
臓器機能の評価方法
【課題】臓器または組織傷害を分析する方法の提供。
【解決手段】 本発明は、以下のステップ:(a)臓器または組織を染料で標識するステップ、(b)前記臓器または組織の生体異物代謝および/または細胞状態を含む多重指標を入手するステップ、ならびに(c)前記指標から前記臓器または組織の傷害を分析するステップ、を含む、臓器または組織傷害を分析する方法を提供する。
【解決手段】 本発明は、以下のステップ:(a)臓器または組織を染料で標識するステップ、(b)前記臓器または組織の生体異物代謝および/または細胞状態を含む多重指標を入手するステップ、ならびに(c)前記指標から前記臓器または組織の傷害を分析するステップ、を含む、臓器または組織傷害を分析する方法を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、臓器機能を評価する方法に関する。さらに詳細には、本発明は、微細解剖配向血管系および/または排泄経路を用いて多重指標を視覚的に分析するための、および/または定量的に決定するための方法に関する。
【背景技術】
【0002】
University of Wisconsin(UW)液の使用は、肝臓移植のための平均保存時間を改善したが、初期移植片機能不全および初期の低機能の発生はなお残っている(1−3)。そのような機能障害の臨床的発生および結果としての移植片生存の損失は、保存時間によって変わる(1−4)。再灌流傷害は、長期冷虚血後の移植片機能不全を引き起こす主要な事象である(5−9)。保存の間、肝細胞は膨潤して、ブレッブを形成する(6−8)。しかしながら再灌流時に、実質細胞におけるこれらの変化は、回復不能な傷害に至ることなく修復された(6−8)。これに対して、類洞内皮細胞は生存能を失い、クッパー細胞は活性化される(6−10)。UW溶液中に保存したラット肝臓移植片を使用した以前の研究によると、類洞細胞の変化が起こるときの臨界保存時間は16時間より長い(6,7)。これらの移植片において、細胞は損傷されて、血小板捕捉(10)、フィブリン沈着(11)、および白血球辺縁趨向(12)を引き起こす。より短い期間に渡って保存された移植片では、肝臓ATP含有率は再灌流後に十分回復することが報告され、実質細胞には生存能力があることを示唆している(13,14)。
【0003】
これらのデータとは別に、肝細胞が、回復不能な傷害を示すことなく、次に全体としての移植片機能を妨害することなく、その機能を失うかどうかは、慎重に調査されていない。胆汁産出量、タウロコール酸およびブロモスルホフタレインのクリアランスは以前の研究で測定されたが(13−15)、保存時間が18時間に延びたときでさえ、それらは肝細胞におけるそのような機能変化を証明できなかった(13)。これらの研究以外には、冷虚血後移植片が胆汁構成要素を分泌する能力の変化に関して、情報はほとんど入手できていない。そのような指標は、肝細胞が胆汁形成のための浸透圧駆動力を生む、そして胆汁塩または有機アニオンを排出する能力を含む。それゆえグルタチオンおよびビリルビン排出は、移植片における早期肝細胞変化を検知するマーカーとして作用できる。有機アニオンを排出する能力は、移植片が生体異物を解毒する効率および臨床移植における同種移植機能障害の危険因子である、冷虚血後高ビリルビン血症の重症度を判定することができる(2,16)。
【発明の開示】
【発明が解決しようとする課題】
【0004】
本発明の目的は、臓器機能を評価する方法を提供することである。上記知見に基づいて、本発明者はさらなる調査を継続した。
【課題を解決するための手段】
【0005】
冷虚血後移植片のそのような能力を判定する試みにおいて、本発明者らは本明細書で胆汁サンプルの構成要素の変化を保存時間の関数として調査し、肝細胞での早期事象としてグルタチオンおよびビリルビンの排出障害を明らかにした。本事象は、有機アニオンの胆汁中排出のためのATP依存性トランスポータである、多剤耐性関連タンパク質2(Mrp2)の細胞質再局在化の結果であることがわかった。本発明者らの結果は、このトランスポータの機能が損なわれるが、これに対して移植片は顕著な肝細胞損傷を示すことなく、その全体のエネルギー電荷を明らかに維持することを示唆している。さらに肝細胞中のそのような変化の機構は、むしろより短期間の、すなわち8時間の冷虚血にさらされた移植片中のクッパー細胞におけるトロンボキサン合成に関与するように思われる。
【0006】
本出願は以下の発明を提供する。
(1)臓器または組織傷害を分析する方法であって、以下のステップ:
(a)臓器または組織を染料で標識するステップ、
(b)前記臓器または組織の生体異物代謝および/または細胞状態を含む多重指標を入手するステップ、ならびに
(c)前記指標から前記臓器または組織の傷害を分析するステップ、
を含む方法。
(2)前記臓器または組織が肝臓、腎臓、肺、膵臓および胃腸管から成る群より選択される少なくとも1つである、(1)に記載の方法。
(3)前記ステップ(b)が血管系および/または排出経路の微細解剖配向を得るステップをさらに含む、(1)または(2)に記載の方法。
(4)前記解析が視覚的におよび/または定量的に実施される、(1)〜(3)のいずれか一項に記載の方法。
(5)前記細胞状態が細胞生存能、細胞傷害、分子輸送、およびミトコンドリア機能から成る群より選択される少なくとも1つである、(1)〜(4)のいずれか一項に記載の方法。
(6)薬剤毒性を評価する方法であって、以下のステップ:
(a)臓器または組織を染料で標識するステップ、
(b)試験薬剤を前記臓器または組織に使用するステップ、
(c)前記臓器または組織の生体異物代謝および/または細胞状態を含む多重指標を入手するステップ、
(d)前記指標から前記臓器または組織の傷害を分析するステップ、ならびに
(e)前記薬剤が前記臓器または組織に対する毒性を有するかどうかを評価するステップ、
を含む方法。
(7)前記臓器または組織が肝臓、腎臓、肺、膵臓および胃腸管から成る群より選択される少なくとも1つである、(6)に記載の方法。
(8)前記ステップ(b)が、血管系および/または排出経路の微細解剖配向を得るステップをさらに含む、(6)または(7)に記載の方法。
(9)前記解析が視覚的におよび/または定量的に実施される、(6)〜(8)のいずれか一項に記載の方法。
(10)前記細胞状態が細胞生存能、細胞傷害、細胞内および細胞周囲での分子の輸送、および生物活性化合物の生成、血流、および組織酸素添加から成る群より選択される少なくとも1つである、(6)〜(9)のいずれか一項に記載の方法。
【発明を実施するための最良の形態】
【0007】
以下、本発明をさらに詳細に説明する。
【0008】
クッパー細胞(KC)は冷虚血後肝細胞傷害において重大な役割を果たしていると考えられているが、非壊死移植片機能障害におけるその役割は未知のままである。本研究は、冷虚血後肝臓移植片におけるKCのそのような役割を解明することを狙っている。KC消耗試薬であるリポソームカプセル化ジクロロメチレンジホスホナートを用いて、または用いずに処理したラット肝臓は、4℃のUniversity of Wisconsin溶液中で8〜24時間保存し、胆汁産出量および構成要素の監視下で再灌流させた。肝細胞が胆汁を排出する能力は、生体内原位置(in situ)でレーザ共焦点マイクロフルオログラフィーによって評価した。冷虚血再灌流移植片は、顕著な細胞傷害なしに8時間にてその胆汁産出量を著しく減少させた。この事象は、グルタチオンおよびビリルビン−IXαの排出障害と一致し、これらの有機アニオンの遅延輸送を示唆している。機構は、多剤耐性関連タンパク質2(Mrp2)の細胞内再局在化を含んでいた。このトランスポータを通じて排出される蛍光プローブである、カルボキシフルオレセインの胆汁中排出の動態解析は、肝細胞から毛細胆管内への染料排出の著しい遅延を明らかにした。KC消耗処理は、細胞質から小管膜へのMrp2の再分布によって、8時間冷虚血移植片中のMrp2によって仲介される胆汁アニオン輸送のこのような低下を著しく減衰させた。さらに機構は、CF排出およびMrp2の細胞質内在化に対するこの酵素を阻害する改善効果がKC消耗移植片において消失したため、KC中にトロンボキサンA2シンターゼを包含するように思われた。要するに、これらの結果は、KC活性化が冷虚血後移植片の解毒能力および有機アニオン代謝の恒常性を脅かす、非壊死肝細胞機能障害の重要な決定要因であることを示唆している。
【0009】
本明細書において、一部の省略形が使用されている:BC、毛細胆管;BR−IXα、ビリルビン−IXα;CF、5−カルボキシフルオレセイン;CFDA、5−カルボキシフルオレセインジアセテート;EHBR、エイザイ高ビリルビン血症ラット;GSH、還元グルタチオン;KC、クッパー細胞;LDD、リポソームカプセル化ジクロロメチレンジホスホナート;Mrp2、多剤耐性関連タンパク質2;TXA2、トロンボキサンA2。
【0010】
本発明の1つの実施形態において、臓器または組織の機能障害および/または傷害を分析する方法が提供される。本方法は以下のステップを含む:
(a)臓器または組織を1つまたは2つの染料で標識するステップ、
(b)前記臓器または組織の分子輸送および排出、生体異物代謝および/または細胞状態を包含する、細胞および/または臓器機能を示す多重指標を入手するステップ、ならびに
(c)前記指標から前記臓器または組織の傷害を分析するステップ。
【0011】
本発明において、臓器または組織の傷害は、壊死、アポトーシスまたは細胞質膜の崩壊を意味する。
【0012】
生活条件下で臓器または組織に標識するために、染料(たとえば蛍光染料、ローダミンまたはフルオレセイン塩またはヨウ化プロピジウム)を使用する従来方法が利用できる。たとえばこれらの染料は、実験動物に静脈内注射または腹腔内注射によって、あるいは肝臓、腎臓、膵臓、胃腸管または肺において隔離された潅流臓器調製物中にその制御濃度での血管内潅流によって、装填できる。
【0013】
多重指標は、細胞生存能、ミトコンドリア膜電位、酸素フリーラジカル、細胞膜を渡るアニオンまたはカチオンの輸送、細胞質膜電位、ならびにアポトーシスまたは壊死を含む細胞死を意味する。多重指標の例は、これに限定されるわけではないが、生体異物代謝および細胞生存能を含む。
【0014】
生体異物代謝は、体内で分解されるあらゆる種類の外因性試薬または薬剤の代謝および/または異化を意味する。
【0015】
細胞状態は、分子輸送(たとえば細胞内および細胞周囲の分子輸送)、膜電位、ミトコンドリア機能、生物活性化合物の生成(生物活性種の生成)、血流および組織酸素添加、ならびにアニオンおよびカチオンの摂取および排出を含む生細胞のあらゆる種類の機能状態、その生存能を失いつつある細胞のあらゆる種類の機能状態、たとえば壊死またはアポトーシスを意味する。細胞状態の例は、これに限定されるわけではないが、細胞生存能および細胞傷害を含む。
【0016】
本発明の1つの実施形態において、「微細解剖配向」という用語は、細胞と、その周囲構造、たとえばリンパ系および/または結合組織(たとえば肝臓における毛細胆管系)を包含する微小血管系および間質腔との間の地理的関係を意味する。それゆえ微細解剖配向の例は、これに限定されるわけではないが肝臓における毛細胆管網目を意味する。
【0017】
「脈管系」という用語は、伝導血管、細動脈、毛細血管および小静脈を包含するマクロおよびミクロ循環系を意味する。
【0018】
「排出経路」という用語は、細胞内分子が細胞外空間に排出される経路を意味する。
【0019】
本発明において、臓器または組織傷害を視覚的または定量的に調査するために、全臓器機能パラメータの監視が利用される。「視覚的に」という用語は、生物活動の定量的情報を含む生体内画像の取り込みを意味する。「定量的に」という用語は、生物活動を示す値を決定することを意味する。全臓器機能パラメータは、これに限定されるわけではないが胆汁産出量、胆汁構成要素、たとえば胆汁酸、リン脂質、コレステロールおよび重炭酸塩の決定を含む。
【0020】
本発明の別の実施形態において、薬剤毒性を評価する方法が提供される。本発明において、以下のステップを含む:
(a)臓器または組織を染料で標識するステップ、
(b)試験薬剤を前記臓器または組織に使用するステップ、
(c)前記臓器または組織の生体異物代謝および/または細胞状態を含む多重指標を入手するステップ、
(d)前記指標から前記臓器または組織の傷害を分析するステップ、ならびに
(e)前記薬剤が前記臓器または組織に対する毒性を有するかどうかを評価するステップ。
【0021】
本発明において、薬剤毒性は、酵素またはトランスポータ機能の阻害を包含する、生細胞に対する試薬または薬剤のあらゆる種類の有害な作用を意味する。薬物の例は、これに限定されるわけではないが、ABCトランスポータの阻害剤(たとえばmrp2の阻害剤としてのプロベネシド、シクロスポリン、アドリアマイシン、シスプラチン、シメチジンなど)を含む。
【0022】
「試験薬剤を使用する」という用語は、臓器または組織に対し目的の薬剤を接触させることを意味する。臓器または組織傷害をある指標から分析することによって薬剤毒性を評価できる。たとえばいずれかのABCトランスポータの機能に関する指標が変化する場合、薬物(たとえばABCトランスポータ)が臓器または組織に対して毒性を有すると評価される。指標が安定に維持されている場合、薬物(たとえばABCトランスポータ)が臓器または組織に対して毒性を有していないと評価される。
【0023】
本発明は、有機アニオンのMrp2依存性排出を実施する肝細胞の機能の障害が、臓器または細胞傷害、たとえば冷虚血とそれに続く短時間の再灌流によって引き起こされた移植片機能障害を示す早期事象を説明するという証拠を提供する。肝細胞でのこのような変化は微小で非壊死性であるが、全移植片のレベルでグルタチオンおよびビリルビンの細胞生成と排出との間の不均衡を引き起こすのに十分に重要である。冷虚血の期間を16時間まで延長すると、その消失および膨張によって特徴付けられる毛細胆管の機能障害を誘発し、それと同時に低温保存の期間が8時間を超えない限り、その多角形網目は無傷のまま維持された。本発明者らの知る限りでは、比較的短期間の冷虚血を受けた肝細胞のそのような非壊死機能障害がKCを包含する類洞細胞の虚血後反応によって仲介できるかどうかは未知のままである。ここで示すように、Mrp2を介して有機アニオンを放出する機能の低下は、KCを消耗させることによって完全に復元され、機能障害の機構へのこの類洞細胞の包含を示唆している。
【0024】
冷虚血8時間後の移植片におけるMrp2仲介輸送の機能障害は、毛細胆管網目の崩壊からでも、トランスポータ単独の酸化的改変からでもなく、小管膜からのこのトランスポータの細胞質再局在化から発生する。このことは、8時間冷虚血および再灌流が肝臓移植片において顕著な酸化的ストレスを一切示さないという本発明者らの以前の観察と一致している(14)。Mrp2の毛細胆管並べ替え(sorting)の決定要因であるcAMPの変化(32,40,41)は、その含有量がKCの存在とかかわりなく変化しなかったため、KC仲介機能障害においてある役割を果たしそうもない。ATPの肝細胞含有量はトランスポータ機能の別の決定要因であるが、8時間冷虚血を受けたKC消耗移植片と対照移植片との間には相違が顕著であったため、機構においては、あるとしてもわずかな役割しか果たしそうもない。KC消耗はMrp2が正常肝臓において有機アニオンを排出する能力を変化させないため、移植片におけるトランスポータ機能の変化は、移植片が冷虚血再灌流を受けない限り引き起こすことができないKCの反応から発生するように思われる。詳細な機構は未知のままであるが、この結果は、KCから放出されるプロスタノイドの主要なクラスであるTXsに関与する酵素である、TXA2シンターゼの関与を示唆する(37,38)。酵素阻害剤の予防効果がKC消耗移植片において完全に無効にされたという観察により、本発明者らは、KCが、肝細胞の細胞質中へのMrp2の内在化を引き起こすTXsの主要な供給源を構成するということを提唱した。TXA2は各種の細胞に強力な生物作用を及ぼすと考えられているが、前の研究は、TXA2の比較的安定な代謝産物であるTXB2が非リソソームプロテイナーゼを活性化することが可能であり、それによって一次培養肝細胞のブレッブ形成を引き起こすという証拠を提供した(42)。それゆえKC由来TXsが肝細胞機能を引き起こすさらなる機構が必要なはずである。
【0025】
CFの制御された量を事前負荷された移植片からの染料排除分析のために新たに開発された方法は、Mrp2の再局在化が肝細胞レベルで発生し、全移植片機能の著しい劣化を引き起こすことを明らかにした。図3に示すように、8時間保管は、あったとしても組織含有量の変化を一切示さずに、胆汁グルタチオン排出を著しく低下させた。この有機アニオンは胆汁酸とは無関係な胆汁形成の浸透圧駆動力を生じる主要な物質として作用するため、胆汁中でのその減少は、産出量の低下を引き起こすことができる。この概念は、8時間保管移植片が産出量の著しい低下を示したという本発明者らの観察とも一致する。
【0026】
この文脈において、移植片におけるBR−IXαの内因性発生と胆汁中排出との不均衡は、現在の研究において非常に興味深い。図2に示すように、対照肝臓は、前の研究と一致して、潅流20分以内に内因性BR−IXαの約75%を胆汁中に排出することができる(24)。これに対して、胆汁色素のそのような迅速な排除は16時間冷虚血移植片では生じなかった。BR−IXαの胆汁中濃度によって判定されるように(図2E)、色素の絶対量は上昇したが、非冷虚血対照移植片と比較して決して低下しなかった。循環中に放出されたBR−IXαの量は無視できるため(データは示さず)、これらの結果は、冷虚血移植片が最初の20分間の再灌流中に、胆汁中に排出するその能力から予測できる量よりも多くの量の色素を合成することを示唆している。この概念は、移植片がヘム分解のストレス誘導型酵素であるヘムオキシゲナーゼ−1を誘発するという本発明者らの観察と良好に一致している(43)。この事象は、冷虚血後移植片の抗酸化ストレス反応の観点から病態生理学的に重要である。本発明者らは最近、低用量ビリルビンが酸化ストレスを改善し、それによって冷虚血後肝臓移植片を保護することが可能であるが、その過剰の用量は明らかに有害であることを報告した(31,43)。冷虚血を受けさせた移植片において、再灌流は、肝臓ビリルビン代謝に大きく影響する2つの重要な事象である、ヘム分解の増加およびMrp2によるBR−IXαの遅延を引き起こすことができる。それゆえこれらの2つの事象の複合作用は、肝細胞を保護するのに十分なこの抗酸化剤の蓄積を引き起こし、さらにその長期効果は再灌流の後の期間に肝細胞損傷および高ビリルビン血症を引き起こす。
【0027】
KCはエイコサノイドの強力なジェネレータとして作用し、その一方肝細胞およびその膜上に発現されたATP結合カセットトランスポータは、その分解および排出をそれぞれ助ける(38,39)。これに対して、抗酸化有機アニオン、たとえばグルタチオンおよびビリルビンは、冷虚血後移植片において胆汁中へのその排出のためにMrpを共有している。それゆえエイコサノイドのKC仲介合成と肝細胞からのその除去との間の均衡は、肝細胞内および肝細胞周囲の抗酸化アニオンの再分布を決定し、それにより肝臓移植の機能的な結果に影響を及ぼす。Mrp仲介有機アニオン輸送のKC仲介改造(remodeling)は、KC産出トロンボキサンが移植片への続いての酸化ストレスに対する早期警告機構として作用できるかどうかを調査するために、グルタチオンの、そしてBR−IXαの細胞内および細胞間動力学に関する定量的情報が利用可能になるという条件で、さらなる研究に値する。
【実施例】
【0028】
以下で本発明を、以下の実施例を参照してさらに詳細に説明する。本発明の技術的範囲はこれらの実施例によって制限されないことに注目すべきである。
【0029】
材料および方法
動物の準備
本明細書に述べた実験プロトコルは、慶應義塾大学医学部の動物実験委員会によって提供された機関指針により承認された。実験用食餌および水道水を自由に摂食させたオスのウィスターラット(220〜260g、CLEA Japan、東京)およびEisai高ビリルビン血症ラット(EHBR)(220〜260g、三共(株)、東京)を、実験前24時間に渡って絶食させた。これらのラットの肝臓を生体外(ex vivo)でベースライン灌水として酸素添加Krebs-Henseleit緩衝液によって灌流し(17,18)、4℃のUniversity of Wisconsin溶液中に所望の期間だけ保存した(14)。必要な場合、本発明者らの前の研究による低温保存のために、ラットは、生体外肝臓灌流の準備の24時間前にリポソームカプセル化ジクロロメチレンジホスホナート(LDD)の静脈内注射で前処理した(14,19)。前に述べたように、この手順は免疫組織化学によって判定されたように、クッパー細胞をほぼ完全に排除した(20)。低温保存の後、移植片を乳酸加リンガー液40mLの経門脈注射によって静かにすすぎ、酸素添加緩衝液を用いて、タウロコール酸ナトリウムの存在または非存在下で、30μmol/Lでの一定流量(32mL/分)にて1回通過方式で灌流した(14,21)。ある実験では、トロンボキサンA2(TXA2)シンターゼの阻害剤であるOKY−046、またはシクロオキシゲナーゼの阻害剤であるインドメタシン(IM)のどちらかを、UW溶液にはもちろんのこと、すすぎ溶液にも所望の濃度で添加した(22)。
【0030】
胆汁および組織構成要素の決定
胆汁サンプルを使用して、全胆汁塩、リン脂質、還元グルタチオン(GSH)およびビリルビン−IXα(BR-IXα)の濃度を決定した(23,24)。BR−IXαは、24G7を使用して酵素結合免疫吸着測定法によって決定した(24)。このモノクローナル抗体は、上述したHO反応によって特異的に生成された最終ヘム分解生成物であるBR−IXαを認識できる(24,25)。乳酸デヒドロゲナーゼ(LDH)の活性は、上述したように測定した(17)。肝臓移植片中のアデノシン三リン酸(ATP)は、別の場所で述べたようにルシフェリン/ルシフェラーゼ法によって決定した(14,21)。移植片中の環状AMPは、酵素結合免疫吸着測定法(Biotrak(商標)system,Amersham Biosciences,バッキンガムシャー)によって決定した。
【0031】
カルボキシフルオレセインの胆汁中排出率の分析
カルボキシフルオレセイン(CF)は、Mrp2によって各種の細胞から排出された有機アニオンである(26,27)。この染料のエステル前駆物質である、CFジアセテート(CFDA)を経門脈的に肝臓内へ、50nmol/Lにて10分間に渡ってMrp2の強力な阻害剤である、1.5mmol/Lプロベネシドの存在下で前負荷した(26,28)。この試薬は肝細胞に進入可能であり、エステル分解酵素によって、胆汁中に排出されるCFに加水分解される(14,17,29)。10分間のCFDA装填の後、胆汁中へのCFの排出を引き起こすために、肝臓を、プロベネシドを含まない緩衝液によって灌流した。冷虚血群において、保存された移植片にCFDA含有緩衝液を10分間に渡ってプロベネシドの存在下で装填し、プロベネシドの除去およびその後の50分間の再灌流を続けた。これらの調製物から胆汁サンプルを収集し、96ウェル多チャネル蛍光分光光度計を使用して蛍光測定を実施するまで、深冷凍結させた。測定は、サンプルのpH値との干渉なしに510nmの蛍光を生じる染料の等吸収波長である、440nmの落射照明で実施した(26)。サンプル中のCFの濃度は、リン酸緩衝生理食塩水に溶解させたCFの既知の濃度を用いて校正した。後の結果でわかるように、CF濃度は時間と共に指数関数的に低下するように見えた。この仮定により、胆汁CF存続期間を指数関数的減衰のT1/2として決定した。それゆえこの方法は、灌流肝臓に装填されたCFの初期量に対して反応しない。
【0032】
肝細胞CF排除の生体内原位置(in situ)描出
上述のプロトコルを使用してCFを装填した肝臓移植片を、上述した生体内レーザ共焦点マイクロフルオログラフィーによって観察した(14,20,30)。後で結果に示すように、CFを、プロベネシドの存在下で肝細胞中に顕著に装填した。試薬の除去時に、染料は肝細胞からただちに排除され、毛細胆管(BC)内に排出されてハニカム網目を示し、最終的に柔組織から消失した。染料排除がMrp2の機能に依存しているかどうかを調査するために、一部の移植片を、1.5mmol/L プロベネシドを含有する緩衝液によって再灌流した。レーザ共焦マイクロフルオログラフは、増強CCDカメラ(C5810,浜松フォトニクス)およびマルチピンホールレーザ共焦プロセッサ(CSU−10,横河電機(株))を装備した倒立型顕微鏡(Diaphot 300, Nikon/Sankei)によって取り込んだ。すべてのマイクロフルオログラフは8ビットグレイレベル画像にデジタル処理した。蛍光強度を校正するために、CFの既知濃度をインビトロで調製して、画像をカメラの同一の光学パラメータの下で取り込んだ。肝細胞のグレーレベルを可変正方形窓(2x2μm2)によって、デジタル画像プロセッサを使用して測定した(18,31)。1回の実験で、対象の顕微鏡視野内で少なくとも10個の異なる肝細胞を分析した。肝臓表面で測定した蛍光強度が溶液中で測定した蛍光強度と同じであると仮定して、グレーレベルを校正線を使用して見かけのCF濃度に変換し、CFappと呼んだ。
【0033】
本発明者らは肝細胞損傷の指標としてのBC網目の構造変化を調査するために、形態計測も実施した。後の結果に示すように、正常に機能している肝細胞は周囲BC中で多角形CF重点を特徴とするが、これに対して損傷を受けた肝細胞は周囲BC網目の部分的消失によって判定される。CFによる完全なBC充填によって包囲されたそのような無傷の肝細胞の数を対象とする範囲においてカウントした。そのような評価のために1回の実験で肝臓表面の約0.05mm2を分析した。
【0034】
細胞内Mrp2分布の免疫組織化学分析
移植片の肝細胞中のMrp2を評価するために、以前の研究に従って、肝臓サンプルの固定、スライスおよびモノクローナル抗体M2III−6による染色を行った(32)。切片上の抗原をフィコエリトリン結合抗マウスIgGによって描出し、別の箇所で述べたように488nmのレーザ共焦マイクロフルオログラフィーによって観察した(20,30)。BC局在化およびMrp2の肝細胞内在化を調査するために、切片を、肝細胞接合部で発現された別のマーカーである、ZO−1に対するモノクローナル抗体によって二重免疫染色した(33)。タンパク質分布の変化を半定量的な方法で決定するために、Mrp2の単染色マイクロフルオログラフをモノクローム8ビット画像として変換した(14)。グレーレベル(1−256)を単一の肝細胞内の細胞質ドメインおよび小管ドメインの両方で測定した。単一の細胞内で各ドメインの少なくとも5つの異なる部位を選択して、細胞質強度の対応する小管強度に対する相対値を計算した。そのような測定を4つの異なる移植片中の40〜60個の肝細胞で実施して、%I−Mrp2(cyt/bc)として定義されるMrp2関連免疫反応性の細胞質強度パーセンテージのヒストグラムを作成した。この指標の上昇は、Mrp2内在化の上昇を示した。ヒストグラムは、対照移植片とTXA2合成酵素阻害剤の存在下または非存在下でKC消耗手順ありまたはなしで冷虚血を受けさせた移植片との間で比較した。
【0035】
群間での全肝臓移植片におけるMrp2発現の相違を調査するために、同じモノクローナル抗体を使用してウェスタンブロット分析を実施した。本発明者らは抗アクロレインモノクローナル抗体(5F6)によるウェスタン分析を追跡するために、抗体M2III−6によってタンパク質を免疫沈降させることによってMrp2の酸化的改変の変化も調査した(34,35)。
【0036】
統計的解析
異なる実験群間のデータの統計的有意差は、一元配置分散分析(ANOVA)およびFischerの多重比較検定によって決定した。P<0.05が有意であると見なされた。
【0037】
結果
肝臓移植片での胆汁産出量の保存時間依存性減少
肝臓移植片の生存能を試験するために、静脈灌水中の乳酸デヒドロゲナーゼの放出を細胞溶解の指標として測定した。表1に見られるように、24時間未満の冷虚血を受けさせた移植片は、乳酸デヒドロゲナーゼの顕著な上昇を示さなかった。
【表1】

データは、開始(0分)および再灌流の開始後60分(0〜24時間;n=5、48時間;n=3)の移植片からの測定値(mIU/分/g肝臓)の平均±SEを表す。他の群の値と比較して*P<0.05。
【0038】
48時間では、乳酸デヒドロゲナーゼ放出は60分間再灌流の開始および終了時に明らかとなり、現在の実験条件下で24時間未満の冷虚血を受けた移植片では壊死性細胞死が検知されないことを示した。図1は、各種の長さの冷虚血を受けた移植片における、再灌流時間の関数としての胆汁産出量の経時変化を示す。タウロコール酸ナトリウムを添加したパネルAで見られるように、8時間虚血を受けた移植片はその産出量を、30分の対照における産出量と匹敵するレベルまで上昇させたが、初期再灌流の50〜60分後にはそれを減少させた。16〜48時間の長期冷虚血を受けた移植片では、産出量のそのような減少はさらに明らかとなった。図1のパネルBは、灌水中のタウロコール酸ナトリウムの非存在下でモニターした胆汁回収の経時変化を示した。図示されているように、16時間を超えて冷虚血によって処置された群は、産出量の著しい減少を示した。
【0039】
冷虚血後肝臓のグルタチオンおよびビリルビンの胆汁排出の変化
16時間及び24時間冷虚血を受けた移植片における胆汁流が著しく減少したことを観察し、本発明者らは胆汁うっ滞変化の原因である胆汁構成成分を決定することができた。図2は、冷虚血に対する保存時間の関数としてプロットした、再灌流開始20分後に測定した胆汁構成成分のデータを示す。図示されているように、胆汁塩の濃度はどの長さの保存時間でも有意な低下を示さなかったが、これに対してリン脂質は、8から24時間冷虚血を受けた群において濃度と流量の両方で顕著な低下を示した(図2Aおよび2B)。リン脂質が主に肝細胞から胆汁コンパートメントに排出されることを考慮すると、これらのデータは8時間を超えて保存した移植片での肝細胞機能障害の存在を示唆する。
【0040】
本発明者らは次に、その組織含有量をモニターしながらGSHの胆汁中排出を調査した(図2C)。GSH中の胆汁中濃度は8時間で有意に低下し、低温保存時間の関数として下降した。本発明者らは次に、GSHの肝臓含有量を調査した。図2Dに示すように、肝臓GSH含有量は20分間の再灌流の前後に変化しなかった。16時間保存した移植片において、含有量はGSHを含有するUniversity of Wisconsin液の使用の結果として明らかに増加した。しかしながら20分間の再灌流時に、GSHが循環から除去されると含有量は対照レベルまで迅速に抑制され、16時間冷虚血後の20分間の再灌流が移植片の基底GSH含有量を変化させないことを示す。これらの結果は、保存時間が16時間より短い限り、冷虚血後肝臓における胆汁GSH排出の減少が移植片でのその減少からではなく、胆汁へのその輸送の障害から生じることを示唆している。GSHはMrp2を通じて排出されるため、本発明者らは次に、同じトランスポータを通じて排出される胆汁色素である、BR−IXαの胆汁中濃度の変化を調査した。図2Eに見られるように、初期20分間の再灌流におけるBR−IXαの胆汁濃度は8時間虚血群で有意に上昇し、16時間保存群で最大となった。最後に24時間冷虚血を受けた移植片において、BR−IXαの初期濃度は突然低下した。図2Fは、移植片が胆汁中への内因性BR−IXαを排除させる機能を示す。対照移植片に示されるように、この胆汁色素の肝臓含有量は初期20分間灌流中に有意に減少した。これに対して、同じ時間の再灌流は、16時間処置移植片におけるそのような減少を引き起こさなかった。これらの結果は、16時間移植片がBR−IXαを新規に(de novo)生成する機能が、色素を胆汁中に排出するその能力を上回ることを示唆している。
【0041】
カルボキシフルオレセイン排除によるMp2機能の全体的および局所的評価
GSHおよびBR−IXαの胆汁中排出の変化は、Mrp2がこれらの有機アニオンを肝細胞から排除させる機能が長期冷虚血を受けた移植片で障害を受ける可能性を上昇させた。しかしながら、グルタチオンおよびBR−IXαの初期量が群間で異なったため、これらの内因性アニオンの胆汁中排出の測定によって、本発明者らは移植片の有機アニオン排出機能の公正な比較を行うことができなかった。この問題を克服するために、移植片に外因性有機アニオンであるCFを装填して、肝細胞から胆汁中へのその排除を調査した。図3Aの左パネルに見られるように、肝細胞CF装填は、異なる長さ(0〜24時間)の冷虚血を受けた移植片間で比較可能に思われた。このことは図3Cで蛍光強度計によって確認され、肝細胞が生存していたことを示している。このことは、LDHの顕著な放出を示さない結果とも一致していた(表1)。プロベネシド除去の直後に、肝細胞に装填されたMrp2の阻害剤、CFは毛細胆管中に迅速に排出されて、10分以内に小葉上にハニカム網目を形成した(図3Aの中間列)。25分(右列)にて、細胞質内でわずかな蛍光があるとすれば、検出されるようになった。冷虚血再灌流を受けた移植片において、胆汁CF排出の2つの大きな変化が起こった。25分での基底蛍光の上昇によって判定された肝細胞染料排除の遅延、および10分にて収集されたマイクログラフに示された毛細胆管網目の消失および変形である。これらの変化は、24時間の長い冷虚血を受けた移植片において明らかとなった(図3Aの下列)。
【0042】
10分にて取り込まれたこれらのマイクロフルオログラフにおける毛細胆管部位の慎重な走査により、小管網目の構造の最小限ではあるが顕著な変化を示した。図3Bに見られるように、CF充填BCによって完全に包囲された肝細胞の数は、冷虚血の期間が長くなるにつれて減少した。そのような多角形の減少は、16時間保管した移植片においてただちに明らかとなった。肝細胞での初期CF装填が0〜24時間の虚血期間の範囲で匹敵していたため、BCの形態的変化は16時間冷虚血で最初に発生するように思われた。言い換えれば、8時間冷虚血を受けた移植片は、BC網目の形態の有意な変化を示さなかった。
【0043】
CF排除の遅延は、移植片の肝細胞における蛍光の時間的変化をモニターすることによって定量的な方向でさらに調査した(図4A)。図示したようにプロベネシドを連続的に灌流させたときに、染料は細胞中にとどまり、小管排出を示さずに、わずかな低下を示した。図4Bにプロットしたように、肝細胞で測定したグレーレベルによって、本発明者らは肝細胞からのCF排除のT1/2を決定した。図4Cは、群間のT1/2値を示す。プロベネシドの非存在下ではT1/2は約6分であったが、これに対してプロベネシドの存在下では、崩壊は実質的に遅延されて、Mrp2輸送のほぼ完全な阻害を示唆した。図示したように、CFを排出する8時間冷虚血移植片のT1/2はさらに著しく大きく、対照値とEHBRの肝臓で測定した値の中間に渡っていた。プロベネシド処置群と比較した、この変異種のより小さいT1/2値は、Mrp3による染料の類洞空間への補償排出によるものであると推測できる(36)。
【0044】
8時間冷虚血移植片における胆汁CF排出に対するKC消耗の効果
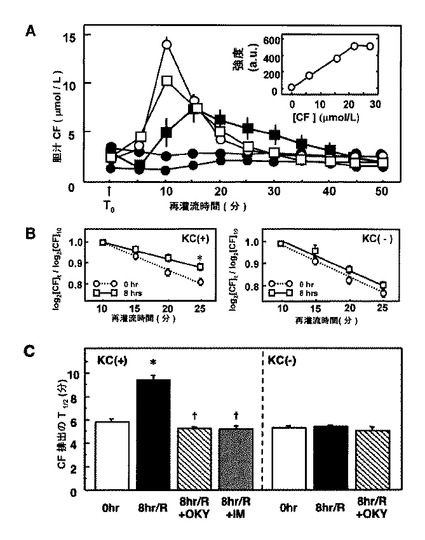
本発明者らは次に、8時間冷虚血移植片の全体としての、CFを排出する機能の評価を試みた。このために、染料排除のT1/2値を決定した(図5)。プロベネシドの除去後(図5AのT0)、胆汁のCF濃度は一時的に上昇して、徐々に基底レベルに戻った。そのような一時的上昇は、プロベネシドの存在下(陰丸)、またはEHBRから単離された移植片(黒丸)のどちらでも観察されず、Mrp2が胆汁CF排出の原因であることを示唆する。8〜16時間保存された全肝臓移植片でCF排除を分析したときに、崩壊は対照においてよりもゆっくりであるように思われた。8時間虚血移植片から収集したデータを使用して、胆汁サンプル中のCF濃度対ピーク時(10分)のCF濃度の対数値を、再灌流時間の関数として再プロットした(図5B)。次に染料排除のT1/2値をKC消耗によって処置した、または処置しなかった移植片間で比較した。図示されているように、LDDによる未処置の肝臓[KC(+)]において、8時間冷虚血は対照よりも長いT1/2値を示した。2つの群間のT1/2のそのような相違は、KC消耗移植片では完全に消失した。注目すべきなのは、非冷虚血対照肝臓においては、KC消耗手順は単独でT1/2値を変化させず、移植片が冷虚血再灌流を受けたときのみに明らかになると示されたことである。
【0045】
8時間冷虚血移植片においてKCがmrp2による長期CF排出を引き起こす機構を解明するために、本発明者らは、冷虚血後条件下で活性化されたKCより放出される主要なプロスタノイドであるトロンボキサンの関与を調査した(37,38)。このために、TXA2合成酵素の阻害剤であるOKY−046の効果を調査した。図5Cに見られるように、保存液およびすすぎ液への240μmol/Lでのこの試薬の使用は、CF排除のT1/2値をほぼ完全に上昇させる。TXA2合成酵素阻害剤は他のプロスタノイド(たとえばPGE2およびPGF2αを合成するための、アラキドン酸の有効性を上昇させるため、本発明者らは、TXsを含むシクロオキシゲナーゼ経路を介して生成されるすべてのプロスタノイドを抑制する阻害剤であるインドメタシン(IM)が、T1/2値を短縮または増大させるかどうかを調査した。図示したように、この阻害剤も長いT1/2値をほぼ完全に短縮した。さらに8時間冷虚血移植片に対するOKY−046の効果は、KCが消耗されたときに消失し、TXsの関与がKC依存性であることを示唆した。
【0046】
8時間冷虚血によるMrp2の細胞内再局在化およびKC消耗によるその減衰
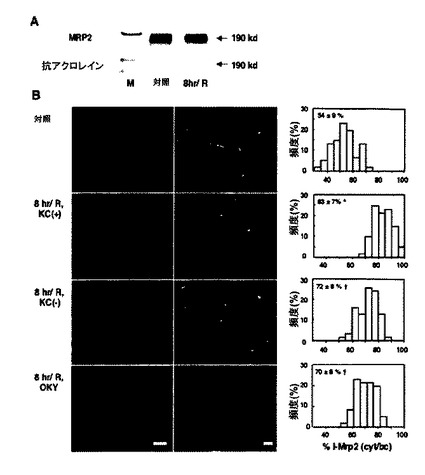
KCが内毒素処置肝臓においてMrp2をダウンレギュレートすることが既知であるため(39)、本発明者らはトランスポータの機能障害の機構にそのような変化が関与するかどうかを調査した。ウェスタンブロット分析によって示されるように、Mrp2タンパク質の量は24時間虚血移植片(データを示さず)と同様に、8時間冷虚血移植片(図6A)において不変であった。本発明者らは、タンパク質が単独で、虚血後酸化的損傷の結果として酸化的改変されたかどうかも調査した。しかしながら、抗アクロレイン抗体を使用した免疫沈降で判定したように、明らかな変化は見られなかった。次にタンパク質の肝細胞局在化が8時間冷虚血移植片において改変されたかどうかを調査した。図6Bに見られるように、BCでのその局在化は顕著に減少したが、これに対して、肝細胞の細胞質におけるバックグラウンド蛍光は8時間虚血再灌流移植片において上昇した。下のパネルに見られるように、ZO−1による二重免疫染色(緑)は、BC中のMrp2の同時局在化(赤)が顕著に妨害されたのに対して、細胞質におけるその強度が上昇したことを明らかにし、タンパク質の内在化を示唆した。そのような変化は、KC消耗によって、またはOXY−046によって処置された移植片では減衰された。%I−Mrp2(cyt/bc)値として、内在化の指標は、8時間冷虚血再灌流がMrp2内在化を有意に上昇させたことと、KC消耗による、またはTXA2合成酵素の遮断による処置がトランスポータの毛細胆管再局在化を改善したことを示した。
【0047】
本発明者らは、8時間冷虚血再灌流後のKC(+)およびKC(−)移植片でのATPおよびcAMPの肝臓含有量の相違を決定したが、2つの群間に顕著な有意性はなかった(それぞれATP含有量では3.4±0.9対2.9±1.1μmol/g肝臓、そしてcAMP含有量では8.9±1.0対9.2±0.5pmol/g肝臓)。これらの結果は、KC消耗によるこのATP結合タンパク質の細胞内回復の改善が、ATPおよびcAMPの組織含有量の変化によって生じなかったことを示唆している。
【0048】
文献
1. Ploeg RJ, D'Alessandro AM, Knechtle SJ, Stegall MD, Pirsch JD, Hoffmann RM, Sasaki T, et al. Risk factors for primary dysfunction after liver transplantation--a multivariate analysis. Transplantation 1993;55:807-813.
2. Deschenes M, Belle SH, Krom RA, Zetterman RK, Lake JR. Early allograft dysfunction after liver transplantation: a definition and predictors of outcome. National Institute of Diabetes and Digestive and Kidney Diseases Liver Transplantation Database. Transplantation 1998;66:302-310.
3. Porte RJ, Ploeg RJ, Hansen B, van Bockel JH, Thorogood J, Persijn GG, Hermans J, et al. Long-term graft survival after liver transplantation in the UW era: late effects of cold ischemia and primary dysfunction. European Multicentre Study Group. Transpl Int 1998;11 Suppl 1:S164-167.
4. Adam R, Bismuth H, Diamond T, Ducot B, Morino M, Astarcioglu I, Johann M, et al. Effect of extended cold ischaemia with UW solution on graft function after liver transplantation. Lancet 1992;340:1373-1376.
5. Jaeschke H. Preservation injury: mechanisms, prevention and consequences. J Hepatol 1996;25:774-780.
6. Caldwell-Kenkel JC, Currin RT, Tanaka Y, Thurman RG, Lemasters JJ. Reperfusion injury to endothelial cells following cold ischemic storage of rat livers. Hepatology 1989;10:292-299.
7. Caldwell-Kenkel JC, Currin RT, Tanaka Y, Thurman RG, Lemasters JJ. Kupffer cell activation and endothelial cell damage after storage of rat livers: effects of reperfusion. Hepatology 1991;13:83-95.
8. Lemasters JJ, Thurman RG. Reperfusion injury after liver preservation for transplantation. Annu Rev Pharmacol Toxicol 1997;37:327-338.
9. Clavien PA. Sinusoidal endothelial cell injury during hepatic preservation and reperfusion. Hepatology 1998;28:281-285.
10. Sindram D, Porte RJ, Hoffman MR, Bentley RC, Clavien PA. Platelets induce sinusoidal endothelial cell apoptosis upon reperfusion of the cold ischemic rat liver. Gastroenterology 2000;118:183-191.
11. Arai M, Mochida S, Ohno A, Fujiwara K. Blood coagulation in the hepatic sinusoids as a contributing factor in liver injury following orthotopic liver transplantation in the rat. Transplantation 1996;62:1398-1401.
12. Clavien PA, Harvey PR, Sanabria JR, Cywes R, Levy GA, Strasberg SM. Lymphocyte adherence in the reperfused rat liver: mechanisms and effects. Hepatology 1993;17:131-142.
13. Vajdova K, Smrekova R, Mislanova C, Kukan M, Lutterova M. Cold-preservation-induced sensitivity of rat hepatocyte function to rewarming injury and its prevention by short-term reperfusion. Hepatology 2000;32:289-296.
14. Kumamoto Y, Suematsu M, Shimazu M, Kato Y, Sano T, Makino N, Hirano KI, et al. Kupffer cell-independent acute hepatocellular oxidative stress and decreased bile formation in post-cold-ischemic rat liver. Hepatology 1999;30:1454-1463.
15. Imamura H, Brault A, Huet PM. Effects of extended cold preservation and transplantation on the rat liver microcirculation. Hepatology 1997;25:664-671.
16. Williams JW, Vera S, Peters TG, Van Voorst S, Britt LG, Dean PJ, Haggitt R, et al. Cholestatic jaundice after hepatic transplantation. A nonimmunologically mediated event. Am J Surg 1986;151:65-70.
17. Suzuki H, Suematsu M, Ishii H, Kato S, Miki H, Mori M, Ishimura Y, et al. Prostaglandin E1 abrogates early reductive stress and zone-specific paradoxical oxidative injury in hypoperfused rat liver. J Clin Invest 1994;93:155-164.
18. Suematsu M, Goda N, Sano T, Kashiwagi S, Egawa T, Shinoda Y, Ishimura Y. Carbon monoxide: an endogenous modulator of sinusoidal tone in the perfused rat liver. J Clin Invest 1995;96:2431-2437.
19. Kyokane T, Norimizu S, Taniai H, Yamaguchi T, Takeoka S, Tsuchida E, Naito M, et al. Carbon monoxide from heme catabolism protects against hepatobiliary dysfunction in endotoxin-treated rat liver. Gastroenterology 2001;120:1227-1240.
20. Goda N, Suzuki K, Naito M, Takeoka S, Tsuchida E, Ishimura Y, Tamatani T, et al. Distribution of heme oxygenase isoforms in rat liver. Topographic basis for carbon monoxide-mediated microvascular relaxation. J Clin Invest 1998;101:604-612.
21. Shiomi M, Wakabayashi Y, Sano T, Shinoda Y, Nimura Y, Ishimura Y, Suematsu M. Nitric oxide suppression reversibly attenuates mitochondrial dysfunction and cholestasis in endotoxemic rat liver. Hepatology 1998;27:108-115.
22. Ishiguro S, Arii S, Monden K, Fujita S, Nakamura T, Niwano M, Harada T, et al. Involvement of thromboxane A2-thromboxane A2 receptor system of the hepatic sinusoid in pathogenesis of cold preservation/reperfusion injury in the rat liver graft. Transplantation 1995; 59: 957-961.
23. Jaeschke H, Smith CV, Mitchell JR. Reactive oxygen species during ischemia-reflow injury in isolated perfused rat liver. J Clin Invest 1988;81:1240-1246.
24. Yamaguchi T, Wakabayashi Y, Tanaka M, Sano T, Ishikawa H, Nakajima H, Suematsu M, et al. Taurocholate induces directional excretion of bilirubin into bile in perfused rat liver. Am J Physiol 1996;270:G1028-1032.
25. Izumi Y, Yamazaki M, Shimizu S, Shimizu K, Yamaguchi T, Nakajima H. Anti-bilirubin monoclonal antibody. II. Enzyme-linked immunosorbent assay for bilirubin fractions by combination of two monoclonal antibodies. Biochim Biophys Acta 1988;967:261-266.
26. Mor-Cohen R, Zivelin A, Rosenberg N, Shani M, Muallem S, Seligsohn U. Identification and functional analysis of two novel mutations in the multidrug resistance protein 2 gene in Israeli patients with Dubin-Johnson syndrome. J Biol Chem 2001;276:36923-36930.
27. van der Kolk DM, de Vries EG, Noordhoek L, van den Berg E, van der Pol MA, Muller M, Vellenga E. Activity and expression of the multidrug resistance proteins P-glycoprotein, MRP1, MRP2, MRP3 and MRP5 in de novo and relapsed acute myeloid leukemia. Leukemia 2001;15:1544-1553.
28. Courtois A, Payen L, Lagadic D, Guillouzo A, Fardel O. Evidence for a multidrug resistance-associated protein 1 (MRP1)-related transport system in cultured rat liver biliary epithelial cells. Life Sci 1999;64:763-774.
29. Suematsu M, Suzuki H, Ishii H, Kato S, Yanagisawa T, Asako H, Suzuki M, et al. Early midzonal oxidative stress preceding cell death in hypoperfused rat liver. Gastroenterology 1992;103:994-1001.
30. Kajimura M, Shimoyama M, Tsuyama S, Suzuki T, Kozaki S, Takenaka S, Tsubota K, et al. Visualization of gaseous monoxide reception by soluble guanylate cyclase in the rat retina. Faseb J 2003;17:506-508.
31. Hayashi S, Takamiya R, Yamaguchi T, Matsumoto K, Tojo SJ, Tamatani T, Kitajima M, et al. Induction of heme oxygenase-1 suppresses venular leukocyte adhesion elicited by oxidative stress: role of bilirubin generated by the enzyme. Circ Res 1999;85:663-671.
32. Mottino AD, Cao J, Veggi LM, Crocenzi F, Roma MG, Vore M. Altered localization and activity of canalicular Mrp2 in estradiol-17beta-D-glucuronide-induced cholestasis. Hepatology 2002;35:1409-1419.
33. Kubitz R, D'Urso D, Keppler D, Haussinger D. Osmodependent dynamic localization of the multidrug resistance protein 2 in the rat hepatocyte canalicular membrane. Gastroenterology 1997;113:1438-1442.
34. Uchida K, Kanematsu M, Sakai K, Matsuda T, Hattori N, Mizuno Y, Suzuki D, et al. Protein-bound acrolein: potential markers for oxidative stress. Proc Natl Acad Sci U S A 1998;95:4882-4887.
35. Tanaka N, Tajima S, Ishibashi A, Uchida K, Shigematsu T. Immunohistochemical detection of lipid peroxidation products, protein-bound acrolein and 4-hydroxynonenal protein adducts, in actinic elastosis of photodamaged skin. Arch Dermatol Res 2001;293:363-367.
36. Ogawa K, Suzuki H, Hirohashi T, Ishikawa T, Meier PJ, Hirose K, Akizawa T, et al. Characterization of inducible nature of MRP3 in rat liver. Am J Physiol Gastrointest Liver Physiol 2000;278:G438-446.
37. Gyenes M, and De Groot H. Prostanoid release by Kupffer cells upon hypoxia-reoxygenation: role of pHi and Ca2+. Am J Physiol 1993; 264: G535-G540.
38. Johnston DE, Peterson MB, Mion F, Berninger RW, Jefferson DM. Synthesis and degradation of eicosanoids in primary rat hepatocyte cultures. Prostaglandins Leukot Essent Fatty Acids 1991; 43: 119-132.
39. Nakamura J, Nishida T, Hayashi K, Kawada N, Ueshima S, Sugiyama Y, Ito T, et al. Kupffer cell-mediated down regulation of rat hepatic CMOAT/MRP2 gene expression. Biochem Biophys Res Commun 1999;255:143-149.
40. Roelofsen H, Soroka CJ, Keppler D, Boyer JL. Cyclic AMP stimulates sorting of the canalicular organic anion transporter (Mrp2/cMoat) to the apical domain in hepatocyte couplets. J Cell Sci 1998;111 ( Pt 8):1137-1145.
41. Kipp H, Arias IM. Intracellular trafficking and regulation of canalicular ATP-binding cassette transporters. Semin Liver Dis 2000;20:339-351.
42. Horton AA,Wood JM. Prevention of thromboxane B2-induced hepatocyte plasma membrane bleb formation by certain prostaglandins and a proteinase inhibitor. Biochim Biophys Acta 1990;1022:319-324
43. Kato Y, Shimazu M, Kondo M, Uchida K, Kumamoto Y, Wakabayashi G, Kitajima M, et al. Bilirubin rinse: A simple protectant against the rat liver graft injury mimicking heme oxygenase-1 preconditioning. Hepatology 2003;38:364-373.
【0049】
すべての文書は、あらゆる目的のために参照により本明細書に組み入れられている。
【産業上の利用可能性】
【0050】
本発明は、その臓器毒性のために以前に不合格になった臨床前試薬の復活に有用である。全体的に、日常的な臨床化学に対して肝臓毒性を示す化合物の毒性の詳細な機構は、大半は未知のままである。そのような化合物を本発明によって詳細に調査し、細胞生存能に影響を及ぼさずにトランスポータ機能のみを遮断することが判明した場合、臨床用途に利用するために化学構造のわずかな改変が有効である。(2)ヒト癌細胞を移植した免疫不全マウスへの本方法の利用によって、本発明者らは、薬物送達の種類の特異性および生体内での腫瘍細胞凝集体または無傷の組織への蓄積について調査できた。トランスポータ、たとえばMrp2の機能は腫瘍の薬物耐性の重要な決定要因であるため、そのような利用は、生体内での移植癌における抗癌試薬の送達および排除を調査するために極めて有用である。抗癌および抗血栓試薬の潜在性をスクリーニングするための臨床前実験の利用。
【図面の簡単な説明】
【0051】
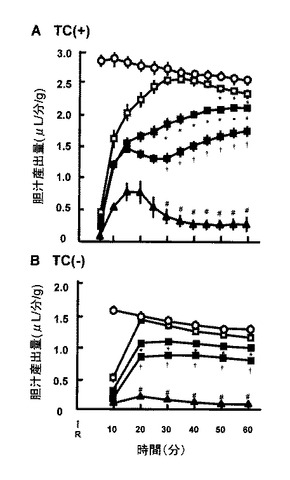
【図1】低温保存、続いて再灌流を受けた肝臓移植片の胆汁産出量の経時変化を示す。白丸は非虚血対照肝臓からのデータを示す。白四角、陰四角および黒四角はそれぞれ、8、16、24時間低温保存を受けた移植片からのデータを示す。黒三角は、48時間保存移植片からのデータを示す。値は、5回の独立した実験の平均±SEである。TC(+)およびTC(−):タウロコール酸ナトリウム 30μmol/Lの存在下または非存在下で収集したデータ。R:再灌流の開始。胆汁塩非依存性および依存性産出量のどちらも、16時間保存移植片で減少したことに注目する。対照からのデータと比較して*P<0.05。16時間移植片からのデータと比較して†P<0.05。24時間移植片からのデータと比較して#P<0.05。
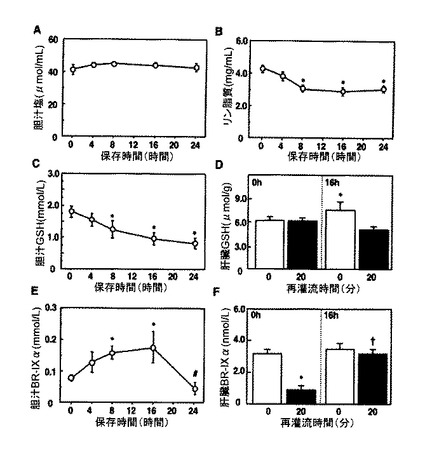
【図2】胆汁構成要素の胆汁中濃度に対する冷虚血の期間の影響を示す。胆汁構成要素のデータは、再灌流開始20分後に収集した。A:胆汁塩(パネルA)。B:リン脂質。C:胆汁中の還元グルタチオン(GSH)の濃度。D:再灌流前および再灌流20分後に測定した対照および16時間保存移植片におけるGSHの肝臓含有量。E:胆汁中のビリルビン(BR)−IXαの濃度。F:再灌流前および再灌流20分後に測定した対照および16時間保存移植片におけるBR−IXαの肝臓含有量。値は、5〜7回の独立した実験の平均±SEである。対照肝臓からのデータと比較して*P<0.05。16時間群からのデータと比較して#P<0.05。20分再灌流を受けた対照移植片からのデータと比較して†P<0.05。
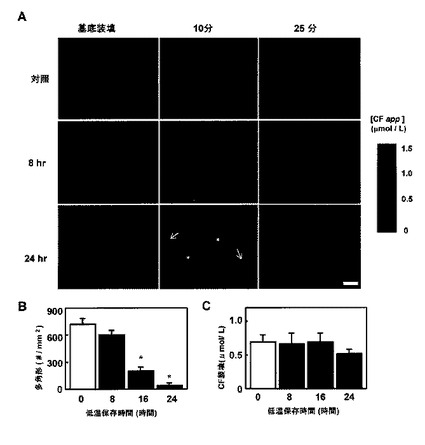
【図3】冷虚血再灌流移植片中の毛細胆管への肝細胞カルボキシフルオレセイン(CF)排出の動態の変化を示す。A:プロベネシド除去前(基底装填)、除去10分後および25分後に取り込まれた小管CF排出の代表的な画像。24時間冷虚血再灌流移植片における毛細胆管網目のハニカムパターンの崩壊に注目する(矢印)。色付きバーは、CFの既知濃度を用いて校正した蛍光強度を示す。バー:30μm。B:各種の冷虚血期間に渡って保管した移植片中のCF充填多角形の密度によって判定された、毛細胆管網目の再灌流誘発崩壊の相違。対照肝臓からのデータと比較して*P<0.05。C:群間の比較CF装填量を示す初期肝細胞CF濃度。値は、5回の独立した実験の平均±SEである。
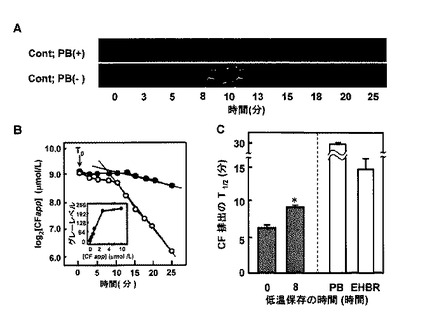
【図4】カルボキシフルオレセイン(CF)の毛細胆管排出を描出することによる、Mrp2機能の生体内定量分析を示す。A:各肝細胞からの染料排出を示す代表的な一連の画像。上;1.5mM プロベネシドの存在下で灌流された移植片からの画像(PB(+))。下、プロベネシド除去後に取り込まれた画像(PB(−))。細胞中の蛍光の時間依存性還元と、それに続く周囲の毛細胆管における染料の濃縮および消失に注目する。細胞中の染料保持に注目すべきである。B:肝細胞CF蛍光の消失。プロベネシドを用いて(黒丸)、または用いずに(白丸)処置した移植片の肝細胞CF濃度を、直線が指数曲線を表すように、時間に対して片対数プロットする。差し込み図:CF濃度と8ビットグレーレベルとの間の関係を示す校正曲線。[CFapp]:CFの見かけの濃度。CF濃度は、3μmol/Lよりも低い濃度でグレーレベルと直線的に関係付けられている(r2=0.996、p<0.05)。C:肝細胞からのCF排除の半減期(T1/2)の相違。値は、各群での5回の独立した実験の平均±SEである。対照肝臓からのデータと比較して*P<0.05。EHBR:Eisai高ビリルビン血症ラットから単離した移植片。PB:1.5mM プロベネシドを用いて灌流した移植片。
【図5】肝臓移植片が冷虚血再灌流時にカルボキシフルオレセイン(CF)を胆汁中に排出する能力の変化を示す。A:各種の長さの冷虚血を受けた移植片における胆汁CF排出の経時変化の相違。白丸:Mrp2阻害剤である1.5mM プロベネシドを除去した直後に灌流させた対照移植片。白四角および陰四角:8および24時間低温保存と、その後のプロベネシドの非存在下での再灌流をそれぞれ受けた移植片。陰丸:プロベネシドの存在下で正常灌流された移植片。黒丸:Eisai高ビリルビン血症ラットから単離された、正常灌流された移植片。値は、5回の独立した実験の平均±SEである。T0:プロベネシドが灌流液から除去された時間。差し込み図:CF濃度と蛍光強度との間の直線関係。B:各種の再灌流期間で収集された胆汁中の相対CF濃度の変化、およびクッパー細胞(KC)の欠乏の効果。左:対照(0時間、破線)と8時間保存肝臓移植片(8時間、実線)との間の胆汁CF排出の減少の相違。右:静脈内リポソームカプセル化ジクロロメチレンジホスホナート(LDD)によるKC消耗の効果。白丸:対照(0時間、破線)。白四角:8時間保存移植片(8時間、実線)。データは、4回の独立した実験による測定値の平均±SEを表す。対照肝臓におけるCF排除の減少と比較して*P<0.05。C:8時間低温保存肝臓におけるT1/2値の延長に対する、LDDおよび/またはトロンボキサンA2合成酵素の阻害剤であるOKY−046(OKY)を用いた処置によるKC消耗[KC(−)]の効果。IM:インドメタシン。保存およびすすぎ溶液中のOKYおよびIMの濃度はそれぞれ、240および28μmol/Lであった。OKYの阻害作用は、KC消耗移植片で消失することに注目する。値は、5〜6回の独立した実験の平均±SEである。対照肝臓からのデータと比較して*P<0.05。8時間保管移植片におけるデータと比較して†P<0.05。
【図6】8時間冷虚血および60分間再灌流(8時間/R)を受けた肝臓移植片におけるMrp2の細胞内分布の崩壊、およびクッパー細胞(KC)欠乏手順[KC(−)]またはOKY−046(OKY)を用いた処置の効果を示す。A:抗アクロレインモノクローナル抗体5F6によるMrp2のウェスタンブロット解析および免疫沈降。M:分子マーカー。B:Mrp2分布の免疫蛍光分析。左;フィコエリトリンによって標識された抗Mrp2モノクローナル抗体(M2III−6)を用いた単染色。バー:30μm。右;FITC標識ZO−1抗体およびフィコエリトリン標識M2III−6抗体を用いた二重免疫染色。バー:10μm。C.肝細胞Mrp2局在化の半定量分析。%I−Mrp2(cyt/bc);毛細胆管にて測定されたものに対する、Mrp2関連免疫反応性の細胞質強度。値は、40〜60個の肝細胞/4個の独立した肝臓からの移植片における測定値の平均±SEである。対照肝臓からのデータと比較して*P<0.05。8時間/R−KC(+)群から収集したデータと比較して†P<0.05。
【技術分野】
【0001】
本発明は、臓器機能を評価する方法に関する。さらに詳細には、本発明は、微細解剖配向血管系および/または排泄経路を用いて多重指標を視覚的に分析するための、および/または定量的に決定するための方法に関する。
【背景技術】
【0002】
University of Wisconsin(UW)液の使用は、肝臓移植のための平均保存時間を改善したが、初期移植片機能不全および初期の低機能の発生はなお残っている(1−3)。そのような機能障害の臨床的発生および結果としての移植片生存の損失は、保存時間によって変わる(1−4)。再灌流傷害は、長期冷虚血後の移植片機能不全を引き起こす主要な事象である(5−9)。保存の間、肝細胞は膨潤して、ブレッブを形成する(6−8)。しかしながら再灌流時に、実質細胞におけるこれらの変化は、回復不能な傷害に至ることなく修復された(6−8)。これに対して、類洞内皮細胞は生存能を失い、クッパー細胞は活性化される(6−10)。UW溶液中に保存したラット肝臓移植片を使用した以前の研究によると、類洞細胞の変化が起こるときの臨界保存時間は16時間より長い(6,7)。これらの移植片において、細胞は損傷されて、血小板捕捉(10)、フィブリン沈着(11)、および白血球辺縁趨向(12)を引き起こす。より短い期間に渡って保存された移植片では、肝臓ATP含有率は再灌流後に十分回復することが報告され、実質細胞には生存能力があることを示唆している(13,14)。
【0003】
これらのデータとは別に、肝細胞が、回復不能な傷害を示すことなく、次に全体としての移植片機能を妨害することなく、その機能を失うかどうかは、慎重に調査されていない。胆汁産出量、タウロコール酸およびブロモスルホフタレインのクリアランスは以前の研究で測定されたが(13−15)、保存時間が18時間に延びたときでさえ、それらは肝細胞におけるそのような機能変化を証明できなかった(13)。これらの研究以外には、冷虚血後移植片が胆汁構成要素を分泌する能力の変化に関して、情報はほとんど入手できていない。そのような指標は、肝細胞が胆汁形成のための浸透圧駆動力を生む、そして胆汁塩または有機アニオンを排出する能力を含む。それゆえグルタチオンおよびビリルビン排出は、移植片における早期肝細胞変化を検知するマーカーとして作用できる。有機アニオンを排出する能力は、移植片が生体異物を解毒する効率および臨床移植における同種移植機能障害の危険因子である、冷虚血後高ビリルビン血症の重症度を判定することができる(2,16)。
【発明の開示】
【発明が解決しようとする課題】
【0004】
本発明の目的は、臓器機能を評価する方法を提供することである。上記知見に基づいて、本発明者はさらなる調査を継続した。
【課題を解決するための手段】
【0005】
冷虚血後移植片のそのような能力を判定する試みにおいて、本発明者らは本明細書で胆汁サンプルの構成要素の変化を保存時間の関数として調査し、肝細胞での早期事象としてグルタチオンおよびビリルビンの排出障害を明らかにした。本事象は、有機アニオンの胆汁中排出のためのATP依存性トランスポータである、多剤耐性関連タンパク質2(Mrp2)の細胞質再局在化の結果であることがわかった。本発明者らの結果は、このトランスポータの機能が損なわれるが、これに対して移植片は顕著な肝細胞損傷を示すことなく、その全体のエネルギー電荷を明らかに維持することを示唆している。さらに肝細胞中のそのような変化の機構は、むしろより短期間の、すなわち8時間の冷虚血にさらされた移植片中のクッパー細胞におけるトロンボキサン合成に関与するように思われる。
【0006】
本出願は以下の発明を提供する。
(1)臓器または組織傷害を分析する方法であって、以下のステップ:
(a)臓器または組織を染料で標識するステップ、
(b)前記臓器または組織の生体異物代謝および/または細胞状態を含む多重指標を入手するステップ、ならびに
(c)前記指標から前記臓器または組織の傷害を分析するステップ、
を含む方法。
(2)前記臓器または組織が肝臓、腎臓、肺、膵臓および胃腸管から成る群より選択される少なくとも1つである、(1)に記載の方法。
(3)前記ステップ(b)が血管系および/または排出経路の微細解剖配向を得るステップをさらに含む、(1)または(2)に記載の方法。
(4)前記解析が視覚的におよび/または定量的に実施される、(1)〜(3)のいずれか一項に記載の方法。
(5)前記細胞状態が細胞生存能、細胞傷害、分子輸送、およびミトコンドリア機能から成る群より選択される少なくとも1つである、(1)〜(4)のいずれか一項に記載の方法。
(6)薬剤毒性を評価する方法であって、以下のステップ:
(a)臓器または組織を染料で標識するステップ、
(b)試験薬剤を前記臓器または組織に使用するステップ、
(c)前記臓器または組織の生体異物代謝および/または細胞状態を含む多重指標を入手するステップ、
(d)前記指標から前記臓器または組織の傷害を分析するステップ、ならびに
(e)前記薬剤が前記臓器または組織に対する毒性を有するかどうかを評価するステップ、
を含む方法。
(7)前記臓器または組織が肝臓、腎臓、肺、膵臓および胃腸管から成る群より選択される少なくとも1つである、(6)に記載の方法。
(8)前記ステップ(b)が、血管系および/または排出経路の微細解剖配向を得るステップをさらに含む、(6)または(7)に記載の方法。
(9)前記解析が視覚的におよび/または定量的に実施される、(6)〜(8)のいずれか一項に記載の方法。
(10)前記細胞状態が細胞生存能、細胞傷害、細胞内および細胞周囲での分子の輸送、および生物活性化合物の生成、血流、および組織酸素添加から成る群より選択される少なくとも1つである、(6)〜(9)のいずれか一項に記載の方法。
【発明を実施するための最良の形態】
【0007】
以下、本発明をさらに詳細に説明する。
【0008】
クッパー細胞(KC)は冷虚血後肝細胞傷害において重大な役割を果たしていると考えられているが、非壊死移植片機能障害におけるその役割は未知のままである。本研究は、冷虚血後肝臓移植片におけるKCのそのような役割を解明することを狙っている。KC消耗試薬であるリポソームカプセル化ジクロロメチレンジホスホナートを用いて、または用いずに処理したラット肝臓は、4℃のUniversity of Wisconsin溶液中で8〜24時間保存し、胆汁産出量および構成要素の監視下で再灌流させた。肝細胞が胆汁を排出する能力は、生体内原位置(in situ)でレーザ共焦点マイクロフルオログラフィーによって評価した。冷虚血再灌流移植片は、顕著な細胞傷害なしに8時間にてその胆汁産出量を著しく減少させた。この事象は、グルタチオンおよびビリルビン−IXαの排出障害と一致し、これらの有機アニオンの遅延輸送を示唆している。機構は、多剤耐性関連タンパク質2(Mrp2)の細胞内再局在化を含んでいた。このトランスポータを通じて排出される蛍光プローブである、カルボキシフルオレセインの胆汁中排出の動態解析は、肝細胞から毛細胆管内への染料排出の著しい遅延を明らかにした。KC消耗処理は、細胞質から小管膜へのMrp2の再分布によって、8時間冷虚血移植片中のMrp2によって仲介される胆汁アニオン輸送のこのような低下を著しく減衰させた。さらに機構は、CF排出およびMrp2の細胞質内在化に対するこの酵素を阻害する改善効果がKC消耗移植片において消失したため、KC中にトロンボキサンA2シンターゼを包含するように思われた。要するに、これらの結果は、KC活性化が冷虚血後移植片の解毒能力および有機アニオン代謝の恒常性を脅かす、非壊死肝細胞機能障害の重要な決定要因であることを示唆している。
【0009】
本明細書において、一部の省略形が使用されている:BC、毛細胆管;BR−IXα、ビリルビン−IXα;CF、5−カルボキシフルオレセイン;CFDA、5−カルボキシフルオレセインジアセテート;EHBR、エイザイ高ビリルビン血症ラット;GSH、還元グルタチオン;KC、クッパー細胞;LDD、リポソームカプセル化ジクロロメチレンジホスホナート;Mrp2、多剤耐性関連タンパク質2;TXA2、トロンボキサンA2。
【0010】
本発明の1つの実施形態において、臓器または組織の機能障害および/または傷害を分析する方法が提供される。本方法は以下のステップを含む:
(a)臓器または組織を1つまたは2つの染料で標識するステップ、
(b)前記臓器または組織の分子輸送および排出、生体異物代謝および/または細胞状態を包含する、細胞および/または臓器機能を示す多重指標を入手するステップ、ならびに
(c)前記指標から前記臓器または組織の傷害を分析するステップ。
【0011】
本発明において、臓器または組織の傷害は、壊死、アポトーシスまたは細胞質膜の崩壊を意味する。
【0012】
生活条件下で臓器または組織に標識するために、染料(たとえば蛍光染料、ローダミンまたはフルオレセイン塩またはヨウ化プロピジウム)を使用する従来方法が利用できる。たとえばこれらの染料は、実験動物に静脈内注射または腹腔内注射によって、あるいは肝臓、腎臓、膵臓、胃腸管または肺において隔離された潅流臓器調製物中にその制御濃度での血管内潅流によって、装填できる。
【0013】
多重指標は、細胞生存能、ミトコンドリア膜電位、酸素フリーラジカル、細胞膜を渡るアニオンまたはカチオンの輸送、細胞質膜電位、ならびにアポトーシスまたは壊死を含む細胞死を意味する。多重指標の例は、これに限定されるわけではないが、生体異物代謝および細胞生存能を含む。
【0014】
生体異物代謝は、体内で分解されるあらゆる種類の外因性試薬または薬剤の代謝および/または異化を意味する。
【0015】
細胞状態は、分子輸送(たとえば細胞内および細胞周囲の分子輸送)、膜電位、ミトコンドリア機能、生物活性化合物の生成(生物活性種の生成)、血流および組織酸素添加、ならびにアニオンおよびカチオンの摂取および排出を含む生細胞のあらゆる種類の機能状態、その生存能を失いつつある細胞のあらゆる種類の機能状態、たとえば壊死またはアポトーシスを意味する。細胞状態の例は、これに限定されるわけではないが、細胞生存能および細胞傷害を含む。
【0016】
本発明の1つの実施形態において、「微細解剖配向」という用語は、細胞と、その周囲構造、たとえばリンパ系および/または結合組織(たとえば肝臓における毛細胆管系)を包含する微小血管系および間質腔との間の地理的関係を意味する。それゆえ微細解剖配向の例は、これに限定されるわけではないが肝臓における毛細胆管網目を意味する。
【0017】
「脈管系」という用語は、伝導血管、細動脈、毛細血管および小静脈を包含するマクロおよびミクロ循環系を意味する。
【0018】
「排出経路」という用語は、細胞内分子が細胞外空間に排出される経路を意味する。
【0019】
本発明において、臓器または組織傷害を視覚的または定量的に調査するために、全臓器機能パラメータの監視が利用される。「視覚的に」という用語は、生物活動の定量的情報を含む生体内画像の取り込みを意味する。「定量的に」という用語は、生物活動を示す値を決定することを意味する。全臓器機能パラメータは、これに限定されるわけではないが胆汁産出量、胆汁構成要素、たとえば胆汁酸、リン脂質、コレステロールおよび重炭酸塩の決定を含む。
【0020】
本発明の別の実施形態において、薬剤毒性を評価する方法が提供される。本発明において、以下のステップを含む:
(a)臓器または組織を染料で標識するステップ、
(b)試験薬剤を前記臓器または組織に使用するステップ、
(c)前記臓器または組織の生体異物代謝および/または細胞状態を含む多重指標を入手するステップ、
(d)前記指標から前記臓器または組織の傷害を分析するステップ、ならびに
(e)前記薬剤が前記臓器または組織に対する毒性を有するかどうかを評価するステップ。
【0021】
本発明において、薬剤毒性は、酵素またはトランスポータ機能の阻害を包含する、生細胞に対する試薬または薬剤のあらゆる種類の有害な作用を意味する。薬物の例は、これに限定されるわけではないが、ABCトランスポータの阻害剤(たとえばmrp2の阻害剤としてのプロベネシド、シクロスポリン、アドリアマイシン、シスプラチン、シメチジンなど)を含む。
【0022】
「試験薬剤を使用する」という用語は、臓器または組織に対し目的の薬剤を接触させることを意味する。臓器または組織傷害をある指標から分析することによって薬剤毒性を評価できる。たとえばいずれかのABCトランスポータの機能に関する指標が変化する場合、薬物(たとえばABCトランスポータ)が臓器または組織に対して毒性を有すると評価される。指標が安定に維持されている場合、薬物(たとえばABCトランスポータ)が臓器または組織に対して毒性を有していないと評価される。
【0023】
本発明は、有機アニオンのMrp2依存性排出を実施する肝細胞の機能の障害が、臓器または細胞傷害、たとえば冷虚血とそれに続く短時間の再灌流によって引き起こされた移植片機能障害を示す早期事象を説明するという証拠を提供する。肝細胞でのこのような変化は微小で非壊死性であるが、全移植片のレベルでグルタチオンおよびビリルビンの細胞生成と排出との間の不均衡を引き起こすのに十分に重要である。冷虚血の期間を16時間まで延長すると、その消失および膨張によって特徴付けられる毛細胆管の機能障害を誘発し、それと同時に低温保存の期間が8時間を超えない限り、その多角形網目は無傷のまま維持された。本発明者らの知る限りでは、比較的短期間の冷虚血を受けた肝細胞のそのような非壊死機能障害がKCを包含する類洞細胞の虚血後反応によって仲介できるかどうかは未知のままである。ここで示すように、Mrp2を介して有機アニオンを放出する機能の低下は、KCを消耗させることによって完全に復元され、機能障害の機構へのこの類洞細胞の包含を示唆している。
【0024】
冷虚血8時間後の移植片におけるMrp2仲介輸送の機能障害は、毛細胆管網目の崩壊からでも、トランスポータ単独の酸化的改変からでもなく、小管膜からのこのトランスポータの細胞質再局在化から発生する。このことは、8時間冷虚血および再灌流が肝臓移植片において顕著な酸化的ストレスを一切示さないという本発明者らの以前の観察と一致している(14)。Mrp2の毛細胆管並べ替え(sorting)の決定要因であるcAMPの変化(32,40,41)は、その含有量がKCの存在とかかわりなく変化しなかったため、KC仲介機能障害においてある役割を果たしそうもない。ATPの肝細胞含有量はトランスポータ機能の別の決定要因であるが、8時間冷虚血を受けたKC消耗移植片と対照移植片との間には相違が顕著であったため、機構においては、あるとしてもわずかな役割しか果たしそうもない。KC消耗はMrp2が正常肝臓において有機アニオンを排出する能力を変化させないため、移植片におけるトランスポータ機能の変化は、移植片が冷虚血再灌流を受けない限り引き起こすことができないKCの反応から発生するように思われる。詳細な機構は未知のままであるが、この結果は、KCから放出されるプロスタノイドの主要なクラスであるTXsに関与する酵素である、TXA2シンターゼの関与を示唆する(37,38)。酵素阻害剤の予防効果がKC消耗移植片において完全に無効にされたという観察により、本発明者らは、KCが、肝細胞の細胞質中へのMrp2の内在化を引き起こすTXsの主要な供給源を構成するということを提唱した。TXA2は各種の細胞に強力な生物作用を及ぼすと考えられているが、前の研究は、TXA2の比較的安定な代謝産物であるTXB2が非リソソームプロテイナーゼを活性化することが可能であり、それによって一次培養肝細胞のブレッブ形成を引き起こすという証拠を提供した(42)。それゆえKC由来TXsが肝細胞機能を引き起こすさらなる機構が必要なはずである。
【0025】
CFの制御された量を事前負荷された移植片からの染料排除分析のために新たに開発された方法は、Mrp2の再局在化が肝細胞レベルで発生し、全移植片機能の著しい劣化を引き起こすことを明らかにした。図3に示すように、8時間保管は、あったとしても組織含有量の変化を一切示さずに、胆汁グルタチオン排出を著しく低下させた。この有機アニオンは胆汁酸とは無関係な胆汁形成の浸透圧駆動力を生じる主要な物質として作用するため、胆汁中でのその減少は、産出量の低下を引き起こすことができる。この概念は、8時間保管移植片が産出量の著しい低下を示したという本発明者らの観察とも一致する。
【0026】
この文脈において、移植片におけるBR−IXαの内因性発生と胆汁中排出との不均衡は、現在の研究において非常に興味深い。図2に示すように、対照肝臓は、前の研究と一致して、潅流20分以内に内因性BR−IXαの約75%を胆汁中に排出することができる(24)。これに対して、胆汁色素のそのような迅速な排除は16時間冷虚血移植片では生じなかった。BR−IXαの胆汁中濃度によって判定されるように(図2E)、色素の絶対量は上昇したが、非冷虚血対照移植片と比較して決して低下しなかった。循環中に放出されたBR−IXαの量は無視できるため(データは示さず)、これらの結果は、冷虚血移植片が最初の20分間の再灌流中に、胆汁中に排出するその能力から予測できる量よりも多くの量の色素を合成することを示唆している。この概念は、移植片がヘム分解のストレス誘導型酵素であるヘムオキシゲナーゼ−1を誘発するという本発明者らの観察と良好に一致している(43)。この事象は、冷虚血後移植片の抗酸化ストレス反応の観点から病態生理学的に重要である。本発明者らは最近、低用量ビリルビンが酸化ストレスを改善し、それによって冷虚血後肝臓移植片を保護することが可能であるが、その過剰の用量は明らかに有害であることを報告した(31,43)。冷虚血を受けさせた移植片において、再灌流は、肝臓ビリルビン代謝に大きく影響する2つの重要な事象である、ヘム分解の増加およびMrp2によるBR−IXαの遅延を引き起こすことができる。それゆえこれらの2つの事象の複合作用は、肝細胞を保護するのに十分なこの抗酸化剤の蓄積を引き起こし、さらにその長期効果は再灌流の後の期間に肝細胞損傷および高ビリルビン血症を引き起こす。
【0027】
KCはエイコサノイドの強力なジェネレータとして作用し、その一方肝細胞およびその膜上に発現されたATP結合カセットトランスポータは、その分解および排出をそれぞれ助ける(38,39)。これに対して、抗酸化有機アニオン、たとえばグルタチオンおよびビリルビンは、冷虚血後移植片において胆汁中へのその排出のためにMrpを共有している。それゆえエイコサノイドのKC仲介合成と肝細胞からのその除去との間の均衡は、肝細胞内および肝細胞周囲の抗酸化アニオンの再分布を決定し、それにより肝臓移植の機能的な結果に影響を及ぼす。Mrp仲介有機アニオン輸送のKC仲介改造(remodeling)は、KC産出トロンボキサンが移植片への続いての酸化ストレスに対する早期警告機構として作用できるかどうかを調査するために、グルタチオンの、そしてBR−IXαの細胞内および細胞間動力学に関する定量的情報が利用可能になるという条件で、さらなる研究に値する。
【実施例】
【0028】
以下で本発明を、以下の実施例を参照してさらに詳細に説明する。本発明の技術的範囲はこれらの実施例によって制限されないことに注目すべきである。
【0029】
材料および方法
動物の準備
本明細書に述べた実験プロトコルは、慶應義塾大学医学部の動物実験委員会によって提供された機関指針により承認された。実験用食餌および水道水を自由に摂食させたオスのウィスターラット(220〜260g、CLEA Japan、東京)およびEisai高ビリルビン血症ラット(EHBR)(220〜260g、三共(株)、東京)を、実験前24時間に渡って絶食させた。これらのラットの肝臓を生体外(ex vivo)でベースライン灌水として酸素添加Krebs-Henseleit緩衝液によって灌流し(17,18)、4℃のUniversity of Wisconsin溶液中に所望の期間だけ保存した(14)。必要な場合、本発明者らの前の研究による低温保存のために、ラットは、生体外肝臓灌流の準備の24時間前にリポソームカプセル化ジクロロメチレンジホスホナート(LDD)の静脈内注射で前処理した(14,19)。前に述べたように、この手順は免疫組織化学によって判定されたように、クッパー細胞をほぼ完全に排除した(20)。低温保存の後、移植片を乳酸加リンガー液40mLの経門脈注射によって静かにすすぎ、酸素添加緩衝液を用いて、タウロコール酸ナトリウムの存在または非存在下で、30μmol/Lでの一定流量(32mL/分)にて1回通過方式で灌流した(14,21)。ある実験では、トロンボキサンA2(TXA2)シンターゼの阻害剤であるOKY−046、またはシクロオキシゲナーゼの阻害剤であるインドメタシン(IM)のどちらかを、UW溶液にはもちろんのこと、すすぎ溶液にも所望の濃度で添加した(22)。
【0030】
胆汁および組織構成要素の決定
胆汁サンプルを使用して、全胆汁塩、リン脂質、還元グルタチオン(GSH)およびビリルビン−IXα(BR-IXα)の濃度を決定した(23,24)。BR−IXαは、24G7を使用して酵素結合免疫吸着測定法によって決定した(24)。このモノクローナル抗体は、上述したHO反応によって特異的に生成された最終ヘム分解生成物であるBR−IXαを認識できる(24,25)。乳酸デヒドロゲナーゼ(LDH)の活性は、上述したように測定した(17)。肝臓移植片中のアデノシン三リン酸(ATP)は、別の場所で述べたようにルシフェリン/ルシフェラーゼ法によって決定した(14,21)。移植片中の環状AMPは、酵素結合免疫吸着測定法(Biotrak(商標)system,Amersham Biosciences,バッキンガムシャー)によって決定した。
【0031】
カルボキシフルオレセインの胆汁中排出率の分析
カルボキシフルオレセイン(CF)は、Mrp2によって各種の細胞から排出された有機アニオンである(26,27)。この染料のエステル前駆物質である、CFジアセテート(CFDA)を経門脈的に肝臓内へ、50nmol/Lにて10分間に渡ってMrp2の強力な阻害剤である、1.5mmol/Lプロベネシドの存在下で前負荷した(26,28)。この試薬は肝細胞に進入可能であり、エステル分解酵素によって、胆汁中に排出されるCFに加水分解される(14,17,29)。10分間のCFDA装填の後、胆汁中へのCFの排出を引き起こすために、肝臓を、プロベネシドを含まない緩衝液によって灌流した。冷虚血群において、保存された移植片にCFDA含有緩衝液を10分間に渡ってプロベネシドの存在下で装填し、プロベネシドの除去およびその後の50分間の再灌流を続けた。これらの調製物から胆汁サンプルを収集し、96ウェル多チャネル蛍光分光光度計を使用して蛍光測定を実施するまで、深冷凍結させた。測定は、サンプルのpH値との干渉なしに510nmの蛍光を生じる染料の等吸収波長である、440nmの落射照明で実施した(26)。サンプル中のCFの濃度は、リン酸緩衝生理食塩水に溶解させたCFの既知の濃度を用いて校正した。後の結果でわかるように、CF濃度は時間と共に指数関数的に低下するように見えた。この仮定により、胆汁CF存続期間を指数関数的減衰のT1/2として決定した。それゆえこの方法は、灌流肝臓に装填されたCFの初期量に対して反応しない。
【0032】
肝細胞CF排除の生体内原位置(in situ)描出
上述のプロトコルを使用してCFを装填した肝臓移植片を、上述した生体内レーザ共焦点マイクロフルオログラフィーによって観察した(14,20,30)。後で結果に示すように、CFを、プロベネシドの存在下で肝細胞中に顕著に装填した。試薬の除去時に、染料は肝細胞からただちに排除され、毛細胆管(BC)内に排出されてハニカム網目を示し、最終的に柔組織から消失した。染料排除がMrp2の機能に依存しているかどうかを調査するために、一部の移植片を、1.5mmol/L プロベネシドを含有する緩衝液によって再灌流した。レーザ共焦マイクロフルオログラフは、増強CCDカメラ(C5810,浜松フォトニクス)およびマルチピンホールレーザ共焦プロセッサ(CSU−10,横河電機(株))を装備した倒立型顕微鏡(Diaphot 300, Nikon/Sankei)によって取り込んだ。すべてのマイクロフルオログラフは8ビットグレイレベル画像にデジタル処理した。蛍光強度を校正するために、CFの既知濃度をインビトロで調製して、画像をカメラの同一の光学パラメータの下で取り込んだ。肝細胞のグレーレベルを可変正方形窓(2x2μm2)によって、デジタル画像プロセッサを使用して測定した(18,31)。1回の実験で、対象の顕微鏡視野内で少なくとも10個の異なる肝細胞を分析した。肝臓表面で測定した蛍光強度が溶液中で測定した蛍光強度と同じであると仮定して、グレーレベルを校正線を使用して見かけのCF濃度に変換し、CFappと呼んだ。
【0033】
本発明者らは肝細胞損傷の指標としてのBC網目の構造変化を調査するために、形態計測も実施した。後の結果に示すように、正常に機能している肝細胞は周囲BC中で多角形CF重点を特徴とするが、これに対して損傷を受けた肝細胞は周囲BC網目の部分的消失によって判定される。CFによる完全なBC充填によって包囲されたそのような無傷の肝細胞の数を対象とする範囲においてカウントした。そのような評価のために1回の実験で肝臓表面の約0.05mm2を分析した。
【0034】
細胞内Mrp2分布の免疫組織化学分析
移植片の肝細胞中のMrp2を評価するために、以前の研究に従って、肝臓サンプルの固定、スライスおよびモノクローナル抗体M2III−6による染色を行った(32)。切片上の抗原をフィコエリトリン結合抗マウスIgGによって描出し、別の箇所で述べたように488nmのレーザ共焦マイクロフルオログラフィーによって観察した(20,30)。BC局在化およびMrp2の肝細胞内在化を調査するために、切片を、肝細胞接合部で発現された別のマーカーである、ZO−1に対するモノクローナル抗体によって二重免疫染色した(33)。タンパク質分布の変化を半定量的な方法で決定するために、Mrp2の単染色マイクロフルオログラフをモノクローム8ビット画像として変換した(14)。グレーレベル(1−256)を単一の肝細胞内の細胞質ドメインおよび小管ドメインの両方で測定した。単一の細胞内で各ドメインの少なくとも5つの異なる部位を選択して、細胞質強度の対応する小管強度に対する相対値を計算した。そのような測定を4つの異なる移植片中の40〜60個の肝細胞で実施して、%I−Mrp2(cyt/bc)として定義されるMrp2関連免疫反応性の細胞質強度パーセンテージのヒストグラムを作成した。この指標の上昇は、Mrp2内在化の上昇を示した。ヒストグラムは、対照移植片とTXA2合成酵素阻害剤の存在下または非存在下でKC消耗手順ありまたはなしで冷虚血を受けさせた移植片との間で比較した。
【0035】
群間での全肝臓移植片におけるMrp2発現の相違を調査するために、同じモノクローナル抗体を使用してウェスタンブロット分析を実施した。本発明者らは抗アクロレインモノクローナル抗体(5F6)によるウェスタン分析を追跡するために、抗体M2III−6によってタンパク質を免疫沈降させることによってMrp2の酸化的改変の変化も調査した(34,35)。
【0036】
統計的解析
異なる実験群間のデータの統計的有意差は、一元配置分散分析(ANOVA)およびFischerの多重比較検定によって決定した。P<0.05が有意であると見なされた。
【0037】
結果
肝臓移植片での胆汁産出量の保存時間依存性減少
肝臓移植片の生存能を試験するために、静脈灌水中の乳酸デヒドロゲナーゼの放出を細胞溶解の指標として測定した。表1に見られるように、24時間未満の冷虚血を受けさせた移植片は、乳酸デヒドロゲナーゼの顕著な上昇を示さなかった。
【表1】
データは、開始(0分)および再灌流の開始後60分(0〜24時間;n=5、48時間;n=3)の移植片からの測定値(mIU/分/g肝臓)の平均±SEを表す。他の群の値と比較して*P<0.05。
【0038】
48時間では、乳酸デヒドロゲナーゼ放出は60分間再灌流の開始および終了時に明らかとなり、現在の実験条件下で24時間未満の冷虚血を受けた移植片では壊死性細胞死が検知されないことを示した。図1は、各種の長さの冷虚血を受けた移植片における、再灌流時間の関数としての胆汁産出量の経時変化を示す。タウロコール酸ナトリウムを添加したパネルAで見られるように、8時間虚血を受けた移植片はその産出量を、30分の対照における産出量と匹敵するレベルまで上昇させたが、初期再灌流の50〜60分後にはそれを減少させた。16〜48時間の長期冷虚血を受けた移植片では、産出量のそのような減少はさらに明らかとなった。図1のパネルBは、灌水中のタウロコール酸ナトリウムの非存在下でモニターした胆汁回収の経時変化を示した。図示されているように、16時間を超えて冷虚血によって処置された群は、産出量の著しい減少を示した。
【0039】
冷虚血後肝臓のグルタチオンおよびビリルビンの胆汁排出の変化
16時間及び24時間冷虚血を受けた移植片における胆汁流が著しく減少したことを観察し、本発明者らは胆汁うっ滞変化の原因である胆汁構成成分を決定することができた。図2は、冷虚血に対する保存時間の関数としてプロットした、再灌流開始20分後に測定した胆汁構成成分のデータを示す。図示されているように、胆汁塩の濃度はどの長さの保存時間でも有意な低下を示さなかったが、これに対してリン脂質は、8から24時間冷虚血を受けた群において濃度と流量の両方で顕著な低下を示した(図2Aおよび2B)。リン脂質が主に肝細胞から胆汁コンパートメントに排出されることを考慮すると、これらのデータは8時間を超えて保存した移植片での肝細胞機能障害の存在を示唆する。
【0040】
本発明者らは次に、その組織含有量をモニターしながらGSHの胆汁中排出を調査した(図2C)。GSH中の胆汁中濃度は8時間で有意に低下し、低温保存時間の関数として下降した。本発明者らは次に、GSHの肝臓含有量を調査した。図2Dに示すように、肝臓GSH含有量は20分間の再灌流の前後に変化しなかった。16時間保存した移植片において、含有量はGSHを含有するUniversity of Wisconsin液の使用の結果として明らかに増加した。しかしながら20分間の再灌流時に、GSHが循環から除去されると含有量は対照レベルまで迅速に抑制され、16時間冷虚血後の20分間の再灌流が移植片の基底GSH含有量を変化させないことを示す。これらの結果は、保存時間が16時間より短い限り、冷虚血後肝臓における胆汁GSH排出の減少が移植片でのその減少からではなく、胆汁へのその輸送の障害から生じることを示唆している。GSHはMrp2を通じて排出されるため、本発明者らは次に、同じトランスポータを通じて排出される胆汁色素である、BR−IXαの胆汁中濃度の変化を調査した。図2Eに見られるように、初期20分間の再灌流におけるBR−IXαの胆汁濃度は8時間虚血群で有意に上昇し、16時間保存群で最大となった。最後に24時間冷虚血を受けた移植片において、BR−IXαの初期濃度は突然低下した。図2Fは、移植片が胆汁中への内因性BR−IXαを排除させる機能を示す。対照移植片に示されるように、この胆汁色素の肝臓含有量は初期20分間灌流中に有意に減少した。これに対して、同じ時間の再灌流は、16時間処置移植片におけるそのような減少を引き起こさなかった。これらの結果は、16時間移植片がBR−IXαを新規に(de novo)生成する機能が、色素を胆汁中に排出するその能力を上回ることを示唆している。
【0041】
カルボキシフルオレセイン排除によるMp2機能の全体的および局所的評価
GSHおよびBR−IXαの胆汁中排出の変化は、Mrp2がこれらの有機アニオンを肝細胞から排除させる機能が長期冷虚血を受けた移植片で障害を受ける可能性を上昇させた。しかしながら、グルタチオンおよびBR−IXαの初期量が群間で異なったため、これらの内因性アニオンの胆汁中排出の測定によって、本発明者らは移植片の有機アニオン排出機能の公正な比較を行うことができなかった。この問題を克服するために、移植片に外因性有機アニオンであるCFを装填して、肝細胞から胆汁中へのその排除を調査した。図3Aの左パネルに見られるように、肝細胞CF装填は、異なる長さ(0〜24時間)の冷虚血を受けた移植片間で比較可能に思われた。このことは図3Cで蛍光強度計によって確認され、肝細胞が生存していたことを示している。このことは、LDHの顕著な放出を示さない結果とも一致していた(表1)。プロベネシド除去の直後に、肝細胞に装填されたMrp2の阻害剤、CFは毛細胆管中に迅速に排出されて、10分以内に小葉上にハニカム網目を形成した(図3Aの中間列)。25分(右列)にて、細胞質内でわずかな蛍光があるとすれば、検出されるようになった。冷虚血再灌流を受けた移植片において、胆汁CF排出の2つの大きな変化が起こった。25分での基底蛍光の上昇によって判定された肝細胞染料排除の遅延、および10分にて収集されたマイクログラフに示された毛細胆管網目の消失および変形である。これらの変化は、24時間の長い冷虚血を受けた移植片において明らかとなった(図3Aの下列)。
【0042】
10分にて取り込まれたこれらのマイクロフルオログラフにおける毛細胆管部位の慎重な走査により、小管網目の構造の最小限ではあるが顕著な変化を示した。図3Bに見られるように、CF充填BCによって完全に包囲された肝細胞の数は、冷虚血の期間が長くなるにつれて減少した。そのような多角形の減少は、16時間保管した移植片においてただちに明らかとなった。肝細胞での初期CF装填が0〜24時間の虚血期間の範囲で匹敵していたため、BCの形態的変化は16時間冷虚血で最初に発生するように思われた。言い換えれば、8時間冷虚血を受けた移植片は、BC網目の形態の有意な変化を示さなかった。
【0043】
CF排除の遅延は、移植片の肝細胞における蛍光の時間的変化をモニターすることによって定量的な方向でさらに調査した(図4A)。図示したようにプロベネシドを連続的に灌流させたときに、染料は細胞中にとどまり、小管排出を示さずに、わずかな低下を示した。図4Bにプロットしたように、肝細胞で測定したグレーレベルによって、本発明者らは肝細胞からのCF排除のT1/2を決定した。図4Cは、群間のT1/2値を示す。プロベネシドの非存在下ではT1/2は約6分であったが、これに対してプロベネシドの存在下では、崩壊は実質的に遅延されて、Mrp2輸送のほぼ完全な阻害を示唆した。図示したように、CFを排出する8時間冷虚血移植片のT1/2はさらに著しく大きく、対照値とEHBRの肝臓で測定した値の中間に渡っていた。プロベネシド処置群と比較した、この変異種のより小さいT1/2値は、Mrp3による染料の類洞空間への補償排出によるものであると推測できる(36)。
【0044】
8時間冷虚血移植片における胆汁CF排出に対するKC消耗の効果
本発明者らは次に、8時間冷虚血移植片の全体としての、CFを排出する機能の評価を試みた。このために、染料排除のT1/2値を決定した(図5)。プロベネシドの除去後(図5AのT0)、胆汁のCF濃度は一時的に上昇して、徐々に基底レベルに戻った。そのような一時的上昇は、プロベネシドの存在下(陰丸)、またはEHBRから単離された移植片(黒丸)のどちらでも観察されず、Mrp2が胆汁CF排出の原因であることを示唆する。8〜16時間保存された全肝臓移植片でCF排除を分析したときに、崩壊は対照においてよりもゆっくりであるように思われた。8時間虚血移植片から収集したデータを使用して、胆汁サンプル中のCF濃度対ピーク時(10分)のCF濃度の対数値を、再灌流時間の関数として再プロットした(図5B)。次に染料排除のT1/2値をKC消耗によって処置した、または処置しなかった移植片間で比較した。図示されているように、LDDによる未処置の肝臓[KC(+)]において、8時間冷虚血は対照よりも長いT1/2値を示した。2つの群間のT1/2のそのような相違は、KC消耗移植片では完全に消失した。注目すべきなのは、非冷虚血対照肝臓においては、KC消耗手順は単独でT1/2値を変化させず、移植片が冷虚血再灌流を受けたときのみに明らかになると示されたことである。
【0045】
8時間冷虚血移植片においてKCがmrp2による長期CF排出を引き起こす機構を解明するために、本発明者らは、冷虚血後条件下で活性化されたKCより放出される主要なプロスタノイドであるトロンボキサンの関与を調査した(37,38)。このために、TXA2合成酵素の阻害剤であるOKY−046の効果を調査した。図5Cに見られるように、保存液およびすすぎ液への240μmol/Lでのこの試薬の使用は、CF排除のT1/2値をほぼ完全に上昇させる。TXA2合成酵素阻害剤は他のプロスタノイド(たとえばPGE2およびPGF2αを合成するための、アラキドン酸の有効性を上昇させるため、本発明者らは、TXsを含むシクロオキシゲナーゼ経路を介して生成されるすべてのプロスタノイドを抑制する阻害剤であるインドメタシン(IM)が、T1/2値を短縮または増大させるかどうかを調査した。図示したように、この阻害剤も長いT1/2値をほぼ完全に短縮した。さらに8時間冷虚血移植片に対するOKY−046の効果は、KCが消耗されたときに消失し、TXsの関与がKC依存性であることを示唆した。
【0046】
8時間冷虚血によるMrp2の細胞内再局在化およびKC消耗によるその減衰
KCが内毒素処置肝臓においてMrp2をダウンレギュレートすることが既知であるため(39)、本発明者らはトランスポータの機能障害の機構にそのような変化が関与するかどうかを調査した。ウェスタンブロット分析によって示されるように、Mrp2タンパク質の量は24時間虚血移植片(データを示さず)と同様に、8時間冷虚血移植片(図6A)において不変であった。本発明者らは、タンパク質が単独で、虚血後酸化的損傷の結果として酸化的改変されたかどうかも調査した。しかしながら、抗アクロレイン抗体を使用した免疫沈降で判定したように、明らかな変化は見られなかった。次にタンパク質の肝細胞局在化が8時間冷虚血移植片において改変されたかどうかを調査した。図6Bに見られるように、BCでのその局在化は顕著に減少したが、これに対して、肝細胞の細胞質におけるバックグラウンド蛍光は8時間虚血再灌流移植片において上昇した。下のパネルに見られるように、ZO−1による二重免疫染色(緑)は、BC中のMrp2の同時局在化(赤)が顕著に妨害されたのに対して、細胞質におけるその強度が上昇したことを明らかにし、タンパク質の内在化を示唆した。そのような変化は、KC消耗によって、またはOXY−046によって処置された移植片では減衰された。%I−Mrp2(cyt/bc)値として、内在化の指標は、8時間冷虚血再灌流がMrp2内在化を有意に上昇させたことと、KC消耗による、またはTXA2合成酵素の遮断による処置がトランスポータの毛細胆管再局在化を改善したことを示した。
【0047】
本発明者らは、8時間冷虚血再灌流後のKC(+)およびKC(−)移植片でのATPおよびcAMPの肝臓含有量の相違を決定したが、2つの群間に顕著な有意性はなかった(それぞれATP含有量では3.4±0.9対2.9±1.1μmol/g肝臓、そしてcAMP含有量では8.9±1.0対9.2±0.5pmol/g肝臓)。これらの結果は、KC消耗によるこのATP結合タンパク質の細胞内回復の改善が、ATPおよびcAMPの組織含有量の変化によって生じなかったことを示唆している。
【0048】
文献
1. Ploeg RJ, D'Alessandro AM, Knechtle SJ, Stegall MD, Pirsch JD, Hoffmann RM, Sasaki T, et al. Risk factors for primary dysfunction after liver transplantation--a multivariate analysis. Transplantation 1993;55:807-813.
2. Deschenes M, Belle SH, Krom RA, Zetterman RK, Lake JR. Early allograft dysfunction after liver transplantation: a definition and predictors of outcome. National Institute of Diabetes and Digestive and Kidney Diseases Liver Transplantation Database. Transplantation 1998;66:302-310.
3. Porte RJ, Ploeg RJ, Hansen B, van Bockel JH, Thorogood J, Persijn GG, Hermans J, et al. Long-term graft survival after liver transplantation in the UW era: late effects of cold ischemia and primary dysfunction. European Multicentre Study Group. Transpl Int 1998;11 Suppl 1:S164-167.
4. Adam R, Bismuth H, Diamond T, Ducot B, Morino M, Astarcioglu I, Johann M, et al. Effect of extended cold ischaemia with UW solution on graft function after liver transplantation. Lancet 1992;340:1373-1376.
5. Jaeschke H. Preservation injury: mechanisms, prevention and consequences. J Hepatol 1996;25:774-780.
6. Caldwell-Kenkel JC, Currin RT, Tanaka Y, Thurman RG, Lemasters JJ. Reperfusion injury to endothelial cells following cold ischemic storage of rat livers. Hepatology 1989;10:292-299.
7. Caldwell-Kenkel JC, Currin RT, Tanaka Y, Thurman RG, Lemasters JJ. Kupffer cell activation and endothelial cell damage after storage of rat livers: effects of reperfusion. Hepatology 1991;13:83-95.
8. Lemasters JJ, Thurman RG. Reperfusion injury after liver preservation for transplantation. Annu Rev Pharmacol Toxicol 1997;37:327-338.
9. Clavien PA. Sinusoidal endothelial cell injury during hepatic preservation and reperfusion. Hepatology 1998;28:281-285.
10. Sindram D, Porte RJ, Hoffman MR, Bentley RC, Clavien PA. Platelets induce sinusoidal endothelial cell apoptosis upon reperfusion of the cold ischemic rat liver. Gastroenterology 2000;118:183-191.
11. Arai M, Mochida S, Ohno A, Fujiwara K. Blood coagulation in the hepatic sinusoids as a contributing factor in liver injury following orthotopic liver transplantation in the rat. Transplantation 1996;62:1398-1401.
12. Clavien PA, Harvey PR, Sanabria JR, Cywes R, Levy GA, Strasberg SM. Lymphocyte adherence in the reperfused rat liver: mechanisms and effects. Hepatology 1993;17:131-142.
13. Vajdova K, Smrekova R, Mislanova C, Kukan M, Lutterova M. Cold-preservation-induced sensitivity of rat hepatocyte function to rewarming injury and its prevention by short-term reperfusion. Hepatology 2000;32:289-296.
14. Kumamoto Y, Suematsu M, Shimazu M, Kato Y, Sano T, Makino N, Hirano KI, et al. Kupffer cell-independent acute hepatocellular oxidative stress and decreased bile formation in post-cold-ischemic rat liver. Hepatology 1999;30:1454-1463.
15. Imamura H, Brault A, Huet PM. Effects of extended cold preservation and transplantation on the rat liver microcirculation. Hepatology 1997;25:664-671.
16. Williams JW, Vera S, Peters TG, Van Voorst S, Britt LG, Dean PJ, Haggitt R, et al. Cholestatic jaundice after hepatic transplantation. A nonimmunologically mediated event. Am J Surg 1986;151:65-70.
17. Suzuki H, Suematsu M, Ishii H, Kato S, Miki H, Mori M, Ishimura Y, et al. Prostaglandin E1 abrogates early reductive stress and zone-specific paradoxical oxidative injury in hypoperfused rat liver. J Clin Invest 1994;93:155-164.
18. Suematsu M, Goda N, Sano T, Kashiwagi S, Egawa T, Shinoda Y, Ishimura Y. Carbon monoxide: an endogenous modulator of sinusoidal tone in the perfused rat liver. J Clin Invest 1995;96:2431-2437.
19. Kyokane T, Norimizu S, Taniai H, Yamaguchi T, Takeoka S, Tsuchida E, Naito M, et al. Carbon monoxide from heme catabolism protects against hepatobiliary dysfunction in endotoxin-treated rat liver. Gastroenterology 2001;120:1227-1240.
20. Goda N, Suzuki K, Naito M, Takeoka S, Tsuchida E, Ishimura Y, Tamatani T, et al. Distribution of heme oxygenase isoforms in rat liver. Topographic basis for carbon monoxide-mediated microvascular relaxation. J Clin Invest 1998;101:604-612.
21. Shiomi M, Wakabayashi Y, Sano T, Shinoda Y, Nimura Y, Ishimura Y, Suematsu M. Nitric oxide suppression reversibly attenuates mitochondrial dysfunction and cholestasis in endotoxemic rat liver. Hepatology 1998;27:108-115.
22. Ishiguro S, Arii S, Monden K, Fujita S, Nakamura T, Niwano M, Harada T, et al. Involvement of thromboxane A2-thromboxane A2 receptor system of the hepatic sinusoid in pathogenesis of cold preservation/reperfusion injury in the rat liver graft. Transplantation 1995; 59: 957-961.
23. Jaeschke H, Smith CV, Mitchell JR. Reactive oxygen species during ischemia-reflow injury in isolated perfused rat liver. J Clin Invest 1988;81:1240-1246.
24. Yamaguchi T, Wakabayashi Y, Tanaka M, Sano T, Ishikawa H, Nakajima H, Suematsu M, et al. Taurocholate induces directional excretion of bilirubin into bile in perfused rat liver. Am J Physiol 1996;270:G1028-1032.
25. Izumi Y, Yamazaki M, Shimizu S, Shimizu K, Yamaguchi T, Nakajima H. Anti-bilirubin monoclonal antibody. II. Enzyme-linked immunosorbent assay for bilirubin fractions by combination of two monoclonal antibodies. Biochim Biophys Acta 1988;967:261-266.
26. Mor-Cohen R, Zivelin A, Rosenberg N, Shani M, Muallem S, Seligsohn U. Identification and functional analysis of two novel mutations in the multidrug resistance protein 2 gene in Israeli patients with Dubin-Johnson syndrome. J Biol Chem 2001;276:36923-36930.
27. van der Kolk DM, de Vries EG, Noordhoek L, van den Berg E, van der Pol MA, Muller M, Vellenga E. Activity and expression of the multidrug resistance proteins P-glycoprotein, MRP1, MRP2, MRP3 and MRP5 in de novo and relapsed acute myeloid leukemia. Leukemia 2001;15:1544-1553.
28. Courtois A, Payen L, Lagadic D, Guillouzo A, Fardel O. Evidence for a multidrug resistance-associated protein 1 (MRP1)-related transport system in cultured rat liver biliary epithelial cells. Life Sci 1999;64:763-774.
29. Suematsu M, Suzuki H, Ishii H, Kato S, Yanagisawa T, Asako H, Suzuki M, et al. Early midzonal oxidative stress preceding cell death in hypoperfused rat liver. Gastroenterology 1992;103:994-1001.
30. Kajimura M, Shimoyama M, Tsuyama S, Suzuki T, Kozaki S, Takenaka S, Tsubota K, et al. Visualization of gaseous monoxide reception by soluble guanylate cyclase in the rat retina. Faseb J 2003;17:506-508.
31. Hayashi S, Takamiya R, Yamaguchi T, Matsumoto K, Tojo SJ, Tamatani T, Kitajima M, et al. Induction of heme oxygenase-1 suppresses venular leukocyte adhesion elicited by oxidative stress: role of bilirubin generated by the enzyme. Circ Res 1999;85:663-671.
32. Mottino AD, Cao J, Veggi LM, Crocenzi F, Roma MG, Vore M. Altered localization and activity of canalicular Mrp2 in estradiol-17beta-D-glucuronide-induced cholestasis. Hepatology 2002;35:1409-1419.
33. Kubitz R, D'Urso D, Keppler D, Haussinger D. Osmodependent dynamic localization of the multidrug resistance protein 2 in the rat hepatocyte canalicular membrane. Gastroenterology 1997;113:1438-1442.
34. Uchida K, Kanematsu M, Sakai K, Matsuda T, Hattori N, Mizuno Y, Suzuki D, et al. Protein-bound acrolein: potential markers for oxidative stress. Proc Natl Acad Sci U S A 1998;95:4882-4887.
35. Tanaka N, Tajima S, Ishibashi A, Uchida K, Shigematsu T. Immunohistochemical detection of lipid peroxidation products, protein-bound acrolein and 4-hydroxynonenal protein adducts, in actinic elastosis of photodamaged skin. Arch Dermatol Res 2001;293:363-367.
36. Ogawa K, Suzuki H, Hirohashi T, Ishikawa T, Meier PJ, Hirose K, Akizawa T, et al. Characterization of inducible nature of MRP3 in rat liver. Am J Physiol Gastrointest Liver Physiol 2000;278:G438-446.
37. Gyenes M, and De Groot H. Prostanoid release by Kupffer cells upon hypoxia-reoxygenation: role of pHi and Ca2+. Am J Physiol 1993; 264: G535-G540.
38. Johnston DE, Peterson MB, Mion F, Berninger RW, Jefferson DM. Synthesis and degradation of eicosanoids in primary rat hepatocyte cultures. Prostaglandins Leukot Essent Fatty Acids 1991; 43: 119-132.
39. Nakamura J, Nishida T, Hayashi K, Kawada N, Ueshima S, Sugiyama Y, Ito T, et al. Kupffer cell-mediated down regulation of rat hepatic CMOAT/MRP2 gene expression. Biochem Biophys Res Commun 1999;255:143-149.
40. Roelofsen H, Soroka CJ, Keppler D, Boyer JL. Cyclic AMP stimulates sorting of the canalicular organic anion transporter (Mrp2/cMoat) to the apical domain in hepatocyte couplets. J Cell Sci 1998;111 ( Pt 8):1137-1145.
41. Kipp H, Arias IM. Intracellular trafficking and regulation of canalicular ATP-binding cassette transporters. Semin Liver Dis 2000;20:339-351.
42. Horton AA,Wood JM. Prevention of thromboxane B2-induced hepatocyte plasma membrane bleb formation by certain prostaglandins and a proteinase inhibitor. Biochim Biophys Acta 1990;1022:319-324
43. Kato Y, Shimazu M, Kondo M, Uchida K, Kumamoto Y, Wakabayashi G, Kitajima M, et al. Bilirubin rinse: A simple protectant against the rat liver graft injury mimicking heme oxygenase-1 preconditioning. Hepatology 2003;38:364-373.
【0049】
すべての文書は、あらゆる目的のために参照により本明細書に組み入れられている。
【産業上の利用可能性】
【0050】
本発明は、その臓器毒性のために以前に不合格になった臨床前試薬の復活に有用である。全体的に、日常的な臨床化学に対して肝臓毒性を示す化合物の毒性の詳細な機構は、大半は未知のままである。そのような化合物を本発明によって詳細に調査し、細胞生存能に影響を及ぼさずにトランスポータ機能のみを遮断することが判明した場合、臨床用途に利用するために化学構造のわずかな改変が有効である。(2)ヒト癌細胞を移植した免疫不全マウスへの本方法の利用によって、本発明者らは、薬物送達の種類の特異性および生体内での腫瘍細胞凝集体または無傷の組織への蓄積について調査できた。トランスポータ、たとえばMrp2の機能は腫瘍の薬物耐性の重要な決定要因であるため、そのような利用は、生体内での移植癌における抗癌試薬の送達および排除を調査するために極めて有用である。抗癌および抗血栓試薬の潜在性をスクリーニングするための臨床前実験の利用。
【図面の簡単な説明】
【0051】
【図1】低温保存、続いて再灌流を受けた肝臓移植片の胆汁産出量の経時変化を示す。白丸は非虚血対照肝臓からのデータを示す。白四角、陰四角および黒四角はそれぞれ、8、16、24時間低温保存を受けた移植片からのデータを示す。黒三角は、48時間保存移植片からのデータを示す。値は、5回の独立した実験の平均±SEである。TC(+)およびTC(−):タウロコール酸ナトリウム 30μmol/Lの存在下または非存在下で収集したデータ。R:再灌流の開始。胆汁塩非依存性および依存性産出量のどちらも、16時間保存移植片で減少したことに注目する。対照からのデータと比較して*P<0.05。16時間移植片からのデータと比較して†P<0.05。24時間移植片からのデータと比較して#P<0.05。
【図2】胆汁構成要素の胆汁中濃度に対する冷虚血の期間の影響を示す。胆汁構成要素のデータは、再灌流開始20分後に収集した。A:胆汁塩(パネルA)。B:リン脂質。C:胆汁中の還元グルタチオン(GSH)の濃度。D:再灌流前および再灌流20分後に測定した対照および16時間保存移植片におけるGSHの肝臓含有量。E:胆汁中のビリルビン(BR)−IXαの濃度。F:再灌流前および再灌流20分後に測定した対照および16時間保存移植片におけるBR−IXαの肝臓含有量。値は、5〜7回の独立した実験の平均±SEである。対照肝臓からのデータと比較して*P<0.05。16時間群からのデータと比較して#P<0.05。20分再灌流を受けた対照移植片からのデータと比較して†P<0.05。
【図3】冷虚血再灌流移植片中の毛細胆管への肝細胞カルボキシフルオレセイン(CF)排出の動態の変化を示す。A:プロベネシド除去前(基底装填)、除去10分後および25分後に取り込まれた小管CF排出の代表的な画像。24時間冷虚血再灌流移植片における毛細胆管網目のハニカムパターンの崩壊に注目する(矢印)。色付きバーは、CFの既知濃度を用いて校正した蛍光強度を示す。バー:30μm。B:各種の冷虚血期間に渡って保管した移植片中のCF充填多角形の密度によって判定された、毛細胆管網目の再灌流誘発崩壊の相違。対照肝臓からのデータと比較して*P<0.05。C:群間の比較CF装填量を示す初期肝細胞CF濃度。値は、5回の独立した実験の平均±SEである。
【図4】カルボキシフルオレセイン(CF)の毛細胆管排出を描出することによる、Mrp2機能の生体内定量分析を示す。A:各肝細胞からの染料排出を示す代表的な一連の画像。上;1.5mM プロベネシドの存在下で灌流された移植片からの画像(PB(+))。下、プロベネシド除去後に取り込まれた画像(PB(−))。細胞中の蛍光の時間依存性還元と、それに続く周囲の毛細胆管における染料の濃縮および消失に注目する。細胞中の染料保持に注目すべきである。B:肝細胞CF蛍光の消失。プロベネシドを用いて(黒丸)、または用いずに(白丸)処置した移植片の肝細胞CF濃度を、直線が指数曲線を表すように、時間に対して片対数プロットする。差し込み図:CF濃度と8ビットグレーレベルとの間の関係を示す校正曲線。[CFapp]:CFの見かけの濃度。CF濃度は、3μmol/Lよりも低い濃度でグレーレベルと直線的に関係付けられている(r2=0.996、p<0.05)。C:肝細胞からのCF排除の半減期(T1/2)の相違。値は、各群での5回の独立した実験の平均±SEである。対照肝臓からのデータと比較して*P<0.05。EHBR:Eisai高ビリルビン血症ラットから単離した移植片。PB:1.5mM プロベネシドを用いて灌流した移植片。
【図5】肝臓移植片が冷虚血再灌流時にカルボキシフルオレセイン(CF)を胆汁中に排出する能力の変化を示す。A:各種の長さの冷虚血を受けた移植片における胆汁CF排出の経時変化の相違。白丸:Mrp2阻害剤である1.5mM プロベネシドを除去した直後に灌流させた対照移植片。白四角および陰四角:8および24時間低温保存と、その後のプロベネシドの非存在下での再灌流をそれぞれ受けた移植片。陰丸:プロベネシドの存在下で正常灌流された移植片。黒丸:Eisai高ビリルビン血症ラットから単離された、正常灌流された移植片。値は、5回の独立した実験の平均±SEである。T0:プロベネシドが灌流液から除去された時間。差し込み図:CF濃度と蛍光強度との間の直線関係。B:各種の再灌流期間で収集された胆汁中の相対CF濃度の変化、およびクッパー細胞(KC)の欠乏の効果。左:対照(0時間、破線)と8時間保存肝臓移植片(8時間、実線)との間の胆汁CF排出の減少の相違。右:静脈内リポソームカプセル化ジクロロメチレンジホスホナート(LDD)によるKC消耗の効果。白丸:対照(0時間、破線)。白四角:8時間保存移植片(8時間、実線)。データは、4回の独立した実験による測定値の平均±SEを表す。対照肝臓におけるCF排除の減少と比較して*P<0.05。C:8時間低温保存肝臓におけるT1/2値の延長に対する、LDDおよび/またはトロンボキサンA2合成酵素の阻害剤であるOKY−046(OKY)を用いた処置によるKC消耗[KC(−)]の効果。IM:インドメタシン。保存およびすすぎ溶液中のOKYおよびIMの濃度はそれぞれ、240および28μmol/Lであった。OKYの阻害作用は、KC消耗移植片で消失することに注目する。値は、5〜6回の独立した実験の平均±SEである。対照肝臓からのデータと比較して*P<0.05。8時間保管移植片におけるデータと比較して†P<0.05。
【図6】8時間冷虚血および60分間再灌流(8時間/R)を受けた肝臓移植片におけるMrp2の細胞内分布の崩壊、およびクッパー細胞(KC)欠乏手順[KC(−)]またはOKY−046(OKY)を用いた処置の効果を示す。A:抗アクロレインモノクローナル抗体5F6によるMrp2のウェスタンブロット解析および免疫沈降。M:分子マーカー。B:Mrp2分布の免疫蛍光分析。左;フィコエリトリンによって標識された抗Mrp2モノクローナル抗体(M2III−6)を用いた単染色。バー:30μm。右;FITC標識ZO−1抗体およびフィコエリトリン標識M2III−6抗体を用いた二重免疫染色。バー:10μm。C.肝細胞Mrp2局在化の半定量分析。%I−Mrp2(cyt/bc);毛細胆管にて測定されたものに対する、Mrp2関連免疫反応性の細胞質強度。値は、40〜60個の肝細胞/4個の独立した肝臓からの移植片における測定値の平均±SEである。対照肝臓からのデータと比較して*P<0.05。8時間/R−KC(+)群から収集したデータと比較して†P<0.05。
【特許請求の範囲】
【請求項1】
臓器または組織傷害を分析する方法であって、以下のステップ:
(a)臓器または組織を染料で標識するステップ、
(b)前記臓器または組織の生体異物代謝および/または細胞状態を含む多重指標を入手するステップ、ならびに
(c)前記指標から前記臓器または組織の傷害を分析するステップ、
を含む方法。
【請求項2】
前記臓器または組織が肝臓、腎臓、肺、膵臓および胃腸管から成る群より選択される少なくとも1つである、請求項1に記載の方法。
【請求項3】
前記ステップ(b)が血管系および/または排出経路の微細解剖配向を得るステップをさらに含む、請求項1または2に記載の方法。
【請求項4】
前記解析が視覚的におよび/または定量的に実施される、請求項1から3のいずれか一項に記載の方法。
【請求項5】
前記細胞状態が細胞生存能、細胞傷害、分子輸送、およびミトコンドリア機能から成る群より選択される少なくとも1つである、請求項1から4のいずれか一項に記載の方法。
【請求項6】
薬剤毒性を評価する方法であって、以下のステップ:
(a)臓器または組織を染料で標識するステップ、
(b)試験薬剤を前記臓器または組織に使用するステップ、
(c)前記臓器または組織の生体異物代謝および/または細胞状態を含む多重指標を入手するステップ、
(d)前記指標から前記臓器または組織の傷害を分析するステップ、ならびに
(e)前記薬剤が前記臓器または組織に対する毒性を有するかどうかを評価するステップ、
を含む方法。
【請求項7】
前記臓器または組織が肝臓、腎臓、肺、膵臓および胃腸管から成る群より選択される少なくとも1つである、請求項6に記載の方法。
【請求項8】
前記ステップ(b)が、血管系および/または排出経路の微細解剖配向を得るステップをさらに含む、請求項6または7に記載の方法。
【請求項9】
前記解析が視覚的におよび/または定量的に実施される、請求項6から8のいずれか一項に記載の方法。
【請求項10】
前記細胞状態が細胞生存能、細胞傷害、細胞内および細胞周囲での分子の輸送、および生物活性化合物の生成、血流、および組織酸素添加から成る群より選択される少なくとも1つである、請求項6から9のいずれか一項に記載の方法。
【請求項1】
臓器または組織傷害を分析する方法であって、以下のステップ:
(a)臓器または組織を染料で標識するステップ、
(b)前記臓器または組織の生体異物代謝および/または細胞状態を含む多重指標を入手するステップ、ならびに
(c)前記指標から前記臓器または組織の傷害を分析するステップ、
を含む方法。
【請求項2】
前記臓器または組織が肝臓、腎臓、肺、膵臓および胃腸管から成る群より選択される少なくとも1つである、請求項1に記載の方法。
【請求項3】
前記ステップ(b)が血管系および/または排出経路の微細解剖配向を得るステップをさらに含む、請求項1または2に記載の方法。
【請求項4】
前記解析が視覚的におよび/または定量的に実施される、請求項1から3のいずれか一項に記載の方法。
【請求項5】
前記細胞状態が細胞生存能、細胞傷害、分子輸送、およびミトコンドリア機能から成る群より選択される少なくとも1つである、請求項1から4のいずれか一項に記載の方法。
【請求項6】
薬剤毒性を評価する方法であって、以下のステップ:
(a)臓器または組織を染料で標識するステップ、
(b)試験薬剤を前記臓器または組織に使用するステップ、
(c)前記臓器または組織の生体異物代謝および/または細胞状態を含む多重指標を入手するステップ、
(d)前記指標から前記臓器または組織の傷害を分析するステップ、ならびに
(e)前記薬剤が前記臓器または組織に対する毒性を有するかどうかを評価するステップ、
を含む方法。
【請求項7】
前記臓器または組織が肝臓、腎臓、肺、膵臓および胃腸管から成る群より選択される少なくとも1つである、請求項6に記載の方法。
【請求項8】
前記ステップ(b)が、血管系および/または排出経路の微細解剖配向を得るステップをさらに含む、請求項6または7に記載の方法。
【請求項9】
前記解析が視覚的におよび/または定量的に実施される、請求項6から8のいずれか一項に記載の方法。
【請求項10】
前記細胞状態が細胞生存能、細胞傷害、細胞内および細胞周囲での分子の輸送、および生物活性化合物の生成、血流、および組織酸素添加から成る群より選択される少なくとも1つである、請求項6から9のいずれか一項に記載の方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図2】
【図3】
【図4】
【図5】
【図6】
【公表番号】特表2007−507690(P2007−507690A)
【公表日】平成19年3月29日(2007.3.29)
【国際特許分類】
【出願番号】特願2006−527774(P2006−527774)
【出願日】平成16年9月28日(2004.9.28)
【国際出願番号】PCT/JP2004/014566
【国際公開番号】WO2005/030264
【国際公開日】平成17年4月7日(2005.4.7)
【出願人】(503185437)株式会社 オキシジェニクス (6)
【Fターム(参考)】
【公表日】平成19年3月29日(2007.3.29)
【国際特許分類】
【出願日】平成16年9月28日(2004.9.28)
【国際出願番号】PCT/JP2004/014566
【国際公開番号】WO2005/030264
【国際公開日】平成17年4月7日(2005.4.7)
【出願人】(503185437)株式会社 オキシジェニクス (6)
【Fターム(参考)】
[ Back to top ]