
自己−集合性ペプチド両親媒性物質および増殖因子送達のための関連する方法
【課題】幹細胞の産生する増殖因子のバイオアベイラビリティに影響を与える両親媒性ペプチド化合物を提供する。
【解決手段】ペプチド成分および疎水性成分を含み、該ペプチド成分はファージディスプレイ法の増殖因子認識産物を含み、該認識産物は該ペプチド成分のほぼN-末端に結合して、1つ以上の対応する増殖因子と結合相互作用するための1つ以上のエピトープ配列を含み、該疎水性成分は該ペプチド成分のほぼC−末端に結合している、両親媒性ペプチド。
【解決手段】ペプチド成分および疎水性成分を含み、該ペプチド成分はファージディスプレイ法の増殖因子認識産物を含み、該認識産物は該ペプチド成分のほぼN-末端に結合して、1つ以上の対応する増殖因子と結合相互作用するための1つ以上のエピトープ配列を含み、該疎水性成分は該ペプチド成分のほぼC−末端に結合している、両親媒性ペプチド。
【発明の詳細な説明】
【技術分野】
【0001】
本願は、全体の内容が参照により本明細書に組み入れられる、2003年12月5日提出の出願番号第60/527,504号の優先権の利益を主張する。
【0002】
米国政府は、エネルギー省(Department of Energy)からノースウェスタン大学(Northwestern University)への付与第DE-FG02-00ER54810号に準じて本発明のある権利を有する。
【背景技術】
【0003】
発明の背景
増殖因子は細胞特殊化において重要な役割を果たし、共に非機能性組織の再生を生じる可能性のある、幹細胞分化を制御するためまたは休止中の生物過程をインビボにおいて再活性化するための有望な因子である。増殖因子は速やかに分解され、細胞部位または損傷から迅速に拡散する。従って、構造または骨格は、制御された様式で周囲の細胞に放出するために、このような部位に増殖因子を保持するのに有用であると思われる。
【0004】
組織再生のために分子的に設計されている材料は高度医療において非常に興味深いものになっている。L-乳酸またはグリコール酸に基づいたポリマーを含む合成ポリマーおよびコラーゲン、フィブリンまたはアルギネートを含むバイオポリマーの骨格が検討されている。増殖因子は、ポリマーなどのヒドロゲルに物理的に捕獲されており、アニオン性ポリマーまたはヘパリンなどの構造物に共有結合または静電気的に結合している。これらのシステムおよび関連するシステムの欠点は、結合している増殖因子の非特異性または望ましい作用を達成するために共有結合の分解の必要性に関する。さらに最近では、受容体に基づいた細胞との相互作用を促進し、細胞接着または分化を促進するために使用されている細胞外タンパク質に見られるペプチドを含有するように天然および合成骨格が改良されている。
【0005】
自己-集合の進歩は、バイオマテリアルの分子設計の新たな機会を提供している。両親媒性分子構築ブロックは、水性環境において集合して、十分に規定されている種々の化学構造と骨格を形成することができる。両親媒性ペプチドおよび一方または両方の末端が疎水性成分(例えば、アルキルテイル)で官能基化されているペプチドを含む種々のクラスのペプチド-系両親媒性物質が文献で報告されている。両親媒性ペプチドは、ナノテープ、リボン、ファイバーおよびツイストリボンのような種々の超分子構造を形成することが示されている。これらの構造は、両親媒性分子のβ-シート形成から生ずる。特別な例は、個々のペプチドブロックの二次構造に強力に依存する特性を有するゲルを形成する両親媒性ペプチドブロックコポリマーの例である。一方の末端にアルキルテイルを有するペプチド両親媒性物質の例には、テイルの長さおよびテイルの数に応じて球状またはディスク-様ミセル構造を形成する三重らせん単位を形成するコラーゲンに見られるペプチドモチーフ由来の両親媒性物質が挙げられる。別のクラスは末端あたり1つの2つのテイルを有する。このクラスの両親媒性物質は、濃度を増加すると、ランダムコイルから繊維状構造を生ずるβ-シート型コンフォーメーションへの遷移を受けるという点においてアミロイド-様挙動を示す。逆平行配列を有する純粋にペプチド性のナノ構造が報告されており、非天然アルキル化四級アンモニウム塩によって誘導される逆平行配列に集合する2つの修飾ペプチド両親媒性に関する報告が1つある。
【0006】
β-シート形成セグメント、溶解のための荷電残基および生物学的エピトープを含むペプチドブロックに結合している直鎖状疎水性テイルを含む1クラスのペプチド両親媒性物質(PA)が2つの同時係属出願の一方または両方に開示されている。アルキルテイルはペプチドのN-末端に結合しており、エピトープセグメントはC-末端に配置されている。pHまたはイオン濃度の変化などのトリガーを適用すると、これらのPA分子は水性媒体中でナノファイバーに自己-集合することができる。アルキル鎖はファイバーのコアに位置し、エピトープは細胞相互作用のために周辺で提示されている。PA分子に組み込まれているエピトープは細胞外マトリックスタンパク質を模倣しており、細胞信号伝達による細胞接着または分化を促進する。異なるエピトープおよび相補的な荷電を有する2つの異なるPA分子が同一のナノファイバーに同時-集合することができることも示されている。各々全体として参照により本明細書に組み入れられる、2003年2月18日提出の同時係属出願第10/368,517号(国際公報番号WO03/070749)(特許文献1)および2002年11月14日提出の出願番号第10/294,114号 (国際公報番号03/054146号)(特許文献2)参照。
【0007】
しかし、このようなPA化合物は、典型的には、固相合成によって製造され、ペプチド成分はC-末端からN-末端方向に作製される。種々のエピトープまたはペプチド配列は生物活性のために遊離のN-末端を必要とすることが多いので、疎水性成分をN-末端に結合すると、得られるミセルナノファイバー集合物の周辺に遊離酸またはアミド基を有するPA化合物が得られ、このようなPA化合物は増殖因子相互作用に無効であると思われる。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】国際公報番号WO03/070749
【特許文献2】国際公報番号03/054146号
【発明の概要】
【0009】
発明の概要
上記を考慮すると、本発明の目的は、ある範囲の増殖因子のバイオアベイラビリティーに影響を与え、それによって上記に概略するものを含む従来技術の種々の欠陥および欠点を克服する両親媒性ペプチド化合物、その集合組成物および/またはそれらを使用するための関連する方法を提供することである。本発明の1つ以上の局面はある目的を満たすことができるが、1つ以上の他の局面は他のある目的を満たすことができることが当業者によって理解される。あらゆる点において、各目的が本発明の全ての局面に等しく適用しない場合がある。従って、以下の目的は、本発明の任意の1つの局面に関して選択的に考慮することができる。
【0010】
本発明の目的は、ファージディスプレイ法の認識産物を含むことができ、それから誘導されうるエピトープである、1つ以上の増殖因子と非共有結合的に相互作用することができるある範囲のエピトープ配列の1つ以上のN-末端に結合しているペプチド成分を含む両親媒性ペプチド化合物を提供することである。
【0011】
本発明の別の目的は、結合しているエピトープ結合配列の提示のための上記の両親媒性ペプチド化合物の1つ以上の集合物を含む組成物であって、このようなエピトープ配列が欠けている他の両親媒性ペプチド化合物を含むことができる組成物を提供することである。
【0012】
本発明の目的は、結合しているエピトープ配列に対応する1つ以上の増殖因子をさらに含む上記の組成物であって、増殖因子送達および放出のために自己-集合することができる組成物の1つ以上を提供することであってもよい。
【0013】
本発明の目的はまた、増殖因子のバイオアベイラビリティーおよび/または幹細胞分化に影響与える上記両親媒性ペプチド化合物の1つ以上の組成物集合物(compositional assemblies)およびそれらの用途を提供することである。
【0014】
本発明の他の目的、特徴、利益および利点はこの概要およびある種の態様の以下の説明から明らかであり、ペプチド組成物および増殖因子送達の骨格または構造の当業者に容易に明らかになる。このような目的、特徴、利益および利点は、添付の実施例、データ、図面およびそれらから引き出される全ての妥当な影響を単独または本明細書に組み入れられている参照文献と併用して考慮すると上記から明らかになる。
【0015】
一部には、本発明は、ペプチド成分および疎水性成分を含み、ペプチド成分がファージディスプレイ法の増殖因子認識産物を含む両親媒性ペプチドに関してもよい。1つ以上のこのような認識産物は、ペプチド成分のN-末端、N-末端付近またはN-末端に近接してペプチド成分に連結または直接結合してもよく、疎水性成分は、C-末端、C-末端付近またはC-末端に近接してペプチド成分と連結または直接結合してもよい。このような認識産物は、増殖因子と1つ以上の結合相互作用を提供するエピトープ配列から選択することができ、このような産物/配列および対応する増殖因子は以下に十分に考察されているものを含むが、これらに限定されない。本発明の化合物および/またはそのペプチド成分は実質的に直鎖状または分岐鎖状であってもよく、分岐鎖状の立体配置およびその製造は、全体の内容が参照により本明細書に組み入れられる、2004年12月6日に同時提出の同時係属出願「Branched Peptide Amphiphiles, Related Epitope Compounds and Self-Assembled Structures Thereof」に開示されている。
【0016】
一部には、本発明は、複数のこのような両親媒性ペプチド化合物を含む組成物に関してもよく、このような化合物各々のペプチド成分は生理的pHにおいて正味荷電を有する。このような組成物は、生理的pHにおいて相補的な正味荷電を有する複数の両親媒性ペプチド化合物をさらに含んでもよく、このような化合物のアミノ酸配列が、認識産物を含む化合物に対応するものより短い長さであるかまたは数が少ないように、増殖因子認識産物が欠ける。結果として、このような化合物のミセル集合物は、適当な培地において、1つ以上の認識産物の提示を増強し、ミセル集合物の周辺全面を越えて延在する。このような提示の増強を使用して、1つ以上の増殖因子のバイオアベイラビリティーに影響を与えるおよび/またはそれを制御することができる。従って、増殖因子が幹細胞によって産生されるても、または上記の組成物の1つ以上と合わせて細胞環境に送達されても、本発明の種々の組成物およびそれに関係する方法は、1つ以上の増殖因子と非共有結合的に相互作用するまたは結合するために幹細胞環境において使用することができる。
【0017】
組成物の用途または適用にかかわらず、本発明のペプチド両親媒性物質は、望ましいフレキシビリティ、荷電および/または分子間相互作用または結合能力に応じて種々の長さまたは配列のペプチド成分を含んでもよい。このような化合物の疎水性成分は異なってもよく(例えば、飽和または不飽和の約C6から約C22超のアルキルまたは置換アルキル等)、このような成分は、得られる両親媒性およびこのような化合物の組成物または集合物に対する影響によってのみ限定される。
【0018】
本発明の一態様は、各々結合エピトープを有するペプチド両親媒性物質と結合エピトープを含有しないフィラーペプチド両親媒性物質の混合物を含む組成物である。混合物は、フィラーペプチド両親媒性物質および結合エピトープを有する自己集合ペプチド両親媒性物質からなる自己集合ナノファイバーまたは他のミセルを形成することができる。組成物中のペプチド両親媒性物質は、アルキルテイル部分、βシート部分および荷電部分を有する。好ましくは、結合エピトープを有するペプチド両親媒性物質のエピトープは、ファージディスプレイ法によって誘導される配列を有し、フィラーペプチド両親媒性物質より長い。
【0019】
本発明の別の態様は、結合エピトープを有するペプチド両親媒性物質およびフィラーペプチド両親媒性物質を含む自己集合ナノファイバーまたは他の自己集合ミセルの組成物またはシステムである。結合エピトープを有するペプチド両親媒性物質は、好ましくは、フィラーペプチド両親媒性物質より長い。自己集合構造物では、結合エピトープを有するペプチド両親媒性物質は、好ましくは、自己集合ナノファイバーまたはミセルの表面から突出している。好ましくは、結合エピトープを有するペプチド両親媒性物質は、非共有結合相互作用により増殖因子または他のペプチド、アミノ酸または核酸と相互作用することができ、よりさらに好ましくは、増殖因子とエピトープの非共有結合相互作用は、増殖因子の細胞外受容体への結合と競合しない。
【0020】
自己集合ナノファイバーまたはミセルの組成物は、結合エピトープを有するナノファイバーのペプチド両親媒性物質に非共有結合している増殖因子をさらに含むことがあるヒドロゲルを含んでもよい。このような増殖因子はインビトロにおいて添加されてもまたはインビボにおいて患者の損傷部位のものであってもよい。自己集合ペプチド両親媒性物質と相互作用する増殖因子には、骨形成タンパク質、トランスフォーミング増殖因子、血管内皮細胞増殖因子、ニューロトロフィンならびにFGF-2、ソニックヘッジホッグおよびWnt-タンパク質のような分裂促進因子を挙げることができるが、これに限定されない。組成物は、ナノファイバーヒドロゲル内に幹細胞などであるが、これに限定されない細胞を含んでもよい。好ましくは、自己集合ペプチド両親媒性物質と非共有結合的に相互作用する増殖因子は、細胞外受容体との相互作用によりまたはナノファイバーマトリックスの分解により周囲組織または細胞に放出される。細胞以外に、他の治療化合物がヒドロゲル内に封入されてもまたはヒドロゲルに結合されてもよい。これらの化合物には、抗-炎症剤、化学療法剤またはナノファイバーヒドロゲル内におけるこれらの併用を挙げることができるが、これに限定されない。好ましくは、自己集合ペプチド両親媒性物質と非共有結合的に相互作用する増殖因子は、細胞外受容体との相互作用によりまたはナノファイバーマトリックスの分解により周囲組織または細胞に放出される。
【0021】
本発明の別の態様は、結合エピトープを有するペプチド両親媒性物質をフィラーペプチド両親媒性物質と混合または組み合わせることによって自己-集合させる段階を含み、結合エピトープを有するペプチド両親媒性物質がフィラーペプチド両親媒性物質より長さが長い、自己集合ペプチド両親媒性ナノファイバーまたはミセルを製造する方法である。ナノファイバーまたはミセルは、多価イオンを添加することによって、相補的荷電ペプチド両親媒性物質を添加することによってまたは脱水によってまたはペプチド両親媒性物質に酸もしくは塩基を添加することによって製造することができる。ヒドロゲルが形成される場合には、好ましくは、ミセルまたはナノファイバーは、多価イオンの添加によってまたは細胞培地もしくは体液にすでに存在するイオンによって形成される。好ましくは、結合エピトープを有するペプチド両親媒性物質は、ファージディスプレイ法によって誘導される配列を有する。
【0022】
本発明の別の態様は、増殖因子に非共有結合するためのエピトープを有するペプチド両親媒性物質を含むナノファイバーまたは他の自己集合ミセルを、組織を再生する必要のある患者の部位に投与する段階を含む組織を治療する方法である。ナノファイバーおよびそれらのヒドロゲルは、幹細胞などであるが、これに限定されない細胞を含んでもよい。細胞以外に、治療化合物がヒドロゲルに封入されてもまたは結合されてもよい。これらの化合物には、抗-炎症剤、化学療法剤またはこれらの併用を挙げることができるが、これに限定されない。好ましくは、自己集合構造に非共有結合する増殖因子は、細胞外受容体との相互作用またはマトリックスの分解によって部位に放出される。好ましくは、増殖因子および/または幹細胞を含有する自己集合ミセルまたはナノファイバーは、骨損傷、軟骨損傷、脊髄損傷、脳組織損傷、神経損傷またはこれらの組み合わせなどであるが、これに限定されない損傷を有する部位または患者に投与される。
【0023】
本発明の別の態様は、第2の末端も有するβ-シート形成ペプチド部分の第1の末端に結合しているアルキルテイル部分を含むペプチド両親媒性物質である。ペプチド両親媒性物質のエピトープペプチド部分は、エピトープが他の分子またはタンパク質との非共有結合相互作用に利用可能であるように、βシート形成ペプチド部分の第2の末端に結合される。好ましくは、エピトープは、ファージディスプレイ法によって誘導される配列を有する。または、ペプチド両親媒性物質は、2つの末端を有し、βシート形成部分の第2の末端とエピトープペプチド部分の間に結合されている荷電部分ペプチドを含んでもよい。
【0024】
本発明の別の態様は、自己集合型ペプチド両親媒性物質から製造される骨格を含む、増殖因子を細胞に放出するための集合物またはシステムを含み、ペプチド両親媒性物質の少なくとも一部は増殖因子と非共有結合するためのペプチドエピトープを含み、増殖因子結合エピトープはナノファイバー面上に突出している。エピトープのアミノ酸配列はファージディスプレイ法から誘導される。このような骨格は幹細胞を含んでもよく、患者において組織を増殖する方法、組織を再生する方法または組織の移植を支持する方法に使用することができる。細胞以外に、他の治療化合物を、骨格のヒドロゲルに封入されてもまたは結合されてもよい。これらの化合物には、抗炎症剤、化学療法剤またはこれらの併用を挙げることができるが、これに限定されない。本発明の方法は、患者の部位に骨格を挿入する段階またはインビボにおいて骨格を形成する段階を含む。骨格は、増殖因子と非共有結合するためのペプチドエピトープを有する自己集合ペプチド両親媒性物質を含み、エピトープは骨格のナノファイバー表面に突出するまたはファージディスプレイ法によって作製され、骨格から周囲細胞に増殖因子を放出する。
【0025】
記載されている増殖因子結合ヒドロゲルは、再生医療および移植医療の分野においていくつかの適用を有する。本発明の組成物、それらを製造する方法およびそれらを使用する方法は、他の増殖因子のためのペプチド両親媒性物質の他のエピトープに、従って多種多様の組織の再生または患者の移植組織の支持に容易に拡大することができる。例えば、BMP-2およびTGF-β1は間葉系幹細胞の造骨および軟骨形成分化に重要な役割を果たし、本発明の自己集合ミセルは骨および軟骨の再生に使用することができる。第二に、BMP-2はまた、脳および脊髄背側部の形成に重要な役割を果たしている。従って、ゲルは、損傷した脊髄の再生のためおよび/または卒中症例の損傷した脳領域を修復するために神経幹細胞と併用して使用してもよい。
【0026】
上記に考察するように、ある態様において、本発明は、選択的非共有結合的相互作用によって増殖因子に結合することができるPAを組み入れるための混合ファイバーの拡張された方法を使用する。この方法では、増殖因子のバイオアベイラビリティーは、結合強度および結合部位の数を調整することによって調節することができる。結合の局所作用の如何または増殖因子を不活性にする結合の影響が支配的であるかどうかに応じて最初の増殖因子濃度は増加または低下される。どちらの場合でも、増殖因子の放出を長期間持続することができる。増殖因子によって認識される結合エピトープを促進するために、エピトープがファイバー表面から延在するようにPAを製造することができる。結合エピトープの必要なアミノ酸配列は、ファージディスプレイを使用して決定することができる。例示目的のために、選択した増殖因子には骨形成タンパク質-2(BMP-2)およびトランスフォーミング増殖因子β1(TGF-β1)が挙げられる。特に、BMP-2は、骨の形成ならびに脳および脊髄背側部の発生に関与するが、TGF-β1は軟骨の形成および平滑筋細胞の分化に関与する。間葉系幹細胞の分化を制御するために対応するゲルを使用することができる。間葉系幹細胞は、インビトロおよびインビボにおいて、骨、軟骨、脂肪、筋肉細胞および心筋に分化することが示されている。BMP-2の影響下における骨系統への分化を以下に示す。
【0027】
ゲルは、間葉系幹細胞の予測される造骨系統への分化を制御する際に培地に増殖因子を添加するより優れている。さらに、結合ゲルは、最初は、全ての他の系統への分化を抑制する。このような挙動は、増殖因子の低いバイオアベイラビリティーによってまたは結合ゲルのこのような増殖因子との細胞の接触がゆっくりであることによって生じると考えられる。最終的に、一部のα-平滑筋発現も結合ゲルに見られる。BMP-2との長期接触または完了までの分化を誘導する因子の欠損は集団の均一性に負の影響を与えることをこれは示唆している。最後に、結合配列がバイオアベイラビリティーを加減するために存在する場合には、内因的に産生されるBMP-2レベルが十分であると思われる。
【0028】
増殖因子結合PAと通常のフィラーPAの同時-集合の成功により、増殖因子に結合し、ゲル状態に維持し、バイオアベイラビリティーを調節することができるヒドロゲルの製造が可能になる。最初に結合PAを含有するゲルにおいて間葉系幹細胞を増殖する場合には、結合PAの非存在下において増殖されるものと比較して、特殊化した細胞のより均一な集団が得られる。適用に応じてゲルの物理的特性を変更するために他のフィラーPAが望ましい場合もあり、結合強度を最適化するために他の結合配列が望ましいこともある。また、本発明によって証明される方法は、多数の結合PAとフィラーの単純な混合によって多数の信号の組み込みを可能にする。おそらく、幹細胞の分化をさらに最適化するためには多数の増殖因子を必要とする。多数の増殖因子に接触させた間葉系幹細胞の分化は現在検討中である。多数の増殖因子系の一時的な放出は、増殖因子の異なる結合定数を有する結合物を選択することによって実施することができる可能性がある。また、本明細書に記載するシステムのインビボにおける検討が実施中である。最後に、本明細書に提供する方法は、細胞動員、特殊化または維持に関与する任意の増殖因子またはタンパク質に容易に拡大することができ、再生医療の有望性の高い方法にしている。
【図面の簡単な説明】
【0029】
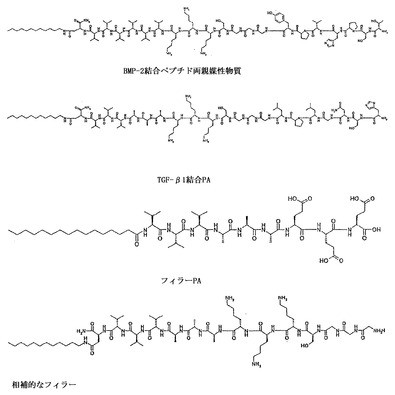
【図1】集合に使用するためのエピトープ配列を含む代表的なPAおよび相補的なPAを例示する。
【発明を実施するための形態】
【0030】
ある種の態様の詳細な説明
本発明は、特異的な非共有結合相互作用によって増殖因子に結合することができる自己集合ペプチド両親媒性物質(PA)に基づいたシステムを記載する。増殖因子は細胞の信号伝達において重要な役割を果たしており、従って、幹細胞の分化を制御するための有望な因子であり、移植した組織の組み込みをサポートするおよび/または非機能組織の再生を生ずる可能性のある休止中の生物学的過程をインビボにおいて分子的または生理的に再-活性化する。増殖因子はインビボにおいて迅速に分解し、損傷部位から迅速に拡散消失する。従って、増殖因子を損傷部位に保持し、増殖因子を周囲の細胞に徐放的に放出する骨格が必要とされる。自己集合ペプチド両親媒性物質を使用しているヒドロゲルは、細胞外マトリックス(ECM)のいくつかの機能を満たす、すなわち、細胞接着を促進し、分化を制御することができることが示されている。本発明は、骨形成タンパク質-2(BMP-2)、トランスフォーミング増殖因子β1(TGF-β1)、VEGFおよびIL-6が挙げられるが、それに限定されるわけではない増殖因子に特異的に結合することができるペプチド配列でこれらのECM置換基を修飾することに関する。これらの増殖因子ペプチド配列はファージディスプレイ技法によって入手することができ、ペプチド両親媒性物質に結合して結合ペプチド両親媒性物質を形成することができる。結合ペプチド両親媒性物質およびフィラーペプチド両親媒性物質は自己集合して、特定の形状に成形され、組織修復の骨格として使用することができるヒドロゲルを形成することができる。または、結合ペプチド両親媒性物質およびフィラーペプチド両親媒性物質はインビボまたはインビトロにおいて細胞試料または組織試料に導入して、試料中でヒドロゲルに自己集合することができる。
【0031】
上記のように、本発明のペプチド両親媒性物質は、アルキルテイル、β-シート形成ペプチド配列および生物活性ペプチド配列を含むことができる。最初の2つのブロックは、ナノファイバーなどであるが、これに限定されないミセルファイバーを形成し、適切な条件下において(多価イオンの添加による中和)、ヒドロゲルまたは溶媒が細孔を満たしている他の湿潤ゲル)を形成する。好ましくは、生物活性なエピトープは溶解のために部分的に荷電しているが、この荷電は、インテグリン-結合配列、向神経活性(neuro-acitive)配列等のような多種多様の配列において異なってもよい。さらに、相補的な荷電のペプチド両親媒性物質は自己集合して混合ファイバーを形成する。この原理を使用して通常のPAおよびファイバー表面から延在している増殖因子結合エピトープ配列を有する長いPAからなるナノファイバーを作製することができる。望ましい増殖因子結合配列は、無作為ペプチドライブラリーを使用するファージディスプレイによって得られ、コンビナトリアル選択過程を使用して強力な結合配列を生ずる。増殖因子の放出は、マトリックスの分解、細胞外受容体への結合またはこれらの組み合わせによって誘導される。
【0032】
既存の技術は、増殖因子をヒドロゲルに捕獲し、それらをポリマーテザーに共有結合する(国際公開公報第03/040336号および米国特許第0020007217号)またはそれらを陰イオンポリマー(国際公開公報第0/13710号)またはヘパリン(国際公開公報第00/64481号)に静電気的に結合する。テザーシステムの欠点は、製造の複雑さおよび増殖因子が細胞外受容体に到達しない可能性である。捕獲されている増殖因子および陰イオンポリマーと複合体形成している増殖因子は比較的迅速に放出され、徐放的な制御は少ない。例えば、陰イオンポリマーに結合している増殖因子は8時間以内に放出される。ヘパリン-結合システムは非特異的であり(例えば、血清に存在する因子の望ましくない結合が生じうる)、増殖因子のサブセットに限定される。有利なことに、本発明は、望ましい増殖因子だけが、結合状態で増殖因子を保持するのに十分に大きい結合強度で複合体形成されるが、細胞外受容体との結合と競合しない程度に弱いように設計される。
【0033】
本明細書に記載する本発明の一局面は、ファージディスプレイ技法を使用して増殖因子を結合するペプチド配列を誘導体化し、その後これらの結合配列を有するペプチド両親媒性物質を製造するために使用することができるということである。ファージ技法によって作製される増殖因子には、骨形成タンパク質、トランスフォーミング増殖因子β1、ニューロトロフィンおよび分裂促進因子FGF-2、ソニックヘッジホッグおよびWnt-3aおよびを挙げることができる。これらの増殖因子タンパク質は、短いペプチド両親媒性物質と結合して、増殖因子ペプチドを有する長いペプチド両親媒性物質を形成する。これらの増殖因子エピトープを有するPAまたは結合ペプチド両親媒性物質は短いフィラーまたは相補的なフィラーPAと同時集合して、結合エピトープが表面から延在するナノファイバーを作製し、その後多価イオンの添加またはpHの変化の結果ヒドロゲルを形成することができる。次いで、得られたヒドロゲルを、ヒドロゲル内部に封入されている幹細胞などであるが、これに限定されない細胞と共に使用してもまたはインビボにおける組織再生のために送達ビヒクルとして単独使用してもよい。細胞以外に、他の治療化合物をヒドロゲルに封入してもまたは結合してもよい。これらの化合物には、抗-炎症剤、化学療法剤またはこれらの併用を挙げることができるが、これに限定されない。
【0034】
ファージディスプレイは、特定の標的に対する分子プローブを作製するためならびにタンパク質-リガンド相互作用を分析および操作するために使用することができる。ファージディスプレイは、アミノ酸、ペプチド、増殖因子、酵素および種々の核酸などであるが、これに限定されない標的分子に結合するアミノ酸配列、ペプチドまたはタンパク質を決定するために使用される。ファージディスプレイは、市販のライブラリーを使用して、マイクロタイターウェルプレートに物理吸着する増殖因子のような標的分子に実施することができる。好ましくは、ライブラリーは、G3コートタンパク質のN-末端が、アミノ酸の全ての可能な組み合わせが存在する方法で、1〜約13以上のランダムアミノ酸が延在しているファージからなる。ライブラリーは増殖因子などの標的分子に接触され、その後非特異的に結合したファージは界面活性剤溶液と共に溶出される。結合ファージの回収後、この集団を例えば、大腸菌(E. coli) において増幅し、溶出条件の緊縮度を増してこの過程を2回以上反復する。最終的な集団を結合ファージに関して濃縮し、クローンのいくつかのDNAを配列決定して、これらのファージ上に提示されているアミノ酸配列の同定を可能にすることができる。その後、ELISAアッセイを実施して、単離されたクローンの相対的な結合強度を推定することができる。固体状態の合成を使用して、同定されたアミノ酸配列を使用して結合ペプチド両親媒性物質を作製するためのペプチドを作製する。
【0035】
本発明のフィラーPAおよび結合エピトープペプチド両親媒性物質のペプチド成分は天然型アミノ酸および人工アミノ酸を含んでもよい。対応する成分がこれに関してペプチド-様であり、ナノファイバーなどのミセルを形成する自己集合であるという効果で、βまたはγアミノ酸などの人工アミノ酸および非天然側鎖を含有する人工アミノ酸および/またはヒドロキシ酸などの他の同様のモノマーの組み込みも考慮されている。
【0036】
本発明の種々のペプチド両親媒性物質は、上記の組み込まれている公報である国際公開公報第03/054146号および全体として参照により本明細書に組み入れられる、Hartgerink, et al. (例えば、J. D. Hartgerink, E. Beniash and S. I. Stupp, Science 294, 1683-1688, 2001参照)によって最初に記載されているものの変法を含む、当業者に周知の製造技法を使用して合成することができる。本発明の合成方法およびエピトープを使用することができる分岐鎖状ペプチド両親媒性物質も本発明において考慮されている。分岐鎖状ペプチド両親媒性物質は、内容が全体として参照により本明細書に組み入れられる、2004年12月6日に本願と同時提出された上記の同時係属出願に開示されている方法および組成物を使用することによって作製することができる。結合エピトープを有するこのような分岐鎖状ペプチド両親媒性物質もナノファイバーに自己集合すると思われる。分岐鎖状ペプチド両親媒性物質は、βシート形成ペプチド部分の第1の末端に結合しているアルキルテイル部分およびエピトープの第2の末端において1つ以上のペプチド分岐鎖を含むペプチド部分を有すると思われる。ペプチドは分岐鎖状アミノ酸および荷電ペプチド部分を含む。分岐鎖状エピトープペプチド部分は、ペプチド両親媒性物質のβシート形成ペプチド部分の第2の末端に結合してもよい。分岐鎖状ペプチドの結合エピトープ配列はファージディスプレイ法から誘導されてもよい。これらの参照文献に記載する合成スキームを本発明に適用することができる。ペプチド両親媒性物質は、完全にプロトン化された形態であっても、部分的にプロトン化された形態であってもまたは酸もしくは塩基付加塩として存在してもよい。一般に、このようなペプチド両親媒性物質は、上記の組み入れられている参照文献に記載されているまたは本明細書に提供する標準的な固相ペプチド化学によって作製することができる。当業者に公知であると思われ、望ましい両親媒性組成物またはペプチド配列に応じて公知の手法および合成技法または直接的な改良を使用して承知しているこれらの合成方法の改良を実施することができる。
【0037】
図1に示す種類のフィラーペプチドは、好ましくは、結合エピトープを有するペプチド両親媒性物質より長さが短く、これらのペプチド両親媒性物質の組み合わせから自己集合するナノファイバーは、増殖因子結合エピトープがナノファイバーの表面から突出する。相補的なフィラーペプチド両親媒性物質は、酸塩基または他の種類の結合によりフィラーペプチドのアミノ酸と相互作用するアミノ酸の側基を有する。
【0038】
ファージディスプレイ法から誘導されるペプチド配列は、増殖因子、酵素、核酸(RNA、DNA)およびアミノ酸を含むが、これらに限定されないペプチドおよび分子に非共有結合するためのペプチド両親媒性物質の結合部位またはエピトープとして使用することができる。ファージディスプレイはエピトープ配列を誘導するために好ましいが、酵母ハイブリッドシステムなどであるが、これに限定されない結合エピトープの配列を同定し、決定する他の相補的な方法を使用することができる。これらの結合ペプチドまたはエピトープをβシート形成ペプチドの遊離端、荷電ペプチドの遊離端またはスペーサーペプチドの遊離端に結合することができる。
【0039】
ペプチド両親媒性物質デザインは、ペプチド両親媒性物質の自己集合のために分子の円錐形状の細長い部分を形成するために働く単純な疎水性テイルを含むことができる。好ましくは、疎水性テイルは、種々のサイズであってもよいが、好ましくは、炭素原子の長さが6を超えるアルキル鎖である。アルキルテイルは、ペプチド両親媒性物質のβシート形成構造セグメントに共有結合されている。
【0040】
βシートペプチド構造セグメントは、テイル基をファージペプチド;または荷電ペプチド、スペーサーペプチドおよびファージペプチド;または荷電ペプチドおよびファージペプチドに共有結合するために使用することができる。βシートペプチド構造セグメントは、一方の末端がテイルに、他方の末端が、種々のペプチドのアミノ酸配列に共有結合する。架橋が望ましい場合には、システインアミノ酸をセグメントのいずれかに使用してもよいが、好ましくは、構造βシートセグメントに使用する。架橋が望ましくない場合には、アラニン、セリンまたはロイシンなどであるが、これに限定されない他の疎水性アミノ酸をこの領域に使用することができる(例えば、本明細書にさらに詳細に記載するSLSLまたはAAAA)。このシステイン-フリーシステムは、ナノファイバーマトリックスの分解速度を制御するために、インビボにおける生物適用にさらに適当となりうる。システムのSLSL改良により、骨格のインビボにおける集合を制御するために使用することができるナノファイバーの自己集合を遅くすることを予想することができる。理論に結びつけたいわけではないが、かさばるロイシン側鎖は、ファイバーへの充填に長い時間を必要とすると考えられる。遅い自己集合は、ナノファイバーの形成が遅いことが有利となることがある、手術室などの機能的なインサイチュー環境における適用性が大きい場合もある。構造的なβシート形成セグメントはグリシンまたは他のアミノ酸を含むフレキシブルなリンカーを含む場合もある。構造セグメントが疎水性アミノ酸を含む場合には、疎水性アミノ酸およびアルキルテイルが疎水性セグメントと考えることができる。構造セグメントが親水性アミノ酸を含む場合には、親水性アミノ酸および親水性ヘッド基が親水性セグメントと考えることができる。
【0041】
β-シート形成単位は、好ましくは、βシートを形成するように相互作用することができ、ペプチド両親媒性物質の全体的な円錐形状を形成する助けをするような疎水性アミノ酸を含む。突出しているペプチド両親媒性物質については、この単位のアミノ酸の数は、ナノファイバーを形成するために使用されるフィラーペプチド両親媒性物質より長く、結合しているペプチド両親媒性物質のペプチドへの接近しやすさを提供するペプチド両親媒性物質を提供するように選択することができる。このセグメントには約4〜約10のアミノ酸が存在してもよく、最も好ましくは、約6のアミノ酸が存在してもよい。β-シート形成セグメントについては、好適なアミノ酸グリシン、アラニン、バリン、ロイシンおよびイソロイシンならびに同様の化学的および構造的な方法でペプチド両親媒性物質において使用することができる他の非天然型アミノ酸には、を挙げることができるが、これに限定されない。水性環境において、好ましくは、患者の部位においてペプチド両親媒性物質の溶解性を提供する荷電セグメントが結合ペプチド両親媒性物質に存在する。荷電ペプチドセグメントには、この溶解性を提供し、自己集合を可能にするようなアミノ酸およびそれらの組み合わせを挙げることができ、ペプチド両親媒性物質の溶解性を改良するためにEまたはKなどの極性アミノ酸およびそれらの組み合わせに限定されない。このセグメントには約2〜約7のアミノ酸(amino)が存在してもよく、好ましくは、約3〜4のアミノ酸が存在する。このセグメントは、第1の末端が構造ペプチドに結合し、第2の末端が、ファージディスプレイ法から誘導されるペプチドに結合するために使用される。スペーサー基ペプチドがペプチド両親媒性物質に含まれてもよい。スペーサー(space)は、SおよびGなどであるが、これに限定されないアミノ酸を含んでもよく、スペーサー(space)は1〜約6のアミノ酸を含んでもよい。ファージ上に提示されているペプチドが遊離N-末端を有する場合には、ペプチド両親媒性物質の典型的な合成スキームを遊離N-末端を提供するように修正することができる。これは、側鎖をドデシルアミンで官能基化することができるアスパラギン酸誘導体を合成することによって実施することができる。このアミノ酸をWang樹脂に結合することにより、N-末端を官能基化する必要なしに、典型的な固相状態のFmoc-化学によってPA合成を完了することができた。遊離PAは、95% TFA/2.5 %水/2.5 %トリイソプロピルシランで処理することにより得ることができる。残存TFAはPAを3 mMのHClに溶解することによって除去することができ、室温において1時間平衡させ、次いで凍結乾燥することができる。これらの分子の合成の成功は、エレクトロスプレー質量分析法を使用して確認することができる。
【0042】
増殖因子エピトープを有するペプチド両親媒性物質は、体液に存在するイオンおよび自己集合を促進するために添加されるイオン/試薬を使用して、インビボにおいて患者の部位において自己集合することができる。または、増殖因子結合エピトープを有する好適なペプチド両親媒性物質の組成物を型に注ぎ、自己集合を使用して、置換または再生される組織または骨の形状の骨格を形成する。成形された骨格を、修復または再生治療を必要としている患者の部位に挿入することができる。組織移植の場合には、増殖因子結合エピトープを有するペプチド両親媒性物質は型で支持構造物またはマトリックスに形成され、組織移植のための患者の支持として使用することができる。ペプチド両親媒性物質ナノファイバーまたは骨格は、幹細胞などであるが、これに限定されない細胞を含んでもよい。他の治療化合物がヒドロゲルに封入されてもまたは結合されてもよい。これらの化合物には、抗炎症剤、化学療法剤またはこれらの組み合わせを挙げることができるが、これに限定されない。
【0043】
増殖因子結合エピトープを有するペプチド両親媒性物質と短いβシート構造ペプチド-荷電ペプチド酸-末端フィラーペプチド両親媒性物質の同時集合を使用して、本発明のナノファイバーおよび骨格を製造することができる。本発明の骨格およびナノファイバーを製造するために、異なる増殖因子結合エピトープを有する1つ以上のペプチド両親媒性物質を使用することができ、種々のフィラーペプチド両親媒性物質を使用することができるが、限定するものではない。フィラーペプチド両親媒性物質と比較した増殖因子結合エピトープを有するペプチド両親媒性物質の量は、本発明の骨格およびナノファイバーを製造する際に異なってもよいが、限定されない。自己集合ミセルおよびナノファイバーは、NOEおよびFT-IR分光分析、円二色性によって特徴づけることができる;ナノファイバーのファイバーネットワークは透過型電子顕微鏡を使用して可視化することができる。
【0044】
結合エピトープを有するペプチド両親媒性物質は、増殖因子またはアミノ酸もしくは核酸などであるが、これに限定されない他のペプチドと非共有結合相互作用によって相互作用することができるエピトープのペプチド配列を選択することによって製造することができる。増殖因子または他のペプチドと結合エピトープを有するペプチド両親媒性物質の非共有結合相互作用の程度は、エピトープが、競合しないまたは細胞外受容体比較して、増殖因子または他のペプチドに対する結合親和性が低いように選択される。受動的な放出実験を使用して、特徴づけ、その後増殖因子結合エピトープを有する種々のナノファイバーの親和性を改良することができる。この特徴づけは、事前ブロックしたマイクロタイタープレート中で増殖因子を含有する自己集合ナノファイバーゲルについて実施することができる。これらのゲルは、適当な比でPAの2%溶液を混合し、次にTBSで1:1希釈することによって作製することができる。次いで、増殖因子を添加し、次に2当量の多価イオンを添加してゲル化および自己集合を誘導することができる。増殖因子含量について上清をアッセイして、このようなゲルの増殖因子結合能を評価することができる。
【0045】
結合エピトープは、近隣の細胞または組織の細胞外受容体より種々のペプチドに対する親和性が弱いことが組織増殖および他の治療に好ましいが、それは、望ましくは、ペプチド、増殖因子、酵素または核酸または体液もしくは細胞試料の他の分子に強力に化学的に結合するペプチド両親媒性物質の結合エピトープであることが考慮されている。この場合には、好適なエピトープを有する結合ペプチド両親媒性を表面で自己集合させて固定して、センサーコーティングまたは除去媒体(removal media)を形成する。これらの強力に結合しているペプチド両親媒性物質の自己集合から形成されるヒドロゲルは、患者の部位に挿入するためまたはろ過システムに使用するために成形してもよい。ヒドロゲルは、患者の関節もしくは腫瘍などの部位または患者の体液からHGFまたはVEGFのような標的ペプチドを除去するために使用してもよい。
【実施例】
【0046】
発明の実施例
以下の限定するものではない実施例およびデータは、増殖因子と非共有結合相互作用をすることができるエピトープ配列を有するまたは有しない特定の範囲の両親媒性ペプチド化合物の合成、このような化合物または組成物の自己集合ならびに増殖因子のバイオアベイラビリティーおよび幹細胞分化に影響を与えるそれらの用途を含む、本発明の化合物、組成物および/または方法に関する種々の局面および特徴を例示している。従来技術と比較すると、本発明の化合物、組成物および/または方法は驚くべきであり、予測されず、従来技術と異なる。本発明の利用性はいくつかの化合物/組成物および集合物を使用することにより例示されているが、同様の結果は、本発明の範囲に相応の種々の他の化合物、組成物および/または集合物で得ることができることが当業者によって理解される。
【0047】
樹脂およびFmoc-1-アミノ酸は全てNovabiochem(San Diego, CA)から入手した。固相合成のための試薬は全て合成用であり、Applied Biosystems(Foster City, CA)から入手した。他の試薬は全てAldrich Chemical Co. (Milwaukee, WI)から入手し、入手したままで使用した。固相ペプチド合成のための溶媒はApplied Biosystemsから入手し、ペプチド合成用であった。他の溶媒はFisher Scientificから入手し、特に記載しない限り、入手したままで使用した。
【0048】
PAは、Applied Biosystems 433A自動ペプチド合成装置を使用して合成した。NMRスペクトルは、Varian Inova 500 MHz分光計で室温において得た。エレクトロスプレー質量スペクトルは、Micromass Quattro II Triple Quadrupole HPLC/MS/MS質量分析計で収集した。CDスペクトルは、Jasco PTC-348WIペルチェ効果温度コントローラー付きJasco J-715分光偏光計で記録した。FT-IRスペクトルは、BioRad FTS-40 FT-IRマシーンで、2 cm-1の分解能で400から4000 nmまで操作した。
【0049】
円二色性分光法. 路程0.1 cmの石英セルを全ての実験に使用した。スキャン速度100 nm/min、反応時間2秒およびバンド幅1 nmで300 nmから190 nmまで各スペクトルを記録し、5回スキャンを平均した。試料は、特に記載しない限り、水で0.1 mg/mLの濃度に調製した。
【0050】
酸-塩基滴定. pKa滴定は、100 mM KClにおいて3.5 mg/mLの濃度において、Fisher Accumet pHメーターを用いてpH 2〜10の範囲のPA 1〜4に実施した。酸性PA 2および4については、低いpHから開始して、0.1 N KOHを1〜5μLずつ添加したが、塩基性PA 1および3については、高いpHから開始して、0.1 N HClを1〜5μLずつ添加した。
【0051】
NMR NOE分光分析. PA1〜6をd6-DMSOに5 mg/mLの濃度で溶解した。NOESYスペクトルは、各PAの10または15 mg/mL濃度、モル比1:1において、ミキシングタイム0.1 s、128スキャンでD2O中で測定した。FT-IR検討については、2重量%の試料を水から凍結乾燥し、次いでKBRペレットに圧縮した。
【0052】
スキーム1. ペプチド両親媒性物質の化学構造

【0053】
実施例1
トリプルリジン配列を有する2つ(1, 3)およびトリプルグルタミン酸配列を有する2つ(2, 4)の4つのPAを製造した(スキーム1)。PA 1および2は、プレロードしておいたWang樹脂を使用し、次にカップリング試薬として2-(1H-ベンゾトリアゾール-1-イル)-1,1,3,3-テトラメチルウロニウムヘキサフルオロホスフェート(HBTU)を用いてパルミチン酸でアルキル化して、標準的なFmoc固相ペプチド技法によって製造した。95%トリフルオロ-酢酸(TFA)、2.5%水および2.5%トリイソプロピルシラン(TIS)の混合物を用いて両親媒性を樹脂から切断した。ペプチド両親媒性物質3および4の合成は、スキーム2によりアミノ酸7を合成した。N-カルボベンジルオキシ-L-アスパラギン酸無水物をドデシルアミンと反応させて、脂肪酸アミノ酸5を得た。次いで、接触水素化によってCBz基を脱離して、6を得、次いでアミンのFmoc-保護を実施した。この合成は、5グラムスケールで容易に進行する。次いで、カップリング試薬としてHBTUを使用して生成物7をリンク樹脂にカップリングした。その後、標準的なFmoc固相技法を使用して、残りのアミノ酸を8に添加した。以前に記載したように、標準的な切断条件により、逆構造を有するPAを得た。1H NMRおよびエレクトロスプレーイオン化質量分析は予想される構造に一致している。
【0054】
スキーム2. 脂肪酸アミノ酸の合成
【0055】
実施例1a
N-ドデシル-2-カルボベンジルオキシアミノ-スクシンアミド酸(5).
N-カルボベンジルオキシ-L-アスパラギン酸無水物(1 mmol)を50 mL塩化メチレンに溶解し、次に1.05 eqのドデシルアミンおよび1.1 eqのトリエチルアミンを添加した。蒸散を防ぐために反応に蓋をし、12時間撹拌した。薄層クロマトグラフィー(TLC)(CH2Cl2, 5% MeOH)によって出発材料がわずかでも検出されなかったら、20 mLの1N塩酸で反応を停止し、次にクロロホルム(5×)で抽出した。有機層を硫酸マグネシウムで乾燥し、7を白色の固体として得た(収率97%)。
【0056】
実施例1b
2-アミノ-N-ドデシル-スクシンアミド酸(6). 100 mlのエタノールに、100 mmolの7を溶解し、PdおよびC(10重量%)を含有する反応容器に移した。次いで、容器を水素雰囲気下(35 Torr)に3時間おいた。反応混合物をセライトでろ過し、減圧下蒸発乾固して、生成物を白色の固体として得た。収率95%。
【0057】
実施例1c
N-ドデシル-2-Fmoc-アミノ-スクシンアミド酸(7). 水/ジオキサン(1:1 v:v)混合物の200 mLに、6.6 mmolの6を1.3 mL(1.5 eq.)のトリエチルアミンと共に溶解し、次に1eq.のFmoc-Oスクシンアミド(Fmoc-OSu)を溶解した。反応をTLC(CH2Cl2, 10% MeOH)でモニターし、2〜3時間後、全てのFmoc-OSuが消費された。反応を酸で停止して、白色の沈殿物を得、ろ過して回収した。収率85%。
【0058】
実施例1d
PA合成および精製. 参照文献3に記載されているように、PA 1〜2を製造した。PA 3〜6は、標準的なリンク樹脂を反応容器に入れ、30%ピペリジンのNMP溶液で3回脱保護し、次いで2eq.の7に終夜カップリングした。ニンヒドリン試験が陰性の結果を示すまで、カップリングを反復した。次いで、この修飾樹脂8を自動合成装置にロードし、PA 1〜2についてと同様にペプチド合成を進行した。自動合成が完了したら、参照文献3に記載されているように、PAを樹脂から切断した。
【0059】
実施例1e
C15H31CONHVal-Val-Val-Ala-Ala-Ala-Lys-Lys-Lys-COOH(1)
【0060】
実施例1f
C15H31CONHVal-Val-Val-Ala-Ala-Ala-Glu-Glu-Glu-COOH(2)
【0061】
実施例1g
Asp(CONHC12)- Val-Val-Val- Val-Val-Val- Lys-Lys-Lys-NH2(3)
【0062】
実施例1h
Asp(CONHC12)- Val-Val-Val- Ala-Ala-Ala-Glu-Glu-Glu--NH2(4)
【0063】
実施例2
PAは、好適な刺激の存在下で水性媒体に分散すると、典型的には、自己集合して高アスペクト比の円筒形ナノファイバーを形成する。以前の研究に基づいて、1つのPA分子または2つのPA分子の混合物からなるファイバーは、β-シート-様水素結合を有する二次構造を示すことが予想されると思われる。負に荷電している2または4とそれぞれ反対に荷電している1または3を混合すると平行なβ-シート-様水素結合配列を生ずると思われるが、1または2とそれぞれ末端アミンが反対に荷電している4または3を混合すると逆平行配列を生ずると思われることが予想された。
【0064】
実施例3
0.1 mMの濃度では、2〜4のCDスペクトルは、主にランダムコイル特性を有するペプチドセグメントを明らかにした。これは、荷電が大きい分子間の静電反発力によって生じる可能性が最も大きい。荷電を中和するようにpHを変化するまたはカルシウムイオンを添加すると、全てのPAはβ-シート構造を示した。一方、1は任意のpHにおいてβ-シートシグニチャーを示す。1は、酸性末端がリジン残基の1つの荷電を中和して、みかけの(formal)正味荷電+2を式に与える可能性があるので、他より荷電が小さい可能性がある。この低い全体荷電は反発力を低下し、ファイバー内にβ-シート水素結合を生じさせる可能性がある。逆に、4は、アミン末端がグルタミン酸残基の1つの荷電を中和すると思われるので、-2のみかけの正味荷電を有し、β-シート相互作用を生じさせると思われる。しかし、4は依然として乱雑なCDシグニチャーを示した。このみかけの矛盾を解決するために、pH 7における集合物の実際の荷電状態およびみかけのpKaを求めて、これらの種々のシステムの自己集合の駆動力をよりよく理解した。
【0065】
実施例4
3 mMの濃度で分子1〜4の集合物のpKa滴定を実施した。自己集合による動態的な影響を回避するために、全ての滴定は、分子がすでに集合した状態にあるpHから開始した。3のpKa滴定だけが、2つのみかけのpKaに関連するシャープな遷移を示し、一方は溶媒接近性の大きいアミン末端の脱プロトン化に起因し、他方はεリジンアミンの1つ以上によって生じた。PA 1、2および4の集合物は複雑な曲線を示し、遷移は広い範囲にわたって生じ、これらの超分子物体のプロトン化/脱プロトン化はゆっくり生じ、酸性度はナノファイバー内の局所的な微小環境により変化することを意味している。文献の最近の報告と矛盾することなく、集合物は酸およびアミン基のみかけのpKaを変化させることがこれらの結果から明らかである。
【0066】
実施例5
荷電相補的PA分子の自己集合により、β-シート水素結合配列を有する2つの成分の混合物を含有するファイバーを生ずるはずである。2をトリプルリジン両親媒性物質3または1と混合すると(2/3または2/1と略す)、得られるCD-スペクトルは純粋なβ-シートに対応する。観察されるβ-シートシグニチャーは、単に2つの成分の個々のCDスペクトルの重ね合わせではないという事実は、2つの分子が1つの集合物構造を形成している混合ナノファイバーの形成を強く示唆している。トリプルグルタミン酸PA4または2と混合したPA1(1/4または1/2)も同様の挙動を示す。同様の荷電の2つのPAを混合すると、1/3リジン混合物はβ-シートを示すが、2/4グルタミン酸混合物は、おそらくグルタミン酸残基間の大きい荷電反発性のために、乱雑な立体配座を示す。
【0067】
実施例6
ファイバー内での分子の自己集合をさらに証明するために、荷電相補的PAから製造した1.5重量%ゲルに核オーバーハウザー(NOE)分光法を実施した。2および3(2/3)の代表的なNOESYは、それぞれ、2のGlu-Hβプロトンと3のLys-HεおよびVal-Hδプロトンの間で観察される密接な接触(<3Å)を示す。考えられるいくつかの他の分子間接触が検出されたが、2/3接触にはっきりと起因させられなかった。これらの結果は、2つのPA分子が同一のナノファイバー内に同時集合されている追加の証拠を提供している。
【0068】
実施例7
両親媒性の自己集合の成功が確立されたら、シングルおよびマルチプルPA由来のシステムのゲル化挙動を検討した。1〜4の1重量パーセント溶液はわずかに不透明で、それぞれ、酸(1、3)または塩基(2、4)の添加によりゲル化することができた。透過型電子顕微鏡は、他のPAにおいて以前に観察されたものと同様の、平均径6.5 nm、平均長さ数百ナノメーターのナノファイバーの形成を明らかにしている。
【0069】
実施例8
ファージディスプレイは、典型的には、受容体-ブロッキングペプチド、すなわち高ブロッキング定数を有する配列を見つけるために使用される。PA結合部位から細胞受容体に増殖因子を移動させるためには、極端に強い結合定数は必要とされず、望ましくない場合もある可能性がある。従って、13-merライブラリーより7-merの市販のファージライブラリーを選択し、中間的な結合強度を得た。増殖因子を96-ウェルプレートに物理吸着させ、その後ファージライブラリーに接触させた。室温において60分インキュベーション後、界面活性剤溶液ですすいで未結合のファージを除去した。その後、増殖因子溶液と共に60分インキュベーションすることによって結合ファージを溶出した。回収したファージを37℃において大腸菌(E. coli) 内で4時間増殖し、パニングを2回以上反復した。その後のパニングラウンドにおいて、界面活性剤濃度を上昇し、ライブラリーへの接触時間を短縮することによって緊縮度を増した。最終的なファージ集団を平板培養し、1つのファージタイプだけを含有するコロニーの10クローンのDNAを配列決定した。次に、ELISAスクリーニングを使用して偽陽性を試験し、選択したクローンの相対的な結合強度を求めた。このプロトコールを使用すると、rh-BMP-2の最良の結合エピトープはYPVHPSTであり、rh-TGF-β1はLPLGNSHであることが見出された(以下の表1参照)。
【0070】
実施例8a
ファージディスプレイは、Ph. D. 7キット(New England Biosystems)を用いて実施した。50マイクロリッターの希釈したM13-ファージライブラリー(2×109ファージおよび可能な全ての7-mer配列を含有する)を96-ウェルマイクロタイタープレートに添加し、0℃において18時間10μg/mlの増殖因子(peprotech)溶液50μlに接触させた。プレートを1時間ブロックし、次いでゆっくり撹拌しながら室温において60分インキュベーションした。TBS/Tween-20(0.1%v/vTween-20)ですすいで未結合のファージを除去した。その後、50μlの増殖因子溶液と共に60分インキュベーションすることによって結合ファージを溶出した。ファージを連続希釈し、その後寒天プレートで大腸菌(E. coli) と共に平板培養することによって結合ファージの存在を判定した。形成されたプラークの数を元の混合物中のファージの数に関連させることができる。その後、ファージを37℃において大腸菌(E. coli) 中で4時間増幅した。その後のパニングラウンドにおいて、Tween-20の濃度を0.5%に上昇し、結合時間および溶出時間をそれぞれ短縮および延長して、選択過程の緊縮度を増した。3ラウンドのパニング後、回収したファージ混合物を希釈し、寒天で細菌と共に平板培養した。その後、1つのDNA配列を含有するプラークを単離する。精製後、10クローンを配列決定した。次に、ELISAスクリーニングを使用して偽陽性を排除し、選択したクローンの相対的な結合強度を求めた。増殖因子をコーティングしたマイクロタイタープレートで各クローンを1時間インキュベーションし、プレートをTBS/Tween-20ですすぎ、その後HRP-結合抗-M13抗体(Amersham Biosciences)を1時間インキュベーションした。ABTSの添加後、30分後に450 nmにおける吸光度を測定し、単離したクローンの相対的な結合強度を求めた。その後、最も強い結合物を選択した。
【0071】
(表1)ファージディスプレイを使用して誘導されるBMP-2およびTGF-β1の結合配列
数値コーディングは、結合強度を最高(1)から最低(4)まで低下する際の選択したクローンの相対的な結合強度を例示している。全ての配列は強く結合している。
【0072】
実施例9
ファージ上で提示されるペプチド配列は遊離N-末端を有するので、PAの典型的な合成経路を使用することができなかった。従って、側鎖をドデシルアミンで修飾したアスパラギン酸をRinkアミド樹脂と反応させた。(上記スキーム2参照)。その後、ペプチドの残りを通常のFmoc-ペプチド合成技法を使用して合成し、逆の極性を有するPAを生じた。選択したペプチド配列は、それぞれ、BMP-2はV6K3SG3YPVHPST(9)であり、TGF-β1はV3A3K3SG3LPLGNSH(10)であった(スキーム3)。K3セグメントは、フィラー2の荷電E3ヘッド基と同時集合するように選択した。SG3配列はスペーサー単位であり、ファージに存在したものと同じスペーサーである。BMP-2のそれぞれYPVHPSTおよびKQALTOTとの結合定数は、蛍光偏光解消によって測定するとき、Ka=1.9±0.9 106 M-1および2.1±0.85 105 M-1であり、結合強度の同調性を例示している。
【0073】
実施例9a
Asp(CONHC12)- Val-Val-Val- Val-Val-Val-Lys-Lys-Lys-Ser-Gly-Gly-Gly-Tyr-Pro-Val-His-Pro-Ser-Thr(9)
【0074】
実施例9b
Asp(CONHC12)- Val-Val-Val-Ala-Ala-Ala- Lys-Lys-Lys- Ser-Gly-Gly-Gly-Leu-Pro-Leu-Gly-Gln-Ser-His-NH2(10)
スキーム3:
【0075】
実施例10
PA 9とフィラーPA 2の同時集合は、円二色性(CD)分光法を使用して証明できると思われる。0.13 mMの濃度において、この濃度における個々のPAナノファイバーのCDスペーサーは主にランダム-コイル特性を有する。しかし、一体として混合すると、CD-スペクトルは純粋なβ-シートを反映し、PAは、1つだけのPA分子からなる個々のホモ-ファイバーを形成するのではなく、1つのファイバーに同時集合することを例示している。同時集合は、KKKおよびEEE部分が複合体を形成する場合には、静電反発力が低下するので、エネルギー的に好ましい。1.5重量%溶液の核オーバーハウザー効果分光法を使用して同時集合の追加の証拠が得られた。EβとKεプロトンの間およびEβとVδプロトンの間に密接な接触(<3Å)が観察され、この距離はホモ-ファイバー間より有意に大きいと思われる。考えられる他の分子間接触は明白に起因させられなかった。最後に、凍結乾燥試料および1重量%溶液のフーリエ変換赤外分光法は、β-シートに典型的な1630 cm-1ピークの存在を示した。同様の結果は10/2混合物で得られた。試料を37℃において24時間アニーリングすることにより、CD効果の強度が有意に増加した。この増加は、完全に結合している発色団のドメインのサイズの増加に起因する。おそらく、最初の混合によって完全でない混合システムの動態的な捕獲を生じ、アニーリングによって、その後両親媒性物質が熱力学的に最も安定な状態に再組織化することが可能になる。水素結合ネットワークおよび脂肪族テイルの予想される融点を十分に上回る80℃における長時間の加熱も超分子キラリティーの任意の顕著な損失を生じず、ナノファイバーの高い安定性を例示している。
【0076】
実施例11
この方法の生理学的重要性を試験するために、異なるレベルの増殖因子を含有するゲルにおいて間葉系幹細胞を培養した。これらの細胞実験では、タンパク質による認識を確実にするために、結合エピトープを50:1(フィラー:結合物)比に希釈した。望ましい量の増殖因子を含有する50μlのプレミックスPA(2重量%)および50μlの間葉系幹細胞懸濁液(100,000細胞)をガラスカバースリップ上で混合し、次に30 mg/mlのCaCl2溶液10μlを添加することによってゲルを調製した。ゲルを1.5時間固化させ、その後1 mlの間葉系幹細胞培地を添加した。この方法では、9/2または2単独からなるおよび0、1または50 ng/mlのBMP-2を含有するゲルが調製された。別の対照として、10,000の幹細胞をガラスカバースリップで平板培養し、同じ濃度のBMP-2を培地に添加した。培地を4日めごとに交換し、培養7日目および21日目にトリプリケート試料を単離した。その後、系統-特異的なマーカーオステオカルシン(骨芽細胞)、アジポネクチン(脂肪細胞)、コラーゲンIIおよびアグリカン(軟骨細胞)ならびにα-平滑筋アクチンおよびデスミン(平滑筋細胞)のタンパク質発現をウェスタンブロットによって評価した。
【0077】
実施例11a
細胞実験のプロトコール:ヒト間葉系幹細胞(Cambrex technologies)を5,000細胞/cm2の密度で4つのT75培養フラスコの15 mlの間葉系幹細胞培地(Cambrex Technologies)に入れ、37℃、5% CO2、湿度90%においてインキュベーションした。集密度90%において、細胞を0.25%トリプシン/EDTA2.5 mlで5分間トリプシン処理した。その後、8 mlの培地を添加して、プロテアーゼ活性を失活させた。細胞懸濁液を合わせて、850 rpmで8分遠心分離した。細胞ペレットを1 mlの培地に再懸濁させ、10μlを細胞計測に使用した。細胞懸濁液を培地で2,000,000細胞/mlの濃度に希釈した。
【0078】
PA 1および3を2重量%の濃度で溶解した。その後、1の50μlを3の1250μlと混合した。混合物を410μlの3部に分割し、さらに3(2重量%)の410μlの3部を調製した。次いで、2μg/ml BMP-2(Peprotech)の20μlを1/3および1の1部に100 ng/ml BMP-2濃度まで添加した。同様に、20 ng/ml BMP-2の41μlを最終濃度2 ng/ml のBMP-2まで添加した。
【0079】
50μlのPA溶液と50μlの細胞懸濁液を電子顕微鏡用ガラスカバースリップ上で混合することによってゲルをトリプリケート(triplo)で調製した。次いで、30 mg/ml CaCl2の10μlを添加し、ゲルを37℃、5% CO2、湿度90%において90分固化させた。得られた濃度は1重量%PAであり、BMP-2では0、1または50 ng/mlである。次に、1 mlの培地を添加した。5μlの細胞懸濁液をガラスカバースリップに置き、1 mlの培地を添加することによって増殖因子の対照溶液を調製した。次いで、2μg/ml BMP-2の25μlおよび2μg/mlのBMP-2の0.5μlを添加して、それぞれ50 ng/mlおよび1 ng/ml濃度のBMP-2溶液を得た。培地の半分を4日目ごとに交換した。
【0080】
実施例12
細胞の生存度は、3週間の培養後に全てのゲルにおいて>95%であることを細胞生存度アッセイは示した。エンドグリンの発現はどの時点においてもウェスタンブロットによって検出することができず、全ての細胞は分化を開始していたことを示している。アジポネクチンおよびアグリカンの発現も任意の時点において検出できなかった。オステオカルシンの発現はゲルと対照間で顕著な差を示した。オステオカルシンは対照では検出できなかったが、全てのゲルは、通常の2〜3週間ではなくすでに7日目に有意なレベルのオステオカルシンを発現したが(図3a)、3週間経過時点において有意に増加しなかった(図3b)。1および3週における同様の発現レベルは、オステオカルシンの発現レベルは培養期間中一定であることを意味している。驚くべきことに、発現はBMP-2濃度に依存しないと思われ、比は、結合PAを含有するゲルと結合PAを含有しないゲルで1:2である。さらに、BMP-2を添加しないゲルも同様のレベルを示す。おそらく、細胞自体によって産生される内因性BMP-2レベルは、骨芽細胞分化を刺激するのに十分であるが、結合ゲルがBMP-2の一部に結合する能力は発現されるオステオカルシン量をわずかに低下する。オステオカルシンの発現はまた免疫細胞化学でも可視化することができた。
【0081】
一方、α-平滑筋アクチンは、9を含有するゲルにおいて検出されなかったが、結合物を含有しないゲルにおいて発現された。しかし、それは3週間後にどちらの場合でも発現されたが(図3b)、9/2のゲルにおける発現は2のゲルよりわずかに低く、対照より有意に低い。さらに、最終分化平滑筋細胞のマーカーであるデスミンの発現を検出することができなかった。
【0082】
実施例12a
ウェスタンブロットプロトコール:培地を除去した後、ゲルを1 mlのリン酸緩衝生理食塩液ですすぎ、その後、200μlの2%ドデシル硫酸ナトリウム、0.08 M Tris、10%グリセロールで細胞を溶解した。3つの溶菌液を合わせ、10μlの溶菌液と200μlのBCAタンパク質アッセイ試薬(Pierce)を混合し、37℃における30分のインキュベーション後BSA標準シリーズに対する562 nmの吸光度を読むことによってタンパク質濃度を測定した。その後、10μgのタンパク質、3μlのβ-メルカプト-エタノール、3μlのブロモフェノールブルーを混合し、水を総容量36μlまで添加した。混合物を5分間煮沸し、4%(エンドグリン、アグリカン)または10〜20%(オステオカルシン、α-平滑筋アクチン、アジポネクチン、デスミン) Novexトリス(グリシン)ゲル(Invitrogen)にロードした。ゲルは、130 Vの定電圧でトリス(グリシン)泳動緩衝液(Invitrogen)で90分泳動した。次いで、タンパク質を、ニトロセルロース膜(Bio-Rad)に移し、190 mAの定電流で90〜120分泳動した。
【0083】
膜を5%脱脂乳(Bio-Rad)中で1時間ブロックし、オステオカルシン(クローン190125, R & D systems)、エンドグリン(クローン166709, R & D systems)、アジポネクチン(クローン166126, R & D systems)、α-平滑筋アクチン(クローン1A4, R & D systems)およびポリクローナルアグリカン(AF1220, R & D systems)に対するモノクローナル抗体を使用して、1%ミルクで1:500希釈のそれぞれのマーカーを2時間探索した。膜を、トリス緩衝生理食塩液/0.1%Tween-20(TTBS)で15分間3回すすいだ。次いで、二次抗体を1%ミルク中で1:3,000希釈で1時間平衡させ、次にTTBSによるすすぎ段階を3回実施した。ECLウェスタンブロット分析溶液(Amersham Biosciences)で1分平衡させることによって膜を形成し、ECLハイパーフィルム(Amersham Biosciences)に接触させた。
【0084】
実施例12b
細胞を2%パラホルムアルデヒド/0.2%グルタルアルデヒド/0.2 Mカルコジレート(calcodylate)ナトリウムで4℃において20分固定した。PBS(2×)ですすいだ後、試料を0.1% Triton-Xと共にPBS溶液中で5分間インキュベーションした。その後、試料をPBSですすいだ(2×)。次に、ヒトオステオカルシンモノクローナル抗体(R & D systems、1:200)の1% BSA/PBS溶液を添加し、試料を4℃において終夜インキュベーションした。PBSで3回すすいだ後、FITC-結合二次抗体の1% BSA/PBSを添加し、2時間インキュベーションした。PBSで3回すすいだ後、Nikon Eclipse TE2000電子顕微鏡で倍率10×で試料を可視化した。
【0085】
実施例13
一連の実験は、TGF-β1の最高濃度が、50 ng/mlでなく20 ng/mlであることを除いて、PA 10、結合TGF-β1、BMP-2実験についてと同様に実施した。試料は10日後に単離した。TGF-β1の存在下における培養により、平滑筋および軟骨系統が分化する。コラーゲンIIの発現は結合ゲルでは高く、TGF-β1の増加により増加すると思われる。驚くべきことに、オステオカルシンおよびコラーゲンXが発現され、この早期時点では予想されない肥大軟骨細胞段階への成熟が生じていることを示している。同様に、α-平滑筋アクチン発現はTGF-β1濃度と共に増加するが、非結合ゲルより結合ゲルにおいて高い。それは対照においてはるかに高いが、デスミンの発現レベルは、これらは主に未熟筋細胞であることを示している。
【0086】
実施例14
7-および12-merファージディスプレイキットを用いて、血管内皮増殖因子(VEGF)、塩基性線維芽細胞増殖因子(FGF-2)、ニューロトロフィン-3(NT-3)およびラミニン-5の数多くの結合配列を決定した;*は最も強い結合物を示す:
【0087】
先の実施例によって例示するように、本発明は、標準的な固相方法に適合する遊離N-末端を有するペプチド両親媒性分子の合成方法を提供する。遊離C-末端PAと混合すると、これらの分子は自己集合して、同じ極性のPAの同時集合物より安定性が高いと思われる熱安定性が高いβ-シート構造を含有するナノファイバーを形成する。表面に遊離N-末端を有する集合物を作製する新規機会により、ペプチド配列のC-末端を露出しているナノ構造物では以前には入手不可能であった生物活性ナノファイバーの設計が可能になる。
【0088】
本発明は、ある好ましい態様を参照してかなり詳細に記載されているが、他の形態も可能である。 例えばファージディスプレイ法によって誘導されるペプチドまたはタンパク質配列を使用して、増殖因子以外のタンパク質に結合して保持する結合ペプチド両親媒性化合物を製造することができる。自己集合して、ナノファイバーヒドロゲルの表面に固定することができる結合ペプチド両親媒性物質を製造するように酵素を結合することができると思われる。このようなヒドロゲルをセンサー適用のコーティングとして使用することができる。または、ペプチド両親媒性物質に結合したファージディスプレイ由来のペプチドの結合相互作用は、HGF(肝細胞増殖因子)またはVEGFなどのペプチドに強力に結合し、それらを除去することによって種々の疾病および状態を治療するように選択してもよい。これらの結合ペプチド両親媒性化合物の自己集合によって形成されるヒドロゲルは患者の部位に挿入するためまたはろ過システムに使用するために成形してもよい。ヒドロゲルは、患者の関節もしくは腫瘍などの部位または患者の体液からHGFまたはVEGFのような標的ペプチドを除去するために使用してもよい。
【技術分野】
【0001】
本願は、全体の内容が参照により本明細書に組み入れられる、2003年12月5日提出の出願番号第60/527,504号の優先権の利益を主張する。
【0002】
米国政府は、エネルギー省(Department of Energy)からノースウェスタン大学(Northwestern University)への付与第DE-FG02-00ER54810号に準じて本発明のある権利を有する。
【背景技術】
【0003】
発明の背景
増殖因子は細胞特殊化において重要な役割を果たし、共に非機能性組織の再生を生じる可能性のある、幹細胞分化を制御するためまたは休止中の生物過程をインビボにおいて再活性化するための有望な因子である。増殖因子は速やかに分解され、細胞部位または損傷から迅速に拡散する。従って、構造または骨格は、制御された様式で周囲の細胞に放出するために、このような部位に増殖因子を保持するのに有用であると思われる。
【0004】
組織再生のために分子的に設計されている材料は高度医療において非常に興味深いものになっている。L-乳酸またはグリコール酸に基づいたポリマーを含む合成ポリマーおよびコラーゲン、フィブリンまたはアルギネートを含むバイオポリマーの骨格が検討されている。増殖因子は、ポリマーなどのヒドロゲルに物理的に捕獲されており、アニオン性ポリマーまたはヘパリンなどの構造物に共有結合または静電気的に結合している。これらのシステムおよび関連するシステムの欠点は、結合している増殖因子の非特異性または望ましい作用を達成するために共有結合の分解の必要性に関する。さらに最近では、受容体に基づいた細胞との相互作用を促進し、細胞接着または分化を促進するために使用されている細胞外タンパク質に見られるペプチドを含有するように天然および合成骨格が改良されている。
【0005】
自己-集合の進歩は、バイオマテリアルの分子設計の新たな機会を提供している。両親媒性分子構築ブロックは、水性環境において集合して、十分に規定されている種々の化学構造と骨格を形成することができる。両親媒性ペプチドおよび一方または両方の末端が疎水性成分(例えば、アルキルテイル)で官能基化されているペプチドを含む種々のクラスのペプチド-系両親媒性物質が文献で報告されている。両親媒性ペプチドは、ナノテープ、リボン、ファイバーおよびツイストリボンのような種々の超分子構造を形成することが示されている。これらの構造は、両親媒性分子のβ-シート形成から生ずる。特別な例は、個々のペプチドブロックの二次構造に強力に依存する特性を有するゲルを形成する両親媒性ペプチドブロックコポリマーの例である。一方の末端にアルキルテイルを有するペプチド両親媒性物質の例には、テイルの長さおよびテイルの数に応じて球状またはディスク-様ミセル構造を形成する三重らせん単位を形成するコラーゲンに見られるペプチドモチーフ由来の両親媒性物質が挙げられる。別のクラスは末端あたり1つの2つのテイルを有する。このクラスの両親媒性物質は、濃度を増加すると、ランダムコイルから繊維状構造を生ずるβ-シート型コンフォーメーションへの遷移を受けるという点においてアミロイド-様挙動を示す。逆平行配列を有する純粋にペプチド性のナノ構造が報告されており、非天然アルキル化四級アンモニウム塩によって誘導される逆平行配列に集合する2つの修飾ペプチド両親媒性に関する報告が1つある。
【0006】
β-シート形成セグメント、溶解のための荷電残基および生物学的エピトープを含むペプチドブロックに結合している直鎖状疎水性テイルを含む1クラスのペプチド両親媒性物質(PA)が2つの同時係属出願の一方または両方に開示されている。アルキルテイルはペプチドのN-末端に結合しており、エピトープセグメントはC-末端に配置されている。pHまたはイオン濃度の変化などのトリガーを適用すると、これらのPA分子は水性媒体中でナノファイバーに自己-集合することができる。アルキル鎖はファイバーのコアに位置し、エピトープは細胞相互作用のために周辺で提示されている。PA分子に組み込まれているエピトープは細胞外マトリックスタンパク質を模倣しており、細胞信号伝達による細胞接着または分化を促進する。異なるエピトープおよび相補的な荷電を有する2つの異なるPA分子が同一のナノファイバーに同時-集合することができることも示されている。各々全体として参照により本明細書に組み入れられる、2003年2月18日提出の同時係属出願第10/368,517号(国際公報番号WO03/070749)(特許文献1)および2002年11月14日提出の出願番号第10/294,114号 (国際公報番号03/054146号)(特許文献2)参照。
【0007】
しかし、このようなPA化合物は、典型的には、固相合成によって製造され、ペプチド成分はC-末端からN-末端方向に作製される。種々のエピトープまたはペプチド配列は生物活性のために遊離のN-末端を必要とすることが多いので、疎水性成分をN-末端に結合すると、得られるミセルナノファイバー集合物の周辺に遊離酸またはアミド基を有するPA化合物が得られ、このようなPA化合物は増殖因子相互作用に無効であると思われる。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】国際公報番号WO03/070749
【特許文献2】国際公報番号03/054146号
【発明の概要】
【0009】
発明の概要
上記を考慮すると、本発明の目的は、ある範囲の増殖因子のバイオアベイラビリティーに影響を与え、それによって上記に概略するものを含む従来技術の種々の欠陥および欠点を克服する両親媒性ペプチド化合物、その集合組成物および/またはそれらを使用するための関連する方法を提供することである。本発明の1つ以上の局面はある目的を満たすことができるが、1つ以上の他の局面は他のある目的を満たすことができることが当業者によって理解される。あらゆる点において、各目的が本発明の全ての局面に等しく適用しない場合がある。従って、以下の目的は、本発明の任意の1つの局面に関して選択的に考慮することができる。
【0010】
本発明の目的は、ファージディスプレイ法の認識産物を含むことができ、それから誘導されうるエピトープである、1つ以上の増殖因子と非共有結合的に相互作用することができるある範囲のエピトープ配列の1つ以上のN-末端に結合しているペプチド成分を含む両親媒性ペプチド化合物を提供することである。
【0011】
本発明の別の目的は、結合しているエピトープ結合配列の提示のための上記の両親媒性ペプチド化合物の1つ以上の集合物を含む組成物であって、このようなエピトープ配列が欠けている他の両親媒性ペプチド化合物を含むことができる組成物を提供することである。
【0012】
本発明の目的は、結合しているエピトープ配列に対応する1つ以上の増殖因子をさらに含む上記の組成物であって、増殖因子送達および放出のために自己-集合することができる組成物の1つ以上を提供することであってもよい。
【0013】
本発明の目的はまた、増殖因子のバイオアベイラビリティーおよび/または幹細胞分化に影響与える上記両親媒性ペプチド化合物の1つ以上の組成物集合物(compositional assemblies)およびそれらの用途を提供することである。
【0014】
本発明の他の目的、特徴、利益および利点はこの概要およびある種の態様の以下の説明から明らかであり、ペプチド組成物および増殖因子送達の骨格または構造の当業者に容易に明らかになる。このような目的、特徴、利益および利点は、添付の実施例、データ、図面およびそれらから引き出される全ての妥当な影響を単独または本明細書に組み入れられている参照文献と併用して考慮すると上記から明らかになる。
【0015】
一部には、本発明は、ペプチド成分および疎水性成分を含み、ペプチド成分がファージディスプレイ法の増殖因子認識産物を含む両親媒性ペプチドに関してもよい。1つ以上のこのような認識産物は、ペプチド成分のN-末端、N-末端付近またはN-末端に近接してペプチド成分に連結または直接結合してもよく、疎水性成分は、C-末端、C-末端付近またはC-末端に近接してペプチド成分と連結または直接結合してもよい。このような認識産物は、増殖因子と1つ以上の結合相互作用を提供するエピトープ配列から選択することができ、このような産物/配列および対応する増殖因子は以下に十分に考察されているものを含むが、これらに限定されない。本発明の化合物および/またはそのペプチド成分は実質的に直鎖状または分岐鎖状であってもよく、分岐鎖状の立体配置およびその製造は、全体の内容が参照により本明細書に組み入れられる、2004年12月6日に同時提出の同時係属出願「Branched Peptide Amphiphiles, Related Epitope Compounds and Self-Assembled Structures Thereof」に開示されている。
【0016】
一部には、本発明は、複数のこのような両親媒性ペプチド化合物を含む組成物に関してもよく、このような化合物各々のペプチド成分は生理的pHにおいて正味荷電を有する。このような組成物は、生理的pHにおいて相補的な正味荷電を有する複数の両親媒性ペプチド化合物をさらに含んでもよく、このような化合物のアミノ酸配列が、認識産物を含む化合物に対応するものより短い長さであるかまたは数が少ないように、増殖因子認識産物が欠ける。結果として、このような化合物のミセル集合物は、適当な培地において、1つ以上の認識産物の提示を増強し、ミセル集合物の周辺全面を越えて延在する。このような提示の増強を使用して、1つ以上の増殖因子のバイオアベイラビリティーに影響を与えるおよび/またはそれを制御することができる。従って、増殖因子が幹細胞によって産生されるても、または上記の組成物の1つ以上と合わせて細胞環境に送達されても、本発明の種々の組成物およびそれに関係する方法は、1つ以上の増殖因子と非共有結合的に相互作用するまたは結合するために幹細胞環境において使用することができる。
【0017】
組成物の用途または適用にかかわらず、本発明のペプチド両親媒性物質は、望ましいフレキシビリティ、荷電および/または分子間相互作用または結合能力に応じて種々の長さまたは配列のペプチド成分を含んでもよい。このような化合物の疎水性成分は異なってもよく(例えば、飽和または不飽和の約C6から約C22超のアルキルまたは置換アルキル等)、このような成分は、得られる両親媒性およびこのような化合物の組成物または集合物に対する影響によってのみ限定される。
【0018】
本発明の一態様は、各々結合エピトープを有するペプチド両親媒性物質と結合エピトープを含有しないフィラーペプチド両親媒性物質の混合物を含む組成物である。混合物は、フィラーペプチド両親媒性物質および結合エピトープを有する自己集合ペプチド両親媒性物質からなる自己集合ナノファイバーまたは他のミセルを形成することができる。組成物中のペプチド両親媒性物質は、アルキルテイル部分、βシート部分および荷電部分を有する。好ましくは、結合エピトープを有するペプチド両親媒性物質のエピトープは、ファージディスプレイ法によって誘導される配列を有し、フィラーペプチド両親媒性物質より長い。
【0019】
本発明の別の態様は、結合エピトープを有するペプチド両親媒性物質およびフィラーペプチド両親媒性物質を含む自己集合ナノファイバーまたは他の自己集合ミセルの組成物またはシステムである。結合エピトープを有するペプチド両親媒性物質は、好ましくは、フィラーペプチド両親媒性物質より長い。自己集合構造物では、結合エピトープを有するペプチド両親媒性物質は、好ましくは、自己集合ナノファイバーまたはミセルの表面から突出している。好ましくは、結合エピトープを有するペプチド両親媒性物質は、非共有結合相互作用により増殖因子または他のペプチド、アミノ酸または核酸と相互作用することができ、よりさらに好ましくは、増殖因子とエピトープの非共有結合相互作用は、増殖因子の細胞外受容体への結合と競合しない。
【0020】
自己集合ナノファイバーまたはミセルの組成物は、結合エピトープを有するナノファイバーのペプチド両親媒性物質に非共有結合している増殖因子をさらに含むことがあるヒドロゲルを含んでもよい。このような増殖因子はインビトロにおいて添加されてもまたはインビボにおいて患者の損傷部位のものであってもよい。自己集合ペプチド両親媒性物質と相互作用する増殖因子には、骨形成タンパク質、トランスフォーミング増殖因子、血管内皮細胞増殖因子、ニューロトロフィンならびにFGF-2、ソニックヘッジホッグおよびWnt-タンパク質のような分裂促進因子を挙げることができるが、これに限定されない。組成物は、ナノファイバーヒドロゲル内に幹細胞などであるが、これに限定されない細胞を含んでもよい。好ましくは、自己集合ペプチド両親媒性物質と非共有結合的に相互作用する増殖因子は、細胞外受容体との相互作用によりまたはナノファイバーマトリックスの分解により周囲組織または細胞に放出される。細胞以外に、他の治療化合物がヒドロゲル内に封入されてもまたはヒドロゲルに結合されてもよい。これらの化合物には、抗-炎症剤、化学療法剤またはナノファイバーヒドロゲル内におけるこれらの併用を挙げることができるが、これに限定されない。好ましくは、自己集合ペプチド両親媒性物質と非共有結合的に相互作用する増殖因子は、細胞外受容体との相互作用によりまたはナノファイバーマトリックスの分解により周囲組織または細胞に放出される。
【0021】
本発明の別の態様は、結合エピトープを有するペプチド両親媒性物質をフィラーペプチド両親媒性物質と混合または組み合わせることによって自己-集合させる段階を含み、結合エピトープを有するペプチド両親媒性物質がフィラーペプチド両親媒性物質より長さが長い、自己集合ペプチド両親媒性ナノファイバーまたはミセルを製造する方法である。ナノファイバーまたはミセルは、多価イオンを添加することによって、相補的荷電ペプチド両親媒性物質を添加することによってまたは脱水によってまたはペプチド両親媒性物質に酸もしくは塩基を添加することによって製造することができる。ヒドロゲルが形成される場合には、好ましくは、ミセルまたはナノファイバーは、多価イオンの添加によってまたは細胞培地もしくは体液にすでに存在するイオンによって形成される。好ましくは、結合エピトープを有するペプチド両親媒性物質は、ファージディスプレイ法によって誘導される配列を有する。
【0022】
本発明の別の態様は、増殖因子に非共有結合するためのエピトープを有するペプチド両親媒性物質を含むナノファイバーまたは他の自己集合ミセルを、組織を再生する必要のある患者の部位に投与する段階を含む組織を治療する方法である。ナノファイバーおよびそれらのヒドロゲルは、幹細胞などであるが、これに限定されない細胞を含んでもよい。細胞以外に、治療化合物がヒドロゲルに封入されてもまたは結合されてもよい。これらの化合物には、抗-炎症剤、化学療法剤またはこれらの併用を挙げることができるが、これに限定されない。好ましくは、自己集合構造に非共有結合する増殖因子は、細胞外受容体との相互作用またはマトリックスの分解によって部位に放出される。好ましくは、増殖因子および/または幹細胞を含有する自己集合ミセルまたはナノファイバーは、骨損傷、軟骨損傷、脊髄損傷、脳組織損傷、神経損傷またはこれらの組み合わせなどであるが、これに限定されない損傷を有する部位または患者に投与される。
【0023】
本発明の別の態様は、第2の末端も有するβ-シート形成ペプチド部分の第1の末端に結合しているアルキルテイル部分を含むペプチド両親媒性物質である。ペプチド両親媒性物質のエピトープペプチド部分は、エピトープが他の分子またはタンパク質との非共有結合相互作用に利用可能であるように、βシート形成ペプチド部分の第2の末端に結合される。好ましくは、エピトープは、ファージディスプレイ法によって誘導される配列を有する。または、ペプチド両親媒性物質は、2つの末端を有し、βシート形成部分の第2の末端とエピトープペプチド部分の間に結合されている荷電部分ペプチドを含んでもよい。
【0024】
本発明の別の態様は、自己集合型ペプチド両親媒性物質から製造される骨格を含む、増殖因子を細胞に放出するための集合物またはシステムを含み、ペプチド両親媒性物質の少なくとも一部は増殖因子と非共有結合するためのペプチドエピトープを含み、増殖因子結合エピトープはナノファイバー面上に突出している。エピトープのアミノ酸配列はファージディスプレイ法から誘導される。このような骨格は幹細胞を含んでもよく、患者において組織を増殖する方法、組織を再生する方法または組織の移植を支持する方法に使用することができる。細胞以外に、他の治療化合物を、骨格のヒドロゲルに封入されてもまたは結合されてもよい。これらの化合物には、抗炎症剤、化学療法剤またはこれらの併用を挙げることができるが、これに限定されない。本発明の方法は、患者の部位に骨格を挿入する段階またはインビボにおいて骨格を形成する段階を含む。骨格は、増殖因子と非共有結合するためのペプチドエピトープを有する自己集合ペプチド両親媒性物質を含み、エピトープは骨格のナノファイバー表面に突出するまたはファージディスプレイ法によって作製され、骨格から周囲細胞に増殖因子を放出する。
【0025】
記載されている増殖因子結合ヒドロゲルは、再生医療および移植医療の分野においていくつかの適用を有する。本発明の組成物、それらを製造する方法およびそれらを使用する方法は、他の増殖因子のためのペプチド両親媒性物質の他のエピトープに、従って多種多様の組織の再生または患者の移植組織の支持に容易に拡大することができる。例えば、BMP-2およびTGF-β1は間葉系幹細胞の造骨および軟骨形成分化に重要な役割を果たし、本発明の自己集合ミセルは骨および軟骨の再生に使用することができる。第二に、BMP-2はまた、脳および脊髄背側部の形成に重要な役割を果たしている。従って、ゲルは、損傷した脊髄の再生のためおよび/または卒中症例の損傷した脳領域を修復するために神経幹細胞と併用して使用してもよい。
【0026】
上記に考察するように、ある態様において、本発明は、選択的非共有結合的相互作用によって増殖因子に結合することができるPAを組み入れるための混合ファイバーの拡張された方法を使用する。この方法では、増殖因子のバイオアベイラビリティーは、結合強度および結合部位の数を調整することによって調節することができる。結合の局所作用の如何または増殖因子を不活性にする結合の影響が支配的であるかどうかに応じて最初の増殖因子濃度は増加または低下される。どちらの場合でも、増殖因子の放出を長期間持続することができる。増殖因子によって認識される結合エピトープを促進するために、エピトープがファイバー表面から延在するようにPAを製造することができる。結合エピトープの必要なアミノ酸配列は、ファージディスプレイを使用して決定することができる。例示目的のために、選択した増殖因子には骨形成タンパク質-2(BMP-2)およびトランスフォーミング増殖因子β1(TGF-β1)が挙げられる。特に、BMP-2は、骨の形成ならびに脳および脊髄背側部の発生に関与するが、TGF-β1は軟骨の形成および平滑筋細胞の分化に関与する。間葉系幹細胞の分化を制御するために対応するゲルを使用することができる。間葉系幹細胞は、インビトロおよびインビボにおいて、骨、軟骨、脂肪、筋肉細胞および心筋に分化することが示されている。BMP-2の影響下における骨系統への分化を以下に示す。
【0027】
ゲルは、間葉系幹細胞の予測される造骨系統への分化を制御する際に培地に増殖因子を添加するより優れている。さらに、結合ゲルは、最初は、全ての他の系統への分化を抑制する。このような挙動は、増殖因子の低いバイオアベイラビリティーによってまたは結合ゲルのこのような増殖因子との細胞の接触がゆっくりであることによって生じると考えられる。最終的に、一部のα-平滑筋発現も結合ゲルに見られる。BMP-2との長期接触または完了までの分化を誘導する因子の欠損は集団の均一性に負の影響を与えることをこれは示唆している。最後に、結合配列がバイオアベイラビリティーを加減するために存在する場合には、内因的に産生されるBMP-2レベルが十分であると思われる。
【0028】
増殖因子結合PAと通常のフィラーPAの同時-集合の成功により、増殖因子に結合し、ゲル状態に維持し、バイオアベイラビリティーを調節することができるヒドロゲルの製造が可能になる。最初に結合PAを含有するゲルにおいて間葉系幹細胞を増殖する場合には、結合PAの非存在下において増殖されるものと比較して、特殊化した細胞のより均一な集団が得られる。適用に応じてゲルの物理的特性を変更するために他のフィラーPAが望ましい場合もあり、結合強度を最適化するために他の結合配列が望ましいこともある。また、本発明によって証明される方法は、多数の結合PAとフィラーの単純な混合によって多数の信号の組み込みを可能にする。おそらく、幹細胞の分化をさらに最適化するためには多数の増殖因子を必要とする。多数の増殖因子に接触させた間葉系幹細胞の分化は現在検討中である。多数の増殖因子系の一時的な放出は、増殖因子の異なる結合定数を有する結合物を選択することによって実施することができる可能性がある。また、本明細書に記載するシステムのインビボにおける検討が実施中である。最後に、本明細書に提供する方法は、細胞動員、特殊化または維持に関与する任意の増殖因子またはタンパク質に容易に拡大することができ、再生医療の有望性の高い方法にしている。
【図面の簡単な説明】
【0029】
【図1】集合に使用するためのエピトープ配列を含む代表的なPAおよび相補的なPAを例示する。
【発明を実施するための形態】
【0030】
ある種の態様の詳細な説明
本発明は、特異的な非共有結合相互作用によって増殖因子に結合することができる自己集合ペプチド両親媒性物質(PA)に基づいたシステムを記載する。増殖因子は細胞の信号伝達において重要な役割を果たしており、従って、幹細胞の分化を制御するための有望な因子であり、移植した組織の組み込みをサポートするおよび/または非機能組織の再生を生ずる可能性のある休止中の生物学的過程をインビボにおいて分子的または生理的に再-活性化する。増殖因子はインビボにおいて迅速に分解し、損傷部位から迅速に拡散消失する。従って、増殖因子を損傷部位に保持し、増殖因子を周囲の細胞に徐放的に放出する骨格が必要とされる。自己集合ペプチド両親媒性物質を使用しているヒドロゲルは、細胞外マトリックス(ECM)のいくつかの機能を満たす、すなわち、細胞接着を促進し、分化を制御することができることが示されている。本発明は、骨形成タンパク質-2(BMP-2)、トランスフォーミング増殖因子β1(TGF-β1)、VEGFおよびIL-6が挙げられるが、それに限定されるわけではない増殖因子に特異的に結合することができるペプチド配列でこれらのECM置換基を修飾することに関する。これらの増殖因子ペプチド配列はファージディスプレイ技法によって入手することができ、ペプチド両親媒性物質に結合して結合ペプチド両親媒性物質を形成することができる。結合ペプチド両親媒性物質およびフィラーペプチド両親媒性物質は自己集合して、特定の形状に成形され、組織修復の骨格として使用することができるヒドロゲルを形成することができる。または、結合ペプチド両親媒性物質およびフィラーペプチド両親媒性物質はインビボまたはインビトロにおいて細胞試料または組織試料に導入して、試料中でヒドロゲルに自己集合することができる。
【0031】
上記のように、本発明のペプチド両親媒性物質は、アルキルテイル、β-シート形成ペプチド配列および生物活性ペプチド配列を含むことができる。最初の2つのブロックは、ナノファイバーなどであるが、これに限定されないミセルファイバーを形成し、適切な条件下において(多価イオンの添加による中和)、ヒドロゲルまたは溶媒が細孔を満たしている他の湿潤ゲル)を形成する。好ましくは、生物活性なエピトープは溶解のために部分的に荷電しているが、この荷電は、インテグリン-結合配列、向神経活性(neuro-acitive)配列等のような多種多様の配列において異なってもよい。さらに、相補的な荷電のペプチド両親媒性物質は自己集合して混合ファイバーを形成する。この原理を使用して通常のPAおよびファイバー表面から延在している増殖因子結合エピトープ配列を有する長いPAからなるナノファイバーを作製することができる。望ましい増殖因子結合配列は、無作為ペプチドライブラリーを使用するファージディスプレイによって得られ、コンビナトリアル選択過程を使用して強力な結合配列を生ずる。増殖因子の放出は、マトリックスの分解、細胞外受容体への結合またはこれらの組み合わせによって誘導される。
【0032】
既存の技術は、増殖因子をヒドロゲルに捕獲し、それらをポリマーテザーに共有結合する(国際公開公報第03/040336号および米国特許第0020007217号)またはそれらを陰イオンポリマー(国際公開公報第0/13710号)またはヘパリン(国際公開公報第00/64481号)に静電気的に結合する。テザーシステムの欠点は、製造の複雑さおよび増殖因子が細胞外受容体に到達しない可能性である。捕獲されている増殖因子および陰イオンポリマーと複合体形成している増殖因子は比較的迅速に放出され、徐放的な制御は少ない。例えば、陰イオンポリマーに結合している増殖因子は8時間以内に放出される。ヘパリン-結合システムは非特異的であり(例えば、血清に存在する因子の望ましくない結合が生じうる)、増殖因子のサブセットに限定される。有利なことに、本発明は、望ましい増殖因子だけが、結合状態で増殖因子を保持するのに十分に大きい結合強度で複合体形成されるが、細胞外受容体との結合と競合しない程度に弱いように設計される。
【0033】
本明細書に記載する本発明の一局面は、ファージディスプレイ技法を使用して増殖因子を結合するペプチド配列を誘導体化し、その後これらの結合配列を有するペプチド両親媒性物質を製造するために使用することができるということである。ファージ技法によって作製される増殖因子には、骨形成タンパク質、トランスフォーミング増殖因子β1、ニューロトロフィンおよび分裂促進因子FGF-2、ソニックヘッジホッグおよびWnt-3aおよびを挙げることができる。これらの増殖因子タンパク質は、短いペプチド両親媒性物質と結合して、増殖因子ペプチドを有する長いペプチド両親媒性物質を形成する。これらの増殖因子エピトープを有するPAまたは結合ペプチド両親媒性物質は短いフィラーまたは相補的なフィラーPAと同時集合して、結合エピトープが表面から延在するナノファイバーを作製し、その後多価イオンの添加またはpHの変化の結果ヒドロゲルを形成することができる。次いで、得られたヒドロゲルを、ヒドロゲル内部に封入されている幹細胞などであるが、これに限定されない細胞と共に使用してもまたはインビボにおける組織再生のために送達ビヒクルとして単独使用してもよい。細胞以外に、他の治療化合物をヒドロゲルに封入してもまたは結合してもよい。これらの化合物には、抗-炎症剤、化学療法剤またはこれらの併用を挙げることができるが、これに限定されない。
【0034】
ファージディスプレイは、特定の標的に対する分子プローブを作製するためならびにタンパク質-リガンド相互作用を分析および操作するために使用することができる。ファージディスプレイは、アミノ酸、ペプチド、増殖因子、酵素および種々の核酸などであるが、これに限定されない標的分子に結合するアミノ酸配列、ペプチドまたはタンパク質を決定するために使用される。ファージディスプレイは、市販のライブラリーを使用して、マイクロタイターウェルプレートに物理吸着する増殖因子のような標的分子に実施することができる。好ましくは、ライブラリーは、G3コートタンパク質のN-末端が、アミノ酸の全ての可能な組み合わせが存在する方法で、1〜約13以上のランダムアミノ酸が延在しているファージからなる。ライブラリーは増殖因子などの標的分子に接触され、その後非特異的に結合したファージは界面活性剤溶液と共に溶出される。結合ファージの回収後、この集団を例えば、大腸菌(E. coli) において増幅し、溶出条件の緊縮度を増してこの過程を2回以上反復する。最終的な集団を結合ファージに関して濃縮し、クローンのいくつかのDNAを配列決定して、これらのファージ上に提示されているアミノ酸配列の同定を可能にすることができる。その後、ELISAアッセイを実施して、単離されたクローンの相対的な結合強度を推定することができる。固体状態の合成を使用して、同定されたアミノ酸配列を使用して結合ペプチド両親媒性物質を作製するためのペプチドを作製する。
【0035】
本発明のフィラーPAおよび結合エピトープペプチド両親媒性物質のペプチド成分は天然型アミノ酸および人工アミノ酸を含んでもよい。対応する成分がこれに関してペプチド-様であり、ナノファイバーなどのミセルを形成する自己集合であるという効果で、βまたはγアミノ酸などの人工アミノ酸および非天然側鎖を含有する人工アミノ酸および/またはヒドロキシ酸などの他の同様のモノマーの組み込みも考慮されている。
【0036】
本発明の種々のペプチド両親媒性物質は、上記の組み込まれている公報である国際公開公報第03/054146号および全体として参照により本明細書に組み入れられる、Hartgerink, et al. (例えば、J. D. Hartgerink, E. Beniash and S. I. Stupp, Science 294, 1683-1688, 2001参照)によって最初に記載されているものの変法を含む、当業者に周知の製造技法を使用して合成することができる。本発明の合成方法およびエピトープを使用することができる分岐鎖状ペプチド両親媒性物質も本発明において考慮されている。分岐鎖状ペプチド両親媒性物質は、内容が全体として参照により本明細書に組み入れられる、2004年12月6日に本願と同時提出された上記の同時係属出願に開示されている方法および組成物を使用することによって作製することができる。結合エピトープを有するこのような分岐鎖状ペプチド両親媒性物質もナノファイバーに自己集合すると思われる。分岐鎖状ペプチド両親媒性物質は、βシート形成ペプチド部分の第1の末端に結合しているアルキルテイル部分およびエピトープの第2の末端において1つ以上のペプチド分岐鎖を含むペプチド部分を有すると思われる。ペプチドは分岐鎖状アミノ酸および荷電ペプチド部分を含む。分岐鎖状エピトープペプチド部分は、ペプチド両親媒性物質のβシート形成ペプチド部分の第2の末端に結合してもよい。分岐鎖状ペプチドの結合エピトープ配列はファージディスプレイ法から誘導されてもよい。これらの参照文献に記載する合成スキームを本発明に適用することができる。ペプチド両親媒性物質は、完全にプロトン化された形態であっても、部分的にプロトン化された形態であってもまたは酸もしくは塩基付加塩として存在してもよい。一般に、このようなペプチド両親媒性物質は、上記の組み入れられている参照文献に記載されているまたは本明細書に提供する標準的な固相ペプチド化学によって作製することができる。当業者に公知であると思われ、望ましい両親媒性組成物またはペプチド配列に応じて公知の手法および合成技法または直接的な改良を使用して承知しているこれらの合成方法の改良を実施することができる。
【0037】
図1に示す種類のフィラーペプチドは、好ましくは、結合エピトープを有するペプチド両親媒性物質より長さが短く、これらのペプチド両親媒性物質の組み合わせから自己集合するナノファイバーは、増殖因子結合エピトープがナノファイバーの表面から突出する。相補的なフィラーペプチド両親媒性物質は、酸塩基または他の種類の結合によりフィラーペプチドのアミノ酸と相互作用するアミノ酸の側基を有する。
【0038】
ファージディスプレイ法から誘導されるペプチド配列は、増殖因子、酵素、核酸(RNA、DNA)およびアミノ酸を含むが、これらに限定されないペプチドおよび分子に非共有結合するためのペプチド両親媒性物質の結合部位またはエピトープとして使用することができる。ファージディスプレイはエピトープ配列を誘導するために好ましいが、酵母ハイブリッドシステムなどであるが、これに限定されない結合エピトープの配列を同定し、決定する他の相補的な方法を使用することができる。これらの結合ペプチドまたはエピトープをβシート形成ペプチドの遊離端、荷電ペプチドの遊離端またはスペーサーペプチドの遊離端に結合することができる。
【0039】
ペプチド両親媒性物質デザインは、ペプチド両親媒性物質の自己集合のために分子の円錐形状の細長い部分を形成するために働く単純な疎水性テイルを含むことができる。好ましくは、疎水性テイルは、種々のサイズであってもよいが、好ましくは、炭素原子の長さが6を超えるアルキル鎖である。アルキルテイルは、ペプチド両親媒性物質のβシート形成構造セグメントに共有結合されている。
【0040】
βシートペプチド構造セグメントは、テイル基をファージペプチド;または荷電ペプチド、スペーサーペプチドおよびファージペプチド;または荷電ペプチドおよびファージペプチドに共有結合するために使用することができる。βシートペプチド構造セグメントは、一方の末端がテイルに、他方の末端が、種々のペプチドのアミノ酸配列に共有結合する。架橋が望ましい場合には、システインアミノ酸をセグメントのいずれかに使用してもよいが、好ましくは、構造βシートセグメントに使用する。架橋が望ましくない場合には、アラニン、セリンまたはロイシンなどであるが、これに限定されない他の疎水性アミノ酸をこの領域に使用することができる(例えば、本明細書にさらに詳細に記載するSLSLまたはAAAA)。このシステイン-フリーシステムは、ナノファイバーマトリックスの分解速度を制御するために、インビボにおける生物適用にさらに適当となりうる。システムのSLSL改良により、骨格のインビボにおける集合を制御するために使用することができるナノファイバーの自己集合を遅くすることを予想することができる。理論に結びつけたいわけではないが、かさばるロイシン側鎖は、ファイバーへの充填に長い時間を必要とすると考えられる。遅い自己集合は、ナノファイバーの形成が遅いことが有利となることがある、手術室などの機能的なインサイチュー環境における適用性が大きい場合もある。構造的なβシート形成セグメントはグリシンまたは他のアミノ酸を含むフレキシブルなリンカーを含む場合もある。構造セグメントが疎水性アミノ酸を含む場合には、疎水性アミノ酸およびアルキルテイルが疎水性セグメントと考えることができる。構造セグメントが親水性アミノ酸を含む場合には、親水性アミノ酸および親水性ヘッド基が親水性セグメントと考えることができる。
【0041】
β-シート形成単位は、好ましくは、βシートを形成するように相互作用することができ、ペプチド両親媒性物質の全体的な円錐形状を形成する助けをするような疎水性アミノ酸を含む。突出しているペプチド両親媒性物質については、この単位のアミノ酸の数は、ナノファイバーを形成するために使用されるフィラーペプチド両親媒性物質より長く、結合しているペプチド両親媒性物質のペプチドへの接近しやすさを提供するペプチド両親媒性物質を提供するように選択することができる。このセグメントには約4〜約10のアミノ酸が存在してもよく、最も好ましくは、約6のアミノ酸が存在してもよい。β-シート形成セグメントについては、好適なアミノ酸グリシン、アラニン、バリン、ロイシンおよびイソロイシンならびに同様の化学的および構造的な方法でペプチド両親媒性物質において使用することができる他の非天然型アミノ酸には、を挙げることができるが、これに限定されない。水性環境において、好ましくは、患者の部位においてペプチド両親媒性物質の溶解性を提供する荷電セグメントが結合ペプチド両親媒性物質に存在する。荷電ペプチドセグメントには、この溶解性を提供し、自己集合を可能にするようなアミノ酸およびそれらの組み合わせを挙げることができ、ペプチド両親媒性物質の溶解性を改良するためにEまたはKなどの極性アミノ酸およびそれらの組み合わせに限定されない。このセグメントには約2〜約7のアミノ酸(amino)が存在してもよく、好ましくは、約3〜4のアミノ酸が存在する。このセグメントは、第1の末端が構造ペプチドに結合し、第2の末端が、ファージディスプレイ法から誘導されるペプチドに結合するために使用される。スペーサー基ペプチドがペプチド両親媒性物質に含まれてもよい。スペーサー(space)は、SおよびGなどであるが、これに限定されないアミノ酸を含んでもよく、スペーサー(space)は1〜約6のアミノ酸を含んでもよい。ファージ上に提示されているペプチドが遊離N-末端を有する場合には、ペプチド両親媒性物質の典型的な合成スキームを遊離N-末端を提供するように修正することができる。これは、側鎖をドデシルアミンで官能基化することができるアスパラギン酸誘導体を合成することによって実施することができる。このアミノ酸をWang樹脂に結合することにより、N-末端を官能基化する必要なしに、典型的な固相状態のFmoc-化学によってPA合成を完了することができた。遊離PAは、95% TFA/2.5 %水/2.5 %トリイソプロピルシランで処理することにより得ることができる。残存TFAはPAを3 mMのHClに溶解することによって除去することができ、室温において1時間平衡させ、次いで凍結乾燥することができる。これらの分子の合成の成功は、エレクトロスプレー質量分析法を使用して確認することができる。
【0042】
増殖因子エピトープを有するペプチド両親媒性物質は、体液に存在するイオンおよび自己集合を促進するために添加されるイオン/試薬を使用して、インビボにおいて患者の部位において自己集合することができる。または、増殖因子結合エピトープを有する好適なペプチド両親媒性物質の組成物を型に注ぎ、自己集合を使用して、置換または再生される組織または骨の形状の骨格を形成する。成形された骨格を、修復または再生治療を必要としている患者の部位に挿入することができる。組織移植の場合には、増殖因子結合エピトープを有するペプチド両親媒性物質は型で支持構造物またはマトリックスに形成され、組織移植のための患者の支持として使用することができる。ペプチド両親媒性物質ナノファイバーまたは骨格は、幹細胞などであるが、これに限定されない細胞を含んでもよい。他の治療化合物がヒドロゲルに封入されてもまたは結合されてもよい。これらの化合物には、抗炎症剤、化学療法剤またはこれらの組み合わせを挙げることができるが、これに限定されない。
【0043】
増殖因子結合エピトープを有するペプチド両親媒性物質と短いβシート構造ペプチド-荷電ペプチド酸-末端フィラーペプチド両親媒性物質の同時集合を使用して、本発明のナノファイバーおよび骨格を製造することができる。本発明の骨格およびナノファイバーを製造するために、異なる増殖因子結合エピトープを有する1つ以上のペプチド両親媒性物質を使用することができ、種々のフィラーペプチド両親媒性物質を使用することができるが、限定するものではない。フィラーペプチド両親媒性物質と比較した増殖因子結合エピトープを有するペプチド両親媒性物質の量は、本発明の骨格およびナノファイバーを製造する際に異なってもよいが、限定されない。自己集合ミセルおよびナノファイバーは、NOEおよびFT-IR分光分析、円二色性によって特徴づけることができる;ナノファイバーのファイバーネットワークは透過型電子顕微鏡を使用して可視化することができる。
【0044】
結合エピトープを有するペプチド両親媒性物質は、増殖因子またはアミノ酸もしくは核酸などであるが、これに限定されない他のペプチドと非共有結合相互作用によって相互作用することができるエピトープのペプチド配列を選択することによって製造することができる。増殖因子または他のペプチドと結合エピトープを有するペプチド両親媒性物質の非共有結合相互作用の程度は、エピトープが、競合しないまたは細胞外受容体比較して、増殖因子または他のペプチドに対する結合親和性が低いように選択される。受動的な放出実験を使用して、特徴づけ、その後増殖因子結合エピトープを有する種々のナノファイバーの親和性を改良することができる。この特徴づけは、事前ブロックしたマイクロタイタープレート中で増殖因子を含有する自己集合ナノファイバーゲルについて実施することができる。これらのゲルは、適当な比でPAの2%溶液を混合し、次にTBSで1:1希釈することによって作製することができる。次いで、増殖因子を添加し、次に2当量の多価イオンを添加してゲル化および自己集合を誘導することができる。増殖因子含量について上清をアッセイして、このようなゲルの増殖因子結合能を評価することができる。
【0045】
結合エピトープは、近隣の細胞または組織の細胞外受容体より種々のペプチドに対する親和性が弱いことが組織増殖および他の治療に好ましいが、それは、望ましくは、ペプチド、増殖因子、酵素または核酸または体液もしくは細胞試料の他の分子に強力に化学的に結合するペプチド両親媒性物質の結合エピトープであることが考慮されている。この場合には、好適なエピトープを有する結合ペプチド両親媒性を表面で自己集合させて固定して、センサーコーティングまたは除去媒体(removal media)を形成する。これらの強力に結合しているペプチド両親媒性物質の自己集合から形成されるヒドロゲルは、患者の部位に挿入するためまたはろ過システムに使用するために成形してもよい。ヒドロゲルは、患者の関節もしくは腫瘍などの部位または患者の体液からHGFまたはVEGFのような標的ペプチドを除去するために使用してもよい。
【実施例】
【0046】
発明の実施例
以下の限定するものではない実施例およびデータは、増殖因子と非共有結合相互作用をすることができるエピトープ配列を有するまたは有しない特定の範囲の両親媒性ペプチド化合物の合成、このような化合物または組成物の自己集合ならびに増殖因子のバイオアベイラビリティーおよび幹細胞分化に影響を与えるそれらの用途を含む、本発明の化合物、組成物および/または方法に関する種々の局面および特徴を例示している。従来技術と比較すると、本発明の化合物、組成物および/または方法は驚くべきであり、予測されず、従来技術と異なる。本発明の利用性はいくつかの化合物/組成物および集合物を使用することにより例示されているが、同様の結果は、本発明の範囲に相応の種々の他の化合物、組成物および/または集合物で得ることができることが当業者によって理解される。
【0047】
樹脂およびFmoc-1-アミノ酸は全てNovabiochem(San Diego, CA)から入手した。固相合成のための試薬は全て合成用であり、Applied Biosystems(Foster City, CA)から入手した。他の試薬は全てAldrich Chemical Co. (Milwaukee, WI)から入手し、入手したままで使用した。固相ペプチド合成のための溶媒はApplied Biosystemsから入手し、ペプチド合成用であった。他の溶媒はFisher Scientificから入手し、特に記載しない限り、入手したままで使用した。
【0048】
PAは、Applied Biosystems 433A自動ペプチド合成装置を使用して合成した。NMRスペクトルは、Varian Inova 500 MHz分光計で室温において得た。エレクトロスプレー質量スペクトルは、Micromass Quattro II Triple Quadrupole HPLC/MS/MS質量分析計で収集した。CDスペクトルは、Jasco PTC-348WIペルチェ効果温度コントローラー付きJasco J-715分光偏光計で記録した。FT-IRスペクトルは、BioRad FTS-40 FT-IRマシーンで、2 cm-1の分解能で400から4000 nmまで操作した。
【0049】
円二色性分光法. 路程0.1 cmの石英セルを全ての実験に使用した。スキャン速度100 nm/min、反応時間2秒およびバンド幅1 nmで300 nmから190 nmまで各スペクトルを記録し、5回スキャンを平均した。試料は、特に記載しない限り、水で0.1 mg/mLの濃度に調製した。
【0050】
酸-塩基滴定. pKa滴定は、100 mM KClにおいて3.5 mg/mLの濃度において、Fisher Accumet pHメーターを用いてpH 2〜10の範囲のPA 1〜4に実施した。酸性PA 2および4については、低いpHから開始して、0.1 N KOHを1〜5μLずつ添加したが、塩基性PA 1および3については、高いpHから開始して、0.1 N HClを1〜5μLずつ添加した。
【0051】
NMR NOE分光分析. PA1〜6をd6-DMSOに5 mg/mLの濃度で溶解した。NOESYスペクトルは、各PAの10または15 mg/mL濃度、モル比1:1において、ミキシングタイム0.1 s、128スキャンでD2O中で測定した。FT-IR検討については、2重量%の試料を水から凍結乾燥し、次いでKBRペレットに圧縮した。
【0052】
スキーム1. ペプチド両親媒性物質の化学構造
【0053】
実施例1
トリプルリジン配列を有する2つ(1, 3)およびトリプルグルタミン酸配列を有する2つ(2, 4)の4つのPAを製造した(スキーム1)。PA 1および2は、プレロードしておいたWang樹脂を使用し、次にカップリング試薬として2-(1H-ベンゾトリアゾール-1-イル)-1,1,3,3-テトラメチルウロニウムヘキサフルオロホスフェート(HBTU)を用いてパルミチン酸でアルキル化して、標準的なFmoc固相ペプチド技法によって製造した。95%トリフルオロ-酢酸(TFA)、2.5%水および2.5%トリイソプロピルシラン(TIS)の混合物を用いて両親媒性を樹脂から切断した。ペプチド両親媒性物質3および4の合成は、スキーム2によりアミノ酸7を合成した。N-カルボベンジルオキシ-L-アスパラギン酸無水物をドデシルアミンと反応させて、脂肪酸アミノ酸5を得た。次いで、接触水素化によってCBz基を脱離して、6を得、次いでアミンのFmoc-保護を実施した。この合成は、5グラムスケールで容易に進行する。次いで、カップリング試薬としてHBTUを使用して生成物7をリンク樹脂にカップリングした。その後、標準的なFmoc固相技法を使用して、残りのアミノ酸を8に添加した。以前に記載したように、標準的な切断条件により、逆構造を有するPAを得た。1H NMRおよびエレクトロスプレーイオン化質量分析は予想される構造に一致している。
【0054】
スキーム2. 脂肪酸アミノ酸の合成
【0055】
実施例1a
N-ドデシル-2-カルボベンジルオキシアミノ-スクシンアミド酸(5).
N-カルボベンジルオキシ-L-アスパラギン酸無水物(1 mmol)を50 mL塩化メチレンに溶解し、次に1.05 eqのドデシルアミンおよび1.1 eqのトリエチルアミンを添加した。蒸散を防ぐために反応に蓋をし、12時間撹拌した。薄層クロマトグラフィー(TLC)(CH2Cl2, 5% MeOH)によって出発材料がわずかでも検出されなかったら、20 mLの1N塩酸で反応を停止し、次にクロロホルム(5×)で抽出した。有機層を硫酸マグネシウムで乾燥し、7を白色の固体として得た(収率97%)。
【0056】
実施例1b
2-アミノ-N-ドデシル-スクシンアミド酸(6). 100 mlのエタノールに、100 mmolの7を溶解し、PdおよびC(10重量%)を含有する反応容器に移した。次いで、容器を水素雰囲気下(35 Torr)に3時間おいた。反応混合物をセライトでろ過し、減圧下蒸発乾固して、生成物を白色の固体として得た。収率95%。
【0057】
実施例1c
N-ドデシル-2-Fmoc-アミノ-スクシンアミド酸(7). 水/ジオキサン(1:1 v:v)混合物の200 mLに、6.6 mmolの6を1.3 mL(1.5 eq.)のトリエチルアミンと共に溶解し、次に1eq.のFmoc-Oスクシンアミド(Fmoc-OSu)を溶解した。反応をTLC(CH2Cl2, 10% MeOH)でモニターし、2〜3時間後、全てのFmoc-OSuが消費された。反応を酸で停止して、白色の沈殿物を得、ろ過して回収した。収率85%。
【0058】
実施例1d
PA合成および精製. 参照文献3に記載されているように、PA 1〜2を製造した。PA 3〜6は、標準的なリンク樹脂を反応容器に入れ、30%ピペリジンのNMP溶液で3回脱保護し、次いで2eq.の7に終夜カップリングした。ニンヒドリン試験が陰性の結果を示すまで、カップリングを反復した。次いで、この修飾樹脂8を自動合成装置にロードし、PA 1〜2についてと同様にペプチド合成を進行した。自動合成が完了したら、参照文献3に記載されているように、PAを樹脂から切断した。
【0059】
実施例1e
C15H31CONHVal-Val-Val-Ala-Ala-Ala-Lys-Lys-Lys-COOH(1)
【0060】
実施例1f
C15H31CONHVal-Val-Val-Ala-Ala-Ala-Glu-Glu-Glu-COOH(2)
【0061】
実施例1g
Asp(CONHC12)- Val-Val-Val- Val-Val-Val- Lys-Lys-Lys-NH2(3)
【0062】
実施例1h
Asp(CONHC12)- Val-Val-Val- Ala-Ala-Ala-Glu-Glu-Glu--NH2(4)
【0063】
実施例2
PAは、好適な刺激の存在下で水性媒体に分散すると、典型的には、自己集合して高アスペクト比の円筒形ナノファイバーを形成する。以前の研究に基づいて、1つのPA分子または2つのPA分子の混合物からなるファイバーは、β-シート-様水素結合を有する二次構造を示すことが予想されると思われる。負に荷電している2または4とそれぞれ反対に荷電している1または3を混合すると平行なβ-シート-様水素結合配列を生ずると思われるが、1または2とそれぞれ末端アミンが反対に荷電している4または3を混合すると逆平行配列を生ずると思われることが予想された。
【0064】
実施例3
0.1 mMの濃度では、2〜4のCDスペクトルは、主にランダムコイル特性を有するペプチドセグメントを明らかにした。これは、荷電が大きい分子間の静電反発力によって生じる可能性が最も大きい。荷電を中和するようにpHを変化するまたはカルシウムイオンを添加すると、全てのPAはβ-シート構造を示した。一方、1は任意のpHにおいてβ-シートシグニチャーを示す。1は、酸性末端がリジン残基の1つの荷電を中和して、みかけの(formal)正味荷電+2を式に与える可能性があるので、他より荷電が小さい可能性がある。この低い全体荷電は反発力を低下し、ファイバー内にβ-シート水素結合を生じさせる可能性がある。逆に、4は、アミン末端がグルタミン酸残基の1つの荷電を中和すると思われるので、-2のみかけの正味荷電を有し、β-シート相互作用を生じさせると思われる。しかし、4は依然として乱雑なCDシグニチャーを示した。このみかけの矛盾を解決するために、pH 7における集合物の実際の荷電状態およびみかけのpKaを求めて、これらの種々のシステムの自己集合の駆動力をよりよく理解した。
【0065】
実施例4
3 mMの濃度で分子1〜4の集合物のpKa滴定を実施した。自己集合による動態的な影響を回避するために、全ての滴定は、分子がすでに集合した状態にあるpHから開始した。3のpKa滴定だけが、2つのみかけのpKaに関連するシャープな遷移を示し、一方は溶媒接近性の大きいアミン末端の脱プロトン化に起因し、他方はεリジンアミンの1つ以上によって生じた。PA 1、2および4の集合物は複雑な曲線を示し、遷移は広い範囲にわたって生じ、これらの超分子物体のプロトン化/脱プロトン化はゆっくり生じ、酸性度はナノファイバー内の局所的な微小環境により変化することを意味している。文献の最近の報告と矛盾することなく、集合物は酸およびアミン基のみかけのpKaを変化させることがこれらの結果から明らかである。
【0066】
実施例5
荷電相補的PA分子の自己集合により、β-シート水素結合配列を有する2つの成分の混合物を含有するファイバーを生ずるはずである。2をトリプルリジン両親媒性物質3または1と混合すると(2/3または2/1と略す)、得られるCD-スペクトルは純粋なβ-シートに対応する。観察されるβ-シートシグニチャーは、単に2つの成分の個々のCDスペクトルの重ね合わせではないという事実は、2つの分子が1つの集合物構造を形成している混合ナノファイバーの形成を強く示唆している。トリプルグルタミン酸PA4または2と混合したPA1(1/4または1/2)も同様の挙動を示す。同様の荷電の2つのPAを混合すると、1/3リジン混合物はβ-シートを示すが、2/4グルタミン酸混合物は、おそらくグルタミン酸残基間の大きい荷電反発性のために、乱雑な立体配座を示す。
【0067】
実施例6
ファイバー内での分子の自己集合をさらに証明するために、荷電相補的PAから製造した1.5重量%ゲルに核オーバーハウザー(NOE)分光法を実施した。2および3(2/3)の代表的なNOESYは、それぞれ、2のGlu-Hβプロトンと3のLys-HεおよびVal-Hδプロトンの間で観察される密接な接触(<3Å)を示す。考えられるいくつかの他の分子間接触が検出されたが、2/3接触にはっきりと起因させられなかった。これらの結果は、2つのPA分子が同一のナノファイバー内に同時集合されている追加の証拠を提供している。
【0068】
実施例7
両親媒性の自己集合の成功が確立されたら、シングルおよびマルチプルPA由来のシステムのゲル化挙動を検討した。1〜4の1重量パーセント溶液はわずかに不透明で、それぞれ、酸(1、3)または塩基(2、4)の添加によりゲル化することができた。透過型電子顕微鏡は、他のPAにおいて以前に観察されたものと同様の、平均径6.5 nm、平均長さ数百ナノメーターのナノファイバーの形成を明らかにしている。
【0069】
実施例8
ファージディスプレイは、典型的には、受容体-ブロッキングペプチド、すなわち高ブロッキング定数を有する配列を見つけるために使用される。PA結合部位から細胞受容体に増殖因子を移動させるためには、極端に強い結合定数は必要とされず、望ましくない場合もある可能性がある。従って、13-merライブラリーより7-merの市販のファージライブラリーを選択し、中間的な結合強度を得た。増殖因子を96-ウェルプレートに物理吸着させ、その後ファージライブラリーに接触させた。室温において60分インキュベーション後、界面活性剤溶液ですすいで未結合のファージを除去した。その後、増殖因子溶液と共に60分インキュベーションすることによって結合ファージを溶出した。回収したファージを37℃において大腸菌(E. coli) 内で4時間増殖し、パニングを2回以上反復した。その後のパニングラウンドにおいて、界面活性剤濃度を上昇し、ライブラリーへの接触時間を短縮することによって緊縮度を増した。最終的なファージ集団を平板培養し、1つのファージタイプだけを含有するコロニーの10クローンのDNAを配列決定した。次に、ELISAスクリーニングを使用して偽陽性を試験し、選択したクローンの相対的な結合強度を求めた。このプロトコールを使用すると、rh-BMP-2の最良の結合エピトープはYPVHPSTであり、rh-TGF-β1はLPLGNSHであることが見出された(以下の表1参照)。
【0070】
実施例8a
ファージディスプレイは、Ph. D. 7キット(New England Biosystems)を用いて実施した。50マイクロリッターの希釈したM13-ファージライブラリー(2×109ファージおよび可能な全ての7-mer配列を含有する)を96-ウェルマイクロタイタープレートに添加し、0℃において18時間10μg/mlの増殖因子(peprotech)溶液50μlに接触させた。プレートを1時間ブロックし、次いでゆっくり撹拌しながら室温において60分インキュベーションした。TBS/Tween-20(0.1%v/vTween-20)ですすいで未結合のファージを除去した。その後、50μlの増殖因子溶液と共に60分インキュベーションすることによって結合ファージを溶出した。ファージを連続希釈し、その後寒天プレートで大腸菌(E. coli) と共に平板培養することによって結合ファージの存在を判定した。形成されたプラークの数を元の混合物中のファージの数に関連させることができる。その後、ファージを37℃において大腸菌(E. coli) 中で4時間増幅した。その後のパニングラウンドにおいて、Tween-20の濃度を0.5%に上昇し、結合時間および溶出時間をそれぞれ短縮および延長して、選択過程の緊縮度を増した。3ラウンドのパニング後、回収したファージ混合物を希釈し、寒天で細菌と共に平板培養した。その後、1つのDNA配列を含有するプラークを単離する。精製後、10クローンを配列決定した。次に、ELISAスクリーニングを使用して偽陽性を排除し、選択したクローンの相対的な結合強度を求めた。増殖因子をコーティングしたマイクロタイタープレートで各クローンを1時間インキュベーションし、プレートをTBS/Tween-20ですすぎ、その後HRP-結合抗-M13抗体(Amersham Biosciences)を1時間インキュベーションした。ABTSの添加後、30分後に450 nmにおける吸光度を測定し、単離したクローンの相対的な結合強度を求めた。その後、最も強い結合物を選択した。
【0071】
(表1)ファージディスプレイを使用して誘導されるBMP-2およびTGF-β1の結合配列
数値コーディングは、結合強度を最高(1)から最低(4)まで低下する際の選択したクローンの相対的な結合強度を例示している。全ての配列は強く結合している。
【0072】
実施例9
ファージ上で提示されるペプチド配列は遊離N-末端を有するので、PAの典型的な合成経路を使用することができなかった。従って、側鎖をドデシルアミンで修飾したアスパラギン酸をRinkアミド樹脂と反応させた。(上記スキーム2参照)。その後、ペプチドの残りを通常のFmoc-ペプチド合成技法を使用して合成し、逆の極性を有するPAを生じた。選択したペプチド配列は、それぞれ、BMP-2はV6K3SG3YPVHPST(9)であり、TGF-β1はV3A3K3SG3LPLGNSH(10)であった(スキーム3)。K3セグメントは、フィラー2の荷電E3ヘッド基と同時集合するように選択した。SG3配列はスペーサー単位であり、ファージに存在したものと同じスペーサーである。BMP-2のそれぞれYPVHPSTおよびKQALTOTとの結合定数は、蛍光偏光解消によって測定するとき、Ka=1.9±0.9 106 M-1および2.1±0.85 105 M-1であり、結合強度の同調性を例示している。
【0073】
実施例9a
Asp(CONHC12)- Val-Val-Val- Val-Val-Val-Lys-Lys-Lys-Ser-Gly-Gly-Gly-Tyr-Pro-Val-His-Pro-Ser-Thr(9)
【0074】
実施例9b
Asp(CONHC12)- Val-Val-Val-Ala-Ala-Ala- Lys-Lys-Lys- Ser-Gly-Gly-Gly-Leu-Pro-Leu-Gly-Gln-Ser-His-NH2(10)
スキーム3:
【0075】
実施例10
PA 9とフィラーPA 2の同時集合は、円二色性(CD)分光法を使用して証明できると思われる。0.13 mMの濃度において、この濃度における個々のPAナノファイバーのCDスペーサーは主にランダム-コイル特性を有する。しかし、一体として混合すると、CD-スペクトルは純粋なβ-シートを反映し、PAは、1つだけのPA分子からなる個々のホモ-ファイバーを形成するのではなく、1つのファイバーに同時集合することを例示している。同時集合は、KKKおよびEEE部分が複合体を形成する場合には、静電反発力が低下するので、エネルギー的に好ましい。1.5重量%溶液の核オーバーハウザー効果分光法を使用して同時集合の追加の証拠が得られた。EβとKεプロトンの間およびEβとVδプロトンの間に密接な接触(<3Å)が観察され、この距離はホモ-ファイバー間より有意に大きいと思われる。考えられる他の分子間接触は明白に起因させられなかった。最後に、凍結乾燥試料および1重量%溶液のフーリエ変換赤外分光法は、β-シートに典型的な1630 cm-1ピークの存在を示した。同様の結果は10/2混合物で得られた。試料を37℃において24時間アニーリングすることにより、CD効果の強度が有意に増加した。この増加は、完全に結合している発色団のドメインのサイズの増加に起因する。おそらく、最初の混合によって完全でない混合システムの動態的な捕獲を生じ、アニーリングによって、その後両親媒性物質が熱力学的に最も安定な状態に再組織化することが可能になる。水素結合ネットワークおよび脂肪族テイルの予想される融点を十分に上回る80℃における長時間の加熱も超分子キラリティーの任意の顕著な損失を生じず、ナノファイバーの高い安定性を例示している。
【0076】
実施例11
この方法の生理学的重要性を試験するために、異なるレベルの増殖因子を含有するゲルにおいて間葉系幹細胞を培養した。これらの細胞実験では、タンパク質による認識を確実にするために、結合エピトープを50:1(フィラー:結合物)比に希釈した。望ましい量の増殖因子を含有する50μlのプレミックスPA(2重量%)および50μlの間葉系幹細胞懸濁液(100,000細胞)をガラスカバースリップ上で混合し、次に30 mg/mlのCaCl2溶液10μlを添加することによってゲルを調製した。ゲルを1.5時間固化させ、その後1 mlの間葉系幹細胞培地を添加した。この方法では、9/2または2単独からなるおよび0、1または50 ng/mlのBMP-2を含有するゲルが調製された。別の対照として、10,000の幹細胞をガラスカバースリップで平板培養し、同じ濃度のBMP-2を培地に添加した。培地を4日めごとに交換し、培養7日目および21日目にトリプリケート試料を単離した。その後、系統-特異的なマーカーオステオカルシン(骨芽細胞)、アジポネクチン(脂肪細胞)、コラーゲンIIおよびアグリカン(軟骨細胞)ならびにα-平滑筋アクチンおよびデスミン(平滑筋細胞)のタンパク質発現をウェスタンブロットによって評価した。
【0077】
実施例11a
細胞実験のプロトコール:ヒト間葉系幹細胞(Cambrex technologies)を5,000細胞/cm2の密度で4つのT75培養フラスコの15 mlの間葉系幹細胞培地(Cambrex Technologies)に入れ、37℃、5% CO2、湿度90%においてインキュベーションした。集密度90%において、細胞を0.25%トリプシン/EDTA2.5 mlで5分間トリプシン処理した。その後、8 mlの培地を添加して、プロテアーゼ活性を失活させた。細胞懸濁液を合わせて、850 rpmで8分遠心分離した。細胞ペレットを1 mlの培地に再懸濁させ、10μlを細胞計測に使用した。細胞懸濁液を培地で2,000,000細胞/mlの濃度に希釈した。
【0078】
PA 1および3を2重量%の濃度で溶解した。その後、1の50μlを3の1250μlと混合した。混合物を410μlの3部に分割し、さらに3(2重量%)の410μlの3部を調製した。次いで、2μg/ml BMP-2(Peprotech)の20μlを1/3および1の1部に100 ng/ml BMP-2濃度まで添加した。同様に、20 ng/ml BMP-2の41μlを最終濃度2 ng/ml のBMP-2まで添加した。
【0079】
50μlのPA溶液と50μlの細胞懸濁液を電子顕微鏡用ガラスカバースリップ上で混合することによってゲルをトリプリケート(triplo)で調製した。次いで、30 mg/ml CaCl2の10μlを添加し、ゲルを37℃、5% CO2、湿度90%において90分固化させた。得られた濃度は1重量%PAであり、BMP-2では0、1または50 ng/mlである。次に、1 mlの培地を添加した。5μlの細胞懸濁液をガラスカバースリップに置き、1 mlの培地を添加することによって増殖因子の対照溶液を調製した。次いで、2μg/ml BMP-2の25μlおよび2μg/mlのBMP-2の0.5μlを添加して、それぞれ50 ng/mlおよび1 ng/ml濃度のBMP-2溶液を得た。培地の半分を4日目ごとに交換した。
【0080】
実施例12
細胞の生存度は、3週間の培養後に全てのゲルにおいて>95%であることを細胞生存度アッセイは示した。エンドグリンの発現はどの時点においてもウェスタンブロットによって検出することができず、全ての細胞は分化を開始していたことを示している。アジポネクチンおよびアグリカンの発現も任意の時点において検出できなかった。オステオカルシンの発現はゲルと対照間で顕著な差を示した。オステオカルシンは対照では検出できなかったが、全てのゲルは、通常の2〜3週間ではなくすでに7日目に有意なレベルのオステオカルシンを発現したが(図3a)、3週間経過時点において有意に増加しなかった(図3b)。1および3週における同様の発現レベルは、オステオカルシンの発現レベルは培養期間中一定であることを意味している。驚くべきことに、発現はBMP-2濃度に依存しないと思われ、比は、結合PAを含有するゲルと結合PAを含有しないゲルで1:2である。さらに、BMP-2を添加しないゲルも同様のレベルを示す。おそらく、細胞自体によって産生される内因性BMP-2レベルは、骨芽細胞分化を刺激するのに十分であるが、結合ゲルがBMP-2の一部に結合する能力は発現されるオステオカルシン量をわずかに低下する。オステオカルシンの発現はまた免疫細胞化学でも可視化することができた。
【0081】
一方、α-平滑筋アクチンは、9を含有するゲルにおいて検出されなかったが、結合物を含有しないゲルにおいて発現された。しかし、それは3週間後にどちらの場合でも発現されたが(図3b)、9/2のゲルにおける発現は2のゲルよりわずかに低く、対照より有意に低い。さらに、最終分化平滑筋細胞のマーカーであるデスミンの発現を検出することができなかった。
【0082】
実施例12a
ウェスタンブロットプロトコール:培地を除去した後、ゲルを1 mlのリン酸緩衝生理食塩液ですすぎ、その後、200μlの2%ドデシル硫酸ナトリウム、0.08 M Tris、10%グリセロールで細胞を溶解した。3つの溶菌液を合わせ、10μlの溶菌液と200μlのBCAタンパク質アッセイ試薬(Pierce)を混合し、37℃における30分のインキュベーション後BSA標準シリーズに対する562 nmの吸光度を読むことによってタンパク質濃度を測定した。その後、10μgのタンパク質、3μlのβ-メルカプト-エタノール、3μlのブロモフェノールブルーを混合し、水を総容量36μlまで添加した。混合物を5分間煮沸し、4%(エンドグリン、アグリカン)または10〜20%(オステオカルシン、α-平滑筋アクチン、アジポネクチン、デスミン) Novexトリス(グリシン)ゲル(Invitrogen)にロードした。ゲルは、130 Vの定電圧でトリス(グリシン)泳動緩衝液(Invitrogen)で90分泳動した。次いで、タンパク質を、ニトロセルロース膜(Bio-Rad)に移し、190 mAの定電流で90〜120分泳動した。
【0083】
膜を5%脱脂乳(Bio-Rad)中で1時間ブロックし、オステオカルシン(クローン190125, R & D systems)、エンドグリン(クローン166709, R & D systems)、アジポネクチン(クローン166126, R & D systems)、α-平滑筋アクチン(クローン1A4, R & D systems)およびポリクローナルアグリカン(AF1220, R & D systems)に対するモノクローナル抗体を使用して、1%ミルクで1:500希釈のそれぞれのマーカーを2時間探索した。膜を、トリス緩衝生理食塩液/0.1%Tween-20(TTBS)で15分間3回すすいだ。次いで、二次抗体を1%ミルク中で1:3,000希釈で1時間平衡させ、次にTTBSによるすすぎ段階を3回実施した。ECLウェスタンブロット分析溶液(Amersham Biosciences)で1分平衡させることによって膜を形成し、ECLハイパーフィルム(Amersham Biosciences)に接触させた。
【0084】
実施例12b
細胞を2%パラホルムアルデヒド/0.2%グルタルアルデヒド/0.2 Mカルコジレート(calcodylate)ナトリウムで4℃において20分固定した。PBS(2×)ですすいだ後、試料を0.1% Triton-Xと共にPBS溶液中で5分間インキュベーションした。その後、試料をPBSですすいだ(2×)。次に、ヒトオステオカルシンモノクローナル抗体(R & D systems、1:200)の1% BSA/PBS溶液を添加し、試料を4℃において終夜インキュベーションした。PBSで3回すすいだ後、FITC-結合二次抗体の1% BSA/PBSを添加し、2時間インキュベーションした。PBSで3回すすいだ後、Nikon Eclipse TE2000電子顕微鏡で倍率10×で試料を可視化した。
【0085】
実施例13
一連の実験は、TGF-β1の最高濃度が、50 ng/mlでなく20 ng/mlであることを除いて、PA 10、結合TGF-β1、BMP-2実験についてと同様に実施した。試料は10日後に単離した。TGF-β1の存在下における培養により、平滑筋および軟骨系統が分化する。コラーゲンIIの発現は結合ゲルでは高く、TGF-β1の増加により増加すると思われる。驚くべきことに、オステオカルシンおよびコラーゲンXが発現され、この早期時点では予想されない肥大軟骨細胞段階への成熟が生じていることを示している。同様に、α-平滑筋アクチン発現はTGF-β1濃度と共に増加するが、非結合ゲルより結合ゲルにおいて高い。それは対照においてはるかに高いが、デスミンの発現レベルは、これらは主に未熟筋細胞であることを示している。
【0086】
実施例14
7-および12-merファージディスプレイキットを用いて、血管内皮増殖因子(VEGF)、塩基性線維芽細胞増殖因子(FGF-2)、ニューロトロフィン-3(NT-3)およびラミニン-5の数多くの結合配列を決定した;*は最も強い結合物を示す:
【0087】
先の実施例によって例示するように、本発明は、標準的な固相方法に適合する遊離N-末端を有するペプチド両親媒性分子の合成方法を提供する。遊離C-末端PAと混合すると、これらの分子は自己集合して、同じ極性のPAの同時集合物より安定性が高いと思われる熱安定性が高いβ-シート構造を含有するナノファイバーを形成する。表面に遊離N-末端を有する集合物を作製する新規機会により、ペプチド配列のC-末端を露出しているナノ構造物では以前には入手不可能であった生物活性ナノファイバーの設計が可能になる。
【0088】
本発明は、ある好ましい態様を参照してかなり詳細に記載されているが、他の形態も可能である。 例えばファージディスプレイ法によって誘導されるペプチドまたはタンパク質配列を使用して、増殖因子以外のタンパク質に結合して保持する結合ペプチド両親媒性化合物を製造することができる。自己集合して、ナノファイバーヒドロゲルの表面に固定することができる結合ペプチド両親媒性物質を製造するように酵素を結合することができると思われる。このようなヒドロゲルをセンサー適用のコーティングとして使用することができる。または、ペプチド両親媒性物質に結合したファージディスプレイ由来のペプチドの結合相互作用は、HGF(肝細胞増殖因子)またはVEGFなどのペプチドに強力に結合し、それらを除去することによって種々の疾病および状態を治療するように選択してもよい。これらの結合ペプチド両親媒性化合物の自己集合によって形成されるヒドロゲルは患者の部位に挿入するためまたはろ過システムに使用するために成形してもよい。ヒドロゲルは、患者の関節もしくは腫瘍などの部位または患者の体液からHGFまたはVEGFのような標的ペプチドを除去するために使用してもよい。
【特許請求の範囲】
【請求項1】
ペプチド成分および疎水性成分を含み、該ペプチド成分はファージディスプレイ法の増殖因子認識産物を含み、該認識産物は該ペプチド成分のほぼN-末端に結合しており、該疎水性成分は該ペプチド成分のほぼC−末端に結合している、両親媒性ペプチド化合物。
【請求項2】
認識産物が、増殖因子BMP-2、TGF-β1、VEGF、FGF-2、NT-3およびラミニン-5との結合相互作用をもたらすエピトープ配列から選択される、請求項1記載の化合物。
【請求項3】
結合するエピトープ配列が、YPVHPSTを含む、請求項2記載の化合物。
【請求項4】
配列が、増殖因子BMP-2との非共有結合相互作用にある、請求項3記載の化合物。
【請求項5】
結合するエピトープ配列が、LPLGNSHを含む、請求項2記載の化合物。
【請求項6】
配列が、増殖因子TGF-β1との非共有結合相互作用にある、請求項5記載の化合物。
【請求項7】
ペプチド成分が、生理的pHにおいて正味荷電を有する、請求項1記載の化合物。
【請求項8】
アラニン、グリシン、ロイシン、システイン、バリンおよびセリンから選択される残基を含む、請求項7記載の化合物。
【請求項9】
疎水性成分が、約C6〜約C22の範囲のアルキル部分を含む、請求項2記載の化合物。
【請求項10】
ペプチド成分が、実質的に直鎖状である、請求項1記載の化合物。
【請求項11】
複数の第1の両親媒性ペプチド化合物を含み、該化合物各々が該化合物のペプチド成分に結合しているファージディスプレイ法の増殖因子認識産物を含み、該ペプチド成分が生理的pHにおいて正味荷電を有し、疎水性成分のほぼC-末端に結合しているペプチド組成物。
【請求項12】
複数の第2の両親媒性ペプチド化合物をさらに含み、該化合物各々は増殖因子認識産物が欠けており、生理的pHにおいて、第1の化合物の正味荷電に相補的な正味荷電を有する、請求項11記載の組成物。
【請求項13】
第2の化合物のペプチド成分のアミノ酸配列が、第1の化合物のペプチドアミノ酸配列の長さより短い長さを有する、請求項12記載の組成物。
【請求項14】
水性媒体にミセル集合物を含む、請求項13記載の組成物。
【請求項15】
非共有結合的に相互作用する少なくとも1つの増殖因子を含む、請求項14記載の組成物。
【請求項16】
増殖因子が、BMP-2、TGF-β1、VEGF、FGF-2、NT-3、ラミニン-5およびそれらの組み合わせから選択される、請求項15記載の組成物。
【請求項17】
幹細胞に接触する、請求項16記載の組成物。
【請求項18】
複数の第2の両親媒性ペプチド化合物をさらに含み、該化合物が第1の化合物のペプチドアミノ酸配列の長さより短い長さを有し、該第2の化合物が増殖因子認識産物が欠けており、生理的pHにおいて該第1の化合物の正味荷電に相補的な正味荷電を有する、請求項11記載の組成物。
【請求項19】
水性媒体にミセル集合物を含む、請求項18記載の組成物。
【請求項20】
幹細胞に接触し、組成物と非共有結合的に相互作用する増殖因子を含む、請求項19記載の組成物。
【請求項21】
以下の段階を含む、両親媒性ペプチド化合物を使用して増殖因子のバイオアベイラビリティーに影響を与える方法:
複数の第1の両親媒性ペプチド化合物および複数の第2の両親媒性ペプチド化合物を含むミセル集合物を提供する段階であって、複数の第1の両親媒性ペプチド化合物の各々が第1の化合物のペプチド成分に結合しているファージディスプレイ法の増殖因子認識産物を含み、該ペプチド成分が生理的pHにおいて正味荷電を有し、疎水性成分のほぼC−末端に結合しており、複数の第2の両親媒性化合物が第1の化合物のペプチド配列の長さより短い長さを有するペプチドアミノ酸配列を含み、増殖因子認識産物が欠けており、生理的pHにおいて該第1の化合物の正味荷電に相補的な正味荷電を有する段階;ならびに
幹細胞に該集合物を接触させる段階。
【請求項22】
増殖因子が、幹細胞によって産生される、請求項21記載の方法。
【請求項23】
増殖因子が、BMP-2、TGF-β1、VEGF、FGF-2、NT-3、ラミニン-5およびそれらの組み合わせから選択される、請求項21記載の方法。
【請求項24】
認識産物が、増殖因子BMP-2およびTGF-β1の1つと非共有結合相互作用をもたらすエピトープ配列から選択される、請求項23記載の方法。
【請求項25】
エピトープ配列が、増殖因子BMP-2と非共有結合的に相互作用するYPVHPSTを含む、請求項24記載の方法。
【請求項26】
エピトープ配列が、増殖因子TGF-β1と非共有結合的に相互作用するLPLGNSHを含む、請求項24記載の方法。
【請求項27】
接触が、幹細胞分化に十分である、請求項21記載の方法。
【請求項28】
集合物が、増殖因子BMP-2を含み、認識産物が、該増殖因子と非共有結合的に相互作用するエピトープ配列を含む、請求項27記載の方法。
【請求項29】
幹細胞が、間葉系幹細胞である、請求項28記載の方法。
【請求項30】
幹細胞が、骨細胞に分化する、請求項29記載の方法。
【請求項1】
ペプチド成分および疎水性成分を含み、該ペプチド成分はファージディスプレイ法の増殖因子認識産物を含み、該認識産物は該ペプチド成分のほぼN-末端に結合しており、該疎水性成分は該ペプチド成分のほぼC−末端に結合している、両親媒性ペプチド化合物。
【請求項2】
認識産物が、増殖因子BMP-2、TGF-β1、VEGF、FGF-2、NT-3およびラミニン-5との結合相互作用をもたらすエピトープ配列から選択される、請求項1記載の化合物。
【請求項3】
結合するエピトープ配列が、YPVHPSTを含む、請求項2記載の化合物。
【請求項4】
配列が、増殖因子BMP-2との非共有結合相互作用にある、請求項3記載の化合物。
【請求項5】
結合するエピトープ配列が、LPLGNSHを含む、請求項2記載の化合物。
【請求項6】
配列が、増殖因子TGF-β1との非共有結合相互作用にある、請求項5記載の化合物。
【請求項7】
ペプチド成分が、生理的pHにおいて正味荷電を有する、請求項1記載の化合物。
【請求項8】
アラニン、グリシン、ロイシン、システイン、バリンおよびセリンから選択される残基を含む、請求項7記載の化合物。
【請求項9】
疎水性成分が、約C6〜約C22の範囲のアルキル部分を含む、請求項2記載の化合物。
【請求項10】
ペプチド成分が、実質的に直鎖状である、請求項1記載の化合物。
【請求項11】
複数の第1の両親媒性ペプチド化合物を含み、該化合物各々が該化合物のペプチド成分に結合しているファージディスプレイ法の増殖因子認識産物を含み、該ペプチド成分が生理的pHにおいて正味荷電を有し、疎水性成分のほぼC-末端に結合しているペプチド組成物。
【請求項12】
複数の第2の両親媒性ペプチド化合物をさらに含み、該化合物各々は増殖因子認識産物が欠けており、生理的pHにおいて、第1の化合物の正味荷電に相補的な正味荷電を有する、請求項11記載の組成物。
【請求項13】
第2の化合物のペプチド成分のアミノ酸配列が、第1の化合物のペプチドアミノ酸配列の長さより短い長さを有する、請求項12記載の組成物。
【請求項14】
水性媒体にミセル集合物を含む、請求項13記載の組成物。
【請求項15】
非共有結合的に相互作用する少なくとも1つの増殖因子を含む、請求項14記載の組成物。
【請求項16】
増殖因子が、BMP-2、TGF-β1、VEGF、FGF-2、NT-3、ラミニン-5およびそれらの組み合わせから選択される、請求項15記載の組成物。
【請求項17】
幹細胞に接触する、請求項16記載の組成物。
【請求項18】
複数の第2の両親媒性ペプチド化合物をさらに含み、該化合物が第1の化合物のペプチドアミノ酸配列の長さより短い長さを有し、該第2の化合物が増殖因子認識産物が欠けており、生理的pHにおいて該第1の化合物の正味荷電に相補的な正味荷電を有する、請求項11記載の組成物。
【請求項19】
水性媒体にミセル集合物を含む、請求項18記載の組成物。
【請求項20】
幹細胞に接触し、組成物と非共有結合的に相互作用する増殖因子を含む、請求項19記載の組成物。
【請求項21】
以下の段階を含む、両親媒性ペプチド化合物を使用して増殖因子のバイオアベイラビリティーに影響を与える方法:
複数の第1の両親媒性ペプチド化合物および複数の第2の両親媒性ペプチド化合物を含むミセル集合物を提供する段階であって、複数の第1の両親媒性ペプチド化合物の各々が第1の化合物のペプチド成分に結合しているファージディスプレイ法の増殖因子認識産物を含み、該ペプチド成分が生理的pHにおいて正味荷電を有し、疎水性成分のほぼC−末端に結合しており、複数の第2の両親媒性化合物が第1の化合物のペプチド配列の長さより短い長さを有するペプチドアミノ酸配列を含み、増殖因子認識産物が欠けており、生理的pHにおいて該第1の化合物の正味荷電に相補的な正味荷電を有する段階;ならびに
幹細胞に該集合物を接触させる段階。
【請求項22】
増殖因子が、幹細胞によって産生される、請求項21記載の方法。
【請求項23】
増殖因子が、BMP-2、TGF-β1、VEGF、FGF-2、NT-3、ラミニン-5およびそれらの組み合わせから選択される、請求項21記載の方法。
【請求項24】
認識産物が、増殖因子BMP-2およびTGF-β1の1つと非共有結合相互作用をもたらすエピトープ配列から選択される、請求項23記載の方法。
【請求項25】
エピトープ配列が、増殖因子BMP-2と非共有結合的に相互作用するYPVHPSTを含む、請求項24記載の方法。
【請求項26】
エピトープ配列が、増殖因子TGF-β1と非共有結合的に相互作用するLPLGNSHを含む、請求項24記載の方法。
【請求項27】
接触が、幹細胞分化に十分である、請求項21記載の方法。
【請求項28】
集合物が、増殖因子BMP-2を含み、認識産物が、該増殖因子と非共有結合的に相互作用するエピトープ配列を含む、請求項27記載の方法。
【請求項29】
幹細胞が、間葉系幹細胞である、請求項28記載の方法。
【請求項30】
幹細胞が、骨細胞に分化する、請求項29記載の方法。
【図1】

【公開番号】特開2012−6951(P2012−6951A)
【公開日】平成24年1月12日(2012.1.12)
【国際特許分類】
【出願番号】特願2011−172533(P2011−172533)
【出願日】平成23年8月8日(2011.8.8)
【分割の表示】特願2006−542795(P2006−542795)の分割
【原出願日】平成16年12月6日(2004.12.6)
【出願人】(596057893)ノースウエスタン ユニバーシティ (35)
【Fターム(参考)】
【公開日】平成24年1月12日(2012.1.12)
【国際特許分類】
【出願日】平成23年8月8日(2011.8.8)
【分割の表示】特願2006−542795(P2006−542795)の分割
【原出願日】平成16年12月6日(2004.12.6)
【出願人】(596057893)ノースウエスタン ユニバーシティ (35)
【Fターム(参考)】
[ Back to top ]