
膜タンパク質の内部移行を検出するための方法
本発明は、細胞の表面に発現される目的の膜貫通タンパク質の内部移行を検出するための方法であって、次の:
a)蛍光金属錯体で目的のタンパク質を標識する段階であって、蛍光金属錯体の寿命が0.1msより長い段階;
b)目的のタンパク質の内部移行を生じることができる組成物を反応媒体に添加する段階;
c)反応媒体に対して以下から選択される調節剤:
a.上記蛍光金属錯体と適合する蛍光又は非蛍光のFRETアクセプター化合物であって、反応媒体中の最終濃度が10−7Mより高く、分子量が50kD未満である化合物;
b.レドックス電位が+0.1V未満、好適には0.25〜0.75Vである還元剤;
c.非共有結合によって、特異的に蛍光金属錯体と結合する薬剤;
d.非蛍光金属錯体を形成するための、希土類と競合する金属イオン、
を添加する段階;
d)蛍光金属錯体の発光波長、及び/又は調節化合物が蛍光アクセプター化合物の場合に調節化合物の発光波長で反応媒体によって放出される発光を測定する段階;
e)段階d)で測定されるシグナルを、段階a)及びc)のみに供された細胞上で測定される基準シグナルと比較する段階、
を含んでなる方法である。
本発明はまた、膜タンパク質の内部移行を検出するための方法である。
a)蛍光金属錯体で目的のタンパク質を標識する段階であって、蛍光金属錯体の寿命が0.1msより長い段階;
b)目的のタンパク質の内部移行を生じることができる組成物を反応媒体に添加する段階;
c)反応媒体に対して以下から選択される調節剤:
a.上記蛍光金属錯体と適合する蛍光又は非蛍光のFRETアクセプター化合物であって、反応媒体中の最終濃度が10−7Mより高く、分子量が50kD未満である化合物;
b.レドックス電位が+0.1V未満、好適には0.25〜0.75Vである還元剤;
c.非共有結合によって、特異的に蛍光金属錯体と結合する薬剤;
d.非蛍光金属錯体を形成するための、希土類と競合する金属イオン、
を添加する段階;
d)蛍光金属錯体の発光波長、及び/又は調節化合物が蛍光アクセプター化合物の場合に調節化合物の発光波長で反応媒体によって放出される発光を測定する段階;
e)段階d)で測定されるシグナルを、段階a)及びc)のみに供された細胞上で測定される基準シグナルと比較する段階、
を含んでなる方法である。
本発明はまた、膜タンパク質の内部移行を検出するための方法である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、膜タンパク質の内部移行を生細胞によって検出するための方法に関し、この内部移行を生じ得る化合物を明らかにするために特に有効である。より詳細には、本発明は、膜貫通タンパク質が内部移行された場合の蛍光金属錯体の発光における変化を測定することに基づいて、蛍光金属錯体と結合された膜貫通タンパク質の内部移行を検出するための方法を説明する。
【背景技術】
【0002】
内部移行:
膜受容体のリガンドによる細胞の活性化の間、膜受容体リガンドは、細胞による内部移行を通常もたらすメカニズムのカスケードへ供される。この方法では、第一に、リガンド、一般にはアゴニスト、の取り込みにより細胞によって受ける刺激の抑制又は減少を可能にし、そして第二に、受容体の脱感作及びこれによる通常のシグナル伝達カスケードの妨害をもたらす。伝達ユニットのこの脱感作は、比較的十分に説明されている複雑な機構である(例えば、Gainetdinov 等, Annu Review Neurosci 2004 v27 p107-44を参照されたい)。
【0003】
この内部移行機構は、Gタンパク質共役型受容体(GPCRs)さらに受容体型チロシンキナーゼを制御するための主要な方法の1つである。
概略的には、アゴニストによるGPCRの活性化は細胞内部における当該受容体のリン酸化をもたらし、この結果一定の細胞内タンパク質、特にクラスリン被覆エンドサイトーシス小胞の形成を生じるアレスチンの補充を誘発する。従って、これらの小胞は、細胞内輸送機構によって処理され、その結果リソソームとの融合によるこれらの分解、又は細胞膜との融合によるこれらの再転換を生じる。
【0004】
GPCRは、治療活性を有する新たな分子の開発に関する選択のターゲットであり、そして候補化合物:アゴニスト、アンタゴニスト又はインバースアゴニストによってこれらの活性化レベルの変化を測定するための複数の技術が存在する。これらの一部は実際に、内部移行に関係する現象を試験することによってGPCRの活性化を検出するという目的を有する。
【0005】
アレスチンは、GPCR内部移行方法に主に関与する細胞内タンパク質であり、以下の記載によって示されるように、内部移行を検出するために一部のアプローチで使用される。
【0006】
WO01/67106は、GPCRとアレスチンの恒常的活性型の変異体との結合の調節を測定することによって、GPCRリガンドを同定することを目的とする方法を説明し、この活性はGPCRのリン酸化レベルに依存しており、当該リガンドは放射性原子、酵素、レポータータンパク質又は蛍光化合物で標識される。
【0007】
WO01/59451は、変異酵素の2つのサブユニットの相補性に基づく「ICAST」テクノロジー(シストロン間相補的分析スクリーニングテクノロジー(Intercistronic Complementary Analysis Screening technology))を使用するGPCRの活性化を検出するための方法を説明し、酵素の各断片は、一方はGPCRと、他方は例えばアレスチンと結合の融合タンパク質の形態で発現される。GPCR−アレスチン相互作用が生じる場合、酵素は相補性によって再構成され、検出可能なシグナルを生じることができるであろう。繰り返して言うが、アレスチンへのこれらの結合の検出によってGPCRの活性化を分析することは重要である。WO01/58923で説明されるように、この技術はGPCR又はアレスチンの変異体を使用することによって改良することができる。
【0008】
WO2005/007822は、化合物が目的の2つのタンパク質間の相互作用を調節するか否かを測定するための方法を説明する。この方法はまた、GPCR−アレスチン結合の検出に基づいている。提示される方法は、特異的なプロテアーゼ感受性のあるペプチド配列を介して転写因子を有する、C末端部で融合したGPCRの使用に基づいている。アレスチンは、この部分でプロテアーゼと融合される。アゴニストリガンドによるGPCRの活性化中、アレスチンはGPCRと相互作用し、これによりプロテアーゼを切断配列に近接させ且つ転写因子の放出を生じ、さらにレポーター遺伝子の転写を活性化でき、この活性は検出することができる。
【0009】
GPCRの活性化に関するマーカーとしてのアレスチン−GPCR結合の使用に付随する問題の1つは、この結合の不安定な性質であり、結合はエンドサイトーシス小胞の形成をもたらす現象のカスケードにおける中間段階にのみ存在する。WO04/065963は、このカスケードのタンパク質パートナーと結合することができない、野生型アレスチンと異なるアレスチン変異体を使用することによってこの問題に対処しており、結果的に受容体に比較的安定して結合した状態である。
Charest等(EMBO Rep. 2005 Apr; 6(4):334-40)は、アレスチンの三次元構造における変化を検出するための技術を開発したが、この変化は活性化GPCRとの相互作用中に生じるものであり、著者等はルシフェラーゼ−アレスチン−YFP(黄色蛍光タンパク質)融合タンパク質を作成し、GPCRの活性化中の分子内BRET(生物発光共鳴エネルギー転移)シグナルの増加を実証した(「2倍輝くアレスチン(double-brilliance arrestin)」として周知の技術)。
【0010】
モルキュラーディバイス社(Molecular Devices)は、特にEP 1 015 608で説明される、「トランスフルオール(Transfluor)(登録商標)」テクノロジーを市販し、このテクノロジーはアレスチン−GFP融合タンパク質の使用に基づいており、この融合タンパク質を発現している細胞の使用により、目的のGPCRの活性化に応じて、適当な画像収集設備を用いて、細胞内でのアレスチン−GFPの転位及びこの再分布の可視化が可能である。
【0011】
膜受容体内部移行を示すための他のアプローチは、GPCR−アレスチン結合又はアレスチンの細胞内転位の検出に基づいておらず、特に以下の文献で説明される。
【0012】
WO00/03246は、GPCR内部移行を誘発することができる化合物を同定するための方法を説明し、当該方法は、GPCR−蛍光タンパク質融合タンパク質を使用して、又はこれらの受容体に結合可能な標識分子を培地に添加することによって、受容体を発光標識することに基づく。この方法は、受容体が内部移行された細胞を同定するために分析しなければならない細胞画像の収集のための段階を含んでなる。
【0013】
アマシャムバイオサイセンス(Amersham Biosciences)社は、商標CypHer5(登録商標)でシアニン誘導体を市販し、特にWO00/75237で説明されるこれらの有機フルオロフォアは、蛍光が中性pHで非常に微弱であり、そして酸性pHで非常に強いという特性を有する。これらのシアニン誘導体は、例えば、GPCRに結合可能な抗体と結合することができ、GPCRへ結合した抗体−Cypher5の内部移行中、Cypher5は中性pHの培地(細胞外培地(extracellular medium))から酸性pHの培地(エンドソーム)まで転移し、この時発光は増加する。
【0014】
ランタニド錯体及び動的FRET(DYNAMIC FRET)による蛍光消光
生物学的現象を研究することに有効な蛍光化合物としての、(以下で「希土類錯体」とも言及する)ランタニド錯体の使用が、1990年代に開発された(例えば、Mathis 等による論文, Clin. Chem. 1995 Sep; 41(9):1391-7を参照されたい)。
FRET現象は、生物学で、特に生物相互作用の研究のために広範囲に渡って使用される。これは、蛍光ドナー化合物(例えば、ランタニド錯体)並びに任意の蛍光アクセプター化合物の使用に基づいており、それぞれ生体分子と結合する。試験対象の生物相互作用によって生体分子が互いに近接し、ドナー化合物が励起されると、ドナーとアクセプターとの間でエネルギー転移が行われ、反応媒体によって放出される発光において変化を生じる。一部の企業は、生物学的な方法を試験するための、このアプローチの実施用の試薬を市販し、例えば希望者には特定の生物学的現象の試験(酵素活性の検出、セカンドメッセンジャーのアッセイ, 等)のためのドナー及びアクセプター化合物並びにキットを準備する。
【0015】
培地中に溶解し自由に拡散している2つのドナーとアクセプター化合物との間の動的エネルギー転移現象は、特にD.Thomas等の、(1978)、PNAS,75:12,5746−5750で説明される。この現象は、頭文字DEFETによってしばしば表示され、これは「拡散増強蛍光エネルギー転移(Diffusion Enhanced Fluorescence Energy Transfer)」を意味する。ドナー及びアクセプター化合物を互いに近づける生物相互作用の発生に基づく、生物学的現象を試験するための通常使用されるエネルギー転移と異なり、動的転移によるFRET又はDEFETは、反応媒体中のFRETパートナーの一方又は両方の自由拡散に基づく。
【0016】
D.Thomas等(1978)の、PNAS,75:12,5746−5750では、溶解したテルビウム錯体(Tb(DPA)3)及びローダミン間のDEFETが説明され、膜小胞の水性溶媒に存在するTb(DPA)3と膜に含有されるエオシン−ホスファチジルコリンの、エオシン発色団との間の最も近い接近距離を測定するためにもまたDEFETが使用された。これらの研究は、特に寿命の長い(2.2 m秒)ドナーフルオロフォアを使用して、「急速拡散限界(rapid diffusion limit)」条件下で行われた。エネルギー転移効率が本質的にドナーとアクセプターとの最も近い接近距離にのみ依存するこれらの条件は、アクセプターの低濃度で特にシグナルの観察を可能にする。
【0017】
GB 2 223 096は、FRETパートナーの拡散定数に依存することが周知の動的FRETの概念にまさに基づく分子(例えば、抗体又は抗原)を検出するための方法を説明する。実際、説明される方法は、ドナー化合物とアクセプター化合物との間の動的FRETの変化を測定するものであり、この変化は、検出される分子との相互作用に続く、培地中のドナー又はアクセプターの拡散の変更に起因する。この方法は、ドナー又はアクセプター化合物の1つが、検出される分子と相互作用が可能なものと結合されることが必要である(例えば、ドナー又はアクセプターは、タンパク質又は抗体と結合する)。
【0018】
Zheng等(J. Am. Chem. Soc. 2005, 127, 16178-16188)は、ランタニド錯体を形成することができ、且つ脂質膜中に非特異的に挿入されることができる親油性大環状分子のクラス(DTPA-PDA-Cn, n=10,12)を、細胞標識の応用及び(ガドリニウム錯体の場合には)核磁気共鳴での使用のための応用のために開発した。著者は、テルビウム錯体とカルセインとの間のエネルギー転移に由来するシグナルを測定することによって、これらの錯体の細胞分布を測定するためにDEFET技術を使用した。著者は、カルセイン(カルセインは細胞膜不透過性である)を細胞外培地に添加する場合に、これらの錯体及びカルセイン間のDEFETを検出することができたが、しかしながら(カルセインの中性エステルの形態で細胞内に導入される)細胞内カルセインを含まず、従ってこれらの脂質ランタニド錯体が細胞膜の外表面上に挿入されると結論付けた。このチームは、これらのランタニド錯体の内部移行の現象におけるシグナル中で生じうる変化を試験しなかった。
【0019】
DEFETはまた、タンパク質の表面で静電ポテンシャルを測定する目的で、エネルギー−アクセプターモチーフを組み込んでいるタンパク質の構造とエネルギー−ドナー化合物とのこれらの相互作用を試験するために、一部のチームによって使用されてきた。
【0020】
ランタニド錯体及びレドックス効果による蛍光消光
ランタニド錯体の発光への一定の還元剤の影響は、文献で説明されている。
【0021】
Kielar等は、例えば、様々な還元剤によって、ユウロピウム及びテルビウム錯体の励起された状態の動的消光を試験し(Org. Biomol. Chem., 2007, 5, 2975-2982)、そして試験した希土類錯体に関して、1Vよりわずかに低い酸化電位を有する化合物が還元により蛍光消光効果を有することができることを示した。これは、例えばヨウ化物イオン(+0.54 V)、尿酸塩(+0.59 V)、アスコルビン酸塩(+0.30 V)及び一定のカテコレート(pH 7で+0.54 V)の場合である。当該著者は、これらの化合物のテルビウム錯体への蛍光消光効果が、同一のキレート構造を有するユウロピウム錯体で観察されたものよりも大きいことをさらに実証した。
【0022】
WO 2008/007089は、テルビウム又はユウロピウム錯体によって還元力を有する分析物をアッセイするための方法に関する。
【0023】
実施することが容易であり且つ高処理の応用に適当であり、目的の膜タンパク質の内部移行の検出に関して感受性のある方法が実際に必要である。このような方法は、特に新規の医薬を検出することを目的としているアプローチにおいて、膜タンパク質の内部移行を生じうる化合物を容易に示すことに関して特に好都合である。さらに、内部移行が可能な受容体の10〜40%のみが実際に内部移行されるので、注目とする方法は、内部移行が生じる場合にシグナル変化の観察を可能にし得ることに十分に敏感であるべきである。
【発明の概要】
【0024】
説明
本発明の対象は、タンパク質の内部移行を検出するための方法であって、次の:
a)ランタニド又はルテニウムを含んでなる蛍光金属錯体で目的のタンパク質を標識する段階であって、蛍光金属錯体の寿命が0.1msより長く、好適には0.5〜6msである段階;
b)目的のタンパク質の内部移行を生じることができる化合物を反応媒体に添加する段階;
c)反応媒体に対して以下から選択される調節剤:
1.上記蛍光金属錯体と適合する蛍光又は非蛍光のFRETアクセプター化合物であって、反応媒体中の最終濃度が10−7Mより高く、好適には10−6M〜10−3Mであり、分子量が50kD未満、好適には0.1〜10kDである化合物;
2.レドックス電位が+0.1V未満、好適には0.25〜0.75Vである還元剤;
3.非共有結合によって、特異的に蛍光金属錯体と結合する薬剤;
4.非蛍光金属錯体を形成するための、ランタニド又はルテニウムと競合する金属イオン、
を添加する段階;
d)蛍光金属錯体の発光波長及び/又は調節剤の発光波長で反応媒体によって放出される発光を測定する段階;
e)段階d)で測定されるシグナルを、段階a)及びc)のみに供した細胞上で測定される基準シグナルと比較する段階であって、基準シグナルと比較される段階d)で測定されたシグナルの違いが目的のタンパク質の内部移行を表す段階、
を含んでなる方法である。
【0025】
これらの必要な段階は、この順番(a, b, c, d, e)で行うことができる。目的のタンパク質の内部移行中、この場合細胞外培地に調節化合物は存在せず、従ってエンドソームに存在する調節化合物のリスクを軽減するので、この実施形態は好適である。
【0026】
本発明の好適な実施形態において、調節因子がFRETアクセプター化合物、又は還元剤である場合、これらの段階はまた、次の順序:a)、c)、b)、d)、e)、で実施することができる。
【0027】
さらに、段階a)とその後の段階との間に洗浄段階を追加することができる。用語「洗浄段階」は、1又は複数回(好適には2又は3回)培地を交換することから成る段階を意味することを目的とする。この洗浄段階は、細胞外培地から目的のタンパク質と結合していない蛍光金属錯体を除去するのに役立つが、本発明は内部移行のシグナルの違いの観察に基づくものであり、この段階は任意である。しかしながら、当該方法の感度を改善するためにこの洗浄を実施することは好適である。
【0028】
本発明の方法は、生細胞及び適当な試薬を含有する反応媒体によって放出されるシグナル、並びに細胞による目的の膜タンパク質の内部移行に従って変化するシグナルの強度の検出に基づいている。
【0029】
より詳細には、本発明の方法では、直接又は間接的に且つ細胞外培地にさらされたドメインのレベルで、目的の膜タンパク質が、蛍光シグナルを放出することができる蛍光金属錯体に結合される。細胞外培地は、(「調節化合物」と呼ばれる)化合物を含み、反応媒体にこの化合物が存在することにより、蛍光金属錯体によって放出されるシグナルが調節される効果を有する。
【0030】
目的の膜タンパク質の内部移行は、蛍光金属錯体の調節化合物への到達性を変更する。蛍光金属錯体はもはや細胞外培地に曝露されないが、しかしながら一定時間後にエンドサイトーシス小胞の内部の培地、又はサイトゾルへ曝露されるであろう。発明者は、調節化合物への蛍光金属錯体の到達性のこの変化が、反応媒体によって放出される発光シグナルに変化をもたらし、内部移行され得る膜貫通受容体の10〜40%だけが実際に内部移行されるという事実にもかかわらず、この変化は検出可能又はさらに任意に定量化できることを発見した。
【0031】
反応媒体によって放出される発光シグナルは、蛍光金属錯体によって放出される発光、或いは調節因子がFRETアクセプター化合物である場合には、調節因子によって放出される発光に対応する。
【0032】
いずれの場合でも、これは、反応媒体で測定される発光の違いにつながる調節化合物への蛍光金属錯体の到達性の違いである。
【0033】
定義
用語「蛍光金属錯体と適合するFRETアクセプター化合物」は、当該蛍光金属錯体とペアのFRETパートナーを形成するFRETアクセプター化合物を意味する。
【0034】
用語「FRETパートナーのペア」は、ドナー蛍光化合物及びアクセプター化合物から成るペアを意味し、これらが互いに近接する場合、及びこれらがドナー蛍光化合物の励起波長で励起される場合、これらの化合物はFRETシグナルを放出する。2つの蛍光化合物がFRETパートナーであるためには、ドナー蛍光化合物の発光がアクセプター化合物の励起スペクトルと部分的に重複しなければならないことは周知である。好適なFRETパートナーペアは、値R0(フォスターディスタンス, エネルギー転移効率が50%である距離)が30 Å以上のものである。
【0035】
「FRETシグナル」は、ドナー蛍光化合物とアクセプター化合物との間のFRETの代表的な、任意の測定可能なシグナルを意味する。従って、FRETシグナルは、アクセプター化合物が蛍光である場合、ドナー蛍光化合物又はアクセプター化合物の発光の強度又は寿命において変化することが可能である。
【0036】
「反応媒体」は、生細胞を試薬と接触させることができる任意の容器、例えばプレートのウェル、試験管等を意味する。
【0037】
「EDTA」は、エチレンジアミン四酢酸を意味する。
「DTPA」は、ジエチレントリアミン五酢酸を意味する。
「TTHA」は、トリエチレンテトラミン−N,N,N’,N’’,N’’’N’’’−六酢酸を意味する。
「DOTA」は、1,4,7,10−テトラアザシクロドデカン−N,N’,N’’,N’’’−四酢酸を意味する。
「NTA」は、ニトリロトリ酢酸を意味する。
「HDTA」は、ヘキサメチレンジアミン四酢酸を意味する。
「DTPP」は、ジエチレントリアミンペンタホスホン酸を意味する。
「EDTP」は、エチレンジニトリロテトラキス(メチルホスホン)酸を意味する。
「NTP」は、ニトリロトリ(メチルホスホン)酸を意味する。
「DOTP」は、1,4,7,10−テトラアザシクロドデカン−N’,N’’,N’’,N’’’−テトラキス(メチルホスホン)酸を意味する。
「DO3A」は、1,4,7,10−テトラアザシクロドデカントリス酢酸を意味する。
「DOTAGA」は、1−(1−カルボキシ−3−カルボキシプロピル)−4,7,10−(カルボキシメチル)−1,4,7,10−テトラアザシクロドデカンを意味する。
「DY647」は、ダイオミクス(Dyomics)社によって市販されるペンタメチン構造を有するフルオロフォアを意味する。
「D2」は、シスビオバイオアッセイ(Cisbio Bioassay)社によって市販される有機蛍光アクセプター化合物を意味する。
「PBS」は、リン酸バッファー溶液を意味する。
「Lumi4−Tb」は、ルミフォール社及びシスビオバイオアッセイ(Cisbio Bioassay)社によって市販されるテルビウム錯体を意味する。
「DMEM」は、「ダルベッコ変法イーグル培地」であり、商業的に入手可能であり且つ多くの応用で使用される細胞培地を意味する。
「NHS」は、N−ヒドロキシスクシンイミドを意味する。
【0038】
本発明の方法で使用される試薬及びこれらの実施の変形もまた、これからより詳細に説明する。
【0039】
A.蛍光金属錯体
一般的に、用語「蛍光金属錯体」は、ランタニド又はルテニウム、及び多座錯体形成剤から成る化合物であって、多座錯体形成剤がすなわち、少なくとも2個、好適には2〜9個の、電子供与体ヘテロ原子、例えばN、O又はSであって、ランタニド又はルテニウムと配位結合を形成している原子を含んでなる剤である化合物を説明することを目的とする。好適には、蛍光金属錯体が芳香族構造から成る1又は複数の発色団を含んでなり、好適には、これらの芳香族構造が、ランタニド又はルテニウム配位原子の役割を果たすN及びOから選択される、1、2又は3個のヘテロ原子を含んでなる。
【0040】
本発明の目的に関して適当な蛍光金属錯体は、錯体形成剤と希土類の会合/解離の面から安定であるべきであり、そしてこの形成定数(Kf)は好適には1010M−1より大きくあるべきである。
【0041】
多くの錯体形成剤が説明されており、そして当業者に周知である。錯体形成剤の例としては、次の化合物:EDTA、DTPA、TTHA、DOTA、NTA、HDTA、DTPP、EDTP、HDTP、NTP、DOTP、DO3A、DOTAGAに言及される。
【0042】
本発明の目的に適当な蛍光金属錯体の実施例は次のものである:
・1又は複数のピリジンユニットを含んでなるランタニドクリプテート。このような希土類クリプテート等は、例えば、EP 0 180 492、EP 0 321 353及びEP 0 601 113並びにWO 01/96 877で説明される。テルビウム(Tb3+)及びユウロピウム(Eu3+)のクリプテートは、本発明の目的に特に適当である。ランタニドクリプテートは、シスビオバイオアッセイ(Cisbio Bioassay)社によって市販される。非限定的な実施例として、以下の式、
【化1】

を有するユウロピウムクリプテートに言及することができ、これらは、反応基例えばNHS基又は反応基R等によって標識することができる化合物と結合することができる。
【0043】
・特にUS4 761 481、US 5 032 677、US 5 055 578、US 5 106 957、US 5 116 989、US 4 761 481、US 4 801 722、US 4 794 191、US 4 637 988、US 4 670 572、US 4 837 169、US 4 859 777で説明されるランタニドキレート。EP 0 403 593、US 5 324 825、US 5 202 423及びUS 5 316 909は、ノナデンテートリガンド、例えばターピリジンから成るキレートを説明する。ランタニドキレートは、パーキンエルマー(PerkinElmer)社によって市販される。
【0044】
・キレート剤から成るランタニド錯体、例えば、芳香環を含んでなる発色団で置換されたテトラアザシクロドデカン、例えばBiomol.Chem,2005,3,1013〜1024の「細胞内(in cellulo)での使用に適当な、高放出で且つ動力学的に安定したランタニド錯体の合成及び評価」でPoole R等によって説明されるものもまた使用することができる。WO2009/10580で説明される錯体もまた使用することができる。
【0045】
・EP 1 154 991及びEP 1 154 990で説明される希土類クリプテートもまた使用することができる。
・シスビオバイオアッセイ(Cisbio Bioassay)社によって市販され、ルミフォール(Lumiphore)社由来の、テルビウムクリプテートLumi4−Tb。
・(反応基、この場合、例えばNHS基によって標識可能な化合物に結合することができる)以下の式:
【化2】
を有するテルビウムクリプテートTb(KR)。この合成は、WO 2008/063721で説明される。
【0046】
・以下の式:
【化3】
を有する(反応基、この場合NCSによって標識される化合物に結合することができる)リサーチオーガニクス(Research Organics)社由来の量子染料。
・ルテニウムキレート、特にルテニウムイオン及び複数のビピリジン、例えばルテニウム(II)トリス(2,2’−ビピリジン)等から成る錯体。
【0047】
・以下の式、
【化4】
を有する(反応基Rによって標識可能な化合物と結合することができる)、インビトロジェン(Invitrogen)社によって市販されるテルビウムキレートDTPA−cs124 Tb。この合成は、US 5,622,821で説明される。
【0048】
ジスプロシウム(Dy3+)、サマリウム(Sm3+)、ネオジム(Nd3+)、イッテルビウム(Yb3+)或いはエルビウム(Er3+)の錯体はまた、本発明の目的に対して適当な希土類錯体であり、ユウロピウム(Eu3+)及びテルビウム(Tb3+)の錯体が特に好適である。
【0049】
非常に多くのランタニド錯体がこれまで説明されており、一部は、特にパーキンエルマー(PerkinElmer)、インビトロジェン(Invitrogen)、モルキュラーデバイシズ(Molecular Devices)及びシスビオバイオアッセイ(Cisbio Bioassay)社によって現在商業的に利用される。図1には、本発明の目的のために使用することができるユウロピウム及びテルビウム錯体の他の例が示される。
【0050】
蛍光金属錯体の使用は本発明の必要な特徴の1つであるが、当該発明は特定のタイプの蛍光金属錯体に限定されず、当業者は、特定の使用のために最も適当であり、0.1ms超、好適には0.5〜6msの寿命を有する蛍光金属錯体から選択が可能である。
【0051】
本発明によれば、蛍光金属錯体は結合される、すなわち、蛍光金属錯体によって目的のタンパク質を直接又は間接的に標識することができるカップリング剤に共有結合される。これらのカップリング剤は、「目的のタンパク質の標識」のセクションで説明され、好適にはリガンド/受容体ペアの1つ、例えば抗タグ抗体、ビオチン、或いは「自殺」酵素の基質、例えばベンジルグアニン、ベンジルシトシン又はクロロアルカンの誘導体である。
【0052】
蛍光金属錯体のカップリング剤との結合は当業者に周知であり、反応基の使用に基づいている。商業的に入手可能な希土類錯体は、カップリング剤へのこの結合を形成することができる反応基を含んでなる、又は既にこのようなカップリング剤を含んでなり、そしていつでも使用できる状態である。
【0053】
B.調節化合物
B.1:調節化合物が、エネルギーアクセプターである。
本発明の方法の一実施形態において、使用は、蛍光金属錯体(エネルギードナー)から調節化合物(エネルギーアクセプター)へのフォルスター共鳴エネルギー転移(FRET)の現象を通じて、蛍光金属錯体によって放出されたシグナルを変更することができる調節化合物から成る。当業者に周知のこの現象は、これらが互いに近接する場合、2つのFRETパートナー化合物間、本願の場合、蛍光金属錯体と反応媒体で自由に拡散する調節化合物との間のエネルギー転移に基づいている。
【0054】
一般的に、生物方法を試験するためのFRET現象の使用は、FRETパートナーのペアのメンバーはそれぞれ、互いに相互作用する化合物へ結合され、よってFRETパートナーは互いに近接され、FRETシグナルを生じることを暗に意味する。本発明の方法において、エネルギーアクセプターは、任意の化合物へ結合されず、反応媒体中で自由に拡散し、一方で蛍光金属錯体は直接又は間接的に目的の膜タンパク質と結合される。2つのFRETパートナー間のエネルギー転移は、DEFET現象に従って、反応媒体中のアクセプター化合物の拡散のみに依存する。
【0055】
発明者は、膜タンパク質と結合された蛍光金属錯体と細胞外培地で拡散しているアクセプターとの間のDEFETの成立に必要な条件を決定し、目的のタンパク質が内部移行される場合に、これらの条件によりシグナルにおける変化の観察も可能となる。
【0056】
この実施形態によれば、目的のタンパク質と蛍光金属錯体の内部移行に依存するDEFETを観察するために使用される実験条件は次の条件である:
・蛍光金属錯体及び調節化合物がFRETパートナーである条件;
・蛍光金属錯体の寿命が、0.1msより長く、好適には0.5〜6msであるべきであるという条件;
・細胞外培地中の調節化合物の濃度が、0.1μM〜1mMであるべきであるという条件。好適にはこの化合物の濃度は、1μM〜100μMであり、これにより急速な拡散限界で機能することができ、そして蛍光金属錯体の濃度に非依存的なDEFETシグナルを測定することが可能である;
・調節化合物は、好適には細胞膜を貫通すべきでないという条件。さらに、この拡散定数が十分高くあるべきである。一般的に、50kD未満、好適には0.1〜10kDの分子量を有する調節化合物が本発明の目的に適当である。
【0057】
本発明の方法のこの実施形態において、調節化合物は蛍光又は非蛍光性であってよい。
【0058】
調節化合物が蛍光エネルギーアクセプター化合物であり、ランタニドがテルビウム又はユウロピウムである場合には、この調節化合物は次の化合物又は化合物ファミリーから選択してよい:シアニン誘導体、DY647、D2、フルオレセイン及びその誘導体、クマリン、ローダミン、カルボピロニン、オキサジン及びその類似体、アレクサフルオールズ(Alexa Fluors)、クリスタルバイオレット、ペリレンビシマイド(bisimide)型のフルオロフォア、スクアレイン、商標ボディピ(BODIPY)(登録商標)で周知のホウ素ジピロメテン誘導体、NBD(ニトロベンズオキサジアゾール)及びその誘導体、DABCYL(4−((4−(ジメチルアミノ)フェニル)アゾ)安息香酸)。
【0059】
アレクサフルオール化合物及びホウ素ジピロメテン誘導体は、インビトロジェン(Invitrogen)社によって市販され、化合物Dy647は、ダイオミクス(Dyomics)社によって市販されるシアニン誘導体であり、シアニン誘導体はまた、特にアマーシャムバイオサイエンス(Amersham Biosciences)社によって市販され、他の化合物は、化学試薬の様々なサプライヤー、例えばシグマ(Sigma)、アルドリッチ(Aldrich)又はアクロス(Acros)社によって市販される。
【0060】
非蛍光化合物もまた、これらの吸収スペクトルが蛍光金属錯体の発光スペクトルと適合するという条件で、使用することができる。次の非蛍光アクセプター:アナスペック社のQXL製品、特にQXL570、QXL610、QXL670及びQXL680、ダイオミクス(Dyomics)社のDYQ660及びDYQ661製品、並びにインビトロジェン(Invitrogen)社のQSY7、QSY9及びQSY21製品、が商業的に入手可能である。
【0061】
反応媒体中で検出されるシグナルは、蛍光金属錯体によって放出される発光、又はアクセプター化合物もまた蛍光の場合にはアクセプター化合物によって放出される発光、これらの強度或いはこれらの寿命である。任意の現象において、これらのシグナルはDEFET現象に依存し、目的の膜貫通タンパク質の内部移行中に、次のように変化する:
・膜貫通タンパク質が内部移行される場合には、発光波長で測定される蛍光金属錯体の発光の強度又は寿命が増加する;
・調節因子が蛍光アクセプター化合物の場合には、タンパク質が内部移行される時に、アクセプターの発光波長で測定されるシグナルの強度又は寿命が変化する。アクセプター化合物由来のシグナルは蛍光金属錯体とのDEFETに依存するが、アクセプター化合物の濃度が高い場合に細胞外培地で生じる、可能性のある分子間の発光−消光効果にも依存することは注目すべきである。この理由のため、内部移行後、アクセプター調節化合物の発光波長で放出されたシグナルは増加又は減少し得る;
・調節因子が蛍光アクセプター化合物である場合、そのシグナルはアクセプターシグナル/ドナーシグナル比を算出することによって補正することができる。FRETシグナルのラジオメトリック補正のこの方法は、広範に使用され文献に説明される。
【0062】
B2:調節化合物が、蛍光金属錯体に関する還元剤である:
本発明者は、蛍光金属錯体及び/又は錯体形成構造のレベルで酸化還元反応によって蛍光金属錯体の発光を減少することができる還元剤が、本発明の方法において好都合に使用することができることを発見した。
【0063】
還元剤は、蛍光金属錯体を還元するのに適当であるレドックス電位を有するべきである。当業者は、選択される蛍光金属錯体に従って、本発明の方法を実施するのにどの還元剤が適当であるかを決定することができるが、レドックス電位が+0.1〜+1.2Vである還元剤が好適である。さらに好適には、還元剤のレドックス電位は+0.25〜+0.75Vである。
【0064】
従って、蛍光金属錯体によって放出されるシグナルを調節する化合物が還元剤である場合、例えば次の還元剤から選択することができる:ヨウ化物、アスコルビン酸塩、尿酸塩、カテコラート、無機化合物、例えばFe(CN)62+。これらのイオンは、対イオン例えばアニオンを有する塩の形態、ナトリウム又はカリウム塩の形態の測定培地へ添加される。
【0065】
本実施形態において、測定されるシグナルは、蛍光金属錯体によって放出され、細胞外培地に存在する還元剤によって調節される発光に相当する:蛍光金属錯体と結合された目的のタンパク質が内部移行される場合、蛍光金属錯体は調節剤と接触又は近接せず、寿命と同様に、この発光は内部移行が生じない場合よりも増大する。
【0066】
この実施形態において、内部移行が存在する場合と内部移行が生じない場合のシグナルの違いを測定することができるように、発明者は蛍光金属錯体/還元剤ペアのシュテルンフォルマー(Stern-Volmer)定数以上の濃度で、反応媒体中に還元剤が存在すべきである、と決定した。この定数は蛍光金属錯体の発光強度の50%の減少を生じるのに必須の還元剤の濃度であると定義される。蛍光金属錯体/還元剤ペアに関するシュテルンフォルマー(Stern-Volmer)定数の例は、特にキラー(Kielar)等による論文(Org. Biomol. Chem., 2007, 5, 2975-2982)に列挙される。
【0067】
従って、実施形態の第一の態様では、これは測定される蛍光金属錯体の発光である。
【0068】
この実施形態の第二の態様では、本発明の方法の第一段階は、測定培地に目的のタンパク質を標識することを目的とする第一の蛍光金属錯体及び第二の蛍光金属錯体を添加することを含んでなる。
【0069】
この第二の蛍光金属錯体は、異なる金属を除いて、第一錯体と同一である。例えば、第一蛍光金属錯体は、ユウロピウム錯体であり、第二金属錯体は同様のキレート構造を含んでなるテルビウム錯体である。そのため、この実施形態において、一定の目的のタンパク質は、潜在的に直接又は間接的に第一蛍光金属錯体に結合する、或いは直接又は間接的に第二蛍光金属錯体に結合する(或いは任意の蛍光金属錯体によって標識されない)。
【0070】
この実施形態において、例えば、第二蛍光金属錯体によって放出される発光の強度との、ランタニド錯体の1つによって放出される発光の割合を形成することによって測定されるシグナルを標準化することが可能である。このタイプの測定により、異なる反応媒体から得られるシグナルを比較することが可能であり、従って特に定量分析を実施するために適当である。
【0071】
B3:調節化合物が、非共有結合によって、特異的に蛍光金属錯体と結合する薬剤である:
調節化合物は、蛍光金属錯体との特異的且つ非共有的な結合を可能とするドメインを有する薬剤であってよく、これらの化合物は実際に蛍光金属錯体の発光において減少を生じることができる。これらの化合物は、蛍光金属錯体−結合ドメインを各々有する、抗体又は抗体断片、ペプチド、或いはアプタマーから選択することができる。
【0072】
これらの薬剤の中で、好適な調節因子は、蛍光金属錯体を認識することができる抗体又は抗体断片である。WO2007/116069(A1)は、希土類錯体に特異的な抗体の調製を説明し、さらに当業者は、これらの自由技術により、このタイプの化合物を生成することが可能である。
【0073】
本発明の方法をこのタイプの調節因子で実施するために、調節剤を添加する前に内部移行を生じる化合物を添加することが望ましく、さもなければ蛍光金属錯体として同時に内部移行される。言い換えれば、この実施形態において、上述の方法の段階は次の順:a)、c)、b)、d)、e)で行われるべきである。
【0074】
B4:調節化合物が、非蛍光金属錯体を形成するするための希土類と競合する金属イオンである:
本発明の方法を実施するのに適当な調節因子の他のタイプは、錯体形成剤を有する非蛍光錯体を形成するための希土類と競合する金属イオンである。従って、希土類キレートの場合には、調節剤はMn2+イオンであってよい。
【0075】
この場合は、適当な対イオンを有する塩の形態、例えばMnCl2の形態で調節剤を細胞外培地に添加する。
【0076】
このタイプの調節因子で本発明の方法を実施するために、調節剤を添加する前に内部移行を生じることができる化合物を添加することが望ましい。言い換えれば、この実施形態において、上述の方法の段階は次の順序:a)、c)、b)、d)、e)で行われるべきである。
【0077】
C.目的のタンパク質の蛍光金属錯体での標識
C.1.発現
内部移行されることができる任意の膜タンパク質が、細胞外部分で蛍光金属錯体と直接又は間接的に結合することができるという条件で、本発明は当該タンパク質を用いて実施することができる。
【0078】
上述のように、本発明は、特にGタンパク質共役型受容体(以下、「GPCR」とする)の内部移行、及び制御機構の1つが内部移行である受容体型チロシンキナーゼ(以下、「RTK」とする)の試験に有効である。それにもかかわらず、内部移行されるか否か確認することが望まれる他の膜タンパク質で本発明を実施することも可能である。
【0079】
目的のタンパク質は細胞膜で当然に発現され、或いは通常の分子生物学技術、特に細胞内に安定的に又は一過性に導入される発現ベクターを使用して発現される。安定に又は一過性に異種DNAを細胞内に導入するための試薬は、商業的に入手可能であり、目的のタンパク質をコードしているDNA配列、特にGPCR及びRTKをコードしているものは、データベース、例えばジェンバンクで入手可能である。目的のタンパク質が細胞によって安定に発現される場合には、非常に多数のGPCRの存在により、細胞毒性現象を観察することができ、この場合、GPCRの発現を制限するために、誘導発現システムを使用することは好都合かもしれない。
【0080】
従って、本発明の方法は目的のタンパク質をコードしている発現ベクターを細胞にトランスフェクトする予備段階を含んでよい。以下に説明されるように、このベクターはまた蛍光金属錯体での目的のタンパク質の共有結合性の標識を可能にする自殺酵素をコードする配列を含んでよい。
【0081】
特定の一実施形態では、第二発現ベクターは目的のタンパク質をコードしているものと同時にトランスフェクトされ、この発現ベクターはβ−アレスチン1をコードしている配列を含んでなる。本発明の実施にこの同時発現は必要ではないが、β−アレスチン1は非特異的に内部移行現象を増幅するので、当該方法の感度の増加を可能にすることができる。
【0082】
C.2.蛍光金属錯体での標識
特に容易な本発明の一実施形態は、目的のタンパク質へ結合することが可能なことが周知の化合物(リガンド)へ蛍光金属錯体を結合することにある:目的のタンパク質への結合により修飾されたリガンドによって、このタンパク質の間接的な標識が可能である。
【0083】
多くの技術はタンパク質を修飾することについて説明されており、当業者は既存ツールをタンパク質を蛍光金属錯体に結合するために使用することが可能である。特に、通常の分子生物学的技術によってタンパク質の一定のドメインを添加、除去又は変更することが可能である。
【0084】
本発明の実施に必要な唯一の特徴は、蛍光金属錯体は細胞外培地に存在するようにそのような方法でタンパク質へ結合され、従って反応媒体によって放出される発光シグナルは実際、調節化合物の細胞外培地における存在に依存的である。
【0085】
目的のタンパク質がGPCR又はRTKである場合、以下に説明される技術の1つに従って直接又は間接的に蛍光金属錯体で標識されることができるように、そのN末端の(細胞外の)ドメインの修飾を介して蛍光金属錯体はこのタンパク質へ結合される。
【0086】
非限定的な説明の目的で、膜貫通タンパク質を蛍光金属錯体に結合するための次の技術に言及することができる:
・間接的に(非共有結合で)目的のタンパク質を蛍光金属錯体と結合すること
【0087】
蛍光金属錯体は、実際は少なくとも一方がタンパク質である、結合パートナーのペアによって、目的のタンパク質と結合されることができる。このアプローチにおいて、目的のタンパク質は、通常の分子生物学技術(タンパク質結合パートナーをコードしているものと融合した目的のタンパク質をコードしているヌクレオチド配列を含んでなる発現ベクターの作成、及び発現ベクターの細胞への導入)によって実際はタンパク質である結合パートナーと融合される。蛍光金属錯体は、明細書中でカップリング剤として言及される他の結合パートナーに共有結合され、続いて細胞外培地に添加される。結合パートナーの認識により、目的のタンパク質を蛍光金属錯体で間接的に標識することができる。
【0088】
本発明を実施するのに特に適当な結合パートナーの非限定的な実施例として以下に言及する:
・Xが任意のアミノ酸であり二ヒ素(biarsenic)化合物である、システイン−システイン−X−X−システイン−システイン配列(配列番号 1)から成るペア。これらの二ヒ素(biarsenic)化合物は、容易にフルオレセイン又はローダミンタイプの有機分子で標識されることができる(当該テクノロジーについて詳述の、B.A.Griffin 等. (1998) Science. 1998, 281, 269-271 及びS.A. Adams 等. (2002) J. Am. Chem. Soc. 2002, 124, 6063-6076 を参照された)。
・ブンガロトキシン(BTX)によって認識される13個のアミノ酸のペプチドから成るBTX(ブンガロトキシン)ペプチドは、蛍光分子と結合されることができる(C. M. McCann 等. (2005), Biotechnique (2005), 38, 945-952を参照されたい)。
・ストレプトアビジン/ビオチンペア:ストレプトアビジン結合配列(SBP-Tag)は、蛍光金属錯体で前標識されることができるビオチンへの高親和性を有する、38個のアミノ酸によって形成される配列である(C. M. McCann 等. (2005), Biotechnique (2005), 38, 945-952を参照されたい)。
・特異的に且つ高親和性を有してリガンド、例えばトリメトプリムを結合する大腸菌ジヒドロ葉酸還元酵素(eDHFR)の配列であり、アクティブモチーフ社からの「リガンドリンクの一般的な標識テクノロジー」として周知のテクノロジーに従って蛍光金属錯体はこれに結合されることができる。
・タグ/抗タグペアは、タンパク質を標識するために頻繁に使用される結合パートナーである。用語「タグ」は、必ずしも非常に短い必要はないが通常非常に短いアミノ酸配列(15個のアミノ酸未満)から成る低分子のタンパク質「標識(label)」を意味し、目的のタンパク質に融合される又はこのタンパク質に当然に存在する。用語「抗タグ」は、当該タグに特異的に結合する抗体を意味する。この実施形態において、抗タグ抗体は、蛍光金属錯体に共有結合される。従って、標識された抗体が細胞外培地に添加される場合、それは目的のタンパク質に結合されるタグに結合され、そしてタグ/抗タグ相互作用によってこのタンパク質の蛍光金属錯体での間接的な標識が可能である。
【0089】
タグ/抗タグペアの非限定的な実施例として、次のペアに言及することができ、これらのメンバーは商業的に入手可能である:GSTが、グルタチオンS−転移酵素又はこの断片を表す、GST/抗GST抗体;6HISが6個のヒスチジンから成るペプチドである、6HIS/抗6HIS抗体;MycがヒトMycタンパク質のアミノ酸410〜419個から成るペプチドであるMyc/抗Myc抗体;FLAGが8個のアミノ酸DYKDDDDK(配列番号 2)を有するペプチドであるFLAG/抗FLAG抗体;HAが9個のアミノ酸YPYDVPFYA(配列番号 3)から成るインフルエンザ赤血球凝集素エピトープであるHA/抗HA抗体。タグの正確な性質は本発明の実施には必要でないことは明らかである。
【0090】
・蛍光金属錯体との目的のタンパク質の直接的な(共有結合性の)結合
このアプローチにおいて、蛍光金属錯体は共有結合によって目的のタンパク質と結合される;一部の技術が説明されており、当該技術を実施するために必要な試薬は、商業的に入手可能である。この結合のために、以下の任意の1つの技術を使用してよい:
・目的のタンパク質上に存在する反応基のレベル、特に次の基の1つのレベルでの共有結合の形成:末端アミノ基、末端カルボキシラート基、アスパラギン酸及びグルタミン酸のカルボキシラート基、リジンのアミン基、アルギニンのグアニジン基、システインのチオール基、チロシンのフェノール基、トリプトファンのインドール環、メチオニンのチオエーテル基、ヒスチジンのイミダゾール基。
【0091】
目的のタンパク質上に存在するこれらの基は、蛍光金属錯体によって保持される反応基と共有結合を形成することができる。
【0092】
適当な反応基は、当業者に周知のものである。マレイミド基で官能性を有する蛍光金属錯体は、例えばタンパク質のシステインによって保持されるチオール基と共有結合が可能であろう。同様に、N−ヒドロキシスクシンイミドエステルを有する蛍光金属錯体は、目的のタンパク質のアミンへ共有結合が可能であろう。
【0093】
・自殺酵素の使用:
用語「自殺酵素」は、特異的突然変異によって修飾された酵素活性を有するタンパク質を意味することを目的とし、当該活性により酵素は急速に共有結合することができる。これらの酵素は各々、ちょうど1つの蛍光分子を結合することができるので、「自殺」と呼ばれ、当該酵素の活性は基質の付着によってブロックされる。これらの酵素は、その結果、1つのタンパク質に対して1つの蛍光分子の割合で目的のタンパク質を特異的に標識するための選択手段を構成する。このアプローチでは、自殺酵素は、通常の分子の生物学的技術によって、膜タンパク質と、好適にはそのN末端部で融合され、そしてドナー/アクセプターに共有結合された酵素の基質は、細胞外培地中に導入される。酵素反応は標識基質の酵素への共有結合をもたらし、従ってドナー又はアクセプターを有する膜タンパク質が標識される。
【0094】
非限定的な実施例として、次の酵素に言及することができる:
−O6−アルキルグアニンDNAアルキル転移酵素(AGT)の変異体。NEB社によって市販される酵素SNAP−tag(Juillerat 等., Chemistry & biology, Vol.10, 313-317 April 2003)及びCLIP−tag(Gautier 等., Chemistry and biology, 15, 128-136, February 2008)は、ヒトAGTの変異体であり、これらの基質は、各々、O6−ベンジルグアニン(以下、BGと略す)及びO2−ベンジルシトシン(以下、BCと略す)である。酵素N−AGT(Gronemeyer 等., Protein engineering, design & selection, vol. 19, No. 7, pp 309-3016, 2006)は、この酵素の他の変異体であり、O6−ベンジルグアニンとのこの反応性は、SNAP−tag酵素のものよりもより良好である;
−脱ハロゲン酵素の変異体(Promegaによって市販されるHaloTag)。これもまた自殺タイプの酵素反応を生じ(WO 04/072232 A2を参照されたい)、この基質の一部はクロロアルカンファミリー、特にユニット−NH−CH2CH2−O−CH2CH2−O−(CH2)6−Clを含んでなるクロロアルカンから構成される。この場合、蛍光金属錯体は、このタイプのユニットに結合される;
−ホスホパンテテイン転移酵素の存在下で、補酵素Aの4’−ホスホパンテテイン残基がACPのセリンに転移される、ACPタンパク質(アシル担体タンパク質)。(N. George 等., Journal of the American Chemical society, 126, (2004) p 8896-8897)。目的のタンパク質を蛍光金属錯体で標識するためにこのアプローチが使用される場合、ホスホパンテテイン転移酵素を反応媒体に添加することが必要である。NEB社は、タンパク質標識のためのACP断片を商標名「ACP−Tag」で市販する。
【0095】
このアプローチが目的のタンパク質を標識するために使用される場合、細胞に自殺酵素及び目的のタンパク質を含んでなる融合タンパク質をコードするDNAを含んでなる発現プラスミドをトランスフェクトする。このプラスミドはまた、これらのタンパク質をコードしているDNA、タグ、例えばFLAGエピトープ、Mycエピトープ、又はインフルエンザ赤血球凝集素(HA)エピトープ等をコードしているDNAの上流を含んでなる。
【0096】
融合タンパク質が細胞膜で発現されることを確実にするために、発現プラスミド中に、目的のタンパク質及び自殺酵素をコードしている配列、膜標的ペプチド、例えばT8シグナルペプチド又はmGluR5受容体のシグナルペプチドをコードしている配列の上流を含むことが有効であり、本目的のためのこの使用は当業者に周知である。最後に、目的のタンパク質をコードしている配列が、目的のタンパク質と自殺酵素の間の結合の翻訳後の切断の対象となりうるいずれの天然の膜標的配列を含んでないことを確実にすることもまた望ましい。この場合には、このドメインを発現プラスミド中に導入しないことが好適である。
【0097】
酵素反応を細胞外培地に存在する酵素の基質(例えばBG−蛍光金属錯体複合体)で生じるために、自殺酵素を細胞外培地にさらすことが必要である。GPCR及びRTKの場合、目的の天然のタンパク質のN末端部が細胞外培地にさらされる時、融合タンパク質が作成され、自殺酵素は融合タンパク質のN末端部で発現されるが、これが存在する場合には常に膜標的ペプチドの下流である。
【0098】
最終的に、蛍光金属錯体で目的のタンパク質を標識するために自殺酵素が使用され、そして目的のタンパク質がGPCR又はRTKである場合、本発明は、融合タンパク質をコードしているDNA配列を含んでなる発現ベクターを細胞にトランスフェクトする予備段階を含んでなり、このN末端部は自殺酵素を含んでなり、そしてC末端部は目的のタンパク質を含んでなる。
【0099】
蛍光金属錯体に結合される酵素の基質の細胞外培地への導入は、目的のタンパク質をこの蛍光金属錯体で標識することとなるであろう。
【0100】
この実施形態において、次の融合タンパク質から選択される融合タンパク質をコードしている発現ベクターを使用することができる:
自殺酵素−目的のタンパク質、又はタグ−自殺酵素−目的のタンパク質、或いは膜標的ペプチド−タグ−自殺酵素−目的のタンパク質。
【図面の簡単な説明】
【0101】
【図1】本発明の目的のために使用することができるユウロピウム及びテルビウム錯体の例を示す。
【図2】バソプレシン濃度に応じた、545nmで測定したドナー発光における変化を示す。
【図3】バソプレシン濃度に応じた、ドナー化合物のΔASR%の変化を示す。
【図4】バソプレシン濃度に応じた、ドナーシグナルのアクセプターシグナルに対する割合の変化を示す。
【図5】EDA9アンタゴニストの濃度に応じた、ドナーシグナルのアクセプターシグナルに対する割合における変化を示す。
【図6】SDF−1の濃度に応じた、ドナーシグナルのアクセプターシグナルに対する割合における変化を示す。
【図7】イソプロテレノールの濃度に応じた、ドナーシグナルのアクセプターシグナルに対する割合における変化を示す。
【実施例】
【0102】
次の実施例は、本発明の好適な実施形態を、すなわち蛍光金属錯体がベンジルグアニン又はベンジルシトシンに共有結合されたユウロピウム又はテルビウムクリプテートであるものを説明しており、この時、目的のタンパク質はアルキルグアニン−DNAアルキル転移酵素変異体を有する融合タンパク質の形態で発現され、そして調節化合物はシアニン誘導体、特にCy5若しくはCy5−COOH、又はフルオレセイン或いはこの誘導体の1つである。ユウロピウムクリプテートが使用される場合、フルオレセインよりもむしろシアニン誘導体を使用することが好適である。
【0103】
実施例1:バソプレシン受容体V2−Rの内部移行の検出
この実施例の目的は、本発明の方法によって、バソプレシンV2受容体を含んでなる融合タンパク質の内部移行、Snap−tag酵素及びHAタグを検出することである。
【0104】
より具体的には、この場合はテルビウムクリプテート(LUMI4-TB)−ベンジルグアニン(BG)複合体がドナー化合物として使用され、この場合にシアニンCy5−COOHはアクセプターとして使用され、そして受容体の内部移行の後に起こるDEFETでの変化に由来するシグナルにおける変化が観察される。
【0105】
使用された試薬及び材料
オプティMEM(OptiMEM)培地(インビトロジェン(Invitrogen)(51985-026))
PBSリン酸バッファー
DMEM woPR(フェノールレッド非含有のDMEM培地、インビトロジェン(Invitrogen))
DMEM+:次のものが添加されたDMEM:1mM L−アラニル/L−グルタミン、1mM Naピルビン酸塩、10%ウシ胎仔血清(インビトロジェン(Invitrogen)SKU#10091-148)、1%ペニシリン−ストレプトマイシン(インビトロジェン(Invitrogen)SKU#15070-063)、2mM HEPES、1%非必須アミノ酸)
DMEM woPR+(フェノールレッドを含まないDMEM+ 培地)
Krebs−Tris+グルコース0.5g/l
BG−LUMI4−TB(シスビオバイオアッセイ(Cisbio Bioassay)によって市販されるベンジルグアニン−テルビウムクリプテート複合体)
BG:O6−ベンジルグアニン(Sigma B2292)
バソプレシン(Bachem)
プラスミドHA−ST−V2:T8膜標的シグナルペプチド、HAエピトープ、SNAPTAG酵素及びV2受容体を含んでなる融合タンパク質をコードしている配列を含んでなるプラスミド。
シアニンCy5−COOH:エラスラボラトリーズ(Eras Laboratories)。
【化5】
【0106】
プレートの処理
ウェルの底への細胞の接着を促進するために、50μlのポリ−L−オルニチン溶液(0.01% 溶液, 分子量 30 000-70 000(SIGMA P4957))を96ウェルプレート(Cellstar, 黒底で黒色)の各ウェル中に分配し、そして、プレートを37℃で30分間インキュベートし、その後100μlのDMEM+で一回リンスした。
【0107】
トランスフェクション
続いて、以下の組成物を有する次のトランスフェクション混合物又はネガティブコントロール混合物を細胞に分配した:
【0108】
【表1】
【0109】
30分間のインキュベーションの後、COS7の細胞を100μl(CIS BIO)、すなわち80 000細胞、を各ウェルに添加し、続いて5%CO2の存在下で37℃で24時間インキュベートした。
【0110】
用量反応曲線
ウェルをDMEM woPR+培地で1回リンスし、その後156nMのBG−LUMI4−TB又は156nMのBG(ネガティブコントロール)溶液をDMEM woPR+培地(100 μl)に添加し、そしてプレートを1時間30分インキュベートした。
【0111】
続いて、各ウェルを100μlのKrebs−トリスバッファー+グルコース(0.5 g/l)で3回洗浄し、その後Krebs−トリスバッファーを添加し、Krebs−トリスバッファーで希釈したバソプレシン(V2 受容体アゴニスト)を添加した(4℃, 10 μl, 可変濃度, プレートプランを参照されたい)。
【0112】
30分間のインキュベーション後、24μMのアクセプター最終濃度を得るために、Krebs−トリスバッファー(10 μl)中の溶液の264 μMのシアニン5−COOHをラインB、D、E〜Hに添加した(プランを参照されたい)。
【0113】
15分間プレートを撹拌し、337nmの窒素レーザー並びに545nm及び665nmの放出フィルターが取り付けられたエンビジョンリーダー(Envision reader)(パーキンエルマー)で読んだ。400μsの遅延及び400〜2900μsの読み取りウィンドウ(window)を有する両チャネルで読み取りを実施した。
【0114】
96ウェルプレートのプランを各ウェルに添加した試薬の量で以下に表す:
【0115】
【表2】
【0116】
ラインA:プラスミドPRK6を有するネガティブコントロールであって、従って標識しているSnaptag(ST)はなく、Cy5アクセプターの非存在下であり、LUMI4−TBドナーの非存在下(A1-A4)、バソプレシンを含まずLUMI4−TBドナーの存在下(A5-A8)又はバソプレシンを含みLUMI4−TBドナーの存在下(A9-A12)で観察されるシグナル。
【0117】
ラインB:プラスミドPRK6を有するネガティブコントロールであって、従って標識しているSnaptag(ST)はなく、Cy5アクセプターの存在下であり、LUMI4−TBドナーの非存在下(B1-B4)、バソプレシンを含まずLUMI4−TBドナーの存在下(B5-B8)又はバソプレシンを含みLUMI4−TBドナーの存在下(B9-B12)で観察されるシグナル。
このラインにより、受容体の標識をすることなくバックグラウンドノイズを測定することが可能であり、測定されたシグナルを補正することが可能である。
【0118】
ラインC:プラスミドHA−ST−V2を有するネガティブコントロールであって、従ってSnaptagの存在下で、バソプレシン又はCy5アクセプターを含まず、非蛍光のBGの存在下(C1-C4)又はLUMI4−TBドナーを有する蛍光のBGの存在下(C5-C12)で観察されるシグナル。
【0119】
ラインD:プラスミドHA−ST−V2を有するネガティブコントロールであって、従ってSnaptagの存在下で、しかしながらバソプレシンを含まず、そしてCy5アクセプターの存在下で;非蛍光のBGの存在下(D1-D4)、又はLUMI4−TBドナーを有する蛍光のBGの存在下(D5-D12)で観察されるシグナル。
このラインにより、BG−LUMI4−TBで標識されるV2受容体の内部移行の非存在下で、基準シグナルを得ることが可能である。
【0120】
ラインE〜Hは、様々な濃度のバソプレシンの存在下で観察されるシグナルに相当する。
【0121】
結果及び考察
図2は、ドナー発光における変化を表し、バソプレシン濃度に応じて、従って受容体の内部移行に応じて、545nmで測定した。
【0122】
ΔAS%は、(バソプレシンを含まず)内部移行を含まずに測定される基準シグナルに対する、内部移行中のドナーシグナルの割合の増加に相当する。
【0123】
より具体的には、バソプレシンの一定の濃度、例えば10pM(ウェルE1〜E6)に対して、ΔAS%は次の方法で算出した。
【0124】
【化6】
【0125】
式中、「[E1−E6]545」、「[D5−D12]545」等は、BG−LUMI4−TBドナーの波長(545 nm)で、ウェルE1〜E6、D5〜D12等において測定されるシグナルの平均値に相当する。
【0126】
分子は、バソプレシンの存在及び非存在下(従って内部移行の非存在下)で測定されるシグナルの違いに相当し、分母は、内部移行の非存在下([D5-D12] 545)における基準シグナルであって、バックグラウンドノイズ(BG-LUMI4-TB ドナーの非存在下で測定される[D1-D4] 545)、さらに非特異的シグナル(Snaptag標識の非存在下での[B5-B8] 545-[B1-B4] 545)に対して補正される基準シグナル相当する。
【0127】
図2は、バソプレシンの増加している濃度の存在下における545でのBG−LUMI4−TBドナー化合物の発光における変化(この場合は増加)を示し、従って膜受容体の内部移行を示すための本発明の方法の使用を検証する。
【0128】
665nm(Cy5)でアクセプターによって放出されるシグナルに焦点を置いているなら、545でのドナーシグナルに対して補正し、同様の結果が得られる(このラジオメトリック補正により、1つのウェルから他のウェルへのシグナル変動(signal variability)を排除することが可能である)。
【0129】
図3は、バソプレシン濃度、従ってV2受容体の内部移行に応じたΔASR%の変化を表す。ΔASR%は、(バソプレシンの非存在下で測定され)内部移行を含まない基準シグナルの割合に相当する。
【0130】
バソプレシンの一定の濃度、例えば10pM(ウェルE1〜E6)に対して、ΔASR%を次の方法で算出した:
【0131】
【化7】
【0132】
バソプレシンの存在下で測定されるシグナル([E1-E6] 665/[E1-E6] 545)及び基準シグナル([D1-D12] 665/[D1-D12] 545)は、バックグラウンドノイズ(BG-LUMI4-TBドナーの非存在下で測定される[D1-D4])及びSnaptag標識の非存在下で測定される非特異的シグナル([B5-B8]-[B1-B4])に対して各々補正される。
【0133】
図3は、バソプレシンの濃度の増加が培地で存在する場合の変化(アクセプター化合物によって放出されるシグナルにおける減少)を示し、従って膜受容体の内部移行を示すための本発明の方法の使用を確認するものである。
【0134】
実施例2:バソプレシン受容体V2−Rの内部移行の検出
本実施例は、V2バソプレシン受容体の内部移行を検出するための本発明の方法の他の実施形態を説明する。特に、フルオレセインをアクセプターとして試験し、そしてV2受容体をコードしている発現プラスミドを、β−アレスチン1をコードしているプラスミドと同時にトランスフェクトした。この同時のトランスフェクション(cotransfection)により、受容体内部移行を促進し、従って本方法の感度を増加することが可能となった。
【0135】
使用した試薬及び材料
オプティMEM(OptiMEM)培地(Invitrogen(51985-026))
Krebs−グルコース:Krebsバッファー+ 0.5g/lグルコース
BG−LUMI4−TB(Cisbio Bioassayによって市販される、ベンジルグアニン−テルビウムクリプテート複合体)
BG:O6−ベンジルグアニン(Sigma B2292)
バソプレシン(Bachem)
プラスミドFLAGST−V2:T8膜標的シグナルペプチド、FLAGエピトープ、SNAPTAG酵素及びV2受容体を含んでなる融合タンパク質をコードしている配列を含んでなるプラスミド。このプラスミドの配列は、配列番号4である。
プラスミドFLAG−アレスチンβ1:FLAGエピトープ及びヒトβ−アレスチン1を含んでなる融合タンパク質をコードしている配列を含んでなるプラスミド。このプラスミドの配列は、配列番号5である。
フルオレセイン(Sigma)。
【0136】
プレートの処理
細胞のウェルの底面への接着を促進するするために、ポリ−L−オルニチンの50μl溶液(0.01% 溶液, 分子量30 000〜70 000 (SIGMA P4957))を96ウェルプレート(Cellstar, 黒底で黒色)の各ウェルに分配し、プレートを37℃で30分間インキュベートした。
【0137】
トランスフェクション
続いて、次のトランスフェクション混合物を各ウェルに分配した:
−0.16μgのプラスミドFLAGST−V2 + 0.16μgのプラスミドFLAG−β1アレスチン
−0.8μlのリポフェクタミン2000
−50μlのオプティMEM(OptiMEM)培地。
【0138】
30分間のインキュベーション後、100 000のCOS7細胞を含有する100μlの懸濁液を各ウェルに添加し、続いて5%のCO2存在下で37℃で24時間インキュベートした。
【0139】
用量反応曲線
トランスフェクション培地を除去し、Krebs−グルコース培地中の100nMのBG−LUMI4−TB溶液を各ウェルに添加した。続いてプレートをCO2の存在下で37℃で1時間インキュベートした。
【0140】
次に、各ウェルを100μlのKrebs−グルコースで4回洗浄し、90μlのKrebs−グルコースバッファー又はバソプレシン溶液を添加した。
【0141】
+37℃及び5%CO2の条件下での30分間のインキュベーション後、アクセプターの24μMの最終濃度を得るために、Krebs−グルコースバッファー中の溶液中の10μlのフルオレセイン240μMをウェルに添加した。
【0142】
その後、プレートを静かに撹拌し、続いて337nmの窒素レーザー並びに620nm及び520nmの放出フィルターが取り付けられたルビスターリーダー(Rubystar reader)(BMG)で読んだ。60μsの遅延及び60〜400μsの読み取りウィンドウを有するアクセプターチャネル(520 nm)、並びに400μsの遅延及び400〜1500μsのウィンドウを有するドナーチャネル(620 nm)に関して読み取りを実施した。
【0143】
結果及び考察
図4は、バソプレシン濃度(AVP)に応じた、(620 nmで測定された)ドナーシグナルの(520 nmで測定された)アクセプターシグナルに対する割合の変化を表す。この割合により、ウェル間の変動を含まないことが可能である。さらに、この図示を容易にするために、これに10 000を乗じる。
【0144】
図4は、バソプレシンの濃度の増加がこの割合における増加を生じることを非常に明確に示し、これはドナー化合物の発光における増加に相当する。
【0145】
実際に、バソプレシンの添加は、BG−LUMI4−Tbドナー複合体が結合するV2受容体の内部移行を生じ、その結果これらはもはや細胞外培地における溶液中のフルオレセインでのDEFET現象に関与しないので、これらの発光で増加を生じる。
【0146】
従って、この実施例から再度、DEFETの減少に相当するドナー化合物のシグナルにおける増加と、そのアゴニスト、バソプレシンの存在下でのV2受容体の内部移行との優れた相関関係が示される。この実施例はまた、目的の受容体のアゴニストである化合物を実証するために本発明の方法を如何に使用することができるかを示す。
【0147】
実施例3:バソプレシン誘発のV2受容体の内部移行へのV2受容体アンタゴニストEDA9の効果
この実施例は、バソプレシンV2受容体アンタゴニストである化合物を実証するための本発明の方法の使用を説明する。この受容体の周知のアンタゴニスト、EDA9(ポリペプチド基)である化合物をこれに使用する。
【0148】
使用される試薬及び材料は同様である。プレートの処理並びにトランスフェクションのための試薬及びプロトコールも同様である。
【0149】
EDA9アンタゴニストの存在下での用量反応曲線
トランスフェクション培地を除去し、Krebs−グルコース培地中のBG−LUMI4−TBの100nM溶液を各ウェルに添加した。続いて、CO2の存在下で37℃で1時間インキュベートした。
【0150】
続いて各ウェルを100μlのKrebs−グルコースで4回洗浄し、その後45μlのKrebs−グルコースバッファー又はEDA9溶液を添加した。
【0151】
+37℃で且つ5%CO2条件下での30分間のインキュベーション後、各ウェルにおいて最終濃度80nMのバソプレシンを得るために、160nMでの45μlのバソプレシン溶液をウェルに添加した。
【0152】
+37℃で且つ5%CO2条件下でのインキュベーション後、最終濃度24μMのアクセプターを得るために、Krebs−グルコースバッファー中240μMで溶解した10μlのフルオレセインをウェルに添加した。
【0153】
続いて、プレートを静かに撹拌し、そして337nm窒素レーザー並びに620nm及び520nm放出フィルターが取り付けられたエンビジョンリーダー(Envision reader)(Perkin Elmer)で読んだ。60μsの遅延及び60〜400μsの読み取りウィンドウを有するアクセプターチャネル(520 nm)、並びに400〜1500μsのウィンドウに関して400μsの遅延を有するドナーチャネル(620 nm)に関して読み取りを実施した。
【0154】
結果及び考察
図5は、EDA9アンタゴニストの濃度に応じ且つ一定濃度のバソプレシン(AVP)の存在下での、(620 nmで測定される)ドナーシグナルの(520 nmで測定される)アクセプターシグナルに対する割合における変化を表す。算出される割合は、この図示を容易にするために10 000を乗じる。
【0155】
図5は、EDA9アンタゴニストの濃度が増加する場合の、当該割合における減少を示す。この減少は、ドナーシグナルにおける減少を説明することができ:EDA9の非存在下、及びバソプレシンの存在下では、V2受容体は内部移行され、ドナーは細胞外培地に存在するアクセプターでのDEFETに関与しない。受容体に結合するバソプレシンと競合するEDA9アンタゴニストの存在下では、V2受容体は活性が低く、従って内部移行も低く、よってドナーは溶液中のアクセプターでのDEFETに関与し、その結果発光はEDA9の非存在下よりも減弱する。
【0156】
従って、この実施例は、受容体の内部移行を検出することを可能にすることによって、本発明の方法から、この受容体のアゴニスト化合物のみならず、アンタゴニスト化合物もまた実証することが可能となることを示す。
【0157】
実施例4:CXCR7ケモカイン受容体の内部移行の検出
この実施例は、アゴニストの1つである、SDF−1(分子名「Stromal Cell-derived factor 1(ストロマ細胞由来因子1)」の頭文字)の作用によって生じるCXCR7受容体の内部移行の検出に関する。この実施例を実施するためのプロトコールは、実施例2と同様である。
【0158】
プラスミドFLAGST−V2の代わりにプラスミドFLAGST−CXCR7(配列番号 6)を使用し、またバソプレシンの代わりにSDF−1(Laboratoire Pasteur, Paris)を使用したことを除いて、使用された試薬及び手順は実施例2と同様である。
【0159】
結果及び考察
図6は、(620 nmで測定される)ドナーシグナルの(520 nmで測定される)アクセプターシグナルに対する割合における変化を、SDF−1の濃度に応じて表す。この割合は、ウェル間で変化しないことが可能である。さらに、この図示を容易にするためにこれに10 000を乗じる。
【0160】
図6は、SDF−1の濃度増加により当該割合における増加を生じ、これはドナー化合物の発光における増加を反映していることを非常に明確に示す。
【0161】
従って、この実施例は、DEFETの減少に相当するドナー化合物のシグナルの増加と、SDF−1アゴニストの存在下におけるCXCR7受容体の内部移行との優れた相関関係を示す。この実施例はまた、活性化が内部移行へつながる全受容体へ、本発明の方法が一般化されうるという事実を支持する。
【0162】
実施例5:β2−アドレナリン受容体の内部移行の検出
この実施例は、アゴニストの1つであるイソプロテレノールの作用によって生じるβ2−アドレナリン受容体の内部移行の検出に注目したことを除いて、実施例2と同様である。
【0163】
使用した試薬及び手順は、プラスミドFLAGST−V2をプラスミドFLAGST−β2AR(配列番号7)に代替し、且つバソプレシンをイソプロテレノール(Tocris)に代替したことを除いて実施例2と同様である。
【0164】
結果及び考察
図7は、イソプロテレノールの濃度に応じた、(620 nmで測定される)ドナーシグナルの(520 nmで測定される)アクセプターシグナルに対する割合における変化を表す。この割合は、ウェル間の変化を生じないことが可能である。さらに、この図示を容易にするために10 000を乗じる。
【0165】
図7は、イソプロテレノールの濃度の増加により、かかる割合において増加が生じ、これはドナー化合物の発光における増加を反映していることを非常に明確に示す。
【0166】
従って、この実施例は、DEFETの減少に相当するドナー化合物のシグナルの増加と、イソプロテレノールの存在下でのβ2−アドレナリン受容体の内部移行の間の優れた相関関係を示す。この実施例はまた、本発明の方法を、活性化が内部移行に繋がるあらゆる受容体へ一般化することができるという事実を支持する。
【技術分野】
【0001】
本発明は、膜タンパク質の内部移行を生細胞によって検出するための方法に関し、この内部移行を生じ得る化合物を明らかにするために特に有効である。より詳細には、本発明は、膜貫通タンパク質が内部移行された場合の蛍光金属錯体の発光における変化を測定することに基づいて、蛍光金属錯体と結合された膜貫通タンパク質の内部移行を検出するための方法を説明する。
【背景技術】
【0002】
内部移行:
膜受容体のリガンドによる細胞の活性化の間、膜受容体リガンドは、細胞による内部移行を通常もたらすメカニズムのカスケードへ供される。この方法では、第一に、リガンド、一般にはアゴニスト、の取り込みにより細胞によって受ける刺激の抑制又は減少を可能にし、そして第二に、受容体の脱感作及びこれによる通常のシグナル伝達カスケードの妨害をもたらす。伝達ユニットのこの脱感作は、比較的十分に説明されている複雑な機構である(例えば、Gainetdinov 等, Annu Review Neurosci 2004 v27 p107-44を参照されたい)。
【0003】
この内部移行機構は、Gタンパク質共役型受容体(GPCRs)さらに受容体型チロシンキナーゼを制御するための主要な方法の1つである。
概略的には、アゴニストによるGPCRの活性化は細胞内部における当該受容体のリン酸化をもたらし、この結果一定の細胞内タンパク質、特にクラスリン被覆エンドサイトーシス小胞の形成を生じるアレスチンの補充を誘発する。従って、これらの小胞は、細胞内輸送機構によって処理され、その結果リソソームとの融合によるこれらの分解、又は細胞膜との融合によるこれらの再転換を生じる。
【0004】
GPCRは、治療活性を有する新たな分子の開発に関する選択のターゲットであり、そして候補化合物:アゴニスト、アンタゴニスト又はインバースアゴニストによってこれらの活性化レベルの変化を測定するための複数の技術が存在する。これらの一部は実際に、内部移行に関係する現象を試験することによってGPCRの活性化を検出するという目的を有する。
【0005】
アレスチンは、GPCR内部移行方法に主に関与する細胞内タンパク質であり、以下の記載によって示されるように、内部移行を検出するために一部のアプローチで使用される。
【0006】
WO01/67106は、GPCRとアレスチンの恒常的活性型の変異体との結合の調節を測定することによって、GPCRリガンドを同定することを目的とする方法を説明し、この活性はGPCRのリン酸化レベルに依存しており、当該リガンドは放射性原子、酵素、レポータータンパク質又は蛍光化合物で標識される。
【0007】
WO01/59451は、変異酵素の2つのサブユニットの相補性に基づく「ICAST」テクノロジー(シストロン間相補的分析スクリーニングテクノロジー(Intercistronic Complementary Analysis Screening technology))を使用するGPCRの活性化を検出するための方法を説明し、酵素の各断片は、一方はGPCRと、他方は例えばアレスチンと結合の融合タンパク質の形態で発現される。GPCR−アレスチン相互作用が生じる場合、酵素は相補性によって再構成され、検出可能なシグナルを生じることができるであろう。繰り返して言うが、アレスチンへのこれらの結合の検出によってGPCRの活性化を分析することは重要である。WO01/58923で説明されるように、この技術はGPCR又はアレスチンの変異体を使用することによって改良することができる。
【0008】
WO2005/007822は、化合物が目的の2つのタンパク質間の相互作用を調節するか否かを測定するための方法を説明する。この方法はまた、GPCR−アレスチン結合の検出に基づいている。提示される方法は、特異的なプロテアーゼ感受性のあるペプチド配列を介して転写因子を有する、C末端部で融合したGPCRの使用に基づいている。アレスチンは、この部分でプロテアーゼと融合される。アゴニストリガンドによるGPCRの活性化中、アレスチンはGPCRと相互作用し、これによりプロテアーゼを切断配列に近接させ且つ転写因子の放出を生じ、さらにレポーター遺伝子の転写を活性化でき、この活性は検出することができる。
【0009】
GPCRの活性化に関するマーカーとしてのアレスチン−GPCR結合の使用に付随する問題の1つは、この結合の不安定な性質であり、結合はエンドサイトーシス小胞の形成をもたらす現象のカスケードにおける中間段階にのみ存在する。WO04/065963は、このカスケードのタンパク質パートナーと結合することができない、野生型アレスチンと異なるアレスチン変異体を使用することによってこの問題に対処しており、結果的に受容体に比較的安定して結合した状態である。
Charest等(EMBO Rep. 2005 Apr; 6(4):334-40)は、アレスチンの三次元構造における変化を検出するための技術を開発したが、この変化は活性化GPCRとの相互作用中に生じるものであり、著者等はルシフェラーゼ−アレスチン−YFP(黄色蛍光タンパク質)融合タンパク質を作成し、GPCRの活性化中の分子内BRET(生物発光共鳴エネルギー転移)シグナルの増加を実証した(「2倍輝くアレスチン(double-brilliance arrestin)」として周知の技術)。
【0010】
モルキュラーディバイス社(Molecular Devices)は、特にEP 1 015 608で説明される、「トランスフルオール(Transfluor)(登録商標)」テクノロジーを市販し、このテクノロジーはアレスチン−GFP融合タンパク質の使用に基づいており、この融合タンパク質を発現している細胞の使用により、目的のGPCRの活性化に応じて、適当な画像収集設備を用いて、細胞内でのアレスチン−GFPの転位及びこの再分布の可視化が可能である。
【0011】
膜受容体内部移行を示すための他のアプローチは、GPCR−アレスチン結合又はアレスチンの細胞内転位の検出に基づいておらず、特に以下の文献で説明される。
【0012】
WO00/03246は、GPCR内部移行を誘発することができる化合物を同定するための方法を説明し、当該方法は、GPCR−蛍光タンパク質融合タンパク質を使用して、又はこれらの受容体に結合可能な標識分子を培地に添加することによって、受容体を発光標識することに基づく。この方法は、受容体が内部移行された細胞を同定するために分析しなければならない細胞画像の収集のための段階を含んでなる。
【0013】
アマシャムバイオサイセンス(Amersham Biosciences)社は、商標CypHer5(登録商標)でシアニン誘導体を市販し、特にWO00/75237で説明されるこれらの有機フルオロフォアは、蛍光が中性pHで非常に微弱であり、そして酸性pHで非常に強いという特性を有する。これらのシアニン誘導体は、例えば、GPCRに結合可能な抗体と結合することができ、GPCRへ結合した抗体−Cypher5の内部移行中、Cypher5は中性pHの培地(細胞外培地(extracellular medium))から酸性pHの培地(エンドソーム)まで転移し、この時発光は増加する。
【0014】
ランタニド錯体及び動的FRET(DYNAMIC FRET)による蛍光消光
生物学的現象を研究することに有効な蛍光化合物としての、(以下で「希土類錯体」とも言及する)ランタニド錯体の使用が、1990年代に開発された(例えば、Mathis 等による論文, Clin. Chem. 1995 Sep; 41(9):1391-7を参照されたい)。
FRET現象は、生物学で、特に生物相互作用の研究のために広範囲に渡って使用される。これは、蛍光ドナー化合物(例えば、ランタニド錯体)並びに任意の蛍光アクセプター化合物の使用に基づいており、それぞれ生体分子と結合する。試験対象の生物相互作用によって生体分子が互いに近接し、ドナー化合物が励起されると、ドナーとアクセプターとの間でエネルギー転移が行われ、反応媒体によって放出される発光において変化を生じる。一部の企業は、生物学的な方法を試験するための、このアプローチの実施用の試薬を市販し、例えば希望者には特定の生物学的現象の試験(酵素活性の検出、セカンドメッセンジャーのアッセイ, 等)のためのドナー及びアクセプター化合物並びにキットを準備する。
【0015】
培地中に溶解し自由に拡散している2つのドナーとアクセプター化合物との間の動的エネルギー転移現象は、特にD.Thomas等の、(1978)、PNAS,75:12,5746−5750で説明される。この現象は、頭文字DEFETによってしばしば表示され、これは「拡散増強蛍光エネルギー転移(Diffusion Enhanced Fluorescence Energy Transfer)」を意味する。ドナー及びアクセプター化合物を互いに近づける生物相互作用の発生に基づく、生物学的現象を試験するための通常使用されるエネルギー転移と異なり、動的転移によるFRET又はDEFETは、反応媒体中のFRETパートナーの一方又は両方の自由拡散に基づく。
【0016】
D.Thomas等(1978)の、PNAS,75:12,5746−5750では、溶解したテルビウム錯体(Tb(DPA)3)及びローダミン間のDEFETが説明され、膜小胞の水性溶媒に存在するTb(DPA)3と膜に含有されるエオシン−ホスファチジルコリンの、エオシン発色団との間の最も近い接近距離を測定するためにもまたDEFETが使用された。これらの研究は、特に寿命の長い(2.2 m秒)ドナーフルオロフォアを使用して、「急速拡散限界(rapid diffusion limit)」条件下で行われた。エネルギー転移効率が本質的にドナーとアクセプターとの最も近い接近距離にのみ依存するこれらの条件は、アクセプターの低濃度で特にシグナルの観察を可能にする。
【0017】
GB 2 223 096は、FRETパートナーの拡散定数に依存することが周知の動的FRETの概念にまさに基づく分子(例えば、抗体又は抗原)を検出するための方法を説明する。実際、説明される方法は、ドナー化合物とアクセプター化合物との間の動的FRETの変化を測定するものであり、この変化は、検出される分子との相互作用に続く、培地中のドナー又はアクセプターの拡散の変更に起因する。この方法は、ドナー又はアクセプター化合物の1つが、検出される分子と相互作用が可能なものと結合されることが必要である(例えば、ドナー又はアクセプターは、タンパク質又は抗体と結合する)。
【0018】
Zheng等(J. Am. Chem. Soc. 2005, 127, 16178-16188)は、ランタニド錯体を形成することができ、且つ脂質膜中に非特異的に挿入されることができる親油性大環状分子のクラス(DTPA-PDA-Cn, n=10,12)を、細胞標識の応用及び(ガドリニウム錯体の場合には)核磁気共鳴での使用のための応用のために開発した。著者は、テルビウム錯体とカルセインとの間のエネルギー転移に由来するシグナルを測定することによって、これらの錯体の細胞分布を測定するためにDEFET技術を使用した。著者は、カルセイン(カルセインは細胞膜不透過性である)を細胞外培地に添加する場合に、これらの錯体及びカルセイン間のDEFETを検出することができたが、しかしながら(カルセインの中性エステルの形態で細胞内に導入される)細胞内カルセインを含まず、従ってこれらの脂質ランタニド錯体が細胞膜の外表面上に挿入されると結論付けた。このチームは、これらのランタニド錯体の内部移行の現象におけるシグナル中で生じうる変化を試験しなかった。
【0019】
DEFETはまた、タンパク質の表面で静電ポテンシャルを測定する目的で、エネルギー−アクセプターモチーフを組み込んでいるタンパク質の構造とエネルギー−ドナー化合物とのこれらの相互作用を試験するために、一部のチームによって使用されてきた。
【0020】
ランタニド錯体及びレドックス効果による蛍光消光
ランタニド錯体の発光への一定の還元剤の影響は、文献で説明されている。
【0021】
Kielar等は、例えば、様々な還元剤によって、ユウロピウム及びテルビウム錯体の励起された状態の動的消光を試験し(Org. Biomol. Chem., 2007, 5, 2975-2982)、そして試験した希土類錯体に関して、1Vよりわずかに低い酸化電位を有する化合物が還元により蛍光消光効果を有することができることを示した。これは、例えばヨウ化物イオン(+0.54 V)、尿酸塩(+0.59 V)、アスコルビン酸塩(+0.30 V)及び一定のカテコレート(pH 7で+0.54 V)の場合である。当該著者は、これらの化合物のテルビウム錯体への蛍光消光効果が、同一のキレート構造を有するユウロピウム錯体で観察されたものよりも大きいことをさらに実証した。
【0022】
WO 2008/007089は、テルビウム又はユウロピウム錯体によって還元力を有する分析物をアッセイするための方法に関する。
【0023】
実施することが容易であり且つ高処理の応用に適当であり、目的の膜タンパク質の内部移行の検出に関して感受性のある方法が実際に必要である。このような方法は、特に新規の医薬を検出することを目的としているアプローチにおいて、膜タンパク質の内部移行を生じうる化合物を容易に示すことに関して特に好都合である。さらに、内部移行が可能な受容体の10〜40%のみが実際に内部移行されるので、注目とする方法は、内部移行が生じる場合にシグナル変化の観察を可能にし得ることに十分に敏感であるべきである。
【発明の概要】
【0024】
説明
本発明の対象は、タンパク質の内部移行を検出するための方法であって、次の:
a)ランタニド又はルテニウムを含んでなる蛍光金属錯体で目的のタンパク質を標識する段階であって、蛍光金属錯体の寿命が0.1msより長く、好適には0.5〜6msである段階;
b)目的のタンパク質の内部移行を生じることができる化合物を反応媒体に添加する段階;
c)反応媒体に対して以下から選択される調節剤:
1.上記蛍光金属錯体と適合する蛍光又は非蛍光のFRETアクセプター化合物であって、反応媒体中の最終濃度が10−7Mより高く、好適には10−6M〜10−3Mであり、分子量が50kD未満、好適には0.1〜10kDである化合物;
2.レドックス電位が+0.1V未満、好適には0.25〜0.75Vである還元剤;
3.非共有結合によって、特異的に蛍光金属錯体と結合する薬剤;
4.非蛍光金属錯体を形成するための、ランタニド又はルテニウムと競合する金属イオン、
を添加する段階;
d)蛍光金属錯体の発光波長及び/又は調節剤の発光波長で反応媒体によって放出される発光を測定する段階;
e)段階d)で測定されるシグナルを、段階a)及びc)のみに供した細胞上で測定される基準シグナルと比較する段階であって、基準シグナルと比較される段階d)で測定されたシグナルの違いが目的のタンパク質の内部移行を表す段階、
を含んでなる方法である。
【0025】
これらの必要な段階は、この順番(a, b, c, d, e)で行うことができる。目的のタンパク質の内部移行中、この場合細胞外培地に調節化合物は存在せず、従ってエンドソームに存在する調節化合物のリスクを軽減するので、この実施形態は好適である。
【0026】
本発明の好適な実施形態において、調節因子がFRETアクセプター化合物、又は還元剤である場合、これらの段階はまた、次の順序:a)、c)、b)、d)、e)、で実施することができる。
【0027】
さらに、段階a)とその後の段階との間に洗浄段階を追加することができる。用語「洗浄段階」は、1又は複数回(好適には2又は3回)培地を交換することから成る段階を意味することを目的とする。この洗浄段階は、細胞外培地から目的のタンパク質と結合していない蛍光金属錯体を除去するのに役立つが、本発明は内部移行のシグナルの違いの観察に基づくものであり、この段階は任意である。しかしながら、当該方法の感度を改善するためにこの洗浄を実施することは好適である。
【0028】
本発明の方法は、生細胞及び適当な試薬を含有する反応媒体によって放出されるシグナル、並びに細胞による目的の膜タンパク質の内部移行に従って変化するシグナルの強度の検出に基づいている。
【0029】
より詳細には、本発明の方法では、直接又は間接的に且つ細胞外培地にさらされたドメインのレベルで、目的の膜タンパク質が、蛍光シグナルを放出することができる蛍光金属錯体に結合される。細胞外培地は、(「調節化合物」と呼ばれる)化合物を含み、反応媒体にこの化合物が存在することにより、蛍光金属錯体によって放出されるシグナルが調節される効果を有する。
【0030】
目的の膜タンパク質の内部移行は、蛍光金属錯体の調節化合物への到達性を変更する。蛍光金属錯体はもはや細胞外培地に曝露されないが、しかしながら一定時間後にエンドサイトーシス小胞の内部の培地、又はサイトゾルへ曝露されるであろう。発明者は、調節化合物への蛍光金属錯体の到達性のこの変化が、反応媒体によって放出される発光シグナルに変化をもたらし、内部移行され得る膜貫通受容体の10〜40%だけが実際に内部移行されるという事実にもかかわらず、この変化は検出可能又はさらに任意に定量化できることを発見した。
【0031】
反応媒体によって放出される発光シグナルは、蛍光金属錯体によって放出される発光、或いは調節因子がFRETアクセプター化合物である場合には、調節因子によって放出される発光に対応する。
【0032】
いずれの場合でも、これは、反応媒体で測定される発光の違いにつながる調節化合物への蛍光金属錯体の到達性の違いである。
【0033】
定義
用語「蛍光金属錯体と適合するFRETアクセプター化合物」は、当該蛍光金属錯体とペアのFRETパートナーを形成するFRETアクセプター化合物を意味する。
【0034】
用語「FRETパートナーのペア」は、ドナー蛍光化合物及びアクセプター化合物から成るペアを意味し、これらが互いに近接する場合、及びこれらがドナー蛍光化合物の励起波長で励起される場合、これらの化合物はFRETシグナルを放出する。2つの蛍光化合物がFRETパートナーであるためには、ドナー蛍光化合物の発光がアクセプター化合物の励起スペクトルと部分的に重複しなければならないことは周知である。好適なFRETパートナーペアは、値R0(フォスターディスタンス, エネルギー転移効率が50%である距離)が30 Å以上のものである。
【0035】
「FRETシグナル」は、ドナー蛍光化合物とアクセプター化合物との間のFRETの代表的な、任意の測定可能なシグナルを意味する。従って、FRETシグナルは、アクセプター化合物が蛍光である場合、ドナー蛍光化合物又はアクセプター化合物の発光の強度又は寿命において変化することが可能である。
【0036】
「反応媒体」は、生細胞を試薬と接触させることができる任意の容器、例えばプレートのウェル、試験管等を意味する。
【0037】
「EDTA」は、エチレンジアミン四酢酸を意味する。
「DTPA」は、ジエチレントリアミン五酢酸を意味する。
「TTHA」は、トリエチレンテトラミン−N,N,N’,N’’,N’’’N’’’−六酢酸を意味する。
「DOTA」は、1,4,7,10−テトラアザシクロドデカン−N,N’,N’’,N’’’−四酢酸を意味する。
「NTA」は、ニトリロトリ酢酸を意味する。
「HDTA」は、ヘキサメチレンジアミン四酢酸を意味する。
「DTPP」は、ジエチレントリアミンペンタホスホン酸を意味する。
「EDTP」は、エチレンジニトリロテトラキス(メチルホスホン)酸を意味する。
「NTP」は、ニトリロトリ(メチルホスホン)酸を意味する。
「DOTP」は、1,4,7,10−テトラアザシクロドデカン−N’,N’’,N’’,N’’’−テトラキス(メチルホスホン)酸を意味する。
「DO3A」は、1,4,7,10−テトラアザシクロドデカントリス酢酸を意味する。
「DOTAGA」は、1−(1−カルボキシ−3−カルボキシプロピル)−4,7,10−(カルボキシメチル)−1,4,7,10−テトラアザシクロドデカンを意味する。
「DY647」は、ダイオミクス(Dyomics)社によって市販されるペンタメチン構造を有するフルオロフォアを意味する。
「D2」は、シスビオバイオアッセイ(Cisbio Bioassay)社によって市販される有機蛍光アクセプター化合物を意味する。
「PBS」は、リン酸バッファー溶液を意味する。
「Lumi4−Tb」は、ルミフォール社及びシスビオバイオアッセイ(Cisbio Bioassay)社によって市販されるテルビウム錯体を意味する。
「DMEM」は、「ダルベッコ変法イーグル培地」であり、商業的に入手可能であり且つ多くの応用で使用される細胞培地を意味する。
「NHS」は、N−ヒドロキシスクシンイミドを意味する。
【0038】
本発明の方法で使用される試薬及びこれらの実施の変形もまた、これからより詳細に説明する。
【0039】
A.蛍光金属錯体
一般的に、用語「蛍光金属錯体」は、ランタニド又はルテニウム、及び多座錯体形成剤から成る化合物であって、多座錯体形成剤がすなわち、少なくとも2個、好適には2〜9個の、電子供与体ヘテロ原子、例えばN、O又はSであって、ランタニド又はルテニウムと配位結合を形成している原子を含んでなる剤である化合物を説明することを目的とする。好適には、蛍光金属錯体が芳香族構造から成る1又は複数の発色団を含んでなり、好適には、これらの芳香族構造が、ランタニド又はルテニウム配位原子の役割を果たすN及びOから選択される、1、2又は3個のヘテロ原子を含んでなる。
【0040】
本発明の目的に関して適当な蛍光金属錯体は、錯体形成剤と希土類の会合/解離の面から安定であるべきであり、そしてこの形成定数(Kf)は好適には1010M−1より大きくあるべきである。
【0041】
多くの錯体形成剤が説明されており、そして当業者に周知である。錯体形成剤の例としては、次の化合物:EDTA、DTPA、TTHA、DOTA、NTA、HDTA、DTPP、EDTP、HDTP、NTP、DOTP、DO3A、DOTAGAに言及される。
【0042】
本発明の目的に適当な蛍光金属錯体の実施例は次のものである:
・1又は複数のピリジンユニットを含んでなるランタニドクリプテート。このような希土類クリプテート等は、例えば、EP 0 180 492、EP 0 321 353及びEP 0 601 113並びにWO 01/96 877で説明される。テルビウム(Tb3+)及びユウロピウム(Eu3+)のクリプテートは、本発明の目的に特に適当である。ランタニドクリプテートは、シスビオバイオアッセイ(Cisbio Bioassay)社によって市販される。非限定的な実施例として、以下の式、
【化1】
を有するユウロピウムクリプテートに言及することができ、これらは、反応基例えばNHS基又は反応基R等によって標識することができる化合物と結合することができる。
【0043】
・特にUS4 761 481、US 5 032 677、US 5 055 578、US 5 106 957、US 5 116 989、US 4 761 481、US 4 801 722、US 4 794 191、US 4 637 988、US 4 670 572、US 4 837 169、US 4 859 777で説明されるランタニドキレート。EP 0 403 593、US 5 324 825、US 5 202 423及びUS 5 316 909は、ノナデンテートリガンド、例えばターピリジンから成るキレートを説明する。ランタニドキレートは、パーキンエルマー(PerkinElmer)社によって市販される。
【0044】
・キレート剤から成るランタニド錯体、例えば、芳香環を含んでなる発色団で置換されたテトラアザシクロドデカン、例えばBiomol.Chem,2005,3,1013〜1024の「細胞内(in cellulo)での使用に適当な、高放出で且つ動力学的に安定したランタニド錯体の合成及び評価」でPoole R等によって説明されるものもまた使用することができる。WO2009/10580で説明される錯体もまた使用することができる。
【0045】
・EP 1 154 991及びEP 1 154 990で説明される希土類クリプテートもまた使用することができる。
・シスビオバイオアッセイ(Cisbio Bioassay)社によって市販され、ルミフォール(Lumiphore)社由来の、テルビウムクリプテートLumi4−Tb。
・(反応基、この場合、例えばNHS基によって標識可能な化合物に結合することができる)以下の式:
【化2】
を有するテルビウムクリプテートTb(KR)。この合成は、WO 2008/063721で説明される。
【0046】
・以下の式:
【化3】
を有する(反応基、この場合NCSによって標識される化合物に結合することができる)リサーチオーガニクス(Research Organics)社由来の量子染料。
・ルテニウムキレート、特にルテニウムイオン及び複数のビピリジン、例えばルテニウム(II)トリス(2,2’−ビピリジン)等から成る錯体。
【0047】
・以下の式、
【化4】
を有する(反応基Rによって標識可能な化合物と結合することができる)、インビトロジェン(Invitrogen)社によって市販されるテルビウムキレートDTPA−cs124 Tb。この合成は、US 5,622,821で説明される。
【0048】
ジスプロシウム(Dy3+)、サマリウム(Sm3+)、ネオジム(Nd3+)、イッテルビウム(Yb3+)或いはエルビウム(Er3+)の錯体はまた、本発明の目的に対して適当な希土類錯体であり、ユウロピウム(Eu3+)及びテルビウム(Tb3+)の錯体が特に好適である。
【0049】
非常に多くのランタニド錯体がこれまで説明されており、一部は、特にパーキンエルマー(PerkinElmer)、インビトロジェン(Invitrogen)、モルキュラーデバイシズ(Molecular Devices)及びシスビオバイオアッセイ(Cisbio Bioassay)社によって現在商業的に利用される。図1には、本発明の目的のために使用することができるユウロピウム及びテルビウム錯体の他の例が示される。
【0050】
蛍光金属錯体の使用は本発明の必要な特徴の1つであるが、当該発明は特定のタイプの蛍光金属錯体に限定されず、当業者は、特定の使用のために最も適当であり、0.1ms超、好適には0.5〜6msの寿命を有する蛍光金属錯体から選択が可能である。
【0051】
本発明によれば、蛍光金属錯体は結合される、すなわち、蛍光金属錯体によって目的のタンパク質を直接又は間接的に標識することができるカップリング剤に共有結合される。これらのカップリング剤は、「目的のタンパク質の標識」のセクションで説明され、好適にはリガンド/受容体ペアの1つ、例えば抗タグ抗体、ビオチン、或いは「自殺」酵素の基質、例えばベンジルグアニン、ベンジルシトシン又はクロロアルカンの誘導体である。
【0052】
蛍光金属錯体のカップリング剤との結合は当業者に周知であり、反応基の使用に基づいている。商業的に入手可能な希土類錯体は、カップリング剤へのこの結合を形成することができる反応基を含んでなる、又は既にこのようなカップリング剤を含んでなり、そしていつでも使用できる状態である。
【0053】
B.調節化合物
B.1:調節化合物が、エネルギーアクセプターである。
本発明の方法の一実施形態において、使用は、蛍光金属錯体(エネルギードナー)から調節化合物(エネルギーアクセプター)へのフォルスター共鳴エネルギー転移(FRET)の現象を通じて、蛍光金属錯体によって放出されたシグナルを変更することができる調節化合物から成る。当業者に周知のこの現象は、これらが互いに近接する場合、2つのFRETパートナー化合物間、本願の場合、蛍光金属錯体と反応媒体で自由に拡散する調節化合物との間のエネルギー転移に基づいている。
【0054】
一般的に、生物方法を試験するためのFRET現象の使用は、FRETパートナーのペアのメンバーはそれぞれ、互いに相互作用する化合物へ結合され、よってFRETパートナーは互いに近接され、FRETシグナルを生じることを暗に意味する。本発明の方法において、エネルギーアクセプターは、任意の化合物へ結合されず、反応媒体中で自由に拡散し、一方で蛍光金属錯体は直接又は間接的に目的の膜タンパク質と結合される。2つのFRETパートナー間のエネルギー転移は、DEFET現象に従って、反応媒体中のアクセプター化合物の拡散のみに依存する。
【0055】
発明者は、膜タンパク質と結合された蛍光金属錯体と細胞外培地で拡散しているアクセプターとの間のDEFETの成立に必要な条件を決定し、目的のタンパク質が内部移行される場合に、これらの条件によりシグナルにおける変化の観察も可能となる。
【0056】
この実施形態によれば、目的のタンパク質と蛍光金属錯体の内部移行に依存するDEFETを観察するために使用される実験条件は次の条件である:
・蛍光金属錯体及び調節化合物がFRETパートナーである条件;
・蛍光金属錯体の寿命が、0.1msより長く、好適には0.5〜6msであるべきであるという条件;
・細胞外培地中の調節化合物の濃度が、0.1μM〜1mMであるべきであるという条件。好適にはこの化合物の濃度は、1μM〜100μMであり、これにより急速な拡散限界で機能することができ、そして蛍光金属錯体の濃度に非依存的なDEFETシグナルを測定することが可能である;
・調節化合物は、好適には細胞膜を貫通すべきでないという条件。さらに、この拡散定数が十分高くあるべきである。一般的に、50kD未満、好適には0.1〜10kDの分子量を有する調節化合物が本発明の目的に適当である。
【0057】
本発明の方法のこの実施形態において、調節化合物は蛍光又は非蛍光性であってよい。
【0058】
調節化合物が蛍光エネルギーアクセプター化合物であり、ランタニドがテルビウム又はユウロピウムである場合には、この調節化合物は次の化合物又は化合物ファミリーから選択してよい:シアニン誘導体、DY647、D2、フルオレセイン及びその誘導体、クマリン、ローダミン、カルボピロニン、オキサジン及びその類似体、アレクサフルオールズ(Alexa Fluors)、クリスタルバイオレット、ペリレンビシマイド(bisimide)型のフルオロフォア、スクアレイン、商標ボディピ(BODIPY)(登録商標)で周知のホウ素ジピロメテン誘導体、NBD(ニトロベンズオキサジアゾール)及びその誘導体、DABCYL(4−((4−(ジメチルアミノ)フェニル)アゾ)安息香酸)。
【0059】
アレクサフルオール化合物及びホウ素ジピロメテン誘導体は、インビトロジェン(Invitrogen)社によって市販され、化合物Dy647は、ダイオミクス(Dyomics)社によって市販されるシアニン誘導体であり、シアニン誘導体はまた、特にアマーシャムバイオサイエンス(Amersham Biosciences)社によって市販され、他の化合物は、化学試薬の様々なサプライヤー、例えばシグマ(Sigma)、アルドリッチ(Aldrich)又はアクロス(Acros)社によって市販される。
【0060】
非蛍光化合物もまた、これらの吸収スペクトルが蛍光金属錯体の発光スペクトルと適合するという条件で、使用することができる。次の非蛍光アクセプター:アナスペック社のQXL製品、特にQXL570、QXL610、QXL670及びQXL680、ダイオミクス(Dyomics)社のDYQ660及びDYQ661製品、並びにインビトロジェン(Invitrogen)社のQSY7、QSY9及びQSY21製品、が商業的に入手可能である。
【0061】
反応媒体中で検出されるシグナルは、蛍光金属錯体によって放出される発光、又はアクセプター化合物もまた蛍光の場合にはアクセプター化合物によって放出される発光、これらの強度或いはこれらの寿命である。任意の現象において、これらのシグナルはDEFET現象に依存し、目的の膜貫通タンパク質の内部移行中に、次のように変化する:
・膜貫通タンパク質が内部移行される場合には、発光波長で測定される蛍光金属錯体の発光の強度又は寿命が増加する;
・調節因子が蛍光アクセプター化合物の場合には、タンパク質が内部移行される時に、アクセプターの発光波長で測定されるシグナルの強度又は寿命が変化する。アクセプター化合物由来のシグナルは蛍光金属錯体とのDEFETに依存するが、アクセプター化合物の濃度が高い場合に細胞外培地で生じる、可能性のある分子間の発光−消光効果にも依存することは注目すべきである。この理由のため、内部移行後、アクセプター調節化合物の発光波長で放出されたシグナルは増加又は減少し得る;
・調節因子が蛍光アクセプター化合物である場合、そのシグナルはアクセプターシグナル/ドナーシグナル比を算出することによって補正することができる。FRETシグナルのラジオメトリック補正のこの方法は、広範に使用され文献に説明される。
【0062】
B2:調節化合物が、蛍光金属錯体に関する還元剤である:
本発明者は、蛍光金属錯体及び/又は錯体形成構造のレベルで酸化還元反応によって蛍光金属錯体の発光を減少することができる還元剤が、本発明の方法において好都合に使用することができることを発見した。
【0063】
還元剤は、蛍光金属錯体を還元するのに適当であるレドックス電位を有するべきである。当業者は、選択される蛍光金属錯体に従って、本発明の方法を実施するのにどの還元剤が適当であるかを決定することができるが、レドックス電位が+0.1〜+1.2Vである還元剤が好適である。さらに好適には、還元剤のレドックス電位は+0.25〜+0.75Vである。
【0064】
従って、蛍光金属錯体によって放出されるシグナルを調節する化合物が還元剤である場合、例えば次の還元剤から選択することができる:ヨウ化物、アスコルビン酸塩、尿酸塩、カテコラート、無機化合物、例えばFe(CN)62+。これらのイオンは、対イオン例えばアニオンを有する塩の形態、ナトリウム又はカリウム塩の形態の測定培地へ添加される。
【0065】
本実施形態において、測定されるシグナルは、蛍光金属錯体によって放出され、細胞外培地に存在する還元剤によって調節される発光に相当する:蛍光金属錯体と結合された目的のタンパク質が内部移行される場合、蛍光金属錯体は調節剤と接触又は近接せず、寿命と同様に、この発光は内部移行が生じない場合よりも増大する。
【0066】
この実施形態において、内部移行が存在する場合と内部移行が生じない場合のシグナルの違いを測定することができるように、発明者は蛍光金属錯体/還元剤ペアのシュテルンフォルマー(Stern-Volmer)定数以上の濃度で、反応媒体中に還元剤が存在すべきである、と決定した。この定数は蛍光金属錯体の発光強度の50%の減少を生じるのに必須の還元剤の濃度であると定義される。蛍光金属錯体/還元剤ペアに関するシュテルンフォルマー(Stern-Volmer)定数の例は、特にキラー(Kielar)等による論文(Org. Biomol. Chem., 2007, 5, 2975-2982)に列挙される。
【0067】
従って、実施形態の第一の態様では、これは測定される蛍光金属錯体の発光である。
【0068】
この実施形態の第二の態様では、本発明の方法の第一段階は、測定培地に目的のタンパク質を標識することを目的とする第一の蛍光金属錯体及び第二の蛍光金属錯体を添加することを含んでなる。
【0069】
この第二の蛍光金属錯体は、異なる金属を除いて、第一錯体と同一である。例えば、第一蛍光金属錯体は、ユウロピウム錯体であり、第二金属錯体は同様のキレート構造を含んでなるテルビウム錯体である。そのため、この実施形態において、一定の目的のタンパク質は、潜在的に直接又は間接的に第一蛍光金属錯体に結合する、或いは直接又は間接的に第二蛍光金属錯体に結合する(或いは任意の蛍光金属錯体によって標識されない)。
【0070】
この実施形態において、例えば、第二蛍光金属錯体によって放出される発光の強度との、ランタニド錯体の1つによって放出される発光の割合を形成することによって測定されるシグナルを標準化することが可能である。このタイプの測定により、異なる反応媒体から得られるシグナルを比較することが可能であり、従って特に定量分析を実施するために適当である。
【0071】
B3:調節化合物が、非共有結合によって、特異的に蛍光金属錯体と結合する薬剤である:
調節化合物は、蛍光金属錯体との特異的且つ非共有的な結合を可能とするドメインを有する薬剤であってよく、これらの化合物は実際に蛍光金属錯体の発光において減少を生じることができる。これらの化合物は、蛍光金属錯体−結合ドメインを各々有する、抗体又は抗体断片、ペプチド、或いはアプタマーから選択することができる。
【0072】
これらの薬剤の中で、好適な調節因子は、蛍光金属錯体を認識することができる抗体又は抗体断片である。WO2007/116069(A1)は、希土類錯体に特異的な抗体の調製を説明し、さらに当業者は、これらの自由技術により、このタイプの化合物を生成することが可能である。
【0073】
本発明の方法をこのタイプの調節因子で実施するために、調節剤を添加する前に内部移行を生じる化合物を添加することが望ましく、さもなければ蛍光金属錯体として同時に内部移行される。言い換えれば、この実施形態において、上述の方法の段階は次の順:a)、c)、b)、d)、e)で行われるべきである。
【0074】
B4:調節化合物が、非蛍光金属錯体を形成するするための希土類と競合する金属イオンである:
本発明の方法を実施するのに適当な調節因子の他のタイプは、錯体形成剤を有する非蛍光錯体を形成するための希土類と競合する金属イオンである。従って、希土類キレートの場合には、調節剤はMn2+イオンであってよい。
【0075】
この場合は、適当な対イオンを有する塩の形態、例えばMnCl2の形態で調節剤を細胞外培地に添加する。
【0076】
このタイプの調節因子で本発明の方法を実施するために、調節剤を添加する前に内部移行を生じることができる化合物を添加することが望ましい。言い換えれば、この実施形態において、上述の方法の段階は次の順序:a)、c)、b)、d)、e)で行われるべきである。
【0077】
C.目的のタンパク質の蛍光金属錯体での標識
C.1.発現
内部移行されることができる任意の膜タンパク質が、細胞外部分で蛍光金属錯体と直接又は間接的に結合することができるという条件で、本発明は当該タンパク質を用いて実施することができる。
【0078】
上述のように、本発明は、特にGタンパク質共役型受容体(以下、「GPCR」とする)の内部移行、及び制御機構の1つが内部移行である受容体型チロシンキナーゼ(以下、「RTK」とする)の試験に有効である。それにもかかわらず、内部移行されるか否か確認することが望まれる他の膜タンパク質で本発明を実施することも可能である。
【0079】
目的のタンパク質は細胞膜で当然に発現され、或いは通常の分子生物学技術、特に細胞内に安定的に又は一過性に導入される発現ベクターを使用して発現される。安定に又は一過性に異種DNAを細胞内に導入するための試薬は、商業的に入手可能であり、目的のタンパク質をコードしているDNA配列、特にGPCR及びRTKをコードしているものは、データベース、例えばジェンバンクで入手可能である。目的のタンパク質が細胞によって安定に発現される場合には、非常に多数のGPCRの存在により、細胞毒性現象を観察することができ、この場合、GPCRの発現を制限するために、誘導発現システムを使用することは好都合かもしれない。
【0080】
従って、本発明の方法は目的のタンパク質をコードしている発現ベクターを細胞にトランスフェクトする予備段階を含んでよい。以下に説明されるように、このベクターはまた蛍光金属錯体での目的のタンパク質の共有結合性の標識を可能にする自殺酵素をコードする配列を含んでよい。
【0081】
特定の一実施形態では、第二発現ベクターは目的のタンパク質をコードしているものと同時にトランスフェクトされ、この発現ベクターはβ−アレスチン1をコードしている配列を含んでなる。本発明の実施にこの同時発現は必要ではないが、β−アレスチン1は非特異的に内部移行現象を増幅するので、当該方法の感度の増加を可能にすることができる。
【0082】
C.2.蛍光金属錯体での標識
特に容易な本発明の一実施形態は、目的のタンパク質へ結合することが可能なことが周知の化合物(リガンド)へ蛍光金属錯体を結合することにある:目的のタンパク質への結合により修飾されたリガンドによって、このタンパク質の間接的な標識が可能である。
【0083】
多くの技術はタンパク質を修飾することについて説明されており、当業者は既存ツールをタンパク質を蛍光金属錯体に結合するために使用することが可能である。特に、通常の分子生物学的技術によってタンパク質の一定のドメインを添加、除去又は変更することが可能である。
【0084】
本発明の実施に必要な唯一の特徴は、蛍光金属錯体は細胞外培地に存在するようにそのような方法でタンパク質へ結合され、従って反応媒体によって放出される発光シグナルは実際、調節化合物の細胞外培地における存在に依存的である。
【0085】
目的のタンパク質がGPCR又はRTKである場合、以下に説明される技術の1つに従って直接又は間接的に蛍光金属錯体で標識されることができるように、そのN末端の(細胞外の)ドメインの修飾を介して蛍光金属錯体はこのタンパク質へ結合される。
【0086】
非限定的な説明の目的で、膜貫通タンパク質を蛍光金属錯体に結合するための次の技術に言及することができる:
・間接的に(非共有結合で)目的のタンパク質を蛍光金属錯体と結合すること
【0087】
蛍光金属錯体は、実際は少なくとも一方がタンパク質である、結合パートナーのペアによって、目的のタンパク質と結合されることができる。このアプローチにおいて、目的のタンパク質は、通常の分子生物学技術(タンパク質結合パートナーをコードしているものと融合した目的のタンパク質をコードしているヌクレオチド配列を含んでなる発現ベクターの作成、及び発現ベクターの細胞への導入)によって実際はタンパク質である結合パートナーと融合される。蛍光金属錯体は、明細書中でカップリング剤として言及される他の結合パートナーに共有結合され、続いて細胞外培地に添加される。結合パートナーの認識により、目的のタンパク質を蛍光金属錯体で間接的に標識することができる。
【0088】
本発明を実施するのに特に適当な結合パートナーの非限定的な実施例として以下に言及する:
・Xが任意のアミノ酸であり二ヒ素(biarsenic)化合物である、システイン−システイン−X−X−システイン−システイン配列(配列番号 1)から成るペア。これらの二ヒ素(biarsenic)化合物は、容易にフルオレセイン又はローダミンタイプの有機分子で標識されることができる(当該テクノロジーについて詳述の、B.A.Griffin 等. (1998) Science. 1998, 281, 269-271 及びS.A. Adams 等. (2002) J. Am. Chem. Soc. 2002, 124, 6063-6076 を参照された)。
・ブンガロトキシン(BTX)によって認識される13個のアミノ酸のペプチドから成るBTX(ブンガロトキシン)ペプチドは、蛍光分子と結合されることができる(C. M. McCann 等. (2005), Biotechnique (2005), 38, 945-952を参照されたい)。
・ストレプトアビジン/ビオチンペア:ストレプトアビジン結合配列(SBP-Tag)は、蛍光金属錯体で前標識されることができるビオチンへの高親和性を有する、38個のアミノ酸によって形成される配列である(C. M. McCann 等. (2005), Biotechnique (2005), 38, 945-952を参照されたい)。
・特異的に且つ高親和性を有してリガンド、例えばトリメトプリムを結合する大腸菌ジヒドロ葉酸還元酵素(eDHFR)の配列であり、アクティブモチーフ社からの「リガンドリンクの一般的な標識テクノロジー」として周知のテクノロジーに従って蛍光金属錯体はこれに結合されることができる。
・タグ/抗タグペアは、タンパク質を標識するために頻繁に使用される結合パートナーである。用語「タグ」は、必ずしも非常に短い必要はないが通常非常に短いアミノ酸配列(15個のアミノ酸未満)から成る低分子のタンパク質「標識(label)」を意味し、目的のタンパク質に融合される又はこのタンパク質に当然に存在する。用語「抗タグ」は、当該タグに特異的に結合する抗体を意味する。この実施形態において、抗タグ抗体は、蛍光金属錯体に共有結合される。従って、標識された抗体が細胞外培地に添加される場合、それは目的のタンパク質に結合されるタグに結合され、そしてタグ/抗タグ相互作用によってこのタンパク質の蛍光金属錯体での間接的な標識が可能である。
【0089】
タグ/抗タグペアの非限定的な実施例として、次のペアに言及することができ、これらのメンバーは商業的に入手可能である:GSTが、グルタチオンS−転移酵素又はこの断片を表す、GST/抗GST抗体;6HISが6個のヒスチジンから成るペプチドである、6HIS/抗6HIS抗体;MycがヒトMycタンパク質のアミノ酸410〜419個から成るペプチドであるMyc/抗Myc抗体;FLAGが8個のアミノ酸DYKDDDDK(配列番号 2)を有するペプチドであるFLAG/抗FLAG抗体;HAが9個のアミノ酸YPYDVPFYA(配列番号 3)から成るインフルエンザ赤血球凝集素エピトープであるHA/抗HA抗体。タグの正確な性質は本発明の実施には必要でないことは明らかである。
【0090】
・蛍光金属錯体との目的のタンパク質の直接的な(共有結合性の)結合
このアプローチにおいて、蛍光金属錯体は共有結合によって目的のタンパク質と結合される;一部の技術が説明されており、当該技術を実施するために必要な試薬は、商業的に入手可能である。この結合のために、以下の任意の1つの技術を使用してよい:
・目的のタンパク質上に存在する反応基のレベル、特に次の基の1つのレベルでの共有結合の形成:末端アミノ基、末端カルボキシラート基、アスパラギン酸及びグルタミン酸のカルボキシラート基、リジンのアミン基、アルギニンのグアニジン基、システインのチオール基、チロシンのフェノール基、トリプトファンのインドール環、メチオニンのチオエーテル基、ヒスチジンのイミダゾール基。
【0091】
目的のタンパク質上に存在するこれらの基は、蛍光金属錯体によって保持される反応基と共有結合を形成することができる。
【0092】
適当な反応基は、当業者に周知のものである。マレイミド基で官能性を有する蛍光金属錯体は、例えばタンパク質のシステインによって保持されるチオール基と共有結合が可能であろう。同様に、N−ヒドロキシスクシンイミドエステルを有する蛍光金属錯体は、目的のタンパク質のアミンへ共有結合が可能であろう。
【0093】
・自殺酵素の使用:
用語「自殺酵素」は、特異的突然変異によって修飾された酵素活性を有するタンパク質を意味することを目的とし、当該活性により酵素は急速に共有結合することができる。これらの酵素は各々、ちょうど1つの蛍光分子を結合することができるので、「自殺」と呼ばれ、当該酵素の活性は基質の付着によってブロックされる。これらの酵素は、その結果、1つのタンパク質に対して1つの蛍光分子の割合で目的のタンパク質を特異的に標識するための選択手段を構成する。このアプローチでは、自殺酵素は、通常の分子の生物学的技術によって、膜タンパク質と、好適にはそのN末端部で融合され、そしてドナー/アクセプターに共有結合された酵素の基質は、細胞外培地中に導入される。酵素反応は標識基質の酵素への共有結合をもたらし、従ってドナー又はアクセプターを有する膜タンパク質が標識される。
【0094】
非限定的な実施例として、次の酵素に言及することができる:
−O6−アルキルグアニンDNAアルキル転移酵素(AGT)の変異体。NEB社によって市販される酵素SNAP−tag(Juillerat 等., Chemistry & biology, Vol.10, 313-317 April 2003)及びCLIP−tag(Gautier 等., Chemistry and biology, 15, 128-136, February 2008)は、ヒトAGTの変異体であり、これらの基質は、各々、O6−ベンジルグアニン(以下、BGと略す)及びO2−ベンジルシトシン(以下、BCと略す)である。酵素N−AGT(Gronemeyer 等., Protein engineering, design & selection, vol. 19, No. 7, pp 309-3016, 2006)は、この酵素の他の変異体であり、O6−ベンジルグアニンとのこの反応性は、SNAP−tag酵素のものよりもより良好である;
−脱ハロゲン酵素の変異体(Promegaによって市販されるHaloTag)。これもまた自殺タイプの酵素反応を生じ(WO 04/072232 A2を参照されたい)、この基質の一部はクロロアルカンファミリー、特にユニット−NH−CH2CH2−O−CH2CH2−O−(CH2)6−Clを含んでなるクロロアルカンから構成される。この場合、蛍光金属錯体は、このタイプのユニットに結合される;
−ホスホパンテテイン転移酵素の存在下で、補酵素Aの4’−ホスホパンテテイン残基がACPのセリンに転移される、ACPタンパク質(アシル担体タンパク質)。(N. George 等., Journal of the American Chemical society, 126, (2004) p 8896-8897)。目的のタンパク質を蛍光金属錯体で標識するためにこのアプローチが使用される場合、ホスホパンテテイン転移酵素を反応媒体に添加することが必要である。NEB社は、タンパク質標識のためのACP断片を商標名「ACP−Tag」で市販する。
【0095】
このアプローチが目的のタンパク質を標識するために使用される場合、細胞に自殺酵素及び目的のタンパク質を含んでなる融合タンパク質をコードするDNAを含んでなる発現プラスミドをトランスフェクトする。このプラスミドはまた、これらのタンパク質をコードしているDNA、タグ、例えばFLAGエピトープ、Mycエピトープ、又はインフルエンザ赤血球凝集素(HA)エピトープ等をコードしているDNAの上流を含んでなる。
【0096】
融合タンパク質が細胞膜で発現されることを確実にするために、発現プラスミド中に、目的のタンパク質及び自殺酵素をコードしている配列、膜標的ペプチド、例えばT8シグナルペプチド又はmGluR5受容体のシグナルペプチドをコードしている配列の上流を含むことが有効であり、本目的のためのこの使用は当業者に周知である。最後に、目的のタンパク質をコードしている配列が、目的のタンパク質と自殺酵素の間の結合の翻訳後の切断の対象となりうるいずれの天然の膜標的配列を含んでないことを確実にすることもまた望ましい。この場合には、このドメインを発現プラスミド中に導入しないことが好適である。
【0097】
酵素反応を細胞外培地に存在する酵素の基質(例えばBG−蛍光金属錯体複合体)で生じるために、自殺酵素を細胞外培地にさらすことが必要である。GPCR及びRTKの場合、目的の天然のタンパク質のN末端部が細胞外培地にさらされる時、融合タンパク質が作成され、自殺酵素は融合タンパク質のN末端部で発現されるが、これが存在する場合には常に膜標的ペプチドの下流である。
【0098】
最終的に、蛍光金属錯体で目的のタンパク質を標識するために自殺酵素が使用され、そして目的のタンパク質がGPCR又はRTKである場合、本発明は、融合タンパク質をコードしているDNA配列を含んでなる発現ベクターを細胞にトランスフェクトする予備段階を含んでなり、このN末端部は自殺酵素を含んでなり、そしてC末端部は目的のタンパク質を含んでなる。
【0099】
蛍光金属錯体に結合される酵素の基質の細胞外培地への導入は、目的のタンパク質をこの蛍光金属錯体で標識することとなるであろう。
【0100】
この実施形態において、次の融合タンパク質から選択される融合タンパク質をコードしている発現ベクターを使用することができる:
自殺酵素−目的のタンパク質、又はタグ−自殺酵素−目的のタンパク質、或いは膜標的ペプチド−タグ−自殺酵素−目的のタンパク質。
【図面の簡単な説明】
【0101】
【図1】本発明の目的のために使用することができるユウロピウム及びテルビウム錯体の例を示す。
【図2】バソプレシン濃度に応じた、545nmで測定したドナー発光における変化を示す。
【図3】バソプレシン濃度に応じた、ドナー化合物のΔASR%の変化を示す。
【図4】バソプレシン濃度に応じた、ドナーシグナルのアクセプターシグナルに対する割合の変化を示す。
【図5】EDA9アンタゴニストの濃度に応じた、ドナーシグナルのアクセプターシグナルに対する割合における変化を示す。
【図6】SDF−1の濃度に応じた、ドナーシグナルのアクセプターシグナルに対する割合における変化を示す。
【図7】イソプロテレノールの濃度に応じた、ドナーシグナルのアクセプターシグナルに対する割合における変化を示す。
【実施例】
【0102】
次の実施例は、本発明の好適な実施形態を、すなわち蛍光金属錯体がベンジルグアニン又はベンジルシトシンに共有結合されたユウロピウム又はテルビウムクリプテートであるものを説明しており、この時、目的のタンパク質はアルキルグアニン−DNAアルキル転移酵素変異体を有する融合タンパク質の形態で発現され、そして調節化合物はシアニン誘導体、特にCy5若しくはCy5−COOH、又はフルオレセイン或いはこの誘導体の1つである。ユウロピウムクリプテートが使用される場合、フルオレセインよりもむしろシアニン誘導体を使用することが好適である。
【0103】
実施例1:バソプレシン受容体V2−Rの内部移行の検出
この実施例の目的は、本発明の方法によって、バソプレシンV2受容体を含んでなる融合タンパク質の内部移行、Snap−tag酵素及びHAタグを検出することである。
【0104】
より具体的には、この場合はテルビウムクリプテート(LUMI4-TB)−ベンジルグアニン(BG)複合体がドナー化合物として使用され、この場合にシアニンCy5−COOHはアクセプターとして使用され、そして受容体の内部移行の後に起こるDEFETでの変化に由来するシグナルにおける変化が観察される。
【0105】
使用された試薬及び材料
オプティMEM(OptiMEM)培地(インビトロジェン(Invitrogen)(51985-026))
PBSリン酸バッファー
DMEM woPR(フェノールレッド非含有のDMEM培地、インビトロジェン(Invitrogen))
DMEM+:次のものが添加されたDMEM:1mM L−アラニル/L−グルタミン、1mM Naピルビン酸塩、10%ウシ胎仔血清(インビトロジェン(Invitrogen)SKU#10091-148)、1%ペニシリン−ストレプトマイシン(インビトロジェン(Invitrogen)SKU#15070-063)、2mM HEPES、1%非必須アミノ酸)
DMEM woPR+(フェノールレッドを含まないDMEM+ 培地)
Krebs−Tris+グルコース0.5g/l
BG−LUMI4−TB(シスビオバイオアッセイ(Cisbio Bioassay)によって市販されるベンジルグアニン−テルビウムクリプテート複合体)
BG:O6−ベンジルグアニン(Sigma B2292)
バソプレシン(Bachem)
プラスミドHA−ST−V2:T8膜標的シグナルペプチド、HAエピトープ、SNAPTAG酵素及びV2受容体を含んでなる融合タンパク質をコードしている配列を含んでなるプラスミド。
シアニンCy5−COOH:エラスラボラトリーズ(Eras Laboratories)。
【化5】
【0106】
プレートの処理
ウェルの底への細胞の接着を促進するために、50μlのポリ−L−オルニチン溶液(0.01% 溶液, 分子量 30 000-70 000(SIGMA P4957))を96ウェルプレート(Cellstar, 黒底で黒色)の各ウェル中に分配し、そして、プレートを37℃で30分間インキュベートし、その後100μlのDMEM+で一回リンスした。
【0107】
トランスフェクション
続いて、以下の組成物を有する次のトランスフェクション混合物又はネガティブコントロール混合物を細胞に分配した:
【0108】
【表1】
【0109】
30分間のインキュベーションの後、COS7の細胞を100μl(CIS BIO)、すなわち80 000細胞、を各ウェルに添加し、続いて5%CO2の存在下で37℃で24時間インキュベートした。
【0110】
用量反応曲線
ウェルをDMEM woPR+培地で1回リンスし、その後156nMのBG−LUMI4−TB又は156nMのBG(ネガティブコントロール)溶液をDMEM woPR+培地(100 μl)に添加し、そしてプレートを1時間30分インキュベートした。
【0111】
続いて、各ウェルを100μlのKrebs−トリスバッファー+グルコース(0.5 g/l)で3回洗浄し、その後Krebs−トリスバッファーを添加し、Krebs−トリスバッファーで希釈したバソプレシン(V2 受容体アゴニスト)を添加した(4℃, 10 μl, 可変濃度, プレートプランを参照されたい)。
【0112】
30分間のインキュベーション後、24μMのアクセプター最終濃度を得るために、Krebs−トリスバッファー(10 μl)中の溶液の264 μMのシアニン5−COOHをラインB、D、E〜Hに添加した(プランを参照されたい)。
【0113】
15分間プレートを撹拌し、337nmの窒素レーザー並びに545nm及び665nmの放出フィルターが取り付けられたエンビジョンリーダー(Envision reader)(パーキンエルマー)で読んだ。400μsの遅延及び400〜2900μsの読み取りウィンドウ(window)を有する両チャネルで読み取りを実施した。
【0114】
96ウェルプレートのプランを各ウェルに添加した試薬の量で以下に表す:
【0115】
【表2】
【0116】
ラインA:プラスミドPRK6を有するネガティブコントロールであって、従って標識しているSnaptag(ST)はなく、Cy5アクセプターの非存在下であり、LUMI4−TBドナーの非存在下(A1-A4)、バソプレシンを含まずLUMI4−TBドナーの存在下(A5-A8)又はバソプレシンを含みLUMI4−TBドナーの存在下(A9-A12)で観察されるシグナル。
【0117】
ラインB:プラスミドPRK6を有するネガティブコントロールであって、従って標識しているSnaptag(ST)はなく、Cy5アクセプターの存在下であり、LUMI4−TBドナーの非存在下(B1-B4)、バソプレシンを含まずLUMI4−TBドナーの存在下(B5-B8)又はバソプレシンを含みLUMI4−TBドナーの存在下(B9-B12)で観察されるシグナル。
このラインにより、受容体の標識をすることなくバックグラウンドノイズを測定することが可能であり、測定されたシグナルを補正することが可能である。
【0118】
ラインC:プラスミドHA−ST−V2を有するネガティブコントロールであって、従ってSnaptagの存在下で、バソプレシン又はCy5アクセプターを含まず、非蛍光のBGの存在下(C1-C4)又はLUMI4−TBドナーを有する蛍光のBGの存在下(C5-C12)で観察されるシグナル。
【0119】
ラインD:プラスミドHA−ST−V2を有するネガティブコントロールであって、従ってSnaptagの存在下で、しかしながらバソプレシンを含まず、そしてCy5アクセプターの存在下で;非蛍光のBGの存在下(D1-D4)、又はLUMI4−TBドナーを有する蛍光のBGの存在下(D5-D12)で観察されるシグナル。
このラインにより、BG−LUMI4−TBで標識されるV2受容体の内部移行の非存在下で、基準シグナルを得ることが可能である。
【0120】
ラインE〜Hは、様々な濃度のバソプレシンの存在下で観察されるシグナルに相当する。
【0121】
結果及び考察
図2は、ドナー発光における変化を表し、バソプレシン濃度に応じて、従って受容体の内部移行に応じて、545nmで測定した。
【0122】
ΔAS%は、(バソプレシンを含まず)内部移行を含まずに測定される基準シグナルに対する、内部移行中のドナーシグナルの割合の増加に相当する。
【0123】
より具体的には、バソプレシンの一定の濃度、例えば10pM(ウェルE1〜E6)に対して、ΔAS%は次の方法で算出した。
【0124】
【化6】
【0125】
式中、「[E1−E6]545」、「[D5−D12]545」等は、BG−LUMI4−TBドナーの波長(545 nm)で、ウェルE1〜E6、D5〜D12等において測定されるシグナルの平均値に相当する。
【0126】
分子は、バソプレシンの存在及び非存在下(従って内部移行の非存在下)で測定されるシグナルの違いに相当し、分母は、内部移行の非存在下([D5-D12] 545)における基準シグナルであって、バックグラウンドノイズ(BG-LUMI4-TB ドナーの非存在下で測定される[D1-D4] 545)、さらに非特異的シグナル(Snaptag標識の非存在下での[B5-B8] 545-[B1-B4] 545)に対して補正される基準シグナル相当する。
【0127】
図2は、バソプレシンの増加している濃度の存在下における545でのBG−LUMI4−TBドナー化合物の発光における変化(この場合は増加)を示し、従って膜受容体の内部移行を示すための本発明の方法の使用を検証する。
【0128】
665nm(Cy5)でアクセプターによって放出されるシグナルに焦点を置いているなら、545でのドナーシグナルに対して補正し、同様の結果が得られる(このラジオメトリック補正により、1つのウェルから他のウェルへのシグナル変動(signal variability)を排除することが可能である)。
【0129】
図3は、バソプレシン濃度、従ってV2受容体の内部移行に応じたΔASR%の変化を表す。ΔASR%は、(バソプレシンの非存在下で測定され)内部移行を含まない基準シグナルの割合に相当する。
【0130】
バソプレシンの一定の濃度、例えば10pM(ウェルE1〜E6)に対して、ΔASR%を次の方法で算出した:
【0131】
【化7】
【0132】
バソプレシンの存在下で測定されるシグナル([E1-E6] 665/[E1-E6] 545)及び基準シグナル([D1-D12] 665/[D1-D12] 545)は、バックグラウンドノイズ(BG-LUMI4-TBドナーの非存在下で測定される[D1-D4])及びSnaptag標識の非存在下で測定される非特異的シグナル([B5-B8]-[B1-B4])に対して各々補正される。
【0133】
図3は、バソプレシンの濃度の増加が培地で存在する場合の変化(アクセプター化合物によって放出されるシグナルにおける減少)を示し、従って膜受容体の内部移行を示すための本発明の方法の使用を確認するものである。
【0134】
実施例2:バソプレシン受容体V2−Rの内部移行の検出
本実施例は、V2バソプレシン受容体の内部移行を検出するための本発明の方法の他の実施形態を説明する。特に、フルオレセインをアクセプターとして試験し、そしてV2受容体をコードしている発現プラスミドを、β−アレスチン1をコードしているプラスミドと同時にトランスフェクトした。この同時のトランスフェクション(cotransfection)により、受容体内部移行を促進し、従って本方法の感度を増加することが可能となった。
【0135】
使用した試薬及び材料
オプティMEM(OptiMEM)培地(Invitrogen(51985-026))
Krebs−グルコース:Krebsバッファー+ 0.5g/lグルコース
BG−LUMI4−TB(Cisbio Bioassayによって市販される、ベンジルグアニン−テルビウムクリプテート複合体)
BG:O6−ベンジルグアニン(Sigma B2292)
バソプレシン(Bachem)
プラスミドFLAGST−V2:T8膜標的シグナルペプチド、FLAGエピトープ、SNAPTAG酵素及びV2受容体を含んでなる融合タンパク質をコードしている配列を含んでなるプラスミド。このプラスミドの配列は、配列番号4である。
プラスミドFLAG−アレスチンβ1:FLAGエピトープ及びヒトβ−アレスチン1を含んでなる融合タンパク質をコードしている配列を含んでなるプラスミド。このプラスミドの配列は、配列番号5である。
フルオレセイン(Sigma)。
【0136】
プレートの処理
細胞のウェルの底面への接着を促進するするために、ポリ−L−オルニチンの50μl溶液(0.01% 溶液, 分子量30 000〜70 000 (SIGMA P4957))を96ウェルプレート(Cellstar, 黒底で黒色)の各ウェルに分配し、プレートを37℃で30分間インキュベートした。
【0137】
トランスフェクション
続いて、次のトランスフェクション混合物を各ウェルに分配した:
−0.16μgのプラスミドFLAGST−V2 + 0.16μgのプラスミドFLAG−β1アレスチン
−0.8μlのリポフェクタミン2000
−50μlのオプティMEM(OptiMEM)培地。
【0138】
30分間のインキュベーション後、100 000のCOS7細胞を含有する100μlの懸濁液を各ウェルに添加し、続いて5%のCO2存在下で37℃で24時間インキュベートした。
【0139】
用量反応曲線
トランスフェクション培地を除去し、Krebs−グルコース培地中の100nMのBG−LUMI4−TB溶液を各ウェルに添加した。続いてプレートをCO2の存在下で37℃で1時間インキュベートした。
【0140】
次に、各ウェルを100μlのKrebs−グルコースで4回洗浄し、90μlのKrebs−グルコースバッファー又はバソプレシン溶液を添加した。
【0141】
+37℃及び5%CO2の条件下での30分間のインキュベーション後、アクセプターの24μMの最終濃度を得るために、Krebs−グルコースバッファー中の溶液中の10μlのフルオレセイン240μMをウェルに添加した。
【0142】
その後、プレートを静かに撹拌し、続いて337nmの窒素レーザー並びに620nm及び520nmの放出フィルターが取り付けられたルビスターリーダー(Rubystar reader)(BMG)で読んだ。60μsの遅延及び60〜400μsの読み取りウィンドウを有するアクセプターチャネル(520 nm)、並びに400μsの遅延及び400〜1500μsのウィンドウを有するドナーチャネル(620 nm)に関して読み取りを実施した。
【0143】
結果及び考察
図4は、バソプレシン濃度(AVP)に応じた、(620 nmで測定された)ドナーシグナルの(520 nmで測定された)アクセプターシグナルに対する割合の変化を表す。この割合により、ウェル間の変動を含まないことが可能である。さらに、この図示を容易にするために、これに10 000を乗じる。
【0144】
図4は、バソプレシンの濃度の増加がこの割合における増加を生じることを非常に明確に示し、これはドナー化合物の発光における増加に相当する。
【0145】
実際に、バソプレシンの添加は、BG−LUMI4−Tbドナー複合体が結合するV2受容体の内部移行を生じ、その結果これらはもはや細胞外培地における溶液中のフルオレセインでのDEFET現象に関与しないので、これらの発光で増加を生じる。
【0146】
従って、この実施例から再度、DEFETの減少に相当するドナー化合物のシグナルにおける増加と、そのアゴニスト、バソプレシンの存在下でのV2受容体の内部移行との優れた相関関係が示される。この実施例はまた、目的の受容体のアゴニストである化合物を実証するために本発明の方法を如何に使用することができるかを示す。
【0147】
実施例3:バソプレシン誘発のV2受容体の内部移行へのV2受容体アンタゴニストEDA9の効果
この実施例は、バソプレシンV2受容体アンタゴニストである化合物を実証するための本発明の方法の使用を説明する。この受容体の周知のアンタゴニスト、EDA9(ポリペプチド基)である化合物をこれに使用する。
【0148】
使用される試薬及び材料は同様である。プレートの処理並びにトランスフェクションのための試薬及びプロトコールも同様である。
【0149】
EDA9アンタゴニストの存在下での用量反応曲線
トランスフェクション培地を除去し、Krebs−グルコース培地中のBG−LUMI4−TBの100nM溶液を各ウェルに添加した。続いて、CO2の存在下で37℃で1時間インキュベートした。
【0150】
続いて各ウェルを100μlのKrebs−グルコースで4回洗浄し、その後45μlのKrebs−グルコースバッファー又はEDA9溶液を添加した。
【0151】
+37℃で且つ5%CO2条件下での30分間のインキュベーション後、各ウェルにおいて最終濃度80nMのバソプレシンを得るために、160nMでの45μlのバソプレシン溶液をウェルに添加した。
【0152】
+37℃で且つ5%CO2条件下でのインキュベーション後、最終濃度24μMのアクセプターを得るために、Krebs−グルコースバッファー中240μMで溶解した10μlのフルオレセインをウェルに添加した。
【0153】
続いて、プレートを静かに撹拌し、そして337nm窒素レーザー並びに620nm及び520nm放出フィルターが取り付けられたエンビジョンリーダー(Envision reader)(Perkin Elmer)で読んだ。60μsの遅延及び60〜400μsの読み取りウィンドウを有するアクセプターチャネル(520 nm)、並びに400〜1500μsのウィンドウに関して400μsの遅延を有するドナーチャネル(620 nm)に関して読み取りを実施した。
【0154】
結果及び考察
図5は、EDA9アンタゴニストの濃度に応じ且つ一定濃度のバソプレシン(AVP)の存在下での、(620 nmで測定される)ドナーシグナルの(520 nmで測定される)アクセプターシグナルに対する割合における変化を表す。算出される割合は、この図示を容易にするために10 000を乗じる。
【0155】
図5は、EDA9アンタゴニストの濃度が増加する場合の、当該割合における減少を示す。この減少は、ドナーシグナルにおける減少を説明することができ:EDA9の非存在下、及びバソプレシンの存在下では、V2受容体は内部移行され、ドナーは細胞外培地に存在するアクセプターでのDEFETに関与しない。受容体に結合するバソプレシンと競合するEDA9アンタゴニストの存在下では、V2受容体は活性が低く、従って内部移行も低く、よってドナーは溶液中のアクセプターでのDEFETに関与し、その結果発光はEDA9の非存在下よりも減弱する。
【0156】
従って、この実施例は、受容体の内部移行を検出することを可能にすることによって、本発明の方法から、この受容体のアゴニスト化合物のみならず、アンタゴニスト化合物もまた実証することが可能となることを示す。
【0157】
実施例4:CXCR7ケモカイン受容体の内部移行の検出
この実施例は、アゴニストの1つである、SDF−1(分子名「Stromal Cell-derived factor 1(ストロマ細胞由来因子1)」の頭文字)の作用によって生じるCXCR7受容体の内部移行の検出に関する。この実施例を実施するためのプロトコールは、実施例2と同様である。
【0158】
プラスミドFLAGST−V2の代わりにプラスミドFLAGST−CXCR7(配列番号 6)を使用し、またバソプレシンの代わりにSDF−1(Laboratoire Pasteur, Paris)を使用したことを除いて、使用された試薬及び手順は実施例2と同様である。
【0159】
結果及び考察
図6は、(620 nmで測定される)ドナーシグナルの(520 nmで測定される)アクセプターシグナルに対する割合における変化を、SDF−1の濃度に応じて表す。この割合は、ウェル間で変化しないことが可能である。さらに、この図示を容易にするためにこれに10 000を乗じる。
【0160】
図6は、SDF−1の濃度増加により当該割合における増加を生じ、これはドナー化合物の発光における増加を反映していることを非常に明確に示す。
【0161】
従って、この実施例は、DEFETの減少に相当するドナー化合物のシグナルの増加と、SDF−1アゴニストの存在下におけるCXCR7受容体の内部移行との優れた相関関係を示す。この実施例はまた、活性化が内部移行へつながる全受容体へ、本発明の方法が一般化されうるという事実を支持する。
【0162】
実施例5:β2−アドレナリン受容体の内部移行の検出
この実施例は、アゴニストの1つであるイソプロテレノールの作用によって生じるβ2−アドレナリン受容体の内部移行の検出に注目したことを除いて、実施例2と同様である。
【0163】
使用した試薬及び手順は、プラスミドFLAGST−V2をプラスミドFLAGST−β2AR(配列番号7)に代替し、且つバソプレシンをイソプロテレノール(Tocris)に代替したことを除いて実施例2と同様である。
【0164】
結果及び考察
図7は、イソプロテレノールの濃度に応じた、(620 nmで測定される)ドナーシグナルの(520 nmで測定される)アクセプターシグナルに対する割合における変化を表す。この割合は、ウェル間の変化を生じないことが可能である。さらに、この図示を容易にするために10 000を乗じる。
【0165】
図7は、イソプロテレノールの濃度の増加により、かかる割合において増加が生じ、これはドナー化合物の発光における増加を反映していることを非常に明確に示す。
【0166】
従って、この実施例は、DEFETの減少に相当するドナー化合物のシグナルの増加と、イソプロテレノールの存在下でのβ2−アドレナリン受容体の内部移行の間の優れた相関関係を示す。この実施例はまた、本発明の方法を、活性化が内部移行に繋がるあらゆる受容体へ一般化することができるという事実を支持する。
【特許請求の範囲】
【請求項1】
細胞の表面で発現される目的の膜貫通タンパク質の内部移行を検出するための方法であって、次の:
a)ランタニド又はルテニウムを含んでなる蛍光金属錯体で目的のタンパク質を標識する段階であって、蛍光金属錯体の寿命が0.1msより長い段階;
b)目的のタンパク質の内部移行を生じることができる化合物を反応媒体に添加する段階;
c)反応媒体に対して以下から選択される調節剤:
1.上記蛍光金属錯体と適合する蛍光又は非蛍光のFRETアクセプター化合物であって、反応媒体中の最終濃度が10−7Mより高く、分子量が50kD未満である化合物;
2.レドックス電位が+0.1V未満である還元剤;
3.非共有結合によって、特異的に蛍光金属錯体と結合する薬剤;
4.非蛍光金属錯体を形成するための、ランタニド又はルテニウムと競合する金属イオン、
を添加する段階;
d)蛍光金属錯体の発光波長、及び/又は調節化合物が蛍光アクセプター化合物の場合に調節化合物の発光波長で反応媒体によって放出される発光を測定する段階;
e)段階d)で測定されるシグナルを、段階a)及びc)のみに供された細胞上で測定される基準シグナルと比較する段階であって、基準シグナルと比較される段階d)で測定されるシグナルの違いが目的のタンパク質の内部移行を表す段階、
を含んでなる方法。
【請求項2】
前記段階が次の順序:a)、b)、c)、d)、e)で行われることを特徴とする、請求項1に記載の方法。
【請求項3】
前記調節剤がFRETアクセプター化合物又は還元剤であり、前記段階が次の順序:a)、c)、b)、d)、e)で行われることを特徴とする、請求項1に記載の方法。
【請求項4】
段階a)とその後の段階との間に洗浄段階を含んでなることを特徴とする、請求項1〜3のいずれか1項に記載の方法。
【請求項5】
前記蛍光金属錯体の寿命が、0.5〜6msであることを特徴とする、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
前記蛍光金属錯体が:
a)ランタニドがユウロピウム、テルビウム、ネオジム、サマリウム及びジスプロシウムから選択されるランタニド錯体;又は
b)ルテニウム錯体、
であることを特徴とする、請求項1〜5のいずれか1項に記載の方法。
【請求項7】
前記蛍光金属錯体がランタニドキレート又はクリプテートであることを特徴とする、請求項6に記載の方法。
【請求項8】
前記蛍光金属錯体が、
(i)N、O及びSから選択される2〜9個の電子供与体ヘテロ原子であって、前記ランタニド又は前記ルテニウムと配位結合を形成している原子、並びに
(ii)N及びOから選択される1、2又は3個のヘテロ原子を含んでなる、芳香族構造を含んでなる1又は複数の発色団であって、ランタニド又はルテニウムの配位原子の役割を果たす発色団、
を含んでなることを特徴とする、請求項7に記載の方法。
【請求項9】
前記調節剤が、前記蛍光金属錯体と適合する蛍光又は非蛍光FRETアクセプター化合物であり、前記反応媒体中の最終濃度が10−7Mより高く、分子量が50kD未満であることを特徴とする、請求項1〜8のいずれか1項に記載の方法。
【請求項10】
反応媒体中のFRETアクセプター化合物の最終濃度が10−6M〜10−3Mであり、分子量が0.1〜10kDであることを特徴とする、請求項9に記載の方法。
【請求項11】
前記調節剤が次の化合物:シアニン誘導体、DY647、D2、CY5、Cy5−COOH、フルオレセイン、クマリン、ローダミン、カルボピロニン、オキサジン及びその類似体、アレクサフルオールフルオロフォア、クリスタルバイオレット、ペリレンビシマイド(bisimide)フルオロフォア、スクアレインフルオロフォア、ホウ素ジピロメテン誘導体、NBD(ニトロベンズオキサジアゾール)及びその誘導体、DABCYL(4−((4−(ジメチルアミノ)フェニル)アゾ)安息香酸)から選択される蛍光アクセプター化合物であることを特徴とする、請求項1〜10のいずれか1項に記載の方法。
【請求項12】
前記調節剤が、次の化合物:フルオレセイン及びその誘導体、並びにシアニン誘導体から選択される蛍光アクセプター化合物であることを特徴とする、請求項1〜11のいずれか1項に記載の方法。
【請求項13】
前記調節剤が、次の化合物:QXL 570、QXL 610、QXL 670及びQXL 680;DYQ660及びDYQ661;QSY7、QSY9及びQSY21、から選択される非蛍光アクセプター化合物であることを特徴とする、請求項1〜12のいずれか1項に記載の方法。
【請求項14】
前記調節化合物が、次の化合物:ヨウ化物、アスコルビン酸塩、尿酸塩、カテコール塩、及び無機種、例えばFe(CN)62+から選択される還元剤であることを特徴とする、請求項1〜8のいずれか1項に記載の方法。
【請求項15】
前記調節剤が還元剤であり、段階a)が第一蛍光金属錯体を添加すること及び第二蛍光金属錯体を添加することを含んでなり、当該蛍光金属錯体が同様のキレート構造を有するが異なる金属と錯体を形成することを特徴とする、請求項14に記載の方法。
【請求項16】
前記第一金属錯体がユウロピウム錯体であり、前記第二金属錯体がテルビウム錯体であり、テルビウム及びユウロピウム錯体が同様のキレート構造を有することを特徴とする、請求項15に記載の方法。
【請求項17】
前記調節化合物が、非共有結合によって、特異的に蛍光金属錯体に結合する薬剤であって、蛍光金属錯体結合ドメインを各々有する次の化合物:抗体、抗体断片、ペプチド又はアプタマー、から選択されることを特徴とする、請求項1または2に記載の方法。
【請求項18】
前記調節化合物が、ランタニド又はルテニウムと競合する金属イオンであり、これにより非蛍光金属錯体が形成されることを特徴とする、請求項1または2に記載の方法。
【請求項19】
前記イオンがマンガンイオンであることを特徴とする、請求項18に記載の方法。
【請求項20】
前記蛍光金属錯体での目的のタンパク質の標識が、結合パートナーのペアを介した間接的な標識であり、ペアのメンバーの1つが蛍光金属錯体に共有結合され、他のメンバーが目的のタンパク質に共有結合されるか又は目的のタンパク質上に当然に存在することを特徴とする、請求項1〜19のいずれか1項に記載の方法。
【請求項21】
前記結合パートナーのペアが、次のペア:配列CCXXCC(配列番号 1)(ここで、C=システイン及びX=任意のアミノ酸)/二ヒ素(biarsenic)化合物;BTXペプチド/ブンガロトキシン;ストレプトアビジン或いはアビジン/ビオチン;eDHFR断片/トリメトプリム;タグ/抗タグ抗体;目的のタンパク質の周知のリガンド/目的のタンパク質、から選択されることを特徴とする、請求項20に記載の方法。
【請求項22】
前記蛍光金属錯体が、目的のタンパク質に対して特異的な抗体;GST、6HIS、Myc、FLAG及びHAから選択されるタグに対して特異的な抗体、から選択される抗タグ抗体に共有結合され、そして目的のタンパク質が対応するタグを含有することを特徴とする、請求項21に記載の方法。
【請求項23】
前記目的のタンパク質を蛍光金属錯体で標識することが、共有結合による直接的な標識であることを特徴とする、請求項1〜19のいずれか1項に記載の方法。
【請求項24】
前記蛍光金属錯体が自殺酵素の基質に結合され、目的のタンパク質が融合タンパク質の形態で発現され、タンパク質の細胞外部分が自殺酵素を含有することを特徴とする、請求項23に記載の方法。
【請求項25】
前記自殺酵素/自殺酵素基質のペアが、次のペア:アルキルグアニン−DNAアルキル転移酵素変異体/ベンジルグアニン;アルキルグアニン−DNAアルキル転移酵素変異体/ベンジルシトシン、脱ハロゲン酵素変異体/クロロアルカン;アシル担体タンパク質/ホスホパンテテイン転移酵素の存在下での補酵素A、から選択されることを特徴とする、請求項24に記載の方法。
【請求項26】
前記蛍光金属錯体が、目的のタンパク質上に存在する第二反応基、特に末端アミノ基、末端カルボキシラート基、アスパラギン酸及びグルタミン酸のカルボキシラート基、リジンのアミン基、アルギニンのグアニジン基、システインのチオール基、チロシンのフェノール基、トリプトファンのインドール環、メチオニンのチオエーテル基、ヒスチジンのイミダゾール基と共有結合を形成することができる第一反応基を含んでなることを特徴とする、請求項23に記載の方法。
【請求項27】
前記蛍光金属錯体が目的のタンパク質のチオール基と反応することを目的とするマレイミド基を含んでなることを特徴とする、請求項26に記載の方法。
【請求項28】
前記蛍光金属錯体が、ベンジルグアニン又はベンジルシトシン或いはこれらの誘導体の1つに共有結合されるユウロピウムクリプテート又はテルビウムクリプテートであり、目的のタンパク質がアルキルグアニン−DNAアルキル転移酵素変異体との融合タンパク質の形態で発現されることを特徴とする、請求項1〜19のいずれか1項に記載の方法。
【請求項29】
前記蛍光金属錯体がベンジルグアニン又はベンジルシトシンに共有結合されるユウロピウムクリプテート又はテルビウムクリプテートであり、目的のタンパク質がアルキルグアニン−DNAアルキル転移酵素変異体との融合タンパク質の形態で発現され、そして調節化合物がシアニン誘導体、特にCy5又はCy5−COOHであることを特徴とする、請求項1〜19のいずれか1項に記載の方法。
【請求項30】
前記蛍光金属錯体が、ベンジルグアニン又はベンジルシトシンに共有結合されるテルビウムクリプテートであり、前記目的のタンパク質がアルキルグアニン−DNAアルキル転移酵素変異体との融合タンパク質の形態で発現され、そして前記調節化合物がフルオレセイン又はその誘導体の1つであることを特徴とする、請求項1〜19のいずれか1項に記載の方法。
【請求項31】
前記目的のタンパク質が、Gタンパク質共役型受容体及び受容体型チロシンキナーゼから選択されることを特徴とする、請求項1〜30のいずれか1項に記載の方法。
【請求項32】
前記目的のタンパク質をコードしているDNA配列を含んでなる発現ベクターを細胞にトランスフェクトする予備段階を含んでなることを特徴とする、請求項1〜31のいずれか1項に記載の方法。
【請求項33】
前記発現ベクターが融合タンパク質をコードしている配列を含んでなり、当該配列のN末端部が自殺酵素を含んでなり、C末端部が目的のタンパク質を含んでなり、目的のタンパク質がGPCR又はRTKであることを特徴とする、請求項32に記載の方法。
【請求項34】
前記トランスフェクション段階がまた、β−アレスチン1をコードしているDNA配列を含んでなるベクターを同時にトランスフェクションすることを含んでなることを特徴とする、請求項32または33に記載の方法。
【請求項1】
細胞の表面で発現される目的の膜貫通タンパク質の内部移行を検出するための方法であって、次の:
a)ランタニド又はルテニウムを含んでなる蛍光金属錯体で目的のタンパク質を標識する段階であって、蛍光金属錯体の寿命が0.1msより長い段階;
b)目的のタンパク質の内部移行を生じることができる化合物を反応媒体に添加する段階;
c)反応媒体に対して以下から選択される調節剤:
1.上記蛍光金属錯体と適合する蛍光又は非蛍光のFRETアクセプター化合物であって、反応媒体中の最終濃度が10−7Mより高く、分子量が50kD未満である化合物;
2.レドックス電位が+0.1V未満である還元剤;
3.非共有結合によって、特異的に蛍光金属錯体と結合する薬剤;
4.非蛍光金属錯体を形成するための、ランタニド又はルテニウムと競合する金属イオン、
を添加する段階;
d)蛍光金属錯体の発光波長、及び/又は調節化合物が蛍光アクセプター化合物の場合に調節化合物の発光波長で反応媒体によって放出される発光を測定する段階;
e)段階d)で測定されるシグナルを、段階a)及びc)のみに供された細胞上で測定される基準シグナルと比較する段階であって、基準シグナルと比較される段階d)で測定されるシグナルの違いが目的のタンパク質の内部移行を表す段階、
を含んでなる方法。
【請求項2】
前記段階が次の順序:a)、b)、c)、d)、e)で行われることを特徴とする、請求項1に記載の方法。
【請求項3】
前記調節剤がFRETアクセプター化合物又は還元剤であり、前記段階が次の順序:a)、c)、b)、d)、e)で行われることを特徴とする、請求項1に記載の方法。
【請求項4】
段階a)とその後の段階との間に洗浄段階を含んでなることを特徴とする、請求項1〜3のいずれか1項に記載の方法。
【請求項5】
前記蛍光金属錯体の寿命が、0.5〜6msであることを特徴とする、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
前記蛍光金属錯体が:
a)ランタニドがユウロピウム、テルビウム、ネオジム、サマリウム及びジスプロシウムから選択されるランタニド錯体;又は
b)ルテニウム錯体、
であることを特徴とする、請求項1〜5のいずれか1項に記載の方法。
【請求項7】
前記蛍光金属錯体がランタニドキレート又はクリプテートであることを特徴とする、請求項6に記載の方法。
【請求項8】
前記蛍光金属錯体が、
(i)N、O及びSから選択される2〜9個の電子供与体ヘテロ原子であって、前記ランタニド又は前記ルテニウムと配位結合を形成している原子、並びに
(ii)N及びOから選択される1、2又は3個のヘテロ原子を含んでなる、芳香族構造を含んでなる1又は複数の発色団であって、ランタニド又はルテニウムの配位原子の役割を果たす発色団、
を含んでなることを特徴とする、請求項7に記載の方法。
【請求項9】
前記調節剤が、前記蛍光金属錯体と適合する蛍光又は非蛍光FRETアクセプター化合物であり、前記反応媒体中の最終濃度が10−7Mより高く、分子量が50kD未満であることを特徴とする、請求項1〜8のいずれか1項に記載の方法。
【請求項10】
反応媒体中のFRETアクセプター化合物の最終濃度が10−6M〜10−3Mであり、分子量が0.1〜10kDであることを特徴とする、請求項9に記載の方法。
【請求項11】
前記調節剤が次の化合物:シアニン誘導体、DY647、D2、CY5、Cy5−COOH、フルオレセイン、クマリン、ローダミン、カルボピロニン、オキサジン及びその類似体、アレクサフルオールフルオロフォア、クリスタルバイオレット、ペリレンビシマイド(bisimide)フルオロフォア、スクアレインフルオロフォア、ホウ素ジピロメテン誘導体、NBD(ニトロベンズオキサジアゾール)及びその誘導体、DABCYL(4−((4−(ジメチルアミノ)フェニル)アゾ)安息香酸)から選択される蛍光アクセプター化合物であることを特徴とする、請求項1〜10のいずれか1項に記載の方法。
【請求項12】
前記調節剤が、次の化合物:フルオレセイン及びその誘導体、並びにシアニン誘導体から選択される蛍光アクセプター化合物であることを特徴とする、請求項1〜11のいずれか1項に記載の方法。
【請求項13】
前記調節剤が、次の化合物:QXL 570、QXL 610、QXL 670及びQXL 680;DYQ660及びDYQ661;QSY7、QSY9及びQSY21、から選択される非蛍光アクセプター化合物であることを特徴とする、請求項1〜12のいずれか1項に記載の方法。
【請求項14】
前記調節化合物が、次の化合物:ヨウ化物、アスコルビン酸塩、尿酸塩、カテコール塩、及び無機種、例えばFe(CN)62+から選択される還元剤であることを特徴とする、請求項1〜8のいずれか1項に記載の方法。
【請求項15】
前記調節剤が還元剤であり、段階a)が第一蛍光金属錯体を添加すること及び第二蛍光金属錯体を添加することを含んでなり、当該蛍光金属錯体が同様のキレート構造を有するが異なる金属と錯体を形成することを特徴とする、請求項14に記載の方法。
【請求項16】
前記第一金属錯体がユウロピウム錯体であり、前記第二金属錯体がテルビウム錯体であり、テルビウム及びユウロピウム錯体が同様のキレート構造を有することを特徴とする、請求項15に記載の方法。
【請求項17】
前記調節化合物が、非共有結合によって、特異的に蛍光金属錯体に結合する薬剤であって、蛍光金属錯体結合ドメインを各々有する次の化合物:抗体、抗体断片、ペプチド又はアプタマー、から選択されることを特徴とする、請求項1または2に記載の方法。
【請求項18】
前記調節化合物が、ランタニド又はルテニウムと競合する金属イオンであり、これにより非蛍光金属錯体が形成されることを特徴とする、請求項1または2に記載の方法。
【請求項19】
前記イオンがマンガンイオンであることを特徴とする、請求項18に記載の方法。
【請求項20】
前記蛍光金属錯体での目的のタンパク質の標識が、結合パートナーのペアを介した間接的な標識であり、ペアのメンバーの1つが蛍光金属錯体に共有結合され、他のメンバーが目的のタンパク質に共有結合されるか又は目的のタンパク質上に当然に存在することを特徴とする、請求項1〜19のいずれか1項に記載の方法。
【請求項21】
前記結合パートナーのペアが、次のペア:配列CCXXCC(配列番号 1)(ここで、C=システイン及びX=任意のアミノ酸)/二ヒ素(biarsenic)化合物;BTXペプチド/ブンガロトキシン;ストレプトアビジン或いはアビジン/ビオチン;eDHFR断片/トリメトプリム;タグ/抗タグ抗体;目的のタンパク質の周知のリガンド/目的のタンパク質、から選択されることを特徴とする、請求項20に記載の方法。
【請求項22】
前記蛍光金属錯体が、目的のタンパク質に対して特異的な抗体;GST、6HIS、Myc、FLAG及びHAから選択されるタグに対して特異的な抗体、から選択される抗タグ抗体に共有結合され、そして目的のタンパク質が対応するタグを含有することを特徴とする、請求項21に記載の方法。
【請求項23】
前記目的のタンパク質を蛍光金属錯体で標識することが、共有結合による直接的な標識であることを特徴とする、請求項1〜19のいずれか1項に記載の方法。
【請求項24】
前記蛍光金属錯体が自殺酵素の基質に結合され、目的のタンパク質が融合タンパク質の形態で発現され、タンパク質の細胞外部分が自殺酵素を含有することを特徴とする、請求項23に記載の方法。
【請求項25】
前記自殺酵素/自殺酵素基質のペアが、次のペア:アルキルグアニン−DNAアルキル転移酵素変異体/ベンジルグアニン;アルキルグアニン−DNAアルキル転移酵素変異体/ベンジルシトシン、脱ハロゲン酵素変異体/クロロアルカン;アシル担体タンパク質/ホスホパンテテイン転移酵素の存在下での補酵素A、から選択されることを特徴とする、請求項24に記載の方法。
【請求項26】
前記蛍光金属錯体が、目的のタンパク質上に存在する第二反応基、特に末端アミノ基、末端カルボキシラート基、アスパラギン酸及びグルタミン酸のカルボキシラート基、リジンのアミン基、アルギニンのグアニジン基、システインのチオール基、チロシンのフェノール基、トリプトファンのインドール環、メチオニンのチオエーテル基、ヒスチジンのイミダゾール基と共有結合を形成することができる第一反応基を含んでなることを特徴とする、請求項23に記載の方法。
【請求項27】
前記蛍光金属錯体が目的のタンパク質のチオール基と反応することを目的とするマレイミド基を含んでなることを特徴とする、請求項26に記載の方法。
【請求項28】
前記蛍光金属錯体が、ベンジルグアニン又はベンジルシトシン或いはこれらの誘導体の1つに共有結合されるユウロピウムクリプテート又はテルビウムクリプテートであり、目的のタンパク質がアルキルグアニン−DNAアルキル転移酵素変異体との融合タンパク質の形態で発現されることを特徴とする、請求項1〜19のいずれか1項に記載の方法。
【請求項29】
前記蛍光金属錯体がベンジルグアニン又はベンジルシトシンに共有結合されるユウロピウムクリプテート又はテルビウムクリプテートであり、目的のタンパク質がアルキルグアニン−DNAアルキル転移酵素変異体との融合タンパク質の形態で発現され、そして調節化合物がシアニン誘導体、特にCy5又はCy5−COOHであることを特徴とする、請求項1〜19のいずれか1項に記載の方法。
【請求項30】
前記蛍光金属錯体が、ベンジルグアニン又はベンジルシトシンに共有結合されるテルビウムクリプテートであり、前記目的のタンパク質がアルキルグアニン−DNAアルキル転移酵素変異体との融合タンパク質の形態で発現され、そして前記調節化合物がフルオレセイン又はその誘導体の1つであることを特徴とする、請求項1〜19のいずれか1項に記載の方法。
【請求項31】
前記目的のタンパク質が、Gタンパク質共役型受容体及び受容体型チロシンキナーゼから選択されることを特徴とする、請求項1〜30のいずれか1項に記載の方法。
【請求項32】
前記目的のタンパク質をコードしているDNA配列を含んでなる発現ベクターを細胞にトランスフェクトする予備段階を含んでなることを特徴とする、請求項1〜31のいずれか1項に記載の方法。
【請求項33】
前記発現ベクターが融合タンパク質をコードしている配列を含んでなり、当該配列のN末端部が自殺酵素を含んでなり、C末端部が目的のタンパク質を含んでなり、目的のタンパク質がGPCR又はRTKであることを特徴とする、請求項32に記載の方法。
【請求項34】
前記トランスフェクション段階がまた、β−アレスチン1をコードしているDNA配列を含んでなるベクターを同時にトランスフェクションすることを含んでなることを特徴とする、請求項32または33に記載の方法。
【図1】

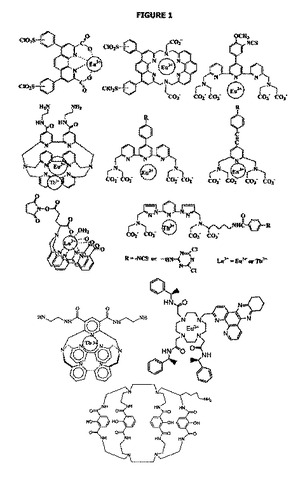
【図2】
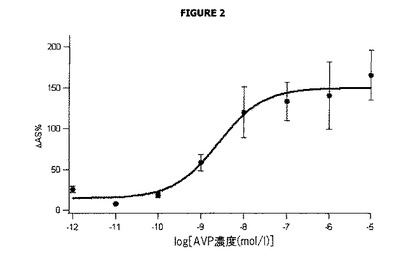
【図3】
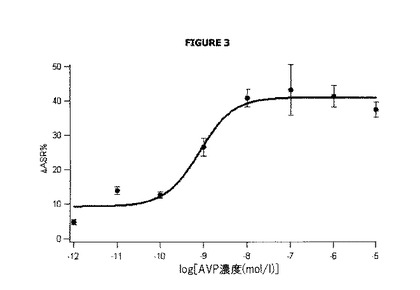
【図4】
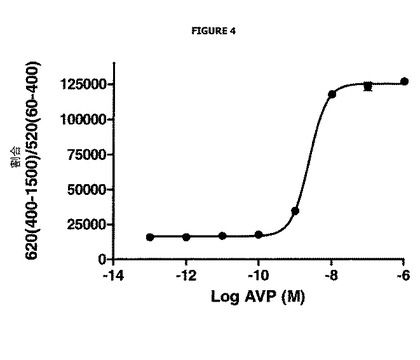
【図5】
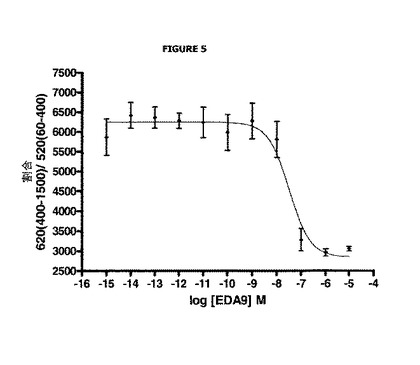
【図6】
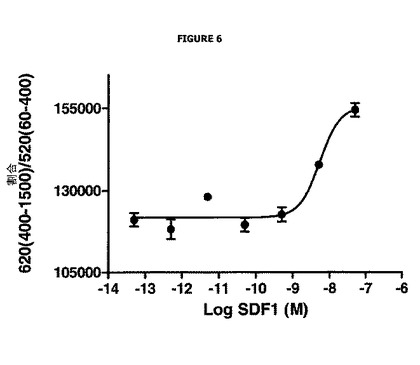
【図7】
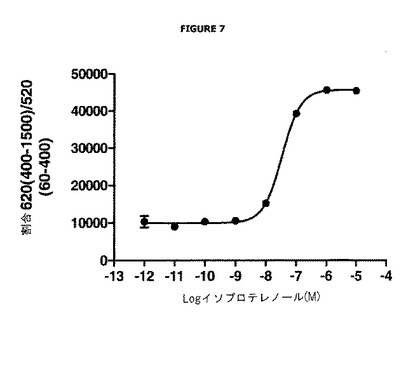
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公表番号】特表2011−529338(P2011−529338A)
【公表日】平成23年12月8日(2011.12.8)
【国際特許分類】
【出願番号】特願2011−520571(P2011−520571)
【出願日】平成21年7月30日(2009.7.30)
【国際出願番号】PCT/FR2009/051538
【国際公開番号】WO2010/012962
【国際公開日】平成22年2月4日(2010.2.4)
【出願人】(507179726)シ ビオ アンテルナショナル (9)
【Fターム(参考)】
【公表日】平成23年12月8日(2011.12.8)
【国際特許分類】
【出願日】平成21年7月30日(2009.7.30)
【国際出願番号】PCT/FR2009/051538
【国際公開番号】WO2010/012962
【国際公開日】平成22年2月4日(2010.2.4)
【出願人】(507179726)シ ビオ アンテルナショナル (9)
【Fターム(参考)】
[ Back to top ]