
膵島細胞の産生およびインスリンの送達
【課題】膵島細胞、特にインスリン産生β細胞の産生による対象へのインスリン送達を提供する。
【解決手段】膵島転写因子または膵島転写因子の発現を誘導する因子をコードする核酸を導入することによって、対象の膵臓において膵島転写因子を発現させることを特徴とし、具体的には膵島転写因子遺伝子および膵島転写因子ポリペプチドを用いて培養中またはインビボにおける細胞分化を改変して、新しいβ細胞を産生する。例えば2型糖尿病の維持および治療に有用である。
【解決手段】膵島転写因子または膵島転写因子の発現を誘導する因子をコードする核酸を導入することによって、対象の膵臓において膵島転写因子を発現させることを特徴とし、具体的には膵島転写因子遺伝子および膵島転写因子ポリペプチドを用いて培養中またはインビボにおける細胞分化を改変して、新しいβ細胞を産生する。例えば2型糖尿病の維持および治療に有用である。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は一般に、膵島細胞、特にインスリン産生β細胞の産生による対象へのインスリン送達の分野に関する。より詳細には、本発明は、膵島細胞の成長および分化に関与する転写因子をコードするヌクレオチド配列の発現調節による膵島細胞の産生に関する。
【背景技術】
【0002】
発明の背景
糖尿病は米国における死因の第3位であり、失明、腎不全、および切断の主要原因である。糖尿病は早発性心臓発作および脳卒中の主要原因でもあり、米国の医療費の15%を占める。アメリカ人のおよそ5%、65歳を超えるアメリカ人の20%が、糖尿病を患っている。
【0003】
糖尿病は、膵臓内分泌部のランゲルハンス島のβ細胞が、代謝要求を満たすのに十分なインスリンを産生できないことに起因する。糖尿病は、1型糖尿病(またはインスリン依存型糖尿病)および2型糖尿病(またはインスリン非依存型糖尿病)という、2つの臨床形態に分類される。1型糖尿病は、インスリン産生β細胞の損失が原因となって起こる。2型糖尿病は1型糖尿病に比べ遺伝病の要素が強く(Zonana およびRimoin、1976 N. Engl. J. Med. 295:603(非特許文献1))、通常は人生のより後期に発症し、米国では糖尿病のおよそ90%の割合を占める。通常、罹患個体は、膵臓のインスリン産生能の低下およびインスリン利用能の欠陥(インスリン抵抗性)の両方を伴う。肥満症によりインスリン抵抗性が引き起こされるが、2型糖尿病を有する個体のおよそ80%が臨床的肥満(標準体重の20%を上回る)である。残念なことに、2型糖尿病に罹患した米国人の約半分は、この疾患を有することに気付いていない。2型糖尿病に関連する臨床症状は疾患の後期まで明らかにならないこともあり、また初期症状が誤診されることも多く、これは治療の遅延および合併症増加の原因になる。2型糖尿病の病因における遺伝の役割は明らかであるが、この疾患に関与する明確な遺伝子の大半が未知である。
【0004】
インスリンは、膵臓のランゲルハンス島のβ細胞によってのみ産生される。発生段階において、β細胞を含む膵島細胞は、成長過程にある膵芽内の未分化前駆細胞から発達する。膵芽が成長するにつれ未分化細胞が菅を形成するが、これが前駆細胞として機能する細胞である。菅細胞は生涯を通じて膵島細胞に分化する能力を維持すると思われ、膵臓が損傷を受けるような状況では、菅細胞から新しい膵島細胞が形成され得る。しかし残念なことに、1型糖尿病で見られるように膵島細胞のみが損傷を受ける場合には、膵島細胞の再生は起こらないようである。
【0005】
この発達過程は、様々な理由で臨床上の問題と関連性がある。まず第一に、膵島細胞および特にβ細胞の形成は、インスリンを産生するためおよびエネルギー代謝を制御するために必須である。β細胞の発達過程が何らかの形で損傷を受けた場合、これは後に個体が糖尿病を発症する素因となる。よってこの過程に関連する遺伝子は、新生児糖尿病、若年発症成人型糖尿病(MODY)、または2型糖尿病に対する候補遺伝子である。これらの遺伝子の配列を用いて、糖尿病を発症する危険性のある個人を同定でき、または糖尿病を予防および治療するための新規な薬理的物質を開発できる。
【0006】
第二に、上記のように、糖尿病を患う個体ではインスリンの産生が損傷される。1型糖尿病ではβ細胞の破壊によりこの障害が生じるが、一方2型糖尿病では、インスリン産生は不十分ではあるが損傷されない。1型糖尿病の治療および大半の2型糖尿病の治療は、β細胞の補充を伴う。β細胞の補充はいくつかの方法によって行われ得るが、前駆細胞からの新しいβ細胞の発生は、培養に行われるかまたは患者内でインビボで行われるのが最も生理的であると考えられる。そのためには、β細胞の分化を制御する分子が必要とされる。
【0007】
したがって糖尿病分野では、膵島前駆細胞を同定する試み、およびインビトロでβ細胞を分化させる方法を開発する試みに多大な努力を払ってきた。しかし今日まで、ほとんど成功していない。本発明は、この問題に取り組むものである。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】Zonana およびRimoin、1976 N. Engl. J. Med. 295:603
【発明の概要】
【0009】
発明の概要
本発明は、例えば膵島転写因子または膵島転写因子の発現を誘導する因子をコードする核酸を導入することにより、対象の膵臓において膵島転写因子の発現を提供することによる、対象における膵島細胞およびインスリンの産生に関し、特にNgn3、NeuroD1、およびMash1が関心対象である。本発明はまた、糖尿病患者を治療するために、膵島転写因子遺伝子および膵島転写因子ポリペプチドを用いて培養中またはインビボにおける細胞分化を改変させて、新しいβ細胞を産生する方法に関する。本発明は、例えば2型糖尿病の維持および治療において有用であると考えられ、かつさらに1型糖尿病を有する個体における自己免疫性破壊によって失われたβ細胞の補充を可能にすると考えられる。
【0010】
本発明の第一の目的は、対象の膵臓において膵島細胞の産生を提供することである。
【0011】
本発明の別の目的は、機能的β細胞形成を誘導することにより、対象においてインスリンの産生を提供することである。
【0012】
本発明の別の目的は、糖尿病患者を治療するために、膵島転写因子遺伝子を用いて培養中またはインビボにおける細胞の分化を改変して新しいβ細胞を産生する方法を提供することである。
【0013】
本発明の別の目的は、エクスビボ治療のための膵島細胞の産生、例えば対象に移植するための膵島転写因子発現細胞の産生を提供することである。
【0014】
本発明の別の目的は、糖尿病等のインスリン関連疾患に罹患した対象を治療する方法を提供することである。
【0015】
本発明のこれらおよび他の目的、利点、および特徴は、以下に示す本発明のより詳細な記載を読むことによって当業者に明らかになるであろう。
【0016】
本発明を以下でさらに詳細に説明する。
【図面の簡単な説明】
【0017】
【図1】CMVプロモーターに機能的に結合されたマウスニューロゲニン3配列を含む、アデノ-X.NGN3ウイルス構築物のマップである。
【図2A】図2Aはアデノ-X.NGN3を注射されたラットの膵臓組織切片におけるインスリンの組織学的染色を高倍率(40×)で示す写真であり、これにより本発明によってインスリンが産生されることが示唆される。矢印はインスリン染色細胞を示す。
【図2B】図2Bは図2Aの連続切片の写真であり、菅細胞のマーカーであるサイトケラチン20について染色したものである。矢印は、連続切片におけるインスリン染色細胞の位置を示す。
【図3】アデノ-XZ.NGN3を注射されたラットの膵臓における、インスリンおよびニューロゲニン3について蛍光同時染色した共焦点像の写真である。図の中心にある細胞では、核においてニューロゲニン3が染色され(矢印で示した濃い灰色の領域)、細胞質においてインスリンが染色される(明るい灰色をした周囲の領域)のが観察される。ニューロゲニン3およびインスリンの産生は、本発明による同じ細胞においても検出され得る。
【図4A】図4Aはストレプトゾトシンにより糖尿病となりかつアデノ-X.NGN3を注射されたラットの膵臓組織切片におけるインスリンの組織学的染色を示す写真であり、これにより、本発明によるインスリン産生細胞の産生が示唆される。白矢印は、個別のおよび小さな凝集塊であるインスリン染色細胞を示す。黒矢印は、ストレプトゾトシン処理後に形成された膵島に残存する残存細胞および変性細胞を示す。
【図4B】図4Bは、対照ウイルス構築物であるアデノLacZを注射されたラットの膵臓組織切片における、インスリンの染色を示す写真である。矢印は、ストレプトゾトシン処理後に形成された膵島において残存する残存細胞および変性細胞を示す。個別のおよび小さな凝集塊であるインスリン染色細胞は全く検出されなかった。
【図5】アデノX.NGN3またはアデノLacZ(対照動物)のいずれかを注射されたストレプトゾトシン処理(糖尿病)動物の血糖値のグラフである。
【図6】図6A〜図6Cは、胚齢12.5日のトランスジェニックマウスにおける膵島ホルモン発現を示す写真である。非トランスジェニック同腹仔(図6A)、およびngn3(図6B)またはneuroD1/BETA2(図6C)を発現するトランスジェニック胎仔の膵臓におけるグルカゴンの免疫組織化学的染色を示す。
【図7】図7Aおよび図7Bは、pdx1プロモーターの制御下でngn3(図7A)またはneuroD1(図7B)を過剰発現する胚齢18.5日のマウス胎仔の腸およびそれに付随する器官を示す写真である。非トランスジェニック同腹仔を各図の右側に示す。矢印は以下の器官を指し示す:肝臓(L)、胃(St)、脾臓(sp)、および膵臓(P)。トランスジェニック動物において、膵臓組織の減少は見られない。
【図8】それぞれCMVプロモーターに機能的に結合したNeuroD1およびMas1配列を含む、アデノX.NeuroD1およびアデノX.MASH1のウイルス構築物のマップである。
【図9】CMVプロモーターに機能的に結合したニューロゲニン3配列を含む、pBAT16.NGN3プラスミドDNA構築物のマップである。
【図10】bHLHタンパク質の系統樹の略図である。h.ngn3はヒトの配列であるが、それ以外はすべてマウスの配列である。
【図11】図11A〜図11Bは、ヒトニューロゲニン3遺伝子プロモーターを示す概略図である。図11Aは、実施例で用いたプロモーター断片の転写開始部位および5'末端の相対的位置を示すプロモーターの略図である。転写因子の結合部位として可能性のあるいくつかの部位を示すが、これにはプロモーターの遠位にある部位クラスターも含まれる。TATAAボックス(実際の配列はGATAA)は、転写開始部位の30 bp上流に示される。図11Bに、クラスター1に対応するプロモーターの-3728位と-3653位の間のDNA配列を示す。ここでも、転写因子の結合部位として可能性のある部位を示す。Hoxおよび関連するクラスのホメオドメインタンパク質の結合部位として機能し得る可能性のある2つの配列を点線で示す。
【図12】細胞株におけるヒトニューロゲニン3プロモーターの機能を示すグラフである。表示した5'末端から+261 bpにわたる配列を含むプロモーター断片をホタルルシフェラーゼ遺伝子の上流に結合し、示した細胞株にトランスフェクションした。レポーター遺伝子活性を、同じ細胞種におけるプロモーターのないルシフェラーゼベクターと相対的に表した。トランスフェクションは少なくとも3連で2回行い、誤差は平均値の±標準誤差として示す。ND、試験せず。
【図13】図13A〜図13C は、インビボでのヒトニューロゲニン3プロモーターの機能を示すグラフである。図13Aでは、5.7 kbのヒトNgn3プロモーターを細菌のβ-ガラクトシダーゼ遺伝子の上流に連結し、トランスジェニックマウス中で発現させた。染色部分(濃い領域)は、標識細胞におけるX-gal基質のβ-ガラクトシダーゼ触媒作用を示す。図13Bおよび図13Cは、E15.5マウス胎仔の膵臓における、β-ガラクトシダーゼ発現の酵素検出による、グルカゴンに対する免疫組織化学染色(ペルオキシダーゼ検出)を示す。β-ガラクトシダーゼの発現は、示したように-5.7 kbまたは-2.6 kbのヒトNgn3プロモーターにより促進される。X-gal染色は、37℃(2.6 kbプロモーター)または室温(5.7 kbプロモーター)で16時間行った。両切片とも倍率10×での比較可能な切片を示す。
【図14】図14A〜図14Cは、ニューロゲニン3の腸での発現を示す写真である。図14Aは、5.7 kbヒトngn3プロモーターによって駆動されるβ-ガラクトシダーゼ導入遺伝子の活性が、E15.5マウス膵臓の腸管上皮内に散在する細胞において検出され得ることを示す(濃い斑点)。図14Bは対照であり、野生型の同腹仔では同様のβ-ガラクトシダーゼ発現がみられないことを示す。図14Cは、E15.5のトランスジェニックマウスの腸における、β-ガラクトシダーゼも同じく発現するいくつかの細胞(黒矢印)における、およびβ-ガラクトシダーゼを発現しないいくつかの細胞(白矢印)におけるNgn3の検出(ペルオキシダーゼ検出)を示す。
【図15】図15A〜図15Bは、HNF3がヒトngn3プロモーターに結合することを示す、電気泳動移動度シフト分析の写真である。図15Aは、ヒトngn3プロモーターのH3-1プローブに対する転写因子HNF3βの結合を実証する電気泳動移動度シフト分析を示す。図15Bは、ヒトNgn3プロモーターにおける近位の部位および遠位の部位がHNF3βへの結合について同等に競合し得ることを実証する電気泳動移動度シフト分析を示す。
【図16】HNF1がヒトngn3プロモーターに結合することを示す、電気泳動移動度シフト分析の写真である。
【図17】図17A〜図17Cは、HES1がニューロゲニン3プロモーターを阻害することを示すグラフである。図17Aは、所定のレポーター構築物でトランスフェクションされ、かつCMVプロモーターの下流に連結されたHES1 cDNAを含む所定の量の発現プラスミド(ベクターpBAT12)で同時トランスフェクションされたNIH3T3細胞を用いた結果を示す。ルシフェラーゼ活性は、レポータープラスミドのみでトランスフェクションされた細胞における活性と相対的に表す。図17Bは、90 ng/106細胞のHES1発現プラスミドとの同時トランスフェクションの存在下または非存在下の、ルシフェラーゼの上流に連結させたNgn3プロモーターの様々な断片およびラウス肉種ウイルス(RSV)プロモーターの相対活性を示す。図17Cは、NIH3T3細胞およびαTC1.6細胞における、同時トランスフェクションしたHES1による、所定のプロモーターからのルシフェラーゼ活性の抑制率を示す。抑制率とは、相対ルシフェラーゼ活性の逆数である。トランスフェクションは三つ組みで少なくとも3回別々に行った。誤差は、平均値の±標準誤差として示す。
【図18】図18Aは、ヒトngn3遺伝子の転写開始部位(+1)のすぐ上流のDNA配列の模式図を示す。HES1結合部位として可能な部位(Nボックス)を示す。図18B〜図18Cは、HES1がヒトngn3プロモーターに結合することを示す電気泳動移動度シフト分析の写真である。図18Bは、細菌的に産生されたHES1が、マウスHES1プロモーター(H1P)の既に特徴づけられている部位だけでなく、近位プロモーターのNボックスにも結合し得ることを実証する電気泳動移動度シフト分析を示す。GST-HES1タンパク質またはGSTタンパク質のいずれかを、表示のプローブと共にインキュベートした。一番右のレーンは、HES1抗血清の添加により、HES1複合体がスーパーシフトすることを示す。図18Cは、GST-HES1タンパク質の結合に関して競合できるHES1結合部位の相対的能力を実証する電気泳動移動度シフト分析を示す。3つのngn3プロモーター部位はすべて、以前に記載されているHES1プロモーター(H1P)のHES1結合部位よりも高い親和性で結合する。一方、200倍過剰量のオリゴヌクレオチドE1(近位プロモーター由来の無関係のEボックス配列)は、複合体形成に効果を及ぼさない。
【図19A】図19A〜図19Cは、β-ガラクトシダーゼ(adCMV-Bgal)、ニューロゲニン3(ヒト)(adCMV-Ngn3)、neuroD1(マウス)(adCMV-ND1)、mash1(マウス)(adCMV-Mash1)、またはmyoD(マウス、筋bHLH遺伝子)(adCMV-MyoD)を発現するアデノウイルスベクターを、2つのマウス膵管細胞株mPAC L20およびmPAC L4S2にトランスフェクションした効果を示すアガロースゲルの写真である。図19Aは、これらのベクターの発現がNgN3、ND1、Mash1、MyoD、およびβ-アクチンRNAレベルに及ぼす影響を示す。
【図19B】図19A〜図19Cは、β-ガラクトシダーゼ(adCMV-Bgal)、ニューロゲニン3(ヒト)(adCMV-Ngn3)、neuroD1(マウス)(adCMV-ND1)、mash1(マウス)(adCMV-Mash1)、またはmyoD(マウス、筋bHLH遺伝子)(adCMV-MyoD)を発現するアデノウイルスベクターを、2つのマウス膵管細胞株mPAC L20およびmPAC L4S2にトランスフェクションした効果を示すアガロースゲルの写真である。図19Bは、これらのベクターの発現がグルカゴン、ソマトスタチン、インスリン、IAPP、グルコキナーゼ、グルコース輸送体2、およびβ-アクチンRNAレベルに及ぼす影響を示す。
【図19C】図19A〜図19Cは、β-ガラクトシダーゼ(adCMV-Bgal)、ニューロゲニン3(ヒト)(adCMV-Ngn3)、neuroD1(マウス)(adCMV-ND1)、mash1(マウス)(adCMV-Mash1)、またはmyoD(マウス、筋bHLH遺伝子)(adCMV-MyoD)を発現するアデノウイルスベクターを、2つのマウス膵管細胞株mPAC L20およびmPAC L4S2にトランスフェクションした効果を示すアガロースゲルの写真である。図19Cは、これらのベクターの発現がPax4、Pax6、Nkx2.2、Nkx6.1、Isl-1、およびPdx-1RNAレベルに及ぼす影響を示す。
【発明を実施するための形態】
【0018】
発明の詳細な説明
膵島細胞およびインスリン産生のための本発明の構成および本方法を説明する前に、本発明は記載した特定の方法、プロトコール、細胞株、ベクター、および試薬に制限されず、当然変更され得ることが理解されねばならない。また、本明細書で使用する専門用語は、特定の態様を説明する目的のためのみに用いられ、添付の特許請求の範囲によってのみ限定される本発明の範囲を制限する意図はないことも理解されねばならない。
【0019】
本明細書および添付の特許請求の範囲で使用する単数形「1つの」、「ある」、および「その」とは、特記する場合を除き、その対象物の複数形も含むことに留意されたい。したがって、例えば「1つの膵島転写因子」についての言及は複数のそのような膵島転写因子およびその機能的同等物を含み、「そのポリヌクレオチド」についての言及は1つまたは複数のポリヌクレオチドおよび当業者に周知のその同等物を含み、以下同様である。
【0020】
特記されない限り、本明細書で使用する専門用語および科学用語はすべて、本発明が属する当技術分野の当業者によって共通に理解されるものと同じ意味をもつ。本発明の実施または試験において、本明細書に記載したものと類似したまたは同等の任意の方法、装置、および材料が使用できるが、本明細書に記載された方法、装置、および材料が好ましい。
【0021】
本明細書で言及したすべての文献は、その文献に記載され本発明に関連して使用される可能性のある、例えば、細胞株、ベクター、および方法を説明および開示する目的で参照として本明細書に組み入れられる。本明細書内で考察される文献は、単に本出願の出願日以前にそれらが開示されているというだけの理由で提供されたものである。本明細書に記載されたいかなるものも、先行発明のせいで本発明者らがこれら開示に先行できないことを認めると解釈されるべきではない。
【0022】
定義
本明細書で用いる「ポリヌクレオチド」とは、オリゴヌクレオチド、ヌクレオチド、およびその断片または一部、ならびにペプチド核酸(PNA)、その断片、部分、またはアンチセンス分子を指し、さらにゲノム由来また合成由来のDNAまたはRNAを指し、これは一本鎖または二本鎖であってよい。「ポリヌクレオチド」とはセンス鎖またはアンチセンス鎖を意味する。ある特定のポリヌクレオチド配列(例えば、Ngn3ポリペプチドをコードするポリヌクレオチド)を指すために「ポリヌクレオチド」という用語を用いる場合、「ポリヌクレオチド」とは、引用されたポリペプチドと機能的に同等なポリペプチド、例えば縮重変種のポリヌクレオチド、または引用されたポリペプチドの生物活性のある変種もしくは断片をコードするポリヌクレオチドを含むことが意図されており、これは本明細書で提供する配列と比較して実質的な配列類似性または配列同一性を有するポリヌクレオチドを含む。同様に、本明細書で用いる「ポリペプチド」とは、オリゴペプチド、ペプチド、またはタンパク質を指す。本明細書で天然型タンパク質分子のアミノ酸配列を指すのに「ポリペプチド」という用語を引用する場合、「ポリペプチド」およびその類語は、アミノ酸配列を、引用されたタンパク質分子と関連する完全な天然アミノ酸配列に限定することを意味してはおらず、それどころか生物活性のある変種または断片をも含むことを意図しており、本明細書で提供されたアミノ酸配列と比較して実質的な配列類似性または配列同一性を有するポリペプチドを含む。
【0023】
本明細書で用いる「縮重変種」とは、所与のポリヌクレオチド配列がコードするのと同じポリペプチドをコードするが、遺伝暗号の縮重によりコード配列の点で異なるようなポリヌクレオチド配列を指す。遺伝暗号は、2つ以上の異なるコドンが同じアミノ酸をコードする場合に「縮重する」。
【0024】
本明細書で用いる「ポリペプチド」とは、i)天然のポリペプチド、ii)生物活性のあるポリペプチド断片、iii)生物活性のある、ポリペプチドのポリペプチド類似体、またはiv)生物活性のあるポリペプチド変種のアミノ酸配列を有する組換えポリペプチドまたは非組換えポリペプチドのアミノ酸配列を指す。本発明で有用なポリペプチドは天然、合成、半合成、または組換えにかかわらず任意の供給源由来の任意の種から取得可能であり、例えば哺乳動物または非哺乳動物(例えば、は虫類、両生類、鳥類(例えばニワトリ))、特にヒトを含む哺乳動物、げっ歯類(例えば、マウスまたはラット)、ウシ、ヒツジ、ブタ、マウス、またはウマ、好ましくはラットまたはヒト等から取得可能である。例えば、「ヒトNgn3ポリペプチド」、「ヒトND1ポリペプチド」、または「ヒトMash1ポリペプチド」は、それぞれヒトに由来する単離ヒトNgn3、ND1、または Mash1ポリペプチドのアミノ酸配列を指し、天然の対立遺伝子変種すべてを含むことを意図するが、引用したタンパク質分子と関連する完全な天然アミノ酸配列のみにアミノ酸配列を限定する意図はない。
【0025】
本明細書で用いる「抗原性アミノ酸配列」とは、単独でまたは担体分子と結合して、哺乳動物において抗体応答を誘発し得るアミノ酸配列を意味する。
【0026】
ポリペプチドの「変種」とは、1つまたは複数のアミノ酸が改変されたアミノ酸配列として定義される。変種は、例えばイソロイシンによるロイシンの置換など、置換されたアミノ酸が同様の構造または化学特性を有するような、「保存的」変化を有する可能性がある。より稀ではあるが、変種は、例えばトリプトファンによるグリシンの置換など、「非保存的な」変化を有する可能性がある。同様の軽微な改変にはまた、アミノ酸の欠失もしくは挿入、またはその両方が含まれうる。生物活性または免疫活性を消失させることなく、どのアミノ酸残基がいくつ置換、挿入、または欠失されうるかを決定する手引きを、例えばDNAStarソフトウェア等の当技術分野で周知のコンピュータープログラムを用いて見出すことができる。
【0027】
「関心対象の核酸」または「関心対象のDNA」とは、哺乳動物対象への投与に望ましいタンパク質または他の分子をコードする任意の核酸(例えばDNA)を意味する。一般に核酸は、プロモーターなどの、発現に必要な他の配列に機能的に結合されている。
【0028】
「欠失」とは、関心対象のの天然ポリペプチドのアミノ酸配列またはヌクレオチド配列と比較して、それぞれ1つまたは複数のアミノ酸またはヌクレオチドの残基を欠いたアミノ酸またはヌクレオチドの配列のいずれかの変化として定義される。
【0029】
「挿入」または「付加」とは、天然ポリペプチドのアミノ酸配列またはヌクレオチド配列と比較して、それぞれ1つまたは複数のアミノ酸またはヌクレオチドの残基の付加をもたらすアミノ酸またはヌクレオチドの配列の変化である。
【0030】
「置換」とは、天然ポリペプチドのアミノ酸配列またはヌクレオチド配列と比較して、1つまたは複数のアミノ酸またはヌクレオチドがそれぞれ別のアミノ酸またはヌクレオチドにより置換されることによって生じる。
【0031】
「生物活性のある」という用語は、例えば、天然の膵島転写因子ポリペプチドの構造機能、制御機能、または生化学的機能を有するヒト膵島転写因子ポリペプチドを指す。同様に「免疫活性のある」とは、天然、組換え、または合成のヒト膵島転写因子ポリペプチド、もしくはその任意のオリゴペプチドが、適当な動物または細胞において特異的免疫応答を誘導して特異的抗体と結合する能力と定義される。
【0032】
本明細書で用いる「誘導体」という用語は、ポリペプチドをコードする核酸または核酸にコードされるポリペプチドの化学的修飾を指す。そのような修飾の実例として、アルキル基、アシル基、またはアミノ基による水素の置換が挙げられるであろう。核酸誘導体は、天然ポリペプチドの本質的な生物学的特徴を保持するポリペプチドをコードする。
【0033】
本明細書で用いる「単離された」という用語は、天然の化合物が存在する環境とは異なる環境にある関心対象の化合物(例えば、ポリヌクレオチドまたはポリペプチドのどちらか)を表すことを意図する。「単離された」とは、関心対象の化合物が十分に濃縮された試料、および/または関心対象の化合物が部分的にまたは十分に精製された試料内に存在する化合物を含むことを意図する。
【0034】
本明細書で用いる「実質的に精製された」または「実質的に遊離した」という用語は、自然環境から取り出されて、これに自然状態で結びついている少なくとも60%、好ましくは75%、最も好ましくは90%の他の化合物を含まないような化合物(例えば、ポリヌクレオチドまたはポリペプチドのどちらか)を指す。
【0035】
「形質転換」または「トランスフェクション」とは、新しい核酸(例えば、細胞にとって外因性のDNAまたはRNA)の取り込み後に細胞に誘導される永続的または一過的の遺伝的変化、好ましくは永続的な遺伝的変化を意味する。遺伝的変化は、新しい核酸を宿主細胞のゲノムに取り込むことによって、またはエピソーム成分として新しいDNAを一過的または安定的に維持することにより達成され得る。
【0036】
「形質転換細胞」とは、組換えDNA技術により、関心対象のタンパク質をコードするDNA分子が導入された細胞(または、DNA分子がその祖先の細胞に導入された細胞)を意味する。
【0037】
「構築物」とは、特異的なヌクレオチド配列を発現する目的で作製された組換え核酸、通常は組換えDNAを意味し、または他の組換えヌクレオチド配列の構築に用いられ得る。
【0038】
「ベクター」とは、関心対象のDNAによる標的細胞の形質転換を促進する、任意の生物的化合物または化学的化合物を意味する。例示的な生物学的ベクターには、ウイルス、特に弱毒化ウイルスおよび/または複製不能ウイルスが含まれる。例示的な化学的ベクターには、脂質複合体および裸のDNA構築物が含まれる。
【0039】
「裸のDNA」または「裸の核酸」またはDNA配列などは、ウイルス粒子、細菌細胞、または標的細胞の細胞質への核酸の送達を促進する他の封入手段に含まれない核酸分子を意味する。標的細胞の部位(例えば、細胞への移動を促進する手段、ヌクレアーゼ分解から核酸を保護する手段等)および/または標的上皮細胞の表面への核酸の送達を促進する手段により裸の核酸を任意に付随(例えば、製剤化)することができる。
【0040】
「プロモーター」とは、転写を指示するのに十分な最小配列を意味する。「プロモーター」はまた、細胞種特異的に制御可能な、組織特異的に制御可能な、または外部のシグナルもしくは薬剤で誘導可能な、プロモーター依存性遺伝子発現に十分なプロモーター成分を含むことが意図される。そのような成分は、天然遺伝子の5'領域または3'領域に位置している可能性がある。
【0041】
「機能的に結合された」とは、適当な分子(例えば、転写活性化タンパク質)が制御配列に結合された時に発現が可能になるように、DNA配列と制御配列が結合していることを意味する。
【0042】
「膵臓」とは、胃の後ろで脾臓と十二指腸の間に横向きに位置する大きく細長いブドウ状の腺を意味する。膵臓は、内分泌部分(内分泌部)および外分泌部分(外分泌部)からなる。ランゲルハンス島を含む内分泌部はインスリンを含むタンパク質を産生し、血流に直接分泌する。外分泌部分は分泌単位を含み、タンパク質の消化に必須の酵素を含む膵液を産生し、十二指腸に分泌する。
【0043】
「正常血糖」または「正常血糖状態」とは、高血糖に関連する疾患または病態を有する対象における血糖値と特に比較して、正常またはほぼ正常な血糖値を伴う状態を意味する。ヒトにおける正常血糖とは、約70 mg/dl〜約110 mg/dlの範囲の血糖値に相当する。「高血糖」とは血糖値の増加を伴う状態を意味し、これは一般に、約120 mg/dl、130 mg/dl、または140 mg/dlを上回る。
【0044】
「前駆細胞」とは膵島細胞に発達することが可能な任意の細胞を意味し、これにはインビボまたはインビトロにおける、胎児膵上皮細胞、成人膵細胞(例えば、膵管細胞、腺房細胞、および膵幹細胞)、腸上皮細胞、腸幹細胞もしくは腺窩細胞、他の組織由来の幹細胞(造血幹細胞または肝幹細胞等)、肝細胞(肝実質細胞および肝管細胞)、唾液腺細胞、神経幹細胞、筋幹細胞、および胚幹細胞等の、哺乳動物不死化細胞株または哺乳動物初代培養細胞が含まれる。
【0045】
「標的細胞」とは転写因子ヌクレオチド配列をコードするDNAを組み込むために選択される任意の細胞を意味し、これにはインビボまたはインビトロにおける、前駆細胞、胎児膵上皮細胞、成人膵細胞(例えば、膵管細胞、腺房細胞、および膵幹細胞)、腸上皮細胞、腸幹細胞もしくは腺窩細胞、他の組織由来の幹細胞(造血幹細胞、皮膚幹細胞、または肝幹細胞等)、肝細胞(肝実質細胞および肝管細胞)、唾液腺細胞、神経幹細胞、筋幹細胞、および胚幹細胞等の、哺乳動物不死化細胞株または哺乳動物初代培養細胞が含まれる。
【0046】
「膵島転写因子」または「膵島因子」または「転写因子」とは、膵島細胞の分化、形成、および/または発達に関与する任意の転写因子を意味し、その発現は膵島細胞の表現型を有する細胞、例えば、インスリンまたは膵島細胞に特徴的な他のマーカー(例えば、グルカゴン、グルコキナーゼ、膵島アミロイドポリペプチド(IAPP)、ソマトスタチン、膵臓ポリペプチド等)を産生する細胞の産生に寄与する。また同様に、機能的に同等な相同体も意味する。特に関心が持たれるのは、膵島細胞の発達に関与するクラスB塩基性ヘリックス・ループ・ヘリックス(bHLH)転写因子であり、これにはニューロゲニン(ニューロゲニン1、ニューロゲニン2、およびニューロゲニン3)、neuroD因子(NeuroD1/BETA2、neuroD2、およびNeuroD4/Math3)、およびMash因子Mash1が含まれる。また同様に、これら転写因子の機能的に同等な相同体も特に関心対象となる。
【0047】
本明細書で用いる「膵島転写因子制御経路」という用語は、活性化事象および非活性化事象のカスケードを介して協調して膵島細胞の発達を制御し、膵島細胞の表現型を決定するシグナル伝達分子および転写因子を含む一連のタンパク質を指す。膵島転写因子経路の正の制御因子は、特定の膵島転写因子またはその下流のエフェクターの発現を促進する制御因子であり、よってこれは膵島細胞表現型の誘導に正の影響を及ぼす。膵島転写因子経路の負の制御因子は、特定の膵島転写因子またはその下流のエフェクターの発現を阻害する制御因子であり、よって膵島細胞の表現型の誘導に負の影響を及ぼす。正の制御因子には、負の制御因子の活性または発現を阻害する因子が含まれる。
【0048】
本明細書で用いる「ニューロゲニン3(Ngn3)制御経路」という用語は、活性化事象および非活性化事象のカスケードを介して協調してNgn3活性を促進し、細胞が膵島細胞となるように導き、膵島細胞表現型の産生または維持を促進するシグナル伝達分子および転写因子を含む一連のタンパク質を指す。Ngn3経路の正の制御因子は、Ngn3またはその下流のエフェクター(例えば、Mash1、NeuroD1等)の発現を促進する制御因子であり、よってこれは膵島細胞の表現型の誘導に正の影響を及ぼす。Ngn3経路の負の制御因子は、Ngn3またはその上流もしくは下流のエフェクター(例えばHES1)の発現を阻害する制御因子であり、よってこれは膵島細胞の表現型の誘導に負の影響を及ぼす。Ngn3経路の正の制御因子には、Ngn3経路の負の制御因子の活性もしくは発現を阻害する因子、またはNgn3の発現を促進する因子が含まれる。Ngn3をコードする核酸は、Ngn3の発現増加を提供しNgn3経路の活性を促進するため、Ngn3経路の正の制御因子の一例である。HNF転写因子(例えば、HNF1、HNF3、およびHNF6)はNgn3の発現を促進する(正に制御する)ため、Ngn3経路の正の制御因子のさらなる一例である。
【0049】
本明細書で用いる「NeuroD1制御経路」という用語は、活性化事象および非活性化事象のカスケードを介して協調してNeuroD1活性を促進し、かつしたがって細胞が膵島細胞となるように導き、膵島細胞の表現型の産生または維持を促進するシグナル伝達分子および転写因子を含む一連のタンパク質を指す。NeuroD1経路の正の制御因子は、NeuroD1またはその下流のエフェクター(例えば、Mash1、Ngn3等)の発現を促進する制御因子であり、よってこれは膵島細胞の表現型の誘導に正の影響を及ぼす。NeuroD1経路の負の制御因子は、NeuroD1またはその上流もしくは下流の正のエフェクターの発現を阻害する制御因子であり、よって膵島細胞の表現型の誘導に負の影響を及ぼす。NeuroD1経路の正の制御因子には、NeuroD1経路の負の制御因子の活性もしくは発現を阻害する因子、またはNeuroD1の発現を促進する因子が含まれる。NeuroD1をコードする核酸は、NeuroD1の発現増加を提供しかつしたがってNeuroD1経路の活性を促進するため、NeuroD1経路の正の制御因子の一例である。正の制御因子には負の制御因子の活性または発現を阻害する因子が含まれ、かつNeuroD1をコードする核酸が含まれる。NeuroD1がNgn3の下流に作用するという条件下で、NeuroD1制御経路はNgn3制御経路と重複することに留意されたい。したがって、Ngn3およびNgn3の正の制御因子もまた、NeuroD1経路の正の制御因子である。
【0050】
本明細書で用いる「Mash1制御経路」という用語は、活性化事象および非活性化事象のカスケードを介して協調してMash1活性を促進し、かつしたがって細胞が膵島細胞となるように導き、膵島細胞の表現型の産生または維持を促進するシグナル伝達分子および転写因子を含む一連のタンパク質を指す。Mash1経路の正の制御因子は、Mash1またはその下流のエフェクター(例えば、Ngn3、NeuroD1等)の発現を促進する制御因子であり、よってこれは膵島細胞の表現型の誘導に正の影響を及ぼす。Mash1経路の負の制御因子は、Mash1もしくはその上流または下流のエフェクターの発現を阻害する制御因子であり、よってこれは膵島細胞の表現型の誘導に負の影響を及ぼす。正の制御因子には、Mash1制御経路の負の制御因子の活性もしくは発現を阻害する因子、またはMash1の発現を促進する因子が含まれる。Mash1をコードする核酸は、Mash1の発現増加を提供してMash1経路の活性を促進するため、Mash1経路の正の制御因子の一例である。Mash1はNeuroD1およびNgn3転写因子の上流に作用するため、Mash1制御経路はNgn3 およびNeuroD1制御経路と重複することに留意されたい。したがって、Mash1はNeuroD1およびNgn3制御経路の正の制御因子である。
【0051】
「膵島細胞」とは、通常膵臓ランゲルハンス島からなるホルモン産生細胞と類似した表現型を有する細胞を意味し、一般に、インスリン、グルカゴン、グルコキナーゼ、ソマトスタチン、膵臓ポリペプチド、または膵島アミロイドポリペプチド(IAPP)等の、膵臓ランゲルハンス島の細胞を他の膵細胞から通常識別するようなマーカーの発現を特徴とする。
【0052】
「β細胞」とは、インスリン、Nkx6.1、またはグルコキナーゼ等の、β細胞を他の膵島細胞から通常識別するマーカーの発現を特徴とする表現型を有する膵島細胞を意味する。
【0053】
「α細胞」とは、プログルカゴンまたはグルカゴン等の、α細胞を他の膵島細胞から通常識別するマーカーの発現を特徴とする表現型を有する膵島細胞を意味する。
【0054】
「標的細胞」とは、例えば、発現について関心対象の核酸の導入が望ましい細胞を意味する。本明細書における「標的細胞」は便宜上用いられているだけであり、かつ例えば、ターゲティング技術(例えば、物質が特定の細胞または細胞種に導入されるように優先的に導く標的分子)の使用を必要とする関心対象の核酸の導入を行うことを意図しているわけではない。
【0055】
「対象」または「患者」または「個体」とは、診断または治療が望ましい任意の哺乳動物対象を意味し、特にヒトを意味する。他の対象には、ウシ、イヌ、ネコ、モルモット、ウサギ、ラット、マウス、ウマ等が含まれ得る。特に関心が持たれるのは、(例えば、膵島転写因子をコードする核酸の対象へのインビボ導入により、または、膵島転写因子発現細胞(例えばβ細胞前駆細胞)の、もしくは膵島転写因子発現細胞から培養されインスリンを産生するほぼ発生し終えたまたは成熟したβ細胞の、対象への移植により)対象の細胞内でいずれかの膵島転写因子をコードする核酸を発現させることによる(例えば、疾患に関連する症状を緩和するための)治療影響を受けやすいインスリン関連疾患を有する対象である。
【0056】
「治療」、「治療すること」、「治療する」などの用語は、本明細書内で一般に、所望の薬理的および/または生理的効果を得ることを指すために用いられる。その効果は、疾病、疾患、もしくはその症状を完全にまたは部分的に防ぐという点で予防的であってよく、かつ/または、疾病や疾患および/もしくは疾病や疾患に起因する有害な影響を部分的にもしくは完全に安定化もしくは治癒させるという点で治療的であってよい。本明細書で用いられる「治療」とは、哺乳動物、特にヒトにおける疾病または疾患の任意の治療を包含し、かつ以下を含む:
(a)疾病、疾患、または症状を起こしやすい可能性があるが、まだそれを有すると診断されていない対象における疾病、疾患、または症状の発達を予防すること;(b)疾病、疾患、もしくは症状を阻害する、すなわちその発症を抑止すること、または、疾病、疾患、もしくは症状を軽減する、すなわち疾病、疾患、もしくは症状の退縮を引き起こすこと。したがって「糖尿病の治療」には、血糖値の低下、インスリン産生の増加等のうち1つまたは複数が含まれる。
【0057】
「インスリン関連疾患」とは、インスリン産生量の変化または対象が、例えば血糖値を調節するためにインスリンを利用する能力の変化によって直接的もしくは間接的に引き起こされる、またはこの変化に伴う、疾病、疾患、もしくは病態を意味する。インスリン関連疾患には、1型糖尿病、2型糖尿病、耐糖能異常、高血糖症、低血糖症等が含まれるが、これらに限定されるわけではない。特に関心が持たれるのは、(例えば、膵島転写因子をコードする核酸をインビボで対象に導入することにより、または、膵島転写因子を発現する細胞(例えばβ細胞前駆細胞)もしくは、膵島転写因子を発現する細胞から培養されかつインスリンを産生するほぼ発達し終えたまたは成熟したβ細胞を対象に移植することにより)対象の細胞内でいずれかの膵島転写因子をコードする核酸を発現させることによる(例えば、疾患に関連する症状を緩和するための)治療の影響を受けやすいインスリン関連疾患である。
【0058】
明瞭にする目的で、以下の表により、本明細書中で用いる転写因子の名称と文献中で用いられる同一因子の名称、および対応するヒト遺伝子の正式名称との相互参照を提供する。

*ヒト遺伝子命名法委員会(Human Gene Nomenclature Committee)により承認されている。
【0059】
発明の概略
本発明は、膵島転写因子をコードするポリヌクレオチド配列を膵細胞または他の適当な細胞に導入することにより、インスリン産生β細胞を含む、膵島細胞の表現型を有する細胞の産生が誘導されるという発見に基づく。この発見そのものは、成熟した膵細胞(非β細胞)内でニューロゲニン3(Ngn3)活性の増加を提供すると、非β膵細胞が、膵β細胞表現型(例えば、インスリンの産生)を有する細胞となるという発見に基づく。
【0060】
膵島転写因子活性の増加は、例えば、膵島転写因子をコードするポリヌクレオチドを細胞に導入して膵島転写因子の発現(これは、細胞における内在性膵島転写因子の発現に追加されうる)を提供することにより、(例えば、膵島転写因子の発現を正に制御する第二の転写因子をコードするポリヌクレオチドを導入することによって、またはさもなくばそのような膵島転写因子の正の制御因子の活性もしくは発現を増加させることによって)膵島転写因子の正の制御因子の発現レベルの増加を提供することにより、(例えば、その発現を阻害することによって)膵島転写因子の発現もしくは活性の負の制御因子または阻害剤の活性を阻害することにより、膵島転写因子によって正に制御される下流のエフェクターの発現を増加させることにより、および、本明細書を読むことによって当業者に容易に明らかになると考えられる他の方法により達成され得る。転写因子の発現または活性の調節(例えば、膵島転写因子活性の増加、または膵島転写因子発現の阻害剤の発現もしくは活性の減少)は、その経路のシグナル伝達分子(受容体、リガンド、細胞内エフェクター)ならびに合成および天然の小分子制御因子の使用によっても達成され得る。
【0061】
1つの態様において、Ngn3活性の増加は、例えば、Ngn3をコードするポリヌクレオチドを細胞に導入してNgn3の発現(これは、細胞における内因性Ngn3発現に追加されうる)を提供することにより、(例えば、Ngn3の発現を正に制御する転写因子(例えば、Mash1、HNF1、HNF3、HNF6等)をコードするポリヌクレオチドを導入することによって、またはさもなくばそのようなNgn3の正の制御因子の活性もしくは発現を増加させることによって)Ngn3の正の制御因子の発現レベルの増加を提供することにより、(例えば、その発現を阻害することによって)Ngn3の発現もしくは活性の負の制御因子または阻害剤の活性を阻害することにより、Ngn3によって正に制御される下流のエフェクター(例えばneuroD1)の発現を増加させることにより、および本明細書を読むことにより当業者に容易に明らかになるであろう他の方法により達成され得る。Ngn3の発現または活性の調節(例えば、Ngn3活性の増加、またはNgn3発現の阻害剤の発現もしくは活性の減少)は、その経路のシグナル伝達分子(受容体、リガンド、細胞内エフェクター)ならびに合成および天然の小分子制御因子の使用によっても達成され得る。
【0062】
別の態様において、NeuroD1活性の増加は、例えば、NeuroD1をコードするポリヌクレオチドを細胞に導入してNeuroD1の発現(これは細胞における内因性NeuroD1発現に追加されうる)を提供することにより、(例えば、NeuroD1の発現を正に制御する転写因子(例えば、Mash1、Ngn3、HNF1、HNF3、HNF6等)をコードするポリヌクレオチドを導入することによって、またはさもなくばそのようなNeuroD1の正の制御因子の活性もしくは発現を増加させることによって)NeuroD1の正の制御因子の発現レベルの増加を提供することにより、(例えば、発現を阻害することによって)NeuroD1の発現もしくは活性の負の制御因子または阻害剤の活性を阻害することにより、NeuroD1によって正に制御される下流のエフェクターの発現を増加させることにより、および本明細書を読むことにより当業者に容易に明らかになるであろう他の方法により達成され得る。NeuroD1の発現または活性の調節(例えば、NeuroD1活性の増加、またはNeuroD1発現の阻害剤の発現もしくは活性の減少)は、その経路のシグナル伝達分子(受容体、リガンド、細胞内エフェクター)ならびに合成および天然の小分子制御因子の使用によっても達成され得る。
【0063】
別の態様において、Mash1活性の増加は、例えば、Mash1をコードするポリヌクレオチドを細胞に導入してMash1の発現(これは、細胞における内因性Mash1発現に追加されうる)を提供することにより、Mash1の正の制御因子の発現レベルの増加を提供することにより(例えば、Mash1の発現を正に制御する転写因子をコードするポリヌクレオチドを導入することによって、またはさもなくばそのようなMash1の正の制御因子の活性もしくは発現を増加させることによって)、(例えば、発現を阻害することによって)Mash1の発現もしくは活性の負の制御因子または阻害剤の活性を阻害することにより、Mash1によって正に制御される下流のエフェクター(例えば、Ngn3、NeuroD1等)の発現を増加させることにより、および本明細書を読むことにより当業者に容易に明らかになるであろう他の方法により達成され得る。Mash1の発現または活性の調節(例えば、Mash1活性の増加、またはMash1発現の阻害剤の発現もしくは活性の減少)は、その経路のシグナル伝達分子(受容体、リガンド、細胞内エフェクター)ならびに合成および天然の小分子制御因子の使用によっても達成され得る。
【0064】
本発明は一般に、ニューロゲニン(ニューロゲニン1/NEUROG1/MATH4C/NeuroD3、ニューロゲニン2/NEUROG2/MATH4A、またはニューロゲニン3/NEUROG3/MATH4B)、neuroD因子(NeuroD1/BETA2/BHF1、NeuroD2/NDRF、MATH2/NEX1/DLX3、NeuroD4/Math3)、Mash因子(Mash1およびMash2)、およびatonal関連因子(MATH1/ATOH1)を含む神経原性の塩基性ヘリックス・ループ・ヘリックス因子(bHLH)から選択される少なくとも1つの膵島転写因子、ならびにそれらの組み合わせまたは他の遺伝子との組み合わせの発現増加を提供し、膵β細胞の誘導を提供することを含む。特に関心が持たれるのはNgn3である。
【0065】
図10は、神経内分泌bHLHタンパク質、ならびにmyoD(筋原性クラスB bHLHタンパク質)およびE47(クラスA(遍在性)bHLHタンパク質)を加えてのアラインメント系図の略図である。ヒトNgn3アミノ酸配列である「h.ngn3」以外はすべて、マウスのアミノ酸配列に基づく。配列は、MacVector6.5.1配列解析プログラム(Oxford Molecular)で提供される多重配列整列アルゴリズムClustal-Wを用いて整列化させた。クラスAおよびBの定義は、Murreら(Murreら、(1989) Cell 58(3)、537-44)の分類に基づく。
【0066】
膵島転写因子は、膵島細胞の分化および発達に関与する。膵島転写因子には、クラスB塩基性ヘリックス・ループ・ヘリックス(bHLH)ファミリーの転写因子に属する転写因子が含まれ、このファミリーの因子は多くの細胞種の成長および分化を制御することが知られている。膵島細胞および発達過程の膵臓は多様なクラスB bHLH遺伝子群を発現するが、中でも最も多く発現されるのはNgn3、NeuroD1/BETA2、Mash1、およびNeuroD4/Math3である。NeuroD1は、膵島細胞の初期分化および膵β細胞におけるインスリン転写の制御に関与することが示されている。ニューロゲニン3は膵臓の発達中にNeuroD1/BETA2を活性化するため、ニューロゲニン3は膵島細胞分化の過程で活性化される膵島転写因子の階層においてneuroD1の上流に位置する。Mash1は膵臓の発達中にNgn3を活性化するため、Mash1は膵島細胞分化の過程で活性化される膵島転写因子の階層においてNgn3の上流に位置する。
【0067】
Ngn3は膵島細胞前駆細胞で発現されて前内分泌遺伝子として機能し、膵島細胞の分化を駆動させる。Ngn3は膵島細胞種4つすべての発達過程において初期に発現され、Pax4、Pax6、Isl1、およびNkx2.2、ならびにNeuroD1/BETA1等の他の膵島転写因子の制御に関与する。Ngn3の初期発現および異所的発現は、膵島細胞の初期分化および異所的分化をもたらしうる。他の膵島転写因子には、例えばPax4、Pax6、Isl1、Nkx2.2、およびNkx6.1といったホメオドメイン因子等の非bHLH因子も含まれる。これらの因子はNgn3のすぐ下流(例えばPax4、Pax6、Isl1、およびNkx2.2)または上流に位置し、膵島細胞の発達に関与する。pou-ホメオドメイン因子HNF1およびwinged-ヘリックス因子HNF3はNgn3の上流に位置し、cut-ホメオドメイン因子HNF6とともに膵島細胞の分化に関与すると見なされており、本発明に基づく膵島転写因子のさらなる例である。
【0068】
β細胞の発達の誘導
膵β細胞は、インビボ(例えば膵管の管腔に核酸を導入することによって例えば膵島転写因子をコードする核酸(例えばRNAまたはDNA)を対象の膵臓に投与することにより)またはインビトロ(例えば、培養中に膵島転写因子をコードする核酸(例えばRNAまたはDNA)を標的細胞(例えば単離した非β膵細胞)に接触させる(次にその細胞を培養して増加させ、対象に移植する)ことにより)のどちらかにおいて、膵細胞における膵島転写因子の産生を提供することにより非β膵細胞から産生され得る。
【0069】
標的細胞における膵島転写因子の発現は、様々な方法により達成され得る。例えば1つの態様において、膵島転写因子の発現は、膵島転写因子をコードする核酸(例えばDNAまたはRNA)を導入し、標的細胞においてコードされる膵島転写因子ポリペプチドの発現を提供することにより達成される。別の態様において、膵島転写因子の発現は、膵島転写因子の発現の誘導を提供するタンパク質をコードする遺伝子の導入(例えば、標的細胞における膵島転写因子発現の「上流の」正の制御因子の発現)により誘導される。別の態様において、膵島転写因子の発現は、膵島転写因子発現の負の制御因子の活性(例えば、機能または発現)を阻害するタンパク質をコードする遺伝子の導入により達成される。別の態様において、膵島転写因子の発現は、膵島転写因子の発現の誘導を提供する小分子(例えば、標的細胞において膵島転写因子の発現を誘導する小分子薬剤)の導入により誘導される。さらに、本発明の膵β細胞の産生は、膵島転写因子により誘導される因子の産生を提供することによっても達成され得る。
【0070】
特に関心の持たれる1つの態様において、β細胞は、標的細胞においてβ細胞の表現型を誘導するのに十分なレベルのニューロゲニン3(Ngn3)の発現を提供することにより産生される。1つの態様において、Ngn3の発現は、Ngn3をコードする核酸(例えばDNAまたはRNA)を導入して、標的細胞においてコードされるNgn3ポリペプチドの発現を提供することにより達成される。別の態様において、Ngn3の発現は、Ngn3発現の誘導を提供するタンパク質をコードする遺伝子の導入(例えば、標的細胞におけるNgn3発現の「上流の」正の制御因子(例えば、Mash1、HNF1、HNF3、HNF6等)の発現)により誘導される。別の態様において、Ngn3の発現は、Ngn3発現の負の制御因子の活性(例えば、機能または発現)を阻害するタンパク質をコードする遺伝子の導入により達成される。別の態様において、Ngn3の発現は、Ngn3の発現の誘導を提供する小分子(例えば、標的細胞においてNgn3の発現を誘導する小分子薬剤)の導入により誘導される。さらに、本発明の膵β細胞の産生は、Ngn3により誘導される因子の産生を提供することによっても達成され得る。
【0071】
別の態様において、β細胞は、標的細胞においてβ細胞の表現型を誘導するのに十分なレベルのNeuroD1の発現を提供することにより産生される。1つの態様において、NeuroD1の発現は、NeuroD1をコードする核酸(例えばDNAまたはRNA)を導入し、標的細胞においてコードされるNeuroD1ポリペプチドの発現を提供することにより達成される。別の態様において、NeuroD1の発現は、NeuroD1の発現の誘導を提供するタンパク質をコードする遺伝子の導入(例えば、標的細胞におけるNeuroD1発現の「上流の」正の制御因子(例えば、Mash1、Ngn3、HNF1、HNF3、HNF6等)の発現)により誘導される。別の態様において、NeuroD1の発現は、NeuroD1発現の負の制御因子の活性(例えば、機能また発現)を阻害するタンパク質をコードする遺伝子の導入により達成される。別の態様において、NeuroD1の発現は、NeuroD1の発現誘導を提供する小分子(例えば、標的細胞においてNeuroD1の発現を誘導する小分子薬剤)の導入により誘導される。さらに、本発明の膵β細胞の産生は、NeuroD1により誘導される因子の産生を提供することによっても達成され得る。
【0072】
別の態様において、β細胞は、標的細胞においてβ細胞の表現型を誘導するのに十分なレベルのMash1の発現を提供することにより産生される。1つの態様において、Mash1の発現は、Mash1をコードする核酸(例えばDNAまたはRNA)を導入し、標的細胞においてコードされるMash1ポリペプチドの発現を提供することにより達成される。別の態様において、Mash1の発現は、Mash1の発現の誘導を提供するタンパク質をコードする遺伝子の導入(例えば、標的細胞におけるMash1発現の「上流の」正の制御因子の発現)により誘導される。別の態様において、Mash1の発現は、Mash1発現の負の制御因子の活性(例えば、機能または発現)を阻害するタンパク質をコードする遺伝子の導入により達成される。別の態様において、Mash1の発現は、Mash1の発現の誘導を提供する小分子(例えば、標的細胞においてNgn3の発現を誘導する小分子薬剤)の導入により誘導される。さらに、本発明の膵β細胞の産生は、Mash1により誘導される因子の産生を提供することによっても達成され得る。
【0073】
本開示を読むことによって当業者に容易に理解されると考えられるように、膵島転写因子の発現は、これらのアプローチの任意の組み合わせを提供することにより達成され得る。例えば、本発明は、膵島転写因子をコードする核酸および内因性膵島転写因子遺伝子の正の制御因子の両方を標的細胞において発現すること、導入した膵島転写因子核酸ならびに内因性膵島転写遺伝子および導入した膵島転写因子配列の負の制御因子の阻害剤を発現すること等も提供する。一般に、例えば(膵島転写因子をコードする核酸の導入によって、または内因性膵島転写因子の発現の提供によって)膵島転写因子自体の発現を提供することにより、および/または、通常は膵島転写因子が発現した結果として産生される膵島転写因子の「下流の」因子の産生を提供することにより、膵島転写因子の活性を提供するようなアプローチの任意の組み合わせは本発明の範囲内である。
【0074】
1つの態様において、本発明は、Ngn3をコードする核酸および内因性Ngn3遺伝子の正の制御因子(例えば、Mash1、HNF1、HNF3、HNF6等)の両方を標的細胞において発現すること、導入したNgn3核酸ならびに内因性Ngn3および導入したNgn3配列の負の制御因子の阻害剤を発現すること等を提供する。一般に、(Ngn3をコードする核酸の導入によって、または内因性Ngn3の発現を提供することによって)例えばNgn3自体の発現を提供することにより、および/または通常はNgn3が発現した結果として産生されるNgn3の「下流の」因子(例えばNeuroD1等)の産生を提供することにより、Ngn3の活性を提供するアプローチの任意の組み合わせは、全般にわたって本発明の範囲内である。Ngn3発現の正の制御因子にはMash1、HNF1、HNF3、およびHNF6が含まれるが、必ずしもこれらに限定されるわけではない。
【0075】
別の態様において、本発明は、NeuroD1をコードする核酸および内因性NeuroD1遺伝子の正の制御因子(例えば、Mash1、Ngn3、HNF1、HNF3、HNF6等)の両方を標的細胞において発現すること、導入したNeuroD1核酸ならびに内因性NeuroD1および導入したNeuroD1配列の負の制御因子の阻害剤を発現すること等を提供する。一般に、(NeuroD1をコードする核酸の導入によって、または内因性NeuroD1の発現を提供することによって)例えばNeuroD1自体の発現を提供することにより、および/または通常はNgn3が発現した結果として産生されるNeuroD1の「下流の」因子の産生を提供することにより、Ngn3の活性を提供するアプローチの任意の組み合わせは、全般にわたって本発明の範囲内である。NeuroD1の正の制御因子にはMash1、Ngn3、HNF1、HNF3、およびHNF6が含まれるが、必ずしもこれらに限定されるわけではない。
【0076】
別の態様において、本発明は、Mash1をコードする核酸および内因性Mash1遺伝子の正の制御因子の両方を標的細胞において発現すること、導入したMash1核酸ならびに内因性Mash1および導入したMash1配列の負の制御因子の阻害剤を発現すること等を提供する。一般に(Mash1をコードする核酸の導入によって、または内因性Mash1の発現を提供することによって)例えばMash1自体の発現を提供することにより、および/または通常はMash1が発現した結果として産生されるMash1の「下流の」因子(例えば、Ngn3、NeuroD1等)の産生を提供することにより、Mash1の活性を提供するアプローチの任意の組み合わせは、全般にわたって本発明の範囲内である。
【0077】
さらに、膵島転写因子経路の活性の誘導は、核酸以外の天然分子または合成分子を用いて達成され得る。例えば、膵島転写因子活性は、例えば膵島転写因子発現の負の制御因子の活性を阻害することによって、その経路に関与する膵島転写因子の発現を促進する合成分子を用いることにより、誘導され得る。
【0078】
1つの態様において、Ngn3制御経路の活性の誘導は、核酸以外の天然または合成分子を用いて達成される。例えば、Ngn3活性は、例えばNgn3発現の負の制御因子の活性を阻害することによって、Ngn3発現を促進する合成分子を用いることにより誘導される。Ngn3発現の抑制的な転写因子にはHES1が含まれるが、必ずしもこれに限定されない。Ngn3発現を阻害する負のシグナル伝達経路にはNotch経路が含まれるが、必ずしもこれに限定されるわけではない。
【0079】
別の態様において、NeuroD1制御経路の活性の誘導は、核酸以外の天然または合成分子を用いて達成される。例えばNeuroD1活性は、例えばNeuroD1発現の負の制御因子の活性を阻害することによって、NeuroD1発現を促進する合成分子を用いることにより誘導される。
【0080】
別の態様において、Mash1制御経路の活性の誘導は、核酸以外の天然または合成分子を用いて達成される。例えばMash1活性は、例えばMash1発現の負の制御因子の活性を阻害することによって、Mash1発現を促進する合成分子を用いることにより誘導される。
【0081】
膵島転写因子核酸
「膵島転写因子遺伝子」という用語は、膵島細胞で発現される転写因子と膵島細胞の発生、分化、または形成に関与する転写因子の両方を表すために用いる。「膵島転写因子遺伝子」という用語はまた、コード領域の範囲を超えて最大約2.5 kb、約5 kb、10 kb、または約15 kbに至る、特定の膵島転写因子ポリペプチドをコードするオープンリーディングフレーム、イントロン、ならびに発現の制御に関与する5'側および3'側の隣接非コードヌクレオチド配列を意味することを意図するが、どちらかの方向にさらに伸長する可能性もある。膵島転写因子をコードするDNA配列は、cDNAまたはゲノムDNAもしくはそれらの断片であってよい。染色体外での維持または宿主への組込みのため、遺伝子は適当なベクターに導入され得る。
【0082】
本明細書で用いる「cDNA」という用語は、天然の成熟RNA種に見られる配列成分の配置を共有する核酸すべてを含むことを意図するが、ここで配列成分とはエキソン(例えば、コードされたポリペプチドのオープンリーディングフレームをコードする配列)ならびに3'側および5'側の非コード領域のことである。通常、mRNA種は、核RNAスプライシングによって介在イントロンが除去された、関心対象のポリペプチドをコードする連続的なオープンリーディングフレームを作製するための連続的エキソンを有する。
【0083】
関心対象の膵島転写因子ゲノム配列は、列挙された配列に定義されるように開始コドンと終止コドンの間の核酸を含み、天然の染色体に通常存在するイントロンのすべてを含む。さらに、成熟mRNAに見られる3'側および5'側の非翻訳領域を含んでもよい。さらに、プロモーター、エンハンサー等のような特定の転写制御配列および翻訳制御配列を含んでもよく、転写領域の5'末端または 3'末端のどちらかに約2.5 kb、約5 kb、約10 kb、または約15 kb、場合によってはさらに長い、隣接ゲノムDNAを含む。ゲノムDNAは、100 kbpもしくはそれ以上の大きな断片、または隣接する染色体配列を実質的に含まない小さな断片として単離され得る。
【0084】
この5'領域の配列、ならびにさらに5'上流の配列および3'下流の配列は、エンハンサー結合部位を含む、膵島転写因子が発現される組織中で発現を提供する
プロモーター成分に利用される可能性がある。本発明の膵島転写因子プロモーター成分の配列は、任意の種(例えば、哺乳動物または非哺乳動物(例えば、は虫類、両生類、鳥類(例えばニワトリ))、特に、ヒトを含む哺乳動物、げっ歯類(例えば、マウスまたはラット)、ウシ、ヒツジ、ブタ、マウス、またはウマ、好ましくは、マウスまたはヒト)のヌクレオチド配列に基づきうり、天然、合成、半合成、もしくは組換えにかかわらず任意の供給源から単離または産生され得る。
【0085】
本発明で用いる核酸組成物は、必要に応じて膵島転写因子ポリペプチドのすべてまたは一部をコードし得るが、通常は少なくとも実質的にそのすべてをコードし得る。DNA配列の断片を、従来法による化学的合成オリゴヌクレオチド、制限酵素消化、PCR増幅等により得ることができる。ほとんどの場合、DNA断片は少なくとも約10個の連続したヌクレオチドからなると考えられ、通常少なくとも約15ヌクレオチド、より通常は少なくとも約18ヌクレオチド〜約20ヌクレオチド、より通常は少なくとも約25ヌクレオチド〜約50ヌクレオチドからなる。そのような小さなDNA断片は、PCRのプライマー、ハイブリダイゼーションスクリーニング等に有用である。より大きなDNA断片、すなわち100ヌクレオチドを上回る断片は、コードされたポリペプチドの産生に有用である。PCR等の増幅反応に利用するために、一対のプライマーが用いられる。プライマー配列の正確な組成は本発明にとって重要ではないが、当技術分野で周知のように、ほとんどの用途においてプライマーはストリンジェントな条件下で対象の配列にハイブリダイズする。少なくとも約50ヌクレオチド、好ましくは約100ヌクレオチドの増幅産物を産生する一対のプライマーを選択することが好ましい。プライマー配列を選択するためのアルゴリズムが周知であり、市販のソフトウェアパッケージで入手できる。増幅プライマーはDNAの相補鎖にハイブリダイズし、互いに向かってプライミングすると考えられる。
【0086】
膵島転写因子遺伝子は、一般的には元の哺乳動物染色体とは別のものとして単離され、実質的に純粋なものとして得られる。通常、DNAは膵島転写因子またはその断片をコードする配列を含まない他の核酸配列を実質的に含まずに取得され、一般には少なくとも約50%、通常は少なくとも約90%の純度であり、これは、典型的な「組換え型」である(すなわち天然の染色体には通常は付随しない1つまたは複数のヌクレオチドが隣接している)。
【0087】
隣接するプロモーター領域およびコード領域を含む膵島転写因子の配列は、当技術分野で周知の様々な方法により変異を導入されうり、プロモーター強度、コードされるタンパク質の配列等においてターゲティングされた変化を生じる。そのような変異を有するDNA配列または産物は本明細書で提供される配列と実質的に類似しており、すなわちそれぞれ少なくとも1つのヌクレオチドまたはアミノ酸が異なっており、かつ少なくとも2つもしくは少なくとも約10個のヌクレオチドまたはアミノ酸が異なっていてもよい。一般に、配列変化とは、置換、挿入、または欠失であってよい。欠失には、ドメインまたはエキソンの欠失のような大きな変化がさらに含まれうる。膵島転写因子配列は主にbHLHドメイン内で保存されているが、このドメインの外側の領域ではそれほど保存されていない可能性があり、かつさらに例えばラット、マウス、およびヒト間ではほとんど保存されていない可能性さえあることに留意されねばならない。したがって、膵島転写因子は、bHLHドメインの外側のヌクレオチドおよびアミノ酸残基のさらなる変化を許容し、かつbHLHドメイン内で起こる変化よりもずっと大きな程度の変化まで機能を保持できる。そのように修飾された膵島転写因子の配列を用いて、例えば、膵島細胞を産生する目的で標的細胞に導入するベクターを作製することができる。
【0088】
クローニングされた遺伝子のインビトロ突然変異誘発を行うための技法は周知である。変異を検出するためのプロトコールの例は、Gustinら、1993 Biotechniques 14:22;Barany、1985 Gene 37:111-23;Colicelliら、1985 Mol Gen Genet 199:537-9;およびPrentkiら、1984 Gene 29:303-13に見出されうる。部位特異的突然変異誘発法は、Sambrookら、1989 Molecular Cloning: A Laboratory Manual、CSH Press、pp. 15.3-15.108;Weinerら、1993 Gene 126:35-41;Sayersら、1992 Biotechniques 13:592-6;JonesおよびWinistorfer、1992 Biotechniques 12:528-30;Bartonら、1990 Nucleic Acids Res 18:7349-55;MarottiおよびTomich、1989 Gene Anal Tech 6:67-70;およびZhu 1989 Anal Biochem 177:120-4に見出され得る。
【0089】
本発明において特に関心の持たれる膵島転写因子はニューロゲニン転写因子ファミリーに属する、例えばニューロゲニン1(Ngn1)、ニューロゲニン2(Ngn2)、ニューロゲニン3(Ngn3)などの転写因子であるが、Ngn3が特に関心対象となる。ヒトNgn3のヌクレオチドおよびアミノ酸の配列は、配列表においてそれぞれ配列番号:1および2として提供される。
【0090】
ヒトNgn1のヌクレオチドおよびアミノ酸の配列は、GenBankアクセッション番号XM_003834(配列表においてそれぞれ配列番号:20および21として提供する)およびNM_006161(配列表においてそれぞれ配列番号:22および23として提供する)で入手可能である。ヒトNgn1のクローニングおよび発現は、例えば、McCormickら、(1996) Mol. Cell. Biol. 16(10)、5792-5800;およびTamimiら、(1997) Genomics 40(2)、355-357 (1997)に記載されている。
【0091】
ヒトNgn2のヌクレオチドおよびアミノ酸の配列は、GenBankアクセッション番号AF303002(配列表においてそれぞれ配列番号:24および25として提供する)およびXM_067897(配列表においてそれぞれ配列番号:26および27として提供する)で入手可能である。ヒトNgn1のクローニングおよび発現は、例えば、Simmonsら、(2001) Dev. Biol. 229(2)、327-339に記載されている。
【0092】
Ngn3の「上流」(したがってNgn3発現を活性化する)またはNgn3の「下流」のどちらかに作用し膵島細胞の表現型の発現を引き起こす転写因子についても、本発明での使用が意図されていることに留意されたい。
【0093】
ニューロゲニン3は、それ自体で十分、未分化の膵上皮細胞を膵島細胞にするように作用する。ニューロゲニン3の発現によりどの前駆細胞が膵島細胞に分化するかが決定されるため、ニューロゲニン3の発現を制御するシグナルもまた膵島細胞の形成に関与する。2.7 kbのngn3プロモーターはトランスジェニックマウスにおいて正しく発現を指示するのに十分であるが、遠位の配列がngn3の発現を大幅に増強することが示されている。この遠位プロモーター領域は、HNF6、HNF1α、およびHNF3βのような膵転写因子の結合部位のクラスターを含む。これらの膵転写因子がngn3遺伝子の発現を制御することが見出されており、したがってこれらも膵島細胞形成の制御に関与する。これらのシグナルは、糖尿病患者に新しい膵島細胞を産生するのに有効となり得る。
【0094】
別の態様において、膵島転写因子はneuroD転写因子ファミリーに属する転写因子であり、例えばNeuroD1、NeuroD2、NeuroD4であるが、NeuroD1が特に関心対象となる。ヒトNeuroD1遺伝子の配列および対応するヒトNeuroD1アミノ酸配列は、GenBankアクセッション番号NM_002500(配列表においてそれぞれ配列番号:28および29として提供する)ならびにXM_002573(配列表においてそれぞれ配列番号:30および31として提供する)およびAF045152(配列表においてそれぞれ配列番号:32および33として提供する)で入手可能である。ヒトNeuroD1のクローニングおよび発現は、例えば、Leeら、(1995) Science 268(5212)、836-844;Tamimiら、(1996) Genomics 34(3)、418-421;Yokoyamaら、(1996) DNA Res. 3(5)、311-320;およびYokoyamaら、(1196) Brain Res. Mol. Brain Res. 42(1)、135-139に記載されている。
【0095】
ヒトNeuroD4遺伝子および対応するヒトNeuroD4アミノ酸配列は、GenBankアクセッション番号AF203901(配列表においてそれぞれ配列番号:34および35として提供する)で入手可能である。
【0096】
NeuroD1の「上流」(したがってNeuroD1の発現を活性化する)またはNeuroD1の「下流」のどちらかに作用して膵島細胞の表現型の発現を引き起こす転写因子についても、本発明での使用が意図されることに留意されたい。
【0097】
別の態様において、膵島転写因子はMash転写因子ファミリーに属する転写因子、例えばMash1、Mash2であるが、Mash1が特に関心対象となる。ヒトMash1遺伝子の配列および対応するMash1アミノ酸配列は、GenBankアクセッション番号XM_006688(配列表においてそれぞれ配列番号:36および37として提供する)およびNM_004316(配列表においてそれぞれ配列番号:38および39として提供する)で入手可能である。ヒトMash1のクローニングおよび発現は、例えば、Ballら、(1993) Proc. Natl. Acad. Sci. U.S.A. 90(12)、5648-5652;Renaultら、(1995) Genomics 30(1)、81-83;Borgesら、(1997) Nature 386(6627)、852-855、およびChenら、(1997) Cell Growth Differ. 8(6)、677-686に記載されている。
【0098】
Mash1の「上流」(したがってMash1の発現を活性化する)またはMash1の「下流」のどちらかに作用して膵島細胞の表現型の発現を引き起こす転写因子についても、本発明での使用が意図されていることに留意されたい。
【0099】
島転写因子核酸を送達するための構築物
送達される膵島転写因子核酸がDNAである場合、関心対象のDNAに機能的に結合されたプロモーター(例えば、真核細胞内で機能するプロモーター)を有する任意の構築物が本発明において使用可能である。本発明に従って使用され得るDNA配列(または対応するRNA配列)を含む構築物は、関心対象のDNAまたRNAの配列を含む任意の真核発現構築物でよい。例えば、プラスミドまたはウイルス性構築物(例えばアデノウイルス)を切断して、連結可能な末端を有する直鎖状DNAを提供し得る。これらの末端を相補様の連結可能な末端を有する外因性DNAに結合させ、完全な複製単位および所望の表現型特性を有する生物学的に機能的な組換えDNA分子を提供する。構築物は真核生物および/または原核生物の宿主において複製可能であることが好ましく(真核生物においてはウイルス、原核生物においてはプラスミド)、そのような構築物は当技術分野で周知であり、市販されている。
【0100】
構築物を、当技術分野で周知の技法を用いて調製することができる。同様に、遺伝的に改変された宿主において外因性のDNA配列またはRNA配列を発現させるための技法は、当技術分野で周知である。(例えば、Kormalら、Proc. Natl. Acad. Sci. USA、84:2150-2154、1987;Sambrookら、Molecular Cloning: a Laboratory Manual、第2版、1989、Cold Spring Harbor Laboratory Press、Cold Spring Harbor、NYを参照のこと。いずれも関心対象のDNAを真核生物で発現させるための方法および組成物に関する参照として、本明細書に組み入れられる。)
【0101】
1つの態様において、DNA構築物はプロモーターを含み、膵細胞内で関心対象のDNAの発現を促進する。プロモーターは、サイトメガロウイルス(CMV)、マウス乳がんウイルス(MMTV)、ラウス肉種ウイルス(RSV)、またはアデノウイルスのプロモーターのような、真核細胞内で機能する強力なウイルスプロモーターであってよい。より具体的には、例示的なプロモーターには、ヒトCMVの前初期遺伝子由来のプロモーター(Boshartら、Cell 41:521-530、1985)およびRSVの長い末端反復配列(LTR)由来のプロモーター(Gormanら、Proc. Natl. Acad. Sci. USA 79:6777-6781、1982)が含まれる。これら2つのプロモーターでは、RSVプロモーターよりもCMVプロモーターを用いた方が発現レベルが高くなるため、ここではCMVプロモーターが好ましい。
【0102】
または、用いるプロモーターは、アクチン遺伝子プロモーターのような強力かつ一般的な真核プロモーターでもよい。1つの態様において、用いるプロモーターは組織特異的プロモーターでもよい。例えば、構築物に用いるプロモーターは、膵臓特異的プロモーター、管細胞特異的プロモーター、または幹細胞特異的プロモーターでよい。本発明の構築物は、プロモーターに加えて標的細胞内で発現を増強する配列も含みうる。
【0103】
別の態様において、プロモーターはテトラサイクリン調節性プロモーターのような調節性プロモーターであり、その発現は外因性の物質(例えばテトラサイクリン)に曝すことにより調節され得る。
【0104】
マーカー等の他の構成成分(例えば、抗生物質耐性遺伝子(アンピシリン耐性遺伝子等)またはβ-ガラクトシダーゼ)は、構築物、細菌細胞における構築物の安定な複製のための複製起点(好ましくは、高コピー数の複製起点)、核局在化シグナル、または、DNA構築物、それにコードされるタンパク質、もしくはその両方の産生を促進するその他の成分を含むおよび/または発現する細胞の選択または同定に役立つ。
【0105】
真核生物において発現させるためには、構築物は関心対象のDNAに機能的に結合された真核生物プロモーターを最低限含むべきであり、関心対象のDNAは次にポリアデニル化シグナル配列に結合させる。ポリアデニル化シグナル配列は、当技術分野で周知の様々なポリアデニル化シグナル配列のいずれかより選択されうる。例示的なポリアデニル化シグナル配列は、SV40初期ポリアデニル化シグナル配列である。適切な場合、特に関心対象のDNAがcDNAである(例えば、天然配列のイントロンを含まない)場合には、構築物は関心対象のDNAの発現レベルを上昇させ得る1つまたは複数のイントロンを含んでもよい。当技術分野で周知の様々なイントロンのうちいずれかのイントロンを使用できる(例えば、ヒトβ-グロビンイントロン、これは関心対象のDNAの5'の位置で構築物に挿入する)。
【0106】
別の態様において、細胞に送達される核酸は、膵島転写因子をコードするRNAである。この態様においては、RNAは標的細胞における発現(すなわちRNAの翻訳)について適合化される。関心対象のタンパク質をコードするRNA(例えばmRNA)の産生方法は当技術分野で周知であり、本発明に有用な膵島転写因子をコードするRNAの産生に容易に応用されうる。
【0107】
膵島転写因子をコードする核酸の送達
膵島転写因子をコードする核酸の送達は、ウイルスベクターまたは非ウイルスベクターを用いて達成され得る。1つの態様において、核酸はアデノウイルスのようなウイルス粒子に入れて送達される。別の態様において、ウイルス粒子(例えばアデノウイルス)またはカチオン性脂質またはリポソームのようなアジュバントと混合された裸のDNAを含む製剤として、核酸を送達する。「アジュバント」とは、それ自体で所望の効果を生じないが、活性化合物の作用を増大させるまたは改善する役割を果たす物質である。使用するベクターおよびベクター製剤の詳細は、導入するDNAの大きさ、利用する送達プロトコール等のようないくつかの因子に依存すると考えられる。例示的な非ウイルスベクターおよびウイルスベクターについて、以下でより詳細に説明する。
【0108】
I. ウイルスベクター
一般に、本発明に従って使用されるウイルスベクターは、複製不能となりかつ本発明による標的細胞での発現用の関心対象の組換え遺伝子を送達するように遺伝的に改変された、天然ウイルス由来のウイルス粒子から構成される。
【0109】
多くのウイルスベクターが当技術分野で周知であり、これには例えば、レトロウイルス、アデノウイルス、アデノ随伴ウイルス、単純ヘルペスウイルス(HSV)、サイトメガロウイルス(CMV)、ワクシニアウイルス、およびポリオウイルスのベクターが含まれる。アデノウイルスおよびAAVは、ゆっくりと複製する細胞および/または最終的に分化した細胞に効率的に感染するため、通常好ましいウイルスベクターである。ウイルスベクターは、標的細胞への選択的感染によって選択されうる。
【0110】
ウイルスベクターとして複製不能ウイルスを使用する場合には、関心対象のDNAに対応するDNAまたはRNAのいずれかを含む感染性ウイルス粒子は、ウイルス複製に必須ながら欠除している成分を提供する組換え細胞株にウイルス構築物を導入することによって産生されうる。1つの態様において、組換えウイルスベクターを用いた組換え細胞株の形質転換は、例えば、組換え細胞株のウイルス配列の、導入されたウイルスベクターへの相同的組換えにより、複製可能なウイルスの産生または実質的な産生をもたらさない。関心対象の核酸を含む複製不能ウイルス粒子の産生方法は当技術分野で周知であり、例えば、Rosenfeldら、Science 252:431-434、1991およびRosenfeld ら、Cell 68:143-155、1992(アデノウイルス)、米国特許第5,139,941号(アデノ随伴ウイルス)、米国特許第4,861,719号(レトロウイルス)、ならびに米国特許第5,356,806号(ワクシニアウイルス)に記載されている。流行性耳下腺炎ウイルスゲノムを操作するための方法と材料、ウイルス融合とウイルス複製に関する流行性耳下腺炎ウイルス遺伝子の特徴づけ、および流行性耳下腺炎ウイルスゲノムの構造と配列は、Tanabayashiら、J. Virol. 67:2928-2931、1993;Takeuchiら、Archiv. Virol.、128:177-183、1993;Tanabayashiら、Virol. 187:801-804、1992;Kawanoら、Virol.、179:857-861、1990;Elangoら、J. Gen. Virol. 69:2893-2900、1988に記載されている。
【0111】
II. 非ウイルスベクター
関心対象の核酸は、非ウイルスベクターを用いて細胞に導入してもよい。本明細書で使用する「非ウイルスベクター」には、裸のDNA(例えば、ウイルス粒子内に含まれず、脂質のような担体分子を含まないDNA)、裸の核酸を含む化学製剤(例えば、DNA(および/またはRNA)とカチオン性化合物(例えば、硫酸デキストラン、カチオン性脂質)との製剤)、およびウイルス粒子のようなアジュバントと混合された裸の核酸(例えば、関心対象のDNAはウイルス粒子内に含まれないが、製剤は裸のDNAとウイルス粒子(例えば、アデノウイルス粒子)との両方からなる)(例えば、Curielら、1992 Am. J. Respir. Cell Mol Biol. 6:247-52を参照のこと)が含まれることを意図する。したがってウイルス粒子が関心対象のDNAをウイルス粒子内に含まない場合にも「非ウイルスベクター」には核酸とウイルス粒子からなるベクターが含まれ得る。
【0112】
1つの態様において、製剤は、投与前に裸のDNA構築物と混合されたウイルス粒子を含む。全量約100μl中で、約108個〜約1010個のウイルス粒子(好ましくは約1×1010個〜約5×1010個、より好ましくは約3×1010個の粒子)を裸のDNA構築物(約5μg 〜50μg DNA、より好ましくは約8μg 〜25μg DNA)と混合する。好ましくは、ウイルス粒子はアデノウイルス粒子である(Curielら、1992、前記)。
【0113】
代替的にまたはさらに、核酸をポリ-L-リジンもしくはDEAC-デキストランのようなポリカチオン性物質、標的リガンド、および/またはDNA結合タンパク質(例えばヒストン)と複合体化することもできる。DNAまたはRNAとリポソームとの複合体製剤は、遺伝物質(DNAまたはRNA)に結合する脂質の混合物を含み、細胞への核酸の送達を促進する。本発明に従って使用できるリポソームには、DOPE(ジオレイルホスファチジルエタノールアミン)、CUDMEDA(N-(5-コレストラム-3-β-オール3-ウレタニル)-N',N'-ジメチルエチレンジアミン)が含まれる。
【0114】
例えば裸のDNAを、約2.5%〜15%(容量:容量)、好ましくは6%〜12%(容量:容量)の範囲にある濃度のリポフェクチン(商標)(LTI/BRL)を含む溶液として投与できる。本発明の方法に基づく送達用のDNA製剤についての好ましい方法および組成は米国特許第5,527,928号に記載されており、その開示は参照として本明細書に組み入れられる。
【0115】
また関心対象の核酸は、宿主細胞の生物学的特性を変化させる目的で、宿主細胞への送達を促進する担体分子(例えば、抗体または受容体リガンド)に結合させたDNAまたはRNAの化学製剤としても投与できる。「化学製剤」という用語は、タンパク質もしくは脂質またはその誘導体のような担体分子に核酸化合物の結合を可能にする、核酸の修飾を意味する。例示的なタンパク質担体分子には、標的膵細胞の細胞に特異的な抗体または受容体リガンド、例えば、標的膵細胞の細胞に結合した受容体と相互作用できる分子が含まれる。
【0116】
膵島転写因子ポリペプチドおよびそのようなポリペプチドに特異的に結合する抗体の産生
Ngn3、NeuroD1、Mash1、または他の膵島転写因子をコードする核酸を用いて、全長ポリペプチドまたはその断片、特に機能的なドメインに対応する断片、DNA結合部位等を合成することができ、他のタンパク質またはその一部への被験ポリペプチドの融合体もこれに含まれる。したがって、本発明での使用に適したポリヌクレオチドおよびポリペプチドには、霊長類、げっ歯類、イヌ、ネコ、ウマ、線虫、酵母などにおいて見出される膵島転写因子ポリペプチドおよびポリヌクレオチド、ならびにそれらの天然変種および非天然変種が含まれるが、これらに限定されるわけではない。
【0117】
膵島転写因子ポリペプチドを用いて抗体を産生することができるが、ここで、短い断片は特定のポリペプチドに特異的な抗体を提供し、より大きな断片またはタンパク質全体ではポリペプチド表面を覆う抗体の産生が可能になる。抗体は、ポリペプチドの野生型または変異型に対して産生され得る。例えば、関心対象のポリペプチドを発現する細胞による免疫、関心対象のポリペプチドが膜に挿入されているリポソームによる免疫等によって、これらのドメインに対応する単離ペプチドまたは天然タンパク質に対して抗体が産生され得る。
【0118】
抗体は従来法に基づいて調製されるが、ここで、発現されたポリペプチドまたはタンパク質は、それ自体でまたは周知の免疫原性担体(例えば、KLH、pre-S HBsAg、他のウイルスまたは真核生物タンパク質等)に結合して、免疫原として使用される。さらなる説明は、Monoclonal Antibodies: A Laboratory Manual、HarlowおよびLane編、Cold Spring Harbor Laboratories、Cold Spring Harbor、New York、1988を参照のこと。
【0119】
膵島転写因子に特異的に結合する抗体は、Ngn3、NeuroD1、またはMash1のような組換え膵島転写因子を発現する細胞の検出に用いることができる(例えば、以下でより詳細に説明するように、移植または体内移植の前など)。
【0120】
転写因子をコードする核酸の発現による膵島細胞の産生
膵島細胞は、本発明に基づいて様々な方法で産生することができる。一般に本発明は、膵島転写因子の産生を刺激することを含む。特に関心の持たれる態様において、本発明は、膵島転写因子をコードする核酸を細胞、通常は膵細胞に導入することによる、膵島転写因子の活性の増強を含む。
【0121】
I. 膵島転写因子をコードする核酸の導入によるインビトロにおける膵島細胞の産生
膵島転写因子をコードする核酸(例えば、Ngn3、MeuroD1、Mash1等)をインビトロで細胞に導入し、細胞内で発現させて、少なくとも一過的に発現させることができる。核酸を導入する細胞は、分化した上皮細胞(例えば、膵細胞、腸細胞、肝細胞、または管細胞)、多能性成人幹細胞もしくは多能性胚幹細胞、またはβ細胞に発達可能な任意の哺乳動物細胞もしくは膵島転写因子をコードする核酸の発現後にインビトロでインスリンの発現が可能な細胞とすることがきる。続いて、膵島細胞置換療法(例えばエクスビボ療法)による治療の影響を受けやすいインスリン関連疾患を有する対象にその細胞を移植する。
【0122】
1つの態様において、膵島転写因子の発現、好ましくはNgn3、MeuroD1、もしくはMash1、特にNgn3の発現、またはNgN3制御経路の他の正の制御因子の発現が提供され、(例えば自家移植を提供するために)対象に移植される宿主細胞は、移植を受ける個体由来の細胞である。例えば、1型糖尿病を有する対象では、多能性幹細胞、肝細胞、腸細胞、または膵細胞を罹患対象から単離し、膵島転写因子をコードするDNAを発現するように細胞を修飾し、細胞を罹患対象に移植してインスリンの産生を提供することができる。または、細胞がインスリン産生β細胞へ発達するのを促進するように形質転換細胞を培養し、その後β細胞を罹患患者に移植することもできる。
【0123】
別の態様において、別の対象(「ドナー」)由来の多能性幹細胞、肝細胞、腸細胞、または膵細胞を修飾して膵島転写因子をコードするDNA、特にNgn3をコードするDNAまたはNgn3制御経路の他の正の制御因子をコードするDNAを発現させ、続いてその細胞を罹患対象に移植して、(例えば、異種移植を提供するために)インスリンの産生を提供する。または、細胞がインスリン産生β細胞へ発達するのを促進するように形質転換細胞を培養し、その後β細胞を罹患患者に移植する。
【0124】
インビトロでの細胞への核酸の導入は、当技術分野で周知の方法に従って(例えば、エレクトロポレーション、マイクロインジェクション、リポフェクション、組換え(好ましくは複製不能)ウイルスによる感染、および当技術分野で周知の他の手段によって)達成され得る。一般に核酸は、所望のレベルのポリペプチド発現を促進するプロモーター(例えば、CMV、SV40、アデノウイルス由来のプロモーター、または組織特異的プロモーターもしくは細胞種特異的プロモーター)に機能的に結合される。組換え核酸を含む形質転換細胞は、例えば、導入した構築物に存在する選択マーカー遺伝子または構築物と同時トランスフェクションした核酸に存在する選択マーカー遺伝子の発現により選択および/または濃縮されうる。一般に、選択マーカーは、テトラサイクリン、ハイグロマイシン、ネオマイシン等のような抗生物質に対する耐性を提供する。そのためマーカーには、チミジンキナーゼ等が含まれ得る。その他のマーカーには、β-ガラクトシダーゼまたは緑色蛍光タンパク質等の、発現細胞を同定するのに使用可能なマーカーが含まれ得る。
【0125】
形質転換細胞における導入した核酸の発現は、当技術分野で周知の様々な方法により評価することができる。例えば、導入した遺伝子の発現は、ノーザンブロット法を用いて関連遺伝子由来のDNAプローブとハイブリダイズするmRNAを検出することにより調べることができる。さらに、所望の遺伝子を発現する細胞を、当技術分野で周知の方法により単離してインビトロで培養し増殖させることができる。形質転換のために選択される宿主細胞は、エクスビボ治療の目的(例えばインスリン産生)、細胞の移植部位、および当業者に理解されうる様々な要因によって変わりうる他の要因によって変化すると考えられる。
【0126】
形質転換細胞を、膵島細胞表現型の発現について調べることもできる。例えば、PCR法、ノーザンブロット法、免疫細胞化学法、ウェスタンブロット法、RIA法、またはELISA法により、インスリン、IAPP、グルコキナーゼ、またはソマトスタチンの発現を検出することができる。または、インスリン遺伝子プロモーター等の膵島特異的プロモーターに機能的に結合された緑色蛍光タンパク質遺伝子または抗生物質耐性遺伝子のようなマーカー遺伝子を使って、分化した膵島細胞を同定または選択することができる。所望の遺伝子産物を発現させるために宿主細胞を操作する方法および操作された細胞を体内移植または移植する方法(例えばエクスビボ療法)は、当技術分野で周知である(例えば、Gilbertら、1993、Cell Transplantation of genetically altered cells on biodegradable polymer scaffolds in syngenic rats、Transplantation 56:423-427を参照のこと)。外因性の細胞または自己細胞における所望遺伝子の発現、および膵臓への細胞(例えば膵島細胞)移植に関しては、例えば、Docherty 1997、Gene Therapy for diabetes mellitus、Clin Sci (Colch) 92:321-330;Hegreら、1976、Transplantation of islet tissue in the rat、Acta Endocrinol Suppl (Copenh) 205:257-281;Sandlerら、1997、Assessment of insulin secretion in vitro from microencapsulated fetal porcine islet-like cell clusters and rat, mouse, and human pancreatic islets、Transplantation 63:1712-1718;Calafiore 1997、Perspectives in pancreatic and islet cell transplantation for the therapy of IDDM、Diabetes Care 20:889-896;Kenyonら、1996、Islet cell transplantation: beyond the paradigms、Diabetes Metab Rev 12:361-372;Sandler;Chickら、1977 Science、Artificial pancreas using living beta cells: effects on glucose homeostasis in diabetic rats、197:780-782を参照されたい。一般に、膵臓、または例えば皮下部位、肝臓、腹膜等の任意の実際的もしくは簡便な部位に、細胞を移植することが可能である。
【0127】
膵島細胞を移植する方法は当技術分野で周知であり、例えば、Hegreら、1976、Transplantation of islet tissue in the rat、Acta Endocrinol Suppl (Copenh) 205:257-281;Sandlerら、1997、Assessment of insulin secretion in vitro from microencapsulated fetal porcine islet-like cell clusters and rat, mouse, and human pancreatic islets、Transplantation 63:1712-1718;Calafiore 1997、Perspectives in pancreatic and islet cell transplantation for the therapy of IDDM、Diabetes Care 20:889-896;Kenyonら、1996、Islet cell transplantation: beyond the paradigms、Diabetes Metab Rev 12:361-372;Sandler;Chickら、1977 Science、Artificial pancreas using living beta cells: effects on glucose homeostasis in diabetic rats、197:780-782を参照されたい。
【0128】
一般に、インビトロで形質転換細胞を増殖させてから、当技術分野で周知の方法により哺乳動物対象にその細胞を移植する。移植される細胞の数は、血糖値を低下させるのに十分なレベルのインスリン発現を提供するのに十分な細胞の数に等しい。インビトロで達成されるポリペプチドの発現レベル、および/または移植の過程で残存する細胞の数等の要因に基づいて、移植する細胞の数が決まり得る。形質転換細胞は、肝臓等の血管新生の盛んな領域に移植され、または対象における外科的処置を最小限にする様式で移植する。形質転換細胞からなる移植細胞の移植は、移植拒絶反応の典型的な兆候、すなわち移植部位における炎症および/または剥離ならびに発熱に関して、哺乳動物対象を試験することにより、および血糖値をモニタリングすることによってモニタリングされる。
【0129】
上記の移植方法は、ニューロゲニン3の発現に限定されない。分化カスケードに存在する他の膵島転写因子、特にNeuroD1およびMash1を、Ngn3とともにまたはNgn3に代えて発現させるように宿主細胞を操作し、インスリン関連疾患を有する対象を治療することもできる。
【0130】
II. インビボでの膵島細胞の発達および膵臓におけるインスリンの産生
膵島転写因子をコードする核酸を対象に直接送達して、標的細胞(例えば、膵細胞、腸細胞、肝細胞、または膵島転写因子の発現およびインスリンの産生が可能な他の臓器の細胞)内で膵島転写因子の発現を提供し、これにより細胞が(例えば膵臓内の)インスリン産生細胞へと発達するのを促進することができ、または対象における膵島転写因子発現の欠損を治療することができる。関心対象の核酸を標的細胞で発現させるためにインビボで送達する方法は、当技術分野で周知である。例えば、インビボで遺伝子送達する方法は、通常、標的細胞にDNAを導入する生物学的手段(例えば、関心対象のDNAを含むウイルス)または標的細胞にDNAを導入する機械的手段(例えば、細胞へのDNAの直接注射、リポソーム融合、または遺伝子銃を用いた気圧注射)のどちらかを利用する。
【0131】
一般に、関心対象のDNAにコードされるタンパク質を発現する形質転換細胞は、治療的に有効な量のタンパク質を産生し、哺乳動物患者において膵島細胞、特に膵β細胞を産生する。1つの態様において、関心対象のDNAは、ニューロゲニン1、ニューロゲニン2、ニューロゲニン3、NeuroD1/BETA2、Mash1、またはNeuroD4/Math3等(特に関心が持たれるのはNgn3である)の膵島転写因子をコードし、関心対象のDNAはプロモーターに機能的に結合されるが、そのプロモーターは転写因子にとって異種性または内在性であってよい。好ましい態様において、関心対象のDNAは、Ngn3、NeuroD1、またはMash1をコードし、特にNgn3をコードする。
【0132】
一般論として、本送達方法は、関心対象のDNAを含むベクターを膵細胞に導入する段階を含む。一例として、関心対象のDNAを含むベクターはウイルスベクターまたは非ウイルスベクター(裸のDNAを含む)のいずれかを含み、菅系を通してインビボで膵臓に導入される。管内投与は、例えば、胃腸管の管腔を介したカニューレの挿入による、外部開口部を介したカニューレの挿入による、または総胆管を介したカニューレの挿入によるカニューレ挿管により、達成され得る。内視鏡的逆行性胆道膵管造影法(ECRP)により、膵臓で逆行性菅投与を行うこともできる。膵臓への菅内送達を達成する例示的な方法は、米国特許第6,004,944号に記載されている。
【0133】
投与する膵島転写因子をコードする核酸の正確な量は、標的細胞の形質転換されやすさ、対象の体長および体重、所望のタンパク質発現のレベル、および治療対象のインスリン関連疾患を含む多くの要素によって大きく変化する。標的組織を感染させ、十分な数の細胞を形質転換し、所望のレベルのインスリン産生を提供するのに効果的な核酸の量および/または感染性ウイルス粒子の数は、インビトロにおける形質転換効率および標的細胞の形質転換されやすさ等の要因に基づいて、容易に決定され得る。例えば、ヒトの膵管に導入するDNA量は、例えば、一般的には約1μg〜約750 mg、好ましくは約500μg〜約500 mg、より好ましくは約10 mg〜約200 mg、最も好ましくは約100mgである。一般に、動物モデルにおける所望の遺伝子の送達と発現に有効なDNA量から、導入されるDNA量を推定できる。例えば、ヒトにおける送達用のDNA量は、ラットにおいて有効なDNA量のおよそ100倍である。
【0134】
特に送達する核酸がDNAでありかつ関心対象のDNAが真核生物の強力なプロモーター(例えば、CMV、MMTV)と機能的に結合している場合、本発明に従って修飾した膵細胞により、関心対象の核酸の十分に高いレベルでの発現が促進される。発現したタンパク質により、膵島細胞の産生およびインスリンの産生が誘導され得る。したがって、本発明の方法は、様々なインスリン関連疾患を有する哺乳動物対象を治療する上で有用である。
【0135】
1つの態様において、コードされるタンパク質は、塩基性ヘリックス・ループ・ヘリックス(bHLH)タンパク質のクラスに由来する膵島転写因子である。例えば、ニューロゲニン3および/またはNeuroD1および/またはMash1の発現により、哺乳動物において膵島細胞の産生およびインスリンの産生が実質的に誘導され得る。
【0136】
関心対象のタンパク質を治療的レベルで得るために必要な形質転換膵細胞の実際の数は、発現対象のタンパク質、形質転換細胞によるタンパク質発現のレベル、タンパク質が膵島細胞産生(特にβ細胞)を誘導する割合、および治療を受ける対象の健康状態を含むいくつかの要因によって変化すると考えられる。
【0137】
膵島転写因子をコードする核酸(または、インビトロで産生される膵島細胞、または、移植後にインビボで膵島細胞に発達するように移植する予定の膵島転写因子核酸を発現する組換え細胞)は、インビボ導入かエクスビボ導入かにかかわらず、他の遺伝子および他の薬剤と組み合わせて投与されうる。
【0138】
治療の評価
本発明の方法によるエクスビボ両方またはインビボ療法の効果を、様々な方法でモニタリングすることができる。一般に、対象の血液試料を、例えば、グルコース、プロインスリン、c-ペプチド、およびインスリンのレベルについてアッセイすることができる。血液試料中のプロインスリン、c-ペプチド、インスリン、およびグルコースを検出する適切なアッセイ法は、当技術分野で周知である。グルタミン酸脱炭酸酵素(GAD)または当技術分野で周知の他の自己抗原などの膵島タンパク質に対する自己反応性T細胞または抗体についてアッセイすることにより、再発性自己免疫の徴候を評価することができる。
【実施例】
【0139】
以下の実施例は本発明を実施するための方法を当業者に完全に開示して説明するために提供されるものであり、本発明者らが本発明だと考える範囲を限定することを意図したものではない。使用する数字(例えば、量、温度等)に関しては正確性を期す努力を行ったが、ある程度の実験誤差および偏差を考慮する必要がある。特記されない限り、割合とは重量割合、分子量とは重量平均分子量、温度とは摂氏、および気圧とは大気圧またはその付近である。
【0140】
実施例1:マウス膵臓におけるNgn3発現の検出
塩基性ヘリックス・ループ・ヘリックス(bHLH)ファミリーの転写因子に属する転写因子は、多くの細胞種の成長および分化を制御する。インスリン遺伝子の発現は、2つのbHLHタンパク質である、偏在的に発現する(クラスA)タンパク質と細胞種特異的(クラスB)パートナーであるNeuroD1/BETA2のヘテロ二量体複合体により活性化される。NeuroD1/BETA2は、β細胞の発達にとっても重要である。マウスのNeuroD1/BETA2遺伝子を標的破壊すると、胎児発生の後期に膵島細胞のアポトーシスが増加することにより、出生時のβ細胞の質量が著しく減少する。しかし、この因子がβ細胞の分化に重要であると考えられているにもかかわらず、β細胞の形成またはインスリン遺伝子の発現には明らかな欠陥は見られない。
【0141】
このように表現型が中程度であるということは、膵臓内分泌部において密接に関連したクラスB bHLHタンパク質が重複して発現していることを反映していると仮定し、本発明者らは、クラスB bHLHタンパク質のbHLHドメインにおいて保存されるアミノ酸配列(Sommerら、1996 Mol. Cell. Neurosci. 8:221)に基づいた縮重オリゴヌクレオチドプライマーを用いて逆転写ポリメラーゼ連鎖反応(RT-PCR)することにより、このファミリーに属するさらなる転写因子を探索した。PCR解析により、膵臓内分泌細胞株および単離した成人膵島細胞は、NeuroD1を発現するだけでなく、Mash1、neuroD2およびneuroD4、ならびにニューロゲニン(ngn)1、ニューロゲニン2、およびニューロゲニン3を含む神経クラスB bHLH遺伝子ファミリーに属するいくつかの他の転写因子も発現することが明らかになった。この著しい重複性により、マウスにおいてNeuroD1/BETA2の喪失が代償されたのであろう。最も共通して増殖していた配列はNeuroD4およびNgn3をコードする2つの配列であったが、マウス膵臓をインサイチューハイブリダイゼーションにより調べたところ、NeuroD1およびNgn3の発現が最も高いことが示された。これらの結果は、免疫組織化学法により確認した。
【0142】
マウスにおいて、Ngn3は胚齢11.5日(e11.5)にまず最初に検出され、e15.5で最大になるまで増加して、e18.5で減少し、成体では染色は見られない。Ngn3は散在する菅細胞および菅周囲の細胞の核において検出されるが、4つの膵島ホルモン(インスリン、グルカゴン、ソマトスタチン、および膵臓ポリペプチド)のいずれとも同時染色されなかった。この時間的かつ空間的な発現パターンは、膵島細胞前駆体のマーカーとしてのNgn3を意味するものである。将来的にはβ細胞の特異的マーカーであるNkx6.1はNgn3陽性細胞の10%〜20%で発現し、これによりさらに膵島細胞前駆細胞のマーカーとしてのNgn3の使用が支持された。e15.5におけるNgn3発現のピークは、胎仔における新しいβ細胞形成のピークとも一致する。Ngn3がNeuroD1/BETA2および他の膵島分化因子の上流に作用して膵島細胞前駆細胞を標識し、最終的に分化する前に消失するというモデルが、本発明者らのデータにより裏付けられる。
【0143】
実施例2:ヒトNgn3ポリペプチドをコードするポリヌクレオチドの単離および配列決定
クローニングされたマウスNgn3遺伝子断片に由来するプローブ(Sommerら、前記)を用いて、ヒトゲノムライブラリーをスクリーニングした。このスクリーニングにより、配列表に配列番号:1として提供するゲノム配列を単離した。マウス胎仔膵臓RNAを用いた5'RACE法によりマウスの開始部位をマッピングした結果に基づくと、ヒトNgn3コード配列の転写開始部位は2643位のヌクレオチド残基である。コード配列は3022位〜3663位のヌクレオチド残基の間にあり、終始部位は3664位〜3666位にある。5'非翻訳領域(UTR)または配列番号:1のコード配列内には、イントロンは存在しない。
【0144】
Ngn3のプロモーターは、マウス、ラット、およびヒトの間で非常によく保存されているという点で特に興味深い。膵臓および膵島細胞の発達におけるNgn3の役割を考えると、Ngn3プロモーターは成熟した膵臓に存在する膵島細胞の数を決定する鍵である可能性が高い。ヒトNgn3プロモーターに対応する調節領域は、ヒトNgn3プロモーター内にある転写開始部位の最大で約500 bp上流に至るまでの配列(例えば、約2144位から、2643位の転写開始部位まで)を含む。FISH法を用いて、Ngn3のヒト染色体上の位置を10q22.1〜22.2上に同定した。
【0145】
実施例3:マウスNgn3ポリペプチドをコードするポリヌクレオチドおよびプロモーターの単離ならびに配列決定
以前に得られたマウスゲノムDNA断片(Sommerら、前記)を配列決定することにより、マウスの全長Ngn3配列およびその5'近接配列が得られたが、これにはマウスNgn3プロモーターも含まれていた。マウスNgn3配列は配列表において配列番号:3として提供されており、コードされるポリペプチドは配列番号:4として提供される。転写開始部位を5'RACE法で決定し、マウス胎仔膵臓由来RNAを用いたRNase保護(protection)法により確認し、719位のヌクレオチド残基であることがわかった。マウスNgn3のコード配列は、1093位のヌクレオチド残基において開始される。プロモーターは、転写開始部位のおよそ500 bp上流の領域を含む。
【0146】
実施例4:ニューロゲニン3をコードするアデノウイルスベクターの構築
マウスの全長ニューロゲニン3コード配列をアデノ-Xウイルスゲノムのサイトメガロウイルス前初期遺伝子プロモーター(PCMV IE)の下流に挿入し、アデノ-XTM発現系についての製造業者(Clontech、カリフォルニア州パロアルト)の説明に従って完全なウイルス粒子を産生した。図1に最終的なアデノX.NGN3構築物のマップを示す。対照構築物をNgn3の代わりにLac Zコード配列を用いて作製し、アデノLacZ構築物とした。以下の実験で使用するため、当技術分野で周知のプロトコールによりアデノウイルスの大量調製を行った。ウイルス濃度は、プラークアッセイ法によってではなく、精製ウイルス保存液のタンパク質濃度に基づいて推定した。ウイルスの同一性は、調製中のPCRにより、および精製保存液に感染した細胞の溶解液を用いたニューロゲニン3についてのウェスタンブロットにより確認した。
【0147】
実施例5:アデノX.NGN3を用いた処置による正常成体ラットにおけるインスリンの誘導
以前に記載された外科的技法(Goldfineら、Nat Biotechnol 15:1378-82、1997)を用いて、体重250 g〜350 gの成体雄性Sprague-Dawleyラットの膵管に、アデノLacZまたはアデノX.NGN3のいずれかを注射した。およそ3×1010個のウイルス粒子(低投与量)またはおよそ3×1011個のウイルス粒子(高投与量)を、各ラットの膵管に注射した。術後回復してから、ラットを通常の食餌に戻した。約48時間後、低投与量のアデノX.NGN3を注射されたラットを屠殺し、膵臓を摘出して固定し、パラフィンに包埋して切片を作製した。確立した免疫組織化学的技法および抗血清(Schwitzgebelら、Development 127:3533-3542、2000)を用いて、膵臓切片をインスリン、グルカゴン、サイトケラチン20(菅のマーカー)、およびマウスニューロゲニン3について染色した。
【0148】
低投与量または高投与量のアデノX.NGN3を注射されたラットは、ランゲルハンス島を形成するグルカゴン染色細胞およびインスリン染色細胞の通常の大きな(およそ1000個の細胞)凝集塊のほかに、個別の、または細胞8個以下からなる小さな凝集塊である、インスリン染色細胞またはグルカゴン染色細胞を有しており、これらは菅の近くに散在して見られた。図2Aは、低投与量(3×1010個)のアデノX.NGN3を注射されたラットの膵臓組織切片におけるインスリンについての組織学的染色の結果を示す。個別のおよび小さな凝集塊であるインスリン染色細胞を矢印で示す。図2Bは同じラットの連続切片を菅細胞のマーカーであるサイトケラチン20について染色した結果を示し、矢印は図2Aにおけるインスリン染色の位置を示す。
【0149】
対照アデノLacZウイルスを注射されたラットでは、ランゲルハンス島の外側に散在するインスリン染色細胞またはグルカゴン染色細胞は見られなかった。高力価のニューロゲニン3発現アデノウイルスを注射されたラットは全て、注射後36時間以内に死亡した。高投与量のラットのうち1匹の死亡前の血糖値は、Glucometer Elite計測器(Bayer Corporation、インディアナ州エルクハート)を用いたグルコースオキシダーゼ法により測定したところ、32 mg/mlであった。絶食ラットの血糖値は通常およそ100 mg/dlであり、70 mg/dlよりも低いことはめったにない。高投与量ラットの血糖値が32 mg/mlであるということは、過剰なインスリンが産生されて、血中の糖を低血糖症レベルにまで低下させたことを示す。Ngn3をコードする高投与量のDNAを注射されたラットは、新たに産生されたβ細胞からインスリンが過剰発現され、これにより低血糖症が誘導されて死亡したと思われる。
【0150】
図3は、高投与量(3×1011個)のアデノX.NGN3に感染させて約24時間後に回収したラットの膵臓における、インスリンおよびニューロゲニン3について蛍光同時染色した共焦点像を示す。図の中心にある細胞では、核がニューロゲニン3について染色され(矢印で示した濃い灰色の領域)、細胞質がインスリンについて染色された(明るい灰色である周辺領域)ことが観察される。対照動物の膵臓では、ニューロゲニン3は検出されなかった。
【0151】
実施例6:マウスNgn3をコードするDNAを膵臓に導入することにより誘導される、糖尿病誘発成体ラットにおける血糖値の正常化
β細胞毒素であるストレプトゾトシン(Sigma:40 mg/kg体重、1 mMクエン酸緩衝液、pH 4.5に溶解)を1日目および2日目に腹腔内注射することにより、体重250 g〜350 gの成体雄性Sprague-Dawleyラットにおいて糖尿病を誘発した。ストレプトゾトシンとは、ラットにおいて糖尿病を誘発するβ細胞毒素である。4日目に、高血糖症を有することが確認されたラット(Glucometer Elite計測器(Bayer Corporation、インディアナ州エルクハート)を用いたグルコースオキシダーゼ法により測定し、300 mg/dlを上回る血糖値を有する)の膵管に、前述の外科的技法によりアデノLacZまたはアデノX.NGN3のいずれかを注射した。およそ3×1010個または3×1011個のウイルス粒子を各ラットの膵管に注射した。
【0152】
術後5日目にラットを屠殺し、膵臓を摘出して固定し、パラフィンに包埋して切片を作製した。インスリン、グルカゴン、サイトケラチン20(菅のマーカー)、およびマウスニューロゲニン3について、膵臓切片を染色した。図4AはアデノX.NGN3を注射されたラットの膵臓組織切片におけるインスリンの組織学的染色の結果であるが、これにより本発明によってインスリンが産生されることが示唆される。個別のおよび小さな凝集塊であるインスリン染色細胞を白矢印で示す。残存するインスリン染色細胞(残りはストレプトゾトシンで破壊された)をわずかに有する膵島を黒矢印で示す。
【0153】
図4Bは、アデノ-LacZウイルスを注射されたラットの膵臓におけるインスリン染色の結果を示す。残存するインスリン染色細胞をわずかに有する2つの膵島を矢印で示す。対照動物において、個別のまたは小さな凝集塊であるインスリン染色細胞は全く検出されなかった。
【0154】
アデノX.NGN3ウイルスを注射されたラットは、アデノLacZを注射された対照ラットよりも有意に低い血糖値を有していた。図5に示すように、アデノX.NGN3を注射された糖尿病ラットにおいて、アデノLacZを注射された糖尿病ラットと比較して、血糖値が有意に減少した。血糖値の減少は、2日目と4日目の両方で観察された。したがってこれらのデータにより、Ngn3をコードするDNAを膵臓に導入することでインスリンが持続的に発現されること、および形質転換膵細胞が発現したインスリンは血流中に分泌され、かつ動物モデルにおける糖尿病を克服するのに十分なレベルで血糖制御のために機能し得ることが示される。
【0155】
本発明は、成熟した膵臓内の細胞、最も適当には菅細胞が、内分泌細胞に分化する能力を保持することを示す。成体菅細胞は、前内分泌bHLH遺伝子、特にNgn3に応答し、これらの遺伝子が他のシグナルと共同して新しいβ細胞の形成を誘導し、その後のインスリン産生を誘導することができる。本発明は2型糖尿病の維持および治療に有用であり、さらに1型糖尿病を有する個体において自己免疫性破壊によって失われたβ細胞の補充を可能にすると考えられる。
【0156】
実施例7:Ngn3トランスジェニック動物におけるNgn3の過剰発現および膵島細胞の産生
以前に記載されたように(Schwitzgebelら、Development 127:3533-3542、2000)、マウスpdx1プロモーター(転写開始部位およびプロモーターを含むマウスPdx1遺伝子の4.4 kbのXbaI-SmaI断片(Apelqvistら、Curr Biol 7:801-804;Wuら、Mol Cell Biol 17:6002-6013 (1997))およびヒトβ-グロビン遺伝子の第一イントロンをpBATポリリンカーの上流に挿入することにより(Germanら、Genes & Dev 6:2165-2176)、ならびにSV40後期遺伝子ポリアデニル化シグナルを挿入することにより、pdx1プロモーターベクターpBAT.PD17を構築した。PCR法によりマウスゲノムニューロゲニン3クローン(Sommerら、Mol Cell Neurosci 8:221-241 (1996))からマウスの全長Ngn3 cDNA をコードする663 bpのDNA断片を取得し、これをpBAT.PD17ポリリンカーに挿入した。
【0157】
このベクターを線状化して精製し、C3Fe/B6親に由来するF1ハイブリッド卵母細胞に前核注射(1.5 ng/μl)することによりトランスジェニックマウスを作製した(Hoganら、(1994) Manipulating the mouse embryo: A Laboratory Manual. New York: Cold Spring Harbour Laboratory Press.)。遺伝子型を、尾部生検試料のゲノムDNAをPCR解析することにより判定した。用いたプライマーは、
(5'側用Pdx1プライマー)および
(3'側用ヒトβ-グロビンイントロン)であった。
【0158】
胚齢12.5日(E12.5)またはE18.5で胎仔を回収した。全部で6匹のpdx1-ngn3マウスをE12.5で、10匹のpdx1-ngn3マウスをE18.5で試験した。トランスジェニック胎仔の大きさには著しい異常は認められなかった。E12.5で回収したトランスジェニック胎仔では、図6Bにおいてグルカゴン染色により示されるように、膵島細胞の数に増加が見られた。対照の非トランスジェニック胎仔は、正常な数のグルカゴン発現細胞を有する(図6A)。E18.5ではトランスジェニック動物の膵臓の大きさが著しく減少しているが、膵島細胞画分は同じ胚齢の対照動物と比較して顕著に増加している(図7A)。
【0159】
これらの実験から、適切な前駆細胞においてニューロゲニン3が膵島細胞の新生を誘導する能力を有することが実証された。
【0160】
実施例8:NeuroD1トランスジェニック動物における膵島細胞の産生
開始コドンから3'UTRにわたるマウスの全長cDNAをコードする1.7 KbのDNA断片(Leeら、(1995) Science 268:836-844)をpBAT.PD17にクローニングすることにより、neuroD1/BETA2ベクターを構築した。
【0161】
このベクターを線状化して精製し、上述のようにC3Fe/B6親に由来するF1ハイブリッド卵母細胞に前核注射(1.5 ng/μl)することによりトランスジェニックマウスを作製した(Hoganら(1994)、前記)。遺伝子型を、尾部生検試料のゲノムDNAをPCR解析することにより決定した。用いたプライマーは、
(5'側用Pdx1プライマー)および
(3'側用ヒトβ-グロビンイントロン)である。
【0162】
胚齢12.5日(E12.5)で、またはE18.5で胎仔を回収した。全部で5匹のpdx1-neuroD1マウスをE12.5で、10匹のpdx1-neuroD1マウスをE18.5で試験した。ニューロゲニン3トランスジェニックマウスと同様に、E12.5で回収されたpdx1-neuroD1トランスジェニック胎仔では、図6Cにおいてグルカゴン染色により示されたように、膵島細胞の数に増加が見られた。E18.5ではトランスジェニックマウスの膵臓の大きさが著しく減少しているが、膵島細胞画分は同じ胚齢の対照マウスと比較して顕著に増加した(図7B)。
【0163】
これらの実験から、neuroD1は、ニューロゲニン3と同様に、適切な前駆細胞において膵島細胞の新生を誘導する能力を有することが実証された。
【0164】
実施例9:neuroD1をコードするアデノウイルスベクターの構築
マウスまたはヒトの全長neuroD1コード配列をアデノウイルスゲノムのサイトメガロウイルス前初期遺伝子プロモーター(PCMV IE)の下流に挿入し、当技術分野で公知の方法によって、例えばアデノ-XTM発現系についての製造業者(Clontech、カリフォルニア州パロアルト)の説明に従って、完全なウイルス粒子を産生した。図8に、そのような例示的アデノ-NeuroD1構築物のマップを示す。
【0165】
実施例10:mash1/ASCL1/ASH1をコードするアデノウイルスベクターの構築
マウスの全長mash1コード配列またはヒトACSL1/ASH1(ヒトにおけるmas1の略号)コード配列を、アデノウイルスゲノムのCMV前初期遺伝子プロモーター(PCMV IE)の下流に挿入し、当技術分野で公知の方法によって、例えばアデノ-XTM発現系についての製造業者(Clontech、カリフォルニア州パロアルト)の説明に従って完全なウイルス粒子を産生した。図8に、そのような例示的アデノ-mash1/ASCL1/ASH1構築物のマップを示す。
【0166】
実施例11:アデノ-NEUROD1を用いた処理による正常成体ラットにおけるインスリン産生β細胞の形成の誘導
以前に記載された外科的技法(例えば、Goldfineら、Nat. Biotechnol 15:1378-82(1997)を参照のこと)を用いて、体重250 g〜350 gの成体雄性Sprague-Dawleyラットの膵管にアデノLacZまたはアデノ-neuroD1(マウスまたはヒト)のいずれかを注射した。およそ3×1010個のウイルス粒子(低投与量)またはおよそ3×1011個のウイルス粒子(高投与量)を、各ラットの膵管に注射した。術後回復してから、ラットを通常の食餌に戻した。約48時間後にラットを屠殺し、膵臓を摘出して固定し、パラフィンに包埋して切片を作製した。確立した免疫組織化学的技法および抗血清(Schwitzgebelら、Development 127:3533-3542、2000)を用いて、膵臓切片をインスリン、グルカゴン、サイトケラチン20(菅のマーカー)、およびneuroD1について染色した。
【0167】
実施例12:アデノ-mash1/ASCL1/ASH1を用いた処理による正常成体ラットにおけるインスリン産生β細胞の形成の誘導
以前に記載された外科的技法(例えば、Goldfineら、Nat. Biotechnol 15:1378-82(1997)を参照のこと)を用いて、体重250 g〜350 gの成体雄性Sprague-Dawleyラットの膵管にアデノLacZまたはアデノ-mash1/ASCL1/ASH1(マウスまたはヒト)のいずれかを注射した。およそ3×1010個のウイルス粒子(低投与量)またはおよそ3×1011個のウイルス粒子(高投与量)を、各ラットの膵管に注射した。術後回復してから、ラットを通常の食餌に戻した。約48時間後にラットを屠殺し、膵臓を摘出して固定し、パラフィンに包埋して切片を作製した。確立した免疫組織化学的技法および抗血清(Schwitzgebelら、Development 127:3533-3542、2000)を用いて、膵臓切片をインスリン、グルカゴン、サイトケラチン20(菅のマーカー)、およびneuroD1について染色した。
【0168】
実施例13:NeuroD1をコードするDNAの膵臓への導入による、糖尿病誘発成体ラットにおけるインスリンの産生
β細胞毒素であるストレプトゾトシン(Sigma:40 mg/kg体重、1 mMクエン酸緩衝液、pH 4.5に溶解)を1日目および2日目に腹腔内注射することにより、体重250 g〜350 gの成体雄性Sprague-Dawleyラットにおいて糖尿病を誘発した。ストレプトゾトシンとは、ラットにおいて糖尿病を誘発するβ細胞毒素である。4日目に、高血糖症を有することが確認されたラット(Glucometer Elite計測器(Bayer Corporation、インディアナ州エルクハート)を用いたグルコースオキシダーゼ法により測定した300 mg/dlを上回る血糖値を有する)の膵管に、前述の外科的技法によりアデノ-LacZまたはアデノ-NEUROD1(マウスまたはヒト)のいずれかを注射した。およそ3×1010個または3×1011個のウイルス粒子を各ラットの膵管に注射した。次に、ラットの血糖値の減少を12時間ごとにモニタリングした。
【0169】
実施例14:mash1/ASCL1/ASH1をコードするDNAの膵臓への導入による糖尿病誘発成体ラットにおけるインスリンの産生
β細胞毒素であるストレプトゾトシン(Sigma社:40 mg/kg体重、1 mMクエン酸緩衝液、pH 4.5に溶解)を1日目および2日目に腹腔内注射することにより、体重250 g〜350 gの成体雄性Sprague-Dawleyラットにおいて糖尿病を誘発した。ストレプトゾトシンとは、ラットにおいて糖尿病を誘発するβ細胞毒素である。4日目に、高血糖症を有することが確認されたラット(Glucometer Elite計測器(Bayer Corporation、インディアナ州エルクハート)を用いたグルコースオキシダーゼ法により測定された、300 mg/dlを上回る血糖値を有する)の膵管に、前述の外科的技法によりアデノ-LacZまたはアデノ-mash1/ASCL1/ASH1(マウスまたはヒト)のいずれかを注射した。およそ3×1010個または3×1011個のウイルス粒子を各ラットの膵管に注射した。次に、ラットの血糖値の減少を12時間ごとにモニタリングした。
【0170】
実施例15:ニューロゲニン3をコードするプラスミドベクターの構築
マウスまたはヒトの全長ニューロゲニン3コード配列cDNAまたはその遺伝子のいずれかを、pBAT16プラスミドベクターのCMV前初期遺伝子プロモーター(PCMV IE)およヒトびβ-グロビン遺伝子イントロンの下流、ならびにSV40ポリアデニル化シグナルの上流に挿入した。このベクターはまた、細菌内での高コピー数の複製のためのcolE1複製起点、およびプラスミド含有細菌を選択するための細菌性アンピシリン耐性遺伝子も含む。精製プラスミドDNAは、当技術分野で周知の方法により作製した。図9に、そのような例示的pBAT16.hNGN3構築物のマップを示す。
【0171】
実施例16:プラスミドDNAベクターpBAT16.hNGN3の膵臓への導入による糖尿病誘発成体ラットにおけるインスリンの産生
β細胞毒素であるストレプトゾトシン(Sigma:40 mg/kg体重、1 mMクエン酸緩衝液、pH 4.5に溶解)を1日目および2日目に腹腔内注射することにより、体重250 g〜350 gの成体雄性Sprague-Dawleyラットにおいて糖尿病を誘発した。ストレプトゾトシンとは、ラットにおいて糖尿病を誘発するβ細胞毒素である。4日目に、高血糖症を有することが確認されたラット(Glucometer Elite計測器(Bayer Corporation、インディアナ州エルクハート)を用いたグルコースオキシダーゼ法により測定された、300 mg/dlを上回る血糖値を有する)の膵管の管腔に、カチオン性脂質Transfast(Promega)等のアジュバントを含むまたは含まない8μg〜25μgのプラスミドDNAを注射した。膵管に注射された場合、カチオン性脂質等のアジュバントを含むまたは含まない裸のDNAが膵細胞において遺伝子を発現する能力は、以前に記載されている(Goldfineら、1997、前記)。その後、ラットの血糖値の減少を12時間ごとにモニタリングした。
【0172】
実施例17:アデノX-NGN3またはプラスミドDNAベクターpBAT16.hNGN3の膵臓への導入による自己免疫性糖尿病マウスにおけるインスリンの産生
ヒトの自己免疫性糖尿病または「1型」糖尿病モデルとして一般的なNODマウスは、自己免疫性糖尿病の形態を発症する(総説として、例えばBachら、The NOD mouse. Research in Immunology、1997 Jun、148(5):285-6を参照のこと)。このマウスは、膵島移植のような糖尿病の自己免疫状態治療の試験モデルとして用いられている(総説として、例えばHahnら、Adv. Exp. Med Biol. 1997 426:411-9;Sutherland Transplantation Proc. 1996、28(4):2131-3を参照のこと)。
【0173】
本実施例では、15週齢を超えた成体マウスを糖尿病(Glucometer Elite計測器(Bayer Corporation、インディアナ州エルクハート)を用いたグルコースオキシダーゼ法により測定された、300 mg/dlを上回る血糖値を有する)について試験し、これを次に前述の外科的技法を用いてマウスの膵管にアデノ-LacZもしくはアデノX-NGN3(マウスまたはヒト)のいずれか、またはプラスミドベクターpBAT16.hNGN3を注射した。およそ3×109個または3×1010個のウイルス粒子、または2μg〜25μgのプラスミドDNAを各マウスの膵管に注射した。その後、マウスの血糖値の減少を12時間毎にモニタリングした。
【0174】
自己免疫の再発および新しく形成されたβ細胞の破壊を防ぐためには、いくつかの方法が当技術分野で周知であり、これには、サイトキサン(cytoxan)もしくはFK506等の免疫系を抑制する薬剤の使用、またはCTLA-4に対する抗体等の同時刺激分子を遮断する試薬の使用(Shapiroら、New Engl. J. Med. 2000 343(4):230-8;Griffinら、J. Immunol、2000 164(9):4433-42)が含まれるが、これらに限定されるわけではない。動物にはDNA注射の2日前から抗自己免疫療法による処置を開始し、実験期間中免疫療法を続ける。
【0175】
実施例18:インビトロでの膵島細胞の形成の誘導
本実施例では、ニューロゲニン3を用いて、インビトロで培養する非膵島細胞から膵島細胞の形成を誘導する。使用する細胞には、膵管細胞、膵腺房細胞、腺窩細胞を含む腸細胞、肝細胞、および唾液腺細胞等の胃腸管系器官由来の細胞、造血幹細胞、神経幹細胞、筋幹細胞、もしくは膵幹細胞等の成体幹細胞、または胚幹細胞を含む、哺乳動物不死化細胞株または初代培養哺乳細胞が含まれるが、必ずしもこれらに限定されるわけではない。細胞は、当技術分野で周知の方法により培養される。ニューロゲニン3は、ウイルスベクター(アデノXZ-NGN3ベクター等)により導入されるか、またはカチオン性脂質トランスフェクション試薬Transfast等のアジュバントと共にpBAT16.hNGN3等のDNAプラスミドベクターを用いた裸のDNAにより導入される。1日から数週間にわたる期間、細胞の培養を続ける。新しく形成された膵島細胞の検出は、ラジオイムノアッセイ法もしくはELISA法を用いて細胞試料から調製された抽出物中でのインスリンもしくはグルカゴン等の膵島ホルモンを測定することにより、または当技術分野で周知の方法を用いて培養液中のホルモンレベルを測定することにより実施される。
【0176】
実施例19:ヒト対象へのNgn3をコードする核酸の送達
本実施例では、糖尿病患者または関連のインスリン欠損を有する患者を、新しい膵島細胞の形成を誘導するという目的で、ニューロゲニン3をコードするDNAを用いて治療する。ニューロゲニン3をコードするDNAは、上述のアデノX-NGN3の例においてと同じくウイルスベクターに含まれることが可能であり、またはカチオン性脂質Transfast等のアジュバントを含むまたは含まない上述のpBAT16.NGN3の例においてと同じく、裸のDNAベクターに含まれることが可能である。ベクターは、膵管を介して膵臓に逆方向に導入される。ベクターがアデノウイルスの場合、注射される量は、血糖値を下げてそれを正常な範囲に維持するのに必要な量によって決まり、約1011個〜約1014個のウイルス粒子であってよい。ベクターが裸のDNAベクターの場合、必要な量は約100μg〜100 mgとなり得る。
【0177】
自己免疫の再発および新しく形成されたβ細胞の破壊を防ぐためにいくつかの方法が当技術分野で周知であり、ヒトにおいて臨床的に用いられているが、これにはサイトキサン、FK506、もしくはシロリムス(sirolimus)の免疫系を抑制する薬剤の使用、またはCTLA-4に対する抗体等の同時刺激分子を遮断する試薬の使用(Shapiroら、New Engl. J. Med. 2000 343(4):230-8;Griffinら、J. Immunol、2000 164(9):4433-42)が含まれるが、これらに限定されるわけではない。患者にはDNA投与の1日前〜2日前から抗自己免疫療法による処置を開始し、その後も免疫療法を続ける。
【0178】
処置後は、1時間ごとに、または連続グルコースモニターの使用により、血糖値を厳密にモニタリングし、必要に応じて皮下注射によるインスリン治療を減らすかまたは中止する。血糖値が外因性インスリン非存在下で完全に正常化されない場合には、処置を繰り返す。再発性自己免疫による新生β細胞の破壊をモニタリングするため、血糖値を定期的にモニタリングする。さらに、自己反応性T細胞、グルタミン酸脱炭酸酵素に対する抗体(抗GAD抗体)または膵島抗原(膵島細胞抗体またはICA)についてアッセイすることにより、新しい細胞に対する免疫応答の直接的徴候を試験することが可能である。再発性自己免疫の徴候は、免疫抑制治療の増加または別の変更により治療されうる。
【0179】
実施例20:ニューロゲニン3プロモーターの特徴づけおよび制御
以下の実験により、Ngn3プロモーターの特徴づけ、およびNgn3発現を制御する調節経路に関与するNgn3の上流と下流の両方の因子の同定を提供する。
【0180】
材料および方法
以下の材料および方法を本実施例において使用した。
【0181】
マウスおよびヒトのngn3遺伝子プロモーターのクローニング
マウスニューロゲニン3オープンリーディングフレームを含むλファージゲノムクローンである、クローン17/6-1-1-2(Sommerら、Mol. Cell. Neurosci. 8:221-241、1996)を使用した。このファージクローンから、オープンリーディングフレームの上流の配列を含む1 kb断片をサブクローニングして配列決定した。ヒトニューロゲニン3ゲノムクローンは、マウスニューロゲニン3ゲノム断片を用いてλDASHヒトゲノムライブラリーをスクリーニングすることにより取得した。最も長い5'近接配列を含むクローン、クローン14Hをサブクローニングして配列決定し、レポーター遺伝子プラスミドの構築に使用した。
【0182】
5'cDNA末端迅速増幅法(5'RACE法)
5'RACE System Version 2.0(GibcoBRL)のプロトコールの変法を用いて、5'RACEによりマウスngn3 cDNAの5'末端を同定した。マウスcDNAについては、2.5 pmolの特異的プライマーJL1
をマウスE15.5膵臓由来の全RNA 1μgとアニールさせた。逆転写は、SuperScript II逆転写酵素(GibcoBRL)を用いて行った。一本鎖cDNA合成の後、元のmRNA鋳型をRNase処理により除去し、次に末端デオキシヌクレオチド転移酵素を用いてcDNAの3'末端にホモポリマーのdCTPテール(tail)を付加した。プライマーとして5'RACE Abridged Anchor Primer(GibcoBRL)およびJL2
を用いた35サイクルのPCRにおいて、これらの産物を鋳型として使用した。入れ子(nested)PCRについては、Abridged Universal Amplification Primer(GibcoBRL)およびJL3
をプライマーとして使用し、35サイクルのPCRを行った。PCR産物をサブクローニングして配列決定した。
【0183】
レポーター遺伝子構築物
レポータープラスミドを構築するため、制限酵素切断によって得たヒトngn3遺伝子の5'領域の断片を、プラスミドpFOXLuc1内のルシフェラーゼ遺伝子の上流、またはプラスミドpFOXLuc1TK内のTK最小プロモーター遺伝子の上流に結合した(Mirmiraら、J Biol Chem 275:14743-51、2000)。
【0184】
細胞培養および一過的トランスフェクション
βTC3細胞、αTC1.6細胞、およびMPAC細胞は、2.5%ウシ胎仔血清および15%ウマ血清を添加したダルベッコ改変イーグル培地(DMEM)で培養した。NIH3T3細胞は、10%子ウシ血清を添加したDMEM培地で培養した。Cos7細胞は、10%ウシ胎仔血清および4 mMグルタミンを添加したDMEM培地で培養した。哺乳動物細胞の一過的トランスフェクションを行うため、トランスフェクションの24時間前に、細胞を6ウェル組織培養プレートにプレーティングした。標準的レポーター遺伝子解析のために、製造業者の説明に従い、TRANSFAST(商標)脂質試薬(Promega)を用いて2μgのルシフェラーゼレポータープラスミドを細胞にトランスフェクションした。HES1の発現がngn3プロモーターに及ぼす影響を評価するためには、所定の量のHES1またはドミナントネガティブなHes1(pcDNA3Hes1)(京都大学、R.Kageyama氏より分与(Sasaiら、Genes Develop. 6:2620-2634、1992))発現プラスミドDNAを2μgのルシフェラーゼレポータープラスミドと共に同時トランスフェクションした。トランスフェクションから48時間後、細胞を回収し、以前に記載されたのと同様の(Germanら、Genes & Dev. 6:2165-2176、1992)ルシフェラーゼアッセイを行った。ルシフェラーゼ活性は、細胞のタンパク質濃度について補正した。レポーター遺伝子解析はすべて少なくとも3回行い、データは平均値±標準誤差として表した。
【0185】
トランスジェニックマウスの作製およびβ-ガラクトシダーゼの検出
ヒトβ-グロビンイントロンの上流の-5.7 kb〜+261 bpおよび-2.6 kb〜+261 bpにわたるヒトngn3プロモーター断片と、細菌のβ-ガラクトシダーゼ遺伝子を連結して、プラスミドpNAT6BおよびpNAT3Bを構築した。それぞれのプラスミドを線状化し、マウス前核に微量注射(1.5 ng/μl)した。注射した胚を偽妊娠した雌性マウスに移植し、e15.5の時点で創始(founder)マウスから胎仔の膵臓を胃および小腸と共に摘出した。組織を4%パラホルムアルデヒド中で4℃で30分間あらかじめ固定し、次にX-gal(400μg/ml)基質中、37℃で(-2.6 kbプロモーター)または室温で(-5.7 kbプロモーター)一晩インキュベートした。その後、組織を再度4%パラホルムアルデヒド中で30分間固定し、パラフィン包埋して5μMで切片を作製した。遺伝子型は、ヒトngn3プロモーター配列に特異的なプライマーを用いたPCRにより判定した。β-ガラクトシダーゼ活性は、-2.6 kbプロモーター構築物を組み込んだ6匹の独立した創始胎仔、および-5.7 kbプロモーター構築物を組み込んだ8匹の独立した創始胎仔で測定した。
【0186】
免疫組織化学法
免疫組織化学法は、以前に記載されたのと同様のパラフィン包埋切片上で行った(Schwitzgebelら、Development 127:3533-3542、2000)。一次抗体は以下の希釈で使用した:モルモット抗インスリン(Linco)、1:5000;モルモット抗グルカゴン(Linco)、1:10000;ウサギ抗ngn3(Schwitzgebelら、Development 127:3533-3542、2000)、1:5000。ビオチン標識二次抗体(Vector)は、ABC Elite免疫ペルオキシダーゼシステム(Vector)で検出した。
【0187】
タンパク質の調製および電気泳動移動度シフト分析法(EMSA)
SP6およびT7 TNT Quick Coupled Lysate System(登録商標)(Promega)を用いて、pGEM-1ラットHNF3β(イリノイ大学シカゴ校、R.Costa氏より分与)およびpcDNA3-HNF1α(ロックフェラー大学、M. Stoffel氏より分与)を鋳型として使用し、HNF3βタンパク質およびHNF1αタンパク質をインビトロで産生した。グルタチオンS転移酵素(GST)を融合したHES1タンパク質は、pGEX2Tプラスミドシステム(Promega)を使用して大腸菌BL21コンピテントセルで産生した。αTC1.6細胞、βTC3細胞、およびNIH3T3細胞の核抽出物は、SadowskiおよびGilman(SadowskiおよびGilman、Nature 362:79-83、1993)により記載されている手順に従って調製した。
【0188】
T4ポリヌクレオチドキナーゼを使用し、ヒトngn3プロモーター内にある配列に対応する1本鎖オリゴヌクレオチドの5'末端を[γ-32P]-ATPで標識した。標識オリゴヌクレオチドをカラム精製し、過剰量の相補鎖にアニールさせた。HNF3βおよびHNF1α結合実験に用いられるEMSA緩衝液および電気泳動条件は、以前に記載されている(Mirmiraら、J Biol Chem 275:14743-51、2000)。HES1結合実験に関しては、ポリ(dI-dC)濃度を15 ng/μlまで減少させる以外は同じ条件である。それぞれの結合反応には、インビトロ反応混合液1μl、または核抽出物2μg、またはGST融合Hes1タンパク質400 ngを使用した。抗体を使用する場合には、ゲル電気泳動を行う前に、各抗体1μlを結合混合液とともに室温で15分間インキュベートした。HNF-3α、HNF-3β、およびHNF-3γに対する抗血清はR.Costa氏(イリノイ大学)から分与されたものであり、HES-1抗血清はY.Jan氏(カリフォルニア大学サンフランシスコ校)から分与されたものである。抗HNF-1α抗血清は、サンタクルーズバイオテクノロジー社(Santa Cruz Biotechnology, Inc.)から購入した。
【0189】
標識プローブまたはEMSA反応における競合物として、以下のオリゴヌクレオチドを使用した(上側の鎖を示す)。
【0190】
ヒトngn3プロモーターのマッピング
ニューロゲニン3遺伝子発現の制御を理解する最初の段階として、マウスおよびヒトのニューロゲニン3プロモーターの配列を決定した(図11A〜図11B)。胚齢15.5日のマウス胎仔の膵臓から精製したRNAを使用し、5'RACE法によりマウスngn3遺伝子の転写開始部位を決定した。5'RACE産物すべてから、推定TATAAボックスから30塩基対下流という、同一の開始部位が同定された(図11B)。開始部位の上流領域は、マウス、ヒト、およびラットにおいて高度に保存されており、マウスとヒトで最も相同性が高い領域はおよそ300 bp上流に及ぶ。CCAAT配列エレメントは、転写開始部位から-85 bpの位置に位置する。可能性のある配列エレメントがいくつか他にも同定されたが、それらを図11Bに示す。
【0191】
細胞株におけるプロモーターの活性
3'末端はそれぞれ+261 bpまでかつ5'側が連続的に欠失された一連のニューロゲニン3プロモーターを、ホタルルシフェラーゼ遺伝子に結合し、これらを細胞株で試験した(図12)。-502 bpまで連続的に欠失させても、インビトロでのプロモーター活性は減少しなかった。驚いたことに、線維芽細胞を含む全被験細胞株において、プロモーターは高レベルで転写を駆動する。最も短い構築物でさえ、全被験細胞株において高い活性を有していることから、この非特異的な高活性はプロモーターの近位にあると思われる。
【0192】
トランスジェニックマウスにおけるプロモーターの活性
細胞株における一過的トランスフェクションによりプロモーター活性の指標がいくつか提供され得るものの、これらの腫瘍細胞は、ニューロゲニン3が通常発現されている発達過程の膵臓の細胞を表さない。したがって、β-ガラクトシダーゼをコードする細菌遺伝子を駆動する、ヒトニューロゲニン3遺伝子から5.7 kb上流または2.6 kb上流の配列を伴う導入遺伝子を保有するマウスを作製した。胚齢15.5日、つまりマウス胎仔の膵臓におけるニューロゲニン3発現の通常のピーク時に(Schwitzgebelら、Development 127:3533-3542、2000)、創始マウスを回収した。
【0193】
5.7 kbの構築物を保有する動物では、発達中の膵臓の中心領域および腸上皮において、すなわち発達中の同時期にニューロゲニン3が通常発現されるのと同じ領域において、β-ガラクトシダーゼが強くかつ選択的に発現されていた(図13A)。2.6 kbの構築物を用いた場合のβ-ガラクトシダーゼの発現レベルは5.7 kb構築物を用いた場合に比べ有意に低いが(図13B、図13C)、β-ガラクトシダーゼ発現の全体的なパターンは同じである。
【0194】
免疫組織化学法を用いて、5.7 kbのプロモーター構築物を保有するトランスジェニックマウスにおいてβ-ガラクトシダーゼを発現する細胞を同定した。β-ガラクトシダーゼ発現細胞は、主に菅に局在する。β細胞において時々はインスリンとβ-ガラクトシダーゼが同時に発現するものの、ほとんどのβ-ガラクトシダーゼ発現細胞は膵島ホルモンを発現しない。
【0195】
β-ガラクトシダーゼ活性およびニューロゲニン3タンパク質発現が発達過程の膵臓および腸の同じ領域に特異的に密接に同時局在するにもかかわらず、完全な一致はみられない。いくつかのβ-ガラクトシダーゼ陽性細胞は高レベルのニューロゲニン3と同時発現するが、多くのβ-ガラクトシダーゼ陽性細胞ではそうではなく、かつ多くのニューロゲニン3発現細胞はβ-ガラクトシダーゼ活性をほとんどまたは全く含まない。おそらくこの矛盾は、遺伝子発現の開始および終結の違いからというよりも、むしろ2つの遺伝子産物の蓄積および分解のタイミングの違いからきている可能性が最も高い。
【0196】
特定の細胞においてそれぞれの遺伝子産物を最初に検出する正確なタイミングは蓄積速度および検出限界に依存するため、これらが同一であると考えるべきではない。さらに、いくつかのニューロゲニン3発現細胞はランダムに導入遺伝子をサイレンシングさせうり、トランスジェニックマウスにおいて多くのプロモーターで観察される現象は理解が不十分である(Graubertら、Nucleic Acids Res 26:2849-58、1998)。前駆細胞でのニューロゲニン3の発現は非常に短期だが大量であることから、mRNAおよびタンパク質は急速に蓄積するが半減期が非常に短いことが示唆される。対照的に、β-ガラクトシダーゼは哺乳動物細胞においてかなり長い半減期を有し(Smithら、J Virol 69:4593-9、1995)、より遅くピークに達し、かつニューロゲニン3がもはや検出できなくなった後まで細胞中に残存すると考えられ得る。したがって、β-ガラクトシダーゼ発現細胞の多くにおいて、ニューロゲニン3遺伝子の産生が終わった後かつホルモン発現が始まる前に、膵島細胞の分化段階が起こる。これらの細胞が多数あることから、この分化の中間段階が初期のニューロゲニン3発現段階よりも長く持続することが示唆される。
【0197】
腸での発現
E15.5の時点から、トランスジェニック胚および非トランスジェニック胚の両方で、腸絨毛の刷子縁に沿って低レベルの内因性β-ガラクトシダーゼ発現が検出され得る。より強いβ-ガラクトシダーゼ活性はトランスジェニックマウスにおいて斑点模様として検出され得り、これは小腸で最も顕著であるが、非トランスジェニック同腹仔には見られない(図13A)。腸の切片を作製すると、このβ-ガラクトシダーゼシグナルが腸管上皮内に散在する細胞に由来することが示される(図14A〜図14C)。このβ-ガラクトシダーゼの発現パターンにより、ニューロゲニン3プロモーターは発達過程の腸にある前駆細胞の亜集団においても活性があることが示唆される。これらは、腸内分泌細胞の前駆細胞であってもよい。膵臓においてと同様に、このβ-ガラクトシダーゼ活性は内因性ニューロゲニン3発現と部分的に重複し、ここでもβ-ガラクトシダーゼの蓄積のピークがニューロゲニン3と比較して遅れることが示唆される(図14C)。
【0198】
複数の因子がニューロゲニン3プロモーターに結合する
ngn3プロモーターに結合する核因子を同定するため、一連のオリゴヌクレオチドを合成した。プロモーター内の重要なDNA結合部位にまたがる可能性のあるオリゴヌクレオチドを、電気泳動移動度シフト分析法(EMSA)により核タンパク質への結合について試験した。
【0199】
HNF3ファミリーの翼状ヘリックス転写因子に属する転写因子は、膵臓の発達および膵島機能と関係している(Wuら、Mol Cell Biol 17:6002-13、1997;Gerrishら、J Biol Chem 275:3485-92、2000;Sharmaら、Mol Cell Biol 17:2598-604、1997;Duncanら、Science 281:692-5、1998;Philippeら、Mol Cell Biol 14:3514-23、1994;Kaestnerら、Genes Dev 13:495-504、1999)。コンセンサスなHNF結合部位(Costaら、Mol Cell Biol 9:1415-25、1989)との類似性に基づくと、5.7 kbヒトニューロゲニン3プロモーター内には潜在的HNF3結合部位がいくつか存在する。最も可能性の高い2つの部位は、-3687 bpおよび-200 bpに位置する。これらの結合部位に関してEMSA試験を行った結果、両部位がインビトロ産生HNF3βに高い親和性で結合することが示された(図15)。βTC3細胞およびαTC1.6細胞の抽出物を用いると、単一の主要な複合体は両部位に結合し、この複合体はHNF3βに対する抗血清により特異的に認識される。さらに、HNF3βを同時発現すると、一過的トランスフェクションされた3T3線維芽細胞においてニューロゲニン3プロモーターが活性化される(該当データ示さず)。
【0200】
-3687 bpの HNF3β結合部位は、公知の膵臓転写因子に対する潜在的DNA結合部位のクラスターの一部を形成し(図11)、これにはhox型ホメオドメイン転写因子ならびにcut-ホメオドメイン転写因子HNF6およびPou-ホメオドメインHNF1因子(Courtoisら、Science 238:688-92、1987)の結合部位として可能な部位が含まれる。ニューロゲニン3プロモーターに結合するHNF6については、以前に実証されている(Jacqueminら、Mol Cell Biol 20:4445-54、2000)。
【0201】
潜在的HNF1結合部位にまたがるオリゴヌクレオチドをEMSAにより試験した結果、インビトロで産生されるHNF-1αに結合し得ることが見出された。さらに、βTC3細胞由来の核抽出物内では、移動度の低い主要な複合体がオリゴヌクレオチドに結合し、HNF1αに対する抗血清により特異的に認識される(図16)。HNF1α結合部位のすぐ下流にある類似hoxホメオドメイン型の結合部位は、HNF1αに結合しないと思われる。
【0202】
ニューロゲニン3プロモーターのHES-1阻害
転写制御因子HES-1を介するNotch受容体のシグナル伝達が、発達過程の膵臓の細胞の小さな亜集団を除くすべての細胞において、ニューロゲニン3の発現を抑制しうることが提唱されている(Jensenら、Nat Genet 24:36-44、2000)。ニューロゲニン3プロモーターを直接阻害するHES-1の能力を試験するため、3T3細胞において、ニューロゲニン3プロモータールシフェラーゼプラスミドと共にCMVプロモーター駆動発現プラスミドからHES-1 cDNAを発現させた(図17A〜図17C)。HES-1は、ニューロゲニン3プロモーターを劇的かつ特異的に阻害する。5'配列を下流に-502 bpまで除去しても、HES-1がプロモーターを阻害する能力は有意に減少しない。
【0203】
HES-1抑制に応答できる配列をさらにマッピングするため、ホタルルシフェラーゼ遺伝子に結合させた-208 bp〜+40 bpのヒトngn3遺伝子プロモーター配列(近位プロモーター(図18A))、またはホタルルシフェラーゼ遺伝子に結合させたヘルペスウイルスチミジンキナーゼ(TK)プロモーターの上流-2.6 kb〜-208 bpの配列(遠位プロモーター)のいずれかを用いてプラスミドを構築した。小さな近位プロモーターはHES-1抑制に関する能力のほとんどを保持しているのに対し、遠位配列はHES-1により弱く抑制される。
【0204】
近位の208 bpのプロモーター内には、HES-1のコンセンサスな結合部位(CTNGTG)(Takebayashiら、J Biol Chem 269:5150-6、1994)およびそのショウジョウバエ相同体hairy/enhancer-of-split(CGCGTC)(Van Dorenら、Genes Dev 8:2729-42、1994;Ohsakoら、Genes Dev 8:2743-55、1994)に基づく、潜在的HES-1結合部位がいくつか存在する(図18A)。ゲル移動度シフト分析法により、これら4つの部位を含む3つのオリゴヌクレオチドを、細菌的に産生されたHES-1タンパク質への結合について調べた(図18B)。以前に記載されたマウスHES-1遺伝子由来の高親和性タンデム部位(Takebayashiら、J Biol Chem 269:5150-6、1994)(図18CではH1と表示)よりも高い親和性で、3つのオリゴヌクレオチドはすべてHES-1に結合する。これら4つの配列はすべて、マウスニューロゲニン3プロモーターに保存されている(図11)。
【0205】
実施例21:膵島転写因子制御経路の特徴づけ
以下の実験により、膵島細胞の形成に関連する転写因子の特徴づけを提供する。
【0206】
簡潔に説明すると、β-ガラクトシダーゼ、ヒトニューロゲニン3(配列番号:1)、マウスNeuroD1(GenBankアクセッション番号BC018241で入手可能でありかつ配列表において配列番号:40として提供される)、マウスMash1(GuillemontおよびJoyner (1993) Mech. Dev. 42(3):171-85)、およびマウスmyoDのコード配列をpACCMV-pLpAプラスミドに挿入し、以前に記載された方法(Bermanら、(1998) J. Biol. Chem. 273:26421-26425;Beckerら、(1994) Methods Cell. Biol. 43:161-189)を用いて組換えアデノウイルスを作製した。β-ガラクトシダーゼ(adCMV-Bgal)、ニューロゲニン3(ヒト)(adCMV-Ngn3)、NeuroD1(マウス)(adCMV-ND1)、mash1(マウス)(adCMV-Mash1)、またはmyoD(マウス、筋bHLH遺伝子)(adCMV-MyoD)を発現するアデノウイルスベクターを用いて、2つのマウス膵管細胞株mPAC L20およびmPAC L4S2を感染させた。
【0207】
48時間後、細胞を回収してRNAを単離し、Ngn3、ND1、Mash1、MyoD、β-アクチン、グルカゴン、ソマトスタチン、インスリン、IAPP、グルコキナーゼ、グルコース輸送体2、Pax4、Pax6、Nkx2.2、Nkx6.1、Isl-1、およびPdx-1についてRT-PCRを行った。RT-PCR産物をアガロースゲル電気泳動により分離してエチジウムブロマイドで染色し、写真撮影した。結果を図19A〜図19Cに示す。PCR産物の同一性は、アガロースゲルから産物を精製し、精製したDNA断片を細菌のプラスミドベクターに挿入し、挿入物を配列決定することにより確認した。これらのデータから、ニューロゲニン3は、膵島細胞の分化および機能に関与する全体の遺伝子群の活性化をともなって内分泌分化を誘導できることが実証された。
【0208】
本発明をその特定の態様に関して説明したが、本発明の精神および範囲に反することなく、様々な変更が行われうりかつ均等物が代用され得ることは当業者に理解されねばならない。さらに、特定の状況、物質、対象組成物、工程、工程段階、または段階を、本発明の目的、精神、および範囲に適合化させるため、多くの改変がなされ得る。そのような修正はすべて、本発明の範囲内に包含されることが意図される。
【技術分野】
【0001】
発明の分野
本発明は一般に、膵島細胞、特にインスリン産生β細胞の産生による対象へのインスリン送達の分野に関する。より詳細には、本発明は、膵島細胞の成長および分化に関与する転写因子をコードするヌクレオチド配列の発現調節による膵島細胞の産生に関する。
【背景技術】
【0002】
発明の背景
糖尿病は米国における死因の第3位であり、失明、腎不全、および切断の主要原因である。糖尿病は早発性心臓発作および脳卒中の主要原因でもあり、米国の医療費の15%を占める。アメリカ人のおよそ5%、65歳を超えるアメリカ人の20%が、糖尿病を患っている。
【0003】
糖尿病は、膵臓内分泌部のランゲルハンス島のβ細胞が、代謝要求を満たすのに十分なインスリンを産生できないことに起因する。糖尿病は、1型糖尿病(またはインスリン依存型糖尿病)および2型糖尿病(またはインスリン非依存型糖尿病)という、2つの臨床形態に分類される。1型糖尿病は、インスリン産生β細胞の損失が原因となって起こる。2型糖尿病は1型糖尿病に比べ遺伝病の要素が強く(Zonana およびRimoin、1976 N. Engl. J. Med. 295:603(非特許文献1))、通常は人生のより後期に発症し、米国では糖尿病のおよそ90%の割合を占める。通常、罹患個体は、膵臓のインスリン産生能の低下およびインスリン利用能の欠陥(インスリン抵抗性)の両方を伴う。肥満症によりインスリン抵抗性が引き起こされるが、2型糖尿病を有する個体のおよそ80%が臨床的肥満(標準体重の20%を上回る)である。残念なことに、2型糖尿病に罹患した米国人の約半分は、この疾患を有することに気付いていない。2型糖尿病に関連する臨床症状は疾患の後期まで明らかにならないこともあり、また初期症状が誤診されることも多く、これは治療の遅延および合併症増加の原因になる。2型糖尿病の病因における遺伝の役割は明らかであるが、この疾患に関与する明確な遺伝子の大半が未知である。
【0004】
インスリンは、膵臓のランゲルハンス島のβ細胞によってのみ産生される。発生段階において、β細胞を含む膵島細胞は、成長過程にある膵芽内の未分化前駆細胞から発達する。膵芽が成長するにつれ未分化細胞が菅を形成するが、これが前駆細胞として機能する細胞である。菅細胞は生涯を通じて膵島細胞に分化する能力を維持すると思われ、膵臓が損傷を受けるような状況では、菅細胞から新しい膵島細胞が形成され得る。しかし残念なことに、1型糖尿病で見られるように膵島細胞のみが損傷を受ける場合には、膵島細胞の再生は起こらないようである。
【0005】
この発達過程は、様々な理由で臨床上の問題と関連性がある。まず第一に、膵島細胞および特にβ細胞の形成は、インスリンを産生するためおよびエネルギー代謝を制御するために必須である。β細胞の発達過程が何らかの形で損傷を受けた場合、これは後に個体が糖尿病を発症する素因となる。よってこの過程に関連する遺伝子は、新生児糖尿病、若年発症成人型糖尿病(MODY)、または2型糖尿病に対する候補遺伝子である。これらの遺伝子の配列を用いて、糖尿病を発症する危険性のある個人を同定でき、または糖尿病を予防および治療するための新規な薬理的物質を開発できる。
【0006】
第二に、上記のように、糖尿病を患う個体ではインスリンの産生が損傷される。1型糖尿病ではβ細胞の破壊によりこの障害が生じるが、一方2型糖尿病では、インスリン産生は不十分ではあるが損傷されない。1型糖尿病の治療および大半の2型糖尿病の治療は、β細胞の補充を伴う。β細胞の補充はいくつかの方法によって行われ得るが、前駆細胞からの新しいβ細胞の発生は、培養に行われるかまたは患者内でインビボで行われるのが最も生理的であると考えられる。そのためには、β細胞の分化を制御する分子が必要とされる。
【0007】
したがって糖尿病分野では、膵島前駆細胞を同定する試み、およびインビトロでβ細胞を分化させる方法を開発する試みに多大な努力を払ってきた。しかし今日まで、ほとんど成功していない。本発明は、この問題に取り組むものである。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】Zonana およびRimoin、1976 N. Engl. J. Med. 295:603
【発明の概要】
【0009】
発明の概要
本発明は、例えば膵島転写因子または膵島転写因子の発現を誘導する因子をコードする核酸を導入することにより、対象の膵臓において膵島転写因子の発現を提供することによる、対象における膵島細胞およびインスリンの産生に関し、特にNgn3、NeuroD1、およびMash1が関心対象である。本発明はまた、糖尿病患者を治療するために、膵島転写因子遺伝子および膵島転写因子ポリペプチドを用いて培養中またはインビボにおける細胞分化を改変させて、新しいβ細胞を産生する方法に関する。本発明は、例えば2型糖尿病の維持および治療において有用であると考えられ、かつさらに1型糖尿病を有する個体における自己免疫性破壊によって失われたβ細胞の補充を可能にすると考えられる。
【0010】
本発明の第一の目的は、対象の膵臓において膵島細胞の産生を提供することである。
【0011】
本発明の別の目的は、機能的β細胞形成を誘導することにより、対象においてインスリンの産生を提供することである。
【0012】
本発明の別の目的は、糖尿病患者を治療するために、膵島転写因子遺伝子を用いて培養中またはインビボにおける細胞の分化を改変して新しいβ細胞を産生する方法を提供することである。
【0013】
本発明の別の目的は、エクスビボ治療のための膵島細胞の産生、例えば対象に移植するための膵島転写因子発現細胞の産生を提供することである。
【0014】
本発明の別の目的は、糖尿病等のインスリン関連疾患に罹患した対象を治療する方法を提供することである。
【0015】
本発明のこれらおよび他の目的、利点、および特徴は、以下に示す本発明のより詳細な記載を読むことによって当業者に明らかになるであろう。
【0016】
本発明を以下でさらに詳細に説明する。
【図面の簡単な説明】
【0017】
【図1】CMVプロモーターに機能的に結合されたマウスニューロゲニン3配列を含む、アデノ-X.NGN3ウイルス構築物のマップである。
【図2A】図2Aはアデノ-X.NGN3を注射されたラットの膵臓組織切片におけるインスリンの組織学的染色を高倍率(40×)で示す写真であり、これにより本発明によってインスリンが産生されることが示唆される。矢印はインスリン染色細胞を示す。
【図2B】図2Bは図2Aの連続切片の写真であり、菅細胞のマーカーであるサイトケラチン20について染色したものである。矢印は、連続切片におけるインスリン染色細胞の位置を示す。
【図3】アデノ-XZ.NGN3を注射されたラットの膵臓における、インスリンおよびニューロゲニン3について蛍光同時染色した共焦点像の写真である。図の中心にある細胞では、核においてニューロゲニン3が染色され(矢印で示した濃い灰色の領域)、細胞質においてインスリンが染色される(明るい灰色をした周囲の領域)のが観察される。ニューロゲニン3およびインスリンの産生は、本発明による同じ細胞においても検出され得る。
【図4A】図4Aはストレプトゾトシンにより糖尿病となりかつアデノ-X.NGN3を注射されたラットの膵臓組織切片におけるインスリンの組織学的染色を示す写真であり、これにより、本発明によるインスリン産生細胞の産生が示唆される。白矢印は、個別のおよび小さな凝集塊であるインスリン染色細胞を示す。黒矢印は、ストレプトゾトシン処理後に形成された膵島に残存する残存細胞および変性細胞を示す。
【図4B】図4Bは、対照ウイルス構築物であるアデノLacZを注射されたラットの膵臓組織切片における、インスリンの染色を示す写真である。矢印は、ストレプトゾトシン処理後に形成された膵島において残存する残存細胞および変性細胞を示す。個別のおよび小さな凝集塊であるインスリン染色細胞は全く検出されなかった。
【図5】アデノX.NGN3またはアデノLacZ(対照動物)のいずれかを注射されたストレプトゾトシン処理(糖尿病)動物の血糖値のグラフである。
【図6】図6A〜図6Cは、胚齢12.5日のトランスジェニックマウスにおける膵島ホルモン発現を示す写真である。非トランスジェニック同腹仔(図6A)、およびngn3(図6B)またはneuroD1/BETA2(図6C)を発現するトランスジェニック胎仔の膵臓におけるグルカゴンの免疫組織化学的染色を示す。
【図7】図7Aおよび図7Bは、pdx1プロモーターの制御下でngn3(図7A)またはneuroD1(図7B)を過剰発現する胚齢18.5日のマウス胎仔の腸およびそれに付随する器官を示す写真である。非トランスジェニック同腹仔を各図の右側に示す。矢印は以下の器官を指し示す:肝臓(L)、胃(St)、脾臓(sp)、および膵臓(P)。トランスジェニック動物において、膵臓組織の減少は見られない。
【図8】それぞれCMVプロモーターに機能的に結合したNeuroD1およびMas1配列を含む、アデノX.NeuroD1およびアデノX.MASH1のウイルス構築物のマップである。
【図9】CMVプロモーターに機能的に結合したニューロゲニン3配列を含む、pBAT16.NGN3プラスミドDNA構築物のマップである。
【図10】bHLHタンパク質の系統樹の略図である。h.ngn3はヒトの配列であるが、それ以外はすべてマウスの配列である。
【図11】図11A〜図11Bは、ヒトニューロゲニン3遺伝子プロモーターを示す概略図である。図11Aは、実施例で用いたプロモーター断片の転写開始部位および5'末端の相対的位置を示すプロモーターの略図である。転写因子の結合部位として可能性のあるいくつかの部位を示すが、これにはプロモーターの遠位にある部位クラスターも含まれる。TATAAボックス(実際の配列はGATAA)は、転写開始部位の30 bp上流に示される。図11Bに、クラスター1に対応するプロモーターの-3728位と-3653位の間のDNA配列を示す。ここでも、転写因子の結合部位として可能性のある部位を示す。Hoxおよび関連するクラスのホメオドメインタンパク質の結合部位として機能し得る可能性のある2つの配列を点線で示す。
【図12】細胞株におけるヒトニューロゲニン3プロモーターの機能を示すグラフである。表示した5'末端から+261 bpにわたる配列を含むプロモーター断片をホタルルシフェラーゼ遺伝子の上流に結合し、示した細胞株にトランスフェクションした。レポーター遺伝子活性を、同じ細胞種におけるプロモーターのないルシフェラーゼベクターと相対的に表した。トランスフェクションは少なくとも3連で2回行い、誤差は平均値の±標準誤差として示す。ND、試験せず。
【図13】図13A〜図13C は、インビボでのヒトニューロゲニン3プロモーターの機能を示すグラフである。図13Aでは、5.7 kbのヒトNgn3プロモーターを細菌のβ-ガラクトシダーゼ遺伝子の上流に連結し、トランスジェニックマウス中で発現させた。染色部分(濃い領域)は、標識細胞におけるX-gal基質のβ-ガラクトシダーゼ触媒作用を示す。図13Bおよび図13Cは、E15.5マウス胎仔の膵臓における、β-ガラクトシダーゼ発現の酵素検出による、グルカゴンに対する免疫組織化学染色(ペルオキシダーゼ検出)を示す。β-ガラクトシダーゼの発現は、示したように-5.7 kbまたは-2.6 kbのヒトNgn3プロモーターにより促進される。X-gal染色は、37℃(2.6 kbプロモーター)または室温(5.7 kbプロモーター)で16時間行った。両切片とも倍率10×での比較可能な切片を示す。
【図14】図14A〜図14Cは、ニューロゲニン3の腸での発現を示す写真である。図14Aは、5.7 kbヒトngn3プロモーターによって駆動されるβ-ガラクトシダーゼ導入遺伝子の活性が、E15.5マウス膵臓の腸管上皮内に散在する細胞において検出され得ることを示す(濃い斑点)。図14Bは対照であり、野生型の同腹仔では同様のβ-ガラクトシダーゼ発現がみられないことを示す。図14Cは、E15.5のトランスジェニックマウスの腸における、β-ガラクトシダーゼも同じく発現するいくつかの細胞(黒矢印)における、およびβ-ガラクトシダーゼを発現しないいくつかの細胞(白矢印)におけるNgn3の検出(ペルオキシダーゼ検出)を示す。
【図15】図15A〜図15Bは、HNF3がヒトngn3プロモーターに結合することを示す、電気泳動移動度シフト分析の写真である。図15Aは、ヒトngn3プロモーターのH3-1プローブに対する転写因子HNF3βの結合を実証する電気泳動移動度シフト分析を示す。図15Bは、ヒトNgn3プロモーターにおける近位の部位および遠位の部位がHNF3βへの結合について同等に競合し得ることを実証する電気泳動移動度シフト分析を示す。
【図16】HNF1がヒトngn3プロモーターに結合することを示す、電気泳動移動度シフト分析の写真である。
【図17】図17A〜図17Cは、HES1がニューロゲニン3プロモーターを阻害することを示すグラフである。図17Aは、所定のレポーター構築物でトランスフェクションされ、かつCMVプロモーターの下流に連結されたHES1 cDNAを含む所定の量の発現プラスミド(ベクターpBAT12)で同時トランスフェクションされたNIH3T3細胞を用いた結果を示す。ルシフェラーゼ活性は、レポータープラスミドのみでトランスフェクションされた細胞における活性と相対的に表す。図17Bは、90 ng/106細胞のHES1発現プラスミドとの同時トランスフェクションの存在下または非存在下の、ルシフェラーゼの上流に連結させたNgn3プロモーターの様々な断片およびラウス肉種ウイルス(RSV)プロモーターの相対活性を示す。図17Cは、NIH3T3細胞およびαTC1.6細胞における、同時トランスフェクションしたHES1による、所定のプロモーターからのルシフェラーゼ活性の抑制率を示す。抑制率とは、相対ルシフェラーゼ活性の逆数である。トランスフェクションは三つ組みで少なくとも3回別々に行った。誤差は、平均値の±標準誤差として示す。
【図18】図18Aは、ヒトngn3遺伝子の転写開始部位(+1)のすぐ上流のDNA配列の模式図を示す。HES1結合部位として可能な部位(Nボックス)を示す。図18B〜図18Cは、HES1がヒトngn3プロモーターに結合することを示す電気泳動移動度シフト分析の写真である。図18Bは、細菌的に産生されたHES1が、マウスHES1プロモーター(H1P)の既に特徴づけられている部位だけでなく、近位プロモーターのNボックスにも結合し得ることを実証する電気泳動移動度シフト分析を示す。GST-HES1タンパク質またはGSTタンパク質のいずれかを、表示のプローブと共にインキュベートした。一番右のレーンは、HES1抗血清の添加により、HES1複合体がスーパーシフトすることを示す。図18Cは、GST-HES1タンパク質の結合に関して競合できるHES1結合部位の相対的能力を実証する電気泳動移動度シフト分析を示す。3つのngn3プロモーター部位はすべて、以前に記載されているHES1プロモーター(H1P)のHES1結合部位よりも高い親和性で結合する。一方、200倍過剰量のオリゴヌクレオチドE1(近位プロモーター由来の無関係のEボックス配列)は、複合体形成に効果を及ぼさない。
【図19A】図19A〜図19Cは、β-ガラクトシダーゼ(adCMV-Bgal)、ニューロゲニン3(ヒト)(adCMV-Ngn3)、neuroD1(マウス)(adCMV-ND1)、mash1(マウス)(adCMV-Mash1)、またはmyoD(マウス、筋bHLH遺伝子)(adCMV-MyoD)を発現するアデノウイルスベクターを、2つのマウス膵管細胞株mPAC L20およびmPAC L4S2にトランスフェクションした効果を示すアガロースゲルの写真である。図19Aは、これらのベクターの発現がNgN3、ND1、Mash1、MyoD、およびβ-アクチンRNAレベルに及ぼす影響を示す。
【図19B】図19A〜図19Cは、β-ガラクトシダーゼ(adCMV-Bgal)、ニューロゲニン3(ヒト)(adCMV-Ngn3)、neuroD1(マウス)(adCMV-ND1)、mash1(マウス)(adCMV-Mash1)、またはmyoD(マウス、筋bHLH遺伝子)(adCMV-MyoD)を発現するアデノウイルスベクターを、2つのマウス膵管細胞株mPAC L20およびmPAC L4S2にトランスフェクションした効果を示すアガロースゲルの写真である。図19Bは、これらのベクターの発現がグルカゴン、ソマトスタチン、インスリン、IAPP、グルコキナーゼ、グルコース輸送体2、およびβ-アクチンRNAレベルに及ぼす影響を示す。
【図19C】図19A〜図19Cは、β-ガラクトシダーゼ(adCMV-Bgal)、ニューロゲニン3(ヒト)(adCMV-Ngn3)、neuroD1(マウス)(adCMV-ND1)、mash1(マウス)(adCMV-Mash1)、またはmyoD(マウス、筋bHLH遺伝子)(adCMV-MyoD)を発現するアデノウイルスベクターを、2つのマウス膵管細胞株mPAC L20およびmPAC L4S2にトランスフェクションした効果を示すアガロースゲルの写真である。図19Cは、これらのベクターの発現がPax4、Pax6、Nkx2.2、Nkx6.1、Isl-1、およびPdx-1RNAレベルに及ぼす影響を示す。
【発明を実施するための形態】
【0018】
発明の詳細な説明
膵島細胞およびインスリン産生のための本発明の構成および本方法を説明する前に、本発明は記載した特定の方法、プロトコール、細胞株、ベクター、および試薬に制限されず、当然変更され得ることが理解されねばならない。また、本明細書で使用する専門用語は、特定の態様を説明する目的のためのみに用いられ、添付の特許請求の範囲によってのみ限定される本発明の範囲を制限する意図はないことも理解されねばならない。
【0019】
本明細書および添付の特許請求の範囲で使用する単数形「1つの」、「ある」、および「その」とは、特記する場合を除き、その対象物の複数形も含むことに留意されたい。したがって、例えば「1つの膵島転写因子」についての言及は複数のそのような膵島転写因子およびその機能的同等物を含み、「そのポリヌクレオチド」についての言及は1つまたは複数のポリヌクレオチドおよび当業者に周知のその同等物を含み、以下同様である。
【0020】
特記されない限り、本明細書で使用する専門用語および科学用語はすべて、本発明が属する当技術分野の当業者によって共通に理解されるものと同じ意味をもつ。本発明の実施または試験において、本明細書に記載したものと類似したまたは同等の任意の方法、装置、および材料が使用できるが、本明細書に記載された方法、装置、および材料が好ましい。
【0021】
本明細書で言及したすべての文献は、その文献に記載され本発明に関連して使用される可能性のある、例えば、細胞株、ベクター、および方法を説明および開示する目的で参照として本明細書に組み入れられる。本明細書内で考察される文献は、単に本出願の出願日以前にそれらが開示されているというだけの理由で提供されたものである。本明細書に記載されたいかなるものも、先行発明のせいで本発明者らがこれら開示に先行できないことを認めると解釈されるべきではない。
【0022】
定義
本明細書で用いる「ポリヌクレオチド」とは、オリゴヌクレオチド、ヌクレオチド、およびその断片または一部、ならびにペプチド核酸(PNA)、その断片、部分、またはアンチセンス分子を指し、さらにゲノム由来また合成由来のDNAまたはRNAを指し、これは一本鎖または二本鎖であってよい。「ポリヌクレオチド」とはセンス鎖またはアンチセンス鎖を意味する。ある特定のポリヌクレオチド配列(例えば、Ngn3ポリペプチドをコードするポリヌクレオチド)を指すために「ポリヌクレオチド」という用語を用いる場合、「ポリヌクレオチド」とは、引用されたポリペプチドと機能的に同等なポリペプチド、例えば縮重変種のポリヌクレオチド、または引用されたポリペプチドの生物活性のある変種もしくは断片をコードするポリヌクレオチドを含むことが意図されており、これは本明細書で提供する配列と比較して実質的な配列類似性または配列同一性を有するポリヌクレオチドを含む。同様に、本明細書で用いる「ポリペプチド」とは、オリゴペプチド、ペプチド、またはタンパク質を指す。本明細書で天然型タンパク質分子のアミノ酸配列を指すのに「ポリペプチド」という用語を引用する場合、「ポリペプチド」およびその類語は、アミノ酸配列を、引用されたタンパク質分子と関連する完全な天然アミノ酸配列に限定することを意味してはおらず、それどころか生物活性のある変種または断片をも含むことを意図しており、本明細書で提供されたアミノ酸配列と比較して実質的な配列類似性または配列同一性を有するポリペプチドを含む。
【0023】
本明細書で用いる「縮重変種」とは、所与のポリヌクレオチド配列がコードするのと同じポリペプチドをコードするが、遺伝暗号の縮重によりコード配列の点で異なるようなポリヌクレオチド配列を指す。遺伝暗号は、2つ以上の異なるコドンが同じアミノ酸をコードする場合に「縮重する」。
【0024】
本明細書で用いる「ポリペプチド」とは、i)天然のポリペプチド、ii)生物活性のあるポリペプチド断片、iii)生物活性のある、ポリペプチドのポリペプチド類似体、またはiv)生物活性のあるポリペプチド変種のアミノ酸配列を有する組換えポリペプチドまたは非組換えポリペプチドのアミノ酸配列を指す。本発明で有用なポリペプチドは天然、合成、半合成、または組換えにかかわらず任意の供給源由来の任意の種から取得可能であり、例えば哺乳動物または非哺乳動物(例えば、は虫類、両生類、鳥類(例えばニワトリ))、特にヒトを含む哺乳動物、げっ歯類(例えば、マウスまたはラット)、ウシ、ヒツジ、ブタ、マウス、またはウマ、好ましくはラットまたはヒト等から取得可能である。例えば、「ヒトNgn3ポリペプチド」、「ヒトND1ポリペプチド」、または「ヒトMash1ポリペプチド」は、それぞれヒトに由来する単離ヒトNgn3、ND1、または Mash1ポリペプチドのアミノ酸配列を指し、天然の対立遺伝子変種すべてを含むことを意図するが、引用したタンパク質分子と関連する完全な天然アミノ酸配列のみにアミノ酸配列を限定する意図はない。
【0025】
本明細書で用いる「抗原性アミノ酸配列」とは、単独でまたは担体分子と結合して、哺乳動物において抗体応答を誘発し得るアミノ酸配列を意味する。
【0026】
ポリペプチドの「変種」とは、1つまたは複数のアミノ酸が改変されたアミノ酸配列として定義される。変種は、例えばイソロイシンによるロイシンの置換など、置換されたアミノ酸が同様の構造または化学特性を有するような、「保存的」変化を有する可能性がある。より稀ではあるが、変種は、例えばトリプトファンによるグリシンの置換など、「非保存的な」変化を有する可能性がある。同様の軽微な改変にはまた、アミノ酸の欠失もしくは挿入、またはその両方が含まれうる。生物活性または免疫活性を消失させることなく、どのアミノ酸残基がいくつ置換、挿入、または欠失されうるかを決定する手引きを、例えばDNAStarソフトウェア等の当技術分野で周知のコンピュータープログラムを用いて見出すことができる。
【0027】
「関心対象の核酸」または「関心対象のDNA」とは、哺乳動物対象への投与に望ましいタンパク質または他の分子をコードする任意の核酸(例えばDNA)を意味する。一般に核酸は、プロモーターなどの、発現に必要な他の配列に機能的に結合されている。
【0028】
「欠失」とは、関心対象のの天然ポリペプチドのアミノ酸配列またはヌクレオチド配列と比較して、それぞれ1つまたは複数のアミノ酸またはヌクレオチドの残基を欠いたアミノ酸またはヌクレオチドの配列のいずれかの変化として定義される。
【0029】
「挿入」または「付加」とは、天然ポリペプチドのアミノ酸配列またはヌクレオチド配列と比較して、それぞれ1つまたは複数のアミノ酸またはヌクレオチドの残基の付加をもたらすアミノ酸またはヌクレオチドの配列の変化である。
【0030】
「置換」とは、天然ポリペプチドのアミノ酸配列またはヌクレオチド配列と比較して、1つまたは複数のアミノ酸またはヌクレオチドがそれぞれ別のアミノ酸またはヌクレオチドにより置換されることによって生じる。
【0031】
「生物活性のある」という用語は、例えば、天然の膵島転写因子ポリペプチドの構造機能、制御機能、または生化学的機能を有するヒト膵島転写因子ポリペプチドを指す。同様に「免疫活性のある」とは、天然、組換え、または合成のヒト膵島転写因子ポリペプチド、もしくはその任意のオリゴペプチドが、適当な動物または細胞において特異的免疫応答を誘導して特異的抗体と結合する能力と定義される。
【0032】
本明細書で用いる「誘導体」という用語は、ポリペプチドをコードする核酸または核酸にコードされるポリペプチドの化学的修飾を指す。そのような修飾の実例として、アルキル基、アシル基、またはアミノ基による水素の置換が挙げられるであろう。核酸誘導体は、天然ポリペプチドの本質的な生物学的特徴を保持するポリペプチドをコードする。
【0033】
本明細書で用いる「単離された」という用語は、天然の化合物が存在する環境とは異なる環境にある関心対象の化合物(例えば、ポリヌクレオチドまたはポリペプチドのどちらか)を表すことを意図する。「単離された」とは、関心対象の化合物が十分に濃縮された試料、および/または関心対象の化合物が部分的にまたは十分に精製された試料内に存在する化合物を含むことを意図する。
【0034】
本明細書で用いる「実質的に精製された」または「実質的に遊離した」という用語は、自然環境から取り出されて、これに自然状態で結びついている少なくとも60%、好ましくは75%、最も好ましくは90%の他の化合物を含まないような化合物(例えば、ポリヌクレオチドまたはポリペプチドのどちらか)を指す。
【0035】
「形質転換」または「トランスフェクション」とは、新しい核酸(例えば、細胞にとって外因性のDNAまたはRNA)の取り込み後に細胞に誘導される永続的または一過的の遺伝的変化、好ましくは永続的な遺伝的変化を意味する。遺伝的変化は、新しい核酸を宿主細胞のゲノムに取り込むことによって、またはエピソーム成分として新しいDNAを一過的または安定的に維持することにより達成され得る。
【0036】
「形質転換細胞」とは、組換えDNA技術により、関心対象のタンパク質をコードするDNA分子が導入された細胞(または、DNA分子がその祖先の細胞に導入された細胞)を意味する。
【0037】
「構築物」とは、特異的なヌクレオチド配列を発現する目的で作製された組換え核酸、通常は組換えDNAを意味し、または他の組換えヌクレオチド配列の構築に用いられ得る。
【0038】
「ベクター」とは、関心対象のDNAによる標的細胞の形質転換を促進する、任意の生物的化合物または化学的化合物を意味する。例示的な生物学的ベクターには、ウイルス、特に弱毒化ウイルスおよび/または複製不能ウイルスが含まれる。例示的な化学的ベクターには、脂質複合体および裸のDNA構築物が含まれる。
【0039】
「裸のDNA」または「裸の核酸」またはDNA配列などは、ウイルス粒子、細菌細胞、または標的細胞の細胞質への核酸の送達を促進する他の封入手段に含まれない核酸分子を意味する。標的細胞の部位(例えば、細胞への移動を促進する手段、ヌクレアーゼ分解から核酸を保護する手段等)および/または標的上皮細胞の表面への核酸の送達を促進する手段により裸の核酸を任意に付随(例えば、製剤化)することができる。
【0040】
「プロモーター」とは、転写を指示するのに十分な最小配列を意味する。「プロモーター」はまた、細胞種特異的に制御可能な、組織特異的に制御可能な、または外部のシグナルもしくは薬剤で誘導可能な、プロモーター依存性遺伝子発現に十分なプロモーター成分を含むことが意図される。そのような成分は、天然遺伝子の5'領域または3'領域に位置している可能性がある。
【0041】
「機能的に結合された」とは、適当な分子(例えば、転写活性化タンパク質)が制御配列に結合された時に発現が可能になるように、DNA配列と制御配列が結合していることを意味する。
【0042】
「膵臓」とは、胃の後ろで脾臓と十二指腸の間に横向きに位置する大きく細長いブドウ状の腺を意味する。膵臓は、内分泌部分(内分泌部)および外分泌部分(外分泌部)からなる。ランゲルハンス島を含む内分泌部はインスリンを含むタンパク質を産生し、血流に直接分泌する。外分泌部分は分泌単位を含み、タンパク質の消化に必須の酵素を含む膵液を産生し、十二指腸に分泌する。
【0043】
「正常血糖」または「正常血糖状態」とは、高血糖に関連する疾患または病態を有する対象における血糖値と特に比較して、正常またはほぼ正常な血糖値を伴う状態を意味する。ヒトにおける正常血糖とは、約70 mg/dl〜約110 mg/dlの範囲の血糖値に相当する。「高血糖」とは血糖値の増加を伴う状態を意味し、これは一般に、約120 mg/dl、130 mg/dl、または140 mg/dlを上回る。
【0044】
「前駆細胞」とは膵島細胞に発達することが可能な任意の細胞を意味し、これにはインビボまたはインビトロにおける、胎児膵上皮細胞、成人膵細胞(例えば、膵管細胞、腺房細胞、および膵幹細胞)、腸上皮細胞、腸幹細胞もしくは腺窩細胞、他の組織由来の幹細胞(造血幹細胞または肝幹細胞等)、肝細胞(肝実質細胞および肝管細胞)、唾液腺細胞、神経幹細胞、筋幹細胞、および胚幹細胞等の、哺乳動物不死化細胞株または哺乳動物初代培養細胞が含まれる。
【0045】
「標的細胞」とは転写因子ヌクレオチド配列をコードするDNAを組み込むために選択される任意の細胞を意味し、これにはインビボまたはインビトロにおける、前駆細胞、胎児膵上皮細胞、成人膵細胞(例えば、膵管細胞、腺房細胞、および膵幹細胞)、腸上皮細胞、腸幹細胞もしくは腺窩細胞、他の組織由来の幹細胞(造血幹細胞、皮膚幹細胞、または肝幹細胞等)、肝細胞(肝実質細胞および肝管細胞)、唾液腺細胞、神経幹細胞、筋幹細胞、および胚幹細胞等の、哺乳動物不死化細胞株または哺乳動物初代培養細胞が含まれる。
【0046】
「膵島転写因子」または「膵島因子」または「転写因子」とは、膵島細胞の分化、形成、および/または発達に関与する任意の転写因子を意味し、その発現は膵島細胞の表現型を有する細胞、例えば、インスリンまたは膵島細胞に特徴的な他のマーカー(例えば、グルカゴン、グルコキナーゼ、膵島アミロイドポリペプチド(IAPP)、ソマトスタチン、膵臓ポリペプチド等)を産生する細胞の産生に寄与する。また同様に、機能的に同等な相同体も意味する。特に関心が持たれるのは、膵島細胞の発達に関与するクラスB塩基性ヘリックス・ループ・ヘリックス(bHLH)転写因子であり、これにはニューロゲニン(ニューロゲニン1、ニューロゲニン2、およびニューロゲニン3)、neuroD因子(NeuroD1/BETA2、neuroD2、およびNeuroD4/Math3)、およびMash因子Mash1が含まれる。また同様に、これら転写因子の機能的に同等な相同体も特に関心対象となる。
【0047】
本明細書で用いる「膵島転写因子制御経路」という用語は、活性化事象および非活性化事象のカスケードを介して協調して膵島細胞の発達を制御し、膵島細胞の表現型を決定するシグナル伝達分子および転写因子を含む一連のタンパク質を指す。膵島転写因子経路の正の制御因子は、特定の膵島転写因子またはその下流のエフェクターの発現を促進する制御因子であり、よってこれは膵島細胞表現型の誘導に正の影響を及ぼす。膵島転写因子経路の負の制御因子は、特定の膵島転写因子またはその下流のエフェクターの発現を阻害する制御因子であり、よって膵島細胞の表現型の誘導に負の影響を及ぼす。正の制御因子には、負の制御因子の活性または発現を阻害する因子が含まれる。
【0048】
本明細書で用いる「ニューロゲニン3(Ngn3)制御経路」という用語は、活性化事象および非活性化事象のカスケードを介して協調してNgn3活性を促進し、細胞が膵島細胞となるように導き、膵島細胞表現型の産生または維持を促進するシグナル伝達分子および転写因子を含む一連のタンパク質を指す。Ngn3経路の正の制御因子は、Ngn3またはその下流のエフェクター(例えば、Mash1、NeuroD1等)の発現を促進する制御因子であり、よってこれは膵島細胞の表現型の誘導に正の影響を及ぼす。Ngn3経路の負の制御因子は、Ngn3またはその上流もしくは下流のエフェクター(例えばHES1)の発現を阻害する制御因子であり、よってこれは膵島細胞の表現型の誘導に負の影響を及ぼす。Ngn3経路の正の制御因子には、Ngn3経路の負の制御因子の活性もしくは発現を阻害する因子、またはNgn3の発現を促進する因子が含まれる。Ngn3をコードする核酸は、Ngn3の発現増加を提供しNgn3経路の活性を促進するため、Ngn3経路の正の制御因子の一例である。HNF転写因子(例えば、HNF1、HNF3、およびHNF6)はNgn3の発現を促進する(正に制御する)ため、Ngn3経路の正の制御因子のさらなる一例である。
【0049】
本明細書で用いる「NeuroD1制御経路」という用語は、活性化事象および非活性化事象のカスケードを介して協調してNeuroD1活性を促進し、かつしたがって細胞が膵島細胞となるように導き、膵島細胞の表現型の産生または維持を促進するシグナル伝達分子および転写因子を含む一連のタンパク質を指す。NeuroD1経路の正の制御因子は、NeuroD1またはその下流のエフェクター(例えば、Mash1、Ngn3等)の発現を促進する制御因子であり、よってこれは膵島細胞の表現型の誘導に正の影響を及ぼす。NeuroD1経路の負の制御因子は、NeuroD1またはその上流もしくは下流の正のエフェクターの発現を阻害する制御因子であり、よって膵島細胞の表現型の誘導に負の影響を及ぼす。NeuroD1経路の正の制御因子には、NeuroD1経路の負の制御因子の活性もしくは発現を阻害する因子、またはNeuroD1の発現を促進する因子が含まれる。NeuroD1をコードする核酸は、NeuroD1の発現増加を提供しかつしたがってNeuroD1経路の活性を促進するため、NeuroD1経路の正の制御因子の一例である。正の制御因子には負の制御因子の活性または発現を阻害する因子が含まれ、かつNeuroD1をコードする核酸が含まれる。NeuroD1がNgn3の下流に作用するという条件下で、NeuroD1制御経路はNgn3制御経路と重複することに留意されたい。したがって、Ngn3およびNgn3の正の制御因子もまた、NeuroD1経路の正の制御因子である。
【0050】
本明細書で用いる「Mash1制御経路」という用語は、活性化事象および非活性化事象のカスケードを介して協調してMash1活性を促進し、かつしたがって細胞が膵島細胞となるように導き、膵島細胞の表現型の産生または維持を促進するシグナル伝達分子および転写因子を含む一連のタンパク質を指す。Mash1経路の正の制御因子は、Mash1またはその下流のエフェクター(例えば、Ngn3、NeuroD1等)の発現を促進する制御因子であり、よってこれは膵島細胞の表現型の誘導に正の影響を及ぼす。Mash1経路の負の制御因子は、Mash1もしくはその上流または下流のエフェクターの発現を阻害する制御因子であり、よってこれは膵島細胞の表現型の誘導に負の影響を及ぼす。正の制御因子には、Mash1制御経路の負の制御因子の活性もしくは発現を阻害する因子、またはMash1の発現を促進する因子が含まれる。Mash1をコードする核酸は、Mash1の発現増加を提供してMash1経路の活性を促進するため、Mash1経路の正の制御因子の一例である。Mash1はNeuroD1およびNgn3転写因子の上流に作用するため、Mash1制御経路はNgn3 およびNeuroD1制御経路と重複することに留意されたい。したがって、Mash1はNeuroD1およびNgn3制御経路の正の制御因子である。
【0051】
「膵島細胞」とは、通常膵臓ランゲルハンス島からなるホルモン産生細胞と類似した表現型を有する細胞を意味し、一般に、インスリン、グルカゴン、グルコキナーゼ、ソマトスタチン、膵臓ポリペプチド、または膵島アミロイドポリペプチド(IAPP)等の、膵臓ランゲルハンス島の細胞を他の膵細胞から通常識別するようなマーカーの発現を特徴とする。
【0052】
「β細胞」とは、インスリン、Nkx6.1、またはグルコキナーゼ等の、β細胞を他の膵島細胞から通常識別するマーカーの発現を特徴とする表現型を有する膵島細胞を意味する。
【0053】
「α細胞」とは、プログルカゴンまたはグルカゴン等の、α細胞を他の膵島細胞から通常識別するマーカーの発現を特徴とする表現型を有する膵島細胞を意味する。
【0054】
「標的細胞」とは、例えば、発現について関心対象の核酸の導入が望ましい細胞を意味する。本明細書における「標的細胞」は便宜上用いられているだけであり、かつ例えば、ターゲティング技術(例えば、物質が特定の細胞または細胞種に導入されるように優先的に導く標的分子)の使用を必要とする関心対象の核酸の導入を行うことを意図しているわけではない。
【0055】
「対象」または「患者」または「個体」とは、診断または治療が望ましい任意の哺乳動物対象を意味し、特にヒトを意味する。他の対象には、ウシ、イヌ、ネコ、モルモット、ウサギ、ラット、マウス、ウマ等が含まれ得る。特に関心が持たれるのは、(例えば、膵島転写因子をコードする核酸の対象へのインビボ導入により、または、膵島転写因子発現細胞(例えばβ細胞前駆細胞)の、もしくは膵島転写因子発現細胞から培養されインスリンを産生するほぼ発生し終えたまたは成熟したβ細胞の、対象への移植により)対象の細胞内でいずれかの膵島転写因子をコードする核酸を発現させることによる(例えば、疾患に関連する症状を緩和するための)治療影響を受けやすいインスリン関連疾患を有する対象である。
【0056】
「治療」、「治療すること」、「治療する」などの用語は、本明細書内で一般に、所望の薬理的および/または生理的効果を得ることを指すために用いられる。その効果は、疾病、疾患、もしくはその症状を完全にまたは部分的に防ぐという点で予防的であってよく、かつ/または、疾病や疾患および/もしくは疾病や疾患に起因する有害な影響を部分的にもしくは完全に安定化もしくは治癒させるという点で治療的であってよい。本明細書で用いられる「治療」とは、哺乳動物、特にヒトにおける疾病または疾患の任意の治療を包含し、かつ以下を含む:
(a)疾病、疾患、または症状を起こしやすい可能性があるが、まだそれを有すると診断されていない対象における疾病、疾患、または症状の発達を予防すること;(b)疾病、疾患、もしくは症状を阻害する、すなわちその発症を抑止すること、または、疾病、疾患、もしくは症状を軽減する、すなわち疾病、疾患、もしくは症状の退縮を引き起こすこと。したがって「糖尿病の治療」には、血糖値の低下、インスリン産生の増加等のうち1つまたは複数が含まれる。
【0057】
「インスリン関連疾患」とは、インスリン産生量の変化または対象が、例えば血糖値を調節するためにインスリンを利用する能力の変化によって直接的もしくは間接的に引き起こされる、またはこの変化に伴う、疾病、疾患、もしくは病態を意味する。インスリン関連疾患には、1型糖尿病、2型糖尿病、耐糖能異常、高血糖症、低血糖症等が含まれるが、これらに限定されるわけではない。特に関心が持たれるのは、(例えば、膵島転写因子をコードする核酸をインビボで対象に導入することにより、または、膵島転写因子を発現する細胞(例えばβ細胞前駆細胞)もしくは、膵島転写因子を発現する細胞から培養されかつインスリンを産生するほぼ発達し終えたまたは成熟したβ細胞を対象に移植することにより)対象の細胞内でいずれかの膵島転写因子をコードする核酸を発現させることによる(例えば、疾患に関連する症状を緩和するための)治療の影響を受けやすいインスリン関連疾患である。
【0058】
明瞭にする目的で、以下の表により、本明細書中で用いる転写因子の名称と文献中で用いられる同一因子の名称、および対応するヒト遺伝子の正式名称との相互参照を提供する。
*ヒト遺伝子命名法委員会(Human Gene Nomenclature Committee)により承認されている。
【0059】
発明の概略
本発明は、膵島転写因子をコードするポリヌクレオチド配列を膵細胞または他の適当な細胞に導入することにより、インスリン産生β細胞を含む、膵島細胞の表現型を有する細胞の産生が誘導されるという発見に基づく。この発見そのものは、成熟した膵細胞(非β細胞)内でニューロゲニン3(Ngn3)活性の増加を提供すると、非β膵細胞が、膵β細胞表現型(例えば、インスリンの産生)を有する細胞となるという発見に基づく。
【0060】
膵島転写因子活性の増加は、例えば、膵島転写因子をコードするポリヌクレオチドを細胞に導入して膵島転写因子の発現(これは、細胞における内在性膵島転写因子の発現に追加されうる)を提供することにより、(例えば、膵島転写因子の発現を正に制御する第二の転写因子をコードするポリヌクレオチドを導入することによって、またはさもなくばそのような膵島転写因子の正の制御因子の活性もしくは発現を増加させることによって)膵島転写因子の正の制御因子の発現レベルの増加を提供することにより、(例えば、その発現を阻害することによって)膵島転写因子の発現もしくは活性の負の制御因子または阻害剤の活性を阻害することにより、膵島転写因子によって正に制御される下流のエフェクターの発現を増加させることにより、および、本明細書を読むことによって当業者に容易に明らかになると考えられる他の方法により達成され得る。転写因子の発現または活性の調節(例えば、膵島転写因子活性の増加、または膵島転写因子発現の阻害剤の発現もしくは活性の減少)は、その経路のシグナル伝達分子(受容体、リガンド、細胞内エフェクター)ならびに合成および天然の小分子制御因子の使用によっても達成され得る。
【0061】
1つの態様において、Ngn3活性の増加は、例えば、Ngn3をコードするポリヌクレオチドを細胞に導入してNgn3の発現(これは、細胞における内因性Ngn3発現に追加されうる)を提供することにより、(例えば、Ngn3の発現を正に制御する転写因子(例えば、Mash1、HNF1、HNF3、HNF6等)をコードするポリヌクレオチドを導入することによって、またはさもなくばそのようなNgn3の正の制御因子の活性もしくは発現を増加させることによって)Ngn3の正の制御因子の発現レベルの増加を提供することにより、(例えば、その発現を阻害することによって)Ngn3の発現もしくは活性の負の制御因子または阻害剤の活性を阻害することにより、Ngn3によって正に制御される下流のエフェクター(例えばneuroD1)の発現を増加させることにより、および本明細書を読むことにより当業者に容易に明らかになるであろう他の方法により達成され得る。Ngn3の発現または活性の調節(例えば、Ngn3活性の増加、またはNgn3発現の阻害剤の発現もしくは活性の減少)は、その経路のシグナル伝達分子(受容体、リガンド、細胞内エフェクター)ならびに合成および天然の小分子制御因子の使用によっても達成され得る。
【0062】
別の態様において、NeuroD1活性の増加は、例えば、NeuroD1をコードするポリヌクレオチドを細胞に導入してNeuroD1の発現(これは細胞における内因性NeuroD1発現に追加されうる)を提供することにより、(例えば、NeuroD1の発現を正に制御する転写因子(例えば、Mash1、Ngn3、HNF1、HNF3、HNF6等)をコードするポリヌクレオチドを導入することによって、またはさもなくばそのようなNeuroD1の正の制御因子の活性もしくは発現を増加させることによって)NeuroD1の正の制御因子の発現レベルの増加を提供することにより、(例えば、発現を阻害することによって)NeuroD1の発現もしくは活性の負の制御因子または阻害剤の活性を阻害することにより、NeuroD1によって正に制御される下流のエフェクターの発現を増加させることにより、および本明細書を読むことにより当業者に容易に明らかになるであろう他の方法により達成され得る。NeuroD1の発現または活性の調節(例えば、NeuroD1活性の増加、またはNeuroD1発現の阻害剤の発現もしくは活性の減少)は、その経路のシグナル伝達分子(受容体、リガンド、細胞内エフェクター)ならびに合成および天然の小分子制御因子の使用によっても達成され得る。
【0063】
別の態様において、Mash1活性の増加は、例えば、Mash1をコードするポリヌクレオチドを細胞に導入してMash1の発現(これは、細胞における内因性Mash1発現に追加されうる)を提供することにより、Mash1の正の制御因子の発現レベルの増加を提供することにより(例えば、Mash1の発現を正に制御する転写因子をコードするポリヌクレオチドを導入することによって、またはさもなくばそのようなMash1の正の制御因子の活性もしくは発現を増加させることによって)、(例えば、発現を阻害することによって)Mash1の発現もしくは活性の負の制御因子または阻害剤の活性を阻害することにより、Mash1によって正に制御される下流のエフェクター(例えば、Ngn3、NeuroD1等)の発現を増加させることにより、および本明細書を読むことにより当業者に容易に明らかになるであろう他の方法により達成され得る。Mash1の発現または活性の調節(例えば、Mash1活性の増加、またはMash1発現の阻害剤の発現もしくは活性の減少)は、その経路のシグナル伝達分子(受容体、リガンド、細胞内エフェクター)ならびに合成および天然の小分子制御因子の使用によっても達成され得る。
【0064】
本発明は一般に、ニューロゲニン(ニューロゲニン1/NEUROG1/MATH4C/NeuroD3、ニューロゲニン2/NEUROG2/MATH4A、またはニューロゲニン3/NEUROG3/MATH4B)、neuroD因子(NeuroD1/BETA2/BHF1、NeuroD2/NDRF、MATH2/NEX1/DLX3、NeuroD4/Math3)、Mash因子(Mash1およびMash2)、およびatonal関連因子(MATH1/ATOH1)を含む神経原性の塩基性ヘリックス・ループ・ヘリックス因子(bHLH)から選択される少なくとも1つの膵島転写因子、ならびにそれらの組み合わせまたは他の遺伝子との組み合わせの発現増加を提供し、膵β細胞の誘導を提供することを含む。特に関心が持たれるのはNgn3である。
【0065】
図10は、神経内分泌bHLHタンパク質、ならびにmyoD(筋原性クラスB bHLHタンパク質)およびE47(クラスA(遍在性)bHLHタンパク質)を加えてのアラインメント系図の略図である。ヒトNgn3アミノ酸配列である「h.ngn3」以外はすべて、マウスのアミノ酸配列に基づく。配列は、MacVector6.5.1配列解析プログラム(Oxford Molecular)で提供される多重配列整列アルゴリズムClustal-Wを用いて整列化させた。クラスAおよびBの定義は、Murreら(Murreら、(1989) Cell 58(3)、537-44)の分類に基づく。
【0066】
膵島転写因子は、膵島細胞の分化および発達に関与する。膵島転写因子には、クラスB塩基性ヘリックス・ループ・ヘリックス(bHLH)ファミリーの転写因子に属する転写因子が含まれ、このファミリーの因子は多くの細胞種の成長および分化を制御することが知られている。膵島細胞および発達過程の膵臓は多様なクラスB bHLH遺伝子群を発現するが、中でも最も多く発現されるのはNgn3、NeuroD1/BETA2、Mash1、およびNeuroD4/Math3である。NeuroD1は、膵島細胞の初期分化および膵β細胞におけるインスリン転写の制御に関与することが示されている。ニューロゲニン3は膵臓の発達中にNeuroD1/BETA2を活性化するため、ニューロゲニン3は膵島細胞分化の過程で活性化される膵島転写因子の階層においてneuroD1の上流に位置する。Mash1は膵臓の発達中にNgn3を活性化するため、Mash1は膵島細胞分化の過程で活性化される膵島転写因子の階層においてNgn3の上流に位置する。
【0067】
Ngn3は膵島細胞前駆細胞で発現されて前内分泌遺伝子として機能し、膵島細胞の分化を駆動させる。Ngn3は膵島細胞種4つすべての発達過程において初期に発現され、Pax4、Pax6、Isl1、およびNkx2.2、ならびにNeuroD1/BETA1等の他の膵島転写因子の制御に関与する。Ngn3の初期発現および異所的発現は、膵島細胞の初期分化および異所的分化をもたらしうる。他の膵島転写因子には、例えばPax4、Pax6、Isl1、Nkx2.2、およびNkx6.1といったホメオドメイン因子等の非bHLH因子も含まれる。これらの因子はNgn3のすぐ下流(例えばPax4、Pax6、Isl1、およびNkx2.2)または上流に位置し、膵島細胞の発達に関与する。pou-ホメオドメイン因子HNF1およびwinged-ヘリックス因子HNF3はNgn3の上流に位置し、cut-ホメオドメイン因子HNF6とともに膵島細胞の分化に関与すると見なされており、本発明に基づく膵島転写因子のさらなる例である。
【0068】
β細胞の発達の誘導
膵β細胞は、インビボ(例えば膵管の管腔に核酸を導入することによって例えば膵島転写因子をコードする核酸(例えばRNAまたはDNA)を対象の膵臓に投与することにより)またはインビトロ(例えば、培養中に膵島転写因子をコードする核酸(例えばRNAまたはDNA)を標的細胞(例えば単離した非β膵細胞)に接触させる(次にその細胞を培養して増加させ、対象に移植する)ことにより)のどちらかにおいて、膵細胞における膵島転写因子の産生を提供することにより非β膵細胞から産生され得る。
【0069】
標的細胞における膵島転写因子の発現は、様々な方法により達成され得る。例えば1つの態様において、膵島転写因子の発現は、膵島転写因子をコードする核酸(例えばDNAまたはRNA)を導入し、標的細胞においてコードされる膵島転写因子ポリペプチドの発現を提供することにより達成される。別の態様において、膵島転写因子の発現は、膵島転写因子の発現の誘導を提供するタンパク質をコードする遺伝子の導入(例えば、標的細胞における膵島転写因子発現の「上流の」正の制御因子の発現)により誘導される。別の態様において、膵島転写因子の発現は、膵島転写因子発現の負の制御因子の活性(例えば、機能または発現)を阻害するタンパク質をコードする遺伝子の導入により達成される。別の態様において、膵島転写因子の発現は、膵島転写因子の発現の誘導を提供する小分子(例えば、標的細胞において膵島転写因子の発現を誘導する小分子薬剤)の導入により誘導される。さらに、本発明の膵β細胞の産生は、膵島転写因子により誘導される因子の産生を提供することによっても達成され得る。
【0070】
特に関心の持たれる1つの態様において、β細胞は、標的細胞においてβ細胞の表現型を誘導するのに十分なレベルのニューロゲニン3(Ngn3)の発現を提供することにより産生される。1つの態様において、Ngn3の発現は、Ngn3をコードする核酸(例えばDNAまたはRNA)を導入して、標的細胞においてコードされるNgn3ポリペプチドの発現を提供することにより達成される。別の態様において、Ngn3の発現は、Ngn3発現の誘導を提供するタンパク質をコードする遺伝子の導入(例えば、標的細胞におけるNgn3発現の「上流の」正の制御因子(例えば、Mash1、HNF1、HNF3、HNF6等)の発現)により誘導される。別の態様において、Ngn3の発現は、Ngn3発現の負の制御因子の活性(例えば、機能または発現)を阻害するタンパク質をコードする遺伝子の導入により達成される。別の態様において、Ngn3の発現は、Ngn3の発現の誘導を提供する小分子(例えば、標的細胞においてNgn3の発現を誘導する小分子薬剤)の導入により誘導される。さらに、本発明の膵β細胞の産生は、Ngn3により誘導される因子の産生を提供することによっても達成され得る。
【0071】
別の態様において、β細胞は、標的細胞においてβ細胞の表現型を誘導するのに十分なレベルのNeuroD1の発現を提供することにより産生される。1つの態様において、NeuroD1の発現は、NeuroD1をコードする核酸(例えばDNAまたはRNA)を導入し、標的細胞においてコードされるNeuroD1ポリペプチドの発現を提供することにより達成される。別の態様において、NeuroD1の発現は、NeuroD1の発現の誘導を提供するタンパク質をコードする遺伝子の導入(例えば、標的細胞におけるNeuroD1発現の「上流の」正の制御因子(例えば、Mash1、Ngn3、HNF1、HNF3、HNF6等)の発現)により誘導される。別の態様において、NeuroD1の発現は、NeuroD1発現の負の制御因子の活性(例えば、機能また発現)を阻害するタンパク質をコードする遺伝子の導入により達成される。別の態様において、NeuroD1の発現は、NeuroD1の発現誘導を提供する小分子(例えば、標的細胞においてNeuroD1の発現を誘導する小分子薬剤)の導入により誘導される。さらに、本発明の膵β細胞の産生は、NeuroD1により誘導される因子の産生を提供することによっても達成され得る。
【0072】
別の態様において、β細胞は、標的細胞においてβ細胞の表現型を誘導するのに十分なレベルのMash1の発現を提供することにより産生される。1つの態様において、Mash1の発現は、Mash1をコードする核酸(例えばDNAまたはRNA)を導入し、標的細胞においてコードされるMash1ポリペプチドの発現を提供することにより達成される。別の態様において、Mash1の発現は、Mash1の発現の誘導を提供するタンパク質をコードする遺伝子の導入(例えば、標的細胞におけるMash1発現の「上流の」正の制御因子の発現)により誘導される。別の態様において、Mash1の発現は、Mash1発現の負の制御因子の活性(例えば、機能または発現)を阻害するタンパク質をコードする遺伝子の導入により達成される。別の態様において、Mash1の発現は、Mash1の発現の誘導を提供する小分子(例えば、標的細胞においてNgn3の発現を誘導する小分子薬剤)の導入により誘導される。さらに、本発明の膵β細胞の産生は、Mash1により誘導される因子の産生を提供することによっても達成され得る。
【0073】
本開示を読むことによって当業者に容易に理解されると考えられるように、膵島転写因子の発現は、これらのアプローチの任意の組み合わせを提供することにより達成され得る。例えば、本発明は、膵島転写因子をコードする核酸および内因性膵島転写因子遺伝子の正の制御因子の両方を標的細胞において発現すること、導入した膵島転写因子核酸ならびに内因性膵島転写遺伝子および導入した膵島転写因子配列の負の制御因子の阻害剤を発現すること等も提供する。一般に、例えば(膵島転写因子をコードする核酸の導入によって、または内因性膵島転写因子の発現の提供によって)膵島転写因子自体の発現を提供することにより、および/または、通常は膵島転写因子が発現した結果として産生される膵島転写因子の「下流の」因子の産生を提供することにより、膵島転写因子の活性を提供するようなアプローチの任意の組み合わせは本発明の範囲内である。
【0074】
1つの態様において、本発明は、Ngn3をコードする核酸および内因性Ngn3遺伝子の正の制御因子(例えば、Mash1、HNF1、HNF3、HNF6等)の両方を標的細胞において発現すること、導入したNgn3核酸ならびに内因性Ngn3および導入したNgn3配列の負の制御因子の阻害剤を発現すること等を提供する。一般に、(Ngn3をコードする核酸の導入によって、または内因性Ngn3の発現を提供することによって)例えばNgn3自体の発現を提供することにより、および/または通常はNgn3が発現した結果として産生されるNgn3の「下流の」因子(例えばNeuroD1等)の産生を提供することにより、Ngn3の活性を提供するアプローチの任意の組み合わせは、全般にわたって本発明の範囲内である。Ngn3発現の正の制御因子にはMash1、HNF1、HNF3、およびHNF6が含まれるが、必ずしもこれらに限定されるわけではない。
【0075】
別の態様において、本発明は、NeuroD1をコードする核酸および内因性NeuroD1遺伝子の正の制御因子(例えば、Mash1、Ngn3、HNF1、HNF3、HNF6等)の両方を標的細胞において発現すること、導入したNeuroD1核酸ならびに内因性NeuroD1および導入したNeuroD1配列の負の制御因子の阻害剤を発現すること等を提供する。一般に、(NeuroD1をコードする核酸の導入によって、または内因性NeuroD1の発現を提供することによって)例えばNeuroD1自体の発現を提供することにより、および/または通常はNgn3が発現した結果として産生されるNeuroD1の「下流の」因子の産生を提供することにより、Ngn3の活性を提供するアプローチの任意の組み合わせは、全般にわたって本発明の範囲内である。NeuroD1の正の制御因子にはMash1、Ngn3、HNF1、HNF3、およびHNF6が含まれるが、必ずしもこれらに限定されるわけではない。
【0076】
別の態様において、本発明は、Mash1をコードする核酸および内因性Mash1遺伝子の正の制御因子の両方を標的細胞において発現すること、導入したMash1核酸ならびに内因性Mash1および導入したMash1配列の負の制御因子の阻害剤を発現すること等を提供する。一般に(Mash1をコードする核酸の導入によって、または内因性Mash1の発現を提供することによって)例えばMash1自体の発現を提供することにより、および/または通常はMash1が発現した結果として産生されるMash1の「下流の」因子(例えば、Ngn3、NeuroD1等)の産生を提供することにより、Mash1の活性を提供するアプローチの任意の組み合わせは、全般にわたって本発明の範囲内である。
【0077】
さらに、膵島転写因子経路の活性の誘導は、核酸以外の天然分子または合成分子を用いて達成され得る。例えば、膵島転写因子活性は、例えば膵島転写因子発現の負の制御因子の活性を阻害することによって、その経路に関与する膵島転写因子の発現を促進する合成分子を用いることにより、誘導され得る。
【0078】
1つの態様において、Ngn3制御経路の活性の誘導は、核酸以外の天然または合成分子を用いて達成される。例えば、Ngn3活性は、例えばNgn3発現の負の制御因子の活性を阻害することによって、Ngn3発現を促進する合成分子を用いることにより誘導される。Ngn3発現の抑制的な転写因子にはHES1が含まれるが、必ずしもこれに限定されない。Ngn3発現を阻害する負のシグナル伝達経路にはNotch経路が含まれるが、必ずしもこれに限定されるわけではない。
【0079】
別の態様において、NeuroD1制御経路の活性の誘導は、核酸以外の天然または合成分子を用いて達成される。例えばNeuroD1活性は、例えばNeuroD1発現の負の制御因子の活性を阻害することによって、NeuroD1発現を促進する合成分子を用いることにより誘導される。
【0080】
別の態様において、Mash1制御経路の活性の誘導は、核酸以外の天然または合成分子を用いて達成される。例えばMash1活性は、例えばMash1発現の負の制御因子の活性を阻害することによって、Mash1発現を促進する合成分子を用いることにより誘導される。
【0081】
膵島転写因子核酸
「膵島転写因子遺伝子」という用語は、膵島細胞で発現される転写因子と膵島細胞の発生、分化、または形成に関与する転写因子の両方を表すために用いる。「膵島転写因子遺伝子」という用語はまた、コード領域の範囲を超えて最大約2.5 kb、約5 kb、10 kb、または約15 kbに至る、特定の膵島転写因子ポリペプチドをコードするオープンリーディングフレーム、イントロン、ならびに発現の制御に関与する5'側および3'側の隣接非コードヌクレオチド配列を意味することを意図するが、どちらかの方向にさらに伸長する可能性もある。膵島転写因子をコードするDNA配列は、cDNAまたはゲノムDNAもしくはそれらの断片であってよい。染色体外での維持または宿主への組込みのため、遺伝子は適当なベクターに導入され得る。
【0082】
本明細書で用いる「cDNA」という用語は、天然の成熟RNA種に見られる配列成分の配置を共有する核酸すべてを含むことを意図するが、ここで配列成分とはエキソン(例えば、コードされたポリペプチドのオープンリーディングフレームをコードする配列)ならびに3'側および5'側の非コード領域のことである。通常、mRNA種は、核RNAスプライシングによって介在イントロンが除去された、関心対象のポリペプチドをコードする連続的なオープンリーディングフレームを作製するための連続的エキソンを有する。
【0083】
関心対象の膵島転写因子ゲノム配列は、列挙された配列に定義されるように開始コドンと終止コドンの間の核酸を含み、天然の染色体に通常存在するイントロンのすべてを含む。さらに、成熟mRNAに見られる3'側および5'側の非翻訳領域を含んでもよい。さらに、プロモーター、エンハンサー等のような特定の転写制御配列および翻訳制御配列を含んでもよく、転写領域の5'末端または 3'末端のどちらかに約2.5 kb、約5 kb、約10 kb、または約15 kb、場合によってはさらに長い、隣接ゲノムDNAを含む。ゲノムDNAは、100 kbpもしくはそれ以上の大きな断片、または隣接する染色体配列を実質的に含まない小さな断片として単離され得る。
【0084】
この5'領域の配列、ならびにさらに5'上流の配列および3'下流の配列は、エンハンサー結合部位を含む、膵島転写因子が発現される組織中で発現を提供する
プロモーター成分に利用される可能性がある。本発明の膵島転写因子プロモーター成分の配列は、任意の種(例えば、哺乳動物または非哺乳動物(例えば、は虫類、両生類、鳥類(例えばニワトリ))、特に、ヒトを含む哺乳動物、げっ歯類(例えば、マウスまたはラット)、ウシ、ヒツジ、ブタ、マウス、またはウマ、好ましくは、マウスまたはヒト)のヌクレオチド配列に基づきうり、天然、合成、半合成、もしくは組換えにかかわらず任意の供給源から単離または産生され得る。
【0085】
本発明で用いる核酸組成物は、必要に応じて膵島転写因子ポリペプチドのすべてまたは一部をコードし得るが、通常は少なくとも実質的にそのすべてをコードし得る。DNA配列の断片を、従来法による化学的合成オリゴヌクレオチド、制限酵素消化、PCR増幅等により得ることができる。ほとんどの場合、DNA断片は少なくとも約10個の連続したヌクレオチドからなると考えられ、通常少なくとも約15ヌクレオチド、より通常は少なくとも約18ヌクレオチド〜約20ヌクレオチド、より通常は少なくとも約25ヌクレオチド〜約50ヌクレオチドからなる。そのような小さなDNA断片は、PCRのプライマー、ハイブリダイゼーションスクリーニング等に有用である。より大きなDNA断片、すなわち100ヌクレオチドを上回る断片は、コードされたポリペプチドの産生に有用である。PCR等の増幅反応に利用するために、一対のプライマーが用いられる。プライマー配列の正確な組成は本発明にとって重要ではないが、当技術分野で周知のように、ほとんどの用途においてプライマーはストリンジェントな条件下で対象の配列にハイブリダイズする。少なくとも約50ヌクレオチド、好ましくは約100ヌクレオチドの増幅産物を産生する一対のプライマーを選択することが好ましい。プライマー配列を選択するためのアルゴリズムが周知であり、市販のソフトウェアパッケージで入手できる。増幅プライマーはDNAの相補鎖にハイブリダイズし、互いに向かってプライミングすると考えられる。
【0086】
膵島転写因子遺伝子は、一般的には元の哺乳動物染色体とは別のものとして単離され、実質的に純粋なものとして得られる。通常、DNAは膵島転写因子またはその断片をコードする配列を含まない他の核酸配列を実質的に含まずに取得され、一般には少なくとも約50%、通常は少なくとも約90%の純度であり、これは、典型的な「組換え型」である(すなわち天然の染色体には通常は付随しない1つまたは複数のヌクレオチドが隣接している)。
【0087】
隣接するプロモーター領域およびコード領域を含む膵島転写因子の配列は、当技術分野で周知の様々な方法により変異を導入されうり、プロモーター強度、コードされるタンパク質の配列等においてターゲティングされた変化を生じる。そのような変異を有するDNA配列または産物は本明細書で提供される配列と実質的に類似しており、すなわちそれぞれ少なくとも1つのヌクレオチドまたはアミノ酸が異なっており、かつ少なくとも2つもしくは少なくとも約10個のヌクレオチドまたはアミノ酸が異なっていてもよい。一般に、配列変化とは、置換、挿入、または欠失であってよい。欠失には、ドメインまたはエキソンの欠失のような大きな変化がさらに含まれうる。膵島転写因子配列は主にbHLHドメイン内で保存されているが、このドメインの外側の領域ではそれほど保存されていない可能性があり、かつさらに例えばラット、マウス、およびヒト間ではほとんど保存されていない可能性さえあることに留意されねばならない。したがって、膵島転写因子は、bHLHドメインの外側のヌクレオチドおよびアミノ酸残基のさらなる変化を許容し、かつbHLHドメイン内で起こる変化よりもずっと大きな程度の変化まで機能を保持できる。そのように修飾された膵島転写因子の配列を用いて、例えば、膵島細胞を産生する目的で標的細胞に導入するベクターを作製することができる。
【0088】
クローニングされた遺伝子のインビトロ突然変異誘発を行うための技法は周知である。変異を検出するためのプロトコールの例は、Gustinら、1993 Biotechniques 14:22;Barany、1985 Gene 37:111-23;Colicelliら、1985 Mol Gen Genet 199:537-9;およびPrentkiら、1984 Gene 29:303-13に見出されうる。部位特異的突然変異誘発法は、Sambrookら、1989 Molecular Cloning: A Laboratory Manual、CSH Press、pp. 15.3-15.108;Weinerら、1993 Gene 126:35-41;Sayersら、1992 Biotechniques 13:592-6;JonesおよびWinistorfer、1992 Biotechniques 12:528-30;Bartonら、1990 Nucleic Acids Res 18:7349-55;MarottiおよびTomich、1989 Gene Anal Tech 6:67-70;およびZhu 1989 Anal Biochem 177:120-4に見出され得る。
【0089】
本発明において特に関心の持たれる膵島転写因子はニューロゲニン転写因子ファミリーに属する、例えばニューロゲニン1(Ngn1)、ニューロゲニン2(Ngn2)、ニューロゲニン3(Ngn3)などの転写因子であるが、Ngn3が特に関心対象となる。ヒトNgn3のヌクレオチドおよびアミノ酸の配列は、配列表においてそれぞれ配列番号:1および2として提供される。
【0090】
ヒトNgn1のヌクレオチドおよびアミノ酸の配列は、GenBankアクセッション番号XM_003834(配列表においてそれぞれ配列番号:20および21として提供する)およびNM_006161(配列表においてそれぞれ配列番号:22および23として提供する)で入手可能である。ヒトNgn1のクローニングおよび発現は、例えば、McCormickら、(1996) Mol. Cell. Biol. 16(10)、5792-5800;およびTamimiら、(1997) Genomics 40(2)、355-357 (1997)に記載されている。
【0091】
ヒトNgn2のヌクレオチドおよびアミノ酸の配列は、GenBankアクセッション番号AF303002(配列表においてそれぞれ配列番号:24および25として提供する)およびXM_067897(配列表においてそれぞれ配列番号:26および27として提供する)で入手可能である。ヒトNgn1のクローニングおよび発現は、例えば、Simmonsら、(2001) Dev. Biol. 229(2)、327-339に記載されている。
【0092】
Ngn3の「上流」(したがってNgn3発現を活性化する)またはNgn3の「下流」のどちらかに作用し膵島細胞の表現型の発現を引き起こす転写因子についても、本発明での使用が意図されていることに留意されたい。
【0093】
ニューロゲニン3は、それ自体で十分、未分化の膵上皮細胞を膵島細胞にするように作用する。ニューロゲニン3の発現によりどの前駆細胞が膵島細胞に分化するかが決定されるため、ニューロゲニン3の発現を制御するシグナルもまた膵島細胞の形成に関与する。2.7 kbのngn3プロモーターはトランスジェニックマウスにおいて正しく発現を指示するのに十分であるが、遠位の配列がngn3の発現を大幅に増強することが示されている。この遠位プロモーター領域は、HNF6、HNF1α、およびHNF3βのような膵転写因子の結合部位のクラスターを含む。これらの膵転写因子がngn3遺伝子の発現を制御することが見出されており、したがってこれらも膵島細胞形成の制御に関与する。これらのシグナルは、糖尿病患者に新しい膵島細胞を産生するのに有効となり得る。
【0094】
別の態様において、膵島転写因子はneuroD転写因子ファミリーに属する転写因子であり、例えばNeuroD1、NeuroD2、NeuroD4であるが、NeuroD1が特に関心対象となる。ヒトNeuroD1遺伝子の配列および対応するヒトNeuroD1アミノ酸配列は、GenBankアクセッション番号NM_002500(配列表においてそれぞれ配列番号:28および29として提供する)ならびにXM_002573(配列表においてそれぞれ配列番号:30および31として提供する)およびAF045152(配列表においてそれぞれ配列番号:32および33として提供する)で入手可能である。ヒトNeuroD1のクローニングおよび発現は、例えば、Leeら、(1995) Science 268(5212)、836-844;Tamimiら、(1996) Genomics 34(3)、418-421;Yokoyamaら、(1996) DNA Res. 3(5)、311-320;およびYokoyamaら、(1196) Brain Res. Mol. Brain Res. 42(1)、135-139に記載されている。
【0095】
ヒトNeuroD4遺伝子および対応するヒトNeuroD4アミノ酸配列は、GenBankアクセッション番号AF203901(配列表においてそれぞれ配列番号:34および35として提供する)で入手可能である。
【0096】
NeuroD1の「上流」(したがってNeuroD1の発現を活性化する)またはNeuroD1の「下流」のどちらかに作用して膵島細胞の表現型の発現を引き起こす転写因子についても、本発明での使用が意図されることに留意されたい。
【0097】
別の態様において、膵島転写因子はMash転写因子ファミリーに属する転写因子、例えばMash1、Mash2であるが、Mash1が特に関心対象となる。ヒトMash1遺伝子の配列および対応するMash1アミノ酸配列は、GenBankアクセッション番号XM_006688(配列表においてそれぞれ配列番号:36および37として提供する)およびNM_004316(配列表においてそれぞれ配列番号:38および39として提供する)で入手可能である。ヒトMash1のクローニングおよび発現は、例えば、Ballら、(1993) Proc. Natl. Acad. Sci. U.S.A. 90(12)、5648-5652;Renaultら、(1995) Genomics 30(1)、81-83;Borgesら、(1997) Nature 386(6627)、852-855、およびChenら、(1997) Cell Growth Differ. 8(6)、677-686に記載されている。
【0098】
Mash1の「上流」(したがってMash1の発現を活性化する)またはMash1の「下流」のどちらかに作用して膵島細胞の表現型の発現を引き起こす転写因子についても、本発明での使用が意図されていることに留意されたい。
【0099】
島転写因子核酸を送達するための構築物
送達される膵島転写因子核酸がDNAである場合、関心対象のDNAに機能的に結合されたプロモーター(例えば、真核細胞内で機能するプロモーター)を有する任意の構築物が本発明において使用可能である。本発明に従って使用され得るDNA配列(または対応するRNA配列)を含む構築物は、関心対象のDNAまたRNAの配列を含む任意の真核発現構築物でよい。例えば、プラスミドまたはウイルス性構築物(例えばアデノウイルス)を切断して、連結可能な末端を有する直鎖状DNAを提供し得る。これらの末端を相補様の連結可能な末端を有する外因性DNAに結合させ、完全な複製単位および所望の表現型特性を有する生物学的に機能的な組換えDNA分子を提供する。構築物は真核生物および/または原核生物の宿主において複製可能であることが好ましく(真核生物においてはウイルス、原核生物においてはプラスミド)、そのような構築物は当技術分野で周知であり、市販されている。
【0100】
構築物を、当技術分野で周知の技法を用いて調製することができる。同様に、遺伝的に改変された宿主において外因性のDNA配列またはRNA配列を発現させるための技法は、当技術分野で周知である。(例えば、Kormalら、Proc. Natl. Acad. Sci. USA、84:2150-2154、1987;Sambrookら、Molecular Cloning: a Laboratory Manual、第2版、1989、Cold Spring Harbor Laboratory Press、Cold Spring Harbor、NYを参照のこと。いずれも関心対象のDNAを真核生物で発現させるための方法および組成物に関する参照として、本明細書に組み入れられる。)
【0101】
1つの態様において、DNA構築物はプロモーターを含み、膵細胞内で関心対象のDNAの発現を促進する。プロモーターは、サイトメガロウイルス(CMV)、マウス乳がんウイルス(MMTV)、ラウス肉種ウイルス(RSV)、またはアデノウイルスのプロモーターのような、真核細胞内で機能する強力なウイルスプロモーターであってよい。より具体的には、例示的なプロモーターには、ヒトCMVの前初期遺伝子由来のプロモーター(Boshartら、Cell 41:521-530、1985)およびRSVの長い末端反復配列(LTR)由来のプロモーター(Gormanら、Proc. Natl. Acad. Sci. USA 79:6777-6781、1982)が含まれる。これら2つのプロモーターでは、RSVプロモーターよりもCMVプロモーターを用いた方が発現レベルが高くなるため、ここではCMVプロモーターが好ましい。
【0102】
または、用いるプロモーターは、アクチン遺伝子プロモーターのような強力かつ一般的な真核プロモーターでもよい。1つの態様において、用いるプロモーターは組織特異的プロモーターでもよい。例えば、構築物に用いるプロモーターは、膵臓特異的プロモーター、管細胞特異的プロモーター、または幹細胞特異的プロモーターでよい。本発明の構築物は、プロモーターに加えて標的細胞内で発現を増強する配列も含みうる。
【0103】
別の態様において、プロモーターはテトラサイクリン調節性プロモーターのような調節性プロモーターであり、その発現は外因性の物質(例えばテトラサイクリン)に曝すことにより調節され得る。
【0104】
マーカー等の他の構成成分(例えば、抗生物質耐性遺伝子(アンピシリン耐性遺伝子等)またはβ-ガラクトシダーゼ)は、構築物、細菌細胞における構築物の安定な複製のための複製起点(好ましくは、高コピー数の複製起点)、核局在化シグナル、または、DNA構築物、それにコードされるタンパク質、もしくはその両方の産生を促進するその他の成分を含むおよび/または発現する細胞の選択または同定に役立つ。
【0105】
真核生物において発現させるためには、構築物は関心対象のDNAに機能的に結合された真核生物プロモーターを最低限含むべきであり、関心対象のDNAは次にポリアデニル化シグナル配列に結合させる。ポリアデニル化シグナル配列は、当技術分野で周知の様々なポリアデニル化シグナル配列のいずれかより選択されうる。例示的なポリアデニル化シグナル配列は、SV40初期ポリアデニル化シグナル配列である。適切な場合、特に関心対象のDNAがcDNAである(例えば、天然配列のイントロンを含まない)場合には、構築物は関心対象のDNAの発現レベルを上昇させ得る1つまたは複数のイントロンを含んでもよい。当技術分野で周知の様々なイントロンのうちいずれかのイントロンを使用できる(例えば、ヒトβ-グロビンイントロン、これは関心対象のDNAの5'の位置で構築物に挿入する)。
【0106】
別の態様において、細胞に送達される核酸は、膵島転写因子をコードするRNAである。この態様においては、RNAは標的細胞における発現(すなわちRNAの翻訳)について適合化される。関心対象のタンパク質をコードするRNA(例えばmRNA)の産生方法は当技術分野で周知であり、本発明に有用な膵島転写因子をコードするRNAの産生に容易に応用されうる。
【0107】
膵島転写因子をコードする核酸の送達
膵島転写因子をコードする核酸の送達は、ウイルスベクターまたは非ウイルスベクターを用いて達成され得る。1つの態様において、核酸はアデノウイルスのようなウイルス粒子に入れて送達される。別の態様において、ウイルス粒子(例えばアデノウイルス)またはカチオン性脂質またはリポソームのようなアジュバントと混合された裸のDNAを含む製剤として、核酸を送達する。「アジュバント」とは、それ自体で所望の効果を生じないが、活性化合物の作用を増大させるまたは改善する役割を果たす物質である。使用するベクターおよびベクター製剤の詳細は、導入するDNAの大きさ、利用する送達プロトコール等のようないくつかの因子に依存すると考えられる。例示的な非ウイルスベクターおよびウイルスベクターについて、以下でより詳細に説明する。
【0108】
I. ウイルスベクター
一般に、本発明に従って使用されるウイルスベクターは、複製不能となりかつ本発明による標的細胞での発現用の関心対象の組換え遺伝子を送達するように遺伝的に改変された、天然ウイルス由来のウイルス粒子から構成される。
【0109】
多くのウイルスベクターが当技術分野で周知であり、これには例えば、レトロウイルス、アデノウイルス、アデノ随伴ウイルス、単純ヘルペスウイルス(HSV)、サイトメガロウイルス(CMV)、ワクシニアウイルス、およびポリオウイルスのベクターが含まれる。アデノウイルスおよびAAVは、ゆっくりと複製する細胞および/または最終的に分化した細胞に効率的に感染するため、通常好ましいウイルスベクターである。ウイルスベクターは、標的細胞への選択的感染によって選択されうる。
【0110】
ウイルスベクターとして複製不能ウイルスを使用する場合には、関心対象のDNAに対応するDNAまたはRNAのいずれかを含む感染性ウイルス粒子は、ウイルス複製に必須ながら欠除している成分を提供する組換え細胞株にウイルス構築物を導入することによって産生されうる。1つの態様において、組換えウイルスベクターを用いた組換え細胞株の形質転換は、例えば、組換え細胞株のウイルス配列の、導入されたウイルスベクターへの相同的組換えにより、複製可能なウイルスの産生または実質的な産生をもたらさない。関心対象の核酸を含む複製不能ウイルス粒子の産生方法は当技術分野で周知であり、例えば、Rosenfeldら、Science 252:431-434、1991およびRosenfeld ら、Cell 68:143-155、1992(アデノウイルス)、米国特許第5,139,941号(アデノ随伴ウイルス)、米国特許第4,861,719号(レトロウイルス)、ならびに米国特許第5,356,806号(ワクシニアウイルス)に記載されている。流行性耳下腺炎ウイルスゲノムを操作するための方法と材料、ウイルス融合とウイルス複製に関する流行性耳下腺炎ウイルス遺伝子の特徴づけ、および流行性耳下腺炎ウイルスゲノムの構造と配列は、Tanabayashiら、J. Virol. 67:2928-2931、1993;Takeuchiら、Archiv. Virol.、128:177-183、1993;Tanabayashiら、Virol. 187:801-804、1992;Kawanoら、Virol.、179:857-861、1990;Elangoら、J. Gen. Virol. 69:2893-2900、1988に記載されている。
【0111】
II. 非ウイルスベクター
関心対象の核酸は、非ウイルスベクターを用いて細胞に導入してもよい。本明細書で使用する「非ウイルスベクター」には、裸のDNA(例えば、ウイルス粒子内に含まれず、脂質のような担体分子を含まないDNA)、裸の核酸を含む化学製剤(例えば、DNA(および/またはRNA)とカチオン性化合物(例えば、硫酸デキストラン、カチオン性脂質)との製剤)、およびウイルス粒子のようなアジュバントと混合された裸の核酸(例えば、関心対象のDNAはウイルス粒子内に含まれないが、製剤は裸のDNAとウイルス粒子(例えば、アデノウイルス粒子)との両方からなる)(例えば、Curielら、1992 Am. J. Respir. Cell Mol Biol. 6:247-52を参照のこと)が含まれることを意図する。したがってウイルス粒子が関心対象のDNAをウイルス粒子内に含まない場合にも「非ウイルスベクター」には核酸とウイルス粒子からなるベクターが含まれ得る。
【0112】
1つの態様において、製剤は、投与前に裸のDNA構築物と混合されたウイルス粒子を含む。全量約100μl中で、約108個〜約1010個のウイルス粒子(好ましくは約1×1010個〜約5×1010個、より好ましくは約3×1010個の粒子)を裸のDNA構築物(約5μg 〜50μg DNA、より好ましくは約8μg 〜25μg DNA)と混合する。好ましくは、ウイルス粒子はアデノウイルス粒子である(Curielら、1992、前記)。
【0113】
代替的にまたはさらに、核酸をポリ-L-リジンもしくはDEAC-デキストランのようなポリカチオン性物質、標的リガンド、および/またはDNA結合タンパク質(例えばヒストン)と複合体化することもできる。DNAまたはRNAとリポソームとの複合体製剤は、遺伝物質(DNAまたはRNA)に結合する脂質の混合物を含み、細胞への核酸の送達を促進する。本発明に従って使用できるリポソームには、DOPE(ジオレイルホスファチジルエタノールアミン)、CUDMEDA(N-(5-コレストラム-3-β-オール3-ウレタニル)-N',N'-ジメチルエチレンジアミン)が含まれる。
【0114】
例えば裸のDNAを、約2.5%〜15%(容量:容量)、好ましくは6%〜12%(容量:容量)の範囲にある濃度のリポフェクチン(商標)(LTI/BRL)を含む溶液として投与できる。本発明の方法に基づく送達用のDNA製剤についての好ましい方法および組成は米国特許第5,527,928号に記載されており、その開示は参照として本明細書に組み入れられる。
【0115】
また関心対象の核酸は、宿主細胞の生物学的特性を変化させる目的で、宿主細胞への送達を促進する担体分子(例えば、抗体または受容体リガンド)に結合させたDNAまたはRNAの化学製剤としても投与できる。「化学製剤」という用語は、タンパク質もしくは脂質またはその誘導体のような担体分子に核酸化合物の結合を可能にする、核酸の修飾を意味する。例示的なタンパク質担体分子には、標的膵細胞の細胞に特異的な抗体または受容体リガンド、例えば、標的膵細胞の細胞に結合した受容体と相互作用できる分子が含まれる。
【0116】
膵島転写因子ポリペプチドおよびそのようなポリペプチドに特異的に結合する抗体の産生
Ngn3、NeuroD1、Mash1、または他の膵島転写因子をコードする核酸を用いて、全長ポリペプチドまたはその断片、特に機能的なドメインに対応する断片、DNA結合部位等を合成することができ、他のタンパク質またはその一部への被験ポリペプチドの融合体もこれに含まれる。したがって、本発明での使用に適したポリヌクレオチドおよびポリペプチドには、霊長類、げっ歯類、イヌ、ネコ、ウマ、線虫、酵母などにおいて見出される膵島転写因子ポリペプチドおよびポリヌクレオチド、ならびにそれらの天然変種および非天然変種が含まれるが、これらに限定されるわけではない。
【0117】
膵島転写因子ポリペプチドを用いて抗体を産生することができるが、ここで、短い断片は特定のポリペプチドに特異的な抗体を提供し、より大きな断片またはタンパク質全体ではポリペプチド表面を覆う抗体の産生が可能になる。抗体は、ポリペプチドの野生型または変異型に対して産生され得る。例えば、関心対象のポリペプチドを発現する細胞による免疫、関心対象のポリペプチドが膜に挿入されているリポソームによる免疫等によって、これらのドメインに対応する単離ペプチドまたは天然タンパク質に対して抗体が産生され得る。
【0118】
抗体は従来法に基づいて調製されるが、ここで、発現されたポリペプチドまたはタンパク質は、それ自体でまたは周知の免疫原性担体(例えば、KLH、pre-S HBsAg、他のウイルスまたは真核生物タンパク質等)に結合して、免疫原として使用される。さらなる説明は、Monoclonal Antibodies: A Laboratory Manual、HarlowおよびLane編、Cold Spring Harbor Laboratories、Cold Spring Harbor、New York、1988を参照のこと。
【0119】
膵島転写因子に特異的に結合する抗体は、Ngn3、NeuroD1、またはMash1のような組換え膵島転写因子を発現する細胞の検出に用いることができる(例えば、以下でより詳細に説明するように、移植または体内移植の前など)。
【0120】
転写因子をコードする核酸の発現による膵島細胞の産生
膵島細胞は、本発明に基づいて様々な方法で産生することができる。一般に本発明は、膵島転写因子の産生を刺激することを含む。特に関心の持たれる態様において、本発明は、膵島転写因子をコードする核酸を細胞、通常は膵細胞に導入することによる、膵島転写因子の活性の増強を含む。
【0121】
I. 膵島転写因子をコードする核酸の導入によるインビトロにおける膵島細胞の産生
膵島転写因子をコードする核酸(例えば、Ngn3、MeuroD1、Mash1等)をインビトロで細胞に導入し、細胞内で発現させて、少なくとも一過的に発現させることができる。核酸を導入する細胞は、分化した上皮細胞(例えば、膵細胞、腸細胞、肝細胞、または管細胞)、多能性成人幹細胞もしくは多能性胚幹細胞、またはβ細胞に発達可能な任意の哺乳動物細胞もしくは膵島転写因子をコードする核酸の発現後にインビトロでインスリンの発現が可能な細胞とすることがきる。続いて、膵島細胞置換療法(例えばエクスビボ療法)による治療の影響を受けやすいインスリン関連疾患を有する対象にその細胞を移植する。
【0122】
1つの態様において、膵島転写因子の発現、好ましくはNgn3、MeuroD1、もしくはMash1、特にNgn3の発現、またはNgN3制御経路の他の正の制御因子の発現が提供され、(例えば自家移植を提供するために)対象に移植される宿主細胞は、移植を受ける個体由来の細胞である。例えば、1型糖尿病を有する対象では、多能性幹細胞、肝細胞、腸細胞、または膵細胞を罹患対象から単離し、膵島転写因子をコードするDNAを発現するように細胞を修飾し、細胞を罹患対象に移植してインスリンの産生を提供することができる。または、細胞がインスリン産生β細胞へ発達するのを促進するように形質転換細胞を培養し、その後β細胞を罹患患者に移植することもできる。
【0123】
別の態様において、別の対象(「ドナー」)由来の多能性幹細胞、肝細胞、腸細胞、または膵細胞を修飾して膵島転写因子をコードするDNA、特にNgn3をコードするDNAまたはNgn3制御経路の他の正の制御因子をコードするDNAを発現させ、続いてその細胞を罹患対象に移植して、(例えば、異種移植を提供するために)インスリンの産生を提供する。または、細胞がインスリン産生β細胞へ発達するのを促進するように形質転換細胞を培養し、その後β細胞を罹患患者に移植する。
【0124】
インビトロでの細胞への核酸の導入は、当技術分野で周知の方法に従って(例えば、エレクトロポレーション、マイクロインジェクション、リポフェクション、組換え(好ましくは複製不能)ウイルスによる感染、および当技術分野で周知の他の手段によって)達成され得る。一般に核酸は、所望のレベルのポリペプチド発現を促進するプロモーター(例えば、CMV、SV40、アデノウイルス由来のプロモーター、または組織特異的プロモーターもしくは細胞種特異的プロモーター)に機能的に結合される。組換え核酸を含む形質転換細胞は、例えば、導入した構築物に存在する選択マーカー遺伝子または構築物と同時トランスフェクションした核酸に存在する選択マーカー遺伝子の発現により選択および/または濃縮されうる。一般に、選択マーカーは、テトラサイクリン、ハイグロマイシン、ネオマイシン等のような抗生物質に対する耐性を提供する。そのためマーカーには、チミジンキナーゼ等が含まれ得る。その他のマーカーには、β-ガラクトシダーゼまたは緑色蛍光タンパク質等の、発現細胞を同定するのに使用可能なマーカーが含まれ得る。
【0125】
形質転換細胞における導入した核酸の発現は、当技術分野で周知の様々な方法により評価することができる。例えば、導入した遺伝子の発現は、ノーザンブロット法を用いて関連遺伝子由来のDNAプローブとハイブリダイズするmRNAを検出することにより調べることができる。さらに、所望の遺伝子を発現する細胞を、当技術分野で周知の方法により単離してインビトロで培養し増殖させることができる。形質転換のために選択される宿主細胞は、エクスビボ治療の目的(例えばインスリン産生)、細胞の移植部位、および当業者に理解されうる様々な要因によって変わりうる他の要因によって変化すると考えられる。
【0126】
形質転換細胞を、膵島細胞表現型の発現について調べることもできる。例えば、PCR法、ノーザンブロット法、免疫細胞化学法、ウェスタンブロット法、RIA法、またはELISA法により、インスリン、IAPP、グルコキナーゼ、またはソマトスタチンの発現を検出することができる。または、インスリン遺伝子プロモーター等の膵島特異的プロモーターに機能的に結合された緑色蛍光タンパク質遺伝子または抗生物質耐性遺伝子のようなマーカー遺伝子を使って、分化した膵島細胞を同定または選択することができる。所望の遺伝子産物を発現させるために宿主細胞を操作する方法および操作された細胞を体内移植または移植する方法(例えばエクスビボ療法)は、当技術分野で周知である(例えば、Gilbertら、1993、Cell Transplantation of genetically altered cells on biodegradable polymer scaffolds in syngenic rats、Transplantation 56:423-427を参照のこと)。外因性の細胞または自己細胞における所望遺伝子の発現、および膵臓への細胞(例えば膵島細胞)移植に関しては、例えば、Docherty 1997、Gene Therapy for diabetes mellitus、Clin Sci (Colch) 92:321-330;Hegreら、1976、Transplantation of islet tissue in the rat、Acta Endocrinol Suppl (Copenh) 205:257-281;Sandlerら、1997、Assessment of insulin secretion in vitro from microencapsulated fetal porcine islet-like cell clusters and rat, mouse, and human pancreatic islets、Transplantation 63:1712-1718;Calafiore 1997、Perspectives in pancreatic and islet cell transplantation for the therapy of IDDM、Diabetes Care 20:889-896;Kenyonら、1996、Islet cell transplantation: beyond the paradigms、Diabetes Metab Rev 12:361-372;Sandler;Chickら、1977 Science、Artificial pancreas using living beta cells: effects on glucose homeostasis in diabetic rats、197:780-782を参照されたい。一般に、膵臓、または例えば皮下部位、肝臓、腹膜等の任意の実際的もしくは簡便な部位に、細胞を移植することが可能である。
【0127】
膵島細胞を移植する方法は当技術分野で周知であり、例えば、Hegreら、1976、Transplantation of islet tissue in the rat、Acta Endocrinol Suppl (Copenh) 205:257-281;Sandlerら、1997、Assessment of insulin secretion in vitro from microencapsulated fetal porcine islet-like cell clusters and rat, mouse, and human pancreatic islets、Transplantation 63:1712-1718;Calafiore 1997、Perspectives in pancreatic and islet cell transplantation for the therapy of IDDM、Diabetes Care 20:889-896;Kenyonら、1996、Islet cell transplantation: beyond the paradigms、Diabetes Metab Rev 12:361-372;Sandler;Chickら、1977 Science、Artificial pancreas using living beta cells: effects on glucose homeostasis in diabetic rats、197:780-782を参照されたい。
【0128】
一般に、インビトロで形質転換細胞を増殖させてから、当技術分野で周知の方法により哺乳動物対象にその細胞を移植する。移植される細胞の数は、血糖値を低下させるのに十分なレベルのインスリン発現を提供するのに十分な細胞の数に等しい。インビトロで達成されるポリペプチドの発現レベル、および/または移植の過程で残存する細胞の数等の要因に基づいて、移植する細胞の数が決まり得る。形質転換細胞は、肝臓等の血管新生の盛んな領域に移植され、または対象における外科的処置を最小限にする様式で移植する。形質転換細胞からなる移植細胞の移植は、移植拒絶反応の典型的な兆候、すなわち移植部位における炎症および/または剥離ならびに発熱に関して、哺乳動物対象を試験することにより、および血糖値をモニタリングすることによってモニタリングされる。
【0129】
上記の移植方法は、ニューロゲニン3の発現に限定されない。分化カスケードに存在する他の膵島転写因子、特にNeuroD1およびMash1を、Ngn3とともにまたはNgn3に代えて発現させるように宿主細胞を操作し、インスリン関連疾患を有する対象を治療することもできる。
【0130】
II. インビボでの膵島細胞の発達および膵臓におけるインスリンの産生
膵島転写因子をコードする核酸を対象に直接送達して、標的細胞(例えば、膵細胞、腸細胞、肝細胞、または膵島転写因子の発現およびインスリンの産生が可能な他の臓器の細胞)内で膵島転写因子の発現を提供し、これにより細胞が(例えば膵臓内の)インスリン産生細胞へと発達するのを促進することができ、または対象における膵島転写因子発現の欠損を治療することができる。関心対象の核酸を標的細胞で発現させるためにインビボで送達する方法は、当技術分野で周知である。例えば、インビボで遺伝子送達する方法は、通常、標的細胞にDNAを導入する生物学的手段(例えば、関心対象のDNAを含むウイルス)または標的細胞にDNAを導入する機械的手段(例えば、細胞へのDNAの直接注射、リポソーム融合、または遺伝子銃を用いた気圧注射)のどちらかを利用する。
【0131】
一般に、関心対象のDNAにコードされるタンパク質を発現する形質転換細胞は、治療的に有効な量のタンパク質を産生し、哺乳動物患者において膵島細胞、特に膵β細胞を産生する。1つの態様において、関心対象のDNAは、ニューロゲニン1、ニューロゲニン2、ニューロゲニン3、NeuroD1/BETA2、Mash1、またはNeuroD4/Math3等(特に関心が持たれるのはNgn3である)の膵島転写因子をコードし、関心対象のDNAはプロモーターに機能的に結合されるが、そのプロモーターは転写因子にとって異種性または内在性であってよい。好ましい態様において、関心対象のDNAは、Ngn3、NeuroD1、またはMash1をコードし、特にNgn3をコードする。
【0132】
一般論として、本送達方法は、関心対象のDNAを含むベクターを膵細胞に導入する段階を含む。一例として、関心対象のDNAを含むベクターはウイルスベクターまたは非ウイルスベクター(裸のDNAを含む)のいずれかを含み、菅系を通してインビボで膵臓に導入される。管内投与は、例えば、胃腸管の管腔を介したカニューレの挿入による、外部開口部を介したカニューレの挿入による、または総胆管を介したカニューレの挿入によるカニューレ挿管により、達成され得る。内視鏡的逆行性胆道膵管造影法(ECRP)により、膵臓で逆行性菅投与を行うこともできる。膵臓への菅内送達を達成する例示的な方法は、米国特許第6,004,944号に記載されている。
【0133】
投与する膵島転写因子をコードする核酸の正確な量は、標的細胞の形質転換されやすさ、対象の体長および体重、所望のタンパク質発現のレベル、および治療対象のインスリン関連疾患を含む多くの要素によって大きく変化する。標的組織を感染させ、十分な数の細胞を形質転換し、所望のレベルのインスリン産生を提供するのに効果的な核酸の量および/または感染性ウイルス粒子の数は、インビトロにおける形質転換効率および標的細胞の形質転換されやすさ等の要因に基づいて、容易に決定され得る。例えば、ヒトの膵管に導入するDNA量は、例えば、一般的には約1μg〜約750 mg、好ましくは約500μg〜約500 mg、より好ましくは約10 mg〜約200 mg、最も好ましくは約100mgである。一般に、動物モデルにおける所望の遺伝子の送達と発現に有効なDNA量から、導入されるDNA量を推定できる。例えば、ヒトにおける送達用のDNA量は、ラットにおいて有効なDNA量のおよそ100倍である。
【0134】
特に送達する核酸がDNAでありかつ関心対象のDNAが真核生物の強力なプロモーター(例えば、CMV、MMTV)と機能的に結合している場合、本発明に従って修飾した膵細胞により、関心対象の核酸の十分に高いレベルでの発現が促進される。発現したタンパク質により、膵島細胞の産生およびインスリンの産生が誘導され得る。したがって、本発明の方法は、様々なインスリン関連疾患を有する哺乳動物対象を治療する上で有用である。
【0135】
1つの態様において、コードされるタンパク質は、塩基性ヘリックス・ループ・ヘリックス(bHLH)タンパク質のクラスに由来する膵島転写因子である。例えば、ニューロゲニン3および/またはNeuroD1および/またはMash1の発現により、哺乳動物において膵島細胞の産生およびインスリンの産生が実質的に誘導され得る。
【0136】
関心対象のタンパク質を治療的レベルで得るために必要な形質転換膵細胞の実際の数は、発現対象のタンパク質、形質転換細胞によるタンパク質発現のレベル、タンパク質が膵島細胞産生(特にβ細胞)を誘導する割合、および治療を受ける対象の健康状態を含むいくつかの要因によって変化すると考えられる。
【0137】
膵島転写因子をコードする核酸(または、インビトロで産生される膵島細胞、または、移植後にインビボで膵島細胞に発達するように移植する予定の膵島転写因子核酸を発現する組換え細胞)は、インビボ導入かエクスビボ導入かにかかわらず、他の遺伝子および他の薬剤と組み合わせて投与されうる。
【0138】
治療の評価
本発明の方法によるエクスビボ両方またはインビボ療法の効果を、様々な方法でモニタリングすることができる。一般に、対象の血液試料を、例えば、グルコース、プロインスリン、c-ペプチド、およびインスリンのレベルについてアッセイすることができる。血液試料中のプロインスリン、c-ペプチド、インスリン、およびグルコースを検出する適切なアッセイ法は、当技術分野で周知である。グルタミン酸脱炭酸酵素(GAD)または当技術分野で周知の他の自己抗原などの膵島タンパク質に対する自己反応性T細胞または抗体についてアッセイすることにより、再発性自己免疫の徴候を評価することができる。
【実施例】
【0139】
以下の実施例は本発明を実施するための方法を当業者に完全に開示して説明するために提供されるものであり、本発明者らが本発明だと考える範囲を限定することを意図したものではない。使用する数字(例えば、量、温度等)に関しては正確性を期す努力を行ったが、ある程度の実験誤差および偏差を考慮する必要がある。特記されない限り、割合とは重量割合、分子量とは重量平均分子量、温度とは摂氏、および気圧とは大気圧またはその付近である。
【0140】
実施例1:マウス膵臓におけるNgn3発現の検出
塩基性ヘリックス・ループ・ヘリックス(bHLH)ファミリーの転写因子に属する転写因子は、多くの細胞種の成長および分化を制御する。インスリン遺伝子の発現は、2つのbHLHタンパク質である、偏在的に発現する(クラスA)タンパク質と細胞種特異的(クラスB)パートナーであるNeuroD1/BETA2のヘテロ二量体複合体により活性化される。NeuroD1/BETA2は、β細胞の発達にとっても重要である。マウスのNeuroD1/BETA2遺伝子を標的破壊すると、胎児発生の後期に膵島細胞のアポトーシスが増加することにより、出生時のβ細胞の質量が著しく減少する。しかし、この因子がβ細胞の分化に重要であると考えられているにもかかわらず、β細胞の形成またはインスリン遺伝子の発現には明らかな欠陥は見られない。
【0141】
このように表現型が中程度であるということは、膵臓内分泌部において密接に関連したクラスB bHLHタンパク質が重複して発現していることを反映していると仮定し、本発明者らは、クラスB bHLHタンパク質のbHLHドメインにおいて保存されるアミノ酸配列(Sommerら、1996 Mol. Cell. Neurosci. 8:221)に基づいた縮重オリゴヌクレオチドプライマーを用いて逆転写ポリメラーゼ連鎖反応(RT-PCR)することにより、このファミリーに属するさらなる転写因子を探索した。PCR解析により、膵臓内分泌細胞株および単離した成人膵島細胞は、NeuroD1を発現するだけでなく、Mash1、neuroD2およびneuroD4、ならびにニューロゲニン(ngn)1、ニューロゲニン2、およびニューロゲニン3を含む神経クラスB bHLH遺伝子ファミリーに属するいくつかの他の転写因子も発現することが明らかになった。この著しい重複性により、マウスにおいてNeuroD1/BETA2の喪失が代償されたのであろう。最も共通して増殖していた配列はNeuroD4およびNgn3をコードする2つの配列であったが、マウス膵臓をインサイチューハイブリダイゼーションにより調べたところ、NeuroD1およびNgn3の発現が最も高いことが示された。これらの結果は、免疫組織化学法により確認した。
【0142】
マウスにおいて、Ngn3は胚齢11.5日(e11.5)にまず最初に検出され、e15.5で最大になるまで増加して、e18.5で減少し、成体では染色は見られない。Ngn3は散在する菅細胞および菅周囲の細胞の核において検出されるが、4つの膵島ホルモン(インスリン、グルカゴン、ソマトスタチン、および膵臓ポリペプチド)のいずれとも同時染色されなかった。この時間的かつ空間的な発現パターンは、膵島細胞前駆体のマーカーとしてのNgn3を意味するものである。将来的にはβ細胞の特異的マーカーであるNkx6.1はNgn3陽性細胞の10%〜20%で発現し、これによりさらに膵島細胞前駆細胞のマーカーとしてのNgn3の使用が支持された。e15.5におけるNgn3発現のピークは、胎仔における新しいβ細胞形成のピークとも一致する。Ngn3がNeuroD1/BETA2および他の膵島分化因子の上流に作用して膵島細胞前駆細胞を標識し、最終的に分化する前に消失するというモデルが、本発明者らのデータにより裏付けられる。
【0143】
実施例2:ヒトNgn3ポリペプチドをコードするポリヌクレオチドの単離および配列決定
クローニングされたマウスNgn3遺伝子断片に由来するプローブ(Sommerら、前記)を用いて、ヒトゲノムライブラリーをスクリーニングした。このスクリーニングにより、配列表に配列番号:1として提供するゲノム配列を単離した。マウス胎仔膵臓RNAを用いた5'RACE法によりマウスの開始部位をマッピングした結果に基づくと、ヒトNgn3コード配列の転写開始部位は2643位のヌクレオチド残基である。コード配列は3022位〜3663位のヌクレオチド残基の間にあり、終始部位は3664位〜3666位にある。5'非翻訳領域(UTR)または配列番号:1のコード配列内には、イントロンは存在しない。
【0144】
Ngn3のプロモーターは、マウス、ラット、およびヒトの間で非常によく保存されているという点で特に興味深い。膵臓および膵島細胞の発達におけるNgn3の役割を考えると、Ngn3プロモーターは成熟した膵臓に存在する膵島細胞の数を決定する鍵である可能性が高い。ヒトNgn3プロモーターに対応する調節領域は、ヒトNgn3プロモーター内にある転写開始部位の最大で約500 bp上流に至るまでの配列(例えば、約2144位から、2643位の転写開始部位まで)を含む。FISH法を用いて、Ngn3のヒト染色体上の位置を10q22.1〜22.2上に同定した。
【0145】
実施例3:マウスNgn3ポリペプチドをコードするポリヌクレオチドおよびプロモーターの単離ならびに配列決定
以前に得られたマウスゲノムDNA断片(Sommerら、前記)を配列決定することにより、マウスの全長Ngn3配列およびその5'近接配列が得られたが、これにはマウスNgn3プロモーターも含まれていた。マウスNgn3配列は配列表において配列番号:3として提供されており、コードされるポリペプチドは配列番号:4として提供される。転写開始部位を5'RACE法で決定し、マウス胎仔膵臓由来RNAを用いたRNase保護(protection)法により確認し、719位のヌクレオチド残基であることがわかった。マウスNgn3のコード配列は、1093位のヌクレオチド残基において開始される。プロモーターは、転写開始部位のおよそ500 bp上流の領域を含む。
【0146】
実施例4:ニューロゲニン3をコードするアデノウイルスベクターの構築
マウスの全長ニューロゲニン3コード配列をアデノ-Xウイルスゲノムのサイトメガロウイルス前初期遺伝子プロモーター(PCMV IE)の下流に挿入し、アデノ-XTM発現系についての製造業者(Clontech、カリフォルニア州パロアルト)の説明に従って完全なウイルス粒子を産生した。図1に最終的なアデノX.NGN3構築物のマップを示す。対照構築物をNgn3の代わりにLac Zコード配列を用いて作製し、アデノLacZ構築物とした。以下の実験で使用するため、当技術分野で周知のプロトコールによりアデノウイルスの大量調製を行った。ウイルス濃度は、プラークアッセイ法によってではなく、精製ウイルス保存液のタンパク質濃度に基づいて推定した。ウイルスの同一性は、調製中のPCRにより、および精製保存液に感染した細胞の溶解液を用いたニューロゲニン3についてのウェスタンブロットにより確認した。
【0147】
実施例5:アデノX.NGN3を用いた処置による正常成体ラットにおけるインスリンの誘導
以前に記載された外科的技法(Goldfineら、Nat Biotechnol 15:1378-82、1997)を用いて、体重250 g〜350 gの成体雄性Sprague-Dawleyラットの膵管に、アデノLacZまたはアデノX.NGN3のいずれかを注射した。およそ3×1010個のウイルス粒子(低投与量)またはおよそ3×1011個のウイルス粒子(高投与量)を、各ラットの膵管に注射した。術後回復してから、ラットを通常の食餌に戻した。約48時間後、低投与量のアデノX.NGN3を注射されたラットを屠殺し、膵臓を摘出して固定し、パラフィンに包埋して切片を作製した。確立した免疫組織化学的技法および抗血清(Schwitzgebelら、Development 127:3533-3542、2000)を用いて、膵臓切片をインスリン、グルカゴン、サイトケラチン20(菅のマーカー)、およびマウスニューロゲニン3について染色した。
【0148】
低投与量または高投与量のアデノX.NGN3を注射されたラットは、ランゲルハンス島を形成するグルカゴン染色細胞およびインスリン染色細胞の通常の大きな(およそ1000個の細胞)凝集塊のほかに、個別の、または細胞8個以下からなる小さな凝集塊である、インスリン染色細胞またはグルカゴン染色細胞を有しており、これらは菅の近くに散在して見られた。図2Aは、低投与量(3×1010個)のアデノX.NGN3を注射されたラットの膵臓組織切片におけるインスリンについての組織学的染色の結果を示す。個別のおよび小さな凝集塊であるインスリン染色細胞を矢印で示す。図2Bは同じラットの連続切片を菅細胞のマーカーであるサイトケラチン20について染色した結果を示し、矢印は図2Aにおけるインスリン染色の位置を示す。
【0149】
対照アデノLacZウイルスを注射されたラットでは、ランゲルハンス島の外側に散在するインスリン染色細胞またはグルカゴン染色細胞は見られなかった。高力価のニューロゲニン3発現アデノウイルスを注射されたラットは全て、注射後36時間以内に死亡した。高投与量のラットのうち1匹の死亡前の血糖値は、Glucometer Elite計測器(Bayer Corporation、インディアナ州エルクハート)を用いたグルコースオキシダーゼ法により測定したところ、32 mg/mlであった。絶食ラットの血糖値は通常およそ100 mg/dlであり、70 mg/dlよりも低いことはめったにない。高投与量ラットの血糖値が32 mg/mlであるということは、過剰なインスリンが産生されて、血中の糖を低血糖症レベルにまで低下させたことを示す。Ngn3をコードする高投与量のDNAを注射されたラットは、新たに産生されたβ細胞からインスリンが過剰発現され、これにより低血糖症が誘導されて死亡したと思われる。
【0150】
図3は、高投与量(3×1011個)のアデノX.NGN3に感染させて約24時間後に回収したラットの膵臓における、インスリンおよびニューロゲニン3について蛍光同時染色した共焦点像を示す。図の中心にある細胞では、核がニューロゲニン3について染色され(矢印で示した濃い灰色の領域)、細胞質がインスリンについて染色された(明るい灰色である周辺領域)ことが観察される。対照動物の膵臓では、ニューロゲニン3は検出されなかった。
【0151】
実施例6:マウスNgn3をコードするDNAを膵臓に導入することにより誘導される、糖尿病誘発成体ラットにおける血糖値の正常化
β細胞毒素であるストレプトゾトシン(Sigma:40 mg/kg体重、1 mMクエン酸緩衝液、pH 4.5に溶解)を1日目および2日目に腹腔内注射することにより、体重250 g〜350 gの成体雄性Sprague-Dawleyラットにおいて糖尿病を誘発した。ストレプトゾトシンとは、ラットにおいて糖尿病を誘発するβ細胞毒素である。4日目に、高血糖症を有することが確認されたラット(Glucometer Elite計測器(Bayer Corporation、インディアナ州エルクハート)を用いたグルコースオキシダーゼ法により測定し、300 mg/dlを上回る血糖値を有する)の膵管に、前述の外科的技法によりアデノLacZまたはアデノX.NGN3のいずれかを注射した。およそ3×1010個または3×1011個のウイルス粒子を各ラットの膵管に注射した。
【0152】
術後5日目にラットを屠殺し、膵臓を摘出して固定し、パラフィンに包埋して切片を作製した。インスリン、グルカゴン、サイトケラチン20(菅のマーカー)、およびマウスニューロゲニン3について、膵臓切片を染色した。図4AはアデノX.NGN3を注射されたラットの膵臓組織切片におけるインスリンの組織学的染色の結果であるが、これにより本発明によってインスリンが産生されることが示唆される。個別のおよび小さな凝集塊であるインスリン染色細胞を白矢印で示す。残存するインスリン染色細胞(残りはストレプトゾトシンで破壊された)をわずかに有する膵島を黒矢印で示す。
【0153】
図4Bは、アデノ-LacZウイルスを注射されたラットの膵臓におけるインスリン染色の結果を示す。残存するインスリン染色細胞をわずかに有する2つの膵島を矢印で示す。対照動物において、個別のまたは小さな凝集塊であるインスリン染色細胞は全く検出されなかった。
【0154】
アデノX.NGN3ウイルスを注射されたラットは、アデノLacZを注射された対照ラットよりも有意に低い血糖値を有していた。図5に示すように、アデノX.NGN3を注射された糖尿病ラットにおいて、アデノLacZを注射された糖尿病ラットと比較して、血糖値が有意に減少した。血糖値の減少は、2日目と4日目の両方で観察された。したがってこれらのデータにより、Ngn3をコードするDNAを膵臓に導入することでインスリンが持続的に発現されること、および形質転換膵細胞が発現したインスリンは血流中に分泌され、かつ動物モデルにおける糖尿病を克服するのに十分なレベルで血糖制御のために機能し得ることが示される。
【0155】
本発明は、成熟した膵臓内の細胞、最も適当には菅細胞が、内分泌細胞に分化する能力を保持することを示す。成体菅細胞は、前内分泌bHLH遺伝子、特にNgn3に応答し、これらの遺伝子が他のシグナルと共同して新しいβ細胞の形成を誘導し、その後のインスリン産生を誘導することができる。本発明は2型糖尿病の維持および治療に有用であり、さらに1型糖尿病を有する個体において自己免疫性破壊によって失われたβ細胞の補充を可能にすると考えられる。
【0156】
実施例7:Ngn3トランスジェニック動物におけるNgn3の過剰発現および膵島細胞の産生
以前に記載されたように(Schwitzgebelら、Development 127:3533-3542、2000)、マウスpdx1プロモーター(転写開始部位およびプロモーターを含むマウスPdx1遺伝子の4.4 kbのXbaI-SmaI断片(Apelqvistら、Curr Biol 7:801-804;Wuら、Mol Cell Biol 17:6002-6013 (1997))およびヒトβ-グロビン遺伝子の第一イントロンをpBATポリリンカーの上流に挿入することにより(Germanら、Genes & Dev 6:2165-2176)、ならびにSV40後期遺伝子ポリアデニル化シグナルを挿入することにより、pdx1プロモーターベクターpBAT.PD17を構築した。PCR法によりマウスゲノムニューロゲニン3クローン(Sommerら、Mol Cell Neurosci 8:221-241 (1996))からマウスの全長Ngn3 cDNA をコードする663 bpのDNA断片を取得し、これをpBAT.PD17ポリリンカーに挿入した。
【0157】
このベクターを線状化して精製し、C3Fe/B6親に由来するF1ハイブリッド卵母細胞に前核注射(1.5 ng/μl)することによりトランスジェニックマウスを作製した(Hoganら、(1994) Manipulating the mouse embryo: A Laboratory Manual. New York: Cold Spring Harbour Laboratory Press.)。遺伝子型を、尾部生検試料のゲノムDNAをPCR解析することにより判定した。用いたプライマーは、
(5'側用Pdx1プライマー)および
(3'側用ヒトβ-グロビンイントロン)であった。
【0158】
胚齢12.5日(E12.5)またはE18.5で胎仔を回収した。全部で6匹のpdx1-ngn3マウスをE12.5で、10匹のpdx1-ngn3マウスをE18.5で試験した。トランスジェニック胎仔の大きさには著しい異常は認められなかった。E12.5で回収したトランスジェニック胎仔では、図6Bにおいてグルカゴン染色により示されるように、膵島細胞の数に増加が見られた。対照の非トランスジェニック胎仔は、正常な数のグルカゴン発現細胞を有する(図6A)。E18.5ではトランスジェニック動物の膵臓の大きさが著しく減少しているが、膵島細胞画分は同じ胚齢の対照動物と比較して顕著に増加している(図7A)。
【0159】
これらの実験から、適切な前駆細胞においてニューロゲニン3が膵島細胞の新生を誘導する能力を有することが実証された。
【0160】
実施例8:NeuroD1トランスジェニック動物における膵島細胞の産生
開始コドンから3'UTRにわたるマウスの全長cDNAをコードする1.7 KbのDNA断片(Leeら、(1995) Science 268:836-844)をpBAT.PD17にクローニングすることにより、neuroD1/BETA2ベクターを構築した。
【0161】
このベクターを線状化して精製し、上述のようにC3Fe/B6親に由来するF1ハイブリッド卵母細胞に前核注射(1.5 ng/μl)することによりトランスジェニックマウスを作製した(Hoganら(1994)、前記)。遺伝子型を、尾部生検試料のゲノムDNAをPCR解析することにより決定した。用いたプライマーは、
(5'側用Pdx1プライマー)および
(3'側用ヒトβ-グロビンイントロン)である。
【0162】
胚齢12.5日(E12.5)で、またはE18.5で胎仔を回収した。全部で5匹のpdx1-neuroD1マウスをE12.5で、10匹のpdx1-neuroD1マウスをE18.5で試験した。ニューロゲニン3トランスジェニックマウスと同様に、E12.5で回収されたpdx1-neuroD1トランスジェニック胎仔では、図6Cにおいてグルカゴン染色により示されたように、膵島細胞の数に増加が見られた。E18.5ではトランスジェニックマウスの膵臓の大きさが著しく減少しているが、膵島細胞画分は同じ胚齢の対照マウスと比較して顕著に増加した(図7B)。
【0163】
これらの実験から、neuroD1は、ニューロゲニン3と同様に、適切な前駆細胞において膵島細胞の新生を誘導する能力を有することが実証された。
【0164】
実施例9:neuroD1をコードするアデノウイルスベクターの構築
マウスまたはヒトの全長neuroD1コード配列をアデノウイルスゲノムのサイトメガロウイルス前初期遺伝子プロモーター(PCMV IE)の下流に挿入し、当技術分野で公知の方法によって、例えばアデノ-XTM発現系についての製造業者(Clontech、カリフォルニア州パロアルト)の説明に従って、完全なウイルス粒子を産生した。図8に、そのような例示的アデノ-NeuroD1構築物のマップを示す。
【0165】
実施例10:mash1/ASCL1/ASH1をコードするアデノウイルスベクターの構築
マウスの全長mash1コード配列またはヒトACSL1/ASH1(ヒトにおけるmas1の略号)コード配列を、アデノウイルスゲノムのCMV前初期遺伝子プロモーター(PCMV IE)の下流に挿入し、当技術分野で公知の方法によって、例えばアデノ-XTM発現系についての製造業者(Clontech、カリフォルニア州パロアルト)の説明に従って完全なウイルス粒子を産生した。図8に、そのような例示的アデノ-mash1/ASCL1/ASH1構築物のマップを示す。
【0166】
実施例11:アデノ-NEUROD1を用いた処理による正常成体ラットにおけるインスリン産生β細胞の形成の誘導
以前に記載された外科的技法(例えば、Goldfineら、Nat. Biotechnol 15:1378-82(1997)を参照のこと)を用いて、体重250 g〜350 gの成体雄性Sprague-Dawleyラットの膵管にアデノLacZまたはアデノ-neuroD1(マウスまたはヒト)のいずれかを注射した。およそ3×1010個のウイルス粒子(低投与量)またはおよそ3×1011個のウイルス粒子(高投与量)を、各ラットの膵管に注射した。術後回復してから、ラットを通常の食餌に戻した。約48時間後にラットを屠殺し、膵臓を摘出して固定し、パラフィンに包埋して切片を作製した。確立した免疫組織化学的技法および抗血清(Schwitzgebelら、Development 127:3533-3542、2000)を用いて、膵臓切片をインスリン、グルカゴン、サイトケラチン20(菅のマーカー)、およびneuroD1について染色した。
【0167】
実施例12:アデノ-mash1/ASCL1/ASH1を用いた処理による正常成体ラットにおけるインスリン産生β細胞の形成の誘導
以前に記載された外科的技法(例えば、Goldfineら、Nat. Biotechnol 15:1378-82(1997)を参照のこと)を用いて、体重250 g〜350 gの成体雄性Sprague-Dawleyラットの膵管にアデノLacZまたはアデノ-mash1/ASCL1/ASH1(マウスまたはヒト)のいずれかを注射した。およそ3×1010個のウイルス粒子(低投与量)またはおよそ3×1011個のウイルス粒子(高投与量)を、各ラットの膵管に注射した。術後回復してから、ラットを通常の食餌に戻した。約48時間後にラットを屠殺し、膵臓を摘出して固定し、パラフィンに包埋して切片を作製した。確立した免疫組織化学的技法および抗血清(Schwitzgebelら、Development 127:3533-3542、2000)を用いて、膵臓切片をインスリン、グルカゴン、サイトケラチン20(菅のマーカー)、およびneuroD1について染色した。
【0168】
実施例13:NeuroD1をコードするDNAの膵臓への導入による、糖尿病誘発成体ラットにおけるインスリンの産生
β細胞毒素であるストレプトゾトシン(Sigma:40 mg/kg体重、1 mMクエン酸緩衝液、pH 4.5に溶解)を1日目および2日目に腹腔内注射することにより、体重250 g〜350 gの成体雄性Sprague-Dawleyラットにおいて糖尿病を誘発した。ストレプトゾトシンとは、ラットにおいて糖尿病を誘発するβ細胞毒素である。4日目に、高血糖症を有することが確認されたラット(Glucometer Elite計測器(Bayer Corporation、インディアナ州エルクハート)を用いたグルコースオキシダーゼ法により測定した300 mg/dlを上回る血糖値を有する)の膵管に、前述の外科的技法によりアデノ-LacZまたはアデノ-NEUROD1(マウスまたはヒト)のいずれかを注射した。およそ3×1010個または3×1011個のウイルス粒子を各ラットの膵管に注射した。次に、ラットの血糖値の減少を12時間ごとにモニタリングした。
【0169】
実施例14:mash1/ASCL1/ASH1をコードするDNAの膵臓への導入による糖尿病誘発成体ラットにおけるインスリンの産生
β細胞毒素であるストレプトゾトシン(Sigma社:40 mg/kg体重、1 mMクエン酸緩衝液、pH 4.5に溶解)を1日目および2日目に腹腔内注射することにより、体重250 g〜350 gの成体雄性Sprague-Dawleyラットにおいて糖尿病を誘発した。ストレプトゾトシンとは、ラットにおいて糖尿病を誘発するβ細胞毒素である。4日目に、高血糖症を有することが確認されたラット(Glucometer Elite計測器(Bayer Corporation、インディアナ州エルクハート)を用いたグルコースオキシダーゼ法により測定された、300 mg/dlを上回る血糖値を有する)の膵管に、前述の外科的技法によりアデノ-LacZまたはアデノ-mash1/ASCL1/ASH1(マウスまたはヒト)のいずれかを注射した。およそ3×1010個または3×1011個のウイルス粒子を各ラットの膵管に注射した。次に、ラットの血糖値の減少を12時間ごとにモニタリングした。
【0170】
実施例15:ニューロゲニン3をコードするプラスミドベクターの構築
マウスまたはヒトの全長ニューロゲニン3コード配列cDNAまたはその遺伝子のいずれかを、pBAT16プラスミドベクターのCMV前初期遺伝子プロモーター(PCMV IE)およヒトびβ-グロビン遺伝子イントロンの下流、ならびにSV40ポリアデニル化シグナルの上流に挿入した。このベクターはまた、細菌内での高コピー数の複製のためのcolE1複製起点、およびプラスミド含有細菌を選択するための細菌性アンピシリン耐性遺伝子も含む。精製プラスミドDNAは、当技術分野で周知の方法により作製した。図9に、そのような例示的pBAT16.hNGN3構築物のマップを示す。
【0171】
実施例16:プラスミドDNAベクターpBAT16.hNGN3の膵臓への導入による糖尿病誘発成体ラットにおけるインスリンの産生
β細胞毒素であるストレプトゾトシン(Sigma:40 mg/kg体重、1 mMクエン酸緩衝液、pH 4.5に溶解)を1日目および2日目に腹腔内注射することにより、体重250 g〜350 gの成体雄性Sprague-Dawleyラットにおいて糖尿病を誘発した。ストレプトゾトシンとは、ラットにおいて糖尿病を誘発するβ細胞毒素である。4日目に、高血糖症を有することが確認されたラット(Glucometer Elite計測器(Bayer Corporation、インディアナ州エルクハート)を用いたグルコースオキシダーゼ法により測定された、300 mg/dlを上回る血糖値を有する)の膵管の管腔に、カチオン性脂質Transfast(Promega)等のアジュバントを含むまたは含まない8μg〜25μgのプラスミドDNAを注射した。膵管に注射された場合、カチオン性脂質等のアジュバントを含むまたは含まない裸のDNAが膵細胞において遺伝子を発現する能力は、以前に記載されている(Goldfineら、1997、前記)。その後、ラットの血糖値の減少を12時間ごとにモニタリングした。
【0172】
実施例17:アデノX-NGN3またはプラスミドDNAベクターpBAT16.hNGN3の膵臓への導入による自己免疫性糖尿病マウスにおけるインスリンの産生
ヒトの自己免疫性糖尿病または「1型」糖尿病モデルとして一般的なNODマウスは、自己免疫性糖尿病の形態を発症する(総説として、例えばBachら、The NOD mouse. Research in Immunology、1997 Jun、148(5):285-6を参照のこと)。このマウスは、膵島移植のような糖尿病の自己免疫状態治療の試験モデルとして用いられている(総説として、例えばHahnら、Adv. Exp. Med Biol. 1997 426:411-9;Sutherland Transplantation Proc. 1996、28(4):2131-3を参照のこと)。
【0173】
本実施例では、15週齢を超えた成体マウスを糖尿病(Glucometer Elite計測器(Bayer Corporation、インディアナ州エルクハート)を用いたグルコースオキシダーゼ法により測定された、300 mg/dlを上回る血糖値を有する)について試験し、これを次に前述の外科的技法を用いてマウスの膵管にアデノ-LacZもしくはアデノX-NGN3(マウスまたはヒト)のいずれか、またはプラスミドベクターpBAT16.hNGN3を注射した。およそ3×109個または3×1010個のウイルス粒子、または2μg〜25μgのプラスミドDNAを各マウスの膵管に注射した。その後、マウスの血糖値の減少を12時間毎にモニタリングした。
【0174】
自己免疫の再発および新しく形成されたβ細胞の破壊を防ぐためには、いくつかの方法が当技術分野で周知であり、これには、サイトキサン(cytoxan)もしくはFK506等の免疫系を抑制する薬剤の使用、またはCTLA-4に対する抗体等の同時刺激分子を遮断する試薬の使用(Shapiroら、New Engl. J. Med. 2000 343(4):230-8;Griffinら、J. Immunol、2000 164(9):4433-42)が含まれるが、これらに限定されるわけではない。動物にはDNA注射の2日前から抗自己免疫療法による処置を開始し、実験期間中免疫療法を続ける。
【0175】
実施例18:インビトロでの膵島細胞の形成の誘導
本実施例では、ニューロゲニン3を用いて、インビトロで培養する非膵島細胞から膵島細胞の形成を誘導する。使用する細胞には、膵管細胞、膵腺房細胞、腺窩細胞を含む腸細胞、肝細胞、および唾液腺細胞等の胃腸管系器官由来の細胞、造血幹細胞、神経幹細胞、筋幹細胞、もしくは膵幹細胞等の成体幹細胞、または胚幹細胞を含む、哺乳動物不死化細胞株または初代培養哺乳細胞が含まれるが、必ずしもこれらに限定されるわけではない。細胞は、当技術分野で周知の方法により培養される。ニューロゲニン3は、ウイルスベクター(アデノXZ-NGN3ベクター等)により導入されるか、またはカチオン性脂質トランスフェクション試薬Transfast等のアジュバントと共にpBAT16.hNGN3等のDNAプラスミドベクターを用いた裸のDNAにより導入される。1日から数週間にわたる期間、細胞の培養を続ける。新しく形成された膵島細胞の検出は、ラジオイムノアッセイ法もしくはELISA法を用いて細胞試料から調製された抽出物中でのインスリンもしくはグルカゴン等の膵島ホルモンを測定することにより、または当技術分野で周知の方法を用いて培養液中のホルモンレベルを測定することにより実施される。
【0176】
実施例19:ヒト対象へのNgn3をコードする核酸の送達
本実施例では、糖尿病患者または関連のインスリン欠損を有する患者を、新しい膵島細胞の形成を誘導するという目的で、ニューロゲニン3をコードするDNAを用いて治療する。ニューロゲニン3をコードするDNAは、上述のアデノX-NGN3の例においてと同じくウイルスベクターに含まれることが可能であり、またはカチオン性脂質Transfast等のアジュバントを含むまたは含まない上述のpBAT16.NGN3の例においてと同じく、裸のDNAベクターに含まれることが可能である。ベクターは、膵管を介して膵臓に逆方向に導入される。ベクターがアデノウイルスの場合、注射される量は、血糖値を下げてそれを正常な範囲に維持するのに必要な量によって決まり、約1011個〜約1014個のウイルス粒子であってよい。ベクターが裸のDNAベクターの場合、必要な量は約100μg〜100 mgとなり得る。
【0177】
自己免疫の再発および新しく形成されたβ細胞の破壊を防ぐためにいくつかの方法が当技術分野で周知であり、ヒトにおいて臨床的に用いられているが、これにはサイトキサン、FK506、もしくはシロリムス(sirolimus)の免疫系を抑制する薬剤の使用、またはCTLA-4に対する抗体等の同時刺激分子を遮断する試薬の使用(Shapiroら、New Engl. J. Med. 2000 343(4):230-8;Griffinら、J. Immunol、2000 164(9):4433-42)が含まれるが、これらに限定されるわけではない。患者にはDNA投与の1日前〜2日前から抗自己免疫療法による処置を開始し、その後も免疫療法を続ける。
【0178】
処置後は、1時間ごとに、または連続グルコースモニターの使用により、血糖値を厳密にモニタリングし、必要に応じて皮下注射によるインスリン治療を減らすかまたは中止する。血糖値が外因性インスリン非存在下で完全に正常化されない場合には、処置を繰り返す。再発性自己免疫による新生β細胞の破壊をモニタリングするため、血糖値を定期的にモニタリングする。さらに、自己反応性T細胞、グルタミン酸脱炭酸酵素に対する抗体(抗GAD抗体)または膵島抗原(膵島細胞抗体またはICA)についてアッセイすることにより、新しい細胞に対する免疫応答の直接的徴候を試験することが可能である。再発性自己免疫の徴候は、免疫抑制治療の増加または別の変更により治療されうる。
【0179】
実施例20:ニューロゲニン3プロモーターの特徴づけおよび制御
以下の実験により、Ngn3プロモーターの特徴づけ、およびNgn3発現を制御する調節経路に関与するNgn3の上流と下流の両方の因子の同定を提供する。
【0180】
材料および方法
以下の材料および方法を本実施例において使用した。
【0181】
マウスおよびヒトのngn3遺伝子プロモーターのクローニング
マウスニューロゲニン3オープンリーディングフレームを含むλファージゲノムクローンである、クローン17/6-1-1-2(Sommerら、Mol. Cell. Neurosci. 8:221-241、1996)を使用した。このファージクローンから、オープンリーディングフレームの上流の配列を含む1 kb断片をサブクローニングして配列決定した。ヒトニューロゲニン3ゲノムクローンは、マウスニューロゲニン3ゲノム断片を用いてλDASHヒトゲノムライブラリーをスクリーニングすることにより取得した。最も長い5'近接配列を含むクローン、クローン14Hをサブクローニングして配列決定し、レポーター遺伝子プラスミドの構築に使用した。
【0182】
5'cDNA末端迅速増幅法(5'RACE法)
5'RACE System Version 2.0(GibcoBRL)のプロトコールの変法を用いて、5'RACEによりマウスngn3 cDNAの5'末端を同定した。マウスcDNAについては、2.5 pmolの特異的プライマーJL1
をマウスE15.5膵臓由来の全RNA 1μgとアニールさせた。逆転写は、SuperScript II逆転写酵素(GibcoBRL)を用いて行った。一本鎖cDNA合成の後、元のmRNA鋳型をRNase処理により除去し、次に末端デオキシヌクレオチド転移酵素を用いてcDNAの3'末端にホモポリマーのdCTPテール(tail)を付加した。プライマーとして5'RACE Abridged Anchor Primer(GibcoBRL)およびJL2
を用いた35サイクルのPCRにおいて、これらの産物を鋳型として使用した。入れ子(nested)PCRについては、Abridged Universal Amplification Primer(GibcoBRL)およびJL3
をプライマーとして使用し、35サイクルのPCRを行った。PCR産物をサブクローニングして配列決定した。
【0183】
レポーター遺伝子構築物
レポータープラスミドを構築するため、制限酵素切断によって得たヒトngn3遺伝子の5'領域の断片を、プラスミドpFOXLuc1内のルシフェラーゼ遺伝子の上流、またはプラスミドpFOXLuc1TK内のTK最小プロモーター遺伝子の上流に結合した(Mirmiraら、J Biol Chem 275:14743-51、2000)。
【0184】
細胞培養および一過的トランスフェクション
βTC3細胞、αTC1.6細胞、およびMPAC細胞は、2.5%ウシ胎仔血清および15%ウマ血清を添加したダルベッコ改変イーグル培地(DMEM)で培養した。NIH3T3細胞は、10%子ウシ血清を添加したDMEM培地で培養した。Cos7細胞は、10%ウシ胎仔血清および4 mMグルタミンを添加したDMEM培地で培養した。哺乳動物細胞の一過的トランスフェクションを行うため、トランスフェクションの24時間前に、細胞を6ウェル組織培養プレートにプレーティングした。標準的レポーター遺伝子解析のために、製造業者の説明に従い、TRANSFAST(商標)脂質試薬(Promega)を用いて2μgのルシフェラーゼレポータープラスミドを細胞にトランスフェクションした。HES1の発現がngn3プロモーターに及ぼす影響を評価するためには、所定の量のHES1またはドミナントネガティブなHes1(pcDNA3Hes1)(京都大学、R.Kageyama氏より分与(Sasaiら、Genes Develop. 6:2620-2634、1992))発現プラスミドDNAを2μgのルシフェラーゼレポータープラスミドと共に同時トランスフェクションした。トランスフェクションから48時間後、細胞を回収し、以前に記載されたのと同様の(Germanら、Genes & Dev. 6:2165-2176、1992)ルシフェラーゼアッセイを行った。ルシフェラーゼ活性は、細胞のタンパク質濃度について補正した。レポーター遺伝子解析はすべて少なくとも3回行い、データは平均値±標準誤差として表した。
【0185】
トランスジェニックマウスの作製およびβ-ガラクトシダーゼの検出
ヒトβ-グロビンイントロンの上流の-5.7 kb〜+261 bpおよび-2.6 kb〜+261 bpにわたるヒトngn3プロモーター断片と、細菌のβ-ガラクトシダーゼ遺伝子を連結して、プラスミドpNAT6BおよびpNAT3Bを構築した。それぞれのプラスミドを線状化し、マウス前核に微量注射(1.5 ng/μl)した。注射した胚を偽妊娠した雌性マウスに移植し、e15.5の時点で創始(founder)マウスから胎仔の膵臓を胃および小腸と共に摘出した。組織を4%パラホルムアルデヒド中で4℃で30分間あらかじめ固定し、次にX-gal(400μg/ml)基質中、37℃で(-2.6 kbプロモーター)または室温で(-5.7 kbプロモーター)一晩インキュベートした。その後、組織を再度4%パラホルムアルデヒド中で30分間固定し、パラフィン包埋して5μMで切片を作製した。遺伝子型は、ヒトngn3プロモーター配列に特異的なプライマーを用いたPCRにより判定した。β-ガラクトシダーゼ活性は、-2.6 kbプロモーター構築物を組み込んだ6匹の独立した創始胎仔、および-5.7 kbプロモーター構築物を組み込んだ8匹の独立した創始胎仔で測定した。
【0186】
免疫組織化学法
免疫組織化学法は、以前に記載されたのと同様のパラフィン包埋切片上で行った(Schwitzgebelら、Development 127:3533-3542、2000)。一次抗体は以下の希釈で使用した:モルモット抗インスリン(Linco)、1:5000;モルモット抗グルカゴン(Linco)、1:10000;ウサギ抗ngn3(Schwitzgebelら、Development 127:3533-3542、2000)、1:5000。ビオチン標識二次抗体(Vector)は、ABC Elite免疫ペルオキシダーゼシステム(Vector)で検出した。
【0187】
タンパク質の調製および電気泳動移動度シフト分析法(EMSA)
SP6およびT7 TNT Quick Coupled Lysate System(登録商標)(Promega)を用いて、pGEM-1ラットHNF3β(イリノイ大学シカゴ校、R.Costa氏より分与)およびpcDNA3-HNF1α(ロックフェラー大学、M. Stoffel氏より分与)を鋳型として使用し、HNF3βタンパク質およびHNF1αタンパク質をインビトロで産生した。グルタチオンS転移酵素(GST)を融合したHES1タンパク質は、pGEX2Tプラスミドシステム(Promega)を使用して大腸菌BL21コンピテントセルで産生した。αTC1.6細胞、βTC3細胞、およびNIH3T3細胞の核抽出物は、SadowskiおよびGilman(SadowskiおよびGilman、Nature 362:79-83、1993)により記載されている手順に従って調製した。
【0188】
T4ポリヌクレオチドキナーゼを使用し、ヒトngn3プロモーター内にある配列に対応する1本鎖オリゴヌクレオチドの5'末端を[γ-32P]-ATPで標識した。標識オリゴヌクレオチドをカラム精製し、過剰量の相補鎖にアニールさせた。HNF3βおよびHNF1α結合実験に用いられるEMSA緩衝液および電気泳動条件は、以前に記載されている(Mirmiraら、J Biol Chem 275:14743-51、2000)。HES1結合実験に関しては、ポリ(dI-dC)濃度を15 ng/μlまで減少させる以外は同じ条件である。それぞれの結合反応には、インビトロ反応混合液1μl、または核抽出物2μg、またはGST融合Hes1タンパク質400 ngを使用した。抗体を使用する場合には、ゲル電気泳動を行う前に、各抗体1μlを結合混合液とともに室温で15分間インキュベートした。HNF-3α、HNF-3β、およびHNF-3γに対する抗血清はR.Costa氏(イリノイ大学)から分与されたものであり、HES-1抗血清はY.Jan氏(カリフォルニア大学サンフランシスコ校)から分与されたものである。抗HNF-1α抗血清は、サンタクルーズバイオテクノロジー社(Santa Cruz Biotechnology, Inc.)から購入した。
【0189】
標識プローブまたはEMSA反応における競合物として、以下のオリゴヌクレオチドを使用した(上側の鎖を示す)。
【0190】
ヒトngn3プロモーターのマッピング
ニューロゲニン3遺伝子発現の制御を理解する最初の段階として、マウスおよびヒトのニューロゲニン3プロモーターの配列を決定した(図11A〜図11B)。胚齢15.5日のマウス胎仔の膵臓から精製したRNAを使用し、5'RACE法によりマウスngn3遺伝子の転写開始部位を決定した。5'RACE産物すべてから、推定TATAAボックスから30塩基対下流という、同一の開始部位が同定された(図11B)。開始部位の上流領域は、マウス、ヒト、およびラットにおいて高度に保存されており、マウスとヒトで最も相同性が高い領域はおよそ300 bp上流に及ぶ。CCAAT配列エレメントは、転写開始部位から-85 bpの位置に位置する。可能性のある配列エレメントがいくつか他にも同定されたが、それらを図11Bに示す。
【0191】
細胞株におけるプロモーターの活性
3'末端はそれぞれ+261 bpまでかつ5'側が連続的に欠失された一連のニューロゲニン3プロモーターを、ホタルルシフェラーゼ遺伝子に結合し、これらを細胞株で試験した(図12)。-502 bpまで連続的に欠失させても、インビトロでのプロモーター活性は減少しなかった。驚いたことに、線維芽細胞を含む全被験細胞株において、プロモーターは高レベルで転写を駆動する。最も短い構築物でさえ、全被験細胞株において高い活性を有していることから、この非特異的な高活性はプロモーターの近位にあると思われる。
【0192】
トランスジェニックマウスにおけるプロモーターの活性
細胞株における一過的トランスフェクションによりプロモーター活性の指標がいくつか提供され得るものの、これらの腫瘍細胞は、ニューロゲニン3が通常発現されている発達過程の膵臓の細胞を表さない。したがって、β-ガラクトシダーゼをコードする細菌遺伝子を駆動する、ヒトニューロゲニン3遺伝子から5.7 kb上流または2.6 kb上流の配列を伴う導入遺伝子を保有するマウスを作製した。胚齢15.5日、つまりマウス胎仔の膵臓におけるニューロゲニン3発現の通常のピーク時に(Schwitzgebelら、Development 127:3533-3542、2000)、創始マウスを回収した。
【0193】
5.7 kbの構築物を保有する動物では、発達中の膵臓の中心領域および腸上皮において、すなわち発達中の同時期にニューロゲニン3が通常発現されるのと同じ領域において、β-ガラクトシダーゼが強くかつ選択的に発現されていた(図13A)。2.6 kbの構築物を用いた場合のβ-ガラクトシダーゼの発現レベルは5.7 kb構築物を用いた場合に比べ有意に低いが(図13B、図13C)、β-ガラクトシダーゼ発現の全体的なパターンは同じである。
【0194】
免疫組織化学法を用いて、5.7 kbのプロモーター構築物を保有するトランスジェニックマウスにおいてβ-ガラクトシダーゼを発現する細胞を同定した。β-ガラクトシダーゼ発現細胞は、主に菅に局在する。β細胞において時々はインスリンとβ-ガラクトシダーゼが同時に発現するものの、ほとんどのβ-ガラクトシダーゼ発現細胞は膵島ホルモンを発現しない。
【0195】
β-ガラクトシダーゼ活性およびニューロゲニン3タンパク質発現が発達過程の膵臓および腸の同じ領域に特異的に密接に同時局在するにもかかわらず、完全な一致はみられない。いくつかのβ-ガラクトシダーゼ陽性細胞は高レベルのニューロゲニン3と同時発現するが、多くのβ-ガラクトシダーゼ陽性細胞ではそうではなく、かつ多くのニューロゲニン3発現細胞はβ-ガラクトシダーゼ活性をほとんどまたは全く含まない。おそらくこの矛盾は、遺伝子発現の開始および終結の違いからというよりも、むしろ2つの遺伝子産物の蓄積および分解のタイミングの違いからきている可能性が最も高い。
【0196】
特定の細胞においてそれぞれの遺伝子産物を最初に検出する正確なタイミングは蓄積速度および検出限界に依存するため、これらが同一であると考えるべきではない。さらに、いくつかのニューロゲニン3発現細胞はランダムに導入遺伝子をサイレンシングさせうり、トランスジェニックマウスにおいて多くのプロモーターで観察される現象は理解が不十分である(Graubertら、Nucleic Acids Res 26:2849-58、1998)。前駆細胞でのニューロゲニン3の発現は非常に短期だが大量であることから、mRNAおよびタンパク質は急速に蓄積するが半減期が非常に短いことが示唆される。対照的に、β-ガラクトシダーゼは哺乳動物細胞においてかなり長い半減期を有し(Smithら、J Virol 69:4593-9、1995)、より遅くピークに達し、かつニューロゲニン3がもはや検出できなくなった後まで細胞中に残存すると考えられ得る。したがって、β-ガラクトシダーゼ発現細胞の多くにおいて、ニューロゲニン3遺伝子の産生が終わった後かつホルモン発現が始まる前に、膵島細胞の分化段階が起こる。これらの細胞が多数あることから、この分化の中間段階が初期のニューロゲニン3発現段階よりも長く持続することが示唆される。
【0197】
腸での発現
E15.5の時点から、トランスジェニック胚および非トランスジェニック胚の両方で、腸絨毛の刷子縁に沿って低レベルの内因性β-ガラクトシダーゼ発現が検出され得る。より強いβ-ガラクトシダーゼ活性はトランスジェニックマウスにおいて斑点模様として検出され得り、これは小腸で最も顕著であるが、非トランスジェニック同腹仔には見られない(図13A)。腸の切片を作製すると、このβ-ガラクトシダーゼシグナルが腸管上皮内に散在する細胞に由来することが示される(図14A〜図14C)。このβ-ガラクトシダーゼの発現パターンにより、ニューロゲニン3プロモーターは発達過程の腸にある前駆細胞の亜集団においても活性があることが示唆される。これらは、腸内分泌細胞の前駆細胞であってもよい。膵臓においてと同様に、このβ-ガラクトシダーゼ活性は内因性ニューロゲニン3発現と部分的に重複し、ここでもβ-ガラクトシダーゼの蓄積のピークがニューロゲニン3と比較して遅れることが示唆される(図14C)。
【0198】
複数の因子がニューロゲニン3プロモーターに結合する
ngn3プロモーターに結合する核因子を同定するため、一連のオリゴヌクレオチドを合成した。プロモーター内の重要なDNA結合部位にまたがる可能性のあるオリゴヌクレオチドを、電気泳動移動度シフト分析法(EMSA)により核タンパク質への結合について試験した。
【0199】
HNF3ファミリーの翼状ヘリックス転写因子に属する転写因子は、膵臓の発達および膵島機能と関係している(Wuら、Mol Cell Biol 17:6002-13、1997;Gerrishら、J Biol Chem 275:3485-92、2000;Sharmaら、Mol Cell Biol 17:2598-604、1997;Duncanら、Science 281:692-5、1998;Philippeら、Mol Cell Biol 14:3514-23、1994;Kaestnerら、Genes Dev 13:495-504、1999)。コンセンサスなHNF結合部位(Costaら、Mol Cell Biol 9:1415-25、1989)との類似性に基づくと、5.7 kbヒトニューロゲニン3プロモーター内には潜在的HNF3結合部位がいくつか存在する。最も可能性の高い2つの部位は、-3687 bpおよび-200 bpに位置する。これらの結合部位に関してEMSA試験を行った結果、両部位がインビトロ産生HNF3βに高い親和性で結合することが示された(図15)。βTC3細胞およびαTC1.6細胞の抽出物を用いると、単一の主要な複合体は両部位に結合し、この複合体はHNF3βに対する抗血清により特異的に認識される。さらに、HNF3βを同時発現すると、一過的トランスフェクションされた3T3線維芽細胞においてニューロゲニン3プロモーターが活性化される(該当データ示さず)。
【0200】
-3687 bpの HNF3β結合部位は、公知の膵臓転写因子に対する潜在的DNA結合部位のクラスターの一部を形成し(図11)、これにはhox型ホメオドメイン転写因子ならびにcut-ホメオドメイン転写因子HNF6およびPou-ホメオドメインHNF1因子(Courtoisら、Science 238:688-92、1987)の結合部位として可能な部位が含まれる。ニューロゲニン3プロモーターに結合するHNF6については、以前に実証されている(Jacqueminら、Mol Cell Biol 20:4445-54、2000)。
【0201】
潜在的HNF1結合部位にまたがるオリゴヌクレオチドをEMSAにより試験した結果、インビトロで産生されるHNF-1αに結合し得ることが見出された。さらに、βTC3細胞由来の核抽出物内では、移動度の低い主要な複合体がオリゴヌクレオチドに結合し、HNF1αに対する抗血清により特異的に認識される(図16)。HNF1α結合部位のすぐ下流にある類似hoxホメオドメイン型の結合部位は、HNF1αに結合しないと思われる。
【0202】
ニューロゲニン3プロモーターのHES-1阻害
転写制御因子HES-1を介するNotch受容体のシグナル伝達が、発達過程の膵臓の細胞の小さな亜集団を除くすべての細胞において、ニューロゲニン3の発現を抑制しうることが提唱されている(Jensenら、Nat Genet 24:36-44、2000)。ニューロゲニン3プロモーターを直接阻害するHES-1の能力を試験するため、3T3細胞において、ニューロゲニン3プロモータールシフェラーゼプラスミドと共にCMVプロモーター駆動発現プラスミドからHES-1 cDNAを発現させた(図17A〜図17C)。HES-1は、ニューロゲニン3プロモーターを劇的かつ特異的に阻害する。5'配列を下流に-502 bpまで除去しても、HES-1がプロモーターを阻害する能力は有意に減少しない。
【0203】
HES-1抑制に応答できる配列をさらにマッピングするため、ホタルルシフェラーゼ遺伝子に結合させた-208 bp〜+40 bpのヒトngn3遺伝子プロモーター配列(近位プロモーター(図18A))、またはホタルルシフェラーゼ遺伝子に結合させたヘルペスウイルスチミジンキナーゼ(TK)プロモーターの上流-2.6 kb〜-208 bpの配列(遠位プロモーター)のいずれかを用いてプラスミドを構築した。小さな近位プロモーターはHES-1抑制に関する能力のほとんどを保持しているのに対し、遠位配列はHES-1により弱く抑制される。
【0204】
近位の208 bpのプロモーター内には、HES-1のコンセンサスな結合部位(CTNGTG)(Takebayashiら、J Biol Chem 269:5150-6、1994)およびそのショウジョウバエ相同体hairy/enhancer-of-split(CGCGTC)(Van Dorenら、Genes Dev 8:2729-42、1994;Ohsakoら、Genes Dev 8:2743-55、1994)に基づく、潜在的HES-1結合部位がいくつか存在する(図18A)。ゲル移動度シフト分析法により、これら4つの部位を含む3つのオリゴヌクレオチドを、細菌的に産生されたHES-1タンパク質への結合について調べた(図18B)。以前に記載されたマウスHES-1遺伝子由来の高親和性タンデム部位(Takebayashiら、J Biol Chem 269:5150-6、1994)(図18CではH1と表示)よりも高い親和性で、3つのオリゴヌクレオチドはすべてHES-1に結合する。これら4つの配列はすべて、マウスニューロゲニン3プロモーターに保存されている(図11)。
【0205】
実施例21:膵島転写因子制御経路の特徴づけ
以下の実験により、膵島細胞の形成に関連する転写因子の特徴づけを提供する。
【0206】
簡潔に説明すると、β-ガラクトシダーゼ、ヒトニューロゲニン3(配列番号:1)、マウスNeuroD1(GenBankアクセッション番号BC018241で入手可能でありかつ配列表において配列番号:40として提供される)、マウスMash1(GuillemontおよびJoyner (1993) Mech. Dev. 42(3):171-85)、およびマウスmyoDのコード配列をpACCMV-pLpAプラスミドに挿入し、以前に記載された方法(Bermanら、(1998) J. Biol. Chem. 273:26421-26425;Beckerら、(1994) Methods Cell. Biol. 43:161-189)を用いて組換えアデノウイルスを作製した。β-ガラクトシダーゼ(adCMV-Bgal)、ニューロゲニン3(ヒト)(adCMV-Ngn3)、NeuroD1(マウス)(adCMV-ND1)、mash1(マウス)(adCMV-Mash1)、またはmyoD(マウス、筋bHLH遺伝子)(adCMV-MyoD)を発現するアデノウイルスベクターを用いて、2つのマウス膵管細胞株mPAC L20およびmPAC L4S2を感染させた。
【0207】
48時間後、細胞を回収してRNAを単離し、Ngn3、ND1、Mash1、MyoD、β-アクチン、グルカゴン、ソマトスタチン、インスリン、IAPP、グルコキナーゼ、グルコース輸送体2、Pax4、Pax6、Nkx2.2、Nkx6.1、Isl-1、およびPdx-1についてRT-PCRを行った。RT-PCR産物をアガロースゲル電気泳動により分離してエチジウムブロマイドで染色し、写真撮影した。結果を図19A〜図19Cに示す。PCR産物の同一性は、アガロースゲルから産物を精製し、精製したDNA断片を細菌のプラスミドベクターに挿入し、挿入物を配列決定することにより確認した。これらのデータから、ニューロゲニン3は、膵島細胞の分化および機能に関与する全体の遺伝子群の活性化をともなって内分泌分化を誘導できることが実証された。
【0208】
本発明をその特定の態様に関して説明したが、本発明の精神および範囲に反することなく、様々な変更が行われうりかつ均等物が代用され得ることは当業者に理解されねばならない。さらに、特定の状況、物質、対象組成物、工程、工程段階、または段階を、本発明の目的、精神、および範囲に適合化させるため、多くの改変がなされ得る。そのような修正はすべて、本発明の範囲内に包含されることが意図される。
【特許請求の範囲】
【請求項1】
哺乳動物膵島細胞を産生する方法であり、膵島転写因子は神経内分泌bHLH転写因子であり、導入により細胞内における膵島転写因子の発現および細胞内における膵島細胞表現型の産生が提供される、哺乳動物膵細胞に膵島転写因子をコードする核酸分子を導入する段階を含む方法。
【請求項2】
神経内分泌bHLH転写因子がニューロゲニン3(Ngn3)、NeuroD1、およびMash1からなる群より選択される、請求項1記載の方法。
【請求項3】
神経内分泌bHLH転写因子がニューロゲニン1、ニューロゲニン2、neuroD2、math2、NeuroD4/Math3、math1/ATOH1、およびmash2からなる群より選択される、請求項1記載の方法。
【請求項4】
bHLH転写因子がニューロゲニン3(Ngn3)制御経路の正の制御因子である、請求項1記載の方法。
【請求項5】
哺乳動物膵島細胞を産生する方法であり、導入により細胞内における膵島表現型の産生が提供される、哺乳動物膵細胞にニューロゲニン3(Ngn3)制御経路の正の制御因子を導入する段階を含む方法。
【請求項6】
正の制御因子が細胞内でNgn3の発現を増加させる、請求項5記載の方法。
【請求項7】
正の制御因子がNgn3をコードする核酸であり、さらに該核酸が細胞におけるNgn3の発現および産生に適合化される、請求項5記載の方法。
【請求項8】
正の制御因子がMash1、NeuroD1、HNF1、HNF2、およびHNF6からなる群より選択される、請求項5記載の方法。
【請求項9】
哺乳動物膵細胞がヒト膵細胞である、請求項1または5記載の方法。
【請求項10】
膵島細胞表現型が膵β細胞の表現型である、請求項1または5記載の方法。
【請求項11】
哺乳動物対象の血流にインスリンを送達する方法であり、導入により膵島細胞によるインスリンの産生および哺乳動物対象の血流へのインスリンの送達が提供される、請求項1または5記載の方法により産生される膵島細胞を哺乳動物対象の膵臓に導入する段階を含む方法。
【請求項12】
哺乳動物対象の血流にインスリンを送達する方法であり、膵島転写因子は神経内分泌bHLH転写因子であり、投与はβ細胞の産生を誘導するのに十分なレベルの膵島転写因子を産生するのに十分な量で行われ、膵島細胞はインスリンを産生し、インスリンは対象の血流に送達される、哺乳動物対象の膵細胞に膵島転写因子をコードする核酸分子を含む製剤を投与する段階を含む方法。
【請求項13】
哺乳動物対象において膵島細胞を産生する方法であり、
製剤は膵島転写因子をコードしかつ膵島転写因子の発現に適合化された核酸分子を含み、膵島転写因子は神経内分泌bHLH転写因子であり、投与は対象の膵臓における膵島転写因子の産生および膵島細胞の産生に十分な量で行われる、哺乳動物対象の膵細胞に製剤を投与する段階を含む方法。
【請求項14】
神経内分泌bHLH転写因子が、ニューロゲニン3(Ngn3)、NeuroD1、およびMash1からなる群より選択される、請求項12または13記載の方法。
【請求項15】
神経内分泌bHLH転写因子が、ニューロゲニン1、ニューロゲニン2、neuroD2、math2、NeuroD4/Math3、math1/ATOH1、およびmash2からなる群より選択される、請求項12または13記載の方法。
【請求項16】
対象がヒトである、請求項12または13記載の方法。
【請求項17】
請求項1、5、または13記載の方法により産生される、単離された組換え哺乳動物膵島細胞。
【請求項1】
哺乳動物膵島細胞を産生する方法であり、膵島転写因子は神経内分泌bHLH転写因子であり、導入により細胞内における膵島転写因子の発現および細胞内における膵島細胞表現型の産生が提供される、哺乳動物膵細胞に膵島転写因子をコードする核酸分子を導入する段階を含む方法。
【請求項2】
神経内分泌bHLH転写因子がニューロゲニン3(Ngn3)、NeuroD1、およびMash1からなる群より選択される、請求項1記載の方法。
【請求項3】
神経内分泌bHLH転写因子がニューロゲニン1、ニューロゲニン2、neuroD2、math2、NeuroD4/Math3、math1/ATOH1、およびmash2からなる群より選択される、請求項1記載の方法。
【請求項4】
bHLH転写因子がニューロゲニン3(Ngn3)制御経路の正の制御因子である、請求項1記載の方法。
【請求項5】
哺乳動物膵島細胞を産生する方法であり、導入により細胞内における膵島表現型の産生が提供される、哺乳動物膵細胞にニューロゲニン3(Ngn3)制御経路の正の制御因子を導入する段階を含む方法。
【請求項6】
正の制御因子が細胞内でNgn3の発現を増加させる、請求項5記載の方法。
【請求項7】
正の制御因子がNgn3をコードする核酸であり、さらに該核酸が細胞におけるNgn3の発現および産生に適合化される、請求項5記載の方法。
【請求項8】
正の制御因子がMash1、NeuroD1、HNF1、HNF2、およびHNF6からなる群より選択される、請求項5記載の方法。
【請求項9】
哺乳動物膵細胞がヒト膵細胞である、請求項1または5記載の方法。
【請求項10】
膵島細胞表現型が膵β細胞の表現型である、請求項1または5記載の方法。
【請求項11】
哺乳動物対象の血流にインスリンを送達する方法であり、導入により膵島細胞によるインスリンの産生および哺乳動物対象の血流へのインスリンの送達が提供される、請求項1または5記載の方法により産生される膵島細胞を哺乳動物対象の膵臓に導入する段階を含む方法。
【請求項12】
哺乳動物対象の血流にインスリンを送達する方法であり、膵島転写因子は神経内分泌bHLH転写因子であり、投与はβ細胞の産生を誘導するのに十分なレベルの膵島転写因子を産生するのに十分な量で行われ、膵島細胞はインスリンを産生し、インスリンは対象の血流に送達される、哺乳動物対象の膵細胞に膵島転写因子をコードする核酸分子を含む製剤を投与する段階を含む方法。
【請求項13】
哺乳動物対象において膵島細胞を産生する方法であり、
製剤は膵島転写因子をコードしかつ膵島転写因子の発現に適合化された核酸分子を含み、膵島転写因子は神経内分泌bHLH転写因子であり、投与は対象の膵臓における膵島転写因子の産生および膵島細胞の産生に十分な量で行われる、哺乳動物対象の膵細胞に製剤を投与する段階を含む方法。
【請求項14】
神経内分泌bHLH転写因子が、ニューロゲニン3(Ngn3)、NeuroD1、およびMash1からなる群より選択される、請求項12または13記載の方法。
【請求項15】
神経内分泌bHLH転写因子が、ニューロゲニン1、ニューロゲニン2、neuroD2、math2、NeuroD4/Math3、math1/ATOH1、およびmash2からなる群より選択される、請求項12または13記載の方法。
【請求項16】
対象がヒトである、請求項12または13記載の方法。
【請求項17】
請求項1、5、または13記載の方法により産生される、単離された組換え哺乳動物膵島細胞。
【図1】

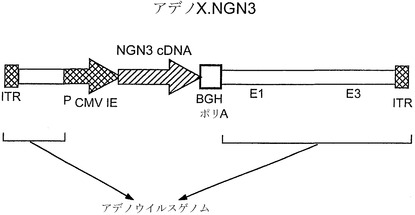
【図2A】

【図2B】
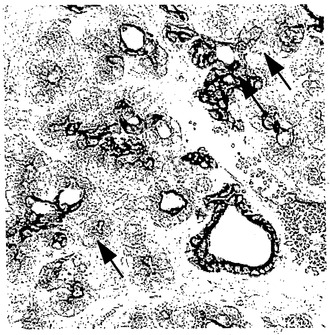
【図3】

【図4A】

【図4B】

【図5】
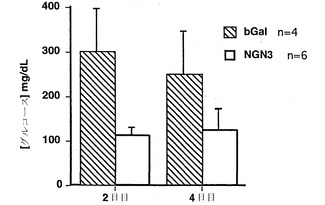
【図6】
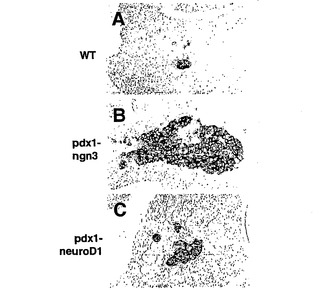
【図7】
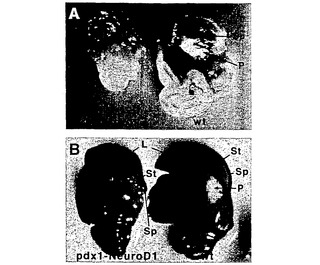
【図8】
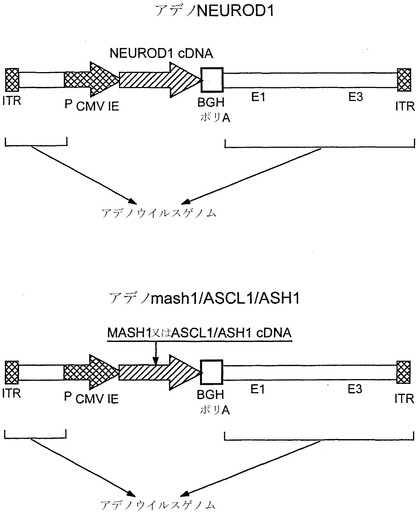
【図9】
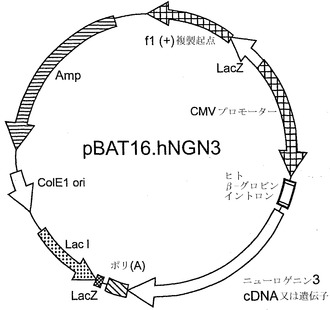
【図10】
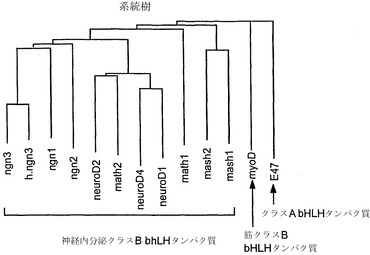
【図11】
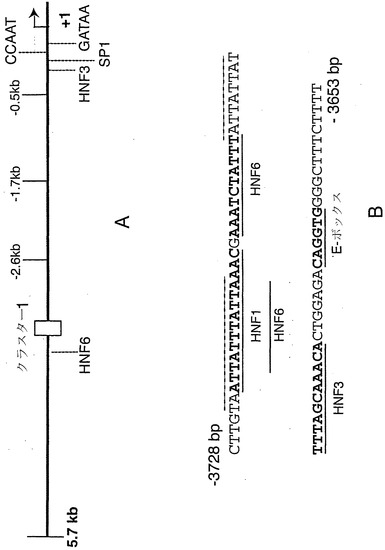
【図12】
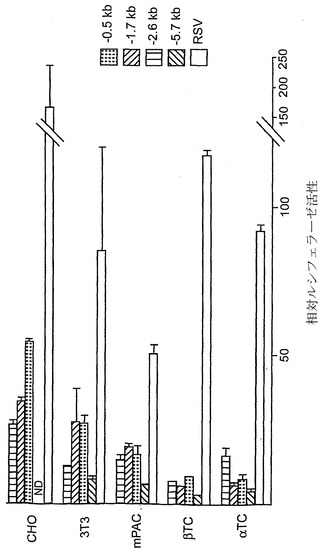
【図13】
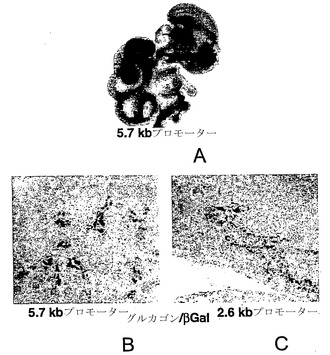
【図14】
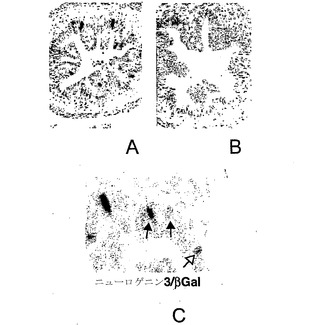
【図15】
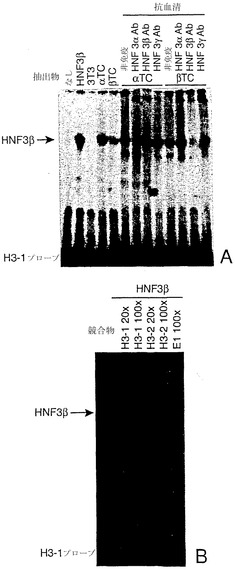
【図16】
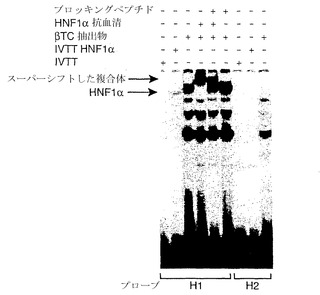
【図17】
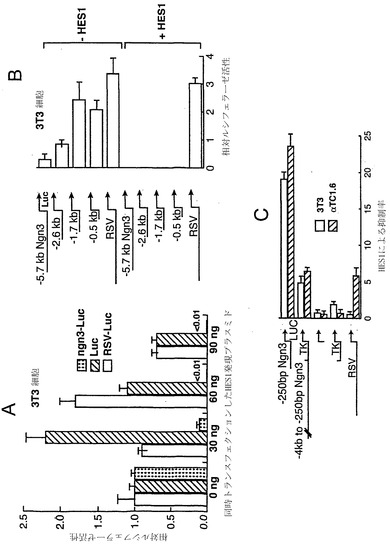
【図18】
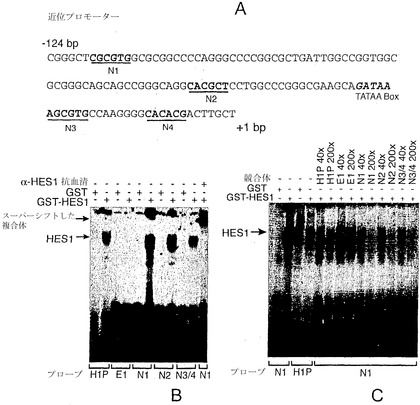
【図19A】
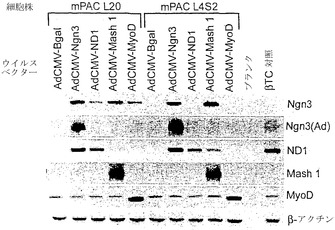
【図19B】
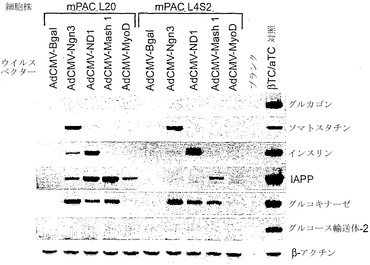
【図19C】
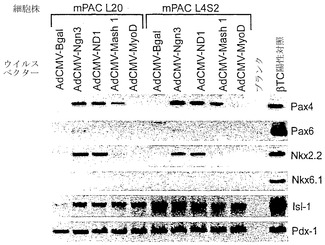
【図2A】
【図2B】
【図3】
【図4A】
【図4B】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19A】
【図19B】
【図19C】
【公開番号】特開2010−148510(P2010−148510A)
【公開日】平成22年7月8日(2010.7.8)
【国際特許分類】
【出願番号】特願2010−11543(P2010−11543)
【出願日】平成22年1月22日(2010.1.22)
【分割の表示】特願2002−572772(P2002−572772)の分割
【原出願日】平成14年3月20日(2002.3.20)
【出願人】(592130699)ザ リージェンツ オブ ザ ユニバーシティ オブ カリフォルニア (364)
【氏名又は名称原語表記】The Regents of The University of California
【Fターム(参考)】
【公開日】平成22年7月8日(2010.7.8)
【国際特許分類】
【出願日】平成22年1月22日(2010.1.22)
【分割の表示】特願2002−572772(P2002−572772)の分割
【原出願日】平成14年3月20日(2002.3.20)
【出願人】(592130699)ザ リージェンツ オブ ザ ユニバーシティ オブ カリフォルニア (364)
【氏名又は名称原語表記】The Regents of The University of California
【Fターム(参考)】
[ Back to top ]