する。これらの結果は組織固有の幹細胞の抗腫瘍作用が同じ組織から形成される腫瘍に対して特異的ではないことを示す。その上、MCF−7乳腺腺癌細胞およびカポジ肉腫細胞の48時間にわたる2、10、15および30μg/mlの、HLSC−6B細胞から由来のMV(図18)が、ビヒクル単独でインキュベートされた対照となる細胞と比べて、アポトーシスを誘発し、その効果も化学療法薬であるドキソルビシンのアポトーシスの誘発と同じくらいである。このことはさらに、組織固有の幹細胞の抗腫瘍作用が同じ組織より生成される腫瘍に対して特異的でないことを確認するものである。
【技術分野】
【0001】
本願発明は腫瘍性疾患の治療的処置に関する。
【背景技術】
【0002】
造血幹細胞移植は、充実性腫瘍の患者にて腫瘍阻害作用を発揮し、ならびに転移性の乳癌、腎臓癌、卵巣癌、前立腺癌および膵臓癌にて抗腫瘍作用を発揮することが知られている。
【0003】
ヒト骨髄間充織幹細胞(BM−MSC)は多種の器官の修復および組織の修復に寄与することが明らかにされ、実証研究はMSCの移植が心臓、肝臓および腎臓を含むいくつかの器官の機能性および構造的回復に有益な効果のある可能性があることを示唆した。
【0004】
幹細胞の微小環境は、増殖を阻害し、分化を促進するシグナルを提供することで発癌を防止するのに不可欠な役割を発揮するようである。
【0005】
しかしながら、幹細胞の治療指標における使用は、かかる幹細胞療法の腫瘍形成能の危険性が報告されていることを考えれば、それほど望ましいものではない(Amariglio Nら、Donor-derived brain tumor following neural stem cell transplantation in an ataxia telangiectasia patient. PLoS Med. 2009 Feb 17;6(2))。
【0006】
細胞由来の微細小胞(MV)は、特徴的な抗原を発現する細胞によって放出される小胞であって、その細胞から膜および細胞質成分を生成して担持し、細胞通信の新たな機構であると記載されている。最近になって、本願発明者らはヒト内皮前駆細胞(EPC)、BM−MSCおよび肝臓幹細胞から由来の微細小胞が、標的となる分化した細胞を内皮細胞、尿細管上皮細胞および肝細胞として再プログラム化し(Deregibus MCら、Endothelial progenitor cell derived microvesicles activate an angiogenic program in endothelial cells by a horizontal transfer of mRNA. Blood. 2007 Oct 1;110(7):2440-8;Bruno Sら、Mesenchymal stem cell-derived microvesicles protect against acute tubular injury. J Am Soc Nephrol. 2009 May;20(5):1053-67;Herrera MBら、Human liver stem cell-derived microvesicles accelerate hepatic regeneration in hepatectomized rats. J Cell Mol Med. 2009 Jul 24)、組織の再生および修復に貢献し得る、遺伝物質(mRNA)の移動のためのビヒクルとして供し得ることを明らかにした。
【発明の概要】
【発明が解決しようとする課題】
【0007】
国際特許出願WO2009/087361は、誘導物質を未分化細胞の第一集団に適用することで分化細胞を産生し、ついで微細小胞をその分化細胞より単離することを開示する。この点については、WO2009/087361の30頁の第1段落のすべてを参照のこと。未分化細胞の第1集団は、例えば、骨髄間充織幹細胞である。しかし、WO2009/087361は未分化細胞より微細小胞を直接得ることを開示していない。
【課題を解決するための手段】
【0008】
本願発明者らは、この度、成体肝細胞、好ましくは骨髄または糸球体間充識幹細胞より、あるいは非楕円形肝臓幹細胞より由来の微細小胞(MV)が、インビトロおよびインビボの両方で顕著な抗腫瘍活性を示し、それにより癌の治療的処置のための対応するすべての幹細胞にわたって別途利点を示しうることを見出した。本願発明の成体幹細胞より由来の微細小胞の抗腫瘍活性は、該微細小胞の、種々のヒト癌細胞株の増殖およびアポトーシスに対する効果、ならびにその毛細血管様構造の形成に対する効果を測定することによりインビボにて明らかにされた。微細小胞の抗腫瘍活性もまた、MV−処置の腫瘍増殖に対する効果を測定することでインビボのマウス実験にて確認された。
【0009】
成体幹細胞より由来の微細小胞の抗腫瘍活性の観察は先行技術では全く予期せぬものである。間充織幹細胞(MSC)の調製物は、実際、いくつかの組織にて再生作用を発揮することが知られている。例えば、骨髄由来のMSCは、可溶性細胞外マトリックス糖蛋白、サイトカインおよび成長因子を含む、多くの栄養性分子を分泌することにより、自然と、血液生成を支持することが知られている。その上、内皮幹細胞より由来の微細小胞は、WO2009/050742より、インビトロおよびインビボの両方で、血管形成およびアポトーシスに対する耐性を促進することが分かっている。WO2009/057165では、幹細胞より由来の微細小胞は、損傷した組織または器官の内皮および上皮再生を誘発することが明らかにされた。
【0010】
WO2009/105044にて、間充織幹細胞より由来の微細小胞は、多数かつ多種の疾患の治療に用いるのに適すると推論している。癌の処置について、実験的証拠はなく、理論的説明もなく、具体的な方法も提供されていない。
【0011】
かくして、本願発明の第1の態様は、腫瘍性疾患の治療的処置に用いるための成体幹細胞より由来の微細小胞である。
【0012】
これに関連して、微細小胞は、それらを投与する同一患者より採取される成体幹細胞に由来する必要はないことを理解すべきである。むしろ、違う対象に由来して調製され、医薬の形態にて維持され、次にその必要とする患者に投与されてもよい。これはいわゆる同種アプローチである。
【0013】
好ましい実施態様において、成体幹細胞はヒト間充織幹細胞またはヒト肝臓幹細胞である。好ましいヒト肝臓幹細胞は、WO2006/126219に開示される間充織および胚幹細胞マーカーの両方を発現するヒト非楕円形肝臓幹細胞(HLSC)である。この細胞株は、成体幹細胞が、肝細胞マーカーを発現し、成熟肝細胞、インスリン産生細胞、骨形成原細胞および上皮細胞への分化能を有する、好ましくはアルブミン、α−フェト蛋白、CK18、CD44、CD29、CD73、CD146、CD105、CD90からなる群より選択されるマーカーを発現し、好ましくはCD133、CD117、CK19、CD34、チトクロムP450からなる群より選択されるマーカーを発現しない、成体組織より単離される、非楕円形ヒト肝臓多能性前駆細胞株である点で特に特徴付けられる。
【0014】
もう一つ別の好ましい実施態様において、ヒト間充織幹細胞は、ヒト成体骨髄(BM−MSC)より誘導される。もう一つ別の好ましい実施態様にて、ヒト間充織幹細胞は、欧州特許出願番号第08425708.8に開示されるように、ヒト成体脱糸球体嚢(Gl−MSC)より誘導される。これらの細胞はさらには、該細胞がCD133陰性、CD146陽性およびCD34陰性であり、有足細胞、内皮細胞およびメサンギウム細胞への分可能を有し、好ましくはまたCD24、Pax−2、CD31、CD29、CD44、CD73、CD90、CD105、CD166、ナノグ、ムサシ、ビメンチン、ネスチンからなる群より選択されるマーカーを発現し、好ましくはα−SMA、Oct−4、CD45、サイトケラチン、CD80、CD86、CD40からなる群より選択されるマーカーを発現しない点で特徴付けられる。
【0015】
本願発明の一の実施態様によれば、腫瘍性疾患は、肝臓腫瘍(例、肝癌)、上皮腫瘍(例、カポシ肉腫)、乳房腫瘍(例、乳腺腺癌)、肺腫瘍、前立腺腫瘍、胃腫瘍、結腸腫瘍および卵巣腫瘍からなる群より選択される。
【0016】
本願発明のもう一つ別の態様は、上記した腫瘍性疾患の治療的処置のための医薬の製造にて、上記の成体幹細胞より誘導される微細小胞を用いることである。
【0017】
一の実施態様にて、該治療的処置は、一または複数の細胞傷害性または細胞増殖抑制性剤を投与することを含む。適当な細胞傷害性および細胞増殖抑制性剤として、例えば、パクリタセル(Paclitaxel)、レナリドミド(Lenalidomide)、ポマリドミド
(Pomalidomide)、エピルビシン(Epirubicin)、5FU、スニチニブ(Sunitinib)、ラ−パチニブ(La-patinib)、カネルチニブ(Canertinib)、シクロホスファミド
(cyclophosphamide)、ドキソルビシン(doxorubicin)、レナリドミデン/デキサメタゾン(Lenalidomiden/Dexamethason)、ポ−マリドミド(Po-malidomide)/デキサメタゾン、カルボプラチン(Carboplatin)、ラパマイシン(Rapamycin)、ミトキサントロン(mitoxantron)、オキサリプラチン(oxaliplatin)、ドキセタセル(docetaxel)、ビノレルビン(vinorelbin)、ビンクリスチン(vincristine)およびその組み合わせが挙げられる。ドキソルビシンおよび/またはビンクリスチンと、成体幹細胞より由来のMVとの組み合わせの投与が極めて好ましい。というのも、かかる組み合わせは相乗作用を発揮することが示されたからである(図13を参照)。
【0018】
微細小胞は、その必要とする患者に、局所または全身投与のいずれかで投与される。局所および全身投与の両方に適する医薬剤形は、例えば、注射可能な剤形である。一例として、微細小胞が、充実性腫瘍で局所的な腫瘍内(i.t.)注射にて、あるいは充実性腫瘍の場合と、転移癌の場合の両方で静脈内注射または注入にて投与される。投与されるべきMVの適当な用量は複数の因子に依存するが、一般に、受容者の体重1kg当たり0.1−200μg、好ましくは受容者の体重1kg当たり1−150μg、さらにより好ましくは受容者の体重1kg当たり3−120μgで構成される。
【0019】
本願明細書にて用いられる「成体幹細胞より由来の微細小胞(MV)」なる語は、少なくとも部分的に成体幹細胞より誘導される、膜粒子に言及する。同様に、「成体幹細胞」なる語は、増殖能(自己再生能)および分化能(塑性能)を有し、それによってライフサイクルの終わりを迎える特化された細胞系統の成熟細胞に取って代わる、未分化または一部未分化の細胞を包含する。本願明細書にて用いられる「成体幹細胞」なる語は、無制限の自己再生能および多能性塑性を有する肝細胞、および多能性塑性を有し、ある場合には制限された自己再生能を有する前駆細胞の両方を包含する。好ましい実施態様において、「成体幹細胞」は、少なくとも2種の、より好ましくは少なくとも3種の異なる型の特化された、十分に分化された成熟細胞への分化能を有する、多能性または複数の能力の塑性を有することを意味する。
【0020】
本願明細書の文脈の中で、「成体幹細胞」なる語は、胚盤胞の内細胞塊より単離される「胚幹細胞」とは異なり、意図的に、成体組織より単離される幹細胞を意味する。成体幹細胞はまた、「体性幹細胞」としても知られている。
【0021】
本願明細書の文脈の中で、「成体幹細胞より由来の微細小胞(MV)」なる表現は、意図的に、微細小胞が未分化の幹細胞より直接誘導されることを意味する。
【0022】
本願明細書の文脈の中で、「直接」なる語は、微細小胞を得る前にいずれの分化工程がなされることも、生じることもなく、微細小胞が未分化の成体幹細胞より誘導されることを意味する。
【0023】
本願発明にて使用される成体幹細胞より誘導される微細小胞は、一般に、形状が球体であり、100nm〜5μm、より典型的には0.2〜1μmの範囲の直径を有する。仮に粒子の形状が球体でない場合、上記した値は該粒子の最大の寸法に言及するものである。
【0024】
本願発明にて使用される微細小胞をもたらす幹細胞は、本願明細書の実験のセクションにて記載されるように単離されてもよい。次に、微細小胞(MV)は、単離された幹細胞の上澄から、例えば本願明細書の実験のセクションに開示される超遠心分離により得られてもよい。ついで、その単離されたMVは、1または複数の抗凍結剤を配合した懸濁液中、超低温、例、−80℃で凍らせることで使用するまで貯蔵されてもよい。適当な抗凍結剤は、例えば、ジメチルスルホキシド(DMSO)およびグリセロールである。DMSOの細胞懸濁液の1容量%濃度での使用は、細胞の良好な保存および患者に再び注入された場合の毒性作用の制限を保証する。抗凍結剤として言及されてもよい他の物質は、いわゆる、密なバリアを細胞表面に形成し、細胞内脱水を減らすように作用する高分子量物質である。
【0025】
本願発明のさらなる目的および利点は、単に例示として用いられる、以下の実施例からさらに明らかになるであろう。実施例において、以下の図面が参照される。
【図面の簡単な説明】
【0026】
【図1】図1AおよびBは、HepG2細胞(図1A)およびKS細胞(図1B)を異なる量の微細小胞(MV)と一緒にインキュベートすることで、ビヒクル単独でインキュベートされた対照となる細胞と比べて、増殖が顕著に阻害されることを示す。HepG2およびKS細胞の増殖は、異なる量のBM−MSCからのMV(1、10および30μg/ml)、またはRNaseで前処理されている、もしくはされていないGl−MSCからのMVと一緒にインキュベーションした48時間後に、BrdU取り込みアッセイで評価された。結果は3回の実験の平均値±SDで表される。
【図2】図2AおよびBは、HepG2およびKS細胞とMVとのインキュベーションが、ビヒクル単独でインキュベートされた対照と、同様にドキソルビシン刺激と比べて、アポトーシスを顕著に促進することを示す。HepG2およびKS細胞のアポトーシスは、BM−MSC(RNaseで前処理されているか、されていない)またはGl−MSCからの異なる量のMVと一緒にインキュベーションした24時間および/または48時間後のアポトーシス細胞の割合としてTunelアッセイで評価された。結果は3回の平均値±SDで表される。
【図3】図3Aおよび3Bは、MCF−7細胞(図3A)およびSKOV−3細胞(図3B)を、BM−MSCからの30μg/mlのMVと一緒に48時間インキュベートすることで、ビヒクル単独でインキュベートされた対照細胞と比べて増殖が顕著に阻害されることを示す。MCF−7およびSKOV−3細胞の増殖をBM−MSCからの、またはGl−MSCからの30μg/mlのMVと一緒にインキュベートした48時間後にBrdU取り込みアッセイにより評価した。結果は3回の実験の平均値±SDで表される。
【図4】BM−MSCからのMVが、特にSKOV−3細胞のG0/G1フェーズにて細胞の増加を誘発することを示す。DNA含量を、10%FCSと共に、FCSを除き(飢餓)、30μg/mlのMVの存在下で24時間培養したSKOV−3細胞にて測定した。MVの存在下、G0/G1フェーズにて細胞数の増加が観察される。
【図5】図5Aおよび5BはMVのインビトロでの血管形成における作用を評価するのに実施された実験の結果を示す。HUVECおよびTECのマトリゲル内での毛細血管様構造を形成する能力について評価した。
【図6】MVのインビトロでのアポトーシスにおける作用を評価するのに実施された実験の結果を示す。HUVECおよびTECのアポトーシスを、異なる量のMVと一緒にインキュベートした48時間後のアポトーシス細胞の割合としてTunelアッセイで評価した。
【図7】MVのTECの増殖に対する作用を評価するのに実施された実験の結果を示す。TECの増殖は、RNaseで前処理されているか、またはされていないBM−MSCからの異なる量のMV(10および30μg/ml)と一緒にインキュベートした48時間後にBrdU取り込みアッセイで評価した。結果は2回の実験の平均値±SDとして表される。
【図8】HepG2異種移植片のSCIDマウスに腫瘍内投与された、RNaseで処理されているか、またはされていないMVのインビボでの抗腫瘍活性を示す。腫瘍塊を毎週該移植片の2つの直交する寸法をキャリパーを用いて測定することにより決定した。結果は腫瘍塊の増大の割合として表され:第1処置(HepG2注射の1週間後)の腫瘍塊を慣例により100%値として固定する。
【図9】BM−MSCから由来のMVがインビボにて腫瘍増殖を減少させることを示す。A)BM−MSCから由来のMVで処置されている(右図)または処理されていない(左図)HepG2腫瘍の代表例;B)BM−MSCからのMVで処置されている(右図)または処理されていない(左図)切除されたHepG2腫瘍の代表例;C)BM−MSCからのMVで処置されている(右図)または処理されていない(左図)HepG2腫瘍のヘマトキシリンおよびエオシン染色。
【図10】MVのRNaseを用いる前処理がBM−MSCから由来のMVの抗腫瘍活性を無効にすることを示す。A)MV−RNaseで処置されている切除されたHepG2腫瘍の代表例;B)MV−RNaseで処置されているHepG2腫瘍のヘマトキシリンおよびエオシン染色。
【図11】HepG2上のBrdUをベースとする増殖アッセイの結果を示すグラフである。HepG2をDMEM単独中に、あるいは異なる量のHLSCより由来のMVを補足したDEMEにて培養した。3日後、HepG2増殖がBrdU取り込みアッセイを用いて定量された。
【図12】HepG2細胞上のアポトーシスアッセイの結果を示すグラフである。HepG2細胞は、DMEM単独中で、または10%FCSを含むDMEM中で、ビンクリスチン(100ng/ml)またはMV−HLSC(30μg/ml)またはMV−HLSC(RNaseで前処理されている、30μg/ml)を補足して培養された。24時間後に分析を行った。
【図13】基底条件下のHepG2細胞、あるいはビトロネクチン、ドキソルビシン、MV−HLSCまたはビトロネクチン+MV−HLSCおよびドキソルビシン+MV−HLSCで処理したHepG2細胞でのアポトーシスアッセイの結果を示すグラフである。
【図14】MV−HLSC(n=3)、ビヒクル(n=2)またはRNase処理のMV−HLSC(n=3)でi.t.処理された後で、マウスを殺した際の、回収されたHepG2腫瘍の体積を測定することで得られたデータを示すグラフである。腫瘍の体積は該移植片の2つの直交する寸法をキャリパーを用いて毎週測定することにより決定された。
【図15】HLSC−MV処理によるインビボでの腫瘍増殖の阻害および腫瘍内アポトーシスの誘発を示す顕微鏡写真を図示する。A)4週間後に回収されたHepG2腫瘍のアポトーシス、PCNAおよびヘマトキシリン&エオシン染色を示す代表的顕微鏡写真;B)MV−処理されたマウスから回収されたHepG2腫瘍のアポトーシス、PCNAおよびヘマトキシリン&エオシン染色を示す代表的顕微鏡写真;C)MV−RNAase処理されたマウスから回収されたHepG2腫瘍のアポトーシス、PCNAおよびヘマトキシリン&エオシン染色を示す代表的顕微鏡写真。
【図16】MCF−7細胞を異なる濃度のHLSC−MVと共にインキュベートすることにより実施されたインビトロでの増殖アッセイの結果を示すグラフである。MCF−7細胞の増殖は、2、10、15および30μg/mlのHLSC細胞から由来のMVと共に48時間インキュベートした後に、BrdU取り込みアッセイにより評価された。該実験を3回重複して行った。P<0.05。
【図17】カポジ細胞(KS)を異なる濃度のHLSC−MVと共にインキュベートすることにより実施されたインビトロでの増殖アッセイの結果を示すグラフである。カポジ細胞の増殖は、2、10、15および30μg/mlのHLSC細胞から由来のMVと共に48時間インキュベートした後に、BrdU取り込みアッセイにより評価された。該実験を3回重複して行った。P<0.05。
【図18】MCF−7細胞およびカポジ細胞をHLSC−MVと共にインキュベートすることにより実施されたインビトロでのアポトーシスアッセイの結果を示すグラフである。アポトーシスは異なる量のMV(2、10、15および30μl/ml)および30μg/mlのRNase処理されたMVと共にインキュベートされた48時間後のアポトーシス細胞の割合としてTUNELアッセイにより評価された。アポトーシス誘発の陽性対照としてドキソルビシンを用いた。陰性対照にて、MCF−7細胞およびカポジ細胞をビヒクル単独で処理した。該実験を3回重複して行った。P<0.05。
【発明を実施するための形態】
【0027】
1.間充織幹細胞(MSC)からの微細小胞(MV)
【0028】
1.1 MSCの単離および特徴付け
【0029】
骨髄細胞をフィコール(Ficoll)グラジエント(密度:1022g/ml;Sigma-Aldrich、St. Louis、MO)上に積層させ、1500rpmで30分間遠心分離に付した。単核細胞を間充織幹細胞基底培地(MSCBM、Lonza)の存在下で培養させた。5日間培養した後、培地を変換した。単離された細胞を拡張させるために、付着した単層を、第1継代について15日目に、その後の継代については7日毎に37℃で5分間トリプシン処理に付して分離した。細胞を10000細胞/cm2の密度で播き、継代6より後の細胞は使用されなかった。
【0030】
糸球体からのMSC集団(Gl−MSC)が、Bruno Sら、Isolation and characterization of resident mesenchymal stem cells in human glomeruli. Stem Cells Dev. 2009;18:867-880に記載されるように、手術で摘出された腎臓の皮質の正常な部分より得られた。皮質の切開した後で、糸球体の懸濁液を標準的な確立された方法を用いて集めた:一連の等級別のメッシュ(60および120メッシュ)に通した後、最上部にある120メッシュのシーブで糸球体を集めた。次に、糸球体を自然沈降(室温で10分間)により円錐管の底部で集め、ボーマン嚢の臓側板を、10mlのピペットを用いて吸引/排出を数回行うことで機械的に、かつコラゲナーゼI(Sigma, St.Louis, MO)を用いて2分間消化することで酵素的に除去した。ついで、細胞およびボーマン嚢を取り出すために、糸球体を自然沈降により円錐管の底部に集め、それをフィブロネクチンを被覆したT25フラスコ(Falcon、BD Bioscience、Two Oak Park、Bedford、MA)に移した。糸球体を間充織幹細胞基礎培地(MSCBM、Lonza)の存在下で培養した。細胞を放置し、継代前に集密とさせた:継代4になるまでの継代間の間隔は変動し(3−7日)、それからは約7日で確立された。
【0031】
各継代の際に、細胞を計数し、細胞蛍光分析および免疫蛍光分析により免疫表現型について分析した。細胞蛍光分析は、次の抗体、すべてのフィコエリトリン(PE)またはフルオレセインイソチオシアネート(FITC)接合体:抗−CD105、−CD29、−CD31、−CD146、−CD44、−CD90(Dakocytomation、Copenhagen、Denmark);−CD73、−CD34、−CD45、−CD80、−CD86、−CD166、HLA−I(Becton Dickinson Biosciences Pharmingen、San Jose、CA);−CD133(Miltenyi Biotec、Auburn);KDR(R&D Systems、Abington、U.K.);−HLA−II(Chemicon International Temecula、CA)、−CD40(Immunotech、Beckman Coulter)、−CD154(Serotec、Raleigh、NC USA)モノクローナル抗体を用いて実施された。マウスIgG同形対照はDakocytomationからのものであった。インキュベーションはすべて、0.1%ウシ血清アルブミンおよび0.1%アジ化ナトリウムを含有する、100μlのリン酸緩衝セイライン(PBS)中、4℃で行われた。各サンプルについて、10000個の細胞をFACSCalibur血球計算器(BD Biosciences Pharmingen)で分析した。ゲーティングは陰性対照に基づいて構成され、行われたすべての分析にて補償対照が含められた。セルクエストソフトウェア(Cell Quest software)(BD Biosciences Pharmingen)を用いて各実験からゲート化された集団についての集団割合および数が生成された。
【0032】
間接免疫蛍光を、チャンバースライド(Nalgen Nunc International、Rochester、NY、USA)上で培養され、2%シュークロースを含有し、必要ならば、ヘペス−トリトンX100緩衝剤(Sigma、St. Louis、MO)で透過処理された、4%パラホルムアルデヒドに固定された、MSCについて行った。次の抗体:マウスモノクローナル抗ビネンチン(Sigma)およびウサギポリクローナル抗−フォン・ヴィレブランド因子(Dakocytomation)を用いた。一次抗体を省略したもの、または非免疫ウサギまたはマウスIgGと置き換えられたものが、適当であるならば、対照として使用された。Alexa Fluor488抗−ウサギおよび 抗−マウスTexas Red(Molecular Probes、Leiden、The Netherlands)が二次抗体として使用された。共焦点顕微鏡分析が、Zeiss LSM 5 Pascal Model Confocal Microscope(Carl Zeiss International、Germany)を用いて実施された。ヘキスト33258染料(Sigma)が核染色のために添加された。
【0033】
BM−MSCおよびGL−MSC調製物は、CD45、CD14およびCD34などの造血マーカーを発現しなかった。該調製物は副刺激性分子(CD80、CD86およびCD40)も内皮マーカー(CD31、フォン・ヴィレブランド因子、KDR)も発現しなかった。異なる培養継代での細胞調製物はすべて典型的なMSCマーカー:CD105、CD73、CD44、CD90、CD166およびCD146を発現した。それらはまたHLAクラスIも発現した。
【0034】
MSCの脂質生成、骨形成および軟骨形成の分化能は、Pittenger MF、Martin BJ、Mesenchymal and their potential as cardiac therapeutics. Circ. Res 2004;95:9-20に記載されるように測定された。簡単には、MSCをAdipogenic Medium(Lonza)で3週間培養した。分化を評価するのに、細胞を4%パラホルムアルデヒド中室温で20分間固定し、メタノール(Sigma)中0.5%のOil Red O(Sigma)を用いて室温で20分間染色した。
【0035】
骨形成分化は、MSCをOsteogenic Medium(Lonza)で培養することにより評価された。培地を週に二回3週間変えた。分化を評価するのに、細胞を4%パラホルムアルデヒド中20分間固定し、Alizarin Red(pH4.1)(Lonza)を用いて室温で20分間染色した。
【0036】
軟骨形成の分化では、2.5x105のMSCを15mlの円錐形ポリプロピレン管(Falcon BD Bioscience)中150gで5分間遠心分離に付し、DMEMで2回洗浄した。該ペレットを10ng/mlのTransforming Growth Factor β3(Lonza)を補足した軟骨培地(Lonza)にて培養した。該培地を28日間で3日毎に変えた。ペレットを4%パラホルムアルデヒド中に一夜固定し、パラフィン包埋切片を0.1%サフラニンO(Sigma)を用いてグリコサミノグリカンについて、および1%アルシャンブルーを用いて硫酸化プロテオグリカンについて染色した。
【0037】
1.2 MSCから由来のMVの単離および特徴付け
【0038】
FCSを除去し、0.5%のBSA(Sigma)を補足したRPMIにて培養された、上記のように得られたBM−MSCまたはGL−MSCの上澄から微細小胞(MV)を得た。2000gで20分間遠心分離に付して残骸を除去した後、無細胞上澄を100000gで超遠心分離器(Beckman Coulter Optima L-90K)を用いて4℃で1時間遠心分離に付し、25mMのN−2−ヒドロキシエチルピペラジン−N’−2−エタンスルホン酸(HEPES)(Sigma)を含有する無血清培地199にて洗浄し、同条件下で第2の超遠心分離に供した。MVのエンドトキシン汚染物を、Limulus試験により、製造業者の使用説明書(Charles River Laboratories、Inc.、Wilmington、MA、USA)に従って排除し、MVを−80℃で貯蔵した。
【0039】
選択される実験においては、MVを1U/mlのRNase(Ambion Inc.、Austin、TX、USA)を用いて37℃で1時間処理し、10U/mlのRNase阻害剤(Ambion Inc.)を添加することで反応を停止させ、超遠心分離によりMVを洗浄した。
【0040】
1.3 MSCから由来のMVを用いて行われるインビトロ実験
【0041】
癌細胞株 ヒト肝臓癌細胞株(HepG2)、ヒト乳腺腺癌細胞株(MCF−7)およびヒト卵巣癌細胞株(SKOV−3)を、10%ウシ胎仔血清(FCS、Euroclone)、100U/mlのペニシリン、100mg/mlのストレプトマイシンおよび1%のグルタミン(すべて、Sigmaより入手)を含有する低グルコースDMEM(Euroclone)中に培養し、5%CO2、37℃の加湿雰囲気下でインキュベーターに維持した。カポジ肉腫細胞(KS細胞)の初代培養を、免疫抑制療法下にある、腎臓同種移植を受けた患者の皮膚病変より得、10%のFCS、100μg/mlのペニシリンおよび100μg/mlのストレプトマイシンを補足したRPMI1640培地にて培養した。
【0042】
腫瘍内皮細胞(TEC):単離および培養 MACSシステム(Miltenyi Biotec、Auburn、CA、USA)を用いる磁気細胞分類により、磁気ビーズに結合した抗−CD105抗体を用いて、明細胞型腎細胞癌の試料よりTECを単離した。TEC細胞株を確立し、上皮細胞増殖因子(10ng/ml)、ヒドロコルチゾン(1mg/ml)、ウシ脳抽出物(すべて、Lonzaより入手)および10%のFCSを補足した内皮基底完全培地(EBM)中に培養を維持した。TECは形態学、vWF抗原、CD105、CD146および血管内皮カドヘリンに対して陽性染色を示し、サイトケラチンおよびデスミン(Bussolati Bら、Altered angiogenesis and survival in endothelial cells derived from renal carcinoma. FASEB J 2003;17:1159-1161)に対して陰性染色を示すことにより、内皮細胞として特徴付けられる。
【0043】
ヒト臍帯静脈内皮細胞(HUVEC):単離および培養 ヒト臍帯静脈内皮細胞(HUVEC)を、以前に記載されるように(Bussolati Bら、Vascular endothelial growth factor receptor-1 modulates vascular endothelial growth factor-mediated angiogenesis via nitric oxide. Am J Pathol. 2001Sep;159(3):993-1008)、臍帯静脈より得、EBMおよび10%FCSに維持された。実験は第2および第3継代のHUVECで行われた。
【0044】
細胞増殖 HepG2、MCF−7、SKOV−3、KS細胞またはHUVECを、2000または4000細胞/ウェルで、FCSを除く、RNaseで前処理されている、または前処理されていない異なる濃度の微細小胞を有するDMEM(Sigma)中の96−ウェルに播種した。DNA合成が、48時間培養した後、5−ブロモ−2’−デオキシ−ウリジン(BrdU)の細胞DNAへの取り込みとして検出された。ついで、細胞を0.5Mのエタノール/HClで固定し、ヌクレアーゼと共にインキュベートし、該DNAを消化した。DNAへのBrdUの取り込みを抗−BrdUペルオキシダーゼ接合のmAbを用いて検出し、可溶性発色基質(Roche Applied Science、Mannheim、Germany)で可視化した。光学密度を405nmの下でELISAを用いて測定した。
【0045】
細胞周期分析 ヒト癌細胞株を、30μg/mlの異なるMVの調製物を用いて24時間刺激し、トリプシンにより分離し、冷80%エタノール中に固定した。細胞を−20℃に少なくとも24時間維持し、次にPBSで洗浄した。ついで、該細胞を室温でヨウ化プロピジウム(50μg/ml)(Sigma)と一緒に1時間インキュベートし、RNase(200μg/ml)(Sigma)および0.5%のNonidetP40(Sigma)含有の溶液中で該DNAを染色した。各サンプルについて、50000個の細胞をFACSCalibur血球計算器(BD Biosciences Pharmingen)で分析した。
【0046】
アポトーシスアッセイ HepG2、MCF−7、SKOV−3、KS細胞、HUVECまたはTECが、8000個の細胞/ウェルで、10%FCSを含み、ドキソルビシン(100ng/ml、Sigma)または異なる濃度(10および30μg/ml)のRNaseで前処理されている、またはされていないMVの存在下で、DMEM(Sigma)中に配置された96−ウェルに播種された。アポトーシスをTUNELアッセイ(ApopTag Oncor、Gaithersburg、MD、USA)により評価した。処理して24または48時間経過した後、該細胞をPBSで洗浄し、1%パラホルムアルデヒド(pH7.4)中に4℃で15分間固定させ、PBSにて2回洗浄し、ついで予め冷却したエタノール−酢酸(2:1)に−20℃で5分間ポスト固定させた。試料を酵素末端デオキシヌクレオチジルトランスフェラーゼ(TdT)で処理した。ついで、該細胞をフルオレセインとの加熱した抗ジゴキシゲニン接合体で処理し、室温で30分間インキュベートした。試料を1μg/mlのヨウ化プロピジウム含有の培地に取り込み、該細胞を免疫蛍光により分析した。結果は赤色蛍光発光性細胞(全細胞)に対する緑色蛍光発光性細胞(アポトーシスを起こした細胞)の割合として表される。
【0047】
インビトロでの血管形成 24ウェルのプレートを成長因子を減少させるマトリゲル(BD Biosciences)を用いて4℃で被覆し、加湿雰囲気下、37℃、5%CO2で30分間インキュベートさせた。HUVECまたはTECを、マトリゲル被覆のウェルの5%FCSを含むRPMIまたはEBM中に、5x104個細胞/ウェルの密度で、異なる濃度の、RNaseで処理されているか、処理されていないMVの存在または不在の下で播種させた。インキュベーションから6時間経過した後、該細胞をニコン倒立顕微鏡(Nikon)で観察し、実験結果を記録した。結果は、管の長さの平均として表され、MicroImage分析システム(Cast Imaging)で測定され、任意単位にて表され、3回の異なる実験からの2つの重複したウェルにて20xの倍率で5個所の異なるフィールドにて評価された。
【0048】
統計分析 異なる実験操作からのデータはすべて平均値±SDとして表される。統計分析は、必要に応じて、Newmann−Keulsのマルチ比較試験を備えたANOVAで行われた。
【0049】
1.4 インビトロ結果
【0050】
1.4.1 BM−MSCおよびGL−MSCより由来のMVの腫瘍細胞株に対するインビトロ生物学的効果
ヒトBM−MSCから由来のMVの抗腫瘍活性を、HepG2、MCF−7、SKOV−3およびKS細胞株に対してその増殖を阻害し、そのアポトーシスを誘発する能力を測定することによりインビトロにて評価した。
【0051】
図1はHepG2細胞(図1A)およびKS細胞(図1B)を異なる量のMVと共に48時間インキュベートすると、ビヒクル単独でインキュベートされた対照細胞と比べて増殖が顕著に阻害されることを示す。
【0052】
図2はHepG2細胞(図2A)およびKS細胞(図2B)をMVと共に24および48時間インキュベートすると、ビヒクル単独でインキュベートされた対照細胞と比べて、ドキソルビシン刺激と同様に、アポトーシスが顕著に促進されることを示す。
【0053】
MVによりシャトルされるRNAの完全な分解を誘発するようにMVをRNaseと一緒にインキュベートした場合、MVにより惹起されるHepG2およびKS細胞に対するアンチ増殖およびプロアポトーシス作用は小さくなった(図1および2)。MVのRNase処理そのものはドキソルビシンにより誘発される癌細胞株のアポトーシスを干渉しなかった(図2)。
【0054】
反対に、MCF−7細胞およびSKOV−3細胞と30μg/mlのBM−MSCからのMVとの48時間に及ぶインキュベーションは、ビヒクル単独でインキュベートされた対照細胞と比べて増殖を顕著に阻害した(図3Aおよび3B)が、アポトーシスを促進しなかった。これら2種の腫瘍細胞株にて、本願発明者らはまた、細胞がG0/G1フェーズにある割合を、細胞がSおよびG2フェーズにある割合と比較して評価するために、細胞周期をヨウ化プロピジウム染色技法を用いて検討した。本願発明者らは、BrdUの取り込みで観察される増殖の阻害を説明しうる、BM−MSCからのMVがG0/G1フェーズにある細胞、特にSKOV−3細胞(図4)の増大を誘発することを観察した。
【0055】
本願発明者らはまた、Gl−MSCから由来のMVの腫瘍細胞株の増殖およびアポトーシスに対する作用を試験した。Gl−MSCはHepG2細胞株の増殖およびアポトーシスに影響を与えなかった。反対に、Gl−MSCから由来のMVはKS細胞の増殖を阻害し、そのアポトーシスを誘発した(図1および2)。その上、MCF−7およびSKOV−3細胞と30μg/mlのGl−MSCから由来のMVとの48時間に及ぶインキュベーションは、ビヒクル単独でインキュベートした対照の細胞と比べて増殖を阻害した(図3)が、アポトーシスを促進しなかった。
【0056】
1.4.2 ヒト線維芽細胞から由来のMV
【0057】
ヒト線維芽細胞から由来のMVは種々の癌細胞株の増殖を阻害せず、そのアポトーシスを誘発しなかった(図示せず)。
【0058】
1.4.3 BM−MSCから由来のMVの内皮細胞に対するインビトロでの効果
【0059】
本願発明者らはまた、BM−MSCから由来のMVのHUVECおよび腫瘍内皮細胞(TEC)の増殖、アポトーシスおよび毛細血管様形成に対するインビトロ作用を検討した。
【0060】
MV処理はHUVECの増殖に影響を及ぼさず(図示せず)、その毛細血管様形成能にも影響しなかった(図5A)。加えて、HUVECの種々の量のMVとの48時間にわたるインキュベーションはアポトーシスを誘発しなかった(図6)。
【0061】
反対に、TECの種々の量のMVとの48時間にわたるインキュベーションは、ビヒクル単独でインキュベートした対照となる細胞と比べて、増殖を顕著に阻害し(図7)、アポトーシスを顕著に促進した(図6)。TECの増殖は、RNaseで前処理されている、またはされていないBM−MSCからの種々の量(10および30μg/ml)のMVと共にインキュベートして48時間経過した後にBrdUの取り込みアッセイにより評価した。図7の結果は2回の実験の平均値±SDとして表される。
【0062】
MVをRNaseと共にインキュベートした場合、MVにより惹起されるTECに対するアポトーシス作用は著しく減少した。マトリゲルに播種されたTECの種々の量のMVとのインキュベーションは、TECの毛細血管様構造をインビトロにて形成する能力を著しく阻害した。MVのRNaseでの前処理は、MVの細管形成に対する阻害作用を失効させた(図5B)。
【0063】
1.5 MSCより由来のMVを用いるインビボ実験
【0064】
腫瘍形成 3x106個のHepG2細胞を集め、SCIDマウス(Charles River、Jackson Laboratories、Bar Harbor、ME)に皮下移植した。トリプシン−EDTAを用いて収穫した培養細胞をPBSで洗浄し、マイクロ血球計算器のチャンバーにて計数し、100μlのDMEMおよび100μlのマトリゲルマトリックス(Becton Dickinson)に再び懸濁させた。細胞を氷冷させ、1mlのシリンジを用いる26−ゲージ針を介してSCIDマウスの左背中に皮下注射した。動物を活動および健康状態について毎日モニター観察し、体重測定および腫瘍塊の測定を3日毎に行った。腫瘍塊は移植片の2つの直交する直径をキャリパー計測により測定し、式:1/2axb2(式中、aは長軸であり、bは短軸である)を用いて計算した(Hou Jら、Experimental therapy of hepatoma with artemisin and its derivatives:in vitro and in vivo activity、chemosensitization and mechanism of action. Clin Cancer Research. 2008;14:5519-5530))。1週間後、移植された腫瘍が約15mm3の体積に達した時に、本願発明者らは、週に一度のMVの腫瘍内注射を開始した。最初の処理は100μgのMV(RNaseで処理された、またはされていない)で、最大容量が20μlであり;その後の腫瘍内注射は50μgのMV(RNaseで処理された、またはされていない)で、最大容量が20μlであった。対照となるマウスには、発明者らは同じ容量のビヒクルを単独で腫瘍内注射した。マウスをランダムに3つの処理群:a)MVを腫瘍内注射した群(n=8);b)RNaseで処理されたMVを腫瘍内注射した群(n=8);およびc)同じ容量のビヒクルを単独で注射した対照群(n=5)に分けた。マトリゲル注射から3週間後に、マウスを殺し、腫瘍を回収し、組織構造について加工処理した。
【0065】
1.6 インビボ結果
【0066】
1.6.1 BM−MSCから由来のMVのHepg2腫瘍増殖に対するインビボでの生物学的作用
【0067】
腫瘍形成および増殖がSCIDマウスにてBM−MSCより由来のMVにより阻害される。BM−MSCより由来のMVのインビボにおける腫瘍形成および増殖に対する効果を測定するために、SCIDマウスに、マトリゲルの存在下にてHepG2を皮下注射した。注射した1週間後に、腫瘍の体積が約15mm3であったならば、発明者らは週に一度のMV(RNaseで処理された、またはされていない)の腫瘍内注射を、20μlの最大容量にて開始した。最初の処理は100μgのMVであり;その後の腫瘍内注射は50μgのMVであった。対照となるマウスには、発明者らは20μlのビヒクルを単独で腫瘍内注射した。
【0068】
マトリゲル注射から3週間経過した後、すべての腫瘍を回収し、分析した。HepG2異種移植片実験にて、MVの腫瘍内注射は腫瘍増殖に対して阻害剤作用を示した(図8)。腫瘍の大きさおよび体積は、MVで処理されたSCIDマウスにて有意に小さく(図9AおよびB)、組織学的分析はMVで処理されたHepG2腫瘍にて壊死の部分を示した(図9C)。RNaseで前処理されたMVを注射された腫瘍は、対照となる腫瘍と大きさおよび組織構造にて差異がなかった(図10AおよびB)。
【0069】
2. 肝臓幹細胞から由来の微細小胞(MV)
【0070】
2.1 成体ヒト肝臓幹細胞(HLSC)の単離および特徴付け
【0071】
HLSCは、L−グルタミン(5mM)、ヘペス(12mM、pH7.4)、ペニシリン(50IU/ml)、ストレプトマイシン(50μg/ml)(すべて、Sigma、St. Louisより入手)、FCS(10%)を補足した、最小必須培地/内皮細胞基底培地−1(α−MEM/EBM)(3:1)(Gibco/Cambrex)にて培養された、Cambrex Bio Science Verviers S.p.r.l.(Verviers、Belgium)より入手の、低温保存された正常なヒト肝細胞より単離された。拡張した細胞をT−75フラスコに移し、それらが集密に達した時に分析された。
【0072】
肝癌細胞株HepG2を10%ウシ胎仔血清(FBS)含有のダルベッコ修飾イーグル培地(DMEM)で培養させた。
【0073】
2.2 HLSCから由来のMVの単離
【0074】
MVを2%ウシ胎仔血清(FBS)補足のMEM−アルファにて培養されたHLSCの上澄より得た。血清無しで一夜インキュベートされた細胞の生存能をトリパンブルーの排除により検出した。2000gで20分間遠心分離に付し、残骸を取り除いた後、無細胞上澄を100000g(Beckman Coulter Optima L-90K超遠心分離器)で4℃で1時間遠心分離に付し、25mMのN−2−ヒドロキシエチルピペラジン−N’−2−エタンスルホン酸(HEPES)(Sigma)含有の無血清培地199で洗浄し、同じ条件下で第2の超遠心分離に供した。インビトロおよびインビボにて、蛍光顕微鏡法またはFACS分析によりMVを追跡するのに、幹細胞由来のMVが赤色蛍光脂肪族発色団を脂質二重層中に挿入するPKH26色素(Sigma)で標識された。標識後、MVを洗浄し、10000gで1時間4℃で超遠心分離に供した。MVペレットを培地199に懸濁させ、蛋白含量をブラッドフォード法(BioRad、Hercules、CA)により定量した。MVの汚染エンドトキシンをリムルス試験により製造者の説明書(Charles River Laboratories、Inc.、Wilmington、MA)に従って排除し、MVを−80℃で貯蔵した。ヨウ化プロピジウムで染色した後のMV懸濁液上でなされた形態学的分析はアポトーシス体の存在を明らかにしなかった。
【0075】
選択された実験にて、HLSCからのMVは1U/mlのRNase(Ambion Inc.、Austin、TX)で1時間37℃で処理され、10U/mlのRNase阻害剤(Ambion Inc.)を添加することで反応を停止させ、MVを超遠心分離で洗浄した。RNase処理の効果を、TRIZOL試薬(Invitrogen、Carlsbad、CA)を用いてRNA抽出を行った後に、抽出された全RNAを分光光度計分析により評価した(未処理:1.3±0.2μg RNA/mgの蛋白MV;RNase処理:<0.2μg RNA/mgの蛋白MV)。加えて、RNAase処理および処理されていないMVより抽出されたRNAをオリゴdTでなされるレトロトランスクリプションにより標識し、0.6%アガロース上で分析し、RNase処理によるRNAの完全な分解を示した。対照として、MVを1U/mlのDNase(Ambion Inc.)で1時間37℃で処理した。
【0076】
2.3 HLSCより単離されたMVを用いて実施されたインビトロ実験
【0077】
増殖アッセイ DNA分析は、製造者の仕様説明書に従って酵素免役吸着測定キット(Chemicon、Temecula、CA)を用いる5−ブロモ−2−デオキシウリジン(BrdU)の細胞DNAへの取り込みとして検出された。簡単には、洗浄した後、細胞を10モル/lのBrdUと一緒に6〜10時間、37℃で、5%CO2にて、加湿雰囲気下でインキュベートさせ、0.5モル/Lのエタノール/HClで固定し、ヌクレアーゼと一緒にインキュベートし、DNAを消化した。DNA中に取り込まれたBrdUを、抗BrdUペルオキシダーゼ接合モノクローナル抗体を用いて検出し、可溶性発色基質で可視化した。光学密度を酵素免疫吸着測定リーダーを用いて405nmで測定した。
【0078】
アポトーシスアッセイ アポトーシスを末端dUTPニックエンド標識アッセイ(ApoTag;Oncor、Gaithersburg、MD)を用いて評価した。細胞(8x103/ウェル)を96ウェルプレートで培養し、リン酸緩衝セイライン(PBS)に懸濁させ、PBS中の1%パラホルムアルデヒド(pH7.4)にて15分間、4℃で、つづいて予め冷却したエタノール/酢酸(2:1)で5分間−20℃で固定させた。細胞を末端デオキシヌクレオチドトランスフェラーゼ酵素で処理し、加湿チャンバーにて1時間37℃でインキュベートし、ついで加温したフルオレセインイソチオシアネート接合のアンチジゴキシゲニンで30分間室温で処理した。洗浄した後、試料を1g/mlのヨウ化プロピジウム含有の培地に固定し、細胞を免疫蛍光により分析した。
【0079】
統計分析 種々の実験操作のデータはすべて平均±SDとして表される。統計分析は、必要に応じて、ニューマン−クールズの多重比較試験のANOVAにより実施された。
【0080】
2.4 インビトロ結果
【0081】
2.4.1 HLSCより由来のMVのHepG2肝癌細胞株の増殖に対する効果
【0082】
本願発明者らはHLSCより由来のMVのHepG2肝癌細胞株の増殖に対する効果を評価した。簡単には、HepG2細胞を種々の量の(10、20および30μg/ml)のHLSC由来のMVそのものと共にインキュベートするか、またはRNaseで3日間処理した。インキュベーションの終わりに、HepG2培養物を計数するか、0.5Mのエタノール/HClに固定し、ヌクレアーゼと一緒にインキュベートし、そのDNAを消化した。該DNAに取り込まれたBrdUを抗BrdUペルオキシダーゼ接合モノクローナル抗体を用いて検出し、可溶性発色基質で可視化した。光学密度をELISAリーダーを用いて405nmで測定した。図11に示されるように、HLSCより由来のMVはHepG2増殖を顕著に阻害しうる。このことはまたRNase処理のMVにも適用する。
【0083】
2.4.2 HLSCより由来のMVのHepG2肝癌細胞株のアポトーシスに対する効果
【0084】
HLSC誘導のMVのHepG2のアポトーシス誘発能を評価した。簡単には、HepG2を8000細胞/ウェルの密度で96−ウェルプレートの10%FCSを補足したDMEMに播種し、FCSの不在下で、癌の化学療法にて使用される2種の有糸分裂阻害剤である、ビンクリスチン(100ng/ml)またはドキソルビシン(50ng/ml)で処理することにより、あるいはMV処理(30μg/ml)で培養してアポトーシスを誘発した。対照として、MVをまた1U/mlのRNase18(Ambion、Austin、TX)で37℃1時間処理し、癌細胞増殖の阻害への寄与がMVによって癌細胞にデリバリーされたmRNAの水平移動に依存するかどうかを評価した。アポトーシスをTUNELアッセイ分析を用いて24および72時間で評価した。図12に示されるように、HLSCより由来のMVは、ビンクリスチンにより誘発されるアポトーシスと比べて、HepG2のアポトーシスを誘発することが可能であった。反対に、RNase処理はアポトーシスを誘発しなかった。加えて、HepG2のビンクリスチン+MV−HLSCまたはドキソルビシン+MV−HLSCでの処理は図13に示されるように相加作用をもたらす。
【0085】
2.5 HLSCより単離されるMVで行われるインビボ実験
【0086】
細胞培養 ヒト肝癌細胞HepG2を、10%ウシ胎仔血清、100μg/mlのペニシリンおよび100μg/mlのストレプトマイシンを補足したDEMEで培養し、5%CO2、37℃の加湿雰囲気のインキュベーター中に維持した。
【0087】
ヒト肝臓幹細胞(HLSC)を、10%ウシ胎仔血清、100μg/mlのペニシリンおよび100μg/mlのストレプトマイシンを補足した、α−MEM/EBM(3:1)にて培養した。EBMはhEGF(ヒト上皮成長因子)、ヒドロコルチゾン、GA(ゲンタマイシン)、BBE(ウシ脳抽出物)で復元された。
【0088】
微細小胞(MV)のHLSCからの単離 MVを2%ウシ胎仔血清を補足したα−MEM培地にて培養したHLSCの上澄より得た。2000gで20分間遠心分離に付して残骸を除去した後、無細胞上澄を100000gで1時間4℃で遠心分離に付し、25mMのN−2−ヒドロキシエチルピペラジン−N’−2−エタンスルホン酸(HEPES)を含有する血清不含培地199で洗浄し、同じ条件下で第2の超遠心分離に供した。MVペレットを0.1%のDMSOの培地199に懸濁させ、蛋白含量をブラッドフォード法を用いて定量した。
【0089】
実験の設計 雄の4ないし5月齢のSCIDをCharles River laboratoriesより得た。すべてのマウスを無菌状態の檻に入れ、1週間保持して順応させた。0日目に、マトリゲル基底膜マトリックスを含む血清不含DMEMに再び懸濁させた3x106のHepG2腫瘍細胞を1:1の割合で2回注射して投与した。細胞懸濁液を合計0.2mlの容量でSCIDマウスの左鼠径部に注射した。すべてのマウスを無作為に3つの処理群:20μlの腫瘍内(i.t.)MV注射(n=3);20μlのi.t.PBS注射(n=2)および20μlのi.t.MV−RNase処理の注射(n=3)に分けた。7日目に、腫瘍細胞移植後の処理を開始した。腫瘍は7日目より触診可能となり;0.1%のDMSOを補足したM199に懸濁させた、50または100μgのMVを腫瘍移植の7、12、14および18日後に注射した。腫瘍の体積が約15mm3になった時に治療を開始した。動物を活性および健康状態について毎日モニター観察し、体重測定および腫瘍塊の測定を3日毎に行った。
【0090】
腫瘍をキャリパーで測定した。腫瘍塊を、キャリパーにより移植片の2つの直交する直径を測定し、式:1/2a x b2(式中、aは長軸であり、bは短軸である)を用いて計算した。28日目に動物を殺し、さらなる分析のために腫瘍を集めた。
【0091】
形態学研究 腫瘍を10%緩衝化中性ホルマリンに固定し、慣用的に加工処理し、パラフィンに包埋し、5μmの薄片にし、顕微鏡試験のためにH&Eで染色した。増殖を検出するための免疫組織化学を抗PCNAモノクローナル抗体を用いて行った。切片を抗マウスHRP二次抗体で遮断して標識化した(1:300希釈)。一次抗体を省略したもの、または非免疫マウスIgGと置き換えられたものが、対照として使用された。アポトーシスをTUNELによりパラフィン包埋組織の切片にて評価した。10個の連続しない切片をアポトーシスの陽性腫瘍細胞について630の倍率で計数した。ヘキスト33258色素を核染色のために添加した。
【0092】
統計分析 種々の実験操作のデータはすべて平均±SDとして表される。統計分析は、必要に応じて、ニューマン−クールズの多重比較試験のANOVAにより実施された。
【0093】
2.6 インビボ結果
【0094】
2.6.1 腫瘍の成長および増殖がSCIDマウスの肝癌異種移植片実験にてHLSCから由来のMVによって阻害された
【0095】
HLSCから由来のMVのインビボでの腫瘍増殖に対する効果を測定するために、SCIDマウスに、ヒト肝細胞癌の細胞株HepG2を皮下移植した。HepG2注射の1および2週間後で、腫瘍の体積が約15mm3であった際に、マウスにMV(50または100μg)を最大20μlの容量で腫瘍内注射して処理した。対照となるマウスにおいては、腫瘍に20μlのビヒクルを単独で注射した。HepG2の注射から3および4週間後に、すべての腫瘍を回収し、分析した。この異種移植片実験にて、MVの腫瘍内注射(図14)は腫瘍成長に対して阻害剤の効果を示した。加えて、組織学的分析はMVで処理された腫瘍にて壊死の領域を示し(図15B)、抗増殖作用がPCNA染色を用いて観察された(図15B)。
【0096】
2.6.2 HLSCより由来のMVによるSCIDマウスにおける肝癌異種移植片実験でのアポトーシスの誘発
【0097】
腫瘍内アポトーシスの効果を測定するために、MVで処理された腫瘍からのパラフィン切片をTUNELで分析した。MV処理はビヒクル単独で処理された腫瘍と比べて腫瘍を誘発した(図15A)。
【0098】
2.7 HLSCから由来のMVの、MCF−7乳腺腺癌およぼカポジ肉腫(KS)細胞に対するインビトロでの生物学的効果
【0099】
1.7.1 材料および方法
【0100】
細胞培養 ヒト非楕円形肝臓幹細胞(HLSC)を、10%ウシ胎仔血清、100μg/mlのペニシリンおよび100μg/mlのストレプトマイシンを補足した、α−MEM/EBM(3:1)にて培養した。EBMはhEGF(ヒト上皮成長因子)、ヒドロコルチゾン、GA(ゲンタマイシン)、BBE(ウシ脳抽出物)で復元された。
【0101】
MCF−7乳腺腺癌細胞株をAmerican Type Culture Collection(Manassas、VA)より入手し、10%FCS、100μg/mlのペニシリンおよび100μg/mlのストレプトマイシンを補足した、DMEMにて培養し、インキュベーター中、5%CO2の加湿雰囲気下、37℃で維持した。
【0102】
カポジ肉腫細胞(KS細胞)の初代培養を、免疫抑制療法を受けている腎同種移植片を有する患者の皮膚病変から入手し、10%FCS、100μg/mlのペニシリンおよび100μg/mlのストレプトマイシンを補足した、RPMI1640培地にて培養した。
【0103】
MVの単離 2%ウシ胎仔血清(FBS)を補足した、MEM−アルファにて18時間培養したHLSCの上澄からMVを得た。選択された実験にて、MVをFBSの不在下で集めた。2%のFBSおよび血清無しで一夜インキュベートされた細胞の生存能をトリパンブルーの排除により検出した(90%より大きい、データは示さず)。2000gで20分間遠心分離に付し、残骸を取り除いた後、無細胞の上澄を100000g(Beckman Coulter Optima L-90K超遠心分離器)で1時間4℃で遠心分離に付し、25mMのN−2−ヒドロキシエチルピペラジン−N'−2−エタンスルホン酸(HEPES)含有の無血清培地199で洗浄し、同じ条件下で第2の超遠心分離に供した。MV細胞を培地199に懸濁させ、蛋白含量をブラッドフォード法で測定した。MVを−80℃で貯蔵した。ヨウ化プロピジウムで染色した後のMV懸濁液上でなされた形態学的分析はアポトーシス体の存在を明らかにしなかった。
【0104】
RNase処理 選択された実験にて、HLSCからのMVを1U/mlのRNaseで1時間37℃で処理した。10U/mlのRNase阻害剤を添加することで反応を停止させ、MVを超遠心分離により洗浄した。
【0105】
細胞増殖 HLSCから由来のMVが種々の腫瘍に対してその抗腫瘍活性を発揮するかどうかを調べるために、MCF−7乳腺腺癌細胞およびカポジ肉腫細胞から由来の細胞株を、各々、8000細胞/ウェルの密度で、種々の濃度のMV(2;10;15;および30μg/mlおよび30μg/mlのRNase処理のMV)を含む、96−ウェルプレートのDMEMおよびRPMIに播種した。DNA合成を、48時間培養した後の、5−ブロモ−2’−デオキシ−ウリジン(BrdU)の細胞DNAへの取り込みとして検出した。次に、細胞を0.5Mのエタノール/HClで固定し、ヌクレアーゼと一緒にインキュベートし、DNAを消化させた。該DNAに取り込まれたBrdUを、抗−BrdUペルオキシダーゼ接合mAbを用いて検出し、可溶性発色基質を用いて可視化した。光学密度は405nmでELISAリーダーを用いて測定された。
【0106】
アポトーシスアッセイ MCF−7およびKS細胞を8000細胞/ウェルの密度で96−ウェルの10%FCSを含み、ドキソルビシン(100ng/ml)または種々の濃度のMV(2;10;15および30μg/mlおよび30μg/mlのRNase処理されたMV)の存在する低グルコースのDMEMに播種した。アポトーシスをTUNELアッセイを用いて評価した。
【0107】
2/7/2 結果:HLSCより由来のMVはMCF−7細胞およびKS細胞のインビトロでの増殖を阻害した
【0108】
MCF−7乳腺腺癌細胞およびカポジ肉腫細胞と、2、10、15および30μg/mlのHLSC−6Bから由来のMV(図16および17)との48時間に及ぶインキュベーションは、ビヒクル単独でインキュベートされた対照細胞と比べて、細胞増殖を顕著に阻害する。これらの結果は組織固有の幹細胞の抗腫瘍作用が同じ組織から形成される腫瘍に対して特異的ではないことを示す。その上、MCF−7乳腺腺癌細胞およびカポジ肉腫細胞の48時間にわたる2、10、15および30μg/mlの、HLSC−6B細胞から由来のMV(図18)が、ビヒクル単独でインキュベートされた対照となる細胞と比べて、アポトーシスを誘発し、その効果も化学療法薬であるドキソルビシンのアポトーシスの誘発と同じくらいである。このことはさらに、組織固有の幹細胞の抗腫瘍作用が同じ組織より生成される腫瘍に対して特異的でないことを確認するものである。
【特許請求の範囲】
【請求項1】
腫瘍性疾患の治療的処置にて使用されるための成体幹細胞から由来の微細小胞。
【請求項2】
成体幹細胞が間充織幹細胞および肝臓幹細胞からなる群より選択される、請求項1記載の微細小胞(MV)。
【請求項3】
成体幹細胞が骨髄から由来のヒト間充織幹細胞(BM−MSC)、脱糸球体嚢から由来のヒト間充織幹細胞(Gl−MSC)およびヒト非楕円形肝臓幹細胞(HLSC)からなる群より選択される、請求項2記載の微細小胞(MV)。
【請求項4】
腫瘍性疾患が肝臓腫瘍、上皮腫瘍、乳房腫瘍、肺腫瘍、前立腺腫瘍、胃腫瘍、結腸腫瘍および卵巣腫瘍からなる群より選択される、請求項1ないし3のいずれかに記載の微細小胞(MV)。
【請求項5】
腫瘍性疾患の治療的処置が、その必要とする患者に、患者の体重1kg当たり0.1−200マイクログラム、好ましくは患者の体重1kg当たり1−150マイクログラム、さらにより好ましくは患者の体重1kg当たり3−120マイクログラムの用量で含まれるMVを投与することを含む、請求項1ないし4のいずれかに記載の微細小胞(MV)。
【請求項6】
局所的または全身的投与に適する医薬剤形の、請求項1ないし5のいずれかに記載の微細小胞(MV)。
【請求項7】
治療的処置が1または複数の細胞傷害性および/または細胞増殖抑制性薬剤を投与することを含む、請求項1ないし6のいずれかに記載の微細小胞(MV)。
【請求項8】
細胞傷害性および/または細胞増殖抑制性薬剤が、パクリタセル、レナリドミド、ポマリドミド、エピルビシン、5FU、スニチニブ、ラ−パチニブ、カネルチニブ、シクロホスファミド、ドキソルビシン、レナリドミド/デキサメタゾン、ポ−マリドミド/デキサメタゾン、カルボプラチン、ラパマイシン、ミトキサントロン、オキサリプラチン、ドキセタセル、ビノレルビン、ビンクリスチンおよびその組み合わせからなる群より選択される、請求項7記載の微細小胞(MV)。
【請求項9】
細胞傷害性および/または細胞増殖抑制性薬剤がドキソルビシンまたはビンクリスチンである、請求項8記載の微細小胞(MV)。
【請求項10】
腫瘍性疾患を治療的に処置する方法であって、その必要とする患者に、成体幹細胞より由来の治療的に効果的な量の微細小胞を投与することを含む、方法。
【請求項11】
成体幹細胞が間充織幹細胞である、請求項10記載の方法。
【請求項12】
成体幹細胞が骨髄から由来のヒト間充織幹細胞(BM−MSC)または脱糸球体嚢から由来のヒト間充織幹細胞(Gl−MSC)である、請求項11記載の方法。
【請求項13】
成体幹細胞が肝臓幹細胞である、請求項10記載の方法。
【請求項14】
成体幹細胞が非楕円形肝臓幹細胞(HLSC)である、請求項13記載の方法。
【請求項15】
腫瘍性疾患が肝臓腫瘍、上皮腫瘍、乳房腫瘍、肺腫瘍、前立腺腫瘍、胃腫瘍、結腸腫瘍および卵巣腫瘍からなる群より選択される、請求項10ないし14のいずれかに記載の方法。
【請求項16】
腫瘍性疾患の治療的処置が、その必要とする患者に、患者の体重1kg当たり0.1−200マイクログラム、好ましくは患者の体重1kg当たり1−150マイクログラム、さらにより好ましくは患者の体重1kg当たり3−120マイクログラムの用量で含まれるMVを投与することを含む、請求項10記載の方法。
【請求項17】
微細小胞(MV)が局所的または全身的経路を介して投与される、請求項16記載の方法。
【請求項18】
治療的処置が1または複数の細胞傷害性および/または細胞増殖抑制性薬剤を投与することを含む、請求項10記載の方法。
【請求項19】
細胞傷害性および/または細胞増殖抑制性薬剤が、パクリタセル、レナリドミド、ポマリドミド、エピルビシン、5FU、スニチニブ、ラ−パチニブ、カネルチニブ、シクロホスファミド、ドキソルビシン、レナリドミド/デキサメタゾン、ポ−マリドミド/デキサメタゾン、カルボプラチン、ラパマイシン、ミトキサントロン、オキサリプラチン、ドキセタセル、ビノレルビン、ビンクリスチンおよびその組み合わせからなる群より選択される、請求項18記載の方法。
【請求項20】
細胞傷害性および/または細胞増殖抑制性薬剤がドキソルビシンまたはビンクリスチンである、請求項19記載の方法。
【請求項1】
腫瘍性疾患の治療的処置にて使用されるための成体幹細胞から由来の微細小胞。
【請求項2】
成体幹細胞が間充織幹細胞および肝臓幹細胞からなる群より選択される、請求項1記載の微細小胞(MV)。
【請求項3】
成体幹細胞が骨髄から由来のヒト間充織幹細胞(BM−MSC)、脱糸球体嚢から由来のヒト間充織幹細胞(Gl−MSC)およびヒト非楕円形肝臓幹細胞(HLSC)からなる群より選択される、請求項2記載の微細小胞(MV)。
【請求項4】
腫瘍性疾患が肝臓腫瘍、上皮腫瘍、乳房腫瘍、肺腫瘍、前立腺腫瘍、胃腫瘍、結腸腫瘍および卵巣腫瘍からなる群より選択される、請求項1ないし3のいずれかに記載の微細小胞(MV)。
【請求項5】
腫瘍性疾患の治療的処置が、その必要とする患者に、患者の体重1kg当たり0.1−200マイクログラム、好ましくは患者の体重1kg当たり1−150マイクログラム、さらにより好ましくは患者の体重1kg当たり3−120マイクログラムの用量で含まれるMVを投与することを含む、請求項1ないし4のいずれかに記載の微細小胞(MV)。
【請求項6】
局所的または全身的投与に適する医薬剤形の、請求項1ないし5のいずれかに記載の微細小胞(MV)。
【請求項7】
治療的処置が1または複数の細胞傷害性および/または細胞増殖抑制性薬剤を投与することを含む、請求項1ないし6のいずれかに記載の微細小胞(MV)。
【請求項8】
細胞傷害性および/または細胞増殖抑制性薬剤が、パクリタセル、レナリドミド、ポマリドミド、エピルビシン、5FU、スニチニブ、ラ−パチニブ、カネルチニブ、シクロホスファミド、ドキソルビシン、レナリドミド/デキサメタゾン、ポ−マリドミド/デキサメタゾン、カルボプラチン、ラパマイシン、ミトキサントロン、オキサリプラチン、ドキセタセル、ビノレルビン、ビンクリスチンおよびその組み合わせからなる群より選択される、請求項7記載の微細小胞(MV)。
【請求項9】
細胞傷害性および/または細胞増殖抑制性薬剤がドキソルビシンまたはビンクリスチンである、請求項8記載の微細小胞(MV)。
【請求項10】
腫瘍性疾患を治療的に処置する方法であって、その必要とする患者に、成体幹細胞より由来の治療的に効果的な量の微細小胞を投与することを含む、方法。
【請求項11】
成体幹細胞が間充織幹細胞である、請求項10記載の方法。
【請求項12】
成体幹細胞が骨髄から由来のヒト間充織幹細胞(BM−MSC)または脱糸球体嚢から由来のヒト間充織幹細胞(Gl−MSC)である、請求項11記載の方法。
【請求項13】
成体幹細胞が肝臓幹細胞である、請求項10記載の方法。
【請求項14】
成体幹細胞が非楕円形肝臓幹細胞(HLSC)である、請求項13記載の方法。
【請求項15】
腫瘍性疾患が肝臓腫瘍、上皮腫瘍、乳房腫瘍、肺腫瘍、前立腺腫瘍、胃腫瘍、結腸腫瘍および卵巣腫瘍からなる群より選択される、請求項10ないし14のいずれかに記載の方法。
【請求項16】
腫瘍性疾患の治療的処置が、その必要とする患者に、患者の体重1kg当たり0.1−200マイクログラム、好ましくは患者の体重1kg当たり1−150マイクログラム、さらにより好ましくは患者の体重1kg当たり3−120マイクログラムの用量で含まれるMVを投与することを含む、請求項10記載の方法。
【請求項17】
微細小胞(MV)が局所的または全身的経路を介して投与される、請求項16記載の方法。
【請求項18】
治療的処置が1または複数の細胞傷害性および/または細胞増殖抑制性薬剤を投与することを含む、請求項10記載の方法。
【請求項19】
細胞傷害性および/または細胞増殖抑制性薬剤が、パクリタセル、レナリドミド、ポマリドミド、エピルビシン、5FU、スニチニブ、ラ−パチニブ、カネルチニブ、シクロホスファミド、ドキソルビシン、レナリドミド/デキサメタゾン、ポ−マリドミド/デキサメタゾン、カルボプラチン、ラパマイシン、ミトキサントロン、オキサリプラチン、ドキセタセル、ビノレルビン、ビンクリスチンおよびその組み合わせからなる群より選択される、請求項18記載の方法。
【請求項20】
細胞傷害性および/または細胞増殖抑制性薬剤がドキソルビシンまたはビンクリスチンである、請求項19記載の方法。
【図1】


【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】

【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【公表番号】特表2013−520971(P2013−520971A)
【公表日】平成25年6月10日(2013.6.10)
【国際特許分類】
【出願番号】特願2012−555384(P2012−555384)
【出願日】平成23年2月28日(2011.2.28)
【国際出願番号】PCT/EP2011/052945
【国際公開番号】WO2011/107437
【国際公開日】平成23年9月9日(2011.9.9)
【出願人】(597075904)フレゼニウス メディカル ケア ドイッチェランド ゲゼルシャフト ミット ベシュレンクテル ハフツング (55)
【Fターム(参考)】
【公表日】平成25年6月10日(2013.6.10)
【国際特許分類】
【出願日】平成23年2月28日(2011.2.28)
【国際出願番号】PCT/EP2011/052945
【国際公開番号】WO2011/107437
【国際公開日】平成23年9月9日(2011.9.9)
【出願人】(597075904)フレゼニウス メディカル ケア ドイッチェランド ゲゼルシャフト ミット ベシュレンクテル ハフツング (55)
【Fターム(参考)】
[ Back to top ]