
臍帯血からの人工多能性幹細胞の生成方法
遺伝子操作された人工多能性幹細胞の生成および使用のための方法ならびに組成物が提供される。

【発明の詳細な説明】
【技術分野】
【0001】
関連出願の相互参照
本出願は、2009年6月19日出願の米国仮出願第61/218,611号に恩典を主張し、その内容は参照により、その全体がすべての目的のために本明細書に組み入れられる。
【背景技術】
【0002】
発明の背景
組込みウイルスの方法を用いた多能性因子および癌遺伝子の異所性発現は、マウスおよびヒト線維芽細胞の両方において多能性を誘導するのに十分である3〜9。しかしながら、この過程は時間がかかり、非効率的で、ゲノム中へのベクターの恒久的な組込みは、治療への適用のためのiPS細胞の使用を制限する1。さらなる研究によって、使用される年齢、起源、および細胞型が再プログラミングの効率に重大な影響を有し、最終的により少ない因子の発現が必要であったり、および/または過程全体の時期が減少したりすることが示されている。最近、レトロウイルスによるヒト角化細胞への形質導入によって、線維芽細胞に比べて100倍も効率が高く、2倍速い多能性への再プログラミングがもたらされることが示された。これらの相違は、出発物の角化細胞集団におけるKLF4およびc-MYCの内在性の発現および/または再プログラミングをより受け入れやすいというエピジェネティックな状態を示す未分化前駆細胞のプールの存在に起因し得るという仮説が立てられた10。この後者の仮説は、マウスにおける他の研究によってさらに裏付けられている11、12。しかしながら、幹細胞は通常は稀であり、大量に入手および単離することは困難である(例えば、神経幹細胞13、14)。
【0003】
人工多能性幹(iPS)細胞は、細胞療法のための潜在的価値を有する患者特異的な前駆体をインビトロで生成できるようにするため、再生医療への関心を引き起こしてきた1。しかしながら、多くの場合において、急性疾患の細胞療法のため、または慢性疾患もしくは加齢の帰結として患者の体細胞が変化した場合等はすぐに使える手段が望ましい。臍帯血(CB)幹細胞は、最小限の遺伝子の変化およびエピジェネティックな変化を有する、生まれたばかりで免疫学的に未成熟な細胞であり、CBバンクの世界的ネットワークを通して数十万の免疫分類されたCBユニットをすぐに利用できるため、この目的のためにはCB幹細胞が理想的に適しているようである2。造血移植のための幹細胞の供給源として骨髄(BM)の代替物と考えられているCB細胞は、ドナーに危険を与えることなく十分な量を採取することができる15。容易に入手できることに加えて、CB細胞は最小限の体細胞変異を有する幼若細胞であるという特徴と新生細胞の免疫学的な未成熟により与えられる長所を兼ね備える15。これらの特性はHLA-ドナー-レシピエント選択のより厳しくない基準を可能にし、移植のための決定的な利点を表す。さらに、臍帯血バンクの世界的で包括的なネットワークは、CB幹細胞の適合ドナーを素早く効果的に検索することを保証する2。最後に、CB CD133+幹細胞はOCT4、SOX2、NANOG、REX1および他の多能性関連マーカーを発現することが示されてきており16〜18、したがって原理上は再プログラミングをより受け入れやすいかもしれない。本明細書において、2つの強力な癌遺伝子(c-MYCおよびKLF4)もしくは追加の化学物質の必要なしに、4つ(OSKM)、3つ(OSK)、およびわずか2つ(OS)の転写因子をレトロウイルスで形質導入することによって、素早く効率的にCB幹細胞を多能性へと再プログラミングすることを含む態様が初めて記載される。
【0004】
本明細書に記載された一定の方法および組成物を用いて、OCT4、SOX2、KLF4、およびc-MYCをレトロウイルスで形質導入することによって、非常に効率的かつ素早い過程においてCB幹細胞を多能性へと再プログラムできる。結果として得られたCB由来のiPS(CBiPS)細胞は、表現型も分子的にもヒト胚性幹(hES)細胞と識別不可能である。さらに、臍帯血iPSの生成はc-MYCおよびKLF4癌遺伝子を使用せずに、OCT4およびSOX2を過剰発現させるだけで効率的に達成できる。本明細書に記載された方法および組成物は、当技術分野における課題を克服し、すぐに使える適用のためのHLA適合CBiPS細胞の包括的バンクの製造のための基礎を築くかもしれない。
【発明の概要】
【0005】
本明細書に提供されているのは、特に、臍帯血から人工多能性幹細胞を作製し、使用するための非常に効率的な方法および組成物である。
【0006】
一つの局面において、人工多能性幹細胞を調製するための方法が提供される。この方法は、OCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成することを含む。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。
【0007】
別の局面において、人工多能性幹細胞を調製するための方法が提供される。この方法は、OCT4タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成することを含む。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。
【0008】
別の局面において、本明細書の方法に従って調製された人工多能性幹細胞が提供される。
【0009】
一つの局面において、OCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を含む臍帯血幹細胞が提供される。
【0010】
別の局面において、OCT4タンパク質をコードする核酸を含む臍帯血幹細胞が提供される。
【0011】
一つの局面において、ヒト体細胞を産生するための方法が提供される。この方法は、細胞増殖因子を人工多能性幹細胞に接触させることを含む。人工多能性幹細胞は分裂させられ、これによりヒト体細胞を形成する。ある態様において、人工多能性幹細胞はOCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程を含む過程によって調製される。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。別の態様において、人工多能性幹細胞はOCT4タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程を含む過程によって調製される。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。
【0012】
別の局面において、組織修復を必要としている哺乳動物を治療するための方法が提供される。この方法は、人工多能性幹を哺乳動物に投与し、哺乳動物において人工多能性幹細胞を分裂および体細胞に分化させ、これにより哺乳動物において組織修復を提供することを含む。ある態様において、人工多能性幹細胞はOCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程を含む過程によって調製される。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。別の態様において、人工多能性幹細胞はOCT4タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程を含む過程によって調製される。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。
【図面の簡単な説明】
【0013】
【図1A】図1A〜1E。OCT4およびSOX2因子のみを用いたCBiPS細胞株の生成。図1A:臍帯血幹細胞再プログラミングの予定表。感染の3日後に、CB CD133+細胞をフィーダーに移動する。9日目頃に小さい接着性のコロニーが観察される。12日後に典型的なhES様コロニーがはっきりと見える。
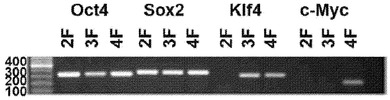
【図1B】図1A〜1E。OCT4およびSOX2因子のみを用いたCBiPS細胞株の生成。図1B:4つ、3つ、および2つのみの導入遺伝子の挿入を確認するゲノムDNAのPCR。
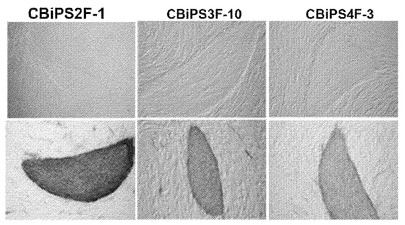
【図1C】図1A〜1E。OCT4およびSOX2因子のみを用いたCBiPS細胞株の生成。図1C:CBiPS2F-1、3F-10、および4F-3細胞株の代表的な位相差画像ならびにアルカリフォスファターゼ(AP)染色。
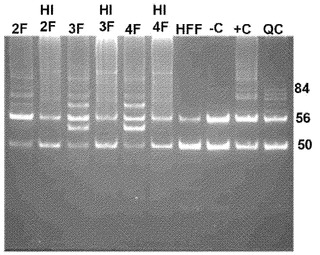
【図1D】図1A〜1E。OCT4およびSOX2因子のみを用いたCBiPS細胞株の生成。図1D:CBiPS2F、3F、および4F細胞株における代表的なテロメラーゼ活性(HI:熱不活化、HFF:ヒト包皮線維芽細胞、−C:陰性対照として溶解緩衝液、+C:陽性対照、およびQC:量的対照)。
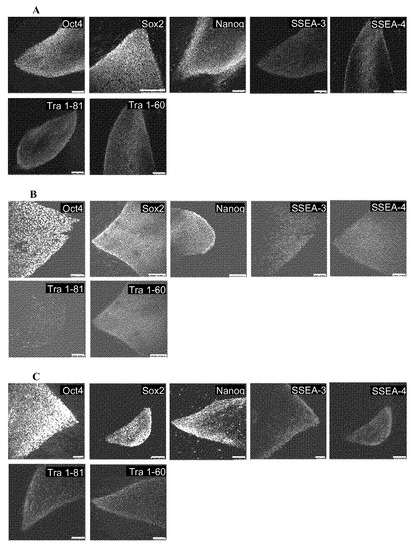
【図1E】図1A〜1E。OCT4およびSOX2因子のみを用いたCBiPS細胞株の生成。図1E:多能性マーカーについてのCBiPS2F-1細胞株の免疫蛍光解析。コロニーは胚性マーカーSSEA-4、SSEA-3、TRA-1-60、TRA-1-81、ならびに転写因子OCT4、SOX2、およびNANOGを発現する。基礎となる線維芽細胞は陰性対照を提供する。スケールバーは250μm。
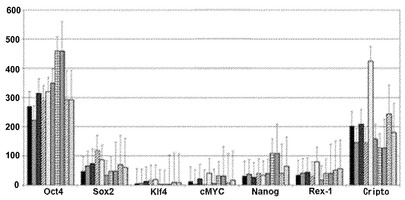
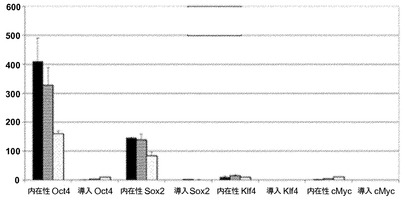
【図2A】図2A〜2G。CBiPS細胞株の特徴付け。図2A:多能性マーカーOCT4、SOX2、NANOG、REX1、CRIPTO、KLF4、およびc-MYCの定量的RT-PCR解析を描写したヒストグラム。新鮮および凍結試料に由来する異なるCBiPS細胞株と一緒にES[2]および角化細胞-iPS(KiPS)細胞株を解析した。エラーバーは、三連より生成されたs.d.(標準偏差)を示す。ヒストグラムの説明(左から右):CBiPS4F-3、CBiPS4F-5、CBiPS3F-10、CBiPS3F-12、CBiPS2F-1、CBiPS2F-2、CBiPSF-1、CBiPSF-5、ES2、およびKiPS。
【図2B】図2A〜2G。CBiPS細胞株の特徴付け。図2B:CBiPS細胞株におけるOCT4、SOX2、KLF4、およびc-MYC導入遺伝子の抑制を示す定量的RT-PCRを描写したヒストグラム。ヒストグラムの説明(左から右):CBiPS4F-3、CBiPS3F-10、およびCBiPS2F-1。
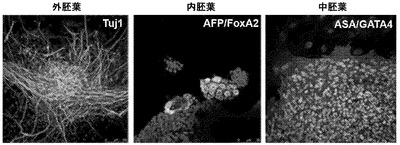
【図2C】図2A〜2G。CBiPS細胞株の特徴付け。図2C:3つの初期生殖細胞層(外肺葉-Tuj1、内胚葉-AFPおよびFOXA2、ならびに中胚葉-ASAおよびGATA4)へのCBiPS2F-1のインビトロ分化。
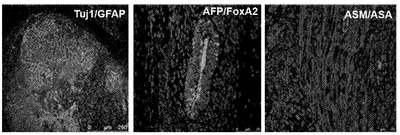
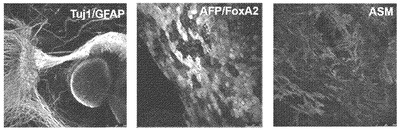
【図2D】図2A〜2G。CBiPS細胞株の特徴付け。図2D:Tuj1/GFAP陽性外胚葉、AFP/FoxA2陽性内胚葉、およびASM/ASA陽性中胚葉を示すCBiPS2F-1の精巣内注射から60日間後の奇形腫切片の免疫蛍光解析。スケールバーは75〜250μm。
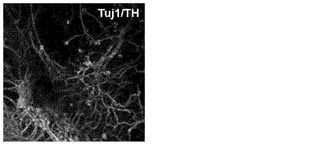
【図2E】図2A〜2G。CBiPS細胞株の特徴付け。図2E:免疫表現型的に成熟したドーパミン作動性ニューロン(Tuj1/THチロシンヒドロキシラーゼ)への、CBiPS2F-1の特異的なインビトロ分化。
【図2F】図2A〜2G。CBiPS細胞株の特徴付け。図2F:免疫表現型的に成熟したドーパミン作動性ニューロン(Tuj1/THチロシンヒドロキシラーゼ)への、CBiPS3F-12の特異的なインビトロ分化。
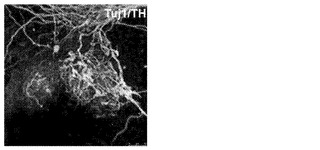
【図2G】図2A〜2G。CBiPS細胞株の特徴付け。図2G:ヒト線維芽細胞およびCD133+細胞において、OCT4、NANOG、HOXB4、およびHOXB5のプロモーター中のK4(H3K4me2)、K27(H3K27me3)、およびK9(H3K9me3)におけるヒストンH3メチル化レベルを比較したクロマチン免疫沈降法を描写したヒストグラム。ヒストグラムの説明:線維芽細胞(整列した点)、CD133+(黒塗り)。
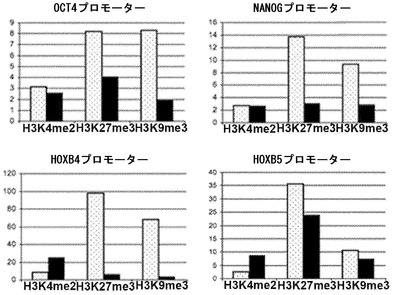
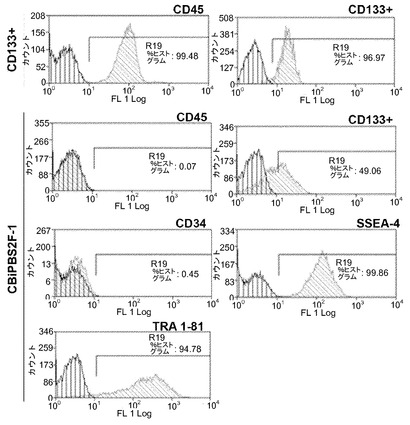
【図3A】図3A〜3C。ヒトCD133+細胞のフローサイトメトリー解析。図3A:免疫選択後のCD133細胞の純度についての代表的なドットプロットを描写する。
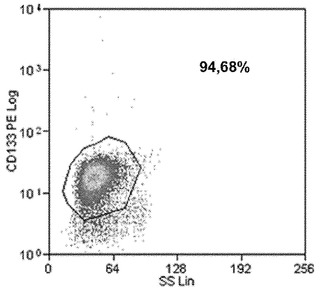
【図3B】図3A〜3C。ヒトCD133+細胞のフローサイトメトリー解析。図3B:感染から3日後の全GFP+細胞および二重陽性GFP/CD133細胞の定量を描写する。
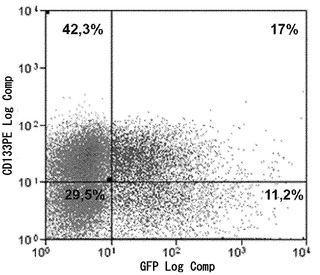
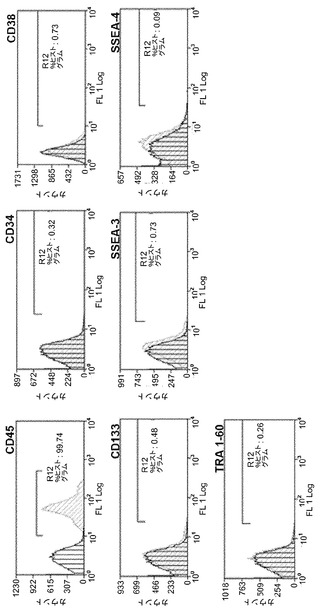
【図3C】図3A〜3C。ヒトCD133+細胞のフローサイトメトリー解析。図3C:ヒストグラムは、hES条件において3週間にわたって培養された形質導入されていないCD133+幹細胞のフローサイトメトリー解析を描写する。造血マーカーCD45、CD34、CD38、およびCD133、ならびにSSEA3、SSEA4、およびTRA-1-60を含む胚性幹細胞マーカーについて細胞を解析した。
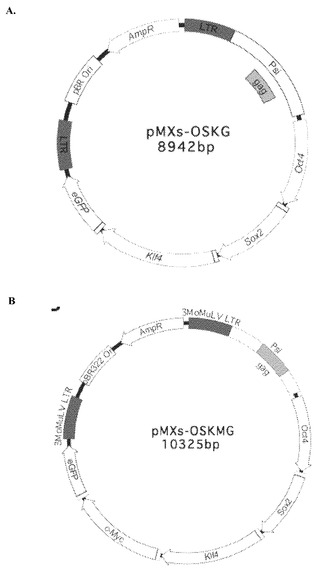
【図4】図4A〜4B。pMXs-OSKMGおよびpMXs-OSKGポリシストロニックレトロウイルスの模式図。図4A:pMXs-OSKGポリシストロニックレトロウイルス。図4B:pMXs-OSKMGポリシストロニックレトロウイルス。
【図5】図5A〜5C。多能性マーカーについての免疫蛍光解析。図5A:CBiPS3F-10。図5B:CBiPS4F-3細胞株はSSEA-4、SSEA-3、TRA-1-60、TRA-1-81を含む他の典型的な多能性マーカー、ならびに転写因子OCT4、SOX2、およびNANOGを発現する。図5C:凍結融解したCBユニットから精製されたCD133+細胞を用いて、OSKレトロウイルスで形質導入した後に生成されたCBiPS凍結(CBiPSFr)-1細胞株は、SSEA-4、SSEA-3、TRA-1-60、TRA-1-81を含む他の典型的な多能性マーカー、ならびに転写因子OCT4、SOX2、およびNANOGを発現する。基礎となる線維芽細胞は陰性対照を提供する。スケールバーは250μm。
【図6】CBiPS2Fのフローサイトメトリー解析。ヒストグラムはCBiPS2F-1細胞がCD45およびCD34等の造血マーカーを喪失し、TRA-1-181およびSSEA-4を含む典型的な多能性マーカーを獲得したことを確認するフローサイトメトリー解析を描写する。
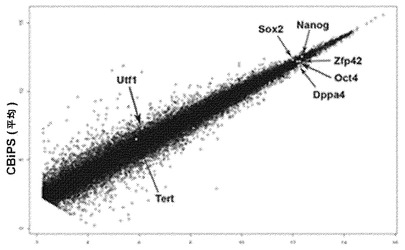
【図7A】図7A〜7B。全般的遺伝子発現解析。図7A:CBiPS(2本の線、各2反復)およびES2(2反復)の間で平均の全般的遺伝子発現パターンを比較したところ、非常に高いレベルの相関を示した。いくつかの多能性遺伝子をプロット中に同定する。
【図7B】図7A〜7B。全般的遺伝子発現解析。図7B:異なる多能性株および各出発物集団のすべての一対比較についての、ゲノム全般の転写プロファイルの相関係数。
【図8】図8A〜8B。レトロウイルスによる導入遺伝子サイレンシング。任意のFLAGタグ付けされたレトロウイルスの転写因子からの導入遺伝子発現のみを検出する特定のFLAG抗体と組み合わせたOCT4に対する免疫蛍光染色。図8A:CBiPS2F-1における内在性OCT4の発現および導入遺伝子のサイレンシング。図8B:陽性対照としてのOCT4およびSOX2に感染させた初代ヒト線維芽細胞。スケールバーは250μm。
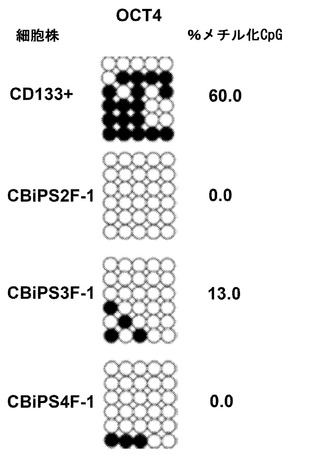
【図9】亜硫酸水素塩ゲノム配列によるメチル化プロモーター解析。すべてのCBiPS細胞株におけるプロモーターの一貫した脱メチル化を確認するOCT4プロモーターのメチル化解析。
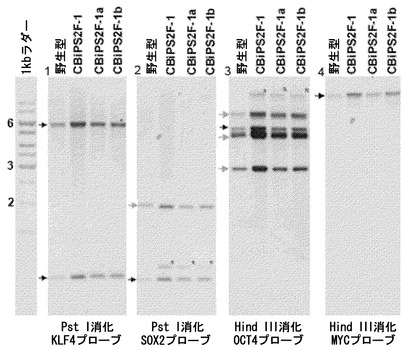
【図10】サザンブロット。CBiPS2F-1株およびサブクローンにおけるレトロウイルスの組込み数を評価するためのサザンブロット。1:KLF4特異的プローブとハイブリダイズした、Pst Iで消化したゲノムDNA。内在性バンド:5.9kbおよび0.9kb(黒い矢頭)。予期されたように、このプローブでは追加のバンドは検出されない。2:SOX2特異的プローブとハイブリダイズした、Pst Iで消化したゲノムDNA。内在性バンド:0.9kb(黒い矢頭)。固有の導入遺伝子の挿入に対応する同じ追加のバンドがCBiPS2F-1、CBiPS2F-1a、およびCBiPS2F-1bに存在する(赤い星印)。3:OCT4特異的プローブとハイブリダイズした、Hind IIIで消化したゲノムDNA。内在性の特異的バンド:4.5kb(黒い矢頭)。内在性の非特異的バンド(灰色の矢頭)。固有の導入遺伝子の挿入に対応する同じ追加のバンドがCBiPS2F-1、CBiPS2F-1a、およびCBiPS2F-1bに存在する(赤い星印)。4:c-MYC特異的プローブとハイブリダイズした、Hind IIIで消化したゲノムDNA。内在性バンド:11kb(黒い矢頭)。予期されたように、このプローブでは追加のバンドは検出されない。
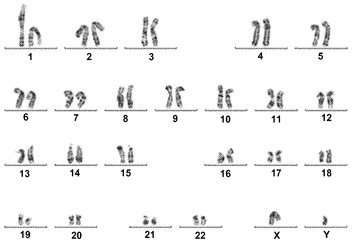
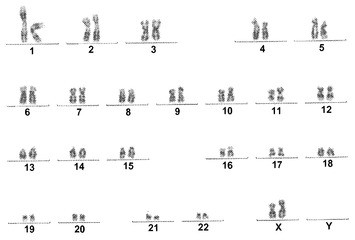
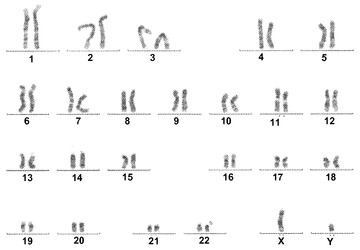
【図11A】図11A〜11C。CBiPS 2F、3F、および4F細胞株についての核型分析。10回の継代後に解析されたCBiPS2F-1(図11A)、CBiPS3F-10(図11B)、およびCBiPS4F-3(図11C)細胞における正常染色体、2倍体染色体、男性染色体、および女性染色体の内容を示す高解像度のGバンド形成核型。
【図11B】図11A〜11C。CBiPS 2F、3F、および4F細胞株についての核型分析。10回の継代後に解析されたCBiPS2F-1(図11A)、CBiPS3F-10(図11B)、およびCBiPS4F-3(図11C)細胞における正常染色体、2倍体染色体、男性染色体、および女性染色体の内容を示す高解像度のGバンド形成核型。
【図11C】図11A〜11C。CBiPS 2F、3F、および4F細胞株についての核型分析。10回の継代後に解析されたCBiPS2F-1(図11A)、CBiPS3F-10(図11B)、およびCBiPS4F-3(図11C)細胞における正常染色体、2倍体染色体、男性染色体、および女性染色体の内容を示す高解像度のGバンド形成核型。
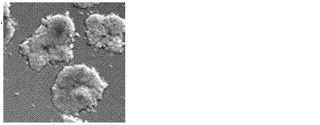
【図12A】図12A〜12F。CBiPS細胞株のインビトロおよびインビボ多能性。図12A:CBiPS2F-1細胞株に由来する胚様体。
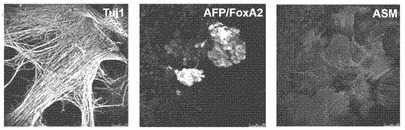
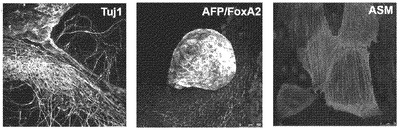
【図12B】図12A〜12F。CBiPS細胞株のインビトロおよびインビボ多能性。CBiPS3F-10(図12B)およびCBiPS4F-3(図12C)はインビトロにおいて、神経(Tuj1/GFAP)、内胚葉(AFP/FoxA2)、および中胚葉(ASM)細胞を含む3つの胚葉に分化できる。
【図12C】図12A〜12F。CBiPS細胞株のインビトロおよびインビボ多能性。CBiPS3F-10(図12B)およびCBiPS4F-3(図12C)はインビトロにおいて、神経(Tuj1/GFAP)、内胚葉(AFP/FoxA2)、および中胚葉(ASM)細胞を含む3つの胚葉に分化できる。
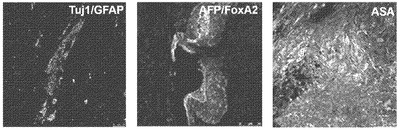
【図12D】図12A〜12F。CBiPS細胞株のインビトロおよびインビボ多能性。図12D:CBiPSFr-1細胞株の神経(Tuj1)、内胚葉(AFP/FoxA2)、および中胚葉(ASM)細胞を含む3つの胚葉へのインビトロ分化。
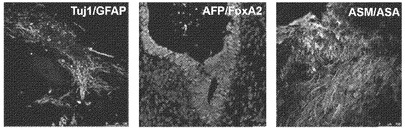
【図12E】図12A〜12F。CBiPS細胞株のインビトロおよびインビボ多能性。CBiPS3F-10(図12E)およびCBiPS4F-3(図12F)のインビボ分化。結果として得られた奇形腫は3つの胚葉のすべて:外胚葉(Tuj1/GFAP)、内胚葉(AFP/FoxA2)、および中胚葉(ASM/ASA)を表す組織を含んでいた。
【図12F】図12A〜12F。CBiPS細胞株のインビトロおよびインビボ多能性。CBiPS3F-10(図12E)およびCBiPS4F-3(図12F)のインビボ分化。結果として得られた奇形腫は3つの胚葉のすべて:外胚葉(Tuj1/GFAP)、内胚葉(AFP/FoxA2)、および中胚葉(ASM/ASA)を表す組織を含んでいた。
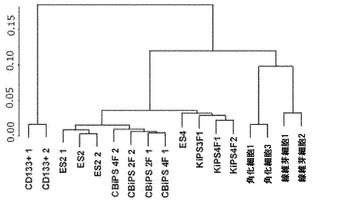
【図13A】図13A〜13D。CB CD133+細胞の遺伝子発現解析。図13A:CD133+細胞は線維芽細胞および角化細胞よりも多能性細胞に近くないことを示す、CD133+、角化細胞、線維芽細胞、ES細胞、KiPS、およびCBiPSについてのゲノム全般の転写プロファイルの階層的クラスタリングを表す系統樹。
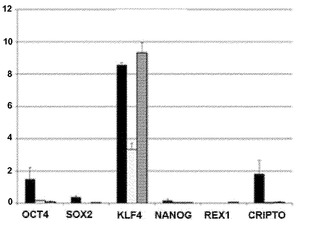
【図13B】図13A〜13D。CB CD133+細胞の遺伝子発現解析。図13B:定量的RT-PCRによる、CD133+細胞、線維芽細胞、および角化細胞における多能性マーカーおよびKLF4の比較遺伝子発現解析を描写したヒストグラム。ヒストグラムの説明(左から右):CD133+(黒色)、線維芽細胞(白色)、および角化細胞(灰色)。
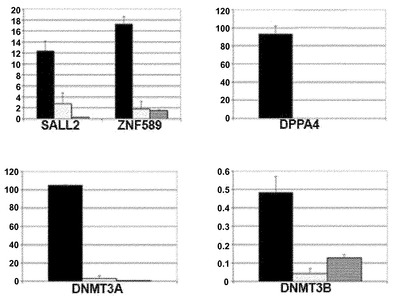
【図13C】図13A〜13D。CB CD133+細胞の遺伝子発現解析。図13C:定量的RT-PCRによる、CD133+細胞におけるSALL2、ZNF589、DPPA4、DNMT3A、およびDNMT3B遺伝子の上方制御の検証。ヒストグラムの説明:図13Bと同様。
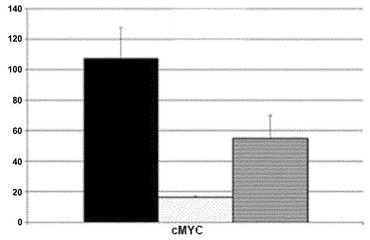
【図13D】図13A〜13D。CB CD133+細胞の遺伝子発現解析。図13D:CD133+細胞、線維芽細胞、および角化細胞におけるc-MYC発現の定量的RT-PCR解析。エラーバーは、三連より生成されたs.d.を示す。ヒストグラムの説明:図13Bと同様。
【発明を実施するための形態】
【0014】
発明の詳細な説明
I.定義
以下の定義は、本明細書中で頻繁に使用される一定の用語の理解を促進するために提供されるものであり、本開示の範囲を限定することを意図しない。
【0015】
「核酸」は、デオキシリボヌクレオチドもしくはリボヌクレオチド、および一本鎖もしくは二本鎖のいずれかの形態におけるそれらのポリマー、ならびにそれらの相補を指す。
【0016】
「相補的」もしくは「相補性」という言葉は、ポリヌクレオチド中の核酸が第二のポリヌクレオチド中の別の核酸と塩基対を形成する能力を指す。例えば、配列A-G-Tは配列T-C-Aと相補的である。相補性は、塩基対合に従って核酸のいくつかだけが一致する部分的なものでもよく、もしくは塩基対合に従ってすべての核酸が一致する完全なものでもよい。
【0017】
「同一の」もしくはパーセント「同一性」という用語は、2以上の核酸との関連では、同じかまたは以下に記載された規定値のパラメーターでBLASTもしくはBLAST 2.0配列比較アルゴリズムを用いて測定するか、または手動の整列および目視検査によって測定すると、特定のパーセントの同じヌクレオチド(すなわち、比較ウィンドウもしくは指定された領域にわたって最大に一致するように比較および整列させた場合に、特定の領域にわたって約60%の同一性、好ましくは65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、もしくはより高い同一性)を有する2以上の配列もしくは部分配列を指す(例えばNCBIウェブサイト等を参照)。このとき、そのような配列は「実質的に同一」と言われる。この定義は試験配列の相補も指すか、もしくは適用されてよい。この定義は欠失および/または付加を有する配列、ならびに置換を有するものも含む。以下に記載されるように、好ましいアルゴリズムはギャップ等を計上できる。好ましくは、同一性は長さが少なくとも約25アミノ酸もしくはヌクレオチド、またはより好ましくは、長さが50〜100アミノ酸もしくはヌクレオチドの領域にわたって存在する。
【0018】
「ストリンジェントなハイブリダイゼーション条件」という語句は、通常は核酸の複合混合物中でプローブがその標的配列とハイブリダイズするが、他の配列とはハイブリダイズしない条件を指す。ストリンジェントな条件は配列依存性であり、異なる状況において異なる。より長い配列は、より高い温度において特異的にハイブリダイズする。核酸のハイブリダイゼーションの広範な手引きはTijssen, Techniques in Biochemistry and Molecular Biology--Hybridization with Nucleic Probes, "Overview of principles of hybridization and the strategy of nucleic acid assays" (1993)に見出される。概して、ストリンジェントな条件は規定されたイオン強度のpHにおいて、特定の配列についての熱融解点(Tm)よりも約5〜10℃低くなるように選択される。Tmは、(規定されたイオン強度、pH、および核の濃度において)標的に相補的なプローブの50%が平衡状態で標的配列にハイブリダイズする温度である(標的配列が過剰に存在するため、Tmでは平衡状態でプローブの50%が占有される)。ストリンジェントな条件は、ホルムアミド等の不安定化剤の添加によっても達成されてよい。選択的もしくは特異的なハイブリダイゼーションのためには、陽性シグナルはバックグラウンドの少なくとも2倍であり、好ましくはバックグラウンドハイブリダイゼーションの10倍である。例示的なストリンジェントなハイブリダイゼーション条件は、次の通りであり得る:50% ホルムアミド、5×SSC、および1% SDSで42℃においてインキュベートするか、もしくは5×SSC、1% SDSで65℃においてインキュベートし、0.2×SSCおよび0.1% SDS中で65℃において洗浄する。
【0019】
核酸ハイブリダイゼーション技術を使用する、特異的なDNAおよびRNAの測定の多岐にわたる方法が当業者に公知である(Sambrook、前出を参照)。いくつかの方法は電気泳動分離を含む(例えば、DNAを検出するためのサザンブロット、およびRNAを検出するためのノザンブロット)が、DNAおよびRNAの測定は電気泳動分離を行わなくても実行できる(例えば、ドットブロットによって)。
【0020】
ハイブリダイゼーション測定法の感度は、検出されている標的核酸を増幅させる核酸増幅システムの使用を通して強化できる。そのようなシステムの例には、ポリメラーゼ連鎖反応(PCR)システムおよびリガーゼ連鎖反応(LCR)システムが含まれる。当技術分野において最近記載された他の方法は、核酸配列に基づく増幅(NASBA、Cangene、Mississauga、Ontario)およびQβレプリカーゼシステムである。PCRもしくはLCRプライマーが、選択配列が存在する場合にのみ伸長もしくは連結するように設計されているところ、これらのシステムは突然変異を直接的に同定するのに使用できる。あるいは、選択された配列は例えば非特異的PCRプライマーを用いて大まかに増幅でき、後に突然変異を示す特異的配列を増幅された標的領域に探索する。増幅反応産物を、例えばリアルタイムで観察するために、Taqman(登録商標)および分子指標プローブを含む様々な検出プローブを使用できることが理解される。
【0021】
「ポリヌクレオチド」という言葉は、ヌクレオチドの直鎖状配列を指す。ヌクレオチドはリボヌクレオチド、デオキシリボヌクレオチド、もしくは両方の混合物であり得る。本明細書において予期されるポリヌクレオチドの例には、一本鎖および二本鎖DNA、一本鎖および二本鎖RNA(miRNAを含む)、ならびに一本鎖および二本鎖DNAおよびRNAの混合物を有するハイブリッド分子を含む。
【0022】
「タンパク質」、「ペプチド」、および「ポリペプチド」という言葉は、アミノ酸ポリマーまたは2つ以上の相互作用する、もしくは結合したアミノ酸ポリマーの組を表示するために互換的に使用される。
【0023】
「遺伝子」という用語は、タンパク質の産生に関与するDNAの断片を意味し、コード領域に先行および後続する領域(リーダーおよびトレーラー)ならびに個々のコード断片(エクソン)の間の介在配列(イントロン)を含む。リーダー、トレーラー、およびイントロンは、遺伝子の転写および翻訳の間に必要な調節要素を含む。さらに、「タンパク質遺伝子産物」は、特定の遺伝子から発現したタンパク質である。
【0024】
「ウイルスベクター」は、別の核酸を細胞中に輸送できるウイルス由来の核酸である。ウイルスベクターは、適切な環境に存在する場合に、ベクターによって運搬される1つ以上の遺伝子にコードされる1つのタンパク質もしくは複数のタンパク質の発現を指示できる。ウイルスベクターの例にはレトロウイルスベクター、アデノウイルスベクター、レンチウイルスベクター、およびアデノ随伴ウイルスベクターが含まれるが、これらに限定されない。
【0025】
「形質移入」または「形質移入する」という用語は、非ウイルス的もしくはウイルスに基づく方法によって核酸分子を細胞に導入する過程として定義される。非ウイルス的な形質移入の方法は、核酸分子を細胞に導入するための送達システムとしてウイルスDNAもしくはウイルス粒子を使用しない任意の適切な形質移入の方法を含む。例示的な非ウイルス的形質移入方法には、リン酸カルシウム形質移入、リポソーム形質移入、ヌクレオフェクション、ソノポレーション、熱ショックによる形質移入、マグネティフェクション、およびエレクトロポレーションが含まれる。ウイルスに基づく形質移入の方法のために、本明細書に記載された方法において任意の有用なウイルスベクターを使用してよい。ウイルスベクターの例にはレトロウイルスベクター、アデノウイルスベクター、レンチウイルスベクター、およびアデノ随伴ウイルスベクターが含まれるが、これらに限定されない。
【0026】
遺伝子に関連して本明細書で使用される「発現」または「発現した」という言葉は、その遺伝子の転写および/または翻訳産物を意味する。細胞におけるDNA分子の発現レベルは、細胞内に存在する対応するmRNAの量もしくは細胞によって産生されるそのDNAにコードされるタンパク質の量のいずれかに基づいて決定してよい(Sambrook et al., 1989 Molecular Cloning: A Laboratory Manual, 18.1-18.88)。
【0027】
「プラスミド」という用語は、遺伝子の発現に必要な遺伝子および/または調節要素をコードする核酸分子を指す。プラスミドからの遺伝子の発現は、シスもしくはトランスで起こり得る。遺伝子がシスで発現する場合は、遺伝子および調節要素は同じプラスミドによってコードされる。トランスでの発現とは、遺伝子および調節要素が別々のプラスミドによってコードされる場合を指す。
【0028】
「エピソームの」という用語は、細胞におけるプラスミドの染色体外の状態を指す。エピソームのプラスミドは、染色体DNAの部分ではなく、染色体DNAとは独立して複製する核酸分子である。
【0029】
「細胞培養物」とは、生物の外側に存在する細胞の集団である。これらの細胞は、任意で細胞バンク、動物、もしくは血液バンクから単離された初代細胞、またはこれらの供給源の一つに由来する二次細胞であり、長寿命のインビトロ培養のために不死化されている。
【0030】
「幹細胞」とは、有糸細胞分裂を通して自己再生する能力および組織もしくは器官に分化する潜在力によって特徴付けされる細胞である。哺乳動物の幹細胞の中で、胚性および成体幹細胞は識別できる。胚性幹細胞は胚盤胞の中に存在し、胚組織を生み出す一方で、成体幹細胞は組織再生および修復のために成体組織中に存在する。
【0031】
「多能性の」もしくは「多能性」という用語は、適切な条件下で3つの胚葉(内胚葉、中胚葉、および外胚葉)から細胞系統に伴う特徴を集団で呈する細胞型に分化を起こすことのできる子孫を生み出す能力を有する細胞を指す。多能性幹細胞は、出生前、出生後、もしくは成体の生物の組織に寄与できる。細胞集団の多能性を確立するために、8〜12週齢のSCIDマウスにおいて奇形腫を形成する能力等の、技術分野において容認された標準的な試験を使用できる。しかしながら、多能性細胞を同定するために、様々な多能性幹細胞の特徴の同定も使用することができる。
【0032】
「多能性幹細胞の特徴」とは、多能性幹細胞を他の細胞と識別する細胞の特徴を指す。分子マーカーの一定の組合せの発現もしくは非発現は、多能性幹細胞の特徴の例である。より具体的には、ヒト多能性幹細胞は次の非限定的な一覧表からのマーカーの少なくともいくつか、任意ですべてを発現してよい:SSEA-3、SSEA-4、TRA-1-60、TRA-1-81、TRA-2-49/6E、ALP、Sox2、E-カドヘリン、UTF-1、Oct4、Lin28、Rex1、およびNanog。多能性幹細胞に伴う細胞の形態も多能性幹細胞の特徴である。
【0033】
「再プログラミング」という用語は、非多能性細胞を多能性幹細胞の特徴を呈する細胞に脱分化させる過程を指す。
【0034】
「治療する」という用語は、治療されている疾患の発症を改善し、抑制し、根絶し、および/または遅延させることを意味する。
【0035】
II.臍帯血から人工多能性幹細胞を調製する方法
一つの局面において、人工多能性幹細胞を調製するための方法が提供される。この方法は、OCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成することを含む。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。
【0036】
「人工多能性幹細胞」とは、非多能性細胞に人工的に由来する多能性幹細胞を指す。「非多能性細胞」は、多能性幹細胞よりも自己再生および分化する潜在力がより低い細胞であり得る。潜在力がより低い細胞とは、成体幹細胞、組織特異的前駆細胞、初代もしくは二次細胞であり得るが、これらに限定されない。成体幹細胞は、胚発生後に体全体に見出される未分化細胞である。成体幹細胞は死に瀕した細胞を補充し、損傷した組織を再生させるために、細胞分裂によって増殖する。成体幹細胞は、分裂して自身に似た別の細胞を製造し、また分裂して自身よりもより分化した細胞を製造する能力を有する。成体幹細胞はRex1、Nanog、Oct4、もしくはSox2等の多能性マーカーの発現を伴ってはいるが、3つのすべての胚葉の細胞型に分化するという多能性幹細胞の能力は有していない。成体幹細胞は、自己再生および独特の細胞型の子孫を生成する限定された潜在力を有する。成体幹細胞は、造血幹細胞、臍帯血幹細胞、間充織幹細胞、上皮幹細胞、皮膚幹細胞、もしくは神経幹細胞であり得るが、これらに限定されない。組織特異的前駆体とは、特定の器官もしくは組織に分化することが約束された、自己再生の潜在力を持たない細胞を指す。初代細胞は、卵細胞、精細胞、および幹細胞を除く成体もしくは胎児の生物の任意の細胞を含む。有用な初代細胞の例は皮膚細胞、骨細胞、血液細胞、内臓の細胞、および結合組織の細胞を含むが、これらに限定されない。二次細胞は初代細胞に由来し、長寿命のインビトロ細胞培養のために不死化されている。
【0037】
「臍帯血幹細胞」とは、臍帯血に存在し、多能性幹細胞よりも自己再生および分化する潜在力がより低いことで特徴付けられる成体幹細胞を指す。
【0038】
「形質移入」または「形質移入する」という用語は、非ウイルス的もしくはウイルスに基づく方法によって核酸分子を細胞に導入する過程として定義される。非ウイルス的な形質移入の方法は、核酸分子を細胞に導入するための送達システムとしてウイルスDNAもしくはウイルス粒子を使用しない任意の適切な形質移入の方法を含む。例示的な非ウイルス的形質移入方法には、リン酸カルシウム形質移入、リポソーム形質移入、ヌクレオフェクション、ソノポレーション、熱ショックによる形質移入、マグネティフェクション、およびエレクトロポレーションが含まれる。ある態様において、核酸分子は当技術分野において周知の標準的な手順に続いてエレクトロポレーションを使用して細胞内に導入される。ウイルスに基づく形質移入の方法のために、本明細書に記載された方法において任意の有用なウイルスベクターを使用してよい。ウイルスベクターの例にはレトロウイルスベクター、アデノウイルスベクター、レンチウイルスベクター、およびアデノ随伴ウイルスベクターが含まれるが、これらに限定されない。ある態様において、核酸分子は当技術分野において周知の標準的な手順に続いてレトロウイルスベクターを使用して細胞内に導入される。
【0039】
形質移入遺伝子の発現は、細胞内で一過的もしくは安定して起こり得る。「一過的発現」の間は、形質移入遺伝子は細胞分裂の間に娘細胞に伝達されない。その発現が形質移入細胞に限定されているため、遺伝子の発現は時間とともに失われる。対照的に、形質移入遺伝子の安定発現は、形質移入細胞に選択の優位性を与える別の遺伝子とその遺伝子を一緒に形質移入したときに起こり得る。そのような選択の優位性は、細胞に提示される一定の毒素に対する抵抗性であってよい。形質移入遺伝子の発現は、宿主ゲノムへのトランスポゾンを介した挿入によって、さらに遂行され得る。トランスポゾンを介した挿入の間に、宿主ゲノムへの挿入および続く切除を行わせる2つのトランスポゾンリンカー配列の間に遺伝子が位置する。
【0040】
本明細書で指すところの「OCT4タンパク質」は、Octomer 4転写因子の任意の天然の形態、またはOct4転写因子活性を(例えば、Oct4と比較して少なくとも50%、80%、90%、95%、96%、97%、98%、99%、もしくは100%の活性の範囲内で)維持しているその変異体を含む。ある態様において、変異体は配列全体もしくは配列の一部分(例えば、50、100、150、もしくは200個の連続したアミノ酸部分)にわたって、天然のOct4ポリペプチド(例えば、配列番号:1、配列番号:2、もしくは配列番号:3)と比較して少なくとも90%、95%、96%、97%、98%、99%、もしくは100%のアミノ酸配列同一性を有する。他の態様において、Oct4タンパク質はアイソフォーム1(配列番号:1)に対応するNCBI参照番号gi:42560248、アイソフォーム2(配列番号:2および配列番号:3)に対応するgi:116235491およびgi:291167755によって同定されるタンパク質である。
【0041】
本明細書で指すところの「Sox2タンパク質」は、Sox2転写因子の任意の天然の形態、またはSox2転写因子活性を(例えば、Sox2と比較して少なくとも50%、80%、90%、95%、96%、97%、98%、99%、もしくは100%の活性の範囲内で)維持しているその変異体を含む。ある態様において、変異体は配列全体もしくは配列の一部分(例えば、50、100、150、もしくは200個の連続したアミノ酸部分)にわたって、天然のSox2ポリペプチド(例えば、配列番号:4)と比較して少なくとも90%、95%、96%、97%、98%、99%、もしくは100%のアミノ酸配列同一性を有する。他の態様において、Sox2タンパク質はNCBI参照番号gi:28195386(配列番号:4)によって同定されるタンパク質である。
【0042】
本明細書で指すところの「KLF4タンパク質」は、KLF4転写因子の任意の天然の形態、またはKLF4転写因子活性を(例えば、KLF4と比較して少なくとも50%、80%、90%、95%、96%、97%、98%、99%、もしくは100%の活性の範囲内で)維持しているその変異体を含む。ある態様において、変異体は配列全体もしくは配列の一部分(例えば、50、100、150、もしくは200個の連続したアミノ酸部分)にわたって、天然のKLF4ポリペプチド(例えば、配列番号:5)と比較して少なくとも90%、95%、96%、97%、98%、99%、もしくは100%のアミノ酸配列同一性を有する。他の態様において、KLF4タンパク質はNCBI参照番号gi:194248077(配列番号:5)によって同定されるタンパク質である。
【0043】
本明細書で指すところの「cMYCタンパク質」は、cMyc転写因子の任意の天然の形態、またはcMyc転写因子活性を(例えば、cMycと比較して少なくとも50%、80%、90%、95%、96%、97%、98%、99%、もしくは100%の活性の範囲内で)維持しているその変異体を含む。ある態様において、変異体は配列全体もしくは配列の一部分(例えば、50、100、150、もしくは200個の連続したアミノ酸部分)にわたって、天然のcMycポリペプチド(例えば、配列番号:6)と比較して少なくとも90%、95%、96%、97%、98%、99%、もしくは100%のアミノ酸配列同一性を有する。他の態様において、cMycタンパク質はNCBI参照番号gi:71774083(配列番号:6)によって同定されるタンパク質である。
【0044】
本明細書で指すところの「NANOGタンパク質」は、Nanog転写因子の任意の天然の形態、またはNanog転写因子活性を(例えば、Nanogと比較して少なくとも50%、80%、90%、95%、96%、97%、98%、99%、もしくは100%の活性の範囲内で)維持しているその変異体を含む。ある態様において、変異体はそれらの配列全体もしくは配列の一部分(例えば、50、100、150、もしくは200個の連続したアミノ酸部分)にわたって、天然のNanogポリペプチド(例えば、配列番号:7)と比較して少なくとも90%、95%、96%、97%、98%、99%、もしくは100%のアミノ酸配列同一性を有する。他の態様において、Nanogタンパク質はNCBI参照番号gi:153945816(配列番号:7)によって同定されるタンパク質である。
【0045】
本明細書で指すところの「LIN28タンパク質」は、Lin28転写因子の任意の天然の形態、またはLin28転写因子活性を(例えば、Lin28と比較して少なくとも50%、80%、90%、95%、96%、97%、98%、99%、もしくは100%の活性の範囲内で)維持しているその変異体を含む。ある態様において、変異体はそれらの配列全体もしくは配列の一部分(例えば、50、100、150、もしくは200個の連続したアミノ酸部分)にわたって、天然のLin28ポリペプチド(例えば、配列番号:8)と比較して少なくとも90%、95%、96%、97%、98%、99%、もしくは100%のアミノ酸配列同一性を有する。他の態様において、Lin28タンパク質はNCBI参照番号gi:13375938(配列番号:8)によって同定されるタンパク質である。
【0046】
形質移入臍帯血幹細胞を分裂させ、これにより人工多能性幹細胞を形成することには、形質移入後に臍帯血幹細胞を増殖させ、任意で形質移入細胞を選択し、多能性幹細胞を同定することを含んでよい。本明細書で使用される増殖には、当技術分野において周知の条件下で、容器中において形質移入臍帯血幹細胞により子孫細胞を産生させることが含まれる。増殖は、適した培地および細胞増殖因子の存在下で起こってもよい。細胞増殖因子は、細胞の遊走、分化、形質転換、もしくは成熟および分裂を引き起こす作用物質である。それらは通常、様々な正常および悪性の哺乳動物細胞型から単離できるポリペプチドである。いくつかの増殖因子は、細菌(大腸菌(E.coli))および酵母等の遺伝子操作された微生物によっても産生され得る。細胞増殖因子は培地に補充してもよく、および/またはそのような細胞増殖因子を分泌する放射線照射された胚線維芽細胞と共培養することによって提供されてもよい。細胞増殖因子の例には、FGF、bFGF2、およびEGFが含まれるが、これらに限定されない。
【0047】
適切な場合には、増殖している形質移入臍帯血幹細胞を選択の過程に供してもよい。選択の過程は、形質移入の際に臍帯血幹細胞に導入した選択マーカーを含んでもよい。選択マーカーは、酵素活性を有するポリペプチドをコードする遺伝子であってもよい。酵素活性は、アセチルトランスフェラーゼおよびフォスフォトランスフェラーゼの活性を含むが、これらに限定されない。ある態様において、選択マーカーの酵素活性はフォスフォトランスフェラーゼの活性である。選択マーカーの酵素活性は、形質移入臍帯血幹細胞に毒素の存在下で増殖する能力を与えてもよい。そのような毒素は通常、細胞増殖を阻害し、および/または細胞死を引き起こす。そのような毒素の例にはハイグロマイシン、ネオマイシン、ピューロマイシン、およびゲンタマイシンが含まれるが、これらに限定されない。ある態様において、毒素はハイグロマイシンである。選択マーカーの酵素活性を通して、毒素はもはや形質移入臍帯血幹細胞の増殖を阻害せず、細胞死を引き起こさない非毒素に変換されてもよい。毒素への暴露の際に、選択マーカーを欠いた細胞は排除され、これにより増殖から除外されてもよい。
【0048】
人工多能性幹細胞の同定は、上述した多能性幹細胞の特徴の評価を含んでもよいが、これに限定されない。そのような多能性幹細胞の特徴には、分子マーカーの一定の組合せの発現もしくは非発現が含まれるが、これにさらに限定されない。さらに、多能性幹細胞に伴う細胞の形態も多能性幹細胞の特徴である。
【0049】
上述したように、本明細書の方法で提供される臍帯血幹細胞はOCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を形質移入されてもよい。ある態様において、臍帯血幹細胞はcMYCタンパク質、LIN28タンパク質、NANOGタンパク質、もしくはKLF4タンパク質をコードする追加の核酸を形質移入されない。
【0050】
ある態様において、OCT4タンパク質をコードする核酸は一つのプラスミドの部分を形成し、SOX2タンパク質をコードする核酸は一つのプラスミドの部分を形成する。別の態様において、OCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸は同じプラスミドの部分を形成する。一つの態様において、OCT4タンパク質をコードする核酸は第一のプラスミドの部分を形成し、SOX2タンパク質をコードする核酸は第二のプラスミドの部分を形成する。
【0051】
別の局面において、人工多能性幹細胞を調製するための方法が提供される。この方法は、OCT4タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成することを含む。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。
【0052】
一つの態様において、臍帯血幹細胞はcMYCタンパク質、LIN28タンパク質、NANOGタンパク質、もしくはKLF4タンパク質をコードする追加の核酸を形質移入されない。
【0053】
ある態様において、本明細書に提供される方法で使用される臍帯血細胞はCD133抗原を発現する。「CD133抗原」は大きさが120キロダルトンの、5つの膜貫通領域を有する糖タンパク質を指す。CD133抗原は成体幹細胞および前駆細胞によって発現されてよい。CD133抗原はPROML1、AC133、造血幹細胞抗原、hProminin、prominin-like 1、prominin、RP41、MCDR2、STGD4、CORD12、もしくはMSTP061としても知られる。ある態様において、CD133抗原はNCBI参照番号gi:225690512(配列番号:9)によって同定される遺伝子にコードされるタンパク質である。
【0054】
ある態様において、本明細書に提供される方法で使用される臍帯血幹細胞は新鮮な臍帯血に由来する。「新鮮な臍帯血」とは新生児の臍帯に由来する血液であって、臍帯が早まってクランプされていない場合には新生児の血液循環に戻される。本明細書で指すところの新鮮な臍帯血は、臍帯から単離された後に凍結保存されていない。「凍結保存」という用語は、液体窒素を用いて臍帯血等の生物学的材料を凍結させ、これにより生物学的材料を長期間にわたって保存する過程を指す。他の態様において、本明細書に提供される方法で使用される臍帯血幹細胞は凍結臍帯血に由来する。凍結臍帯血とは、本明細書に提供される方法に従って処理される前に凍結保存されていた新生児の臍帯に由来する血液である。
【0055】
III.人工多能性幹細胞
別の局面において、本明細書の方法に従って調製された人工多能性幹細胞が提供される。「臍帯血から人工多能性幹細胞を調製する方法」と題された節において上述された方法は、本明細書で提供される人工多能性幹細胞に対して同じように適用できる。
【0056】
IV.臍帯血幹細胞
一つの局面において、OCT4タンパク質をコードする核酸(例えば、OCT4タンパク質をコードする外来性の核酸もしくはOCT4タンパク質をコードする組換え核酸)およびSOX2タンパク質をコードする核酸(例えば、SOX2タンパク質をコードする外来性の核酸もしくはSOX2タンパク質をコードする組換え核酸)を含む臍帯血幹細胞が提供される。本明細書で使用されるタンパク質をコードする核酸に関連した「外来性」という用語は、それが見出される細胞(例えば、臍帯血細胞)において天然ではないことを意味する。ある態様において、OCT4タンパク質をコードする核酸は一つのプラスミドの部分を形成し、SOX2タンパク質をコードする核酸は一つのプラスミドの部分を形成する。別の態様において、OCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸は同じプラスミドの部分を形成する。一つの態様において、OCT4タンパク質をコードする核酸は第一のプラスミドの部分を形成し、SOX2タンパク質をコードする核酸は第二のプラスミドの部分を形成する。ある態様において、臍帯血幹細胞はcMYCタンパク質をコードする核酸(例えば、cMYCタンパク質をコードする外来性の核酸もしくはcMYCタンパク質をコードする組換え核酸)、LIN28タンパク質をコードする核酸(例えば、LIN28タンパク質をコードする外来性の核酸もしくはLIN28タンパク質をコードする組換え核酸)、NANOGタンパク質をコードする核酸(例えば、NANOGタンパク質をコードする外来性の核酸もしくはNANOGタンパク質をコードする組換え核酸)、および/またはKLF4タンパク質をコードする核酸(例えば、KLF4タンパク質をコードする外来性の核酸もしくはKLF4タンパク質をコードする組換え核酸)等のiPS細胞形成において有用であることが知られている他の転写因子をコードする核酸を含まない。
【0057】
他の態様において、臍帯血幹細胞はOCT4タンパク質をコードする核酸(例えば、OCT4タンパク質をコードする外来性の核酸もしくはOCT4タンパク質をコードする組換え核酸)およびSOX2タンパク質をコードする核酸(例えば、SOX2タンパク質をコードする外来性の核酸もしくはSOX2タンパク質をコードする組換え核酸)から本質的になる。臍帯血幹細胞がOCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸「から本質的になる」場合は、臍帯血幹細胞はcMYCタンパク質をコードする核酸(例えば、cMYCタンパク質をコードする外来性の核酸もしくはcMYCタンパク質をコードする組換え核酸)、LIN28タンパク質をコードする核酸(例えば、LIN28タンパク質をコードする外来性の核酸もしくはLIN28タンパク質をコードする組換え核酸)、NANOGタンパク質をコードする核酸(例えば、NANOGタンパク質をコードする外来性の核酸もしくはNANOGタンパク質をコードする組換え核酸)、および/またはKLF4タンパク質をコードする核酸(例えば、KLF4タンパク質をコードする外来性の核酸もしくはKLF4タンパク質をコードする組換え核酸)等のiPS細胞形成において有用であることが知られている他の転写因子をコードする核酸を含まない。ある態様において、臍帯血幹細胞は他の転写因子をコードする核酸(例えば、転写因子をコードする他の外来性の核酸もしくは転写因子をコードする他の組換え核酸)を含まない。他の態様において、臍帯血幹細胞は他のタンパク質を発現する遺伝子をコードする核酸(例えば、タンパク質をコードする他の外来性の核酸もしくはタンパク質をコードする他の組換え核酸)を含まない。
【0058】
別の局面において、OCT4タンパク質をコードする核酸(例えば、OCT4タンパク質をコードする外来性の核酸もしくはOCT4タンパク質をコードする組換え核酸)を含む臍帯血幹細胞が提供される。他の態様において、臍帯血幹細胞はOCT4タンパク質をコードする核酸(例えば、OCT4タンパク質をコードする外来性の核酸もしくはOCT4タンパク質をコードする組換え核酸)から本質的になる。臍帯血幹細胞がOCT4タンパク質をコードする核酸「から本質的になる」場合は、臍帯血幹細胞はcMYCタンパク質をコードする核酸(例えば、cMYCタンパク質をコードする外来性の核酸もしくはcMYCタンパク質をコードする組換え核酸)、LIN28タンパク質をコードする核酸(例えば、LIN28タンパク質をコードする外来性の核酸もしくはLIN28タンパク質をコードする組換え核酸)、NANOGタンパク質をコードする核酸(例えば、NANOGタンパク質をコードする外来性の核酸もしくはNANOGタンパク質をコードする組換え核酸)、および/またはKLF4タンパク質をコードする核酸(例えば、KLF4タンパク質をコードする外来性の核酸もしくはKLF4タンパク質をコードする組換え核酸)等のiPS細胞形成において有用であることが知られている他の転写因子をコードする核酸を含まない。ある態様において、臍帯血幹細胞は他の転写因子をコードする核酸(例えば、転写因子をコードする他の外来性の核酸もしくは転写因子をコードする他の組換え核酸)を含まない。他の態様において、臍帯血幹細胞は他のタンパク質を発現する遺伝子をコードする核酸(例えば、タンパク質をコードする他の外来性の核酸もしくはタンパク質をコードする他の組換え核酸)を含まない。
【0059】
ある態様において、臍帯血幹細胞はCD133抗原を発現する。他の態様において、臍帯血幹細胞は新鮮な臍帯血に由来する。ある態様において、臍帯血幹細胞は凍結臍帯血に由来する。
【0060】
V.フットプリントを含まない、ヒト人工多能性幹細胞からヒト体細胞を産生するための方法
一つの局面において、ヒト体細胞を産生するための方法が提供される。この方法は、細胞増殖因子を人工多能性幹細胞に接触させることを含む。人工多能性幹細胞は分裂させられ、これによりヒト体細胞を形成する。人工多能性幹細胞は適切な培地および細胞増殖因子の存在下で分裂させられる。細胞増殖因子の例にはSCF、GMCSF、FGF、TNF、IFN、EGF、IGF、およびインターロイキンファミリーのメンバーが含まれるが、これらに限定されない。人工多能性幹細胞は、本発明によって提供される方法に従って調製される。ある態様において、人工多能性幹細胞はOCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程を含む過程によって調製される。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。別の態様において、人工多能性幹細胞はOCT4タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程を含む過程によって調製される。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。
【0061】
別の局面において、組織修復を必要としている哺乳動物を治療する方法が提供される。この方法は、人工多能性幹を哺乳動物に投与し、哺乳動物において人工多能性幹細胞を分裂および体細胞に分化させ、これにより哺乳動物において組織修復を提供することを含む。ある態様において、人工多能性幹細胞はOCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程を含む過程によって調製される。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。別の態様において、人工多能性幹細胞はOCT4タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程を含む過程によって調製される。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。
【実施例】
【0062】
CB幹細胞を単離するために、免疫磁気選択法を用いてCBユニットからCD133+細胞を精製し、純度の範囲が90〜94%の細胞の集団を取得した(図3A)。静止状態のCD133+幹細胞の増殖を促進するために、細胞を幹細胞因子(SCF)、トロンボポエチン(TPO)、Flt ligand 3(Flt3)、およびインターロイキン6(IL-6)の存在下で24時間にわたって培養した。いくつかの修正を加えた接着細胞のための手順に従って10、これらの細胞に感染させた。手短には、以前に記載したように19、ウイルス粒子で前もって前吸着させた、レトロネクチンのコートされたプレートの上に細胞を播種した。対照実験において、構成的GFPレトロウイルスを用いて12時間ごとに精製CD133+集団を感染の3サイクルに供し、結果として得られた集団をフローサイトメトリーを用いて感染の3日後に解析した。図3Bは、典型的な実験結果を示す:全細胞の28%がGFP陽性であり、58%がCD133抗原に対して依然として陽性であった。GFP陽性集団のうちの17%がCD133+/GFP+であった一方で、11%がCD133−/GFP+であった。
【0063】
OSKM因子の任意の単独もしくは組合せを用いてCD133+細胞の再プログラムが試みられた。形質導入の3日後に、放射線照射されたヒト包皮線維芽細胞(HFF-1)フィーダー細胞の上に細胞を播種し、bFGF含有hES培地中で培養した。単なるES様培養条件によって再プログラミング自体を誘導できる可能性を排除するために、形質導入されていないCD133+幹細胞もhES細胞条件において3週間にわたって培養した。これらのプレートでは、コロニー形成は観察されなかった。これらの細胞のフローサイトメトリー解析によって、これらの細胞が幹細胞マーカーCD133、CD34、およびCD38をもはや発現しておらず、造血マーカーCD45については陽性のままであるが、胚性マーカーSSEA-3、SSEA-4、もしくはTRA1-60は獲得していないことが明らかになり、全体として、hES細胞条件において培養すると形質導入されていないCD133+細胞は成熟造血細胞に分化することが示唆された(図1C)。
【0064】
感染の約9日後に、OSKM、OSK、もしくはOSで形質導入された細胞において小さいコロニーが出現し始めた。感染の12〜15日後に、コロニーのいくつかははっきりとした境界を有する典型的なhES細胞の形態を呈し、大きい核および明確に視認できる核小体を有する小さい密集した細胞集団から構成されていた(図1A)。8×104個のCD133+細胞の標準的な感染から4〜5個のhES様コロニーが観察され、CBiPSと名付けられた。コロニーを手作業のピッキングで増殖させ、各因子の組合せ(CBiPS 4F-1、CBiPS 3F-1、CBiPS 2F-1)からのCBiPS株をさらなる特徴付けのために増幅させた。各レトロウイルス導入遺伝子の存在をPCR遺伝子型決定によって確認し、CBiPS 4F-1、CBiPS 3F-1、CBiPS 2F-1のそれぞれにおいて予期された4、3、もしくは2個の転写因子の挿入が明らかにされた(図1B)。
【0065】
3つのCBiPS株はすべてアルカリフォスファターゼにより陽性に染色され(図1C)、免疫蛍光染色で評価されたように、多能性マーカーOCT4、SOX2、TRA-1-81、TRA-1-60、SSEA3、SSEA4、およびNANOGを発現した(図1D)。加えて、フローサイトメトリーで測定されたように、再プログラムされたCBiPS株は造血幹細胞マーカーCD45、CD34、およびCD38について陰性であった。しかしながら、これらの株は造血および胚性幹細胞の共通マーカーCD133について陽性であった(図1E)。以前の免疫特徴付けと一致して、リアルタイムPCR解析により、3つのCBiPS株はすべてOCT4、SOX2、NANOG、CRIPTO、およびREXを含む多数の多能性遺伝子を発現していることが示され、他のiPS10およびhES [2]細胞株20に匹敵する遺伝子発現プロファイルが明らかにされた(図2A)。加えて、レトロウイルス導入遺伝子の発現は適切に停止され、CBiPS株におけるOCT4、SOX2、KLF4、およびc-MYCの発現が対応する内在性遺伝子によって推進された(図2B)。このことは、FLAGタグ付けされた遺伝子導入因子に対して特異的な抗体を用いた免疫蛍光染色によっても確認された(図8A〜8B)。
【0066】
線維芽細胞および角化細胞は内在性のOCT4、NANOG、SOX2、REX1、およびCRIPTOを発現しなかったが、CD 133+幹細胞は低いレベルで発現したため(図2C)、迅速な再プログラミングが可能な、より可塑的でエピジェネティックな状態に向けられた。さらに、CD133+細胞におけるOCT4およびNANOGのプロモーターは線維芽細胞に比べてヒストン抑制の特徴(H3K7およびH3K9のメチル化)をより低いレベルで有することも見出され(図2D)、過剰発現した因子によるこれらの遺伝子の結合および転写活性化を助けるかもしれない、より許容的なクロマチン構成の存在が示唆された。加えて、線維芽細胞および角化細胞と比較して、CB CD133+幹細胞におけるKLF4およびc-MYCの高いレベルの組合せは(図2C、図2E)、これらの因子の内在性の発現が、これらの細胞のより迅速および/または促進された再プログラミングを可能にするかもしれないことをさらに示しているのかもしれない10。
【0067】
細胞遺伝学的解析によって、3つの細胞株は10回の継代後も正常な46XYの核型を維持していることが示された。加えて、男性染色体の内容は、再プログラムされた細胞が最初の臍帯血試料中に存在することが知られている混入した母の細胞の小さい画分に起因するという可能性を排除する(図11A)。次に、インビトロ胚様体形成によるCBiPS細胞株の分化の潜在力を評価した。すべての細胞株は胚様体(EBs)を高い効率で形成でき(図12A)、EBsはFoxA2およびα-アクチニン陽性中胚葉、GFAPおよびTuj1陽性外胚葉、ならびにα-フェトプロテイン陽性内胚葉を含む3つのすべての胚性胚葉の細胞型に分化できた(図12B〜12F)。この結果によって、CBiPS 4F-1、CBiPS 3F-1、CBiPS 2F-1細胞株が他のhiPSおよびhES細胞株と類似した状態にまで転写的に再プログラムされており、核型的に安定で、多能性と一致したインビトロの発生潜在力を見せることが確認される。
【0068】
体細胞の再プログラミングは、多能性因子および癌遺伝子の組合せの発現を通して遂行されてきた。たった2週間で2個の因子を用いてCB CD133+幹細胞を再プログラムできたという事実は、再生医療のために臨床的に適したiPS細胞の発生に向けた体細胞の理想的な供給源としてのCB細胞の潜在力を強調する。これには、非組込みもしくは半組込み手段の使用21〜23、および小分子によるOCT4もしくはSOX2の置換24、25を含んでもよい。他の受け入れやすい体細胞の供給源に関する限りでは、動員された末梢血(mPB)細胞もiPS誘導のための効果的な供給源を表し得ることが最近示された26。しかしながら、新生児のCB幹細胞と比較して、成体のmPB細胞は加齢の結果としてもしくは特定の疾患の直接的な帰結としてゲノムの変異が蓄積される可能性がより高い。さらに、成体造血幹細胞の区画を動員するために使用される薬物治療はドナーの健康上の危険性を表す:ドナーの小さいけれどかなり多い割合において、この手順は脾破裂27を含む激しい反応を引き起こし得る。
【0069】
CB幹細胞はこれらの課題を克服し、容易に入手可能で、その起源が早いおかげでまだ免疫学的に未成熟なため、HLA-ドナー-レシピエント選択のためのより厳しくない基準を可能にする15。今日までに、臍帯血バンクの包括的なネットワークにおいて400,000を超えるCBユニットが世界的に入手可能であり、CBiPS生成のための適合ドナーの素早く効果的な検索を促進する2。患者に特異的なiPS株の生成が、理論上は理想的な臨床の選択肢として繰り返し唱えられてきたにもかかわらず、実務的および費用対効果の局面から、この手段は多くの場合において実現不可能かもしれない。公的に入手可能なネットワークにおける様々なHLAハプロタイプを表すCBiPS株の大規模な産生およびバンクへの保存は、ヒトiPS細胞の基礎研究および将来的な臨床への適用のための貴重な手段を表すであろう。
【0070】
VI.材料および方法
試料の採取
臍のCB試料は、Banc de Sang i Teixits, Hospital Duran i Reynals, Barcelonaより取得した。
【0071】
CD133+細胞の精製
Lympholyte-H(Cederlane, Ontario, CA)密度勾配遠心を用いて、単核細胞(MNC)をCBから単離した。Mini-Macs免疫磁気分離システム(Miltenyi Biotec, Bergisch Gladbach, Germany)を用いて、CD133+細胞を陽性選択した。CD133-フィコエリトリン(PE;Miltenyi Biotec, Bergisch Gladbach, Germany)抗体で染色し、フローサイトメトリー解析によって精製効率を検証した。
【0072】
構築物およびレトロウイルス産生
RT-PCRによって、ES[4]の全RNAからOCT4およびSOX2のヒトcDNAを増幅した。IMAGEクローン5111134からヒトKLF4を増幅し、Luciano Di Croceよりご提供いただいたDNA鋳型から変異ヒトc-MYCT58Aを増幅した。N末のFLAGタグ付けタンパク質を発現できる修正pMSCVpuroベクターのEcoR I/Cla Iサイトに増幅したcDNAをクローン化した。pMXs-OSKMGを次のように構築した:Oct4終止コドンを排除してBspE Iサイトを追加した逆方向プライマーを用いてマウスOct4 cDNAを増幅し、pCRII(Invitrogen)にクローン化してpCRII-Oct4-Bsp(Not I-5'cDNA3'-Acc65 Iの向き)を得た。Age Iサイトに続くP2Aペプチド配列を含む順方向プライマーおよびSox2終止コドンを排除してBspE Iサイトを含んだ逆方向プライマーを用いてマウスSox2 cDNAを増幅した。この断片をpCRIIにクローン化してpCRII-Age-Sox2-Bsp(Not I-5'cDNA3'-Acc65 Iの向き)を得た。pCRII-Age-Sox2-BspをAge IおよびAcc65 Iで切断し、BspE I-Acc65 Iで切断したpCRII-Oct4-Bspにクローン化してpCRII-Oct4-P2A-Sox2-BspE Iを産生した。マウスKlf4およびeGFP(pCRII-OSKGを産生)もしくはマウスKlf4、c-Myc、およびeGFP(pCRII-OSKMGを産生)を組み入れるために、同じクローン化の手段を2回繰り返した。最後に、pCRII-OSKGおよびpCRII-OSKMGをEcoR Iで切断し、レトロウイルス空ベクターpMXsの唯一のEcoR Iサイトにクローン化してpMXs-OSKGおよびpMXs-OSKMGを産生した。製造業者の指示に従って、Fugene 6試薬(Roche)を用いてPhoenix Amphotropic細胞株に形質移入した後に、4個の因子についてのレトロウイルスを独立して産生した。24時間後に培地を置換し、細胞を32℃で培養して、12時間ごとにウイルス上清を回収した。
【0073】
CD133+細胞の形質導入
SCF(50ng/ml)+Flt3(50ng/ml)+TPO(10ng/ml)+IL-6(10ng/ml)(PeproTech)の存在下で、10%のFBSを添加したDMEM中において24時間にわたってCB CD133+細胞(1mlにつき1×105細胞)を前刺激した。マルチウェル非組織培養処理プレートをフィブロネクチン断片CH-296(15mg/cm2)であるレトロネクチン(Takara, Otsu, Japan, www.takara-bio.com)でコートし、OCT4、SOX2、KLF4、およびc-MYC因子についてのレトロウイルス上清の濾過した1:1:1:1混合物とともに2,500RPMで30分間にわたってプレートを遠心することにより前処置した。DMEM+10% FBSおよび上述したサイトカイン混合物の存在下で約80,000個のCD133+細胞を播種した。12時間ごとに、サイトカイン混合物を含んだ新鮮なウイルス上清で培地の半分を置換し、37℃、5% CO2で培養して、3回の感染サイクルを実行した。3日目に細胞を回収し、放射線照射されたヒト線維芽細胞ならびに、20% KO-Serum Replacement(GIBCO)を添加したKO-DMEM培地(Invitrogen)、非必須アミノ酸(Lonza)、2-β-メルカプトエタノール(GIBCO)、ペニシリン/ストレプトマイシン(GIBCO)、GlutaMAX(商標)(Invitrogene)、および10ng/ml bFGF(Peprotech)からなるES培地を含む6ウェルプレートに移動させた。放射線照射されたヒト線維芽細胞の上でCBiPS細胞を培養し、機械的にピッキングした。
【0074】
全RNAの精製および定量的RT-PCR
入手可能な細胞数に基づいて、Trizol(登録商標)試薬(Invitrogen, Carlsbad, CA)もしくはRNAqueous(登録商標)-Microキット(Ambion Inc., Austin TX)のいずれかを用いてCB CD133+幹細胞、hES[2]細胞、KiPS細胞(14)、およびCBiPSからの全RNAの単離を実行した。残存するいかなるゲノムDNAも除去するために、すべての試料をTURBO DNase 阻害剤(Ambion)で処理し、Invitrogen SuperScript(商標) II Reverse Transcriptaseキットを用いて、1μgのRNAを使用してcDNAを合成した。以前に記載したように10、プライマーを用いた定量的RT-PCRによって、25ngのcDNAを使用して遺伝子発現を定量した。
【0075】
GeneChip(登録商標)発現解析
Institute for Research in Biomedicine(Barcelona, Spain)において、製造業者の手順(Affymetrix, Santa Clara, CA)に従って、Functional Genomica CoreによりGeneChip(登録商標)マイクロアレイ処理を実行した。25ngの出発物のRNAを用いて、Nugenの手順に示されたように増幅および標識の処理を行った。各試料について3.75μgのssDNAを標識し、Affymetrix HG-U133 Plus 2.0チップにハイブリダイズさせた。Affymetrix GeneChip Scanner(7Gアップグレード)上で発現シグナルを走査した。Affymetrix GCOSソフトウェア v.1.4によってデータ抽出を行った。R Project for Statistical ComputingからのプログラムRを用いてデータの統計解析を実行した。最初に、Rに実装されたgcRMAアルゴリズムを用いて生データを正規化し、正規化されたデータについてピアソンの相関係数を用いた階層的クラスタリングを実行した。2つの異なる実験(我々の角化細胞再プログラミング(GEOアクセス番号:GSE12583)、および本実験(GSE16694))で取得されたデータセットを組み込むために、RにおいてgcRMAアルゴリズムを用いて生のCELファイルを一緒に正規化し、その後、当技術分野において公知であるように、ComBatアルゴリズムを用いてバッチ効果を補正した。例えば、Johnson et al., 2007, Biostatistics 8:118-127を参照。
【0076】
サザンブロット
製造業者の指針に従って、All Prep DNA/RNAカラム(Qiagen)を用いて各細胞株からのゲノムDNAを単離した。サザンブロットの各レーンは、40UのPst IもしくはHind III制限酵素(New England Biolabs)のいずれかで消化し、1% アガロースゲル上で電気泳動し、中性ナイロン膜(Hybond(商標)-N, Amersham)に転写し、PCR DIG Probe Synthesis Kit(Roche Diagnostics)を用いてPCRにより生成されたDIG-dUTP標識プローブとハイブリダイズさせた4μgのゲノムDNAに相当する。化学発光のための基質としてCDP-Star(Sigma-Aldrich)を用いて、AP結合DIG-Antibody(Roche Diagnostics)によりプローブを検出した。条件は製造業者の指示通りであった。プローブは以下のプライマー(F、順方向;R、逆方向)とともにSOX2、OCT4、KLF4、およびc-MYC cDNAを鋳型として用いて生成した:
。
【0077】
免疫蛍光解析およびAP解析
プラスチックのカバースライド容器上でCBiPSを増殖させ、4% パラホルムアルデヒド(PFA)で固定した。以下の抗体を使用した:TRA-1-60(MAB4360、1:200)、TRA-1-81(MAB4381、1:200)、SOX2(AB5603、1:500)、すべてChemicon、SSEA-4(MC-813-70、1:2)、SSEA-3(MC-631、1:2)、すべてIowa、Tuj1(1:500;Covance)、α-フェトプロテイン(1:400;Dako)、α-アクチニン(1:100;Sigma)、OCT4(C-10、SantaCruz、sc-5279、1:100)、NANOG(Everest Biotech EB06860、1:100)、GATA4(1:50、SantaCruz)、平滑筋アクチン(1:400、Sigma)、FoxA2(1:50、R&D System)、GFAP(1:1000、Dako)、α-筋節アクチン(1:400、Sigma)、抗Flag(Sigma M2)。Leica SP5共焦点顕微鏡を用いて画像を撮影した。製造業者の指針に従って、Alkaline Phosphatase Blue/Red Membrane基質溶液キット(Sigma)を用いて直接的なAP活性を解析した。
【0078】
インビトロ分化
機械的に採取され、その後hES培地の存在下で24時間にわたって懸濁液中に維持されたコロニー断片からEBsの形成を誘導した。我々は超低吸着プレートの上で3つの異なる分化培地中にEBsが維持される前条件培養を2〜3日間にわたって実行した。特に、内胚葉分化のために、10% ウシ胎仔血清(FBS)、2mM L-グルタミン、0.1mM 2-β-メルカプトエタノール、非必須アミノ酸、およびペニシリン-ストレプトマイシンを添加したKO-DMEM培地の存在下でEBsを培養した。中胚葉分化のために、我々は上述したのと同じであるが、アスコルビン酸(0.5mM)を追加した培地を使用した。外胚葉誘導のために、N2/B27培地中でEBsを培養した。前条件の工程の後に、0.1% ゼラチンのコートされたプラスチック容器スライドに内胚葉および中胚葉条件のEBsを移動し、それぞれ分化培地およびアスコルビン酸(0.5nM)添加分化培地において2週間にわたって培養した。外胚葉分化のために、N2/B27培地の存在下で間質細胞株PA6の上にEBsを2週間にわたって移動した。各条件のための培地を1日おきに交換した。
【0079】
クロマチン免疫沈降
製造業者の指示に従って、Magnetic Low cell ChIP Kit from Diagenodeを用いて、免疫沈降ごとに15,000細胞を使用してクロマチン免疫沈降実験を実行した。Millipore 07-440(抗H3K27me3)、07-030(抗H3K4me2)、および17-625(抗H3K9me3)からの抗体を使用した。
【0080】
プロモーターメチル化解析
QIA AMP DNA Mini Kit(Qiagen)を用いて、約500,000個のCD133+およびCBiPS細胞の試料によってゲノムDNAを抽出した。製造業者の仕様書に従って、Epitect Bisulfite Kit(Qiagen)で2μgの精製DNAに突然変異を起こさせた。以前に記載したプライマーを用いて10、続く2回のPCRによって関心対象のプロモーター配列を増幅させた。結果として得られた増幅産物をpGEM T Easyプラスミドにクローン化し、TOP10細胞中で増幅させ、精製および配列決定を行った。
【0081】
奇形腫の形成
重症複合免疫不全(SCID)ベージュマウス(Charles River Laboratories)を麻酔し、20〜40μlのhES培地に再懸濁した約0.5×106個のCBiPS細胞を精巣に注射した。細胞注射の6〜8週間後にマウスを安楽死させ、従来型の免疫組織化学の手順(マッソントリクローム染色)および免疫蛍光に従って腫瘍を処理および解析した。
【0082】
VII.参考文献
【技術分野】
【0001】
関連出願の相互参照
本出願は、2009年6月19日出願の米国仮出願第61/218,611号に恩典を主張し、その内容は参照により、その全体がすべての目的のために本明細書に組み入れられる。
【背景技術】
【0002】
発明の背景
組込みウイルスの方法を用いた多能性因子および癌遺伝子の異所性発現は、マウスおよびヒト線維芽細胞の両方において多能性を誘導するのに十分である3〜9。しかしながら、この過程は時間がかかり、非効率的で、ゲノム中へのベクターの恒久的な組込みは、治療への適用のためのiPS細胞の使用を制限する1。さらなる研究によって、使用される年齢、起源、および細胞型が再プログラミングの効率に重大な影響を有し、最終的により少ない因子の発現が必要であったり、および/または過程全体の時期が減少したりすることが示されている。最近、レトロウイルスによるヒト角化細胞への形質導入によって、線維芽細胞に比べて100倍も効率が高く、2倍速い多能性への再プログラミングがもたらされることが示された。これらの相違は、出発物の角化細胞集団におけるKLF4およびc-MYCの内在性の発現および/または再プログラミングをより受け入れやすいというエピジェネティックな状態を示す未分化前駆細胞のプールの存在に起因し得るという仮説が立てられた10。この後者の仮説は、マウスにおける他の研究によってさらに裏付けられている11、12。しかしながら、幹細胞は通常は稀であり、大量に入手および単離することは困難である(例えば、神経幹細胞13、14)。
【0003】
人工多能性幹(iPS)細胞は、細胞療法のための潜在的価値を有する患者特異的な前駆体をインビトロで生成できるようにするため、再生医療への関心を引き起こしてきた1。しかしながら、多くの場合において、急性疾患の細胞療法のため、または慢性疾患もしくは加齢の帰結として患者の体細胞が変化した場合等はすぐに使える手段が望ましい。臍帯血(CB)幹細胞は、最小限の遺伝子の変化およびエピジェネティックな変化を有する、生まれたばかりで免疫学的に未成熟な細胞であり、CBバンクの世界的ネットワークを通して数十万の免疫分類されたCBユニットをすぐに利用できるため、この目的のためにはCB幹細胞が理想的に適しているようである2。造血移植のための幹細胞の供給源として骨髄(BM)の代替物と考えられているCB細胞は、ドナーに危険を与えることなく十分な量を採取することができる15。容易に入手できることに加えて、CB細胞は最小限の体細胞変異を有する幼若細胞であるという特徴と新生細胞の免疫学的な未成熟により与えられる長所を兼ね備える15。これらの特性はHLA-ドナー-レシピエント選択のより厳しくない基準を可能にし、移植のための決定的な利点を表す。さらに、臍帯血バンクの世界的で包括的なネットワークは、CB幹細胞の適合ドナーを素早く効果的に検索することを保証する2。最後に、CB CD133+幹細胞はOCT4、SOX2、NANOG、REX1および他の多能性関連マーカーを発現することが示されてきており16〜18、したがって原理上は再プログラミングをより受け入れやすいかもしれない。本明細書において、2つの強力な癌遺伝子(c-MYCおよびKLF4)もしくは追加の化学物質の必要なしに、4つ(OSKM)、3つ(OSK)、およびわずか2つ(OS)の転写因子をレトロウイルスで形質導入することによって、素早く効率的にCB幹細胞を多能性へと再プログラミングすることを含む態様が初めて記載される。
【0004】
本明細書に記載された一定の方法および組成物を用いて、OCT4、SOX2、KLF4、およびc-MYCをレトロウイルスで形質導入することによって、非常に効率的かつ素早い過程においてCB幹細胞を多能性へと再プログラムできる。結果として得られたCB由来のiPS(CBiPS)細胞は、表現型も分子的にもヒト胚性幹(hES)細胞と識別不可能である。さらに、臍帯血iPSの生成はc-MYCおよびKLF4癌遺伝子を使用せずに、OCT4およびSOX2を過剰発現させるだけで効率的に達成できる。本明細書に記載された方法および組成物は、当技術分野における課題を克服し、すぐに使える適用のためのHLA適合CBiPS細胞の包括的バンクの製造のための基礎を築くかもしれない。
【発明の概要】
【0005】
本明細書に提供されているのは、特に、臍帯血から人工多能性幹細胞を作製し、使用するための非常に効率的な方法および組成物である。
【0006】
一つの局面において、人工多能性幹細胞を調製するための方法が提供される。この方法は、OCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成することを含む。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。
【0007】
別の局面において、人工多能性幹細胞を調製するための方法が提供される。この方法は、OCT4タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成することを含む。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。
【0008】
別の局面において、本明細書の方法に従って調製された人工多能性幹細胞が提供される。
【0009】
一つの局面において、OCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を含む臍帯血幹細胞が提供される。
【0010】
別の局面において、OCT4タンパク質をコードする核酸を含む臍帯血幹細胞が提供される。
【0011】
一つの局面において、ヒト体細胞を産生するための方法が提供される。この方法は、細胞増殖因子を人工多能性幹細胞に接触させることを含む。人工多能性幹細胞は分裂させられ、これによりヒト体細胞を形成する。ある態様において、人工多能性幹細胞はOCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程を含む過程によって調製される。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。別の態様において、人工多能性幹細胞はOCT4タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程を含む過程によって調製される。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。
【0012】
別の局面において、組織修復を必要としている哺乳動物を治療するための方法が提供される。この方法は、人工多能性幹を哺乳動物に投与し、哺乳動物において人工多能性幹細胞を分裂および体細胞に分化させ、これにより哺乳動物において組織修復を提供することを含む。ある態様において、人工多能性幹細胞はOCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程を含む過程によって調製される。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。別の態様において、人工多能性幹細胞はOCT4タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程を含む過程によって調製される。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。
【図面の簡単な説明】
【0013】
【図1A】図1A〜1E。OCT4およびSOX2因子のみを用いたCBiPS細胞株の生成。図1A:臍帯血幹細胞再プログラミングの予定表。感染の3日後に、CB CD133+細胞をフィーダーに移動する。9日目頃に小さい接着性のコロニーが観察される。12日後に典型的なhES様コロニーがはっきりと見える。
【図1B】図1A〜1E。OCT4およびSOX2因子のみを用いたCBiPS細胞株の生成。図1B:4つ、3つ、および2つのみの導入遺伝子の挿入を確認するゲノムDNAのPCR。
【図1C】図1A〜1E。OCT4およびSOX2因子のみを用いたCBiPS細胞株の生成。図1C:CBiPS2F-1、3F-10、および4F-3細胞株の代表的な位相差画像ならびにアルカリフォスファターゼ(AP)染色。
【図1D】図1A〜1E。OCT4およびSOX2因子のみを用いたCBiPS細胞株の生成。図1D:CBiPS2F、3F、および4F細胞株における代表的なテロメラーゼ活性(HI:熱不活化、HFF:ヒト包皮線維芽細胞、−C:陰性対照として溶解緩衝液、+C:陽性対照、およびQC:量的対照)。
【図1E】図1A〜1E。OCT4およびSOX2因子のみを用いたCBiPS細胞株の生成。図1E:多能性マーカーについてのCBiPS2F-1細胞株の免疫蛍光解析。コロニーは胚性マーカーSSEA-4、SSEA-3、TRA-1-60、TRA-1-81、ならびに転写因子OCT4、SOX2、およびNANOGを発現する。基礎となる線維芽細胞は陰性対照を提供する。スケールバーは250μm。
【図2A】図2A〜2G。CBiPS細胞株の特徴付け。図2A:多能性マーカーOCT4、SOX2、NANOG、REX1、CRIPTO、KLF4、およびc-MYCの定量的RT-PCR解析を描写したヒストグラム。新鮮および凍結試料に由来する異なるCBiPS細胞株と一緒にES[2]および角化細胞-iPS(KiPS)細胞株を解析した。エラーバーは、三連より生成されたs.d.(標準偏差)を示す。ヒストグラムの説明(左から右):CBiPS4F-3、CBiPS4F-5、CBiPS3F-10、CBiPS3F-12、CBiPS2F-1、CBiPS2F-2、CBiPSF-1、CBiPSF-5、ES2、およびKiPS。
【図2B】図2A〜2G。CBiPS細胞株の特徴付け。図2B:CBiPS細胞株におけるOCT4、SOX2、KLF4、およびc-MYC導入遺伝子の抑制を示す定量的RT-PCRを描写したヒストグラム。ヒストグラムの説明(左から右):CBiPS4F-3、CBiPS3F-10、およびCBiPS2F-1。
【図2C】図2A〜2G。CBiPS細胞株の特徴付け。図2C:3つの初期生殖細胞層(外肺葉-Tuj1、内胚葉-AFPおよびFOXA2、ならびに中胚葉-ASAおよびGATA4)へのCBiPS2F-1のインビトロ分化。
【図2D】図2A〜2G。CBiPS細胞株の特徴付け。図2D:Tuj1/GFAP陽性外胚葉、AFP/FoxA2陽性内胚葉、およびASM/ASA陽性中胚葉を示すCBiPS2F-1の精巣内注射から60日間後の奇形腫切片の免疫蛍光解析。スケールバーは75〜250μm。
【図2E】図2A〜2G。CBiPS細胞株の特徴付け。図2E:免疫表現型的に成熟したドーパミン作動性ニューロン(Tuj1/THチロシンヒドロキシラーゼ)への、CBiPS2F-1の特異的なインビトロ分化。
【図2F】図2A〜2G。CBiPS細胞株の特徴付け。図2F:免疫表現型的に成熟したドーパミン作動性ニューロン(Tuj1/THチロシンヒドロキシラーゼ)への、CBiPS3F-12の特異的なインビトロ分化。
【図2G】図2A〜2G。CBiPS細胞株の特徴付け。図2G:ヒト線維芽細胞およびCD133+細胞において、OCT4、NANOG、HOXB4、およびHOXB5のプロモーター中のK4(H3K4me2)、K27(H3K27me3)、およびK9(H3K9me3)におけるヒストンH3メチル化レベルを比較したクロマチン免疫沈降法を描写したヒストグラム。ヒストグラムの説明:線維芽細胞(整列した点)、CD133+(黒塗り)。
【図3A】図3A〜3C。ヒトCD133+細胞のフローサイトメトリー解析。図3A:免疫選択後のCD133細胞の純度についての代表的なドットプロットを描写する。
【図3B】図3A〜3C。ヒトCD133+細胞のフローサイトメトリー解析。図3B:感染から3日後の全GFP+細胞および二重陽性GFP/CD133細胞の定量を描写する。
【図3C】図3A〜3C。ヒトCD133+細胞のフローサイトメトリー解析。図3C:ヒストグラムは、hES条件において3週間にわたって培養された形質導入されていないCD133+幹細胞のフローサイトメトリー解析を描写する。造血マーカーCD45、CD34、CD38、およびCD133、ならびにSSEA3、SSEA4、およびTRA-1-60を含む胚性幹細胞マーカーについて細胞を解析した。
【図4】図4A〜4B。pMXs-OSKMGおよびpMXs-OSKGポリシストロニックレトロウイルスの模式図。図4A:pMXs-OSKGポリシストロニックレトロウイルス。図4B:pMXs-OSKMGポリシストロニックレトロウイルス。
【図5】図5A〜5C。多能性マーカーについての免疫蛍光解析。図5A:CBiPS3F-10。図5B:CBiPS4F-3細胞株はSSEA-4、SSEA-3、TRA-1-60、TRA-1-81を含む他の典型的な多能性マーカー、ならびに転写因子OCT4、SOX2、およびNANOGを発現する。図5C:凍結融解したCBユニットから精製されたCD133+細胞を用いて、OSKレトロウイルスで形質導入した後に生成されたCBiPS凍結(CBiPSFr)-1細胞株は、SSEA-4、SSEA-3、TRA-1-60、TRA-1-81を含む他の典型的な多能性マーカー、ならびに転写因子OCT4、SOX2、およびNANOGを発現する。基礎となる線維芽細胞は陰性対照を提供する。スケールバーは250μm。
【図6】CBiPS2Fのフローサイトメトリー解析。ヒストグラムはCBiPS2F-1細胞がCD45およびCD34等の造血マーカーを喪失し、TRA-1-181およびSSEA-4を含む典型的な多能性マーカーを獲得したことを確認するフローサイトメトリー解析を描写する。
【図7A】図7A〜7B。全般的遺伝子発現解析。図7A:CBiPS(2本の線、各2反復)およびES2(2反復)の間で平均の全般的遺伝子発現パターンを比較したところ、非常に高いレベルの相関を示した。いくつかの多能性遺伝子をプロット中に同定する。
【図7B】図7A〜7B。全般的遺伝子発現解析。図7B:異なる多能性株および各出発物集団のすべての一対比較についての、ゲノム全般の転写プロファイルの相関係数。
【図8】図8A〜8B。レトロウイルスによる導入遺伝子サイレンシング。任意のFLAGタグ付けされたレトロウイルスの転写因子からの導入遺伝子発現のみを検出する特定のFLAG抗体と組み合わせたOCT4に対する免疫蛍光染色。図8A:CBiPS2F-1における内在性OCT4の発現および導入遺伝子のサイレンシング。図8B:陽性対照としてのOCT4およびSOX2に感染させた初代ヒト線維芽細胞。スケールバーは250μm。
【図9】亜硫酸水素塩ゲノム配列によるメチル化プロモーター解析。すべてのCBiPS細胞株におけるプロモーターの一貫した脱メチル化を確認するOCT4プロモーターのメチル化解析。
【図10】サザンブロット。CBiPS2F-1株およびサブクローンにおけるレトロウイルスの組込み数を評価するためのサザンブロット。1:KLF4特異的プローブとハイブリダイズした、Pst Iで消化したゲノムDNA。内在性バンド:5.9kbおよび0.9kb(黒い矢頭)。予期されたように、このプローブでは追加のバンドは検出されない。2:SOX2特異的プローブとハイブリダイズした、Pst Iで消化したゲノムDNA。内在性バンド:0.9kb(黒い矢頭)。固有の導入遺伝子の挿入に対応する同じ追加のバンドがCBiPS2F-1、CBiPS2F-1a、およびCBiPS2F-1bに存在する(赤い星印)。3:OCT4特異的プローブとハイブリダイズした、Hind IIIで消化したゲノムDNA。内在性の特異的バンド:4.5kb(黒い矢頭)。内在性の非特異的バンド(灰色の矢頭)。固有の導入遺伝子の挿入に対応する同じ追加のバンドがCBiPS2F-1、CBiPS2F-1a、およびCBiPS2F-1bに存在する(赤い星印)。4:c-MYC特異的プローブとハイブリダイズした、Hind IIIで消化したゲノムDNA。内在性バンド:11kb(黒い矢頭)。予期されたように、このプローブでは追加のバンドは検出されない。
【図11A】図11A〜11C。CBiPS 2F、3F、および4F細胞株についての核型分析。10回の継代後に解析されたCBiPS2F-1(図11A)、CBiPS3F-10(図11B)、およびCBiPS4F-3(図11C)細胞における正常染色体、2倍体染色体、男性染色体、および女性染色体の内容を示す高解像度のGバンド形成核型。
【図11B】図11A〜11C。CBiPS 2F、3F、および4F細胞株についての核型分析。10回の継代後に解析されたCBiPS2F-1(図11A)、CBiPS3F-10(図11B)、およびCBiPS4F-3(図11C)細胞における正常染色体、2倍体染色体、男性染色体、および女性染色体の内容を示す高解像度のGバンド形成核型。
【図11C】図11A〜11C。CBiPS 2F、3F、および4F細胞株についての核型分析。10回の継代後に解析されたCBiPS2F-1(図11A)、CBiPS3F-10(図11B)、およびCBiPS4F-3(図11C)細胞における正常染色体、2倍体染色体、男性染色体、および女性染色体の内容を示す高解像度のGバンド形成核型。
【図12A】図12A〜12F。CBiPS細胞株のインビトロおよびインビボ多能性。図12A:CBiPS2F-1細胞株に由来する胚様体。
【図12B】図12A〜12F。CBiPS細胞株のインビトロおよびインビボ多能性。CBiPS3F-10(図12B)およびCBiPS4F-3(図12C)はインビトロにおいて、神経(Tuj1/GFAP)、内胚葉(AFP/FoxA2)、および中胚葉(ASM)細胞を含む3つの胚葉に分化できる。
【図12C】図12A〜12F。CBiPS細胞株のインビトロおよびインビボ多能性。CBiPS3F-10(図12B)およびCBiPS4F-3(図12C)はインビトロにおいて、神経(Tuj1/GFAP)、内胚葉(AFP/FoxA2)、および中胚葉(ASM)細胞を含む3つの胚葉に分化できる。
【図12D】図12A〜12F。CBiPS細胞株のインビトロおよびインビボ多能性。図12D:CBiPSFr-1細胞株の神経(Tuj1)、内胚葉(AFP/FoxA2)、および中胚葉(ASM)細胞を含む3つの胚葉へのインビトロ分化。
【図12E】図12A〜12F。CBiPS細胞株のインビトロおよびインビボ多能性。CBiPS3F-10(図12E)およびCBiPS4F-3(図12F)のインビボ分化。結果として得られた奇形腫は3つの胚葉のすべて:外胚葉(Tuj1/GFAP)、内胚葉(AFP/FoxA2)、および中胚葉(ASM/ASA)を表す組織を含んでいた。
【図12F】図12A〜12F。CBiPS細胞株のインビトロおよびインビボ多能性。CBiPS3F-10(図12E)およびCBiPS4F-3(図12F)のインビボ分化。結果として得られた奇形腫は3つの胚葉のすべて:外胚葉(Tuj1/GFAP)、内胚葉(AFP/FoxA2)、および中胚葉(ASM/ASA)を表す組織を含んでいた。
【図13A】図13A〜13D。CB CD133+細胞の遺伝子発現解析。図13A:CD133+細胞は線維芽細胞および角化細胞よりも多能性細胞に近くないことを示す、CD133+、角化細胞、線維芽細胞、ES細胞、KiPS、およびCBiPSについてのゲノム全般の転写プロファイルの階層的クラスタリングを表す系統樹。
【図13B】図13A〜13D。CB CD133+細胞の遺伝子発現解析。図13B:定量的RT-PCRによる、CD133+細胞、線維芽細胞、および角化細胞における多能性マーカーおよびKLF4の比較遺伝子発現解析を描写したヒストグラム。ヒストグラムの説明(左から右):CD133+(黒色)、線維芽細胞(白色)、および角化細胞(灰色)。
【図13C】図13A〜13D。CB CD133+細胞の遺伝子発現解析。図13C:定量的RT-PCRによる、CD133+細胞におけるSALL2、ZNF589、DPPA4、DNMT3A、およびDNMT3B遺伝子の上方制御の検証。ヒストグラムの説明:図13Bと同様。
【図13D】図13A〜13D。CB CD133+細胞の遺伝子発現解析。図13D:CD133+細胞、線維芽細胞、および角化細胞におけるc-MYC発現の定量的RT-PCR解析。エラーバーは、三連より生成されたs.d.を示す。ヒストグラムの説明:図13Bと同様。
【発明を実施するための形態】
【0014】
発明の詳細な説明
I.定義
以下の定義は、本明細書中で頻繁に使用される一定の用語の理解を促進するために提供されるものであり、本開示の範囲を限定することを意図しない。
【0015】
「核酸」は、デオキシリボヌクレオチドもしくはリボヌクレオチド、および一本鎖もしくは二本鎖のいずれかの形態におけるそれらのポリマー、ならびにそれらの相補を指す。
【0016】
「相補的」もしくは「相補性」という言葉は、ポリヌクレオチド中の核酸が第二のポリヌクレオチド中の別の核酸と塩基対を形成する能力を指す。例えば、配列A-G-Tは配列T-C-Aと相補的である。相補性は、塩基対合に従って核酸のいくつかだけが一致する部分的なものでもよく、もしくは塩基対合に従ってすべての核酸が一致する完全なものでもよい。
【0017】
「同一の」もしくはパーセント「同一性」という用語は、2以上の核酸との関連では、同じかまたは以下に記載された規定値のパラメーターでBLASTもしくはBLAST 2.0配列比較アルゴリズムを用いて測定するか、または手動の整列および目視検査によって測定すると、特定のパーセントの同じヌクレオチド(すなわち、比較ウィンドウもしくは指定された領域にわたって最大に一致するように比較および整列させた場合に、特定の領域にわたって約60%の同一性、好ましくは65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、もしくはより高い同一性)を有する2以上の配列もしくは部分配列を指す(例えばNCBIウェブサイト等を参照)。このとき、そのような配列は「実質的に同一」と言われる。この定義は試験配列の相補も指すか、もしくは適用されてよい。この定義は欠失および/または付加を有する配列、ならびに置換を有するものも含む。以下に記載されるように、好ましいアルゴリズムはギャップ等を計上できる。好ましくは、同一性は長さが少なくとも約25アミノ酸もしくはヌクレオチド、またはより好ましくは、長さが50〜100アミノ酸もしくはヌクレオチドの領域にわたって存在する。
【0018】
「ストリンジェントなハイブリダイゼーション条件」という語句は、通常は核酸の複合混合物中でプローブがその標的配列とハイブリダイズするが、他の配列とはハイブリダイズしない条件を指す。ストリンジェントな条件は配列依存性であり、異なる状況において異なる。より長い配列は、より高い温度において特異的にハイブリダイズする。核酸のハイブリダイゼーションの広範な手引きはTijssen, Techniques in Biochemistry and Molecular Biology--Hybridization with Nucleic Probes, "Overview of principles of hybridization and the strategy of nucleic acid assays" (1993)に見出される。概して、ストリンジェントな条件は規定されたイオン強度のpHにおいて、特定の配列についての熱融解点(Tm)よりも約5〜10℃低くなるように選択される。Tmは、(規定されたイオン強度、pH、および核の濃度において)標的に相補的なプローブの50%が平衡状態で標的配列にハイブリダイズする温度である(標的配列が過剰に存在するため、Tmでは平衡状態でプローブの50%が占有される)。ストリンジェントな条件は、ホルムアミド等の不安定化剤の添加によっても達成されてよい。選択的もしくは特異的なハイブリダイゼーションのためには、陽性シグナルはバックグラウンドの少なくとも2倍であり、好ましくはバックグラウンドハイブリダイゼーションの10倍である。例示的なストリンジェントなハイブリダイゼーション条件は、次の通りであり得る:50% ホルムアミド、5×SSC、および1% SDSで42℃においてインキュベートするか、もしくは5×SSC、1% SDSで65℃においてインキュベートし、0.2×SSCおよび0.1% SDS中で65℃において洗浄する。
【0019】
核酸ハイブリダイゼーション技術を使用する、特異的なDNAおよびRNAの測定の多岐にわたる方法が当業者に公知である(Sambrook、前出を参照)。いくつかの方法は電気泳動分離を含む(例えば、DNAを検出するためのサザンブロット、およびRNAを検出するためのノザンブロット)が、DNAおよびRNAの測定は電気泳動分離を行わなくても実行できる(例えば、ドットブロットによって)。
【0020】
ハイブリダイゼーション測定法の感度は、検出されている標的核酸を増幅させる核酸増幅システムの使用を通して強化できる。そのようなシステムの例には、ポリメラーゼ連鎖反応(PCR)システムおよびリガーゼ連鎖反応(LCR)システムが含まれる。当技術分野において最近記載された他の方法は、核酸配列に基づく増幅(NASBA、Cangene、Mississauga、Ontario)およびQβレプリカーゼシステムである。PCRもしくはLCRプライマーが、選択配列が存在する場合にのみ伸長もしくは連結するように設計されているところ、これらのシステムは突然変異を直接的に同定するのに使用できる。あるいは、選択された配列は例えば非特異的PCRプライマーを用いて大まかに増幅でき、後に突然変異を示す特異的配列を増幅された標的領域に探索する。増幅反応産物を、例えばリアルタイムで観察するために、Taqman(登録商標)および分子指標プローブを含む様々な検出プローブを使用できることが理解される。
【0021】
「ポリヌクレオチド」という言葉は、ヌクレオチドの直鎖状配列を指す。ヌクレオチドはリボヌクレオチド、デオキシリボヌクレオチド、もしくは両方の混合物であり得る。本明細書において予期されるポリヌクレオチドの例には、一本鎖および二本鎖DNA、一本鎖および二本鎖RNA(miRNAを含む)、ならびに一本鎖および二本鎖DNAおよびRNAの混合物を有するハイブリッド分子を含む。
【0022】
「タンパク質」、「ペプチド」、および「ポリペプチド」という言葉は、アミノ酸ポリマーまたは2つ以上の相互作用する、もしくは結合したアミノ酸ポリマーの組を表示するために互換的に使用される。
【0023】
「遺伝子」という用語は、タンパク質の産生に関与するDNAの断片を意味し、コード領域に先行および後続する領域(リーダーおよびトレーラー)ならびに個々のコード断片(エクソン)の間の介在配列(イントロン)を含む。リーダー、トレーラー、およびイントロンは、遺伝子の転写および翻訳の間に必要な調節要素を含む。さらに、「タンパク質遺伝子産物」は、特定の遺伝子から発現したタンパク質である。
【0024】
「ウイルスベクター」は、別の核酸を細胞中に輸送できるウイルス由来の核酸である。ウイルスベクターは、適切な環境に存在する場合に、ベクターによって運搬される1つ以上の遺伝子にコードされる1つのタンパク質もしくは複数のタンパク質の発現を指示できる。ウイルスベクターの例にはレトロウイルスベクター、アデノウイルスベクター、レンチウイルスベクター、およびアデノ随伴ウイルスベクターが含まれるが、これらに限定されない。
【0025】
「形質移入」または「形質移入する」という用語は、非ウイルス的もしくはウイルスに基づく方法によって核酸分子を細胞に導入する過程として定義される。非ウイルス的な形質移入の方法は、核酸分子を細胞に導入するための送達システムとしてウイルスDNAもしくはウイルス粒子を使用しない任意の適切な形質移入の方法を含む。例示的な非ウイルス的形質移入方法には、リン酸カルシウム形質移入、リポソーム形質移入、ヌクレオフェクション、ソノポレーション、熱ショックによる形質移入、マグネティフェクション、およびエレクトロポレーションが含まれる。ウイルスに基づく形質移入の方法のために、本明細書に記載された方法において任意の有用なウイルスベクターを使用してよい。ウイルスベクターの例にはレトロウイルスベクター、アデノウイルスベクター、レンチウイルスベクター、およびアデノ随伴ウイルスベクターが含まれるが、これらに限定されない。
【0026】
遺伝子に関連して本明細書で使用される「発現」または「発現した」という言葉は、その遺伝子の転写および/または翻訳産物を意味する。細胞におけるDNA分子の発現レベルは、細胞内に存在する対応するmRNAの量もしくは細胞によって産生されるそのDNAにコードされるタンパク質の量のいずれかに基づいて決定してよい(Sambrook et al., 1989 Molecular Cloning: A Laboratory Manual, 18.1-18.88)。
【0027】
「プラスミド」という用語は、遺伝子の発現に必要な遺伝子および/または調節要素をコードする核酸分子を指す。プラスミドからの遺伝子の発現は、シスもしくはトランスで起こり得る。遺伝子がシスで発現する場合は、遺伝子および調節要素は同じプラスミドによってコードされる。トランスでの発現とは、遺伝子および調節要素が別々のプラスミドによってコードされる場合を指す。
【0028】
「エピソームの」という用語は、細胞におけるプラスミドの染色体外の状態を指す。エピソームのプラスミドは、染色体DNAの部分ではなく、染色体DNAとは独立して複製する核酸分子である。
【0029】
「細胞培養物」とは、生物の外側に存在する細胞の集団である。これらの細胞は、任意で細胞バンク、動物、もしくは血液バンクから単離された初代細胞、またはこれらの供給源の一つに由来する二次細胞であり、長寿命のインビトロ培養のために不死化されている。
【0030】
「幹細胞」とは、有糸細胞分裂を通して自己再生する能力および組織もしくは器官に分化する潜在力によって特徴付けされる細胞である。哺乳動物の幹細胞の中で、胚性および成体幹細胞は識別できる。胚性幹細胞は胚盤胞の中に存在し、胚組織を生み出す一方で、成体幹細胞は組織再生および修復のために成体組織中に存在する。
【0031】
「多能性の」もしくは「多能性」という用語は、適切な条件下で3つの胚葉(内胚葉、中胚葉、および外胚葉)から細胞系統に伴う特徴を集団で呈する細胞型に分化を起こすことのできる子孫を生み出す能力を有する細胞を指す。多能性幹細胞は、出生前、出生後、もしくは成体の生物の組織に寄与できる。細胞集団の多能性を確立するために、8〜12週齢のSCIDマウスにおいて奇形腫を形成する能力等の、技術分野において容認された標準的な試験を使用できる。しかしながら、多能性細胞を同定するために、様々な多能性幹細胞の特徴の同定も使用することができる。
【0032】
「多能性幹細胞の特徴」とは、多能性幹細胞を他の細胞と識別する細胞の特徴を指す。分子マーカーの一定の組合せの発現もしくは非発現は、多能性幹細胞の特徴の例である。より具体的には、ヒト多能性幹細胞は次の非限定的な一覧表からのマーカーの少なくともいくつか、任意ですべてを発現してよい:SSEA-3、SSEA-4、TRA-1-60、TRA-1-81、TRA-2-49/6E、ALP、Sox2、E-カドヘリン、UTF-1、Oct4、Lin28、Rex1、およびNanog。多能性幹細胞に伴う細胞の形態も多能性幹細胞の特徴である。
【0033】
「再プログラミング」という用語は、非多能性細胞を多能性幹細胞の特徴を呈する細胞に脱分化させる過程を指す。
【0034】
「治療する」という用語は、治療されている疾患の発症を改善し、抑制し、根絶し、および/または遅延させることを意味する。
【0035】
II.臍帯血から人工多能性幹細胞を調製する方法
一つの局面において、人工多能性幹細胞を調製するための方法が提供される。この方法は、OCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成することを含む。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。
【0036】
「人工多能性幹細胞」とは、非多能性細胞に人工的に由来する多能性幹細胞を指す。「非多能性細胞」は、多能性幹細胞よりも自己再生および分化する潜在力がより低い細胞であり得る。潜在力がより低い細胞とは、成体幹細胞、組織特異的前駆細胞、初代もしくは二次細胞であり得るが、これらに限定されない。成体幹細胞は、胚発生後に体全体に見出される未分化細胞である。成体幹細胞は死に瀕した細胞を補充し、損傷した組織を再生させるために、細胞分裂によって増殖する。成体幹細胞は、分裂して自身に似た別の細胞を製造し、また分裂して自身よりもより分化した細胞を製造する能力を有する。成体幹細胞はRex1、Nanog、Oct4、もしくはSox2等の多能性マーカーの発現を伴ってはいるが、3つのすべての胚葉の細胞型に分化するという多能性幹細胞の能力は有していない。成体幹細胞は、自己再生および独特の細胞型の子孫を生成する限定された潜在力を有する。成体幹細胞は、造血幹細胞、臍帯血幹細胞、間充織幹細胞、上皮幹細胞、皮膚幹細胞、もしくは神経幹細胞であり得るが、これらに限定されない。組織特異的前駆体とは、特定の器官もしくは組織に分化することが約束された、自己再生の潜在力を持たない細胞を指す。初代細胞は、卵細胞、精細胞、および幹細胞を除く成体もしくは胎児の生物の任意の細胞を含む。有用な初代細胞の例は皮膚細胞、骨細胞、血液細胞、内臓の細胞、および結合組織の細胞を含むが、これらに限定されない。二次細胞は初代細胞に由来し、長寿命のインビトロ細胞培養のために不死化されている。
【0037】
「臍帯血幹細胞」とは、臍帯血に存在し、多能性幹細胞よりも自己再生および分化する潜在力がより低いことで特徴付けられる成体幹細胞を指す。
【0038】
「形質移入」または「形質移入する」という用語は、非ウイルス的もしくはウイルスに基づく方法によって核酸分子を細胞に導入する過程として定義される。非ウイルス的な形質移入の方法は、核酸分子を細胞に導入するための送達システムとしてウイルスDNAもしくはウイルス粒子を使用しない任意の適切な形質移入の方法を含む。例示的な非ウイルス的形質移入方法には、リン酸カルシウム形質移入、リポソーム形質移入、ヌクレオフェクション、ソノポレーション、熱ショックによる形質移入、マグネティフェクション、およびエレクトロポレーションが含まれる。ある態様において、核酸分子は当技術分野において周知の標準的な手順に続いてエレクトロポレーションを使用して細胞内に導入される。ウイルスに基づく形質移入の方法のために、本明細書に記載された方法において任意の有用なウイルスベクターを使用してよい。ウイルスベクターの例にはレトロウイルスベクター、アデノウイルスベクター、レンチウイルスベクター、およびアデノ随伴ウイルスベクターが含まれるが、これらに限定されない。ある態様において、核酸分子は当技術分野において周知の標準的な手順に続いてレトロウイルスベクターを使用して細胞内に導入される。
【0039】
形質移入遺伝子の発現は、細胞内で一過的もしくは安定して起こり得る。「一過的発現」の間は、形質移入遺伝子は細胞分裂の間に娘細胞に伝達されない。その発現が形質移入細胞に限定されているため、遺伝子の発現は時間とともに失われる。対照的に、形質移入遺伝子の安定発現は、形質移入細胞に選択の優位性を与える別の遺伝子とその遺伝子を一緒に形質移入したときに起こり得る。そのような選択の優位性は、細胞に提示される一定の毒素に対する抵抗性であってよい。形質移入遺伝子の発現は、宿主ゲノムへのトランスポゾンを介した挿入によって、さらに遂行され得る。トランスポゾンを介した挿入の間に、宿主ゲノムへの挿入および続く切除を行わせる2つのトランスポゾンリンカー配列の間に遺伝子が位置する。
【0040】
本明細書で指すところの「OCT4タンパク質」は、Octomer 4転写因子の任意の天然の形態、またはOct4転写因子活性を(例えば、Oct4と比較して少なくとも50%、80%、90%、95%、96%、97%、98%、99%、もしくは100%の活性の範囲内で)維持しているその変異体を含む。ある態様において、変異体は配列全体もしくは配列の一部分(例えば、50、100、150、もしくは200個の連続したアミノ酸部分)にわたって、天然のOct4ポリペプチド(例えば、配列番号:1、配列番号:2、もしくは配列番号:3)と比較して少なくとも90%、95%、96%、97%、98%、99%、もしくは100%のアミノ酸配列同一性を有する。他の態様において、Oct4タンパク質はアイソフォーム1(配列番号:1)に対応するNCBI参照番号gi:42560248、アイソフォーム2(配列番号:2および配列番号:3)に対応するgi:116235491およびgi:291167755によって同定されるタンパク質である。
【0041】
本明細書で指すところの「Sox2タンパク質」は、Sox2転写因子の任意の天然の形態、またはSox2転写因子活性を(例えば、Sox2と比較して少なくとも50%、80%、90%、95%、96%、97%、98%、99%、もしくは100%の活性の範囲内で)維持しているその変異体を含む。ある態様において、変異体は配列全体もしくは配列の一部分(例えば、50、100、150、もしくは200個の連続したアミノ酸部分)にわたって、天然のSox2ポリペプチド(例えば、配列番号:4)と比較して少なくとも90%、95%、96%、97%、98%、99%、もしくは100%のアミノ酸配列同一性を有する。他の態様において、Sox2タンパク質はNCBI参照番号gi:28195386(配列番号:4)によって同定されるタンパク質である。
【0042】
本明細書で指すところの「KLF4タンパク質」は、KLF4転写因子の任意の天然の形態、またはKLF4転写因子活性を(例えば、KLF4と比較して少なくとも50%、80%、90%、95%、96%、97%、98%、99%、もしくは100%の活性の範囲内で)維持しているその変異体を含む。ある態様において、変異体は配列全体もしくは配列の一部分(例えば、50、100、150、もしくは200個の連続したアミノ酸部分)にわたって、天然のKLF4ポリペプチド(例えば、配列番号:5)と比較して少なくとも90%、95%、96%、97%、98%、99%、もしくは100%のアミノ酸配列同一性を有する。他の態様において、KLF4タンパク質はNCBI参照番号gi:194248077(配列番号:5)によって同定されるタンパク質である。
【0043】
本明細書で指すところの「cMYCタンパク質」は、cMyc転写因子の任意の天然の形態、またはcMyc転写因子活性を(例えば、cMycと比較して少なくとも50%、80%、90%、95%、96%、97%、98%、99%、もしくは100%の活性の範囲内で)維持しているその変異体を含む。ある態様において、変異体は配列全体もしくは配列の一部分(例えば、50、100、150、もしくは200個の連続したアミノ酸部分)にわたって、天然のcMycポリペプチド(例えば、配列番号:6)と比較して少なくとも90%、95%、96%、97%、98%、99%、もしくは100%のアミノ酸配列同一性を有する。他の態様において、cMycタンパク質はNCBI参照番号gi:71774083(配列番号:6)によって同定されるタンパク質である。
【0044】
本明細書で指すところの「NANOGタンパク質」は、Nanog転写因子の任意の天然の形態、またはNanog転写因子活性を(例えば、Nanogと比較して少なくとも50%、80%、90%、95%、96%、97%、98%、99%、もしくは100%の活性の範囲内で)維持しているその変異体を含む。ある態様において、変異体はそれらの配列全体もしくは配列の一部分(例えば、50、100、150、もしくは200個の連続したアミノ酸部分)にわたって、天然のNanogポリペプチド(例えば、配列番号:7)と比較して少なくとも90%、95%、96%、97%、98%、99%、もしくは100%のアミノ酸配列同一性を有する。他の態様において、Nanogタンパク質はNCBI参照番号gi:153945816(配列番号:7)によって同定されるタンパク質である。
【0045】
本明細書で指すところの「LIN28タンパク質」は、Lin28転写因子の任意の天然の形態、またはLin28転写因子活性を(例えば、Lin28と比較して少なくとも50%、80%、90%、95%、96%、97%、98%、99%、もしくは100%の活性の範囲内で)維持しているその変異体を含む。ある態様において、変異体はそれらの配列全体もしくは配列の一部分(例えば、50、100、150、もしくは200個の連続したアミノ酸部分)にわたって、天然のLin28ポリペプチド(例えば、配列番号:8)と比較して少なくとも90%、95%、96%、97%、98%、99%、もしくは100%のアミノ酸配列同一性を有する。他の態様において、Lin28タンパク質はNCBI参照番号gi:13375938(配列番号:8)によって同定されるタンパク質である。
【0046】
形質移入臍帯血幹細胞を分裂させ、これにより人工多能性幹細胞を形成することには、形質移入後に臍帯血幹細胞を増殖させ、任意で形質移入細胞を選択し、多能性幹細胞を同定することを含んでよい。本明細書で使用される増殖には、当技術分野において周知の条件下で、容器中において形質移入臍帯血幹細胞により子孫細胞を産生させることが含まれる。増殖は、適した培地および細胞増殖因子の存在下で起こってもよい。細胞増殖因子は、細胞の遊走、分化、形質転換、もしくは成熟および分裂を引き起こす作用物質である。それらは通常、様々な正常および悪性の哺乳動物細胞型から単離できるポリペプチドである。いくつかの増殖因子は、細菌(大腸菌(E.coli))および酵母等の遺伝子操作された微生物によっても産生され得る。細胞増殖因子は培地に補充してもよく、および/またはそのような細胞増殖因子を分泌する放射線照射された胚線維芽細胞と共培養することによって提供されてもよい。細胞増殖因子の例には、FGF、bFGF2、およびEGFが含まれるが、これらに限定されない。
【0047】
適切な場合には、増殖している形質移入臍帯血幹細胞を選択の過程に供してもよい。選択の過程は、形質移入の際に臍帯血幹細胞に導入した選択マーカーを含んでもよい。選択マーカーは、酵素活性を有するポリペプチドをコードする遺伝子であってもよい。酵素活性は、アセチルトランスフェラーゼおよびフォスフォトランスフェラーゼの活性を含むが、これらに限定されない。ある態様において、選択マーカーの酵素活性はフォスフォトランスフェラーゼの活性である。選択マーカーの酵素活性は、形質移入臍帯血幹細胞に毒素の存在下で増殖する能力を与えてもよい。そのような毒素は通常、細胞増殖を阻害し、および/または細胞死を引き起こす。そのような毒素の例にはハイグロマイシン、ネオマイシン、ピューロマイシン、およびゲンタマイシンが含まれるが、これらに限定されない。ある態様において、毒素はハイグロマイシンである。選択マーカーの酵素活性を通して、毒素はもはや形質移入臍帯血幹細胞の増殖を阻害せず、細胞死を引き起こさない非毒素に変換されてもよい。毒素への暴露の際に、選択マーカーを欠いた細胞は排除され、これにより増殖から除外されてもよい。
【0048】
人工多能性幹細胞の同定は、上述した多能性幹細胞の特徴の評価を含んでもよいが、これに限定されない。そのような多能性幹細胞の特徴には、分子マーカーの一定の組合せの発現もしくは非発現が含まれるが、これにさらに限定されない。さらに、多能性幹細胞に伴う細胞の形態も多能性幹細胞の特徴である。
【0049】
上述したように、本明細書の方法で提供される臍帯血幹細胞はOCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を形質移入されてもよい。ある態様において、臍帯血幹細胞はcMYCタンパク質、LIN28タンパク質、NANOGタンパク質、もしくはKLF4タンパク質をコードする追加の核酸を形質移入されない。
【0050】
ある態様において、OCT4タンパク質をコードする核酸は一つのプラスミドの部分を形成し、SOX2タンパク質をコードする核酸は一つのプラスミドの部分を形成する。別の態様において、OCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸は同じプラスミドの部分を形成する。一つの態様において、OCT4タンパク質をコードする核酸は第一のプラスミドの部分を形成し、SOX2タンパク質をコードする核酸は第二のプラスミドの部分を形成する。
【0051】
別の局面において、人工多能性幹細胞を調製するための方法が提供される。この方法は、OCT4タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成することを含む。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。
【0052】
一つの態様において、臍帯血幹細胞はcMYCタンパク質、LIN28タンパク質、NANOGタンパク質、もしくはKLF4タンパク質をコードする追加の核酸を形質移入されない。
【0053】
ある態様において、本明細書に提供される方法で使用される臍帯血細胞はCD133抗原を発現する。「CD133抗原」は大きさが120キロダルトンの、5つの膜貫通領域を有する糖タンパク質を指す。CD133抗原は成体幹細胞および前駆細胞によって発現されてよい。CD133抗原はPROML1、AC133、造血幹細胞抗原、hProminin、prominin-like 1、prominin、RP41、MCDR2、STGD4、CORD12、もしくはMSTP061としても知られる。ある態様において、CD133抗原はNCBI参照番号gi:225690512(配列番号:9)によって同定される遺伝子にコードされるタンパク質である。
【0054】
ある態様において、本明細書に提供される方法で使用される臍帯血幹細胞は新鮮な臍帯血に由来する。「新鮮な臍帯血」とは新生児の臍帯に由来する血液であって、臍帯が早まってクランプされていない場合には新生児の血液循環に戻される。本明細書で指すところの新鮮な臍帯血は、臍帯から単離された後に凍結保存されていない。「凍結保存」という用語は、液体窒素を用いて臍帯血等の生物学的材料を凍結させ、これにより生物学的材料を長期間にわたって保存する過程を指す。他の態様において、本明細書に提供される方法で使用される臍帯血幹細胞は凍結臍帯血に由来する。凍結臍帯血とは、本明細書に提供される方法に従って処理される前に凍結保存されていた新生児の臍帯に由来する血液である。
【0055】
III.人工多能性幹細胞
別の局面において、本明細書の方法に従って調製された人工多能性幹細胞が提供される。「臍帯血から人工多能性幹細胞を調製する方法」と題された節において上述された方法は、本明細書で提供される人工多能性幹細胞に対して同じように適用できる。
【0056】
IV.臍帯血幹細胞
一つの局面において、OCT4タンパク質をコードする核酸(例えば、OCT4タンパク質をコードする外来性の核酸もしくはOCT4タンパク質をコードする組換え核酸)およびSOX2タンパク質をコードする核酸(例えば、SOX2タンパク質をコードする外来性の核酸もしくはSOX2タンパク質をコードする組換え核酸)を含む臍帯血幹細胞が提供される。本明細書で使用されるタンパク質をコードする核酸に関連した「外来性」という用語は、それが見出される細胞(例えば、臍帯血細胞)において天然ではないことを意味する。ある態様において、OCT4タンパク質をコードする核酸は一つのプラスミドの部分を形成し、SOX2タンパク質をコードする核酸は一つのプラスミドの部分を形成する。別の態様において、OCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸は同じプラスミドの部分を形成する。一つの態様において、OCT4タンパク質をコードする核酸は第一のプラスミドの部分を形成し、SOX2タンパク質をコードする核酸は第二のプラスミドの部分を形成する。ある態様において、臍帯血幹細胞はcMYCタンパク質をコードする核酸(例えば、cMYCタンパク質をコードする外来性の核酸もしくはcMYCタンパク質をコードする組換え核酸)、LIN28タンパク質をコードする核酸(例えば、LIN28タンパク質をコードする外来性の核酸もしくはLIN28タンパク質をコードする組換え核酸)、NANOGタンパク質をコードする核酸(例えば、NANOGタンパク質をコードする外来性の核酸もしくはNANOGタンパク質をコードする組換え核酸)、および/またはKLF4タンパク質をコードする核酸(例えば、KLF4タンパク質をコードする外来性の核酸もしくはKLF4タンパク質をコードする組換え核酸)等のiPS細胞形成において有用であることが知られている他の転写因子をコードする核酸を含まない。
【0057】
他の態様において、臍帯血幹細胞はOCT4タンパク質をコードする核酸(例えば、OCT4タンパク質をコードする外来性の核酸もしくはOCT4タンパク質をコードする組換え核酸)およびSOX2タンパク質をコードする核酸(例えば、SOX2タンパク質をコードする外来性の核酸もしくはSOX2タンパク質をコードする組換え核酸)から本質的になる。臍帯血幹細胞がOCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸「から本質的になる」場合は、臍帯血幹細胞はcMYCタンパク質をコードする核酸(例えば、cMYCタンパク質をコードする外来性の核酸もしくはcMYCタンパク質をコードする組換え核酸)、LIN28タンパク質をコードする核酸(例えば、LIN28タンパク質をコードする外来性の核酸もしくはLIN28タンパク質をコードする組換え核酸)、NANOGタンパク質をコードする核酸(例えば、NANOGタンパク質をコードする外来性の核酸もしくはNANOGタンパク質をコードする組換え核酸)、および/またはKLF4タンパク質をコードする核酸(例えば、KLF4タンパク質をコードする外来性の核酸もしくはKLF4タンパク質をコードする組換え核酸)等のiPS細胞形成において有用であることが知られている他の転写因子をコードする核酸を含まない。ある態様において、臍帯血幹細胞は他の転写因子をコードする核酸(例えば、転写因子をコードする他の外来性の核酸もしくは転写因子をコードする他の組換え核酸)を含まない。他の態様において、臍帯血幹細胞は他のタンパク質を発現する遺伝子をコードする核酸(例えば、タンパク質をコードする他の外来性の核酸もしくはタンパク質をコードする他の組換え核酸)を含まない。
【0058】
別の局面において、OCT4タンパク質をコードする核酸(例えば、OCT4タンパク質をコードする外来性の核酸もしくはOCT4タンパク質をコードする組換え核酸)を含む臍帯血幹細胞が提供される。他の態様において、臍帯血幹細胞はOCT4タンパク質をコードする核酸(例えば、OCT4タンパク質をコードする外来性の核酸もしくはOCT4タンパク質をコードする組換え核酸)から本質的になる。臍帯血幹細胞がOCT4タンパク質をコードする核酸「から本質的になる」場合は、臍帯血幹細胞はcMYCタンパク質をコードする核酸(例えば、cMYCタンパク質をコードする外来性の核酸もしくはcMYCタンパク質をコードする組換え核酸)、LIN28タンパク質をコードする核酸(例えば、LIN28タンパク質をコードする外来性の核酸もしくはLIN28タンパク質をコードする組換え核酸)、NANOGタンパク質をコードする核酸(例えば、NANOGタンパク質をコードする外来性の核酸もしくはNANOGタンパク質をコードする組換え核酸)、および/またはKLF4タンパク質をコードする核酸(例えば、KLF4タンパク質をコードする外来性の核酸もしくはKLF4タンパク質をコードする組換え核酸)等のiPS細胞形成において有用であることが知られている他の転写因子をコードする核酸を含まない。ある態様において、臍帯血幹細胞は他の転写因子をコードする核酸(例えば、転写因子をコードする他の外来性の核酸もしくは転写因子をコードする他の組換え核酸)を含まない。他の態様において、臍帯血幹細胞は他のタンパク質を発現する遺伝子をコードする核酸(例えば、タンパク質をコードする他の外来性の核酸もしくはタンパク質をコードする他の組換え核酸)を含まない。
【0059】
ある態様において、臍帯血幹細胞はCD133抗原を発現する。他の態様において、臍帯血幹細胞は新鮮な臍帯血に由来する。ある態様において、臍帯血幹細胞は凍結臍帯血に由来する。
【0060】
V.フットプリントを含まない、ヒト人工多能性幹細胞からヒト体細胞を産生するための方法
一つの局面において、ヒト体細胞を産生するための方法が提供される。この方法は、細胞増殖因子を人工多能性幹細胞に接触させることを含む。人工多能性幹細胞は分裂させられ、これによりヒト体細胞を形成する。人工多能性幹細胞は適切な培地および細胞増殖因子の存在下で分裂させられる。細胞増殖因子の例にはSCF、GMCSF、FGF、TNF、IFN、EGF、IGF、およびインターロイキンファミリーのメンバーが含まれるが、これらに限定されない。人工多能性幹細胞は、本発明によって提供される方法に従って調製される。ある態様において、人工多能性幹細胞はOCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程を含む過程によって調製される。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。別の態様において、人工多能性幹細胞はOCT4タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程を含む過程によって調製される。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。
【0061】
別の局面において、組織修復を必要としている哺乳動物を治療する方法が提供される。この方法は、人工多能性幹を哺乳動物に投与し、哺乳動物において人工多能性幹細胞を分裂および体細胞に分化させ、これにより哺乳動物において組織修復を提供することを含む。ある態様において、人工多能性幹細胞はOCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程を含む過程によって調製される。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。別の態様において、人工多能性幹細胞はOCT4タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程を含む過程によって調製される。形質移入臍帯血幹細胞は分裂させられ、これにより人工多能性幹細胞を形成する。
【実施例】
【0062】
CB幹細胞を単離するために、免疫磁気選択法を用いてCBユニットからCD133+細胞を精製し、純度の範囲が90〜94%の細胞の集団を取得した(図3A)。静止状態のCD133+幹細胞の増殖を促進するために、細胞を幹細胞因子(SCF)、トロンボポエチン(TPO)、Flt ligand 3(Flt3)、およびインターロイキン6(IL-6)の存在下で24時間にわたって培養した。いくつかの修正を加えた接着細胞のための手順に従って10、これらの細胞に感染させた。手短には、以前に記載したように19、ウイルス粒子で前もって前吸着させた、レトロネクチンのコートされたプレートの上に細胞を播種した。対照実験において、構成的GFPレトロウイルスを用いて12時間ごとに精製CD133+集団を感染の3サイクルに供し、結果として得られた集団をフローサイトメトリーを用いて感染の3日後に解析した。図3Bは、典型的な実験結果を示す:全細胞の28%がGFP陽性であり、58%がCD133抗原に対して依然として陽性であった。GFP陽性集団のうちの17%がCD133+/GFP+であった一方で、11%がCD133−/GFP+であった。
【0063】
OSKM因子の任意の単独もしくは組合せを用いてCD133+細胞の再プログラムが試みられた。形質導入の3日後に、放射線照射されたヒト包皮線維芽細胞(HFF-1)フィーダー細胞の上に細胞を播種し、bFGF含有hES培地中で培養した。単なるES様培養条件によって再プログラミング自体を誘導できる可能性を排除するために、形質導入されていないCD133+幹細胞もhES細胞条件において3週間にわたって培養した。これらのプレートでは、コロニー形成は観察されなかった。これらの細胞のフローサイトメトリー解析によって、これらの細胞が幹細胞マーカーCD133、CD34、およびCD38をもはや発現しておらず、造血マーカーCD45については陽性のままであるが、胚性マーカーSSEA-3、SSEA-4、もしくはTRA1-60は獲得していないことが明らかになり、全体として、hES細胞条件において培養すると形質導入されていないCD133+細胞は成熟造血細胞に分化することが示唆された(図1C)。
【0064】
感染の約9日後に、OSKM、OSK、もしくはOSで形質導入された細胞において小さいコロニーが出現し始めた。感染の12〜15日後に、コロニーのいくつかははっきりとした境界を有する典型的なhES細胞の形態を呈し、大きい核および明確に視認できる核小体を有する小さい密集した細胞集団から構成されていた(図1A)。8×104個のCD133+細胞の標準的な感染から4〜5個のhES様コロニーが観察され、CBiPSと名付けられた。コロニーを手作業のピッキングで増殖させ、各因子の組合せ(CBiPS 4F-1、CBiPS 3F-1、CBiPS 2F-1)からのCBiPS株をさらなる特徴付けのために増幅させた。各レトロウイルス導入遺伝子の存在をPCR遺伝子型決定によって確認し、CBiPS 4F-1、CBiPS 3F-1、CBiPS 2F-1のそれぞれにおいて予期された4、3、もしくは2個の転写因子の挿入が明らかにされた(図1B)。
【0065】
3つのCBiPS株はすべてアルカリフォスファターゼにより陽性に染色され(図1C)、免疫蛍光染色で評価されたように、多能性マーカーOCT4、SOX2、TRA-1-81、TRA-1-60、SSEA3、SSEA4、およびNANOGを発現した(図1D)。加えて、フローサイトメトリーで測定されたように、再プログラムされたCBiPS株は造血幹細胞マーカーCD45、CD34、およびCD38について陰性であった。しかしながら、これらの株は造血および胚性幹細胞の共通マーカーCD133について陽性であった(図1E)。以前の免疫特徴付けと一致して、リアルタイムPCR解析により、3つのCBiPS株はすべてOCT4、SOX2、NANOG、CRIPTO、およびREXを含む多数の多能性遺伝子を発現していることが示され、他のiPS10およびhES [2]細胞株20に匹敵する遺伝子発現プロファイルが明らかにされた(図2A)。加えて、レトロウイルス導入遺伝子の発現は適切に停止され、CBiPS株におけるOCT4、SOX2、KLF4、およびc-MYCの発現が対応する内在性遺伝子によって推進された(図2B)。このことは、FLAGタグ付けされた遺伝子導入因子に対して特異的な抗体を用いた免疫蛍光染色によっても確認された(図8A〜8B)。
【0066】
線維芽細胞および角化細胞は内在性のOCT4、NANOG、SOX2、REX1、およびCRIPTOを発現しなかったが、CD 133+幹細胞は低いレベルで発現したため(図2C)、迅速な再プログラミングが可能な、より可塑的でエピジェネティックな状態に向けられた。さらに、CD133+細胞におけるOCT4およびNANOGのプロモーターは線維芽細胞に比べてヒストン抑制の特徴(H3K7およびH3K9のメチル化)をより低いレベルで有することも見出され(図2D)、過剰発現した因子によるこれらの遺伝子の結合および転写活性化を助けるかもしれない、より許容的なクロマチン構成の存在が示唆された。加えて、線維芽細胞および角化細胞と比較して、CB CD133+幹細胞におけるKLF4およびc-MYCの高いレベルの組合せは(図2C、図2E)、これらの因子の内在性の発現が、これらの細胞のより迅速および/または促進された再プログラミングを可能にするかもしれないことをさらに示しているのかもしれない10。
【0067】
細胞遺伝学的解析によって、3つの細胞株は10回の継代後も正常な46XYの核型を維持していることが示された。加えて、男性染色体の内容は、再プログラムされた細胞が最初の臍帯血試料中に存在することが知られている混入した母の細胞の小さい画分に起因するという可能性を排除する(図11A)。次に、インビトロ胚様体形成によるCBiPS細胞株の分化の潜在力を評価した。すべての細胞株は胚様体(EBs)を高い効率で形成でき(図12A)、EBsはFoxA2およびα-アクチニン陽性中胚葉、GFAPおよびTuj1陽性外胚葉、ならびにα-フェトプロテイン陽性内胚葉を含む3つのすべての胚性胚葉の細胞型に分化できた(図12B〜12F)。この結果によって、CBiPS 4F-1、CBiPS 3F-1、CBiPS 2F-1細胞株が他のhiPSおよびhES細胞株と類似した状態にまで転写的に再プログラムされており、核型的に安定で、多能性と一致したインビトロの発生潜在力を見せることが確認される。
【0068】
体細胞の再プログラミングは、多能性因子および癌遺伝子の組合せの発現を通して遂行されてきた。たった2週間で2個の因子を用いてCB CD133+幹細胞を再プログラムできたという事実は、再生医療のために臨床的に適したiPS細胞の発生に向けた体細胞の理想的な供給源としてのCB細胞の潜在力を強調する。これには、非組込みもしくは半組込み手段の使用21〜23、および小分子によるOCT4もしくはSOX2の置換24、25を含んでもよい。他の受け入れやすい体細胞の供給源に関する限りでは、動員された末梢血(mPB)細胞もiPS誘導のための効果的な供給源を表し得ることが最近示された26。しかしながら、新生児のCB幹細胞と比較して、成体のmPB細胞は加齢の結果としてもしくは特定の疾患の直接的な帰結としてゲノムの変異が蓄積される可能性がより高い。さらに、成体造血幹細胞の区画を動員するために使用される薬物治療はドナーの健康上の危険性を表す:ドナーの小さいけれどかなり多い割合において、この手順は脾破裂27を含む激しい反応を引き起こし得る。
【0069】
CB幹細胞はこれらの課題を克服し、容易に入手可能で、その起源が早いおかげでまだ免疫学的に未成熟なため、HLA-ドナー-レシピエント選択のためのより厳しくない基準を可能にする15。今日までに、臍帯血バンクの包括的なネットワークにおいて400,000を超えるCBユニットが世界的に入手可能であり、CBiPS生成のための適合ドナーの素早く効果的な検索を促進する2。患者に特異的なiPS株の生成が、理論上は理想的な臨床の選択肢として繰り返し唱えられてきたにもかかわらず、実務的および費用対効果の局面から、この手段は多くの場合において実現不可能かもしれない。公的に入手可能なネットワークにおける様々なHLAハプロタイプを表すCBiPS株の大規模な産生およびバンクへの保存は、ヒトiPS細胞の基礎研究および将来的な臨床への適用のための貴重な手段を表すであろう。
【0070】
VI.材料および方法
試料の採取
臍のCB試料は、Banc de Sang i Teixits, Hospital Duran i Reynals, Barcelonaより取得した。
【0071】
CD133+細胞の精製
Lympholyte-H(Cederlane, Ontario, CA)密度勾配遠心を用いて、単核細胞(MNC)をCBから単離した。Mini-Macs免疫磁気分離システム(Miltenyi Biotec, Bergisch Gladbach, Germany)を用いて、CD133+細胞を陽性選択した。CD133-フィコエリトリン(PE;Miltenyi Biotec, Bergisch Gladbach, Germany)抗体で染色し、フローサイトメトリー解析によって精製効率を検証した。
【0072】
構築物およびレトロウイルス産生
RT-PCRによって、ES[4]の全RNAからOCT4およびSOX2のヒトcDNAを増幅した。IMAGEクローン5111134からヒトKLF4を増幅し、Luciano Di Croceよりご提供いただいたDNA鋳型から変異ヒトc-MYCT58Aを増幅した。N末のFLAGタグ付けタンパク質を発現できる修正pMSCVpuroベクターのEcoR I/Cla Iサイトに増幅したcDNAをクローン化した。pMXs-OSKMGを次のように構築した:Oct4終止コドンを排除してBspE Iサイトを追加した逆方向プライマーを用いてマウスOct4 cDNAを増幅し、pCRII(Invitrogen)にクローン化してpCRII-Oct4-Bsp(Not I-5'cDNA3'-Acc65 Iの向き)を得た。Age Iサイトに続くP2Aペプチド配列を含む順方向プライマーおよびSox2終止コドンを排除してBspE Iサイトを含んだ逆方向プライマーを用いてマウスSox2 cDNAを増幅した。この断片をpCRIIにクローン化してpCRII-Age-Sox2-Bsp(Not I-5'cDNA3'-Acc65 Iの向き)を得た。pCRII-Age-Sox2-BspをAge IおよびAcc65 Iで切断し、BspE I-Acc65 Iで切断したpCRII-Oct4-Bspにクローン化してpCRII-Oct4-P2A-Sox2-BspE Iを産生した。マウスKlf4およびeGFP(pCRII-OSKGを産生)もしくはマウスKlf4、c-Myc、およびeGFP(pCRII-OSKMGを産生)を組み入れるために、同じクローン化の手段を2回繰り返した。最後に、pCRII-OSKGおよびpCRII-OSKMGをEcoR Iで切断し、レトロウイルス空ベクターpMXsの唯一のEcoR Iサイトにクローン化してpMXs-OSKGおよびpMXs-OSKMGを産生した。製造業者の指示に従って、Fugene 6試薬(Roche)を用いてPhoenix Amphotropic細胞株に形質移入した後に、4個の因子についてのレトロウイルスを独立して産生した。24時間後に培地を置換し、細胞を32℃で培養して、12時間ごとにウイルス上清を回収した。
【0073】
CD133+細胞の形質導入
SCF(50ng/ml)+Flt3(50ng/ml)+TPO(10ng/ml)+IL-6(10ng/ml)(PeproTech)の存在下で、10%のFBSを添加したDMEM中において24時間にわたってCB CD133+細胞(1mlにつき1×105細胞)を前刺激した。マルチウェル非組織培養処理プレートをフィブロネクチン断片CH-296(15mg/cm2)であるレトロネクチン(Takara, Otsu, Japan, www.takara-bio.com)でコートし、OCT4、SOX2、KLF4、およびc-MYC因子についてのレトロウイルス上清の濾過した1:1:1:1混合物とともに2,500RPMで30分間にわたってプレートを遠心することにより前処置した。DMEM+10% FBSおよび上述したサイトカイン混合物の存在下で約80,000個のCD133+細胞を播種した。12時間ごとに、サイトカイン混合物を含んだ新鮮なウイルス上清で培地の半分を置換し、37℃、5% CO2で培養して、3回の感染サイクルを実行した。3日目に細胞を回収し、放射線照射されたヒト線維芽細胞ならびに、20% KO-Serum Replacement(GIBCO)を添加したKO-DMEM培地(Invitrogen)、非必須アミノ酸(Lonza)、2-β-メルカプトエタノール(GIBCO)、ペニシリン/ストレプトマイシン(GIBCO)、GlutaMAX(商標)(Invitrogene)、および10ng/ml bFGF(Peprotech)からなるES培地を含む6ウェルプレートに移動させた。放射線照射されたヒト線維芽細胞の上でCBiPS細胞を培養し、機械的にピッキングした。
【0074】
全RNAの精製および定量的RT-PCR
入手可能な細胞数に基づいて、Trizol(登録商標)試薬(Invitrogen, Carlsbad, CA)もしくはRNAqueous(登録商標)-Microキット(Ambion Inc., Austin TX)のいずれかを用いてCB CD133+幹細胞、hES[2]細胞、KiPS細胞(14)、およびCBiPSからの全RNAの単離を実行した。残存するいかなるゲノムDNAも除去するために、すべての試料をTURBO DNase 阻害剤(Ambion)で処理し、Invitrogen SuperScript(商標) II Reverse Transcriptaseキットを用いて、1μgのRNAを使用してcDNAを合成した。以前に記載したように10、プライマーを用いた定量的RT-PCRによって、25ngのcDNAを使用して遺伝子発現を定量した。
【0075】
GeneChip(登録商標)発現解析
Institute for Research in Biomedicine(Barcelona, Spain)において、製造業者の手順(Affymetrix, Santa Clara, CA)に従って、Functional Genomica CoreによりGeneChip(登録商標)マイクロアレイ処理を実行した。25ngの出発物のRNAを用いて、Nugenの手順に示されたように増幅および標識の処理を行った。各試料について3.75μgのssDNAを標識し、Affymetrix HG-U133 Plus 2.0チップにハイブリダイズさせた。Affymetrix GeneChip Scanner(7Gアップグレード)上で発現シグナルを走査した。Affymetrix GCOSソフトウェア v.1.4によってデータ抽出を行った。R Project for Statistical ComputingからのプログラムRを用いてデータの統計解析を実行した。最初に、Rに実装されたgcRMAアルゴリズムを用いて生データを正規化し、正規化されたデータについてピアソンの相関係数を用いた階層的クラスタリングを実行した。2つの異なる実験(我々の角化細胞再プログラミング(GEOアクセス番号:GSE12583)、および本実験(GSE16694))で取得されたデータセットを組み込むために、RにおいてgcRMAアルゴリズムを用いて生のCELファイルを一緒に正規化し、その後、当技術分野において公知であるように、ComBatアルゴリズムを用いてバッチ効果を補正した。例えば、Johnson et al., 2007, Biostatistics 8:118-127を参照。
【0076】
サザンブロット
製造業者の指針に従って、All Prep DNA/RNAカラム(Qiagen)を用いて各細胞株からのゲノムDNAを単離した。サザンブロットの各レーンは、40UのPst IもしくはHind III制限酵素(New England Biolabs)のいずれかで消化し、1% アガロースゲル上で電気泳動し、中性ナイロン膜(Hybond(商標)-N, Amersham)に転写し、PCR DIG Probe Synthesis Kit(Roche Diagnostics)を用いてPCRにより生成されたDIG-dUTP標識プローブとハイブリダイズさせた4μgのゲノムDNAに相当する。化学発光のための基質としてCDP-Star(Sigma-Aldrich)を用いて、AP結合DIG-Antibody(Roche Diagnostics)によりプローブを検出した。条件は製造業者の指示通りであった。プローブは以下のプライマー(F、順方向;R、逆方向)とともにSOX2、OCT4、KLF4、およびc-MYC cDNAを鋳型として用いて生成した:
。
【0077】
免疫蛍光解析およびAP解析
プラスチックのカバースライド容器上でCBiPSを増殖させ、4% パラホルムアルデヒド(PFA)で固定した。以下の抗体を使用した:TRA-1-60(MAB4360、1:200)、TRA-1-81(MAB4381、1:200)、SOX2(AB5603、1:500)、すべてChemicon、SSEA-4(MC-813-70、1:2)、SSEA-3(MC-631、1:2)、すべてIowa、Tuj1(1:500;Covance)、α-フェトプロテイン(1:400;Dako)、α-アクチニン(1:100;Sigma)、OCT4(C-10、SantaCruz、sc-5279、1:100)、NANOG(Everest Biotech EB06860、1:100)、GATA4(1:50、SantaCruz)、平滑筋アクチン(1:400、Sigma)、FoxA2(1:50、R&D System)、GFAP(1:1000、Dako)、α-筋節アクチン(1:400、Sigma)、抗Flag(Sigma M2)。Leica SP5共焦点顕微鏡を用いて画像を撮影した。製造業者の指針に従って、Alkaline Phosphatase Blue/Red Membrane基質溶液キット(Sigma)を用いて直接的なAP活性を解析した。
【0078】
インビトロ分化
機械的に採取され、その後hES培地の存在下で24時間にわたって懸濁液中に維持されたコロニー断片からEBsの形成を誘導した。我々は超低吸着プレートの上で3つの異なる分化培地中にEBsが維持される前条件培養を2〜3日間にわたって実行した。特に、内胚葉分化のために、10% ウシ胎仔血清(FBS)、2mM L-グルタミン、0.1mM 2-β-メルカプトエタノール、非必須アミノ酸、およびペニシリン-ストレプトマイシンを添加したKO-DMEM培地の存在下でEBsを培養した。中胚葉分化のために、我々は上述したのと同じであるが、アスコルビン酸(0.5mM)を追加した培地を使用した。外胚葉誘導のために、N2/B27培地中でEBsを培養した。前条件の工程の後に、0.1% ゼラチンのコートされたプラスチック容器スライドに内胚葉および中胚葉条件のEBsを移動し、それぞれ分化培地およびアスコルビン酸(0.5nM)添加分化培地において2週間にわたって培養した。外胚葉分化のために、N2/B27培地の存在下で間質細胞株PA6の上にEBsを2週間にわたって移動した。各条件のための培地を1日おきに交換した。
【0079】
クロマチン免疫沈降
製造業者の指示に従って、Magnetic Low cell ChIP Kit from Diagenodeを用いて、免疫沈降ごとに15,000細胞を使用してクロマチン免疫沈降実験を実行した。Millipore 07-440(抗H3K27me3)、07-030(抗H3K4me2)、および17-625(抗H3K9me3)からの抗体を使用した。
【0080】
プロモーターメチル化解析
QIA AMP DNA Mini Kit(Qiagen)を用いて、約500,000個のCD133+およびCBiPS細胞の試料によってゲノムDNAを抽出した。製造業者の仕様書に従って、Epitect Bisulfite Kit(Qiagen)で2μgの精製DNAに突然変異を起こさせた。以前に記載したプライマーを用いて10、続く2回のPCRによって関心対象のプロモーター配列を増幅させた。結果として得られた増幅産物をpGEM T Easyプラスミドにクローン化し、TOP10細胞中で増幅させ、精製および配列決定を行った。
【0081】
奇形腫の形成
重症複合免疫不全(SCID)ベージュマウス(Charles River Laboratories)を麻酔し、20〜40μlのhES培地に再懸濁した約0.5×106個のCBiPS細胞を精巣に注射した。細胞注射の6〜8週間後にマウスを安楽死させ、従来型の免疫組織化学の手順(マッソントリクローム染色)および免疫蛍光に従って腫瘍を処理および解析した。
【0082】
VII.参考文献
【特許請求の範囲】
【請求項1】
人工多能性幹細胞を調製するための方法であって、以下の工程を含む方法:
(i) OCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程、および
(ii) 形質移入臍帯血幹細胞を分裂させ、これにより人工多能性幹細胞を形成する工程。
【請求項2】
cMYCタンパク質、LIN28タンパク質、NANOGタンパク質もしくはKLF4タンパク質をコードする追加の核酸を臍帯血幹細胞に形質移入しない、請求項1記載の方法。
【請求項3】
人工多能性幹細胞を調製するための方法であって、以下の工程を含む方法:
(i) OCT4タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程、および
(ii) 形質移入臍帯血幹細胞を分裂させ、これにより人工多能性幹細胞を形成する工程。
【請求項4】
cMYCタンパク質、LIN28タンパク質、NANOGタンパク質もしくはKLF4タンパク質をコードする追加の核酸を臍帯血幹細胞に形質移入しない、請求項3記載の方法。
【請求項5】
臍帯血幹細胞がCD133抗原を発現する、請求項1または3記載の方法。
【請求項6】
臍帯血幹細胞が新鮮な臍帯血に由来する、請求項1または3記載の方法。
【請求項7】
臍帯血幹細胞が凍結臍帯血に由来する、請求項1または3記載の方法。
【請求項8】
請求項1〜7の一項記載の方法に従って調製された人工多能性幹細胞。
【請求項9】
OCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を含む臍帯血幹細胞。
【請求項10】
OCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸から本質的になる、請求項9記載の臍帯血幹細胞。
【請求項11】
OCT4タンパク質をコードする核酸を含む臍帯血幹細胞。
【請求項12】
OCT4タンパク質をコードする核酸から本質的になる、請求項11記載の臍帯血幹細胞。
【請求項13】
CD133抗原を発現する、請求項9、10、11または12記載の臍帯血幹細胞。
【請求項14】
新鮮な臍帯血に由来する、請求項9、10、11または12記載の臍帯血幹細胞。
【請求項15】
凍結臍帯血に由来する、請求項9、10、11または12記載の臍帯血幹細胞。
【請求項16】
ヒト体細胞を産生するための方法であって、以下の工程を含む方法:
(i) 細胞増殖因子を人工多能性幹細胞に接触させる工程;および
(ii) 人工多能性幹細胞を分裂させ、これによりヒト体細胞を形成する工程;
ここで、人工多能性幹細胞は以下の工程を含む過程によって調製される:
(i) OCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程、および
(ii) 形質移入臍帯血幹細胞を分裂させ、これにより人工多能性幹細胞を形成する工程;
もしくは、ここで、人工多能性幹細胞は以下の工程を含む過程によって調製される:
(i) OCT4タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程、および
(ii) 形質移入臍帯血幹細胞を分裂させ、これにより人工多能性幹細胞を形成する工程。
【請求項17】
組織修復を必要としている哺乳動物を治療する方法であって、以下の工程を含む方法:
(i) 人工多能性幹を哺乳動物に投与する工程、
(ii) 哺乳動物において人工多能性幹細胞を分裂および体細胞に分化させ、これにより哺乳動物において組織修復を提供する工程;
ここで、人工多能性幹細胞は以下の工程を含む過程によって調製される:
(i) OCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程、および
(ii) 形質移入臍帯血幹細胞を分裂させ、これにより人工多能性幹細胞を形成する工程;
もしくは、ここで、人工多能性幹細胞は以下の工程を含む過程によって調製される:
(i) OCT4タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程、および
(ii) 形質移入臍帯血幹細胞を分裂させ、これにより人工多能性幹細胞を形成する工程。
【請求項1】
人工多能性幹細胞を調製するための方法であって、以下の工程を含む方法:
(i) OCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程、および
(ii) 形質移入臍帯血幹細胞を分裂させ、これにより人工多能性幹細胞を形成する工程。
【請求項2】
cMYCタンパク質、LIN28タンパク質、NANOGタンパク質もしくはKLF4タンパク質をコードする追加の核酸を臍帯血幹細胞に形質移入しない、請求項1記載の方法。
【請求項3】
人工多能性幹細胞を調製するための方法であって、以下の工程を含む方法:
(i) OCT4タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程、および
(ii) 形質移入臍帯血幹細胞を分裂させ、これにより人工多能性幹細胞を形成する工程。
【請求項4】
cMYCタンパク質、LIN28タンパク質、NANOGタンパク質もしくはKLF4タンパク質をコードする追加の核酸を臍帯血幹細胞に形質移入しない、請求項3記載の方法。
【請求項5】
臍帯血幹細胞がCD133抗原を発現する、請求項1または3記載の方法。
【請求項6】
臍帯血幹細胞が新鮮な臍帯血に由来する、請求項1または3記載の方法。
【請求項7】
臍帯血幹細胞が凍結臍帯血に由来する、請求項1または3記載の方法。
【請求項8】
請求項1〜7の一項記載の方法に従って調製された人工多能性幹細胞。
【請求項9】
OCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を含む臍帯血幹細胞。
【請求項10】
OCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸から本質的になる、請求項9記載の臍帯血幹細胞。
【請求項11】
OCT4タンパク質をコードする核酸を含む臍帯血幹細胞。
【請求項12】
OCT4タンパク質をコードする核酸から本質的になる、請求項11記載の臍帯血幹細胞。
【請求項13】
CD133抗原を発現する、請求項9、10、11または12記載の臍帯血幹細胞。
【請求項14】
新鮮な臍帯血に由来する、請求項9、10、11または12記載の臍帯血幹細胞。
【請求項15】
凍結臍帯血に由来する、請求項9、10、11または12記載の臍帯血幹細胞。
【請求項16】
ヒト体細胞を産生するための方法であって、以下の工程を含む方法:
(i) 細胞増殖因子を人工多能性幹細胞に接触させる工程;および
(ii) 人工多能性幹細胞を分裂させ、これによりヒト体細胞を形成する工程;
ここで、人工多能性幹細胞は以下の工程を含む過程によって調製される:
(i) OCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程、および
(ii) 形質移入臍帯血幹細胞を分裂させ、これにより人工多能性幹細胞を形成する工程;
もしくは、ここで、人工多能性幹細胞は以下の工程を含む過程によって調製される:
(i) OCT4タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程、および
(ii) 形質移入臍帯血幹細胞を分裂させ、これにより人工多能性幹細胞を形成する工程。
【請求項17】
組織修復を必要としている哺乳動物を治療する方法であって、以下の工程を含む方法:
(i) 人工多能性幹を哺乳動物に投与する工程、
(ii) 哺乳動物において人工多能性幹細胞を分裂および体細胞に分化させ、これにより哺乳動物において組織修復を提供する工程;
ここで、人工多能性幹細胞は以下の工程を含む過程によって調製される:
(i) OCT4タンパク質をコードする核酸およびSOX2タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程、および
(ii) 形質移入臍帯血幹細胞を分裂させ、これにより人工多能性幹細胞を形成する工程;
もしくは、ここで、人工多能性幹細胞は以下の工程を含む過程によって調製される:
(i) OCT4タンパク質をコードする核酸を臍帯血幹細胞に形質移入して、形質移入臍帯血幹細胞を形成する工程、および
(ii) 形質移入臍帯血幹細胞を分裂させ、これにより人工多能性幹細胞を形成する工程。
【図1A】


【図1B】
【図1C】
【図1D】
【図1E】

【図2A】
【図2B】
【図2C】
【図2D】
【図2E】
【図2F】
【図2G】
【図3A】
【図3B】
【図3C】
【図4】
【図5】
【図6】
【図7A】
【図7B】

【図8】

【図9】
【図10】
【図11A】
【図11B】
【図11C】
【図12A】
【図12B】
【図12C】
【図12D】
【図12E】
【図12F】
【図13A】
【図13B】
【図13C】
【図13D】
【図1B】
【図1C】
【図1D】
【図1E】
【図2A】
【図2B】
【図2C】
【図2D】
【図2E】
【図2F】
【図2G】
【図3A】
【図3B】
【図3C】
【図4】
【図5】
【図6】
【図7A】
【図7B】
【図8】
【図9】
【図10】
【図11A】
【図11B】
【図11C】
【図12A】
【図12B】
【図12C】
【図12D】
【図12E】
【図12F】
【図13A】
【図13B】
【図13C】
【図13D】
【公表番号】特表2012−530497(P2012−530497A)
【公表日】平成24年12月6日(2012.12.6)
【国際特許分類】
【出願番号】特願2012−516345(P2012−516345)
【出願日】平成22年6月18日(2010.6.18)
【国際出願番号】PCT/US2010/039222
【国際公開番号】WO2010/148334
【国際公開日】平成22年12月23日(2010.12.23)
【出願人】(502207839)ザ・ソーク・インスティチュート・フォー・バイオロジカル・スタディーズ (2)
【出願人】(511286300)
【Fターム(参考)】
【公表日】平成24年12月6日(2012.12.6)
【国際特許分類】
【出願日】平成22年6月18日(2010.6.18)
【国際出願番号】PCT/US2010/039222
【国際公開番号】WO2010/148334
【国際公開日】平成22年12月23日(2010.12.23)
【出願人】(502207839)ザ・ソーク・インスティチュート・フォー・バイオロジカル・スタディーズ (2)
【出願人】(511286300)
【Fターム(参考)】
[ Back to top ]