
腫瘍発生を制御する方法および腫瘍発生のリスクを診断する方法
本発明は、腫瘍発生を予防、阻害、停止または逆行させることを含む、腫瘍発生を制御する方法に関する。本発明は、転移性癌などの癌を含む、新生芽腫の治療、予防および診断の方法も提供する。1つの方法では、発癌性になる素因を有する1つまたは複数の細胞を同定する。該1つまたは複数の細胞において、Rab結合タンパク質の量の変化、細胞内局在の変化、または活性の変化を検出する。少なくとも1つの発現レベルで新生物を発症するリスクを診断する方法では、Rab結合タンパク質の活性レベルおよび細胞内局在を決定する。Rab結合タンパク質と複合体を形成することが可能な化合物を同定するin vitro方法では、各複合体を形成する構成要素を互いに接触させる。新生物が細胞内カルシウムレベルの変化に感受性であるかどうかを決定する方法では、Rab結合タンパク質の発現および活性の少なくとも1つを決定する。Rab結合タンパク質の発現の変化または活性の変化は、新生物が細胞内カルシウムレベルの変化に感受性であることを示す。

【発明の詳細な説明】
【技術分野】
【0001】
関連出願の相互参照
本出願は、米国特許商標局に2007年4月13日に出願され、シリアル番号60/911,573が割り当てられた「Preventing,inhibiting,arresting or reversing tumorigenesis by modulation of a rab binding protein and related uses of a rab binding protein」の出願を参照し、かつその優先権を主張するものである。2007年4月13日に出願された前記出願の内容は、PCTの規則4.18に準じて、本願明細書に含まれないPCTの規則20.5(a)で言及された記載事項、特許請求の範囲または図面の任意の要素または部分の援用を含め、すべての目的で本願明細書に援用する。
【0002】
本発明は、腫瘍発生を制御する方法および腫瘍発生のリスクを診断する方法に関する。これらの方法は、Rab結合タンパク質の量の変化および/または活性の変化を利用するものである。本発明は、Rab結合タンパク質と複合体を形成することが可能な化合物、およびRab結合タンパク質の量、発現および/または活性を調節することが可能な化合物を同定する方法にも関する。本発明は、新生物が細胞内カルシウムレベルの変化に感受性であるかどうかを決定する方法も提供する。
【背景技術】
【0003】
癌は世界的に主たる死因であり、先進国では2番目の主な死因であり、さらに例えばオーストラリア、日本、韓国、シンガポールにおいて、ならびに英国およびスペインの男性人口では、第1番目の死因である。毎年癌を発症する人の数は増大している。それにもかかわらず、癌療法は最近30年間で癌死亡率を低下させることができていない。
【0004】
組織浸潤および転移をもたらす自律的細胞増殖は、すべての悪性新生物の顕著な特徴である。癌は必ずしも加速的速度の細胞増殖の結果としてのみ生じるわけではなく、癌は、一方では細胞周期進行(細胞分裂)および細胞増殖(細胞増大)の速度と、他方ではプログラムされた細胞死(アポトーシス)の速度との間の不均衡の結果である。
【0005】
腫瘍発生および転移増殖の発癌誘導因子の同一性を解明することは、癌の病理生物学を理解するためだけでなく、患者を治療するためにも重要である。というのは、このような遺伝子は、治療的有効性(ヘルセプチン、アバスチン、グリーベック)のためにますます活用されているからである。ヒトの癌では、ゲノム増幅、配列突然変異またはプロモーターの低メチル化から生じる異常な過剰発現によって癌遺伝子が活性化されることが多い。アンプリコンとして知られる、癌において増幅されることが多い染色体上の遺伝子座は、1つまたは複数の癌遺伝子によって誘導されると考えられ、多くの反復アンプリコンが観察されているが、それらの癌遺伝子決定因子は依然として大部分が知られていない。これらの遺伝子座は、コピー数増大の程度および頻度を評価する比較ゲノムハイブリダイゼーション(CGH)などの細胞遺伝学的技法によって伝統的に調べられており、この戦略は考慮対象の多くの遺伝子候補を与えるが、遺伝子リストが広範囲であり、かつ機能情報がわずかであるため、癌遺伝子の発見に対する進展は限られている。それよりも、大抵は異なる実験背景における遺伝子機能の累積的観察結果から導かれる仮説の生成によって決まるため、癌遺伝子の発見は緩慢で実験プロセスとなることが多い。
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明の目的は、腫瘍発生の予防を含めた腫瘍発生を制御する手段、および発癌性になる素因を有する細胞を同定する手段を提供することである。
【課題を解決するための手段】
【0007】
第1の態様において、本発明は、腫瘍発生を予防、阻害、停止または逆行させることを含む、腫瘍発生を制御する方法を提供する。本発明は、転移性癌などの癌を含む、新生芽腫の治療、予防および診断の方法も提供する。
【0008】
1つのこのような方法は、Rab結合タンパク質、特にRab11FIPタンパク質(Rab11ファミリー相互作用タンパク質)の量または活性を調節することを含む。
本発明のさらなる方法は、発癌性になる素因を有する1つまたは複数の細胞を同定することに関する。方法は、1つまたは複数の細胞において、Rab結合タンパク質、特にRab11FIPタンパク質の量の変化、細胞内局在の変化、または活性の変化を検出することを含む。
【0009】
本発明は、新生物を発症するリスクを診断する方法も提供する。方法は、Rab結合タンパク質、特にRab11FIPタンパク質の発現レベル、活性レベルおよび細胞内局在の少なくとも1つを決定することを含む。
【0010】
本出願は、Rab結合タンパク質、特にRab11FIPタンパク質の活性を調節することができ、したがって例えば抗癌剤として作用することができる、適切な化合物の同定にも関する。
【0011】
これに関し、本発明は、Rab結合タンパク質、特にRab11FIPタンパク質と複合体を形成することが可能な化合物を同定するin vitro方法を提供する。方法は、各複合体を形成する構成要素を互いに接触させることを含む。
【0012】
さらなる態様において、本発明は、in vivoでRab結合タンパク質、特にRab11FIPタンパク質の活性を調節することが可能な化合物を同定するための方法を提供する。方法は、Rab結合タンパク質またはその機能断片を発現する微生物を提供することを含む。方法は、微生物に化合物を加えることをさらに含む。方法は、Rab結合タンパク質の細胞表現型または活性の変化をモニターすることも含む。
【0013】
他の態様において、本発明は、Rab結合タンパク質、特にRab11FIPタンパク質の量または活性を調節することが可能な化合物を同定する方法を提供する。
1つのこのような方法は、宿主生物を提供することを含む。宿主生物は、癌細胞を収容し増殖させることができる。方法は癌細胞を宿主生物に導入することも含む。さらに方法は、Rab結合タンパク質の発現および/または活性を調節し得ると考えられる化合物を投与することを含む。方法は、宿主生物中の任意の腫瘍(複数の腫瘍を含む)の増殖をモニターすることをさらに含む。
【0014】
さらなるこのような方法は、宿主生物を提供することも含む。宿主生物は癌細胞を収容し増殖させることができる。方法は癌細胞を宿主生物に導入することも含む。癌細胞は、Rab結合タンパク質の量または活性を調節し得ると考えられる化合物を含む。方法は、宿主生物中の任意の腫瘍(複数の腫瘍を含む)の増殖をモニターすることをさらに含む。
【0015】
他のさらなる態様において本発明は、新生物が細胞内カルシウムレベルの変化に感受性であるかどうかを決定する方法を提供する。方法は、Rab結合タンパク質の発現および/または活性を決定することを含む。Rab結合タンパク質の発現の増大または活性の増大は、新生物が細胞内カルシウムレベルの変化に感受性であることを示す。
【0016】
本発明は、限定されない実施例および添付の図面と併せて検討した場合、詳細な説明に関してさらに理解されるであろう。
【図面の簡単な説明】
【0017】
【図1】TRIAGE方法論の複数工程の略図。
【図2】統合されたマイクロアレイのデータセットを構築するために使用された乳癌コホートの臨床的特徴を示す表。
【図3】乳癌の癌ゲノムのLSVD分析の図。固有値(y軸)を、各染色体(x軸)の長さに沿ってプロットする。最も高いスコアを得た主固有ピークのゲノムの位置を強調してある。
【図4A】図4は、増殖サインおよび無遠隔転移生存率(DMFS)を示す。図4Aは、ウプサラ(Uppsala)コホート(n=251)におけるDMFS不良(P<0.05;尤度比検定)と相関した全プローブセット(n=2,377)が、階層的にクラスタ化されたことを示す図。優性遺伝子のクラスタは、増殖サインを反映する。増殖サインの中核遺伝子を、クラスタ化の順番に以下の実施例において名付ける。クラスタグラムの下に、転移した腫瘍(白マル)および転移しなかった腫瘍(黒マル)を時間の関数として示す。
【図4B】ウプサラ(Uppsala)コホート(n=251)におけるDMFS不良(P<0.05;尤度比検定)と相関した全プローブセット(n=2,377)が、階層的にクラスタ化されたことを示す図。優性遺伝子のクラスタは、増殖サインを反映する。増殖サインの中核遺伝子を、クラスタ化の順番に以下の実施例において名付ける。クラスタグラムの下に、移した腫瘍(白マル)および転移しなかった腫瘍(黒マル)を時間の関数として示す。
【図4C】DMFS不良(n=2,377)に関連するプローブのセットを、P値に従ってビン分割し、>0.4の増殖サインに相関するプローブセットの分画を白で示し、各ビンのパーセントを示す図。全DMFS関連プローブセットのおよそ35%が、>0.4の増殖サインと相関する。
【図5A】図5は、17q12および8p11のアンプリコンのTRIAGE分析を示す。図5Aは、17q12における推定発現アンプリコン(PEA)の発現ヒートマップを示す図。腫瘍をカラムに配置し、LSVDにより同定した遺伝子を列の中で体系化する(染色体に沿った順番で)。腫瘍を、固有重量の絶対値によりランク付けする(ヒートマップの下に記載)。平均発現より上を白で示す。TRIAGEの第3工程において、生存率に有意に関連する遺伝子を右に示す(図1を参照)。白い文字で示された遺伝子(TCAPおよびERBB2)は、TRIAGEによる、最も有力な癌遺伝子候補を示す。
【図5B】8p11における推定発現アンプリコン(PEA)の発現ヒートマップを示す図。腫瘍をカラムに配置し、LSVDにより同定した遺伝子を列の中で体系化する(染色体に沿った順番で)。腫瘍を、固有重量の絶対値によりランク付けする(ヒートマップの下に記載)。平均発現より上を白で示す。TRIAGEの第3工程において、生存率に有意に関連する遺伝子を右に示す(図1を参照)。白い文字で示された遺伝子(RAB11FIP1およびWHSC1L1)は、TRIAGEにより、最も有力な癌遺伝子候補を示す。
【図5C】図5Aと同じデータを、平均発現より下を白く示して表した図。
【図5D】図5Bと同じデータを、平均発現より下を白く示して表した図。
【図5E】得られたデータを要約した図。NSは、「有意差なし」を示唆する。アスタリスクは、増殖サインとの相関が>0.4であることを示す。パネルAのヒートマップの上の黒色の垂直なバーは、免疫組織化学によるERBB2(HER2/neu)陽性を表す。
【図6A】図6は、MCF10A細胞においてRCPの過剰発現が癌表現型を誘導することを示す。図6Aは、MCF10Aおよび乳癌の細胞系において、ウェスタンブロットにより評価した内因性RCPのレベルを示す図。
【図6B】RCP過剰発現の、細胞形態学における影響の図(スケールバー、100μm)。
【図6C】免疫蛍光法による、細胞間接合部のE−カドヘリンの局在化(上)、F−アクチンの組織化(中)および血清飢餓下のフィブロネクチンの発現(下)の、RCP過剰発現の影響の図(スケールバー、50μm)。
【図6D】WSTアッセイによるMCF10A細胞の成長因子非依存性増殖に関するRCP過剰発現の影響の図(平均+/−標準偏差)。
【図6E】軟寒天における、足場非依存性コロニー形成に関するRCP過剰発現の影響の図(倍率、×5)。
【図6F】創傷治癒(擦過傷)アッセイによる細胞遊走に関するRCP過剰発現の影響の図(スケールバー、100μm)。
【図7A】図7は、RCPの作用機序を示している。図7Aは、フローサイトメトリーによる、血清飢餓MCF10A−RCP細胞および対照細胞の細胞周期分析の図。
【図7B】ウェスタンブロットにより示された、RCP過剰発現(MCF10A細胞)に反応したErkおよびAktのリン酸化状態およびRCP(MCF7細胞)のRNAi仲介ノックダウンの図。
【図7C】MAPK阻害剤であるU0126によるRCP仲介Erkリン酸化阻害の図(上のパネル)およびRCP増殖表現型のU0126仲介阻害の図(下のパネル、平均+/−標準偏差)。
【図8A】図8は、乳癌細胞系におけるRCPの癌遺伝子表現型を示す。図8Aは、WSTアッセイによる、細胞増殖に関する、RCP過剰発現およびRCPのRNAi仲介ノックダウンの影響の図。左のパネル、形質移入6日後の結果。中のパネル、MCF10A細胞におけるRCP過剰発現の増殖時間経過。右のパネル、2種の異なるRNAi RCP構築体および(スクランブル配列の)対照を使用する、MCF7細胞におけるRCP阻害の増殖時間経過。
【図8B】軟寒天における、足場非依存性コロニー形成に関するRCPの過剰発現および阻害の影響の図。
【図8C】マトリゲルを介した、細胞浸潤に関するRCPの過剰発現および阻害の影響の図。
【図8D】細胞遊走に関するRCPの過剰発現および阻害の影響の図。すべてのエラーバーは、平均+/−標準偏差*、p<0.05、**、p<0.01および***p<0.001、スチューデントt検定(両側)から計算した。
【図9A】図9は、RCPが、ヌードマウスにおいて腫瘍形成を制御することを示している。図9Aは、MCF7細胞を使用した、in vivoの腫瘍成長に関するRCP阻害の影響の図。時間の関数としてプロットした、平均腫瘍容量を表す(平均+/−s.e.m.)。
【図9B】MCF7細胞を使用した、in vivoの腫瘍成長に関するRCP阻害の影響の図。5週(MCF7)および7週(MB231)における重量をプロットした腫瘍を表す。平均重量は実線により示した。
【図9C】MCF7細胞を使用した、in vivoの腫瘍成長に関するRCP阻害の影響の図。切除された全腫瘍を示す。
【図9D】MB231細胞を使用した、in vivoの腫瘍成長に関するRCP阻害の影響の図。時間の関数としてプロットした、平均腫瘍容量を表す(平均+/−s.e.m.)。
【図9E】MB231細胞を使用した、in vivoの腫瘍成長に関するRCP阻害の影響の図。5週(MCF7)および7週(MB231)における重量をプロットした腫瘍を表す。平均重量は実線により示した。
【図9F】MB231細胞を使用した、in vivoの腫瘍成長に関するRCP阻害の影響の図。切除された全腫瘍を示す。
【図9G】腫瘍試料中のRCPの免疫組織化学的染色を表す図(スケールバー、100μm)。
【図9H】TUNEL染色による、血清飢餓MB231細胞のRCP阻害のアポトーシス効果を示す図(スケールバー、100μm)。
【図10】増殖サインの由来を表す図。細胞周期制御遺伝子(ヒーラ細胞において)として、ウィットフィールドら(Whitfield et al.)((2002)Mol Biol Cell、13、1977−2000)により同定された1,134IMAGEクローンを、2,113 Affymetrixプローブセット(U133AおよびU133B)に対応させて、884の UniGene ID(build #187)にマッピングした。これらのプローブセットを使用して、ウプサラ腫瘍コホート(n=251)の発現プロフィールを、階層的にクラスタ化し(ピアソンの相関、平均連結法)、固有クラスタグラムのヒートマップにおいて視覚化した(左側)。図10Aにおける白いシグナルは、平均発現より上を示し、図10Bにおける白いシグナルは、平均発現より下を表す。明るさのそれぞれの程度は、発現の大きさを反映している。(紙面に限りがあるため、遺伝子の系統樹は省略した)。暗いバーは、ヒト乳癌の増殖に関連するとして、ウィットフィールド(Whitfield)およびペロー(Perou)によってすでに同定されたG2/Mにおいて大部分が発現する遺伝子のクラスタを示す。増殖クラスタの67の遺伝子に対応する78のプローブのセットは、>0.8の平均相関(白いバーで示されている)を有し、正しい階層的クラスタ化が示されている。これらのプローブセット(右側)は、コア増殖サイン遺伝子として示されており、全プローブセットと、増殖サインとのピアソン相関を導くために平均した(すなわち、TRIAGEの第6の工程)。
【発明を実施するための形態】
【0018】
本発明は、Rab結合タンパク質、および特にRab11FIPタンパク質(Rab11ファミリー相互作用タンパク質)が、腫瘍、および特に癌を含む、新生物の形成を促進することができるという、驚くべき発見に基づく。
【0019】
本発明者らは、反復増幅のゲノム部位の遺伝子の病態生理学的関与は、ゲノム位置、遺伝子発現および臨床データの統合および三角測量によって高い解明能力で推測することができ、三角測量は遺伝子発現と腫瘍攻撃性の臨床測定の間の統計学的関係を引き出すことができると仮定した。この目的のために、多段階のデータマイニング戦略を開発し、癌遺伝子としてのRab11FIPタンパク質の発見に至った。本発明の方法はこの発見に基づく。
【0020】
本発明者らのこの発見は、RAB1lFIPのタンパク質をコードする遺伝子の局在と、癌の確率の増大との関係に関して報告されたデータの説明も示す。例えば、ヒトRAB11FIP1遺伝子(Ensemblデータベースのアクセッション番号ENSG00000156675)は、第8染色体上、p12領域中、位置37、835、628〜37、876、161に見られる。同様に、マカクザルRAB11FIP1遺伝子(Ensemblデータベースのアクセッション番号ENSMMUG00000003213)は、第8染色体上、位置38、309、752〜38、349、824に見られる。イヌRAB11FIP1遺伝子(Ensemblデータベースのアクセッション番号ENSCAFG00000006176)は、第16染色体上、位置30、456、385〜30、484、441に見られる。ニワトリRAB11FIP1遺伝子(Ensemblデータベースのアクセッション番号ENSGALG00000003129)は、第22染色体上、位置2、258、738〜2、267、830に見られる。いくつかの分析済みの乳癌において、ヒト染色体領域8p12が増幅されることが以前に示されている(レテジアー、エーら(Letessier,A.,et al.)、(2006)BMC Cancer(2006)6、1〜13ページ、http://www.biomedcentral.com/content/pdf/1471−2407−6−245.pdfで入
手可能)。第8染色体の短いアームの消失、増大、増幅および再配列も、1組の乳、結腸および膵臓癌細胞系中で見られることが示されている(ポール、ジェイ シー エムら(Pole,J.C.M.,et al.)、(2006)Oncogene 25、5693〜5706ページ)。外科的切除を施された患者における、ヒト染色体領域8p12の変化と肺癌の間の相関関係が観察されてきている(キム、ティー−エムら(Kim,T.−M.,et al.)、(2005)Clinical Cancer Research、11、23、8235〜8242ページ)。比較ゲノムハイブリダイゼーションを使用して、遺伝子の遺伝子座における増幅および欠失が、膵臓および肺癌亜型において起こることも示された(トノン、ジーら(Tonon,G.,et al.)、(2005)Proc Natl Acad Sci USA、102、27、9625〜9630ページ)。同じ遺伝子座におけるコピー数の消失は、複数の患者から得た結腸癌試料においても観察された(ヤギヌマ、ワイら(Yaginuma,Y.,et al.)、(2006)Journal of International Medical Research、34、4、390〜396ページ)。
【0021】
Rab11ファミリー相互作用タンパク質(FIP)タンパク質は、C末端またはその近辺に位置する相同的Rab11結合ドメインを共有する(例えば、ヘイルズ、シー エムら(Hales,C.M.,et al.)、J.Biol.Chem.(2001)276、42、39067〜39075ページを参照)。それらのドメイン構造に基づいて、Rab11FIPタンパク質は、クラスI(FIP5/Rip11、FIP1およびFIP2)およびクラスII(FIP3/エフェリン、およびFIP4)の2つのサブファミリーに分けられている。したがって、本明細書で使用する用語「Rab11FIPタンパク質」は、Rab11ファミリー相互作用タンパク質のファミリーの任意のメンバー、特にRab11FIP1、Rab11FIP2、Rab11FIP3(エフェリン)、Rab11FIP4、Rab11FIP5(Rip11)、およびそれらのアイソフォームを指す。
【0022】
Rab11FIP1タンパク質の例には、SwissProtアクセッション番号Q05A58(Q05A58_マウス)、Q8K028(Q8K028_マウス)、Q6WKZ4(RFIP1_ヒト)、Q9D620(RFIP1_マウス)、Q3B7T9(RFIP1_ラット)、Q6KAN5(Q6KAN5_マウス)、Q307T1(Q307T1_ヒト)、Q28BL8(Q28BL8_XENTR)、(ニシツメガエル由来)、A4IHL6(A4IHL6_XENTR)(ニシツメガエル由来)の、例えばSwissProtアクセッション番号ENSG00000156675(ヒト)、ENSMUSG00000031488(マウス)、ENSPTRG00000020164(チンパンジー)、ENSPPYG00000018515(オランウータン)、ENSMMUG00000003213(アカゲザル)、ENSCAFG00000006176(イヌ)、ENSGALG00000003129(ニワトリ)、ENSMODG00000010813(ハイイロジネズミオポッサム)、ENSMLUG00000007472(ホオヒゲコウモリ属)、ENSMICG00000010174(ハイイロショウネズミキツネザル)、ENSOCUG00000004502(ウサギ)、ENSCPOG00000008298(モルモット)、ENSETEG00000007044(マダガスカルモヘアハリネズミ)、ENSECAG00000016134(ウマ)、ENSOANG00000003030(カモノハシ)、ENSOPRG00000010860(アメリカナキウサギ)、ENSSTOG00000005565(ジュウサンセンジリス)、ENSDNOG00000008299(ココノオビアルマジロ)、ENSLAFG00000004250(アフリカサバンナゾウ)、ENSOGAG00000005737(コビトガラゴ)の遺伝子、例えばSwissProtアクセッション番号DQ236342(ヒト)、BC125400(マウス)、BC132094(マウス)、BC034201(マウス)、BC077720(ヒト)、CX078607(アカゲザル)、D
R005339(ヒト)、BC135580(ニシツメガエル)の核酸配列を有し、SwissProtアクセッション番号AAH34201(マウス)、AAI25401(マウス)、AAI32095(マウス)、ABB43161(ヒト)、AAH77720(ヒト)、EAW63349(ヒト)、EAW63347(ヒト)、EAW63348(ヒト)、AAI35581(ニシツメガエル)およびCAJ81459(ニシツメガエル)のコード配列を含むタンパク質があるが、これらだけには限られない。
【0023】
Rab11FIP2タンパク質の例には、SwissProtアクセッション番号Q5HZI0(Q5HZI0_マウス)、Q7L804(RFIP2_ヒト)、Q3U366(Q3U366_マウス)の、例えばSwissProtアクセッション番号ENSMUSG00000040022(マウス)、ENSG00000107560(ヒト)、ENSGALG00000009304(ニワトリ)、ENSPTRG00000002982(チンパンジー)、ENSMODG00000009385(ハイイロジネズミオポッサム)、ENSXETG00000009541(ニシツメガエル)、ENSMMUG00000021805(アカゲザル)、ENSCAFG00000011991(イヌ)、ENSPPYG00000002692(オランウータン)、ENSEEUG00000006651(ナミハリネズミ)、ENSMLUG00000015200(ホオヒゲコウモリ属)、ENSMICG00000001976(ハイイロショウネズミキツネザル)、ENSOCUG00000009241(ウサギ)、ENSCPOG00000000453(モルモット)、ENSETEG00000019294(マダガスカルモヘアハリネズミ)、ENSECAG00000019791(ウマ)、ENSSARG00000001794(ヨーロッパトガリネズミ)、IGI00014376(Integr8データベース、ヒト)、IGI00031145(Integr8データベース、マウス)およびIGI02699308(Integr8データベース、ウシ)の遺伝子、および例えばSwissProtアクセッション番号BC089010(マウス)、BC075073(ヒト)、BC075074(ヒト)およびDN994254(ヒト)の核酸配列を有し、SwissProtアクセッション番号AAH89010(マウス)、EAW49423(ヒト)、AAH75073(ヒト)およびAAH75074(ヒト)のコード配列を含むタンパク質があるが、これらだけには限られない。
【0024】
Rab11FIP3タンパク質の例には、SwissProtアクセッション番号Q75154(RFIP3_ヒト)、Q8JZT3(Q8JZT3_マウス)、A2IDE0(A2IDE0_ヒト)、A2A2G1(A2A2G1_ヒト)、A2IDD9(A2IDD9_ヒト)、A2A2G0(A2A2G0_ヒト)、およびQ8CHD8(Q8CHD8_マウス)の、例えばSwissProtアクセッション番号ENSG00000090565(ヒト)、ENSMUSG00000037098(マウス)、ENSPTRG00000007543(チンパンジー)、ENSMMUG00000000613、IGI00010332(Integr8データベース、ヒト)およびIGI00087092(Integr8データベース、ニワトリ)、IGI00026472(Integr8データベース、マウス)の遺伝子、例えばSwissProtアクセッション番号AL049542(ヒト)、Z98882(ヒト)、BC023364(マウス)、BC037132(マウス)、BC051360(ヒト)、およびAL023881(ヒト)の核酸配列を有し、SwissProtアクセッション番号AAH23364(マウス)、AAH37132(マウス)、CAI95788(マウス)、CAI95591(ヒト)、CAI95593(ヒト)、CAM26382(ヒト)、CAM26383(ヒト)、CAQ09643(ヒト)、CAQ11003(ヒト)、CAM28374(ヒト)、AAH51360(ヒト)、EAW85805(ヒト)、EAW85806(ヒト)、EAW85807(ヒト)、EAW85808(ヒト)、EAW85809(ヒト)、およびEAW85810(ヒト)のコード配列を含むタンパク質があるが、これらだけには限られない。
【0025】
Rab11FIP4タンパク質の例には、SwissProtアクセッション番号Q6DGI2(Q6DGI2_BRARE)、Q86YS3(RFIP4_ヒト)、Q8BQP8(RFIP4_マウス)、Q5SYH4(Q5SYH4_マウス)、Q3LGD4(Q3LGD4_BRARE)、Q3U4R6(Q3U4R6_マウス)およびQ3LGD3(Q3LGD3_BRARE)の、例えばSwissProtアクセッション番号ENSDARG00000053855(ゼブラフィッシュ)、ENSG00000131242(ヒト)、ENSMUSG00000017639(マウス)、ENSMODG00000019209(ハイイロジネズミオポッサム)、ENSPTRG00000008974(チンパンジー)、ENSMMUG00000008127(アカゲザル)、ENSOCUG00000007443(ウサギ)、ENSMLUG00000016407(ホオヒゲコウモリ属)、ENSCPOG00000011634(モルモット)、IGI00013025(Integr8データベース、ヒト)、IGI00029750(Integr8データベース、マウス)、IGI02693035(Integr8データベース、ゼブラフィッシュ)およびIGI00051410(Integr8データベース、ゼブラフィッシュ)の遺伝子、例えばSwissProtアクセッション番号BC076363(ゼブラフィッシュ)、BC093914(ヒト)、BC101517(ヒト)、AL591174(マウス)、BC135989(ニシツメガエル)およびDR000701(ヒト)の核酸配列を有し、SwissProtアクセッション番号AAH76363(ゼブラフィッシュ)、AAI35990(ニシツメガエル)、CAI24833(マウス)、CAI24836(マウス)、EAW80265(ヒト)、AAI01518(ヒト)およびAAH93914(ヒト)のコード配列を含むタンパク質があるが、これらだけには限られない。
【0026】
Rab11FIP5タンパク質の例には、SwissProtアクセッション番号Q6ZQ33(Q6ZQ33_マウス)、Q9BXF6(RFIP5_ヒト)、Q8R361(RFIP5_マウス)およびQ3UM85_マウスの、例えばSwissProtアクセッション番号ENSG00000135631(ヒト)、ENSMUSG00000051343(マウス)、ENSGALG00000016092(ニワトリ)、ENSBTAG00000006162(ウシ)、ENSXETG00000011781(ニシツメガエル)、ENSOCUG00000000840(ウサギ)、NSPTRG00000012057(チンパンジー)、ENSMMUG00000002301(アカゲザル)、ENSOGAG00000009150(コビトガラゴ)、ENSECAG00000007084(ウマ)、IGI00035642(Integr8データベース、マウス)およびIGI00006075(Integr8データベース、ヒト)の遺伝子、例えばSwissProtアクセッション番号DQ890991(ヒトクローンの合成構築体)、DQ894168(ヒトクローンの合成構築体)、BC035013(ヒト)、BC026473(マウス)、BC044833(マウス)、BC051063(マウス)、DR770942(ベンガルザル)の核酸配列を有し、SwissProtアクセッション番号AAH26473(マウス)、AAH44833(マウス)、AAH51063(マウス)、ABM81917(合成構築体)、ABM85094(合成構築体)、AAH35013(ヒト)、EAW99750(ヒト)、EAW99749(ヒト)およびEAW99751(ヒト)のコード配列を含むタンパク質があるが、これらだけには限られない。
【0027】
Rabファミリーの低分子GTPアーゼタンパク質は、細胞内膜輸送、シグナル変換、受容体リサイクリングおよびエフェクタータンパク質の細胞膜への動員を含めた、細胞活性の制御において重要な役割を有することが知られている。したがって同様の役割が、Rab11ファミリー相互作用タンパク質などのRabファミリー相互作用タンパク質によるものと考えられている。Rab11−FIP3は例えば、エンドソーム循環区画の機能
において重要な役割を果たすことが示されている(例えば、ホーガン、シー ピーら(Horgan,C.P.,et al.)、(2007)、Traffic、8、414〜430ページを参照)。
【0028】
Rabタンパク質は、細胞骨格編成および細胞形態、細胞質分裂、小胞輸送および分泌、細胞周期進行および細胞分化を含めた広範囲の基本的細胞機能を制御する、Ras関連低分子Gタンパク質(Rho関連タンパク質を含む)のさらに大きなファミリーに属する。いくつかのこれらのタンパク質ならびにそれらのレギュレーターおよびエフェクターは、発癌現象および癌の進行に関与している。これらのうち、乳癌および卵巣癌において頻繁に増幅されるRAB25は、in vitroで増殖およびアポトーシス耐性を高め、かつin vivoで腫瘍形成を高めることが示されている。RABファミリーのタンパク質は膜輸送およびオルガネラ区画化において中心的役割を果たし、RAB11、RCPの主な結合パートナーは、膜タンパク質のリサイクリングに特に関係している。RAB11は細胞移動に関与しており、RAB11経路の混乱は異なる細胞型において移動を害する(ジョーンズ、エム シーら(Jones,M.C.,et al.)、(2006)Curr Opin Cell Biol 18、549〜557ページ;ポウェルカ、エー エムら(Powelka,A.M.,et al.)、(2004)Traffic 5、20〜36ページ;ユーン、エス オーら(Yoon,S.O.,et al.)、(2005)Cancer Res 65、2761〜2769ページ)。さらに、RAB11機能の拮抗的阻害は乳癌細胞の低酸素誘導侵襲性を低下させ(ユーンら(Yoon,et al.)、2005、上記)、インテグリンα6β4、EGFR、CXCR2およびPKCαなどの侵襲と関係がある膜受容体の輸送を変える(ジョーンズら(Jones,et al.)、2006、上記)。RAB遺伝子発現の異常調節は多くのヒト腫瘍の一般的要素として関与する。しかしながら、腫瘍進行中のRab相互作用タンパク質の役割は研究されていない。
【0029】
本発明者らは、Rab11FIP1の発現と関係がある広範囲の癌促進効果を確認している。非癌性乳房上皮細胞では、Rab11FIP1の過剰発現は、悪性形質転換、ならびに増大した細胞死亡率および移動をもたらした。乳癌細胞系では、RNA阻害による内因性Rab11FIP1の弱化は、増殖、足場非依存性増殖、侵襲およびアポトーシス耐性を低減し、ヌードマウスにおいて腫瘍形成を低下させた。生理学上、Rab11FIP1は主にRab11と相互作用して、管状エンドソームにおけるタンパク質の選別を制御すると考えられる(ピーデン、エー エーら(Peden,A.A.,et al.)、(2004)Mol Biol Cell 15、3530〜3541ページ)。Rab11−Rab11FIP1複合体は、トランスフェリン受容体をリソソーム(分解経路)からリサイクリングエンドソーム(リサイクリング経路)にチャンネル移動させ、それによって膜結合受容体のレベルの相対的増大を促進することが示されている(ピーデンら(Peden et al.)、2004、上記)。推測的ではあるが、Rab11FIP1の形質転換効果はリサイクリング経路へのこの移動によって仲介され、癌遺伝子的役割を有するインテグリンまたは増殖因子受容体などの、特定の膜タンパク質の膜局在(したがって機能)を延長する可能性があることは妥当である。本発明者らが観察したRab11FIP1の発現とERKのリン酸化の間の強い相関関係はこの可能性を反映し得る。というのは、それは、Rab11FIP1(以下ではRCPとも名付ける)と増殖因子受容体仲介MAPK活性化の間の特異的関係を示し得るからである。
【0030】
理論によって縛られるわけではないが、本発明者らによって得られた免疫沈降データ(示さず)は、Rab11FIP1と低分子GTPアーゼRasの間に直接的相互作用が存在することを示し、この2つのタンパク質間の複合体の形成を示す。Rasタンパク質は、アポトーシス、細胞増殖、および癌におけるそれらの中心的役割が当該技術分野で知られている低分子モノマータンパク質のファミリーである。少なくとも11個の異なる機能
クラスのタンパク質が、Rasタンパク質のエフェクターであることが示されている。したがって、Rasタンパク質は複合体シグナル相互作用ネットワークの中心に位置し、その中でRasタンパク質は、少なくとも3つの重複および共同タンパク質キナーゼカスケード、すなわちRaf/MEK/ERK、RalGTPアーゼ、およびPI3K/Aktを活性化すると考えられる。1つのランダムに選択したさらなる例示例として、それらはRb/E2FおよびDnmt−1を介して作用し、それによって腫瘍細胞増殖を活性化するp16の不活性化などのメチル化による後成的変化を引き起こす。次いでrasタンパク質は複雑な制御機構に曝される。点突然変異したRas遺伝子は様々な腫瘍の3分の1で頻繁に発見され、Ras遺伝子は癌遺伝子であることが知られている。
【0031】
本発明によるいくつかの方法は、腫瘍発生を制御する方法である。これらの方法は、腫瘍発生を予防、阻害、停止または逆行させる個々の方法を含む。腫瘍発生は例えば、悪性型の癌腫の形成を含めた発癌現象であってよい。したがって、例えば、本発明の方法は、癌などの増殖性疾患または障害の治療または予防に含まれ得る。
【0032】
本発明の方法を使用して、例えば哺乳動物、魚類、両生類、鳥類または微生物を含めた任意の生物において、腫瘍発生を予防、阻害、停止または逆行させることができる。いくつかの実施形態では、それぞれの微生物は細胞である。このような細胞は、直接採取またはそこからの単離を含めて、哺乳動物などの例えば前述の例の1つなどの生物から得ることができる。適切な哺乳動物の例には、ラット、マウス、ウサギ、モルモット、オポッサム、イヌ、ネコ、チンパンジー、アカゲザル、マカクザル、オランウータン、ウシ(メウシ)、マーモセット、アメリカナキウサギ、ガラゴ(「コビトガラゴ」)、リス、ココノオビアルマジロ、ゾウおよびヒトがあるが、これらだけには限られない。細胞は、従来の細胞系、例えば癌細胞系の細胞であってもよい。いくつかの実施形態では、細胞を培養する。他の実施形態では、それぞれの細胞は、例えば前述の例の1つなどの生物の一部分に含まれ得る。
【0033】
本発明による方法のいくつかの実施形態は、Rab11FIPタンパク質などのRab結合タンパク質の量を調節することを含む。Rab結合の量の調節は、細胞中、試験管などのin vitro設定中、または生物中に存在するRab結合タンパク質分子の全数の一部分を分解状態にすることを含むことができる。Rab結合タンパク質の分解は、例えば(カルシウム活性化プロテアーゼまたはカルパインとも呼ばれる)カルパインプロテアーゼなどのプロテアーゼによって実施することができる。例示例として、Rab11FIP1は、カルパインの標的として働きin vivoでカルパインによって切断される、3つのPEST(Pro、GlyおよびSer/Thr多量)モチーフを含むことが開示されている(ニコラス、エムら(Nicolas,M.,et al.)、(2005)Biochem.J.、389、223〜231ページ)。Rab結合タンパク質の量の調節は、Rab11FIPタンパク質の発現を調節することも含むことができる。これは例えば、Rab11FIPタンパク質の発現を調節する化合物を投与することによって実施することができる。Rab11FIPタンパク質の発現は、増大させてもよいし低減させてもよい。例示例として、Rab11FIPタンパク質の発現は、非コード核酸分子などの核酸分子によって低減することが可能である。
【0034】
本明細書で使用する用語「核酸分子」は、一本鎖、二本鎖またはそれらの組合せなどの、任意の考えられる形状の任意の核酸を指す。核酸は、例えばDNA分子(例えば、cDNAまたはゲノムDNA)、RNA分子(例えば、mRNA)、ヌクレオチド類似体の使用または核酸化学の使用によって作製したDNAまたはRNAの類似体、およびPNA(タンパク質核酸)を含む。DNAまたはRNAはゲノム起源または合成起源であってよく、一本鎖または二本鎖であってよい。本発明の方法では、典型的には、ただし必ずではないが、RNAまたはDNA分子を使用する。このような核酸は、例えばmRNA、cRN
A、合成RNA、ゲノムDNA、cDNA、合成DNA、DNAとRNAのコポリマー、オリゴヌクレオチドなどであってよい。それぞれの核酸は非天然ヌクレオチド類似体をさらに含むことができ、および/または親和性タグまたは標識と連結することができる。いくつかの実施形態では、核酸分子を単離、濃縮、または精製することができる。核酸分子は、例えばcDNAクローニングまたはサブトラクティブハイブリダイゼーションによって天然源から単離することができる。天然源は、ヒト、血液、精液、または組織などの哺乳動物由来であってよい。核酸は、例えばトリエステル法によって、または自動DNA合成装置を使用することによって合成することもできる。
【0035】
多くのヌクレオチド類似体が知られており、本発明の方法中で使用する核酸およびオリゴヌクレオチドにおいて、それらを使用することができる。ヌクレオチド類似体は、例えば塩基、糖、またはホスフェート部分に修飾を含むヌクレオチドである。塩基部分における修飾には、A、C、G、およびT/U、異なるプリンまたはピリミジン塩基、ウラシル−5−イル、ヒポキサンチン−9−イル、および2−アミノアデニン−9−イルなど、ならびに非プリンまたは非ピリミジンヌクレオチド塩基の天然および合成修飾がある。他のヌクレオチド類似体は共通塩基として働く。共通塩基には3−ニトロピロールおよび5−ニトロインドールがある。共通塩基は、任意の他の塩基と塩基対を形成することができる。塩基の修飾は、例えば2’−O−メトキシエチルなどの糖の修飾と頻繁に組合せて、例えば二本鎖安定性の増大などの特有の性質を得ることができる。
【0036】
本発明の方法のいくつかの実施形態では、核酸分子は、例えばアプタマーまたはSpiegelmer(登録商標)(WO01/92655中に記載)などの非コード核酸分子である。非コード核酸分子はnc−RNA分子であってもよい(例えば、天然nc−RNA分子の導入に関するコスタ、エフエフ(Costa,FF)、Gene(2005)、357、83〜94ページを参照)。nc−RNA分子の例には、アンチセンスRNA分子、L−RNASpiegelmer(登録商標)、サイレンサーRNA分子(二本鎖ニューロン限定サイレンサーエレメントなど)、マイクロRNA(miRNA)分子、低分子ヘアピンRNA(shRNA)分子、低分子干渉RNA(siRNA)分子、反復配列関連低分子干渉RNA(rasiRNA)またはPiwiタンパク質と相互作用するRNA(piRNA)(簡単な概説に関しては、例えばリン、エイチ(Lin,H.)、Science(2007)316、397を参照)があるが、これらだけには限られない。適切なshRNAの例示例は、以下の実施例中に配列番号3および配列番号4として示す。
【0037】
低分子干渉RNAの使用は、特定の遺伝子を「ノックダウン」するためのツールとなっている。それは、転写後レベルで起こりmRNA分解を含むRNA干渉(RNAi)を介した、遺伝子サイレンシングまたは遺伝子抑制を利用する。RNA干渉はゲノムを保護する細胞機構となる。siRNA分子は、いわゆるRNA誘導型サイレンシング複合体(RISC)を形成するためのsiRNAと複合酵素複合体の結合によって、それらの相補的RNAの分解を仲介する。siRNAはRISCの一部分となり、相補的RNA種を標的化し、それは次いで切断される。これはそれぞれの遺伝子の発現の消失をもたらす(簡単な概説に関しては、(ザモア、ピーディーおよびハーレー、ビー(Zamore,PD,& Haley,B)(2005)Science 309、1519〜1524ページを参照)。この技法は例えば、米国特許出願2005/0191618中に開示されたHIV RNAの切断などの、寄生虫DNA配列のサイレンシングに施されている。
【0038】
本発明のこのようなsiRNAの典型的実施形態は、10〜35ヌクレオチド、いくつかの実施形態では15〜25ヌクレオチドのin vitroまたはin vivo合成分子を含む。それぞれのsiRNA分子は、(微生物および動物の一部分である細胞を含めた)対象の細胞内で直接合成することができる。siRNA分子をそれぞれの細胞に導
入する、および/またはそれぞれの細胞に送達することもできる。in vivoでの選択細胞へのsiRNA分子の送達の例示例は、重鎖抗体断片(Fab)と核酸結合タンパク質プロタミンの融合タンパク質との、その非共有結合である(ソング、イーら(Song,E.et al.)、(2005)、Nature Biotech.23、6、709〜717ページ)。本発明の1実施形態では、siRNA分子を使用して、Rab結合タンパク質の(例えばin vivo)合成と関係がある1つまたは複数の転写因子をコードするmRNA分子の分解を誘導する。
【0039】
本発明による本発明の方法のいくつかの実施形態は、Rab結合タンパク質、例えばRab11FIPタンパク質の活性を調節することを含む。Rab結合タンパク質の活性の調節は、Rab結合タンパク質と化合物の間の複合体を形成することを含むことができる。これは例えば、Rab結合タンパク質の活性を調節する化合物を投与することによって実施することができる。Rab結合タンパク質(例えばRab11FIPタンパク質)の活性は増大または低減することができる。
【0040】
それぞれの化合物は任意の適切な手段によって投与することができる。宿主生物が哺乳動物である場合、化合物は例えば経口または非経口(経腸)投与することができる。哺乳動物への投与の典型的実施形態では、例えば化合物の調製物の経口、静脈内投与によって、または吸入によって、その施用は血液および肝臓への送達を確実にする。経口施用のための調製物の例は、錠剤、ピルまたは飲料溶液であり、静脈内投与のための調製物の例は注射または輸液であり、吸入による投与のための調製物の例はエアロゾル混合物またはスプレーである。宿主生物が組換え微生物である場合、投与の例は、微生物の環境への化合物の注射または添加である。微生物が単細胞である場合、微生物を改変する技法と組合せて、後者の形の投与をおそらく実施することができる。このような技法は、細胞膜のエレクトロポレーションまたは透過処理を含むことができる。
【0041】
Rab結合タンパク質、例えばRab11FIPタンパク質の活性の調節は、Rab結合タンパク質の翻訳後修飾のパターンの改変を含めた、Rab結合タンパク質の翻訳後修飾の改変を含むことができる。いくつかの実施形態では、Rab11FIPタンパク質などのRab結合タンパク質の翻訳後修飾の改変は、Rab結合タンパク質の翻訳後修飾のパターンを改変する化合物の使用を含む。それぞれの翻訳後修飾の例示例は、リン酸化、脱リン酸化、アセチル化、およびユビキチン化である。それぞれの翻訳後修飾、例えばリン酸化の結果として、Rab結合タンパク質の活性が変わる。例示例として、Rab11−FIP2機能はセリン227におけるリン酸化によって制御されることが示されている(デゥチャルメ、エヌ エーら(Ducharme,N.A.,et al.)、Molecular Biology of the Cell(2006)7、8、3625〜3637ページ)。キナーゼMARK2/EMK1/Par−1B(MARK2)によってリン酸化されると、Rab11−FIP2は上皮細胞の極性を確立する際に働く(上記)。
【0042】
Rab11FIPタンパク質などのRab結合タンパク質の活性、発現、量または細胞内局在を調節する化合物は、任意の化合物であってよい。例には、核酸(上記参照)、ペプチド、ペプトイド、無機分子および小有機分子があるが、これらだけには限られない。ペプトイドは、ペプチドよりはるかに高い細胞透過性を有し得る(例えば、クウォン、ワイ ユーおよびコダデック、(2007)ティー(Kwon,Y.−U,and Kodadek,(2007)T.)、J.Am.Chem.Soc.129、1508〜1509ページ参照)。ペプチドは合成起源であってよく、または当該技術分野でよく知られている方法によって天然源から単離することができる。天然源はヒトなどの哺乳動物、血液、精液、または組織であってよい。ポリペプチドを含めたペプチドは、例えば自動ポリペプチド合成装置を使用して合成することができる。ポリペプチドの例示例は、抗体、抗
体断片、および抗体様機能を有するタンパク質結合分子である。(組換え)抗体断片の例は、Fab断片、Fv断片、単鎖Fv断片(scFv)、ダイアボディまたはドメイン抗体である(ホルト、エル ジェイら(Holt,L.J.,et al.)、(2003)Trends Biotechnol.、21、11、484〜490ページ)。抗体様機能を有するタンパク質結合分子の1例は、リポカリンファミリーのポリペプチドに基づくムテインである(WO03/029462、ベステら(Beste et al.)、(1999)Proc.Natl.Acad.Sci.U.S.A.、96、1898〜1903ページ)。ビリン結合タンパク質、ヒト好中球ゼラチナーゼ結合性リポカリン、ヒトアポリポタンパク質Dまたはグリコデリンなどのリポカリンは、それらがハプテンとして知られる選択した小さなタンパク質領域と結合するように修飾することができる天然リガンド結合部位を有する。他のタンパク質結合分子の例は、いわゆるグルボディ(WO96/23879参照)、アンキリン骨格(モザビ、エル ケイら(Mosavi,L.K.,et al.)、(2004)Protein Science 13、6、1435〜1448ページ)または結晶骨格(WO01/04144)に基づくタンパク質、スケラ(Skerra)、(2000)J.Mal.Recognit.13、167〜187ページ中に記載されたタンパク質、アドネクチン、テトラネクチン、およびアビマーである。アビマーは、いくつかの細胞表面受容体において一連の多数のドメインとして存在する、いわゆるA−ドメインを含む(シルバーマン、ジェイら(Silverman,J.,et al.)、(2005)Nature Biotechnology 23、1556〜1561ページ)。ヒトフィブロネクチンのドメインに由来するアドネクチンは、標的との免疫グロブリン様結合用に工学処理することができる3個のループを含む(ギル、ディー エスおよびダムレ、エヌ ケイ(Gill,D.S.& Damle,N.K.)、(2006)Current Opinion in Biotechnology 17、653〜658ページ)。それぞれのヒトホモ三量体タンパク質に由来するテトラネクチンは、C型レクチンドメイン中に所望の結合用に工学処理することができるループ領域を同様に含む(上記)。タンパク質リガンドとして作用することができるペプトイドは、側鎖がα炭素原子ではなくアミド窒素と結合している点でペプチドと異なる、オリゴ(N−アルキル)グリシンである。ペプトイドは典型的にはプロテアーゼおよび他の修飾酵素に耐性があり、ペプチドよりはるかに高い細胞透過性を有し得る(例えば、クウォン、ワイ ユーおよびコダデック、(2007)ティー(Kwon,Y.−U,and Kodadek,(2007)T.)、J.Am.Chem.Soc.129、1508〜1509ページ参照)。所望の場合、任意のまたは特定の形、クラスなどの標的物質に対するそれぞれの部分の親和性をさらに増大させる修飾作用物質を使用してもよい。
【0043】
それぞれの化合物は、(Rab11−FIPタンパク質などの)Rab結合タンパク質の翻訳後修飾を改変することもできる。化合物は、例えば、Rab結合タンパク質を含めた細胞要素のリン酸化状態を変えることができる。それぞれの化合物は、Rab結合タンパク質と異なるタンパク質を含めた細胞内タンパク質の、リン酸化の程度の調節物質であってもよい。このようなタンパク質は、したがってRab結合タンパク質の翻訳後修飾を改変することができる。それぞれの化合物の例示例はチロシンキナーゼ阻害剤である。チロホスチン、キナゾリン、キノキサリン、キノリン、2−フェニルアミノピリミジン、フラボノイド、ベンゾキノイド、アミノサリチル酸またはスチルベン(これらは例えばWO9618738、WO03035621およびその中に引用された参照文献中に記載されており、例えばそれらの実験による同定の例に関しては、例えば米国特許第6,740,665号を参照)などの多数のチロシンキナーゼ阻害剤が市販されている。チロホスチンの例は、AG213、AG490、AG879、AG1295、AG1478、AG1517、AGL2043、チロホスチン46およびメチル2,5−ジヒドロキシケイ皮酸である。キナゾリンは、例えばPD153035、PD156273、ゲフィチニブまたはラパチニブであり、キノキサリンは、例えばPD153035またはZD1839である
。キノリンの1例は5−メチル−5H−インドール[2,3−β]キノリンであり、2−フェニルアミノピリミジンの1例はイマチニブであり、フラボノイドの例はゲニステインまたはクエルセチンであり、ベンゾキノイドの1例はハービマイシンAであり、アミノサリチル酸の1例はラベンドウスチンAであり、スチルベンの1例はピセアタノールである。他の適切な化合物は、チロホスチンエルブスタチンなどの受容体チロシンキナーゼ阻害剤、WHI−P97またはチロホスチンAG592などのEGFR特異的受容体チロシンキナーゼ阻害剤、アウリントリカルボン酸などのチロシンリン酸化刺激剤、または過バナジン酸ナトリウムまたはイソキサゾールカルボン酸などのチロシンホスファターゼ阻害剤を含むことができる。
【0044】
タンパク質キナーゼの機能を阻害することが報告されている化合物のさらなる例には、ビス単環式、二環式または複素環式アリール化合物(国際特許出願WO92/20642)ビニレン−アザインドール誘導体(PCTWO94/14808)、1−シクロプロピル−4−ピリジル−キノロン(米国特許第5,330,992号)、スチリル化合物(米国特許第5,217,999号)、スチリル置換ピリジル化合物(米国特許第5,302,606号)、特定のキナゾリン誘導体(欧州特許出願No.0566266A1)、セレオインドールおよびセレニド(国際特許出願WO94/03427)、三環式多価化合物(国際特許出願WO92/21660)、およびベンジル−ホスホン酸化合物(国際特許出願WO91/15495)があるが、これらだけには限られない。調節物質として使用することができるさらに他の化合物には、1996年8月23日に出願された米国特許出願第08/702,232号中に記載されたオキシンドリノン(oxindolinone)などのオキシンドリノンがある。
【0045】
Rab結合タンパク質の翻訳後修飾を調節する化合物のさらなる例は、タンパク質キナーゼまたはタンパク質ホスファターゼの制御を誘導することができる細胞表面分子に対するアゴニストまたはアンタゴニストである。
【0046】
Rab11FIPタンパク質などのRab結合タンパク質の活性を調節する化合物の他の例は、例えばビタミンD3またはビタミンD4などのビタミンDのメンバーである。ビタミンD3(コレカルシフェロール、カルシオールまたはアラキトールとも呼ばれ、IUPAC名(1S,3Z)−3−[(2E)−2−[(1R,3aS,7aR)−1−[(1R)−1,5−ジメチルヘキシル]オクタヒドロ−7a−メチル−4H−インデン−4−イリデン]エチリデン]−4−メチレン−シクロヘキサノール、(3β、5Z、7E)−9,10−セココレスタ−5,7,10(19)−トリエン−3−オールとも名付けられる)は、その活性形で、乳癌を含めた様々な起源の悪性細胞において、増殖停止、分化、および細胞死を誘導する能力を有する。この観察結果が、考えられる抗癌剤としてのビタミンD類似体の開発をもたらしている(例えば、バウデット、シーら(Baudet,C.,et al.)(1996)Cancer Lett.100、3〜10ページ;ヒジー、ティー、およびウー、ジェイ エム(Hsieh,T.,& Wu,J.M.)、(1997)Biochem.Biophys.Res.Commun.235、539〜544ページ;ジェームス、エス ワイら(James,S.Y.,et al.)、(1999)Gen.Pharmacol.32、143〜154ページ;マティアセン、アイ エスら(Mathiasen,I.S.,et al.)、(1999)Cancer Res.59、4848〜4856ページを参照)。化合物EB1089は1つのこのような類似体であり、現在癌の治療に関する第3相臨床試験中である(例えば、マティアセン、アイ エスら(Mathiasen,I.S.,et al.)、(1993)J.Steroid Biochem.Mol.Biol.46、365〜371ページ;コルストン、ケイ ダブリュら(Colston,K.W.,et al.)、(1992)Biochem.Pharmacol.44、2273〜2280ページ;ハンセン、シー エムら(Hansen,C.M.,et al.)、(2000)Cu
rr.Pharm.Des.6、803〜828ページを参照)。ビタミンD化合物の抗癌活性は細胞内遊離カルシウム([Ca2+])に対するその影響と直接関係がある。ビタミンD化合物で処理した細胞中の遊離Ca2+の上昇は、乳癌細胞におけるアポトーシスの誘導と相関関係があり(サージーブ、アイ エヌら(Sergeev,I.N.,et al.)、(2000)Vitamin D Endocrine System:Structural、Biological、Genetic and Clinical Aspects中(ノーマン、エー ダブリュ、ボイロン、アール、およびトーマセット、エム(Norman,A.W.,Bouillon,R.,and Thomasset,M.)編)、399〜402ページ、University of California、Riverside、CA;バンデバーレ、ビーら(Vandewalle,B.,et al.)(1995)Int.J.Cancer 61、806〜8112ページ;サージーブ、アイ エヌおよびローテン、ダブリュ ビー(Sergeev,I.N.,& Rhoten,W.B.)(1998)Endocrine 9、321〜327ページ)、カルパインとしても知られるカルシウム依存性システインプロテアーゼの活性化における主な機構であることが示されている(ナカガワ、ティーおよびユーアン、ジェイ(Nakagawa,T.,& Yuan,J.)、(2000)J.Cell Biol.150、887〜894ページ;レイ、エス ケイら(Ray,S.K.,et al.)(2000)Brain Res.852、326〜334ページ;イシハラ、アイら(Ishihara,I.,et al.)(2000)Neurosci.Lett.279、97〜100ページ)。
【0047】
理論によって縛られることを望まないが、乳癌細胞における試験からの最近の証拠は、カルパインがビタミンD3およびEB1089によって誘導されるアポトーシス死における主な実行プロテアーゼであり得ることを示唆している(マティアセン、アイ エスら(Mathiasen,I.S.,et al.)、(2002)J.Bio.Chem.277、30738〜30745ページ)。カルパインのプロテアーゼ(タンパク質分解)機能は、カルパインのタンパク質標的のペプチド配列中に存在するPEST(Pro、Gly、およびSer/Thr多量)アミノ酸モチーフに依存してる(ダイス、ジェイ エフ(Dice,J.F.)(1987)FASEB J.1,349〜357ページ;トンパ、ピーら(Tompa,P.,et al.)、(2004)J.Biol.Chem.279、20775〜20785ページ;レヒシュタイナー、エム、およびロジャース、エス ダブリュ(Rechsteiner,M.,& Rogers,S.W)(1996)Trends Biochem.Sci.21、267〜271ページ;シャムウェイ、エス ディーら(Shumway,S.D.,et al.)、(1999)J.Biol.Chem.274、30874〜30881ページ;フクダ、エムおよびイトー、ティー(Fukuda,M.,& Itoh,T)(2004)J.Biol.Chem.279、22314〜22321ページ)。RAB11FIP1は、例えばC2ドメインとRBD(Rab結合ドメイン)の間に位置する3個のこのようなPESTモチーフを含む。近年の試験において、マリーら(Marie,et.al.)は、RAB11FIP1中のPESTモチーフは、カルパインによる制御型分解に関してそれを標的化するかどうか決定しようと努めた(マリー、エヌら(Marie,N.,et.al.)(2005)Biochem.J.389、223〜231ページ)。マリーらは、高濃度のカルシウムを用いた細胞の処理はRAB11FIP1の分解を誘導し、この分解は特異的カルパイン阻害剤を加えることにより無効にすることができることを観察した。さらに、このカルシウム制御型分解におけるPEST配列の関与は、その中のPEST配列を除去したRAB11FIP1突然変異体を使用して立証した。したがってマリーらは、RAB11FIP1のPEST配列は、おそらく細胞中の生理的代謝回転の機構として、カルパイン仲介型分解に関してタンパク質を標的化すると結論付けた。例えばRAB11FIP1がカルパイン作用に感受性である活性PESTドメインを含み、カルパインがビタミンD3およびEB1089などのその化学療法剤類似体によってタンパク質分解活性
に関して活性化されることを考慮して、本発明の方法は、ビタミンD3およびEB1089を含めたその関連類似体に基づく化学療法レジメン、および細胞内遊離カルシウムレベルを上昇させるため、またはカルパインを活性化するために生化学的に作用する任意の他の治療レジメンに関して患者を選択するためのバイオマーカーとして、RAB11FIP1のmRNAまたはタンパク質レベルを使用することを可能にする。
【0048】
前に記したように、細胞における腫瘍発生を制御する方法、例えば細胞における腫瘍発生を予防、停止または逆行させる方法のいくつかの実施形態では、Rab結合タンパク質のレベル、発現および/または活性を分析する。細胞は、(例えば単離した)個々の細胞、癌組織を有し得る生物の細胞、癌組織を含む組織の細胞、または細胞培養物の細胞であってよい。癌細胞は、例えば神経、グリア、肺、肝臓、脳、乳房、膀胱、血液、白血病、結腸、子宮内膜、胃、皮膚、卵巣、脂肪、骨、頸部、食道、膵臓、前立腺、腎臓、または甲状腺細胞であってよい。いくつかの態様では、癌は、星状細胞腫、急性骨髄性白血病、乳癌、膀胱癌、子宮頸癌、結腸直腸癌、子宮内膜癌、食道扁平上皮細胞癌、グリオーマ、グリア芽腫、胃癌、肝細胞癌、ホジキンリンパ腫、白血病、脂肪腫、メラノーマ、マントル細胞リンパ腫、粘液線維肉腫、多発性骨髄腫、神経芽細胞腫、非ホジキンリンパ腫、肺癌、非小細胞肺癌、卵巣癌、食道癌、骨肉腫、膵臓癌、前立腺癌、頭部または頸部の扁平細胞癌、甲状腺癌および尿路上皮癌を含むが、これらだけには限られない。
【0049】
細胞におけるRab結合タンパク質の量の変化、発現の変化または活性の変化は、ほぼ平均レベルまたは一般に見られるレベルに対応するレベルに調整することができる。細胞におけるRab結合タンパク質の量の変化は、プロテアーゼの活性または発現の調節によって実施することができる。いくつかの実施形態では、このようなプロテアーゼはカルシウム依存性プロテアーゼ(カルパイン)、例えばカルパインIまたはIIである。いくつかの実施形態では、カルパインプロテアーゼの活性および/または発現を、細胞内カルシウムレベル、例えば遊離細胞内カルシウムの変化によって調節する。
【0050】
いくつかの実施形態では、カルパインプロテアーゼの活性および/または発現を、それらの施用を含めた、ビタミンDファミリーのメンバー、このようなビタミンDファミリーのメンバーのプロドラッグまたはこのようなビタミンDファミリーのメンバーの類似体の使用によって調節、例えば活性化する。それぞれのビタミンD、その類似体またはプロドラッグは、例えばそれぞれの細胞と接触させることが可能であり、またはそれぞれの対象を含めた、それぞれの生物に投与することができる。プロテアーゼの活性および/または発現は、シグナル伝達カスケードによって次いで調節されるプロテアーゼ調節物質などの、他の因子によって調節することもできる。多数のプロテアーゼが、それらの活性とそれらの発現の両方の点で制御を受けることは知られている(例えば、トゥルク、ビー(Turk,B.)、(2006)Nature Reviews Drug Discovery、5、9、785〜799ページ参照)。アポトーシスプロテアーゼ活性化因子−1またはメタロプロテイナーゼの組織阻害剤は例示例として役立つ可能性がある。
【0051】
ビタミンDファミリーのメンバーまたはビタミンDファミリーのメンバーの類似体の「プロドラッグ」は、in vivoで(例えば段階的または急速に)転換されて、酵素作用または一般的な酸または塩基加溶媒分解のいずれかによる、例えば血液中での加水分解または酸化による代謝を含めた、例えば宿主中での代謝によって親ビタミンD/ビタミンD類似体化合物を生成し、活性成分を形成する化合物であることは理解されている。典型的にはプロドラッグは、in vivoで使用すると化学的または生理的工程によって元の化合物を放出する、親ビタミンD/ビタミンD類似体の誘導体である。プロドラッグは例えば、生理的pHに達すると、(1つまたは複数の)自発性化学反応、(1つまたは複数の)酵素触媒化学反応、光分解、および/または(1つまたは複数の)代謝化学反応によって、親ビタミンD/ビタミンD類似体に変換され得る。一般的な例は、カルボン酸部
分を有する活性形を有するエステルおよびアミド形の化合物だけには限られないが、これらを含めた、親化合物の官能基部分に生物学的に不安定な保護基を有する化合物である。さらなる例には、酸化、還元、アミノ化、脱アミノ化、エステル化、脱エステル化、アルキル化、脱アルキル化、アシル化、脱アシル化、リン酸化、脱リン酸化、光分解、加水分解することが可能な化合物の部分、またはプロドラッグにおける化学結合の形成または破壊に関する他の官能基の変化または変換がある。それぞれのプロドラッグは、その治療活性または利用可能形にin vivoで変換されるまで、完全に活性状態または生理的に利用可能でないプロドラッグでよい。「プロドラッグ」は、それぞれの薬剤分子(この場合、親ビタミンD/ビタミンD類似体)と担体部分の間の共有結合または非共有結合によって、in vitroまたは生理的に形成される生成物も含み得る。特定の化合物は、溶媒和および非溶媒和形、例えば水和形で存在し得ることも理解されよう。溶媒和形は単に「溶媒和物」とも呼ばれる。
【0052】
前で他に記したように、発癌性になる素因を有する細胞を同定する方法のいくつかの実施形態では、(例えば単離した)個々の細胞、生物の細胞または細胞培養物の細胞であってよい細胞におけるRab結合タンパク質の量の変化、発現の変化または活性の変化を決定することができる。同様に、対象、例えば患者における新生物を発症するリスクを診断する方法のいくつかの実施形態では、Rab結合タンパク質の量の変化、発現の変化または活性の変化を決定することができる。このような場合、それぞれの変化の決定は、それぞれの新生物または腫瘍が対象の1つまたは複数の細胞におけるプロテアーゼの活性および/または量の変化に感受性であることを示す可能性がある。いくつかの実施形態では、このようなプロテアーゼは、カルパインプロテアーゼカルパインIまたはII(カルシウム活性化中性プロテアーゼIおよびIIとも呼ばれる)としても知られるカルシウム依存性プロテアーゼCDPIまたはCDPIIなどの、カルシウム依存性プロテアーゼ(カルパイン)である。いくつかの実施形態では、Rab結合タンパク質の活性および/または発現の変化の決定は、それぞれの新生物または腫瘍が細胞内カルシウムレベルの変化に感受性であることを示す可能性がある。
【0053】
この点において、各方法は、対象から試料を得ること、および試料から核酸を単離することをさらに含むことができる。核酸を増幅し、標識することができる。選択したタンパク質の量を決定する方法は、当該技術分野でよく知られている。各方法は、例えばプロテオミクス系の技法に依存することができる。それは、それぞれのRab結合タンパク質と結合する、抗体、抗体断片、または抗体様機能を有するタンパク質結合分子の使用を含むことができる。タンパク質の発現タンパク質を分析する方法は、当該技術分野で十分確立している。mRNAの発現は、例えばノーザンブロッティングおよびin situハイブリダイゼーション、RNAse保護アッセイおよびPCR系の方法を使用して定量化することができる。さらなる例として、DNA二本鎖、RNA二本鎖、およびDNA−RNAハイブリッド二本鎖またはDNA−タンパク質二本鎖を含めた特異的二本鎖を認識することができる、抗体、抗体断片、またはタンパク質結合分子を利用することができる。塩基配列決定に基づく遺伝子発現分析の方法の例示例には、遺伝子発現の連続分析(SAGE)、および超並列型サイン塩基配列決定(MPSS)による遺伝子発現分析がある。
【0054】
(組換え)抗体断片の例は、Fab断片、Fv断片、単鎖Fv断片(scFv)、ダイアボディ、トリアボディ(イリアデ、ピーら(Iliades,P.,et al.)、FEBS Lett(1997)409、437〜441ページ)、デカボディ(ストーン、イーら(Stone,E.,et al.)、Journal of Immunological Methods(2007)318、88〜94ページ)、および他のドメイン抗体である(ホルト エル ジェイら(Holt,L.J.,et al.)、Trends Biotechnol.、(2003)21、11、484〜490ページ)である。単鎖Fv断片は、例えば免疫グロブリン分子の1本の重鎖と1本の軽鎖由来
の可変領域の融合体である。抗体様機能を有するタンパク質結合分子の1例は、リポカリンファミリーのポリペプチドに基づくムテインである(WO2003/029462;WO2005/019254;WO2005/019255;WO2005/019256;ベステら(Beste et al.)、Proc.Natl.Acad.Sci.U.S.A.(1999)、96、1898〜1903ページ)。ビリン結合タンパク質、ヒト好中球ゼラチナーゼ結合性リポカリン、ヒトアポリポタンパク質D、ヒト涙液リポカイン、またはグリコデリンなどのリポカリンは、それらがハプテンとして知られる選択した小さなタンパク質領域と結合するように修飾することができる天然リガンド結合部位を有する。さらなるタンパク質結合分子の他の非制限的な例は、いわゆるグルボディ(WO96/23879参照)、アンキリン骨格(モザビ、エル ケイら(Mosavi,L.K.,et al.)、Protein Science(2004)13、6、1435〜1448ページ)または結晶骨格(WO2001/04144)に基づくタンパク質、スケラ(Skerra)(J.Mal.Recognit.(2000)13、167〜187ページ)中に記載されたタンパク質、アドネクチン、テトラネクチン、アビマー、およびペプトイドである。アビマーは、いくつかの細胞表面受容体において一連の多数のドメインとして存在する、いわゆるA−ドメインを含む(シルバーマン、ジェイら(Silverman,J.,et al.)、Nature Biotechnology(2005)23、1556〜1561ページ)。ヒトフィブロネクチンのドメインに由来するアドネクチンは、標的との免疫グロブリン様結合用に工学処理することができる3個のループを含む(ギル、ディー エスおよびダムレ、エヌ ケイ(Gill,D.S.& Damle,N.K.)、Current Opinion in Biotechnology(2006)17、653〜658ページ)。それぞれのヒトホモ三量体タンパク質に由来するテトラネクチンは、C型レクチンドメイン中に所望の結合用に工学処理することができるループ領域を同様に含む(上記)。タンパク質リガンドとして作用することができるペプトイドは、側鎖がα炭素原子ではなくアミド窒素と結合している点でペプチドと異なる、オリゴ(N−アルキル)グリシンである。ペプトイドは典型的にはプロテアーゼおよび他の修飾酵素に耐性があり、ペプチドよりはるかに高い細胞透過性を有し得る(例えば、クウォン、ワイ ユーおよびコダデック、ティー(Kwon,Y.−U,and Kodadek,T.)、J.Am.Chem.Soc.(2007)129、1508〜1509ページ参照)。所望の場合、任意のまたは特定の形、クラスなどの標的物質に対するそれぞれの部分の親和性をさらに増大させる修飾作用物質を使用することができる。
【0055】
PCR(ポリメラーゼ連鎖反応)は、細胞由来またはメラノーマ試料などの組織試料由来の転写産物を、増幅および検出するのに有用な標準的な技法である。逆転写酵素PCR(RT−PCR)は例えば、それを使用して異なる試料(例えば、非転移性および転移性メラノーマ試料、または良性皮膚およびメラノーマ試料)中のmRNAレベルを比較し、遺伝子発現サインを調べることができる感度の良い定量法である。RT−PCRを実施するために、mRNAを試料(例えば、ヒトメラノーマ試料から単離した全RNA)から単離する。mRNAは例えば、凍結または保存済みパラフィン包埋および固定(例えば、ホルマリン固定)組織試料から抽出することができる。RNA単離用の精製キットは市販されている。RT−PCR技法は、二重標識蛍光プローブによってPCR産物の蓄積を測定する、リアルタイム定量PCRの実施形態中で使用することもできる。リアルタイムPCRは、それぞれの標的配列用の内部競合剤を標準化に使用する定量競合的PCRと、試料内に含まれる標準化遺伝子、またはRT−PCR用のハウスキーピング遺伝子を使用する定量比較PCRとの両方に適合する。さらなる例として、競合的PCR設計は、おそらく自動、高スループットマトリクス支援レーザー脱離イオン化法飛行時間型(MALDI−TOF)質量分析検出法および核酸分子の定量化を含めた、遺伝子発現分析に使用することができる。遺伝子発現分析に適したPCR系の方法中に含めることができる技法のさらなる例は、ディファレンシャルディスプレイ、増幅断片長多型(iAFLP)法、Bea
dAmiy(商標)技術、遺伝子発現の検出用のBeadsArvay(BADGE)および広範囲の発現プロファイリング(HiCEP)分析だけには限られないが、これらを含むことができる。さらなる例として、組織試料の遺伝子発現の決定は、マイクロアレイを用いて実施することもできる。
【0056】
いくつかの実施形態では、Rab結合タンパク質の活性および/または発現の変化、特にRab結合タンパク質の活性および/または発現の増大の決定は、それぞれの新生物または腫瘍がビタミンDのメンバー、その類似体またはプロドラッグを用いた治療に感受性であることを示す可能性がある。この点において本発明は、(腫瘍、例えば癌を含めた)新生物がプロテアーゼの活性および/または量の変化に感受性であるかどうかを決定する方法も提供する。新生物中、特にその細胞中で発現されるプロテアーゼを含めて、プロテアーゼが存在してよい。いくつかの実施形態では、方法は、新生物がビタミンDファミリーのメンバー、その類似体またはプロドラッグに感受性であるかどうかを決定する方法である。
【0057】
本発明による方法に使用することができる、ビタミンD3類似体を含めた多数のビタミンD類似体が、当該技術分野で知られている(概説に関しては、例えば、クボデラ、エヌ(Kubodera,N.)(2006)Current Bioactive Compounds、2(3)、301〜315ページを参照されたい)。例えばカルシトリオール、マクサカルシトール、タカルシトール、セカルシフェロール、アルファカルシドールまたはセオカルシトール(EB1089)を含めた適切な例は、例えば国際特許出願WO2007/082542中に挙げられている。セオカルシトールは、乳癌腫瘍細胞中の電離放射線に対する応答性を高めることが以前に示されており、これはDNA損傷の初期誘導の細胞シグナル伝達経路下流の変化の結果であることが示唆されている(デマスターズ、ジーら(DeMasters,G.,et al.)、(2006)Mol Cancer Ther、5、11、2786〜2797ページ)。さらなる最近の例は、その形成がシミズら(Shimizu et al.)、((2006)Bioorganic & Medicinal Chemistry Letters、16、3323〜3329ページおよびその中に挙げられた参照文献)によって記載されている、SMD−429などの16−エン−22−オキサビタミンD3類似体、例えばリーら(Lee et al.)、((2007)、Cancer Research、67(24)、11840〜11847ページ)によって記載された、1α,25−ジヒドロキシ−20S,21(3−ヒドロキシ−3−メチルブチル)−23−イン−26,27−ヘキサフルオロコル−カルシフェロール、またはヨーンら(Yoon et al.)、((2008)International Journal of Oncology、23、387〜396ページ)によって表された1α,25−ジヒドロキシビタミンD3類似体、1α−ヒドロキシ−5,6−トランス−ビタミンD3の19官能性誘導体(ボジットキーレビッズ、エー(Wojtkielewicz,A)、(2007)Steroids、72(6〜7)、552〜558ページ)、オノら(Ono et al.)、((2007)、Journal of Combinatorial Chemistry9(4)、711〜716ページ)によって記載された延長側鎖を有する類似体、またはポスナーら(Posner et al.)、((2007)、Journal of Steroid Biochemistry & Molecular Biology、103、213〜221ページ)によって記載されたジフルオロ類似体である。
【0058】
本発明のいくつかの実施形態用に、ライブラリーの形で化合物を使用することができる。このようなライブラリーの例は、モデル化合物として化学的に合成した様々な有機小分子、または多数の配列変異体を含む核酸分子の集合である。
【0059】
いくつかの実施形態では、本発明の方法は、例えば生物またはその組織もしくは細胞に
おける、Rab11−FIPタンパク質の発現、細胞内局在および/または活性を検出することを含む。例示例として、Rab11−FIPタンパク質の細胞内量、発現、細胞内局在および/または活性を一定時間モニターすることができる。
【0060】
本発明の方法は、Rab11−FIPタンパク質の細胞内量、発現、細胞内局在および/または活性を測定した結果と、対照測定(または「参照」測定)の結果を比較することをさらに含むことができる。対照測定は、Rab11FIPタンパク質の細胞内量、発現、細胞内局在および/または活性を調節しない条件の使用を含むことができる。細胞内量、発現、および/または活性の比較において、検出レベルを例えば対照レベルと比較することができる。本明細書で使用する用語「対照レベル」は、例えばRab11FIPタンパク質の細胞、mRNAまたはタンパク質発現レベルでの、それぞれのタンパク質の分子の数、および対照試料中のRab11FIPタンパク質の活性レベルを指す。したがってこの用語は、正常対照レベルと癌の対照レベルの両方を含む(これも以下参照)。この用語は1つの参照測定値、または複数の参照測定値を指すことができる。いくつかの実施形態では、対照レベルは以前に実施した測定からの発現または活性値のデータベースであってよい。用語「通常レベル」は、正常、健康な個体中、または癌を含めた新生物に罹患していないことが知られている個体の集団中で検出される、Rab11FIPタンパク質の発現のレベルまたはRab11FIPタンパク質の活性レベルを指す。正常な個体は、それぞれの新生物の臨床症状がない個体である。
【0061】
本発明によれば、例えば細胞中の、遺伝子発現レベルまたは活性レベルまたはタンパク質の量は、対照レベルと比較して遺伝子発現/活性/量が約10%、約25%、約50%、約75%、約100%、またはそれより多く増大または低下するとき、「変化した」または「異なる」とみなされる。あるいは、発現レベルまたは活性レベルは、対照レベルと比較して遺伝子発現/または活性が少なくとも約0.1、少なくとも約0.2、少なくとも約1、少なくとも約2、少なくとも約5、または少なくとも約10またはそれより多くの倍数増大または低下するとき、「増大した」または「低下した」とみなされる。
【0062】
本発明の方法は、細胞外シグナル制御キナーゼ(ERK)の活性を変えることを含むことができる。いくつかの実施形態では、方法はERKの活性化を予防、阻害、停止または逆行させることを含む。典型的実施形態では、Rab11−FIPタンパク質の量または活性の調節は、ERKの活性の調節をもたらす可能性がある。
【0063】
本発明のさらなる方法は、発癌現象を含めて発癌性になる素因を有する細胞を同定する方法である。方法は、細胞におけるRab11FIPタンパク質の量の変化、発現の変化、細胞内局在の変化または活性の変化を検出することを含む。関連法に関して前ですでに説明したように、方法は、Rab11−FIPタンパク質の細胞内量、発現、細胞内局在および/または活性を測定することを含むことができる。いくつかの実施形態では、方法は、Rab11−FIPタンパク質の細胞内量、発現、細胞内局在および/または活性を測定した結果と、対照測定の結果を比較することも含むことができる(上記を参照)。
【0064】
本発明は、対象において癌を含む腫瘍などの新生物を発症するリスクを診断する方法にも関する。それぞれの腫瘍は、例えば乳房腫瘍、肺腫瘍、結腸直腸腫瘍、膀胱の腫瘍または卵管(卵巣管とも呼ばれる)の腫瘍であってよい。同様に、それぞれの癌は、対応する癌腫の1つを含めた、例えば乳房癌、肺癌、結腸直腸癌、膀胱の癌または卵管(卵巣管とも呼ばれる)の癌であってよい。肺癌の癌腫の例示例は、非小細胞肺癌腫である。方法は、Rab11FIPタンパク質の量、発現レベル、活性レベルおよび細胞内局在の少なくとも1つを決定することを含む。測定はいくつかの実施形態では、対象由来の組織試料または細胞試料などの試料において実施することができる。いくつかの実施形態では、方法は、Rab11−FIPタンパク質の細胞内量、発現、細胞内局在および/または活性を
測定した結果と、対照測定の結果を比較することも含むことができる(上記を参照)。それぞれの対照測定用に、Rab結合タンパク質の発現および/または活性が通常(「正常」)レベルである試料を使用することができる。典型的実施形態では、対照測定と比較したRab結合タンパク質の量、発現レベル、活性レベルまたは細胞内局在の変化は、対象が新生物に罹患しているかまたは新生物を発症するリスクがあることを示す。
【0065】
本発明のさらなる方法は、それぞれの化合物を同定するin−vivo法とin−vitro法の両方の方法である。化合物は発癌現象を含めた腫瘍発生を予防、阻害、停止または逆行させることができる。化合物は、Rab11FIPタンパク質などのRab結合タンパク質と複合体を形成することができる。典型的にはこれらの方法は、in−vitroまたはin−vivoであれこの複合体の構成要素を互いに曝すことを含む。1つのこのような方法は、複合体を形成する、または複合体を形成すると考えられる構成要素を互いに接触させることを含むin−vitro法である。例えばRab11FIPタンパク質と、Rab11FIPタンパク質と複合体を形成すると考えられる薬剤候補分子を、互いに接触させることが可能である。いくつかの実施形態では、方法は、複合体の形成を検出することをさらに含む。
【0066】
複合体の形成を検出する任意の適切な方法を使用することができる。検出法は、例えば電気泳動法、HPLC、フローサイトメトリー、蛍光相関分光法またはこれらの技法の改変型を含むことができる。他の技法は生物分子の結合自体の測定を含む。このような測定は、例えば分光的、光化学的、光度分析的、蛍光分析的、放射線学的、酵素的または熱力学的手段、または細胞効果に基づくことができる。分光的検出法の1例は、蛍光相関分光法である(トンプソン、エヌ エルら(Thompson,N.L.,et al.)、Curr.Opin.Struct.Biol.(2002)12(5)、634〜641ページ)。光化学的方法は、例えば光化学的架橋である(スティーン、エイチ、ジェンセン、オー エヌ(Steen,H.,Jensen,O.N.)、Mass Spectrom Rev.(2002)21、3、163〜182ページ)。それぞれ光活性、蛍光、放射性または酵素標識の使用(概説に関しては、:リッペ、アール エーら(Rippe,R.A.,et al.)、Methods Mol.Biol.(2001)160、459〜479ページ参照)は、光度分析的、蛍光分析的、放射線学的および酵素的検出法の例である。熱力学的検出法の1例は、等温滴定型熱量計である(ITC、概説に関しては、:ベラツクエツ−カンポイ、エーら(Velazquez−Campoy,A.,et al.)、Methods Mol.BioL(2004)261、35〜54ページ参照)。細胞効果を使用する方法の1例は、その酵素的検出または細胞複製を含めた細胞増殖または細胞生存の測定である。
【0067】
さらなる関連法は微生物を提供することを含むin vivo法である。微生物はRab結合タンパク質を発現し、該Rab結合タンパク質と共に化合物は複合体を形成し得ると考えられ、またはRab結合タンパク質の発現を化合物が変え得る(例えば、抑制し得る)と考えられる。いくつかの実施形態では、微生物はRab結合タンパク質を内生的に発現することができる。いくつかの実施形態では、微生物は組換え細胞またはトランスジェニック微生物である。いくつかの実施形態では、微生物は、乳癌細胞または肺癌細胞を含めた癌細胞などの単細胞である。いくつかの実施形態では、微生物は生物から得られる細胞である。それぞれの生物は、例えば哺乳動物、魚類、両生類または鳥類であってよい。細胞は典型的には、皮膚組織または乳房組織などの、それぞれの生物の組織に由来する。いくつかの実施形態では、微生物は、乳癌細胞系または肺癌細胞系を含めた、癌細胞系などの細胞系の細胞である。適切な乳癌細胞系の例には、HMEC細胞系、3D、Bcap−37、BT−20、BT−474、BT474M1、Hs578T、Hs578Bst、JIMT−1、MCF−7、M231、MDA−231、MDA−MB−134、MDA−MB−175、MDA−MB−231、MDA−MB−361、MDA−MB−3
61DYT2、MDA−MB−435、MDAMB−435HM、MDA−MB−468、MB231、MW1、PMC42、SK−BR−3、SK−BR−7、SUM149、SUM159、SUM1315、SUM−52PE、T−47D、ZR−75、ZR−75−1およびZR−75−30があるが、これらだけには限られない。肺癌細胞系は、例えば腺癌細胞系、扁平上皮細胞癌細胞系または小細胞癌細胞系であってよい。適切な肺癌細胞系の例には、ABC−1、A549、Calu−6、EBC−1、H322、H446、H460、HCC2279、HTB−56R、HUT29、IMEC−2、KU−T1、L9981、LCD、LCOK、LK−2、LC−1/sq、Lu135、MS−1、NCI−H69、NCI−H82、NCI−H209、NCI−H446、NCI−H1395、NCI−H1437、NCI−H1770、NCI−H2009、NCI−H2087、NCI−H2122、NCI−H2126、NCI−H2171、NCI−N226、NCI−N231、PC1、PC3、PC7、PC9、PC10、PC14、PLA−801D、QG56、RERF−LCAI、RERF−LCMS、RERF−LCKJ、SBC3、SPC−A1、SQ5およびYTLMC−90があるが、これらだけには限られない。多数の他の癌細胞系、例えばポールら(Pole et al.)(2006、上記)中で命名された追加的細胞系が当該技術分野で知られている。
【0068】
いくつかの実施形態では、微生物は、結腸直腸癌細胞系、膀胱癌細胞系および卵管癌細胞系の細胞である。適切な結腸直腸癌細胞系の例には、Colo−320、HCT116、HT−29、L0、L0−200、L0−500、L0−1000、LoVo、LS1034、LST−R1、RKO、SW480、およびWiDr、ならびにドウグラら(Douglas et al.)((2004)Cancer Research 64、14、4817〜4825ページ)によって言及されたさらなる結腸直腸癌細胞系があるが、これらだけには限られない。適切な膀胱癌細胞系の例には、BIU−87、EJ、KoTCC−1、MBT−2、RT4、ScaBER、T24およびUM−UC−2があるが、これらだけには限られない。卵管癌細胞系の2つの例示例は、FT−MZ−1(ルンネバウム、アイ ビーら(Runnebaum,I.B.,et al.)、(1994)FEBS Lett.、353、1、29〜32ページ)およびHEPFT(イシワタ、アイら(Ishiwata,I.,et al.)、(2007)Human Cell、20、119〜130ページを参照)である。
【0069】
いくつかの実施形態では、微生物は、Rab11FIP、例えばRab11FIP1などのRab結合タンパク質を過剰発現する乳癌上皮細胞系MCF10Aなどの非癌性組織細胞系を含めた、非腫瘍性の細胞である。同様に、任意の細胞−細胞系の細胞、個々の細胞、多細胞生物内の細胞、(直接入手および単離を含めて)多細胞生物から入手した細胞であろうと選択して、Rab11FIP1などのRab結合タンパク質を過剰発現させることによって癌細胞に形質転換することができる。以下の実施例および例えば図6は、細胞がどのようにして癌細胞に形質転換され得るかを然るべく例示する。この点において、本発明の方法は癌細胞を入手する方法も提供し、それは細胞中のRab結合タンパク質の発現を調節する、特に増大させる方法を含む。それによって細胞は癌細胞に形質転換される。それぞれのRab結合タンパク質は、例えばRab11FIP1などのRab11結合タンパク質であってよい。Rab結合タンパク質は、内因性または異種タンパク質、すなわち細胞中に本来存在するコード核酸配列を有するタンパク質、またはコード核酸配列がそれぞれの細胞中に導入される/導入されているタンパク質であってよい。本発明は、in vivoでの対応するRab結合タンパク質の発現を調節することが可能な化合物を同定するための、このような細胞の使用にも関する(以下参照)。
【0070】
本発明の方法は、Rab結合タンパク質と複合体を形成し得ると考えられる化合物、またはその発現を変えると考えられる化合物を微生物に加えることを含む。各方法のいくつかの実施形態では、Rab結合タンパク質、例えばRab11FIP結合タンパク質の発
現をモニターする。各方法のいくつかの実施形態では、Rab結合タンパク質の活性をモニターする。いくつかの実施形態では、Rab結合タンパク質の細胞表現型の変化をモニターする。いくつかの実施形態では、このような方法は対照測定を含む(上記も参照)。対照測定の結果は化合物を使用して得た結果と比較する。対照測定は、例えば、Rab結合タンパク質の発現または活性に影響を与えないことが知られている化合物の使用を含むことができる。
【0071】
さらに他の関連法は、宿主生物を提供することを含むin−vivo法である。任意の望ましい宿主生物を、それが癌細胞を収容し増殖させることが可能である限り提供することができる。宿主生物の例には、哺乳動物、魚類、両生類、および鳥類があるが、これらだけには限られない。適切な哺乳動物の例には、ラット、ウサギ、モルモット、オポッサム、イヌ、およびネコがあるが、これらだけには限られない。
【0072】
任意の望ましい癌細胞をこの目的で使用することができる(例えば上記を参照)。方法は、宿主生物に癌細胞を導入することをさらに含む。さらに方法は、先に記載した化合物、すなわちRab結合タンパク質と複合体を形成し得ると考えられる化合物、Rab結合タンパク質の量を調節し得ると考えられる化合物、またはRab結合タンパク質の活性を調節し得ると考えられる化合物の使用を含む。いくつかの実施形態では、癌細胞は化合物を含む。したがって、宿主生物に癌細胞を導入する前に、化合物を癌細胞に導入することができる。いくつかの実施形態では、宿主生物への癌細胞の導入の前、後、または同時に、化合物を宿主生物に投与する。典型的には、方法の特定の段階で化合物を癌細胞に導入する。方法は、宿主生物中の腫瘍の増殖をモニターすることをさらに含む。
【0073】
本発明を容易に理解し実務上の効果をもたらすことを可能にするために、個々の実施形態をここで以下の非制限的な実施例によって記載する。
【実施例】
【0074】
本発明者らが開発したTRIAGE法の、特に関連する態様および特徴は、以下のように要約できる(図1も参照されたい)。
【0075】
【表1】
【0076】
細胞および薬剤治療
マイクロアレイデータおよび腫瘍試料。4種の前述の乳癌マイクロアレイデータセットを、この研究に使用し、全発現プロフィールを、原典であるウプサラコホート(ミラー、エル ディーら(Miller,L.D.et al.)(2005)Proc Natl Acad Sci USA 102、13550〜13555)、ストックホルムコホート(パウィタン、ワイら(Pawitan,Y.et al.)(2005)Breast Cancer Res 7、(R953〜964)、ベルギーコホート(ルイ、エスら(Loi,S.et al.)(2007)J Clin Oncol 25、1239〜1246)およびシンガポールコホート(イブシナ、エー ブイら(Ivshina,A.V.et al.)(2006)Cancer Res 66、10292〜10301)に記載されているように、AffymetrixU133AおよびU133
Bの遺伝子チップを使用して発生させた。全AffymetrixCELファイルを、Gene Expression Omnibus(http://www.ncbi.nlm.nih.gov/geo/)から検索し、グローバル平均法(ミラーら(Miller et al.)、2005、上記)を使用して標準化し、プローブセットのシグナル強度を自然対数変換し、平均強度をlog 500の標的シグナル値に調整することによって尺度化した。コホート依存性アレイのバイアスを取り除くために、それぞれのコホート内のアレイを、それらを1つのデータセットに組み込む前に平均化した。すべての患者および腫瘍試料の臨床的特徴を、図2に要約した。
【0077】
局所的特異値分解(LSVD)。マイクロアレイのプローブセットを、UCSD Genome Browserに従ってゲノム座標にマッピングした。候補PEAを、対応する腫瘍のサブセット中の遺伝子の、局在座標化された過剰発現として同定した。第1に、遺伝子を、試料中の遺伝子の発現が、全試料にわたったその発現中央値から調整された中央絶対偏差(aMAD)の2.5倍を超えた場合に、試料中の過剰発現として定義した(図1、第1工程)。第2に、局所座標化された過剰発現を、50のプローブのスライディングウィンドウに適用したSVDにより得られた主固有値によって定量化した。同じ遺伝子に対応するプローブ(UniGene ID、build #177に対応する)を平均し、単一遺伝子の測定値を表した。候補PEAを、ベースライン閾値を上回る局所主固有値を有する連続した染色体領域として同定した。候補PEAにスコアを付け、主固有値のピークによりランク付けした。最も高いスコアを得た候補PEAを、ピーク構造をさらに解明するために(すなわち、複数のサブピークから1つを識別するために)、10のプローブのウィンドウサイズを使用して、LSVDの第2ラウンドに供した。最後に、最も高いスコアを得たサブピークを、個人の遺伝子および腫瘍に関する固有重量を導くために、その後SVDに供した。固有重量は、主固有ピークの大きさに対する、遺伝子および腫瘍の相対的寄与率を反映する。得られたピークに関して、非ゼロの固有重量を有する遺伝子は、PEAの成分であると考えられる。中央値+2標準偏差を超える固有重量を有する腫瘍を、アンプリコン含有腫瘍として分類した。(TRIAGE、第4工程)。
【0078】
再発性ゲノム増幅を同定するための局所特異値分解(LSVD)
腫瘍試料のコホートに由来する遺伝子発現プロフィールのLSVD分析は、推定発現アンプリコン(PEA)の同定における鍵となる第1の工程であり、これにより、転移に寄与する癌遺伝子を、TRIAGE(Clinico−Genomic Intersectsを介した癌遺伝子三角検証(Triangulating Oncogenes through Clinico−Genomic Intersects))方法論により発見できる。LSVDは、再発性アンプリコンが、腫瘍試料のサブセット中にゲノム的に局在化された遺伝子の座標化された過剰発現として現れるという仮説に基づき、PEAを同定する。LSVDは、特異値分解(SVD)(ストラング、ジー(Strang,G.)、(1998)「Introduction to Linear Algebra」、Section 6.7.第3版、ウェルズリ−ケンブリッジプレス社(Wellesley−Cambridge Press.)ISBN 0−9614088−5−5)を中心に構築されており、染色体ごとのSVDの局所適用により得られる、局所主固有値(LPEV)のプロットである、座標化された局所過剰発現プロットのピークとしてPEAを予測する。LSVDは、ゲノム上の各ゲノムの位置に関する局所発現行列を導き、バイナリ連結行列に変換し、バイナリ行列にSVDを適用することにより達成される。このような適用で得られた主固有値は、その遺伝子座における遺伝子の座標化された過剰発現の基準である(クルーガー、ワイら(Kluger,Y.et al.)、(2003)Genome Research、13、703〜716;クラインバーグ、ジェイ エム(Kleinberg,J.M.)、(1999)ACMのJl、46、5、604〜632)。LPEV対ゲノム位置のプロット(LPEVプロット)における特定のベース閾値を上回る、固有値の「ピーク」は、PEAの境界を定める。LSVDの手順の詳細
な説明は、以下に提供する。
【0079】
Ecを、染色体Ccの遺伝子発現行列とし、Ecij(注:EcijはPCT出願時英文ではベクトルのため太字)を、腫瘍Tjの中の位置liにおける遺伝子の発現のlog2とし、i=1、2、...、Nc、およびj=1、2、...、Mである。Cc上の推定発現アンプリコン(PEA)は、LSVDを使用してEcを分析することによって予測され、LSVDは以下の順序の工程:(1)Ec(注:EcはPCT出願時英文ではベクトルのため太字)を、バイナリ連結行列Ac(注:AcはPCT出願時英文ではベクトルのため太字)に変換する工程、(2)染色体局在化行列Apc(注:ApcはPCT出願時英文ではベクトルのため太字)を導く工程、(3)ApcにSVDを適用する工程、および(4)PEAを同定する工程、を有する。
【0080】
1.Ecのバイナリ連結行列Acへの変換:Ecを、以下のようなブールの離散化定理を介して、バイナリ連結行列Acへ変換する(注:Ec、AcはPCT出願時英文ではベクトルのため太字)。
【0081】
Ecij>μi+x*νiの場合Acij=1
Ecij≦μi+x*νiの場合Acij=0
μiおよびνiが中央値であり、調整中央絶対偏差(aMAD)が、Eci={Eci1、Eci2、...、EciM}である場合、aMADはMADの1.4826倍である。xは、粗いマップの第1のパスに関しては2.5であり、第2のパスにおいて得られた、より詳細なマップに関しては2である。
【0082】
2.局所行列Apcの導き(注:ApcはPCT出願時英文ではベクトルのため太字)。
位置Lpcにおける局所行列Apcは、染色体Cc上の位置p−wからp+wまでの遺伝子を使用してAcから導き、wはウィンドウサイズであり、所定のパラメーターである。wは、第1のパスの粗いマップに関しては50であり、第2のパスのより詳細なマップに関しては10である。
【0083】
3.特異値分解(SVD):SVDのApcへの適用は、PEAを予測するための重要な工程である。ApcのSVDは、Apcを3つの行列、Upc、Σpc、Vpc(注:Upc、Σpc、VpcはPCT出願時英文ではベクトルのため太字)の積、すなわちApc=Upc×Σpc×VpcTに分解する。UpcおよびVpcは、それぞれ、(2w+1)×(2w+1)およびM×Mの行列であり、一方Σpcは、(2w+1)×Mの対角行列である。UpcおよびVpcの列ベクトルは、それぞれ、ApcApcTおよびApcTApc(注:ApcApcTおよびApcTApcはPCT出願時英文ではベクトルのため太字)の固有ベクトルであり、一方Σpc(注:ΣpcはPCT出願時英文ではベクトルのため太字)の対角成分は、各固有値である。最も高い固有値を、主固有値(λpcで表す)と呼び、各固有ベクトルを主固有ベクトルと呼ぶ。位置Lpcにおける腫瘍Tjの固有重量はTpcjで表され、行列ApcTApcの主固有ベクトルのjth成分の絶対値として定義する。同様に、Gpcjで表される、位置Lpcにおける遺伝子Giの固有重量は、行列ApcApcTの主固有ベクトルのjth成分の絶対値である。
【0084】
真のPEAシグナルとバックグラウンドとの差異を改善するために、本発明者らのLSVDの実施において、本発明者らは、SVDを、Apcについて使用する代わりに(ApcApcT)3について使用し、λpcを見出した。上記の定義に従って、腫瘍および遺伝子の固有重量が、それぞれ(ApcTApc)4および(ApcApcT)4の主固有ベクトルから得られた。ここで、λpcは、(ApcApcT)4対M4の主固有値4乗根との比であり、遺伝子および腫瘍の固有重量は、それぞれ、(ApcApcT)4/M
4および(ApcTApc)4/M4の主固有ベクトルの各成分の4乗根である(注:ApcおよびApcTはPCT出願時英文ではベクトルのため太字)。
【0085】
4.PEAの予測:
λpc対Lpcをプロットすることにより、LPEVのプロットが得られ、これをLPEVプロットと呼ぶ。このプロットのピークは、PEAを示す。ピークにおけるλpcの値が高いほど、Lpcの周りのPEAの信頼性が高い。最終的に、PEAは、λpc>Tλ=20を含む、連続した領域を対象とする範囲を有するピークおよびこのピークを中心として報告される。
【0086】
TRIAGE遺伝子生存率分析。各プローブセットに関して、平均発現レベルを、所与のコホートに関してまず計算した。その後、患者を、平均より下の群と平均より上の群とに階層化し、コックス比例ハザード回帰モデルをこのデータに適合させ、無遠隔転移生存率(DMFS)に基づくハザード比を計算した。帰無仮説は、2つの群は再発の危険性が同じであろうと言うものであり、尤度比検定のp値はこの仮説に対する証拠として計算した。コックスモデルの適合および統計的計算を、Rでサバイバルパッケージ(the survival package)を使用して実施した。同じ遺伝子にマッピングされた複数のプローブセットの場合、最も有意に関与するプローブセットの結果が報告された。DMFSを、外科手術から最初の遠隔再発事象または最後の追跡調査の日までの時間間隔として定義した。両側および対側の癌を有するすべての患者ならびに再発したまたは診断後10年を超えて疾患特異的な死を迎えた人を、事象に関して体系的に審査した。ウプサラコホートは最も長く、最も完全に患者を追跡調査したコホートに基づく、本発明者らの最も大きい集団であったので、それを使用し、TRIAGEにおける生存率データを作成した。
【0087】
増殖サインおよび遺伝子相関。cDNAマイクロアレイを使用して、ウィットフィールドら(Whitfield et al)は850を超える細胞周期調節遺伝子を、ヒーラ子宮頸癌細胞においてすでに同定していた(ウィットフィールド、エム エルら(Whitfield,M.L.et al.)、(2002)Mol Biol Cell、13、1977〜2000)。著者らは、乳房の腫瘍における高度に相関した発現を示し、ペローら(Perou et al)の乳癌データセット(ペロー、シー エムら(Perou,C.M.et al.)(2000)Nature 406、747〜752)の腫瘍細胞増殖に関与した、これらの遺伝子の分画を発見した。この増殖サインの近似値を求めるために、本発明者らは、細胞周期調節遺伝子(ウィットフィールド(Whitfield)の研究における1,134 IMAGEクローンに対応する)を、Uni Gene build #187を介してAffymetrixプローブセットにマッピングし、ウプサラコホートの腫瘍を階層的にクラスタ化した。増殖関連遺伝子を含む独特のクラスタが現れ(図10を参照されたい)、組織学的悪性度、分裂指数およびKI67スコアを含むウプサラコホートにおける増殖のマーカーと高度に相関することが見出された(データ非掲載)。>0.8の平均相関を有する、このクラスタの中心となるプローブセット(n=78のプローブセットが67の遺伝子に対応する(図10を参照されたい))を、「コア増殖サイン」遺伝子に指定し、それらの平均発現を使用して、増殖サインと、TRIAGE分析の第7工程のための他のすべてのプローブセットとのピアソン相関を計算した。増殖と癌の再発との臨床的関連性次第では、特に、この工程は、他の癌型のTRIAGE分析には必須ではないことがある。
【0088】
RAB11FIP1(RCP)の選択およびクローン化。RCPの3種のスプライス変異体:変異体1(NM_025151)、変異体2(NM_001002233)および変異体3(NM_001002814)を、GenBankで同定した。これらの中で、変異体1および3だけが、腫瘍試料および細胞系においてRT−PCRにより一貫して検
出でき(データ非掲載)、変異体1だけが、ポリクロナールRCP抗体(GenWay)を用いたウェスタンブロットにより観察できた。したがって、RCPの完全長変異体1は、正常なヒト血液からRT−PCRにより増幅され(順方向プライマー:5’−ACCATGTCCCTAATGGTCTCGGCT−3’(配列番号1)、逆方向プライマー:5’−TGCTGATTTACATCTTTCCTCT−3’)(配列番号2))、配列を確認し、この研究においてRCPの機能を調査するために使用した。
【0089】
細胞系および一過性形質移入。ヒト乳房細胞系、MCF10A、MCF7およびMDA−MB231を、American Type Culture Collection(ATCC)から入手し、37℃、5%CO2で、ATCCにより推奨されている成長培地で維持した。完全長RCPをpcDNA3.1のBamHlおよびEcoR1部位に連結した。2種のRCP特異的RNAi構築体を、先に記載したように(レイノルズ、エーら(Reynolds,A.et al.)、(2004)Nat Biotechnol、22、326〜330)設計し、Ambionにより作製した。MCF7について、一過性プラスミドおよびRNAiの形質移入を、Lipofectamine 2000(インビトロジェン社(Invitrogen))を使用し、製造業者のプロトコルに従って実施した。
【0090】
レンチウィルスに感染させることによる安定細胞系の作製。shRNAを安定して過剰発現する細胞系を作成するために、標的配列(順方向:5’−TCGATAAGCAAGAAGGAGTTTTCAAgAgAAACTCCTTCTTGCTTATCGTTTTTTC−3’(配列番号3)および逆方向:5’−TCGAGAAAAAACGATAAGCAAGAAGGAGTTTCTCTTGAAAACTCCTTCTTGCTTATCGA−3’)(配列番号4)をコードするオリゴヌクレオチドを、LentiLox pLL3.7ベクターにアニーリングし、クローン化した。対照shRNAのために、非標的「スクランブル」配列を、pLL3.7.(順方向:5’−TGAACGGCATCAAGGTGAACttcaagagaGTTCACCTTGATGCCGTTCTTTTTTC−3’(配列番号5)および逆方向:5’−TCGAGAAAAAAGAACGGCATCAAGGTGAACTCTCTTGAAGTTCACCTTGATGCCGTTCA−3’)(配列番号6)内にクローン化した。レンチウィルスによるRCPの過剰発現のために、RCPをまず、Invitrogen gatewayエントリーベクターpENTR3Cにクローン化し、さらに、レンチウィルスベクターpLenti6/V5−DEST(インビトロジェン社(Invitrogen))に、製造業者のプロトコルに従ってLR組換え反応により組換えた。レンチウィルスの作製のために、pLL3.7またはpLenti6/V5−DESTを、パッケージングベクターを用いて、293FT細胞に共形質移入し、48時間後に上清を収穫した。ウィルスを、Beckman SW28ローター内で、25,000rpmで2時間、超遠心分離器にかけて濃縮し、リン酸緩衝食塩水に再懸濁した。力価を、濃縮ウィルスの段階希釈液を用いて、NIH/3T3細胞に感染させることによって確定した。標準的な調製用に、力価は、pLL3.7に関してはおよそ1〜5×108粒子であり、pLenti6/V5−DESTベクターに関しては4−10×107粒子であった。MCF10A、MCF7およびMB231細胞の感染のために、2×105細胞を、1×107粒子および8μg/mlポリブレンの懸濁液中で、3時間、37℃においてインキュベートした。細胞をその後、再度播種し、前記のように培養した。RCPノックダウン安定細胞の純粋な集団を得るために、GFP陽性細胞を、培養4日後に選り分けた。RCP過剰発現安定系の作製のために、細胞を、ブラストサイジンを用いて感染2週間後に選択した。
【0091】
ウェスタンブロット分析。すべての細胞溶解物を、RIPAバッファーを使用して調製した。タンパク質を、SDS−PAGEにより分離し、Hybond−P PVDF メンブラン(ジーイーヘルスケア社(GE Healthcare))に転写した。RCP
(ジェンウェイ社(Genway)、カタログ番号(CN):15−288−21574A)、β−アクチン(シグマ社(Sigma)、CN:A5441)、ホスホ−Akt(セルシグナリングテクノロジー社(Cell Signaling Technology)、CN:9271)、Akt(セルシグナリングテクノロジー社(Cell Signaling Technology)、CN:9272)、ホスホ−Erk(セルシグナリングテクノロジー社(Cell Signaling Technology)、CN:9101)およびErk(セルシグナリング社(Cell Signalling)、CN:9102)に対する抗体を使用して、メンブランを調査し、抗体−タンパク質複合体を、HRP標識抗体およびECL(アマシャムバイオサイエンス社(Amersham Biosciences))により検出した。
【0092】
細胞増殖およびコロニー形成。細胞を、96ウェルプレートに、5000細胞/ウェルの濃度で播種し、細胞増殖を、WST−1(ロッシュ社(Roche))を使用し、製造業者のプロトコルに従って4重に測定した(すなわち、条件当たり4ウェル)。MAPK阻害アッセイのために、96ウェルプレートに再度播種する前に、細胞を、MEK阻害剤U0126(プロメガ社(Promega))を用いて、0.5%の血清を含む培地で一晩処理した。足場非依存性コロニー形成に関するRCPの作用を試験するために、細胞を、25%FBSを含む完全培地に溶解した0.3%寒天(シグマ社(Sigma))250μlに再懸濁し、500μlの0.6%寒天ベースであらかじめコーティングした24ウェルプレートに4重(すなわち、条件当たり4ウェル)に播種した。コロニー形成の効率を、播種の21日またはそれ以後に、ヨードニトロテトラゾリウムクロリド(シグマ社(Sigma))を用いて染色することによって検査した。コロニーの大きさが50umを超えるコロニーをLeica QWinソフトウェアを使用して計測した。
【0093】
in vitroの浸潤および遊走。トランスウェルの遊走および浸潤アッセイを、孔サイズ8μmのFalcon FluoroBlok 24マルチウェルインサート(ビーディーバイオサイエンス社(BD Biosciences))を使用して実施した。浸潤アッセイのために、インサートを、80μLの、無血清成長培地中の、20μgのMatrigel(ビーディーバイオサイエンス社(BD Biosciences))でコーティングした。両方のアッセイのために、200μL無血清成長培地中の5000細胞を、10%ウシ胎児血清を含む750μL完全成長培地を、下方のチャンバーに入れた、各トランスウェルインサートに入れた。24時間後、インサートの孔を介して遊走または浸潤した細胞を、3.7%ホルムアルデヒドで固定し、2.5μg/mL Hoechst 33342(インビトロジェン社(Invitrogen))を用いて15分間染色し、PBSで洗浄し、Target Activation Bioapplicationを使用してArrayScan VTI(セロミクス社(Cellomics))上で計測した。フィールドサイズは1mm2であった。浸潤アッセイのために、実験を4〜5倍の複製を用いて実施し、実験ごとに10〜16個のフィールドをスキャンした。遊走アッセイのために、実験を3倍の複製を用いて実施し、実験ごとに10〜16個のフィールドをスキャンした。
【0094】
細胞の免疫蛍光顕微鏡検査。細胞培養物を、4%パラホルムアルデヒドを用いて固定し、0.25%Triton X−100を用いて透過処理し、その後1%ヤギ血清を用いてPBS中でブロッキングした。細胞を、抗E−Cadherin(ビーディーファルミンゲン社(BD Pharmingen)、CN:610404)および1:500希釈のフィブロネクチン抗体(ビーディーファルミンゲン社(BD Pharmingen)、CN:610077)を用いて染色し、その後、マウスおよびウサギのIGgを検出する適切な2次抗体を、それぞれAlexa Fluor 594または488(モレキュラープローブ社(Molecular Probes))に接合した。F−アクチンは、Texas Red−X phalloidin(モレキュラープローブ社(Molec
ular Probes))を用いて標識した。DAPIを、核の染色に使用した。画像を共焦点顕微鏡、(LSM 510 META、ツァイス社(Zeiss))を用いて獲得した。
【0095】
細胞周期分析。MCF10A細胞を、1%血清中で48時間飢餓状態にした。細胞をその後、75%エタノールで固定し、RNase A(0.25mg/ml)を用いて処理し、ヨウ化プロピジウム(10μg/ml)を用いて染色し、LDR2フローサイトメーター(ビーディーバイオサイエンス社(BD Biosciences))で分析した。分類した細胞を、FACS DiVaソフトウェア(ビーディーバイオサイエンス社(BD Biosciences))を使用して分析した。
【0096】
アポトーシスアッセイ。MB231細胞を、48時間、完全培地で維持または1%血清を用いて飢餓状態にした。アポトーシス細胞を、In Situ Cell Death
Detection Kit(ロッシュ社(Roche))を用いて検出した。DNA鎖の壊れた細胞を、TMR(赤)を用いて、TUNEL(TdT−mediated dUTP nick end labeling)反応により標識し、蛍光顕微鏡検査およびフローサイトメトリーにより分析した。
【0097】
ヌードマウスにおける腫瘍原性。メスのBALB/c−nu/nu(ヌード)マウス、6〜8週齢を、無菌条件下で、温度調節室において、12/12時間の明/暗スケジュールで、食餌および水は好きな時に取らせて飼育した。動物およびそれらの世話に関するすべての手順は、国内的および国際的規格に従った。MCF−7腫瘍形成実験のために、1.7mgの17β−エストラジオールのペレット(90日放出、イノベイティブリサーチオブアメリカ社(Innovative Research of America))を、腫瘍細胞の注入1週間前に各マウスに移植し、MCF−7細胞のエストロゲン要求性を補った。実験群は、MCF7−RNAi RCP(安定RCPノックダウン)、MCF7−RNAi Ctrl(対照)、MB231−RNAi RCP、MB231−RNAi Ctrlであった。各実験群に関して、8匹のマウスの皮下(右および左の後脚上部)に2.0×106の細胞を注入した。皮下腫瘍を、デジタル計測器を用いて、週に1回測定した。腫瘍の堆積は、ab2/2としてmm3で計算し、aおよびbは、それぞれ直交する、腫瘍の最も長い径および最も短い径である。腫瘍重量は終了時に測定し、シンガポール動物飼育規則に従って死亡させた。
【0098】
免疫組織化学。4μmの厚さのパラフィン包埋腫瘍組織切片を、ABC免疫組織化学試薬およびアビジン−ビオチンブロッッキング試薬(ベクターラボラトリーズ社(Vector Laboratories))を使用して、製造業者のプロトコルに従って染色した。スライドを、1:500希釈の抗RCP抗体と共にインキュベートし、ヘマトキシリンを用いて対比染色した。
【0099】
結果
上記のデータにより、ゲノム、転写および臨床的な情報の座標分析が、転移性再発および患者の死に寄与していると思われる癌の新規遺伝子の発見を良好に対象とでき、その結果、下流の機能研究のための遺伝子の理論的優先順位付けが容易になることが実証された。ヒト乳癌における、複数機能の頻繁に発生する癌遺伝子としてのRAB11FIP1の例示的検証は、治療標的としてのその重要性を強調し、発癌におけるエンドサイトーシス再循環経路の新たな役割に新しい洞察を提供する。
【0100】
例示目的で、ヒト乳癌形成において複数の病理学的役割を有する新規な癌遺伝子として染色体8p11に位置する、RAB11FIP1の機能的検証を本明細書で提供する。RAB11FIP1は、Rab4およびRab11の結合タンパク質としてすでに同定され
ており、Rabカップリングタンパク質(RCP)とも呼ばれ、これはエンドソームの再循環および受容体の分類において機能する(リンゼイ、エー ジェイら(Lindsay,A.J.et al.)(2002)J Biol Chem 277、12190〜12199;ペデン、エー エーら(Peden,A.A.et al.)(2004)Mol Biol Cell 15、3530〜3541)。RAB11およびRAB25の異常調節を介して、発現は、多くのヒトの腫瘍の一般的要素として示され(例えば、チェン、ケイ ダブリュら(Cheng,K.W.et al.)(2004)Nat Med 10、1251〜1256;チェン、ケイ ダブリュら(Cheng,K.W.et al.)(2005)Methods Enzymol 403、202〜215;ゲブハルド、シーら(Gebhardt,C.et al.)(2005)Am JPathol 167、243〜253;ゴールデンリング、ジェイ アールら(Goldenring,J.R.et al.)(1999)Yale JBiol Med 72、113〜120;ヨーン、エス オーら(Yoon,S.O.et al.)(2005)Cancer Res 65、2761〜2769を参照されたい)、腫瘍進行におけるRab結合タンパク質の役割については記載されていない。本発明者らは、臨床的およびゲノム的交差を介して癌形成の関連癌遺伝子に三角検証を用いる、統合分析戦略を考案し、これは、発現の頻度、アンプリコンにおける共発現、患者の無病生存率との関連を考慮し、腫瘍増殖の偶発的関連は考慮に入れない。この多段階工程のデータ検索研究法である、略してTRIAGE(Triangulating Oncogenes through Clinico−Genomic Intersects、以下を参照されたい)を使用して、本発明者らは、RAB11FIP1を癌遺伝子として同定した。
【0101】
TRIAGEによる癌遺伝子の推論
進行性乳癌に寄与する癌遺伝子を同定するために、このような遺伝子の特定のサブセットは、転写学的、ゲノム的および臨床的情報の三角検証を介して発見できるという仮説を立てた。特に、本発明者らは、1)再発性増幅の領域に位置し、2)遺伝子の異常調節により活性化し、3)癌の転移性播種に関連する癌遺伝子を明らかにするよう探求した。この目標のために、本発明者らは、TRIAGE(Triangulating Oncogenes through Clinico−Genomic Intersects)と呼ばれる、複数工程のデータ検索研究法を開発し、これは、発現マイクロアレイデータにおける再発性ゲノムアンプリコンの発現フットプリントを同定し、遺伝子発現を、転移性再発の危険性と関連付け、生存率相関および比較mRNA動学に基づく遺伝子の立候補を評価する(図1)。
【0102】
第1の工程において、737例の原発性乳房腫瘍のマイクロアレイ発現プロフィール(図2)を、局所特異値分解(Local Singular Value Decomposition)(LSVD)により分析し、再発性ゲノムアンプリコンの存在および位置を推測した。この研究は、再発性アンプリコンは、関連遺伝子の過剰発現を座標化することによって検出できるという前提に基づき(チン、ケーら(Chin,K.et al.)、(2006)Cancer Cell 10、529〜541;レアル、エフら(Reyal,F.et al.)(2005)Cancer Res 65、1376〜1383)、乳癌において高度に増幅した遺伝子の大部分が同時過剰発現であるという証拠に支持されている(ポラック、ジェイ アールら(Pollack,J.R.et al.)、(2002)Proc Natl Acad Sci USA、99、12963〜12968)。LSVDは、遺伝子の過剰発現が濃縮されたゲノム座および「遺伝子座特異的」過剰発現を生じる腫瘍分画の同時同定を可能にする。アウトプットは、遺伝子座特異的過剰発現の程度および関連する腫瘍数の両方を反映する、主固有ピークとしてみなされる。全ゲノムLSVDにより発見された、最も高いスコアを得た主固有ピークは、再発性コピー数増加の公知の領域に対応する遺伝子座に優勢にマッピングされた(図3を参照されたい)。LSVDによりすべてのピークが同定されたわけではないが、再発性ア
ンプリコン事象を表すことが期待され、公知の再発性アンプリコンと、非常に大きく、正確なゲノム重複を有するものを推定発現アンプリコン(PEA)と呼び、TRIAGE手順によりさらに調査した。
【0103】
対象となる癌遺伝子の発現レベルは、患者の予後不良に相関するだろうという指針に基づき、PEA内の遺伝子を、遠隔再発関連の有意性に関して評価した。各遺伝子のために、腫瘍(および対応する患者)を、低発現群(平均より低い)または高発現群(平均より高い)に、2つに分け、コックス比例ハザード回帰により、無遠隔転移生存率(DMFS)に関して分析した(図1、第2および第3工程)。得られたハザード比の有意性を反映する尤度比検定のP値を、PEAの遺伝子の発癌性の初期基準として使用した(図1、第3工程、任意抽出の生存率)。しかし、再発性ゲノムアンプリコンは、患者の生存率不良に相関し得るので、アンプリコン上に物理的には存在する良性の「パッセンジャー遺伝子」もまた、腫瘍の侵攻に機能的に寄与することなくDMFS不良に相関でき、一方、ドライビング癌遺伝子(コピー数増加以外の機序により癌において活性化できる)は、アンプリコンを欠いた腫瘍においてDMFS不良との相関を維持することが予想され、パッセンジャー遺伝子は予想されない。したがって、この分析から審査される、遺伝子発現と、含有アンプリコン腫瘍のDMFSとの相関を再検査した。(図1、第4工程、任意抽出)。この方法では、増幅事象に非依存性の、DMFS不良に確実に関連するそれらの候補癌遺伝子のみが分かってくるであろう。
【0104】
生存率分析の両方の工程において有意な候補癌遺伝子を、mRNA発現パターンの比較を介してさらに精査した(図1、第5および6工程)。ドライビング癌遺伝子は、アンプリコン含有腫瘍において比較的高度に一貫して発現されるはずであるという仮定に従って、PEA内の遺伝子発現レベルを平均し、その次に遺伝子を平均発現によりランク付けした(ランク1が、最も高い平均発現レベルを表す)。この方法では、発現のランキングは、所与の遺伝子座における発癌性のさらなる相対的尺度として機能する。
【0105】
高増殖性乳房腫瘍は、再発する可能性が高く、増殖に関連する遺伝子は、多くの場合DMFS不良に相関している。このことは、細胞周期機能を濃縮する、何百の高度関連遺伝子からなる増殖サインにおいて実証され、これは多くの癌型の再発に正に相関することが見出された(ダイ、エイチら(Dai,H.et al.)(2005)Cancer Res 65、4059〜4066;イブシナ、エー ブイら(Ivshina,A.V.et al.)(2006)Cancer Res 66、10292〜10301;ペロー、シー エムら(Perou,C.M.et al.)(2000)Nature
406、747〜752;ローゼンウォルド、エーら(Rosenwald,A.et
al.)(2003)Cancer Cell 3、185〜197;ウィットフィールド、エム エルら(Whitfield,M.L.et al.)(2002)Mol
Biol Cell 13、1977〜2000;ウィットフィールド、エム エルら(Whitfield,M.L.et al.)(2006)Nat Rev Cancer6、99〜106)。この調査は、増殖との関連とは非依存性の、発癌に寄与する固有の遺伝子に関し、TRIAGEの最終工程において、候補遺伝子の発現パターンを増殖サインとの相関について試験し、生存率との関連が腫瘍の増殖率により説明できる程度を検討した。DMFS不良に相関する、ウプサラ乳房腫瘍コホートのプローブセット(n=251腫瘍)を図4に表す。DMFS不良(p<0.05)に有意に相関することが見出された、2,377のプローブセットの約35%が、ピアソン相関が、>0.4でこの増殖サインのメンバーであることが見出された(図4)。増殖サインの中核コア遺伝子は、クラスタ化の順に、B.222958_s_(DEPDC1)、A.204170_s_(CKS2)、B.222680_s_(DTL)、A.209680_s_(KIFC1)、A.202503_s_(CSNKIG1)、A.201291_s_(TOP2A)、A.201292_(TOP2A)、A.204033_(TRIP13)、B.
225687_(FAM83D)、A.200853_(H2AFZ)、A.213911_s_(H2AFZ)、A.203362_s_(MAD2L1)、B.228729_(CCNB1)、A.214710_s_(CCNB1)、A.203755_(BUB1B)、B.228323_(CASC5)、A.204825_(MELK)、A.204822_(TTK)、A.207828_s_(未知)、A.202870_s_(CDC20)、A.222039_(LOC146909)、A.204962_s_(CENPA)、B.228273_(未知)、A.209408_(KIF2C)、A.202705_(CCNB2)、A.222077_s_(RACGAP1)、A.218542_(CEP55)、A.203213_(CDC2)、A.203214_x_(CDC2)、A.210559_s_(CDC2)、A.218039_(NUSAPI)、A.218009_s_(PRC1)、A.204444_(KIF11)、A.210052_s_(TPX2)、A.219918_s_(未知)、A.203764_(DLG7)、B.223307_(CDCA3)、A.202095_s_(BIRC5)、A.202954_(UBE2C)、A.202580_x_(FOXM1)、A.218726_(DKFZp762E1312)、A.203554_x_(PTTGI)、A.209642_(BUB1)、A.209773_s_(RRM2)、A.201890_(RRM2)、A.204641_(NEK2)、A.218355_(KIF4A)、A.208079_s_(未知)、A.204092_s_(AURKA)、A.38158_(SP1)およびA.218755_(KIF20A)。乳癌において、DMFS不良に有意に相関することが見出されたプローブセットのおよそ1/3が、増殖サインのメンバーであることが同定されたので、生存率不良関連遺伝子のおよそ3つに1つが、非依存性発癌機序ではなく、増殖との関連により最もよく説明できる。したがって、発明者らは、選択過程において、生存率に有意に関連し、さらに(>0.4の)増殖サインと相関する候補遺伝子の優先順位を下げた、または考慮に入れなかった。
【0106】
この分析研究の妥当性を試験するために、TRIAGEを、全ゲノムのLSVDにより同定された最も大きな主固有ピークに用いた。これは、約30の遺伝子を包含する染色体17q12の1Mb領域に対応する。このPEAの遺伝子成分を、EGFR−関連癌遺伝子ERBB2(HER2/neu)に収めた、乳癌の症例の約20〜30%に生じることが公知である、十分に特徴づけられた17q12アンプリコンに正確にマッピングした。特に、このアンプリコンの発現フットプリントは、すでにマイクロアレイ研究において観察されており、アレイ−CGHにより判別されるように、物理的アンプリコンの境界とよく相関することが示されている(チンら(Chin et al.)、2006、上記;ポラックら(Pollack et al.)、2002、上記)。このPEAを示す腫瘍において、本発明者らは、免疫組織化学によりHER2陽性の強い濃縮を見出し(P<1.0×10−26;カイ二乗検定)、このPEAが物理的17gl2アンプリコンに対応する証拠と一致する(図5A、図5C、黒色の垂直のバー)。初期の(任意抽出の)遺伝子生存率分析により、DMFSに有意に関連するこのPEAの中の6つの遺伝子が同定された(図5A)。PEAを有する腫瘍を有する患者の不在下での生存率(すなわち選択された生存率)の再分析では、DMFSに有意に関連する2つの遺伝子:TCAPおよびERBB2のみが残った。どちらも増殖サインと強く相関しないが、ERBB2は最も強い発現ランキングを有した(図5E)。したがって、TRIAGEにより、ERBB2(受容体チロシン−タンパク質キナーゼ、erbB−2の前駆体をコードする)を、転移促進癌遺伝子および乳癌における治療標的としてのその公知の役割とは無関係に、17q12遺伝子座における発癌ドライバーの「トップ候補」として同定した。
【0107】
LSVDにより同定された第2の大きな主固有ピークを、1.5Mbにわたる領域で22の遺伝子を包含する、第8染色体のp腕にマッピングした。このPEAは、乳癌の10〜25%に発生することが公知の、すでに記載された8p11−12アンプリコンと遺伝子レベルで正確に重複することが見出された。(ガルシア、エム ジェイら(Garci
a,M.J.et al.)(2005)Oncogene24、5235〜5245;ジェルシ−ボイヤーら(Gelsi−Boyer et al.)、(2005)Mol
Cancer Res 3、655〜667;ルテシア、エーら(Letessier,A.et al.)(2006)BMC Cancer 6、245;レイ、エム イーら(Ray,M.E.et al.)(2004)Cancer Res 64、40〜47)。このアンプリコンの発癌ドライバーは、十分に特徴づけられていない。TRIAGEによって、本発明者らは、遠隔転移の危険性の増加に有意に関連するこのアンプリコン上の3つの遺伝子:RAB11FIP1、EIF4EBP1およびWHSC1LIを特定した(図5B)。選択された遺伝子−生存率の相関の分析において、遠隔転移に有意に関連する、わずか2つの遺伝子、RAB11FIP1およびWHSC1LI(ヒストン−リシンN−メチルトランスフェラーゼNSD3をコードする)が残り、RAB11FIP1は、遠隔再発との最も有意な関連および最も高い発現ランクを有した(図5E)。この発見に基づいて、本発明者らは、乳癌の発癌特性に関するRAB11FIP1の機能的影響を調査した。
【0108】
RCP過剰発現は、MCF10A細胞を形質転換させる
RAB11FIP1(本明細書ではRCPとも称する)が、乳腺腫瘍においてタンパク質レベルで増幅するかどうかを確認するために、腫瘍進行の様々な悪性度の細胞系のパネルにおいて、市販の抗体を使用して、ウェスタンブロット分析を実施した。RCPは非腫瘍発生(正常な)乳腺上皮細胞系MCF10Aにおいて最も低いが、乳癌細胞系、CAMA−1、MDA−MB231、MCF7およびT47Dにおいて4〜10倍高いレベルで発現した(図6A)。
【0109】
次に、乳癌におけるRCP増幅の役割を、完全長RCP(MCF10A−RCP)(Genbankアクセス番号:NM_025151)または対照ベクターを安定して過剰発現するMCF10A細胞系を確立することによって調査した。対照細胞は、平板な形態および正常な上皮の単層成長の特徴を示すが(図6B、左のパネル)、RCP過剰発現体は、紡錘様の突起および所々に、形質転換線維芽細胞を連想させる病巣様構造がちりばめられ散在した成長パターンを含む劇的な形態の変化をもたらした(図6B、右のパネル)。一方、対照MCF10A細胞は、平板な形態および正常な上皮においてより特徴的な上皮型島における単層成長を示した。さらに、コンフルエントで、MCF10ARCP過剰発現細胞は、接触阻止の損失を示し、連続的な多層成長を示した。細胞の免疫蛍光染色により、RCP過剰発現が、細胞間の接合部におけるE−カドヘリンの欠失およびβアクチン破壊に関連したことが明らかになった。その上、血清飢餓状態において、対照細胞より、MCF10A−RCP細胞により多くのフィブロネクチンが観察された(図6C)。細胞間接触部位におけるE−カドヘリン(上皮マーカー)の欠失およびフィブロネクチン(間葉細胞マーカー)の発現増加は、両方ともRCP−依存性上皮間葉移行(EMT)と一致する。
【0110】
RCPの作用をさらに評価するために、本発明者らは、血清および追加成長因子に対する、MCF10A細胞の厳しい要求を活用し、それらの増殖を支えた。安定系を、成長因子の存在下および不在下の両方において維持した。対照細胞と比較して、MCF10A−RCP細胞は両方の培養条件において増殖率の有意な増加を示した(図6Dおよび図8A)。最も興味深いことに、追加成長因子(EGF、インスリン)を含まない、1%FBSの最も厳しい培地において、対照系は6日以内に死亡したが、一方MCF10A−RCP細胞は、1カ月を超えて保持でき、成長因子依存性のRCP−仲介の欠失を示した(図6D)。ヨウ化プロピジウム染色による細胞周期分析により、血清および成長因子の飢餓の48時間後、対照細胞の93%がG1期で停止していたのに対してG2M期の細胞はわずか3%であったが、一方、MCF10A−RCP細胞において、細胞の約80%がG1期にあり、細胞の約12%はG2M期にあることが明らかになり、細胞周期進行の調節にお
けるRCPの役割が確認された(図7A)。
【0111】
次に、軟寒天でのコロニー形成アッセイにおいて、MCF10A細胞の足場非依存性成長を推進するRCPの能力を試験した。予想通り、対照細胞は、軟寒天においてコロニー形成ができなかった。しかし、それとは大いに異なり、MCF10A−RCP細胞は4週間後に非常に多くのコロニーを形成し、RCPが形質転換表現型を誘導できることが実証された(図6Eおよび図8B)。その上、本発明者らは、創傷治癒アッセイ(図6F)およびトランスウェルアッセイ(図8D)により実証されるように、RCPの過剰発現がMCF10A細胞の遊走能力を増強していることを観察した。
【0112】
RCPがその癌表現型に影響を与える、1つまたは複数の機序を見抜くために、RCPの発現が、発癌性成長を制御する主要経路の2つであるERKまたはAKTのどちらかを介して成長シグナル伝達を活性化できるかどうかを試験した。MCF10A−RCP細胞は、ERKの強い活性化を示したが、AKTの活性化は示さなかった(図7B)。RCP誘導性Erk活性化がMCF10A−RCP細胞の増殖の有利性と関連があることを試験するために、MCF10A細胞を、活性および不活性の両方のMEK1/2の強力な阻害因子であるU0126で処理した。対照細胞は、U0126濃度の範囲にわたってERKのリン酸化が低いことを示したのに対して、MCF10A−RCP細胞におけるERKのリン酸化状態は、濃度依存性であった(図7C)。30μMのU0126により、ERKのRCP誘導性リン酸化は、ベースラインまで減少し(対照と同じレベル)、一方、RCPのレベルは変化なかった。重要なのは、30uMのU0126により、ERKのリン酸化が低減したMCF10A−RCP細胞が、RCPの発現が高いにもかかわらず、対照レベルまで増殖の減少を示したことである(図7D)。RCP増殖表現型がERKのリン酸化を介してMAPK経路の活性化に依存していることが示唆される。
【0113】
RCPは、多数の乳癌細胞系における腫瘍原性を増強する。
MCF7(エストロゲン受容体陽性、p53−野生型)乳癌細胞におけるRCPの過剰発現は、細胞増殖率、軟寒天におけるコロニー形成活性、および浸潤能を中程度に増加させた(図8A〜C)。MDA−MB−231(MB231;エストロゲン受容体陰性、p53陰性)細胞では、RCPの過剰発現は、コロニー形成活性を増加させたが、細胞増殖率、浸潤および遊走能は増加させなかった(図8A〜D)。さらに腫瘍原性の調節における内因性RCPの役割を検証するために、本発明らは、RNAi技術を用いて内因性RCPの発現を低減した。RCPを、siRNA構築体の一過性形質移入、またはRCPを標的とするshRNAを発現するレンチウィルスベクターの安定形質導入のどちらかによって「ノックダウン」させた。スクランブル配列を有するRNAi構築体を、対照として使用した。RCPを対象とする非依存性かつ非重複性の、2つのsiRNAは、MCF7細胞において、タンパク質レベルの40〜80%を減少させ、細胞増殖を有意に減少させた(図8A、右のパネル)。MCF7細胞におけるRCPのノックダウンもまた、軟寒天におけるコロニー形成を有意に低下させた(図8B)。MB231細胞において、RCPは、コロニー形成の阻害を低下させたが、細胞増殖率は低下させなかった(図8A、B)。両方の細胞系において、RCPノックダウンは、浸潤および遊走を著しく低減した(図8C、D)。さらに、MCF10A細胞における観察と一致して、MCF7細胞におけるRCPの減少もまた、ERKの脱リン酸化をもたらす(図7B)。これらの結果を合わせると、腫瘍細胞における内因性RCPの発現が、増殖、遊走および浸潤の能力の維持にとって重要であり、RCP機能の標的低減は、実行可能な治療戦力を表し得ることを示唆している。
【0114】
RCPのノックダウンはin vivoにおける腫瘍形成を阻害する。
in vivoでの癌細胞におけるRCP低減の作用を調査するために、本発明者らは、RCP(RNAi RCP)に対するshRNAを安定して発現するMCF7およびM
B231細胞の腫瘍形成能と、ヌードマウスにおける対照(RNAi Ctrl)の腫瘍形成能とを比較した。各細胞型を、8匹のマウスの左および右の後脚上部の皮下に埋め込んだ。MCF7は、追加のエストロゲンの存在下で、ヌードマウスにおいて中程度の悪性腫瘍を形成する、エストロゲン依存性の細胞系である。MCF7−RNAi Ctrl(n=8)またはMCF7−RNAi RCP(n=8)を埋め込んだすべてのエストロゲン追加マウスが明らかな腫瘍を発生したが、RCP阻害細胞は、有意により小さいサイズの腫瘍を発生した(P=0.00007;スチューデントt検定)(図9A、図9B、図9C)。一方で、MB231は、ヌードマウスにおいて高度悪性かつ転移性の腫瘍を形成する、エストロゲン非依存性乳癌細胞系である。MB231−RNAi Ctrl細胞を注入されたすべてのマウスは、大型の腫瘍を発生したが、MB231−RNAi RCP細胞を注入された8匹のマウスではそのうちわずか2匹しか腫瘍を発生しなかった。(すなわち、1匹のマウスに2つの腫瘍および別のマウスに1つの腫瘍)(図9D、図9E、図9F)。特に、これらの腫瘍は対照マウスの腫瘍より非常に小型であり、平均重量は、対照のわずか1%であった。これらの腫瘍におけるRCPのノックダウンを、免疫組織化学により確認した(図9G)。
【0115】
MB231細胞におけるRCPのノックダウンがin vitroの増殖に作用しないという事実は、in vivoのRCPの阻害に関して観察された強い抗腫瘍作用が、血清の喪失下での細胞死の誘導に関連し得ることを示唆しており、皮下に埋め込みの腫瘍の血管新生が現れるには数日かかることを示している。したがって、本発明者らは、MB231安定細胞系の細胞死およびアポトーシスへの感受性を、様々な培養条件下で調査した。完全培地において、MB231−RNAi RCPおよび対照系は、どちらの系においてもアポトーシス細胞がほとんどないことにより証明されたように、細胞死において差がないことを示した。しかし、血清飢餓条件下において、対照は、細胞死に対する耐性を維持していたが、RNAi RCP細胞は維持していなかった。有意によりRCPを低減した細胞は、血清飢餓の結果として死亡し、その中でもアポトーシスが約13%検出できた(図9H)。このことは、RCPの減少が、栄養不良環境に対するMB231細胞の脆弱性を増強することを示唆し、これによりin vivoのRCPノックダウンの劇的な抗腫瘍作用が説明できる。興味深いことに、本発明者らは、in vivoのRCP過剰発現に関連する腫瘍原性の増加を観察せず(データ非掲載)、おそらくRCPの内因性レベルにより達成される腫瘍形成に関する最大の病理学的作用を反映する。全体的に見て、結果は、RCPの標的ノックダウンにより、エストロゲン依存性および非依存性の乳房腫瘍の両方のin vivo成長が抑制され、RCPが、治療介入の有益な新規の標的候補として関係があることを示している。
【0116】
考察
原発性腫瘍のゲノムワイドマイクロアレイ分析により、遺伝子発現の固有のパターンにより定義された、新規な臨床的に意義のある腫瘍亜型の発見が可能となった。しかし、より最近、この概念の逆が、腫瘍発生において機能的役割を有する遺伝子亜型を同定しようとするボトムアップ分析戦略を介して調査された。これらの戦略は、遺伝子発現ならびに癌の臨床的特徴およびゲノムの特徴の間の条件関係を解析する。アドラー(Adler)および仲間は、遺伝子連鎖研究(マイクロアレイサインのステップワイズ連鎖分析)を使用して、乳癌における予測的「創傷サイン」の発現を推進する転写機序を明らかにした(アドラー、エー エスおよびチャン、エイチ ワイ(Adler,A.S. and Chang,H.Y.)、(2006)Cell Cycle 5、1148〜1151)。創傷サインに関して腫瘍陽性または腫瘍陰性に有意に関連した、コピー数および遺伝子発現の両方の変化を同定することにより、著者らは、MYC(8q24)およびCSN5(8q13)が機能的に相互作用し、創傷応答遺伝子の転写を制御することを推論し、生化学的に確認した。本研究において、本発明者らは、データ統合および機能的発見上にこの概念を築き、癌進行における機序的役割を有する遺伝子を推定する、相関に基づく新規
な戦略を開発した。
【0117】
TRIAGEと呼ばれる上記の研究は、遺伝子発現の特徴と位置情報(ゲノムの位置)、臨床データ(患者の無遠隔転移生存率)および予測腫瘍生物学(増殖能)とを組み合わせ、過剰発現し、生存率に相関する再発性増幅の遺伝子部位を突き止める。TRIAGEは、設計により機能的癌遺伝子のすべての型を検出するものではないが、ヒト患者において遠隔転移に寄与する悪性過程を推進する、機能的癌遺伝子を解明する可能性を有する。TRIAGEはまた、癌において高い頻度で活性化される癌遺伝子および日常の療法への耐性を伴う癌遺伝子の発見を最適化する。例えば、癌遺伝子に関する本発明者らの調査は、LSVD手順により同定された主固有ピークの大きさにより優先順位を付ける。この大きさは、増幅事象の頻度と正に関連するので、高頻度のアンプリコン(ピークの高さに対応する)は、高い優先順位が得られる。LSVDは、腫瘍発現プロフィールの任意の包括的コレクションに、TRIAGEのロバスト適用を可能にするが、留意すベき固有の限界を有する。LSVDは、所与の増幅事象を有する腫瘍の近似値のみを提供し、実際のアンプリコンの境界の解明は十分ではない。高解像度のアレイ−CGHにより提供されるような、対応するコピー数のデータは、アンプリコン決定の精度および正確さを改善することによって、TRIAGEの性能を実質的に増強できる。
【0118】
TRIAGEの性能に影響を与える追加の因子には、コホートサイズ、患者の追跡の期間および治療の特徴が含まれ、これらのすべては、候補遺伝子の選択に影響を与える。本発明者らの遺伝子−生存率の関連は、本発明者らの最も大きな集団に基づくコホートであるウプサラコホート(n=251例の患者)を使用して計算され、これはさらに、最も長く、最も完全な患者の追跡データを有する。重要なことには、このコレクションの中の腫瘍は、侵攻型の範囲(図2)が、早期乳癌から(小型径、リンパ節陰性、未処理)後期乳癌(大型径、リンパ節陽性、侵攻的に処理された)までの範囲にわたる。したがって、このコホートは、おそらく様々な癌遺伝子により推進されている様々な癌病態生理学を反映している。しかし、患者の混合された治療バックグラウンドは、治療の型に非依存性の生存率関連遺伝子の同定を有利にする。したがって、治療に依存性または2機能の生存率関連癌遺伝子(アンドレ、エフら(Andre,F et al.)(2007)Clin
Cancer Res 13、2061〜2067)は、このコホートでは見分けられないが、他においては同定される可能性がある。
【0119】
本発明者らの統合乳房腫瘍データセットのLSVD分析は、8p11−12の頻繁に増幅される領域に位置する遺伝子群に集中する大きな固有ピークの発見をもたらした。この遺伝子座の増幅は、乳房腫瘍症例の約10〜25%において観察され(アドナン、ジェイら(Adnane,J.et al.)(1991)Oncogene 6、659〜663;ガルシア、エム ジェイら(Garcia,M.J.et al.)(2005)Oncogene 24、5235〜5245;ルテシアら(Letessier et
al.)2006上記、テイレット、シーら(Theillet,C.et al.)(1993)Genes Chromosomes Cancer 7、219〜226)、多数の非依存性研究において、患者の生存率不良および遠隔転移の短い間隔に関連していた(チンら(Chin et al.)、2006、上記、;キュニー、エムら(Cuny,M.et al.)(2000)Cancer Res 60、1077〜1083;ジェルシ−ボイヤー、ブイら(Gelsi−Boyer,V.et al.)(2005)Mol Cancer Res 3、655〜667;ルテシアら(Letessier et al.)2006、上記)。近年、このアンプリコンは、原発性乳房腫瘍および細胞系に関する、いくつかの集中的な機能的ゲノム研究の焦点であった(ガルシアら(Garcia et al.)、2005、上記;ジェルシ−ボイヤーら(Gelsi−Boyer et al.)、2005、上記;レイら(Ray et al.)、2004、上記)。染色体8pに特異的な高解析度BACマイクロアレイを使用して、
ジェルシ−ボイヤー(Gelsi−Boyer)および仲間は(2005、上記)、8p11−12アンプリコンが、実際は境界の明確なA1、A2、A3およびA4と呼ばれる重複が最小の4つのサブアンプリコンを決定した。この研究および他の研究の著者は、遺伝子増幅とmRNAの過剰発現との間の強い相関に基づき、この領域の数多くの候補癌遺伝子を提示してきた。(ガルシアら(Garcia et al.)、2005、上記;ジェルシ−ボイヤーら(Gelsi−Boyer et al.)、2005、上記;レイら(Ray et al.)、2004、上記)。累積的に、これらの研究は、ZNF703(FLJ14299)、ERLIN2(SPFH2)、PROSC、BRF2、RAB11FIP1(RCP)およびLSM1を、増幅と過剰発現との間に最も強い相関を有するとして同定し、LSM1以外のすべては、1−Mb「最小アンプリコン」に局在し(ガルシアら(Garcia et al.)、2005、上記)、A1サブアンプリコンと正確に重複する(ジェルシ−ボイヤーら(Gelsi−Boyer et al.)、2005、上記)。特に、本発明者らの研究においてLSVDにより同定された遺伝子は、特にAlおよびA2サブアンプリコンからなり、したがって、これら早期の研究により記載された、すべての「最も優れた候補」癌遺伝子を含む。
【0120】
TRIAGEの8p11−12への適用は、患者における播種性転移を潜在的に推進する、このアンプリコンにおける最も強い候補癌遺伝子として、RAB11FIP1(RCP)の同定をもたらした。この領域内の他の候補癌遺伝子の中でも、乳房上皮細胞において、いくつかの形質転換特性を有することがすでに示されている。(シュトライヒャー、ケイ エルら(Streicher,K.L.et al.)、(2007)Oncogene 26、2104〜2114;ヤン、ゼット キューら(Yang,Z.Q.et
al.)、(2006)Cancer Res 66、11632〜11643)、LSM1、BAG4およびC8orf4(TCI)はすべて、A2およびA3のサブアンプリコン上に主に存在する(ジェルシ−ボイヤーら(Gelsi−Boyer et al.)、2005、上記)。反対に、RAB11FIP1(RCP)は、1−Mb最小アンプリコン上のA1アンプリコンのコアのより近くに位置し、(ガルシアら(Garcia
et al.)、2005、上記)、癌遺伝子として機能的に特徴づけられたこの領域において第1である。
【0121】
それらの新規な研究を使用して、本発明者らはRCPの発現に連結した広範囲の癌促進作用を発見した。非癌性の乳房上皮細胞において、RCPの過剰発現は、悪性の形質転換をもたらし、細胞の運動および遊走を増加させる。乳癌細胞系において、RNA阻害による内因性RCPの低減は、増殖、足場非依存性成長、浸潤およびアポトーシス耐性を減少させ、ヌードマウスにおいて腫瘍形成を減退させた。生理的に、RCPは、主にRab11と相互作用し、管状エンドソームにおいてタンパク質選別を制御すると考えられている(ペデンら(Peden et al.)、2004、上記)。Rab11−RCP複合体は、トランスフェリン受容体をリソソームから離し(分解経路)エンドソームの再循環(再循環経路)へ導き、その結果膜結合受容体のレベルの相対的増加を促進することが示されている(ペデンら(Peden et al.)、2004、上記)。推論ではあるが、RCPの形質転換作用が再循環経路へのこの推移により仲介され得、インテグリンなどの特定の膜タンパク質または癌遺伝子の役割を有する成長因子受容体の膜への局在(およびこのような機能)を引き延ばすことは妥当と思われる。RCP発現とERKリン酸化の間の、本発明者らが観察した強い相関は、RCPと成長因子受容体仲介MAPK活性化との間の特定の連結を示し得るので、この可能性を反映し得る。上記のデータは、RAB11FIP1が、8p11−12アンプリコンの再発性の選択の陰の鍵となる推進力を表し得、この増幅領域およびRAB11FIP1それ自体の発現に関連する、乳癌の転移拡散に機能的に寄与し得ることを示している。このアンプリコンが、非小細胞肺癌(バルサラ、ビー アールら(Balsara,B.R.et al.)、(1997)Cancer Res 57、2116〜2120)、結腸直腸癌(ナカオ、ケーら(Nakao
,K.et al.)、(2004)Carcinogenesis 25、1345〜1357)および膀胱の腫瘍(サイモン、アールら(Simon,R.et al.)、(2001)Cancer Res 61、4514〜4519)およびファローピウス管(スナイデルス、エー エムら(Snijders,A.M.et al.)、(2003)Oncogene 22、4281〜4286)を含む他の固体腫瘍に頻繁に観察されていることは、RAB11FIP1が単独または近隣の癌遺伝子との相乗効果において作用する、癌の進行へのありふれた経路を表し得ることを示唆している。
【0122】
この明細書における、公知文献のリストまたは考察は、必ずしもかかる公知文献が技術水準の一部である、または一般的常識であると認めるものとしてとらえるべきではない。
本発明は、本明細書中に広範にかつ一般的に記載してある。包括的開示の範囲内に入る、より狭い種および亜属の分類もまた、それぞれ本発明を形成する。これは、削除された材料を本明細書中に具体的に列挙するかどうかにかかわらず、条件付き、または任意の対象を属から除去する否定的限定を伴う、本発明の包括的記載を含む。
【0123】
当業者は、本発明が、目標を達成すること、内在する目的および利点と同様に本明細書で言及した目的および利点を獲得するのに適していることを、容易に理解するであろう。さらに、様々な置き換えおよび変形が、本発明の範囲および精神から逸脱することなく、本明細書に開示した発明に対して実施できることは、当業者には容易に理解されるであろう。本明細書に記載された分子複合体および方法、手順、処理、分子、本明細書に記載された特定の化合物は、好ましい実施形態を例示する目下の典型であり、本発明の範囲を限定する意図のものではない。当業者は、特許請求の範囲により定義される本発明の精神に包含される、その変更および他の使用が思い浮かぶであろう。
【0124】
本明細書において適切に例示的に記載された発明は、任意の、1つまたは複数の要素、本明細書に具体的に開示されなかった、1つまたは複数の限定が存在しない状況下においても実施できる。したがって、例えば、本明細書における各事例において、「有する」、「本質的に〜からなる」および「からなる」などの任意の用語は、広い意味で、限定することなく理解すべきであり、直接参照される列挙された成分だけに限定されず、他の明記されていない成分および要素もまた含む。このように、それらは相互に交換することもできる。さらに、用いた用語および表現は説明の用語として使用し、限定するものではなく、示し、記載した特徴の任意の同等物またはそれらの一部を排除したような用語または表現の使用に意図はなく、しかし、様々な変形が本発明の特許請求の範囲内で可能であることが認識される。
【0125】
他の実施形態は以下の特許請求の範囲内である。さらに、本発明の特徴または態様が、Markush群に関して記載されている場合、当業者には、本発明がMarkush群の任意の個々のメンバーまたは該メンバーのサブグループに関しても記載されていることが理解されるであろう。
【図3−1】
【図3−2】
【図3−3】
【図3−4】
【図10A】
【図10B】
【技術分野】
【0001】
関連出願の相互参照
本出願は、米国特許商標局に2007年4月13日に出願され、シリアル番号60/911,573が割り当てられた「Preventing,inhibiting,arresting or reversing tumorigenesis by modulation of a rab binding protein and related uses of a rab binding protein」の出願を参照し、かつその優先権を主張するものである。2007年4月13日に出願された前記出願の内容は、PCTの規則4.18に準じて、本願明細書に含まれないPCTの規則20.5(a)で言及された記載事項、特許請求の範囲または図面の任意の要素または部分の援用を含め、すべての目的で本願明細書に援用する。
【0002】
本発明は、腫瘍発生を制御する方法および腫瘍発生のリスクを診断する方法に関する。これらの方法は、Rab結合タンパク質の量の変化および/または活性の変化を利用するものである。本発明は、Rab結合タンパク質と複合体を形成することが可能な化合物、およびRab結合タンパク質の量、発現および/または活性を調節することが可能な化合物を同定する方法にも関する。本発明は、新生物が細胞内カルシウムレベルの変化に感受性であるかどうかを決定する方法も提供する。
【背景技術】
【0003】
癌は世界的に主たる死因であり、先進国では2番目の主な死因であり、さらに例えばオーストラリア、日本、韓国、シンガポールにおいて、ならびに英国およびスペインの男性人口では、第1番目の死因である。毎年癌を発症する人の数は増大している。それにもかかわらず、癌療法は最近30年間で癌死亡率を低下させることができていない。
【0004】
組織浸潤および転移をもたらす自律的細胞増殖は、すべての悪性新生物の顕著な特徴である。癌は必ずしも加速的速度の細胞増殖の結果としてのみ生じるわけではなく、癌は、一方では細胞周期進行(細胞分裂)および細胞増殖(細胞増大)の速度と、他方ではプログラムされた細胞死(アポトーシス)の速度との間の不均衡の結果である。
【0005】
腫瘍発生および転移増殖の発癌誘導因子の同一性を解明することは、癌の病理生物学を理解するためだけでなく、患者を治療するためにも重要である。というのは、このような遺伝子は、治療的有効性(ヘルセプチン、アバスチン、グリーベック)のためにますます活用されているからである。ヒトの癌では、ゲノム増幅、配列突然変異またはプロモーターの低メチル化から生じる異常な過剰発現によって癌遺伝子が活性化されることが多い。アンプリコンとして知られる、癌において増幅されることが多い染色体上の遺伝子座は、1つまたは複数の癌遺伝子によって誘導されると考えられ、多くの反復アンプリコンが観察されているが、それらの癌遺伝子決定因子は依然として大部分が知られていない。これらの遺伝子座は、コピー数増大の程度および頻度を評価する比較ゲノムハイブリダイゼーション(CGH)などの細胞遺伝学的技法によって伝統的に調べられており、この戦略は考慮対象の多くの遺伝子候補を与えるが、遺伝子リストが広範囲であり、かつ機能情報がわずかであるため、癌遺伝子の発見に対する進展は限られている。それよりも、大抵は異なる実験背景における遺伝子機能の累積的観察結果から導かれる仮説の生成によって決まるため、癌遺伝子の発見は緩慢で実験プロセスとなることが多い。
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明の目的は、腫瘍発生の予防を含めた腫瘍発生を制御する手段、および発癌性になる素因を有する細胞を同定する手段を提供することである。
【課題を解決するための手段】
【0007】
第1の態様において、本発明は、腫瘍発生を予防、阻害、停止または逆行させることを含む、腫瘍発生を制御する方法を提供する。本発明は、転移性癌などの癌を含む、新生芽腫の治療、予防および診断の方法も提供する。
【0008】
1つのこのような方法は、Rab結合タンパク質、特にRab11FIPタンパク質(Rab11ファミリー相互作用タンパク質)の量または活性を調節することを含む。
本発明のさらなる方法は、発癌性になる素因を有する1つまたは複数の細胞を同定することに関する。方法は、1つまたは複数の細胞において、Rab結合タンパク質、特にRab11FIPタンパク質の量の変化、細胞内局在の変化、または活性の変化を検出することを含む。
【0009】
本発明は、新生物を発症するリスクを診断する方法も提供する。方法は、Rab結合タンパク質、特にRab11FIPタンパク質の発現レベル、活性レベルおよび細胞内局在の少なくとも1つを決定することを含む。
【0010】
本出願は、Rab結合タンパク質、特にRab11FIPタンパク質の活性を調節することができ、したがって例えば抗癌剤として作用することができる、適切な化合物の同定にも関する。
【0011】
これに関し、本発明は、Rab結合タンパク質、特にRab11FIPタンパク質と複合体を形成することが可能な化合物を同定するin vitro方法を提供する。方法は、各複合体を形成する構成要素を互いに接触させることを含む。
【0012】
さらなる態様において、本発明は、in vivoでRab結合タンパク質、特にRab11FIPタンパク質の活性を調節することが可能な化合物を同定するための方法を提供する。方法は、Rab結合タンパク質またはその機能断片を発現する微生物を提供することを含む。方法は、微生物に化合物を加えることをさらに含む。方法は、Rab結合タンパク質の細胞表現型または活性の変化をモニターすることも含む。
【0013】
他の態様において、本発明は、Rab結合タンパク質、特にRab11FIPタンパク質の量または活性を調節することが可能な化合物を同定する方法を提供する。
1つのこのような方法は、宿主生物を提供することを含む。宿主生物は、癌細胞を収容し増殖させることができる。方法は癌細胞を宿主生物に導入することも含む。さらに方法は、Rab結合タンパク質の発現および/または活性を調節し得ると考えられる化合物を投与することを含む。方法は、宿主生物中の任意の腫瘍(複数の腫瘍を含む)の増殖をモニターすることをさらに含む。
【0014】
さらなるこのような方法は、宿主生物を提供することも含む。宿主生物は癌細胞を収容し増殖させることができる。方法は癌細胞を宿主生物に導入することも含む。癌細胞は、Rab結合タンパク質の量または活性を調節し得ると考えられる化合物を含む。方法は、宿主生物中の任意の腫瘍(複数の腫瘍を含む)の増殖をモニターすることをさらに含む。
【0015】
他のさらなる態様において本発明は、新生物が細胞内カルシウムレベルの変化に感受性であるかどうかを決定する方法を提供する。方法は、Rab結合タンパク質の発現および/または活性を決定することを含む。Rab結合タンパク質の発現の増大または活性の増大は、新生物が細胞内カルシウムレベルの変化に感受性であることを示す。
【0016】
本発明は、限定されない実施例および添付の図面と併せて検討した場合、詳細な説明に関してさらに理解されるであろう。
【図面の簡単な説明】
【0017】
【図1】TRIAGE方法論の複数工程の略図。
【図2】統合されたマイクロアレイのデータセットを構築するために使用された乳癌コホートの臨床的特徴を示す表。
【図3】乳癌の癌ゲノムのLSVD分析の図。固有値(y軸)を、各染色体(x軸)の長さに沿ってプロットする。最も高いスコアを得た主固有ピークのゲノムの位置を強調してある。
【図4A】図4は、増殖サインおよび無遠隔転移生存率(DMFS)を示す。図4Aは、ウプサラ(Uppsala)コホート(n=251)におけるDMFS不良(P<0.05;尤度比検定)と相関した全プローブセット(n=2,377)が、階層的にクラスタ化されたことを示す図。優性遺伝子のクラスタは、増殖サインを反映する。増殖サインの中核遺伝子を、クラスタ化の順番に以下の実施例において名付ける。クラスタグラムの下に、転移した腫瘍(白マル)および転移しなかった腫瘍(黒マル)を時間の関数として示す。
【図4B】ウプサラ(Uppsala)コホート(n=251)におけるDMFS不良(P<0.05;尤度比検定)と相関した全プローブセット(n=2,377)が、階層的にクラスタ化されたことを示す図。優性遺伝子のクラスタは、増殖サインを反映する。増殖サインの中核遺伝子を、クラスタ化の順番に以下の実施例において名付ける。クラスタグラムの下に、移した腫瘍(白マル)および転移しなかった腫瘍(黒マル)を時間の関数として示す。
【図4C】DMFS不良(n=2,377)に関連するプローブのセットを、P値に従ってビン分割し、>0.4の増殖サインに相関するプローブセットの分画を白で示し、各ビンのパーセントを示す図。全DMFS関連プローブセットのおよそ35%が、>0.4の増殖サインと相関する。
【図5A】図5は、17q12および8p11のアンプリコンのTRIAGE分析を示す。図5Aは、17q12における推定発現アンプリコン(PEA)の発現ヒートマップを示す図。腫瘍をカラムに配置し、LSVDにより同定した遺伝子を列の中で体系化する(染色体に沿った順番で)。腫瘍を、固有重量の絶対値によりランク付けする(ヒートマップの下に記載)。平均発現より上を白で示す。TRIAGEの第3工程において、生存率に有意に関連する遺伝子を右に示す(図1を参照)。白い文字で示された遺伝子(TCAPおよびERBB2)は、TRIAGEによる、最も有力な癌遺伝子候補を示す。
【図5B】8p11における推定発現アンプリコン(PEA)の発現ヒートマップを示す図。腫瘍をカラムに配置し、LSVDにより同定した遺伝子を列の中で体系化する(染色体に沿った順番で)。腫瘍を、固有重量の絶対値によりランク付けする(ヒートマップの下に記載)。平均発現より上を白で示す。TRIAGEの第3工程において、生存率に有意に関連する遺伝子を右に示す(図1を参照)。白い文字で示された遺伝子(RAB11FIP1およびWHSC1L1)は、TRIAGEにより、最も有力な癌遺伝子候補を示す。
【図5C】図5Aと同じデータを、平均発現より下を白く示して表した図。
【図5D】図5Bと同じデータを、平均発現より下を白く示して表した図。
【図5E】得られたデータを要約した図。NSは、「有意差なし」を示唆する。アスタリスクは、増殖サインとの相関が>0.4であることを示す。パネルAのヒートマップの上の黒色の垂直なバーは、免疫組織化学によるERBB2(HER2/neu)陽性を表す。
【図6A】図6は、MCF10A細胞においてRCPの過剰発現が癌表現型を誘導することを示す。図6Aは、MCF10Aおよび乳癌の細胞系において、ウェスタンブロットにより評価した内因性RCPのレベルを示す図。
【図6B】RCP過剰発現の、細胞形態学における影響の図(スケールバー、100μm)。
【図6C】免疫蛍光法による、細胞間接合部のE−カドヘリンの局在化(上)、F−アクチンの組織化(中)および血清飢餓下のフィブロネクチンの発現(下)の、RCP過剰発現の影響の図(スケールバー、50μm)。
【図6D】WSTアッセイによるMCF10A細胞の成長因子非依存性増殖に関するRCP過剰発現の影響の図(平均+/−標準偏差)。
【図6E】軟寒天における、足場非依存性コロニー形成に関するRCP過剰発現の影響の図(倍率、×5)。
【図6F】創傷治癒(擦過傷)アッセイによる細胞遊走に関するRCP過剰発現の影響の図(スケールバー、100μm)。
【図7A】図7は、RCPの作用機序を示している。図7Aは、フローサイトメトリーによる、血清飢餓MCF10A−RCP細胞および対照細胞の細胞周期分析の図。
【図7B】ウェスタンブロットにより示された、RCP過剰発現(MCF10A細胞)に反応したErkおよびAktのリン酸化状態およびRCP(MCF7細胞)のRNAi仲介ノックダウンの図。
【図7C】MAPK阻害剤であるU0126によるRCP仲介Erkリン酸化阻害の図(上のパネル)およびRCP増殖表現型のU0126仲介阻害の図(下のパネル、平均+/−標準偏差)。
【図8A】図8は、乳癌細胞系におけるRCPの癌遺伝子表現型を示す。図8Aは、WSTアッセイによる、細胞増殖に関する、RCP過剰発現およびRCPのRNAi仲介ノックダウンの影響の図。左のパネル、形質移入6日後の結果。中のパネル、MCF10A細胞におけるRCP過剰発現の増殖時間経過。右のパネル、2種の異なるRNAi RCP構築体および(スクランブル配列の)対照を使用する、MCF7細胞におけるRCP阻害の増殖時間経過。
【図8B】軟寒天における、足場非依存性コロニー形成に関するRCPの過剰発現および阻害の影響の図。
【図8C】マトリゲルを介した、細胞浸潤に関するRCPの過剰発現および阻害の影響の図。
【図8D】細胞遊走に関するRCPの過剰発現および阻害の影響の図。すべてのエラーバーは、平均+/−標準偏差*、p<0.05、**、p<0.01および***p<0.001、スチューデントt検定(両側)から計算した。
【図9A】図9は、RCPが、ヌードマウスにおいて腫瘍形成を制御することを示している。図9Aは、MCF7細胞を使用した、in vivoの腫瘍成長に関するRCP阻害の影響の図。時間の関数としてプロットした、平均腫瘍容量を表す(平均+/−s.e.m.)。
【図9B】MCF7細胞を使用した、in vivoの腫瘍成長に関するRCP阻害の影響の図。5週(MCF7)および7週(MB231)における重量をプロットした腫瘍を表す。平均重量は実線により示した。
【図9C】MCF7細胞を使用した、in vivoの腫瘍成長に関するRCP阻害の影響の図。切除された全腫瘍を示す。
【図9D】MB231細胞を使用した、in vivoの腫瘍成長に関するRCP阻害の影響の図。時間の関数としてプロットした、平均腫瘍容量を表す(平均+/−s.e.m.)。
【図9E】MB231細胞を使用した、in vivoの腫瘍成長に関するRCP阻害の影響の図。5週(MCF7)および7週(MB231)における重量をプロットした腫瘍を表す。平均重量は実線により示した。
【図9F】MB231細胞を使用した、in vivoの腫瘍成長に関するRCP阻害の影響の図。切除された全腫瘍を示す。
【図9G】腫瘍試料中のRCPの免疫組織化学的染色を表す図(スケールバー、100μm)。
【図9H】TUNEL染色による、血清飢餓MB231細胞のRCP阻害のアポトーシス効果を示す図(スケールバー、100μm)。
【図10】増殖サインの由来を表す図。細胞周期制御遺伝子(ヒーラ細胞において)として、ウィットフィールドら(Whitfield et al.)((2002)Mol Biol Cell、13、1977−2000)により同定された1,134IMAGEクローンを、2,113 Affymetrixプローブセット(U133AおよびU133B)に対応させて、884の UniGene ID(build #187)にマッピングした。これらのプローブセットを使用して、ウプサラ腫瘍コホート(n=251)の発現プロフィールを、階層的にクラスタ化し(ピアソンの相関、平均連結法)、固有クラスタグラムのヒートマップにおいて視覚化した(左側)。図10Aにおける白いシグナルは、平均発現より上を示し、図10Bにおける白いシグナルは、平均発現より下を表す。明るさのそれぞれの程度は、発現の大きさを反映している。(紙面に限りがあるため、遺伝子の系統樹は省略した)。暗いバーは、ヒト乳癌の増殖に関連するとして、ウィットフィールド(Whitfield)およびペロー(Perou)によってすでに同定されたG2/Mにおいて大部分が発現する遺伝子のクラスタを示す。増殖クラスタの67の遺伝子に対応する78のプローブのセットは、>0.8の平均相関(白いバーで示されている)を有し、正しい階層的クラスタ化が示されている。これらのプローブセット(右側)は、コア増殖サイン遺伝子として示されており、全プローブセットと、増殖サインとのピアソン相関を導くために平均した(すなわち、TRIAGEの第6の工程)。
【発明を実施するための形態】
【0018】
本発明は、Rab結合タンパク質、および特にRab11FIPタンパク質(Rab11ファミリー相互作用タンパク質)が、腫瘍、および特に癌を含む、新生物の形成を促進することができるという、驚くべき発見に基づく。
【0019】
本発明者らは、反復増幅のゲノム部位の遺伝子の病態生理学的関与は、ゲノム位置、遺伝子発現および臨床データの統合および三角測量によって高い解明能力で推測することができ、三角測量は遺伝子発現と腫瘍攻撃性の臨床測定の間の統計学的関係を引き出すことができると仮定した。この目的のために、多段階のデータマイニング戦略を開発し、癌遺伝子としてのRab11FIPタンパク質の発見に至った。本発明の方法はこの発見に基づく。
【0020】
本発明者らのこの発見は、RAB1lFIPのタンパク質をコードする遺伝子の局在と、癌の確率の増大との関係に関して報告されたデータの説明も示す。例えば、ヒトRAB11FIP1遺伝子(Ensemblデータベースのアクセッション番号ENSG00000156675)は、第8染色体上、p12領域中、位置37、835、628〜37、876、161に見られる。同様に、マカクザルRAB11FIP1遺伝子(Ensemblデータベースのアクセッション番号ENSMMUG00000003213)は、第8染色体上、位置38、309、752〜38、349、824に見られる。イヌRAB11FIP1遺伝子(Ensemblデータベースのアクセッション番号ENSCAFG00000006176)は、第16染色体上、位置30、456、385〜30、484、441に見られる。ニワトリRAB11FIP1遺伝子(Ensemblデータベースのアクセッション番号ENSGALG00000003129)は、第22染色体上、位置2、258、738〜2、267、830に見られる。いくつかの分析済みの乳癌において、ヒト染色体領域8p12が増幅されることが以前に示されている(レテジアー、エーら(Letessier,A.,et al.)、(2006)BMC Cancer(2006)6、1〜13ページ、http://www.biomedcentral.com/content/pdf/1471−2407−6−245.pdfで入
手可能)。第8染色体の短いアームの消失、増大、増幅および再配列も、1組の乳、結腸および膵臓癌細胞系中で見られることが示されている(ポール、ジェイ シー エムら(Pole,J.C.M.,et al.)、(2006)Oncogene 25、5693〜5706ページ)。外科的切除を施された患者における、ヒト染色体領域8p12の変化と肺癌の間の相関関係が観察されてきている(キム、ティー−エムら(Kim,T.−M.,et al.)、(2005)Clinical Cancer Research、11、23、8235〜8242ページ)。比較ゲノムハイブリダイゼーションを使用して、遺伝子の遺伝子座における増幅および欠失が、膵臓および肺癌亜型において起こることも示された(トノン、ジーら(Tonon,G.,et al.)、(2005)Proc Natl Acad Sci USA、102、27、9625〜9630ページ)。同じ遺伝子座におけるコピー数の消失は、複数の患者から得た結腸癌試料においても観察された(ヤギヌマ、ワイら(Yaginuma,Y.,et al.)、(2006)Journal of International Medical Research、34、4、390〜396ページ)。
【0021】
Rab11ファミリー相互作用タンパク質(FIP)タンパク質は、C末端またはその近辺に位置する相同的Rab11結合ドメインを共有する(例えば、ヘイルズ、シー エムら(Hales,C.M.,et al.)、J.Biol.Chem.(2001)276、42、39067〜39075ページを参照)。それらのドメイン構造に基づいて、Rab11FIPタンパク質は、クラスI(FIP5/Rip11、FIP1およびFIP2)およびクラスII(FIP3/エフェリン、およびFIP4)の2つのサブファミリーに分けられている。したがって、本明細書で使用する用語「Rab11FIPタンパク質」は、Rab11ファミリー相互作用タンパク質のファミリーの任意のメンバー、特にRab11FIP1、Rab11FIP2、Rab11FIP3(エフェリン)、Rab11FIP4、Rab11FIP5(Rip11)、およびそれらのアイソフォームを指す。
【0022】
Rab11FIP1タンパク質の例には、SwissProtアクセッション番号Q05A58(Q05A58_マウス)、Q8K028(Q8K028_マウス)、Q6WKZ4(RFIP1_ヒト)、Q9D620(RFIP1_マウス)、Q3B7T9(RFIP1_ラット)、Q6KAN5(Q6KAN5_マウス)、Q307T1(Q307T1_ヒト)、Q28BL8(Q28BL8_XENTR)、(ニシツメガエル由来)、A4IHL6(A4IHL6_XENTR)(ニシツメガエル由来)の、例えばSwissProtアクセッション番号ENSG00000156675(ヒト)、ENSMUSG00000031488(マウス)、ENSPTRG00000020164(チンパンジー)、ENSPPYG00000018515(オランウータン)、ENSMMUG00000003213(アカゲザル)、ENSCAFG00000006176(イヌ)、ENSGALG00000003129(ニワトリ)、ENSMODG00000010813(ハイイロジネズミオポッサム)、ENSMLUG00000007472(ホオヒゲコウモリ属)、ENSMICG00000010174(ハイイロショウネズミキツネザル)、ENSOCUG00000004502(ウサギ)、ENSCPOG00000008298(モルモット)、ENSETEG00000007044(マダガスカルモヘアハリネズミ)、ENSECAG00000016134(ウマ)、ENSOANG00000003030(カモノハシ)、ENSOPRG00000010860(アメリカナキウサギ)、ENSSTOG00000005565(ジュウサンセンジリス)、ENSDNOG00000008299(ココノオビアルマジロ)、ENSLAFG00000004250(アフリカサバンナゾウ)、ENSOGAG00000005737(コビトガラゴ)の遺伝子、例えばSwissProtアクセッション番号DQ236342(ヒト)、BC125400(マウス)、BC132094(マウス)、BC034201(マウス)、BC077720(ヒト)、CX078607(アカゲザル)、D
R005339(ヒト)、BC135580(ニシツメガエル)の核酸配列を有し、SwissProtアクセッション番号AAH34201(マウス)、AAI25401(マウス)、AAI32095(マウス)、ABB43161(ヒト)、AAH77720(ヒト)、EAW63349(ヒト)、EAW63347(ヒト)、EAW63348(ヒト)、AAI35581(ニシツメガエル)およびCAJ81459(ニシツメガエル)のコード配列を含むタンパク質があるが、これらだけには限られない。
【0023】
Rab11FIP2タンパク質の例には、SwissProtアクセッション番号Q5HZI0(Q5HZI0_マウス)、Q7L804(RFIP2_ヒト)、Q3U366(Q3U366_マウス)の、例えばSwissProtアクセッション番号ENSMUSG00000040022(マウス)、ENSG00000107560(ヒト)、ENSGALG00000009304(ニワトリ)、ENSPTRG00000002982(チンパンジー)、ENSMODG00000009385(ハイイロジネズミオポッサム)、ENSXETG00000009541(ニシツメガエル)、ENSMMUG00000021805(アカゲザル)、ENSCAFG00000011991(イヌ)、ENSPPYG00000002692(オランウータン)、ENSEEUG00000006651(ナミハリネズミ)、ENSMLUG00000015200(ホオヒゲコウモリ属)、ENSMICG00000001976(ハイイロショウネズミキツネザル)、ENSOCUG00000009241(ウサギ)、ENSCPOG00000000453(モルモット)、ENSETEG00000019294(マダガスカルモヘアハリネズミ)、ENSECAG00000019791(ウマ)、ENSSARG00000001794(ヨーロッパトガリネズミ)、IGI00014376(Integr8データベース、ヒト)、IGI00031145(Integr8データベース、マウス)およびIGI02699308(Integr8データベース、ウシ)の遺伝子、および例えばSwissProtアクセッション番号BC089010(マウス)、BC075073(ヒト)、BC075074(ヒト)およびDN994254(ヒト)の核酸配列を有し、SwissProtアクセッション番号AAH89010(マウス)、EAW49423(ヒト)、AAH75073(ヒト)およびAAH75074(ヒト)のコード配列を含むタンパク質があるが、これらだけには限られない。
【0024】
Rab11FIP3タンパク質の例には、SwissProtアクセッション番号Q75154(RFIP3_ヒト)、Q8JZT3(Q8JZT3_マウス)、A2IDE0(A2IDE0_ヒト)、A2A2G1(A2A2G1_ヒト)、A2IDD9(A2IDD9_ヒト)、A2A2G0(A2A2G0_ヒト)、およびQ8CHD8(Q8CHD8_マウス)の、例えばSwissProtアクセッション番号ENSG00000090565(ヒト)、ENSMUSG00000037098(マウス)、ENSPTRG00000007543(チンパンジー)、ENSMMUG00000000613、IGI00010332(Integr8データベース、ヒト)およびIGI00087092(Integr8データベース、ニワトリ)、IGI00026472(Integr8データベース、マウス)の遺伝子、例えばSwissProtアクセッション番号AL049542(ヒト)、Z98882(ヒト)、BC023364(マウス)、BC037132(マウス)、BC051360(ヒト)、およびAL023881(ヒト)の核酸配列を有し、SwissProtアクセッション番号AAH23364(マウス)、AAH37132(マウス)、CAI95788(マウス)、CAI95591(ヒト)、CAI95593(ヒト)、CAM26382(ヒト)、CAM26383(ヒト)、CAQ09643(ヒト)、CAQ11003(ヒト)、CAM28374(ヒト)、AAH51360(ヒト)、EAW85805(ヒト)、EAW85806(ヒト)、EAW85807(ヒト)、EAW85808(ヒト)、EAW85809(ヒト)、およびEAW85810(ヒト)のコード配列を含むタンパク質があるが、これらだけには限られない。
【0025】
Rab11FIP4タンパク質の例には、SwissProtアクセッション番号Q6DGI2(Q6DGI2_BRARE)、Q86YS3(RFIP4_ヒト)、Q8BQP8(RFIP4_マウス)、Q5SYH4(Q5SYH4_マウス)、Q3LGD4(Q3LGD4_BRARE)、Q3U4R6(Q3U4R6_マウス)およびQ3LGD3(Q3LGD3_BRARE)の、例えばSwissProtアクセッション番号ENSDARG00000053855(ゼブラフィッシュ)、ENSG00000131242(ヒト)、ENSMUSG00000017639(マウス)、ENSMODG00000019209(ハイイロジネズミオポッサム)、ENSPTRG00000008974(チンパンジー)、ENSMMUG00000008127(アカゲザル)、ENSOCUG00000007443(ウサギ)、ENSMLUG00000016407(ホオヒゲコウモリ属)、ENSCPOG00000011634(モルモット)、IGI00013025(Integr8データベース、ヒト)、IGI00029750(Integr8データベース、マウス)、IGI02693035(Integr8データベース、ゼブラフィッシュ)およびIGI00051410(Integr8データベース、ゼブラフィッシュ)の遺伝子、例えばSwissProtアクセッション番号BC076363(ゼブラフィッシュ)、BC093914(ヒト)、BC101517(ヒト)、AL591174(マウス)、BC135989(ニシツメガエル)およびDR000701(ヒト)の核酸配列を有し、SwissProtアクセッション番号AAH76363(ゼブラフィッシュ)、AAI35990(ニシツメガエル)、CAI24833(マウス)、CAI24836(マウス)、EAW80265(ヒト)、AAI01518(ヒト)およびAAH93914(ヒト)のコード配列を含むタンパク質があるが、これらだけには限られない。
【0026】
Rab11FIP5タンパク質の例には、SwissProtアクセッション番号Q6ZQ33(Q6ZQ33_マウス)、Q9BXF6(RFIP5_ヒト)、Q8R361(RFIP5_マウス)およびQ3UM85_マウスの、例えばSwissProtアクセッション番号ENSG00000135631(ヒト)、ENSMUSG00000051343(マウス)、ENSGALG00000016092(ニワトリ)、ENSBTAG00000006162(ウシ)、ENSXETG00000011781(ニシツメガエル)、ENSOCUG00000000840(ウサギ)、NSPTRG00000012057(チンパンジー)、ENSMMUG00000002301(アカゲザル)、ENSOGAG00000009150(コビトガラゴ)、ENSECAG00000007084(ウマ)、IGI00035642(Integr8データベース、マウス)およびIGI00006075(Integr8データベース、ヒト)の遺伝子、例えばSwissProtアクセッション番号DQ890991(ヒトクローンの合成構築体)、DQ894168(ヒトクローンの合成構築体)、BC035013(ヒト)、BC026473(マウス)、BC044833(マウス)、BC051063(マウス)、DR770942(ベンガルザル)の核酸配列を有し、SwissProtアクセッション番号AAH26473(マウス)、AAH44833(マウス)、AAH51063(マウス)、ABM81917(合成構築体)、ABM85094(合成構築体)、AAH35013(ヒト)、EAW99750(ヒト)、EAW99749(ヒト)およびEAW99751(ヒト)のコード配列を含むタンパク質があるが、これらだけには限られない。
【0027】
Rabファミリーの低分子GTPアーゼタンパク質は、細胞内膜輸送、シグナル変換、受容体リサイクリングおよびエフェクタータンパク質の細胞膜への動員を含めた、細胞活性の制御において重要な役割を有することが知られている。したがって同様の役割が、Rab11ファミリー相互作用タンパク質などのRabファミリー相互作用タンパク質によるものと考えられている。Rab11−FIP3は例えば、エンドソーム循環区画の機能
において重要な役割を果たすことが示されている(例えば、ホーガン、シー ピーら(Horgan,C.P.,et al.)、(2007)、Traffic、8、414〜430ページを参照)。
【0028】
Rabタンパク質は、細胞骨格編成および細胞形態、細胞質分裂、小胞輸送および分泌、細胞周期進行および細胞分化を含めた広範囲の基本的細胞機能を制御する、Ras関連低分子Gタンパク質(Rho関連タンパク質を含む)のさらに大きなファミリーに属する。いくつかのこれらのタンパク質ならびにそれらのレギュレーターおよびエフェクターは、発癌現象および癌の進行に関与している。これらのうち、乳癌および卵巣癌において頻繁に増幅されるRAB25は、in vitroで増殖およびアポトーシス耐性を高め、かつin vivoで腫瘍形成を高めることが示されている。RABファミリーのタンパク質は膜輸送およびオルガネラ区画化において中心的役割を果たし、RAB11、RCPの主な結合パートナーは、膜タンパク質のリサイクリングに特に関係している。RAB11は細胞移動に関与しており、RAB11経路の混乱は異なる細胞型において移動を害する(ジョーンズ、エム シーら(Jones,M.C.,et al.)、(2006)Curr Opin Cell Biol 18、549〜557ページ;ポウェルカ、エー エムら(Powelka,A.M.,et al.)、(2004)Traffic 5、20〜36ページ;ユーン、エス オーら(Yoon,S.O.,et al.)、(2005)Cancer Res 65、2761〜2769ページ)。さらに、RAB11機能の拮抗的阻害は乳癌細胞の低酸素誘導侵襲性を低下させ(ユーンら(Yoon,et al.)、2005、上記)、インテグリンα6β4、EGFR、CXCR2およびPKCαなどの侵襲と関係がある膜受容体の輸送を変える(ジョーンズら(Jones,et al.)、2006、上記)。RAB遺伝子発現の異常調節は多くのヒト腫瘍の一般的要素として関与する。しかしながら、腫瘍進行中のRab相互作用タンパク質の役割は研究されていない。
【0029】
本発明者らは、Rab11FIP1の発現と関係がある広範囲の癌促進効果を確認している。非癌性乳房上皮細胞では、Rab11FIP1の過剰発現は、悪性形質転換、ならびに増大した細胞死亡率および移動をもたらした。乳癌細胞系では、RNA阻害による内因性Rab11FIP1の弱化は、増殖、足場非依存性増殖、侵襲およびアポトーシス耐性を低減し、ヌードマウスにおいて腫瘍形成を低下させた。生理学上、Rab11FIP1は主にRab11と相互作用して、管状エンドソームにおけるタンパク質の選別を制御すると考えられる(ピーデン、エー エーら(Peden,A.A.,et al.)、(2004)Mol Biol Cell 15、3530〜3541ページ)。Rab11−Rab11FIP1複合体は、トランスフェリン受容体をリソソーム(分解経路)からリサイクリングエンドソーム(リサイクリング経路)にチャンネル移動させ、それによって膜結合受容体のレベルの相対的増大を促進することが示されている(ピーデンら(Peden et al.)、2004、上記)。推測的ではあるが、Rab11FIP1の形質転換効果はリサイクリング経路へのこの移動によって仲介され、癌遺伝子的役割を有するインテグリンまたは増殖因子受容体などの、特定の膜タンパク質の膜局在(したがって機能)を延長する可能性があることは妥当である。本発明者らが観察したRab11FIP1の発現とERKのリン酸化の間の強い相関関係はこの可能性を反映し得る。というのは、それは、Rab11FIP1(以下ではRCPとも名付ける)と増殖因子受容体仲介MAPK活性化の間の特異的関係を示し得るからである。
【0030】
理論によって縛られるわけではないが、本発明者らによって得られた免疫沈降データ(示さず)は、Rab11FIP1と低分子GTPアーゼRasの間に直接的相互作用が存在することを示し、この2つのタンパク質間の複合体の形成を示す。Rasタンパク質は、アポトーシス、細胞増殖、および癌におけるそれらの中心的役割が当該技術分野で知られている低分子モノマータンパク質のファミリーである。少なくとも11個の異なる機能
クラスのタンパク質が、Rasタンパク質のエフェクターであることが示されている。したがって、Rasタンパク質は複合体シグナル相互作用ネットワークの中心に位置し、その中でRasタンパク質は、少なくとも3つの重複および共同タンパク質キナーゼカスケード、すなわちRaf/MEK/ERK、RalGTPアーゼ、およびPI3K/Aktを活性化すると考えられる。1つのランダムに選択したさらなる例示例として、それらはRb/E2FおよびDnmt−1を介して作用し、それによって腫瘍細胞増殖を活性化するp16の不活性化などのメチル化による後成的変化を引き起こす。次いでrasタンパク質は複雑な制御機構に曝される。点突然変異したRas遺伝子は様々な腫瘍の3分の1で頻繁に発見され、Ras遺伝子は癌遺伝子であることが知られている。
【0031】
本発明によるいくつかの方法は、腫瘍発生を制御する方法である。これらの方法は、腫瘍発生を予防、阻害、停止または逆行させる個々の方法を含む。腫瘍発生は例えば、悪性型の癌腫の形成を含めた発癌現象であってよい。したがって、例えば、本発明の方法は、癌などの増殖性疾患または障害の治療または予防に含まれ得る。
【0032】
本発明の方法を使用して、例えば哺乳動物、魚類、両生類、鳥類または微生物を含めた任意の生物において、腫瘍発生を予防、阻害、停止または逆行させることができる。いくつかの実施形態では、それぞれの微生物は細胞である。このような細胞は、直接採取またはそこからの単離を含めて、哺乳動物などの例えば前述の例の1つなどの生物から得ることができる。適切な哺乳動物の例には、ラット、マウス、ウサギ、モルモット、オポッサム、イヌ、ネコ、チンパンジー、アカゲザル、マカクザル、オランウータン、ウシ(メウシ)、マーモセット、アメリカナキウサギ、ガラゴ(「コビトガラゴ」)、リス、ココノオビアルマジロ、ゾウおよびヒトがあるが、これらだけには限られない。細胞は、従来の細胞系、例えば癌細胞系の細胞であってもよい。いくつかの実施形態では、細胞を培養する。他の実施形態では、それぞれの細胞は、例えば前述の例の1つなどの生物の一部分に含まれ得る。
【0033】
本発明による方法のいくつかの実施形態は、Rab11FIPタンパク質などのRab結合タンパク質の量を調節することを含む。Rab結合の量の調節は、細胞中、試験管などのin vitro設定中、または生物中に存在するRab結合タンパク質分子の全数の一部分を分解状態にすることを含むことができる。Rab結合タンパク質の分解は、例えば(カルシウム活性化プロテアーゼまたはカルパインとも呼ばれる)カルパインプロテアーゼなどのプロテアーゼによって実施することができる。例示例として、Rab11FIP1は、カルパインの標的として働きin vivoでカルパインによって切断される、3つのPEST(Pro、GlyおよびSer/Thr多量)モチーフを含むことが開示されている(ニコラス、エムら(Nicolas,M.,et al.)、(2005)Biochem.J.、389、223〜231ページ)。Rab結合タンパク質の量の調節は、Rab11FIPタンパク質の発現を調節することも含むことができる。これは例えば、Rab11FIPタンパク質の発現を調節する化合物を投与することによって実施することができる。Rab11FIPタンパク質の発現は、増大させてもよいし低減させてもよい。例示例として、Rab11FIPタンパク質の発現は、非コード核酸分子などの核酸分子によって低減することが可能である。
【0034】
本明細書で使用する用語「核酸分子」は、一本鎖、二本鎖またはそれらの組合せなどの、任意の考えられる形状の任意の核酸を指す。核酸は、例えばDNA分子(例えば、cDNAまたはゲノムDNA)、RNA分子(例えば、mRNA)、ヌクレオチド類似体の使用または核酸化学の使用によって作製したDNAまたはRNAの類似体、およびPNA(タンパク質核酸)を含む。DNAまたはRNAはゲノム起源または合成起源であってよく、一本鎖または二本鎖であってよい。本発明の方法では、典型的には、ただし必ずではないが、RNAまたはDNA分子を使用する。このような核酸は、例えばmRNA、cRN
A、合成RNA、ゲノムDNA、cDNA、合成DNA、DNAとRNAのコポリマー、オリゴヌクレオチドなどであってよい。それぞれの核酸は非天然ヌクレオチド類似体をさらに含むことができ、および/または親和性タグまたは標識と連結することができる。いくつかの実施形態では、核酸分子を単離、濃縮、または精製することができる。核酸分子は、例えばcDNAクローニングまたはサブトラクティブハイブリダイゼーションによって天然源から単離することができる。天然源は、ヒト、血液、精液、または組織などの哺乳動物由来であってよい。核酸は、例えばトリエステル法によって、または自動DNA合成装置を使用することによって合成することもできる。
【0035】
多くのヌクレオチド類似体が知られており、本発明の方法中で使用する核酸およびオリゴヌクレオチドにおいて、それらを使用することができる。ヌクレオチド類似体は、例えば塩基、糖、またはホスフェート部分に修飾を含むヌクレオチドである。塩基部分における修飾には、A、C、G、およびT/U、異なるプリンまたはピリミジン塩基、ウラシル−5−イル、ヒポキサンチン−9−イル、および2−アミノアデニン−9−イルなど、ならびに非プリンまたは非ピリミジンヌクレオチド塩基の天然および合成修飾がある。他のヌクレオチド類似体は共通塩基として働く。共通塩基には3−ニトロピロールおよび5−ニトロインドールがある。共通塩基は、任意の他の塩基と塩基対を形成することができる。塩基の修飾は、例えば2’−O−メトキシエチルなどの糖の修飾と頻繁に組合せて、例えば二本鎖安定性の増大などの特有の性質を得ることができる。
【0036】
本発明の方法のいくつかの実施形態では、核酸分子は、例えばアプタマーまたはSpiegelmer(登録商標)(WO01/92655中に記載)などの非コード核酸分子である。非コード核酸分子はnc−RNA分子であってもよい(例えば、天然nc−RNA分子の導入に関するコスタ、エフエフ(Costa,FF)、Gene(2005)、357、83〜94ページを参照)。nc−RNA分子の例には、アンチセンスRNA分子、L−RNASpiegelmer(登録商標)、サイレンサーRNA分子(二本鎖ニューロン限定サイレンサーエレメントなど)、マイクロRNA(miRNA)分子、低分子ヘアピンRNA(shRNA)分子、低分子干渉RNA(siRNA)分子、反復配列関連低分子干渉RNA(rasiRNA)またはPiwiタンパク質と相互作用するRNA(piRNA)(簡単な概説に関しては、例えばリン、エイチ(Lin,H.)、Science(2007)316、397を参照)があるが、これらだけには限られない。適切なshRNAの例示例は、以下の実施例中に配列番号3および配列番号4として示す。
【0037】
低分子干渉RNAの使用は、特定の遺伝子を「ノックダウン」するためのツールとなっている。それは、転写後レベルで起こりmRNA分解を含むRNA干渉(RNAi)を介した、遺伝子サイレンシングまたは遺伝子抑制を利用する。RNA干渉はゲノムを保護する細胞機構となる。siRNA分子は、いわゆるRNA誘導型サイレンシング複合体(RISC)を形成するためのsiRNAと複合酵素複合体の結合によって、それらの相補的RNAの分解を仲介する。siRNAはRISCの一部分となり、相補的RNA種を標的化し、それは次いで切断される。これはそれぞれの遺伝子の発現の消失をもたらす(簡単な概説に関しては、(ザモア、ピーディーおよびハーレー、ビー(Zamore,PD,& Haley,B)(2005)Science 309、1519〜1524ページを参照)。この技法は例えば、米国特許出願2005/0191618中に開示されたHIV RNAの切断などの、寄生虫DNA配列のサイレンシングに施されている。
【0038】
本発明のこのようなsiRNAの典型的実施形態は、10〜35ヌクレオチド、いくつかの実施形態では15〜25ヌクレオチドのin vitroまたはin vivo合成分子を含む。それぞれのsiRNA分子は、(微生物および動物の一部分である細胞を含めた)対象の細胞内で直接合成することができる。siRNA分子をそれぞれの細胞に導
入する、および/またはそれぞれの細胞に送達することもできる。in vivoでの選択細胞へのsiRNA分子の送達の例示例は、重鎖抗体断片(Fab)と核酸結合タンパク質プロタミンの融合タンパク質との、その非共有結合である(ソング、イーら(Song,E.et al.)、(2005)、Nature Biotech.23、6、709〜717ページ)。本発明の1実施形態では、siRNA分子を使用して、Rab結合タンパク質の(例えばin vivo)合成と関係がある1つまたは複数の転写因子をコードするmRNA分子の分解を誘導する。
【0039】
本発明による本発明の方法のいくつかの実施形態は、Rab結合タンパク質、例えばRab11FIPタンパク質の活性を調節することを含む。Rab結合タンパク質の活性の調節は、Rab結合タンパク質と化合物の間の複合体を形成することを含むことができる。これは例えば、Rab結合タンパク質の活性を調節する化合物を投与することによって実施することができる。Rab結合タンパク質(例えばRab11FIPタンパク質)の活性は増大または低減することができる。
【0040】
それぞれの化合物は任意の適切な手段によって投与することができる。宿主生物が哺乳動物である場合、化合物は例えば経口または非経口(経腸)投与することができる。哺乳動物への投与の典型的実施形態では、例えば化合物の調製物の経口、静脈内投与によって、または吸入によって、その施用は血液および肝臓への送達を確実にする。経口施用のための調製物の例は、錠剤、ピルまたは飲料溶液であり、静脈内投与のための調製物の例は注射または輸液であり、吸入による投与のための調製物の例はエアロゾル混合物またはスプレーである。宿主生物が組換え微生物である場合、投与の例は、微生物の環境への化合物の注射または添加である。微生物が単細胞である場合、微生物を改変する技法と組合せて、後者の形の投与をおそらく実施することができる。このような技法は、細胞膜のエレクトロポレーションまたは透過処理を含むことができる。
【0041】
Rab結合タンパク質、例えばRab11FIPタンパク質の活性の調節は、Rab結合タンパク質の翻訳後修飾のパターンの改変を含めた、Rab結合タンパク質の翻訳後修飾の改変を含むことができる。いくつかの実施形態では、Rab11FIPタンパク質などのRab結合タンパク質の翻訳後修飾の改変は、Rab結合タンパク質の翻訳後修飾のパターンを改変する化合物の使用を含む。それぞれの翻訳後修飾の例示例は、リン酸化、脱リン酸化、アセチル化、およびユビキチン化である。それぞれの翻訳後修飾、例えばリン酸化の結果として、Rab結合タンパク質の活性が変わる。例示例として、Rab11−FIP2機能はセリン227におけるリン酸化によって制御されることが示されている(デゥチャルメ、エヌ エーら(Ducharme,N.A.,et al.)、Molecular Biology of the Cell(2006)7、8、3625〜3637ページ)。キナーゼMARK2/EMK1/Par−1B(MARK2)によってリン酸化されると、Rab11−FIP2は上皮細胞の極性を確立する際に働く(上記)。
【0042】
Rab11FIPタンパク質などのRab結合タンパク質の活性、発現、量または細胞内局在を調節する化合物は、任意の化合物であってよい。例には、核酸(上記参照)、ペプチド、ペプトイド、無機分子および小有機分子があるが、これらだけには限られない。ペプトイドは、ペプチドよりはるかに高い細胞透過性を有し得る(例えば、クウォン、ワイ ユーおよびコダデック、(2007)ティー(Kwon,Y.−U,and Kodadek,(2007)T.)、J.Am.Chem.Soc.129、1508〜1509ページ参照)。ペプチドは合成起源であってよく、または当該技術分野でよく知られている方法によって天然源から単離することができる。天然源はヒトなどの哺乳動物、血液、精液、または組織であってよい。ポリペプチドを含めたペプチドは、例えば自動ポリペプチド合成装置を使用して合成することができる。ポリペプチドの例示例は、抗体、抗
体断片、および抗体様機能を有するタンパク質結合分子である。(組換え)抗体断片の例は、Fab断片、Fv断片、単鎖Fv断片(scFv)、ダイアボディまたはドメイン抗体である(ホルト、エル ジェイら(Holt,L.J.,et al.)、(2003)Trends Biotechnol.、21、11、484〜490ページ)。抗体様機能を有するタンパク質結合分子の1例は、リポカリンファミリーのポリペプチドに基づくムテインである(WO03/029462、ベステら(Beste et al.)、(1999)Proc.Natl.Acad.Sci.U.S.A.、96、1898〜1903ページ)。ビリン結合タンパク質、ヒト好中球ゼラチナーゼ結合性リポカリン、ヒトアポリポタンパク質Dまたはグリコデリンなどのリポカリンは、それらがハプテンとして知られる選択した小さなタンパク質領域と結合するように修飾することができる天然リガンド結合部位を有する。他のタンパク質結合分子の例は、いわゆるグルボディ(WO96/23879参照)、アンキリン骨格(モザビ、エル ケイら(Mosavi,L.K.,et al.)、(2004)Protein Science 13、6、1435〜1448ページ)または結晶骨格(WO01/04144)に基づくタンパク質、スケラ(Skerra)、(2000)J.Mal.Recognit.13、167〜187ページ中に記載されたタンパク質、アドネクチン、テトラネクチン、およびアビマーである。アビマーは、いくつかの細胞表面受容体において一連の多数のドメインとして存在する、いわゆるA−ドメインを含む(シルバーマン、ジェイら(Silverman,J.,et al.)、(2005)Nature Biotechnology 23、1556〜1561ページ)。ヒトフィブロネクチンのドメインに由来するアドネクチンは、標的との免疫グロブリン様結合用に工学処理することができる3個のループを含む(ギル、ディー エスおよびダムレ、エヌ ケイ(Gill,D.S.& Damle,N.K.)、(2006)Current Opinion in Biotechnology 17、653〜658ページ)。それぞれのヒトホモ三量体タンパク質に由来するテトラネクチンは、C型レクチンドメイン中に所望の結合用に工学処理することができるループ領域を同様に含む(上記)。タンパク質リガンドとして作用することができるペプトイドは、側鎖がα炭素原子ではなくアミド窒素と結合している点でペプチドと異なる、オリゴ(N−アルキル)グリシンである。ペプトイドは典型的にはプロテアーゼおよび他の修飾酵素に耐性があり、ペプチドよりはるかに高い細胞透過性を有し得る(例えば、クウォン、ワイ ユーおよびコダデック、(2007)ティー(Kwon,Y.−U,and Kodadek,(2007)T.)、J.Am.Chem.Soc.129、1508〜1509ページ参照)。所望の場合、任意のまたは特定の形、クラスなどの標的物質に対するそれぞれの部分の親和性をさらに増大させる修飾作用物質を使用してもよい。
【0043】
それぞれの化合物は、(Rab11−FIPタンパク質などの)Rab結合タンパク質の翻訳後修飾を改変することもできる。化合物は、例えば、Rab結合タンパク質を含めた細胞要素のリン酸化状態を変えることができる。それぞれの化合物は、Rab結合タンパク質と異なるタンパク質を含めた細胞内タンパク質の、リン酸化の程度の調節物質であってもよい。このようなタンパク質は、したがってRab結合タンパク質の翻訳後修飾を改変することができる。それぞれの化合物の例示例はチロシンキナーゼ阻害剤である。チロホスチン、キナゾリン、キノキサリン、キノリン、2−フェニルアミノピリミジン、フラボノイド、ベンゾキノイド、アミノサリチル酸またはスチルベン(これらは例えばWO9618738、WO03035621およびその中に引用された参照文献中に記載されており、例えばそれらの実験による同定の例に関しては、例えば米国特許第6,740,665号を参照)などの多数のチロシンキナーゼ阻害剤が市販されている。チロホスチンの例は、AG213、AG490、AG879、AG1295、AG1478、AG1517、AGL2043、チロホスチン46およびメチル2,5−ジヒドロキシケイ皮酸である。キナゾリンは、例えばPD153035、PD156273、ゲフィチニブまたはラパチニブであり、キノキサリンは、例えばPD153035またはZD1839である
。キノリンの1例は5−メチル−5H−インドール[2,3−β]キノリンであり、2−フェニルアミノピリミジンの1例はイマチニブであり、フラボノイドの例はゲニステインまたはクエルセチンであり、ベンゾキノイドの1例はハービマイシンAであり、アミノサリチル酸の1例はラベンドウスチンAであり、スチルベンの1例はピセアタノールである。他の適切な化合物は、チロホスチンエルブスタチンなどの受容体チロシンキナーゼ阻害剤、WHI−P97またはチロホスチンAG592などのEGFR特異的受容体チロシンキナーゼ阻害剤、アウリントリカルボン酸などのチロシンリン酸化刺激剤、または過バナジン酸ナトリウムまたはイソキサゾールカルボン酸などのチロシンホスファターゼ阻害剤を含むことができる。
【0044】
タンパク質キナーゼの機能を阻害することが報告されている化合物のさらなる例には、ビス単環式、二環式または複素環式アリール化合物(国際特許出願WO92/20642)ビニレン−アザインドール誘導体(PCTWO94/14808)、1−シクロプロピル−4−ピリジル−キノロン(米国特許第5,330,992号)、スチリル化合物(米国特許第5,217,999号)、スチリル置換ピリジル化合物(米国特許第5,302,606号)、特定のキナゾリン誘導体(欧州特許出願No.0566266A1)、セレオインドールおよびセレニド(国際特許出願WO94/03427)、三環式多価化合物(国際特許出願WO92/21660)、およびベンジル−ホスホン酸化合物(国際特許出願WO91/15495)があるが、これらだけには限られない。調節物質として使用することができるさらに他の化合物には、1996年8月23日に出願された米国特許出願第08/702,232号中に記載されたオキシンドリノン(oxindolinone)などのオキシンドリノンがある。
【0045】
Rab結合タンパク質の翻訳後修飾を調節する化合物のさらなる例は、タンパク質キナーゼまたはタンパク質ホスファターゼの制御を誘導することができる細胞表面分子に対するアゴニストまたはアンタゴニストである。
【0046】
Rab11FIPタンパク質などのRab結合タンパク質の活性を調節する化合物の他の例は、例えばビタミンD3またはビタミンD4などのビタミンDのメンバーである。ビタミンD3(コレカルシフェロール、カルシオールまたはアラキトールとも呼ばれ、IUPAC名(1S,3Z)−3−[(2E)−2−[(1R,3aS,7aR)−1−[(1R)−1,5−ジメチルヘキシル]オクタヒドロ−7a−メチル−4H−インデン−4−イリデン]エチリデン]−4−メチレン−シクロヘキサノール、(3β、5Z、7E)−9,10−セココレスタ−5,7,10(19)−トリエン−3−オールとも名付けられる)は、その活性形で、乳癌を含めた様々な起源の悪性細胞において、増殖停止、分化、および細胞死を誘導する能力を有する。この観察結果が、考えられる抗癌剤としてのビタミンD類似体の開発をもたらしている(例えば、バウデット、シーら(Baudet,C.,et al.)(1996)Cancer Lett.100、3〜10ページ;ヒジー、ティー、およびウー、ジェイ エム(Hsieh,T.,& Wu,J.M.)、(1997)Biochem.Biophys.Res.Commun.235、539〜544ページ;ジェームス、エス ワイら(James,S.Y.,et al.)、(1999)Gen.Pharmacol.32、143〜154ページ;マティアセン、アイ エスら(Mathiasen,I.S.,et al.)、(1999)Cancer Res.59、4848〜4856ページを参照)。化合物EB1089は1つのこのような類似体であり、現在癌の治療に関する第3相臨床試験中である(例えば、マティアセン、アイ エスら(Mathiasen,I.S.,et al.)、(1993)J.Steroid Biochem.Mol.Biol.46、365〜371ページ;コルストン、ケイ ダブリュら(Colston,K.W.,et al.)、(1992)Biochem.Pharmacol.44、2273〜2280ページ;ハンセン、シー エムら(Hansen,C.M.,et al.)、(2000)Cu
rr.Pharm.Des.6、803〜828ページを参照)。ビタミンD化合物の抗癌活性は細胞内遊離カルシウム([Ca2+])に対するその影響と直接関係がある。ビタミンD化合物で処理した細胞中の遊離Ca2+の上昇は、乳癌細胞におけるアポトーシスの誘導と相関関係があり(サージーブ、アイ エヌら(Sergeev,I.N.,et al.)、(2000)Vitamin D Endocrine System:Structural、Biological、Genetic and Clinical Aspects中(ノーマン、エー ダブリュ、ボイロン、アール、およびトーマセット、エム(Norman,A.W.,Bouillon,R.,and Thomasset,M.)編)、399〜402ページ、University of California、Riverside、CA;バンデバーレ、ビーら(Vandewalle,B.,et al.)(1995)Int.J.Cancer 61、806〜8112ページ;サージーブ、アイ エヌおよびローテン、ダブリュ ビー(Sergeev,I.N.,& Rhoten,W.B.)(1998)Endocrine 9、321〜327ページ)、カルパインとしても知られるカルシウム依存性システインプロテアーゼの活性化における主な機構であることが示されている(ナカガワ、ティーおよびユーアン、ジェイ(Nakagawa,T.,& Yuan,J.)、(2000)J.Cell Biol.150、887〜894ページ;レイ、エス ケイら(Ray,S.K.,et al.)(2000)Brain Res.852、326〜334ページ;イシハラ、アイら(Ishihara,I.,et al.)(2000)Neurosci.Lett.279、97〜100ページ)。
【0047】
理論によって縛られることを望まないが、乳癌細胞における試験からの最近の証拠は、カルパインがビタミンD3およびEB1089によって誘導されるアポトーシス死における主な実行プロテアーゼであり得ることを示唆している(マティアセン、アイ エスら(Mathiasen,I.S.,et al.)、(2002)J.Bio.Chem.277、30738〜30745ページ)。カルパインのプロテアーゼ(タンパク質分解)機能は、カルパインのタンパク質標的のペプチド配列中に存在するPEST(Pro、Gly、およびSer/Thr多量)アミノ酸モチーフに依存してる(ダイス、ジェイ エフ(Dice,J.F.)(1987)FASEB J.1,349〜357ページ;トンパ、ピーら(Tompa,P.,et al.)、(2004)J.Biol.Chem.279、20775〜20785ページ;レヒシュタイナー、エム、およびロジャース、エス ダブリュ(Rechsteiner,M.,& Rogers,S.W)(1996)Trends Biochem.Sci.21、267〜271ページ;シャムウェイ、エス ディーら(Shumway,S.D.,et al.)、(1999)J.Biol.Chem.274、30874〜30881ページ;フクダ、エムおよびイトー、ティー(Fukuda,M.,& Itoh,T)(2004)J.Biol.Chem.279、22314〜22321ページ)。RAB11FIP1は、例えばC2ドメインとRBD(Rab結合ドメイン)の間に位置する3個のこのようなPESTモチーフを含む。近年の試験において、マリーら(Marie,et.al.)は、RAB11FIP1中のPESTモチーフは、カルパインによる制御型分解に関してそれを標的化するかどうか決定しようと努めた(マリー、エヌら(Marie,N.,et.al.)(2005)Biochem.J.389、223〜231ページ)。マリーらは、高濃度のカルシウムを用いた細胞の処理はRAB11FIP1の分解を誘導し、この分解は特異的カルパイン阻害剤を加えることにより無効にすることができることを観察した。さらに、このカルシウム制御型分解におけるPEST配列の関与は、その中のPEST配列を除去したRAB11FIP1突然変異体を使用して立証した。したがってマリーらは、RAB11FIP1のPEST配列は、おそらく細胞中の生理的代謝回転の機構として、カルパイン仲介型分解に関してタンパク質を標的化すると結論付けた。例えばRAB11FIP1がカルパイン作用に感受性である活性PESTドメインを含み、カルパインがビタミンD3およびEB1089などのその化学療法剤類似体によってタンパク質分解活性
に関して活性化されることを考慮して、本発明の方法は、ビタミンD3およびEB1089を含めたその関連類似体に基づく化学療法レジメン、および細胞内遊離カルシウムレベルを上昇させるため、またはカルパインを活性化するために生化学的に作用する任意の他の治療レジメンに関して患者を選択するためのバイオマーカーとして、RAB11FIP1のmRNAまたはタンパク質レベルを使用することを可能にする。
【0048】
前に記したように、細胞における腫瘍発生を制御する方法、例えば細胞における腫瘍発生を予防、停止または逆行させる方法のいくつかの実施形態では、Rab結合タンパク質のレベル、発現および/または活性を分析する。細胞は、(例えば単離した)個々の細胞、癌組織を有し得る生物の細胞、癌組織を含む組織の細胞、または細胞培養物の細胞であってよい。癌細胞は、例えば神経、グリア、肺、肝臓、脳、乳房、膀胱、血液、白血病、結腸、子宮内膜、胃、皮膚、卵巣、脂肪、骨、頸部、食道、膵臓、前立腺、腎臓、または甲状腺細胞であってよい。いくつかの態様では、癌は、星状細胞腫、急性骨髄性白血病、乳癌、膀胱癌、子宮頸癌、結腸直腸癌、子宮内膜癌、食道扁平上皮細胞癌、グリオーマ、グリア芽腫、胃癌、肝細胞癌、ホジキンリンパ腫、白血病、脂肪腫、メラノーマ、マントル細胞リンパ腫、粘液線維肉腫、多発性骨髄腫、神経芽細胞腫、非ホジキンリンパ腫、肺癌、非小細胞肺癌、卵巣癌、食道癌、骨肉腫、膵臓癌、前立腺癌、頭部または頸部の扁平細胞癌、甲状腺癌および尿路上皮癌を含むが、これらだけには限られない。
【0049】
細胞におけるRab結合タンパク質の量の変化、発現の変化または活性の変化は、ほぼ平均レベルまたは一般に見られるレベルに対応するレベルに調整することができる。細胞におけるRab結合タンパク質の量の変化は、プロテアーゼの活性または発現の調節によって実施することができる。いくつかの実施形態では、このようなプロテアーゼはカルシウム依存性プロテアーゼ(カルパイン)、例えばカルパインIまたはIIである。いくつかの実施形態では、カルパインプロテアーゼの活性および/または発現を、細胞内カルシウムレベル、例えば遊離細胞内カルシウムの変化によって調節する。
【0050】
いくつかの実施形態では、カルパインプロテアーゼの活性および/または発現を、それらの施用を含めた、ビタミンDファミリーのメンバー、このようなビタミンDファミリーのメンバーのプロドラッグまたはこのようなビタミンDファミリーのメンバーの類似体の使用によって調節、例えば活性化する。それぞれのビタミンD、その類似体またはプロドラッグは、例えばそれぞれの細胞と接触させることが可能であり、またはそれぞれの対象を含めた、それぞれの生物に投与することができる。プロテアーゼの活性および/または発現は、シグナル伝達カスケードによって次いで調節されるプロテアーゼ調節物質などの、他の因子によって調節することもできる。多数のプロテアーゼが、それらの活性とそれらの発現の両方の点で制御を受けることは知られている(例えば、トゥルク、ビー(Turk,B.)、(2006)Nature Reviews Drug Discovery、5、9、785〜799ページ参照)。アポトーシスプロテアーゼ活性化因子−1またはメタロプロテイナーゼの組織阻害剤は例示例として役立つ可能性がある。
【0051】
ビタミンDファミリーのメンバーまたはビタミンDファミリーのメンバーの類似体の「プロドラッグ」は、in vivoで(例えば段階的または急速に)転換されて、酵素作用または一般的な酸または塩基加溶媒分解のいずれかによる、例えば血液中での加水分解または酸化による代謝を含めた、例えば宿主中での代謝によって親ビタミンD/ビタミンD類似体化合物を生成し、活性成分を形成する化合物であることは理解されている。典型的にはプロドラッグは、in vivoで使用すると化学的または生理的工程によって元の化合物を放出する、親ビタミンD/ビタミンD類似体の誘導体である。プロドラッグは例えば、生理的pHに達すると、(1つまたは複数の)自発性化学反応、(1つまたは複数の)酵素触媒化学反応、光分解、および/または(1つまたは複数の)代謝化学反応によって、親ビタミンD/ビタミンD類似体に変換され得る。一般的な例は、カルボン酸部
分を有する活性形を有するエステルおよびアミド形の化合物だけには限られないが、これらを含めた、親化合物の官能基部分に生物学的に不安定な保護基を有する化合物である。さらなる例には、酸化、還元、アミノ化、脱アミノ化、エステル化、脱エステル化、アルキル化、脱アルキル化、アシル化、脱アシル化、リン酸化、脱リン酸化、光分解、加水分解することが可能な化合物の部分、またはプロドラッグにおける化学結合の形成または破壊に関する他の官能基の変化または変換がある。それぞれのプロドラッグは、その治療活性または利用可能形にin vivoで変換されるまで、完全に活性状態または生理的に利用可能でないプロドラッグでよい。「プロドラッグ」は、それぞれの薬剤分子(この場合、親ビタミンD/ビタミンD類似体)と担体部分の間の共有結合または非共有結合によって、in vitroまたは生理的に形成される生成物も含み得る。特定の化合物は、溶媒和および非溶媒和形、例えば水和形で存在し得ることも理解されよう。溶媒和形は単に「溶媒和物」とも呼ばれる。
【0052】
前で他に記したように、発癌性になる素因を有する細胞を同定する方法のいくつかの実施形態では、(例えば単離した)個々の細胞、生物の細胞または細胞培養物の細胞であってよい細胞におけるRab結合タンパク質の量の変化、発現の変化または活性の変化を決定することができる。同様に、対象、例えば患者における新生物を発症するリスクを診断する方法のいくつかの実施形態では、Rab結合タンパク質の量の変化、発現の変化または活性の変化を決定することができる。このような場合、それぞれの変化の決定は、それぞれの新生物または腫瘍が対象の1つまたは複数の細胞におけるプロテアーゼの活性および/または量の変化に感受性であることを示す可能性がある。いくつかの実施形態では、このようなプロテアーゼは、カルパインプロテアーゼカルパインIまたはII(カルシウム活性化中性プロテアーゼIおよびIIとも呼ばれる)としても知られるカルシウム依存性プロテアーゼCDPIまたはCDPIIなどの、カルシウム依存性プロテアーゼ(カルパイン)である。いくつかの実施形態では、Rab結合タンパク質の活性および/または発現の変化の決定は、それぞれの新生物または腫瘍が細胞内カルシウムレベルの変化に感受性であることを示す可能性がある。
【0053】
この点において、各方法は、対象から試料を得ること、および試料から核酸を単離することをさらに含むことができる。核酸を増幅し、標識することができる。選択したタンパク質の量を決定する方法は、当該技術分野でよく知られている。各方法は、例えばプロテオミクス系の技法に依存することができる。それは、それぞれのRab結合タンパク質と結合する、抗体、抗体断片、または抗体様機能を有するタンパク質結合分子の使用を含むことができる。タンパク質の発現タンパク質を分析する方法は、当該技術分野で十分確立している。mRNAの発現は、例えばノーザンブロッティングおよびin situハイブリダイゼーション、RNAse保護アッセイおよびPCR系の方法を使用して定量化することができる。さらなる例として、DNA二本鎖、RNA二本鎖、およびDNA−RNAハイブリッド二本鎖またはDNA−タンパク質二本鎖を含めた特異的二本鎖を認識することができる、抗体、抗体断片、またはタンパク質結合分子を利用することができる。塩基配列決定に基づく遺伝子発現分析の方法の例示例には、遺伝子発現の連続分析(SAGE)、および超並列型サイン塩基配列決定(MPSS)による遺伝子発現分析がある。
【0054】
(組換え)抗体断片の例は、Fab断片、Fv断片、単鎖Fv断片(scFv)、ダイアボディ、トリアボディ(イリアデ、ピーら(Iliades,P.,et al.)、FEBS Lett(1997)409、437〜441ページ)、デカボディ(ストーン、イーら(Stone,E.,et al.)、Journal of Immunological Methods(2007)318、88〜94ページ)、および他のドメイン抗体である(ホルト エル ジェイら(Holt,L.J.,et al.)、Trends Biotechnol.、(2003)21、11、484〜490ページ)である。単鎖Fv断片は、例えば免疫グロブリン分子の1本の重鎖と1本の軽鎖由来
の可変領域の融合体である。抗体様機能を有するタンパク質結合分子の1例は、リポカリンファミリーのポリペプチドに基づくムテインである(WO2003/029462;WO2005/019254;WO2005/019255;WO2005/019256;ベステら(Beste et al.)、Proc.Natl.Acad.Sci.U.S.A.(1999)、96、1898〜1903ページ)。ビリン結合タンパク質、ヒト好中球ゼラチナーゼ結合性リポカリン、ヒトアポリポタンパク質D、ヒト涙液リポカイン、またはグリコデリンなどのリポカリンは、それらがハプテンとして知られる選択した小さなタンパク質領域と結合するように修飾することができる天然リガンド結合部位を有する。さらなるタンパク質結合分子の他の非制限的な例は、いわゆるグルボディ(WO96/23879参照)、アンキリン骨格(モザビ、エル ケイら(Mosavi,L.K.,et al.)、Protein Science(2004)13、6、1435〜1448ページ)または結晶骨格(WO2001/04144)に基づくタンパク質、スケラ(Skerra)(J.Mal.Recognit.(2000)13、167〜187ページ)中に記載されたタンパク質、アドネクチン、テトラネクチン、アビマー、およびペプトイドである。アビマーは、いくつかの細胞表面受容体において一連の多数のドメインとして存在する、いわゆるA−ドメインを含む(シルバーマン、ジェイら(Silverman,J.,et al.)、Nature Biotechnology(2005)23、1556〜1561ページ)。ヒトフィブロネクチンのドメインに由来するアドネクチンは、標的との免疫グロブリン様結合用に工学処理することができる3個のループを含む(ギル、ディー エスおよびダムレ、エヌ ケイ(Gill,D.S.& Damle,N.K.)、Current Opinion in Biotechnology(2006)17、653〜658ページ)。それぞれのヒトホモ三量体タンパク質に由来するテトラネクチンは、C型レクチンドメイン中に所望の結合用に工学処理することができるループ領域を同様に含む(上記)。タンパク質リガンドとして作用することができるペプトイドは、側鎖がα炭素原子ではなくアミド窒素と結合している点でペプチドと異なる、オリゴ(N−アルキル)グリシンである。ペプトイドは典型的にはプロテアーゼおよび他の修飾酵素に耐性があり、ペプチドよりはるかに高い細胞透過性を有し得る(例えば、クウォン、ワイ ユーおよびコダデック、ティー(Kwon,Y.−U,and Kodadek,T.)、J.Am.Chem.Soc.(2007)129、1508〜1509ページ参照)。所望の場合、任意のまたは特定の形、クラスなどの標的物質に対するそれぞれの部分の親和性をさらに増大させる修飾作用物質を使用することができる。
【0055】
PCR(ポリメラーゼ連鎖反応)は、細胞由来またはメラノーマ試料などの組織試料由来の転写産物を、増幅および検出するのに有用な標準的な技法である。逆転写酵素PCR(RT−PCR)は例えば、それを使用して異なる試料(例えば、非転移性および転移性メラノーマ試料、または良性皮膚およびメラノーマ試料)中のmRNAレベルを比較し、遺伝子発現サインを調べることができる感度の良い定量法である。RT−PCRを実施するために、mRNAを試料(例えば、ヒトメラノーマ試料から単離した全RNA)から単離する。mRNAは例えば、凍結または保存済みパラフィン包埋および固定(例えば、ホルマリン固定)組織試料から抽出することができる。RNA単離用の精製キットは市販されている。RT−PCR技法は、二重標識蛍光プローブによってPCR産物の蓄積を測定する、リアルタイム定量PCRの実施形態中で使用することもできる。リアルタイムPCRは、それぞれの標的配列用の内部競合剤を標準化に使用する定量競合的PCRと、試料内に含まれる標準化遺伝子、またはRT−PCR用のハウスキーピング遺伝子を使用する定量比較PCRとの両方に適合する。さらなる例として、競合的PCR設計は、おそらく自動、高スループットマトリクス支援レーザー脱離イオン化法飛行時間型(MALDI−TOF)質量分析検出法および核酸分子の定量化を含めた、遺伝子発現分析に使用することができる。遺伝子発現分析に適したPCR系の方法中に含めることができる技法のさらなる例は、ディファレンシャルディスプレイ、増幅断片長多型(iAFLP)法、Bea
dAmiy(商標)技術、遺伝子発現の検出用のBeadsArvay(BADGE)および広範囲の発現プロファイリング(HiCEP)分析だけには限られないが、これらを含むことができる。さらなる例として、組織試料の遺伝子発現の決定は、マイクロアレイを用いて実施することもできる。
【0056】
いくつかの実施形態では、Rab結合タンパク質の活性および/または発現の変化、特にRab結合タンパク質の活性および/または発現の増大の決定は、それぞれの新生物または腫瘍がビタミンDのメンバー、その類似体またはプロドラッグを用いた治療に感受性であることを示す可能性がある。この点において本発明は、(腫瘍、例えば癌を含めた)新生物がプロテアーゼの活性および/または量の変化に感受性であるかどうかを決定する方法も提供する。新生物中、特にその細胞中で発現されるプロテアーゼを含めて、プロテアーゼが存在してよい。いくつかの実施形態では、方法は、新生物がビタミンDファミリーのメンバー、その類似体またはプロドラッグに感受性であるかどうかを決定する方法である。
【0057】
本発明による方法に使用することができる、ビタミンD3類似体を含めた多数のビタミンD類似体が、当該技術分野で知られている(概説に関しては、例えば、クボデラ、エヌ(Kubodera,N.)(2006)Current Bioactive Compounds、2(3)、301〜315ページを参照されたい)。例えばカルシトリオール、マクサカルシトール、タカルシトール、セカルシフェロール、アルファカルシドールまたはセオカルシトール(EB1089)を含めた適切な例は、例えば国際特許出願WO2007/082542中に挙げられている。セオカルシトールは、乳癌腫瘍細胞中の電離放射線に対する応答性を高めることが以前に示されており、これはDNA損傷の初期誘導の細胞シグナル伝達経路下流の変化の結果であることが示唆されている(デマスターズ、ジーら(DeMasters,G.,et al.)、(2006)Mol Cancer Ther、5、11、2786〜2797ページ)。さらなる最近の例は、その形成がシミズら(Shimizu et al.)、((2006)Bioorganic & Medicinal Chemistry Letters、16、3323〜3329ページおよびその中に挙げられた参照文献)によって記載されている、SMD−429などの16−エン−22−オキサビタミンD3類似体、例えばリーら(Lee et al.)、((2007)、Cancer Research、67(24)、11840〜11847ページ)によって記載された、1α,25−ジヒドロキシ−20S,21(3−ヒドロキシ−3−メチルブチル)−23−イン−26,27−ヘキサフルオロコル−カルシフェロール、またはヨーンら(Yoon et al.)、((2008)International Journal of Oncology、23、387〜396ページ)によって表された1α,25−ジヒドロキシビタミンD3類似体、1α−ヒドロキシ−5,6−トランス−ビタミンD3の19官能性誘導体(ボジットキーレビッズ、エー(Wojtkielewicz,A)、(2007)Steroids、72(6〜7)、552〜558ページ)、オノら(Ono et al.)、((2007)、Journal of Combinatorial Chemistry9(4)、711〜716ページ)によって記載された延長側鎖を有する類似体、またはポスナーら(Posner et al.)、((2007)、Journal of Steroid Biochemistry & Molecular Biology、103、213〜221ページ)によって記載されたジフルオロ類似体である。
【0058】
本発明のいくつかの実施形態用に、ライブラリーの形で化合物を使用することができる。このようなライブラリーの例は、モデル化合物として化学的に合成した様々な有機小分子、または多数の配列変異体を含む核酸分子の集合である。
【0059】
いくつかの実施形態では、本発明の方法は、例えば生物またはその組織もしくは細胞に
おける、Rab11−FIPタンパク質の発現、細胞内局在および/または活性を検出することを含む。例示例として、Rab11−FIPタンパク質の細胞内量、発現、細胞内局在および/または活性を一定時間モニターすることができる。
【0060】
本発明の方法は、Rab11−FIPタンパク質の細胞内量、発現、細胞内局在および/または活性を測定した結果と、対照測定(または「参照」測定)の結果を比較することをさらに含むことができる。対照測定は、Rab11FIPタンパク質の細胞内量、発現、細胞内局在および/または活性を調節しない条件の使用を含むことができる。細胞内量、発現、および/または活性の比較において、検出レベルを例えば対照レベルと比較することができる。本明細書で使用する用語「対照レベル」は、例えばRab11FIPタンパク質の細胞、mRNAまたはタンパク質発現レベルでの、それぞれのタンパク質の分子の数、および対照試料中のRab11FIPタンパク質の活性レベルを指す。したがってこの用語は、正常対照レベルと癌の対照レベルの両方を含む(これも以下参照)。この用語は1つの参照測定値、または複数の参照測定値を指すことができる。いくつかの実施形態では、対照レベルは以前に実施した測定からの発現または活性値のデータベースであってよい。用語「通常レベル」は、正常、健康な個体中、または癌を含めた新生物に罹患していないことが知られている個体の集団中で検出される、Rab11FIPタンパク質の発現のレベルまたはRab11FIPタンパク質の活性レベルを指す。正常な個体は、それぞれの新生物の臨床症状がない個体である。
【0061】
本発明によれば、例えば細胞中の、遺伝子発現レベルまたは活性レベルまたはタンパク質の量は、対照レベルと比較して遺伝子発現/活性/量が約10%、約25%、約50%、約75%、約100%、またはそれより多く増大または低下するとき、「変化した」または「異なる」とみなされる。あるいは、発現レベルまたは活性レベルは、対照レベルと比較して遺伝子発現/または活性が少なくとも約0.1、少なくとも約0.2、少なくとも約1、少なくとも約2、少なくとも約5、または少なくとも約10またはそれより多くの倍数増大または低下するとき、「増大した」または「低下した」とみなされる。
【0062】
本発明の方法は、細胞外シグナル制御キナーゼ(ERK)の活性を変えることを含むことができる。いくつかの実施形態では、方法はERKの活性化を予防、阻害、停止または逆行させることを含む。典型的実施形態では、Rab11−FIPタンパク質の量または活性の調節は、ERKの活性の調節をもたらす可能性がある。
【0063】
本発明のさらなる方法は、発癌現象を含めて発癌性になる素因を有する細胞を同定する方法である。方法は、細胞におけるRab11FIPタンパク質の量の変化、発現の変化、細胞内局在の変化または活性の変化を検出することを含む。関連法に関して前ですでに説明したように、方法は、Rab11−FIPタンパク質の細胞内量、発現、細胞内局在および/または活性を測定することを含むことができる。いくつかの実施形態では、方法は、Rab11−FIPタンパク質の細胞内量、発現、細胞内局在および/または活性を測定した結果と、対照測定の結果を比較することも含むことができる(上記を参照)。
【0064】
本発明は、対象において癌を含む腫瘍などの新生物を発症するリスクを診断する方法にも関する。それぞれの腫瘍は、例えば乳房腫瘍、肺腫瘍、結腸直腸腫瘍、膀胱の腫瘍または卵管(卵巣管とも呼ばれる)の腫瘍であってよい。同様に、それぞれの癌は、対応する癌腫の1つを含めた、例えば乳房癌、肺癌、結腸直腸癌、膀胱の癌または卵管(卵巣管とも呼ばれる)の癌であってよい。肺癌の癌腫の例示例は、非小細胞肺癌腫である。方法は、Rab11FIPタンパク質の量、発現レベル、活性レベルおよび細胞内局在の少なくとも1つを決定することを含む。測定はいくつかの実施形態では、対象由来の組織試料または細胞試料などの試料において実施することができる。いくつかの実施形態では、方法は、Rab11−FIPタンパク質の細胞内量、発現、細胞内局在および/または活性を
測定した結果と、対照測定の結果を比較することも含むことができる(上記を参照)。それぞれの対照測定用に、Rab結合タンパク質の発現および/または活性が通常(「正常」)レベルである試料を使用することができる。典型的実施形態では、対照測定と比較したRab結合タンパク質の量、発現レベル、活性レベルまたは細胞内局在の変化は、対象が新生物に罹患しているかまたは新生物を発症するリスクがあることを示す。
【0065】
本発明のさらなる方法は、それぞれの化合物を同定するin−vivo法とin−vitro法の両方の方法である。化合物は発癌現象を含めた腫瘍発生を予防、阻害、停止または逆行させることができる。化合物は、Rab11FIPタンパク質などのRab結合タンパク質と複合体を形成することができる。典型的にはこれらの方法は、in−vitroまたはin−vivoであれこの複合体の構成要素を互いに曝すことを含む。1つのこのような方法は、複合体を形成する、または複合体を形成すると考えられる構成要素を互いに接触させることを含むin−vitro法である。例えばRab11FIPタンパク質と、Rab11FIPタンパク質と複合体を形成すると考えられる薬剤候補分子を、互いに接触させることが可能である。いくつかの実施形態では、方法は、複合体の形成を検出することをさらに含む。
【0066】
複合体の形成を検出する任意の適切な方法を使用することができる。検出法は、例えば電気泳動法、HPLC、フローサイトメトリー、蛍光相関分光法またはこれらの技法の改変型を含むことができる。他の技法は生物分子の結合自体の測定を含む。このような測定は、例えば分光的、光化学的、光度分析的、蛍光分析的、放射線学的、酵素的または熱力学的手段、または細胞効果に基づくことができる。分光的検出法の1例は、蛍光相関分光法である(トンプソン、エヌ エルら(Thompson,N.L.,et al.)、Curr.Opin.Struct.Biol.(2002)12(5)、634〜641ページ)。光化学的方法は、例えば光化学的架橋である(スティーン、エイチ、ジェンセン、オー エヌ(Steen,H.,Jensen,O.N.)、Mass Spectrom Rev.(2002)21、3、163〜182ページ)。それぞれ光活性、蛍光、放射性または酵素標識の使用(概説に関しては、:リッペ、アール エーら(Rippe,R.A.,et al.)、Methods Mol.Biol.(2001)160、459〜479ページ参照)は、光度分析的、蛍光分析的、放射線学的および酵素的検出法の例である。熱力学的検出法の1例は、等温滴定型熱量計である(ITC、概説に関しては、:ベラツクエツ−カンポイ、エーら(Velazquez−Campoy,A.,et al.)、Methods Mol.BioL(2004)261、35〜54ページ参照)。細胞効果を使用する方法の1例は、その酵素的検出または細胞複製を含めた細胞増殖または細胞生存の測定である。
【0067】
さらなる関連法は微生物を提供することを含むin vivo法である。微生物はRab結合タンパク質を発現し、該Rab結合タンパク質と共に化合物は複合体を形成し得ると考えられ、またはRab結合タンパク質の発現を化合物が変え得る(例えば、抑制し得る)と考えられる。いくつかの実施形態では、微生物はRab結合タンパク質を内生的に発現することができる。いくつかの実施形態では、微生物は組換え細胞またはトランスジェニック微生物である。いくつかの実施形態では、微生物は、乳癌細胞または肺癌細胞を含めた癌細胞などの単細胞である。いくつかの実施形態では、微生物は生物から得られる細胞である。それぞれの生物は、例えば哺乳動物、魚類、両生類または鳥類であってよい。細胞は典型的には、皮膚組織または乳房組織などの、それぞれの生物の組織に由来する。いくつかの実施形態では、微生物は、乳癌細胞系または肺癌細胞系を含めた、癌細胞系などの細胞系の細胞である。適切な乳癌細胞系の例には、HMEC細胞系、3D、Bcap−37、BT−20、BT−474、BT474M1、Hs578T、Hs578Bst、JIMT−1、MCF−7、M231、MDA−231、MDA−MB−134、MDA−MB−175、MDA−MB−231、MDA−MB−361、MDA−MB−3
61DYT2、MDA−MB−435、MDAMB−435HM、MDA−MB−468、MB231、MW1、PMC42、SK−BR−3、SK−BR−7、SUM149、SUM159、SUM1315、SUM−52PE、T−47D、ZR−75、ZR−75−1およびZR−75−30があるが、これらだけには限られない。肺癌細胞系は、例えば腺癌細胞系、扁平上皮細胞癌細胞系または小細胞癌細胞系であってよい。適切な肺癌細胞系の例には、ABC−1、A549、Calu−6、EBC−1、H322、H446、H460、HCC2279、HTB−56R、HUT29、IMEC−2、KU−T1、L9981、LCD、LCOK、LK−2、LC−1/sq、Lu135、MS−1、NCI−H69、NCI−H82、NCI−H209、NCI−H446、NCI−H1395、NCI−H1437、NCI−H1770、NCI−H2009、NCI−H2087、NCI−H2122、NCI−H2126、NCI−H2171、NCI−N226、NCI−N231、PC1、PC3、PC7、PC9、PC10、PC14、PLA−801D、QG56、RERF−LCAI、RERF−LCMS、RERF−LCKJ、SBC3、SPC−A1、SQ5およびYTLMC−90があるが、これらだけには限られない。多数の他の癌細胞系、例えばポールら(Pole et al.)(2006、上記)中で命名された追加的細胞系が当該技術分野で知られている。
【0068】
いくつかの実施形態では、微生物は、結腸直腸癌細胞系、膀胱癌細胞系および卵管癌細胞系の細胞である。適切な結腸直腸癌細胞系の例には、Colo−320、HCT116、HT−29、L0、L0−200、L0−500、L0−1000、LoVo、LS1034、LST−R1、RKO、SW480、およびWiDr、ならびにドウグラら(Douglas et al.)((2004)Cancer Research 64、14、4817〜4825ページ)によって言及されたさらなる結腸直腸癌細胞系があるが、これらだけには限られない。適切な膀胱癌細胞系の例には、BIU−87、EJ、KoTCC−1、MBT−2、RT4、ScaBER、T24およびUM−UC−2があるが、これらだけには限られない。卵管癌細胞系の2つの例示例は、FT−MZ−1(ルンネバウム、アイ ビーら(Runnebaum,I.B.,et al.)、(1994)FEBS Lett.、353、1、29〜32ページ)およびHEPFT(イシワタ、アイら(Ishiwata,I.,et al.)、(2007)Human Cell、20、119〜130ページを参照)である。
【0069】
いくつかの実施形態では、微生物は、Rab11FIP、例えばRab11FIP1などのRab結合タンパク質を過剰発現する乳癌上皮細胞系MCF10Aなどの非癌性組織細胞系を含めた、非腫瘍性の細胞である。同様に、任意の細胞−細胞系の細胞、個々の細胞、多細胞生物内の細胞、(直接入手および単離を含めて)多細胞生物から入手した細胞であろうと選択して、Rab11FIP1などのRab結合タンパク質を過剰発現させることによって癌細胞に形質転換することができる。以下の実施例および例えば図6は、細胞がどのようにして癌細胞に形質転換され得るかを然るべく例示する。この点において、本発明の方法は癌細胞を入手する方法も提供し、それは細胞中のRab結合タンパク質の発現を調節する、特に増大させる方法を含む。それによって細胞は癌細胞に形質転換される。それぞれのRab結合タンパク質は、例えばRab11FIP1などのRab11結合タンパク質であってよい。Rab結合タンパク質は、内因性または異種タンパク質、すなわち細胞中に本来存在するコード核酸配列を有するタンパク質、またはコード核酸配列がそれぞれの細胞中に導入される/導入されているタンパク質であってよい。本発明は、in vivoでの対応するRab結合タンパク質の発現を調節することが可能な化合物を同定するための、このような細胞の使用にも関する(以下参照)。
【0070】
本発明の方法は、Rab結合タンパク質と複合体を形成し得ると考えられる化合物、またはその発現を変えると考えられる化合物を微生物に加えることを含む。各方法のいくつかの実施形態では、Rab結合タンパク質、例えばRab11FIP結合タンパク質の発
現をモニターする。各方法のいくつかの実施形態では、Rab結合タンパク質の活性をモニターする。いくつかの実施形態では、Rab結合タンパク質の細胞表現型の変化をモニターする。いくつかの実施形態では、このような方法は対照測定を含む(上記も参照)。対照測定の結果は化合物を使用して得た結果と比較する。対照測定は、例えば、Rab結合タンパク質の発現または活性に影響を与えないことが知られている化合物の使用を含むことができる。
【0071】
さらに他の関連法は、宿主生物を提供することを含むin−vivo法である。任意の望ましい宿主生物を、それが癌細胞を収容し増殖させることが可能である限り提供することができる。宿主生物の例には、哺乳動物、魚類、両生類、および鳥類があるが、これらだけには限られない。適切な哺乳動物の例には、ラット、ウサギ、モルモット、オポッサム、イヌ、およびネコがあるが、これらだけには限られない。
【0072】
任意の望ましい癌細胞をこの目的で使用することができる(例えば上記を参照)。方法は、宿主生物に癌細胞を導入することをさらに含む。さらに方法は、先に記載した化合物、すなわちRab結合タンパク質と複合体を形成し得ると考えられる化合物、Rab結合タンパク質の量を調節し得ると考えられる化合物、またはRab結合タンパク質の活性を調節し得ると考えられる化合物の使用を含む。いくつかの実施形態では、癌細胞は化合物を含む。したがって、宿主生物に癌細胞を導入する前に、化合物を癌細胞に導入することができる。いくつかの実施形態では、宿主生物への癌細胞の導入の前、後、または同時に、化合物を宿主生物に投与する。典型的には、方法の特定の段階で化合物を癌細胞に導入する。方法は、宿主生物中の腫瘍の増殖をモニターすることをさらに含む。
【0073】
本発明を容易に理解し実務上の効果をもたらすことを可能にするために、個々の実施形態をここで以下の非制限的な実施例によって記載する。
【実施例】
【0074】
本発明者らが開発したTRIAGE法の、特に関連する態様および特徴は、以下のように要約できる(図1も参照されたい)。
【0075】
【表1】
【0076】
細胞および薬剤治療
マイクロアレイデータおよび腫瘍試料。4種の前述の乳癌マイクロアレイデータセットを、この研究に使用し、全発現プロフィールを、原典であるウプサラコホート(ミラー、エル ディーら(Miller,L.D.et al.)(2005)Proc Natl Acad Sci USA 102、13550〜13555)、ストックホルムコホート(パウィタン、ワイら(Pawitan,Y.et al.)(2005)Breast Cancer Res 7、(R953〜964)、ベルギーコホート(ルイ、エスら(Loi,S.et al.)(2007)J Clin Oncol 25、1239〜1246)およびシンガポールコホート(イブシナ、エー ブイら(Ivshina,A.V.et al.)(2006)Cancer Res 66、10292〜10301)に記載されているように、AffymetrixU133AおよびU133
Bの遺伝子チップを使用して発生させた。全AffymetrixCELファイルを、Gene Expression Omnibus(http://www.ncbi.nlm.nih.gov/geo/)から検索し、グローバル平均法(ミラーら(Miller et al.)、2005、上記)を使用して標準化し、プローブセットのシグナル強度を自然対数変換し、平均強度をlog 500の標的シグナル値に調整することによって尺度化した。コホート依存性アレイのバイアスを取り除くために、それぞれのコホート内のアレイを、それらを1つのデータセットに組み込む前に平均化した。すべての患者および腫瘍試料の臨床的特徴を、図2に要約した。
【0077】
局所的特異値分解(LSVD)。マイクロアレイのプローブセットを、UCSD Genome Browserに従ってゲノム座標にマッピングした。候補PEAを、対応する腫瘍のサブセット中の遺伝子の、局在座標化された過剰発現として同定した。第1に、遺伝子を、試料中の遺伝子の発現が、全試料にわたったその発現中央値から調整された中央絶対偏差(aMAD)の2.5倍を超えた場合に、試料中の過剰発現として定義した(図1、第1工程)。第2に、局所座標化された過剰発現を、50のプローブのスライディングウィンドウに適用したSVDにより得られた主固有値によって定量化した。同じ遺伝子に対応するプローブ(UniGene ID、build #177に対応する)を平均し、単一遺伝子の測定値を表した。候補PEAを、ベースライン閾値を上回る局所主固有値を有する連続した染色体領域として同定した。候補PEAにスコアを付け、主固有値のピークによりランク付けした。最も高いスコアを得た候補PEAを、ピーク構造をさらに解明するために(すなわち、複数のサブピークから1つを識別するために)、10のプローブのウィンドウサイズを使用して、LSVDの第2ラウンドに供した。最後に、最も高いスコアを得たサブピークを、個人の遺伝子および腫瘍に関する固有重量を導くために、その後SVDに供した。固有重量は、主固有ピークの大きさに対する、遺伝子および腫瘍の相対的寄与率を反映する。得られたピークに関して、非ゼロの固有重量を有する遺伝子は、PEAの成分であると考えられる。中央値+2標準偏差を超える固有重量を有する腫瘍を、アンプリコン含有腫瘍として分類した。(TRIAGE、第4工程)。
【0078】
再発性ゲノム増幅を同定するための局所特異値分解(LSVD)
腫瘍試料のコホートに由来する遺伝子発現プロフィールのLSVD分析は、推定発現アンプリコン(PEA)の同定における鍵となる第1の工程であり、これにより、転移に寄与する癌遺伝子を、TRIAGE(Clinico−Genomic Intersectsを介した癌遺伝子三角検証(Triangulating Oncogenes through Clinico−Genomic Intersects))方法論により発見できる。LSVDは、再発性アンプリコンが、腫瘍試料のサブセット中にゲノム的に局在化された遺伝子の座標化された過剰発現として現れるという仮説に基づき、PEAを同定する。LSVDは、特異値分解(SVD)(ストラング、ジー(Strang,G.)、(1998)「Introduction to Linear Algebra」、Section 6.7.第3版、ウェルズリ−ケンブリッジプレス社(Wellesley−Cambridge Press.)ISBN 0−9614088−5−5)を中心に構築されており、染色体ごとのSVDの局所適用により得られる、局所主固有値(LPEV)のプロットである、座標化された局所過剰発現プロットのピークとしてPEAを予測する。LSVDは、ゲノム上の各ゲノムの位置に関する局所発現行列を導き、バイナリ連結行列に変換し、バイナリ行列にSVDを適用することにより達成される。このような適用で得られた主固有値は、その遺伝子座における遺伝子の座標化された過剰発現の基準である(クルーガー、ワイら(Kluger,Y.et al.)、(2003)Genome Research、13、703〜716;クラインバーグ、ジェイ エム(Kleinberg,J.M.)、(1999)ACMのJl、46、5、604〜632)。LPEV対ゲノム位置のプロット(LPEVプロット)における特定のベース閾値を上回る、固有値の「ピーク」は、PEAの境界を定める。LSVDの手順の詳細
な説明は、以下に提供する。
【0079】
Ecを、染色体Ccの遺伝子発現行列とし、Ecij(注:EcijはPCT出願時英文ではベクトルのため太字)を、腫瘍Tjの中の位置liにおける遺伝子の発現のlog2とし、i=1、2、...、Nc、およびj=1、2、...、Mである。Cc上の推定発現アンプリコン(PEA)は、LSVDを使用してEcを分析することによって予測され、LSVDは以下の順序の工程:(1)Ec(注:EcはPCT出願時英文ではベクトルのため太字)を、バイナリ連結行列Ac(注:AcはPCT出願時英文ではベクトルのため太字)に変換する工程、(2)染色体局在化行列Apc(注:ApcはPCT出願時英文ではベクトルのため太字)を導く工程、(3)ApcにSVDを適用する工程、および(4)PEAを同定する工程、を有する。
【0080】
1.Ecのバイナリ連結行列Acへの変換:Ecを、以下のようなブールの離散化定理を介して、バイナリ連結行列Acへ変換する(注:Ec、AcはPCT出願時英文ではベクトルのため太字)。
【0081】
Ecij>μi+x*νiの場合Acij=1
Ecij≦μi+x*νiの場合Acij=0
μiおよびνiが中央値であり、調整中央絶対偏差(aMAD)が、Eci={Eci1、Eci2、...、EciM}である場合、aMADはMADの1.4826倍である。xは、粗いマップの第1のパスに関しては2.5であり、第2のパスにおいて得られた、より詳細なマップに関しては2である。
【0082】
2.局所行列Apcの導き(注:ApcはPCT出願時英文ではベクトルのため太字)。
位置Lpcにおける局所行列Apcは、染色体Cc上の位置p−wからp+wまでの遺伝子を使用してAcから導き、wはウィンドウサイズであり、所定のパラメーターである。wは、第1のパスの粗いマップに関しては50であり、第2のパスのより詳細なマップに関しては10である。
【0083】
3.特異値分解(SVD):SVDのApcへの適用は、PEAを予測するための重要な工程である。ApcのSVDは、Apcを3つの行列、Upc、Σpc、Vpc(注:Upc、Σpc、VpcはPCT出願時英文ではベクトルのため太字)の積、すなわちApc=Upc×Σpc×VpcTに分解する。UpcおよびVpcは、それぞれ、(2w+1)×(2w+1)およびM×Mの行列であり、一方Σpcは、(2w+1)×Mの対角行列である。UpcおよびVpcの列ベクトルは、それぞれ、ApcApcTおよびApcTApc(注:ApcApcTおよびApcTApcはPCT出願時英文ではベクトルのため太字)の固有ベクトルであり、一方Σpc(注:ΣpcはPCT出願時英文ではベクトルのため太字)の対角成分は、各固有値である。最も高い固有値を、主固有値(λpcで表す)と呼び、各固有ベクトルを主固有ベクトルと呼ぶ。位置Lpcにおける腫瘍Tjの固有重量はTpcjで表され、行列ApcTApcの主固有ベクトルのjth成分の絶対値として定義する。同様に、Gpcjで表される、位置Lpcにおける遺伝子Giの固有重量は、行列ApcApcTの主固有ベクトルのjth成分の絶対値である。
【0084】
真のPEAシグナルとバックグラウンドとの差異を改善するために、本発明者らのLSVDの実施において、本発明者らは、SVDを、Apcについて使用する代わりに(ApcApcT)3について使用し、λpcを見出した。上記の定義に従って、腫瘍および遺伝子の固有重量が、それぞれ(ApcTApc)4および(ApcApcT)4の主固有ベクトルから得られた。ここで、λpcは、(ApcApcT)4対M4の主固有値4乗根との比であり、遺伝子および腫瘍の固有重量は、それぞれ、(ApcApcT)4/M
4および(ApcTApc)4/M4の主固有ベクトルの各成分の4乗根である(注:ApcおよびApcTはPCT出願時英文ではベクトルのため太字)。
【0085】
4.PEAの予測:
λpc対Lpcをプロットすることにより、LPEVのプロットが得られ、これをLPEVプロットと呼ぶ。このプロットのピークは、PEAを示す。ピークにおけるλpcの値が高いほど、Lpcの周りのPEAの信頼性が高い。最終的に、PEAは、λpc>Tλ=20を含む、連続した領域を対象とする範囲を有するピークおよびこのピークを中心として報告される。
【0086】
TRIAGE遺伝子生存率分析。各プローブセットに関して、平均発現レベルを、所与のコホートに関してまず計算した。その後、患者を、平均より下の群と平均より上の群とに階層化し、コックス比例ハザード回帰モデルをこのデータに適合させ、無遠隔転移生存率(DMFS)に基づくハザード比を計算した。帰無仮説は、2つの群は再発の危険性が同じであろうと言うものであり、尤度比検定のp値はこの仮説に対する証拠として計算した。コックスモデルの適合および統計的計算を、Rでサバイバルパッケージ(the survival package)を使用して実施した。同じ遺伝子にマッピングされた複数のプローブセットの場合、最も有意に関与するプローブセットの結果が報告された。DMFSを、外科手術から最初の遠隔再発事象または最後の追跡調査の日までの時間間隔として定義した。両側および対側の癌を有するすべての患者ならびに再発したまたは診断後10年を超えて疾患特異的な死を迎えた人を、事象に関して体系的に審査した。ウプサラコホートは最も長く、最も完全に患者を追跡調査したコホートに基づく、本発明者らの最も大きい集団であったので、それを使用し、TRIAGEにおける生存率データを作成した。
【0087】
増殖サインおよび遺伝子相関。cDNAマイクロアレイを使用して、ウィットフィールドら(Whitfield et al)は850を超える細胞周期調節遺伝子を、ヒーラ子宮頸癌細胞においてすでに同定していた(ウィットフィールド、エム エルら(Whitfield,M.L.et al.)、(2002)Mol Biol Cell、13、1977〜2000)。著者らは、乳房の腫瘍における高度に相関した発現を示し、ペローら(Perou et al)の乳癌データセット(ペロー、シー エムら(Perou,C.M.et al.)(2000)Nature 406、747〜752)の腫瘍細胞増殖に関与した、これらの遺伝子の分画を発見した。この増殖サインの近似値を求めるために、本発明者らは、細胞周期調節遺伝子(ウィットフィールド(Whitfield)の研究における1,134 IMAGEクローンに対応する)を、Uni Gene build #187を介してAffymetrixプローブセットにマッピングし、ウプサラコホートの腫瘍を階層的にクラスタ化した。増殖関連遺伝子を含む独特のクラスタが現れ(図10を参照されたい)、組織学的悪性度、分裂指数およびKI67スコアを含むウプサラコホートにおける増殖のマーカーと高度に相関することが見出された(データ非掲載)。>0.8の平均相関を有する、このクラスタの中心となるプローブセット(n=78のプローブセットが67の遺伝子に対応する(図10を参照されたい))を、「コア増殖サイン」遺伝子に指定し、それらの平均発現を使用して、増殖サインと、TRIAGE分析の第7工程のための他のすべてのプローブセットとのピアソン相関を計算した。増殖と癌の再発との臨床的関連性次第では、特に、この工程は、他の癌型のTRIAGE分析には必須ではないことがある。
【0088】
RAB11FIP1(RCP)の選択およびクローン化。RCPの3種のスプライス変異体:変異体1(NM_025151)、変異体2(NM_001002233)および変異体3(NM_001002814)を、GenBankで同定した。これらの中で、変異体1および3だけが、腫瘍試料および細胞系においてRT−PCRにより一貫して検
出でき(データ非掲載)、変異体1だけが、ポリクロナールRCP抗体(GenWay)を用いたウェスタンブロットにより観察できた。したがって、RCPの完全長変異体1は、正常なヒト血液からRT−PCRにより増幅され(順方向プライマー:5’−ACCATGTCCCTAATGGTCTCGGCT−3’(配列番号1)、逆方向プライマー:5’−TGCTGATTTACATCTTTCCTCT−3’)(配列番号2))、配列を確認し、この研究においてRCPの機能を調査するために使用した。
【0089】
細胞系および一過性形質移入。ヒト乳房細胞系、MCF10A、MCF7およびMDA−MB231を、American Type Culture Collection(ATCC)から入手し、37℃、5%CO2で、ATCCにより推奨されている成長培地で維持した。完全長RCPをpcDNA3.1のBamHlおよびEcoR1部位に連結した。2種のRCP特異的RNAi構築体を、先に記載したように(レイノルズ、エーら(Reynolds,A.et al.)、(2004)Nat Biotechnol、22、326〜330)設計し、Ambionにより作製した。MCF7について、一過性プラスミドおよびRNAiの形質移入を、Lipofectamine 2000(インビトロジェン社(Invitrogen))を使用し、製造業者のプロトコルに従って実施した。
【0090】
レンチウィルスに感染させることによる安定細胞系の作製。shRNAを安定して過剰発現する細胞系を作成するために、標的配列(順方向:5’−TCGATAAGCAAGAAGGAGTTTTCAAgAgAAACTCCTTCTTGCTTATCGTTTTTTC−3’(配列番号3)および逆方向:5’−TCGAGAAAAAACGATAAGCAAGAAGGAGTTTCTCTTGAAAACTCCTTCTTGCTTATCGA−3’)(配列番号4)をコードするオリゴヌクレオチドを、LentiLox pLL3.7ベクターにアニーリングし、クローン化した。対照shRNAのために、非標的「スクランブル」配列を、pLL3.7.(順方向:5’−TGAACGGCATCAAGGTGAACttcaagagaGTTCACCTTGATGCCGTTCTTTTTTC−3’(配列番号5)および逆方向:5’−TCGAGAAAAAAGAACGGCATCAAGGTGAACTCTCTTGAAGTTCACCTTGATGCCGTTCA−3’)(配列番号6)内にクローン化した。レンチウィルスによるRCPの過剰発現のために、RCPをまず、Invitrogen gatewayエントリーベクターpENTR3Cにクローン化し、さらに、レンチウィルスベクターpLenti6/V5−DEST(インビトロジェン社(Invitrogen))に、製造業者のプロトコルに従ってLR組換え反応により組換えた。レンチウィルスの作製のために、pLL3.7またはpLenti6/V5−DESTを、パッケージングベクターを用いて、293FT細胞に共形質移入し、48時間後に上清を収穫した。ウィルスを、Beckman SW28ローター内で、25,000rpmで2時間、超遠心分離器にかけて濃縮し、リン酸緩衝食塩水に再懸濁した。力価を、濃縮ウィルスの段階希釈液を用いて、NIH/3T3細胞に感染させることによって確定した。標準的な調製用に、力価は、pLL3.7に関してはおよそ1〜5×108粒子であり、pLenti6/V5−DESTベクターに関しては4−10×107粒子であった。MCF10A、MCF7およびMB231細胞の感染のために、2×105細胞を、1×107粒子および8μg/mlポリブレンの懸濁液中で、3時間、37℃においてインキュベートした。細胞をその後、再度播種し、前記のように培養した。RCPノックダウン安定細胞の純粋な集団を得るために、GFP陽性細胞を、培養4日後に選り分けた。RCP過剰発現安定系の作製のために、細胞を、ブラストサイジンを用いて感染2週間後に選択した。
【0091】
ウェスタンブロット分析。すべての細胞溶解物を、RIPAバッファーを使用して調製した。タンパク質を、SDS−PAGEにより分離し、Hybond−P PVDF メンブラン(ジーイーヘルスケア社(GE Healthcare))に転写した。RCP
(ジェンウェイ社(Genway)、カタログ番号(CN):15−288−21574A)、β−アクチン(シグマ社(Sigma)、CN:A5441)、ホスホ−Akt(セルシグナリングテクノロジー社(Cell Signaling Technology)、CN:9271)、Akt(セルシグナリングテクノロジー社(Cell Signaling Technology)、CN:9272)、ホスホ−Erk(セルシグナリングテクノロジー社(Cell Signaling Technology)、CN:9101)およびErk(セルシグナリング社(Cell Signalling)、CN:9102)に対する抗体を使用して、メンブランを調査し、抗体−タンパク質複合体を、HRP標識抗体およびECL(アマシャムバイオサイエンス社(Amersham Biosciences))により検出した。
【0092】
細胞増殖およびコロニー形成。細胞を、96ウェルプレートに、5000細胞/ウェルの濃度で播種し、細胞増殖を、WST−1(ロッシュ社(Roche))を使用し、製造業者のプロトコルに従って4重に測定した(すなわち、条件当たり4ウェル)。MAPK阻害アッセイのために、96ウェルプレートに再度播種する前に、細胞を、MEK阻害剤U0126(プロメガ社(Promega))を用いて、0.5%の血清を含む培地で一晩処理した。足場非依存性コロニー形成に関するRCPの作用を試験するために、細胞を、25%FBSを含む完全培地に溶解した0.3%寒天(シグマ社(Sigma))250μlに再懸濁し、500μlの0.6%寒天ベースであらかじめコーティングした24ウェルプレートに4重(すなわち、条件当たり4ウェル)に播種した。コロニー形成の効率を、播種の21日またはそれ以後に、ヨードニトロテトラゾリウムクロリド(シグマ社(Sigma))を用いて染色することによって検査した。コロニーの大きさが50umを超えるコロニーをLeica QWinソフトウェアを使用して計測した。
【0093】
in vitroの浸潤および遊走。トランスウェルの遊走および浸潤アッセイを、孔サイズ8μmのFalcon FluoroBlok 24マルチウェルインサート(ビーディーバイオサイエンス社(BD Biosciences))を使用して実施した。浸潤アッセイのために、インサートを、80μLの、無血清成長培地中の、20μgのMatrigel(ビーディーバイオサイエンス社(BD Biosciences))でコーティングした。両方のアッセイのために、200μL無血清成長培地中の5000細胞を、10%ウシ胎児血清を含む750μL完全成長培地を、下方のチャンバーに入れた、各トランスウェルインサートに入れた。24時間後、インサートの孔を介して遊走または浸潤した細胞を、3.7%ホルムアルデヒドで固定し、2.5μg/mL Hoechst 33342(インビトロジェン社(Invitrogen))を用いて15分間染色し、PBSで洗浄し、Target Activation Bioapplicationを使用してArrayScan VTI(セロミクス社(Cellomics))上で計測した。フィールドサイズは1mm2であった。浸潤アッセイのために、実験を4〜5倍の複製を用いて実施し、実験ごとに10〜16個のフィールドをスキャンした。遊走アッセイのために、実験を3倍の複製を用いて実施し、実験ごとに10〜16個のフィールドをスキャンした。
【0094】
細胞の免疫蛍光顕微鏡検査。細胞培養物を、4%パラホルムアルデヒドを用いて固定し、0.25%Triton X−100を用いて透過処理し、その後1%ヤギ血清を用いてPBS中でブロッキングした。細胞を、抗E−Cadherin(ビーディーファルミンゲン社(BD Pharmingen)、CN:610404)および1:500希釈のフィブロネクチン抗体(ビーディーファルミンゲン社(BD Pharmingen)、CN:610077)を用いて染色し、その後、マウスおよびウサギのIGgを検出する適切な2次抗体を、それぞれAlexa Fluor 594または488(モレキュラープローブ社(Molecular Probes))に接合した。F−アクチンは、Texas Red−X phalloidin(モレキュラープローブ社(Molec
ular Probes))を用いて標識した。DAPIを、核の染色に使用した。画像を共焦点顕微鏡、(LSM 510 META、ツァイス社(Zeiss))を用いて獲得した。
【0095】
細胞周期分析。MCF10A細胞を、1%血清中で48時間飢餓状態にした。細胞をその後、75%エタノールで固定し、RNase A(0.25mg/ml)を用いて処理し、ヨウ化プロピジウム(10μg/ml)を用いて染色し、LDR2フローサイトメーター(ビーディーバイオサイエンス社(BD Biosciences))で分析した。分類した細胞を、FACS DiVaソフトウェア(ビーディーバイオサイエンス社(BD Biosciences))を使用して分析した。
【0096】
アポトーシスアッセイ。MB231細胞を、48時間、完全培地で維持または1%血清を用いて飢餓状態にした。アポトーシス細胞を、In Situ Cell Death
Detection Kit(ロッシュ社(Roche))を用いて検出した。DNA鎖の壊れた細胞を、TMR(赤)を用いて、TUNEL(TdT−mediated dUTP nick end labeling)反応により標識し、蛍光顕微鏡検査およびフローサイトメトリーにより分析した。
【0097】
ヌードマウスにおける腫瘍原性。メスのBALB/c−nu/nu(ヌード)マウス、6〜8週齢を、無菌条件下で、温度調節室において、12/12時間の明/暗スケジュールで、食餌および水は好きな時に取らせて飼育した。動物およびそれらの世話に関するすべての手順は、国内的および国際的規格に従った。MCF−7腫瘍形成実験のために、1.7mgの17β−エストラジオールのペレット(90日放出、イノベイティブリサーチオブアメリカ社(Innovative Research of America))を、腫瘍細胞の注入1週間前に各マウスに移植し、MCF−7細胞のエストロゲン要求性を補った。実験群は、MCF7−RNAi RCP(安定RCPノックダウン)、MCF7−RNAi Ctrl(対照)、MB231−RNAi RCP、MB231−RNAi Ctrlであった。各実験群に関して、8匹のマウスの皮下(右および左の後脚上部)に2.0×106の細胞を注入した。皮下腫瘍を、デジタル計測器を用いて、週に1回測定した。腫瘍の堆積は、ab2/2としてmm3で計算し、aおよびbは、それぞれ直交する、腫瘍の最も長い径および最も短い径である。腫瘍重量は終了時に測定し、シンガポール動物飼育規則に従って死亡させた。
【0098】
免疫組織化学。4μmの厚さのパラフィン包埋腫瘍組織切片を、ABC免疫組織化学試薬およびアビジン−ビオチンブロッッキング試薬(ベクターラボラトリーズ社(Vector Laboratories))を使用して、製造業者のプロトコルに従って染色した。スライドを、1:500希釈の抗RCP抗体と共にインキュベートし、ヘマトキシリンを用いて対比染色した。
【0099】
結果
上記のデータにより、ゲノム、転写および臨床的な情報の座標分析が、転移性再発および患者の死に寄与していると思われる癌の新規遺伝子の発見を良好に対象とでき、その結果、下流の機能研究のための遺伝子の理論的優先順位付けが容易になることが実証された。ヒト乳癌における、複数機能の頻繁に発生する癌遺伝子としてのRAB11FIP1の例示的検証は、治療標的としてのその重要性を強調し、発癌におけるエンドサイトーシス再循環経路の新たな役割に新しい洞察を提供する。
【0100】
例示目的で、ヒト乳癌形成において複数の病理学的役割を有する新規な癌遺伝子として染色体8p11に位置する、RAB11FIP1の機能的検証を本明細書で提供する。RAB11FIP1は、Rab4およびRab11の結合タンパク質としてすでに同定され
ており、Rabカップリングタンパク質(RCP)とも呼ばれ、これはエンドソームの再循環および受容体の分類において機能する(リンゼイ、エー ジェイら(Lindsay,A.J.et al.)(2002)J Biol Chem 277、12190〜12199;ペデン、エー エーら(Peden,A.A.et al.)(2004)Mol Biol Cell 15、3530〜3541)。RAB11およびRAB25の異常調節を介して、発現は、多くのヒトの腫瘍の一般的要素として示され(例えば、チェン、ケイ ダブリュら(Cheng,K.W.et al.)(2004)Nat Med 10、1251〜1256;チェン、ケイ ダブリュら(Cheng,K.W.et al.)(2005)Methods Enzymol 403、202〜215;ゲブハルド、シーら(Gebhardt,C.et al.)(2005)Am JPathol 167、243〜253;ゴールデンリング、ジェイ アールら(Goldenring,J.R.et al.)(1999)Yale JBiol Med 72、113〜120;ヨーン、エス オーら(Yoon,S.O.et al.)(2005)Cancer Res 65、2761〜2769を参照されたい)、腫瘍進行におけるRab結合タンパク質の役割については記載されていない。本発明者らは、臨床的およびゲノム的交差を介して癌形成の関連癌遺伝子に三角検証を用いる、統合分析戦略を考案し、これは、発現の頻度、アンプリコンにおける共発現、患者の無病生存率との関連を考慮し、腫瘍増殖の偶発的関連は考慮に入れない。この多段階工程のデータ検索研究法である、略してTRIAGE(Triangulating Oncogenes through Clinico−Genomic Intersects、以下を参照されたい)を使用して、本発明者らは、RAB11FIP1を癌遺伝子として同定した。
【0101】
TRIAGEによる癌遺伝子の推論
進行性乳癌に寄与する癌遺伝子を同定するために、このような遺伝子の特定のサブセットは、転写学的、ゲノム的および臨床的情報の三角検証を介して発見できるという仮説を立てた。特に、本発明者らは、1)再発性増幅の領域に位置し、2)遺伝子の異常調節により活性化し、3)癌の転移性播種に関連する癌遺伝子を明らかにするよう探求した。この目標のために、本発明者らは、TRIAGE(Triangulating Oncogenes through Clinico−Genomic Intersects)と呼ばれる、複数工程のデータ検索研究法を開発し、これは、発現マイクロアレイデータにおける再発性ゲノムアンプリコンの発現フットプリントを同定し、遺伝子発現を、転移性再発の危険性と関連付け、生存率相関および比較mRNA動学に基づく遺伝子の立候補を評価する(図1)。
【0102】
第1の工程において、737例の原発性乳房腫瘍のマイクロアレイ発現プロフィール(図2)を、局所特異値分解(Local Singular Value Decomposition)(LSVD)により分析し、再発性ゲノムアンプリコンの存在および位置を推測した。この研究は、再発性アンプリコンは、関連遺伝子の過剰発現を座標化することによって検出できるという前提に基づき(チン、ケーら(Chin,K.et al.)、(2006)Cancer Cell 10、529〜541;レアル、エフら(Reyal,F.et al.)(2005)Cancer Res 65、1376〜1383)、乳癌において高度に増幅した遺伝子の大部分が同時過剰発現であるという証拠に支持されている(ポラック、ジェイ アールら(Pollack,J.R.et al.)、(2002)Proc Natl Acad Sci USA、99、12963〜12968)。LSVDは、遺伝子の過剰発現が濃縮されたゲノム座および「遺伝子座特異的」過剰発現を生じる腫瘍分画の同時同定を可能にする。アウトプットは、遺伝子座特異的過剰発現の程度および関連する腫瘍数の両方を反映する、主固有ピークとしてみなされる。全ゲノムLSVDにより発見された、最も高いスコアを得た主固有ピークは、再発性コピー数増加の公知の領域に対応する遺伝子座に優勢にマッピングされた(図3を参照されたい)。LSVDによりすべてのピークが同定されたわけではないが、再発性ア
ンプリコン事象を表すことが期待され、公知の再発性アンプリコンと、非常に大きく、正確なゲノム重複を有するものを推定発現アンプリコン(PEA)と呼び、TRIAGE手順によりさらに調査した。
【0103】
対象となる癌遺伝子の発現レベルは、患者の予後不良に相関するだろうという指針に基づき、PEA内の遺伝子を、遠隔再発関連の有意性に関して評価した。各遺伝子のために、腫瘍(および対応する患者)を、低発現群(平均より低い)または高発現群(平均より高い)に、2つに分け、コックス比例ハザード回帰により、無遠隔転移生存率(DMFS)に関して分析した(図1、第2および第3工程)。得られたハザード比の有意性を反映する尤度比検定のP値を、PEAの遺伝子の発癌性の初期基準として使用した(図1、第3工程、任意抽出の生存率)。しかし、再発性ゲノムアンプリコンは、患者の生存率不良に相関し得るので、アンプリコン上に物理的には存在する良性の「パッセンジャー遺伝子」もまた、腫瘍の侵攻に機能的に寄与することなくDMFS不良に相関でき、一方、ドライビング癌遺伝子(コピー数増加以外の機序により癌において活性化できる)は、アンプリコンを欠いた腫瘍においてDMFS不良との相関を維持することが予想され、パッセンジャー遺伝子は予想されない。したがって、この分析から審査される、遺伝子発現と、含有アンプリコン腫瘍のDMFSとの相関を再検査した。(図1、第4工程、任意抽出)。この方法では、増幅事象に非依存性の、DMFS不良に確実に関連するそれらの候補癌遺伝子のみが分かってくるであろう。
【0104】
生存率分析の両方の工程において有意な候補癌遺伝子を、mRNA発現パターンの比較を介してさらに精査した(図1、第5および6工程)。ドライビング癌遺伝子は、アンプリコン含有腫瘍において比較的高度に一貫して発現されるはずであるという仮定に従って、PEA内の遺伝子発現レベルを平均し、その次に遺伝子を平均発現によりランク付けした(ランク1が、最も高い平均発現レベルを表す)。この方法では、発現のランキングは、所与の遺伝子座における発癌性のさらなる相対的尺度として機能する。
【0105】
高増殖性乳房腫瘍は、再発する可能性が高く、増殖に関連する遺伝子は、多くの場合DMFS不良に相関している。このことは、細胞周期機能を濃縮する、何百の高度関連遺伝子からなる増殖サインにおいて実証され、これは多くの癌型の再発に正に相関することが見出された(ダイ、エイチら(Dai,H.et al.)(2005)Cancer Res 65、4059〜4066;イブシナ、エー ブイら(Ivshina,A.V.et al.)(2006)Cancer Res 66、10292〜10301;ペロー、シー エムら(Perou,C.M.et al.)(2000)Nature
406、747〜752;ローゼンウォルド、エーら(Rosenwald,A.et
al.)(2003)Cancer Cell 3、185〜197;ウィットフィールド、エム エルら(Whitfield,M.L.et al.)(2002)Mol
Biol Cell 13、1977〜2000;ウィットフィールド、エム エルら(Whitfield,M.L.et al.)(2006)Nat Rev Cancer6、99〜106)。この調査は、増殖との関連とは非依存性の、発癌に寄与する固有の遺伝子に関し、TRIAGEの最終工程において、候補遺伝子の発現パターンを増殖サインとの相関について試験し、生存率との関連が腫瘍の増殖率により説明できる程度を検討した。DMFS不良に相関する、ウプサラ乳房腫瘍コホートのプローブセット(n=251腫瘍)を図4に表す。DMFS不良(p<0.05)に有意に相関することが見出された、2,377のプローブセットの約35%が、ピアソン相関が、>0.4でこの増殖サインのメンバーであることが見出された(図4)。増殖サインの中核コア遺伝子は、クラスタ化の順に、B.222958_s_(DEPDC1)、A.204170_s_(CKS2)、B.222680_s_(DTL)、A.209680_s_(KIFC1)、A.202503_s_(CSNKIG1)、A.201291_s_(TOP2A)、A.201292_(TOP2A)、A.204033_(TRIP13)、B.
225687_(FAM83D)、A.200853_(H2AFZ)、A.213911_s_(H2AFZ)、A.203362_s_(MAD2L1)、B.228729_(CCNB1)、A.214710_s_(CCNB1)、A.203755_(BUB1B)、B.228323_(CASC5)、A.204825_(MELK)、A.204822_(TTK)、A.207828_s_(未知)、A.202870_s_(CDC20)、A.222039_(LOC146909)、A.204962_s_(CENPA)、B.228273_(未知)、A.209408_(KIF2C)、A.202705_(CCNB2)、A.222077_s_(RACGAP1)、A.218542_(CEP55)、A.203213_(CDC2)、A.203214_x_(CDC2)、A.210559_s_(CDC2)、A.218039_(NUSAPI)、A.218009_s_(PRC1)、A.204444_(KIF11)、A.210052_s_(TPX2)、A.219918_s_(未知)、A.203764_(DLG7)、B.223307_(CDCA3)、A.202095_s_(BIRC5)、A.202954_(UBE2C)、A.202580_x_(FOXM1)、A.218726_(DKFZp762E1312)、A.203554_x_(PTTGI)、A.209642_(BUB1)、A.209773_s_(RRM2)、A.201890_(RRM2)、A.204641_(NEK2)、A.218355_(KIF4A)、A.208079_s_(未知)、A.204092_s_(AURKA)、A.38158_(SP1)およびA.218755_(KIF20A)。乳癌において、DMFS不良に有意に相関することが見出されたプローブセットのおよそ1/3が、増殖サインのメンバーであることが同定されたので、生存率不良関連遺伝子のおよそ3つに1つが、非依存性発癌機序ではなく、増殖との関連により最もよく説明できる。したがって、発明者らは、選択過程において、生存率に有意に関連し、さらに(>0.4の)増殖サインと相関する候補遺伝子の優先順位を下げた、または考慮に入れなかった。
【0106】
この分析研究の妥当性を試験するために、TRIAGEを、全ゲノムのLSVDにより同定された最も大きな主固有ピークに用いた。これは、約30の遺伝子を包含する染色体17q12の1Mb領域に対応する。このPEAの遺伝子成分を、EGFR−関連癌遺伝子ERBB2(HER2/neu)に収めた、乳癌の症例の約20〜30%に生じることが公知である、十分に特徴づけられた17q12アンプリコンに正確にマッピングした。特に、このアンプリコンの発現フットプリントは、すでにマイクロアレイ研究において観察されており、アレイ−CGHにより判別されるように、物理的アンプリコンの境界とよく相関することが示されている(チンら(Chin et al.)、2006、上記;ポラックら(Pollack et al.)、2002、上記)。このPEAを示す腫瘍において、本発明者らは、免疫組織化学によりHER2陽性の強い濃縮を見出し(P<1.0×10−26;カイ二乗検定)、このPEAが物理的17gl2アンプリコンに対応する証拠と一致する(図5A、図5C、黒色の垂直のバー)。初期の(任意抽出の)遺伝子生存率分析により、DMFSに有意に関連するこのPEAの中の6つの遺伝子が同定された(図5A)。PEAを有する腫瘍を有する患者の不在下での生存率(すなわち選択された生存率)の再分析では、DMFSに有意に関連する2つの遺伝子:TCAPおよびERBB2のみが残った。どちらも増殖サインと強く相関しないが、ERBB2は最も強い発現ランキングを有した(図5E)。したがって、TRIAGEにより、ERBB2(受容体チロシン−タンパク質キナーゼ、erbB−2の前駆体をコードする)を、転移促進癌遺伝子および乳癌における治療標的としてのその公知の役割とは無関係に、17q12遺伝子座における発癌ドライバーの「トップ候補」として同定した。
【0107】
LSVDにより同定された第2の大きな主固有ピークを、1.5Mbにわたる領域で22の遺伝子を包含する、第8染色体のp腕にマッピングした。このPEAは、乳癌の10〜25%に発生することが公知の、すでに記載された8p11−12アンプリコンと遺伝子レベルで正確に重複することが見出された。(ガルシア、エム ジェイら(Garci
a,M.J.et al.)(2005)Oncogene24、5235〜5245;ジェルシ−ボイヤーら(Gelsi−Boyer et al.)、(2005)Mol
Cancer Res 3、655〜667;ルテシア、エーら(Letessier,A.et al.)(2006)BMC Cancer 6、245;レイ、エム イーら(Ray,M.E.et al.)(2004)Cancer Res 64、40〜47)。このアンプリコンの発癌ドライバーは、十分に特徴づけられていない。TRIAGEによって、本発明者らは、遠隔転移の危険性の増加に有意に関連するこのアンプリコン上の3つの遺伝子:RAB11FIP1、EIF4EBP1およびWHSC1LIを特定した(図5B)。選択された遺伝子−生存率の相関の分析において、遠隔転移に有意に関連する、わずか2つの遺伝子、RAB11FIP1およびWHSC1LI(ヒストン−リシンN−メチルトランスフェラーゼNSD3をコードする)が残り、RAB11FIP1は、遠隔再発との最も有意な関連および最も高い発現ランクを有した(図5E)。この発見に基づいて、本発明者らは、乳癌の発癌特性に関するRAB11FIP1の機能的影響を調査した。
【0108】
RCP過剰発現は、MCF10A細胞を形質転換させる
RAB11FIP1(本明細書ではRCPとも称する)が、乳腺腫瘍においてタンパク質レベルで増幅するかどうかを確認するために、腫瘍進行の様々な悪性度の細胞系のパネルにおいて、市販の抗体を使用して、ウェスタンブロット分析を実施した。RCPは非腫瘍発生(正常な)乳腺上皮細胞系MCF10Aにおいて最も低いが、乳癌細胞系、CAMA−1、MDA−MB231、MCF7およびT47Dにおいて4〜10倍高いレベルで発現した(図6A)。
【0109】
次に、乳癌におけるRCP増幅の役割を、完全長RCP(MCF10A−RCP)(Genbankアクセス番号:NM_025151)または対照ベクターを安定して過剰発現するMCF10A細胞系を確立することによって調査した。対照細胞は、平板な形態および正常な上皮の単層成長の特徴を示すが(図6B、左のパネル)、RCP過剰発現体は、紡錘様の突起および所々に、形質転換線維芽細胞を連想させる病巣様構造がちりばめられ散在した成長パターンを含む劇的な形態の変化をもたらした(図6B、右のパネル)。一方、対照MCF10A細胞は、平板な形態および正常な上皮においてより特徴的な上皮型島における単層成長を示した。さらに、コンフルエントで、MCF10ARCP過剰発現細胞は、接触阻止の損失を示し、連続的な多層成長を示した。細胞の免疫蛍光染色により、RCP過剰発現が、細胞間の接合部におけるE−カドヘリンの欠失およびβアクチン破壊に関連したことが明らかになった。その上、血清飢餓状態において、対照細胞より、MCF10A−RCP細胞により多くのフィブロネクチンが観察された(図6C)。細胞間接触部位におけるE−カドヘリン(上皮マーカー)の欠失およびフィブロネクチン(間葉細胞マーカー)の発現増加は、両方ともRCP−依存性上皮間葉移行(EMT)と一致する。
【0110】
RCPの作用をさらに評価するために、本発明者らは、血清および追加成長因子に対する、MCF10A細胞の厳しい要求を活用し、それらの増殖を支えた。安定系を、成長因子の存在下および不在下の両方において維持した。対照細胞と比較して、MCF10A−RCP細胞は両方の培養条件において増殖率の有意な増加を示した(図6Dおよび図8A)。最も興味深いことに、追加成長因子(EGF、インスリン)を含まない、1%FBSの最も厳しい培地において、対照系は6日以内に死亡したが、一方MCF10A−RCP細胞は、1カ月を超えて保持でき、成長因子依存性のRCP−仲介の欠失を示した(図6D)。ヨウ化プロピジウム染色による細胞周期分析により、血清および成長因子の飢餓の48時間後、対照細胞の93%がG1期で停止していたのに対してG2M期の細胞はわずか3%であったが、一方、MCF10A−RCP細胞において、細胞の約80%がG1期にあり、細胞の約12%はG2M期にあることが明らかになり、細胞周期進行の調節にお
けるRCPの役割が確認された(図7A)。
【0111】
次に、軟寒天でのコロニー形成アッセイにおいて、MCF10A細胞の足場非依存性成長を推進するRCPの能力を試験した。予想通り、対照細胞は、軟寒天においてコロニー形成ができなかった。しかし、それとは大いに異なり、MCF10A−RCP細胞は4週間後に非常に多くのコロニーを形成し、RCPが形質転換表現型を誘導できることが実証された(図6Eおよび図8B)。その上、本発明者らは、創傷治癒アッセイ(図6F)およびトランスウェルアッセイ(図8D)により実証されるように、RCPの過剰発現がMCF10A細胞の遊走能力を増強していることを観察した。
【0112】
RCPがその癌表現型に影響を与える、1つまたは複数の機序を見抜くために、RCPの発現が、発癌性成長を制御する主要経路の2つであるERKまたはAKTのどちらかを介して成長シグナル伝達を活性化できるかどうかを試験した。MCF10A−RCP細胞は、ERKの強い活性化を示したが、AKTの活性化は示さなかった(図7B)。RCP誘導性Erk活性化がMCF10A−RCP細胞の増殖の有利性と関連があることを試験するために、MCF10A細胞を、活性および不活性の両方のMEK1/2の強力な阻害因子であるU0126で処理した。対照細胞は、U0126濃度の範囲にわたってERKのリン酸化が低いことを示したのに対して、MCF10A−RCP細胞におけるERKのリン酸化状態は、濃度依存性であった(図7C)。30μMのU0126により、ERKのRCP誘導性リン酸化は、ベースラインまで減少し(対照と同じレベル)、一方、RCPのレベルは変化なかった。重要なのは、30uMのU0126により、ERKのリン酸化が低減したMCF10A−RCP細胞が、RCPの発現が高いにもかかわらず、対照レベルまで増殖の減少を示したことである(図7D)。RCP増殖表現型がERKのリン酸化を介してMAPK経路の活性化に依存していることが示唆される。
【0113】
RCPは、多数の乳癌細胞系における腫瘍原性を増強する。
MCF7(エストロゲン受容体陽性、p53−野生型)乳癌細胞におけるRCPの過剰発現は、細胞増殖率、軟寒天におけるコロニー形成活性、および浸潤能を中程度に増加させた(図8A〜C)。MDA−MB−231(MB231;エストロゲン受容体陰性、p53陰性)細胞では、RCPの過剰発現は、コロニー形成活性を増加させたが、細胞増殖率、浸潤および遊走能は増加させなかった(図8A〜D)。さらに腫瘍原性の調節における内因性RCPの役割を検証するために、本発明らは、RNAi技術を用いて内因性RCPの発現を低減した。RCPを、siRNA構築体の一過性形質移入、またはRCPを標的とするshRNAを発現するレンチウィルスベクターの安定形質導入のどちらかによって「ノックダウン」させた。スクランブル配列を有するRNAi構築体を、対照として使用した。RCPを対象とする非依存性かつ非重複性の、2つのsiRNAは、MCF7細胞において、タンパク質レベルの40〜80%を減少させ、細胞増殖を有意に減少させた(図8A、右のパネル)。MCF7細胞におけるRCPのノックダウンもまた、軟寒天におけるコロニー形成を有意に低下させた(図8B)。MB231細胞において、RCPは、コロニー形成の阻害を低下させたが、細胞増殖率は低下させなかった(図8A、B)。両方の細胞系において、RCPノックダウンは、浸潤および遊走を著しく低減した(図8C、D)。さらに、MCF10A細胞における観察と一致して、MCF7細胞におけるRCPの減少もまた、ERKの脱リン酸化をもたらす(図7B)。これらの結果を合わせると、腫瘍細胞における内因性RCPの発現が、増殖、遊走および浸潤の能力の維持にとって重要であり、RCP機能の標的低減は、実行可能な治療戦力を表し得ることを示唆している。
【0114】
RCPのノックダウンはin vivoにおける腫瘍形成を阻害する。
in vivoでの癌細胞におけるRCP低減の作用を調査するために、本発明者らは、RCP(RNAi RCP)に対するshRNAを安定して発現するMCF7およびM
B231細胞の腫瘍形成能と、ヌードマウスにおける対照(RNAi Ctrl)の腫瘍形成能とを比較した。各細胞型を、8匹のマウスの左および右の後脚上部の皮下に埋め込んだ。MCF7は、追加のエストロゲンの存在下で、ヌードマウスにおいて中程度の悪性腫瘍を形成する、エストロゲン依存性の細胞系である。MCF7−RNAi Ctrl(n=8)またはMCF7−RNAi RCP(n=8)を埋め込んだすべてのエストロゲン追加マウスが明らかな腫瘍を発生したが、RCP阻害細胞は、有意により小さいサイズの腫瘍を発生した(P=0.00007;スチューデントt検定)(図9A、図9B、図9C)。一方で、MB231は、ヌードマウスにおいて高度悪性かつ転移性の腫瘍を形成する、エストロゲン非依存性乳癌細胞系である。MB231−RNAi Ctrl細胞を注入されたすべてのマウスは、大型の腫瘍を発生したが、MB231−RNAi RCP細胞を注入された8匹のマウスではそのうちわずか2匹しか腫瘍を発生しなかった。(すなわち、1匹のマウスに2つの腫瘍および別のマウスに1つの腫瘍)(図9D、図9E、図9F)。特に、これらの腫瘍は対照マウスの腫瘍より非常に小型であり、平均重量は、対照のわずか1%であった。これらの腫瘍におけるRCPのノックダウンを、免疫組織化学により確認した(図9G)。
【0115】
MB231細胞におけるRCPのノックダウンがin vitroの増殖に作用しないという事実は、in vivoのRCPの阻害に関して観察された強い抗腫瘍作用が、血清の喪失下での細胞死の誘導に関連し得ることを示唆しており、皮下に埋め込みの腫瘍の血管新生が現れるには数日かかることを示している。したがって、本発明者らは、MB231安定細胞系の細胞死およびアポトーシスへの感受性を、様々な培養条件下で調査した。完全培地において、MB231−RNAi RCPおよび対照系は、どちらの系においてもアポトーシス細胞がほとんどないことにより証明されたように、細胞死において差がないことを示した。しかし、血清飢餓条件下において、対照は、細胞死に対する耐性を維持していたが、RNAi RCP細胞は維持していなかった。有意によりRCPを低減した細胞は、血清飢餓の結果として死亡し、その中でもアポトーシスが約13%検出できた(図9H)。このことは、RCPの減少が、栄養不良環境に対するMB231細胞の脆弱性を増強することを示唆し、これによりin vivoのRCPノックダウンの劇的な抗腫瘍作用が説明できる。興味深いことに、本発明者らは、in vivoのRCP過剰発現に関連する腫瘍原性の増加を観察せず(データ非掲載)、おそらくRCPの内因性レベルにより達成される腫瘍形成に関する最大の病理学的作用を反映する。全体的に見て、結果は、RCPの標的ノックダウンにより、エストロゲン依存性および非依存性の乳房腫瘍の両方のin vivo成長が抑制され、RCPが、治療介入の有益な新規の標的候補として関係があることを示している。
【0116】
考察
原発性腫瘍のゲノムワイドマイクロアレイ分析により、遺伝子発現の固有のパターンにより定義された、新規な臨床的に意義のある腫瘍亜型の発見が可能となった。しかし、より最近、この概念の逆が、腫瘍発生において機能的役割を有する遺伝子亜型を同定しようとするボトムアップ分析戦略を介して調査された。これらの戦略は、遺伝子発現ならびに癌の臨床的特徴およびゲノムの特徴の間の条件関係を解析する。アドラー(Adler)および仲間は、遺伝子連鎖研究(マイクロアレイサインのステップワイズ連鎖分析)を使用して、乳癌における予測的「創傷サイン」の発現を推進する転写機序を明らかにした(アドラー、エー エスおよびチャン、エイチ ワイ(Adler,A.S. and Chang,H.Y.)、(2006)Cell Cycle 5、1148〜1151)。創傷サインに関して腫瘍陽性または腫瘍陰性に有意に関連した、コピー数および遺伝子発現の両方の変化を同定することにより、著者らは、MYC(8q24)およびCSN5(8q13)が機能的に相互作用し、創傷応答遺伝子の転写を制御することを推論し、生化学的に確認した。本研究において、本発明者らは、データ統合および機能的発見上にこの概念を築き、癌進行における機序的役割を有する遺伝子を推定する、相関に基づく新規
な戦略を開発した。
【0117】
TRIAGEと呼ばれる上記の研究は、遺伝子発現の特徴と位置情報(ゲノムの位置)、臨床データ(患者の無遠隔転移生存率)および予測腫瘍生物学(増殖能)とを組み合わせ、過剰発現し、生存率に相関する再発性増幅の遺伝子部位を突き止める。TRIAGEは、設計により機能的癌遺伝子のすべての型を検出するものではないが、ヒト患者において遠隔転移に寄与する悪性過程を推進する、機能的癌遺伝子を解明する可能性を有する。TRIAGEはまた、癌において高い頻度で活性化される癌遺伝子および日常の療法への耐性を伴う癌遺伝子の発見を最適化する。例えば、癌遺伝子に関する本発明者らの調査は、LSVD手順により同定された主固有ピークの大きさにより優先順位を付ける。この大きさは、増幅事象の頻度と正に関連するので、高頻度のアンプリコン(ピークの高さに対応する)は、高い優先順位が得られる。LSVDは、腫瘍発現プロフィールの任意の包括的コレクションに、TRIAGEのロバスト適用を可能にするが、留意すベき固有の限界を有する。LSVDは、所与の増幅事象を有する腫瘍の近似値のみを提供し、実際のアンプリコンの境界の解明は十分ではない。高解像度のアレイ−CGHにより提供されるような、対応するコピー数のデータは、アンプリコン決定の精度および正確さを改善することによって、TRIAGEの性能を実質的に増強できる。
【0118】
TRIAGEの性能に影響を与える追加の因子には、コホートサイズ、患者の追跡の期間および治療の特徴が含まれ、これらのすべては、候補遺伝子の選択に影響を与える。本発明者らの遺伝子−生存率の関連は、本発明者らの最も大きな集団に基づくコホートであるウプサラコホート(n=251例の患者)を使用して計算され、これはさらに、最も長く、最も完全な患者の追跡データを有する。重要なことには、このコレクションの中の腫瘍は、侵攻型の範囲(図2)が、早期乳癌から(小型径、リンパ節陰性、未処理)後期乳癌(大型径、リンパ節陽性、侵攻的に処理された)までの範囲にわたる。したがって、このコホートは、おそらく様々な癌遺伝子により推進されている様々な癌病態生理学を反映している。しかし、患者の混合された治療バックグラウンドは、治療の型に非依存性の生存率関連遺伝子の同定を有利にする。したがって、治療に依存性または2機能の生存率関連癌遺伝子(アンドレ、エフら(Andre,F et al.)(2007)Clin
Cancer Res 13、2061〜2067)は、このコホートでは見分けられないが、他においては同定される可能性がある。
【0119】
本発明者らの統合乳房腫瘍データセットのLSVD分析は、8p11−12の頻繁に増幅される領域に位置する遺伝子群に集中する大きな固有ピークの発見をもたらした。この遺伝子座の増幅は、乳房腫瘍症例の約10〜25%において観察され(アドナン、ジェイら(Adnane,J.et al.)(1991)Oncogene 6、659〜663;ガルシア、エム ジェイら(Garcia,M.J.et al.)(2005)Oncogene 24、5235〜5245;ルテシアら(Letessier et
al.)2006上記、テイレット、シーら(Theillet,C.et al.)(1993)Genes Chromosomes Cancer 7、219〜226)、多数の非依存性研究において、患者の生存率不良および遠隔転移の短い間隔に関連していた(チンら(Chin et al.)、2006、上記、;キュニー、エムら(Cuny,M.et al.)(2000)Cancer Res 60、1077〜1083;ジェルシ−ボイヤー、ブイら(Gelsi−Boyer,V.et al.)(2005)Mol Cancer Res 3、655〜667;ルテシアら(Letessier et al.)2006、上記)。近年、このアンプリコンは、原発性乳房腫瘍および細胞系に関する、いくつかの集中的な機能的ゲノム研究の焦点であった(ガルシアら(Garcia et al.)、2005、上記;ジェルシ−ボイヤーら(Gelsi−Boyer et al.)、2005、上記;レイら(Ray et al.)、2004、上記)。染色体8pに特異的な高解析度BACマイクロアレイを使用して、
ジェルシ−ボイヤー(Gelsi−Boyer)および仲間は(2005、上記)、8p11−12アンプリコンが、実際は境界の明確なA1、A2、A3およびA4と呼ばれる重複が最小の4つのサブアンプリコンを決定した。この研究および他の研究の著者は、遺伝子増幅とmRNAの過剰発現との間の強い相関に基づき、この領域の数多くの候補癌遺伝子を提示してきた。(ガルシアら(Garcia et al.)、2005、上記;ジェルシ−ボイヤーら(Gelsi−Boyer et al.)、2005、上記;レイら(Ray et al.)、2004、上記)。累積的に、これらの研究は、ZNF703(FLJ14299)、ERLIN2(SPFH2)、PROSC、BRF2、RAB11FIP1(RCP)およびLSM1を、増幅と過剰発現との間に最も強い相関を有するとして同定し、LSM1以外のすべては、1−Mb「最小アンプリコン」に局在し(ガルシアら(Garcia et al.)、2005、上記)、A1サブアンプリコンと正確に重複する(ジェルシ−ボイヤーら(Gelsi−Boyer et al.)、2005、上記)。特に、本発明者らの研究においてLSVDにより同定された遺伝子は、特にAlおよびA2サブアンプリコンからなり、したがって、これら早期の研究により記載された、すべての「最も優れた候補」癌遺伝子を含む。
【0120】
TRIAGEの8p11−12への適用は、患者における播種性転移を潜在的に推進する、このアンプリコンにおける最も強い候補癌遺伝子として、RAB11FIP1(RCP)の同定をもたらした。この領域内の他の候補癌遺伝子の中でも、乳房上皮細胞において、いくつかの形質転換特性を有することがすでに示されている。(シュトライヒャー、ケイ エルら(Streicher,K.L.et al.)、(2007)Oncogene 26、2104〜2114;ヤン、ゼット キューら(Yang,Z.Q.et
al.)、(2006)Cancer Res 66、11632〜11643)、LSM1、BAG4およびC8orf4(TCI)はすべて、A2およびA3のサブアンプリコン上に主に存在する(ジェルシ−ボイヤーら(Gelsi−Boyer et al.)、2005、上記)。反対に、RAB11FIP1(RCP)は、1−Mb最小アンプリコン上のA1アンプリコンのコアのより近くに位置し、(ガルシアら(Garcia
et al.)、2005、上記)、癌遺伝子として機能的に特徴づけられたこの領域において第1である。
【0121】
それらの新規な研究を使用して、本発明者らはRCPの発現に連結した広範囲の癌促進作用を発見した。非癌性の乳房上皮細胞において、RCPの過剰発現は、悪性の形質転換をもたらし、細胞の運動および遊走を増加させる。乳癌細胞系において、RNA阻害による内因性RCPの低減は、増殖、足場非依存性成長、浸潤およびアポトーシス耐性を減少させ、ヌードマウスにおいて腫瘍形成を減退させた。生理的に、RCPは、主にRab11と相互作用し、管状エンドソームにおいてタンパク質選別を制御すると考えられている(ペデンら(Peden et al.)、2004、上記)。Rab11−RCP複合体は、トランスフェリン受容体をリソソームから離し(分解経路)エンドソームの再循環(再循環経路)へ導き、その結果膜結合受容体のレベルの相対的増加を促進することが示されている(ペデンら(Peden et al.)、2004、上記)。推論ではあるが、RCPの形質転換作用が再循環経路へのこの推移により仲介され得、インテグリンなどの特定の膜タンパク質または癌遺伝子の役割を有する成長因子受容体の膜への局在(およびこのような機能)を引き延ばすことは妥当と思われる。RCP発現とERKリン酸化の間の、本発明者らが観察した強い相関は、RCPと成長因子受容体仲介MAPK活性化との間の特定の連結を示し得るので、この可能性を反映し得る。上記のデータは、RAB11FIP1が、8p11−12アンプリコンの再発性の選択の陰の鍵となる推進力を表し得、この増幅領域およびRAB11FIP1それ自体の発現に関連する、乳癌の転移拡散に機能的に寄与し得ることを示している。このアンプリコンが、非小細胞肺癌(バルサラ、ビー アールら(Balsara,B.R.et al.)、(1997)Cancer Res 57、2116〜2120)、結腸直腸癌(ナカオ、ケーら(Nakao
,K.et al.)、(2004)Carcinogenesis 25、1345〜1357)および膀胱の腫瘍(サイモン、アールら(Simon,R.et al.)、(2001)Cancer Res 61、4514〜4519)およびファローピウス管(スナイデルス、エー エムら(Snijders,A.M.et al.)、(2003)Oncogene 22、4281〜4286)を含む他の固体腫瘍に頻繁に観察されていることは、RAB11FIP1が単独または近隣の癌遺伝子との相乗効果において作用する、癌の進行へのありふれた経路を表し得ることを示唆している。
【0122】
この明細書における、公知文献のリストまたは考察は、必ずしもかかる公知文献が技術水準の一部である、または一般的常識であると認めるものとしてとらえるべきではない。
本発明は、本明細書中に広範にかつ一般的に記載してある。包括的開示の範囲内に入る、より狭い種および亜属の分類もまた、それぞれ本発明を形成する。これは、削除された材料を本明細書中に具体的に列挙するかどうかにかかわらず、条件付き、または任意の対象を属から除去する否定的限定を伴う、本発明の包括的記載を含む。
【0123】
当業者は、本発明が、目標を達成すること、内在する目的および利点と同様に本明細書で言及した目的および利点を獲得するのに適していることを、容易に理解するであろう。さらに、様々な置き換えおよび変形が、本発明の範囲および精神から逸脱することなく、本明細書に開示した発明に対して実施できることは、当業者には容易に理解されるであろう。本明細書に記載された分子複合体および方法、手順、処理、分子、本明細書に記載された特定の化合物は、好ましい実施形態を例示する目下の典型であり、本発明の範囲を限定する意図のものではない。当業者は、特許請求の範囲により定義される本発明の精神に包含される、その変更および他の使用が思い浮かぶであろう。
【0124】
本明細書において適切に例示的に記載された発明は、任意の、1つまたは複数の要素、本明細書に具体的に開示されなかった、1つまたは複数の限定が存在しない状況下においても実施できる。したがって、例えば、本明細書における各事例において、「有する」、「本質的に〜からなる」および「からなる」などの任意の用語は、広い意味で、限定することなく理解すべきであり、直接参照される列挙された成分だけに限定されず、他の明記されていない成分および要素もまた含む。このように、それらは相互に交換することもできる。さらに、用いた用語および表現は説明の用語として使用し、限定するものではなく、示し、記載した特徴の任意の同等物またはそれらの一部を排除したような用語または表現の使用に意図はなく、しかし、様々な変形が本発明の特許請求の範囲内で可能であることが認識される。
【0125】
他の実施形態は以下の特許請求の範囲内である。さらに、本発明の特徴または態様が、Markush群に関して記載されている場合、当業者には、本発明がMarkush群の任意の個々のメンバーまたは該メンバーのサブグループに関しても記載されていることが理解されるであろう。
【図3−1】
【図3−2】
【図3−3】
【図3−4】
【図10A】
【図10B】
【特許請求の範囲】
【請求項1】
Rab結合タンパク質の量または活性を調節する工程からなる、細胞における腫瘍発生を予防、阻害、停止または逆行させる方法。
【請求項2】
前記Rab結合タンパク質がRab11結合タンパク質である、請求項1に記載の方法。
【請求項3】
前記Rab11結合タンパク質がRab11FIPタンパク質(Rab11ファミリー相互作用タンパク質)である、請求項2に記載の方法。
【請求項4】
細胞における前記Rab結合タンパク質の量を調節する工程が、前記Rab結合タンパク質の発現を調節することからなる、請求項1乃至3のいずれか1項に記載の方法。
【請求項5】
細胞における前記Rab結合タンパク質の量を調節する工程が、前記Rab結合タンパク質を分解することからなる、請求項1乃至3のいずれか1項に記載の方法。
【請求項6】
細胞における前記Rab結合タンパク質の活性を調節する工程が、前記Rab結合タンパク質と化合物との複合体を形成することからなる、請求項1乃至3のいずれか1項に記載の方法の方法。
【請求項7】
前記化合物が、ペプチド、ペプトイド、無機分子および小有機分子からなる群から選択される、請求項6に記載の方法。
【請求項8】
前記Rab結合タンパク質の分解がカルパインプロテアーゼによって実施される、請求項5に記載の方法。
【請求項9】
前記腫瘍発生が発癌である、請求項1乃至8のいずれか1項に記載の方法。
【請求項10】
増殖性疾患または障害の治療または予防に含まれる、請求項1乃至8のいずれか1項に記載の方法。
【請求項11】
治療される前記増殖性疾患または障害が癌である、請求項10に記載の方法。
【請求項12】
Rab結合タンパク質の発現および活性のうちの少なくとも一方を調節する化合物を投与する工程を含む、請求項1乃至11のいずれか1項に記載の方法。
【請求項13】
前記Rab結合タンパク質が、自身の量または活性を調節することが可能なRab結合タンパク質である、請求項12に記載の方法。
【請求項14】
前記Rab結合タンパク質の発現を核酸分子によって低減する、請求項1乃至13のいずれか1項に記載の方法。
【請求項15】
前記核酸分子が非コード核酸分子である、請求項14に記載の方法。
【請求項16】
前記核酸分子がRNAまたはDNAである、請求項14または15に記載の方法。
【請求項17】
前記核酸分子が、アプタマー、アンチセンスRNA分子、サイレンサーRNA分子、低分子ヘアピンRNA(shRNA)分子、マイクロRNA(miRNA)分子、低分子干渉RNA(siRNA)分子、および反復配列関連低分子干渉RNA(rasiRNA)からなる群から選択される、請求項15または16に記載の方法。
【請求項18】
哺乳動物、魚類、両生類および鳥類からなる群から選択される生物に含まれる細胞における腫瘍発生を予防、阻害、停止または逆行させる方法である、請求項1乃至17のいずれか1項に記載の方法。
【請求項19】
哺乳動物、魚類、両生類および鳥類からなる群から選択される生物から得られた細胞における腫瘍発生を予防、阻害、停止または逆行させる方法である、請求項1乃至17のいずれか1項に記載の方法。
【請求項20】
前記細胞が培養される、請求項19に記載の方法。
【請求項21】
前記哺乳動物が、ラット、マウス、ウサギ、モルモット、オポッサム、リス、ココノオビアルマジロ、イヌ、ネコ、ゾウ、チンパンジー、アカゲザル、マカクザル、オランウータン、ウシ(メウシ)、マーモセット、アメリカナキウサギ、ガラゴ、およびヒトからなる群から選択される、請求項18乃至20のいずれか1項に記載の方法。
【請求項22】
細胞外シグナル制御キナーゼ(ERK)の活性化を予防、阻害、停止または逆行させる工程を含む、請求項1乃至21のいずれか1項に記載の方法。
【請求項23】
前記Rab結合タンパク質の発現および活性のうちの少なくとも一方を検出する工程をさらに含む、請求項1乃至22のいずれか1項に記載の方法。
【請求項24】
前記発現および活性のうちの少なくとも一方を検出する工程が、該発現および活性のうちの少なくとも一方を一定時間測定することからなる、請求項23に記載の方法。
【請求項25】
得られた結果を対照測定の結果と比較する工程をさらに含む、請求項23または24に記載の方法。
【請求項26】
前記対照測定がRab結合タンパク質の発現および活性のうちの少なくとも一方を調節しない条件の使用からなる、請求項25に記載の方法。
【請求項27】
細胞におけるRab結合タンパク質の量の変化、発現の変化、細胞内局在の変化、または活性の変化を検出する工程からなる、発癌性になる素因を有する細胞を同定する方法。
【請求項28】
前記Rab結合タンパク質がRab11結合タンパク質である、請求項27に記載の方法。
【請求項29】
前記Rab11結合タンパク質がRab11FIPタンパク質である、請求項28に記載の方法。
【請求項30】
Rab結合タンパク質の発現の変化および活性の変化の少なくとも1つが決定され、該Rab結合タンパク質の発現の増大または活性の増大が、各腫瘍が細胞内カルシウムレベルの変化に感受性であることを示す、請求項27乃至29のいずれか1項に記載の方法。
【請求項31】
前記細胞内カルシウムレベルの増大が、ビタミンDのメンバー、該ビタミンDのメンバーのプロドラッグおよび該ビタミンDのメンバーの類似体の使用によって達成可能である、請求項30に記載の方法。
【請求項32】
前記ビタミンDのメンバーがビタミンD3である、請求項32に記載の方法。
【請求項33】
Rab結合タンパク質の発現レベル、活性レベル、および細胞内局在の少なくとも1つを決定する工程からなる、対象において新生物を発症するリスクを診断する方法。
【請求項34】
前記Rab結合タンパク質がRab11結合タンパク質である、請求項33に記載の方法。
【請求項35】
前記Rab11結合タンパク質がRab11FIPタンパク質である、請求項34に記載の方法。
【請求項36】
前記新生物が腫瘍である、請求項33に記載の方法。
【請求項37】
前記腫瘍が、乳房腫瘍、肺腫瘍、結腸直腸腫瘍、膀胱の腫瘍および卵管の腫瘍の1つである、請求項36に記載の方法。
【請求項38】
前記腫瘍が癌である、請求項36または37に記載の方法。
【請求項39】
Rab結合タンパク質の発現レベル、活性レベル、および細胞内局在の少なくとも1つを決定する工程を前記対象から得た試料において実施する、請求項33に記載の方法。
【請求項40】
得られた結果を対照測定の結果と比較する工程をさらに含む、請求項33に記載の方法。
【請求項41】
前記対照測定が、前記Rab結合タンパク質の発現および活性のうちの少なくとも一方が通常レベルである試料の使用からなる、請求項40に記載の方法。
【請求項42】
前記対照測定と比較したRab結合タンパク質の発現レベル、活性レベル、または細胞内局在の変化が、前記対象が新生物に罹患しているかまたは新生物を発症するリスクがあることを示す、請求項41に記載の方法。
【請求項43】
Rab結合タンパク質の発現の変化および活性の変化の少なくとも1つが決定され、前記Rab結合タンパク質の発現の増大または活性の増大が、各新生物が細胞内カルシウムレベルの変化に感受性であることを示す、請求項33乃至42のいずれか1項に記載の方法。
【請求項44】
前記細胞内カルシウムレベルの増大が、ビタミンDのメンバー、該ビタミンDのメンバーのプロドラッグおよび該ビタミンDのメンバーの類似体の使用によって達成可能である、請求項43に記載の方法。
【請求項45】
前記ビタミンDのメンバーがビタミンD3である、請求項44に記載の方法。
【請求項46】
前記Rab結合タンパク質が、Rab11FIP1、Rab11FIP2、Rab11FIP3(エフェリン)、Rab11FIP4、Rab11FIP5(Rip11)、およびそれらのアイソフォームからなる群から選択されるRab11FIPタンパク質である、請求項1乃至45のいずれか1項に記載の方法。
【請求項47】
複合体を形成する構成要素を互いに接触させる工程からなる、Rab結合タンパク質と複合体を形成することが可能な化合物を同定するin vitro方法。
【請求項48】
前記Rab結合タンパク質がRab11結合タンパク質である、請求項47に記載の方法。
【請求項49】
前記Rab11結合タンパク質がRab11FIPタンパク質である、請求項48に記載の方法。
【請求項50】
(a)試験管中に前記Rab結合タンパク質を提供する工程、
(b)前記Rab結合タンパク質と複合体を形成可能であると考えられる化合物を該試験管に加える工程、および
(c)該複合体形成を検出する工程を含む、請求項47乃至49のいずれか1項に記載の方法。
【請求項51】
前記検出工程が、適切な分光的、光化学的、光度分析的、蛍光分析的、放射線学的、酵素的または熱力学的方法によって実施されるか、または細胞効果に基づいている、請求項50に記載の方法。
【請求項52】
腫瘍発生を予防、阻害、停止または逆行させることができる化合物を同定する方法である、請求項49乃至51のいずれか1項に記載の方法。
【請求項53】
in vivoでの該Rab結合タンパク質の活性を調節することが可能な化合物を同定する方法であって、
(a)Rab結合タンパク質またはその機能断片を発現する微生物を提供する工程、
(b)該微生物に化合物を加える工程、および
(c)Rab結合タンパク質の細胞表現型または活性の変化をモニターする工程
からなる方法。
【請求項54】
前記Rab結合タンパク質がRab11結合タンパク質である、請求項53に記載の方法。
【請求項55】
前記Rab11結合タンパク質がRab11FIPタンパク質である、請求項54に記載の方法。
【請求項56】
得られた結果を対照測定の結果と比較する工程をさらに含む、請求項53乃至55のいずれか1項に記載の方法。
【請求項57】
前記対照測定が、前記Rab11FIPタンパク質の発現および活性のうちの少なくとも一方を調節しない化合物の使用からなる、請求項56に記載の方法。
【請求項58】
in vivoでの該Rab結合タンパク質の発現を調節することが可能な化合物を同定する方法であって、
(a)Rab11FIPタンパク質またはその機能断片を発現する微生物を提供する工程、
(b)該微生物に化合物を加える工程、および
(c)該Rab結合タンパク質の発現をモニターする工程
からなる方法。
【請求項59】
前記Rab結合タンパク質がRab11結合タンパク質である、請求項58に記載の方法。
【請求項60】
前記Rab11結合タンパク質がRab11FIPタンパク質である、請求項59に記載の方法。
【請求項61】
得られた結果を対照測定の結果と比較する工程をさらに含む、請求項58乃至60のいず
れか1項に記載の方法。
【請求項62】
前記対照測定が、前記Rab結合タンパク質の発現を調節しない化合物の使用からなる、請求項61に記載の方法。
【請求項63】
前記微生物が癌細胞である、請求項53乃至62のいずれか1項に記載の方法。
【請求項64】
前記癌細胞が、乳房癌細胞、肺癌細胞、結腸直腸癌細胞、膀胱の癌細胞および卵管の癌細胞の1つである、請求項63に記載の方法。
【請求項65】
前記微生物が、乳房組織、肺組織、結腸直腸組織、膀胱組織および卵管の組織の1つに由来する細胞である、請求項53乃至64のいずれか1項に記載の方法。
【請求項66】
前記細胞が、HMEC細胞系、3D、ABC−1、A549、Bcap−37、BIU−87、BT−20、BT−474、BT474M1、Calu−6、EBC−1、Colo−320、EJ、FT−MZ−1、H322、H446、H460、HCC2279、HCT116、HEPFT、Hs578T、Hs578Bst、HT−29、HTB−56R、HUT29、IMEC−2、JIMT−1、KoTCC−1、KU−T1、L0、L0−200、L0−500、L0−1000、L9981、LC−1/sq、LCD、LCOK、LK−2、LoVo、LS1034、LST−R1、Lu135、MBT−2、MCF−7、M231、MDA−231、MDA−MB−134、MDA−MB−175、MDA−MB−231、MDA−MB−361、MDA−MB−361DYT2、MDA−MB−435、MDA−MB−435HM、MDA−MB−468、MB231、MS−1、MW1、NCI−H69、NCI−H82、NCI−H209、NCI−H446、NCI−H1395、NCI−H1437、NCI−H1770、NCI−H2009、NCI−H2087、NCI−H2122、NCI−H2126、NCI−H2171、NCI−N226、NCI−N231、PC1、PC3、PC7、PC9、PC10、PC14、PMC42、PLA−801D、QG56、RERF−LCAI、RERF−LCMS、RERF−LCKJ、RKO、RT4、SBC3、ScaBER、SK−BR−3、SK−BR−7、SPC−A1、SQ5、SUM149、SUM159、SUM1315、SUM−52PE、SW480、T24、T−47D、UM−UC−2、WiDr、YTLMC−90、ZR−75、ZR−75−1、およびZR−75−30からなる群から選択される細胞系の細胞である、請求項63に記載の方法。
【請求項67】
前記細胞が、Rab結合タンパク質を過剰発現させることによって非癌性細胞から得られるものである、請求項63に記載の方法。
【請求項68】
前記細胞がRAB11FIP1を過剰発現している、請求項63に記載の方法。
【請求項69】
前記微生物が組換え細胞またはトランスジェニック微生物である、請求項55乃至64のいずれか1項に記載の方法。
【請求項70】
Rab結合タンパク質の量または活性を調節することが可能な化合物を同定する方法であって、
(a)癌細胞を収容し増殖させることができる宿主生物を提供する工程、
(b)癌細胞を該宿主生物に導入する工程、
(c)Rab結合タンパク質の発現および活性のうちの少なくとも一方を調節し得ると考えられる化合物を投与する工程、および
(c)該宿主生物中の腫瘍の増殖をモニターする工程
からなる方法。
【請求項71】
前記Rab結合タンパク質がRab11結合タンパク質である、請求項70に記載の方法。
【請求項72】
前記Rab11結合タンパク質がRab11FIPタンパク質である、請求項71に記載の方法。
【請求項73】
得られた結果を対照測定の結果と比較する工程をさらに含む、請求項70乃至72のいずれか1項に記載の方法。
【請求項74】
前記対照測定が、前記Rab11FIPタンパク質の発現および活性のうちの少なくとも一方を調節しない化合物の使用からなる、請求項73に記載の方法。
【請求項75】
Rab結合タンパク質の量または活性を調節することが可能な化合物を同定する方法であって、
(a)癌細胞を収容し増殖させることができる宿主生物を提供する工程、
(b)Rab結合タンパク質の量または活性を調節し得ると考えられる化合物からなる癌細胞を該宿主生物に導入する工程、および
(c)該宿主生物中の腫瘍の増殖をモニターする工程
からなる方法。
【請求項76】
前記Rab結合タンパク質がRab11結合タンパク質である、請求項75に記載の方法。
【請求項77】
前記Rab11結合タンパク質がRab11FIPタンパク質である、請求項76に記載の方法。
【請求項78】
得られた結果を対照測定の結果と比較する工程をさらに含む、請求項75乃至77のいずれか1項に記載の方法。
【請求項79】
前記対照測定が、前記Rab11FIPタンパク質の発現および活性のうちの少なくとも一方を調節しない化合物の使用からなる、請求項78に記載の方法。
【請求項80】
前記宿主生物が、マウス、ラット、ゾウ、ウサギ、モルモット、オポッサム、リス、ココノオビアルマジロ、イヌ、ネコ、アメリカナキウサギ、ガラゴ、チンパンジー、アカゲザル、マカクザル、オランウータン、ウシ(メウシ)およびマーモセットからなる群から選択される、請求項70乃至79のいずれか1項に記載の方法。
【請求項81】
Rab結合タンパク質の発現および活性の少なくとも1つを決定する工程からなり、該Rab結合タンパク質の発現の変化または活性の変化が、新生物が細胞内カルシウムレベルの変化に感受性であることを示す、該新生物が細胞内カルシウムレベルの変化に感受性であるかどうかを決定する方法。
【請求項82】
前記新生物が哺乳動物中に含まれる、請求項81に記載の方法。
【請求項83】
前記新生物が腫瘍である、請求項81または82に記載の方法。
【請求項84】
前記腫瘍が、乳房腫瘍、肺腫瘍、結腸直腸腫瘍、膀胱の腫瘍および卵管の腫瘍の1つである、請求項83に記載の方法。
【請求項85】
前記腫瘍が癌である、請求項81または82に記載の方法。
【請求項86】
前記細胞内カルシウムレベルの変化が遊離細胞内カルシウムの増大である、請求項81乃至85のいずれか1項に記載の方法。
【請求項87】
前記遊離細胞内カルシウムの増大が、ビタミンDファミリーのメンバー、該ビタミンDのメンバーのプロドラッグおよび該ビタミンDのメンバーの類似体の1つの使用によって達成可能であり、前記Rab結合タンパク質の発現の増大または活性の増大が、前記新生物が該ビタミンDのメンバー、そのプロドラッグまたは類似体を用いた治療に感受性であることを示す、請求項86に記載の方法。
【請求項88】
前記ビタミンDのメンバーがビタミンD3である、請求項87に記載の方法。
【請求項1】
Rab結合タンパク質の量または活性を調節する工程からなる、細胞における腫瘍発生を予防、阻害、停止または逆行させる方法。
【請求項2】
前記Rab結合タンパク質がRab11結合タンパク質である、請求項1に記載の方法。
【請求項3】
前記Rab11結合タンパク質がRab11FIPタンパク質(Rab11ファミリー相互作用タンパク質)である、請求項2に記載の方法。
【請求項4】
細胞における前記Rab結合タンパク質の量を調節する工程が、前記Rab結合タンパク質の発現を調節することからなる、請求項1乃至3のいずれか1項に記載の方法。
【請求項5】
細胞における前記Rab結合タンパク質の量を調節する工程が、前記Rab結合タンパク質を分解することからなる、請求項1乃至3のいずれか1項に記載の方法。
【請求項6】
細胞における前記Rab結合タンパク質の活性を調節する工程が、前記Rab結合タンパク質と化合物との複合体を形成することからなる、請求項1乃至3のいずれか1項に記載の方法の方法。
【請求項7】
前記化合物が、ペプチド、ペプトイド、無機分子および小有機分子からなる群から選択される、請求項6に記載の方法。
【請求項8】
前記Rab結合タンパク質の分解がカルパインプロテアーゼによって実施される、請求項5に記載の方法。
【請求項9】
前記腫瘍発生が発癌である、請求項1乃至8のいずれか1項に記載の方法。
【請求項10】
増殖性疾患または障害の治療または予防に含まれる、請求項1乃至8のいずれか1項に記載の方法。
【請求項11】
治療される前記増殖性疾患または障害が癌である、請求項10に記載の方法。
【請求項12】
Rab結合タンパク質の発現および活性のうちの少なくとも一方を調節する化合物を投与する工程を含む、請求項1乃至11のいずれか1項に記載の方法。
【請求項13】
前記Rab結合タンパク質が、自身の量または活性を調節することが可能なRab結合タンパク質である、請求項12に記載の方法。
【請求項14】
前記Rab結合タンパク質の発現を核酸分子によって低減する、請求項1乃至13のいずれか1項に記載の方法。
【請求項15】
前記核酸分子が非コード核酸分子である、請求項14に記載の方法。
【請求項16】
前記核酸分子がRNAまたはDNAである、請求項14または15に記載の方法。
【請求項17】
前記核酸分子が、アプタマー、アンチセンスRNA分子、サイレンサーRNA分子、低分子ヘアピンRNA(shRNA)分子、マイクロRNA(miRNA)分子、低分子干渉RNA(siRNA)分子、および反復配列関連低分子干渉RNA(rasiRNA)からなる群から選択される、請求項15または16に記載の方法。
【請求項18】
哺乳動物、魚類、両生類および鳥類からなる群から選択される生物に含まれる細胞における腫瘍発生を予防、阻害、停止または逆行させる方法である、請求項1乃至17のいずれか1項に記載の方法。
【請求項19】
哺乳動物、魚類、両生類および鳥類からなる群から選択される生物から得られた細胞における腫瘍発生を予防、阻害、停止または逆行させる方法である、請求項1乃至17のいずれか1項に記載の方法。
【請求項20】
前記細胞が培養される、請求項19に記載の方法。
【請求項21】
前記哺乳動物が、ラット、マウス、ウサギ、モルモット、オポッサム、リス、ココノオビアルマジロ、イヌ、ネコ、ゾウ、チンパンジー、アカゲザル、マカクザル、オランウータン、ウシ(メウシ)、マーモセット、アメリカナキウサギ、ガラゴ、およびヒトからなる群から選択される、請求項18乃至20のいずれか1項に記載の方法。
【請求項22】
細胞外シグナル制御キナーゼ(ERK)の活性化を予防、阻害、停止または逆行させる工程を含む、請求項1乃至21のいずれか1項に記載の方法。
【請求項23】
前記Rab結合タンパク質の発現および活性のうちの少なくとも一方を検出する工程をさらに含む、請求項1乃至22のいずれか1項に記載の方法。
【請求項24】
前記発現および活性のうちの少なくとも一方を検出する工程が、該発現および活性のうちの少なくとも一方を一定時間測定することからなる、請求項23に記載の方法。
【請求項25】
得られた結果を対照測定の結果と比較する工程をさらに含む、請求項23または24に記載の方法。
【請求項26】
前記対照測定がRab結合タンパク質の発現および活性のうちの少なくとも一方を調節しない条件の使用からなる、請求項25に記載の方法。
【請求項27】
細胞におけるRab結合タンパク質の量の変化、発現の変化、細胞内局在の変化、または活性の変化を検出する工程からなる、発癌性になる素因を有する細胞を同定する方法。
【請求項28】
前記Rab結合タンパク質がRab11結合タンパク質である、請求項27に記載の方法。
【請求項29】
前記Rab11結合タンパク質がRab11FIPタンパク質である、請求項28に記載の方法。
【請求項30】
Rab結合タンパク質の発現の変化および活性の変化の少なくとも1つが決定され、該Rab結合タンパク質の発現の増大または活性の増大が、各腫瘍が細胞内カルシウムレベルの変化に感受性であることを示す、請求項27乃至29のいずれか1項に記載の方法。
【請求項31】
前記細胞内カルシウムレベルの増大が、ビタミンDのメンバー、該ビタミンDのメンバーのプロドラッグおよび該ビタミンDのメンバーの類似体の使用によって達成可能である、請求項30に記載の方法。
【請求項32】
前記ビタミンDのメンバーがビタミンD3である、請求項32に記載の方法。
【請求項33】
Rab結合タンパク質の発現レベル、活性レベル、および細胞内局在の少なくとも1つを決定する工程からなる、対象において新生物を発症するリスクを診断する方法。
【請求項34】
前記Rab結合タンパク質がRab11結合タンパク質である、請求項33に記載の方法。
【請求項35】
前記Rab11結合タンパク質がRab11FIPタンパク質である、請求項34に記載の方法。
【請求項36】
前記新生物が腫瘍である、請求項33に記載の方法。
【請求項37】
前記腫瘍が、乳房腫瘍、肺腫瘍、結腸直腸腫瘍、膀胱の腫瘍および卵管の腫瘍の1つである、請求項36に記載の方法。
【請求項38】
前記腫瘍が癌である、請求項36または37に記載の方法。
【請求項39】
Rab結合タンパク質の発現レベル、活性レベル、および細胞内局在の少なくとも1つを決定する工程を前記対象から得た試料において実施する、請求項33に記載の方法。
【請求項40】
得られた結果を対照測定の結果と比較する工程をさらに含む、請求項33に記載の方法。
【請求項41】
前記対照測定が、前記Rab結合タンパク質の発現および活性のうちの少なくとも一方が通常レベルである試料の使用からなる、請求項40に記載の方法。
【請求項42】
前記対照測定と比較したRab結合タンパク質の発現レベル、活性レベル、または細胞内局在の変化が、前記対象が新生物に罹患しているかまたは新生物を発症するリスクがあることを示す、請求項41に記載の方法。
【請求項43】
Rab結合タンパク質の発現の変化および活性の変化の少なくとも1つが決定され、前記Rab結合タンパク質の発現の増大または活性の増大が、各新生物が細胞内カルシウムレベルの変化に感受性であることを示す、請求項33乃至42のいずれか1項に記載の方法。
【請求項44】
前記細胞内カルシウムレベルの増大が、ビタミンDのメンバー、該ビタミンDのメンバーのプロドラッグおよび該ビタミンDのメンバーの類似体の使用によって達成可能である、請求項43に記載の方法。
【請求項45】
前記ビタミンDのメンバーがビタミンD3である、請求項44に記載の方法。
【請求項46】
前記Rab結合タンパク質が、Rab11FIP1、Rab11FIP2、Rab11FIP3(エフェリン)、Rab11FIP4、Rab11FIP5(Rip11)、およびそれらのアイソフォームからなる群から選択されるRab11FIPタンパク質である、請求項1乃至45のいずれか1項に記載の方法。
【請求項47】
複合体を形成する構成要素を互いに接触させる工程からなる、Rab結合タンパク質と複合体を形成することが可能な化合物を同定するin vitro方法。
【請求項48】
前記Rab結合タンパク質がRab11結合タンパク質である、請求項47に記載の方法。
【請求項49】
前記Rab11結合タンパク質がRab11FIPタンパク質である、請求項48に記載の方法。
【請求項50】
(a)試験管中に前記Rab結合タンパク質を提供する工程、
(b)前記Rab結合タンパク質と複合体を形成可能であると考えられる化合物を該試験管に加える工程、および
(c)該複合体形成を検出する工程を含む、請求項47乃至49のいずれか1項に記載の方法。
【請求項51】
前記検出工程が、適切な分光的、光化学的、光度分析的、蛍光分析的、放射線学的、酵素的または熱力学的方法によって実施されるか、または細胞効果に基づいている、請求項50に記載の方法。
【請求項52】
腫瘍発生を予防、阻害、停止または逆行させることができる化合物を同定する方法である、請求項49乃至51のいずれか1項に記載の方法。
【請求項53】
in vivoでの該Rab結合タンパク質の活性を調節することが可能な化合物を同定する方法であって、
(a)Rab結合タンパク質またはその機能断片を発現する微生物を提供する工程、
(b)該微生物に化合物を加える工程、および
(c)Rab結合タンパク質の細胞表現型または活性の変化をモニターする工程
からなる方法。
【請求項54】
前記Rab結合タンパク質がRab11結合タンパク質である、請求項53に記載の方法。
【請求項55】
前記Rab11結合タンパク質がRab11FIPタンパク質である、請求項54に記載の方法。
【請求項56】
得られた結果を対照測定の結果と比較する工程をさらに含む、請求項53乃至55のいずれか1項に記載の方法。
【請求項57】
前記対照測定が、前記Rab11FIPタンパク質の発現および活性のうちの少なくとも一方を調節しない化合物の使用からなる、請求項56に記載の方法。
【請求項58】
in vivoでの該Rab結合タンパク質の発現を調節することが可能な化合物を同定する方法であって、
(a)Rab11FIPタンパク質またはその機能断片を発現する微生物を提供する工程、
(b)該微生物に化合物を加える工程、および
(c)該Rab結合タンパク質の発現をモニターする工程
からなる方法。
【請求項59】
前記Rab結合タンパク質がRab11結合タンパク質である、請求項58に記載の方法。
【請求項60】
前記Rab11結合タンパク質がRab11FIPタンパク質である、請求項59に記載の方法。
【請求項61】
得られた結果を対照測定の結果と比較する工程をさらに含む、請求項58乃至60のいず
れか1項に記載の方法。
【請求項62】
前記対照測定が、前記Rab結合タンパク質の発現を調節しない化合物の使用からなる、請求項61に記載の方法。
【請求項63】
前記微生物が癌細胞である、請求項53乃至62のいずれか1項に記載の方法。
【請求項64】
前記癌細胞が、乳房癌細胞、肺癌細胞、結腸直腸癌細胞、膀胱の癌細胞および卵管の癌細胞の1つである、請求項63に記載の方法。
【請求項65】
前記微生物が、乳房組織、肺組織、結腸直腸組織、膀胱組織および卵管の組織の1つに由来する細胞である、請求項53乃至64のいずれか1項に記載の方法。
【請求項66】
前記細胞が、HMEC細胞系、3D、ABC−1、A549、Bcap−37、BIU−87、BT−20、BT−474、BT474M1、Calu−6、EBC−1、Colo−320、EJ、FT−MZ−1、H322、H446、H460、HCC2279、HCT116、HEPFT、Hs578T、Hs578Bst、HT−29、HTB−56R、HUT29、IMEC−2、JIMT−1、KoTCC−1、KU−T1、L0、L0−200、L0−500、L0−1000、L9981、LC−1/sq、LCD、LCOK、LK−2、LoVo、LS1034、LST−R1、Lu135、MBT−2、MCF−7、M231、MDA−231、MDA−MB−134、MDA−MB−175、MDA−MB−231、MDA−MB−361、MDA−MB−361DYT2、MDA−MB−435、MDA−MB−435HM、MDA−MB−468、MB231、MS−1、MW1、NCI−H69、NCI−H82、NCI−H209、NCI−H446、NCI−H1395、NCI−H1437、NCI−H1770、NCI−H2009、NCI−H2087、NCI−H2122、NCI−H2126、NCI−H2171、NCI−N226、NCI−N231、PC1、PC3、PC7、PC9、PC10、PC14、PMC42、PLA−801D、QG56、RERF−LCAI、RERF−LCMS、RERF−LCKJ、RKO、RT4、SBC3、ScaBER、SK−BR−3、SK−BR−7、SPC−A1、SQ5、SUM149、SUM159、SUM1315、SUM−52PE、SW480、T24、T−47D、UM−UC−2、WiDr、YTLMC−90、ZR−75、ZR−75−1、およびZR−75−30からなる群から選択される細胞系の細胞である、請求項63に記載の方法。
【請求項67】
前記細胞が、Rab結合タンパク質を過剰発現させることによって非癌性細胞から得られるものである、請求項63に記載の方法。
【請求項68】
前記細胞がRAB11FIP1を過剰発現している、請求項63に記載の方法。
【請求項69】
前記微生物が組換え細胞またはトランスジェニック微生物である、請求項55乃至64のいずれか1項に記載の方法。
【請求項70】
Rab結合タンパク質の量または活性を調節することが可能な化合物を同定する方法であって、
(a)癌細胞を収容し増殖させることができる宿主生物を提供する工程、
(b)癌細胞を該宿主生物に導入する工程、
(c)Rab結合タンパク質の発現および活性のうちの少なくとも一方を調節し得ると考えられる化合物を投与する工程、および
(c)該宿主生物中の腫瘍の増殖をモニターする工程
からなる方法。
【請求項71】
前記Rab結合タンパク質がRab11結合タンパク質である、請求項70に記載の方法。
【請求項72】
前記Rab11結合タンパク質がRab11FIPタンパク質である、請求項71に記載の方法。
【請求項73】
得られた結果を対照測定の結果と比較する工程をさらに含む、請求項70乃至72のいずれか1項に記載の方法。
【請求項74】
前記対照測定が、前記Rab11FIPタンパク質の発現および活性のうちの少なくとも一方を調節しない化合物の使用からなる、請求項73に記載の方法。
【請求項75】
Rab結合タンパク質の量または活性を調節することが可能な化合物を同定する方法であって、
(a)癌細胞を収容し増殖させることができる宿主生物を提供する工程、
(b)Rab結合タンパク質の量または活性を調節し得ると考えられる化合物からなる癌細胞を該宿主生物に導入する工程、および
(c)該宿主生物中の腫瘍の増殖をモニターする工程
からなる方法。
【請求項76】
前記Rab結合タンパク質がRab11結合タンパク質である、請求項75に記載の方法。
【請求項77】
前記Rab11結合タンパク質がRab11FIPタンパク質である、請求項76に記載の方法。
【請求項78】
得られた結果を対照測定の結果と比較する工程をさらに含む、請求項75乃至77のいずれか1項に記載の方法。
【請求項79】
前記対照測定が、前記Rab11FIPタンパク質の発現および活性のうちの少なくとも一方を調節しない化合物の使用からなる、請求項78に記載の方法。
【請求項80】
前記宿主生物が、マウス、ラット、ゾウ、ウサギ、モルモット、オポッサム、リス、ココノオビアルマジロ、イヌ、ネコ、アメリカナキウサギ、ガラゴ、チンパンジー、アカゲザル、マカクザル、オランウータン、ウシ(メウシ)およびマーモセットからなる群から選択される、請求項70乃至79のいずれか1項に記載の方法。
【請求項81】
Rab結合タンパク質の発現および活性の少なくとも1つを決定する工程からなり、該Rab結合タンパク質の発現の変化または活性の変化が、新生物が細胞内カルシウムレベルの変化に感受性であることを示す、該新生物が細胞内カルシウムレベルの変化に感受性であるかどうかを決定する方法。
【請求項82】
前記新生物が哺乳動物中に含まれる、請求項81に記載の方法。
【請求項83】
前記新生物が腫瘍である、請求項81または82に記載の方法。
【請求項84】
前記腫瘍が、乳房腫瘍、肺腫瘍、結腸直腸腫瘍、膀胱の腫瘍および卵管の腫瘍の1つである、請求項83に記載の方法。
【請求項85】
前記腫瘍が癌である、請求項81または82に記載の方法。
【請求項86】
前記細胞内カルシウムレベルの変化が遊離細胞内カルシウムの増大である、請求項81乃至85のいずれか1項に記載の方法。
【請求項87】
前記遊離細胞内カルシウムの増大が、ビタミンDファミリーのメンバー、該ビタミンDのメンバーのプロドラッグおよび該ビタミンDのメンバーの類似体の1つの使用によって達成可能であり、前記Rab結合タンパク質の発現の増大または活性の増大が、前記新生物が該ビタミンDのメンバー、そのプロドラッグまたは類似体を用いた治療に感受性であることを示す、請求項86に記載の方法。
【請求項88】
前記ビタミンDのメンバーがビタミンD3である、請求項87に記載の方法。
【図1】

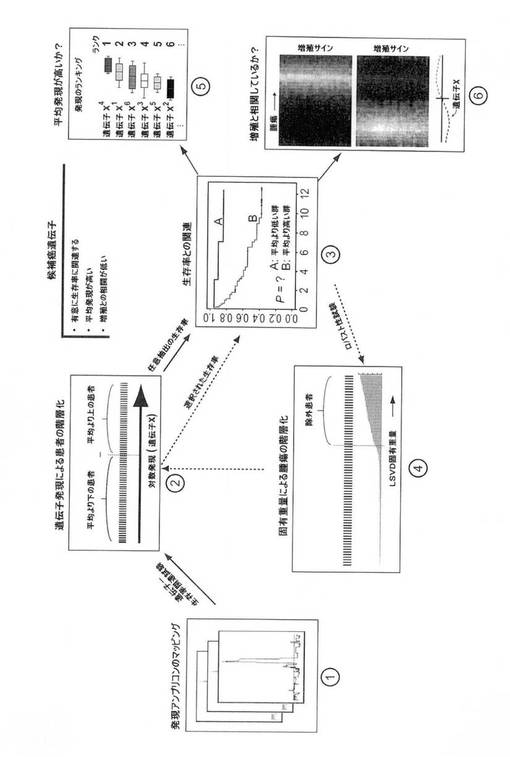
【図2】
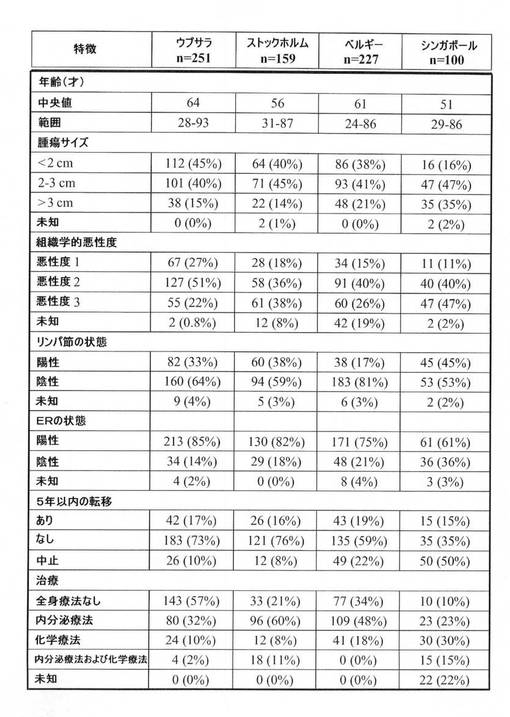
【図4A】

【図4B】

【図4C】
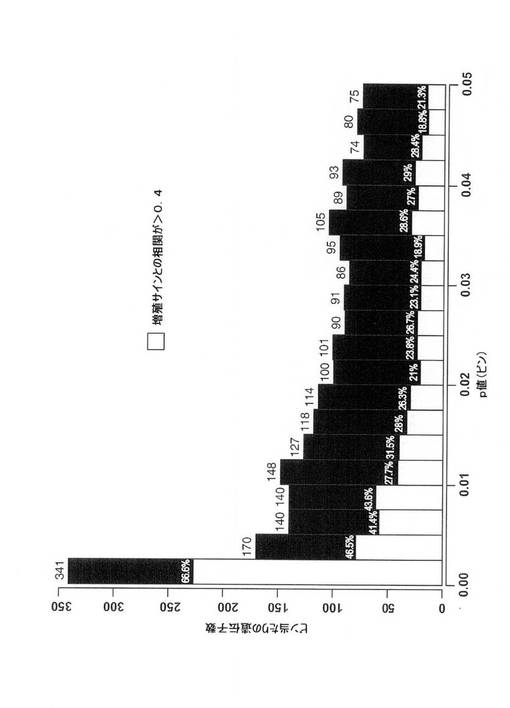
【図5A】
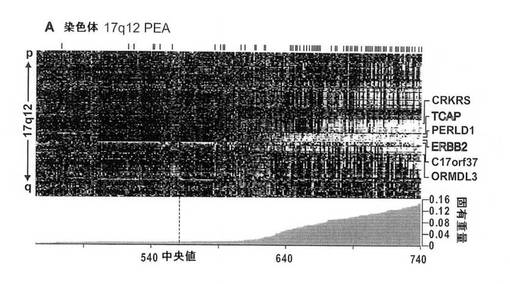
【図5B】
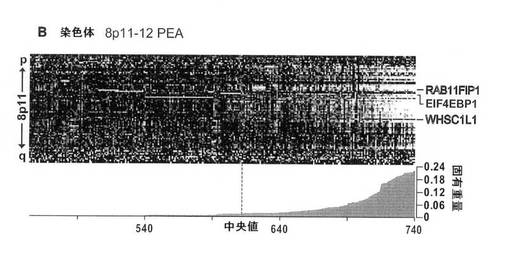
【図5C】

【図5D】

【図5E】
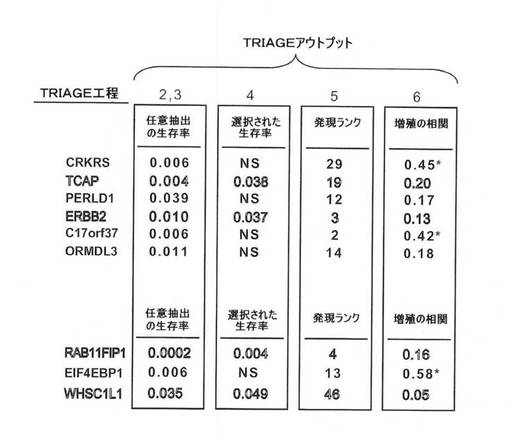
【図6A】
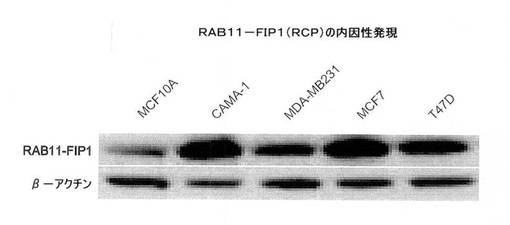
【図6B】
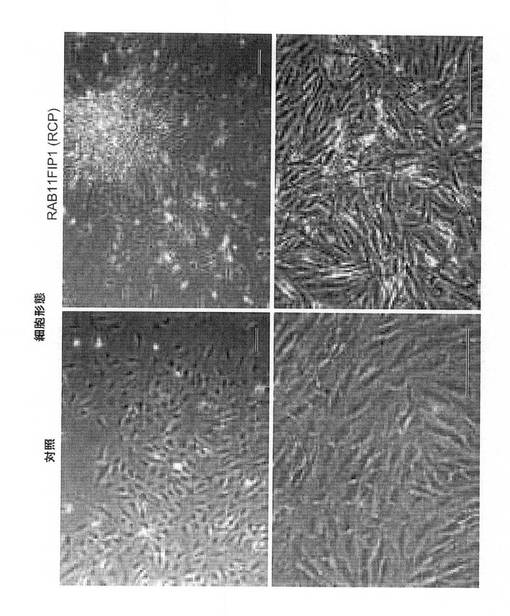
【図6C】
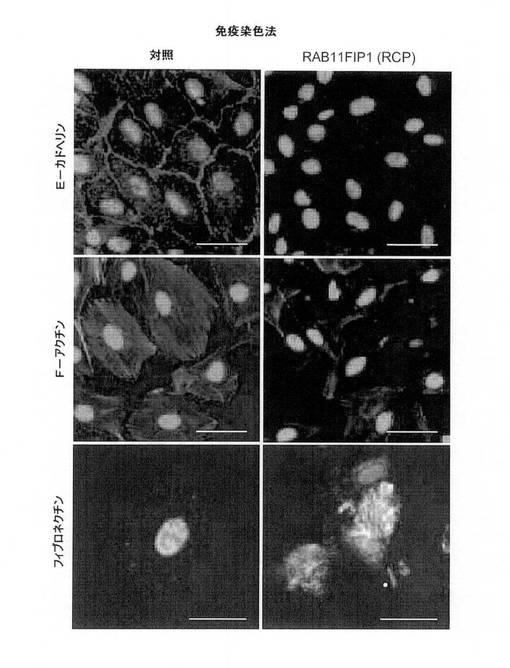
【図6D】
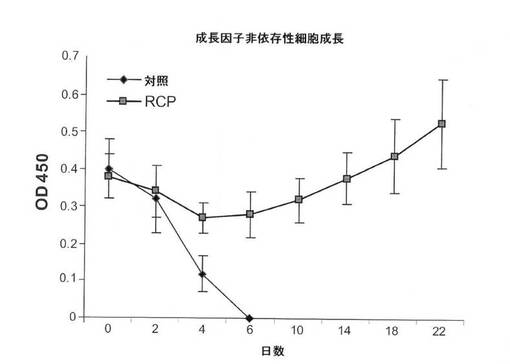
【図6E】
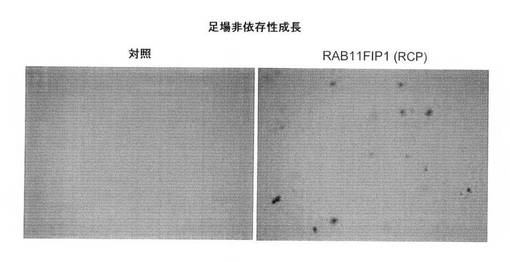
【図6F】
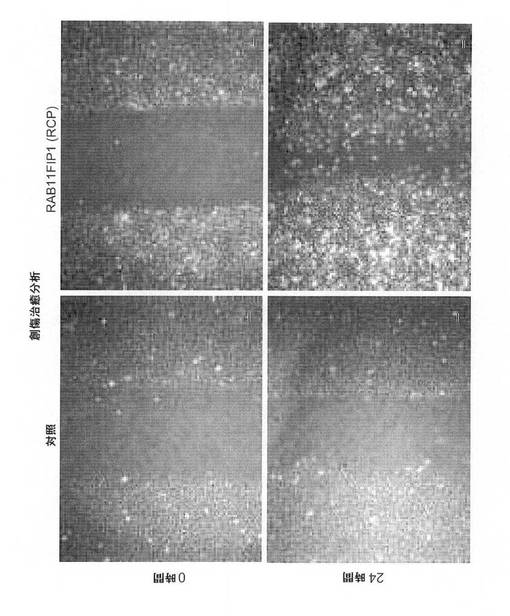
【図7A】
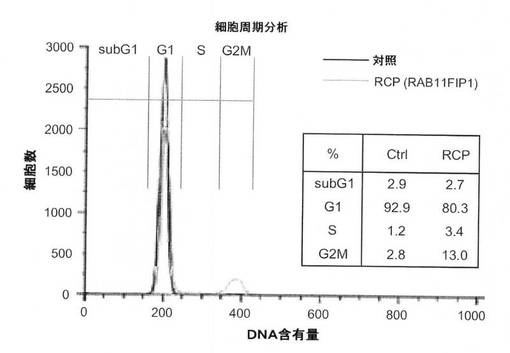
【図7B】
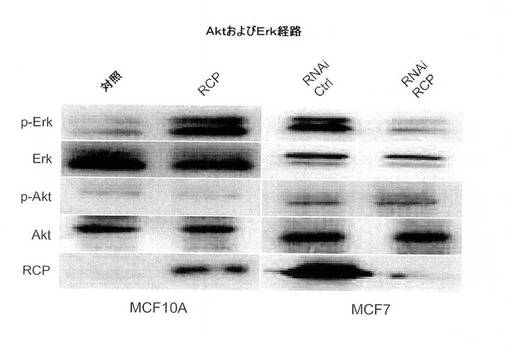
【図7C】

【図7D】
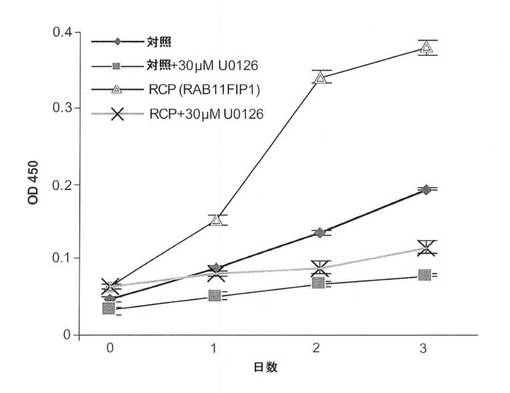
【図8A】
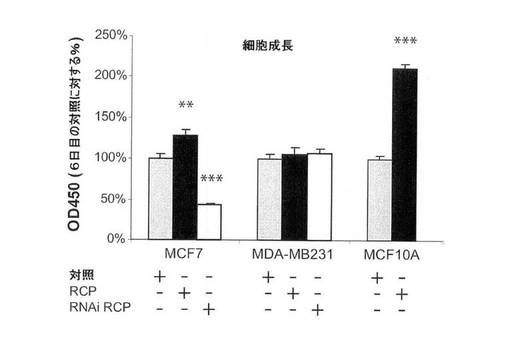
【図8B】
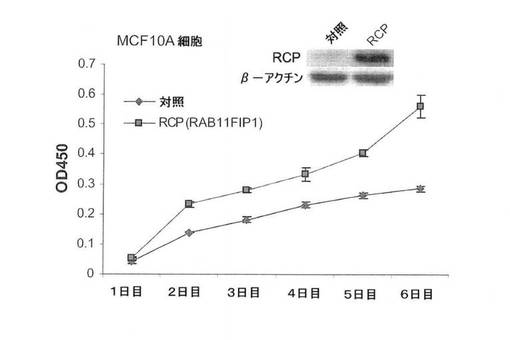
【図8C】
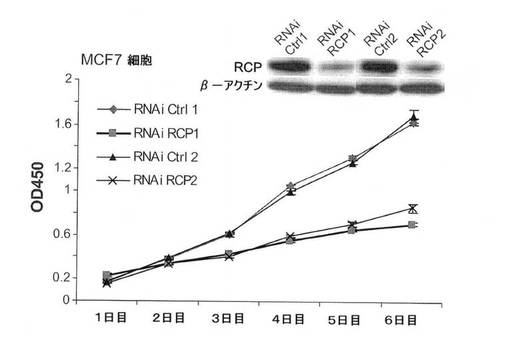
【図8D】
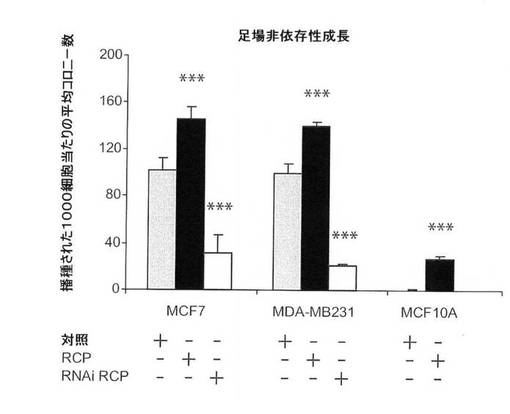
【図8E】
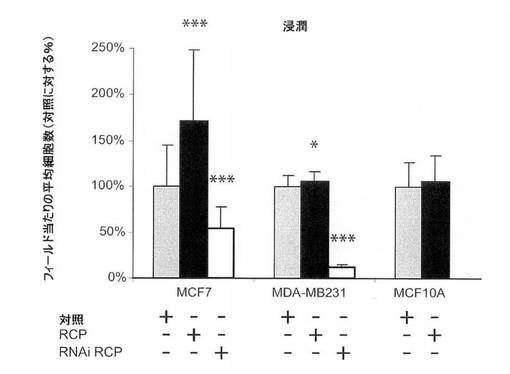
【図8F】
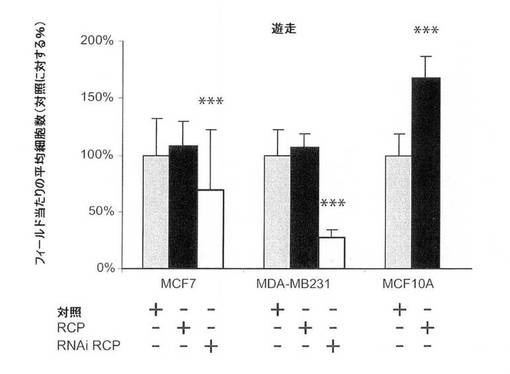
【図9A】
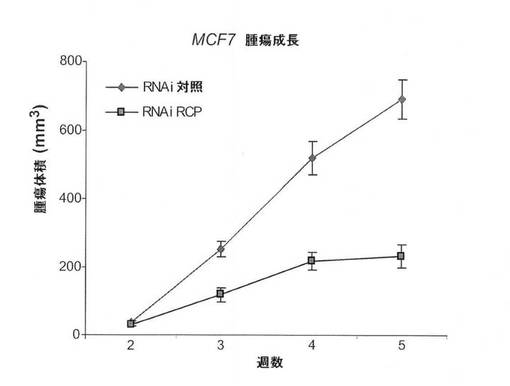
【図9B】
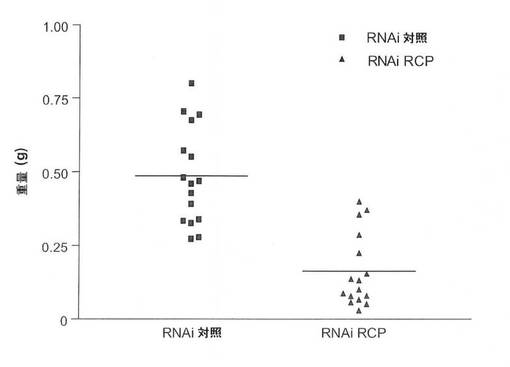
【図9C】
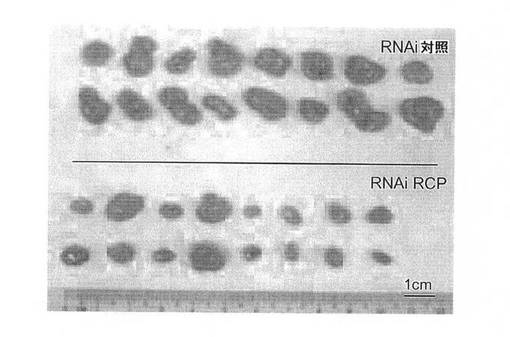
【図9D】
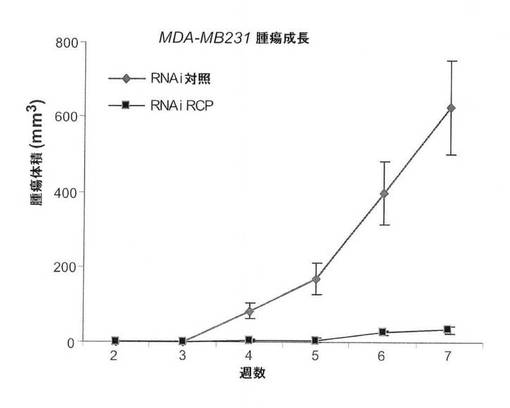
【図9E】
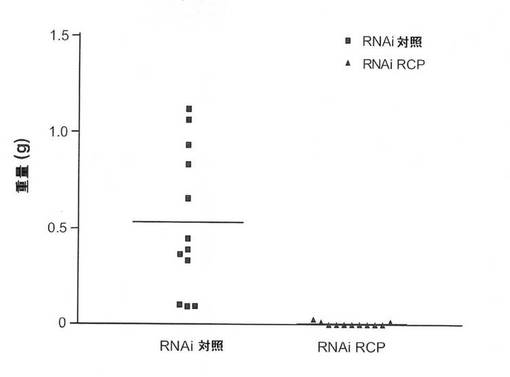
【図9F】
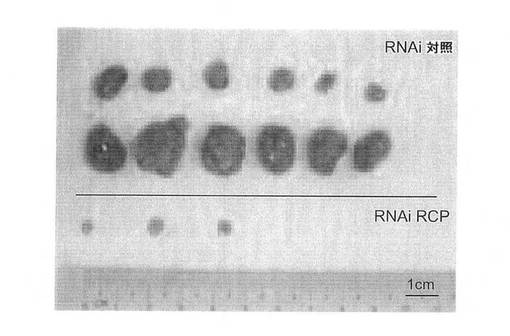
【図9G】
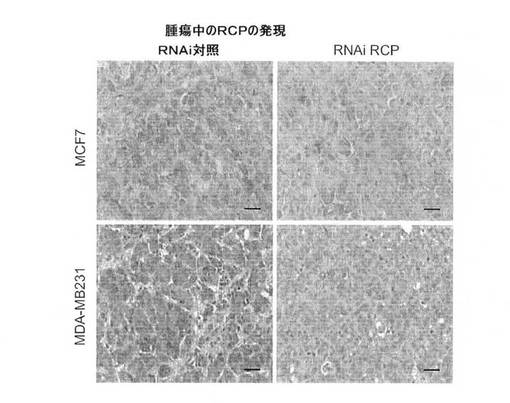
【図9H】
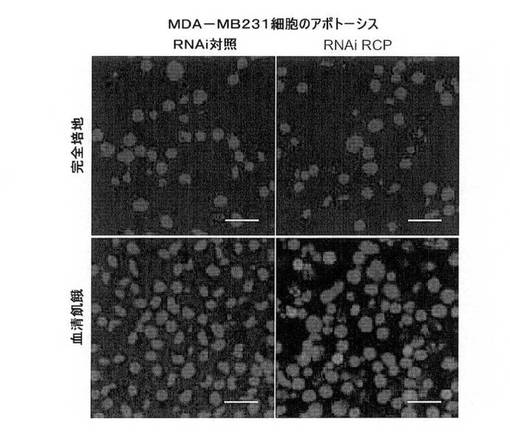
【図2】
【図4A】
【図4B】
【図4C】
【図5A】
【図5B】
【図5C】
【図5D】
【図5E】
【図6A】
【図6B】
【図6C】
【図6D】
【図6E】
【図6F】
【図7A】
【図7B】
【図7C】
【図7D】
【図8A】
【図8B】
【図8C】
【図8D】
【図8E】
【図8F】
【図9A】
【図9B】
【図9C】
【図9D】
【図9E】
【図9F】
【図9G】
【図9H】
【公表番号】特表2010−523652(P2010−523652A)
【公表日】平成22年7月15日(2010.7.15)
【国際特許分類】
【出願番号】特願2010−502978(P2010−502978)
【出願日】平成20年4月11日(2008.4.11)
【国際出願番号】PCT/SG2008/000116
【国際公開番号】WO2008/127199
【国際公開日】平成20年10月23日(2008.10.23)
【出願人】(503231882)エージェンシー フォー サイエンス,テクノロジー アンド リサーチ (179)
【Fターム(参考)】
【公表日】平成22年7月15日(2010.7.15)
【国際特許分類】
【出願日】平成20年4月11日(2008.4.11)
【国際出願番号】PCT/SG2008/000116
【国際公開番号】WO2008/127199
【国際公開日】平成20年10月23日(2008.10.23)
【出願人】(503231882)エージェンシー フォー サイエンス,テクノロジー アンド リサーチ (179)
【Fターム(参考)】
[ Back to top ]